no slide title · •giardia lamblia. ... el complejo i de la cadena respiratoria no sería...
TRANSCRIPT

PROTOZOARIOS
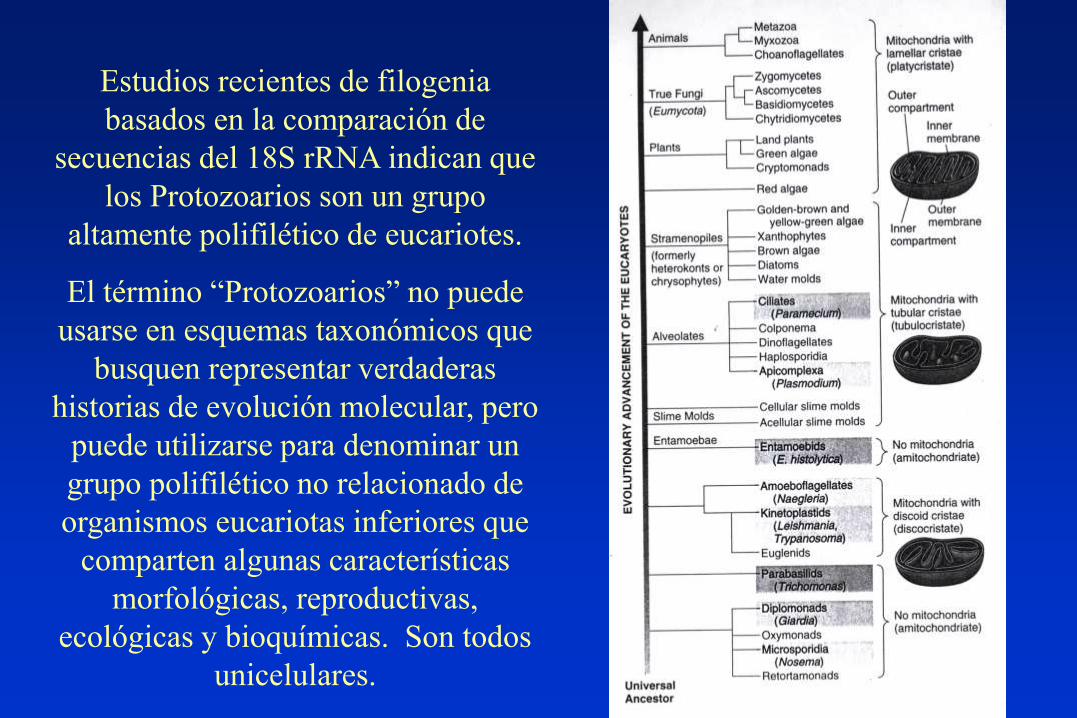
Estudios recientes de filogenia
basados en la comparación de
secuencias del 18S rRNA indican que
los Protozoarios son un grupo
altamente polifilético de eucariotes.
El término “Protozoarios” no puede
usarse en esquemas taxonómicos que
busquen representar verdaderas
historias de evolución molecular, pero
puede utilizarse para denominar un
grupo polifilético no relacionado de
organismos eucariotas inferiores que
comparten algunas características
morfológicas, reproductivas,
ecológicas y bioquímicas. Son todos
unicelulares.
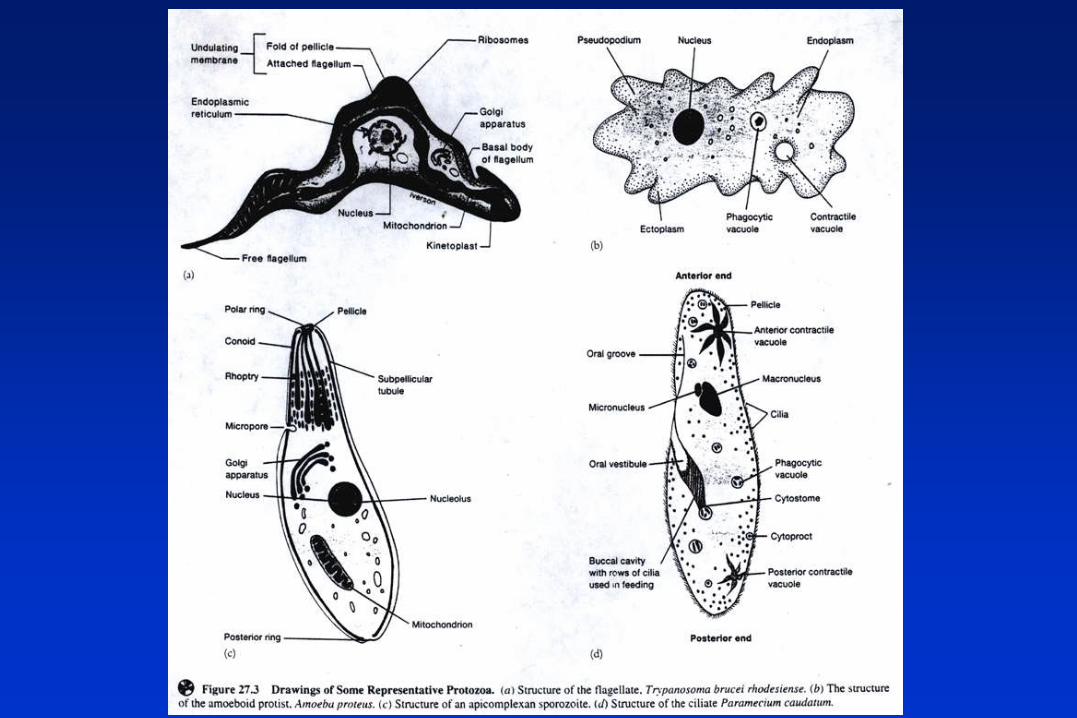
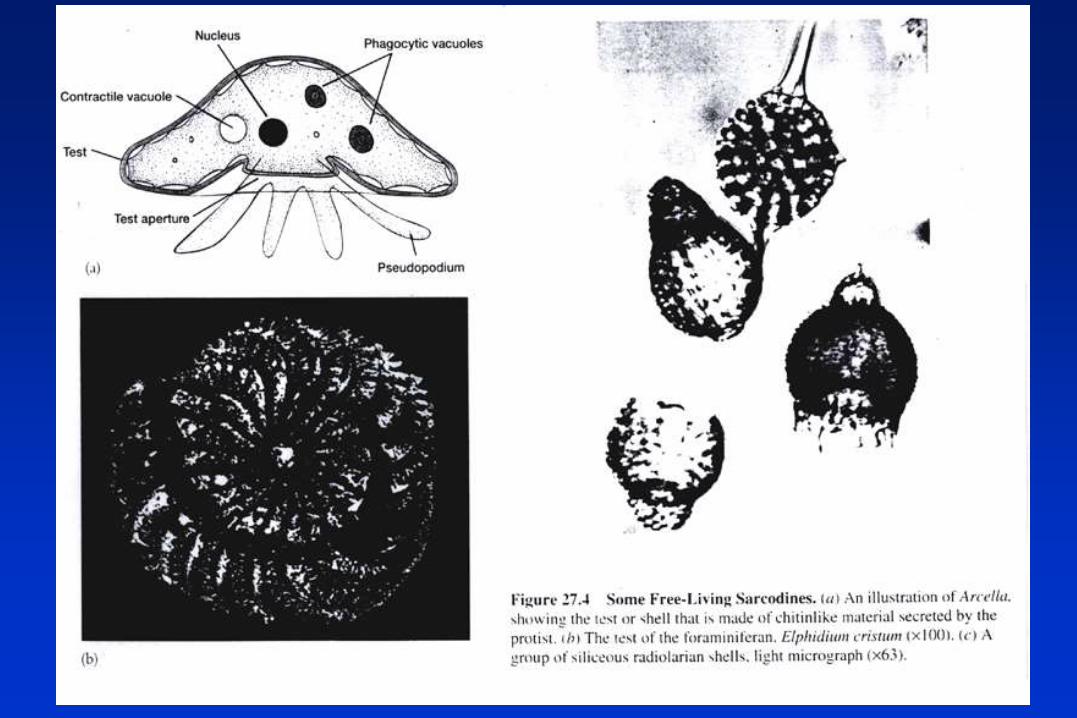



PROTOZOARIOS PARASITOS.
45.000 especies actuales; 10.000 de ellas son parásitos de animales y, en algunos casos, de plantas.
REINO PROTISTA: Organismos eucarióticos unicelulares.
Grupo 1: Flagelados (locomoción por flagelos).
• Trypanosoma cruzi, Trypanosoma brucei, Leishmania spp.
• Giardia lamblia.
• Trichomonas vaginalis.
Grupo 2: Amebas (locomoción por pseudópodos).
• Entamoeba histolytica.
Grupo 3: Esporozoarios (Apicomplexa) (sin medios visibles de locomoción).
• Plasmodium falciparum.
• Toxoplasma gondii.
Grupo 4: Ciliados (locomoción por cilias).
• Balantidium coli.

PARASITOS INTRACELULARES
Salvo en los casos de los bdellovibrios y los Microsporidia, que invaden
células por penetración directa, los parásitos intracelulares penetran a
través de un proceso que deja la membrana celular intacta y resulta en la
formacion de un fagosoma. El ciclo del parásito puede continuar en uno
de tres compartimentos:
1) Intrafagosomal, sin fusión lisosomal y generalmente sin acidificación:
Legionella pneumophila, Mycobacterium tuberculosis, Toxoplasma gondii.
2) Intrafagolisosomal, es decir después de la fusión con lisosomas:
Yersinia pestis, Leishmania spp.
3) Intracitosólico (rompen la vacuola y escapan al citosol): Rickettsia
spp., Listeria monocytogenes, Shigella flexneri, Babesia bovis, Theileria
parva, Trypanosoma cruzi.

