utilizaci6n de papas desarrollo transgenicas resistentes a enfermedades...
TRANSCRIPT

Desarrollo y Utilizaci6n de Papas Transgenicas Resistentes
a Enfermedades Bacterianas Taller 24 al 26 de Noviembre, 1998
Lima y San Ramon - Peru
Centro Internacional de la Papa
I lllilll llll l il/1 111111111111111111111 111111111111 •I60029CJ,
CIP G4 C45.1
Deutsche Gesellschaft fur Technische Zusammenarbeit (GTZ)
Centro Internacional de la Papa

Citacion correcta de las Memorias:
Centro Internacional de la Papa (CIP). 2000. Desarrollo y Utilizacion de Papas Transgenicas Resistentes a Enfermedades Bacterianas. 24 al 26 de noviembre, 1998. Memorias del Taller. Lima y San Ramon, Peru.
Desarrollo y Utilizaci6n de Papas Transgenicas Resistentes a Enfermedades Bacterianas
Memorias del Taller Lima y San Ramon, Peru . 1998
Coordinacion tecnica :
Procesado por
Coordinacion
Edicion
Dr. Marc Ghislain Big. Luis Nopo
Departamento de Capacitacion y Comunicaciones, CIP
Dr. Patricio Malagamba Big. Margarita Lopez
Big. Edda Echeandfa
Estas Memorias han sido financiadas con los recurses de Deutsche Gesellschaft fUr Technische Zusammenarbeit (GTZ)
CENTRO INTERNACIONAL DE LA PAPA {CIP) Apartado 1558 - Lima, Peru
Tel.: (51-1) 349-5783; 349-6017. Fax: (51-1) 349-5638. Correo-E: [email protected]
Participl
l . Lorentz Bulow 2. Fausto Buitron 3. Jorge Benavides 4. Daniel Chirito 5. Cesar Giron 6. Ricardo Gutierrez 7. Patricio Arce-Johnson 8. Eduardo Perez 9. Jorge Tenorio l 0. Marc Ghislain l l . Cesar Aguilar

CtP ~'t C45.A TABLA DE CONTENIDO
Centro lnt~rMr.ional de la Papa NOTA DEL EDITOR ••.•....• ··········I:a··MO"~mei··lima·········· ............ 3
OBJETIVOS •••••••••••••••••••• ·········1··5··'!1f.AY1.l""?UOO""""""""" ..•.••••.••• 5
PROGRAMA ••••••••..••.•••••••••••••••.•••.•..••••••••••••••.•.••••••••••••••••.•••• 7
PREAMBULO BIBLIOTECA /\4ericleth Bonierhale •••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••• 9
RESISTENCIA A ENFERMEDADES DE PAPA MEDIADA POR PROTEiNAS ANTIMICROBIANAS Marc Ghislain •••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••• 11 /,
EVALUACION DE TUBERCULOS DE PAPA TRANSGENICA A LA PUDRICION CAUSADA POR Erwinia carotovora SUBSP. carotovora ,~vlvie Priou ••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••• 21
MEJORAMIENTO PARA OBTENER RESISTENCIA A MARCHITEZ BACTERIANA ,~1·/vie Priou ••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••• 25
DESARROLLO DE ESTRATEGIAS DE INGENIERIA GENETICA PARA LA RESISTENCIA A BACTERIAS FITOPATOGENICAS Petra Porsch •••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••• 39
EXPERIMENTOS CON PAPAS TRANSGENICAS EN INVERNADERO Y CAMPO EN EL PERU
I
/
/
Ali Gohnirzaie ••••••••..••..•••.•••••••••••••••••••••••••••••.•.•.••.•••.•.••••..••••••• 45 /

TRANSFORMACION DE PLANTAS DE PAPA CON GENES DE LISOZIMA, A'fACI~ ~fCECROPINA PARA CONFERIR RESISTENCIA. A:SAc:;T-=~IA;S· Patricio Arce-Johnson ................................................................ 53 /
HIBRIDIZACION ENTRE Y DENTRO DE VARIEDADES CULTIVADAS Y SILVESTRES DE PAPA Bodo Trognitz ............................................................................ 57
INVESTIGACION EN BIOSEGURIDAD EN ALEMANIA: PAPAS ._ •1
CON LISOZIMA T 4 Y SUS IMPLICACIONES EN LA COMUNIDAD MICROBIANA COMO UN CASO DE ESTUDIO Andreas Mahn .......................................................................... 63 I
ROL DEL SENASA EN LA SANIDAD VEGETAL DEL PERU Enrique Carranza ...................................................................... 69
't· -·
'c6t.1o LIBERAR PLANTAS TRANSGENICAS RESISTENTES A PLAGAS EN CAMPO Jorge Benavides •••.•••••••••••••.••••.•.•.•••.•••.••...•••••••••••••.••••.••••..•.••.•. 73
EXPERIMENTO EN CAMPO: PAPAS RESISTENTES AL ATAQUE DE BACTERIAS Luis Nopo ................................................................................ 77
EXPERIMENTO DE CAMPO: MEDIDAS DE BIOSEGURIDAD EN EL CENTRO INTERNACIONAL DE LA PAPA Jorge Tenorio ..•.•.••.•.•..•••.•.•..•••.•.•••...•..••.•••.••..•••.••...•.•.•..••....••.. 83
RECOMENDACIONES Y CONCLUSIONES .............................. 89
LISTA DE PARTICIPANTES ............................................... 91

NOTA DEL EDITOR ,·\i·;•.:" ·
A note 10 lhe fe'?ider ~· ·
Desde el inicio de la agricultura, el hombre ha estado preocupado por mejorar las cultivos con la finalidad de obtener variedades mas productivas y resistentes a las enfermedades que las atacan. Con el desarrollo de las tecnicas de transformacion de plantas en el afio 1983, el mejoramiento de las cultivos de importancia agrfcola se via impulsado. Se ofreda una alternativa rapida para introducir genes foraneos en plantas, versus la metodologfa clasica.
Actualmente, el mejoramiento genetico de plantas se hace utilizando la biotecnologfa no solo con fines agrfcolas y nutricionales, sino tambien para obtener plantas de mejores caracterfsticas morfol6gicas (ornamentales) y/o coma fabrica de productos de inte.res industrial, farmaceutico, qufmico, agro-alimenticio, entre otros.
Durante mas de 28 anos, desde su fundaci6n, el Centro Internacional de la Papa (CIP) ha realizado avances importantes en el mejoramiento genetico, no solo de la planta de papa, sino tambien de otros cultivos andinos coma el camote y recientemente en las cultivos denominados "rafces y tuberosas andinas". Con este fin, constantemente se incorporan a la metodologfa empleada las tecnicas que han ido apareciendo dentro de esta actividad cientffica.
En 1989 el CIP empezo a elaborar las primeros lineamientos para el desarrollo de la introduccion de genes foraneos en el genoma de la papa, con la finalidad de conferir resistencia a pat6genos coma Erwinia sp. y Ralstonia sp., bacterias que causan enfermedades en este cultivo y que incluso lo eliminan. Asf, el CIP se convirtio en el pionero para la aplicacion de esta tecnica en el Peru.
El uso de Agrobacterium tumefaciens coma mediador de la transferencia de genes permitio a las cientfficos realizar estudios en
3

los ambientes de laboratorio, bajo condiciones controladas, para comprobar al utilicjad ~de este sistema ante las distintas variaciones climaticas y de suelo~· .. :~.
Agrobacterium tumefaciens result6 ser una herramienta tan util, que pronto empezaron los trabajos de mejoramiento con el fin de disminuir la incidencia de enfermedades de gran importancia a nivel mundial como por ejemplo la "podredumbre blanda" y la "pierna negra".
En la actualidad estos trabajos se siguen realizando a traves de programas especiales respaldados por una experiencia cientffica de cooperaci6n internacional. En estos programas se vigila el cumplimiento de las normas de bioseguridad aprobadas en el ano 1994 por el gobierno peruano para llevar a cabo estudios de esta naturaleza sin causar cambios en el lugar en el que se realicen (campos de cultivo rurales, ambiente de los diferentes nichos ecol6gicos ).
Gracias al apoyo del Centro Federal para la Investigaci6n del Mejoramiento en Plantas Cultivadas (Bundesanstalt fi.ir ZOchtungsforschung an Kulturpflanzen, BMZ, Alemania), el CIP ha crefdo conveniente la organizaci6n del taller "Desarrollo y utilizaci6n de papas transgenicas: Resistencia a enfermedades bacterianas", con la finalidad de exponer las bases del mejoramiento biotecnol6gico a las instituciones educativas y gubernamentales locales, para lo cual se ha irivitado a investigadores entendidos en el area, tanto del CIP como de institutes colaboradores en el extranjero.
4
Edda M. Echeandfa Chiappe Editora

• f ' • ~
OBJETIVOS: Objectives
Objetivos Generales
Revision de temas estrategicos relacionados al desarrollo y uso de papas transgenicas en el control de enfermedades.
Establecer recomendaciones para el empleo apropiado de las variedades transgenicas de papa.
Objetivos Especificos
Revision de experiencias relacionadas con la introduccion de papas transgenicas tolerantes a enfermedades bacterianas y proponer estrategias de manejo para maximizar las beneficios y minimizar las riesgos.
Preguntas espedficas a responder:
•!• lCuales son las beneficios de la papas mejoradas par ingenJerfa genetica con tolerancia a enfermedades bacterianas?
•!• lCual es la probabilidad de una transferencia horizontal de genes desde las papas transgenicas hacia las microorganismos en el suelo?
•!• lCuales son las riesgos de la hibridizacion entre las papas transgenicas con tolerancia a enfermedades bacterianas y papas no transformadas con especies silvestres?
•!• lEs importante realmente el origen? lSon acaso los genes de defensa y resistencia "natural" mejores que los genes quimericos o sinteticos?
5

•!• lCuales son las medidas de seguridad para manejar apropiadamente plantas transgenicas con tolerancia a enfermedades bacterianas en el Peru?
•!• lQue otros experimentos y nuevas informaciones deben desarrollarse?

PROGRAMA Program
Martes 24 de noviembre Lima
08:00 08:30 09:00
09:30 09:45 10:30 11:15
12:00 13:30 14:00
14:45 15:00
15:45
16:15
Asuntos administrativos Inauguracion del taller Resistencia a enfermedades de la papa mediante protefnas antimicrobianas Cafe Marchitez bacteriana y su control Mejoramiento para resistencia a enfermedades Desarrollo de estrategias de ingenierfa genetica para la resistencia a bacterias fitopatogenicas Almuerzo Procedimientos de bioseguridad en el CIP Experimentos con papas transgenicas en invernaderos y campo en Peru Cafe Hibridizacion entre y dentro de papas cultivadas y especies silvestres Transformacion de plantas de papa con genes de lisozima, atacina y cecropina Discusion general
Miercoles 25 de noviembre San Ramon
07:00 12:00
13:00
Viaje a San Ramon Visita al campo seguida por una discusion sabre la adecuacion de los procedimientos de bioseguridad Almuerzo
N. Espinoza M. Bonierbale M. Ghislain
S. Priou S. Priou P. Persch L. Bulow
M. Ghislain A. Golmirzaie
B. Trognitz
P. Arce
M. Ghislain
P. Aley

14:00 Investigaci6n sabre bioseguridad en Alemania: Papas con el gen T4 de la lisozima y su implicancia en la comunidad microbiana coma un caso de estudio
15:00 Protocolos de bioseguridad en el Peru SENASA 15:45 Cafe 16:00 Experimento de campo: Papas resistentes a
insectos 16:30 Experimento de campo: Papas resistentes a
bacterias 17:00 Experimento de campo: Medidas de bioseguridad
Jueves 26 de noviembre Lima
08:00 11:00
12:00 12:30
Regreso a Lima Desarrollo de recomendaciones sabre bioseg u rid ad Clausura del taller Almuerzo
,":. ':; .·.
A. Mahn
E. Carranza
J. Benavides
L. Nopo
J. Tenorio
M. Ghislain

PREAMBULO Foreword
Merideth Bonierbale *
La Biotecnologfa y la Bioseguridad nos ofrecen una serie de avances que nos ayudan en la actualidad a extender las fronteras del mejoramiento genetico y de la Biologfa en general.
Par un lado, utilizando las herramientas de la Biotecnologfa, ahora es posible enriquecer las acervos geneticos de nuestros cultivos con informaci6n de variedades nativas y para fines espedficos, coma par ejem plo:
•!• La defensa y/o resistencia de plantas ante estreses abi6ticos y bi6ticos
•!• Aumentar la nutrici6n y/o calidad de los alimentos •!• Mejorar la productividad
Y por otro lado, el concepto de Bioseguridad en el manejo de las plantas transgenicas ha sido siempre una preocupaci6n de los biotecn61ogos conscientes del equilibria ecol6gico de nuestro media. Casi en forma simultanea, muchas autoridades a nivel mundial han tornado interes en estas actividades cientfficas. Ahora el Peru empieza a formar parte del grupo de pafses que desarrollan nuevas tecnologfas en una manera responsable.
En la practica, la Bioseguridad aplicada a los transgenes comprende la evaluaci6n de los posibles riesgos inherentes a la experimentaci6n en laboratorio, invernadero o campo, utilizando un conjunto de medidas o procedimientos especfficos para cada caso.
Actualmente, existe literatura abundante que trata sabre las aspectos concernientes a la experimentaci6n con plantas transgenicas en el ambiente. Varios talleres han sido llevados a cabo a nivel
* Centro Internacional de la Papa (CIP). Estacion Central, La Molina. Lima - Peru 9

mundial, inclusive en America del Sur (Iguazu, Argentina), donde se analizaron los posibles riesgos de la liberacion de papas transgenicas en la zona andina (Riesgos ambientales de las plantas transgenicas en centros de diversidad: la papa como modelo, Parque Nacional Iguazu, Argentina, junio de 1995). En el caso espedfico de Peru dependiendo del tipo de mejora que se realiza, por ejemplo, la introduccion de una planta transgenica de papa con un gen de resistencia a bacterias implica distintos riesgos y por ende distintas medidas de seguridad que la introduccion de una planta transgenica de soya con un gen de resistencia a herbicidas, puesto que dicho cultivo es originario de esta region andina.
Otro concepto que ha evolucionado paralelamente con el de Bioseguridad es la Bioetica. Varios pafses de la Union Europea ya han desarrollado comites consultivos sobre Bioetica, lo que refleja una madurez en la opinion publica de la biotecnologfa, asf como en las clases polfticas encargadas de la regulacion de los avances cientfficos.
La Bioseguridad, tal como se discutira en el presente taller, esta relacionada con los riesgos y precauciones que deberan considerarse para el manejo responsable de plantas transgenicas de papa con genes de resistencia a enfermedades bacterianas en el Peru. Como un ejemplo de un caso espedfico.
•!• lQue clases de riesgos existen? •!• lCual es la probabilidad que dichos riesgos se lleven a cabo? •!• lCuales de estos riesgos son economicamente y/o ecologicamente
inaceptables? •!• LQue medidas deberan tomarse en cuenta? •!• El SENASA y el CIP han elaborado reglamentos. lSon adecuados? •!• lQue fallas podrfan detectarse en nuestros procedimientos
actuales?
Estos aspectos seran revisados durante este taUer, y esperamos llegar al final a recomendaciones y conclusiones que puedan ser utiles para el desarrollo y establecimiento de procedimientos ajustados a nuestra realidad, estructurados con una solida base cientffica. De esta manera, esperamos contribuir con las autoridades gubernamentales encargadas de la regulacion de la investigacion con plantas tra nsgen icas.
10

RESISTENCIA A ENFERMEDADES DE PAPA MEDIADA POR PROTEiNAS
ANTIMICROBIANAS Potato Disease Resistance Mediated by
Anti-microbial proteins
Marc Ghislain *
INTRODUCCION
El presente taller forma parte del programa de un proyecto realizado en colaboracion con el "Federal Center for Breeding Research on Cultivated Plants" en Quedlinburg, Alemania.
Muches son los factores que afectan la produccion de papa, entre estos se encuentran las ~ y_ enfermedades.
Se estima que hay una perdida de 21 % de la produccion total de papa que esta relacionada a enfermedades propias del cultivo. Para disminuir esta perdida, es necesario desarrollar resistencia a plagas y enfermedades en papa para garantizar la seguridad alimentaria de la poblacion, disminuir los costos de produccion, mejorar la seguridad y salud de los productores y su familia, disminuir la contaminacion ambiental y bajar la concentracion de residues qufmicos en la alimentacion, para beneficio del consumidor.
A continuacion presentamos un resumen de las actividades de este proyecto relacionadas al taller y que se estan llevando a cabo con el fin de contribuir en la obtencion de mejores variedades de cultivos de importancia.
Comencemos mencionando los objetivos del proyecto:
EL OBJETIVO PRINCIPAL es la manipulacion de la expresion de genes que codifican para protef nas de accion antimicrobiana con la finalidad de aumentar la resistencia a la marchitez bacteriana y al tizon tardfo en papa.
* Centro Internacional de la Papa (CIP). Estacion Central, La Molina. Lima - Peru 11

Para cumplir con este objetivo, se estan considerando estas dos enfermedades dentro del proyecto: Ambas son las enfermedades de mayor importancia en el cultivo de la papa en los pafses en vfas de desarrollo. Por esta raz6n, el CIP, esta dedicando sus mayores esfuerzos a la investigaci6n en este campo, a pesar de las dificultades para mejorar los niveles de resistencia a estas enfermedades.
Varios fueron /os OBJETIVOS ESPECIFICOS definidos para este proyecto:
•!• Estrategias utilizadas para combatir al Tizon Tardio
Aislamiento del gen codificante, en variedades de papas nativas, de la protefna osmotina, conocida como factor inhibidor de crecimiento del hongo responsable del Tizon Tardio. Una vez obtenido el gen, se desarrolla una construccion genica que codifique para la expresi6n constitutiva, inducida y con secreci6n a nivel vacuolar de la protefna osmotina. Ademas, come una alternativa, se esta trabajando en el desarrollo de construcciones di- y tri- genicas ( que incluirfan osmotina y/o lisozima y/o glucanasa) para obtener un efecto sinergico durante el proceso de defensa al Tizon Tardio. Estes genes quimericos pueden ser transferidos a variedades de papa susceptibles coma: Achirana INTA, Amarilis y Tomasa Condemayta. Finalmente, se realizan ensayos de campo para determinar el nivel de resistencia alcanzado.
•!• Estrategias utilizadas para combatir la Marchitez Bacteriana
En el caso de la Marchitez Bacteriana, se busca la transferencia del gen que codifica para la lisozima del bacteriofago T4 a variedades susceptibles de papa como: Achirana INTA, Amarilis y Tomasa Condemayta. Primera, se desarrollan construcciones genicas con expresi6n dirigida al espacio intercelular de tejidos espedficos ( como los haces vasculares) e inducida por estres abiotico (presencia de heridas) o condiciones anaer6bicas. Estas construcciones incluiran el gen de interes del bacteriofago T4. Posteriormente se realizara la transferencia de estas construcciones a variedades de papa susceptibles coma: Achirana INTA, Amarilis y Tomasa Condemayta. En este case, tambien se realizan ensayos de campo para determinar el nivel de resistencia alcanzado.
12

