“variación estacional de la actividad antimicótica tesis...
TRANSCRIPT

UNIVERSIDAD AUTONOMA METROPOLITANA
“Variación estacional de la actividad antimicótica
de algunas macroalgas de las costas
del Estado de Veracruz, México.”
TESIS
Que para obtener el grado de
M a e s t r o e n B i o l o g í a
PRESENTA
A b e l a r d o I n c l á n S á n c h e z
DICIEMBRE 2004


ii
LA MAESTRIA EN BIOLOGÍA DE LA
UNIVERSIDAD AUTONOMA METROPOLITANA
PERTENECE AL PADRÓN DE EXCELENCIA DEL CONACyT.

iii
El jurado designado por laDivisión de Ciencias Biológicas y de la Salud
De la Unidad Iztapalapa aprobó la tesis que presentó
Hidrobiól. Abelardo Inclán Sánchez
El día 02 de Diciembre del año 2004
Comité Tutoral
Firma_________________________________________________________________
Tutor: Dra. Graciela De Lara Isassi_________________________________________________________________
Asesor: Dr. Abel Sentiés Granados_________________________________________________________________
Asesor: M. En B. E. Sergio Alvárez Hernández_________________________________________________________________
Sinodal: Dra. Alejandra Hernádez Santoyo_________________________________________________________________
Sinodal: Dr. Francisco Cruz Sosa_________________________________________________________________

iv
DEDICATORIA
A mi esposa amada Norma, motor incansable de motivación constante en la realización
de este proyecto, agradezco tu compañía durante mis desvelos, tus palabras de aliento
cuando mi cansancio derrumbaba mi ser, tu compresión por las horas de dedicación que
parecían interminables y restaba tiempo para lo nuestro. Por todos y cada uno de los
valiosos elementos que aportaste para cumplir con la meta.
¡Y sobre todo agradezco a Dios la vida tu existencia, mil gracias!
Abelardo Inclán Sánchez

v
AGRADECIMIENTOS
Agradezco al Comité Tutoral las sugerencias realizadas para llevar a cabo mi proyecto
de tesis, a la Dra. Graciela De Lara Isassi, al Dr. Abel Sentiés Granados y al M. en B. E.
Sergio Alvárez Hernández. También, gracias por las recomendaciones y atenciones de la
Dra. Alejandra Hernández Santoyo y Dr. Francisco Cruz Sosa. A todos los compañeros
del Laboratorio de Ficología Aplicada, le sdoy gracias por su comprensión y apoyo
durante el trabajo experimental. A CONACYT por el apoyo económico y a todos
aquellos que de alguna manera contribuyeron en la realización de una meta.

vi
CONTENIDO
RESUMEN.__________________________________________________________ vii
ABSTRACT. ________________________________________________________ viii
1. INTRODUCCION. ___________________________________________________ 1
2. ANTECEDENTES. __________________________________________________ 11
3. JUSTIFICACION. __________________________________________________ 19
4. OBJETIVOS. ______________________________________________________ 21
5. AREA DE ESTUDIO.________________________________________________ 22
6. MATERIAL Y METODOS.___________________________________________ 24
6.1. Trabajo en campo. _____________________________________________________ 24
6.2. Trabajo en laboratorio. _________________________________________________ 24
6.3. Pruebas antimicóticas. __________________________________________________ 26
6.3.1. Técnica de difusión.______________________________________________________ 27
6.3.2. Técnica de dilución.______________________________________________________ 29
7. RESULTADOS. ____________________________________________________ 31
7.1. Variación estacional ____________________________________________________ 45
7.2. Efectividad de los solventes ______________________________________________ 49
8. DISCUSION _______________________________________________________ 51
8.1. Variación estacional ____________________________________________________ 58
9. CONCLUSIONES___________________________________________________ 68
10. LITERATURA CITADA.____________________________________________ 70
11. ANEXO_____________________________________________________________ 82

vii
RESUMEN
En México, la investigación con macroalgas marinas dedicada a la búsqueda de alternativas
antimicóticas es prácticamente nula. El presente trabajo es pionero dentro del campo de la
Ficología aplicada con fines de investigación, a partir del cual se puede afirmar que existe un
gran porcentaje de especies algales provenientes de las zonas de La Mancha y Costa de Oro
del estado de Veracruz, México, cuyo potencial antimicótico resultó evidente. Los extractos de
37 algas fueron probados en contra de seis cepas de hongos. El 95 % del total de las especies
mostró actividad en contra de por lo menos alguna de las cepas de Aspergillus niger, Rhizopus
nigricans y Candida albicans. Del total de los extractos probados, ninguno reflejó actividad en
contra de Rhyzopus oryzae, Alternaria alternata y Beauveria bassiana. En las pruebas de
detección se emplearon las técnicas de difusión y de dilución, con la segunda se obtuvieron los
mejores resultados. Para la obtención de los extractos se utilizaron cuatro diferentes mezclas
de solventes: cloroformo-metanol-agua (C-M), cloroformo-alcohol etílico-agua (C-OH), éter-
metanol-agua (E-M) y éter-alcohol etílico-agua (E-OH). La mezcla de E-OH reportó el mayor
porcentaje de extractos activos (29 %), para las mezclas de E-M, C-OH y C-M, los extractos
con actividad antimicótica fueron de 28 %, 22% y 21 %, respectivamente. Las especies con
actividad antimicótica pertenecientes a la División Rhodophyta representaron el 49 %;
mientras que para la División Phaeophyta fue del 34 % y en la División Chlorophyta, sólo el
17 % del total de especies fueron activas. Las algas Digenea simplex, Padina boergesenii y
Cymopolia barbata colectadas en La Mancha, así como Ulva fasciata obtenida en Costa de
Oro reportaron variación estacional de la actividad antimicótica. Con nuestros resultados se
puede concluir que algunos extractos tienen la capacidad de inhibir el crecimiento de los
hongos utilizados, por esta razón se deben establecer colectas programadas de algas para
obtener nuevos compuestos para el control de las micosis.

viii
ABSTRACT
There is no research in Mexico looking for antimicotic activity on seaweeds. This is the first
work in this field of applied phycology with pharmaceutical purposes. With the results of this
research we find that there are several algal species with antimicotic potential from La Mancha
and Costa de Oro, Veracruz. Extracts of thirty seven algal species were tested against six fungi
strains. The ninety five percent of the analized species showed activity against at least of one
of the strains of Aspergillus niger, Rhizopus nigricans and Candida albicans. None of the
tested extracts showed activity against Rhyzopus oryzae, Alternaria alternata and Beauveria
bassiana. In order to detect antimicotic activity of the extracts, diffusion and dilution
techniques were used being more useful the second. The extracts were made with four
different mixtures of solvents: chlorophorm-methanol-water (C-M), chlorophorm-ethanol-
water (C-OH), ether-methanol-water (E-M) and ether-ethanol-water (E-OH). The percentage
of active extracts were: 29 % with E-OH, 28 % with E-M, 22 % with C-OH and 21% with
C-M mixture. The active species were 49% from the Rhodophyta, 34 % from Phaeophyta and
17 % from Chlorophyta. The antimicotic activity of Digenea simplex, Padina boergesenii and
Cymopolia barbata from La Mancha and Ulva fasciata collected at Costa de Oro showed
seasonal variation. This is the first report on antimicotic activity of Digenea simplex and
Padina boergesenii. With the results obtained we can concluded that some algae have
substances with the capacity of inhibit growth of the fungi used, for this reason we have to
stablish programmed collections of algae in order to obtain new compounds for the control of
mycosis.

1
1. INTRODUCCIÓN.
El conocimiento del potencial farmacológico que presentan las algas, tuvo sus orígenes en la
investigación con las microalgas, específicamente en cianofitas, de las cuales la especie
dulceacuícola Nostoc punctiforme fue una de las primeras en ser examinada con dichos fines
(Harder, 1917). Los primeros trabajos para conocer la actividad antimicrobiana de las
macroalgas los realizaron Mautner et al. (1953) con Rhodomela larix, Martínez–Nadal (1958)
con Sargassum natans y Chondria littoralis, y Martínez–Nadal et al. (1966) con Cymopolia
barbata. Desde entonces, el desarrollo de la investigación en este campo a escala mundial, se
ha enfocado a realizar trabajos experimentales con el fin de conocer la actividad
antibacteriana, antimicótica y antiviral de las algas, predominando los trabajos con microalgas
(Zajic y Knetting, 1970; Hoppe, 1979; Biard et al., 1980; Pesando, 1990; Sridhar y
Vidyavathi, 1991).
La mayoría de estudios encaminados hacia el conocimiento de la actividad antimicrobiana de
las algas, aprovechan las propiedades que presentan los disolventes para la obtención de
extractos, cuyo contenido es rico en metabolitos, los cuales de acuerdo a Karp (1998) son
moléculas o productos intermediarios originados en las rutas metabólicas que en último
término conducen a la formación de un producto final con una función específica.
De acuerdo a Fenical (1982), existen dos tipos de metabolitos, los primarios y los secundarios,
al primer grupo pertenecen los carotenoides y los esteroles. Los carotenoides son formados en
las vías metabólicas de síntesis de los terpenos, y son considerados metabolitos primarios. En

2
las algas marinas suelen encontrarse formas carotenoideas muy diversas, por el contrario en
las plantas terrestres, las formas son muy similares entre sí, ocurre lo mismo para los esteroles,
encontrándose en las algas marinas gran diversidad de dichos compuestos. Por lo que respecta
a los metabolitos secundarios, al igual que sucede en las plantas terrestres, las estructuras son
modificadas debido a los elementos presentes en el medio marino, tales como los cloruros,
bromuros y yoduros. Dichos elementos son incorporados a los organismos marinos y
participan en las síntesis de compuestos halogenados, vía común en el medio marino, mientras
que en el terrestre es un proceso raro.
Los tres principales tipos de metabolitos secundarios de acuerdo a Fenical (1982) son
terpenos, acetogeninos y los metabolitos nitrogenados. Los terpenos son los metabolitos
secundarios más importantes en el medio marino, se forman vía condensación de unidades de
isopentenil de pirofosfato y son metabolitos de peso molecular alto como los diterpenos. Por el
contrario, en las plantas terrestres, es común la obtención de terpenos de menor peso
molecular como los monoterpenos responsables del aceite aromático de las plantas. En la
biosíntesis de terpenos marinos participan los halógenos, especialmente los bromuros, a través
de reacciones cíclicas produciendo haloterpenos.
El segundo grupo de metabolitos secundarios es el de los acetogeninos, compuestos lineales
producidos a través del precursor acetil CoA, este patrón es el responsable de la síntesis de
lípidos como los ácidos grasos. A este grupo pertenecen los halometanos, considerados como
potentes biocidas, como ejemplo se tiene al ácido acético halogenado. Finalmente, los
metabolitos nitrogenados son comunes en el medio terrestre y marino, en éste último, existen

3
las condiciones que propician la síntesis de una gran cantidad de aminoácidos como algunos
derivados de la tirosina. Por otro lado, el metabolismo del aminoácido triptofano es un proceso
que genera residuos indol, como los polihaloindoles de Laurencia brongniartii y Rhodophyllis
membranaceae. En general, dichas moléculas son las que confieren a las algas su actividad
antimicrobiana. El aislamiento de los metabolitos dependerá en primer lugar del tipo de
solvente usado en la obtención del extracto, es decir, del grado de afinidad polar entre los
metabolitos y los solventes utilizados; y en segundo lugar, de la metodología o técnica de
separación de los metabolitos del extracto.
En forma alterna a la investigación de la actividad antimicrobiana de las algas, existe el interés
por conocer cuáles son los elementos que confieren dichas propiedades, por lo que se han
aislado algunos compuestos como el caso de los lípidos simples no saponificables que son
insolubles en agua y se extraen por medio de solventes no polares como el cloroformo, éter y
benceno, entre otros. (Lehninger, 1984). A este grupo pertenecen los terpenos, los cuales ya
han probado actividad y se ha logrado su separación (Fenical y Paul, 1984; Reichelt y
Borowitzka, 1984; Tringali et al., 1986).
Sobre los lípidos, se sabe que forman parte de la estructura de las membranas; sirven de
transporte y almacenamiento de sustancias; forman una cubierta protectora sobre la superficie
de muchos organismos y, son componentes de la superficie celular relacionados con el
reconocimiento de las células, la especificidad de especie y la inmunidad de los tejidos
(Lehninger, 1984). Por otro lado Spoehr et al. (1949), establecieron que en las plantas
vasculares existe una mezcla de ácidos grasos, en la cual los compuestos más insaturados son

4
convertidos por fotoxidación hacia derivados antibióticamente activos. Desde este punto de
vista, es factible establecer que en las algas ocurra un proceso semejante dado el grado de
parentesco que existe entre ambos grupos, apuntando a las algas como antecesoras de las
plantas vasculares (Scagel et al., 1991).
De acuerdo con Pesando (1990), las algas marinas bentónicas producen una gran variedad de
metabolitos biológicamente activos. En general, las algas rojas son ricas en ácido acrílico,
terpenoides halogenados, cetonas halogenadas, polisulfurados y aldehídos. En contraste con
las rodofitas, en las clorofitas y en las feofitas no se conocen metabolitos halogenados. Las
clorofitas son ricas en ácido acrílico, aldehídos y terpenoides. Los metabolitos de las feofitas,
son principalmente el ácido acrílico y los terpenos (Martínez-Nadal et al., 1966; Glombitza,
1970; Fenical et al., 1973; Fenical, 1974; Robertson y Fenical, 1977; Carter et al., 1978;
Caccamese y Azzolina, 1979; Finer et al., 1979; Enoki et al., 1983; Baker, 1984; Fenical y
Paul, 1984; Reichelt y Borowitzka, 1984; Tringali et al., 1986; Perry et al., 1991; Gunasekera
et al., 1995).
Dentro del campo de la investigación química, una de las técnicas para el aislamiento de los
componentes de interés ha sido la cromatografía, de la cual, la de capa fina aprovecha las
características polares de los metabolitos, facilitando su separación. A partir de ésta técnica se
han logrado aislar de los extractos de macroalgas, varios componentes que han demostrado su
actividad en contra de bacterias y hongos (Perry et al., 1991; Gunasekera et al., 1995; Robles
y Ballantine, 1999).

5
El desarrollo de la economía agrícola a escala mundial ha generado una importante fuente de
alimentos, por ello es importante el control de enfermedades de las plantas causadas por
hongos, los cuales llegan a mermar la producción con las consecuentes pérdidas de los
cultivos. Actualmente existen cerca de 8,000 especies de hongos susceptibles de causar
enfermedades, afortunadamente el control de muchos de ellos es fácil. Los hongos poseen
características como el tener alta supervivencia, rápido crecimiento y reproducción explosiva,
debido a esto es que su diseminación y efectos destructores sobre los cultivos han resultado, en
ocasiones, catastróficos (Romero, 1988).
Los hongos son microorganismos eucarióticos quimiorganotróficos, se reproducen de manera
natural por medio de esporas, no tienen clorofila, sus cuerpos son alargados o filamentosos
con pared celular quitinosa o de celulosa, o ambas. Las esporas se reproducen de dos maneras
en casi todas las clases de hongos: a) sexuales, estas tienen núcleo derivado de las células
originales y ambas son haploides que al fundirse forman el zigoto diploide. b) Las esporas
asexuales se producen por simple diferenciación del talo en crecimiento, se conoce como talo
a todo el hongo en forma individual, incluidas las porciones vegetativas o no sexuales y todas
las estructuras especializadas. El talo de los hongos se compone de filamentos tubulares
microscópicos llamados hifas y al conjunto de hifas se le llama micelio (Pelczar et al., 1990).
Los hongos se adaptan a condiciones más severas que otros microorganismos, toleran y se
desarrollan en concentraciones de acidez elevada, soportan pH entre 2.0 a 9.0, siendo el
óptimo de 5.6. Pueden sobrevivir en ambientes deshidratados, cuando el medio se deseca,
producen esporas o pasan al estadio de resistencia. Se desarrollan en condiciones de

6
temperatura muy variadas, siendo la óptima de 22 a 30 °C, algunos hongos pueden crecer a
0 °C por lo que pueden dañar los alimentos refrigerados (Pelczar et al., 1990).
Todos los hongos se encuentran en la división Mycota, la cual presenta dos subdivisiones: a)
Myxomycotina que comprende a los hongos plasmodiales; y, b) Eumycotina que incluye a los
hongos típicamente filamentosos. Su clasificación de acuerdo con Pelczar et al. (1990) se
muestra en la figura 1.
Figura 1. Clasificación de la División Mycota (según Pelczar et al., 1990).
División Subdivisión Clase OrdenMyxomycotina Myxomycetes
ChytridialesChytridiomycetes Blastocladiales
Monoblepharidales
Hyphochytridiomycetes HyphochytridalesMycota
Plasmodiophoromycetes Plasmodiophoraceae
SaprolegnialesOomycetes Leptomitales
LegnidialesPernosporales
Eumycotina MucoralesZygomycetes Entomophtnorales
Zoopagales
HemiascomycetidaeAscomycetes Euascomycetidae
Loculoascomycetidae
Basidiomycetes HeterobasidiomycetidaeHomobasidiomycetidae
SphaeropsidalesDeutoromycetes Melanconiales
MonilialesMycelia Sterilia

7
Los principales grupos de hongos pertenecientes a la subdivisión Eumycotina son:
Chytridiomycetes, los cuales presentan células que se mueven por flagelos, viven en medios
acuáticos y algunos en el suelo.
Los Hyphochytridiomycetes, son especies acuáticas parásitas de algas y de otros hongos.
Los Oomycetes, producen enfermedades a los peces y causan la roya de la papa.
Plasmodiophoromycetes, se encuentran los mohos limosos que parasitan a las plantas
vasculares, causan la hinchazón de la raíz de la col y la roya polvosa de la papa.
Los Zygomycetes, son organismos que parasitan a las frutas y vegetales en crecimiento, otros
atacan a las frutas almacenadas. Dentro de ésta clase se encuentra el orden de los Mucorales,
principalmente a los géneros Rhyzopus y Mucor, algunos de sus miembros ocasionan
enfermedades nerviosas al hombre.
Los Ascomycetes, son organismos que se caracterizan por poseer un micelio bien desarrollado
(excepto las levaduras), la mayor parte de ellos son benéficos y de uso frecuente en la
industria, los que son dañinos producen enfermedad al olmo americano, la costra de las
manzanas y el mildio polvoso.

8
Los Basidiomycetes, producen enfermedades como el añublo y el tizón que destruyen los
graneros de trigo.
Deuteromycetes, se reproducen a través de conidios o esporas, carecen de fase sexual, son
llamados hongos anamorfos. Como ejemplos típicos del grupo se tiene a los géneros
Aspergillus, Penicillium, Fusarium, Trichophyton, principialmente.
La utilización de las macroalgas como agentes antimicóticos, es escasa por lo que se hace
necesario dedicar esfuerzos encaminados hacia este campo de la ficología. Los resultados
obtenidos hasta hoy indican que existe un gran potencial en las macroalgas para ser usadas en
el control de las enfermedades producidas por hongos que afectan a las plantas, animales y por
supuesto al hombre.
En la tabla 1 se presenta un resumen de las principales enfermedades que producen los hongos
a las plantas y en la tabla 2 algunos hongos causantes del deterioro de la salud del hombre. En
ambos cuadros se resaltan las especies que se utilizaron en este estudio.

