identificaciÓn de macroinvertebrados fÓsiles de...
TRANSCRIPT

Edificio Escuelas Facultad de Ciencias · Campus Isla Teja· Valdivia · Chile
Casilla 567 · Fono-Fax: 56 63 221581 · email:[email protected] · www.uach.cl
Profesor Patrocinante:
Dr. Sven N. Nielsen
Instituto de Ciencias Ambientales y Evolutivas
Facultad de Ciencias – Universidad Austral de Chile
IDENTIFICACIÓN DE MACROINVERTEBRADOS FÓSILES
DE LA CUESTA SANTO DOMINGO, FORMACIÓN SANTO
DOMINGO, MIOCENO INFERIOR, CHILE.
Tesis de Grado presentada como parte
de los requisitos para optar al grado de
Licenciado en Biología Marina y
Título Profesional de Biólogo Marino.
FRANCISCA BELÉN FLORES NASS
VALDIVIA - CHILE
2014

I
ÍNDICE DE FIGURAS ......................................................................................................... II
ÍNDICE DE TABLAS .......................................................................................................... III
ÍNDICE DE LÁMINAS ....................................................................................................... IV
RESUMEN ............................................................................................................................. 1
SUMMARY ........................................................................................................................... 2
1-. INTRODUCCIÓN ............................................................................................................ 3
1.1-. MIOCENO MARINO DEL CENTRO Y SUR DE CHILE ....................................... 3
1.2-. FORMACIÓN SANTO DOMINGO ......................................................................... 5
1.2.1-. Sedimentología .................................................................................................... 6
1.2.2-. Microfauna ........................................................................................................... 7
1.2.3-. Macrofauna .......................................................................................................... 8
1.2.4-. Icnofósiles ............................................................................................................ 8
1.2.5-. Paleoflora ............................................................................................................. 9
1.3-. HIPÓTESIS ................................................................................................................ 9
1.4-. OBJETIVO ................................................................................................................. 9
1.4.1-. OBJETIVOS ESPECÍFICOS ............................................................................ 10
2-. MATERIALES Y MÉTODOS ....................................................................................... 10
