tema 5.3 replicaciÓn del...
TRANSCRIPT

TEMA 5.3
REPLICACIÓN DEL DNA
1. CARACTERÍSTICAS GENERALES DE LA REPLICACIÓN 2. LAS DNA POLIMERASAS 2.1. DNA polimerasas de E.coli 2.1.1. La DNApol I 2.1.2. La DNApol II 2.1.3. La DNApol III 2.2. DNA polimerasas eucariotas 3.3. Otras DNA polimerasas 3. ELEMENTOS PARTICIPANTES EN LA REPLICACIÓN 3.1. Sitios específicos de origen de la replicación 3.1. Actividad Primasa 3.2. Actividad DNA-ligasa 3.3. Actividad Helicasa, Topoisomerasa (girasa) y proteínas fijadoras de monohebra (SSB) 4. EL COMPLEJO DE REPLICACIÓN: MODELOS DE REPLICACIÓN. 4.1. E.coli como modelo básico de para replicación del DNA: modelo de lazo 4.1.1. Inicio y progresión de la síntesis de DNA 4.1.2. Terminación de la síntesis de DNA 4.2. Modelo de replicación de círculo rodante 5. LA REPLICACIÓN DEL DNA EN EUCARIOTAS 5.1. Papel de las DNApolimerasas eucarioticas 5.2. El origen de replicación en eucariotas 5.3. Replicación del DNA en el extremo de cromosomas lineales 5.3.1. Telomerasas eucariotas

El metabolismo del DNA comprende el proceso mediante el cual se hacen copias ‘fieles’ de las moléculas de DNA (REPLICACIÓN), junto a los procesos que afectan a la estructura inherentes a la información (reparación y recombinación) Cada una de las cadenas es complementaria de la otra. Cada cadena molde parental genera una cadena ‘hija’
cuya secuencia es predecible y complementaria
Necesidad de alto grado de precisión (los errores cometidos pueden tener funestas consecuencias: el cambio es hereditario) y velocidad (moléculas de millones de bases) Desafío: Separación de hebras en lugar determinado y soportar la tensión (superenrollamiento que provoca) Determinados procesos requieren ‘intercambio de material entre moléculas parentales’: recombinación: generación de diversidad en secuencias de anticuerpos, integración de genoma viríco en DNA hospedador, …

CARACTERÍSTICAS GENERALES DE LA REPLICACIÓN
1).- Complementariedad entre bases del DNA (DNA molde): Chargaff. 2).- Carácter semiconservativo. 3).- Síntesis de replicación 5´→3´. 4).- Necesidad de Cebador o Primer: primasas. 5).- Carácter secuencial de la replicación a partir del origen: procesividad de la DNApolimerasa. 6).- Origen fijo de replicación: replicones. 7).- Replicación bidireccional: horquillas replicativas. 8).- Replicación semidiscontinua: fragmentos de Okazaki.

1).- Complementariedad entre bases del DNA (DNA molde): Chargaff. 2).- Carácter semiconservativo. 3).- Síntesis de replicación 5´→3´. 4).- Necesidad de Cebador o Primer: primasas. 5).- Carácter secuencial de la replicación a partir del origen: procesividad de la DNApolimerasa. 6).- Origen fijo de replicación: replicones. 7).- Replicación bidireccional: horquillas replicativas. 8).- Replicación semidiscontinua: fragmentos de Okazaki.
CARACTERÍSTICAS GENERALES DE LA REPLICACIÓN

TIPOS HIPOTÉTICOS DE REPLICACIÓN

EXPERIMENTO DE MESELSON Y STAHL: REPLICACIÓN SEMICONSERVATIVA
Cultivo celular en medio pesado (15N)
Transfieren las células a medio ligero (14N)

