la partÍcula viral -...
TRANSCRIPT

1 Virología – Modulo II – Lic. Daniel Rensetti
Área de Virología – FCV - UNCPBA
Módulo II: Los virus como partículas extracelulares.
Rensetti Daniel
LA PARTÍCULA VIRAL
La partícula viral madura, denominada virión consiste básicamente de un
bloque de material genético rodeado de proteínas que lo protegen del medio
ambiente y le sirven como vehículo para permitir su transmisión de una célula a otra.
En estado extracelular el virión es inerte metabólicamente, siendo su única función
transportar el ácido nucleico viral desde la célula en la que se ha reproducido hasta
otra célula en la que se pueda reproducir. La morfología de los virus se refiere a este
estado, en el que constan de ácido nucleico, una estructura que los recubre y enzimas.
Los virus pueden alternar entre dos estados distintos, uno intracelular y otro
extracelular, cuyas características fundamentales son su composición simple y su
forma de multiplicación especial, siendo ambas propiedades determinantes de su
parasitismo intracelular obligado.
Los virus más pequeños y simples están constituidos únicamente por ácido
nucleico y proteínas. El ácido nucleico (genoma viral) está ubicado en el interior de la
partícula viral, y puede ser ADN o ARN. Debido al escaso espacio para codificar la
información, los virus han evolucionado para lograr la máxima eficiencia replicativa
con el menor número de genes. Generalmente, el genoma, está asociado con un
número pequeño de moléculas proteicas que pueden tener actividad enzimática o
cumplir alguna función estabilizadora para el plegamiento del ácido nucleico y armado
de la partícula viral. Este conjunto, de genoma y proteínas íntimamente asociadas, es
llamado "core", núcleo, nucleoproteína o nucleoide. Este núcleo central está rodeado
por una cubierta proteica, la cápside, que junto con el genoma constituye la
nucleocápside. Las cápsides virales están formadas por un gran número de
subunidades polipeptídicas que se ensamblan adoptando una simetría de tipo
helicoidal (nucleocápside en forma de bastón) o icosaédrica (partículas casi esféricas).
En algunos virus más complejos, por fuera de la cápside se encuentra otra cubierta, la
envoltura, que es una estructura membranosa constituida por lípidos provenientes de
la célula hospedadora y glicoproteínas (Fig. 1).

2 Virología – Modulo II – Lic. Daniel Rensetti
CÁPSIDE:
La cápside es la cubierta proteica que aloja en su interior al ácido nucleico.
Tiene como misión proteger al ácido nucleico y reconocer los lugares de unión y
penetración a las células, en el caso de los virus que carecen de envoltura. Está
formada por proteínas globulares llamadas capsómeros. La variedad de proteínas que
forman estos capsómeros es baja, aunque su número es elevado. Para evaluar la
importancia que tiene la constitución de la cápside hay que tener en cuenta que los
virus, debido a su proceso de reproducción, están obligados a ser parásitos
intracelulares; necesitan una célula hospedadora donde replicar su ácido nucleico y
utilizar la maquinaria metabólica para reproducirse completamente y transmitirse a
otras células. En este proceso, donde el metabolismo del organismo afectado se altera,
la cápside del virus protege a éste del exterior, pero a la vez es una parte fundamental
que el organismo invadido reconoce para crear anticuerpos. Además, la morfología de
la cápside con protuberancias y hendiduras, que se corresponden con la estructura
poliédrica, determinan las zonas de mayor o menor accesibilidad para los anticuerpos
que el sistema inmunológico del hospedador puede crear. Las subunidades de la
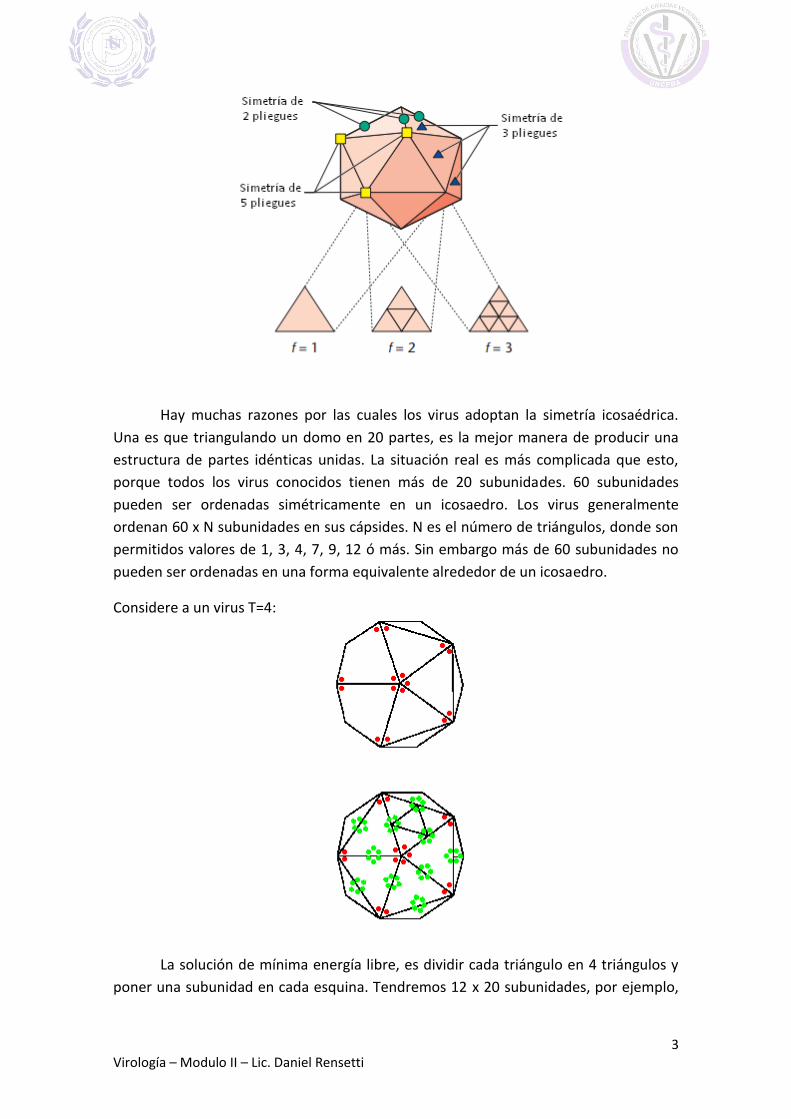
cápside se ubican alrededor de los vértices o cara de un icosaedro. Un icosaedro tiene
20 triángulos equiláteros dispuestos alrededor de la cara de una esfera. Está definido
por tener 2, 3 y 5 ejes de simetría.
