impacto de las plantaciones de palma aceitera,
TRANSCRIPT

1

2
Impacto de las plantaciones de palma aceitera,
Elaeis guineensis Jacq., sobre la regeneración natural de los
bosques en la cuenca del río Curbaradó (Chocó)
Estudiante:
Johanna Ramírez Herrera
c.c. 32220684
ASESORES:
Cristina López Gallego
Bióloga PhD.
Profesora UdeA
César A. Acosta
Ingeniero Agrónomo, especialista en Desarrollo Rural
Investigador del Centro de Investigación y Educación Popular -CINEP-
Estela Quintera Vallejo
Bióloga Msc.
Los productos académicos del proyecto de investigación tendrán como autora principal a la
estudiante Johanna Ramírez Herrera, y los asesores serán los autores secundarios si es
del caso.
UNIVERSIDAD DE ANTIOQUIA
FACULTAD DE CIENCIAS EXACTAS Y NATURALES
INSTITUTO DE BIOLOGÍA
MEDELLÍN
2009

3
…y ese misterio tan sencillo:
en la semilla de un árbol se encierra todo el espacio y el tiempo del árbol
incluyendo las semillas de ese árbol
todo el tamaño de su edad,
su frondosidad con frescos pájaros y sus años,
incluyendo sus semillas, y las semillas de sus semillas.
Cántico Cósmico. Ernesto Cardenal
El lugar donde muchos de mis sueños volvieron a tener sentido…
En aquel tiempo les dijo esta parábola:
si el óvulo fecundado y maduro no cae en tierra,
abre el tegumento que lo cubre
y salen la raicilla y la yemecilla con sus cotiledones
mientras se pudre el tegumento,
permanecerá solo.
Cántico Cósmico. Ernesto Cardenal

4
AGRADECIMIENTOS
A mi familia, por su paciente e inconmensurable amor, por ser ese refugio
donde empecé a amar la vida, y al que siempre vuelvo sin que estén ausentes
las cosas que quiero.
A los parceros del alma, esos hermanos que la vida me dio la oportunidad de
elegir y compartir con ellos el camino. A la Naty, quien conspira y construye
conmigo sueños a seis manos, también las de la Esperanza. A Dieguito, La
Vale, Andrés, Sandy, la Deysi, el César y Cris por compartir las “…locuras para
la esperanza, también las locuras del dolor, las locuras de allá donde el cuerdo
no alcanza, locuras de otro color…”
A los profes, de la universidad y de afuera, y a los del herbario, que me
compartieron su pasión por la maravilla de la vida.
A la familia Rentería Mosquera, y todos los relacionados, incluido Narciso, por
dejarme entrar en sus vidas y por entrar en la mía, por enseñarme que hay que
defender tercamente la dignidad de la condición humana y devolverme la fe en
la humanidad.
A los niños de Cetino, esa promesa de que un mañana diferente es posible.
Esta investigación fue posible gracias entre muchas otras cosas, al apoyo
económico de la ONG OXFAM-GB, y del Comité de para el Desarrollo de la
Investigación de la Universidad de Antioquia-CODI.

5
RESUMEN
Los bosques tropicales están sometidos, en mayor o menor medida, a
perturbaciones naturales y de origen antrópico que interactúan con
características bióticas y abióticas para influir en las dinámicas poblacionales y
de las comunidades de estos ecosistemas. Una vez la perturbación ha cesado,
se inicia un proceso de sucesión natural de los ecosistemas y su curso
dependerá, entre otras cosas, de las condiciones iniciales de los sitios. En este
estudio, evaluamos el impacto que tienen los monocultivos de palma aceitera
sobre la regeneración natural de los bosques en la cuenca del río Curbaradó
(Chocó). Nos enfocamos en tres procesos, la lluvia de semillas, la germinación
y supervivencia inicial de plántulas, y la regeneración natural en estados
iniciales de la sucesión (comunidad de plántulas y juveniles) en tres unidades
de paisaje: Bosque, Rastrojo y Plantación. Dispusimos trampas para
caracterizar la lluvia de semillas, realizamos siembras de semillas, y trazamos
transectos para describir la comunidad de plántulas y juveniles en cada unidad
de paisaje. Encontramos que la riqueza de especies de la lluvia de semillas y
de la comunidad de plántulas es menor en las plantaciones de palma. Además,
la composición de especies de la lluvia de semillas en las plantaciones fue
ligeramente diferente a la de los bosques, mientras que para la comunidad de
plántulas las plantaciones no se diferencian claramente de las otras unidades
de paisaje. Las plantaciones mostraron una menor variabilidad en la riqueza de
especies de la lluvia de semillas y la comunidad de plántulas, y tuvieron una
mayor similitud en la composición de especies entre ellas que la mostrada por
los bosques y los rastrojos. Estos resultados sugieren que estas plantaciones,
que como perturbación tienen una baja intensidad de uso previo, no
representan una barrera significativa para la lluvia de semillas y el
establecimiento de plántulas en los estados iniciales de la sucesión, sin
embargo los efectos observados sobre la riqueza de especies, la variabilidad
de la riqueza y la similitud de la composición de especies en las plantaciones, a
largo plazo podrían incidir sobre el curso de la sucesión natural, lo que debe
tenerse en cuenta al momento de diseñar un plan de restauración.

6
TABLA DE CONTENIDO
1. INTRODUCCIÓN ....................................................................................... 10
Impactos de los monocultivos de palma aceitera sobre la biodiversidad ...... 10
Restauración ecológica y barreras a la sucesión natural de bosques .......... 15
Las plantaciones de palma aceitera como potenciales barreras para la
regeneración del bosque ............................................................................... 18
2. MATERIALES Y MÉTODOS ...................................................................... 22
Área de estudio ............................................................................................. 22
Experimentos ................................................................................................ 24
Lluvia de semillas .......................................................................................... 25
Germinación y supervivencia inicial de plántulas .......................................... 28
Regeneración natural: establecimiento de plántulas ..................................... 29
3. RESULTADOS .......................................................................................... 32
Lluvia de semillas .......................................................................................... 32
Germinación y supervivencia inicial de plántulas. ......................................... 39
Regeneración natural: establecimiento de plántulas ..................................... 40
4. DISCUSIÓN ............................................................................................... 46
BIBLIOGRAFÍA ................................................................................................ 52

7
LISTA DE TABLAS
Tabla 1. Especies compartidas entre las unidades de paisaje. ........................ 33
Tabla 2. Prueba U de Mann-Whitney para la riqueza promedio de especies de
la lluvia de semillas (gl=1). ............................................................................... 33
Tabla 3. Matriz de distancia entre las unidades de paisaje de según el índice de
similitud de Hamman de acuerdo a la composición de especies de la lluvia de
semillas (valores entre 0 y 1). ........................................................................... 35
Tabla 4. Porcentajes de germinación de 4 especies de árboles en tres unidades
de paisaje. ........................................................................................................ 39
Tabla 5. Porcentajes de supervivencia de 4 especies de árboles en tres
unidades de paisaje. ........................................................................................ 40
Tabla 6. Especies compartidas entre las unidades de paisaje. ........................ 40
Tabla 7. Prueba U de Mann-Whitney para la riqueza promedio de especies de
plántulas (gl=1). ................................................................................................ 41
Tabla 8. Matriz de distancia entre las unidades de paisaje de según el índice de
similitud de Hamman de acuerdo a la composición especies de la comunidad
de plántulas (valores entre 0 y 1). .................................................................... 43

8
LISTA DE FIGURAS
Figura 1. Municipio Carmen del Darién (Curbaradó), Chocó……………………21
Figura 2. Parte media de la cuenca del río Curbaradó…………………………..21
Figura 3. Unidad de paisaje Bosque……………………………………………….22
Figura 4. Trampas de caída de semillas…………………………………………..24
Figura 5.Llenado de las trampas de semillas.…………………………………….24
Figura 6. Lavado del sustrato de las trampas de semillas………………………25
Figura 7. Secado del sustrato de las trampas de semillas………………………25
Figura 8. Parcelas de germinación…………………………………………………27
Figura 9. Plántulas presentes en el censo de germinación. Bálsamo a la
izquierda, algarrobo a la derecha…………………………………………………..27
Figura 10. Riqueza de especies (media-error estándar) en la lluvia semillas por
unidad de paisaje…………………………………………………………………….31
Figura 11. Agrupación de las unidades de paisaje de acuerdo a la composición
de especies en la lluvia de semillas………………………………………………..33
Figura 12. Análisis de ordenación de las unidades de paisaje de acuerdo a la
composición de especies en la lluvia de semillas………………………………...35
Figura 13. Distribución de especies de la lluvia de semillas de acuerdo a su
estado sucesional…………………………………………………………………….36
Figura 14. Distribución de especies de la lluvia de semillas de acuerdo a su
hábito de crecimiento…………………………………………………………..........37
Figura 15. Riqueza de especies (media-error estándar) de plántulas por unidad
de paisaje…………………………………………………………………………......40
Figura 16. Agrupación de las unidades de paisaje de acuerdo a la composición
de especies en la comunidad de plántulas………………………………………..41

9
Figura 17. Análisis de ordenación de las unidades de paisaje de acuerdo a la
composición de especies en la comunidad de plántulas………………………...43
Figura 18. Distribución de especies de la comunidad de plántulas de acuerdo a
su estado sucesional. La especie introducida se refiere a la palma aceitera…44
Figura 19. Distribución de especies de la comunidad de plántulas de acuerdo a
su hábito de crecimiento. La especie introducida se refiere a la palma
aceitera………………………………………………………………………………..45

10
1. INTRODUCCIÓN
Los bosques tropicales están sometidos constantemente a diversas
perturbaciones de origen natural o antrópico (Brown y Lugo, 1990; Chazdon,
2003). Con la intensificación de las relaciones hombre-naturaleza y el
crecimiento de poblaciones humanas, la presión sobre los bosques para la
extracción de recursos naturales es una de las principales causas de pérdida
de la biodiversidad. Entre los años 1990 y 2000, 10 millones de hectáreas de
bosque en regeneración fueron documentadas en los trópicos del mundo
comparadas a los 142 millones de hectáreas de bosques convertidos a usos no
boscosos del suelo, así, por cada hectárea de bosque en recuperación natural
en los trópicos 14,2 hectáreas están siendo destruidas. (Chazdon, 2003).
Según el informe de la FAO “Global Forest Resources Assessment 2005” la
expansión de la frontera agrícola fue la principal causa de la pérdida de 13
millones de hectáreas anuales de bosques tropicales (FAO, 2006). Si se tiene
en cuenta que en la actualidad, además de la producción de alimentos, el
sector agrícola incluye la producción de materias primas para la agroindustria
de los “biocombustibles”, especialmente caña de azúcar, maíz, soya y palma
aceitera, el impacto de la agricultura se incrementa sustancialmente.
Impactos de los monocultivos de palma aceitera sobr e la biodiversidad
Con la problemática del cambio climático y especialmente con la reducción de
las reservas de combustibles de origen fósil, ha aumentado la demanda de
“biocombustibles” causando la expansión del área dedicada a la siembra de
palma aceitera (la producción global de palma aceitera incrementó en 55%
entre 2001 y 2006)1 y la aparición de nuevos países productores. Las
plantaciones industriales de palma aceitera, Elaeis guineensis Jacq., fueron
desarrolladas inicialmente en Indonesia y Malasia, donde se han documentado
graves impactos sobre la biodiversidad. Entre 1990 y 2005 el área de palma
1 http://faostat.fao.org. Consultada en diciembre de 2010.

