iib alberto sols memoria de la...
TRANSCRIPT

MEMORIA IIB ALBERTO SOLS 1
Alberto Maurel
4-8 julio 2016
IIB Alberto Sols
Memoria de la estancia
Lunes
Hemos visitado el laboratorio de cultivos y preparado un gel de acrilamida.
Cultivos celulares
El medio utilizado para mantener nuestras células es el DMEM. Uno de los componentes
de este medio es el rojo fenol. Este indicador permite detectar de visu, gracias a que es un
indicador colorimétrico, el estado del medio. A medida que las células van consumiendo el
medio, provocan que el rojo fenol vire del rojo intenso inicial hacia un rojo pálido,
anaranjado y posteriormente naranja amarillento, indicándonos estos últimos que es
necesario renovar el medio.

MEMORIA IIB ALBERTO SOLS 2
Las células que vamos a cultivar están adheridas a la placa petri. Por ello, quitamos el
medio a través de una bomba de vacío y una pipeta. Las células muertas se separan de la
placa, por lo que también las eliminamos.
Posteriormente, añadimos de nuevo DMEM para que las células sigan reproduciéndose.
Y añadimos también puromicina para que destruya aquellas células que no contengan el
plásmido transfectado (que contiene un gen de resistencia para este antibiótico).

MEMORIA IIB ALBERTO SOLS 3
Y ya tenemos nuestras células vivas listas para seleccionar y seguir reproduciéndose.

MEMORIA IIB ALBERTO SOLS 4
Otro de los cultivos está ya listo para cambiarlo de placa. De esta forma, evitamos que la
confluencia de las células sea excesiva, y permitimos que sigan creciendo. Para separar las
células en primer lugar eliminamos el DMEM (llevándonos al mismo tiempo las células
muertas).
Posteriormente las limpiamos con PBS (solución salina).
Tras esto, añadimos tripsina, que se encarga de separar las células vivas de la placa, y lo
incubamos. Al cabo de unos cinco minutos, las células ya están separadas.

MEMORIA IIB ALBERTO SOLS 5
Trasladamos esta disolución a un falcon para
centrifugarla. Tras 5 minutos a 1200 rpm, queda en el
sobrenadante la tripsina y en el pellet las células vivas.
Retiramos con cuidado la tripsina por medio de la bomba de vacío y trasladamos de
nuevo las células a una placa petri con DMEM. Las células ya están listas para seguir
reproduciéndose. !Y por supuesto rotulamos todo para no confundirnos!
Ruta de señalización Hedgehog (Hh)
La ruta de señalización Hedgehog está implicada en el desarrollo embrionario. El motivo de su estudio en
este caso es su relación con el síndrome de Ellis-van Creveld, causante de lesiones en los dientes, polidactilia y

MEMORIA IIB ALBERTO SOLS 6
desaparición del tabique entre aurículas. Este síndrome está causado por la mutación de las proteínas Evc y
Evc2, que forman parte de la ruta Hh.
La proteína IFT74 es una proteína que transporta unidades de tubulina de la base del cilio primario a la
punta. Dicha proteína interacciona con la Evc, y es la relación entre estas dos proteínas la que se trata de
dilucidar mediante este experimento.
Se ha descubierto que en genotipos IFT74 +/- (con solo una copia del gen que codifica para la proteína) se
forma menos IFT74 y Evc, formándose además cilio primario. En el caso de genotipos IFT74 -/- (sin copias
del gen para esta proteína) no se forma IFT74, pero se forma más Evc que en genotipos IFT74 +/+.
Además, no se forma cilio primario.
La hipótesis que se baraja para estos resultados es que la proteína IFT74 codifica para la formación del
cilio primario, estructura en la que se produce la degradación de la proteína Evc. En los genotipos IFT74 -/-
no se forma el cilio primario y, por ende, no se produce la degradación de la Evc.
En este experimento se infecta con retrovirus una célula IFT74 -/-. Dicho retrovirus contiene una
construcción con el gen que codifica la proteína IFT74. Si la hipótesis es acertada, en la célula infectada se
comenzaría a producir IFT74, se formaría de nuevo el cilio primario y se restablecerían los valores normales de
Evc.
Para comprobar la veracidad de esta hipótesis, tras introducir el gen se emplea el Western Blot para
cuantificar las cantidades de Evc e IFT74 sintetizadas, y microscopía de fluorescencia para detectar, por medio
de fluoróforos, la formación del cilio primario.
Preparación del gel de acrilamida para realizar el Western Blot
En primer lugar, montamos el molde para los geles.

MEMORIA IIB ALBERTO SOLS 7
Posteriormente, siguiendo el protocolo preparamos gel para dos moldes. Primero
cargamos el gel separador, cubriéndolo de agua mientras solidifica.
Cuando ha solidificado (lo podemos comprobar gracias a que en el falcon la disolución
sobrante ha solidificado), cargamos el gel concentrador, sobre el cual colocamos el peine para
formar los pocillos.

