trabajando juntos para reducir -...
TRANSCRIPT

41
Avances recientes en calidad de leche y control de mastitis en explotaciones lecherasnº 40
Incluso el mejor tratamiento antibiótico de vaca seca deja la puerta abierta a nuevas infecciones intramamarias.
OrbeSeal, el sellador interno del pezón, proporciona una protección de larga duracióncerrando la entrada a la ubre de nuevas infecciones.
La administración de OrbeSeal en combinación con el antibiótico de secadopreviene nuevas infecciones intramamarias durante el periodo seco
dando como resultado vacas más sanasy mayor rentabilidad de la explotación.
Trabajando juntos para reducirdrásticamente la mamitis
ORBESEAL®. Composición cuantitativa y cualitativa: Ingrediente(s) activo(s): Subnitrato de Bismuto, denso (2,6 g). Indicaciones de uso: Prevención de nuevas infecciones intramamarias durante el periodo de secado. Contraindicaciones: No usar en vacas en lactación. En vacas con mamitis subclínicas al secado no debe utilizarse solo el producto. No utilizar en vacas con mamitis clínica al secado. Efectos adversos (frecuencia y gravedad): No se conocen. Precauciones especiales de uso: Es una buena práctica observar regularmente las vacas secas para detectar signos de mamitis clínica. Si un cuarto sellado desarrolla mamitis clínica, deberá retirarse el sello manualmente del cuarto afectado antes de establecer una terapia antimicrobiana adecuada. Para reducir riesgos de contaminación, no introducir la jeringa en agua. Utilizar la jeringa una sola vez. No administrar ningún otro producto intramamario después de la administración de Orbeseal. Uso durante la gestación y la lactancia: No deberá administrarse Orbeseal durante la lactación. Orbeseal puede utilizarse en animales gestantes, dado que el producto no se absorbe después de la administración intramamaria. En el parto, el sello puede ser ingerido por el ternero. La ingestión de Orbeseal por el ternero es segura y no produce efectos adversos. Tiempo de espera: Carne: 0 días; Leche: 0 días. Precauciones especiales que deberá tomar la persona que administre el producto a los animales: Lavarse las manos después de su uso.
A-OrbeSeal-1/10
Anuncio Cria y Salud 225x320mm.indd 1 30/06/11 10:02CYS 40.indd 41 26/12/11 10:48
Año 8
• n
º42
Revista de Medicina VETERINARIA
Programas de alimentación en Porcino en función del sexo: efectos de la castración quirúrgica y la inmunocastración
cambios en el esPermatozoide desde la eyaculación hasta la fecundación
factores que afectan la Pérdida de la gestación en el Periodo fetal temPrano. una aProximación teraPéutica
Patología del sistema resPiratorio Para veterinarios de camPo
Ahora síguenos en:

41
Avances recientes en calidad de leche y control de mastitis en explotaciones lecherasnº 40
Incluso el mejor tratamiento antibiótico de vaca seca deja la puerta abierta a nuevas infecciones intramamarias.
OrbeSeal, el sellador interno del pezón, proporciona una protección de larga duracióncerrando la entrada a la ubre de nuevas infecciones.
La administración de OrbeSeal en combinación con el antibiótico de secadopreviene nuevas infecciones intramamarias durante el periodo seco
dando como resultado vacas más sanasy mayor rentabilidad de la explotación.
Trabajando juntos para reducirdrásticamente la mamitis
ORBESEAL®. Composición cuantitativa y cualitativa: Ingrediente(s) activo(s): Subnitrato de Bismuto, denso (2,6 g). Indicaciones de uso: Prevención de nuevas infecciones intramamarias durante el periodo de secado. Contraindicaciones: No usar en vacas en lactación. En vacas con mamitis subclínicas al secado no debe utilizarse solo el producto. No utilizar en vacas con mamitis clínica al secado. Efectos adversos (frecuencia y gravedad): No se conocen. Precauciones especiales de uso: Es una buena práctica observar regularmente las vacas secas para detectar signos de mamitis clínica. Si un cuarto sellado desarrolla mamitis clínica, deberá retirarse el sello manualmente del cuarto afectado antes de establecer una terapia antimicrobiana adecuada. Para reducir riesgos de contaminación, no introducir la jeringa en agua. Utilizar la jeringa una sola vez. No administrar ningún otro producto intramamario después de la administración de Orbeseal. Uso durante la gestación y la lactancia: No deberá administrarse Orbeseal durante la lactación. Orbeseal puede utilizarse en animales gestantes, dado que el producto no se absorbe después de la administración intramamaria. En el parto, el sello puede ser ingerido por el ternero. La ingestión de Orbeseal por el ternero es segura y no produce efectos adversos. Tiempo de espera: Carne: 0 días; Leche: 0 días. Precauciones especiales que deberá tomar la persona que administre el producto a los animales: Lavarse las manos después de su uso.
A-OrbeSeal-1/10
Anuncio Cria y Salud 225x320mm.indd 1 30/06/11 10:02CYS 40.indd 41 26/12/11 10:48
Año 8
• n
º42
Revista de Medicina VETERINARIA
Programas de alimentación en Porcino en función del sexo: efectos de la castración quirúrgica y la inmunocastración
cambios en el esPermatozoide desde la eyaculación hasta la fecundación
factores que afectan la Pérdida de la gestación en el Periodo fetal temPrano. una aProximación teraPéutica
Patología del sistema resPiratorio Para veterinarios de camPo
Ahora síguenos en:

3
nº 42
Yo fui uno de esos estudiantes de veterinaria que disfrutó de una universidad con instalaciones obsoletas, pocas prácticas y menos medios. Por eso ahora leo con tristeza como proyectos que se desarrollaron con la intención de cambiar esta situación y equipararnos a los estudiantes de veterinaria de otros países europeos están en peligro. Este es el caso del Hospital Clínico Veterinario de Castilla y León (HCV).
El reciente anuncio por parte de la Junta de Castilla y León de suprimir la Fundación del Hospital Clínico Veterinario encargada de la financiación del hospital, pone en serios apuros a la facultad de Veterinaria. El fin primordial de esta fundación, formada por la Universidad de León y las consejerías de Agricultura y Ganadería y Educación en el año 2005, era impulsar el hospital clínico veterinario y convertirlo en un centro de referencia. El director-gerente del HCV, Norberto González insiste en que el hospital veterinario no ha sido un agujero para la Universidad y que cuando se creó la fundación la previsión de aportaciones por parte de la Junta estaban fechadas hasta el ejercicio 2020. En el contexto actual, parece que la única salida sea que la Universidad de León, estrangulada por lo recortes, decida asumir su gestión en un futuro.
Creo que ha llegado el momento de recordar a nuestros políticos un viejo proverbio chino: “Si haces planes para un año, siembra arroz. Si los haces por dos lustros, planta árboles. Si los haces para toda la vida, educa una persona”.
Redacción
Editorial
editorial.indd 3 14/05/12 10:54

4
EDITA:
DIRECTOR EJECUTIVO: Enrique Marcos [email protected]
DIRECTORA DE MÁRKETING: Valle Garcí[email protected]
COORDINADOR DE PROYECTOS GRÁFICOS: Miguel Ángel Gá[email protected]
DISEñO Y MAqUETACIóN: José GilJavier Pérez
FOTOGRAFÍA: Eva García Martín (Veterinaria)
COORDINADORA DE REDACCIóN: Ana del Cura (Veterinaria)[email protected]
DEPARTAMENTO DE SUSCRIPCIONES: Alicia [email protected]
DEPARTAMENTO COMERCIAl:Fco. David Muñ[email protected]
REDACCIóN, PUBlICIDAD Y ADMINISTRACIóN: Calle Dulcinea 42 (4º B)28020 MadridTél.: 902 36 39 34Fax: 91 628 92 77 [email protected]
IMPRIME: everyone+
DEPóSITO lEGAl: M-14245-2005ISSN 1889-2094
COlABORA:
DIFUSIÓN GRATUÍTA
Queda totalmente prohibida cualquier reproducción total o parcial, ya sea por medios mecánicos, electrónicos y expresamente por fotocopias de todos los textos, figuras, fotografías dibujos incluidos en nuestra revista CRIA Y SALUD. Las opiniones expresadas en cada trabajo o artícu-lo son de la exclusiva responsabilidad del autor,pudiendo la empresa editora no compartir la opinión de los mismos.
CONSEJO DE REDACCIÓNde Cría y Salud en Bovino y PorcinoDra. Susana AstizInvestigadora Titular Dpto. de Reproducción (INIA).Prof. María Alcaide Licenciada en Veterinaria.Prof. Dr. Alex BachDirector de la Unidad de Rumiantes del IRTA.Doctor en Veterinaria por la Universidad de Minnesota.Dr. Joaquín BaucellsCentro Veterinario Tona.Prof. Dr. Sergio CalsamigliaDepartamento de Patología y Producción Animal.Facultad de Veterinaria. Universidad Autónoma de Barcelona.Prof. Dr. Javier Cañón Catedrático de Genética.Universidad Complutense de Madrid. Facultad de Veterinaria de Madrid.Prof. Dr. Pere Costa-BatlloriVeterinario. Universidad Politécnica de Cataluña.Prof. Dr. Carlos FernándezDepartamento de Producción Animal y Ciencia de los Alimentos.Facultad de Ciencias Experimentales y de la Salud. Veterinaria.Universidad Cardenal Herrera CEU.Dra. Eva Mª FronteraDra. en Veterinaria.Dr. Julio de la FuenteDepartamento de Reproducción Animal y Conservación de Recursos Zoogenéticos. Ministerio de Ciencia y Tecnología. I.N.I.A.Prof. Dr. Antonio GázquezCatedrático de Histología y Anatomía Patológica de la Facultad de Veterinaria de Extremadura.D. Ignacio R. García Gómez Director Veterinario.Albeitares Consultores S.L.D. Javier Gil PascualVeterinario Asesor en Porcino.Prof. Dr. Juan Vicente González Dipl. ECBHM. Prof. Titular Dto. Medicina y Cirugía Animal, UCMTRIALVET Asesoria e Investigación Veterinaria SL.Prof. Dr. Gonzalo González Departamento de Producción Animal.Escuela Técnica Superior de Ingenieros Agrónomos.Universidad Politécnica de Madrid.Prof. Dr. Xavier MantecaUnidad de Fisiología. Facultad de Veterinaria.Universidad Autónoma de Barcelona.D. Juan C. Marco MeleroJefe del Laboratorio de Salud Pública.Departamento de Sanidad. Gobierno Vasco.Prof. Dr. Francisco MazzucchelliJefe del Servicio de Clínica Bovina. Hospital Clínico Veterinario.Universidad Complutense de Madrid.Dr. Alfonso MongeDirector de AMASVET. Veterinario especialista en vacuno.Prof. Dr. Antonio Muñoz Catedrático de Producción Animal.Facultad de Veterinaria. Universidad de Murcia.Prof. Dr. Antonio Palomo Departamento de Mediciona y Cirugía Animal.Facultad de Veterinaria. Universidad Complutense de Madrid.Prof. Gabriel Parrilla Veterinario. Profesor del Hospital Clínico de la F.V. de Madrid.
Dr. José Pérez Tapia Servicios Veterinarios.Prof. Manuel Pizarro Departamento de Medicina y Cirugía Animal.Universidad Complutense de Madrid. Facultad de Veterinaria de Madrid.Dr. David Reina Dr. en Veterinaria.Prof. Dr. Elías Rodríguez Catedrático de Microbiología e Inmunología.Departamento de Sanidad Animal. Facultad de Veterinaria de León.Prof. Dr. Manuel Rodríguez Catedrático de la Universidad Complutense de Madrid.Vicerrector de la Facultad de Veterinaria de Madrid.Prof. Dr. Pedro RubioDepartamento de Sanidad Animal.Facultad de Veterinaria. Universidad de León.Prof. Gregorio Salcedo Profesor de Nutrición Animal y Análisis Químico Agrícola.
S u m a r i o
Ahora síguenos en:
creditos y sumario.indd 4 21/05/12 10:34

5
Agenda �������������������������������������������������������������������������������������������������������������������������������6
Noticias ������������������������������������������������������������������������������������������������������������������������������8
Pfizer promueve el intercambio de experiencias profesionales entre Italia y España en Ovino de Leche �����������������������������������������������������������16
Jefo y Beldem celebran 10 años de colaboración entorno a Belfeed ���18
Sección Innovación Tecnológica: Avances en el tratamiento de la artritis equina ����������������������������������������������������������������������������������������������������22
Programas de alimentación en porcino en función del sexo: efectos de la castración quirúrgica y la inmunocastración� ��������������������������������������������������������������������������������������������28
Cambios en el espermatozoide desde la eyaculación hasta la fecundación� �����������������������������������������������������������������������������������������������38
Factores que afectan la pérdida de la gestación en el periodo fetal temprano� Una aproximación terapéutica �����������������52
Patología del sistema respiratorio para veterinarios de campo� ������������60
5
Cría y saludS U m A r I O
creditos y sumario.indd 5 14/05/12 10:58

6
F E R I A S• EDF CONGRESS 2012
27-29/06/2012. Hasselt, Bélgicau http://www.dairyfarmer.net/
• SEPOR 2012 17-20/09/2012. Lorca, Murcia, España http://www.seporlorca.com/
• VIV CHINA 2012Beijing, China. 23-25/09/2012http://www.vivchina.nl/en/Bezoeker.aspx
• FIGAP/VIV MÉXICO 2012Guadalajara, México. 17-19/10/2012http://www.vivamericalatina.nl/en/Bezoeker.aspx
C u R S O S y C O N G R E S O S
FERIAS • CONGRESOS • CONCURSOS • FERIAS • CONGRESOS • CONCURSOS
• 12TH INTERNATIONAL SyMPOSIuM ON DIGESTIVE PHySIOLOGy IN PIGSKeystone, Colorado, Usa. 29/05-01/06/2012http://www.dppigs.org/2012/default.asp
• 22ND INTERNATIONAL PIG VETERINARy SOCIETy CONGRESSJeju, Korea. 10-13/06/2012http://www.ipvs2012.kr/
• PORCIAMÉRICAS 2012Periera, Colombia. 17-19/07/2012http://www.bhrgrupoestrategico.com/PORCIAMERICAS2012/index.php
• XI CONGRESO NACIONAL DE PRODuCCIÓN PORCINA, (XI CNPP)Salta, Argentina. 14-17/08/2012http://www.congresoporcino.com/
• PORKEXPO 2012 & VI FÓRuM INTERNACIONAL DE SuINOCuLTuRACuritiba, Brasil. 26-28/09/2012http://www.porkexpo.com.br/
• XI ENCuENTRO NACIONAL DE PORCICuLTuRACancún, México. 10-13/10/2012http://www.encuentroporcicultura.com.mx/
• ITALPIGCremona, Italia. 25-28/10/2012http://www.italpig.it
BOvINO
PORCINO
CYS42 noticias/AE/agenda.indd 6 14/05/12 10:14

7
nº 42
FERIAS • CONGRESOS • CONCURSOS • FERIAS • CONGRESOS • CONCURSOS
• XXVIIWORLDBUIATRICSCONGRESSLisboa, Portugal. 03-08/06/2012http://wbc-2012.com/
• 12ºCONGRESOPANAMERICANODELALECHEAsunción, Paraguay. 05-07/06/2012http://www.fepale.org/es/proximos-eventos/136-12congreso.html
• WORLDDAIRYEXPO2012Madison, Wisconsin, USA. 02-06/10/2012http://www.worlddairyexpo.com/
• IIJORNADASTÉCNICASSOBRELACALIDADDE LECHE - SERIVET ASTuR-GALAICORibadeo, Lugo, España. 26-27/10/2012http://www.serivet.com/
BOvINO
O T R O S C O N G R E S O S• II CONGRESO INTERNACIONAL DE PODOLOGÍA EQuINA
Segovia, España. 24-26/05/2012http://www.colvema.org
• WORLDBIOENERGY2012Jönköping, Suecia. 29-31/05/2012http://www.elmia.se/en/worldbioenergy/
• IV CONFERENCIA ANuAL 2012 DE LA PLATAFORMA TECNOLÓGICA ESPAÑOLA DE SANIDAD ANIMALMadrid, España. 30/05/2012http://www.vetmasi.es
• MEAT CONGRESSParis, Francia. 04-06/06/2012http://www.worldmeatcongress2012.com
• INTERNATIONAL SyMPOSIuM ON EMISSIONS OF GAS AND DuST FROM LIVESTOCKSaint-Malo, Francia. 10-13/06/2012https://colloque4.inra.fr/emili2012
• CONFERENCIAFAO/OIESOBRECONTROLDELLAFIEBREAFTOSABangkok, Tailandia. 27-29/06/2012http://www.oie.int/eng/A_FMD2012/Background.html
• 17TH INTERNATIONAL CONGRESS ON ANIMAL REPRODuCTION (ICAR)vancouver, Canadá. 29/07-02/08/2012http://www.icar2012.com/
• 5 CONGRESO INTERNACIONAL AuTOCONTROL y SEGuRIDAD ALIMENTARIABarcelona, España. 17-19/10/2012.http://www.kausal.cat/
CYS42 noticias/AE/agenda.indd 7 14/05/12 10:14

8
Noticias
Reconocimiento de las organizaciones
de productores
El Ministerio de Agricultura francés ha pu-blicado un decreto que establece las condi-ciones para el reconocimiento de las orga-
nizaciones de productores (OP) de leche de vaca (Decreto 2012/512 del 19 de abril de 2012, publi-cado el pasado el viernes). Establece un reconoci-miento de tipo general y otro para el caso de pro-ducción de leche bajo alguna norma de calidad.
En España, el reconocimiento de las organi-
zaciones de productores de leche (no solo de vaca, sino también de oveja y cabra) se reguló hace un año, con el RD 460/2011. Según dicho RD, la producción mínima comercializable de las OP de leche de vaca es de 200.000 tn para el caso general y 10.000 tn en el caso de las Islas Baleares, Canarias, ganadería ecológica, gana-dería integrada y fabricación de productos bajo IGP, DOP y ETG. El compromiso mínimo de per-manencia exigido a los productores miembro es de 3 años. El reconocimiento de las OP es muy importante, teniendo en cuenta que el pasado 30 de marzo se publicó el “Paquete leche” (Re-glamento 261/2012). Uno de sus puntos clave es permitir a las OP que puedan negociar los precios de la leche cruda para los productores a los que representen, tanto si cuentan como si no, con la transferencia de la propiedad de la leche cruda de los ganaderos y tanto si el precio negociado es el mismo o no, para la producción conjunta de algunos o todos los miembros.
CargosdirectivosdeANEMBE
El 9 de mayo tuvo lugar en Madrid la pri-mera reunión de la nueva junta directiva de la Asociación Nacional de Especialistas
en Medicina Bovina de España (ANEMBE), ele-gida en Santander durante su último congreso. Tal y como fi gura en sus estatutos, el primer punto del orden del día fue la elección de los cargos directivos de la Asociación. El nuevo or-ganigrama de ANEMBE es:
Presidente: Luis Miguel Cebrián (vocal de relaciones externas y deontología)Vicepresidente: Luis Miguel Jiménez (vo-cal de calidad de leche)Secretario: Gorka Aduriz (vocal de seguri-dad alimentaria y bienestar animal)
Tesorero: Francisco González (vocal de cebo)Vocales electos:Luis Miguel Ortega (vocal de investigación y di-vulgación y presidente del comité científi co)Susana Astiz (vocal de reproducción)Roger Guix (vocal de gestión de granjas e instalaciones)
Los vocales cuyas vocalías quedaron de-siertas en las últimas elecciones continua-rán hasta la próxima asamblea general:
Juan vicente González (vocal de medicina, cirugía y podología)Adrián González (vocal de nutrición)
El Consejo de Ministros aprobó el pasado mes un Real Decreto por el que se modifica la normativa vigen-
te sobre el Programa Nacional de con-servación, Mejora y Fomento de las razas ganaderas en materia de concurrencia de asociaciones para la gestión del libro genealógico de una raza. Por ello, y reco-nociendo la posibilidad de que haya más de una asociación que legítimamente pretenda gestionar un libro genealógico
de una misma raza, la nueva normativa elimina el número mínimo de animales y ganaderos afiliados a una segunda aso-ciación, como condición para conceder o denegar el reconocimiento de la misma. Se toma así en cuenta el hecho de que no todas las razas se encuentran en las mismas circunstancias censales, territo-riales y genéticas, que pueden determi-nar las actuaciones para su mejor regu-lación y fomento.
Nuevas condiciones de reconocimiento de asociaciones de criadores de animales de raza
Según el informe “El sector de la carne de cerdo en cifras. Principales indicadores económicos en 2011”, publicado re-
cientemente por el Ministerio de Agricultura, la producción porcina supuso el 34,2% de la producción fi nal ganadera, con una disminu-ción del 1% respecto a 2010 y el 12,4% de la producción fi nal agraria española (un 0,4% más que en 2010).
El informe abarca otros aspectos como el número de explotaciones, la producción de carne o las exportaciones. En relación con el número de explotaciones destaca la impor-tante disminución de las explotaciones tota-les, en particular intensivas, y el claro aumen-to de las explotaciones mixtas. En relación con el censo de animales, hay un leve descenso de la cabaña porcina respecto al año anterior y Cataluña y Aragón se mantienen como princi-pales zonas productoras.
En el 2011 la producción porcina supuso el 34,2% de la producción final
El Consejo de Ministros ha apro-bado un Real Decreto por el que se modifi ca la normativa
vigente sobre control ofi cial del ren-dimiento lechero para la evaluación genética en las especies bovina, ovi-na y caprina.
En esta norma se definen los pa-rámetros para la realización de los controles de producción láctea de las hembras de animales de raza pura de esas especies. Con las nue-vas modificaciones se flexibiliza la parte técnica relativa al control lechero oficial para adaptarlo a los avances tecnológicos alcanzados en los últimos años en el ámbito de la comprobación de rendimientos del ganado. Así, se ajusta el con-tenido de la norma vigente a las
directrices del Comité Internacio-nal para el Control del Rendimiento Animal (ICAR). Esta organización -de la que forma parte España como miembro fundador a través del Magrama- armoniza los dife-rentes métodos de control de los principales parámetros productivos del ganado para que sean compa-rables con los utilizados en otros países.
La aplicación de las recomen-daciones ICAR se llevará a cabo de acuerdo con las indicaciones esta-blecidas por la Comisión nacional del control lechero oficial, que se harán públicas mediante Resolución de la Dirección General de Produc-ciones y Mercados Agrarios, que se publicará en el BOE.
Modificada la norma española sobre el control oficial del rendimiento lecheroModificada la norma española sobre el control oficial del rendimiento lecherocontrol oficial del rendimiento lechero
CYS42 noticias/AE/agenda.indd 8 14/05/12 10:14

9
nº 42
El virus de Schmallenberg afecta ya a más de 3.300 explotaciones en la uE
La presencia del virus de Schmallenberg (SBv) se ha detecta-do ya en 3.316 explotaciones ganaderas europeas, según han confi rmado los servicios veterinarios de Alemania, Holanda,
Bélgica, Gran Bretaña, Francia, Luxemburgo, Italia y España, paí-ses afectados hasta la fecha.
En su conjunto, se han confi rmado un total de 2.407 focos en explotaciones de ovino, 842 en ganaderías de bovino y 67 en granjas de caprino. Alemania es el país que más focos ha loca-lizado (1.191 confi rmaciones), con la detección de 841 casos en explotaciones de ovino, 305 de bovino y 45 de caprino, según los datos del Instituto Friedrich-Loeffl er, principal encargado del seguimiento de esta epizootia.
En Holanda, el Ministerio de Asuntos Económicos, Agricultura e Innovación ha comunicado que este nuevo mal del ganado ha sido declarado ya en 272 explotaciones, de las cuales 107 son de ovino, 159 de vacuno y 6 de caprino.
En el caso de Bélgica, la Agencia Federal para la Seguridad en la Cadena Alimentaria (Afsca) ha reportado ya la presencia del SBv en 364 granjas del país, con 167 focos en ovino, 195 en bovino y dos en caprino.
En Gran Bretaña, el Departamento de Medio Ambiente, Ali-mentación y Asuntos Rurales (Defra) ha confi rmado que el virus de Schmallenberg ha llegado ya a 243 explotaciones, con 215 fo-cos declarados en ovino y 28 más en ganaderías de vacuno. En Francia continúan aumentando los focos verifi cados del virus y el Ministerio de Agricultura, Alimentación, Pesca y Desarrollo Rural y Regional ha reconocido ya la presencia del SBv en 1.232 ex-plotaciones, mayoritariamente de ovino (1.070), pero también en granjas de vacuno (149) y caprino (13).
Luxemburgo e Italia están entre los últimos países en constatar la presencia del SBv, con seis granjas de ovino y seis de bovino en el pri-mero, y otro en una explotación de caprino, en el segundo.
En España, veterinarios de la Consejería de Agricultura y Pes-ca de la Junta de Andalucía detectaron el pasado 6 de marzo un aborto de un animal con malformaciones -y síntomas clínicos de la enfermedad- en una ganadería mixta de ganado ovino y caprino de la comarca de Hinojosa del Duque (Córdoba). El Laboratorio Central de veterinaria de Algete (Madrid) del Minis-terio de Agricultura, Alimentación y Medio Ambiente confi rmó el resultado positivo, en lo que fue el primer caso detectado en España.
E l Comité Permanente de la Cadena Alimentaria y de Sanidad Animal (CPCASA) apoyó un paquete pro-
puesto por la Comisión Europea para ac-tualizar las normas relativas a la lengua azul. La propuesta desarrolla la nueva normativa que entró en vigor el 22 de marzo de 2012, que permitía el uso de vacunas inactivadas contra la lengua azul en la U.E. La propuesta de modifi -cación del Reglamento (CE) nº 1266/2007 permite una mayor fl exibilidad para que los países elaboren programas de vigilan-cia de la fi ebre catarral ovina y reduzcan la carga para los agricultores y las autori-dades centrales. También fueron aproba-das enmiendas a las normas que regulan los requisitos aplicables a la importación de ganado bovino, ovino y caprino en la U.E. en lo referente a la lengua azul. Así, las normas de importación de la U.E. para los rumiantes domésticos (vacas, ovejas y cabras) se han actualizado para tener en cuenta los avances científi cos y estar más en línea con las normas de movimiento dentro de la Unión.
Se actualizan las normas comunitarias sobre la lengua azul
No habrá moratoria ni plan de acción para la adaptación a la
normativa de bienestar
En el marco de una jornada técnica or-ganizada por la Asociación de veteri-narios de Porcino de Aragón el pasado
mes de abril en Zaragoza, Juan Antonio Agüero Monedero, veterinario del Minis-terio de Agricultura, Alimentación y Medio Ambiente, fue contundente respecto el cumplimiento de la normativa europea de bienestar animal. En su ponencia, dejó cla-ro que si bien la intención de las autorida-des estatales y autonómicas es facilitar al máximo la adaptación de explotaciones de ganado porcino a las condiciones estipula-das, se va a ser estricto a la hora de com-probar la idoneidad de las instalaciones, y en ningún caso se contempla la posibilidad de solicitar una prórroga a la Unión Euro-pea o establecer un plan de acción (como en el caso de las explotaciones de gallinas ponedoras, que pueden desviar los produc-tos procedentes de naves no adaptadas a canales de distribución alternativos). Así que, para aquellas granjas que no cumplan los requisitos el día 1 de enero, la única sa-lida es el cese de su actividad.
Las estimaciones actuales hablan de que en España hay más de un 53% de las explotaciones que cumplen con la nor-mativa europea de bienestar animal, y se prevé que el 100% lo hagan el 1 de enero de 2013. Según Agüero, hay un 37% de granjas que tienen que adaptarse, mien-tras que es probable que el 10% restante, de pequeño tamaño, acabe por cerrar.
Según el informe publicado por la in-terprofesional porcina del Reino Unido (BPEX), la entrada en vigor de la nueva ley de bienestar el próximo 1 de enero de 2013, que obligará a alojar a las cer-das gestantes en grupo, puede conllevar una reducción de la producción porcina europea de entre el 5 al 10% así como un aumento sustancial de los precios para procesadores y minoristas. Algunos Estados miembros han indicado que es-peran que un “número signifi cativo” de productores porcinos dejen este tipo de producción ya que no pueden o no quie-ren cumplir con la nueva legislación.
CYS42 noticias/AE/agenda.indd 9 14/05/12 10:14

10
Actualidad de empresa
MMerial Laboratorios convoca a los ve-terinarios de las principales distribui-doras de animales de producción, con
el objetivo de llevar a cabo una actualización técnica de los productos principales, noveda-des y herramientas de marketing.
Con este fin, Merial ha inaugurado un encuentro anual a través del cual potenciará el trabajo en equipo y el intercambio de ex-periencias entre las distintas zonas de España.
Córdoba y Burgos, puntos de encuentro
Para estrenar este tipo de encuentros, Me-rial Laboratorios ha organizado dos jornadas en el territorio nacional. El pasado 27 y 28 de marzo, Córdoba reunió a un grupo de veteri-narios comerciales en torno a los productos más destacados de las ramas de porcino y de rumiantes. A ella asistieron veterinarios de las distribuidoras de zona centro-sur.
El segundo evento se realizará los días 3 y 4 de mayo en Burgos, donde se abordaron las novedades más destacadas de las tres
ramas de animales de producción y a la que asistirán los contactos de las distribuidoras del resto de España.
Merial Laboratorios afianza su compromiso con las principales distribuidoras
Boehringer Ingelheim lanza un nuevo enva-se de Ingelvac CircoFLEX (100 ml), la única vacuna autorizada para su uso combinado
con Ingelvac MycoFLEX para proteger frente a la circovirosis y neumonía enzooótica en 1 inyección de 2 ml. Su adyuvante ImpranFLEX, de naturaleza totalmente acuosa, asegura un rápido inicio de in-munidad y una protección completa durante todo el ciclo productivo incluso en cerdos sacrificados a los 9 meses*.
Además de eficaz, su elevado perfil de seguridad asegura la protección sin comprometer el arran-que de los lechones durante el destete. Ingelvac CircoFLEX, disponible en España desde 2009, ha demostrado su eficacia bajo distintos sistemas de explotación y formas de enfermedad, y se ha con-vertido en la vacuna de elección global con más de 500 millones de cerdos vacunados en todo el mundo.
* Terreni et al. (2009)
Ingelvac CircoFLEX, ahora disponible en 100 ml
Grupo Copese 40 años El el sector cárnico porcino
Cerca de 40 años de actividad en el mercado del sector cár-nico del porcino avalan la experiencia de Grupo Copese. La empresa, constituida en 1976, como negocio dedicado a la
fabricación y comercialización de piensos y complementos alimen-ticios para el ganado porcino, con sede en Segovia, es hoy una en-tidad consolidada en la producción y transformación de productos del cerdo blanco Duroc e ibérico de cebo, que comercializa bajo su marca “ERESMA”, referente de calidad, con especial presencia en Castilla y León. Con un incremento de su facturación de un 8,4 %, con respecto a la cifra de 2010, Grupo Copese alcanzó los 35,85 millones de euros en 2011.
Sin duda uno de los factores que determinan la personalidad de Grupo Copese es su Proceso Integrado de Producción (P.I.P.), en el que se integran todas las fases de la cadena de producción, desde la granja hasta el punto de venta, incluida la fabricación –en su propia instalación- de piensos y correctores con los que alimentan a su ca-baña porcina.
Esta empresa de carácter familiar ha apostado desde sus oríge-nes por destinar una parte importante de su inversión a investigación y desarrollo, que la han situado en una posición de liderazgo en la provincia de Segovia, y entre las primeras de Castilla y León. Son
numerosos los proyectos desarrollados en el ámbito de la Selección Porcina por Grupo Copese, que cuenta con un departamento propio dedicado a esta labor que realiza con el know-how que le proporciona su participación como socio de GENE+ (organización especializada en Ingeniería porcina para la producción de cerdos blancos e ibéricos de calidad diferenciada).
CYS42 noticias/AE/agenda.indd 10 21/05/12 10:23

11
nº 42
El viernes día 27 de abril tuvo lugar la entrega del PREMIO SYvA 2012. Este galardón, que alcanza ya su 15ª edición, premia cada año a la mejor tesis doctoral en sanidad animal y tiene una do-
tación económica de 15.000 euros.
El tribunal del premio SYvA 2012 ha decidido que la tesis ganadora de esta edición sea la presentada por el Dr. Álvaro Hidalgo Uña, titulada
“Brachyspira hyodysenteriae: sensibilidad antibiótica y epidemio-logía molecular- Brachyspira spp. en perros.”
EL ACTO DE ENTREGA DEL PREMIO tuvo lugar en el Paraninfo de la Facultad de veterinaria. El evento tuvo en esta edición una relevancia espe-cial, ya que intervinieron Carmen vela Olmo, Secretaria de Estado de I+D+I del Ministerio de Economía y Competitividad y Jorge Blanco Alvarez, Direc-tor del LREC (Laboratorio de Referencia de E.coli) de la Universidad de San-tiago de Compostela, que pronunció una magnífica conferencia titulada: “Emergencia de clones de E. coli altamente virulentos y multirresistentes”.
La primera intervención fue la de Luis Bascuñan Herrera, Director General de SYvA, que comenzó hablando de la importante difusión y el reconocido prestigio alcanzado por este galardón que alcanza ya su 15ª edición. Destacó asimismo la elevada calidad científica de las 25 tesis presentadas al PREMIO SYvA en esta convocatoria y alabó especialmente la calidad del trabajo premiado y el extraordinario cu-rriculum del Dr. Hidalgo Uña.
Finalmente explicó que se había elegido en esta ocasión como sede la Facultad de veterinaria, por la relación indisoluble del PRE-MIO SYvA y la Sanidad Animal.
A continuación, Luisa de Arriba Martín, Secretaria del Tribunal, procedió a la lectura del acta de la reunión del Tribunal calificador para dar paso seguidamente a la intervención de Alvaro Hidalgo Uña, autor de la Tesis Premiada, que agradeció la concesión del PREMIO a Laboratorios SYvA y a la Universidad de León.
Carmen vela Olmo, intervino después para agradecer a Labo-ratorios SYvA y la Universidad de León la oportunidad de volver a tener contacto con el mundo de la sanidad animal, en el que ha trabajado a lo largo de su trayectoria profesional en la empresa privada. Habló sobre la reducción del presupuesto en su Secretaría de estado y planteó que la excelencia será el criterio que prime a la hora de asignar esos recursos, abogando además por la estrecha colaboración entre equipos de investigación, Administración y em-presa privada.
Jorge Blanco Alvarez, pronunció una brillante conferencia sobre la emergencia de nuevas cepas de E. coli altamente virulentas y multirresistentes , como la que causó hace un año la famosa “crisis de los pe-pinos” en Alemania. Destacó la importancia de seguir investigando en este campo desde el punto de vista de sanidad animal y de salud humana y hacerlo de manera conjunta y coordinada con los diferentes ex-pertos internacionales.
Cerró el acto José Angel Hermida Alonso, Rector de la Universidad de León, que calificó el Premio SYvA como el más importante en sanidad animal en nuestro país y destacó la colaboración entre SYvA y la Universidad de León durante to-dos estos años plasmada, no sólo en el PREMIO SYvA, sino también en los proyectos de investiga-ción conjuntos o en la contratación de personal por parte de SYvA entre los licenciados y doctores de la Universidad de León.
La entrega del PREMIO contó con una amplia cobertura de los medios de comunicación, tanto generales como especializados (pe-riódicos, revistas, televisiones y radios).
RESuMEN DE LA TESIS GANADORA
Dr. Álvaro Hidalgo Uña
“Brachyspira hyodysenteriae: sensibilidad antibiótica y epidemio-logía molecular- Brachyspira spp. en perros.”
Las espiroquetas del género Brachyspira son bacterias anaerobias que colonizan el intestino de aves y mamíferos. Una de las especies más relevantes de este género es Brachyspira hyodysenteriae, agente etiológico de la disentería porcina. Esta enfermedad afecta especial-mente a los cerdos de cebo y se caracteriza, en su forma clínica más clásica, por la presencia de diarrea sanguinolenta. La disentería origina elevados costes económicos en las explotaciones porcinas, derivados sobre todo del aumento del índice de conversión, de la disminución de la ganancia media diaria y del incremento en gastos de medicación.
El tratamiento y el control de la disentería porcina se basan prin-
cipalmente en la utilización de antibióticos. De todos ellos, los que más se han empleado en los últimos años son la tilosina, la tiamulina, la valnemulina y la lincomicina. Recientemente, se ha aprobado el uso de la tilvalosina en España para tratar esta enfermedad.
Esta tesis doctoral, basada en cinco publicaciones científicas, describe la sensibilidad de aislados españoles de B. hyodysenteriae a estos antibióticos durante la última década e investiga la base gené-tica de sus resistencias. Asimismo, profundiza en la caracterización fenotípica y molecular de B. hyodysenteriae y en las posibles relacio-nes epidemiológicas existentes entre distintos clones, con especial referencia a los aislados españoles. Además, se estudió la presen-cia de espiroquetas del género Brachyspira en perros, tanto por su potencial patógeno en esta especie como por su posible papel de reservorio epidemiológico.
Una tesis doctoral sobre disentería porcina ganadora de la XV edición del PREMIO SYVA
CYS42 noticias/AE/agenda.indd 11 14/05/12 10:14

12
Actualidad de empresa
Pfi zer Salud Animal, a través de vet Sup-port, programa de Servicios de valor Añadido, ha fi rmado un acuerdo con
la prestigiosa escuela de idiomas vaughan System, para ofrecer a sus clientes de por-cino la oportunidad de asistir a cursos resi-denciales con los que poder mejorar su nivel de inglés de una manera práctica, entrete-nida y rápida.
El método vaughan Town se caracte-riza por su dinamismo y rápido aprendiza-je, consiguiendo mejorar el nivel de inglés profesional y de negocios en un entorno exclusivo y con atención personalizada. Un total de 30 veterinarios especializados en porcino han asistido a tres cursos intensi-vos, desarrollados durante el mes de mayo en diferentes centros vaughan de nuestro país, lo que les ha permitido interactuar con otros profesionales en inglés adquiriendo la fl uidez necesaria para su actividad profesio-nal diaria.
Se trata de un programa de inmersión lingüística en inglés testado y reconocido a
nivel internacional en el que, durante 5 días, y sin salir de España, conviven participan-tes españoles con nativos angloparlantes procedentes de todas partes del mundo. Comprende actividades como las conver-saciones one-to-one (sesión individual de conversación con un angloparlante), sesio-nes telefónicas y conferencias simuladas, la preparación de presentaciones y conversa-ciones en grupo, todo ello con el objetivo de ganar confi anza y comunicarse con decisión en inglés, aumentando además la compren-sión auditiva.
vet Support es la plataforma líder de servicios de valor añadido de Pfi zer Salud Animal que ofrece formación teórico prác-tica para veterinarios, además de la posibi-lidad de realizar estancias con especialistas acreditados. vet Support pone igualmente a disposición de sus clientes servicios ex-clusivos como los Jornadas de Formación Avanzada y los Servicios de Consultoría y Atención personalizada.
Vet Support colabora con Vaughan System en el diseño de cursos de inglés específicos para veterinarios de porcino
La presentación de STARTvAC reunió 200 veterinarios expertos en calidad de la leche en Izmir, Turquía.
El Dr. Ynte Schukken (Universidad de Cornell, EEUU), el Dr. valerio Bronzo (Uni-versidad de Milano, Italia), y el Dr. Bastan (Universidad de Ankara, Turquía) fueron invitados por HIPRA para realizar la se-
sión de apertura. Entre los temas trata-dos por los ponentes destacan el manejo de la salud del rebaño, la importancia del biofilm bacteriano y el coste de la mas-titis bovina.
HIPRA refuerza de esta manera, su compromiso con la Prevención en la Salud Animal a nivel internacional.
HIPRA lanza la 1ª vacuna HIPRA lanza la 1ª vacuna HIPRA lanza la 1ª vacuna europea contra la mastitis europea contra la mastitis europea contra la mastitis bovina en turquía bovina en turquía bovina en turquía
En el próximo mes de Junio, HIPRA participará en el 27º Congreso Mundial de Buiatría, que se realiza-
rá en Lisboa, Portugal, del 3 al 8 de Junio de 2012.
Como Premium sponsor HIPRA dis-pondrá de un stand de 100 m2 donde se realizaran diferentes actividades relacio-nales.
Durante el Congreso, HIPRA también patrocinará un exclusivo workshop de-dicado al manejo en la sala de ordeño que contará con la participación del Dr. Ynte Schukken de la Universidad de Cornell, EEUU.
Con este tipo de actividades, HIPRA refuerza su compromiso con la Preven-ción en la Salud Animal a nivel interna-cional.
Hipra Premium sponsor en el Congreso Mundial de Buiatria’12 en Lisboa
ElXVIICongresodeANEMBEsehacelebrado,durantelosdías18,19 y 20 de abril, en la ciudad de Santander.
Laboratorios SyVA ha participado, como empresa patrocinadora, en el XVII Congreso de la Asociación Nacional de Especialistas en MedicinaBovinadeEspaña,quehareunidoaungrannúmerodeprofesionales del sector en el Palacio de Congresos y Exposiciones de Santander.
Enestecongresosehanpresentadotrabajosclínicosydeinves-tigación aplicada, tanto por parte de los socios como de los diversos organismos(Universidades,OPIs,etc.)quetrabajanenelámbitodelabuiatría.Elprogramaseestructuróencincograndesbloquesgenera-les:Sanidad,Reproducción,Nutrición,BienestaryMercados.
Paralelamente al Congreso, se realizó una exposición comercial ycientíficadondeseexpusieronlosúltimosproductosfarmacéu-ticosyen laqueLaboratorioSYVAparticipómedianteunstandcomercialenelquerecibiónumerosasvisitasyenelquepromo-cionó, entre otros, los siguientes productos para ganado bovino: ACTIONIS,ceftiofurquepresentalamayorfacilidaddeaplicaciónlo que permite la inyección demás animales enmenos tiempo,PARAMECTIN, ivermectina inyectable y GONAySL-LuTEOSyL, el tratamiento hormonal completo para ganado vacuno.
Para más informaciónLaboratorios SyVA
www.syva.es
Syva participa en el XVII Congreso Internacional ANEMBE de Medicina BovinaSyva participa en el XVII Congreso Internacional ANEMBE de Medicina BovinaSyva participa en el XVII Congreso Internacional ANEMBE de Medicina Bovina
CYS42 noticias/AE/agenda.indd 12 14/05/12 10:14