11
aceitera en Malasia incrementó de 1,8 a 4,2 millones de hectáreas, mientras
que se perdieron 1,1 millones de hectáreas de bosque, y en Indonesia el área
sembrada de palma incrementó de 4,4 a 6,1 millones de hectáreas, mientras
que el total de bosque perdido fue de 28,1 millones de hectáreas (FAO, 2006;
Koh y Wilcove, 2008).
La expansión de los cultivos industriales de palma aceitera podría en principio
contribuir a la deforestación de 4 maneras a menudo indistinguibles: (i) como el
motivo primario para la tala de bosques intactos; (ii) por reemplazo de los
bosques previamente degradados por extracción de madera o fuego; (iii) como
parte de una empresa económica combinada, usando otros beneficios como la
extracción de madera o pulpa de papel para compensar los costos de
establecimiento de las plantaciones; o (iv) indirectamente, a través de mejorar
vías de acceso a los bosques previamente inaccesibles o desplazando otros
cultivos a los bosques (Fitzherbert et al., 2008).
La eliminación de bosques tropicales para el establecimiento de monocultivos
como los de palma aceitera, deja la capa orgánica del suelo expuesta,
causando la erosión del suelo y la contaminación de fuentes de agua. Además,
los hábitats naturales de un diverso rango de animales son eliminados y las
especies que sobreviven llegan a ser plagas para los productores de palma,
induciendo al uso de pesticidas químicos que dañan la biodiversidad, las
fuentes de agua y la salud de los trabajadores y comunidades locales
(Fitzherbert et al, 2008; Holt, 2007; Koh y Wilcove 2008).
Hay grandes emisiones de gases de efecto invernadero asociadas con el
establecimiento de plantaciones de palma aceitera en reemplazo de los
bosques, la desecación de suelos orgánicos, y el uso de combustibles fósiles
para la cosecha de los cultivos, procesamiento y transporte (Danielsen et al,
2008; Holt, 2007). Sólo si las plantaciones de palma aceitera son establecidas
en potreros degradados con bajo contenido de carbono es probable que ellas
lleguen a ser sumideros de carbono (Danielsen et al, 2008; Holt, 2007; IAvH,
2000). Es probable que el impacto más grande provenga de la polución,

12
contaminantes potenciales incluyen los desechos industriales (POME, por sus
siglas en inglés), fertilizantes, insecticidas, rodenticidas y herbicidas (Danielsen
et al, 2008; Fitzherbert et al., 2008; Holt, 2007; Koh y Wilcove, 2008).
Tomará décadas o siglos que las emisiones de carbono desde combustibles
fósiles evitadas con el uso de “biodiesel” compensen las emisiones liberadas
cuando los bosques o suelos orgánicos son convertidos a plantaciones de
palma aceitera (Danielsen et al, 2008). Hasta que sea demostrado que los
cultivos de oleaginosas no están reemplazando bosques, es probable que el
uso de palma y otros aceites vegetales que son materias primas de
biocombustibles exacerbe el cambio climático, conduzca a un aumento de los
precios de los alimentos y acelere la pérdida de biodiversidad (Danielsen et al,
2008; Fitzherbert et al., 2008; Holt, 2007; IAvH, 2000; Koh y Wilcove, 2008).
Una vez establecidas las plantaciones de palma, como sucede con otros
monocultivos, la respuesta de la biodiversidad al cambio en la cobertura de la
tierra depende del alcance al cual las características del hábitat natural son
replicadas y de la variación en las sensibilidades de las especies al cambio
(Fischer y Lindenmayer, 2007). Las plantaciones de palma aceitera son
estructuralmente menos complejas que los bosques naturales, con una
estructura uniforme de edad de los árboles, dosel más bajo, escasas hierbas,
microclima menos estable y mayores perturbaciones antrópicas (Danielsen et
al, 2008; Fitzherbert et al., 2008; Koh y Wilkove, 2008).
De acuerdo con investigaciones realizadas por el Instituto de Investigación de
Recursos Biológicos Alexander von Humboldt, “la palma africana no se integra
en el ambiente existente, lo remplaza con el suyo, lo que quiere decir que
destruye las especies de fauna y flora que no son compatibles con ella o las
transforma” (IAvH, 2000). Fitzherbert et al., (2008) encontraron que sólo el 15%
de las especies registradas en bosque primario fueron también encontradas en
las plantaciones de palma en Malasia e Indonesia. También encontraron que la
palma aceitera es particularmente pobre sustituto para bosques primarios o
degradados porque las plantaciones mantienen incluso mucho menos especies

13
de bosque que la mayoría de otras opciones agrícolas (Hevea brasiliensis
(Willd. ex A. Juss.) Müll. Arg., Theobroma cacao (Mill.) Bernoulli, y Acacia
mangium Willd.).
Aunque el cultivo extensivo de palma africana con fines comerciales en
Colombia empezó en 1945 con la instalación de una plantación en el
departamento de Magdalena y un vivero en Calima, Valle, por parte de la
compañía estadounidense United Fruit Company (Mignorance et al., 2004),
sólo a partir de la administración del ex presidente Álvaro Uribe la agroindustria
palmícola se considera como el sector estratégico que redimirá el agro
colombiano, y la expansión del área sembrada se convirtió en un propósito
nacional. Tal como lo ha declarado públicamente en diversas ocasiones el ex
presidente, el país debe alcanzar por lo menos 6 millones de hectáreas
sembradas con palma. Como resultado del impulso que se ha dado en
Colombia a esta agroindustria tenemos que de 157.327 hectáreas sembradas
con palma en el año 2000 se pasó a 301.810 hectáreas al primero de febrero
de 20072.
En el Chocó, específicamente en las cuencas de los ríos Curbaradó y
Jiguamiandó, en el municipio Carmen del Darién, la siembra de palma africana
fue la principal causa de la deforestación de aproximadamente 3.000
hectáreas, la desviación o desecación de cauces de agua y la pérdida de por lo
menos 24 especies de plantas de hábito arbustivo, 23 especies de plantas de
hábitos epífitos, 43 especies de peces, 23 especies de anfibios, 40 especies de
reptiles, 90 especies de aves y 33 especies de mamíferos (IIAP, 2008). Al
impacto de esta práctica de agricultura intensiva deben sumarse las prácticas
de ganadería extensiva y la extracción de madera, muchas veces impulsadas
por las mismas empresas palmeras.
2 http://www.presidencia.gov.co/prensa_new/sne/2007/febrero/01/12012007.htm. Consultada en enero de 2011.

14
Esta agroindustria llegó a la zona del Urabá chocoano luego del
desplazamiento de por lo menos 15.000 pobladores debido a la agudización del
conflicto armado (Defensoría del Pueblo, 2005). Estos territorios son propiedad
colectiva de las comunidades que los han habitado ancestralmente, tal como lo
reconocen la Ley 70 de 1993 y las resoluciones 02809 y 02801 del año 2000
por medio de las cuales el Instituto Colombiano de Desarrollo Rural -
INCODER- otorga el título colectivo al Consejo Comunitario del Río Curbaradó
sobre 46.084 ha con 50 m2 y al Consejo Comunitario del Río Jiguamiandó
sobre 54.974 ha con 8.368 m2.
Las comunidades conformadas principalmente por afrodescendientes y en
menor medida por indígenas y mestizos, que se encontraban en situación de
desplazamiento, decidieron retornar a sus territorios en el 2006,
estableciéndose bajo las figuras de “Zona Humanitaria” y “Zona de
Biodiversidad”. Paralelo a ello organizaron sus Consejos Comunitarios y
emprendieron acciones de diversa índole para la restitución legal de sus
territorios. Ante la gravedad de las denuncias sobre la violación de derechos
humanos relacionada con el impulso del megaproyecto palmícola, el
INCODER, por medio de la resolución 2424 del 10 de septiembre de 2007,
deslindó el Consejo Comunitario del Río Curbaradó con el fin de establecer
cuáles de los territorios sembrados con palma aceitera pertenecen al título
colectivo del Consejo Comunitario, y decretó la restitución de 25.273 hectáreas
a las comunidades, que sólo hasta ahora se está materializando (Defensoría
del Pueblo, 2005).
El gobierno colombiano, a través del Ministerio de Agricultura, condiciona la
restitución de los territorios a la implementación de diversas propuestas
agroindustriales, a saber, continuar con la industria de la palma como socios, o
la producción a gran escala de caucho, banano, cacao o teca, para la oferta
agroexportadora. Las comunidades no están conformes con estas propuestas y
para restablecer las relaciones desarrolladas ancestralmente con sus territorios
han decidido recuperar la biodiversidad de sus bosques, de los que depende su

15
identidad cultural e incluso su existencia como comunidades3. Con el retorno
de las comunidades a la zona gran parte de los cultivos de palma han sido
abandonados, dejando en la cuenca del río Curbaradó un paisaje fragmentado
compuesto por grandes áreas sembradas con palma, potreros provenientes de
terrenos que no alcanzaron a ser sembrados o que estuvieron dedicados a
ganadería, y parches de bosque intervenido debido a la extracción de madera.
Restauración ecológica y barreras a la sucesión nat ural de bosques
El nivel de avance de la regeneración del bosque en un sitio previamente
perturbado varía según el tipo e intensidad de la práctica previa, el tiempo
transcurrido desde el abandono, y la existencia de vegetación remanente (Aide
et al, 1996; Brown y Lugo, 1990; Castillo y Stevenson, 2010; Chazdon, 2003;
Cubiña y Aide, 2001; Guevara et al, 1996; Wijdeven y Kuzee, 2000). Además,
con la aparición de barreras que limitan la continuidad del proceso sucesional,
este se ralentiza o incluso puede estancarse (Shono et al., 2007; Guariguata,
1999), resultando en sitios con diferente composición y estructura vegetal.
Aunque la regeneración natural puede producir un bosque secundario similar
en estructura a los bosque previamente establecidos entre 40 y 70 años
aproximadamente (Aide et al, 1996; Aide et al, 2000; Capers et al, 2005;
Chazdon, 2003), es deseable acelerar la recuperación para el control de la
erosión, incrementar la biodiversidad, o promover otras interacciones
biológicas. En los sitios sometidos a perturbaciones de origen natural o
antrópico, el proceso de sucesión natural está influenciado por la compleja
interacción de varios factores incluyendo condiciones edáficas, cantidad y
composición de especies de la lluvia de semillas, y niveles de depredación de
3 Para estas comunidades “el territorio tiene su significado en el marco de una relación particular entre comunidad, ser humano y naturaleza, en la cual esta no es simplemente el entorno que rodea al ser humano, sino que tanto el individuo como la comunidad son sujetos que hacen parte de la naturaleza” Jesús Alfonso Flórez y Constanza Millán Echeverría. 2007. “Derecho a la Alimentación y al Territorio en el Pacífico Colombiano”. Editado por Diócesis de Quibdó, pág 39.

