sistemática y evolución de los anfibios de chile
TRANSCRIPT

Arch. Biol. Med. Exper. 12:59-70, 1979
Sistemática y evolución de los anfibios de Chi le 1
Systematics and Evolution of Amphibians from Chile
NELSON F . DÍAZ*, ALBERTO VELOSO M.**
' D e p a r t a m e n t o de Bio logía . Facultad de Cienc ias . Sede Oriente , Univers idad de Ch i l e . Sant iago Cas i l la Ó53,
Sant iago , Chi le .
* * D e p a r t a m e n t o de Bio log ía Celu lar y Genét ica . Facultad de M e d i c i n a Sede Norte . Univers idad de Chi l e , San
t iago.
(Rec ib ido para publ icación el 3 de dic iembre de 1 9 7 7 )
D Í A Z , N . F . , VELOSO, M.A. Sistemática y evolución de los anfibios de Chile. (Systematics and evolution of amphibians from Chile). Arch. Med. Exper. 12:59-70, 1979.
A review of previous taxonomic arrangements, together with chromosomes and molecular evidences are used to give an up to date classification of Chilean amphibians. Thirty three species are recognized belonging to two major families, Leptodactylidae and Bufonidae. These species are grouped in the following genera: Caudiver-bera (one species) Telmatobufo (two species),
A M P H I B I A N S
El conocimiento de la fauna de Anfibios Chilenos ha recibido aportes significativos por las contribuciones de Molina, Bell, Guichenot, Gün-ther, Philippi, Schneider y otros herpetólogos que, desde 1780 hasta comienzos de este siglo realizaron descripciones tipológicas de numerosas especies.
El conocimiento de los límites distribucio-nales, de la existencia de poblaciones estables en determinados ambientes biocenóticos, de información relativa a la variación interpo-blacional de caracteres, de datos de la fisiología y reproducción, así como una tesonera labor de recopilación biliográfica y estudio de tipos y colecciones en museos del país y del extranjero permitieron a Cei (10) hacer una actualización de estos conocimientos.
Sin embargo, la casi inexistencia de colecciones locales y el hecho frecuente del depósito
Batrachyla (three species), Eupsophus (four species), Hylorina (one species), Alsodes (four species), Telmatobius (seven species), Insuetophrynus (one species), Pleurodema (three species), Rhinoderma (two species) and Bufo (five species). Intergeneric and intra-generic relationships between these taxa are pointed out.
E V O L U T I O N
de tipos en instituciones extranjeras dificultan los estudios taxonómicos. A esto se agregan características peculiares de esta fauna, como su alto endemismo, y en numerosas especies un alto grado de especialización ecológica que determina tamaños poblacionales y distribuciones geográficas restringidas. Esto puede explicar las descripciones de nuevas especies basadas en escaso número de ejemplares y los frecuentes cambios de ubicación genérica que se encuentran en las contribuciones sistemáticas de autores locales y extranjeros que se han ocupado del tema en los últimos quince años.
CRITERIOS DE ANÁLISIS Y SISTEMAS DE
CLASIFICACIÓN
En el estudio de los Anfibios Chilenos, los criterios taxonómicos utilizados han estado en corres-
S Y S T E M A T I C S
' F i n a n c i a d o parc ia lmente por los Proyectos 4 2 9 7 - R y B 7 0 5 - 7 9 1 , de la Univers idad de Ch i l e .

60 D Í A Z y V E L O S O
pondencia con la información biológica disponible. Una primera etapa, caracterizada por el interés de los taxónomos en descripciones de naturaleza tipológica, es superada con el advenimiento de los criterios poblacionales que centran el interés de los naturalistas en el estudio de la variación de caracteres. Sin embargo, hay ausencia de un método de análisis explicitado para construir los esquemas de clasificación.
Una tercera etapa es reconocible con la aplicación de elementos y conceptos de la Sistemática Filogenética (23). Las principales diferencias en las clasificaciones propuestas y actualmente utilizadas resultan de los criterios aplicados a los caracteres para la obtención de ciado-gramas (24, 30).
Esta aproximación cuenta sin embargo con serios detractores en una escuela diferente de clasificación representada por Mayr, Simp-son y otros sistemáticos evolucionistas, cuyos esquemas de clasificación difieren en los postulados básicos para la construcción de los modelos filogenéticos. Estos modelos constituyen hipótesis que sólo pueden verse reafirmadas o rechazadas de manera indirecta mediante el aporte de nuevas evidencias (13).
Los principales propósitos de este trabajo son la entrega de una actualización detallada del ordenamiento taxonómico de las especies Chilenas de Anfibios y la discusión de esquemas de clasificación y evolución introduciendo antecedentes cromosómicos y moleculares.
MATERIALES Y M É T O D O S
El anál i s i s c romosómico se efectuó a dos n ive les : estudio
comparat ivo de los c o m p l e m e n t o s diploides de 26 espec ies ,
y anál is is comparat ivo de car iot ipos bandeados en la Sub
familia T e l m a t o b i i n a e . Los c r o m o s o m a s se obtuvieron en
an imales metamorfoseados inyectados con colchicina al
1%. por aplastado de epite l io de córnea y / o suspens ión celu
lar de bazo y médula ósea. Las t inc iones ut i l i zadas fueron
Orce ína acética y G i e m s a Se real izaron medic io
nes de los c r o m o s o m a s metafás icos sobre sus reproduccio
nes fotográficas y se construyeron id iogramas . La n o m e n
clatura para denominar los c r o m o s o m a s es la propuesta
por Levan y col. ( 2 8 ) . El bandeo cromosómico se realizó
adaptando y es tandar izando técnicas descritas para M a m í
feros, al uso en Anfibios anuros ( 5 0 ) .
