metabolismo - profesora maribel arnes · el metabolismo, por regla general, representa la suma de...
TRANSCRIPT

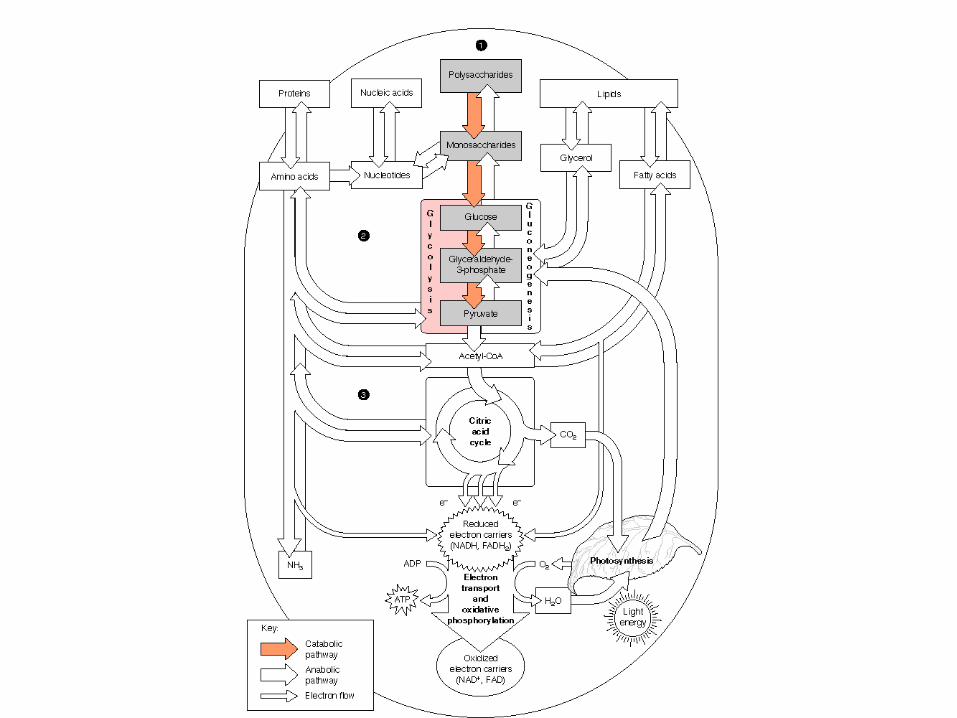
METABOLISMO

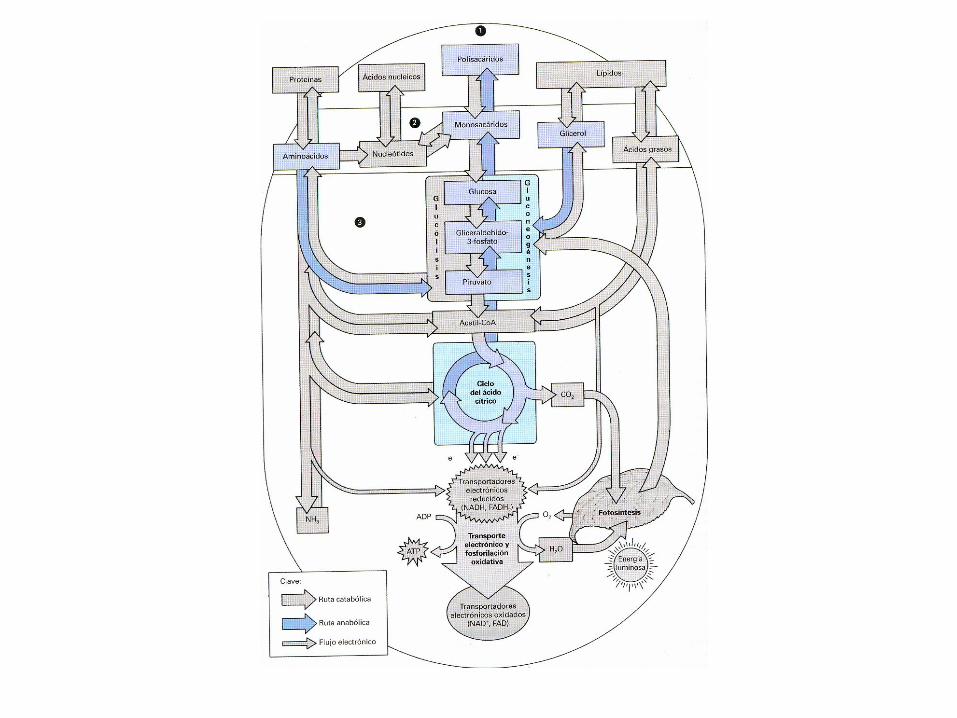
• El metabolismo tiene dos propósitosfundamentales: la generación de energíapara poder realizar funciones vitales para elorganismo y la síntesis de moléculasbiológicas
• El metabolismo es el proceso general por elcual los sistemas vivos adquieren y utilizanla energía libre que necesitan para realizarlas diversas funciones que ocurren dentro deellos. Y lo consiguen acoplando lasreacciones exoergónicas de la oxidación delos nutrientes a los procesos endoergónicosrequeridos para mantener los sistemasvivos.

