modelización del desarrollo postembrionario de
TRANSCRIPT

Bol. San. Veg. Plagas, 25: 241-253, 1999
Modelización del desarrollo postembrionario deDociostaurus maroccanus (Thunberg, 1815)
P. BARRANCO, F. PASCUAL Y T. CABELLO
Se realiza el ajuste de los datos del desarrollo en campo de una población de Docios-taurus maroccanus del Cabo de Gata (Almería) durante los años 1994, 1995 y 1996. Lasimulación se hace mediante la aplicación de tres modelos logísticos autónomos clási-cos: Verhuslt-Pearl, Gomperz y Richards; y uno no autónomo: modelo de Carreño.
Los ajustes siempre fueron altamente significativos (P < 0.01). En el 80% de loscasos r2 >0.99 y en ningún caso r2 es inferior a 0.9. Por lo que cualquiera de los modeloselegidos es válido para la simulación del desarrollo de la langosta en sus diferentes esta-dios.
Se establece la duración de cada estadio aplicando estos modelos, siendo la duraciónen días para cada uno de los estadios 7-10, 6-7, 4-8, 4-12 y 7-11, y la duración total deldesarrollo 37-44 días.
P. BARRANCO Y T. CABELLO: Dpto. Biología Aplicada. Escuela Politécnica Superior.Universidad de Almería. 04120 La Cañada (Almería), España, [email protected],[email protected].
F. PASCUAL: Dpto. Biología Animal y Ecología. Facultad de Ciencias. Universidad deGranada. 14071 Granada, España, [email protected].
Palabras clave: Dociostaurus maroccanus, desarrollo postembrionario, modelosmatemáticos, Almería
INTRODUCCIÓN
Los modelos matemáticos contribuyen almanejo y esclarecimiento de la dinámica delas poblaciones y revisten especial interéscomo herramientas predictivas en las espe-cies plaga, siendo cada vez más frecuenteslos trabajos en los que se utilizan los modelosanalíticos aplicados a la ecología de pobla-ciones.
La modelización del desarrollo postem-brionario se puede realizar mediante el ajustede la fenología en días o en grados-día acu-mulados (que establece una tasa de creci-miento en función del tiempo y temperatura).Los datos de campo contienen valores de dis-tintos estadios simultáneamente, cuyas pro-
porciones van variando en cada muestra. Elmodelo fenológico se expresa como un con-junto de ecuaciones predictivas, donde cadaecuación describe la proporción de insectosque han alcanzado un determinado estadioconforme pasa el tiempo (HUDES & SHOEMA-KER, 1988). Además, al ajustar independien-temente cada estadio se pueden aplicar tasasde crecimiento diferentes para cada uno deellos (DENNIS et al, 1986).
Al expresar el modelo fenológico como unconjunto de ecuaciones predictivas, dondecada una describe la proporción de insectosque han alcanzado un determinado estadio,se consiguen una serie de propiedades. Porun lado, la variabilidad en la duración de losestadios es considerada; se permite la acó-
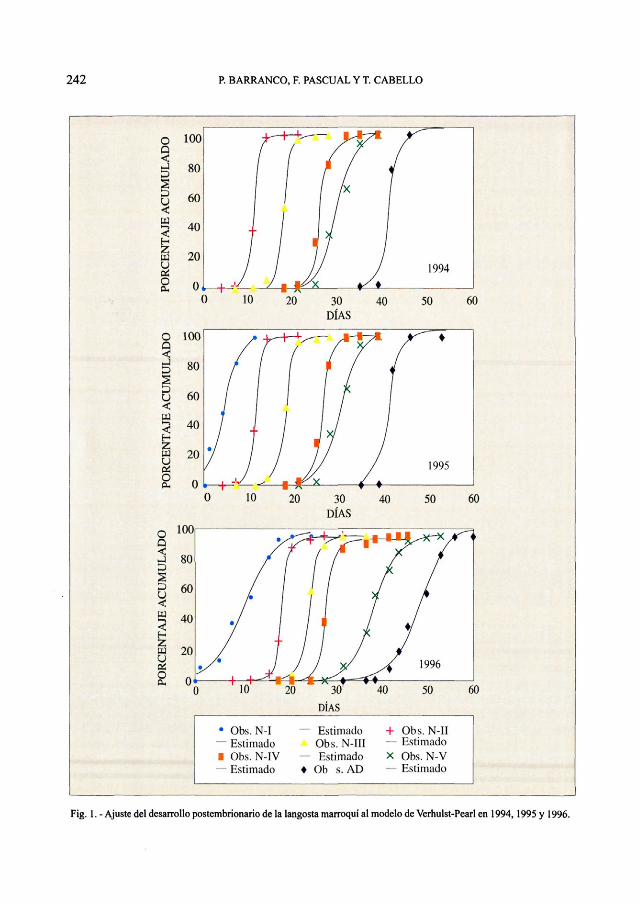
Fig. 1. - Ajuste del desarrollo postembrionario de la langosta marroquí al modelo de Verhulst-Pearl en 1994,1995 y 1996.