Se estudiaron las i s o e n z i m a s de Lactato D e s h i d r o g e n a s a
( L D H ) de h o m o g e n e i z a d o s de cr is ta l inos , s eparadas elec-
troforéticamente en ge les de pol iacri lamida. L o s geles se
prepararon de acuerdo a Dav i s ( 1 4 ) , modif icando la con
centración del gel separador al 6%. Se real izaron evaluaciones
dens i tométr icas de los ge les . Se ut i l izaron an imales perte
nec ientes a dieciocho especies , en la mayoría de las cuales
se efectuó para le lamente la separación de i s o e n z i m a s fos-
forilantes de g lucosa presentes en el h ígado , mediante cro
matografía de intercambio iónico ( 4 1 ) .
RESULTADOS
Los resultados que se entregan comprenden un conjunto de treinta y tres especies, incluyendo las descritas con posterioridad a la revisión de Cei (10) y una entidad aun no descrita que incluimos con la denominación Alsodes sp. (ver ** en Tabla II).
RESULTADOS CITOGENÉTICOS
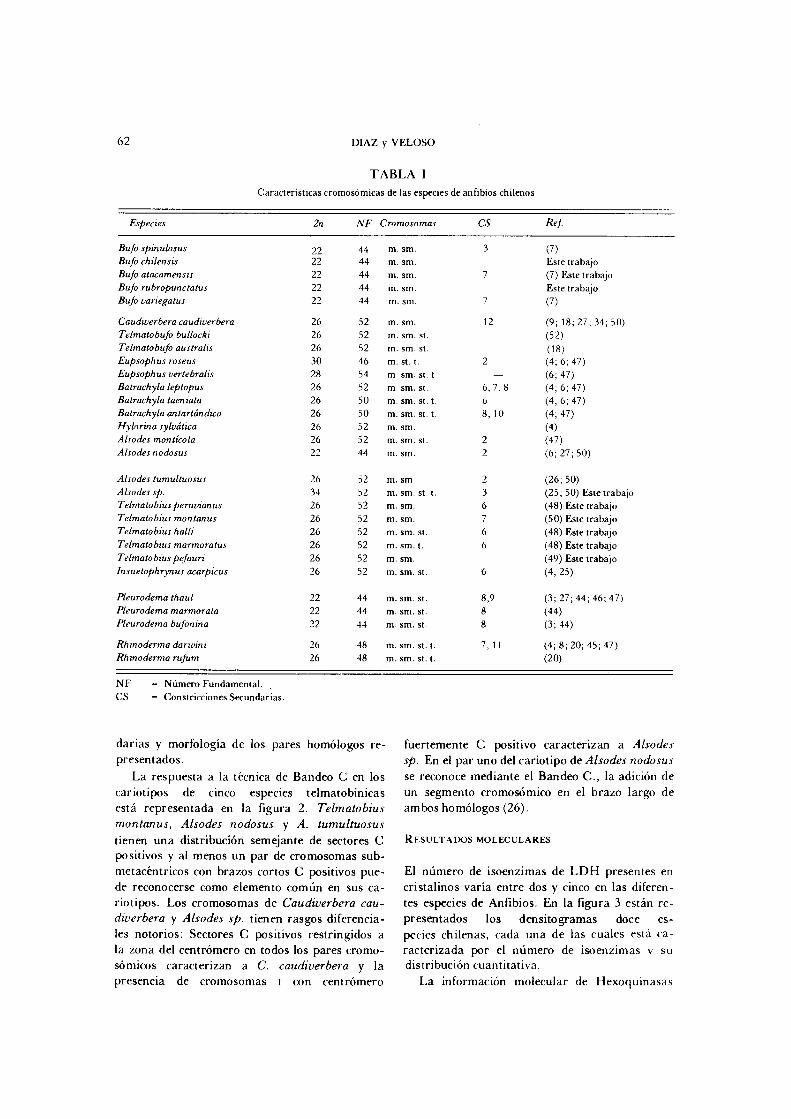
La Tabla I contiene un resumen de características de los complementos diploides conocidos para las especies de Anfibios Chilenos, tanto de la información bibliográfica disponible como de nuestros resultados. En la figura 1 se muestran los idiogramas de 26 especies. Los complementos cromosómicos de Eupsophus coppingeri, E. vanzolim, Telmatobius laevis y T. grandisonae se desconocen. Estas son especies endémicas, de distribución muy restringida de acuerdo a las descripciones originales, y que probablemente en razón de tamaños poblacionales reducidos no han sido recientemente colectadas.
La existencia de un número diploide 2n =22 en nueve especies de la muestra, cinco de las cuales pertenecen al Género Bufo, tres al Género Pleurodema de la Subfamilia Leptodactylinae y Alsodes nodosus de la Subfamilia Telmatobiinae, determina que este no sea un carácter útil para reconocer agrupaciones naturales de estas especies. Quince especies de la Subfamilia Telma tobiinae tienen un cariotipo de 26 cromosomas, número que aparece como el más generalizado, también representado en el Género Rhinoderma de la Subfamilia Rhinodermatinae. Las tres especies restantes tienen números cromosómicos de 28, 30 y 34, que son los mas altos en este grupo, y en el caso de Alsodes sp., uno de los más altos reportados en Leptodactylidae. Estos tres cario-tipos pueden caracterizarse además, por la disminución gradual en el tamaño de los cromosomas y por la presencia de cromosomas telocén-tricos, numerosos en Eupsophus roseus y Alsodes sp. La variación cromosómica en el conjunto de las especies se refiere a diferencias en el número diploide, proporción de cromosomas grandes y pequeños, localización de constricciones secun-

A N F I B I O S C H I L E N O S 61
S S 8 a B U F O s p i n u ' o s u s
8 a a a B U F O a t a c a m e n s i s
8 8 8 S
8. 8 8
t a d B a a a a a a H B 11
B U F O v a n e g a t u s
11 P L E U R O D E M A t h a u l
í B 8 B 0 8 8 u B a 8 B 11
P L E U R O D E M A m a r m o r a l a
í u u u O B 8 B B 8 B B 11
P L E U R O D E M A b u i o n i n »
II A L S O O E S n o d o s u s
8 8 8 8 8 s s 11 13
A L S O O E S t u f n u l t u o t u s
8 8 8 8 8 8 8 11 13
A L S O O E S m o n t í c o l a
8 8 8 8 8 8 8 11 13
C A U D I V E R B E R A c a u d i v « r b « r a
0 8 8 8 8 8 a s s
8 S 8 8 S
E U P S O P H U S « e r t e b r a l i s 11 13 M
O D 0 8 S S O Q 8 D 8 11 13 H 15 E U P S O P H U S r o s e u s
1 * -
'Í8 8 L U U S U U U Ü U Q B Ü D ú a S B o ' 2 3 4 S 6 7 > 9 10 11 12 13 14 15 1» 17
| A L S O O E S SO