•!• Organizaci6n de talleres sobre manejo de papas transgenicas en el medio ambiente
Se realizaran dos talleres especializados en el manejo de papas transgenicas con resistencia a enfermedades bacterianas y al Tizon Tardio en las cuales se trataran las siguientes temas :
1. Identificaci6n de las riesgos ( Lconsecuencias?)
2. Evaluaci6n de estos riesgos (Lconsecuencias?)
3. Experimentos que deben realizarse antes de liberar papas transgenicas
4. Medidas a seguir para el manejo eficiente de papas transgenicas
5. Revision de la situaci6n actual de las reglas y normas en el Peru para el manejo de papas transgenicas.
•!• Uso de la ingenieria genetica para la resistencia a enfermedades bacterianas en papa
El mejoramiento convencional para aumentar la resistencia a la Marchitez Bacteriana no logr6 hasta ahora altos niveles de resistencia. Generalmente se cree que hay poca resistencia par lo menos dentro del germoplasma cultivado. Es par eso que el mejoramiento mediante el uso de la ingenierfa genetica, tambien llamado mejoramiento molecular, se ha convertido en un nuevo campo de investigaci6n con promisorios resultados. Antes de definir cuales son las estrategias para el mejoramiento molecular de la resistencia a enfermedades bacterianas en papa, es importante resaltar varios aspectos del cultivo de la papa:
(a) La papa es el cuarto cultivo en importancia en el mundo despues del trigo, arroz y mafz.
(b) Tiene un alto valor nutritivo y gran potencial en rendimiento.
(c) Constituye aproximadamente la mitad del volumen de la producci6n mundial de rafces y tuberculos.
(d) Forma parte de la dieta de media bi116n de consumidores en pafses en vfas de desarrollo.
( e) La producci6n global para la decada de los 60 era de 29 millones de toneladas, se preve que llegara a superar las 100 millones de toneladas para fines de milenio.
13

(f) El rendimiento par pafs fluctua entre las 7 t/ha, en pafses de la region del sub-Sahara, y las 27 t/ha caracterfsticos de la provincia de Shandong en la China. El rendimiento promedio mundial esta en 13 t/ha en pafses en vfas de desarrollo.
(g) En pafses desarrollados, la mayor produccion se presenta en Holanda, llegando hasta 42 t/ha.
(h) En condiciones 6ptimas de crecimiento, se estima que la producci6n podrfa sobrepasar las 100 t/ha.
4!Que ventajas tiene la papa en la Transformacion Genetica?
Se ha realizado porque:
1. Fue el primer cultivo alimenticio en ser transformado a principios de la decada de las 80, la tecnolog fa en pa pa ya esta bien establecida y durante su desarrollo no se report6 ningun efecto indeseable.
2. La transferencia de genes es muy eficiente y esta mediada par el sistema natural caracterfstico de Agrobacterium tumefaciens llamado "agroinfeccion".
3. Existe la posibilidad de aplicar el sistema a larga escala en pafses en vfa de desarrollo sin mayor costo.
4. La agroinfeccion favorece la insercion de algunas copias del transgen en la planta.
5. La multiplicaci6n clonal caracterfstica de esta planta permite mantener el caracter agregado en forma estable.
4!Cuales son las estrategias antimicrobianas en la papa transgenica?
(a) Los genes codificantes para cecropina B (y sus analogos artificiales coma Shiva y 58-37) y atacina E, aislados de celulas del insecto Hya/ophora cecropia fueron introducidos en plantas de tabaco, papa y manzana. Se han presentado reportes con resultados contradictorios en trabajos realizados con cecropina para determinar la expresi6n de este gen foraneo y el nivel de resistencia adquirido.
14

(b) El gen que codifica para la sarcotoxina ( del mismo ti po que la cecropina), aislado del insecto Sarcophaga peregrina e introducido en papa. Este material esta actualmente en proceso de evaluacion en el campo en San Ramon (proyecto de colaboracion entre el Weizmann Institute of Science, Israel y el CIP).
( c) Uso de genes codificantes de tioninas (peso de 5000 da, ricas en cistefnas) presentes en trigo y cebada. Fueron transferidos a plantas de tabaco (de expresi6n intracelular). Se observe un aumento en la resistencia a P. syringae en tamizados de hojas infiltradas, pero no en condiciones de campo. Otras pruebas con tioninas se hicieron en tomate y papas sin mayor exito.
( d) El gen de un peptido antimicrobiano, tachyplesin, aislado de un cangrejo asiatico, fue expresado en papas con secrecion dirigida hacia el espacio intercelular. Una ligera disminucion del grade de pudricion fue observada luego de la inoculacion con Erwinia spp.
(e) Protefnas de transferencia no espedfica de lfpidos, nsLTP, (aisladas de plantas de cebolla) fueron introducidas en tabaco para conferir resistencia a la presencia de Pseudomonas syringae, resistencia que se manifesto en la forma de una menor presencia de sfntomas en las hojas infectadas.
(f) El gen que codifica para la enzima glucosa oxidasa del hongo Aspergillus niger fue introducido en papas. Esta enzima promueve la produccion de peroxide de hidrogeno (H20 2), molecula altamente toxica para las microbios, pero tambien involucrada en la reaccion local de hipersensibilidad y coma sefial de defensa celular. Se observaron altos niveles de resistencia a Erwinia carotovora. Sin embargo, debe evaluarse el efecto de la expresion de este gen sabre la salud humana por la toxicidad del H20 2 en eucariotes.
(g) Genes de lisozima fueron tambien introducidos exitosamente en papas para aumentar la resistencia a enfermedades bacterianas. Se trata de las genes que codifican la lisozima del bacteri6fago T4,
de la clara de huevo de gallina, y de bovino.
15

Posibles mejoras en la expresion de los genes antimicrobianos.
Expresi6n controlada de la lisozima L en !2BJ29.
• La expresi6n constitutiva del gen de lisozima T4 y su posterior secreci6n hacia el espacio intercelular permiti6 la reducci6n del grado de pudrici6n observado ante la presencia de Erwinia carotovora en el cultivo.
• Par la naturaleza del proceso de infecci6n se ha centrado el estudio en la construcci6n de genes quimericos. Asf, se busca que la lisozima se acumule en el momenta yen el tejido oportuno.
Expresi6n diriqida en teiidos
• El uso del promotor pat del gen de patatina que se expresa en tuberculos, podrfa ser muy util para aumentar la resistencia a enfermedades bacterianas causadas par Erwinia spp.
• El uso del promotor grpl.8 que se expresa en los haces vasculares durante el proceso de diferenciaci6n, podrfa ser especialmente util para R. solanacearum.
Expresi6n controlada QQ[ factores abi6ticos
• Uso de genes promotores coma el mas y el wunl que son activados por heridas en el tejido vegetal. Estas heridas constituyen una de las vfas de ingreso mas utilizadas par las bacterias durante el proceso de infecci6n. Con este gen quimerico se busca la resistencia a R. so/anacearum.
• Uso del gen promoter GapC4, inducido bajo condiciones anaer6bicas, condiciones que prevalecen durante el proceso de pudrici6n del tuberculo. Este gen podrfa ser util para incrementar la resistencia a Erwinia spp.
(h) Protefnas tipo hevein (aisladas de amaranto) que se caracterizan par su afinidad a la quitina, compuesto que forma parte de la membrana celular de varios pat6genos.
(i) Protef nas del ti po knottin ( aisladas de Mirabilis jalapa) que tienen las mismos ranges de actividad antimicrobiana. Estas protefnas se transfirieron a tabaco pero nose observ6 ningun efecto in vivo de actividad antimicrobiana.
16

(j) Defensinas, protelnas aisladas en semillas de rabanito. Fueron utilizadas exitosamente en tabaco para incrementar su defensa ante la presencia de hongos.
c!Otras estrategias en camino para el futuro?
• La identificacion de nuevas protef nas de accion antimicrobiana en rafces y tuberculos.
• Expresion en plantas del gen de origen bacteriano que codifica la oligo-galacturonido liasa, enzima que convierte los compuestos resultantes de la degradacion de la membrana vegetal por las enzimas de la bacteria patogenica y que actuan como inductores de patogenicidad, en compuestos inactivos.
• Expresion en plantas de anticuerpos especfficos que bloqueen los factores de patogenicidad.
• Determinar un mecanismo de muerte celular inducida por un gen suicida activado por el pat6geno.
PERSPECTIVAS
Las construcciones gerncas que se utilizaron podrfan conferir resistencia a variedades susceptibles a marchitez bacteriana.
Se probaran los genes en cultivares como Achirana INTA, Amarilis, Tomasa Condemayta y Desiree.
La o las construcciones que demuestren conferir cierto grado de resistencia a marchitez bacteriana seran utilizadas conjuntamente para buscar un efecto sinergico de los genes quimericos sobre la resistencia bacteriana.
POSIBLES LIMITACIONES
• La opinion publica esta todavfa poco preparada a aceptar los alimentos obtenidos mediante el uso de la biotecnologfa. Se trata, en mi opinion mayormente de una falta de informacion sobre la naturaleza de los cambios geneticos producidos por este metodo en comparaci6n con los metodos convencionales que tambien generan modificaciones geneticas. En realidad, el termino genenco de Organismos Modificados Geneticamente, ya revela la mala informacion del publico,
17

porque cualquier producto del mejoramiento es una modificacion genetica que nose encuentra en la naturaleza. Tambien existe en la opinion publica la creencia de que no existe ningun tipo de regulacion de los cultivos transgenicos. Esto tampoco es verdad, al igual que las otros productos obtenidos par mejoramiento, estos se analizan con respecto a varios caracteres conocidos coma dafiinos para la salud humana. Sin embargo, algunos riesgos ambientales necesitan una regulaci6n espedfica que en muchos casos todavia no existe en los paises en desarrollo.
La desconfianza del consumidor hacia los cultivos geneticamente modificados podrfa interferir con la inversion que se realiza en este tipo de investigaci6n, atrasando el desarrollo de esta tecnologfa en los pafses donde se venden estos productos y afectando a los productores envueltos en esta actividad.
Tambien, un efecto perverse de esta actitud podrfa ser que se promuevan los productos que no hayan sido modificados geneticamente y mantener el sistema actual de producci6n asistido por numerosos aportes en productos fitosanitarios. Todos sabemos que estos contaminan el media ambiente, son peligrosos para la salud humana, tanto para el agricultor coma para el consumidor, y mantienen una dependencia del agricultor hacia las empresas de agroqufmicos.
• La regulacion de la bioseguridad referente a los cultivos transgenicos es tambien un factor limitante para el desarrollo de esta tecnologia. En general, la obligaci6n de establecer normas nacionales de bioseguridad conduce a una demora para el desarrollo de este tipo de productos en los pafses en desarrollo. Par esta razon, es muy importante que en pafses coma el Peru se establezca a la brevedad posible normas y regulaciones, que permitan el desarrollo de esta tecnologfa bajo la supervision de instancias nacionales donde las sectores industriales, de proteccion del media ambiente y de la investigacion agrfcola esten representados.
Los derechos de propiedad intelectual constituyen un tercer tipo de posibles limitaciones para el desarrollo de esta tecnologfa. Existe un peligro real que concentra la tecnologfa en unos pocos "duefios". Compafifas transnacionales, que pueden comprar patentes de otros y defender sus patentes por la gran capacidad financiera que tienen. Las pequenas empresas o el sector publico generalmente no tienen el
18

respaldo financiero suficiente como para entablar acci6n judicial en contra de estas companfas. Entonces, los derechos de propiedad intelectuales podrfan limitar esta tecnologfa fuera de pafses en vfa de desarrollo por carecer estos de legislaciones adecuadas para protegerlos.
Tabla 1. Costas adicionales en el cultivo de papa ocasionados par la presencia de las 'plagas mayores'.
Plagas y Area Perdidas Proteccion Valor de Total Enfermedades afectada enla del cultivo perdida {$US)
{ha)1 produccion {$US)1 del cultivo {x106)
{x103) {$US)1 {x106
) {$US)2
{x106) {x106
)
Tizon Tardf o 1,960 2,430 742 3,172
Marchitez 1,150 720 720 Bacteriana
Virus 1,405 705 705
Insectos 746 655 655
1 Estimado a partir de Walker y Collion, 1997 2 Estimado en base a la depreciacion y las costos par insecticidas para gorgojo de las
Andes de la papa, mosca minadora y polilla de papa. Tornado de Walker y Collion, 1997
19


" ,I
EVALUACION DE TUBERCULOS DE PAPA TRANSGENICA A LA PUDRICION
CAUSADA POR Erwinia carotovora subsp. carotovora
Evaluation of transgenic tubers against bacterial wilt due to Erwinia carotovora
subsp. carotovora
Sylvie Priou *
MATERIALES Y METODOS
Se evaluaron 16 clones transformados con el gen de lisozima C2, 9 con el gen de lisozima T4 y 32 con el gen Sarcotoxina IA para resistencia a la pudrici6n blanda causada por Erwinia carotovora subsp. carotovora bajo condiciones de laboratorio y empleando de 5 a 10 tuberculos por don. Los tuberculos fueron cortados por la mitad e inoculados con la cepa CIP-400. Se usaron dos concentraciones diferentes de ineculo (5x108 y 2.Sx 109 ufc/ml) para cada mitad. La inoculacien se realize por el metodo del sacabocado. Se utilize 20 ~d de la suspension bacteriana que fue depositada en el hoyo de 2 mm de profundidad por 3 mm de diametro hecha en la parte central de la mitad del tuberculo con un sacabocados N°3. Los tuberculos fueron incubados en una camara humeda a 25°C durante dos dfas, al cabo de los cuales se evalu6 el peso (g) de tejido podrido. Luego se calcul6 el porcentaje relative de pudrici6n blanda (PRPB):
Peso to ta I - Peso sa no PRPB = x 100
Peso total
* Centro Internacional de la Papa (CIP). Estacion Central, La Molina. Lima - Peru 21

Los datos fueron transformados a:
Arc Sena --J PRPB
para ser analizados por SAS (programa de computacion utilizado para el analisis estadfstico).
RESULTADOS
Todos los clones evaluados desarrollaron pudricion, sin embargo, en los clones Desiree con el gen lisozima C2 se encontr6 que los clones Desiree/Lys-C2/ AA-09 y Desiree/Lys-C2/ AA-15 son significativamente menos susceptibles que el testigo Desiree (sin el gen lisozima C2). En clones con el gen de lisozima T4 se encontr6 que los clones Desiree/Lys-T4/DL-10 y Desiree/Lys-T4/DL-12 tambien son significativamente menos susceptibles que el testigo Desiree (Tabla 1).
En el case de los clones con el gen Sarcotoxina IA se encontr6 que los clones Achirana - INTA / Saree - IA/PS - 12, TS-10 / Sarco -IA/ PS - 8, Achirana - INTA/Sarco-IA/PS - 1, Desiree/Saree - IA/TS -14, TS-10 /Saree - IA/ PS - 9 y TS - 10/Sarco - IA/PS - 6 son significativamente mejores con respecto a los testigos Desiree y TS -10 sin transformar pero mas susceptibles que el cultivar Yana Puna (testigo moderadamente resistente; Tabla 2).
Tabla 1. Porcentaje relativo de pudrici6n blanda (PRPB) en clones de papa transgenicas (genes C2 y T4 Lisozima) inoculadas mediante sacabocado con dos concentraciones de Erwinia carotovora subsp. carotovora.
DL12 (T4)
WIKAA15 (C2)
WIKAA09 (C2)
DLlO (T4)
Desiree cs, NT) Dunnett, P = 0.05 S = Susceptible; NT = Clan no transgenico.
22
Concentraci6n de inoculo 5 x 108 ufc/ml 2.5 x 109 ufc/ml
2.5 a 3.7 a 3.2 b 3.9 a 3.9 b 4.1 a 5.2 c
10.8 d
4.8 b
12.8 c

Tabla 2. Porcentaje relativo de pudricion blanda (PRPB) en clones de papa transgenicas (Sarcotoxina IA) inoculadas por el metodo del sacabocado con dos concentraciones de Etwinia carotovora subsp. carotovora.
Concentracion de inoculo
Clon 5 x 108 ufc/ml
PS-12 (Achirana INTA) 2.6* b PS-8 (TS-10) 2.9 b
PS-1 (Achirana INTA) 3.6 c
TS-14 (Desiree) 4.0 c PS-9 (TS-10) 4.1 c
PS-6 (TS-10) 4.2 c
Desiree (S, NT) 9.7 d Yana Puna (MR, NT) 0.12 a
TS-10 (S, NT) 10.8 e Dunnett, P = 0.05 MR = Moderadamente resistente; S = Susceptible; NT= Clon no transgenico.
2.5 x 109 ufc/ml
6.7 e
3.8 b
4.8 c 3.9 b
5.6 d
4.7 c
10.5 f
0.49 a 14.2 g
23