Fig. 1: Virus desnudo y virus envuelto.
3 Virología – Modulo II – Lic. Daniel Rensetti
Hay muchas razones por las cuales los virus adoptan la simetría icosaédrica.
Una es que triangulando un domo en 20 partes, es la mejor manera de producir una
estructura de partes idénticas unidas. La situación real es más complicada que esto,
porque todos los virus conocidos tienen más de 20 subunidades. 60 subunidades
pueden ser ordenadas simétricamente en un icosaedro. Los virus generalmente
ordenan 60 x N subunidades en sus cápsides. N es el número de triángulos, donde son
permitidos valores de 1, 3, 4, 7, 9, 12 ó más. Sin embargo más de 60 subunidades no
pueden ser ordenadas en una forma equivalente alrededor de un icosaedro.
Considere a un virus T=4:
La solución de mínima energía libre, es dividir cada triángulo en 4 triángulos y
poner una subunidad en cada esquina. Tendremos 12 x 20 subunidades, por ejemplo,

4 Virología – Modulo II – Lic. Daniel Rensetti
240 con 12 pentámeros y 30 hexámeros. Obviamente no son equivalentes, pero 180 y
los restantes 60 hacen contacto. Se dice que son casi equivalentes.
¿PORQUÉ UNA CONSTRUCCIÓN EN SUBUNIDADES?
Necesidad: Los genomas virales son los más pequeños que se conocen. Pueden
codificar pocas proteínas, pero pueden producirlas repetidamente con alta eficiencia.
En consecuencia, la construcción en subunidades es la solución más simple y práctica.
En el mejor de los casos, un ácido nucleico puede codificar un 15% de su peso en
proteína. Como los virus están compuestos del 50 al 90% de su peso en proteína, debe
haber más de una proteína y la construcción en subunidades, por lo tanto, es esencial.
Fidelidad: La síntesis de ADN, ARN y proteínas, es un proceso que puede sufrir errores
ocasionales. Al tratarse de genomas y proteínas pequeños, hay poca oportunidad de
que ocurra un error.
Economía: Una estructura correcta puede ser formada con el mínimo de gasto
energético, ya que si una unidad es sintetizada incorrectamente, entonces, solo esa
unidad es descartada.
AUTOENSAMBLE:
Los virus son construidos de acuerdo con principios de diseño eficientes y, por
lo tanto, son capaces de autoensamblarse sin la participación de ningún factor
organizador externo. Esto es posible debido a la formación de un gran número de
uniones débiles cuando los componentes del virus son colocados en la configuración
adecuada por medio de movimientos al azar propiciados por factores termodinámicos.
La repulsión entre las cargas electrostáticas negativas de los fosfatos presentes en las
proteínas dificultan el plegado de las mismas. Los virus contrarrestan este problema,
empaquetando moléculas con carga positiva como iones K+, Na+ y Mg++, poliaminas o
proteínas unidas al ácido nucleico. Algunas de estas últimas son codificadas por el
virus. Por ejemplo, si se trata al Virus del Mosaico del Tabaco con una solución
concentrada de urea, ocurre una descomposición del virus en subunidades de proteína
y ARN. Si se elimina la urea y son incubadas de nuevo las subunidades de proteína en
presencia del ARN viral, las macromoléculas se agregan en forma espontánea dando
origen a partículas virales infecciosas. Sin embargo, cuando se omite el ARN viral
durante el proceso de repolimerización, se obtienen cilindros de proteína, pero en este
caso las subunidades están apiladas en forma de discos en lugar de tener una
disposición helicoidal. Además, estos cilindros son menos estables que la partícula viral
completa, lo que señala la importancia del ácido nucleico en la estabilización de la
estructura viral.
5 Virología – Modulo II – Lic. Daniel Rensetti
En el caso de algunos virus icosaédricos, ha sido posible realizar también
experimentos de reconstrucción o reconstitución in vitro (o sea, en el tubo de ensayo).
Sin embargo, modificando las condiciones de estos experimentos, se han obtenido
estructuras tubulares y otras variables morfológicas. Esto indica que las propiedades
de empaquetamiento de las proteínas son las que en última instancia determinan la
estructura de la partícula viral.
