gucolisis - uncp - ii semestre
TRANSCRIPT

UNIVERSIDAD NACIONAL DEL CENTRO DEL PERÚ
FACULTAD DE ZOOTECNIA
GLUCOLISIS
Presentado por:Kely Shirley Quispe Pariona
Asignatura:Bioquímica
Docente:
Ing. Rafael Pantoja Huancayo, noviembre del 2013

INTRODUCCIÓN
El presente trabajo tiene por objetivo conocer el funcionamiento de nuestro organismo para lo cual uno de los mas importantes es la glucolisis que es el metabolismo anaerobio (no requiere oxígeno) de la glucosa, que tiene por función obtener energía a partir de la conversión de una molécula de glucosa en dos moléculas de piruvato todo este proceso ayudara a que nuestro cuerpo se encuentre en buen estado.
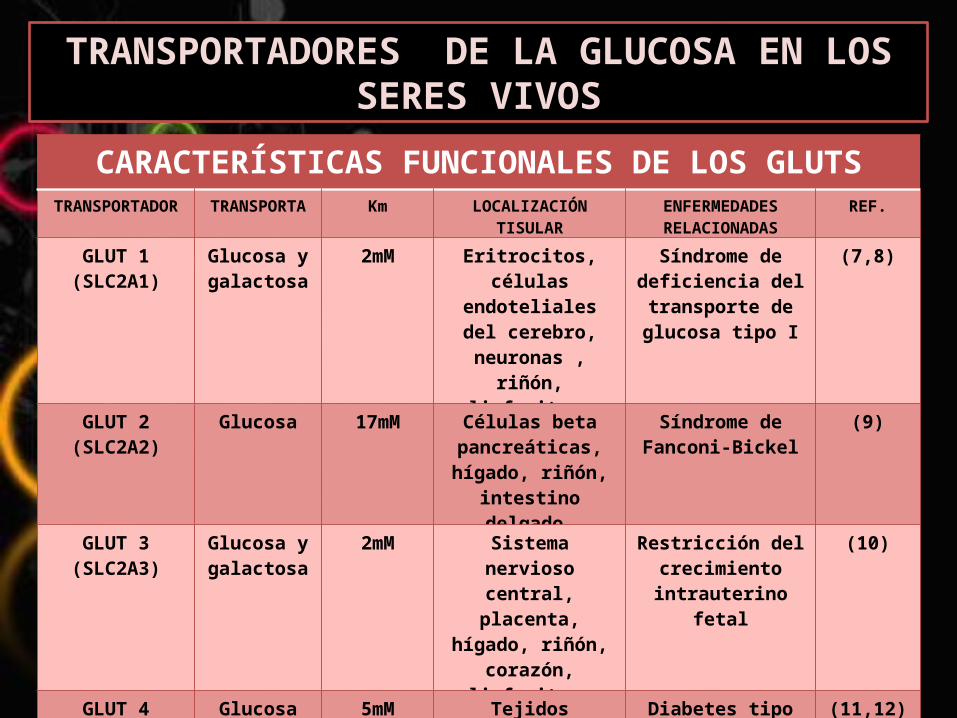
TRANSPORTADORES DE LA GLUCOSA EN LOS SERES VIVOS
CARACTERÍSTICAS FUNCIONALES DE LOS GLUTSTRANSPORTAD
ORTRANSPORT
AKm LOCALIZACIÓN
TISULARENFERMEDADES RELACIONADAS
REF.
GLUT 1(SLC2A1)
Glucosa y galactosa
2mM Eritrocitos, células
endoteliales del cerebro,
neuronas , riñón, linfocitos
Síndrome de deficiencia del transporte de glucosa tipo I
(7,8)
GLUT 2(SLC2A2)
Glucosa 17mM Células beta pancreáticas, hígado, riñón,
intestino delgado.
Síndrome de Fanconi-Bickel
(9)
GLUT 3(SLC2A3)
Glucosa y galactosa
2mM Sistema nervioso central,
placenta, hígado, riñón,
corazón, linfocitos.
Restricción del crecimiento intrauterino
fetal
(10)
GLUT 4(SLC2A4)
Glucosa 5mM Tejidos sensibles a la
insulina linfocitos.
Diabetes tipo II (11,12)
GLUT 5(SLC2A5)
Fructosa 10 Mm Intestino delgado,
testículo, riñón.