El metabolismo, por regla general, representa la suma de todoslos cambios químicos que convierten los nutrientes, losmateriales de partida utilizables por los organismos, enenergía y productos celulares químicamente complejo, esdecir, consiste literalmente en cientos de reaccionesenzimáticas organizadas en rutas características.
• Obtener energía química a partir de la energía solar
o degradando nutrientes del medioambiente.
• Convertir nutrientes en moléculas propias de la
célula.
• Polimerizar moléculas pequeñas en
macromoléculas (proteínas, ácidos nucleicos y
polisacáridos).
• Sintetizar y degradar biomoléculas necesarias para
funciones especificas de la célula.
RUTAS
METABOLICAS
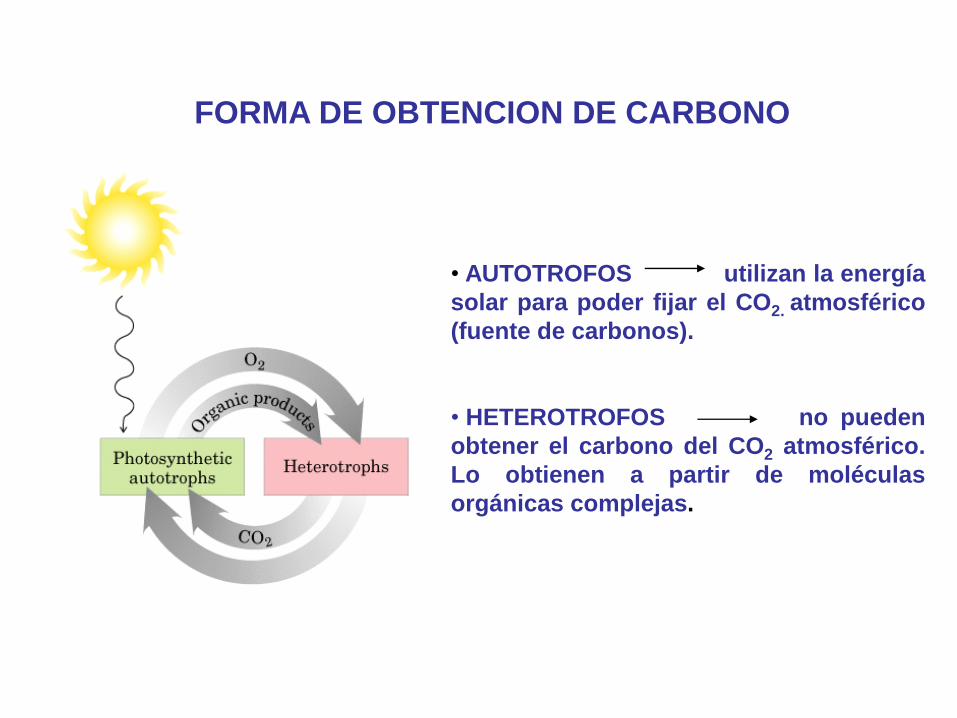
FORMA DE OBTENCION DE CARBONO
• AUTOTROFOS utilizan la energía
solar para poder fijar el CO2. atmosférico
(fuente de carbonos).
• HETEROTROFOS no pueden
obtener el carbono del CO2 atmosférico.
Lo obtienen a partir de moléculas
orgánicas complejas.
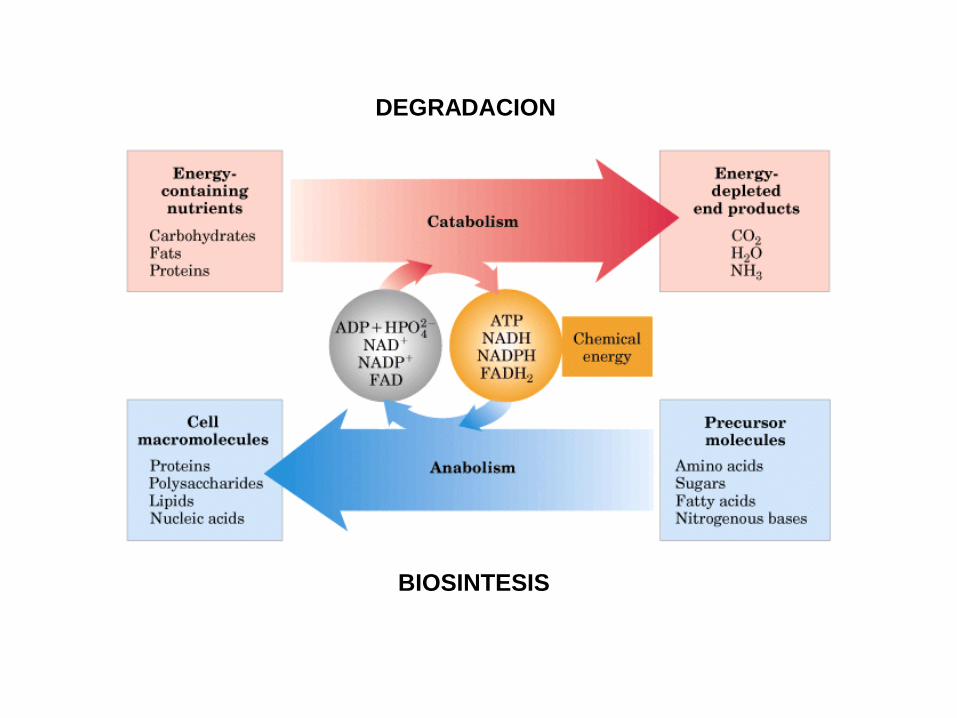
DEGRADACION
BIOSINTESIS


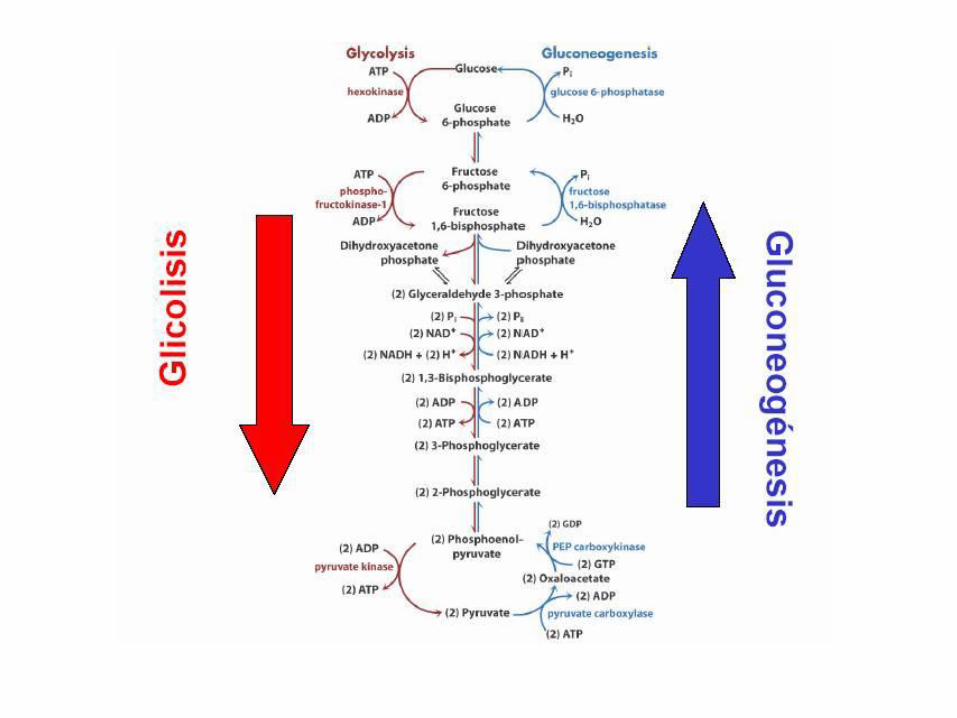
GLUCÓLISIS
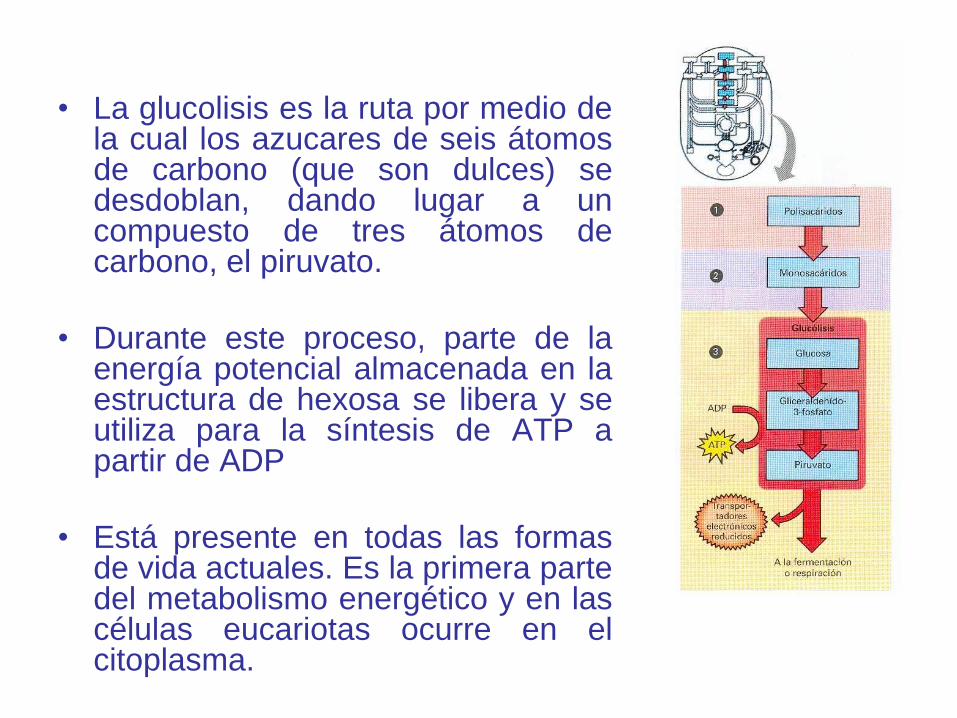
• La glucolisis es la ruta por medio dela cual los azucares de seis átomosde carbono (que son dulces) sedesdoblan, dando lugar a uncompuesto de tres átomos decarbono, el piruvato.
• Durante este proceso, parte de laenergía potencial almacenada en laestructura de hexosa se libera y seutiliza para la síntesis de ATP apartir de ADP
• Está presente en todas las formasde vida actuales. Es la primera partedel metabolismo energético y en lascélulas eucariotas ocurre en elcitoplasma.