9
Tabla 1. Principales micosis que afectan a las plantas (Pelczar et al., 1990; Herrera y Ulloa, 1998).
Clase ENFERMEDADChytridiomycetesOlpidium brassicae Pudrición de las raíces de las coles y del lino.Synchytrium endobioticum Roña negra o verruga de la papa.
PlasmodiophoromycetesPlasmodiophora brassicae Hernia de las raíces de la col, del nabo y otras crucíferas.
Spongospora subterranea Sarna negra o verrugosa de la papa
OomycetesAphanomyces Podredumbre de las raíces vegetales
Pythium debaryanum Ahogamiento de las plántulas de la papa, jitomate, chícharo, maíz y caña de azúcar.
Phytophtora infestans Tizón tardío del follaje y tubérculos de la papa y follaje y fruto del jitomate.
Albugo candica Roya blanca del follaje de la col, rábano y otras crucíferas.
Plasmopara viticola Roya vellosa del follaje y fruto de la vid.
ZygomycetesRhizopus nigricans Pudrición húmeda de los frutos de la fresa, plátano y otras frutas.
Ascomycetes:Erysiphe graminis Roya pulverulenta o cenicilla del follaje del trigo, maíz y otras gramíneas.Ceratocystis ulmi Enfermedad del olmo Holandés
Claviceps purpurea Cornezuelo de las espigas del centeno, trigo y otras gramíneas.
Mycosphaerella Nódulos en las hojas de muchas plantas
Sclerotium rolfsii Pudrición de la corona de especies hortícolas y ornamentales.
Basidiomycetes:Tilletia caries Carbón cubierto del trigo
Ustilago nuda Carbón descubierto del trigo y cebada.
Cronartium ribicola Roya del pino blanco y de la planta de grosella.
Puccinia graminis Roya del tallo y follaje de diversas gramíneas.
Uromyces phaseoli Roya del frijol.
Deuteromycetes:Colletotrichum gloeosporioides Antracnosis del naranjo, limonero, aguacatero y de cientos de plantas.
Alternaria solani Podredumbre de la papa y del follaje y fruto del jitomate.
Aspergillus niger Pudrición de gran variedad de granos y semillas almacenadas.
Botrytis cinerea Pudrición de los frutos de la vid, fresa y de otras plantas y flores ornamentales.
Fusarium oxysporum Marchitamiento de las plantas de diversas especies.
Penicillium italicum Podredumbre de cítricos.

10
Tabla 2. Principales micosis que afectan al hombre (Pelczar et al., 1990; Herrera y Ulloa, 1998).
ESPECIE OCURRENCIA Y ENFERMEDAD
Epidermophyton floccosum Moho dermatofito que produce dermatofitosis, micosis cutánea en pies (tiña
de los pies pie de atleta), uñas y otras partes del cuerpo.
Microsporum canis Infección de la piel y pelo en perros y gatos, moho dermatofito productor de
dermatofitosis, tiña de cabeza en los niños y tiña del cuerpo (piel glabra).
Microsporum. gypseum Moho dermatofito que ocasiona la tiña en cuero cabelludo y pelo, así como
la piel glabra.
Trichophyton subgrupo gypseum:
T. mentagrophytes Moho dermatofito, parásito primario de cabello piel u uñas (tiñas).
T. rubrum Dermatofitosis por moho dermatofito, produce tiña en muchas partes del
cuerpo, infecta el pelo y cuero cabelludo
T. tonsurans Dermatofito que infecta el pelo y el cuero cabelludo
Trichophyton subgrupo faviforme:
T schoenleini
T. violaceum Causan infecciones de piel, cuero cabelludo y la piel lampiña en el hombre
T. ferrugineum
T. concentricum
T. verrucossum
Nocardia asteroides, N.. brasiliensis,N. caviae
Moho que produce la enfermedad nocardiosis de los pelos axilares y
púbicos, de tipo subcutánea o sintética.
Aspergillus clavatus, A. niger, A. flavus Producen aspergilosis u otomicosis, son cutáneas, pueden afectar el
sistema respiratorio
Penicillium commune, P. marneffei Causan peniciliosis, oportunistas, inicialmente pulmonares y después
sistémicas.
Mucor circinelloides, M. pusillus,M. racemosus
Provocan mucormicosis, subcutánea localizada, principalmente
rinocerebral; gastrointestinal, y diseminada o sistémica.
Rhizopus oryzae, Rh. arrhizus Producen mucormicosis, subcutánea localizada, principalmente
rinocerebral; gastrointestinal, y diseminada o sistémica.
Beauveria bassiana En forma ocasional ha producido beauveriosis, afectando pulmones.
Candida albicans Levadura que provoca cistitis (inflamación de la vejiga) y endometritis
(inflamación endometrial)

11
2. ANTECEDENTES.
Existe un amplio grupo de organismos acuáticos que sintetizan una gran cantidad de
compuestos químicos con actividad antimicrobiana que pueden ser examinados en la búsqueda
de nuevas alternativas (Faulkner, 1986; Metting y Pyne, 1986). Dentro de este grupo, algunas
especies de algas marinas han sido reportadas con propiedades antibióticas (Hornsey y Hide,
1974; Glombitza, 1979; Henriquez et al., 1979; Accorinti, 1983; Reichelt y Borowitzka, 1984;
Caccamese et al., 1985; De Lara-Isassi et al. 1989; De Lara-Isassi, 1991; De Lara-Isassi y
Alvarez-Hernández, 1994), aunque la gran mayoría de los estudios se han enfocado a la
búsqueda de actividad antibacteriana, también se han obtenido, aunque en menor grado,
resultados de actividad antimicótica (Accorinti, 1983; Moreau et al., 1984; Pesando y Caram,
1984; Calvo et al., 1986; Padmakumar y Ayyakkannu, 1997; Pesando, 1990; Melo et al.,
1997; Robles y Ballantine, 1999; Zamora y Ballantine, 2000).
De los primeros estudios con el objeto de conocer el efecto de la actividad antimicótica se
encuentran los reportes de Mauter et al. (1953), quienes observaron actividad con extractos
etéreos de Rhodomela larix en contra de los hongos Candida albicans, Trichophyton
mentagrophytes y T. rubrum. Welch (1962), probó extractos acuosos algales de 35 especies
marinas de las divisiones Rhodophyta, Chlorophyta y Phaeophyta, destacando por su
actividad: Amphiroa fragilissima, Hormothamnion enteromorphoides, Laurencia obtusa,
Halimeda opuntia, Udotea flabellum, Dictyosphaeria favulosa, Dictyota bartayresii, Dictyota
divaricata, Dictyota indica y Padina gymnospora, dichas especies fueron probadas en contra

12
de los hongos Aspergillus niger, Mucor racemosus, Rhizopus oryzae, Candida albicans,
Cryptococcus neoformans y una especie de Penicillum sp.
Olesen et al. (1964) observaron la inhibición del crecimiento de Candida albicans con
extractos etanólicos de Wrangelia argus y W. bicuspidata; extractos etéreos y clorofórmicos
de Laurencia obtusa y, extractos acetónicos de Falkenbergia hillebrandii y L. obtusa. Por otro
lado, Martínez-Nadal et al. (1966) encontraron actividad antimicótica en contra de
C. albicans, Penicillium chrysogenum, Aspergillus niger, Torulopsis utilis y Saccharomyces
cerevisiae con extractos etéreos de Cymopolia barbata.
El estudio de la actividad antimicótica de las macroalgas no sólo se ha quedado en el
conocimiento general de cuáles especies son las que registran actividad, sino que ha
trascendido hacia investigaciones más finas, con el objeto de conocer las características del
elemento o metabolito contenido en las algas y así poderlo extraer con diversos solventes para
después con técnicas cromatográficas poder separarlo y posteriormente aislarlo.
De los metabolitos con capacidad antimicótica se tienen a las hidroquinonas zonarol e
isozonarol, ambos aislados de la feofita Dictyopteris zonaroides; el cicloendesmol, del tipo de
los ciclopropanos, el cual actúa en contra de Candida albicans y que fue aislado del alga
rodofita Chondria oppositoclada (Fenical et al., 1973; Fenical, 1974; Mori y Komatsu, 1986).
Robertson y Fenical (1977) aislaron el paquidictiol, un diterpeno obtenido de Dictyota
flabellata con actividad antibiótica. Otros metabolitos como la naftoquinona y derivados
extraídos de la feofita Landsburgia quercifolia, mostraron actividad en contra de C. albicans y

13
Trichophyton mentagrophytes (Perry et al., 1991), así como un compuesto fenólico obtenido
de Sporochnus pedunculatus con propiedades antimicóticas contra C. albicans (Gunasekera et
al., 1995).
Khaleafa et al. (1975) probaron extractos de 18 feofitas de las costas egipcias en el
Mediterráneo, resultando sólo cinco con actividad que inhibió el crecimiento de algunos
hongos. En Sri Lanka se observó que Ulva fasciata presentó una considerable inhibición del
crecimiento de la levadura Candida albicans (Sachithananthan y Sivapalan, 1975). Codomier
et al. (1977) estudiaron las propiedades antibacterianas y antimicóticas de las rodofitas
Asparagopsis armata y Falkenbergia rufolanosa encontrando actividad en contra de los
dermatofitos Microsporum gypseum y Trichophyton mentagrophytes.
Caccamese y Azzolina (1979) trabajaron con extractos obtenidos con una mezcla de tolueno-
metanol (1:3) de algas de la costa de Sicilia, encontrando actividad en contra de la levadura
Saccharomyces cerevisiae y del hongo Penicillium digitatum; contra este último, presentaron
actividad las algas: Dictyota dichotoma, D. dichotoma var. intricata, Dilophus fasciola,
Cystoseira compressa, C. elegans, C. stricta, Gracilaria bursa-pastoris, Stypocaulon
scoparium, Nereia filiformis, Laurencia obtusa, Rytiphloea tinctoria, Phyllophora nervosa,
Liagora viscida, Pterocladia pinnata y Caulerpa prolifera. Se registró actividad en contra de
la levadura con los extractos de C. elegans, L. obtusa, G. bursa-pastoris y C. prolifera.
Biard et al. (1980), con algas provenientes de las costas francesas, probaron extractos a partir
de éter y etanol, encontrando actividad antimicótica de las especies Asparagopsis armata y

14
Falkenbergia rufolanosa en contra de Candida albicans, Trichophyton tonsurans y
Epidermophyton floccosum, mientras que Polysiphonia thuyoides y Laminaria digitata sólo
inhibieron el crecimiento de E. floccosum.
Moreau et al. (1984), trabajaron con extractos obtenidos con hexano y dicloruro de metileno
de las especies Dictyota dichotoma, Dictyota lineraris, Dilophus fasciola y Dilophus spiralis,
provenientes de las costas del Mediterráneo, encontraron que existe una fuerte relación entre la
variación de la actividad antimicótica de dichas macroalgas y los cambios estacionales. Los
resultados obtenidos con las especies señaladas probaron inhibir del crecimiento de la levadura
C. albicans, los mohos Aspergillus fumigatus y Sporothrix schenckii, así como los
dermatofitos Trichophyton schoenleini, T. rubrum, T. mentagrophytes, Microsporum gypseum,
M. audouini, M. fulvum, M. canis y Epidermophyton floccosum.
En 1984, Pesando y Caram trabajaron con extractos etánolicos de macroalgas del
Mediterráneo, registrando actividad antimicótica las feofitas Dictyota dichotoma, Dictyota
linearis, Dilophus fasciola, Nereia filiformis y Sporochnus pedunculatus y las rodofitas
Hypnea musciformis, Falkenbergia rufolanosa y Laurencia obtusa, en contra del crecimiento
de los dermatofítos Trichophyton mentagrophytes y Microsporum fulvum y de los mohos
Sporothrix schnenkii y Aspergillus fumigatus.
Padmakumar y Ayyakkannu (1986), realizaron estudios de antimicosis con el alga Gracilaria
verrucosa de las costas índicas, los extractos obtenidos a partir de una mezcla de tolueno-
metanol fueron sometidos a las pruebas antimicóticas, encontrando inhibición del crecimiento

15
del hongo Aspergillus fumigatus. Moreau et al. (1988), realizaron un estudio anual con las
macroalgas Dictyota dichotoma, D. dichotoma var. implexa y Dilophus spiralis colectadas en
las costas del Mediterráneo, a partir de ellas se obtuvieron los extractos con hexano y se
efectuaron las pruebas antimicóticas, encontrando un amplio espectro de actividad, el cual
varió dependiendo de las estaciones de colecta. Esta actividad fue en contra de Candida
albicans, C. troicalis, C. parapsilosis, C. pseudotropicalis, Cryptococcus neoformans,
Torulopsis glabrata, Aspergillus fumigatus, A. niger, Sporothrix schenckii, Epidermophyton
floccosum, Microsporum audouini, M. canis, M. fulvum, M. gypseum, Trichophyton
mentagrophytes, T. rubrum, T. schoenleinii, Fusarium oxysporum albedinis, F. oxysporum
dianthi, F. oxysporum lycopersici, Colletotrichium acutatum, Phialophora cinerescens y
Phoma tracheiphila.
Perry et al. (1991), trabajaron con Landsburgia quercifolia, especie endémica de Nueva
Zelanda, de la cual obtuvieron un extracto con metanol-cloruro de metilo (3:1) y aislaron un
metabolito derivado de la naftoquinona que mostró actividad en contra de Candida albicans y
de Trichophyton mentagrophytes. Por su parte, Tariq (1991) trabajó con extractos de
macroalgas de rodofitas Irlandesas y encontró que Dilsea carnosa, Laurencia pinnatifida,
Odonthalia dentata y Polysiphonia lanosa redujeron la extensión de las colonias de los
dermatofitos Microsporum canis y Trichophyton verrucosum, además observaron claras
diferencias con relación a las variaciones estacionales.
En 1992, Ballesteros et al. probaron la actividad antimicrobiana de 71 especies del
Mediterráneo con extractos obtenidos con la mezcla metanol-tolueno (3:1), de las cuales el

16
70% reportaron inhibición de crecimiento de las colonias de Candida albicans y Aspergillus
niger, de estas las especies más activas fueron Corallina elongata, Lithophyllum lichenoides,
Phyllophora crispa, Halimeda tuna, Valonia utricularis, Cystoseira spp, Halopteris spp y
Codium spp.
Gunasekera et al. (1995) de la feofita Sporochnus pedunculatus colectada en las costas de las
Islas Canarias, obtuvieron extractos etanólicos, posteriormente aislaron y probaron un
compuesto con grupo fenol presente, el cual que inhibió el crecimiento de Candida albicans.
Vora y colaboradores (1995), colectaron macroalgas de las costas de la India y realizaron
pruebas antimicóticas con extractos a partir de cloroformo-metanol para detectar su actividad,
encontrando que las colonias de Saccharomyces cerevesciae y C. albicans fueron inhibidas
por Chnoospora fastigiata.
Padmakumar y Ayyakkannu en 1997 analizaron algas de la costa sur de la India, hicieron
extractos etanólicos y llevaron a cabo un estudio para conocer la actividad antimicrobiana, del
total de especies probadas sólo el 27.5% mostró actividad antimicótica, predominando las
algas de la división Rhodophyta seguido de las Phaeophyta y finalmente las Chlorophyta. Los
extractos de las algas rojas que presentaron inhibición de las colonias de Candida albicans,
Helminthosporium oryzae, Trichophyton mentagrophytes, Aspergillus terreus, Fusarium
oxysporium y Trichoderma viridae fueron las especies Centroceras clavulatum, Gelidiella
acerosa, Gracilaria corticata, Halymenia floresia, Hypnea musciformis y Sarconema
filiforme. Del mismo estudio, las feofitas que tuvieron actividad en contra de Candida
albicans, Aspergillus terreus, Fusarium oxysporium, Helminthosporium oryzae, Trichophyton

17
mentagrophytes y Trichoderma viridae fueron las algas Dictyopteris delicatulata, Dyctyota
bartayresiana, Dyctiota dichotoma, Padina gymnospora, Sargassum plagiophyllum,
Spatoglossum asperum y Stoechospermum marginatum. Finalmente, sólo las algas verdes
Caulerpa racemosa y Codium arabicum mostraron actividad en contra de Aspergillus terreus,
Helminthosporium oryzae y Trichophyton mentagrophytes.
En Sudáfrica, Barreto et al. (1997) obtuvieron extractos etanólicos de las clorofitas Caulerpa
filiformis y Ulva rigida, de la feofita Zonaria tournefortii y de las rodofitas Hypnea spicifera,
Gelidium abottiorum y Osmundaria serrata, los cuales provocaron una inhibición del 50 % en
el crecimiento de las colonias de Rhizoctonia solani. Por otra parte, los extractos de las
rodofitas Spyridia cupressina y Beckerela pinnatifida mostraron ligera actividad antimicótica.
En un estudio con aglutininas provenientes de Hypnea musciformis colectada en la costa de
Brasil, Melo et al. (1997) obtuvieron una excelente actividad antimicótica contra Trichophyton
rubrum.
Robles y Ballantine (1999), realizaron cultivos controlados de la rodofíta Spyridia filamentosa
proveniente de las costas de Puerto Rico, de la cual los gametofitos femeninos, masculinos y
tetrasporofitos fueron sometidos a diferentes intensidades de radiación luminosa. Con una
mezcla cloroformo metanol (2:1) se obtuvieron los extractos algales y se realizaron las
pruebas antimicrobianas en contra de Candida albicans y se concluyó que existían diferencias
en la actividad con respecto a los diferentes estados del ciclo de vida y de la intensidad
luminosa; en donde los gametofitos masculinos sometidos a radiaciones mayores a 30 µmol

18
fotón/m2s y los tetrasporofitos con radiaciones superiores a 70 µmol fotón/m2s, produjeron
inhibición del crecimiento de la levadura, con los gametofitos femeninos no se presentó
actividad.
Por su parte Zamora y Ballantine (2000), al probar la actividad antimicótica de extractos
obtenidos a partir de la mezcla cloroformo y metanol (2:1) de Spyridia filamentosa y probarlos
en contra de Candida albicans encontraron resultados positivos, posteriormente en un análisis
de cromatografía de alta resolución (HPLC), reveló la presencia de diversas fracciones, las
cuales de acuerdo a los autores, cada una representa a un compuesto, por lo que la observación
de diversos picos les llevó a concluir que el extracto de S. filamentosa tiene cualidades de
múltiple actividad antibiótica. Pese a que en dicho estudio no se aislaron e identificaron los
compuestos responsables de la actividad antibiótica, se puede suponer de acuerdo a Pesando
(1990) que la alga roja S. filamentosa puede ser rica en metabolitos biológicamente activos del
tipo del ácido acrílico, terpenoides halogenados, cetonas halogenadas, polisulfurados o
aldehídos.

19
3. JUSTIFICACIÓN.
La mayoría de los medicamentos tienen su origen en productos provenientes del medio
natural, del cual las plantas terrestres son la base principal en la industria farmacéutica; sin
embargo, debemos de dirigir nuestro interés hacia los recursos acuáticos, en especial al medio
marino ya que existen organismos con alto potencial para ser aprovechados, previa
investigación por parte de los especialistas en el ramo y a partir de ella generar las bases para
la creación de nuevos fármacos. Uno de estos recursos acuáticos son las macroalgas marinas,
que si bien ya se han venido aprovechando como fuente generadora de materia prima para las
industrias agroquímica y alimenticia, hace falta mayor esfuerzo encaminado hacia la
investigación farmacéutica para el control de microorganismos.
El presente estudio pretende contribuir al conocimiento de las propiedades antimicóticas de
las macroalgas de México, específicamente de las provenientes del litoral del estado de
Veracruz; además, este trabajo es uno de los primeros en realizarse dentro del campo de la
Ficología aplicada en nuestro país y revela la presencia de macroalgas marinas en México que
sintetizan moléculas capaces de inhibir el crecimiento de hongos patógenos y fitopatógenos, y
pretende coadyuvar en el control de las enfermedades producidas por hongos que van en
detrimento del bienestar de plantas, animales y del hombre.

20
Por otro lado, se prestende conocer cómo pueden afectar los cambios estacionales a la
expresión de la actividad antimicótica de las macroalgas. A partir de los resultados obtenidos,
se pudo conocer algunas de las especies con actividad antimicótica y tener la certeza de
disponer de un recurso durante todo el año para aquellas especies en las cuales su efecto
antimicótico no se vio afectado por los cambios estacionales en las zonas de colecta.
Por lo que respecta con las metas a mediano plazo, sería importante realizar estudios de
separación y aislamiento de las sustancias responsables de la actividad antimicótica de las
macroalgas que resultaron activas; y, por otro lado desarrollar la tecnología apropiada, para
realizar el cultivo controlado de las especies con propiedades antimicóticas y de esta manera
asegurar la disponibilidad del recurso algal con estas características.

21
4. OBJETIVOS.
OBJETIVO GENERAL.
Ø Determinar la actividad antimicótica de algunas macroalgas marinas de las Divisiones
Rhodophyta, Phaeophyta y Chlorophyta, colectadas en la costa del estado de Veracruz y
estudiar su posible variación estacional.
OBJETIVOS PARTICULARES.
Ø Obtener extractos algales de diferente polaridad.
Ø Realizar pruebas específicas en contra de cepas de hongos patógenos para determinar sí
presentan actividad.
Ø Determinar sí existe variación estacional en la actividad antimicótica.
Ø Determinar sí existen diferencias en la eficacia de los solventes para obtener
extractos algales con fines antimicóticos.