Algunas células cancerígenas, HPTG, HPINS
(14)
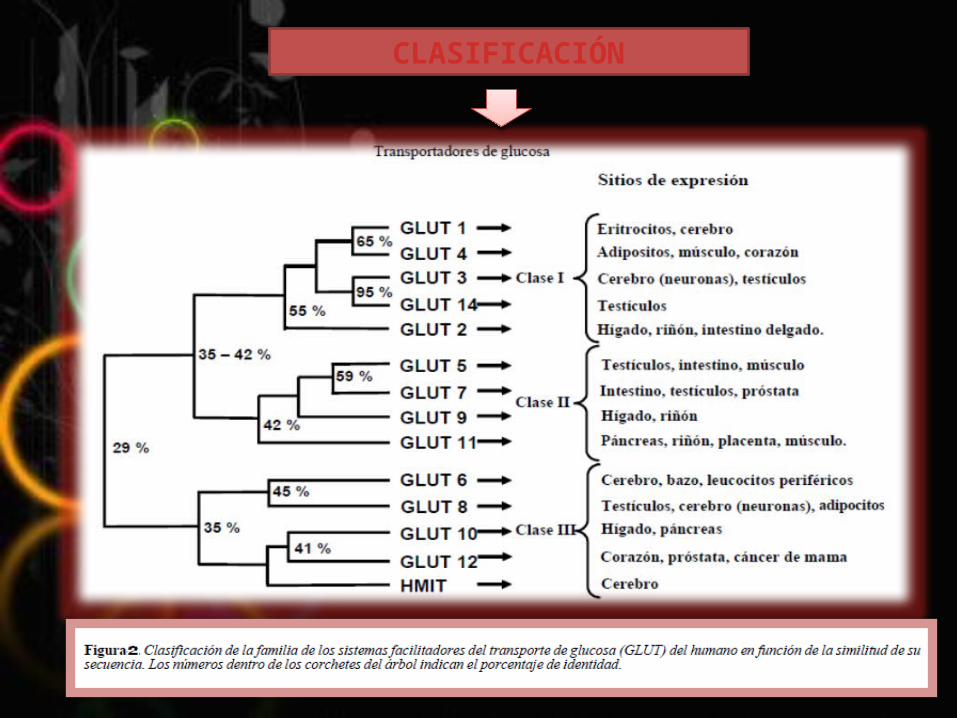
CLASIFICACIÓN
TRANSPORTADORES DE GLUCOSA GLUT 1 Y GLUT 2
TRANSPORTADORES DE GLUCOSA GLUT 1 Y GLUT 3

IMPORTANCIA DE LAS RUTAS O DESTINOS CATABÓLICOS DEL
PIRUVATO
DESTINO
A
FERMENTACIÓN HOMOLÁCTICA
En el músculo, en periodos de alta actividad en los cuales la demanda de ATP es alta y el oxígeno ha sido consumido, la lactato deshidrogenasa (LDH) cataliza la reducción del piruvato a L-lactato
Glu + 2 NAD+ + 2 Pi 2- + 2 ADP 3-
2 Pir - + 2 NADH + 2 H+
Þ
Þ
2 Pir - + 2 ATP 4- + 2 NADH + 2 H+ + 2 H2O
2 lactato - + 2 NAD+
Glu + 2 Pi 2- + 2 ADP 3-
Þ
2 lactato - + 2 H+ + 2 ATP 4- + 2 H2O
Reacción global.