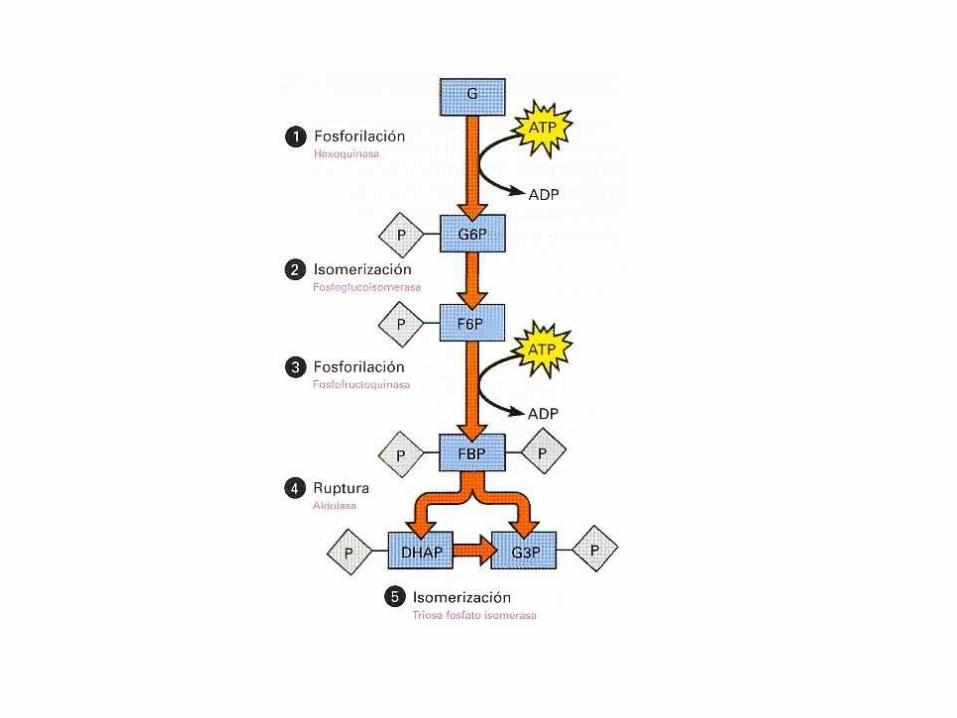
Primera fase
• Las cinco primeras reacciones constituyen una fase de
inversión de energía, en la que se sintetizan azúcares-
fosfato a costa de la conversión de ATP en ADP, y el
sustrato de seis carbonos se desdobla en dos azúcares-
fosfato de tres carbonos.