MEMORIA IIB ALBERTO SOLS 8
Martes
Tras hablar sobre la historia de la secuenciación del ADN, hemos acudido al Instituto de
Genética Médica y Molecular (INGEMM) para ver las máquinas y técnicas de secuenciación.
Osteogénesis imperfecta
Este laboratorio del IIB está especializado en las alteraciones del desarrollo óseo, concretamente en dos: el
síndrome de Ellis-van Creveld (del que ya hemos hablado) y el síndrome de osteogénesis imperfecta.
Esta enfermedad está causada en un 90% de los casos por mutaciones que afectan a los genes Col1a1 y
Col1a2, y en un 10% de los casos por mutaciones en los genes que regulan el plegamiento de la proteínas en el
retículo.
El colágeno es una proteína formada por una triple hélice. Dos de las hebras son alfa 1, codificada por el
gen Col1a1, y la otra es de alfa 2, codificada por el gen Col1a2.
Tras formarse en los polirribosomas, el procolágeno no funcional pasa al retículo endoplasmático. Allí se
cortan los propéptidos (se encuentran en los extremos del procolágeno) y se entrecruzan las dos cadenas alfa 1
con otra alfa 2 (a este proceso afectaría el 10% de las mutaciones previamente mencionadas).
En esta enfermedad podemos distinguir además varias clases de mutaciones que la originan:
1. Puede aparecer una delección en una de las copias del ADN. Esto provoca que se produzca tan solo la
mitad del colágeno, lo que ya origina la enfermedad.
2. Puede existir una mutación, que cause una modificación en los aminoácidos (aa) de la cadena. Las
cadenas están formadas por 338 repeticiones de un trímero de aminoácidos, de los cuales el primero siempre
es la glicina. La modificación de estos aa puede conducir a que todo el colágeno que se forme sea defectuoso.
Historia de la secuenciación
Inicios
En sus inicios, para localizar los genes causantes de las enfermedades monogénicas se empleaban zonas
polimórficas. Estas zonas se localizan en los intrones, y son repeticiones de determinadas secuencias de
nucleótidos, y que presentan varias variantes en la población. Si no se realiza recombinación entre la zona
polimórfica y la enfermedad, sabemos que están próximos en el cromosoma.
Para realizar un mapeo del gen requiere tener por un lado un mapa con las zonas polimórficas y por otro un
árbol genealógico grande con el que hacer el estudio (y ver de padres a hijos con que zonas polimórficas
recombinan los genes causantes de la enfermedad y con cuáles no).