Reacción global:
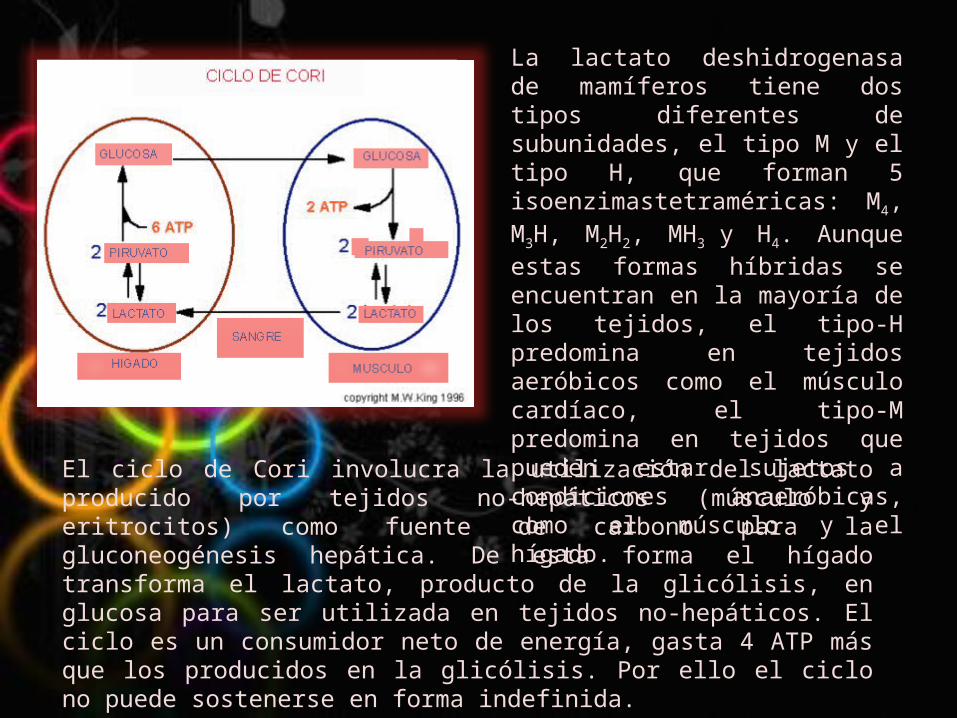
La lactato deshidrogenasa de mamíferos tiene dos tipos diferentes de subunidades, el tipo M y el tipo H, que forman 5 isoenzimastetraméricas: M4, M3H, M2H2, MH3 y H4. Aunque estas formas híbridas se encuentran en la mayoría de los tejidos, el tipo-H predomina en tejidos aeróbicos como el músculo cardíaco, el tipo-M predomina en tejidos que pueden estar sujetos a condiciones anaeróbicas, como el músculo y el hígado.
El ciclo de Cori involucra la utilización del lactato producido por tejidos no-hepáticos (músculo y eritrocitos) como fuente de carbono para la gluconeogénesis hepática. De esta forma el hígado transforma el lactato, producto de la glicólisis, en glucosa para ser utilizada en tejidos no-hepáticos. El ciclo es un consumidor neto de energía, gasta 4 ATP más que los producidos en la glicólisis. Por ello el ciclo no puede sostenerse en forma indefinida.
DESTINO
B
FERMENTACIÓN ALCOHÓLICA
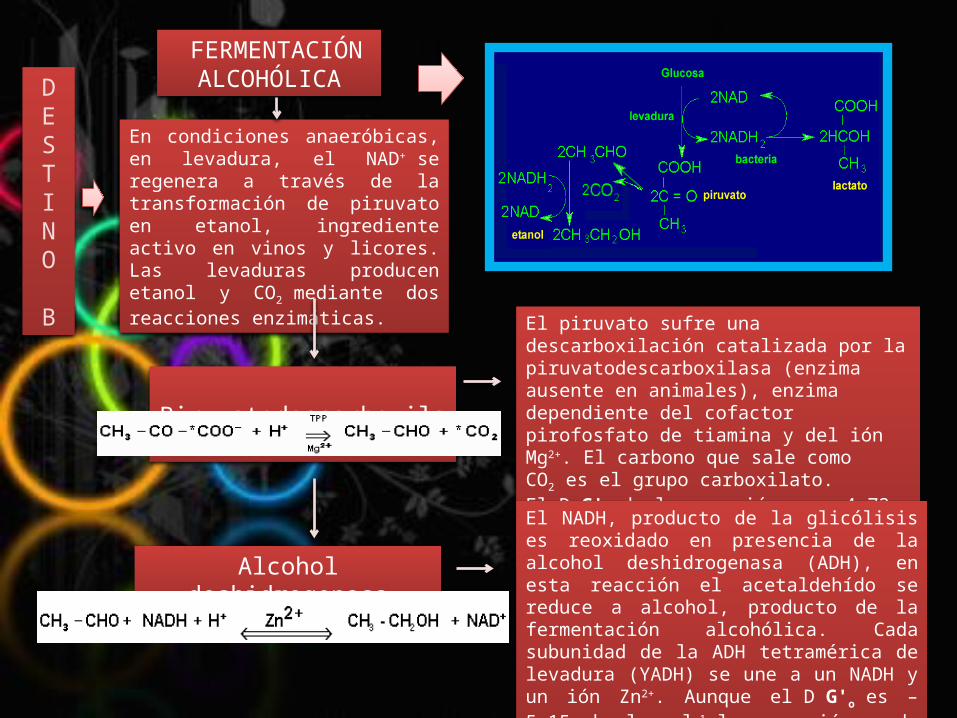
En condiciones anaeróbicas, en levadura, el NAD+ se regenera a través de la transformación de piruvato en etanol, ingrediente activo en vinos y licores. Las levaduras producen etanol y CO2 mediante dos reacciones enzimáticas.