1. Primera inversión del ATP
• En esta etapa la glucosa es fosforilada mediante un
ATP, esta reacción es catalizada por la hexoquinasa
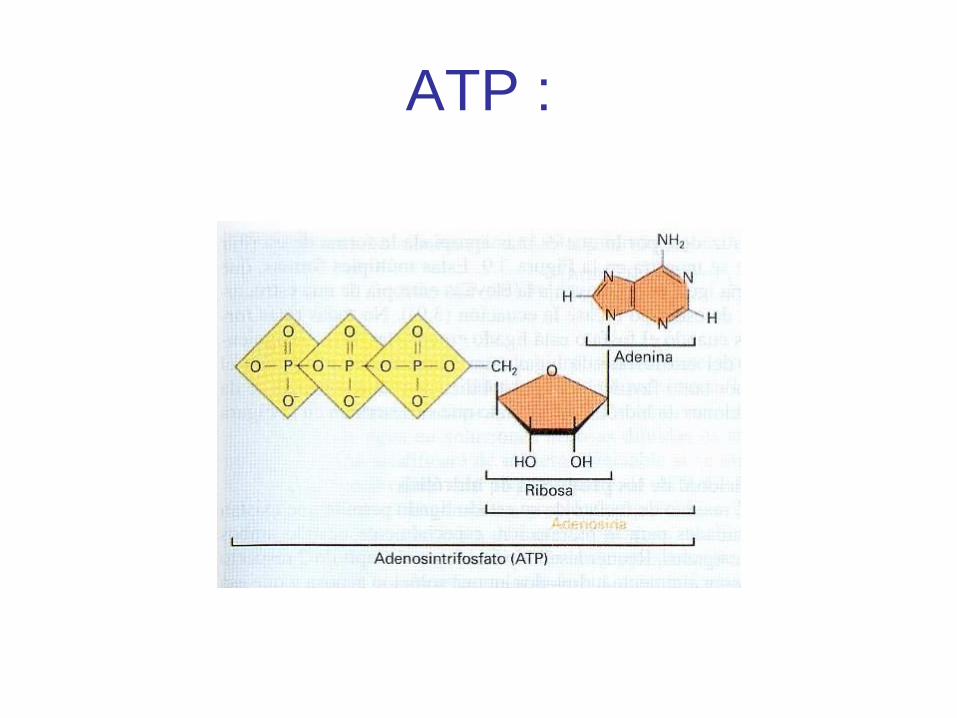
ATP :
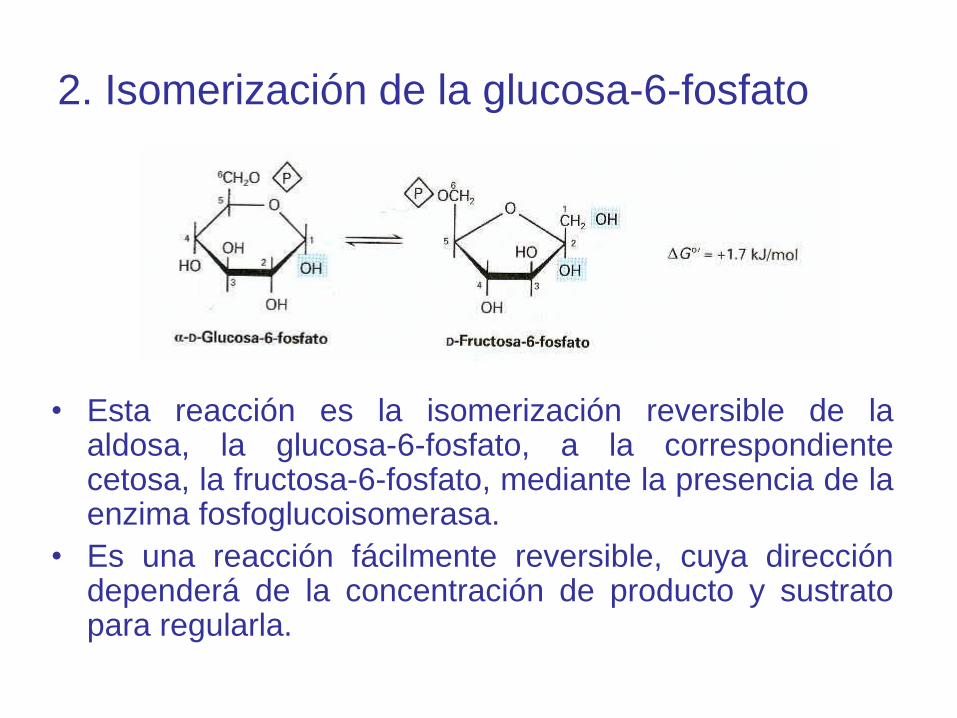
2. Isomerización de la glucosa-6-fosfato
• Esta reacción es la isomerización reversible de laaldosa, la glucosa-6-fosfato, a la correspondientecetosa, la fructosa-6-fosfato, mediante la presencia de laenzima fosfoglucoisomerasa.
• Es una reacción fácilmente reversible, cuya direccióndependerá de la concentración de producto y sustratopara regularla.
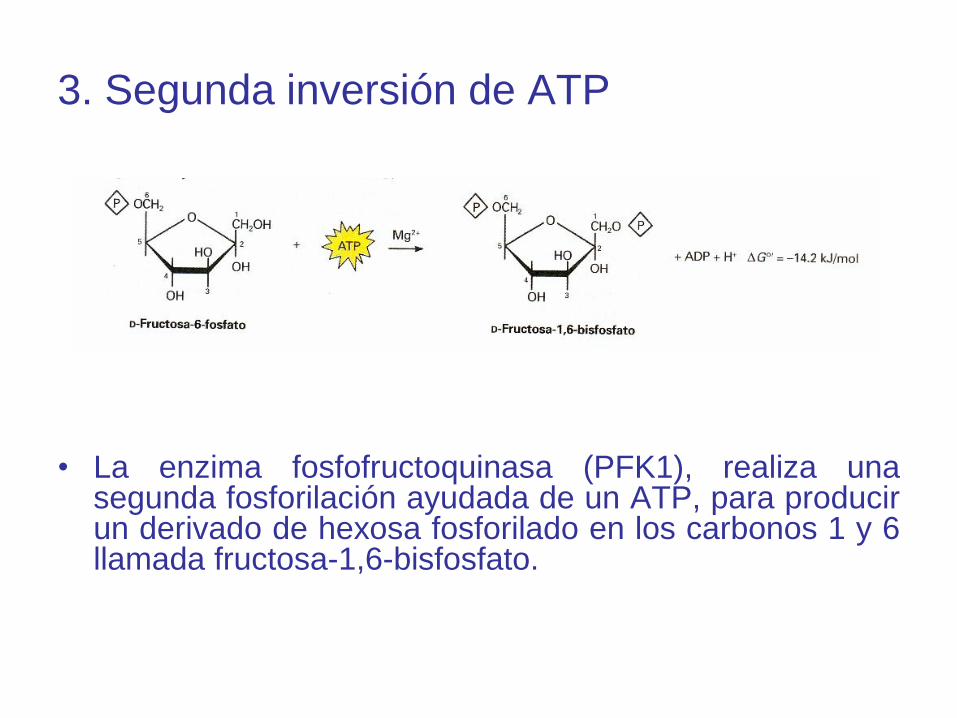
3. Segunda inversión de ATP
• La enzima fosfofructoquinasa (PFK1), realiza unasegunda fosforilación ayudada de un ATP, para producirun derivado de hexosa fosforilado en los carbonos 1 y 6llamada fructosa-1,6-bisfosfato.
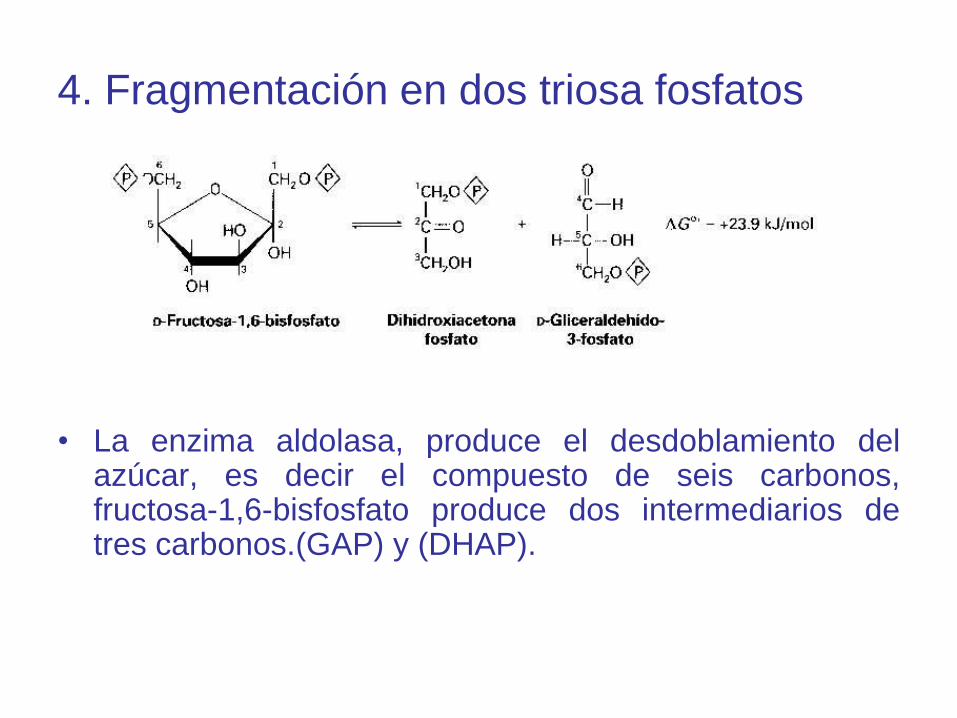
4. Fragmentación en dos triosa fosfatos
• La enzima aldolasa, produce el desdoblamiento delazúcar, es decir el compuesto de seis carbonos,fructosa-1,6-bisfosfato produce dos intermediarios detres carbonos.(GAP) y (DHAP).
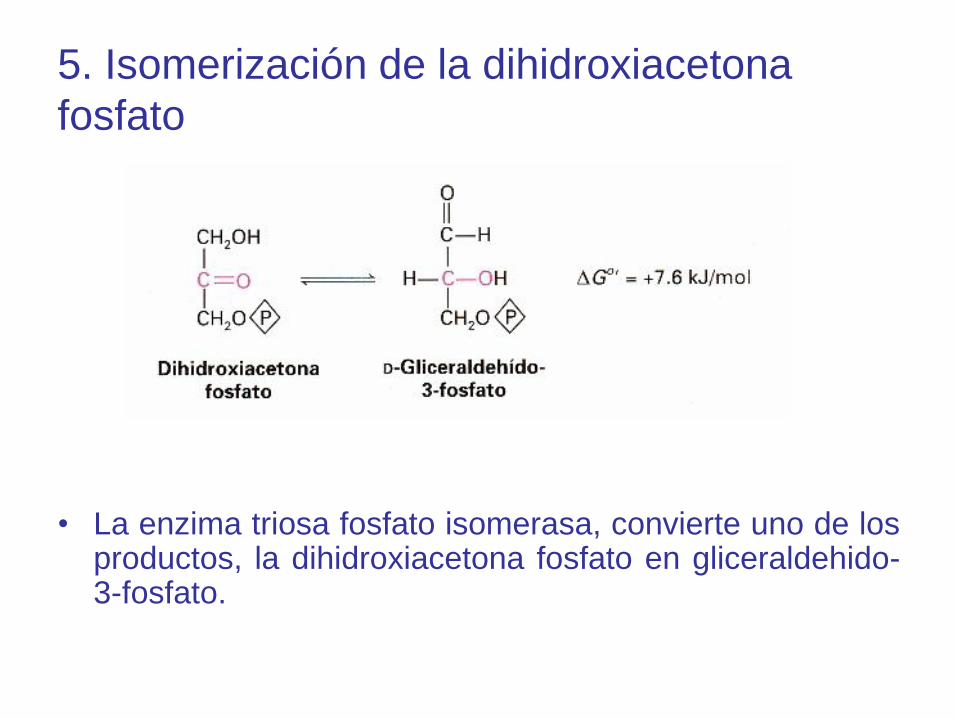
5. Isomerización de la dihidroxiacetona
fosfato
• La enzima triosa fosfato isomerasa, convierte uno de losproductos, la dihidroxiacetona fosfato en gliceraldehido-3-fosfato.

Segunda fase
• Las cinco últimas reacciones corresponden a una fase
de generación de energía, en esta fase, las triosas-
fosfato se convierten en compuestos ricos en energía,
que transfieren fosfato al ADP, dando lugar a la síntesis
de ATP.