'7
6oos~~~~~~~~~~~-----, MEJORAMIENTO PARA OBTENER
RESISTENCIA A MARCHITEZ BACTERIA NA
Breeding for Resistance to Bacterial Wilt
Sylvie Priou *
HISTORIA DEL MEJORAMIENTO GENETICO DE LA PAPA EN EL CIP
En la decada de los 50 las doctores F. Haynes y L. Nielsen de la Universidad de Carolina del Norte evaluaron mas de 9000 cultivares de papa para la resistencia a marchitez bacteriana.
Muy pocos cultivares demostraron ser resistentes, el mejor de estos fue Prisca. Posteriormente se hallaron otros tres cultivares resistentes: Achat, Ontario y Cruza 148. Prisca es un cultivar aleman que ya no es cultivado en Europa. Este cultivar fue hallado en Madagascar por personal del CIP y anadido al PTL (Pathogen TestedList Collection, un grupo de cultivares libres de pat6genos que se conserva en el CIP, bajo condiciones in vitro) coma el numero 800966. Achat es una variedad que se cultiva en gran cantidad en Brasil debido al hecho de que es tolerante a la marchitez bacteriana, pero el poseer caracterfsticas culinarias pobres y carecer de periodo de floraci6n, llev6 a EMBRAPA - CNHP a iniciar en 1987, un programa de mejoramiento en resistencia a marchitez bacteriana en colaboraci6n con el CIP.
Desde 1987 hasta 1995 se recibieron progenies TPS de los ultimas cruces realizados en CIP de los que se habfan seleccionado contra la raza 130 clones cuyo desempeno era igual o mejor que Achat (Lopes et al., 1998).
Cruza 148 es un cv. de Costa Rica de pedigrf desconocido que ha sido utilizado ampliamente en programas de mejoramiento en el CIP, CNPH-Brasil y CAAS-China y cultivado en muchos pafses por su resistencia a la marchitez bacteriana (Tabla 1). Sin embargo, sus tuberculos poseen caracterfsticas de plantas silvestres (anillo morado,
* Centro Internacional de la Papa (CIP). Estacion Central, La Molina. Lima - Peru 25

alto contenido de alcaloides), valor de comercializacion pobre y sus tuberculos son infectados en estadfo de latencia por Ralstonia so/anacearum (Rs).
El programa de mejoramiento para la resistencia a marchitez bacteriana fue iniciado en 1972 en la Universidad de Wisconsin por Rowe y Sequeira despues de la identificaci6n de la resistencia en los clones colombianos de So/anum phureja (Sequeira y Rowe, 1969).
Toda el material evaluado en el Peru durante los primeros ocho anos del CIP vino del programa de mejoramiento de Wisconsin.
De 1972 a 1976 (French y Sequeira)
Poblaciones basadas en los clones colombianos resistentes de S. phureja (CCC) fueron cruzadas con 5. tuberosum subsp. tuberosum (provistas por el programa de mejoramiento de Wisconsin). Los hfbridos resultantes fueron luego cruzados con clones mejicanos resistentes al tiz6n tardfo (LB) ( derivados de la variedad silvestre hexaploide de S. demissum): los c6digos utilizados fueron BR, MS, PSP y PSW (Figura 1). A partir de este material, muchas variedades se han convertido en nuevas variedades (coma Cajamarca y Molinera) o han sido cultivadas en grandes cantidades durante los ultimas anos de la decada de los 70 (Tabla 1). Se han incorporado nuevas fuentes de resistencia a la primera poblacion derivada de las especies silvestres 5. chacoense y S. raphanifolium.
La mayorfa de estas eran de maduracion tardfa. Mas aun, existfa una interaccion altamente especifica hospedero - lfnea - media ambiente y la resistencia se expresaba solamente bajo condiciones de temperatura frfas. Ademas, la base genetica para la poblaci6n que llevaba la resistencia tuvo que ser ampliada y adaptada a condiciones de temperatura calidas junta con la adici6n de la caracterfstica de maduracion temprana.
De 1977 a 1979 (Mendoza)
En 1977 los clones BR fueron cruzados con clones de maduraci6n temprana (series 3778 del CIP, Grupo I).
Los clones del Grupo I fueron cruzados en 1979 con genotipos adaptados a condiciones tropicales para obtener los clones que constituyen el Grupo II (series 3796 del CIP).
26

El Grupo III (serie 3805 del CIP) fue producida entre 1979 y 1980 para restaurar la resistencia a marchitez bacteriana y tiz6n tardfo que se perdieron cuando las Grupos I y II fueron creados. La autopolinizaci6n se convirti6 en un procedimiento imprescindible debido a las restricciones de material que surge de la necesidad de prevenir el uso de plantas infectadas con PS1V.
De 1980 a 1985 (Schmiediche)
Los resultados obtenidos en Sri Lanka confirmaron que la expresi6n de la resistencia a marchitez bacteriana esta en funci6n a la adaptabilidad al calor. Con el fin de aumentar la base genetica para la resistencia, se genera el Grupo IV (series 3810 CIP) a partir del clan AVRDC-1287.19 proveniente de Taiwan (resistente a marchitez bacteriana y tolerante al calor) cuya fuentes de resistencia a marchitez bacteriana son S. chacoense y S. raphanifolium y Serrana INTA ( cv. de Argentina, resistente a PLRV y PVY e inmune a PVX). Los niveles de resistencia eran superiores a las del cv. variedad Phureja en las Filipinas.
El Grupo V fue generado en 1982 cruzando las clones con mejor resistencia a marchitez bacteriana con Cruza 148 (resistente a marchitez bacteriana y tiz6n tardfo) e India 853 ( resistente a tiz6n tardfo) para introducir nuevas fuentes de resistencia a marchitez bacteriana, tiz6n tardfo y maduraci6n temprana (y para superar la tendencia a la autofecundaci6n).
De 1986 a 1995 (Anguiz, Mendoza y Chujoy)
Una poblaci6n diploide llamada MBN fue producida, derivada de una poblaci6n de S. sparsipilum ( combinando resistencia al nude de la rafz causada par nematodos), chacoense, microdontum y phureja. Esta poblaci6n fue cruzada con una poblaci6n diploide cultivada que fue derivada de S. stenotomum, phureja y gonioca/yx (del Dr. Haynes en la Universidad de Carolina del Norte). La poblaci6n diploide final fue cruzada con el germoplasma tetraploide mas avanzado resistente a marchitez bacteriana (TD). Los genotipos TD fueron finalmente cruzados con las mejores genotipos de las Grupos VI y V obteniendose una poblaci6n con un alto grado de heterocigocidad y te6ricamente con un amplio espectro de resistencia a marchitez bacteriana ( combinando, par lo menos, cuatro fuentes especfficas de resistencia; Figura 2). Fueron clones de alta productividad cuya forma de tuberculo es similar a S. tuberosum, sin embargo, tenfan alga de
27

maduracion tardfa. Esos clones fueron enviados a las Filipinas de los cuales se seleccionaron 27 (series BP) como resistentes a la marchitez bacteriana con un nivel mas alto de resistencia que los clones Phureja (entraron en el PTL del CIP). Sin embargo, estos clones eran susceptibles al tiz6n tardfo.
Cruces realizados luego con los materiales que mejor se adaptaban a las altas temperaturas (materiales de tropicos de tierras bajas [LT]) y de maduracion temprana y tambien con material resistente al tizon tardfo, y/o Hneas resistentes a RKN y/o clones inmunes a PVX y PVY llevaron a obtener tres poblaciones que aun se encuentran en evaluaci6n en la ciudad de Carhuaz, Peru (contra raza 3):
•!• MB raza 3, PVX, PVY, LB; •!• MB raza 1, PVX, PVY, LT, maduraci6n temprana media y buena
prod uctividad; •!• MB raza 1, LT, RKN y maduraci6n temprana.
CARACTERISTICAS DE LA RESISTENCIA DE PAPA A LA MARCHITEZ BACTERIANA
1. Tipo poligenico y cuantitativo (con genes aditivos y no aditivos);
2. !nestable debido a una fuerte interacci6n genotipo - patogeno, genotipo - medio ambiente;
3. Bajo nivel de resistencia en los clones mas resistentes y si no todas son potencialmente susceptibles a la infecci6n latente;
4. Se requiere una amplia adaptacion del genotipo hospedero para que haya una expresion estable de resistencia;
5. No se ha evidenciado una relacion gen por gen entre Rs y papa, pero la resistencia es especffica del linaje (no es especffica de la raza o cu ltiva r);
6. La alta variabilidad genetica en (Rs) requiere de un ti po de resistencia de amplia base para que pueda manejarse;
7. la base genetica es diferente para cada especie silvestre de Solanum (algunas de ellas de tipo poligenico, genes mayores y menores) ademas el concepto de resistencia de amplio espectro es realfstico.
28

CONSECUENCIAS DE LAS ESTRATEGIAS DE MEJORAMIENTO
La magnitud relativa de los componentes y rangos de heredabilidad de las varianzas geneticas sugieren que la seleccion clonal serfa exitosa (pero con un progreso lento en la poblacion). La resistencia a la marchitez bacteriana es mas estable cuando se le relaciona con la tolerancia al calor o con alguna adaptacion relativamente amplia dentro de genotipo. La selecci6n clonal puede ser exitosa para condiciones de crecimiento especffico con prevalencia de lfneas especificas de Rs (la limpieza y elecci6n de los cultivares debe realizarse en el lugar donde se utilizara el cv.). Por ejemplo: en las Filipinas, los clones BP que combinan maduraci6n temprana, adaptaci6n al calor y alguna resistencia a marchitez bacteriana de especies silvestres, presentaron resistencia exitosa (R) a la marchitez bacteriana (razas 1 y 3). Las lfneas altamente virulentas de Rs que mantienen patogenicidad por encima de un amplio rango de condiciones ambientales serfan las mas apropiadas para ser elegidas coma portadoras de una resistencia estable.
RESISTENCIA A LA MARCHITEZ BACTERIANA EN OTROS HOSPEDEROS
Materiales resistentes han sido encontrados en tomate, pimientos, berenjena, tabaco y manf. No se ha evidenciado inmunidad alguna y en ninguno de los casos, las plantas resistentes estan infectadas de manera latente con Rs. La resistencia no es estable de una localidad y/o lfnea a otra, y se quiebra ante la presencia de altas poblaciones de Me/oidogyne sp. en el suelo.
Se han encontrado dos tipos de tomate resistentes (Prior et al., 1994):
(a) tipo poligenico (CRA66);
(b) tipo de herencia simple monogenica y dominante (coma Hawai 7996) que sigue las leyes de la segregacion Mendeliana (en Guadeloupe, Francia), pero recientemente caracterizada por el mapeo genetico a partir del cual se hallo que es controlado por un loci resistente cuantitativo ubicado en el cromosoma 6. En este cromosoma tambien se encuentra el gen Mi que lleva la resistencia a Meloidogyne sp. (Deberdt y Prior, 1998). Estudios de mapeo posteriores demostraron que pueden existir dos QTLs, uno controlando la resistencia adaptativa y el otro controlando la
29

resistencia constitutiva (e.g., no dependiente del media ambiente; Wang et al., 1998).
Sin embargo, todos las materiales resistentes de tomate tienen frutos pequenos (gen localizado en cromosoma 2). En un experimento reciente, coordinado por AVRDC para comparar el comportamiento de 35 llneas de tomate resistentes (variedades comerciales, llneas para mejoramiento y especies silvestres) mas de 11 palses confirmaron que hay no inmunidad. Pero las resultados mostraron que hay 7 entradas que mostraron mas de 90% de supervivencia en las diferentes localidades. Ademas, parece que la estabilidad de la resistencia en tomate esta disponible y que el desarrollo de variedades para una distribuci6n mas amplia es posible (Wang et al., 1998).
VARIABILIDAD GENETICA DE Ralstonia solanacearum
Se utilizan- dos sistemas de clasificaci6n para Rs. las sistemas de raza y de biovar.
El sistema de razas esta basado en el rango de hospederos bajo condiciones de campo. A partir de esto, se pueden distinguir cuatro razas (Hayward, 1991):
1. Raza 1: afecta un amplio rango de cultivos solanaceos, incluyendo la papa, varias malas hierbas y bananas diploides. Algunas llneas pueden afectar al manly al jengibre. La mayorla de las veces se le halla en lugares de poca elevaci6n, en las tr6picos y subtr6picos.
2. Raza 2: afecta plantas de la familia de las musaceae, coma la banana triploide, el platano y la He/iconia spp. en las tr6picos.
3. Raza 3: afecta principalmente papa y ocasionalmente tomate junta con otros cultivos solanaceos y malas hierbas. Se la haya comunmente en lugares de mayor elevaci6n o latitudes ( climas frfos).
4. Raza 4: afecta a la mora y se halla solamente en China.
El sistema biovar consiste en pruebas bioqulmicas basadas en la habilidad de la bacteria para utilizar tres disacaridos y/u oxidar tres alcoholes de hexosa.
30

La clasificacion de las biovares se basa en lo siguiente (Hayward, 1991): 1. Bv 1 = utilizacion de disacaridos negativa, oxidacion de alcoholes
negativa; 2. Bv 2 = utilizacion de disacaridos positiva, oxidacion de alcoholes
negativa; 3. Bv 3 = utilizacion de disacaridos positiva, oxidacion de alcoholes
positiva; 4. Bv 4 = utilizacion de disacaridos negativa, oxidacion de alcoholes
positiva; 5. Bv 5 = utilizacion de disacaridos positiva, oxidacion solo de
manitol.
Hay alguna equivalencia entre la raza y el biovar: la raza 3 coincide con el biovar 2A; las lfneas de raza 1 son del biovar 1, 3 o 4, el biovar 1 es el mas comun en papa. No se ha establecido una raza para el biovar 2T, se le haya principalmente en las tierras bajas de la cuenca del Amazonas. Tiene muchas lfneas y un rango amplio de hospederos coma en el caso de la raza 1; sin embargo, el biovar 2T son menos agresivas. Las lfneas de la raza 2 son las biovares 1 y 3, y toda la raza 4 son el biovar 5 (Hayward, 1991).
De hecho, el sistema de biovares es el mas utilizado en la actualidad debido a la poca importancia que tiene la raza: no hay especificidad del hospedero que haya sido evidenciada y ademas se ha presentado superposicion entre las razas. Rs es extremadamente adaptable desde el momenta en el que puede afectar 200 especies de plantas diferentes.
Muchos estudios realizados con el fin de acceder a la diversidad genetica de Rs han llevado a tener una imagen mas clara de SU
distribucion y evolucion geografica. Rs es una especie muy heterogenea. Estudios en RFLP utilizando sondas de 8 ADNs dividieron las lfneas de Rs en 38 genotipos multilocus agrupados en dos divisiones mayores (Cook y Sequeira, 1994): (a) Division 1: contiene las biovares 3, 4 y 5 (razas 1 y 4); (b) Division 2: reagrupa las biovares 1, 2A y 2T (razas 1, 2 y 3).
La caracterfstica que ha sido correlacionada de manera mas estrecha con las datos obtenidos del RFLP fue el origen geografico de las lfneas. Los dos grupos mayores que se han podido distinguir provienen o del Viejo Mundo o del Nuevo Mundo. Cerca del 92% de las lfneas de la Division I provienen de Australia y Asia, y el 97% de
31

las lfneas de la Division II son de las Americas. Al final, la lfneas de los biovar 1 y 2 fueron separadas en dos grupos. Dentro del grupo del biovar 2 se separaron aquellas lfneas que proceden del biovar 2A de las tierras altas de los Andes de aquellas que proceden del biovar 2T indfgenas de los valles de la tierras bajas en Peru y Brasil lo que es consistente con la especializacion de la papa dentro del biovar 2A.
Al secuenciar el ADNr 165, la division 1 fue dividida en dos subdivisiones (Fegan et al., 1998):
i. Division 2a: lfneas de los biovares 1, 2A y 2T provenientes de Indonesia; y Ralstonia syzygii (agente de la enfermedad del clavo de Sumatra) y Pseudomonas ce/ebensis ( agente la enfermedad bacteriana de la sangre del banana presente en Asia), dos patogeno muy estrechamente relacionados a Rs,
ii. Division 2b: todas las otras lfneas de los biovares 1, 2A y 2T.
Estas tecnicas moleculares han confirmado las conclusiones a las que han llegado patologos luego de estudiar los fenotipos y la patogenicidad. Los subgrupos mas homogeneos fenotfpica y genotfpicamente de Rs son: iii. Uneas de la Raza 3 / biovar 2A (grupos RFLP 26, 27, 34) que
causan marchitez bacteriana / pudricion marron de papa; iv. Uneas de la Raza 2 I biovar 1 (grupos RFLP 24, 25 y 28) que
causan la enfermedad de Bugtok del platano y la enfermedad Moko de la banana.
Todas la lfneas Europeas pertenecen al grupo 26 de RFLP, pero al aplicarse la tecnicas de AFLP y RC-PFGE se encontro cierto polimorfismo: tres grupos mostraron evidencias de tener varias lfneas clonales presentes en Europa. No habfa correlacion entre la distribucion geografica o la adaptaci6n ecol6gica y los perfiles que podrfan obtenerse (Van der Wolf et al., 1998).
PROSPECTOS: MAYORES EVALUACIONES DE PAPA Y Solanum spp. SILVESTRE EN EL CIP
La infeccion latente no esta relacionada con los sfntomas fuera del suelo. Casi nada se conoce acerca de la genetica de la resistencia a la infecci6n latente, este aspecto fue descartado como objetivo en la investigacion realizada en favor del mejoramiento. En un esfuerzo mayor por el mejoramiento en el CIP, se pondra enfasis tanto en la expresion de la enfermedad como en la infeccion latente en tuberculos (utilizando el kit post-enrichment NCM-ELISA del CIP).
32

Actualmente se realizan las siguientes actividades en el CIP relacionadas con la investigacion en resistencia a Marchitez Bacteriana:
1. Evaluacion de campo (campo de raza 3 infectada) de la ultima poblacion que tiene al menos cuatro fuentes de resistencia a Marchitez Bacteriana combinada con la inmunidad a PVY y PVX y/o resistencia a RKN, madurez prematura y tolerancia al calor (Figura 2).
2. Pruebas en el invernadero con las accesiones mas resistentes de las especies silvestres evaluadas por Sequeira en 1986. A partir de 1573 accesiones de la coleccion Internacional probadas en Sturgeon Bay, Wisconsin, 25 accesiones mostraron menos de 10% plantas marchitas despues de la inoculacion con Rs. La progenies resistentes fueron de las siguientes especies: 5. demissum, S. acaule, S. po!ytrichon, S. raphanifolium, S. commersonii y S. boliviense. Estas accesiones seran evaluadas nuevamente en los invernaderos del CIP. Se ran inoculadas con varias lfneas de Rs de la razas 1 y 3, y las progenies mas resistentes seran clonadas y evaluadas para infeccion latente en tuberculos despues de la inoculacion del suelo.
3. Pruebas en el invernadero (siguiendo la misma estrategia descrita anteriormente) del germoplasma silvestre que aun no haya sido probado, algunos de ellos colectados en la areas endemicas de Marchitez Bacteriana de Latinoamerica: 5. a!bicans, S. bukasovii, S. multiinterruptum, S. wittmackii.
4. Pruebas en el invernadero de especies cultivadas de Solanum incluyendo 5. tuberosum ssp. andigena.
5. Evaluacion en el campo de plantas transgenicas (genes de sarcotoxina y lisozima) en San Ramon (cam po infectado con la raza 1).
6. Caracterizacion de la resistencia a Marchitez Bacteriana (mas lfneas, infeccion latente ... ) de los hfbridos somaticos de haploides de S. tuberosum ssp. tuberosum y las especies diploides silvestres de S. commersonii recientemente obtenidas en Wisconsin (Laferriere et al., 1998).
33