Piruvatodescarboxilasa.
Alcohol deshidrogenasa
El piruvato sufre una descarboxilación catalizada por la piruvatodescarboxilasa (enzima ausente en animales), enzima dependiente del cofactor pirofosfato de tiamina y del ión Mg2+. El carbono que sale como CO2 es el grupo carboxilato. El D G'o de la reacción es – 4,72 kcal mol-1, es una reacción irreversible.
El NADH, producto de la glicólisis es reoxidado en presencia de la alcohol deshidrogenasa (ADH), en esta reacción el acetaldehído se reduce a alcohol, producto de la fermentación alcohólica. Cada subunidad de la ADH tetramérica de levadura (YADH) se une a un NADH y un ión Zn2+. Aunque el D G'o es – 5,15 kcal mol-1 la reacción pude realizarse en ambas direcciones.
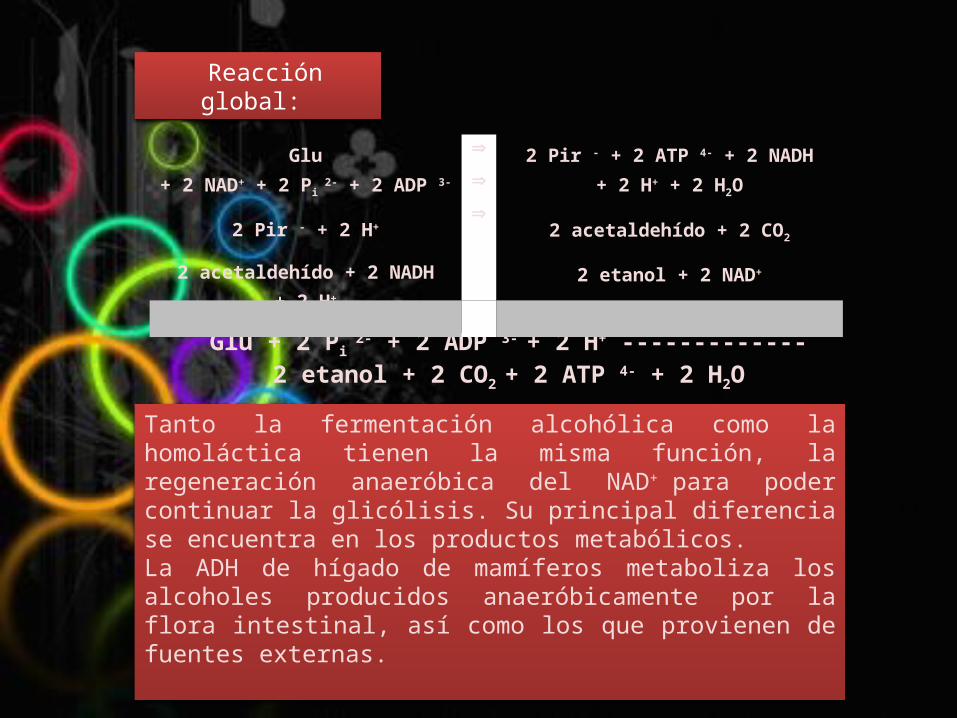
Glu + 2 Pi 2- + 2 ADP 3- + 2 H+ ------------- 2 etanol + 2 CO2
+ 2 ATP 4- + 2 H2O
Glu + 2 NAD+ + 2 Pi 2- + 2 ADP 3-
2 Pir - + 2 H+
2 acetaldehído + 2 NADH + 2 H+
Þ
Þ
Þ
2 Pir - + 2 ATP 4- + 2 NADH + 2 H+ + 2 H2O
2 acetaldehído + 2 CO2
2 etanol + 2 NAD+
Tanto la fermentación alcohólica como la homoláctica tienen la misma función, la regeneración anaeróbica del NAD+ para poder continuar la glicólisis. Su principal diferencia se encuentra en los productos metabólicos.La ADH de hígado de mamíferos metaboliza los alcoholes producidos anaeróbicamente por la flora intestinal, así como los que provienen de fuentes externas.