1).- Complementariedad entre bases del DNA (DNA molde): Chargaff. 2).- Carácter semiconservativo. 3).- Síntesis de replicación 5´→3´. 4).- Necesidad de Cebador o Primer: primasas. 5).- Carácter secuencial de la replicación a partir del origen: procesividad de la DNApolimerasa. 6).- Origen fijo de replicación: replicones. 7).- Replicación bidireccional: horquillas replicativas. 8).- Replicación semidiscontinua: fragmentos de Okazaki.
CARACTERÍSTICAS GENERALES DE LA REPLICACIÓN
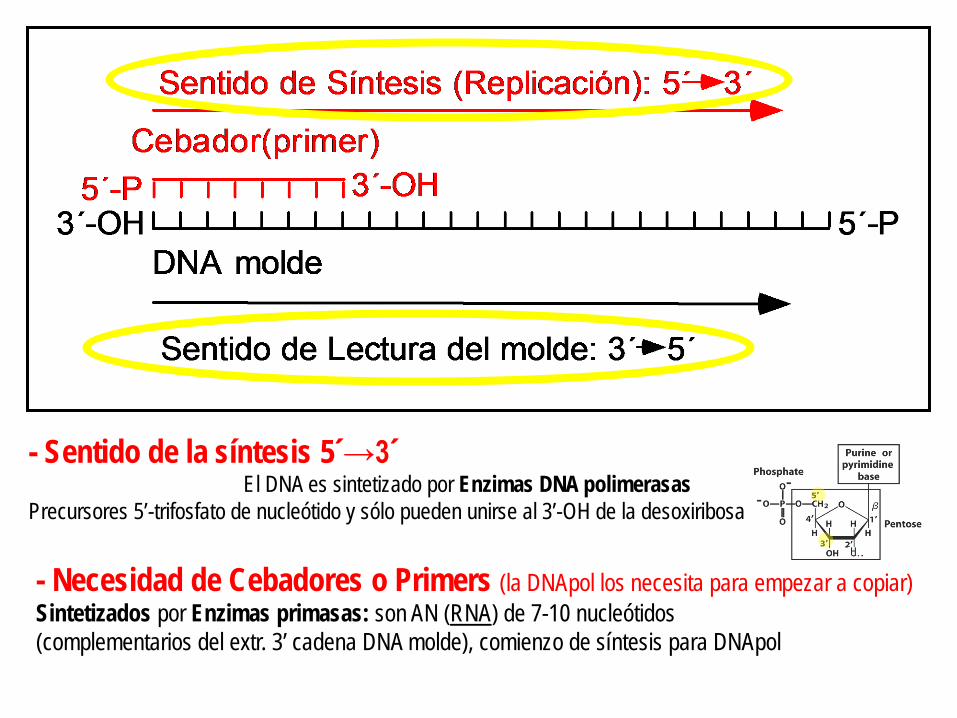
- Sentido de la síntesis 5´→3´ El DNA es sintetizado por Enzimas DNA polimerasas
Precursores 5’-trifosfato de nucleótido y sólo pueden unirse al 3’-OH de la desoxiribosa
- Necesidad de Cebadores o Primers (la DNApol los necesita para empezar a copiar) Sintetizados por Enzimas primasas: son AN (RNA) de 7-10 nucleótidos (complementarios del extr. 3’ cadena DNA molde), comienzo de síntesis para DNApol
H

1).- Complementariedad entre bases del DNA (DNA molde): Chargaff. 2).- Carácter semiconservativo. 3).- Síntesis de replicación 5´-3´. 4).- Necesidad de Cebador o Primer: primasas. 5).- Carácter secuencial de la replicación a partir del origen: procesividad de la DNApolimerasa. 6).- Origen fijo de replicación: replicones. 7).- Replicación bidireccional: horquillas replicativas. 8).- Replicación semidiscontinua: fragmentos de Okazaki.
CARACTERÍSTICAS GENERALES DE LA REPLICACIÓN
Una vez iniciada la replicación desde el origen, los genes codificados se replican siempre en un orden determinado (uno tras otro),
NO al azar Procesividad: nº medio de nucleótidos que son añadidos antes que la DNApol se disocie

1).- Complementariedad entre bases del DNA (DNA molde): Chargaff. 2).- Carácter semiconservativo. 3).- Síntesis de replicación 5´→3´. 4).- Necesidad de Cebador o Primer: primasas. 5).- Carácter secuencial de la replicación a partir del origen: procesividad de la DNApolimerasa. 6).- Origen fijo de replicación: replicones. 7).- Replicación bidireccional: horquillas replicativas. 8).- Replicación semidiscontinua: fragmentos de Okazaki.
CARACTERÍSTICAS GENERALES DE LA REPLICACIÓN
Los lazos de replicación siempre se originan en un punto único denominado origen
Trozo DNA que se replica con un único origen de replicación: replicón