El autoensamble resulta económico para los virus, pues no requieren de
información genética específica para lograrlo, y además proporciona un método
sencillo y eficiente para eliminar subunidades defectuosas producidas durante la
replicación de los componentes virales.
Según su simetría, las cápsides se pueden clasificar en:
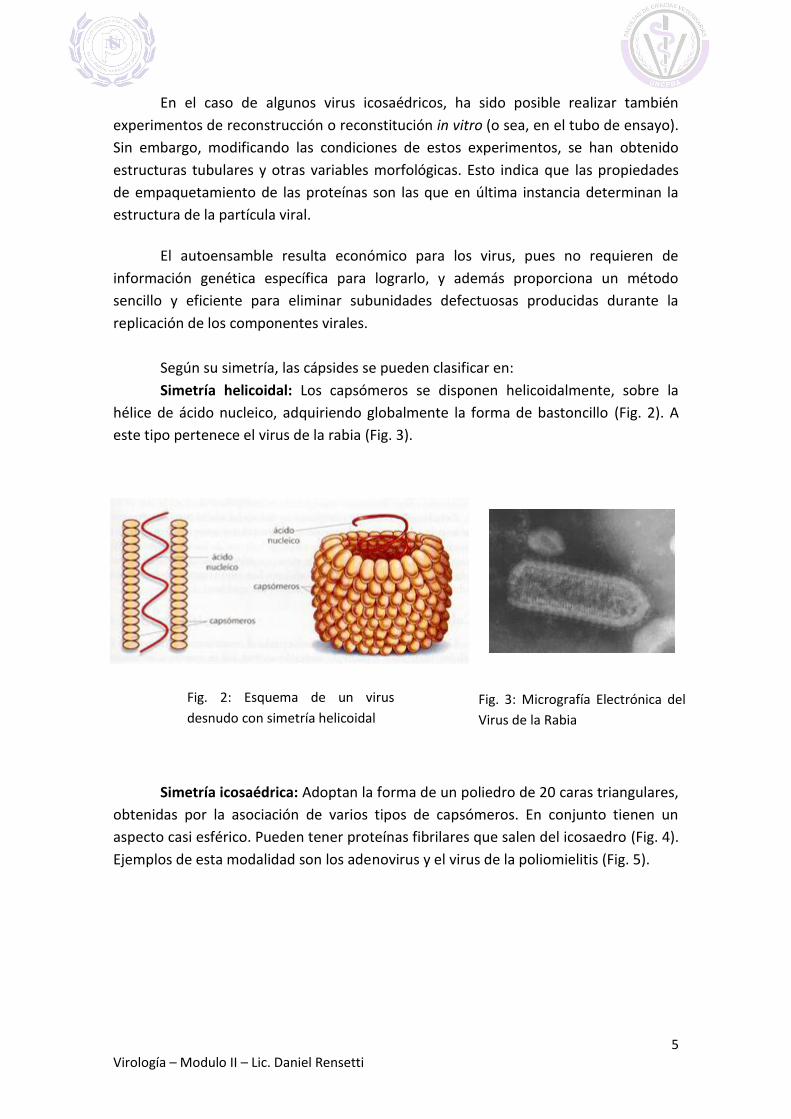
Simetría helicoidal: Los capsómeros se disponen helicoidalmente, sobre la
hélice de ácido nucleico, adquiriendo globalmente la forma de bastoncillo (Fig. 2). A
este tipo pertenece el virus de la rabia (Fig. 3).
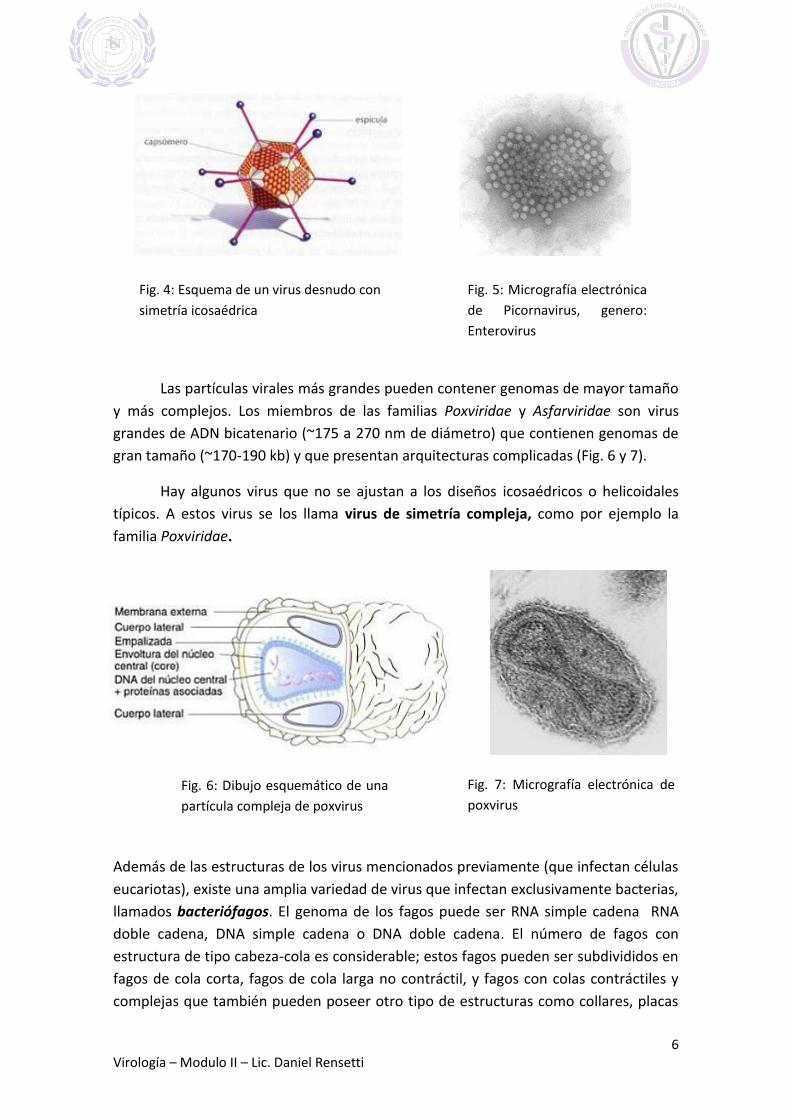
Simetría icosaédrica: Adoptan la forma de un poliedro de 20 caras triangulares,
obtenidas por la asociación de varios tipos de capsómeros. En conjunto tienen un
aspecto casi esférico. Pueden tener proteínas fibrilares que salen del icosaedro (Fig. 4).