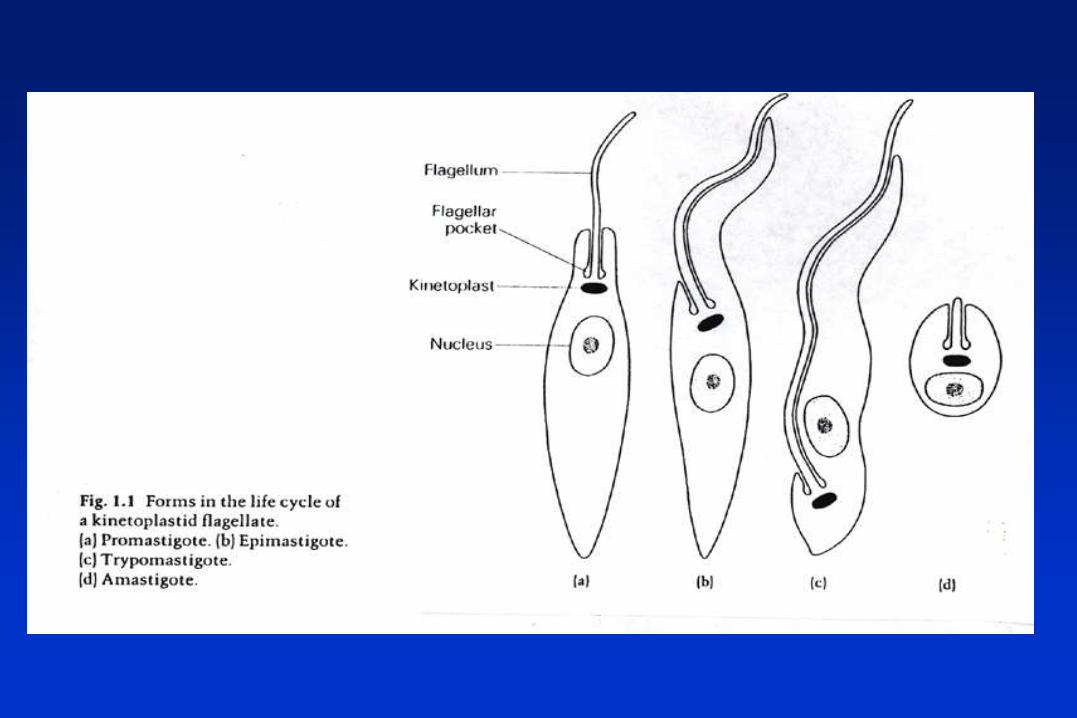
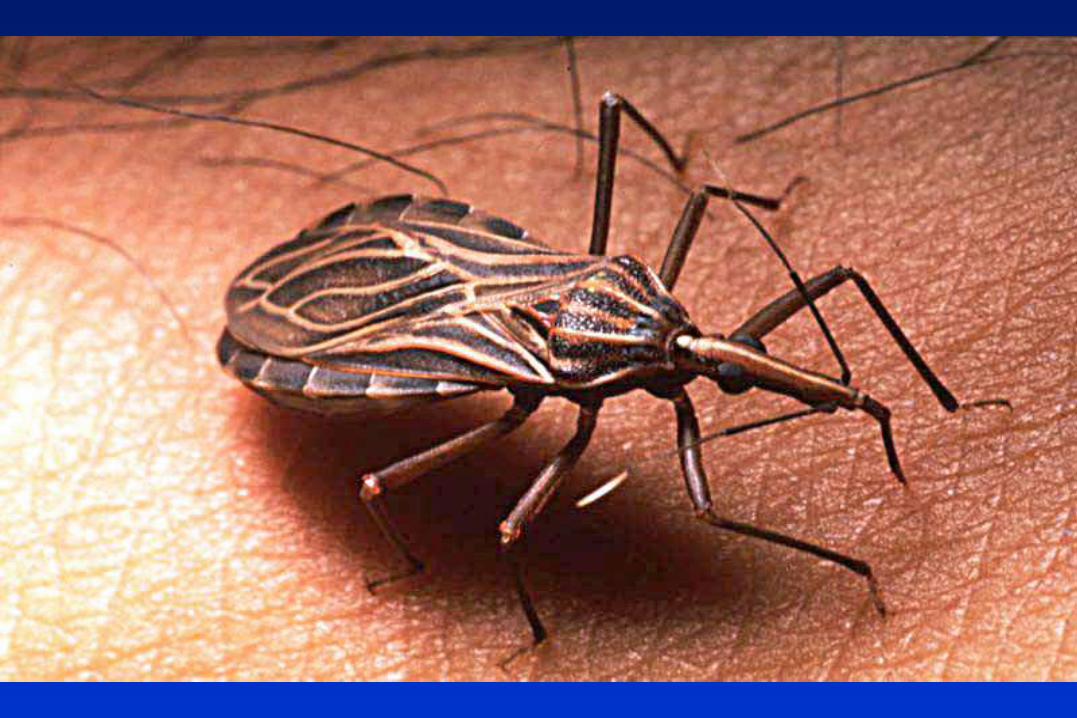
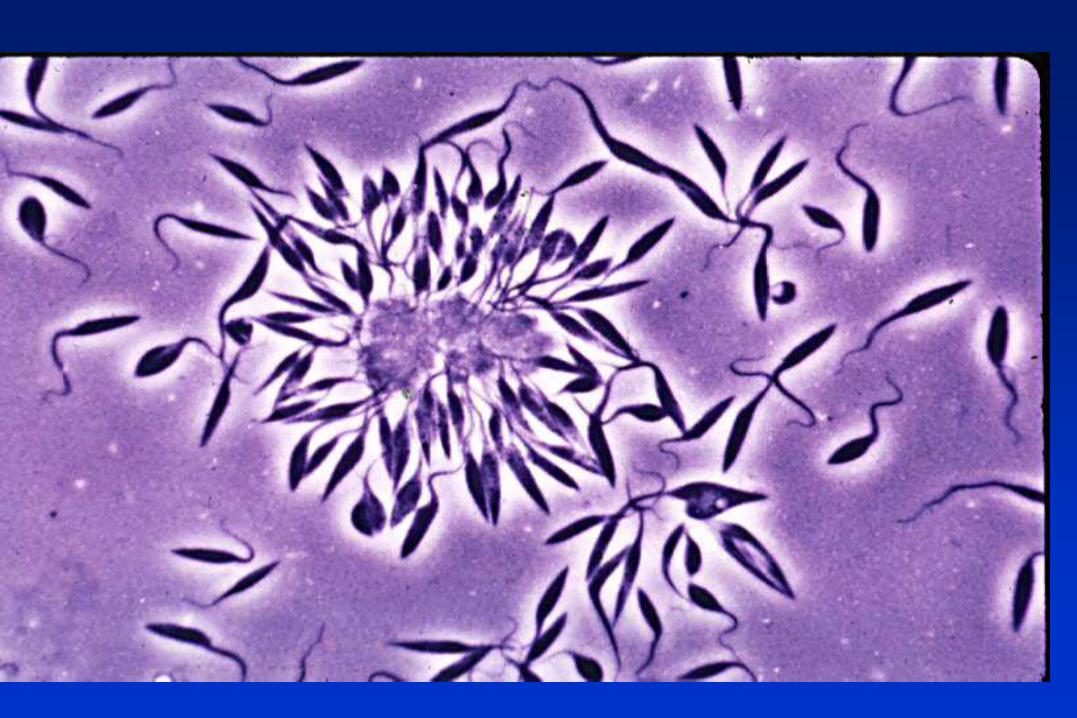
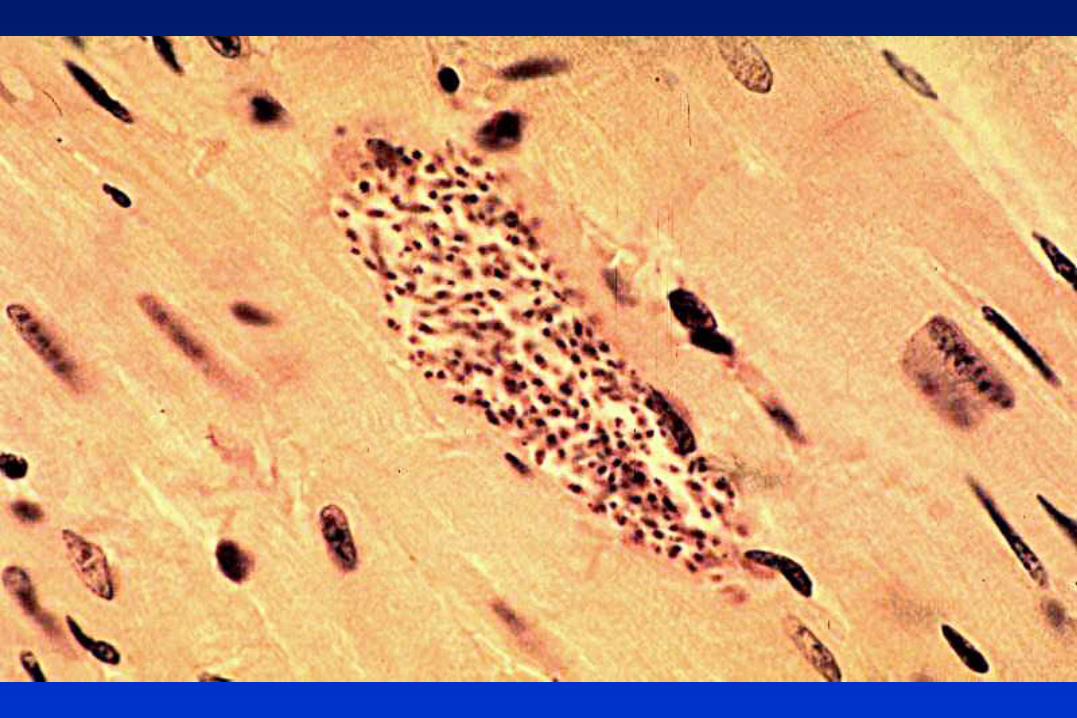
TRIPANOSOMÁTIDOS
Protozoarios flagelados, pertenecientes al Orden Kinetoplastida,
caracterizado por la presencia del complejo kinetoplasto-mitocondria.
Parásito Vector Enfermedad
Trypanosoma cruzi Triatomino Enfermedad de Chagas
Trypanosoma brucei
T. b. gambiense
T. b. rhodesiense Mosca Tse-Tse Enfermedad del Sueño
T. b. brucei Mosca Tse-Tse Nagana (ganado)
Leishmania mexicana Flebótomo Leishmanisasis cutánea
Phytomonas spp. Hemípteros Enfermedades de
fitófagos Plantas.

EL TRYPANOSOMA
CRUZI Y LA
ENFERMEDAD DE
CHAGAS

Carlos
Ribeiro
Justiniano
das Chagas
(1879 – 1934).


En 2012 se contabilizaban 21 países endémicos en
América Latina. El número de personas infectadas se
estima en 6 a 8 millones, estando 65 millones en riesgo
de infección.
Se estiman 28,000 casos nuevos por año. Y alrededor
de 12,000 muertes anuales causadas por la enfermedad.
Por las migraciones, se encuentran enfermos
chagásicos en muchos países no endémicos, como
Estados Unidos (se calculan unos 300,000), España,
Suecia, Japón, y otros.

Transmisión de la Enfermedad de Chagas
1) Transmisión vectorial (vinchuca)
2) Transmisión congénita (de madre infectada a hijo, por infección
transplacentaria o connatal).
3) Transmisión oral, por alimentos o bebidas contaminados.
4) Transmisión transfusional (sangre infectada).
5) Transplante de órganos infectados.
6) Accidentes de laboratorio.
Fases de la Enfermedad de Chagas.
1) Fase aguda. 2) Fase indeterminada 3) Fase crónica (20 a 30
% de casos con sintomatología cardíaca o digestiva).

