informe proy invest 2005 - sappi.ipn.mxsappi.ipn.mx/cgpi/archivos_anexo/20050153_2609.pdf28...
TRANSCRIPT

27
Anexo del informe final del proyecto Determinación de B lactamasas de cepas de P. aeruginosa resistentes
a antibióticos B lactámicos, con clave SIP 20050153.
Responsable: Dr. Gerardo Aparicio Ozores
INTRODUCCIÓN P. aeruginosa es uno de los principales agentes involucrados en enfermedades intrahospitalarias con una
significativa incidencia en nuestro país; presentando una tasa alta de mortalidad entre los pacientes. Esta
bacteria por sus características de resistencia extrínseca hacia diversos antibióticos ha tomado gran interés
y se ha convertido en un importante problema para el tratamiento; que no resulta fácil, por la limitada
acción de las penicilinas, las cefalosporinas de la primera, la segunda generación y de amplio espectro
debido a la producción de β-lactamasas de espectro extendido Las bacterias adquieren los factores que
confieren la resistencia a los antibióticos a través de la transferencia genética por los mecanismos de
conjugación, transformación, transducción y/o transposición. Los genes adquiridos pueden ser detectados
por técnicas de biología molecular como son la PCR y la hibridación. En nuestro país, se ha demostrado el
carácter de multirresistencia de cepas de P. aeruginosa recuperadas de pacientes con algún tipo de
infección intrahospitalaria; sin embargo, no existen reportes que asocien la producción de ESBLs. Todo lo
anterior expresa la necesidad de detectar este tipo de enzimas en nuestro país y así contar con datos que
fundamenten a los programas de control y manejo de antibióticos que son implementados en los
hospitales.
H. influenzae es una de las principales especies bacterianas que causa infecciones en niños. Las cepas
capsuladas son responsables de enfermedades invasoras, de entre las que destaca la meningitis; aunque
también pueden causar epiglotitis, artritis, neumonía y celulitis. Las cepas no capsuladas causan
generalmente otitis. Esta bacteria puede llegar a formar parte de la biota de nasofaringe de niños,
pudiendo alcanzar hasta un 60%. Sin embargo, el nivel de colonización varia de acuerdo a diferentes
factores como la edad, contagio intrafamiliar y condiciones de vida. Además se deben considerar factores
de las cepas, como la resistencia que presentan a diferentes antibióticos, así como que presenten cápsula
del tipo b; aunque para esto último, la vacunación con el antígeno Hib en la población ha llevado a una
disminución en la colonización de las cepas capsuladas del tipo b. Es necesario conocer el nivel de
colonización y las características que presentan las cepas de H. influenzae que están distribuidas entre la
población infantil mexicana. Por lo anterior, en este estudio se presentarán datos orientados a monitorear
el tipo de cepas de H. influenzae que colonizan a niños que asisten a guarderías en diferentes ciudades de
la república mexicana; así como conocer las características de resistencia a varios antibióticos y la
presencia de β-lactamasas que presentan dichas cepas.
Hasta hace poco se consideró a los SCN como bacterias contaminantes en muestras de origen clínico; sin
embargo, con el uso y abuso descontrolado de los antibióticos en el tratamiento de infecciones, estas
bacterias han adquirido la capacidad de resistir a la acción de los antibióticos; transformándose así en
potenciales agentes etiológicos de infecciones en humanos; sobre todo en aquellas de origen nosocomial
en las que S. epidermidis y S. saprophyticus son las dos especies aisladas con mayor frecuencia. Las
diferentes especies de SCN son capaces de producir infecciones tales como endocarditis, bacteriemia,

28
infecciones en vías urinarias entre otras; lo anterior se debe a la facilidad de estas bacterias de colonizar
dispositivos permanentes tales como catéteres intravenosos y para hemodiálisis, válvulas cardiacas,
prótesis y marcapasos; además de considerar el estado de inmunosupresión del paciente. El monitoreo y
vigilancia de los Staphylococcus en los nosocomios ha tomado importancia, dado que además de casos
aislados, llegan a causar brotes. Considerando todo lo anterior, es de interés conocer los patrones de
resistencia a los antibióticos que presentan los SCN, además de detectar la presencia de elementos
genéticos que son los plásmidos y que pueden asociarse con la transferencia de los determinantes de
resistencia en las cepas, para así proponer recomendaciones sobre el impacto que está resultando del uso
y abuso de los y así colaborar a la interrupción de la diseminación de las cepas multirresistentes en los
hospitales.
V. cholerae es el agente causal del cólera, entidad clínica con posibilidad de presentación epidémica que
se caracteriza por una diarrea profusa de tipo secretora que puede llevar a la muerte a los individuos al
producirse acidosis y choque hipovulémico. Este patógeno ha acompañado al hombre a lo largo de la
historia debido a que se transmite principalmente por agua y alimentos contaminados, sin embargo no se
tiene del todo claro en que parte de la naturaleza se encuentra la bacteria antes de ser consumida por el
hombre. Se ha observado que V. cholerae ha desarrollado tres estadios fisiológicos diferentes, los cuales
le pueden ayudar a mantenerse en el medio ambiente: viable y cultivable, que es la forma recuperada de
muestras clínicas; rugoso, que puede aislarse de muestras clínicas y del medio ambiente; y viable no
cultivable, que se ha obtenido en el laboratorio aunque no se ha demostrado su presencia en la naturaleza,
pero que debido a sus características se propone que sea una forma de persistencia en el medio ambiente.
Para mantener la viabilidad, las bacterias tienen que contar con mecanismos de generación de energía y
así llevar las actividades fisiológicas que permitan su crecimiento. La forma más eficiente de generación de
energía la llevan a cabo a través de la cadena transportadora de electrones, de las cuales existen
diferentes versiones, directa y alterna, como se ha descrito en Escherichia coli. Sin embargo, en el caso de
V. cholerae sólo se conoce que presenta la citocromo oxidasa, por lo que se desconocen los otros
elementos que pudieran participar en el transporte de los electrones. Con base en lo anterior y
considerando la presión ambiental a la que es sometido este organismo, se sugiere que puede presentarse
alguna vía alterna del transporte electrónico, como una medida de sobrevivencia y adaptación al medio
ambiente, lo cual tampoco ha sido establecido para esta especie. Este estudio pretende contribuir en el
conocimiento de los elementos que forman las cadenas transportadoras de electrones de V. cholerae.

29
MÉTODOS
CEPAS EMPLEADAS: Cepas de S. aureus empleadas
ESPECIE
CLAVE ASIGNADA
ORIGEN
OBSERVACIONES
CEPAS CLÍNICAS S. aureus S. aureus S. aureus S. aureus S. aureus S. aureus S. aureus S. aureus S. aureus S. aureus S. aureus
SA1 SA2 SA3 SA4 SA5 SA6 SA7 SA8 SA9 SA10 SA11
Hemocultivo Secreción de herida
Hemocultivo Punta de catéter
Hemocultivo Líquido de diálisis
Secreción de herida Líquido peritoneal Líquido peritoneal Punta de catéter
Secreción de herida
HICMR HICMR HICMR HICMR HICMR HICMR HICMR HICMR HICMR HICMR HICMR
CEPAS CONTROL S. aureus
ATCC 25923
Para realizar el control de calidad de susceptibilidad a los antibióticos
Pseudomonas aeruginosa
ATCC 27853
Cepas de referencia
Para el método de CEFINASE
E. coli
DH5�
Con plásmido de resistencia a la ampicilina pIAGO
E. coli
DH5�
Cepas proporcionadas por M en C Juan Castelán Vega
Con plásmido de resistencia a la tobramicina pKC
E. coli
IJ133
Cepa proporcionada por MSP Elizabeth Fernández R
Receptora de conjugación.