22
5. ÁREA DE ESTUDIO.
El área de estudio comprendió dos localidades del estado de Veracruz, La Mancha y Costa de
Oro (Figura 2).
Figura 2. Área de estudio. Localidades de La Mancha y Costa de Oro, Veracruz.
La Mancha se localiza en el litoral del Golfo de México en la parte central del estado de
Veracruz, aproximadamente a 30 km al noroeste de Ciudad José Cardel, perteneciente al
municipio de Actopan. Su vegetación circundante es predominantemente agregada a selva baja
subperennifolia, selva baja caducifolia, vegetación de dunas costeras, selva baja perennifolia
inundable, tular, ceibal y asociaciones de macroalgas marinas (Contreras, 1993)
N
COSTA DE ORO
LA MANCHA
VeracruzLa Mancha
Costa de Oro
100 km

23
Costa de Oro se encuentra en el centro del estado de Veracruz, es considerada una
prolongación de las playas de Mocambo, pertenece al municipio de Boca del Río, se localiza
al sureste de la Isla Sacrificios, aproximadamente a 15 km al sur de la ciudad de Veracruz y
aproximadamente a 30 km al norte de la ciudad de Antón Lizardo. La playa está formada por
sustrato arenoso modificado por escolleras artificiales, mar adentro se alcanza una profundidad
máxima de 2 m aproximadamente. Su vegetación es predominantemente de dunas costeras
(Blanco y Ramírez, 1993).
Para el área de estudio, Castillo y Carabias (1982) dividen al año en tres estaciones climáticas
bien establecidas, la primera llamada de “secas” que abarca de marzo a mayo con
temperaturas en promedio de 24 ºC, velocidades del viento de 6.1 m/s y precipitación mínima
de 6.0 mm. Durante la época de “lluvias”, que comprende de junio a octubre, se registran las
temperaturas más altas, siendo en promedio de 26.8 ºC, velocidades del viento mínimas de
alrededor de 3.8 m/s y con las máximas precipitaciones en promedio de 200 a 400 mm.
Finalmente, se tiene a la temporada de “nortes” de noviembre a febrero, caracterizada por
temperaturas bajas con promedios de 20 ºC, velocidades del viento altas de 7 m/s y
precipitación alta de 24.0 mm en promedio

24
6. MATERIAL Y METODOS.
6.1. Trabajo en campo. Con el objeto de tener representadas las características climáticas de
las zonas de estudio, para el año 2002 y principios del 2003 se efectuaron colectas
programadas durante las temporadas de “secas” y de “lluvias”
El trabajo de campo inició con la colecta de las muestras de forma manual en la zona
intermareal, posteriormente se limpiaron quitando los organismos epífitos, se lavaron con agua
marina para retirar el exceso de arena y se colocaron en forma individual en bolsas de plástico
con numeración progresiva. En la bitácora de campo se registraron los siguientes datos:
número de muestra, fecha de colecta, localidad, hora, temperatura del agua, tipo de sustrato
(arenoso o rocoso), profundidad de la columna de agua y observaciones pertinentes. Una vez
llenada la bitácora y enumeradas las bolsas, se colocaron en una hielera con hielo seco con el
objeto de congelar las muestras y preservar el principio activo, evitando al máximo la
degradación enzimática y se trasladadaron al laboratorio de Ficología Aplicada de la
Universidad Autónoma Metropolitana Iztapalapa para su estudio.
6.2. Trabajo en laboratorio. Las muestras se descongelaron, se lavaron perfectamente con la
finalidad de retirar todos los organismos epífitos de sus superficie, así como partículas de
arena y otros residuos.
La obtención de los extractos se realizó a partir de muestras húmedas, de acuerdo a las
técnicas propuestas por Bernard y Pesando (1989) y Bligh y Dyer (1959).

25
En la figura 3 se muestra el proceso general de la obtención de los extractos algales.
TÉCNICA DE EXTRACCIÓN
Se probaron cuatro diferentes mezclas de solventes a una proporción de 2:2:1.8 v/v/v, las
mezclas utilizadas fueron: 1) cloroformo-metanol-agua (C-M), 2) cloroformo-alcohol etílico-
agua (C-OH), 3) éter-metanol-agua (E-M) y 4) éter-alcohol etílico-agua (E-OH). En total se
obtuvieron seis diferentes extractos por cada muestra procesada. Cuando la biomasa de una
muestra era escasa, sólo se utilizaron las mezclas de C-M y E-M.
MUESTRA LIMPIAPESAR 15 g
MACERAR EN MORTERO
** UTILIZAR MEZCLA DE SOLVENTES (2 : 2 :1.8 V/V/V)*
POR SEPARADO, CONCENTRAR AMBASFASES A PRESIÓN REDUCIDA A 45 °C
RECONSTITUIR LA FASE ORGANICA CON ACETONA (1 mL)Y LA FASE ACUOSA CON AGUA DESTILADA (1 mL)
FILTRAR LA MUESTRA Y CENTRIFUGAR A 3500 rpmDURANTE 20 min
FASE ACUOSA FASE ORGANICA
Figura 3. Diagrama de flujo para la obtención de los extractos algales.**Mezclas de solventes: C-M, C-OH, E-M y E-OH.

26
Para obtener los extractos se utilizaron 15 g de muestra húmeda. La fase inicial de la
extracción consistió en triturar en un mortero la muestra con la mezcla de solventes
seleccionada (1, 2, 3 ó 4). Una vez que quedó perfectamente triturada la muestra, la solución
se decantó y se sometió a centrifugación a una velocidad de 3500 rpm durante 20 min. Para las
mezclas en que se utilizó cloroformo (C-M y C-OH), se obtuvieron dos fases, una acuosa y
otra orgánica u oleoso. En las mezclas en que se utilizó éter, sólo se formó una fase de aspecto
oleoso.
Posteriormente, ambas fases fueron secadas en su totalidad en matraces tarados, dentro de un
rotoevaporador a 45 °C con presión reducida. Una vez secas las fases, los recipientes
conteniéndolas se pesaron nuevamente y por diferencia de pesos, se obtuvo el peso en mg de
cada fase. Para reconstituir cada una de ellas, se adicionaron 1 mL de acetona y 1 mL de agua
a las fases orgánicas y acuosas, respectivamente.
6.3. Pruebas antimicóticas.
Se utilizaron las técnicas de difusión y dilución para conocer la actividad antimicótica de los
extractos algales en contra de las cepas: Rhizopus oryzae y Rhizopus nigricans (Clase
Zygomycetes), Aspergillus niger, Alternaria alternata y Beuaveria bassiana (Clase
Deuteromycetes), y Candida albicans (Clase Blastomycetes).

27
6.3.1. Técnica de difusión.
Se usaron cajas de Petri estériles desechables de 15 mL con medio sólido agar dextrosa
Sabouraud. Las pruebas con Candida albicans se llevaron a cabo utilizando la técnica de
estriado. Por lo que respecta a los hongos filamentosos, se realizó la técnica de inoculación
directa en el medio. El medio de cultivo fue preparado de acuerdo con las indicaciones
descritas por el fabricante. Las pruebas se realizaron por duplicado.
Para cada prueba se usaron dos discos de papel filtro Wathman No. 1 estéril de 6.0 mm de
diámetro. Para concentrar los discos se utilizó una micropipeta, el primero se concentró con el
extracto de características oleosas y el segundo disco con el extracto acuoso y se dejaron secar
a temperatura ambiente. Dependiendo de la muestra procesada, fueron obtenidos extractos con
diferentes concentraciones, por lo que también varió la concentración de los discos de prueba,
siendo de entre 4.8 a 8.0 mg/disco. Se utilizaron dos discos control, uno se concentró con el
antimicótico comercial miconazol a una concentración de 10 mg/mL y el otro únicamente con
acetona.
La siguiente etapa consistió en colocar dentro de la caja de Petri con medio conteniendo al
microorganismo, los discos impregnados con los extractos así como los discos control. Se
incubaron por 72 h a una temperatura de 27 °C, transcurrido este tiempo se revisaron las cajas,
en caso de apreciarse escaso o nulo crecimiento se dejaron en incubación por 24 h más, al
final, se registró la presencia o ausencia de halos de inhibición, en caso de obtenerse un
28
resultado positivo, se midió en mm el halo formado, considerando para ello el promedio entre
el diámetro mayor y el menor.
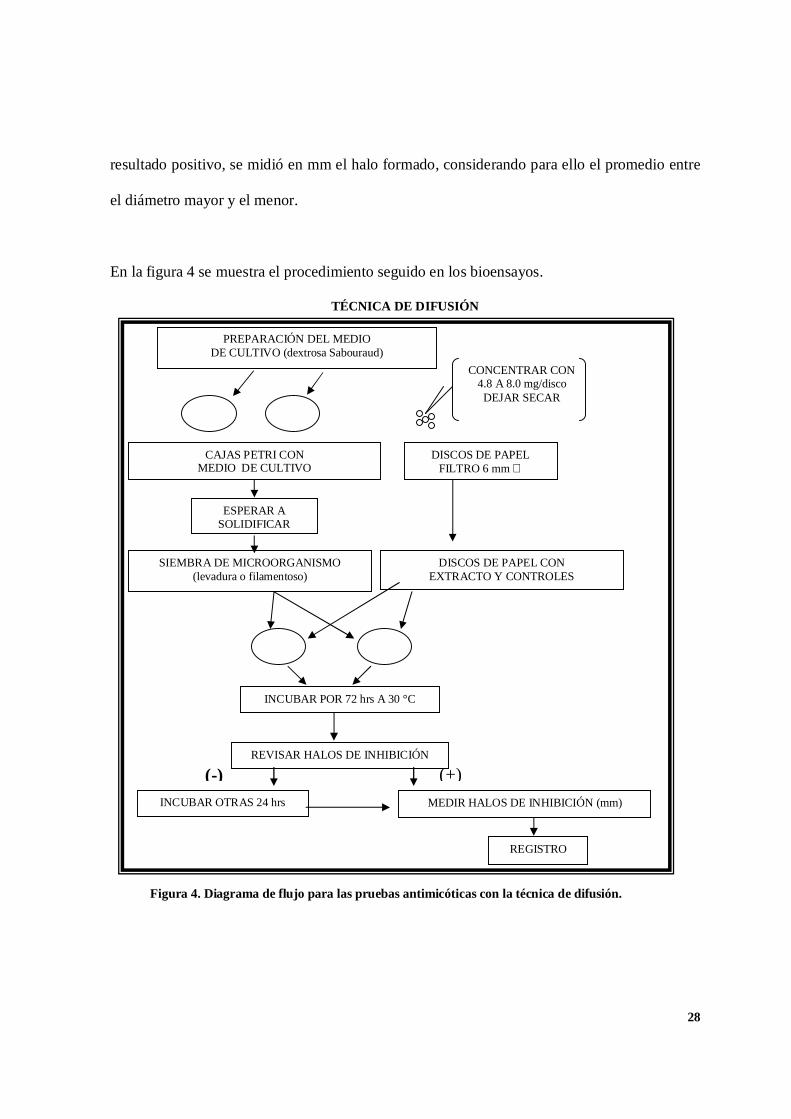
En la figura 4 se muestra el procedimiento seguido en los bioensayos.
TÉCNICA DE DIFUSIÓN
PREPARACIÓN DEL MEDIODE CULTIVO (dextrosa Sabouraud)
DISCOS DE PAPELFILTRO 6 mm ∅
CAJAS PETRI CONMEDIO DE CULTIVO
CONCENTRAR CON4.8 A 8.0 mg/discoDEJAR SECAR
ESPERAR ASOLIDIFICAR
SIEMBRA DE MICROORGANISMO(levadura o filamentoso)
DISCOS DE PAPEL CONEXTRACTO Y CONTROLES
INCUBAR POR 72 hrs A 30 °C
REVISAR HALOS DE INHIBICIÓN
MEDIR HALOS DE INHIBICIÓN (mm)INCUBAR OTRAS 24 hrs
REGISTRO
(-) (+)
Figura 4. Diagrama de flujo para las pruebas antimicóticas con la técnica de difusión.

29
6.3.2. Técnica de dilución.
Se utilizó la técnica propuesta por Atlas et al. (1988) y Kim et al. (2000), para lo cual se
realizó una suspensión de esporas utilizando una solución de 1 ml de Tween 80 en 10 mL de
agua destilada, posteriormente se prepararon los microcultivos conteniendo 75 µL de caldo
maltosa Sabouraud, con 25 µL de suspensión de esporas y 35 µL del extracto algal.
Se prepararon dos testigos, para el primero se llevó a acabo el mismo proceso, sólo que en
lugar de extracto algal, se utilizó el mismo volumen de acetona, para el segundo testigo sólo se
utilizó agua estéril. Ambos fueron empleados como referencias, el primero tuvo la función de
observar el posible efecto del solvente sobre las esporas y el testigo de agua se utilizó para
observar la viabilidad de las esporas.
El microcultivo se mantuvo durante 48 h a 27 °C, posteriormente se realizó la siembra en
placas con medio sólido de dextrosa Sabouraud, para ello se colocaron 15 µL del microcultivo
y se mantuvieron en incubación por espacio de 5 días reportándose la inhibición o no del
crecimiento de las cepas (Figura 5).
Es importante mencionar que con el objeto de conocer la susceptibilidad de las esporas en
suspensión de las cepas en contra de la acetona, se realizó una curva con diferentes volúmenes
del solvente, para ello se realizaron microcultivos con 75 µL de caldo maltosa Sabouraud y
25 µL de suspensión de esporas. El volumen de acetona probado fue de 5, 10, 15, 20, 25, 30,
30
35, 40, 45 y 50 µL. Los resultados de la curva indicaron que a 35 µL de acetona, no se observó
afectación en el crecimiento de las cepas, por lo cual este fue el volumen utilizado para las
pruebas de detección antimicótica.
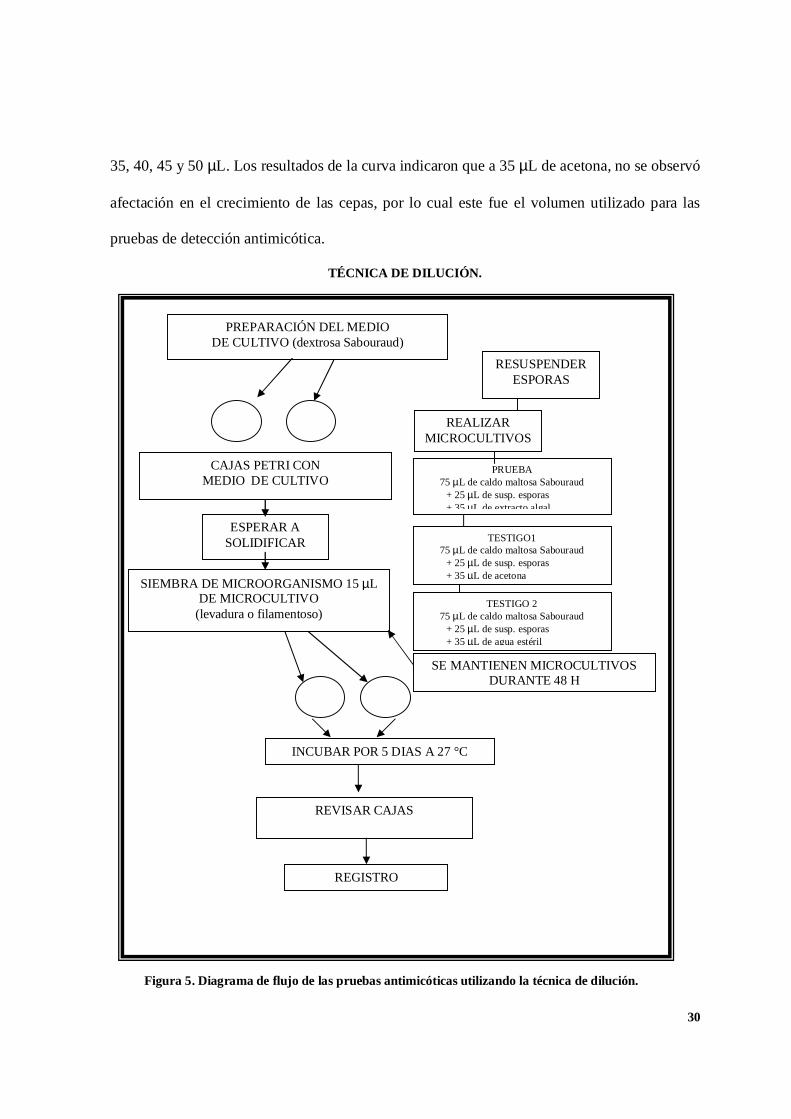
TÉCNICA DE DILUCIÓN.
Figura 5. Diagrama de flujo de las pruebas antimicóticas utilizando la técnica de dilución.
PREPARACIÓN DEL MEDIODE CULTIVO (dextrosa Sabouraud)
REALIZARMICROCULTIVOS
CAJAS PETRI CONMEDIO DE CULTIVO
ESPERAR ASOLIDIFICAR
SIEMBRA DE MICROORGANISMO 15 µLDE MICROCULTIVO
(levadura o filamentoso)
SE MANTIENEN MICROCULTIVOSDURANTE 48 H
INCUBAR POR 5 DIAS A 27 °C
REVISAR CAJAS
REGISTRO
RESUSPENDERESPORAS
PRUEBA75 µL de caldo maltosa Sabouraud
+ 25 µL de susp. esporas+ 35 µL de extracto algal
TESTIGO175 µL de caldo maltosa Sabouraud
+ 25 µL de susp. esporas+ 35 µL de acetona
TESTIGO 275 µL de caldo maltosa Sabouraud
+ 25 µL de susp. esporas+ 35 µL de agua estéril

31
7. RESULTADOS.
En el presente estudio se reportan los resultados obtenidos con los extractos de algas
provenientes de tres colectas llevadas a cabo en dos localidades costeras del estado de
Veracruz, la Mancha y Costa de Oro. Las dos primeras se realizaron durante mayo y agosto de
2002 y la tercera se realizó en marzo-abril del 2003.
En la tabla 3 se presentan las 37 especies colectadas en ambas localidades, distribuidas por
División, 18 Rhodophyta, 13 Phaeophyta y 6 Chlorophyta. Se colectaron ocho especies
comunes para los dos sitios de estudio, las cuales fueron: Acanthophora spicifera, Gracilaria
caudata, G. cervicornis e Hypnea musciformis (Rhodophyta); Dictyoya ciliolata y Padina
pavonica (Phaeophyta); Cymopolia barbata y Ulva fasciata (Chlorophyta).
Tabla 3. Especies colectadas en las localidades de la Mancha y Costa de Oro, 2002 y 2003.
Rhodophyta (18) Phaeophyta (13) Chlorophyta (6)
Acantophora spiciferaBryothamnium triquetumCentroceras clavulatumChondracantus acicularisChondrophycus papillosusDasya collinsianaDigenea simplexGalaxaura obtusataGanonema farinosumGracilaria caudataGracilaria cervicornisGracilaria domingensisGracilariopsis lemaneiformisHaliptilon subulatumHydropuntia corneaHypnea musciformisHypnea spinellaLiagora dendroidea
Colpomenia sinuosaDictyopteris delicatulaDictyota cervicornisDictyota ciliolataPadina boergeseniiPadina pavonicaSargassum fluitansSargassum natansSargassum platycarpumSargassum polyceratiumSargassum vulgareSpatoglossum schroederiStypopodium zonale
Caulerpa racemosa var. lamourouxiiCladophora proliferaCodium simplexCymopolia barbataHalimeda scabraUlva fasciata

32
En la figura 6 se presenta el porcentaje de especies por División que resultaron activas de las
tres colectas, sólo Hydropuntia cornea y Stypopodium zonale no reportaron actividad.
Rhodophyta49%
Phaeophyta34%
Chlorophyta17%
Durante mayo del 2002 se recolectaron y probaron 11 especies de ambas localidades, 5
pertenecieron a la División Rhodophyta, 4 a la División Phaeophyta y 2 a la División
Chlorophyta. Del total de especies probadas, 7 mostraron ser activas, de ellas al menos uno de
sus extractos mostró actividad antimicótica (Tabla 4). En términos porcentuales, el 64 % del
total de las especies mostró efectos de inhibición del crecimiento de las cepas de Aspergillus
niger, Rhyzopus nigricans y Candida albicans.
Tabla 4. Especies colectadas en dos localidades del Estado de Veracruz durante mayo del 2002,a las que se les evaluó su actividad antimicótica.
División La Mancha Costa de Oro
Rhodophyta Digenea simplexGracilaria caudataHypnea musciformis
Gracilaria domingensisHydropuntia cornea *Hypnea musciformis
Phaeophyta Padina boergeseniiSargassum vulgare
Sargassum fluitansStypopodium zonale *
Chlorophyta Cymopolia barbata * Ulva fasciata *Cymopolia barbata *
* Especies sin actividad.
Figura 6. Porcentaje de especies activa por División (2002 y 2003)

33
De la recolecta efectuada durante el mes de agosto del 2002, 17 especies fueron estudiadas, de
las cuales 8 pertenecen a la División Rhodophyta, 3 a la División Phaeophyta y 6 a la División
Chlorophyta (Tabla 5). Las pruebas antimicóticas mostraron que el 100% de las especies
colectadas en esta época resultaron ser activas al menos en uno de sus extractos en contra de
las cepas de Aspergillus niger, Rhyzopus nigricans y Candida albicans.
Tabla 5. Especies colectadas en dos localidades del Estado de Veracruz durante agosto del 2002,a las que se les evaluó su actividad antimicótica.
División La Mancha Costa de Oro
Rhodophyta Chondrophycus papillosusDigenea simplexGalaxaura obtusataGracilaria cervicornisLiagora dendroidea.
Acantophora spiciferaGracilaria caudataGracilaria cervicornisGracilariopsis lemaneiformis
Phaeophyta Dictyota cervicornis.Padina boergesenii.Sargassum platycarpum
Chlorophyta Caulerpa racemosa var. LamourouxiiCodium simplexCymopolia barbataHalimeda scabraUlva fasciata
Cladophora proliferaUlva fasciata
Por lo que respecta a la recolecta llevada a cabo en marzo-abril del 2003, 23 especies fueron
estudiadas, perteneciendo 13 a la División Rhodophyta, 7 a la Phaeophyta y 3 a la
Chlorophyta (Tabla 6). El 96 % de las especies colectadas mostraron extractos con actividad
antimicótica en contra de las cepas de Aspergillus niger, Rhyzopus nigricans y Candida
albicans.