MEMORIA IIB ALBERTO SOLS 9
Además, se construían bibliotecas genómicas, en las que en el interior de una bacteria (BAC) o de una
levadura (YAC) se guardaban fragmentos del ADN. Tras acotar por el procedimiento previamente mencionado
la posición del gen, se podía trabajar con esa región del ADN, acotándola aún más.
Secuenciación Sanger
La secuenciación Sanger es una técnica de secuenciación del ADN que se lleva a cabo en unos aparatos
llamados secuenciadores, y que permite secuencias fragmentos de tamaño variable dependiendo de la longitud del
capilar (unos 600 pb para 36 cm de capilar). Para llevar a cabo esta técnica se parte de una copia de ADN
molde, la cual se amplifica por medio de una PCR. Posteriormente, se introduce el fragmento amplificado en
una mezcla que contiene DNTPs, didesoxirribonucleótidos marcados con fluorocromos (cada tipo de base
nitrogenada está marcada con un fluorocromo distinto), ADN polimerasas, cebadores y factores de duplicación
(esencialmente cationes como el Mg 2+).
La ADN polimerasa comienza a sintetizar copias de la cadena amplificada. De forma aleatoria la ADN
polimerasa inserta o bien un desoxirribonucleótido o bien un didesoxirribonucleótido. En el caso de que inserte
un desoxirribonucleótido, se sigue sintetizando la copia de ADN. En el caso de insertar un
didesoxirribonucleótido, no puede continuar sintetizando la copia porque carece del 3OH, el cual debería formar
el enlace fosfodiéster.
Después, la disolución con el ADN sintetizado con diferentes longitudes se hace pasar por un capilar con un
diámetro que solo permite que pase una copia de ADN a la vez. En el interior del capilar se encuentra un gel de
acrilamida. En el extremo opuesto del capilar se halla una carga positiva (el ADN posee carga negativa). Los
fragmentos más pequeños correrán por el gel a mayor velocidad. En un lugar del capilar se encuentra un lector
de fluorescencia, que permite leer las bases de los didesoxirribonucleótidos marcadas con fluorocromos. De esta
forma, se obtiene la secuencia de bases del fragmento.
Arrays
Sirven para detectar mutaciones (delecciones, inserciones...) de tamaño superior a unas cuantas bases o para
realizar mapeos de homocigosidad. En un chip se sitúan las variantes de oligonucleótidos (SNPS) que se
emplean como marcadores. Posteriormente, se marca el ADN humano, se híbrida el ADN con los
oligonucleótidos del array y este chip se puede leer con un escáner.
Entre los datos que nos arroja esta técnica, destacan dos:
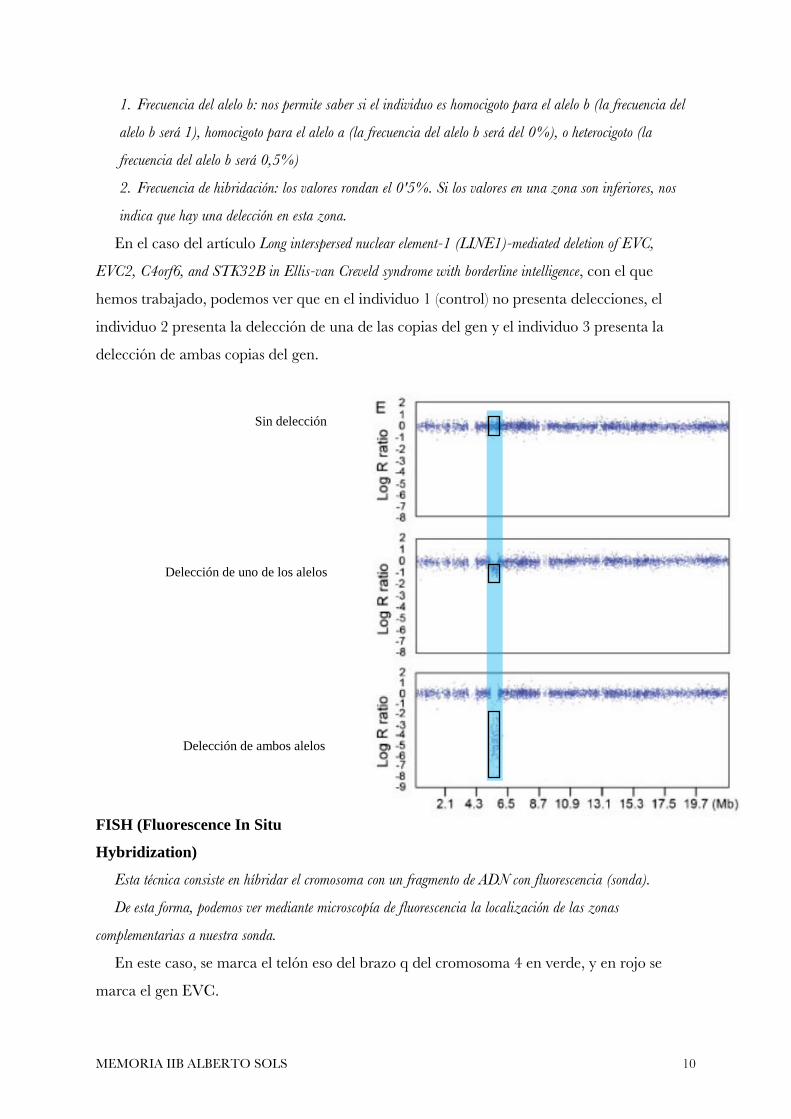
MEMORIA IIB ALBERTO SOLS 10
1. Frecuencia del alelo b: nos permite saber si el individuo es homocigoto para el alelo b (la frecuencia del
alelo b será 1), homocigoto para el alelo a (la frecuencia del alelo b será del 0%), o heterocigoto (la
frecuencia del alelo b será 0,5%)
2. Frecuencia de hibridación: los valores rondan el 0'5%. Si los valores en una zona son inferiores, nos
indica que hay una delección en esta zona.
En el caso del artículo Long interspersed nuclear element-1 (LINE1)-mediated deletion of EVC,
EVC2, C4orf6, and STK32B in Ellis-van Creveld syndrome with borderline intelligence, con el que
hemos trabajado, podemos ver que en el individuo 1 (control) no presenta delecciones, el
individuo 2 presenta la delección de una de las copias del gen y el individuo 3 presenta la
delección de ambas copias del gen.
FISH (Fluorescence In Situ
Hybridization)
Esta técnica consiste en híbridar el cromosoma con un fragmento de ADN con fluorescencia (sonda).
De esta forma, podemos ver mediante microscopía de fluorescencia la localización de las zonas
complementarias a nuestra sonda.
En este caso, se marca el telón eso del brazo q del cromosoma 4 en verde, y en rojo se
marca el gen EVC.
Sin delección
Delección de uno de los alelos
Delección de ambos alelos
MEMORIA IIB ALBERTO SOLS 11
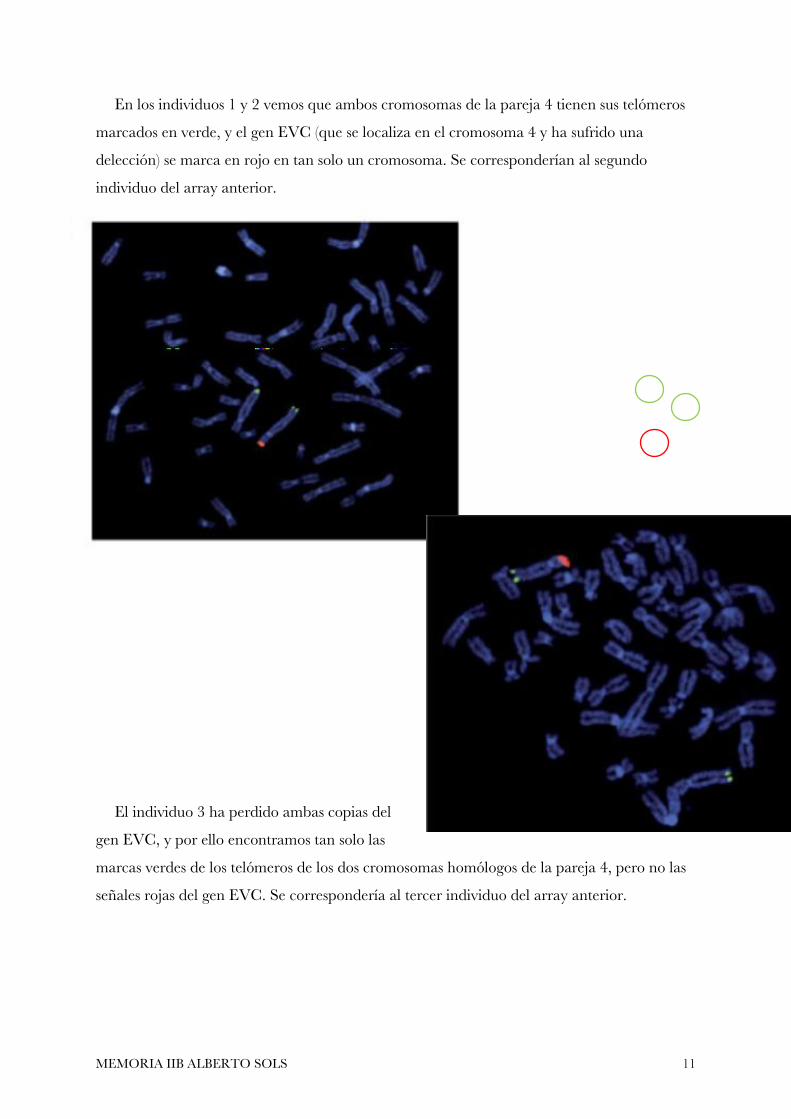
En los individuos 1 y 2 vemos que ambos cromosomas de la pareja 4 tienen sus telómeros
marcados en verde, y el gen EVC (que se localiza en el cromosoma 4 y ha sufrido una
delección) se marca en rojo en tan solo un cromosoma. Se corresponderían al segundo
individuo del array anterior.
El individuo 3 ha perdido ambas copias del
gen EVC, y por ello encontramos tan solo las
marcas verdes de los telómeros de los dos cromosomas homólogos de la pareja 4, pero no las
señales rojas del gen EVC. Se correspondería al tercer individuo del array anterior.
MEMORIA IIB ALBERTO SOLS 12
Secuenciación masiva o NGS (Next Generation Sequency)
A diferencia de la secuenciación Sanger, que tan solo
permite secuenciar una secuencia de bases corta, la NGS
permite secuenciar un conjunto de exones es concreto, el
exoma o el genoma completo. Este proceso consta de 4
pasos:
A. Preparación de los fragmentos de ADN
que queremos secuenciar.
B. Fijación de los fragmentos al clúster por
medio de adaptadores (uno a cada extremo) y
posterior replicación de los fragmentos.
C. Secuenciación de los fragmentos gracias a
nucleótidos marcados con fluorocromos
D. Alineación de los miles de fragmentos secuencia
dos y análisis de las secuencias obtenidas.