16
semillas y plántulas, lo que dificulta determinar la cantidad de tiempo necesaria
para que el bosque sea restaurado (Aide y Cavelier, 1994; Brown y Lugo, 1990;
Capers et al, 2005; Shono et al., 2007). Estos factores interactúan fuertemente
con la edad desde el abandono y la proximidad de las áreas a parches de
bosque remanentes para influir procesos sucesionales (Aide y Cavelier, 1994;
Chazdon, 2003).
Mientras más intenso o duradero es el uso del sitio previo al abandono, la
recuperación de la diversidad vegetal y la estructura de la vegetación a través
de la sucesión es más lenta debido, parcialmente, a la reducción del número de
posibles mecanismos de colonización: lluvia de semillas, banco de semillas,
rebrote de vegetación remanente, etc (Guariguata, 1999; (Wijdeven y Kuzee,
2000)). Las tasas de recuperación son aceleradas si la intensidad del uso
previo de la tierra es baja, si los suelos están fértiles, y si hay áreas cercanas
de bosque remanente (Aide y Cavelier, 1994; Chazdon, 2003; Guariguata y
Ostertag, 2001); en contraste, la recuperación natural en áreas sujetas a
efectos antropogénicos intensivos es muy lenta debido a la degradación del
suelo, perturbaciones recurrentes, y aislamiento de bosques intactos (Chazdon,
2003; Shono et al; 2007). En consecuencia, la elección de una estrategia de
restauración apropiada depende del nivel de degradación, la tasa deseada de
recuperación y la similitud a la composición de especies de bosque nativo
deseado (Aide et al, 2000).
Mientras que algunas propiedades de las comunidades, tales como la densidad
de especies parecen converger con el tiempo en bosques tropicales, la
convergencia de otras propiedades, incluyendo la composición de especies,
puede depender de las características de los sitios individuales tales como la
proximidad al bosque maduro y el grado de disturbio (Capers et al, 2005). La
recuperación de la estructura del bosque, las reservas de nutrientes en el
suelo, y la riqueza de especies es mucho más rápida que la recuperación de la
composición de especies, a menudo debido a una dispersión inadecuada de
especies de sucesión tardía (Chazdon, 2003). Es improbable que la
composición de especies alcance los niveles pre-disturbio dentro de un período

17
de menos de 200 años, particularmente en suelos de baja fertilidad
(Saldarriaga et al, 1988).
Aunque la riqueza de especies en bosques secundarios se acerca o iguala a la
de bosques maduros dentro de 40-70 años, la composición de la comunidad
permanece diferente por mucho más tiempo, posiblemente por siglos (Aide, et
al, 2000; Finegan, 1996; Capers et al, 2005). En un estudio de cronosecuencias
de regeneración de bosque en potreros abandonados en Puerto Rico, Aide et
al., (2000) encontraron que las características estructurales de los pastizales
abandonados en Puerto Rico (p.e. área basal, altura de los árboles, densidad
de tallos, riqueza de especies) estuvieron dentro del rango de valores de los
bosques más antiguos después de aproximadamente 40 años de regeneración,
sin embargo, la composición de especies permaneció diferente.
En un estudio sobre las dinámicas sucesionales de comunidades de plántulas
de bosques secundarios en Costa Rica, Capers et al., (2005) encontraron que
comunidades de plántulas divergen en cuanto a sus trayectorias sucesionales
debido a una colonización altamente estocástica, a la variedad de sitios fuentes
de semillas que circundan el área a restaurar y a las condiciones iniciales de
estas áreas (tipo e intensidad de la perturbación). Una vez abandonadas, estas
condiciones inhiben la recuperación del bosque original y pueden resultar en un
ecosistema distinto (Aide y Cavelier, 1994; Cavelier et al, 1998). En este caso,
si la meta de un proyecto de restauración es restaurar la composición de
especies similar a la de bosque maduro, será necesario realizar siembras de
enriquecimiento. Sin embargo, si interacciones tales como polinización,
dispersión, y depredación se han perdido, entonces la siembra no será
suficiente para asegurar el funcionamiento ecológico a largo plazo de estos
bosques (Aide et al, 2000).
Muchas de las especies de bosques maduros existen en el paisaje pero no
están colonizando los bosques secundarios derivados de pastizales
abandonados. Estas especies a menudo son tolerantes a la sombra, de
manera que el enriquecimiento por siembra de plántulas puede ser suficiente

18
para su establecimiento (Aide et al, 2000). La vegetación remanente cumple un
papel crítico en la recuperación del bosque, promoviendo rápido incremento en
la riqueza de especies, densidad de árboles y biomasa arriba del suelo
(Guariguata & Ostertag 2001; Wijdeven y Kuzee, 2000). Las especies pioneras
se establecen abundantemente en pastizales y campos de cultivo
abandonados, pero las especies colonizadoras particulares dependerán de la
fertilidad, drenaje y compactación del suelo, la presencia o ausencia de un
banco de semillas, y de vegetación remanente periférica (Chazdon, 2003).
La colonización exitosa parece deberse a diferentes combinaciones de
características tales como: semillas grandes, semillas dispersadas por el viento
(Wijdeven y Kuzee, 2000), crecimiento vegetativo, capacidad para rebrotar,
entre otras (Aide et al, 2000). Si estas especies colonizadoras logran
establecerse temprano por germinación de semillas o siembra de plántulas,
pueden ser capaces de acelerar la regeneración del bosque al proveer hábitat y
alimento para los dispersores de otras especies de bosque (Castillo y
Stevenson, 2010; Cole et al, 2010; Fuentes, 2000; Holl et al, 2000; Wunderlee,
1997) y un microhábitat apropiado para el establecimiento de plantas (Aide et
al, 2000; Guevara et al, 1986; Holl et al, 2000; Wijdeven y Kuzee, 2000;
Wunderle, 1997), de esta manera el establecimiento de árboles pioneros fija el
curso de la sucesión secundaria (Chazdon, 2003).
Las plantaciones de palma aceitera como potenciales barreras para la regeneración del bosque
Algunos monocultivos de palma aceitera establecidos en la cuenca del río
Curbaradó fueron abandonados seis años luego de ser establecidos, y su
intensidad de manejo fue alta, pero comparado con otras actividades agrícolas
y de ganadería donde la perturbación ha tenido duraciones de más de 20 años,
la intensidad del uso previo del área de estudio la consideramos baja en
términos generales. A partir de entonces se inició un proceso de sucesión
natural cuyo curso está determinado por la presencia de las plantaciones según

19
la manera como estas afecten los mecanismos que regulan el proceso
sucesional, especialmente la lluvia de semillas y el establecimiento de
plántulas.
En muchos sitios en el trópico, la dispersión pobre de semillas (Aide y Cavelier,
1994; Cole et al, 2010; Holl, 1999; Wunderlee, 1997), la competencia con
vegetación herbácea, la compactación y pobreza de nutrientes del suelo
(Chazdon, 2003), o la baja disponibilidad lumínica pueden inhibir el
establecimiento inicial de especies leñosas (Aide et al, 2000), creando barreras
al proceso sucesional relacionadas con la lluvia de semillas o el
establecimiento de plántulas. De acuerdo con esto, el primer paso para
restaurar los ecosistemas es evaluar cuáles de estas barreras limitan la
sucesión natural. Las principales barreras que causas un retardo en la
velocidad de este proceso son: físicas (poco agua, poca luz, compactación);
químicas (baja fertilidad o exceso de compuestos tóxicos en el suelo); y
biológicas (falta de semillas dispersadas naturalmente, defoliación por insectos,
amplia cobertura por pastos y malezas) (Aide, et al, 2000; Cavelier et al, 1996;
Fuentes, 2000; Guariguata, 1999; Holl et al, 2000). Una vez identificadas las
barreras, los esfuerzos de restauración deben enfocarse en solucionarlas de
forma tal que la sucesión proceda (Chazdon, 2003; Shono et al, 2007).
La restauración ecológica acelera la sucesión por remoción o reducción de
barreras a la regeneración natural del bosque: la competencia con hierbas es
reducida, las perturbaciones son prevenidas, microclimas inadecuados son
mejorados por el crecimiento acelerado de pioneras establecidas naturalmente,
y la dispersión de semillas dentro de los sitios por aves y animales es
potenciada por la restauración del hábitat boscoso (Shono et al, 2007). Sin
embargo, cuando el proceso sucesional ha empezado pueden aparecer otro
tipo de barreras. Por ejemplo, las especies herbáceas (pastos intolerantes a la
sombra y helechos) pueden formar ecosistemas que se auto perpetúan,
bloqueando efectivamente la regeneración natural del bosque por décadas e
incluso siglos (Shono et al, 2007; Chazdon, 2003). Por otro lado, las

20
plantaciones que forman parte de una matriz de paisaje pueden actuar como
una barrera a los movimientos de animales (Fitzherbert et al, 2008).
El establecimiento de especies de árboles de rápido crecimiento, de manera
espontánea o por siembras de enriquecimiento, particularmente leguminosas
que fijan nitrógeno, pueden incrementar la materia orgánica en el suelo,
prevenir la erosión, y potenciar el ciclaje de nutrientes (Chazdon, 2003).
Además, los árboles tienen una función crítica en atraer aves y murciélagos
dispersores de semillas a las áreas en restauración (Castillo y Stevenson,
2010; Fuentes, 2000; Holl et al, 2000; Wunderle, 1997). El establecimiento de
especies exitosas en la colonización de pastizales (pioneras) puede excluir por
la sombra a especies herbáceas, abriendo el sotobosque para la colonización
de especies leñosas dispersadas al sitio. La colonización exitosa parece
deberse a diferentes combinaciones de características tales como: semillas
grandes, semillas dispersadas por el viento, crecimiento vegetativo, capacidad
para rebrotar, entre otras (Aide et al, 2000). Si estas especies colonizadoras
pueden establecerse temprano por germinación de semillas o siembra de
plántulas pueden ser capaces de acelerar la regeneración del bosque al
proveer hábitat y alimento para los dispersores de otras especies de bosque y
un microhábitat apropiado para su establecimiento (Aide et al, 2000).
Para asegurar el éxito de cualquier proyecto de restauración además de la
parte técnica es fundamental el aspecto social. Los bosques proveen una red
segura para las poblaciones rurales alrededor del mundo al proveer alimento,
medicina, y otros materiales vegetales necesarios para la comunidad. El
aumento de conciencia, la capacidad de construcción, y la promoción de
procesos participativos en el manejo de los recursos del bosque pueden ser un
componente integral en la propuesta de Regeneración Natural Asistida -RNA-
(Shono et al, 2007).
Una vez un bosque secundario es establecido, este puede ser manejado para
proporcionar los productos y servicios deseados de acuerdo a los objetivos de
manejo específicos y a la disponibilidad de recursos (Brown y Lugo, 1990;

21
Shono et al, 2007). Según la Sociedad Internacional para la Restauración
Ecológica (SER International, por sus siglas en inglés) la restauración ecológica
fomenta, y quizás hasta depende de la participación a largo plazo de la
población local. Para SER International, lo que hace única a la restauración
ecológica es su capacidad inherente de proveer a las comunidades con la
oportunidad no sólo de reparar el daño ecológico, sino también de mejorar la
condición humana. En muchas instancias, la restauración ecológica ha sido
capaz de renovar las oportunidades económicas, renovar las prácticas
culturales tradicionales y enfocar las aspiraciones de las comunidades locales
(SER International, 2004).
Con el fin de acompañar el proceso de sucesión se debe elaborar un plan de
restauración ecológica del bosque a largo plazo, y definir las metas y
estrategias de restauración. Para esto un primer paso es evaluar si las
plantaciones podrían ser una barrera para algunos procesos característicos de
la regeneración inicial del bosque. En este sentido, nuestro proyecto evalúa si
la presencia de plantaciones industriales de palma aceitera constituye una
barrera para la regeneración natural del bosque. Si las plantaciones industriales
de la palma aceitera afectan la regeneración natural de los bosques porque
restringen la dispersión de semillas, disminuyen la germinación y la
supervivencia inicial de plántulas, y afectan el establecimiento de plántulas;
entonces la riqueza de especies de la lluvia de semillas y de la comunidad de
plántulas y juveniles será menor en las plantaciones comparadas con los
rastrojos y los bosques, la composición de especies será distinta entre las
unidades de paisaje, y los porcentajes de germinación y supervivencia inicial
serán más bajos en las plantaciones que en los rastrojos y los bosques. Para
probar estas predicciones realizamos experimentos con trampas de caída para
caracterizar la lluvia de semillas, siembras de semillas, y trasectos para
describir la comunidad de plántulas y juveniles en unidades de paisaje de
bosques y rastrojos y plantaciones abandonadas, en estados iniciales de
sucesión del bosque.

