introducciÓn - instituto politécnico...
TRANSCRIPT

INTRODUCCIÓN
Hoy en día, existe una gran demanda del uso de enzimas para llevar a cabo
procesos industriales, lo cual, se ha tratado de solucionar buscando nuevas
fuentes para su obtención, siendo las de origen microbiano las más socorridas por
ofrecer mejores ventajas sobre otras como la vegetal o animal, además de
comprobar que su empleo permite a la industria obtener productos de menor
costo, mayor estabilidad y calidad. Lo anterior ha podido sustentar que el
desarrollo de las industrias que basan sus procesos en el empleo de enzimas,
depende fuertemente de los microorganismos y su capacidad de producción
enzimática.
Son muchos los productos que se obtienen del procesamiento de algunos
azúcares, principalmente de la manipulación de polisacáridos como el almidón, por
lo que el empleo de enzimas capaces de hidrolizar los enlaces glucosídicos resulta
ser fundamental. Hasta ahora las enzimas de microorganismos más empleados
para llevar esta actividad son las provenientes de hongos filamentosos y las
bacterias; sin embargo, existen varios reportes en donde se establece que las
levaduras también pueden jugar un papel importante en cuanto a la producción
enzimática y su empleo industrial.
En la actualidad, la ingeniería genética y la biotecnología han demostrado
sustentar las herramientas necesarias para lograr la caracterización genética y
proteica, es por ello que en nuestro estudio se hará uso exhaustivo de éstas para
la búsqueda del gen o genes involucrados en llevar a cabo esta actividad, así
como el análisis de la regulación de su expresión, y de esta forma, ofrecer a la
industria el empleo de un nuevo microorganismo con actividad enzimática bien
caracterizada.
Las enzimas microbianas poseen mejorías sobre las obtenidas de origen
animal y vegetal debido a las ventajas técnicas y económicas que poseen.
La principal ventaja económica de las enzimas microbianas es su
producción a gran escala. La cantidad de producto que se puede obtener en una

superficie pequeña en poco tiempo, supera muy de lejos a la que se puede
conseguir de enzimas animales o vegetales. La segunda ventaja es la facilidad de
extracción. Una gran proporción de las enzimas industriales de origen microbiano,
por ejemplo las amilasas son extracelulares y se obtienen por tanto del medio de
cultivo, obviándose así los problemas de extracción.
Hay cuatro aspectos por los que los microorganismos son más ventajosos
como productores de enzimas que los animales o las plantas.
La primera es la enorme variedad de vías metabólicas (y, por tanto, de
enzimas) que existen.
La segunda ventaja de los microorganismos está en que crecen en un
amplio rango de condiciones ambientales.
La tercera ventaja radica en la mayor flexibilidad genética de los
microorganismos, lo que supone una mayor facilidad para su manipulación con el
fin de incrementar el rendimiento de enzima.
La ventaja final es el corto tiempo de generación de los microorganismos.
La propuesta del uso de levaduras con alta actividad enzimática resulta ser
relevante, es por ello que en nuestro grupo de trabajo se ha puesto gran empeño
en lograr la caracterización de los genes codificantes de enzimas con actividad
amilolítica y proteolítica en levaduras aisladas de diversas fuentes naturales que
pertenecen al laboratorio de Microbiología Industrial de la ENCB – IPN.
Así mismo resulta interesante buscar las condiciones ambientales que
favorezcan la producción de las mismas, además de conocer los mecanismos que
regulan su biosíntesis.
Dentro de las levaduras que han mostrado mayor actividad de estas
enzimas se encuentran Yarrowia lipolytica y Wickerhamia sp.
Características de Whickerhamia sp.
Este género no ha reportado ninguna levadura amilolítica, solo se sabe que
pertenece al grupo de levaduras tipo “GRAS” (Generaly Reported As Safe), es por
ello que se usa en el tratamiento de pielonefritis, y de la hiperlipemia al igual que
otras especies de Saccharomyces y Schizosaccharomyces.

No se han descrito usos relevantes para Wickerhamia desde el punto de vista
biotecnológico; de tal manera que es atractiva en el ámbito de la producción de
enzimas con alta demanda a nivel industrial.
Se tienen resultados preliminares de esta levadura en relación con la producción
de amilasas. Con base en los estudios cinéticos de crecimiento y producción se
determinó que la amilasa extracelular que produce es un metabolito asociado al
crecimiento.
Estudios previos de caracterización bioquímica muestran que el pH óptimo de
actividad y estabilidad es 6.0, es una enzima termoestable ya que su temperatura
optima es a 50°C y su estabilidad se mantiene al 100% a 40°C.
Características deYarrowia lipolytica
Las cepas de Yarrowia lipolytica son aisladas fácilmente de productos comunes
tales como queso, yogurt y salchichas (sustratos que contienen proteínas). Su
incapacidad para sobrevivir en condiciones anaeróbicas permiten su eliminación
de productos diarios, esta especie no se considera patógena, debido en parte a
que su temperatura máxima de crecimiento rara vez excede los 32 o 34ºC.
Y. lipolytica es una levadura dimórfica, cuyo genoma en contraste a otras
levaduras comparte propiedades con eucariotes superiores, como la dispersión de
los genes 5S RNA y un mayor tamaño de snRNA . Y. lipolytica es heterotálica,
sus aislados son en su mayoría haploides, evocativo de los hongos filamentosos
A pesar de las características que esta levadura comparte con eucariotes
superiores y hongos filamentosos, la filogenia la coloca inequívocamente entre las
levaduras hemiascomicetales
Y. lipolytica es un hongo dimórfico natural, que forma células levaduriformes,
pseudohifas e hifas septadas.

Y. lipolytica secreta grandes cantidades de proteasa alcalina extracelular
(AEP), alrededor de 1-2 g/L cuando es crecida en un medio rico a pH de 6.8. La
AEP es codificada por el gen XPR2, que ha sido clonado y secuenciado, fue
identificado entre al menos 11 genes que controlan la síntesis, secreción y
actividad de AEP.
La regulación de la proteasa es compleja y depende de la fuente de carbono,
nitrógeno, sulfuro y pH entre otros.
La proteasa ácida extracelular es detectada en medio rico YPD en pH
ácido, los niveles más altos de mRNA del gen estructural (AXP) que codifica esta
proteasa ácida extracelular, han sido reportados a pH 5.5, mientras que en pH
arriba de 7.0 no se obtiene expresión.
Se han reportado la presencia de tres proteasas: aminopeptidasa yylAPE,
carboxipeptidasa yylCP y una dipeptidil aminopeptidasa yylDAP, se ha observado
que el nivel más alto de enzimas proteolíticas intracelulares se obtienen en medio
con peptona durante la fase estacionaria de crecimiento.
Las tres enzimas se ven afectadas por inhibidores de las serín proteasas y los
agentes quelantes afectan principalmente la actividad APE. La aminopeptidasa fue
purificada, encontrándose que pesa 97 kDa, tiene un pH y temperatura óptimas de
7.0 y 37 ºC respectivamente.
En cuanto a la DAP, sólo se ha intentado describir su papel en la maduración
de la AEP; sin embargo, el gen codificante de esta enzima no ha sido
caracterizado.
MATERIALES Y MÉTODOS
MICROORGANISMOS
Para el desarrollo de este trabajo se emplearon cepas de levadura aisladas de
diversas fuentes naturales como Wickerhamia s, Y.lipolytica y otras que aún no
han sido identificadas. Todas ellas pertenecen al cepario del laboratorio de
Microbiología Industrial del Departamento de Microbiología de la E. N. C. B.
También se utilizaron bacterias que sirvieron como cepas de referencia en algunos
experimentos o bien para las diversas técnicas de biología molecular.

La cepas se conservaron en tubos con agar inclinado, usando diversos medios:
Sabouraud, BHI, Medio de Castañeda-almidón o caseína a 4°C o bien en glicerol
al 50% a -60°C.
MEDIOS DE CULTIVO
Se utilizó la base de Castañeda-Agulló, que utiliza los siguientes componentes
COMPUESTO CONCENTRACIÓN(g/L)
(NH4)2C6H6O7
NaCl
KH2PO4
MgSO4.7H2O
Na2CO3
0.625
0.250
0.375
0.125
0.375
Dependiendo de los experimentos, se utilizaron diversos sustratos como almidón,
caseína, o bien sustratos comerciales amiláceos o proteicos, a diferentes
concentración para los estudios cinéticos relacionados con la producción y
regulación de las enzimas.
Para los estudios genéticos se utilizaron
• YEPD (medio rico de mantenimiento): Extracto de levadura 1%, peptona
2%, dextrosa 2%.
• YNB (medio mínimo): Base nitrogenada de levadura sin aminoácidos y sin
sulfato de amonio 0.17%, dextrosa 2%, sulfato de amonio 0.5%.
• YNB sin fuente de carbono: Base nitrogenada de levadura sin aminoácidos
y sin sulfato de amonio 0.17%, sulfato de amonio 0.5%.
• YNB con distintas fuentes de carbono: Base nitrogenada de levadura sin
aminoácidos y sin sulfato de amonio 0.17%, sulfato de amonio 0.5%, las
fuentes de carbono al 2% en cada caso.