MEMORIA IIB ALBERTO SOLS 13
Esta técnica permite secuenciar de una forma relativamente barata (1000€ ) el genoma de un paciente, para
así poder detectar el origen de su patología. En el caso de la osteogénesis imperfecta, son 17 los genes en los que
suele residir la mutación, por lo que mediante la secuenciación de esos exones concretos podemos conocer su
origen.
Instituto de Genética Médica y Molecular (INGEMM)
El INGEMM está situado en el Hospital de La Paz. En él, entre otros, encontramos un
servicio de secuenciación Sanger, arrays y secuenciación masiva, que se emplea para detectar
el origen de patologías con origen genético
Secuenciación Sanger
Se llevan a cabo en estos aparatos llamados secuenciadores

MEMORIA IIB ALBERTO SOLS 14
La muestra se coloca en una placa de 96 pocillos, y se va haciendo correr por los capilares
para realizar la secuenciación
Arrays
En un portaobjetos con pocillos se colocan las muestras de ADN
Y posteriormente se cubre con otro portaobjetos que tiene los oligonucleótidos fijados.
Tras fijarse el ADN en el segundo portaobjetos, se lee el resultado en esta máquina
Obteniendo en el ordenador algo similar a lo mostrado en el apartado teórico
Capilares
P96

MEMORIA IIB ALBERTO SOLS 15
Secuenciación masiva
Esta máquina es la encargada de
realizar la NGS.
Las muestras se preparan de forma automatizada gracias a este robot