El número de casos agudos debidos a la transmisión vectorial
es bastante bajo (7 casos informados a las autoridades de
Salud Pública en 2009), pero los expertos del Programa
estiman que el número real de casos debe ser mayor.
La transmisión accidental por transfusiones de sangre es muy
poco frecuente, dado que la sangre a transfundir es
chequeada sistemáticamente buscando anticuerpos. La
transmisión congénita, sin embargo, se está transformando en
la causa principal de infección. Las autoridades sanitarias
estiman alrededor de 1.300 casos por año; 690 recién nacidos
fueron informados como diagnosticados y tratados en 2009.
La prevalencia más alta es en las Provincias norteñas de Salta
y Formosa, vecinas de Bolivia.
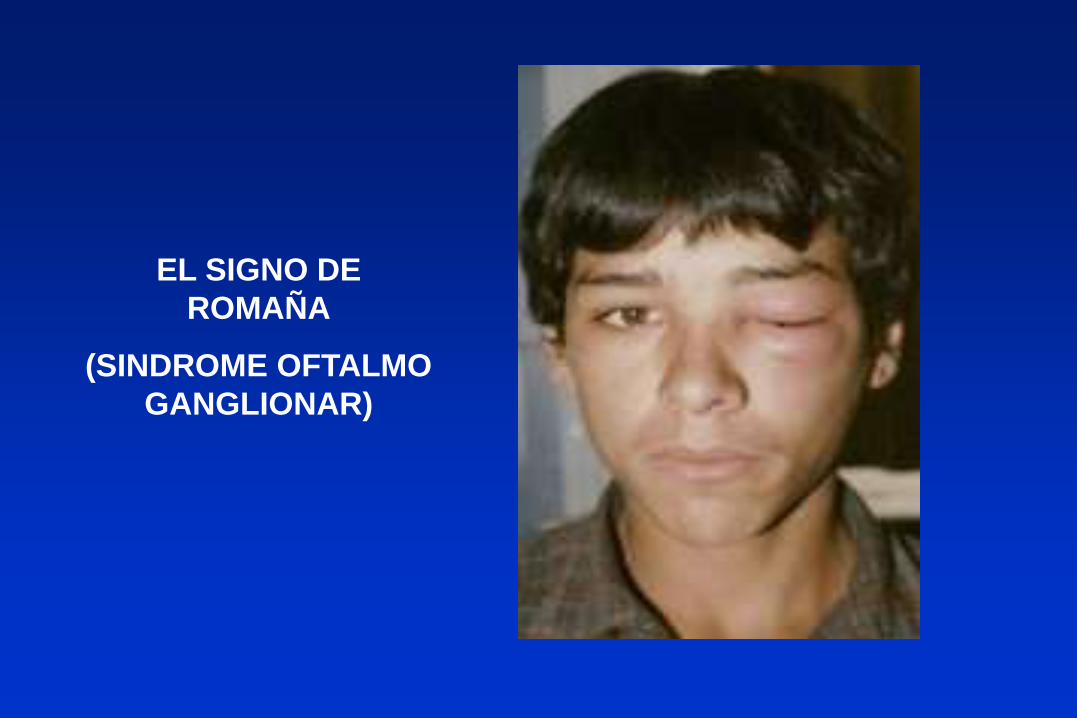
EL SIGNO DE
ROMAÑA
(SINDROME OFTALMO
GANGLIONAR)
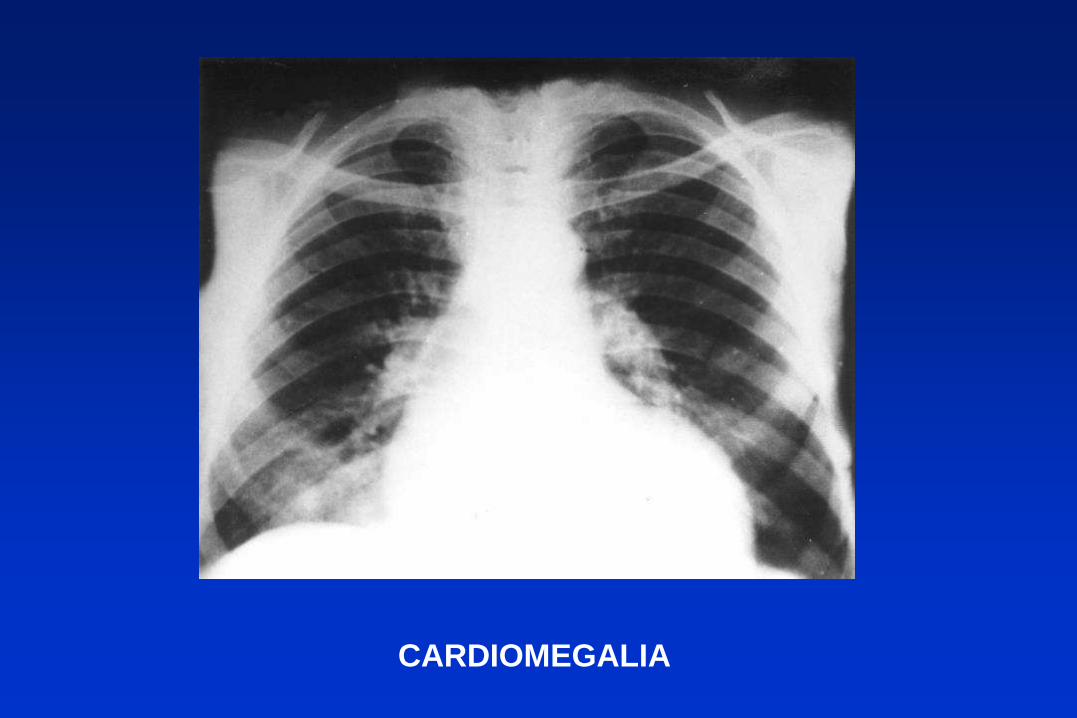
CARDIOMEGALIA

MEGAESÓFAGO
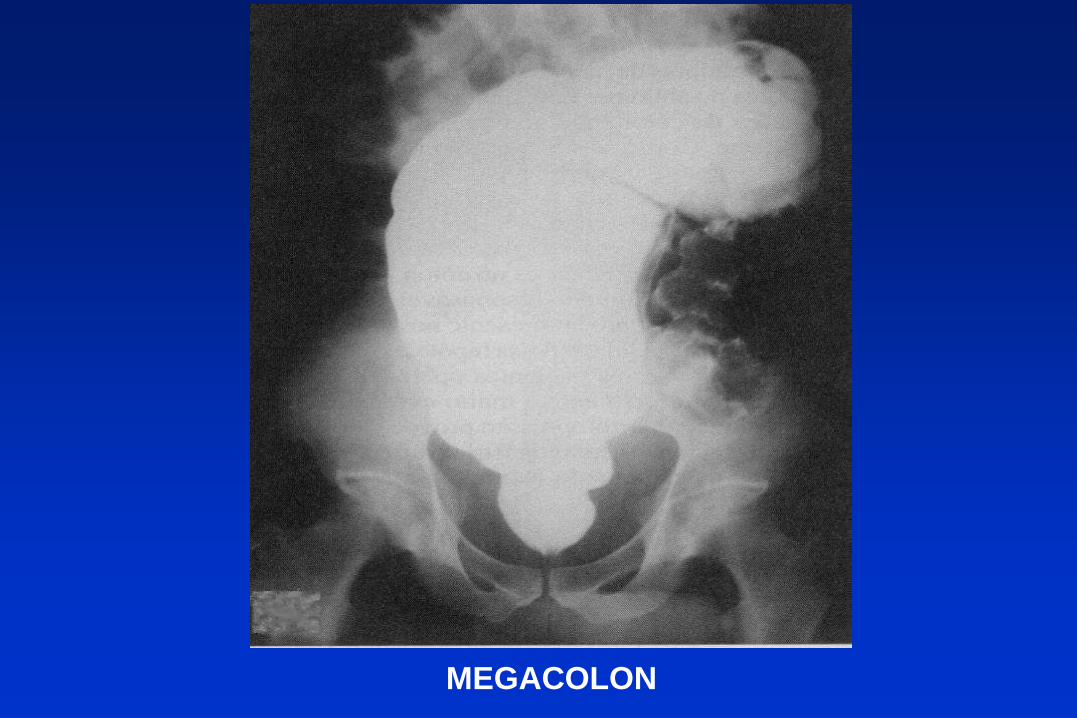
MEGACOLON

El Programa Nacional de Chagas de Argentina
comenzó en 1962.
Argentina no está aún libre de transmisión de la enfermedad, pese a los
esfuerzos para controlar al vector y mejorar la vivienda.
Triatoma infestans está aún presente en casas pobres del area conocida
como el Gran Chaco, que ocupa regiones de Argentina (62 % del area
total, con 3.8 millones de habitantes), Bolivia y Paraguay.
La resistencia a Deltametrina ha aparecido en el Norte de Argentina, y
está presente in Bolivia.

Las últimas estimaciones de casos (OPS, 2015) indican que en Argentina
habría 7.300.000 personas expuestas, 1.505.235 infectadas, de las cuales
376.309 presentarían cardiopatías de origen chagásico. En el año 2014 se
notificaron un total de 271.766 embarazadas estudiadas cuya seroprevalencia
para T. cruzi fue de 2,57% y la transmisión congénita sobre los niños
estudiados y seguidos hasta el año de vida, fue de 5,72% (Iniciativa del Cono
Sur para controlar e eliminar la enfermedad de Chagas (INCOSUR) , 2014).
La prevalencia media de infección en niños menores de 14 años fue de 1,5%
en 2009 y de 1.17 % en 2012. Afortunadamente, 9 de cada 10 niños tratados
en fase aguda y 7 de cada 10 tratados en fase crónica, se curan.
Sobre las actividades de control y vigilancia vectorial, durante el 2014 se
evaluaron 266.987 viviendas por búsqueda activa del vector (triatominos), por
personal técnico de los programas de Chagas, municipal y agentes de
Atención Primaria de Salud. Se trataron 49.077 viviendas en áreas con ataque
químico y bajo vigilancia. De las acciones de control realizadas se obtuvo un
índice de infestación intradomiciliaria promedio, de 1,55% y para la Unidad
Domiciliaria de 3,17% para el año 2014 (INCOSUR, 2014).

Actualmente, el escenario nacional de la enfermedad de Chagas es el
siguiente:
Situación de alto riesgo para la transmisión vectorial
Las provincias de Chaco, Catamarca, Formosa, Santiago del Estero, San Juan y Mendoza
presentan una re-emergencia de la transmisión vectorial de Chagas debido a un aumento de
la infestación domiciliaria y a una alta seroprevalencia en grupos vulnerables.
Situación de riesgo moderado para la transmisión vectorial
Las provincias de Córdoba, Corrientes, La Rioja, Salta, y Tucumán, muestran una situación
de riesgo intermedio con un índice de re-infestación mayor al 5% en algunos departamentos,
e insuficiente cobertura de vigilancia en algunos casos.
Situación de bajo riesgo para la transmisión vectorial
Comprende las provincias certificadas como libres de transmisión vectorial.
Situación de riesgo universal
Las 24 jurisdicciones poseen riesgo de transmisión congénita derivado de las migraciones
internas e internacionales de países vecinos con elevada endemicidad.

Evolución del área de interrupción de transmisión vectorial de
T. cruzi
Hasta el 2015 se alcanzó la certificación por la interrupción de la
transmisión vectorial domiciliaria de Trypanosoma cruzi por
Triatoma infestans en un total de 9 provincias. Durante 2001-2004
lograron la certificación las provincias de Jujuy, Río Negro, Entre
Ríos, La Pampa y Neuquen. En el año 2011 certifica la provincia
de Misiones. Durante el año 2012 se realizó la re-certificación de
las provincias y se certificaron 9 departamentos de Santiago del
Estero. Entre el 2014 y 2015 la Comisión Internacional visitó
áreas endémicas de la Provincia de San Luis y Tucumán, con la
finalidad de evaluar la situación epidemiológica y de prevención,
control y atención de la Enfermedad de Chagas otorgando el
status de interrupción de la transmisión vectorial domiciliaria de
Trypanosoma cruzi en ambas provincias.

PORQUE NECESITAMOS NUEVAS DROGAS CONTRA LA
ENFERMEDAD DE CHAGAS
Las drogas en uso, Benznidazol y Nifurtimox, tienen malos efectos
colaterales y no son efectivas en todos los casos.
El tratamiento de los pacientes es necesario, particularmente en los
casos agudos y congénitos, aunque actualmente los casos crónicos
están siendo tambien tratados. Tanto Médicos sin Fronteras como
la Drugs for Neglected Diseases Initiative (DNDi) opinan que todos
los pacientes chagásicos deben ser tratados.
Las nuevas drogas deberían ser mas efectivas, menos tóxicas y lo
suficientemente baratas como para poder ser adquiridas por
pacientes pobres, en países o regiones pobres.