E. coli
ES1
Transconjugante obtenida en este estudio
Obtenida de SA1
E. coli
ES3
Transconjugante obtenida en este estudio
Obtenida de SA3
HICMR= Hospital de Infectología del Centro Médico la Raza
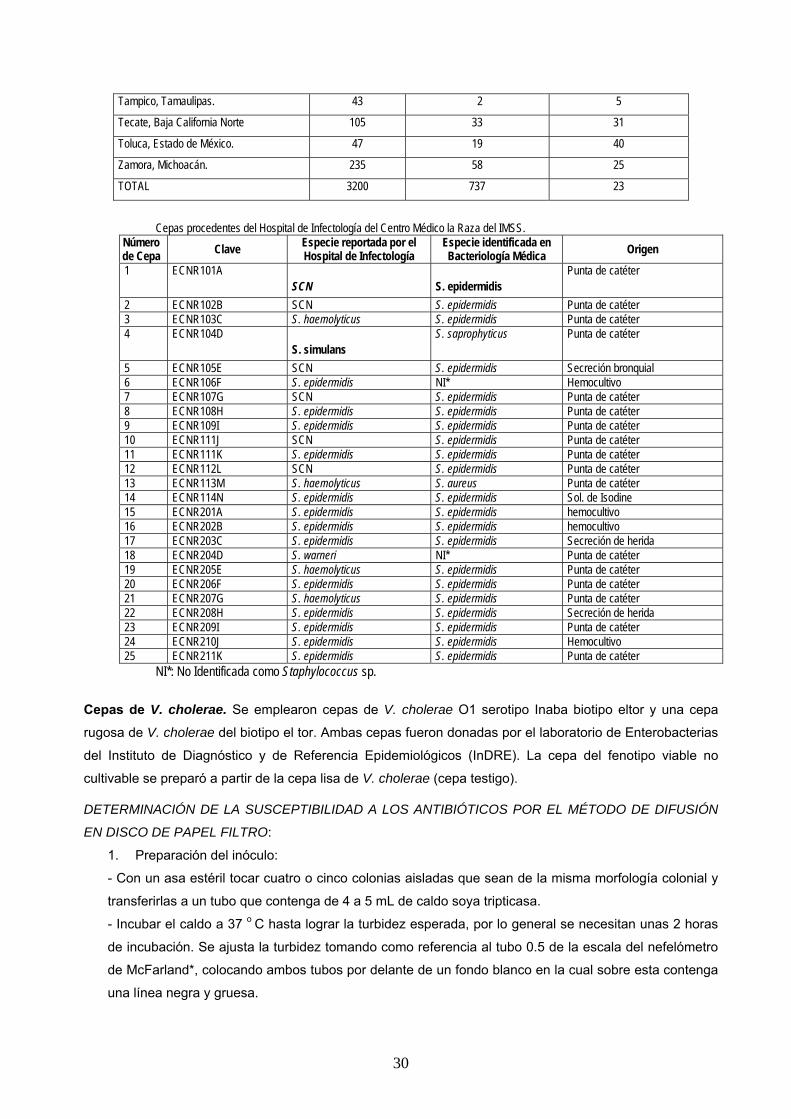
Incidencia de H. influenzae en niños que asistieron a guarderías de 15 ciudades de la república mexicana.
CIUDAD NÚMERO DE MUESTRAS
POR CIUDAD
AISLAMIENTOS DE
H. influenzae
FRECUENCIA DE RECUPERACIÓN
(%)
Ciudad Madero, Tamaulipas 183 66 36
Distrito Federal 626 254 41
Ensenada, Baja California Norte 282 47 17
Guadalajara, Jalisco. 155 19 12
Jalapa, Veracruz. 112 30 27
León, Guanajuato. 295 31 11
Mexicali, Baja California Norte 82 2 2
Monterrey, Nuevo León. 248 16 6
Oaxaca, Oaxaca. 354 73 21
Pachuca, Hidalgo. 218 66 30
San Luís Potosí, San Luís Potosí.
215 21 10
30
Tampico, Tamaulipas. 43 2 5
Tecate, Baja California Norte 105 33 31
Toluca, Estado de México. 47 19 40
Zamora, Michoacán. 235 58 25
TOTAL 3200 737 23
Cepas procedentes del Hospital de Infectología del Centro Médico la Raza del IMSS.
Número de Cepa Clave Especie reportada por el
Hospital de Infectología Especie identificada en Bacteriología Médica Origen
1 ECNR101A SCN S. epidermidis
Punta de catéter
2 ECNR102B SCN S. epidermidis Punta de catéter 3 ECNR103C S. haemolyticus S. epidermidis Punta de catéter 4 ECNR104D
S. simulans S. saprophyticus Punta de catéter
5 ECNR105E SCN S. epidermidis Secreción bronquial 6 ECNR106F S. epidermidis NI* Hemocultivo 7 ECNR107G SCN S. epidermidis Punta de catéter 8 ECNR108H S. epidermidis S. epidermidis Punta de catéter 9 ECNR109I S. epidermidis S. epidermidis Punta de catéter 10 ECNR111J SCN S. epidermidis Punta de catéter 11 ECNR111K S. epidermidis S. epidermidis Punta de catéter 12 ECNR112L SCN S. epidermidis Punta de catéter 13 ECNR113M S. haemolyticus S. aureus Punta de catéter 14 ECNR114N S. epidermidis S. epidermidis Sol. de Isodine 15 ECNR201A S. epidermidis S. epidermidis hemocultivo 16 ECNR202B S. epidermidis S. epidermidis hemocultivo 17 ECNR203C S. epidermidis S. epidermidis Secreción de herida 18 ECNR204D S. warneri NI* Punta de catéter 19 ECNR205E S. haemolyticus S. epidermidis Punta de catéter 20 ECNR206F S. epidermidis S. epidermidis Punta de catéter 21 ECNR207G S. haemolyticus S. epidermidis Punta de catéter 22 ECNR208H S. epidermidis S. epidermidis Secreción de herida 23 ECNR209I S. epidermidis S. epidermidis Punta de catéter 24 ECNR210J S. epidermidis S. epidermidis Hemocultivo 25 ECNR211K S. epidermidis S. epidermidis Punta de catéter
NI*: No Identificada como Staphylococcus sp.
Cepas de V. cholerae. Se emplearon cepas de V. cholerae O1 serotipo Inaba biotipo eltor y una cepa
rugosa de V. cholerae del biotipo el tor. Ambas cepas fueron donadas por el laboratorio de Enterobacterias
del Instituto de Diagnóstico y de Referencia Epidemiológicos (InDRE). La cepa del fenotipo viable no
cultivable se preparó a partir de la cepa lisa de V. cholerae (cepa testigo).
DETERMINACIÓN DE LA SUSCEPTIBILIDAD A LOS ANTIBIÓTICOS POR EL MÉTODO DE DIFUSIÓN
EN DISCO DE PAPEL FILTRO:
1. Preparación del inóculo:
- Con un asa estéril tocar cuatro o cinco colonias aisladas que sean de la misma morfología colonial y
transferirlas a un tubo que contenga de 4 a 5 mL de caldo soya tripticasa.
- Incubar el caldo a 37 o C hasta lograr la turbidez esperada, por lo general se necesitan unas 2 horas
de incubación. Se ajusta la turbidez tomando como referencia al tubo 0.5 de la escala del nefelómetro
de McFarland*, colocando ambos tubos por delante de un fondo blanco en la cual sobre esta contenga
una línea negra y gruesa.

31
- *Este estándar se prepara agregando 0.05 mL de cloruro de bario hidratado (BaCl2 H2O) al
1% (0.048 M) a 9.95 mL de ácido sulfúrico puro y frío al 1% (0.36 N). Esta suspensión
corresponde a 1.5 X 108 bacterias/mL (19).
2. Siembra masiva en placas de gelosa Müeller Hinton y colocación de los sensidiscos.
- Sembrar en placas de Müeller Hinton usando un hisopo a partir de la suspensión ajustada.
- Humedecer el hisopo en la suspensión bacteriana quitándole el exceso de caldo presionando y
girando sobre la pared interna de tubo por arriba del nivel del caldo.
- Pasar el hisopo varias veces sobre la superficie de la gelosa (siembra masiva). Dejar
reposar la placa cinco minutos a temperatura ambiente para que se seque.
- Colocar los sensidiscos sobre la gelosa y con ayuda de una pinza estéril, presionarlos ligeramente
hacia abajo para asegurar un contacto con la superficie, que no sean más de seis en cada caja de
Petri que mida 10 cms de diámetro y deben ser distribuidos uniformemente de tal manera que se
pueda evitar una sobreposición de los halos de inhibición.
3. Lectura de los diámetros de las zonas de inhibición.
- Medir los halos de inhibición en mm con Vernier o regla por debajo de la caja Petri e interpretar las
categorías de susceptible, intermedio o resistente de acuerdo a las tablas incluidas en la publicación
del NCCLS.
4. Control de calidad de la prueba utilizando a la cepa Pseudomonas aeruginosa ATCC 27853.
- El control de calidad de la prueba se realiza exactamente como se describió anteriormente de los
puntos 1 al 3, los halos de inhibición para el crecimiento obtenidos, se compararon con los valores
contenidos en las tablas publicadas por el NCCLS para la parte de control de calidad.
Los antibióticos utilizados fueron comprados a la compañía Beckton and Dickinson, en la tabla 4 se
muestra una clasificación de dichos antibióticos agrupados en familias.