6. Generación del primer compuesto de alta
energía
• Esta reacción la cataliza la gliceraldehído-3-fosfatodeshidrogenasa, para producir 1,3-Bifosfoglicerato y unamolécula de NADH (dinucleótido de nicotinamida y adenina) y H+.
• El fosfato se ha introducido sin utilizar ATP, sinoaprovechando la energía producida por la reacciónredox.
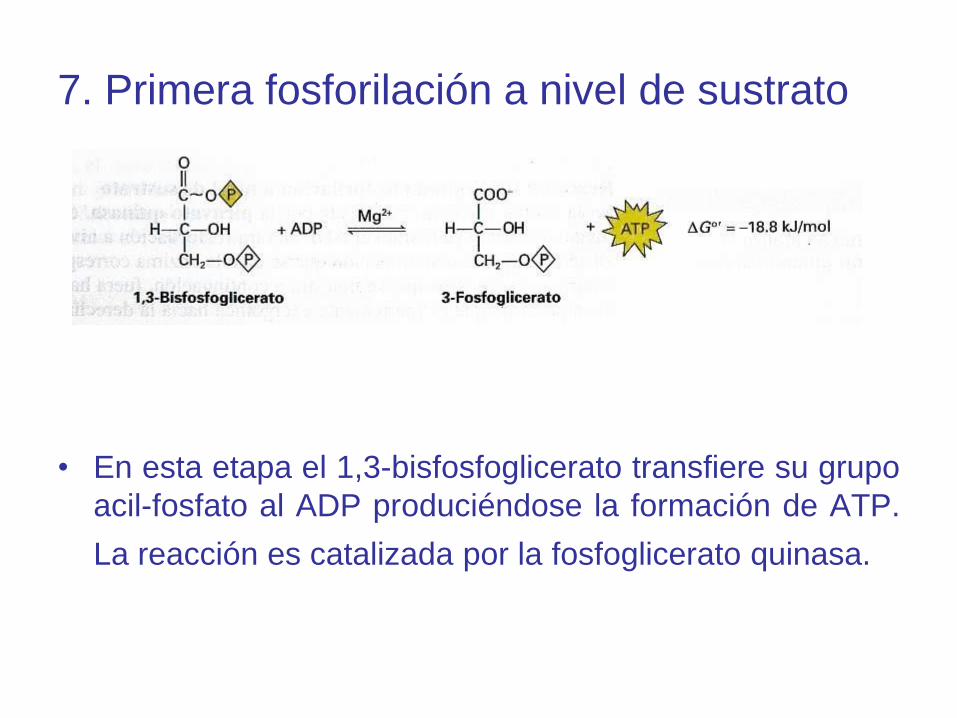
7. Primera fosforilación a nivel de sustrato
• En esta etapa el 1,3-bisfosfoglicerato transfiere su grupo
acil-fosfato al ADP produciéndose la formación de ATP.
La reacción es catalizada por la fosfoglicerato quinasa.
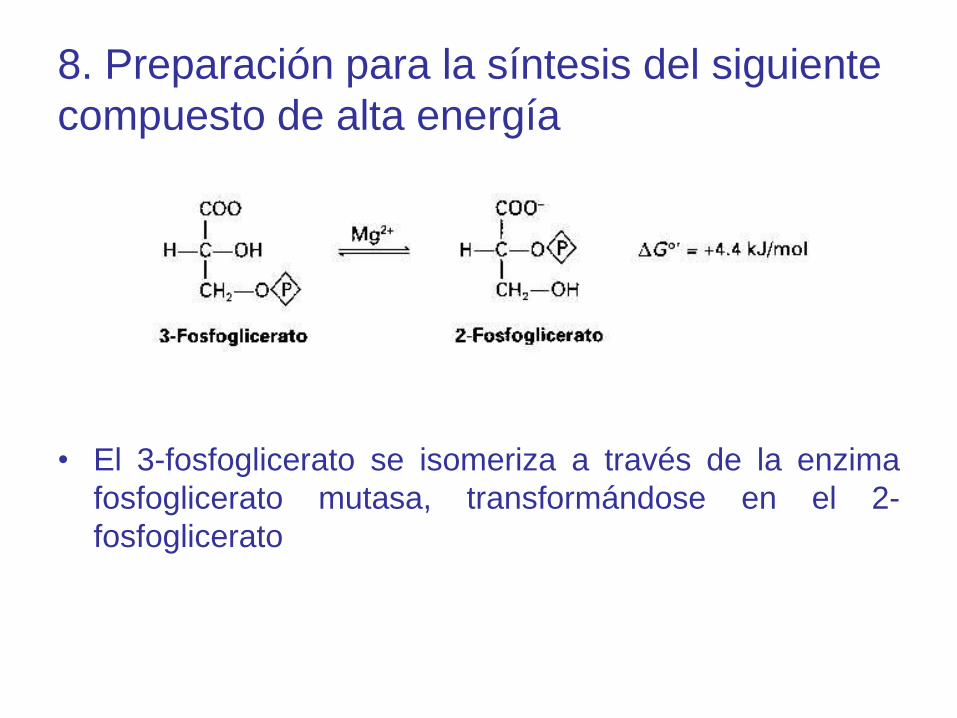
8. Preparación para la síntesis del siguiente
compuesto de alta energía
• El 3-fosfoglicerato se isomeriza a través de la enzima
fosfoglicerato mutasa, transformándose en el 2-
fosfoglicerato

9. Síntesis del segundo compuesto de alta
energía
• En esta reacción ocurre una deshidratación simple del 3-
fosfoglicerato para dar el fosfoenolpiruvato bajo la
acción de la enzima enolasa.

10. Segunda fosforilación a nivel de sustrato
• Desfosforilación del Fosfoenolpiruvato, obteniéndose
piruvato y ATP. Reacción irreversible mediada por la
Piruvato quinasa.



El rendimiento total de la glucólisis es de 2 ATP y 2 NADH.
Glucosa + 2ADP + 2Pi + 2 NAD+ 2 Piruvato + 2ATP + 2NADH + 2H+ + 2H2O
∆G°’= -73,3 KJ/mol
Consume ATP Hexoquinasa
Fosfofructoquinasa
Produce ATP Fosfoglicerato quinasa
Piruvato quinasa
Produce NADH Gliceraldehido 3 P deshidrogenasa

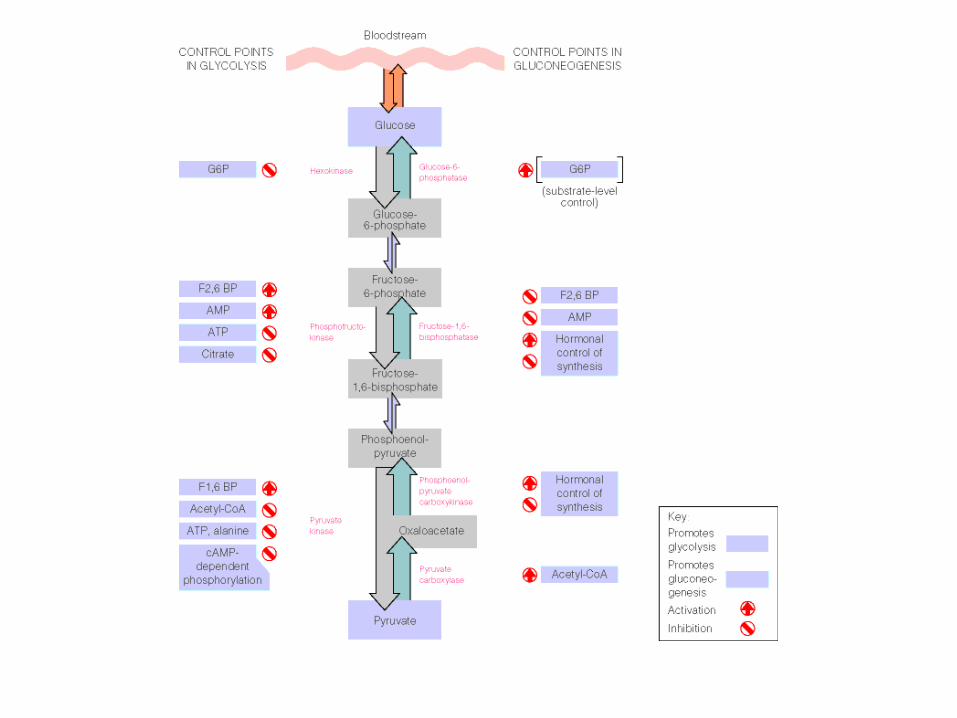
Regulación de la glucólisis
• La glucólisis se regula enzimáticamente en los
tres puntos irreversibles de esta ruta, esto es, en
la primera reacción (G -- >G-6P), por medio de
la Hexoquinasa; en la tercera reacción (F-6P -->
F-1,6-BP) por medio de la PFK1 y en el último
paso (PEP --> Piruvato) por la Piruvatoquinasa.