MEMORIA IIB ALBERTO SOLS 16
Y posteriormente se introducen en esta placa, llamada flow cell.
Miércoles
Realizamos una PCR con electroforesis y visitamos el citómetro de flujo.
Relación entre metabolismo del colesterol y la respuesta inflamatoria en la
proteína LXR
El LXR es una proteína que posee dos isotipos: alfa y beta. Esta proteína actúa como un receptor nuclear,
ya que se encuentra disuelta en el núcleo. Este receptor se activa con los oxisteroles. Al activarse cumple dos
funciones. Por un lado se activan genes que se encargan de metabolizar y degradar el colesterol. Por otro lado
promueve la transcripción de genes implicados en la regulación de la inflamación.
Esta función (la de reducir la inflamación) es llevada a cabo el LXR en los macrófagos, de ahí que estas
células sean el objeto de este experimento.
En el experimento se emplean dos clases de ratones:
- DKO (doble ko): carece de ambas copias de los alelos que codifican para los isotipos alfa y beta.
- KO para los alelos que codifican el LXR beta y carecen de uno de los alelos que codifica para el LXR alfa,
que está sustituido por dos genes. Uno que codifica para una proteína rojo fluorescente (DSRed), y el otro
para sintetizar el receptor para la toxina de la difteria humana (DTR).
El KO se logra introduciendo un virus que codifica para una enzima de restricción que corta el gen deseado.
Citometería de flujo
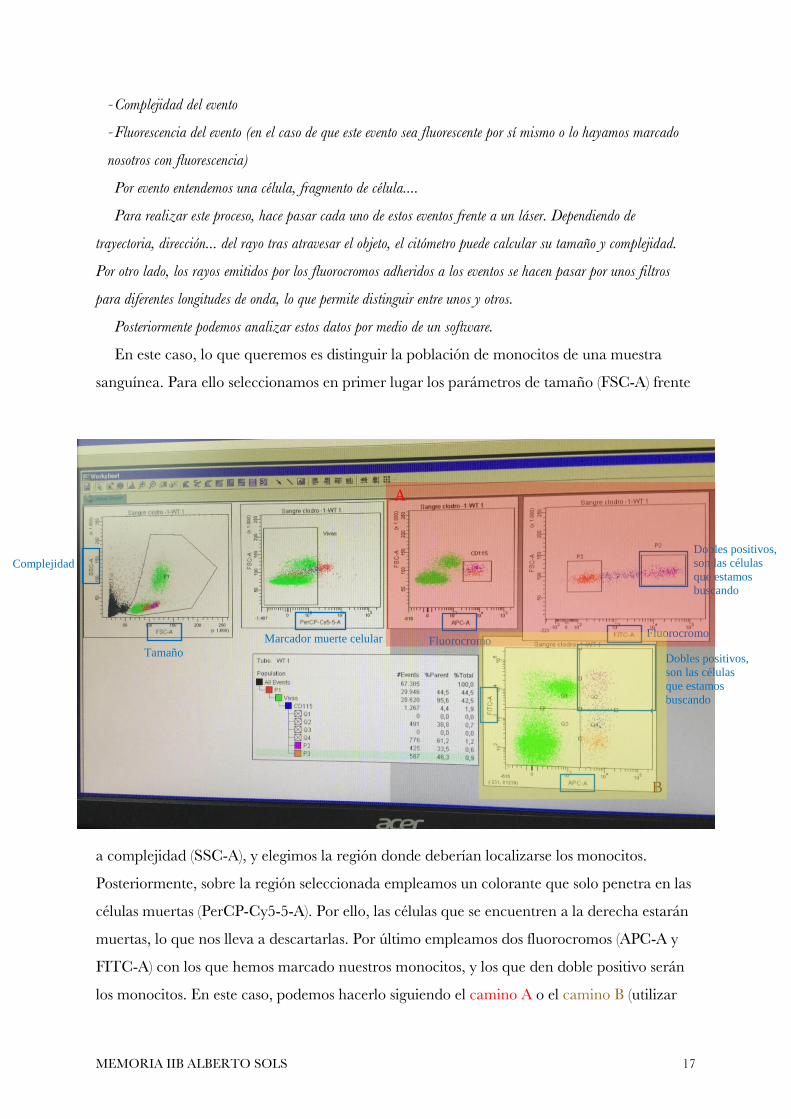
El citómetro de flujo es un aparato que permite medir de forma cuantitativa 3 parámetros:
- Tamaño del evento
MEMORIA IIB ALBERTO SOLS 17
- Complejidad del evento
- Fluorescencia del evento (en el caso de que este evento sea fluorescente por sí mismo o lo hayamos marcado
nosotros con fluorescencia)
Por evento entendemos una célula, fragmento de célula....
Para realizar este proceso, hace pasar cada uno de estos eventos frente a un láser. Dependiendo de
trayectoria, dirección... del rayo tras atravesar el objeto, el citómetro puede calcular su tamaño y complejidad.
Por otro lado, los rayos emitidos por los fluorocromos adheridos a los eventos se hacen pasar por unos filtros
para diferentes longitudes de onda, lo que permite distinguir entre unos y otros.
Posteriormente podemos analizar estos datos por medio de un software.
En este caso, lo que queremos es distinguir la población de monocitos de una muestra
sanguínea. Para ello seleccionamos en primer lugar los parámetros de tamaño (FSC-A) frente
a complejidad (SSC-A), y elegimos la región donde deberían localizarse los monocitos.
Posteriormente, sobre la región seleccionada empleamos un colorante que solo penetra en las
células muertas (PerCP-Cy5-5-A). Por ello, las células que se encuentren a la derecha estarán
muertas, lo que nos lleva a descartarlas. Por último empleamos dos fluorocromos (APC-A y
FITC-A) con los que hemos marcado nuestros monocitos, y los que den doble positivo serán
los monocitos. En este caso, podemos hacerlo siguiendo el camino A o el camino B (utilizar
Complejidad
Tamaño
Marcador muerte celular Fluorocromo Fluorocromo
Dobles positivos,
son las células
que estamos
buscando
Dobles positivos,
son las células
que estamos
buscando
A
B

MEMORIA IIB ALBERTO SOLS 18
los dos es redundante, simplemente estamos colocando los parámetros en los ejes de forma
distinta). Los nombres entre paréntesis (PerCP-Cy5-5-A, APC-A, …) son el nombre del canal
en el que se encuentran esos parámetros, no el nombre de los colorantes.
PCR (Reacción en cadena de la polimerasa)
La PCR es una técnica que nos permite amplificar un fragmento de DNA. Para ello, se introduce una
mezcla que, además de contener el fragmento a amplificar, contiene también DNTPs, ADN polimerasas,
factores de duplicación y dos clases de cebadores (cada uno de
ellos es complementario a uno de los extremos del fragmento a
amplificar).
Esta técnica se lleva a cabo en un aparato llamado
termociclador, que repite de forma cíclica 3 pasos:
- Desnaturalización: se separan las hebras del ADN
- Cebado: se une el primer a la hebra monocatenaria de ADN
- Elongación: la DNApolimerasa sintetiza la cadena de ADN
La repetición de estos tres pasos permite duplicar de forma
exponencial el ADN.
En este caso, se amplifica ADN de un trozo de oreja
de ratón para identificar su genotipo.
Electroforesis
La electroforesis es una técnica que se emplea a continuación de la PCR y que permite cuantificar los
resultados. En primer lugar se fabrica el gel de agarosa en el que se van a correr las muestras. Para ello se
mezcla agarosa, buffer y RedSafe (agente que favorece la desnaturalización del ADN) y se deja solidificar.
Eppendorf de 200 µl, especial para el termociclador

MEMORIA IIB ALBERTO SOLS 19
Posteriormente, se carga en el gel con una micropipeta el ADN amplificado, y se conecta a la corriente. La carga
negativa del ADN hace que se desplace hacia el polo positivo, separándose los fragmentos de ADN de acuerdo a
su tamaño y complejidad.
Y al finalizarla, se ve el gel en el transiluminador para poder
observar las bandas de ADN.
En esta ocasión se emplea la electroforesis para ver si el ADN de los ratones amplificados con
la PCR pertenecía a ratones WT o heterocigotos.