REFERENCIAS
Anguiz R.J. & Mendoza H.A. 1997. General and specific combining abilities for resistance to bacterial wilt (Pseudomonas solanacearum E.F. Smith) in PVX and PVY immune autotetraploid potatoes. Fitopatologia 32(1): 71-80.
Beverdt P. & Prior P. 1998. Mi introgression lines as tools for the genetic analysis of bacterial wilt resistance in tomato. In Bacteria/ wilt disease: molecular and ecological aspects (eds. Prior, Ph., Allen, C. and Elphinstone, J.) p. 255-262. INRA edn, Springer Verlag, Berlin, Germany.
Cook D. & Sequeira L. 1994. Strain differentiation of Pseudomonas solanacearum by molecular genetic methods. In Bacteria/ wilt: the disease and its causative agent Pseudomonas solanacearum (eds. Hayward, A. C. and Hartman, G. L.) p. 77-93. CAB International, Wallingford, UK.
Fegan M., Taghavi M., Sly L. I. & Hayward A. C. 1998. Phylogeny, diversity and molecular diagnostics of Ralstonia so/anacearum. In Bacterial wilt disease: molecular and ecological aspects (eds. Prior, Ph., Allen, C. and Elphinstone, J.) pp. 19-33. INRA edn, Springer Verlag, Berlin (Germany).
French E.R. & Sequeira L. 1987. Additional sources of resistance to bacterial wilt. In Bacterial diseases of the potato. Planning Conference on Bacterial Diseases of the Potato, pp 29-33, Lima, Peru.
French E.R., Anguiz R. & Aley P. 1997. The usefulness of potato resistance to Ralstonia solanacearum, for the integrated control of bacterial wilt. In Bacteria/ wilt disease: Molecular and ecological aspects (eds. Prior P., Allen C. and Elphinstone J. eds.) p. 381-385. INRA edn, Springer Verlag, Berlin, Germany.
Hayward, A.C. 1991. Biology and epidemiology of bacterial wilt caused by Pseudomonas so/anacearum. Annu. Rev. Phytopathol. 29: 65-87.
Jatala P., Martin C. & Mendoza, H.A. 1987. Role of nematodes in disease expression by Pseudomonas so/anacearum and strategies for screening and breeding for combined resistance. In Bacterial diseases of the potato. Planning Conference on Bacterial Diseases of the Potato, p. 35-37, Lima, Peru.
34

Laferriere L. T., Hegelson J. P. and Allen C. 1998. Solanum tuberosum - S. commersonii somatic hybrids are resistant to brown rot caused by Ralstonia so!anacearum. In Bacterial wilt disease: Molecular and ecological aspects (Prior P., Allen C. and Elphinstone J. eds.) p. 316-320. INRA edn, Springer Verlag, Berlin (Germany).
Lopes C. A., Quezada-Soares A. M., Busa J. A. and Melo P. E. 1998. Breeding for resistance to bacterial wilt of potatoes in Brazil. In Bacterial wilt disease: Molecular and ecological aspects (Prior P., Allen C. and Elphinstone J. eds.) p. 290-293. INRA edn, Springer Verlag, Berlin (Germany).
Prior P., Grimault V. and Schmit J. 1994. Resistance to bacterial wilt (Pseudomonas solanacearum) in tomato: Present status and prospects. In Bacterial wilt: the disease and its causative agent Pseudomonas solanacearum (Hayward, A. C. and Hartman, G. L. eds.) p. 209-223. CAB International, Wallingford (UK).
Sequeira L. and Rowe P. R. 1969. Selection and utilization of Solanum phureja clones with high resistance to different strains of Pseudomonas solanacearum. Am. Potato J. 46:451-462.
Schmiediche P. 1985. Breeding potatoes for resistance to bacterial wilt caused by Pseudomonas solanacearum. In ACIAR Proceedings N° 13, p. 105-111 (ed. Persley, G.J.), Canberra (Australia).
Schmiediche P. 1988. Breeding for resistance to Pseudomonas so!anacearum. In Bacteria! diseases of the potato. In Bacterial diseases of the potato. Planning Conference on Bacterial Diseases of the Potato, p. 19-27, Lima (Peru).
Schmiediche P. and Martin C. 1986. The use of wild species in breeding for resistance to bacterial wilt ( Pseudomonas so!anacearum). American Potato Journa/63:453.
Tung P.X. and Schmiediche P. 1995. Breeding potato for resistance to bacterial wilt (Pseudomonas solanacearum): looking for stable resistance? In Integrated management of bacteria/ wilt, Proceedings of the International Workshop (eds. Hardy B. and French E.R.) p. 173-176. International Potato Center (CIP) and Indian Council of Agricultural Research, New Delhi (India).
35

Van Der Wolf J. M., Bonants P. J. M., Smith J. J., Hagenaar M., Nijhuis E., Van Beckhoven J. R. C. M., Saddler G. S., Trigalet A. and Feuillade R. 1998. Genetic diversity of Ralstonia solanacearum race 3 in Western Europe determined by AFLP, RC-PFGE and Rep-PCR. In Bacteria/ wilt disease: molecular and ecological aspects (eds. Prior, Ph., Allen, C. and Elphinstone, J.) p. 44-49. INRA edn, Springer Verlag, Berlin (Germany).
Wang J-F., Thoquet P., Olivier J. and Grimsley N. 1998. Genetic analysis of quantitative resistance loci (QRL) of tomato variety Hawaii 7996 in Tawain. In Bacteria/ wilt disease: molecular and ecological aspects (eds. Prior, Ph., Allen, C. and Elphinstone, J.) p. 245-249. INRA edn, Springer Verlag, Berlin (Germany).
Wang J-F., Hanson P. and Barnes J. A. 1998. Worldwide evaluation of an interantional set of resistant sources to bacterial wilt in tomato. In Bacterial wilt disease: molecular and ecological aspects (eds. Prior, Ph., Allen, C. and Elphinstone, J.) pp. 269-275. INRA edn, Springer Verlag, Berlin (Germany).
36
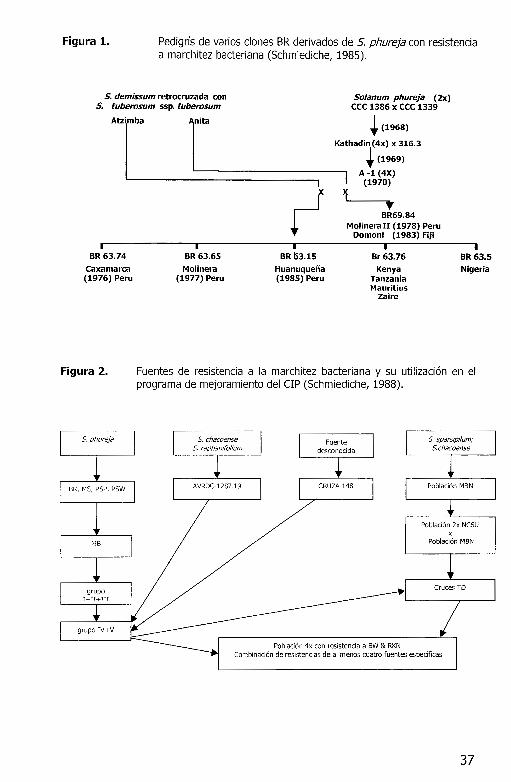
Figura 1. Pedigrfs de varios clones BR derivados de S. phureja con resistencia a marchitez bacteriana (Schmiediche, 1985).
S. demissum retrocruzada con S. tuberosum ssp. tuberosum
So/anum phureja (2x) CCC 1386 x CCC 1339
i (1968) Atzilmba 1.n-ita ------.
Kathadin(4x) x 316.3
~ (1969)
I A -1 (4X) I (1970)
BR 63.74
Caxamarca (1976) Peru
Figura 2.
S. phureja
BR63.65
Molinera (1977) Peru
't BR69.84 r
~
Molinerall (1978) Peru Domoni (1983) Fiji
BR 63.15
Huanuquefia (1985) Peru
Br 63.76
Kenya Tanzania Mauritius
Zaire
BR 63.S
Nigeria
Fuentes de resistencia a la marchitez bacteriana y su utilizacion en el programa de mejoramiento del CIP (Schmiediche, 1988).
S. chacoense S. raphamfolium
AVRDC-1287.19
Fuente desconocida
CRUZA 148
Poblacion 4x con resistencia a BW & RKN
S. sparsipilum; S.chacoense
Poblaci6n 2x NCSU x
Poblaci6n MBN
Combinacion de resistencias de al menos cuatro fuentes especificas
37


INTRODUCCION
DESARROLLO DE ESTRA TEGIAS DE INGENIERIA GENETICA PARA LA
RESISTENCIA A BACTERIAS FITOPATOGENICAS
Development of genetic engineering strategies for resistance to phytopathogenic bacteria
Petra Porsch *
La podredumbre blanda causada por Erwinia chrysantemi y Erwinia carotovora es una de las enfermedades bacterianas de papa de mayor importancia. Ya que el control qufmico no es posible, es preferible mejorar geneticamente la resistencia de la planta. La mejora genetica mediante el metodo clasico es diffcil de lograr ya que hay escasez de variedades resistentes. Es por eso que la ingenierfa genetica se ha convertido en la alternativa mas prometedora.
Nuestra investigaci6n se basa en dos ideas:
1. Transformaci6n de papa con el gen para lisozima de bacteri6fago
T4 2. Producci6n de anticuerpos contra diferentes enzimas peptolfticas
de Erwinia - transformaci6n de papa con las construcciones de anticuerpos mas exitosas.
* Federal Center for Breeding Research on Cultivated Plants (BAZ), Quedlinburg, Germany
39

LISOZIMAS
Las lisozimas hidrolizan espedficamente el peptidoglicano componente de la pared celular de las bacterias llamado murelna.
En la mayorla de las plantas, la actividad de la lisozima es relativamente baja y esta localizada en la vacuola. Nuestro fin fue integrar una lisozima foranea a los genes de papa donde deba ser secretada a los espacios intercelulares, en el lugar de la infecci6n y multiplicaci6n.
Nuestro grupo eligi6 la lisozima de bacteri6fago T4 para la transformaci6n de papa porque ha sido reportada como la mas activa contra bacterias gram (-) y gram ( + ).
En lugar de utilizar la lisozima natural, utilizamos el mutante M6K que tiene un aminoacido cambiado en la posici6n 6 de metionina a lisina. Mientras la actividad muramidasa de este mutante es la misma que la de la enzima natural, la actividad bactericida ( que sera explicada mas adelante) es cuatro veces la actividad normal.
En todas las construcciones, el gen para lisozima T4 esta enlazado con un secuencia para un peptide senal (de cebada, oc -
amilasa) de localizaci6n extracelular. En dos de las construcciones (pSR 8-36 y pSR 8-40) el gen de lisozima esta dirigido por el promoter 35s del CaMV para lograr su expresi6n constitutiva. En pSR 8-40 la unidad de lisozima esta flanqueada por regiones asociadas a la matriz que han sido reportadas como regiones que aumentan la expresi6n del transgen y reducen la variaci6n.
Mediante la evaluaci6n densitometrica de los Western blots el nivel de expresi6n de lisozima en plantas in vitro fue calculada entre 0.0002 y 0.0006% del total de protelna soluble.
Se prob6 la susceptibilidad ante Erwinia en ensayos con discos de tuberculos. Algunas llneas transgenicas especialmente DL 11 y DL 13 presentaron menor laceraci6n.
40

Dos lfneas (DL 4 y DL 5) fueron finalmente cultivadas en campo en dos localidades: en Quedlinburg en Alemania Central y en GroB Lusewitz al norte de Alemania en los afios 1997 y 1998.
Para poder probar los 'tuberculos-semillas' ante pierna negra y podredumbre blanda, tuvimos que infectarlos con Eca. Dos metodos fueron utilizados: infiltraci6n al vado e infecci6n por sacabocados. Para el ultimo metodo se hizo un agujero profundo en el tuberculo con un sacabocado y se llen6 el agujero con una suspension de Erwinia.
Mientras que la mayorfa de los tuberculos brotaron despues de la infiltraci6n al vado, varios tuberculos no lo hicieron despues de la infeccion con sacabocado y se pudrieron en el suelo.
El porcentaje de tuberculos infectados con Erwinia que no brotaron fue de 70% de la lfnea DL 4, 40-60% del Desiree no transformado y 25-40% de la lfnea DL 5. Se supone que el pesimo resultado con DL 4 es una consecuencia de su desarrollo tardfo, probablemente causado por una variaci6n somaclonal en la etapa de cultivo de tejidos.
Las plantas fueron evaluadas para sfntomas de pierna negra una vez al mes. No se pudo encontrar diferencias entre las lfneas en Quedinburg, pero si se detectaron marcadas diferencias en GroBLusewitz. DL 4 present6 mayor cantidad de sf ntomas mientras que y DL 5 fue se mostro coma la mejor planta.
Despues de la cosecha se evalu6 la susceptibilidad de las tuberculos ante podredumbre blanda. Para esto se utilizaron diferentes metodos: 1) Infecci6n de mitades de tuberculos con suspension de Erwinia y 2) Una prueba de almacenamiento en balsas plasticas selladas par varios meses a temperatura ambiente. En el ensayo de las tuberculos en mitades, DL 4 y DL 5 mostraron una maceraci6n significativamente menor que las If neas control (transformadas y no transformadas ). La pudrici6n en los tuberculos dentro de las balsas selladas no mostraron diferencias entre las lfneas.
41

En una segunda fase nuestro proyecto, el promotor constitutivo Ubi3 fue utilizado junta con otros promotores para obtener la expresi6n temporal o localizada de lisozima: el promotor Mas de Agrobacterium (inducido par heridas), y el promotor Wun de papa; el promotor GapC4 de mafz (inducido par anaerobi6sis) y el promotor Pat de papa especffico de tuberculos.
El promotor Ubi presenta un nivel de expresion comparable al del promotor 35s. Los ensayos de Western Blot confirmaron una inducci6n de la lisozima luego de hacer heridas en hojas de papa que llevaban el promotor Mas, mientras que no se pudo evidenciar ninguna expresi6n en plantas con el promotor Wun. En un primer intento, los Western blots de plantas transformadas con la construccion Pat-lisozima dieron resultados negativos. Ya que la expresi6n de estas construcciones esta prevista a suceder en las capas mas externas del tuberculo, se debio repetir el experimento solo con este tejido. El promotor GapC4 sera discutido par separado.
Al realizar las analisis de las plantas con expresion de la lisozima T4, se detect6 que estas plantas no solo presentaban resistencia ante bacterias fitopat6genas, sino tambien ante hongos fitopat6genos. Esto no se puede explicar ni la actividad muramidasa, ni con la actividad quitinasa, ya que el hongo Phytophthora infestans implicado en el proceso no contiene quitina en su estructura. Esto nos llev6 a investigar los posibles mecanismos de accion de la lisozima. Un analisis de computadora revel6 varias regiones amfipaticas en la parte terminal C de la lisozima T4 en donde las aminoacidos basicos se encuentran agrupados a un lado de la molecula y los aminoacidos hidrof6bicos se hallan en el otro. De acuerdo con la secuencia en esta region, se han sintetizado dos peptidos que posteriormente fueron evaluados en sus acciones bactericida y fungicida.
Para cumplir con este objetivo, se introdujeron dos sistemas de evaluacion. La acci6n bactericida fue medida par media de un ensayo de conteo en placa (count plating assay). Celulas de E coli fueron incubadas con lisozima T4 par una hara a 37°C. Las soluciones fueron luego sembradas en placas y se contaron las colonias crecidas para el dfa siguiente. La supervivencia de celulas se redujo en una proporci6n muy grande luego de la incubaci6n con lisozima natural, lisozima desnaturalizada par temperatura, lisozima mutante M6K y un peptido (A4). El segundo peptido (A23) no tuvo efecto en la supervivencia de las bacterias. Las medidas de la acci6n fungicida
42

fueron realizadas en el MPI en Cologne en el grupo de Werner Gieffers. Se midi6 el largo de las tubas germinativos de las hongos luego de un periodo de incubacion de varias horas de conidias con lisozima T4 • En el caso de Phytophthora nicotianae/ Fusarium oxysporum y Botiytis cinerea las tubas germinativos fueron mas cortos.
En un ensayo de discos de hoja con Phytophthora infestans razas 1 - 11, algunas llneas transgenicas portando lisozima T4 fueron mas resistentes que las plantas control.
Los resultados con las peptides muestran que la actividad antimicrobiana de T4 reside en la region amfipatica. Se supone que helice cargada positivamente interactua con las componentes de membrana (de carga negativa) de las bacterias u hongos y posiblemente se inserta en la membrana llevando a la muerte celular. Se conoce un caso similar con HEWL.
Estos resultados son la base para la construccion de nuevas protefnas o peptides para aumentar la accion antibacteriana y la estabilidad estructural.
ANTICUERPOS
Se hicieron anticuerpos por media de la tecnica tradicional de hibridomas; los anticuerpos de cadena simple fueron hechos por el metodo de muestreo de fagos (phage display). Las cadenas simples estan constituidas par las regiones variables de las cadenas livianas y pesadas combinadas con un peptido de union. Los anticuerpos fueron probados contra 5 enzimas de pared celular de Erwinia: liasas pecticas C y D, poligalacturonasa y celulasa de Ecc, y contra PME de Ech. En los ensayos de disco de tuberculos, las celulas de Erwinia fueron mezcladas con una cierta cantidad de anticuerpo y se determine el grado de maceraci6n. Hemos encontrado anticuerpos contra la poligalacturonasa y anticuerpos de cadena simple contra las Hasas pecticas Cy D logrando asf inhibir la maceracion.
43
. ~
I