22
2. MATERIALES Y MÉTODOS
Área de estudio
El trabajo de campo de lo realizamos entre marzo de 2009 y mayo de 2010 en
la parte baja de la Cuenca del río Curbaradó (7°09, 598N y 76°58,505E),
tributario del río Atrato, municipio Carmen del Darién, departamento de Chocó
(Figura 1). El sitio de estudio específicamente se localiza en predios del
territorio colectivo de las comunidades afro-descendientes de Cetino y La
Iguana que comprenden un área aproximada de 8000 hectáreas,
pertenecientes al Consejo Mayor de Curbaradó (Figura 2). La zona se
encuentra a una altura de 12 msnm, tiene una temperatura promedio anual de
26°C (alcanzando un máximo de 28°C en diciembre y u n mínimo de 24°C en
febrero), una precipitación anual de 5523 mm/año (varía desde 684 mm/mes en
octubre hasta 14,7 mm/mes en enero) (Codechocó, 2001), y el bosque está
clasificado como Bosque muy Húmedo Tropical (BmH-T) (Holdridge, 1971).
Gran parte de estos territorios fue deforestada en el año 2002 para la
implementación de monocultivos de palma aceitera. Del área deforestada,
3.864 hectáreas fueron sembradas con palma (IIAP, 2008), y otra parte, cuya
extensión se desconoce, se dejó en potreros usados de diferentes maneras.
Debido a regulaciones ambientales parte del bosque no se taló, y actualmente
existen sólo tres fragmentos de bosque de un área menor de 100 hectáreas
cada uno. Estos cultivos fueron abandonados en el año 2008, dejando un
paisaje fragmentado compuesto básicamente por tres unidades: Bosque,
Rastrojo y Plantación. Al momento en que iniciamos esta investigación, este
mosaico llevaba 2 años de abandono.

23
Figura 1. Municipio Carmen del Darién (Curbaradó), Chocó.
Fuente: www.colombiassh.org
Figura 2. Parte media de la cuenca del río Curbaradó.
Fuente: www.colombiassh.org

24
Experimentos
Para evaluar las hipótesis empleamos tres réplicas de cada unidad de paisaje
denominadas así: Bosque 1, Rastrojo 1, Plantación 1; Bosque 2, Rastrojo 2,
Plantación 2 y Bosque 3, Rastrojo 3, Plantación 3. Para tratar de minimizar los
efectos que pudieran tener la calidad del suelo, la distancia al ecosistema de
referencia o fuente de semillas, y el uso previo (ganadería o terrenos en
preparación para la siembra de palma) sobre la dispersión de semillas y el
establecimiento de plántulas (Chazdon, 2003), elegimos sitios con
características similares para los siguientes criterios: distancia del bosque,
edad de abandono, uso previo y topografía.
Los bosques remanentes son bosques secundarios, altamente perturbados
debido a la extracción maderera con fines comerciales y en menor medida para
la construcción, y ocupan un área muy pequeña respecto a la matriz de
plantaciones donde se encuentran. Entre ellos el más conservado es el Bosque
3, donde se encuentran algunos individuos de Prioria copaifera Griseb. (cativo),
Carapa guianensis Aubl. (güino), Tabebuia rosea (Bertol.) A. DC. (roble) y
Dipteryx oleifera Benth. (choibá). Probablemente esto se deba a que el lugar se
encuentra alejado de caminos, cultivos alimenticios o casas. Por el contrario, el
Bosque 2 es el más degradado, por quedar más cerca del río y de caseríos,
además está atravesado por una vía de balastro y una trocha por la que se
extienden cables de polea empleadas para el transporte del corozo de la
palma. El Bosque 2 tiene numerosos individuos de varias especies de Cecropia
sp. (yarumos) en el borde que limita con la carretera (Figura 3).
Figura 3. Unidad de paisaje Bosque 2.

25
Los rastrojos tienen su origen en potreros que serían destinados a ganadería o
a la siembra de palma aceitera y que por diversos motivos no se usaron para
estos fines. En esta unidad de paisaje se encuentran abundantes individuos de
varias especies de Urera sp. (pringamosa), Cecropia sp. (yarumos), Calathea
lutea Schult. (hoja blanca) y Carludovica palmata Ruiz & Pav. (iraca). En el
área de estudio también se encuentran rastrojos derivados del abandono de
cultivos de pan coger como yuca, plátano, arroz o maíz, ya fuera para el
autoabastecimiento o para su comercialización. Estos últimos no los incluimos
en el estudio porque el tiempo transcurrido desde su abandono es menor, lo
que podría causar ruido en los resultados por el efecto del tiempo de abandono
sobre la regeneración natural (Aide et al, 1996; Brown y Lugo, 1990; Castillo y
Stevenson, 2010; Chazdon, 2003; Cubiña y Aide, 2001; Guevara et al, 1996;
Wijdeven y Kuzee, 2000). De los tres rastrojos, el más conservado es el
Rastrojo 3 porque está alejado de viviendas y caminos. El Rastrojo 1 tiene un
mayor grado de intervención por estar en medio de una matriz de plantaciones
de plátano, y a la mitad del desarrollo de esta investigación encontramos este
sitio rozado y sembrado con maíz.
Entre las plantaciones, al igual que con las otras dos unidades de paisaje, la
menos intervenida fue la Plantación 3. Las otras dos réplicas tuvieron un grado
similar de intervención, y en el segundo muestreo de la lluvia de semillas los
sitios se encontraron rozados y sembrados con plátano y maíz. En las tres
plantaciones se encuentran abundantes individuos de especies de Cecropia sp.
(yarumos), Calathea lutea (hoja blanca) y Trema micrantha (L.) Blume
(guayuyos). En términos generales, las plantaciones de palma aceitera fueron
asistidas durante poco tiempo o, en el caso de la Plantación 3, el cultivo nunca
recibió mantenimiento de fertilización, fumigación o eliminación de “malezas”.
Lluvia de semillas
Para monitorear la lluvia de semillas por el período de un año ubicamos en
cada sitio dos trampas de caída que consistían en cajones de 0,5 x 0,5 m

26
(Figura 4), abarcando un área de muestreo de 1,5 m2 para cada unidad de
paisaje. Los cajones fueron ubicados de tal manera que distaran por lo menos
50 m del borde del bosque y evitando claros de bosque. En cada cajón
vertimos suelo tomado del bosque a 1,5m de profundidad donde se asume que
la presencia de semillas es insignificante (Figura 5) (Wijdeven y Kuzee, 2000).
Instalamos las trampas en marzo de 2009, e hicimos los muestreos en junio y
octubre de 2009 y febrero de 2010. En cada muestreo removimos el sustrato, lo
diluimos en agua para pasarlo por un tamiz # 64 (Figura 6), se secó al sol
(Figura 7) y se almacenó en bolsas plásticas para su posterior procesamiento.
Luego de remover el sustrato de cada cajón se vertió más sustrato para el
siguiente período de muestreo.
Figura 4. Trampas de caída de semillas.
Figura 5. Llenado de las trampas de semillas.

27
Figura 6. Lavado del sustrato de las trampas de semillas.
Figura 7. Secado del sustrato de las trampas de semillas.
Procesamos las muestras en el laboratorio pasando el sustrato por un juego de
4 tamices de diferentes tamaños, de los cuales el menor fue el #64, por lo tanto
las semillas de un tamaño menor no se consideraron en este estudio. Vertimos
el contenido de cada tamiz en una caja de petri y separamos las semillas
presentes con la ayuda de un estereoscopio. El conjunto de semillas de cada
muestra fue separado por morfotipos para estimar la riqueza de especies y
establecer la composición de la lluvia de semillas en cada unidad de paisaje.

28
Identificamos los morfotipos con la mayor resolución taxonómica posible por
comparación con una colección de referencia de plantas con frutos colectadas
durante todo el estudio a las cuales extrajimos las semillas. Los ejemplares de
herbario fueron depositados en la colección del Herbario de la Universidad de
Antioquia (HUA). También empleamos colecciones de referencia de otros
estudios realizados en bosques húmedos tropicales disponibles en el Herbario
de la Universidad de Antioquia; y usamos fotografías del libro Seed Dispersal
by Bats in the Neotropics (Lobova, 2009) y de bases de datos disponibles en
internet (http://striweb.si.edu/esp/tesp/plant_images_a.htm de Smithsonian
Tropical Research Institute y http://nt.ars-
grin.gov/sbmlweb/SeedsFruits/rptSeedsFruitsFam.cfm de United States
Department of Agriculture, USDA). Con esta información estimamos la riqueza
de especies y establecimos la composición de la lluvia de semillas en cada
unidad de paisaje. Los morfotipos identificados se caracterizaron de acuerdo a
su hábito de crecimiento en árbol, arbusto, palma, hierba, pasto, trepadora y
helecho; además, los árboles y arbustos se clasificaron de acuerdo a su estado
sucesional en pioneras y no pioneras de acuerdo con la literatura (Aide y
Cavelier, 1994; Aide et al, 1995; Aide et al, 2000; Cavelier et al, 1998; Cole et
al, 2000; Cubiña y Aide, 2001; Guevara et al, 1986; Olea-Wagner, 2007;
Wijdeven y Kuzee, 2000) y con la ayuda de expertos del Herbario de la
Unversidad de Antioquia.
Germinación y supervivencia inicial de plántulas
Escogimos cuatro especies de árboles de acuerdo a su presencia en los
bosques antes de la siembra de palma y su importancia para la comunidad
(usos tradicionales o comerciales): Ceratonia siliqua L. (algarrobo),
Centrolobium paraense Tul. (guayacán hobo), Couroupita guianensis Aubl.
(cocuelo) y Myroxylon balsamum (L.) Harms (bálsamo). En cada réplica de
unidad de paisaje establecimos una parcela de germinación en junio de 2009
(para un total de nueve parcelas) en la que sembramos 80 semillas de cada
especie. Hicimos un censo en octubre de 2009 para determinar el porcentaje

29
de germinación de cada especie en cada unidad de paisaje (Figura 8).
Finalmente hicimos un censo en febrero de 2010 para establecer el porcentaje
de supervivencia de cada especie en cada unidad de paisaje (Figura 9).
Figura 8. Parcelas de germinación.
Figura 9. Plántulas presentes en el censo de germinación. Bálsamo a la
izquierda, algarrobo a la derecha
Regeneración natural: establecimiento de plántulas
Para evaluar la regeneración natural en cada réplica de unidad de paisaje
hicimos un transecto de 50 x 2 m para registrar la presencia de todas las
plántulas de arbustos, árboles y hierbas. En el muestreo sólo consideramos las

30
plántulas y los juveniles con una altura menor de dos metros para garantizar
que fuesen plantas establecidas luego del abandono de los sitios. Se
colectaron muestras botánicas de cada una de los morfotipos separados en
campo para su posterior identificación taxonómica en el Herbario de la
Universidad de Antioquia.
Identificamos los morfotipos de las plántulas y juveniles con la mayor resolución
taxonómica posible empleando claves para plántulas propuestas por Barrera
(1985, 1986), Duke (1965) y del Amo (1979) y con la ayuda de de expertos del
Herbario de la Universidad de Antioquia. Con la información de los nombres
comunes proporcionados por el ayudante de campo y habitantes de la
comunidad de Cetino, consultamos el Diccionario de los Nombres Comunes de
las Plantas de Colombia, del Instituto de Ciencias Naturales de la Universidad
Nacional de Colombia (http://www.biovirtual.unal.edu.co/diccionario/), y
corroboramos esta información con los especímenes de herbario. Con esta
información estimamos la riqueza de especies y establecimos la composición
de la comunidad de plántulas en cada unidad de paisaje. Cada plántula
identificada se caracterizó según su hábito de crecimiento en árbol, arbusto,
palma, hierba, pasto, trepadora y helecho, y los árboles y arbustos se
clasificaron de acuerdo a su estado sucesional en especies pioneras y no
pioneras igual que con las semillas.
Análisis estadístico
Para evaluar si existen diferencias significativas en la riqueza de especies de la
lluvia de semillas y de la regeneración natural, entre las tres unidades de
paisaje realizamos la prueba no paramétrica de Kruskal-Wallis, dado el bajo
número de réplicas. Posteriormente hicimos pruebas post-hoc usando la
prueba U de Mann-Whitney para comparar la riqueza de especies por pares de
unidad de paisaje. Para estos análisis utilizamos el programa estadístico SPSS
17.0 (SPSS, 2008).