Ejemplos de esta modalidad son los adenovirus y el virus de la poliomielitis (Fig. 5).
Fig. 3: Micrografía Electrónica del
Virus de la Rabia
Fig. 2: Esquema de un virus
desnudo con simetría helicoidal
6 Virología – Modulo II – Lic. Daniel Rensetti
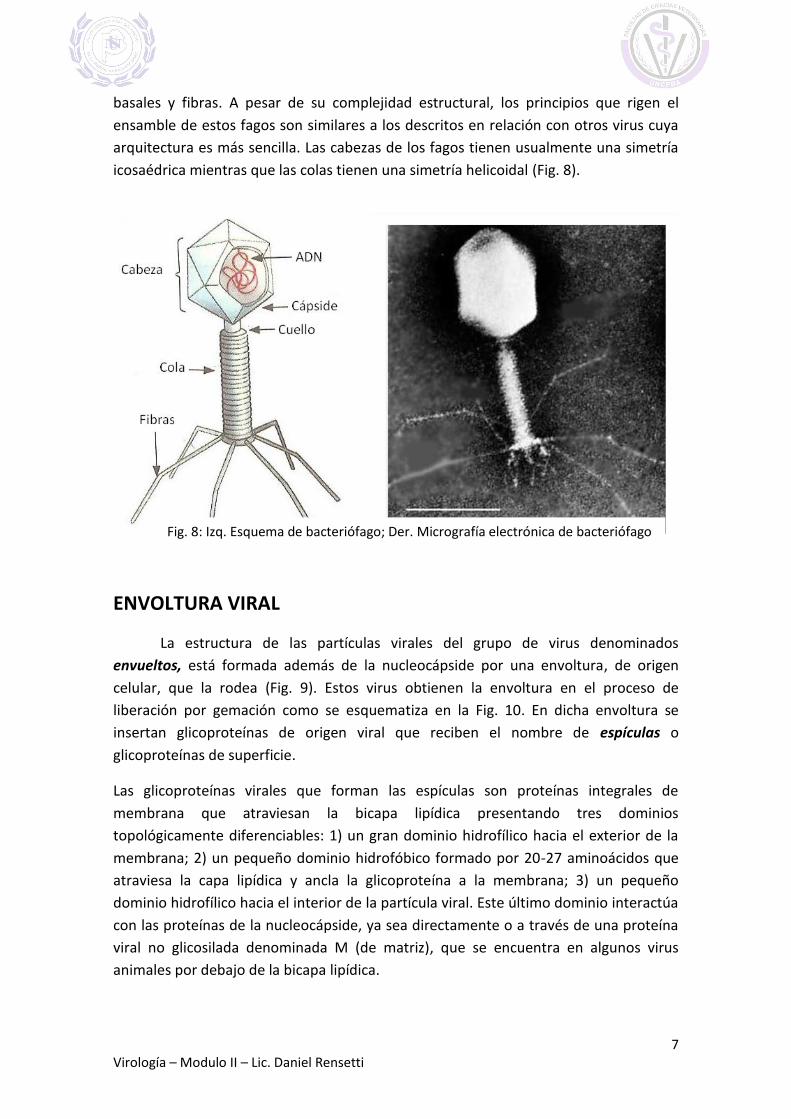
Las partículas virales más grandes pueden contener genomas de mayor tamaño
y más complejos. Los miembros de las familias Poxviridae y Asfarviridae son virus
grandes de ADN bicatenario (~175 a 270 nm de diámetro) que contienen genomas de
gran tamaño (~170-190 kb) y que presentan arquitecturas complicadas (Fig. 6 y 7).
Hay algunos virus que no se ajustan a los diseños icosaédricos o helicoidales
típicos. A estos virus se los llama virus de simetría compleja, como por ejemplo la
familia Poxviridae.