'\ 0
INTRODUCCION
EXPERIMENTOS CON PAPAS TRANSGENICAS EN INVERNADERO Y
CAMPO EN EL PERU Experiments with transgenic potato plants at
field and greenhouses in Peru
Ali Golmirzaie *
El CIP cuenta con tres estaciones experimentales donde mantiene colecciones de germoplasma de papa, camote y, rafces y tuberosas andinas (RTAs). En estas estaciones tambien se realizan pruebas con las materiales mejorados obtenidos par metodos convencionales y/o par biotecnologfa. Las estaciones estan ubicadas en cada una de las regiones agroecologicas mas importantes del Peru. La estaci6n principal esta en la costa, Lima (La Molina); la segunda estaci6n esta en la sierra, en Huancayo, a 3,200 msnm, y la tercera en la ceja de selva, en San Ramon, a 800 msnm.
La investigacion en el CIP esta dirigida hacia la producci6n de papas mejoradas que contribuyan en la soluci6n de las problemas de hambre en el mundo. Para lograr este objetivo, desde 1986, CIP esta aplicando tecnicas de cultivo in vitro que junta con la transformacion genetica se han convertido en herramientas primordiales en el fin de incorparar y/o aumentar la resistencia natural a enfermedades y plagas en las cultivas de papa y camate.
Durante el periodo 1986-1997, se han realizado cerca de 25,000 ensayos con plantas cultivadas in vitro y plantas transgenicas en campas de cultivo a nivel mundial. El numero de cultivas probados en cerca 45 pafses diferentes es aproximadamente 60. Del total de las 25,000 ensayos, aproximadamente el 60% (15,000) se han realizado
* Centro Internacional de la Papa (CIP). Estacion Central, La Molina. Lima - Peru 45
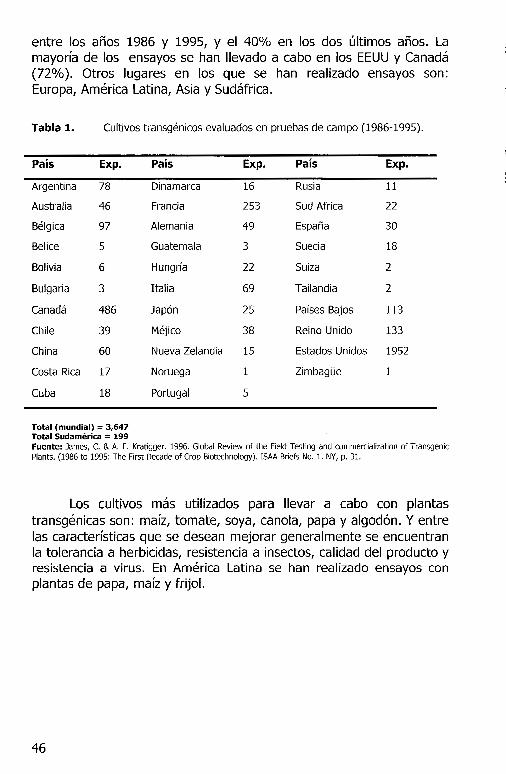
entre los a nos 1986 y 1995, y el 40% en los dos ultimas anos. La mayorfa de los ensayos se han llevado a cabo en los EEUU y Canada (72%). Otros lugares en los que se han realizado ensayos Europa, America Latina, Asia y Sudafrica.
Tabla 1. Cultivos transgenicos evaluados en pruebas de campo (1986-1995).
Pais Exp.
Argentina 78
Australia 46
Belgica 97
Be lice 5
Bolivia 6
Bulgaria 3
Canada 486
Chile 39
China 60
Costa Rica 17
Cuba 18
Total (mundial) = 3,647 Total Sudamerica = 199
Pais
Dinamarca
Francia
Alemania
Guatemala
Hungrfa
Italia
Japan
Mejico
Nueva Zelandia
Noruega
Portugal
Exp. Pais Exp.
16 Rusi a 11
253 Sud Africa 22
49 Espana 30
3 Suecia 18
22 Suiza 2
69 Tailandia 2
25 Pafses Bajos 113
38 Reino Unido 133
15 Estados Unidos 1952
1 ZimbagOe 1
5
son:
Fuente: James, C. & A. F. Kratigger. 1996. Global Review of the Field Testing and commercialization of Transgenic Plants. (1986 to 1995: The First Decade of Crop Biotechnology). !SAA Briefs No. 1. NY, p. 31.
Los cultivos mas utilizados para llevar a cabo con plantas transgenicas son: mafz, tomate, soya, canola, papa y algod6n. Y entre las caracterfsticas que se desean mejorar generalmente se encuentran la tolerancia a herbicidas, resistencia a insectos, calidad del producto y resistencia a virus. En America Latina se han realizado ensayos con plantas de papa, mafz y frijol.
46

REQUISITOS PARA El MANEJO DE PLANTAS TRANSGENICAS EN PERU
1. Establecimiento de las reg/as de bioseguridad: Antes de iniciar trabajos con plantas transgenicas a nivel de invernadero y campo, el CIP desarroll6 un Reglamento de Bioseguridad para el manejo de material transgenico que, luego de ser revisado por el Ministerio de Agricultura, fue aprobado por el Gobierno Peruano.
2. Acuerdo sabre derechos de propiedad inte!ectual con los donantes de genes.· Para poder empezar con los trabajos de transformaci6n genetica, el CIP estableci6 acuerdos con las instituciones donantes de genes en los que se delineaban las reglas del uso de los genes donados y de las derechos de uso de las plantas posiblemente mejoradas resultantes del proceso.
3. Autorizaci6n def Ministerio de Agricu/tura de/ Gobierno Peruano: NingCm experimento de campo puede realizarse sin la previa autorizacic:Sn del Gobierno Peruano, otorgada a traves de su oficina , competente, SENASA.
Sin embargo, existen · algunas restricciones para conducir experimentos con plantas transgenicas:
No debe realizarse experimentos de campo que signifiquen un riesgo de contaminacicSn de media ambiente del lugar donde se desea montar un estudio. En caso de que las pruebas en el invernadero demuestren que dicho riesgo no existe, el experimento puede llevarse a cabo bajo medidas estrictas de bioseguridad.
Por ejemplo: Plantas transformadas con protefnas de cubierta de virus, en el CIP los experimentos con plantas transgenicas para evaluar la resistencia al viroide PSTV, el patcSgeno que causa las mayores perdidas en los cultivos de papa, solo se realizan a nivel de i nvernadero.
47
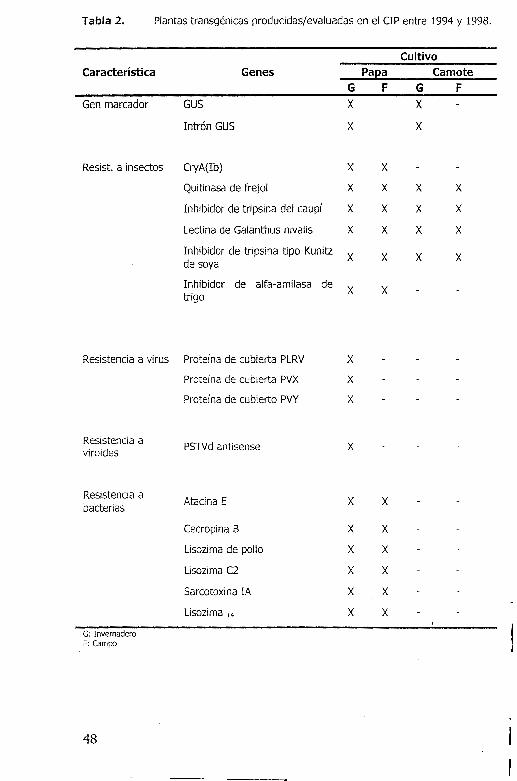
Tabla 2. Plantas transgenicas producidas/evaluadas en el CIP entre 1994 y 1998.
Cultivo
Caracteristica Genes Pa ea Ca mote
G F G F Gen marcador GUS x x
Intr6n GUS x x
Resist. a insectos CryA(Ib) x x Quitinasa de frejol x x x x Inhibidor de tripsina de! caupf x x x x Lectina de Galanthus nivalis x x x x Inhibidor de tripsina tipo Kunitz x x x x de soya
Inhibidor de alfa-amilasa de x x trigo
Resistencia a virus Protef na de cubierta PLRV x Protefna de cubierta PVX x Protef na de cubierto PVY x
Resistencia a PSTVd antisense x
viroides
Resistencia a Atacina E x x
bacterias
Cecropina B x x Lisozima de pollo x x
Lisozima C2 x x Sarcotoxina IA x x Lisozima T4 x x
G: Invernadero F: Campo
48

Para poder llevar a cabo experimentos en campo y/o invernadero se deben tener en cuenta algunas reglas de seguridad basicas:
1. Aislamiento de las plantas con las que se realizara el trabajo. 2. Realizar un manejo cuidadoso del material en evaluacion. 3. Remocion de yemas florales del material transformado. 4. Incineracion de las plantas despues de su evaluaci6n.
En el caso en que se realicen pruebas en el campo, se deben tener en cuenta las siguientes cuidados adicionales:
1. Utilizar barreras o cercos en las bordes del campo, par ejemplo, mafz.
2. Realizar la irrigacion par goteo. 3. Rotacion de cultivo.
TRABAJOS REALIZADOS PARA OBTENER RESISTENCIA A VIRUS
Se han realizado diferentes ensayos para evaluar la resistencia a enfermedades causadas por virus. Todos estos ensayos se han realizado solo en invernadero "Proyectos de Colaboracion" con diferentes instituciones de pafses desarrollados:
(a) Virus del enrollamiento de la hoja de la papa (PLRV): Resistencia a PLRV con el gen de la protefna de cubierta de PLRV (CP-PLRV) proporcionado por CSIRO (Australia). Resultado: dos lfneas transgenicas resistentes.
(b) Virus Y de papa (PVY), Virus X de papa (PVX) : Resistencia a PVY y PVX con genes proporcionados por MONSANTO (USA). Para el ensayo de resistencia a PVY en invernadero se utilizaron plantas de 8 cultivares diploides y 1 tetraploide que fueron inoculados por injerto con plantas infectadas (Solanum tuberosum vr. Rosita) con PVY. Resultado: 5 lfneas diploides resistentes y 4 lfneas tetraploides resistentes. Para el ensayo de resistencia a PVX en invernadero, se utilizaron plantas de 8 cultivares diploides y 1 tetraploide que fueron inoculados mecanicamente con PVX. I Resultado: 6 lfneas diploides resistentes y 2 lfneas tetraploides resistentes. ·
(c) Resistencia a PSTVd: Este trabajo fue realizado bajo un proyecto en colaboraci6n con la Universidad de Lousiana (EEUU). Las
49

construcciones geneticas fueron preparadas por LSU y el CIP realiz6 la transformaci6n genetica y las pruebas de resistencia a nivel de invernadero. Durante las pruebas de resistencia a PSTV se guardan todos los cuidados necesarios para evitar contaminaci6n con este viroide. Resultado: 2 If neas tolerantes a PSTV. Estas lfneas no pueden ser utilizadas porque se pueden comportar como portadores sanos de este pat6geno.
TRABAJOS REALIZADOS PARA OBTENER RESISTENCIA A BACTERIAS
Se estan realizando diferentes proyectos con el objetivo de obtener resistencia a Pseudomonas so/anacearum y Erwinia carotovora. Se estan utilizando genes de peptidos lfticos:
(a) Gen Lisozima T4 (origen: fago T4) provisto por el Inst. For .Breeding Methods in Vegetables (BAZ-Alemania).
(b) Gen de Lisozima C2 ( origen bovine) provisto por Smart Plant Institute (USA).
( c) Gen Sarcotoxina ( origen: mosca de la came - insecto) provisto par Weizmann Institute of Science. En los resultados del ensayo en invernadero de pruebas de resistencia a Erwinia carotovora con el Gen de Sarcotoxina (gen de la mosca de la came) se encontr6 que las plantas transgenicas muestran cierto nivel de resistencia a Erwinia, aun asf se requieren mas ensayos para confirmar los resultados mencionados.
TRABAJOS REALIZADOS PARA OBTENER RESISTENCIA A GORGOJO DE LOS ANDES (APW)
Proyecto realizado en colaboracion con AXIS Genetics ( compafifa inglesa privada). Se realize un experimento en invernadero con la lfnea PWG-70 conteniendo el gen inhibidor de Alpha-amilasa de trigo y una lectina de Galanthus niva/is (gorgojo de los andes). Esta lfnea result6 resistente al ataque del gorgojo, despues de dos meses de inoculaci6n.
Otro experimento fue realizado en campo con plantas transgenicas de papa, transformadas con genes inhibidores de proteasas. Esta prueba fue en San Ramon en 1995.
50

TRABAJOS REALIZADOS PARA OBTENER RESISTENCIA A PTM
Proyecto realizado en colaboraci6n Plant Genetic System entre 1993 y 1997. Se realiz6 la evaluaci6n de este material en su nivel de resistencia a la polilla del tuberculo de papa (Potato Tuber Moth, PTM).
La lfnea transgenica LT-8 (Costanera) mostr6 resistencia a PTM en una prueba de laboratorio (Bioensayo ).
En 1996, en lea, la cosecha de la lfnea transgenica resistente a PTM comparada con cosecha de material control no diferla en sus caracterfsticas agron6micas. Para liberar esta If nea transgenica co mo variedad se estan realizando otros ensayos de campo en Tacna.
51


RESUMEN
TRANSFORMACION DE PLANTAS DE PAPA CON GENES DE LISOZIMA, ATACINA Y
CECROPINA PARA CONFERIR RESISTENCIA A BACTERIAS
Potato Plants Transformation for Anti-Bacterial Resistance with Lysozyme/ Attacin and Cecropine
Genes
Dr. Patricio Arce-Johnson*
La pudrici6n blanda y el pie negro son dos importantes enfermedades que afectan al tuberculo en almacen y a la planta de papa cultivada en el campo respectivamente en Chile. Estas enfermedades son producidas por bacterias del genera Erwinia spp. siendo principalmente Erwinia carotovora subsp. atroseptica la responsable de los mayores danos en nuestro pals. Con el objeto de conferir resistencia a bacterias en este cultivo, transformamos plantas transgenicas de papa de la variedad Desiree para que porten de manera independiente los genes de lisozima de pollo ( chly), atacina acida de Hyalophora cecropia (att), o un peptide analogo a cecropina (sb-37). Mas de 200 lfneas transgenicas de papa resultaron del proceso de transformaci6n vfa Agrobacterium tumefaciens.
Aquellas plantas que manifestaron variaci6n somaclonal (30% aproximadamente, que se reflej6 en cambios en la coloraci6n o la forma de los tuberculos o tambien en la forma de crecimiento de la plantas), fueron descartadas. Aquellas lfneas que mantuvieron las caracterfsticas fenotfpicas de la variedad Desiree, fueron analizadas para verificar la presencia y expresi6n del gen de interes mediante la evaluaci6n de su resistencia a Erwinia spp.
Aproximadamente, el 5% de las If neas analizadas de lisozima, el 7% de las lfneas de atacina, y el 13% de las lfneas de cecropina mostraron
* Departamento de Genetica Molecular y Microbiologla, Fae. de Ciencias Biol6gicas, 53 Pontificia Universidad Cat61ica de Chile. Apartado 114-D, Santiago, Chile

alta resistencia al pie negro o a la pudrici6n blanda en ensayos de invernadero y laboratorio.
PRESENTACION
En Chile se producen anualmente importantes perdidas econ6micas en los cultivos de papa a consecuencia de las enfermedades bacterianas causadas par Erwinia spp. Estas bacterias infectan el tuberculo en almacen pudriendolo completa o parcialmente causando lo que se llama "pudrici6n blanda". En el campo, las plantas afectadas desarrollan el "pie negro", que se caracteriza par un empardecimiento y posterior necrosis en la base del tallo. Ambas afecciones se presentan principalmente en el sur de nuestro pals en zonas caracterizadas por alta humedad y bajas temperaturas, siendo Erwinia carotovora subsp. atroseptica/ la principal bacteria responsable de dichas enfermedades.
Con el prop6sito de obtener resistencia a Erwinia spp. en papas, aplicamos tecnicas de ingenierfa genetica a este cultivo. Mediante transformaci6n binaria via Agrobacterium tumefaciens, produjimos 214 llneas transgenicas de papa que portan uno de los siguientes genes: lisozima de polio ( chly) con su correspondiente peptide senal o atacina acida de Hya/ophora cecropia (att) o un peptide analogo a cecropina (sb-37). Todas las construcciones estan bajo el control transcripcional del promotor 355 del virus mosaico de la coliflor, y la secuencia terminadora del gen de la nopalina sintetasa de Agrobacterium spp. (nosT). Ademas, portan el gen de neomicina fosfotransferasa II (nptll) bajo el mismo promoter, que confiere a la planta resistencia al antibi6tico kanamicina. Todas las plantas que presentaron variaci6n somaclonal (63 llneas, equivalentes al 30% ), que se reflej6 principalmente en deformaci6n y cambios en la coloraci6n de los tuberculos o cambios en la apariencia de la planta o de su habito de crecimiento, fueron descartadas. El resto de las llneas fueron analizadas mediante las tecnicas de Southern-blot para verificar la presencia y numero de copias del transgen, y de Northern-blot para evaluar el nivel de expresi6n de los ARN mensajeros de las genes bactericidas, ademas se les hizo un analisis de resistencia a bacterias en tuberculos y plantas.
Para evaluar la resistencia a Erwinia spp. en papa se montaron dos tipos de ensayo. El primero consisti6 en evaluar susceptibilidad a
54

desarrollar pie negro en esquejes de plantas transgenicas de papas tratadas con Erwinia spp. en concentraciones de 5.8xl04 y 5.8x106
unidades formadoras de colonias par mililitro de cultivo (ufc/ml).
Con este fin, se depositaron los esquejes en vasos que contenfan una mezcla de perlita/tierra y posteriormente se les reg6 una vez con la solucion de bacterias. Se evaluo a los 4 y 6 dfas despues de la infeccion para determinar si hubo desarrollo de sfntomas, de acuerdo a un fndice de dano entre 1 y 5. En este fndice, el valor 1 correspondfa a esquejes que no presentaron sfntomas de la enfermedad, y el valor 5 correspondla a esquejes completamente necrosados. Valores intermedios correspondlan a daifos porcentuales en las esquejes.
Para comparar el grado de resistencia entre las lfneas transgenicas con el control, regado con agua, se determino un coeficiente de resistencia. El coeficiente de resistencia resulta de la division del lndice de resistencia promedio de las plantas controles, entre el lndice de resistencia promedio de las plantas transgenicas. Con este tipo de analisis en invernadero, el 16% de las plantas que portaban el gen de la lisozima, el 35% que portaba el gen de la atacina, y el 45% que portaba el gen de la cecropina, presentaron algCm grado de resistencia a Erwinia spp. Sin embargo, solo unas pocas lfneas presentaron alto nivel de resistencia a Erwinia carotovora subsp. atroseptica.
El segundo ensayo se realiz6 para evaluar la resistencia en tuberculos a la pudricion blanda. Para ello, utilizamos mitades de tuberculos que fueron tratadas con una solucion de 106 ufc/ml de Erwinia carotovora subsp. atroseptica.
Cada mitad de tuberculo fue depositada en el fondo de un vaso plastico transparente y cubierta con tierra esteril. Con este procedimiento fue posible ir examinado la evolucion de las sf ntomas en las tuberculos en el tiempo, sin destruir o remover el ensayo. Se evaluo la capacidad de brote de las tuberculos tratados con bacterias o con agua (en el caso del control) a lo largo del tiempo. De acuerdo a este ensayo, se obtuvo un nivel de resistencia media o alto en 12 de 69 llneas transformadas analizados de lisozima. De la misma manera, se obtuvo un grado de resistencia a la pudrici6n blanda superior al control en 11 de 35 lfneas transformadas analizados de atacina, y en 14 de 31 lfneas transformadas analizados de cecropina.
55