Reacción global:
DESTINO
C
CONDICIONES AERÓBICAS
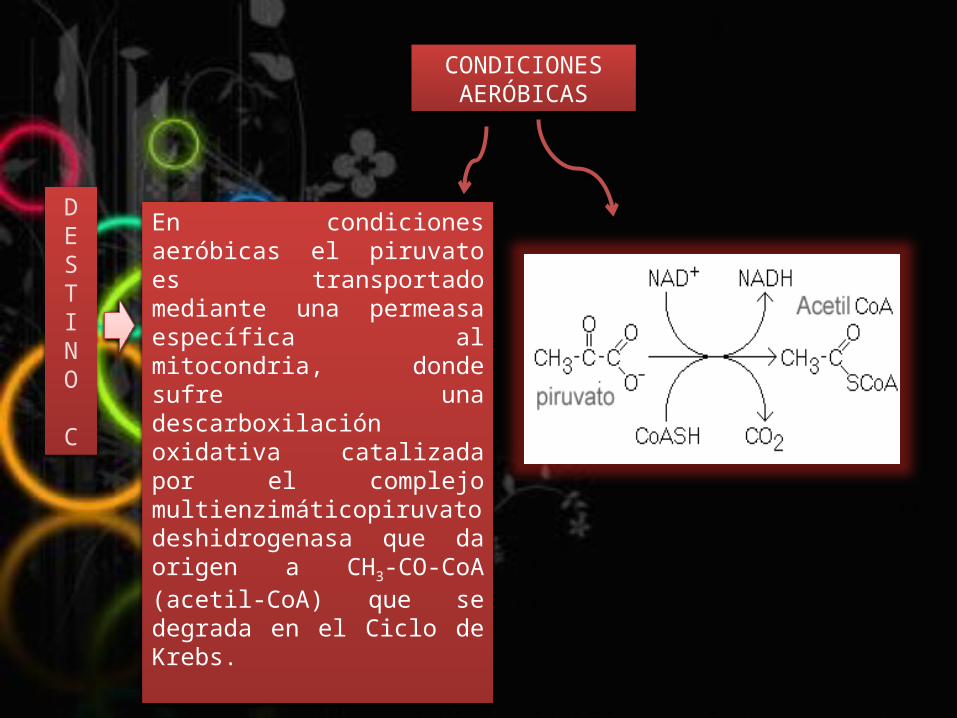
En condiciones aeróbicas el piruvato es transportado mediante una permeasa específica al mitocondria, donde sufre una descarboxilación oxidativa catalizada por el complejo multienzimáticopiruvato deshidrogenasa que da origen a CH3-CO-CoA (acetil-CoA) que se degrada en el Ciclo de Krebs.

MECANISMOS DE REGULACIÓN DE LA GLUCOLISIS
LA FOSFOFRUCTOQUINAS
A
Es el elemento de control más importante en la vía glucolítica en mamíferos. Su actividad está regulada principalmente por el nivel de energía: cuando se observan niveles altos de ATP, el enzima se inhibe alostéricamente disminuyendo la afinidad del enzima por la fructosa 6-Fosfato. La relación inhibidora del ATP se contraresta por el AMP, de manera que cuanto menor sea la relación ATP/AMP, mayor será la actividad del enzima.
También se ve afectada por el pH, esto es para evitar la acumulación excesiva de lactato y la caída brusca del pH sanguíneo provocando acidosis.