• YNB sin fuente de nitrógeno: Base nitrogenada de levadura sin
aminoácidos y sin sulfato de amonio 0.17%, dextrosa 2%.
• YNB con distintas fuentes de nitrógeno: Base nitrogenada de levadura sin
aminoácidos y sin sulfato de amonio 0.17%, dextrosa 2%, las fuentes de
nitrógeno al 0.5% (peptona 2%) en cada caso.
• * Medio de Luria (LB) bajo en sal: Bacto Triptona 10%, extracto de levadura
5%, NaCl 5%, pH 7.5.
• * Medio de Luria (LB): Bacto Triptona 1%, extracto de levadura 0.5%, NaCl
1%, pH 7. Añadir ampicilina 100μg/ml. Al medio sólido añadir además IPTG,
para seleccionar las colonias que contengan el vector con el inserto.
Análisis Bioinformático
• Análisis teórico de genes codificantes de α-amilasa en hongos y levaduras .
• Se buscaron las secuencias nucleotídicas y proteicas de genes que
codifican para α-amilasas en hongos filamentosos y levaduras, en la base
de datos del NCBI (http://www.ncbi.nim.nih.gov) y con éstas se realizó un
análisis tipo BLAST en línea, con la finalidad de enriquecer el número de
secuencias. Se realizó un alineamiento múltiple de las secuencias
nucleotídicas y aminoacídicas encontradas, utilizando en software
CLUSTALX versión 1.81 y se ubicaron las regiones conservadas de las
secuencias.
• Diseño de oligonucleótidos.
• Se diseño un par de oligonucleótidos degenerados, a partir de las
secuencias de nucleótidos de las regiones conservadas.
• A partir de la secuencia obtenida del o los genes putativos de α-amilasa de
Wickerhamia sp., se diseñaron dos oligonucleótidos específicos para la
búsqueda de los extremos 5´y 3´del ORF. Una vez obtenida esta secuencia
se diseñó un par de iniciadores específicos para la amplificación de ORF.

Técnicas de biología molecular
• Extracción de DNA genómico y RNA.
• Para obtener DNA genómico de Wickerhamia sp. se utiliza el método
descrito por Hoffman y Winston (1987). El RNA se obtendrá por el método
descrito por Sherman et al., 1986.
• Para valorar la concentración de RNA, se determinará
espectrofotométricamente y se cuantificará mediante la formula A260 X
factor de dilución X 40 μg/ mL = μg/ mL de RNA X 1000 = mg/ μL de
RNA, la relación A260/A280 debe ser como mínimo de 2.0.
• Electroforesis de DNA en geles de agarosa.
• El DNA genómico y los productos de amplificación son evidenciados por
• electroforesis en geles de agarosa al 1%, teñidos con bromuro de etidio y
observados con luz UV.
• Amplificación de los fragmentos del gen o genes codificantes para la α-
amilasa
• Con los iniciadores degenerados y el DNA genómico de Wickerhamia sp.,
se amplificaron los fragmentos codificantes, utilizando la técnica de PCR
bajo las siguientes condiciones:
Mezcla de reacción para amplificar un fragmento de DNA por PCR
Mezcla de reacción
MgCl2 50 mM 1.0 µL
Regulador para PCR 10X 2.5µL
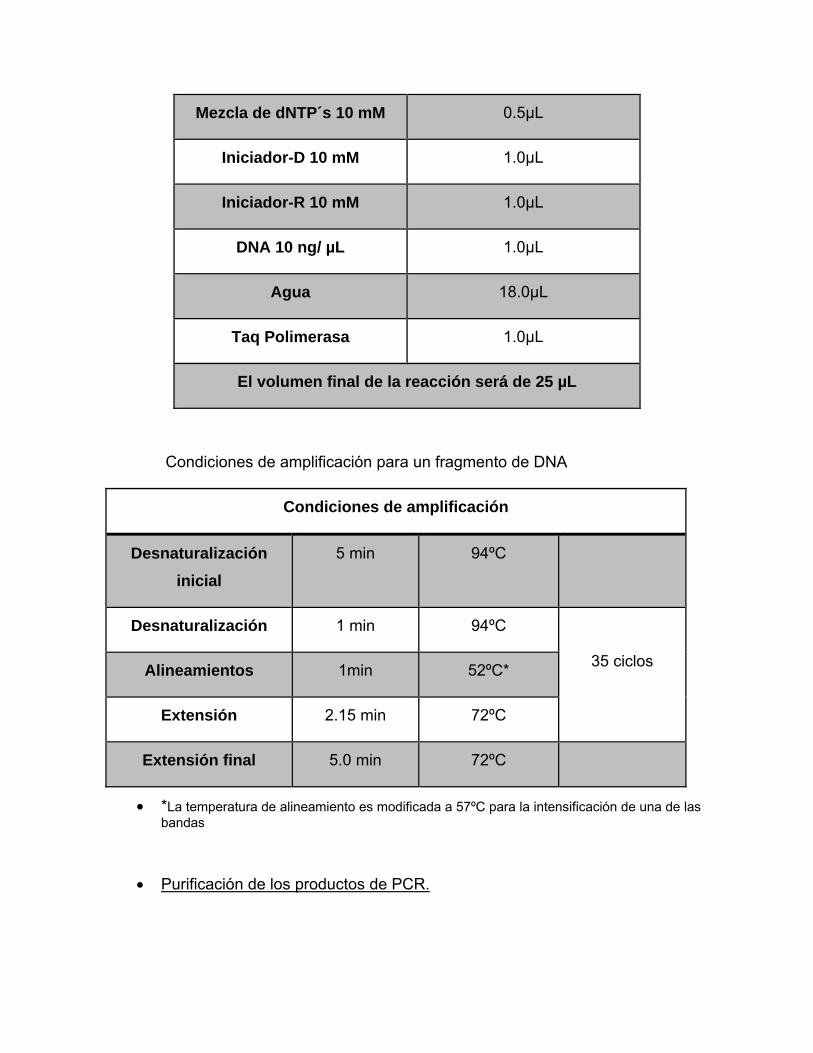
Mezcla de dNTP´s 10 mM 0.5µL
Iniciador-D 10 mM 1.0µL
Iniciador-R 10 mM 1.0µL
DNA 10 ng/ µL 1.0µL
Agua 18.0µL
Taq Polimerasa 1.0µL
El volumen final de la reacción será de 25 µL
Condiciones de amplificación para un fragmento de DNA
Condiciones de amplificación
Desnaturalización inicial
5 min 94ºC
Desnaturalización 1 min 94ºC
35 ciclos Alineamientos 1min 52ºC*
Extensión 2.15 min 72ºC
Extensión final 5.0 min 72ºC
• *La temperatura de alineamiento es modificada a 57ºC para la intensificación de una de las bandas
• Purificación de los productos de PCR.