Clasificación en familias de los antibióticos usados
I. Penicilinas semisintéticas.
1.1 Carboxipenicilinas. Carbecilina (CB100).
Ticarcilina (TIC75).
1.2 Ureidopenicilinas. Azlocilina (AZ75).
Mezlocilina (MZ75).
Piperacilina (PIP100).
II. Otras β-Lactamasas. Imipenem (IPM10), Meropenem (MEM10), Aztreonam (ATM30).
Aminoglucósidos: Amikacina (AN30), Tobramicina (NN10), Gentamicina (GM10), Netilmicina (NET30).
Cefalosporinas:
1. Amplio espectro (tercera generación). Cefoperazona (CFP75), Ceftazidima (CAZ30), ceftriaxona (CRO30), ceftizoxima (ZOX30).

32
2. Espectro extendido (cuarta generación). Cefepima (FEP30).
Quinolonas:
1. Amplio espectro (segunda generación). Ciprofloxacina (CIP5), levofloxacina (LVX5), ofloxacina (OFX5), norfloxacina (NOR10).
• DETERMINACIÓN DE LA CONCENTRACIÓN MÍNIMA INHIBITORIA (MIC) POR EL MÉTODO DE
DILUCIONES EN AGAR.
El procedimiento que se describe a continuación se realizó tanto en la cepa de referencia empleada para
llevar a cabo el control de calidad del procedimiento, así como en las cepas clínicas proporcionadas.
PREPARACIÓN DE LAS CAJAS CONTENIENDO DIFERENTES CONCENTRACIONES DEL
ANTIBIÓTICO
1.-Se utilizó la sal pura del antibiótico β lactámico, imipenem.
2.-Se disolvió y diluyó en agua desionizada. La concentración de abasto correspondió a 64,000 µg/ml y a
partir de la cual se realizó una serie de diluciones considerando la preparación de diluciones dobles en 16
ml de agar con una concentración de 64 µg/ml hasta 0.25 µg/ml.
3.- La serie de diluciones con volumen de 8 ml se llevó a 16 ml agregando 8 ml de agar Müeller Hinton en
doble concentración y después de mezclar gentilmente para evitar la formación de burbujas se vertió el
contenido en cajas de Petri colocadas sobre una superficie plana sin desniveles
PREPARACIÓN DEL INÓCULO
1.-Con un asa se tocaron de cuatro o cinco colonias aisladas del mismo tipo morfológico y se inocularon en
un tubo con caldo Müeller- Hinton.
2.-Se incubó el caldo Müeller-Hinton a 37ºC hasta que apareció una leve opacidad (2 a 5 h) con el
propósito de ajustar el inóculo de prueba.
3.-Se ajustó la opacidad tomando como referencia el tubo 0.5 de la escala del nefelómetro de McFarland,
colocando el tubo sobre un fondo blanco bien iluminado que contenía una línea negra gruesa.
4.-Se realizó una dilución 1:10 de la suspensión previamente ajustada, para obtener una densidad
bacteriana de 107 y se cargo el replicador Steers.
INOCULACIÓN DE LAS PLACAS QUE CONTENÍAN LAS DIFERENTES CONCENTRACIONES DEL
ANTIBIÓTICO
1.-Se colocó la placa de puntas del replicador de Steers en su posición.
2.-Se colocó la base del replicador de Steers que contenía las cepas ajustadas en su sitio.
3.-Se inocularon con el replicador de Steers una a una las cajas de petri que contenían las diferentes
concentraciones del antibiótico
4.-Las cajas se incubaron a 35oC por 18 h y después se determino la concentración mínima que inhibió el
desarrollo de cada una de las cepas
5.-Se determinaron variaciones en el nivel de resistencia considerando una diferencia de más de dos
diluciones (Leannette y col., 1985)

33
DETERMINACIÓN DE LA PRODUCCIÓN DE �-LACTAMASAS POR EL MÉTODO DE CEFINASE:
MÉTODO DE LA CEFALOSPORINA COLORIDA (CEFINASE) ® - A partir del crecimiento obtenido en placa se tomaron asadas en distintas colonias y se colocaron sobre
discos impregnados con la cefalosporina colorida.
- Se observó una coloración rosa en los discos que se habían colocado las cepas que presentaron �-
lactamasas.
MÉTODO DE LA INHIBICIÓN DEL ÁCIDO CLAVULÁNICO Y/O EDTA-Con un asa se tocaron cuatro o
cinco colonias aisladas del mismo tipo morfológico y se inocularon en un tubo con caldo Müeller- Hinton.
2.-Se incubó el caldo Müeller-Hinton a 37ºC hasta que apareció una leve opacidad (2 a 5 h) con el
propósito de ajustar el inóculo de prueba.
3.-Se ajustó la opacidad tomando como referencia el tubo 0.5 de la escala del nefelómetro de McFarland,
colocando el tubo sobre un fondo blanco bien iluminado que contenía una línea negra gruesa
4.-Se colocó el disco que contenía amoxicilina-clavulanato y en otra placa el disco que contenía EDTA y a
una distancia de 15mm (centro a centro), se colocaron dos discos de los antibióticos ceftazidima o
cefotaxima y ceftazidima respectivamente. Un tercer par de discos con ceftazidima y cefotaxima
respectivamente se colocaron en el extremo opuesto de la caja usándolo como control de la técnica
5.-Se incubó a 37ºC durante 24 h.
6.-Interpretación: la formación de un halo de inhibición ampliado en la región en la que hacen sinergia los
dos discos se consideró como una prueba positiva, es decir la cepa produjo ESBLs; mientras que la
ausencia de una ampliación en la inhibición se consideró como una prueba negativa (Jarlier y col., 1988;
Arakawa y col 2003; Tankhiwale y col., 2000; Sader y col., 2005).
Cepa control positivo E. coli SHV.
Cepa control negativo P. aeruginosa ATCC 27853.
AISLAMIENTO DE PLÁSMIDO POR EL MÉTODO DE LISIS ALCALINA 1.- Se parte de un cultivo líquido (3ml de caldo soja y tripticaseína) de 24 h, el cual se vacía en un tubo
eppendorf y se centrifuga a 13,400Xg. El sobrenadante se elimina y se vuelve a repetir el mismo paso con
el resto del cultivo.
2.- Se resuspende el sedimento con vórtex después se le añade 100 �l de solución I fría (NaCl 0.1M,
tris.Cl 10mM pH 8.0 1mM EDTA pH 8.0), se agita en vórtex y se deja reposar durante 5 minutos a
temperatura ambiente.
3.- Pasado el tiempo de incubación se le añade 200 �l de solución II (NaOH 0.2 N, SDS 1%) y se agita por
inversión diez veces, inmediatamente se debe de introducir en hielo durante 5 minutos.
4.- Añadir 150 �l de solución III (Acetato de potasio 3M pH 4.8) y agitar manualmente 10veces. Efectuado
esto se deja reposar 5 minutos en hielo. Después del tiempo de incubación requerido se centrifuga 5
minutos.
5.-.- Se toma aproximadamente 400 �l de sobrenadante sin sedimento y se coloca en otro tubo eppendorf.
En este tubo es en donde se lleva la extracción con el fenol-cloroformo (100-200 �l), se debe de agitar por
inversión (15 veces), después se centrifuga durante 5 minutos.

34
6.- Se precipita la fase acuosa con isopropanol a temperatura ambiente vol/vol (400 �l) durante toda la
noche. Una vez efectuado este paso se vuelve a centrifugar durante 5 minutos y el tubo se coloca dentro
del speed vac (desecador) para que se seque el DNA.
7.- Finalmente se resuspende el DNA obtenido en 30-50 �l de regulador TE y se corre sobre un gel de
agarosa al 0.8% en regulador TBX a 80-100 voltios.
ELECTROFORESIS DE DNA PLASMÍDICO EN GEL DE AGAROSA AL 0.8%. 1.- Se funde la agarosa a 50oC. Se coloca un molde sobre una superficie plana (usando una burbuja), y se
pone el peine de modo que quede separado de la base del molde al menos el espesor de una hoja de
papel.
2.- Se vacía la agarosa fundida dentro del molde y deja solidificar a temperatura ambiente. Se quita el
peine y posteriormente se introduce el molde dentro de la cámara de electroforesis.
3.- Se llena la cámara con el regulador TBE con una concentración de 0.5-1 X hasta que se cubra el gel
(250 mL aproximadamente).
4.- La preparación de las muestras y la colocación de éstas dentro de cada pozo incluyendo al DNA del
fago Lambda digerido con EcoRI y Hind III se explica a continuación:
• Se mezcla 2 �L de densificante + 3 �L de muestra sobre el papel parafilm.