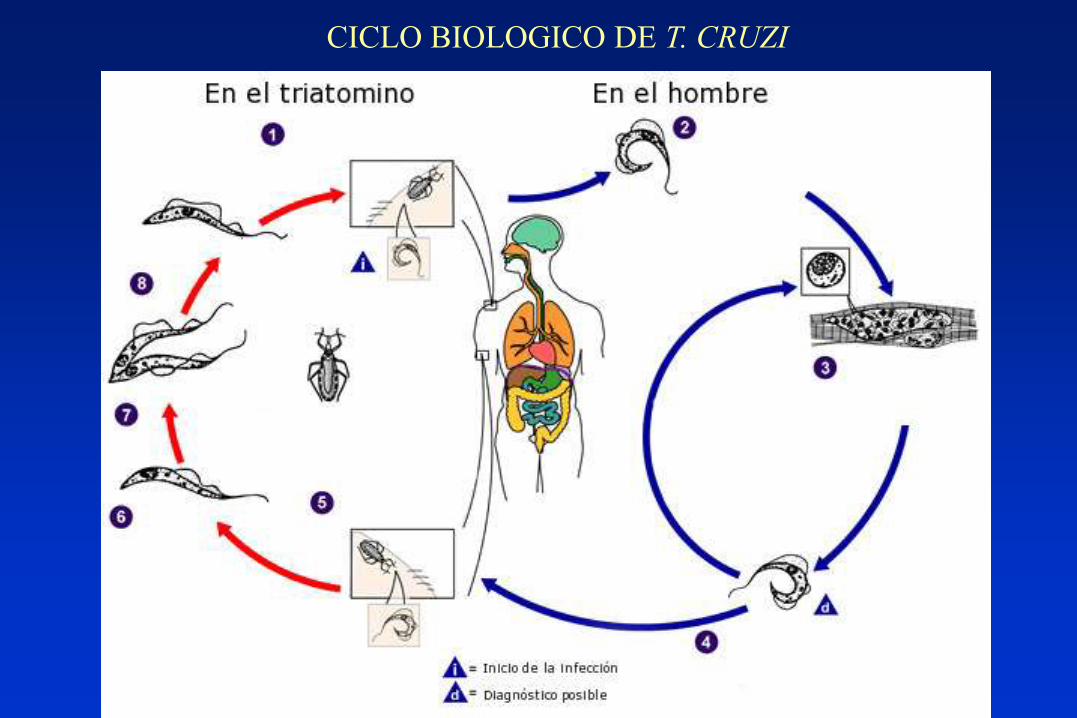
CICLO BIOLOGICO DE T. CRUZI
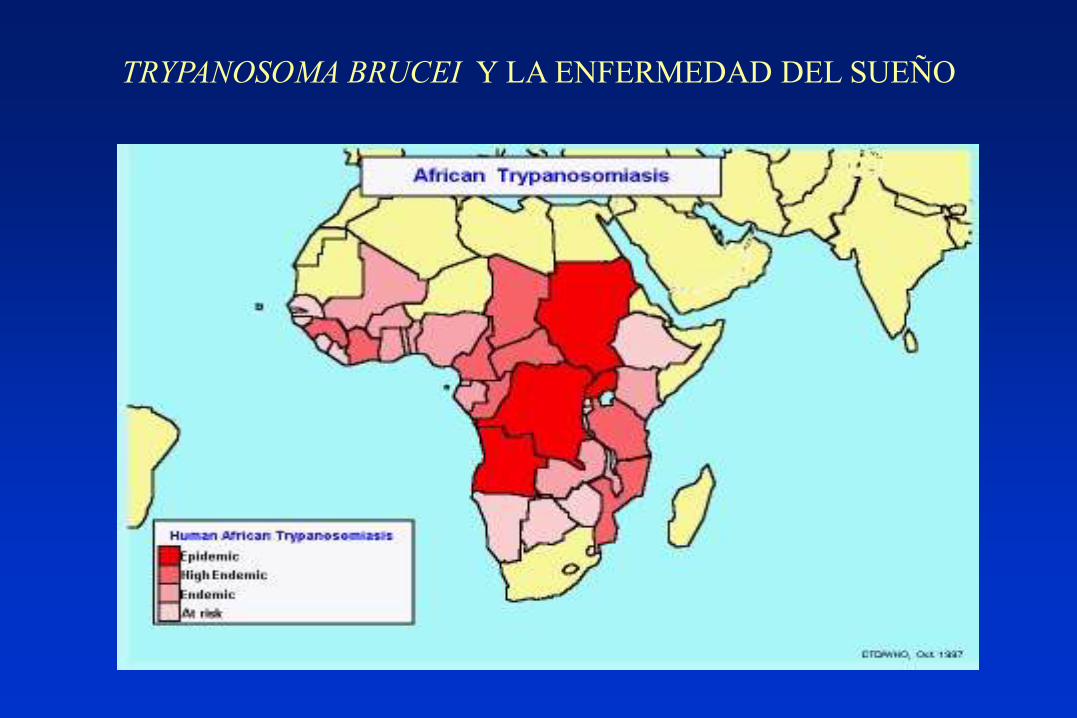
TRYPANOSOMA BRUCEI Y LA ENFERMEDAD DEL SUEÑO
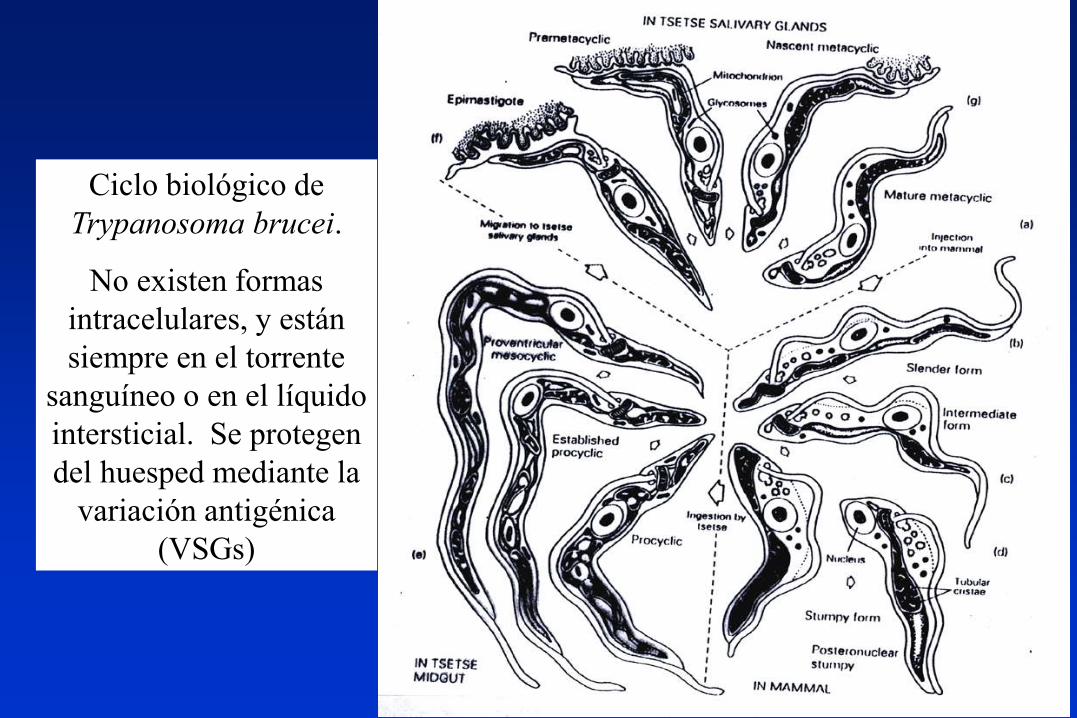
Ciclo biológico de
Trypanosoma brucei.
No existen formas
intracelulares, y están
siempre en el torrente
sanguíneo o en el líquido
intersticial. Se protegen
del huesped mediante la
variación antigénica
(VSGs)

La variación antigénica en Trypanosoma brucei.
T. brucei se encuentra expuesto continuamente al sistema inmune del
hospedador, pues no tiene un estadio intracelular, como T. cruzi y las
Leishmanias. Su solución al problema ha sido el desarrollo de la
variación antigénica.
Las formas del parásito expuestas al sistema inmune del hospedador
tienen su membrana plasmática recubierta por una capa formada por
una única glicoproteína de superficie, llamada VSG (variant surface
glycoprotein). Esta proteína es muy antigénica, y genera una
respuesta que destruye al parásito. Pero éste tiene alrededor de 1000
genes que codifican para diferentes variantes de la VSG, que difieren
en la parte N-terminal, expuesta al medio, y algunos parásitos
comienzan a expresar otra variante, que les permite sobrevivir y
multiplicarse hasta que se genera la respuesta inmune contra ella, y
así sucesivamente.
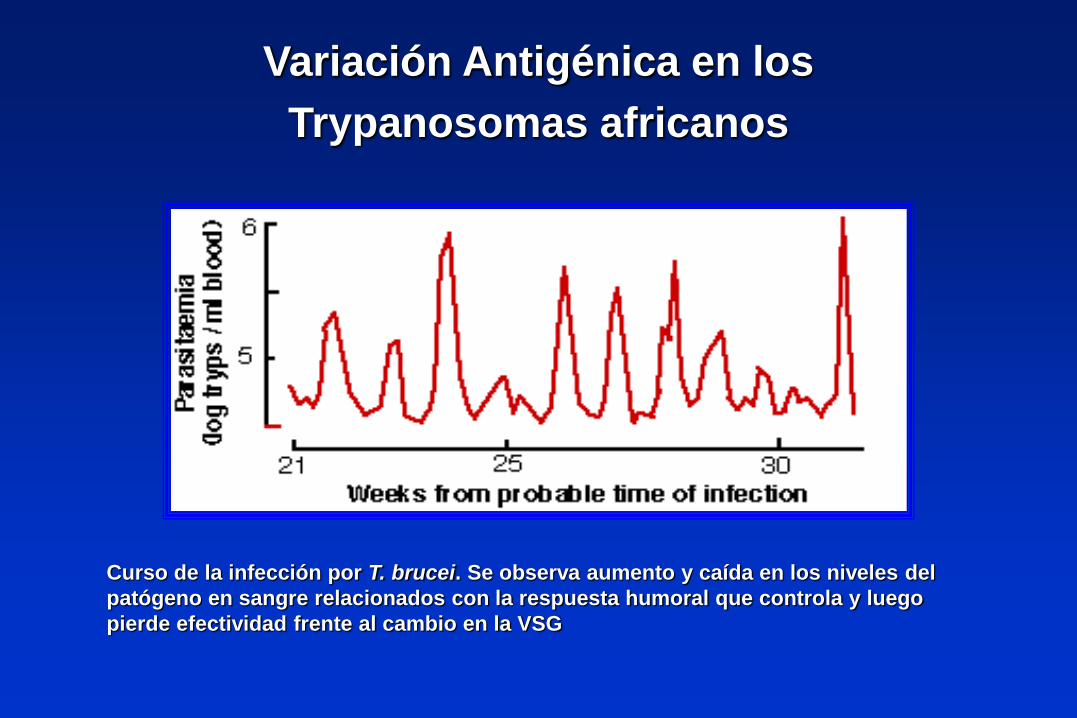
Variación Antigénica en los
Trypanosomas africanos
Curso de la infección por T. brucei. Se observa aumento y caída en los niveles del
patógeno en sangre relacionados con la respuesta humoral que controla y luego
pierde efectividad frente al cambio en la VSG