modación de la correlación entre los distintosestadios, siguiendo su ritmo normal de desa-rrollo; y no es necesario conocer la tasa demortalidad al asumir la supervivencia comouna función temporal (HUDES & SHOEMA-KER, 1988). Estos autores comparan los ajus-tes realizados de la fenología de un lepidóp-tero plaga en días y tiempo fisiológico, sien-do mejor el ajuste en días en determinadosestadios y en otros a tiempo fisiológico.Cuando las tasas de crecimiento de los insec-tos no son funciones estrictamente linealesde la temperatura las escalas simples de tiem-po fisiológico no dan resultados exactos(STINNERétfa/., 1974).
METODOLOGIA
La modelización del desarrollo postem-brionario se ha realizado en días, utilizandolos datos de seguimiento en campo. Para ellose tomaron muestras cada tres días desde elcomienzo de la avivación de la langosta has-ta que todos los individuos alcanzaban elestado adulto. En cada uno de las fechas seobtuvo el porcentaje de individuos que seencontraba en los distintos estadios. De estemodo se estableció una estructura de edadesa lo largo del tiempo para las poblaciones delHaza de la Veedora durante los años 1994 y1995 y la de los alrededores de Fernán-Pérezen 1996.
Se utilizan los modelos matemáticos logís-ticos como herramienta para el conocimientode la dinámica poblacional. Estos modelos sebasan en la tasa de crecimiento de las pobla-ciones sometidas a una capacidad de cargaque viene determinada por las limitacionesfísicas y biológicas del ecosistema.
Se aplican a los datos de desarrollo pos-tembrionario de las poblaciones de langostade Almería una serie de modelos logísticospara conocer su ajuste a una determinadacurva y comparar los distintos ajustes. Seemplean dos tipos de modelos: los clásicos yautónomos, y uno no autónomo. Los autóno-mos consideran el tiempo como una funciónperiódica que se repite y que se presenta en la
naturaleza con pequeñas variaciones y noestá recogido en sus ecuaciones diferencia-les. Los modelos no autónomos, por el con-trario, si consideran el tiempo.
Los modelos logísticos clásicos exigenque exista uniformidad entre los individuos yque la tasa de crecimiento de la poblacióncambie instantáneamente con su densidad. Elmodelo logístico no autónomo de CARREÑO(1996) utilizado, establece que el comporta-miento dinámico de la población está deter-minado por el tamaño de ésta y por el tiempo.Se han aplicado tres modelos logísticos clási-cos de los más usuales señalados por MORE-NO (1994): VERHULST-PEARL, GOMPERZ yRICHARDS. Estos cuatro modelos y sus curvassolución están recogidos en CARREÑO et al.(1997b).
Para poder aplicar a los datos los modelosmatemáticos se han utilizado los siguientesprogramas informáticos: STATISTIC, Ver. 4.1, Analytical Software, 1994; STATGRAP-HICS, Ver. 7.1, Statistical Graphics Corpora-tion, 1994 y Table Curve 2D™, JANDELSCIENTIFIC, AISN Software, 1994.
RESULTADOS Y DISCUSIÓN
Los resultados de estos ajustes están reco-gidos en los Cuadros 1,2, y 3; y en las Figu-ras 1,2,3 y 4.
El desarrollo postembrionario de la lan-gosta sigue una función logística que se ajus-ta de forma significativa a todos los modelosutilizados (Figs. 1-4). En el 80% de los casosr2 > 0.99 y en ningún caso r2 es inferior a 0.9siendo la significación siempre alta (P <0.001) (Cuadros 1-4). Por lo que cualquierade los modelos elegidos es válido para lasimulación del desarrollo de la langosta porestadios. Sin embargo, no todos los modelosse comportan de igual modo. En el 83% de loscasos, el modelo de Carreño ha presentadouna r2 mayor, aunque este modelo no es logís-tico, y si continuamos la función al infinito,no se mantiene estable, sino que dependiendodel parámetro que actúa de freno de la fun-ción n ésta continúa creciendo o se extingue.