31
Para tomar la decisión de rechazar la hipótesis nula en las pruebas de hipótesis
que evalúan el efecto de la unidad del paisaje sobre la riqueza de especies
decidimos tomar como nivel de significancia un valor de α=0.1. Al incrementar
la probabilidad de cometer error tipo I en la prueba de hipótesis hasta un 10%
en nuestro análisis incrementamos el poder de la prueba para detectar un
efecto del factor sobre la riqueza de especies y por tanto se disminuye la
probabilidad de cometer error tipo II. En escenarios de estudios que pretenden
hacer recomendaciones para el manejo y conservación de ecosistemas esta es
una aproximación razonable, dados los mayores riesgos que representa
cometer error tipo II y dejar de reconocer la importancia del factor en el proceso
ecológico de interés y el reducido número de réplicas que pueden obtenerse en
estudios a escala de paisaje (Feisinger, 2001)
Para observar el patrón de agrupación entre las unidades de paisaje de
acuerdo con la similitud en su composición especies en la lluvia de semillas y
de especies de plántulas y juveniles en la regeneración natural, hicimos un
análisis de agrupamiento. Para este análisis empleamos el método de
centroides y como medida de similitud el índice de Hamann por ser una medida
binaria, pues nuestros datos son de presencia/ausencia de especies en cada
unidad de paisaje. Este índice de similitud se estima como la probabilidad de
que las especies tengan el mismo estado en dos réplicas (presente o ausente
en los dos) menos la probabilidad de que tengan estados diferentes en las dos
réplicas, y toma valores entre cero (similitud baja) y uno (similitud alta). Para
estos análisis y para comparar las riquezas promedios de especies de cada
sitio, utilizamos el programa estadístico SPSS 17.0 (SPSS, 2008).
Además del análisis de agrupamiento realizamos un análisis de ordenación tipo
DCA (Detrended Correspondence Analysis, por sus siglas en inglés) para
facilitar la interpretación de los resultados del agrupamiento según la
composición de especies de la lluvia de semillas y de la regeneración natural.
En el DCA es posible explorar la similitud en la composición de especies entre
sitios y además visualizar cuáles especies son importantes para agrupar los
sitios con base en su similitud, y cuáles especies son importantes para aislar

32
unos sitios de otros al ser exclusivas o poco comunes. Para hacer el DCA
empleamos el programa estadístico PAST (Hammer et al, 2008). Con los
valores de las coordenadas del DCA para cada especie de semilla y plántula
identificada de la cual sabíamos su hábito de crecimiento y estado sucesional,
hicimos una gráfica donde pudiéramos visualizar cómo se distribuyen estas
especies entre las unidades de paisaje. Debido a que numerosas especies
presentaron el mismo valor para ambos ejes en el DCA, se dejó sólo un dato
de aquellas que tenían el mismo hábito de crecimiento y/o estado sucesional
para evitar puntos superpuestos que pudieran ocultar otros puntos referentes
hábitos o estados sucesionales diferentes, de esta manera, los puntos en esta
gráfica no corresponden al número de especies para cada característica.
3. RESULTADOS
Lluvia de semillas
Al monitorear la lluvia de semillas por el período de un año se encontraron 113
morfoespecies de las cuales 38 se identificaron hasta especie, 29 sólo hasta
género, 11 sólo hasta familia y 35 no se identificaron. En la unidad de paisaje
Bosque se encontraron 84 especies (15 no pioneras y 29 pioneras), en
Rastrojo 63 (8 no pioneras y 27 pioneras) y en Plantación 62 (5 no pioneras y
29 pioneras). Las unidades Bosque y el Rastrojo compartieron un mayor
número de especies, la mayoría de las cuales fueron especies pioneras (Tabla
1). La riqueza promedio de especies de la lluvia de semillas fue mayor en
Bosque, en Rastrojo fue intermedia y fue menor en Plantación (Figura 9), y sólo
fue significativamente diferente entre las unidades de paisaje Bosque y
Plantación (Tabla 2), y la variabilidad fue menor para las plantaciones (Figura
10).

33
Tabla 1. Especies compartidas entre las unidades de paisaje.
Unidades de paisaje N° de especies compartidas
Total Pioneras No pioneras Sin caracterizar Bosque-Rastrojo 46 21 4 21 Bosque-Plantación 41 22 1 18 Rastrojo-Plantación 43 22 1 20 Bosque-Rastrojo-Plantación 34 19 1 14
Tabla 2. Prueba U de Mann-Whitney para la riqueza promedio de especies de la lluvia de semillas (gl=1).
Unidades de Paisaje comparadas U P Bosque- Rastrojo 1,000 0,127 Bosque- Plantación ≤0,001 0,050* Rastrojo-Plantación 3,500 0,658 *diferencia significativa
Figura 10. Riqueza de especies (media-error estándar) en la lluvia semillas por unidad de paisaje.

34
Al realizar el análisis de agrupamiento se formaron dos conjuntos (Figura 11).
Dentro del primer grupo, el Rastrojo 3 y la Plantación 3 forman un subconjunto,
y el Bosque 1 y 3 forman otro subconjunto. En el segundo grupo la Plantación 1
y el Rastrojo 1 están más relacionados entre sí que cualquiera de ellos con el
subconjunto formado por la Plantación 2 y el Rastrojo 2. En medio de estos dos
conjuntos se encuentra el Bosque 2, estando igual de relacionando con
cualquiera de los dos grupos. Al observar con más detalle la matriz de
distancias que se emplea para la construcción del dendrograma según los
índices de similitud de Hamann (Tabla 3) encontramos que una similitud muy
alta dentro de las unidades de paisaje Plantación, muy bajas entre las réplicas
de la unidad de paisaje Bosque, e intermedias dentro de las réplicas de
Rastrojo.
Figura 11. Agrupación de las unidades de paisaje de acuerdo a la composición de especies en la lluvia de semillas.

35
Tabla 3. Matriz de distancia entre las unidades de paisaje según el índice de similitud de Hamman de acuerdo a la composición de especies de la lluvia de semillas (valores entre 0 y 1).
Unidad de
Paisaje
Bosque Rastrojo Plantación
1 2 3 1 2 3 1 2 3
Bosque 1 1.000 0.150 0.381 0.363 0.221 0.327 0.239 0.221 0.363
2 0.150 1.000 0.239 0.150 0.221 0.221 0.133 0.221 0.186
3 0.381 0.239 1.000 0.345 0.310 0.487 0.363 0.345 0.487
Rastrojo
1 0.363 0.150 0.345 1.000 0.327 0.398 0.628 0.434 0.363
2 0.221 0.221 0.310 0.327 1.000 0.504 0.487 0.398 0.363
3 0.327 0.221 0.487 0.398 0.504 1.000 0.487 0.398 0.646
Plantación
1 0.239 0.133 0.363 0.628 0.487 0.487 1.000 0.522 0.416
2 0.221 0.221 0.345 0.434 0.398 0.398 0.522 1.000 0.363
3 0.363 0.186 0.487 0.363 0.363 0.646 0.416 0.363 1.000
En el DCA se observa un patrón parecido al encontrado en el análisis de
agrupamiento en la similitud entre las unidades de paisaje, pero aquí el Bosque
2 se encuentra más cercano a los otros dos bosques (Figura 12). Al separar las
especies de la lluvia de semillas según su estado sucesional se observa que
las especies que son compartidas por las unidades de paisaje son especies
pioneras muy comunes en la sucesión en bosques húmedos de tierras bajas.
Estas pioneras incluyen especies como: Cecropia peltata L., Cecropia insignis
Liebm., Cecropia engleriana Snethl., Vismia baccifera (L.) Triana & Planch.,
Cecropia goudotiana Trécul, Ficus insipida Willd., Trema integerrima (Beurl.)
Standl, Piper asperiusculum Kunth y Piper cyrtopodon C. DC. (agrupadas
dentro del óvalo punteado en la Figura 13). Las especies más cercanas a las
unidades Plantación 1 y Rastrojo 1 son las especies pioneras Trema micrantha
(L.) Blume y Ochroma pyramidale (Cav. ex Lam.) Urb. (óvalo sólido, Figura 4).

36
Figura 12. Análisis de ordenación de las unidades de paisaje de acuerdo a la composición de especies en la lluvia de semillas.
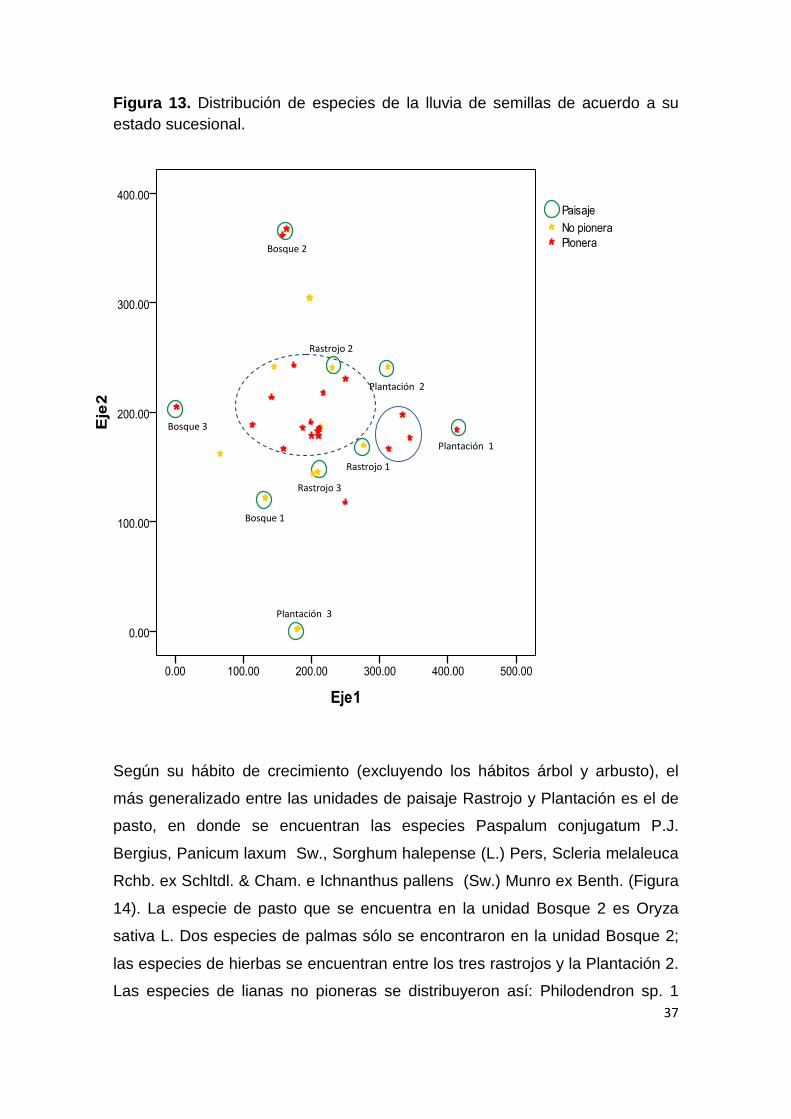
37
Figura 13. Distribución de especies de la lluvia de semillas de acuerdo a su estado sucesional.
Según su hábito de crecimiento (excluyendo los hábitos árbol y arbusto), el
más generalizado entre las unidades de paisaje Rastrojo y Plantación es el de
pasto, en donde se encuentran las especies Paspalum conjugatum P.J.
Bergius, Panicum laxum Sw., Sorghum halepense (L.) Pers, Scleria melaleuca
Rchb. ex Schltdl. & Cham. e Ichnanthus pallens (Sw.) Munro ex Benth. (Figura
14). La especie de pasto que se encuentra en la unidad Bosque 2 es Oryza
sativa L. Dos especies de palmas sólo se encontraron en la unidad Bosque 2;
las especies de hierbas se encuentran entre los tres rastrojos y la Plantación 2.
Las especies de lianas no pioneras se distribuyeron así: Philodendron sp. 1
Bosque 1
Bosque 3
Bosque 2
Plantación 1
Plantación 3
Plantación 2
Rastrojo 3
Rastrojo 1
Rastrojo 2