• Para purificar los productos de PCR se utilizo el equipo de reactivos
comerciales Zymoclean™ Gel DNA Recovery (ZYMO RESEARCH), según
las instrucciones del proveedor.
• Clonación de los productos de PCR en un vector de clonación.
• Se utilizó el equipo de reactivos comerciales CloneJET™ PCR Cloning y el
vector linearizado pJET1.2 blunt (Fermentas).
• pJET1.2 blunt es un vector de clonación linearizado, que tiene un gen letal
el cual es interrumpido cuando un amplificado de PCR es ligado al sitio de
clonación, como resultado, sólo las células con el plásmido recombinante
son propagadas. Además el vector tiene un marcador de selección que
confiere resistencia a ampicilina.
• Las células de E. coli DH10b competentes fueron transformadas con las
construcciones del vector de clonación.
• Después de transformar, las células se recuperan con caldo Luria bajo en
sales, se sembraron por espatulado en agar LB suplementado con 40µL de
ampicilina (50 mg/mL) y 40µL de IPTG (100mM). Se Incubaron durante 24h,
las colonias crecidas se seleccionan como transformantes y se siembran
en 5mL de medio LB adicionado con 10 µL ampicilina (50mg/mL) toda la
noche para extraer el DNA plasmídico.
• Extracción de DNA plamídico de E. coli.
• Se hace por el método de lisis alcalina de Birnboim y Doly.
Confirmación del inserto en el plásmido pCR- por restricción y PCR.
• La confirmación de la inserción de los productos de amplificación en el
vector se determinó por la digestión del DNA plasmídico con las enzimas
XhoI y XbaI, evidenciando por medio de electroforesis en gel de agarosa.
• Se realizó un análisis de restricción, para verificar si no existían sitios de
corte en los amplificados clonados con las dos enzimas anteriores,
realizando una digestión con 3 enzimas de alta frecuencia (HindIII, PstI,

PvuII) posteriormente se realizó una electroforesis con los productos de la
digestión.
• Secuenciación del fragmento del gen o genes que codifican para la α-
amilasa en las transformantes seleccionadas y a partir de los productos de
PCR purificados.
• Se secuenciaron aquellas transformantes que mostraron tener el vector con
el inserto deseado.
• Se secuenciaron los purificados obtenidos a partir de productos de PCR.
• Búsqueda del ORF por RACE.
• De acuerdo a las recomendaciones del kit Invitrogen (GeneRacerTM cDNA).
Análisis de la expresión del gen o genes codificantes para la α-amilasa
• Construcción de sondas para el gen o los genes codificantes de α-amilasa
en Wickerhamia sp.
• Con los iniciadores específicos de cada gen, se amplificarán sondas por
PCR, para marcarlas mediante la técnica de Random Primed usando el DIG
DNA Labeling Kit (Roche). Las sondas marcadas se almacenan a -20ºC.
• Análisis de la expresión del o los genes codificantes de α-amilasa en
Wickerhamia sp. por análisis Northern blot.
• Se elaborará una curva de crecimiento en caldo Castañeda 2% almidón, a
28ºC/150rpm, para determinar la fase de crecimiento logarítmica, las
lecturas se realizarán a los 0, 15, 30, 60 min, 3, 6, 12, 24, 48, 72 h, leyendo
a una OD600.
• Inocular medio Castañeda con un preinóculo en fase logarítmica
proveniente de medio Castañeda con almidón. Los parámetros a evaluar

serán: fuentes de carbono, intervalo de temperaturas, e intervalo de pH, los
cuales serán establecidos posteriormente.
• Tratamiento del RNA con DNAsa 1
• Se llevó a cabo como se describe en el Kit Deoxyribonuclease I,
Amplification Grade, Invitrogen.
• Electroforesis de RNA en geles de agarosa en condiciones
desnaturalizantes
• Las muestras de RNA serán sometidas a una electroforesis en geles de
agarosa al 1.2% en condiciones desnaturalizantes (Lerach et al., 1977). Con formaldehído 37% como agente desnaturalizante, MOPS 0.04 M a un
voltaje máximo de 5 V/cm de distancia entre los electrodos (90 V).
• Se cargarán en cada carril 20 μg de RNA con 2 μL de regulador MOPS 5X,
3.3 μL de formaldehído, 10 µL de formamida, 2µL de regulador de carga
(glicerol al 50%, EDTA 1 mM pH 8.0, azul de bromocresol 0.25% y xylen
cyanol 0.25 %), y 1 μL de bromuro de etidio. Se desnaturalizarán las
muestras a 55ºC por 15 min para asegurar que el RNA migrará en función
de su tamaño molecular.
• Al finalizar la electroforesis el gel se visualizará con luz UV, la presencia de
los RNA ribosomales indica que el RNA no sufrió una degradación externa
(Sambrook et al., 1989).
• Transferencia e hibridación
• La transferencia del RNA sometido a electroforesis en gel de agarosa a
una membrana de Nylon, se realizará usando un puente salino y 10X SSC
como regulador de transferencia (Sambrook et al., 1989).
• Una vez efectuada la transferencia del RNA, éste se fijará a la membrana
de Nylon con luz ultravioleta, colocando la parte de la membrana que
estuvo en contacto con el gel hacia la lámpara.

• La membrana se prehibridará durante 3 h a 42ºC, la hibridación se
efectuará con 3 μg de la sonda marcada durante toda la noche a la misma
temperatura; los reguladores de prehibridación e hibridación serán
preparados de acuerdo al instructivo del “DIG Nucleic Acid Detection Kit”
(Roche). Se realizarán dos lavados, el primero en condiciones de baja
astringencia con un regulador de 2X SSC y 0.1% de SDS, y el segundo,
con un regulador de moderada astringencia que contenía 1X SSC y 0.1%
de SDS, ambos a 42°C durante 30 min cada uno (Sambrook et al., 1989).
El tratamiento para el revelado se realizará de acuerdo a las
especificaciones del “DIG Nucleic Acid Detection Kit” (Roche).
Posteriormente la membrana se pondrá en contacto con la película para
realizar la autoradiografía.
• Análisis de resultados para los Northern blot
• Una vez efectuada la autoradiografía las películas se digitalizarán, se
estimará el tamaño molecular de las señales de expresión, así como la
intensidad de las bandas obtenidas (en píxeles inversos) con ayuda del
programa Sigmagel (Versión 1.0, Jadel Corporation 1994, 1995).
• Se efectuará la normalización de las intensidades de las bandas de
expresión del gen problema, en relación a la señal del gen 18S rDNA, que
se utiliza como testigo de los niveles constitutivos de expresión.
Normalización
• La normalización se realizó utilizando el programa SigmaGel, calculando el
cociente de pixeles invertidos del gen DAP1 o XPR2 / pixeles invertidos del
gen ACT1
RESULTADOS
Para describir los resultados, se presentan los diversos objetivos que tuvo el
proyecto:

Objetivo 1
Realizar un estudio sobre la regulación de la síntesis de la enzima α amilasa de
Wickerhamia sp por análisis de la producción y actividad en respuesta a cambios
de sustratos en cultivo por lote.
DETERMINACIÓN DE AZÚCARES REDUCTORES Y ACTIVIDAD ENZIMÁTICA
EN ALMIDONES GRADO REACTIVO Y COMERCIALES.
Se encuentran ciertas similitudes entre las actividades de el almidón grado
reactivo Baker (patrón) y las harinas comerciales de trigo y arroz, siendo el
almidón patrón el de más baja actividad.
Para la actividad de el almidón grado reactivo Monterrey (patrón) se encuentra que
la harina de maíz tiene cierta semejanza en el comportamiento; siendo que el
almidón grado reactivo Monterrey es el que muestra mejor actividad para
producción de α- amilasa de Wicherhamia sp.
Por último se nota que tanto almidones de grado reactivo y los almidones
comerciales su actividad máxima coincide dentro del periodo de fase acelerada del
crecimiento; por lo que decimos que la producción de α- amilasa está asociada al
crecimiento.
DETERMINACIÓN DE AZÚCARES REDUCTORES Y ACTIVIDAD ENZIMÁTICA
EN MALTOSA.
Debido a que la α- amilasa deja como ultima unidad a la maltosa o dextrinas como
la iso-maltosa o malto-triosa estos podrían ser componentes que provoquen
represión catabólica. Anteriormente se observo que los azúcares no se
acumulaban demasiado y aunque están presentes no parecen tener un efecto
negativo en la producción de α- amilasa, lo que indicaría que posiblemente el
efecto de represión catabólica en el microorganismo no esta tan acentuado.

