“purificación a homogeneidad de la sustancia tipo
TRANSCRIPT

Profesor Patrocinante Marcia Costa Lobo Instituto de Ciencia y Tecnología de los Alimentos Facultad de Ciencias Agrarias
“Purificación a homogeneidad de la sustancia tipo bacteriocina de la cepa BALB considerando como parámetro
la actividad inhibitoria frente Listeria monocytogenes ”
Tesis de Grado presentada como parte de los requisitos para optar al grado de Licenciado en Bioquímica y Título Profesional de Bioquímico
CLAUDIA NATALIA HUANQUILEF ARRIAGADA VALDIVIA-CHILE
2010

ii
AGRADECIMIENTOS
Este trabajo se realizó bajo la dirección y apoyo de la Ing Civil Bioquímico. Marcia
Costa Lobo y de los integrantes que conforman su laboratorio a quienes agradezco su
entrega y compromiso; extiendo este agradecimiento a mis profesores informantes
Dr. Juan Guillermo Cárcamo, Tec. Med. M. sc. Renate Schöbitz por aceptar formar
parte de la comisión evaluadora y aportar en la corrección de esta tesis. Debo
mencionar a los que con su conocimiento y gran calidad humana me apoyaron en el
desarrollo de este trabajo de titulo; Dr. Joel Asenjo, Dr. Ociel Muñoz, Ing. Alimento
Yanina Nahuelquin, BQ. Tatiana Pérez. Una especial mención a mis compañeros de
trabajo, Mónica, don José, don Ramón, Genny, por estar conmigo, apoyarme y
escucharme cada vez que lo necesité, y también a esos amigos incondicionales que
forman parte de este proceso y de mi vida personal mis amigos Jorge, Mabel y Wallys.
Finalmente esta mi familia mi gran fortaleza, los que siempre estarán; gracias a mis
padres por su amor y apoyo, a mis hermanas por su compañerismo, a mis sobrinos
por su alegría.
Quiero agradecer en especial a mi cuñado Cristián Castañeda por el cariño y apoyo
entregado durante estos años de universidad.
Esta tesis fue financiada por los aportes otorgados por el Fondo de Desarrollo
Científico y Tecnológico FONDEF D04i 1153.

iii
A mi familia y a Cristián por el cariño y el apoyo entregado en estos años.

iv
INDICE DE CONTENIDOS
Páginas
1. RESUMEN 1
1.1 SUMMARY 3
2. INTRODUCCIÓN 5
2.1 Bacterias ácido lácticas 7
2.2 Bacteriocinas 8
2.2.1 Clasificación 9
2.2.2 Estructura tridimensional de las bacteriocinas 11
2.2.3 Biosíntesis y organización genética 15
2.2.4 Bacteriocina ν/s antibiótico 19
2.2.5 Mecanismo de acción 21
2.3 Bacterias Gram positivas resistentes a péptidos antimicrobianos 24
2.4 Aplicación de las bacteriocinas en la industria alimentaria 24
2.5 Género Listeria spp 25
2.6 Hipótesis y objetivos 27
2.6.1 Hipótesis 27
2.6.2 Objetivo general 27
2.6.3 Objetivos específicos 28
3. MATERIALES Y MÉTODOS 29
3.1 Materiales 29
3.1.1 Reactivos 29

v
3.1.2 Equipos 31
3.1.3 Soluciones 32
3.2 Métodos 34
3.2.1 Cepa bacteriana 34
3.2.2 Cepa indicadora 34
3.2.3 Medios de cultivos y crecimientos de la cepa BALB 34
3.2.4 Determinación de la actividad antimicrobiana de BALB 35
3.2.5 Purificación a homogeneidad de la cepa BALB 38
3.2.6 Liofilización de las muestras 42
3.2.7 Geles Tris-tricina 44
3.2.8 Determinación de proteína mediante el método de Bradford 45
3.2.9 Determinación de la composición aminoácidica de STB 45
4. RESULTADOS 48
4.1 Cromatografía de filtración en gel sephadex G-25 Hip-Trap 54
4.2 Ultrafiltración en tubos Centriplus 54
4.3 Cromatografía de intercambio catiónico 55
4.4 Cromatografía de fase reversa sobre soporte solido C18 57
4.5 Cromatografía líquida de alta resolución HPLC 59
4.6 Electroforesis en geles de poliacrilamida Tris-tricina
en condiciones desnaturantes 61
4.7 Determinación de la composición aminoacídica de la STB 63

vi
5. DISCUSIÓN 65
6. CONCLUSIÓN 72
7. SUGERENCIA 73
8. BIBLIOGRAFÍA 74
9. ANEXOS 86
9.1 Cromatografía de filtración en mini columna Hi-Trap desalting 86
9.2 Cuantificación de proteínas por el método de Bradford 87
9.3 Determinación de la actividad inhibitoria con relación al volumen 90
9.4 Determinación del peso molecular de la STB 91
9.5 Determinación de aminoácidos por el método PITC 94
9.6 Fotografía cepa BALB y extracto crudo de la STB 97
9.7 Fotografía de las precipitaciones de la STB 98

vii
INDICE DE FIGURAS
Figura Páginas
1 Ilustración tridimensional de bacteriocina de la clase IIa 12
2 Orientación de las bacteriocinas de la clase IIa en la membrana
de la célula Blanco 14
3 Representación esquemática de la síntesis de las bacteriocinas
clase IIa 18
4 Esquemas de los diferentes mecanismos de acción propuestos para
los péptidos Antimicrobiano 23
5 Técnicas “gota sobre césped” 37
6 Metodología empleada para la purificación de la STB 43
7 Reacción de derivatización de los aá libres PITC 47
8 Actividad inhibitoria detectada por el ensayo “gota sobre césped”
utilizando Como M.O indicador Lm 53 9 Perfil de elución de la STB obtenido con por SP-Fast Flow 56
10 Halos de inhibición presente en las fracciones VIa y VIb 58
11 Cromatogramas de la fracción VIIa obtenido por HPLC 60
12 Geles Tris-tricina de las fracciones VIa y VIb 62

viii
13 Cromatogramas de la derivatización de aminoácidos obtenido
por HPLC 64

ix
INDICE DE TABLAS
Tabla Páginas
I Principales diferencias entre bacteriocinas y antibióticos 20 II Purificación parcial de STB concentrado con acetona 50
III Purificación parcial de STB concentrado con sulfato de amonio 51

x
LISTAS DE ABREVIATURAS
BAL B Cepa Bacteriana.
BSA Albúmina Sérica de Bovino.
EC Extracto Crudo.
EDTA Ácido Etilendiaminotetracético.
FPLC Cromatografía Líquida Rápida de Proteínas.
HPLC Cromatografía Líquida de alta Presión.
PBS Tampón Fosfato Salino.
PITC Fenilisotiacianato.
SDS Dodecil Sulfato Sódico.
STB Sustancia Tipo Bacteriocinas.
TFA Ácido Trifluoroacético.
TRIS Tris (hidroximetil) amino-metano.
UA Unidades formadores de colonias.

1
1. RESUMEN
En las últimas décadas, en la industria alimentaria, se ha detectado la presencia de
diversos microorganismos “emergentes”, causante de una gran variedad de
enfermedades, asociadas al consumo de alimentos contaminados. Dentro de este
grupo se encuentra la Listeria monocitogenes, un microorganismo capaz de sobrevivir
y crecer a temperatura de refrigeración, convirtiéndose en una amenaza permanente
para la salud de los consumidores y para la industria alimentaria. En la industria
salmonera para reducir y/o eliminar la presencia de Listeria monocytogenes en las
líneas de producción, se ha propuesto el uso de un biocontrolador natural, formulado a
base de un extracto crudo que contiene sustancias inhibitorias, las cuales son
liberadas al medio extracelular por diferentes cepas lácticas (BALA, BALB, BALC).
Estas sustancias inhibitorias actúan en la membrana celular de las células sensibles
provocando un aumento en la permeabilidad y posterior muerte celular.
En el presente trabajo, se purificó a homogeneidad una Sustancia Antimicrobiana Tipo
Bacteriocina (STB) extraída de la cepa BALB. El procedimiento de purificación se
logró, a través, de las siguientes etapas: a) Precipitación del extracto con acetona y
sulfato de amonio independientemente, b) Cromatografía Sephadex G-25,
c) Ultrafiltración con Centriplus (YM30, YM10, YM3), d) Cromatografía intercambio iónico
SP-Fast Flow, y e) RP-HPLC C18.
Se obtuvo un péptido catiónico de peso molecular aproximado de 3 kDa con actividad
inhibitoria frente Listeria monocytogenes.

2
Se obtuvo un bajo rendimiento en la purificación utilizando ambas metodologías, no
obstante la purificación parcial con acetona resulto ser ligeramente superior con
respecto a la precipitación con sulfato de amonio.

3
1.1
During the last decades, in the food industry, it has been detected the presence of a
variety of emerging microorganisms, which cause a wide variety of illnesses that are
associated to the consumption of contaminated food. Inside this group it is found the
Listeria monocytogenes, which is a microorganism capable of living and developing at
cooling temperature, that is why it has become a permanent threat to not only the
consumers health but to the food industry as well. The salmon industry for reducing or
eliminate the presence of Listeria Monocytogenes from in the production lines has
been proposed implemented the use of a natural biocontroller, formulated based on a
crude extract which contains inhibitory substances that are released to the extracellular
environment for different lactic strains (BALA, BALB, BALC). These antimicrobial
substances act on the cellular membranes from the sensitive cells causing a rise in the
permeability and subsequent cellular death.
In the following work, an inhibitory substance of the bacteriocin type (STB) extracted
from the LABB strain. The purification procedure was accomplished through the
following stages: a) Precipitation of the extract with acetone and ammonium sulfate
independently, b) Cromatography Sephadex G-25 c) ultrafiltration with Centriplu (YM30,
YM10, YM3), d) Ion exchange chromatography SP-Fast Flow, y e) RP-HPLC C18.
It was obtained a cationic peptide of approximate molecular weight 3 kDa with
inhibitory activity against Listeria monocytogenes.

4
Using both methodologies is obtained a low little performance in the purification,
despite the partial purification with acetone turned out to be slightly superior respect to
the ammonium sulfate precipitation.

5
2. INTRODUCCION
En las últimas décadas, se han detectados la presencia de diversos microorganismos
“emergentes”, causantes de una gran variedad de enfermedades asociadas al
consumo de alimentos contaminados. Uno de ellos es la Listeria monocytogenes (Lm),
patógeno intracelular, capaz de sobrevivir y de desarrollarse a temperatura de
refrigeración, (Doyle et al., 2001), siendo una amenaza permanente, para los
consumidores y la industria alimentaria.
Listeria monocytogenes, se encuentra ampliamente distribuida en la naturaleza;
dejando de manifiesto su extraordinaria adaptación a condiciones ambientales
adversas, pudiendo crecer en un amplio rango de pH, así como temperaturas, en
presencia de altas concentraciones salinas y a baja actividad del agua aw, teniendo
además la capacidad de formar biofilm, (Gandhi y Chikindas, 2007).
La enfermedad producida por este agente patógeno se denomina listeriosis, la cual es
considerada un serio problema de salud pública, registrándose un aumento de esta en
los últimos años.
La dosis infectiva de Lm no ha sido determinada aún, pero se considera un número
superior a 100 células, pudiendo variar según la cepa y la susceptibilidad de las
personas.
La población más vulnerable para contraer esta enfermedad son las mujeres
embarazadas, inmunodeprimidos, lactantes y adultos mayores.
Las manifestaciones clínicas en humanos son usualmente asintomáticas, sin embargo,
estas pueden ser variables, desde un cuadro gastrointestinal leve o en casos extremos

6
de alta mortalidad por meningitis, encefalitis, septicemias y abortos espontáneos
atribuibles a este patógeno.
Para obtener una línea segura de producción, las industrias alimentarias se han visto
en la necesidad de implementar nuevas técnicas de control de calidad, para poder
reforzar sus puntos críticos de contaminación, y así cumplir con las normas sanitarias
impuesta por el mercado internacional, garantizando de esta manera la seguridad e
inocuidad de los productos ofrecidos.
En el Instituto de Ciencia y Tecnología de los Alimentos, (ICYTAL), de la Universidad
Austral de Chille, se ha desarrollado un biocontrolador natural, elaborado a partir de los
extractos crudos, provenientes de los cultivos de diferentes bacterias lácticas, en
proceso de identificación y a la fecha denominadas como cepas (BALA, BALB, BALC)
aisladas de carne al vacío y de salmón. Este grupo de bacterias son consideradas,
como “microorganismo seguros” GRAS (Generally Recognized As Safe) por la FDA
(Food and Drug Administration), lo que permite ser utilizadas con un alto potencial
biotecnológico en la preservación de los alimentos
Este biocontrolador se ha desarrollado con la finalidad de introducir su uso en las
plantas de procesos de la industria del salmón, con la finalidad de reducir los riesgos
asociados a la posible presencia de Lm.