38
está entre los bosques 1 y 3, Philodendron sp. 2 sólo se encontró en la unidad
Bosque 3, Philodendron sp. 3 sólo en Rastrojo 2, y Paullinia fuscescens Kunth
sólo se encontró en la Plantación 3, y Cissus erosa Rich., una liana pionera, se
encontró en los bosques 1 y 2.
Figura 14. Distribución de especies de la lluvia de semillas de acuerdo a su
hábito de crecimiento.
Bosque 1
Bosque 3
Bosque 2
Plantación 1
Plantación 3
Plantación 2
Rastrojo 3
Rastrojo 1
Rastrojo 2

39
Germinación y supervivencia inicial de plántulas.
Los datos de los experimentos de una réplica de bosque, una de rastrojo y dos
de plantaciones no se tomaron porque estos sitios fueron rozados por colonos
para la siembra de plátano y maíz, o para continuar con la extracción del
corozo de la palma aceitera. Por lo tanto, los resultados que se muestran no
tienen un número suficiente de réplicas para realizar análisis estadísticos, pues
se tenían sólo dos réplicas para Bosque, dos para Rastrojo y una para
Plantación.
De las cuatro especies empleadas en el experimento de germinación, sólo
germinaron semillas de dos especies, y sus porcentajes de germinación fueron
muy bajos. Para Ceratonia siliqua el porcentaje de germinación fue mayor en
Rastrojo, en Plantación fue intermedio y en Bosque fue menor. Para Myroxylon
balsamum, el porcentaje de germinación fue mayor en Plantación, seguido por
Rastrojo, y en Bosque no hubo germinación (Tabla 4). El porcentaje de
supervivencia de las plántulas de Ceratonia siliqua fue mayor en Bosque,
seguida por Rastrojo y menor en Plantación. El porcentaje de supervivencia de
Myroxylon balsamum, fue mayor en Plantación que en Rastrojo (Tabla 5).
Tabla 4. Porcentajes de germinación de 4 especies de árboles en tres unidades de paisaje.
Especie Nombre común % de germinación
Bosque Rastrojo Plantación Ceratonia siliqua Algarrobo 5,95% 23,21% 15,12% Centrolobium paraense Guayacán hobo 0,00% 0,00% 0,00% Couroupita guianensis Cocuelo 0,00% 0,00% 0,00% Myroxylon balsamum Bálsamo 0,00% 3,01% 13,25%

40
Tabla 5. Porcentajes de supervivencia de 4 especies de árboles en tres unidades de paisaje.
Especie Nombre común % de supervivencia
Bosque Rastrojo Plantación Ceratonia siliqua Algarrobo 100% 93,75% 84,62% Centrolobium paraense Guayacán hobo 0,00% 0,00% 0,00% Couroupita guianensis Cocuelo 0,00% 0,00% 0,00% Myroxylon balsamum Bálsamo 0,00% 66,66% 100%
Regeneración natural: establecimiento de plántulas
Se registraron 177 morfoespecies de plántulas de las cuales 83 fueron
identificadas hasta el nivel de especies, 71 sólo hasta género, 16 sólo hasta
familia y 6 no se pudieron identificar. En la unidad de paisaje Bosque se
encontraron 112 especies, en Rastrojo 92, y en Plantación 61. El Bosque y el
Rastrojo comparten muchas más especies las demás unidades, la mayoría de
las cuales son no pioneras (Tabla 6). La riqueza promedio de especies de la
comunidad de plántulas fue mayor en la unidad de paisaje Bosque, seguida por
Rastrojo y por último está la Plantación (Figura 15), y sólo fue
significativamente diferente entre las unidades de paisaje Bosque y Plantación
(Tabla 7). La riqueza de especies tuvo una mayor varianza en Rastrojo, seguida
por Bosque y finalmente se encuentra la Plantación (Figura 15).
Tabla 6. Especies compartidas entre las unidades de paisaje.
Unidades de paisaje N° de especies comp artidas
Total Pioneras No pioneras Sin caracterizar Bosque-Rastrojo 53 9 36 8 Bosque-Plantación 30 7 18 5 Rastrojo-Plantación 27 8 16 3 Bosque-Rastrojo-Plantación 22 5 14 3

41
Tabla 7. Prueba U de Mann-Whitney para la riqueza promedio de especies de plántulas (gl=1).
Unidades de Paisaje comparadas U P
Bosque- Rastrojo 2,000 0,275
Bosque- Plantación ≤0,001 0,046*
Rastrojo-Plantación 2,000 0,268
*diferencia significativa
Figura 15. Riqueza de especies (media-error estándar) de plántulas por unidad de paisaje.
Al realizar el análisis de agrupamiento se formaron dos grupos (Figura 16).
Dentro del primero, las plantaciones 1 y 2 están más relacionadas que
cualquiera de ellas con un subgrupo conformado por el Rastrojo 3 y la

42
Plantación 3 por un lado, y el Bosque 1 y el Rastrojo 2 por el otro. En el
segundo grupo el Bosque 2 está más relacionado con el Rastrojo 1 que
cualquiera de ellos con el Bosque 3. Las unidades Bosque y Plantación nunca
se agrupan, mientras que la unidad Rastrojo se agrupa con ambas. Al observar
con más detalle la matriz de distancias que se emplea para la construcción del
dendrograma según los índices de similitud de Hamann (Tabla 8) encontramos
que en general, dentro de la unidad de paisaje Plantación las réplicas tienen
índices de similitud mucho más altos que las réplicas de la unidad de paisaje
Bosque y que Rastrojo.
Figura 16. Agrupación de las unidades de paisaje de acuerdo a la composición de especies en la comunidad de plántulas.
43
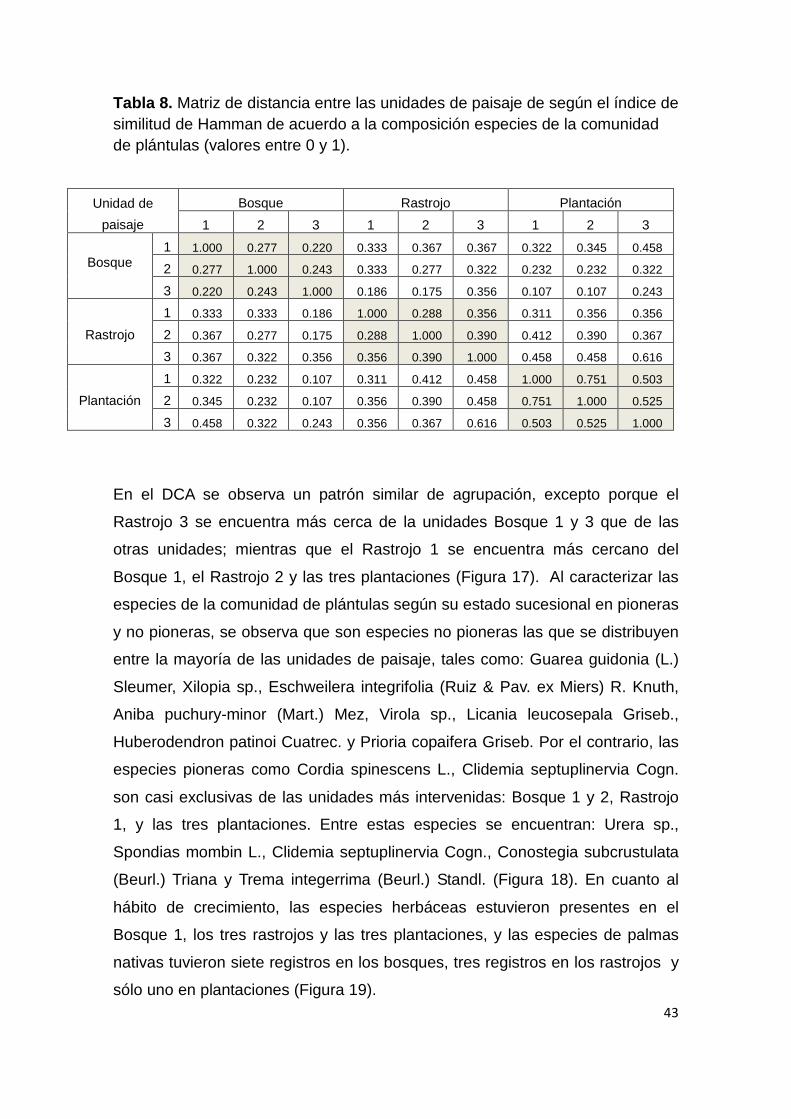
Tabla 8. Matriz de distancia entre las unidades de paisaje de según el índice de similitud de Hamman de acuerdo a la composición especies de la comunidad de plántulas (valores entre 0 y 1).
Unidad de
paisaje
Bosque Rastrojo Plantación
1 2 3 1 2 3 1 2 3
Bosque 1 1.000 0.277 0.220 0.333 0.367 0.367 0.322 0.345 0.458
2 0.277 1.000 0.243 0.333 0.277 0.322 0.232 0.232 0.322
3 0.220 0.243 1.000 0.186 0.175 0.356 0.107 0.107 0.243
Rastrojo
1 0.333 0.333 0.186 1.000 0.288 0.356 0.311 0.356 0.356
2 0.367 0.277 0.175 0.288 1.000 0.390 0.412 0.390 0.367
3 0.367 0.322 0.356 0.356 0.390 1.000 0.458 0.458 0.616
Plantación
1 0.322 0.232 0.107 0.311 0.412 0.458 1.000 0.751 0.503
2 0.345 0.232 0.107 0.356 0.390 0.458 0.751 1.000 0.525
3 0.458 0.322 0.243 0.356 0.367 0.616 0.503 0.525 1.000
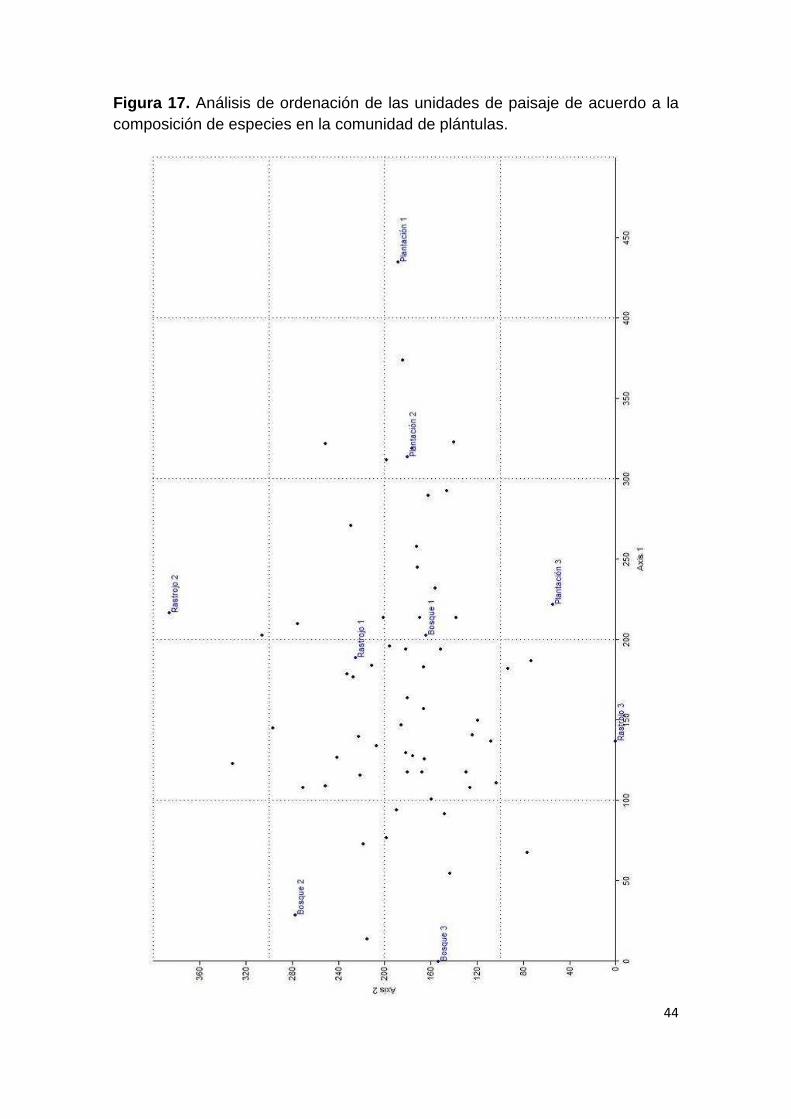
En el DCA se observa un patrón similar de agrupación, excepto porque el
Rastrojo 3 se encuentra más cerca de la unidades Bosque 1 y 3 que de las
otras unidades; mientras que el Rastrojo 1 se encuentra más cercano del
Bosque 1, el Rastrojo 2 y las tres plantaciones (Figura 17). Al caracterizar las
especies de la comunidad de plántulas según su estado sucesional en pioneras
y no pioneras, se observa que son especies no pioneras las que se distribuyen
entre la mayoría de las unidades de paisaje, tales como: Guarea guidonia (L.)
Sleumer, Xilopia sp., Eschweilera integrifolia (Ruiz & Pav. ex Miers) R. Knuth,
Aniba puchury-minor (Mart.) Mez, Virola sp., Licania leucosepala Griseb.,
Huberodendron patinoi Cuatrec. y Prioria copaifera Griseb. Por el contrario, las
especies pioneras como Cordia spinescens L., Clidemia septuplinervia Cogn.
son casi exclusivas de las unidades más intervenidas: Bosque 1 y 2, Rastrojo
1, y las tres plantaciones. Entre estas especies se encuentran: Urera sp.,
Spondias mombin L., Clidemia septuplinervia Cogn., Conostegia subcrustulata
(Beurl.) Triana y Trema integerrima (Beurl.) Standl. (Figura 18). En cuanto al
hábito de crecimiento, las especies herbáceas estuvieron presentes en el
Bosque 1, los tres rastrojos y las tres plantaciones, y las especies de palmas
nativas tuvieron siete registros en los bosques, tres registros en los rastrojos y
sólo uno en plantaciones (Figura 19).
44
Figura 17. Análisis de ordenación de las unidades de paisaje de acuerdo a la composición de especies en la comunidad de plántulas.