1).- Complementariedad entre bases del DNA (DNA molde): Chargaff. 2).- Carácter semiconservativo. 3).- Síntesis de replicación 5´→3´. 4).- Necesidad de Cebador o Primer: primasas. 5).- Carácter secuencial de la replicación a partir del origen: procesividad de la DNApolimerasa. 6).- Origen fijo de replicación: replicones. 7).- Replicación bidireccional: horquillas replicativas. 8).- Replicación semidiscontinua: fragmentos de Okazaki.
CARACTERÍSTICAS GENERALES DE LA REPLICACIÓN

Replicación de un cromosoma circular: exp de timidina marcada con tritio (3H) E.coli
Replicación bidireccional: horquillas replicativas El DNA bicatenario de las 2 hebras debe desenrollarse y separarse antes de ser replicado: requiere la intervención de enzimas Helicasas y Topoisomerasas Replicación evoluciona a ambos lados originando las burbujas u horquillas de replicación Los dos extremos tienen replicación activa: bidireccional

1).- Complementariedad entre bases del DNA (DNA molde): Chargaff. 2).- Carácter semiconservativo. 3).- Síntesis de replicación 5´→3´. 4).- Necesidad de Cebador o Primer: primasas. 5).- Carácter secuencial de la replicación a partir del origen: procesividad de la DNApolimerasa. 6).- Origen fijo de replicación: replicones. 7).- Replicación bidireccional: horquillas replicativas. 8).- Replicación semidiscontinua: fragmentos de Okazaki.
CARACTERÍSTICAS GENERALES DE LA REPLICACIÓN

Replicación semidiscontinua: fragmentos de Okazaki
Okazaki (1968) (experimentos de Pulso y Caza) Cadena continua o conductora (leading strand) síntesis transcurre en la misma dirección que el movimiento de la horquilla (y continua). La otra cadena se sintetiza en fragmentos cortos ‘de Okazaki’ (cadena discontinua o rezagada “lagging strand”) Fragmentos de Okazaki: -En E.coli de 1000 a 2000 nucleótidos -En eucariotas de 150-200 nucleótidos
Necesidad de Ligasas y Primasas
Hebra rezagada
Dirección del movimiento de la orquilla de replicación
Hebra Conductora o líder Las 2 cadenas DNA son antiparalelas y
Las 2 son sintetizadas en la dirección 5’→3’ ¡¿cómo simultáneamente?!

DNA-POLIMERASAS
Kornberg (1958): primera actividad DNApol de E.coli: DNApol I Existen más enzimas DNA pol ≠ en E. coli DNApol II, DNApol III, … (al menos 5 DNApol)
Reacción fundamental:
(dNMP)n + dNTP → (dNMP)n+1 + PPi
La DNA polimerasa es una enzima que cataliza la síntesis de DNA a partir de desoxirribonucleótidos y de una molécula de DNA molde
Requiere un “molde” y un “cebador”

DNA polimerasa: requerimientos
Los 4 dNTPs como sustrato
-Molécula de DNA molde (según reglas de apareamiento) -Cebador: segmento de cadena (RNA complementario del molde) con extremo 3’-OH libre: extremo cebador o

Los puentes de hidrógeno especifican el apareamiento y la geometría común entre bases complementarias. El centro activo de la DNApol acomoda solamente pares de bases con esa geometría Medida in vitro indican error cada 104-105 nucleótidos
La tasa de error disminuye gracias a mecanismos enzimáticos adicionales
La replicación es muy precisa E. Coli un error cada 109-1010 nucleótidos (cromosoma de 4,6×106 pb)→ un error cada 1000-10000 replicaciones