7
2.1 Bacterias ácido lácticas
Las bacterias ácido lácticas (BAL) comprenden un número elevado de bacterias Gram-
positivas, que participan en la elaboración y maduración de una gran variedad de
alimentos fermentados, otorgándoles las características organolépticas típicas como
son el sabor y la textura a los alimentos.
Las BAL han sido reportadas en numerosos géneros de microorganismos, como
Aerococcus, Alloiococcus, Carnobacterium, Dolosigranulun, Enterococcus, Globicatella,
Lactobacillus, Lactococcus, Lactosphaera, Leuconostoc, Oenococcus, Pediococcus,
Streptococcus, Tetragenococcus, Vagococcus y Weissella (Axelsson, et al., 1998;
Cintas et al., 2000a).
Las BAL poseen una morfología bacilar o cocoide, son generalmente inmóviles, no
esporulados, no pigmentados y no reductores de nitratos, además son anaerobio
facultativos, carentes de citocromo y catalasa, presentando en el ADN un porcentaje de
G+C inferior al 50 %. Las BAL se caracterizan principalmente por producir ácido láctico
como producto mayoritario de la fermentación de los carbohidratos. Las BAL sólo
pueden obtener energía a partir de azúcares y otros compuestos relacionados, debido
a su limitada capacidad biosintética, razón por la cual necesitan una gran variedad de
factores de crecimiento como aminoácidos, vitaminas, purinas y pirimidinas (Dellagio et
al., 1994), para poder desarrollarse y es por eso que se suele encontrar en un medio
tan rico nutritivamente como la leche.
Los principales mecanismos antimicrobianos que poseen las BAL son la competencia
por nutrientes, y la producción de ácidos láctico y acético, los cuales permiten preservar

8
el valor nutritivo y la calidad higiénica de los alimentos fermentados (Kandler, 1983;
Daeschel, 1989; Lindgren y Dobrogosz, 1990). No obstante, estas bacterias también
producen otras sustancias antimicrobianas, como etanol, dióxido de carbono, diacetilo,
acetaldehído, peróxido de hidrogeno, ácido benzoico, reuterinas y bacteriocinas (Piard
y Desmazeaud, 1992; Requena y Peláez, 1995). Destacando de estas últimas el alto
potencial biotecnológico en la preservación de los alimentos y tratamientos biomédicos
(López et al., 2008).
2.2 Bacteriocinas
Las bacteriocinas son péptidos evolutivos conservados, producidos por la mayoría de
los géneros bacterianos, como un mecanismo de defensa y de competencia frente a
otros microorganismos. En general las bacteriocinas de bacterias Gram positivas se
definen como un grupo heterogéneo de péptidos antimicrobiano de síntesis ribosomal,
secretados extracelularmente y que poseen en común el tamaño molecular, la
naturaleza catiónica y anfipática. Su actividad antimicrobiana frente a otros
microorganismos, es en general relacionadas taxonómicamente con la cepa
productora, (Hancock et al., 1995; Nissen-Meyer, 1997, Cotter et al., 2005).
Las primeras bacteriocinas descritas, fueron las colicinas, producidas por Escherichia
coli (Gratia, 1925) y posteriormente, en bacterias Gram positivas producidas por las
cepas Lactococcus lactis, (Jacob et al., 1953; Tagg et al., 1976).
Actualmente, las cepas productoras de bacteriocinas poseen un alto potencial en la
bioconservación, de los alimentos, al inhibir el crecimiento de patógenos alimentarios,

9
como Bacillus cereus. Clostridium perfringes, Staphylococcus aureus y Listeria
monocytogenes, los cuales presentan un alto riesgo para la salud de los consumidores.
2.2.1 Clasificación
Las bacteriocinas han sido agrupadas según sus diferentes características, como
tamaño molecular, mecanismo de acción, estructura química, estabilidad térmica,
organización genética, presencia de aminoácidos modificados, etc, (Nes et al., 1996;
Moll et al., 1999). Existen en la actualidad tres clases de bacteriocinas bien definidas:
Clase I: Son bacteriocinas de tamaño molecular inferior a 5 kDa, que presentan
modificaciones post-traduccionales y son térmicamente estables. Éstos péptidos
poseen aminoácidos inusuales en su estructura primaria como lantionina, β-metil-
lantionina, deshidroalanina (DHA), y deshidrobutirina (DHB) (Bierbaum et al., 1996; Moll
et al., 1999) los cuales son originados por la deshidratación de la serina y la treonina
respectivamente.
A este grupo de bacteriocinas se les conocen como lantibióticos estos se subdividen en
dos grupos Ia y Ib. Los lantibióticos Ia son bacteriocinas alargadas y flexibles poseen
una carga neta positiva y un peso molecular entre 2-5 kDa, son anfipáticos y
formadores de poros. Un ejemplo de lantibiótico Ia es la Nisina. Los lantibióticos Ib
poseen una forma globular, carecen de carga neta y su peso molecular es de 2 kDa,
también son anfipáticos y actúan como inhibidores de enzimas (Sahl et al., 1995).

10
Clase II: Son péptidos pequeños, con un tamaño molecular inferior de 10 kDa, éste
grupo de bacteriocinas no poseen aminoácidos modificados en su estructura primaria,
son termoestables y no sufren modificaciones post-traduccionales. Estas bacteriocinas
tienen en común una secuencia péptido líder Gly-Gly y comparten un alto contenido de
aminoácidos catiónicos. Se subdividen en cuatro subclases IIa, IIb, IIc, IId. Las
bacteriocinas de la subclase IIa se caracterizan por ser particularmente activas frente al
género Listeria (Muriana, 1996). Sus secuencias aminoacídicas muestran un elevado
grado de homología (40-70 %), (Aymerich et al, 1996). Todos los miembros de esta
subclase comparten la siguiente secuencia de consenso, YYGNGV(X)C(X)4C(X)V(X)4A
(X significa cualquier aminoácidos) ubicada en el extremo amino-terminal (Cleveland et
al., 2001; Ennahar et al., 2000; Klaenhammer et al., 1993). Los aminoácidos resaltados
en negrita muestran prácticamente un 100 % de conservación (Nes y Holo, 2000), y en
el otro extremo en el motivo C-terminal poseen aminoácidos hidrofóbicos que
interaccionan con la membrana de las células diana originando los poros de inhibición
(Chikindas et al., 1993). Otra característica de esta subclase es que contienen uno o
dos puentes disulfuro en la parte N-terminal de la molécula (Cleveland et al., 2001). La
mayoría de estos péptidos son secretados mediante transportadores específicos de
tipo ABC, sin embargo, se ha observado que la enterocina P utiliza el sistema sec
dependiente para ser transportada (Cintas et al., 1997). Esta subclase es la más
estudiada dentro de las bacteriocinas de la clase II.
La subclase IIb requiere la acción combinada de dos péptidos para ser una molécula
biológicamente activa, ya que estos péptidos no muestran actividad inhibitoria

11
individual. La lactococcina G y M de L Lactis es un miembro representativo de esta
subclase (Moll et al., 1996).
Las bacteriocinas de la subclase IIc poseen en común una estructura cíclica como
resultado de la unión covalente entre los extremos amino-terminal y C-terminal. Un
representante de esta subclase es la enterocina AS-48 de Enterococcus faecalis
(Sánchez et al., 2003).
La subclase IId está formada por los péptidos de la clase II que no han podido ser
clasificados en ninguno de estos subgrupos, siendo la lactocoquina A de L lactis. Un
ejemplo de esta subclase (Holo et al., 1991)
Clase III: Está formada por bacteriocinas de un tamaño molecular mayor a 30 kDa, son
termolábiles a temperaturas mayores a 60 ºC. Se desconoce el mecanismo de acción
de este grupo de bacteriocinas, sólo se sabe que no actúan a nivel de membrana. Esta
clase de bacteriocinas ha sido poco estudiada, presentando un bajo interés en los
investigadores dada su termolabilidad y escasa aplicación en la industria alimentaria.
2.2.2 Estructura tridimensional de las bacteriocinas
En los últimos años se ha conocido la estructura tridimensional (3D) de cuatro
bacteriocinas como carnobacteriocina B2 (Wang et al., 1999), curvacina A (Haugen et
al., 2005), leucocina A (Fregeau Gallagher et al., 1997) y sakacina P (176), todas ellas
producidas por la Clase IIa. Estas estructuras han sido bien dilucidadas por resonancia
magnética nuclear, como se muestra en la figura 1.

12
Figura 1. Ilustración Tridimensional de Bacteriocinas de la
Clase IIa. Determinadas por resonancia magnética nuclear.
Fuente: Drider et al., 2006.

13
En general todas las bacteriocinas producidas por la Clase IIa presentan en su extremo
N-terminal una estructura lámina β, estabilizada por el puente de disulfuro altamente
conservado. En el extremo C-terminal de estos péptidos adoptan una conformación α
hélices (Figura 2), la sakacina P presenta un puente disulfuro extra con respecto a
carnbateriocina B2, curvacina A, y leucocina, lo cual podría jugar un importante rol en la
estabilización de la estructura tridimensional del extremo C-terminal y la
termoestabilidad que presentan este tipo de bacteriocinas (Fimland et al., 2005), esto
se pudo comprobar cuando se incorporó un segundo puente de sulfuro a una variante
sintética de sakacina P, se comprobó que poseía una termoestabilidad superior a las
de las bacteriocinas originales.

14
Figura 2. Orientación de las bacteriocinas de la Clase IIa en la
membrana de la célula blanco.
Fuente: Johnsen et al., 2005.

15
2.2.3 Biosíntesis y Organización Genética
Los genes que codifican a las proteínas necesarias para la síntesis de las bacteriocinas
están organizados en uno o dos operones, siendo localizados en el cromosoma y en
los plásmidos (Klaenhammer, 1993; Quadri et al., 1995; Kanatani et al., 1995). Para
llevar a cabo las síntesis de estas bacteriocinas, se necesita a lo menos de la
participación de 4 genes: i) gen estructural, que codifica la prebacteriocina, la cual
posee una secuencia señal amino terminal de longitud variable entre 14 a 30
aminoácidos, ii) gen de inmunidad que codifica a la proteína de inmunidad la cual
protege a la célula frente al efecto antimicrobiano de las bacteriocinas, iii) gen
transportador que codifica a una proteína transportadora del tipo ABC necesario para la
secreción de la bacteriocina, iv) gen codificador que codifica a la proteína accesoria de
función aún desconocida.
En general, las bacteriocinas son sintetizadas como pre-péptidos inactivos que
contienen un péptido señal en el extremo N-terminal altamente conservado, esta señal
las mantiene de forma inactiva dentro de la célula productora. Esta pre-secuencia es
removida proteolíticamente durante la exportación del pre-péptido hacia el espacio
periplásmico por las mismas proteínas transportadoras (transportador ABC
dependientes de ATP, que también puede contener un dominio proteolítico) o por
proteínas serina-proteasa presentes en la parte externa de la membrana celular, lo cual
con lleva la liberación del péptido biológicamente activo al espacio extracelular
(Havarstein et al., 1995; Cotter et al., 2005; Ennahar et al., 2000). Esta pre-secuencia
parece jugar una doble función en la biosíntesis de la bacteriocina., teniendo un rol

16
protector en la cara citosólica de la membrana, manteniendo la bacteriocina inactiva y
evitando así ser atacado por síntesis de nuevas bacteriocinas, también pueden jugar un
rol como una señal de reconocimiento durante la exportación de la bacteriocina.
Las bacterias productoras de bacteriocinas han desarrollado un mecanismo de
inmunidad que evita su autodestrucción cuando es provocado por sus propios péptidos.
Se desconocen los mecanismos moleculares exactos por los que estas proteínas les
confieren protección a las bacterias productoras, sin embargo, se han propuesto dos
sistemas de protección, siendo la más interesante, la que postula la presencia de una
proteínas de inmunidad, la cual se une al receptor de la bacteriocina, provocando
cambios, en su estructura conformacional, impidiendo de esta manera la acción
antimicrobiana de las bacteriocinas, tal receptor sería la proteína transmembrana
manosa permeasa, para las bacteriocinas subclase IIa (Dalet et al., 2001; Héchard et
al., 2001). No se conocen en detalle los mecanismos de inmunidad presentes en las
células productoras. Sólo se han purificado las proteínas de inmunidad de la
lactococina A-LciA (Nissen-Meyer et al., 1993) y de la carnobacteriocina B2–CiB2
(Quadri et al., 1995).
La regulación de la síntesis de algunas bacteriocinas está mediada por dos sistemas de
transducción de señales constituidos por dos o tres componentes. Siendo estos
activados, como respuestas a diferentes modificaciones ambientales, como es la
temperatura, pH o por un mecanismo de “censado poblacional” o “quorom sensing”. El
sistema de tres componentes está constituido de una molécula señal o feromona (IF),
de una proteína histidín-quinasa (HPK) y de una proteína reguladora de respuesta (RR)
(Nes et al., 1996; Kleerebezem et al., 1997).

17
La proteína histidina-quinasa, contiene un dominio N-terminal, con una función sensora,
localizado en el lado periplásmico de la membrana y el dominio C-terminal posee
actividad autoquinasa y contiene un residuo de histidina absolutamente conservado
que constituye el sitio de fosforilación. La proteína RR (“Respanse Regulador”) actúa
como un regulador de respuesta citoplasmática, mediando una respuesta adaptativa
(Nes et al., 1996). El factor de inducción IF, constituye la señal inicial del sistema de
tres componentes, cuya presencia es detectada por la proteína HPK, dando inicio a la
transcripción de los genes involucrados en la síntesis de las bacteriocinas (Figura 3).
Por otra parte, las bacteriocinas que poseen sistemas de dos componentes, presentan
una función dual, actuando como antimicrobiano e inductor de su propia síntesis, un
representante de este sistema es la nisina de L. lactis (Kleerebezem, 2004).

18
Figura 3. Representación esquemática de la síntesis de las
bacteriocinas clase IIa: El péptido inductor (IP) interacciona con la
proteína Histidina-quinasa (HPK) provocando la fosforilación de esta
proteína a expensa de ATP. El grupo fosfato es transferido a la proteína
reguladora (RR), provocando la activación de los genes de la
prebacteriocinas y de otros genes asociados. El procesamiento y secreción
de la prebacteriocina y del IP, a través, de un transportador ABC.
Fuente: Drieder et al., 2006.

19
2.2.4 Bacteriocinas v/s antibióticos
Las bacteriocinas fueron consideradas antibióticos por algunos investigadores en los
inicios de su investigación. Sin embargo se diferencian de los antibióticos
(Hurst, 1981; Hansen et al., 1994), en su estructura molecular, su vía de síntesis,
espectro de acción, entre otras propiedades, como se detalla en la Tabla 1. Las
ventajas que presentan las bacteriocinas respecto de los antibióticos, radica
principalmente en que son sustancias inocuas para la salud de las personas, distinto es
el caso de los antibióticos que pueden producir microorganismos resistente, toxicidad y
efectos secundarios en las personas.

20
Tabla 1. Principales diferencias entre bacteriocinas y antibióticos
BACTERIOCINA
ANTIBIOTICO
Aplicación
Fundamentalmente en
Alimentación
Clínica
Síntesis
Ribosómica
Metabolitos secundarios
Actividad
Espectro reducido
Espectro variable
Inmunidad del
productor
Sí
No
Mecanismo de
resistencia
Adaptación
relacionada
con la composición de
membrana
Modificación genética
Modo de
acción
Principalmente,
formación de poros
Dianas intra o
Extracelulares
Toxicidad
No descritos
Sí
Fuente: Cleveland, 2001.