s s a s s s I N S U E T O P H R V N U S a c a r p . c u s
8 8 8 S 8 S 8 S M Y L O R I N A s y l v a t r c a
8 § 8 S S s o R H I N O D E R M A d a r w i m
Í O f l O B R H I N O D E R M A r u f u m
t 8 B 8 8
8 8 Ü 8 8 8 S O
8 8 8 8 8 8 s T E L M A T O B U F O a u s l r a l i s
8 8 8 8 8 S S T E L M A T O B U F O b u l l o c k i
^ 8 8 8 8 3 8 8 S 8 2 8 8 o 13
B A T R A C H Y LA a n t a r t a n d i c a
Í B 8 8 8 8 § 1 1 s 8 s s B B A T R A C H Y L A l e p t o p u s
S 8 8 8 8 8 Ü S B A T R A C H Y L A l a e n i a l a
B 8 8 8 S 8 8 8 T E L M A T O B I U S h a l l
5 8 8 8 8 8 8 8 T E L M A T O B I U S p w u v i a n u s
8 8 8 8 8 8 8 8 T E L M A T O B I U S m a r m o r a l u s
1 2 3 4 5 1 T E L M A T O B I U S m o n t a n u
Ü G D I 8 8 8 8 8 8 4 5 6 7 1 9 10 11 1 2 1 3
Fig. 7. Id iogramas . La escala permite determinar el % de cada par cromosómico en relación al c o m p l e m e n t o haplo ide .
62 D I A Z y V E L O S O
TABLA I Característ icas c r o m o s ó m i c a s de las espec ies de anfibios ch i lenos
Especies 2n NF Cromosomas cs Ref.
Bufo spinulosus 22 44 m. sm. 3 (7) Bufo chilensis 22 44 m. sm. Este trabajo Bufo atacamensis 22 44 m. sm. 7 (7) Este trabajo Bufo rubropunctatus 22 44 m. sm. Este trabajo Bufo variégatus 22 44 m. sm. 7 (7)
Caudwerbera caudiverbera 26 52 m. sm. 12 (9; 18; 2 7 ; 34 ; 5 0 ) Telmatobufo butlocki 26 52 m. sm. st. ( 52 ) Telmatobufo australis 26 52 m. sm. St. (18) Eupsophus roseus 3 0 4 6 m. st. t 2 (4 ; 6; 4 7 ) Eupsophus vertebralis 28 54 m. sm. St. t. — (6 ; 4 7 ) Batrachyla leptopus 26 52 m. sm. St. 6 , 7 , 8 (4; 6; 47) Batrachyla taen iala 26 5 0 m. sm. St . t . 6 (4; 6; 47) Batrachyla arxlartándica 26 5 0 m. sm. St . t. 8 , 10 (4; 47) Hylorina sylvática 26 52 m. sm. (4) Alsodes montícola 26 52 m. sm. St. 2 (47 ) Alsodes nodosus 22 44 m. sm. 2 (6; 2 7 ; 5 0 )
Alsodes tumultuosus 26 52 m. sm. 2 ( 2 6 ; 50) Alsodes sp. 34 52 m. sm. st. t. 3 ( 2 5 ; 50) Este trabajo Telmatobius peruvianus 26 52 m. sm. 6 ( 4 8 ) Este trabajo Telmatobius montanus 26 52 m. sm. 7 ( 5 0 ) Este trabajo Telmatobius halli 26 52 m. sm. st. 6 (48 ) Este trabajo Telmatobius marmoratus 26 52 m. sm. t. 6 (48 ) Este trabajo Telmato bius pefauri 26 52 m. sm. ( 4 9 ) Este trabajo lnsuetophrynus acarpicus 26 52 m. sm. st. 6 ( 4 ; 2 5 )
Pleurodema thaul 22 4 4 m. sm. st. 8 ,9 ( 3 ; 2 7 ; 4 4 ; 4 6 ; 4 7 ) Pleurodema marmorata 22 44 m. sm. st. 8 ( 4 4 ) Pleurodema bufonina 22 44 m. sm. st. 8 (3 ; 4 4 )
Rhinoderma darwini 26 48 m. sm. st. t. 7, 11 (4; 8; 2 0 ; 4 5 ; 4 7 ) Rhino derma rufum 26 48 m. sm. st. t. ( 2 0 )
N F = N ú m e r o Fundamenta l .
C S = Constr icc iones Secundar ias .
darías y morfología de los pares homólogos representados.
La respuesta a la técnica de Bandeo C en los cariotipos de cinco especies telmatobínicas está representada en la figura 2. Telmatobius montanus, Alsodes nodosus y A. tumultuosus tienen una distribución semejante de sectores C positivos y al menos un par de cromosomas sub-metacéntricos con brazos cortos C positivos puede reconocerse como elemento común en sus cariotipos. Los cromosomas de Caudwerbera caudiverbera y Alsodes sp. tienen rasgos diferenciales notorios; Sectores C positivos restringidos a la zona del centrómero en todos los pares cromo-sómicos caracterizan a C. caudwerbera y la presencia de cromosomas t con centrómero
fuertemente C positivo caracterizan a Alsodes sp. En el par uno del cariotipo de Alsodes nodosus se reconoce mediante el Bandeo C , la adición de un segmento cromosómico en el brazo largo de ambos homólogos (26).