• Se agrega la mezcla a los pozos del gel
6.- Por último se tapa la cámara y se conecta a una fuente de poder. El voltaje debe de ser a 80-100v. La
cámara debe estar nivelada sobre una superficie plana horizontal.
OBTENCIÓN DE DNA TOTAL BACTERIANO
Para cada cepa se recuperaron las semillas, las cuales se mantuvieron congeladas a -70°C. Cada bacteria
se transfirió a un tubo conteniendo 10 ml de caldo soya y tripticaseína. Se extrajo el DNA genómico con
(Fenol-Cloroformo), para lo cual se centrifugaron 10 ml. de la suspensión bacteriana de un cultivo de toda
la noche a 37°C, a 6,000 x g durante 10 min Se removió el sobrenadante, y el paquete bacteriano se lavó
con 100µl de una solución STE más lisozima a una concentración de 10µg/ml hecha previamente, se
incubó a 37°C durante 45 min, se adicionó 300µl de una solución de Tris-HCL 50mM de EDTA 1mM, SDS
1%-Proteinasa K 50µg/ml, se incubó durante 60 min a 50°C. Al término de la incubación se llevó a cabo la
extracción con 200µl de fenol saturado y equilibrado con Tris a pH de 8.0 se centrifugó 1 min a 13,000 x g,
luego se separó la fase acuosa y la interfase es decir. la fase orgánica donde se encontraba el DNA
genómico, se extrajo con fenol-cloroformo (100µl +100µl), se mezcló vigorosamente y se centrifugó a
13,000 x g durante 1 min, se volvió a separar la fase acuosa y finalmente se llevó a cabo la extracción con
200µl de cloroformo, se mezcló vigorosamente y se centrifugó durante 2 min a 13,000 x g. Se separó la
fase acuosa y se adicionó 1µl de RNasa, se incubó a 37°C durante 30 min; al DNA precipitado se le
adicionó 1 volumen de acetato de sodio 3M y 2 volúmenes de etanol absoluto frío y se dejó precipitando
toda la noche a -20°C al siguiente día se centrifugó durante 10 min a 13,000 x g y se desechó el etanol
absoluto y se lavó el DNA con 300 µl de etanol al 70%. Finalmente el DNA se recuperó a 13,000 x g
durante 5 min se eliminó el etanol secándolo en un equipo Speed Vac y el DNA se disolvió en 50 μl de
agua bidestilada y estéril. La integridad del DNA se analizó en un gel de agarosa al 0.8%, diluyendo la
muestra 4:1 en regulador de muestra. Se sometió a electroforesis en gel de agarosa al 0.8% con una
35
corriente de 90 Voltios durante 90 min El gel se tiñó con bromuro de etidio. Las bandas del DNA genómico
se visualizaron y digitalizaron.
REACCIÓN EN CADENA DE LA POLIMERASA (PCR)
Con el objeto de identificar que tipo de genes están asociados a la producción de las metalo-β-Lactamasas
VIM, IMP, SPM y GIM, se realizó la reacción en cadena de la polimerasa utilizando como molde el DNA
genómico y los iniciadores específicos. Las condiciones se establecieron para la amplificación de cada
región génica.
IMP
Los oligonucleótidos que se utilizaron para amplificar el gene blaIMP por la técnica de la PCR se diseñaron
con base en un análisis de la secuencia reportada por Senda y col. (1996). Se utilizaron los iniciadores, en
sentido 5´ CTA CGG CAG CAG AGT CTT TG 3´ y anti-sentido 5´ AAC CAG TTT TGC CTT ACC AT 3´. De
igual forma se diseñaron oligonucleótidos degenerados para tratar de amplificar diferentes tipos de genes
blaIMP; utilizando los iniciadores en sentido 5’ AGC RTT KCT ACC GCA GSA G 3’ y anti sentido 5’AAC
AAC HAG KTT TGC HTT MCC A 3’. El tamaño esperado del amplificado del gene blaIMP es de 587 pb. Las
secuencias de los oligonucleótidos fueron analizadas con los programas Primer 3, CODEHOP y
CLUSTALX versión 1.8(URL1, 2 y 5) para comprobar que la secuencia de oligonucleótidos era la más
adecuada para la amplificación del gen.
Los siguientes parámetros específicos de amplificación con los 2 pares de iniciadores consistieron en:
Nota: estas condiciones se
emplearon para ambos juegos de
iniciadores
a) Condiciones de reacción cuando se utilizaron los iniciadores descritos por Senda y col. (1996). Una fase
de desnaturalización de 2 min a 94°C, durante un ciclo, 1 min a 94°C para desnaturalización, 1 min a 55°C
para alineamiento y 1.5 min a 72°C para extensión durante 30 ciclos de reacción y finalmente un ciclo a
72°C durante 10 min.
DNA molde (1:100) 1.0 μl.
Buffer PCR 10X 2.5 μl
MgCl2 50mM 0.75μl.
dNTP (2.0mM) 2.5 μl.
Oligonucleótidos (30pmoles/µl 0.5 μl
Agua estéril 17 μl.
Taq polimerasa (5 unidades/μl) 0.25 μl.
Volumen total 25 μl.

36
b) Condiciones de reacción cuando se utilizaron los iniciadores degenerados: la reacción se realizó
utilizando 5 min a 95°C para desnaturalización, durante un ciclo, 0.50 min a 95°C para desnaturalización, 1
min a 50°C para alineamiento y 1.5 mins a 72°C para extensión durante 30 ciclos de reacción. Y finalmente
un ciclo a 72°C durante 10 min El producto de PCR se analizó en un gel de agarosa al 1.5% a 90 Voltios
durante 90min.
Las muestras se analizaron mediante electroforesis en gel de agarosa al 1.5% a 90 voltios durante 1 h y se
utilizó como marcador de tamaño molecular el DNA de λ Pst I (2-23.3 kpb) y la escalera de 100 pb, ambos
de Invitrogen.
Para llevar acabo el corrimiento electroforético de los productos de PCR primero se preparo la agarosa al
1.5% en TBE 0.5X, una vez fundida se coloco en la placa de la cámara de electroforesis y se dejó gelificar
durante 30 min la cámara se llenó con regular TBE 0.5X y se agregó la mezcla de 1 μl. de azul de
bromofenol con 5 μl. de producto de PCR en el pozo correspondiente. En el primer pozo, se agregaron 5
μl. de los marcadores de peso molecular. El corrimiento electroforético se realizó durante 90 min con un
voltaje de 90 (fuente de poder Bio-Rad, modelo 200/2.0, Richmond, CA), posteriormente el gel se retiró de
la cámara y se sumergió en regular TBE 1X con bromuro de etidio, durante 15 min La imagen se digitalizó
en el equipo ChemiImager 4000 Low Light Imaging System (San Leandro, CA).
GIM
Los oligonucleótidos que se utilizaron para amplificar el gene blaGIM por la técnica de PCR se diseñaron
con base en un análisis de la secuencia reportada por Casthaneria y col. (2004). Para amplificar el gen
blaGIM se utilizaron los iniciadores, en sentido 5´ CTA CGG CAG CAG AGT CTT TG 3´ y anti-sentido 5´
AAC CAG TTT TGC CTT ACC AT 3´. El tamaño esperado del amplificado del gene blaGIM sería de 753 pb.
Las secuencias de los oligonucleótidos fueron analizadas con los programas Primer 3, CODEHOP y
CLUSTALX versión 1.8(URL1, 2 y 5) para comprobar que la secuencia de oligonucleótidos era la más
adecuada para la amplificación del gen.
a) Las condiciones de reacción fueron. Una
fase de desnaturalización de
5 min a 95°C, durante un ciclo, 1 min a 95°C
para desnaturalización, 40
s a 40°C para alineamiento y 1 min
a 68°C para extensión durante 30
ciclos de reacción y finalmente un ciclo a
68°C durante 5 min
VIM
Los oligonucleótidos que se utilizaron para amplificar el gene blaVIM por la técnica de PCR fueron los
descritos por Nordmann & Poirel (2002). Para amplificar el gen blaVIM se utilizaron los iniciadores, en
sentido 5´ ATG GTG TTT GGT CGC ATA TC 3´ y anti-sentido 5´ GAT CTG GCT GAA TGG CCC A 3´. El
tamaño esperado del amplificado del gene blaVIM sería de 490 pb. Las secuencias de los oligonucleótidos
DNA molde (1:100) 1.0 μl.
Buffer PCR 10X 2.5 μl
MgCl2 50mM 0.75μl.
dNTP (2.0mM) 2.5 μl.