Para poder ver de qué magnitud es el efecto de la represión catabólica se hicieron
cinéticas en maltosa a dos concentraciones diferentes (1 y 2%) para observar el
comportamiento de la levadura; a diferencia de las metodologías anteriores para el
desarrollo del cultivo este se desarrollo a partir de un inóculo de maltosa para
observar si la actividad se abatía y el posterior efecto que ocasionaría.
Para la actividad en el medio con maltosa al 1%, al principio hay una actividad de
58 UA se considera que es alta; aunque podría ser remanente de la actividad que
ocurre en almidón ya que la cepa se conserva en medio inclinado de almidón
grado reactivo Baker.
Para comprobar dicho comportamiento se realizo otra determinación de azúcares
y actividad con maltosa al 2%, mostrando una cantidad de actividad mayor (68
UA) y más cantidad de azúcares (45 mM/mL) al transcurrir el tiempo esta
actividad aumenta aún más (75 UA), lo que indica que la maltosa no está
ejerciendo un efecto de represión en la producción de α- amilasa de Wicherhamia
sp; o bien la cepa es insensible a la represión catabólica por maltosa, aunque la
maltosa se acumule no va a afectar a la producción.
DETERMINACIÓN DE AZÚCARES REDUCTORES Y ACTIVIDAD ENZIMÁTICA,
ACTIVIDAD ESPECÍFICA, CRECIMIENTO Y pH EN ALMIDÓN GRADO
REACTIVO MONTERREY.
Como se observó en los experimentos anteriores el almidón de grado reactivo
Monterrey fue el que obtuvo mayor actividad, por ello se realizó la cinética sobre
dicho almidón; partiendo de un inóculo de almidón Monterrey.
Ésta se realizó en un reactor de 1L, el cual se agitó magnéticamente y se mantuvo
con aireación por medio de una bomba a la que se adaptó un filtro con fibra de
vidrio para esterilizar el aire, así también se realizó la conexión de tubería de
forma que, por medio de una jeringa se tomara la muestra y por las conexiones
fluyera aire estéril para evitar una contaminación, se adaptó un embudo de

separación el cual ayudó a proporcionar agua estéril ya que el medio se
evaporaba. El montaje del reactor se puede observar en la Figura 1.
A las muestras recolectadas se les realizó azúcares reductores, actividad
enzimática, pH, crecimiento y proteínas para realizar el cálculo de actividad
específica.
En la curva de crecimiento se observa una fase de adaptación, una de crecimiento
acelerado (10 a 28 h) y finalmente la fase estacionaria. En cuanto a las curvas de
azúcares reductores y actividad se asemejan notándose que la cantidad de
azúcares no se ve acumulada, teniendo una máxima actividad de 150 UA a las 14
h. Finalmente la actividad especifica que se observa es muy alta (500 µM/mgp*h)
comenzando a las 12 h permaneciendo constante hasta cerca de las 24 h, esto es
debido a que la cantidad de proteínas que se detectaron por el método de Lowry
eran muy pequeñas ya que se pierden proteínas que no son importantes para la
actividad, indicando que la proteína presente es la enzima de interés, en este caso
la α-amilasa revelando que la enzima esta en un estado más puro, de esta manera
mientras menor cantidad de proteína exista la actividad específica será mayor y se
hallará debido a la presencia de la enzima de interés.
En este sistema a diferencia de el matraz por las actividades observadas
encontramos que se mejoró el sistema de aireación y agitación, además de que
fueron mejor controlados.

DETERMINACIÓN DE AZÚCARES REDUCTORES, ACTIVIDAD ENZIMÁTICA Y
ACTIVIDAD ESPECÍFICA EN MEDIO DE CASTAÑEDA-GLUCOSA-ALMIDÓN EN
CULTIVO SUMERGIDO EN REACTOR.
En este experimento se trabajo en un reactor de 1L de igual forma que la cinética
de Almidón grado reactivo Monterrey. Lo que se hizo fue partir de un inoculo de
glucosa para ver los efectos que este sustrato podía tener en el sistema del
microorganismo.
Existe una fase de adaptación muy larga (0 a 10 h) la actividad es muy baja
(menor de 50 UA), notándose que la actividad se empieza a recuperar a las 12 h
se nota la actividad enzimática máxima (80 UA) la cual permaneció constante
hasta el final de la cinética y fue hasta cerca de las 24 h donde se obtiene la
mayor actividad específica (360 µM/mgp*h) ya que la glucosa es un sustrato de
fácil asimilación, así el microorganismo creció mejor (mayor cantidad de biomasa),
por lo tanto al ser un número considerable de microorganismos la actividad se ve
incrementada.
Además en cinéticas con almidón se observa la mayor actividad alrededor de las
12 h, y en este experimento se observa una fase con poca actividad (0 a 10 h) la
cual nos indica un retraso, por lo que se supone que la glucosa tiene un efecto de
represión catabólica.
DETERMINACIÓN DE AZÚCARES REDUCTORES, ACTIVIDAD ENZIMÁTICA Y
ACTIVIDAD ESPECÍFICA EN MEDIO DE CASTAÑEDA-ALMIDON-GLUCOSA-
GLUCOSA.
A diferencia del experimento anterior en este se manejo un pre-inoculo de
almidón, un inoculo en glucosa y la cinética se realizo en glucosa al 2%.

Para esto el microorganismo al estar creciendo en almidón viene en forma
inducida (está produciendo la enzima), por ello no es suficiente un solo pase a
glucosa para abatir la actividad, por lo tanto se hicieron dos pases en glucosa
(inoculo y cinética en reactor) para observar el efecto.
Se realizó una cinética en reactor, la actividad que se encuentra al principio es
considerablemente alta (250 UA y 170 µM/mgp*h), lo que nos indica que la enzima
que viene del almidón permanece y la actividad también.
Posteriormente la actividad específica aumenta a las 12 h (410 µM/mgp*h) pero no
se explicaría como una producción, mejor dicho se considera una permanencia de
la actividad, probablemente en este momento la glucosa ya es muy poca en el
medio y aunque la enzima no se está produciendo si se mantiene en el cultivo.
Se tiene la hipótesis de que se manejara almidón después la actividad se
recuperaría.
Para comprobar los resultados se realizó la misma cinética sólo que en matraz,
encontrando resultados semejantes solo que en matraz hay menor actividad por
las condiciones de aireación y agitación
DETERMINACIÓN DE AZÚCARES REDUCTORES, ACTIVIDAD ENZIMÁTICA Y
ACTIVIDAD ESPECÍFICA EN MEDIO DE CASTAÑEDA-ALMIDON-MALTOSA-
MALTOSA.
En experimentos realizados anteriormente donde se uso maltosa, se encontró que
probablemente es un inductor.
Para comprobar que la maltosa es un inductor se hizo un pre-inoculo en almidón,
un inoculo en maltosa y la cinética sobre maltosa al 2%, esto para tratar de abatir
el efecto del almidón.

La cinética en reactor (Figura 5) notamos que la actividad es más constante y se
mantiene por más tiempo, disminuyendo considerablemente hasta después de las
24 h, esta disminución no se atribuye a que la actividad desaparezca sino a que
sustrato se está agotando.
La fluctuación de la curva de actividad específica puede deberse a la diferencia de
concentración de proteínas que el microorganismo va excretando, pero a pesar de
la oscilación permanece dentro de un rango de actividad.
Para corroborar resultados la misma cinética se realizo en matraz, encontrado
resultados semejantes, sólo que en la cinética de matraz las curvas se ven más
constantes y permanecen a lo largo del tiempo que se mantuvo la cinética, y se
atribuye a las condiciones de agitación y aireación que se manejaron en los dos
sistemas.
La maltosa funciona en dado caso como sustrato de mantenimiento.
DETERMINACIÓN DE AZÚCARES REDUCTORES, ACTIVIDAD ENZIMÁTICA Y
ACTIVIDAD ESPECIFICA EN MEDIO DE CASTAÑEDA-ALMIDON-MALTOSA-
MALTOSA Y CASTAÑEDA-ALMIDON-GLUCOSA-GLUCOSA CON INÓCULO
LAVADO Y SIN LAVAR.
Dado que en los experimentos anteriores donde se uso Maltosa y Glucosa se
observo una alta actividad de la enzima al principio de la cinética, se propuso
realizar los dos experimentos anteriores con el mismo sistema, solo que usando
para una misma cinética un inoculo lavado con solución salina al 0.85 % y un
inoculo sin lavar, de esta forma se tuvieron 2 cinéticas de Maltosa y 2 cinéticas de
Glucosa.
Se tiene como hipótesis que al realizar lavado a uno de los inóculos este
provocaría una disminución de la actividad remanente proveniente del pre-inoculo

de almidón y así corroborar el efecto real de la glucosa y la maltosa sobre el
sistema de Wickerhamia sp
Para las cinéticas sobre Maltosa, se observa que la actividad específica para la
cinética realizada con inoculo lavado (MIL) es más alta que para la cinética con el
inoculo sin lavar (M) lo que indica que la maltosa tiene cierto grado de inducción
en el sistema de Wickerhamia sp
En Glucosa se supone una fuerte inducción (en almidón) y esta señal permanece
en las células, es muy posible que si haya represión pero el sistema ya inducido
continua expresándose, o sea que el sistema de inducción es fuerte. El promotor
es fuerte, tiene una alta afinidad por la RNA pol y aunque existan bajos niveles de
cAMP continua la trascripción.
En ambos casos observamos que las actividades en los inóculos lavados y sin
lavar no difieren mucho, tal vez la solución salina no fue suficiente para quitar el
remanente de actividad y hubiera sido mejor realizar los lavados con detergente.
OBJETIVO 2
Producción en cultivo por lote y continuo de amilasas a partir de Wickerhamia sp.
Crecimiento
Las cinéticas se realizaron en medio base Castañeda-almidón grado reactivo con
almidón marca Baker, en matraz de 250 ml, como ya fue referido en materiales y
métodos.
Como se observa se obtuvo un máximo de 5 g/l de biomasa en un tiempo de 48
horas.
La velocidad específica máxima (µ) de crecimiento en almidón grado reactivo
Baker fue de 0.2511 h¯¹. En este caso se tendría que ver la forma de alargar la

