relaciÓn amonio/nitrato en la producciÓn de … · es el principal proveedor de albahaca verde,...
TRANSCRIPT

Agricultura Técnica en México Vol. 35 Núm.1 1 de enero-31 de marzo 2009 p. 5-11
RELACIÓN AMONIO/NITRATO EN LA PRODUCCIÓN DE HIERBAS AROMÁTICAS EN HIDROPONÍA*
AMMONIUM/NITRATE RATIO IN THE PRODUCTION OF AROMATIC HERBS IN HYDROPONICSJosé Luis González García1, María de las Nieves Rodríguez Mendoza1§, Prometeo Sánchez García1 y Elda Araceli Gaytán Acuña2
1Posgrado en Edafología-Nutrición Vegetal, Colegio de Postgraduados. ([email protected]), ([email protected]). 2Posgrado en Fruticultura-Recursos Genéticos y Productividad. Colegio de Postgraduados, Campus. Montecillo, km 36.5 carretera México-Texcoco. Montecillo, Estado de México. C. P. 56230. ([email protected]). §Autora para correspondencia: [email protected].
* Recibido: Enero, 2008 Aceptado: Febrero, 2009
RESUMEN
La producción de hierbas aromáticas puede ser una alternativa rentable bajo condiciones de invernadero. El objetivo de esta investigación fue evaluar el efecto de diferentes relaciones NH4
+ /NO3- en la solución nutritiva sobre el crecimiento y
producción de tres hierbas aromáticas: cebollín (Allium schoenoprasum), albahaca (Ocimum basilicum L.) y eneldo (Anethum graveolens L.). El experimento se realizó en el Colegio de Postgraduados Campus Montecillo, Estado de México, México, durante los meses de enero-mayo de 2005 bajo condiciones de invernadero. Las hierbas fueron cultivadas en un sistema hidropónico con riego por goteo, el sustrato utilizado fue una mezcla de tezontle y fibra de coco (60:40), la solución nutritiva universal Steiner se modificó para obtener cuatro diferentes relaciones de NH4
+/NO3-
(0/100, 20/80, 40/60 y 100/0) con las que fueron regadas las hierbas. Las variables determinadas fueron: altura de planta, área foliar y biomasa total. La relación 0/100, incrementó significativamente el área foliar y la producción de biomasa total en cebollín. En albahaca, la mayor altura, área foliar y biomasa total se obtuvieron con la relación 20/80. En eneldo no se observaron diferencias por efecto de las relaciones NH4
+ / NO3- estudiadas.
Palabras clave: Allium schoenoprasum, Anethum graveolens L., Ocimum basilicum L., relación NH4
+/NO3-.
ABSTRACT
The production of aromatic herbs could be a profitable alternative for greenhouse production. The objective of this research was to determine the effects of different NH4
+ /NO3- ratios on the growth and production of three
aromatic herbs: chives (Allium schoenoprasum), basil (Ocimum basilicum L.) and dill (Anethum graveolens L.). The experiment was conducted in the Colegio de Postgraduados at Montecillo, Estado de Mexico, Mexico, from January to May 2005, under greenhouse conditions. The herbs were cultivated in a hydroponic system with drip irrigation. A mixture of volcanic rock, locally called “tezontle” and coconut fiber (60:40) was used as substrate; the universal Steiner’s nutritive solution was modified to obtain four different NH4
+/NO3- ratios:
0/100, 20/80, 40/60 and 100/0. The studied parameters were: plant height, leaf area and total biomass. The 0/100-ratio significantly increased leaf area and total biomass production of chives. The greater height, leaf area and total biomass production of basil were observed with the 20/80 ratio. For dill, no significant differences were observed for the effects of the NH4
+
/NO3- ratios studied.
Key words: All ium schoenoprasum , Anethum graveolens L., Ocimum basilicum L., NH4
+/NO3- ratio.

6 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
José Luis González García et al.
INTRODUCCIÓN
La producc ión de h ie rbas f inas (cu l ina r ias y aromáticas), pueden ser una alternativa de producción bajo condiciones de invernadero, especialmente en sistemas hidropónicos. Cabe mencionar que México es el principal proveedor de albahaca verde, cilantro (Coriandrum sat ivum ) y perej i l (Petrosel inum sativum) a E.E. U.U. (Minero, 2004).
El nitrógeno es el cuarto elemento más abundante que se encuentra en el tejido vegetal después del carbono, oxígeno e hidrogeno, además éste es parte importante de un gran número de los constituyentes de las plantas, proteínas y clorofila, entre otros. Las plantas pueden aprovechar el nitrógeno en forma de NO3
- o NH4+, por lo que en hidroponía es posible
utilizar nitrato y amonio en las soluciones nutritivas. Se ha argumentando que en cualquiera de las dos formas es benéfico o de igual forma puede causar desbalances nutrimentales en la solución nutritiva. Se ha demostrado que un adecuado balance entre el amonio y el nitrato es benéfico para el crecimiento de las plantas pero sólo bajo ciertas circunstancias y este efecto benéfico varia entre cultivos (Mengel y Kirkby, 1987).
En muchos cultivos se ha observado que la combinación de NO3
- con bajas cantidades de NH4+ produce un
mayor crecimiento; sin embargo, la proporción óptima probablemente difiere entre las distintas especies y podría cambiar con la edad de la planta (Haynes, 1986). Por otro lado, Mengel y Kirkby (1987) reportaron que muchas especies vegetales crecen mejor cuando el nitrógeno se suministra en forma de NO3
- comparado con NH4+.
El nitrógeno en forma NO3- es preferentemente
absorbido por la mayoría de las plantas vasculares, mientras que la forma NH4
+ resulta tóxica para muchas de ellas, incluso en bajas concentraciones cuando ésta es la única fuente de nitrógeno o en combinación con N-NO3
- (Salsac et al., 1987).
El objetivo de la presente investigación fue determinar el efecto de cuatro relaciones NH4
+/NO3- en soluciones
nutritivas, en la producción de tres hierbas aromáticas (cebollín, albahaca y eneldo).
MATERIALES Y MÉTODOS
La investigación se realizó durante los meses de enero a mayo de 2005, en un invernadero tipo túnel, con cubierta de polietileno UVII-720, y estructura de acero galvanizado, con ventilación lateral, presentándose una temperatura máxima de 38 ºC y 9 ºC la mínima, con una intensidad luminosa de 653.43 mmol m-2 s-1, localizado en el Colegio de Postgraduados, Campus Montecillo, el cual se ubica a 19º 29’ latitud oeste y 98º 53’ latitud norte a 2 250 m, en Montecillo, Estado de México. Para la germinación de las semillas de los tres cultivos se emplearon charolas de plástico de 200 cavidades (27.94 cm de ancho por 54.61 cm de largo). El trasplante se efectuó a las seis semanas después de la emergencia, cuando las plántulas tenían una altura de 5-7 cm, en charolas de fibra de vidrio con una superficie de 1 m2 (1 m de largo x 1m de ancho) y 20 cm de altura; el sustrato utilizado fue una mezcla de 60% tezontle (granulometría de 1-3 mm) y 40% de fibra de coco. Las plántulas de las tres especies se colocaron a una distancia entre hileras de 15 cm y una separación entre plantas de 15 cm, en un arreglo de tresbolillo, obteniéndose un total de 45 plantas por charola. Se utilizó un sistema de riego por goteo, constituido por cuatro recipientes de 100 L cada uno (en los que se prepararon las cuatro diferentes soluciones nutritivas correspondientes a cada uno de los tratamientos), tubería de conducción de 16 mm de diámetro y estacas tipo gotero de 2 L h-1 regulados por un distribuidor de cuatro salidas con un gasto de 8 L h-1. El diseño experimental utilizado fue un factorial 4 X 3 distribuido como parcelas divididas; la parcela grande correspondió a las relaciones de NH4
+/NO3- y las
parcelas chicas estuvieron constituidas por las tres especies; de esta combinación resultaron 12 tratamientos, cada uno de los tratamientos se constituyó por 45 plantas, cada planta se tomó como una unidad experimental, obteniéndose un total de 540 unidades experimentales.
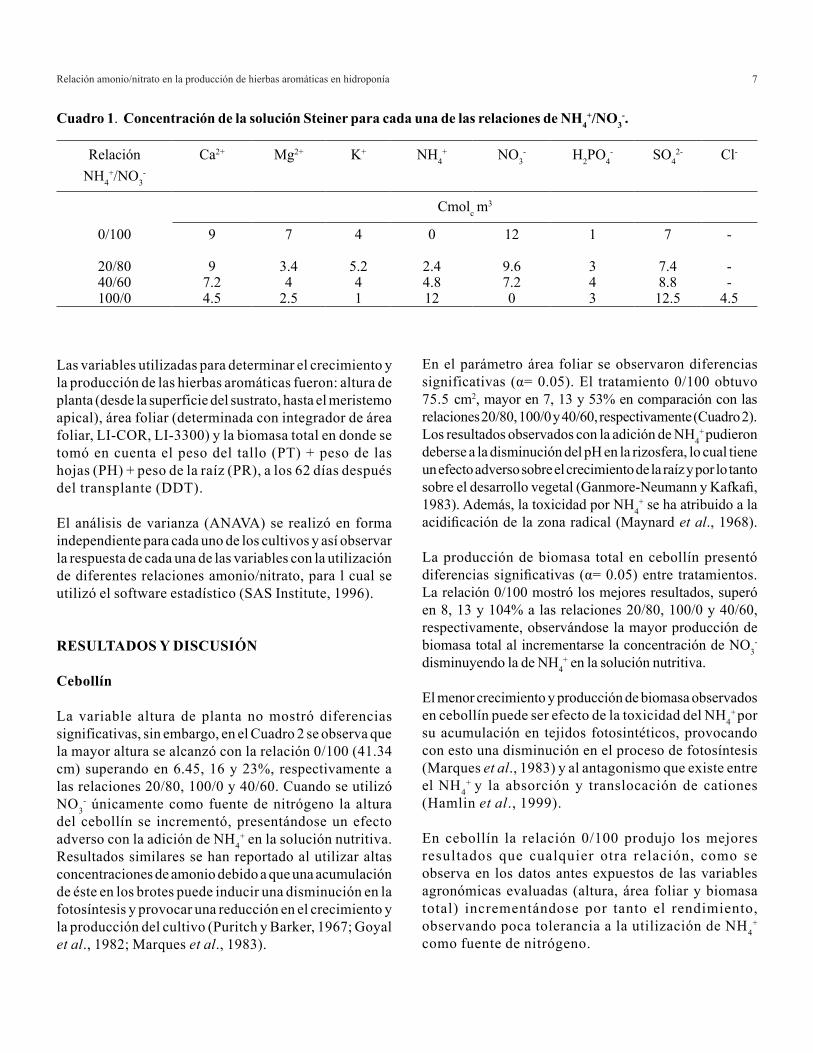
La solución nutritiva base utilizada en el experimento fue la solución universal Steiner (Steiner, 1961) modificada. Las soluciones nutritivas se prepararon en tanques de 100 L de capacidad, en donde se agregaron las cantidades de fertilizantes requeridas de acuerdo al tratamiento, se mezcló y ajustó el pH en el rango de 5.5-6.0. La solución fue renovada cada ocho días para evitar deficiencias nutrimentales. El Cuadro 1 muestra, en general, los balances de cada nutrimento en relación con las concentraciones de NH4
+ y NO3-.
Relación amonio/nitrato en la producción de hierbas aromáticas en hidroponía 7
Las variables utilizadas para determinar el crecimiento y la producción de las hierbas aromáticas fueron: altura de planta (desde la superficie del sustrato, hasta el meristemo apical), área foliar (determinada con integrador de área foliar, LI-COR, LI-3300) y la biomasa total en donde se tomó en cuenta el peso del tallo (PT) + peso de las hojas (PH) + peso de la raíz (PR), a los 62 días después del transplante (DDT).
El análisis de varianza (ANAVA) se realizó en forma independiente para cada uno de los cultivos y así observar la respuesta de cada una de las variables con la utilización de diferentes relaciones amonio/nitrato, para l cual se utilizó el software estadístico (SAS Institute, 1996).
RESULTADOS Y DISCUSIÓN
Cebollín
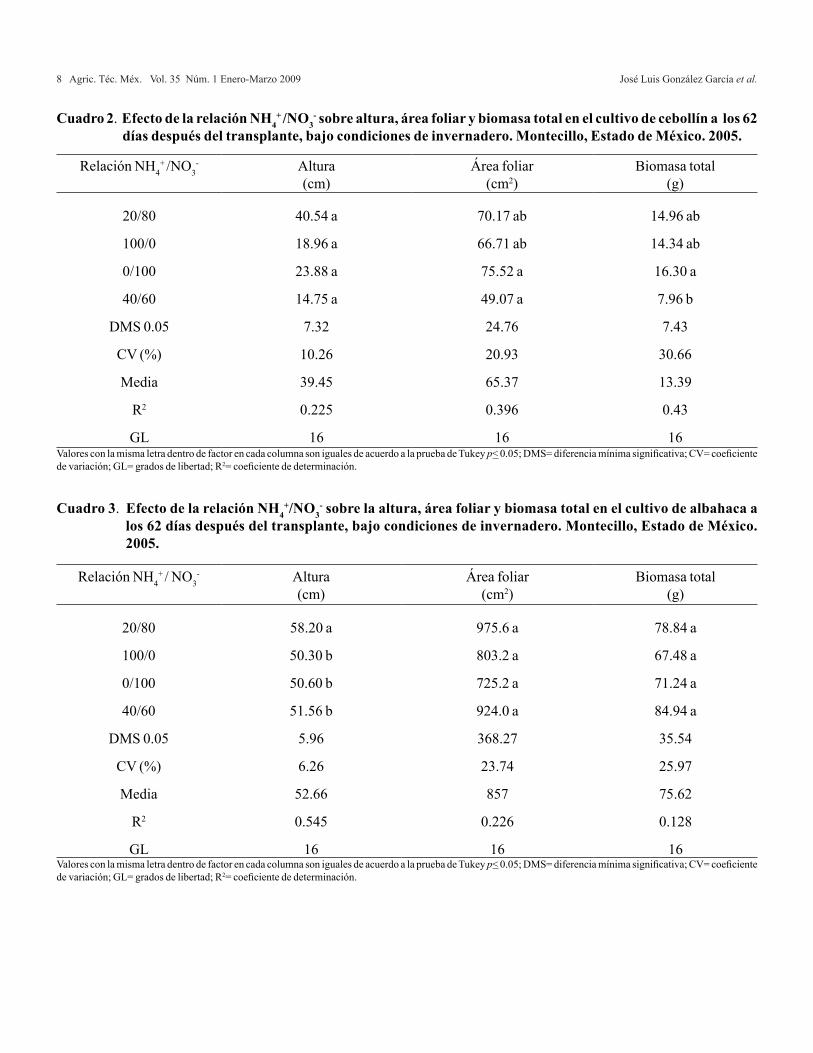
La variable altura de planta no mostró diferencias significativas, sin embargo, en el Cuadro 2 se observa que la mayor altura se alcanzó con la relación 0/100 (41.34 cm) superando en 6.45, 16 y 23%, respectivamente a las relaciones 20/80, 100/0 y 40/60. Cuando se utilizó NO3
- únicamente como fuente de nitrógeno la altura del cebollín se incrementó, presentándose un efecto adverso con la adición de NH4
+ en la solución nutritiva. Resultados similares se han reportado al utilizar altas concentraciones de amonio debido a que una acumulación de éste en los brotes puede inducir una disminución en la fotosíntesis y provocar una reducción en el crecimiento y la producción del cultivo (Puritch y Barker, 1967; Goyal et al., 1982; Marques et al., 1983).
En el parámetro área foliar se observaron diferencias significativas (α= 0.05). El tratamiento 0/100 obtuvo 75.5 cm2, mayor en 7, 13 y 53% en comparación con las relaciones 20/80, 100/0 y 40/60, respectivamente (Cuadro 2). Los resultados observados con la adición de NH4
+ pudieron deberse a la disminución del pH en la rizosfera, lo cual tiene un efecto adverso sobre el crecimiento de la raíz y por lo tanto sobre el desarrollo vegetal (Ganmore-Neumann y Kafkafi, 1983). Además, la toxicidad por NH4
+ se ha atribuido a la acidificación de la zona radical (Maynard et al., 1968).
La producción de biomasa total en cebollín presentó diferencias significativas (α= 0.05) entre tratamientos. La relación 0/100 mostró los mejores resultados, superó en 8, 13 y 104% a las relaciones 20/80, 100/0 y 40/60, respectivamente, observándose la mayor producción de biomasa total al incrementarse la concentración de NO3
-
disminuyendo la de NH4+ en la solución nutritiva.
El menor crecimiento y producción de biomasa observados en cebollín puede ser efecto de la toxicidad del NH4
+ por su acumulación en tejidos fotosintéticos, provocando con esto una disminución en el proceso de fotosíntesis (Marques et al., 1983) y al antagonismo que existe entre el NH4
+ y la absorción y translocación de cationes (Hamlin et al., 1999).
En cebollín la relación 0/100 produjo los mejores resultados que cualquier otra relación, como se observa en los datos antes expuestos de las variables agronómicas evaluadas (altura, área foliar y biomasa total) incrementándose por tanto el rendimiento, observando poca tolerancia a la utilización de NH4
+
como fuente de nitrógeno.
RelaciónNH4
+/NO3-
Ca2+ Mg2+ K+ NH4+ NO3
- H2PO4- SO4
2- Cl-
Cmolc m3
0/100 9 7 4 0 12 1 7 -
20/80 9 3.4 5.2 2.4 9.6 3 7.4 -40/60 7.2 4 4 4.8 7.2 4 8.8 -100/0 4.5 2.5 1 12 0 3 12.5 4.5
Cuadro 1. Concentración de la solución Steiner para cada una de las relaciones de NH4+/NO3
-.
8 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
José Luis González García et al.
Relación NH4+ / NO3
- Altura(cm)
Área foliar(cm2)
Biomasa total(g)
20/80 58.20 a 975.6 a 78.84 a
100/0 50.30 b 803.2 a 67.48 a
0/100 50.60 b 725.2 a 71.24 a
40/60 51.56 b 924.0 a 84.94 a
DMS 0.05 5.96 368.27 35.54
CV (%) 6.26 23.74 25.97
Media 52.66 857 75.62
R2 0.545 0.226 0.128
GL 16 16 16
Relación NH4+ /NO3
- Altura(cm)
Área foliar(cm2)
Biomasa total(g)
20/80 40.54 a 70.17 ab 14.96 ab
100/0 18.96 a 66.71 ab 14.34 ab
0/100 23.88 a 75.52 a 16.30 a
40/60 14.75 a 49.07 a 7.96 b
DMS 0.05 7.32 24.76 7.43
CV (%) 10.26 20.93 30.66
Media 39.45 65.37 13.39
R2 0.225 0.396 0.43
GL 16 16 16
Cuadro 2. Efecto de la relación NH4+ /NO3
- sobre altura, área foliar y biomasa total en el cultivo de cebollín a los 62 días después del transplante, bajo condiciones de invernadero. Montecillo, Estado de México. 2005.
Valores con la misma letra dentro de factor en cada columna son iguales de acuerdo a la prueba de Tukey p< 0.05; DMS= diferencia mínima significativa; CV= coeficiente de variación; GL= grados de libertad; R2= coeficiente de determinación.
Cuadro 3. Efecto de la relación NH4+/NO3
- sobre la altura, área foliar y biomasa total en el cultivo de albahaca a los 62 días después del transplante, bajo condiciones de invernadero. Montecillo, Estado de México. 2005.
Valores con la misma letra dentro de factor en cada columna son iguales de acuerdo a la prueba de Tukey p< 0.05; DMS= diferencia mínima significativa; CV= coeficiente de variación; GL= grados de libertad; R2= coeficiente de determinación.

Relación amonio/nitrato en la producción de hierbas aromáticas en hidroponía 9
Relación NH4+ / NO3
- Altura(cm)
Área foliar(cm2)
Biomasa total(g)
20/80 58.20 a 975.6 a 78.84 a
100/0 50.30 b 803.2 a 67.48 a
0/100 50.60 b 725.2 a 71.24 a
40/60 51.56 b 924.0 a 84.94 a
DMS 0.05 5.96 368.27 35.54
CV (%) 6.26 23.74 25.97
Media 52.66 857 75.62
R2 0.545 0.226 0.128
GL 16 16 16
Albahaca
Los resultados observados en altura de planta fueron significativamente diferentes (α=0.05). El tratamiento 20/80 (58.2 cm) superó los tratamientos 40/60, 0/100 y 100/0 en 12, 15 y 14%, respectivamente (Cuadro 3); así mismo, se observó una mayor respuesta de la planta al ser nutrida con ambas fuentes de N (NH4
+ y NO3- ) a diferencia
de los valores obtenidos en plantas tratadas con soluciones nutritivas que únicamente contenían alguna de la fuente individual de nitrógeno; esto concuerda con lo reportado con Zornoza et al. (1988) para el cultivo de pimiento, en donde la utilización de una relación 80/20 (NO3
- / NH4+)
presentó un incremento de 10% en la altura de planta en comparación con plantas tratadas con la solución nutritiva que contenía NO3
- como fuente única de N. Otros autores coinciden en que el crecimiento es mayor en plantas nutridas con ambas formas de nitrógeno de manera conjunta en comparación con la utilización de NH4
+ o NO3
- de forma independiente (Greidanus et al., 1972; Goyal et al., 1982; Xu et al.,1992).
Los resultados observados por el efecto de las relaciones de amonio/nitrato no presentaron diferencias significativas en el área foliar producida en el cultivo de albahaca; sin embargo, los valores más altos se obtuvieron al utilizar la relación 20/80 con un incremento de 5, 21 y 34% con respecto a las relaciones 40/60, 100/0 y 0/100 (Cuadro 3). Los más altos valores de área foliar se observaron con la utilización de ambas fuentes de nitrógeno en comparación con el uso individual de NH4
+ o NO3- . De la misma forma, Kyunghwan
y Yongbeom, (2004) reportaron un incremento en la producción de biomasa en albahaca al utilizar relaciones de amonio:nitrato 30:70 y 15:85. Estos resultados coinciden con lo reportado por Miyoung y Byoungryong (2001) quienes obtuvieron resultados similares con la utilización de una relación (NH4
+/NO3-) 25/75 en el cultivo de petunia.
De igual manera a los efectos producidos con la utilización de diferentes relaciones NH4
+ /NO3- en los resultados
de área foliar, la producción de biomasa total no se vio modificada por los tratamientos; sin embargo, al igual que en las otras variables la presencia de ambas formas de nitrógeno en la solución nutritiva incrementó la biomasa total en comparación con aquellos obtenidos en plantas tratadas con NH4
+ o NO3- de forma separada, obteniéndose
la mayor producción de biomasa total con la utilización de la relación 40/60, en contraste con las relaciones 20/80, 0/100 y 100/0 (Cuadro 3). Muñoz (1993) reportó una producción
de biomasa total en albahaca de 10,000 a 15,000 kg ha-1, la cual fue superada en esta investigación, ya que a los 62 DDT la producción fue de 35,478 kg ha-1 al utilizar la relación 20/80. Esta relación, aunque no alcanzó la mayor producción de biomasa total, presentó las mejores características físicas para su consumo, en comparación con las plantas tratadas con la relación 40/60 las cuales presentaron un amarillamiento en las hojas, probablemente provocado por toxicidad de amonio.
En general, el cultivo de albahaca respondió mejor a la nutrición con la relación 20/80 obteniéndose los mejores resultados en las variables agronómicas evaluadas (altura, área foliar y biomasa total) que al utilizar NH4
+ y NO3- de
forma separada (0/100 y 100/0); estos resultados coinciden con lo reportado por Cao y Tibbitts (1993) que al utilizar ambas fuentes de nitrógeno en la solución nutritiva con pequeñas proporciones de NH4
+ (8 a 20%), promovieron el crecimiento y desarrollo de tubérculos en el cultivo de papa, en comparación a la utilización de NH4
+ o NO3- de forma
separada. Sandoval et al. (1992) reportaron en el cultivo de trigo (Triticum aestivum L.), que se puede obtener una mayor producción de grano y de materia seca al utilizar NH4
+ en porcentajes menores o iguales a 50% de la cantidad de nitrógeno aplicado que cuando la planta es suministrada exclusivamente con NO3
-.
Eneldo
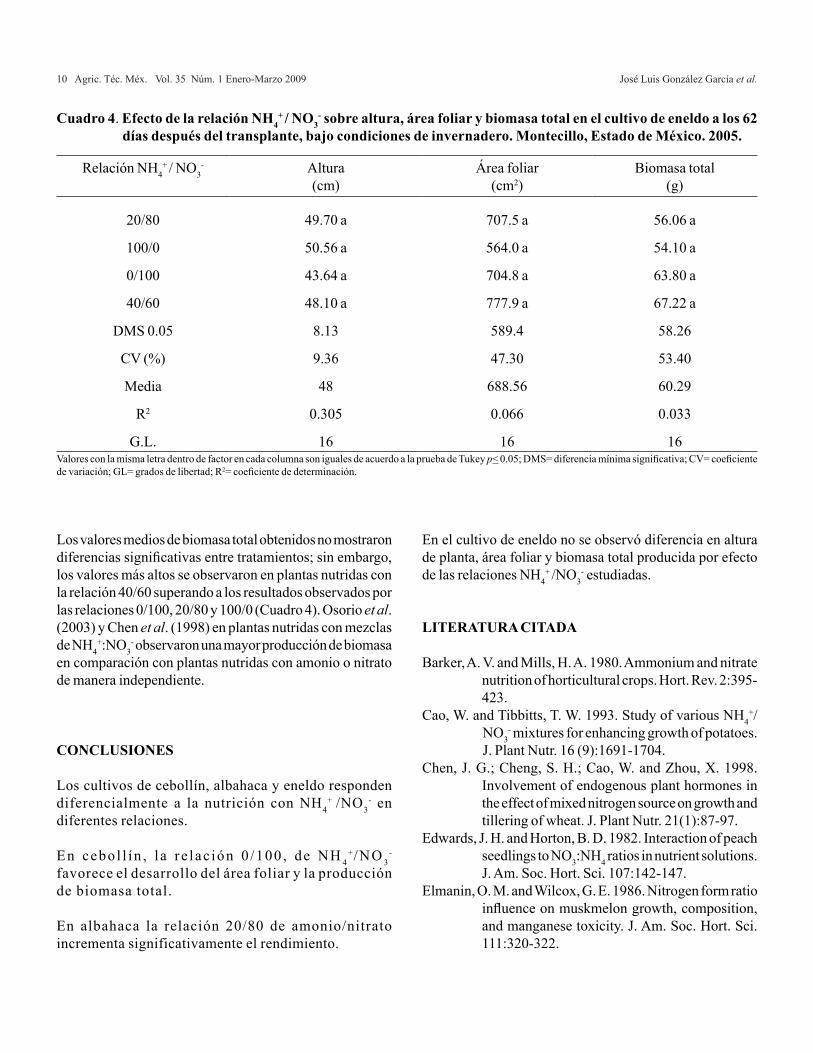
La altura de las plantas tratadas con las diferentes relaciones amonio/nitrato no presentaron diferencias significativas; no obstante, la mayor altura de planta se observó con la relación 100/0 en comparación con las relaciones en las cuales hubo presencia de NO3
- en cualquier proporción (20/80, 40/60 y 0/100) (Cuadro 4). En cuanto a el área foliar en este cultivo tampoco se observó diferencia significativa entre tratamientos; sin embargo, se incrementó cuando se utilizó conjuntamente NH4
+ y NO3- en
la solución nutritiva, en comparación con la utilización de NH4+
o NO3- de manera independiente (0/100 y 100/0). Resultados
similares fueron reportados por Kraus et al. (2002) en rudbeckia (Rudbeckia hirta), quienes al suministrar diferentes relaciones de NH4
+/NO3- (75/25, 50/50, 25/75) observaron mayor área foliar
en comparación a la obtenida únicamente con NO3- como fuente
de nitrógeno. Otros autores han observado en varios cultivos que cuando el amonio y el nitrato son utilizados de manera conjunta, el crecimiento y la producción mejoran en comparación cuando se adicionan éstos de manera separada (Barker y Mills, 1980; Edwards y Horton, 1982; Elmanin y Wilcox, 1986; Errebhi y Wilcox, 1990).
10 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
José Luis González García et al.
Los valores medios de biomasa total obtenidos no mostraron diferencias significativas entre tratamientos; sin embargo, los valores más altos se observaron en plantas nutridas con la relación 40/60 superando a los resultados observados por las relaciones 0/100, 20/80 y 100/0 (Cuadro 4). Osorio et al. (2003) y Chen et al. (1998) en plantas nutridas con mezclas de NH4
+:NO3- observaron una mayor producción de biomasa
en comparación con plantas nutridas con amonio o nitrato de manera independiente.
CONCLUSIONES
Los cultivos de cebollín, albahaca y eneldo responden diferencialmente a la nutrición con NH4
+ /NO3- en
diferentes relaciones.
En cebo l l ín , l a re lac ión 0 /100 , de NH 4+/NO 3
-
favorece el desarrollo del área foliar y la producción de biomasa total.
En albahaca la relación 20/80 de amonio/nitrato incrementa significativamente el rendimiento.
En el cultivo de eneldo no se observó diferencia en altura de planta, área foliar y biomasa total producida por efecto de las relaciones NH4
+ /NO3- estudiadas.
LITERATURA CITADA
Barker, A. V. and Mills, H. A. 1980. Ammonium and nitrate nutrition of horticultural crops. Hort. Rev. 2:395-423.
Cao, W. and Tibbitts, T. W. 1993. Study of various NH4+/
NO3- mixtures for enhancing growth of potatoes.
J. Plant Nutr. 16 (9):1691-1704.Chen, J. G.; Cheng, S. H.; Cao, W. and Zhou, X. 1998.
Involvement of endogenous plant hormones in the effect of mixed nitrogen source on growth and tillering of wheat. J. Plant Nutr. 21(1):87-97.
Edwards, J. H. and Horton, B. D. 1982. Interaction of peach seedlings to NO3:NH4 ratios in nutrient solutions. J. Am. Soc. Hort. Sci. 107:142-147.
Elmanin, O. M. and Wilcox, G. E. 1986. Nitrogen form ratio influence on muskmelon growth, composition, and manganese toxicity. J. Am. Soc. Hort. Sci. 111:320-322.
Relación NH4+ / NO3
- Altura(cm)
Área foliar(cm2)
Biomasa total(g)
20/80 49.70 a 707.5 a 56.06 a
100/0 50.56 a 564.0 a 54.10 a
0/100 43.64 a 704.8 a 63.80 a
40/60 48.10 a 777.9 a 67.22 a
DMS 0.05 8.13 589.4 58.26
CV (%) 9.36 47.30 53.40
Media 48 688.56 60.29
R2 0.305 0.066 0.033
G.L. 16 16 16
Cuadro 4. Efecto de la relación NH4+ / NO3
- sobre altura, área foliar y biomasa total en el cultivo de eneldo a los 62 días después del transplante, bajo condiciones de invernadero. Montecillo, Estado de México. 2005.
Valores con la misma letra dentro de factor en cada columna son iguales de acuerdo a la prueba de Tukey p< 0.05; DMS= diferencia mínima significativa; CV= coeficiente de variación; GL= grados de libertad; R2= coeficiente de determinación.

Relación amonio/nitrato en la producción de hierbas aromáticas en hidroponía 11
Errebi, M. and Wilcox, G. E. 1990. Plant species response to ammonium-nitrate concentration ratios. J. Plant Nutr. 13(8):1017-1029.
Ganmore-Neumann, R. and Kafkafi, U. 1983. Root temperature and percentage NO3
-/NH4+ effect
on strawberry plants. I Growth, flowering and root development. Agron. J. 75:941-947.
Goyal, S. S.; Huffaker, R. C. and Lorenz, O. A. 1982. Inhibitory effects of ammoniacal nitrogen on growth of radish plants. II. Investigations on the possible causes of ammonium toxicity to radish plants and its reversal by nitrate. J. Am. Soc. Hort. Sci. 107:130-135.
Greidanus, T.; Peterson, L. A.; Schrader, L. E. and Dana, M. N. 1972. Essentiality of ammonium for cranberry nutrition. J. Am. Soc. Hort. Sci. 97:272-277.
Hamlin, R. L.; Mills, H. A. and Randle W., N. 1999. Growth and nutrition of pansy as influenced by N-form ratio and temperature. J. Plant Nutr. 22(10):1637-1650.
Haynes, R. J. 1986. Uptake and assimilation of mineral nitrogen by plants, 303-358. In: Mineral Nitrogen in the Soil Plant System. Haynes, R. J.; Cameron, K. C.; Goh, K. M.; Sherlock, R. R. (eds.). Academic Press, Inc. Florida, USA.
Kyunghwan, Y. and Yongbeom, L. 2004. The effect of NO3
-N and NH4+ -N ratio in the nutrient
solution on growth and quality of sweet basil. Korean J. Hort. Sce & Techn. 22(1):29:36.
Kraus, H. T.; Warren, S. L. and Anderson, C. E. 2002. Nitrogen form affects growth, mineral nutrient content, and root anatomy of cotoneaster and rudbeckia. HortScience 37(1):126-129.
Marques, Y. A.; Oberholzer, M. J. and Erismann K., H. 1983. Effects of different nitrogen sources on photosyntetic carbon metabolism in primary leaves of non-nodulated Phaseolus vulgaris L. Plant Physiol. 71:555-561.
Maynard, N. D.; Barker, A. V. and Lachman, W. H. 1968. Influence of potassium on the utilization of ammonium by tomato plants. Proc. Amer. Soc. Hort. Science 92:537-542.
Mengel, K. and Kirkby, E. A. 1987. Nitrogen, pp. 347-374. In: Principles of plant nutrition. Mengel, K. and Kirkby, E. A. (eds.). 4th edition. International Potash Ins t i tu te . Wor ldblaufenBern / Switzerland. 593p.
Minero, A. A. 2004. Mercados nichos: hierbas finas. Productores de Hortalizas 13(10):24-31.
Miyoung, L. and Byoungryong, J. 2001. Effect of N concentration and NH4
+:NO3- ratio in nutrient
solution on growth and flowering of Petunia hybrida ‘Midnight’ and ‘Romeo’. J. Korean Soc. Hort. Sce. 42:748-751.
Muñoz, F. 1993. Plantas medicinales y aromáticas: estudio, cultivo y procesado. Ed. Mundi Prensa. Madrid España.
Osorio, N. W.; Shuai, X.; Miyasaka, S.; Wang, B.; Shirley, R. L. and Wigmore, W. J. 2003. Nitrogen level and form affect taro growth and nutrition. HortScience 38(1):36-40.
Puritch, G. S. and Barker, A. V. 1967. Structure and function of tomato leaf chloroplasts during ammonium toxicity. Plant Physiol. 42(9):1229-1238.
Salsac, L.; Chaillou, S.; Morot, J. F.; Lesaint, C. and Jolivet, E. 1987. Nitrate and ammonium nutrition in plants. Plant Physiol. Biochem. 25(6):805-812.
Sandoval V. M., G. Alcántar G., J.L. Tirado T. and A. Aguilar S. 1992. Effect of the NH4
+/NO3- ratio on GS and
PEP Case activities and on dry matter production in wheat. J. Plant Nutr. 15(11):2545-2557.
Satatistical Analysis System Institute (SAS Institute) 1996. SAS/ STAT. Guide for personal computers. Version V8.1. Cary, N. C. USA. 1028 p.
Steiner, A. A. 1961. A universal method for preparing nutrient solutions of a certain desired composition. Plant and Soil 15:134-154.
Xu, Q. F.; Tsai, C. L. and Tsai, C. Y. 1992. Interaction of potassium with the form and amount of nitrogen nutrition on growth and nitrogen uptake of maize. J. Plant Nutr. 15(1):23-33.
Zornoza, P.; Caselles, J. and Carpena, O. 1988. Influence of Light and NO3
-:NH4+ ratio on nutrient
uptake by pepper plant in sand culture. Soilless Culture 4:65-74.

Agricultura Técnica en México Vol. 35 Núm.1 1 de enero-31 de marzo 2009 p. 12-22
ESTIMACIÓN DEL RENDIMIENTO DE CEBADA (Hordeum vulgare L.) MALTERA CON EL MÉTODO FAO*
ESTIMATION OF MALTING BARLEY (Hordeum vulgare L.) YIELD BY THE FAO METHODRené Gómez Mercado1§ , Carlos Alberto Ortiz Solorio5, Mauro Zamora Díaz2, Jesús Soria Ruíz3, Antonio Trinidad Santos4 y Aquiles Carballo Carballo5
1INIFAP-Hidalgo, km 3.6 carretera Pachuca - Cd. Sahagún, Centro Comercial El Saucillo, Torre Norte 1er. piso despacho 111, Mineral de la Reforma, Hidalgo, México. Tel. 01 771 7136387. 2Campo Experimental Valle de México, INIFAP, km 18.5 carretera Los Reyes-Lechería C. P. 56230, Chapingo, Texcoco, Estado de México, Tel. 01 595 9542277 Ext. 126, ([email protected]). 3Laboratorio de Geomática, Campo Experimental Toluca, INFIAP, km 4.5, carretera Toluca-Zitacuaro, Zinancantepec C. P. 52176, Estado de México, México. Tel. 01 722 2784339, ([email protected]). 4Posgrado en Edafología, IRENAT. Colegio de Postgraduados, km 36.5 carretera México-Texcoco, Montecillo, Estado de México, C. P. 56230. Tel. 01 595 9511474 y 9520200 Ext. 1241, ([email protected]), ([email protected]). 5Posgrado en Producción de Semillas. IREGEP. Colegio de Postgraduados, km 36.5 carretera México-Texcoco, Montecillo, Estado de México, C. P. 56230. Tel. 01 595 9520200 Ext. 1552, ([email protected]) §Autor para correspondencia: [email protected].
RESUMEN
Al emplear modelos para predecir rendimientos de cultivos, con frecuencia existen limitantes por las variables que utilizan. De ahí, que resulte la necesidad de elegir al que contenga el menor número de variables o bien que éstas puedan estimarse con algún procedimiento conocido y fácil de realizar. La metodología propuesta por el proyecto de zonas agroecológicas de la Organización para la Alimentación y la Agricultura (FAO) presenta esta alternativa, por su sencillez, pero a la vez con fundamentos ecofisiológicos que soportan los procesos que tienen los cultivos para producir biomasa. El uso de ese método permite validar, calibrar y hacer el ajuste, de acuerdo con sus variables, mismas que pueden calcularse en campo, de manera experimental o por un procedimiento de muestreo que evalúe una superficie o región, para corroborar que el método estima el rendimiento de manera aceptable (alta precisión y bajo sesgo). Para el caso de estudio en el cultivo de la cebada, el método no permite captar la variación local del potencial de rendimiento, pero a nivel regional produce una media aceptable, por lo que se sugiere el ampliar el estudio, para explorar la diferencia entre regiones y genotipos. Se propone un factor de ajuste a la
metodología de la FAO, para que la estimación se aproxime a la realidad, tanto por región como por genotipo. Se presenta una relación entre factor de ajuste del rendimiento y la densidad de espigas por hectárea.
Palabras clave: Hordeum vulgare L., índice de área foliar, índice de cosecha, modelos de predicción.
ABSTRACT
When using models to predict crop yields, with frequency there are limitations due to the variables fed into those models. Hence, the need to select the model with either the smallest number of variables or variables that can be estimated by some well known procedures that are also easy to fit. The methodology provided by the Agro-ecological Zones Project, proposed by the Food and Agriculture Organization (FAO) constitutes a good alternative, not only due to its simplicity, but also for the eco-physiological principles that support the processes for biomass production by crops.
* Recibido: Enero, 2008
Aceptado: Marzo, 2009

Estimación del rendimiento de cebada (Hordeum vulgare L.) maltera con el método FAO 13
The use of this method allows for validating, calibrateing and fitting the model, according to the needed variables, these must be calculated in the field, experimentally or by a sampling procedure that evaluates an area or a region to make sure that the method estimates crop yields in an acceptable form (highly accurate and low bias). For the studied case in the barley crop, the method does not allow for the detection of the local variation in yield potential, but at a regional scale the method produces a reasonable average yield, therefore we suggest the increase of the study to explore differences among regions and genotypes. The results indicate the need for an adjustment factor to FAO's methodology, so that the estimated yield approaches reality for both, region and genotype.. A relationship between the fitting factor of crop yield and spikes density per hectare, is presented.
Key words: Hordeum vulgare L., harvest index, leaf area index, prediction models.
INTRODUCCIÓN
El cultivo de la cebada de temporal se siembra principalmente, para la elaboración de malta, y en consecuencia requiere índices de calidad industrial, que estén determinados por las cualidades genéticas, manejo, suelo y cantidad y distribución de la precipitación. Las características genéticas y el manejo agronómico son variables que pueden controlarse, pero algunas propiedades del suelo y lluvia son factores incontrolables, que resultan determinantes para obtener un buen rendimiento y una buena calidad de grano para malta (Gómez et al., 2001).
La diferencia climática y de suelo para producir cultivos, hace más complejo el obtener tecnología y en consecuencia crea modelos de simulación que tengan un mayor ajuste y estimen los rendimientos de acuerdo a la fluctuación que existe entre los diferentes ambientes (Ortiz et al., 2005). Bajo este principio, el
cultivo de la cebada no escapa a esa variación, además de tener un ciclo de cultivo corto y requerimientos hídricos menores que otros cultivos, como maíz, trigo y avena (Castañeda et al., 2004).
De manera general, los modelos de simulación de rendimientos se clasifican en simples y complejos. Los primeros se relacionan con el clima, la biomasa y el índice de área foliar (Tinoco et al., 2003; Gómez et al., 2003); los segundos, involucran estas variables, pero incluyen aspectos genéticos y del suelo (Ovalle et al., 2005).
Para simular y predecir el rendimiento, se han utilizado desde modelos simples empíricos hasta los complejos (Ortiz et al., 2005; Castellano et al., 2006). Sin embargo, cuando se trata simular el rendimiento de cultivos no existe información para cubrir los requerimientos de la mayoría de los modelos, que explican de manera teórica, los fenómenos que influyen en el rendimiento. Los modelos empíricos son más simples, pero no aclaran los procesos que ocurren para obtener rendimientos. La información no disponible se refiere al tipo de cultivar (fenología, fisiología y genética) y cómo interactúa con el clima y los suelos (Barrales et al., 2004).
El propósito de este trabajo fue determinar un modelo que simule adecuadamente el rendimiento de cebada maltera para el estado de Hidalgo, con base en variables de planta y clima, sustentado en el método propuesto por la Organización para la Alimentación y la Agricultura (FAO, 2000).
MATERIALES Y MÉTODOS
La presente investigación se realizó en tres regiones del estado de Hidalgo donde se siembra cebada maltera; en cada región se instaló un experimento que permitiera explorar diferentes ámbitos de producción durante el ciclo primavera-verano 2006 (Cuadro 1).
Región Localidad Latitud Longitud Altitud (m) Temperatura
1 Almoloya 19° 45’ 14.4” 98º 21’ 57” 2 730 12.2 2 Apan 19° 41’ 16.6” 98º 23’ 33” 2 547 14.1
3 Villa de Tezontepec 19° 54’ 16.0” 98º 53’ 4” 2 355 15.4
Cuadro 1. Características de las regiones de estudio del estado de Hidalgo, México.