Oligonucleótidos (30pmoles/µl) 0.5 μl
Agua estéril 17 μl.
Taq polimerasa (5 unidades/μl) 0.25 μl.
Volumen total 25 μl.
37
fueron analizadas con los programas Primer 3, CODEHOP y CLUSTALX versión 1.8(URL1, 2 y 5) para
comprobar que la secuencia de oligonucleótidos era la más adecuada para la amplificación del gen.
a) Las condiciones fueron. Una fase de desnaturalización de 5 min a 94°C, durante un ciclo, 30 s a 94°C
para desnaturalización, 40 s a 52°C para alineamiento y 50 s a 72°C para extensión durante 30 ciclos de
reacción y finalmente un ciclo a 72°C durante 6 min
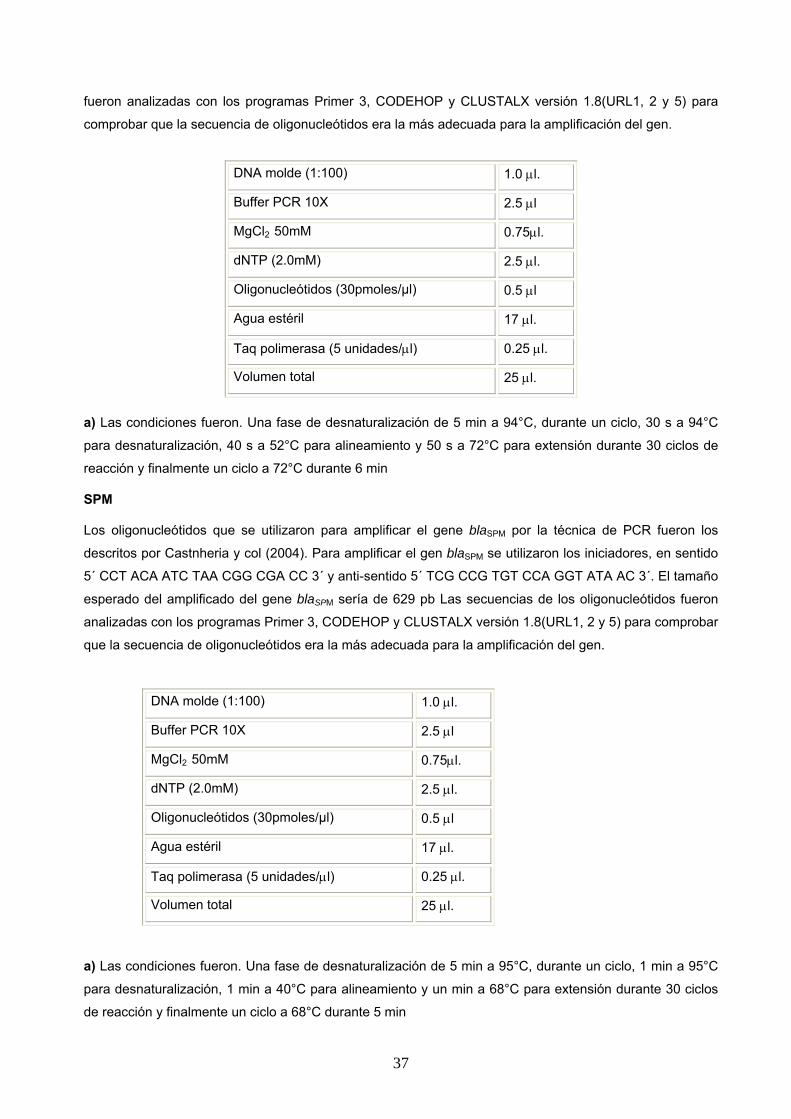
SPM
Los oligonucleótidos que se utilizaron para amplificar el gene blaSPM por la técnica de PCR fueron los
descritos por Castnheria y col (2004). Para amplificar el gen blaSPM se utilizaron los iniciadores, en sentido
5´ CCT ACA ATC TAA CGG CGA CC 3´ y anti-sentido 5´ TCG CCG TGT CCA GGT ATA AC 3´. El tamaño
esperado del amplificado del gene blaSPM sería de 629 pb Las secuencias de los oligonucleótidos fueron
analizadas con los programas Primer 3, CODEHOP y CLUSTALX versión 1.8(URL1, 2 y 5) para comprobar
que la secuencia de oligonucleótidos era la más adecuada para la amplificación del gen.
a) Las condiciones fueron. Una fase de desnaturalización de 5 min a 95°C, durante un ciclo, 1 min a 95°C
para desnaturalización, 1 min a 40°C para alineamiento y un min a 68°C para extensión durante 30 ciclos
de reacción y finalmente un ciclo a 68°C durante 5 min
DNA molde (1:100) 1.0 μl.
Buffer PCR 10X 2.5 μl
MgCl2 50mM 0.75μl.
dNTP (2.0mM) 2.5 μl.
Oligonucleótidos (30pmoles/µl) 0.5 μl
Agua estéril 17 μl.
Taq polimerasa (5 unidades/μl) 0.25 μl.
Volumen total 25 μl.
DNA molde (1:100) 1.0 μl.
Buffer PCR 10X 2.5 μl
MgCl2 50mM 0.75μl.
dNTP (2.0mM) 2.5 μl.
Oligonucleótidos (30pmoles/µl) 0.5 μl
Agua estéril 17 μl.
Taq polimerasa (5 unidades/μl) 0.25 μl.
Volumen total 25 μl.

38

39
RESULTADOS
Resultados de la transferencia de material genético por conjugación, utilizando como cepas donadoras S. aureus y como cepa receptora E. coli IJ133
ANTIBIÓTICO E. coli IJ133
(antes de conjugar)
SA1
ES1
SA3
ES3
AMC R R R R R Penicilinas IPM S S S R S