34
Tabla 6. Especies colectadas en dos localidades del Estado de Veracruz durante marzo-abril del 2003,a las que se les evaluó su actividad antimicótica.
División La Mancha Costa de Oro
Rhodophyta Acantophora spiciferaBryothamnion triquetrumCentroceras clavulatumChondrophycus papillosusDasya collinsianaDigenea simplesGanonema farinosumGracilaria caudataGracilaria cervicornis *Haliptilon subulatumHypnea musciformisHypnea spinella
Acantophora spiciferaChondracanthus acicularisGracilaria caudataHypnea musciformis
Phaeophyta Colpomenia sinuosaDictyopteris delicatulaDictyota ciliolataPadina pavonicaSargassum poltycceratiumSpatoglossum schroederi
Dictyota ciliolataPadina pavonicaSargassum natans
Chlorophyta Codium simplexCymopolia barbataUlva fasciata
Ulva fasciata
* Especie sin actividad.
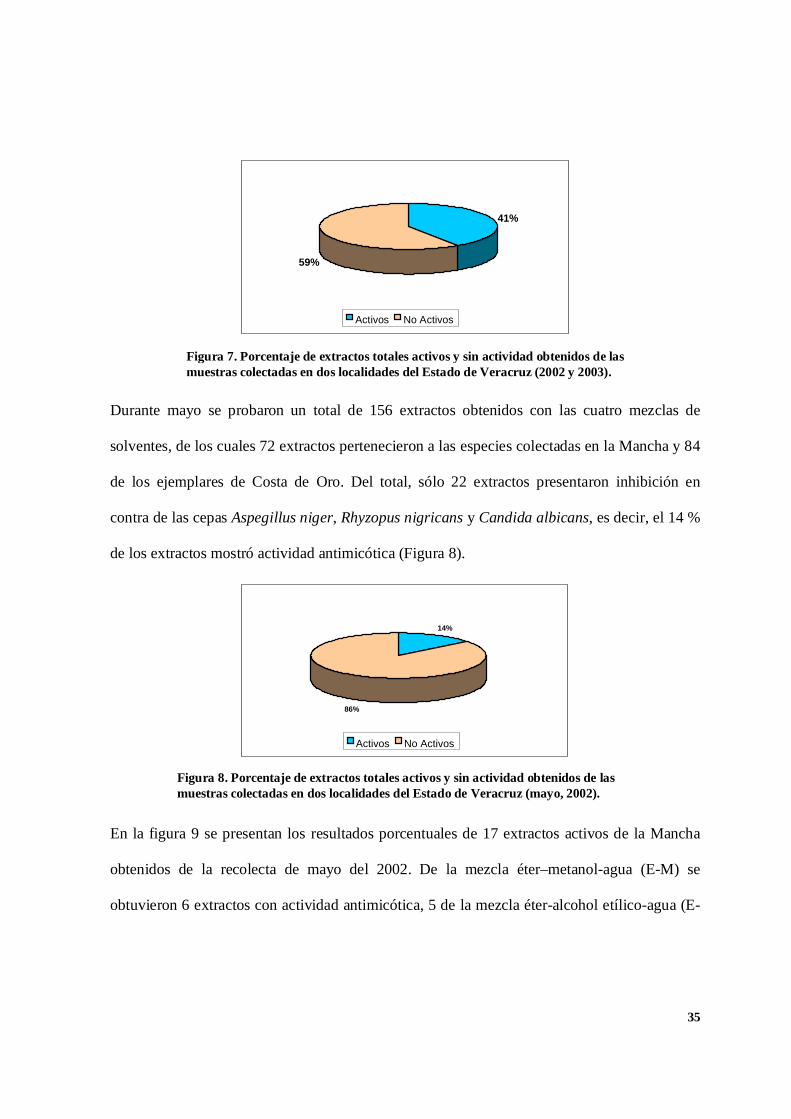
Durante el desarrollo de la fase experimental, se estudiaron 639 extractos algales de las
especies obtenidas en las tres colectas usando las mezclas de solventes: cloroformo-metanol-
agua (C-M), cloroformo-alcohol etílico-agua (C-OH), éter-metanol-agua (E-M) y éter-alcohol
etílico-agua (E-OH). Del total, 261 extractos inhibieron el crecimiento de las cepas Aspergillus
niger, Rhyzopus nigricans y Candida albicans. Es decir, alrededor del 41 % de los extractos
probados fueron activos. En contra de las cepas de Rhyzopus oryzae, Alternaria alternata y
Beauveria bassiana no se observaron efectos inhibitorios (Figura 7).
35
41%
59%
Activos No Activos
Durante mayo se probaron un total de 156 extractos obtenidos con las cuatro mezclas de
solventes, de los cuales 72 extractos pertenecieron a las especies colectadas en la Mancha y 84
de los ejemplares de Costa de Oro. Del total, sólo 22 extractos presentaron inhibición en
contra de las cepas Aspegillus niger, Rhyzopus nigricans y Candida albicans, es decir, el 14 %
de los extractos mostró actividad antimicótica (Figura 8).
14%
86%
Activos No Activos
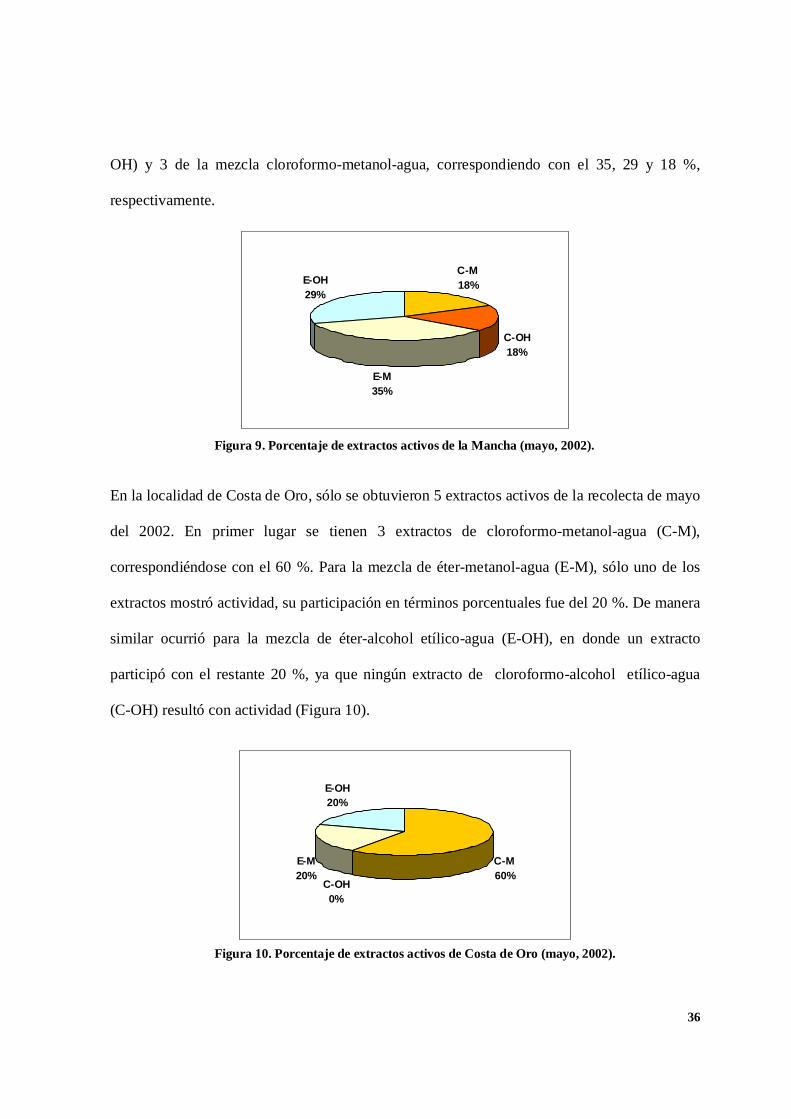
En la figura 9 se presentan los resultados porcentuales de 17 extractos activos de la Mancha
obtenidos de la recolecta de mayo del 2002. De la mezcla éter–metanol-agua (E-M) se
obtuvieron 6 extractos con actividad antimicótica, 5 de la mezcla éter-alcohol etílico-agua (E-
Figura 7. Porcentaje de extractos totales activos y sin actividad obtenidos de lasmuestras colectadas en dos localidades del Estado de Veracruz (2002 y 2003).
Figura 8. Porcentaje de extractos totales activos y sin actividad obtenidos de lasmuestras colectadas en dos localidades del Estado de Veracruz (mayo, 2002).
36
OH) y 3 de la mezcla cloroformo-metanol-agua, correspondiendo con el 35, 29 y 18 %,
respectivamente.
C-M18%
C-OH18%
E-M35%
E-OH29%
En la localidad de Costa de Oro, sólo se obtuvieron 5 extractos activos de la recolecta de mayo
del 2002. En primer lugar se tienen 3 extractos de cloroformo-metanol-agua (C-M),
correspondiéndose con el 60 %. Para la mezcla de éter-metanol-agua (E-M), sólo uno de los
extractos mostró actividad, su participación en términos porcentuales fue del 20 %. De manera
similar ocurrió para la mezcla de éter-alcohol etílico-agua (E-OH), en donde un extracto
participó con el restante 20 %, ya que ningún extracto de cloroformo-alcohol etílico-agua
(C-OH) resultó con actividad (Figura 10).
C-M60%
C-OH0%
E-M20%
E-OH20%
Figura 9. Porcentaje de extractos activos de la Mancha (mayo, 2002).
Figura 10. Porcentaje de extractos activos de Costa de Oro (mayo, 2002).

37
Durante agosto se estudiaron un total de 159 extractos algales obtenidos de las cuatro mezclas
de solventes utilizadas. De ellos, 111 extractos produjeron inhibición en contra de Aspergillus
niger, Rhyzopus nigricans y Candida albicans, es decir, el 70 % de los extractos presentó
actividad antimicótica. De los 159 extractos, 105 se obtuvieron de las especies de la Mancha,
de los cuales 77 resultaron activos. 54 extractos fueron obtenidos de las especies de Costa de
Oro, de los cuales 34 presentaron actividad antimicótica (Figura 11).
70%
30%
Activos No Activos
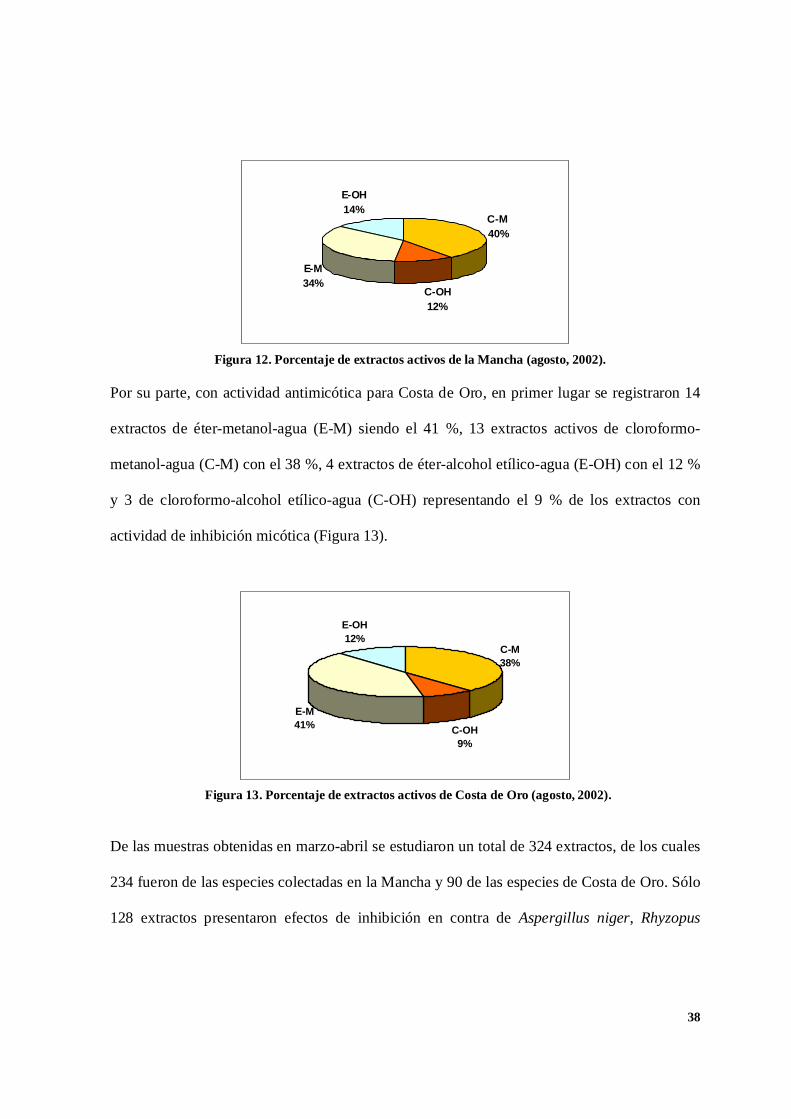
En la figura 12 se presentan, para la Mancha, los extractos activos obtenidos de la recolecta
realizada en agosto del 2002. Con la mezcla de cloroformo-metanol-agua (C-M) se obtuvieron
31 extractos, correspondiéndose con el 40 %. En segundo lugar, a partir de la mezcla de éter-
metanol-agua (E-M) se obtuvieron 26 extractos activos, lo que significó el 34 %. En orden de
importancia, le siguen los extractos obtenidos de la mezcla de éter-alcohol etílico-agua
(E-OH), contabilizándose un total de 11 extractos activos, representando el 14 %. Finalmente,
con el 12 % se reportaron 9 extractos con actividad para la mezcla de cloroformo-alcohol
etílico-agua (C-OH).
Figura 11. Porcentaje de extractos totales activos y sin actividad obtenidos de lasmuestras colectadas en dos localidades del Estado de Veracruz (agosto, 2002).
38
C-M40%
C-OH12%
E-M34%
E-OH14%
Por su parte, con actividad antimicótica para Costa de Oro, en primer lugar se registraron 14
extractos de éter-metanol-agua (E-M) siendo el 41 %, 13 extractos activos de cloroformo-
metanol-agua (C-M) con el 38 %, 4 extractos de éter-alcohol etílico-agua (E-OH) con el 12 %
y 3 de cloroformo-alcohol etílico-agua (C-OH) representando el 9 % de los extractos con
actividad de inhibición micótica (Figura 13).
C-M38%
C-OH9%
E-M41%
E-OH12%
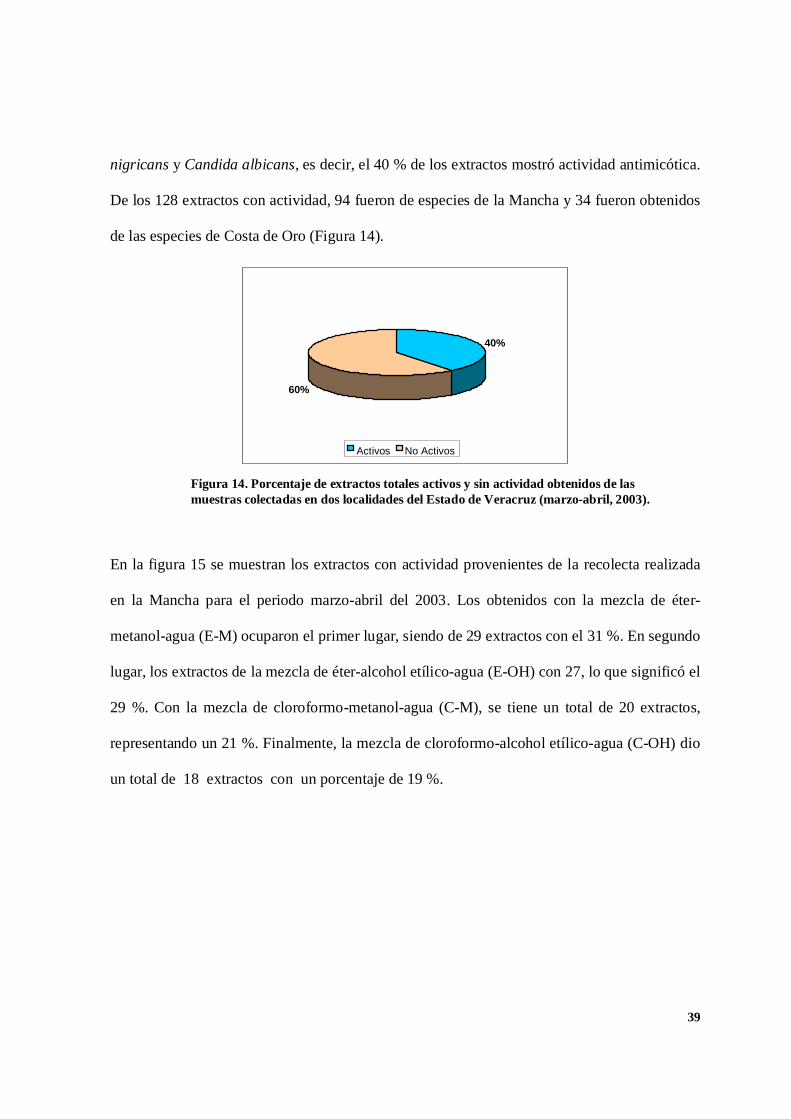
De las muestras obtenidas en marzo-abril se estudiaron un total de 324 extractos, de los cuales
234 fueron de las especies colectadas en la Mancha y 90 de las especies de Costa de Oro. Sólo
128 extractos presentaron efectos de inhibición en contra de Aspergillus niger, Rhyzopus
Figura 12. Porcentaje de extractos activos de la Mancha (agosto, 2002).
Figura 13. Porcentaje de extractos activos de Costa de Oro (agosto, 2002).
39
nigricans y Candida albicans, es decir, el 40 % de los extractos mostró actividad antimicótica.
De los 128 extractos con actividad, 94 fueron de especies de la Mancha y 34 fueron obtenidos
de las especies de Costa de Oro (Figura 14).
40%
60%
Activos No Activos
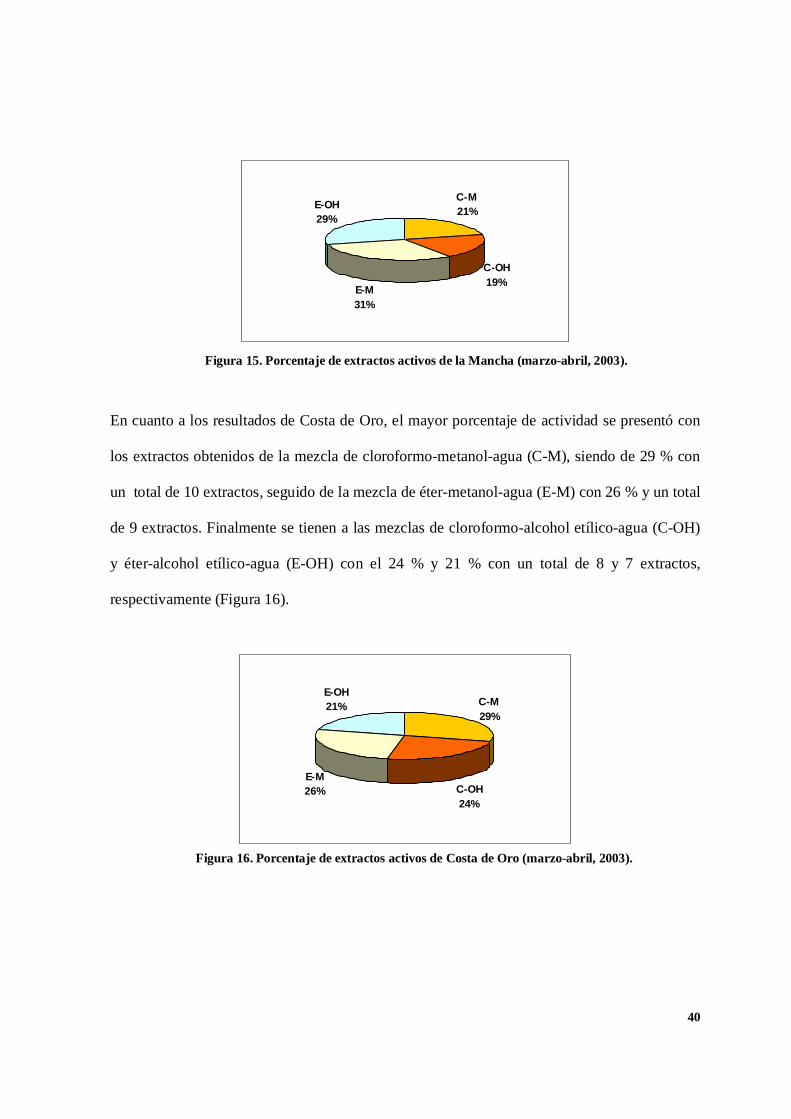
En la figura 15 se muestran los extractos con actividad provenientes de la recolecta realizada
en la Mancha para el periodo marzo-abril del 2003. Los obtenidos con la mezcla de éter-
metanol-agua (E-M) ocuparon el primer lugar, siendo de 29 extractos con el 31 %. En segundo
lugar, los extractos de la mezcla de éter-alcohol etílico-agua (E-OH) con 27, lo que significó el
29 %. Con la mezcla de cloroformo-metanol-agua (C-M), se tiene un total de 20 extractos,
representando un 21 %. Finalmente, la mezcla de cloroformo-alcohol etílico-agua (C-OH) dio
un total de 18 extractos con un porcentaje de 19 %.
Figura 14. Porcentaje de extractos totales activos y sin actividad obtenidos de lasmuestras colectadas en dos localidades del Estado de Veracruz (marzo-abril, 2003).
40
C-M21%
C-OH19%
E-M31%
E-OH29%
En cuanto a los resultados de Costa de Oro, el mayor porcentaje de actividad se presentó con
los extractos obtenidos de la mezcla de cloroformo-metanol-agua (C-M), siendo de 29 % con
un total de 10 extractos, seguido de la mezcla de éter-metanol-agua (E-M) con 26 % y un total
de 9 extractos. Finalmente se tienen a las mezclas de cloroformo-alcohol etílico-agua (C-OH)
y éter-alcohol etílico-agua (E-OH) con el 24 % y 21 % con un total de 8 y 7 extractos,
respectivamente (Figura 16).
C-M29%
C-OH24%
E-M26%
E-OH21%
Figura 15. Porcentaje de extractos activos de la Mancha (marzo-abril, 2003).
Figura 16. Porcentaje de extractos activos de Costa de Oro (marzo-abril, 2003).