45
Figura 18. Distribución de especies de la comunidad de plántulas de acuerdo a su estado sucesional. La especie introducida se refiere a la palma aceitera.
Bosque 1 Bosque 3
Bosque 2
Rastrojo 1
Rastrojo 2
Rastrojo 3
Plantación 3
Plantación 2
Plantación 1

46
Figura 1. Distribución de especies de la comunidad de plántulas de acuerdo a su hábito de crecimiento. La especie introducida se refiere a la palma aceitera.
4. DISCUSIÓN
Con el presente estudio buscamos explorar el efecto del cambio en el uso del
suelo de bosques naturales a plantaciones industriales de palma aceitera sobre
el proceso de sucesión natural una vez los cultivos son abandonados. Para
esto evaluamos tres procesos críticos en la sucesión: la lluvia de semillas, la
germinación, y el establecimiento de plántulas (Aide y Cavelier, 1994; Brown y
Lugo, 1990; Capers et al, 2005; Shono et al., 2007). Encontramos que los
monocultivos de palma aceitera tienden a mostrar una menor riqueza de
especies en comparación con los bosques para la lluvia de semillas y el
Bosque 1 Bosque 3
Bosque 2 Rastrojo 1
Rastrojo 2
Rastrojo 3
Plantación 3
Plantación 2
Plantación 1

47
establecimiento de plántulas en los estados tempranos de sucesión (2 años de
abandono), por lo menos para plantaciones con una intensidad de manejo baja.
En términos de la composición de especies de la lluvia de semillas las
plantaciones son similares a algunos de los rastrojos, y las unidades de bosque
fueron similares a las unidades de rastrojo y no tanto a las plantaciones. Por
otro lado, no hubo un efecto marcado de las plantaciones sobre la composición
de especies de la comunidad de plántulas, pues no hay un patrón claro
respecto a la similitud de las unidades de paisaje entre sí. Finalmente,
encontramos que para la lluvia de semillas y la comunidad de plántulas la
variabilidad en la riqueza de especies es menor entre las réplicas de
plantaciones comparadas con las otras dos unidades de paisaje, y la similitud
en términos de la composición de especies entre las plantaciones es mayor
entre las plantaciones que entre los rastrojos y entre los bosques.
Lluvia de semillas
La riqueza de especies promedio en la lluvia de semillas fue mayor en la
unidad de paisaje Bosque comparada con la Plantación, mientras que la unidad
Rastrojo no se diferenció en su riqueza de especies de las otras dos unidades
de paisaje. Las plantaciones tienen un dosel dominado principalmente por
palma aceitera, lo cual podría representar una menor oferta de recursos para
los dispersores de semillas (Danielsen, 2008; Fitzherbert et al, 2008). Esto
podría explicar en parte por qué hay una menor riqueza de especies en la lluvia
de semillas en plantaciones comparadas con los bosques, (Aide et al, 2000;
Castillo y Stevenson, 2010; Cole et al, 2010; Fuentes, 2000; Holl et al, 2000;
Wunderlee, 1997). La riqueza en la lluvia de semillas en los rastrojos tiende a
ser menor que en los bosques (aunque las medias no son significativamente
diferentes debido a la alta variabilidad en los rastrojos) y similar a la de las
plantaciones. Es probable que en los rastrojos la atracción de dispersores sea
menor que en el bosque, pero para esto se requiere un estudio más detallado
sobre la dispersión de semillas.

48
Respecto a la composición de especies en la lluvia de semillas el efecto de las
plantaciones de palma no es tan marcado como ocurre con la riqueza de
especies, pero las unidades de Bosque nunca se agrupan directamente con las
unidades de Plantaciones en el análisis de agrupamiento. Los bosques se
agrupan con unidades de rastrojo, lo que puede deberse a que hay una réplica
de bosque (Bosque 2) con un grado mayor de intervención antrópica, y que una
unidad de plantación y otra de rastrojo (Plantación 3 y Rastrojo 3) se
encuentran en un estado sucesional más avanzado porque han sido menos
intervenidos que las demás réplicas. Específicamente, el Bosque 2 comparte
especies con todas las unidades de paisaje, la mayoría de las cuales son
especies pioneras muy comunes en estos ecosistemas. Además, cerca del
70% de las especies de semillas en las plantaciones y en los rastrojos se
encuentran también en los bosques, y corresponden principalmente a especies
pioneras, como se ha reportado en otros estudios (Fitzherbert et al, 2008;
Danielsen et al, 2008). De acuerdo a las condiciones particulares del nuestro
sitio de estudio, esta similitud en la composición por compartir un gran número
de especies pioneras puede deberse en parte al alto grado de intervención de
los bosques, y que los principales vectores de semillas de áreas abiertas, aves
y murciélagos, generalmente sólo dispersan semillas pequeñas, livianas de
especies pioneras, opuesto a las semillas grandes y pesadas de especies de
bosques maduros (Wijdeven y Kuzee, 2000). En lo que se refiere a los hábitos,
las especies de pastos fueron comunes entre las plantaciones y los rastrojos y
no entre los bosques, mientras que las especies de lianas no pioneras se
encontraron en las unidades de paisaje más conservadas.
Germinación y supervivencia
A pesar de que el experimento de germinación no arrojó suficientes datos, la
evidencia disponible no refleja un patrón claro del efecto de las plantaciones
sobre el porcentaje de germinación y supervivencia inicial de plántulas. Es
posible que haya ausencia de un efecto de las plantaciones sobre la
germinación y supervivencia de plántulas porque el manejo de las plantaciones

49
luego de la remoción del bosque no fue muy intensivo en estos ecosistemas. Si
el suelo no fue profundamente degradado en las plantaciones respecto a los
potreros abandonados es probable que no haya una barrera para la
regeneración del bosque en términos de la germinación de semillas y
supervivencia inicial de las plántulas.
Regeneración natural
Hay un efecto de las plantaciones de palma aceitera sobre la regeneración
natural (comunidad de plántulas y juveniles) en términos de la riqueza de
especies, pero no para la composición de especies. Igual que ocurrió con la
lluvia de semillas la riqueza de especies de la comunidad de plántulas sólo fue
significativamente diferente entre Bosque y Plantación. En las plantaciones la
riqueza de la comunidad de plántulas fue menor que en los bosques,
posiblemente como consecuencia de una lluvia de semillas con menos
especies. Además, la comunidad de plántulas en las plantaciones está
compuesta principalmente por especies pioneras muchas de las cuales tienen
altas tasas de mortalidad en un sotobosque con una intensidad lumínica tan
baja como la de las plantaciones (Danielsen, 2008; Fitzherbert et al, 2008).
El efecto de las plantaciones sobre la composición de especies de la
comunidad de plántulas es mucho menos evidente, porque las unidades de
paisaje no se separan claramente en el análisis de agrupamiento. La
comunidad de plántulas es muy semejante entre bosques y rastrojos, lo cual se
explica probablemente porque estos bosques han sido severamente
intervenidos y contienen pocas especies de bosque maduro; las especies no
pioneras que comparte son especies que pueden establecerse en estados
tempranos de la sucesión (Aide et al, 2000; Chazdon, 2003). En cuanto a las
plantaciones, estas no se separan marcadamente de las otras dos unidades de
paisaje, pues en su sotobosque se han establecido algunas plántulas de
especies pioneras con cierto grado de tolerancia a la sombra que también se