Reparación mediada por actividad Exonucleasas de la DNApol I: -Exonucleasa 3´→5´: “Comprobación de Lectura ó Corrección de pruebas” (proof reading) -Exonucleasa 5´→3´: “ Traslado de mella” (Nick translation)
Segunda comprobación, quita aquellos nucleótidos que por fallo de la propia actividad polimerasa se hallan mal apareados: corrige y la polimerasa ‘vuelve a empezar’. Mejora la precisión en factor 102-103 En DNApol I actividades polimerización y corrección de pruebas en centros activos diferentes
Reparación de apareamientos incorrectos: quita aquellos nucleótidos que se hallan apareados por delante de la actividad polimerasa (RNA o DNA) Si existe una mella ‘nick’ en la cadena (enlace P cortado), lo desplaza en la dirección de avance de la polimerasa: simultáneamente degrada 5´→3´ y reemplaza por actividad polimerasa Precisión adicional en factor 102-103

Exonucleasa 3´→5´: “Comprobación de Lectura ó Corrección de pruebas”
Una base mal apareada impide la translocación de la DNApol I al sitio siguiente. Deslizándose hacia atrás el enzima corrige el error con su actividad exonucleasa 3’→5’ y, a continuación, remprende su actividad polimerasa en la dirección 5’→3’
centros activos diferentes
Comprobación 1 a 1

ERROR
Exonucleasa 3’→5’: “Comprobación de Lectura ó Corrección de pruebas” (proof reading)

Exonucleasa 3’→5’: “Comprobación de Lectura ó Corrección de pruebas” (proof reading)
ERROR

Exonucleasa 5´→3´: “Traslado de mella” (Nick translation)
Importantes en la reparación del DNA y eliminación de cebadores de RNA durante la replicación La síntesis de DNA comienza en una mella (fosfodiester con 3´-OH, y fosfato en 5’ libre) La DNApol I extiende la hebra que no hace de molde y desplaza la mella a lo largo del DNA hasta el sitio de disociación, donde quedará una mella que sellará otro enzima
El AN a eliminar en verde

Exonucleasa 5’→3’: “Traslado de mella” (Nick translation)
Traslada la MELLA

Dominios estructurales de la DNApol I de E.coli Mediante tratamiento proteolítico suave se pueden separar dos fragmentos: *de Klenow ( 2 centros activos independientes) *exonucleasa 5’→3’ o N-terminal (no presente en otras DNApol)
Fragmento Klenow
Los dominios de los dedos (fingers) y pulgar (trumb) se enrollan alrededor del DNA y lo mantienen sujeto al centro activo (residuos de la palma) Fragmento de Klenow incluye el centro activo exonucleasa 3’→5’
La reacción polimerasa depende de dos iones metálicos (Mg2+)

-FUNCIÓN y CINETICA DE LA DNApol I * Es LENTA (Velocidad=600-1000 nucleotidos/min) * PROCESIVIDAD muy BAJA (20 nucleotidos) Nucleótidos añadidos antes de la disociación de la polimerasa * NUMERO DE COPIAS (400 moléculas/célula) Existe Cepa bacteriana con el gen de la DNApol I alterado (enzima inactivo) es VIABLE
(anormalmente sensible a los agentes que dañan el DNA)
DNApol I NO ES LA REPLICASA DE E.coli SU FUNCIÓN ES REPARADORA de fragmentos cortos de DNA
Incluidos fragmentos de Okazaki
En la replicación intervienen muchos genes, y por tanto muchas proteínas
DNApol I NO puede ser la encargada de replicar todo el DNA DNApol I de E.coli no es la replicasa del DNA

DNA polimerasa II -LENTA (50 nucleotidos/min) -PROCESIVIDAD BAJA -Con EXONUCLEASA 3´→5´ (Sin EXONUCLEASA 5´→3’)
Actividad DNA polimerasa III:
-VELOCIDAD ALTA (9000 nucleótidos/min) -PROCESIVIDAD ALTA. -CON EXO 3´→5´ (sin EXONUCLEASA 5´→3’) -NUMERO DE COPIAS (10-20 moléculas/célula)
-Núcleo o Core: subunidades α (polimerasa), ε (corrección pruebas) y θ (estructural). -Resto: subunidades β,δ,γ,τ,.. aumentan la procesividad y velocidad del core (15000 nucleotidos/min)
Función REPARADORA
DNA pol III forma parte de un complejo con multimérico denominado HOLOENZIMA pol III: verdadera REPLICASA de E.coli
Subunidad catalítica
Actividad Exonucleasa 3’-5’
DNA polimerasa IV y V (1999) están implicadas en un tipo de reparación poco habitual