RESULTADOS MOLECULARES
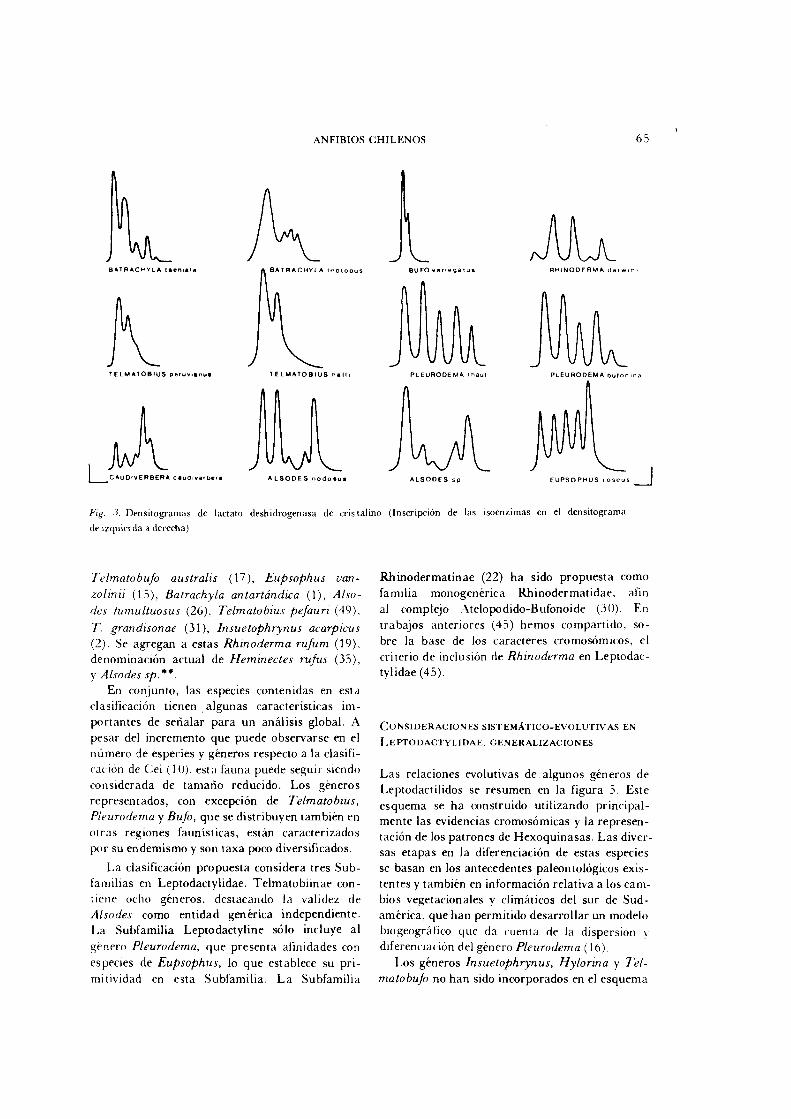
El número de isoenzimas de L D H presentes en cristalinos varía entre dos y cinco en las diferentes especies de Anfibios. En la figura 3 están representados los densitogramas doce especies chilenas, cada una de las cuales está caracterizada por el número de isoenzimas y su distribución cuantitativa.
La información molecular de Hexoquinasas

A N F I B I O S C H I L E N O S 63
hepáticas se resume de la siguiente manera: en el hígado de Anfibios pueden encontrarse cuatro Hexoquinasas, aunque en ninguna de las especies estudiadas existen simultáneamente más de tres, y sólo se encuentran las combinaciones de isoenzimas A.B.D. o C.B.D. (42). En la figura 4 se muestran los patrones cromatográfi-cos de estas isoenzimas en doce especies chilenas de anuros, pertenecientes a ocho géneros: Pleurodema, Rhinoderma, Bufo, Telmatobius y Alsodes (part.) tiene un cromatograma A.B.D.; Alsodes (part.), Eupsophus, Batrachyla y Cau-diverbera tienen un cromatograma C.B.D.
DISCUSIÓN
Los géneros agrupados en Telmatobiinae constituyen de acuerdo a la evidencia fósil disponible, representantes actuales de una fauna de Leptodactylidos ya diversificados en el Terciario Inferior de la Patagonia (Oligoceno Inferior) (38). Los análisis cladísticos realizados en Leptodactylidae (16), (24), (30) y los planteamientos sistemático-evolutivos en Bu-fonide (5), junto con los datos fósiles señalados para Telmatobiinae y los aportados para Bufo (40), contribuyen a señalar un carácter holo-filético a cada una de estas agrupaciones. En el ordenamiento propuesto por Cei en 1962 se considera a Bufo en Chile representado por B. variegatus y cuatro subespecies del Grupo de Bufo spinulosus. Con posterioridad se han incorporado criterios moleculares por estudio de aminas biógenas así como de relaciones inmu-nológicas para señalar el carácter específico de cada una de estas entidades (11). El holofile-tismo del Grupo spinulosus se reafirma por los estudios de compatibilidad genética (5) y particularmente por las evidencias cromosómicas (6), (7). La divergencia de Bufo variegatus respecto al Grupo spinulosus se refuerza por sus caracteres osteológicos (32). La discusión sistemática se centra fundamentalmente en Leptodactylidae; las proposiciones sistemáticas de Lynch (29), (30) incluyen en Telmatobiinae un total de veinticinco géneros. A estos debe agregarse el Género Alsodes (Bell, 1843), revalidado por Gallardo (21).