21
2.2.5 Mecanismo de acción
Los péptidos antimicrobianos presentan una serie de pasos que conducen a la muerte
celular que ocurren con independencia del tiempo y/o la especificidad del mecanismo
de acción (Brogden, 2005; Hale y Hancock, 2007).
El primer paso consiste en la interacción electroestática entre los péptidos catiónicos y
la membrana citoplasmática las cuales poseen en sus superficies lipopolisacáridos
(LPS) en el caso de las bacterias Gram-negativas y ácidos teicoicos y lipoteicoicos en
las bacterias Gram positivas, el segundo paso consiste en la adhesión de estos
péptidos de forma paralela sobre la superficie de la membrana y cuando la razón
péptido/lípido aumenta los péptidos se orientan de forma perpendicular, este
reordenamiento provoca la inserción y por consiguiente la permeabilización de estos
péptidos en la membrana para culminar con la formación de poros transmembrana
(Yang et al., 2001).
El mecanismo de acción de los péptidos antimicrobianos, presentan en general algunos
elementos comunes como las característica a nivel molecular, los cuales permiten
agruparlos según sus diferentes propiedades (Ganz y lehrer, 1999; Chmiel, 2001;
Brogden, 2005; Kamysz et al., 2003, Hale y Hancock, 2007).
Los diferentes mecanismos de acción se representan en diversos modelos propuestos
en la actualidad, los cuales se describen a continuación:
Modelo en barril: También conocido como modelo de haces de hélices
transmembrana (Transmembrane helical-hundle model), los péptidos helicoidales
orientados en paralelo, se insertan perpendicularmente en la membrana de modo que

22
las regiones hidrofóbicas de los péptidos se alinean con la región del core lipídico de la
membrana, dejando las regiones hidrofílica hacia el lúmen central, figura 4A. Se forma
así un poro transmembrana muy similar a un barril sin fondo cuyas paredes son los
péptidos helicoidales (Bechinger, 1997; Shai, 1999).
Modelo de carpeta: Los péptidos se acumulan en la superficie de la bicapa cubriendo
la superficie de la membrana celular como una carpeta. En altas concentraciones estos
péptidos interrumpen la continuidad de la membrana insertándose en la membrana en
paralelo a la bicapa lipídica, mediantes sus regiones hidrofóbicas. Las regiones polares
de los péptidos quedan expuestas al exterior, de modo que actúan de manera similar a
los detergentes, como se observa en la figura 4B (Shai, 1999; Ladokhin y White, 2001).
Modelo de poro toroidal: Los péptidos se acumulan en la superficie de la membrana
celular y se insertan en la membrana induciendo la distorsión de la monocapa de
lípidos a curvarse a través del poro, de modo que las regiones polares de ambas capas
llegan a unirse. Así, las paredes del poro quedan formadas por las cabezas polares de
los lípidos de membrana y por los péptidos insertados en ella, como se observa en la
figura 4C (Bechinger, 1997).

23
Figura 4. Esquema de los diferentes mecanismos de acción
propuestos para los péptidos antimicrobianos. (A) Modelo de barril
(B) Modelo de carpeta (C) Modelo toroidal. Las regiones hidrofílica del
péptido se observa de color rojo y las hidrofóbicas en azul.
Fuente: Brogden, 2005.

24
2.3 Bacterias Gram positivas resistentes a péptidos antimicrobianos
La resistencia bacteriana, puede presentarse de forma natural (resistencia intrínseca) o
bien puede originarse como resultado de una exposición continua del péptido
(resistencia adquirida).
Se han registrado cepas mutantes para L.monocytogenes, L.innocua, Streptococcus
pneumoniae, las cuales han mostrado cambios en la pared y en la membrana celular,
observándose en algunos casos variación en el grosor y fluidez en la membrana. Es así
como, en las bacteriocinas de la Clase IIa, se ha observado la disminución de la
expresión del probable receptor manosa permeasa (Dalet et al., 2001)
2.4 Aplicación de las bacteriocinas en la industria alimentaria
Como se planteó anteriormente las bacteriocinas cumplen un rol inhibitorio contra
microorganismos patógenos presentes en los alimentos, además de presentar una
amplia estabilidad térmica y ser activas en amplios rangos de pH.
La utilización de las bacteriocinas, como bioconservador, es una alternativa segura
para los consumidores, al ser inactivadas fácilmente por las proteasas intestinales.
En la actualidad la nisina y la pediocina PA1, se utilizan de manera comercial como
bioconservantes de alimentos. La nisina se encuentra comercialmente en la forma de
“nisaplina” (Danisco) y la pediocina PA1 en la forma de “ALTA 2431” (Quest). A pesar
de ser las únicas comerciables en el mercado, no cabe duda que en corto plazo
aparecerán nuevas formulaciones de nuevas bacteriocinas, para poder abastecer un

25
mercado demandante de nuevas tecnologías que impidan el crecimiento de patógenos
que alteran la calidad de los alimentos.
2.5 Género Listeria spp
El género Listeria spp es un bacilo Gram positivo, catalasa positivo, anaerobio
facultativo, no esporulado y móvil por poseer flagelos perítricos cuando son incubadas
a 20-25 ºC. Son capaces de crecer en un rango de pH extremo. Tienen una
temperatura óptima de crecimiento entre los 30 ºC y 37 ºC, su crecimiento se restringe
entre 1 ºC y los 45 ºC.
El género Listeria comprende seis especies: L. monocytogenes, L. ivanovii, L. seeligeri,
L. innocua, L. welshimeri, L. grayi subsp. murrayi y L. grayi subsp. grayi. Solamente
cuatro de estas especies han sido reportadas en enfermedades causadas al hombre o
en animales (L. monocytogenes, L.ivanovii, L. seeligeri y L. innocua). Siendo la especie
L. monocytogenes, la más peligrosa por ser el agente causal de la listeriosis (Duffes et
al., 1999; Eklund et al., 1995 y Stanier et al., 1996). Este microorganismo intracelular es
capaz de sobrevivir en los macrófagos e invadir numerosos tipos de células no
fagocíticas, como células epiteliales, hepatocitos y células endoteliales, permitiendo de
esta forma diseminarse dentro de las células sin exponerse al ambiente extracelular,
protegiéndose del sistema inmune humoral. La Lm es un enteropatógeno que mide 0,5
x 1,5 µm y capaz de sobrevivir a temperaturas de refrigeración, por lo que la infección
puede ser transmitida incluso a través de alimentos refrigerados o congelados (Gandhi
y Chikindas, 2007). Sin embargo, Lm es destruido a través de la pasteurización y por la
mayoría de los agentes desinfectantes. La continua aparición de este patógeno en las

26
plantas de procesos se debe a la formación de biofilm, siendo esta otra característica
de Lm.
Se han descrito 17 serotipos de Lm en base a sus antígenos somáticos y flagelares.
Corresponden a 15 antígenos somáticos (I-XV) y 4 flagelares (A-D). Sin embargo, los
aislamientos de muestras clínicas pertenecen mayoritariamente a los serotipos 4b y
1/2a (Liu, 2006), siendo el serotipo 4b el responsable de la mayoría de los brotes
asociados al consumo de alimentos. La Lm es el agente infeccioso causal de la
listeriosis en humanos y animales cuyas manifestaciones clínicas pueden ser variables
según el estado inmune del paciente, ya que puede presentarse como un cuadro
gastrointestinal leve o bien como un cuadro severo y de alta mortalidad, como
meningitis, abscesos cerebrales y sepsis, en pacientes inmunocomprometidos
(Doganay, 2003).

27
En base a estos antecedentes, la presente investigación tiene como finalidad purificar a
homogeneidad un péptido tipo bacteriocina producido por la cepa BALB que posee
actividad inhibitoria contra Lm, para lo cual se pretende lograr a través de dos
metodologías diferentes la purificación a homogeneidad del extracto crudo obtenida de
la cepa BALB. Éste extracto crudo será incorporado a futuro en la industria del salmón
para contribuir en la calidad higiénicas de sus productos alimenticios.
Esta investigación plantea la siguiente hipótesis
2.6 Hipótesis y objetivos
2.6.1 Hipótesis
La sustancia tipo bacteriocina (STB) producida por la cepa láctica BALB conservará su
acción inhibitoria frente Lm después de utilizar diferentes métodos experimentales para
su aislamiento y purificación a homogeneidad.
2.6.2 Objetivo general
Evaluar dos metodologías para la purificación a homogeneidad de STB producida por
la cepa BALB en caldo D-MRS, considerando como parámetro la actividad inhibitoria
frente Lm.

28
2.6.3 Objetivo específicos
Obtener un extracto crudo con actividad inhibitoria sobre Lm luego de precipitar con
acetona.
Obtener un extracto crudo con actividad inhibitoria sobre Lm luego de precipitar con
sulfato de amonio.
Purificar a homogeneidad cada uno de los extractos crudos obtenidos mediante el
empleo de columnas cromatográficas de intercambio catiónico, RP-HPLC y en geles
Tris-tricina, tomando como parámetro la actividad inhibitoria frente Lm en cada etapa.
Analizar cada una de los procedimientos considerando los rendimientos de cada
etapa.

29
3. MATERIALES Y MÉTODOS
3.1 Materiales
3.1.1 Reactivos
Acetonitrilo, C2H3N/CH3CN, (Merck).
Acetona, CH3(CO)CH3, (Merck).
Aceite de inmersión, (Winkler).
Acido acético glacial, C2H4O2, (Merck).
Acido clorhídrico, HCl, (Merck).
Ácido etilendiamino tetracético, EDTA, C10H16N2O8, (Riedel-de Haën).
Agar, (Oxoid).
Agua ultra pura HPLC, H2O, (Merck).
Alcohol Isopropìlico, H3C-HCOH-CH3, (Baker Chemical Co).
Azul de bromofenol, C19H10Br4O5S, (Merck).
Albúmina de suero bovino, BSA, (Winkler).
Azul de coomassie G-250, (Winkler).
Bisacrilamida, C3H5NO + C7H10N2O2 + H2O, (Winkler).
β-mercaptoetanol, HOCH2CH2SH, (Winkler).
Caldo Soya Tripticasa, ST, (Difco).
Citrato de amonio, (NH4)2C6H6O7, (Winkler).
Cloruro de sodio, NaCl, (Winkler).
Columna Pico Tag 3,9 x 150 mm, (Waters).
Columna Symmetry C18 4,6 x 250 mm, (Waters–Micromass).

30
Dodecil sulfato sódico, SDS, C12H25NaO4S, (Winkler).
Etanol, CH3-CH2-OH, (Winkler).
Extracto de carne, fosfato di potásico, (Bacto).
Extracto de levadura, (Bacto).
Fosfato monosódico, NaH2PO4, (Winkler).
Fosfato disódico, Na2HPO4, (Winkler).
Fosfato dipotásico, K2HPO4, (Winkler).
Glicerol, C3H8O3, (Winkler).
Glicina, C2H5NO2, (Winkler).
Glucosa, C6H12O6, (Winkler).
Hidróxido de sodio, NaOH, (Winkler).
Marcador de peso molecular proteínas, (Sigma).
Metanol, CH3OH, (Merck).
Persulfato de amonio, (NH4)2S2O8, (Merck).
Proteína peptona, (Bacto).
Tricina, C6H13NO5, (Merck).
Tris-base, (HOCH2)3CNH2, (Winkler).
TFA, C2HF3O2, (Merck).
Sacarosa, C12H22O11, (Merck).
Soya tripticasa, (Bacto).
Sep-pak C18, (Waters).
SP-fast flow, 3-acetyl-1-(3-sulfopropyl), (GE Healthcare Bio-Sciencies AB).

31
Sulfato de amonio, (NH4)2 SO4, (Merck).
3.1.2 Equipos
Agitador magnético, VELP.
Agitador orbital, STUART.
Auto clave, TEMPERA.
Balanza analítica, AND GR-200.
Baño termorregulador, MEMMERT.
Centrífuga, Hitachi CR21GII.
Cámara de electroforesis, Bio-Rad.
Cámara flujo laminar, LABCONCO.
Espectrofotómetro, Spectronic Genesys 5.
Estufa, BINDER.
Fuente de poder, Bio-rad (Power Pac Basic 300 V, 400.A, 75 W).
FPLC, GE Healthcare.
HPLC, Fisher.
Liofilizador, LABCONCO.
Pipetas, Hamilton 20µl, 100µl, 1000µl.
pH metro, HANNA INSTRUMENTS HI 932.
Incubadora Shaker, MA 410/CF, MARCONI.
Tubos ultrafiltración, Centriplus YM30, YM10 e YM3, MILLIPORE.
Ultrasonicador S30 H, ELMA SONIC.

32
3.1.3 Soluciones
Agar semi-sólido 0,75 %: Cada 100 ml contiene 0,5 g de agar-agar y 3,0 g de
soya tripticasa. Autoclavado.
Agar placa 1,5 %: Cada 100 ml contiene 1,5 g de agar-agar y 3,0 g soya de
tripticasa. Autoclavado
Buffer fosfato de sodio 0,05 M pH 8,0: Cada 100 ml contiene 0,6 g de NaH2P04 x
H2O; 0,7 g de Na2HPO4 ·2 H2O. Autoclavado.
Caldo de cultivo D-MRS: Proteína peptona 10 g/l; extracto de carne 10 g/l;
extracto de levadura 5 g/l; citrato de amonio 2 g/l, K2HPO4 2 g/l, 20 g sacarosa.
* El litro de solución contiene un buffer fosfato pH 8,0 (NaH2PO4 0,56 g/l; K2HPO4
18 g/l). Autoclavado.
Caldo soya tripticasa: Cada 100 ml contiene 3 g de soya tripticasa. Autoclavado.
Solución de equilibrio para Columna Sep-pack C18: Cada 100 ml contiene 0,154 g
de acetato de amonio 20 mM.
Solución de elución para columna Sep-pack C18: 5ml de acetato de amonio +
10 % de acetonitrilo; 5 ml de acetato de amonio + 20 % de acetonitrilo: 5 ml de
acetato amonio + 30 % acetonitrilo; 10 ml de acetato de amonio + 80 % de
acetonitrilo.
Solución gel Tris-tricina 5 %: 2,75 ml de H2O; 0,51 ml de Tris-HCl 1 M pH 6,8; 0,66
ml Acril:bis (30:0,8 %); 0,3 % de SDS; 0,03 ml de PSA 10 %; 7 µl de Temed.

33
Solución gel Tris-tricina 10 %: 1,84 ml de H2O destilada; 1,33 ml de Tris-HCl 3 M
pH 8,9; 0,8 ml acril:bis (48:1,5 %); 0,3 % de SDS; 0,018 ml de PSA 10 %; 4 µl de
temed.
Solución gel Tris tricina 17 %: 2,43 ml de agua destilada; 3,3 ml de Tris-HCl 3 M
pH 8,9; 3,3 ml de acril:bis (51:0,5 %); 1,1 ml de glicerol; 0,3% de SDS; 0,033 ml
de PSA 10 %; 3 µl de temed.
Solución de carga de muestra: 20 % de glicerol, 4µl de azul de bromofenol 1 %;
Beta-mercaptoetanol 5 %.
Tampón ánodo geles Tris-tricina : Tris-HCl 0,2 M, pH 8,9
Tampón Cátodo geles Tris-tricina: Tris-HCl 0,1 M, tricina 0,1 M y SDS 0,1 % (p/v).
Solución de fijación: Isopropanol 25 % (v/v), ácido acético 10 % (v/v), por 4 h.
Solución de teñido: Metanol 40 % (v/v), ácido acético 10 % (v/v), azul de
coomassie.
Solución de desteñido para geles de poliacrilamida: Metanol 35 % (v/v), ácido
acético 10 % (v/v).
Solución para HPLC: 225 ml de agua para HPLC; 25 ml de acetonitrilo; 2,5 ml de
ácido trifluoroacético. La solución es desgasificada con un sonicador durante
10 min.