1. La hexoquinasa es un punto de regulación pocoimportante, ya que se inhibe cuando hay mucho G-6Pen músculo. Es un punto poco importante ya que el G-6P se utiliza para otras vías.
HQ: Inhibe G-6P

2. La PFK1 es la enzima principal de la regulación de laglucólisis, si está activa cataliza muchas reacciones y seobtiene más Fructosa 1,6 bifosfato, lo que permitirá a lasenzimas siguientes transformar mucho piruvato. Si estáinhibida, se obtienen bajas concentraciones de productoy por lo tanto se obtiene poco piruvato.
Esta enzima es controlada por regulación alostéricamediante: Por un lado se activa gracias a nivelesenergéticos elevados de ADP y AMP, inhibiéndose enabundancia de ATP y citrato, y por otro se activa enpresencia de un metabolito generado por la PFK2 quees la Fructosa-2,6-Bisfosfato (F-2,6-BP)

La lógica de la inhibición y activación son las siguientes:
– ATP: inhibe esta enzima pues si hay una altaconcentración de ATP entonces la célula no necesitagenerar más.
– Citrato: si hay una alta concentración de citratoentonces, se está llevando a cabo el ciclo del ácidocítrico (o ciclo de Krebs) y este ciclo aporta muchaenergía, entonces no se necesita realizar glucólisispara obtener más ATP, ni piruvato.
– AMP, ADP: la baja concentración de estas moléculasimplica que hay una carencia de ATP, por lo que esnecesario realizar glucólisis, para generar piruvato yenergía.

PFK1: Inhibe: ATP - Activa: ADP, AMP y F-2,6-BP.
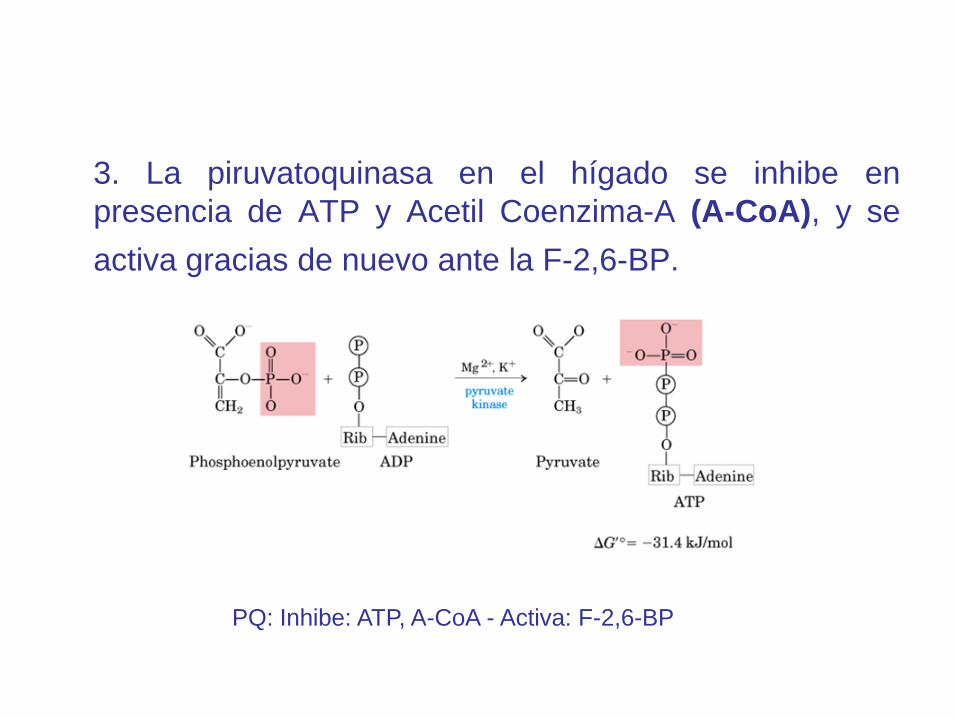
3. La piruvatoquinasa en el hígado se inhibe en
presencia de ATP y Acetil Coenzima-A (A-CoA), y se
activa gracias de nuevo ante la F-2,6-BP.
PQ: Inhibe: ATP, A-CoA - Activa: F-2,6-BP


Sustratos de la gluconeogénesis
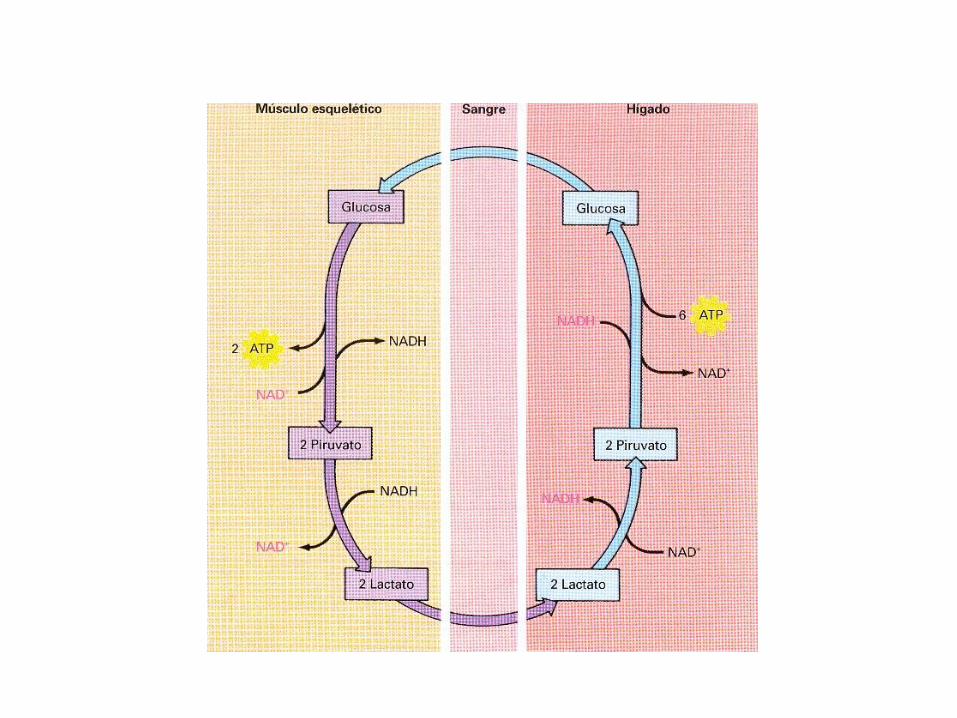
1.- Lactato, producido fundamentalmente mediante laglucólisis en el músculo esquelético y los eritrocitos
Durante el ejercicio intenso se movilizan las reservas deglucógeno y la glucosa-6-fosfato se convierte enpiruvato con mayor rapidez de lo que pueda alcanzar suulterior matabolismo a través de la piruvatodeshidrogenasa y del ciclo de ácido cítrico. Como elpiruvato es muy abundante la enzima lactatodeshidrogenasa lo reduce rápidamente a lactato. Esteproceso se denomina Ciclo de Cori.

2.- El aminoácido específico alanina,producido en el músculo mediante el cicloglucosa alanina.
En un proceso paralelo, denominado Cicloglucosa-alanina, el piruvato de los tejidosperiféricos experimenta unatransaminación a alanina que se devuelveal hígado y se utiliza para lagluconeogénesis.