El. APORTE DE LOS CRITERIOS CROMOSÓMICOS
El número diploide, la morfología y las cons
tricciones secundarias, junto con la distribución de heterocromatina constitutiva (bandas C) son los parámetros citogenéticos considerados en el análisis cromosómico. Los idiogramas de la figura 1 muestran la escasa variación intra-genérica de las especies de Telmatobius, Telma-tobufo, Rhinoderma y Pleurodema. Las especies de Batrachyla difieren en las constricciones secundarias. Además, el cariotipo de B. leptopus está constituido solamente por cromosomas bibraquiados, en tanto que en B. antartándica el par 11 y en B. taeniata el par 12 son telocéntri-cos. La mayor variación cromosómica entre los Leptodactílidos analizados ocurre en los géneros Eupsophus y Alsodes. En Telmatobiinae los números diploies 2n = 22 de Alsodes nodosus y 2n = 3 4 de Alsodes sp. constituyen excepciones. Sin embargo, la unidad genérica de Alsodes ha sido demostrada parcialmente mediante el análisis de cariotipos bandeados (50). De acuerdo a la evidencia disponible, en el Género Alsodes concurren posiblemente dos mecanismos responsables de la modificación del N F (NF = Número Fundamental de brazos cromosómicos (33)). Los cromosomas grandes de A. nodosus, en particular el par uno, presentan en sus brazos largos evidencias de traslocaciones (banda C intercalar en el brazo largo de ambos homólogos); este mecanismo de reordenamiento podría dar cuenta de la disminución del número de pares pequeños a partir de un cariotipo 2n = 26 o de mayor número, similar al que presenta A. tumultuosus. Otro mecanismo participante de la modificación del N F puede ser el crecimiento de brazos por adición de heterocromatina constitutiva, como ha sido propuesto en Mamíferos (39).
La distribución de heterocromatina constitutiva en Telmatobius montanus (Fig. 2) muestra, al igual que en las especies de Alsodes, brazos completos C positivos. Una modificación del N F por mecanismos semejantes en especies de Alsodes y Telmatobius avala una relación intergenérica. Las posibles relaciones intergenéricas de Alsodes y Eupsophus aún no han sido determinadas mediante bandeo. Sin embargo, la relación telmatobínica de estas especies puede sugerirse si se comparan los cariotipos con altos números cromosómicos y presencia de pares telocéntricos de Eupsophus vertebralis, E. ro-seus y Alsodes sp.

64 D Í A Z y V E L O S O
C A U D I V E R B E H A c a u d i v e f b e r a
SI.... .... t a * » « T E L M A T O B I U S m o n t a n u s
A L S O 0 E S n o d o s u s
4 4i í . i * y M #l" »<* *•* ft» HI »* A L S O D E S t u m u l l u o s u s
* # # t * * A t S O O E S S » .
r7/^. 2. Distr ibución de Heterocromat ina const i tut iva en especies de te lmatobi inae .
CRITERIOS MOLECULARES
Los resultados de nuestro estudio comparativo de LDH de cristalinos pueden ser un aporte taxonómico ya que cada especie estudiada presenta un patrón característico (Fig. 3), dando al sistema un alto grado de resolución. Otro aspecto importante resulta de considerar las similitudes al nivel supraespecífico; dos especies de Pleurodema tienen densitogramas muy similares, sólo diferenfiables si se considera la expresión cuantitativa de cada isoenzi-ma. Dos especies del Genero Batrachyla aparecen diferentes en cuanto a este fenotipo, en concordancia con la divergencia cariológica del género. Dos especies de Telmatobius tienen patrones muy similares (T. peruvianus no se muestra), en tanto que T. montanus diverge mostrando un fenotipo que tiene similitud al que presentan las especies de Alsodes. Esta situación concuerda con las afinidades cariológicas de T. montanus. El fenotipo de LDH también agrupa naturalmente a las especies de Alsodes reforzando la identidad del género frente a Eupsophus. Bufo variegatus, B. atacamensis (también B.
chilensis y B. spinulosus que no se muestran) tienen entre sí una alta identidad del patrón electroforético. Ambas especies del Género Rhinoderma (R. rufum no se muestra) son mas similares entre sí que a otras categorías. Caudi-verbera presenta también un fenotipo característico no compartido.
Los resultados obtenidos en el enfoque comparativo de Hexoquinasas hepáticas no tienen el mismo valor taxonómico que asignamos al sistema de LDH. Sólo es posible reconocer dos tipos de cromatogramas en las especies estudiadas. Sin embargo, algunas afinidades genéri cas pueden discutirse a este nivel. Las especies de los géneros Alsodes y Telmatobius, con excepción de Alsodes sp. tienen un cromatograma del tipo A.B.D. diferente al encontrado en Eup sophus y Caudwerbera.
Basados en las evidencias eromosómicas y moleculares presentadas, que interpretan la validez de algunas agrupaciones supraespecífi-cas, proponemos la Clasificación que se señala en la tabla II. En esta tabla se indican (*) las especies descritas con posterioridad a 1962:
A N F I B I O S C H I L E N O S
T E L M A T O B I U S p e r u v m n u » T E L M A T O B I U S h « l h
Fig. 3. D e n s i t o g r a m a s de lactato desh idrogenasa de cr is ta l
de izquierda a derecha) .
Telmatobufo australis (17), Eupsophus van-zolinii (15), Batrachyla antartándica (1), Alsodes tumultuosus (26), Telmatobius pefauri (49), T. grandisonae (31), Insuetophrynus acarpicus (2). Se agregan a estas Rhinoderma rufum (19), denominación actual de Heminectes rufus (35), y Alsodes sp.**.
En conjunto, las especies contenidas en esta clasificación tienen algunas características importantes de señalar para un análisis global. A pesar del incremento que puede observarse en el número de especies y géneros respecto a la clasificación de Cei (10), esta fauna puede seguir siendo considerada de tamaño reducido. Los géneros representados, con excepción de Telmatobius, Pleurodema y Bufo, que se distribuyen también en otras regiones faunísticas, están caracterizados por su endemismo y son taxa poco diversificados.
La clasificación propuesta considera tres Subfamilias en Leptodactylidae. Telmatobiinae contiene ocho géneros, destacando la validez de Alsodes como entidad genérica independiente. La Subfamilia Leptodactyline sólo incluye al género Pleurodema, que presenta afinidades con especies de Eupsophus, lo que establece su primitividad en esta Subfamilia. La Subfamilia
ino (Inscripción de las i soenz imas en el dens i tograma
Rhinodermatinae (22) ha sido propuesta como familia monogenérica Rhinodermatidae, afin al complejo Atelopodido-Bufonoide (30). En trabajos anteriores (45) hemos compartido, sobre la base de los caracteres cromosómicos, el criterio de inclusión de Rhinoderma en Leptodactylidae (45).