34
3.2 Métodos
3.2.1 Cepa bacteriana
La bacteria ácido láctica, cepa BALB, fue aislada a partir de una muestra de carne
envasada al vacio, descrita por Schöbitz (1990). La cepa BALB se conservó en caldo
D-MRS con glicerol al 1 % a –20 ºC hasta iniciar su empleo.
3.2.2 Cepa indicadora
El microorganismo indicador utilizado para evaluar la actividad inhibitoria presente en la
sustancia tipo bacteriocina (STB) fue Lm Lsa 4/00, la cual fue aislada de salmón
ahumado en frío. La cepa Lsa 4/00 se conservó en caldo Soya Tripticasa, con glicerol
al 1% a -20 ºC.
3.2.3 Medios de cultivos y crecimiento de la cepa BALB
Para el crecimiento de la cepa BALB, se empleó un medio de cultivo rico en nutrientes
como es el caldo D-MRS. El cual es una variante del caldo MRS (Man Rogosa Sharpe),
al no presentar acetato de amonio entre sus ingredientes y la dextrosa también es
reemplazado por sacarosa (Shillinger et al., 1998)
Para obtener la STB, se cultivaron 1,7 litros de caldo D-MRS fraccionados en diferentes
matraces de 1 litro respectivamente, siendo estos inoculados al 1 % con la cepa BALB,
para posteriormente ser incubados a 25 ºC a 180 rpm por 24 h. El caldo D-MRS
empleado no contiene Tween 80 por ser considerado un agente interferente para las

35
etapas posteriores a la purificación (MØrtvedt et al, 1991; Muriana y Klaenhammer,
1991).
Finalizada la incubación de estos, se realizaron tinciones de Gram para confirmar la
pureza y la morfología de los cultivos. Una vez verificada la pureza de los cultivos se
recogieron los sobrenadantes por centrifugación a 9000 rpm por 15 min a 4 ºC
(Anexo 9.5).
A continuación los sobrenadantes obtenidos fueron ajustados a pH 8 con una solución
de NaOH 3 N. Este paso es considerado como el punto de partida de la purificación.
Seguidamente los sobrenadantes fueron filtrados en membranas de 0,45 y 0,22 µm
respectivamente, con la finalidad de obtener una solución estéril libre de biomasa, a
esta solución se le denominó extracto crudo (EC) o Fracción I.
3.2.4 Determinación de la actividad inhibitoria de STB
La determinación de la actividad inhibitoria, se llevó a cabo mediante la técnica “gota
sobre césped”, descrita por (Barefoot y Klaenhammer, 1983), empleando un césped de
Lm, Lsa 4/002, como microorganismo indicador.
La Fracción I, fue diluída seriadamente (1:1, 1:2, 1:4, 1:8, 1:16, 1:32) en buffer fosfato
0,05 mM a pH 7,0. A continuación se extrajo de cada dilución 20 µl para luego ser
inoculadas sobre una placa de agar soya tripticasa que contiene el césped indicador,
dejando como control el EC sin diluir. Una vez secadas las gotas sobre el césped
fueron incubadas a 25 ºC ± 2 ºC por 18 h, finalizado el tiempo de incubación, se
procedió a detectar la actividad inhibitoria, la cual fue registrada con la aparición de
halos de inhibición presente en la placa de agar, la unidad de actividad (AU) inhibitoria

36
se determinó como el inverso de la dilución más alta que origina un halo de inhibición
presente en 20 µl. el ensayo de actividad inhibitoria se observa en la figura 5.

37
Figura 5. Técnica “gota sobre césped” (1) 20 µl de la sustancia tipo
bacteriocina son inoculados sobre una placa de agar con la cepa
indicadora “Lm”, la placa de agar es incubada por 18 h a 25 °C. La
actividad inhibitoria se detecta con la aparición de halos de inhibición (2)
Determinación de la actividad inhibitoria presente en el extracto crudo
(E.C) detectada por el ensayo “gota sobre césped”.
Fuente: Barefoot y Klaenhammer, 1983

38
3.2.5 Purificación a homogeneidad de la cepa BALB
La purificación a homogeneidad de la sustancia tipo bacteriocina (STB), se realizó a
través de diferentes técnicas cromatográficas, lo cual permiten aislar y caracterizar un
péptido, en función de sus diferentes propiedades físico-químicas, como por ejemplo, el
tamaño molecular, su carga electroestática neta y su hidrofobicidad.
Los procedimientos empleados para la purificación con la metodología con acetona
presentaron algunas modificaciones a los descritos por zendo, 2006 al incluir nuevas
etapas de purificación, como los tubos centriplus. La precipitación con sulfato de
amonio, se llevo a cabo según lo descrito por Jack, 1996.
El procedimiento de purificación son descritos a continuación:
1) La purificación a homogeneidad de la STB comienza, cuando el cultivo de la cepa BALB
es cultivado en el caldo D-MRS, como se describe en la sección 3.2.3.
2) Los sobrenadantes del cultivo productor fueron recogidos, cuando el cultivo se
encontraba en el inicio de la fase estacionaria. Registrándose en esta fase la mayor
secreción de la STB, hacia el medio extracelular, de acuerdo a lo descrito por Lamas
(2002).
Una vez obtenidos los sobrenadantes libres de células fueron congelados a –20 ºC
hasta su empleo. Estos sobrenadantes se les denominaron extracto crudo (EC) o
Fracción I.
3) A continuación, se llevó a cabo la precipitación de la Fracción I con acetona y sulfato de
amonio respectivamente.

39
Precipitación con acetona: 250 ml de la Fracción I fueron concentradas con 750 ml
de acetona. La incorporación de acetona fue realizada gradualmente y en agitación
constante a 4 ºC. Posteriormente la solución fue almacenada a -20 ºC por 18 h (Zendo
et al, 2006). Una vez finalizado este tiempo de precipitación, se procedió a centrifugar
la solución a 9000 rpm a 4 ºC por 15 min, con la finalidad de obtener un precipitado
homogéneo libre de acetona, seguido este paso se procedió a eliminar los restos de
acetona presentes en la solución, para lo cual se utilizó nitrógeno gaseoso. Una vez
obtenido el precipitado libre de acetona, fue solubilizado en 18 ml de buffer fosfato
pH 5,6. La muestra obtenida en esta etapa se denominó Fracción IIa (Anexo 9.6).
Precipitación con sulfato de amonio: 516 g de sulfato de amonio (saturación al 65
%), fueron incorporados de forma gradual y en agitación constante a 4 ºC para 1 litro
de EC o Fracción I. La muestra se mantuvo en agitación constante durante 3 h a 4 ºC.
Finalizado este tiempo de agitación, se procedió a centrifugar la solución a 9000 rpm a
4 ºC por 15 min. El precipitado obtenido fue solubilizado en 18 ml de buffer fosfato
0,05 mM pH 5,6. Obteniéndose de esta etapa la Fracción IIb. (Anexo 9.6).
4) La Fracción IIa y IIb fueron desaladas mediante cromatografía de filtración en gel
empleando mini columnas de polipropileno Hi-Trap desalting, (GE Healthcare) pre-
empaquetadas con 1 ml de Sephadex G-25 en un equipo de FPLC (GE Healthcare).
Para ello se depositaron 1,5 ml de la Fracción IIa y IIb en la columna Hi-trap
previamente equilibrada con 30 ml de buffer fosfato pH 5,6. Todas las fracciones
desaladas se juntaron y formaron la Fracción IIIa y Fracción IIIb respectivamente
(Anexo 9.1).

40
5) Las fracciones anteriores fueron ultrafiltradas en tubos de Centriplus a través de
membranas de 30, 10 y 3 kDa de diámetro nominal de corte (YM30, YM10, YM3)
siguiendo las instrucciones señaladas por el fabricante (Millipore). Los extractos
concentrados con actividad inhibitoria fueron congelados a -20 °C, formando las
Fracciones IVa y IVb.
6) Posteriormente, las Fracciones IVa y IVb, fueron sometidas a una cromatografía de
intercambio catiónico SP-Fast Flow* (GE Healthcare) empleando una columna de
1,5 × 18 cm. La columna fue equilibrada con 70 ml de solución buffer fosfato 50 mM pH
5,6. Una vez equilibrada la columna, se procedió a aplicar lentamente, con una pipeta
pasteur 500 µl de la fracción IVa y IV b. Una vez ingresada la muestra en la resina la
columna fue lavada con 50 ml de buffer fosfato pH 5,6. La muestra fue eluída en un
gradiente escalonado de sal de 0,1 a 1 M de NaCl a un flujo moderado de 1 ml/min. Las
fracciones eluídas con actividad antimicrobiana, fueron recolectadas en una sola
fracción, formando las Fracción Va y Fracción Vb. Una vez obtenidas estas fracciones
se ajustaron a pH 8,0 con NaOH 3 N. Las fracciones Va y Vb fueron liofilizadas por
24 h, para luego ser resuspendidas en 2 ml de buffer fosfato pH 5,6, formando las
Fracciones VIa y VIb.
* La Cromatografía de intercambio iónico está basada en la atracción entre iones de
soluto y de los grupos cargados que existen en la fase estacionaria de la resina. En el
caso de intercambiadores catiónicos, los grupos cargados negativamente en la fase
estacionaria atraen a los cationes presentes en el soluto o muestra a purificar.

41
7) Las Fracciones VIa y VIb proveniente de la etapa anterior, fueron repurificadas
mediante cromatografía de fase reversa utilizando cartuchos rellenos de C18*. Estas
minicolumnas o cartuchos fueron lavados con 10 ml de etanol (99%) y con 5 ml de
acetonitrilo y para posteriormente ser equilibrados con 10 ml de acetato de amonio
20 mM. Para ello, se depositaron 2 ml de la Fracciones VIa y VIb respectivamente en la
columna C18 a un flujo moderado de 1 ml/min, lavando el material no adsorbido con
5 ml de acetato de amonio 20 mM. La muestra retenida se recuperó al eluir la muestra
en un gradiente de acetato de amonio 20 mM, que contienen diferentes porcentajes de
acetonitrilo (10 %, 20 %, 30 %, 80 %). Las fracciones con actividad fueron recolectadas
en una sola fracción, formando las Fracciones VIIa y VIIb, respectivamente.
* Las minicolumnas de fase reversas Sep-pack C18, son columnas que vienen
preempaquetadas con un soporte de sílica, donde llevan unidas covalentemente
cadenas hidrocarbonadas de 18 átomos de carbono. El principio de separación de
molécula de la fase reversa se basa en las interacciones hidrofóbicas entre las
moléculas de soluto en la fase móvil y el ligando hidrofóbico inmovilizado en la fase
estacionaria.
8) Las Fracciones VIIa y Fracciones VIIb obtenidas en la etapa anterior fueron
repurificadas a homogeneidad mediante una cromatografía liquida de fase reversa (RP-
HPLC*) utilizando una columna Simmestry C18 de 4,6 x 250 mm, con tamaño de
partícula de 5 µm (Waters), las muestras fueron monitoreadas a 210 nm. Las fracciones
VIIa y VIIb retenidas en la columna fueron eluídas utilizando un método socrático a un
flujo de 1 ml/minuto. La fase móvil se describe en la sección 3.1.3 de materiales y

42
métodos. Las fracciones VIIIa y VIIIb obtenidas en esta etapa, fueron recolectadas y
guardadas a -20 ºC.
* Todas las fracciones obtenidas en las diferentes etapas de la purificación fueron
recolectadas a 4 ºC, con la finalidad de preservar la actividad inhibitoria presente en
estas fracciones.
* La cromatografía líquida de alta eficacia o HPLC, es el método más sofisticado y
moderno de cromatografía líquida, esta técnica proporciona retención y selectividad
óptimas, cuando las muestras tienen carácter predominantemente alifático o aromático.
La HPLC ofrece una mayor variedad de fases estacionarias, lo que permite una mayor
gama de estas interacciones selectivas y más posibilidades para la separación.
3.2.6 Liofilización de las muestras
Todas las fracciones obtenidas durante las diferentes etapas cromatográficas
(FPLC, SP-Fast Flow, Sep-pak y RP-HPLC), fueron todas ellas concentradas por
liofilización con nitrógeno líquido en un equipo LABCONCO, con el objeto de obtener
un sólido de baja densidad.
En la figura 6, se detalla la metodología empleada para llevar a cabo la purificación a
homogeneidad de la STB.

43
Figura 6. Metodología empleada para la purificación de la STB

44
3.2.7 Geles de Tris-tricina
El gel de Tris-tricina descrito por Schägger y Von Jagow (1987), están constituido por
tres fragmentos de geles, los cuales poseen diferentes densidades de poliacrilamida: el
primer gel superior o concentrador, el gel intermedio o espaciador y el gel inferior o
separador. Los geles de Tris-tricina son muy resolutivos para la separación de
proteínas o péptidos en función de su tamaño. La primera parte del gel mide 6,5 cm
conteniendo una relación del 48:3 % (p/v) de acrilamida/bisacrilamida. La segunda
parte del gel mide aproximadamente 2 cm de alto, presentando una relación de un
48:15 % (p/v) de acrilamida/bisacrilamida. La tercera parte que corresponde al gel
espaciador, mide aproximadamente 2 cm de alto con una relación de 30:0,8 %
acrilamida/bisacrilamida.
Las muestras se hicieron migrar aplicando una corriente de 22 mA por 5 h
aproximadamente. El tampón ánodo se coloca al interior de la cámara electroforética y
el tampón de cátodo afuera. Finalizada la electroforesis, el gel fue fijado en la solución
de fijación durante al menos 4 horas y posteriormente, se tiñó en una solución de
tinción azul de Coomassie por 1 h. Para desteñir el gel se utilizó una mezcla de
metanol y ácido acético, permaneciendo en esta solución hasta la aparición de las
bandas de proteínas y eliminación del fondo. Las soluciones electroforéticas se
describen en la sección 3.2.1.
Para evaluar la actividad inhibitoria presente en el gel Tris-tricina, se procedió a lavar el
gel durante 30 min en agua desionizada estéril, con la finalidad de eliminar los restos
de buffer presente en el gel. Una vez terminado el tiempo de lavado se procedió a

45
colocar el gel sobre una placa de agar soya tripticasa, para después agregar sobre el
gel una capa de agar semi-sólido (7 ml), conteniendo la cepa indicadora Lm (0,1 ml).
Esta placa fue incubada a 25 ºC durante 16-18 h, la aparición de una zona de
inhibición, es indicadora de la actividad inhibitoria frente a Lm.
3.2.8 Determinación de proteína mediante el método de Bradford
La determinación cuantitativa de proteínas se llevó a cabo por el método descrito por
Bradford (1976). El colorante azul de Commassie se une a la proteína de modo
inespecífico. El reactivo colorante fue diluido 5 veces en buffer fosfato 0,05 M pH 5,6
hasta obtener una concentración 1X, se tomaron 2 ml de la solución 1X y se mezclaron
con 10 µl de muestra. Las diferentes muestras obtenidas en las diferentes etapas de la
purificación fueron incubadas a temperatura ambiente por 10 min, para posteriormente
leer la absorbancia a 595 nm en un espectrofotómetro Genesys 5.
La curva de calibración se preparó a diferentes concentraciones de BSA de 0,4375;
0,875; 1,313, 1,750; 2,188; 2,625; 3,063; 3,238; 3,325 mg/ml. Finalmente, la
concentración de las muestras se calculó por interpolación de la absorbancia obtenida
en la curva de calibración (Anexo 9.2).
3.2.9 Determinación de la composición aminoacídica de la STB
El análisis de aminoácidos fue realizado por HPLC por medio de la derivatización de los
aminoácidos libres con fenilisotiocianato (PITC), el cual permite identificar y cuantificar
los derivados feniltiocarbamilo-aminoácidos a través de un cromatógrafo líquido de alta

46
resolución, integrado a un detector UV, empleando una columna Pico Tag, diámetro de
partícula de 4 µm, con una dimensión de 3,9 x 150 mm (Waters). Las muestras
provenientes de la Fracción VIIIa y Fracción VIIIb descritas en la sección 3.2.5, fueron
hidrolizadas con HCl 6 N y fenol 1 %, por 24 h, a 110 ºC (Anexo 9.4) para obtener los
aminoácidos libres que componen la STB. Una vez realizado el tratamiento se procedió
analizar la mezcla obtenida a través de una columna de HPLC, la derivatización de los
aminoácidos fue detectada a 254 nm, teniendo como referencia el tiempo de retención
para la identificación de los aminoácidos que lo componen para ser comparados con la
solución patrón.
En la figura 7 se presenta la reacción de derivatización de los aminoácidos libres con
PITC.