14 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
René Gómez Mercado et al.
T 5 10 15 20 25 30 35 40 45Pm 5 15 20 20 15 5 0 0 0
Cuadro 2. Fotosíntesis máxima (Pm) kg ha-1 h-1 de CH2O, como función de temperatura (T) media (oC) del ciclo para el cultivo de cebada (FAO, 2000).
Se parte del supuesto que tiene la metodología de la FAO para estimar rendimiento, es decir, se considera que no existen limitaciones en cuanto a la humedad del suelo.
La metodología adoptada para realizar la presente investigación constó de tres etapas:
1) determinación de componentes de rendimiento de cebada en las tres regiones del estado. Para ello, se condujeron tres experimentos (Cuadro 1), con tres genotipos (Esmeralda, Adabella y M-152A) para obtener su biomasa neta (Bn), índice de cosecha (Hi), densidad de espigas por hectárea (DsE) y rendimiento de grano. El diseño experimental fue de bloques al azar, con 10 repeticiones de cada genotipo (30 parcelas experimentales). Cada unidad experimental constó de cuatro surcos a 0.30 m de ancho por 5 m de largo (6 m2).
Se obtuvo la Bn con base en raíces, tallos, hojas y espigas; Hi se calculó con la siguiente relación: Hi= rendimiento de grano/Bn total; el rendimiento de grano se calculó de acuerdo con Gómez et al., 2003; se cuantificó la densidad de espigas por hectárea (DsE); y el índice de área foliar (IAF) se calculó con base a Bn de hojas y el área foliar específica (AFE) para el cultivo de cebada (FAO, 2000), de acuerdo con la siguiente ecuación: IAF= (Bn de hojas)(AFE/10 000). El IAF es una variable determinante en los modelos de predicción de rendimiento que permite la estimar el rendimiento de cultivos (Loke y Soós, 2002).
2) aplicación del método FAO. La información climática se obtuvo directamente de las estaciones agroclimáticas automatizadas que se encuentran en cada localidad de estudio, localizadas de 2 a 5 km de distancia en línea recta donde se establecieron los experimentos de campo. Este método se basa en principios ecofisiológicos que consisten en obtener la cantidad total de biomasa neta (Bn) generada por el cultivo y multiplicarla por el índice de cosecha (Hi), para obtener el rendimiento estimado (Y), esto es: Y= Bn(Hi). Para tener la Bn se calcula la diferencia entre biomasa total (Bg) y las pérdidas por respiración (R), por lo que es necesario calcular la tasa de producción de biomasa neta (bn), con la tasa de producción de biomasa gruesa (bg) y la tasa de respiración (r). Es decir, bn= bg - r.
Se asume que la Bn, durante el ciclo, sigue una curva sigmoidal perfecta y que bn una curva normal, lo que resulta en una tasa promedio diaria de producción de biomasa neta de la mitad de la tasa de biomasa neta máxima (0.5bnm), con lo que se calcula Bn= 0.5bnm(N), donde: N es la duración en días del ciclo del cultivo. Es necesario generar la tasa máxima de producción (bgm), la cual se relaciona con la fotosíntesis máxima (Pm), ésta a su vez depende de la temperatura media, el tipo de cultivo (C3 o C4) y la concentración de CO2 en la atmósfera (Ortiz et al., 2005).
Se calculó bgm con base a la producción de biomasa en días despejados (bc), en días nublados (bo) y la fracción del día en la que el ciclo está nublado (F): bgm= F(bo) + (1 - F)bc, donde: F= (Ac - 0.5Rg)/0.8Ac; Rg= radiación global y Ac= radiación fotosintéticamente activa.
Los valores de Ac, bc y bo se calcularon con modelos que emplean valores de radiación teórica (RA), se consideró que si se tiene una Pm= 20 kg ha-1 h-1 de CH2O, entonces bgm= F(bo) + (1 - F)bc; y si es menor que 20, la fórmula cambia a: bgm= F(bo)((100 - (x/2))/100) + (1 - F)(bc)((100 - x)/100); donde: x= |(Pm - 20)/20|(100).
Por otra parte, la respiración involucra dos procesos: crecimiento y mantenimiento, que se expresa por la ecuación Bn= 0.36bgm/((1/N) + 0.25Ct); donde Ct es el coeficiente de respiración y se obtiene de la ecuación Ct= C30(0.044 + 0.0019T + 0.001T2), con C30= 0.0283 para leguminosas y 0.0108 para no leguminosas, T es la temperatura (oC) media mensual del ciclo; y N es el número de días del ciclo del cultivo. La expresión matemática de Bn es válida para un IAF > 5; cuando el IAF es < 5, se multiplica por un factor de corrección (L), que se estima como: L= 0.3424 + 0.9051(log10IAF).
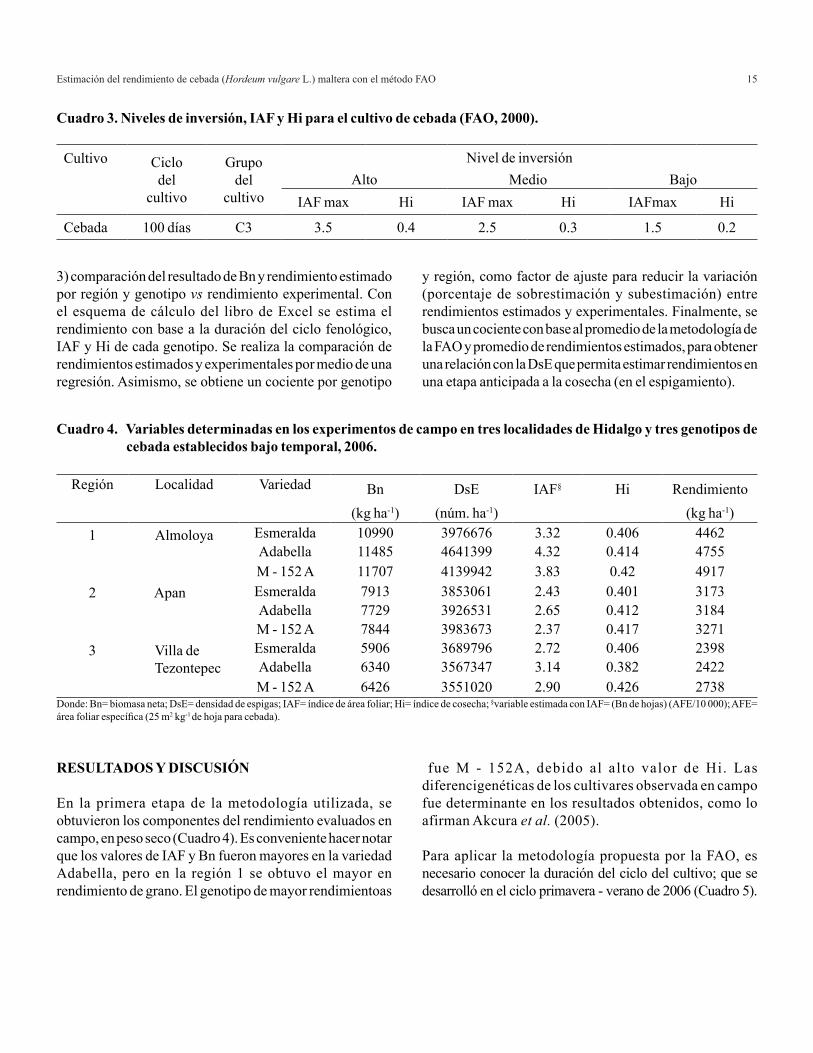
La radiación global (Rg) y la temperatura media se obtuvieron de las estaciones agroclimáticas automatizadas. Para el cálculo de Pm, se utilizó datos determinados por FAO (2000) (Cuadro 2). Los diversos niveles de inversión empleados para la estimación de rendimientos genera diferentes índices de cosecha (Hi) e índices de área foliar máxima (IAF), las cuales se muestran en el Cuadro 3 (FAO, 2000).
Estimación del rendimiento de cebada (Hordeum vulgare L.) maltera con el método FAO 15
3) comparación del resultado de Bn y rendimiento estimado por región y genotipo vs rendimiento experimental. Con el esquema de cálculo del libro de Excel se estima el rendimiento con base a la duración del ciclo fenológico, IAF y Hi de cada genotipo. Se realiza la comparación de rendimientos estimados y experimentales por medio de una regresión. Asimismo, se obtiene un cociente por genotipo
RESULTADOS Y DISCUSIÓN
En la primera etapa de la metodología utilizada, se obtuvieron los componentes del rendimiento evaluados en campo, en peso seco (Cuadro 4). Es conveniente hacer notar que los valores de IAF y Bn fueron mayores en la variedad Adabella, pero en la región 1 se obtuvo el mayor en rendimiento de grano. El genotipo de mayor rendimientoas
y región, como factor de ajuste para reducir la variación (porcentaje de sobrestimación y subestimación) entre rendimientos estimados y experimentales. Finalmente, se busca un cociente con base al promedio de la metodología de la FAO y promedio de rendimientos estimados, para obtener una relación con la DsE que permita estimar rendimientos en una etapa anticipada a la cosecha (en el espigamiento).
fue M - 152A, debido al alto valor de Hi. Las diferencigenéticas de los cultivares observada en campo fue determinante en los resultados obtenidos, como lo afirman Akcura et al. (2005).
Para aplicar la metodología propuesta por la FAO, es necesario conocer la duración del ciclo del cultivo; que se desarrolló en el ciclo primavera - verano de 2006 (Cuadro 5).
Región Localidad Variedad Bn DsE IAF§ Hi Rendimiento(kg ha-1) (núm. ha-1) (kg ha-1)
1 Almoloya Esmeralda 10990 3976676 3.32 0.406 4462Adabella 11485 4641399 4.32 0.414 4755M - 152 A 11707 4139942 3.83 0.42 4917
2 Apan Esmeralda 7913 3853061 2.43 0.401 3173Adabella 7729 3926531 2.65 0.412 3184M - 152 A 7844 3983673 2.37 0.417 3271
3 Villa de Tezontepec
Esmeralda 5906 3689796 2.72 0.406 2398Adabella 6340 3567347 3.14 0.382 2422M - 152 A 6426 3551020 2.90 0.426 2738
Cultivo Ciclodel
cultivo
Grupodel
cultivo
Nivel de inversiónAlto Medio Bajo
IAF max Hi IAF max Hi IAFmax Hi
Cebada 100 días C3 3.5 0.4 2.5 0.3 1.5 0.2
Cuadro 3. Niveles de inversión, IAF y Hi para el cultivo de cebada (FAO, 2000).
Cuadro 4. Variables determinadas en los experimentos de campo en tres localidades de Hidalgo y tres genotipos de cebada establecidos bajo temporal, 2006.
Donde: Bn= biomasa neta; DsE= densidad de espigas; IAF= índice de área foliar; Hi= índice de cosecha; §variable estimada con IAF= (Bn de hojas) (AFE/10 000); AFE= área foliar específica (25 m2 kg-1 de hoja para cebada).
16 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
René Gómez Mercado et al.
Cálculo de la tasa bruta o gruesa (bgm) de producción de biomasa, biomasa neta (Bn) y rendimiento de grano de localidad
Se determinó el valor promedio para el ciclo del cultivo de: la radiación global (Rg); radiación teórica (RA); radiación fotosintéticamente activa (Ac); la producción fotosintética para días despejados (bc) y para días nublados (bo); y la temperatura media. Con base a la temperatura media del ciclo se determinó la fotosíntesis máxima (Pm) kg ha-1 h-1 de CH2O (Cuadros 6, 9 y 12). Se obtuvo la bgm con base a Ac, bc, bo, Rg y Pm de cada localidad en estudio (Cuadros 7, 10 y 13).
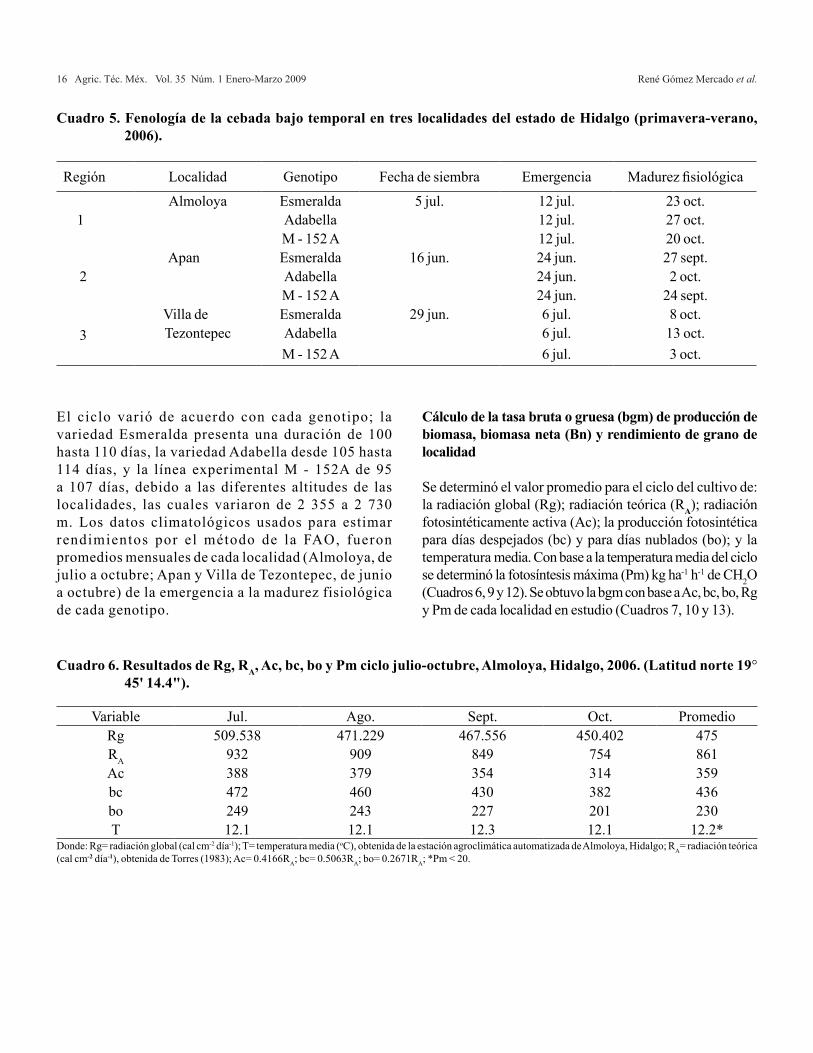
El ciclo varió de acuerdo con cada genotipo; la variedad Esmeralda presenta una duración de 100 hasta 110 días, la variedad Adabella desde 105 hasta 114 días, y la línea experimental M - 152A de 95 a 107 días, debido a las diferentes altitudes de las localidades, las cuales variaron de 2 355 a 2 730 m. Los datos climatológicos usados para estimar rendimientos por el método de la FAO, fueron promedios mensuales de cada localidad (Almoloya, de julio a octubre; Apan y Villa de Tezontepec, de junio a octubre) de la emergencia a la madurez fisiológica de cada genotipo.
Región Localidad Genotipo Fecha de siembra Emergencia Madurez fisiológica
1Almoloya Esmeralda 5 jul. 12 jul. 23 oct.
Adabella 12 jul. 27 oct.M - 152 A 12 jul. 20 oct.
2 Apan Esmeralda 16 jun. 24 jun. 27 sept.
Adabella 24 jun. 2 oct.M - 152 A 24 jun. 24 sept.
3 Villa de Esmeralda 29 jun. 6 jul. 8 oct.
Tezontepec Adabella 6 jul. 13 oct.M - 152 A 6 jul. 3 oct.
Variable Jul. Ago. Sept. Oct. PromedioRg 509.538 471.229 467.556 450.402 475RA 932 909 849 754 861Ac 388 379 354 314 359bc 472 460 430 382 436bo 249 243 227 201 230T 12.1 12.1 12.3 12.1 12.2*
Cuadro 5. Fenología de la cebada bajo temporal en tres localidades del estado de Hidalgo (primavera-verano, 2006).
Cuadro 6. Resultados de Rg, RA, Ac, bc, bo y Pm ciclo julio-octubre, Almoloya, Hidalgo, 2006. (Latitud norte 19° 45' 14.4").
Donde: Rg= radiación global (cal cm-2 día-1); T= temperatura media (oC), obtenida de la estación agroclimática automatizada de Almoloya, Hidalgo; RA= radiación teórica (cal cm-2 día-1), obtenida de Torres (1983); Ac= 0.4166RA; bc= 0.5063RA; bo= 0.2671RA; *Pm < 20.
Estimación del rendimiento de cebada (Hordeum vulgare L.) maltera con el método FAO 17
IAF Hi bgm L Ct Bn Rendimiento(kg ha-1)*
3.5 0.4 307.89 0.83483599 0.00209445 9496.44264 38502.5 0.3 307.89 0.7025755 0.00209445 7991.95058 24301.5 0.2 307.89 0.5017802 0.00209445 5707.85992 1735
Variable Jun. Jul. Ago. Sept. PromedioRg 585.9 532.0 521.9 495.0 534RA 933 931 909 850 906Ac 389 388 379 354 377bc 472 471 460 430 459bo 249 249 243 227 242T 14.1 14.2 14.0 14.1 14.1*
Ac bc bo Rg F x bgm
359 436 230 475 0.422896845 15 307.89
Ac bc bo Rg F x bgm377 459 242 534 0.365998361 5 363.33
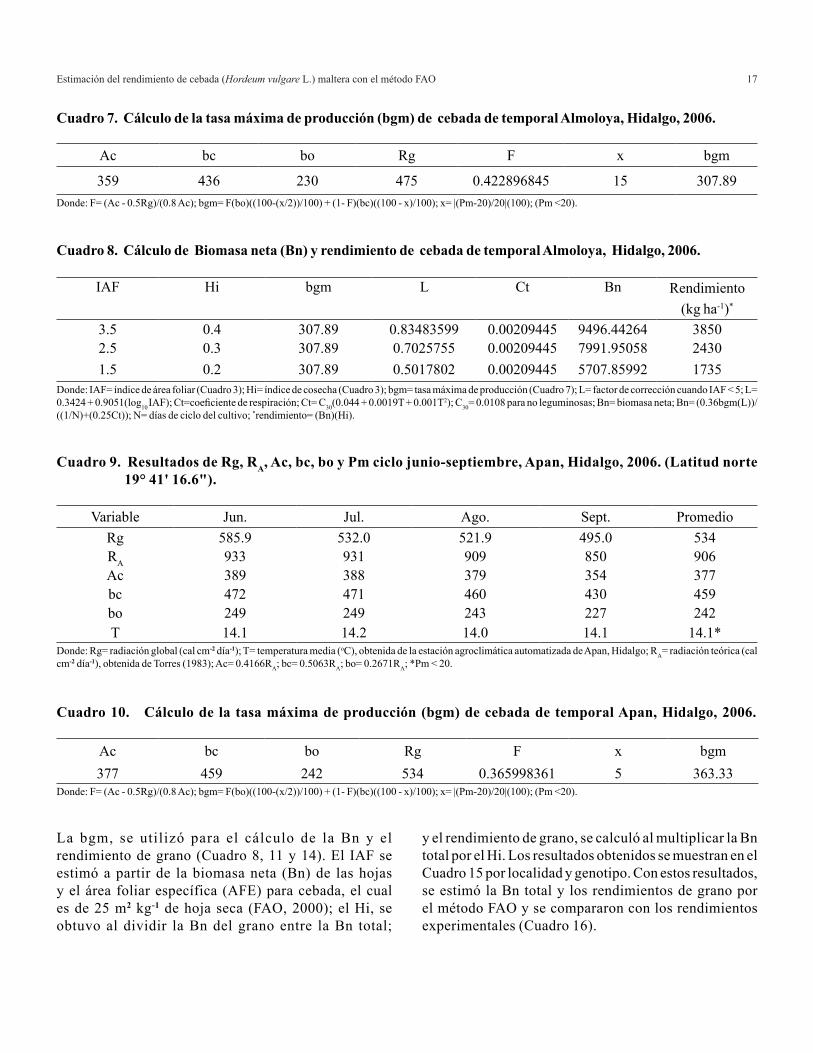
Cuadro 7. Cálculo de la tasa máxima de producción (bgm) de cebada de temporal Almoloya, Hidalgo, 2006.
Donde: F= (Ac - 0.5Rg)/(0.8 Ac); bgm= F(bo)((100-(x/2))/100) + (1- F)(bc)((100 - x)/100); x= |(Pm-20)/20|(100); (Pm <20).
Cuadro 9. Resultados de Rg, RA, Ac, bc, bo y Pm ciclo junio-septiembre, Apan, Hidalgo, 2006. (Latitud norte 19° 41' 16.6").
Donde: Rg= radiación global (cal cm-2 día-1); T= temperatura media (oC), obtenida de la estación agroclimática automatizada de Apan, Hidalgo; RA= radiación teórica (cal cm-2 día-1), obtenida de Torres (1983); Ac= 0.4166RA; bc= 0.5063RA; bo= 0.2671RA; *Pm < 20.
Cuadro 8. Cálculo de Biomasa neta (Bn) y rendimiento de cebada de temporal Almoloya, Hidalgo, 2006.
Donde: IAF= índice de área foliar (Cuadro 3); Hi= índice de cosecha (Cuadro 3); bgm= tasa máxima de producción (Cuadro 7); L= factor de corrección cuando IAF < 5; L= 0.3424 + 0.9051(log10 IAF); Ct=coeficiente de respiración; Ct= C30(0.044 + 0.0019T + 0.001T2); C30= 0.0108 para no leguminosas; Bn= biomasa neta; Bn= (0.36bgm(L))/((1/N)+(0.25Ct)); N= días de ciclo del cultivo; *rendimiento= (Bn)(Hi).
Cuadro 10. Cálculo de la tasa máxima de producción (bgm) de cebada de temporal Apan, Hidalgo, 2006.
Donde: F= (Ac - 0.5Rg)/(0.8 Ac); bgm= F(bo)((100-(x/2))/100) + (1- F)(bc)((100 - x)/100); x= |(Pm-20)/20|(100); (Pm <20).
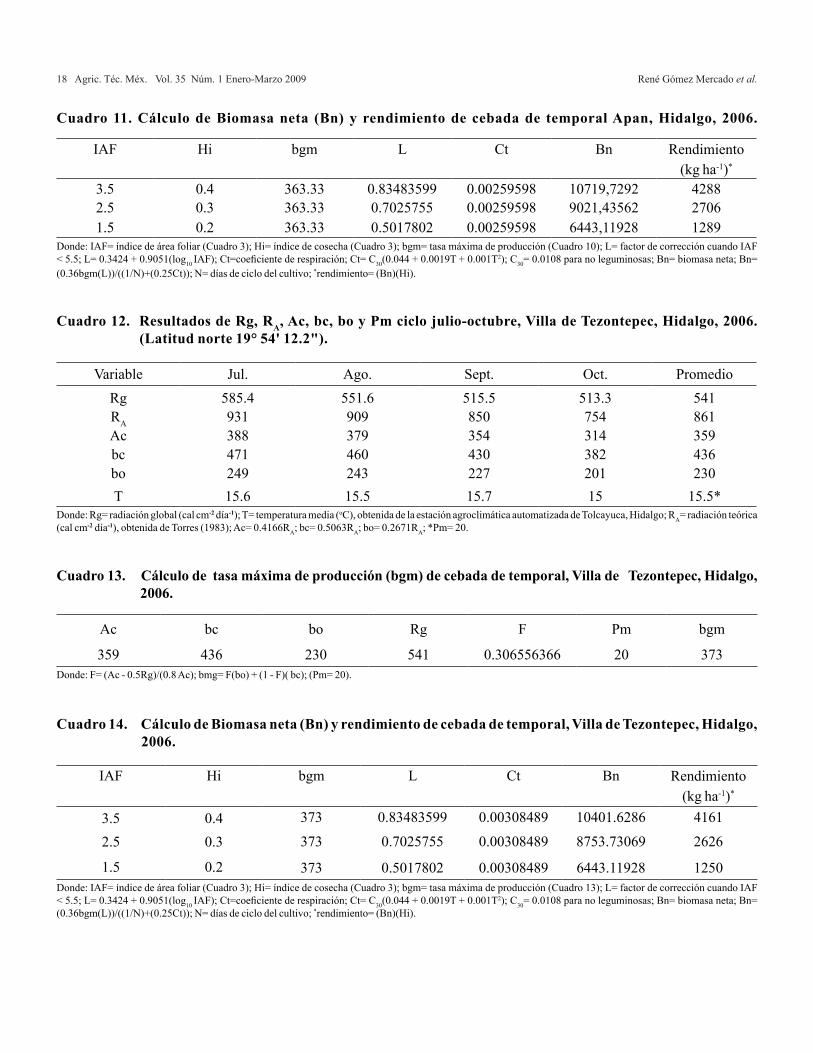
La bgm, se utilizó para el cálculo de la Bn y el rendimiento de grano (Cuadro 8, 11 y 14). El IAF se estimó a partir de la biomasa neta (Bn) de las hojas y el área foliar específica (AFE) para cebada, el cual es de 25 m2 kg-1 de hoja seca (FAO, 2000); el Hi, se obtuvo al dividir la Bn del grano entre la Bn total;
y el rendimiento de grano, se calculó al multiplicar la Bn total por el Hi. Los resultados obtenidos se muestran en el Cuadro 15 por localidad y genotipo. Con estos resultados, se estimó la Bn total y los rendimientos de grano por el método FAO y se compararon con los rendimientos experimentales (Cuadro 16).
18 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
René Gómez Mercado et al.
IAF Hi bgm L Ct Bn Rendimiento(kg ha-1)*
3.5 0.4 373 0.83483599 0.00308489 10401.6286 4161
2.5 0.3 373 0.7025755 0.00308489 8753.73069 2626
1.5 0.2 373 0.5017802 0.00308489 6443.11928 1250
Ac bc bo Rg F Pm bgm
359 436 230 541 0.306556366 20 373
Variable Jul. Ago. Sept. Oct. PromedioRg 585.4 551.6 515.5 513.3 541RA 931 909 850 754 861Ac 388 379 354 314 359bc 471 460 430 382 436bo 249 243 227 201 230T 15.6 15.5 15.7 15 15.5*
IAF Hi bgm L Ct Bn Rendimiento(kg ha-1)*
3.5 0.4 363.33 0.83483599 0.00259598 10719,7292 42882.5 0.3 363.33 0.7025755 0.00259598 9021,43562 27061.5 0.2 363.33 0.5017802 0.00259598 6443,11928 1289
Cuadro 11. Cálculo de Biomasa neta (Bn) y rendimiento de cebada de temporal Apan, Hidalgo, 2006.
Donde: IAF= índice de área foliar (Cuadro 3); Hi= índice de cosecha (Cuadro 3); bgm= tasa máxima de producción (Cuadro 10); L= factor de corrección cuando IAF < 5.5; L= 0.3424 + 0.9051(log10 IAF); Ct=coeficiente de respiración; Ct= C30(0.044 + 0.0019T + 0.001T2); C30= 0.0108 para no leguminosas; Bn= biomasa neta; Bn= (0.36bgm(L))/((1/N)+(0.25Ct)); N= días de ciclo del cultivo; *rendimiento= (Bn)(Hi).
Cuadro 12. Resultados de Rg, RA, Ac, bc, bo y Pm ciclo julio-octubre, Villa de Tezontepec, Hidalgo, 2006. (Latitud norte 19° 54' 12.2").
Donde: Rg= radiación global (cal cm-2 día-1); T= temperatura media (oC), obtenida de la estación agroclimática automatizada de Tolcayuca, Hidalgo; RA= radiación teórica (cal cm-2 día-1), obtenida de Torres (1983); Ac= 0.4166RA; bc= 0.5063RA; bo= 0.2671RA; *Pm= 20.
Cuadro 13. Cálculo de tasa máxima de producción (bgm) de cebada de temporal, Villa de Tezontepec, Hidalgo, 2006.
Donde: F= (Ac - 0.5Rg)/(0.8 Ac); bmg= F(bo) + (1 - F)( bc); (Pm= 20).
Cuadro 14. Cálculo de Biomasa neta (Bn) y rendimiento de cebada de temporal, Villa de Tezontepec, Hidalgo, 2006.
Donde: IAF= índice de área foliar (Cuadro 3); Hi= índice de cosecha (Cuadro 3); bgm= tasa máxima de producción (Cuadro 13); L= factor de corrección cuando IAF < 5.5; L= 0.3424 + 0.9051(log10 IAF); Ct=coeficiente de respiración; Ct= C30(0.044 + 0.0019T + 0.001T2); C30= 0.0108 para no leguminosas; Bn= biomasa neta; Bn= (0.36bgm(L))/((1/N)+(0.25Ct)); N= días de ciclo del cultivo; *rendimiento= (Bn)(Hi).
Estimación del rendimiento de cebada (Hordeum vulgare L.) maltera con el método FAO 19
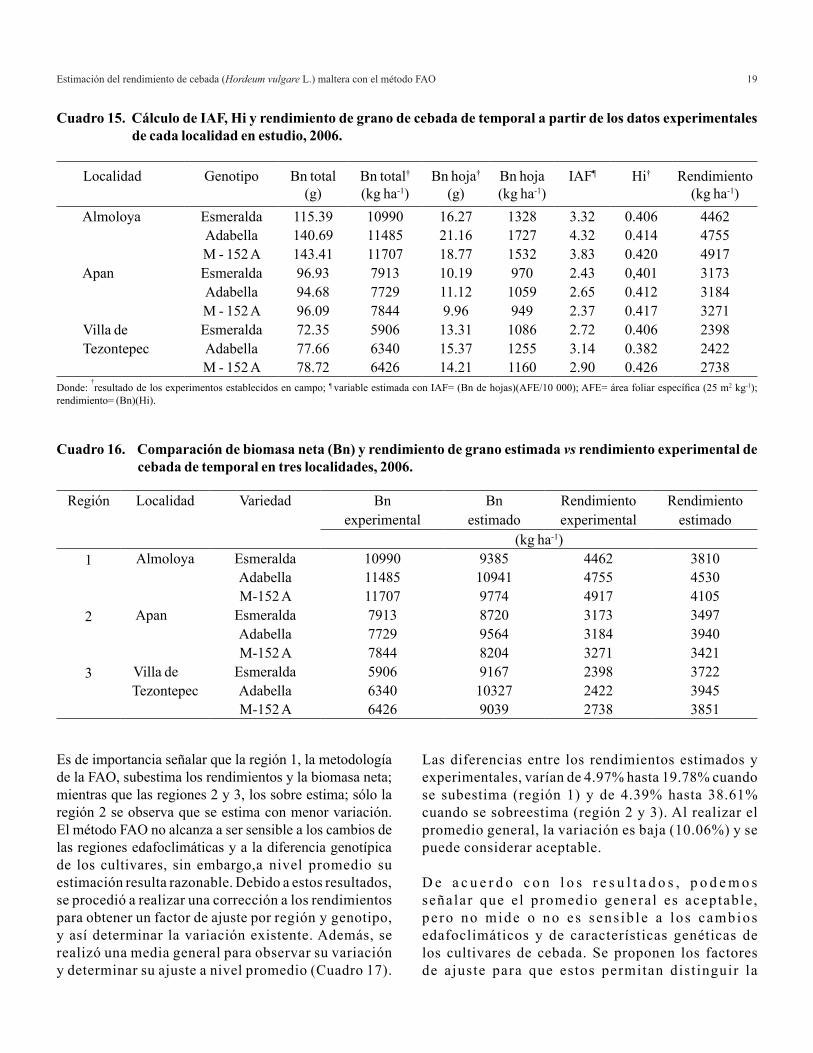
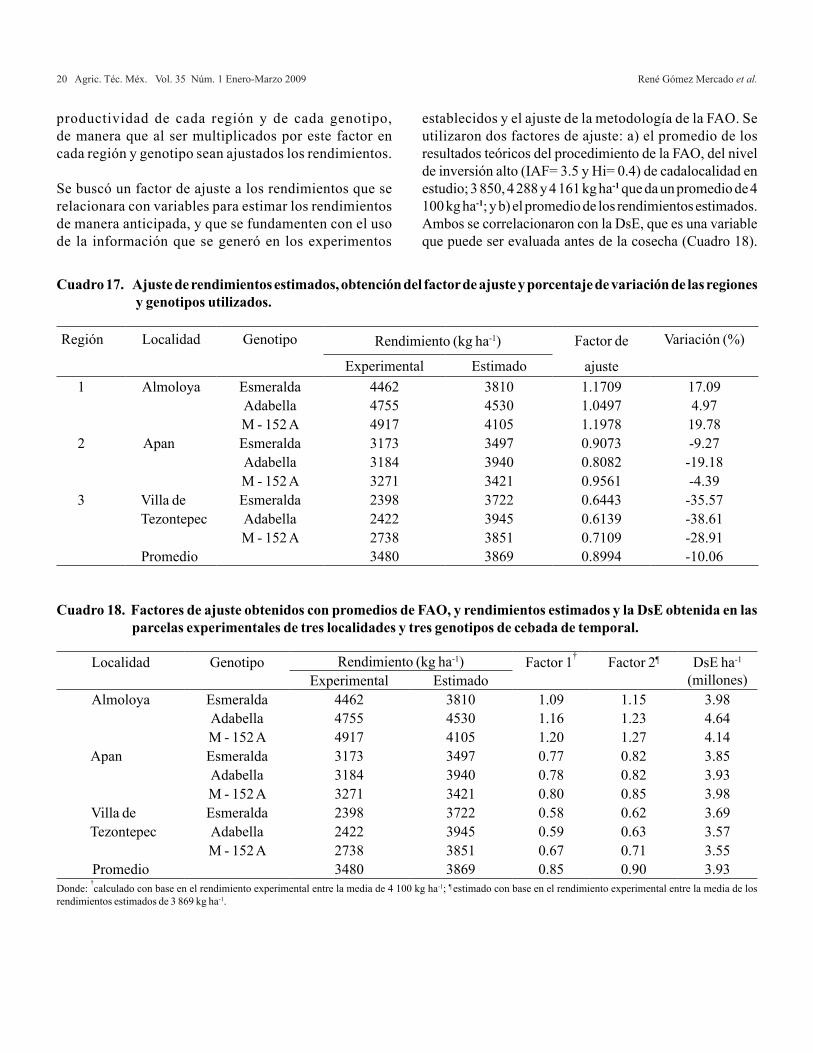
Las diferencias entre los rendimientos estimados y experimentales, varían de 4.97% hasta 19.78% cuando se subestima (región 1) y de 4.39% hasta 38.61% cuando se sobreestima (región 2 y 3). Al realizar el promedio general, la variación es baja (10.06%) y se puede considerar aceptable.
D e a c u e r d o c o n l o s r e s u l t a d o s , p o d e m o s señalar que el promedio general es aceptable , pe ro no mide o no es sens ib le a los cambios edafoclimáticos y de características genéticas de los cultivares de cebada. Se proponen los factores de ajuste para que estos permitan dist inguir la
Región Localidad Variedad Bn Bn Rendimiento Rendimiento experimental estimado experimental estimado
(kg ha-1)1 Almoloya Esmeralda 10990 9385 4462 3810
Adabella 11485 10941 4755 4530M-152 A 11707 9774 4917 4105
2 Apan Esmeralda 7913 8720 3173 3497Adabella 7729 9564 3184 3940M-152 A 7844 8204 3271 3421
3 Villa de Esmeralda 5906 9167 2398 3722Tezontepec Adabella 6340 10327 2422 3945
M-152 A 6426 9039 2738 3851
Localidad Genotipo Bn total(g)
Bn total†
(kg ha-1)Bn hoja†
(g)Bn hoja(kg ha-1)
IAF¶ Hi† Rendimiento(kg ha-1)
Almoloya Esmeralda 115.39 10990 16.27 1328 3.32 0.406 4462Adabella 140.69 11485 21.16 1727 4.32 0.414 4755M - 152 A 143.41 11707 18.77 1532 3.83 0.420 4917
Apan Esmeralda 96.93 7913 10.19 970 2.43 0,401 3173Adabella 94.68 7729 11.12 1059 2.65 0.412 3184M - 152 A 96.09 7844 9.96 949 2.37 0.417 3271
Villa de Esmeralda 72.35 5906 13.31 1086 2.72 0.406 2398 Tezontepec Adabella 77.66 6340 15.37 1255 3.14 0.382 2422
M - 152 A 78.72 6426 14.21 1160 2.90 0.426 2738
Cuadro 15. Cálculo de IAF, Hi y rendimiento de grano de cebada de temporal a partir de los datos experimentales de cada localidad en estudio, 2006.
Donde: †resultado de los experimentos establecidos en campo; ¶ variable estimada con IAF= (Bn de hojas)(AFE/10 000); AFE= área foliar específica (25 m2 kg-1);
rendimiento= (Bn)(Hi).
Cuadro 16. Comparación de biomasa neta (Bn) y rendimiento de grano estimada vs rendimiento experimental de cebada de temporal en tres localidades, 2006.
Es de importancia señalar que la región 1, la metodología de la FAO, subestima los rendimientos y la biomasa neta; mientras que las regiones 2 y 3, los sobre estima; sólo la región 2 se observa que se estima con menor variación. El método FAO no alcanza a ser sensible a los cambios de las regiones edafoclimáticas y a la diferencia genotípica de los cultivares, sin embargo,a nivel promedio su estimación resulta razonable. Debido a estos resultados, se procedió a realizar una corrección a los rendimientos para obtener un factor de ajuste por región y genotipo, y así determinar la variación existente. Además, se realizó una media general para observar su variación y determinar su ajuste a nivel promedio (Cuadro 17).
20 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
René Gómez Mercado et al.
productividad de cada región y de cada genotipo, de manera que al ser multiplicados por este factor en cada región y genotipo sean ajustados los rendimientos.
Se buscó un factor de ajuste a los rendimientos que se relacionara con variables para estimar los rendimientos de manera anticipada, y que se fundamenten con el uso de la información que se generó en los experimentos
establecidos y el ajuste de la metodología de la FAO. Se utilizaron dos factores de ajuste: a) el promedio de los resultados teóricos del procedimiento de la FAO, del nivel de inversión alto (IAF= 3.5 y Hi= 0.4) de cadalocalidad en estudio; 3 850, 4 288 y 4 161 kg ha-1 que da un promedio de 4 100 kg ha-1; y b) el promedio de los rendimientos estimados. Ambos se correlacionaron con la DsE, que es una variable que puede ser evaluada antes de la cosecha (Cuadro 18).
Región Localidad Genotipo Rendimiento (kg ha-1) Factor de
ajuste
Variación (%)
Experimental Estimado 1 Almoloya Esmeralda 4462 3810 1.1709 17.09
Adabella 4755 4530 1.0497 4.97M - 152 A 4917 4105 1.1978 19.78
2 Apan Esmeralda 3173 3497 0.9073 -9.27Adabella 3184 3940 0.8082 -19.18M - 152 A 3271 3421 0.9561 -4.39
3 Villa de Esmeralda 2398 3722 0.6443 -35.57 Tezontepec Adabella 2422 3945 0.6139 -38.61
M - 152 A 2738 3851 0.7109 -28.91 Promedio 3480 3869 0.8994 -10.06
Cuadro 17. Ajuste de rendimientos estimados, obtención del factor de ajuste y porcentaje de variación de las regiones y genotipos utilizados.
Donde: †calculado con base en el rendimiento experimental entre la media de 4 100 kg ha-1; ¶ estimado con base en el rendimiento experimental entre la media de los
rendimientos estimados de 3 869 kg ha-1.
Localidad Genotipo Rendimiento (kg ha-1) Factor 1† Factor 2¶ DsE ha-1
(millones)Experimental EstimadoAlmoloya Esmeralda 4462 3810 1.09 1.15 3.98
Adabella 4755 4530 1.16 1.23 4.64M - 152 A 4917 4105 1.20 1.27 4.14
Apan Esmeralda 3173 3497 0.77 0.82 3.85Adabella 3184 3940 0.78 0.82 3.93M - 152 A 3271 3421 0.80 0.85 3.98
Villa de Esmeralda 2398 3722 0.58 0.62 3.69 Tezontepec Adabella 2422 3945 0.59 0.63 3.57
M - 152 A 2738 3851 0.67 0.71 3.55Promedio 3480 3869 0.85 0.90 3.93
Cuadro 18. Factores de ajuste obtenidos con promedios de FAO, y rendimientos estimados y la DsE obtenida en las parcelas experimentales de tres localidades y tres genotipos de cebada de temporal.
Estimación del rendimiento de cebada (Hordeum vulgare L.) maltera con el método FAO 21
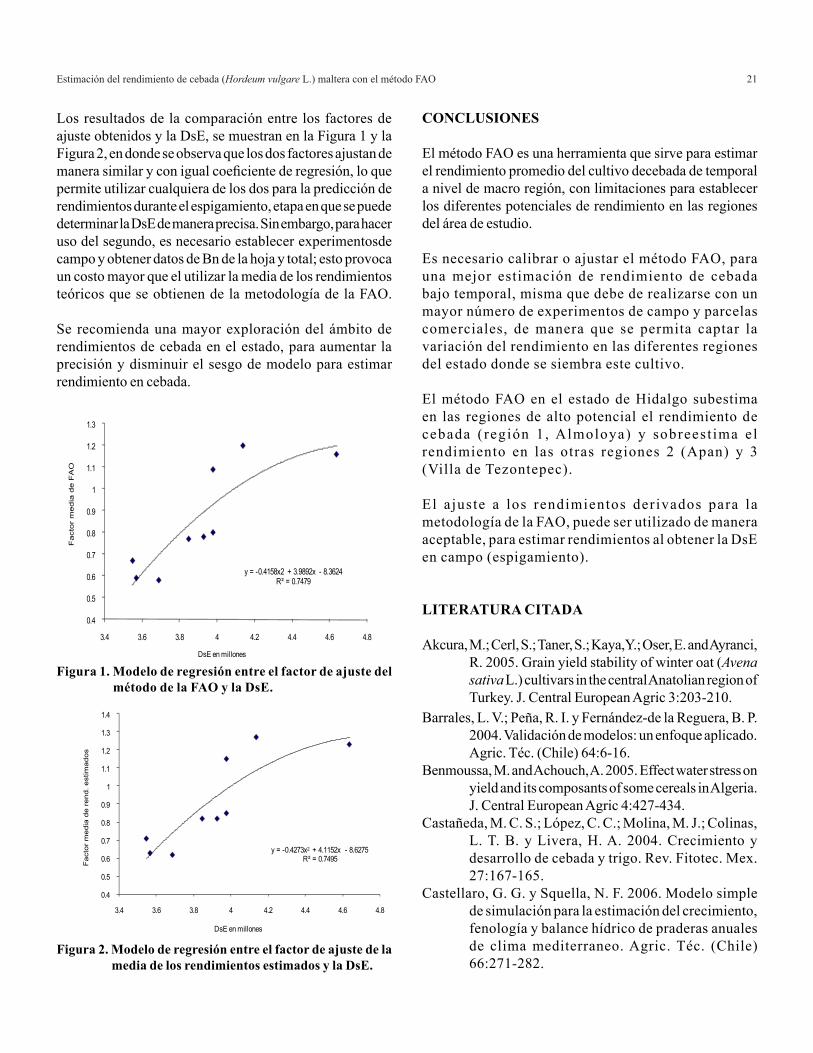
Los resultados de la comparación entre los factores de ajuste obtenidos y la DsE, se muestran en la Figura 1 y la Figura 2, en donde se observa que los dos factores ajustan de manera similar y con igual coeficiente de regresión, lo que permite utilizar cualquiera de los dos para la predicción de rendimientos durante el espigamiento, etapa en que se puede determinar la DsE de manera precisa. Sin embargo, para hacer uso del segundo, es necesario establecer experimentosde campo y obtener datos de Bn de la hoja y total; esto provoca un costo mayor que el utilizar la media de los rendimientos teóricos que se obtienen de la metodología de la FAO.
Se recomienda una mayor exploración del ámbito de rendimientos de cebada en el estado, para aumentar la precisión y disminuir el sesgo de modelo para estimar rendimiento en cebada.
CONCLUSIONES
El método FAO es una herramienta que sirve para estimar el rendimiento promedio del cultivo decebada de temporal a nivel de macro región, con limitaciones para establecer los diferentes potenciales de rendimiento en las regiones del área de estudio.
Es necesario calibrar o ajustar el método FAO, para una mejor estimación de rendimiento de cebada bajo temporal, misma que debe de realizarse con un mayor número de experimentos de campo y parcelas comerciales, de manera que se permita captar la variación del rendimiento en las diferentes regiones del estado donde se siembra este cultivo.
El método FAO en el estado de Hidalgo subestima en las regiones de alto potencial el rendimiento de cebada (región 1, Almoloya) y sobreest ima el rendimiento en las otras regiones 2 (Apan) y 3 (Villa de Tezontepec).
El ajuste a los rendimientos derivados para la metodología de la FAO, puede ser utilizado de manera aceptable, para estimar rendimientos al obtener la DsE en campo (espigamiento).
LITERATURA CITADA
Akcura, M.; Cerl, S.; Taner, S.; Kaya,Y.; Oser, E. and Ayranci, R. 2005. Grain yield stability of winter oat (Avena sativa L.) cultivars in the central Anatolian region of Turkey. J. Central European Agric 3:203-210.
Barrales, L. V.; Peña, R. I. y Fernández-de la Reguera, B. P. 2004. Validación de modelos: un enfoque aplicado. Agric. Téc. (Chile) 64:6-16.
Benmoussa, M. and Achouch, A. 2005. Effect water stress on yield and its composants of some cereals in Algeria. J. Central European Agric 4:427-434.
Castañeda, M. C. S.; López, C. C.; Molina, M. J.; Colinas, L. T. B. y Livera, H. A. 2004. Crecimiento y desarrollo de cebada y trigo. Rev. Fitotec. Mex. 27:167-165.
Castellaro, G. G. y Squella, N. F. 2006. Modelo simple de simulación para la estimación del crecimiento, fenología y balance hídrico de praderas anuales de clima mediterraneo. Agric. Téc. (Chile) 66:271-282.
Figura 1. Modelo de regresión entre el factor de ajuste del método de la FAO y la DsE.
Figura 2. Modelo de regresión entre el factor de ajuste de la media de los rendimientos estimados y la DsE.
y = -0.4158x2 + 3.9892x - 8.3624R² = 0.7479
0.4
0.5
0.6
0.7
0.8
0.9
1
1.1
1.2
1.3
3.4 3.6 3.8 4 4.2 4.4 4.6 4.8
Fa
cto
r m
ed
ia d
e F
AO
y = -0.4273x2 + 4.1152x - 8.6275R² = 0.7495
0.4
0.5
0.6
0.7
0.8
0.9
1
1.1
1.2
1.3
1.4
3.4 3.6 3.8 4 4.2 4.4 4.6 4.8
Fac
tor
med
ia d
e re
nd. e
stim
ados
DsE en millones
y = -0.4273x2 + 4.1152x - 8.6275R² = 0.7495
0.4
0.5
0.6
0.7
0.8
0.9
1
1.1
1.2
1.3
1.4
3.4 3.6 3.8 4 4.2 4.4 4.6 4.8
Fac
tor
med
ia d
e re
nd. e
stim
ados
DsE en millones

22 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
René Gómez Mercado et al.
Gómez, M. R.; Turrent, F. A.; Ortiz, S. C. A. y Peña, O. B. 2001. Productividad en cebada maltera. I. Uso de factoriales 2k en el estudio integrado de factores controlables e incontrolables. Agric. Téc. Méx. 27:84-94.
Gómez, M. R.; Turrent, F. A.; Ortiz, S. C. A. y Peña, O. B. 2001. Productividad en cebada maltera. II. Análisis de las interacciones de seis factores de la producción. Agric. Téc. Méx. 27:95-105.
Gómez, M. R.; Gómez, C. J. L. y Tiscareño, L. M. 2003. Predicción de rendimientos de maíz de riego en el Valle del Mezquital, Hidalgo. In: Simposio Binacional de Modelaje y Sensores Remotos en Agricultura Ed. Alba, A. A., L. Reyes M. y M. Tiscareño L. Aguascalientes, Aguascalientes, México. 3 y 4 de junio de 2003.
Loke, Zs. and Soós, G. 2002. Determination of leaf area and plant cover by using digital image processing. J. Central European Agric 4:343-352.
Ortiz, S. C. A.; Gutiérrez, C. M. C. y Nieves, F. J. 2005. Estimación de rendimientos de maíz con el método FAO en el ejido de Atenco, Estado de México. Rev. Geografía Agríc. 35:57-65.
Ovalle, C. M.; Pozo, L. A. del; Arredondo, S. S. y Cavaría, M. J. 2005. Crecimiento y producción de nuevas leguminosas forrajeras anuales en la zona mediterránea de Chile. I. Comportamiento de las especies en la Cordillera Andina. Agric. Téc. (Chile) 65:35-47.
Theo, C. C.; Benjamin, E. and Okechukwu, N. 2005. Relation between global radiation and food production in a humid tropical climate of West Africa. J. Central European Agric. 2:179-184.
Tinoco, A. C. A.; Tiscareño, L. M. y Baéz, G. A. 2003. Predicción de cosechas de maíz: experiencias generadas en el sur de Veracruz, México. In: Simposio Binacional de Modelaje y Sensores Remotos en Agricultura. Ed. Alba, A. A., L. Reyes M. y M. Tiscareño L. Aguascalientes, Aguascalientes, México 3 y 4 de junio de 2003.
Torres, B. E.; Mejía, S. E.; Cortés, B. J.; Palacios, V. E. y Exebio, G. A. 2005. Adaptación de un modelo de simulación hidrológica a la cuenca del río Laja, Guanajuato, México. Agrociencia 39:481-490.

Agricultura Técnica en México Vol. 35 Núm.1 1 de enero-31 de marzo 2009 p. 23-35
ESTUDIO DE CASO DEL IMPACTO DE LA TRANSFERENCIA DE TECNOLOGÍA EN TRIGO DEL INIFAP*
STUDY OF CASE OF THE IMPACT OF THE TRANSFERENCE OF TECHNOLOGY IN WHEAT OF THE INIFAP
Dora M. de J. Sangerman-Jarquín1§, Eduardo Espitia Rangel2, Héctor Eduardo Villaseñor Mir1, Benito Ramírez Valverde3 y Pilar Alberti Manzanares4
1Campo Experimental Valle de México, INIFAP. Km 18.5 carretera Los Reyes- Lechería, A. P. 10, C. P. 56230 Chapingo, Texcoco, Estado de México. Tel. y Fax. 01 595 9542964. 2Campo Experimental Bajío, INIFAP. Km. 6.5 carretera Celaya-San Miguel de Allende. C. P. 38110, Celaya, Guanajuato. Tel. 01 461 6177508, ([email protected]). 1Programa de Trigo, Campo Experimental Valle de México. Km 18.5 carretera Los Reyes- Lechería, A. P. 10, C. P. 56230 Chapingo, Texcoco, Estado de México. Tel. 01 595 9542877 Ext. 127, (villaseñ[email protected]). 3Posgrado en Estrategias para el Desarrollo Agrícola Regional, Campus- Puebla, Colegio de Postgraduados en Ciencias Agrícolas. Km 125.5 carretera Federal México- Puebla C. P. 72130, Puebla, Puebla, A. P. 2-12, Colonia Libertad, Tel. 01 222 28514 42, ([email protected]). 4Posgrado en Desarrollo Rural- Estudios de Género, Colegio de Postgraduados en Ciencias Agrícolas. Km 35.5, carretera México- Texcoco, C. P. 56230, Montecillo, Texcoco, Estado de México. Tel. 01 595 9520200 Ext. 1878. 1§Autora para correspondencia: [email protected].
RESUMEN
El objetivo de esta investigación fue identificar los factores que favorecieron o limitaron el uso de las variedades de trigo generadas por el Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP). El estudio de campo se realizó en 2006 en el municipio de Nanacamilpa, Tlaxcala, México; región donde el trigo representa un componente de importancia económica familiar. La información base se obtuvo de una muestra simple aleatoria, la cual consistió en aplicar un cuestionario a 50 agricultores con preguntas cerradas y abiertas. Los resultados más destacados fueron. 1) las variedades de mayor impacto fueron: Temporalera M87 y Romoga F96. Además de haberse sembrado Zacatecas VT74, México M82, Batán F96, Náhuatl F2000 y Tlaxcala F2000, todas generadas por el INIFAP, 2) 60% de los agricultores sabe que el INIFAP genera semilla de trigo de las variedades mejoradas, 3) los productores indicaron que usar semillas mejoradas les proporciona beneficios como: altos rendimientos y mayor calidad de semilla para siembra, 4) los principales problemas de producción en el cereal fueron: malezas, plagas y enfermedades en las diversas etapas del desarrollo
del cultivo, y 5) la generación de tecnología en la propia comunidad es un factor importante en la transferencia y adopción de la misma.
Palabras clave: agricultores, transferencia de tecnología,trigo, variedades.
ABSTRACT
The goal of this research was to identify the factors that favor or limit the use of bred wheat cultivars that have been developed by the breeding program of the National Research Institute for Forestry, Agriculture and Livestock (INIFAP). The study was conducted during 2006 in the Nanacamilpa county, Tlaxcala, Mexico, where wheat cultivation is an important component in the families’ economy. The base information was a simple random sample consisting in the application of a questionnaire with open and close questions to 50 farmers. The most important results were: 1) the
* Recibido: Diciembre, 2007
Aceptado: Marzo, 2009

24 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
Dora M. de J. Sangerman-Jarquín et al.
cultivars of great impact are Temporalera M87 and Romaga F96. In addition, Zacatecas VT74, Mexico M82, Batan F96, Náhuatl F2000 and Tlaxcala F2000, all developed by INIFAP, were also used, 2) 60% of the farmers knows that INIFAP produces wheat seeds of improved cultivars, 3) farmers whom had used seeds from INIFAP indicated that the use of improved cultivars is beneficial due to higher yields and superior seed quality and, 4) the main production problems identified in the cereal were: weeds, pests and diseases throughout crop development, 5) the generation of technology in their own community is an important factor in its transference and adoption.
Key words: farmers, technology transference, cultivars, wheat.
INTRODUCCIÓN
En el contexto de las políticas neoliberales y de ajuste estructural en América Latina y el Caribe, durante los últimos quince años se han dado cambios e innovaciones en gran número de sistemas y procesos de transferencia tecnológica. En general, las reformas se han caracterizado por una descentralización de la transferencia y adopción de tecnología como un bien público. La dinámica compuesta por múltiples redes organizativas, económicas, políticas, sociales y tecnológicas se presenta con los productores que cultivan trigo. Ante este contexto, se planteo el escenario de Nanacamilpa, Tlaxcala. En el estado de Tlaxcala, se cultivaron alrededor de 5 mil hectáreas de trigo durante la década de los setenta; en los años ochenta se llegaron a sembrar más de 50 000 hectáreas y durante los noventa la superficie alcanzó un incremento entre 30 y 80 000 hectáreas, para 2000 la superficie fue de 90 000 hectáreas (SIAP, 2006). Los incrementos por área sembrada se debe a que este cereal representa una alternativa para las áreas donde el maíz tiene de mediana a baja productividad, debido a que su ciclo biológico es más corto (Villaseñor, 2000). Asimismo, es un cultivo altamente mecanizado y necesita menos insumos; por lo que es más rentable en las tierras limitativas para la producción de maíz (Villaseñor y Espitia, 2000). El objetivo planteado en este estudio fue analizar el papel de la generación y transferencia de tecnología del INIFAP en 2006, como factor impulsor de las estrategias para el desarrollo agrícola en Nanacamilpa.
El INIFAP, genera y valida la transferencia de conocimientos e innovaciones tecnológicas como soporte al desarrollo sostenible de los procesos productivos forestales, agrícolas y pecuarios sin deterioro de los recursos naturales. Estas tecnologías permiten mejorar los niveles de productividad, rentabilidad, competitividad y sostenibilidad de la producción nacional en sus diversas regiones agroecológicas. El Campo Experimental “Valle de México” (CEVAMEX) uno de los 30 Campos Experimentales que tiene el INIFAP diseminados en el país, inició sus actividades en 1943 como Campo Experimental “El Horno”, en 2003 cumplió 60 años de existencia, durante estos años se han conjugado como resultado aportaciones científicas y tecnológicas que contribuyen al desarrollo agrícola, pecuario y forestal de México (Islas, 2003). En los períodos de 1945-1985 los investigadores de la oficina de estudios especiales, hoy INIFAP, liberaron a partir de 1960, 633 variedades con más de 24 especies de cultivos básicos como: trigo, maíz y frijol. De 1986-1996 se agregaron 71 variedades, para totalizar en los años de 1945-1996, 704 variedades. El programa de mejoramiento genético de trigo de temporal en México se inició en 1969 por el INIA, institución que posteriormente pasó a ser el INIFAP. Durante 1969-2000 se liberaron 18 variedades específicas para las diferentes áreas agroecológicas del país, con el objetivo de que el productor aumente los rendimientos. En el año 2000 se liberó la variedad Tlaxcala F2000, para la disposición de los productores de la región de Tlaxcala con características ideales y específicas del lugar (Villaseñor y Espitia, 2000).
MARCO TEÓRICO
No podemos entender ni analizar los modelos de transferencia de tecnología que un país impulsa para la población rural, si no conocemos el contexto socioeconómico y político en el que se inserta; es decir, hay que enmarcarlo en la corriente de desarrollo que impera en él. Para ello consideramos el desarrollo como una estrategia para el impulso de cambios que, al operar en la población producen cambios sociales (Muro, 1999; Rodríguez 2000; Reijntjes, 2006). El desarrollo es un proceso que debe analizarse en perspectiva histórica y es un fenómeno complejo, que incluye múltiples aspectos a considerar, entre los que se encuentran: 1) territorio, 2) población, 3) conjunto de asentamientos, y 4) conjunto de instituciones públicas y privadas (Boisier, 1999; Ceña, 2000; Rolls, 2003).

Estudio de caso del impacto de la transferencia de tecnología en trigo del INIFAP 25
La transferencia de tecnología se ha concebido de diversas maneras: como un flujo o movimientos de conocimientos o como un proceso donde dicho flujo tiene lugar; como un método de hacer algo con orden y de manera sistemática (Medellín y Bocanegra, 2002). Estos autores refieren que la transferencia de tecnología se circunscribe como: a) un flujo de conocimientos, b) un método ordenado y sistemático de transmisión de saberes tecnológicos, y c) como transmisión de conocimientos estructurados, que requiere y exige una solución organizacional. Diversos autores (De Janvry, 1998; Rosengrant y Svenden, 1999; Simmons, 2003; Unnevehr, 2003) indican que la tecnología que incorpora los bienes de capital puede ser: ahorradora de trabajo, neutral o ahorradora de tierra, de acuerdo con el sentido de la tecnología. Estos autores van clasificando, cada uno a la tecnología en cuatro categorías: 1) mecánicas, 2) biológicas, 3) químicas, y 4) prácticas agronómicas culturales y de manejo.
Una tecnología es una combinación de todas las prácticas de manejo para producir un cultivo o una mezcla de cultivos, o para almacenar los productos agrícolas provenientes de
éstos. Cada práctica se define por el tiempo, y por la cantidad y el tipo de componentes tecnológicos utilizados, tales como la preparación de la cama de siembra, uso de fertilizantes, el deshierbe, etc.
MARCO REGIONAL DE LA ZONA DE ESTUDIO
Esta región fue seleccionada para el estudio por las siguientes razones: a) casi 30 años de trabajos de investigación realizados por el INIFAP, b) la importancia del trigo en la economía regional, c) siembra de trigo en condiciones de temporal, y d) productores minifundistas y con escasos recursos. Nanacamilpa, Tlaxcala, se localiza en el Altiplano central mexicano a 2 720 m, el municipio de Nanacamilpa se sitúa en las coordenadas geográficas de 19o 29’ latitud norte y 98o 32’ longitud oeste. Éste municipio colinda al norte y poniente con el municipio de Calpulalpan, al sur con el estado de Puebla y al oriente se establecen linderos con el municipio de Sanctórum de Lázaro Cárdenas (García, 2004; INEGI, 2002) (Figura1).
Figura 1. Localización geográfica del municipio de Nanacamilpa, Tlaxcala.
2160000
2140000
2120000
2180000
2160000
2140000
2120000
580000 580000 600000 620000 640000
540000 560000 580000 600000 620000 640000
Estado de Tlaxcala
Municipio Nanacamilpa
Sistema de Proyección: Universal Transversa de Mercator Zona UTM: 14 Norte Datum: NAD 27 Esferoide: Clarke 1886
ESCALA GRÁFICA
0 100000 Metros 200000
SIMBOLOGÍA
DATOS DE CONSTRUCCIÓN
N
S
O E

26 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
Dora M. de J. Sangerman-Jarquín et al.
MATERIALES Y MÉTODOS
Para la realización de este estudio, se utilizó un muestreo cualitativo. La ecuación para calcular el tamaño de muestra es la siguiente:
donde, N= tamaño de la población; Zα/2= confiabilidad; pn= proporción de unidades con la característica de interés; qn= corresponde a la proporción que no posee la característica, y d= precisión de la muestra.
El número productores en la comunidad fue de 227. Con una precisión de 90% y utilizando la variable conocimiento de la tecnología recomendada por el INIFAP. Donde, pn= 0.6 y consecuentemente; qn= 0.4. La precisión fue de 10%, por lo que el tamaño de muestra quedó definido en 50 productores.
Como parte de la metodología, se procedió a entrevistar a informantes claves, entre los que se localizaron investigadores, funcionarios del Distrito de Desarrollo Rural y del CADER, autoridades municipales y ejidales, productoras y productores. En el análisis estadístico se utiliza la prueba de t de Student, exacta de Fisher, chi - cuadrada de Pearson y además de estadística descriptiva como cuadros de cruce de información, tablas de frecuencia, gráficos, etc.
RESULTADOS Y DISCUSIÓN
Estudio de campo
El programa de mejoramiento genético de trigo del INIFAP, específicamente en la región de estudio, ha alcanzado impactos, logros y aportaciones en el cultivo de trigo, que por su importancia son: resistencia a plagas y enfermedades, potencial de rendimiento, calidad industrial, áreas de recomendación, entre otras. El funcionamiento de una estrategia, que ponga de relieve el alto rendimiento de las variedades liberadas por el INIFAP, en la región de Nanacamilpa se pueden considerar las siguientes: 1) región definida de alto potencial para siembras comerciales, así como alto potencial en rendimiento (Villaseñor y Espitia, 2000), y 2) las variedades deben reunir las condiciones y características que los productores de la región requieren,
según los datos observados en campo son: rendimiento, resistencia a enfermedades y al acame, grano y planta grande, que sea del gusto de los compradores y la industria harinera.
Porcentaje de productores con y sin asistencia técnica
Los resultados muestran que 52% de los productores recibió asistencia técnica, y el restante 48% no recibió este servicio.
Edad de los productores
La edad de los productores es una característica que ha sido analizada con insistencia en la literatura sobre desarrollo agrícola y rural como un factor que debe tomarse en cuenta, sobre todo para fomentar procesos de cambio tecnológico e introducir nuevas actividades productivas. Al comparar el promedio de edad de los padres de familia que recibieron asistencia técnica y aquellos productores que no recibieron, se observa que mediante prueba de t, no existe diferencia estadística entre estos grupos, es decir, se presentan condiciones similares de edad y esto puede observarse al revisar los promedios de edad, donde se encontró que los que recibieron asistencia técnica tienen un promedio de 54.73 años con un mínimo de 27 y máximo 87 años de edad, y el grupo que no recibió posee en promedio de 55.08 años con un mínimo de 30 y un máximo de 75 años, como se muestra en el Cuadro 1. Se puede concluir entonces que los productores son en términos generales de edad madura y que la edad no es determinante en el empleo de asistencia técnica en la región.
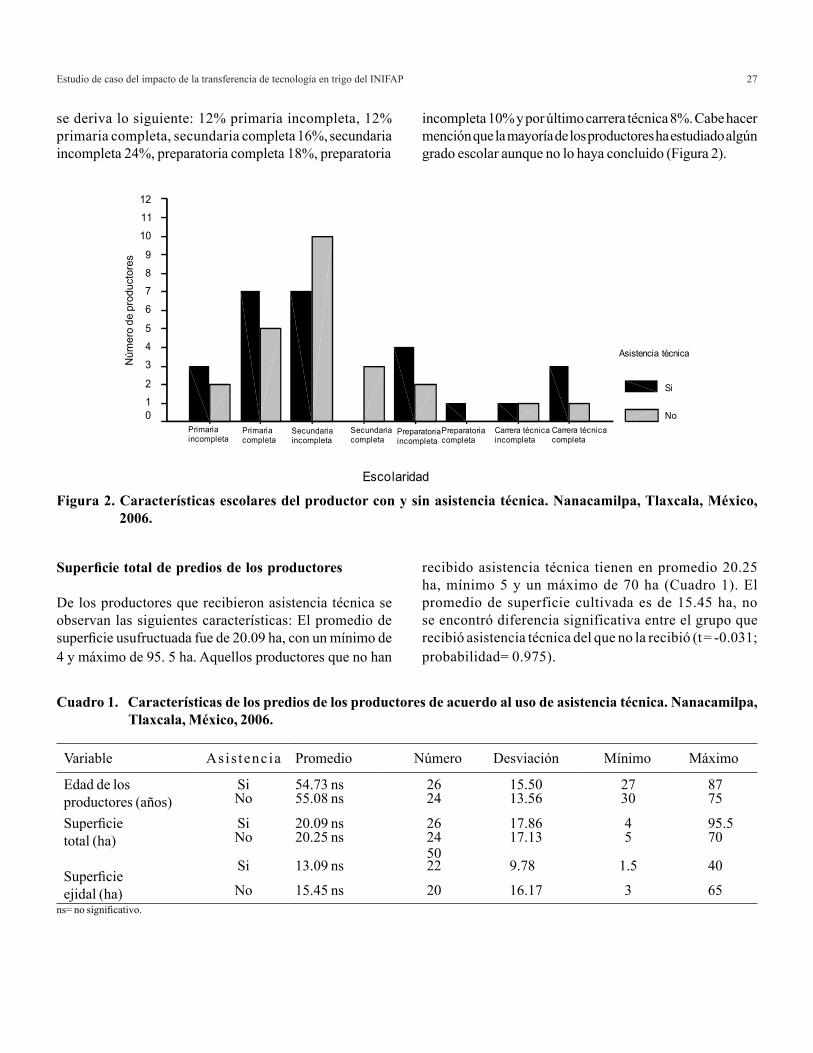
Escolaridad de los productores
Los productores que han participado en la adopción de transferencia de tecnología del INIFAP, han mostrado características escolares medio y bajo (primarias, secundaria y en un menor porcentaje preparatoria), los productores más jóvenes ostentan un mejor nivel educativo formal (secundaria, preparatoria, y los menos alguna carrera técnica). En cuanto a los porcentajes de escolaridad de los productores con asistencia técnica, 16% tienen primaria terminada, 8% primaria incompleta, 18% secundaria completa, 23%, preparatoria completa, 20% preparatoria incompleta, y 15% carrera técnica, para los productores que no recibieron asistencia técnica
nn2/22
nn2/2
qp Z+ d NqpZN
= nα
α
Estudio de caso del impacto de la transferencia de tecnología en trigo del INIFAP 27
se deriva lo siguiente: 12% primaria incompleta, 12% primaria completa, secundaria completa 16%, secundaria incompleta 24%, preparatoria completa 18%, preparatoria
Superficie total de predios de los productores
De los productores que recibieron asistencia técnica se observan las siguientes características: El promedio de superficie usufructuada fue de 20.09 ha, con un mínimo de 4 y máximo de 95. 5 ha. Aquellos productores que no han
incompleta 10% y por último carrera técnica 8%. Cabe hacer mención que la mayoría de los productores ha estudiado algún grado escolar aunque no lo haya concluido (Figura 2).
recibido asistencia técnica tienen en promedio 20.25 ha, mínimo 5 y un máximo de 70 ha (Cuadro 1). El promedio de superficie cultivada es de 15.45 ha, no se encontró diferencia significativa entre el grupo que recibió asistencia técnica del que no la recibió (t = -0.031; probabilidad= 0.975).
Variable Asis tencia Promedio Número Desviación Mínimo Máximo
Edad de los productores (años)
Si 54.73 ns 26 15.50 27 87No 55.08 ns 24 13.56 30 75
Superficietotal (ha)
Si 20.09 ns 26 17.86 4 95.5No 20.25 ns 24 17.13 5 70
50Superficieejidal (ha)
Si 13.09 ns 22 9.78 1.5 40
No 15.45 ns 20 16.17 3 65
Figura 2. Características escolares del productor con y sin asistencia técnica. Nanacamilpa, Tlaxcala, México, 2006.
Cuadro 1. Características de los predios de los productores de acuerdo al uso de asistencia técnica. Nanacamilpa, Tlaxcala, México, 2006.
ns= no significativo.
Núm
ero
de p
rodu
ctor
es
Escolaridad
Carrera técnica completa
Carrera técnica incompleta
Preparatoria completa
Preparatoria incompleta
Secundaria completa
Secundaria incompleta
Primariacompleta
Primariaincompleta
12
11
10
9
8
7
6
5
4
3
2
10
Asistencia técnica
Si
No
28 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
Dora M. de J. Sangerman-Jarquín et al.
Promedio superficie ejidal de temporal (SET) en los predios de los productores
De las familias entrevistadas con asistencia técnica, reportan una superficie ejidal de temporal (SET), con un promedio de 13.09 ha, un mínimo de 1.50 y máximo de 40 ha. Para aquellos productores que no han recibido asistencia técnica el porcentaje de (SET) es de 15.45 ha mínimo de 3 y máximo 65 ha. El Cuadro 1, también muestra, que no se encontró diferencia significativa entre el grupo que recibió asistencia técnica del que no la recibió (t = -0.578; probabilidad= 0.566). En la región de estudio la superficie ejidal usufructuada por los productores, no es determinante para tener asistencia técnica.
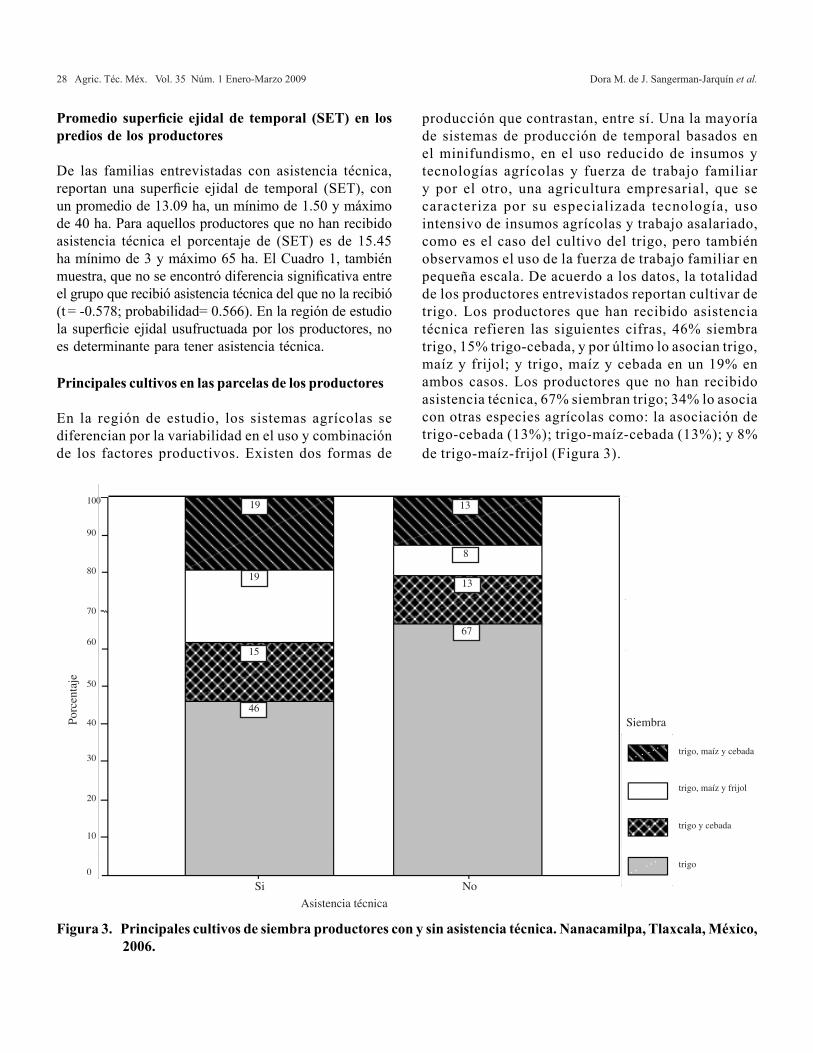
Principales cultivos en las parcelas de los productores
En la región de estudio, los sistemas agrícolas se diferencian por la variabilidad en el uso y combinación de los factores productivos. Existen dos formas de
producción que contrastan, entre sí. Una la mayoría de sistemas de producción de temporal basados en el minifundismo, en el uso reducido de insumos y tecnologías agrícolas y fuerza de trabajo familiar y por el otro, una agricultura empresarial, que se caracteriza por su especializada tecnología, uso intensivo de insumos agrícolas y trabajo asalariado, como es el caso del cultivo del trigo, pero también observamos el uso de la fuerza de trabajo familiar en pequeña escala. De acuerdo a los datos, la totalidad de los productores entrevistados reportan cultivar de trigo. Los productores que han recibido asistencia técnica refieren las siguientes cifras, 46% siembra trigo, 15% trigo-cebada, y por último lo asocian trigo, maíz y frijol; y trigo, maíz y cebada en un 19% en ambos casos. Los productores que no han recibido asistencia técnica, 67% siembran trigo; 34% lo asocia con otras especies agrícolas como: la asociación de trigo-cebada (13%); trigo-maíz-cebada (13%); y 8% de trigo-maíz-frijol (Figura 3).
Figura 3. Principales cultivos de siembra productores con y sin asistencia técnica. Nanacamilpa, Tlaxcala, México, 2006.
100
90
80
70
60
50
40
30
20
10
0
trigo, maíz y cebada
trigo, maíz y frijol
trigo y cebada
trigo
Porc
enta
je
Asistencia técnicaSi No
Siembra
19
19
15
46
13
8
13
67

Estudio de caso del impacto de la transferencia de tecnología en trigo del INIFAP 29
Variedades de trigo Con asistencia técnica Sin asistencia técnicaProbabilidad
Frecuencia (%) Frecuencia (%)Temporalera M87 13 50 12 50 1.00 ns1
Romoga F96 12 46.2 14 58.3 0.389 nsTlaxcala F2000, Náhuatl F2000 y Romoga F96 10 38.5 9 37.5 0.944 nsTlaxcala F2000 5 19.2 6 25 0.623 nsNáhuatl F2000 y Tlaxcala F2000 10 38.5 4 16.7 0.086 nsNáhuatl F2000 14 53.8 11 45.8 0.571 nsJuchitepec F2000 11 42.3 4 16.7 0.048 *Zacatecas 74 10 38.5 4 16.7 0.086 nsZacatecas 74 y Náhuatl F2000 5 19.2 8 33.3 0.256 nsTemporalera M87, Mixteco S82 y Romoga F96 8 30.8 7 29.2 0.902 nsNáhuatl F2000 y Romoga F96 3 11.5 7 29.2 0.164 nsTemporalera M87 y Pavón F76 0 0 3 12.5 0.103 nsCleopatra y Romoga F96 10 38.5 10 41.7 0.817 nsPavón y Romoga F96 14 53.8 11 45.8 0.571 nsGálvez M87 y Mixteco S82 0 0 3 12.5 0.103 ns
La totalidad de los productores siembra trigo, aunque en el caso de los que tienen asistencia técnica 46.2% siembra solo trigo y el resto con otra especie. Para el caso de los que no cuentan con este servicio la mayoría (66.7%) siembra sólo trigo. Los productores que no tienen asesoría técnica tienden a sembrar únicamente trigo mientras que los que reciben asesoría técnica tienen mayor diversificación.
Uso de variedades mejoradas del INIFAP por los productores
Los productores remarcan que las variedades han jugado un papel importante y en el futuro será mayor, ya que indican que esta problemática detiene sequías cada vez másfrecuentes, suelos degradados,
y las heladas tempranas que se presentan con mayor continuidad. En el cultivo de trigo ha sido más intensa la labor de transferencia de tecnología, ya que desde mediados de la década de los setenta el INIFAP, a través del Campo Experimental “Valle de México”, el programa de mejoramiento genético ha realizado trabajos de investigación, para estudiar el comportamiento de líneas y variedades, así como lotes demostrativos de variedades. Con relación a la región de estudio (Cuadro 2), presenta las variedades de trigo que predominan en las siembras comerciales como: Temporalera M87, variedad única, pero ha sido sembrada con la variedad Romoga F96, variedades liberadas hace 17 y 8 años respectivamente. Otras variedades sembradas continuamente por los productores son Tlaxcala F2000, Náhuatl F2000, éstas fueron liberadas en 2000.
recibieron este año asistencia técnica como los que no yla explicación a lo anterior se encuentra en el hecho de que el INIFAP ha realizado trabajos de investigación, durante muchos años en la región; generando nuevas variedades y empleando tecnologías adecuadas.
Beneficios en el uso de semillas del INIFAP
Los productores encuestados fueron consultados con respecto a los beneficios que tendrían al usar semillas de
Cuadro 2. Uso de variedades mejoradas del INIFAP por los productores. Nanacamilpa, Tlaxcala, México, 2006.
1ns= no significativo; *= significativo (α= 0.05).
El análisis estadístico se realizó mediante prueba de chi-cuadrada y prueba exacta de Fisher de acuerdo a las características de la tabla de 2 x 2 y la significancia se muestra en la última columna (Cuadro 2), como puede observarse, existe un componente similar en los dos grupos respecto al uso de variedades, excepto en la variedad Juchitepec 2000, que fue liberada por el INIFAP en el año 2000, donde se aprecia mejor uso en los productores que este año recibieron asistencia técnica. Los productores usan variedades recientemente puestas a disposición por el INIFAP. Tanto los que

30 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
Dora M. de J. Sangerman-Jarquín et al.
las variedades mejoradas del INIFAP, los datos se indican en el Cuadro 3. Los resultados indican que 64% de los agricultores(as) hacen referencia al mejoramiento de
las semillas que sembrará, 56% refieren aplicar bien los agroquímicos en las semillas, 52% mejor rendimiento, 42% aprender nuevas formas de siembra, entre las principales.
Ventajas que el agricultor mencionó Frecuencia (%)
Mejoramiento de las semillas que sembrará 32 64Aplicar bien los agroquímicos en las semillas 28 56Mejor rendimiento 26 52Aprender nuevas formas de siembra 21 42Calidad de la semilla que se va a usar 21 42Sembrar nuevas variedades 19 38Tendría semilla que no se conoce hasta ahora 19 38Conocer nuevas variedades 18 36Ser un buen productor de trigo 17 34Adquirir nuevos conocimientos del trigo 15 30Que sea una mejor variedad para la comunidad 14 28Tener buenos resultado de las nuevas variedades 13 26Que le compren a mejor precio su semilla 13 26Controlar mejor malezas, plagas y enfermedades 13 26Aprender nuevas técnicas del trigo 11 22Que les dejen las semillas 9 18Beneficio para el campo 8 16Quieren asistencia sobre manejo de suelos 7 14Para ver que semilla es mejor en rendimiento 7 14Agradecimiento a los ingenieros por su experiencia con estas semillas 6 12
Cuadro 3. Beneficios en el uso semillas del INIFAP. Nanacamilpa, Tlaxcala, México, 2006.
Nota: las frecuencias son sobre 100%. Los encuestados señalaron de 1-5 ventajas.
Fertilizantes utilizados en las parcelas
La tecnología de producción generada por el INIFAP ha sido fundamental para impulsar la superficie sembrada y productividad del trigo. Las variedades han jugado un papel importante y en el futuro serán mayores, ya que la problemática biótica y abiótica es cada vez más compleja, como por ejemplo: enfermedades más agresivas, las sequías son más frecuentes año con año, el suelo se degradan conforme son cultivados excesivamente, heladas con mayor continuidad, y mayor aplicación de los fertilizantes, ésta se ha cuestionado por la contaminación al medio ambiente, que representa en su utilización.
En el Cuadro 4, señala que el uso de seis fertilizantes frecuentemente utilizados por los productores, que recibieron asistencia técnica fueron: superfosfato de calcio triple 42.3%, cloruro de potasio 38.5%, urea 30.8%, triple 17 26.9% y fullmina 15.4%. Cabe mencionar, que la utilización de abono orgánico representa al igual que
el superfosfato de calcio triple 42.3% el fertilizante más utilizado. El empleo del abono orgánico ha sido una práctica ancestral que los productores han hecho común en sus parcelas, además los productores están conscientes del deterioro ambiental y desean utilizarlo en mayor medida. Los productores que han no han recibido asistencia mencionan en primer lugar el uso de la fullmina 45.8%, cloruro de calcio triple y triple 17 en 33.3%, cloruro de potasio 33.3%, urea 29.2% y por último abono orgánico 16.7%.
La utilización de urea en la fertilización del cultivo es 69.2% y 70.8%, para productores con y sin asistencia técnica, respectivamente. Como puede notarse en el Cuadro 4, ambos grupos de productores utilizan este fertilizante, casi en la misma proporción. Para reforzar lo anterior, la prueba de chi - cuadrada, nos indica que no existe diferencia estadística (χ2= 0.015; probabilidad= 0.902), es decir, ambos grupos de productores lo utiliza en la misma proporción.
Estudio de caso del impacto de la transferencia de tecnología en trigo del INIFAP 31
El Cuadro 4, muestra la utilización del fertilizante superfosfatado, en ambos grupos de productores es 37.7% y 58.3% con y sin asistencia técnica, respectivamente. Por lo que la prueba de chi- cuadrada, nos indica que no existe diferencia estadística. (χ2= 0.002; probabilidad= 0.963), es decir, el superfosfato, es utilizado en la misma proporción en ambos grupos de productores.
La utilización de cloruros en el cultivo de trigo, es 61.5% y 66.7% en productores con y sin asistencia técnica, respectivamente. Como puede observarse, en el Cuadro 4, ambos grupos utilizan cloruro casi en la misma proporción, lo anterior lo refuerza la prueba de chi - cuadrada, misma que nos indica que no existe diferencia estadística en la utilización de cloruro en el cultivo del trigo, en ambos grupos de productores (χ2= 0.142; probabilidad= 0.706).
Para el grupo de productores con asistencia técnica 84.6% utiliza fullmina, mientras que 15.4% no lo utiliza. Ahora en el grupo de productores que no han adoptado asistencia. écnica, se analizó que 54.2%, utiliza la fullmina y el resto no (Cuadro 4). Se puede notar que existe una mayor tutilización de fullmina en productores con asistencia técnica en comparación con el otro grupo. Pero la prueba de chi - cuadrada, nos demuestra que existe diferencia estadística en la utilización de este agroquímico (fullmina) en ambos grupos de productores (χ2= 5.510; probabilidad= 0.019).
Costos de fertilización y t ha-1 utilizados por productores. Nanacamilpa, Tlaxcala, México, 2006.
En el Cuadro 5, se realizó una comparación de medias en cuanto a la utilización de fertilizantes t ha-1, donde se demostró que no existe diferencia estadística en la utilización de fertilizantes t ha-1 (t= 1.369; probabilidad=
0.178). Se identificó que no existe diferencia estadística en el costo del fertilizante por hectárea en ambos grupos de productores (t= 1.474; probabilidad= 0.147). El costo de este fertilizante es de $ 33 656 00 en promedio. Cabe aclarar, que los datos de los fertilizantes por hectárea y costo del fertilizante son generalizados, por los diferentes tipos de fertilizantes utilizados por los productores.
Instituciones que dieron asistencia técnica a los productores. Nanacamilpa, Tlaxcala, México, 2006.
La estrategia de investigación agrícola adoptada por el INIFAP y sus antecesores, se fundamenta metodológica y operativamente en el reconocimiento y delimitación de las regiones agrícolas, como ha sido la región de Nanacamilpa, las cuales se convierten en el sustrato metodológico para la generación y transferencia de tecnología, en ésta región de estudio los investigadores en trigo han tenido una permanencia por casi 30 años, validando y generando variedades de trigo aptas para la región y del gusto de los productores. En el Cuadro 6, se hace referencia a las instituciones que han dado asistencia técnica en la región de estudio a través del tiempo y no necesariamente este año. El 47.7% el INIFAP, como institución que dio asistencia técnica, la Secretaría de Agricultura, Desarrollo Rural, Pesca y Alimentación (SAGARPA) 25.9%, los Distritos de Desarrollo Rural (DDR), 12.7%, Universidad Autónoma Chapingo (UACH) 7.8% y por último el Colegio de Postgraduados en Ciencias Agrícolas (CP) 5.9%.
Con estos resultados se fortalece el papel que las instituciones de enseñanza e investigación han realizado en la región, especialmente destaca el papel del INIFAP, no sólo en la generación de variedades, sino también en la transferencia de la tecnología.
Fertilizantes Asistencia técnica Nivel de significanciaSi No Total
F1 (%) F (%) F (%)Triple 17 7 26.9 10 41.7 17 34 0.272 ns2
Urea 8 30.8 7 29.2 15 30 0.902 nsSuperfosfato de calcio triple 11 42.3 10 41.7 21 42 0.963 nsCloruro de potasio 11 42.3 10 41.7 21 42 0.963 nsFullmina 4 15.4 11 45.8 15 30 0.019*Abono orgánico 11 42.3 4 16.7 15 30 0.048**
Cuadro 4. Resumen de los fertilizantes utilizados. Nanacamilpa, Tlaxcala, México, 2006.
1F= frecuencia; 2ns= no significativo; * significativo; ** significancia con la prueba exacta de Fisher.
32 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
Dora M. de J. Sangerman-Jarquín et al.
y mientras que el grupo que no recibió asistencia técnica es 34.6%. Por otro lado, con la prueba de chi -cuadrada, se encontró que no existe diferencia estadística en cuanto al conocimiento del establecimiento de parcelas demostrativas (χ2= 0.045; probabilidad= 0.832), quiere decir, que ambos grupos de productores tiene conocimiento similar del establecimiento de estas parcelas.
Años que los productores recuerdan haber participado en días demostrativos
A través de las encuestas realizadas, se observó que los productores que participaron en días demostrativos durante los años de investigación, están mejor informados de los avances tecnológicos del cultivo, de tal manera, se espera que ellos utilizaran las nuevas variedades aptas para la región.
Instituciones participantes Porcentaje
Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP) 47.7SAGARPA 25.9Distritos de Desarrollo Rural (DDR) 12.7Universidad Autónoma Chapingo (UACH) 7.8Colegio de Postgraduados en Ciencias Agrícolas (CP) 5.9Total 100
Cuadro 6. Instituciones que dieron asistencia técnica. Nanacamilpa, Tlaxcala, México, 2006.
Participación de los productores en el establecimiento de parcelas demostrativas en la comunidad
En el estado de Tlaxcala es donde han sido más notorios lo incrementos de trigo, debido, que la siembra de variedades mejoradas para temporal, ha sido más seguro y rentable que el maíz, pero también gracias a la participación de los agricultores en el establecimiento de parcelas demostrativas en la comunidad, por parte del INIFAP a través del Campo Experimental “Valle de México” y del programa de trigo. Difundir in situ las innovaciones generadas en el cultivo es materia de comunicación para la transferencia de tecnología.
Asimismo, los productores que tienen conocimiento del establecimiento de parcelas demostrativas en la región, el grupo de productores con asistencia técnica es 65.4%,
Cuadro 5. Cantidad y costo de fertilizante empleado por productores con y sin asistencia técnica. Nanacamilpa, Tlaxcala, México, 2006.
Asistencia técnica Cantidad de fertilizante (t ha-1)
Costo de fertilizante ($ ha-1)
Si Media 4.0435 3 873.9615N 26 26Desviación estándar 3.9632 3 466.2715Mínimo 0.20 540Máximo 14 12 810
No Media 2.6313 2 564.3542N 24 24Desviación estándar 3.2653 2737.0451Mínimo 0.20 312Máximo 12 10 980
Total Media 3.3656 3245.35N 50 50Desviación estándar 3.6778 3175.4242Mínimo 0.20 312Máximo 14 12 810

Estudio de caso del impacto de la transferencia de tecnología en trigo del INIFAP 33
También se realizó una comparación de medias, para observar si ambos grupos de productores conocen del establecimiento de las parcelas demostrativas implementadas por el INIFAP en la región de estudio, se percibió que no existe diferencia estadística entre productores con y sin asistencia técnica (t= 0.358; probabilidad= 0.725). Para el primer grupo de productores tienen mínimo de un año y un máximo de 10 y para los que no tienen asistencia técnica un mínimo de 2 y máximo de 10.
DISCUSIÓN
En la región de Nanacamilpa, Tlaxcala en los últimos 30 años el INIFAP ha sido una de las instituciones soporte en la validación, generación y transferencia de tecnología, ha realizado trabajos de investigación en una forma constante y en comunicación directa con los productores de la región. De los productores entrevistados, 67.8% recibió asistencia técnica y el resto no, pero cabe aclarar, que estos últimos han conocido de la transferencia de tecnología del INIFAP, ya que los productores transmiten el conocimiento a través de comunicación informal, y de observación directa que hacen en los terrenos de los productores que han aceptado el proceso de transferencia, y adoptado las tecnologías del INIFAP. Los productores de la región conocen y han utilizado variedades generadas por el INIFAP, por factores propicios como: mayores rendimientos, variedades resistentes a plagas y enfermedades, facilidad en el control químico, variedades recientes, etc., que han permitido conocer en gran medida cómo producir el cereal.
Es importante mencionar, que los productores que han recibido asistencia técnica y aquellos que no han recibido el servicio, sobre el uso de tecnologías; Viana y Villar (2001), refieren que los factores que intervienen en el uso son: patrón de difusión de variedades mejoradas, formas de adquisición de la semilla y variedades mejoradas utilizadas por los productores; situación que se reafirma en el análisis realizado en la región, además, se asocian otros factores como: superficie cultivada, tenencia y renta de tierras, son componentes esenciales para la alta rentabilidad del cereal.
Estudios han reportado, que productores que han adoptado paquetes tecnológicos de variedades de trigo en la India y Bangladesh, usualmente son de paquetes
tecnológicos demostrados con semilla mejorada, insumos y bajo sus propias circunstancias (Biggs, 1997; Power y Doran, 2001; Nesbitt y Samuel, 2006). En el presente caso, se suma la categoría de días demostrativos que se realiza en la comunidad, con la participación de los productores es uno de los factores más importantes en la toma de decisiones del productor para el uso de variedades mejoradas del cereal.
Damián (2004), señala en este sentido, en un estudio realizado en el estado de Tlaxcala, sobre apropiación de tecnología agrícola, que los productores encuestados con respecto a la escolaridad la apropiación se basa en la edad, el nivel de escolaridad tienen una alta apropiación, acceden a folletos y revistas técnicas con información agrícola. Los productores refieren que el gobierno cuenta con programas de apoyo al campo, de estos conocen: Programas de Alianza para el Campo (PROCAMPO), PROGRESA, Kilo x Kilo, Apoyos y Servicios a la Comercialización Agropecuaria (ACERCA), SOFOA- SAGARPA, Diesel y Alianza y Capacitación para el Desarrollo Rural (CADER), aunque no todos son elegibles por estos, ya que indicaron que estos programas son insuficientes y que tener acceso a ellos es difícil, en el sentido de que existe mucha burocracia (les hacen dar muchas vueltas), como refieren estudios realizados en México, Syria e India (Galindo et al., 2000; Jenkyn et al., 2001; Álvarez et al., 2006).
Otros factores que Ramírez (2004) plantea, es el nivel de apropiación de tecnología de los agricultores se relaciona con la importancia de las recomendaciones que a nivel regional, específicamente en el estado de Tlaxcala, realiza el INIFAP y otras instituciones de enseñanza e investigación. Villaseñor (com. pers. 2004), refiere a que la comunicación informal da relevancia a este proceso, toda vez, que los productores, se acercan a los terrenos donde existen experimentos, para preguntar de una forma constante sobre el cultivo. Ante esta situación la obtención de conocimientos es precisamente la recomendación dada por otro agricultor.
CONCLUSIONES
La totalidad de los productores siembra trigo, aunque existen asociaciones con otros cultivos, lo cual demuestra la importancia económica del cultivo en la región.
Las variedades empleadas en la zona de estudio son: Temporalera M87 y Romoga F96, las cuales tienen 17 y

34 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
Dora M. de J. Sangerman-Jarquín et al.
8 años de haberse liberado, pero siguen en la preferencia de los agricultores. Además de haber sembrado Zacatecas T74, México M82, Batán F96, Náhuatl F2000 y Tlaxcala F2000. La totalidad de las variedades sembradas en la región han sido generadas por el INIFAP.
El papel del INIFAP en la región fue determinante no sólo en la generación de tecnología, sino también en la difusión de este conocimiento.
Las ventajas al usar semilla del INIFAP son: mayores rendimientos, mejoramiento y calidad de la semilla que siembra el agricultor, semilla de nuevas variedades, control de malezas, plagas y enfermedades entre otras. Lo cual constata la presencia que el INIFAP tiene en la región.
La mayoría de los productores reconocen que la tecnología generada por el INIFAP ha tenido un impacto positivo en la producción de trigo de la región.
AGRADECIMIENTO
A Doña Gloria Jarquín Vda. de Sangerman. Eres una madre extraordinaria, gracias por tu inmenso amor.
LITERATURA CITADA
Álvarez, D. P. A.; Luna, F. M.; Hernández, M. J.; Lara, H. A.; Salas, L. M. A. y Cabañas, C. B. 2006. Sistemas de producción de cebada maltera (Hordeum vulgare L.) en el estado de Zacatecas. Agric. Téc. Méx. 33:181-190.
Biggs, S. 1997. Resource-poor farmer participation in research: a synthesis of experience in nine national agricultural research systems. OFCOR. Comparative Study Paper N° 3. ISNAR. The Hague.
Boisier, S. 1999. La articulación Estado-Región, clave del desarrollo regional. In: Ávila, H. S. (comp.) Lecturas de desarrollo rural en México y América Latina. Chapingo, México. Universidad Autónoma Chapingo, p. 309-335.
Ceña, E. 2000. El desarrollo rural en un sentido amplio. In: El Desarrollo Rural Andaluz a las puertas del Siglo XXI. Congreso y Jornadas. Andalucía, España. Núm. 32. p. 14-17.
Damián, H. M. A. 2004. Apropiación de tecnología por actividades del ciclo agrícola del maíz. In: apropiación de tecnología agrícola. Damián, H. M. A (coord.) Benemérita Universidad Autónoma de Puebla- CONACYT-Siza- H. Congreso del Estado Tlaxcala, LVII Legislatura, Puebla, Puebla.
De Janvry, A. 1998. The political economy of rural development in Latin America. An interpretation. In: America Journal of Agricultural Economics, 57:67.
Galindo, G. G.; Gómez, A. G. y Tabares, R. W. C. 2000. Caracterización de los extensionistas del programa elemental de asistencia técnica en Zacatecas. Rev. Fitotec. Mex. 23:307-320.
García, E. 2004. Modificaciones al sistema de clasificación de Köppen. 5ª. Ed. Instituto de Geografía- Universidad Nacional Autónoma de México. México, D. F. 90 p.
Instituto Nacional de Estadística Geografía e Informática. INEGI. 2002. Censo general de población y vivienda. Información de cabeceras municipales. México, D. F.
Islas, G. J. 2003. Reseña Histórica Campo Experimental Valle de México “El Horno”. 60 años de Investigación al servicio de México. Día de campo CEVAMEX, 3ª Expo Nacional de Maquinaria Agrícola, 25 años de Investigación Forestal. Región Centro, septiembre. p.5. (Publicación Especial Núm. 1).
Jenkyn. J. F.; Chiristian, D. G.; Bacon, E. T.; Gutteridge, R. J. and Todd, A. D. 2001. Effects of incorporating different amounts of straw on growth, diseases and yield of consecutive crops of winter wheat grown on contrasting soil types. J. Agric. Sci. 136:1-14.
Medellín, C. E y Bocanegra, G. C. 2002. Un caso de transferencia de tecnología. Centro de innovación tecnológica. Universidad Nacional Autónoma de México (UNAM). 54 p.
Muro, B. P. 1999. Problemas del campesinado y desarrollo rural alternativo. Universidad Autónoma Chapingo. Chapingo, México. 125 p.
Nesbitt, M. And Samuel, D. 2006. ¿From staple to crop extinction?. The archaeology and history of the wheats. In: Padulosi, S., K. Hammer and J. Heller (eds) “Hulled wheats”. Proceeding of the firs international workshop on hulled wheats. Italy, Rome. p. 41-100.