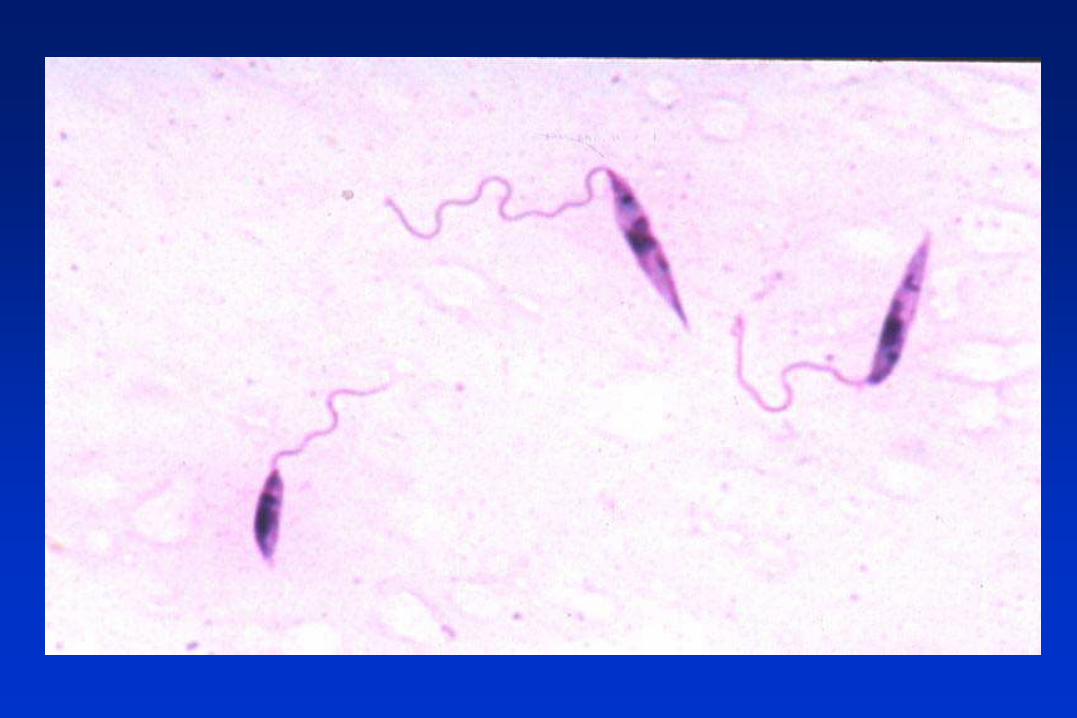
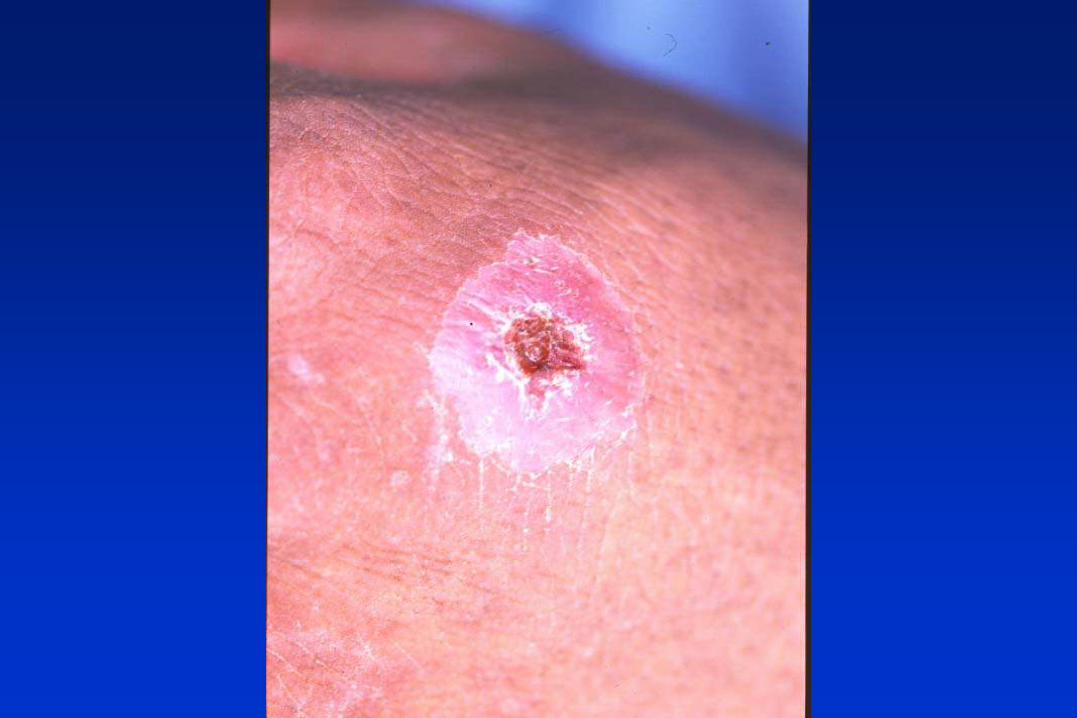
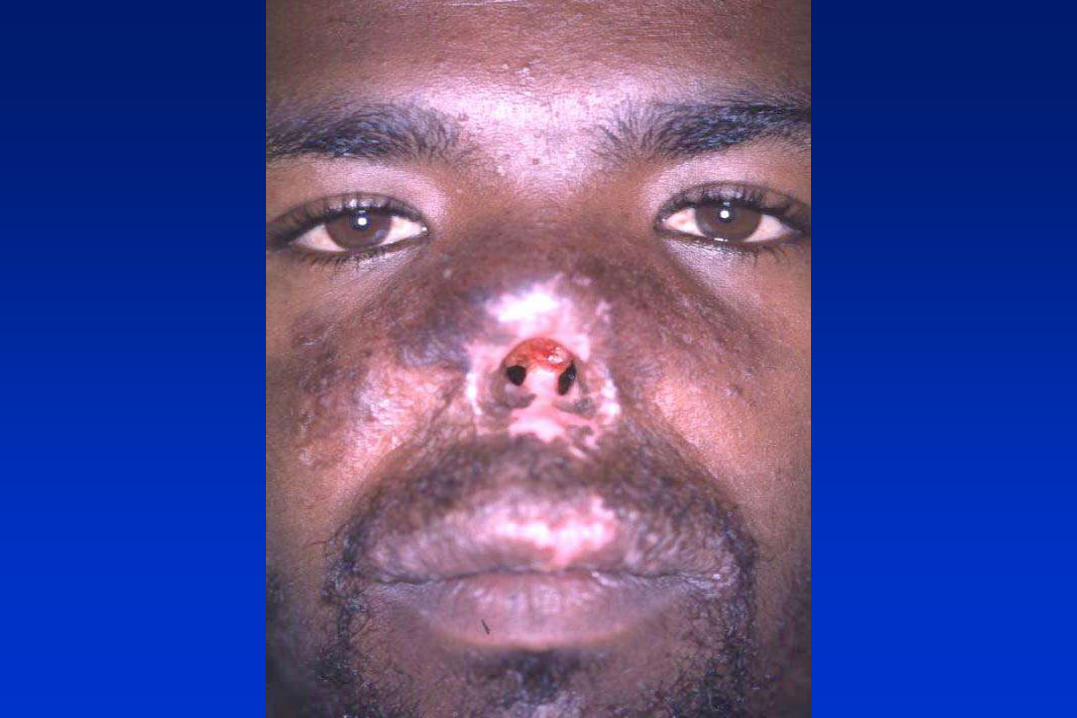
LEISHMANIA spp. Y LAS LEISHMANIASIS


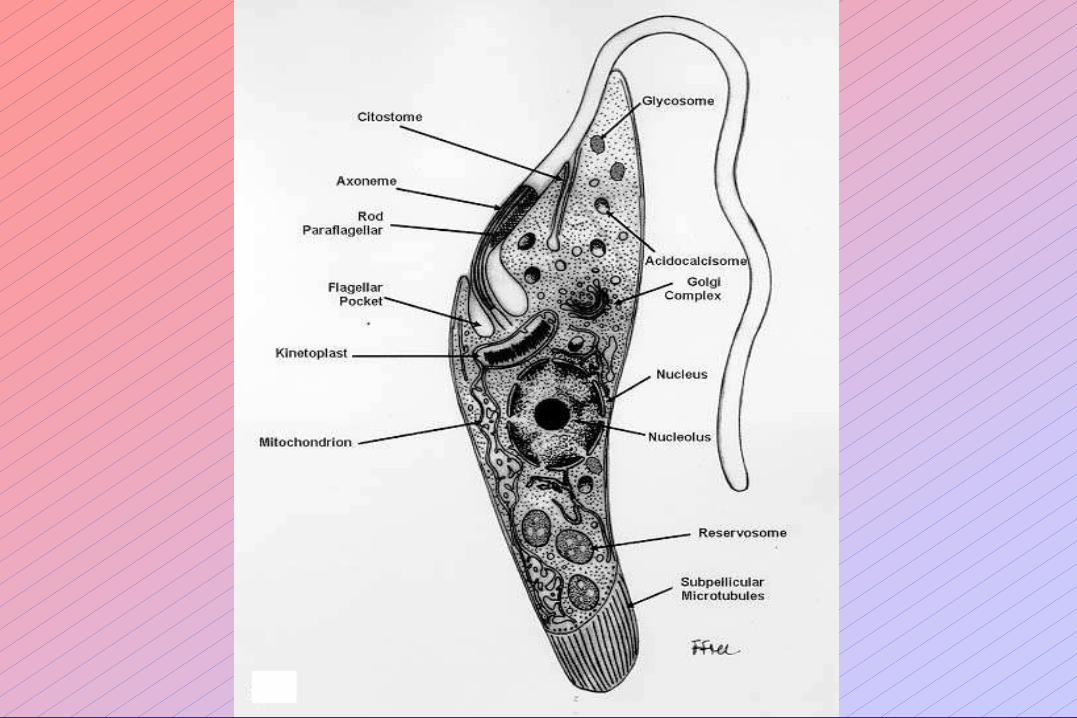
ULTRAESTRUCTURA Y METABOLISMO.
Los distintos grupos de Protozoarios parásitos presentan diferencias metabólicas considerables
entre ellos y con sus huéspedes.
Conocer estas diferencias es importante para poder explotarlas para el diseño de agentes
quimioterápicos (= medicamentos) que ataquen específicamente al parásito y no a su
hospedador.
Las diferencias metabólicas se relacionan en algunos casos con claras diferencias en la
ultraestructura de las células de los Protozoarios.
• Glicosoma de los Trypanosomas y Leishmanias: Metabolismo muy intenso,
aunque incompleto, de la glucosa.
• Cobertura antigénica variable de Trypanosoma brucei: Evasión de la respuesta
inmune del hospedador.
• Carencia de mitocondrias en Entamoeba, Giardia y Trichomonas: Metabolismo
fermentativo obligado; microaerofilia.
• Hidrogenosoma de Trichomonas vaginalis: Producción de H2.
• Apicoplasto (cloroplasto degenerado) en Apicomplexa: Presencia de enzimas y
vías metabólicas características de plantas
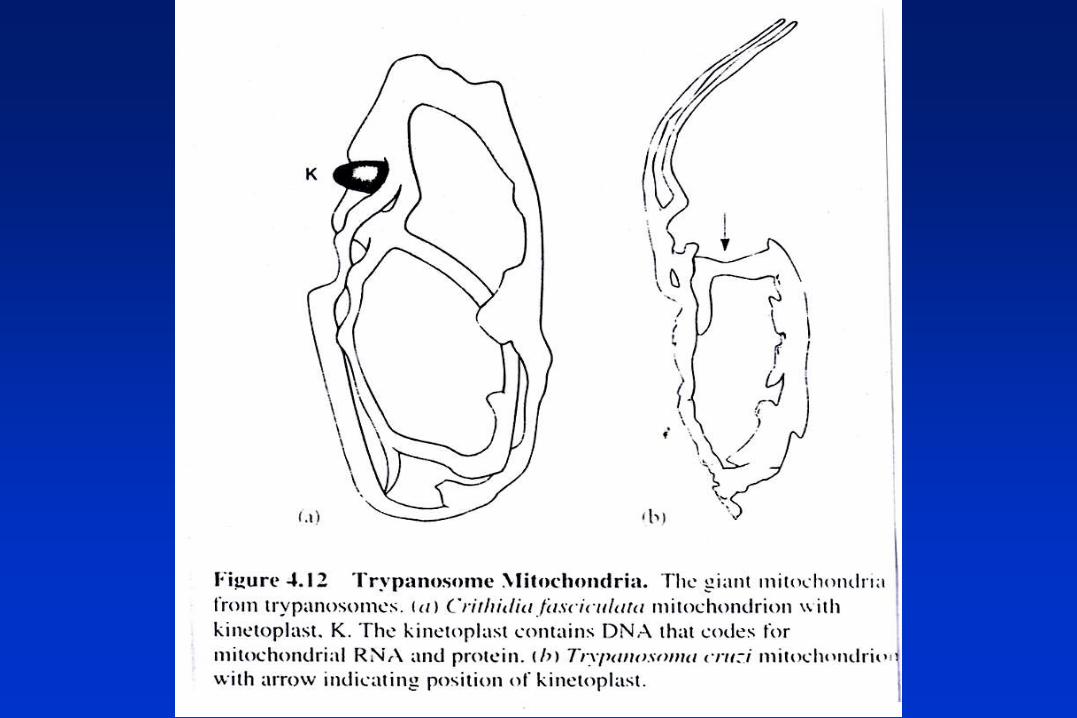
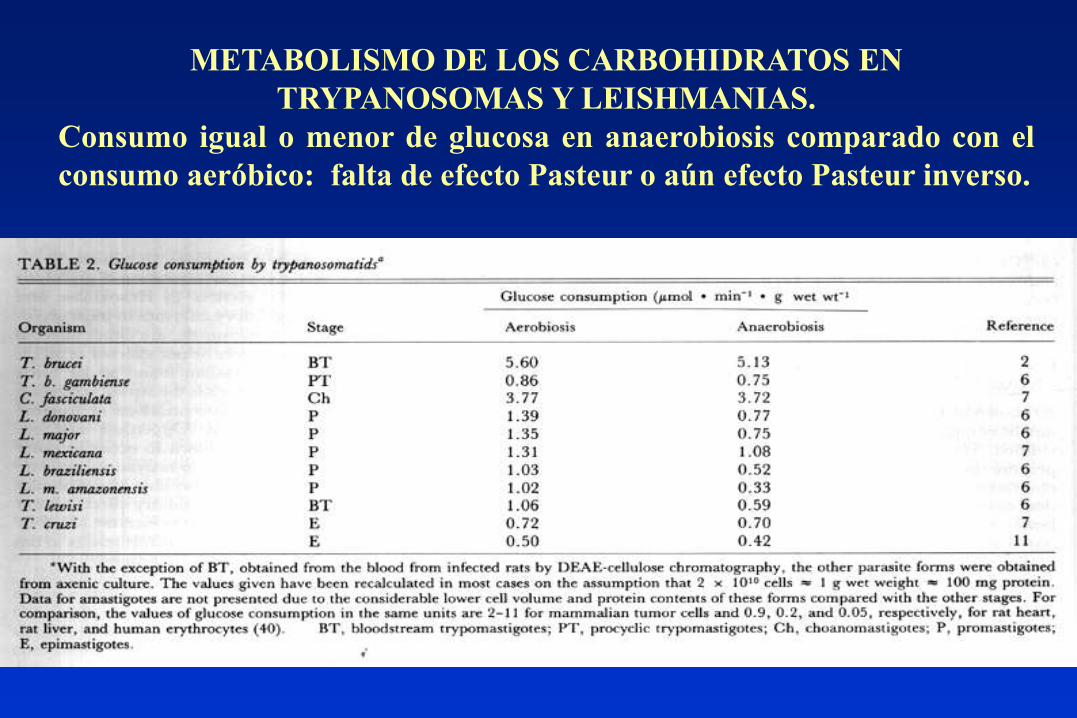
METABOLISMO DE LOS CARBOHIDRATOS EN
TRYPANOSOMAS Y LEISHMANIAS.
Consumo igual o menor de glucosa en anaerobiosis comparado con el
consumo aeróbico: falta de efecto Pasteur o aún efecto Pasteur inverso.