Además de las estructuras de los virus mencionados previamente (que infectan células
eucariotas), existe una amplia variedad de virus que infectan exclusivamente bacterias,
llamados bacteriófagos. El genoma de los fagos puede ser RNA simple cadena RNA
doble cadena, DNA simple cadena o DNA doble cadena. El número de fagos con
estructura de tipo cabeza-cola es considerable; estos fagos pueden ser subdivididos en
fagos de cola corta, fagos de cola larga no contráctil, y fagos con colas contráctiles y
complejas que también pueden poseer otro tipo de estructuras como collares, placas
Fig. 4: Esquema de un virus desnudo con
simetría icosaédrica
Fig. 5: Micrografía electrónica
de Picornavirus, genero:
Enterovirus
Fig. 7: Micrografía electrónica de
poxvirus
Fig. 6: Dibujo esquemático de una
partícula compleja de poxvirus
7 Virología – Modulo II – Lic. Daniel Rensetti
basales y fibras. A pesar de su complejidad estructural, los principios que rigen el
ensamble de estos fagos son similares a los descritos en relación con otros virus cuya
arquitectura es más sencilla. Las cabezas de los fagos tienen usualmente una simetría
icosaédrica mientras que las colas tienen una simetría helicoidal (Fig. 8).
ENVOLTURA VIRAL
La estructura de las partículas virales del grupo de virus denominados
envueltos, está formada además de la nucleocápside por una envoltura, de origen
celular, que la rodea (Fig. 9). Estos virus obtienen la envoltura en el proceso de
liberación por gemación como se esquematiza en la Fig. 10. En dicha envoltura se
insertan glicoproteínas de origen viral que reciben el nombre de espículas o
glicoproteínas de superficie.
Las glicoproteínas virales que forman las espículas son proteínas integrales de
membrana que atraviesan la bicapa lipídica presentando tres dominios
topológicamente diferenciables: 1) un gran dominio hidrofílico hacia el exterior de la
membrana; 2) un pequeño dominio hidrofóbico formado por 20-27 aminoácidos que
atraviesa la capa lipídica y ancla la glicoproteína a la membrana; 3) un pequeño
dominio hidrofílico hacia el interior de la partícula viral. Este último dominio interactúa
con las proteínas de la nucleocápside, ya sea directamente o a través de una proteína
viral no glicosilada denominada M (de matriz), que se encuentra en algunos virus
animales por debajo de la bicapa lipídica.
Fig. 8: Izq. Esquema de bacteriófago; Der. Micrografía electrónica de bacteriófago

8 Virología – Modulo II – Lic. Daniel Rensetti
Las glicoproteínas virales cumplen diversas funciones biológicas durante el ciclo de
vida de un virus, siendo esenciales para la infectividad, ya que actúan: 1) en la
adsorción a la célula huésped; 2) en el proceso de fusión que permite la entrada de la
nucleocápside viral al citoplasma de la célula; 3) en la gemación, que permite la salida
del virus envuelto a partir de la célula infectada. Además las glicoproteínas son el
blanco de reacción para el sistema inmune tanto en la respuesta humoral como celular
PROTEINAS VIRALES:
La transcripción viral conduce a la síntesis del ARN mensajero que codifica proteínas
virales tempranas y tardías. Los genomas de los virus animales codifican de tan sólo
una proteína a más de 100. Las proteínas tempranas, también llamadas proteínas
funcionales, son elementos no estructurales generalmente con actividad enzimática,
que participan en la ensamble de la partícula, la replicación del genoma, o en la
modificación de la respuesta innata del huésped a la infección.
Las proteínas tardías, llamadas también proteínas estructurales, forman parte de los
componentes estructurales de la partícula viral.
ÁCIDOS NUCLEICOS VIRALES Los virus se caracterizan, a diferencia de los otros organismos, por presentar una única
especie de ácido nucleico constitutiva que puede ser ADN o ARN, monocatenario o
bicatenario con estructura de doble hélice.
Fig. 9: Esquema de virus envuelto Fig. 10: Esquema del proceso de
gemación virus envuelto

9 Virología – Modulo II – Lic. Daniel Rensetti
Tipos de ADN virales
La mayoría de los virus ADN presentan un genoma bicatenario, con excepción de los
parvovirus y circovirus, constituidos por ADN monocatenario. Además las moléculas de
ADN viral pueden ser lineales o circulares.
La conformación circular que presentan los virus de las familias
Papovaviridae y Hepadnaviridae, confiere una serie de ventajas al ácido nucleico
respecto de la estructura lineal, otorgándole protección frente al ataque de
exonucleasas, facilitando la replicación completa de la molécula y su posible
integración al ADN celular. En el caso de los papovavirus, el ADN puede presentar tres
conformaciones: la forma I corresponde a la molécula circular covalentemente cerrada
y superenrollada sobre sí misma. Si se produce una ruptura en una unión en una de las
cadenas, la doble hélice se desenrolla y resulta una molécula circular relajada (forma
II). Por último, la forma III es el resultado de una ruptura en la otra cadena que origina
una molécula bicatenaria lineal.
El ADN doble circular incompleto de los hepadnavirus tiene una estructura muy
peculiar y de características únicas dentro de los ADN virales: una de las cadenas (S,
corta) es incompleta, de manera que el 15-50% de la molécula es monocatenaria; la
otra cadena (L, larga) posee una proteína covalentemente unida a su extremo 5´. Los
genomas de los hepadnavirus tienen tres (avihepadnavirus) o cuatro
(orthohepadnavirus) marcos de lectura abiertos.
Tipos de ARN virales
Los ARN de los virus animales son en su gran mayoría de cadena simple,
siendo Reoviridae y Birnaviridae las únicas familias que presentan como genoma ARN
bicatenario. En algunos grupos de virus, el ARN genómico está segmentado en varios
fragmentos, cuyo número es característico de cada familia.