CFP S I R R R CXM S R R R R CAZ S R R R R CRO S R R R R FEP S I S R S
Cefalosporinas
ZOX S R R R R AN S R S R S NN S R R R R
Aminoglicósidos
NET S I I S I Macrólidos E R I R R R
CIP S R R R R LVX S R R R R NOR S R R R R
Quinolonas OFX S R R R R
Otros β-lactámicos MEM S R S R S D S R S R S
RA R S R R R

40
TE S R S S S SA1 y SA3 = Cepas donadoras de S. aureus; ES1 y ES3 = Cepas de E. coli transconjugantes
41
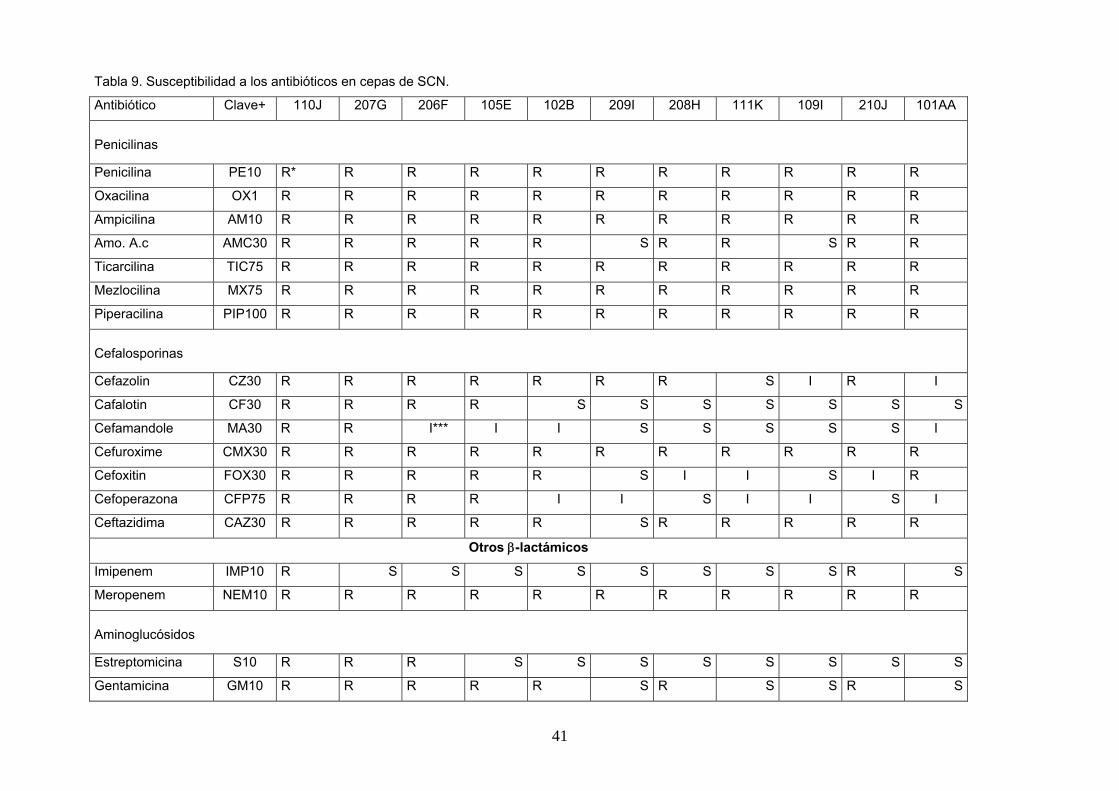
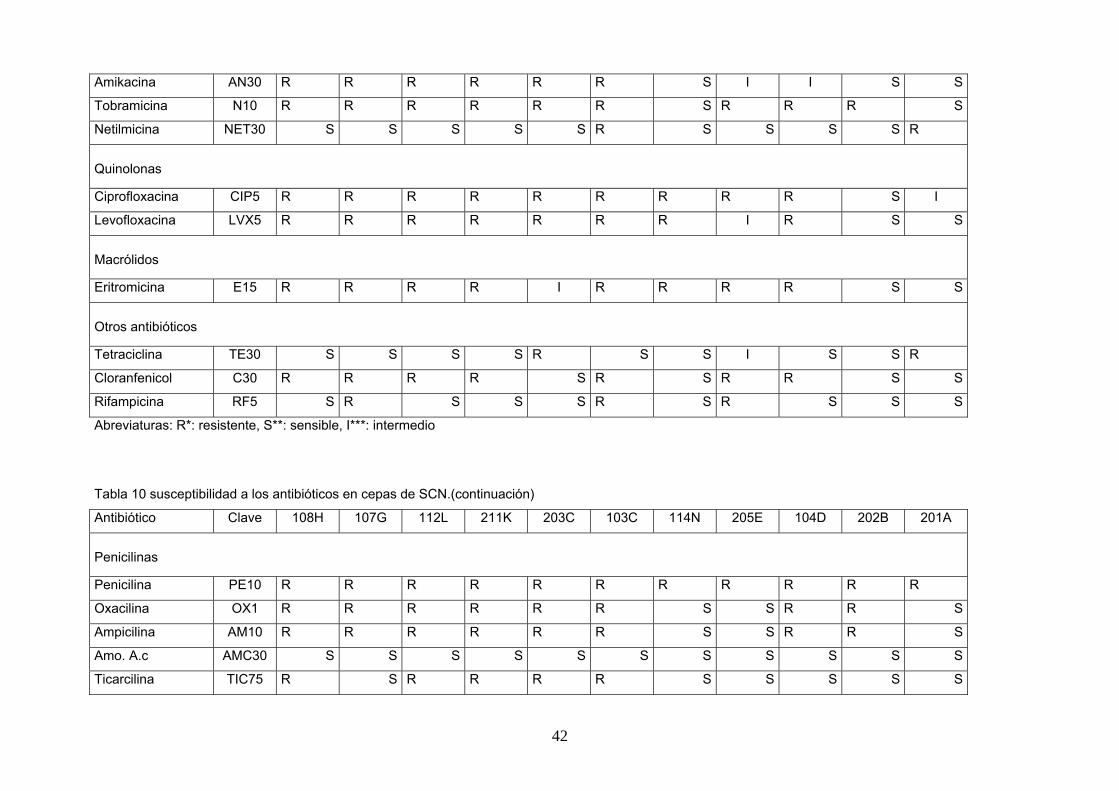
Tabla 9. Susceptibilidad a los antibióticos en cepas de SCN.
Antibiótico Clave+ 110J 207G 206F 105E 102B 209I 208H 111K 109I 210J 101AA
Penicilinas
Penicilina PE10 R* R R R R R R R R R R
Oxacilina OX1 R R R R R R R R R R R
Ampicilina AM10 R R R R R R R R R R R
Amo. A.c AMC30 R R R R R S R R S R R
Ticarcilina TIC75 R R R R R R R R R R R
Mezlocilina MX75 R R R R R R R R R R R
Piperacilina PIP100 R R R R R R R R R R R
Cefalosporinas
Cefazolin CZ30 R R R R R R R S I R I
Cafalotin CF30 R R R R S S S S S S S
Cefamandole MA30 R R I*** I I S S S S S I
Cefuroxime CMX30 R R R R R R R R R R R
Cefoxitin FOX30 R R R R R S I I S I R
Cefoperazona CFP75 R R R R I I S I I S I
Ceftazidima CAZ30 R R R R R S R R R R R
Otros β-lactámicos
Imipenem IMP10 R S S S S S S S S R S
Meropenem NEM10 R R R R R R R R R R R
Aminoglucósidos
Estreptomicina S10 R R R S S S S S S S S
Gentamicina GM10 R R R R R S R S S R S
42
Amikacina AN30 R R R R R R S I I S S
Tobramicina N10 R R R R R R S R R R S
Netilmicina NET30 S S S S S R S S S S R
Quinolonas
Ciprofloxacina CIP5 R R R R R R R R R S I
Levofloxacina LVX5 R R R R R R R I R S S
Macrólidos
Eritromicina E15 R R R R I R R R R S S
Otros antibióticos
Tetraciclina TE30 S S S S R S S I S S R
Cloranfenicol C30 R R R R S R S R R S S
Rifampicina RF5 S R S S S R S R S S S
Abreviaturas: R*: resistente, S**: sensible, I***: intermedio
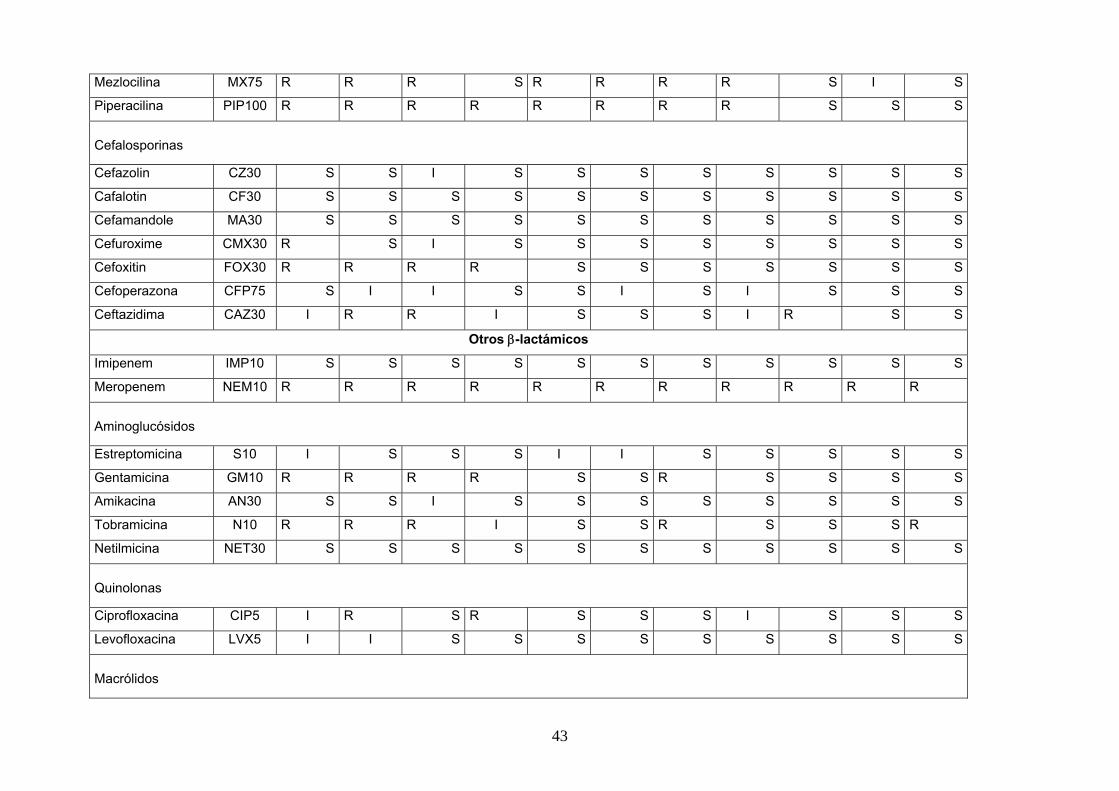
Tabla 10 susceptibilidad a los antibióticos en cepas de SCN.(continuación)
Antibiótico Clave 108H 107G 112L 211K 203C 103C 114N 205E 104D 202B 201A
Penicilinas
Penicilina PE10 R R R R R R R R R R R
Oxacilina OX1 R R R R R R S S R R S
Ampicilina AM10 R R R R R R S S R R S
Amo. A.c AMC30 S S S S S S S S S S S