Producción y liberación al medio de catabolitos de la glucosa parcialmente
reducidos (= fermentación) aún en presencia de oxígeno: succinato, acetato,
glicerol, L-alanina, etanol. (= fermentación aeróbica de la glucosa).
Carecen de reserva de glucógeno, al igual que otros hemoparásitos. Las Leishmanias
contienen un polímero de manosa, el manógeno, que cumple una función similar.
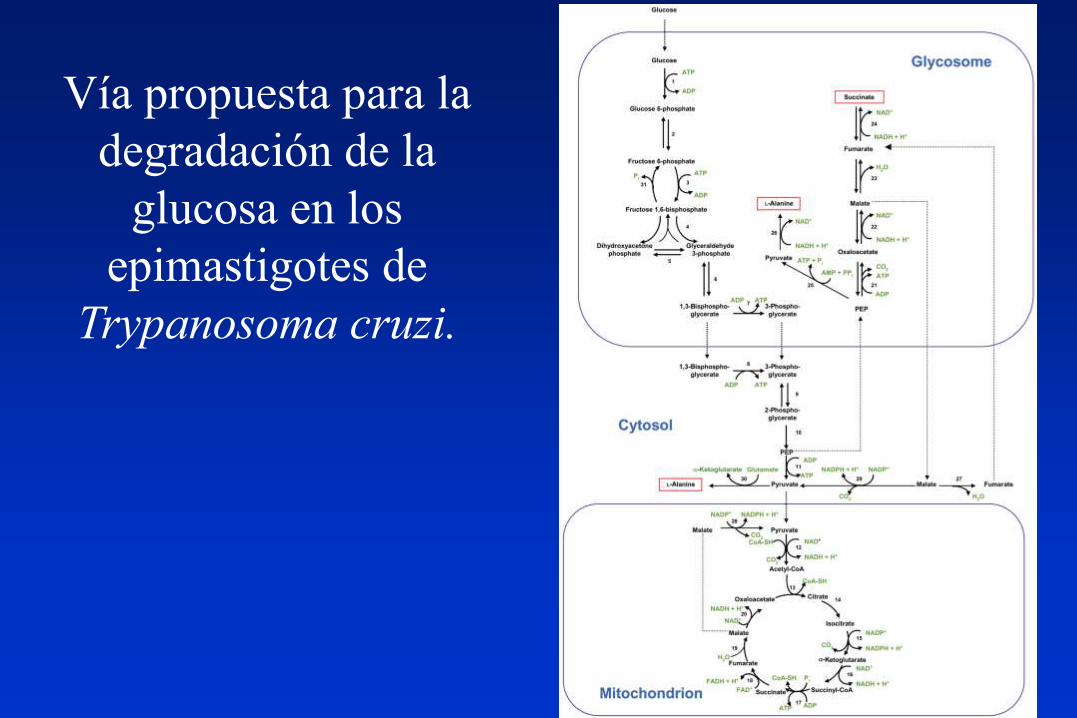
Vía propuesta para la
degradación de la
glucosa en los
epimastigotes de
Trypanosoma cruzi.
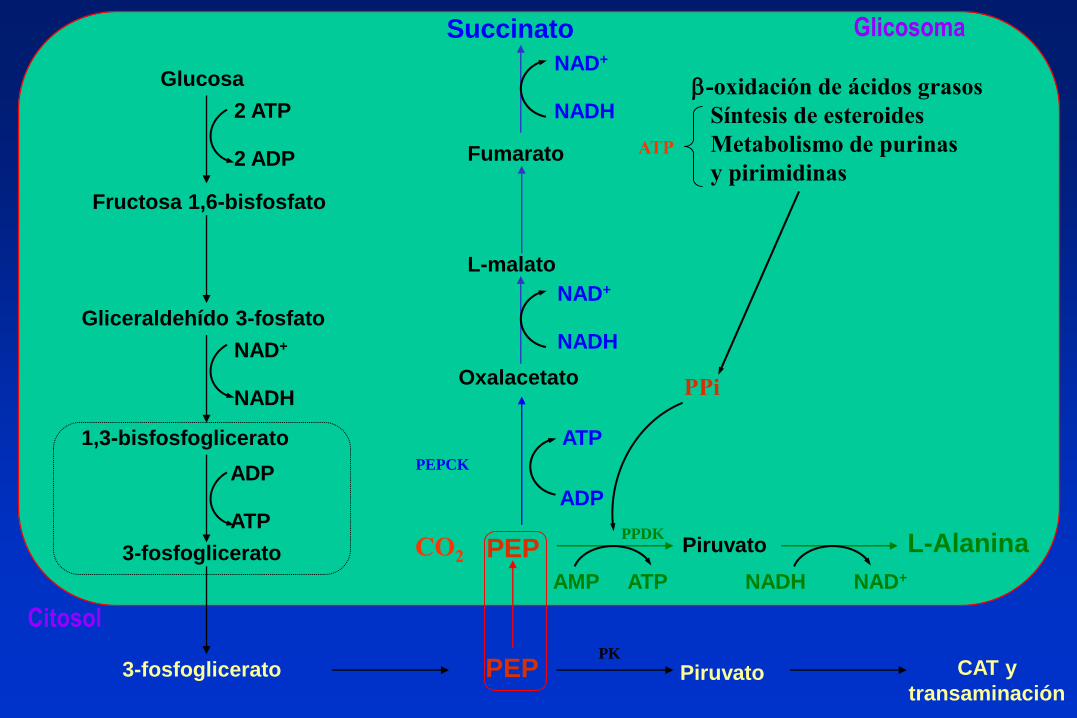
PEP Piruvato
PEP Piruvato L-Alanina
Succinato
Fumarato
L-malato
Oxalacetato
CAT y
transaminación
Glucosa
Fructosa 1,6-bisfosfato
Gliceraldehído 3-fosfato
1,3-bisfosfoglicerato
3-fosfoglicerato
3-fosfoglicerato
2 ATP
2 ADP
ADP
ATP
NAD+
NADH
NADH
NAD+
NADH
NAD+
ADP
ATP
AMP ATP NADH NAD+
b-oxidación de ácidos grasos
Síntesis de esteroides
Metabolismo de purinas
y pirimidinas
PPi
ATP
PK
PPDK
Glicosoma
Citosol
PEPCK
CO2

Resumen del Metabolismo Energético en epimastigotes de T. cruzi.
1) Se ha estudiado el transporte de la glucosa en epimastigotes y trypomastigotes
sanguíneos; los amastigotes intracelulares no transportan glucosa y consumirían
esencialmente aminoácidos, y talvez lípidos.
2) Los productos finales del catabolismo de la glucosa por los epimastigotes son el
succinato y la L-alanina; el acetato no proviene de la glucosa. Una concentración alta
de CO3H- favorece la producción de succinato, y la inhibición de la PEPCK la
producción de L-alanina.
3) La falta del efecto Pasteur se debería a la falta del control de la HK (que no es inhibida
por G-6-P) y de la PFK (que no es inhibida por ATP, citrato o Pi y es activada por AMP).
4) Otras dos diferencias con otros organismos son que la PFK, siendo dependiente de
ATP, tiene la máxima similitud de secuencia con las PFKs dependientes de PPi, y que la
PK es activada por F 2,6-bisP, que contrarresta su inhibición por ATP.
5) No hay una prueba definitiva de que el Ciclo de Krebs sea completamente funcional.
6) El Complejo I de la cadena respiratoria no sería funcional, y la oxidación del NADH se
haría a través de la fumarato reductasa dependiente de NADH, dando succinato que
entraría por el Complejo II, funcional, como los complejos III y IV y la F0F1-ATP sintasa.

Trypanosoma
brucei,
forma sangu’nea.
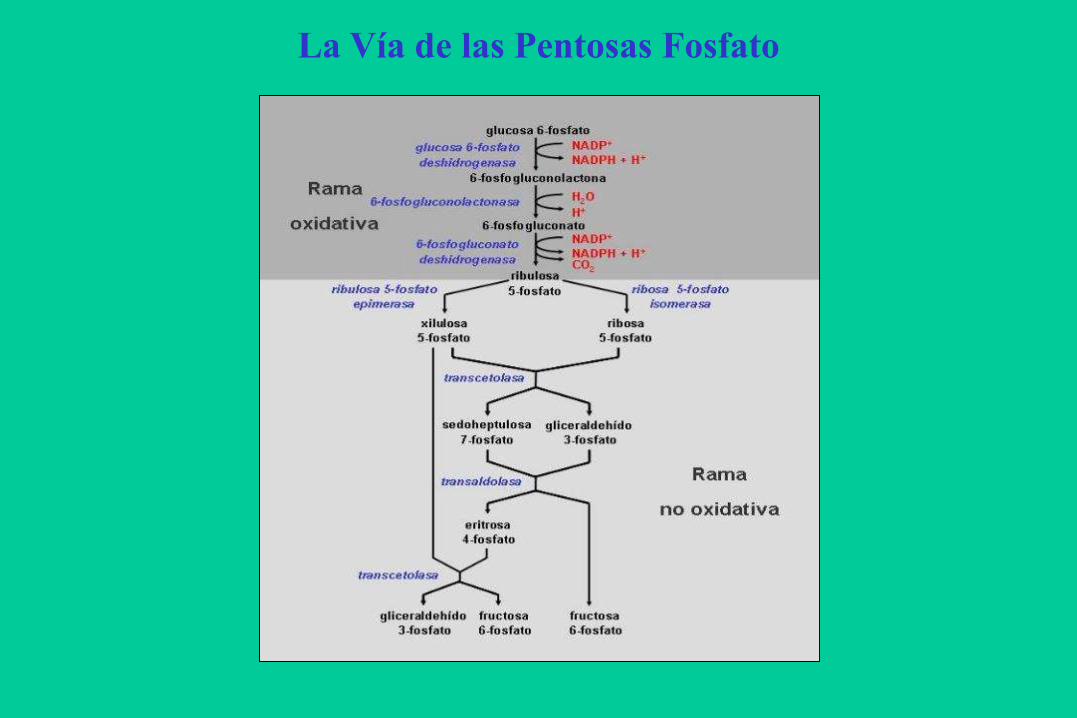
La Vía de las Pentosas Fosfato

METABOLISMO DE PROTEINAS, AMINOACIDOS, PURINAS Y PIRIMIDINAS.
PROTEINASAS.
Las proteinasas se clasifican, según su mecanismo de reacción, en cistein
proteinasas, serin proteinasas, treonin proteinasas (el proteasoma), aspartil
proteinasas y metaloproteinasas. Todas estas clases han sido encontradas en
Protozoarios parásitos, aunque no todas en todos ellos. Son en general importantes
no sólo para la nutrición del parásito, sino que en muchos casos participan
activamente en el mecanismo de patogénesis, y puede considerarselas factores de
virulencia.
En Trypanosomas, Leishmanias, Entamoeba, Giardia y Trichomonas,
predominan las cisteín proteinasas. Las Leishmanias poseen tambien una
metaloproteinasa de superficie cuantitativamente muy importante.
En Plasmodium falciparum la digestión de la hemoglobina del glóbulo rojo
parasitado se hace por la acción concertada de cisteín proteinasas (las falcipaínas) y
aspartil proteinasas (las plasmepsinas).
La inhibición de cistein proteinasas de Trypanosoma cruzi (la cruzipaína) y de
Plasmodium falciparum mata al parásito o impide etapas de diferenciación en su ciclo
biológico. Se están ensayando inhibidores de la cruzipaína como eventuales agentes
quimioterápicos contra la enfermedad de Chagas.