41
Ninguno de los extractos de las especies colectadas presentó actividad antimicótica en contra
de Rhyzopus oryzae, Alternaria alternata y Beauveria bassiana. Los resultados obtenidos con
los extractos de las especies colectadas se presentan en las tablas 7, 8 y 9, donde se presentó la
actividad observada con los extractos probados. Cuando algún extracto mostró inhibición total
o bien el nulo crecimiento de colonias de la cepa utilizada en la prueba de detección, el
extracto fue señalado como activo y con ello la actividad antimicótica fue indicada con el
signo (+).
Por el contrario, cuando en alguno de los extractos se observó la aparición de colonias de la
cepa y además el medio de cultivo era cubierto por completo, dicho extracto fue registrado con
nula actividad o carente de actividad antimicótica, en las tablas se indicó con el signo (-).
Por último, cuando en el medio de cultivo se observaron algunas colonias que resultaron
fácilmente cuantificables, el extracto fue considerado con actividad parcial y se reportaron
los datos del número de colonias observadas, utilizando para ello las siglas UFC (unidades
formadoras de colonias).
42
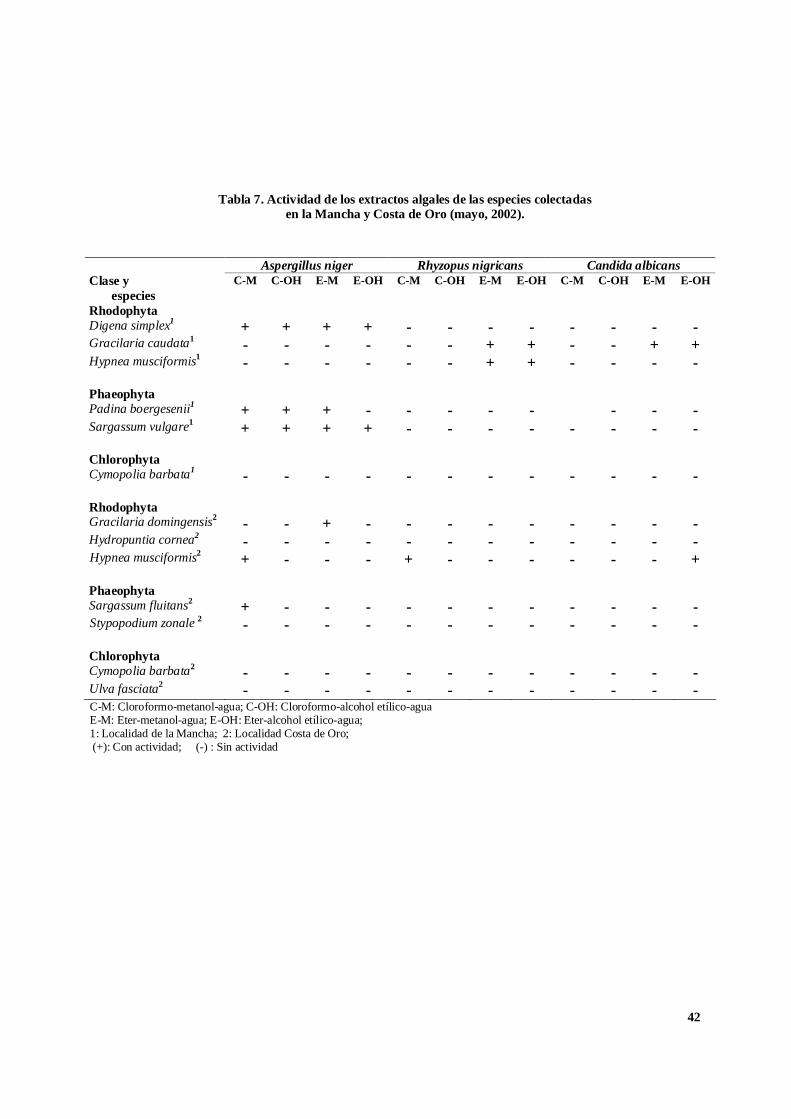
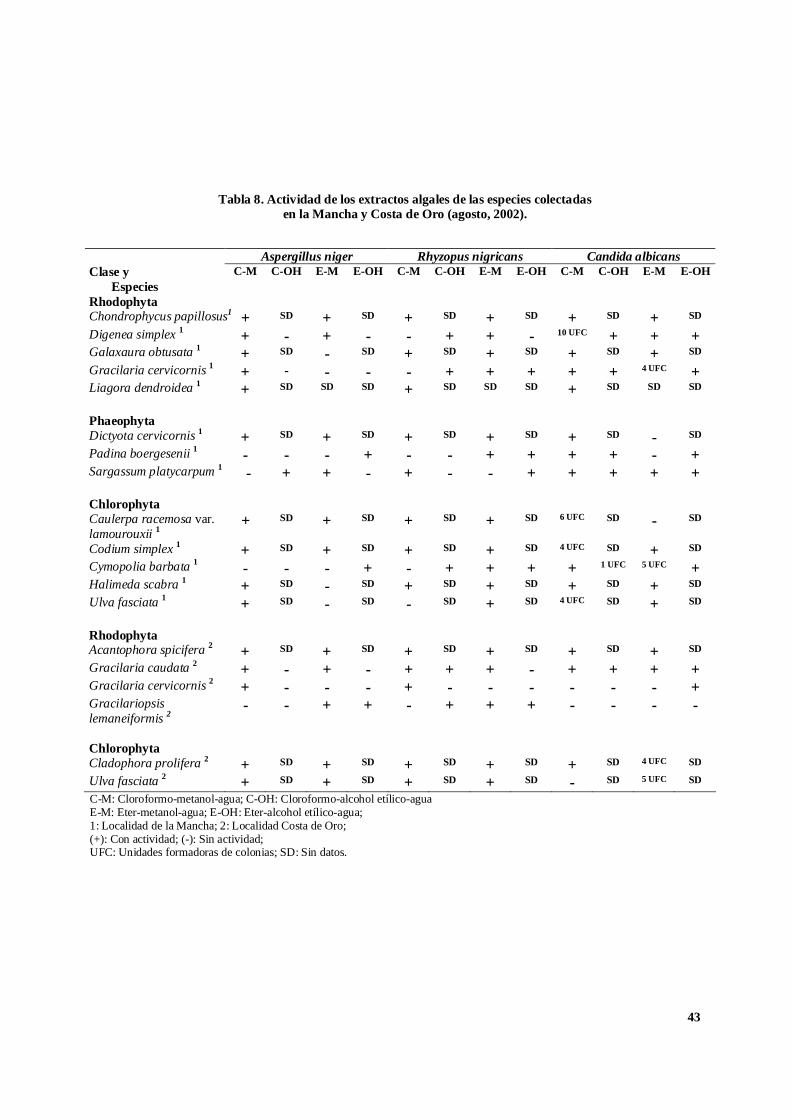
Tabla 7. Actividad de los extractos algales de las especies colectadasen la Mancha y Costa de Oro (mayo, 2002).
Aspergillus niger Rhyzopus nigricans Candida albicansClase y
especiesC-M C-OH E-M E-OH C-M C-OH E-M E-OH C-M C-OH E-M E-OH
RhodophytaDigena simplex1 + + + + - - - - - - - -Gracilaria caudata1 - - - - - - + + - - + +Hypnea musciformis1 - - - - - - + + - - - -
PhaeophytaPadina boergesenii1 + + + - - - - - - - -Sargassum vulgare1 + + + + - - - - - - - -
ChlorophytaCymopolia barbata1 - - - - - - - - - - - -
RhodophytaGracilaria domingensis2 - - + - - - - - - - - -Hydropuntia cornea2 - - - - - - - - - - - -Hypnea musciformis2 + - - - + - - - - - - +
PhaeophytaSargassum fluitans2 + - - - - - - - - - - -Stypopodium zonale 2 - - - - - - - - - - - -
ChlorophytaCymopolia barbata2 - - - - - - - - - - - -Ulva fasciata2 - - - - - - - - - - - -C-M: Cloroformo-metanol-agua; C-OH: Cloroformo-alcohol etílico-aguaE-M: Eter-metanol-agua; E-OH: Eter-alcohol etílico-agua;1: Localidad de la Mancha; 2: Localidad Costa de Oro; (+): Con actividad; (-) : Sin actividad
43
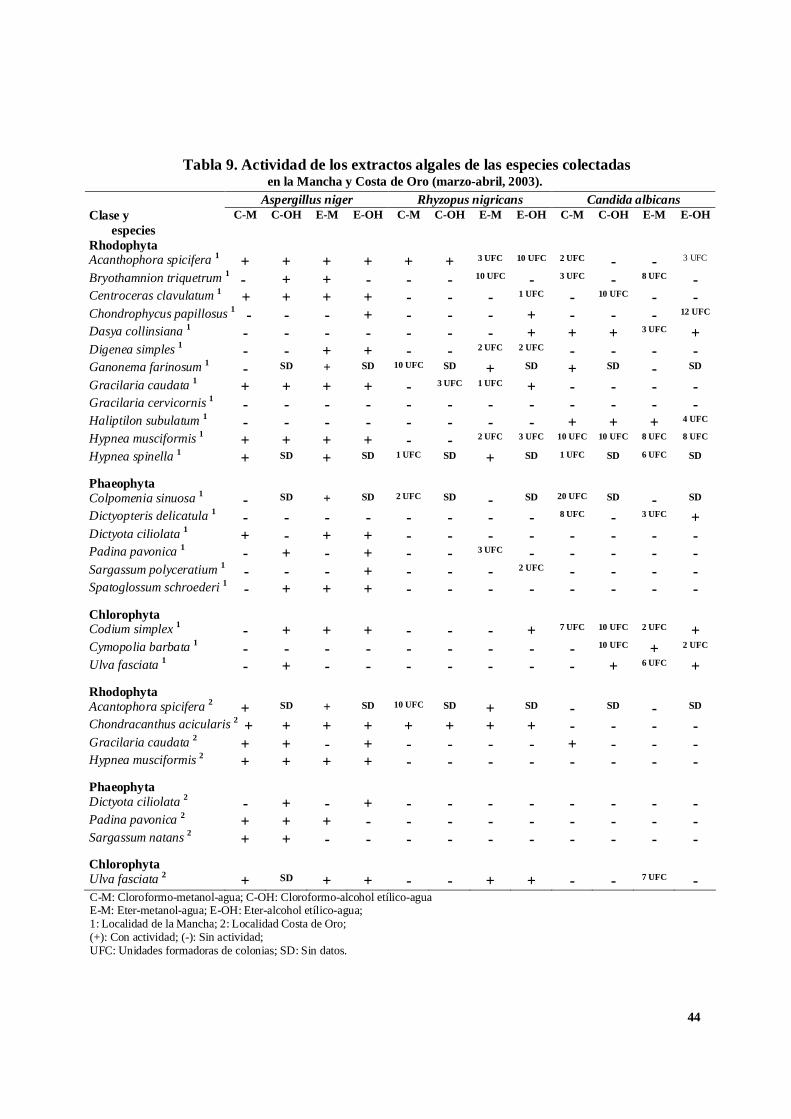
Tabla 8. Actividad de los extractos algales de las especies colectadasen la Mancha y Costa de Oro (agosto, 2002).
Aspergillus niger Rhyzopus nigricans Candida albicansClase y
EspeciesC-M C-OH E-M E-OH C-M C-OH E-M E-OH C-M C-OH E-M E-OH
RhodophytaChondrophycus papillosus1 + SD + SD + SD + SD + SD + SD
Digenea simplex 1 + - + - - + + - 10 UFC + + +Galaxaura obtusata 1 + SD - SD + SD + SD + SD + SD
Gracilaria cervicornis 1 + - - - - + + + + + 4 UFC +Liagora dendroidea 1 + SD SD SD + SD SD SD + SD SD SD
PhaeophytaDictyota cervicornis 1 + SD + SD + SD + SD + SD - SD
Padina boergesenii 1 - - - + - - + + + + - +Sargassum platycarpum 1 - + + - + - - + + + + +
ChlorophytaCaulerpa racemosa var.lamourouxii 1
+ SD + SD + SD + SD 6 UFC SD - SD
Codium simplex 1 + SD + SD + SD + SD 4 UFC SD + SD
Cymopolia barbata 1 - - - + - + + + + 1 UFC 5 UFC +Halimeda scabra 1 + SD - SD + SD + SD + SD + SD
Ulva fasciata 1 + SD - SD - SD + SD 4 UFC SD + SD
RhodophytaAcantophora spicifera 2 + SD + SD + SD + SD + SD + SD
Gracilaria caudata 2 + - + - + + + - + + + +Gracilaria cervicornis 2 + - - - + - - - - - - +Gracilariopsislemaneiformis 2
- - + + - + + + - - - -
ChlorophytaCladophora prolifera 2 + SD + SD + SD + SD + SD 4 UFC SD
Ulva fasciata 2 + SD + SD + SD + SD - SD 5 UFC SD
C-M: Cloroformo-metanol-agua; C-OH: Cloroformo-alcohol etílico-aguaE-M: Eter-metanol-agua; E-OH: Eter-alcohol etílico-agua;1: Localidad de la Mancha; 2: Localidad Costa de Oro;(+): Con actividad; (-): Sin actividad;UFC: Unidades formadoras de colonias; SD: Sin datos.
44
Tabla 9. Actividad de los extractos algales de las especies colectadasen la Mancha y Costa de Oro (marzo-abril, 2003).
Aspergillus niger Rhyzopus nigricans Candida albicansClase y
especiesC-M C-OH E-M E-OH C-M C-OH E-M E-OH C-M C-OH E-M E-OH
RhodophytaAcanthophora spicifera 1 + + + + + + 3 UFC 10 UFC 2 UFC - - 3 UFC
Bryothamnion triquetrum 1 - + + - - - 10 UFC - 3 UFC - 8 UFC -Centroceras clavulatum 1 + + + + - - - 1 UFC - 10 UFC - -Chondrophycus papillosus 1 - - - + - - - + - - - 12 UFC
Dasya collinsiana 1 - - - - - - - + + + 3 UFC +Digenea simples 1 - - + + - - 2 UFC 2 UFC - - - -Ganonema farinosum 1 - SD + SD 10 UFC SD + SD + SD - SD
Gracilaria caudata 1 + + + + - 3 UFC 1 UFC + - - - -Gracilaria cervicornis 1 - - - - - - - - - - - -Haliptilon subulatum 1 - - - - - - - - + + + 4 UFC
Hypnea musciformis 1 + + + + - - 2 UFC 3 UFC 10 UFC 10 UFC 8 UFC 8 UFC
Hypnea spinella 1 + SD + SD 1 UFC SD + SD 1 UFC SD 6 UFC SD
PhaeophytaColpomenia sinuosa 1 - SD + SD 2 UFC SD - SD 20 UFC SD - SD
Dictyopteris delicatula 1 - - - - - - - - 8 UFC - 3 UFC +Dictyota ciliolata 1 + - + + - - - - - - - -Padina pavonica 1 - + - + - - 3 UFC - - - - -Sargassum polyceratium 1 - - - + - - - 2 UFC - - - -Spatoglossum schroederi 1 - + + + - - - - - - - -ChlorophytaCodium simplex 1 - + + + - - - + 7 UFC 10 UFC 2 UFC +Cymopolia barbata 1 - - - - - - - - - 10 UFC + 2 UFC
Ulva fasciata 1 - + - - - - - - - + 6 UFC +RhodophytaAcantophora spicifera 2 + SD + SD 10 UFC SD + SD - SD - SD
Chondracanthus acicularis 2 + + + + + + + + - - - -Gracilaria caudata 2 + + - + - - - - + - - -Hypnea musciformis 2 + + + + - - - - - - - -PhaeophytaDictyota ciliolata 2 - + - + - - - - - - - -Padina pavonica 2 + + + - - - - - - - - -Sargassum natans 2 + + - - - - - - - - - -ChlorophytaUlva fasciata 2 + SD + + - - + + - - 7 UFC -C-M: Cloroformo-metanol-agua; C-OH: Cloroformo-alcohol etílico-aguaE-M: Eter-metanol-agua; E-OH: Eter-alcohol etílico-agua;1: Localidad de la Mancha; 2: Localidad Costa de Oro;(+): Con actividad; (-): Sin actividad;UFC: Unidades formadoras de colonias; SD: Sin datos.