13
nº 42
Pfizer Salud Animal, en colaboración con Elasa, su distribui-dor en la zona, organizó el pasado 22 de marzo en Sego-via, una Jornada titulada “Futuro del Ovino” que reunió a
más de 200 productores y veterinarios y en el que se trataron temas de gran importancia para el sector y la perspectiva del mismo ante la nueva reforma de la PAC.
Las enfermedades parasitarias en pequeños rumiantes centraron gran parte de la jornada, en la que Emilio Legaz, gerente de Marketing de Rumiantes de Pfizer Salud Animal, habló de su importancia y expuso el programa de control destinado a evitar su problemática; y por su parte, Manuela Jimenez, Feed Industrial Manager de Pfizer Salud Animal, se centró en la coccidiosis que afecta en la recría de corderas, y que puede ocasionar grandes repercusiones económicas en la
rentabilidad de las granjas. También intervino Antonio Torres, ingeniero Agrónomo y director Técnico de APAG (Asociación Provincial de Agricultores y Ganaderos de Guadalajara), quien trató la situación actual de la PAC (Política Agraria Común) y el futuro de las ayudas en el horizonte de 2020.
Más de 200 ganaderos y veterinarios acuden a la Jornada Pfizer sobre el futuro del ovino
FICHA TECNICA PORCILIS PRRS
Vacuna viva atenuada contra la infección con virus PRRS.
Liofilizado y disolvente para suspensión inyectable. Vía intramuscular o intradérmica.
COMPOSICIÓN POR DOSIS (2 ml (administración intramuscular) o 0,2 ml (administración intradérmica) de vacuna reconstituida): Fracción liofilizada: Sustancia activa: Virus PRRS cepa DV vivo atenuado: 104,0 – 106,3 TCID50*. Fracción disolvente: Adyuvante: acetato de dl-α-tocoferilo: 75 mg/ml. *Dosis infectiva de cultivo tisular 50%.
INDICACIONES Y ESPECIES DE DESTINO: Porcino: Inmunización activa de cerdos clínica-mente sanos en un ambiente contaminado con virus de PRRS para reducir la viremia causada por la infección con cepas europeas del virus PRRS.
CONTRAINDICACIONES: No usar en explotaciones donde la prevalencia de virus de PRRS europeo no haya sido establecida mediante métodos de diagnóstico fiables.
PRECAUCIONES: Porcilis PRRS debe utilizarse solamente en explotaciones contaminadas con virus de PRRS, donde se haya establecido la prevalencia de virus de PRRS europeo mediante mé-todos de diagnóstico virológico fiables. No se dispone de datos sobre la seguridad de la vacuna para el rendimiento reproductivo en verracos. No utilizar en explotaciones en las que se haya adoptado un programa de erradicación de PRRS basado en la serología. Deben tomarse precau-ciones para evitar la introducción de la cepa vacunal en un área en la que no esté presente el virus de PRRS. El virus vacunal puede transmitirse a cerdos en contacto durante 5 semanas después de la vacunación. La vía de transmisión más común es el contacto directo, pero no puede excluirse la transmisión a través de objetos contaminados o a través del aire. Deben tomarse precauciones para evitar la transmisión del virus vacunal de animales vacunados a animales no vacunados (es decir, cerdas gestantes sin inmunidad) que deben permanecer libres de virus de PRRS. No utilizar en verracos donantes de semen para explotaciones seronegativas, puesto que el virus de PRRS puede ser excretado en el semen durante muchas semanas. En caso de autoinyección accidental, consulte con un médico inmediatamente y muéstrele el prospecto o la etiqueta.
Las cerdas adultas y nulíparas sin inmunidad frente a virus de PRRS no deben ser vacunadas durante la gestación, puesto que esto puede tener efectos negativos. La vacunación durante la gestación es segura cuando se lleva a cabo en cerdas adultas y nulíparas ya inmunizadas frente al virus de PRRS europeo mediante la vacunación o por infección de campo. La vacuna puede ser utilizada durante la lactancia.
Utilizar jeringas y agujas estériles o un equipo intradérmico limpio.
Existe información disponible sobre la seguridad y la eficacia de la inyección intramuscular en cerdos de cebo a partir de 4 semanas, que demuestran que esta vacuna puede mezclarse con Porcilis M Hyo. Se debe consultar también la información sobre el medicamento Porcilis M Hyo antes de la administración de los medicamentos mezclados. No existe información disponible so-bre la seguridad y la eficacia del uso de esta vacuna con cualquier otro medicamento veterinario excepto el medicamento mencionado anteriormente. La decisión sobre el uso de esta vacuna antes o después de la administración de cualquier otro medicamento veterinario se deberá realizar caso por caso. No existe información disponible sobre la seguridad y la eficacia del uso de Porcilis PRRS mezclado con Porcilis M Hyo en animales destinados a la cría o durante la gestación. No mezclar con ningún otro medicamento veterinario, excepto el disolvente suministrado con el medicamento o con Porcilis M Hyo.
Precauciones especiales de conservación: Fracción liofilizada o caja combinada: con-servar en nevera (entre 2ºC y 8ºC). Proteger de la luz. Fracción disolvente: conservar a temperatura inferior a 25ºC.
TIEMPO DE ESPERA: Cero días.
Uso Veterinario - medicamento sujeto a prescripción veterinaria.
Instrucciones completas en el prospecto.
Mantener fuera del alcance y la vista de los niños.
Formatos: Formatos para administración por vía intradérmica: Caja con 5 viales de 100 dosis de liofilizado y 5 viales de PET de 20 ml de disolvente. Formatos para administración por vía intramuscular: Caja con una caja de 10 viales de 25 dosis de liofilizado y 10 viales de PET de 50 ml de disolvente, caja con una caja de 10 viales de 50 dosis de liofilizado y 10 viales de PET de 100 ml de disolvente, caja con una caja de 10 viales de 100 dosis de liofilizado y 10 viales de PET de 200 ml de disolvente.
Reg Nº: 1361 ESPIntervet International B.V.Wim de Körverstraat 35
5831 AN Boxmeer, Países Bajos
El Patronato del Centre de Recerca en Sa-nitat Animal (CReSA), reunido el 17 de abril de 2012, ha nombrado al Dr. Joaquim
Segalés nuevo director del centro, e iniciará su mandato a partir del próximo 1 de mayo de 2012. Sustituye así al Dr. Mariano Domingo Álvarez, quien ha dirigido el CReSA desde su creación, hace ya casi 12 años.
El CReSA es una fundación pública creada por la Universitat Autònoma de Barcelona (UAB) y el Institut de Recerca i Tecnologia Agroalimen-tàries (IRTA) para la investigación en sanidad ani-mal, y es uno de los centros de investigación del sistema CERCA de la Generalitat de Catalunya. Actualmente, el centro cuenta con 118 trabaja-
dores altamente especializados de los cuales 38 son investigadores.
El Dr. Segalés es Profesor Titular del De-partament de Sanitat i d’Anatomia Animals de la Facultat de Veterinària de la UAB e inves-tigador adscrito al CReSA. Asimismo, durante los dos últimos años ha ocupado el cargo de responsable del subprograma de investigación del CReSA sobre enfermedades víricas endé-micas. Tras haberse doctorado en Ciencias Ve-terinarias por la UAB (1996), el Dr. Segalés se diplomó por el European College of Veterinary Pathologists (2000) y posteriormente por el European College of Porcine Health and Ma-nagement (2004).
Su amplia experiencia en investigación se ha centrado principalmente en el estudio de las enfermedades víricas porcinas, especialmente en infecciones por circovirus porcino tipo 2 (PCV2). El Dr. Segalés cuenta con más de 200 publicacio-nes científicas (SCI), más de 240 comunicaciones y 50 conferencias en congresos nacionales e in-ternacionales, ha dirigido 11 tesis doctorales y ha recibido 3 premios de investigación en sanidad animal. Además, imparte docencia en diferentes cursos y másteres, es revisor en múltiples revistas científicas y en proyectos nacionales, y miembro de diversos comités científicos.
Dr. Segalés: [email protected]
El Dr. Joaquim Segalés Coma, nuevo Director del CReSA
CYS42 noticias/AE/agenda.indd 13 21/05/12 10:24

14
Actualidad de empresa
Continúan las Jornadas de Nutrición Porcina Vet Support en La Coruña
Vet Support, programa de Servicios de Valor Añadido de Pfi zer Salud Animal, sigue adelante con su compromiso de formación para sus principales clientes, celebrando una nueva Jornada
de Nutrición Porcina que tuvo lugar en El Centro de Investigaciones Agrarias de Mabegondo (La Coruña), el día 18 de abril, y que contó con la asistencia de más de 30 veterinarios especialistas en porcino.
Impartida por Gonzalo Bruna, Josep Gasa, Antonio Palomo e Ig-nasi Riu, profesionales especialistas en nutrición porcina de recono-cido prestigio y con amplia experiencia, tiene por objetivo actualizar los conocimientos en una materia clave para la producción porcina moderna, favoreciendo la obtención de altos rendimientos y una me-jora en la rentabilidad económica de la granja de cerdos.
Con estas jornadas la compañía quiere aportar más que produc-tos y ofrecer soluciones a medida para los problemas que surgen en el día a día de la explotación
El pasado mes de abril Santander acogió la XVII Congreso Internacional
Anembe de Medicina Bovina. Bajo el lema “Abriendo cami-no al conocimiento”, el evento contó con el apoyo de Merial Laboratorios, en calidad de co-laborador y de Zactran como patrocinador.
30 años Ivomec®
Merial Laboratorios apro-vechó el marco del evento de referencia para los veterina-rios especializados en medi-cina bovina, para poner en escena el 30º aniversario de Ivomec® y la nueva campaña
de Zactran®, “Respira con-fianza”.
De este modo, Merial dio a conocer a los profesionales las campañas que tiene previsto desarrollar a lo largo del año 2012 para conmemorar los 30 años de Ivomec®, el producto que mayor impacto ha tenido en la historia de la sanidad animal.
Avalado por la tradición pero mirando de frente al fu-turo, Merial Laboratorios pone en marcha diversas acciones enfocadas a los estudiantes que se encuentran en la recta fi nal de sus licenciaturas. Así,
a lo largo del año, los futuros veterinarios podrán entrar a www.30aniversarioivomec.com y acceder a la colección de cuadernos de campo digi-talizados a través de códigos de descarga. Además, tendrán acceso a la versión digital del Atlas de patología parasitaria en rumiantes. Este compen-dio, que consta de más de 400 páginas, recoge los procesos parasitarios de mayor frecuen-cia e importancia patógena en el ganado vacuno, ovino y caprino. La iniciativa ha sido desarrollada por un equipo de docentes liderado por Magda-lena Garijo Toledo, profesora de Parasitología y Enfermeda-
des Parasitarias en la Facultad de Veterinaria de la Univer-sidad CEU-Cardenal Herrera (Valencia).
Estas iniciativas refl ejan el compromiso con la formación que siempre ha distinguido la comunidad Ivomec® y que ahora, en una fecha conme-morativa, da un paso adelante para presentarse ante aquellos que representan el futuro de la profesión.
Merial conmemora los 30 años de Ivomec® en el Congreso Anembe
El español Guillermo Romero nombrado vicepresidente de Mercados Emergentes en Pfizer Animal Health
Guillermo Romero Jiménez ha sido nom-brado Vicepresidente de Mercados Emer-gentes para Europa, África y Oriente
Medio (EuAf ME) de la multinacional de sanidad animal, Pfi zer Animal Health (PAH), un área es-tratégica para el crecimiento de la compañía que engloba 80 países de cuatro regiones (Rusia, Eu-ropa del Este y Asia Central; Turquía, Israel, Irán y Magreb; Egipto y Oriente Medio; y África SubSa-hariana) y cuenta con un potencial de mercado estimado para 2012 de 2.3 billones $US.
En palabras de Alejandro Bernal, pre-sidente del Área EuAf ME de PAH, “esta promoción refl eja los logros de Guiller-mo Romero, no solo por haber liderado el crecimiento de Pfi zer Animal Health en los mercados emergentes, sino también por el talento, conocimiento del mercado y capa-cidad de liderazgo demostrado en su ex-
tensa carrera dentro de la compañía”.Guillermo Romero se incorporó a la
compañía en 1992, siendo nombrado cua-tro años más tarde Director de la División de PAH en España y Portugal, cargo que ocupó hasta el año 2006, cuando pasó a ser Director General de Pfi zer Salud Animal en España.
Tras la adquisición de Fort Dodge por parte de PAH en 2009, Romero fue pro-mocionado como Director Senior de Mer-cados Emergentes, trasladando su resi-dencia a París, ciudad en la que se sitúan las oficinas centrales de Pfizer Animal Health- EuAfME.
En estos años PAH ha pasado de tener un 7% de la cuota de mercado (año 2010) en estos países, a contar en la actualidad
con un 11%, encabezando el ranking de empresas de sanidad animal en esta área.
Biológicos de Pfizer
Pfizer cuenta con una selección de productos biológicospara el ganado ovinodestinados a la prevención de enfermedadescuyo impacto económicoes significativo.
la protección SÍ importa
¿Te vas a arriesgar?
www.pfizer-salud-animal.es
Bedsa-Vac
Gangrevac
Capri-Vac
Covexin-10
Covexin-8
PFIZER_Ad_Biologicos_Ovino_A4-Abr2012-1.indd 1 07/05/12 10:48CYS42 noticias/AE/agenda.indd 14 21/05/12 10:24

15
nº 42nº 42
Biológicos de Pfizer
Pfizer cuenta con una selección de productos biológicospara el ganado ovinodestinados a la prevención de enfermedadescuyo impacto económicoes significativo.
la protección SÍ importa
¿Te vas a arriesgar?
www.pfizer-salud-animal.es
Bedsa-Vac
Gangrevac
Capri-Vac
Covexin-10
Covexin-8
PFIZER_Ad_Biologicos_Ovino_A4-Abr2012-1.indd 1 07/05/12 10:48CYS42 noticias/AE/agenda.indd 15 21/05/12 10:24

16
Pfizer Salud Animal reunió el 4 de mayo en Salamanca a más de 70 especialistas del sector de ovino de leche procedentes de Italia y España, con el fin de intercambiar experiencias profesionales y compartir conocimientos para
aportar soluciones a los problemas que surgen en el día a día de cada explotación.
Emilio Legaz, gerente de Marketing de la Unidad de Rumiantes de Pfizer Salud Animal, fue el encargado de dar la bienvenida a los asistentes y recalcó la importancia de la jornada para conocer la perspectiva del sector desde el punto de vista de ambos países y poder intercambiar experiencias al respecto.
Para comenzar, Luis Fernando de la Fuente (Departamento de Producción Animal de La Universidad de León) expuso la situación de la producción láctea de ovino en España. “Durante los últimos años nos encontramos en una regresión en el censo ovino debida principalmente a la desaparición de explotaciones por el no relevo generacional y la falta de rentabilidad motivada por el encareci-miento de los piensos, por la subida del precio de los cereales, y el estancamiento de los precios de la leche y la carne”, indicó.
A continuación Gian Lorenzo d’Alterio (veterinario clínico de la región italiana de Lazio) transmitió su experiencia personal como veterinario en las zonas de Lazio y Toscana. Tras presentar la raza ovina italiana, la Sarda - “una raza rústica con aptitud de alta producción láctea y de fuerte dependencia al pastoreo” - , habló de la producción típica en Italia- “el queso Pecorino Romano” - , el mercado de la leche de ovino y las características de la produc-ción ovina en Italia, donde a diferencia con España, predomina la producción extensiva.
Giuseppe Argiolas (veterinario clínico de la región italiana de Cerdeña) centró su ponencia en la gestión de la reproducción con el objetivo de maximizar la rentabilidad de la explotación. Propuso un plan de actuación comenzando con un análisis de la situación de la granja para a continuación gestionar la fertilidad, valorando los animales vacíos o retrasados, que serán los que mayores pérdi-das ocasionarán en la rentabilidad económica.
En la misma línea continuó Ángel Ruiz Mantecón (CSIC – Consejo Superior de Investigaciones Científicas – de León) quien planteó dife-rentes matices de los sistemas de producción ovina de leche en España. Habló de la evolución del sector, indicando el cambio racial que ha tenido lugar con el paso de los años, pasando de la raza Churra y Castellana a predominar la Assaf y Lacaune, y el cambio en la intensificación y el desarrollo tecnológico, predominando las explotaciones intensivas de mayor tamaño y mayor producción láctea. Además recalcó la impor-tancia de realizar una adecuada gestión de los gastos de alimentación, especialmente en épocas de crisis, o cuando los precios de las materias primas son elevados, evitando compensar esta situación, como así ocu-rre, con una reducción de los gastos zoosanitarios que puede derivar en problemas productivos a medio y largo plazo.
De izqda a dcha.- Giuseppe Argiolas, Gian Lorenzo d’Alterio, Ángel Ruiz, Luis Fernando de la Fuente, y Jesús Matilla.
La jornada contó con la asistencia de más de 70 especialistas.
Pfizer promueve el intercambio de experiencias profesionales entre Italia y España en Ovino de Leche
rep pfizer.indd 16 21/05/12 10:23

17
nº 42
Jesus Matilla (OCEVA Veterinarios) expu-so diferentes factores a tener en cuenta para rentabilizar las explotaciones. La importancia de ligar la explotación a la tierra, la capacita-ción técnica y cultural de los ganaderos, una alimentación sana y equilibrada, el manejo y organización de la reproducción, la importan-cia de unas buenas instalaciones, y la gestión de residuos, fueron, entre otros, algunos de los factores en los que se centró Matilla.
Antonio Scala (Departamento de Parasitología y Enfermedades Parasitarias de la Universidad de Sassari) habló de la iden-tificación, control y perspectiva futura de los problemas parasitológicos de la oveja de leche en Italia. Concretamente se centró en la región de Cerdeña donde se crían más de 3’5 millones de ovejas de raza Sarda, una raza muy dependiente del pastoreo, siendo éste el principal factor de riesgo para las parasitosis.
Finalmente, Giuseppe Cringoli (Dep. de Parasitología y Enfermedades Parasitarias de la Universidad de Nápoles) mostró las nuevas estrategias en el diagnóstico parasitario en ovino y caprino, en concreto un innovador pro-ducto, Flotac, desarrollado por la Universidad de Nápoles para promover el correcto diag-nóstico parasitológico, como herramienta necesaria para el uso racional y eficaz de los antihelmínticos.
Emilio Legaz.
Giuseppe Argiolas. Ángel Ruiz.
Jesús Matilla. Antonio Scala. Giuseppe Cringoli.
Luis Fernando de la Fuente. Gian Lorenzo d’Alterio.
La reducción de gastos zoosanitarios puede deivar en problemas productivos a medio y largo plazo
“”
rep pfizer.indd 17 21/05/12 10:23

18
Jefo, división Europa/África especialista de la nutrición animal, y Beldem, filial del grupo Puratos, especialista en producción de ingredientes para la alimentación animal y humana, nos invitaron a celebrar en madrid los 10 años que
llevan colaborando. En la jornada celebrada el pasado 8 de marzo presentaron distintos trabajos de investigación sobre Belfeed, la única xilanasa de origen bacteriano autorizada en el mercado europeo en lechones, cerdos de cebo, pollos, pavos, ponedoras y patos. Hubo espacio también para el intercambio de impresiones en la comida, la actividad, y la cena posterior, que sirvieron de colofón a un interesante día.
inauguraron las jornadas Sabrina Vandeplas, Jefe de pro-ducto de Enzimas de Beldem, y Sergio merinero, responsable Comercial de Jefo en nuestro país. Tras recordar cómo surgió la colaboración entre las dos empresas en el año 1999, repasaron la cronología del desarrollo de Belfeed, la primera xilanasa bacteriana autorizada en Europa. Belfeed se lanzó en Francia en el año 2000 para su uso en pollos, posteriormente esta autori-
zación se fue ampliando -lechones, cerdos de cebo, pavos, etc.- hasta que en el año 2009 se obtuvo la autorización permanente de la Unión Europea para su uso en todas las especies. También explicaron cómo paralelamente diferentes estudios validaron su eficacia en piensos en base a maíz, DDGS (tanto de trigo como de maíz), mandioca y sorgo.
Después intervino Jérôme Fouquet, Director de Jefo, para expo-ner el programa de la jornada, pero antes, teniendo en cuenta la fecha, felicitó a las mujeres que se encontraban en la sala.
El Dr. Joaquin Brufau (irTA) inició el ciclo de conferencias explicando el concepto de xilanasa y cuáles son sus ventajas a nivel de resultados zootécnicos. Esta enzima aumenta la diges-tibilidad de los NSP, sobre todo en piensos en base a maíz y trigo, y en aves consigue optimizar los principales parámetros productivos -la ganancia media diaria, el consumo medio diario, el índice de conversión, el % de puesta- además de aumentar el peso del huevo y mejorar su digestibilidad. Asimismo aporta otros efectos adicionales como la mejora del ratio agua/pienso, el incremento en la materia seca de la excreta, menos heces adheridas a la cloaca o la disminución del peso de las vísceras en los pollos que llegan al matadero. Analizar su efecto en cerdos es algo más complicado, pero la disminución de la vis-cosidad aumenta la disponibilidad de los aminoácidos a nivel intestinal, lo que tiene una gran importancia en el crecimiento. Para finalizar su intervención expuso cómo las xilanasas pue-den jugar un papel estratégico en la prevención del síndrome Coccidiosis-Enteritis Necrótica.
Después, Sergio merinero y Anna Fernández oller (Jefe de producto Avicultura) presentaron las especificidades de la enzima de Beldem. Belfeed tiene una actividad del tipo de las endo-xilanasas sobre la viscosidad, una actividad importante sobre los arabinoxilanos solubles, pero sobre todo sobre los ara-binoxilanos insolubles. Gracias a esta propiedad Belfeed ofrece resultados estables en numerosos perfiles de materias primas incluyendo los piensos a base de maíz ricos en arabinoxilanos insolubles. Además, gracias a su origen bacteriano, Belfeed tiene una sensibilidad reducida a los inhibidores de xilanasas (TAxi y
Jérôme Fouquet, Director de Jefo, exponiendo el programa de la jornada.
Sabrina Vandeplas, Jefe de producto de Enzimas de Beldem.
Jefo y Beldem celebran 10 años de colaboración entorno a Belfeed
CYS 42.indb 18 14/05/12 10:10

19
nº 42
xiP). otras características destacables son su resistencia al tra-tamiento térmico, su afinidad por un pH neutro y la estabilidad en piensos y premix.
Tras una pausa, la Dra. Anna maría Pérez Vendrell (irTA) subrayó el impacto de los inhibidores de xilanasas sobre los cereales y los piensos para monogástricos. La presencia de inhi-bidores de xilanasa varía entre cereales, y no está relacionada con el contenido en arabinoxilanos. Extractos de trigo, centeno o cebada presentan actividades inhibidoras contra xilanasas de diferentes microorganismos, especialmente hongos; sin embar-go la xilanasa de Bacillus (Belfeed) ha sido muy poco inhibida. Los inhibidores de xilanasas también se encuentran en subpro-
ductos de cereales y son más estables que los propios enzimas xilanasas en la fabricación de piensos. Para apoyar estos datos mostró los resultados de distintos ensayos in vivo en los que se valoraba la actividad de tres enzimas diferentes sobre dietas de trigo tratadas (pasadas por autoclave) y sin tratar. Si se traba-jaba con el trigo sin tratar, es decir con inhibidores presentes, los mejores resultados se obtenían con Bacillus. En el caso de la cebada estos ensayos pusieron de manifiesto una interacción clara entre el tipo de cebada y la enzima; la energía metaboliza-ble había aumentado más en aquella variedad con un contenido de inhibidores menor. Del mismo modo, cuando trabajamos con variedades de avena o centeno donde los inhibidores no están presentes, o su presencia es pequeña, el incremento del peso vivo, la GmD y el iC siempre mejoran.
Dr. Joaquin Brufau (IRTA).
Anna Fernández Oller, Jefe de producto Avicultura de Jefo.
Dra. Anna María Pérez Vendrell (IRTA).
Belfeed es la única xilanasa de origen bacteriano cuyo uso está autorizado en Europa. La dosis recomendada es de 100 ppm para todas las especies
“ˮJosé Luis Fernández (Delegado Extremadura y Andalucía Jefo.
Jefo y Beldem celebran 10 años de colaboración entorno a Belfeed
CYS 42.indb 19 14/05/12 10:10

20
Sabrina Vandeplas volvió a intervenir para resumir la acción de Belfeed, xilanasa producida por Bacillus subtilis, sobre la flora intestinal. Por su efecto Belfeed permite reducir los problemas de digestión en producción, mejorar el estatus sanitario de los animales y actuar en contra de la disminución de rendimientos asociados a los problemas de digestión. Los polisacáridos no amiláceos no digestibles presentes en las raciones disminuyen la cantidad de sustrato disponible en el intestino delgado y aumentan la viscosidad del contenido intes-tinal. Esto provoca que la velocidad de tránsito se reduzca y de
Sergio Merinero, Responsable Comercial de Jefo en España.
Jonathan Rastel (Jefe de producto porcino Jefo).
Jean-Christophe Bodin (Director técnico Jefo).
Todas las xilanasas no producen el mismo tipo de AXOS. Belfeed, a diferencia de las xilanasas fúgicas, no produce xilosa que origina diarreas osmóticas
“ˮ
Belfeed permite reducir los problemas de digestión en producción, mejorar el estatus sanitario y evitar la disminución de rendimientos asociados a los problemas de digestión
“ˮ
jefo.com
BELFEED®
LA ENZIMA INCOMPARABLE
Aditivos para cada especie
JEFE DE PRODUCTO PORCINO : Sergio MERINERO I [email protected] I 616 47 34 69
JEFE DE PRODUCTO AVICULTURA : Anna FERNÁNDEZ I [email protected] I 618 60 21 43
La única xilanasa de origen bacteriano autorizada en el mercado europeo en pollos, pavos, ponedoras, patos, lechones y cerdos de cebo.
Alta eficacia en todo tipo de dietas : trigo, cebada, maíz...
BELFEED también tiene una sensibilidad reducida a los inhibidores de xilanasa (TAXI), lo que permite una homogeneidad de respuesta zootécnica durante todo el año (en cereales de “nueva cosecha” o almacenados, de campañas anteriores).Está disponible bajo las formas polvo y líquido y tiene una dosis de uso única.No duden en ponerse en contacto con nuestros jefes de producto para tener más información sobre el producto.
BELFEED.indd 1 09/05/12 09:15
más tiempo a que proliferen ciertas bacterias: Bifidobacterium y Lactobacillus. Esta última disminuye la digestibilidad de las materias grasas. Además el aumento de viscosidad favorece a las bacterias mucolíticas (Clostridium perfringes en aves y Helicobacter pylori en cerdos). El efecto de las xilanasas puede ser asociado a cambios en la cantidad y composición de la flora microbiana intestinal. Los arabinoxilo-olgosacáridos (AxoS)
producidos por la actividad xilanásica tienen un potencial pre-biótico, lo que significa que no son digeridos por los enzimas del animal, e implica que pueden ser potencialmente fermentados de manera específica por bacterias endógenas beneficiosas (bacterias sacarolíticas que estimulan la inmunidad, producen vitaminas y tienen actividad antibacteriana) que compiten con las patógenas, lo que resulta positivo para el animal. Sabrina Vandeplas destacó además como Belfeed, a diferencia de otras xilanasas fúngicas, no produce xilosas (un tipo de AxoS) origen de diarreas osmóticas.
Las dos últimas intervenciones estuvieron dedicadas a la Valoración de Belfeed. José Luis Fernández (Delegado Extremadura y Andalucía Jefo), tomando como ejemplo un perfil típico usado en cerdo ibérico, demostró con el nuevo software de formulación Jefo que la valoración de Belfeed permite optimizar las fórmulas de manera tangible y, como consecuencia, ahorrar sobre el coste de la fórmula.
Para terminar, Jean-Christophe Bodin (Director técnico Jefo) y Jonathan rastel (Jefe de producto porcino Jefo) nos instruyeron acerca de un concepto innovador desarrollado por Jefo y Beldem para tener más precisión en sus recomen-daciones. Este sistema está basado en una recopilación de casi 40 pruebas zootécnicas por especie, de proporciones de Arabinoxilanos (solubles e insolubles) en las materias primas y de modelos matemáticos. Ambos especialistas explicaron como distintas pruebas de campo permitieron validar la aplicación zootécnica del concepto para un uso aplicado de manera rutinaria. Así, este nuevo sistema permite precisar la valoración aplicada a Belfeed basándose en la cantidad de Arabinoxilanos.
CYS 42.indb 20 14/05/12 10:10

21
nº 42
jefo.com
BELFEED®
LA ENZIMA INCOMPARABLE
Aditivos para cada especie
JEFE DE PRODUCTO PORCINO : Sergio MERINERO I [email protected] I 616 47 34 69
JEFE DE PRODUCTO AVICULTURA : Anna FERNÁNDEZ I [email protected] I 618 60 21 43
La única xilanasa de origen bacteriano autorizada en el mercado europeo en pollos, pavos, ponedoras, patos, lechones y cerdos de cebo.
Alta eficacia en todo tipo de dietas : trigo, cebada, maíz...
BELFEED también tiene una sensibilidad reducida a los inhibidores de xilanasa (TAXI), lo que permite una homogeneidad de respuesta zootécnica durante todo el año (en cereales de “nueva cosecha” o almacenados, de campañas anteriores).Está disponible bajo las formas polvo y líquido y tiene una dosis de uso única.No duden en ponerse en contacto con nuestros jefes de producto para tener más información sobre el producto.
BELFEED.indd 1 09/05/12 09:15jefo.indd 21 14/05/12 11:01

22
nº 42
Las artropatías traumáticas agudas pertenecen a las causas más frecuentes de cojeras en el caballo.
El creciente uso y abuso del equino se asocia a un incre-mento de patologías del aparato locomotor, que obligan a la inactividad del paciente. La interrupción de la actividad física por reconvalescencia puede durar hasta varios meses, con la consiguiente frustración en cuanto a sus consecuencias laborales, deportivas o económicas.
Las articulaciones de las extremidades son estructuras sometidas a diferentes y grandes tensiones y presiones bio-mecánicas. Junto a las anomalías de conformidad anatómica como causas predisponentes, los traumatismos únicos o repe-tidos son la causa más común de inflamación y dolor articular. Debido a la complejidad e interrelación estructural y funcional de las articulaciones, muy frecuentemente quedan afectadas varias estructuras a la vez. Así, son hallazgos muy comunes la sinovitis y capsulitis asociadas. También se pueden encontrar daños en los ligamentos, meniscos, cartílago articular o placa ósea subcondral. Dependiendo de la intensidad de la lesión inicial, y sobre todo, de cuándo y cómo se inicia el tratamiento, cualquiera de estas condiciones por si sola puede progresar a la osteoartritis o enfermedad articular degenerativa, ocasio-nando daños irreversibles en el cartílago articular.
Uno de los componentes naturales de las estructuras articulares es el hialuronato sódico. Es un glucosamino-glicano formado principalmente por los sinoviocitos y condrocitos, y que en la membrana sinovial y en la matriz cartilaginosa se encuentra formando un entramado tridi-mensional. Su propiedad altamente higroscópica contribuye a la viscocidad y capacidad lubricante del líquido sinovial, en el que se encuentra en altas concentraciones. En cuanto al cartílago articular, contribuye a darle su característica firme-za tensioelástica y, mediante ello, optimiza la distribución de fuerzas y presiones mecánicas recibidas.
Básicamente, la articulación sinovial se compone de:
a. Tejidos blandos (membrana sinovial, cápsula, meniscos, ligamentos colaterales o cruzados, y tendones).
b. Tejidos duros (cartílago y placa ósea subcondral).
Tejidos blandos de la articulación
La localización primaria de una artropatía traumática aguda puede darse en cada uno de los componentes de tejido blando. La membrana sinovial, abundantemente irrigada por vasos sanguíneos, se compone básicamente de una delgada mem-brana de sinoviocitos, tejido conectivo y hialuronato sódico, contribuyendo éste a la regulación de la composición físico-química del líquido sinovial. Generalmente es la responsable del inicio de la cascada de inflamación, con la disminución o pérdida de sus múltiples funciones en la homeostasis articular.
La capsulitis también contribuye a la liberación de media-dores de la inflamación en el líquido sinovial. No tratada correctamente, la progresión de la afección lleva a la for-mación de tejido cicatrizal, pudiendo reducirse la movilidad articular y alterarse la estabilización articular. La ruptura fibrilar ligamentosa puede llevar tanto a una inflamación local, como a una desestabilización articular, con el consiguiente incremento de presión mecánica focal en el cartílago articular.
Tejidos duros de la articulación
La matriz del cartílago hialino se compone básicamente de un entramado de fibras de colágeno tipo ii, entre el cual se encuentran moléculas de hialuronato sódico y de proteogli-canos. Las patologías articulares van asociadas a una pérdida de hialuronatos y proteoglicanos. La deficiente capacidad autoregenerativa del cartílago se caracteriza porque las
Avances en el tratamiento de la artritis equina
Hyonate®, el innovador antiinflamatorio y
condroprotector de Bayer
CYS 42.indb 22 14/05/12 10:10

23
fibras de colágeno de tipo ii no se renuevan, reemplazándose por fibras de colágeno de tipo i, típicas de tejido fibroso. Esto se asocia a una pérdida de las características morfológicas y funcionales del cartílago articular.
Artritis traumática aséptica
Un traumatismo articular agudo afecta a las estructuras articulares en forma directa, por daño mecánico, o en forma indirecta, por liberación de múltiples mediadores de la infla-mación. La acción catabólica de diversas enzimas inflamatorias sobre la membrana sinovial y el cartílago articular disminuye también el hialuronato sódico presente en sus estructuras. La destrucción del entramado de hialuronato sódico de la membrana sinovial conlleva a una pérdida de su capacidad homeostática, lo que facilita el ingreso de células inflamatorias al líquido sinovial, incrementándose la liberación de mediado-res de la inflamación, la fagocitosis y destrucción celular. Por otra parte, el catabolismo de los componentes de la matriz cartilaginosa reduce también el contenido de hialuronato sódi-co, con la consiguiente disminución de la firmeza y elasticidad cartilaginosa, y del aporte de nutrientes.
En resumen, un traumatismo articular agudo implica la pérdida de hialuronato sódico naturalmente presente en la membrana sinovial y en el cartílago articular, lo que puede desencadenar un peligroso círculo vicioso. De no mediar un tratamiento efectivo y rápido, la consecuencia frecuente-mente son patologías articulares degenerativas crónicas, con la consiguiente reducción del período de vida útil del caballo.
Hyonate® es un innovador antiinflamatorio y condro-protector a base de hialuronato sódico de muy alto grado de pureza. Es el único que se obtiene a partir de un proceso biotecnológico de fermentación bacteriana selectiva, lo que evita la contaminación con moléculas extrañas. Por ello, se puede administrar en forma segura tanto por vía intraveno-sa, como también por vía intraarticular.
Numerosos estudios han confirmado la seguridad y eficacia clínica de Hyonate® en el tratamiento de las artri-tis agudas asépticas de los equinos, asociadas al complejo sinovitis/capsulitis. El efecto antiinflamatorio de Hyonate® consiste en la inhibición tanto de la proliferación, migración y fagocitosis leucocitaria, como también en la reducción de prostaglandinas inflamatorias, interleucinas y radicales libres (Kawcak et al., 1996; White et al, 1996, Kawcak et al., 1997).
La mayor ventaja de Hyonate®, sin embargo, radica en su administración intravenosa. El hialuronato sódico admi-nistrado por vía intravenosa entra directa y más rápidamente en contacto con los sinoviocitos. Esto se debe al estrecho contacto de las membranas basales de los vasos sanguíneos con los de la membrana sinovial, facilitándose así la difusión de moléculas.
Las ventajas de la administración intravenosa de Hyonate® son:
• Fácil, cómoda y rápida aplicación.
• Caballos inquietos pueden ser tratados en forma más segura.
Sinovioc.
Leucocitos
Líquido sinovial
Enzimas degradantes
Sinovitis
Alteraciones ante un traumatismo
Enzimas lisosomales Prostaglandinas
Interleucina-1 Radicales libres
Trauma
Inflamación de la membrana
Lesión directa de sinoviocitos y penetración
de leucocitos
Degradación de HS, proteoglicanos, y colágeno
Reducción del contenido de hyaluronato en el
líquido sinovial
Sinovioc.
Leucocitos
Líquido sinovial
Enzimas degradantes
Artritis
CYS 42.indb 23 14/05/12 10:10