CATABOLISMO DE AMINOACIDOS. Los protozoarios parásitos toman en general los aminoácidos del hospedador, y sus capacidades
sintéticas son limitadas. Los utilizan para la biosíntesis proteica, para otros procesos
biosintéticos y para la generación de energía por oxidación de la cadena carbonada (en los casos
en que hay un Ciclo de Krebs y una cadena respiratoria funcionales).
Algunas de las vías metabólicas son apreciablemente diferentes de las presentes en el
hospedador. Por ejemplo, el catabolismo de los aminoácidos aromáticos (fenilalanina, tirosina y
triptofano) en los Trypanosomas es mucho mas simple que en los mamíferos, y resulta en la
liberación al medio de los derivados aromáticos del lactato, a través de solo dos reacciones
enzimáticas, una transaminación y una reducción.
BIOSINTESIS Y RECUPERACION DE PIRIMIDINAS Y PURINAS.Los parásitos tienden a simplificar su metabolismo, tomando del hospedador tantos componentes
esenciales ya formados como sea posible. Esto es particularmente claro en el caso de las bases
púricas, las cuales no son sintetizadas, sino que los organismos las recuperan del medio. En el
caso de las bases pirimidínicas, además de recuperación del medio, la generalidad de los
Protozoarios parásitos son capaces de sintetizarlas.
HEMOLos Trypanosomas y las Leishmanias son totalmente incapaces de sintetizar el hemo necesario
para sus citocromos y otras hemoproteínas, y deben obtenerlo del medio.
Los Plasmodios tienen la necesidad de evitar los efectos tóxicos de la acumulación de hemo
proveniente de la hemoglobina digerida, y para ello lo precipitan en forma de pigmento malárico o
hemozoína.

PARASITOS ANAEROBIOS
(MICROAEROFILOS)
Trichomonas vaginalis - Tritrichomonas
foetus
Entamoeba histolytica
Giardia intestinalis (Giardia lamblia)
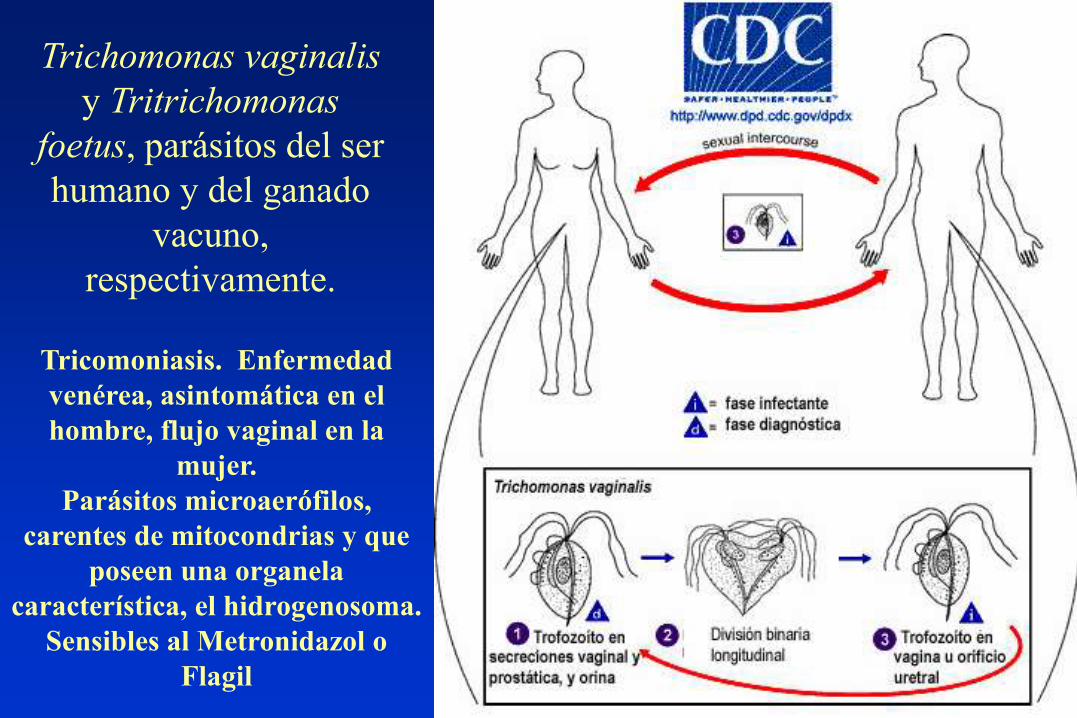
Trichomonas vaginalis
y Tritrichomonas
foetus, parásitos del ser
humano y del ganado
vacuno,
respectivamente.
Tricomoniasis. Enfermedad
venérea, asintomática en el
hombre, flujo vaginal en la
mujer.
Parásitos microaerófilos,
carentes de mitocondrias y que
poseen una organela
característica, el hidrogenosoma.
Sensibles al Metronidazol o
Flagil
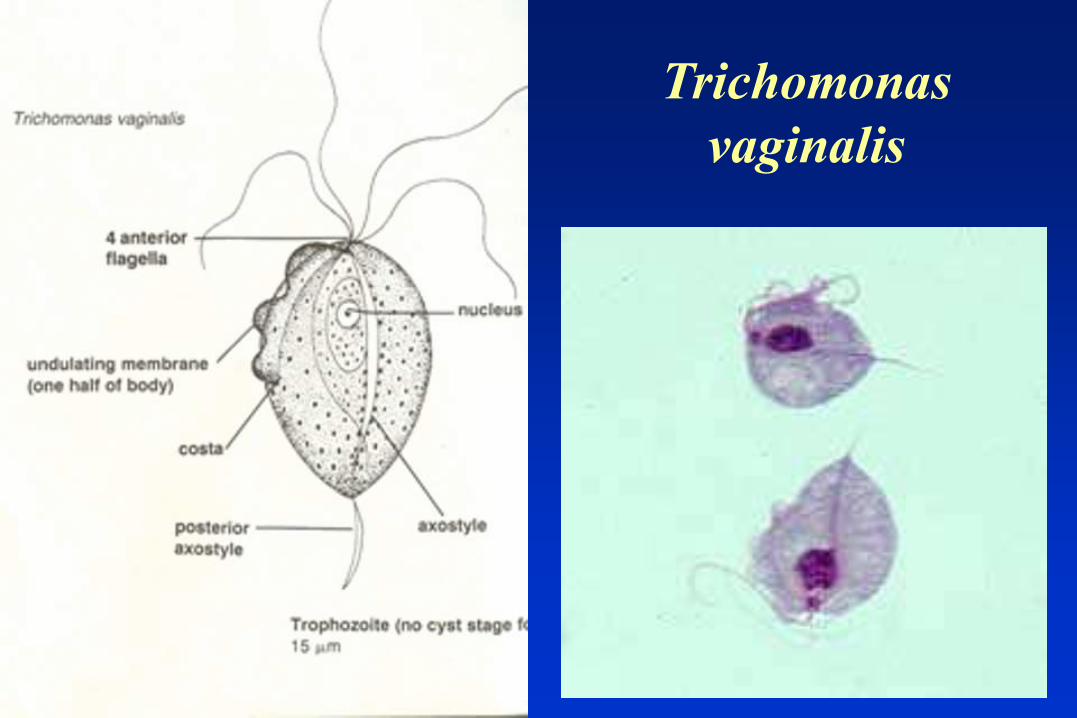
Trichomonas
vaginalis
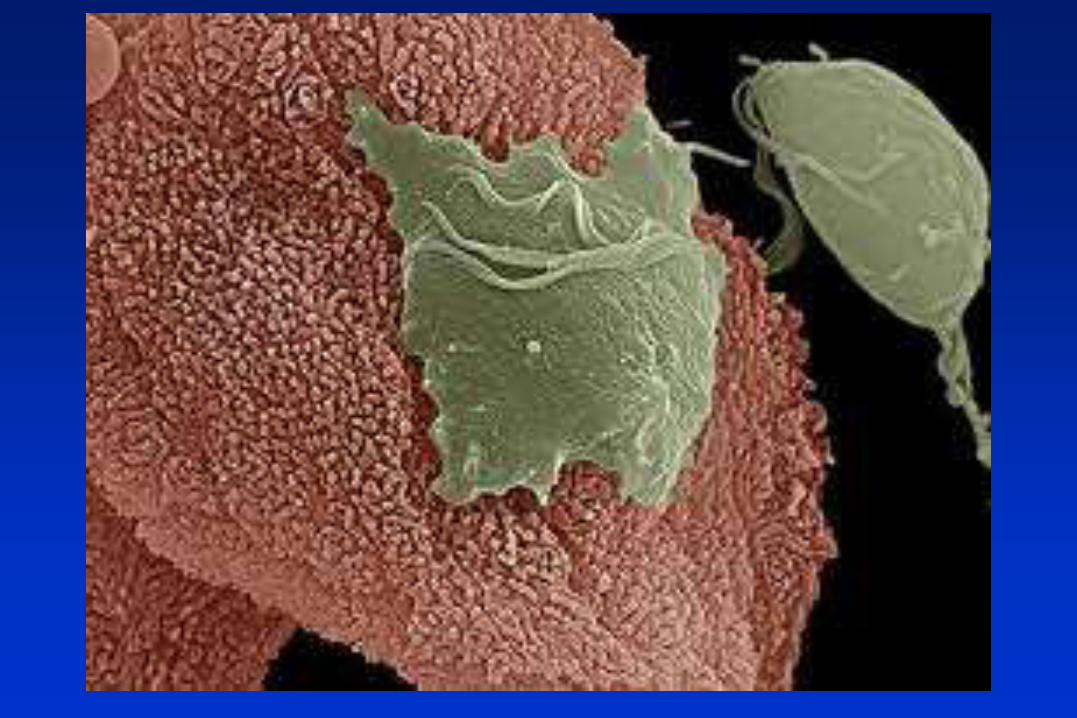

Trichomonas vaginalis : el Hidrogenosoma
.
METABOLISMO DE LA GLUCOSA
POR TRICHOMONAS VAGINALIS
La glucólisis tiene lugar en el citosol, y
es similar a la de otros organismos,
salvo que la PFK es dependiente de PPi
y no de ATP.
La produción de hidrógeno tiene lugar
sólo en anaerobiosis estricta. Los
demás productos finales: glicerol,
lactato y acetato, son iguales en
aerobiosis o en anaerobiosis.
El hidrogenosoma no es exclusivo de T.
vaginalis o Tritrichomonas foetus. Se
encuentra tambien en Protistas muy
distantes evolutivamente, como ciliados
del rumen, que son anaerobios
estrictos, y tambien en algunos
hongos..