Fig. 2. - Ajuste del desarrollo postembrionario de la langosta marroquí al modelo de Gomperz en 1994, 1995 y 1996.

Fig. 3. - Ajuste del desarrollo postembrionario de la langosta marroquí al modelo de Richards en 1994, 1995 y 1996.

Fig. 4. - Ajuste del desarrollo postembrionarío de la langosta marroquí al modelo de Carreño en 1994, 1995 y 1996.

Cuadro 1 - Valores de los parámetros obtenidos del ajuste y significación de los distintos modelosmatemáticos para el año 1994
Su estabilidad depende de que \i sea suficien-temente pequeño, y cuando se hace cero, estemodelo se convierte en el de Richards.
Por otro lado, a pesar de que en la mayoríade los casos el modelo de Carreño es elmejor, su interpretación biológica al igualque en el de Richards es difícil, pues apartede los parámetros ecológicos reconocidos, K(capacidad de carga del sistema) y r (tasa decrecimiento de la población), los otros dosparámetros, v y ji, tienen un sentido única-
mente matemático. Además, en el modelo deCarreño la tasa de crecimiento tiene valoresdemasiado altos, comparados con los otrosajustes, para los estadios quinto y adulto en1994, así como para el tercero y adulto en1995.
Esta primera aproximación a la modeliza-ción del crecimiento de la langosta ha de ser-vir de base para futuros trabajos en campo,para los que previamente en laboratorio sedeterminen los umbrales térmicos de desa-
Verhulst-PearlGompertz
Richards
Carreño
Verhulst-Pearl
Gompertz
Richards
Carreño
Verhulst-Pearl
Gompertz
Richards
Carreño
Verhulst-PearlGompertzRichardsCarreño
Verhulst-Pearl
Gompertz
Richards
Carreño
Verhulst-Pearl
Gompertz
Richards
Carreño
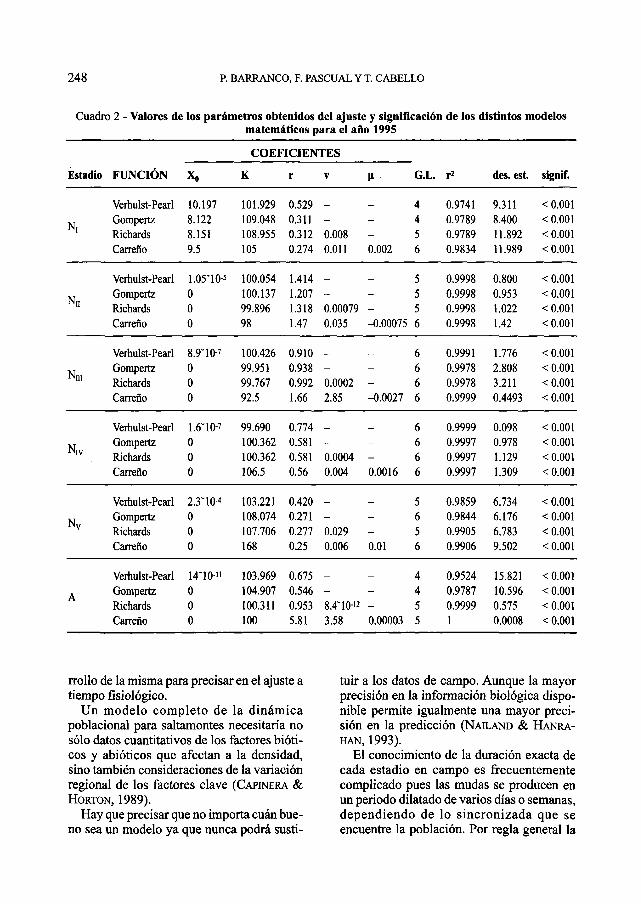
Cuadro 2 - Valores de los parámetros obtenidos del ajuste y significación de los distintos modelosmatemáticos para el año 1995
rrollo de la misma para precisar en el ajuste atiempo fisiológico.
Un modelo completo de la dinámicapoblacional para saltamontes necesitaría nosólo datos cuantitativos de los factores bióti-cos y abióticos que afectan a la densidad,sino también consideraciones de la variaciónregional de los factores clave (CAPINERA &HORTON, 1989).