24
nº 42
• No hay riesgos de daño articular iatrogénico (ej: lesión mecánica; artritis séptica).
• Efecto dual rápido y directo en el tejido diana.
• Una inyección hace efecto en más de una articulación a la vez.
• Efectividad similar o superior a la administración intra-articular.
La terapia por vía intravenosa logra resultados clínicos similares a aquellos por vía intraarticular o, más frecuentemen-te, resultados clínicos superiores (Niederhofer, 2002). mediante la administración intravenosa secuencial de Hyonate®, los sinoviocitos son estimulados a formar hialuronato endógeno, logrando una prolongación del efecto antiinflamatorio de hasta 45 días tras la última de tres aplicaciones intravenosas (Kawcak y cols., 1995). Con Hyonate® se reducen los riesgos de la administración por vía intraarticular al mínimo necesario.
Hyonate® se presenta en unidades de dos viales de 2 ml cada uno. La dosis de 4 ml (vía intravenosa) se administra hasta tres veces a intervalos semanales, según el cuadro clínico. La dosis de 2 ml corresponde a la administración por vía intraarticular. Como en todo tratamiento de afecciones traumáticas del aparato locomotor, además del tratamiento médico es muy importante someter al paciente a un período de reposo, seguido por una reanudación paulatina del ejercicio físico en función de la intensi-dad del cuadro clínico inicial y de la recuperación posterior.
Dr. ralf Ebertmarketing y Ventas iberia. Bayer HealthCare
División Sanidad Animal. E-mail: [email protected]
Bibliografía
• C.E. KAWCAK, D.D. FriSBiE, C.W. mCiLLWrAiTH, G.W. TroTTEr. JoiNT
DiSEASE iN THE HorSE. C.W. mCiLLWrAiTH, G.W. TroTTEr; W.B.
SAUNDErS ComPANY, PHiLADELPHiA, 264-265 (1996).
• G.W. WHiTE ET AL. EFFiCACY oF SiSTEmiCALLY ADmiNiSTErED ANTiAr-
THriTiC DrUGS iN AN iNDUCED EqUiNE CArPiTiS moDEL. ProC. AAEP
42, 135-138 (1996).
• C.E. KAWCAK, DVm, mS; D.D. FriSBiE, DVm, mS; G.W. TroTTEr, DVm, mS;
C.W. mCiLLWrAiTH, BVSC, PHD; S.m. GiLLETE, DVm, PHD; B.E. PoWErS,
DVm, PHD; r.m. WALToN, DVm. EFFECTS oF iNTrAVENoUS ADmiNiS-
TrATioN oF SoDiUm HYALUroNATE oN CArPAL JoiNTS iN ExErCiSiNG
HorSES AFTEr ArTHoSCoPiC SUFGErY AND oSTEoCHoNDrAL FrAG-
mENTATioN. AJVr, VoL 58, Nº 10 (1997).
• m. NiEDErHoFEr. WirKSAmKEiT UND VErTräGLiCHKEiT EiNES 1%iGEN
NATriUmHYALUroNATES BEi DEr BEHANDLUNG AKUTEr ArTHriTiDEN
DES PFErDES: VErGLEiCH DEr iNTrAVENöSEN UND iNTrAArTiKULärEN
APPLiKATioN. SoNDErDrUCK AUS TiErärzTLiCHE UmSCHAU, 57.
JAHrGANG / Nr. 9; 441-487, (2002).
-
Leucocitos
Líquido sinovial
Efectos de la administración de Hyaluronato sódico
Enzimas lisosomales Prostaglandinas
Interleucina-1 Radicales libres
Mediadores de la inflamación
Inhibe la producción de enzimas lisosomales
Inhibe la Interleucina 1
Inhibe la producción de radicales libres
Suprime la producción de
prostaglandinas
Hyaluronato sódico
- -
- -
Previene que los leucocitos atraviesen los capilares
PosologíaIV: 4 ml
iA: 2 mlTres inyecciones a intervalos semanales
1 semana 1 semana
5 días
CYS 42.indb 24 14/05/12 10:10

25
CYS 42.indb 25 14/05/12 10:10

26
CYS 42.indb 26 14/05/12 10:10

27
Cría y saludPORCINO
42
Programas de alimentación en porcino en función del sexo: efectos de la castración quirúrgica y la inmunocastración
Cambios en el espermatozoide desde la eyaculación hasta la fecundación
C I E N T Í F I C O S
CYS 42.indb 27 14/05/12 10:10

28
Bravo, F.
❱❱ Introducción
Debido a la actual situación económica, los elevados precios de las materias primas, y la alta presión de la oferta de carne de cerdo que ha hecho que la competitividad haya aumentado drásticamente, uno de los retos más importantes de la producción porcina moderna es mejorar la eficiencia de produc-ción. Además, cada vez existen más regula-ciones y normativas que hay que cumplir. Estas regulaciones afectan a todos los esla-bones de la cadena de producción (inclusión de ciertos aditivos o materias primas en el pienso, instalaciones y proceso productivo de acuerdo con la Ley Europea de Bienestar Animal en su Directiva 2008/120/CE, control de la seguridad alimentaria), aumentando mucho la competitividad.
Esta nueva situación requiere profe-sionales cualificados que sean capaces de adaptar los recursos disponibles a las demandas del mercado. Por lo tanto, la producción porcina europea en general, y española en particular, continuará en los próximos años con su proceso de moderni-zación, en el que se aplicarán nuevas técni-cas de manejo y de alimentación adaptadas al tipo de animal presente hoy en día en nuestras granjas y a la legislación vigente, y enfocados a una mejora de la eficiencia productiva con el fin de disminuir costes y así aumentar los márgenes para el ganadero.
Una de las prácticas que provocan un mayor debate es la castración. La castración tiene muchas implicaciones tanto desde el punto de vista del bienestar animal como del de la eficiencia productiva. La prohibición de esta práctica va a suponer nuevos retos para los productores que tradicionalmente han producido cerdo castrado, y que tendrán que adaptarse a los nuevos tiempos para seguir siendo competitivos.
En este trabajo se pretende describir lo que supondrá la desaparición de la cas-tración desde el punto de vista productivo; se describe una alternativa a la castración que cada vez está más en uso, la inmuno-castración; y se analizan las alternativas aplicables en la alimentación de los cerdos para sacarles el mayor rendimiento posible.
Programas de alimentación en porcino en función del sexo: efectos de la castración quirúrgica y la inmunocastración
F. Bravo de laguna OrtegaNutreco Swine research Centre. Países Bajos
Ponencia Presentada en el XXVii curso de esPecialización Fedna
Fotos Axón Comunicación
CYS 42.indb 28 14/05/12 10:10

29
nº 42Programas de alimentación en porcino en función del sexo: efectos de la castración quirúrgica y la inmunocastración
❱❱ Castración
Castración física o quirúrgica
1.1. Normativa
Como ya se ha mencionado, los aspectos legales alrededor de la castración están regulados a nivel europeo, en la Directiva 2008/120/CE, relativa a las normas mínimas para la protección de cerdos. En ella, se indica que “la castración puede provocar un dolor prolongado que es más fuerte si se produce un desgarro de los tejidos [ ... ] Por consiguiente, deben establecerse normas para garantizar unas mejores prácticas”.
El Anexo 1 indica, en su Capítulo 1, Punto 8, que se prohíben todos los procedimientos que “provoquen lesiones, o la pérdida de una parte sensible del cuerpo o la alteración de la estructura ósea”. No obstante, señala algunas excepciones, entre ellas “la castración de los cerdos macho por medios que no sean el desgarre de tejidos”. Además solo se autoriza a un veterinario o una persona formada a llevar a cabo la castración. Si ésta se lleva a cabo a partir del séptimo día de vida, “se utilizará anestesia y una analgesia prolongada practi-cada por un veterinario”. Cada Estado miembro puede ser más restrictivo si lo considera. En el caso de España, el real Decreto 1135/2002 relativo a las normas mínimas para la protec-ción de cerdos ratifica la normativa europea en cuanto a castración de los machos.
Con vistas al futuro, hay organizacio-nes del sector porcino que han suscrito la “Declaración europea sobre alternativas a la castración quirúrgica de los cerdos” (2010), en el que se comprometen a la apli-cación de anestesia y analgesia prolongada desde el 1 de enero de 2012, y a poner fin voluntariamente a la castración quirúrgica antes del 1 de enero de 2018.
La EFSA (2004) indica que más de 600 millones de lechones son castrados al año en el mundo. En Europa se sacrifican anualmen-te unos 250 millones de cerdos, de los que la mitad son machos. De ellos, el 80% son cas-trados, de los cuales solo el 3% (3 millones), son castrados con anestesia (Frederiksen et al., 2009). En cuanto a la castración de hem-bras, el real Decreto 1221/2009 por el que se establecen normas básicas de ordenación de las explotaciones de ganado porcino exten-sivo, señala que por motivos justificados, la castración de una hembra “deberá ser certificada y realizada por un veterinario, con anestesia y analgesia prolongada”.
1.2. Aspectos generales
La castración de lechones ha sido y sigue siendo una práctica común en muchos paí-ses europeos, que se lleva a cabo después del nacimiento, casi siempre durante la primera semana de vida. No obstante, el porcentaje de cerdos que llegan enteros a matadero está creciendo cada vez más, y debido a las nuevas normativas sobre bienestar animal, dicha práctica tiende a desaparecer (Stevenson, 2000). En la figura 1 se repre-senta el porcentaje de los cerdos producidos que son castrados en distintos países euro-peos. La tendencia al abandono del uso de la castración va a suponer en la mayor parte de Europa un cambio drástico en la manera de producir cerdos, teniendo que adaptar tanto la alimentación como en el manejo.
En España también se presenta la pro-blemática de la castración en cerdo ibérico. Debido al elevado peso al sacrificio de estos animales, la práctica de la castración en cerdo ibérico se lleva a cabo tanto en machos como en hembras, siendo en estas últimas donde se genera un mayor debate en cuanto al bienestar animal, ya que nor-malmente se realiza alrededor de los 20 kg.
¿Por qué castramos? La castración es habitual por varios motivos: controlar mejor el comportamiento de los animales, ya que los machos enteros son más activos y agresivos que los castrados, y para prevenir el olor sexual de la carne, y así mejorar la calidad organolép-tica de la misma. El olor sexual se produce por la acumulación de androstenona y escatol en el tejido adiposo del animal, produciendo un olor y un sabor desagradables para el consumidor. La androstenona se forma en los testículos al alcanzar los animales la pubertad. En cambio, el escatol procede de la fermentación anaeró-bica bacteriana del triptófano en el intestino grueso (Lundstrom y zamaratskaia, 2006) de tanto machos como de hembras, aunque en mayor concentración en los machos enteros, ya que ciertos esteroides testiculares inhiben su degradación en el hígado, llegando al intes-tino y estando disponible para la degradación bacteriana. Yokohama y Carlson (1979) des-cribieron su proceso de producción (figura 2). Este proceso se da en el ciego y colon de los cerdos, y también se ha detectado en el rumen de las vacas (Jensen, 2006).
Lundstrom y zamaratskaia (2006) estudiaron los factores que afectan a la producción de escatol y androstenona en machos enteros (figura 3). Los que afectan directamente a la producción de ambos compuestos son: el metabolismo del ani-mal, la capacidad de síntesis, el estado hormonal y la genética. El fotoperíodo parece afectar solo a la producción de androstenona, mientras que el ambiente y la dieta afectan más a la síntesis de escatol.
Como hemos mencionado anterior-mente, la androstenona se forma en los
Figura 2. Formación de escatol a partir de L-Triptófano (Yokoyama y Carlson, 1979).
CH2-C-CooH
CH2-C-CooH
NH2
N
- -
NH
NH
CH3
NH
L-TRYPTOPHAN
INDOLEAcETIc AcID
3-METHYLINDOLE (SkATOLE)
100-
90-
80-
70-
60-
50-
40-
30-
20-
10-
0-
Aust
ria
Bélg
ica
Fran
cia
Alem
ania
Pais
es B
ajos
Suiz
a
repú
blic
a Ch
eca
Eslo
veni
a
Hun
gría
P olo
nia
Eslo
vaqu
ia
Dina
mar
ca
Esto
nia
Finl
andi
a
rein
o U
nido
irlan
da
Leto
nia
Litu
ania
Nor
uega
Suec
ia
Chip
re
Gre
cia
italia
Port
ugal
Espa
ña
Figura 1. Porcentaje de cerdos castrados por país de la UE-25 (datos 2007-2008) (Fredriksen et al., 2009).
CYS 42.indb 29 14/05/12 10:10

30
Bravo, F.
testículos, y el nivel que alcanza está espe-cialmente influido por la genética (figura 4). Según Bonneau (2006) las líneas genéti-cas que alcanzan la madurez a edades más tempranas son más propensas a producirla. El nivel máximo de androstenona para que la carne se mantenga en buenas condi-ciones de olor y sabor ha sido determina-do experimentalmente por Zamaratskaia (2004), y se sitúa entre 0,5-1 ppm.
El peso que tiene el escatol en el olor sexual es mayor que el de la androstenona. Su mecanismo de distribución se muestra en la figura 5 (Font i Furnols, 2002). El triptófa-no no metabolizado en el hígado es trans-formado en escatol en el intestino grueso y pasa al torrente sanguíneo, desde donde es depositado en diversos tejidos corporales, sobre todo en el adiposo, ya que tiene una naturaleza afín a las grasas (Jensen, 2006).
Hay autores que han estudiado cómo minimizar el contenido de escatol en grasa. Bonneau y Squire (2000), concluyeron que algunas estrategias podrían ser: tener un acceso ilimitado al agua, la cría de los ani-males sobre slat, y el uso de alimentación líquida. En cuanto a otros factores que afec-tan al nivel de escatol: un pH bajo y condi-ciones anaeróbicas estimulan su formación, según Kemmer et al. (1997). También se ha reportado la edad del animal como factor. En cerdos sacrificados entre 90 y 100 kg se encontraron niveles significativamente más bajos de escatol en grasa dorsal que en cerdos sacrificados a los 110 kg (Andersson et al., 2005). Lawlor (2010) determinó que cerdos sacrificados con menos de 105 kg es poco probable que presenten olor sexual. Aunque esto dependerá de la genética, ya que la variación en el nivel de escatol a lo largo de la vida del animal difiere entre razas (Lundstrom et al., 2004). Estos auto-res observaron que el nivel de escatol en machos enteros incrementaba a los 180-200 días de vida, y que posteriormente empezó a reducirse entre los 240-260 días de vida en cerdos Yorkshire y Landrace, y entre los 310-360 en cerdos Hampshire y Duroc.
Otro de los factores influyentes ha resul-tado ser el tamaño de los órganos sexuales. Zamaratskaia et al (2005) observaron que cerdos con un peso de los testículos <565 g no producían grandes cantidades de escatol. Respecto al efecto del ambiente, Hansen et al. (1994) concluyeron que cerdos criados en ambientes limpios acumulaban menos escatol que los criados en corrales más sucios. Otra consideración es que en verano el nivel de escatol es mayor que en invierno (Lundstrom y Zamaratskaia, 2005).
No todas las personas son capaces de percibir el olor sexual de la carne, la per-cepción de estos compuestos está deter-minada genéticamente y además difiere entre países. En España hay un porcentaje más alto de mujeres que de hombres que lo detectan (Font i Furnols et al., 2009).
Además de lo discutido sobre el olor sexual, la castración es importante en España para la producción de ciertos pro-ductos cárnicos, como por ejemplo algunos embutidos. Hay ciertas indicaciones geo-gráficas protegidas, denominaciones de origen o tipo de producto, que imponen ciertos requisitos en los animales de los
cuales se van a producir dichos embuti-dos. Estos requisitos conciernen a la raza (sobrasada de Mallorca de cerdo negro, productos ibéricos, jamón de Teruel), al peso al sacrificio (productos ibéricos, jamón de Teruel), al tipo de alimentación (cho-rizo de Cantimpalo, Ibérico de bellota), a su composición en cuanto a ingredientes (chosco de Tineo), a su tiempo de curación (jamón ibérico) al sexo (jamón de hembra) o a la castración (longaniza de Vic). Para la producción de estos productos, en oca-siones como hemos dicho es necesario un mínimo de grasa, ya sea por requerimientos legales, ya sea por facilitar el proceso de preparado y curación. El contenido graso es mayor en machos castrados que en hem-bras y en machos enteros, por este orden.
❱❱ Alternativa a la castración quirúrgica: inmunocastración
2.1. Estatus legal
Actualmente existe la Decisión de Ejecución de la Comisión 2011/C 243/06 del 19 de
Figura 5. Proceso de distribución del escatol (Font i Furnols, 2002).
Bile Rectum
Breathing Urine
Feeding (trp) IntestineSkatole Faeces
Hepatic metabolism
Adipose tissue
Other tissues
Excretion
Lungs
blood
Figura 3. Factores que afectan la producción de androstenona y escatol en machos enteros (Lundstrom y Zamaratskaia, 2006).
Hormonal status
Genetic background
Androstenone SkatolePhotoperiod
MetabolismSynthesis
Enviroment
Dietary ingredients
Figura 4. Influencia de la genética y el ambiente en la producción de androstenona (Bonneau, 2006).
Genetic
16 androstenes
16 androstenes
16 androstenes
androgensProgestagens
estrogens
androstenes
Sexual maturity
LHTestis Blood Fat
Salivary glands
Andien- synthase
Catabolism
Environment Nutrition
alimentación en porcino.indd 30 21/05/12 10:27

31
nº 42Programas de alimentación en porcino en función del sexo: efectos de la castración quirúrgica y la inmunocastración
agosto, por la que se adopta un programa de trabajo para la financiación de las acti-vidades de la Unión sobre alternativas a la castración quirúrgica de los cerdos. Este programa constará de “medidas técnicas, científicas y educativas encaminadas a buscar y adoptar alternativas a la castra-ción quirúrgica”. Entre otras cosas, en su consideración número 4 indica que “para evitar el olor sexual en la carne de porcino se suele recurrir a 3 medidas: el sacrificio temprano, la ablación de los testículos (castración quirúrgica) o la inmunocastra-ción”, y que “la castración quirúrgica de los cerdos constituye una preocupación de bienestar animal”. Teniendo en cuenta los compromisos adquiridos en la Declaración europea sobre alternativas a la castración quirúrgica de los cerdos (2010) mencio-nados anteriormente al hablar sobre la normativa de la castración, la Decisión de la Comisión indica en su consideración número 8 que «conviene que la Unión invierta en estudios sobre alternativas a la castración quirúrgica de los cerdos».
Este programa incluye:
• el desarrollo de métodos de referencia reconocidos por la UE para la detección y medición de los compuestos respon-sables del olor sexual del cerdo.
• estudios sobre la aceptación por parte de los consumidores de carne y pro-ductos procedentes de cerdos enteros.
• estudios sobre métodos de detección rápida del olor sexual que se usen o desa-rrollen en plantas de sacrificio de la UE.
• estudio sobre cómo reducir los com-puestos responsables del olor sexual por medio de técnicas de reproducción, alimentación y manejo.
• estudio económico sobre costes y beneficios de abandonar la castración quirúrgica.
• crear un sitio web que proporcione edu-cación y formación sobre alternativas a las castración quirúrgica de los cerdos.
En cuanto al estatus de la aplicación de la inmunocastración en España, la vacuna que existe en el mercado (de la que se
habla más adelante) está incorporada en la Agencia Española de medicamentos y Productos Sanitarios, y su uso está autori-zado en machos. Para su uso en hembras (especialmente ibéricas, aunque en italia también hay interés por la producción de jamón de Parma), está amparado por el real Decreto 1132/2010, que en su artículo 82 señala que “cuando no existan medicamen-tos veterinarios autorizados para una enfer-medad, el veterinario podrá […], para evitar sufrimientos inaceptables, tratar al animal o animales afectados de una explotación concreta administrando un medicamento veterinario, con similar efecto terapéutico al deseado, autorizado para su uso en otra especie, o para tratar otra enfermedad de la misma especie, por la Agencia Española de medicamentos y Productos Sanitarios.
2.2. Aspectos generales
Debido a las limitaciones que presenta la castración quirúrgica en cuanto a bienestar animal y a la pérdida de rendimientos pro-ductivos y calidad de canal con respecto a los machos enteros (este aspecto se discute en el apartado 3 del presente trabajo), se ha trabajado en encontrar técnicas alter-nativas con las que se consigan los mismos efectos sin necesidad de la aplicación de la castración quirúrgica. Uno de estas alter-nativas consiste en una vacunación que
desencadena una serie de procesos que inhiben el desarrollo de los órganos sexua-les. Es la inmunocastración.
La inmunocastración consiste en una vacuna que estimula el sistema inmunitario del animal para la producción de anticuer-pos que inhiban la secreción de la hormona liberadora de gonadotropina (GnrH), y así la proliferación de los tejidos masculinos (Pfizer Animal Health; figura 6). Así, se inhibe el desarrollo de los testículos, y con-siguientemente la producción de los esteroi-des que allí se producen, como testosterona y androstenona; esta última se ha indicado anteriormente como uno de los causantes directos del olor sexual de la carne.
La inmunocastración actúa directamen-te sobre el eje hipotálamo-hipófisiariogo-nadal, que es el que controla la síntesis de las hormonas luteinizante (LH), y folículo estimulante (FSH, figura 6): en el hipotálamo se libera la GnrH, que se une a receptores específicos de la hipófisis para la secreción de LH y FSH. En el macho, la LH induce en los testículos la producción de testosterona, que es, junto con la FSH, la desencadenante de la fabricación de los espermatozoides. Así, con la vacuna se consigue:
• inhibir la producción de espermatozoi-des, es decir, esterilizar al animal.
• inhibir la producción de androstenona dis-minuyendo así el olor sexual de la carne.
La vacuna consiste en dos dosis, y los animales inmunizados comienzan a comportarse como castrados a partir de la segunda dosis. Hasta ese momento crece como un entero. El efecto de la inmuno-castración es temporal, por lo que el animal
Figura 6. Efecto de la inmunocastración en la producción de androstenona y escatol (Pfizer Animal Health, 2011).
Hypothalamus
Brain
Pituitary
Testes
LH FSH
G rF
many esteroids
Skatole
P tiliLibBeiourSindronenone
Cuadro 1. Ventajas e inconvenientes de la castración.
Ventajas Inconvenientes• reducción de olor sexual• Disminuyen las agresiones• Disminuyen las montas• Deposición de grasa necesaria para
producción de ciertos productos cárnicos
• menor eficiencia del macho castrado• Efecto negativo sobre el sitema inmune y el
crecimiento• Aumenta la mortalidad• Aumenta el uso de antibióticos• Dolor y estrés asociado
CYS 42.indb 31 14/05/12 10:10

32
Bravo, F.
vuelve a su estado de entero tras un perío-do aproximado de 10 semanas. Además, al tratarse de una vacuna, no es necesario un período de retirada antes del sacrificio. Los efectos observados tras la inmunocastra-ción se sintetizan en el cuadro 3.
Uno de los obstáculos para su acepta-ción es el temor de los productores y de las empresas a una depreciación del produc-to final. Esta técnica precisa de estudios para demostrar que otras características relacionadas con la calidad de la carne no se ven afectadas, y que no repercute negativamente en la seguridad alimentaria. La EFSA (2004) señala en sus conclusiones una serie de incertidumbres alrededor de la inmunocastración:
• la aceptabilidad que va a tener entre la población.
• problemas de seguridad alimentaria debi-do a la presencia de residuos en la carne.
• problemas de seguridad del personal que la aplica, al poder inyectarse a sí mismos, con los mismos efectos que en los animales.
❱❱ Efecto de la castración en los rendimientos productivos
Son muchos los factores que se han repor-tado que influyen en los rendimientos productivos de los cerdos: genética, estatus sanitario, factores ambientales, alimenta-ción, manejo... En este trabajo nos vamos a
centrar en el efecto del sexo, considerando que en producción porcina tenemos 3 sexos: macho entero, macho castrado y hembra (el caso de la hembra castrada, común en producción de cerdo ibérico, no se tratará). Entre los machos castrados dis-tinguiremos entre castrados físicamente e inmunizados, ya que se ha observado expe-rimentalmente que no se comportan igual a partir de la segunda dosis de la vacuna.
Se ha observado que los machos cas-trados comen más y crecen más rápido que los enteros. Dunshea (2010) llevó a cabo un meta-análisis en el que revi-só diversos estudios en que se utiliza-ron machos enteros y machos castrados durante la última fase del cebo (cuadro 4), desde las 4 ó 5 semanas previas al sacri-ficio. observó que los castrados tenían un crecimiento (GmD) más rápido (+31 g/d), un consumo (CmD) más alto (+467 g/d), un índice de conversión (iC) peor (-0,48 kg/d), un espesor de grasa dorsal (EGD) más alto (+4,9 mm), y se sacrificaron a mayor peso (+2,14 kg).
Dunshea (2010) también estudió por el mismo método de meta-análisis, el efec-to en los rendimientos productivos de machos inmunizados, entre la segunda vacunación y el sacrificio, comparado con los rendimientos de los enteros (cuadro 5). El sacrificio en todos los estudios utilizados para este meta-análisis se produjo 4 ó 5 semanas tras la segunda dosis. Como se observa, a partir de la segunda dosis los animales inmunizados empiezan a com-portarse como castrados y se diferencian de los enteros con una GmD más alta (149 g/d), un mayor CmD (512 g/d), un peor iC (0,07 kg/d), un mayor EGD (1,2 mm), y un mayor peso canal (1,48 kg).
Al comparar los valores de los cuadros 4 y 5, teniendo en cuenta que ambos metaanálisis han recogido datos corres-pondientes al mismo período de tiempo pre-sacrificio, se puede deducir que a partir de la segunda vacunación los animales inmunizados se comportan como castra-dos. Además, aunque el espesor de la grasa dorsal aumenta, no llega al nivel que el de los castrados.
En la misma línea que estos resultados, en un estudio llevado a cabo en Nutreco Canadá Agresearch (2009), se compararon los rendimientos productivos de machos enteros, machos castrados físicamente y machos inmunizados (cuadro 6).
Los machos inmunizados presentaron una eficiencia intermedia entre los enteros y los castrados físicamente. Esto se debe al hecho de que hasta la segunda vacunación se desarrollan como enteros. A partir de este momento su metabolismo se asemeja más al de un castrado, e incluso parece que intenta compensar, como se intuye de los datos de GmD y CmD desde la segunda dosis hasta el sacrificio. Parece como si el cerdo deseara tener una composición corporal para cada estado metabólico y en cada momento. En el momento en que el cerdo es inmunizado, tiene que compensar la deposición de grasa de un castrado físi-camente, y para igualarse lo hace aumen-tando el consumo de pienso.
Hay numerosos estudios que reportan una mayor eficiencia de los machos ente-ros que los castrados durante la fase final del cebo, también resultan más eficientes si miramos el período completo (Campbell y Tavemer, 1988; Hennessy et al., 2009). Si sólo miramos el período post-destete (de 28 a 63 días de edad), quiniou et al. (2010) observaron un menor CmD en machos enteros (720 g/d) que en castrados o hem-bras (760 g/d y 740 g/d respectivamente), lo que produjo una mayor eficiencia en los enteros, aunque estas diferencias no fueron significativas. Pero es a partir de
Cuadro 3. Efectos de la inmunocastración.
• El olor sexual de la carne procedente de estos animales se reduce a un nivel imperceptible: ya que disminuye la producción de testorena, lo que hace que dismunuya la androstenona, causando un aumento del metabolismo hepático y provocado una mayor eliminación de escatol.
• Disminución del tamaño de los testiculos tras la vacunación.
• menos montas y agresividad en la cría en grupos.
• menos espesor de grasa dorsal al secrificio que el macho castrado quirúrgicamente.
Cuadro 4. Diferencias de rendimiento entre enteros y castrados (Dunshea, 2010).
Enteros vs. Castrados N
GmD ((g/d) -31 8
CmD (g/d) -467 7
iC (Kg/kg) -0,48 7
EGD (mm) -4,9 10
Peso canal (kg) -2,14 11
N = número de estudios incluídos en el meta-análisis.
Cuadro 5.Diferencias de rendimiento entre inmuno-castrados y enteros (Dunshea, 2010).
Inmunocastrados vs. Castrados N
GmD ((g/d) 149 14
CmD (g/d) 512 10
iC (Kg/kg) 0,07 16
EGD (mm) 1,2 16
Peso canal (kg) 1,48 16
N = número de estudios incluídos en el meta-análisis.
Cuadro 6. rendimientos de machos enteros, castrados fisicamente e inmunizados (Nutreco Canada Agresearch, 2009).
2º dosis-matadero Cebo completo (27-127 kg)GMD CMD IC GMD CMD IC EGD
Entero 1,240 3,050 2,46 1,097 2,302 2,10 18,8
Inmunocastrado 1,362 3,712 2,72 1,142 2,539 2,22 20,2
castrado 1,087 3,452 3,16 1,091 2,677 2,45 22,1
Diferencia entre Inmunocastrado vs. Entero y castrado quirúrgicamente
Entero +122 +662 +0,26 +45 +237 +0,12 +14
castrado +275 +260 -0,44 +51 -138 -0,23 -1,9
CYS 42.indb 32 14/05/12 10:10

33
nº 42Programas de alimentación en porcino en función del sexo: efectos de la castración quirúrgica y la inmunocastración
los 63 días de edad (aproximadamente 25 kg de peso) cuando se empiezan a observar diferencias significativas. En el trabajo mencionado de quiniou et al. (2010), la mayor GmD se observó en los machos, independientemente de si eran castrados o enteros. mirando en detalle las dos mitades del período de cebo, se observó cómo desde los 25 kg hasta los 70 kg, los castrados crecieron significativa-mente más que los enteros y las hembras. A partir de 70 kg fueron los enteros los que tuvieron mayor crecimiento que los castrados y las hembras. Hay que tener en cuenta que en este estudio se utilizó un pienso único, y que existe un desajuste entre necesidades y consumo durante gran parte del cebo, lo que perjudica sig-nificativamente el crecimiento.
Se han llevado a cabo diversos estudios sobre el efecto de la castración en la com-posición de la canal. Suster et al. (2006) compararon machos enteros y castrados (cuadro 7) a los 122 y a los 150 días de edad. Como se observa en el cuadro, los enteros presentaron más tejido magro y menos tejido graso que los castrados, ade-más de un menor EGD. Con estudios de sacrificio comparado se puede determinar la curva de deposición de proteína de los distintos sexos (figura 7). Estos datos pue-den utilizarse para la creación de modelos matemáticos de simulación, con los que predecir el desarrollo de los animales.
En el gráfico se aprecian las curvas de deposición de proteína de machos enteros, castrados físicamente, e inmunocastrados, según el modelo Watson®, desarrollado en Nutreco Canadá. Como se observa, el animal inmunocastrado es un macho entero hasta el momento de la segunda vacunación, cuando empieza a comportarse como un castrado. Estas diferencias son las que hacen que las necesidades de un macho entero y de un macho castrado sean distintas, que es lo que se analiza en el apartado 4.
❱❱ Recomendaciones nutricionales
Para el diseño de un programa de alimen-tación óptimo, es necesario conocer: i) las necesidades de los animales a los que va dirigido dicho programa de alimentación, y ii) la capacidad de consumo de estos ani-males. Es en base a estos dos parámetros como se pueden hacer recomendaciones más precisas para los niveles de inclusión de los distintos nutrientes en las dietas.
Energía
La técnica más utilizada para el cálculo de las necesidades de energía es el método factorial. Este método consiste en sumar la energía necesaria para cada una de las actividades de los animales. En el caso de la fase de cebo, que es la que nos ocupa, la energía es la nece-saria para mantenimiento, termorregulación, y crecimiento. En el caso del mantenimiento y la termorregulación, no existen diferencias entre sexos. Los gastos de mantenimiento se calculan en función del peso metabólico del animal (Peso vivo 0,60), mediante la ecuación:
Ecuación 1: EMm (kcal/d) = 206 x PV 0,60 (FEDNA, 2006)
Los gastos de termorregulación se cal-culan en función de la diferencia entre la temperatura ambiente (T) y la temperatura crítica inferior (TCi), mediante las ecuaciones:
Ecuación 2: TCI = 26 - 0,061 x PV (FEDNA, 2006)
Ecuación 3: EMt (kcal/d) = 46-134 x (TCI- T) (FEDNA, 2006)
En cambio, los gastos de crecimiento se calculan en función de la naturaleza de dicho crecimiento, básicamente cantidad de grasa y de proteína depuesta. Cada uno de los sexos tratados muestra una composición corporal distinta y una composición del crecimiento distintas. Por este motivo, las necesidades de energía serán diferentes para cada sexo. Las necesidades se calculan mediante la ecuación:
Ecuación 4: EMc (kcal) = 12,8 x gramo grasa + 12,1 x gramo proteína (FEDNA, 2006)
En definitiva, habiendo estimado la composición corporal de cada sexo, FEDNA estima las siguientes necesidades de creci-miento, para un crecimiento de 700 g/d. En el cuadro 8 se muestra la composición del crecimiento de machos enteros, hembras y castrados, y las necesidades de energía para crecimiento de los mismos. Hay que tener en cuenta que los castrados crecen más que las hembras y que los machos enteros, por lo que las diferencias que se observan en el cuadro, se verían incremen-tadas algo más en condiciones reales.
❱❱ Proteína
Las necesidades de proteína están direc-tamente relacionadas con la capacidad de deposición de proteína y síntesis de tejido muscular que tenga el animal. Entre los factores que influyen en esta capacidad,
Figura 7. Deposición de proteína diaria de machos enteros, castrados físicamente e inmunocastrados (Modelo Watson®, Nutreco).
190-
180-
170-
160-
150-
140-
130-
120-
110-
100-
Edad (días)
Deposición de proteina
Depo
sici
ón (g
/d)
63 119 14191 133 15377 127 145105 137 159
EnteroCastradoinmunocastrado
Cuadro 7. Características de la canal en machos enteros y castrados, criados en grupo (Suster et al., 2006).
Enteros Castrados
Peso vivo (kg)
122 76,8 78,3
150 103 108
Peso magro (kg)
122 51,4 50,2
150 69,1 66,2
Peso grasa (kg)
122 11,9 13,4
150 18,2 23,8
EGD (mm)
150 15,1 17,1
Cuadro 8. Necesidades de crecimiento en la fase de cebo en función del sexo.
Sexo Proteína (% crecimiento) Grasa (% crecimiento) kcal EMC/d*Macho entero 16,1 20,3 3182
Hembra < 95 kg 15,3 24,1 3455
Hembra > 95 kg 15,3 28,0 3805
castrado < 95 kg 15,3 29,0 3894
castrado > 95 kg 15,3 32,0 4163
*recomendaciones FEDNA (2006) basadas en una GmD de 700 g/d.
CYS 42.indb 33 14/05/12 10:10

34
Bravo, F.
están el peso vivo, la genética y el sexo (Lizaso, 1994). En la figura 8 vemos cómo varía la retención de proteína con el peso vivo. Como se observa, los machos enteros tienen un mayor potencial de deposición de proteína para un mismo peso que las hembras y que los machos castrados.
No obstante, hay que tener en cuenta que el animal alcanza su potencial genético siempre y cuando no exista otro tipo de limitación. Una de estas posibles limitaciones es la energía disponible. Para que el proceso de deposición de proteína se dé, es necesaria una cantidad mínima de energía. Esto ha sido estudiado por distintos autores. Uno de los modelos teóricos más aceptados es el desa-rrollado por el NrC (1998; figura 9), que con-siste en un modelo “lineal-plateau”. El modelo “lineal-plateau” consiste en que la respuesta de los cerdos a un incremento en el consumo de energía, aumenta de manera lineal hasta un punto a partir del cual un aumento de la energía consumida no produce un aumento en la deposición proteica. Esto ocurre siempre y cuando el nivel de proteína no sea limitante. En la figura 9 se aprecia que para cada peso vivo existe una capacidad de deposición de proteína máxima. Para cada genética y cada sexo tendríamos un gráfico distinto (figura 10). En la figura se observa como con el mismo peso vivo, los machos son capaces de retener más nitrógeno, y por consiguiente depositar más proteína, que las hembras.
En el Swine research Centre se ha rea-lizado un estudio en el que se han recogido datos productivos de machos enteros y de hembras, y se ha observado su respuesta a niveles crecientes de aminoácidos. En los cuadros 9 y 10 se muestran los datos corres-pondientes al período de cebo completo. A ambos sexos se les alimentó con las mismas dietas y durante los mismos intervalos de peso. Se observó que la GmD de las hembras fue mayor que la de los machos. En cuanto al
iC, en los niveles más bajos de aminoácidos, los machos presentaron una peor conversión, pero esta fue mejorando con niveles más altos de aminoácidos, resultando ser, en el nivel más óptimo mejor que en las hembras. Esto sugiere que en machos, con un mayor potencial de deposición de proteína, un défi-cit de aminoácidos penaliza más que en hem-bras. Por último, se observa como las hembras presentaron un mayor EGD que los machos.
El programa de alimentación fue de 4 fases: de los 25 kg a los 40 kg, de los 40 kg a los 55 kg, de los 55 kg a los 75 kg, y de los 75 kg hasta el sacrificio a los 110 kg. Las dietas experimentales se formularon de acuerdo al concepto de proteína ideal, con diferentes niveles de lisina digestible aparente (AiD lys) y el resto de aminoácidos se incluyeron a un contenido fijo en relación a la lisina. En las dos primeras fases no se observaron diferencias entre machos y hembras. Es a partir de los 55 kg cuando se empieza a observar una respuesta distinta a niveles de aminoácidos entre los dos sexos. En cada fase y para cada sexo se dibujaron las curvas de dosis-respuesta a los diferentes niveles de aminoácidos tanto para la GmD como para el iC, aplicando el modelo de “lineal-plateau”.
Como se observa en la figura 11, la res-puesta del iC de los machos enteros ante un nivel creciente de lisina digestible se alargó más que en las hembras, debido al mayor potencial de deposición de proteína. Esto quiere decir que para aprovechar todo el potencial de machos y hembras, y ser econó-micamente óptimo al mismo tiempo, habría que alimentar separadamente a machos
Figura 8. Predicción de la retención de proteína máxima en machos enteros, castrados y hembras (Miller, 2002).
0,25-
0,20-
0,15-
0,10-
0,05-
0-
Livewight (kg) (6 times protein mass)
max
imum
rat
e of
pro
tein
ret
entio
n (k
g/d)
0 150100 200 25050 300
maleFemaleCastrate
Figura 9. Relación entre deposición de proteína y consumo de energía digestible (NRC, 1998).
|
0|
1|
2|
3|
4|
5|
6|
7|
8|
9|
10|
11|
12
Digestible Energy intake (mcal/d)
Who
le B
ody
Prot
ein
Gai
n (g
/d)
Pig Weight (kg)
200 -
150 -
100 -
50 -
0 -
-50 -
-100 -
5255075100150
Figura 10. Efecto del sexo sobre el nitrógeno retenido (Lizaso, 1994).
|
1|
2|
3|
4|
5|
6|
7|
8|
9|
10|
11|
12
Em ingerida mcal/d
N r
eten
ido
g/d
30 -
25 -
20 -
15 -
10 -
5 -
Hembras 40 kg PV
machos 40 kg PV
AÑO 7
• n
º39
Ganadería sostenible
Revista de Medicina VETERINARIA
OPCIONES PARA EL CONTROL DE LAS ENFERMEDADES COMPARTIDAS CON LA FAUNA SILVESTRE
ENFERMEDADES ENTÉRICAS EN LECHONES LACTANTES
COMPOSICIÓN Y VALOR NUTRITIVO DEL GUISANTEEN LA ALIMENTACIÓN PORCINA
TRATAMIENTO Y CONTROL DEL SÍNDROME DE DISGALAXIA POSTPARTO EN CERDAS
ESTRATEGIAS DE MANEJO DE LOTES DE VACAS DURANTE EL PERÍODO DE TRANSICIÓN
HIDROCEFALIA EN UN TERNERO
INDIGESTIÓN VAGAL: CASO CLÍNICO
ColimutinaSolución inyectable
La mejor combinación sinérgica para porcinoTiamulina + Colistina
ComposiciónTiamulina (fumarato ácido)...... 10 g(equivalente a 12,35 g de fumarato ácido de tiamulina)Colistina...... 18.000.000 U.l.Disolvente c.s.p. ...... 100 mlEspecies de destino e indicacionesPorcino: Tratamiento de la disentería procina producida por B. hyodysenteriae. Neumonía enzoótica producida por M. hyopneumoniae. Tratamiento de artritis producida por M. hyosynoviae. Colibacilosis. Ileitis.Bovino y ovino: Micoplasmosis y colibacilosis.Posología y modo de administraciónVía intramuscular.Tratamiento de la disentería procina, micoplasmosis bovina y ovina y colibacilosis: 1 ml de COLIMUTINA INYECTABLE / 10 kg p.v. / día, durante 3 días.Neumonía enzoótica y artritis micoplásmica: 1,5 ml de COLIMUTINA INYECTABLE / 10 kg p.v. / día, durante 3 días.En animales pequeños, especialmente recomendamos diluir el producto en suero gluco-sado para no sobrepasar las dosis indicadas.Tiempo de esperaCarne: 21 díasRegistro n.º: 9.134Envase de 100 y 250 ml
Ctra. Reus-Vinyols Km. 4,1 - Ap. Correos, 60 - Teléfono 977 850 170* - Fax 977 850 405 - 43330 RIUDOMS (Tarragona)
www.spveterinaria.com
LA SOLUCIÓN CON EL ESPECTRO MÁS COMPLETO
EN PROBLEMAS RESPIRATORIOS Y
DIGESTIVOS EN GANADO PORCINO
an_colimutina.indd 1 28/04/11 12:08 Cubierta CyS39.indd 1 16/11/11 12:19CYS 42.indb 34 14/05/12 10:10