Estudio de caso del impacto de la transferencia de tecnología en trigo del INIFAP 35
Power, J. F. and Doran, J. W. 2001. Role of crop residue management in nitrogen cycling and use. In: “Cropping strategies for efficient use of water and nitrogen” Madison, WI., Am. Soc. Agron. 56:101-113.
Reijntjes, C. 2006. Farming for the future: an introduction to low- external- input and sustainable agriculture, Londres, Mc. Millan Press. p.133-157.
Ramírez, V. B. 2004. Apropiación de tecnología por actividades del ciclo agrícola del maíz. In: Apropiación de tecnología agrícola. Damían-Huato, M. A (coord.) Benemérita Universidad Autónoma de Puebla- CONACYT-Siza- H. Congreso del Estado Tlaxcala, LVII Legislatura, Puebla, Puebla.
Rodríguez, C. A. 2000. Bases y componentes de una estrategia para el desarrollo rural regional. In: Nueva Época. Textual. Análisis del medio rural. Núm. 36. Universidad Autónoma Chapingo. México. p. 86-87.
Rolls, M. 2003. The dimensions of Rural Extension In: Jones Gwyn (ed.). Investing in rural extension: strategies and goals. Elsevier Applied Science. Publishers London. p. 167-174.
Rosegrant, M. and Svenden, M. 1999. Asian food production in the 90's: irrigation investment and management policy. Food Policy. 32:815-819.
Servicio de Información Agroalimentaria y Pesquera (SIAP). 2006. Estadísticas sistema-producto-trigo.www.siap.sagarpa.gob.mx. (consultado 27 de febrero 2006).
Simmons, S. R. 2003. Growth, development and Physiology. In: Heyne, E. G. (ed) “Wheat and wheat improvement”. Madison, Wl., USA, Am. Soc. Agron. p. 3(3):77-113.
Unnevehr, L. 2003. “Food safety, food security and food trade”. Washington. D. C. IFPRI 2020. p. 4-14.
Viana, A. y Villar, B. 2001. Adopción de variedades mejoradas de frijol en la región de la Frailesca, Chiapas, México. Secretaría de Agricultura Ganadería y Desarrollo Rural. Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias. Centro de Investigación Regional del Pacífico Sur. Campo Experimental, Centro de Chiapas. División Agrícola. México, D. F. p. 1-25 (Libro Técnico Núm. 1).
Villaseñor, M. E. 2000. Importancia del trigo. Características de las áreas productoras de trigo de temporal: problemática y condiciones de producción”. In: Villaseñor, H. E y Espitia, R. E (eds.). El trigo de temporal en México. Secretaría de Agricultura Ganadería y Desarrollo Rural, Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias, Centro de Investigación Regional del Centro, Campo Experimental Valle de México, División Agrícola. México. p. 7-21 (Libro Técnico Núm. 1).
Villaseñor, M. H. E. y Espitia, R. E. 2000. Características de las áreas productoras de trigo de temporal: problemática y condiciones de producción. In: Villaseñor, M. H. E y Espitia, R. E (eds.). El trigo de temporal en México. Secretaría de Agricultura Ganadería y Desarrollo Rural, Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias, Centro de Investigación Regional del Centro, Campo Experimental Valle de México, División Agrícola. México. p. 85-98 (Libro Técnico Núm. 1).

Agricultura Técnica en México Vol. 35 Núm.1 1 de enero-31 de marzo 2009 p. 36-45
INFLUENCIA DE LA SUBDIVISIÓN DEL CALLO EMBRIOGÉNICO EN LA FORMACIÓN DE EMBRIONES SOMÁTICOS DE COCOTERO*
INFLUENCE OF EMBRYOGENIC CALLUS SUBDIVISION ON COCONUT SOMATIC EMBRYO FORMATION
Alfonso Azpeitia Morales1§, José Luis Chan2, Luis Sáenz Carbonell2 y Carlos Oropeza Salin2
1Programa de Biotecnología, Campo Experimental Huimanguillo, INIFAP. Km 1, carretera Huimanguillo-Cárdenas, A. P. Núm. 17, C. P. 86400. Tel. 01 917 3750396 Huimanguillo, Tabasco. 2Unidad de Biotecnología, Centro de Investigación Científica de Yucatán, CICY. Calle 43 No. 130 Col. Chuburna de Hidalgo, C. P. 97200 Mérida, Yucatán. ([email protected]), ([email protected]), ([email protected]). Tel. 01 999 9813966. §Autor para correspondencia: [email protected].
* Recibido: Febrero, 2008
Aceptado: Marzo, 2009
RESUMEN El presente estudio tuvo como objetivo determinar la influencia de la subdivisión del callo embriogénico primario de cocotero en la formación de embriones somáticos. El estudio se llevó a cabo en el laboratorio de biotecnología del Centro de Investigaciones Científicas (CICY-Yucatán), México. Se utilizaron callos embriogénicos de tres meses de cultivo procedentes del cultivo de plúmulas en condiciones de obscuridad por tres meses del genotipo de cocotero Enano Malayo Verde, cultivados previamente en un medio Y3 y adicionado con 0.55 mM de 2,4-D. Los resultados permitieron observar que con callos embriogénicos subdivididos en cuatro partes se incrementa en 3.36 veces el número de embriones somáticos tipo torpedo con respecto al control en los primeros 15 días de cultivo. En un segundo ensayo en el que se utilizó mayor número de muestras la respuesta fue similar, aunque en menor cantidad (2.7 veces) y el peso fresco se incrementó 2.36 veces en subdivisiones de callo embriogénico cultivadas sin fitohormonas, con respecto al control con fitohormonas durante los primeros 15 días de cultivo en condiciones de luminosidad. Los resultados obtenidos son prometedores y establecen las bases para incrementar el número de embriones somáticos
y eventualmente la multiplicación de callos embriogénicos en cocotero, lo cual podrá facilitar la propagación a gran escala de palmas híbridas o palmas élite.
Palabras clave: Cocos mucifera L., embriogénesis somática, subdivisión de callo.
ABSTRACT
The objective of this study was to determine the influence of primary embryogenic callus subdivision on the formation of somatic embryos. The research was carried out at the biotechnology laboratory of the Centro de Investigación Científica de Yucatán, Mexico. Three months old embryogenic callus from plumules cultivated during three months under dark conditions in Y3 medium added with 0.55 mM of 2,4-D of the cultivar Enano Malayo Verde, were used. The subdivision in four parts of the embryogenic callus showed after fifteen days in culture a 3.36 times increase in the number of torpedo type somatic embryos compared to the control. In a second experiment, using a

Influencia de la subdivisión del callo embriogénico en la formación de embriones somáticos de cocotero 37
larger number of samples, the results were similar although in smaller amount (2.7 times more somatic embryos) and 2.36 times increase in fresh weight with the subdivision of callus cultured without phytohormones in relation to the control with phytohormones after fifteen days under light culture conditions. These results are promising and could be useful to increase the number of somatic embryos and eventually facilitate large-scale propagation of hybrids or elite coconut palms.
Key words: Cocos mucifera L., somatic embryogenesis, callus subdivision.
INTRODUCCIÓN
En los últimos años la demanda de palmas de cocotero resistentes a enfermedades se ha incrementado en México, debido a que las plantas de grandes superficies estan siendo afectadas y devastadas principalmente por la enfermedad conocida como “amarillamiento letal”. Esta enfermedad ha ocasionado la desaparición de aproximadamente 15 mil ha en la península de Yucatán y su avance continua hacia los estados del Golfo de México (Tabasco y Veracruz) y del Pacífico (Chiapas, Oaxaca, Guerrero). Aunado a este problema el 80% de las plantaciones son improductivas porque rebasan los 40 años de establecidas (Piña, 1998), por lo que es necesario establecer nuevas plantaciones para lo cual se requiere producir alrededor de 50 millones de nueces de cocotero para propagar 32 millones 200 mil palmas. La escasez de material, hace necesario contar con un método de multiplicación masiva de palmas seleccionadas o mejoradas, lo cual podría efectuarse a través de la micropropagación.
Desde los años setenta se han realizado intentos para clonar in vitro el cocotero, en ellos se han evaluado diferentes tipos de explantes: hojas inmaduras, raíces, embriones e inflorescencias inmaduras con diferentes grados de éxito, siendo posible la inducción de la embriogénesis y regeneración a partir de inflorescencias inmaduras (Verdeil y Buffard-Morel, 1995) y de plúmula (Hornung, 1995; Oropeza y Chan, 1995).
Con la finalidad de mejorar la eficiencia en el proceso de propagación in vitro de esta especie se han realizado estudios sobre diferentes aspectos que incluyen: la asimilación de nutrimentos en callos de cocotero (Dussert et al., 1995; Magnaval et al., 1997), estudios sobre condiciones de cultivo (Chan et al., 1998), evaluación de los cambios en los niveles
del 2,4-D en el medio de cultivo en presencia de carbón activado y la respuesta en el crecimiento en explantes de inflorescencias (Ebert y Taylor, 1990). Recientemente, se ha demostrado que la respuesta embriogénica en cocotero varía con el uso de diferentes fuentes de carbón activado (Herrera, 2002).
En el caso particular de plúmula es posible obtener embriones somáticos y regeneración de plantas en forma reproducible y con una eficiencia mayor a la alcanzada con otros tipos de explantes (Chan et al., 1998), pero su magnitud es todavía baja en relación a lo necesario para una aplicación práctica. En general, la baja capacidad embriogénica de esta especie se ha asociado a diversas causas: intenso necrosamiento del tejido, heterogeneidad en la respuesta, fuerte capacidad de enraizamiento, alta formación de tejido tipo haustorial (Verdeil y Buffard Morel, 1995), así como la formación anormal de embriones somáticos (ES). De acuerdo a Blake (1990), las anormalidades en el desarrollo de los ES, son provocados probablemente por niveles sub óptimos o supra óptimos de las auxinas y/o citocininas.
El cocotero es una especie recalcitrante debido a que presenta una baja capacidad embriogénica atribuida a diversas causas. En esta especie, la formación de embriones somáticos a partir de callos embriogénicos (CE) ocurre cuando el callo es cultivado íntegramente sin sufrir fragmentación o división. Los ES crecen fusionados y su desarrollo no es sincronizado, lo que limita la individualización de éstos así como su desarrollo independiente hasta la formación de una planta completa. Una práctica común en algunas especies como Begonia, Saintpaulia y Streptocarpus es la de causar heridas en la superficie de las hojas o éstas son cortadas en pequeñas piezas, conduciendo a un significativo incremento de brotes adventicios (George, 1993). Los embriones cigóticos de zanahoria inducen a la formación de ES cuando se les causa una herida y son cultivados en un medio libre de fitohormonas (Smith y Krikorian, 1989). Una observación similar fue hecha en Dysosma pleiantha por Meng-Jin y Wei-Chin (1987), quienes a partir de embriones cigóticos heridos, indujeron callos embriogénicos mientras que los embriones intactos no respondieron.
En el protocolo de embriogénesis somática a partir de plúmula es de uso común la auxina 2, 4-diclorofenoxiacético (2,4-D) y la citocinina bencil aminopurina (BAP) en fase de inducción de callo embriogénico y en la formación de embriones somáticos (100 veces menor cantidad) (Chan et al., 1998). No obstante, probablemente la utilización del

38 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
Alfonso Azpeitia Morales et al.
2,4-D en esta segunda fase no sea necesaria, dado que puede ejercer un efecto negativo en la embriogénesis somática. En zanahoria por ejemplo, el proceso de ES se inhibe cuando se adiciona 2,4-D después del estado de corazón por lo que los embriones somáticos se deforman. Se postula que la adición de la auxina al medio de cultivo interrumpe el gradiente endógeno adquirido en el embrión y por dicha razón se inhibe su desarrollo (Kawahara y Komamine, 1995). La regeneración in vitro de cocotero es un proceso relativamente largo, además de que la producción de ES es baja, por lo anterior el objetivo de la presente investigación fue determinar la influencia de la subdivisión de callos embriogénicos en la formación de embriones somáticos,
así como determinar si la adición de las fitohormonas 2,4-D y la citocinina BAP favorecen la formación de embriones somáticos.
MATERIALES Y MÉTODOS
Material vegetal. En este estudio se utilizaron callos embriogénicos de tres meses de cultivo procedentes del cultivo de plúmulas en condiciones de obscuridad por tres meses del genotipo de cocotero Enano Malayo Verde (EMV), cultivados previamente en un medio Y3 (Eeuwens, 1976) adicionado con 0.55 mM de 2,4-D (Figura 1).
Figura 1. Estrategia utilizada para incrementar la eficiencia de regeneración de cocotero in vitro mediante embriogénesis somática.
1Estrategia en desarrollo por el grupo de cocotero en el CICY; 2estrategias utilizada en esta investigación.
Estrategias Proceso Tiempo Etapas
Etapa I:
1 Em
brio
géne
sis s
ecun
dari
a
Explante 0 mMedio I:
Y3 + carbón activado
2.5 g l-1 + gelrite 3 g l-1
+ 2,4-D 0.55 mM. Callo 1 m
Subdivisión de CE2 Callo
embriogénico 3 mEtapa II:
Embriones
somáticos
5 m Medio II:
Y3 + carbón activado
2.5 g l-1 + gelrite 3 g l-1
+ 2,4-D 6 µM +
6-BAP 300 µM.
Brotes 8 m
Plántulas 12 m

Influencia de la subdivisión del callo embriogénico en la formación de embriones somáticos de cocotero 39
Condiciones de cultivo. Se utilizaron dos condiciones de cultivo: a) cultivo de callos embriogénicos (CE) con un peso de entre 0.1 y 0.14 g, y b) de los CE se realizaron subdivisiones (SCE) con ayuda de un bisturí con porciones de un peso de entre 0.1 y 0.14 g. Para determinar el número de subdivisiones adecuado para la formación de embriones somáticos se estableció un experimento, con cuatro tratamientos: 1) cultivo de CE (Control), 2) subdivisión del CE en 2 secciones (SCE), 3) CE subdividido en 4 SCE, y 4) CE subdividido en 8 SCE; y todos cultivados en un frasco de cultivo, con cinco repeticiones por tratamiento. Este experimento se desarrollo en etapa II, utilizando un medio Y3 con 2,4-D (6 µM) y BAP (300 µM), así como condiciones II: fotoperíodo de 16 h de iluminación (45-60 µmol m-2s-1
PPFD)/ 8 h de oscuridad a 27 ± 2 ºC y subcultivo cada dos meses (Figura 1). En un segundo experimento se utilizó el mejor tratamiento anterior e incluyó las fitohormonas 2,4-D y BAP y un control, sin fitohormonas, los cuatro tratamientos fueron: a) callo embriogénico + fitohormonas (CE+F) como testigo, b) callo embriogénico sin fitohormonas (CE-F), y c) CE subdividido en 4 SCE + F y d) CE subdividido en 4 SCE-F. Se utilizaron cuatro repeticiones con 10 frascos de cultivo. Los cultivos se establecieron en etapa II en un medio de acuerdo a Chan et al. (1998) conteniendo las fitohormonas 2,4-D (6 µM) y BAP (300 µM) y un testigo sin estas (Figura 1). Las variables a medir fueron: a) número de embriones somáticos, y b) peso fresco. En ambos experimentos las variables se registraron a los 15 y 30 días de cultivo.
Histología. El procedimiento histológico fue realizado de acuerdo a Buffard Morel et al. (1992), con modificaciones. Las muestras de tejido fueron fijadas en paraformaldehido 4% en buffer fosfato (pH 7.2) por 24 h bajo presión
negativa. Las muestras fueron deshidratadas con etanol al 30, 50, 70, 80, 90, 95 y 100% durante 60 min cada una. Posteriormente se realizó la impregnación con resina JB-4 (Poliscience, USA). De los tejidos impregnados en resina se hicieron cortes histológicos de 3 micras y fueron teñidos con PAS-Naphtol blue black.
Análisis estadístico. Los experimentos se establecieron bajo un diseño experimental completamente al azar con un arreglo factorial y los resultados de las variables se sometieron a análisis de varianza (ANAVA) y la prueba de medias diferencia mínima significativa (p<0.05) para diferenciar los tratamientos, para lo cual se utilizó el paquete de diseños experimentales FEUANL, versión 2.5 (Olivares, 1994).
RESULTADO Y DISCUSIÓN
Influencia del número de subdivisiones en la formación de embriones somáticos. En el análisis de varianza se observó significancia entre tratamientos. El análisis realizado para el factor A formado por 15 y 30 días se encontraron diferencias estadísticas (DMS p<0.05= 1.19) entre períodos, siendo superior 15 días, con valor promedio de 8.48 ES contra 6.40 ES a 30 días. En cuanto al factor B correspondiente al número de subdivisiones de callo embriogénico, se observó que el mayor número de ES se obtuvo con el tratamiento con cuatro subdivisiones de callo embriogénico (SCE)/frasco con 13.58 ES tipo torpedo, mientras que con 2 y 8 SCE el número de ES fue de 7.16 y 5.01 respectivamente. El número de ES en SCE fue mayor a los observados en el tratamiento control, en el cual solamente se presentaron 4.01 ES del tipo globular principalmente (Cuadro 1).
Factor A (días de cultivo)
Factor B (núm. de subdivisiones)
Control 2 4 8
15 4.56 8.0 15.66 8.48
30 3.46 6.33 11.50 6.40
Valor promedio 4.01 c 7.16 b 13.58 a 5.01 c
Cuadro 1. Número de embriones somáticos observados en función del número de veces que el callo embriogénico fue subdividido a los 15 y 30 días de cultivo (n= 5, cada repetición se estableció con cinco callos embriogénicos).
Los valores con la misma letra son iguales estadísticamente, DMS p<0.05= 1.69.

40 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
Alfonso Azpeitia Morales et al.
En todos los tratamientos se observó a los 30 días una pérdida de ES ocasionada principalmente por una fuerte desdiferenciación del tejido embriogénico; sin embargo, durante este período de cultivo, se continuó observando un efecto favorable cuando el CE es subdividido respecto al control, sobresaliendo con 11 ES el tratamiento con 4 SCE por frasco, mientras que el control solamente presento 3.8 ES por CE. La interacción de ambos factores no fue estadísticamente significativa.
De acuerdo con Chan et al. (1998) la formación de embriones somáticos generados a partir de plúmula parte de la formación de callos iniciales que conducen a la formación de callos embriogénicos, estos CE pueden ser cul t ivados ín tegramente s in sufr i r fragmentación o división posterior. Los ES crecen
fusionados y su desarrollo no es sincronizado, lo cual limita su individualización, así como su desarrollo independiente hasta la formación de una planta completa, por lo que se decidió evaluar la posibilidad de que al subdividir callos embriogénicos (CE) se pudiera promover la formación de embriones somáticos (ES), tomando en cuenta el aumento de la superficie en contacto con el medio de cultivo. Los resultados permitieron observar que cuando un callo embriogénico es subdividido en 4 partes, el número de ES fue favorecido y se incrementó en 3.36 veces con respecto al control en los primeros 15 días de cultivo (Figura 1) y cuando este ensayo se efectuó de nuevo con mayor número de repeticiones, la respuesta fue similar aunque en una cantidad menor, obteniendo 2.7 veces más ES y 2.36 veces su peso fresco en SCE -F con respecto al control (CE +F) durante los primeros 15 días en condiciones II (Cuadro 2).
Factor A(días de cultivo)
Factor B
Control CE+F Control CE -F SCE +F SCE -F15 4.0 6.50 9.0 12.3530 3.70 5.80 8.80 9.90
Valor promedio 3.85 d 6.15 c 8.90 b 11.12 z
Cuadro 2. Número de embriones somáticos por callo embriogénico a los 15 y 30 días en cultivo en condiciones II n= 4, donde cada repetición se estableció con 40 callos embriogénicos y CE subdividido.
Factor B= callos embriogénicos en medio de cultivo con fitohormonas; CE +F= desprovisto de éstas; CE-F= subdivisiones de callo embriogénico; SCE= en medio con fitohormonas; +F= sin fitohormonas; -F. Valores con la misma letra son estadísticamente iguales DMS p<0.05= 2.0.
Cultivo de callos embriogénicos y subdivisión de callos embriogénicos en un medio con y sin fitohormonas 2,4-D (6 µM) y BAP (300 µM)
Número de embriones somáticos. El análisis de varianza mostró significancia en los tratamientos y en la prueba de comparación de medias DMS (p<0.05) para el factor A correspondiente a los 15 y 30 días, éstos fueron iguales estadísticamente con 7.96 ES a los 15 días, mientras que el valor promedio a los 30 días fue de 7.0 (DMS p<0.05= 2.30). En cuanto al factor B correspondiente al número de subdivisiones de callo embriogénico se encontraron diferencias estadísticas entre tratamientos. El mayor número de ES se observó en los tratamientos con subdivisiones de callo embriogénico con fitohormonas (SCE +F) y subdivisiones de callo embriogénico sin fitohormonas (SCE -F) presentando mayor número de ES (12.35 ES) en comparación a los controles (CE + F y CE - F). Sin embargo,
es importante señalar que cuando los CE y las SCE son cultivados en un medio desprovisto de fitohormonas fue mayor el número de ES (Cuadro 2).
Durante los 15 días de cultivo se presentó mayor número de ES en todos los tratamientos, sobresalió el tratamiento con SCE -F con 12.35 ES que en su mayoría fueron del tipo torpedo (EST); con 9.0 ES en el SCE +F y 6.50 ES en el tratamiento control -F, mientras que el control con fitohormonas (CE + F) se observaron 4.0 ES del tipo globular y muy pocos EST. A los 30 días se observó una disminución de ES en todos los tratamientos, siendo sobresaliente el SCE -F con 9.90 ES, superior al control (CE + F) que indujo 3.70 ES (Cuadro 2). En la interacción de ambos factores, no se observó significancia estadística.
Peso fresco. El análisis de varianza para esta variable fue significativo y en la prueba de comparación de medias

Influencia de la subdivisión del callo embriogénico en la formación de embriones somáticos de cocotero 41
DMS (p<0.05) se encontraron diferencias entre tratamientos. El factor A (días de cultivo) fue superior a los 30 días con un incremento de 0.62 g contra 0.26 g a los 15 días. Los tratamiento con SCE con + F y - F presentaron mayor peso fresco y fueron iguales estadísticamente y superiores a los tratamientos con
CE + F y - F (Cuadro 3). Se observó un incremento en 10 veces su peso fresco inicial en el tratamiento con SCE -F y un incremento de 2.36 veces respecto al control (CE + F, a los 30 días). La interacción entre los factores A (días de cultivo) y B (CE o SCE con o sin fitohormonas) no mostró significancia.
Factor A
(días de cultivo)
Factor B
Control CE+F Control CE -F SCE +F SCE –F
15 0.18 0.16 0.30 0.4030 0.39 0.40 0.80 0.90
Valor promedio 0.28 b 0.28 b 0.55 a 0.65 a
Cuadro 3. Peso fresco en los tratamientos con cultivo de callos embriogénicos (CE) y subdivisiones de callo embriogénico (SCE) en medio conteniendo fitohormonas (+F) y sin fitohormonas (-F) a los 15 y 30 días de cultivo (n= 4, cada repetición se formó con 4 CE y CE subdividido.
Factor B= callos embriogénicos en medio de cultivo con fitohormonas; CE + F= sin ellas; CE - F= subdivisiones de callo embriogénico; SCE= en medio con fitohormonas; +F= sin -F. Valores con la misma letra son iguales estadísticamente (DMS 0.05).
Con la inclusión o exclusión de las fitohormonas en etapa II, se observó que no es necesario incluir las fitohormonas 2,4-D (6 µM) y BAP (300 µM) durante los primeros 30 días de cultivo como fue reportado por Chan et al. (1998); sin embargo, Chan (com. pers.) mencionó que es más conveniente incluirlas debido a que su exclusión tiende a afectar el proceso de germinación.
Se ha propuesto que la inducción de heridas puede incrementar principalmente la formación de fitohormonas, mayor accesibilidad de nutrimentos así como la promoción de la división celular (Mariotti y Arcioni, 1983; Park y Son, 1988; Ambrozic-Dolinsek et al., 2001). Es importante mencionar que los CE así como las SCE mostraron diferencias en la formación de ES. Los CE hasta los 30 días presentaron ES tipo globular y esporádicamente los ES tipo torpedo, mientras que al utilizar SCE a los 15 y 30 días se observaron ES tipo torpedo y mayor sincronización entre ellos, además se observó un desarrollo y diferenciación más acelerado con respecto al CE. La SCE también estimuló una mayor ganancia en peso fresco, lo cual puede deberse a que la herida estimula la división celular (Mariotti y Arcioni, 1983) y una mayor síntesis endógena de fitohormonas en el explante (Park y Son, 1988). Asimismo, se ha reportado que la inducción de heridas en un tejido vegetal pueden estimular la organogénesis en condiciones in vitro (Ambrozic-Dolinsek et al., 2001); por ejemplo, cuando las hojas de Platycerium bifurcatum son heridas se
incrementa significativamente el potencial de formación de gametofitos en comparación con hojas intactas. Los gametofitos se originaron de áreas entre las heridas (Ambrozic-Dolinsek et al., 2001). Los autores mencionan que la síntesis o un cambio en el sitio de localización de síntesis de las fitohormonas dentro de la hoja herida podrían contribuir a la inducción de los gametofitos.
Por otra parte en cotiledones de Pyrus malus previamente seccionados a la mitad, en la región del corte se formó callo que dio origen a múltiples brotes adventicios (Browning et al., 1987), mientras que lo mismo no ocurrió con los callos embriogénicos subdivididos de cocotero. En las partes del corte del callo embriogénico no se observaron formaciones de embriones somáticos, estos ES se formaron en la periferia de las subdivisiones de los callos embriogénicos, zonas donde se forman normalmente aun sin realizar subdivisiones.
El aumento de ES en SCE cultivados en medio sólido, puede estar asociado a un mayor contacto con el medio de cultivo, dado que existe una mayor entrada de nutrimentos a las subdivisiones del CE. En trébol (Lotus corniculatus L.) se ha reportado que cuando un callo embriogénico es homogeneizado (macerado) se incrementan los embriones globulares en comparación con callos no homogeneizados (Orshinsky et al., 1983), por lo que se presume un aumento del contacto de las células maceradas con el medio de cultivo.
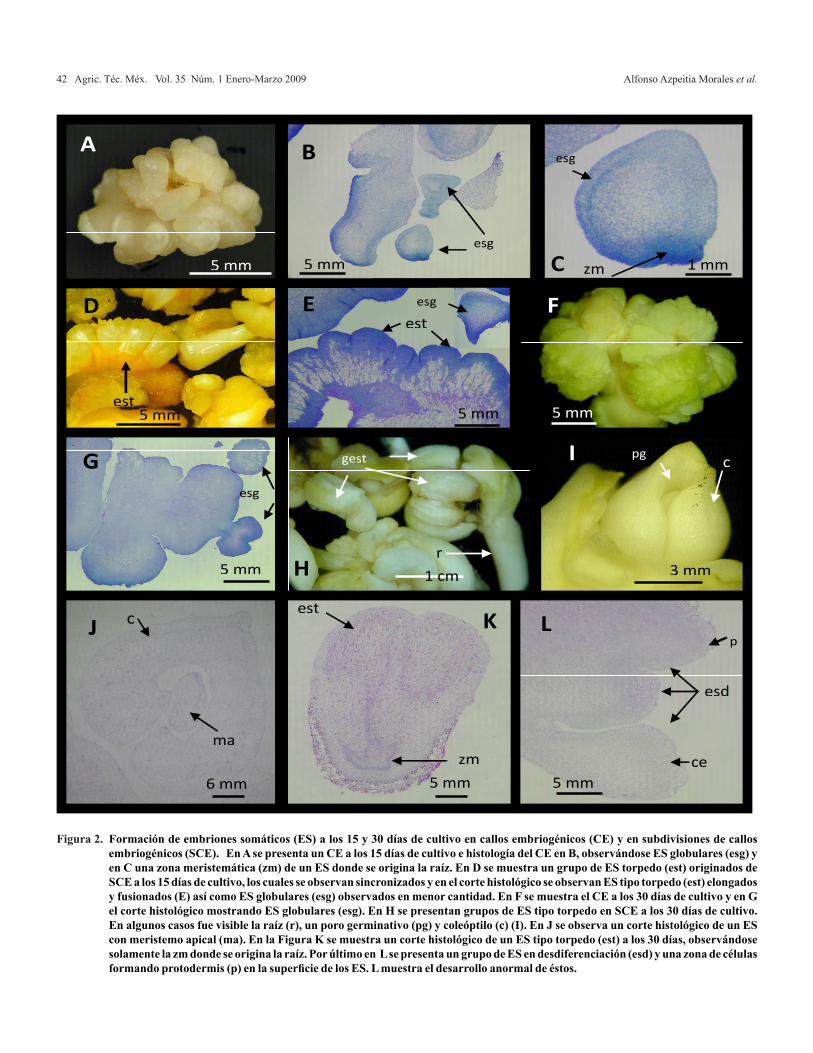
42 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
Alfonso Azpeitia Morales et al.
Formación de embriones somáticos (ES) a los 15 y 30 días de cultivo en callos embriogénicos (CE) y en subdivisiones de callos embriogénicos (SCE). En A se presenta un CE a los 15 días de cultivo e histología del CE en B, observándose ES globulares (esg) y en C una zona meristemática (zm) de un ES donde se origina la raíz. En D se muestra un grupo de ES torpedo (est) originados de SCE a los 15 días de cultivo, los cuales se observan sincronizados y en el corte histológico se observan ES tipo torpedo (est) elongados y fusionados (E) así como ES globulares (esg) observados en menor cantidad. En F se muestra el CE a los 30 días de cultivo y en G el corte histológico mostrando ES globulares (esg). En H se presentan grupos de ES tipo torpedo en SCE a los 30 días de cultivo. En algunos casos fue visible la raíz (r), un poro germinativo (pg) y coleóptilo (c) (I). En J se observa un corte histológico de un ES con meristemo apical (ma). En la Figura K se muestra un corte histológico de un ES tipo torpedo (est) a los 30 días, observándose solamente la zm donde se origina la raíz. Por último en L se presenta un grupo de ES en desdiferenciación (esd) y una zona de células formando protodermis (p) en la superficie de los ES. L muestra el desarrollo anormal de éstos.
Figura 2.
A B
C
E
L
I
esg
esg
est
gest c
ma
r
zm ce
zm 5 mm 5 mm 1 mm
5 mm
1 cm
5 mm 5 mm
3 mm
J
pg
6 mm
c K
esd
est
5 mm
esg
G esg
5 mm
D F
H
p
est 5 mm

Influencia de la subdivisión del callo embriogénico en la formación de embriones somáticos de cocotero 43
Evaluación histológica y morfológica de los callos embriogénicos y subdivisiones
Observaciones a los 15 días de cultivo. Los callos embriogénicos mostraron sobre su superficie embriones somáticos del tipo globular (sólo visibles bajo microscópico estereoscópico) y en los cortes histológicos se observaron embriones somáticos en formación y del tipo globular (esg) sin presentarse el meristemo apical, pero si se observó una zona meristemática (zm) donde se origina la raíz (Figura 2 A, B y C). En cuanto a las subdivisiones de callo embriogénico (SCE) se observaron secciones con ES con un estado de desarrollo sincronizado con ES tipo torpedo (est) como se puede observar en la Figura 2D, estos ES fueron visibles a simple vista; sin embargo, los cortes histológicos mostraron formación de ES fusionados en desarrollo y también del tipo globular (Figuras 2 E). Durante este período de cultivo no fue visible el meristemo apical en los cortes histológicos de los ES originados de las SCE.
Observaciones a los 30 días de cultivo. Durante este período de cultivo los CE se tornaron de color verde claro (Figura 2F) y no se observaron embriones tipo torpedo, pero si un crecimiento de tejido haustorial. En los cortes histológicos se observaron pocos ES globulares (Figura 2G) y tejido desdiferenciado conocido generalmente como haustorio con ES fusionados y poco definidos. Además, es importante mencionar que fue raro observar embriones somáticos tipo torpedo. En cuanto a las SCE, éstas presentaron grupos de ES tipo torpedo, con aparente buena conformación (Figura 2H) con color blanco y en pocos casos con sistema radicular (r). En algunos casos se encontraron ES con poro germinativo (Figura 2 I y J) observándose un coleóptilo (c) protegiendo al meristemo apical (ma). La mayor parte de los ES fueron bien definidos sin observarse el meristemo apical (Figuras 2 K), solamente fue posible observar una zona meristemática donde se origina la raíz. Además, los cortes histológicos también mostraron grupos de ES fusionados (Figura 2 L) con una capa de células bien definida formando protodermis en la periferia, zona donde se observó tejido haustorial por lo que estas células localizadas en esta zona dan origen a este tipo de tejido.
Como ha sido indicado, el cocotero es una especie recalcitrante debido a que presenta baja capacidad embriogénica (Verdeil y Buffard Morel, 1995) y por lo tanto poca respuesta morfogenética. En este estudio se presentaron principalmente dos respuestas morfogenéticas:
1) diferenciación de embriones somáticos de 15 a los 30 días, y 2) desdiferenciación de ES después de los 30 días, con crecimiento de callo tipo haustorial en la periferia de los ES formados por lo que esta desdiferenciación del CE, lo que ocasiona la disminución de ES. Esta disminución de ES ocurre después de 30 días de cultivo en callos embriogénicos así como en subdivisiones de callo embriogénico, lo anterior es debido a que se promueve una proliferación rápida de células en la superficie de los ES o del CE, que conduce a la formación de callo o tejido tipo haustorial. En los cortes histológicos realizados fue posible observar una capa de células formando protodermis bien definida en la periferia de los ES, los cuales presentaron un ligero crecimiento de tejido tipo haustorial, ya que estas células dan origen a este tipo de tejido (Figura 2 L).
Aunque fue posible observar ES bien formados (SCE -F) a los 15 días de cultivo, en los cortes histológicos no se observó el meristemo apical (Figura 2 F), sólo en algunos casos los ES mostraron las partes características que conforman un embrión como: poro germinativo, coleóptilo y meristemo apical, pero sólo después de los 30 días de cultivo (Figura 2 J). En la actualidad se desconocen las causas que influye en la fuerte heterogeneidad presentada en el desarrollo de los ES formados de cocotero. De acuerdo a Blake (1990), estas anormalidades en el desarrollo de ES son atribuidas a diversos factores, pero probablemente provocadas por niveles sub o supra óptimos de auxinas y/o citocininas.
Indudablemente, la formación del meristemo apical en un embrión somático es importante, debido a que las células no diferenciadas de esta estructura son las responsables de la iniciación de órganos en las plantas superiores (Clark et al., 1997). La ausencia del meristemo apical en los ES formados esta probablemente asociado, a cambios en el nivel de expresión de genes, por ejemplo, se conoce que la expresión de los genes cdc2 y cdc2a son un factor critico en la regulación de la actividad meristemática y el establecimiento de la competencia proliferativa en Arabidopsis thaliana y pueden contribuir a la regulación temporal y espacial de la división celular en plantas (Martínez et al., 1992; Hemerly et al., 1993).
CONCLUSIONES
La subdivisión del callo embriogénico del cocotero favoreció la obtención de embriones somáticos tipo torpedo a los 15 días de cultivo en medio Y3 sin fitohormonas en condiciones de luminosidad.

44 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
Alfonso Azpeitia Morales et al.
Los resultados son prometedores y establecen las bases para incrementar el número de embriones somáticos y eventualmente la multiplicación de callos embriogénicos en cocotero, lo cual podrá permitir la propagación a gran escala de palmas híbridas o palmas élite.
AGRADECIMIENTOS
Alfonso Azpeitia Morales agradece al Centro de Investigación Científica de Yucatán (CICY) por haber llevado a cabo en sus instalaciones el trabajo de investigación de la tesis de doctorado, así como al Instituto Nacional de Investigaciones Agrícolas, Forestales y Pecuarias (INIFAP) y al CONACYT (beca 119335) por el apoyo económico para su manutención. El presente artículo es parte de la tesis doctoral. Este proyecto fue financiado por el CONACYT-SISIERRA (proyecto número 990130).
LITERATURA CITADA
Ambrozic-Dolinsek, J. M.; Camloh, B. B. and Zel, J. 2001. Apospory in leaf culture of staghorn fern (Platycerium bifurcatum). Plant Cell Reports (on line), file://A:\Plant%20Cell%20Rep%20-%20Full%20Text%20DOI%2010_1007-s00299-001-.
Blake, J. 1990. Coconut (Cocos nucifera L.): Micropropagation. In: Bajaj, Y. P. S (ed.). Biotechnology in agriculture and forestry. Springer-Verlag Berlin Heidelberg. Vol. 10, pp. 538-554.
Browning, G. V. O.; Passey, A. J. and James, D. J. 1987. Multiple Shoot and root regeneration from pear embryo cotyledon explants in vitro. J. Hortic. Sci. 62:305-311.
Buffard-Morel, J.; Verdeil, J. L. and Pannetier, C. 1992. Embryogenèse somatique du cocotier (Cocos nucifera L.) à partir de tissus foliaires: étude histologique. Can J. Bot. 70:735-741.
Clark, E. S.; Robert, W. W. and Meyerowitz, E. M. 1997. The CLAVATA1 gene encodes a putative receptor kinase that controls shoot and floral meristem size in Arabidopsis. Cell 89:575-585.
Chan, L. J.; Saénz, L.; Talavera, C.; Hornung, R.; Robert, M. and Oropeza, C. 1998. Regeneration of coconut (Cocos nucifera L.) from plumule explants through somatic embryogenesis. Plant Cell Reports. 17:515-521.
Dussert, S.; Verdeil, J. L.; Rival, A. M. N. and Buffard-Morel, J. 1995. Nutrient uptake and growth of in vitro coconut (Cocos nucifera L.) calluses. Plant Sci. 106:185-193.
Eeuwens, C. J. 1976. Mineral requirements for growth and callus initiation of tissue explants excised from mature coconut (Cocos nucifera) and date (Phoenix dactylifera) palms cultured in vitro. Physiol. Plant. 36:23-28.
Ebert, A. and Taylor, H. F. 1990. Assessment of the changes of 2,4-dichlorophenoxyacetic acid concentrations in plant tissue culture media in the presence of activated charcoal. Plant Cell, Tissue and Organ Culture. 20:165-172.
George, F. E. 1993. Plant growth regulators. Micropropagation in practice. In: George, E. F. (ed.) Plant Propagation by Tissue Culture (Part 2). pp. 834-1122.
Hemerly, A. S.; Ferreira, P.; De Almeida, J. E.; Van Montagu, M.; Engler, G. and Inzé, D. 1993. cdc2a expression in Arabidopsis is linked with competence for cell division. Plant Cell. 5:1711-1723.
Herrera, H. G. A. 2002. Caracterización fisicoquímica del carbón activado y su efecto en el cultivo in vitro de Cocos nucifera L. Tesis de Licenciatura. Instituto Tecnológico Agropecuario Núm. 2. 49 p.
Hornung, R. 1995. Micropropagation of Cocos nucifera L. from plumular tissue excised from mature zygotic embryos. Plantations, Recherche, Development. 2:38-41.
Kawahara, R. and Komamine, A. 1995. Molecular basis of somatic embryogenesis. In: Bajaj, Y. P. S. (ed). Biotechnology in agriculture and forestry, Vol. 30, Somatic embryogenesis and synthetic seed I. Spriger-Verlag, Berlin Heidelberg, Alemania. pp. 30-39.
Magnaval, C.; Noirot, M.; Verdeil, J. L.; Blattes, A.; Huet, C.; Grosdemange, F.; Beule, T. and Buffard-Morel, J. 1997. Specific nutritional requirements of coconut calli (Cocos nucifera L.) during somatic embryogenesis induction. J. Plant Physiol. 150:719-728.
Mariotti, D. and Arcioni, S. 1983. Coronilla varia L. (Crown vetch): plant regeneration through somatic embryogenesis. Plant Cell, Tissue and Organ Culture. 2:103-110.
Martinez, C. M.; Jan-Elo, J.; Lawton, M. A.; Lamb, C. J. and Doerner, P. W. 1992. Spatial pattern of cdc2 expression in relation to meristem activity and cell proliferation during plant development. Proc. Natl. Acad. Sci. 89:7360-7364.

Influencia de la subdivisión del callo embriogénico en la formación de embriones somáticos de cocotero 45
Meng-Jin, C. and Wei-Chin, C. 1987. Somatic embryogenesis and plant regeneration in callus culture derived from immature seeds and mature zygotic embryos of Dysosma pleiantha (Hance) Woodson. Plant Cell Reports. 6:484-485.
Olivares, S. E. 1994. Paquete de diseños experimentales FEUANL. Versión 2.5. Facultad de Agronomía Universidad Autónoma de Nuevo León (UANL). Marín, Nuevo León, México.
Oropeza, C. and Chan. J. L. 1995. Protocol for the formation of somatic embryos from coconut plumule explants. In: Compendium of protocols. Verdeil, J. L. (ed.). STD3 (ERTBTS3*CT940298). ORSTOM-CIRAD, Montpellier, Francia 280 p.
Orshinsky, B. R.; Swanson, E. B. and Tomes, D. T. 1983. Enhanced shoot regeneration from homogenized callus cultures of birdsfoot trefoil (Lotus corniculatus L.). Plant Cell Tissue and Organ Culture. 2:341-347.
Park, Y. G. and Son, S. H. 1988. In vitro organogenesis and somatic embryogenesis from punctured leaf of Populus nigra x P. maximowiezii. Plant Cell Tissue and Organ Culture. 15:95-105.
Piña, R. J. 1998. Programa nacional de investigación del cocotero. Memoria de la 1ra. Reunión Nacional de Palma de Coco, Acapulco , Guerrero, México. pp 5-17.
Smith, D. L. and Krikorian, A. D. 1989. Release of somatic embryogenic potential from excised zygotic embryos of carrot and maintenance of polyembryonic cultures in hormone-free medium. Plant Sci. 58:1832-1843.
Verdeil, J-L. and Buffard-Morel, J. 1995. Somatic embryogenesis in coconut (Cocos nucifera L.): In: Bajaj, Y. P. S. (ed) Biotechnology in agriculture and forestry, Vol. 30. Somatic embryogenesis and synthetic seed I. Spriger-Verlag, Berlin Heidelberg, Alemania pp. 299-317.

Agricultura Técnica en México Vol. 35 Núm.1 1 de enero-31 de marzo 2009 p. 46-57
PATOTIPOS DE Colletotrichum lindemuthianum EN OAXACA Y SAN LUIS POTOSÍ, MÉXICO, Y RESISTENCIA EN GENOTIPOS DE FRIJOL*
Colletotrichum lindemuthianum PATHOTYPES IN OAXACA AND SAN LUIS POTOSI, MEXICO, AND RESISTANCE IN COMMON BEAN
Bertha María Sánchez-García1, Alberto Flores-Olivas1, Abiel Sánchez-Arizpe1, Susana Pineda-Rodríguez2, Gabriela López-Jiménez2, Saúl Fraire-Velásquez3, Eduardo Raymundo Garrido-Ramírez4, Jorge Alberto Acosta-Gallegos2, June Simpson Williamson5 y Raúl Rodríguez-Guerra6§
1Universidad Autónoma Agraria Antonio Narro, Departamento de Parasitología Agrícola, A. P. 342, Buenavista, Saltillo, Coahuila, México. C. P. 25315. Tel. 01 844 4110226, ([email protected]), ([email protected]), ([email protected]). 2Campo Experimental Bajío, INIFAP, km 6.5 carretera Celaya-San Miguel de Allende, A. P. 112, Celaya, Guanajuato, México. C.P. 38000. Tel. 01 461 6115323, ([email protected]), ([email protected]), ([email protected]). 3Unidad Académica de Biología Experimental, Universidad Autónoma de Zacatecas, Av. Revolución Mexicana S/N, Col. Tierra y Libertad, Guadalupe, Zacatecas, México C. P. 98600. Tel. 01 492 8993420, ([email protected]). 4Campo Experimental Centro de Chiapas, INIFAP, km 3.0 carretera Ocozocoautla-Cintalapa, Ocozocoautla, Chiapas, México, C. P. 29140. Tel. 01 968 6882915, ([email protected]). 5Centro de Investigación y Estudios Avanzados del Instituto Politécnico Nacional-Unidad Irapuato, A. P. 629, Irapuato, Guanajuato, México C. P. 36500. Tel. 01 462 6239667, ([email protected]). 6Campo Experimental General Terán, INIFAP, km. 31 carretera Montemorelos-China, A. P. Núm. 3, General Terán, Nuevo León, México, C. P. 67400. Tel. 01 826 2670539. §Autor para correspondencia: [email protected].
* Recibido: Enero, 2008
Aceptado: Marzo, 2009
RESUMEN
La antracnosis del frijol (Phaseolus vulgaris L.), causada por Colletotrichum lindemuthianum (Sacc. et Magn.) Scrib., es una enfermedad ampliamente distribuida en las regiones productoras del mundo. Este hongo muestra una gran diversidad de patotipos identificados alrededor del mundo, cuyo conocimiento es importante para el desarrollo de variedades resistentes. En México se conocen 54 patotipos; sin embargo, en algunos estados se desconocen los patotipos existentes y la frecuencia con que ocurren. En esta investigación, se caracterizaron patogénicamente 23 cepas de C. lindemuthianum de Oaxaca (15) y San Luis Potosí (8) con base en la reacción de 12 variedades diferenciales de 2005-2007. Asimismo, el nivel de resistencia de 24 materiales nativos de San Luis Potosí, 115 de Guanajuato y 55 variedades mejoradas, fue registrado por su reacción a los patotipos 64 y 320 (presentes en ambos estados) que
infectan a variedades diferenciales de origen mesoamericano. En Oaxaca y San Luis Potosí se identificaron 12 y 5 patotipos, respectivamente, siete de los cuales (69, 100, 260, 324, 325, 356 y 485) son reportados por primera vez en México. Lo anterior indica que la variabilidad de este patógeno es mayor que la reportada anteriormente y que nuevas combinaciones de genes de avirulencia están presentes o desarrollándose en México. De los materiales de San Luis Potosí, Guanajuato y variedades mejoradas de frijol 12, 76 y 28 respectivamente, fueron resistentes a los patotipos 64 y 320; estos genotipos podrían utilizarse como fuentes de resistencia a C. lindemuthianum.
Palabras clave: Glomerella lindemuthiana (Sacc. et Magn.) Scrib., frecuencia y distribución de patotipos, fuentes de resistencia.

Patotipos de Colletotrichum lindemuthianum en Oaxaca y San Luis Potosí, México, y resistencia en genotipos de frijol 47
ABSTRACT
Anthracnose, a disease of common bean caused by Colletotrichum lindemuthianum (Sacc. et Magn.) Scrib., is worldwide distributed. C. lindemuthianum displays a significant pathogenic diversity which is important to know for the development of resistant cultivars. In Mexico 54 pathotypes are known; however, some of the important bean producing states have not been thoroughly sampled. In this research 23 strains of C. lindemuthianum from Oaxaca (15) and San Luis Potosí (8) were pathotypically characterized using twelve differential cultivars, during 2005-2007. In addition, 24 bean landraces from San Luis Potosí, 115 from Guanajuato, as well as 55 bread cultivars, were challenged with the pathotypes 64 and 320 (which occur in both states) that infect differential cultivars from Mesoamerican origin, in the search for sources of resistance. Twelve and five pathotypes were identified in Oaxaca and San Luis Potosí, respectively, with seven of them (69, 100, 260, 324, 325, 356 and 485) reported for the first time in Mexico. Results indicate higher pathogenic diversity than previously reported and that newly form or undetected combinations of avirulence genes are present in Mexico. From the 24 bean landraces from San Luis Potosí, 115 from Guanajuato, and the 55 bred cultivars challenged, 12, 76 and 28, respectively, were resistant to both races; these genotypes could be used as sources of resistance against C. lindemuthianum.
Key words: Glomerella lindemuthiana (Sacc. et Magn.) Scrib., frequency and distribution of pathotypes, sources of resistance.
INTRODUCCIÓN
La antracnosis del frijol, ocasionada por el hongo Colletotrichum lindemuthianum (Sacc. et Magn.) Scrib. (teleomorfo Glomerella lindemuthiana), es de amplia distribución en el mundo y puede ser devastadora si se siembran variedades susceptibles y las condiciones climáticas son favorables (temperaturas entre 13 y 26 ºC y humedad superior a 92%) para el desarrollo de la enfermedad (Pastor-Corrales y Tu, 1989). En México, la enfermedad esta presente en todas las regiones productoras de frijol, con excepción de regiones donde se cultiva bajo condiciones de riego durante la época seca en el invierno (González-Chavira et al., 2004; Rodríguez-Guerra et al., 2006). Entre las diversas estrategias para combatirla está la utilización de cultivares resistentes. Sin embargo, para el
desarrollo de cultivares con resistencia a la antracnosis, es importante conocer los patotipos presentes en las regiones para las cuales se desarrollarán los nuevos cultivares; esto es considerando que la interacción entre C. lindemuthianum y Phaseolus vulgaris es del tipo gen por gen, que considera que por cada gen de resistencia en el hospedero existe un gen de avirulencia en el patógeno (Flor, 1942; Flor, 1956), aunque se ha reportado evidencia de resistencia parcial conferida por varios genes (Geffroy et al., 2000). También, se ha demostrado una estrecha relación coevolutiva entre patotipos y el germoplasma originado en los centros de origen del frijol (andino y mesoamericano; Gepts y Debouck, 1991), lo que significa que patotipos de un origen son capaces de infectar preferentemente genotipos de frijol del mismo origen (Pastor-Corrales, 1996; Araya, 2003).
El hongo de la antracnosis muestra una gran diversidad de patotipos, a fines del siglo pasado se habían reportado 93 patotipos en el mundo (Hernández-G. et al., 1998); actualmente son más de 100 y de éstos, 54 están presentes en México (Rodríguez-Guerra et al., 2003; González-Chavira et al., 2004; Rodríguez-Guerra et al., 2006). Los aislados de C. lindemuthianum provenientes de trece estados de México, incluyendo los principales estados que dedican mayor superficie al cultivo del frijol (Chihuahua, Durango, Zacatecas, Jalisco, Michoacán, Guerrero, Hidalgo, Morelos, México, Tlaxcala, Puebla, Veracruz, Chiapas) han sido caracterizados patogénicamente (Rodríguez-Guerra et al., 2006). Pero otros estados han sido escasamente estudiados o en estos ningún estudio de este tipo se ha realizado. El estado con mayor número de patotipos es Jalisco (Rodríguez-Guerra et al., 2006), siendo los patotipos 448, 256 y 0 los más frecuentes en México (González-Chavira et al., 2004; Rodríguez-Guerra et al., 2006); y junto con el 320, son los más ampliamente distribuidos, y ocurren en la mayoría de los estados donde se han realizado estudios sobre la identificación de patotipos de C. lindemuthianum (Rodríguez-Guerra et al, 2006).
El conocimiento de la diversidad patogénica de C. lindemuthianum en México, ha permitido identificar fuentes de resistencia a patotipos virulentos, ampliamente distribuidos y capaces de infectar incluso a genotipos de origen andino usados como variedades diferenciales para la identificación de patotipos (González-Chavira et al., 2004). La búsqueda de fuentes de resistencia se ha extendido a frijoles criollos, silvestres y a especies afines al frijol común (Mahuku et al., 2002; Ferreira et al., 2003; Sartorato y Arraes, 2003). Recientemente se ha sugerido que el uso

48 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
Bertha María Sánchez-García et al.
mediante inoculaciones artificiales de un reducido número de patotipos, que en conjunto sean capaces de vencer la resistencia de la mayoría de las variedades diferenciales, puede permitir la identificación y selección de fuentes de resistencia a la mayoría de los patotipos presentes en México (López-Jiménez et al., 2006).
Los objetivos de esta investigación fueron determinar los patotipos de C. lindemuthianum a partir de aislados provenientes de los estados de Oaxaca y San Luis Potosí, en donde se desconoce la variabilidad patogénica del hongo, e identificar fuentes de resistencia a dos patotipos que ocurren en común en ambos estados.
MATERIALES Y MÉTODOS
Muestreo y aislamiento de cepas
Durante el ciclo primavera-verano 2005 y 2006 se hicieron recorridos en campos de producción de frijol de los estados de San Luis Potosí y Oaxaca, ocho y catorce, respectivamente. Se colectaron vainas de frijol con síntomas evidentes de antracnosis. El aislamiento del hongo se realizó como fue descrito previamente (González et al., 1998). Se obtuvo una suspensión de conidios a partir de lesiones esporuladas y fue transferida y dispersada en caja de Petri conteniendo
el medio de cultivo papa dextrosa agar (PDA) acidificado (200 μL de acido láctico al 85% incorporado después de la esterilización). Para desarrollar cepas monoconidiales de cada campo muestreado se extrajo un conidio germinado entre las 24-48 h y se transfirió a una nueva caja Petri con medio de cultivo. Se obtuvieron 23 cultivos monoconidiales, uno por cada campo muestreado, sólo en un campo de Oaxaca se obtuvieron dos. Las cepas fueron duplicadas en PDA para mantenerlas en activo a 4 ºC y ser usadas en la determinación de patotipos e identificación de fuentes de resistencia. Los cultivos monoconidiales se conservaron a 4 ºC por periodos prolongados en tubos de transporte conteniendo PDA y cubiertas con glicerol al 50%.
Determinación de patotipos
Los patotipos de los aislados fueron determinados utilizando un grupo de 12 variedades diferenciales de frijol y la nomenclatura binaria descrita por Pastor-Corrales (1991). En este sistema de denominación de patotipos cada diferencial (nueve de origen mesoamericano y tres de origen andino) posee un valor asignado (Cuadro 1), y el nombre o número del patotipo corresponde a la suma de los valores de cada variedad que reacciona como susceptible a algún aislado en cuestión. En caso de que ninguna variedad diferencial reaccione como susceptible a algún aislado, éste será designado como patotipo cero.
Núm. Variedad diferencial Valor asignado Origen1 Gen de resistencia conocido1 Michelite (20)= 1 Mesoamericano2 Michigan Dark Red Kidney (21)= 2 Andino Co-13 Perry Marrow (22)= 4 Andino Co-13
4 Cornell 49242 (23)= 8 Mesoamericano Co-25 Widusa (24)= 16 Mesoamericano Co-15, Co-93
6 Kaboon (25)= 32 Andino Co-12
7 Mexico 222 (26)= 64 Mesoamericano Co-38 PI 207262 (27)= 128 Mesoamericano Co-43, Co-99 To (28)= 256 Mesoamericano Co-410 Tu (29)= 512 Mesoamericano Co-511 AB 136 (210)= 1024 Mesoamericano Co-6, Co-812 G 2333 (211)= 2048 Mesoamericano Co-42, Co-5, Co-7
Cuadro 1. Características de las variedades de frijol utilizadas actualmente como diferenciales para la identificación de patotipos de Colletotrichum lindemuthianum (Pastor-Corrales, 1991).
1Se refiere al acervo genético del cual proviene cada diferencial; el andino se originó en los andes sudamericanos, mientras que el mesoamericano comprende principalmente América Central y México (Gepts y Debouck, 1991).