50
han establecido en las otras dos unidades de paisaje. La mitad de las especies
encontradas en las plantaciones estuvieron también en los bosques, pero a
diferencia de lo observado con las especies de la lluvia de semillas, en este
caso son principalmente especies no pioneras. En cuanto a los hábitos de
crecimiento, rastrojo y plantaciones compartieron especies herbáceas, y
bosques y rastrojos tienen palmas aunque de diferentes especies.
Probablemente la comunidad de plántulas de las plantaciones no se diferencia
marcadamente de las otras unidades de paisaje porque el suelo no fue
severamente degradado, y las plantaciones tienen un sotobosque sombreado
permiten el establecimiento de las plántulas (Aide et al, 2000; Chazdon, 2003).
Además de la tendencia observada para las medias, encontramos un patrón en
cuanto a la variabilidad de la riqueza de especies y los índices de similitud en la
composición en la lluvia de semillas y la comunidad de plántulas. Respecto a la
riqueza de especies de la lluvia de semillas la variabilidad fue muy baja en las
plantaciones comparadas con la variabilidad de los bosques y la variabilidad en
los rastrojos fue intermedia. En el mismo sentido, los índices de similitud
respecto a la composición de especies fueron mayores entre las réplicas de
plantaciones que entre los bosques e intermedios para los rastrojos. En la
comunidad de plántulas, hay una alta variabilidad en la riqueza de especies en
bosque y rastrojos y es muy baja en las plantaciones, y los índices de similitud
en la composición son muy bajos para bosques, intermedios para rastrojos y
altos para las plantaciones. Esto sugiere la homogenidad en la estructura y
composición de estas plantaciones (reportado por Danielsen et al, 2008;
Fitzherbert, 2008; IAvH, 2000 en plantaciones que con una intensidad de
manejo mayor) afecta procesos como la lluvia de semillas y el establecimiento
de plántulas. Esta homogenidad estructural puede tener consecuencias a largo
plazo para la sucesión natural afectando la velocidad a la que ocurre la
regeneración y conduciendo a que la composición de los bosques de estados
sucesionales más avanzados diverja de aquellos bosques naturales
característicos de ese ecosistema (Aide y Cavelier, 1994; Capers et al, 2005;
Chazdon, 2007; Cavelier et al, 1998; Guariguata, 1999; Shono et al., 2007)

51
A pesar de que se han documentado graves impactos del establecimiento de
plantaciones de palma aceitera sobre la biodiversidad, especialmente porque
se han establecido en lugares donde antes habían bosques (Danielsen et al,
2008; Fitzherbert, 2008; IAvH, 2000; Koh y Wilcove, 2008), al momento no se
ha publicado ningún estudio sobre el proceso de recuperación natural que
siguen estos sitios una vez han sido abandonados. Los resultados de este
estudio muestran que para el caso de las plantaciones de palma en la cuenca
del río Curbaradó, en donde la intensidad del uso previo fue baja, la
implementación de estos monocultivos afecta la riqueza de especies que llegan
al sitio en la lluvia de semillas y las plántulas que se establecen, pero tiene
poco efecto sobre la composición de especies en estados tempranos de la
sucesión. Este estudio también sugiere que las plantaciones podrían afectar la
homogenidad en la riqueza de especies y la similitud en la composición de
especies de la lluvia de semillas y las plántulas, lo cual podría tener efectos a
largo plazo sobre la trayectoria que sigue la sucesión del bosque, porque las
plantas establecidas pueden llegar a dominar el paisaje y excluir a otras por
competencia (Danielsen et al, 2008; Chazdon, 2003; Shono et al., 2007). Esta
investigación se constituye en el primer paso de una investigación sobre
potenciales barreras y descripción de la dinámica de la sucesión que puede
generar recomendaciones para emprender un proceso de restauración de los
bosques de la cuenca del río Curbaradó.

52
BIBLIOGRAFÍA
Aide TM, y Cavelier J. 1994. Barriers to Tropical Lowland Forest Restoration in
the Sierra Nevada de Santa Marta, Colombia. Restoration Ecology, 2:219-229.
Aide TM, Zimmerman JK, Rosario M, y Marcano H . 1996. Forest Recovery in
Abandoned Cattle Pastures Along an Elevational Gradient in Northeastern
Puerto Rico. Biotropica 28(4):537-548
AIDE, T. M., J. K. Ziemmerman, J. B. Pascarella, L. Rivera, y H. Marcano-
Vega.2000. Forest Regeneratios in a Chronosequence of Tropical Abandoned
Pastures: Implications for Restoration Ecology. Restoration Ecology. 8(4):328-
338.
Barrera E. 1985. Identificación de Plántulas de Algunas Especies Arbóreas del
Bosque de Niebla. Pérez-Arbelaezia, 1(1):39-95.
Barrera E. 1986. Identificación de Plántulas de Algunas Especies Arbóreas del
Bosque de Niebla. Pérez-Arbelaezia, 1(2):165-209.
Brown S y Lugo AE. 1990. Tropical secondary forest. Journal of Tropical
Ecology 6:1-32.
Capers R, Chazdon R, Redondo A, y Vilchez B . 2005. Succesional Dynamics
of Woody Seedling Communities in Wet Tropical Secondary Forests. Journal of
Ecology, 93:1071-1084.
Castillo LS y Stevenson PR . 2010. Relative Importance of Seed-bank and
Post-disturbance Seed Dispersal on Early Gap Regeneration in a Colombian
Amazon Forest. Biotropica, 42(4): 488–492.

53
Cavelier, J, Aide TM, Santos C, Eusse AM, y Dupuy J M. 1998. The
Savannization of Moist Forests in the Sierra Nevada de Santa Marta, Colombia.
Journal of Biogeography, 25:901-912
Chazdon RL. 2003. Tropical forest recovery: legacies of human impact and
natural disturbances. Perspectives in Plant Ecology, Evolution and Systematics,
6:51-71.
Codechocó. 2002. Plan de Ordenamiento Territorial de las Tierras Colectivas
de las Comunidades Negras del Bajo Atrato: Municipios De Riosucio, Carmen
Del Darien y Belén De Bajirá. Ríosucio, Chocó.
Cole RJ, Holl KD y Zahawi RA . 2010. Seed rain under tree islands planted to
restore degraded lands in a tropical agricultural landscape. Ecological
Applications, 20(5): 1255–1269
Cooke SJ y Suski CD. 2008. Ecological Restoration and Physiology: An
Overdue Integration. BioScience, 58(10):957.
Cubiña A y Aide TM. 2001. The Effect of Distance from Forest Edge on Seed
Rain and Soil Seed Bank in a Tropical Pasture. Biotropica, 33(2): 260–267.
Danielsen F, Beukema H, Burgess ND, Parish F, Brühl CA, Donald PF,
Murdiyarso D, Phalan B, Reijnders L, Struebig M y F itzherbert EB. 2008.
Biofuel Plantations on Forested Lands: Double Jeopardy for Biodiversity and
Climate. Conservation Biology, 23(2):348–358.
Defensoría del Pueblo. 2005. Violación de los Derechos Humanos por
Siembra de Palma Africana en Territorios Colectivos de Jiguamiandó y
Curbaradó-Chocó. Resolución defensorial No. 39. Bogotá, Colombia

54
Del Amo S. 1979. Clave para Plántulas y Estados Juveniles de Especies
Primarias de una Selva Alta Perennifolia en Veracruz, México. Biótica, 4(2):59-
108.
Dodds WK, Wilson KC, Rehmeier RL, Knight GL, Wiggam S, Falke JA,
Dalgleish HJ y Bertrand KN. 2008. Comparing Ecosystem Goods and
Services Provided by Restored and Native Lands. BioScience, 58(9), p.837.
FAO- Food and Agriculture Organization of the Unite d Nations. 2006.
Evaluación de los Recursos Forestales Mundiales 2005.
Feisinger P . 2001. Designing field studies for biodiversity conservation. The
Nature Conservancy & Island Press. Washington DC
Finegan B. 1996. Pattern and process in neotropical secondary rain forests: the
first 100 years of succession. Trends in Ecology & Evolution, 11(3):119-124.
Fischer J y Lindenmayer DB. 2007. Landscape modification and habitat
fragmentation: a synthesis. Global Ecology and Biogeography, 16:265–280.
Fitzherbert E, Struebig MJ, Morel A, Danielsen F, B rühl C, Donald P y
Phalan B . 2008. ¿How will oil palm expansion affect biodiversity? Trends in
Ecology and Evolution, 23(10).
Fuentes M . 2000. Frugivory, Seed Dispersal and plant Community Ecology.
Trends in Ecology and Evolution, 15(12):487-488.
Guariguata MR. 1999. Bases Ecológicas Generales para el Seguimiento de
proyectos de Restauración de Bosques. Memorias del Seminario de
Restauración Ecológica y Reforestación. Colombia. pág 83-95.

55
Guariguata MR y Ostertag R. 2001. Neotropical secondary forest succession:
changes in structural and functional characteristics. Forest Ecology and
Management, 148(1-3):185-206.
Guevara S, Purata SE y Van der Maarel E . 1986. The role of remnant forest
trees in tropical secondary succession. Vegetatio, 66: 77-84.
Hammer O, Harper DAT y Ryan PD. 2008. PAST: Paleontological statistics.
version 1.77.
Holdridge LR, Grenke WC, Hatheway WH, Liany T y Tos i JA. 1971. Forest
Environments in Tropical Lifezones. Pergamon Press, New York, New York,
USA.
Holl KD . 1999. Factors Limiting Tropical Rain Forest Regeneration in
Abandoned Pasture: Seed Rain , Seed Germination , Microclimate , and Soil.
Biotropica, 31(2):229-242.
Holl KD, Loik ME, Lin E y Samuels IA . 2000. Tropical Montane Forest
Restoration in Costa Rica: Overcoming Barriers to Dispersal and Establishment.
Restoration Ecology, 8(4):339-349.
Holt E . 2007. Biofuels: Myths of the Agro-fuel Transition. Backgrounder 13(2).
Publicación del Institute for Food and Development Policy-Food First.
Disponible en <www.foodfirst.org>. Fecha de consulta: 19 de febrero de 2009.
IAvH-Instituto de Investigacion de Recursos Biológi cos Alexander von
Humboldt . 2000. Biosíntesis, boletín N° 21. Bogotá, Colombi a.
IIAP, Instituto de Investigaciones Ambientales del Pacífico. 2008.
Evaluación Interdimensional de los Daños Ambientales Ocasionados por el
Cultivo de la Palma Aceitera y la Ganadería en Jiguamiandó y Curvaradó,
Chocó-Colombia. Sin publicar.

56
Koh, LP. y Wilcove, DS . 2008. Is oil palm agriculture really destroying tropical
biodiversity? Conservation. Letters. 1(2): 60–64.
Lobova TA, Geiselman CK, Mori SA. 2009. Seed dispersal by bats in the
Neotropics. Memoirs of the New York Botanical Garden, Vol 101.
Mingorance F, Minelli F y Le Du H. 2004. “El Cultivo de la Palma Africana en
el Chocó – Legalidad Ambiental, Territorial y Derechos Humanos”. Human
Rights Everywhere, Diócesis de Quibdó. Bogotá, Colombia.
Olea-Wagner A, Lorenzo C, Naranjo E, Ortiz D y León -Paniagua L. 2007.
Diversidad de frutos que consumen tres especies de murciélagos (Chiroptera:
Phyllostomidae) en la selva lacandona, Chiapas, México. Revista Mexicana de
Biodiversidad, 78:191-200
Saldarriaga JC, West DC, Tharp ML & Uhl C. 1988. Longterm
chronosequence of forest succession in the upper Rio Negro of Colombia and
Venezuela. Journal of Ecology, 76: 939–958.
Shono K, Cadaweng E, y Durst P. 2007. Application of Assisted Natural
Regeneration to Restore Degraded Tropical Forestlands. Restoration Ecology,
15(4):620–626.
SPSS, 2008. SPSS, Statistical Package for the Social Sciences, version 17.0.
Widjeven SM, y Kuzee M. 2000. Seed Availability as a Limiting Factor in
Forest Recovery Processes in Costa Rica. Restoration Ecology 8(4):414-424.
White, R. et al., 2009. Developing an integrated conceptual framework to
understand biodiversity conflicts. Land Use Policy, 26(2), pp.242-253.

57
Wunderle, JM. 1997. The Role of Animal Seed Dispersal in Accelerating Native
forest Regeneration on Degraded Tropical Lands. Forest Ecology and
Management, 99:223-225.