35
nº 42Programas de alimentación en porcino en función del sexo: efectos de la castración quirúrgica y la inmunocastración
AÑO 7
• n
º39
Ganadería sostenible
Revista de Medicina VETERINARIA
OPCIONES PARA EL CONTROL DE LAS ENFERMEDADES COMPARTIDAS CON LA FAUNA SILVESTRE
ENFERMEDADES ENTÉRICAS EN LECHONES LACTANTES
COMPOSICIÓN Y VALOR NUTRITIVO DEL GUISANTEEN LA ALIMENTACIÓN PORCINA
TRATAMIENTO Y CONTROL DEL SÍNDROME DE DISGALAXIA POSTPARTO EN CERDAS
ESTRATEGIAS DE MANEJO DE LOTES DE VACAS DURANTE EL PERÍODO DE TRANSICIÓN
HIDROCEFALIA EN UN TERNERO
INDIGESTIÓN VAGAL: CASO CLÍNICO
ColimutinaSolución inyectable
La mejor combinación sinérgica para porcinoTiamulina + Colistina
ComposiciónTiamulina (fumarato ácido)...... 10 g(equivalente a 12,35 g de fumarato ácido de tiamulina)Colistina...... 18.000.000 U.l.Disolvente c.s.p. ...... 100 mlEspecies de destino e indicacionesPorcino: Tratamiento de la disentería procina producida por B. hyodysenteriae. Neumonía enzoótica producida por M. hyopneumoniae. Tratamiento de artritis producida por M. hyosynoviae. Colibacilosis. Ileitis.Bovino y ovino: Micoplasmosis y colibacilosis.Posología y modo de administraciónVía intramuscular.Tratamiento de la disentería procina, micoplasmosis bovina y ovina y colibacilosis: 1 ml de COLIMUTINA INYECTABLE / 10 kg p.v. / día, durante 3 días.Neumonía enzoótica y artritis micoplásmica: 1,5 ml de COLIMUTINA INYECTABLE / 10 kg p.v. / día, durante 3 días.En animales pequeños, especialmente recomendamos diluir el producto en suero gluco-sado para no sobrepasar las dosis indicadas.Tiempo de esperaCarne: 21 díasRegistro n.º: 9.134Envase de 100 y 250 ml
Ctra. Reus-Vinyols Km. 4,1 - Ap. Correos, 60 - Teléfono 977 850 170* - Fax 977 850 405 - 43330 RIUDOMS (Tarragona)
www.spveterinaria.com
LA SOLUCIÓN CON EL ESPECTRO MÁS COMPLETO
EN PROBLEMAS RESPIRATORIOS Y
DIGESTIVOS EN GANADO PORCINO
an_colimutina.indd 1 28/04/11 12:08 Cubierta CyS39.indd 1 16/11/11 12:19CYS 42.indb 35 14/05/12 10:10
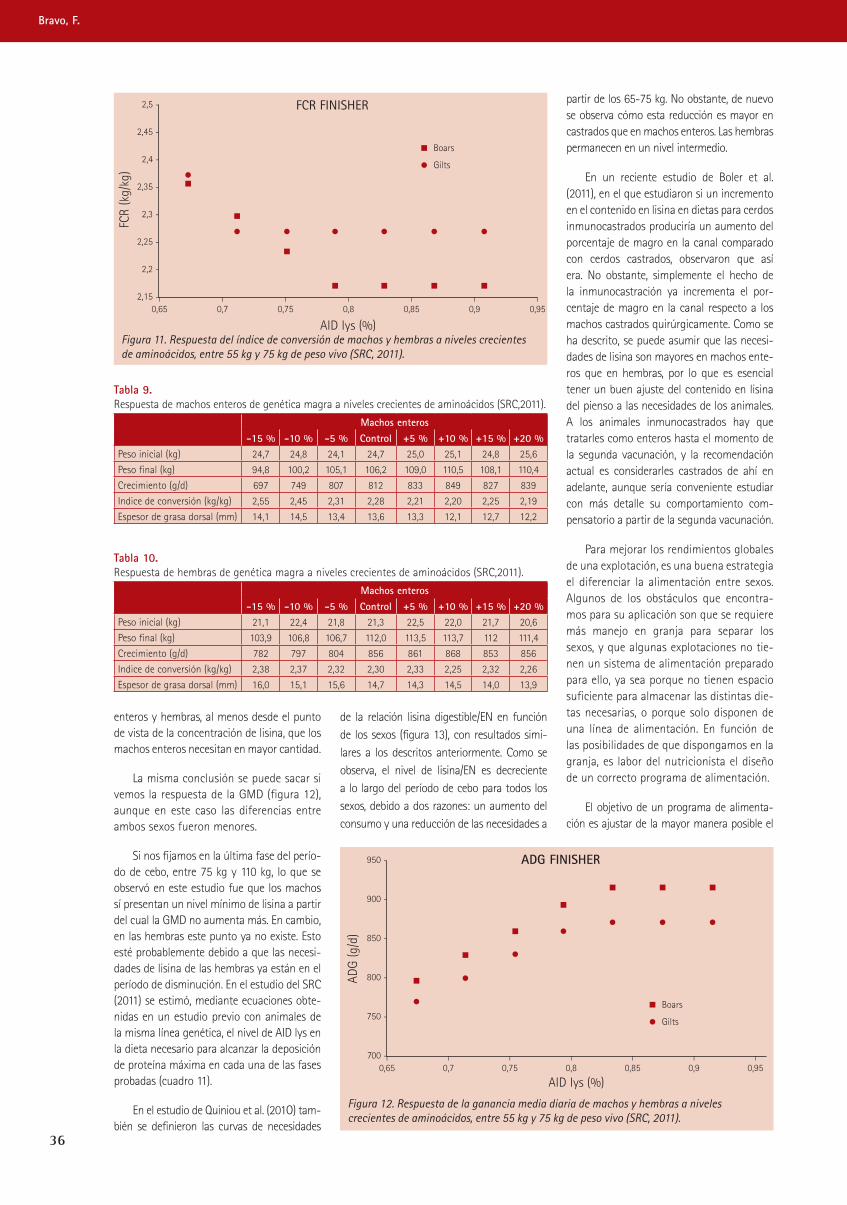
36
enteros y hembras, al menos desde el punto de vista de la concentración de lisina, que los machos enteros necesitan en mayor cantidad.
La misma conclusión se puede sacar si vemos la respuesta de la GmD (figura 12), aunque en este caso las diferencias entre ambos sexos fueron menores.
Si nos fijamos en la última fase del perío-do de cebo, entre 75 kg y 110 kg, lo que se observó en este estudio fue que los machos sí presentan un nivel mínimo de lisina a partir del cual la GmD no aumenta más. En cambio, en las hembras este punto ya no existe. Esto esté probablemente debido a que las necesi-dades de lisina de las hembras ya están en el período de disminución. En el estudio del SrC (2011) se estimó, mediante ecuaciones obte-nidas en un estudio previo con animales de la misma línea genética, el nivel de AiD lys en la dieta necesario para alcanzar la deposición de proteína máxima en cada una de las fases probadas (cuadro 11).
En el estudio de quiniou et al. (201o) tam-bién se definieron las curvas de necesidades
de la relación lisina digestible/EN en función de los sexos (figura 13), con resultados simi-lares a los descritos anteriormente. Como se observa, el nivel de lisina/EN es decreciente a lo largo del período de cebo para todos los sexos, debido a dos razones: un aumento del consumo y una reducción de las necesidades a
partir de los 65-75 kg. No obstante, de nuevo se observa cómo esta reducción es mayor en castrados que en machos enteros. Las hembras permanecen en un nivel intermedio.
En un reciente estudio de Boler et al. (2011), en el que estudiaron si un incremento en el contenido en lisina en dietas para cerdos inmunocastrados produciría un aumento del porcentaje de magro en la canal comparado con cerdos castrados, observaron que así era. No obstante, simplemente el hecho de la inmunocastración ya incrementa el por-centaje de magro en la canal respecto a los machos castrados quirúrgicamente. Como se ha descrito, se puede asumir que las necesi-dades de lisina son mayores en machos ente-ros que en hembras, por lo que es esencial tener un buen ajuste del contenido en lisina del pienso a las necesidades de los animales. A los animales inmunocastrados hay que tratarles como enteros hasta el momento de la segunda vacunación, y la recomendación actual es considerarles castrados de ahí en adelante, aunque sería conveniente estudiar con más detalle su comportamiento com-pensatorio a partir de la segunda vacunación.
Para mejorar los rendimientos globales de una explotación, es una buena estrategia el diferenciar la alimentación entre sexos. Algunos de los obstáculos que encontra-mos para su aplicación son que se requiere más manejo en granja para separar los sexos, y que algunas explotaciones no tie-nen un sistema de alimentación preparado para ello, ya sea porque no tienen espacio suficiente para almacenar las distintas die-tas necesarias, o porque solo disponen de una línea de alimentación. En función de las posibilidades de que dispongamos en la granja, es labor del nutricionista el diseño de un correcto programa de alimentación.
El objetivo de un programa de alimenta-ción es ajustar de la mayor manera posible el
Bravo, F.
Tabla 9.respuesta de machos enteros de genética magra a niveles crecientes de aminoácidos (SrC,2011).
Machos enteros-15 % -10 % -5 % Control +5 % +10 % +15 % +20 %
Peso inicial (kg) 24,7 24,8 24,1 24,7 25,0 25,1 24,8 25,6
Peso final (kg) 94,8 100,2 105,1 106,2 109,0 110,5 108,1 110,4
Crecimiento (g/d) 697 749 807 812 833 849 827 839
indice de conversión (kg/kg) 2,55 2,45 2,31 2,28 2,21 2,20 2,25 2,19
Espesor de grasa dorsal (mm) 14,1 14,5 13,4 13,6 13,3 12,1 12,7 12,2
Tabla 10. respuesta de hembras de genética magra a niveles crecientes de aminoácidos (SrC,2011).
Machos enteros-15 % -10 % -5 % Control +5 % +10 % +15 % +20 %
Peso inicial (kg) 21,1 22,4 21,8 21,3 22,5 22,0 21,7 20,6
Peso final (kg) 103,9 106,8 106,7 112,0 113,5 113,7 112 111,4
Crecimiento (g/d) 782 797 804 856 861 868 853 856
indice de conversión (kg/kg) 2,38 2,37 2,32 2,30 2,33 2,25 2,32 2,26
Espesor de grasa dorsal (mm) 16,0 15,1 15,6 14,7 14,3 14,5 14,0 13,9
Figura 12. Respuesta de la ganancia media diaria de machos y hembras a niveles crecientes de aminoácidos, entre 55 kg y 75 kg de peso vivo (SRC, 2011).
|
0,65|
0,7|
0,75|
0,8|
0,85|
0,9|
0,95
950 -
900 -
850 -
800 -
750 -
700 -
AiD iys (%)
ADG
(g/d
)
Gilts
Boars
ADG FINISHER
Figura 11. Respuesta del índice de conversión de machos y hembras a niveles crecientes de aminoácidos, entre 55 kg y 75 kg de peso vivo (SRC, 2011).
|
0,65|
0,7|
0,75|
0,8|
0,85|
0,9|
0,95
2,5 -
2,45 -
2,4 -
2,35 -
2,3 -
2,25 -
2,2 -
2,15 -
AiD iys (%)
FCr
(kg/
kg) Gilts
Boars
FcR FINISHER
CYS 42.indb 36 14/05/12 10:11
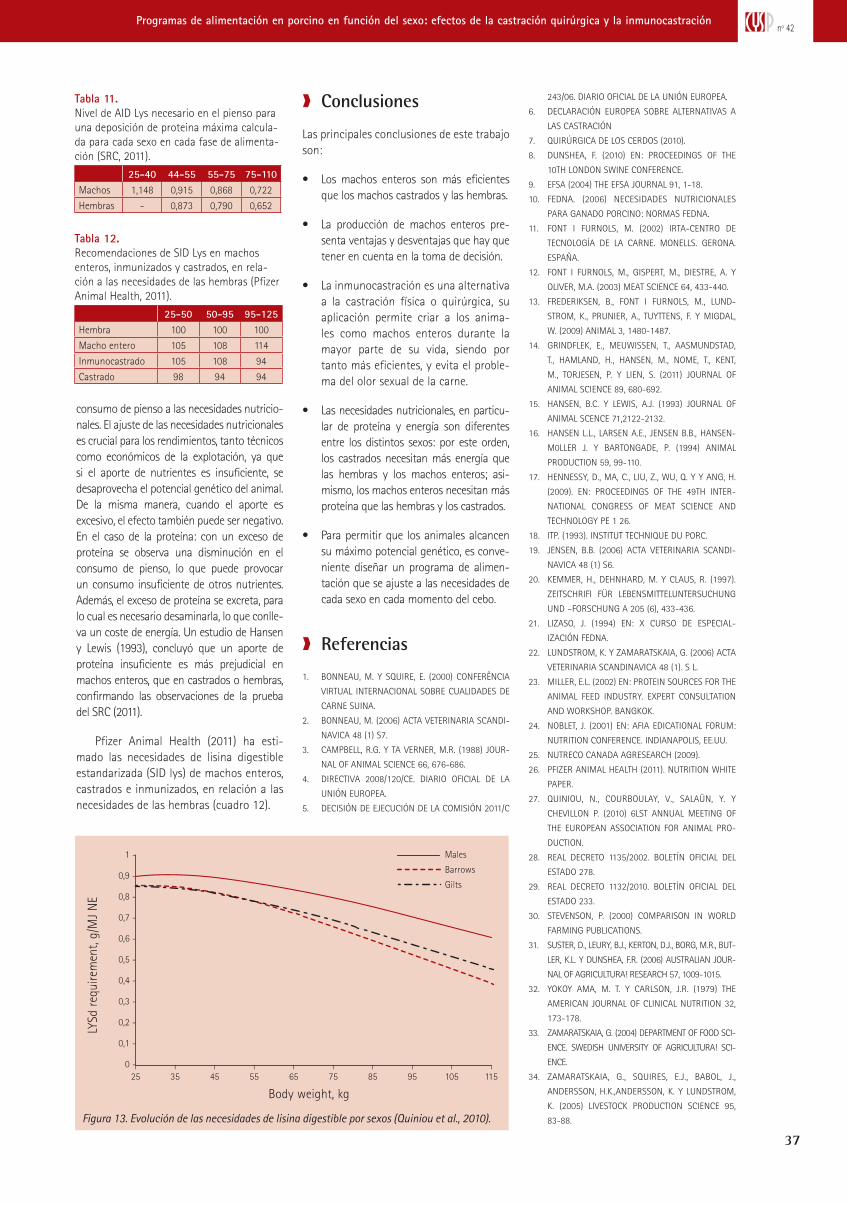
37
nº 42Programas de alimentación en porcino en función del sexo: efectos de la castración quirúrgica y la inmunocastración
Figura 13. Evolución de las necesidades de lisina digestible por sexos (Quiniou et al., 2010).
|
25|
35|
45|
55|
65|
75|
85|
95|
105|
115
Body weight, kg
LYSd
req
uire
men
t, g/
mJ
NE
1 -
0,9 -
0,8 -
0,7 -
0,6 -
0,5 -
0,4 -
0,3 -
0,2 -
0,1 -
0 -
males
Barrows
Gilts
Tabla 11. Nivel de AiD Lys necesario en el pienso para una deposición de proteina máxima calcula-da para cada sexo en cada fase de alimenta-ción (SrC, 2011).
25-40 44-55 55-75 75-110machos 1,148 0,915 0,868 0,722
Hembras - 0,873 0,790 0,652
Tabla 12.recomendaciones de SiD Lys en machos enteros, inmunizados y castrados, en rela-ción a las necesidades de las hembras (Pfizer Animal Health, 2011).
25-50 50-95 95-125Hembra 100 100 100
macho entero 105 108 114
inmunocastrado 105 108 94
Castrado 98 94 94
consumo de pienso a las necesidades nutricio-nales. El ajuste de las necesidades nutricionales es crucial para los rendimientos, tanto técnicos como económicos de la explotación, ya que si el aporte de nutrientes es insuficiente, se desaprovecha el potencial genético del animal. De la misma manera, cuando el aporte es excesivo, el efecto también puede ser negativo. En el caso de la proteína: con un exceso de proteína se observa una disminución en el consumo de pienso, lo que puede provocar un consumo insuficiente de otros nutrientes. Además, el exceso de proteína se excreta, para lo cual es necesario desaminarla, lo que conlle-va un coste de energía. Un estudio de Hansen y Lewis (1993), concluyó que un aporte de proteína insuficiente es más prejudicial en machos enteros, que en castrados o hembras, confirmando las observaciones de la prueba del SrC (2011).
Pfizer Animal Health (2011) ha esti-mado las necesidades de lisina digestible estandarizada (SiD lys) de machos enteros, castrados e inmunizados, en relación a las necesidades de las hembras (cuadro 12).
❱❱ Conclusiones
Las principales conclusiones de este trabajo son:
• Los machos enteros son más eficientes que los machos castrados y las hembras.
• La producción de machos enteros pre-senta ventajas y desventajas que hay que tener en cuenta en la toma de decisión.
• La inmunocastración es una alternativa a la castración física o quirúrgica, su aplicación permite criar a los anima-les como machos enteros durante la mayor parte de su vida, siendo por tanto más eficientes, y evita el proble-ma del olor sexual de la carne.
• Las necesidades nutricionales, en particu-lar de proteína y energía son diferentes entre los distintos sexos: por este orden, los castrados necesitan más energía que las hembras y los machos enteros; asi-mismo, los machos enteros necesitan más proteína que las hembras y los castrados.
• Para permitir que los animales alcancen su máximo potencial genético, es conve-niente diseñar un programa de alimen-tación que se ajuste a las necesidades de cada sexo en cada momento del cebo.
❱❱ Referencias
1. BoNNEAU, m. Y SqUirE, E. (2000) CoNFErêNCiA
VirTUAL iNTErNACioNAL SoBrE CUALiDADES DE
CArNE SUiNA.
2. BoNNEAU, m. (2006) ACTA VETEriNAriA SCANDi-
NAViCA 48 (1) S7.
3. CAmPBELL, r.G. Y TA VErNEr, m.r. (1988) JoUr-
NAL oF ANimAL SCiENCE 66, 676-686.
4. DirECTiVA 2008/120/CE. DiArio oFiCiAL DE LA
UNióN EUroPEA.
5. DECiSióN DE EJECUCióN DE LA ComiSióN 2011/C
243/06. DiArio oFiCiAL DE LA UNióN EUroPEA.
6. DECLArACióN EUroPEA SoBrE ALTErNATiVAS A
LAS CASTrACióN
7. qUirÚrGiCA DE LoS CErDoS (2010).
8. DUNSHEA, F. (2010) EN: ProCEEDiNGS oF THE
10TH LoNDoN SWiNE CoNFErENCE.
9. EFSA (2004) THE EFSA JoUrNAL 91, 1-18.
10. FEDNA. (2006) NECESiDADES NUTriCioNALES
PArA GANADo PorCiNo: NormAS FEDNA.
11. FoNT i FUrNoLS, m. (2002) irTA-CENTro DE
TECNoLoGíA DE LA CArNE. moNELLS. GEroNA.
ESPAñA.
12. FoNT i FUrNoLS, m., GiSPErT, m., DiESTrE, A. Y
oLiVEr, m.A. (2003) mEAT SCiENCE 64, 433-440.
13. FrEDEriKSEN, B., FoNT i FUrNoLS, m., LUND-
STrom, K., PrUNiEr, A., TUYTTENS, F. Y miGDAL,
W. (2009) ANimAL 3, 1480-1487.
14. GriNDFLEK, E., mEUWiSSEN, T., AASmUNDSTAD,
T., HAmLAND, H., HANSEN, m., NomE, T., KENT,
m., TorJESEN, P. Y LiEN, S. (2011) JoUrNAL oF
ANimAL SCiENCE 89, 680-692.
15. HANSEN, B.C. Y LEWiS, A.J. (1993) JoUrNAL oF
ANimAL SCENCE 71,2122-2132.
16. HANSEN L.L., LArSEN A.E., JENSEN B.B., HANSEN-
m0LLEr J. Y BArToNGADE, P. (1994) ANimAL
ProDUCTioN 59, 99-110.
17. HENNESSY, D., mA, C., LiU, z., WU, q. Y Y ANG, H.
(2009). EN: ProCEEDiNGS oF THE 49TH iNTEr-
NATioNAL CoNGrESS oF mEAT SCiENCE AND
TECHNoLoGY PE 1 26.
18. iTP. (1993). iNSTiTUT TECHNiqUE DU PorC.
19. JENSEN, B.B. (2006) ACTA VETEriNAriA SCANDi-
NAViCA 48 (1) S6.
20. KEmmEr, H., DEHNHArD, m. Y CLAUS, r. (1997).
zEiTSCHriFi Fur LEBENSmiTTELUNTErSUCHUNG
UND –ForSCHUNG A 205 (6), 433-436.
21. LizASo, J. (1994) EN: x CUrSo DE ESPECiAL-
izACióN FEDNA.
22. LUNDSTrom, K. Y zAmArATSKAiA, G. (2006) ACTA
VETEriNAriA SCANDiNAViCA 48 (1). S L.
23. miLLEr, E.L. (2002) EN: ProTEiN SoUrCES For THE
ANimAL FEED iNDUSTrY. ExPErT CoNSULTATioN
AND WorKSHoP. BANGKoK.
24. NoBLET, J. (2001) EN: AFiA EDiCATioNAL ForUm:
NUTriTioN CoNFErENCE. iNDiANAPoLiS, EE.UU.
25. NUTrECo CANADA AGrESEArCH (2009).
26. PFizEr ANimAL HEALTH (2011). NUTriTioN WHiTE
PAPEr.
27. qUiNioU, N., CoUrBoULAY, V., SALAuN, Y. Y
CHEViLLoN P. (2010) 6LST ANNUAL mEETiNG oF
THE EUroPEAN ASSoCiATioN For ANimAL Pro-
DUCTioN.
28. rEAL DECrETo 1135/2002. BoLETíN oFiCiAL DEL
ESTADo 278.
29. rEAL DECrETo 1132/2010. BoLETíN oFiCiAL DEL
ESTADo 233.
30. STEVENSoN, P. (2000) ComPAriSoN iN WorLD
FArmiNG PUBLiCATioNS.
31. SUSTEr, D., LEUrY, B.J., KErToN, D.J., BorG, m.r., BUT-
LEr, K.L. Y DUNSHEA, F.r. (2006) AUSTrALiAN JoUr-
NAL oF AGriCULTUrA! rESEArCH 57, 1009-1015.
32. YoKoY AmA, m. T. Y CArLSoN, J.r. (1979) THE
AmEriCAN JoUrNAL oF CLiNiCAL NUTriTioN 32,
173-178.
33. zAmArATSKAiA, G. (2004) DEPArTmENT oF FooD SCi-
ENCE. SWEDiSH UNiVErSiTY oF AGriCULTUrA! SCi-
ENCE.
34. zAmArATSKAiA, G., SqUirES, E.J., BABoL, J.,
ANDErSSoN, H.K.,ANDErSSoN, K. Y LUNDSTrom,
K. (2005) LiVESToCK ProDUCTioN SCiENCE 95,
83-88.
CYS 42.indb 37 14/05/12 10:11

38
luño lázaro, Victoria y Matas Parra, Carmen
Cambios en el espermatozoide desde la eyaculación hasta la fecundación
Victoria luño lázaro1 y Carmen Matas Parra2
1Dpto. Patología Animal (Área de reproducción y obstetricia). Facultad de Veterinaria. Universidad de zaragoza
2Dpto. Fisiología (Fisiología Veterinaria).Facultad de Veterinaria. Universidad de murcia. [email protected]. http://www.um.es/grupo-fisiovet
Foto Axón Comunicación
❱❱ Introducción
Los espermatozoides tras ser eyaculados ascienden por el tracto genital femenino, donde solo una pequeña proporción es capaz de unirse a las células epiteliales del oviducto en el denominado reservorio espermático. Aquí, permanecen hasta el momento de la ovulación en el que son liberados secuencialmente gracias a modi-ficaciones endocrinas, que conllevan una serie de cambios a nivel de membrana plas-mática del epitelio oviductal o en el fluido intraluminal (Fazeli et al., 1999) (Figura 1).
El oviducto juega un papel modulador importante en el proceso reproductivo, controlando y sincronizando a los gametos hasta su unión. La interacción entre los espermatozoides y las células oviductales durante dicho ascenso permite que los espermatozoides adquieran la capacidad fecundante a través de un proceso denomi-nado capacitación, el cual fue descrito por primera vez simultáneamente por Austin y Chang (1951), y le confiere a los esper-matozoides la capacidad de penetrar al ovocito (Figura 2).
Las modificaciones producidas por este proceso tienen varias consecuencias en el gameto. Entre ellas podemos encontrarnos la modificación en el patrón de movimien-to, la generación de radicales libres, el incremento del nivel de calcio, o la modi-ficación de lípidos de membrana (matas et al., 2010). No obstante, parece ser que una de las modificaciones más importantes que se producen es el aumento los niveles de fosforilación en tirosina de las proteínas (Figura 3).
En varias especies de mamíferos la fosforilación en residuos en tirosina ha sido correlacionada con el estado de capa-citación del espermatozoide. (Visconti et al., 1995; 1999; Tadiff et al., 2001; Pommer et al., 2003; Grasa et al., 2006). éste se plantea como uno de los principales criterios para la unión entre las células oviductales y los espermatozoides, representando un meca-nismo de selección de la población esper-mática más competente para la fecunda-ción (Fazeli et al., 1999).
CYS 42.indb 38 14/05/12 10:11

39
nº 42Cambios en el espermatozoide desde la eyaculación hasta la fecundación
NBc: Cotransportador de Na+/HCo3, SAcY: Adenilato Ciclasa Soluble; PkA: Protein kinasa A (Adaptado de Visconti, 2009).
In vitro se han desarrollado diversas técnicas para capacitar a los espermato-zoides intentando imitar las condiciones fisiológicas. Entre ellas se encuentran, los lavados con albúmina, lavados en gradien-tes de diferente densidad o simplemente centrifugación e incubación en un medio rico en calcio y bicarbonato (matás et al., 2003). De la misma forma, median-te cultivos in vitro de células oviductales porcinas (PoEC) se ha intentado simular las condiciones que los espermatozoides encuentran en el tracto genital femenino. Se ha determinado un mayor porcentaje de
espermatozoides unidos con un nivel bajo de fosforilación en tirosina y de concen-tración de calcio intracelular en diferen-tes especies, representando la población más viable destinada a la fecundación (Petrunkina et al., 2001; 2003a; zumoffen et al., 2010). otra posibilidad es el uso de explantes procedentes de oviductos de hembras recién ovuladas en el momento del sacrificio en mataderos, o la utilización de oviductos de hembras que han sido inseminadas bajo condiciones de campo. También se ha visualizado los esperma-tozoides in situ tras inseminaciones en cerda y vaca, demostrándose interacciones entre el epitelio y la membrana plasmática espermática en la zona caudal del istmo (Hunter, 1991).
❱❱ El espermatozoide. Generalidades
El espermatozoide (del griego esperma-semilla, y zoon-animal) es una célula haploide que constituye el gameto mascu-lino y cuya función es la formación de un cigoto totipotente, resultado de la fusión de su núcleo con el del gameto femenino, fenómeno que dará lugar, posteriormente, al embrión y al feto.
La célula espermática tiene 3 regio-nes altamente diferenciadas y especia-lizadas: cabeza, cuello y cola o flagelo (Figura 4). En el caso del verraco la longitud total del espermatozoide es
Figura 1. Transporte espermático desde la eyaculación hasta la fecundación.
Eyaculación
Progesterona
Útero
Adquisición de la motilidad progresiva Supresión de la motilidad progresiva,
seguida de la reactivación coincidente con la ovulación
Hiperactivación espermática
Ovario
Útero
Istmo
AmpollaAmpolla
Figura 2. Secuencia de las modificaciones que experimenta el espermatozoide en el útero y oviducto: 1. Activación 2. Capacitación 3. Hiperactivación 4. Reconocimiento entre gametos 5. Reacción acrosómica 6. Adhesión y 7. Fusión (Olivera et al., 2006).
CYS 42.indb 39 14/05/12 10:11

40
Bravo, F.
de 50-60 µm (Pesch y Bergmann, 2006). A continuación vamos a describir cada una de las regiones que componen el espermatozoide: Ver la figura 4.
Cabeza
Es una estructura de morfología más o menos piriforme en mamíferos. En el verraco posee una longitud de 8.84-8.95 µm y un perímetro de 26.33-27.65 µm (quintero-moreno et al., 2004). En ella podemos encontrar el acrosoma y el núcleo, encontrándose ambas envueltas por la membrana plasmática (Figura 5).
Además, estructuralmente, la cabeza, se puede subdividir en 4 segmentos: apical, pre-ecuatorial, ecuatorial y post-ecuatorial (Figura 6).
• Acrosoma: El acrosoma cubre aproxi-madamente los dos primeros tercios de la cabeza del espermatozoide. Posee una membrana interna y una exter-na que delimita su contenido (Pesch y Bergmann, 2006) (Figura 6). Entre el acrosoma y el núcleo hay un pequeño espacio sub-acrosomal cuya función, aún discutida, parece ser una capa adhesiva que tiende a unir el acrosoma al núcleo subyacente (Fawcett, 1970). El acrosoma contiene enzimas hidrolíticas como la acrosina, hialorunidasa y muchas otras hidrolasas y esterasas. Estas enzimas son esenciales para la lisis de la zona pelúcida y la penetración de las células que rodean al ovocito. Cuando la membrana acroso-mal externa se fusiona con la membrana plasmática, los enzimas son liberados. Este proceso se conoce como reacción acrosómica (rA) (Figura 6).
• Núcleo: El núcleo espermático está formado por ADN y proteínas nuclea-res, todo ello rodeado por la membra-na nuclear. Estas proteínas nucleares tienen funciones tanto estructurales (compactación del ADN) como enzi-máticas (reparación). Aparentemente, la estructura y/o composición de la cromatina en el núcleo de los esper-matozoides puede ser muy variable dependiendo de la especie.
El núcleo espermático se encuentra rodeado por una estructura conocida como teca perinuclear (TP) que es com-primida por al menos 6 proteínas distin-tas (oko y maravei, 1994), y tiene como función la unión de membranas esper-máticas (Korley et al., 1997) y la preser-vación de su integridad (Watson, 2000).
Cuello
El cuello es un segmento de unión corto (1 µm) entre el flagelo y la cabeza esper-mática, compuesto de columnas segmen-tadas y una estructura densa y fibrosa denominada capitulum. Seguido del capi-tulum se encuentran los centriolos proxi-mal y distal. Este último está ausente en el espermatozoide maduro. El cuello sirve como conector y pieza articular (Pesh y Bergmann, 2006).
Cola
La cola es la parte más larga del esper-matozoide; sus componentes estructura-les principales son: el axonema, la vaina mitocondrial, las fibras externas densas y la vaina fibrosa. Puede dividirse en diversas porciones: pieza media, pieza principal y pieza terminal (Figura 7).
La pieza media (Figura 7) tiene una longitud en la especie porcina de 10 µm (Smollich y michel, 1992), y está caracteri-zada por la presencia de una gran cantidad de mitocondrias concentradas en una vaina helicoidal, que rodea al complejo axonema y las fibras densas.
La pieza principal (Figura 7) es el seg-mento más largo (entre 30-50 µm), siendo esta longitud en la especie porcina de 30 µm. La función principal es la de propor-cionar movilidad al espermatozoide (Pesch y Bergmann, 2006).
Figura 3. Esquema de las modificaciones en el espermatozoide durante la capacitación espermática que inducen la fosforilación de la tirosina en proteínas.
Figura 5. A) Partes principales del espermato-zoide: 1) Cabeza, 2) Cola. B) Corte transversal de la cabeza espermática: 3) Acrosoma, 4) Membrana plasmática y 5) Núcleo (http://www.oocities.org/ar/biolembrio/).
Figura 4. Estructura del espermatozoide porcino.
Cabeza
Cuello
Flagelo
Pieza terminal
Cola
Pieza principal
mitocondrias
CentriolosNúcleo
Acrosoma
Segmento intermedio
CYS 42.indb 40 14/05/12 10:11

41
nº 42Cambios en el espermatozoide desde la eyaculación hasta la fecundación
La pieza terminal (Figura 7) es la parte distal del flagelo, es una porción corta y tiene una longitud de 3 µm.
❱❱ Eyaculación
La eyaculación es el reflejo de expulsión del semen fuera del tracto reproductivo a través de la uretra mediante contracciones musculares coordinadas. El cerdo eyacula un volumen que varía entre 200 y 250 ml y se encuentra compuesto por una sus-pensión de espermatozoides en un fluido denominado plasma seminal.
Se diferencian tres fracciones en el eyaculado en función de la procedencia de las mismas (Garner y Hafez, 1996):
1. Fracción pre-espermática: formada por las secreciones procedentes de la prós-tata, las vesículas seminales y de las glándulas bulbouretrales.
2. Fracción espermática: presenta una concentración elevada de espermato-zoides junto con secreciones de la próstata y de las vesículas seminales.
3. Fracción post-espermática: tiene una baja concentración espermática y está constituida por las secreciones de la próstata y de las glándulas bul-bouretrales. éstas últimas producen abundantes corpúsculos gelatinosos denominados comúnmente “tapio-ca”, que van a formar una especie de tapón para sellar el cérvix de la hembra, evitando en la monta natural el reflujo del eyaculado del verraco (Geisert, 1999).
❱❱ Plasma seminal
El plasma seminal es el fluido en el que se encuentran inmersos los espermato-zoides de los mamíferos domésticos. Está compuesto por una mezcla compleja de secreciones procedentes de los testícu-los, el epidídimo y las glándulas sexuales accesorias (vesículas seminales, próstata y glándulas bulbouretrales). El conteni-do de estas glándulas es específico de cada especie y muy variable incluso entre individuos de la misma especie o entre eyaculados del mismo animal. Entre las causas más importantes de esta variación, aparecen diversos procesos patológicos, la estación del año, el estado fisiológico del animal o las variaciones de temperatura (Pérez-Pé et al., 2001; Cardozo et al., 2006; Colenbrander y Kemp, 1990). Entre los compuestos secretados encontramos dis-tintos iones como por ejemplo, el zn2+ que tiene propiedades antibacterianas (Strzezek y Cieresko, 1987) y que en el caso del plasma seminal de verraco alcanza concen-traciones muy altas (0.3-0.7 nm) (Arver y Eliasoson, 1980); y el Ca2+, que participa en los procesos de capacitación espermática y reacción acrosómica (Visconti et al., 2002). También vamos a encontrar distintos sus-tratos energéticos para el metabolismo de la célula, como fructosa, inositol, ácido cítrico y ácido ascórbico (Frei et al., 1990), aminoácidos (ácido glutámico, carnitina, taurina, hipotaurina) que servirán como fuente de energía y de protección contra sustancias oxígeno reactivas y distintas enzimas (proteasas, acrosina, nucleasas, fosfatasa ácida y alcalina y superóxido dismutasa) que intervendrán en la licue-facción seminal, penetración del ovocito
por parte del espermatozoide, digestión de espermatozoides muertos y dañados, y protección contra sustancias oxígeno reactivas (Strzezek, 2002). Por tanto, el papel del plasma seminal no es el de ser un líquido meramente nutritivo y de trans-porte, sino que también posee funciones de protección, regulación de la motilidad y capacitación de los espermatozoides, reconocimiento y unión entre gametos (Johnson et al., 2000), (Figura 9).
De todos los componentes del plasma seminal, parece ser que son las proteínas las que contribuyen de forma más impor-tante en el proceso reproductivo (Töpfer-Petersen, 1999). Además, el plasma seminal va a contener tanto factores que van a prevenir reacciones acrosómicas espontá-neas o prematuras (factores decapacitan-tes) como otros factores que van a propi-ciar la fertilización de los espermatozoides (Thérien et al., 1995). Estudios recientes sugieren que la acción concertada de los factores seminales reguladores del plas-ma, modulan el estado de capacitación de los espermatozoides de las especies de mamíferos. Los efectos biológicos de estos factores del plasma en la funcionalidad espermática, no están hoy en día del todo
Figura 7. A) Dibujo esquemático de un espermatozoide. B) Sección longitudinal de un espermatozoide; C) y H) secciones transversales de la cola del espermato-zoide: (C) pieza media proximal, (D) pieza media distal, E) y F) pieza principal, G) y H) pieza terminal. (1) Membrana plasmá-tica, (2) acrosoma, (3) núcleo, (4) vacuola nuclear, (5) centriolo proximal, (6) mito-condria, (7) fibras externas densas (ODF), (8) túbulo exterior doble, (9) vaina o envol-tura fibrosa; (←) unión de la pieza media con la hélice mitocondrial y la pieza prin-cipal con la vaina fibrosa (Imagen tomada de Pesch y Bergmann, 2006).
Figura 6. A) Esquema de la estructura del espermatozoide. B) Corte sagital de la célula espermática: 1: Membrana espermática, 2: Membrana acrosomal externa, 3: Contenido acrosomal, 4: Membrana acrosomal interna, 5: Núcleo celular, 6: Contenido de ADN altamente condensado, 7: Anillo posterior o placa basal, 8: Pieza media, 9: Mitocondria, 10: Anillo anular, 11: Flagelo. C) 12: Vesícula (fusión de la membrana plasmática con la membrana acrosomal externa, 13: Secreción acrosomal. D) 14: Estructura en horquilla (Imagen de Flesch y Gadella 2000).
1234
5
6
7
8
9
1
1
C.
D.
E.
F.
G.
H.1
1
1
1
2
6
6
3
3
1
4
4
4
4
2
8
7
1
8
9
4
A. B.
CYS 42.indb 41 14/05/12 10:11