DNA polimerasas Eucarióticas DNA pol α: replicasa nuclear, Sin Exo 3´→5´, procesividad media. Actividad fundamental PRIMASA de Eucariotas. Actua en cad retardada?
DNA pol δ: verdadera REPLICASA nuclear, comparable a la DNApol III. Exo 3´-5´y sin actividad Primasa. Muy procesiva en presencia de la proteína del antígeno nuclear (PCNA ) Papel ~subunidades procesivas de DNApol III
DNA pol β: comparable a DNApol I, Sin Exo 3´→5´, Poco procesiva. No Primasa. Actividad REPARACIÓN.
DNA pol ε: algunos autores proponen actividad ~DNApol I, Exo 3´→5´, muy procesiva. Sin actividad primasa. Función REPARACIÓN Lesiones U.V.
DNA pol γ: replicasa mitocondrial, poco conocida.

Otras DNA polimerasas -DNA polimerasas de bacterias termófilas: Taq polimerasa → PCR
-DNA polimerasas de fagos: diversas estrategias
M13 y φx174: DNApolIII de E.coli modificada por factores propios. T4: DNApolimerasa monomérica con exonucleasa 3´→5´ muy fuerte → Biología molecular T7 ó secuenasa DNA polimerasa monomerica con un nivel de fidelidad de copia muy alto → Secuenciación -DNA polimerasas RNA dependiente ó Transcriptasa reversa: Copia un molde de RNA para dar una copia de DNA

La replicación de DNA requiere muchos enzimas y factores proteicos
o Topoisomerasa II

Proteínas que intervienen en la horquilla de replicación
o Topoisomerasa

ELEMENTOS PARTICIPANTES EN LA REPLICACIÓN

Sitios específicos de origen de la replicación (E. coli)
En eucariotas varios orígenes fijos de replicación En E. coli uno único para todo el genoma: oriC (un solo replicón)
oriC: 245 pb, con 3 secuencias casi idénticas en tandem (13-mer) y cuatro lugares de unión (9-mer) para la dnaA (proteína de iniciación cuya unión inicia secuencia sucesos)
13-mer Secuencia de 13 p.b.
9-mer

La replicación necesita un cebador

ACTIVIDAD PRIMASA (RNA polimerasa especializada): síntesis de PRIMERS o cebadores.
-Primasa de Eucariotas: es una de las replicasas la DNA pol α. -Primasa de E. Coli: es un enzima especial exclusivo para sintetizar PRIMERS. Es POCO PROCESIVA (20 nucleotidos max) y puede usar dNTPs y NTPs (usa más los segundos sólo por su abundancia)→RNA No actúa aislado sino como SUBUNIDAD DE UN COMPLEJO llamado PRIMOSOMA (Primasa + 7 subunidades más, incluyendo helicasa)
Necesidad PRIMASAS: Para asegurar ‘fidelidad’ DNApol comprueba la validez del par de bases precedentes antes de formar un nuevo enlace fosfodiester. Las RNApol NO son tan restrictivas y pueden comenzar la síntesis de novo sin comprobaciones → mayor tasa de error, pero NO problema porque es provisional (RNA cebador se elimina por exonucleasa 5’→3’)
Otros modelos: -Primasa de fago M13: emplea la RNA polimerasa de E.coli Como PRIMASA en el paso de la forma infectiva del fago (ss), a la forma replicativa (Rf).
-Primasa de φx174: aunque es también fago de E.coli, sintetiza un enzima multimérico propio especial para la actividad PRIMASA.