fase de producción, lo cual puede lograrse modificando el tipo de cultivo; por
ejemplo, se podría establecer un cultivo continuo.
Al analizar la actividad enzimática y crecimiento en almidón grado reactivo marca
Baker, observamos que en las primeras 12 horas de fermentación que son las de
mayor actividad, ésta se encuentra asociada al crecimiento.
Cinética de producción de la amilasa extracelular de Wickerhamia sp. en dos
almidones de grado reactivo.
El experimento se realizó con almidón Baker y almidón Monterrey para conocer la
actividad que presenta la enzima sobre dos diferentes almidones grado reactivo .
La máxima actividad del almidón grado reactivo Baker fue de 77.11 UA siendo
ésta a las 12 horas, después de este tiempo la actividad comienza a disminuir
hasta permanecer constante hasta las 41 horas.
También se obtuvo que la enzima tiene una mayor actividad sobre el almidón tipo
Monterrey que sobre el tipo Baker; siendo la máxima actividad de 112.481 UA a
las 12 horas permaneciendo esta constante hasta las 24 horas, después de este
tiempo comienza el descenso de la actividad enzimática. En cuanto a los azúcares
se puede apreciar que en ambos casos permanecen en concentraciones bajas.
Producción de la amilasa extracelular con diferentes sustratos.
Se realizaron cinéticas con la levadura probando diferentes sustratos amiláceos
con el propósito de conocer cuál de ello se obtenía una mayor producción de la
enzima. Es importante señalar que debido a las propiedades de las harinas no se
pudo determinar crecimiento debido a que existía una gran turbidez en éstas y
afectaban los métodos tanto el de peso seco y como el de densidad óptica, siendo
el más afectado.
Una de las harinas probadas fue la de arroz esta contiene 75% de almidón, del
cual un 15-35% es amilosa y un 63-85% es amilopectina. La acción de la enzima

sobre el sustrato de la harina de arroz alcanza su máximo a las 12 horas con
88.593 UA después de esta hora la actividad comienza a disminuir y permanece
constante.También se puede apreciar que cuando la actividad enzimática es
máxima los azúcares disminuyen y posteriormente vuelven a permanecer
constantes.
El siguiente sustrato estudiado fue la harina de maíz , esta harina contiene 75% de
almidón, 25% de amilosa y 75% de Amilopectina. La máxima actividad de la
enzima sobre el sustrato ocurre a las 24 horas siendo ésta de 112.781UA después
de este tiempo la actividad enzimática disminuye considerablemente.
A continuación se probó la harina de papa ésta contiene 30% de almidón, y a su
vez 23% de amilosa y 77% de amilopectina., es importante señalar que esta
harina presento una viscosidad muy alta. Es por esta razón que consume
rápidamente el sustrato debido a que existe poco almidón como sustrato en el
medió siendo la máxima actividad a las 12 horas, en cuanto a los azúcares se
aprecia como después de las 12 horas empieza a existir una acumulación y a las
24 horas empiezan a disminuir nuevamente.
Finalmente se probó la harina de trigo esta contiene 75% de almidón, 20% de
amilosa y 80% de amilopectina. .
Al realizar la comparación de los sustratos utilizados en todas las cinéticas harina
de maíz, harina de trigo, harina de arroz, fécula de papa y almidón soluble como
referencia (almidón grado reactivo marca Baker y Monterrey). Se obtuvo que con
el almidón grado reactivo Monterrey se obtienen los mayores niveles de actividad
(118.481 UA). La harina de maíz presenta un comportamiento similar con respecto
al almidón de referencia (118 UA))
En cuanto a la harina de arroz, harina de trigo y fécula de papa presentaron
valores menores de actividad en cuanto al almidón de referencia (Monterrey).
Al ser el almidón grado reactivo Baker uno de los almidones que dió una de las
actividades más bajas se decidió para los siguientes experimentos trabajar con el
almidón grado reactivo Monterrey. También es importante mencionar que la mayor
actividad enzimática que ejerce la enzima sobre cualquier almidón alcanza su
máximo a las 12 horas este tiempo será determinante ya que se desea realizar
otro tipo de producción es este caso un cultivo continuo y este tiempo junto con la
velocidad máxima de crecimiento (0.2511 h) serán indispensables.
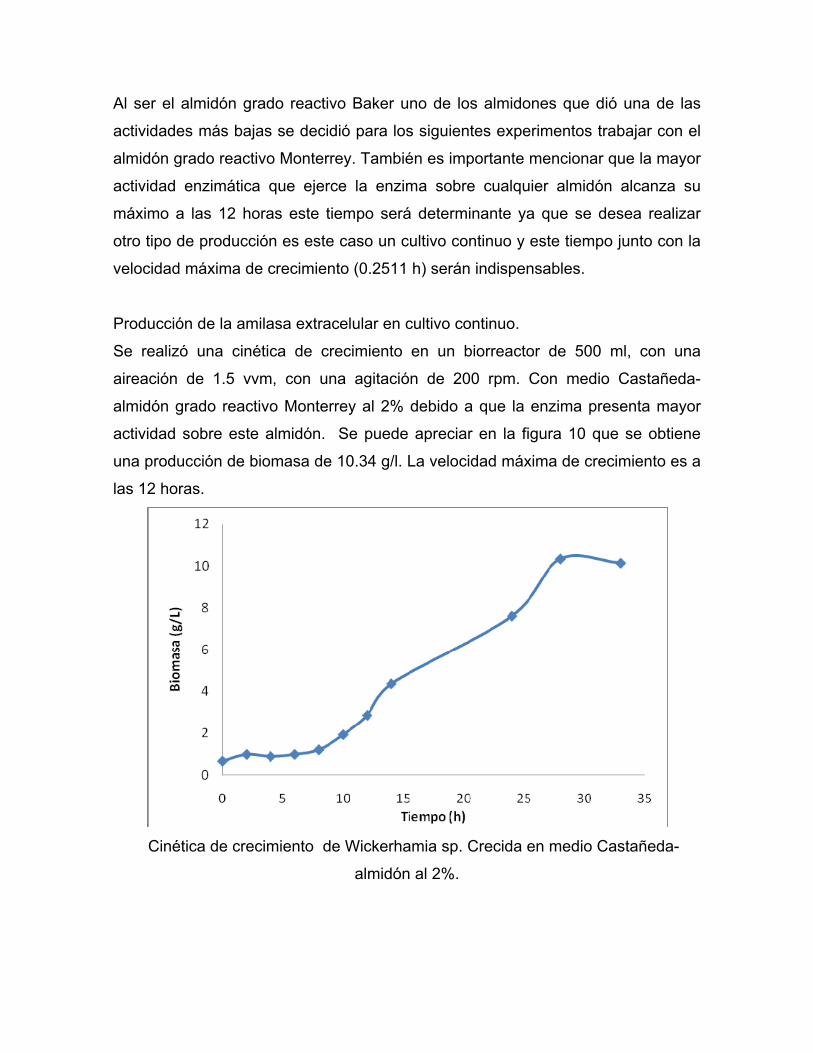
Producción de la amilasa extracelular en cultivo continuo.
Se realizó una cinética de crecimiento en un biorreactor de 500 ml, con una
aireación de 1.5 vvm, con una agitación de 200 rpm. Con medio Castañeda-
almidón grado reactivo Monterrey al 2% debido a que la enzima presenta mayor
actividad sobre este almidón. Se puede apreciar en la figura 10 que se obtiene
una producción de biomasa de 10.34 g/l. La velocidad máxima de crecimiento es a
las 12 horas.
Cinética de crecimiento de Wickerhamia sp. Crecida en medio Castañeda-
almidón al 2%.