47
Figura 7. Reacción de derivatización de los aminoácidos libres con
PITC.
Fuente: White y Hart, 1992a.

48
4. RESULTADOS
Las bacteriocinas son un variado grupo de péptidos antimicrobianos, que poseen
algunas características comunes entre sí, como el tamaño molecular, la carga neta, y
su hidrofobicidad, lo que permitió adoptar metodologías generales de purificación.
Para la purificación a homogeneidad de la STB, se utilizaron diferentes metodologías
comprendidas en el protocolo descrito por (Zendo et al., 2006) con algunas
modificaciones.
Como se indicó anteriormente, se tomó como punto de partida los sobrenadantes libres
de biomasa, denominándoles extractos crudos. El crecimiento bacteriano de la cepa
BALB se detuvo cuando el cultivo se encontraba en la fase exponencial tardía,
momento en el cual se registra una mayor producción de la STB.
En las Tablas II y III se presentan los pasos seguidos para dicha purificación: Centriplus
YM30, cromatografía de intercambio catiónico (Sp-Fast Flow), cromatografía de fase
reversa sobre soporte sólido empleando matrices de C18 y finalmente repurificación
mediante cromatografía líquida en fase reversa de alta resolución (HPLC) sobre C18.
En cada unos de estos pasos se evalúo la actividad inhibitoria como un parámetro de
detección de la STB.
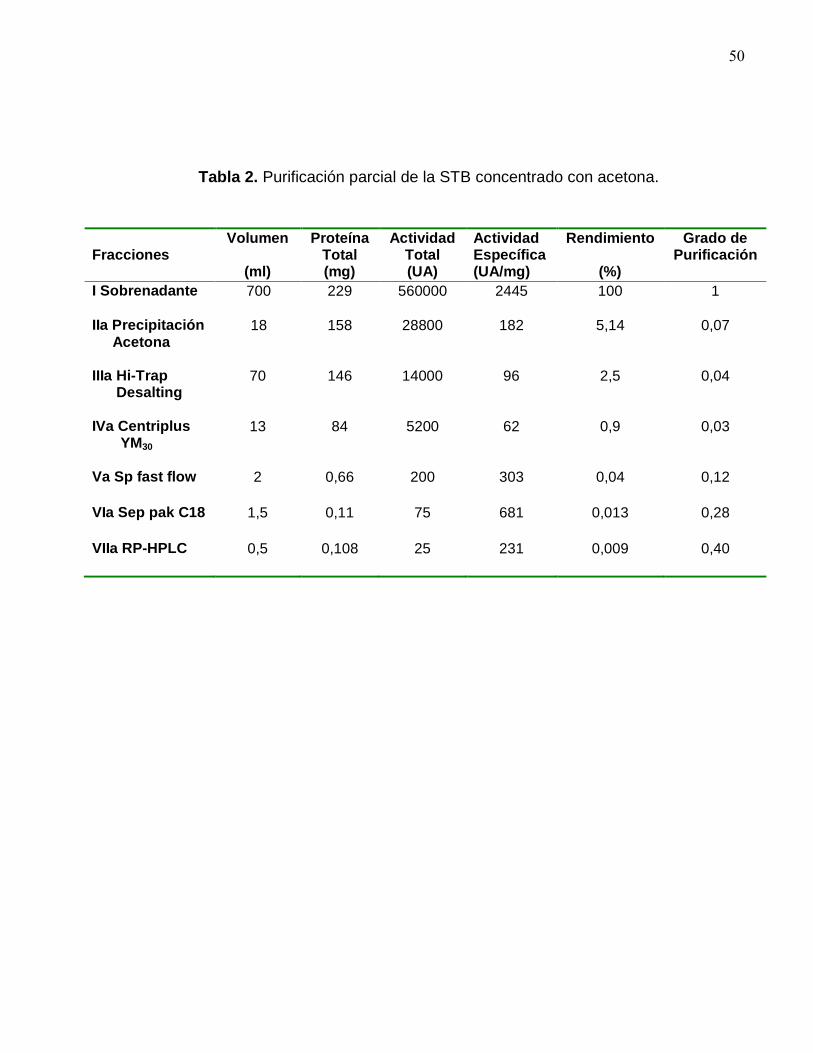
La actividad específica del extracto crudo o Fracción I fue de 24445 UA/mg.
A continuación la Fracción I fue concentrada por precipitación con acetona (Fracción Ia)
y sulfato de amonio (Fracción Ib) respectivamente. Los sedimentos obtenidos fueron
resuspendido en un buffer fosfato 50 mM pH 5,6. El rendimiento de este paso fue del
5,14 % para la precipitación con sulfato de amonio y de un 18 % para la precipitación

49
con acetona. La actividad específica para la Fracción Ia y Ib fue de 182 UA/mg y 118
UA/mg respectivamente.
En las Tablas 2 y 3, se muestra el proceso de purificación de la STB, donde se detallan
entre otros resultados la actividad inhibitoria específica y el grado de purificación
presente en cada uno de estas etapas.
50
Tabla 2. Purificación parcial de la STB concentrado con acetona.
Fracciones
Volumen
(ml)
Proteína Total (mg)
Actividad Total (UA)
Actividad Específica (UA/mg)
Rendimiento
(%)
Grado de Purificación
I Sobrenadante 700 229 560000 2445 100 1
IIa Precipitación Acetona
18 158 28800 182 5,14 0,07
IIIa Hi-Trap Desalting
70 146 14000 96 2,5 0,04
IVa Centriplus YM30
13 84 5200 62 0,9 0,03
Va Sp fast flow 2 0,66 200 303 0,04 0,12
VIa Sep pak C18 1,5 0,11 75 681 0,013 0,28
VIIa RP-HPLC 0,5 0,108 25 231 0,009 0,40

51
Tabla 3. Purificación parcial de la STB concentrado con sulfato de amonio.
Fracciones
Volumen
(ml)
Proteina Total (mg)
Actividad Total (UA)
Actividad Especifica (UA/mg)
Rendimiento
(%)
Grado de Purificación
I Sobrenadante 1000 475 1600000 3368 100 1
IIb Precipitación (NH4)2SO4
18 244 28800 118 18 0,035
IIIb Hi-Trap Desalting
70 66 56000 85 3,5 0,252
IVb Centriplus YM30
15 3,85 6000 1558 3,8 0,462
Vb Sp fast flow 2 1,03 200 194 0,13 0,058
VIb Sep pak C18 1,5 0,318 75 235 0,05 0,070
VIIb RP-HPLC 0,5 0,050 - - - -

52
El grado de purificación se calculó como la actividad específica de la Fracción
X/Actividad específica de la Fracción I
El rendimiento se calculó como la actividad total de la Fracción X/Actividad total
de la Fracción I, multiplicado por 100.
En la figura 8 se observa la actividad inhibitoria de la STB frente a Lm, durante las
primeras etapas de la purificación, Fracción I, Fracción Ia y Fracción Ib.

53
Figura 8. Actividad inhibitoria detectada por el ensayo “gota sobre
césped” utilizando como microorganismo indicador Lm. (A)
Actividad inhibitoria presente en el extracto crudo. (B) Actividad
inhibitoria presente en el precipitado con acetona. (C) Actividad
inhibitoria presente en el precipitado con sulfato de amonio.
A
B C

54
4.1 Cromatografía de filtración en gel Sephadex G-25 Hip-Trap
Para desalinizar las fracciones Ia y Ib provenientes de las diferentes precipitaciones, se
utilizaron columnas de filtración en gel Hi-TrapTM (Desalting Columnm, GE Healthcare).
Las primeras ocho fracciones estaban libres de sales (Anexo 9.1), siendo estas
recolectadas en una sola fracción (fracción IIa y IIb respectivamente). Las fracciones IIa
y IIb fueron liofilizadas con el objeto de obtener fracciones más concentradas. La
actividad específica de esta etapa fue 96 AU/mg para la Fracción IIa y 85 AU/mg para
la fracción IIb.
4.2 Ultrafiltración en tubos Centriplus
Las fracciones obtenidas en el paso anterior fueron ultrafiltradas, en tubos Centriplus,
con diferentes cortes de membranas como se describe sección 2.4.6 de materiales y
método. La ultrafiltración de las fracciones IIa y IIb, tuvo como objetivo la depuración de
la STB, donde las fracciones fueron separadas según el tamaño molecular presente en
la solución. Una vez finalizada la ultrafiltración se procedió a determinar la actividad
antimicrobiana presentes en cada una fracciones obtenidas en los tubos Centriplus
(retenido y permeado).
En un principio todas las fracciones de retenidos y permeados de los tubos Centriplus
presentaron actividad inhibitoria frente a Lm, pero la actividad no perduró en todos los
tubos de Centriplus. Los únicos que conservaron la actividad antimicrobiana fueron los
tubos con membrana de corte de 30 kDa. El resto de las fracciones fueron descartadas
al no presentar actividad antimicrobiana frente a Lm.

55
4.3 Cromatografía de intercambio catiónico
Las fracciones IIIa y IIIb fueron sometidas a una cromatografía de intercambio iónico
SP Fast Flow, como se detalla en la sección 3.2.5. La actividad inhibitoria de la STB fue
eluida con 0,6 M de NaCl (Figura 9). Las fracciones entre 52-60, presentaron actividad
inhibitoria frente Lm. La actividad específica de la fracción IVa fue de 303 AU/mg y 194
UA/mg para la Fracción IVb.

56
Figura 9. Cromatograma y actividad inhibitoria obtenido por
SP–Fast Flow. (A) Perfil de elución de la STB. B) Actividad inhibitoria
de la Fracción Va obtenida en las diferentes fracciones cromatográficas.
(C) Actividad inhibitoria de la Fracción Vb en las diferentes fracciones
cromatográficas. Las fracciones con actividad inhibitoria frentes Lm
fueron recolectadas, para continuar con el protocolo de purificación.
B C
A

57
4.4 Cromatografía de fase reversa sobre soporte sólido C18
Las fracciones Va y Vb provenientes de la cromatografía intercambio catiónico, fueron
sometidas a cromatografía de fase reversa utilizando cartuchos rellenos de C18, como
se describe en la sección 2.3.4 materiales y métodos. Para ello, las muestras se
filtraron a través de C18 a un flujo moderado de 1 ml/min. El material retenido se
recuperó (STB) eluyendo con distintos porcentajes (entre el 30-80 %) del solvente de
elución como se indica en la sección 2.3.4. La actividad inhibitoria de las muestras
recogidas fueron ensayadas mediante la técnica gota sobre césped frente a Lm antes
de ser congeladas a -20 ºC. La actividad específica de este paso fue de 681 AU/mg y
235 AU/mg, para las fracciones VIa y VIb respectivamente. Una vez recolectadas las
fracciones con actividad inhibitoria (Figura 10), fueron liofilizadas con la finalidad de
concentrar, para continuar con el siguiente paso de la purificación.

58
Figura 10. Halos de inhibición presente en las Fracciones VIa y
VIb. Ensayadas sobre un césped de Lm.
Fracción VIa
Fracción VIb

59
4.5 Cromatografía liquida de alta resolución HPLC
Las fracciones con actividad antimicrobiana obtenidas del paso anterior fueron
sometidas a una cromatografía líquida de fase reversa RP-HPLC, en una columna
Simmetry C18, con un tamaño de partícula de 5 µm y una dimensión de 4,6x 250 mm.
En este paso se obtuvo una purificación a homogeneidad de la STB para ambas
metodologías descritas en la sección 3.2.5 de materiales y métodos. Los perfiles se
observan en la (Figura 11).

60
Figura 11. Cromatogramas obtenidos por RP-HPLC. (A) Perfil
cromatográfico de la Fracción VIIa, el perfil destacado con rojo presenta
actividad inbitoria frente Lm. (B) Perfil cromatográficos de la Fracción
VIIIb.

61
4.6 Electroforesis en geles de Tris-tricina en condiciones desnaturantes
Para determinar el peso molecular de la STB, los extractos semi-purificados de
fracciones VIIIa y VIIIB obtenidas por RP-HPLC, como se describe en la sección 3.2.5,
fueron sometidas a electroforesis en geles de Tris-tricina en condiciones desnaturantes,
como se describe en la sección 3.2.7 de materiales y métodos.
Se realizaron dos geles bajos las mismas condiciones electroforéticas, el primer gel fue
fijado con gluteraldehido y teñido con azul de Coomassie, el segundo gel se evaluó la
actividad inhibitoria presentes en las fracciones VIIIa y VIIIb como se describe en la
sección 3.2.7.
La fracción VIIIb no presentó ninguna banda en el gel de poliacrilamida y no se detecto
la actividad inhibitoria frente Lm.
El peso molecular aproximado de la sustancia inhibitoria de la fracción VIIIa fue de
3 kDa, como se observa en la figura 13. El peso molecular fue obtenido por la
interpolación de los pesos moleculares (Anexo 9.3) y la actividad inhibitoria fue
detectada en la fracción VIIIa, como se observa en la (Figura 12).

62
Figura 12. (A) Gel Tris-tricina de las Fracciones VIa y VIb; Carril 1:
estándar de peso molecular (sigma).- 26600, 17000, 14200, 6500, 3496,
1060 Da. Carril 2: Fracción VIIa. Carril 3: Fracciones VIIb. (B) Detección de
la actividad inhibitoria sobre L. monocitogenes de las fracciones VIIIa y
VIIIb.
Fracción VIIIa

63
4.7 Determinación de la Composición aminoacídica de la STB
El análisis de aminoácidos fue realizado en un sistema PICO TAG (Waters), donde la
STB fue sometida a una hidrólisis ácida, por 24 h a 110 ºC. Como se indica en la
sección 7.4. La identificación de los aminoácidos fue realizada a través de una
cromatografía en HPLC. Los aminoácidos libres provenientes de la hidrólisis ácida, son
derivatizados con fenilisotiocianato PITC. La sustancia tipo bacteriocina posee los
siguientes aminoácidos: ácido aspártico, ácido glutámico, glicina, histidina,treonina,
alanina, prolina, valina, tirosina, metionina, isoleucina, fenilalanina y lisina, siendo estos
comparados con el tiempo de retardo, de la solución patrón (Anexo 9.4).
Los perfiles cromatográficos de la sustancia tipo bacteriocina, como la solución patrón
de los aminoácidos se muestran en la (Figura 13).