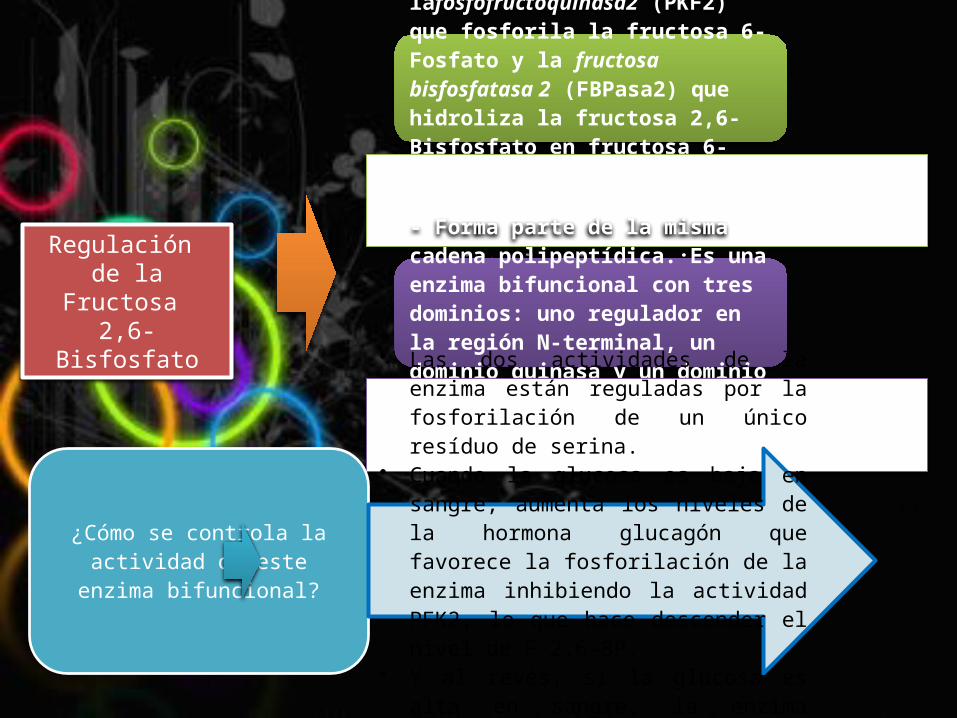
Regulación de la Fructosa 2,6-Bisfosfato
- La concentración de fructosa 2,6-Bisfosfato está regulada por dos enzimas: lafosfofructoquinasa2 (PKF2) que fosforila la fructosa 6-Fosfato y la fructosa bisfosfatasa 2 (FBPasa2) que hidroliza la fructosa 2,6-Bisfosfato en fructosa 6-Fosfato. Pero esto no es del todo correcto; ya que la PKF2 yla FBPasa2 son en realidad la misma enzima. - Forma parte de la misma cadena polipeptídica. Es una enzima bifuncional con tres dominios: uno regulador en la región N-terminal, un dominio quinasa y un dominio fosfatasa.
¿Cómo se controla la actividad de este enzima
bifuncional?
• Las dos actividades de la enzima están reguladas por la fosforilación de un único resíduo de serina.
• Cuando la glucosa es baja en sangre, aumenta los niveles de la hormona glucagón que favorece la fosforilación de la enzima inhibiendo la actividad PFK2, lo que hace descender el nivel de F-2,6-BP.
• Y al revés, si la glucosa es alta en sangre, la enzima pierde el fosfato unido activando la PFK2.

Hexoquinasa y
Glucoquinasa
- La hexoquinasa cataliza la primera etapa de la glicólisis y se inhibe con su producto: la glucosa 6-fosfato. Altas concentraciones de esta indican que la célula no precisa de más glucosa.
- En el hígado se encuentra una isozima especializada de la hexoquinasa: laglucoquinasa, la cual no se inhibe por la glucosa 6-fosfato. Pero esta isozima de lahexoquinasa sólo es activa a altas concentraciones de glucosa (tiene una afinidad 50 veces menor que la hexoquinasa).
- Su función es suministrar glucosa 6-fosfato para la síntesis de glocógeno como medio de almacenamiento de la glucosa.