En el caso de las llneas transgenicas de lisozima, cecropina y atacina que presentaron alto nivel de resistencia a Erwinia, encontramos generalmente una alta correlaci6n con el nivel de expresi6n del transgen. En aquellos cases en los que esto no ocurri6, el nivel de resistencia fue atribuido a las diferencias fisiol6gicas propias de las plantas y tuberculos, que son dificiles de controlar cuando se realiza el ensayo de resistencia.
A modo de resumen general, actualmente contamos con aproximadamente 6 diferentes clones transgenicos que portan alguno de los genes bactericidas descritos y que en ensayos de laboratorio e invernadero presentan alta resistencia a Erwinia spp.
Recientemente, hemos producido plantas transgenicas de papa que portan conjuntamente el gen de la lisozima de pollo y el gen ap-24 que corresponde a una protelna relacionada con patogenesis (PRS), del grupo de la osmotina tipo taumatina. Este gen afecta la membrana plasmatica de la celula, par lo que esperamos un efecto sinergico al de la lisozima en la actividad bactericida. Las Hneas transgenicas producidas con estas construcciones dobles las evaluaremos en laboratorio y terreno durante el ano 1999.
56

HIBRIDIZACION ENTRE Y DENTRO DE VARIEDADES CULTIVADAS Y
SILVESTRES DE PAPA Hibridization within and between cultivated
and wild species
Bodo Trognitz *
Las plantas transgenicas se han hecho cada vez mas disponibles para la experimentaci6n y uso en la agricultura. Este desarrollo ha aumentado la preocupaci6n por la protecci6n de la biodiversidad frente a la posible infiltraci6n de genes foraneos (extranos a la papa). Un investigador cuidadoso debe probar la hip6tesis de que un transgen puede ser transmitido en forma creciente desde el material modificado geneticamente hacia sus parientes silvestres y cultivados. El mecanismo mas importante que puede llevar a este tipo de transferencia es la recombinacion meiotica luego de una polinizacion cruzada accidental y una fertilizaci6n exitosa subsecuente en los parientes silvestres o cultivados que puedan hallarse creciendo al lado del campo transgenico. Este escenario podrfa llevar a la suposicion de que cualquier hfbrido potencialmente resultante podrfa transmitir el transgen a poblaciones de especies relacionadas.
El objetivo del presente estudio de factibilidad de cruce entre la papa cultivada y salvaje es resaltar los procesos que influyen en el "outcrossing" y la formacion de organismos hfbridos, evaluar las oportunidades que un transgen podrfa incorporarse accidentalmente en el contenido genetico de una especie relacionada.
Las condiciones necesarias para que se lleve a cabo una fertilizaci6n cruzada exitosa implica la ruptura de ambas barreras, espacial y biol6gica, que normalmente previenen que este evento suceda (Tabla 1).
* Centro Internacional de la Papa (CIP). Estacion Central, La Molina. Lima - Peru 57

Tabla 1. Barreras de la polinizaci6n cruzada, entre especies de papa relacionadas. Las barreras estan enumeradas en orden jerarquico de arriba a abajo. Basta que una de las cuatro barreras permanezca intacta para prevenir la hibridaci6n.
Complejidad Barreras Componentes
Espacial Distancia Insectos polinizadores Baja propensi6n del polen de papa a ser transportado por el aire
Tiempo de floraci6n Tiempo de vida largo del polen
Biol6gica Pre cig6tica Fertilidad (pol en, 6vulo) Incompatibilidad estilar
Post cig6tica Incompatibilidad gen6mica Incompatibilidad citoplasmatica Ploidfa
Se ve claramente que el aislamiento espacial de las plantas transgenicas puede ser llevado a cabo en su totalidad bajo condiciones experimentales. Sin embargo, el aislamiento de un cultivo transgenico en condiciones agrfcolas regulares no puede ser garantizado bajo todas las circunstancias, ya que el aislamiento espacial del cultivo no puede alcanzarse completamente.
Consecuentemente, el potencial de las barreras biol6gicas para prevenir un flujo genetico accidental, debe ser discutido.
Se les llama barreras pre cig6ticas a las procesos que evitan la fertilizaci6n. La fertilidad es un requerimiento basico para la hibridizaci6n. Los sistemas de esterilidad citoplasmatica masculina (Grun, 1979) pueden ser utilizados eficientemente para reducir la fertilidad del polen de un genotipo utilizado para transformaci6n genetica. Estos sistemas pueden causar tambien la esterilidad de una progenie putativa en caso de que ocurra una hibridizaci6n con un ancestro silvestre, ademas reduce la posibilidad de multiplicaci6n de un transgen.
La incompatibilidad de estilos es un mecanismo importante que dirige o previene la hibridizaci6n entre dos plantas fertiles de papa. Las dos formas, la autocompatibilidad y la incongruencia unilateral ( o incompatibilidad interespedfica), estan condicionadas en parte par el
58

mismo loci genetico (Bernacchi y Tanksley, 1997). El polen de genotipos diploides autoincompatibles frecuentemente puede crecer con exito dentro de los estilos de una planta autocompatible, mientras que el cruce redproco usualmente no llega a su fin (Trognitz y Schmiediche, 1993). Los estilos de papas tetraploides (5. tuberosum ssp. tuberosum y ssp. andigena) frecuentemente no apoyan el desarrollo de los tubes de polinizaci6n de sus parientes silvestres (Tabla 2). Sin embargo, los experimentos en polinizaci6n han demostrado que el polen de papa comun puede crecer hacia el ovario a lo largo de los estilos de varies parientes tetraploides (Figura 1).
Para saber los grades de crecimiento de los tubes poll nicos, ver Tabla 2. B: formaci6n de la baya; no P, no hay polen remanente en el estigma despues de la polinizaci6n.
Las barreras postcig6ticas actuan durante y despues de la fertilizaci6n, llevan a aberraciones del embri6n y del desarrollo de la semilla y previenen de esta manera la formaci6n de hfbridos. Los factores gen6micos hipoteticos que evitan el desarrollo del embri6n debido a la malformaci6n del endospermo se conocen como los factores del numero de balance endospermico, EBN (Johnston y Hanneman, 1980). Los valores de EBN entre 1 y 4 han sido asignados a varias especies de so/anum que forman tuberculos (Johnston et al., 1980) y la hibridaci6n solo puede ser exitosa entre parejas con numero EBN identico. En este caso, el entrecruzamiento de la papa comun con un valor EBN de 4 se vera reducido a especies relacionadas con el mismo valor, los que representa menos del 20% de todas las especies conocidas.
La habilidad de apareamiento, mei6tico y mit6tico, de los cromosomas hom61ogos tambien contribuye a la reducci6n de la hibridaci6n exitosa. Varias combinaciones de los cinco genomas de papa y sus parientes: A, B, C, D y E (Matsubayashi, 1991) llevaron a distorsiones en el emparejamiento y recombinaci6n. Sin embargo, estos problemas puede ser sobrepasados per algunas plantas con la union de gametes no reducidos, que llevan en sf un genoma diploide en lugar de un equivalente al genoma haploide y resultan en embriones alopoliploides con emparejamiento normal de los cromosomas hom61ogos.
59

Tabla 2. Grado de crecimiento del tuba polfnico de un diploide silvestre y polinizadores tetraploides de papa dentro de los estilos de cultivares de papa.
Fem.
Cruz a 148
Molinera Perricholi
Fem.
Cruz a 148
Acg abz amb 7* 12 3
3 3 1 1
oxc Phu pcs 10 10 3
Molinera 10 2 16 9 Perricholi 16 3
Masculino bue ber Chq chm cmm Cnd 9 2 15 3 10
9 1 2
9 15
Masculino
2 5
10 1
pur sol uru San mol 18 16 10 10 6
9 9
10 9 12 2
3 9
2
1 3
lpt 3
2
lxs 2
12 3
mcc 15
10 3
brd etb 12 13
13 13 11
Grados de crecimiento del tubo polfnico entre 1-3 son suficientes para la fertilizacion y el desarrollo de la semilla, los valores entre 4-9 raramente resultan en fertilizacion y valore entre 9-18 previenen eficazmente el contacto de los tubos polfnicos con el ovario, y asf excluyendo la posibilidad de polinizacion. Para las abreviaciones de los nombres de la especies, ver Huaman y Ross (1985).
Este mismo mecanismo puede tambien llevar a la formaci6n de embriones fertiles (euploides) en un cruce entre especies de diferente nivel de ploidla. Por ejemplo, el cruce de una planta de papa tetraploide con un pariente diploide resulta normalmente en hfbridos triploides esteriles, que se hallan sexualmente aislados de sus progenitores y de cualquiera de los parientes euploides. Sin embargo, cuando el progenitor diploide de tal cruce forma gametes no reducidos, diploides, un cruce de estos con una papa tetraploide, con gametes diploides regulares, resultarfa en una progenie tetraploide fertil.
Finalmente, las combinaciones de tipos espedficos de citoplasma con tipos particulares de genoma, pueden resultar en una progenie esteril o no viable (Lossl, 1996).
Los cruces interespedficos que ocurren dependen de un sistema de circunstancias, como la presencia de genotipos y condiciones que lleven a la hibridaci6n. Varias barreras para la hibridizaci6n se hayan virtualmente en cualquier situaci6n y la presencia de solo una o algunas de ellas es suficiente para hacer la hibridaci6n imposible. Sin embargo, pueden existir situaciones aisladas en las que te6ricamente ocurra la hibridaci6n. Asf, el riesgo de un escape accidental de un transgen debe estar completamente eliminado bajo condiciones de experimentaci6n apropiadas y la liberaci6n de cultivos de papa alterados geneticamente, especialmente para la
60

producci6n, en el area de distribuci6n de parientes del cultivo debe ser considerada cuidadosamente bajo un sistema de estudio de las riesgos caso a caso.
REFERENCIAS
Bernacchi D, SD Tanksley 1997. An interspecific backcross of Lycopersicon esculentum x L. hirsutum: linkage analysis and QTL study of sexual compatibility factors and floral traits. Genetics 147: 861-877.
Grun P 1979. Evolution of the cultivated potato: a cytoplasmic analysis. In: The Biology and Taxonomy of the Solanaceae. Linnean Society Symposium Series, No.7. Eds. Hawkes JG; RN Lester AE Skelding, London, Academic Press Inc. :655-665.
Huaman Z, H Ross, 1985. Updated listing of potato species names, abbreviations and taxonomic status. Amer Potato J 62: 629-641.
Johnston SA, RE Hanneman Jr. 1980. Support of the endosperm balance number hypothesis utilizing some tuber-bearing Solanum species. Amer Potato J 57(1):7-14.
Johnston SA, TPM den Nijs, SJ Peloquin, RE Hanneman Jr. 1980. The significance of genie balance to endosperm development in interspecific crosses. Theor Appl Gen 57: 5-9.
Lossl A 1996. Analyse der DNA variabilitat in Zellorganellen somatischer Fusionshybriden der Kartoffel und deren phanotypische Auswirkungen. Ph. D. thesis, Dissertations Druck Darmstadt GmbH, Darmstadt, Germany: 111 p.
Matsubayashi M, 1991. Phylogenetic relationships in the potato and its related species. In: Tsuchiya T, PK Gupta (eds.), p. 93-118. Chromosome engineering in plants: genetics, breeding, evolution. Part B. Elsevier Science Publishers BV, Amsterdam.
Trognitz BR, PE Schmiediche 1993. A new look at incompatibility relationships in higher plants. Sex Plant Reprod 6: 183-190.
61

Figura 1.
rrtrn 10-13
a::i 10-13
tt.qO 13
Jxt 13
h.q2 11
i:x:;i
a1J 13
trr
62
Incompatibilidad unilateral entre especies solanaceas tetraploides, expresada por el grado de crecimiento del tubo polf nico dentro de un estilo.
Yalaes:
1-3 irtibid6n &bil o ma, r:ffibilic:Ecl c:E fertilizadOn.
9-18 irtiad6n fu:rte en la p:irte SLµrior cl2I estigra,
sin r:ffialic:Ecl c:E fertilizad6n.
Especiesde .2>'.nmErtreouzaclas:
:d sdisii
oxc oxyGJrpun
rrtrn rmt:amn
cd craie
h.q tup:rrense
Jxt jB:uia (a 4x hbicb)
JXj {J3Uiju;µn
a1J arrig:na trr lliErrELm

INVESTIGACION EN BIOSEGURIDAD EN ALEMAN IA: PAPAS CON LISOZIMA T 4 Y SUS
IMPLICACIONES EN LA COMUNIDAD MICROBIANA COMO UN CASO DE ESTUDIO
Biosafety Researh in Germany: T,, Lysozyme Potatoes and Their Implications on the Microbial Community
as a Case Study.
Andreas Mahn *
INTRODUCCION
En 1990, el Ministerio Aleman para la Educacion y la investigacion estableci6 el programa llamado "Biotecnologfa 2000", con los siguientes objetivos:
•!• Fortalecer la investigaci6n como base para las provisiones de salud, ambiente y energfa.
•!• Desarrollar el potencial de la biotecnologfa para su aplicacion en la economfa.
En las anos 1995 y 1996 se destino un fondo promedio de 150 millones de marcos alemanes anuales para la promocion de proyectos de investigacion. Las siguientes areas de investigacion recibieron mayor importancia:
•!• Desarrollo de metodos para la biotecnologfa (DM 4 millones) •!• Metodos alternativos para experimentos con animales
(DM 7.5 millones) •!• Bioseguridad y evaluaci6n de riesgos (DM 7.5 millones) •!• Investigacion en biologfa celular y protefnas (DM 10 millones) •!• Investigacion neurobiologica (DM 4 millones) •!• Investigacion genomica (OM 35 millones)
* Federal Center for Breeding Research on Cultivated Plants (BAZ), Quedlinburg, Germany
63

•!• Mejoramiento de plantas y desarrollo de pesticidas biol6gicos (DM 9.5 millones)
•!• Investigaci6n molecular en sustancias naturales (DM 20 millones) •!• Bases biol6gicas de las tecnologfas destinadas a la reducci6n de la
poluci6n ambiental (DM 5 millones)
En 1997, el programa "Biotecnologla 2000", y especialmente el campo de la bioseguridad y la evaluaci6n del riesgo, se complement6 con el programa "BioMonitor", promocionando las siguientes aspectos de investigaci6n:
•!• Ecologfa y monitorizaci6n de plantas transgenicas en campo. 'r Entrecruzamiento de transgenes durante el cultivo de plantas
tolerantes a herbicidas Y Influencia de las plantas resistentes a virus en la emergencia
de poblaciones de pat6genos alteradas ";;- Efectos de la expresi6n de la toxina Bt en plantas transgenicas
sabre la poblaci6n de insectos 'r Efectos de la resistencia a pat6genos geneticamente
aumentada en plantas asociadas a bacterias y hongos >-- Efectos de las alteraciones metab61icas en las caracterfsticas
fitopatol6gicas de las plantas transgenicas •!• Ecologfa molecular de los microbios •!• Ingenierfa genetica y alimentos •!• Vectores para la terapia genetica somatica
PROYECTO: "Efecto del cultivo de papas transgenicas (con lisozima T 4 )
sobre Los microorganismos en el campo"
Este proyecto de investigaci6n es resultante de la colaboraci6n entre las siguientes instituciones:
a) Federal Centre for Breeding Research on Cultivated Plants, Quedlinberg. Colaboradores: Klaus DUring, Andreas Mahn, Ina Hempbel.
64
•!• Producci6n de material vegetal y organizaci6n de ensayos de cam po
•!• Producci6n de lisozima T4
•!• Efecto de la lisozima T4 en celulas eucari6ticas •!• Efecto de la micorrizaci6n en rafces de papa

b) Federal Biological Research Centre for Agriculture and Forestry, Braunschweig. Colaboradores: Cornelia Samalla, Holger Heuer. •!• Efectos en las comunidades bacterianas de la riz6sfera
(incluyendo bacterias no cultivables) c) Universidad de Oldenburg. Colaboradores: Wilfried Wackernagel,
Johann de Vries, Klaus Harms. •!• Liberacion de la lisozima T4 y su actividad en el suelo •!• Liberacion del AON vegetal y su persistencia en el suelo •!• Transferencia genetica horizontal
d) University of Restock. Colaborador: Inge Broer •!• Efectos en Rhizobia. Nodulaci6n y fijaci6n de nitr6geno en
leguminosas e) University of Restock. Colaboradores: Gabriele Berg, Jana
Lottmann. •!• Efectos de las bacterias beneficiosas y sus antagonistas.
Las investigaciones de estos grupos han permitido obtener importantes hallazgos respecto al uso potencial de la lisozima T4 coma agente bactericida. A continuacion se resumen algunos aspectos:
•!• Se produjeron varias lfneas transgenicas de papa del cultivar Desiree, conteniendo tanto el gen de la lisozima T4 coma el gen marcador NPTII de resistencia al antibi6tico kanamicina. De la misma manera, se obtuvo plantas transgenicas del mismo cultivar, pero conteniendo solamente el gen NPTII.
•!• La accion bactericida de la lisozima T4 no solo es debida a la actividad enzimatica sino tambien a la actividad sabre la membrana celular. Para demostrarlo, se realiz6 una prueba con protoplastos de papa utilizando SYTOX, un sistema de tincion verde-fluorescente de acido nucleico; este colorante es impermeable a la membrana y tine solo a las protoplastos con la membrana danada. Despues de aplicar lisozima de huevo de gallina y T4, a un grupo de protoplastos, pudo apreciarse que muchos protoplastos tuvieron la membrana danada, en comparacion a las controles con PBS ("Phosphate Buffered Saline") o BSA ("Bovine Serum Albumin").
•!• En un ensayo de hemolisis, la lisozima T4 no mostr6 reacci6n.
•!• Se redujo la micorrizacion de las rafces de las lfneas transgenicas que expresaron lisozima T4 •
65

66
•!• Se tomaron muestras bacterianas de la riz6sfera de plantas transgenicas y controles sembrados en campo. Se aisl6 una fracci6n bacteriana y se establecieron cultivos axenicos. Para investigar aquellas bacterias que pudieron cultivarse, se aisl6 la fracci6n total del ADN bacteriano y los fragmentos 16S del rADN fueron amplificados por PCR. El uso de DGGE (Denaturating Gradient Gel Electrophoresis) evidenci6 un "fingerprint" caracterfstico; el secuenciamiento posterior de las bandas halladas no mostr6 diferencias entre las plantas transgenicas y los controles, pero sf una disminuci6n de los acti nom icetos.
•!• Utilizando placas con medias de cultivo, se estudi6 el efecto de la lisozima T4 sobre diferentes bacterias pat6genas de plantas asf como del suelo. Esto permiti6 apreciar que la lisozima T4 tuvo mayor efecto sobre las bacterias grampositivas Bacillus subtilis y Clavibacter michiganense, y sabre la bacteria gram-negativa Xanthomonas campestris. Ademas, una prueba espectrofotometrica empleando B. subti!is en 0.3X PBS y extractos de tuberculos de papa, mostr6 que dicho extracto proveniente de papas transgenicas expresando lisozima T4 causaron una disminuci6n significativa de la densidad 6ptica por minuto, respecto a los controles. Sin embargo, la actividad de esta enzima estuvo disminuida drasticamente en el entorno qufmico del suelo (tierra, arena, podzol, loamy soil), permitiendo un mayor crecimiento de las poblaciones de B. subti!is. Par otro lado, ha sido posible la detecci6n de la lisozima T4 a traves de un Western blot realizado con una soluci6n de crecimiento de plantas transgenicas, demostrando que la lisozima T4 es excretada de la planta transformada.
•!• La expresi6n de lisozima T4 en rafces transgenicas de Vicia hirsuta, una planta hospedera de Rhizobia, redujo el numero de n6dulos inducidos por esta bacteria. Sin embargo, el experimento de campo mostr6 un efecto distinto. Se sembraron plantas de Vicia hirsuta en los alrededores de plantas transgenicas de papa y luego fueron cosechadas las primeras, contandose el numero de n6dulos formados asf como la tasa de fijaci6n de nitr6geno mediante cromatograffa de gas, no encontrandose diferencias respecto a los experimentos con papas control.