64
Figura 13. Cromatogramas de la derivatización de aminoácidos obtenido
por HPLC (A) Perfil cromatográfico del producto feniltiocarbomil aminoácido
proveniente de la hidrólisis ácida de la Fracción VIIIa. (B) Perfil cromatográfico
del producto feniltiocarbomil, proveniente de la hidrólisis ácida, de la solución
patrón. La fracción VIIIb, no se obtuvo un perfil cromatográfico.
A
B

65
5. DISCUSION
Los avances en el campo de la biotecnología han requerido del uso de técnicas y
metodologías más eficaces para la separación y purificación de péptidos
antimicrobianos, como las bacteriocinas. El empleo de diferentes técnicas
cromatográficas, han permitido la separación de estos a partir de medios de cultivos
complejos. En la actualidad se han aislado y caracterizado una amplia variedad de
bacteriocinas siendo todas ellas clasificadas según sus diferentes propiedades
moleculares.
La biosíntesis de la STB está estrechamente relacionada con la fase de crecimiento del
microorganismo productor, como lo describe Lamas, (2002). Detectándose la presencia
de la STB desde el inicio de la fase exponencial, alcanzando un máximo de producción
justo antes que el cultivo entre a la fase estacionaria. También en esta fase
estacionaria se ha registrado la disminución de la actividad inhibitoria, hasta
desaparecer por completo al cabo de 24 h. Esta inactivación, ha sido descrita por
varios autores (Geis et al., 1983; Piard et al., De Vuys et al., 1992), quienes explican
que la posible inhibición se deba a la acción de enzimas proteolíticas liberadas por el
microorganismo productor o por el aumento de la acidez registrada en esta fase del
crecimiento. Estos resultados concuerdan con el crecimiento de otras bacteriocinas
descritas hasta la fecha.
Además se ha demostrado que la STB es estable a un intervalo de pH 5 a 8, lo cual fue
demostrado por Lamas (2002) detectándose la mayor actividad inhibitoria de la STB,
cuando el medio de cultivo es alcalino. En este trabajo se obtuvo la mayor unidad de

66
actividad cuando el pH era 8. Esto se puede deber a que las bacteriocinas se
encuentran cargadas positivamente a pH fisiológico, debido a la presencia de
aminoácidos catiónicos, como Lis y His. El pH de amortiguación afecta a la ionización
de las cadenas laterales provocando la desprotonación de los residuos aminoácidicos,
favoreciendo la unión de la bacteriocina a la membrana (Chen y Montaville, 1995).
El análisis de la estructura tridimensional 3D ha demostrado que las bacteriocinas, que
no poseen puentes de disulfuro en el extremo C- terminal pierden parte de la estructura
helicoidal a altas temperaturas y son significativamente menos activas a 37 ºC que a
25 ºC, dejando en evidencia que la forma helicoidal es crítica para la actividad de las
bacteriocinas (kaur et al., 2004). Estos resultados explicarían lo observado durante la
purificación de la STB, ya que cada vez que se expuso una muestra a temperaturas
superiores a 25 ºC la actividad de la STB disminuía o desaparecía, por completo.
El medio de cultivo empleado para obtener la STB fue el caldo D-MRS por ser el medio
de cultivo con mejores resultados de actividad inhibitoria descrito por Lamas (2002), el
caldo D-MRS empleado en este trabajo no contaba con Tween 80 por ser considerado
un agente interferente en las etapas posteriores a la purificación, en especial en la
secuenciación de péptidos (MØrtvedt et al., 1991; Muriana y Klaenhammer, 1991).
También se ha registrado que interfieren en la producción y/o detección de la actividad
inhibitoria, como es el caso de la enterocina A y B, que en presencia de Tween 80
disminuye su producción (Aymerich et al., 2000). Sin embargo, Tween 80 se
caracteriza por ser un agente antiagregante, impidiendo a la formación de agregados,
también favorece a la secreción de las bacteriocinas (Nissen-meyer, et al., 1993;
Aymerich et al., 2000).

67
Antes de proceder a la purificación se evaluaron las condiciones óptimas para el
crecimiento de la cepa BALB, con el fin de aislar la sustancia activa del resto de las
otras proteínas y péptidos presentes en del medio de cultivo productor.
Para la realización de este trabajo se planteó como objetivo general evaluar dos
metodologías de purificación que difieren solamente en la etapa de la concentración de
la STB, realizándose con acetona y sulfato de amonio respectivamente, registrándose
en cada una de estas etapas la actividad inhibitoria frente Lm.
Debido a la heterogeneidad de las bacteriocinas, los protocolos de purificación
utilizados, se han diseñado empíricamente para cada uno de ellos. La mayoría de las
metodologías empleadas para llevar a cabo una purificación a homogeneidad,
incorporan precipitaciones con sulfato de amonio, solventes orgánicos entre otros,
cromatografía de intercambio iónico y/o interacción hidrofóbica, y finalmente HPLC en
fase inversa.
A continuación, la purificación a homogeneidad de la (STB), se realizó según la
metodología descrita por (Zendo. et al., 2006), con algunas modificaciones. El primer
paso de la purificación consistió en la precipitación con acetona y sulfato de amonio
respectivamente, este procedimiento tuvo como objetivo concentrar la STB presente en
los sobrenadantes de los cultivos productores. Este paso demostró que la STB resistía
al proceso de la precipitación y que ambos precipitados presentaban una alta unidad de
actividad de inhibición.
El segundo paso de la purificación consistió en desalinizar las muestras provenientes
de las precipitaciones. La precipitación con acetona no necesitaba pasar por la etapa
de desalinización, ya que se precipitaba con un solvente orgánico, sin embargo, una

68
vez precipitado el sobrenadante de BALB se observó un precipitado blanco sugiriendo
la presencia de sales provenientes del medio de cultivo. Por lo cual se procedió a
desalinizar en las columnas Hi-Trap desalting, como se describe en la sección 3.2.5,
donde se muestran los cromatogramas obtenidos, destacándose la alta conductividad
obtenida para ambas precipitaciones (Anexo 9.1).
El tercer paso de purificación se basó en la ultrafiltración de las Fracciones IIIa y IIIb,
utilizándose tubos Centriplus, con diferentes cortes de membrana, este procedimiento
tuvo como objeto la separación de moléculas presentes en estas fracciones IVa y IVb
de acuerdo a su tamaño molecular. Sin embargo, este paso presenta desventajas, ya
que se registró una pérdida de la actividad inhibitoria después de 24 h realizada la
ultrafiltración en los tubos con membrana de corte YM10 y YM3, siendo estos
descartados para la continuidad de la purificación.
El cuarto paso de la purificación se realizó una cromatografía de intercambio catiónico,
utilizando como resina SP-Fast Flow descrita en la sección 3.2.5. Una de las ventajas
que presenta este tipo de cromatografía es su alta capacidad de resolución, la alta
capacidad de procesamiento de muestras y la elevada concentración de componentes
activos (Karlsson et al., 1989). El resultado principal obtenido en esta etapa fue la unión
de la STB al grupo sulfopropil de la resina, uniéndose fuertemente a ésta ya que la STB
eluyó a una alta concentración de sal (0,7 M de NaCl), el cual indicó que la STB posee
una alta carga neta positiva.
Esta característica de la STB aumentó el poder resolutivo de esta cromatografía,
obteniéndose un grado de purificación del 0,12 % para la metodología desarrollada con
acetona y de un 0,058% desarrollada para la metodología con sulfato de amonio y, es

69
en esta etapa donde se registró una disminución en la concentración de los
componentes que conforman las fracciones con actividad inhibitoria de la STB. La
actividad específica de la STB no aumentó al final de esta etapa, lo cual podría indicar,
que existe una baja secreción de la STB hacia el medio de cultivo de la cepa
productora.
Por otra parte, el perfil cromatográfico obtenido por HPLC como se observa en la
figura 11, la STB fue detectada a los 3 minutos una vez iniciada la corrida. Se obtuvo
un pico cromatográfico mayoritario, estrecho y simétrico. Éste resultado obtenido indica
que se está en presencia de una molécula altamente hidrofílica, debido a que
prácticamente no fue retenido en la matriz C18 empleada. La ventaja principal de esta
cromatografía en fase reversa aún cuando el inhibidor se retiene poco, es el tiempo de
retención en la columna el cual fue suficiente para eluir completamente las sales. Sin
embargo, la actividad inhibitoria presente en las fracciones provenientes de HPLC, sólo
fue detectada en esta etapa la metodología iniciada con acetona, una vez realizado el
gel Tris-tricina. La actividad inhibitoria en esta etapa no fue detectada cuando se realizó
la técnica gota sobre césped, aun cuando se duplicó y triplicó el volumen de
inoculación.
El peso molecular aproximado de la STB fue de 3 kDa, el cual fue obtenido, a través,
de geles de Tris-tricina, observándose en este gel una banda muy débil para la
metodología iniciada con acetona. Estos resultados fueron confirmados cuando se
realizó la determinación de la composición aminoacídica de la STB.
En la figura 13 se puede observar el perfil cromatográfico obtenido por la metodología
con acetona, donde se observa la presencia de 14 aminoácidos para la STB, pero en la

70
metodología con sulfato de amonio, no se observa ningún perfil cromatográfico, y por
ende ninguna banda en los geles de Tris-tricina. Una vez comparado con el perfil
patrón de aminoácidos, se puede concluir, que no existen aminoácidos inusuales en la
estructura primaria de la STB.
Al comparar los resultados obtenidos con la precipitación con acetona y sulfato de
amonio (Tabla 2 y 3) se pudo observar que en términos generales se obtiene una
escasa purificación con ambas técnicas, siendo la purificación parcial con acetona
ligeramente superior a la metodología empleada con sulfato de amonio, al conservar la
actividad inhibitoria de la STB hasta las últimos pasos de la purificación. En general, las
diferencias entre las dos metodologías no son tan marcadas, el rendimiento obtenido
es muy bajo para ambas de un 0,4 % para acetona y 0,07% para sulfato de amonio y la
actividad específica de la STB no aumentó al final del proceso, lo cual indicaría, que
existe una baja concentración de la STB en el sobrenadante de cultivo de la cepa
productora. La única probable ventaja que presentaba la precipitación con acetona era
que se obviaba el procedimiento de diálisis, sin embargo, en la práctica no fue así, ya
que en ambas precipitaciones tuvieron que ser desalinizadas, no obstante la
metodología con acetona es bastante más corta y fácil de llevar a cabo.
De acuerdo a la hipótesis planteada la STB producida por la cepa BALB logra mantener
su actividad inhibitoria frente Lm durante todas las etapas de la purificación realizada.

71
La aplicación de la STB en la industria alimentaria presenta un gran potencial como un
biocontrolador natural, para prevenir el crecimiento de microorganismos patógenos,
como Listeria monocytogenes que provocan alteraciones en los alimentos
contaminados.
Estos tipos de inhibidores provenientes de las BAL presentan una alternativa segura
para ser implementadas, como nueva tecnología de control de la calidad. Sin embargo,
la aplicación de la STB debe ser aplicada en combinaciones con otras STB o
bacteriocinas producidas por otros géneros que componen al variado grupo de las
bacterias ácido lácticas, para impedir la aparición de microorganismos resistentes.

72
6. CONCLUSIÓN
De los resultados obtenidos en el presente trabajo se han extraído las siguientes
conclusiones:
Se ha identificado y caracterizado parcialmente un péptido catiónico tipo
bacteriocina de peso molecular aproximado de 3 kDa con actividad inhibitoria
contra Lm
Este péptido catiónico no presentaría en su estructura primaria aminoácidos
inusuales como lantionina, lo cual permite inferir con los resultados obtenidos
que este péptido se comporta como una bacteriocina de la subclase IIa, por
presentar un tamaño molecular inferior a 10 kDa y por su alta actividad inhibitoria
frente a Lm. Sin embargo, no se puede asegurar esta afirmación hasta una vez
obtenida la secuencia aminoacídica, para poder llegar a esta conclusión.
La precipitación con acetona permite la purificación hasta homogeneidad, pero
con bajos rendimientos.

73
7. SUGERENCIAS
Para lograr mejores resultados en la purificación sugiero realizar las siguientes
modificaciones:
1) Realizar un cultivo de varios litros (5 lt), para poder garantizar una alto rendimiento
en los cultivos de la cepa productora.
2) Una vez obtenido los sobrenadantes realizar una etapa de ultrafiltración, para
eliminar las proteínas mayores de 100 kDa
3) La precipitación se realizaría con acetona, según los resultados obtenidos
4) Utilizar una columna analítica de exclusión molecular Superdex 75 HR 10/30, por
separar moléculas que poseen un peso molecular entre 3-70 kDa.
5) Al utilizar la columna de exclusión molecular no se logra separa la STB, se debería
realizar la última separación en una columna C18, para lograr de esta forma una
mayor purificación total.

74
8. BIBLIOGRAFÌA
Aymerich, M.T., Artigas, M. Garriga, J. Monfort, M. y Hugas, M. (2000). Effect of sausage
ingredients and additives on the production of enterocins A and B by Enterococcus
faecium CTC492. Optimization of in vitro production and anti-listeria effect in dry
fermented sausages. J. Appl. Microbiol., 88, 686–694.
Aymerich, T., Holo, H. Håvarstein, L.S. Hugas, M. Garriga, M. y Nes, I.F. (1996).
Biochemical and genetic characterization of enterocin A from Enterococcus faecium, a
new antilisterial bacteriocin in the pediocin family of bacteriocins. Appl. Environ.
Microbiol., 62, 1676-1682.
Axelsson, L., Katla, T. Bjornslett, M. Eijsink, V.G.H. y Holck, A. (1998). A system for
heterologous expression of bacteriocins in Lactobacillus sake. FEMS Microbiol. Lett.,
168, 137-143.
Barefoot, S., y Klaenhammer, T. (1983). Detection and activity of lactacin B, a bacteriocin
produced by Lactobacillus acidophilus. Appl. Environ. Microbiol., 45, 1808–1815
Bechinger, B., (1997). Structure and function of channel-forming peptides: magainins,
cecropins, melittin and alamethicin. J. Membrane Biol., 156, 197-211.
Bierbaum, G., Szekat, C. Josten, M. Hedrich, C. Kempter, J. y Sahl, H.G. (1996).
Engineering of a novel thiother bridge and role of modifies residues in the lantibiotic
Pep5. Appl. Environ. Microbial., 62, 385-392.