Patotipos de Colletotrichum lindemuthianum en Oaxaca y San Luis Potosí, México, y resistencia en genotipos de frijol 49
Núm. Variedad diferencial Valor asignado Origen1 Gen de resistencia conocido1 Michelite (20)= 1 Mesoamericano2 Michigan Dark Red Kidney (21)= 2 Andino Co-13 Perry Marrow (22)= 4 Andino Co-13
4 Cornell 49242 (23)= 8 Mesoamericano Co-25 Widusa (24)= 16 Mesoamericano Co-15, Co-93
6 Kaboon (25)= 32 Andino Co-12
7 Mexico 222 (26)= 64 Mesoamericano Co-38 PI 207262 (27)= 128 Mesoamericano Co-43, Co-99 To (28)= 256 Mesoamericano Co-410 Tu (29)= 512 Mesoamericano Co-511 AB 136 (210)= 1024 Mesoamericano Co-6, Co-812 G 2333 (211)= 2048 Mesoamericano Co-42, Co-5, Co-7
La semilla de las variedades diferenciales fue tratada con hipoclorito de sodio al 1% por un min y lavada en dos pasos de agua estéril. Posteriormente se sembró en macetas de plástico con capacidad de 0.5 L conteniendo vermiculita, y permanecieron en invernadero durante 10 a 15 días hasta que las hojas primarias estuvieron completamente expandidas para ser inoculadas.
Las cepas monoconidiales mantenidas en PDA acidificado se transfirieron a cajas de Petri conteniendo el medio de cultivo harina de maíz-agar (infusión de 20 g de harina de maíz en un L de agua por una h a 60 ºC, 2 g de glucosa, 1 g de extracto de levadura y 18 g de agar por litro de medio) para inducir abundante esporulación. Las cajas inoculadas se mantuvieron a 25-27 ºC por 10 días y los conidios fueron obtenidos al agregar agua estéril a las cajas, mediante el raspado de la superficie con una varilla de vidrio. Finalmente se obtuvo una suspensión con una concentración de 1.5 x 106 conidios mL-1, a la cual se le agregaron 2 gotas de Tween 20 por cada 100 mL de la suspensión.
Diez días después de la emergencia, cinco plantas de cada uno de los 12 cultivares diferenciales fueron inoculadas asperjando la suspensión conidial en el envés de las hojas primarias. Las plantas inoculadas fueron cubiertas con plástico transparente para mantener una humedad aproximada al 95%, a una temperatura de 22 ºC, por tres días. Diez días después de la inoculación los síntomas fueron evaluados usando una escala de cinco valores (Garrido-Ramírez y Romero-Cova, 1989). La reacción de las plantas fue clasificada como: 0= síntomas no visibles, 1= pequeñas lesiones en las nervaduras principales visibles sólo en el envés de las hojas, 2= lesiones en haz y envés de las hojas, 3= defoliación de la planta y esporulación del hongo, 4= planta muerta. Las plantas con reacción de 0 a 2 se consideraron como resistentes y las plantas con reacción de 3 y 4 se consideraron susceptibles. Cuando plantas distintas de una misma variedad diferencial mostraron reacciones distintas de resistencia o susceptibilidad a algún aislado, nuevas plantas de la variedad fueron inoculadas con el aislado en cuestión hasta obtener una reacción consistente de las plantas.
Considerando la reacción que causaron los patotipos sobre las 12 variedades diferenciales, se obtuvo el índice de resistencia (IR) el cual se estima mediante la siguiente relación= núm. de patotipos que no causan susceptibilidad a la variedad en cuestión/núm. de patotipos inoculados sobre la variedad x 100) de cada una, y el (IV) el cual
se representa por: índice de virulencia (IV= núm. de variedades susceptibles al patotipo en cuestión/núm. total de variedades inoculadas X 100) de los patotipos (Antunes et al., 2003). Ambos índices pueden tomar valores de 0 a 100, valores elevados del IR implica que las diferenciales son resistentes a un mayor número de patotipos y valores elevados del IV indican que los patotipos son capaces de causar enfermedad a un mayor número de variedades diferenciales. Para ambos estados se determinó el índice de diversidad de patotipos utilizando la fórmula: H= -Σpi lnpi;
donde, pi= representa la frecuencia de cada patotipo (Shannon y Weaver, 1963). Este índice permite obtener valores de cero (ausencia de variabilidad) o superiores (mientras el valor sea más elevado la diversidad de patotipos es mayor), y también fue aplicado a los datos de patotipos de los estados de Chihuahua, Durango, Zacatecas, Jalisco, Michoacán (González et al., 1998), Hidalgo, Tlaxcala, México, Puebla (González-Chavira et al., 2004) para fines de análisis y comparación.
Fuentes de resistencia en cultivares nativos y mejorados
Se evaluaron 24 materiales nativos de San Luis Potosí y 115 de Guanajuato pertenecientes al banco de germoplasma del Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP) y 55 variedades mejoradas liberadas por el INIFAP, por su reacción a los patotipos 64 (aislado CoSLP-8) y 320 (aislado CoOax-9B) de C. lindemuthianum. Los patotipos fueron seleccionados por ocurrir en los dos estados considerados en el estudio, infectar variedades diferenciales de origen mesoamericano y para determinar si la estrategia presentada por López-Jiménez et al. (2006) permite demostrar que los genotipos resistentes al patotipo 320, también lo son al 64. La semilla fue tratada como se mencionó anteriormente, al igual que la elaboración del inóculo de los patotipos. Al menos cinco plantas de cada material fueron inoculadas con cada uno de los patotipos; el procedimiento de inoculación utilizado fue el mismo que en la caracterización de patotipos y la reacción de los materiales se determinó con la escala de cinco valores antes mencionada. Cuando plantas individuales mostraron reacciones de resistencia o susceptibilidad dentro de los materiales nativos o mejorados de frijol, nuevas inoculaciones fueron realizadas para confirmar la reacción de los materiales.
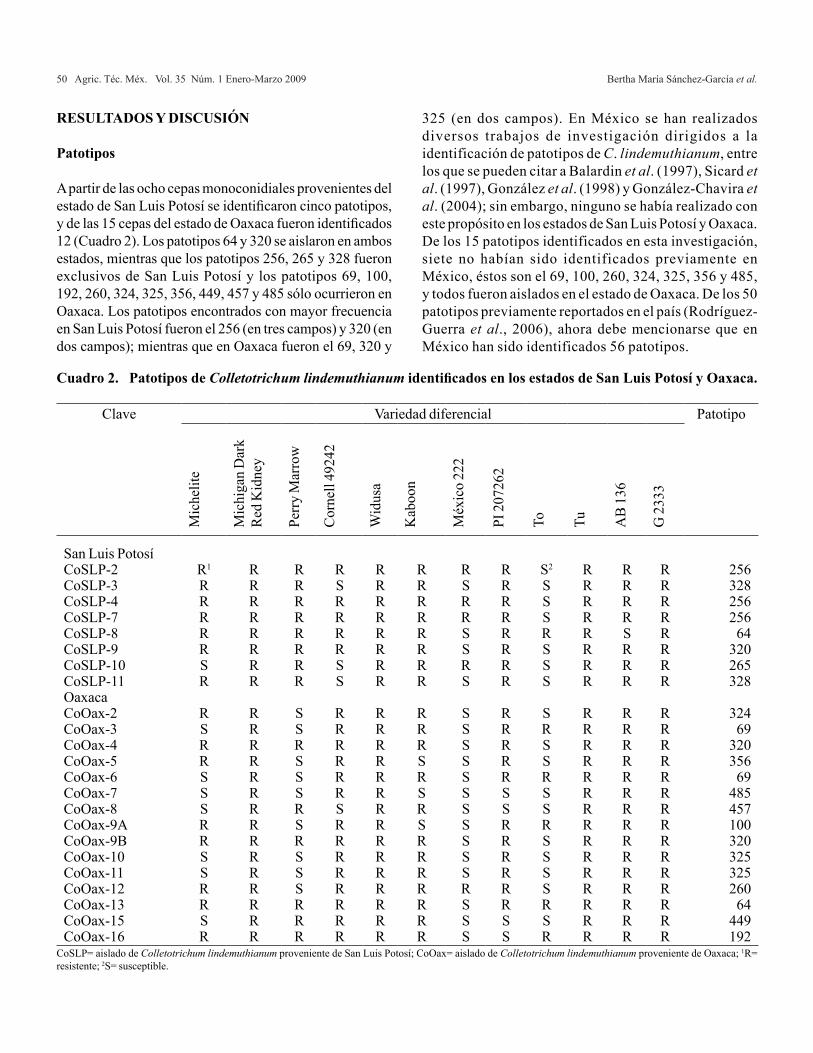
50 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
Bertha María Sánchez-García et al.
325 (en dos campos). En México se han realizados diversos trabajos de investigación dirigidos a la identificación de patotipos de C. lindemuthianum, entre los que se pueden citar a Balardin et al. (1997), Sicard et al. (1997), González et al. (1998) y González-Chavira et al. (2004); sin embargo, ninguno se había realizado con este propósito en los estados de San Luis Potosí y Oaxaca. De los 15 patotipos identificados en esta investigación, siete no habían sido identificados previamente en México, éstos son el 69, 100, 260, 324, 325, 356 y 485, y todos fueron aislados en el estado de Oaxaca. De los 50 patotipos previamente reportados en el país (Rodríguez-Guerra et al., 2006), ahora debe mencionarse que en México han sido identificados 56 patotipos.
RESULTADOS Y DISCUSIÓN
Patotipos
A partir de las ocho cepas monoconidiales provenientes del estado de San Luis Potosí se identificaron cinco patotipos, y de las 15 cepas del estado de Oaxaca fueron identificados 12 (Cuadro 2). Los patotipos 64 y 320 se aislaron en ambos estados, mientras que los patotipos 256, 265 y 328 fueron exclusivos de San Luis Potosí y los patotipos 69, 100, 192, 260, 324, 325, 356, 449, 457 y 485 sólo ocurrieron en Oaxaca. Los patotipos encontrados con mayor frecuencia en San Luis Potosí fueron el 256 (en tres campos) y 320 (en dos campos); mientras que en Oaxaca fueron el 69, 320 y
Clave Variedad diferencial Patotipo
Mic
helit
e
Mic
higa
n D
ark
Red
Kid
ney
Perr
y M
arro
w
Cor
nell
4924
2
Wid
usa
Kab
oon
Méx
ico
222
PI 2
0726
2
To Tu AB
136
G 2
333
San Luis PotosíCoSLP-2 R1 R R R R R R R S2 R R R 256CoSLP-3 R R R S R R S R S R R R 328CoSLP-4 R R R R R R R R S R R R 256CoSLP-7 R R R R R R R R S R R R 256CoSLP-8 R R R R R R S R R R S R 64CoSLP-9 R R R R R R S R S R R R 320CoSLP-10 S R R S R R R R S R R R 265CoSLP-11 R R R S R R S R S R R R 328OaxacaCoOax-2 R R S R R R S R S R R R 324CoOax-3 S R S R R R S R R R R R 69CoOax-4 R R R R R R S R S R R R 320CoOax-5 R R S R R S S R S R R R 356CoOax-6 S R S R R R S R R R R R 69CoOax-7 S R S R R S S S S R R R 485CoOax-8 S R R S R R S S S R R R 457CoOax-9A R R S R R S S R R R R R 100CoOax-9B R R R R R R S R S R R R 320CoOax-10 S R S R R R S R S R R R 325CoOax-11 S R S R R R S R S R R R 325CoOax-12 R R S R R R R R S R R R 260CoOax-13 R R R R R R S R R R R R 64CoOax-15 S R R R R R S S S R R R 449CoOax-16 R R R R R R S S R R R R 192
Cuadro 2. Patotipos de Colletotrichum lindemuthianum identificados en los estados de San Luis Potosí y Oaxaca.
CoSLP= aislado de Colletotrichum lindemuthianum proveniente de San Luis Potosí; CoOax= aislado de Colletotrichum lindemuthianum proveniente de Oaxaca; 1R= resistente; 2S= susceptible.

Patotipos de Colletotrichum lindemuthianum en Oaxaca y San Luis Potosí, México, y resistencia en genotipos de frijol 51
Clave Variedad diferencial Patotipo
Mic
helit
e
Mic
higa
n D
ark
Red
Kid
ney
Perr
y M
arro
w
Cor
nell
4924
2
Wid
usa
Kab
oon
Méx
ico
222
PI 2
0726
2
To Tu AB
136
G 2
333
San Luis PotosíCoSLP-2 R1 R R R R R R R S2 R R R 256CoSLP-3 R R R S R R S R S R R R 328CoSLP-4 R R R R R R R R S R R R 256CoSLP-7 R R R R R R R R S R R R 256CoSLP-8 R R R R R R S R R R S R 64CoSLP-9 R R R R R R S R S R R R 320CoSLP-10 S R R S R R R R S R R R 265CoSLP-11 R R R S R R S R S R R R 328OaxacaCoOax-2 R R S R R R S R S R R R 324CoOax-3 S R S R R R S R R R R R 69CoOax-4 R R R R R R S R S R R R 320CoOax-5 R R S R R S S R S R R R 356CoOax-6 S R S R R R S R R R R R 69CoOax-7 S R S R R S S S S R R R 485CoOax-8 S R R S R R S S S R R R 457CoOax-9A R R S R R S S R R R R R 100CoOax-9B R R R R R R S R S R R R 320CoOax-10 S R S R R R S R S R R R 325CoOax-11 S R S R R R S R S R R R 325CoOax-12 R R S R R R R R S R R R 260CoOax-13 R R R R R R S R R R R R 64CoOax-15 S R R R R R S S S R R R 449CoOax-16 R R R R R R S S R R R R 192
Los patotipos presentes en San Luis Potosí ya habían sido reportados previamente en estados aledaños (González et al., 1998), y nuevos patotipos fueron identificados en Oaxaca. Aunque ambos estados están separados, los patotipos 64 y 320 (ampliamente dis t r ibuido en México) ocurr ieron en los dos , mientras que el resto de los patotipos de cada estado fue exclusivo de cada uno. Tanto la ocurrencia de patotipos comunes en diferentes estados como la ocurrencia exclusiva de patotipos en algunos estados o regiones de México, ha sido reportado previamente (Rodríguez-Guerra et al., 2006). Aun no es posible establecer las causas precisas de la distribución de estos patotipos, pero es posible que se deba a la base genética del hospedero, prácticas de cultivo y factores climáticos (González et al., 1998), entre otros.
Los estados de San Luis Potosí y Oaxaca son contrastantes geográficamente, y también en cuanto al tipo de frijol que se cultivan en cada uno. San Luis Potosí se ubica en el centro del país y ahí se cultivan principalmente variedades de hábito de crecimiento indeterminado arbustivo, mientras que Oaxaca se localiza al sur y ahí principalmente se cultiva frijol criollo de hábito indeterminado postrado y trepador y variedades nativas de otras especies de frijol como son P. coccineus y P. dumosus. Esto puede explicar las diferencias en patotipos, índice de diversidad de patotipos y origen de las variedades diferenciales que reaccionan como susceptibles a los patotipos identificados en estos estados. Por otra parte, el hecho de que se hayan aislado los patotipos 64 y 320 en ambos estados, quizás tiene que ver con la costumbre de
los productores de usar semilla producida en localidades distintas que pudieran portar patotipos ausentes en algunas áreas de producción, lo que promovería su dispersión, como ha sido referido por Kelly et al. (1994).
Es posible que las la base genética de los cultivares de frijol y las condiciones climáticas influyan en la diversidad de patotipos presentes en estados particulares de México, como ha sido sugerido en estudios similares (González et al., 1998; González-Chavira et al., 2004; Rodríguez-Guerra et al., 2006). El elevado índice de diversidad de patotipos observado en esta investigación en el estado de Oaxaca (2.43), muestra que las poblaciones del patógeno presentes en estados más al sur del país, con valles intermontanos y mayor
Estado Fuente1 Núm. de aislados Núm. de patotipos H2
Chihuahua 1 22 1 0.00Durango 1 9 3 0.85Zacatecas 1 7 4 1.28San Luis Potosí Esta investigación 8 5 1.49Jalisco 1 15 3 1.08Michoacán 1 6 5 1.56México 2 6 3 1.01Hidalgo 2 2 2 0.69Tlaxcala 2 2 2 0.69Puebla 2 7 4 1.15Oaxaca Esta investigación 15 12 2.43
11= González et al., 1998; 12= González-Chavira et al., 2004; H2= índice de diversidad de patotipos (Shannon y Weaver, 1963).
Cuadro 3. Número de patotipos de Colletotrichum lindemuthianum reportados por varios autores e índices de diversidad de patotipos en varios estados de México.
El índice de diversidad de patotipos fue de 1.49 en San Luis Potosí y de 2.43 en Oaxaca, lo que indica que la mayor diversidad en patotipos ocurre en este último estado. Sin embargo, los índices de diversidad son altos en ambos estados con relación a otros estados del país (Cuadro 3). La ocurrencia de cinco patotipos en San Luis Potosí y 12 en Oaxaca a partir de ocho y 15 aislados respectivamente, muestra que C. lindemuthianum muestra gran variabilidad patogénica en ambos estados respecto a otros estados del país donde previamente se han caracterizado patogénicamente aislados del hongo (González et al., 1998; González-Chavira et al., 2004). Esto es apoyado por los altos índices de diversidad que muestran San Luis Potosí y Oaxaca (Cuadro 3). Sin embargo, es posible que nuevos patotipos puedan ser identificados al determinar el patotipo de más aislados en el futuro.

52 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
Bertha María Sánchez-García et al.
diversidad climática, son más variables patogénicamente que las poblaciones que ocurren del centro hacia al norte de México, como ha sido comparada la diversidad de patotipos en estados del centro y norte de México (González-Chavira et al., 2004). Esto también puede ser debido al cultivo de una gran diversidad de materiales nativos de frijol con una amplia variabilidad genética en estados del sur, que no es común en estados del centro-norte de México donde cultivares mejorados uniformes son comúnmente utilizados.
Los resultados anteriores sugieren que existe cierto grado de aislamiento entre las poblaciones del patógeno que ocurren en ambos estados. Esto también es apoyado por la ocurrencia en Oaxaca de un gran número de patotipos capaces de infectar variedades diferenciales de origen andino que no ocurren en San Luis Potosí y otros estados del norte de México, pero si en estados vecinos de Oaxaca como Puebla, Tlaxcala y México (González et al., 1998; González-Chavira et al., 2004). Aunque Balardin et al.
(1997) también reportaron patotipos presentes en el país capaces de infectar diferenciales de origen mesoamericano y andino, se desconoce el origen geográfico de ellos. Aunque se ha considerado que la ocurrencia de estos patotipos que infectan variedades diferenciales de ambos acervos no es raro en México (González-Chavira et al., 2004), no se haplanteado una posible explicación de esto. Considerando que existe coevolución entre las poblaciones andinas y mesoamericanas del patógeno y los acervos genéticos andinos y mesoamericanos del frijol, respectivamente (Pastor-Corrales, 1996; Araya, 2003), es posible la existencia de cultivares de origen andino (tipos canarios y cacahuates) en los estados del centro y sur de México cuya interacción con el patógeno ha permitido el desarrollo de patotipos que afectan genotipos de ambos acervos genéticos; sin embargo, también la introducción a México de patotipos que infectan genotipos andinos de frijol pudiera explicar su ocurrencia en el país, entre otras posibles causas.
Patotipo Variedad diferencial IV
Mic
helit
e
Mic
higa
n D
ark
Red
Kid
ney
Perr
y M
arro
w
Cor
nell
4924
2
Wid
usa
Kab
oon
Méx
ico
222
PI 2
0726
2
To Tu AB
136
G 2
333
64 R1 R R R R R S2 R R R R R 8.369 S R S R R R S R R R R R 25
100 R R S R R S S R R R R R 25192 R R R R R R S S R R R R 16.7256 R R R R R R R R S R R R 8.3260 R R S R R R R R S R R R 16.7265 S R R S R R R R S R R R 25320 R R R R R R S R S R R R 16.7324 R R S R R R S R S R R R 25325 S R S R R R S R S R R R 33.3328 R R R S R R S R S R R R 25356 R R S R R S S R S R R R 33.3449 S R R R R R S S S R R R 33.3457 S R R S R R S S S R R R 41.7485 S R S R R S S S S R R R 50
IR 60 100 53.3 80 100 80 20 73.3 26.7 100 100 100
Cuadro 4. Índice de resistencia (IR) de las variedades diferenciales e índice de virulencia (IV) de los patotipos de Colletotrichum lindemuthianum identificados en San Luis Potosí y Oaxaca.
1R= resistente; 2S= susceptible.

Patotipos de Colletotrichum lindemuthianum en Oaxaca y San Luis Potosí, México, y resistencia en genotipos de frijol 53
Sólo siete de las variedades diferenciales fueron suscep t ib les a a lguno de los pa to t ipos de C. l indemuthianum (Cuadro 4) . México 222 y To fueron susceptibles a la mayoría de los patotipos, mostrando IR bajos de 20.0 y 26.7, respectivamente. Rodríguez-Guerra et al. (2006) también reportó que ambas variedades diferenciales muestran bajos IR a 50 patotipos previamente reportados en México. Las variedades diferenciales Michigan Dark Red Kidney, Widusa, Tu, AB 136 y G 2333 fueron resistentes a los patotipos identificados en los estados bajo estudio, pero pueden reaccionar como susceptibles a otros patotipos presentes en México (Rodríguez-Guerra et al., 2006).
Ningún patotipo identificado en San Luis Potosí fue capaz de producir una reacción de susceptibilidad sobre variedades diferenciales de origen andino, mientras que seis (50%) de los patotipos identificados en Oaxaca (69, 100, 260, 324, 325, 356) causaron una reacción de susceptibilidad a alguna diferencial de ese acervo genético (Cuadro 2 y 4). Los patotipos más virulentos fueron el 457 (causa susceptibilidad a cinco variedades diferenciales) y 485 (causa susceptibilidad a seis variedades diferenciales) de l es tado de Oaxaca , con IV de 41 .7 y 50 .0 respectivamente (Cuadro 4), ubicándolos entre los más virulentos en el país. Sin embargo, patotipos más virulentos (1 869, 3 785, 3 993, 3 995, 4 027, 4 077) han sido identificados en el estado de Chiapas, México, capaces de infectar de siete a diez variedades diferenciales (Rodríguez-Guerra et al . , 2006).
Cinco variedades diferenciales (Michigan Dark Red Kidney, Widusa, Tu, AB 136, G 2333) mostraron resistencia a los patotipos identificados en San Luis Potosí y Oaxaca, y pueden ser utilizadas como fuentes de resistencia contra los patotipos presentes en ambos estados. Entre éstas, la variedad diferencial Tu (porta el gen de resistencia Co-5) continua siendo una importante fuente de resistencia a la mayoría de los patotipos presentes en México (González-Chavira et al., 2004).
Fuentes de resistencia
Entre los materiales nativos de San Luis Potosí y Guanajuato ocurrieron cuatro grupos de reacción: I) resistencia a ambos patotipos, II) resistencia al
patotipo 64 y susceptibilidad al 320, III) resistencia a l patot ipo 320 y suscept ibi l idad al 64, y IV) susceptibilidad a ambos patotipos. Mientras que en los cultivares mejorados ninguno mostró reacción de resistencia al patotipo 320 con susceptibilidad al 64 (grupo de reacción III). La mayoría de los materiales nativos de San Luis Potosí (12) y de Guanajuato (76) fueron resistentes a ambos patotipos (grupo de reacción I), mientras que cinco y 15 materiales fueron resistentes a uno u otro patotipo (grupos de reacción II y III) (Cuadro 5). Los restantes materiales nativos fueron susceptibles a los patotipos 320 y 64. De los 55 cultivares mejorados, 21 fueron susceptibles a los patotipos 320 y 64, seis susceptibles al primero pero resistentes al 64, y 28 resistentes a ambos patotipos (Cuadro 6). Con anterioridad se identificaron fuentes de resistencia a patotipos de C. lindemuthianum presentes en México en un reducido grupo elite de cultivares de frijol del INIFAP (González-Chavira et al., 2004); sin embargo, ningún esfuerzo se había realizado con el propósito de identificar fuentes de resistencia en materiales nativos de frijol en México. La búsqueda de fuentes de resistencia en materiales nativos de frijol ha permitido identificar fuentes de resistencia contra patotipos altamente virulentos presentes en Colombia (Santana y Mahuku, 2002).
En esta investigación, la mayoría de los materiales evaluados por su reacción a los patotipos 320 y 64 fueron resistentes a ambos (grupo de reacción I). Es posible que la resistencia de los materiales nativos y mejorados que reaccionaron como resistentes al patotipo 64 pero fueron susceptibles al 320, se deba al menos a un gen de resistencia similar al presente en la variedad diferencial To, la cual es susceptible a este último patotipo, o se deba a otro gen de resistencia que esta ausente en México 222 (susceptible al patotipo 64).
López-Jiménez et al. (2006) sugieren que el uso sistemático de un reducido y selecto grupo de patotipos de C. lindemuthianum puede ser útil para identificar fuentes de resistencia contra el patógeno. Esta estrategia considera que los patotipos seleccionados deben causar una reacción de susceptibilidad a la mayoría de las variedades diferenciales, y que los materiales resistentes a ellos también deben ser resistentes a patotipos que infectan a cualquiera de las diferenciales que son infectadas en conjunto por los patotipos utilizados.

54 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
Bertha María Sánchez-García et al.
México 222 y To) y 64 (causa reacción de susceptibilidad a la diferencial México 222), como debería esperarse según la estrategia de López-Jiménez et al. (2006).
En esta investigación se esperaba que los materiales nativos y cultivares mejorados que reaccionaron como resistentes al patotipo 320, también lo fueran al 64. Lo anterior ocurrió en 116 materiales nativos de los 125 que fueron resistentes al patotipo 320; los otros nueve materiales fueron susceptibles al patotipo 64. Sin embargo, resultados contradictorios a los esperados con la estrategia de López-Jiménez et al. (2006) y de manera similar a lo observado con esos nueve materiales, se han reportado en investigaciones realizadas previamente
Grupo de reacción Estado/Accesión Tipo de semilla representado
San Luis PotosíI (resistencia a los patotipos 320 y 64)
852, 853, 854, 855, 857, 859, 860, 861, 862, 3905, 3983, 4002 Negro, Flor de Mayo, Ojo de Cabra, Tejano
I I ( r e s i s t e n c i a a l pa to t ipo 64 y susceptibilidad al 320)
858, 2564, 3942, 4031 Canario, Tejano, Bayo
II I ( res is tente a l p a t o t i p o 3 2 0 y susceptible al 64)
4011 Canario
IV (susceptible a los patotipos 320 y 64)
2563, 2566, 2568, 3924, 3930, 3935, 3990 Bayo, Café, Cacahuate, Negro, Flor de Mayo
GuanajuatoI (resistencia a los patotipos 320 y 64) 149, 153, 176, 179, 189, 196, 203, 207, 208, 233, 234, 239, 1513,
2921, 2922, 2925, 2931, 2932, 2938, 2939, 2942, 2943, 2944, 2947, 2948, 2949, 2953, 2954, 2955, 2950, 2957, 2958, 2959, 2960, 2961, 2962, 2963, 2965, 2967, 2968, 2969, 2970, 2971, 2972, 2973, 2974, 2975, 2977, 2979, 2981, 2982, 2984, 2985, 2986, 2987, 2988, 2989, 2990, 2993, 2994, 2995, 2996, 2997, 2998, 2999, 3000, 3002, 3003, 3005, 3006, 4804, 4807, 1514, 2976, 2978, 2917
Canario, Bayo, Pardo, Café, Negro, Pinto, Ojo de Cabra
I I ( r e s i s t e n c i a a l pa to t ipo 64 y susceptibilidad al 320)
150, 180, 190, 204, 219, 222, 236, Negro, Ojo de Liebre
I I I ( res is tente a l p a t o t i p o 3 2 0 y susceptible al 64)
173, 182, 186, 221, 238, 245, 1518, 2920 Flor de Mayo, Café, C a n a r i o y B a y o
IV (susceptible a los patotipos 320 y 64)
151, 152, 157, 174, 175, 192, 197, 211, 213, 215, 217, 218, 219, 223, 227, 228, 230, 231, 232, 2929, 2934, 2935, 2941, 2945
Bayo, Negro, Ojo de Liebre, Flor de Mayo, Canario, Ojo de Cabra , Pinto
Cuadro 5. Grupos de reacción de materiales nativos de frijol común (P. vulgaris) del estado de San Luis Potosí y Guanajuato a dos patotipos de Colletotrichum lindemuthianum.
Lo anterior ha ocurrido con siete cultivares elite inoculados con el patotipo 1 472 (infecta a las variedades diferenciales México 222, PI 207262, To y AB 136), los cuales fueron resistentes a ese patotipo y también al 448 (que al igual que el patotipo 1472, infecta en común variedades diferenciales México 222, PI 207262 y To) (González-Chavira et al., 2004); así como con la variedad mejorada Bayo Mecentral, reportada por González-Chavira et al. (2004) como resis tentes a l patot ipo 448 (causa reacción de susceptibilidad a las diferenciales México 222, PI 207262 y To), la cual en esta investigación fue resistente a los patotipos 320 (causa reacción de susceptibilidad a las diferenciales
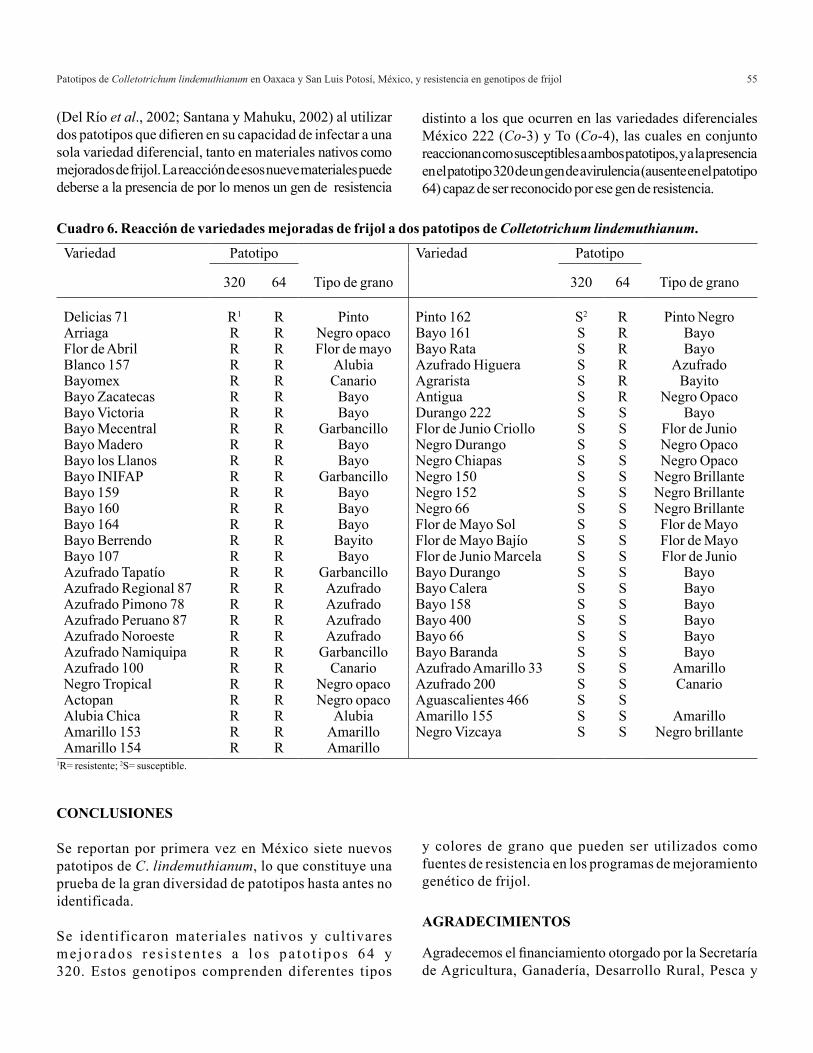
Patotipos de Colletotrichum lindemuthianum en Oaxaca y San Luis Potosí, México, y resistencia en genotipos de frijol 55
(Del Río et al., 2002; Santana y Mahuku, 2002) al utilizar dos patotipos que difieren en su capacidad de infectar a una sola variedad diferencial, tanto en materiales nativos como mejorados de frijol. La reacción de esos nueve materiales puede deberse a la presencia de por lo menos un gen de resistencia
distinto a los que ocurren en las variedades diferenciales México 222 (Co-3) y To (Co-4), las cuales en conjunto reaccionan como susceptibles a ambos patotipos, y a la presencia en el patotipo 320 de un gen de avirulencia (ausente en el patotipo 64) capaz de ser reconocido por ese gen de resistencia.
y colores de grano que pueden ser utilizados como fuentes de resistencia en los programas de mejoramiento genético de frijol.
AGRADECIMIENTOS
Agradecemos el financiamiento otorgado por la Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y
Variedad Patotipo Variedad Patotipo
320 64 Tipo de grano 320 64 Tipo de grano
Delicias 71 R1 R Pinto Pinto 162 S2 R Pinto NegroArriaga R R Negro opaco Bayo 161 S R BayoFlor de Abril R R Flor de mayo Bayo Rata S R BayoBlanco 157 R R Alubia Azufrado Higuera S R AzufradoBayomex R R Canario Agrarista S R BayitoBayo Zacatecas R R Bayo Antigua S R Negro OpacoBayo Victoria R R Bayo Durango 222 S S BayoBayo Mecentral R R Garbancillo Flor de Junio Criollo S S Flor de JunioBayo Madero R R Bayo Negro Durango S S Negro OpacoBayo los Llanos R R Bayo Negro Chiapas S S Negro OpacoBayo INIFAP R R Garbancillo Negro 150 S S Negro BrillanteBayo 159 R R Bayo Negro 152 S S Negro BrillanteBayo 160 R R Bayo Negro 66 S S Negro BrillanteBayo 164 R R Bayo Flor de Mayo Sol S S Flor de MayoBayo Berrendo R R Bayito Flor de Mayo Bajío S S Flor de MayoBayo 107 R R Bayo Flor de Junio Marcela S S Flor de JunioAzufrado Tapatío R R Garbancillo Bayo Durango S S BayoAzufrado Regional 87 R R Azufrado Bayo Calera S S BayoAzufrado Pimono 78 R R Azufrado Bayo 158 S S BayoAzufrado Peruano 87 R R Azufrado Bayo 400 S S BayoAzufrado Noroeste R R Azufrado Bayo 66 S S BayoAzufrado Namiquipa R R Garbancillo Bayo Baranda S S BayoAzufrado 100 R R Canario Azufrado Amarillo 33 S S AmarilloNegro Tropical R R Negro opaco Azufrado 200 S S CanarioActopan R R Negro opaco Aguascalientes 466 S SAlubia Chica R R Alubia Amarillo 155 S S AmarilloAmarillo 153 R R Amarillo Negro Vizcaya S S Negro brillanteAmarillo 154 R R Amarillo
Cuadro 6. Reacción de variedades mejoradas de frijol a dos patotipos de Colletotrichum lindemuthianum.
1R= resistente; 2S= susceptible.
CONCLUSIONES
Se reportan por primera vez en México siete nuevos patotipos de C. lindemuthianum, lo que constituye una prueba de la gran diversidad de patotipos hasta antes no identificada.
Se identificaron materiales nativos y cultivares m e j o r a d o s r e s i s t e n t e s a l o s p a t o t i p o s 6 4 y 320. Estos genotipos comprenden diferentes tipos

56 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
Bertha María Sánchez-García et al.
Alimentación (SAGARPA)-Consejo Nacional de Ciencia y Tecnología (CONACYT) (convenio 2004-C01-28) para la realización de esta investigación y al CONACYT por la beca otorgada a Bertha María Sánchez García, para realizar los estudios de la Maestría en Ciencias.
LITERATURA CITADA
Antunes, I. J.; Santin, R. de C. M.; Mastrantonio, J. J. da S. and Bonemann, C. 2003. New sources of resistance, race identification and virulence and resistance indexes in anthracnose research. Ann. Rep. Bean Improv. Coop. 46:179-180.
Araya, C. M. 2003. Coevolución de interacciones hospedan te -pa tógeno en f r i jo l común . Fitopatología Brasileira 28:221-228.
Balardin, R. S.; Jarosz, A. M. and Kelly, J . D. 1997. Virulence and molecular diversity in Colletotrichum lindemuthianum from South, Central, and North America. Phytopathology 87:1184-1191.
Del Río, L. E.; Lamppa, R. S. and Gross, P. L. 2002. Reaction of dry bean cultivars of the Northern Plains to anthracnose. Ann. Rep. Bean Improv. Coop. 45:72-73.
Ferreira, J. J.; Rodríguez, C.; Pañeda, A. and Giradles, R. 2003. Evaluation of Phaseolus vulgaris germplasm for resistance to five anthracnose races isolated in Northern Spain. Ann. Rep. Bean Improv. Coop. 46:171-172.
Flor, H. H. 1942. Inheritance of pathogenicity in Melampsora lini. Phytopathology 32:653-669.
Flor, H. H. 1956. The complementary genetic system in flax and flax rust. Advances in Genetics 8:29-54.
Garrido-Ramírez, E. R. y Romero-Cova, S. 1989. Identificación de razas de Colletotrichum lindemuthianum (Sacc. & Magn.) Scrib. En México y búsqueda de resistencia genética a este hongo. Agrociencia 77:139-156.
Geffroy, V.; Sévignac, M.; De Oliveira, J. C. F.; Fouilloux, G.; Skroch, P.; Thoquet, P.; Gepts, P.; Langin, T. and Dron, M. 2000. Inheritance of parcial resistance against Colletotrichum lindemuthianum in Phaseolus vulgaris and co-localization of quantitative trait loci with genes involved in specific resistance. MPMI 13:287-296.
Gepts, P. and Debouck, D. G. 1991. Origin, Domestication and Evolution of the Common Bean (Phaseolus vulgaris L.) pp. 7-53. In: Schoonhoven, van A. and Voysest, O. (eds.). Common Beans: Research for Crop Improvement. CAB. Inter. Wallingford, UK and CIAT, Cali, Colombia.
González, M.; Rodríguez, R.; Zavala, M. E.; Jacobo, J. L.; Hernández, F.; Acosta, J.; Martínez, O. y Simpson, J. 1998. Characterization of Mexican isolates of Colletotrichum lindemuthianum by using differential cultivar and molecular markers. Phytopathology. 88:292-299.
González-Chavira, M.; Rodríguez-Guerra, R.; Hernández-Godínez, F.; Acosta-Gallegos, J. A.; Martínez-de la Vega, O. and Simpson, J. 2004. Analysis of pathotypes of Colletotrichum lindemuthianum found in the central region of Mexico and resistance in elite germplasm of Phaseolus vulgaris. Plant Disease. 88:152-156.
Hernández-G., F.; González-Ch., M.; Rodríguez-G., R.; Acosta-G., J. y Simpson, J. 1998. La variación patogénica de Colletotrichum lindemuthianum y su importancia en los programas de mejoramiento genético del frijol. Rev. Mex. Fitopatol. 16(S1):63.
Kelly, J. D.; Afanador, L. and Cameron, L. S. 1994. New races of Colletotrichum lindemuthianum in Michigan and implications in dry bean resistance breeding. Plant Disease 78:892-894.
López-Jiménez, G.; Pineda-Rodríguez, S.; Sánchez-García, B. M.; Acosta-Gallegos, J. A.; Simpson, J. y Rodríguez-Guerra, R. 2006. Estrategia para identificar fuentes de resistencia en germoplasma de frijol común a razas de Colletotrichum lindemuthianum presentes en México. XXXIII Congreso Nacional de Fitopatología/VIII Congreso Internacional de Fitopatología. Resumen C100.
Mahuku, G. S.; Jara, C. E.; Cajiao, C. and Beebe, S. 2002. Sources of resistance to Colletotrichum lindemuthianum in the secondary gene pool of Phaseolus vulgaris and crosses of primary and secondary gene pools. Plant Disease 86:1383-1387.
Pastor-Corrales M. A. 1991. Estandarización de variedades diferenciales y de designación de razas de Colletotrichum lindemuthianum. (Abstract) Phytopathology 81:694.

Patotipos de Colletotrichum lindemuthianum en Oaxaca y San Luis Potosí, México, y resistencia en genotipos de frijol 57
Pastor-Corrales, M. A. and Tu, J. C. 1989. Anthracnose. In: Schwartz, H. F. and Pastor-Corrales, M. A. (eds). Bean Production Problems in the Tropics. Centro Internacional de Agricultura Tropical, Colombia. p. 77-104.
Pastor-Corrales, M. A. 1996. Tradicional and molecular confirmation of the coevolution of beans and pathogens in Latin America. Ann. Rep. Bean Improv. Coop. 39:46-47.
Shannon, C. E. and Weaver, W. 1963. Mathematical theory of comunication. Univ. Illinois Press, USA. 117p.
Rodríguez-Guerra, R.; Ramírez-R., M.T.; Martínez de la Vega, O. and Simpson, J. 2003. Variation in genotype, pathotype and anastomosis groups of Colletotrichum lindemuthianum isolates from Mexico. Plant Pathology 52:228-235.
Rodríguez-Guerra, R.; Acosta-Gallegos, J. A.; González-Chavira, M. M. y Simpson, J. 2006. Patotipos de Colletotrichum lindemuthianum y su implicación en la generación de cultivares resistentes de frijol. Agri. Téc. Méx. 32:99-112.
Santana, G. E. and Mahuku, G. 2002. Diversidad de razas de Colletotrichum lindemuthianum en Antioquia y evaluación de germoplasma de frijol crema-rojo por resistencia a antracnosis. Agronomía Mesoamericana 13:95-103.
Sartorato, A. and Arraes, P. A. 2003. Wild beans as source of resistance to Colletotrichum lindemuthianum. Ann. Rep. Bean Improv. Coop. 46:9-10.
Sicard, D.; Michalakis, Y.; Dron, M. and Neema, C. 1997. Genetic diversity and pathogenic variation of Colletotrichum lindemuthianum in three centers of diversity of its host, Phaseolus vulgaris. Phytopathology 87:807-813.

Agricultura Técnica en México Vol. 35 Núm.1 1 de enero-31 de marzo 2009 p. 58-69
ASOCIACIÓN DE HEMIPTERA: TRIOZIDAE CON LA ENFERMEDAD ‘PERMANENTE DEL TOMATE’ EN MÉXICO*
ASSOCIATION OF HEMIPTERA: TRIOZIDAE WITH THE DISEASE ‘PERMANENTE DEL TOMATE’ IN MEXICO
José Antonio Garzón-Tiznado3§, Oralia Guadalupe Cárdenas-Valenzuela3, Rafael Bujanos-Muñiz2, Antonio Marín-Jarillo2, Alicia Becerra-Flora6, Sixto Velarde-Felix1, Cuauhtémoc Reyes-Moreno3, Mario González-Chavira4 y José Luis Martínez-Carrillo5
1Unidad de Biotecnología del Noroeste, Campo Experimental Valle de Culiacán, INIFAP, carretera Culiacán-El Dorado, km 16.5, Culiacán, Sinaloa, México. Tel. 01 667 8461013, ([email protected]). 2Laboratorio de Taxonomía de Insectos, Campo Experimental del Bajío, INIFAP, carretera Celaya-San Miguel Allende, km 6.5, Celaya, Guanajuato, México. Tel. 01 461 016115323, ([email protected].), ([email protected]). 3Programa Regional del Noroeste para el Doctorado en Biotecnología, Universidad Autónoma de Sinaloa. Tel. 01 667 7157641, ([email protected]). 4Unidad de Biotecnología del Bajío, Campo Experimental del Bajío, INIFAP. Celaya, Guanajuato, México. Tel. 01 461 6115323, ([email protected]). 5Programa Nacional de Protección Vegetal, Campo Experimental Valle del Yaqui, INIFAP, Ciudad Obregón, Sonora, México. Tel. 01 644 4143173, ([email protected]). 6Instituto Potosino de Investigación Científica y Tecnológica, A. C. Carretera a la Presa. Tel. 01 444 8342000, ([email protected]). §Autor para correspondencia: [email protected].
RESUMEN
La enfermedad conocida como permanente del tomate causa grandes daños en regiones donde el tomate se siembra en el ciclo primavera-verano. La etiología de la enfermedad se ha relacionado con organismos tipo bacteria y su transmisión con el psílido Bactericera (=Paratrioza) cockerelli (Sulc) (Hemiptera: Triozidae). El objetivo de esta investigación fue elucidar la asociación entre el psílido y la enfermedad. La obtención de la fuente de inoculo y los bioensayos de transmisión se realizaron en jaulas entomológicas bajo condiciones de invernadero. La transmisión planta de tomate a planta de tomate fue por injerto a partir de plantas que mostraban los síntomas característicos en campo. El tejido de plantas enfermas y del psílido fueron analizados por PCR, hibridación y secuenciación de productos de PCR clonados. Los resultados mostraron la reproducción de los síntomas de la enfermedad en las plantas de tomate injertadas y en las expuestas a B. cockerelli. Los bioensayos de transmisión del patógeno por el insecto fueron positivos, los peróidos de adquisición fueron: 15 min, 2, 3 y 48 h para
ninfas y 30 min, 2, 3, 8 y 48 h para adultos, con períodos de transmisión de 15 min, 2 y 24 h para el caso de adultos y 24 h de incubación. El producto de PCR de B. cockerelli fue clonado y secuenciado parcialmente e ingresado al gene bank con el numero de accesión DQ355020, esta secuencia presentó una similaridad de 99% con las accesiones DQ355018 y DQ355019 obtenidas de plantas de tomate con permanente del tomate, lo cual evidencia la asociación entre el insecto y la enfermedad.
Palabra clave: Bactericera (=Paratrioza) cockerelli (Sulc) (Hemiptera: Triozidae), enfermedad permanente del tomate, psílido.
ABSTRACT
The disease known as ‘permanente del tomate’ produces the greatest damage in those regions where tomato
* Recibido: Noviembre, 2007
Aceptado: Febrero, 2009

Asociación de Hemiptera: Triozidae con la enfermedad permanente del tomate en México 59
plantings are established in the spring-summer season. The etiology of this disease has been related to non-cultured type bacteria and its transmission with the psillid Bactericera (=Paratrioza) cockerelli (Sulc) (Hemiptera: Triozidae). The objective of this research was to discern the association between the psillid and the disease. The obtention of inoculum and the bioassays for transmission were performed in entomological cages under greenhouse conditions. Transmission from tomato plant to tomato plant was performed through grafting from plants that showed the characteristic symptoms of the disease in the field. Tissue from diseased plants and from the psillid was analyzed by PCR, hybridization and sequenciation of cloned products. The reproduction of the disease symptoms was observed in tomato plants grafted and in plants exposed to B. cockerelli. Bioassays of transmission of this pathogen by the insect were positive, and the acquisition periods were: 15 min, 2, 3 and 48 h for nymphs and 30 min, 2, 3, 8 and 48 h for adults, with transmission periods of 15 min, 2 and 24 h for adults and 24 h of incubation. The product of PCR of B cockerelli was cloned and partially sequenced and registered in the gene bank with the number DQ355020, this sequence presented 99% similarity with accessions DQ355018 and DQ355019 obtained from diseased tomato plants with permanente del tomate, which confirms the association between the insect and the disease.
Key words: Bactericera (=Paratrioza) cockerelli (Sulc) (Hemiptera: Triozidae), disease permanente del tomate, psillid.
INTRODUCCIÓN
En la región de El Bajío, México, se reportó por primera vez la enfermedad denominada permanente del tomate (PT) (Garzón-Tiznado, 1984). Es la enfermedad más dañina en las siembras de tomate (Solanum lycopersicum L.) del ciclo primavera-verano.
La etiología de esta enfermedad no es muy clara, ya que observaciones al microscopio electrónico, describen la presencia de cuerpos pleomórficos en el floema de plantas enfermas; sin embargo, el análisis de alineamiento de secuencias de los productos amplificados del gen 16S rRNA, de estas plantas, presentó sólo 80% de similaridad con fitoplasmas, lo cual no es un elemento para indicar su relación con este tipo de patógenos (Ing. Ming Lee, com.
pers.); no obstante, con algunas bacterias no cultivables su similaridad fue mayor al 98%, de ahí que se haya decidido denominarlas temporalmente como “organismo tipo bacteria no cultivable”.
Este patógeno se ha asociado al insecto Bactericera cockerelli en plantaciones de tomate donde la enfermedad está presente (Becerra-Flora, 1986 y Garzón-Tiznado et al., 2005) y consistentemente se ha mencionado que B. cockerelli es inductor de una enfermedad no contagiosa conocida como “amarillamiento de la papa”, la cual se describe como un producto de la toxina secretada por las ninfas durante el proceso de alimentación en las plantas (Richards y Blood, 1933). B. cockerelli pertenece a la familia Psylloidea, que junto con la familia Cicadellidea y Fulgoridea, se han descrito como vectores de procariotes (Jensen et al., 1964; Kaloostian y Jones, 1968; Harris, 1980; Kaloostian, 1980; Kummert y Rufflard, 1997; Kawakita et al., 2000; Palermo et al., 2001; Pilkington et al., 2004).
En la familia Psylloidea se han mencionado el menor número de especies vectores y dentro de estos, sólo dos géneros, Cacopsyla con varias especies y Bactericera trigonica (Font et al., 1999). Los del primer género, se han reportado como transmisores de fitoplasmas asociados a enfermedades en árboles f ru ta les , en donde se descr iben a Cacopsy l lapyri L. y C. pyricola (Forster), transmisores del ‘Pear decline’ phytoplasma (Davies et al., 1992), del grupo 16SrX-C (Kummert y Rufflard, 1997; Lee et al., 1998); C. melanoneura (Forster) y C. costales; Cacopsylla picta transmisores del ‘Apple proliferation’ phytoplasma (Alma et al., 1997; Frisinghelli et al., 2000; Jarausch et al., 2003), grupo 16SrX-A (Lee et al., 1998), y C. pruni como transmisor del European ‘stone fruit yellow’ phytoplasma (Carraro et al., 1998), grupo 16SrX-B (Lee et al., 1998) y Bactericera t r i gon i ca como t r ansmiso r de un f i t op l a sma asociado al grupo del ‘Stolbur ’ (Sr16XII) en el cultivo de zanahoria (Font et al., 1999; Weintraub y Beanland, 2006).
Por otro lado, se han descri to dos especies de bacterias no cultivables restringidas al floema, que causan una enfermedad en cítricos descrita como “Huanglongbing”, pertenecientes a la subdivisión α-Proteobacteria y previamente propuestas como “Candidatus Liberobacter africanum” y “Candidatus
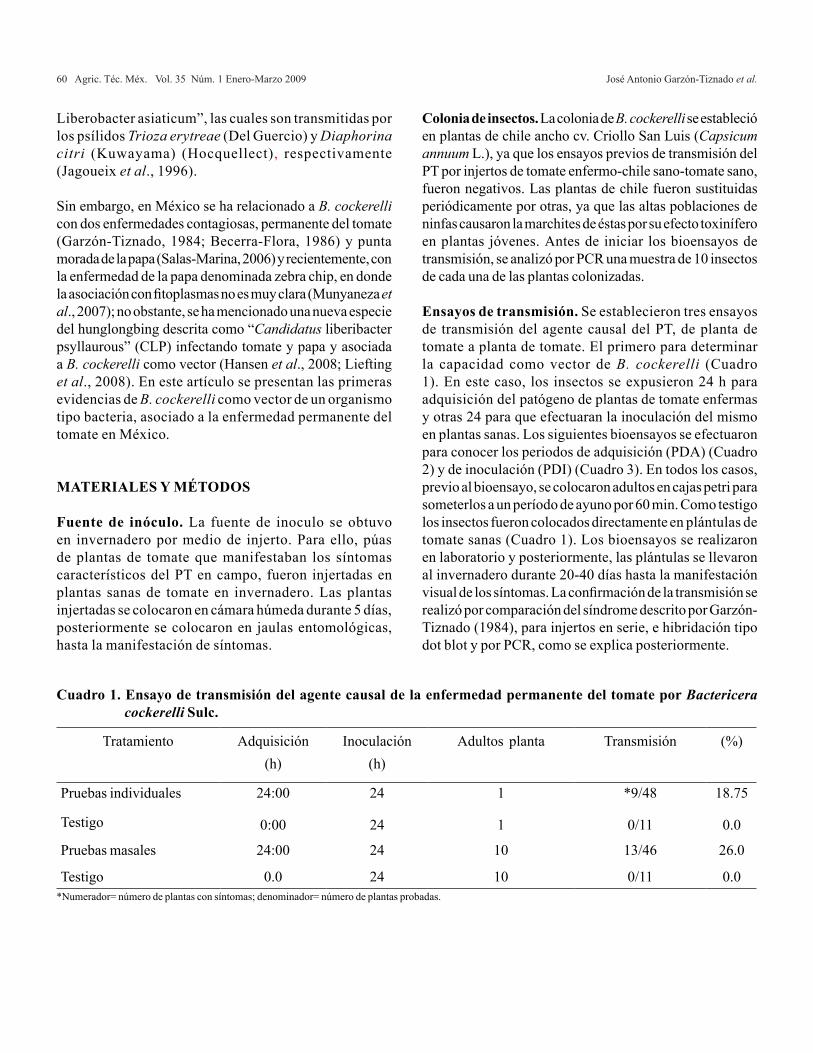
60 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
José Antonio Garzón-Tiznado et al.
Liberobacter asiaticum”, las cuales son transmitidas por los psílidos Trioza erytreae (Del Guercio) y Diaphorina citri (Kuwayama) (Hocquellect), respectivamente (Jagoueix et al., 1996).
Sin embargo, en México se ha relacionado a B. cockerelli con dos enfermedades contagiosas, permanente del tomate (Garzón-Tiznado, 1984; Becerra-Flora, 1986) y punta morada de la papa (Salas-Marina, 2006) y recientemente, con la enfermedad de la papa denominada zebra chip, en donde la asociación con fitoplasmas no es muy clara (Munyaneza et al., 2007); no obstante, se ha mencionado una nueva especie del hunglongbing descrita como “Candidatus liberibacter psyllaurous” (CLP) infectando tomate y papa y asociada a B. cockerelli como vector (Hansen et al., 2008; Liefting et al., 2008). En este artículo se presentan las primeras evidencias de B. cockerelli como vector de un organismo tipo bacteria, asociado a la enfermedad permanente del tomate en México.
MATERIALES Y MÉTODOS
Fuente de inóculo. La fuente de inoculo se obtuvo en invernadero por medio de injerto. Para ello, púas de plantas de tomate que manifestaban los síntomas característicos del PT en campo, fueron injertadas en plantas sanas de tomate en invernadero. Las plantas injertadas se colocaron en cámara húmeda durante 5 días, posteriormente se colocaron en jaulas entomológicas, hasta la manifestación de síntomas.
Colonia de insectos. La colonia de B. cockerelli se estableció en plantas de chile ancho cv. Criollo San Luis (Capsicum annuum L.), ya que los ensayos previos de transmisión del PT por injertos de tomate enfermo-chile sano-tomate sano, fueron negativos. Las plantas de chile fueron sustituidas periódicamente por otras, ya que las altas poblaciones de ninfas causaron la marchites de éstas por su efecto toxinífero en plantas jóvenes. Antes de iniciar los bioensayos de transmisión, se analizó por PCR una muestra de 10 insectos de cada una de las plantas colonizadas.
Ensayos de transmisión. Se establecieron tres ensayos de transmisión del agente causal del PT, de planta de tomate a planta de tomate. El primero para determinar la capacidad como vector de B. cockerelli (Cuadro 1). En este caso, los insectos se expusieron 24 h para adquisición del patógeno de plantas de tomate enfermas y otras 24 para que efectuaran la inoculación del mismo en plantas sanas. Los siguientes bioensayos se efectuaron para conocer los periodos de adquisición (PDA) (Cuadro 2) y de inoculación (PDI) (Cuadro 3). En todos los casos, previo al bioensayo, se colocaron adultos en cajas petri para someterlos a un período de ayuno por 60 min. Como testigo los insectos fueron colocados directamente en plántulas de tomate sanas (Cuadro 1). Los bioensayos se realizaron en laboratorio y posteriormente, las plántulas se llevaron al invernadero durante 20-40 días hasta la manifestación visual de los síntomas. La confirmación de la transmisión se realizó por comparación del síndrome descrito por Garzón-Tiznado (1984), para injertos en serie, e hibridación tipo dot blot y por PCR, como se explica posteriormente.
Tratamiento Adquisición(h)
Inoculación(h)
Adultos planta Transmisión (%)
Pruebas individuales 24:00 24 1 *9/48 18.75
Testigo 0:00 24 1 0/11 0.0
Pruebas masales 24:00 24 10 13/46 26.0
Testigo 0.0 24 10 0/11 0.0
Cuadro 1. Ensayo de transmisión del agente causal de la enfermedad permanente del tomate por Bactericera cockerelli Sulc.
*Numerador= número de plantas con síntomas; denominador= número de plantas probadas.
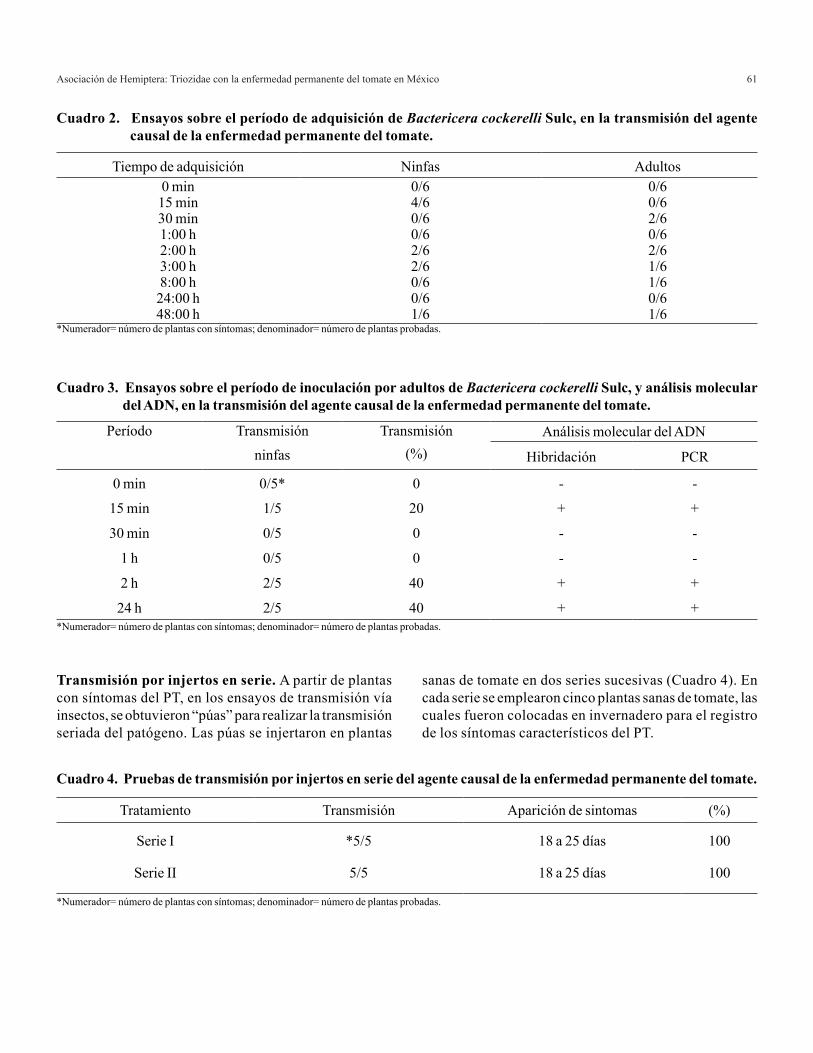
Asociación de Hemiptera: Triozidae con la enfermedad permanente del tomate en México 61
Transmisión por injertos en serie. A partir de plantas con síntomas del PT, en los ensayos de transmisión vía insectos, se obtuvieron “púas” para realizar la transmisión seriada del patógeno. Las púas se injertaron en plantas
sanas de tomate en dos series sucesivas (Cuadro 4). En cada serie se emplearon cinco plantas sanas de tomate, las cuales fueron colocadas en invernadero para el registro de los síntomas característicos del PT.
Tratamiento Transmisión Aparición de sintomas (%)
Serie I *5/5 18 a 25 días 100
Serie II 5/5 18 a 25 días 100
Período Transmisión
ninfas Transmisión
(%)Análisis molecular del ADN
Hibridación PCR
0 min 0/5* 0 - -
15 min 1/5 20 + +
30 min 0/5 0 - -
1 h 0/5 0 - -
2 h 2/5 40 + +
24 h 2/5 40 + +
Tiempo de adquisición Ninfas Adultos0 min 0/6 0/615 min 4/6 0/630 min 0/6 2/61:00 h 0/6 0/62:00 h 2/6 2/63:00 h 2/6 1/68:00 h 0/6 1/624:00 h 0/6 0/648:00 h 1/6 1/6
Cuadro 2. Ensayos sobre el período de adquisición de Bactericera cockerelli Sulc, en la transmisión del agente causal de la enfermedad permanente del tomate.
*Numerador= número de plantas con síntomas; denominador= número de plantas probadas.
*Numerador= número de plantas con síntomas; denominador= número de plantas probadas.
Cuadro 3. Ensayos sobre el período de inoculación por adultos de Bactericera cockerelli Sulc, y análisis molecular del ADN, en la transmisión del agente causal de la enfermedad permanente del tomate.
Cuadro 4. Pruebas de transmisión por injertos en serie del agente causal de la enfermedad permanente del tomate.
*Numerador= número de plantas con síntomas; denominador= número de plantas probadas.