3.- Los aminoácidos generados a partir de
proteínas de la dieta o a partir de la degradación
de las proteínas musculares durante la
inanición.
Los aminoácidos capaces de convertirse en
glucosa se denominan glucogénicos. Durante el
ayuno, el catabolismo de las proteínas
musculares constituye la principal fuente de
mantenimiento de las concentraciones normales
de glucosa en sangre

4.- El propianato, procedente de la degradación
de algunos ácidos grasos y aminoácidos
En todos los organismos la propionil-CoA se
genera, bien a partir de la degradación de
algunos aminoácidos o bien a partir de la
oxidación de los ácidos grasos que tienen un
número impar de átomos de carbono. Esta
propionil-CoA entra en la gluconeogénesis a
través de su conversión en succinil-CoA y de
esta a oxalacetato.

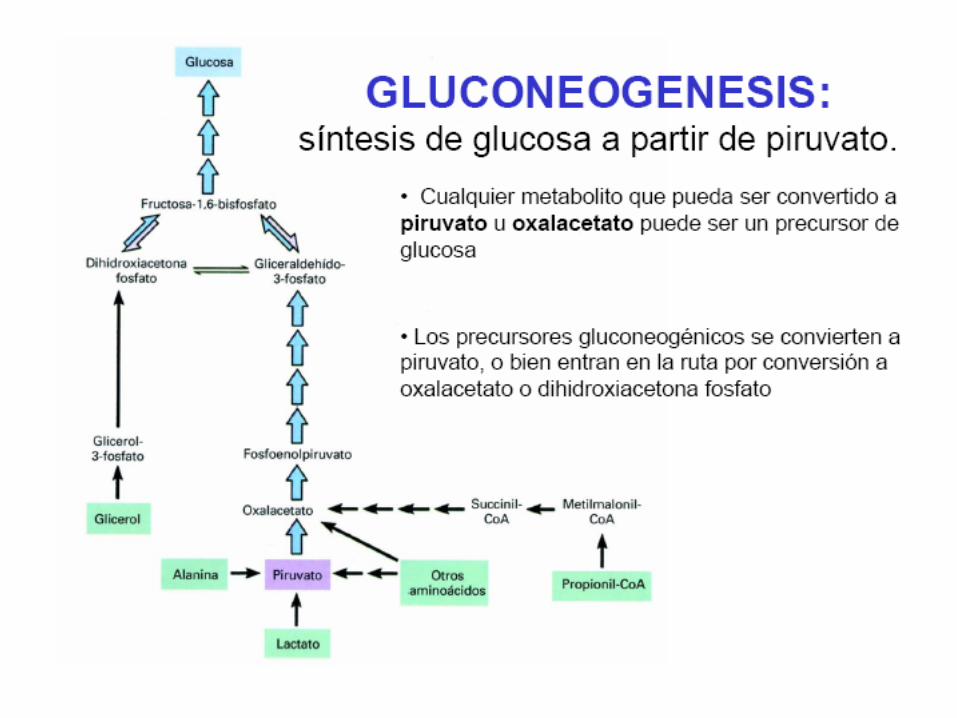
5.- El glicerol procedente del catabolismo de las grasas.
Es muy importante señalar que los lípidos son malosprecursores gluconeogénicos. El catabolismo de lostriacilgliceridos produce ácidos grasos y glicerol. Losácidos grasos sufren una -oxidación para produciracetil-CoA el cual en los animales no puede convertirseen piruvato ni en ningún otro precursor gluconeogénico,por lo tanto no pueden experimentar una conversiónneta en hidratos de carbono. En consecuencia en ladegradación de las grasas el único producto que puedeentrar en la gluconeogénesis es el glicerol. Esteexperimenta una fosforilación, seguida dedeshidrogenación para producir dihidroxicetona fosfato

Etapas de la gluconeogénesis
• Para que una ruta metabólica pueda darse sin dificultadsu G’ debe ser fuertemente negativo para el conjuntode la ruta. En el caso de la glucólisis, desde la glucosahasta el piruvato el G’ es aproximadamente –96KJ/mol, dado que tres de sus reacciones de la rutaglucolítica son muy negativas (exergónicas)presentándose en forma irreversible. Estas son lacatalizada por la hexoquinasa, la fosfofructoquinasa y lapiruvato quinasa.
• Para que la gluconeogénesis pueda desarrollarse senecesitan distintas enzimas que puedan evitar las tresreacciones irreversibles catalizándolas en la dirección dela síntesis de glucosa
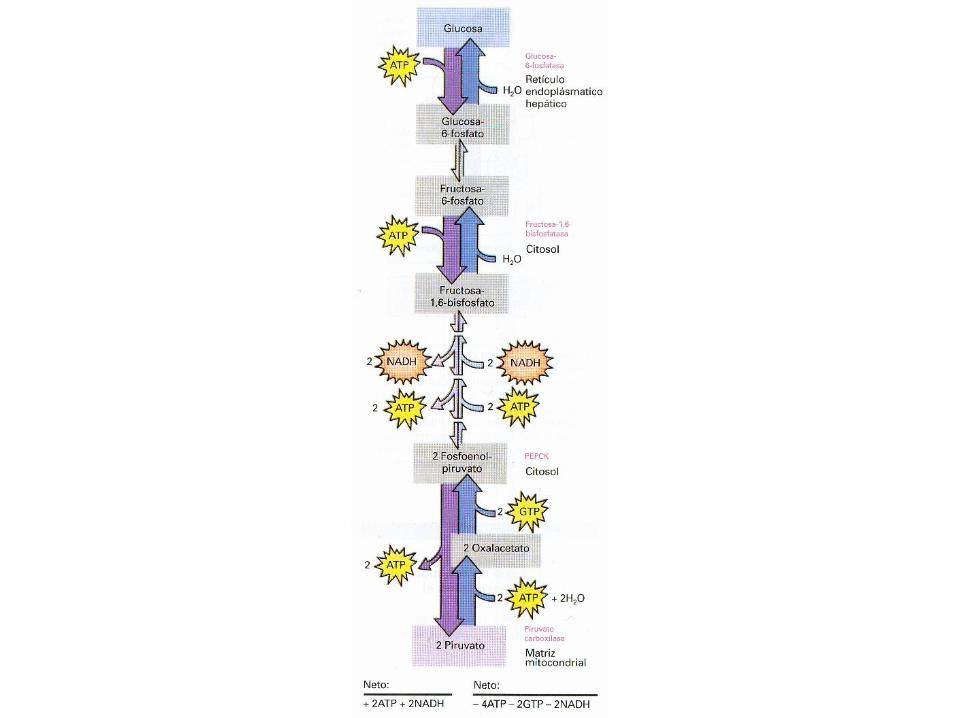

Paso 1 Conversión del piruvato en fosfoenolpiruvato
A.- La enzima piruvato carboxilasa cataliza la
conversión dependiente de ATP y de Biotina,
del piruvato en oxalacetato
Piruvato + CO2 +H2O + ATP Oxalacetato + ADP + Pi + 2H+
Gº’ = -2,1 KJ/mol
La enzima requiere de manera absoluta
acetil-CoA como activador alostérico.