Ticarcilina TIC75 R S R R R R S S S S S
43
Mezlocilina MX75 R R R S R R R R S I S
Piperacilina PIP100 R R R R R R R R S S S
Cefalosporinas
Cefazolin CZ30 S S I S S S S S S S S
Cafalotin CF30 S S S S S S S S S S S
Cefamandole MA30 S S S S S S S S S S S
Cefuroxime CMX30 R S I S S S S S S S S
Cefoxitin FOX30 R R R R S S S S S S S
Cefoperazona CFP75 S I I S S I S I S S S
Ceftazidima CAZ30 I R R I S S S I R S S
Otros β-lactámicos
Imipenem IMP10 S S S S S S S S S S S
Meropenem NEM10 R R R R R R R R R R R
Aminoglucósidos
Estreptomicina S10 I S S S I I S S S S S
Gentamicina GM10 R R R R S S R S S S S
Amikacina AN30 S S I S S S S S S S S
Tobramicina N10 R R R I S S R S S S R
Netilmicina NET30 S S S S S S S S S S S
Quinolonas
Ciprofloxacina CIP5 I R S R S S S I S S S
Levofloxacina LVX5 I I S S S S S S S S S
Macrólidos

44
Eritromicina E15 S I I S I I S R S S S
Otros antibióticos
Tetraciclina TE30 R R S S R S R S S S S
Cloranfenicol C30 S S S S S S S S S S S
Rifampicina RF5 S S S S R S S S S S S
Abreviaturas: R*: resistente, S**: sensible, I***: intermedio
45
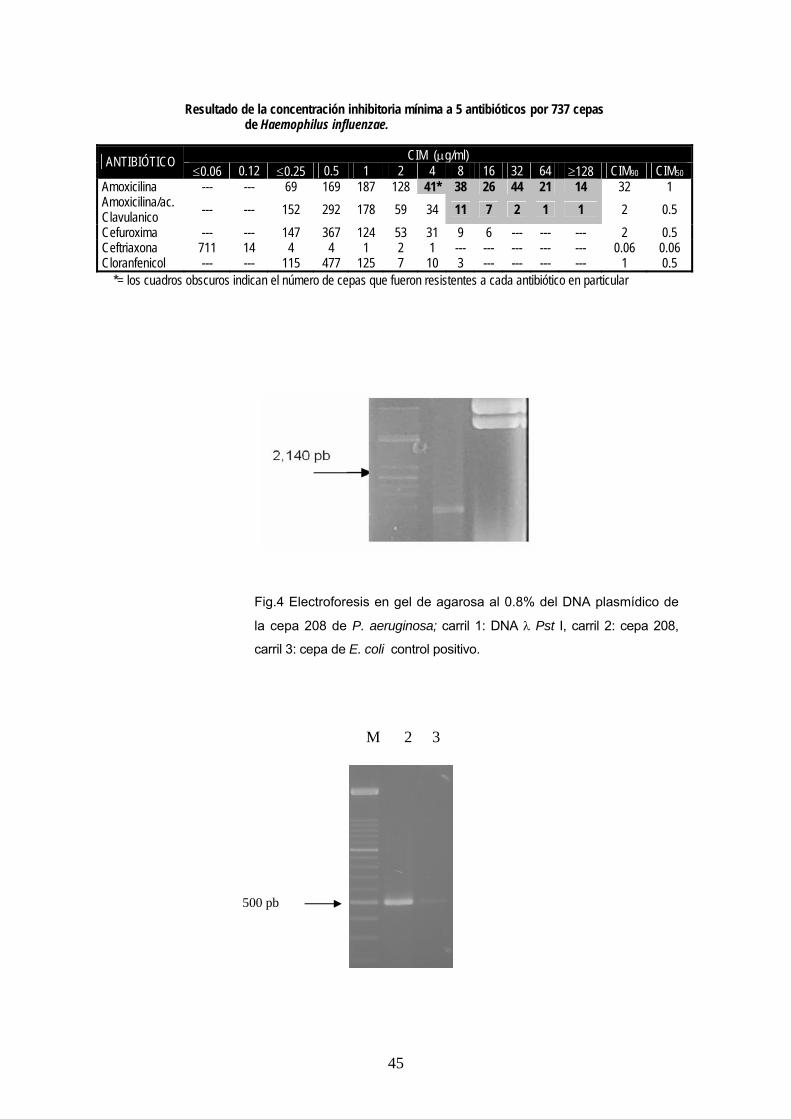
Resultado de la concentración inhibitoria mínima a 5 antibióticos por 737 cepas de Haemophilus influenzae.
*= los cuadros obscuros indican el número de cepas que fueron resistentes a cada antibiótico en particular
Fig.4 Electroforesis en gel de agarosa al 0.8% del DNA plasmídico de
la cepa 208 de P. aeruginosa; carril 1: DNA λ Pst I, carril 2: cepa 208,
carril 3: cepa de E. coli control positivo.
CIM (μg/ml) ANTIBIÓTICO ≤0.06 0.12 ≤0.25 0.5 1 2 4 8 16 32 64 ≥128 CIM90 CIM50
Amoxicilina --- --- 69 169 187 128 41* 38 26 44 21 14 32 1 Amoxicilina/ac. Clavulanico --- --- 152 292 178 59 34 11 7 2 1 1 2 0.5 Cefuroxima --- --- 147 367 124 53 31 9 6 --- --- --- 2 0.5 Ceftriaxona 711 14 4 4 1 2 1 --- --- --- --- --- 0.06 0.06 Cloranfenicol --- --- 115 477 125 7 10 3 --- --- --- --- 1 0.5
M 2 3
500 pb
46
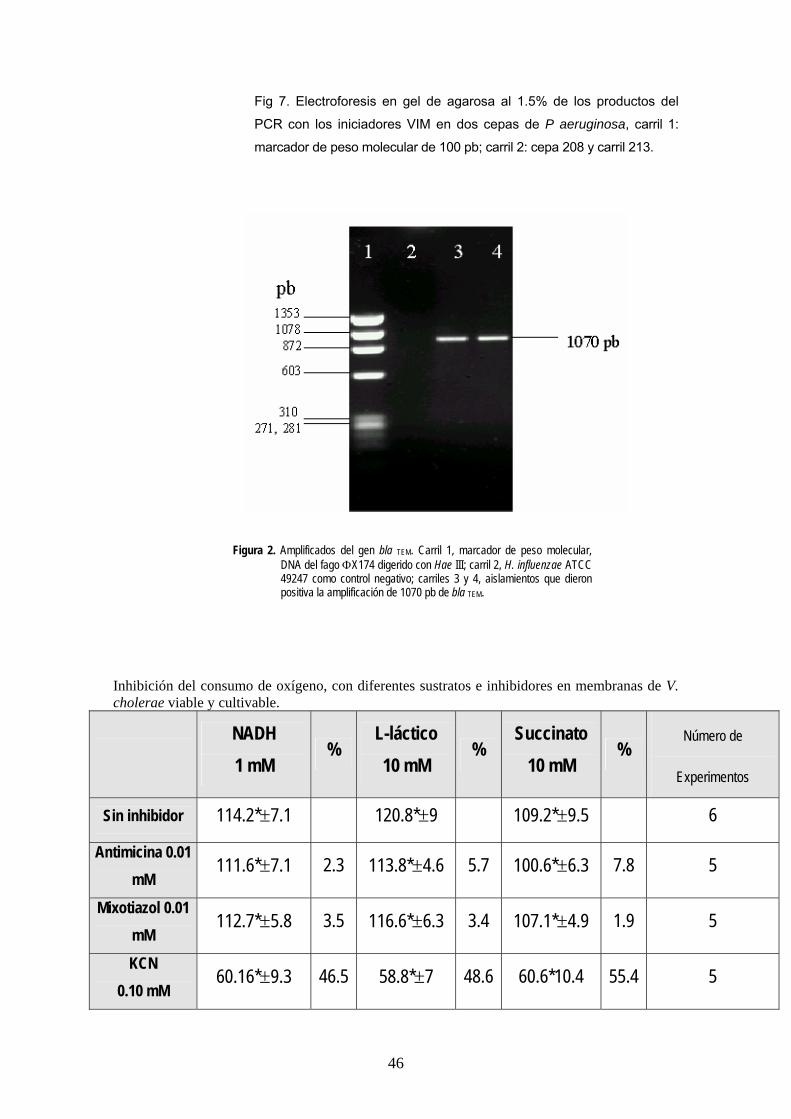
Fig 7. Electroforesis en gel de agarosa al 1.5% de los productos del
PCR con los iniciadores VIM en dos cepas de P aeruginosa, carril 1:
marcador de peso molecular de 100 pb; carril 2: cepa 208 y carril 213.