En resumen, los datos obtenidos de los experimentos de campo han permitido demostrar que no siempre estos resultados coinciden con aquellos obtenidos bajo condiciones controladas de laboratorio.
67


lQUE ES EL SENASA?
ROL DEL SENASA EN LA SANIDAD VEGETAL DEL PERU
SENASA: It's role on Peruvian agriculture health
Enrique Carranza Hernandez *
El Servicio Nacional de Sanidad Agraria, SENASA, es el organismo responsable de desarrollar y promover la participacicSn de la actividad privada y de las instituciones publicas, para la ejecucicSn de los Planes y Programas de PrevencicSn, Control y Erradicacion de Plagas y Enfermedades que inciden con mayor significacion socioeconomica en la actividad agraria. A su vez es la institucion responsable de cautelar la seguridad sanitaria del Agro Nacional.
El SENASA ha sido creado por Decreto Ley N° 25902 el 27 de noviembre de 1992, es un organismo publico descentralizado del Ministerio de Agricultura, con personerfa jurfdica de Derecho Publico Interno, autonomfa tecnica, administrativa, economica y financiera; y se rige por su Reglamento de OrganizacicSn y Funciones, aprobado por Decreto Supremo N° 24-95-AG.
lCUAL ES LA FUNCION DEL SENASA?
La principal funcion del SENASA, es la de preservar la sanidad en el campo agrfcola, a traves de la ejecuci6n de planes y programas de prevenci6n, control y erradicaci6n de plagas y enfermedades que afecten o puedan afectar el normal desarrollo de la actividad agropecuaria en el pafs.
El SENASA promueve y desarrolla la participacion de la actividad publica y privada en la busqueda y consolidacion de un marco de seguridad sanitaria, que garantice, ayude al mantenimiento y logro de ventajas competitivas del sector agrfcola y pecuario en los mercados i nternaciona les.
* Programa de Marchitez Bacteriana. Servicio Nacional de Sanidad Agraria (SENASA) Ministerio de Agricultura. Lima, Peru 69

PRINCIPALES FUNCIONES
•!• Controlar y supervisar el estado sanitario de animales, vegetales y de productos e insumos agrarios, en el comercio nacional y en el de importaci6n o exportaci6n que realice nuestro pafs.
•!• Normar las aspectos sanitarios dentro de las actividades de importaci6n, exportaci6n, comercializaci6n, transito interno de animales y vegetales; asl coma productos e insumos agrarios.
•!• Proponer al Ministerio de Agricultura las normas de alcance nacional y regional en relaci6n a las actividades de vigilancia, inspecci6n, registro, control, supervision y evaluaciones sanitarias del agro.
•!• Planear y organizar programas y proyectos de sanidad agraria a desarrollarse en el pals, a traves de entes estatales o privados, con recursos nacionales y apoyo de la Cooperaci6n Tecnica Internacional.
•!• Realizar analisis de riesgo e impacto sanitaries, a fin de generar la informaci6n tecnico econ6mica necesaria para predecir la magnitud de las efectos potenciales o reales; coma consecuencia de la comercializaci6n de determinadas plantas, animales e insumos agrarios.
•!• Organizar y conducir el sistema de cuarentena nacional, evitando la introducci6n de nuevos problemas sanitarios y la dispersion de las existentes hacia otras regiones o pafses libres de ellos.
Debido a las adelantos tecnol6gicos en el campo cientffico para lograr desarrollar resistencia en las cultivos a plagas y enfermedades mediante la utilizaci6n de productos transgenicos, con la intenci6n de elevar el nivel socioecon6mico de las agricultores al obtener mejores rendimientos en sus cosechas; y debido a las polfticas existentes de libre comercio, el Servicio Nacional de Sanidad Agraria - SENASA, responsable de velar par las importaciones de origen vegetal que ingresen al palsy sabre todo de aquellas que puedan poner en riesgo la conservaci6n y explotaci6n sostenible de la diversidad biol6gica y la salud humana, actua en forma responsable al ser integrante del Grupo Nacional de Trabajo en Bioseguridad, en donde actualmente se
70

elaboro un Proyecto de Ley de Seguridad de la Biotecnologfa que ya ha sido elevado al Congreso de la Republica para su revision.
Actualmente solo se esta permitiendo la importaci6n de material transgenico al Centro Internacional de la Papa, par ser una instituci6n de investigaci6n seria y de prestigio internacional que acata las disposiciones reglamentarias del pafs. Se cuenta con la RESOLUCION MINISTERIAL N° 0682-94-AG; en esta se resuelve:
Artfculo 1°. Aprobar las Normas Internas de Biotecnologfa y Bioseguridad del Centro Internacional de la Papa (CIP) para la experimentacion y utilizacion de Organismos Modificados Geneticamente (GMO) que contiene 16 paginas y 4 formularios de procedimientos, las mismas que coma anexo forman parte integrante de la presente Resoluci6n.
Artfculo 2°. El Servicio Nacional de Sanidad Agraria - SENASA, mediante la Direcci6n General de Sanidad Vegetal, sera la entidad oficial encargada de supervisar, normar y evaluar la movilizaci6n de material biologico nacional resultante de la experimentacion de GMO de importaci6n, cuarentena post - entrada y autorizaciones para estudios de campo y distribuci6n par la Red Internacional del Centro Internacional de la Papa - CIP para las cultivos de papa, camote y, tuberculos y rafces andinas.
Artfculo 3°. El Servicio Nacional de Sanidad Agraria, designara a un representante especializado para integrar el Comite de Bioseguridad, quien informara permanentemente de las avances y alternativas alcanzados, para su utilizacion en la agricultura nacional, asf coma recomendara las medidas requeridas para la mutua colaboraci6n institucional.
El procedimiento para la importaci6n de este tipo de material, es el siguiente:
1. Solicitud de los requisitos fitosanitarios para la importacion. 2. Solicitud del Formato "O", en el cual se alcanza al SENASA las datos
acerca de la ubicacion del predio de post - entrada. 3. Comunicacion a la Coordinacion a cargo del lugar especificado en el
Formato "O", para la verificacion del predio en donde se llevara a cabo la cuarentena post - entrada.
71

4. Determinaci6n del Predio (Aprobaci6n o no aprobaci6n). 5. Emisi6n de los requisites fitosanitarios para la importaci6n. 6. Autarizaci6n par parte de SENASA de las estudias de campa y
distribuci6n de las cultivas. 7. El Camite de Biaseguridad informara peri6dicamente al SENASA
acerca de las avances y alternativas alcanzadas en la siembra de estas cultivas.
72

INTRODUCCION
COMO LIBERAR PLANTAS TRANSGENICAS RESISTENTES A
PLAGAS EN CAMPO How to Set Free in Held Transgenic Plants
Resistant to Pests
Jorge Benavides *
La polilla de la papa, Phthorimaea operculella, es la plaga mas importante de papa en zonas tropicales y subtropicales a nivel mundial. Los intentos de control biologico clasico han tenido exitos solo en limitadas producciones, y la resistencia genetica serfa el principal componente en un manejo integrado de plagas. La resistencia genetica obtenida hasta ahora par mejoramiento clasico no ha podido proveer suficiente resistencia contra la plaga.
La produccion de lfneas transgenicas de importantes variedades de papa con resistencia a la polilla ha sido alcanzadas exitosamente en el laboratorio y en ensayos de invernadero, las cuales han sido introducidas en pruebas de campo bajo condiciones naturales.
PROYECTO COLABORATIVO
En 1992, el Centro Internacional de la Papa (CIP), y la empresa privada belga llamada "Plant Genetic Systems" (PGS), iniciaron un proyecto en colaboracion para desarrollar variedades transgenicas de papa con resistencia a la polilla en el campo y en almacen. Esta estrategia consiste en introducir el gen de Bacillus thuringiensis (Bt) que codifica una protefna insecticida, en el genoma de la papa.
* Centro Internacional de la Papa (CIP). Estacion Central, La Molina. Lima - Peru 73

RESULTADOS
Diez importantes variedades de papa, (Tabla 1) han sido transformadas utilizando el sistema de transformaci6n vfa Agrobacterium tumefaciens llevando el gen Bt. Las lfneas transgenicas han mostrado alta resistencia contra el ataque de la larva de la polilla en hojas y tuberculos, en comparaci6n con plantas de la misma variedad no transformadas. Mediante pruebas moleculares y bioquimicas hemos comprobado la inserci6n y la expresi6n del gen Bt en estas plantas transgenicas.
Un total de 81 lfneas transgenicas de las variedades Costanera, Sangema y Cruza-148 han sido evaluadas en campo (Tabla 2), siguiendo las medidas de bioseguridad establecidas en la Gu fa de Bioseguridad del CIP.
De 48 lfneas transgenicas evaluadas en campo de la variedad Costanera, 9 han sido seleccionadas par sus caracterfsticas agron6micas y de resistencia a la plaga, las cuales han sido evaluadas nuevamente en Tacna. Actualmente, se estan evaluando en campo, el restante de lfneas transgenicas de las variedades Sangema y Cruza-148.
El manejo de resistencia de la polilla a plantas transgenicas resistentes en campo, se ha analizado en el Taller con el ejemplo reportado para la principal plaga del algod6n en las Estados Unidos que es la He/iotis virences. El manejo propuesto para evitar el aumento de poblaci6n resistente en la plaga serfa de colocar un refugio de plantas no transformadas para mantener la poblaci6n susceptible, con la que puedan cruzarse con la poblaci6n resistente sobreviviente en las plantas transgenicas y producir una poblacion heterocigotico susceptible. Se ha propuesto que sembrando plantas no transformadas (refugio) hasta 4% del total de area sembradas con plantas transgenicas retardarfa la resistencia en par lo menos 10 afios.
74

Tabla 1. Relacion de plantas transgenicas de papa par variedad.
Pais Variedad Origen Atributos #
Lineas transg.
Revoluci6n Peru Peru, Bolivia 27 Perricholi CIP Peru, Bolivia, Guatemala, LB, FT 2
Burundi, Etiopfa, Uganda
Cruza-148 Mexico Burundi, Ruanda, LB, BW 115 Uganda, Zaire
Achirana lnta Argentina China, Nepal, Kenia, PLRV, LB, H 83 Madagascar
Sangema Rwanda Burundi, Ruanda, Zaire LB 46
Parda Colombia 125 Pastuza
Costanera CIP Peru PVX, PVY, 80 TPS
Marfa CIP Peru LMF 90 Tambena Atzimba Mexico Costa Rica, Zaire 43
Desiree Egipto, Tunez, 60 Marruecos, Chile
Tabla 2. Uneas transgenicas de papa evaluadas en campo, 1994-1998.
Variedad Lineas Selecci6n Luga res
Costanera 48 (3X) 9 (3X) lea Ta en a
Sangema 4 (3X) ND lea 10 (lX)
Cruza-148 19 (2X) ND Arequipa
75


, C\
EXPERIMENTO EN CAMPO: PAPAS RESISTENTES AL ATAQUE
DE BACTERIAS Field Trial:
Potato plants resistant to bacterial attack
Luis Nopo *
OBJETIVO
Evaluar plantas transgenicas de papa bajo condiciones de campo.
EXPERIMENTOS EN LABORATORIO E INVERNADERO
En el laboratorio se realiza el proceso de transformaci6n genetica de papa. Para llevarlo a cabo, se toman segmentos de hojas o entrenudos de plantulas cultivadas in vitro. Estos segmentos de tejido (llamados explantes), son puestos en contacto con una suspension de Agrobacterium tumefaciens conteniendo un gen foraneo y un gen marcador que seran introducidos en la planta. La bacteria A. tumefaciens transfiere dicho gen foraneo al genoma de la planta. Los explantes son trasladados a un media de cultivo semisolido conteniendo la sustancia selectora (la mas empleada hasta la fecha es el antibi6tico kanamicina), lo cual permite la regeneraci6n exclusiva de plantas que han asimilado el gen deseado. Este proceso se puede realizar tambien con trozos de microtuberculos.
Las plantas asf obtenidas (llneas transgenicas) son rotuladas en laboratorio y propagadas in vitro. Un grupo de plantas de cada llnea es sometida a la extracci6n de ADN para determinar en laboratorio si se realize la inserci6n del gen en el genoma vegetal mediante la realizaci6n de la prueba Southern Blot con una sonda espedfica para el gen de importancia.
77 * Centro Internacional de la Papa (CIP). Estacion Central, La Molina. Lima - Peru

Por otro lado, en invernadero se producen tuberculos de cada una de las lfneas transgenicas y de los respectivos controles (plantas no transformadas). Todos estos tuberculos son infectados experimentalmente con cepas bacterianas aisladas en laboratorio, para evaluar el nivel de resistencia al pat6geno, Erwinia carotovora o Ralstonia solanacearum segun se evalue el dafio.
EXPERIMENTO EN CAMPO
Aquellas Hneas que resultaron transgenicas en las pruebas moleculares y que mostraron diferencias significativas en la infecci6n bacteriana en laboratorio e invernadero con respecto a los controles, son evaluadas en campo.
El experimento en campo se inicia con un diseno experimental con base estadlstica que incluye una apropiada distribuci6n de las plantas en el area donde se realizara la prueba (latices o bloques completes al azar). Ademas, es necesario realizar pruebas previas al suelo del campo con la finalidad de estimar la poblaci6n bacteriana infectante, asl coma durante y/o despues de la cosecha para conocer la presi6n patogenica a la que se ha expuesto el cultivo. Asl, el material transgenico y el control suele distribuirse aleatoriamente en el campo, empleandose par lo general tres repeticiones de siembra, lo cual minimiza las errores en caso de tenerse una distribuci6n heterogenea de la poblaci6n bacteriana en el suelo, o de estar presentes otros factores que pudieran influir en el experimento. Dos semanas antes de la siembra de papa, el campo a utilizarse es aislado del resto mediante la siembra perimetrica de cinco filas de un cultivo apropiado segun la region agroecol6gica de experimentaci6n: malz (Zea mays) en zonas de clima de selva o tarwi en zona andina.
Es preferible la siembra de plantas brotadas a partir de tuberculos de papa; se garantiza que se realice el experimento con plantas mas vigorosas.
Luego de dos semanas de haber sido sembrado el campo, se realiza el aporque, y de allf en adelante, se realiza la evaluaci6n quincenal de los danos en las plantas. Como es evidente, aquellas llneas transgenicas o controles susceptibles a la marchitez bacteriana,
78

no sobreviviran al ataque de Pseudomonas, Ralstonia o Erwinia. Como norma de bioseguridad, cada dos dfas se realiza la eliminaci6n manual y posterior autoclavado de todos las botones florales que pudieran desarrollarse coma medida de protecci6n al germoplasma natural y compatible que pudiera encontrarse cerca al area experimental, y coma prevenci6n de bioseguridad hasta que se demuestre la ausencia de riesgo al ecosistema.
Llegado el momenta de la cosecha, se realiza la evaluaci6n final de la performance en campo, la cual incluye no solo al follaje sino tambien al desarrollo y apariencia de las tuberculos. Los tuberculos que representan el menor o ningun dano (idealmente), son separados para estudiar su infecci6n latente en invernadero. Es decir, son conservados en contenedores plasticos y en suelo durante 3 meses, para determinar si las bacterias presentes en la cascara de las tuberculos llegan a causarles dano.
Toda el material excedente de la cosecha, es autoclavado o incinerado, dependiendo del volumen. Ademas, el campo no es utilizado para la siembra de papa en la siguiente campafia agrfcola, o se siembra otro cultivo. Si hubieran brotes posteriores de papa, seran inmediatamente eliminados. Toda esto se realiza con la finalidad de impedir el escape del material transgenico en experimentaci6n.
79
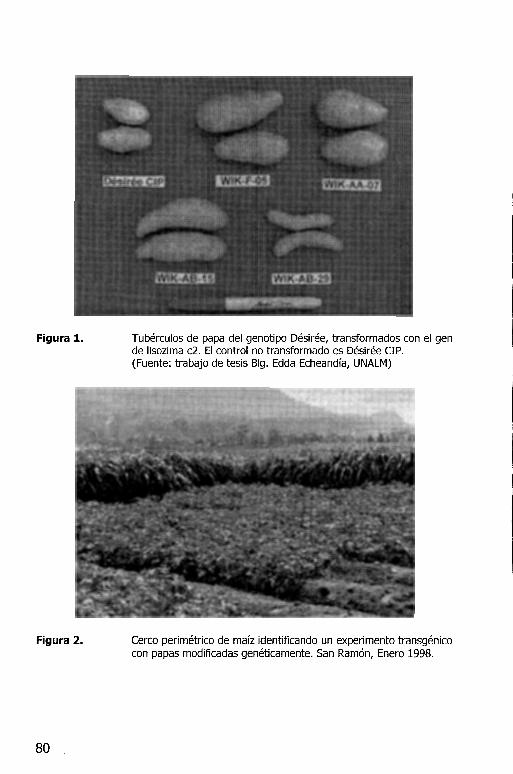
Figura 1.
Figura 2.
80
Tuberculos de papa del genotipo Desiree, transformados con el gen de lisozima c2. El control no transformado es Desiree CIP. (Fuente: trabajo de tesis Big. Edda Echeandfa, UNALM)
Cereo perimetrico de mafz identificando un experimento transgenico con papas modificadas geneticamente. San Ramon, Enero 1998.