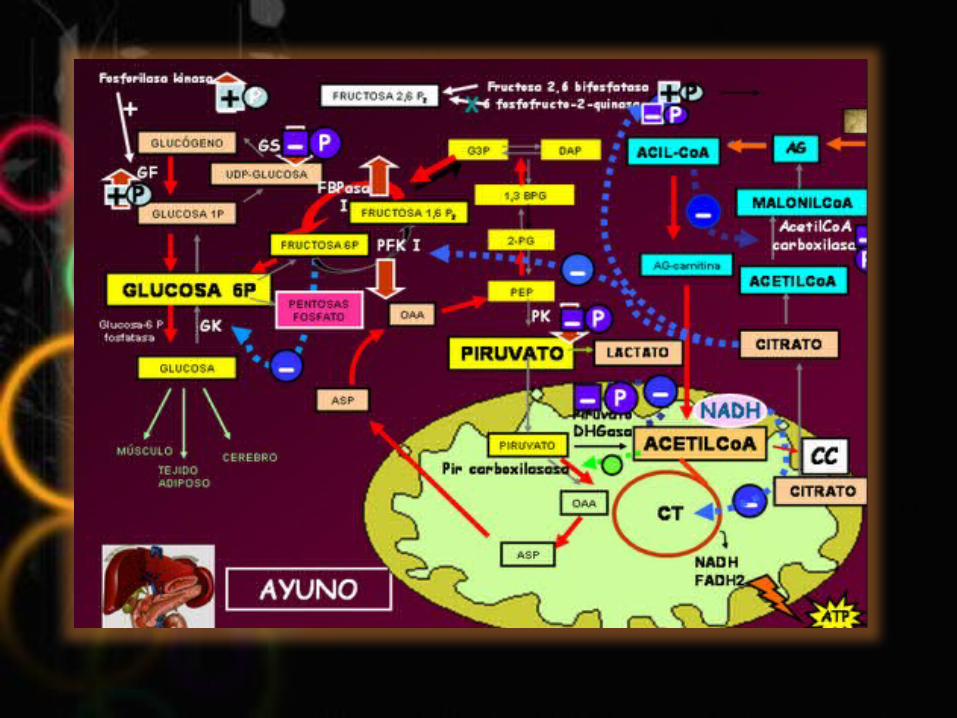
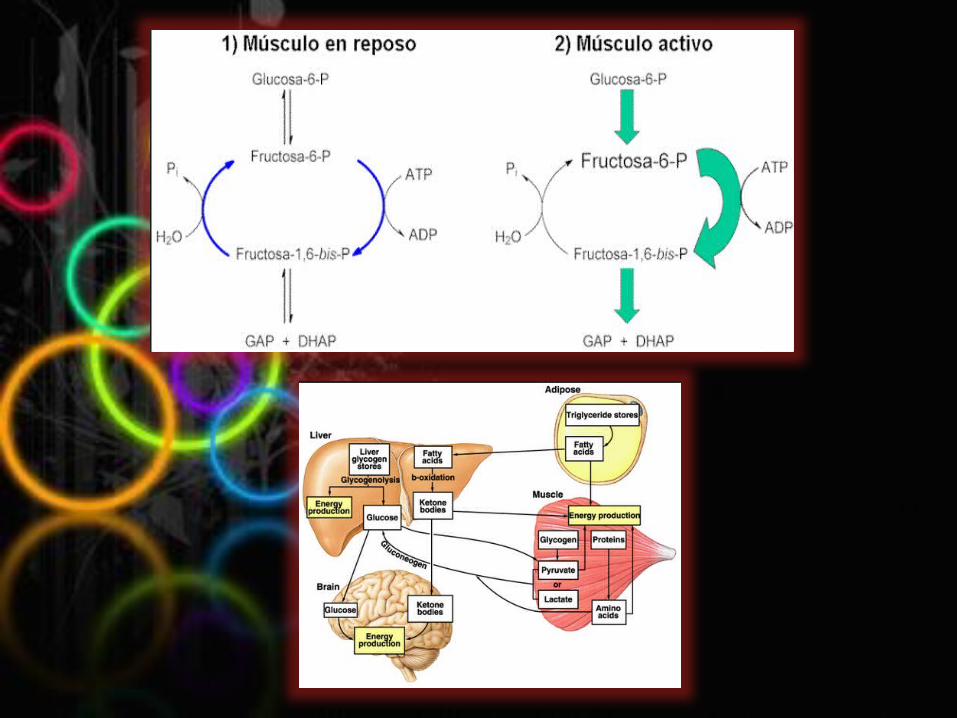

CONCLUSIONES Pudimos llegar a la conclusión que por
metabolismo entendemos :• Definición: Es el nombre que recibe
el metabolismo anaerobio (no requiere oxígeno) de la glucosa.
• Función: Obtener energía a partir de la conversión de una molécula de glucosa en dos moléculas de piruvato.
• Balance energético neto: por cada molécula de glucosa convertida en dos de piruvato se obtiene 2 ATP.
• nº de reacciones de las que consta: 10 agrupadas en 3 bloques de 3, 1 y 5 reacciones respectivamente, cada una de ellas mediada por una enzima específica.
• Regulación: La regulación de este proceso está dirigida por tres enzimas: la fosfofructoquinasa , la fructosa 2,6-Bisfosfato y la hexoquinasa las cuales manifestarán un comportamiento u otro en función de unos factores específicos como pueden ser el pH o la concentración de una determinada sustancia.