MEMORIA IIB ALBERTO SOLS 20
Jueves
Realizamos un citospyn y teñimos una muestra de macrófagos para observar mañana.
Citospyn
El citospyn es una técnica que se emplea para concentrar la muestra líquida en un punto de un portaobjetos.
Esto facilita el recuento diferencial.
Para ello en primer lugar montamos nuestro portaobjetos en el compartimento del citospyn. Lo cubrimos con un
cartón que deja tan solo un círculo del portaobjetos libre, donde se concentrará la muestra.
Por último, pipetamos en el círculo la disolución a concentrar.
Tras esto, introducimos el montaje en la máquina y lo tenemos a 600 rpm durante 4 minutos.Al sacar la
muestra, la disolución se ha secado y la tenemos ya en el portaobjetos.
Tinción azul de metileno según Giemsa
Con esta tinción teñimos los núcleos de la célula de color azul y el citoplasma de color rosáceo.

MEMORIA IIB ALBERTO SOLS 21
El protocolo es el siguiente. Tras realizar el citospyn cubrimos la muestra con azul de metileno . Tras 1
minuto añadimos la misma cantidad de agua y mezclamos soplando y agitando el colorante con el agua. 3
minutos después se lava el portaobjetos para retirar el exceso de colorante.
Como tenemos que observar la muestra a gran aumento, necesitamos emplear un objetivo de inmersión. Por
ello es necesario fijar un cubreobjetos sobre nuestra muestra. Para este proceso se emplea un pegamento
denominado DPX. Primero colocamos el DPX sobre nuestra muestra y posteriormente dejamos caer suavemente
el cubreobjetos encima. Si se forman burbujas aplastamos
cuidadosamente el cubreobjetos para
eliminarlas.
Esto nos permite ver los monocitos seleccionados mediante un sorter al
microscopio.
MEMORIA IIB ALBERTO SOLS 22
Viernes
Realizamos una herida y visitamos el servicio de microscopía del IIB.
Realización de una herida
Consiste en trazar una línea en una placa de Petri confluente (se puede trazar con
una punta de micropipeta). Las células de la placa, si tienen capacidad de movimiento, van a
moverse para rellenar la línea vacía de células que ha quedado. Tras trazarla, vamos
tomando fotos cada cierto intervalo de tiempo para poder medir la velocidad de movimiento
las células. Este estudio será evidentemente así solo cuando las células no estén en división.
La forma más fácil de realizarlo es introducir nuestra herida en el cell observer, microscopio
que será explicado posteriormente.
Microscopía
Los microscopios ópticos nos permiten observar muestras de pequeño tamaño. Dentro de
los microscopios encontramos múltiples variedades. Hoy vemos microscopios de campo claro,
de fluorescencia, el cell observer y con focal.
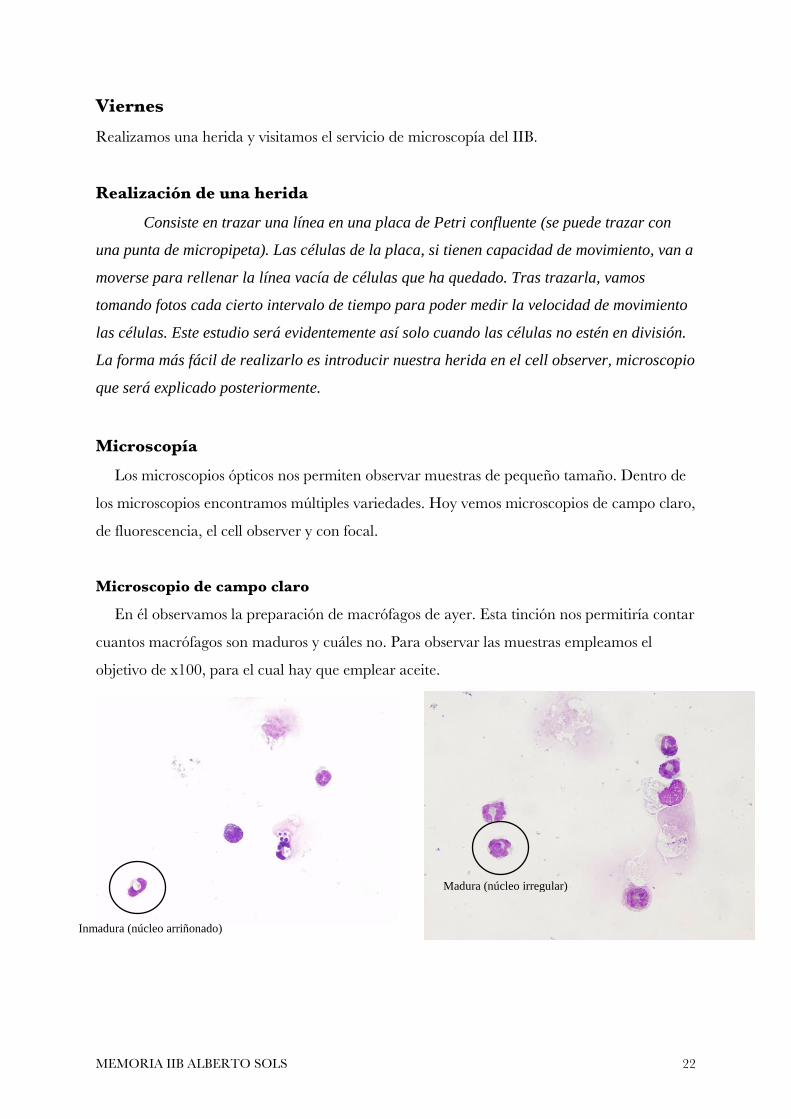
Microscopio de campo claro
En él observamos la preparación de macrófagos de ayer. Esta tinción nos permitiría contar
cuantos macrófagos son maduros y cuáles no. Para observar las muestras empleamos el
objetivo de x100, para el cual hay que emplear aceite.
Inmadura (núcleo arriñonado)
Madura (núcleo irregular)