Hay que precisar que no importa cuan bue-no sea un modelo ya que nunca podrá susti-
tuir a los datos de campo. Aunque la mayorprecisión en la información biológica dispo-nible permite igualmente una mayor preci-sión en la predicción (NAILAND & HANRA-HAN, 1993).
El conocimiento de la duración exacta decada estadio en campo es frecuentementecomplicado pues las mudas se producen enun periodo dilatado de varios días o semanas,dependiendo de lo sincronizada que seencuentre la población. Por regla general la
Verhulst-PearlGompertzRichardsCarreño
Verhulst-PearlGompertzRichardsCarreño
Verhulst-PearlGompertzRichardsCarreño
Verhulst-PearlGompertzRichardsCarreño
Verhulst-PearlGompertzRichardsCarreño
Verhulst-PearlGompertzRichardsCarreño
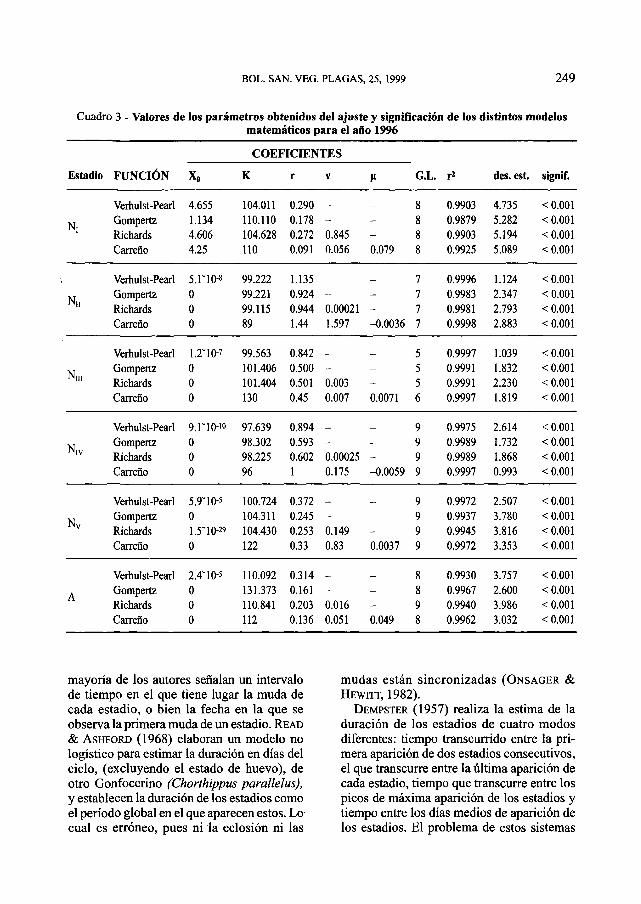
Cuadro 3 - Valores de los parámetros obtenidos del ajuste y significación de los distintos modelosmatemáticos para el año 1996
mayoría de los autores señalan un intervalode tiempo en el que tiene lugar la muda decada estadio, o bien la fecha en la que seobserva la primera muda de un estadio. READ& ASHFORD (1968) elaboran un modelo nologístico para estimar la duración en días delciclo, (excluyendo el estado de huevo), deotro Gonfocerino (Chorthippus parallelus),y establecen la duración de los estadios comoel período global en el que aparecen estos. Locual es erróneo, pues ni la eclosión ni las
mudas están sincronizadas (ONSAGER &HEWITT, 1982).
DEMPSTER (1957) realiza la estima de laduración de los estadios de cuatro modosdiferentes: tiempo transcurrido entre la pri-mera aparición de dos estadios consecutivos,el que transcurre entre la última aparición decada estadio, tiempo que transcurre entre lospicos de máxima aparición de los estadios ytiempo entre los días medios de aparición delos estadios. El problema de estos sistemas
Verhulst-Pearl
Gompertz
Richards
Carreño
Verhulst-PearlGompertzRichardsCarreño
Verhulst-Pearl
Gompertz
Richards
Carreño
Verhulst-Pearl
Gompertz
Richards
Carreño
Verhulst-Pearl
Gompertz
Richards
Carreño
Verhulst-PearlGompertzRichardsCarreño

Fig. 5. - Ninfas de Dociostaurus maroccanus tomandoel sol. (Foto: P. Barranco).