Figura 2. Amplificados del gen bla TEM. Carril 1, marcador de peso molecular, DNA del fago ΦX174 digerido con Hae III; carril 2, H. influenzae ATCC 49247 como control negativo; carriles 3 y 4, aislamientos que dieron positiva la amplificación de 1070 pb de bla TEM.
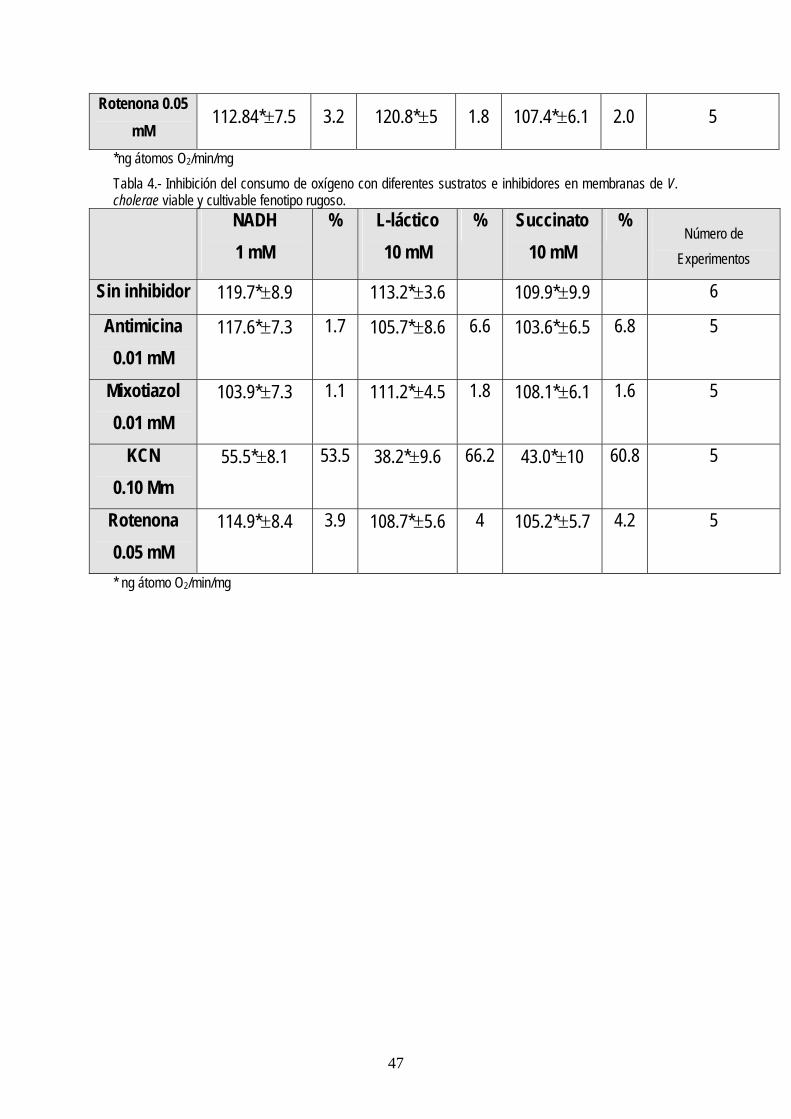
Inhibición del consumo de oxígeno, con diferentes sustratos e inhibidores en membranas de V. cholerae viable y cultivable.
NADH 1 mM
% L-láctico 10 mM
% Succinato
10 mM %
Número de
Experimentos
Sin inhibidor 114.2*±7.1 120.8*±9 109.2*±9.5 6
Antimicina 0.01 mM
111.6*±7.1 2.3 113.8*±4.6 5.7 100.6*±6.3 7.8 5
Mixotiazol 0.01 mM
112.7*±5.8 3.5 116.6*±6.3 3.4 107.1*±4.9 1.9 5
KCN 0.10 mM
60.16*±9.3 46.5 58.8*±7 48.6 60.6*10.4 55.4 5
47
Rotenona 0.05 mM
112.84*±7.5 3.2 120.8*±5 1.8 107.4*±6.1 2.0 5
*ng átomos O2/min/mg Tabla 4.- Inhibición del consumo de oxígeno con diferentes sustratos e inhibidores en membranas de V. cholerae viable y cultivable fenotipo rugoso.
NADH 1 mM
% L-láctico 10 mM
% Succinato 10 mM
% Número de
Experimentos
Sin inhibidor 119.7*±8.9 113.2*±3.6 109.9*±9.9 6
Antimicina 0.01 mM
117.6*±7.3 1.7 105.7*±8.6 6.6 103.6*±6.5 6.8 5
Mixotiazol 0.01 mM
103.9*±7.3 1.1 111.2*±4.5 1.8 108.1*±6.1 1.6 5
KCN 0.10 Mm
55.5*±8.1 53.5 38.2*±9.6 66.2 43.0*±10 60.8 5
Rotenona 0.05 mM
114.9*±8.4 3.9 108.7*±5.6 4 105.2*±5.7 4.2 5
* ng átomo O2/min/mg

48
IMPACTO
• Se realizó por primera vez en nuestro país, la detección del gene blavim en cepas
panresistentes de P. aeruginosa.
• Únicamente dos de las once cepas de S. aureus presentaron multirresistencia a la acción
de 34 antibióticos de diferentes familias.
• Todas las cepas fueron resistentes a penicilina y ampicilina y fueron sensibles a
cloranfenicol y nitrofurantoina.
• Todas las cepas de S. aureus fueron productoras de β-lactamasas, las cuales se sugiere
que probablemente sean del tipo penicilinasas.
• Se logró realizar la conjugación entre las cepas Gram positivas de S. aureus (SA1 y SA3,
donadoras) y la cepa Gram negativa E. coli (receptora).
• Por el método de lisis alcalina se encontraron plásmidos en las cepas transconjugantes
ES1 y ES3 que por lo tanto posiblemente están implicados en el mecanismo de resistencia
de S. aureus (SA1 y SA3).
• Las cepas de S. aureus a las cuales no se les realizó un proceso de conjugación tienen
β-lactamasas de reducido espectro y se desconoce si estas son de origen plasmídico o
cromosómico.
• Se determinó que el porcentaje global de aislamientos de H. influenzae en las entidades de
la Republica Mexicana fue 23%.
• Sólo 5% (38/737) de los aislados demostraron ser capsulados, siendo el serotipo f el más
frecuente (29%); en tanto que el b se presentó en 16%.
• Los aislados de H. influenzae mostraron resistencia a amoxicilina en 25% (184/737). De
estas 16% permanecieron resistentes a amoxicilina, aún en combinación con ácido
clavulánico.
• La resistencia al resto de los antibióticos probados no superó el 1%, observándose que las
cepas resistentes a estos se encontraban dentro del grupo de las resistentes a amoxicilina.
• En nuestro estudio se encontró que 16% (118/737) de los aislamientos de H. influenzae
fueron productores de ß-lactamasas y todos ellos mostraron resistencia a la amoxicilina.
• El 84% de las cepas productoras de ß-lactamasa, presentaron el gen bla TEM , mientras
que el 16% mostraron poseer el gen bla ROB.
• Todas las cepas de Staphylococcus coagulasa negativa empleadas en este estudio presentaron resistencia a algún tipo de antibiótico β-lactámico y produjeron algún tipo de β-lactamasa
• 10 de las 22 cepas de Staphylococcus coagulasa negativa en estudio, presentaron plásmidos, los cuales podrían estar relacionados con la resistencia a los antibióticos

49
• La integridad observada en las membranas de los tres fenotipos de Vibrio cholerae
obtenidos sugiere que los organismos se encontraban viables
• Vibrio cholerae presenta en su cadena transportadora de electrones enzimas como la
NADH deshidrogenasa, succinato deshidrogenasa y l-latico deshidrogenasa además de
citocromos como el b, el c y el o.
• No se encontró diferencia significativa entre las quinonas y menaquinonas de los tres
fenotipos de Vibrio cholerae
• Vibrio cholerae presenta en los tres diferentes fenotipos citocromo c oxidasa
• Las oxidasas terminales en la cadena transportadora de electrones en Vibrio cholerae son
la citocromo oxidasa co y la citocromo oxidasa bo