62 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
José Antonio Garzón-Tiznado et al.
Análisis del ADN por PCR. En el ensayo de transmisión descrito en el Cuadro 3 se analizó por PCR el ADN extraído de plantas de tomate que manifestaron los síntomas del PT y de aquellas que no los manifestaron. En otro experimento, se analizó tejido de una planta de tomate con síntomas del PT, colectada en el municipio de Comonfort, Guanajuato, así como de huevecillos, ninfas y adultos de B. cockerelli que colonizaban esta planta. Por otro lado, en Dolores Hidalgo, Guanajuato, fueron colectados adultos en plantas de chile (Capsicum annuum L.) y otros en una especie silvestre conocida como “jarilla” (Figura 1). La extracción del ADN de tejido vegetal se realizó a partir de la metodología previamente descrita (Dellaporta et al., 1983; Almeida-León, 2001) y la de insectos por el método de Doyle (Doyle y Doyle, 1990). Para el análisis por PCR se empleó el método “anidado” reportado por Gunderson y Lee (1996). Se utilizaron dos pares de iniciadores que amplifican la región correspondiente al gen 16S Rrna y el espacio intergénico 16S-23S.
En la primera reacción se utilizó el par P1/P7 y para la segunda amplificación o “anidado” el par R16MF2/R16R2, con lo que se genera un fragmento predicho de 1.4 kb aproximadamente. En la mezcla de reacción se
empleó el amortiguador 1X, a base de Tris HCl 10 mM, pH 8.3, KCl 50 mM, MgCl2 1.0 mM, desoxinucleotidos 0.1 mm (Datp, Dctp, Dgtp, Dttp), iniciadores 5 mΜ, Taq ADN polimeraza 0.6 U y 25 ng de ADN genómico. Se agregó agua libre de nucleasas hasta completar un volumen de 15 mL. Para la segunda reacción se sustituyó el ADN por un mL de la suspensión de la primera reacción de amplificación, el par de iniciadores empleado fue el R16MF2/R16R2, y se llevó a un volumen total de 25 mL con agua libre de nucleasas. La reacción de amplificación se llevó a cabo en un termociclador Eppendorf programado para 35 ciclos con temperatura de desnaturalización de 1 min a 94 ºC, alineamiento 2 min a 55 ºC, polimerización 3 min a 72 ºC, con un precalentamiento a 94 ºC durante 1 min y una extensión final de 10 min a 72 ºC. Los productos de PCR se analizaron en un gel de agarosa al 1.5%, teñido con bromuro de etidio (0.5ug/mL) y se visualizaron en un transiluminador de luz ultravioleta.
Análisis por hibridación tipo dot blot. La hibridación tipo dot blot se utilizó en el análisis del ADN extraído del ensayo de PDI (Cuadro 3), para ello, un fragmento de ADN amplificado por PCR correspondiente al ADN de plantas de tomate enfermas colectadas en un invernadero comercial, en Celaya, Guanajuato, fue empleado como sonda en el análisis de hibridación. Este ADN fue aislado y purificado según las especificaciones del proveedor (QUANTUM PREP GEL SLICE, BIO-RAD®).
El ADN amplificado fue marcado con Fluoresceína-Dutp según las recomendaciones (GENE IMAGES RANDOM PRIME LABELLING MODULE, AMERSHAM PHARMACIA BIOTECH®). La sonda se almacenó en alícuotas de 10 µL, a -12 °C para su uso en la hibridación molecular. Esta se desarrolló de acuerdo al método descrito por la empresa (GENE IMAGES CDP-STAR DETECTION MODULE, AMERSHAM PHARMACIA BIOTECH®).
Para ello, una vez confirmada la presencia del ADN purificado en agarosa, se procedió a desnaturalizar y neutralizarlo para su transferencia a una membrana de Nylon, Hybond-N+ (AMERSHAM PHARMACIA BIOTECH®). El ADN se fijó a la membrana por exposición durante 5 min a luz UV, y se almacenó en refrigeración. Cada membrana fue prehibridada durante 30 min a 60 °C, en una solución SSC 5X, SDS 0.1% (p/v) + líquido de bloqueo 20X y Dextrán sulfato 5%, para de ahí hibridarse en 10 µL la sonda marcada a 60 °C durante
1 2 3 4 5 6 7 8 9
Figura 1. Gel de agarosa de productos de PCR anidado. Carriles 1 y 9, marcadores de tamaño molecular (escalera de 1-Kb); carriles= 2, huevecillos de B. cockerelli; 3= ninfas; 4= adultos; 5= planta de tomate con síntomas de PT donde se colectaron los insectos; 6= adultos colectados en planta de chile; 7= adultos colectados en “jarilla”; 8= planta de tomate sana.
1400 bp
1 2 3 4 5 6 7 8 9

Asociación de Hemiptera: Triozidae con la enfermedad permanente del tomate en México 63
toda la noche y con agitación constante. Terminado el tiempo, se realizaron los lavados en solución SSC 1X/SDS 0.1% a 60 °C/15 min y un segundo lavado en solución de SSC 0.1X/SDS 0.1% bajo las condiciones anteriores. Cada membrana se lavó con Tween 20 (0.3%) y posteriormente se le adicionó el reactivo de detección y se dejó incubando por 5 min, para ser expuesta a una película de rayos X (Hyperfilm, Amersham), durante 15 min.
Clonación y secuenciación del ADN de B. cockerelli. Un fragmento amplificado de 1.4 Kb a partir del ADN de ninfas de B. cockerelli (1194-2), fue clonado empleando el paquete TOPO TA Cloning (Invitrogen Life Technologies) siguiendo la reacción indicada en el protocolo del proveedor y secuenciado según el método 2',triphosphates 3'-dideoxynucleotide (ddNTPs) (Tanne et al., 2001), utilizando el Secuenciador Automático de ADN LI-COR® (LI-COR Inc., Lincoln, NE, USA 68504) modelo 4200 Global Edition IR2 System. La secuenciación vía PCR del vector PCR®4-TOPO® (Invitrogen™ Corp., Carlsbad, CA, USA 92008) con el inserto de interés se realizó mediante iniciadores universales marcados fluorescentemente IRD800 (heptamethine, λ 795 nm) e IRD700 (pentamethine carbocyanine, λ 685 nm), y los protocolos específicos para Termo Sequenase® y SequiTherm EXCEL™ II. El proceso de electroforésis y captura de datos de la Secuencias de ADN fue a través del programa e-Seq™ versión 3.0.1 y Gene ImagIR™ Software para Windows y LI-COR® (LI-COR Inc., Lincoln, NE, USA 68504).
Análisis del alineamiento de secuencias (clustal W). Las secuencias obtenidas a partir del ADN de B. cockerelli en esta investigación (1194-2), las obtenidas de plantas enfermas de tomate, depositadas en el gene bank con los números DQ355019 y DQ355018, así como las secuencias descritas para fitoplasmas: L33760 (tomato big bug mycoplasma-like organism, 16SrVI), AF222064 (tomato big bud phytoplasma-16SrVI), AF494529 (tomato big bud phytoplasma-16SrVI), DQ375328 (tomato little leaf phytoplasma) y DQ370434 (tomato yellows phytoplasma) (Figura 2), fueron alineadas base-a-base con el software MultAlin (Multiple Sequence Alignment, http://www.jouy.inra.fr) del programa Clustal W (Corpet, 1988). Con el método "Blast 2 sequences” se obtuvieron los porcentajes de identidad entre las secuencias (Tanne, 2001).
RESULTADOS
Colonia de insectos. El insecto B. cockerelli fue identificado de acuerdo a las características morfológicas descritas
previamente (Marín-Jarillo et al., 1995) y las colonias en plantas de chile se desarrollaron en forma satisfactoria; sin embargo, antes del inicio de la floración las plantas se marchitaban y morían, tal vez por el efecto de las toxinas secretadas por las ninfas que se desarrollaban en éstas (Richards y Blood, 1933), por lo cual se sustituyeron en forma frecuente por nuevas plantas. Los ADN’s extraídos de ninfas y adultos colectados de estas plantas fueron analizados por PCR y sólo se emplearon insectos de colonias que resultaron negativas.
Ensayos de transmisión. En el Cuadro 1 se describen los resultados de transmisión del PT, en donde nueve de 48 plantas manifestaron los síntomas de la enfermedad en el ensayo con un insecto por planta, lo que mostró 18.75% de plantas con síntomas. En el ensayo con 10 insectos por planta, 13 de 46 plantas manifestaron los síntomas de la enfermedad, con lo que se obtuvo 26% de plantas con síntomas. Para el caso de los testigos correspondientes, no se detectaron plantas con síntomas. En todos los casos, los síntomas característicos de la enfermedad se manifestaron entre 25 a 30 días posteriores a la transmisión.
Transmisión por injertos en serie. El 100% de las plantas injertadas en forma seriada, a partir de púas obtenidas de plantas de tomate empleadas en la transmisión porinsectos, presentaron los síntomas característicos del PT entre los 18 a 25 días posteriores a la realización del injerto (Cuadro 4).
Período de adquisición. Se detectaron plantas con síntomas del PT, en los PDA por las ninfas a los 15 min, 2:00, 3:00 y 48:00 h, con cuatro, dos dos y una planta, respectivamente (Cuadro 2), no se detectó transmisión a partir de insectos que permanecieron durante 30 min, y 1:00, 8:00 y 24:00 h, mientras que para los adultos, los síntomas se detectaron en dos, dos, una, una y una planta a partir de los PDA de 30 min, 2:00, 3:00, 8:00 y 48 :00 h, respectivamente, siendo negativos los tiempos de 15 min, 1:00 y 24:00 h (Cuadro 2). La manifestación de síntomas en ambos casos se dio a los los 25 días después del inicio del bioensayo.
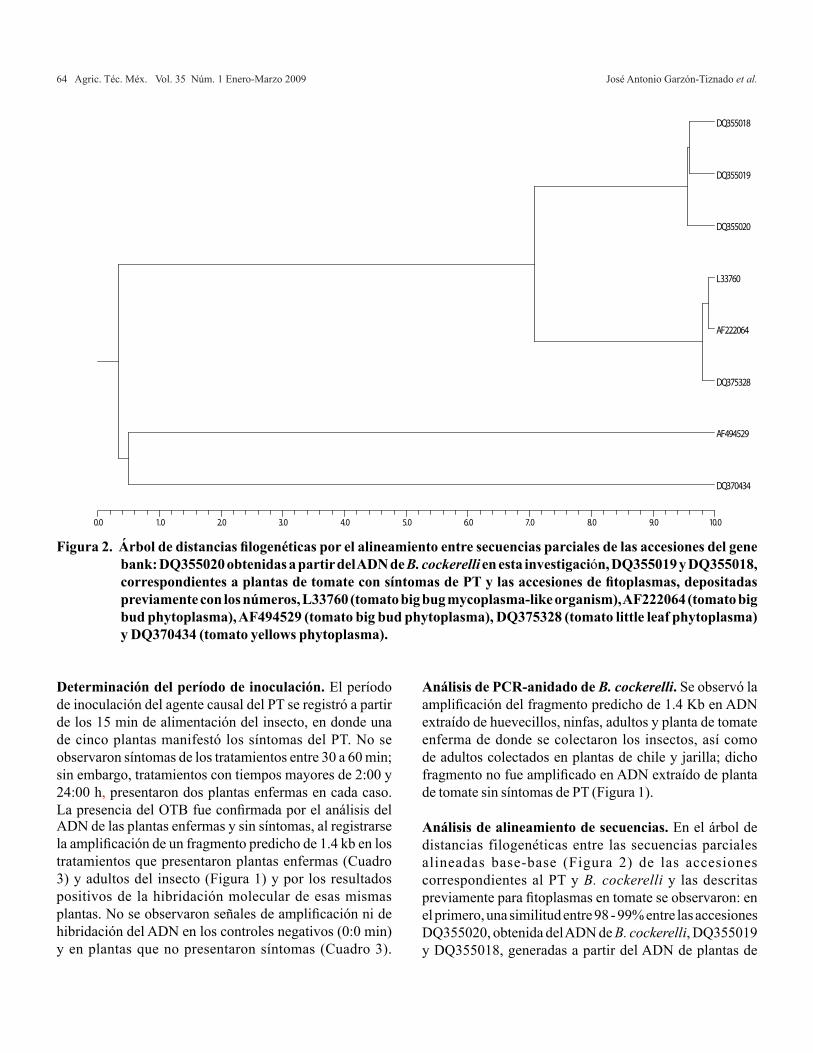
64 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
José Antonio Garzón-Tiznado et al.
Análisis de PCR-anidado de B. cockerelli. Se observó la amplificación del fragmento predicho de 1.4 Kb en ADN extraído de huevecillos, ninfas, adultos y planta de tomate enferma de donde se colectaron los insectos, así como de adultos colectados en plantas de chile y jarilla; dicho fragmento no fue amplificado en ADN extraído de planta de tomate sin síntomas de PT (Figura 1).
Análisis de alineamiento de secuencias. En el árbol de distancias filogenéticas entre las secuencias parciales alineadas base-base (Figura 2) de las accesiones correspondientes al PT y B. cockerelli y las descritas previamente para fitoplasmas en tomate se observaron: en el primero, una similitud entre 98 - 99% entre las accesiones DQ355020, obtenida del ADN de B. cockerelli, DQ355019 y DQ355018, generadas a partir del ADN de plantas de
0.0 1.0 2.0 3.0 4.0 5.0 6.0 7.0 8.0 9.0 10.0
DQ355018
DQ355019
DQ355020
L33760
AF222064
DQ375328
AF494529
DQ370434
Figura 2. Árbol de distancias filogenéticas por el alineamiento entre secuencias parciales de las accesiones del gene bank: DQ355020 obtenidas a partir del ADN de B. cockerelli en esta investigación, DQ355019 y DQ355018, correspondientes a plantas de tomate con síntomas de PT y las accesiones de fitoplasmas, depositadas previamente con los números, L33760 (tomato big bug mycoplasma-like organism), AF222064 (tomato big bud phytoplasma), AF494529 (tomato big bud phytoplasma), DQ375328 (tomato little leaf phytoplasma) y DQ370434 (tomato yellows phytoplasma).
Determinación del período de inoculación. El período de inoculación del agente causal del PT se registró a partir de los 15 min de alimentación del insecto, en donde una de cinco plantas manifestó los síntomas del PT. No se observaron síntomas de los tratamientos entre 30 a 60 min; sin embargo, tratamientos con tiempos mayores de 2:00 y 24:00 h, presentaron dos plantas enfermas en cada caso. La presencia del OTB fue confirmada por el análisis del ADN de las plantas enfermas y sin síntomas, al registrarse la amplificación de un fragmento predicho de 1.4 kb en los tratamientos que presentaron plantas enfermas (Cuadro 3) y adultos del insecto (Figura 1) y por los resultados positivos de la hibridación molecular de esas mismas plantas. No se observaron señales de amplificación ni de hibridación del ADN en los controles negativos (0:0 min) y en plantas que no presentaron síntomas (Cuadro 3).

Asociación de Hemiptera: Triozidae con la enfermedad permanente del tomate en México 65
tomate con síntomas del PT (Cardenas-Valenzuela et al., en proceso); en un segundo grupo, con una similitud de 80% al primero, correspondientes a las accesiones descritas para fitoplasmas, L33760 (tomato big bug mycoplasma-like organism), AF222064 (tomato big bud phytoplasma), DQ375328 (tomato little leaf phytoplasma) con una similitud 99% entre ellas, y un tercer grupo muy distante correspondiente a las accesiones AF494529 (tomato big bud phytoplasma) y DQ370434.
DISCUSIÓN
La planta de chile (Capsicum annuum L.), fue un buen hospedante para el desarrollo de B. cockerelli; sin embargo, fue necesario utilizar plantas entre la primera y la segunda flor, ya que las jóvenes en estado de seis hojas, morían después de 20 días de estar colonizadas por ninfas del insecto, lo mismo sucedió con plantas jóvenes de tomate; esta situación puede estar relacionada con el efecto toxinifero de las ninfas, las cuales se ha reportado que llegan a causar la muerte de plantas de papa o tomate (Richards y Blood, 1933).
La fuente de inóculo del PT, se estableció en invernadero por medio de injerto y posteriormente, después del primer ensayo de transmisión con B. cockerelli (Cuadro 1), el agente infeccioso fue transmitido en dos series de injerto (Cuadro 4), esto indica que la enfermedad puede establecerse sin necesidad del vector, por lo que deducimos que existe un agente causal del PT, al que temporalmente denominamos “organismo tipo bacteria” (OTB), por la alta identidad entre este agente y las secuencias alineadas con bacterias (98%) y por la baja similitud detectada en alineamientos con fitoplasmas (82%) (Cardenas-Valenzuela et al., en proceso). Los resultados posteriores de transmisión del OTB descritos en los Cuadros 2 y 3, fueron consistentes con el Cuadro 1, al permitir la reproducibilidad de los síntomas del PT en plantas de tomate.
Este tipo de experimentos, tal y como lo describen algunos autores (Lefol et al., 1993; Vega et al., 1993; Vega et al., 1994), son básicos en el proceso de búsqueda del vector de un agente infeccioso. Para esto, inicialmente se emplearon ninfas en 3° y 4° estadio de desarrollo por ser fáciles de manejar y para mejorar la eficiencia en la transmisión del OTB de acuerdo a resultados previos
en este estudio (Cuadro 2); no obstante, se utilizaron adultos para reducir la confusión o “enmascaramiento” de los síntomas de amarillamiento inducidos por las ninfas (Richards y Blood, 1933), con los inducidos por este OTB (Garzón, 1984).
Con los resultados de transmisión por injerto y por el uso de adultos de B. cockerelli, descritos en los Cuadros 1, 2, 3 y 4, se presentan los primeros indicios de que la enfermedad permanente del tomate en México, es causada por un agente infeccioso asociado al insecto B. cockerelli y elimina la posibilidad de que estos síntomas hayan sido inducidos por la toxina que produce éste insecto. Evidencia adicional se observó con el análisis molecular realizado a partir del ADN extraído del insecto y de tejido de plantas de tomate enfermas, ya que ambos fueron positivos por hibridación tipo dot blot con el uso de un fragmento amplificado de ADN, extraído de plantas de tomate con síntomas del PT, y confirmado posteriormente con la amplificación tipo PCR anidado de este agente en las plantas de tomate con síntomas; en todos los casos el control fue negativo. En lo que respecta al tipo de transmisión, los resultados indicaron que la relación B. cockerelli/ PT, tiende a ser persistente, ya que el patógeno logró transmitirse después de 24 h de incubación en el insecto, con PDA entre los 15 a 30 min y PDI entre los 15 a 30 min para ninfas y adultos, respectivamente (Cuadros 2 y 3). Además, se observó que a medida que el período de adquisición fue mayor, se registró un ligero incrementó en la eficiencia de transmisión.
Algunos de los resultados anteriores difieren parcialmente con lo mencionado en la literatura, como en el caso de los fitoplasmas en su vector las chicharritas, ya que el ciclo completo “adquisición-incubación-transmisión” de estos ha sido reportado desde unos pocos hasta 80 días (Moya-Raygoza y Nault, 1998; Murral et al., 1996), tiempo mayor que el requerido por B. cockerelli, para transmitir el OTB, cuyo período de latencia en nuestro estudio fue de sólo 24 h y de minutos en adquisición y transmisión; sin embargo, el período de adquisición que va de minutos hasta varias horas en chicharritas (Purcell, 1982), fue similar a lo obtenido en este bioensayo. Asimismo se establece que B. cockerelli logró adquirir y transmitir el OTB a partir de plantas de tomate y la transmisión fue directa a tomate, a diferencia de los fitoplasmas transmitidos por chicharritas los cuales tienen que ser adquiridos a partir de especies silvestres antes de ser transmitidos al cultivo (Bosco et al., 1997).

66 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
José Antonio Garzón-Tiznado et al.
En lo que se refiere a la edad fisiológica de los insectos como vectores, no obstante que se ha mencionado que ésta no influye en su capacidad para transmitir el fitoplasma (Chiykowski y Sinha, 1998), estudios recientes describen que ninfas recién eclosionadas de Euscelidius variegatus fueron menos eficaces en adquirir al CY (amarillamiento del crisantemo) que ninfas del quinto estadio (Palermo et al., 2001), y en ciertos casos las ninfas son más eficientes para adquirir al fitoplasma que los adultos (Nagaich et al., 1974; Murral, 1996). Este último reporte coincide con los observados en este estudio, en donde las ninfas de B. cockerelli requirieron de menor tiempo (15 min) que los adultos (30 min) para adquirir al agente causal del PT y en general el número de plantas que resultaron positivas en el bioensayo fue mayor con el uso de ninfas, con nueve plantas infectadas y siete con adultos (Cuadro 2).
No obstante que pocos agentes fitopatógenos u otros procariotes simbióticos pueden ser transmitidos en forma transovárica por su vector, se consideraba que los fitoplasmas no se podían transmitir de esta manera; sin embargo, algunas investigaciones han reportado este tipo de transmisión para Hishimonoides sellatiformis, vector del fitoplasma mulberry dwarf (grupo 16SrI) (Kawakita et al., 2000), y en Schaphoideus titanus, transmisor del aster yellow (grupo 16SrI), los cuales se han detectado en huevecillo y ninfas mediante PCR (Alma et al., 1997), lo que sugiere la posible transmisión transovarial de estos patógenos. Estudios recientes han asociado a B. cockerelli con síntomas de amarillamiento de hojas en papa y de “zebra chip” (síntomas de manchado del tubérculo) en papa (Munyanesa et al., 2007) y más específicamente, otros asocian a “Candidatus Liberibacter psyllaurous” (CLP) con zebra chip en papa, tomate y B. cockerelli (Liefting et al., 2008; Hansen et al., 2008), coincidiendo con nuestros resultados, estos últimos investigadores, mencionaron que por medio de PCR, lograron detectar a CLP, en huevecillos y ninfas de B. cockerelli colonizando plantas de papa y tomate.
Coincidentemente con estos autores, en este estudio, se amplificó un fragmento de 1.4 Kb en el ADN correspondiente a huevecillos de este insecto, hospedando plantas de tomate con síntomas del PT, así como en ninfas y adultos, correspondientes a esta colonia de insectos (Figura 2), lo que representa un indicio más de que B. cockerelli puede estar transmitiendo un agente infeccioso en forma transovárica en tomate. No obstante, se ha mencionado que los fitoplasmas en el caso de las chicharritas puede penetrar
intracelularmente a través del epitelio celular del intestino, pasar al intestino medio, de ahí al hemocele e infectar otros tejidos como tubos de Malpighi (Lherminier et al., 1990) y sólo si llegan a los órganos reproductivos (Kawakita et al., 2000), pueden pasar de ahí a los huevecillos. Por lo anterior, la confirmación transovárica se obtendrá cuando se realicen los ensayos biológicos de transmisibilidad a plantas de tomate (Vega et al., 1994).
En lo que respecta a la relación filogenética establecida por el análisis de las secuencias parciales de las accesiones DQ355020, DQ355019 y DQ355018, correspondientes al ADN de ninfas de B. cockerelli y plantas de tomate con síntomas del PT, respectivamente, el análisis reveló una identidad de 98 a 99% entre ellas, con lo cual se evidencia la elevada relación genética entre las tres secuencias asociadas a la enfermedad del PT y por otro lado, una relación filogenética muy lejana con fitoplasmas, en el sentido de que el agente causal del PT está mas relacionado con un OTB que con un fitoplasma. Por otro lado, en publicaciones recientes, se ha descrito a CLP asociado a B. cockerelli (Hansen et al., 2008; Liefting et al., 2008), lo que coincide con nuestros resultados. Sin embargo, aún y cuando existen similitudes en varios aspecto relacionados con la transmisión, como es la detección del agente infeccioso en huevecillos y ninfas de B. cockerelli del CLP y OTB (nuestro estudio), así como en el tiempo de expresión de síntomas que va de 3 a 4 semanas, es posible que los síntomas no coincidan exactamente, ya que los descritos para el CLP son un amarillamiento en tomate, mientras que para el OTB asociado al PT son de aborto de flor, hojas quebradizas, de un color verde intenso, aunque finalmente la planta llega a amarillarse y morir.
Los resultados aquí descritos representan la evidencia molecular y biológica de que B. cockerelli está asociada a la transmisión del agente causal del PT; dicha transmisión parece ocurrir de manera persistente y se presentan los primeros indicios de que su forma de transmisión es transovárica.
CONCLUSIONES
El agente causal de la enfermedad PT, se logró transmitir por injerto.
El psílido B. cockerelli participa como transmisor del agente causal de la enfermedad permanente del tomate.

Asociación de Hemiptera: Triozidae con la enfermedad permanente del tomate en México 67
La transmisión del agente causal de la enfermedad permanente del tomate, se logró de planta de tomate a otra de tomate.
La adquisición y transmisión del patógeno por B. cockerelli, indican que la relación es persistente y posiblemente transovárica.
Se reporta por primera vez a B. cockerelli como vector de un organismo tipo bacteria en plantas de tomate.
LITERATURA CITADA
Almeida-León, I. H.; Rocha-Peña, M. A.; Piña-Razo, J. and Martínez-Soriano, J. P. 2001. The use of polymerase chain reaction and molecular hybridization for detection of phytoplasms in different plant species in Mexico. Rev. Mex. de Fitopatol. 19:1-9.
Alma, A.; Bosco, D.; Danielli, A.; Bertaccini, A. and Vibio, M. 1997. Identification of phytoplasms in eggs, nymphs and adults of Scaphoideus titanus Ball reared on healthy plants. Insect Mol. Biol, 6:115-121.
Becerra-Flora, A. 1986. Biología de Paratrioza cockerelli (Sulc) y su relación con la enfermedad permanente del tomate en el Bajío. Tesis de Licenciatura. Universidad Autónoma de Querétaro, México. Facultad de Química. 55 p.
Bosco, D.; Minucci, C.; Boccardo, G. and Conti, M., 1997. Differential acquisition of chrysanthemum yellows phytoplasm by three leafhopper species. Entomol. Exp. Appl. 83:219-224.
Carraro, L.; Osler, R.; Loi, N.; Ermacora, P. and Refati, E., 1999. Transmission of European stone fruit yellows phytoplasm by Cacopsylla pruni. J. Plant Pathol. 80:233-239.
Chiykowski, L. N. and Sinha, R. C. 1998. Some factors affecting the transmission of eastern peach X-mycoplasmalike organism by the leafhopper Paraphlepsius irroratus. Can. J. Plant Pathol. 10:85-92.
Davies, D. L.; Guise, C. M.; Clark, M. F. and Adams, A. N. 1992. Parry’s disease of pears is similar to pear declaine and is associated with mycoplasm-like organisms transmitted by Cacopsylla pyricola. Plant Pathol. 41:195-203.
Dellaporta, S. L.; Wood, J. and Hicks, J. B. 1983. A plant DNA minipreparation: version II. Plant Mol Biol Rep 1:19-21.
Doyle, J. J. and Doyle, J. L. 1990. Isolation of plant DNA from fresh tissue. Focus 12:13-15.
Font, I.; Abad, P.; Albinana, M.; Espino, A. I. y Dally, E. L. 1999. Amarilleos y enrojecimientos en zanahoria: una enfermedad a diagnóstico. Bol. San. Veg. Plagas 25:405-415.
Frisinghelli, C.; Delaiti, L.; Grando, M. S.; Forti D. and Vendimian, M. E. 2000. Cacopsylla costalis (Flor 1861), as a vector of apple proliferation in Trentino. J. Phytopathol. 148:425-431.
Garzon-Tiznado, J . A. , 1984. Enfermedad del “Permanente” del j i tomate (Lycopersicon esculentum Mill.) en Celaya, Guanajuato XI Congreso Nacional de Fitopatologia. San Luis Potosí, S. L. P. 1984. Resúmenes, Soc. Mex. Fitopatología 138 p.
Garzón-Tiznado, J. A.; Garzón-Ceballos, J. A.; Velarde-Felix, S.; Marin-Jarillo, A. y Cardenas-Valenzuela, O. G. 2005. Ensayos de transmisión del fitoplasma asociado al “permanente del tomate” por el psílido Bactericera cockerelli Sulc en México. Entomología Mexicana 4:672-675.
Gundersen, D. E. and Lee, I. M. 1996. Ultrasensitive detection of phytoplasms by nested-PCR assays using two universal primer pairs. Phtopathol Medditerr 35:144-151.
Hansen, A. K.; Trumble, J. T.; Stouthamer, T. and Paine, T. D., 2008. A new huanglongbing species, “Candidatus Liberibacter psyllaurous”, found to infect tomato and potato, is vectored by the psyllid Bactericera cockerelli Sulc. Appl. Environ. Microbiol 74(18):5862-5865.
Harris, K. F. 1980. Aphids, leafhoppers and planthoppers. Pags, 1-13 In: Vectors of plant pathogens. Harris, K. F. and Maramorosch, K. (editors). 1980. Nueva York, Academic Press. 467 p.
Jensen, D. D.; Griggs, W. H.; Gonzales, C. Q. and Scheneider, H. 1964. Pear decline virus transmission by pear psylla. Phytopathol. 54:1346-1351.
Jarausch, B.; Schwind, N.; Jarausch, W.; Kresal, G.; Dickler, E. and SeemÜller E. 2003. First report of Cacopsylla picta as a vector of apple proliferation phytoplasma in Germany. Plant Dis. 87:101.

68 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
José Antonio Garzón-Tiznado et al.
Jagoueix, S.; Bové, J. M. and Garnier M. 1996. PCR detection of the two ‘Candidatus’ liberobacter species associated with greening disease of citrus. Molecular and Cellular Probes 10:43-50.
Kaloostian, G. H. and Jones, L. S. 1968. Pear leaf curl virus transmitted by pear psylla. Plant Dis. Rep. 52:924-925.
Kaloostian, G. H. 1980. Psyllids. Pags. 87-91 In: Vectors of plant pathogens. Harris, K. F. and Maramorosch, K. (editors). 1980. New York, Academic Press. 467 p.
Kawakita, H.; Saiki, T.; Wei, W.; Mitsuhashi, W. and Watanabe, K. 2000. Identification of mulberry dwarf phytoplasms in the genital organs and eggs of leafhopper Hishimonoides sellatiformis. Phytopathology 90:909-914.
Kummert, J. and Rufflard, G. 1997. A preliminary report on the detection of phytoplasms by PCR. Biochemica 1:22.
Lee, I. M.; Gundersen-Rindal, D. E. and Bertaccini, A. 1998. Phytoplasm: Ecology and genomic diversity. Phytopathol. 88:1359-1366.
Lefol, C.; Caudwell, A.; Lherminier. J. and Larue, J. 1993. Attachment of the flavescence dorée pathogen (MLO) to leafhopper vectors and other insects. Ann. Appl. Biol. 123:611-622.
Lherminier, J.; Prensier, G.; Boudon-Padieu, E. and Caudwell, A. 1990. Immunolabeling of grapevine flavescence dorée MLO in salivary glands of Euscelidius variegatus: light and electron microscopy study. J. Histochem & Cytochem. 38:79-86.
Liefting L. W.; Perez-Egusquiza, Z.C.; Clover, G. R. G. and Anderson, J. A. D. 2008. A New ‘Candidatus Liberibacter’ species in Solanum tuberosum in New Zealand. Plant Dis. 92 (10):1474..
Marín-Jarillo, A., Garzón-Tiznado, J. A.; Becerra-Flora, A.; Mejia-Avila, C.; Bujanos-Muñiz, R. y Byerly-Murphuy, K. F. 1995. Ciclo biológico y morfología del "salerillo" Paratrioza cockerelli Sulc. (Homoptera-Psillidae), vector de la enfermedad permanente del jitomate en el Bajío. CATIE, Turrialba, Costa Rica. Revista Manejo Integrado de Plagas 38:25-32.
Moya-Raygoza, G. and Nault, L. R. 1998. Transmission biology of mayze bushy stunt phytoplasm by the corn leafhopper (Homoptera-Vicadellidae). Ann. Entomol. Soc. Am. 91:668-676.
Munyaneza, J. E.; Crosslin, J. M. and Upton, J. E. 2007. Association of Bactericera cockerelli (Homoptera: Psyllidae) with “zebra chip”, a new potato disease in southwestern United State and Mexico. J. Econ. Entom. 100 (3) 656-663.
Murral, D. J.; Nault, L. R.; Hoy, C. W.; Madden, L. V. and Miller, S. A. 1996. Effects of temperature and vector age on transmission of two Ohio strains of aster yellows phytoplasm by the aster leafhopper (Homoptera: Cicadellidae). J. Econ. Entomol. 89:1223-1232.
Nagaich, B. B.; Puri, B. K.; Sinha, R. C.; Dhingra, M. K. and Bhardwaj V. P. 1974. Mycoplasm like organisms in plants affected with purple top-roll, marginal flavescence and witches broom diseases of potatoes. Phytopathol. Z. 81:273-379.
Palermo, S. ; Arzone, A. and Bosco, D. 2001. Vector-pathogen-host plant relationships of chrysanthemum yellows (CY) phytoplasma and the vector leafhoppers Macrosteles quadripunctulatus and Euscelidius variegatus. Entomol. Exp. Appl. 99:347-354.
Pilkington, L.; Gurr, G. M.; Fletcher, M. Y.; Nikandrow, A. and Elliott, E. 2004. Vector status of three leafhopper species for Australian Lucerne yellows phytoplasm. Aust. J. Entomol. 43:366-373.
Purcell, A. H. 1982. Insect vector relation-ships with prokaryotic plant pathogens. Annu. Rev. Phytopathol. 20:397-417.
Richards, B. L. and Blood, H. L. 1933. Psyllid yellows of the potato. J. Agr. Res. 46:189-216.
Salas-Marina, M. A., 2006. Eficiencia de insectos vectores en la transmisión de fitoplasma de la punta morada de la papa. Tesis de Maestría. UAAAN. Saltillo, Coahuila, México. 49 p.
Tanne, E.; Boudon-Padieu, E.; Clair, D.; Davidovich, M.; Melamed, S. and Klein M. 2001. Detection of phytoplasm by polimerase chain reaction of insect feeding medium and its use in determining vectoring ability. Phytopathol. 91:741-746.

Asociación de Hemiptera: Triozidae con la enfermedad permanente del tomate en México 69
Vega, F. E.; Davis, R. E.; Dally, E. L.; Barbosa, P. and Purcell. A. H. 1994. Use of a biotinylated DNA probe for detection of the aster yellows mycoplasmlike organism in Dalbulus maidis and Macrosteles fascifrons (Homoptera : Cicadellidae). Fla. Entomol. 77:330-334.
Vega, F. E.; Davis, R. E.; Barbosa, P.; Dally, E. L. and Purcell, A. H. 1993. Detection of a plant pathogen in a nonvector insect species by the polymerase chain reaction. Phytopathol. 83:621-624.
Weintraub, G. P. and Beanland, L. 2006. Insect vectors of phytoplasms. Annu. Rev. Entomol. 51:91-111.

Agricultura Técnica en México Vol. 35 Núm.1 1 de enero-31 de marzo 2009 p. 70-77
ALTAS DENSIDADES CON DESPUNTE TEMPRANO EN RENDIMIENTO Y PERÍODO DE COSECHA EN CHILE PIMIENTO*
HIGH PLANT STAND WITH EARLY PRUNING ON YIELD AND HARVEST PERIOD IN BELL PEPPER
Nicacio Cruz Huerta1§, Felipe Sánchez del Castillo2, Joaquín Ortiz Cereceres1 y Ma. del Carmen Mendoza Castillo1
1Posgrado en Recursos Genéticos y Productividad, Colegio de Postgraduados, carretera México-Texcoco, km 35.5 Montecillo, Texcoco, Estado de México, México. 56230.([email protected]), ([email protected]). 2Departamento de Fitotecnia, Universidad Autónoma Chapingo, carretera México-Texcoco, km 38.8 Chapingo, Estado de México, México. ([email protected]). §Autor para correspondencia: [email protected].
RESUMEN
El presente estudio se realizó con el objetivo de determinar el efecto de la densidad de población combinado con el despunte temprano de las yemas terminales de plantas de pimiento dulce sobre el rendimiento y el período de cosecha. El experimento se llevó a cabo en hidroponía bajo condiciones de invernadero en Texcoco, Estado de México, México en 2004; se utilizó el cv. Ariane. Se establecieron tres tratamientos: 3.3 plantas m2 (testigo sin poda), 8 plantas m2 con despunte arriba de la cuarta bifurcación del tallo y 14 plantas m2 con despunte arriba de la tercera bifurcación. Se utilizó un diseño en bloques al azar con cuatro repeticiones. Se determinó el rendimiento de fruto comercial por planta y por m2 e índice de área foliar (IAF). El rendimiento de fruto comercial por planta fue 873 g en el testigo a 604 y 404 g en las densidades 8 y 14 plantas m2, respectivamente; mientras que el rendimiento por unidad de superficie fue mayor en las altas densidades de población con despunte temprano (5.63 y 4.83 kg m2 en 14 y 8 plantas m2, respectivamente) que en el testigo (2.88 kg m-2). El sistema de producción con alta densidad de población con despunte temprano de las plantas incrementó significativamente el rendimiento de chile pimiento dulce por unidad de superficie y mostró potencial para reducir el período de cosecha para aprovechar ventanas específicas de mercado.
Palabras clave: Capsicum annuum L., hidroponía, invernadero.
ABSTRACT
The present research was carried out with the aim of determining the effect of plant stand combined with early pruning of the terminal buds on yield and length of harvest period in bell pepper. The experiment was carried out under soil-less culture conditions in a greenhouse at Texcoco, Estado de Mexico, Mexico, from 2004. The cv. Ariane was utilized. Three treatments were studied: 3.3 plants m2 with no pruning (control), 8 plants m2
with pruning above the fourth stem bifurcation and 14 plants m2 with pruning above the third stem bifurcation. A randomized block design with three replications was used. Marketable fruit yield per plant and per m2 and LAI were recorded. Fruit yield was 873 g per plant in the control as compared to 604 and 404 g at 8 and 14 plants m2, respectively; however, yield per area basis was greater under high plant stands (5.63 and 4.83 kg m2 at 14 and 8 plants m2, respectively) as compared to the control (2.88 kg m-2). A high plant stand combined with early
* Recibido: Octubre, 2007
Aceptado: Marzo, 2009

Altas densidades con despunte temprano en rendimiento y período de cosecha en chile pimiento 71
pruning of the terminal buds, significantly increased yield per unit area in bell pepper and showed potential to reduce the harvest period for specific market windows.
Key words: Capsicum annuum L., hydroponic, greenhouse.
INTRODUCCIÓN
La agricultura bajo condiciones de invernadero implica una alta inversión inicial y por tanto requiere producir cultivos de alto valor comercial y económicamente rentables, como hortalizas y flores. En las hortalizas de fruto encontramos tomate y pimiento, en las cuales el período de cosecha en un ciclo de producción bajo invernadero es largo (4 a 7 meses) y se da en un contexto donde el precio del producto oscila constantemente en el mercado. Los precios más altos se presentan en ventanas de tiempo específicas, generalmente de corta duración por lo que diseñar sistemas de producción para concentrar la cosecha en estos períodos, favorece directamente al productor.
La producción de pimiento bajo invernadero en el norte de Europa y de América generalmente se basa en la práctica de poda tipo “Holandés”, que consiste en el uso de cultivares de ciclo indeterminado que se conducen a dos tallos hasta alcanzar 2 a 3 m de altura. Con esta práctica se logran rendimientos altos de 100 a 200 t ha-1 año-1; sin embargo, el ciclos de cultivo es de 9 a 11 meses y el período de inicio a fin de cosecha de 6 a 7 meses (Challinor, 1996; Nuez et al., 1996), lo que hace difícil programar la producción para obtenerla en los períodos cortos de alto precio en el mercado.
En Almería, España, el pimiento se maneja sin poda o con un mínimo de podas de tallos y flores o frutos, para obtener un flujo de producción concentrada. Esta práctica detiene en forma temporal el crecimiento vegetativo y la caída de flores ocasionados por la excesiva demanda de los frutos; no obstante, una vez que inicia la cosecha se recupera el crecimiento que, en condiciones climáticas favorables, puede llevar a otro flujo de producción dos a tres meses después (Posadas, 1993; Nuez et al., 1996); sin embargo, también los ciclos son largos y el rendimiento por unidad de superficie es menor (50 a 80 t ha-1 año-1). Por la altura que alcanzan las plantas con este tipo de manejo, se utilizan densidades de población que oscilan de 2 a 4 plantas m2.
Mediante trasplantes tardíos y despuntes tempranos (un racimo por planta) en el cultivo de tomate (Lycopersicon esculentum Mill.) es posible, manejar hasta 180 mil plantas ha, reducir el ciclo de cultivo de 11 a 2.5 meses y concentrar la cosecha en un período menor a 15 días. Este tipo de manejo permite 3-4 ciclos por año y por lo tanto, el rendimiento potencial anual supera al del sistema convencional que se práctica en invernaderos europeos y norteamericanos (Sánchez y Ponce, 1998; Sánchez et al., 1999).
En pimiento morrón también es posible lograr altos rendimientos por unidad de superficie por medio de ciclos de cultivo más cortos y períodos de cosecha concentrados, mediante el manejo combinado de trasplante de plántulas de mayor edad (60 a 70 días), despunte temprano de los puntos de crecimiento para desarrollar plantas compactas con pocos frutos y altas densidades de población para compensar el menor rendimiento por planta. El despunte temprano y la poda dan como resultado plantas de menor área foliar y altura. El índice de área foliar (IAF) óptimo para máxima tasa de crecimiento se logra con alta densidad de población pero en ciclos más cortos (Cruz et al., 2005).
Dentro de ciertos límites, el aumento en la densidad de población causa disminución en el rendimiento por planta, pero incrementa el rendimiento por unidad de superficie (Batal y Smittle, 1981; Stoffella y Bryan, 1988; Decoteau y Graham, 1994; Cebula, 1995; Lorenzo y Castilla, 1995). La relación que existe entre la densidad de población y el rendimiento por planta no siempre es la misma; en pimiento y tomate, el incremento en la densidad de población disminuye el número de frutos por planta sin afectar el peso medio del fruto, (Stoffella y Bryan, 1988; Cebula, 1995; Sánchez et al., 1998). Sin embargo, no se ha estudiado el potencial productivo del cultivo de pimiento en densidades de población mayores de 10 plantas m2, bajo un sistema intensivo de producción en un medio no restrictivo como el propiciado por invernaderos e hidroponía, como lo reportaron Sánchez et al. (1999). Además, es de interés para el productor cosechar en los períodos con menor oferta para obtener mejor precio. Por esta razón, se requiere generar tecnología que permita obtener altos rendimientos por unidad de superficie, concentrar la cosecha en períodos cortos y mantener la calidad del producto.
El objetivo del presente trabajo fue determinar el efecto de alta densidad de población y despunte temprano de los brotes terminales sobre el rendimiento, período de cosecha y calidad comercial del fruto de pimiento bajo condiciones

72 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
Nicacio Cruz Huerta et al.
de la cuarta bifurcación y densidad alta, 14 plantas m2, con despunte arriba de la tercera bifurcación. Los despuntes se realizaron entre los 60 y 70 días después del transplante. La unidad experimental fue de 5.7 m2 de invernadero para el testigo y de 3.8 m2 para las otras densidades. Se utilizó un diseño de bloques completos al azar con cuatro repeticiones.
Los frutos fueron cosechados cuando al menos 50% de su superficie había cambiado de color verde a naranja. Se realizaron 11 cortes de frutos maduros durante 61 días de cosecha. El primer corte se efectuó a los 97 días después de transplante y los siguientes conforme maduraron los frutos. Los frutos cosechados en cada corte fueron clasificados en forma visual en comerciales y no comerciales, con base en la uniformidad, sanidad y tamaño. Los frutos comerciales se separaron visualmente en tres tamaños: grandes de primera calidad, medianos y chicos. Los frutos no comerciales incluyeron frutos muy pequeños (25-70 g), deformes, o dañados.
El rendimiento acumulado en los primeros 15 días de cosecha fue considerado como rendimiento temprano y el acumulado durante todo el período de la cosecha como rendimiento final. Se analizaron el rendimiento por planta y por metro cuadrado de invernadero. El IAF se determinó a los 120 dds con Un ceptometer (Lincoln, NE).
Se realizó un análisis de varianza en todas las variables usando el paquete SAS® (SAS Institute, Inc. Cary, NC). En los casos en que el efecto de tratamientos fue significativo se realizó la comparación de medias por el método de Tukey (0.05).
RESULTADOS Y DISCUSIÓN
El rendimiento y el número de frutos por planta d i sminuyeron s ign i f i ca t ivamente (p<0.05) a l aumentar la densidad y despuntar la planta por arriba de la tercera o cuarta bifurcación respecto al tratamiento testigo sin despunte en baja densidad. Por el contrario, el rendimiento y número de frutos por unidad de superficie fueron significativamente (p<0.05) mayores en los tratamientos de alta densidad despuntados (Cuadro 1).
de invernadero. Bajo las hipótesis de que con el incremento de la densidad de población combinada con el despunte temprano de yemas terminales se incrementa el rendimiento por unidad de superficie sin afectar significativamente la calidad de fruto y se acorta el ciclo de cultivo lo que permite obtener mayor productividad por unidad de tiempo.
MATERIALES Y MÉTODOS
Se utilizó el híbrido de chile pimiento cv. ‘Ariane’, resistente al virus del mosaico del tabaco, de crecimiento rápido y follaje denso; el fruto es verde hasta la madurez fisiológica y posteriormente se torna naranja; posee 3 a 4 lóculos y paredes gruesas y madura en alrededor de 70 días después de antesis.
Las semillas se germinaron en charolas de poliestireno expandido rellenas con una mezcla comercial de turba orgánica y vermiculita (Ball Seed®, Geo J. Ball Inc.®, USA), se regaron con agua durante los primeros 20 días, luego con solución nutritiva a la mitad de su concentración durante los siguientes 10 días y con concentración completa hasta el fin del ciclo. La concentración de cada nutrimento (mg L-1) en la solución nutritiva completa fue: N:200, P:60, K:250, Ca:250, S:200, Mg:60, Fe: 3.0, Mn:0.5, Bo:0.5, Cu:0.5 y Zn:0.5.
Las plántulas fueron transplantadas a los 55 días después de la siembra (dds) en tinas de 0.3 m de profundidad y 1.2 m de ancho, que contenían arena de tezontle rojo con partículas de 1 a 4 mm de diámetro en un invernadero de vidrio en Texcoco, Estado de México (2 251 m, 19° 29’ latitud norte y 98° 53’ longitud oeste), en 2005. Las plantas fueron regadas con la solución nutritiva antes descrita por medio de un sistema de cintas de goteo (4 L m-2 d-1), se aplicaron en dos riegos por día en promedio en función del estado de desarrollo y las condiciones ambientales. El control químico de plagas y enfermedades y otras prácticas se realizaron de acuerdo con lo descrito por Cruz et al. (2005).
A partir de la antesis de las primeras flores, se provocaron corrientes de aire dos veces por semana con un ventilador a la altura de las plantas para mejorar la polinización. Se permitió el desarrollo de todos los frutos que amarraron, incluso el de la yema terminal.
Se aplicaron tres tratamientos: testigo, 3.3 plantas m2 sin despunte cosechando sólo el primer flujo de fructificación, densidad intermedia, 8 plantas m2; con despunte arriba
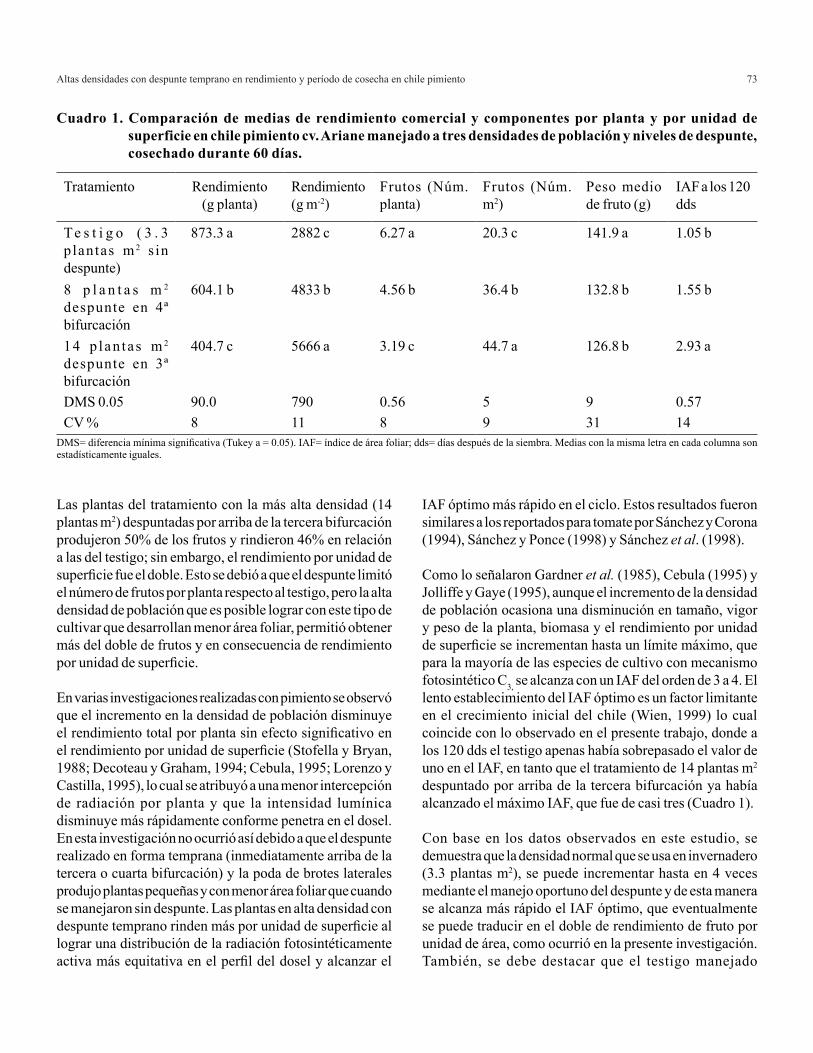
Altas densidades con despunte temprano en rendimiento y período de cosecha en chile pimiento 73
Las plantas del tratamiento con la más alta densidad (14 plantas m2) despuntadas por arriba de la tercera bifurcación produjeron 50% de los frutos y rindieron 46% en relación a las del testigo; sin embargo, el rendimiento por unidad de superficie fue el doble. Esto se debió a que el despunte limitó el número de frutos por planta respecto al testigo, pero la alta densidad de población que es posible lograr con este tipo de cultivar que desarrollan menor área foliar, permitió obtener más del doble de frutos y en consecuencia de rendimiento por unidad de superficie.
En varias investigaciones realizadas con pimiento se observó que el incremento en la densidad de población disminuye el rendimiento total por planta sin efecto significativo en el rendimiento por unidad de superficie (Stofella y Bryan, 1988; Decoteau y Graham, 1994; Cebula, 1995; Lorenzo y Castilla, 1995), lo cual se atribuyó a una menor intercepción de radiación por planta y que la intensidad lumínica disminuye más rápidamente conforme penetra en el dosel. En esta investigación no ocurrió así debido a que el despunte realizado en forma temprana (inmediatamente arriba de la tercera o cuarta bifurcación) y la poda de brotes laterales produjo plantas pequeñas y con menor área foliar que cuando se manejaron sin despunte. Las plantas en alta densidad con despunte temprano rinden más por unidad de superficie al lograr una distribución de la radiación fotosintéticamente activa más equitativa en el perfil del dosel y alcanzar el
IAF óptimo más rápido en el ciclo. Estos resultados fueron similares a los reportados para tomate por Sánchez y Corona (1994), Sánchez y Ponce (1998) y Sánchez et al. (1998).
Como lo señalaron Gardner et al. (1985), Cebula (1995) y Jolliffe y Gaye (1995), aunque el incremento de la densidad de población ocasiona una disminución en tamaño, vigor y peso de la planta, biomasa y el rendimiento por unidad de superficie se incrementan hasta un límite máximo, que para la mayoría de las especies de cultivo con mecanismo fotosintético C3, se alcanza con un IAF del orden de 3 a 4. El lento establecimiento del IAF óptimo es un factor limitante en el crecimiento inicial del chile (Wien, 1999) lo cual coincide con lo observado en el presente trabajo, donde a los 120 dds el testigo apenas había sobrepasado el valor de uno en el IAF, en tanto que el tratamiento de 14 plantas m2 despuntado por arriba de la tercera bifurcación ya había alcanzado el máximo IAF, que fue de casi tres (Cuadro 1).
Con base en los datos observados en este estudio, se demuestra que la densidad normal que se usa en invernadero (3.3 plantas m2), se puede incrementar hasta en 4 veces mediante el manejo oportuno del despunte y de esta manera se alcanza más rápido el IAF óptimo, que eventualmente se puede traducir en el doble de rendimiento de fruto por unidad de área, como ocurrió en la presente investigación. También, se debe destacar que el testigo manejado
Cuadro 1. Comparación de medias de rendimiento comercial y componentes por planta y por unidad de superficie en chile pimiento cv. Ariane manejado a tres densidades de población y niveles de despunte, cosechado durante 60 días.
DMS= diferencia mínima significativa (Tukey a = 0.05). IAF= índice de área foliar; dds= días después de la siembra. Medias con la misma letra en cada columna son estadísticamente iguales.
Tratamiento Rendimiento(g planta)
Rendimiento (g m-2)
Frutos (Núm. planta)
Frutos (Núm. m2)
Peso medio de fruto (g)
IAF a los 120 dds
Te s t i g o ( 3 . 3 p lantas m 2 s in despunte)
873.3 a 2882 c 6.27 a 20.3 c 141.9 a 1.05 b
8 p l a n t a s m 2 despunte en 4ª bifurcación
604.1 b 4833 b 4.56 b 36.4 b 132.8 b 1.55 b
14 p lan tas m 2 despunte en 3ª bifurcación
404.7 c 5666 a 3.19 c 44.7 a 126.8 b 2.93 a
DMS 0.05 90.0 790 0.56 5 9 0.57CV % 8 11 8 9 31 14
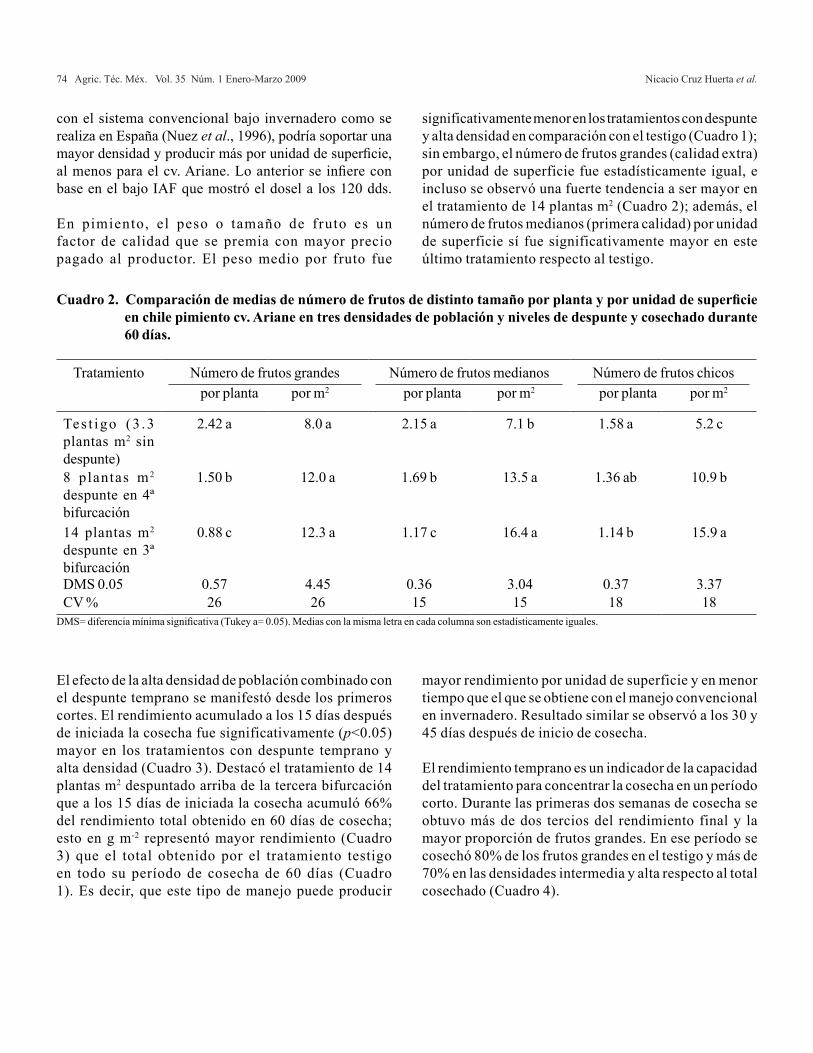
74 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
Nicacio Cruz Huerta et al.
con el sistema convencional bajo invernadero como se realiza en España (Nuez et al., 1996), podría soportar una mayor densidad y producir más por unidad de superficie, al menos para el cv. Ariane. Lo anterior se infiere con base en el bajo IAF que mostró el dosel a los 120 dds.
En pimiento, el peso o tamaño de fruto es un factor de calidad que se premia con mayor precio pagado al productor. El peso medio por fruto fue
El efecto de la alta densidad de población combinado con el despunte temprano se manifestó desde los primeros cortes. El rendimiento acumulado a los 15 días después de iniciada la cosecha fue significativamente (p<0.05) mayor en los tratamientos con despunte temprano y alta densidad (Cuadro 3). Destacó el tratamiento de 14 plantas m2 despuntado arriba de la tercera bifurcación que a los 15 días de iniciada la cosecha acumuló 66% del rendimiento total obtenido en 60 días de cosecha; esto en g m-2 representó mayor rendimiento (Cuadro 3) que el total obtenido por el tratamiento testigo en todo su período de cosecha de 60 días (Cuadro 1). Es decir, que este tipo de manejo puede producir
significativamente menor en los tratamientos con despunte y alta densidad en comparación con el testigo (Cuadro 1); sin embargo, el número de frutos grandes (calidad extra) por unidad de superficie fue estadísticamente igual, e incluso se observó una fuerte tendencia a ser mayor en el tratamiento de 14 plantas m2 (Cuadro 2); además, el número de frutos medianos (primera calidad) por unidad de superficie sí fue significativamente mayor en este último tratamiento respecto al testigo.
mayor rendimiento por unidad de superficie y en menor tiempo que el que se obtiene con el manejo convencional en invernadero. Resultado similar se observó a los 30 y 45 días después de inicio de cosecha.
El rendimiento temprano es un indicador de la capacidad del tratamiento para concentrar la cosecha en un período corto. Durante las primeras dos semanas de cosecha se obtuvo más de dos tercios del rendimiento final y la mayor proporción de frutos grandes. En ese período se cosechó 80% de los frutos grandes en el testigo y más de 70% en las densidades intermedia y alta respecto al total cosechado (Cuadro 4).
Tratamiento Número de frutos grandes por planta por m2
Número de frutos medianospor planta por m2
Número de frutos chicos por planta por m2
Tes t igo (3 .3 plantas m2 sin despunte)
2.42 a 8.0 a 2.15 a 7.1 b 1.58 a 5.2 c
8 plantas m2 despunte en 4ª bifurcación
1.50 b 12.0 a 1.69 b 13.5 a 1.36 ab 10.9 b
14 plantas m2 despunte en 3ª bifurcación
0.88 c 12.3 a 1.17 c 16.4 a 1.14 b 15.9 a
DMS 0.05 0.57 4.45 0.36 3.04 0.37 3.37CV % 26 26 15 15 18 18
Cuadro 2. Comparación de medias de número de frutos de distinto tamaño por planta y por unidad de superficie en chile pimiento cv. Ariane en tres densidades de población y niveles de despunte y cosechado durante 60 días.
DMS= diferencia mínima significativa (Tukey a= 0.05). Medias con la misma letra en cada columna son estadísticamente iguales.
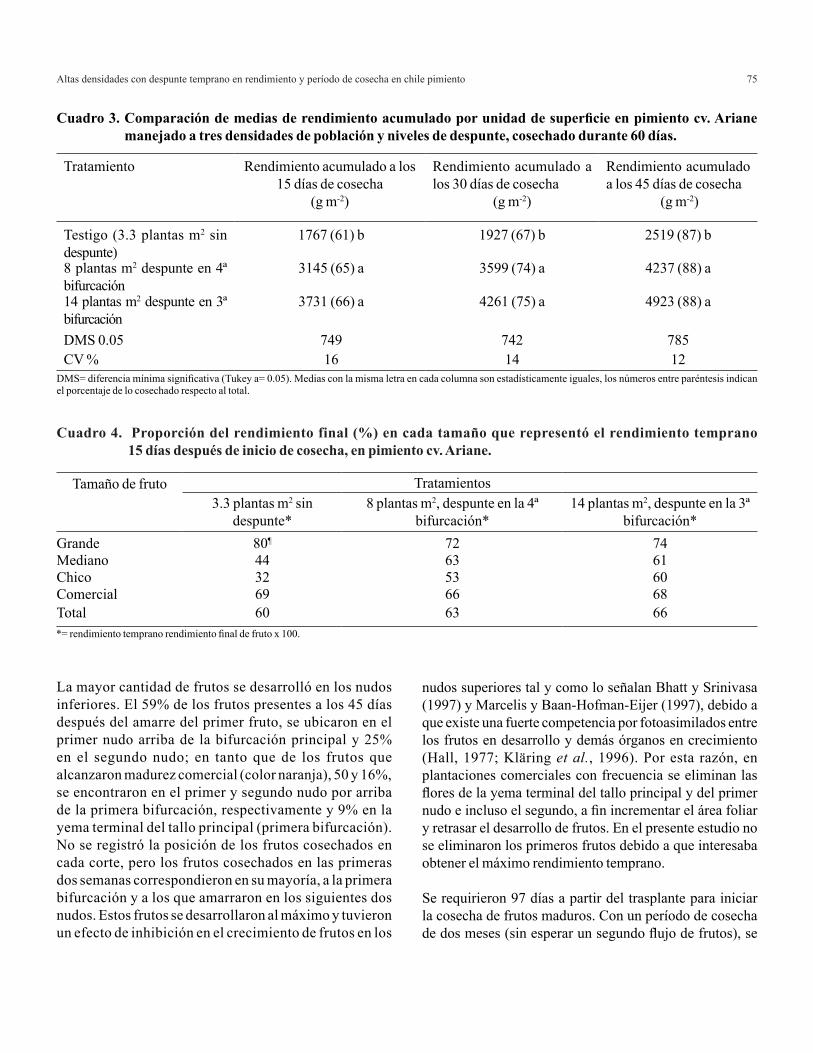
Altas densidades con despunte temprano en rendimiento y período de cosecha en chile pimiento 75
Tamaño de fruto Tratamientos3.3 plantas m2 sin
despunte*8 plantas m2, despunte en la 4ª
bifurcación*14 plantas m2, despunte en la 3ª
bifurcación*Grande 80¶ 72 74Mediano 44 63 61Chico 32 53 60Comercial 69 66 68Total 60 63 66
Cuadro 3. Comparación de medias de rendimiento acumulado por unidad de superficie en pimiento cv. Ariane manejado a tres densidades de población y niveles de despunte, cosechado durante 60 días.
Tratamiento Rendimiento acumulado a los 15 días de cosecha
(g m-2)
Rendimiento acumulado a los 30 días de cosecha
(g m-2)
Rendimiento acumulado a los 45 días de cosecha
(g m-2)
Testigo (3.3 plantas m2 sin despunte)
1767 (61) b 1927 (67) b 2519 (87) b
8 plantas m2 despunte en 4ª bifurcación
3145 (65) a 3599 (74) a 4237 (88) a
14 plantas m2 despunte en 3ª bifurcación
3731 (66) a 4261 (75) a 4923 (88) a
DMS 0.05 749 742 785CV % 16 14 12
DMS= diferencia mínima significativa (Tukey a= 0.05). Medias con la misma letra en cada columna son estadísticamente iguales, los números entre paréntesis indican el porcentaje de lo cosechado respecto al total.
La mayor cantidad de frutos se desarrolló en los nudos inferiores. El 59% de los frutos presentes a los 45 días después del amarre del primer fruto, se ubicaron en el primer nudo arriba de la bifurcación principal y 25% en el segundo nudo; en tanto que de los frutos que alcanzaron madurez comercial (color naranja), 50 y 16%, se encontraron en el primer y segundo nudo por arriba de la primera bifurcación, respectivamente y 9% en la yema terminal del tallo principal (primera bifurcación). No se registró la posición de los frutos cosechados en cada corte, pero los frutos cosechados en las primeras dos semanas correspondieron en su mayoría, a la primera bifurcación y a los que amarraron en los siguientes dos nudos. Estos frutos se desarrollaron al máximo y tuvieron un efecto de inhibición en el crecimiento de frutos en los
Cuadro 4. Proporción del rendimiento final (%) en cada tamaño que representó el rendimiento temprano 15 días después de inicio de cosecha, en pimiento cv. Ariane.
*= rendimiento temprano rendimiento final de fruto x 100.
nudos superiores tal y como lo señalan Bhatt y Srinivasa (1997) y Marcelis y Baan-Hofman-Eijer (1997), debido a que existe una fuerte competencia por fotoasimilados entre los frutos en desarrollo y demás órganos en crecimiento (Hall, 1977; Kläring et al., 1996). Por esta razón, en plantaciones comerciales con frecuencia se eliminan las flores de la yema terminal del tallo principal y del primer nudo e incluso el segundo, a fin incrementar el área foliar y retrasar el desarrollo de frutos. En el presente estudio no se eliminaron los primeros frutos debido a que interesaba obtener el máximo rendimiento temprano.
Se requirieron 97 días a partir del trasplante para iniciar la cosecha de frutos maduros. Con un período de cosecha de dos meses (sin esperar un segundo flujo de frutos), se

76 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
Nicacio Cruz Huerta et al.
necesitaron 158 días para completar un ciclo (transplante a finde cosecha), por lo que, bajo condiciones similares de radiación solar y temperatura del invernadero se podrían obtener 2.3 ciclos por año. Si se toman como base los rendimientos por unidad de superficie obtenidos (Cuadro 1) y se multiplican por ese factor de 2.3, con un manejo en baja densidad y sin despunte (testigo) se podrían obtener 6.63 kg m-2 año-1, mientras que con densidad de 8 plantas m2 despuntadas arriba de la cuarta bifurcación se obtendrían 11.12 kg m-2 año-1 y con 14 plantas m2 en plantas despuntadas arriba de la tercera bifurcación se tendrían 13.03 kg m-2 año-1.
Si el período de cosecha se acotara a los primeros 15 días en el que se cosechó más de 60% del rendimiento total final y más de 70% de frutos de primera calidad (grandes), el ciclo se reduce a 112 días (de transplante a fin de cosecha), lo que permitiría aumentar el número de ciclos a 3.2 por año y potencial de rendimiento de fruto comercial sería de 3.145 y 3.731 kg m-2 por ciclo (Cuadro 4), lo que equivale a 10.06 y 11.94 kg m-2 año-1 para 8 y 14 plantas m2, respectivamente.
Lo anterior indica que este sistema de manejo intensivo del pimiento dulce tiene un alto potencial para incrementar la productividad anual por unidad de superficie; sin embargo, se requiere investigación adicional para concentrar la cosecha en 2 ó 3 semanas, de tal manera que se reduzca el ciclo, mejorar la calidad y aprovechar ventanas de mercado favorables.
CONCLUSIONES
El aumento de la densidad de población de 3.3 plantas m2 sin despuntar a 14 plantas m2 con despunte inmediatamente arriba de la tercera bifurcación disminuye el rendimiento por planta, debido a un menor peso y número frutos por planta, pero incrementa significativamente el rendimiento de fruto comercial de pimiento dulce por unidad de superficie.
Alta densidad de plantación con despunte reduce en 46 días el ciclo de cultivolo que permite producir de 2.3 hasta 3.2 ciclos por año con rendimiento potencial de 13.03 kg m-2 año-1.
LITERATURA CITADA
Batal, K. M. and Smittle, D. A. 1981. Response of bell pepper to irrigation, nitrogen, and plant-population. J. Am. Soc. Hort. Sci. 106:259-262.
Bhatt, R. M. and Srinivasa-Rao, N. K. 1997. Growth and photosynthesis in bell-pepper as affected by sink manipulation. Biol. Plantarum 39:437-439.
Cebula, S. 1995. Optimization of plant and shoot spacing in greenhouse production of sweet pepper. Acta Hort. 412:321-329.
Challinor, P. F. 1996. Producción de pimiento en climas fríos. In: Pimientos. Namesny, V. (ed). Ed. de Horticultura. Reus, España. p. 41-44.
Cruz-Huerta, N.; Ortiz-Cereceres, J.; Sanchez-Del-Castillo, F. y Mendoza-Castillo, M. C. 2005. Biomasa e indices fisiológicos en chile morrón cultivado en altas densidades. Rev. Fitotec. Mex. 28:287-293.
Decoteau, D. R. and Graham, H. A. H. 1994. Plant spatial arrangement affects growth, yield, and pod distribution of cayenne peppers. Hortsci. 29:149-151.
Gardner, F. P.; Pearce, R. B. and Mitchell, R. L. 1985. Physiology of crop plants. Iowa State University Press. Ames. USA. 327 p.
Hall, A. J. 1977. Assimilate source-sink relationships in Capsicum Annuum L. 1. Dynamics of growth in fruiting and deflorated plants. Aust. J. Plant Physiol- 4:623-636.
Jolliffe, P. A. and Gaye, M. M. 1995. Dynamics of growth and yield components of bell peppers (Capsicum annuum L.) to row covers and population density. Sci. Hort. 62:153-164.
Kläring, H. P.; Heiβner, A. and Fink, M. 1996. Growth of a sweet pepper crop: measurements for modelling. Acta Hort. 417:107-112.
Lorenzo, P. y Castilla, N. 1995. Bell pepper yield response to plant density and radiation in unheated plastic greenhouse. Acta Hort . 412:330-334.
Marcelis, L. F. and Baan-Hofman-Eijer, L. R. 1997. Effects of seed number on competition and dominance among fruits in Capsicum annuum L. Ann. Bot. 79:687-693.
Nuez, V. F.; Gil, R. O. y Costa, J. G. 1996. El cultivo de pimientos, chiles y ajíes. Ed. Mundi-Prensa. Madrid, España. 607 p.
Posadas, S. F. 1993. El cultivo del pimiento en sistemas hidropónicos. In: cultivos sin suelo. Martínez, J. C. F. Díaz, A. (eds). FIAPA. Almería, España. p. 313-317.