Al realizar la comparación de producción de biomasa entre el matraz (5 g/l) y el
biorreactor (10.34 g/L) en el biorreactor se aumentó la producción de biomasa un
48.36% esto se puede atribuir a una mejor aireación en biorreactor, por lo tanto el
microorganismo creció rápidamente.
En cuanto a la actividad enzimática en el biorreactor se alcanzaron 168UA,
existiendo conforme transcurría el tiempo una disminución del pH.
En cuanto a la actividad específica de la enzima se alcanza su máximo a las 14
horas., mientras que la actividad ocurre al mismo tiempo como se puede apreciar
en el cuadro 5.
Para la realización del cultivo continuo se calculó la velocidad específica máxima
(µ max) de crecimiento en almidón Monterrey que fue de 0.2554 h¯¹ (ver cuadro 6).
Como en cultivo continuo la velocidad específica máxima de crecimiento (µ max)
es igual a la velocidad de dilución critica (Dc)., se decidió probar dos velocidades
más aparte de la crítica una al 60% , la otra al 80% de la Dc.
También se calculó la productividad en cultivo por lote siendo la máxima a las 12
horas, como se puede apreciar en el cuadro 7.Se observa que ésta es baja y solo
dura por dos horas., pues después de este tiempo la productividad decae
considerablemente.
Una manera de aumentar la productividad por más tiempo es realizando un cultivo
continuo debido a que se alargará la fase en donde se está produciendo el
metabolito de interés industrial y su única limitante seria una contaminación o
algún desperfecto en el equipo.
El tiempo de duplicación del Wickerhamia sp. es de 2.71 horas, este tiempo se
deberá considerar para tomar en cuenta por lo menos tres tiempos de residencia.
Se calculó la velocidad de flujo, que es el volumen de medio que pasa a través del
reactor por hora siendo éste de 100ml/h cuando se utilizó una velocidad de
dilución de 0.2 h-1

Se inició un cultivo por lote mantenido por 12 horas, a partir de este momento se
inició la alimentación con medio estéril, probándose diferentes velocidades de
dilución, lográndose extender hasta por 100 horas el cultivo
La primera velocidad de dilución probada fue de 0.24 h¯¹ en donde se puede
apreciar que se mantiene la actividad de la enzima sobre el almidón teniendo
hasta 170 UA, se esperaron tres tiempos de residencia y se procedió a cambiar la
velocidad de dilución esta vez a 0.20 h¯¹ se puede observar un período en el cual
el cultivo se desestabiliza, esto es debido al cambio de la velocidad de dilución,
una vez pasado el tiempo de transición el cultivo se vuelve a estabilizar en cuanto
a actividad, habiendo tenido una disminución del 23% con respecto a la velocidad
de dilución anterior, como se había mencionado anteriormente, la enzima durante
el crecimiento exponencial está asociada al crecimiento, por lo tanto si
disminuimos la velocidad de dilución (velocidad de crecimiento ) la actividad
disminuirá.
Cuando se probó la velocidad de dilución de 0.15 h¯¹ se puede apreciar
nuevamente un estado de transición y también se puede apreciar una disminución
de la actividad que tiene la enzima sobre el sustrato.
Se puede apreciar completamente el efecto que tiene la velocidad de dilución
sobre la actividad enzimática en Wickerhamia sp. Mostrando mayor actividad
cuando se trabaja a velocidades de dilución más altas, cabe mencionar que no se
encuentra una diferencia significativa en las velocidades de dilución de 0.20 h¯¹ y
0.15 h¯¹.
El problema de mantener la velocidad de dilución de 0.24 h-1 sería el riesgo de que
se presente un lavado del reactor, debido a que se está trabajando muy cerca de
la velocidad de dilución crítica.

OBJETIVO 3
Caracterizar el gen o los genes codificantes de la(s) amilasa(s) extracelular(es)
de Wickerhamia sp.
RESULTADOS
Diseño de oligonucleótidos.
Se realizó un análisis tipo BLAST a partir de secuencias aminoacídicas y
nucleotídicas de genes codificantes de amilasas en hongos filamentosos y
levaduras, que se encuentran reportados en la base de datos del NCBI
Se realizó un alineamiento de las secuencias aminoácidicas seleccionadas y
de las correspondientes secuencias nucleóticas, utilizando el programa
CLUSTALX 8.1, en donde se ubicaron 4 regiones conservadas, que son
características de las α-amilasa.
A partir de la ubicación de las regiones conservadas en la secuencia
nucleótidica, se diseñó un par de oligonicleótidos degenerados para la
amplificación parcial del o los genes codificantes de una α-amilasa.
A cada oligonucleótido se le asignó un nombre para identificarlo: Amyder para
el iniciador derecho, y Amyrev para el iniciador reverso. Las regiones para el
diseño de dichos iniciadores, fueron elegidas ubicando dentro de ellas a las 4
regiones conservadas; las secuencias nucleótidicas se muestran en la tabla 1.
Tabla 1. Secuencias nucleótidicas de los oligonucleótidos degenerados,
diseñados para la amplificación parcial del gen o genes codificantes de una α-
amilasa
Oligonucleótido Secuencias*
Amyder 5’ GGN ATG GGN TTC ACW GC 3’
Amyrev 5’ GGA TAG TAN AHD GGR TAG TT 3’
*Degeneraciones: N= G A T ó C W= A ó T H= A C ó T D= A G ó T R= G ó A

Se determinó la temperatura media (Tm) de cada uno de los iniciadores para
establecer la temperatura de alineamiento dentro de las condiciones de PCR,
siendo ésta de 52ºC.
Amplificación parcial del o los genes codificantes de una α-amilasa.
Después de establecidas las condiciones de PCR, se procedió a realizar
extracciones de DNA llevándolo a la concentración indicada, se procedió a
montar la reacción de PCR, teniendo como resultado 5 amplificados con
tamaños aproximados de 1500 pb, 1200pb, 900pb, 700pb y 650pb.
Se realizaron ensayos con diferentes temperaturas de alineamiento,
observando que a 57ºC se intensificaba el producto de 1500pb, es por ello que
en las condiciones del programa de PCR se manejaron temperaturas de 52ºC y
57ºC, con la finalidad de obtener el purificado de cada una de las bandas.
Purificación de los amplificados de PCR
Siguiendo las indicaciones del kit de purificación Zymoclean™ Gel DNA
Recovery (ZYMO RESEARCH), se logró obtener 3 purificados: el de 1500 pb,
1200pb y el tercero, correspondiente a los productos de 700 y 650pb.
Secuenciación de los productos de PCR
Una vez obtenidos los purificados, se procedió a secuenciarlos directamente
como producto de PCR. A cada una de las secuencias se le realizó un análisis
tipo BLAST en la base de datos del NCBI, para la búsqueda de similitud con
secuencias codificantes de α amiasas de microorganismos ya reportados. El
purificado correspondiente a los productos de 700 y 650 pb, nombrado como
3Amyder fue el único que mostró una alta similitud con lo reportado en el NCBI.
La secuencia obtenida de 3Amyder fue de 575 pb, la cual posiblemente
corresponda a la secuencia parcial de uno o del único gen codificante de α
amilasa en Wickerhamia sp.

A partir del análisis tipo BLAST de 3Amyder con los otros microorganismos
(que fueron sólo hongos filamentosos y levaduras) se realizó un alineamiento
con el programa CLUSTALX 8.1. Se observó un mejor alineamiento entre las
secuencias de levaduras, así que para el alineamiento final no se incluyó a
hongos filamentosos. Se procedió a buscar las 4 regiones conservadas en
3Amyder, localizando a 3 de ellas: Región A, B y B’ (Fig 1)
Región A
Región B’
Región B
Región A
Figura 1. Ubicación de 3 regiones conservadas: A, B, y B’ en el alineamiento
de 3Amyder. Los microorganismos elegidos para dicho alineamiento
corresponden a Debariomyces occidentalis, Saccharomyces fibuligera, Pichia
burtonii, y Lipomyces kononenkoae, que se muestran de forma abreviada.