Fig. 6. - Adultos de Dociostaurus maroccanus. (Foto:P. Barranco).
de estima, a excepción del tercero (Cuadro5), es que se basan únicamente en días sintener en cuenta la tasa de desarrollo de lapoblación.
En este estudio, se ha optado por estable-cer la duración de cada estadio como el tiem-po que transcurre desde que el 50% de lapoblación está en determinado estadio hastaque ese porcentaje alcanza el siguiente(ONSAGER & HEWITT, op. cit. y DENNIS &KEMP, 1988). Para ello se han utilizado losajustes a los modelos matemáticos, despejan-do la / en cada modelo y haciendo x = 50%.En el caso del modelo de Carreño, no es posi-ble despejar el tiempo, por lo que se ha cal-culado la duración del estadio medianteinterpolación. La duración estimada resul-tante para cada estadio y la total del desarro-llo se recogen en el Cuadro 4. Para cada añola duración total del desarrollo apenas varíade un ajuste a otro, lo que pone de manifiestola idoneidad de todos los modelos utilizados(CARREÑO, etal, 1997). La duración del cre-cimiento es semejante en 1995 y 96 y algomás largo en 1994. La variación máximaentre años es de casi ocho días y se produceentre 1994 y 1996. Las causas que ocasionanestas diferencias son las señaladas en el pun-to anterior.
La duración media de cada estadio es sinembargo muy variable entre años, como seaprecia en el Cuadro 4. Aunque dentro de unmismo año los ajustes proporcionan estimasmuy semejantes, a excepción del modelo deRichads que establece duraciones anómalaspara los estadios IV y V en 1994, y I en 1996.
En el Cuadro 5 se han recogido los datosofrecidos por los distintos autores sobre laduración de los estadios ninfales de la lan-gosta marroquí, incluyendo en último lugarlos promedios de los obtenidos a partir de losajustes de nuestros datos. Se puede apreciarque la variabilidad es muy amplia, además seignora cómo se han calculado en la mayoríade los casos, a excepción de los DEMPSTER(1957), ya mencionado. Y precisamente esteautor realiza, en el caso recogido, una estimasemejante a la nuestra. De todos los datosexpuestos, son lógicamente, los de las zonaseuropeas los más próximos a los del Cabo deGata: La Baume (1918), Melis (1934), Jan-none (1934) y Dempster (1957).
Es importante señalar, que en los datostomados de otros autores, los límites delintervalo que indica la duración total deldesarrollo embrionario, son producto de lasuma de los valores máximos y mínimospara cada estadio. Se considera que este

Cuadro 4. - Duración estimada en días para cada estadio y total del desarollo postembrionario enDociostaurus maroccanus en condiciones de campo en el sureste de España
Cuadro 5. - Duración de los distintos estadios ninfales y total del desarrollo postembrionarioen D. maroccanus según los distintos autores
(*) Tomado de PAOLI (1937)(+) Tomado de LATCHININSKY y LAUNOIS-LUONG (1992).
Verhulst-PearlGomperzRichardsCarreño
Verhulst-PearlGomperzRichardsCarreño
Verhulst-PearlGomperzRichardsCarrefio-
LABAUME, 1918*
MELIS, 1934*JANNONE, 1934
ZHDANOV, 1934+TARBINSKI, 1940+EVSTROPOV, 1948+SVIRIDENKO, 1924+
BOUSHOÍEV, 1905+SIÍAZOV, 1912+
PLOTNIKOV, 1917+
POUKHOV, 1925+POUDOVKINE, 1931 +
IVANOV, 1946+
TOKGAÍEV, 1966+
SHAMONINE, 1964+LATCHININSKY, 1992
BODENHEIMER, 1944
DEMPSTER, 1957

modo es incorrecto, puesto que en condicio-nes de campo, si un estadio tiene una dura-ción breve por determinadas causas, el restono tiene por qué estar sometido a esas cir-
cunstancias. Por eso en nuestro caso esemodo no se cumple, porque se han conside-rado los valores totales independientementede los parciales.
ABSTRACT
BARRANCO R, F. PASCUAL y T. CABELLO, 1999: Modelling postembrionary developmentof Dociostaurus maroccanus (Thunberg, 1815). Bol. San Veg. Plagas, 25 (2): 241-253
Postembrionary development field data of a population of Moroccan locust fromCabo de Gata (Almería, SE Spain) during 1994, 1995 and 1996 are modelled.Simulation is made by three clasic and autonomous logistic models: Verhuslt-Pearl,Gomperz and Richards, and one not autonomous: model of Carreño. All adjust had ahight significance (P < 0.01). In 80% of the cases r2 > 0.99 and never was lower than0.9. So any of the modeis chosen is valid to simulate the development of locust in eachinstar.