Figura 3.
Figura 4.
Extracci6n de botones florales de las plantas transgenicas. (Fuente: trabajo de tesis Big. Edda Echeandfa, UNALM)
Papas transgenicas (WIK-AB-17 y WIK-AB-25) vs. papas control (Desiree). San Ramon, Febrero 1998. (Fuente: trabajo de tesis Big. Edda Echeandfa, UNALM)
81

Figura 5.
82
Quema post-cosecha de desechos transgenicos en un campo sembrado con papa en el anexo de La Libertad, Huancayo, a 3,800 msnm. Marzo 1998.

(' ,.) f ~QL· .----~~~~~~~~~~~~~~~~~ \Q \_
EXPERIMENTO DE CAMPO: MEDIDAS DE BIOSEGURIDAD EN EL
CENTRO INTERNACIONAL DE LA PAPA Field Experiment.·
Biosafety at the International Potato Center
Jorge Tenorio *
Experimentaci6n en el campo
La experimentaci6n en el campo de las plantas transgenicas tiene coma fin evaluar el comportamiento agron6mico, recuperar la variedad original y analizar las caracterfsticas incorporadas. Un requisite previo para la realizacion en forma segura de estos experimentos consiste en reunir la informacion apropiada sabre el ambiente donde se van a realizar los experimentos de campo. Esta selecci6n es particularmente importante cuando se requiere de aislamiento. Puede incluir experimentos preliminares a pequefia escala con plantas no transformadas para verificar que el lugar reline las condiciones necesarias para el estudio del genotipo transformado.
1. Aorobaci6n del ensayo de campo
•!• Antes de iniciar un experimento en campo con plantas modificadas geneticamente, debera someterse el formulario de Bioseguridad BSC-2 (formulario para experimentos en cam po) para su aprobacion por el Presidente del Comite de Seguridad.
•!• La solicitud para realizar el ensayo en el campo debera enviarse a las autoridades locales pertinentes (SENASA en el Peru). Los ensayos con plantas transgenicas solo se iniciaran una vez que se haya recibido la autorizaci6n escrita por parte de las autoridades.
* Centro Internacional de la Papa (CIP). Estacion Central, La Molina. Lima - Peru 83

2. Autorizacion especial para realizar experimentos fuera del CIP
Cuando los experimentos con plantas transgenicas desarrolladas en el CIP no se van a realizar en nuestras estaciones experimentales, aquella instituci6n que va a llevar a cabo el experimento (una instituci6n nacional, una extension del Ministerio de Agricultura o una universidad) debera firmar un acuerdo con el Comite de Bioseguridad del CIP para realizar el experimento y hacerle un seguimiento. Se utilizara material transgenico proveniente del CIP, seg(m las normas de Bioseguridad Nacional.
3. Documentacion en el campo
Cada clon debe se identificado con letreros especiales que indiquen variedad/ transgen/ N° de clon y en caso de existir una nomenclatura espedfica, tambien debe utilizarse .
Ejemplos:
Desiree/Lys/DL13
Achirana INTA/Bt2/1-19
Jewel/CTI/13
La variedad de papa (Desiree) ha sido transformada con un gen Lisozima (Lys) dando como resultado el clon (DL 1 3)
La variedad de papa (Achirana INTA) ha sido transformada con un gen de Bacillus thuringiensis (Bt2) dando coma resultado el clon (1-19)
La variedad de camote (Jewel) ha sido transformada con el gen inhibidor de tripsina del caupf (CTI) dando como resultado el clon (13)
•!• El formulario de Bioseguridad BSC-2 debe acompanar al libro del campo y guardarse en el archive bajo la responsabilidad del encargado de la estaci6n respectiva.
84

•!• Todos los campos deben identificarse con un panel pequefio que indique el material de la planta y el nombre del investigador principal.
4. Diseiio del campo
•!• El investigador principal esta a cargo del disefio del campo. No existe ningun disefio especifico del campo para las plantas transgenicas; sin embargo, se deben tomar precauciones espedficas durante su manipulacion.
•!• Los ensayos de campo transgenicos y no transgenicos para el mismo cultivo deben estar separados por una distancia minima de 20 m en todas las direcciones.
•!• Los campos transgenicos pueden separarse con barreras ( cercos, filas de malz, etc.) en el caso que se justifique el aislamiento fisico. La justificacion de estas barreras debera notificarse en el formularlo BSC-2.
5. Dispersion del polen
•!• Las flares deben retirarse para evitar la polinizacion entre plantas del mismo cultivo o de la parcela aledafia. Se debe impedir que el polen llegue a las especies afines que se sepa esten presentes en el ambiente, o a otros clones del mismo cultivo.
•!• Si las flares son necesarias para una futura experimentacion, se debe cubrir la inflorescencia antes de la maduracion. En estos casos, se debe aislar la parcela para evitar las transmision de polen hacia otras parcelas que tengan clones compatibles o especies afines.
6. Manipulaci6n de los materiales transgenicos cosechados
•:• Para transportar el material cosechado, debe ser empacado en balsas de malla para prevenir perdidas accidentales en el camino entre el campo y los de almacenamiento.
85

•!• El almacenamiento del material transgenico cosechado no requiere de condiciones especificas; sin embargo, debe rotularse segun el protocolo del CIP: variedad/ transgen/ N° de don.
•!• La evaluaci6n del material cosechado proveniente de la experimentaci6n en el campo se debe realizar bajo un examen selectivo y cuidadoso para detectar las fenotipos anormales, basado en las descriptores para el cultivo respective y el analisis de las caracterlsticas incorporadas.
7. Eliminaci6n de los restos transgenicos
•!• Durante la cosecha, todos las restos de la planta, incluyendo el material que podrla haber sido fertilizado accidentalmente (es decir controles no transgenicos), deben incinerarse en las areas designadas.
8. Monitoreo despues de la cosecha
•!• Se debe monitorear el campo despues de la cosecha. Todas las plantas "voluntarias" que aparezcan despues de la cosecha en las parcelas expuestas tambien deberan esterilizarse en autoclave o incinerarse.
•!• Esto debe incluir una rotaci6n de cultivos sistematica en todos las lugares donde se haya sembrado material transgenico (es decir: la papa transgenica, el perlodo de descanso, el camote transgenico ).
9. Informe del ensayo de campo
•!• Se debe enviar un "Informe del Ensayo" al Comite de Bioseguridad dentro de los 30 dlas siguientes a las ensayos de campo. Posteriormente, el Comite de Bioseguridad lo enviara a las autoridades locales pertinentes.
•!• El investigador principal, o en todo caso, el funcionario de Bioseguridad asignado, debera tomar parte en cualquier pregunta o decision relacionada al material transgenico.
86

Referencias:
1. "Biotecnologla y Bioseguridad en el CIP". Reglamento Interno. Centro Internacional de la Papa, junio de 1993.
2. Reglamento para la "Manipulacion de Plantas Modificadas Geneticamente en el Centro internacional de la Papa". Reglamento interno, 1998.
87


RECOMENDACIONES Y CONCLUSION ES
Los participantes al taller hicieron especial enfasis en los siguientes puntos:
•!• Las papas transgenicas con genes de resistencia a enfermedades bacterianas, lisozima, atacina, cecropina, sarcotoxina, no presentan riesgos conocidos para el media ambiente donde ocurre esta enfermedad.
•!• Las medidas de bioseguridad establecidas antes de la realizaci6n de este taller y discutidas en el transcurso del evento son adecuadas para el manejo de este material en zonas agro ecol6gicas del tipo de San Ramon (ceja de selva = "lowland tropics") donde se presentan las enfermedades bacterianas.
•!• La autoridad competente en la materia en el Peru es el SENASA. Se deben fortalecer y desarrollar las herramientas internas con apoyo cientffico y capacitacion por parte de instituciones expertas coma el CIP.
•!• El personal involucrado directamente con el manejo y/o monitoreo de experimentos en campo debe tener el conocimiento suficiente de los riesgos potenciales de los cultivos transgenicos para el media ambiente y la salud humana.
•!• Las medidas de bioseguridad, el equitaje, el cerco de malz, el panel, y las permisos de ensayos en campo, son disenados para ayuda r en la tom a de conciencia de las ca racterf sticas de este nuevo material, por lo tanto, es importante que este disponible la informaci6n en los lugares donde se desarrollan los ensayos de cam po.
•!• La dispersion del polen y por ende, del transgen no fue considerada un riesgo dado que no ocurre en las especies silvestres de las zonas agroecol6gicas donde existen estas
89

90
enfermedades bacterianas. Aun asf, serfa deseable que se llevara a cabo un experimento de flujo de polen hacia especies silvestres en los lugares donde las hibridizaciones interespecfficas podrfan ocurrir con el fin de establecer las normas de bioseguridad para estas areas.

Big. Jorge Alcantara
LISTA DE PARTICIPANTES Participants
Programa Nacional de Investigacion en Recursos Geneticos y Biotecnologfa (PRONARGEB) Av. La Universidad s/n, La Molina, Lima, Peru Tel. +(51-1) 349-5646 Correo electronico: [email protected]
Ing. Agr. Pedro Aley Centro Internacional de la Papa (CIP) Av. La Universidad 795, La Molina, Lima, Peru Tel. +(51-1) 349-6017, anexo 2052 Correo electr6nico: [email protected]
Dr. Patricio Arce Johnson Pontificia Universidad Catolica de Chile Departamento de Genetica Molecular y Microbiologfa Chile Tel. +(56-2) 686 2897 Correo electronico: [email protected]
Big. Jorge Benavides Centro Internacional de la Papa (CIP) Av. La Universidad 795, La Molina, Lima, Peru Tel. +(51-1) 349-6017, anexo 2113 Correo electronico: [email protected]
Ora. Merideth Bonierbale Centro Internacional de la Papa (CIP) Av. La Universidad 795, La Molina, Lima, Peru Tel. +(51-1) 349-6017, anexo 3051 Correo electr6nico: [email protected]
91

Ing. Agr. Fausto Buitron Agrotec Jose Marfa Eguren 156, Magdalena, Lima, Peru Tel. +(51-1) 4616871
Sr. Lorentz Buelow Bundesanstalt fi.ir ZOchtungsforschung an Kulturpflanzen, BMZ (Centro Federal para la Investigaci6n del Mejoramiento en Plantas Cultivadas) Nuer Weg 22/23 D-06484, Quedlinburg, Alemania Tel. +(03946) 47201 Correo electr6nico: [email protected]
Bach. Ing. Agr. Vilma Gutarra Servicio Nacional de Sanidad Agraria (SENASA) Av. La Universidad s/n, La Molina, Lima, Peru Tel. +(51-1) 433-2851 Correo electr6nico: [email protected]
Lie. Big. Enrique Carranza Servicio Nacional de Sanidad Agraria (SENASA) Pasaje Francisco de Zela s/n, Jesus Marfa Edif. Min. Agricultura, piso 10 SENASA Tel. +(51-1) 423-4682 Correo electr6nico: [email protected]
Sr. Daniel Chirito Universidad Nacional Agraria La Molina (UNALM) Av. La Universidad s/n, La Molina, Lima, Peru Tel. +(51-1) 349-5647
Ing. Agr. Noemi Flores Institute Nacional de Investigaci6n Agraria (INIA) Av. La Universidad s/n, La Molina, Lima, Peru Tel. +(51-1) 349-5626 Correo electr6nico: [email protected]
92

Dr. Marc Ghislain Centro Internacional de la Papa (CIP) Av. La Universidad 795, La Molina, Lima, Peru Tel. +(51-1) 349-6017 Correo electronico: [email protected]
Big. Cesar Giron Servicio Nacional de Sanidad Agraria (SENASA) Programa Mosca de la Fruta Av. La Universidad s/n, La Molina, Lima, Peru Tel. +(51-1) 349-5756 Email: [email protected]
Dr. Ali Golmirzaie Direcci6n actual: College of Agriculture Department of Horticulture University of Arkansas 316 Plant Sciences Building Fayetteville Arkansas 72701, U. S. A. Phone: +(501) 575-8780 Email: [email protected]
Big. Ricardo Gutierrez Instituto Nacional de Recursos Naturales (INRENA) Calle 17 # 355, Urb. El Palomar, Lima, Peru Tel. +(51-1) 225-1112 Correo electronico: [email protected]
Big. Carmen Herrera Centro Internacional de la Papa (CIP) Av. La Universidad 795, La Molina, Lima, Peru Tel. +(51-1) 349-6017 Anexo 2124 Correo electronico: [email protected]
Sr. Andreas Mahn Bundesanstalt fi.ir ZUchtungsforschung an Kulturpflanzen, BMZ (Centro Federal para la Investigaci6n del Mejoramiento en Plantas Cultivadas) Nuer Weg 22/23, D-06484, Quedlinburg, Alemania Tel. +(03946) 47201
93

Big. Luis Nopo Centro Internacional de la Papa (CIP) Av. La Universidad 795, La Molina, Lima, Peru Tel. +(51-1) 349-6017, anexo 2124 Correo electr6nico: [email protected]
Ing. Eduardo Perez Sandoval Director de Investigaci6n en Recurses Naturales Consejo Nacional de Ciencia y Tecnologla (CONCYTEC) Calle del Comercio 197, San Borja , Lima, Peru Tel. +(51-1) 225-1150 Correo electr6nico: [email protected]
Srta. Petra Porsch Bundesanstalt fOr ZOchtungsforschung an Kulturpflanzen, BMZ (Centro Federal para la Investigaci6n del Mejoramiento en Plantas Cultivadas) Nuer Weg 22/23, D-06484, Quedlinburg, Alemania Tel. +(03946) 47201 Correo electr6nico: [email protected]
Ora. Sylvie Priou Centro Internacional de la Papa (CIP) Av. La Universidad 795, La Molina, Lima, Peru Tel. +(51-1) 349-6017 Anexo 3049 Correo electr6nico: [email protected]
M. Sc. Big. Jorge Tenorio Centro Internacional de la Papa (CIP) Av. La Universidad 795, La Molina, Lima, Peru Tel. +(51-1) 349-6017, anexo 2024 Correo electronico: [email protected]
Dr. Bodo Trognitz Centro Internacional de la Papa (CIP) Av. La Universidad 795, La Molina, Lima, Peru Tel. +(51-1) 349-6017, anexo 3055 Correo electronico: [email protected] Correo electr6nico: [email protected]
94

Unidad de Capacitaci6n, Centro Internacional de la Papa
Dr. Patricio Malagamba Centro Internacional de la Papa (CIP) .. Av. La Universidad 795, La Molina, Lima,· P~~ru Tel. +(51-1) ~49-6017, anexo 3099 ':'",'C:•fH·<'~ Correo electronico: p.malagamba@~gia~.org
Big. Nelson Espinoza Centro Internacional de la Papa (CIP) Av. La Universidad 795, La Molina, Lima, Peru Tel. +(51-1) 349-6017, anexo 3102 Correo electr6nico: [email protected]
95

Unidad de Capacitaci6n, Centro Internacional de la Papa
Dr. Patricio Malagamba Centro Internacional de la Papa (CIP) , .... ,, Av. La Universidad 795, La Molina, Lima, Pefru Tel. +(51-1) ~49-6017, anexo 3099 ':··,,, .. ,~k''~ Correo electronico: p.malagamba@~9ia~.org
Big. Nelson Espinoza Centro Internacional de la Papa (CIP) Av. La Universidad 795, La Molina, Lima, Peru Tel. +(51-1) 349-6017, anexo 3102 Correo electr6nico: [email protected]
/ }"?
95

-: las Memorias:
1al de la Papa (CIP). 2000. Desarrollo y .. s Transgenicas Resistentes a Enfermedades 26 de noviembre, 1998. Memorias del Taller . . Peru.
Utilizaci6n de Papas Transgenicas ermedades Bacterianas
r . Peru .1998
ca : Dr. Marc Ghislain Big. Luis Nopo
Departamento de Capacitaci6n y Comunicaciones, CIP
Dr. Patricio Malagamba Big . Margarita Lopez
Big . Edda Echeandfa
1an sido financiadas con las recurses de chaft fi.ir Technische Zusammenarbeit
ERNACIONAL DE LA PAPA (CIP) Apartado 1558 - Lima, Peru
349-5783; 349-6017. Fax: (51-1) 349-5638. Correo-E: [email protected]
l
Participantes
l . Lorentz Bulow 2. Fausto Buitron 3. Jorge Benavides 4. Daniel Chirito 5. Cesar Giron 6. Ricardo Gutierrez 7. Patricio Arce-Johnson 8. Eduardo Perez 9. Jorge Tenorio l 0. Marc Ghislain l l . Cesar Aguilar
12. Petra Porsch 13. Nelson Espinoza l 4. Pedro Aley 15. Jorge Alcantara l 6. Enrique Carranza l 7. Ali Golmirzaie 18. Sylvie Priou 19. Luis Nopo 20. Carmen Herrera 21 . Vilma Gutarra 22. Noemi Flores

Desarrollo y Utilizaci Transgenicas R1
a Enfermedades Taller 24 al 26 de tt
Lima y
Centro Internacional de la Papa
I lllilll l/111 1111111111111111111111111111111111111 I A)6002 9 CI A
CIP G4 C45.1
Deutsche Gesellschaft fUr Technische Zusammenarbeit (GTZ)