45
7.1 Variación estacional de la actividad de las algas colectadas en mayo y agosto del 2002.
Al comparar los resultados de las pruebas antimicóticas de las especies provenientes de las
colectas realizadas durante mayo y agosto del 2002, se observaron diferencias en la actividad
en las siguientes especies: Digenea simplex, Padina boergesenii y Cymopolia barbata de la
Mancha y Ulva fasciata de Costa de Oro.
De las muestras obtenidas en mayo, los extractos de Digenea simplex con las mezclas C-M, C-
OH, E-M y E-OH provocaron la inhibición completa del crecimiento de Aspergillus niger, con
los ejemplares colectados en agosto sólo los extractos obtenidos con C-M y E-M dieron
efectos similares, es decir, en estos dos tipos de extractos no existió variación estacional en la
actividad de inhibición de la cepa. Si se observó variación de los efectos inhibitorios de los
extractos obtenidos con C-OH y E-OH, ya que si los presentaron los ejemplares colectados en
mayo y no los de agosto.
En las pruebas con los extractos de Digenea simplex de la misma localidad en contra de
Rhizopus nigricans, en el mes de mayo no se observó actividad con ninguno de sus extractos;
con los ejemplares del mes de agosto, sólo los extractos de C-OH y E-M presentaron
inhibición en el crecimiento de la cepa. De tal manera que entre éstos extractos se presentaron
diferencias en la actividad inhibitoria con los ejemplares colectados en ambas épocas.
Al realizar las pruebas en contra de Candida albicans con los extractos de Digenea simplex, se
observaron diferencias estacionales de la actividad, siendo negativa para el mes de mayo en

46
todos sus extractos y positiva con los cuatro extractos del mes de agosto, de estos el extracto
obtenido con C-M presentó una inhibición parcial.
Con Padina boergesenii también pudimos observar diferencias en la actividad antimicótica
entre los ejemplares colectados en mayo y los colectados en agosto. Ninguno de los extractos
de las especies colectadas en mayo reportó inhibición en contra de Aspergillus niger, mientras
que el extracto obtenido con la mezcla E-OH de los ejemplares colectados en agosto sí
presentó actividad.
Los extractos de Padina boergesenii utilizados con las cepas de Rhizopus nigricans mostraron
variación estacional en su actividad para las mezclas de C-OH, E-M y E-OH, en donde para
los ejemplares colectados en el mes de mayo ninguna de las mezclas presentaron inhibición de
la cepa y por el contrario para agosto, se observó actividad positiva con los extractos
señalados.
En la prueba en contra de Candida albicans, los extractos de Padina boergesenii también
mostraron cambios estacionales en sus efectos de inhibición. Ninguno de los extractos hechos
con los ejemplares de mayo presentaron actividad antimicótica y por el contrario con los de
agosto, se observó inhibición de la cepa, siendo positiva para las mezclas de C-M, C-OH y E-
OH. El extracto de E-M de las muestras dc ambas épocas de colecta no mostró actividad.
Con Cymopolia barbata se observaron cambios en la actividad antimicótica entre los meses
señalados. En el extracto de E-OH del mes de mayo, no se observó actividad, mientras que

47
para el obtenido de la muestra de agosto, se registró inhibición de la cepa de Aspergillus niger,
los tres extractos restantes no tuvieron actividad.
Para la prueba en contra de Rhizopus nigricans, los extractos de C-OH, E-M y E-OH
presentaron variación estacional. Los extractos de las muestras de mayo, no mostraron
inhibición y por el contrario, los del mes de agosto, si tuvieron efectos inhibitorios. Para el
extracto con C-M, no existieron diferencias en la actividad, siendo nula para ambos meses.
En la prueba de detección en contra de Candida albicans, los extractos de Cymopolia barbata
presentaron un comportamiento similar, para mayo no se reportó actividad en ninguno de sus
extractos. Por el contrario, para agosto existió variación, ya que en todos los extractos se
observó la inhibición de la cepa, siendo total con los extractos C-M y E-OH y parcial con los
extractos C-OH y E-M.
Por lo que respecta a las especies colectadas en Costa Oro, sólo Ulva fasciata presentó
cambios de la actividad antimicótica entre las muestras de las dos épocas de colecta. Para las
pruebas de detección en contra de Aspergillus niger y Rhyzopus nigricans, los extractos
obtenidos con las mezclas de C-M y E-M de los ejemplares colectados en mayo, no
presentaron actividad. Los obtenidos en la colecta de agosto si la presentaron. Para la prueba
en contra de Candida albicans, existió variación sólo para el extracto de E-M, siendo nula la
actividad para mayo y de forma parcial para el extracto de agosto.
48
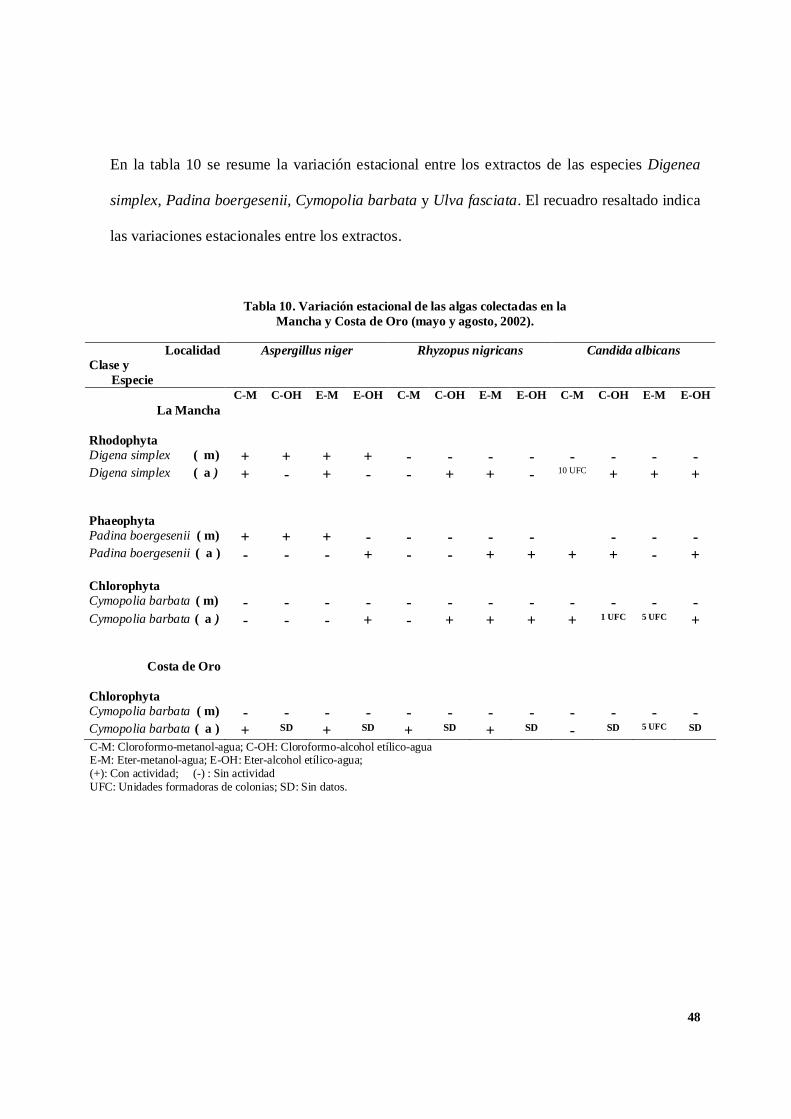
En la tabla 10 se resume la variación estacional entre los extractos de las especies Digenea
simplex, Padina boergesenii, Cymopolia barbata y Ulva fasciata. El recuadro resaltado indica
las variaciones estacionales entre los extractos.
Tabla 10. Variación estacional de las algas colectadas en laMancha y Costa de Oro (mayo y agosto, 2002).
LocalidadClase y
Especie
Aspergillus niger Rhyzopus nigricans Candida albicans
C-M C-OH E-M E-OH C-M C-OH E-M E-OH C-M C-OH E-M E-OHLa Mancha
RhodophytaDigena simplex ( m) + + + + - - - - - - - -Digena simplex ( a ) + - + - - + + - 10 UFC + + +
PhaeophytaPadina boergesenii ( m) + + + - - - - - - - -Padina boergesenii ( a ) - - - + - - + + + + - +
ChlorophytaCymopolia barbata ( m) - - - - - - - - - - - -Cymopolia barbata ( a ) - - - + - + + + + 1 UFC 5 UFC +
Costa de Oro
ChlorophytaCymopolia barbata ( m) - - - - - - - - - - - -Cymopolia barbata ( a ) + SD + SD + SD + SD - SD 5 UFC SD
C-M: Cloroformo-metanol-agua; C-OH: Cloroformo-alcohol etílico-aguaE-M: Eter-metanol-agua; E-OH: Eter-alcohol etílico-agua;(+): Con actividad; (-) : Sin actividadUFC: Unidades formadoras de colonias; SD: Sin datos.

49
7. 2. Efectividad de las mezclas de solventes.
Con la finalidad de evaluar, cuál mezcla de solventes resultó ser más efectiva en el proceso de
extracción de los compuestos químicos con actividad antimicótica, fueron considerados sólo
los extractos que conformaron series de cuatro. Es decir, se tomaron en cuenta a las especies
de algas cuya biomasa resultó suficiente para utilizar las cuatro mezclas de solventes y así
obtener la serie de cuatro extractos (C-M, C-OH, E-M y E-OH).
De esta manera, el total de extractos entre las tres colectas es de 552, distribuidos por colecta
de la siguiente manera: para el mes de mayo del 2002 se contabilizaron 156 extractos; para
agosto del mismo año, el total fue de 96 extractos; y, 300 extractos para la colecta realizada
durante marzo-abril del 2003.
De los 552 extractos, sólo resultaron 188 extractos con actividad antimicótica, los cuales de
acuerdo a la mezcla de solventes, tenemos lo siguiente: de la colecta obtenida en el mes de
mayo, los extractos de C-M fueron 6, 3 extractos de la mezcla de C-OH, los de la mezcla de E-
M resultaron 7 y de la mezcla E-OH, se reportaron 6 extractos (Figura 16). En la colecta de
agosto, el total de extractos de las mezclas de C-M, C-OH, E-M y E-OH fue de 13, 12, 15 y
15, respectivamente (Figura 17). Finalmente, el total de extractos de la colecta efectuada en
marzo-abril fue de 21, 26, 30 y 34 de las mezclas de C-M, C-OH, E-M y E-OH,
respectivamente (Figura 18).
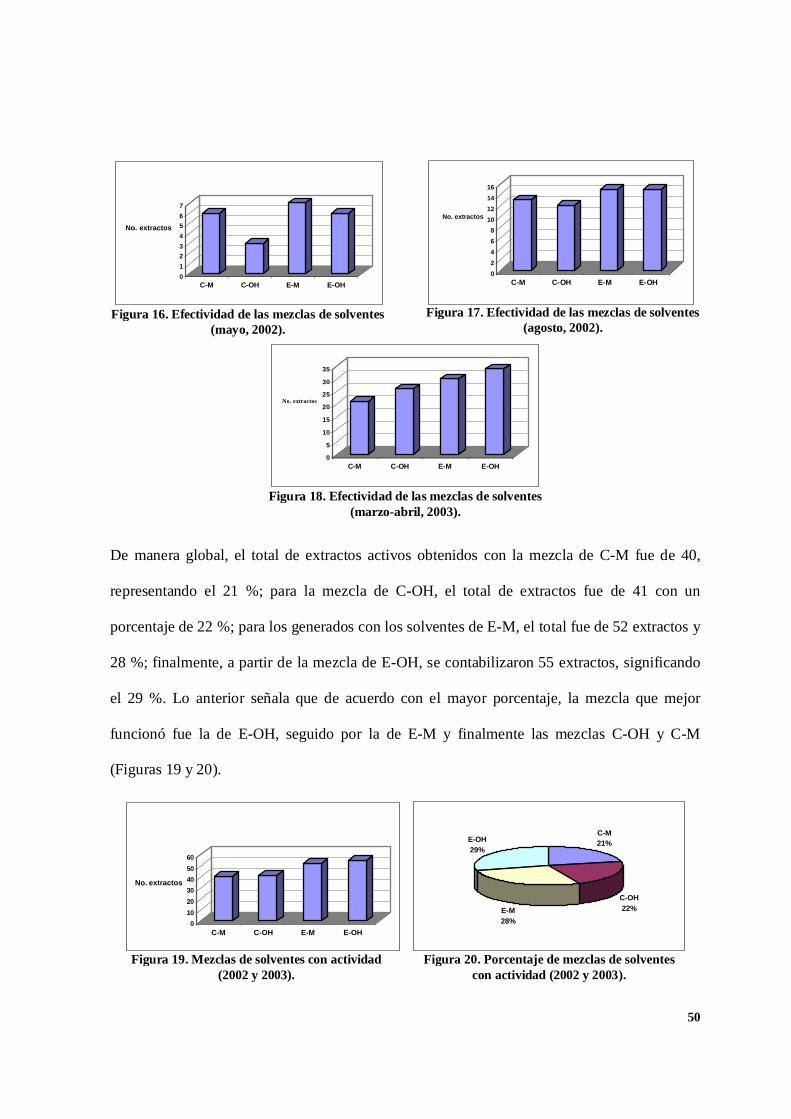
50
01234567
No. extractos
C-M C-OH E-M E-OH02468
10121416
No. extractos
C-M C-OH E-M E-OH
0
5
10
15
20
25
30
35
No. extractos
C-M C-OH E-M E-OH
De manera global, el total de extractos activos obtenidos con la mezcla de C-M fue de 40,
representando el 21 %; para la mezcla de C-OH, el total de extractos fue de 41 con un
porcentaje de 22 %; para los generados con los solventes de E-M, el total fue de 52 extractos y
28 %; finalmente, a partir de la mezcla de E-OH, se contabilizaron 55 extractos, significando
el 29 %. Lo anterior señala que de acuerdo con el mayor porcentaje, la mezcla que mejor
funcionó fue la de E-OH, seguido por la de E-M y finalmente las mezclas C-OH y C-M
(Figuras 19 y 20).
0102030405060
No. extractos
C-M C-OH E-M E-OH
C-M21%
E-M28%
E-OH29%
C-OH22%
Figura 16. Efectividad de las mezclas de solventes(mayo, 2002).
Figura 17. Efectividad de las mezclas de solventes(agosto, 2002).
Figura 18. Efectividad de las mezclas de solventes(marzo-abril, 2003).
Figura 19. Mezclas de solventes con actividad(2002 y 2003).
Figura 20. Porcentaje de mezclas de solventescon actividad (2002 y 2003).

51
8. DISCUSIÓN.
Al inicio de la fase experimental se consideró utilizar el método de difusión, en el cual se
utilizan discos de papel estériles concentrados con el extracto de prueba y posteriormente son
colocados en las placas de agar sólido conteniendo a los microorganismos. De esta manera, la
potencia del extracto es medida por el diámetro de la zona de inhibición. Este método fue
empleado para conocer la posible variación estacional de los extractos de las especies
colectadas durante los meses de mayo y agosto del 2002. Dicha técnica fue utilizada debido a
que existen antecedentes de actividad antimicótica como el que se realizó con material seco
(200 a 500 g) por Padmakumar y Ayyakkannu (1997). En este estudio se obtuvieron extractos
etanólicos de ochenta algas marinas provenientes de las costas de la India. Ellos encontraron
actividad en contra de las ceepas de Candida albicans, Aspergillus terreus, Fusarium
oxysporum, Helminthosporium oryzae, Trichophyton mentagrophytes y Trichoderma viridae.
Los resultados obtenidos en el presente estudio al compararse con los obtenidos por
Padmakumar y Ayyakkannu en 1997, fueron nulos debido a que se deduce la existencia de una
baja concentración de las sustancias químicas que pudieran ser responsables de la actividad
antimicótica, ya que en el presente estudio, los extractos fueron obtenidos a partir de 15 g de
peso húmedo.
Por otro lado, y que es de suma importancia, es el hecho de que técnicamente se dificultó la
labor de concentración, ya que los discos se saturaban y duraban más tiempo en secarse,
inclusive, una gran parte del extracto se quedaba impregnado en la superficie de trabajo con la

52
consecuente pérdida de concentración, es decir, los discos al final contenían menor cantidad de
extracto del que realmente se pretendía que tuvieran.
De acuerdo con Hay et al. (1987) se sabe que en los estudios en donde se realizan pruebas
para detectar la presencia de compuestos antimicrobianos, los investigadores suelen
enfrentarse a problemas de carácter metodológico, básicamente en el proceso de obtención de
los extractos, ya que durante la extracción, raramente ésta se lleva de forma completa, es decir,
existe una proporción de las sustancias que se quedan en el material algal y no logran ser
recuperadas. Por otro lado, durante las primeras fases de un proceso de aislamiento de las
sustancias activas, de manera gradual parte de dichos compuestos se va perdiendo en cada una
de las etapas; y además, hay que considerar el hecho de que una vez obtenidos, pueden ser
sujetos por cuestiones simplemente ambientales, a procesos de degradación en forma completa
o parcial durante la secuencia.
Con respecto al desarrollo de las pruebas antimicóticas llevadas a cabo en éste estudio con la
técnica de difusión, se observaron diferencias en la forma del crecimiento del micelio. En
ocasiones se presentaba un crecimiento en forma rápida cubriendo toda la superficie del medio
de cultivo, incluidos los discos de prueba. En otros casos, el crecimiento del micelio llegaba
justo al borde del disco, de manera que el micelio podía crecer alrededor de todo el contorno
del disco, posteriormente, aparecían algunas hifas sobre el disco y finalmente el micelio
terminaba cubriendo todo el disco de prueba, dicho situación obligó a reconsiderar la
posibilidad de que se tenía una baja concentración de sustancias activas en el extracto algal,

53
probablemente a consecuencia de utilizar muy poco material algal (15 g peso húmedo), o bien,
los resultados se debían a problemas de carácter técnico durante la metodología empleada.
Además, se observaron crecimientos miceliales llegando al disco de prueba y redondeándolo
de forma similar al caso anterior, sólo que en ningún momento el disco fue invadido por
alguna hifa. Para el caso de la levadura, de forma similar a los hongos filamentosos, sus
colonias también mostraron diferencias en su crecimiento, en ocasiones las colonias
invadieron el disco y en otras nunca aparecieron colonias sobre el disco de prueba. Debido a lo
anterior se evaluó el método de prueba señalado, por lo que se utilizaron discos control
concentrados con miconazol, un antimicótico comercial, el resultado demostró que la técnica
de difusión con el uso de discos de papel estériles, sí funcionó para la levadura y los hongos
filamentosos usados en este estudio, ya que se formaron halos de inhibición de forma
irregular, lo cual comprobó el efecto inhibitorio del miconazol y la eficacia de la técnica.
Otro factor a considerar y que resulta de importancia durante las pruebas de actividad
antimicótica, es el hecho de que las paredes celulares de los hongos están constituidas
principalmente por quitina en combinación con diversos polisacáridos como la celulosa y
lignina (Karp, 1998). A través de la pared, se da la absorción de nutrientes y se secretan
enzimas extracelulares para degradar los polímeros que no pueden absorber. Esta característica
es la que les permite resistir los efectos de sustancias que pudieran afectar su permeabilidad.
La función de los antimicóticos comerciales como el miconazol, es degradar la pared y
ocasionar la pérdida de elementos como el sodio y potasio, con la consecuente disminución de
la permeabilidad y muerte del hongo (Karp, 1998).