75
Bradford, M.M., (1976). A rapid and sensitive for the quanttation of microgram
qunatitiesutilizing the principle-dye binding. Anal. Biochem., 72, 248-254.
Brogden, K.A., (2005). Antimicrobial peptides: pore formers or metabolic inhibitors in
bacteria? Nat. Rev. Microbiol., 3, 238-250
Chen, Y., y Montville, T.J. (1995). Efflux of ions and ATP depletion induced by pediocin
PA-1 are concomitant with cell death in Listeria monocitogenes Scott A. J. Appl.
Bacteriol., 79, 684-690.
Chikindas, M. L., García-Garcerá, A. J. M. Driessen, A. M. Ledeboer, J. Nissen-Meyer, I.
F. Nes, T. Abee, W.N. Konings y G. Venema. (1993). Pediocin PA-1, a bacteriocin from
Pediococcus acidilactici PAC1.0, forms hydrophilic pores in the cytoplasmic membrane of
target cells. Appl. Environ. Microbiol., 59, 3577-3584.
Chile, Ministerio de salud., (2009). Departamento de Epidemiología, Informe Listeriosis
http://epi.minsal.cl/epi/html/bolets/reportes/Listeriosis/Informe%20brote%20Listeria.pdf,
Chmiel, D., (2001). Mode of action of antimicrobial peptides. Adv. Cell. Biol., 16, 261-73
Cintas, L.M., Casaus, M.P. Herranz, C. Nes, I.F. Hernández, P.E. (2001). Review:
Bacteriocins of lactic acid bacteria. Int. J. Food Sci. Technol., 7, 281-305.
Cintas, L.M., Casaus, P. y Hernández. P (2000a). Actividad antimicrobiana de las
bacterias lácticas (I). Ácidos orgánicos, metabolitos del oxígeno y productos finales del
catabolismo. Alim. Equip. Tecnol., 83-90.

76
Cintas, L.M., Casaus, P., Håvarstein, L.S., Hernández, P.E. y Nes, I.F. (1997).
Biochemical and genetic characterization of enterocin P, a novel secdependent
bacteriocin from Enterococcus faecium P13 with a broad antimicrobial spectrum. Appl.
Environ. Microbiol., 63, 4321-4330.
Cleveland, J., Montville, T.J. Nes, I.F. y Chikindas, M.L. (2001). Bacteriocins: safe,
natural antimicrobials for food preservation. Int. J. Food Microbiol., 71, 1-20.
Cotter, P.D., Hill, C. y Ross, R.P. (2005). Bacteriocins: developing innate immunity for
food Nature, Microbiology Reviews., 3, 777-788.
Dalet, K., Cenatiempo. y Cossart, P. (2001) the European Listeria Genome Consortium, y
Héchard. Y. A σ54dependent PTS permease of the mannose family is responsible for
sensitivity of Listeria monocytogenes to mesentericin Y105. Microbiology., 147, 3263–
3269.
Daeschel, M.A., (1989). Antimicrobial substances from lactic acid bacteria for use as food
preservatives. Int .J. Food Technol., 43, 164-167.
Dellagio, F., de Roissart, H. Toniani, S. Curk, M.C. y Janssens, D. (1994).
“Caractèristiques gènèrales des bactèries lactiques” En Bacteries Lactiques. H. de
Roissart y F.M. Luquet., 25-116.
De Vuyst, L., y Vandamme E.J. (1992). Influence of the carbon source on nisin
production in
Lactococcus lactis subsp. lactis batch fermentations. J. Gen. Microbiol., 138, 571-578.

77
Dalet K., Cenatiempo Y., Cossart P., Hechard Y. (2001). A σ54-dependent PTS
permease of the mannose family is responsable for sensitivity of Listeria monocytogenes
to mesentericin Y105. Microbiology, 147:3263-3269.
Drider, D., Gunnar, F. Yann, H. Lynn, M. y Hervé, P. (2006). The Continuing Story of
Class IIa Bacteriocins. Microbiol Mol Biol Rev., 70, 564–582.
Doganay, M., (2003). Listeriosis: clinical presentation. FEMS Immunol Med Microbiol.,
35, 173-175.
Doyle, M.E., Mazzota, AS. Wang, T. Wiseman, D. y Scott, V. (2001). Heat resistance of
Listeria monocitogenes. J. of Food Protection., 3, 410-429.
Duffes, F., Correo, C. H, Leroi, F. Dousset, X. y Boyaval, P. (1999). Inhibition of Listeria
monocytogenes by In Situ Produced and Semipurified Bacteriocins of Carnobacterium
spp. on Vacuum-Packed, Refrigerated Cold-Smoked Salmon. J. Food Prot., 62, 1394-
1403.
Eklund, W., Poysky, T. Paranjpye, N. Lashibrook, C. Peterson, E. y Pelroy, A. (1995).
Incidence and Sources of Listeria monocytogenes in Cold-Smoked Fishery Products and
Processing Plants. J. Food Prot., 58, 502-508.
Ennahar, S., Sashihara, K. Sonomoto, A. (2000). Class IIa bacteriocins: biosynthesis,
structure and activity. FEMS Microbiol. Rev., 24, 85-106

78
Fimland, G., Johnsen, L. Dalhus, B. y Nissen-Meyer, J. (2005). Pediocinlike antimicrobial
peptides (class IIa bacteriocins) and their immunity proteins: biosynthesis, structure and
mode of action. J. Pept. Sci., 11, 688–696.
Fregeau Gallagher, N.L., Sailer, M. Niemczura, W. Nakashima, T.T Stiles, M. E. y
Vederas, J.C. (1997). Three-dimensional structure of leucocin A in trifluoroethanol and
dodecylphosphocholine micelles: spatial location of residues critical for biological activity
in type IIa bacteriocins from lactic acid bacteria. Biochemistry., 36, 15062–15072.
Gandhi, M., Chinkindas, M.L. (2007). Listeria: A foodborne pathogen that knows how to
survive. Int J Food Microbiol., 113, 1-15.
Ganz, T. and Lehrer, R. (1999). Antibiotic peptides from higher eukaryotes: biology and
applications. Mol. Med., Today., 5, 292-7
Gandhi, M., y Chikindas, M. (2007). Listeria. A foodborne pathogen that knows how to
survive, International J. of Food Microbilogy., 113, 1-15.
Geis, A., Singh, J. y Teuber, M. (1983). “Potential of lactic streptococci to produced
bacteriocins”. Appl. Environ. Microbiol., 45, 205-211.
Gratia, A., (1925). Sur un remarquable exemple d’antagonisme entre deux souches de
colibacille. C.R.Soc. Biol. Fill., 93, 1040-1041.
Hale, J.D., Hancock, R.E. (2007). Alternative mechanusms of action of cationic
antimicrobial peptides on bacteria. Expert. Rex. Anti Infect., 5, 951-959.

79
Hancock, R.E.W., Falla, T. y. Brown, M. (1995). Cationic bactericidal peptides. Adv.
Microbiol. Physiol., 37, 135-175.
Hansen, J. N. (1994). Nisin as a model food preservative. Crit. Rev. Food Sci. Nutr., 34,
69-93.
Haugen, H. S., G. Fimland, J. Nissen-Meyer, y P. E. Kristiansen. (2005). Three-
dimensional structure in lipid micelles of the pediocin-like antimicrobial peptide curvacin
A. Biochemistry., 44, 16149–16157.
Havarstein, L.S., Diep, D.B. y Nes, I.F. (1995). A family of bacteriocin ABC transporters
carry out proteolytic processing of their substrates concomitant with export. Mol.
Microbiol., 16, 229-240.
Héchard, Y., Pelletier, C., Cenatiempo, Y. y Frere, J. (2001). Analysis of (54)-dependent
genes in Enterococcus faecalis: A mannose PTS permease (EIIMan) is involced in
sensitivity to a bacteriocin mesentericin Y105. Microbiology., 147, 1575-1580.
Holo, H., Nilssen O., Nes I.F. (1991). Lactococcin A, a new bacteriocin from Lactococcus
lactis subsp. cremoris isolation and characterization of the protein and its gene. Journal
of Bacteriology., 173, 3879-3887.
Hurst, A., (1981). Nisin. Adv. Appl. Microbiol., 27, 85-123.
Jack W. R., WAN, J. Gordon, J. Harmark, J. Davison, B. Hillier, A. Wettenhall, R. Hickey,
M y Coventry, J. (1996). Characterization of the Chemical and Antimicrobial Properties of
Piscicolin 126, a Bacteriocin Produced by Carnobacterium piscicola JG126. Scientific

80
and Industrial Research Organization, Division of Food Science and Technology. 62:
2897-2903
Jacob, F., Lwoff, A., Siminovitch, A. y Wollman, E.L. (1953). Définition de quelques
termes relatifs à la lysogénie. Ann. Inst. Pasteur., 84, 222-224
Johnsen, L., Dalhus, B. Leiros, I. y Nissen-Meyer, J (2005). 1.56 Å cristal structure of
entA-im: a bacterial immunity protein conferring immunity to the antimicrobial activity of
the pediocin-like bacteriocin enterocin A. J. Biol. Chem., 280, 19045–19050.
Kamysz, W., Okruj, M. y Lukasiak, J.(2003). Novel properties of antimicrobial peptides.
Acta Biochim., 50, 461-469.
Kanatani, K., Oshimura, M. y Sano, K. (1995). Isolation and characterization of acidocin
A and cloning of the bacteriocin gene from Lactobacillus acidophilus. Appl. Environ.
Microbiol., 61, 1061-1067.
Kandler, O., (1983). Carbohydrate metabolism in lactic acid bacteria. Antonie van
Leeuwenhoek. Microbiol. Serol., 42, 209-224.
Karlsson, E., Ryden, L. Brewer, J. (1989). Ion Exchange Chromatography. En: Janson
JC, Ryden L, eds. Protein Purification, Principles, High resolution methods and
application. New York. Publishers Inc., 107-48.
Kaur, K., Andrew, L. Wishart, D.S. y Vederas, J.C (2004). Dynamic Relationships among
Type IIa Bacteriocins: Temperature Effectes on Antimicrobial Activity and on Structure of

81
The C-Terminal Amphipathic α Helix as a Receptor-Binding Region. Biochemistry., 43,
9009-9020.
Klaenhammer, T.R., (1993). Genetics of bacteriocin produced by lactic acid bacteria.
FEMS Microbiol. Rev., 12, 39-86.
Kleerebezem M., (2004). Quorum sensing control of lantibiotic production; nisin and
subtilin autoregulate their own biosynthesis. Peptides., 25, 1405-1414.
Kleerebezem, M., Beerthuyzen M, Vaughan E. (1997). Controlled gene expression
systems for lactic acid bacteria: transferable nisin-inducible expression cassettes for
Lactococcus, Leuconostoc and Lactobacillus spp. Applied and Environmental.
Microbiology., 63, 4581-4584.
Lamas, P., (2002). Cracaterización de una bacteriocina producida por una bacteria ácido
láctica de carne envasda al vació. Tesis de pregrado, Escuela de Ingeniería en
alimentos, Facultad de Ciencias Agrarias., Univ. Austral de Chile, 30-50 pp.
Liu., (2006). Identification, subtyping and virulence determination of Listeria
monocytogenes, an important foodborne pathogen. J Med Microbiol., 55, 645-59.
Lindgren, E., y Dobrogosz, J. (1990). Antagonistic activies of Iactic acid bacteria in food
and feed fermentations. F.E.M.S. Microbiol. Rey., BZ, 149-164.
Ladokin, A., y White, H. (2001). Detergent-like permeabilization of anionic lipid vesicles
by melittin. Biochim. Biophys., 1514, 253-260.

82
López, J., Ochoa, A. Santoyo, G. Anaya, J. Medina, E. Martínez, M. Loeza, P. (2008)
Bacteriocins of Gram positive bacteria a potencial source of new biomedical tratments.
Revista Mexicana de ciencias farmacéutica., 39, 49-57.
Moll, G.N., Konings, W.N. y Driessen, A.J.M. (1999). Bacteriocins: mechanism of
membrane insertion and pore formation. Antonie van Leeuwenkoek., 76, 185-198.
Moll, G., Ubbink, K. T. Hildeng, H. H. Nissen, M.J. Nes, I. F. Konings, W. Driessen, A.
(1996). Lactococcin G is a potassium ion-conducting, two-component bacteriocin.
Journal of Bacteriology., 178, 600-605
Mørtvedt, C.I., Nissen-Meyer, J., Sletten, K. y Nes, I.F. (1991). “Purification and amino
acid sequence of lactocin S, a bacteriocin produced by Lactobacillus sake L45”. Appl.
Environ. Microbiol., 57, 1829-1834.
Muriana, P.M., y Klaenhammer, T. R. (1991). Purification and partial characterization of
lactacin F, a bacteriocin produced by Lacrobacillus acidophilus. AppI. Environ. Microbiol.,
57, 114-121.
Muriana, P.M., (1996). Bacteriocins for control of Listeria spp. in food. J. Food Prot., 54-
63.
Nes, I.F., Diep, D.B. Havarstein, L.S. Brurberg, M.B. Eijsinnk, V. y Holo, H. (1996).
Biosynthesis of bacteriocins in lactic acid bacteria. Antoine Van leeuwenhoek., 70, 113-
128.

83
Nes, I.F., y Holo, H. (2000). Class II antimicrobial peptides from lactic acid bacteria.
Biopoly., 50, 50-61.
Nissen-Meyer, J., e I.F. Nes (1997). Ribosomally synthesized antimicrobial peptides: their
function, structure, biogenesis, and mechanism of action. Arch. Microbiol., 167, 67-77.
Nissen-Meyer, J., Håvarstein, L.S. Holo, H. Sletten, K. y Nes, I. F (1993). Association of
the lactococcin A immunity factor with the cell membrane: purification and
characterization of the immunity factor. J. Gen. Microbiol., 139, 1503–1509.
Piard, J.C., y Desmazeaud, M. (1992). Inhibiting Factors produced by Lactic Acid
Bacteria. Bacteriocins and others antibacterial substances. Letters Applied International
Technology., 72, 113-142.
Quadri, L.E.N., Kleerebezem, M. Kuipers, O. P. de Vos, W. M. Roy, K.L J.C. Vederas y
Stiles M. E (1997). Characterization of a locus from Carnobacterium piscicola LV17B
involved in bacteriocin production and immunity: evidence for global inducer-mediated
transcriptional regulation. J. Bacteriol., 179, 6163-6171
Quadri, L.E.N., Sailer, M. Terebiznik, M.R. Roy, K.L. Vederas, J.C. y Stiles, M.E. (1995).
Characterization of the protein conferring immunity to the antimicrobial peptide
carnobacteriocin B2 and expression of carnobacteriocins B2 and BM1. J. Bacteriol., 177,
1144-1151.