B.- El oxalacetato a través de la acción de la
fosfoenolpiruvato carboxiquinasa (PEPCK)
da fosfoenolpiruvato
Oxalacetato + GTP Fosfoenolpiruvato + CO2 + GDP
Gº’ = +2,9 KJ/mol

• La reacción global es la siguiente
Piruvato + ATP + GTP + H2O
Fosfoenolpiruvato + ADP + GDP +Pi + 2H+
Gº’ = +0,8 KJ/mol

Paso2. Conversión de la fructosa 1,6-bisfosfato en
fructosa 6-fosfato
• La reacción que tiene lugar en este pasoconsiste en una simple reacciónhidrolítica, catalizada por la fructosa-1,6-bisfosfatasa.
Fructosa-1,6-bisfosfato + H2O
Fructosa-6-fosfato + Pi
Gº’ = -16,3 KJ/mol

Paso 3. Conversión de la glucosa-6-fosfato en glucosa
• A través de la enzima glucosa-6-fosfatasa
se produce una reacción de hidrólisis
Glucosa-6-fosfato + H2O Glucosa + Pi
Gº’ = -12,1 KJ/mol
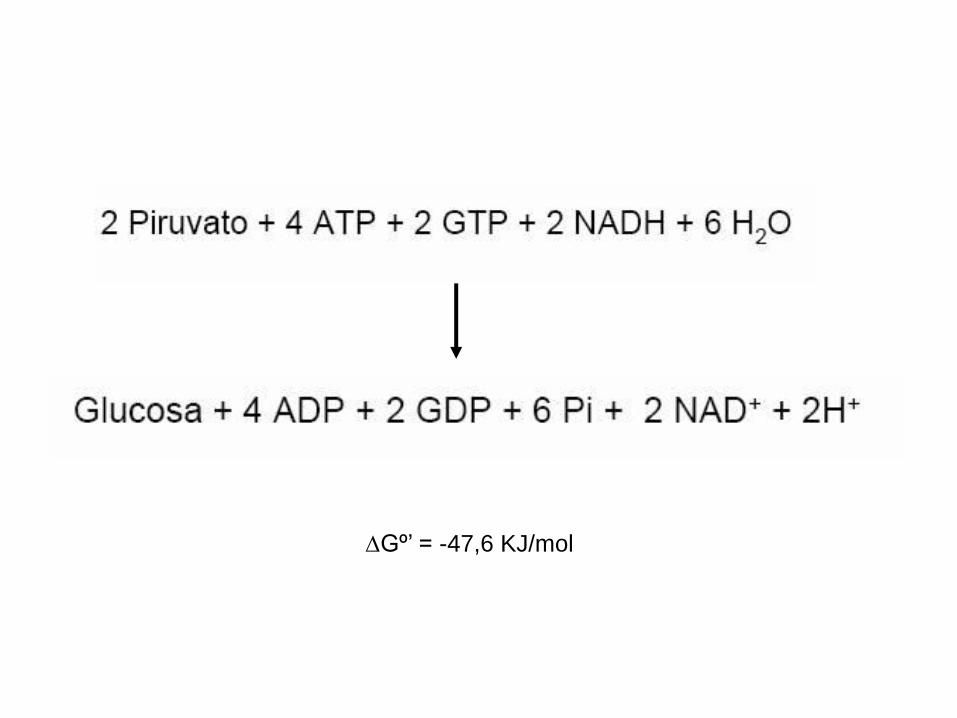
Gº’ = -47,6 KJ/mol

Regulación de la gluconeogésis
La glicólisis y la gluconeogénesis están
reguladas de forma coordinada y
reciproca.
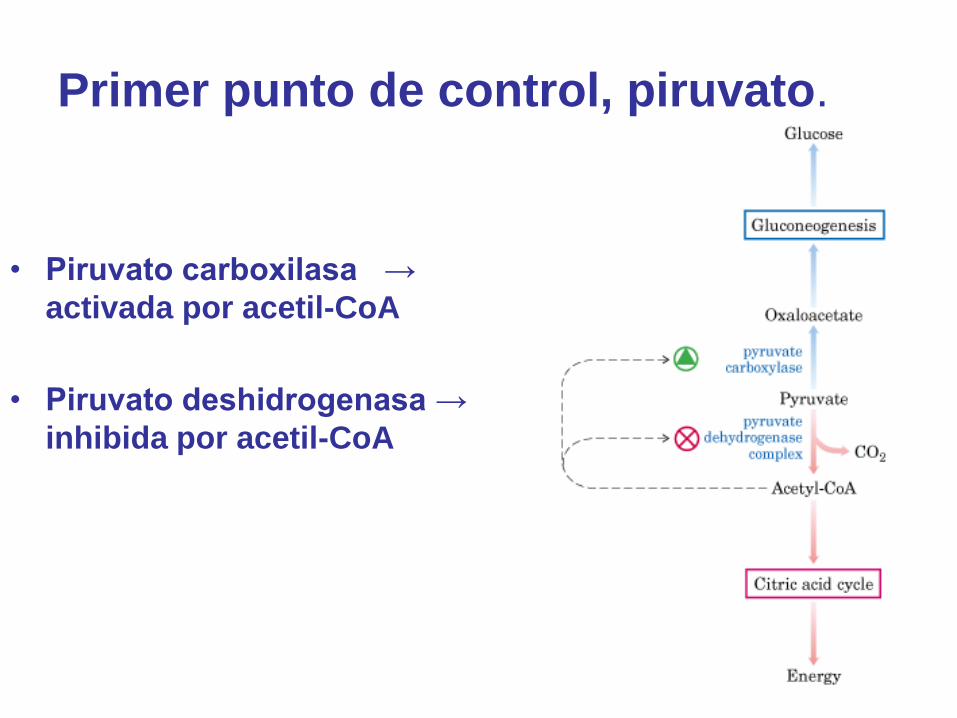
Primer punto de control, piruvato.
• Piruvato carboxilasa →
activada por acetil-CoA
• Piruvato deshidrogenasa →
inhibida por acetil-CoA
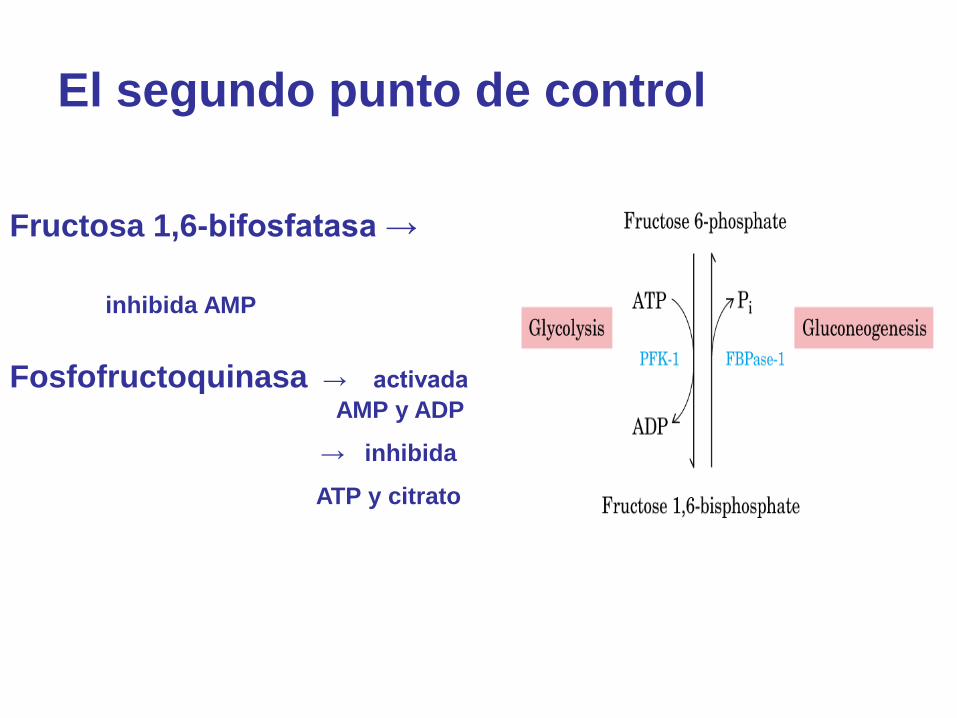
El segundo punto de control
Fructosa 1,6-bifosfatasa →
inhibida AMP
Fosfofructoquinasa → activada
AMP y ADP
→ inhibida
ATP y citrato

El tercer punto de control
• La glucosa-6-fosfatasa no tiene un control
alostérico conocido
• La actividad intracelular se controla por la
concentración del sustrato glucosa-6-
fosfato