Además de las características físicas y químicas mencionadas, la polaridad o sentido de
la cadena de ARN es una propiedad fundamental utilizada para definir los distintos
tipos de ARN viral. Se parte de definir como polaridad positiva la secuencia de bases
correspondiente al ARNm y polaridad negativa a la secuencia complementaria a la del
ARNm. Un virus es de cadena positiva cuando su ARN genómico tiene la polaridad que
le permite actuar como ARNm, o sea ser traducido en proteínas, inmediatamente
después de haber entrado a la célula.
Por el contrario, en los virus de polaridad negativa el ARN genómico tiene la secuencia
complementaria al ARNm viral; por lo tanto, cuando se produce la infección y el ARN
viral entra en la célula debe sintetizar la cadena complementaria que será el ARNm.
Para ello, los virus de polaridad negativa llevan en el virión, asociada a su genoma, una
ARN polimerasa dependiente de ARN, enzima denominada transcriptasa, que efectúa
la transcripción del ARN mensajero a partir del ARN genómico.

10 Virología – Modulo II – Lic. Daniel Rensetti
CLASIFICACIÓN DE LOS VIRUS
NOMENCLATURA
El nombre de los virus obedece a distintas consideraciones. Algunas veces se debe a la
enfermedad que ellos producen, por ejemplo el virus polio se llama así porque
produce la poliomielitis. También puede deberse al nombre de los descubridores como
el virus del Epstein-Barr, o a características estructurales de los mismos como los
coronavirus. Algunos poseen un nombre derivado del lugar donde se los halló por
primera vez, tal es el caso del virus Coxsackie o Norwalk.
El ICTV (International Committee on taxonomy of viruses) ha propuesto un sistema
universal de clasificación viral. El sistema utiliza una serie de taxones como se indica a
continuación: Por ejemplo, el virus de la rinotraqueitis infecciosa bovina se clasifica de
la siguiente manera
(Fuente: http://www.ictvonline.org/)
CLASIFICACIÓN DE BALTIMORE:
Los virus se clasifican o se agrupan de acuerdo con criterios como el tamaño, la
simetría de la cápside, si son desnudos o envueltos, el modo de transmisión, el
huésped que infectan, el tipo de ácido nucleico que contienen o las enfermedades que
causan. David Baltimore (Premio Nobel de Fisiología o Medicina en 1975) propuso un
sistema para clasificar a los virus de acuerdo con el tipo de ácido nucleico que
contienen y su mecanismo de replicación. El esquema se basa en el contexto de que
todos los virus deben generar ARNm simple cadena sentido positivo (+) a partir de sus
genomas con el fin de generar proteínas y replicarse. El mecanismo preciso por el cual

11 Virología – Modulo II – Lic. Daniel Rensetti
esto se logra es diferente para cada familia de virus. Baltimore dividió a los genomas
virales en 7 grupos o clases:
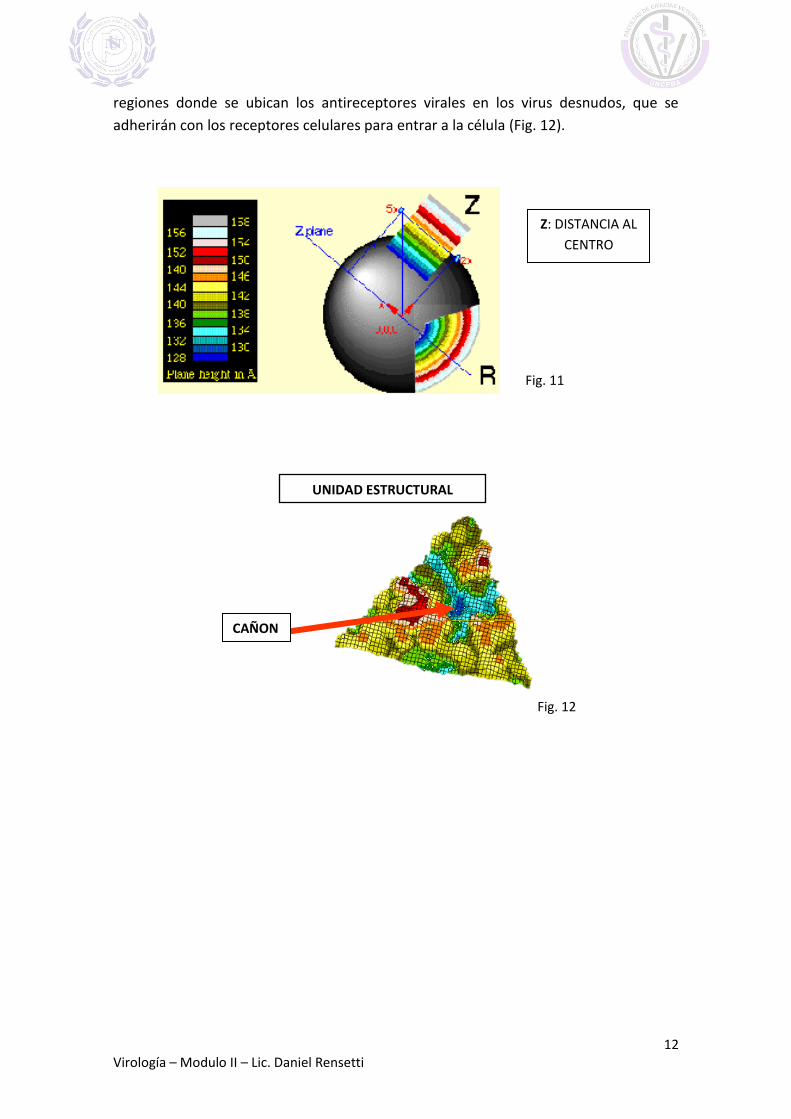
LOS MAPAS TOPOGRÁFICOS
Los mapas topográficos representan 1 de las 60 unidades estructurales del
icosaedro. La topografía se ilustra coloreando las regiones según su distancia al centro
del virus. Esta distancia es el valor “Z” (Fig.11).