Mecanismo del PRIMOSOMA de E.coli sobre la hebra rezagada (síntesis de fragmentos de Okazaki)
Primosoma sintetiza el primer y luego avanza en sentido contrario sobre la
hebra parental retrasada (5’→ 3’) hasta encontrar ‘nuevos’ sitios de ‘inicio de
fragmento’ donde se produce otro primer

ACTIVIDAD DNA-LIGASA: une los fragmentos de Okasaki unión extremos de dos cadenas de DNA
3´-OH + 5´-P → 3´-P-5´ + H2O
Reacción energéticamente desfavorable Requiere un Donador de energía -Donador ATP: Ligasa de fago T4 y Ligasa de Eucariotas -Donador NAD: Ligasa de E.coli
LIGASA

Etapas de la actividad DNA-ligasa:
1) Activación de la DNA-ligasa 2) Reacción nucleofílica 3) Liberación de AMP
La actividad ligasa puede ser REVERSIBLE Reducir enrollamiento introduciendo un nick

Comparación entre
Ligasa de E.coli
Ligasa de T4 (Biología Molecular)

OTRAS ACTIVIDADES: Helicasa y SSB(P)
Helicasa: va al frente de la horquilla replicativa, separando las 2 hebras mediante ruptura puentes hidrógeno: requiere ATP
SSB: proteínas de alta afinidad por el DNA monohebra. Una vez ‘separadas’ las hebras por helicasa estas son inestables (tienden a aparearse): se estabilizan, hacen que permanezcan separadas por unión SSB
En E. coli se han descrito varias: helicasa II, dnaB, proteína Rep

OTRAS ACTIVIDADES: Topoisomerasa
Topoisomerasa II (girasa): introduce superenrollamiento (-) para relajar el superenrollamiento (+), generado por la helicasa ATP dependiente Topoisomerasa I: relaja estructuras superenrolladas por otros mecanismos (escisión 1 sola hebra) NO es dependiente de ATP
Etapas generales y comunes: 1) escisión hebras 2) paso del segundo segmento DNA a través del hueco 3) reparación corte
En E. coli
Cambio conformacional
Unión topoisomerasa a duplex DNA

Topoisomerasa II o DNA girasa → blanco de varios antibióticos: *NOVOBIOCINA impide unión de ATP a la girasa *Ácido Nalidíxico y CIPROFLOXACINA dificultan rotura y empalme de cadenas de DNA
Tratamiento de infecciones de las vías urinarias al inhibir la replicación bacteriana

EL COMPLEJO DE REPLICACIÓN (En procariotas)
Topoisomerasa
Pero… ¿Cómo se replican simultáneamente las 2 hebras?

Horquilla de replicación procariotica replisoma de E.coli
Holoenzima pol III actua como un dímero: replica las 2 hebras simultáneamente
(Cerrar Nicks) (abrazadera de deslizamiento)
Formando un bucle

Coordinación en la síntesis de la hebra guía y la hebra retardada del DNA
Hebra retardada se sintetiza en fragmentos Crecimiento 3’→5’ y Polimerización 5’→3’
Bucle del molde lo sitúa en posición para que tenga lugar la polimerización en sentido 5’→3’ (orientado de manera favorable al avance de la horquilla de replicación) Al avanzar va tirando del lazo, haciéndolo más grande. DNApol III abandona el molde y se forma nuevo bucle.
Huecos entre fragmentos se rellena por DNApol I, que también elimina el RNA cebador (actividad exonucleasa 5’-3’)
DNA ligasa empalma los fragmentos
~1000 nucleótidos en E. coli