CONSIDERACIONES SISTEMÁTICO-EVOLUTIVAS EN
LEPTODACTYLIDAE. GENERALIZACIONES
Las relaciones evolutivas de algunos géneros de Leptodactílidos se resumen en la figura 5. Este esquema se ha construido utilizando principalmente las evidencias cromosómicas y la representación de los patrones de Hexoquinasas. Las diversas etapas en la diferenciación de estas especies se basan en los antecedentes paleontológicos existentes y también en información relativa a los cambios vegetación ales y climáticos del sur de Sud-américa, que han permitido desarrollar un modelo biogeográfico que da cuenta de la dispersión y diferenciación del género Pleurodema (16).
Los géneros Insuetophrynus, Hylorina y Telmatobufo no han sido incorporados en el esquema

66 D I A Z y V E L O S O
TABLA II
Clasif icación de los anfibios ch i lenos
Familia Bufomdae
Género Bufo Laurent i , 1768
Bufo spinulosus W i e g m a n , 1835
Bufo chilensis T s c h u d i , 1838
Bufo rubropunctatus Guichenot , 1854
Bufo atacamensis Cei , 1961
Bufo variegatus Giinther , 1 8 7 0
Familia Leptodactylidae Subfamilia Telmatobiinae Género caudiverbera Laurent i , 1768
Caudiverbera caudiverbera L in naeus , 1758
Género Telmatobufo Schmidt , 1952
Telmatobufo bullocki Schmidt , 1952
Telmatobufo australis Formas , 1 9 7 2 *
Género Eusophus Fitzinger, 1843
Eupsophus roseus Dumeri l y B ibron , 1841
Eupsophus coppingen Gunther , 1881
Eupsophus vertebralis G r a n d i s o n , 1961 Eupsophus vanzolinü D o n o s o - B a r r o s , 1 9 7 4 *
Género Batrachyla Bel l , 1843 Batrachyla leptopus Bell , 1843
Batrachyla taeniata Girard, 1854
Batrachyla antartándica Barrio, 1 9 6 7 *
Género Hylorina Bell , 1843
Hylorina sylvática Bell , 1843
Género Alsodes Bell , 1843
Alsodes nodosus Dumer i l y Bibron, 1841
Alsodes montícola Bel l , 1843 Alsodes tumultuosus V e l o s o y c o l . , 1 9 7 7 *
Alsodes sp. ** V e l o s o y c o l . , 1 9 7 7 *
Género Telmatobius W i e g m a n , 1835
Telmatobius peruvianus W i e g m a n , 1835
Telmatobius marmoralus Dumer i l y Bibron, 1841
Telmatobius montanus Lataste , 1897
Telmatobius laevis Phi l ippi , 1902
Telmatobius halli N o b l e , 1938
Telmatobius grandísonae Lynch, 1 9 7 5 * Telmatobius pefauri V e l o s o y T r u e b , 1 9 7 6 *
Género Insuetophrynus Barrio, 1970
Insuetophrynus acarpicus Barrio, 1 9 7 0 * Subfamilia Leptodactylinae Género Pleurodema T s c h u d i , 1838
Pleurodema thaul Garnot y L e s s o n , 1826
Pleurodema marmorata Dumer i l y B ibron , 1841
Pleurodema bufonina Bell , 1 8 4 3
Subfamilia Rhinodermatinae Género Rhinoderma Dumeri l y B ibron , 1841
Rhinoderma darwini Dumer i l y B ibron , 1841
Rhinoderma rufum Phi l ippi , 1 9 0 2 *
' E s p e c i e s descritas con posterioridad a 1962 .
" E s t a especie , rec ientemente colectada ( N a h u e l b u t a ) , y cuya descripción formal está en e tapas de re
dacción, se agrega a la lista por considerarla de interés en el anál i s i s .

A N F I B I O S C H I L E N O S 67
por carecer de información relacionada con sus respectivos sistemas de Hexoquinasas y la diferenciación longitudinal de sus cromosomas. Insue-tophrynus acarpicus, a pesar de su condición fir-misternia, presenta desarrollo de caracteres sexuales secundarios en los machos y una estructura del oído medio (atrofia), semejante a las especies de Alsodes. Una posible relación intergenérica entre Alsodes e Insuetophrynus también se ve avalada por la distribución geográfica de este último, restringida al bosque de Nothofagus y por sus cromosomas, cuyo número y morfología no difieren de Alsodes (2n = 26). La condición fir-misternia de Insuetophrynus acarpicus es una excepción en Telmatobiinae, cuyas restantes especies son todas arcíferas y con los epicoracoides superpuestos. La fusión de los epicoracoides sería una condición derivada para el caso particular de esta especie.
Las relaciones intergenéricas de Telmatobufo en Telmatobiinae han sido sugeridas en relación con las semejanzas del cariotipo, con Caudiver-bera y Telmatobius (18), (30), (52). Sin embargo, hemos señalado que las especies de estos dos últimos géneros difieren tanto por sus patrones de Hexoquinasas como por la modalidad de distribución de heterocromatina constitutiva. En ausencia de estos antecedentes para Telmatobufo, si bien su adscripción a Telmatobiinae no se discute, una posible estrecha relación con Caudiver-bera y o Telmatobius requiere de mayores antecedentes.
El análisis de conjunto de los cariotipos sugiere que la posesión de un complejo cromosómico común de 2n = 26 es indicativo de la "raigambre telmatobínica" de estas especies (4). Esta afirmación no considera la heterocromatina constitutiva como parámetro citogenético. En especies de los géneros de Telmatobinos, con la excepción posiblemente de Eupsophus y Caudiverbera, la distribución de la heterocromatina constitutiva muestra la existencia de un mecanismo de adición de material C positivo responsable del crecimiento de nuevos brazos cromosómicos y consiguiente modificación del NF.