Con programas informáticos de alta complejidad y modelos virales, es posible ilustrar
con distintos colores las zonas proteicas según su profundidad. Por ej., las zonas
profundas se ven azules y corresponden a los denominados “cañones”, que son las
12 Virología – Modulo II – Lic. Daniel Rensetti
regiones donde se ubican los antireceptores virales en los virus desnudos, que se
adherirán con los receptores celulares para entrar a la célula (Fig. 12).
Z: DISTANCIA AL
CENTRO
Fig. 11
Fig. 12
CAÑON
http://
onlineli
brary.w
iley.co
m/doi/
10.111
1/cmi.1
2224/p
df
UNIDAD ESTRUCTURAL

13 Virología – Modulo II – Lic. Daniel Rensetti
ANEXO
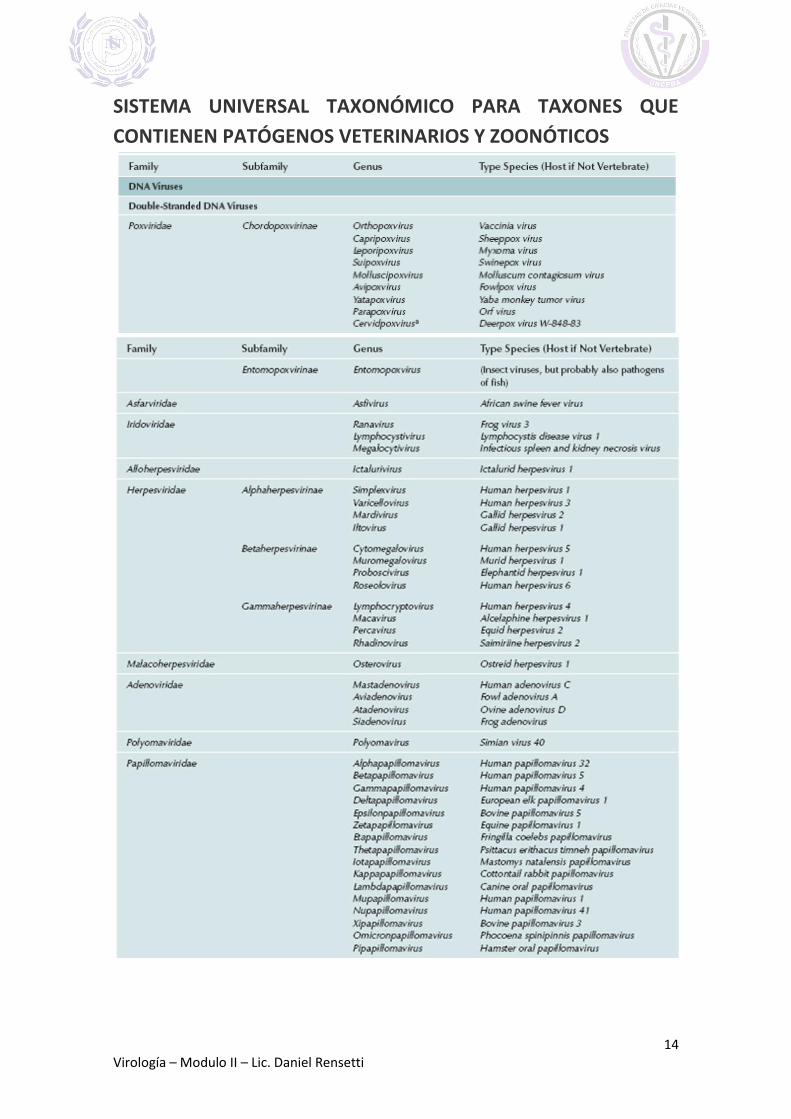
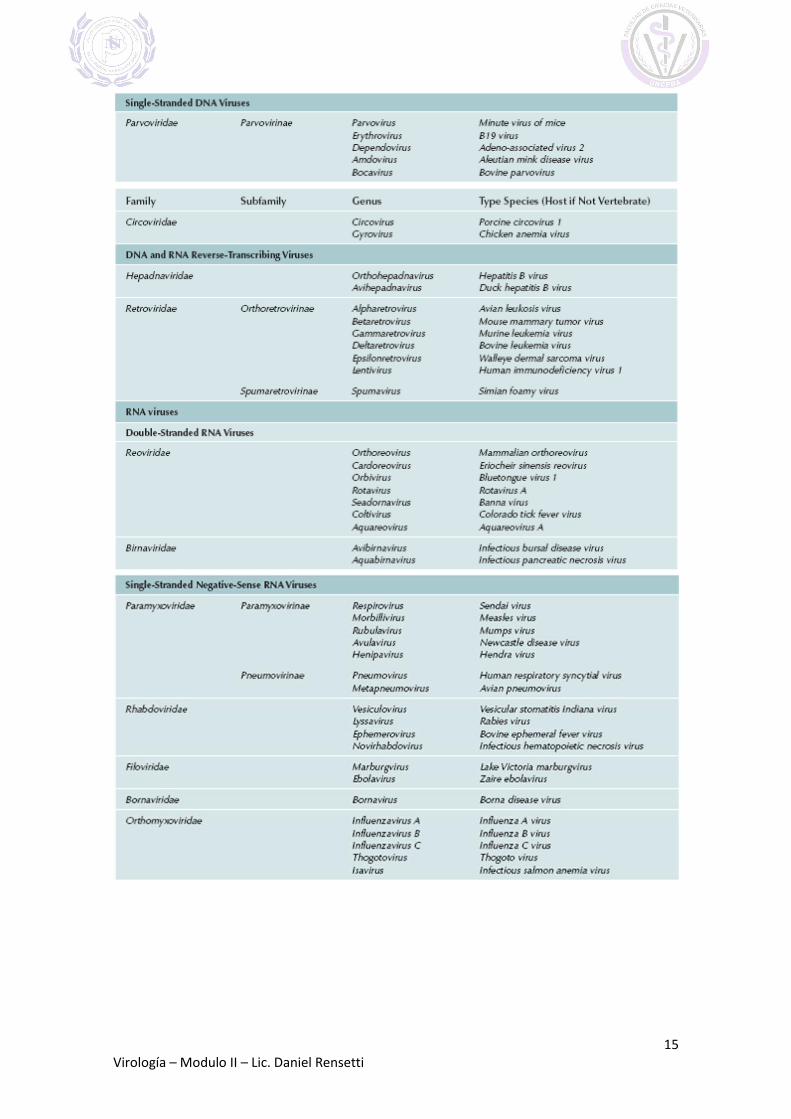