MEMORIA IIB ALBERTO SOLS 23
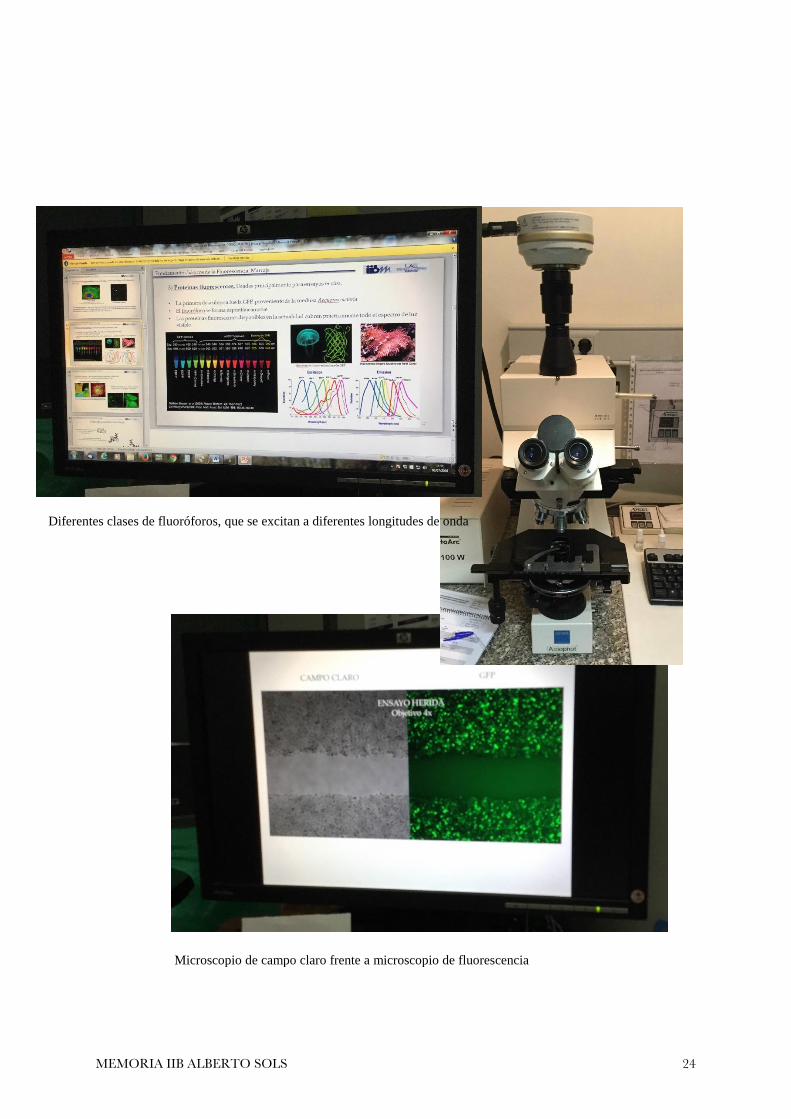
Microscopía de fluorescencia
Permite observar estructuras marcadas con fluoróforos. Su funcionamiento es el siguiente:
1. Una lámpara de mercurio, de haluros metálicos o led emiten un rayo de luz a con una longitud
determinada.
2. Un espejo dicroico es aquel que deja pasar prácticamente todas las longitudes de onda, excepto una, que
es reflejada.
3. Esta longitud reflejada incide sobre la muestra, que excita el fluoróforo.
4. El fluoróforo excitado emite energía. En la transacción energética se pierde energía, por lo que la
energía liberada es menor que la incidida. Siguiendo la ley de Plank
5. La frecuencia será mayor y por tanto, la longitud de onda emitida será mayor que la incidida. Al ser
luz en el aire, v ≈ c
6. Al ser una mayor longitud de onda, el espejo dicroico deja pasar el rayo, que va al ocular y permite al
observador ver la fluorescencia de la muestra.
Funcionamiento de un microscopio de
fluorescencia
Longitudes de onda a la que el espejo deja pasar la luz
Longitudes de onda a la
que el espejo refleja la luz
MEMORIA IIB ALBERTO SOLS 24
Microscopio de campo claro frente a microscopio de fluorescencia
Diferentes clases de fluoróforos, que se excitan a diferentes longitudes de onda