42
luño lázaro, Victoria y Matas Parra, Carmen
claros debido en parte a la alta variabilidad en la composición del plasma seminal entre especies, entre machos dentro de la misma especie y entre eyaculados de un mismo macho (Killian et al., 1993; zhu et al., 2000).
❱❱ Ascenso por el tracto genital femenino
Tras la monta o la inseminación artificial, alrededor de 3000 millones de espermato-zoides son depositados en el cérvix uterino, de los cuales solo una pequeña proporción es capaz de alcanzar el oviducto (Figura 9).
Durante el ascenso la mayoría de los espermatozoides (>70%) son eliminados en la luz uterina, parte de ellos mediante el reflujo retrógrado (35%), por lo que solo unos pocos son capaces de atravesar la unión útero-tubárica (rodriguez-.martinez et al., 2005). También se produce una fago-citosis, mediada principalmente por leu-cocitos que migran al lumen procedentes del endometrio en los primeros 30 minu-tos tras la monta (matthijs et al., 2003). Hay varios factores implicados que inician esta repuesta, como la propia distensión uterina, los espermatozoides o el plasma seminal (Bischof et al., 1994). Proteínas como el heterodímero glicosilado PSP-i/
PSP-ii posee actividad inmuno-estimula-dora, y es capaz de inducir la migración de leucocitos (Leshin et al., 1998). Ese periodo de latencia hasta que se produce la migración al lumen uterino, permite que cierta población espermática atraviese el útero y no sea fagocitada (rodríguez-.martínez et al., 2005). Durante dicho ascenso se produce también la progresiva y coordinada modificación de ciertos fac-tores procedentes del plasma seminal que están unidos a la superficie plasmática del espermatozoide (rodríguez-.martínez et al., 2004). Estos, previenen capacitaciones prematuras, intervienen en la tolerancia inmunitaria, a la vez que están involucra-dos en la unión con las células oviductales (rodríguez-martínez, 2009). Finalmente solo una pequeña proporción atravesará la unión útero-tubárica.
Selección de la población espermática
Existen diferentes mecanismos a lo largo del tracto genital femenino que permiten una selección progresiva de la población más apta para fecundar, estableciéndose diferentes subpoblaciones espermáticas. Estas, son parcial o totalmente deficientes en uno o más aspectos fenotípicos o fun-cionales necesarios para participar en los diferentes aspectos de la fecundación. Si inseminamos con la misma concentración de espermatozoides, el número capaz de acceder al oviducto varía de un individuo a otro (Satake et al., 2006), ya que cada macho produce su particular proporción de subpoblaciones (Holt, 2009).
Existen variaciones fenotípicas que determinan diferentes tipos de esperma-tozoides, éstas van desde la forma de la cabeza, la motilidad o la amplitud fla-gelar (Thurston et al., 2001), a otras que involucran la eficiencia de las señales de transducción (Holt y Harrison 2002) y la integridad de la información genética y epigenética que transmite al ovocito (miller, 2007).
El paso por la (UTJ) es uno de los prin-cipales mecanismos de selección (Figura 9), ya que alrededor del 96% de los esper-matozoides localizados en el oviducto se encuentran allí (mburu et al., 1996). Es atravesada por gametos que presentan una motilidad progresiva y una determinada composición bioquímica a nivel de mem-brana. La integridad molecular de la super-ficie del espermatozoide es determinante para su entrada, por lo que sucesos como la capacitación que producen modificacio-nes estructurales, disminuyen la capacidad del espermatozoide de llegar al oviducto y
Figura 8. Representación esquemática de la membrana plasmática bajo distintas circuns-tancias: en el epidídimo, en contacto con el plasma seminal (eyaculado) y en el genital de la hembra (capacitado).
Figura 9. Transporte espermático en el genital femenino y eventos que acontecen en las distintas secciones del tracto reproductivo de la hembra (Modificado de Seger 1999).
4. OviductoCapacitación completada
Hiperactividad
3. Úteroinicio de capacitación
Fagocitos
2. cérvixBarrera espermática
reservorio espermático
1. Transporte inmediatoPérdida retrógrada
FagocitosEntrada en cérvix y útero
5. Fecundaciónreacción acrosómica
Penetración espermáticaFormación de pronúcleos
UT
Lumen
CYS 42.indb 42 14/05/12 10:11

43
nº 42Cambios en el espermatozoide desde la eyaculación hasta la fecundación
unirse a las células oviductales (Fazeli et al., 1999). Se ha determinado que aproxi-madamente el 70 % de los espermatozoi-des en el oviducto están no capacitados (Tienthai et al., 2004). El daño del material genético presente en el espermatozoide ha sido correlacionado con el incremento de la ubiquitinación de la superficie espermática (Sutovsky et al., 2002), lo cual produciría alteraciones en la membrana similares a las ocurridas en procesos como la capacita-ción, lo que disminuiría su paso por la UTJ.
Podemos concluir que la selección de la población espermática que llega a oviducto (Figura 9) está determinada por diferentes factores clasificados como:
• Tipo 1 (factores fijos): específicos del testículo, están determinados gené-ticamente o afectados por las con-diciones como elevada temperatura testicular.
• Tipo 2 (factores variables): influencia-dos por tratamientos como el lavado, la congelación…La motilidad se incluye en ambos porque está determinada por aspectos genéticos y puede ser modifi-cada su expresión.
• Tipo 3 (factores variables): determinada por señales endocrinas dentro del trac-to genital femenino.
Oviducto
El oviducto tiene un papel significativo en el proceso reproductivo de los mamíferos (Figura 10), ya que proporciona un ambiente apropiado para el transporte y maduración de gametos, la fecundación y el desarrollo embrionario temprano (Hunter, 2005). Está dividido en cuatro segmentos funcionales: el infundíbulo, la ampolla, el istmo y la unión útero-tubárica, cada una de ellas con una función determinada (Hunter ,1994). El infundíbulo es el encargado de captar al ovocito en el momento de la ovulación gracias a la estructura que presenta en forma de dedos. De este modo, el gameto es capaz de llegar hasta la ampolla donde tendrá lugar la fecundación. En el istmo se produce la formación del reservorio espermático (Suarez, 2002), que permite el mantenimiento de la población viable apara la fecundación. Por último la unión útero-tubárica representa la conexión con el cuerno uterino y permite regu-lar y controlar el transporte espermático (Hunter, 1991).
Histológicamente el oviducto consta de tres capas, una serosa externa, una muscular media y una mucosa interna.
El epitelio está formado por células cilia-das y secretoras, distribuidas en diferentes proporciones según la región del oviducto (Gandolfi, 1995). Así, en el infundíbulo y la ampolla se encuentran en mayor pro-porción las células ciliadas y en menor las secretoras, mientras que en la región del istmo ocurre lo contrario (Abe, 1996). Se ha demostrado la presencia de receptores para estrógenos y progesterona a lo largo de todo el oviducto (Amso et al., 1994), que modulan la cantidad celular e intensidad de movimiento ciliar según la fase del ciclo estral. Las células secretoras del istmo acu-mulan gránulos secretores durante la fase folicular, los cuales son eliminados al fluido folicular tras la ovulación (Abe, 1996). Estas células vierten al lumen diferentes macro-moléculas (Abe, 1996), que proporcionan el ambiente idóneo para las diferentes fases del proceso de fecundación.
Fluido oviductal
El fluido oviductal representa el medio acuo-so en el que gametos y embriones están sus-pendidos durante su recorrido por el oviduc-to. Está formado por una secreción específica de origen epitelial y un exudado procedente del plasma sanguíneo (Leese et al., 2001). Sin embargo, posee una concentración iónica, pH, osmolaridad y contenido macromolecular diferente a la que se encuentra en el plasma sanguíneo (Leese, 1988). El pH, determinado principalmente por la concentración de bicar-bonato, varían según el momento del ciclo estral y la región oviductal (zhou et al., 2005). En la cerda, hacia mitad del ciclo, el pH fluc-túa entre 0,3 y 0,7 unidades entre la ampolla y el istmo respectivamente, aumentando en el momento de la ovulación hasta 0,4 uni-dades (Nichol et al., 1997). Estas variaciones pueden tener un gran significado, dado que
ese pH alcalino podría influenciar en la moti-lidad y capacitación espermática cuando está en la proximidad el gameto femenino.
El fluido oviductal también varía en volumen y composición según el momento del ciclo, bajo influencia hormonal (Leese et al., 2001). Está compuesto por una varia-da cantidad de proteínas, entre las que destacan un grupo de glicoproteínas espe-cificas del oviducto (oGP) u oviductinas (Killian, 2004), que podrían estar involucra-das en modificaciones estructurales de la zP del ovocito (Hunter, 1994). También posee un alto contenido en glicosaminoglicanos (GAG), sulfatados (acido hialurónico) y no sulfatados (Tienthai et al., 2000). Las con-centraciones varían según el momento del ciclo estral y entre las regiones oviductales. Estas se incrementan durante los momentos preovulatorios, principalmente a nivel del istmo en el cerdo (Tienthai et al., 2000). Parece que están involucrados en la super-vivencia espermática la capacitación y la unión-liberación del reservorio espermático (rodríguez-martínez et al., 2001; 2005).
Reservorio espermático
En la porción caudal del istmo oviductal se produce la formación del denomina-do reservorio oviductal (Figura10) durante el periodo pre y peri-ovulatorio (Suarez et al., 1991; Töpfer-Petersen et al., 2002). La permanencia de los espermatozoides depende del momento del ciclo estral, variando entre 36 y 48 horas (Hunter, 1984), situándose tanto en el lumen como unidos al epitelio oviductal, principalmente a las células ciliadas (mburu et al., 1996).
Tiene carácter multifuncional, ya que modula la capacitación espermática, man-
Figura 10. Esquema del estado de los espermatozoides en las distintas secciones del oviducto.
CYS 42.indb 43 14/05/12 10:11

44
Luño Lázaro, Victoria y Matas Parra, Carmen
tiene la viabilidad del gameto masculino, sincroniza el tiempo de maduración esper-mática con el momento de la ovulación y selecciona los espermatozoides con mejor morfología (Töpfer-Petersen, 1999). La unión con las células oviductales permite que un número adecuado de gametos viables y fértiles estén disponibles para la fecundación en el momento que tiene lugar la ovulación, y por otro reducir el riesgo de polispermia al permitir que un número reducido de espermatozoides asciendan a la vez hasta el lugar de fecundación.
Presenta una capacidad de unión selec-tiva, escogiendo la población espermática más competente para la fecundación, la cual presenta determinadas características relacionadas con la morfología, motili-dad, integridad de membrana, nivel de calcio citosólico o estado de capacitación y fosforilación en tirosina de las pro-teínas (Ellington et al., 1998; Fazeli et al., 1999; Petrunkina et al., 2001; Gualtieri y Talevi, 2003).
Varios factores pueden estar involucra-dos en la formación del reservorio esper-mático. La unión se produce por contacto directo mediante una interacción ligando-receptor entre las moléculas presentes en la cara rostral del espermatozoide y car-bohidratos de la membrana plasmática de las células oviductales de manera especie-especifico (Lefebvre y Suarez, 1996; Suarez et al., 1998). Dicha unión es un proce-so reversible que involucra en todas las especies estudiadas a diferentes azúcares (Dobrinski et al., 1996a, Suarez et al., 1998). La relativa contribución de otros factores como las secreciones mucosas, las propie-dades químicas del fluido oviductal o los gradientes de temperatura, podría contri-buir en diferente grado a la formación del reservorio.
En el porcino las espermadesinas, en especial AQN1, se encuentran íntimamen-te unidas a través de fosfolípidos a la
Cambios en los patrones de movimiento: hiperactivación
El espermatozoide adquiere la capacidad de mover el flagelo en su tránsito por el epidí-dimo, pero el movimiento empieza después de la eyaculación. Este proceso es conocido como “activación del espermatozoide”. El movimiento del flagelo es característico y consiste en un batido simétrico de la cola que hace que el espermatozoide se despla-ce de forma progresiva. El semen pasa rápi-damente a través del cuello y el útero de la hembra; a los 10 minutos de su deposición encontramos espermatozoides en la unión útero-tubárica. De aquí, los espermatozoi-des pasan al reservorio del oviducto (istmo) caracterizado por cilios luminares epitelia-les y plegamientos o criptas de la mucosa oviductal (Figura 11).
Los espermatozoides son retenidos en las criptas oviductales y allí pierden los fac-tores decapacitantes que habían aportado las glándulas accesorias, experimentando la capacitación.
In vivo, los espermatozoides de verra-co se acumulan en el istmo del oviducto uniéndose a la membrana de las célu-las del epitelio (Suarez, 2002; Tienthai et al., 2004). Después, estos espermatozoides pueden ser liberados de este lugar cuan-do son capacitados (Fazeli et al., 1999; Rodríguez-Martínez et al., 2001). El proceso de capacitación espermática está asociado al desarrollo de un patrón de movimiento diferente conocido con el nombre de hipe-ractivación y que se observa en el momen-to cercano a la fecundación. Se considera que la hiperactivación es crítica para la fecundación (Stauss et al., 1995) porque permite al espermatozoide desprenderse de la pared del oviducto, nadar en el lumen del oviducto o penetrar las células del cumulus oophorus para alcanzar el ovocito. Además, se va a considerar como uno de los pará-metros más importantes en la valoración de la capacidad fecundante de un eyacula-do (Flowers, 1997). Este movimiento apare-ce en diferentes condiciones físicas y varía según especies, pero básicamente consiste en un movimiento no lineal de la cabeza espermática, con aumento de la amplitud de batido de la cola y movimientos asimé-tricos (Yanagimachi, 1994), una marcada trayectoria en giro y baja progresión del movimiento (Figura 12).
La hiperactividad de los espermato-zoides ha sido observada in vitro en varias especies de mamíferos, sobre todo si se induce la capacitación de los espermatozoi-des con Ca2+ y HCO3
- (Yanagimachi, 1994). Para ello se han utilizado sistemas de aná-lisis de imagen computerizado (CASA) que
membrana espermática del espermatozoi-de (Calvete et al., 1997), reconociendo diferentes carbohidratos como manosa y galactosa (Ekhlasi-Hundriesen et al., 2005). La adición de AQN1 inhibe in vitro la unión del espermatozoide a las células epitelia-les de explantes oviductales de manera dosis dependiente (Ekhlasi-Hundriesen et al., 2005). Además, los sitios de unión están localizados en la región rostral de la cabeza del espermatozoide, perdiéndose duran-te la capacitación espermática (Ekhlasi-Hundriesen et al., 2005).
Mediante el análisis de las células de la superficie oviductal se ha deter-minado la interacción de la membrana periacrosomal del espermatozoide con una proteína presente en las células epi-teliales del istmo (Marini y Cabada 2003), la proteína de unión espermática (SBG). Está localizada en la cara apical, cerca del lumen oviductal (Pérez et al., 2006), exponiendo el disacárido o-unido Galβ1-3GalNac, el cual tiene capacidad de unión con la espermadesina AQN1 (Marini y Cabada 2003). SBG podría estar involu-crada en una selección espermática nega-tiva, ya que esta proteína solo se une a espermatozoides capacitados, alterando la integridad acrosomal y suprimiendo la motilidad (Teijeiro et al., 2008). Además produce fosforilación en tirosina de una proteína con peso molecular de 97 KDa, que podría estar involucrada en el meca-nismo de acción de la proteína (Marini y Cabada 2003). Las anexinas aisladas de las células del oviducto porcino también presentan afinidad por proteínas de la membrana periacrosomal. En particular, la anexina A2, localizada en la cara apical del epitelio oviductal puede estar involucrada en la formación del reservorio espermá-tico a través de la interacción con AQN1 (Teijeiro et al., 2009). Otras proteínas pre-sentes en la membrana apical del epitelio oviductal, como HSPA8 tienen capacidad de unión a los glicolípidos sulfatados a la región rostral del espermatozoide, además de aumentar la capacidad de superviven-cia del espermatozoide (Elliot et al., 2009).
Figura 11. Esquema de los diferentes tipos de movimiento del espermatozoide según su localización: a) epidídimo, b) eyaculado y c) en el oviducto (hiperactivado).
a b c Figura 12. Representación esquemática del movimiento de los espermatozoides capacitados y no capacitados.
Parámetros de movilidad regulares
Alta frecuenciaBaja amplitud
Baja frecuenciaGran amplitud
Parámetros de movilidad post-capacitación
Porcino semen del cerdo.indd 44 14/05/12 11:10

45
nº 42Cambios en el espermatozoide desde la eyaculación hasta la fecundación
permiten evaluar tanto el tipo de movi-miento como la velocidad de las células (Figura 13).
Presumiblemente, el inicio de la hipe-ractivación se debe a una señal que se pro-duce en el oviducto en un momento exacto, pero todavía no ha sido identificada.
Fosforilación en tirosina de las proteínas espermáticas
La fosforilación de las proteínas es un evento implicado en la regulación de una gran variedad de funciones celulares como el crecimiento, el control del ciclo celular, cambios en el citoesqueleto, exocitosis, etc. (Pawson, 2004). La adición y/o elimina-ción de los grupos fosfato a aminoácidos de distintas proteínas es un mecanismo que tiene como resultado la producción de cambios conformacionales de dichas proteínas, conduciendo a su activación o inactivación. La fosforilación puede ocurrir en los residuos de serina, treonina, y tiro-sina de las proteínas. Sin embargo, la más ampliamente descrita y estudiada ha sido la fosforilación en tirosina, ya que puede ser el primer indicador de las vías de seña-lización de transducción en las células (Naz y Rajesh, 2004).
En espermatozoides maduros se pro-ducen fenómenos de fosforilación/desfos-forilación de proteínas en la regulación de fenómenos importantes, como la capa-citación espermática, la hipermotilidad y la reacción acrosómica que van a ser indispensables par a que el espermatozoi-de pueda alcanzar y penetrar al ovocito (Galantino- Homer et al., 1997).
En varias especies de mamíferos la fosforilación en residuos en tirosina ha sido correlacionada con el estado de capa-citación del espermatozoide. (Visconti et al., 1995; 1999; Tadiff et al., 2001; Pommer et al., 2003; Grasa et al., 2006). Dichos autores observaron un incremento tiem-po-dependiente en la fosforilación de la tirosina de varias proteínas específicas en
rangos moleculares de 40-120 kDa, con el estado de capacitación de los espermato-zoides. Evidenciaron que la fosforilación de la tirosina es un evento importante en la modulación de los cambios bioquímicos asociados a la capacitación.
Se han descrito numerosas proteínas en las que se produce la fosforilación en tirosina en las diferentes especies animales. En la cerda se ha descrito la fosforilación de la tirosina de 2 sets de proteínas de la membrana plasmática, las mayores (27 kDa, 37 kDa y 40 kDa) y las menores (34 kDa, 47 kDa y 55 kDa) (Flesch et al, 1999) (Figura 14).
La fosforilación en tirosina de las pro-teínas espermáticas ha sido localizada principalmente a nivel del flagelo, estando relacionada la fosforilación de la pieza principal con la adquisición de la hiper-motilidad (Nassar et al., 1999; Petrunkina et al., 2003). Además, la localización de determinadas proteínas fosforiladas en el área acrosomal pueden estar relacionadas con la interacción con la zona pelúcida y la penetración (Dube et al., 2005). También se ha descrito la aparición de un área triangular dentro del segmento ecuatorial en la unión con la zona postacrosomal, el subsegmento ecuatorial (Figura 15). Éste, presenta proteínas fosforiladas en residuos en tirosina, las cuales podrían jugar un
papel clave como centro organizador de ensamblaje de complejos multi-molecula-res mediando en la fusión de gametos y la reacción acrosómica (Jones et al., 2008).
Modulación de la capacitación espermática en el oviducto
Tras la ascensión por el tracto genital feme-nino, los gametos van a quedar retenidos en el reservorio espermático donde completarán el periodo de capacitación (Figura 2). Como es un proceso de desestabilización gradual controlado, que reduce la vida fértil del espermatozoide (Harrison, 1996), el manteni-miento de la viabilidad durante cierto tiempo y el control de la capacitación son pro-cesos que están íntimamente relacionados (Töper-Petersen 1999; Hunter y Rodríguez-Marínez, 2004). Se produce una sincroni-zación entre el momento de ovulación y la adquisición de la capacidad fecundante por parte del espermatozoide, gracias a la unión con las células oviductales en el reser-vorio espermático (Dobrinski et al., 1996b; Petrunkina et al., 2007) (Figura 16).
El estado de capacitación se plan-tea como uno de los principales criterios para la unión entre las células oviducta-les y los espermatozoides, representando un mecanismo de selección de la pobla-ción espermática más competente para la fecundación (Fazeli et al., 1999). También se determinan otros aspectos relacionados con el estado o morfología del espermato-zoide. Existe una correlación negativa entre el porcentaje de espermatozoides unidos y la presencia de alteraciones morfológi-cas o gotas citoplasmáticas (Petrunkina et al., 2001). Es necesario cierto estado de maduración o integridad morfológica para que se produzca la interacción. Además los espermatozoides unidos presentan bajas concentraciones de calcio (Petrunkina et al., 2001), lo que se correlaciona con estadíos de baja capacitación. Se produce una supresión de la capacitación hasta el momento de la ovulación (Rodríguez-Martínez et al., 2001).
Figura 13. Sistema de análisis computerizado de la motilidad espermática. a) microscopio de contraste de fases con placa térmica adaptado a un ordenador, b) imagen del software empleado para la valoración del movimiento espermático mediante análisis de imagen (CASA).
a b
Figura 14. Imagen 1: espermatozoides lavados con proteínas fosforiladas en tirosina evaluados mediante inmunofluorescencia indirecta (1a microscopia de contraste de fases, 1b microscopia de fluorescencia). En la imagen 2 se observan diferencias en las proteínas fosforiladas en tirosina de los espermatozoides lavados (L) y eyaculados (E) sin tratar evaluados mediante western blot.
1a 1b 2
Porcino semen del cerdo.indd 45 14/05/12 11:10
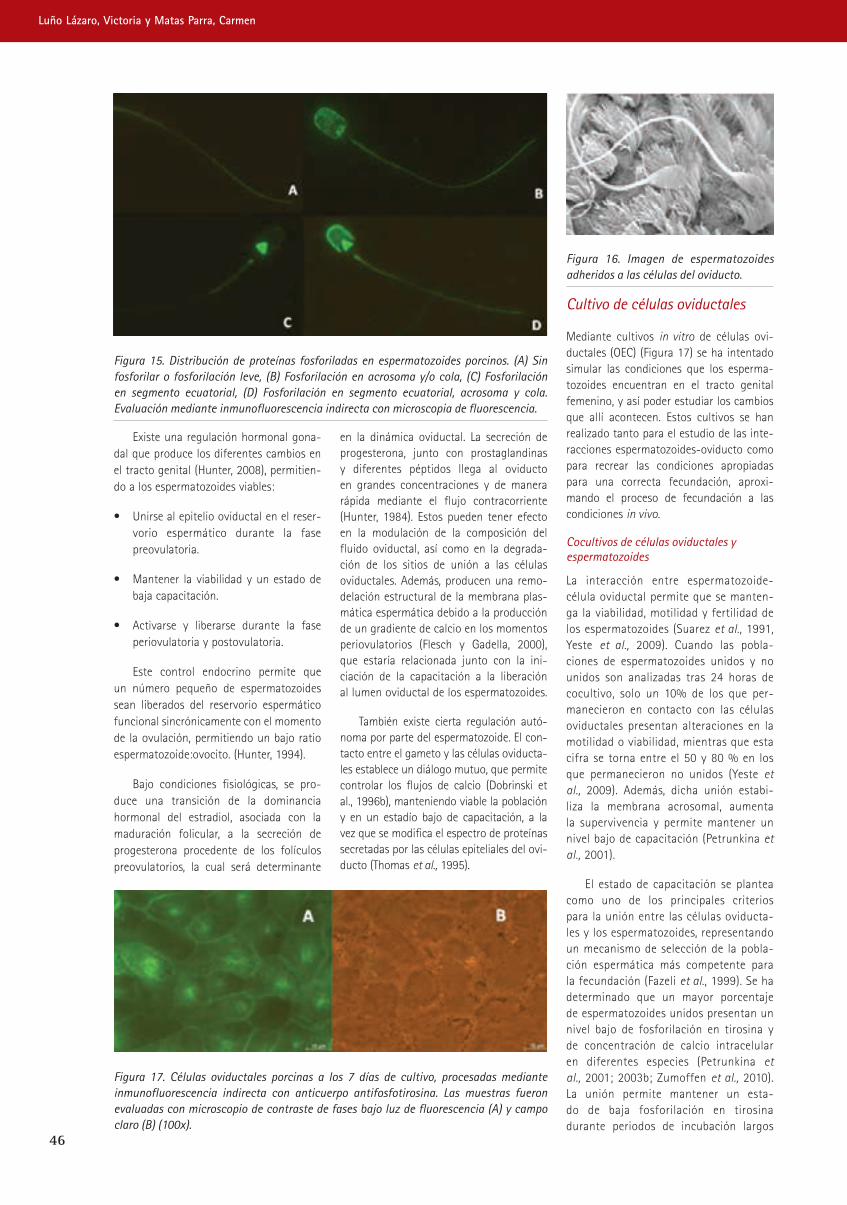
46
luño lázaro, Victoria y Matas Parra, Carmen
Existe una regulación hormonal gona-dal que produce los diferentes cambios en el tracto genital (Hunter, 2008), permitien-do a los espermatozoides viables:
• Unirse al epitelio oviductal en el reser-vorio espermático durante la fase preovulatoria.
• mantener la viabilidad y un estado de baja capacitación.
• Activarse y liberarse durante la fase periovulatoria y postovulatoria.
Este control endocrino permite que un número pequeño de espermatozoides sean liberados del reservorio espermático funcional sincrónicamente con el momento de la ovulación, permitiendo un bajo ratio espermatozoide:ovocito. (Hunter, 1994).
Bajo condiciones fisiológicas, se pro-duce una transición de la dominancia hormonal del estradiol, asociada con la maduración folicular, a la secreción de progesterona procedente de los folículos preovulatorios, la cual será determinante
en la dinámica oviductal. La secreción de progesterona, junto con prostaglandinas y diferentes péptidos llega al oviducto en grandes concentraciones y de manera rápida mediante el flujo contracorriente (Hunter, 1984). Estos pueden tener efecto en la modulación de la composición del fluido oviductal, así como en la degrada-ción de los sitios de unión a las células oviductales. Además, producen una remo-delación estructural de la membrana plas-mática espermática debido a la producción de un gradiente de calcio en los momentos periovulatorios (Flesch y Gadella, 2000), que estaría relacionada junto con la ini-ciación de la capacitación a la liberación al lumen oviductal de los espermatozoides.
También existe cierta regulación autó-noma por parte del espermatozoide. El con-tacto entre el gameto y las células oviducta-les establece un diálogo mutuo, que permite controlar los flujos de calcio (Dobrinski et al., 1996b), manteniendo viable la población y en un estadío bajo de capacitación, a la vez que se modifica el espectro de proteínas secretadas por las células epiteliales del ovi-ducto (Thomas et al., 1995).
Cultivo de células oviductales
mediante cultivos in vitro de células ovi-ductales (oEC) (Figura 17) se ha intentado simular las condiciones que los esperma-tozoides encuentran en el tracto genital femenino, y así poder estudiar los cambios que allí acontecen. Estos cultivos se han realizado tanto para el estudio de las inte-racciones espermatozoides-oviducto como para recrear las condiciones apropiadas para una correcta fecundación, aproxi-mando el proceso de fecundación a las condiciones in vivo.
Cocultivos de células oviductales y espermatozoides
La interacción entre espermatozoide-célula oviductal permite que se manten-ga la viabilidad, motilidad y fertilidad de los espermatozoides (Suarez et al., 1991, Yeste et al., 2009). Cuando las pobla-ciones de espermatozoides unidos y no unidos son analizadas tras 24 horas de cocultivo, solo un 10% de los que per-manecieron en contacto con las células oviductales presentan alteraciones en la motilidad o viabilidad, mientras que esta cifra se torna entre el 50 y 80 % en los que permanecieron no unidos (Yeste et al., 2009). Además, dicha unión estabi-liza la membrana acrosomal, aumenta la supervivencia y permite mantener un nivel bajo de capacitación (Petrunkina et al., 2001).
El estado de capacitación se plantea como uno de los principales criterios para la unión entre las células oviducta-les y los espermatozoides, representando un mecanismo de selección de la pobla-ción espermática más competente para la fecundación (Fazeli et al., 1999). Se ha determinado que un mayor porcentaje de espermatozoides unidos presentan un nivel bajo de fosforilación en tirosina y de concentración de calcio intracelular en diferentes especies (Petrunkina et al., 2001; 2003b; zumoffen et al., 2010). La unión permite mantener un esta-do de baja fosforilación en tirosina durante periodos de incubación largos
Figura 15. Distribución de proteínas fosforiladas en espermatozoides porcinos. (A) Sin fosforilar o fosforilación leve, (B) Fosforilación en acrosoma y/o cola, (C) Fosforilación en segmento ecuatorial, (D) Fosforilación en segmento ecuatorial, acrosoma y cola. Evaluación mediante inmunofluorescencia indirecta con microscopia de fluorescencia.
Figura 17. Células oviductales porcinas a los 7 días de cultivo, procesadas mediante inmunofluorescencia indirecta con anticuerpo antifosfotirosina. Las muestras fueron evaluadas con microscopio de contraste de fases bajo luz de fluorescencia (A) y campo claro (B) (100x).
Figura 16. Imagen de espermatozoides adheridos a las células del oviducto.
· Aumenta los beneficios en cerdos de engorde*.
· Mejora los parámetros reproductivos en cerdas**.
· La vacunación frente a PRRS de cerdas y lechones multiplica el beneficio total de la granja.
R e p o s i c i ó n L e c h o n e sR e p r o d u c t o r a s
* M
arte
lli 1
7º IP
VS
Con
gres
s 20
02 *
* A
lexo
pou
lus
Vet
Mic
200
5
Consulte con nuestro Departamento Técnico para un programa personalizado
A4 PUBLICIDAD 2 PORCILIS PRRS.indd 1 13/03/12 12:19CYS 42.indb 46 14/05/12 10:11

47
nº 42Cambios en el espermatozoide desde la eyaculación hasta la fecundación
· Aumenta los beneficios en cerdos de engorde*.
· Mejora los parámetros reproductivos en cerdas**.
· La vacunación frente a PRRS de cerdas y lechones multiplica el beneficio total de la granja.
R e p o s i c i ó n L e c h o n e sR e p r o d u c t o r a s
* M
arte
lli 1
7º IP
VS
Con
gres
s 20
02 *
* A
lexo
pou
lus
Vet
Mic
200
5
Consulte con nuestro Departamento Técnico para un programa personalizado
Consulte con nuestro Departamento Técnico para un programa personalizado
A4 PUBLICIDAD 2 PORCILIS PRRS.indd 1 13/03/12 12:19CYS 42.indb 47 14/05/12 10:11

48 luño lázaro, Victoria y Matas Parra, Carmen
(Figura 18), permitiendo prolongar la viabilidad espermática y disminuir el estado de capacitación para sincronizar la liberación con el momento exacto de la ovulación.
❱❱ Bibliografía1. ABE H. THE mAmmALiAN oViDUCTAL EPiTHELiUm:
rEGioNAL VAriATioNS iN CYToLoGiCAL AND FUNC-TioNAL ASPECTS oF THE oViDUCTAL SECrETorY CELLS. HiSToL HiSToPATHoL 1996; 11: 743-768.
2. AmSo NN, CroW J, SHAW rW. ComPArATiVE immUNoHiSToCHEmiCAL STUDY oF oESTroGEN AND ProGESTEroNE rECEPTorS iN THE FALLoPiAN TUBE AND UTErUS AT DiFFErENT STAGES oF THE mENSTrUAL CYCLE AND THE mENoPAUSE. HUm rEProD 1994; 9: 1027-1037.
3. ArVEr S, ELiASSoN r. ziNC AND mAGNESiUm iN BULL AND BoAr SPErmATozoA. J rEProD FErTiL 1980; 60: 481-484.
4. AUSTiN Cr. oBSErVATioNS oN THE PENETrATioN oF THE SPErm iN THE mAmmALiAN EGG. AUST J SCi rES B 1951; 4: 581-596.
5. BiSCHoF rJ, LEE CS, BrANDoN mr, mEEUSEN E. iNFLAmmATorY rESPoNSE iN THE PiG UTErUS iNDUCED BY SEmiNAL PLASmA. J rEProD immUNoL 1994; 26: 131-146.
6. CALVETE JJ, ENSSLiN m, mBUrU J, iBorrA A, mArTíNEz P, ADErmANN K, WABErSKi D, SANz L, TöPFEr-PETErSEN E, WEiTzE KF, EiNArSSoN S, roDríGUEz-mArTíNEz H. moNoCLoNAL ANTi-BoDiES AGAiNST BoAr SPErm zoNA PELLUCiDA-BiNDiNG ProTEiN AWN-1.CHArACTErizATioN oF A CoNTiNUoUS ANTiGENiC DETErmiNANT AND immUNoLoCALizATioN oF AWN EPiToPES iN iNSEmiNATED SoWS. BioL rEProD 1997; 57: 735-742.
7. CArDozo JA, FErNÁNDEz-JUAN m, ForCADA F, ABECiA A, mUiño-BLANCo T, CEBriÁN-PérEz JA. moNTHLY VAriATioNS iN oViNE SEmiNAL PLASmA ProTEiNS ANALYzED BY TWo-DimEN-SioNAL PoLYACrYLAmiDE GEL ELECTroPHorESiS. THErioGENoLoGY 2006; 66: 841-850.
8. CHANG mC. FErTiLiziNG CAPACiTY oF SPErmATo-zoA DEPoSiTED iNTo THE FALLoPiAN TUBES. NATUrE 1951; 168: 697-698.
9. CoLENBrANDEr B, KEmP B. FACTorS iNFLUENCiNG SEmEN qUALiTY iN PiGS. J rEProD FErTiL SUPPL 1990; 40: 105-115.
10. DoBriNSKi i, iGNoTz GG, THomAS PG, BALL BA. roLE oF CArBoHYDrATES iN THE ATTACHmENT oF EqUiNE SPErmATozoA To UTEriNE TUBAL (oViDUC-TAL) EPiTHELiAL CELLS iN ViTro. Am J VET rES 1996A; 57: 1635-1639.
11. DoBriNSKi i, SUArEz SS, BALL BA. iNTrACELLULAr CALCiUm CoNCENTrATioN iN EqUiNE SPErmATo-zoA ATTACHED To oViDUCTAL EPiTHELiAL CELLS iN ViTro. BioL rEProD 1996B; 54: 783-788.
12. DUBé C, LECLErC P, BABA T, rEYES-morENo C, BAiLEY JL. THE ProACroSiN BiNDiNG ProTEiN, SP32, iS TYroSiNE PHoSPHorYLATED DUriNG CAPACiTATioN oF PiG SPErm. J ANDroL 2005; 26: 519-528.
13. EKHLASi-HUNDriESEr m, GoHr K, WAGNEr A, TSoLoVA m, PETrUNKiNA A, TöPFEr-PETErSEN E. SPErmADHESiN AqN1 iS A CANDiDATE rECEPTor moLECULE iNVoLVED iN THE FormATioN oF THE oViDUCTAL SPErm rESErVoir iN THE PiG. BioL rEProD 2005; 73: 536-545.
14. ELLiNGToN JE, EVENSoN DP, FLEmiNG JE, BriSBoiS rS, HiSS GA, BroDEr SJ, WriGHT rW. CoCULTUrE oF HUmAN SPErm WiTH BoViNE oViDUCT EPiTHE-LiAL CELLS DECrEASES SPErm CHromATiN STrUC-TUrAL CHANGES SEEN DUriNG CULTUrE iN mEDiA ALoNE. FErTiL STEriL 1998; 69: 643-649.
15. ELLioTT rm, LLoYD rE, FAzELi A, SoSTAriC E, GEorGioU AS, SATAKE N, WATSoN PF, HoLT WV. EFFECTS oF HSPA8, AN EVoLUTioNAriLY CoNSErVED oViDUCTAL ProTEiN, oN BoAr AND BULL SPErmA-TozoA. rEProDUCTioN 2009; 137: 191-203.
16. FAWCETT DW. A ComPArATiVE ViEW oF SPErm ULTrASTrUCTUrE. BioL rEProD 1970; 2: SUPPL 2:90-127.
17. FAzELi A, DUNCAN AE, WATSoN PF, HoLT WV. SPErm-oViDUCT iNTErACTioN: iNDUCTioN oF CAPACiTATioN AND PrEFErENTiAL BiNDiNG oF UNCAPACiTATED SPErmATozoA To oViDUCTAL EPi-THELiAL CELLS iN PorCiNE SPECiES. BioL rEProD 1999; 60: 879-886.
18. FLESCH Fm, CoLENBrANDEr B, VAN GoLDE Lm, GADELLA Bm. CAPACiTATioN iNDUCES TYroSiNE PHoSPHorYLATioN oF ProTEiNS iN THE BoAr SPErm PLASmA mEmBrANE. BioCHEm BioPHYS rES CommUN. 1999; 262(3):787-92.
19. FLESCH Fm, GADELLA Bm. DYNAmiCS oF THE mAm-mALiAN SPErm PLASmA mEmBrANE iN THE Pro-CESS oF FErTiLizATioN. BioCHim BioPHYS ACTA 2000; 1469: 197-235.
20. FLoWErS WL. mANAGEmENT oF BoArS For EFFi-CiENT SEmEN ProDUCTioN. J rEProD FErTiL SUPPL 1997; 52: 67-78.
21. FrEi B, SToCKEr r, ENGLAND L, AmES BN. ASCorBATE: THE moST EFFECTiVE ANTioxiDANT iN
HUmAN BLooD PLASmA. ADV ExP mED BioL 1990; 264: 155-163.
22. GALANTiNo-HomEr HL, ViSCoNTi PE, KoPF GS. rEGULATioN oF ProTEiN TYroSiNE PHoSPHorY-LATioN DUriNG BoViNE SPErm CAPACiTATioN BY A CYCLiC ADENoSiNE 3’5’-moNoPHoSPHATE-DEPENDENT PATHWAY. BioL rEProD 1997; 56: 707-719.
23. GANDoLFi F. FUNCTioNS oF ProTEiNS SECrETED BY oViDUCT EPiTHELiAL CELLS. miCroSC rES TECH 1995; 32: 1-12.
24. GArNEr DL, HAFEz ESE (1996) ESPErmATozoiDES Y PLASmA SEmiNAL. EN: rEProDUCCióN E iNSEmi-NACióN ArTiFiCiAL EN ANimALES. HAFEz ESE (ED.) iNTErAmEriCANA mCGrAW-HiLL (méxiCo). PP 158-179.
25. GEiSErT rD (1999) PiGS. EN: ENCYCLoPEDiA oF rEProDUCTioN. KNoBiL E, NEiLL JD (EDS.) ACADEmiC PrESS (SAN DiEGo, USA). PP 792-799.
26. GrASA P, CEBriÁN-PérEz JA, mUiño-BLANCo T. SiGNAL TrANSDUCTioN mECHANiSmS iNVoLVED iN iN ViTro rAm SPErm CAPACiTATioN. rEProDUCTioN 2006; 132: 721-732.
27. GUALTiEri r, TALEVi r. SELECTioN oF HiGHLY FErTiLizATioN-ComPETENT BoViNE SPErmATozoA THroUGH ADHESioN To THE FALLoPiAN TUBE EPiTHELiUm iN ViTro. rEProDUCTioN 2003; 125: 251-258.
28. HArriSoN rA. CAPACiTATioN mECHANiSmS, AND THE roLE oF CAPACiTATioN AS SEEN iN EUTHEriAN mAmmALS. rEProD FErTiL DEV 1996; 8: 581-594.
29. HoLT WV, HArriSoN rA. BiCArBoNATE STimULA-TioN oF BoAr SPErm moTiLiTY ViA A ProTEiN KiNASE A-DEPENDENT PATHWAY: BETWEEN-CELL AND BETWEEN-EJACULATE DiFFErENCES ArE NoT DUE To DEFiCiENCiES iN ProTEiN KiNASE A ACTi-VATioN. J ANDroL 2002; 23: 557-565.
30. HoLT WV. iS SEmEN ANALYSiS USEFUL To PrEDiCT THE oDDS THAT THE SPErm WiLL mEET THE EGG? rEProD DomEST ANim 2009; 44 SUPPL 3: 31-38.
31. HUNTEr rH. PrE-oVULATorY ArrEST AND PEri-oVULATorY rEDiSTriBUTioN oF ComPETENT SPEr-mATozoA iN THE iSTHmUS oF THE PiG oViDUCT. J rEProD FErTiL 1984; 72: 203-211.
32. HUNTEr rH, FLéCHoN B, FLéCHoN JE. DiSTriBUTioN, morPHoLoGY AND EPiTHELiAL iNTErACTioNS oF BoViNE SPErmATozoA iN THE oViDUCT BEForE AND AFTEr oVULATioN: A SCANNiNG ELECTroN miCroSCoPE STUDY. TiSSUE CELL 1991; 23: 641-656.
33. HUNTEr rH. oVAriAN rEGULATioN oF SPErm ProGrESSioN iN THE FALLoPiAN TUBES. zYGoTE 1994; 2: 363-366.
34. HUNTEr rH, roDriGUEz-mArTiNEz H. CAPACiTATioN oF mAmmALiAN SPErmATozoA iN ViVo, WiTH A SPECiFiC FoCUS oN EVENTS iN THE FALLoPiAN TUBES. moL rEProD DEV 2004; 67: 243-250.
35. HUNTEr rH. THE FALLoPiAN TUBES iN DomESTiC mAmmALS: HoW ViTAL iS THEir PHYSioLoGiCAL ACTiViTY? rEProD NUTr DEV 2005; 45: 281-290.
36. HUNTEr rH. SPErm rELEASE From oViDUCT EPi-THELiAL BiNDiNG iS CoNTroLLED HormoNALLY BY PEri-oVULATorY GrAAFiAN FoLLiCLES. moL rEProD DEV 2008; 75: 167-174.
37. JoHNSoN LA, WEiTzE KF, FiSEr P, mAxWELL Wm. STorAGE oF BoAr SEmEN. ANim rEProD SCi 2000; 62: 143-172.
38. JoNES r, JAmES PS, oxLEY D, CoADWELL J, SUzUKi-ToYoTA F, HoWES EA. THE EqUAToriAL SUBSEG-mENT iN mAmmALiAN SPErmATozoA iS ENriCHED iN TYroSiNE PHoSPHorYLATED ProTEiNS. BioL rEProD 2008; 79: 421-431.
39. KiLLiAN GJ, CHAPmAN DA, roGoWSKi LA. FErTiLiTY-ASSoCiATED ProTEiNS iN HoLSTEiN BULL SEmiNAL PLASmA. BioL rEProD 1993; 49: 1202-1207.
40. KiLLiAN GJ. EViDENCE For THE roLE oF oViDUCT SECrETioNS iN SPErm FUNCTioN, FErTiLizATioN AND EmBrYo DEVELoPmENT. ANim rEProD SCi 2004; 82-83: 141-153.
41. KorLEY r, PoUrESmAEiLi F, oKo r. ANALYSiS oF THE ProTEiN ComPoSiTioN oF THE moUSE SPErm PEriNUCLEAr THECA AND CHArACTErizATioN oF iTS mAJor ProTEiN CoNSTiTUENT. BioL rEProD 1997; 57: 1426-1432.
42. LEESE HJ. THE FormATioN AND FUNCTioN oF oVi-DUCT FLUiD. J rEProD FErTiL 1988; 82: 843-856.
Figura 18. Espermatozoides unidos a las células oviductales in vitro. La imagen A, muestra una fosforilación en tirosina de proteínas en la región la ecuatorial y el flagelo. La ima-gen B presenta espermatozoides fosforilados en la región ecuatorial y sin fosforilar (*). Muestras evaluadas mediante inmunofluorescencia indirecta. Imagenes obtenidas con microscopio de contraste de fases bajo luz de fluorescencia y campo claro (100x).
A
B
CYS 42.indb 48 14/05/12 10:11