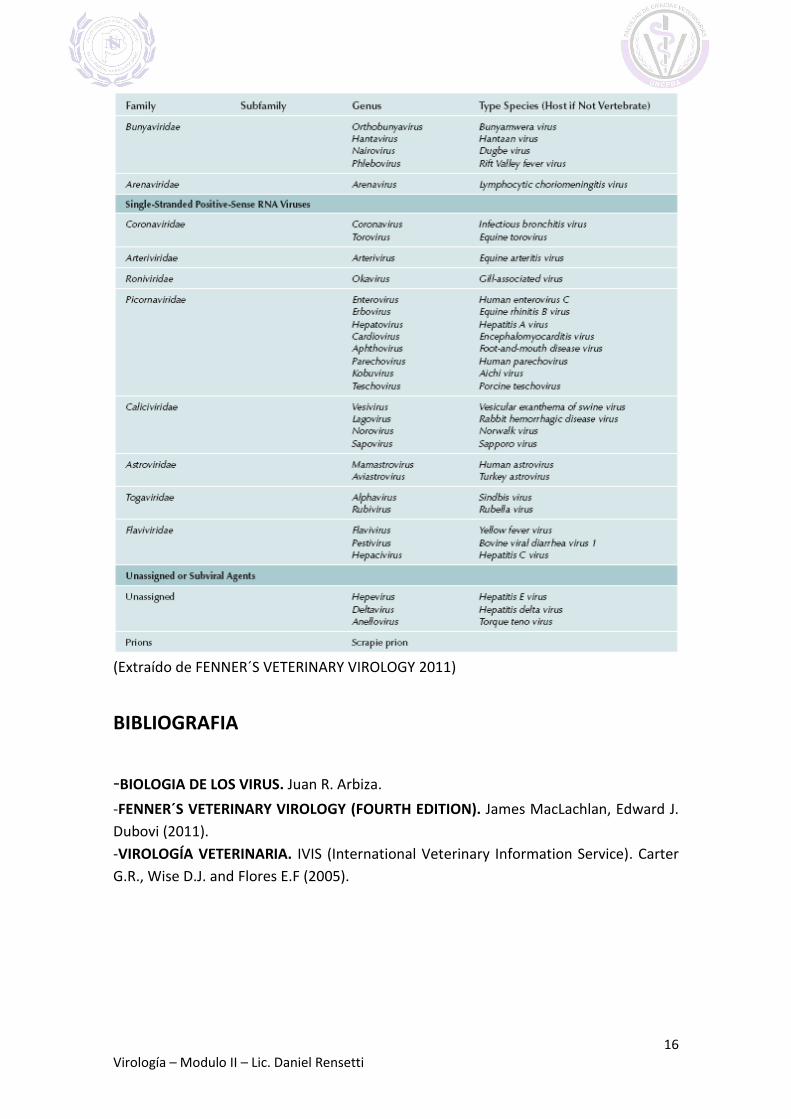
FAMILIAS Y GENEROS DE VIRUS QUE INFECTAN VERTEBRADOS
(Extraído de FENNER´S VETERINARY VIROLOGY 2011)
14 Virología – Modulo II – Lic. Daniel Rensetti
SISTEMA UNIVERSAL TAXONÓMICO PARA TAXONES QUE
CONTIENEN PATÓGENOS VETERINARIOS Y ZOONÓTICOS
15 Virología – Modulo II – Lic. Daniel Rensetti
16 Virología – Modulo II – Lic. Daniel Rensetti
(Extraído de FENNER´S VETERINARY VIROLOGY 2011)
BIBLIOGRAFIA
-BIOLOGIA DE LOS VIRUS. Juan R. Arbiza.
-FENNER´S VETERINARY VIROLOGY (FOURTH EDITION). James MacLachlan, Edward J.
Dubovi (2011).
-VIROLOGÍA VETERINARIA. IVIS (International Veterinary Information Service). Carter
G.R., Wise D.J. and Flores E.F (2005).