54
Por todo lo señalado anteriormente así como lo observado en el presente estudio y de acuerdo
con Atlas et al. (1988), Kim et al. (2000), Koh et al. (2002), se decidió utilizar la técnica de
dilución, para conocer los posibles efectos antimicóticos de las algas de este estudio. El
método de dilución resultó ser más efectivo para llevar a cabo pruebas de detección
antimicótica con hongos filamentosos, esta técnica permite poner en suspensión a las esporas
dándoles homogeneidad y con ello mantenerlas uniformemente expuestas a los extractos.
Además, con dicho método se obtuvieron excelentes resultados con Candida albicans, cuya
cepa no es del tipo filamentoso, ya que se trata de una levadura, por lo tanto el procedimiento
en cuestión, se recomienda para éste tipo de estudios de farmacognosia antimicótica.
Los resultados obtenidos no son de carácter cuantitativo, debido a que sólo se registró la
inhibición el crecimiento de la cepa en turno o bien la nula actividad antimicótica del extracto,
esto sin conocer su magnitud del efecto de inhibición observado. Por lo que de acuerdo con los
resultados obtenidos en este estudio y de manera porcentual, las algas rojas, fueron las que
presentaron el mayor número de especies con actividad (46 %) en contra de las cepas de
hongos que fueron usadas (Aspergillus niger, Rhizopus nigricans y Candida albicans). En
segundo lugar, las algas café con el 32 % y finalmente, las algas verdes con 16 %. Esta
proporción resultó similar a la obtenida por Padmakummar y Ayyakkannu en 1997 con
extractos etanólicos de algas del sur de la India con la técnica de difusión. A pesar de que las
proporciones de actividad son similares para ambos estudios, es posible establecer con toda
seguridad que la proporción encontrada en el presente estudio, en realidad se debe a que se
recolectó una mayor proporción de las algas rojas y con ello se probaron los extractos de igual

55
número de especies de este grupo de algas, en comparación con las algas de los otros dos
grupos.
De acuerdo a los antecedentes dentro del campo de la Ficología experimental, existen otros
estudios que han mostrado actividad antimicótica en contra de las cepas de hongos utilizadas
en el presente trabajo. Uno de estos casos es el de Cymopolia barbata, cuyos extractos a través
de la técnica de dilución reportaron inhibición del crecimiento de las cepas de
Aspergillus niger, Rhyzopus nigricans y Candida albicans. Dichos resultados confirman
que los extractos de la especie C. barbata contienen compuestos químicos antimicóticos,
ya que en un estudio llevado a cabo por Martínez-Nadal en 1966 con el método de difusión,
únicamente encontró actividad de extractos etanólicos en contra de A. niger y C. albicans. Por
lo que a través de la técnica de dilución, fue posible ampliar el campo de detección de
actividad, encontrando que dicha especie también tiene cualidades de poder inhibir el
crecimiento de la cepa de R. nigricans.
En el actual estudio se observó actividad antimicótica con dos especies del género Dictyota, la
especie D. cervicornis inhibió las cepas de Aspergillus niger, Rhyzopus nigricans y Candida
Albicans y por su parte, D. ciliolata sólo inhibió el crecimiento de A. niger. Por otro lado,
Moreau et al. en 1988 trabajando con el método de difusión, obtuvieron actividad de los
extractos con hexano de Dictyota dichotoma y de D. linearis en contra de A. niger y C.
albicans. Con estos resultados, pesar de que las pruebas se realizaron con técnicas diferentes
(difusión y dilución), se corrobora la actividad para el género Dictyota y se está en la

56
posibilidad de mencionar que existan otras especies del mismo grupo cuyos extractos
presenten condiciones para inhibir el crecimiento de algunas cepas de hongos.
Existen otros estudios más recientes con el método de difusión como el de Ballesteros et al.
(1992) en el Mediterráneo, los cuales encontraron actividad antimicótica en extractos
obtenidos con mezclas de metanol-tolueno y el de Padmakumar y Ayyakkannu (1997) en la
India, quienes sólo emplearon etanol. Los primeros señalan los efectos en contra de
Aspergillus niger y Candida albicans, mientras que los segundos sólo en contra de
C. albicans. Los extractos fueron obtenidos de especies como Halimeda tuna, varias especies
del género Codium, Dictyopteris delicatula, Dictyota dichotoma, Stoechospermum
marginatum, Gelidiela acerosa, Gracilaria loliifera y Halimenia floresia. Al comparar los
resultados obtenidos con la técnica de dilución en el presente estudio utilizando especies
recolectadas en Veracruz, con algunos de los géneros señalados, se observó que la especie D.
delicatula fue común confirmándose actividad en contra de C. albicans. Por otro lado,
comparando especies de los mismos géneros, los extractos de las especies Dictyota ciliolata y
Gracilaria dominguensis sólo mostraron inhibición en contra de A. niger. Para los extractos de
las especies Halimeda scabra, Codium simplex, Dictyota cervicornis, Gracilaria caudata y G.
cervicornis se reportaron efectos similares, inhibiendo las cepas de A. niger y C. albicans
además de mostrar actividad en contra de R. nigricans.
A partir de los resultados obtenidos, se comprobó lo señalado por Atlas et al. (1988) y Kim et
al. (2000), acerca de la técnica de dilución. En las pruebas se trabajó con cepas de
características filamentosas, además de una levadura, obteniéndose resultados que permitieron

57
conocer la actividad de los extractos algales, por lo que de acuerdo con éstos autores, dicha
técnica se recomienda para este tipo de estudios ya que la de difusión, también utilizada en el
estudio, resultó ser de alcances limitados, a menos que se disponga de una buena cantidad de
biomasa para las pruebas.
De acuerdo con Koh et al. (2002) el método de dilución tiene la ventaja de conocer los valores
absolutos o la concentración a la cual algún extracto resulta efectivo contra los
microorganismos de prueba. Otra ventaja del método es la homogeneidad que se obtiene en las
pruebas, ya que las esporas en suspensión son uniformemente expuestas a los extractos, lo que
se traduce en resultados más exactos.
Es importante resaltar que en los estudios señalados existieron especies o en su caso géneros
que no tuvieron actividad antimicótica. En éste estudio debido con toda seguridad al empleo
de la técnica de dilución, la cual probó ser más efectiva para las pruebas antimicóticas, fue
posible evidenciar a 35 especies algales que anteriormente no habían sido señaladas con
actividad hacia las cepas de Aspergillus niger, Rhyzopus nigricans y Candida albicans. Las
especies Centroceras clavulatum, Hypnea musciformis, Dictyopteris delicatula y Caulerpa
racemosa ya habían sido reportadas por Padmakumar y Ayyakkannu (1997) con actividad en
contra de Aspergillus terreus, Trichophyton mentagrophytes, Trichoderma viridae, Sporothrix
schenkii y Helminthosporium oryzae, pero no en contra de A. niger, R. nigricans y C.
albicans.

58
8.1. Variación estacional.
La variación estacional observada en la actividad antimicótica de los extractos de las algas
Digenea simplex, Padina boergesenii y Cymopolia barbata colectadas en la Mancha y Ulva
fasciata proveniente de Costa de Oro, es debida principalmente a factores ambientales
imperantes en las zonas de colecta, uno de éstos factores a considerar es la luz, este factor
afecta la actividad antimicótica, de acuerdo con Robles y Ballantine (1999) quienes realizaron
cultivos controlados de Spyridia filamentosa, sometiéndola a diferentes intensidades de
radiación luminosa desde 5 a 70 µmol fotón/m2s, posteriormente obtuvieron y probaron sus
extractos en contra de Candida albicans del estudio concluyeron que hubo diferencias en la
actividad con respecto a los diferentes estados del ciclo de vida y de la intensidad luminosa, en
donde los gametofitos masculinos sometidos a radiaciones mayores a 30 µmol fotón/m2s y los
tetrasporofitos con radiaciones superiores a 70 µmol fotón/m2s, produjeron inhibición del
crecimiento de la cepa de C. albicans. Claramente se observa que a mayor intensidad de
radiación luminosa, la formación de compuestos químicos antimicóticos se ve favorecida y
con ello, los extractos inhiben el desarrollo de los hongos. Por lo que éste factor es uno de los
condicionantes hacia las respuestas fisiológicas y bioquímicas de las algas y por ello es fuente
de origen de los metabolitos responsables de la actividad antimicótica.
Lo anterior está directamente relacionado con los resultados obtenidos, ya que además de
acuerdo con Castillo y Carabias (1982) se precisa que para la zona de estudio existen

59
temporadas climáticas bien establecidas en donde las condiciones físicas están regidas por los
diferentes cambios atmosféricos (Tabla 11).
Tabla 11. Estacionalidad en las zonas de La Mancha y Costa de Oro,Veracruz,Ver. (Castillo y Carabias, 1982).
SECASMarzo-Mayo
LLUVIASJunio-Octubre
NORTESNoviembre-Febrero
Temp. ambiental=24 ºC;Vel viento= 6.1 m/sPrecipitación= 6.0 mm
Temp. ambiental=26.8 ºC;Vel. viento= 3.8 m/sPrecipitación= 200-400 mm
Temp. ambiental=20 ºC;Vel. viento= 7.0 m/sPrecipitación= 24.0 mm
Complementando la información anterior sobre los factores ambientales, también es posible
conocer la tendencia de la precipitación (Contreras, com. per, 2004), ya que se dispone de
registros promedio de lluvia a lo largo de varios años, al graficar claramente se observan las
oscilaciones en forma cíclica (Figura 21), donde en los primeros meses del año, es decir de
enero a principios de mayo, la frecuencia de las precipitaciones es baja; posteriormente,
durante el mes de mayo se da un incremento considerable en el volumen de la precipitación
pluvial, con un ligero descenso para el mes de agosto y de nuevo un repunte para el mes de
septiembre, a partir de aquí de nueva cuenta se presenta una disminución en la intensidad de
las lluvias hasta el mes de diciembre.
Figura 21. Promedio mensual de precitación 1998 a 2002.Contreras com. per, 2004 Estación Mareográfica Veracruz,Ver.

60
Con relación a lo señalado anteriormente, se observa que la primera colecta fue realizada en el
mes de mayo, se llevó a cabo justo al final de la temporada de “secas” y la colecta de agosto
fue realizada prácticamente a la mitad de la época de “lluvias”, es evidente que en las especies
colectadas en el mes de agosto, las sustancias producidas en sus rutas metabólicas,
probablemente se encontraban en un punto máximo ya que dichos compuestos fueron el
resultado de los procesos bioquímicos llevados a cabo por efecto del paso de la temporada de
secas, en donde influyeron factores ambientales como la alta velocidad del viento (6.1 m/s)
favoreciendo las turbulencias así como temperaturas relativamente elevadas (24 ºC), además
de esto se debe de considerar el propio efecto del cambio de estación, el cual por sí sólo debió
generar efectos “estresantes” en las comunidades algales de las zonas de estudio.
El mayor número de especies con compuestos químicos antimicóticos encontrado para el mes
de agosto, se debió a que dichos compuestos, se formaron en la temporada de “secas”, a esto
se debe sumar el efecto del cambio de la temporada de “secas” a “lluvias”, lo cual marcó la
variación de la actividad antimicótica estacional para las especies Padina boergesenni,
Digenea simplex, Cymopolia barbata de la localidad de la Mancha y Ulva fasciata de Costa de
Oro
El nivel de las mareas es otro factor que está directamente relacionado con la formación de los
metabolitos responsables de la actividad antimicótica. Para ello las observaciones realizadas
en campo durante la recolecta permitieron suponer que éste factor tendría consecuencias en la
comunidad algal de los sitios de interés, ya que se presentaron niveles de la altura del agua
relativamente bajos, incluso por debajo de 1 metro, especialmente para la localidad de la

61
Mancha, en donde éste fenómeno ocasionó que las algas quedaran expuestas por periodos más
prolongados a la incidencia de los rayos solares en forma más directa.
Existen estudios en donde se indica que las comunidades instaladas en las zonas litorales y que
están expuesta a los cambios en los niveles de marea, las especies se han adaptado
fisiológicamente dando como resultado incrementos en las tasas fotosintéticas, debido a que el
CO2 se difunde del aire hacia la película ocasionando que penetre en mayor medida hacia las
células (Johnson et al. 1974; Soeder y Stengel, 1974; Brinkhuis et al. 1976; Quadir et al.
1979). En relación con lo señalado anteriormente, Dring y Brown (1982) mencionan que
existen especies que se ubican a lo largo de la zona litoral y que sufre los efectos de la
desecación, pueden mantener activa la fotosíntesis aún con un bajo contenido de agua celular
y sus tasas fotosíntéticas, después del periodo de exposición son incrementadas drásticamente.
Por otro lado, de acuerdo con Lüning (1990), la relación entre la superficie y el volumen de las
algas, es el principal factor que controla la velocidad de los procesos de desecación, entre
menor sea la relación expuesta a la intemperie, mayor será el tiempo que transcurra para que el
alga pierda agua, por el contrario, si la proporción expuesta es mayor, la pérdida de agua será
de forma más acelerada. Dawes (1991), señala que dichas especies, además de ser
fotosintéticamente más activas cuando pasan por periodos de exposición y posteriormente al
quedar nuevamente cubiertas por el agua, son estimuladas para favorecer su crecimiento, por
lo que este factor no puede ser considerado como una condición adversa.

62
Además, Lobban y Harrison (1997), señalan que durante los factores estresantes de la
exposición a la intemperie, las tasas fotosintéticas se ven disminuidas drásticamente aún antes
de que la desecación ocurra, esto es debido a que se agota el aporte de carbón inorgánico,
quedando en su superficie una película de agua con una pequeña cantidad de bicarbonato
disponible para la fotosíntesis.
Para conocer el comportamiento de las mareas en la zona de estudio, se consultó la Base de
datos del nivel del mar (2004) del Servicio Mareográfico del Instituto de Geofísica de la
UNAM, a partir de ésta información, al igual que el análisis realizado para los datos de
precipitación, fueron graficados los últimos cinco años disponibles para conocer la tendencia
de las variaciones del nivel de las mareas en la zona de Veracruz (Figura 22).
Al igual que para la precipitación, existen oscilaciones cíclicas que se repiten año con año, es
decir, durante el inicio del año se mantienen niveles alrededor de los 140 cm con un ligero
incremento hasta los 155 cm aproximadamente hasta llegar al mes de mayo, a partir de aquí se
observa un descenso del nivel de la marea durante casi tres meses, es decir de mayo a julio,
posteriormente sobreviene un incremento de manera considerable hasta llegar a los 170 a 180
cm aproximadamente para el mes de octubre, a partir de entonces de nueva cuenta se observa
un descenso drástico de manera constante hasta el mes de diciembre.

63
Las observaciones realizadas en el campo coincidieron con la tendencia del nivel de este
parámetro, lo cual al igual que el razonamiento realizado para la precipitación, el mayor
número de especies con actividad antimicótica se debe precisamente a estas oscilaciones de las
mareas, ya que como se ha mencionado, un gran porcentaje de especies activas fue reportado
de la colecta de agosto, en comparación con el porcentaje de la colecta de mayo.
Este comportamiento se comprueba con los resultados provenientes de la recolecta de marzo-
abril del 2003, en donde también se reporta a un gran porcentaje de especies con actividad
antimicótica. Para éste periodo, observando la gráfica y corroborando con lo observado en el
campo, tenemos que justo al principio del año el nivel de la marea es muy bajo, lo que
Figura 22. Promedio mensual de altura de marea de 1998 a 2002.Estación Mareográfica Veracruz, Ver. Servicio Mareográfico, Instituto deGeofísica, UNAM. Base de datos del nivel del mar (2004, en preparación).

64
significa que al igual que para la colecta de agosto, en la colecta del periodo marzo-abril
también las algas fueron afectadas por los factores ambientales.
Además se debe considerar que durante esta colecta, la zona de estudio se encontraba justo en
plena época de secas, con escasas lluvias, reportándose promedios de 6.0 mm de precipitación.
Por el contrario, el escaso porcentaje de especies con actividad antimicótica encontradas para
la recolecta de mayo, se debe a que las especies provenientes de dicha colecta atravesaron por
un periodo de “estabilidad”, en donde se presentó un incremento en los niveles de la altura de
la columna del agua ocasionada por el cambio en el nivel de la marea.
La variación estacional por causas ambientales, registrada con las especies recolectadas en las
localidades de la Mancha y Costa de Oro, coincide con el estudio de Moreau et al. (1984),
quienes también encontraron una relación entre la variación de la actividad antimicótica de las
algas Dictyota dichotoma, Dictyota lineraris, Dilophus fasciola y Dilophus spiralis
provenientes del Mediterráneo, y los cambios estacionales de las épocas en que se realizaron
las colectas. Las especies señaladas probaron inhibir del crecimiento de la levadura Candida
albicans y de otros mohos. De igual manera, en el estudio anual de Moreau et al. (1988) con
las macroalgas Dictyota dichotoma, D. dichotoma var. implexa y Dilophus spiralis
recolectadas en el Mediterráneo, encontraron un amplio espectro de actividad, el cual también
varió dependiendo de los factores ambientales de las estaciones de recolecta. Esta actividad se
detectó en contra de C. albicans y de otras cepas diferentes a las utilizadas en el presente
estudio.

65
Por lo que se refiere a las diferencias en la actividad antimicótica reportada por otros autores o
bien la nula actividad, éstas probablemente se debieron a los diferentes tipos de solventes
usados. Este trabajo no es el primero en considerar la combinación de solventes para la
obtención de extractos algales, al respecto existen estudios como el de Caccamese y Azzolina
(1979), Padmakumar y Ayyakkannu (1986), Perry et al. (1991), Ballesteros et al. (1992), Vora
et al. (1995), Robles y Ballantine (1999) y Zamora y Ballantine (2000). Dichos autores
realizaron sus pruebas de detección antimicótica con extractos algales producidos a través de
combinar dos o más solventes, por lo cual y de manera similar se considera que la gran
cantidad de especies algales detectadas en el presente estudio fue debida principalmente a la
variante en la técnica de obtención de los extractos a partir de la mezcla de los solventes
propuestos (C-M, C-OH, E-M y E-OH), lo cual probablemente no hubiera sido posible con el
uso de un sólo solvente.
A este respecto, existe una gran cantidad de solventes con diferente polaridad que se ubican a
lo largo de una escala desde los no polares hasta los muy polares, que permiten extraer un
grupo de moléculas bien definido de acuerdo a su carga, a su vez, algunas moléculas que no
tienen cualidades para comportarse con características de bioactividad, pueden ser excluidas
durante el proceso de extracción y quedarse en la fase acuosa, la cual en las pruebas
antimicóticas comprobó carecer de actividad. Además, se debe de considerar que existen
diferencias en las propiedades de los compuestos químicos responsables de la actividad
antimicótica presentes en las algas (terpenos, fenoles, compuestos halogenados, etcétera). De
la buena elección del conjunto de dichos factores en el proceso de extracción, dependerá la

66
calidad de los extractos en lo que se refiere a la concentración de componentes activos que
actúan en contra de los microorganismos de prueba.
Otro factor responsable de las discrepancias entre los diferentes estudios, es el que tiene que
ver con el momento en el cual se programan las recolectas en una determinada época del año,
las cuales pudieran haberse llevado a cabo durante el periodo de inactividad relativa de las
algas, sin que esto implique la nula actividad o ausencia de compuestos químicos responsables
de la actividad antimicrobiana. Un punto que se debe de tomar en cuenta es el que se refiere a
la preservación de las muestras, vale destacar que desde el momento en que las algas son
extraídas de su medio natural, se dan procesos enzimáticos que aceleran la descomposición de
las muestras, lo cual puede llevar a obtener resultados poco confiables durante las pruebas de
detección de actividad antimicótica (Hay et al. 1987). Sin embargo, debido a que la
metodología utilizada en la recolección de las muestras consideró el empleo de hielo seco para
preservarlas en congelación, este último factor no influyó en la obtención de los resultados.
Al realizar un simple análisis porcentual de los resultados obtenidos con la mezclas de
solventes, se encontró de manera general que las mezclas de éter-alcohol etílico-agua (E-OH)
y éter-metanol-agua (E-M), fueron las que dieron mejores resultados.
Con la técnica de extracción y al considerar cuatro mezclas de solventes, se abrieron las
posibilidades de obtener resultados positivos de actividad antimicótica, lo cual al parecer no
hubiera sido posible sí sólo se hubiera contemplado utilizar una mezcla de solventes o en su
caso un sólo solvente, ya que se observó que cuando alguno de los extractos obtenidos de

67
cualquiera de las mezclas no presentaba actividad, en alguno de los otros extractos, la
actividad se hacía evidente. Por lo anterior, no es posible poder determinar con certeza cuál o
cuáles de las mezclas pueden tener un mayor poder para extraer los compuestos activos.
La ventaja del presente estudio en la búsqueda de especies algales con propiedades
antimicóticas fue la de utilizar mezclas de solventes, con lo cual es altamente probable obtener
una extracción más selectiva de los componentes que pudieran ser responsables de la
actividad. Esto es debido a que los compuestos bioactivos varían ampliamente en sus
polaridades químicas, de otra manera la utilización de un solo solvente pudo haber repercutido
en la obtención de resultados que indicaran la inactividad en los extractos, lo cual aunado a la
utilización del método de suspensión de esporas para las pruebas antimicóticas, nos ha llevado
a considerar el actual estudio como todo un éxito dentro de la investigación ficológica con
fines de investigación farmacéutica.
Los antecedentes ficológicos encaminados hacia la búsqueda de algas con potenciales
antimicóticos, son escasos, ya que han aparecido relativamente pocos estudios, al respecto
para México, prácticamente se puede considerar como nulos, por lo cual el presente trabajo es
considerado si no el pionero, uno de los primeros dentro de este campo. Los resultados hasta el
momento han demostrado que las algas colectadas en la Mancha y Costa de Oro, representan
un potencial para la búsqueda de metabolitos con efectos antimicóticos.