49
nº 42Cambios en el espermatozoide desde la eyaculación hasta la fecundación
43. LEESE HJ, TAY Ji, rEiSCHL J, DoWNiNG SJ. FormATioN oF FALLoPiAN TUBAL FLUiD: roLE oF A NEGLECTED EPiTHELiUm. rEProDUCTioN 2001; 121: 339-346.
44. LEFEBVrE r, SUArEz SS. EFFECT oF CAPACiTA-TioN oN BULL SPErm BiNDiNG To HomoLoGoUS oViDUCTAL EPiTHELiUm. BioL rEProD 1996; 54: 575-582.
45. LESHiN LS, rAJ Sm, SmiTH CK, KWoK SC, KrAELiNG rr, Li Wi. immUNoSTimULATorY EFFECTS oF PiG SEmiNAL ProTEiNS oN PiG LYmPHoCYTES. J rEProD FErTiL 1998; 114: 77-84.
46. mAriNi PE, CABADA mo. oNE STEP PUriFiCA-TioN AND BioCHEmiCAL CHArACTErizATioN oF A SPErmATozoA-BiNDiNG ProTEiN From PorCiNE oViDUCTAL EPiTHELiAL CELLS. moL rEProD DEV 2003; 66: 383-390.
47. mATTHiJS A, ENGEL B, WoELDErS H. NEUTroPHiL rECrUiTmENT AND PHAGoCYToSiS oF BoAr SPEr-mATozoA AFTEr ArTiFiCiAL iNSEmiNATioN oF SoWS, AND THE EFFECTS oF iNSEmiNATE VoLUmE, SPErm DoSE AND SPECiFiC ADDiTiVES iN THE ExTENDEr. rEProDUCTioN 2003; 125: 357-367.
48. mATÁS C, CoY P, romAr r, mArCo m, GADEA J, rUiz S. EFFECT oF SPErm PrEPArATioN mETHoD oN iN ViTro FErTiLizATioN iN PiGS. rEProDUCTioN 2003; 125: 133-141.
49. mATÁS C, SANSEGUNDo m, rUiz S, GArCíA-VÁzqUEz FA, GADEA J, romAr r, CoY P. SPErm TrEATmENT AFFECTS CAPACiTATioN PArAm-ETErS AND PENETrATioN ABiLiTY oF EJACU-LATED AND EPiDiDYmAL BoAr SPErmATozoA. THErioGENoLoGY 2010; 74: 1327-1340.
50. mBUrU JN, EiNArSSoN S, LUNDEHEim N, roDriGUEz-mArTiNEz H. DiSTriBUTioN, NUmBEr AND mEmBrANE iNTEGriTY oF SPErmATozoA iN THE PiG oViDUCT iN rELATioN To SPoNTANEoUS oVULATioN. ANim rEProD SCi 1996; 45: 109-121.
51. miLLEr D. SPErmATozoAL rNA AS rESErVoir, mArKEr AND CArriEr oF EPiGENETiC iNFor-mATioN: imPLiCATioNS For CLoNiNG. rEProD DomEST ANim 2007; 42 SUPPL 2: 2-9.
52. NAz rK, rAJESH PB. roLE oF TYroSiNE PHoS-PHorYLATioN iN SPErm CAPACiTATioN / ACro-SomE rEACTioN. rEProD BioL ENDoCriNoL 2004; 2: 75.
53. NiCHoL r, HUNTEr rH, CooKE Gm. oViDUCT FLUiD PH iN iNTACT AND UNiLATErALLY oVAriECTo-mizED PiGS. CAN J PHYSioL PHArmACoL 1997; 75: 1069-1074.
54. oLiVErA m, rUiz T, TArAzoNA A, GirALDo C (2006) EL ESPErmATozoiDE, DESDE LA EYACU-LACióN HASTA LA FErTiLizACióN. rEV CoL CiENC PEC. 19:426-436.
55. oKo r, mArAVEi D. ProTEiN ComPoSiTioN oF THE PEriNUCLEAr THECA oF BULL SPErmATozoA. BioL rEProD 1994; 50: 1000-1014.
56. PAWSoN T. SPECiFiCiTY iN SiGNAL TrANSDUCTioN: From PHoSPHoTYroSiNE-SH2 DomAiN iNTErAC-TioNS To ComPLEx CELLULAr SYSTEmS. CELL 2004; 116: 191-203.
57. PESCH S, BErGmANN m. STrUCTUrE oF mAm-mALiAN SPErmATozoA iN rESPECT To ViABiLiTY, FErTiLiTY AND CrYoPrESErVATioN. miCroN 2006; 37: 597-612.
58. PETrUNKiNA A, FriEDriCH J, DrommEr W, BiCKEr G, WABErSKi D, TöPFEr-PETErSEN E. KiNETiC CHArACTErizATioN oF THE CHANGES iN ProTEiN TYroSiNE PHoSPHorYLATioN oF mEmBrANES, CYToSoLiC CA2+ CoNCENTrATioN AND ViABiL-iTY iN BoAr SPErm PoPULATioNS SELECTED BY BiNDiNG To oViDUCTAL EPiTHELiAL CELLS. rEProDUCTioN 2001; 122: 469-480.
59. PETrUNKiNA A, SimoN K, GuNzEL-APEL A, TöPFEr-PETErSEN E. rEGULATioN oF CAPACiTATioN oF CANiNE SPErmATozoA DUriNG Co-CULTUrE WiTH HETEroLoGoUS oViDUCTAL EPiTHELiAL CELLS. rEProD DomEST ANim 2003A; 38: 455-463.
60. PETrUNKiNA Am, SimoN K, GuNzEL-APEL Ar, TöPFEr-PETErSEN E. SPECiFiC orDEr iN THE APPEArANCE oF ProTEiN TYroSiNE PHoSPHorY-LATioN PATTErNS iS FUNCTioNALLY CoorDiNATED WiTH DoG SPErm HYPErACTiVATioN AND CAPACi-TATioN. J ANDroL 2003B; 24: 423-437.
61. PETrUNKiNA Am, WABErSKi D, GuNzEL-APEL Ar, TöPFEr-PETErSEN E. DETErmiNANTS oF SPErm
qUALiTY AND FErTiLiTY iN DomESTiC SPECiES. rEProDUCTioN 2007; 134: 3-17.
62. PommEr AC, rUTLLANT J, mEYErS SA. PHoSPHorYLATioN oF ProTEiN TYroSiNE rESi-DUES iN FrESH AND CrYoPrESErVED STALLioN SPErmATozoA UNDEr CAPACiTATiNG CoNDi-TioNS. BioL rEProD 2003; 68: 1208-1214.
63. PérEz FA, romA Sm, CABADA mo, mAriNi PE. SPErm BiNDiNG GLYCoProTEiN iS DiFFErENTiALLY PrESENT SUrroUNDiNG THE LUmEN oF iSTH-mUS AND AmPULLA oF THE PiG’S oViDUCT. ANAT EmBrYoL (BErL) 2006; 211: 619-624.
64. PérEz-Pé r, BArrioS B, mUiño-BLANCo T, CEBriÁN-PérEz JA. SEASoNAL DiFFErENCES iN rAm SEmiNAL PLASmA rEVEALED BY PArTiTioN iN AN AqUEoUS TWo-PHASE SYSTEm. J CHromAToGr B BiomED SCi APPL 2001; 760: 113-121.
65. qUiNTEro-morENo A, riGAU T, roDríGUEz-GiL JE. rEGrESSioN ANALYSES AND moTiLE SPErm SUBPoPULATioN STrUCTUrE STUDY AS imProV-iNG TooLS iN BoAr SEmEN qUALiTY ANALYSiS. THErioGENoLoGY 2004; 61: 673-690.
66. roDriGUEz-mArTiNEz H, TiENTHAi P, SUzUKi K, FUNAHASHi H, EKWALL H, JoHANNiSSoN A. iNVoLVEmENT oF oViDUCT iN SPErm CAPACiTA-TioN AND ooCYTE DEVELoPmENT iN PiGS. rEProD SUPPL 2001; 58: 129-145.
67. roDríGUEz-mArTíNEz H, SArAViA F, WALLGrEN m, TiENTHAi P, JoHANNiSSoN A, VÁzqUEz Jm, mArTíNEz E, roCA J, SANz L, CALVETE JJ. BoAr SPErmATozoA iN THE oViDUCT. THErioGENoLoGY 2005; 63: 514-535.64.
68. roDríGUEz-mArTíNEz H, KViST U, SArAViA F, WALLGrEN m, JoHANNiSSoN A, SANz L, PEñA FJ, mArTíNEz EA, roCA J, VÁzqUEz Jm, CALVETE JJ. THE PHYSioLoGiCAL roLES oF THE BoAr EJACU-LATE. SoC rEProD FErTiL SUPPL 2009; 66: 1-21.
69. SATAKE N, ELLioTT rm, WATSoN PF, HoLT WV. SPErm SELECTioN AND ComPETiTioN iN PiGS mAY BE mEDiATED BY THE DiFFErENTiAL moTiL-iTY ACTiVATioN AND SUPPrESSioN oF SPErm SUBPoPULATioNS WiTHiN THE oViDUCT. J ExP BioL 2006; 209: 1560-1572.
70. SENGEr PL. PATHWAYS To PrEGNANCY AND PArTUriTioN.1ST EDiTioN.CUrrENT CoNCEPTioNS, iNC, 1999, 1997. WASHiNGToN STATE UNiVErSiTYrESEArCH & TECHNoLoGY PArK. 1615NE EASTGATE BLVD PULLmAN, WA 99163-5607.
71. SmoLLiCH A, miCHEL G. miCroSCoPiCAL ANAT-omY oF DomESTiC ANimALS. GUSTAV FiSCHEr, (STUTTGArT) 1992. PP 245.
72. STAUSS Cr, VoTTA TJ, SUArEz SS. SPErm moTiL-iTY HYPErACTiVATioN FACiLiTATES PENETrATioN oF THE HAmSTEr zoNA PELLUCiDA. BioL rEProD 1995; 53: 1280-1285.
73. STrzEzEK J, CiErESzKo A. HETEroGENEiTY oF ASPArTATE AmiNoTrANSFErASE (AAT) iN BULL SEmEN. ComP BioCHEm PHYSioL B 1987; 86: 373-375.
74. STrzEzEK J. SECrETorY ACTiViTY oF BoAr SEmiNAL VESiCLE GLANDS. rEProD BioL 2002; 2: 243-266.
75. SUArEz SS, KATz DF, oWEN DH, ANDrEW JB, PoWELL rL. EViDENCE For THE FUNCTioN oF HYPErACTiVATED moTiLiTY iN SPErm. BioL rEProD 1991; 44: 375-381.
76. SUArEz SS, rEVAH i, Lo m, KöLLE S. BULL SPErm BiNDiNG To oViDUCTAL EPiTHELiUm iS mEDi-ATED BY A CA2+-DEPENDENT LECTiN oN SPErm THAT rECoGNizES LEWiS-A TriSACCHAriDE. BioL rEProD 1998; 59: 39-44.
77. SUArEz SS. FormATioN oF A rESErVoir oF SPErm iN THE oViDUCT. rEProD DomEST ANim 2002; 37: 140-143.
78. SUToVSKY P, NEUBEr E, SCHATTEN G. UBiqUiTiN-DEPENDENT SPErm qUALiTY CoNTroL mECHA-NiSm rECoGNizES SPErmATozoA WiTH DNA DEFECTS AS rEVEALED BY DUAL UBiqUiTiN-TUNEL ASSAY. moL rEProD DEV 2002; 61: 406-413.
79. TArDiF S, DUBé C, CHEVALiEr S, BAiLEY JL. CAPACiTATioN iS ASSoCiATED WiTH TYroSiNE PHoSPHorYLATioN AND TYroSiNE KiNASE-LiKE ACTiViTY oF PiG SPErm ProTEiNS. BioL rEProD 2001; 65: 784-792.
80. TEiJEiro Jm, CABADA mo, mAriNi PE. SPErm BiND-iNG GLYCoProTEiN (SBG) ProDUCES CALCiUm AND BiCArBoNATE DEPENDENT ALTErATioN oF
ACroSomE morPHoLoGY AND ProTEiN TYro-SiNE PHoSPHorYLATioN oN BoAr SPErm. J CELL BioCHEm 2008; 103: 1413-1423.
81. TEiJEiro Jm, iGNoTz GG, mAriNi PE. ANNExiN A2 iS iNVoLVED iN PiG (SUS SCroFA) SPErm-oViDUCT iNTErACTioN. moL rEProD DEV 2009; 76: 334-341.
82. THomAS PG, iGNoTz GG, BALL BA, BriNSKo SP, CUrriE WB. EFFECT oF CoCULTUrE WiTH STALLioN SPErmATozoA oN DE NoVo ProTEiN SYNTHESiS AND SECrETioN BY EqUiNE oViDUCT EPiTHELiAL CELLS. Am J VET rES 1995; 56: 1657-1662.
83. THUrSToN Lm, WATSoN PF, miLEHAm AJ, HoLT WV. morPHoLoGiCALLY DiSTiNCT SPErm SUBPoPULA-TioNS DEFiNED BY FoUriEr SHAPE DESCriPTorS iN FrESH EJACULATES CorrELATE WiTH VAriA-TioN iN BoAr SEmEN qUALiTY FoLLoWiNG CrYo-PrESErVATioN. J ANDroL 2001; 22: 382-394.
84. THériEN i, BLEAU G, mANJUNATH P. PHoSPHATiDYLCHoLiNE-BiNDiNG ProTEiNS oF BoViNE SEmiNAL PLASmA moDULATE CAPACiTA-TioN oF SPErmATozoA BY HEPAriN. BioL rEProD 1995; 52: 1372-1379.
85. TiENTHAi P, KJELLéN L, PErToFT H, SUzUKi K, roDriGUEz-mArTiNEz H. LoCALizATioN AND qUANTiTATioN oF HYALUroNAN AND SUL-FATED GLYCoSAmiNoGLYCANS iN THE TiSSUES AND iNTrALUmiNAL FLUiD oF THE PiG oViDUCT. rEProD FErTiL DEV 2000; 12: 173-182.
86. TiENTHAi P, JoHANNiSSoN A, roDriGUEz-mArTiNEz H. SPErm CAPACiTATioN iN THE Por-CiNE oViDUCT. ANim rEProD SCi 2004; 80: 131-146.
87. TöPFEr-PETErSEN E. moLECULES oN THE SPErm’S roUTE To FErTiLizATioN. J ExP zooL 1999; 285: 259-266.
88. TöPFEr-PETErSEN E, WAGNEr A, FriEDriCH J, PETrUNKiNA A, EKHLASi-HUNDriESEr m, WABErSKi D, DrommEr W. FUNCTioN oF THE mAmmALiAN oViDUCTAL SPErm rESErVoir. J ExP zooL 2002; 292: 210-215.
89. ViSCoNTi P, BAiLEY J, moorE G, PAN D, oLDS-CLArKE P, KoPF G. CAPACiTATioN oF moUSE SPErmATozoA. i. CorrELATioN BETWEEN THE CAPACiTATioN STATE AND ProTEiN TYroSiNE PHoSPHorYLATioN. DEVELoPmENT 1995; 121: 1129-1137.
90. ViSCoNTi PE, STEWArT-SAVAGE J, BLASCo A, BATTAGLiA L, mirANDA P, KoPF GS, TEzóN JG. roLES oF BiCArBoNATE, CAmP, AND ProTEiN TYroSiNE PHoSPHorYLATioN oN CAPACiTATioN AND THE SPoNTANEoUS ACroSomE rEACTioN oF HAmSTEr SPErm. BioL rEProD 1999; 61: 76-84.
91. ViSCoNTi PE, WESTBrooK VA, CHErTiHiN o, DEmArCo i, SLEiGHT S, DiEKmAN AB. NoVEL SiG-NALiNG PATHWAYS iNVoLVED iN SPErm ACqUi-SiTioN oF FErTiLiziNG CAPACiTY. J rEProD immUNoL 2002; 53: 133-150.
92. ViSCoNTi PE. UNDErSTANDiNG THE moLECU-LAr BASiS oF SPErm CAPACiTATioN THroUGH KiNASE DESiGN. ProC NATL ACAD SCi U S A. 2009; 106(3):667-668.
93. WATSoN PF. THE CAUSES oF rEDUCED FErTiLiTY WiTH CrYoPrESErVED SEmEN. ANim rEProD SCi 2000; 60-61: 481-492.
94. YANAGimACHi r. FErTiLiTY oF mAmmALiAN SPEr-mATozoA: iTS DEVELoPmENT AND rELATiViTY. zYGoTE 1994; 2: 371-372.
95. YESTE m, LLoYD rE, BADiA E, Briz m, BoNET S, HoLT WV. DirECT CoNTACT BETWEEN BoAr SPErmA-TozoA AND PorCiNE oViDUCTAL EPiTHELiAL CELL (oEC) CULTUrES iS NEEDED For oPTimAL SPErm SUrViVAL iN ViTro. ANim rEProD SCi 2009; 113: 263-278.
96. zHoU Cx, WANG xF, CHAN HC. BiCArBoNATE SECrETioN BY THE FEmALE rEProDUCTiVE TrACT AND iTS imPACT oN SPErm FErTiLiziNG CAPACiTY. SHENG Li xUE BAo 2005; 57: 115-124.
97. zHU J, xU x, CoSGroVE Jr, FoxEroFT Gr. EFFECTS oF SEmEN PLASmA From DiFFErENT FrACTioNS oF iNDiViDUAL EJACULATES oN iVF iN PiGS. THErioGENoLoGY 2000; 54: 1443-1452.
98. zUmoFFEN Cm, CAiLLE Am, mUNUCE mJ, CABADA mo, GHErSEViCH SA. ProTEiNS From HUmAN oViDUCTAL TiSSUE-CoNDiTioNED mEDiUm moDU-LATE SPErm CAPACiTATioN. HUm rEProD 2010; 25: 1504-1512.
CYS 42.indb 49 14/05/12 10:11

44
índice bovino.indd 44 14/05/12 11:11

45
Cría y saludBOVINO
42
Factores que afectan a la pérdida de la gestación en el periodo fetal temprano. Una aproximación terapeútica
Patología del sistema respiratorio para veterinarios de campo
C I E N T Í F I C O S
índice bovino.indd 45 14/05/12 11:11

52
lópez-Gatius F.
En este artículo revisamos una serie de factores de naturaleza no infecciosa relacionados con la pérdida de la gestación durante el periodo fetal temprano. Parte de la información deriva de una compilación de nuestros estudios en explotaciones de ganado vacuno lechero de alta producción en el noreste español, la zona limítrofe sur de las provincias de Lérida y Huesca,
incluyendo 15525 gestaciones. Aspectos a resaltar son el fuerte efecto negativo sobre el mantenimiento de la gestación que pueden tener la edad (vacas respecto novillas), el toro
utilizado en la inseminación artificial (IA), el periodo cálido del año (mayo a septiembre) y la gestación doble. En contraste, la presencia de un cuerpo lúteo adicional (un número mayor de cuerpos lúteos que el de embriones) se ha relacionado con la reducción drástica de la pérdida de la gestación. El tratamiento con progesterona o GnRH en el momento del diagnóstico de la gestación (28-34 días post-IA) tiene la capacidad de reducir la incidencia de la pérdida de
gestación en vacas sin cuerpo lúteo adicional con gestación sencilla o doble, respectivamente, en explotaciones con una alta incidencia de pérdidas. Desde un punto de vista práctico, se
sugiere la confirmación de la gestación los días 60 y 90 post-IA.
Factores que afectan la pérdida de la gestación en el periodo fetal temprano. Una aproximación terapeútica
F. lópez-GatiusDepartamento de Producción Animal, Universidad de Lleida
Ponencia Presentada en el XV congreso internacional aneMBe de Medicina BoVina
Fotos Axón Comunicación
CYS 42.indb 52 14/05/12 10:11

53
nº 42Factores que afectan la pérdida de la gestación en el periodo fetal temprano. Una aproximación terapeútica
❱❱ Introducción
El diálogo inmuno-endocrino implicado en la biología reproductiva de la hembra cíclica varía enormemente cuando ocurre la gestación. Es necesario que la mucosa uterina tolere el embrión, elemento antigé-nicamente extraño, sin reducir la capacidad de la respuesta uterina a una posible con-taminación microbiana. Tanto el delicado proceso de inmuno-modulación endocrina de la gestación como la serie de interaccio-nes embriomaternas en los micro-ambien-tes de los órganos genitales tubulares son muy sensibles a diferentes tipos de estrés. Procesos como la regulación de la función ovárica (transformación de un cuerpo lúteo cíclico en uno de gestación), reconocimien-to materno de la gestación, implantación y placentación pueden verse seriamente comprometidos, y así la gestación, cuando son sometidos a una serie de factores de carácter no infeccioso relacionados con la vida productiva de la vaca.
En la vaca, el periodo embrionario de la gestación se extiende desde la fecundación hasta el fin del estadio de diferenciación, sobre los 45 días, y el periodo fetal desde el día 45 hasta el parto (Committee on Bovine reproductive Nomenclature, 1972). La placentación finaliza antes del día 60 de gestación, periodo en el que se considera que la gestación está firmemente estable-cida y el riesgo de pérdidas se reduce (Ball, 1997). En ganado vacuno lechero, aunque la mayor parte de las pérdidas de la gesta-ción ocurren durante el periodo embrionario temprano (Peters, 1996; Hanzen et al., 1999), el riesgo de pérdida fetal temprana pare-ce incrementar en condiciones de manejo intensivo (Forar et al., 1995; Hanzen et al., 1999; LópezGatius et al., 2008). Este tipo de pérdida escapa a menudo a un control clínico y se asume que es un problema de origen multifactorial (Vanroose et al., 2000). Una pérdida fetal de un 10% se asume como normal (Thurmond et al., 1990; Vanroose et al., 2000; Santos et al., 2004b; López-Gatius et al., 2008). En nuestra área geográfica, localizada en el noreste español, en la zona limítrofe sur de las provincias de Lérida y Huesca (Labernia et al., 1996; López-Gatius et al., 1996, 2002, 2004a), y en otros lugares (Silke et al., 2002; mcDougall et al., 2005; ]ousan et al., 2005; Grimard et al., 2006; Gábor et al., 2008; Santos et al., 2009; rhinehart et al., 2009), factores de naturale-za no infecciosa relacionados con la vaca y su manejo se han descrito como factores de riesgo afectando la pérdida fetal temprana. Además, en países cálidos, como es el caso de España, el estrés térmico calórico no sólo afecta a la fertilidad (Labernia et al., 1998; López-Gatius, 2003; López-Gatius et
al., 2005a; García-ispierto et al., 2007) sino también a la pérdida fetal (López-Gatius et al., 2004a-c; García-ispierto et al., 2006). De hecho, la pérdida fetal temprana, que ocurre principalmente entre los 40 y 50 días de la gestación (López-Gatius et al., 2004a; Santos et al., 2004b) se está convirtiendo en el desorden más frecuente del periodo gestacional en nuestra zona. más del 90% de las pérdidas tras un diagnóstico de ges-tación positivo ocurren antes del día 90 de gestación (López-Gatius et al., 2002, 2004a).
En este artículo, que no pretende ser una revisión exhaustiva sobre el tema, consideramos una serie de factores de naturaleza no infecciosa relacionados con la pérdida de la gestación durante el periodo fetal temprano. Parte de la información deriva de una compilación de nuestros estudios en explotaciones de ganado vacuno lechero de alta producción en el noreste español incluyendo 15525 gestaciones (López-Gatius et al., 2008). Todos los diagnósticos de gestación fueron realizados por el autor, bien por palpación rectal o por ecografía. Consideraremos una serie de factores de riesgo, sin efecto y preventivos de la pérdida fetal. Finalmente, expondremos un acercamiento terapéutico al problema.
❱❱ Factores de riesgo
Edad
Agrupando la información de dos estudios (Labernia et al., 1996; López-Gatius et al., 2004b), la incidencia de pérdidas en novi-llas (2,6%: 47/1801) fue inferior a la de vacas lactantes (9%: 469/5184). Estos valo-res son comparables a los obtenidos entre 1949 y 1955: 2,5% para las novillas y 13% para vacas multíparas (mares et al., 1961) y a los de la compilación reciente sobre varios estudios de Santos et al. (2004b): 4,2% para las novillas y 10,7% para vacas lactantes. Estos resultados sugieren que la selección genética para la alta producción no ha afectado a la incidencia de pérdidas en las últimas décadas.
Toro
En tres estudios incluyendo 2141 gestacio-nes pudimos registrar un riego relativo de pérdida relacionado con el uso del semen de un toro determinado de 2,1 a 21,7 veces superior al de otros toros (López-Gatius et al., 2002, 2004a, 2007b). En cambio, no pudimos encontrar este efecto en otros tres estudios sobre 3095 gestaciones (López-Gatius et al., 2004c; García-ispierto et al., 2006; Bech-Sabat et al., 2009). La asocia-
ción de determinados toros utilizados en la lA con la pérdida de la gestación se ha demostrado en varios estudios (Bulman, 1979; Pegorer et al., 2007), incluyendo uno epidemiológico sobre 58048 gestaciones (markusfeld-Nir, 1997). Posibles razones para explicar este efecto negativo sobre la gestación incluyen defectos anatómicos y genéticos de algunos toros (Ball, 1997), pero el uso de semen contaminado no debiera ser excluido.
Aunque la atención principal en el con-trol de los toros dentro de los programas de iA está dirigida a la fertilidad, una vez la vaca queda gestante, el semen utilizado de determinados toros parece emerger como un importante factor de riesgo de la pér-dida de la gestación. Dado que los niveles plasmáticos de proteínas asociadas a la gestación (PAGs), marcadores del bienestar placentario y fetal, varían marcadamente para cada toro (López-Gatius et al., 2007a, 2007b), bien pudieran ser un elemento de medida para informarnos del posible efecto de un toro sobre el mantenimiento de la gestación. Nuevos estudios debieran corroborar este punto.
Desórdenes durante el periodo post-parto previo a la gestación
La incidencia de pérdidas de gestación fue 2,6 y 1,8 veces superior para las vacas que durante el periodo post-parto previo a la
Distintos estudios señalan que la incidencia de pérdidas en novillas es inferior a la de vacas lactantes.
CYS 42.indb 53 14/05/12 10:11

54
López-Gatius F.
gestación sufrieron piómetra y retención pla-centaria, respectivamente, respecto las que no sufrieron estos desórdenes (López-Gatius et al., 1996). Vacas con un pérdida de con-dición corporal de 1 punto (escala de 1 a 5) durante los primeros 30 días del periodo post-parto tuvieron un riesgo de pérdida fetal 2,4 veces más alto que las que tuvieron una pérdida inferior a 1 punto (López-Gatius et al., 2002). Resultados similares relacionados con los valores de la condición corporal desde el parto a la IA se han descrito recientemente (Santos et al., 2009). Dado que ambos tipos de desórdenes, metabólicos y uterinos, fue-ron un factor de riesgo independiente del intervalo parto-gestación (López-Gatius et al., 1996, 2002), estos resultados sugieren un prolongado efecto de los estreses sufridos durante el periodo postparto.
Mastitis y cojeras
La asociación entre mastitis y muerte embrio-naria se ha propuesto como ejemplo de cómo las sutiles interacciones entre los sistemas inmune y endocrino pueden afectar el éxito reproductivo (Hansen et al., 2004). Los pro-cesos inflamatorios implicados en la mastitis o cojeras agudas pueden incrementar la secreción de prostaglandinas, causar luteo-lisis y estar relacionados con la pérdida fetal (Graham et al., 1995). De hecho, la mastitis clínica (Schirick et al., 2001; Santos et al., 2004a; Chebel et al., 2004; McDougall et al., 2005) y el recuento de células somáticas (Jousan et al., 2005; Moore et al., 2005) se han relacionado con la pérdida de la ges-tación. Solo en un estudio (López-Gatius et al., 2002) incluimos estas patologías como posibles factores de riesgo y no afectaron a la pérdida fetal, posiblemente debido a que durante el estudio se aplicó a las vacas enfermas, entre otros productos, flunixin meglumine, un potente anti-inflamatorio no
esteroideo que inhibe la síntesis de prosta-glandinas (Kindahl et al., 1999).
Estación (período cálido)
En seis estudios incluyendo 7341 gestacio-nes (López-Gatius et al., 2004a-c; García-Ispierto et al., 2006; Bech-Sabat et al., 2008, 2009), el riesgo de pérdida fetal fue de 1,6 a 5,4 veces más alto en las vacas que quedaron gestantes durante el periodo cálido (mayo a septiembre) respecto al periodo frío (octubre a abril). La proporción más alta de pérdidas (54%) se registró en las gestaciones dobles durante el período cálido (López-Gatius et al., 2004a). Además, se registró una fuerte relación entre el estrés térmico, medido por el índice tem-peratura-humedad (THI), sufrido durante el periodo peri-implantacional (días 21 a 30 de gestación) y la posterior pérdida fetal (García-Ispierto et al., 2006).
La introducción de sistemas anti-estrés, tales como ventiladores y aspersores, a finales de los noventa redujo sustancial-mente la incidencia de pérdidas (datos no publicados). Sin embargo, el porcentaje de pérdidas se ha ido incrementando desde entonces junto a la producción de leche. Es obvio que necesitamos nuevos plan-teamientos, tanto de medidas del estrés térmico a nivel de explotación, como de mejora de los sistemas anti-estrés. Quizás debamos comenzar a estudiar posibles sistemas de refrigeración, más que mejorar los actuales sistemas que dependen, sobre todo, del movimiento forzado de aire.
Gestaciones gemelares
Quizás la gestación gemelar sea el princi-pal factor relacionado con la pérdida fetal. Utilizando la gestación sencilla (con un
embrión) como referencia, el riesgo rela-tivo mínimo de pérdida fue de 3,1 veces para las gestaciones gemelares, inclu-yendo datos de cinco estudios con 4018 gestaciones (López-Gatius y Hunter, 2005; López-Gatius et al. 2002, 2004a, 2006; García-Ispierto et al., 2006). Como se ha apuntado antes, se registraron hasta un 54% de pérdidas en gestaciones dobles durante el periodo cálido (López-Gatius et al., 2004a). En un estudio epidemiológico sobre 52362 lactaciones se relacionó una mayor incidencia de partos gemelares con una mayor producción de leche (Kinsel et al., 1998). En vacas de alta producción, la tasa de doble ovulación puede exceder el 20% (Fricke y Wiltbank, 1999) o incluso el 25%, si se trata de vacas en su tercera lac-tación o más (López-Gatius et al., 2005b). La genética parece ser un factor deter-minante en el incremento de los partos dobles. En un estudio sobre 37174 toros y 1324678 partos se demostró cómo la descendencia de los toros nacidos después de 1990 sufría una mayor incidencia de partos dobles que la de los nacidos antes de 1980 (Johanson et al., 2001 ). Es razo-nable sugerir que el incremento de partos dobles los últimos años esté relacionado con la selección hacia el incremento de producción de leche.
Pero, paralelas al progreso genético, mejoras en la nutrición y en las prácticas de manejo han favorecido también el incre-mento de la producción. Probablemente, el manejo necesario para una alta producción vaya acompañado de un descenso de las pérdidas de gestación en el caso de ges-taciones gemelares. De una forma u otra, podemos esperar un incremento de los par-tos dobles acompañando al incremento de producción de leche en los próximos años.
La gestación gemelar es no deseable en ganado vacuno lechero, no sólo por el riesgo de pérdida de la gestación durante el periodo fetal temprano, sino también por sus efectos negativos como riesgo de abor-to, distocia, retención de placenta, muerte perinatal, freemartinismo, costes terapéu-
El riesgo de pérdida fetal fue de 1,6 a 5,4 veces más alto en las vacas que quedaron gestantes
durante el periodo cálido.
En ganado vacuno lechero, aunque la mayor parte de las pérdidas de la gestación ocurren durante el periodo embrionario temprano, el riesgo de pérdida fetal temprana parece incrementarse en condiciones de manejo intensivo. Este tipo de pérdida (hasta de un 10%) escapa a menudo a un control clínico y se asume que es un problema de origen multifactorial
pérdida de la gestación.indd 54 14/05/12 11:14

55
nº 42Factores que afectan la pérdida de la gestación en el periodo fetal temprano. Una aproximación terapeútica
TEMA: Seguro de Explotación de Reproductores Bovinos de Aptitud Cárnica
Mediante este seguro, el ganadero cubre la muerte de los animales, por causas ajenas a la voluntad humana, con el límite del capital ase-gurado, el cual se fija en el 100 por ciento del valor asegurado de la explotación, calculado a partir del valor base medio entre el máximo y el mínimo fijado por el Ministerio para cada tipo de animal, según tipos de razas, y su condición de puras o no puras. Es importante recordar que todos los animales de una explotación deben asegurarse al mismo porcentaje respecto del valor unitario máximo que les corresponda por tipo.
Existen unas garantías básicas, con tres opciones encadenadas y unas garantías adicionales, independiente de la opción elegida, tal y como se especifica a continuación:
Garantías básicas Riesgos cubiertos
Opción 1
Muerte de al menos 2 ó 3 animales y pérdida de producción por muerte masiva de reproductores
Muerte, sacrificio o decomiso por EEB
Muerte o sacrificio necesario por Fiebre aftosa e inmovilización de animales por Fiebre aftosa
Opción 2Cubre lo mismo que en la opción 1
más el exceso de mortandad de reproductoras y recría en la explotación
Opción 3Cubre lo mismo que en la opción 2
más la muerte de las crías desde el parto hasta un mes de edad
Garantías adicionales Riesgos cubiertos
Saneamiento ganadero
Saneamiento ganadero
Saneamiento ganadero extra
Pastos estivales y Pastos invernales
El Ministerio de Agricultura, Alimentación y Medio Ambiente, a través de ENESA, subvenciona al ganadero parte del coste neto del seguro. Esta subvención se obtiene mediante la suma de los distintos porcentajes y depende, entre otros aspectos, de las características del asegurado, siendo los siguientes:
Tipo de Subvención Porcentajes
Subvención base aplicable a todos los asegurados 14%
Subvención por contratación colectiva 5%
Subvención adicional según las condiciones del asegurado 14% - 16% *
Subvención por renovación de contrato según se hayan asegurado uno o dos años anteriores 6% ó 9%
Subvención por pertenecer a una Agrupación de Defensa Sanitaria (ADSG) 5%
(*) En el caso de una ganadera joven o ganadera profesional, la subvención adicional según las condiciones del asegurado es del 16%.
Las Comunidades Autónomas también pueden subvencionar este seguro, acumulándose a la subvención que aporta el Ministerio.
El ganadero interesado en este seguro puede solicitar más información a la ENTIDAD ESTATAL DE SEGUROS AGRARIOS C/ Miguel Angel 23-5ª planta 28010 MADRID con teléfono: 913475001, fax: 913085446 y correo electrónico: [email protected] y a través de la página web www.enesa.es. Y sobre todo contactando con su Tomador del Seguro o con su Mediador, ya que éstos se encuentran más próximos y le pueden aclarar cuantas dudas se le planteen antes de realizar la póliza y posteriormente asesorarle en caso de siniestro.
MINISTERIO DE AGRICULTURA, ALIMENTACIÓN Y MEDIO AMBIENTE
“ENESA Informa”
pérdida de la gestación.indd 55 14/05/12 11:14

56
lópez-Gatius F.
ticos post-parto y un retraso en el retorno a la ciclicidad tras el parto (Nielen et al., 1989). Además, un incremento de los partos dobles se ha descrito en los meses de verano (De rensis y Scaramuzzi, 2003), agudizando su efecto en regiones de clima cálido. Los efectos negativos de la gestación gemelar deberían ser menores si intentáramos redu-cir el número de embriones.
métodos de reducción embrionaria se utilizan en la reproducción asistida humana (mansour et al., 1999) y en el tratamiento de la gestación gemelar en yeguas (macpherson y reimer, 2000). Un importante éxito (75%) se registró tras la ruptura manual de la vesícula amnió-tica de un gemelo antes del día 30 de gestación en yegua (Pascoe et al., 1987). intentamos el mismo procedimiento en la vaca (López-Gatius, 2005). La ruptura del amnios sin tratamiento adicional resultó en un 100% de pérdidas (11/11), mien-tras que el procedimiento de ruptura acompañado de un tratamiento con pro-gesterona permitió la reducción geme-lar: 4 de 11 animales mantuvieron la gestación. Sin embargo, en una de estas gestaciones sobrevivientes, el embrión sobrevivió a la ruptura del amnios y la vaca parió gemelos. La aspiración trans-vaginal del embrión guiada por ecografía podría ser una alternativa a la ruptura manual, tal como se ha propuesto en la yegua (mari et al., 2004) y en la especie humana (Coffler et al., 1999).
❱❱ Factores sin efecto
La condición quística previa a la gesta-ción no pudo ser relacionada con la pér-dida de la misma en dos estudios inclu-
yendo 3623 gestaciones (López-Gatius et al., 1996, 2002). Tampoco se pudo relacionar la producción de leche en el momento del diagnóstico de gestación ni el intervalo parto-gestación con la pérdida fetal, agrupando 5138 gesta-ciones de cinco estudios (López-Gatius et al., 2004a, 2004c; García-ispierto et al., 2006; Bech-Sabat et al., 2009). La calidad del manejo relacionada con la alta producción probablemente reduce al mínimo los posibles efectos negativos de estos factores. referentes a la pro-ducción de leche, nuestros resultados están de acuerdo con otros estudios en los que se ha observado escaso o nulo efecto de la producción de leche sobre el incremento del riesgo de pérdida de la gestación (Santos et al., 2004b; Chebel et al., 2004; ]ousan et al., 2005; mcDougall et al., 2005).
❱❱ Factor preventivo
Cuerpo lúteo adicional
registrado en gestaciones con un número de cuerpos lúteos superior al número de los embriones, el cuerpo lúteo adicional ha mostrado ser un potente factor favore-cedor del mantenimiento de la gestación. Agrupando los datos de cinco estudios (López-Gatius et al., 2002, 2004a, 2006; García-ispierto et al., 2006; Bech-Sabat et al., 2008), de un total de 363 vacas gestantes con cuerpo lúteo adicional, 1,7% sufrieron pérdida fetal, mientras que un 9,9% de pérdidas se registraron en las 3643 vacas restantes que no lo tenían. Se da el caso que, en tres de los cinco estudios, que sumaron un total de 126 vacas con cuerpo lúteo adicional y con un único embrión,
ninguna de estas vacas sufrió pérdida fetal (López-Gatius et al., 2002, 2006; Bech-Sabat et al., 2008).
❱❱ Aproximación terapéutica
Tratamiento con GnRH o hCG
La progesterona es la hormona clave para el mantenimiento de la gestación (Spencer et al., 2004), estando el mantenimiento de la gestación positivamente correlacionado con los niveles plasmáticos de progeste-rona en la semana quinta de la gestación (Starbuck et al., 2004). Además, las con-centraciones de progesterona influyen en la actividad secretora del trofoblasto e hipófisis durante el primer trimestre de la gestación (Ayad et al., 2007). Sin embargo, una de las consecuencias de la alta producción es la alta tasa metabó-lica asociada a una mayor capacidad de ingestión de materia seca. Este proceso reduce los niveles plasmáticos de hor-monas esteroideas, como es el caso de la progesterona (Sangsritavong et al., 2002), con lógico impacto sobre la gestación. De hecho, la producción láctea puede afectar negativamente los niveles plasmáticos de progesterona al inicio del periodo fetal (Bech-Sabat et al., 2008; rhinehart et al., 2009). Por lo tanto, parece razonable suponer que una de las causas de la pér-dida fetal temprana en vacas de alta pro-ducción de leche podría ser una concen-tración subóptima de progesterona, bien debido a un catabolismo incrementado de la progesterona, a una sub-función luteal, o a ambos. En este sentido, estrategias que inducen la formación de un cuerpo lúteo adicional deberían incrementar los niveles plasmáticos de progesterona y favorecer el mantenimiento de la gesta-ción. Sin embargo, aunque el tratamiento con GnrH en el momento de la iA (López-Gatius et al., 2006) o con GnrH o hCG en el momento del diagnóstico de gestación (Bartolomé et al., 2006; Stevenson et al., 2008) indujo cuerpos lúteos adicionales, el tratamiento no redujo la incidencia de pérdidas fetales en ninguno de los estudios.
Tratamiento con progesterona
Para constatar la hipótesis que niveles sub-óptimos de progesterona pueden compro-meter el desarrollo de la gestación, aplica-mos con un dispositivo intra-vaginal (PriD) progesterona en el momento del diagnós-tico de la gestación (días 36-42 post-iA). El dispositivo se mantuvo durante 28 días y el
Los procesos inflamatorios implicados en las cojeras agudas pueden incrementar la secreción de prostaglandinas, causar luteolisis y estar relacionados con la pérdida fetal.
Sin título-1 1 20/03/2012 9:54:13
Si quieres más información [email protected]
Sin título-14 1 28/03/12 11:54CYS 42.indb 56 14/05/12 10:11