Altas densidades con despunte temprano en rendimiento y período de cosecha en chile pimiento 77
Stoffella, P. J. and Bryan, H. H. 1988. Plant population influences growth and yields of bell pepper. J. Am. Soc. Hort. Sci. 113:835-839.
Sánchez, Del C. F. y Corona, S. T. 1994. Evaluación de cuatro variedades de tomate (Lycopersicon esculentum Mill.) bajo un sistema hidropónico de producción a base de despuntes y altas densidades. Rev. Chapingo, Serie Horticultura 1:109-114.
Sánchez, Del C. F.; Ortiz, C. J.; Mendoza, C. M. C.; González, H. V. A. y Bustamante, O. J. 1998. Physiological and agronomical parameters of tomato in two new production systems. Fitotec. Mex. Vol. 21(1):1-13.
Sánchez, Del C. F. y Ponce, O. J. 1998. Densidades de población y niveles de despunte en tomate (Lycopersicum esculentum Mill.) cultivado en hidroponía. Rev. Chapingo, Serie Horticultura. 4:89-94.
Sánchez, Del C. F.; Ortíz, C. J.; Mendoza, C. M. C.; González, H. V. A. y Colinas, L. M. T. 1999. Características morfológicas asociadas con un arquetipo de tomate apto para un ambiente no restrictivo. Agrociencia. 33:21-29.
Wien, H. C. 1999. Peppers. In: the physiology of vegetable crops. Wien, H. C. (ed). CABI Publishing. Cambridge, UK. p. 259-293.

Agricultura Técnica en México Vol. 35 Núm.1 1 de enero-31 de marzo 2009 p. 78-84
ANÁLISIS DE LA DIVERSIDAD GENÉTICA DE ‘CAÑA FLECHA’ Gynerium sagittatum Aubl. UTILIZANDO LA TÉCNICA DE AFLP*
ANALYSIS OF THE GENETIC DIVERSITY OF ‘CAÑA FLECHA’ Gynerium sagittatum Aubl. USING THE AFLP TECHNIQUE
Hernando Javier Rivera Jiménez1§, Isidro Elias Suárez Padron2 y Juan Diego Palacio Mejía3
1Laboratorio de Biotecnología Vegetal, Facultad de Ciencias Agrícolas. Universidad de Córdoba. A. A 354. Montería, Córdoba. Colombia. Tel. 00 57 4 7908023. 2Departamento de Ingeniería Agronómica y Desarrollo Rural. Universidad de Córdoba. A. A 354. Montería, Córdoba. Colombia. Tel. 5747908023, ([email protected]). 3Laboratorio Central Interinstitucional de Detección y Monitoreo de Organismos Genéticamente Modificados, Instituto Colombiano Agropecuario -ICA, km 14 vía Mosquera ICA- Tibaitatá. Mosquera, Cundinamarca, Colombia. Tel. 00 57 1 4227371 Ext. 1829, ([email protected]). §Autor para correspondencia: [email protected].
* Recibido: Febrero, 2008
Aceptado: Febrero, 2009
RESUMEN
La fibra de la caña flecha Gynerium sagittatum (Aubl.) es materia prima para la elaboración de artesanías (Zorro y Prieto, 1999), entre ellas el sombrero vueltiao, símbolo de identidad cultural colombiana. El objetivo de este estudio fue la caracterización molecular de 25 accesiones de G. sagittatum (Aubl.) mediante la técnica de AFLP utilizando cuatro combinaciones de cebadores para determinar como se distribuye la variabilidad genética entre las poblaciones de distintas regiones de Colombia considerando criterios geográficos y morfológicos. El análisis de correspondencia múltiple discriminó tres grupos, donde se identifican características de importancia artesanal (comercial) y atributos agronómicos deseables para la elaboración de artesanías. Se observó una baja correlación entre la distancia geográfica y nivel de diferenciación genética de la especie, lo cual indicó un flujo del mismo material por su carácter de reproducción asexual a las diferentes regiones del país, consecuencia de un traslado de cultivares por acción antrópica.
Palabras clave: Gynerium sagittatum (Aubl.), recursos genéticos, agrupamiento, evaluación.
ABSTRACT
The fiber wild cane Gynerium sagittatum (Aubl.) is used as raw material for the elaboration of crafts (Zorro y Prieto, 1999), among them the hat “vueltiao”, symbol of Colombian cultural identity. The objective of this study was the characterization molecular of 25 accessions of G. sagittatum (Aubl.) by means of AFLP to determine the distribution of the genetic variation among populations from different regions in Colombia considering geographic and morphological criteria. The analyses of multiple correspondences discriminated three groups, where desirable characteristics of artisan importance (commercial) and agronomic attributes for the elaboration of crafts are identified. A low correlation was observed between the geographic distance and the level of genetic differentiation of the species, which indicated a flow of the same material by its character of asexual reproduction to the different regions in the country, as a consequence of the transfer of valuable material by entropic action.
Key words: Gynerium sagittatum (Aubl.), genetic resources, clustering pattern, evaluation.

Análisis de la diversidad genética de caña flecha Gynerium sagittatum Aubl. utilizando la técnica de AFLP 79
INTRODUCCIÓN
La fibra de caña flecha Gyneryum sagitatum Aubl., es la materia prima para la elaboración de artesanías, donde se destaca el sombrero “vueltiao” símbolo de identidad cultural colombiana (Zorro y Prieto, 1999). La caña flecha es nativa del oeste de la India y se distribuye desde el sur de México a través de América Central y Suramérica hasta el Paraguay (Howard, 1979). En Colombia se distribuye en gran parte de la geografía nacional localizándose espontáneamente a lo largo de riberas de los ríos y zonas de altas precipitaciones (Zorro y Prieto, 1999). El diagnóstico más reciente sobre el estado de la biodiversidad de la caña flecha en Colombia indica que varios de sus ecosistemas se encuentran seriamente amenazados, con alto riesgo de pérdida de este recurso genético producto de la destrucción del hábitat debido actividades como la ganadería y la agricultura está conduciendo a la reducción de la variabilidad genética (Aramendiz et al., 2005). Actualmente, no se conocen estudios de diversidad genética mediante técnicas moleculares de las diferentes accesiones de caña flecha. La carencia de semilla de caña flecha de óptima calidad, acorde con la demanda, amerita realizar ésta investigación con el fin de generar conocimientos para apoyar tecnológicamente el cultivo y así incrementar los ingresos de los productores.
El banco de germoplasma de caña flecha de la Universidad de Córdoba el cual contiene 25 introducciones, está caracterizado mediante la descripción de la variación morfológica, particularmente características de interés directo para los usuarios (Aramendiz et al., 2005). Este enfoque aunque es muy útil, tiene ciertas limitaciones: son características de baja heredabilidad frecuentemente muestra poca variación para muchos materiales estudiados y la expresión de las características está sujeta a variación ambiental y puede ser difícil de medir (Rivera et al., 2007). Una manera de mitigar estas limitaciones y complementar la caracterización de esta colección es con el empleo de técnicas moleculares que analizan el polimorfismo del ADN (Karpa, 1997).
El objetivo de esta investigación fue determinar la variabilidad genética de 25 accesiones de caña flecha del banco de germoplasma de la Universidad de Córdoba mediante la caracterización molecular con marcadores AFLP (Voz et al., 1995), que permiten establecer el índice de diferenciación genética de G. sagittatum Aubl. y conocer como se distribuye la variabilidad genética entre las poblaciones de caña flecha de distintas regiones de Colombia.
MATERIALES Y MÉTODOS
Material y condiciones experimentalesLa investigación se realizó en el período enero 2006 a diciembre 2007, en el laboratorio de biología molecular del Instituto de Investigación de Recursos Biológicos "Alexander von Humboldt" utilizando 25 accesiones de caña flecha entre cultivadas y silvestres, procedentes de diferentes regiones de los departamentos de Córdoba, Antioquia y Caldas, Colombia. Las accesiones pertenecen al banco de germoplasma de la Universidad de Córdoba, Montería, Colombia.
Se tomó tejido foliar del las diferentes accesiones, sin nervadura central con un período no mayor de seis h después de la colecta, se almacenó la muestra en una bolsa plástica con 50 g de Sílica gel, Posteriormente se maceró con nitrógeno líquido y almacenó a -196 °C para la conservación del material vegetal. La extracción de ADN se realizó en base en los protocolos del manual Dnaeasy Plant Mini Kit (Qiagen, 2006) con algunas modificaciones, reduciendo temperaturas de incubación y tiempo de centrifugado según Rivera et al. (2007). La calidad del ADN se visualizó mediante electroforesis en gel de agarosa al 1% teñido con bromuro de etidio; la cuantificación se hizo mediante el uso del marcador de peso de ADN Lambda 20-bp para la cantidad de ADN total, adicionalmente se hicieron electroforesis en gel de agarosa al 1% teñido con bromuro de etidio, y visualización en luz UV.
Se utilizó la metodología descrita por Vos et al., (1995) modificada por Invitrogen (2003), y se utilizó el Kit de AFLP invitrogen starter primer kit, siguiendo las instrucciones del fabricante. Para la amplificación del ADN se empleó un termociclador MJ Research PTC-100. Después de haber medido la concentración de ADN se procedió a realizar la digestión y ligación de adaptadores de los 25 ADNs de toda la población. Se evaluaron ocho combinaciones de cebadores de los cuales cuatro fueron utilizados en todas la poblaciones. Las reacciones de amplificación fueron visualizadas en un gel de poliacrilamida al 6% en una cámara de electroforesis vertical BIO-RAD® y se utilizó una fuente de poder BIO-RAD Power PAC 3000.
Análisis de datos
Para realizar los análisis de diversidad genética se realizó el análisis de agrupamiento a partir de matrices
80 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
Hernando Javier Rivera Jiménez et al.
de similitud mediante la presencia (1) o ausencia (0) de bandas específicas originadas en la amplificación de los fragmentos digeridos que se transformaron en una matriz binaria. Con la matriz de datos presencia/ausencia se calculó el índice de similitud desarrollado por Dice (1945) y adaptado por Nei y Li (1979) para datos moleculares. El índice promedia los valores de similitud por par de individuos mediante la siguiente ecuación: Sij= 2a / (2a+b+c); donde, Sij= similitud entre los individuos i y j; a= número de loci compartidos por i y j; b= número de loci presentes en i pero ausentes en j, y c= número de loci presentes en j pero ausentes en i.
Las matrices y los dendrogramas de similitud se calcularon con el programa NTSYS-PC, versión 2.02i (Rohlf, 1998) mediante el método UPGMA y el agrupamiento SAHN respectivamente. También se analizaron las relaciones entre
individuos mediante el análisis de correspondencia múltiple (ACM) con toda la población para obtener una representación grafica de la distancia entre las introducciones.
RESULTADO Y DISCUSIÓN
La reacción de amplificación mostró que la combinación de cebadores en los primeros cuatros tratamientos presentó mayor polimorfismo (Cuadro 1). La calidad del ADN fue buena en la segunda elusión (Figura 1a), presentándose así digestión completa con las enzimas de restricción EcoRI y MseI del kit del sistema de análisis I de AFLP de Invitrogen, (2003), (Figura 1b). La reacción de preamplificación de AFLP, presentó buena amplificación durante la PCR, confirmando así el buen estado de los ADNs obtenidos.
Tratamiento Combinación de cebadores
Núm. de loci †
(%) de polimorfismo ¥
P= polimórfico y M= monomórficoTotal P M
1 E-ACC/M-CAG 43 17 26 39.532 E-ACA/M-CAT 52 20 32 38.463 E-ACA/M-CTA 72 23 49 31.944 E-ACT/M-CAA 56 15 41 25.80Total 223 75 148 33.63 §Promedio 37
Cuadro 1. Grado del polimorfismo detectado en las introducciones con diversas combinaciones de cebadores AFLP.
† P= polimórfico y M= monomórfico; ¥= determinado en base del número de loci polimórficos fuera del número total de loci amplificadas por una combinación de cebadores a través de todas las variedades; §= polimorfismo promedio.
Figura 1. a) ADN total de caña flecha, y b) ADN digerido con enzimas EcoRI y MseI.
18.7555.75
MP 21 22 23 24 25
b
1- elusión de ADN
2- elusión de ADN
a

Análisis de la diversidad genética de caña flecha Gynerium sagittatum Aubl. utilizando la técnica de AFLP 81
La combinación de cebadores (E-ACC/M-CAG) y (E-ACA/M-CAT) presentaron el mayor porcentaje de polimorfismo. Se obtuvieron 223 loci y se extendieron desde 43 (E-ACC/M-CAG) hasta 72 loci (E-ACA/M-CTA), con un promedio de 55.75 loci por cebador. En el estudio, 75 loci fueron polimórficos (Cuadro 1). El tratamiento con la combinación de cebadores E-ACC/M-CAG, mostró mayor porcentaje de polimorfismo. Los fragmentos de ADN amplificados estuvieron en un rango de 330 y 20 pb., obteniéndose 43 loci, 17 de los cuales mostraron polimorfismo. Utilizando la combinación de cebadores E-ACA/M-CAT se obtuvo 38.46% de fragmentos de ADN amplificados, los fragmentos estuvieron en un rango de 330 y 20 pb obteniéndose 52 loci las cuales 20 loci mostraron polimorfismo. Las combinaciones E-ACA/M-CTA y la combinación E-ACT/M-CAA mostraron un total de 72 y 56 loci, de las cuales 23 y 15 loci respectivamente eran polimórficas, obteniendo así un porcentaje de polimorfismo de 31.94% y 25.80% (Cuadro 1), los fragmentos de ADN amplificados estuvieron en un rango de 300 y 20 pb, respectivamente. Cada una de estas combinaciones presentaron perfiles de loci diferentes, por lo tanto estas combinaciones de cebadores lograron tener un alto escrutinio del genoma, identificando así la variabilidad genética en la especie.
Parte de las combinaciones de cebadores utilizados en este estudio se tomaron de trabajos recientes en caña de azúcar (Saccharum spp.), divulgados por Selvi et al. (2005), donde se realizaron estudios poblacionales de diversidad genética. Estos datos son similares a los encontrados en caña flecha con respecto al grado de polimorfismo de las combinación de cebadores utilizada para Saccharum spp. Besse et al. (1998) realizaron estudios en la misma especie obteniendo resultados similares a los descritos por Selvi et al. (2005), Otros estudios de similaridad genética en colecciones de cultivares brasileños de la caña de azúcar usando los marcadores de AFLP revelaron promedios altos de polimorfismo (Lima et al., 2002).
El análisis de correspondencia múltiple reveló que los marcadores de AFLP eran informativos y reflejó las relaciones de similitud genética existentes entre las accesiones. Al aumentar el número de cebadores se hizo más selectivo y discriminatorio el estudio permitiendo un análisis más detallado. Al analizar la matriz de datos moleculares de todos los individuos de la población de caña flecha uniendo las matrices de las cuatros combinaciones de cebadores en un ACM bidimensional se presentó una tendencia de distribución de tres grupos (Figura 2).
Esta tendencia de agrupamiento fue similar a los grupos reportados por Aramendiz et al. (2005). El segundo grupo donde se encuentran las accesiones: 1, 13, 20 b, 23, 5, 14, 22, 2, 6, 8, 16 15 y 24, presentan atributos como fibra de textura suave, pubescencia escasa de la vaina verde y pared del tallo gruesa, asimismo se asocia con el área útil de la nervadura inferior y diámetro del tallo delgado y fueron colectados en los municipios de San Andrés de Sotavento, Sahagún, Ciénaga de Oro y Montelibano (departamento de Córdoba-Colombia), recibiendo el nombre de “criolla”. Los individuos de los grupos uno y tres, tienden a conformar el segundo grupo a los reportados por Aramendiz et al. (2005), caracterizándose por textura gruesa, abundante pubescencia en la base de la hoja y vaina, pared delgada del tallo, diámetro del tallo delgado y ligeramente grueso, área útil de la nervadura inferior a intermedia y ángulo de la hoja agudo. Este grupo contiene en su mayoría introducciones provenientes de Tierralta y San Andrés de Sotavento (Córdoba); Caucasia (Antioquia) y Chinchina (Caldas), recibiendo el nombre de caña flecha, caña brava y martinera, por tener estos cultivares una textura áspera, no apta para elaborar artesanías finas.
El análisis de similaridad utilizando el coeficiente de DICE, formó tres grupos (Figura 3). De acuerdo con la evidencia, la agrupación de las introducciones de caña flecha se puede deber a efectos influenciados por su constitución genética y
Figura 2. Análisis de correspondencia múltiple de acuerdo con los resultados de análisis AFLP en las accesiones de caña flecha (Gyneryum sagitatum Aubl.).
-0.36 -0.21 -0.06 0.09 0.24 Dim-1
0.31
0.16
0.02Dim-2
-0.13
-0.27
Grupo 3Textura gruesa
Grupo 1 Textura gruesa
Grupo 2Textura finapo
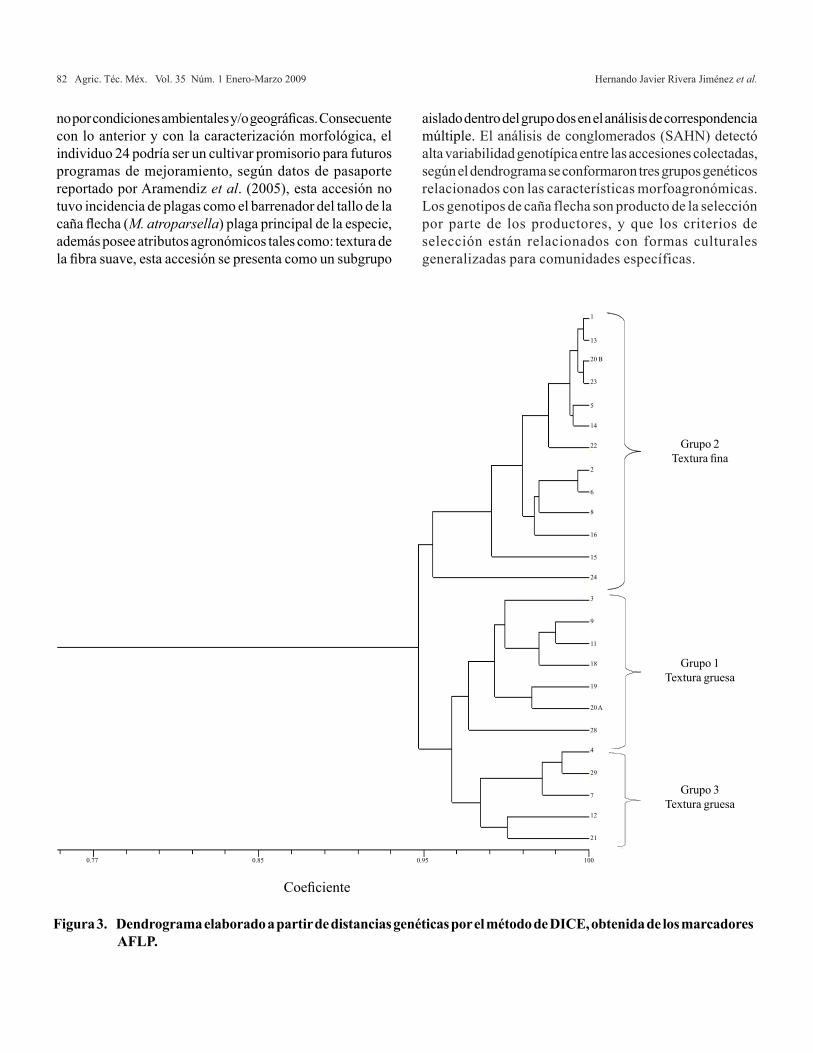
82 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
Hernando Javier Rivera Jiménez et al.
aislado dentro del grupo dos en el análisis de correspondencia múltiple. El análisis de conglomerados (SAHN) detectó alta variabilidad genotípica entre las accesiones colectadas, según el dendrograma se conformaron tres grupos genéticos relacionados con las características morfoagronómicas. Los genotipos de caña flecha son producto de la selección por parte de los productores, y que los criterios de selección están relacionados con formas culturales generalizadas para comunidades específicas.
no por condiciones ambientales y/o geográficas. Consecuente con lo anterior y con la caracterización morfológica, el individuo 24 podría ser un cultivar promisorio para futuros programas de mejoramiento, según datos de pasaporte reportado por Aramendiz et al. (2005), esta accesión no tuvo incidencia de plagas como el barrenador del tallo de la caña flecha (M. atroparsella) plaga principal de la especie, además posee atributos agronómicos tales como: textura de la fibra suave, esta accesión se presenta como un subgrupo
Figura 3. Dendrograma elaborado a partir de distancias genéticas por el método de DICE, obtenida de los marcadores AFLP.
0.77 0.85 0.95 100
1
13
20 B
23
5
14
22
2
6
8
16
15
24
3
9
11
18
19
20 A
28
4
29
7
12
21
Grupo 2Textura fina
Grupo 1Textura gruesa
Grupo 3Textura gruesa
Coeficiente

Análisis de la diversidad genética de caña flecha Gynerium sagittatum Aubl. utilizando la técnica de AFLP 83
La variación molecular detectada por medio de la técnica AFLP en G. sagittatum (Aubl.) detecta claramente tres grupos, diferenciados por sus características artesanales (comercial) y atributos agronómicos deseables tales como textura de la fibra, grosor del tallo y área útil de la nervadura inferior. El grado de diferenciación genética existente entre las introducciones de G. sagittatum (Aubl.) es alto, presentando baja correlación entre la distancia geográfica y el nivel de diferenciación genética.
CONCLUSIONES
La variación molecular detectada por medio de la técnica AFLP mostró en Gynerium sagittatum Aubl. fragmentos específicos para cada una de las accesiones identificando individuos genéticamente diferentes centrados claramente en tres grupos, diferenciados por sus características artesanales (comercial) y atributos agronómicos deseables tales como textura de la fibra, grosor del tallo y área útil de la nervadura inferior.
La diferenciación de los tres grupos genéticos muestra que no existe agrupamiento de acuerdo a la ubicación geográfica de las introducciones, más bien un agrupamiento de acuerdo a su parentesco teniendo en cuenta similitud de caracteres cualitativos y cuantitativos de tipo genético tales como textura de la fibra, grosor del tallo y área útil de la nervadura inferior. Lo cual indica un flujo del mismo material por su carácter de reproducción asexual a las diferentes regiones del país, es consecuencia de un traslado de cultivares por acción antrópica.
Esta variabilidad genética puede ser utilizada en un programa de mejoramiento utilizando genotipos elites que permitan obtener variedades resistentes al barrenador del tallo de la caña flecha (M. atroparsella) plaga principal de la especie, además poseer atributos agronómicos tales como: textura de la fibra suave, que posee una alta demanda en el mercado por sus atributos artesanales.
AGRADECIMIENTOS
Los autores expresan sus agradecimientos a la Universidad de Córdoba por el financiamiento de esta investigación a través del proyecto “Caracterización molecular,
propagación in vitro de caña flecha (Gynrium sagittatum Aubl.) del Caribe Colombiano”, al personal del laboratorio de biotecnología vegetal y el grupo del laboratorio de fitomejoramiento de la Universidad de Córdoba, Montería, Córdoba - Colombia, así como del laboratorio de biología molecular del Instituto de Investigación de recursos biológicos "Alexander von Humboldt". CIAT. Palmira, Valle del Cauca - Colombia.
LITERATURA CITADA
Aramendiz, H.; Espitia, M. y Robles, J. 2005. Colección, caracterización morfoagronómica y producción de semilla de caña flecha (Gynrium sagittatum Aubl) del Caribe Colombiano. Informe final, Universidad de Córdoba, Facultad de Ciencias Agrícolas, Programa de Ingeniería Agronómica, Montería. p. 52-54.
Besse, P. G.; Tailor, B.; Carroll, N.; Berding, D.; Burner, and McIntyre, C. L. 1998. Assessing genetic diversity in a sugarcane germplasm collection using an automated AFLP analysis. Genetics (The Hague) 104:143-153.
Dice , L . R. 1945. Measures of the amount of ecological associat ion between species . Ecology 26:297-302.
Howard, R. A. 1979. Flora of the lesser antilles, leeward and windward islands. Vol. 3. Arnold Arboretum, Harvard University, Jamaica Plain, MA. 586 p.
Invitrogen. Life technologies. 2003. Instruction manual, AFLP(R) Analysis System I and AFLP(R) Starter primer Kit, Version B: 12 p.
Karpa, K.; Bhat, K. V.; Ayad, W. G. and Hodgkin, T. 1997. Molecular tools in plant genetic resources conservation: a guide to the technologies. IPGRI. Technical Bulletin No. 2. International Plant Genetic Resources Institute, Rome, Italy. p. 23 - 27.
Lima, M. L. A.; Garcia, A. A. F.; Oliveira, K. M.; Matsuoka, S.; Arizono, H.; De Souza, J. R. C. L. and De Souza A. P. 2002. Analysis of genetic similarity detected by AFLP and coeffcient of parentage among genotypes of cane Saccharum spp. Theor. Appl. Genet. 104:30-38.
Nei, M. and Li, W. H. 1979. Mathematical model for studying genetic variation in terms of restriction endo nucleases. Proceedings of the National Academy of Sciences of the United States of America 76(10):5269-5273.

84 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
Hernando Javier Rivera Jiménez et al.
Qiagen, 2006. Protocol: purification of total DNA from plant tissue (mini protocol), In: DNeasy plant mini kit for miniprep purification of total cellular DNA from plant cells and tissues, or fungi. p. 19-21.
Rivera, H. J.; Vallejo, F. A.; Suárez, I. y Palacio, J. D. 2007. Estandarización de la técnica AFLP para la caracterización de accesiones colombianas de caña flecha (Gynerium sagitatum Aubl); In: resúmenes XI Congreso “Asociación Colombiana de Fitomejoramiento y Producción de Cultivos”. Universidad de Nariño. San Juan de Pasto. Colombia. p. 36.
Rohlf, F. J. 1998. NTSYS-PC, Numerical taxonomy and multivariate analysis system. Version 1.60. Applied Biostatistics, New York.
Selvi, A. J. L.; Nair, N. K; Noyer, N.; Singh, K. C.; Balasundaram, K. R.; Bansal, K. and Mohapatra, T. 2005. Genomic constitution and genetic relationship among the tropical and subtropical indian sugarcane cultivars revealed by AFLP. Crop Science 45:1750-1757.
Vos, P.; Hogers, R.; Bleeker, M.; Van Der Lee, T.; Hornes, M.; Frijters, A; Pot, J.; Peleman, J.; Kuiper, M. and Zabeau, M. 1995. AFLP: a new technique for DNA fingerprinting. Nucleic Acids Res. 23:4407-4414
Zorro, W. A. y Prieto F. V. 1999. Aproximación a la problemática económica productiva de la comunidad indígena Zenú. Tesis de grado. Facultad de Medicina Veterinaria y Zootecnia. Universidad Nacional de Colombia. Santa fé de Bogotá. 369 p.

Agricultura Técnica en México Vol. 35 Núm.1 1 de enero-31 de marzo 2009 p. 85-91
ÓPTIMOS ANALÍTICOS Y ECONÓMICOS DE MODELOS APLICADOS EN EXPERIMENTACIÓN AGRÍCOLA*
OPTIMAL ANALYTICAL AND ECONOMIC OF MODELS APPLIED IN AGRICULTURAL EXPERIMENTATION
Florencio Briones Encinia1§, Sergio Castro Nava1, José Alberto López Santillán1 y Antonio Trinidad Santos2 1Unidad Académica Multidisciplinaria Agronomía y Ciencias, Universidad Autónoma de Tamaulipas. Centro Universitario Adolfo López Mateos. C. P. 87149. Ciudad Victoria, Tamaulipas. Tel. y Fax. 01 834 3181721 Ext. 2120. ([email protected]), ([email protected]). 2Posgrado de Recursos Naturales, Colegio de Postgraduados, km. 36.5 carretera México-Texcoco. C. P. 56230 Montecillo, Texcoco, Estado de México. Tel. 01 595 9520200 Ext. 1241. ([email protected]). §Autor para correspondencia: [email protected].
* Recibido: Diciembre, 2007
Aceptado: Enero, 2009
RESUMEN
En investigaciones relacionadas con dosis de fertilización de cultivos, es frecuente la aplicación de un modelo pseudocuadrático como una aproximación de una superficie de respuesta. Sin embargo, la determinación del rendimiento óptimo analítico o económico, puede presentar dificultades, particularmente cuando los óptimos generados corresponden a valores que están fuera del intervalo de exploración, se obtiene un punto silla o cuando no es posible llegar a una solución analítica de las ecuaciones. La obtención del óptimo económico en un modelo pseudocuadrático, requiere la aplicación de métodos de cálculo numérico y el acceso a un sistema de cómputo estadístico. Los objetivos del presente trabajo fueron: presentar un procedimiento para el cálculo de los óptimos analíticos o económicos en un modelo pseudocuadrático general con uno y dos factores. El modelo se desarrolló en la Unidad Académica Multidisciplinaria Agronomía y Ciencias de la Universidad Autónoma de Tamaulipas durante 2006 y se aplicó en datos experimentales obtenidos de estudios de fertilización en maíz de temporal efectuados en dos localidades del estado de Puebla, México. Para un factor de estudio, el rendimiento óptimo económico determinado fue 4 569.45 kg ha-1 de maíz con una dosis
de 138.95 kg ha-1 de nitrógeno. Para dos factores, el rendimiento óptimo (2 295.20 kg ha-1) se observó en las dosis de 110 kg ha-1 de nitrógeno y 0 kg ha-1 de fósforo.
Palabras clave: económico, óptimo analítico y modelo pseudocuadrático.
ABSTRACT
In research on crop fertilization it is frequent the application of pseudocuadratic models as a response surface approach. However, the determination of analytic and economic optimum yields may present difficulties, particularly when the corresponding values lie out of the exploration range, when a saddle point is obtained or when is impossible to reach an analytic solution for the equation. To obtain the economic optimums in a pseudocuadratic model, the use of numerical calculation methods and a statistic computer system, is required. The objectives of this work were: to present a procedure for calculating the analytical and economic optimum yields on a pseudocuadratic general model with one and two factors and to test it with real experimental data. The model was developed at the Unidad

86 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
Florencio Briones Encinia et al.
Académica Multidisciplinaria Agronomía y Ciencias de la Universidad Autónoma de Tamaulipas, in 2006 and was applied on experimental data obtained from maize fertilization trials carried out under rainfed conditions at two sites in the state of Puebla, Mexico. For one factor, the optimal economic yield (4 569.45 kg ha-1) was obtained with 138.95 kg ha-1 of nitrogen. For two factors, the optimal yield was 2 295.20 kg ha-1 with 110 kg ha-1 of nitrogen and 0 kg ha-1 of phosphorous.
Key words : analytic and economic optimum, pseudocuadratic model.
INTRODUCCIÓN
A través del tiempo los polinomios de orden bajo (menor o igual a 3) son los que más se han utilizado para estimar la superficie de respuesta (González et al., 2000; Colmenares et al., 2002; Hernández et al., 2006), debido principalmente a su fácil interpretación y cálculo de los estimadores. Con el desarrollo tecnológico y científico, en los últimos años se ha desarrollado equipos y programas de cómputo con capacidad para realizar cálculos de mayor magnitud, precisión y rapidez, lo cual permite ajustar modelos más complejos. Con base en lo anterior, es posible proponer modelos alternativos con mayor ajuste a la superficie de respuesta y expliquen la relación entre las variables de estudio utilizadas en experimentación agrícola. Los modelos pseudocuadráticos representan una alternativa y comprenden al modelo raíz cuadrada que se ha utilizado ampliamente en experimentos relacionados con dosis de fertilización.
Los modelos pseudocuadráticos han sido objeto de estudio en trabajos orientados a determinar algunas características del ajuste de la superficie, como eficiencia de estimación de los coeficientes de regresión y varianza, así como sesgo de la respuesta estimada (Díaz et al., 1991; Castillo et al., 1996; Briones y Martínez, 2002). En general, el propósito de la estimación de una superficie de respuesta es obtener los niveles de los factores que optimicen la respuesta en las variables de interés. En ocasiones, la dificultad que se presenta en el caso de los modelos pseudocuadráticos es el procedimiento de cálculo para determinar los puntos extremos o la estimación de los coeficientes de regresión.
Los objetivos del presente trabajo fueron: presentar un procedimiento para el cálculo de los óptimos analíticos o económicos en un modelo pseudocuadrático general con uno y dos factores y aplicar el procedimiento generado a los datos obtenidos en dos experimentos de fertilización en maíz bajo condiciones de temporal.
La metodología de superficies de respuesta involucra en sus propósitos, determinar un modelo que aproxime una relación funcional desconocida η=f(ξ1,ξ2,...,ξp), esto es, entre una variable respuesta η, y p variables explicativas ξ1, ξ2,..., ξp. En esta investigación se asumió que η se puede aproximar en alguna región de interés por medio de un modelo pseudocuadrático en términos de p variables x1,x2,...xp, las cuales son funciones lineales simples de las variables explicativas:
(1)
donde, las β's son los parámetros a estimar, xij es el nivel del factor j para la observación i, yi es la respuesta de la observación i experimental de η, y εi es el error de ajuste del modelo, correspondiente a la observación i, con E((εi)=0, E(εi
2)=σ2, E((εi εj)=0 si i≠j, y aj y ak están en el intervalo (0,1). Cuando en el modelo (Ecuación 1) aj y ak son diferentes de uno, se obtiene la superficie de respuesta pseudocuadrática (Briones y Martínez, 2002). En esta investigación se implementó y se aplicó un procedimiento para determinar los óptimos analíticos y económicos de la respuesta estimada para cada uno de los dos casos, p= 1 y p= 2, uno y dos factores.
Los cálculos se realizaron con el paquete Mathematica (1999), que contiene un programa computacional que manipula operaciones matemáticas en forma simbólica y el paquete SAS (1998). El procedimiento para identificar los puntos óptimos analíticos u óptimos económicos cuando se ajusta un modelo como la Ecuación 1, comprende el uso del criterio de optimización de la segunda derivada (Bers y Karal, 1985).
Para un factor:
1) se identifica el modelo pseudocuadrático:
yi=f(xi1,xi2,...xip) = ,...,n,,iåxxâxâxââ ika
i k
p
k
ja
i jj k
p
jkj
ja
i j
p
j j j
p
j
ja
i jj
21????0 11
2
11=++++
==<
==
,...,n,,iåxxâxâxââ ika
i k
p
k
ja
i jj k
p
jkj
ja
i j
p
j j j
p
j
ja
i jj
21????0 11
2
11=++++
==<
==
β0 β β
β εi, i ,...,n,,iåxxâxâxââ ika
i k
p
k
ja
i jj k
p
jkj
ja
i j
p
j j j
p
j
ja
i jj
21????0 11
2
11=++++
==<
==
nixxy i
a
i
aii ,...,2,1,1 110
2=+++= εββββ0 β1 β11

Óptimos analíticos y económicos de modelos aplicados en experimentación agrícola 87
con mayor ajuste a la variable respuesta (yi) en función del factor o variable independiente (xi). Este modelo pseudocuadrático es no lineal en a. Para determinar el mejor ajuste se requiere de un equipo de cómputo con un programa específico para ajustar este tipo de modelos, como el procedimiento NLIN de SAS (SAS, 1998), que permite ajustar modelos no lineales.
Para determinar el mejor ajuste de ese modelo; primero, se calcula el estimador del exponente a, para lo cual se puede utilizar el procedimiento NLIN de SAS mediante el programa siguiente:
DATA MAIZ;INPUT x y;CARDS;DATOSPROC NLIN METHOD=GAUSS;PARAMETERS =-8201.59, =2129.57 =-88.75 a=0.5;
MODEL y= + *x**a + *x**(2*a);
DER. =1;
DER. =x**a;
DER. =x**(2*a);
DER.a= *x**a*LOG(x) +2* *x**(2*a)*LOG(x);
RUN;
La opción METHOD comprende los métodos: GAUSS, MARQUARDT y DUD; los dos primeros requieren del comando DER, el cual implica que se deben de incluir las derivadas de los parámetros involucrados en el modelo ( , y a parámetros, para el modelo en consideración). El método DUD no requiere de dichas derivadas.
El comando PARAMETERS requiere que se especifiquen los valores iniciales de cada parámetro del modelo por ajustar. Un criterio que se puede aplicar en experimentos con fertilizantes, es asignar el valor de 0.5 al parámetro a y estimar los parámetros lineales por medio de regresión lineal y los estimadores obtenidos usarlos como valores iniciales; la propuesta anterior se justifica, debido a que se han realizado trabajos con fertilizantes en los que el modelo pseudocuadrático con valores de a´s igual a 0.5 son los mejores; resultados similares que se observaron en este trabajo.
2) una vez obtenido el mejor ajuste, calcule:
Al evaluar f´´(x) se obtiene como resultado uno de los siguientes casos:
a) que el valor obtenido sea mayor que cero, entonces se tiene un mínimo relativo en x0;b) que el valor obtenido sea menor que cero, entonces se tiene un máximo relativo en x0;c) que el valor obtenido sea igual a cero, entonces no se puede concluir. En este caso y cuando ocurra que el valor de x0 esté fuera del intervalo de exploración, se debe encontrar la mejor respuesta en la región de exploración.El procedimiento anterior conduce a la determinación del óptimo analítico. Para determinar el óptimo económico se debe resolver la ecuación:
(2)
donde, Px y Py son: el costo por unidad del factor x aplicado y el precio de venta por unidad del producto y respectivamente (Colmenares et al., 2002). La solución para x en la Ecuación 2, en general, se dificulta por métodos algebraicos ordinarios, por lo que se puede emplear un proceso iterativo como es el método de Newton-Rampson para lo cual se requiere un sistema de cómputo que contenga este procedimiento. Con el paquete Mathematica es relativamente fácil encontrar la solución para x, mediante la siguiente instrucción:
β0 β1 β2
β0
β1 β11β0
β1
β2
β1 β11
β0 β1, β11
β0, β1 y β11, El modelo pseudocuadrático es no lineal en a, pero es lineal en los parámetros por lo que con base en los resultados obtenidos, se sugiere sólo estimar a con un procedimiento no lineal y determinar los estimadores lineales por regresión lineal, después de estimar a. Al aplicarlo de esta manera a los datos del ejemplo utilizado para ilustrar el procedimiento y determinar los óptimos analíticos y los óptimos económicos, se logró disminuir el cuadrado medio del error.
a xa-1+2a x2a-1=
y
x
PP
3) luego evalúe la segunda derivada f`´´(x)=(a-1)a xa-
2+2a(2a-1) x2a-2, en x0 del punto anterior.
x0=
a1
1 12
1
∧
∧
β
β1
∧
β
β111
∧
β
β1
β1 β11
β1 β11

88 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
Florencio Briones Encinia et al.
(3)
El procedimiento de Newton-Rampson requiere de valores iniciales de x; ya que en general, los óptimos analítico y económico están cercanos. Se pueden dar como valores iniciales de x, los valores determinados en el óptimo analítico.
En caso de que el valor de x obtenido con la instrucción dada en Ecuación 3, no se encuentre en el intervalo de exploración, aplicar el paso 3 c.
Para dos factores:
1) identifique el modelo pseudocuadrático:
que mejor ajuste la variable respuesta (yi) en función de los factores o variables independientes (x1 y x2). El ajuste se puede obtener con el procedimiento NLIN de SAS en forma similar al caso de un factor.
2) calcule los óptimos analíticos obtenidos de la primera derivada:
3) obtenga el Hessiano o matriz de segundas derivadas (Bers y Karal, 1985):
y defina D=f11(x01,x02)f22(x01,x02)-f22 1 (x01,x02). Entonces, sí:
a) D<0, yi tiene un punto silla en (x01,x02)b) D>0 y f11(x01,x02)<0, yi tiene un punto máximo relativo en (x01,x02)
c) D>0 y f11(x01,x02)>0, yi tiene un punto mínimo relativo en (x01,x02)
En el caso a), y cuando el punto (x01,x02) esté fuera del intervalo de exploración, se debe determinar la mejor respuesta en la región de exploración.
Igual que en el caso de un factor, los pasos anteriores conducen a la determinación de un óptimo analítico. El cálculo del óptimo económico implica resolver el siguiente sistema de ecuaciones:
donde, Px 1, Px 2
y Py son, respectivamente, el costo por unidad de los factores x1 y x2 aplicados y el precio de venta del producto y. También, como en el caso de un factor, la solución al sistema de ecuaciones anterior requiere de un proceso iterativo, como el del método de Newton-Rampson. Con el paquete Mathematica se resuelve mediante la siguiente instrucción:
Para dirigir la búsqueda en la solución al sistema anterior, como se comentó para el caso de un factor, conviene asignar como valores iniciales a x1 y x2, los valores determinados en el óptimo analítico.
Cuando para un factor resulte 3c) o para dos factores se obtenga un punto silla, o los valores de x1 o x2 se ubiquen fuera de la región de exploración, ya sea con, uno o dos factores, el procedimiento a seguir consiste en aplicar un método numérico para determinar la mayor respuesta en la región de exploración de los factores en estudio. El método consiste en tomar una rejilla de puntos en la región de exploración y evaluar cada punto en el modelo ajustado hasta encontrar la mayor respuesta. El procedimiento puede efectuarse mediante de una hoja de cálculo o un paquete estadístico.
yi= β0+ β1xa1 + β2xa2 + β11x2a1+ β22x2a2 +β12xa1xa2 + i=1,2,...,n
i1 i2 i1 i2 i1 i2
FindRoot[a xa-1+2a x
2a-1== ,{x,x0}] β1 β11
x01=
1
1
2
2 12 21 14-
2 12-2 212
a
∧
+
∧∧
∧∧∧∧
βββ
ββββ
β1
β2 β12
β22 -
β22
β11 2
β12-4 +
x02=
2
1
2
2 1-2 21 14
2 1121 12-
a
∧∧∧
∧∧
+
∧∧
βββ
ββββ β11
β2 + β1
β12
4 - β11 β22
2
β12
a1 x1-
11a +2a1 x
1-21
1a +a1 x1-
11a x 2
2a = β1
β12
β11
a2 x1-
22a +2a2 x
1-22
2a +a2 x1-
22a x 1
1a =
β2
β22
β12
FindRoot[{a1 x1-
11a +2a1 x
1-21
1a +a1 x1-
11a x 2
2a = ,
β11
β
12
β1
a2 x1-
22a +2a2 x
1-22
2a +a2 x1-
22a x 1
1a = β
2
β
22
β12
y
x
P
P
εi
^β12
f11(x01,x02)= jaia
iiia
xiiia
ii xxaaaaxaa 2 0
2-
1 0
2-2
1 0
2-
1 0 2 1)1-(1 1)1-2(21)1-(∧
+∧
+∧
βββjaia
iiia
xiiia
ii xxaaaaxaa 2 0
2-
1 0
2-2
1 0
2-
1 0 2 1)1-(1 1)1-2(21)1-(∧
+∧
+∧
βββ
β1
^β11
jaiaii
iaxii
iaii xxaaaaxaa 2 0
2-
1 0
2-2
1 0
2-
1 0 2 1)1-(1 1)1-2(21)1-(∧
+∧
+∧
βββ β12
jaiaii
iaxii
iaii xxaaaaxaa 2 0
2-
1 0
2-2
1 0
2-
1 0 2 1)1-(1 1)1-2(21)1-(∧
+∧
+∧
βββjaia
iiia
xiiia
ii xxaaaaxaa 2 0
2-
1 0
2-2
1 0
2-
1 0 2 1)1-(1 1)1-2(21)1-(∧
+∧
+∧
βββ
jaiaii
iaxii
iaii xxaaaaxaa 2 0
2-
1 0
2-2
1 0
2-
1 0 2 1)1-(1 1)1-2(21)1-(∧
+∧
+∧
βββ
f22(x01,x02)= 22
2 0
2
2 01 0
2
2 0 2 2)12(22 1)1(2)1(−−− ∧
−+∧
−+∧
−ja
jjjaia
xjjja
jj xaaxaaxaa ββββ2
22
2 0
2
2 01 0
2
2 0 2 2)12(22 1)1(2)1(−−− ∧
−+∧
−+∧
−ja
jjjaia
xjjja
jj xaaxaaxaa βββ22
2 0
2
2 01 0
2
2 0 2 2)12(22 1)1(2)1(−−− ∧
−+∧
−+∧
−ja
jjjaia
xjjja
jj xaaxaaxaa βββ
β12
22
2 0
2
2 01 0
2
2 0 2 2)12(22 1)1(2)1(−−− ∧
−+∧
−+∧
−ja
jjjaia
xjjja
jj xaaxaaxaa βββ β22
22
2 0
2
2 01 0
2
2 0 2 2)12(22 1)1(2)1(−−− ∧
−+∧
−+∧
−ja
jjjaia
xjjja
jj xaaxaaxaa βββ22
2 0
2
2 01 0
2
2 0 2 2)12(22 1)1(2)1(−−− ∧
−+∧
−+∧
−ja
jjjaia
xjjja
jj xaaxaaxaa βββ
f12(x01,x02)=1
2 0
1
1 02 1−−
∧jaia
ji xxaa β1
2 0
1
1 02 1−−
∧jaia
ji xxaa β
y
x
P
P1
y
x
P
P2
y
x
P
P1
y
x
P
P2 },{x1, x01},{ x2, x02}]
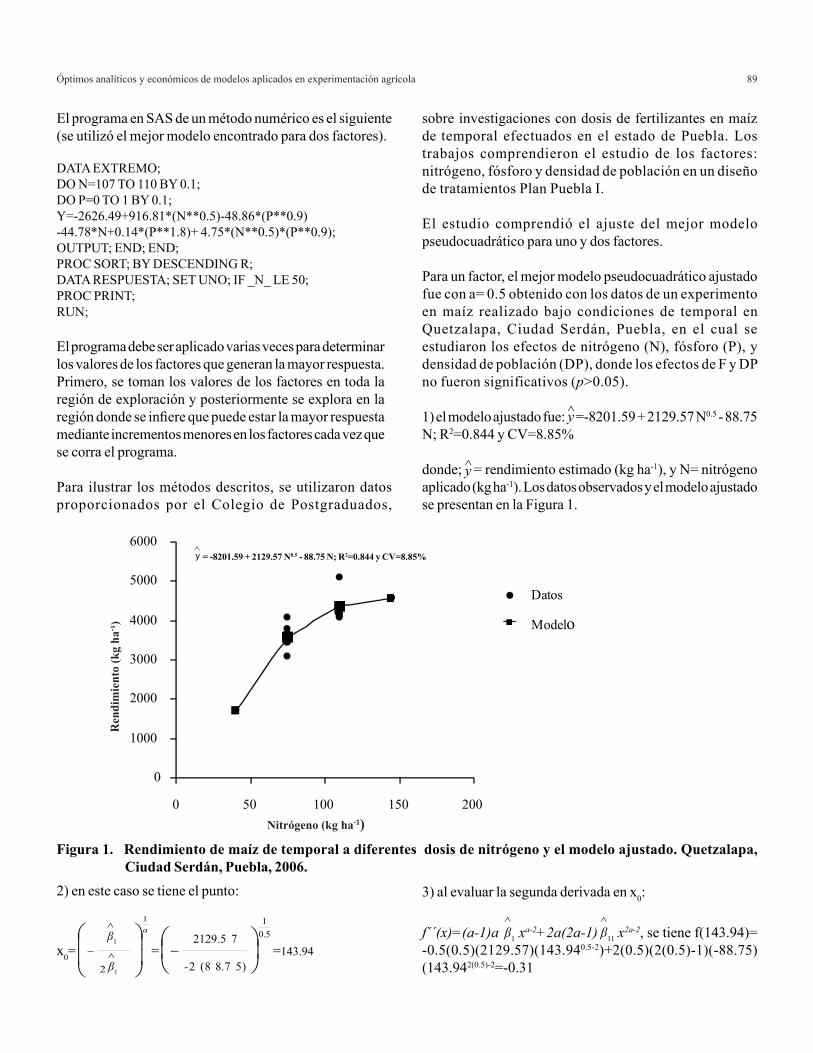
Óptimos analíticos y económicos de modelos aplicados en experimentación agrícola 89
El programa en SAS de un método numérico es el siguiente (se utilizó el mejor modelo encontrado para dos factores).
DATA EXTREMO;DO N=107 TO 110 BY 0.1;DO P=0 TO 1 BY 0.1;Y=-2626.49+916.81*(N**0.5)-48.86*(P**0.9)-44.78*N+0.14*(P**1.8)+ 4.75*(N**0.5)*(P**0.9);OUTPUT; END; END;PROC SORT; BY DESCENDING R;DATA RESPUESTA; SET UNO; IF _N_ LE 50;PROC PRINT;RUN;
El programa debe ser aplicado varias veces para determinar los valores de los factores que generan la mayor respuesta. Primero, se toman los valores de los factores en toda la región de exploración y posteriormente se explora en la región donde se infiere que puede estar la mayor respuesta mediante incrementos menores en los factores cada vez que se corra el programa.
Para ilustrar los métodos descritos, se utilizaron datos proporcionados por el Colegio de Postgraduados,
2) en este caso se tiene el punto:
sobre investigaciones con dosis de fertilizantes en maíz de temporal efectuados en el estado de Puebla. Los trabajos comprendieron el estudio de los factores: nitrógeno, fósforo y densidad de población en un diseño de tratamientos Plan Puebla I.
El estudio comprendió el ajuste del mejor modelo pseudocuadrático para uno y dos factores.
Para un factor, el mejor modelo pseudocuadrático ajustado fue con a= 0.5 obtenido con los datos de un experimento en maíz realizado bajo condiciones de temporal en Quetzalapa, Ciudad Serdán, Puebla, en el cual se estudiaron los efectos de nitrógeno (N), fósforo (P), y densidad de población (DP), donde los efectos de F y DP no fueron significativos (p>0.05).
1) el modelo ajustado fue: =-8201.59 + 2129.57 N0.5 - 88.75 N; R2=0.844 y CV=8.85%
donde; = rendimiento estimado (kg ha-1), y N= nitrógeno aplicado (kg ha-1). Los datos observados y el modelo ajustado se presentan en la Figura 1.
3) al evaluar la segunda derivada en x0:
f´´(x)=(a-1)a xa-2+2a(2a-1) x2a-2, se tiene f(143.94)=-0.5(0.5)(2129.57)(143.940.5-2)+2(0.5)(2(0.5)-1)(-88.75)(143.942(0.5)-2=-0.31
0
1000
2000
3000
4000
5000
6000
0 50 100 150 200Nitrógeno (kg ha ¹)
Ren
dim
ient
o (k
g ha
¹)
Datos
Modelo
Figura 1. Rendimiento de maíz de temporal a diferentes dosis de nitrógeno y el modelo ajustado. Quetzalapa, Ciudad Serdán, Puebla, 2006.
= -8201.59 + 2129.57 N0.5 - 88.75 N; R2=0.844 y CV=8.85% ∧y ∧y
x0=
a1
112
1
∧
∧
−
β
β=
5.01
)5788(2
752129
−
. -
.=143.94
β1
β1
^β1
Ren
dim
ient
o (k
g ha
- ¹)
Nitrógeno (kg ha-1)
y
y
^β11

90 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
Florencio Briones Encinia et al.
Esto implica que para N=143.94 kg ha-1 de Nitrógeno se obtiene el rendimiento máximo de maíz,
= -8201.59 + 2129.57(143.940.5)-88.75(143.94)=4573.26 kg ha-1.
Lo anterior no implica que sea la mejor opción, pues la mejor sería el óptimo económico. Éste se obtiene al resolver para N la ecuación:
a Na-1 +2a N2a-1= , sustituyendo valores y se tiene:
0.5(2129.57) N0.5-1+2(0.5)(-88.75)N2(0.5)-1=
Al despejar N se encuentra el valor de 138.95 kg ha-1 de N lo cual implica que con esta dosis se obtiene el rendimiento óptimo económico. Al sustituir este valor en el modelo dado en (Ecuación 1), se obtiene el rendimiento óptimo económico de 4 569.45 kg ha-1 de maíz.
Para dos factores, se tiene:
El mejor modelo pseudocuadrático ajustado fue para a1= 0.5 y a2= 0.9, obtenido con los datos de un experimento realizado en San José Guerrero, Ciudad Serdán, Puebla, en el cual se probaron diferentes dosis de nitrógeno, fósforo y densidad de población; ésta última no mostró efecto significativo (α > 0.05). El modelo fue:
= -2626.49+916.81N0.5-48.86P0.9 4.66N+0.14P1.8+4.75N0.5P0.9; R2= 0.91, CV=6.8%
Calcule:
Obtenga:
=0.5(0.5-1)(916.81)(105.273)0.5-2+2(0.5)(2(0.5)-1)(-44.78)(105.2737)0.5-2
+0.5(0.5-1)(4.75) 17(105.273)0.5-2(0.4036)0.9=-0.2117
= 0 . 9 ( 0 . 9 - 1 ) ( - 4 8 . 8 6 ) ( 0 . 4 0 3 6 ) 0 . 9 - 2 + 0 . 9 ( 0 . 9 - 1 ) ( 4 . 7 5 )(105.2687)0.5(0.4036)0.9-2 +2(0.9)(2(0.9)-1) (0.14)(0.4036)2(0.9)-2=0.2719
Como el valor de D es negativo, se infiere que en el punto (105.273, 0.4036) el modelo ajustado tiene un punto silla. En estos casos se debe aplicar un método de exploración numérico, como el que se específica en Materiales y Métodos hasta determinar los niveles de los factores en la región de exploración que producen el mayor rendimiento.
El mayor rendimiento determinado con el programa en SAS, fue para 110 kg ha-1 de nitrógeno y 0 kg ha-1 de fósforo, con rendimiento de = 2 295.2 kg ha-1 de maíz. Colmenares et al. (2003) ajustaron un modelo pseudocuadrático con a1= 0.7 y a2= 0.9, ambos fijos, a datos de un estudio sobre dosis de nitrógeno y cantidad de semilla sobre el rendimiento maíz y encontraron la dosis óptima económica de nitrógeno de 118.97 kg ha-1; con los mismos datos pero ajustando un polinomio de segundo grado, estos autores encontraron una dosis de nitrógeno mayor a 120 kg ha-1, lo cual refleja la influencia del modelo propuesto.
AGRADECIMIENTOS
El autor principal agradece al Dr. Antonio Trinidad Santos, por haber proporcionado los datos experimentales de campo para ilustrar los métodos descritos en este documento.
CONCLUSIONES
El modelo pseudocuadrático permite obtener una solución única para los óptimos analíticos de una superficie de respuesta, con valores de los exponentes en (0,1).
β2 β1
9.1
3
x01=
ia1
2
2 12 21 14-
2 12-2 212
∧+
∧∧
∧∧∧∧
βββ
ββββ=
5.01
24.7514)-44.78)(0.(4-
.75)(-48.86)(4-)4 1.0 ( )1 8.916(2
+=105.2687
β1
β222 - β2
β12
-4 + β11 β22
2
β12
2
x02=
ja1
2
2 1-2 21 14
2 1121 12-
∧∧∧
∧∧+
∧∧
βββ
ββββ=
9.01
24.75-14)-44.78)(0.(4
75)916.81)(4.)44.78- ( )6 8.-48(2-
+
=0.4036 β11
β2-2 + β1
β12
β22 β12
2
β124 -2
β12 β22
y
y
y
x
P
P
y
f11(x01,x02) = jaia
iiia
iiia
ii xxaaxaaxaa2 0
2
1 0
2
1 0
2
1 0 2 1)1(1 1)12(21)1( −−−∧
−+∧
−+∧
− βββ
jaia
iiia
iiia
ii xxaaxaaxaa2 0
2
1 0
2
1 0
2
1 0 2 1)1(1 1)12(21)1( −−−∧
−+∧
−+∧
− βββ
β1 β11
β12
jaia
iiia
iiia
ii xxaaxaaxaa2 0
2
1 0
2
1 0
2
1 0 2 1)1(1 1)12(21)1( −−−∧
−+∧
−+∧
− βββ
f22(x01,x02) = 22
2 0
2
2 01 0
2
2 0 2 2)12(22 1)1(2)1(−−−
∧−+
∧−+
∧− ja
jj
jaiaxjj
ja
jj xaaxaaxaa βββ β2
22
2 0
2
2 01 0
2
2 0 2 2)12(22 1)1(2)1(−−−
∧−+
∧−+
∧− ja
jj
jaiaxjj
ja
jj xaaxaaxaa βββ22
2 0
2
2 01 0
2
2 0 2 2)12(22 1)1(2)1(−−−
∧−+
∧−+
∧− ja
jj
jaia
jj
ja
jj xaaxxaaxaa βββ22
2 0
2
2 01 0
2
2 0 2 2)12(22 1)1(2)1(−−−
∧−+
∧−+
∧− ja
jj
jaia
jj
ja
jj xaaxxaaxaa βββ
f12(x01,x02)=1
2 0
1
1 02 1−−
∧jaia
ji xxaa β
=0.5(0.9)(4.75)(105.2687)0.5-1
(0.4036)0.9-1=0.2281
β12
1
2 0
1
1 02 1−−
∧jaia
ji xxaa β
D = f11(x01,x02)f22(x01,x02)-f2
2 1(x01,x02) = -0.2117(0.2719)
-0.22812=-0.1096

Óptimos analíticos y económicos de modelos aplicados en experimentación agrícola 91
La utilidad práctica del modelo aplicado para un factor mostró que en Quetzalapa, Puebla, la aplicación de 138.95 kg ha-1 de nitrógeno produce un rendimiento óptimo económico de 4 569.45 kg ha-1 de maíz. Para dos factores en San José Guerrero, Puebla, se determinó que sólo el nitrógeno tuvo efecto en el rendimiento de maíz y que la aplicación de 110 kg ha-1 produce el mayor rendimiento de 2 295.2 kg ha-1.
LITERATURA CITADA
Bears, L. y Karal, F. 1985. Cálculo. 2a Edición, traducción del inglés; Agut, A. V. 2a Edición. Interamericana. México. 746 p.
Briones, E. F. y Martínez G., A. 2002. Eficiencia de algunos diseños experimentales en la estimación de una superficie de respuesta. Agrociencia 36:201-210.
Castillo, V. M.; Martínez, G. A.; Castillo, M. A. y Santizo, R. J. A. 1996. Comparación de diseños a estimar superficies de respuesta. Agrociencia 30:75-81.
Colmenares, C. B.; Martínez, G. A; Martínez, D. A. y Ramírez, G. M. E. 2002. Una región confidencial para óptimos económicos. Agrociencia 36:337:344.
Colmenares, C. B.; Martínez, G. A; Martínez, D. A.; Ramírez, G. M. E. y González, C. F. 2003. Región confidencial para óptimos económicos en modelos pseudocuadráticos. Agrociencia 37:177-185.
Díaz, G. J. A.; Martínez, G. A.; González, C. F.; Castillo, M. A. y Santizo, R. J. A. 1991. La eficiencia de los diseños de tratamientos empleados en la investigación agrícola, considerando el ajuste de modelos pseudocuadráticos. Agrociencia 3:27-47.
González, E. D. R.; Alcalde, B. S.; Ortiz, C. J. y Castillo, M. A. 2000. Dinámica de la acumulación de potasio por trigo cultivado en diferentes ambientes. Agrociencia 34:1-11.
Hernández, S. J.; Cuca, G. M.; Pró, M. A; González, A. M. y Becerril, P. C. 2006. Nivel óptimo biológico y económico de calcio en gallinas leghorn blancas de segundo ciclo de postura. Agrociencia 40:49-57.
Mathematica. 1999. Users´s Guide; release 4.01 Edition. Wolfram Research Inc. Champaign, IL. USA. 1470 p.
Statistical Analysis System (SAS). 1998. SAS/STAT Users´s Guide; release 6.03 Edition. SAS Institute, Cary, N. C. USA. 1028 p.