The duration of the different intars is calculated by these models, and it is for eachone: 7-10, 6-7, 4-8, 4-12 and 7-11 days. The whole postembrionary developtment is37-44 days.
Key words: Docioslaunis maroccanus, postembrionary development, mathematicalmodels, Almería, Spain.
REFERENCIAS
BODENHEIMER, F. S., 1944: Studies on the ecology andcontrol of the Moroccan locust (Dociostaurus maroc-canus) in 'Iraq. I. Results of a mission of the 'IraqDepartment of Agriculture to N. 'Iraq in spring 1943.Bull. Dir. Gen. Agrie. Iraq., 29: 121 pp.
CAPINERA, J. L. & HORTON, D. R., 1989: Geographicvariation in effects of weather on grasshopper infesta-tion. Environ. Entomol, 18(1): 8-14.
CARREÑO, R., 1996: Modelos logísticos. Aplicaciones ala agronomía. Tesis doctoral. Universidad de Alme-ría. 168 pp.
CARREÑO, R.; BARRANCO, P.; GÁMEZ, M. y CABELLO, T.,1997a: Modelos logísticos aplicados a la biología deArtrópodos. Resúmenes del I Congreso Internacionalsobre «Modelos y métodos matemáticos aplicados ala Biología y Medicina», Alicante, 62-64.
CARREÑO, R.; GÁMEZ, M.; BARRANCO, P.; BELDA, J. yCABELLO, T., 1997b: Modelos analíticos aplicados a ladinámica de población de especies plagas. Actas de laVI Conferecia Española de Biometría, Córdoba,291-304.
DEMPSTER, J. S., 1957: The population dynamics of themoroccan locust {Dociostaurus maroccanus Thun-berg) in Cyprus. Anti-Locust Bull, London, 27:60 pp.
DENNIS, B., KEMP, W. P., 1988: Further stadistical infe-rence methods for a stochastic model of insect pheno-logy. Environ. Entomol, 17(5): 887-893.
DENNIS, B.; KEMP, W. P. & BECKWITH, R. C , 1986: Sto-chastic model of insect phenology: estimation andtesting. Environ. Entomol, 15: 540-546.
HUDES, E. S. & SHOEMAKER, C. S., 1988: Inferentialmethod for modeling insect phenology and its appli-cation to spruce budworm (Lepidoptera: Tortricidae).Environ. Entomol, 17(1): 97-108.
LATCHININSKY, A.V. y LAUNOIS-LUONG, M. H., 1992: LeCriquet marocain, Dociostaurus maroccanus (Thun-berg, 1815), dans la partie orientale de son aire dedistribution. Etude monographique relative àI'ex-URSS et aux pays proches. CIRAD-GERDAT-PRI-FAS, Montpellier, / VIZR, Saint-Pétersbourg, 270 pp.
MORENO, R., 1994: El papel de la epidemiología en laprotección fitosanitaria. En: R. MORENO (Ed.): Semi-

dad vegetal en la horticultura protegida. Consejeríade Agricultura y Pesca, Junta de Andalucía, Sevilla,263-276.
NAILAND, P. & HANRAHAN, S. A., 1993: Modellingbrown locust, Locus tana pardalina (Walker), outbre-aks in the Karoo. S. Afr. J. Sel, 89: 420-424.
ONSAGER, J. A. & HEWITT, G. B., 1982: Rangelandgrasshoppers: average longevity rate of mortalityamong six species in Nature. Environ. Entomol, 11:127-133.
PAOLI, G., 1937: Studi sulle cavallette di foggia (Docios-taurus maroccanus Thnb.) e sui loro ofagi (DitteriBombiliidi e Coleotteri Meloidi) ed acari ectofagi(Eritreii e Trombidiidi). Redia, 23: 27-206.
READ, K. L. Q. & ASHFORD, J. R., 1968: A system ofmodels for the life cycle of a biological organism.Biometrika, 55(1): 211-221.
STINNER, R. E., GUTIÉRREZ, A. P. & BUTLER, G. D., 1974:An algorithm for temperature-dependent growth ratesimulation. Can. Ent., 106: 519-524.
(Recepción: 9 abril 1999)(Aceptación: 13 julio 1999)