57
nº 42Factores que afectan la pérdida de la gestación en el periodo fetal temprano. Una aproximación terapeútica
Sin título-1 1 20/03/2012 9:54:13
Si quieres más información [email protected]
Sin título-14 1 28/03/12 11:54CYS 42.indb 57 14/05/12 10:11

58
lópez-Gatius F.
estudio se realizó con 1098 vacas (López-Gatius et al., 2004c). El riesgo relativo de pérdida de la gestación fue 2,4 veces más alto para las vacas no tratadas. En un estu-dio más reciente en el que el tratamiento con progesterona se aplicó el día 28 post-iA se obtuvieron resultados semejantes (Bech-Sabat et al., 2007). Estos resultados soportan la hipótesis que las concentraciones sub-óptimas de progesterona pueden compro-meter la gestación. En estas condiciones, el tratamiento con progesterona puede reducir la incidencia de la pérdida de gestación durante el periodo fetal temprano.
El caso de las gestaciones gemelares
En un estudio reciente en dos explota-ciones sufriendo una alta incidencia de pérdidas, se establecieron dos grupos de tratamiento: GnrH o progesterona (Bech-Sabat et al., 2009). En el caso de vacas con un único cuerpo lúteo, la incidencia de pérdidas se redujo en un factor de 0,51 para el grupo que recibió el tratamiento con progesterona, comparado al grupo GnrH. Sin embargo, en vacas con dos o más cuerpos lúteos, el tratamiento con progesterona incrementó la probabilidad de pérdida por un factor de 3, comparado al tratamiento con GnrH. Además, en vacas gestantes de gemelos, el porcentaje de reducción gemelar fue superior en el grupo que recibió GnrH (36%), respec-to del que recibió progesterona (16%). Probablemente el efecto anti-luteolítico del tratamiento con GnrH (Thatcher et al., 1989) redujo la tasa de pérdidas en el caso de reducción de gemelos, proceso que suele resultar en una alta incidencia de pérdida de los dos gemelos en vacas sin tratamiento (López-Gatius y Hunter, 2005). Estos resultados sugieren que en el momento del diagnóstico de gestación,
por ejemplo entre los días 28 y 34, es tan importante determinar el número de embriones como el de cuerpos lúteos. Las implicaciones prácticas de estos resul-tados es que en explotaciones con alta incidencia de pérdidas de gestación de carácter no infeccioso, el tratamiento con progesterona a las vacas con un único cuerpo lúteo y con GnrH a las vacas con gemelos, debería reducir el porcentaje de las pérdidas.
❱❱ Conclusiones
Una vez que la vaca se ha diagnostica-do gestante, la pérdida de la gestación durante el periodo fetal temprano se está convirtiendo en el problema más fre-cuente de la gestación en explotaciones de alta producción de leche. Los principa-les factores que favorecen la pérdida son la edad (vacas que han parido respecto de novillas), algunos toros utilizados en la inseminación, el periodo cálido y las gestaciones gemelares, mientras que la presencia de un cuerpo lúteo adicional se ha identificado como un potente factor favorecedor del mantenimiento de la gestación. El tratamiento con pro-gesterona y GnrH puede reducir el nivel de pérdidas aplicado a vacas con uno o con dos o más cuerpos lúteos, respecti-vamente, en explotaciones con alto nivel de pérdidas.
❱❱ Referencias
• AYAD A, SoUSA Nm, SULoN J, HorNiCK JL, WATTS
J, LóPEz-GATiUS F, iGUEr-oUADA m, BECKErS JF.
LNFLUENCE oF ProGESTEroNE CoNCENTrATioNS
oN SECrETorY FUNCTioNS oF TroPHoBLAST
AND PiTUiTArY DUriNG THE FirST TrimESTEr oF
PrEGNANCY iN DAirY CATTLE. THErioGENoLoGY
2007; 67: 1503-11.
• BALL PJH. LATEr EmBrYo AND EArLY FETAL
morTALiTY iN THE CoW. ANim BrEED ABS 1997;
65: 167-75.
• BArToLomE JA, KAmimUrA S, SiLVESTrE F,
ArTECHE ACm, TriGG T, THATCHEr WW. THE
USE oF A DESLorELiN imPLANT (GNrH AGoNiST)
DUriNG THE LATE EmBrYoNiC PErioD To rEDUCE
PrEGNANCY LoSS. THErioGENoLoGY 2006; 65:
1443-53.
• BECH-SABAT G, GArCíA-iSPiErTo i, YÁNiz
JL, LóPEz-GATiUS F. HiGH miLK ProDUCTioN
LNCrEASES THE riSK oF EArLY FETAL LoSS iN
DAirY CoWS. rEProD DomEST ANim 2007;
42(S2): 122.
• BECH-SABAT G, LóPEz-GATiUS F, YÁNiz JL,
GArCíA-iSPiErTo i, SANToLAriA P, SErrANo B,
SULoN J, DE SoUSA Nm, BECKErS JF. F ACTorS
AFFECTiNG PLASmA ProGESTEroNE iN THE EArLY
FETAL PErioD iN HiGH ProDUCiNG DAirY CoWS.
THErioGENoLoGY 2008; 69: 426-32.
• BECH-SABAT G, LóPEz-GATiUS F, GArCíA-iSPiErTo
i, SANToLAriA P, SErrANo B, NoGArEDA C, DE
SoUSA Nm, BECKErS JF, YÁNiz J. PrEGNANCY
PATTErNS DUriNG THE EArLY FETAL PErioD iN
HiGH ProDUCiNG DAirY CoWS TrEATED WiTH
GNrH or ProGESTEroNE. THErioGENoLoGY
2009; 71: 920-9.
• BULmAN DC. A PoSSiBLE iNFLUENCE oF THE BULL
oN THE iNCiDENCE oF EmBrYoNiC morTALiTY iN
DAirY CATTLE. VET rEC 1979; 105: 420-2.
• CHEBEL rC, SANToS JEP, rEYNoLDS JP, CErri rLA,
JUCHEm So, oVErToN m. FACTorS AFFECTiNG
CoNCEPTioN rATE AFTEr ArTiFiCiAL iNSEmiNA-
TioN AND PrEGNANCY LoSS iN LACTATiNG DAirY
CoWS. ANim rEProD SCi 2004; 84: 239-55.
• CoFFLEr mS, KoL S, DrUGAN A, iTSKoViTz-EiDor
). EArLY TrANSVAGiNAL EmBrYo ASPirATioN:
A SAFEr mETHoD For SELECTiVE rEDUCTioN
iN HiGH orDEr mULTiPLE GESTATioNS. HUmAN
rEProD 1999; 14: 1875-8.
• CommiTTEE oN BoViNE rEProDUCTiVE
NomENCLATUrE, 1972: rECommENDATioNS For
STANDArDiziNG BoViNE rEProDUCTiVE TErmS.
CorNELL VET 1972; 62: 216-37.
• DE rENSiS F, SCArAmUzzi rJ. HEAT STrESS AND
SEASoNAL EFFECTS oN rEProDUCTioN iN DAirY
CoW- A rEViEW. THErioGENoLoGY 2003; 60:
1139-51.
• ForAr AL, GAY Jm, HANCoCK DD. THE FrEqUENCY
oF ENDEmiC FETAL LoSS iN DAirY CATTLE: A
rEViEW. THErioGENoLoGY 1995; 43: 989-1000.
• FriCKE Pm, WiLTBANK mC. EFFECT oF miLK Pro-
DUCTioN oN THE iNCiDENCE oF DoUBLE oVULA-
TioN iN DAirY CoWS. THErioGENoLoGY 1999;
52: 1133-43.
• GÁBor G, TóTH F, ózSVÁri L, ABoNYi-TóTH zS,
SASSEr rG. FACTorS iNFLUENCiNG PrEGNANCY
rATE AND LATE EmBrYoNiC LoSS iN DAirY CAT-
TLE. rEProD DomEST ANim 2008; 43: 53-8.
• GArCíA-iSPiErTo i, LóPEz-GATiUS F, SANToLAriA
P, YÁNiz JL, NoGArEDA C, LóPEz-BéJAr m,
DE rENSiS F. rELATioNSHiP BETWEEN HEAT
STrESS DUriNG THE PEri-imPLANTATioN PEri-
oD AND EArLY FETAL LoSS iN DAirY CATTLE.
THErioGENoLoGY 2006; 65: 799-807.
• GArCíA-iSPiErTo i, LóPEz-GATiUS F, BECH-
SABAT G, SANToLAriA P, YÁNiz JL, NoGArEDA
C, DE rENSiS F, LóPEzBéJAr, m. CLimATE FAC-
TorS AFFECTiNG THE FErTiLiTY oF HiGH Pro-
DUCiNG DAirY CoWS iN NorTHEASTErN SPAiN.
THErioGENoLoGY 2007; 67: 1379-85.
• GrAHAm TW, Giri SN, DAELS P F, CULLor
JS, KEEN CL, THUrmoND mC, DELLiNGEr JD,
STABENFELDT GH, oSBUrN BL. ASSoCiATioNS
AmoNG ProSTAGLANDiN F2AiPHA, PLAS-
mA ziNC, CoPPEr AND iroN CoNCENTrA-
TioNS AND FETAL LoSS iN CoWS AND mArES.
THErioGENoLoGY 1995; 44: 379-90.
• GrimArD B, FrErET S, CHEVALLiEr A, PiNTo A,
PoNSArT C, HUmBLoT P. GENETiC AND ENViroN-
mENTAL FACTorS iNFLUENCiNG FirST SErViCE
CoNCEPTioN rATE AND LATE EmBrYoNiC/FoETAL
morTALiTiY iN LoW FErTiLiTY DAirY HErDS. ANim
rEProD SCi 2006; 91: 31-44.
• HANSEN PJ, SoTo P, NATzKE rP. mASTiTiS AND
FErTiLiTY iN CATTLE - PoSSiBLE iNVoLVEmENT
oF iNFLAmmATioN or immUNE ACTiVATioN iN
EmBrYoNiC morTALiTY. Am J rEProD LmmUNoL
2004; 51: 294-301.
• HANzEN CH, DrioN PV, LoUTriE o, DEPiErrEUx
C, CHriSTiANS E. EmBrYoNiC morTALiTY. PArT
L. CLiNiCAL ASPECTS AND ETioLoGiCAL FACTorS
iN THE BoViNE SPECiES. ANN mED VET 1999; 143:
La progesterona es la hormona clave para el mantenimiento de la gestación,
estando el mantenimiento de la gestación positivamente correlacionado con los
niveles plasmáticos de progesterona en la semana quinta de la gestación.
CYS 42.indb 58 14/05/12 10:11

59
nº 42Factores que afectan la pérdida de la gestación en el periodo fetal temprano. Una aproximación terapeútica
91-118.
• JoHANSoN Jm, BErGEr PJ, KirKPATriCK BW,
DENTiNES mr. TWiNNiNG rATES For NorTH
AmEriCAN HoLSTEiN SirES. J DAirY SCi 2001;
84: 2081-8.
• JoUSAN FD, DroST m, HANSEN PJ. FACTorS
ASSoCiATED WiTH EArLY AND miD-To-LATE
FETAL LoSS iN LACTATiNG AND NoN-LACTATiNG
HoLSTEiN CATTLE iN A HoT CLimATE. J ANim SCi
2005; 83: 1017-22.
• KiNDAHL H, BEKAmA m, KASK K, KoNiGSSoN
K, GuSTAFSSoN H, oDENSViK K. ENDoCriNE
ASPECTS oF UTEriNE iNVoLUTioN iN THE CoW.
rEProD DomEST ANim 1999; 34: 261-8.
• KiNSEL mL, mArSH WE, rUEGG PL, ETHEriNGToN
WG. riSK FACTorS For TWiNNiNG iN DAirY
CoWS. J DAirY SCi 1998; 81: 989-93.
• LABErNiA J, LóPEz-GATiUS F, SANToLAriA P,
LóPEz-BéJAr m, rUTLLANT J. LNFLUENCE oF
mANAGEmENT FACTorS oN PrEGNANCY ATTri-
TioN iN DAirY CATTLE. THErioGENoLoGY 1996;
45: 1247-53.
• LABErNiA J, LóPEz-GATiUS F, SANToLAriA P,
LóPEz-BéJAr m, rUTLLANT J. LNFLUENCE oF
CALViNG SEASoN oN THE iNTErACTioNS AmoNG
rEProDUCTiVE DiSorDErS oF DAirY CoWS.
ANim SCi 1998; 67: 387-93.
• LóPEz-GATiUS F. iS FErTiLiTY DECLiNiNG iN DAirY
CATTLE? A rETroSPECTiVE STUDY iN NorTHEAST-
ErN SPAiN. THErioGENoLoGY 2003; 60: 89-99.
• LóPEz-GATiUS F. THE EFFECT oN PrEGNANCY
rATE oF ProGESTEroNE ADmiNiSTrATioN AFTEr
mANUAL rEDUCTioN oF TWiN EmBrYoS iN DAirY
CATTLE. J VET mED A PHYSioL PATHoL CLiN mED
2005; 52: 199-201.
• LóPEz-GATiUS F, HUNTEr rHF. SPoNTANEoUS
rEDUCTioN oF ADVANCED TWiN EmBrYoS: iTS
oCCUrrENCE AND CLiNiCAL rELEVANCE iN DAirY
CATTLE. THErioGENoLoGY 2005; 63: 118-25.
• LóPEz-GATiUS F, LABErNiA J, SANToLAriA P,
LóPEz-BEJAr m, rUTLLANT J. EFFECT oF rEPro-
DUCTiVE DiSorDErS PrEVioUS To CoNCEPTioN
oN PrEGNANCY ATTriTioN iN DAirY CoWS.
THErioGENoLoGY 1996; 46: 643-8.
• LóPEz-GATiUS F, SANToLAriA P, YANiz J,
rUTLLANT J, LóPEz-BéJAr m. FACTorS AFFECTiNG
PrEGNANCY LoSS From GESTATioN DAY 38 To 90
iN LACTATiNG DAirY CoWS From A SiNGLE HErD.
THErioGENoLoGY 2002; 57: 1251-61.
• LóPEz-GATiUS F, SANToLAriA P, YÁNiz JL,
GArBAYo Jm, HUNTEr rHF. TimiNG oF EArLY
FoETAL LoSS For SiNGLE AN TWiN PrEGNANCiES
iN DAirY CATTLE. rEProD DomEST ANim 2004A;
39: 429-33.
• LóPEz-GATiUS F, PABoN m, ALmEríA S. NEoSPorA
CANiNUm iNFECTioN DoES NoT AFFECT EArLY
PrEGNANCY iN DAirY CATTLE. THErioGENoLoGY
2004B; 62: 606-L 3.
• LóPEz-GATiUS F, SANToLAriA P, YÁNiz JL,
HUNTEr rHF. ProGESTEroNE SUPPLEmENTATioN
DUriNG THE EArLY FETAL PErioD rEDUCES PrEG-
NANCY LoSS iN HiGH-YiELDiNG DAirY CATTLE.
THErioGENoLoGY 2004C; 62: 1529-35.
• LóPEz-GATiUS F, SANToLAriA P, ALmEríA
S. NEoSPorA CANiNUm iNFECTioN DoES
NoT AFFECT THE FErTiLiTY oF DAirY CoWS iN
HErDS WiTH HiGH iNCiDENCE oF NEoSPoro-
ASSoCiATED ABorTioNS. J. VET. mED. B LNFECT.
DiS. VET. PUBLiC HEALTH, 2005A; 52: 51-3.
• LóPEz-GATiUS F, LóPEz-BéJAr m, FENECH m,
HUNTEr rHF. oVULATioN FAiLUrE AND DoUBLE
oVULATioN iN DAirY CATTLE: riSK FACTorS AND
EFFECTS. THErioGENoLoGY 2005B; 63: 1298-307.
• LóPEz-GATiUS F, SANToLAriA P, mArTiNA A,
DELéTANG F, DE rENSiS F. THE EFFECTS oF GNrH
TrEATmENT AT THE TimE oF Ai AND 12 DAYS LATEr
oN rEProDUCTiVE PErFormANCE oF HiGH Pro-
DUCiNG DAirY CoWS DUriNG THE WArm SEA-
SoN iN NorTHEASTErN SPAiN. THErioGENoLoGY
2006; 65: 820-30.
• LóPEz-GATiUS F, GArBAYo Jm, SANToLAriA P,
YÁNiz J, AYAD A, DE SoUSA Nm, BECKErS JF.
miLK ProDUCTioN CorrELATES NEGATiVELY WiTH
PLASmA LEVELS oF PrEGNANCY-ASSoCiATED
GLYCoProTEiN (PAG) DUriNG THE EArLY FETAL
PErioD iN HiGH ProDUCiNG DAirY CoWS WiTH
LiVE FETUSES. DomEST ANim ENDoCriNo! 2007A;
32: 29-42.
• LóPEz-GATiUS F, HUNTEr rH, GArBAYo Jm,
SANToLAriA P, YANiz], SErrANo B, AYAD A, DE
SoUSA Nm, BECKErS JF. PLASmA CoNCENTrA-
TioNS oF PrEGNANCY-ASSoCiATED GLYCoPro-
TEiN-1 (PAG-1) iN HiGH ProDUCiNG DAirY CoWS
SUFFEriNG EArLY FETAL LoSS DUriNG THE WArm
SEASoN. THErioGENoLoGY 2007B; 67: 1324-30.
• LóPEz-GATiUS F, SzENCi o, BECH-SABAT G,
GArCiA-iSPiErTo i, SErrANo B, SANToLAriA P,
YÁNiz J. FACTorS oF NoNiNFECTioUS NATUrE
AFFECTiNG EArLY FoETAL LoSS iN HiGH Pro-
DUCiNG DAirY HErDS iN NorTHEASTErN
SPAiN. 2008; xxV WorLD BUiATriC CoNGrESS,
BooKLET oF THE WorKSHoP: FACTorS AFFECTiNG
rEProDUCTiVE PErFormANCE iN THE CoW. EDS.
SzENCi o, BAJCSY AES, PP 118-29.
• mACPHErSoN mL, rEimEr Jm. TWiN rEDUCTioN
iN THE mArE: CUrrENT oPTioNS. ANim rEProD
SCi, 2000; 60-61: 233-44.
• mANSoUr rT, ABoULGHAr mA, SEroUr Gi,
SATTAr mA, KAmAL A, AmiN Ym. mULTiFETAL
PrEGNANCY rEDUCTioN: moDiFiCATioN oF THE
TECHNiqUE AND ANALYSiS oF THE oUTComE.
FErTiL STEriL 1999; 71: 380-4.
• mArES SE, mENGE AC, TYLEr WJ, CASi DA LE.
GENETiC FACTorS AFFECTiNG CoNCEPTioN rATE
AND EArLY PrEGNANCY LoSS iN HoLSTEiN CAT-
TLE. J DAirY SCi 1961; 44: 96-103.
• mAri G, iACoNo E, mErLo B, CASTAGNETTi C.
rEDUCTioN oF TWiN PrEGNANCY iN THE mArE
BY TrANSVAGiNAL ULTrASoUND-GUiDED ASPi-
rATioN. rEProD DomEST ANim 2004; 39: 434-7.
• mArKUSFELD-Nir o. EPiDEmioLoGY oF BoViNE
ABorTioNS iN iSrAELi DAirY HErDS. PrEV VET
mED 1997; 31: 245-55.
• mCDoUGALL S, rHoDES Fm, VErKErK GA.
PrEGNANCY LoSS iN DAirY CATTLE iN THE
WAiKATo rEGioN oF NEW zEALAND. Nz VET J
2005; 53: 279-87.
• moorE DA, oVErToN mW, CHEBEL rC, TrUSCoTT
mL, BoNDUrANT rH. EVALUATioN oF FACTorS
THAT AFFECT EmBrYoNiC LoSS iN DAirY CATTLE. J
Am VET mED ASSoC 2005; 226: 1112-8.
• NiELEN m, SCHUKKEN YH, SCHoLL DT, WiLBriNK
HJ, BrAND A. TWiNNiNG iN DAirY CATTLE: A STUDY
oF riSK FACTorS AND EFFECTS. THErioGENoLoGY
1989; 32: 845-62.
• PASCoE D, PASCoE r, HUGHES J, STABENFELDT
G, KiNDAHL H. ComPAriSoN oF TWo TECH-
NiqUES AND THrEE HormoNE THErAPiES For
mANAGEmENT oF TWiN CoNCEPTUSES BY mAN-
UAL EmBrYoNiC rEDUCTioN. J rEProD FErTiL
1987;SUPPL 35: 701-2.
• PEGorEr mF, VASCoNCELoS JLm, TriNCA LA,
HANSEN PJ, BArroS Cm. LNFLUENCE oF SirE AND
SirE BrEED (GYrVErSUS HoLSTEiN) oN ESTAB-
LiSHmENT oF PrEGNANCY AND EmBrYoNiC LoSS
iN LACTATiNG HoLSTEiN CoWS DUriNG SUmmEr
HEAT STrESS. THErioGENoLoGY 2007; 67: 692-7.
• PETErS Ar. EmBrYo morTALiTY iN THE CoW.
ANim BrEED ABS 1996; 64: 587-98.
• rHiNEHArT JD, STArBUCK-CiEmmEr mJ, FLorES
JA, miLVAE rA, YAo J, PooLE DH, LNSKEEP EK. LoW
PEriPHErAL ProGESTEroNE AND LATE EmBrY-
oNiC/EArLY FETAL LoSS iN SUCKLED BEEF AND
LACTATiNG DAirY CoWS. THErioGENoLoGY 2009;
71: 480-90.
• SANGSriTAVoNG S, ComBS DK, SArTori r,
ArmENTANo LE, WiLTBANK mC. HiGH FEED iNTAKE
iNCrEASES LiVEr BLooD FLoW AND mETABoLiSm
oF ProGESTEroNE AND ESTrADioL-1 7 iN DAirY
CATTLE. J DAirY SCi 2002; 85: 2831-42.
• SANToS JEP, CErri rLA, BALLoU mA,
HiGGiNBoTHAm GE, KirK JH. EFFECT oF TimiNG
oF FirST CLiNiCAL mASTiTiS oCCUrrENCE oN
LACTATioNAL AND rEProDUCTiVE PErFormANCE
oF HoLSTEiN DAirY CoWS. ANim rEProD SCi
2004A; 80: 31-45.
• SANToS JEP, THATCHEr WW, CHEBEL rC, CErri
rLA, GALVAo, KN. THE EFFECT oF EmBrYoNiC
DEATH rATES iN CATTLE oN THE EFFiCACY oF
ESTrUS SYNCHroNizATioN ProGrAmS. ANim
rEProD SCi 2004B; 82-83: 513-35.
• SANToS JEP, rUTiGLiANo Hm, SA mF. riSK FAC-
TorS For rESUmPTioN oF PoSTPArTUm ESTroUS
CYCLES AND EmBrYoNiC SUrViVAL iN LACTAT-
iNG DAirY CoWS. ANim rEProD SCi 2009; 110:
207-21.
• SCHiriCK FN, HoCKETT mE, SAxToN Am, LEWiS
mJ, DoWLEN HH, oLiVEr SP. LNFLUENCE oF SUB-
CLiNiCAL mASTiTiS DUriNG EArLY LACTATioN oN
rEProDUCTiVE PArAmETErS. J DAirY SCi 2001;
84: 1407-12.
• SiLKE V, DiSKiN mG, KENNY DA, BoLAND mP,
DiLLoN P, mEE JF, SrEENAN Jm. ExTENT, PATTErN
AND FACTorS ASSoCiATED WiTH LATE EmBrYoNiC
LoSS iN DAirY CoWS. ANim rEProD SCi 2002;
71: 1-12.
• SPENCEr TE, JoHNSoN GA, BUrGHArT rC, BAzEr
FW. ProGESTEroNE AND PLACENTAL HormoNE
ACTioNS oN THE UTErUS: iNSiGHTS From
DomESTiC ANimALS. BioL rEProD 2004; 71: 2-1o.
• STArBUCK mJ, DAiLEY rA, LNSKEEP EK. FACTorS
AFFECTiNG rETENTioN oF EArLY PrEGNANCY
iN DAirY CATTLE. ANim rEProD SCi 2004; 84:
27-39.
• STEVENSoN JS, TiFFANY Sm, LNSKEEP EK.
mAiNTENANCE oF PrEGNANCY iN DAirY CATTLE
AFTEr TrEATmENT WiTH HUmAN CHorioNiC
GoNADoTroPiN or GoNADoTroPiN-rELEASiNG
HormoNE. J DAirY SCi 2008; 91: 3092-101.
• THATCHEr WW, mACmiLLAN KL, HANSEN PJ,
DroST m. CoNCEPTS For rEGULATioN oF Cor-
PUS LUTEUm FUNCTioN BY THE CoNCEPTUS
AND oVAriAN FoLLiCLES To imProVE FErTiLiTY.
THErioGENoLoGY 1989; 31: 149-64.
• THUrmoND mC, PiCANSo JP, JAmESoN Cm.
CoNSiDErATioNS For USE oF DESCriPTiVE
EPiDEmioLoGY To iNVESTiGATE FETAL LoSS iN
DAirY CoWS. J Am VET mED ASSoC 1990; 197:
1305-12.
• VANrooSE G, DE KrUiF A, VAN Soom A.
EmBrYoNiC morTALiTY AND EmBrYo-PATHoGEN
iNTErACTioNS. ANim rEProD SCi 2000; 60-61:
131-43.
CYS 42.indb 59 14/05/12 10:11

60
lópez Mayagoitia A.
Patología del sistema respiratorio para veterinarios de campo
Dr. Alfonso lópez MayagoitiaUniversity of Prince Edward / Slano, CanadáLoPEz@UPEi. CA
Fotos Axón Comunicación
❱❱ Morfofisiología del aparato respiratorio del bovino
Para comprender los procesos patológicos del sistema respiratorio es conveniente dividirlo en tres compartimentos estruc-turales ya que cada uno de ellos posee sus propios mecanismos de defensa y forma particular de responder al daño celular. Estos compartimentos son: a- Sistema de conducción, que incluye desde fosas nasa-les hasta los bronquios intra-pulmonares; b- Sistema de transición, compuesto exclu-sivamente por los bronquiolos; y e- Sistema de intercambio, constituido por los alveolos pulmonares.
El pulmón de los bovinos, al igual que el de los porcinos, tiene importantes diferencias anatómicas en relación a otras especies animales. Una de estas diferencias es la marcada individualización de los lobulillos pulmonares por la abundancia de
tejido conectivo en sus paredes interlobuli-llares. Esta separación interlobulillar hace al pulmón bovino particularmente vulnerable a procesos de inflamación y reparación, así como también al enfisema observado en neumonías de tipo intersticial.
❱❱ Flora normal del aparato respiratorio
Como cualquier otra membrana mucosa en contacto con el medio ambiente, la muco-sa del aparato respiratorio de los bovinos posee su propia flora o microbiota normal. Si se introduce un hisopo en condiciones asépticas hasta las partes profundas de la cavidad nasal y se envía al laboratorio crecerán varios géneros y especies de bac-terias. Estas bacterias constituyen la flora nasal del aparato respiratorio y habitan únicamente en la porción proximal del sistema de conducción, o sea la cavidad nasal, nasofaringe y laringe. La tráquea, bronquios, bronquiolos (sistema de tran-
CYS 42.indb 60 14/05/12 10:11

61
nº 42Patología del sistema respiratorio para veterinarios de campo
sición) y los alvéolos (sistema de inter-cambio), en comparación, son membranas esencialmente estériles. En estas regiones anatómicas las bacterias son típicamente eliminadas o destruidas por los mecanis-mos de defensa.
Las bacterias de la flora nasal varían considerablemente entre las especies ani-males, así como también varían de acuerdo al medio ambiente y a las aéreas geo-gráficas donde se crían los animales. Es importante destacar que algunas bacterias presentes en la flora normal del sistema de conducción en rumiantes pueden, bajo ciertas condiciones, invadir el pulmón y producir infecciones respiratorias severas. Un ejemplo es la Mannheimia haemolytica, un habitante normal de la mucosa nasal capaz de producir la fiebre de embarque o mannheimiosis neumónica. Un error en el diagnóstico clínico es interpretar equi-vocadamente un aislamiento positivo atri-buyéndole a Mannhemia haemolytica la causa de la enfermedad.
La flora nasal no es constante sino dinámica, el número de algunos microor-ganismos aumenta mientras el de otros disminuye. Estudios microbiológicos en bovinos de engorde demuestran un aumento notable en Mannhemia haemo-lytica cuando los animales son someti-dos al estrés prolongado del transporte. Estudios experimentales también indican que el aire traqueal de los bovinos en con-diciones normales acarrea hasta los pul-mones bacterias normalmente presentes en flora normal, incluyendo desde luego a la Mannheimia haemolytica. Además de este constante acarreo de bacterias, los pulmo-nes de los bovinos y ovicaprinos son tam-bién hostigados por partículas de polvo, polen, fibras, gases tóxicos (S02, N02, H2S) y vapores como el amoniaco. Estas partícu-las, gases y vapores están presentes en el medio ambiente de un rumiante, sea éste criado en estabulación, pastizal o corral de engorde. A pesar de la constante exposi-ción a bacterias, micoplasmas, partículas, vapores y gases, el pulmón se mantiene sano gracias a sus eficientes mecanismos de defensa del aparato respiratorio.
❱❱ Mecanismos de defensa del aparato respiratorio
El principal mecanismo de defensa del sis-tema de conducción lo forma la alfombra o escalera mucociliar. Esta alfombra está constituida por epitelio pseudoestratificado ciliado y las células caliciformes productoras de moco. Cada célula ciliada tiene alrededor de 250 cilios pulsando a una frecuencia
estimada de 1000 pulsaciones/minuto y produciendo un movimiento longitudinal promedio de 20 mm/minuto. muchas par-tículas y gases inhalados son atrapados y eliminados en el moco. Las turbulencias de aire generadas por las conchas nasa-les dentro de la cavidad nasal facilitan la retención de partículas. Se estima que las partículas mayores de 10 µm son atrapadas en el moco de las conchas nasales mientras que las partículas entre 3-10 µm son atra-padas en las bifurcaciones de los bronquios debido al cambio súbito en la dirección del aire inhalado. Las partículas atrapadas en el moco son eliminadas por el movimiento mucociliar hacia la faringe en donde son finalmente deglutidas.
El moco de la alfombra mucociliar tam-bién juega un papel en los mecanismos de defensa de los rumiantes contra los gases tóxicos. Los gases hidrosolubles se disuel-ven en el moco reduciéndose así la concen-tración tóxica del gas que llega al pulmón. El moco contiene lgA, inmunoglobulina que inhibe la adherencia de microorganismos a las células de la alfombra mucociliar.
Sólo las partículas menores a las dos micras (<2.0 µm) llegan hasta los bron-quiolos y alvéolos. Los mecanismos de defensa en los bronquiolos son una com-binación de los encontrados en los sis-temas de conducción y de intercambio. Las secreciones de células bronquiolares protegen esta región que, por naturale-za, es altamente vulnerable al daño. Esta
vulnerabilidad de la región bronquiolar se manifiesta en algunas enfermedades respi-ratorias de los bovinos como es el caso de la infecciones por virus respiratorios (rBi, VrSB y Pi3) y daño toxico como el causado por el 3-metil indol en el llamado “edema y enfisema pulmonar bovino.”
Los alvéolos carecen de cilios y moco, por lo que esta región pulmonar tiene un mecanismo de defensa diferente para protegerse de las partículas y patóge-nos inhalados. El principal mecanismo de defensa en el alveolo lo constituyen los macrófagos alveolares, células altamente fagocíticas originadas en la médula ósea. De la medula ósea se originan los mono-citos sanguíneos, los cuales viajan por el sistema circulatorio hasta llegar al pulmón, en cuyo intersticio maduran y se convier-ten en macrófagos. Durante el tránsito en el intersticio pulmonar los macrófagos adquieren la capacidad de fagocitar en un medio aeróbico. El número de macrófagos alveolares es proporcional al número de partículas inhaladas que llegan al pulmón, o sea que el animal expuesto a un número alto de partículas tendrá mayor número de macrófagos en los alvéolos. Las secreciones alveolares poseen lgG y lgm; estas inmuno-globulinas juegan un papel importante en la opsonización y fagocitosis por macrófa-gos alveolares. Las secreciones alveolares, particularmente el surfactante, favorecen la fagocitosis y actúan como antioxidantes previniendo el daño celular causado por el estrés oxidativo.
Se deben enviar muestras frescas o refrigeradas de pulmón para bacteriología cuando sea necesario, sobre todo cuando haya lesiones con exudado purulento y/o abscesos.
CYS 42.indb 61 14/05/12 10:11

62
lópez Mayagoitia A.
Los mecanismos de defensa del aparato respiratorio de los bovinos son notable-mente eficientes atrapando, destruyendo, eliminando agentes patógenos y detoxifi-cando gases tóxicos. Estudios experimen-tales con aerosoles han demostrado que terneros normales son capaces de recibir dosis masivas de Mannhemia haemolytica (3x106 bacterias/gr de pulmón) sin que estas bacterias logren colonizar el pulmón o causar enfermedad. Sin embargo, cuando los mecanismos de defensa son deprimidos, como sucede durante infecciones virales, las bacterias fácilmente pueden colonizar el pulmón y causar neumonías bacterianas.
Durante muchos años se ha sabido que en epidemias de influenza muchas perso-nas mueren de neumonía secundaria bac-teriana. Esto mismo sucede en los bovinos durante brotes de Herpes bovino-1 (iBr), Virus respiratorio Sincitial Bovino (BrSV), Parainfluenza-3 (Pi-3) y Coronavirus respiratorio Bovino. Estudios experimenta-
les con aerosoles han demostrado que las curvas de eliminación de Mannhemia hae-molytica en terneros se ven notablemente reducidas 5 a 7 días después de una infec-ción viral. Una vez deprimidos los meca-nismos de defensa, las bacterias de la flora nasal fácilmente colonizan el pulmón. Estas infecciones mixtas virales y bacterianas son frecuentemente el resultado del llamado sinergismo virus-bacteria. o sea, las lesio-nes causadas por la combinación del virus y Mannhemia haemolytica son mucho más severas que la suma de las lesiones cau-sadas por estos dos patógenos de forma independiente. Es importante enfatizar que la mayoría de las infecciones por iBr o BrSV causan únicamente una “neumonía bronco intersticial” leve y transitoria. Esto explica el porqué muchos bovinos sanos que nunca han sido vacunados pueden poseer anticuerpos contra estos virus. Por razones todavía desconocidas algunos ter-neros mueren por una infección puramente viral especulándose que esto se debe al
fenómeno de daño alveolar difuso (DAD) presente en el Síndrome de Disfunción respiratoria Aguda (AroS en inglés).
Además de las infecciones virales, otros factores como el edema pulmonar, uremia, amoniaco, deshidratación y estrés pueden inhibir los mecanismos de defensa en el pulmón. El edema pulmonar frecuente-mente observado en rumiantes deprime los mecanismos de defensa y predispone al pulmón a infecciones bacterianas secunda-rias. La deshidratación es otro factor inhibi-torio de los mecanismos de defensa por lo que no es sorprendente encontrar neumo-nía bacteriana en terneros o corderos con diarreas neonatales. Se ha postulado que la deshidratación aumenta la viscosidad del moco interfiriendo así con el movimiento de los cilios en la alfombra mucociliar. De la misma forma, se conoce que factores como la acidosis, hipoxia y estrés también faci-litan la colonización del pulmón por bac-terias patógenas, sobre todo Mannhemia haemolytica, Histophilus somni y posi-blemente también Mycoplasma bovis y Arcanobacterium pyogenes.
❱❱ Examen del aparato respiratorio en la necropsia
La cavidad nasal se evalúa en la necropsia con cortes transversales o longitudinales de la cabeza; la laringe, tráquea y bron-quios se evalúan en cortes longitudinales. La mucosa de estos órganos debe ser cuidadosamente observada determinando la presencia de exudados, parásitos, erosio-nes, úlceras, o presencia de fluido, espuma, o material inhalado.
Los pulmones se observan primero y luego se palpan para detectar cambios en su textura como elasticidad, consolidación, nódulos, crepitación. Todos los reportes de necropsia deben incluir la siguiente infor-mación 1.- Distribución de las lesiones (focal/multifocal, lobar, difusa, localmente extensiva). 2.- magnitud de la lesión expre-sada en porcentaje (%) de pulmón afectado. No debemos olvidar examinar los vasos san-guíneos pulmonares investigando la posible presencia de trombos, particularmente en bovinos con abscesos cerca de la vena cava. Antes de cortar los pulmones es importante tomar fotografías digitales de las lesiones.
Para histopatología se recomienda enviar al laboratorio no solo muestras de pulmón anormal sino también pulmón nor-mal. Es importante que la fijación de tejidos para histopatología sea correcta debiéndose utilizar suficiente fijador (1:10 volúmenes de tejido: formalina) y cortando trozos de
Los pulmones se observan primero y luego se palpan para detectar cambios en su textura como elasticidad, consolidación, nódulos, crepitación.
CYS 42.indb 62 14/05/12 10:11