Diseño de oligonucleótidos específicos.
Después de localizadas las regiones conservadas, se procedió al diseño de
iniciadores específicos para la búsqueda del ORF y al diseño de la sonda para
llevar a cabo el análisis de expresión por Northern blot,, tomados a partir de las
secuencias que corresponden a las regiones conservadas; este trabajo se lleva
a cabo en este momento..
Clonación en el vector pJET1.2 blunt.
La clonación se realizó como se describe en materiales y métodos. Se intentó
obtener clonas de E. coli con los amplificados de 1500pb y 1200pb, de los que
no se logró obtener una secuencia clara a partir de secuenciación directa de
productos de PCR. Se obtuvieron 10 transformantes, a las que se les extrajo el
plásmido para realizarles un análisis con las enzimas de restricción XhoI, y
XbaI, por separado.
XhoI, y XbaI tienen un sólo sitio de corte en el sitio de clonación del vector
(ver Fig 2 de materiales y métodos), quedando linearizada la construcción (si
es que el amplificado clonado no tuviera sitios de corte para éstas dos
enzimas); de tal manera que se espera fragmentos de 4500pb, 4200pb y
3000pb (plásmido vacío).
De las 10 transformantes obtenidas, dos mostraron tener el inserto, una con el
de 1500pb, y la otra con el de 1200pb.
Las dos clonas obtenidas se secuenciarán para posteriormente someterlas a un análisis de homología con una α amilasa. OBJETIVO 4
Análisis de los genes codificantes de la dipeptidil aminopeptidasa (DAP) y la
proteasa alcalina extracelular (AEP) de Yarrowia lipolytica y las proteínas
deducidas. Las secuencias codificantes para la actina, dipeptidil aminopeptidasa (DAP) y la
proteasa alcalina extracelular (AEP) se localizaron en el genoma de Yarrowia
lipolytica utilizando el sitio de la NCBI (http://www.ncbi.nlm.nih.gov/) y

éstas fueron verificadas en Génolevures: Genomic Exploration of the
Hemiascomycete Yeasts (http://cbi. labri.fr/Genolevures/).
El gen XPR2 codificante de la proteasa alcalina extracelular, está compuesto
por 1278 bp, la proteína deducida consta de 425 aminoácidos que constituyen
al precursor no activo.
Con respecto al gen DAP1 codificante de la dipeptidil aminopeptidasa presenta
una secuencia nucleotídica de 2607 bp, la proteína deducida consta de 868
aminoácidos, peso molecular de 97.8 kDa, punto isoeléctrico de 4.9 y no
presenta intrones. La secuencia aminoacídica deducida se analizó con el
programa PSORT II y se encontró que aparentemente no presenta péptido
señal en el extremo amino terminal y que presenta una posible región
transmembranal de 23 aminoácidos que va de los residuos 74-96 (Figura 1), la
topología membranal la ubica como una proteína de membrana tipo II, es decir,
que presenta el extremo amino terminal dirigido hacia el citoplasma. Así mismo
el programa sugiere que la posible localización subcelular de la proteína es
retículo endoplásmico (44.4%).
Se realizó la búsqueda de secuencias motifs en la secuencia aminoacídica de
la dipeptidil aminopeptidasa encontrándose los motivos Pfam00930.8 (del
residuo 209-573) y Pfam00326.8 (del residuo 659-866), que pertenecen a las
DAPs tipo IV y a las peptidasas respectivamente, de la misma manera se
encontró la secuencia DWVYEEE de los residuos 301-307, que es
característica de las DAPs tipo IV (Figura 1).
Figura 1. Representación esquemática de la dipeptidil aminopeptidasa (DAP) de Y. lipolytica. Características de la proteína deducida del gen DAP1.

Análisis bioinformático de las secuencias reguladoras de los genes DAP1 y
XPR2.
Una vez que se localizaron e identificaron los genes de interés, las secuencias
reguladoras de ambos genes se analizaron in silico utilizando el paquete
bioinformático MatInspector.
En la región reguladora del gen DAP1 que codifica para una dipeptidil
aminopeptidasa y se compone de 1131 pb, se ubicaron 30 posibles sitios de
unión a factores de transcripción, de los cuales sólo los de mayor interés fueron
representados en la Figura 2. De igual manera río arriba del gen XPR2
codificante de una proteasa alcalina extracelular, se tiene la región reguladora
que consta de 1161 pb, el resultado marcó la existencia de 41 sitios hipotéticos
de unión a factores de transcripción, los de mayor interés fueron representados
en la Figura 3.
Las características de los factores de transcripción que afectan la expresión de
los genes en estudio se establecen en la Tabla 1.
Figura 2. Gen que codifica para la dipeptidil aminopeptidasa de Y. lipolytica (YALI0B02838g ) y su región reguladora.
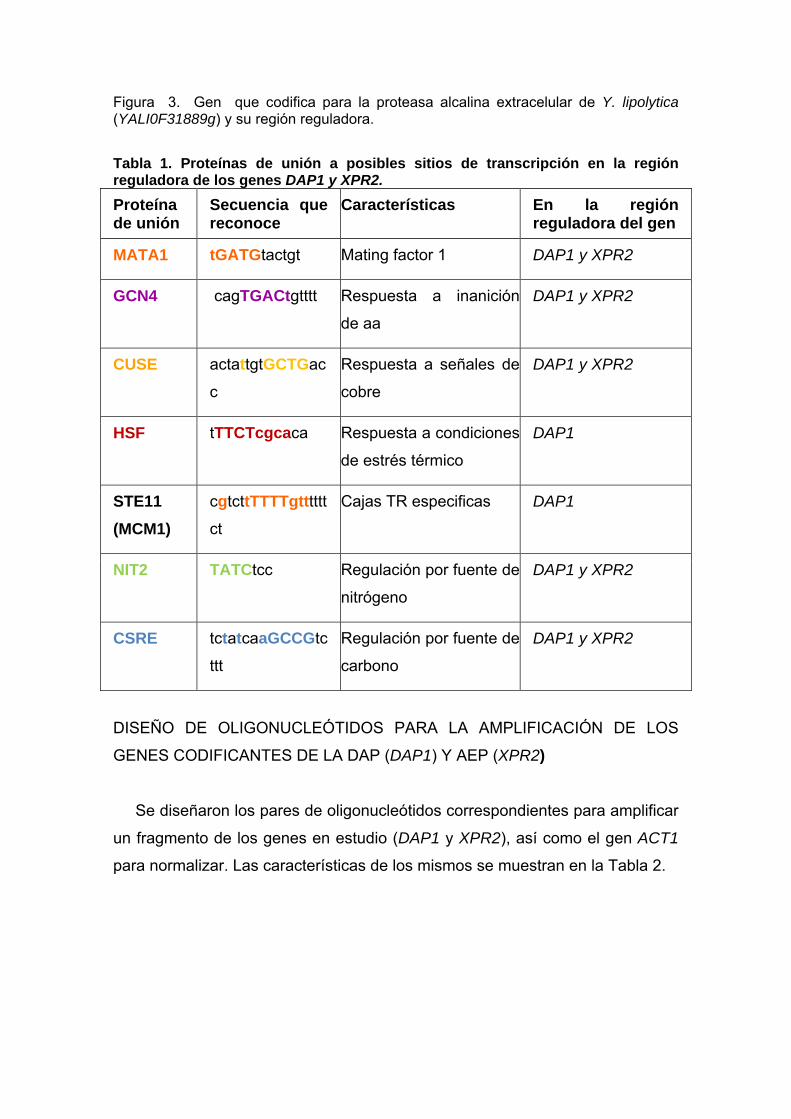
Figura 3. Gen que codifica para la proteasa alcalina extracelular de Y. lipolytica (YALI0F31889g) y su región reguladora. Tabla 1. Proteínas de unión a posibles sitios de transcripción en la región reguladora de los genes DAP1 y XPR2. Proteína de unión
Secuencia que reconoce
Características En la región reguladora del gen
MATA1 tGATGtactgt Mating factor 1 DAP1 y XPR2
GCN4 cagTGACtgtttt Respuesta a inanición
de aa
DAP1 y XPR2
CUSE actattgtGCTGac
c
Respuesta a señales de
cobre
DAP1 y XPR2
HSF tTTCTcgcaca Respuesta a condiciones
de estrés térmico
DAP1
STE11 (MCM1)
cgtcttTTTTgttttttct
Cajas TR especificas DAP1
NIT2 TATCtcc Regulación por fuente de
nitrógeno
DAP1 y XPR2
CSRE tctatcaaGCCGtc
ttt
Regulación por fuente de
carbono
DAP1 y XPR2
DISEÑO DE OLIGONUCLEÓTIDOS PARA LA AMPLIFICACIÓN DE LOS
GENES CODIFICANTES DE LA DAP (DAP1) Y AEP (XPR2)
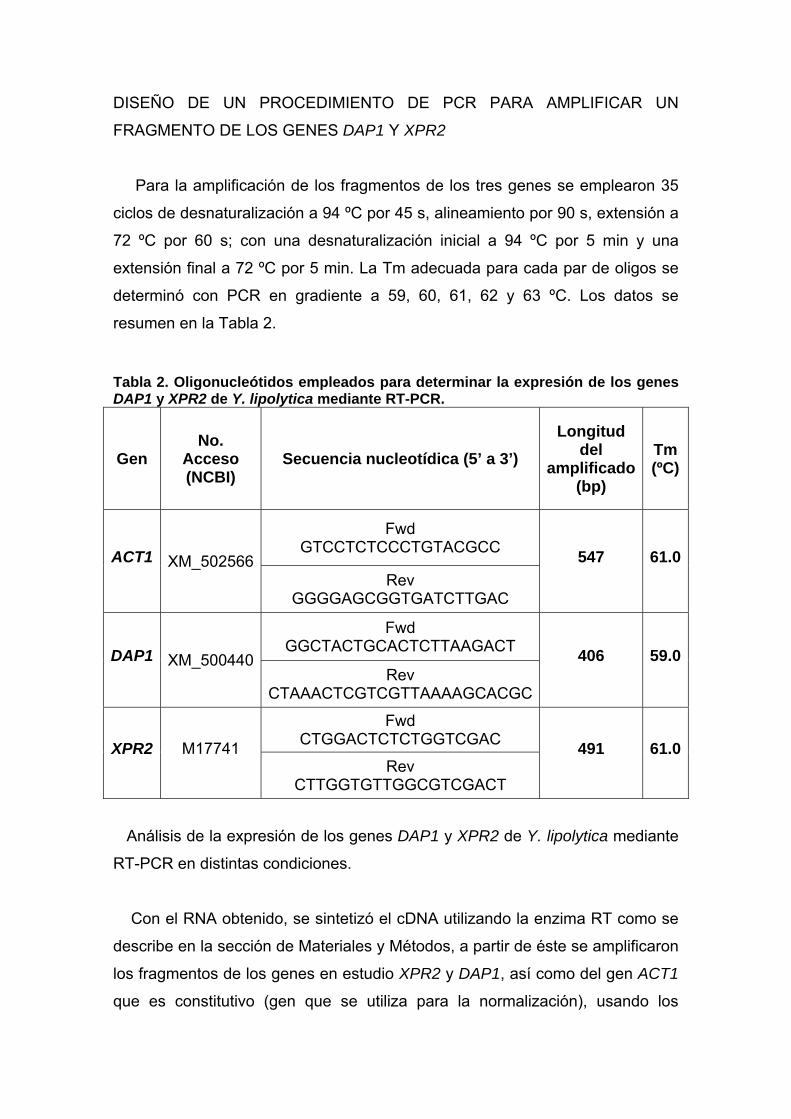
Se diseñaron los pares de oligonucleótidos correspondientes para amplificar
un fragmento de los genes en estudio (DAP1 y XPR2), así como el gen ACT1
para normalizar. Las características de los mismos se muestran en la Tabla 2.
DISEÑO DE UN PROCEDIMIENTO DE PCR PARA AMPLIFICAR UN
FRAGMENTO DE LOS GENES DAP1 Y XPR2
Para la amplificación de los fragmentos de los tres genes se emplearon 35
ciclos de desnaturalización a 94 ºC por 45 s, alineamiento por 90 s, extensión a
72 ºC por 60 s; con una desnaturalización inicial a 94 ºC por 5 min y una
extensión final a 72 ºC por 5 min. La Tm adecuada para cada par de oligos se
determinó con PCR en gradiente a 59, 60, 61, 62 y 63 ºC. Los datos se
resumen en la Tabla 2. Tabla 2. Oligonucleótidos empleados para determinar la expresión de los genes DAP1 y XPR2 de Y. lipolytica mediante RT-PCR.
Gen No.
Acceso (NCBI)
Secuencia nucleotídica (5’ a 3’) Longitud
del amplificado
(bp)
Tm (ºC)
ACT1 XM_502566
Fwd GTCCTCTCCCTGTACGCC 547 61.0
Rev GGGGAGCGGTGATCTTGAC
DAP1 XM_500440
Fwd GGCTACTGCACTCTTAAGACT 406 59.0
Rev CTAAACTCGTCGTTAAAAGCACGC
XPR2 M17741
Fwd CTGGACTCTCTGGTCGAC 491 61.0
Rev CTTGGTGTTGGCGTCGACT
Análisis de la expresión de los genes DAP1 y XPR2 de Y. lipolytica mediante
RT-PCR en distintas condiciones.
Con el RNA obtenido, se sintetizó el cDNA utilizando la enzima RT como se
describe en la sección de Materiales y Métodos, a partir de éste se amplificaron
los fragmentos de los genes en estudio XPR2 y DAP1, así como del gen ACT1
que es constitutivo (gen que se utiliza para la normalización), usando los