Agricultura Técnica en México Vol. 35 Núm.1 1 de enero-31 de marzo 2009 p. 92-106
ÍNDICE DE ESTRÉS HÍDRICO COMO UN INDICADOR DEL MOMENTO DE RIEGOEN CULTIVOS AGRÍCOLAS*
WATER STRESS INDEX AS AN INDICATOR OF IRRIGATION TIMINGIN AGRICULTURAL CROPS
Rutilo López López1§, Ramón Arteaga Ramírez2, Mario Alberto Vázquez Peña3, Irineo López Cruz2 e Ignacio Sánchez Cohen4
1Campo Experimental Huimanguillo, km 1 carretera Huimanguillo-Cárdenas, Huimanguillo, Tabasco, México. C. P. 86400, Tel. 01 917 3750516 Fax: 9173750397. 2Posgrado en Ingeniería Agrícola y Uso Integral del Agua, Universidad Autónoma Chapingo, km. 38.5, carretera México-Texcoco, Chapingo, México. C. P. 56230, Tel. 01 595 9521551, Fax. 5959521551, ([email protected]) ([email protected]). 3Departamento de Irrigación, Universidad Autónoma Chapingo, km. 38.5, carretera México-Texcoco, Chapingo, México. C. P. 56230, Tel. 01 595 9521551, Fax. 5959521551, ([email protected]). 4Centro Nacional de Investigación Disciplinaria en Relación Agua-Suelo-Planta-Atmósfera, INIFAP, km. 6.5 Canal Sacramento Zona industrial Gómez Palacio Durango, México. Tel. 01 871 7191076 y 7191077, Fax: 8717191134, ([email protected]). *Autor para correspondencia: [email protected].
* Recibido: Marzo, 2008
Aceptado: Marzo, 2009
de estrés hídrico de los cultivos, así como la utilidad práctica que tiene en la calendarización y optimización del agua de riego en los cultivos agrícolas.
Palabras clave: estrés hídrico, predicción de rendimiento, programación del riego.
ABSTRACT
Crop water stress index (CWSI) is based on the determination of the actual crop transpiration rate by means of the measurements of canopy temperature and vapor pressure deficit. The objectives of this assay were: to present theoretical and methodological aspects of CWSI and its practical application for irrigation scheduling. The CWSI method has been of practical use in arid and semiarid regions mainly because the required equipment to calculate it is easy to use. This method can also be utilized in humid regions where irrigation is needed during the dry season and there is high solar radiation. The CWSI is based on the theoryof energy balance on a surface, the linear functions of the
RESUMEN
El índice de estrés hídrico del cultivo (CWSI), se basa en la determinación de la tasa de transpiración actual de un cultivo mediante la medición de la temperatura del dosel y el déficit de presión de vapor. Los objetivos de este ensayo fueron: presentar los aspectos teóricos y metodológicos del CWSI y la utilidad práctica que tiene para la programación del riego en los cultivos agrícolas. El método CWSI ha tenido uso práctico en regiones áridas y semiáridas debido a que el equipo requerido para su implementación es de fácil manejo. Este método, también se puede utilizar en regiones con clima húmedo, donde los cultivos requieren riego en la época seca y la radiación solar es alta. El CWSI se basa en la teoría del balance de energía en una superficie, las funciones lineales del diferencial de temperatura entre el cultivo y el aire y el déficit de presión de vapor, las cuales dependen del cultivo y las condiciones atmosféricas. La termometría infrarroja es una herramienta sencilla, práctica y confiable para la estimación del estrés hídrico, la programación del riego y la predicción de rendimiento. El objetivo de este ensayo fue presentar los aspectos teóricos y metodológicos del índice

Índice de estrés hídrico como un indicador del momento de riego en cultivos agrícolas 93
differential temperature between the crop and the air and the vapor pressure deficit; which depends on the crop water status and the weather conditions. Infrared thermometry is a simple, practical and reliable tool to estimate crop water stress, irrigation scheduling and yield prediction.
Key words: water stress, irrigation timing, yield prediction.
INTRODUCCIÓN
En la programación del riego se han utilizado diversos métodos: determinación de la humedad del suelo, mediciones del estado energético del agua en el suelo, estimaciones de la demanda atmosférica y determinaciones del potencial hídrico de las plantas (Buchner et al., 1994) o temperatura del dosel por medio de termometría infrarroja (Giuliani et al., 2001). Otros métodos utilizados en la programación del riego, se basan en mediciones directas del crecimiento de algunos órganos de la planta con diferentes tipos de dendrómetros, asociados al uso de sensores de flujo de savia (basándose en el método del pulso de calor), para determinar el momento y la intensidad con la cual la disponibilidad de agua limita la transpiración del cultivo.
Las limitaciones críticas en el registro continuo del contenido de humedad en el suelo han sido resumidas recientemente por Van Leeuwen et al. (2001), en comparación con el monitoreo directo de las micro variaciones en el diámetro del tallo y del fruto, que han sido reconocidos por Ton et al. (2001) como indicadores más apropiados del déficit hídrico de poca intensidad y duración, producidos en condiciones de campo. El fitomonitoreo es una técnica basada en la microelectrónica y la informática, que permite conocer en tiempo real el estado hídrico de la planta de una forma no destructiva, a través del registro de las respuestas anatómicas y fisiológicas de diversos órganos de las plantas, frente a las condiciones medioambientales y de manejo agronómico (Novak, 1997).
El método del índice de estrés hídrico del cultivo (CWSI, por sus siglas en ingles), es una medida de la transpiración que ocurre en una planta en el tiempo de medición, mediante el registro de la temperatura de la planta y el déficit de presión de vapor del agua. Este enfoque metodológico es necesario para la correcta
programación del riego en cultivos agrícolas en campo abierto. Las mediciones del estrés hídrico de los cultivos combinado con sistemas eficientes de riego permiten maximizar el rendimiento a través de un manejo eficiente del riego (Garrot et al., 1990).
El CWSI ha tenido un importante uso práctico dentro de la programación del riego en regiones áridas y semiáridas (Calado et al., 1990; Itier et al., 1993; Anconelli et al., 1994; Jones, 1999; Orta et al., 2003; Yuan et al., 2004; Şimşek et al., 2005; Erdem et al., 2005); esto se debe principalmente a que el equipo requerido es de fácil manejo. En la agricultura de riego, el costo económico y ecológico del agua es alto, si se considera la incertidumbre en su disponibilidad acrecentada por el cambio climático, de ahí que el costo del equipo para cuantificar las variables climáticas y el estrés hídrico puede justificar la inversión (Feldhake et al., 1997). Los objetivos del presente ensayo bibliográfico fueron presentar los aspectos teóricos y metodológicos del índice de estrés hídrico de los cultivos, así como la utilidad práctica que tiene en la calendarización y optimización del agua de riego en los cultivos agrícolas.
Teoría del índice de estrés hídrico del cultivo
El agua evaporada por una superficie vegetal funciona al mismo tiempo como un estabilizador de la temperatura de las hojas ante la demanda evapotranspirativa de la atmósfera. A partir de esto, Jackson et al. (1981) presentaron la teoría del balance energético que separa la radiación neta en calor sensible del aire y calor latente que incide en la transpiración. Cuando el cultivo se somete a estrés hídrico, los estomas se cierran, la transpiración decrece y la temperatura de la hoja aumenta. Cuando una planta transpira sin estrés hídrico, la temperatura de la hoja es entre 1 - 4 °C menor que la temperatura ambiental, en este caso el índice de estrés hídrico del cultivo (CWSI) es 0. Cuando la transpiración decrece, la temperatura de la hoja asciende y puede alcanzar de 4 a 6 °C más que la temperatura del aire. En este caso, el déficit hídrico es alto, la transpiración de las hojas se ve drásticamente reducida con el incremento de la temperatura foliar; cuando la planta está muerta o no transpira durante mucho tiempo el CWSI es 1.
Idso et al. (1981) desarrollaron el método empírico (CWSI) para la cuantificación de la humedad atmosférica y de la planta en regiones áridas, la cual depende de la

94 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
Rutilo López López et al.
determinación “de las líneas base sin estrés hídrico” que facilitan la normalización de cambios en la temperatura del dosel para condiciones ambientales. Sin embargo, estas líneas base son específicas del cultivo y están influenciadas por el clima (Bucks et al., 1985). Además, Jackson et al. (1981) modificaron el CWSI para incluir una predicción más teórica de los efectos del clima sobre la temperatura del dosel que incluye explícitamente el déficit de presión del vapor de agua, la radiación neta y la resistencia aerodinámica. Esta aproximación teórica para determinar la humedad atmosférica que se utiliza en el CWSI es más precisa que la aproximación empírica, particularmente en climas más húmedos (Keener y Kircher 1983).
El índice de estrés hídrico del cultivo según Idso (1981) y Jackson et al. (1981) está definido como:
donde, Tc= temperatura del cultivo; Ta= temperatura del aire. El subíndice m denota la diferencia medida entre las dos temperaturas, el subíndice li (límite inferior) denota la diferencia entre las dos temperaturas cuando la evapotranspiración no está restringida por disponibilidad del agua, de modo que la Tc es el valor más bajo para las condiciones del clima, y el subíndice ls (límite superior) denota la diferencia hipotética entre las dos temperaturas cuando la evapotranspiración es cero, como resultado de falta de disponibilidad de agua en el sistema suelo planta atmósfera.
Se espera que el CWSI varíe entre 0 a 1 cuando las plantas van de una condición bien regada a una condición totalmente estresada. Puesto que no es normalmente factible medir simultáneamente la temperatura del cultivo sin estrés y un cultivo con estrés, los valores del límite inferior y superior de un dosel se pueden calcular mediante el análisis de balance de energía en la superficie. Este balance energético se puede expresar como:
(2)
donde, Rn= radiación neta (W m-2); G= flujo del calor en la superficie del suelo (W m-2); H= flujo del calor sensible del aire (W m-2); λE= flujo de calor latente (W m-2). Los
términos H y λE en la Ecuación 2 son una función de los gradientes de la temperatura y de la presión de vapor respectivamente y se pueden expresar como:
(3)
y
(4)
donde, ρa= densidad del aire (kg m-3); Cp= calor especifico del aire (J kg-1 °C); es= presión de vapor de agua a saturación (kPa) a Tc; ea= presión de vapor de agua actual del aire; γ= constante psicrométrica (kPa °C-1); ra= resistencia aerodinámica (s m-1); rc= resistencia del dosel al flujo de vapor de agua (s m-1).
La Ecuación 2 puede ser simplificada asumiendo que G= 0.lRn (Feldhake et al., 1996) y por lo tanto, definiendo Ic como coeficiente de intercepción de la radiación igual a 0.9, de modo que la Ecuación 2 se convierte en:
(5)
Cuando un volumen de aire se encuentra retenido sobre una superficie evaporante de agua, se alcanza un equilibrio entre las moléculas de agua que se incorporan al aire y las que regresan a la fuente de agua. En ese momento, se considera que el aire está saturado puesto que no puede incorporar ninguna molécula de agua adicional. La presión correspondiente se llama presión de vapor de agua a saturación (es). La cantidad de moléculas de agua que pueden ser incorporadas al aire depende de la temperatura (T). Mientras más alta es la temperatura del aire, más alta es la capacidad de incorporar vapor de agua y más alta es la presión de vapor a saturación (Figura 1). La pendiente de la curva cambia exponencialmente con la temperatura. A temperaturas bajas, la pendiente es pequeña y varía levemente con el incremento de temperatura. A temperaturas altas, la pendiente es mayor y pequeños cambios de temperatura producen grandes cambios en la pendiente. La pendiente de la curva de presión de vapor de saturación (∆), es un parámetro importante para la descripción de la evaporación y se utiliza en el cálculo de la evapotranspiración de referencia (ET0) con datos climáticos.
EHRI nc λ+=
EHGRn λ++=
[(Tc –Ta )m – (Tc – Ta )li ]
(1)CWSI = [(Tc –Ta )ls – (Tc – Ta )li ]
H= ρa Cp (TC - Ta )
ra
λE= ρaCp (es - ea )
(ra + rc )γ
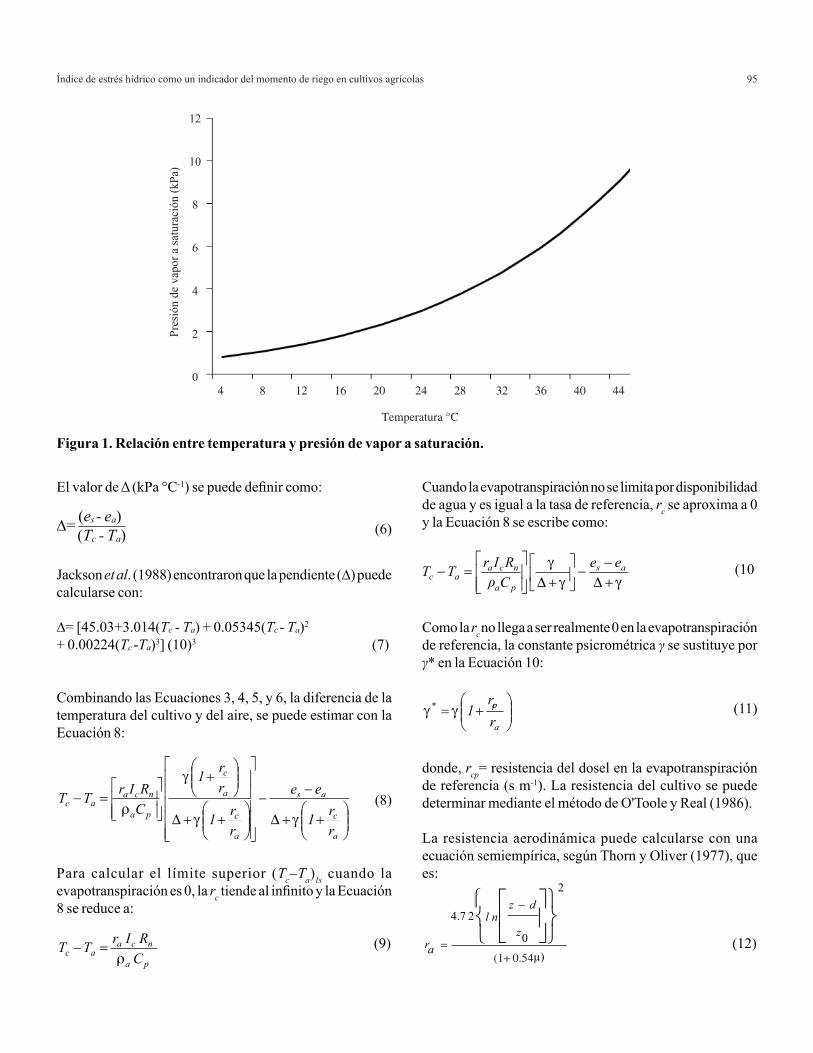
Índice de estrés hídrico como un indicador del momento de riego en cultivos agrícolas 95
El valor de Δ (kPa °C-1) se puede definir como:
(6)
Jackson et al. (1988) encontraron que la pendiente (∆) puede calcularse con:
Combinando las Ecuaciones 3, 4, 5, y 6, la diferencia de la temperatura del cultivo y del aire, se puede estimar con la Ecuación 8:
(8)
Para calcular el límite superior (Tc–Ta) ls cuando la evapotranspiración es 0, la rc tiende al infinito y la Ecuación 8 se reduce a:
(9)
Cuando la evapotranspiración no se limita por disponibilidad de agua y es igual a la tasa de referencia, rc se aproxima a 0 y la Ecuación 8 se escribe como: (10
Como la rc no llega a ser realmente 0 en la evapotranspiración de referencia, la constante psicrométrica γ se sustituye por γ* en la Ecuación 10:
(11)
donde, rcp= resistencia del dosel en la evapotranspiración de referencia (s m-1). La resistencia del cultivo se puede determinar mediante el método de O'Toole y Real (1986).
La resistencia aerodinámica puede calcularse con una ecuación semiempírica, según Thorn y Oliver (1977), que es:
(12)
Figura 1. Relación entre temperatura y presión de vapor a saturación.
++
−−
++
+
=−
a
c
as
a
c
a
c
pa
ncaac
rr1
ee
rr1
rr1
CRIrTT
γ∆γ∆
γ
ρ
pa
ncaac C
RIrTTρ
=−
+=
a
cp*
rr
1γγ
Pres
ión
de v
apor
a sa
tura
ción
(kPa
)
12
10
8
6
4
2
0 4 8 12 16 20 24 28 32 36 40 44
Temperatura °C
∆= (es - ea) (Tc - Ta)
∆= [45.03+3.014(Tc - Ta) + 0.05345(Tc - Ta)2 + 0.00224(Tc -Ta)3] (10)3 (7)
γ∆γ∆γ
ρ +−
−
+
=− as
pa
ncaac
eeCRIrTT
ρ
( )µ4 5.01
2
0
72.4
+
−
=
z
dznl
ar (1+ 0.54µ)( )µ4 5.01
2
0
2 7.4
+
−
=
z
dznl
ar ( )µ4 5.01
2
0
72.4
+
−
=
z
dznl
ar

96 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
Rutilo López López et al.
donde, z= altura de referencia (m); d= altura de desplazamiento (m); z0= longitud de la rugosidad (m); μ= velocidad del viento (m s-1). Los términos z0 y d se pueden calcular a partir de la altura de la planta (h), en cultivos con cobertura completa, estos parámetros se calculan con:
z0= 0.13h (13) y
(14)
Otra forma de desarrollar la ecuación de balance de energía para predecir la diferencia de la temperatura del cultivo y la temperatura del aire (Tc-Ta, oC), es ordenando los términos del balance de energía superficial (Jackson et al., 1981):
(15a)
(15b)
(15c)
(15d)
De esta manera, la Ecuación 15b es igual a la Ecuación 9 y pueden ser utilizadas para obtener el límite superior (dTls) del CWSI, donde la resistencia del cultivo (rc) se aproxima al infinito. Las Ecuaciones 15c y 15d se utilizan en el caso de un cultivo sin estrés hídrico (límite inferior), donde rc se asume igual a cero.
El CWSI también se puede expresar en términos de la evapotranspiración, según Jackson et al. (1981):
pEE1CWSI −=
(16)
donde, E= evapotranspiración real del cultivo; Ep= evapotranspiración de referencia; sustituyendo sus valores se tiene:
(17)
La relación rc/ra se expresa como:
(18)
donde, rcp= resistencia del dosel de un cultivo bien regado; rc= resistencia real del dosel; ra= resistencia aerodinámica al transporte de calor sensible del aire.
La relación de la evapotranspiración real (E) y la de referencia (Ep) se aproxima más a los valores teóricos de Jackson (1982), debido a que no depende tanto de las condiciones de velocidad del viento como en la forma clásica según Idso et al. (1981). Al remplazar las estimaciones del DPV a partir de la temperatura mínima (Idso, 1982) por las mediciones de temperatura del aire de la radiación directa en las parcelas de riego, no sólo aumenta la correlación con la evapotranspiración real, además, corrige parcialmente la influencia de la temperatura superficial del suelo en valores pequeños del índice de área foliar (IAF). Por lo tanto, la exactitud de las estimaciones de la evapotranspiración por medio del CWSI se da por la relación de la evapotranspiración real (E) y el potencial hídrico de la hoja (Ep) en el amanecer (Itier et al., 1993).
Aspectos metodológicos del índice de estrés hídrico en los cultivos
La evapotranspiración medida o real (E) dividida por la evapotranspiración de referencia (Ep), definida en la Ecuación 16 al despejar E/Ep queda como: E/Ep = 1- CWSI, debido a las diferencias en las mediciones de Tc-Ta vs DPV, el cultivo no requiere el riego hasta que el CWSI alcance un
hd 3 6.0=
321ac XXXTT −=−
( )
++
−=
a
c
as3
rr1
eeXγ∆
++
+
=
a
c
a
c
2
rr1
rr1
Xγ∆
γ
++
+−
+
=
a
c
a
cp
a
c
rr1
rr
1rr1
CWSIγ∆
γγ
X1 = ra (Rn - G) (ρa Cp )
(es - ea)
( )( ) ( )
( )
−−
−−+−−
=
p
naac
asacp
a
a
c
CRr
TT
eeTTCRr
rr
ργ
γ∆ρ (Tc - Ta)(∆ + γ) - (es - ea)ρ
(Tc - Ta)ρ

Índice de estrés hídrico como un indicador del momento de riego en cultivos agrícolas 97
) (
) (
iTdsTd
iTdTdCWSI
−
−=
valor umbral, el cual puede ser de 0.1 a 0.2, dependiendo del cultivo. En este tiempo el cultivo transpira en una tasa menor que el óptimo y el rendimiento del cultivo comienza a decrecer. El límite inferior de un cultivo en un lugar específico puede determinarse dos días después de que se aplique una lámina máxima de riego en el cultivo.
Los termómetros infrarrojos o buscadores térmicos se usan para medir la temperatura superficial del cultivo (Figura 2). Miden la cantidad de radiación de onda larga emitida de una superficie y se describe por la ley del cuerpo negro de Stefan-Boltzman en función de la temperatura.
(19)
donde, I= radiación emitida por la superficie (W m-2); σ= constante de Stefan-Boltzman (5.674 x 10-8 W m-2 K-4); ε= energía que emite un cuerpo a una temperatura dada, para el cuerpo negro es 1 y para otros es menor que 1; T= temperatura en la superficie (°K).
Este método utiliza los datos de temperatura para el momento del riego y otra forma de expresarlo, reduciendo términos de la Ecuación 1), se escribe como: (20)
donde, dT= diferencia medida entre la temperatura del aire y del cultivo; dT= límite superior de las temperaturas del aire menos la temperatura del follaje (cultivo sin transpiración); dT= límite inferior de la temperatura del aire menos la temperatura del follaje (cultivo bien regado).
Para determinar el límite superior e inferior en la ecuación del CWSI, se usa el método desarrollado por Idso et al. (1981) que consideran los cambios en los límites superior e inferior debido a la variación del déficit de presión de vapor de agua (DPV). El DPV es la diferencia entre la presión de saturación y la presión actual de vapor (Ecuación 21) y es un indicador preciso de la capacidad real evaporante del aire.
as eeDPV −= (21)
donde, es= presión de vapor de agua a saturación a una temperatura del aire dada; ea= presión de vapor de agua actual (presión parcial de vapor de agua en la atmósfera). Cuando el aire no se satura, la presión actual de vapor será más baja que la presión de vapor de saturación.
La presión de vapor de agua a saturación (es), en kPa, es la máxima cantidad de vapor de agua que puede contener el aire a una temperatura (T en °C) dada y se calcula con la siguiente ecuación: (22)
La presión de vapor de agua actual ea se puede obtener de la Ecuación 23 utilizando la humedad relativa (HR) y la temperatura del cultivo medidas con el termómetro de rayos infrarrojos:
(23)
Un DPV igual a cero indica que el aire sostiene el máximo vapor de agua posible (esto corresponde a una humedad relativa de 100%). El límite inferior del CWSI cambia en función de la presión de vapor del agua debido al DPV. El CWSI varía entre 0 y 1, cuando las plantas se someten a condiciones apropiadas de riego y hasta condiciones totalmente de estrés hídrico. Idso (1982) demostró que el límite inferior del CWSI es una función lineal del DPV para diversos cultivos y localidades como se puede observar en el Cuadros 1 y 2.
4TI σε=
Figura 2. Medición de los parámetros en el cálculo del índice de estrés hídrico con termómetro de rayos infrarrojos en tomate de cáscara (Physalis ixocarpa Brot).
( )
+=
3.237
7 2 .7 1exp611.0
T
TTse (T )
100 se
aeRH =
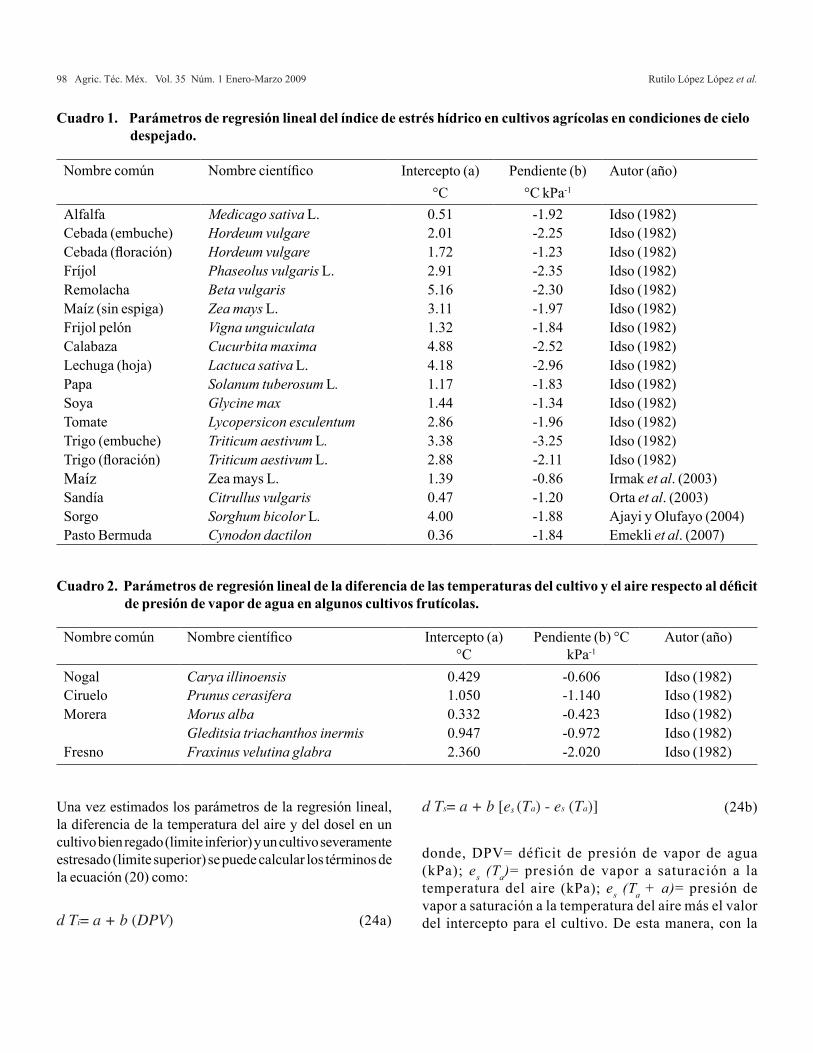
98 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
Rutilo López López et al.
(24b)
donde, DPV= déficit de presión de vapor de agua (kPa); es (Ta)= presión de vapor a saturación a la temperatura del aire (kPa); es (Ta + a)= presión de vapor a saturación a la temperatura del aire más el valor del intercepto para el cultivo. De esta manera, con la
Nombre común Nombre científico Intercepto (a) Pendiente (b) Autor (año)°C °C kPa-1
Alfalfa Medicago sativa L. 0.51 -1.92 Idso (1982)Cebada (embuche) Hordeum vulgare 2.01 -2.25 Idso (1982)Cebada (floración) Hordeum vulgare 1.72 -1.23 Idso (1982)Fríjol Phaseolus vulgaris L. 2.91 -2.35 Idso (1982)Remolacha Beta vulgaris 5.16 -2.30 Idso (1982)Maíz (sin espiga) Zea mays L. 3.11 -1.97 Idso (1982)Frijol pelón Vigna unguiculata 1.32 -1.84 Idso (1982)Calabaza Cucurbita maxima 4.88 -2.52 Idso (1982)Lechuga (hoja) Lactuca sativa L. 4.18 -2.96 Idso (1982)Papa Solanum tuberosum L. 1.17 -1.83 Idso (1982)Soya Glycine max 1.44 -1.34 Idso (1982)Tomate Lycopersicon esculentum 2.86 -1.96 Idso (1982)Trigo (embuche) Triticum aestivum L. 3.38 -3.25 Idso (1982)Trigo (floración) Triticum aestivum L. 2.88 -2.11 Idso (1982)Maíz Zea mays L. 1.39 -0.86 Irmak et al. (2003)Sandía Citrullus vulgaris 0.47 -1.20 Orta et al. (2003)Sorgo Sorghum bicolor L. 4.00 -1.88 Ajayi y Olufayo (2004)Pasto Bermuda Cynodon dactilon 0.36 -1.84 Emekli et al. (2007)
Cuadro 1. Parámetros de regresión lineal del índice de estrés hídrico en cultivos agrícolas en condiciones de cielo despejado.
Nombre común Nombre científico Intercepto (a) °C
Pendiente (b) °C kPa-1
Autor (año)
Nogal Carya illinoensis 0.429 -0.606 Idso (1982)Ciruelo Prunus cerasifera 1.050 -1.140 Idso (1982)Morera Morus alba 0.332 -0.423 Idso (1982)
Gleditsia triachanthos inermis 0.947 -0.972 Idso (1982)Fresno Fraxinus velutina glabra 2.360 -2.020 Idso (1982)
Cuadro 2. Parámetros de regresión lineal de la diferencia de las temperaturas del cultivo y el aire respecto al déficit de presión de vapor de agua en algunos cultivos frutícolas.
Una vez estimados los parámetros de la regresión lineal, la diferencia de la temperatura del aire y del dosel en un cultivo bien regado (limite inferior) y un cultivo severamente estresado (limite superior) se puede calcular los términos de la ecuación (20) como:
(24a)
d Ts= a + b [es (Ta) - es (Ta)]
d Ti= a + b (DPV)
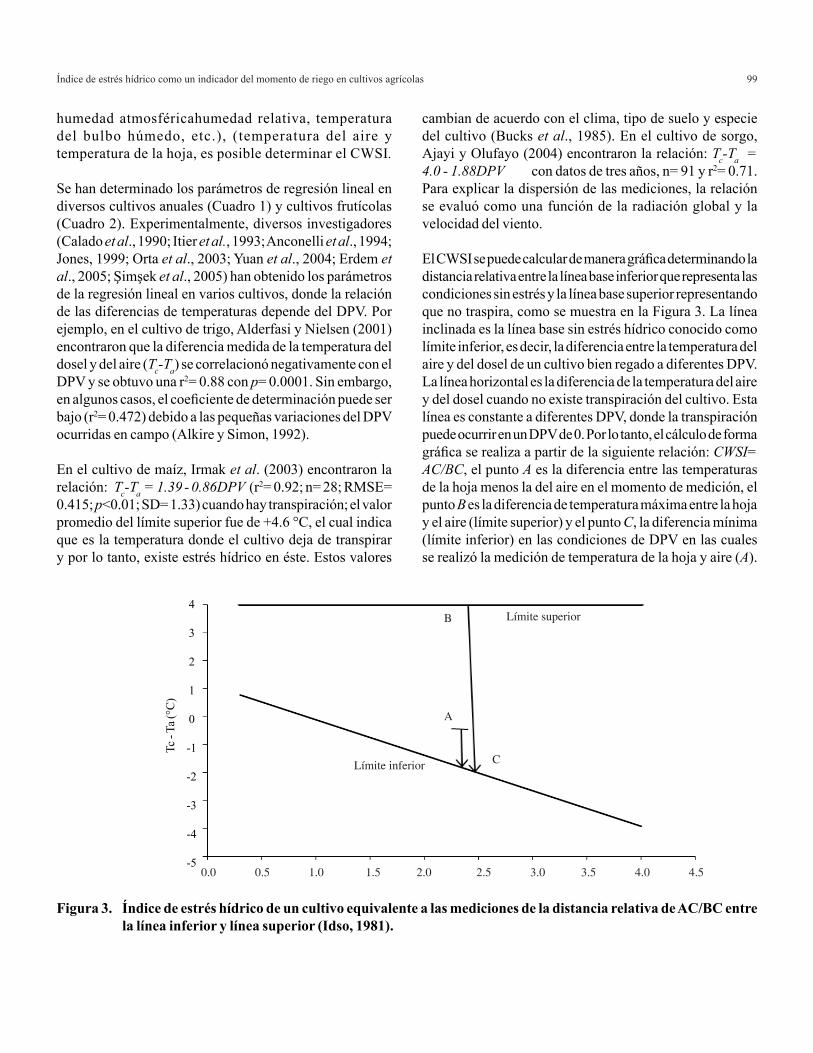
Índice de estrés hídrico como un indicador del momento de riego en cultivos agrícolas 99
humedad atmosféricahumedad relativa, temperatura del bulbo húmedo, etc.), (temperatura del aire y temperatura de la hoja, es posible determinar el CWSI.
Se han determinado los parámetros de regresión lineal en diversos cultivos anuales (Cuadro 1) y cultivos frutícolas (Cuadro 2). Experimentalmente, diversos investigadores (Calado et al., 1990; Itier et al., 1993; Anconelli et al., 1994; Jones, 1999; Orta et al., 2003; Yuan et al., 2004; Erdem et al., 2005; Şimşek et al., 2005) han obtenido los parámetros de la regresión lineal en varios cultivos, donde la relación de las diferencias de temperaturas depende del DPV. Por ejemplo, en el cultivo de trigo, Alderfasi y Nielsen (2001) encontraron que la diferencia medida de la temperatura del dosel y del aire (Tc-Ta) se correlacionó negativamente con el DPV y se obtuvo una r2= 0.88 con p= 0.0001. Sin embargo, en algunos casos, el coeficiente de determinación puede ser bajo (r2= 0.472) debido a las pequeñas variaciones del DPV ocurridas en campo (Alkire y Simon, 1992).
En el cultivo de maíz, Irmak et al. (2003) encontraron la relación: Tc-Ta = 1.39 - 0.86DPV (r2= 0.92; n= 28; RMSE= 0.415; p<0.01; SD= 1.33) cuando hay transpiración; el valor promedio del límite superior fue de +4.6 °C, el cual indica que es la temperatura donde el cultivo deja de transpirar y por lo tanto, existe estrés hídrico en éste. Estos valores
cambian de acuerdo con el clima, tipo de suelo y especie del cultivo (Bucks et al., 1985). En el cultivo de sorgo, Ajayi y Olufayo (2004) encontraron la relación: Tc-Ta = 4.0 - 1.88DPV con datos de tres años, n= 91 y r2= 0.71. Para explicar la dispersión de las mediciones, la relación se evaluó como una función de la radiación global y la velocidad del viento.
El CWSI se puede calcular de manera gráfica determinando la distancia relativa entre la línea base inferior que representa las condiciones sin estrés y la línea base superior representando que no traspira, como se muestra en la Figura 3. La línea inclinada es la línea base sin estrés hídrico conocido como límite inferior, es decir, la diferencia entre la temperatura del aire y del dosel de un cultivo bien regado a diferentes DPV. La línea horizontal es la diferencia de la temperatura del aire y del dosel cuando no existe transpiración del cultivo. Esta línea es constante a diferentes DPV, donde la transpiración puede ocurrir en un DPV de 0. Por lo tanto, el cálculo de forma gráfica se realiza a partir de la siguiente relación: CWSI= AC/BC, el punto A es la diferencia entre las temperaturas de la hoja menos la del aire en el momento de medición, el punto B es la diferencia de temperatura máxima entre la hoja y el aire (límite superior) y el punto C, la diferencia mínima (límite inferior) en las condiciones de DPV en las cuales se realizó la medición de temperatura de la hoja y aire (A).
Figura 3. Índice de estrés hídrico de un cultivo equivalente a las mediciones de la distancia relativa de AC/BC entre la línea inferior y línea superior (Idso, 1981).
Tc -
Ta (°
C)
4
3
2
1
0
-1
-2
-3
-4
-50.0 0.5 1.0 1.5 2.0 2.5 3.0 3.5 4.0 4.5
Límite superior
Límite inferior
B
A
C

100 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
Rutilo López López et al.
También se puede utilizar la conductancia estomática de la hoja estimada a través de la termometría infrarroja en la programación del riego, Jones (1999) desarrolló un procedimiento para determinar el CWSI con el valor teórico de la temperatura de la hoja o temperatura superficial de referencia estimada mediante un reordenamiento estándar del balance de energía de la hoja, expresando el CWSI como:
(25)
donde, Tl –Ta= diferencia de la temperatura de la hoja y del aire; rlw= resistencia de la hoja al transporte de vapor de agua (se asume igual a la resistencia estomática); raw= resistencia de la capa límite al vapor de agua; Rni= radiación neta isotérmica (radiación que recibe una superficie equivalente a la temperatura del aire); rHR= resistencia paralela al calor y transporte radiactivo. Para el análisis de la resistencia (raH) se estima con la siguiente ecuación:
(26)
donde, raH= resistencia de la capa límite de la hoja al transporte de calor (s m-1); d= dimensión característica de la hoja (m); µ= velocidad del viento (m s-1).
En una superficie seca, se tienen las mismas propiedades radiactivas y aerodinámicas, la pérdida de calor sensible es igual que la radiación neta absorbida, usando el concepto de radiación neta isotérmica se puede estimar la temperatura de una superficie seca (Td) a partir de la temperatura máxima:
(27)
La temperatura correspondiente a la superficie húmeda (Tw) puede calcularse a partir de la Ecuación 25 estableciendo rlw igual que cero, la cual resulta:
(28)
Restando la Ecuación 28 de la Ecuación 27 y dividiendo por la diferencia de temperaturas entre las Ecuaciones 25 y 27, se reduce a:
(29)
La Ecuación 29 puede ser reordenada para obtener una expresión simple de la resistencia de la hoja:
(30)
Una importante característica de la Ecuación 30 es que rlw puede determinarse únicamente a partir de una combinación de mediciones de temperaturas de la hoja, de equivalencias de humedad y superficies secas y un término que depende sólo de la resistencia para calor y pérdida de humedad a través de la capa límite de la hoja. Es notable que este factor, en la ecuación no depende de la radiación neta absorbida o del déficit de presión de vapor del aire, sólo depende de las temperaturas. Debido a que el término (Td – Tl) en esta ecuación tiende a ser muy pequeño y es la diferencia entre dos cantidades de diversas variables en el campo, el comportamiento puede ser inestable, generalmente es preferible usar su inverso, la conductancia de la hoja (clw).
(31)
La tasa de transpiración está estrechamente relacionada con la conductancia, mm s-1, también se puede encontrar en la literatura como unidades molar (mol m-2 s-1) y se obtiene simplemente multiplicando por 0.04, este factor es ligeramente sensible a la temperatura.
Una posibilidad de usar un índice de estrés de forma análoga en el CWSI, es sustituyendo Td por la temperatura máxima (Tmax) y Tw por la temperatura del límite inferior (dTi).
(32)
Tl –Ta = rHR (raw + rlw ) γ Rni -
rHR DPV ρa Cp [γ(raw + rlw) + ∆ rHR] γ(raw + rlw) + ∆ rHR
Td – Ta = rHR Rni
ρa Cp
TW – Ta = rHR raw + rlw γ Rni -
rHR DPV ρa Cp [γ (raw) + ∆ rHR] γ raw+ ∆ rHR
( )( )ld
wlRHwawl TT
TTrrr−−
∆+= γ
(Tl –TW )
(Td – Tl )
RHrwar
RHrwarwlr
lTdT
wTdT
∆
+
∆++
=−
−
γ
γ
( )( )
∆
+−
−=
RHrwarwTlT
lTdTwlc
γ
(Td – Tl )
(Tl –TW )
=µ
dHar 100
(Tl –TW ) (Td – TW )
( )( )wTdT
wTlTCWSI
−
−=22

Índice de estrés hídrico como un indicador del momento de riego en cultivos agrícolas 101
Una alternativa para definir CWSI3, que es proporcional a la resistencia estomática, por lo tanto, a partir de la ecuación (30) se puede escribir como:
(33)
Finalmente, a partir de la ecuación (31) se puede definir un índice (CWSI4) el cual es proporcional a la conductancia estomática.
(34)
Por otra parte, De Lorenzi et al. (1993) probaron las tres versiones del CWSI: la aproximación teórica (Ecuación 1) difundida por Jackson et al. (1981), una modificación propuesta por Clawson et al. (1989), y el desarrollo original que considera el valor real de la resistencia del dosel en un cultivo bien regado. Los tres métodos fueron considerados útiles para calcular el estrés hídrico en cultivo de chile dulce a campo abierto. Concluyeron que los valores de CWSI fueron influenciados fuertemente por la adaptación rápida de las plantas a la escasez de agua.
Yuan et al. (2004) evaluaron en trigo de invierno, el modelo empírico de Idso et al. (1981) (Ecuación 1), el modelo teórico de Jackson et al. (1981) (Ecuaciones 15a, 15b, 15c y 15d) y el modelo de Alves (citado por los mismos autores), que sustituye la temperatura superficial radiométrica por una temperatura superficial del “bulbo húmedo”, de tal forma que evita la medición de la resistencia superficial del cultivo. Los resultados muestran que el CWSI basado en la definición de Jackson y Alves son mejores que el método empírico. Ambos métodos fueron herramientas útiles para evaluar el estrés hídrico, pero el CWSI basado en el concepto de Alves es más práctico, mientras que el CWSI basado en el concepto de Jackson es más aceptable en la cuantificación del estrés hídrico.
Aplicaciones del índice de estrés hídrico en la programación de riegos
Según Idso (1982), el CWSI no es apropiado para regiones húmedas, debido a que en estas condiciones, generalmente no se presentan condiciones de sequía. Sin embargo, en
climas cálidos húmedo, como el sureste de México, se presentan de tres a cuatro meses con períodos de sequías, principalmente en el ciclo otoño invierno, período donde la cantidad de radiación solar es mayor y los cultivos agrícolas requieren el agua de riego para satisfacer sus necesidades hídricas. Jones et al. (1997) señalan que la humedad relativamente alta en climas húmedos y la variación de la velocidad del viento y de la radiación solar pueden limitar seriamente el uso del método original, pero demostraron que la sensibilidad del método mejora cuando se considera la temperatura de la hoja y no la temperatura del aire (como en el método de CWSI original), es decir, la temperatura de las “hojas modelo” expuestas dentro del cultivo.
Diversos investigadores han desarrollado el método del CWSI en la programación del riego (Howell et al., 1984; O'Toole et al., 1984; Reginato y Howe, 1985; Reginato y Garrett, 1987; Wanjura et al., 1990). Sin embargo, otra de las desventajas de este método es que los valores del CWSI son difíciles de obtener al inicio del ciclo de los cultivos a causa de la cobertura parcial del dosel. Al comienzo del ciclo, cuando las plantas son pequeñas o tienen escasa población, una parte de la superficie del suelo puede ser considerada por la pistola de rayos infrarrojos cuando se efectúa la medición.
Nielsen (1990) utilizó diferentes umbrales de CWSI (0.2, 0.3, 0.4 y 0.5) en la programación del riego en el cultivo de soya (Glycine max L. Merrill.) con riego por goteo, aplicándose láminas de riego de 180, 181, 174 y 145 mm y los rendimientos obtenidos fueron 2 656, 2 566, 2 430, y 2 189 kg ha-1 respectivamente. Los rendimientos se incrementaron ligeramente cuando aumentó la lámina de riego en 25 y 51 mm adicionales, y presentó las mismas tendencias cuando se incrementó el umbral de CWSI.
Anconelli et al. (1994) encontraron en el cultivo de tomate (Lycopersicon esculentum Mill.), la respuesta en rendimiento de tres valores de CSWI, usados como umbrales para el momento del riego, los cuales fueron comparados con un control (sin riego). Con el riego se obtuvieron aumentos significativos en el rendimiento de frutos. Mientras que con el tratamiento sin riego se obtuvieron 35 t ha-1 de fruto, los tratamientos de riegos con umbrales alcanzados de 0.6, 0.35 y 0.1 de CWSI se obtuvieron rendimientos de frutos de 51, 57 y 60 t ha-1 respectivamente. Las características organolépticas (sabor y aroma) se alteraron por el riego, pero se obtuvo una mejora en la proporción de sacarosa. No se observaron diferencias significativas en los umbrales de
(Tl –TW ) (Td – Tl )
( )( )wTlT
lTdTCWSI
−
−=4
( )( )lTdT
wTlTCWSI
−
−=3
(Td –T l ) (Tl –TW )

102 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
Rutilo López López et al.
CWSI de 0.1 y 0.35, concluyeron que el cultivo es capaz de soportar valores 0.35 de índice de estrés sin que haya disminución de la producción y calidad del fruto.
Calado et al. (1990) estudiaron los efectos de disminución del uso del agua en los indicadores de estrés hídrico en la producción de tomates con riego por goteo. Los tratamientos de riego fueron distribuidos en un diseño de bloques al azar, con cinco niveles de láminas de riego: 0.40 ETc, 0.60 ETc, 0.80 ETc, 1.0 ETc y 1.20 ETc. El riego fue controlado automáticamente por una computadora usando la evaporación de tanque clase “A” adaptada a un sensor electrónico. Los parámetros de suelo y planta fueron seguidos sistemáticamente en forma manual y por computadora. Los resultados indicaron que: i) los sensores del potencial matricial del suelo (SMPS) pueden utilizarse en la programación y control del riego. Las mediciones simultáneas (suelo y planta) permite tomar decisiones correctas basadas en condiciones normales, ii) el método del CWSI es una herramienta en la programación o control del riego para caracterizar el estrés hídrico en plantas de jitomate. Se determinó una variabilidad significativa en este método, y iii) se encontraron algunas correlaciones significativas estadísticas entre los cambios del diámetro del tallo y potencial hídrico de la hoja.
Con base en los resultados de Garrot et al. (1990), Orta et al. (2003) determinan el índice de estrés hídrico en la programación de riego en el cultivo de sandía (Citrullus vulgaris) producida con riego por goteo, estudiaron los efectos de cinco niveles de láminas de riego (reposición de 100, 75, 50, 25 y 0% de la humedad consumida del suelo a partir de la profundidad de 0.90 m del suelo) en el rendimiento de frutos. El rendimiento más alto y uso eficiente del agua se obtuvo en condiciones de riego completo (reposición de 100% de la humedad consumida del suelo) durante dos años. El CWSI fue calculado a partir de mediciones de las temperaturas del cultivo, temperatura ambiental del aire y de los valores del déficit de presión de vapor para cinco niveles de láminas de riego, obteniéndose el siguiente limite inferior: Tc-Ta= -1.2DPV+0.47 con r2= 0.52 y desviación estándar de 0.57. Las tendencias en valores de CWSI fueron constantes con los contenidos de humedad del suelo inducidos por el déficit del riego; el CWSI aumentó con el déficit creciente del agua en el suelo. Un promedio de CWSI alrededor de 0.41 antes del riego produjo un aumento en el rendimiento máximo, éste se correlacionó
directamente con valores medios de CWSI y se obtuvo la ecuación lineal: Y= 91.143- 66.077(CWSI), la cual se puede utilizar en la predicción del rendimiento.
Posteriormente, Erdem et al. (2005) a partir de los resultados anteriores evaluaron diferentes valores de umbral del CWSI para programar el riego en el cultivo de sandía (Citrullus vulgaris) producida con riego por goteo, los riegos se iniciaron cuando los valores de CWSI alcanzaron: 0.2, 0.4, 0.6, 0.8 y 1.0 (sin riego). La cantidad total aplicada de láminas de riegos fueron: 342, 280, 248 y 193 y 0 mm respectivamente. La evapotranspiración máxima estacional fue de 412 mm, medida a partir del tratamiento de 0.2 de CWSI. Concluyeron que los niveles de láminas de riego afectaron significativamente el rendimiento de frutos y la producción más alta (76.3 t ha-1) fue obtenida con el tratamiento de 0.2 de CWSI, los tratamientos 0.4 y 0.6 de CWSI fueron estadísticamente iguales comparado con el tratamiento de 0.2. La eficiencia máxima del uso del agua y la eficiencia del uso del agua de riego se obtuvieron a partir del tratamiento 0.6 de CWSI con 22.1 y 13.3 kg m-3, respectivamente. Con base en estos resultados, el valor de 0.6 de CWSI puede utilizarse en la programación del riego en el cultivo de sandía.
Şimşek et al. (2005) realizaron estudios para determinar los efectos de diferentes láminas de riego por goteo en el rendimiento del cultivo de pepino (Cucumbis sativus L.) y sus componentes, y para determinar un valor de umbral del CWSI basado en la programación del riego. Aplicaron cuatro tratamientos de riego: 50% (T-50), 75% (T-75), 100% (T-100) y 125% (T-125) de láminas de riego basado en la evaporación de tanque “A” acumulada cada tercer día. Los valores de la evapotranspiración del cultivo (ETc) fueron 633, 740, 815 y 903 mm en el primer año y 679, 777, 875 y 990 mm en el segundo año para T-50, T-75, T-100 y T-125, respectivamente. La producción comercial máxima del fruto se obtuvo con el tratamiento T-100 con 76.65 t ha-1 en 2002 y 68.13 t ha-1 en 2003. La producción de frutos se redujo notablemente cuando disminuyó la lámina de riego. La eficiencia del uso del agua varió de 7.37 a 9.40 kg m-3 y 6.32 a 7.79 kg m-3 en 2002 y 2003, respectivamente, mientras que las eficiencias del uso del agua del riego estuvieron entre 7.02 y 9.93 kg m-3 en 2002 y entre 6.11 y 8.82 kg m-3 en 2003. Cuando disminuyó la lámina de riego, la tasa de transpiración del cultivo disminuyó dando por resultado aumentos de temperaturas del dosel del cultivo y del CWSI, esto resultó en una disminución en el rendimiento. Un CWSI igual que

Índice de estrés hídrico como un indicador del momento de riego en cultivos agrícolas 103
0.2 se considera como el valor umbral para el inicio del riego en el cultivo de pepino producido a campo abierto en condiciones semiáridas.
En el cultivo de algodón (Gossypium hirsutum) Barbosa y Ramana (2005) concluyeron que el riego se debe iniciar cuando el CWSI se aproxima a 0.3. Después de un segundo evento de estrés, el cultivo requiere de tres días para su tasa normal de transpiración. La respuesta de CWSI con respecto de la temperatura del cultivo (Tc), rc/ra y Rn es evidente y probablemente es la fuente de error más importante en la determinación del CWSI.
Hutmacher et al . (1991) en el cultivo de alfalfa (Medicago sativa L.) encontraron que los valores de CWSI fueron altamente y linealmente correlacionados con el potencial hídrico del tallo. Concluyeron que el uso de la línea base sin estrés en el período vegetativo para calcular los valores durante la floración y llenado de vaina resultaron valores de 0.1 a 0.2 más altos que los determinados usando líneas base que depende de la etapa de crecimiento. En la programación del riego, sugieren el uso de dos líneasbase, una para el período vegetativo y otra línea base determinada mediante una base de datos de los períodos de floración y llenado de vaina.
En estudios realizados con pastos, Feldhake et al. (1997) encontraron que la relación entre CWSI y la relación E/Ep depende de la especie. Además que el CWSI predice más rápidamente el estrés hídrico en Festuca arundinacea
Carcova et al. (1998) validaron el método de Idso (1981) para determinar el CWSI y describieron el comportamiento del CWSI en tres híbridos de maíz (Zea mays L.) que crecieron en diferentes ambientes y calidad del suelo. Determinaron el efecto de la calidad del suelo y de los híbridos en relación con el CWSI y el agua disponible del suelo. El CWSI aumentó durante el período de crecimiento en todos los ambientes y híbridos. La relación entre CWSI y agua disponible del suelo fue significativo para los tres híbridos y los diferentes ambientes (r2= 0.52, n= 51, p< 0.001). Un umbral mayor que 0.25, que se relacionó con 60% de agua disponible en el suelo, el cultivo presentó síntomas de estrés hídrico. Irmak et al. (2000) encontraron que cuando el valor promedio del CWSI en cualquier etapa de crecimiento es mayor que 0.22, el rendimiento de grano del cultivo de maíz disminuye. El polinomio cuadrático determinado para predecir el rendimiento del cultivo de maíz (Y) a partir del CWSI fue: Y= -4.38CWSI2+1.58CWSI+0.46 con r2= 0.98.
En el Cuadro 3 se presenta un resumen de valores umbrales de CWSI en algunos cultivos agrícolas para el momento del riego. La mayoría de los autores señalan que una vez alcanzado el umbral del CWSI es recomendable programar el riego para evitar daños por estrés hídrico y se disminuya el rendimiento de los cultivos.
Schreb., que en Dactylis glomerata (E/Ep de 0.84 vs 0.70). Encontraron que la relación entre CWSI y E/Ep no es lineal, la relación es más sensible a niveles más altos de índice de estrés. Más tarde, Al-Faraj et al. (2001) estudiaron el
Cuadro 3. Valores umbrales del índice de estrés hídrico para la programación del riego en algunos cultivos agrícolas.
Autor (año) Cultivo Umbral CWSI
Nielsen (1990) Soya (Glycine max) 0.2Garrot et al. (1990) y Orta et al. (2003) Sandía (Citrullus vulgaris L.) 0.41Hutmacher et al. (1991) Alfalfa (Medicago sativa L.) 0.1-0.2Gardner et al. (1992) Trigo (Triticum aestivum L.) 0.2-0.3Anconelli et al. (1994) Tomate (Lycopersicum esculentum) 0.35Anconelli y Battilani (2000) Vid (Vitis vinifera) 0.4Irmak et al. (2003) Maíz (Zea mays L.) 0.22Barbosa y Ramana (2005) Algodón (Gossipium hirsutum) 0.3Erdem et al. (2005) Sandía (Citrullus vulgaris L.) 0.6Simsek et al. (2005) Pepino (Cucumbis sativus) 0.2Emekli et al. (2007) Pasto Bermuda (Cynodon dactilon) 0.1

104 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
Rutilo López López et al.
CWSI y las líneas base con el aumento de la tensión de humedad del suelo en el pasto Festuca arundinacea Schreb. Se midieron continuamente con termómetros infrarrojos las temperaturas del dosel del pasto, junto con la evapotranspiración del cultivo, medido con mini-lisímetros electrónicos. Concluyeron que la diferencia de la temperatura del cultivo-aire (Tc-Ta) aumentó con la disminución del contenido de humedad del suelo. La Tc-Ta aumentó cuando la radiación neta fue mayor, independiente de la tensión de humedad del suelo. La temperatura del dosel de plantas bien regadas disminuyó el índice en 2.4 °C por cada disminución de 1 kPa del DPV para todos los niveles de la radiación neta. Por cada aumento de 100 W m-2 de la radiación neta, la temperatura del dosel de las plantas bien regadas se incrementó en un índice de 0.6 °C y mostró alta correlación (línea base sin estrés) con el DPV. El incremento de la temperatura del dosel junto con la disminución de la tasa de la transpiración fueron los signos del incremento del estrés hídrico. Sin embargo, las relaciones de la Tc-Ta y la línea base del DPV tuvieron una correlación baja para las condiciones de estrés moderadas y severas sin importar los niveles de la radiación neta. Además, en ambiente controlado, al aumentar la precisión y número de repeticiones, el límite inferior de la línea base del estrés hídrico fue muy variable.
CONCLUSIONES
De los aspectos metodológicos y teóricos que han utilizado las diferentes formas como se ha expresado el CWSI, todos se basan en la teoría del balance de energía en el dosel del cultivo, las funciones lineales de la temperatura diferencial (Tc-Ta) y el déficit de presión de vapor que dependen del cultivo y las condiciones atmosféricas.
La termometría infrarroja es una herramienta sencilla, práctica, no destructiva y confiable para la estimación del estrés hídrico de los cultivos, la cual se ha aplicado en una diversidad de cultivos agrícolas en la programación del riego y es útil para la predicción de rendimientos.
El método del CWSI es oportuno y eficaz en la programación del riego cuando se determina el valor umbral en las diferentes etapas fenológicas del cultivo para evitar daños por estrés hídrico y disminuir el rendimiento de los cultivos.
LITERATURA CITADA
Alderfasi, A. A. and Nielsen D. 2001. Use of crop water stress index for monitoring water status and scheduling irrigation in wheat. Agricultural water management. 47:69-75.
Al-Faraj, A.; Meyer, E. G. and Horst, L. G. 2001. A crop water stress index for tall fescue (Festuca arundinacea Schreb.) irrigation decision-making a traditional method. Computers and Electronics in Agriculture. 31:107-124.
Alkire, B. H., and Simon, J. E. 1992. Use of infrared thermometry to monitor water s tatus in peppermint (Mentha x Piperita L.). Acta Hort. 306:385-396.
Anconelli, S.; Mannini, P. and Battilani, A. 1994. CWSI and baseline studies to increase quality of processing tomatoes. Acta Hort. 376:303-306.
Anconelli, S. and Battilani, A. 2000. Use of leaf temperature to evaluate grapevine (Vitis vinifera) yield and quality response to irrigation. Acta Hort. 537:124-138.
Ajayi, A. E. and Olufayo, A. A. 2004. Evaluation of two temperature stress indices to estimate grain sorghum yield and evapotranspiration. Agron. J. 96:1282-1287.
Barbosa da Silva, B., and Ramana, R. T. A. 2005. The CWSI variations of a cotton crop in a semi-arid region of Northeast Brazil. J. Arid Environ. 62:649-659.
Bucks, D. A.; Nakavamma, F. S.; French, O. F.; Regard, W. W.; and Alexander, W. L. 1985. Irrigated guayule evapotranspiration and plant water s t ress . Agr icul tura l Water Management 10:61-79.
Buchner, R. P., Goldhamer, D. A. and Shaw, D. A. 1994. Irrigation scheduling in kiwifruit growing and handling, In: Hasey J. K.; Johnson, R. S.; Grant, J. A. and Reil, W. O. (eds.). University of California Publication 3344. p. 43-49.
Calado, A. M.; Monzon, A.; Clark, D. A.; Phene, C. J.; Ma, C.; and Wang, Y. 1990. Monitoring and control of plant water stress in processing tomatoes. Acta Hort. 277:129-136.
Cárcova, J.; Maddonni A. G.; and Ghersa, C. M. 1998. Crop water stress index of three maize hybrids grown in soils with different quality. Field Crops Res. 55:165-174.

Índice de estrés hídrico como un indicador del momento de riego en cultivos agrícolas 105
De Lorenzi, F. ; Stanghellini, C. and Pitacco, A. 1993. Water shortage sensing through infrared canopy temperature: timely detection is imperative. Acta Hort. 335:373-380.
Emekli, Y.; Bastug, R.; Buyuktas, D. and Yasemin, N. E. 2007. Evaluation of a crop water stress index for irrigation scheduling of bermudagrass. Akdeniz University, Faculty of Agriculture, Department of Farm Structures and Irrigation, Antalya, Turkey.
Erdem, Y.; Erdem, A. T.; Orta H.; and Okursoy, H. 2005. Irrigation scheduling for watermelon with crop water stress index (CWSI). J. Central European Agric. 6:449-460.
Feldhake, C. M.; Glenn, D. M.; and Peterson, D. L. 1996. Effect of drought on the thermal environment of a pasture soil surface. Agron. J. 88:652-656.
Feldhake, C. M.; Glenn, D. M.; Edwards, W. M. and Peterson, D. L. 1997. Quantifying drought for humid, temperate pastures using the crop water stress index (CWSI). N. Z. J. Agric. Res. 40:17-23.
Garrot Jr, D. J.; Ray, D.T.; Livingston, M.; Fangmeier, D. D. and Husman, S. H. 1990. Watermelon field irrigation management using the crop water stress index. Acta Hort. 278:755-762.
Giuliani, R.; Magnanini, A. and Flore, J. A. 2001. Potential use of infrared thermometry for the detection of water deficit in apple and peach orchards. Acta Hort. 557:38-43.
Howell, T. A.; Hatfield, J. L.; Yamada, H. and Davis, K. R. 1984. Evaluation of cotton canopy temperature to detect crop water stress. Trans. Amer. Soc. Agric. Eng. 27:84-88.
Hutmacher, R. B.; Steiner, J. J.; Vail, S. S. and Ayars, J. E. 1991. Crop water stress index for seed alfalfa: influences of within-season changes in plant morphology. Agricultural Water Management. 19:135-149.
Idso, S.B.; Jackson, R. D.; Pinter, P. J.; Reginato, R. J. and Hatfield, J. L. 1981. Normalizing the stress-degree-day parameter for environmental variability. Agric. Meteorol. 24:45-55.
Idso, S. B. 1982. Non-water-stressed baselines: a key to measuring and interpreting plant water stress. Agric. Meteorol. 27:59-70.
Irmak, S.; Hamman D. Z., and Bastug, R. 2000. Determination of crop water stress index for irrigation timing and yield estimation of corn. Agron. J. 92:1221-1227.
Itier, B.; Flura, D. and Belabbes, K. 1993. An alternative way for CWSI. Calcula t ion to improve relative evapotranspiration estimates-results of an experiment over soybean-. Acta Hort. 335:333-340.
Jackson, R. D.; Idso, S. B., Reginato, R. J. and Pinter Jr, P. J. 1981. Canopy temperature as a crop water stress indicator. Water Resour. Res. 17:1133.
Jackson, R. D. 1982. Canopy temperature and crop water stress. In: advances in irrigation. Volume 1. Hillel, D. I. Editor. Academic Press. p. 43-85.
Jackson, R. D.; Kustas, W. P.; and Choudhury, B. J. 1988. A reexamination of the crop water stress index. Irrig. Sci. 9:309-317.
Jones, H. G.; Aikman, D. and McBurney, T. A. 1997. Improvements to infra-red thermometry for irrigation scheduling in humid climates. Acta Hort. 449:259-266.
Jones, H. G. 1999. Use of infrared thermometry for estimation of stomatal conductance as a possible aid to irrigation scheduling. Agric. & For. Meteorol. 95:139-149.
Keener, M. E. and Kircher, P. L. 1983. The use of canopy temperature as an indicator of drought stress in humid regions. Agric. Meteorol. 28:339-349.
Nielsen, D. C. 1990. Scheduling irrigations for soybeans with the crop water stress index (CWSI). Field Crops Res. 23:103-116.
Novak, V. A. 1997. Phitomonitoring in plant physiology. Arrangement and possibilities. Russian J. Plant Physiol. 44:121-128.
Orta, A. H.; Erdem Y.; and Erdem T. 2003. Crop water stress index for watermelon. Sci. Hort. 98:121-130.
O’Toole, J. C.; Turner, N. C.; Namuco, O. P.; Dingkukn, M.; and Gomez, K. A. 1984. Comparison of some crop water stress measurement methods. Crop Sci. 24:1121-1128.
O’Toole, J. C., and Real, J. G. 1986. Estimation of aerodynamic and crop resistances from canopy temperature. Agron. J. 78:305-310.
Reginato, R. J., and Garrot Jr., D. J. 1987. Irrigation scheduling with the crop water stress index. p. 7-10. In: western cotton production conf. Summary Proc., Phoenix, AZ. Cotton Growers Assoc., Memphis, TN.
Reginato, R. J. and Howe, J. 1985. Irrigation scheduling using crop indicators. J. Irrigation Drainage Engineering. 3:125-133.

106 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
Rutilo López López et al.
Stangehellini, C. and De-Lorenzi, F. 1994. A comparison of soil and canopy temperature-based methods for early detection of water stress in a simulated patch of pasture. Irrig. Sci. U. 8:141:153.
Şimşek, M.; Tonkaz T.; Kaçıra, M.; Çömlekçioğlu, N.; and Doğan, Z. 2005. The effects of different irrigation regimes on cucumber (Cucumbis sativus L.) yield and yield characteristics under open field conditions. Agricultural Water Management. 73:240-252.
Ton, Y.; Kopyt, M. K.; and Nilov, N. 2001. Phytomonitoring technique for tuning irrigation of vineyards. Proc. International symposium on irrigation and water relations in Grapevine and Fruit Trees. Mendoza, Argentina. p. 147-168.
Thorn, A. S., and Oliver, H. R. 1977. On Penman´s equation for estimating regional evaporation. Quaterly J. Royal Meteorol. Soc. 103:345-357
Van Leeuwen, C.; Chone, X.; Tregoat, O.; and Gaudillere, J. P. 2001. The use of physiological indicators to assess vine water uptake and to manage vineyard irrigation. The Australian Grapegrower and Winemaker. 449:18-24.
Wanjura, D. F.; Hatfield, J. L.; and Upchurch, D. R. 1990. Crop water stress index relationship with crop productivity. Irrig. Sci. 11:93-99.
Yuan, G.; Luo, Y.; Sun, X.; and Tang, D. 2004. Evaluation of a crop water stress index for detecting water stress in winter wheat in the North China Plain. Agricultural Water Management. 64:29-40.