Las diferencias cromosómicas de C. caudiverbera aportan un antecedente más en relación con su aislamiento genérico en Telmatobiinae (Fig. 2), como también es evidente al analizar su osteología (36), sus relaciones inmunológicas (12) y su comportamiento agresivo (51). La inclusión de este género monofilético en Telmatobiinae resta
uniformidad a esta Subfamilia y sólo es posible considerarla si se acepta la aparición de una línea filética 2n = 26 en una etapa muy primitiva de la diferenciación del grupo y posiblemente en correspondencia con una primera radiación adapta-tiva. Por los antecedentes fósiles que se disponen, esta primera radiación adaptativa ya se habría completado en el Terciario Inferior (Oligoceno). En esta primititiva radiación, correspondiente a un tronco leptodactílido ancestral, también estarían representadas especies del género Eupsophus y posiblemente Batrachyla. A diferencia de Caudiverbera, estos dos últimos géneros son diversificados y lo mismo ocurre con los géneros andinos Telmatobius y Alsodes. Para estos últimos no hay registro fósil, si bien Neopro-coela (38) ha sido considerado afín a Telmatobius (30).
En atención a los números cromosómicos representados, la diversificación de los géneros analizados se habría producido de manera secuen-cial. El proceso de reducción cromosómica mostraría una progresión a partir de especies con alto número cromosómico a números diploides 2n =26 y 2n = 2 2 (Figs. 4 y 5).
En cuanto a la distribución del patrón de Hexoquinasas en Vertebrados, se ha señalado (41) que el patrón C.B.D. está presente en diversas familias de anuros, comprendiendo las más antiguas y también en Leptodactylidae (part.). Esto determinaría el reconocimiento en las especies actuales de estos grupos, de un carácter ancestral en la filogenia del Orden. El patrón A.B.D. de las especies de algunos géneros de Leptodactylidae y de las especies del género Bufo, representaría una condición derivada.
Los cuatro grupos mayores considerados en este trabajo: Leptodactylinae, Telmatobiinae, Rhino-dermatinae y Bufo, comparten la ocurrencia del patrón ABD. El patrón de Hexoquinasas en Pleu-rodema, constituye un antecedente más de la
estrecha relación de este género primitivo de Leptodactylinae (24) con el grupo telmatobino con patrón ABD, como ha sido señalado con anterioridad (16), (30). La historia de la diferenciación y dispersión de Pleurodema puede ser trazada a partir del Oligoceno. El antecedente que permite suponer la existencia de Pleurodema en el Terciario Inferior es la existencia de fósiles de Eupsophus, telmatobino que osteológicamente se relaciona con Pleurodema (16), (30).

D Í A Z y V E L O S O
WOLUMtM 0« ILUCtOM IBU
4. C r o m a t o g r a m a s de H e x o q u i n a s a s hepát icas . Las flechas muestran el comienzo de la gradiente de e luc ión .
5 Re lac iones intergenéricas en Leptodacty l idae .
y los n ú m e r o s 1 al 4 indican cambios cromosómicos a
n = 26 , 2n = 30, 2n = 22 y 2n = 28 respect ivamente .
El único género de Telmatobiinae en que ambos fenotipos de Hexoquinasas están presentes es Alsodes, (Alsodes sp. con fenotipo CBD) lo que posiblemente señala que la transición del patrón C B D a ABD en Leptodactylidae puede haber tenido lugar en Alsodes.
El patrón ABD también está presente en Rhinoderma y Bufo. El significado de que en estos géneros concurra un patrón similar de Hexoquinasas podría reforzar la relación de Rhinoderma con el complejo Atelopódido-Bufonoide. Además, la concurrencia simultánea de este carácter tanto en Leptodactylidae como en Bufonidae estaría en concordancia con la proposición, generalmente aceptada, que los Bufonidos constituyen una rama colateral originada a partir de un grupo lepto-dactiloideo primitivo.
SUMMARY
Since Cei's (1962) Batracios de Chile, no other attempt to propose a taxonomic arrangement of Chilean amphibian species has bee done. During last fifteen years, new species has been described and new biological information concerning these taxa become available.

ANFIBIOS CHILENOS 6 9
The aim of this work is to give a review of the taxonomic status of Chilean amphibians. Chromosomal and molecular evidence are proposed for a first time as valid criteria to pointed out inter-generic and intrageneric relationships of these particular fauna.
Diploid number, chromosome morphology and structural features of karyotypes, such as secondary constrictions and distribution of constitutive heterochromatin (C bands) of certain species, were the chromosomal parameters considered. LDH and Hexokinase isoenzyme patterns were the molecular evidences.
Chromosome and L D H , show variability at the specific-level. Most species has its own karyotype features and L D H isoenzyme pattern, giving these systems a high degre of taxonomic resolution. Hexokinase isoenzyme pattern is less variable. Only two types different chromatograms are represented. In Abodes genus (Telmatobiinae) both chromatograms are present.
Thirty three amphibian species are recognized. Bufonidae is represented by genus Bufo with five species. Three suprageneric arrangements of Leptodactylidae, give account of remainig species.
They are Leptodactylinae, genus Pleurodema with three species, Telmatobiinae, represented by eight genera, Alsodes, Eupsophus, Batrachyla, Telmatobufo, Hylorina; Telmatobius, Insueto-phrynus and Caudiverbera, with a total of twenty five species and Rhinodermatinae, genus Rhino-derma with two species.
REFERENCIAS
1. BARRIO, A., Physis 27(74):101, 1967. 2. BARRIO, A. Physis 28:95, 1970.
3. BARRIO, A., RINALDI DECHIERI, P. Physis 30:309, 1970.
4. BARRIO, A. RINALDI DE CHIERI, P. Physis 39(81):673,
1971. 5. BLAIR, W.F. en: Evolution of the Genus Bufo. W.F.
Blair Ed. p. 91-101. Univ. of Texas, Press, 1972. 6. BOGART, J.P., Cytogenetics 9.369, 1970. 7 BOGART, J.P., en: Evolution of the Genus Bufo. W.F.
Blair Ed. p. 171-195. Univ. of Texas Pess, 1972. 8. BOGART, J.P., en: Evolutionary Biology of the Anurans.
James Vial Ed. p. 337-349. Univ. of Missouri Press, 1973.
9. BRUM-ZORRILLA, N., SAEZ, F.A., Experiencia 24:969,
1968. 10. CEI, J.M., Batracios de Chile, Ediciones de la Univer
sidad de Chile. Santiago, 1962.