INICIACIÓN de replicación en E.coli
1. El complejo con la dnaA se une a la secuencia 9-mer (caja dnaA) dando una conformación plegada especial en OriC (colabora proteína Hu)
2. La conformación plegada en OriC promueve desapareamiento y apertura de las zonas repetidas de 13 pb (13-mer) ricas en A-T
3. Un complejo formado por las proteínas de actividad helicasa dnaB y dnaC comienza a abrir las 2 hebras en colaboración con la topoisomerasa II
13-mer secuencia rica en A-T
9-mer Cajas dnaA
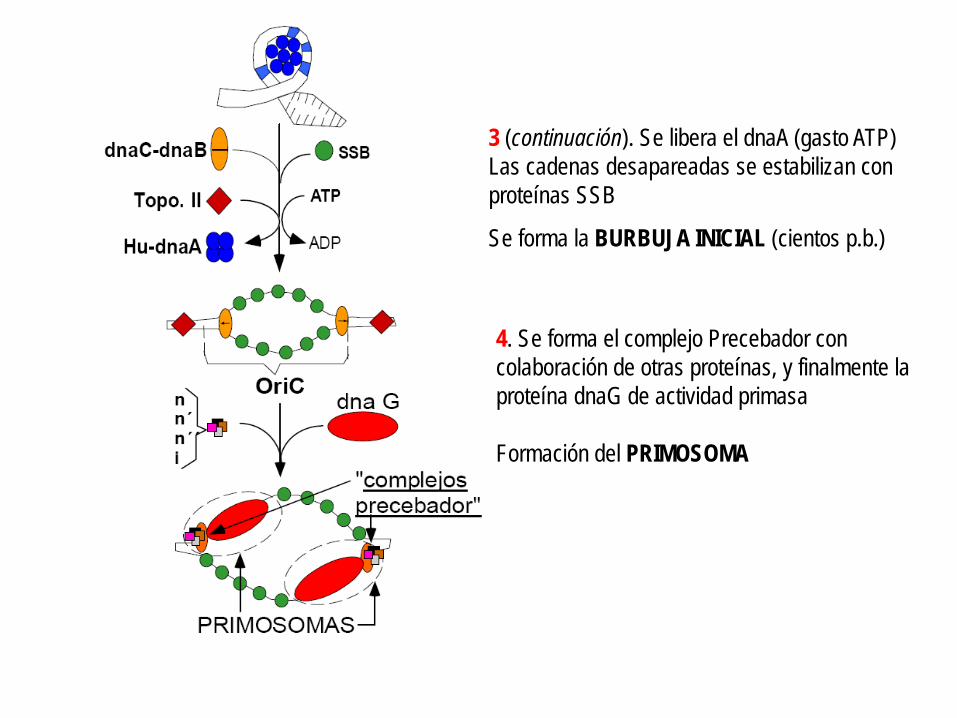
3 (continuación). Se libera el dnaA (gasto ATP) Las cadenas desapareadas se estabilizan con proteínas SSB
Se forma la BURBUJA INICIAL (cientos p.b.)
4. Se forma el complejo Precebador con colaboración de otras proteínas, y finalmente la proteína dnaG de actividad primasa Formación del PRIMOSOMA

5. El Primosoma se une a las 2 hebras de DNA molde e inicia la síntesis de primers de RNA (en cada hebra, bidireccional) 6. Unión de DNApol III y de la prot rep (Helicasa) convierte el primosoma en un REPLISOMA

Resumen: Proteínas requeridas para iniciar la REPLICACIÓN en E. coli

Terminación:
Las dos horquillas de replicación del cromosoma circular de E. coli se encuentran en una región terminal que contiene múltiples copias de una secuencia de 20 pb (Ter) Trampa de donde no puede ‘escapar’ la horquilla de replicación Secuencia Ter sitios de unión de proteína Tus (contrahelicasa). Complejo Ter-Tus puede detener la horquilla de replicación en una única dirección pocos centenares de bases que quedan entre grandes complejos proteicos se replican por mecanismo desconocido Resultado final dos cromosomas circulares ligados (catenados) se separan mediado por topoisomerasa IV (del tipo II)
Está regulado, por un mecanismo no del todo conocido, que sólo haya una replicación en cada ciclo celular

REPLICACIÓN EN EUCARIOTAS
Proceso parecido pero MÁS COMPLEJO que en Procariotas *tamaño GENOMA Eucariot (hum) 6000×106 pb Procar (E coli) 4,8×106 pb *nº cromosomas 23 parejas lineales 1 circular

Papel de las DNA polimerasas Eucarióticas DNA pol α: PRIMASA de los eucariotas. También polimerasa de iniciación. Primers de cadena Lider y de los fragmentos de la cadena rezagada. Algún autor la considera la Replicasa de los Fragmentos de Okazaki (en discusión)
DNA pol δ: verdadera REPLICASA de la cadena Lider según autores también de la rezagada en colaboración con la DNApol ε (en discusión)
DNA pol β: comparable a DNApol I, Sin Exo 3´-5´,
DNA pol ε: función REPARACIÓN de lesiones U.V. y rellenado de huecos dejados por los Primers en los fragmentos de Okazaki. Según autores colaboraría con la DNApol δ, en replicar la hebra rezagada (en discusión)
Tabla comparativa tipos DNApol procariotas y eucariotas