Agricultura Técnica en México Vol. 35 Núm.1 1 de enero-31 de marzo 2009 p. 107-116
ASOCIACIÓN DE ESPECIES ARBÓREAS FORRAJERAS PARA MEJORAR LA PRODUCTIVIDAD Y EL RECICLAJE DE NUTRIMENTOS*
FODDER TREE SPECIES IN ASSOCIATION TO IMPROVE PRODUCTIVITY AND NUTRIENTS CYCLING
Judith Petit Aldana1§, Fernando Casanova Lugo1 y Francisco Javier Solorio Sánchez1
1Cuerpo Académico de Producción Animal en Agroecosistemas Tropicales. Campus - Ciencias Biológicas y Agropecuarias, Universidad Autónoma de Yucatán, México. Carretera Mérida-Xmatkuil, km. 15.5 C. P. 97100, Mérida, Yucatán, México. Tel. 01 99 99 42 32 00 y 9423205, ([email protected]) ([email protected]). §Autora para correspondencia: [email protected].
* Recibido: Enero, 2008
Aceptado: Marzo, 2009
RESUMEN
Se llevo a cabo una revisión bibliográfica relacionada con la asociación de especies arbóreas forrajeras para mejorar la productividad y el reciclaje de nutrimentos en agroecosistemas. Los temas tratados abarcan la situación actual de los sistemas de producción y las alternativas para mejorar su eficiencia. Asimismo, se discute acerca de la contribución de los sistemas agroforestales al ciclo de nutrimentos y su mejoramiento a través de estrategias de manejo, también se analizan algunos trabajos realizados en plantaciones forestales mixtas y su aporte en la fertilidad de los suelos, reciclaje de nutrimentos y fijación biológica de nitrógeno. Se concluye que, la asociación de especies arbóreas forrajeras puede contribuir a la productividad y sostenibilidad de los agroecosistemas, ya que estas adquieren elementos de alto valor nutricional que ayudan a mejorar la fertilidad del suelo; producen materia orgánica de alta calidad para un eficiente reciclaje de nutrimentos, lo que podría incrementar la productividad. De igual forma, se fundamenta que las asociaciones con leguminosas arbóreas establecen simbiosis con microorganismos que mejoran la disponibilidad de elementos en el suelo. Por otra parte, lo anterior no sólo depende de las especies, si no también de los recursos disponibles en el sitio y del manejo del agroecosistema. Sin embargo, aun existen vacíos de
investigación en este tema, puesto que la mayoría de trabajos se han realizado para especies arbóreas maderables en plantaciones forestales.
Palabras clave: forraje, producción, reciclaje de nutrimentos.
ABSTRACT
A bibliographical review was made related to the association of fodder tree species for improving the productivity and nutrients recycling in agroecosystems. The covered topics include the situation of actual production systems and strategies to improve their efficiency. Also, the contribution of agroforestry systems in nutrient cycling and their improvement through management is discussed. Documents on mixed forest plantations and their role in soil fertility, nutrient cycling and biological nitrogen fixation, were also analyzed. It was concluded that fodder tree species in association can contribute to the productivity and sustainability of agroecosystems, since they take up nutrients of high nutritional value and contribute towards the improvement of soil fertility, they produce litter of high-

108 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
Judith Petit Aldana et al.
quality in an efficient nutrient recycling, that can improve productivity. Furthermore associations of legumes trees species in symbiosis with microorganisms upgrade the availability of soils nutrients. On the other hand, the above-mentioned facts do not depend exclusively on the species, but on the resources available and the management of the agroecosystem. Moreover, there is a lack of research studies in this topic, since most research has been conducted in wooden species in forest plantations.
Key words: fodder, production, nutrient cycling.
INTRODUCCIÓN
La asociación de especies leñosas forrajeras para la alimentación animal es una práctica antigua. Sin embargo, recientemente ha cobrado mayor atención debido a la creciente necesidad de buscar alternativas locales, que reduzcan la dependencia de insumos externos y minimicen daños sobre los recursos naturales.
Algunas leguminosas producen alimentos ricos en proteínas. En ciertas circunstancias los árboles pueden producir más forraje que los cereales, caso del algarrobo de las zonas secas del norte de África. Los árboles y los arbustos pueden contribuir a asegurar una dieta nutritiva para el ganado. Muchos árboles forrajeros son originarios de zonas áridas y semiáridas, donde las condiciones del medio son difíciles para el cultivo de los pastos tradicionales; y en estos casos los animales no podrían sobrevivir, sin embargo el pastoreo depende exclusivamente de estos árboles (National Academy of Sciences, 1979).
En muchas zonas tropicales donde la estacionalidad del clima concentra las labores de arado y siembra en la estación lluviosa, se necesitan muchos más animales de los que pueden mantenerse con los alimentos producidos dentro del sistema agrícola. Esto hace que el agricultor tenga acceso a pastos o forrajes fuera de la explotación agrícola, en este caso los bosques y matorrales son a menudo la principal fuente suplementaria de alimentos, y es allí donde los árboles forrajeros toman un papel fundamental en la alimentación animal (Petit,1994).
Por lo tanto, cada vez se presta más atención a las relaciones existentes entre las actividades forestales comunitarias y la obtención de forraje: el potencial de
aumentar los suministros de forraje derivado de árboles y bosques, y la necesidad de no interrumpir los suministros actuales de forraje, así como las interacciones entre agricultura y ganadería cuando la tierra se dedica a la silvicultura (National Academy of Sciences, 1979).
Lo anterior, forma parte del panorama mundial del manejo y uso de la tierra, en donde el papel del árbol como estabilizador de suelos no ha sido tomado en cuenta. Muchos árboles restablecen al suelo los nutrientes que habían absorbido los cultivos, y como protegen el humus, salvaguardan los suministros de fertilizantes naturales (Petit, 1994).
En muchas zonas de los trópicos subhúmedos y semiáridos, la producción ganadera se ve seriamente limitada por la escasez y la baja calidad de forraje durante la estación seca. El bajo contenido de proteína cruda es la limitación más común de la producción ganadera de los pastos nativos y algunos sistemas se han desarrollado para complementar o mejorar la ingesta de proteína cruda de los animales que pastorean en pastos nativos mediante el acceso, ya sea por temporadas o todo el año. Para ello se establecen áreas sembradas de leguminosas arbóreas tropicales que forman lo que se conoce como un banco de proteínas. Esta forma de disposición de una alta calidad de forraje para el ganado se ha utilizado con éxito utilizando Leucaena leucocephala como suplemento alimenticio para el ganado en pastos nativos (Solorio y Solorio, 2002).
En África y la India, los ganaderos confían excesivamente en el pastoreo y el ramoneo como suplemento alimenticio, particularmente en los últimos meses de la larga estación seca que conlleva la escasez de alimentos. Por ejemplo, se estimó para Botswana que 25% de la dieta anual del ganado fue compuesta por ramoneos en árboles y arbustos, también se observó que durante los meses de mayor sequía del año, este consumo se eleva a un 45% y representa esencialmente toda la proteína adquirida por los animales (National Academy of Sciences, 1979).
Esta revisión discute los aspectos más importantes de los sistemas asociados de especies arbóreas, los cuales requieren de estrategias de manejo adecuadas que puedan interactuar conjuntamente para lograr un nivel equilibrado de productividad. Por ello, es oportuno explicar las técnicas de manejo eficiente de los nutrimentos, que se emplean como indicadores para

Asociación de especies arbóreas forrajeras para mejorar la productividad y el reciclaje de nutrimentos 109
determinar la productividad y sustentabilidad de los sistemas agropecuarios. En este sentido, la búsqueda y fomento de prácticas tendientes a maximizar la productividad y eficiencia en el reciclaje de nutrimentos se hace indispensable.
Situación actual de los sistemas de producción y alternativas
El continuo crecimiento poblacional y la demanda por alimento han sobrepasado la capacidad de los suelos para mantener la productividad de manera constante, lo que ha ocasionado la degradación ambiental poniendo en riesgo la estabilidad productiva de los sistemas agropecuarios (Craswell et al., 2004). El problema de la fertilidad de suelos se encuentra estrechamente relacionado con un balance negativo de nutrimentos y es uno de los principales problemas a los que se enfrentan los sistemas de producción agropecuarios (Craswell et al., 2004). El balance de nutrimentos, es decir, la diferencia entre las entradas y salidas de un sistema, es negativo en casi todos los sistemas de producción actuales tradicionales (Powell et al., 2004).
La remoción excesiva y constante de vegetación en el caso de la ganadería resulta en un desbalance de nutrimentos dejando los suelos sin suficientes reservas para mantenerse productivos. El problema se agudiza cuando los nutrientes “cosechados” no son, aunque sea en parte, regresados a su lugar de origen tal es el caso de la ganadería extensiva con base en pastizales. Estos aunque se mantienen productivos los primeros años son de mala calidad, principalmente deficientes en nitrógeno (N) e insuficientes en la época de secas, ya que declinan drásticamente su productividad. Numerosos estudios en los trópicos han concluido que la proteína y la energía son los principales elementos que limitan la producción de los sistemas agropecuarios basados en pastos (Ku-Vera et al., 1999).
Es de primordial importancia mantener la productividad de los sistemas incorporando estrategias tendientes a mejorar la fertilidad del suelo por medio de un reciclaje más eficiente de nutrimentos entre los diferentes componentes de un sistema agroforestal. Estos sistemas deben tener el potencial de aportar nutrimentos a los sistemas de producción animal principalmente de proteína, ya que es considerada el factor principal que limita la producción animal en los trópicos (Nair et al., 1999).
Las especies arbustivas juegan un papel principal en la fertilidad de los suelos ya que incorporan N atmosférico y reciclan cantidades significativas de nutrientes por medio de producción e incorporación de hojarasca al suelo (Chikowo et al., 2006; Sileshi y Mafongoya, 2007). Estudios recientes indican que las arbustivas cuando crecen en callejones producen aproximadamente 20 t por ha de MS al año conteniendo alrededor de 358 kg de N, 28 kg de P, 232 kg de K y 144 kg de Ca (Palm, 1995). Las leguminosas como Leucaena (Huaxim) tienen la capacidad de liberar en un tiempo corto más de 50% del contenido total de nutrimentos como el N, K y el P cuando se incorporan al suelo (Bossa et al., 2005), lo cual indica la excelente calidad de la biomasa que estas especies poseen para ser utilizadas en la recuperación de suelos degradados.
El asociar especies podría ser una estrategia para mejorar la fertilidad del suelo como es el caso de asociar especies fijadoras de N atmosférico con especies no leguminosas. Datos exploratorios indican que la mezcla del follaje de especies de diferente calidad con una relación C/N baja puede favorecer la descomposición el follaje de otra especie de baja calidad, adicionalmente fomentan significativamente el reciclaje de nutrimentos en sistemas de cultivos asociados (Forrester et al., 2005).
Sin embargo, la dinámica de estos nutrimentos y como pueden estar relacionadas estas diferencias en tasa descomposición son aún desconocidas, ya que los estudios de producción y descomposición de materia orgánica (MO) de especies que crecen asociadas han sido escasos (Forrester et al., 2006) pero con resultados muy prometedores en cuanto a la calidad y pertinencia en la liberación e incorporación de nutrimentos al suelo ya que ocurre un mayor flujo de estos nutrimentos causados principalmente por las diferencias en la composición química y propiedades físicas de la hojarasca originadas por las distintas especies (Hättenschwiler et al., 2005).
Adicionalmente, las arbustivas como L. leucocephala y Moringa oleífera son especies de rápido crecimiento que producen considerables cantidades de biomasa de excelente calidad para alimentar animales (rumiantes y monogástricos), principalmente como suplemento de pasturas en la época de escasez (Gutteridge y Shelton, 1994). Leucaena puede fijar aproximadamente 70% del

110 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
Judith Petit Aldana et al.
N que necesita (Van Kessel et al., 1994). Si se considera de que en sistemas asociados se incrementa los niveles de N por consiguiente también se incrementa el área foliar y la captura de luz y fotosíntesis y por tanto se incrementa la calidad y producción total del sistema (Smethurst et al., 2003). Estudios agroforestales han mostrado que una gran proporción del N fijado puede ser transferido a las plantas no leguminosas asociadas (Crews y Peoples, 2005); sin embargo, los procesos que explican dichas interacciones entre especies arbóreas cuando estas se encuentran asociadas no han sido dilucidados del todo (Moyer-Henry et al., 2006).
En el sistema suelo-planta, los nutrimentos de la planta están en un estado de continua transferencia dinámica. Las plantas toman los alimentos del suelo y los utilizan para los procesos metabólicos. Algunas de las partes de la planta tales como hojas y raíces muertas vuelven al suelo durante el crecimiento vegetal, y dependiendo del tipo de utilización del suelo y de la naturaleza de las plantas, las partes de la planta son adicionadas al suelo cuando son cosechadas (Nair, 1993).
La hojarasca o biomasa agregada se descompone a través de los microorganismos del suelo, y los nutrimentos que han sido confinados en las partes de las plantas son liberados al suelo donde llegan a estar una vez más disponibles para ser tomados de nuevo por las plantas. En sentido limitado, el ciclo de nutrientes se refiere a esta continua transferencia de nutrimentos del suelo a la planta y de vuelta al suelo. En un sentido amplio el ciclo de nutrientes implica la continua transferencia de nutrimentos dentro de los diferentes componentes del ecosistema e incluye procesos como: la meteorización de los minerales, actividades de la biota del suelo, y otras transformaciones que ocurren en la biosfera, atmósfera, litosfera e hidrosfera (Jordan, 1985).
Los sistemas agroforestales y otros sistemas basados en árboles son reconocidos como más eficientes en el ciclo de nutrientes que muchos otros sistemas, porque la presencia del componente leñoso sugiere efectos beneficiosos sobre el suelo. Estas leñosas perennes tienen, teóricamente, sistemas radiculares más extensos y profundos que las plantas herbáceas,
puesto que tienen un potencial para capturar y reciclar una gran cantidad de nutrimentos. La contribución de la hojarasca a la superficie del suelo es probablemente mayor que aquella de las plantas herbáceas (Nair, 1993).
Contribución de los sistemas agroforestales al ciclo de nutrimentos
Una representación esquemática del patrón general del ciclo de nutrientes en un sistema agroforestal en comparación con un sistema agrícola y un sistema forestal se muestra en la Figura 1. Se puede observar que los ciclos del N, P, K y otros elementos, varían considerablemente, y deben ser considerados en forma individual. No obstante, todos ellos tienen algunas características comunes tal como se indica en el modelo. El ciclo consiste de entradas (ganancias), salidas (pérdidas) y transferencia y retorno dentro del sistema. Las rutas de estas ganancias, pérdidas y transferencias también son similares: las entradas a través de fertilizantes, precipitación, polvo, materiales orgánicos fuera del sistema, y fijación de N, así como meteorización de las rocas (por otros elementos); las principales salidas se derivan de la erosión, percolación, y de la cosecha de los cultivos (para todos los nutrimentos), desnitrificación y volatización (por N) y combustión (por N y S).
Los ecosistemas forestales representan ciclo de nutrientes en sistemas cerrados y eficientes, esto significa que estos sistemas tienen altas tasas de retorno y bajas tasas de pérdidas (así como también de ganancias), en otras palabras, son autosostenibles. Por otra parte, los sistemas agrícolas a menudo son abiertos o permeables, lo que significa que el retorno dentro del sistema es relativamente bajo y las pérdidas así como las ganancias son comparativamente altas. El ciclo de nutrientes en sistemas agroforestales está entre estos dos extremos, más nutrimentos en el sistema son reusados por las plantas antes de que se pierdan del mismo. La principal diferencia entre sistemas agroforestales y otros usos de la tierra yace en la transferencia o retorno de los nutrimentos dentro del sistema de un componente a otro, y de la posibilidad del manejo del sistema o sus componentes para facilitar el aumento de las tasas de retorno sin afectar la productividad total del mismo (Nair,1993).

Asociación de especies arbóreas forrajeras para mejorar la productividad y el reciclaje de nutrimentos 111
Mejoramiento de la eficiencia del ciclo de nutrientes a través del manejo
Los sistemas agroforestales proporcionan una oportunidad para modificar el ciclo de nutrientes a través del manejo, cuando se comparan con sistemas agrícolas, el cual resulta en el uso más eficiente de los nutrimentos del suelo, sin agregados externos (tal como los fertilizantes) o la disponibilidad a través de procesos naturales (e.g., la meteorización). Los mecanismos fundamentales que contribuyen a un eficiente ciclaje de nutrientes, así como también a otras consideraciones en sistemas agroforestales, las resume Nair (1993) a como sigue:
1) hay un potencial para mejorar la absorción de nutrimentos desde los horizontes más profundos del suelo. 2) las ganancias de la fijación simbiótica de N por los árboles pueden ser mejoradas a través de la selección de especies de árboles y las mezclas de estos. 3) los nutrimentos liberados desde la biomasa del árbol pueden ser sincronizados con los requerimientos del cultivo al controlar la calidad, cantidad, tiempo y método
de aplicación de las podas, como el abono, especialmente en cultivos en callejones.4) prácticas de manejo que conducen a mejorar el estado de materia orgánica en el suelo, inevitablemente conducirán a mejorar el ciclo de nutrientes y la productividad del suelo.5) otra consideración principal de manejo en agroforestería es la posibilidad de reducir la pérdida de nutrimentos a través de métodos de conservación de suelos.
El papel de las plantaciones mixtas
Las plantaciones mixtas son plantaciones con dos o más especies, que pueden estar constituidas tanto por varias especies principales como por una o más especie(s) principal(es) asociadas a una o más especie(s) secundaria(s), sean éstas arbóreas o arbustivas.
Las plantaciones mixtas tienen el potencial de aumentar la producción de biomasa y el secuestro de carbono (Kaye et al., 2000; Resh et al., 2002; Binkley et al., 2003; Bauhus et al., 2004), así como otros beneficios, entre ellos la mejora de la fertilidad del suelo y el ciclaje de nutrientes (Binkley
Figura 1. Representación esquemática de las relaciones de nutrimentos y ventajas de un sistema agroforestal “ideal”en comparación con sistemas forestales y agrícolas. Fuente: Nair (1993).
Ecosistema Forestal (común)
Superficiedel suelo
Pérdida pequeña del sistema (producción)
Pequeña erosión por escorrentía
Granexportación de nutrientes
Gra
n ex
port
ació
n co
mpe
nsad
a po
r la
circ
ulac
ión
dent
ro d
elsi
stem
a y
el u
so e
ficie
nte
Gran volumen de canopia
Adi
ción
por
cal
idad
de h
ojar
asca
Microclima favorable Caída de hojarasca
Asociacióncon micorrizas
Extracción porlas raíces más profundas
Lixiviaciónligera
Liberación debidoa la descomposición de raíces
--
---
Sistema Agrícola (común)
Poco o nulo reciclaje por las plantasPoco aporte de hojarascaPoca lixiviación de follaje
Nutrientes en lalluvia no captada
LL
UV
IA
Acumulación superficial de raíces
Fuerte lixiviación, especialmente durante la época sin cultivo
Liberación por intemperización
Pérdida considerable del sistema (producción)
Aportación de fertilizantes
Erosión severa por escorrentía
Árboles de forma y sistemas radicales deseables y con manejo apropiado
LL
UV
IAA
port
ació
n de
mat
eria
or
gáni
ca: p
odas
Extracción de las capas más profundasdel suelo
Liberación por intemperización
Pérdida pequeña del sistema (producción)
--
Uso complementariode nutrientes
---
Poca erosión
Ecosistema Agroforestal (ideal)
Liberación por la descomposición de raíces

112 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
Judith Petit Aldana et al.
et al., 2000; Montagnini, 2000), la protección contra plagas y enfermedades, y la conservación de agua (FAO, 1992; Montagnini, 2000). Además, pueden funcionar como un sistema silvicultural para la producción de madera de alto valor y una gama más amplia de productos (Keenan et al., 1995; Montagnini et al., 1995; De Bell et al., 1997). Por lo tanto, las plantaciones mixtas, puede ser utilizada para satisfacer una amplia gama de objetivos económicos, silvicultura, y sostenibilidad. El éxito de una plantación mixta dependerá de los objetivos específicos del rodal. Por otra parte es difícil predecir qué combinaciones de especies conducirán a los aumentos en productividad en las plantaciones mixtas cuando no existe ninguna información. Esto depende no sólo de las cualidades de la especie, sino también de factores del sitio tales como disponibilidad del agua y de alimento (Forrester et al ., 2005).
La mayoría de los suelos pobres en el mundo son deficitarios en N y en P (Hussein Zahran, 1999). La fijación biológica de N (FBN) puede ser utilizada como una herramienta de mejora en esos casos. Las hojas de los árboles fijadores de N (AFN) se suelen usar para mejorar la fertilidad de suelos pobres ya sea en forma de mantillo o de abono verde; esta hojarasca suele ser de rápida descomposición aunque existen grandes diferencias entre géneros (Binkley y Giardina, 1997).
Cuando se utilizan AFN en sistemas agroforestales, la liberación de nutrientes desde la hojarasca debe estar en sincronía con la demanda del cultivo para obtener el beneficio buscado (Powell, 1995). En la rehabilitación de suelos agotados por actividades agrícola-ganaderas intensivas se han utilizado pasturas y cultivos perennes o anuales (Francis et al., 1999) y también la reforestación con AFN como Acacia mangium, Alnus spp., Casuarina spp., L. leucocephala, Mimosa scabrella y Paraserianthe falcataria (Halenda, 1990; Halenda y Ting, 1993; Powell, 1995). El mayor crecimiento de los AFN respecto de los no fijadores puede generar en aquellos una mayor demanda de nutrientes del suelo, especialmente de N, P y Mg (Binkley y Giardina, 1997), situación que puede limitar su crecimiento.
En suelos infértiles de Malasia, abandonados por agricultura migratoria, Acacia mangium creció más que L. leucocephala y mostró una buena producción de hojarasca que contribuyó a estabilizar el sitio contra la erosión (Halenda, 1988). Por el contrario, experimentos realizados en suelos de India empobrecidos por décadas de deforestación y mal uso
de la tierra, permitieron concluir que no hay suficiente evidencia de que los árboles fijadores de N crezcan mejor que los no fijadores (Puri y Naugraiya, 1998, citado por Ferrari y Wall, 2004), aunque no se verificó si los árboles tenían nódulos o si estaban fijando N.
En suelos deficientes en N, la selección de genotipos puede ser importante dada la diversidad genética observada en algunas especies en cuanto a la FBN; en un experimento con Casuarina spp. las plantas de distintos orígenes inoculadas con Frankia tuvieron crecimiento más variable que las plantas bien fertilizadas con N mineral (Sanginga et al., 1990).
Los AFN, pueden estimular el crecimiento de la vegetación vecina debido a la descomposición de sus hojas, a la exudación radicular de N fijado y a la intervención de las micorrizas (Binkley y Giardina, 1997).
En Puerto Rico, se realizó un estudio en plantaciones mixtas, entre Casuarina equisetifolia, L. leucocephala y Eucalyptus robusta. La mezcla Casuarina-Leucaena tuvo la mayor biomasa tras cuatro años de crecimiento mientras que la biomasa de Eucalyptus en la mezcla con Leucaena fue menor que en el monocultivo (Parrotta et al., 1994), indicando que el beneficio de una mejor nutrición de N puede ser mitigado por competencia por otros nutrientes del suelo (Binkley y Giardina, 1997).
Los suelos de Hawai presentan limitaciones en N para el crecimiento de Eucalyptus, por lo cual se ensayan plantaciones mixtas con AFN. En la costa húmeda de Hamakua en Hawai (4 600 mm anuales de lluvia), Eucalyptus saligna creció más en cinco años en mezclas 50:50 con Acacia melanoxylon (acacia australiana) o con Paraserianthes falcataria que en la plantación pura. En otro experimento, la mezcla Eucalyptus-Paraserianthes (50:50) sin fertilización durante seis meses creció más que una plantación pura de Eucalyptus fertilizada cada 6 meses con N durante los tres primeros años. En cambio en la costa sudeste más seca (1 700 mm anuales) la mezcla no funcionó tan bien. Se concluye que en la costa húmeda la necesidad de fertilizar Eucalyptus con N después del primer año pudo ser reemplazada por plantaciones mixtas 50:50 con Paraserianthes falcataria (Schubert et al., 1988; citado por Ferrari y Wall, 2004).
Otros experimentos realizados en Hawai, referidos por Binkley y Giardina (1997), demostraron que la productividad de las plantaciones mixtas dependería

Asociación de especies arbóreas forrajeras para mejorar la productividad y el reciclaje de nutrimentos 113
de complejas interacciones entre el suministro de recursos y la eficiencia de uso de los mismos, así como de características propias de cada sitio.
Los tres tipos más importantes de interacciones que pueden presentarse en plantaciones mixtas son: la competencia, reducción de la competencia, y facilitación (Vandermeer, 1989; Kelty, 1992). La competencia se produce cuando las plantas interactúan y al menos una especie ejerce un efecto negativo sobre la otra (Vandermeer, 1989). La reducción de la competencia se produce cuando la competencia ínterespecífica por la limitación de recursos en la asociación es más débil que la competencia intra-específica en los monocultivos (Kelty y Cameron, 1995). Esto ocurre a menudo, cuando hay una repartición de los recursos, ya sea superficial (luz) o subterráneo (agua o nutrimentos) (Kelty y Cameron, 1995; Casanova et al., 2007).
La facilidad se produce cuando una especie tiene un efecto positivo en la otra especie (Vandermeer, 1989), por ejemplo, cuando las especies que fijan el N aumentan el crecimiento de otra especie. Las plantaciones de especies mixtas serán más product ivas que los monocultivos, en el caso de que la facili tación y reducción de la interacción por competencia superen las interacciones competitivas.
Uno de los objetivos principales de la plantación de especies fijadoras de N con árboles como Eucalyptus spp, es aumentar la disponibilidad de N a través de la fijación simbiótica y la aceleración de las tasas del ciclaje de nutrientes. En un ensayo de plantaciones mixtas de Eucalyptus globulus y Acacia mearnsii, el aumento en la disponibilidad de nutrientes surgió a los 25 meses, cuando los incrementos en las concentraciones de N en el follaje senescente de E. globulus se produjo en las plantaciones mezcladas en comparación con las plantaciones puras (Khanna, 1997).
Existen varios mecanismos mediante los cuales la disponibilidad de N se haya incrementado para E. globulus en asociación con A. mearnsii: 1) las especies que no fijan N pueden utilizarlo después que la planta y los tejidos microbianos mueren y se descomponen expulsando el N para así completar su ciclo en el ecosistema (Van Kessel et al., 1994; May y Attiwill, 2003); 2) también el N puede ser transferido entre las plantas vía exudación de la raíz o por las conexiones de las
micorrizas entre los sistemas radiculares, cuando ambas especies forman simbiosis con las mismas micorrizas. Eucalyptus spp. y Acacia spp. pueden formar simbiosis con los hongos de la micorriza arbuscular (Bellei et al., 1992; Adjoud-Sadadou y Halli-Hargas, 2000).
Sin embargo, no hay evidencias que este proceso ocurra en mezclas de eucaliptos y de acacias; 3) porque las plantas fijadoras de N, a menudo dependen mucho del N fijado, el cual puede estar entre 10% hasta aproximadamente 100% del usado por la planta fijadora (Binkley y Giardina, 1997; Khanna, 1998; Fisher y Binkley, 2000; May y Attiwill, 2003) y así, más N puede estar disponible en el suelo para las plantas antes que N fijado sea ciclado y transferidos a los árboles de Eucalyptus globulus.
Las tasas de fijación de N son muy variables y en general son causadas por los mismos factores que afectan el crecimiento de las plantas (Danso et al., 1992; Binkley y Giardina, 1997; Fisher y Binkley, 2000). Las altas tasas de fijación de N (>50 kg ha-1 año-1) se han estimado para muchas especies, por ejemplo, L. leucocephala y Facaltaria molucanna (Binkley, 1992; Binkley y Giardina, 1997; Khanna, 1998; Fisher y Binkley, 2000; May y Attiwill, 2003). Sin embargo, algunas especies fijadoras de N como A. melanoxylon parecen tener bajas tasas de fijación de N (<10 kg ha-1 año-1), y pueden no ser capaces de contribuir de manera significativa a la nutrición de las especies no fijadoras de N. La capacidad de facilitación de estas especies dependerá de sus habilidades para realizar el ciclo de nutrientes.
Además de los aumentos en la disponibilidad de N a través de la fijación simbiótica, mediciones en hojarasca mostraron que las tasas de ciclaje de N y P fueron significativamente mayores en las parcelas que contienen sólo 25% A. mearnsii, en comparación con E. globulus en plantaciones puras (Forrester et al., 2004). Las parcelas que contienen sólo A. mearnsii también tuvieron altas tasas de descomposición de la hojarasca en comparación con las plantaciones puras de E. globulus. Incrementos similares en el ciclo de nutrientes o en la disponibilidad de N y P de especies fijadoras de N han sido observados en otros estudios, tanto en plantaciones y bosques nativos, incluyendo algunas comunidades con mezcla de especies que contienen Eucalyptus (Binkley et al., 2000; May y Attiwill, 2003). Es importante señalar que los aumentos en el ciclo del N y en la disponibilidad de plantas fijadoras de N, pueden influir en la disponibilidad de otros nutrientes (Binkley et al., 2000; Kaye et al., 2000).

114 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
Judith Petit Aldana et al.
El efecto de las plantas fijadoras de N sobre el ciclo de nutrientes en plantaciones mixtas depende de la cantidad de N (y de otros nutrimentos) en la hojarasca y de la velocidad a la que estos nutrimentos son liberados a través de la descomposición. Para aumentar efectivamente las tasas del ciclo de nutrientes en plantaciones mixtas, las especies fijadoras de N deben producir grandes cantidades de hojarasca fácilmente descomponible. Incluso especies con bajas tasas de fijación de N (<10 kg-1 ha-1 año-1), pero con alto contenido de nutrientes foliares pueden mejorar el ciclo de nutrientes en rodales mixtos. En plantaciones mixtas de 11 años de edad, en mezclas 1:1 de E. globulus y A. mearnsii, cayeron 44 kg de N ha-1año-1 de hojas en el mantillo en comparación con plantaciones puras E. globulus que aportaron 14 kg de N ha-1 año-1 y las tasas de descomposición fueron del 0.56 y 0.32 año-1, respectivamente (Forrester et al., 2004).
Pocos estudios han examinado los mecanismos responsables del crecimiento resultante de una alta disponibilidad de N en rodales mixtos. Esa alta disponibilidad puede acrecentar el crecimiento por el aumento y la eficiencia del área foliar (Cromer y Eldridge, 2001).
CONCLUSIONES
Esta revisión, evidencia que la asociación de especies arbóreas forrajeras puede contribuir a la productividad y sostenibilidad de los agroecosistemas.
Es de resaltar que las especies arbóreas poseen elementos de alto valor nutricional que ayudan a reducir problemas de fertilidad en los suelos; incorporando hojarasca de buena calidad para un eficiente reciclaje de nutrimentos y de esta manera aumentar la posibilidad de mejorar la productividad de la especie(s) asociada(s).
Del mismo modo, las leguminosas arbustivas pueden establecer simbiosis con bacterias y hongos, que mejoran la disponibilidad de elementos limitantes para su crecimiento.
La asociación de especies arbóreas, puede utilizarse para satisfacer una amplia gama de objetivos productivos, ecológicos y económicos. Sin embargo, esto no sólo depende de las características de las especies, sino también en
gran medida de los recursos disponibles del sitio, tales como; luz, agua y nutrimentos del suelo, así como también del manejo del agroecosistema.
Es importante señalar que existen vacíos de investigación sobre este tema en la literatura, específicamente con especies arbóreas forrajeras, puesto que la mayoría de trabajos se han realizado para especies arbóreas maderables en plantaciones forestales.
LITERATURA CITADA
Adjoud-Sadadou, D. and Halli-Hargas, R. 2000. Occurrence of arbuscular mycorrhiza on aged Eucalyptus. Mycorrhiza 9:287-290.
Bauhus, J.; van Winden, A. P. and Nicotra, A. B. 2004. Above-ground interactions and productivity in mixed-species plantations of Acacia mearnsii and Eucalyptus globulus. Can. J. Forest. Res. 34:686-694.
Bellei, M. M.; Garbaye, J. and Gil, M. 1992. Mycorrhizal succession in young Eucalyptus viminalis plantations in Santa Catarina (southern Brasil). Forest. Ecol. Managem. 54:205-213.
Binkley, D. 1992. Mixtures of nitrogen-fixing and non-nitrogen-fixing tree species. In: Cannell, M.G.R., Malcolm, D. C., Robertson, P. A. (Eds.). The ecology of mixed species stands of trees. Blackwell Scientific, London, p. 99-123.
Binkley, D. and Giardina, C. 1997. Nitrogen fixation in tropical forest plantations. In: Nambiar, E. K. S., Brown, A. G. (Eds.), Management of Soil, Nutrients and Water in Tropical Plantation Forests. Australian Centre for International Agricultural Research, Canberra. p. 297-337.
Binkley, D.; Giardina, C. and Bashkin, M. A. 2000. Soil phosphorous pools and supply under the influence of Eucalyptus saligna and nitrogen-fixing Albizia facaltaria. Forest. Ecology and Management. 128:241-247.
Binkley, D.; Senock, R.; Bird, S. and Cole, T. G. 2003. Twenty years of stand development in pure and mixed stands of Eucalyptus saligna and N-fixing Facaltaria moluccana. Forest Ecol. Managem. 182:93-102.
Bossa, J. R.; Adams, J. F.; Shannon, D. A. and Mullins, G. L. 2005. Phosphorus and potassium release pattern from leucaena leaves in three environments of Haiti. Nutr. Cycl. Agroecosys. 73:25-35.

Asociación de especies arbóreas forrajeras para mejorar la productividad y el reciclaje de nutrimentos 115
Casanova, L. F.; Ramírez, A. L. and Solorio, S. F. J. 2007. Interacciones radiculares en sistemas agroforestales: mecanismos y opciones de manejo. Avances en Investigación Agropecuaria. 11(3):41-52.
Chicowo, R.; Mapfumo, P.; Leffelaar, P. A. and Giller, K. E. 2006. Integrating legumes to improve N cycling on smallholder farms in sub-humid Zimbabwe: resource quality, biophysical and environmental limitations. Nutr. Cycl. Agroecosys. 76:219-231.
Craswell, E. T.; Grote, U.; Henao, J. and Vlek, P. L. G. 2004. Nutrient flows in agricultural production and international trade: ecological and policy issues. Discussion papers on development policy, Center for Development Research (ZEF) Bonn, Germany.
Crews, T. E. and Peoples, M. B. 2005. Can the synchrony of nitrogen supply and crop demand be improved in legume and fertilizer-based agroecosystems? A review. Nutr. Cycl. Agroecosys. 72:101-120.
Danso, S. K. A.; Bowen, G. D. and Sanginga, N. 1992. Biological nitrogen-fixation in trees in agroecosystems. Plant. Soil. 141:177-196.
De Bell, D. S.; Cole, T. C. and Whitesell, C. D. 1997. Growth, development, and yield of pure and mixed stands of Eucalyptus and Albizia. For. Sci. 43:286-298.
Ferrari, A. E. y Wall, L. G. 2004. Utilización de árboles fijadores de nitrógeno para la revegetación de suelos degradados. Rev. Fac. Agron. 105 (2):63-87.
Fisher, R. F. and Binkley, D. 2000. Ecology and management of forest soils. Wiley, New York.
Food and Agriculture Organization of the United Nations (FAO). 1992. Mixed and pure forest plantations in the tropics and subtropics. FAO. Forestry paper 103 (based on the work of T. J. Wormald). FAO of the UN, Rome, Italy.
Forrester, D. I.; Bauhus, J. and Cowie, A. L. 2005. On the success and failure of mixed-species tree plantations: lessons learned from a model system of Eucalyptus globules and Acacia mearnsii. For. Ecol. Managem. 209:147-155.
Forrester, D. I.; Bauhus, J.; Cowie, A. L. and Vanclay, J. 2006. Mixed-species plantations: of Eucalyptus globules and Acacia mearnsii with nitrogen-fixing trees: a review. For. Ecol. Managem. 233:211-230.
Francis, G. S.; Tabley, F. J. and White, K. M. 1999. Restorative crops for the amelioration of degraded soil conditions in New Zealand. Aust. J. Soil Res. 37:1017-1034.
Gutteridge, R. C. and Shelton, H. M. 1994. Forage tree legumes in tropical agriculture. CAB International, Wallingford, UK.
Halenda, C. 1988. Performance of Acacia mangium Willd. and Leucaena leucocephala (Lam.) de Wit. at Niah forest reserve, Sarawak. Nitrogen Fixing Tree Research Reports 6:15-17.
Halenda, C. J. 1990. Growth rates of three leguminous tree species on degraded acidic soils. Nitrogen Fixing Tree Research Reports 8:40.
Halenda, C. J. and Ting, S. P. 1993. Performance of three legume tree species on degraded acidic soils. Nitrogen Fixing Tree Research Reports 11:29.
Hättenschwiler, S.; Tiunov, A. V. and Scheu, S. 2005. Biodiversity and li t ter decomposition in terrestrial ecosystems. Annu. Rev. Ecol. Evol. Syst. 36:191-218.
Hussein-Zahran, H. 1999. Rhizobium-legume symbiosis and nitrogen fixation under severe conditions and in an arid climate. Microbiol. Molec. Biol. 63:968-989.
Jordan, C. F. 1985. Nutrient cycling in tropical forest ecosystems. John Wiley & Sons. Chichester. 189 p.
Kaye, J. P.; Resh, S. C.; Kaye, M. W. and Chimmer, R. A. 2000. Nutrient and carbon dynamics in a replacement series of Eucalyptus and Albizia trees. Ecology. 81:3267-3273.
Keenan, R.; Lamb, D. and Sexton, G. 1995. Experience with mixed species rainforest plantations in North Queensland. Commun. For. Rev. 74:315-321.
Kelty, M. J. 1992. Comparative productivity of monocultures and mixed-species stands. In: Kelty, M. J., Larson, B. C., Oliver, C. D. (Eds.). The ecology and silviculture of mixed-species forests. Kluwer Academic Publishers, Dordrecht. p. 125-141.
Kelty, M. J. and Cameron, I. R. 1995. Plot designs for the analysis of species interactions in mixed stands. Commun. For. Rev. 74:322-332.
Khanna, P. K. 1997. Comparison of growth and nutrition of young monocultures and mixed stands of Eucalyptus globulus and Acacia mearnsii. For. Ecol. Managem. 94:105-113.
Khanna, P. K. 1998. Nutrient cycling under mixed-species tree systems in Southeast Asia. Agrofores. Systems. 38:99-120.

116 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
Judith Petit Aldana et al.
Parrota, J. A.; Baker, D. D. and Fried, M. 1994. Application of 15 N-enrichment methodologies to estimate nitrogen fixation in Casuarina equisetifolia. Canadian J. Forest. Research. 24:201-207.
Petit, A. J. 1994. Árboles y arbustos forrajeros. Instituto Forestal Latinoamericano. Mérida, Venezuela 174 p.
Powell, J. M.; Pearson, A. R. and Hiernaux, P. H. 2004. Crop-Livestock Interactions in the West African Drylands. Review and Interpretation. Agron. J. 96:469-483.
Powell, M. H. 1995. Nitrogen fixing trees and shrubs for acid soils-An overview. In: Nitrogen fixing trees for fodder production. Daniel J. N. & J. M. Roshetko, Ed. Winrock International, Morrilton, p.73-81.
Resh, S. C.; Binkley, D. Parrotta, J. A. 2002. Greater soil carbon sequestration under nitrogen-fixing trees compared with Eucalyptus species. Ecosystems 5:217-231.
Sanginga, N.; Danso, S. K. A. and Bower, G. D. 1989. Nodulation and growth response of Allocasuarina and Casuarina species to phosphorus fertiliaztion. Plant. Soil. 118:125-132.
Solorio, S. F. J and Solorio, S. B. 2002. Integrating fodder trees in to animal production systems in the tropics. Tropical & Subtropical Agroeosystems. 1:1-11.
Sileshi, G. and Mafongoya, P. L. 2007. Quantity and quality of organic inputs from coppicing leguminous trees influence abundance of soil macrofauna in maize crops in eastern Zambia. Biology Fertility Soils. 43:333-340.
Smethurst, P.; Baillie, C.; Cherry, M. and Holz, G. 2003. Fertilizer effects on LAI and growth of four Eucalyptus nitens plantations. For. Ecol. Managem.176:531-542.
Van Kessel, C.; Farrel, R. E.; Roskoski, J. P. and Keane, K. M. 1994. Recycling of the naturally occurring 15N in an established stand of Leucaena leucocephala. Soil Biology Biochemistry. 26:757-762.
Vandermeer, J. 1989. The ecology of intercropping. Cambridge University Press, New York.
Ku-Vera, J. C.; Ramírez, A. L.; Jiménez, F. G.; Alayon, J. A. and Ramírez, C. L. 1999. Árboles y arbustos para la producción animal en el trópico. In: Sánchez, M. D. y Rosales, M. M. (eds). Agroforestería para la producción animal en América Latina. Roma, Italia. p. 231-258.
May, B. M. and Attiwill, P. M. 2003. Nitrogen-fixation by Acacia dealbata and changes in soil properties 5 years after mechanical disturbance or slash-burning following timber harvest. For. Ecol. Managem. 181:339-355.
Montagnini, F. 2000. Accumulation in above-ground biomass and soil storage of mineral nutrients in pure and mixed plantations in a humid tropical lowland. For. Ecol. Managem. 134:257-270.
Montagnini, F.; Gonzales, E. and Porras, C. 1995. Mixed and pure forest plantations in the humid neotropics: a comparison of early growth, pest damage and establishment costs. Commun. For. Rev. 74:306-314.
Moyer-Henry, K. A.; Burton, J. W.; Israel, D. W. and Rufty, T. W. 2006. Nitrogen transfer between plants: A 15N natural abundance study with crop and weed species. Plant. Soil. 282:7-20.
Nair, P. K. R.; Buresh, D. N.; Mugendi, D. N. and Latt, C. R. 1999. Nutrient Cycling in tropical agroforestry systems: Myths and science. In: Buck L. E., Lassoie J. P. and Fernandez E. C. M. (ed) Agroforesty in sustainable agricultural systems. CRC Press, Lewis Publ., Boca Raton, FL.
Nair, P. K. R. 1993. An introduction to agroforestry. Kluwer Academic Publisher. Dordrecht, The Netherlands. 499 p.
National Academic of Sciences. 1979. Tropical legumes: resources for the future . NAS Washington D.C. 331 p.
Palm, C. A. 1995. Contribution of agroforestry trees to nutrient requirements of intercropped plants. Agrofores. Systems. 30:105-124.

Agricultura Técnica en México Vol. 35 Núm.1 1 de enero-31 de marzo 2009 p. 117-123
PSILOIDEOS Y CICADÉLIDOS EN EL CULTIVO DE LA PAPA EN EL BAJÍO, GUANAJUATO, MÉXICO*
PSYLLOIDS AND LEAFHOPPERS ON THE POTATO CROP IN EL BAJIO, GUANAJUATO, MEXICO
Antonio Marín Jarillo1§, Rafael Bujanos Muñiz1 y Felipe Delgadillo Sánchez1
1Entomología, Campo Experimental Bajío, INIFAP, km 6.5 carretera Celaya-San Miguel de Allende, Celaya, Guanajuato C. P. 38010 Celaya, Guanajuato. México. Tel. 01 461 6115323. §Autor para correspondencia: [email protected], [email protected] y [email protected].
El objetivo de este estudio fue identificar los psiloideos y chicharritas presentes en una localidad de la región productora de papa del estado de Guanajuato, México. La investigación se realizó durante dos ciclos agrícolas del cultivo en 2003, se utilizaron charolas amarillas (trampas) con agua. Los géneros y especies de psílidos colectados fueron: Bactericera cockerelli (Sulcer.), Paracarsidara gigantea (Crawford), Paratrioza maculipenis (Crawford), Heteropsilla texana (Crawford), y Rhinopsylla prob antennata (Crawford). Los géneros y especies de chicharritas colectadas fueron: Carneocephala sp., Empoasca kraemeri Ross & Moore, E. mexara Ross & Moore y E. solana DeLong, Elymana sp., Oncopsis sp., Phera centrolineata (Signoret), Scaphytopius sp., Agallia barretti Ball, Paraphlepsius sp., Agalliopsis sp., Aceratagallia sp. y Draeculacephala minerva Ball.
Palabras clave: chicharritas, papa, psiloideos.
ABSTRACT
The aim of this study was to identify the psylloids and leafhoppers present at one locality in the potato production region of the state of Guanajuato, Mexico. The work was
carried out during two crop cycles in 2003, using yellow traps with water. The genera and species of psylloids collected were: Bactericera cockerelli (Sulcer.), Paracarsidara gigantea (Crawford), Paratrioza maculipenis Crawford, Heteropsilla texana Crawford, and Rhinopsylla prob antennata (Crawford).The genera and species of leafhoppers collected were: Carneocephala sp., Empoasca kraemeri Ross & Moore, E. mexara Ross & Moore and E. solana DeLong., Elymana sp., Oncopsis sp., Phera centrolineata (Signoret), Scaphytopius sp., Agallia barretti Ball, Paraphlepsius sp., Agalliopsis sp., Aceratagallia sp. and Draeculacephala minerva Ball.
Key words: leafhoppers, potato, psylloids.
En México el pulgón saltador del tomate/papa Bactericera (=Paratrioza) cockerelli (Sulc.) transmite el fitoplasma causante del “permanente del tomate” (Garzón et al., 2005) y con base en los avances reportados por el mismo autor, se infiere que el síntoma “punta morada” de la papa es causado por un fitoplasma, el cual es transmitido por B. cockerelli (Garzón, 2002). En los de Estados Unidos de América, México y Centro América fue detectado un defecto en las papas al momento de freírlas al que se le conoce como “zebra
* Recibido: Enero, 2007
Aceptado: Febrero, 2009
RESUMEN

118 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
Antonio Marín Jarillo et al.
chips” (ZC) cuyos síntomas foliares se asemejan a los de la enfermedad conocida como “punta morada de la papa” o “enfermedad del amarillamiento por psílidos”. A la fecha, el agente(s) causal(s) y el vector(s) de ZC son desconocidos; sin embargo, una inspección de los insectos asociados al cultivo de papa en el estado de Texas, EUA y en los estados productores de México indicaron que el psílido de la papa Bactericera cockerelli (Sulc.), fue el insecto más común y abundante en todos los campos afectados con los síntomas de “zebra chips” por lo que se concluyó que existe una fuerte asociación entre el pulgón saltador de la papa (B. cockerelli) y la enfermedad “zebra chips” (Munyaneza et al., 2007 y Munyaneza y Crosslin, 2008).
Poblaciones de formas adultas e inmaduras (huevecillos y ninfas) de esta plaga son abundantes en cultivos de papa, jitomate, tomate de cáscara y chile, además de otras solanáceas silvestres, no así las poblaciones de chicharritas, las cuales son bajas y sólo se encuentran sus formas adultas ya que debido a la constante aplicación de plaguicidas y a los hábitos de vida de estos insectos, la colonización en los cultivos antes mencionados es casi imposible, principalmente en el cultivo de la papa.
Las chicharritas son insectos comunes en diversos cultivos de importancia económica en México, varias de sus especies causan daños económicos a cultivos como frijol, maíz, betabel, jitomate, remolacha, alfalfa, aguacate, melón, tabaco, vid, algodón, henequén y arroz entre otros. Su daño directo (alimentación) es mínimo comparado con la transmisión de agentes patógenos causantes de diversas enfermedades como el virus del enrollamiento de la punta, amarillamiento del aster, punta morada y el agente virescente (BLTVA) transmitido por la chicharrita del betabel (Munyaneza, 2003). Al igual que los insectos antes mencionados, las poblaciones de formas adultas e inmaduras (huevecillos y ninfas) del pulgón saltador del tomate/papa Bactericera (=Paratrioza) cockerelli (Sulc.) son abundantes en los cultivos de papa, jitomate, tomate de cáscara y chile, además de otras solanáceas silvestres.
Los psílidos y chicharritas tienen un papel importante en la transmisión de agentes fitopatógenos y en México pocos son los trabajos sobre taxonomía, biología, comportamiento, distribución, etc. de estos grupos de importancia económica, por lo cual se estableció como objetivo del presente trabajo el conocer las especies de estos insectos que están reportadas sobre el cultivo de papa en una localidad de León, Guanajuato, México, en 2003.
Se tomó como base la encuesta realizada por la Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO), 1995, en la que se enlistan las instituciones nacionales que cuentan con colecciones entomológicas, en búsqueda de los registros de ejemplares de la familia Cicadellidae y la superfamilia Psylloidea o especialistas en la identificación de estos grupos.
En campo, los especímenes utilizados para la realización del presente estudio fueron colectados en charolas amarillas (trampas) con agua con una pizca de jabón, colocadas fuera y dentro del cultivo de papa siempre a una altura en la cual sobrepasaran el follaje de estas plantas durante dos ciclos agrícolas de 2003, en “La Escondida” localizado en el municipio de León, Guanajuato a 1 785 m y coordenadas 21o 01’ 41.8’’ longitud oeste y 101o 43.6’ 51’’ latitud norte, la localidad antes mencionada fue referenciada geográficamente con un GPS de 12 canales XL marca GARMIN.
El material colectado se almacenó en frascos que contenían solución de alcohol al 70% y en el laboratorio de entomología del Campo Experimental Bajío, se realizó la separación de las chicharritas y psílidos, así como la aclaración de genitales masculinos útiles en la identificación de las especies de estos grupos; y con ayuda de claves dicotómicas de diversos autores (Osborn, 1928; Medler, 1942; DeLong, 1948; Nielson, 1968 y Sterling, 1982) se realizó la identificación de subfamilias, géneros y especies de la familia Cicadellidae y para la identificación de ejemplares de Psylloidea se utilizaron claves de Crawford 1910a, Crawford 1910b, Crawford 1911a, Crawford 1911b, Crawford 1911c; Crawford 1914; Caldwell y Martorell, 1952 y Brown y Hodkinson, 1988. Las observaciones fueron realizadas con un microscopio estereoscópico marca ZEISS Stemi 2000-C. Los ejemplares referidos en esta investigación se encuentran depositados en la colección nacional de insectos del Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP), localizada en el Campo Experimental Bajío (Celaya, Guanajuato).
En la consulta realizada se encontraron 21 colecciones de insectos de diversas instituciones nacionales, de estas sólo dos cuentan con ejemplares de la familia Cicadellidae; la colección del Colegio de Postgraduados, Campus Tabasco y la colección nacional de insectos (CNI) del INIFAP; la primera cuenta con 530 ejemplares sin identificar y la segunda con 1 514 ejemplares identificados y montados en seco que representan a 88 géneros y 205 especies así

Psiloideos y cicadélidos en el cultivo de la papa en el Bajío, Guanajuato, México 119
como con 5 823 especímenes almacenados en alcohol al 70% representados por 27 géneros y 44 especies las cuales fueron identificadas en el Departamento de Agricultura de los Estados Unidos (USDA) por el Dr. J. P. Kramer de 1977 a 1987. En cuanto a colecciones de psiloideos sólo la CNI del INIFAP cuenta con ejemplares de la superfamilia Psylloidea, la cual contempla 1 624 ejemplares que representan a 10 géneros y 12 especies.
En campo durante el ciclo agrícola iniciado en febrero de 2003, el número máximo de chicharritas adultas colectadas fue de 11 y 9 ejemplares en plantas arvenses y en el cultivo de papa respectivamente, ambos en el mes de marzo.
En el segundo ciclo iniciado en septiembre 2003, el número máximo de chicharritas colectados en cultivo de papa fue de 20 especímenes atrapados en los primeros días de la segunda quincena de noviembre; en plantas arvenses
el número fue de 24 ejemplares colectados a finales de diciembre y principios de enero 2004; esto debido a que en esa fecha el follaje del cultivo había sido cortado y las poblaciones de chicharritas se refugiaron en las plantas arvenses.
En los dos ciclos agrícolas en que se llevó a cabo esta investigación se identificaron 11 géneros de chicharritas (Cuadro 1). Los géneros Carneocephala, Empoasca, Oncopsis, Phera, y Agallia, fueron colectados durante los dos ciclos agrícolas tanto en cultivo de papa como en plantas arvenses. Los géneros Paraphlepsius, Aceratagallia y Draeculacephala sólo se encontraron en plantas arvenses. La importancia de la presencia de estos géneros radica en que algunas especies de los mismos pueden ser transmisoras de diversas enfermedades de tipo viral y representan una amenaza permanente en el cultivo de papa.
un sólo ejemplar se capturó en plantas arvenses; en estas, la especie P. gigantea se presentó de manera constante pero en poblaciones muy bajas lo que sucedió de igual manera en el cultivo de papa (Figura 1). Tanto en el cultivo de papa como en arvenses se presentaron otras especies de psílidos las cuales se encuentran en proceso de identificación.
Género I Ciclo agrícola- meses II - V II Ciclo agrícola- meses IX - XIIPapa Arvenses Papa Arvenses
Núm. ejemplares
(%) Núm. ejemplares
(%) Núm. ejemplares
(%) Núm. ejemplares
(%)
Carneocephala 3 6 1 2 1 2 4 5Empoasca 24 51 14 28 22 37 21 27Elymana 2 4 0 0 1 2 6 7Oncopsis 3 6 19 38 2 3 13 16Phera 1 2 1 2 4 7 4 5Scaphytopius 1 2 0 0 1 2 1 1Agallia 14 29 13 26 15 26 9 11Paraphlepsius 0 0 2 4 0 0 1 1Agallopsis 0 0 0 0 12 21 20 25Aceratagallia 0 0 0 0 0 0 1 1Draeculacephala 0 0 0 0 0 0 1 1
Cuadro 1. Géneros de chicharritas identificados en dos ciclos agrícolas del cultivo de papa en “La Escondida”, León, Guanajuato. 2003.
Las especies identificadas en esta investigación fueron: Empoasca kraemeri Ross & Moore, E. mexara Ross & Moore y E. solana DeLong, Phera centrolineata (Signoret), Agallia barretti Ball, y Draeculacephala minerva Ball.
En el monitoreo de psiloideos iniciado en febrero, se detectaron las especies Bactericera cockerelli (Sulcer), Paracarsidara gigantea Crawford y otras especies de psiloideos. El número máximo de adultos de la primer especie (15) se obtuvo en las trampas colocadas dentro del cultivo de papa no obstante al uso intensivo de plaguicidas y
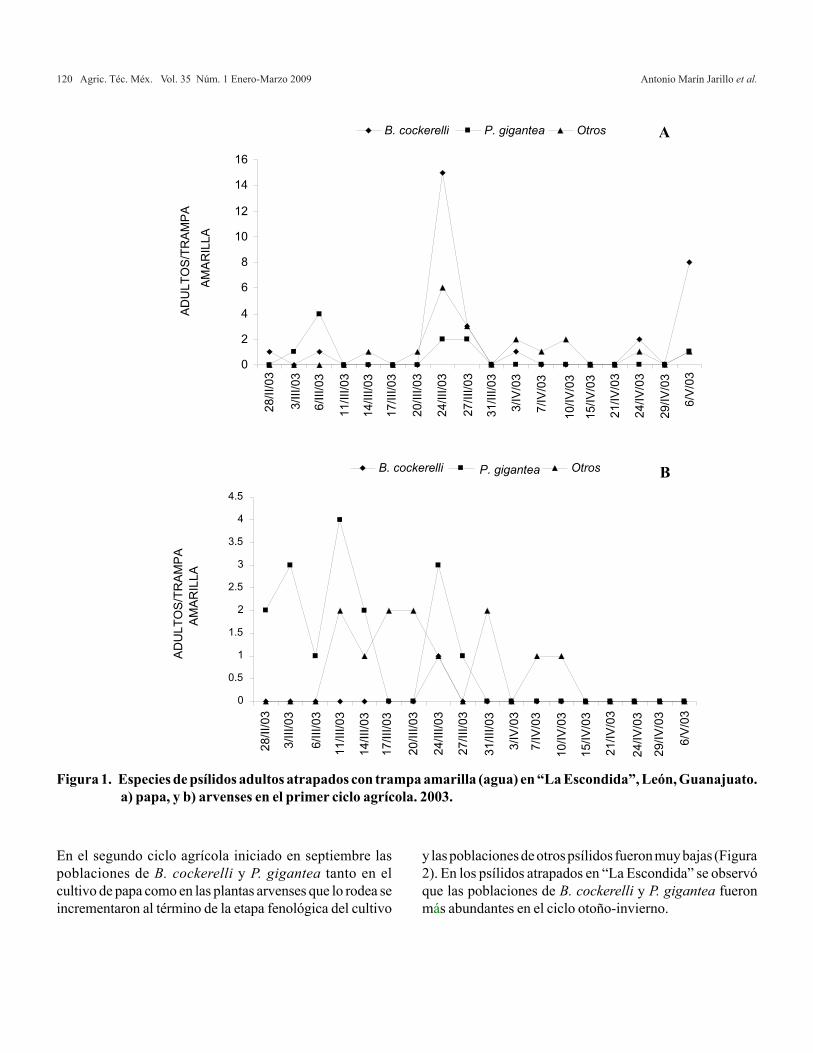
120 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
Antonio Marín Jarillo et al.
En el segundo ciclo agrícola iniciado en septiembre las poblaciones de B. cockerelli y P. gigantea tanto en el cultivo de papa como en las plantas arvenses que lo rodea se incrementaron al término de la etapa fenológica del cultivo
y las poblaciones de otros psílidos fueron muy bajas (Figura 2). En los psílidos atrapados en “La Escondida” se observó que las poblaciones de B. cockerelli y P. gigantea fueron más abundantes en el ciclo otoño-invierno.
0
2
4
6
8
10
12
14
16
28/II
/03
3/III
/03
6/III
/03
11/II
I/03
14/II
I/03
17/II
I/03
20/II
I/03
24/II
I/03
27/II
I/03
31/II
I/03
3/IV
/03
7/IV
/03
10/IV
/03
15/IV
/03
21/IV
/03
24/IV
/03
29/IV
/03
6/V
/03
AD
ULT
OS
/TR
AM
PA
A
MA
RIL
LA
B. cockerelli P. gigantea Otros
Figura 1. Especies de psílidos adultos atrapados con trampa amarilla (agua) en “La Escondida”, León, Guanajuato. a) papa, y b) arvenses en el primer ciclo agrícola. 2003.
B
A
0
0.5
1
1.5
2
2.5
3
3.5
4
4.5
28/II
/03
3/III
/03
6/III
/03
11/II
I/03
14/II
I/03
17/II
I/03
20/II
I/03
24/II
I/03
27/II
I/03
31/II
I/03
3/IV
/03
7/IV
/03
10/IV
/03
15/IV
/03
21/IV
/03
24/IV
/03
29/IV
/03
6/V
/03
AD
ULT
OS
/TR
AM
PA
A
MA
RIL
LA
B. cockerelli P. gigantea Otros
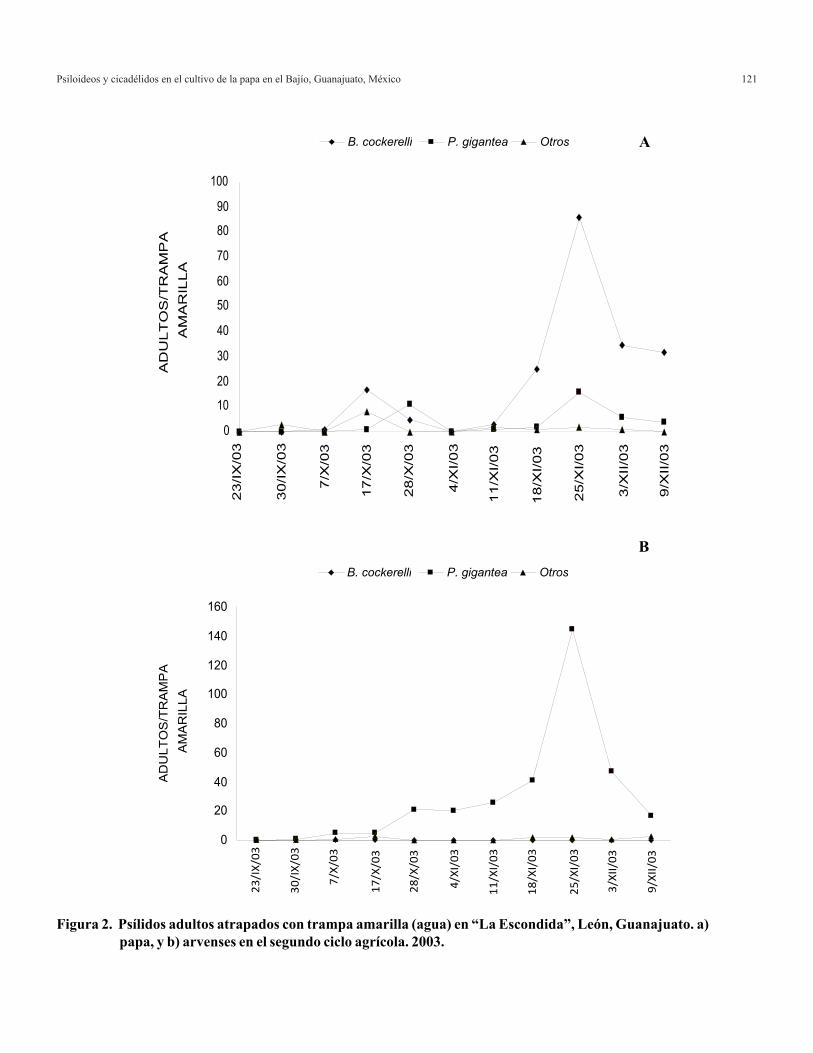
Psiloideos y cicadélidos en el cultivo de la papa en el Bajío, Guanajuato, México 121
Figura 2. Psílidos adultos atrapados con trampa amarilla (agua) en “La Escondida”, León, Guanajuato. a) papa, y b) arvenses en el segundo ciclo agrícola. 2003.
A
B
0 10 20 30 40 50 60 70 80 90
100
23/IX
/03
30/IX
/03
7/X
/03
17/X
/03
28/X
/03
4/X
I/03
11/X
I/03
18/X
I/03
25/X
I/03
3/X
II/0
3
9/X
II/0
3
AD
ULT
OS
/TR
AM
PA
A
MA
RIL
LA
B. cockerelli P. gigantea Otros
0
20
40
60
80
100
120
140
160
23/I
X/03
30/I
X/03
7/X/
03
17/X
/03
28/X
/03
4/XI
/03
11/X
I/03
18/X
I/03
25/X
I/03
3/XI
I/03
9/XI
I/03
AD
ULT
OS
/TR
AM
PA
A
MA
RIL
LA
B. cockerelli P. gigantea Otros
0
2
4
6
8
10
12
14
16
28/II
/03
3/III
/03
6/III
/03
11/II
I/03
14/II
I/03
17/II
I/03
20/II
I/03
24/II
I/03
27/II
I/03
31/II
I/03
3/IV
/03
7/IV
/03
10/IV
/03
15/IV
/03
21/IV
/03
24/IV
/03
29/IV
/03
6/V
/03
AD
ULT
OS
/TR
AM
PA
A
MA
RIL
LA
B. cockerelli P. gigantea Otros
0
2
4
6
8
10
12
14
16
28/II
/03
3/III
/03
6/III
/03
11/II
I/03
14/II
I/03
17/II
I/03
20/II
I/03
24/II
I/03
27/II
I/03
31/II
I/03
3/IV
/03
7/IV
/03
10/IV
/03
15/IV
/03
21/IV
/03
24/IV
/03
29/IV
/03
6/V
/03
AD
ULT
OS
/TR
AM
PA
A
MA
RIL
LA
B. cockerelli P. gigantea Otros

122 Agric. Téc. Méx. Vol. 35 Núm. 1 Enero-Marzo 2009
Antonio Marín Jarillo et al.
Las especies de psílidos colectadas dentro y fuera del cultivo de la papa durante los dos ciclos agrícolas se presentan en el Cuadro 2. B. cockerelli estuvo presente en los dos ciclos agrícolas del cultivo de papa y en las plantas arvenses que lo rodea sólo se atrapó un ejemplar de P. gigantea en los meses de febrero a mayo. Las otras especies de psiloidea presentes en las trampas amarillas fueron Paratrioza maculipenis Crawford, Heteropsilla texana Crawford y Rhinopsylla prob. antennata (Crawford) la primera es muy parecida a B. cockerelli y sólo se presentó en los meses de febrero a mayo; esta especie desarrolla su ciclo de vida (huevecillo, ninfas y adultos) sobre la jarilla amarilla (Senecio salignusDC.) la cual es su hospedera, esta es una planta arvense de hábito
ruderal que crece en terrenos abandonados (Terroneet al., 2004); esta planta se localiza fácilmente a orillas de carreteras y terracerías así como en los alrededores de diversos cultivos en la región de El Bajío. La especie Heteropsila texana sólo fue colectada en trampas colocadas dentro del cultivo de papa lo que no ocurrió con Rhinopsylla prob. antennata la cual sólo se presentó en plantas arvenses. R. prob. antennata pertenece a un grupo monofilético junto con Bactericera Puton y Paratrioza Crawford las cuales tienen cápsulas genitales, antenas y venación muy similares a tal grado que pudieran llegar a ser sinónimos (Brown & Hodkinson, 1988). A esta lista de especies se deben agregar otras 10 especies de Psylloidea las cuales están en proceso de identificación.
Género y/o especie I Ciclo agrícola- meses II - V II Ciclo agrícola- meses IX - XII Papa Arvenses Papa Arvenses
Núm. ejemplares
(%) Núm. ejemplares
(%) Núm. ejemplares
(%) Núm. ejemplares
(%)
Bactericera cockerelli
31 52 1 3 204 78 0 0
Paracarsidara gigantea
10 17 16 56 41 16 328 96
Otros 18 31 12 41 17 6 12 4
Cuadro 2. Especies de psiloidea colectados en trampa amarilla (con agua) en “La Escondida”, León, Guanajuato. 2003.
CONCLUSIONES
Se identificaron cinco especies de psiloideos y 13 especies de chicharritas asociados a papa cultivada y arvenses aledañas en La Escondida, León, Guanajuato, México.
LITERATURA CITADA
Brown, R. G. and Hodkinson, I. D. 1988. Taxonomy and ecology of the jumping plant-lice of Panama. Entomonograph Vol. 9. Ed. L. Lyneborg. E. J. Brill/Scandinavian Science Press Ltd. New York. 304 p.
Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO). 1995. Colecciones científicas. Información de las colecciones mexicanas. (Disponible en línea en http://www.conabio.gob.mx).
Caldwell, J. S. and Martorell, L. F. 1952. A brief review of the psyllidae of Puerto Rico. Ann. Ent. Soc. Amer. 44:603-613.
Crawford, D. L. 1910a. American Psyllidae I (Triozinae). Pomona College Journal of Entomology. 2:228-237.
Crawford, D. L. 1910b. American Psyllidae II (Triozinae). Pomona College Journal of Entomology. 2:347-362.
Crawford, D. L. 1911a. American Psyllidae III (Triozinae). Pomona College Journal of Entomology. 3:422-453.
Crawford, D. L. 1911b. American Psyllidae IV. Pomona College Journal of Entomology. 3:480-503.
Crawford, D. L. 1911c. American Psyllidae V. Pomona College Journal of Entomology. 3:628-632.
Crawford, D. L. 1914. A monograph of the jumping plant-lice or psyllidae of the new worldBulletin of the United States National Museum. 85:1-186.

Psiloideos y cicadélidos en el cultivo de la papa en el Bajío, Guanajuato, México 123
DeLong, D. M. 1948. The leafhoppers, or Cicadellidae, of Illinois (Eurymelinae-Balcluthinae). Bull. Illinois Nat. History Survey 24: 2:97-376.
Garzón-Tiznado, J. A. 2002. Asociación de Paratrioza cockerelli Sulc. con enfermedades en papa (Solanum tuberosum) y Tomate (Lycopersicon lycopersicum Mil. Ex Fawnl) en México. In: Avilés, G. M. A., Gálvez R. J. B. y Garzón T. J. A. (Eds). Memoria del Taller sobre Paratrioza cockerelli Sulc. Culiacán, Sin., México. Primera Edición.
Garzón-Tiznado, J. A.; Garzón-Ceballos, J. A.; Velarde-Felix, S.; Marín-Jarillo, A. y Cárdenas Valenzuela, O. G. 2005. Ensayos de transmisión del fitoplasma asociado al “Permanente del Tomate” por el psílido Bactericera cockerelli Sulc. en México. In: Morales M. A.; Mendoza, E. A., Ibarra, G. M. P. y Stanford, C. S. (Eds). Entomología Mexicana 4:672-675.
Medler, J. T. 1942. The leafhoppers of Minnesota Homoptera:Cicadellidae. Agric. Exp. Station. University of Minnesota. 196 p.
Munyaneza, J. E.; Crosslin, J. and Upton, J. E. 2007. Association of Bactericera cockerelli (Homoptera: Psyllidae) with 'zebra chip', a new potato disease in southwestern United States and Mexico. J. Econ. Entomol. 100(3):656-663.
Munyaneza, J. E. 2003. Leafhopper identification and biology. Pacific Northwest Vegetable Association Proceedings. 89-91.
Munyaneza, J. E. and Crosslin, J. 2008. Zebra chip, a new potato disease in North and Central America, is associated with the potato psyllid. pp. 124-127. In: Chiru, S.; Olteanu, G.; Aldea, C. and Badarau, C. (eds.), "potato for a changing world". Transilvania University of Brasov Publishing House, Brasov, Romania.
Nielson, M. W. 1968. The leafhopper vectors of phytopathogenic viruses (Homoptera:Cicadellidae) taxonomy, biology and virus transmission. Bull. Tech. No. 1382 Agric. Res. Serv., U.S. Dep. Agric. Washington. 386 p.
Osborn, H. 1928. The leafhoppers of Ohio. Vol. III: No. 4. Bulletin No. 14. 374 p.
Sterling, P. S. 1982. A taxonomic study of the leafhopper genus Empoasca (Homoptera:Cicadellidae) in eastern Peru. Tech. Bull. 272. North Carolina Agric. Res. Serv. 194 p.
Terrones, R. T. del R. L.; González, S. C. y Ríos, R. S. A. 2004. Arbustivas nativas de uso múltiple en Guanajuato. Instituto Nacional de Investigaciones Forestales, Agricolas y Pecuarias, Campo Experimental Bajío, Celaya, Guanajuato, México 216 p. (Libro Técnico Núm. 2).