84
Requena, T., y Peláez, C. (1995). Actividad antimicrobiana de bacterias lácticas.
Producción de bacteriocinas. Revista Española de Ciencia y Tecnología de los
Alimentos., 35 (1): 19-44.
Sahl, H.G., Jack, R.W. y Bierbaum, G. (1995). Latibiotics: biosynthesis and biological
activities of peptides with unique posttranslational modification. Eur. J. Biochem., 230,
827-853.
Sánchez B.M.J., Martínez R. M., Gálvez A., Valdivia E., Maqueda M., Cruz V., Albert A.
(2003). Structure of bacteriocin AS-48: From soluble state to membrane bound state.
Journal of Molecular Biology., 334, 541-549.
Schägger, H., y von Jagow, G. (1987). Tricine-sodium dodecyl sulfate-polyacrylamide gel
electrophoresis for the separation of proteins in the range from 1 to 100 kDa. Anal.
Biochem., 166, 368-397.
Shai, Y., (1999). Mechanism of the binding, insertion and destabilization of phosphoslipid
bilayer membranes by α-helical antimicrobial and cell non-selective membrane-lytic
peptides. Biochimica et Biophysica Acta., 1462, 55-70.
Schillinger., U. y Stiles, M. (1993). Bacteriocin production by Carnobacterium piscicola LV
61. J. Food Microbiol. 20: 131-141.
Schöbitz, R., Raddatz, M., Tamayo, R. y Miranda, C. (1990). Actividad antibaceriana de
cepas lácticas aisladas a partir de carne envasada al vacío. Fleischwirtsch., 2, 12-19.

85
Stainer R., Ingraham, J. Wheelis, M. y Painter, P. (1996). Microbiología. Segunda
edición. Ed. Reverté., 750.
Tagg, J.R., Dajani, A.S. y Wannamaker, L.W. (1976). Bacteriocins of Gram-positive
bacteria. Bacteriol. Rev., 40, 722-756.
Wang, Y., Henz, M.E. Gallagher, N. L, Chai, S. Gibbs, A.C. Yan, Z. Stiles, M. E D.
Wishart, S. y Vederas. J. C (1999). Solution structure of carnobacteriocin B2 and
implications for structure-activity relationships among type IIa bacteriocins from lactic acid
bacteria. Biochemistry., 38, 15438–15447.
White, A., Hart, J. (1992a). Derivatization methods for liquid chromatographic separation of amino
acids. En: Analysis by HPLC. Nollet, L.M.L. Ed. pp. 53-74.
Yang, L., Harroun, T. A. Weiss, T. M. Ding, L. y Huang, H. W. (2001). Barrel-stave model
or toroidal model? A case study on melittin pores. Biophys. J., 81, 1475–1485
Zendo, T., Koga, S., Shigeri, Y. Nakayam, J. y Sonomoto, K. (2006). Lactococcin Q, a
novel two-peptide Bacteriocin Procuded by Lactococus lactis QU 4. Applied and
environmental microbiology., 3383–3389

86
9. ANEXOS
9.1 Cromatografía de filtración en mini columna Hi-Trap desalting
Figura 14. Perfil cromatográficos de la STB obtenidos por Hi-Trap
desalting. (A) Fracciones IIa y (B) Fracciones IIb obtenidos por mini
columnas Hi-trap desalting. Se recolectaron los primeros 8 tubos libres
de sales.
A
B

87
9.2 Cuantificación de proteínas por el método de Bradford
La cuantificación de proteínas se dterminó por el método de Bradford (1976). Para la
cuantificación de proteínas presente en los diferenteds pasos de purificación, se debió
construir una curva de calibración, usando una solución estándar de 3,5 mg/ml de
albúmina sérica de bovino, en la tabla 4 se muestran los datos de la curva de
calibración.

88
Tabla 4. Preparación de las muestras para construir la curva de calibración
Tubos Estándar (µl)
Agua (µl)
Absorbancia 595 nm
Concentración mg/ml
K
1 ------ 40 0,000 0,000 0,000
2 5 35 0,429 0,438 0,98
3 10 30 0,788 0,875 0.90
4 15 25 1,050 1,313 0,80
5 20 20 1,260 1,750 0,72
6 25 15 1,386 2,188 0,66
7 30 10 1,523 2,625 0,58
8 35 5 1,654 3,063 0,54
9 37 3 1,781 3,238 0,55
10 38 2 1,729 3,325 0,52

89
Figura 15. Curva de calibración por el método de Bradford; para
determinar la concentración de proteinas presentes en los diferentes
pasos de la purificación.
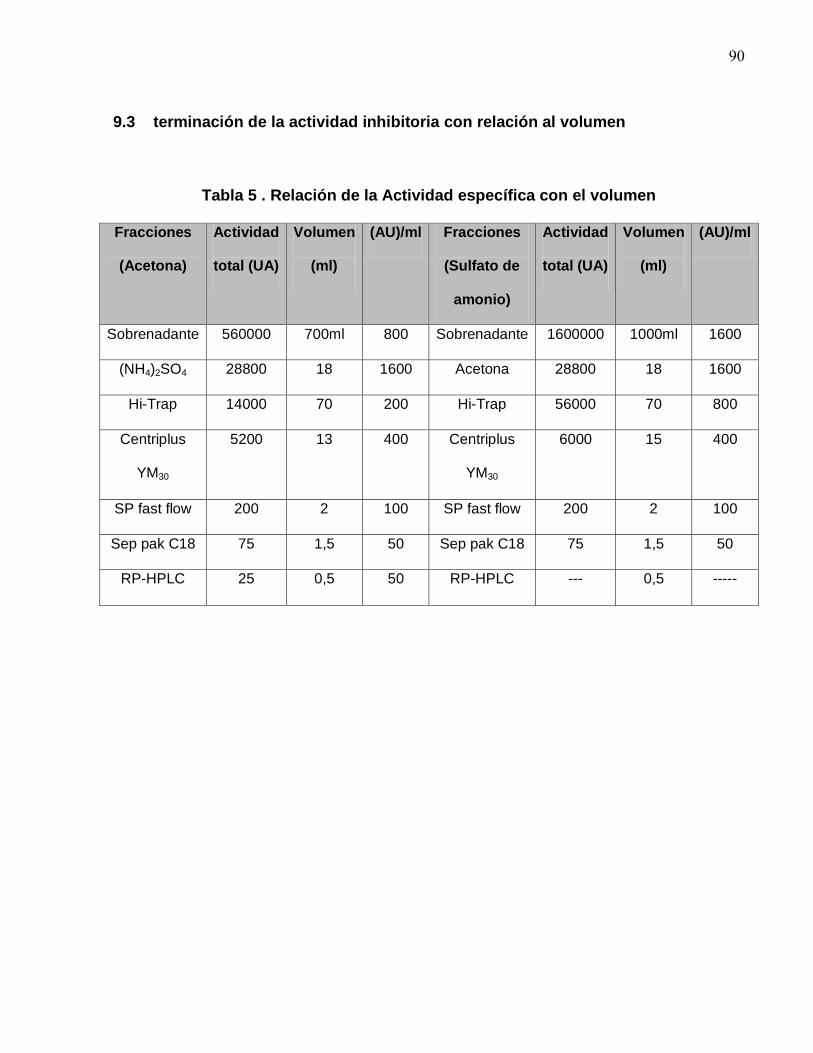
90
9.3 terminación de la actividad inhibitoria con relación al volumen
Tabla 5 . Relación de la Actividad específica con el volumen Fracciones
(Acetona)
Actividad
total (UA)
Volumen
(ml)
(AU)/ml Fracciones
(Sulfato de
amonio)
Actividad
total (UA)
Volumen
(ml)
(AU)/ml
Sobrenadante 560000 700ml 800 Sobrenadante 1600000 1000ml 1600
(NH4)2SO4 28800 18 1600 Acetona 28800 18 1600
Hi-Trap 14000 70 200 Hi-Trap 56000 70 800
Centriplus
YM30
5200 13 400 Centriplus
YM30
6000 15 400
SP fast flow 200 2 100 SP fast flow 200 2 100
Sep pak C18 75 1,5 50 Sep pak C18 75 1,5 50
RP-HPLC 25 0,5 50 RP-HPLC --- 0,5 -----

91
9.4 Determinación del peso molecular de la STB
La determinación del peso moleculara de la STB se logró a traves de la interpolación
en la gráfica del logaritmo de sus pesos moleculares estandar versus la distancia de
migración.
La distancia migrada de STB fue de 3 cm, la cual fue interpolada en la curva dando
como peso molecular de 3 Kda.

92
Tabla 6. Datos necesarios para construir la curva de calibración
Estandar (Da) Log Estándar Distancia migrada (cm)
26600 4,42 0,65
17000 4,23 1,20
14200 4,15 1,70
6500 3,81 2,20
3496 3,54 2,90
1060 3,03 3,70
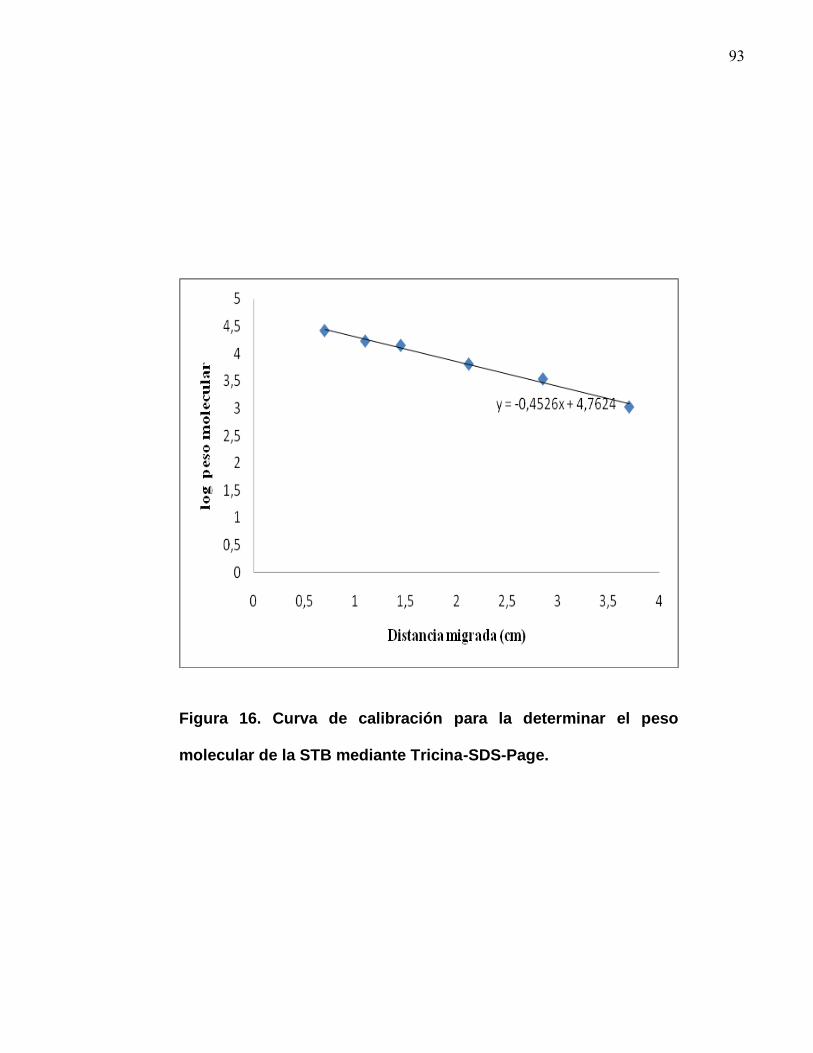
93
Figura 16. Curva de calibración para la determinar el peso
molecular de la STB mediante Tricina-SDS-Page.

94
9.5 Determinacion de aminoácidos por el método PITC
Preparacion de la muestra Se utilizo 20 µl de la STB proveniente de la fracción VIIa y se almacenó a -20ºC,
hasta su uso.
Preparación de los tubos de reacción Para utilizar los tubos de reacción fueron limpiados con 20 ml de HCL 6N y
lavados con abundante agua de calidad Milli Q.luego s adicionaron 5ml de etanol
100% y los tubos fueron secados a vacío .
Preparacion de los patrones de aminoácidos Se disolviéron 0,2 g de cada aminoácidos en 50 ml de HCL 0,1 N. Se tomó 0,5 ml
de cada patrón (15) y se introducieron en un tubo de 10 ml.
Prepararción solución de resecado (solución A) La solución de resecado(1ml), esta constituida de etanol, agua y trietilamina; en
relacion 2:2:1.
Preparación solución de derivatización La solución de derivatización ( 1ml), esta constituida de etanol, agua, trietilamina y
fenilisotiocianato; en relación 7:1:1:1.

95
Procedimiento:
I. Se utilizó 20 µl de las fracciones VIIa y VIIb, respectivamente. Los tubos
con las muestras fueron secado a través de una bomba de vacío durante
30 min.
II. A continuación se procedio a hidrolizar las muestras con HCL 6N por 24 h
a 110ºC.
III. Una vez hidrolizadas las muestra se continuó con el proceso de resecado,
donde se adicionaron 10 µl de la solución A, para cada tubo y se secaron
las muestras al vació durante 30 min.
IV. Trancurrido este tiempo se agregó 20 µl de la solución B acada tub o y se
secó al vacío duarnte 30 min. Las muestras fueron alamcenadas a -20ºC,
hasta su determinación por HPLC.
V. La solución estandar fue sometida al mismo procedimiento, para ser
analizadas en el equipo de HPLC.

96
Tabla 7. Tiempo de retención del estándar de aminoácidos y de la STB
NOMBRE GENERAL
SÍMBOLO TIEMPO DE RETENCION ESTÁNDAR
AREA [mV.s]
ESTÁNDAR
TIEMPO DE RETENCION
AREA [mV.s]
Aspartato D 5,893 2541,783 5,797 307,418
Glutamato E 6,560 2320,969 7,180 2890,385
Serina S 13,073 2686,81 12,247 332,778
Glicina G 13,760 2709,949 12,820 338,960
Histidina H 14333 2717,613 13,313 344,795
Arginina R 15,483 2786,04 14,007 357,274
Alanina A 15,923 2488,91 14,453 672,695
Prolina P 16,173 2826,331 15,020 319,449
Valina V 16,503 2841,761 19,620 372,569
Cistina C 21,133 2941,66 20,013 303,490
Isoleucina I 21,690 2621,512 21,340 342,142
Leucina L 22,553 2096,577 22,323 193,739
Fenilalanina F 23,327 1685,176 22,480 209,983
Lisina K 23590 1873,324 22,630 206,491

97
Figura 17. (A) Cultivo de la Cepa BALB en caldo D-MRS. (B)
extracto crudo (E.C) o Fracción I.
A B
9.6 Fotografías Cepa BALB y extracto crudo de la STB

98
Figura 18. Precipitación de la STB (A) Fracción Ia (precipitación
con acetona). (B) Fracción Ib (precipitación con sulfato de amonio).
A B
9.7 Fotografías de las precipitaciones de la STB