oligonucleótidos diseñados para tales fines. La normalización de los datos se
realizó utilizando el programa Sigma Gel.
Se estudió la expresión de los genes sometiendo a la levadura Y. lipolytica a
diferentes condiciones fisiológicas, en todos los casos las células provenían de
medio YEPD y se encontraban en la fase logarítmica de crecimiento.
En inanición por fuente de carbono, se observó que la DAP1, aumenta su
expresión en contraste a XPR2, que a partir de los 30 minutos cesa su
expresión.
Cuando la levadura fue sometida a un medio conteniendo glucosa como
fuente de carbono, ambas expresiones tuvieron un comportamiento similar, al
anterior, donde la expresión de XPR2 deja de verse a los 60 minutos.
Si Y. lipolytica se somete a condiciones de estrés por ausencia de fuente de
nitrógeno, no deja de expresarse XPR2, pero sí decrece, por su parte DAP1
aparece manteniendo bajos niveles de expresión. En un medio que contenga
peptona, se obtiene la sobreexpresión del gen XPR2, mientras que DAP1 por
el contrario baja totalmente su expresión después de tener un pequeño
incremento en la misma.
Cuando se utiliza un medio con sulfato de amonio como fuente de nitrógeno,
DAP1 eleva su expresión y la mantiene, mientras que XPR2 la aumenta y
posteriormente decrece. OBJETIVO 5 Producción de Proteasas por microorganismos aislados de fuentes naturales
empleando diferentes sistemas de reacción
Mediante técnicas de selección primaria
La selección secundaria de levaduras ha permitido recuperar 2 cepas de Y.
lipolytica con alta actividad de proteasas; éstas han sido probadas en medios

de tipo industrial formulados a base de harinas y desechos pesqueros. Los
resultados han mostrado que la harina de trigo y la caseína son los mejores
sustratos de fermentación. Actualmente los estudios se realizan en un sistema
de cultivo continuo para ver cómo influye el estado fisiológico en la síntesis de
estas enzimas
Los estudios de caracterización revelan que la proteasa extracelular de Y.
lipolytica es termoestable y de tipo alcalino, lo que sugiere que puede ser
utilizada como formulante de detergentes o bien para el tratamiento en los
procesos de curtiduría.
PERSPECTIVAS DE TRABAJO
Hasta el momento se ha logrado obtener la secuencia parcial (575 pb) de un
gen codificante para una α amilasa en Wickerhamia sp, desconociendo si
exista más de una copia en su genoma, es por ello que a partir de esta
secuencia se diseñará una sonda para realizar análisis por Northern blot, que si
bien se llevará a cabo para hacer estudios de regulación de expresión, también
nos permitirá conocer el número de copias existentes; a su vez, se diseñará un
nuevo par de oligonucleótidos específicos para la amplificación del ORF para
tratar de obtener la secuencia nucleotídica del genoma de Wickerhamia sp,
ubicando en éste la o las secuencias codificantes para la actividad amilolítica.
En cuanto a los análisis de expresión que se llevarán a cabo, éstos tendrán
como objetivo determinar qué genes son los que se transcriben para la
expresión de la actividad amilolítica cuando Wickerhamia sp es crecida bajo
ciertas condiciones ambientales y nutricionales, lo que nos permitirá saber cual
es la mejor condición para la máxima expresión de la proteína.
Con respecto a las condiciones de producción de estas enzimas se tienen las
bases para poder ensayar diferentes sistemas de reacción y poder simular un
modelo de producción a mayor escala. Los resultados de los estudios de
regulación presentan aparentemente a una enzima que es resistente a
represión por catabolito, lo cual la hace atractiva para su producción a mayor

escala; sin embargo, esto tiene que ser confirmado haciendo estudios de
regulación bajo diferentes estados fisiológicos en un sistema de cultivo
continuo y probando diferentes inductores y represores de la síntesis.
Se tienen identificados los genes que codifican para la expresión de la proteasa
específica DAP y las condiciones que regulan su expresión, lo cual permitirá
clonarlos en otra levadura que pueda sobreproducirla y secretarla al medio, lo
que simplificaría el proceso de producción para esta proteasa que es utilizada
en la industria de los alimentos.