MEMORIA IIB ALBERTO SOLS 25
Cell observer
Es un microscopio de campo claro (que además posee fluorescencia). Tiene la particularidad de que las
muestras pueden mantenerse a una temperatura y concentración de CO2 constante, lo que permite observar las
células de un cultivo mientras este prolifera. También permite tomar fotos de forma automatizada,
algo muy útil para observar, por ejemplo, como se desplazan las células en el cultivo en el que
hemos hecho anteriormente la herida.
Campana que mantiene la temperatura
Membrana que mantiene el CO2
Herida a medio regenerar
MEMORIA IIB ALBERTO SOLS 26
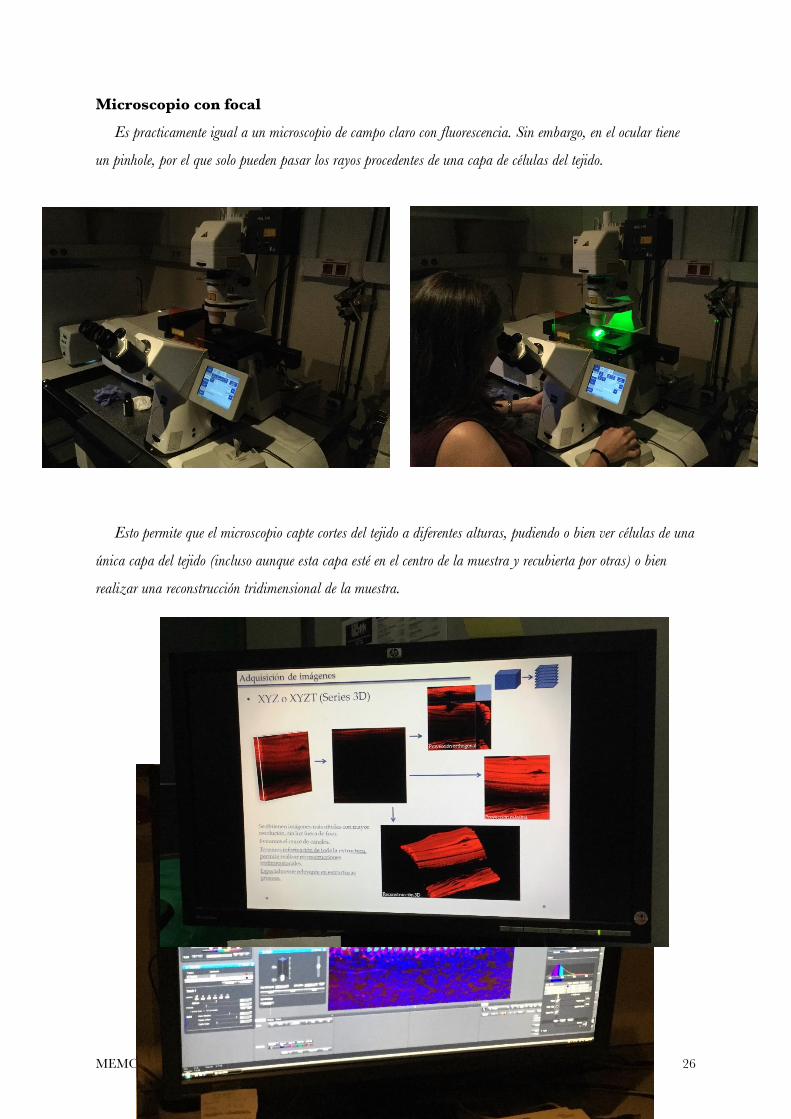
Microscopio con focal
Es practicamente igual a un microscopio de campo claro con fluorescencia. Sin embargo, en el ocular tiene
un pinhole, por el que solo pueden pasar los rayos procedentes de una capa de células del tejido.
Esto permite que el microscopio capte cortes del tejido a diferentes alturas, pudiendo o bien ver células de una
única capa del tejido (incluso aunque esta capa esté en el centro de la muestra y recubierta por otras) o bien
realizar una reconstrucción tridimensional de la muestra.
MEMORIA IIB ALBERTO SOLS 27
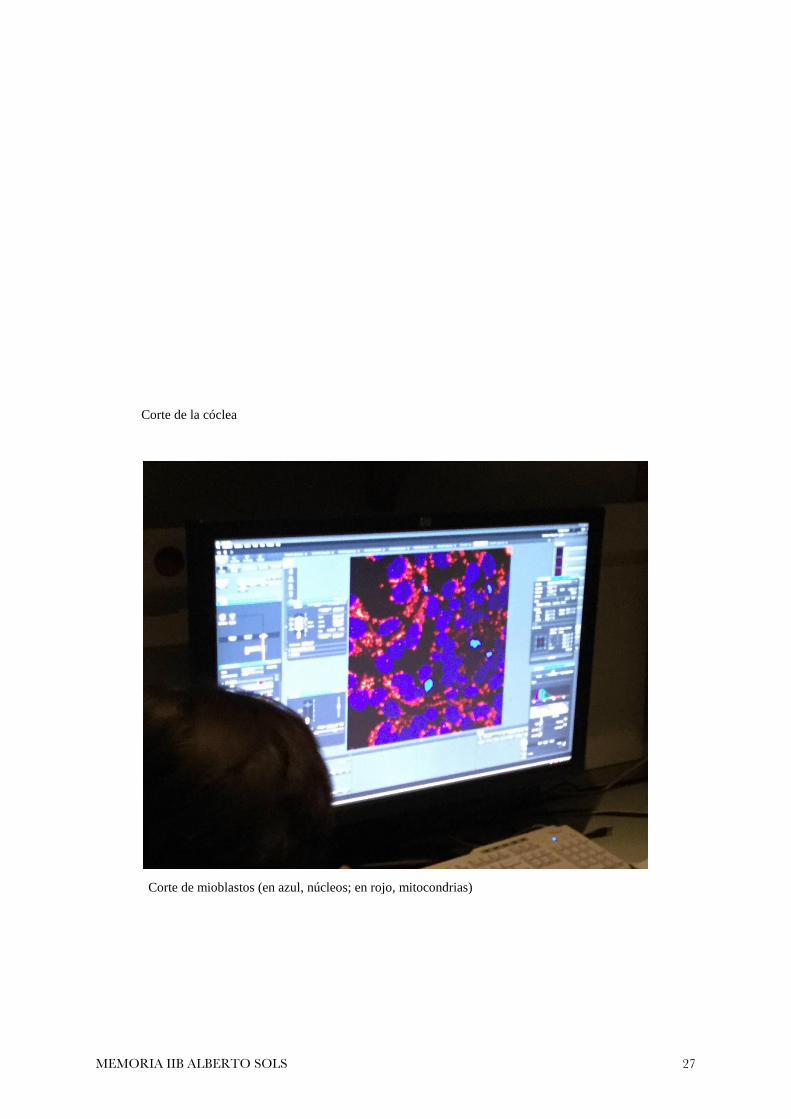
Corte de la cóclea
Corte de mioblastos (en azul, núcleos; en rojo, mitocondrias)