APICOMPLEXA
Toxoplasma gondii
Plasmodium spp.
Piroplasmas: Babesia spp. y Theileria spp.
Parásitos de mamíferos, en particular el
ganado
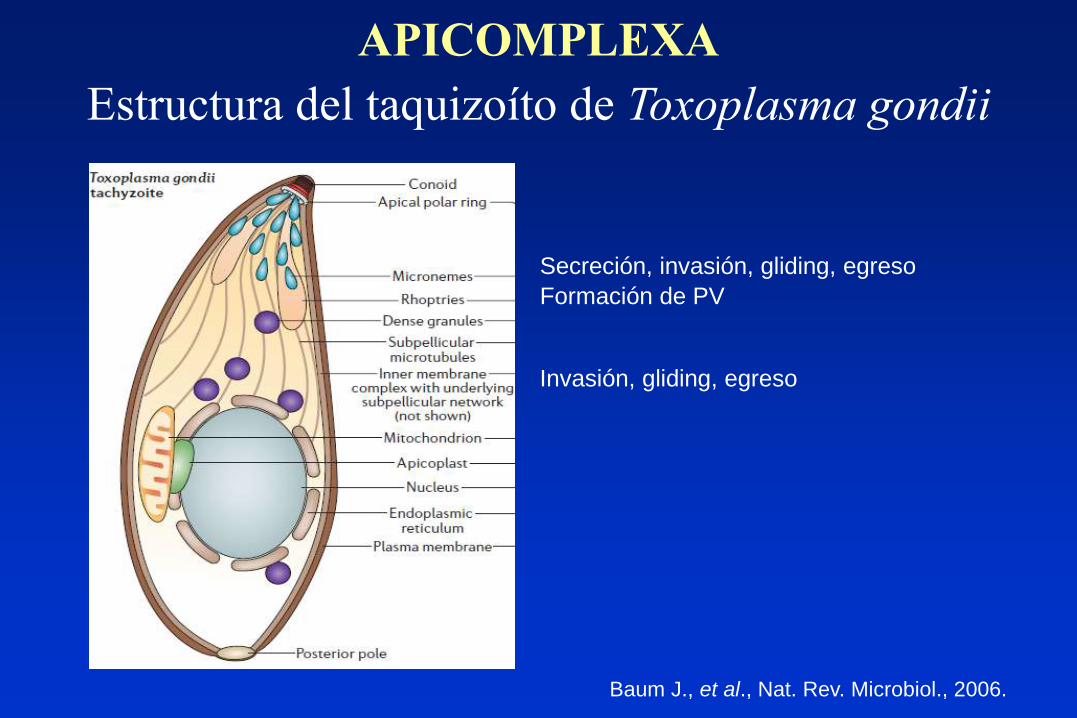
Estructura del taquizoíto de Toxoplasma gondii
Formación de PV
Secreción, invasión, gliding, egreso
Invasión, gliding, egreso
Baum J., et al., Nat. Rev. Microbiol., 2006.
APICOMPLEXA
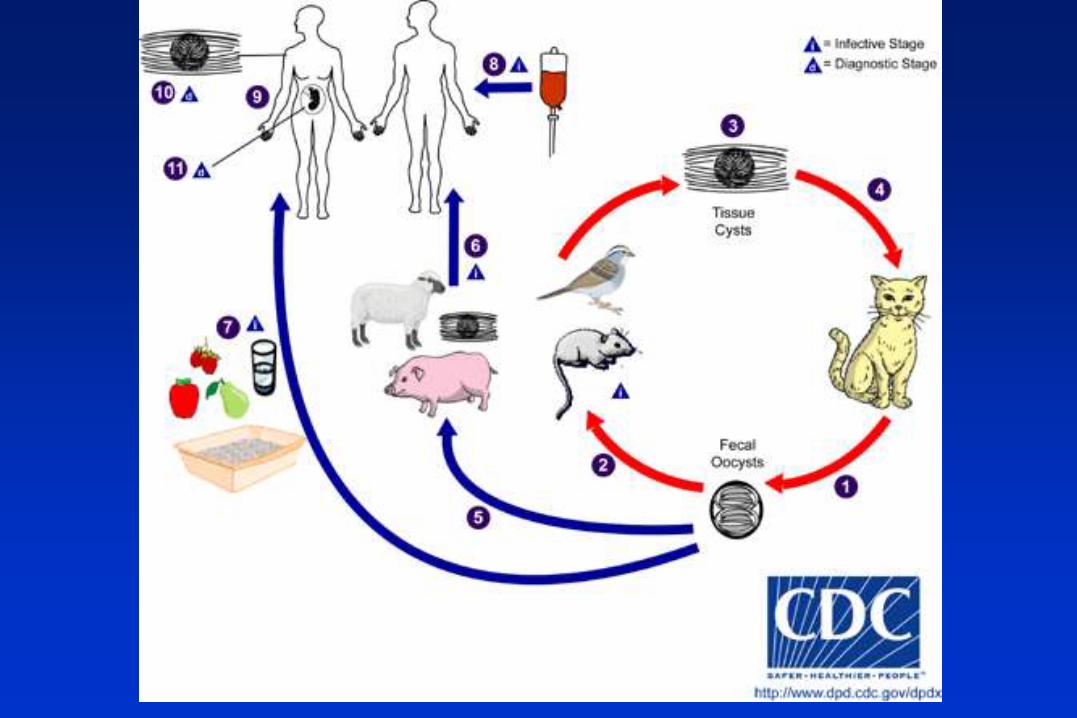

Los únicos huéspedes definitivos de Toxoplasma gondii son miembros de la familia Felidae (gatos
domésticos y otros félidos). Los oocistos no esporulados se eliminan en la materia fecal del gato.
Aunque los oocistos se eliminan sólo durante 1-2 semanas, pueden eliminarse en gran número. Los
oocistos tardan 1-5 días para esporular en el ambiente y hacerse infectivos. Los huéspedes
intermedios naturales (que incluyen aves y roedores) se infectan después de ingerir tierra, agua o
material vegetal contaminado con oocistos. Los oocistos se transforman en taquizoítos poco después
de la ingestión. Estos taquizoítos se localizan en tejido muscular y neural y se desarrollan como
bradizoítos en quistes tisulares.
Los gatos se infectan después de consumir huéspedes intermedios que contienen quistes tisulares. Los
gatos pueden también infectarse directamente por ingestión de oocistos esporulados. Los animales
criados para consumo humano o salvajes pueden también infectarse con quistes tisulares después de
la ingestión de oocistos esporulados en el ambiente.
Los humanos pueden infectarse por una de varias rutas:
Comiendo carne mal cocida de animales que contienen quistes tisulares.
Consumiendo comida o agua contaminada con heces de gato o por muestras ambientales
contaminadas (por ejemplo tierra contaminada o al cambiar el contenido de la caja de un gato
mascota).
Por transfusión de sangre o transplante de órganos.
Transplacentariamente, de la madre al feto.
En el huésped humano, los parásitos forman quistes tisulares, comúnmente en el músculo esquelético,
miocardio, cerebro y ojos; estos quistes pueden permanecer a lo largo de toda la vida del huésped. El
diagnóstico se hace usualmente por serología, aunque los quistes tisulares pueden ser observados en
espcímenes teñidos de biopsias. El diagnóstico de la infección congénita puede lograrse detectando
DNA de T. gondii en el flúido amniótico usando métodos moleculares como la PCR.

Causada por parásitos sanguíneos del género Plasmodium. Hay
aproximadamente 156 especies de Plasmodium que pueden infectar
varias especies de vertebrados. Son muy específicos de la especie que
invaden. Cuatro especies son consideradas verdaderos parásitos de
humanos, pues utilizan humanos casi exclusivamente como un
hospedador intermedio natural: P. falciparum (terciana maligna), P.
malariae (cuartana), P. vivax (terciana benigna), P. ovale (terciana
ovale). Sin embargo, hay informes periódicos de plasmodios de simios
encontrados en humanos, en la mayoría de los casos implicando a P.
knowlesi. No se ha determinado aún si P. knowlesi puede ser
transmitido naturalmente de humano a humano por el mosquito, sin
intervención del hospedador intermedio natural (macacos, género
Macaca). Por lo tanto, P. knowlesi es aún considerado como una
malaria zoonótica.
PLASMODIUM spp. Y LA MALARIA
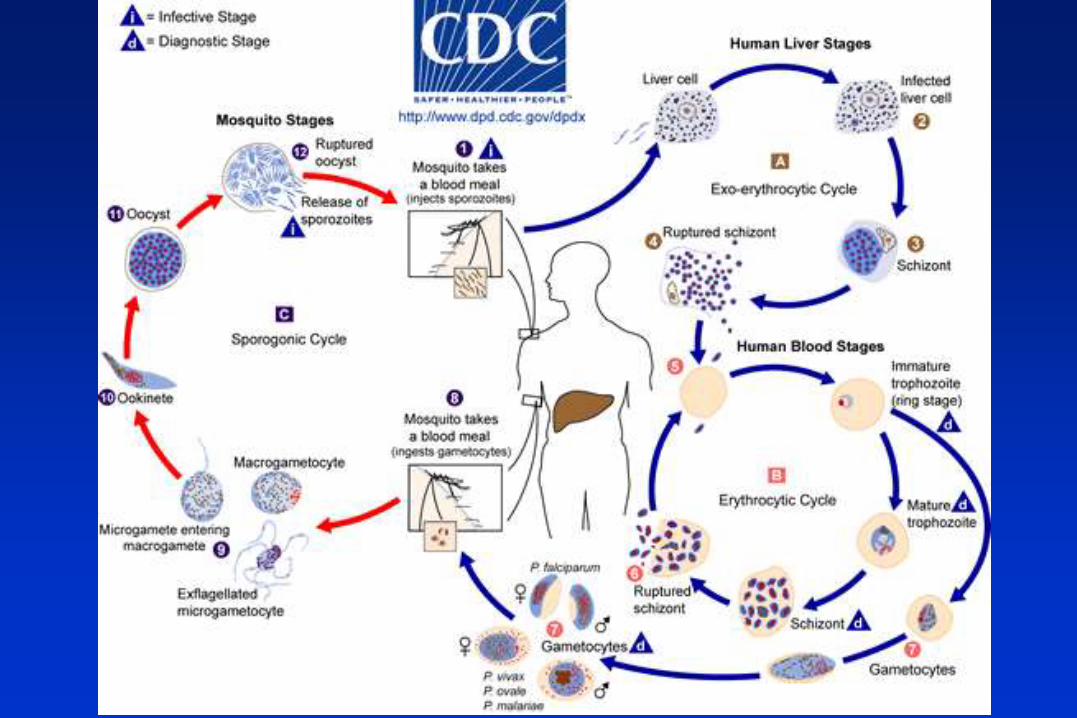

El ciclo biológico del parásito de la malaria involucra dos huéspedes. Durante su
alimentación con sangre, una hembra infectada del mosquito Anopheles inocula
esporozoitos al hospedador humano. Los esporozoitos infectan hepatocitos y maduran
dando esquizontes, que se rompen liberando merozoitos. (En los casos de P. vivax y P.
ovale un estadio durmiente [hipnozoitos] puede persistir en el hígado y causar relapsos
invadiendo la sangre semanas, o aún años, después de la infección.) Después de la
replicación inicial en el hígado (esquizogonia exo-eritrocítica), los parásitos se replican
asexualmente en los eritrocitos (esquizogonia eritrocítica). Los merozoitos infectan los
glóbulos rojos y se transforman en trofozoítos en estadío de anillo, los cuales luego
maduran dando esquizontes, que se rompen liberando merozoitos. Algunos parásitos se
diferencian a estadíos sexuales eritrocíticos (gametocitos). El estadío sanguíneo de los
parásitos es responsable de las manifestaciones clínicas de la enfermedad.
Los gametocitos, masculinos (microgametocitos) y femeninos (macrogametocitos), son
ingeridos por un mosquito Anopheles mientras se alimenta con la sangre del humano
infectado. La multiplicación de los parásitos en el mosquito se denomina ciclo
esporogónico. En el estómago del mosquito las microgametas penetran en las
macrogametas generando zigotas. Las zigotas adquieren motilidad y se elongan
transformándose en oocinetos, los cuales invaden la pared del intestino medio del
mosquito, donde se desarrollan como oocistos. Los oocistos crecen, se rompen, y liberan
esporozoitos, los que se dirigen a las glándulas salivales. La inoculación de los
esporozoitos a un nuevo hospedador humano perpetúa el ciclo biológico de la malaria.