Origen de replicación en Eucarióticas
En Levaduras se denominan Secuencias de replicación autónoma (ARS), 100-120 pb:
1-Las ARS constan de una región central de 11pb rica en A-T (análoga a secuencias de 13pb rica en A-T de procariotas), denominada ”Secuencia consenso ARS”
2-Complejo de 6 proteínas reconocen las zonas flanqueantes de la región central de rica en A-T y se unen a ellas formando el “complejo de Orígen de replicación (ORC)”, que promueve la apertura de la zona rica en A-T.
helicasa extiende la burbuja inicial
3-También se unen unas proteínas de unión a monohebra (equivalentes a las SSB procariot), denominadas Proteínas de replicación A (RPA), estabilizando la zona abierta monohebra y permitiendo entrada de DNApol α (primasa)
4-Tras añadir el cebador (activ. primasa) y ~20 desoxinucleótidos más (activ. polimerasa), el Factor de replicación C (RFC) desplaza a la DNApol α y atrae al Antígeno nuclear de células proliferantes (PCNA) que se une a la DNApol δ (cambio de polimerasa).
SON MÚLTIPLES y en organismos superiores poco conocidos En humanos ~30.000 orígenes, cada cromosoma cientos

Cromatina en Eucarióticas: empaquetamiento El DNA eucariótico se enrolla alrededor de proteínas Histonas, formando los Nucleosomas -Modelo hipotético de Replicación de los nucleosomas:
Las histonas no llegarían a separarse físicamente del DNA durante la replicación, los nucleosomas ‘viejos’ permanecerían en la hebra conductora. Tras la replicación, se incorporarían nuevas histonas dando lugar a ‘nucleosomas hijos’.

-Mecanismos de replicación en cromosomas lineales: Problema de ‘terminación’ replicación en cromosomas lineales: resulta difícil replicar por completo los extremos porque las polimerasas actúan únicamente en dirección 3’→5’
Le hebra retardada presentaría un extremo 5’ incompleto tras la eliminación del DNA
cebador. Cada ronda de replicación acortaría aún más el cromosoma.
ESTO NO es un problema gracias a los TELÓMEROS (DNA no codificante de los extremos de los cromosomas que mantienen la integridad del cromosoma). El DNA de los telómeros contiene cientos de repeticiones en tandem secuencia 6 nucleótidos (en humanos hasta 2000 copias de 3’ AGGGTT 5’). Una de las hebras enriquecida en G extremo 3’ y ligeramente más larga. Forma bucle, con la hebra sencilla creando un duplex de DNA con forma de lazo con otra de las zonas de secuencia repetida
Protegen y enmascaran el extremo del cromosoma
de lectura

Mecanismo Telomerasas Eucariota Los telómeros se replican mediante la telomerasa, polimerasa especializada
que lleva su propio molde de RNA (ribonucleoproteína)
Cuando un cebador que termina en GGTT se añade al enzima telomerasa (en humanos) se generan secuencias más largas: GGTTAGGGTT y GGTTAGGGTTAGGGTT y más largas La telomerasa contiene una molécula de RNA que actúa como molde para la elongación de la hebra enriquecida en G el propio enzima tiene le secuencia para generar las secuencias del telómero Componente proteico de las telomerasas relacionado con las transcriptasas inversas de los retrovirus
Telomerasa es un transcriptasa inversa especializada que lleva su propio molde

Telomerasa “alarga” extremo 3’

Finalmente DNApol α con su actividad primasa introduce un primer en el extremo recién alargado y replica el fragmento

RECOMBINACIÓN Reordenamiento de la información genética dentro y entre moléculas de DNA
En Eucariotas -Meiosis: intercambio limitado entre cromosomas emparejados genera diversidad genética en la población (entrecruzamiento) -Transposición del DNA: tipo diferente recombinación, en que un segmento corto de DNA presenta una notable capacidad de trasladarse de un lugar a otro del cromosoma -Generación de diversidad molecular en anticuerpos (y algunas otras moléculas inmunitarias) -Algunos virus para integrar su material genético en el DNA cel huesped
En el laboratorio herramienta para manipular genes, por ej. Ratones knockout

Flujo de información en los retrovirus: del RNA al DNA