11. CEI, J.M., ERSPAMER, V. Actas III Congreso Latinoamericano de Zoología. Santiago, Chile, 1965.
12. CEI, J .M., Acta Zoológica Lilloana 27:181, 1970. 13. CRACRAFT, J.,Syst. Zool. 23:71, 1974. 14. DAVIS, B.J., Annals N. York Acad. Sci. 121 Art. 2:404,
1964. 15. DONOSOBARROS, R , Bol. Soc. Biol. Concepción 48:
217, 1974. 16. DUELLMAN, W . E , VELOSO, A., Ocass. Papers of the
Mus. Nat. Hist. The Univ. of Kansas, 64:1, 1977. 17. FORMAS,! .R. ,J .o fHerpeto logy ,6( l ) : l , 1972. 18. FORMAS, J.R., ESPINOZA, N.D. , Herpetológica, 37:
(4):429, 1975. 19. FORMAS, R., PUGIN, E., JORQUERA, E., Physis, 34(89):
147, 1975. 20. FORMAS, J R . , Experientia, 32:1000, 1976. 21. GALLARDO, J.M., Neotrópica, 76:73, 1970. 22. GRIFFITH, I., Proc. Zool. Soc., 732:457, 1959. 23. HENNING, W . , Elementos de una Sistemática Filo-
genética, EUDEBA, 1968.
24. HEYER, W . R . , Smithsonian Constributions to Zoology N° 199, 1975.
25. ITURRA, P., VELOSO, A. Res XVIII Reunión Anual Soc. Biol. Chile. 71 R25 , 1975.
26. ITURRA, P., VELOSO, A., Res. Ill Congreso Latinoameri
cano de Genética, p. 39 (Abstract), 1977. 27. KURAMOTO, M., Bull. Inknoka Univ. 27:133, 1971. 28. LEVAN, A., FREDGA, K., SANDBERG, A.A., Hereditas
52:201, 1964. 29. LYNCH, J.D. , Herpetológica, 24(3):25S, 1968. 30. LYNCH, J.D. , Univ. of Kansas Publ. Mus. Nat. Hist.,
53:1, 1971. 31. LYNCH, J .D. , Bull. South California Acad. Sci.,
7 4 ( 3 ) : 1 6 0 , 1975.
32. MARTIN, R.F., en: Evolution of the Genus Bufo. W . F . Blair Ed. p. 37-70, 1972.
33. MATTHEY, R., Experientia, 7:50, 1945.
34. MORESCALCHI, A., en: Cytotaxonomy and Vertebrate Evolution. A.B. Chiarelli and E. Capanna Eds. p. 233-347, 1973.
35. PHILIPPI, R A . , Suplemento a los Batraquios Chilenos descritos en la Historia Física y Política de Chile de don Claudio Gay, Stgo. Chile, 1902.
36. REIG, O., Actas y Trab. I Congreso Sudamericano de Zoología, 4:113, 1959.
37. SAEZ, F.A., VENEGAS, W . , Chromosome Information
Service N° 13, 1972. 38. SCHAEFFER, B., Bull. Amer. Nat. Hist., 93:47, 1949. 39. STOCK, A., Hsu, T.C., Chromosoma, 43:211, 1973.
2(5:69, 1973.
40. TIHEN, J., Amere. Midi. Nat., <57:(1):157, 1962. 41. URETA, T., en: Isozymes III Developmental Biology
Academic Press, p. 575-601, 1975.
42. URETA, T., RADOJKOVIC, J., UIAZ, N., SLEBE, J.C., LOZANC
C , Arch of Biochem. and Biophys., 7«6(2):235-247, 1978.
43. VELOSO, A., GALLEGUILLOS, R., DÍAZ, N., Res. Ill Jornadas
Argentinas de Zoología, p. 24-25 (Abstract), Mendoza, 1972.
44. VELOSO, A., DIAZ, N., GALLEGUILLOS, R. Anales Mus.
Hist. Nat. Valpso., 6:49, 1973.

70 D I A Z y V E L O S O
4 5 . VELOSO, A . , DÍAZ, GALLEGUILLOS, R., A n a l e s M u s . His t .
N a t . V a l p s o . , 6 : 5 7 , 1 9 7 3 .
4 6 . VELOSO, A . , GALLEGUILLOS, R., DIAZ, M . , Caryo log ia ,
2 6 . 6 9 , 1 9 7 3 .
4 7 . VELOSO, A . , GALLEGUILLOS, R. DÍAZ, N . , Ana le s M u s .
His t . N a t . , V a l p s o . , 7 : 2 9 7 , 1 9 7 4 .
4 8 . VELOSO, A. , ITURRA, P. , Res . X V I I R e u n i ó n A n u a l Soc.
Biol . Chi l e . 1 1 4 R 7 3 , 1 9 7 4 .
4 9 . VELOSO, A . , T R U E B , L . , O c c a s . Papers of the M u s .
of N a t . H i s t . K a n s a s , 6 2 : 1 , 1 9 7 6 .
5 0 . VELOSO, A . , Procc. S e m . y III C o n g r e s o Lat inamer ica-
no de Genét i ca , M o n t e v i d e o , 1 8 8 , 1 9 7 7 .
5 1 . VELOSO, A . , Herpeto lóg ica , 3 3 : 4 3 4 , 1 9 7 7 .
5 2 . VENEGAS, W . , Bol . Soc. Biol . C o n c e p c i ó n , 4 9 . 7 1 , 1 9 7 5 .
C o n posterioridad a la entrega de este manuscr i to , F o r m a s ha descrito Eupsophus migueli, nueva espec ie de M e h u i n -
Vald iv ia . ( F o r m a s , J . R . Studies o n Neotropica l F a u n a and Env ironment 13:1, 1 9 7 8 ) . Entre los a n i m a l e s uti
l i zados para estudiar L D H de cr i s ta l inos se inc luyó Eupsophus de M e h u í n , por lo q u e el d e n s i l o g r a m a que en la
F igura 3 se atr ibuye a E. roseus corresponde a E. migueli.