68
9. CONCLUSIONES.
♦ El potencial antimicótico que representan las especies algales de la Mancha y Costa de Oro
es significativo, ya que alrededor del 95 % de las especies probadas mostraron actividad en
contra de por lo menos alguna de las cepas de Aspergillus niger, Rhizopus nigricans y
Candida albicans.
♦ El mayor porcentaje de especies activas fue de la División Rhodophyta con 49 %; para la
División Phaeophyta se reportaron el 34 % y la División Chlorophyta con el 17 %.
♦ Se encontró variación estacional en la actividad antimicótica en las especies de algas
Digenea simplex, Padina boergesenii, y Cymopolia barbata colectadas en la Mancha y en
la especie Ulva fasciata colectada en Costa de Oro. La importancia del estudio radica en
que se pueden establecer programas de recolecta de las especies con una visión dentro del
campo de la industria farmacéutica con el objetivo de obtener nuevos compuestos para el
control de las enfermedades producidas por hongos.

69
♦ De manera porcentual es posible señalar a la mezcla de E-OH como la más efectiva, sin
embargo, debido a que los porcentajes de las demás mezclas se encuentran entre 21 al
28 %, no existen diferencias que indiquen cual de ellas es más efectiva para extraer los
compuestos antimicóticos.
♦ Del total de los extractos probados, ninguno reflejó actividad en contra de Rhyzopus
oryzae, Alternaria alternata y Beauveria bassiana.
♦ Las algas provenientes de las zonas de la Mancha y Costa de Oro del estado de Veracruz,
representan un gran potencial en la búsqueda de nuevas alternativas en el área
farmacológica que permitan obtener compuestos para el control de enfermedades
producidas por las cepas Aspergillus niger, Rhizopus nigricans y Candida albicans.

70
10. LITERATURA CITADA.
Accorinti, J. 1983. Antifungal product from algal origin (Review). Rev. Int. Oceanogr. Med.,
72: 45-53.
Atlas, R. M., A. E. Brown, K. W. Dobra y Ll. Miller. 1988. Experimental microbiology.
Fundamentals and applications. 2° Ed. Mcmillan Publishing Company. New York. 618
p.
Baker, J. T. 1984. Seaweeds in pharmaceutical studies and applications. Hydrobiologia,
116/117: 29-40.
Ballesteros, E., D. Martin y M. J. Uriz. 1992. Biological activity of extracts from some
Mediterranean macrophytes. Bot. Mar., 35 (6): 481-485.
Barreto, M., C. J. Straker y A. T. Critchley. 1997. Short note on the effects of ethanolic
extracts of selected South African seaweeds on the growth of commercially important
plant pathogens, Rhizoctonia solani Kuehn and Verticillum sp. S. African J. Bot., 63 (6):
521-523.
Base de datos del nivel del mar 2004. Servicio Mareográfico del Instituto de Geofísica.
UNAM. México

71
Bernard, P. y D. Pesando. 1989. Antibacterial and antifungal activity of extracts from the
rhizomes of the Mediterranean seagrass Posidonia oceanica (L.) Delile. Bot. Mar., 32:
85-88.
Biard, J. F., J. F. Verbist, J. Le Boterff, G. Ragas y M. M. Lecocq. 1980. Algues fixées de la
côte atlantique franςaise contenant des substances antibacterérinnes et antifungiques. Pl.
Med., suppl., 136-151.
Blanco, P. R. Y R. Ramírez. 1993. Algas marinas en el Golfo de México 1. Escolleras de
Mocambo, Veracruz. Ficología III. Congreso Latinoamericano. 1° Reunión
Iberoamericana. I. Congreso Mexicano. México: 167-171.
Bligh, E. G. y W. J. Dyer. 1959. A rapid method of total lipid extraction and purification. Can.
J. Biochem. Physiol. 37: 911-917.
Brinkhuis, B. H., N. R. Tempel y R. F. Jones. 1976. Photosynthesis and respiration of exposed
salt-marhs fucoids. Mar. Biol. 34: 349-359.
Caccamese, S. y R. Azzolina. 1979. Screenig for antimicrobial activities in marine algae from
eastern Sicily. Pl. Med., 37: 333-339.

72
Caccamese, S., R. M. Toscano, G. Furnari y M. Cormaci. 1985. Antimicrobial activities of red
an brown algae from southern Italy coast. Bot. Mar., 28: 505-508.
Calvo, M. A., F. J. Cabanes y L. Abarca. 1986. Antifungal activity of some Mediterranean
algae. Mycopathol., 93: 61-63.
Carter, G. T., K. L. Rinehart, L. H. Li, S. L. Kuentzel y J. L. Connor. 1978. Brominated
indoles from Laurencia brongniartii. Tetrahedron Lett., 46: 4479-4482.
Castillo, S. Y J. Carabias. 1982. Ecología de la vegetación de dunas y costeras: fenología.
Biotica 7: 551-568.
Codomier, L., G. Bruneau, G. Cambaut y J. Teste. 1977. Etude biologique et chimique d´
Asparagopsis armata et de Falkenbergia rufolanosa (Rhodophyce´es,
Bonnemalsonniales). C. R. Acad. Sc. Paris. Serie D. 284-1163-1165.
Contreras, F. 1993. Ecosistemas costeros mexicanos. U. A. M. Iztapalapa. México. 415 pp.
Dawes, C. J. 1991. Botánica marina. Edit. Limusa. México. 673 pp.

73
De Lara-Isassi, G., A. Sobrino-Figueroa, C. Lozano-Ramírez, M. E. Ponce-Marquez y K.
Dreckman-Estay. 1989. Evaluación de la actividad antibiótica de las macroalgas de las
costas de Michoacán, México. Bol. Inst. Oceanogr. Venezuela, Univ. Oriente. 28 (1
& 2): 99-104.
De Lara-Isassi, G. 1991. Propiedades antibióticas de algunas especies de algas marinas
bentónicas. Hidrobiólogica 1 (2): 21-28.
De Lara-Isassi, G. y S. Alvarez-Hernández. 1994. Actividad biológica de las macroalgas
marinas mexicanas. Rev. Soc. Mex. Hist. Nat. 45: 51-60.
Dring, M. J. y F. A Brown. 1982. Photosynthesis of intertidal brown algae during and after
periods of emersion: a renewed search for physiological causes of zonation. Mar. Ecol.
Prog. Ser. 8: 301-308.
Enoki, N., K. Tsuzuki, S. Omura, R. Ishida y T. Matsumoto. 1983. New antimicrobial
diterpenes, dictyol F, anti-epidictyol F, from the brown alga Dictyota dichotoma. Chem.
Lett., 290: 1627-1630.
Faulkner, D. J. 1986. Marine natural products. Nat. Prod. Rep., 3: 1-33.
Fenical, N. 1974. Cycloendesmol, an antibiotic cyclopropane containing sesquiterpene from
the marine alga, Chondria oppositocloda Dawson. Tetrahedron Lett., 13: 1137-1140.

74
Fenical, W. 1982. Natural products chemistry in the marine environment. Science, 215 (4534):
923-928.
Fenical, W. y V. J. Paul. 1984. Antimicrobial and cytotoxic terpenoids from tropical green
algae of the Family Udoteaceae. Hydrobiologia, 116/117: 137-140.
Fenical, W. F., J. J. Sims, D. Squatrito, R. Wing y P. Radlick. 1973. Zonarol and isozonarol.
fungitoxic hydroquinones from the brown seaweed Dictyopteris zonarioides. J. Org.
Chem., 38: 2383.
Finer, J., J. Clardy, S. Fenical, L. Minale, R. Riccio, J. Battaile, M. Kirkup y R. E. Moore.
1979. Structure of dictyodial and dictyolactone, unusual marine diterpenoid. J. Org.
Chem., 44: 2044-2047.
Glombitza, K. W. 1970. Antimicrobial contituents in algae. Quantitative determination of
acrylic acid in sea-algae. Pl. Med., 18: 210-221.
Glombitza, K. W. 1979. Antibiotics from algae. In: H. A. Hoppe, T. Levring, Y. Tanaka.
(Eds.). Marine Algae Pharmaceutical Science. Walter de Gruyter & Co. Germany. p.
303-342.

75
Gunasekera, L. S., A. E. Wrigth, S. P. Gunasekera, P. McCarthy y J. Reed. 1995.
Antimicrobial constituent of the brown alga Spochnus pedunculatus. Int. J.
Pharmacognosy, 33 (3): 253-255.
Harder, R. 1917. Ernährungsphysiologische untersuchungen an Cyanophyceen, hauptsächlich
dem endophytischen Nostoc puctiforme. Z. Bot., 9:145-242.
Hay, M. E., J. E. Duffy y C. A. Pfister. 1987. Chemical defense against different marine
herbivores are amphipods insects equivalents?. Ecology, 68 (6): 1567-1580.
Henriquez, P., A. Candia, P. Norambuena, M. Silva y R. Zemelman. 1979. Antibiotic
properties of marine algae II. Screning of chilean marine algae for antimicrobial activity.
Bot. Mar., 22: 451-453.
Herrera, T. y M. Ulloa. 1998. El reino de los hongos. Micología básica y aplicada. UNAM.
Fondo de Cultura Económica. México. 552 pp.
Hoppe, H. A. 1979. Marine algae and their products and constituents in pharmacy. In: H. A.
Hoppe, R. Levring y Y. Tanaka (Eds.). Marine Algae in Pharmaceutical Science. Walter
de Gruyter, Berlin. p. 25-119.
Hornsey, I. S. y D. Hide. 1974. The production of antimicrobial compounds by British marine
algae. I. Antibiotic producing marine algae. Br. Phycol. J., 9: 353-394.

76
Johnson, W. S., A. Gigon, S. L. Gulmon y H. A. Mooney. 1974. Comparative photosynthetic
capacities of intertidal algae under exposed and submerged conditions. Ecology, 55:
450-453.
Karp, G. 1998. Biología celular y molecular. McGraw-Hill Interamericana Editores. México.
805 pp.
Khaleafa, A. F., A. M. Kharboush, A. Metwalli, A. F. Mohsen y A. Serwi. 1975. Antibiotic
fungicidal action from extracts of some seaweeds. Bot. Mar., 18: 163-165.
Kim, K., C. D. Harvell, P. D. Kim, G. W. Smith y S. M. Merkel. 2000. Fungal disease
resistance of Caribbean sea fan corals (Gorgonia spp.). Mar. Biol., 136: 259-267.
Koh, T. K. Tan, L. M. Chou y N. K. C. Goh. 2002. Antifungal properties of Singapore
gorgonians: a preliminary study. J. Exp. Mar. Biol. Ecol., 273: 121-130.
Lehninger, A. 1984. Bioquímica. Las bases moleculares de la estructura y función celular.
Ediciones Omega. Barcelona.
Lobban, Ch. S. y P. J. Harrison. 1997. Seaweed ecology and physiology. Cambridge
University Press. USA. 366 p.

77
Lüning, K. 1990. Seaweeds. Their environment, biogeography and ecophysiology. A Wiley
Interscience Publication. John Wiley & Sons, Inc. USA. 527 p.
Martínez-Nadal, N. G. 1958. Information Bulletin 1958-59. University of Puerto Rico. Puerto
Rico.
Martínez-Nadal, N. G. M., C. M. Casillas-Chapel, L. V. Rodríguez, J. R. Rodríguez-Perazza y
L. Torres-Vera. 1966. Antibiotic properties of marine algae. III. Cymopolia barbata.
Bot. Mar., 9:21-26.
Mautner, H. C., Gardner, G. M. y R. Pratt. 1953. Antibiotic activity of seaweed extracts. II.
Rhodomela larix. J. Amer. Pharm. Ass. Sci. Ed., 42: 294-296.
Melo, V. M. M., D. A. Medeiros, F. J. B. Rios, L. T. M. Castelar y F. F. U. Carvalho. 1997.
Antifungal properties of proteins (agglutinins) from the red alga Hypnea musciformis
(Wolfen) Lamouroux. Bot. Mar., 40: 281-284.
Metting, B. y J. W. Pyne. 1986. Biologically active compounds from microalgae. Enzime
Microb. Technol., 8: 381-386.
Moreau, J., D. Pesando y B. Caram. 1984. Antifungal and antibacterial screening of
dyctiotales from the French Mediterranean coast. Hydrobiologia, 116/117: 521-524.

78
Moreau, J., D. Pesando, P. Bernard, B. Caram y J. C. Pionnat. 1988. Seasonal variations in the
production of antifungal substances by some dictyotales (brown algae) from the French
Mediterranean coast. Hydrobiologia , 16(2): 157-162.
Mori, K. y M. Komatsu. 1986. Synthesis and absolute configuration of zonarol. A fungitoxic
hydroquinone from the brown seaweed Dictyopteris zonaroides. Bull. Soc. Chim. Belg.,
95(9-10): 771-782.
Olesen, P. E., A. Maretzki y L. A. Almondovar. 1964. An investigation of antimicrobial
substances from marine algae. Bot. Mar., 6: 224-232.
Padmakumar, K. y K. Ayyakkannu. 1986. Antimicrobial activity of some marine algae of
Porto Novo and Pondicherry waters, east coast of India. Ind. J. of Mar. Sci. New Delhi,
15 (3): 187-188.
Padmakumar, K. y K. Ayyakkannu. 1997. Seasonal variation of antibacterial and antifungal
activities of the extracts of marine algae from southern coast of India. Bot. Mar., 40:
507-515.
Pelczar, M. J., R. D. Reid y E. C. S. Chan. 1990. Microbiología. 4° edición. Editorial
McGraw-Hill. México. 826 pp.

79
Perry, N. B., J. W. Blunt y M. H. G. Munro 1991. A cytotoxic and antifungal 1,4
naphthoquinone and related compounds from a New Zeland brown alga, Landsburgia
quercifolia. J. Nat. Prod., 54(4): 978-985.
Pesando, D. y B. Caram. 1984. Screening of marine algae from the french Mediterranean coast
for antibacterial y antifungal activity. Bot. Mar, 27: 381-386
Pesando, D. 1990. Antibacterial and antifungal activities of marine algae. In: I. Akatsuda (Ed.)
Introduction to Applied Phycology. pp. 3-26. SPB Academic Publishing. The Hague,
The Netherlands.
Quadir, A., P. J. Harrison y R. E. De Wreede. 1979. The effects of emergence and
submergence on the photosynthesis and respiration of marine macrophytes. Phycologia
18: 83-88.
Reichelt, J. L. y M. A. Borowitzka. 1984. Antimicrobial activity from marine algae: Results of
a large scale screening programme. Hydrobiologia, 116/117: 158-168.
Robertson, K. J. y W. Fenical. 1977. Pachydictyol-a epoxide, a new diterpene from the brown
seaweed Dictyota flabellata. Phytochemestry, 16: 1071-1073.

80
Robles, C. P. O. y D. L. Ballantine. 1999. Effects of culture conditions on production of
antibiotically active metabolites by the marine alga Spyridia filamentosa (Ceramiaceae,
Rhodophyta). I. Light. J. Appl. Phycol. 10: 453-460.
Romero, S. 1988. Hongos Fitopatógenos. Universidad Autónoma de Chapingo. México. 360
pp.
Sachithananthan, K. y A. Sivapalan. 1975. Antibacterial properties of some marine algae of
Sri Lanka. Bull. Fish. Res. Stn., Sri Lanka (Ceylan), 26 (1-2): 5-9.
Scagel, R. F., R. J. Bandoni, J. R. Maze, G. E. Rouse, W.B. Schofield y J. R. Stein. 1991.
Plantas no vasculares. Ediciones Omega. Barcelona. 548 pp.
Soeder, C. y E. Stengel. 1974. Physico-chemical factors affecting metabolism and growth rate.
In: W. D. Stewart (Ed.). Algal physiology and biochemistry. University of California
Press. Great Britain. p. 714-740.
Spoehr, H. A., J. H. C. Smith, H. H. Strain, H. W. Milner y G. J. Hardin. 1949. Fatty acid
antibacterials from plants. Carnegie Inst. Washington Pub., 586: 1-67.
Sridhar, K. R. y N. Vidyavathi. 1991. Antimicrobial activity of seaweeds. Acta Hydrochim.
Hydrobiol., 19 (5): 455-496.

81
Tariq, V. N. 1991. Antifungal activity in crude extracts of marine red algae. Micol. Res., 95
(12): 1433-1440.
Tringali, C. M. Piatelli, G. Nicolosi y K. Hostettmann. 1986. Molluscicidal and antifungal
activity of diterpenoids from brown algae of the family Dictyotaceae. Pl. Med. 5: 404-
405.
Vora, J. D., J. M. Babu, R. Sridharan, G. K. Trivedi y H. H. Mathur. 1995. Antimicrobial
activities of marine algae from the Indian coast. J. Mar. Biol. Assoc. India, 36(1-2): 322-
325.
Welch. A. M. 1962. Preliminary survey of fungistatic properties of marine algae. J. Bacteriol.,
83: 97-99.
Zajic, J. E. y E. Knetting. 1970. Antibiotics and medicinal uses. In: J. E. Zajic (Ed.).
Properties and Products of Algae. Proc. Symp. Culture of Algae, N. Y. City, 7-12, Sep.,
1969. Plenum press, N,.Y. 154 p.
Zamora T. C. y D. L. Ballantine. 2000. Multiple antimicrobial activities of the marine alga
Spyridia filamentosa (Ceramiacea, Rhodophyta). Bot.. Mar., 43: 233-238.

82
11. ANEXOS.
A. Medio de cultivo agar dextrosa Sabouraud.
Se utiliza medio de cultivo de la marca BD Bioxon con pH final de 5.6. Suspender 65 g del
medio en un litro de agua destilada. Mezclar perfectamente y calentar con agitación de manera
frecuente, dejar hervir durante un minuto hasta que se haya disuelto el polvo. Finalmente
llevar a una autoclave y esterilizar a 121 °C durante 15 minutos a 15 libras de presión. Dejar
entibiar y verter a cajas Petrí estériles desechables de 100 x 15 mm y dejar solidificar. Las
cajas con medio sólido se deben dejar por lo menos 36 horas para verificar que no exista
contaminación por parte de microorganismos no deseados.
B. Caldo de cultivo Maltosa Sabouraud.
Se utiliza medio de cultivo de la marca Difco con un pH final de 5.6. Para hidratarlo, utilizar
50 g del polvo y disolver en 1000 mL de agua destilada. Esterilizar en autoclave, por 15
minutos a 121 °C a 15 libras de presión. Se deja enfriar y posteriormente es utilizado para los
microcultivos.