63
nº 42Patología del sistema respiratorio para veterinarios de campo
no más de 1 cm de ancho (la formalina no penetra adecuadamente si la muestra es más ancha). Se recomienda no dejar los tejidos en formol más de 5 días pues esto entorpece la inmunohistoquímica. En casos sospechosos de infección viral es recomen-dable congelar pulmón. Se deben enviar muestras frescas o refrigeradas de pulmón para bacteriología cuando sea necesario, sobre todo cuando haya lesiones con exu-dado purulento y/o abscesos. Los resultados microbiológicos deben interpretarse cuida-dosamente tomando en cuenta la historia clínica y los hallazgos a la necropsia.
Congestión o hiperemia nasal son fre-cuentes debido a la gran vascularización de la mucosa del sistema de conducción. Estos cambios vasculares pueden deberse a timpanismo, toxemia, trauma, agonía (fallo circulatorio) o inhalación de gases irritantes y no necesariamente a inflamación o rinitis. Las rinitis y sinusitis virales son comunes en los rumiantes y generalmente aparecen en la necropsia como un enrojecimiento severo de la mucosa. De acuerdo al tipo de exudado, las rinitis y sinusitis pueden designarse como catarral, fibrinosa (diftéri-ca), purulenta, o granulomatosa. De acuer-
do a la apariencia de la inflamación, estas pueden ser también ulcerativas, necróticas, o polipoides. Los senos paranasales tienen un drenaje deficiente y en casos crónicos van frecuentemente acompañadas por una notable acumulación de moco (mucocele) o exudado purulento (empiema). La epistaxis es una secuela común a neoplasias nasales, rinitis o presencia de cuerpo extraño, parti-cularmente en ovicaprinos.
A diferencia del aparato digestivo en el cual se puede situar anatómicamente el origen de la hemorragia de acuerdo al color de las heces, en el aparato res-piratorio la apariencia de la sangre es la misma en hemorragia nasal y en hemo-rragia pulmonar. Hemoptisis en ganado bovino está también asociada a ruptura de vasos pulmonares o tromboembolismos agudos por ruptura de abscesos hepáticos hacia la vena cava. En rumiantes con his-toria clínica de emaciación progresiva no específica debe investigarse la posibilidad de un carcinoma nasal enzoótico.
La laringitis necrótica o difteria de los terneros es una infección secundaria causada por Fusobacterium necrophorum generalmente precedida por una infección
viral o trauma en la mucosa de la laringe, como en el caso de iBr o excesiva voca-lización. Exuberantes placas de exudado fibrino-necrótico se observan en la mucosa. En ocasiones este exudado en terneros es aspirado, lo que causa una neumonía por aspiración, o puede diseminarse sistémica-mente causando toxemia o fusobacteremia.
El pulmón es un tejido comúnmente afectado por diferentes enfermedades de los bovinos por lo que es imprescindible que el clínico revise detalladamente estos órganos. Las anomalías congénitas como hipoplasia o hamartoma pulmonar, aunque raras, deben investigarse en terneros y corderos con problemas respiratorios. La melanosis pulmonar es un hallazgo inci-dental sin ninguna relevancia patológica que ocasionalmente es interpretado equi-vocadamente.
Cambios circulatorios en pulmón son muy frecuentes dada la enorme red capilar presente en estos órganos. La congestión pulmonar o acumulación pasiva de san-gre se asocia a falla cardiaca congestiva (corazón izquierdo) la cual puede progresar a edema pulmonar con hemorragias intra-alveolares. Es imperativo en estos casos
No pierda ni una gota
* Autorizado en la Unión Europea en bovinos destinados a la producción de leche y de carne, ovejas y cabras de leche, corderos y caballos (E1711/4a1711/4b1711).
- R
CS
Lal
lem
and
405
720
194
- 0
6/20
11.
Levucell SC valoriza su ración y maximiza los Ingresos sobre los Costes de Alimentación (IOFC)
Eficacia probada de Saccharomyces cerevisiae CNCM I-1077, la cepa específica para rumiantes seleccionada conjuntamente con el INRA:
• Rendimiento lechero: +1,2* a 2,5 litros/vaca/día• Eficacia alimentaria: +50g* a 120g de leche
por kg de MSI• Optimiza el pH del rumen (menos acidosis) y
mejora la digestibilidad de las fibras.
* Meta-análisis ADSA, USA 2009 probado con UNA cepa (I-1077), UNA dosis recomendada (10 mil millones/día).
Levadura Específica Rumiantes€
LALLEMAND BIO, SL
Telf: +34 93 241 33 80 Email: [email protected]
www.levucellsc.com
CYS 42.indb 63 14/05/12 10:11

64
lópez Mayagoitia A.
revisar el corazón poniendo especial aten-ción a cardiopatías congénitas como son el defecto de septo ventricular, y la hipoplasia o displasia de válvulas, entre otras muchas.
Causas comunes de hemorragia pul-monar incluyen: falla cardiaca congestiva (ver congestión), trauma (penetración o laceración pulmonar), coagulopatías (e.g., intoxicación con dicumarol), trombo embo-lismos pulmonares, septicemias, coagula-ción intravascular diseminada, embolismos, o ruptura de vasos sanguíneos por aneu-rismas pulmonares. Los tromboembolis-mos pulmonares tienen poca importancia clínica y sus causas más comunes son endocarditis y trombosis de venas yugu-lares en cateterización prolongada. Los infartos pulmonares también tienen poca importancia clínica y aparecen general-mente en la necropsia como focos de color rojo oscuro en los márgenes del pulmón.
El edema pulmonar es una lesión fre-cuente en rumiantes, particularmente en
los ovinos. Se caracterizada por acumula-ción de líquido en el intersticio pulmonar o alvéolos. En condiciones normales los pul-mones producen trasudados (líquido bajo en proteína) el cual es absorbido por los vasos linfáticos. Si la producción de líquido excede la remoción, ya sea por exceso en producción o por fallo en la absorción, el líquido se acumula causando edema pul-monar. El edema pulmonar es causa común de muerte, sin embargo, en muchos casos es puramente una lesión terminal que ocurre durante la agonía o por el uso de barbitúricos en la eutanasia. Es importante interpretar correctamente el significado del edema pulmonar en una necropsia y con frecuencia es imprescindible la histopato-logía para establecer un diagnóstico causal.
El enfisema pulmonar es una lesión particularmente común en los bovinos
dada su marcada separación interlobu-lillar. Al igual que el edema pulmonar, el enfisema pulmonar requiere de una correcta interpretación para evitar errores en el diagnóstico. Esta lesión puede ser un hallazgo incidental o parte de la enferme-dad conocida como el “edema y enfisema pulmonar bovino.” La histopatología es imprescindible en todos estos casos y es un error hacer diagnósticos únicamente macroscópicos.
❱❱ Neumonias en rumiantes
No existe una clasificación universal de neumonías en medicina veterinaria. El valor de cualquier clasificación depende funda-mentalmente de su utilidad práctica. Existe una clasificación morfológica sencilla que permite predecir con buen grado de con-fiabilidad la probable etiología y la ruta de entrada del patógeno durante la necropsia. Esta clasificación se basa en el tipo y loca-lización de la inflamación macroscópica en el pulmón y consta de cinco formas morfológicas:
1. Bronconeumonía supurativa; 2. Bronconeumonía fibrinosa;3. Neumonía intersticial; 4. Neumonía embólica; y 5. S. Neumonía granulomatosa.
1. La bronconeumonía supurativa tiene distribución craneoventral y la textura del pulmón afectado es firme (conso-lidado). La vía de entrada del patógeno
en este tipo de neumonía es aeró-gena y generalmente es causada por bacterias como Pasteurella multocida, Actinomyces pyogenes, Mycoplasma bovis. Para que un animal muera de bronconeumonía supurativa es nece-sario que por lo menos el 60% de los pulmones estén afectados. El pulmón afectado está consolidado, tiene una apariencia lobulillar acentuada y el color varía de rojo intenso en casos agudos (hiperemia) a gris pálido en casos crónicos (inflamación, atelecta-sia, fibrosis). Típicamente, los bronquios contienen exudado purulento el cual es evidente al comprimir el parénquima pulmonar. Habitualmente al exprimir en superficie de corte un pulmón afec-tado, pus o moco sale de los bronquios. En los casos crónicos el exudado se vuelve mucoide debido a la hiperplasia de células caliciformes. En la histopa-tología se observan siempre un gran número de neutrófilos y macrófagos tanto en bronquios como en bron-quiolos y alveolos. Entre los ejemplos de enfermedades que se manifiestan como una bronconeumonía supurativa figuran las neumonías enzoóticas de terneros y corderos. Entre las secuelas más comunes de la bronconeumonía supurativa figuran los abscesos pulmo-nares y la bronquiectasia, esta última es una dilatación permanente de un bronquio debido a la destrucción de sus paredes por el proceso inflamatorio.
2. La bronconeumonía fibrinosa, también llamada pleuroneumonía o neumonía lobar, es similar a la bronconeumonía supurativa pero se diferencia macros-cópicamente por la presencia de fibrina en la pleura. La textura es dura y la vía de entrada de los patógenos invo-lucrados es aerógena. Estos agentes generalmente bacterianos producen un daño severo al tejido pulmonar y su pronóstico es menos favorable que en la bronconeumonía supurati-va. La Mannheimia haemolytica y el Mycoplasma mycoides sub mycoides son dos ejemplos de agentes causantes de neumonía fibrinosa en rumiantes. El Histophilus somni también puede producir esta lesión. El animal puede morir con 50% o menos de pulmón afectado debido a que este tipo de bronconeumonía va generalmente acompañada de necrosis pulmonar y toxemia. Al abrir la cavidad torácica se observa efusión pleural con abundante fibrina sobre las superficies pleurales. En los casos más agudos la pleura tiene una apariencia de vidrio moli-do. En superficie de corte el pulmón
La neumonía intersticial es la más difícil de diagnosticar en la necropsia y es siempre necesario realizar un análisis histopatológico
Sin embargo, cuando los mecanismos de defensa son deprimidos, como sucede durante infecciones virales, las bacterias fácilmente pueden colonizar el pulmón y causar neumonías bacterianas. .fácilmen-te pueden colonizar el pulmón y causar neumonías bacterianas
CYS 42.indb 64 14/05/12 10:11

65
nº 42Patología del sistema respiratorio para veterinarios de campo
afectado muestra áreas irregulares de necrosis comúnmente referidas como “marmoleado.” microscópicamente el tejido pulmonar muestra ensancha-miento de los septos inter-lobulillares, perdida de los espacios aéreos y con mucha frecuencia se presentan áreas de necrosis coagulativa las cuales están rodeadas de un típico anillo de leuco-citos degenerados (células de avena). Entre las secuelas más importantes en los animales que sobreviven a la bronconeumonía fibrinosa se encuen-tran secuestros pulmonares, los cuales consisten en grandes áreas de tejido necrótico rodeado de tejido conectivo y abundantes adherencias pleurales.
3. La neumonía intersticial, también conocida como neumonitis, tiene una distribución pulmonar difusa o sea que afecta a todos los lóbulos pulmona-res. Su textura es típicamente elástica. Estos cambios morfológicos de color y textura son poco marcados y difí-ciles de diagnosticar en la necropsia, por lo que se requiere siempre de la histopatología para su confirmación. Las neumonías intersticiales van fre-cuentemente acompañadas de edema y enfisema. La vía de entrada puede ser hematógena, como en septicemia, viremia y toxemia, o aerógena, como en infecciones por virus respiratorios (BrSV, rBi, Pl-3), reacciones a alérge-nos, o inhalación de gases tóxicos. En la necropsia se observa que los pul-mones no se colapsan cuando se abre la cavidad torácica y que hay claras impresiones costales en la pleura vis-ceral. El color de los pulmones varía de rojo en casos agudos, a color pálido en casos crónicos. microscópicamente, la lesión está centrada en la pared alveo-lar, la cual está engrosada. Ejemplos de enfermedades que se manifiestan con una neumonía intersticial incluyen las infecciones por los virus de iBr, BrSV y Pl-3, al igual que el “edema y enfisema pulmonar bovino”, una alveo-litis extrínseca alérgica causada por la hipersensibilidad a esporas de hongos en malos ensilados o hipersensibilidad a proteínas de parásitos pulmonares. En términos generales la neumonía intersticial es la más difícil de diag-nosticar en la necropsia y es siempre necesario que el clínico envíe muestras al laboratorio para histopatología. Se recomienda investigar la presencia de virus respiratorios al mismo tiempo mediante muestras remitidas a labora-torios de virología.
4. La neumonía embólica tiene una dis-tribución típicamente multifocal carac-terizada por la presencia de numero-sos focos de inflamación afectando todos los lóbulos pulmonares. Su vía de entrada es obviamente hematógena donde pequeños émbolos sépticos o bacterias circulantes logran adherirse a la pared de los capilares pulmonares y evadir la fagocitosis de los macrófagos intravasculares. En términos generales, los infartos pulmonares y la neumonía embólica son similares, excepto que en esta última los émbolos son general-mente sépticos y el foco de infección se extiende centrífugamente. Ejemplos de enfermedades que se manifiestan con una neumonía embólica en rumiantes incluyen la endocarditis vegetativa en el lado derecho del corazón (válvula tricúspide o de la arteria pulmonar), émbolos originados por la ruptura de abscesos hepáticos hacia la vena cava en ganado bovino y onfaloflebitis en terneros o corderos recién nacidos. En la necropsia se observan un número variable de pequeños focos de infla-mación. En los casos más agudos se aprecia un centro blanquecino rodea-do de un halo hiperémico; con el tiempo estas lesiones maduran hasta formar pequeños abscesos pulmonares en casos más crónicos. Las secuelas comunes de neumonía embólica inclu-yen abscesos pulmonares distribuidos al azar en todos los lóbulos pulmona-res. microscópicamente existen focos de necrosis en las paredes alveolares rodeadas de una intensa reacción infla-matoria con edema y neutrófilos en los estadios iniciales y posteriormente con macrófagos y tejido conectivo en estadios más crónicos. Es frecuente ver en los focos de necrosis e inflamación colonias de bacterias demostrables tanto en tinciones de HE o en tincio-nes de Gram. En casos de neumonía embólica es imperativo que el clínico de campo haga una inspección deta-llada para localizar el foco de infección poniendo especial atención a las pare-des y válvulas de corazón.
5. La neumonía granulomatosa se caracteriza por numerosos nódulos de tamaño variable y en ocasiones confluyentes afectando uno a varios lóbulos pulmonares. Este tipo de neumonía puede tener una vía de entrada tanto aerógena como hema-tógena. Las lesiones son multifocales caracterizadas por la presencia de nódulos o granulomas pulmonares. muy frecuentemente, las lesiones van acompañadas por nódulos similares
en otros órganos, particularmente en los linfonodos. Ejemplos de enferme-dades que se manifiestan típicamen-te por una neumonía granulomatosa incluyen tuberculosis, las micosis sistémicas (profundas) causadas por hongos como el Coccidiodes immitis, infecciones por algas como la Prototheca spp o Chlorella spp, algunos parásitos como Muellerius capillaris en pequeños rumiantes, Fasciola hepática, larvas migratorias aberrantes, y partículas vegetales aspiradas a pulmón. Nódulos firmes distribuidos en el parénquima pul-monar. Las neumonías granuloma-tosas pueden ser fácilmente confun-didas con neoplasias requiriéndose del uso de la histopatología para su confirmación. microscópicamente, los nódulos están constituidos por un centro necrótico infiltrado en su periferia por macrófagos, células gigantes y tejido conectivo infiltrado con linfocitos y células plasmáticas. Es interesante notar que en las neu-monías granulomatosas casi siempre es posible observar microscópica-mente el agente causal por lo que es recomendable mandar muestras de pulmón y linfonodos al laboratorio para estudios histopatológicos.
En conclusión, para llegar a un diag-nóstico correcto es importante que el clínico realice necropsias de campo y se apoye en los laboratorios de diagnós-tico. El significado de un aislamiento bacteriano debe siempre ir acorde con la historia clínica y hallazgos en la necrop-sia. En algunos casos una necropsia bien hecha es suficiente para formalizar un buen diagnóstico tentativo a nivel de campo. Esta necropsia debe ser sis-temática pues, como comúnmente se dice en diagnóstico veterinario, “más se pierde por no buscar que por no saber.” Se requiere de tiempo y esfuerzo para llegar a buen diagnóstico clínico o patológico. En casos de neumonías, es importante comenzar diferenciando los tipos de inflamación para así poder seleccionar las muestras apropiadas y enviarlas oportunamente al laborato-rio. Es un error tratar de hacer siempre diagnósticos etiológicos sin apoyo del laboratorio. Un estudio reciente mostró que cerca de la mitad de las neumonías intersticiales diagnosticadas en necrop-sias eran equivocadas por no contar con el apoyo de histopatología.
CYS 42.indb 65 14/05/12 10:11

¡¡¡Oferta web!!50 € [email protected]
Consíguelo en www.axoncomunicacion.net
Imprescindible en tu clínica
4-Mastocitoma cutáneo canino
112 113
Elena Martínez de Merlo5 - Tumores de tejidos blandos
Incluyen una amplia variedad de tipos histológicos, que presentan unas ca-racterísticas comunes:
• Alta capacidad de invasión local: Son tumores de márgenes poco defi ni-dos y alto potencial infi ltrativo; aunque aparentemente están encapsula-dos, realmente se trata de una seudocápsula constituida por las propias células tumorales y los tumores presentan numerosas prolongaciones, en forma de dedos de guante, hacia tejidos profundos.
• Por su grado de infi ltración, su potencial de recidiva es muy elevado
• Si desarrollan metástasis, la mayor parte de los casos se diseminan por vía hematógena, por lo que los ganglios regionales no suelen estar afectados
• Baja respuesta a la quimioterapia.
Los STB se clasifi can en función de su grado histológico, determinado, fundamentalmente por su similitud con el tejido de procedencia, su índice mitótico y su porcentaje de necrosis. Este grado histológico es el factor que
1: Lipoma de grandes dimensiones en la región axilar de un Labrador. Previo a la cirugía se realizó un estudio con RM para determinar la extensión del tumor
ecografía es una excelente técnica diagnóstica, ya que la grasa presenta una ecogenicidad característica que le permite diferenciarlos de otras lesiones, infl amatorias o tumorales. El tratamiento, cuando se propone (masas de gran tamaño, que se localicen en zonas donde puedan presionar otras estructuras, como los plexos nerviosos, o por motivos estéticos), es exclusivamente qui-rúrgico (Foto 1).
Otros tumores de tejidos blandos benignos son los fi bromas, hemangio-mas o mixomas. Son poco frecuentes y se presentan también como masas encapsuladas y móviles. Desde el punto de vista diagnóstico, la citología no resulta de utilidad, ya que se obtienen escasas células o sangre (en el caso del hemangioma); sin embargo es muy recomendable realizarla para descartar otros diferenciales. El tratamiento es exclusivamente quirúrgico.
Tumores de tejidos blandos malignos
Los sarcomas de tejidos blandos (STB) son tumores muy frecuentes en la especie canina. Constituyen el 36,8% de todos los tumores de tejidos blan-dos de presentación subcutánea y un 17% de todos los tumores cutáneos.
Tabla 1: Tipos de tumores de tejidos blandos y distribución de su incidencia en tejido subcutáneo en la especie canina
Tejido de origen Benigno Maligno
Pericitos Hemangiopericitoma (14,6%)
Nervios Schwanoma/neurofi broma (4,8%)
Fibroso Fibroma (1%) Fibrosarcoma (3,6%)
Mixomatoso Mixoma (0,2%) Mixosarcoma (0,7%)
Fibroso/ histiocitos Histiocitoma fi broso maligno (0,2%)
Adiposo Lipoma (58%) Liposarcoma (1,7%)
Células sinoviales Sinovioma Sarcoma sinovial (0,2%)
MúsculoLeiomioma
Rabdomioma
Leiomiosarcoma
Rabdomiosarcoma (0,5%)
Endotelio vascular Hemangioma (3,8%) Hemangiosarcoma (4,8%)
Indiferenciado Sarcoma indiferenciado (5,7%)
ecografía es una excelente técnica diagnóstica, ya que la grasa presenta una ecogenicidad característica que le permite diferenciarlos de otras lesiones, infl amatorias o tumorales. El tratamiento, cuando se propone (masas de gran tamaño, que se localicen en zonas donde puedan presionar otras estructuras, como los plexos nerviosos, o por motivos estéticos), es exclusivamente qui-
Otros tumores de tejidos blandos benignos son los fi bromas, hemangio-mas o mixomas. Son poco frecuentes y se presentan también como masas encapsuladas y móviles. Desde el punto de vista diagnóstico, la citología no resulta de utilidad, ya que se obtienen escasas células o sangre (en el caso del hemangioma); sin embargo es muy recomendable realizarla para descartar otros diferenciales. El tratamiento es exclusivamente quirúrgico.
Tumores de tejidos blandos malignos
Tabla 1: Tipos de tumores de tejidos blandos y distribución de su incidencia en tejido subcutáneo en la especie canina
Hemangiopericitoma (14,6%)
Schwanoma/neurofi broma (4,8%)
Fibrosarcoma (3,6%)
Mixosarcoma (0,7%)
Histiocitoma fi broso maligno (0,2%)
Liposarcoma (1,7%)
Sarcoma sinovial (0,2%)
Leiomiosarcoma
Rabdomiosarcoma (0,5%)
Hemangiosarcoma (4,8%)
Sarcoma indiferenciado (5,7%)
4-Mastocitoma cutáneo canino
80 81
Elena Martínez de Merlo3 - Tumores cutáneos
sejable mantener a los gatos susceptibles dentro de casa, aunque hay que tener la precaución de aplicar fi ltros anti-radiación en los cristales. El empleo de cremas de protección solar y tatuajes en las áreas sensibles es controver-tido. Los derivados sintéticos de la vitamina A se han probado como medida preventiva y para revertir o limitar el crecimiento de las lesiones preneoplá-sicas (etretinato: 0,5-2 mg/kg/día); no son efi caces en lesiones avanzadas.
Melanoma cutáneo
Los melanomas son más frecuentes en la especie canina que en la feli-na, afectando con mayor frecuencia a razas pigmentadas.
La mayoría (> 85%) de los melanomas localizados en áreas cubiertas de pelo y párpados tienen un comportamiento benigno. Son lesiones muy pigmentadas, de crecimiento lento, menores de 2 cm, bien delimitadas y no adheridas (Foto 21).
19: Citología de un carcinoma de células escamosas, en la que se observan células poligonales, de gran tamaño, de citoplasma basófi lo, con maduración asincrónica de núcleos y vacuolización perinuclear (May-Grünwald Giemsa, 100x)
En las lesiones nodulares caninas, la citología permite una buena aproximación al diagnóstico, aunque también es habitual que exista una in-tensa reacción infl amatoria que puede complicar la interpretación. Exfolian abundantes células aisladas o en pequeños grupos con escasa cohesión; son células grandes, globosas o poligonales, de citoplasma fuertemente basófi lo. Además de otros criterios citológicos de malignidad, las células presentan maduración asincrónica (falta de correlación entre el tamaño nuclear y la cantidad de citoplasma, de forma que aparecen células grandes con abun-dante citoplasma que retienen un núcleo grande, funcional, no picnótico) y/o vacuolización perinuclear (Foto 19).
El abordaje terapéutico de los CCE es diferente en función de la localiza-ción del tumor, su grado de invasión y su extensión (Tabla 4). En las lesiones pequeñas y superfi ciales, pueden elegirse diferentes modalidades terapéu-ticas que, en general, tienen buenos resultados, de forma que se consiguen tiempos libres de enfermedad prolongados (superiores a 12 meses); sin em-bargo, las lesiones profundas o múltiples se tratan con difi cultad y, en general, las respuestas son peores, aunque las cirugías agresivas de plano nasal en gatos pueden proporcionar buenos resultados (Foto 20a y 20b).
La aplicación de medidas preventivas no es demasiado efi caz, por la di-fi cultad de limitar la exposición al sol de las áreas despigmentadas. Es acon-
18: Carcinoma de células escamosas en el plano nasal de una Dogo Alemán
3 - Tumores cutáneos
En las lesiones nodulares caninas, la citología permite una buena aproximación al diagnóstico, aunque también es habitual que exista una in-tensa reacción infl amatoria que puede complicar la interpretación. Exfolian abundantes células aisladas o en pequeños grupos con escasa cohesión; son células grandes, globosas o poligonales, de citoplasma fuertemente basófi lo. Además de otros criterios citológicos de malignidad, las células presentan maduración asincrónica (falta de correlación entre el tamaño nuclear y la cantidad de citoplasma, de forma que aparecen células grandes con abun-dante citoplasma que retienen un núcleo grande, funcional, no picnótico) y/o vacuolización perinuclear (Foto 19).
El abordaje terapéutico de los CCE es diferente en función de la localiza-ción del tumor, su grado de invasión y su extensión pequeñas y superfi ciales, pueden elegirse diferentes modalidades terapéu-ticas que, en general, tienen buenos resultados, de forma que se consiguen tiempos libres de enfermedad prolongados (superiores a 12 meses); sin em-bargo, las lesiones profundas o múltiples se tratan con difi cultad y, en general, las respuestas son peores, aunque las cirugías agresivas de plano nasal en
18: Carcinoma de células escamosas en el plano nasal de una Dogo Alemán 18: Carcinoma de células escamosas en el plano nasal de una Dogo Alemán
4-Mastocitoma cutáneo canino
122 123
Elena Martínez de Merlo5 - Tumores de tejidos blandos
un AINE (preferiblemente anticox2 selectivo) a las dosis habituales. Se ha comprobado que la quimioterapia metronómica prolonga el tiempo libre de enfermedad en STB de bajo grado.
3. Quimioterapia intralesional: se basa en la administración local de 5 fl uo-racilo (solo o mezclado con aceite de sésamo) o cisplatino (solo, mezclado con aceite de sésamo o en implantes) en la cicatriz. Un estudio prelimi-nar describe una forma sencilla de practicar este tipo de quimioterapia: administrar 150 mg/m2 de 5 fl uoracilo (sin exceder los 150 mg) a lo largo de la cicatriz o en los bordes de la herida quirúrgica una vez a la semana durante un mínimo de 6 sesiones (que se pueden prolongar mientras no se observen signos de recidiva). El tratamiento debe iniciarse lo antes posible (preferiblemente a los 7-14 días de la intervención); la administra-ción con una aguja de 22 g ha demostrado que no requiere sedación del paciente. Esta técnica no tiene, prácticamente, efectos secundarios.
El pronóstico de los pacientes con STB depende del grado histológico, que condiciona los tiempos de supervivencia en función de su potencial me-tastático. El tiempo medio de supervivencia de los STB de bajo grado prácti-camente duplica al de los STB de alto grado (460 días vs 241). Los STB de bajo grado presentan un alto índice de recidivas si no es posible realizar una ciru-gía lo sufi cientemente agresiva; incluso en casos con márgenes libres de cé-lulas tumorales se describe un 30% de recidivas. Es importante destacar que, normalmente, las recidivas de los STB de bajo grado son lentas, pudiéndose retrasar hasta 2 años. Los STB de alto grado tienen mal pronóstico, tanto por la capacidad de recidiva como por la aparición de enfermedad metastásica, lo que condiciona menores tiempos de supervivencia. En general, hay que tener en cuenta que la agresividad de las recidivas es mayor, incrementándose el potencial metastásico.
Sarcomas felinos asociados a puntos de inyección
En 1991 y 1992, en Estados Unidos, se publicaron las primeras eviden-cias de que se estaba produciendo un incremento de la incidencia de fi brosar-comas localizados en los puntos habituales de inyección en el gato (espacio interescapular, costado, región lumbar), coincidiendo con las aplicación de las leyes obligatorias de vacunación frente a la rabia en esta especie. Estudios posteriores demostraron que existe una relación epidemiológica entre la apa-rición de diferentes tipos de sarcomas y las vacunas de leucemia felina y rabia (predominantemente). Más recientemente, se ha reconocido que otros productos inyectados también pueden originar el desarrollo de sarcomas, por lo que antigua denominación de sarcomas asociados a la vacunación se ha modifi cado a sarcomas asociados a puntos de inyección (SAPI).
111a y 11b: STB en la cara de un Husky. La RM permitió valorar su extensión real
Pub oncología.indd 1 19/04/12 16:25
Completa tu biblioteca
CYS 42.indb 66 14/05/12 10:11

ColimutinaSolución inyectable
La mejor combinación sinérgica para porcinoTiamulina + Colistina
ComposiciónTiamulina (fumarato ácido)...... 10 g(equivalente a 12,35 g de fumarato ácido de tiamulina)Colistina...... 18.000.000 U.l.Disolvente c.s.p. ...... 100 mlEspecies de destino e indicacionesPorcino: Tratamiento de la disentería procina producida por B. hyodysenteriae. Neumonía enzoótica producida por M. hyopneumoniae. Tratamiento de artritis producida por M. hyosynoviae. Colibacilosis. Ileitis.Bovino y ovino: Micoplasmosis y colibacilosis.Posología y modo de administraciónVía intramuscular.Tratamiento de la disentería procina, micoplasmosis bovina y ovina y colibacilosis: 1 ml de COLIMUTINA INYECTABLE / 10 kg p.v. / día, durante 3 días.Neumonía enzoótica y artritis micoplásmica: 1,5 ml de COLIMUTINA INYECTABLE / 10 kg p.v. / día, durante 3 días.En animales pequeños, especialmente recomendamos diluir el producto en suero gluco-sado para no sobrepasar las dosis indicadas.Tiempo de esperaCarne: 21 díasRegistro n.º: 9.134Envase de 100 y 250 ml
Ctra. Reus-Vinyols Km. 4,1 - Ap. Correos, 60 - Teléfono 977 850 170* - Fax 977 850 405 - 43330 RIUDOMS (Tarragona)
www.spveterinaria.com
LA SOLUCIÓN CON EL ESPECTRO MÁS COMPLETO
EN PROBLEMAS RESPIRATORIOS Y
DIGESTIVOS EN GANADO PORCINO
an_colimutina.indd 1 28/04/11 12:08
S í n d r o m e R e s p i r a t o r i o B o v i n o
Sé responsablePiensa en rapidez, actúa con efi cacia
NUEVO
Tratamiento antibiótico de las infecciones causadas por Pasteurellaceae en una sola dosis por vía IM
Dosis: 10 ml por 160 kg p.v.
Cortos tiempos de espera: • Carne: 5 días • Leche: 48 horas
T
FORCYL 160 mg/ml SOLUCIÓN INYECTABLE PARA BOVINO. Marbofl oxacino. Cada ml contiene: Sustancia activa: Marbofl oxacino 160 mg; Excipiente: Alcohol bencílico (E 1519) 15 mg. Indicaciones de uso: Tratamiento terapéutico de infecciones respiratorias en bovino causadas por cepas sensibles a Pasteurella multocida y Mannheimia haemolytica. Contraindicaciones: No utilizar en animales con hipersensibilidad conocida a las fl uoroquinolonas o a algún excipiente. No utilizar en caso de patógenos resistentes a otras fl uoroquinolonas (resistencias cruzadas). Precauciones especiales para su uso en animales: Cuando se use este medicamento se deben tener en cuenta las recomendaciones ofi ciales sobre el uso de antimicrobianos. El uso de fl uoroquinolonas debe ser reservado para el tratamiento de aquellos casos clínicos que hayan respondido pobremente, o se espera que respondan pobremente, a otras clases de antimicrobianos. Siempre que sea posible las fl uoroquinolonas deben ser usadas después de realizar un test de sensibilidad. El uso del medicamento en condiciones distintas a las recomendadas en la Ficha Técnica puede incrementar la pre-valencia de bacterias resistentes a las fl uoroquinolonas y disminuir la efi cacia del tratamiento con otras quinolonas debido a las resistencias cruzadas. Precauciones específi cas que debe tomar la persona que administre el medicamento a los animales: Las personas con hipersensibilidad conocida a las (fl uoro) quinolonas deberán evitar cualquier contacto con el medicamento. Si el medicamento entrara en contacto con la piel o los ojos, lávese abundantemente con agua. En caso de autoinyección accidental, consulte con un médico inmediatamente. En caso de auto inyección accidental puede producir irritación leve. Reacciones adversas (frecuencia y gravedad): La administración por vía intramuscular puede causar reacciones locales transitorias como dolor e hinchazón en el punto de inyección que pueden persistir hasta 7 días después de la inyección. Se sabe que las fl uoroquinolonas inducen a artropatías. En bovinos, tales lesiones fueron observadas después de un tratamiento de 3 días con solución de marbofl oxacino al 16 %. Estas lesiones no indujeron a signos clínicos y deben ser reversibles, particularmente si fueron observadas después de una administración única. No fueron observadas otras reacciones adversas en bovinos. Uso durante la gestación o la lactancia: Los estudios en animales de laboratorios (ratas, conejos) no mostraron evidencias de efectos teratógenicos, embriotóxicos o maternotóxicos asociados al uso de marbofl oxacino. La seguridad del medicamento a dosis de 10 mg/kg no se ha demostrado en vacas en gestación ni en terneros lactantes cuando se utiliza en vacas. Por tanto, se debe utilizar según la evaluación benefi cio/riesgo realizada por el veterinario responsable. Posología y vía de administración: La dosis recomendada es de 10 mg/kg de peso vivo, es decir, 10 ml /160kg de peso vivo en una única inyección intramuscular. Si el volumen a inyectar es mayor de 20 ml, la dosis debe dividirse en dos o más puntos de inyección. Para garantizar una dosis correcta, debe determinarse el peso corporal con la mayor precisión posible para evitar infradosifi cación. En caso de presentar ligera turbidez o partículas visibles, dicha turbidez o partículas desaparecerán al agitar el frasco antes del uso. Tiempo de espera: Carne: 5 días; Leche: 48 horas. Formatos: Caja con vial de 100 ml y Caja con vial de 250 ml. VETOQUINOL ESPECIALIDADES VETERINARIAS, S.A. Carretera de Fuencarral, nº 24, Edifi cio Europa I 28108 Madrid-España. NÚMERO DE LA AUTORIZACIÓN DE COMERCIALIZACIÓN 2319 ESP. Medicamento sujeto a prescripción veterinaria. Administración bajo control o supervisión del veterinario.
www.vetoquinol.es
FIeeslvlmaetúLe1tvvdmdLVAb
DIMENSIÓN DEANTIBIOTERAPIA
DOLOR
CARD IO -RENAL

ColimutinaSolución inyectable
La mejor combinación sinérgica para porcinoTiamulina + Colistina
ComposiciónTiamulina (fumarato ácido)...... 10 g(equivalente a 12,35 g de fumarato ácido de tiamulina)Colistina...... 18.000.000 U.l.Disolvente c.s.p. ...... 100 mlEspecies de destino e indicacionesPorcino: Tratamiento de la disentería procina producida por B. hyodysenteriae. Neumonía enzoótica producida por M. hyopneumoniae. Tratamiento de artritis producida por M. hyosynoviae. Colibacilosis. Ileitis.Bovino y ovino: Micoplasmosis y colibacilosis.Posología y modo de administraciónVía intramuscular.Tratamiento de la disentería procina, micoplasmosis bovina y ovina y colibacilosis: 1 ml de COLIMUTINA INYECTABLE / 10 kg p.v. / día, durante 3 días.Neumonía enzoótica y artritis micoplásmica: 1,5 ml de COLIMUTINA INYECTABLE / 10 kg p.v. / día, durante 3 días.En animales pequeños, especialmente recomendamos diluir el producto en suero gluco-sado para no sobrepasar las dosis indicadas.Tiempo de esperaCarne: 21 díasRegistro n.º: 9.134Envase de 100 y 250 ml
Ctra. Reus-Vinyols Km. 4,1 - Ap. Correos, 60 - Teléfono 977 850 170* - Fax 977 850 405 - 43330 RIUDOMS (Tarragona)
www.spveterinaria.com
LA SOLUCIÓN CON EL ESPECTRO MÁS COMPLETO
EN PROBLEMAS RESPIRATORIOS Y
DIGESTIVOS EN GANADO PORCINO
an_colimutina.indd 1 28/04/11 12:08
S í n d r o m e R e s p i r a t o r i o B o v i n o
Sé responsablePiensa en rapidez, actúa con efi cacia
NUEVO
Tratamiento antibiótico de las infecciones causadas por Pasteurellaceae en una sola dosis por vía IM
Dosis: 10 ml por 160 kg p.v.
Cortos tiempos de espera: • Carne: 5 días • Leche: 48 horas
T
FORCYL 160 mg/ml SOLUCIÓN INYECTABLE PARA BOVINO. Marbofl oxacino. Cada ml contiene: Sustancia activa: Marbofl oxacino 160 mg; Excipiente: Alcohol bencílico (E 1519) 15 mg. Indicaciones de uso: Tratamiento terapéutico de infecciones respiratorias en bovino causadas por cepas sensibles a Pasteurella multocida y Mannheimia haemolytica. Contraindicaciones: No utilizar en animales con hipersensibilidad conocida a las fl uoroquinolonas o a algún excipiente. No utilizar en caso de patógenos resistentes a otras fl uoroquinolonas (resistencias cruzadas). Precauciones especiales para su uso en animales: Cuando se use este medicamento se deben tener en cuenta las recomendaciones ofi ciales sobre el uso de antimicrobianos. El uso de fl uoroquinolonas debe ser reservado para el tratamiento de aquellos casos clínicos que hayan respondido pobremente, o se espera que respondan pobremente, a otras clases de antimicrobianos. Siempre que sea posible las fl uoroquinolonas deben ser usadas después de realizar un test de sensibilidad. El uso del medicamento en condiciones distintas a las recomendadas en la Ficha Técnica puede incrementar la pre-valencia de bacterias resistentes a las fl uoroquinolonas y disminuir la efi cacia del tratamiento con otras quinolonas debido a las resistencias cruzadas. Precauciones específi cas que debe tomar la persona que administre el medicamento a los animales: Las personas con hipersensibilidad conocida a las (fl uoro) quinolonas deberán evitar cualquier contacto con el medicamento. Si el medicamento entrara en contacto con la piel o los ojos, lávese abundantemente con agua. En caso de autoinyección accidental, consulte con un médico inmediatamente. En caso de auto inyección accidental puede producir irritación leve. Reacciones adversas (frecuencia y gravedad): La administración por vía intramuscular puede causar reacciones locales transitorias como dolor e hinchazón en el punto de inyección que pueden persistir hasta 7 días después de la inyección. Se sabe que las fl uoroquinolonas inducen a artropatías. En bovinos, tales lesiones fueron observadas después de un tratamiento de 3 días con solución de marbofl oxacino al 16 %. Estas lesiones no indujeron a signos clínicos y deben ser reversibles, particularmente si fueron observadas después de una administración única. No fueron observadas otras reacciones adversas en bovinos. Uso durante la gestación o la lactancia: Los estudios en animales de laboratorios (ratas, conejos) no mostraron evidencias de efectos teratógenicos, embriotóxicos o maternotóxicos asociados al uso de marbofl oxacino. La seguridad del medicamento a dosis de 10 mg/kg no se ha demostrado en vacas en gestación ni en terneros lactantes cuando se utiliza en vacas. Por tanto, se debe utilizar según la evaluación benefi cio/riesgo realizada por el veterinario responsable. Posología y vía de administración: La dosis recomendada es de 10 mg/kg de peso vivo, es decir, 10 ml /160kg de peso vivo en una única inyección intramuscular. Si el volumen a inyectar es mayor de 20 ml, la dosis debe dividirse en dos o más puntos de inyección. Para garantizar una dosis correcta, debe determinarse el peso corporal con la mayor precisión posible para evitar infradosifi cación. En caso de presentar ligera turbidez o partículas visibles, dicha turbidez o partículas desaparecerán al agitar el frasco antes del uso. Tiempo de espera: Carne: 5 días; Leche: 48 horas. Formatos: Caja con vial de 100 ml y Caja con vial de 250 ml. VETOQUINOL ESPECIALIDADES VETERINARIAS, S.A. Carretera de Fuencarral, nº 24, Edifi cio Europa I 28108 Madrid-España. NÚMERO DE LA AUTORIZACIÓN DE COMERCIALIZACIÓN 2319 ESP. Medicamento sujeto a prescripción veterinaria. Administración bajo control o supervisión del veterinario.
www.vetoquinol.es
FIeeslvlmaetúLe1tvvdmdLVAb
DIMENSIÓN DEANTIBIOTERAPIA
DOLOR
CARD IO -RENAL