2.1-. ÁREA DE MUESTREO .......................................................................................... 10
2.2-. METODOLOGÍA ..................................................................................................... 10
3.- RESULTADOS ............................................................................................................... 11
3.1.- SISTEMÁTICA PALEONTOLÓGICA .................................................................. 11
4-. DISCUSIÓN ................................................................................................................... 24
5-. CONCLUSIONES .......................................................................................................... 27
6-. BIBLIOGRAFIA ............................................................................................................. 28
7-. ANEXOS ......................................................................................................................... 34
7.1-. FIGURAS ................................................................................................................. 34
7.2-. TABLAS ................................................................................................................... 38
7.3-. LÁMINAS ................................................................................................................ 43

II
ÍNDICE DE FIGURAS
Figura 1. Columna estratigráfica definida por Pino y Beltrán (1979) en un área cercana al
sitio del presente estudio dentro de la Cuesta Santo Domingo. En azul, se muestra la
posición los niveles desde los cuales se extrajo sedimento para su análisis......................... 34
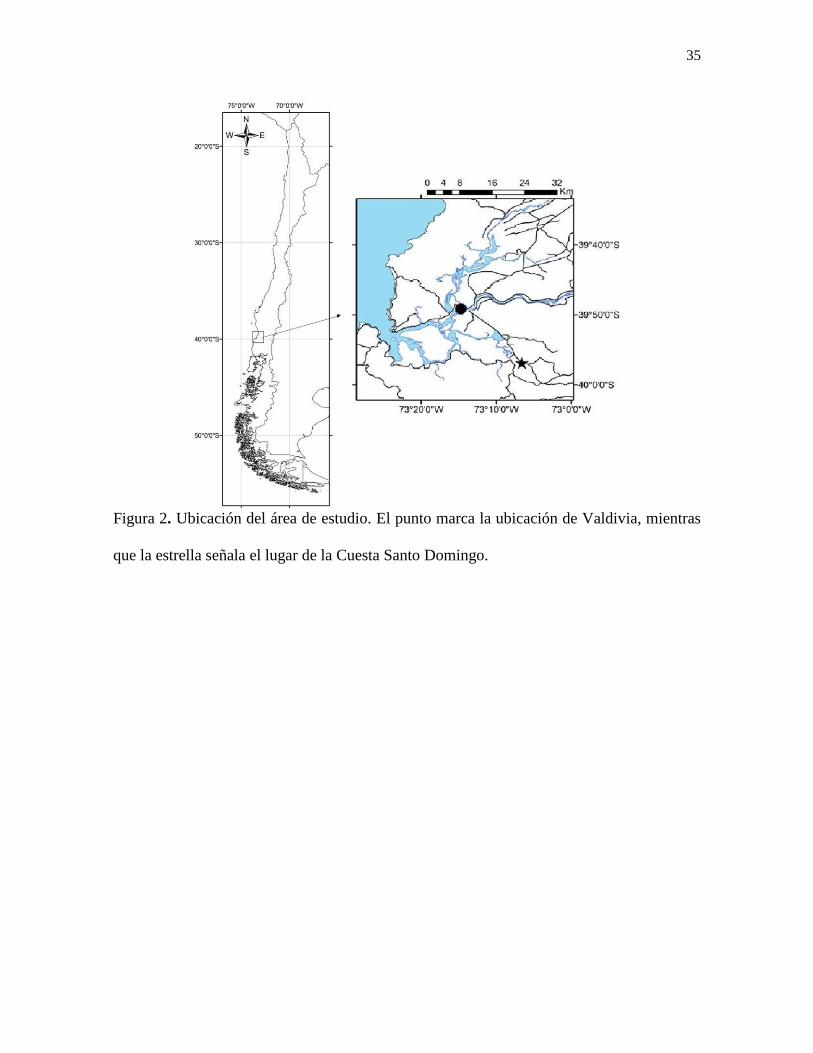
Figura 2. Ubicación del área de estudio. El punto marca la ubicación de Valdivia, mientras
que la estrella señala el lugar de la Cuesta Santo Domingo. ................................................ 35
Figura 3. Posición de los 6 niveles de muestro en la columna de estudio. ........................... 36
Figura 4. Período Neógeno dividido en época y edades....................................................... 37

III
ÍNDICE DE TABLAS
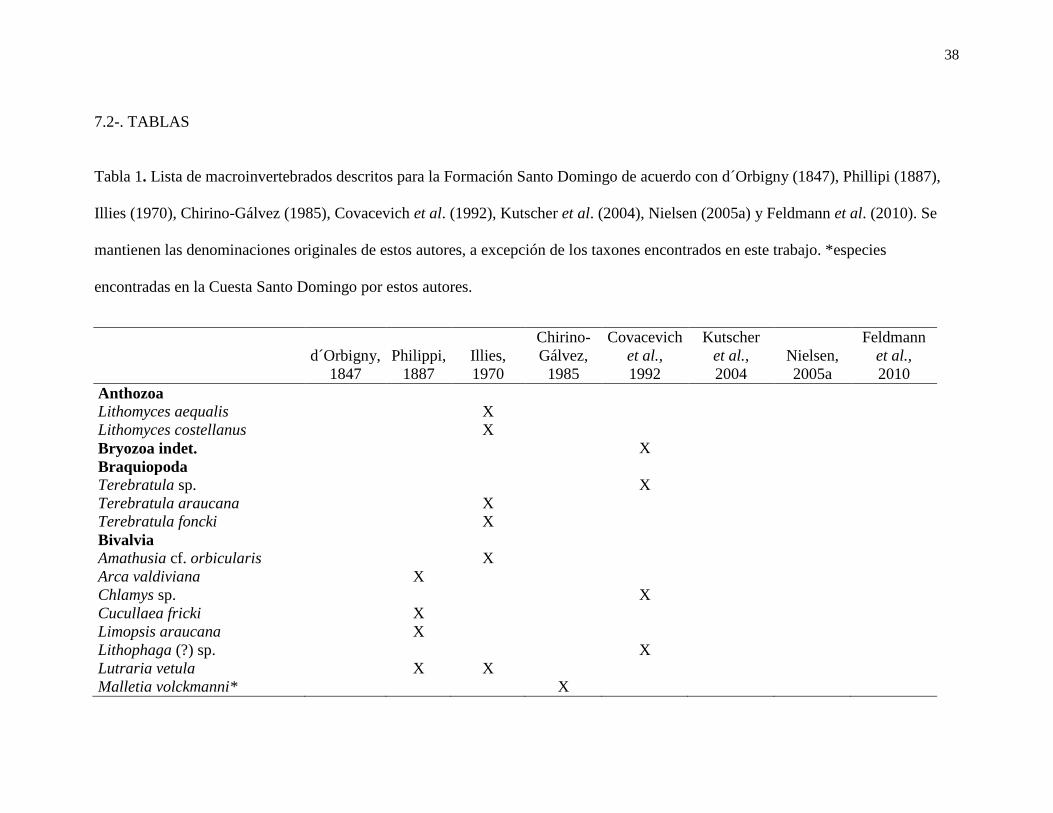
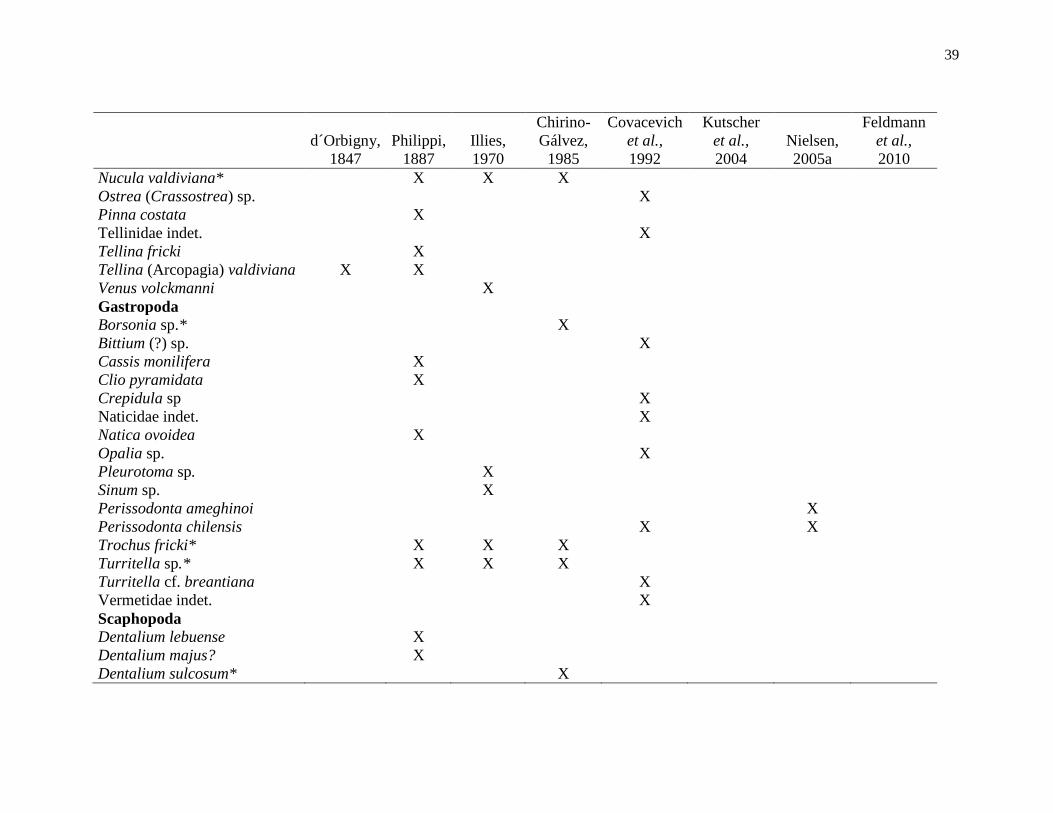
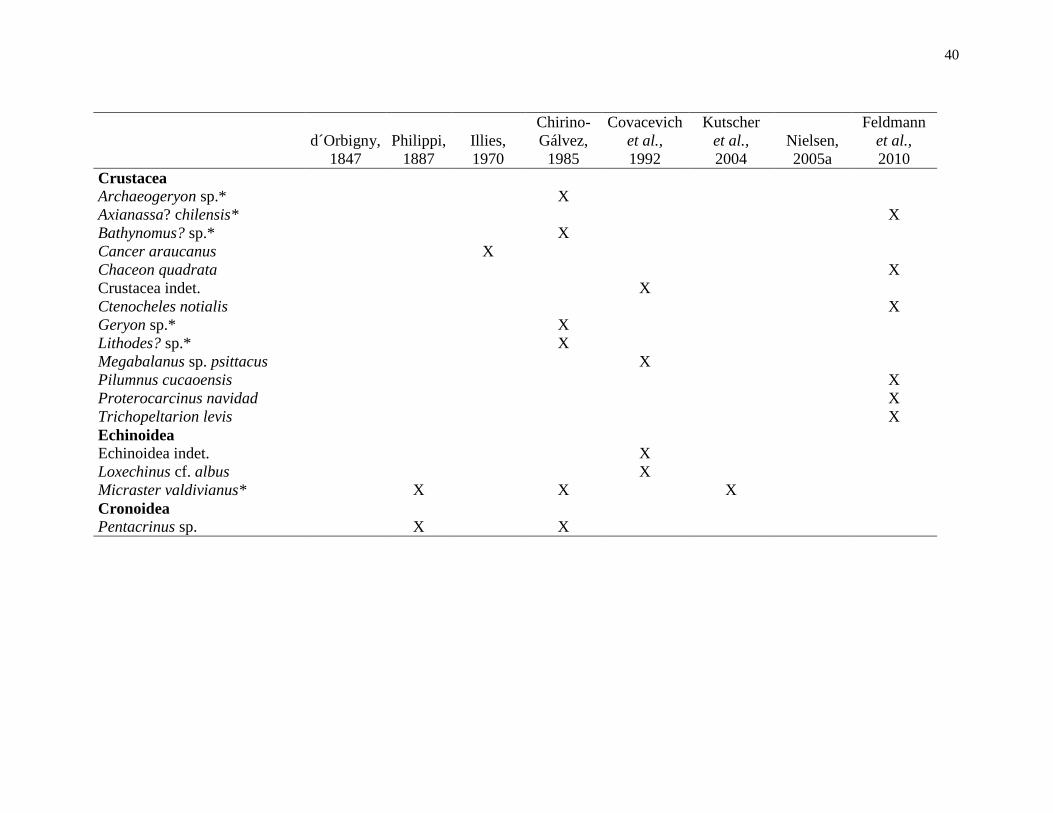
Tabla 1. Lista de macroinvertebrados descritos para la Formación Santo Domingo de
acuerdo con d´Orbigny (1847), Phillipi (1887), Illies (1970), Chirino-Gálvez (1985),
Covacevich et al. (1992), Kutscher et al. (2004), Nielsen (2005a) y Feldmann et al. (2010).
Se mantienen las denominaciones originales de estos autores, a excepción de los taxones
encontrados en este trabajo. *especies encontradas en la Cuesta Santo Domingo............... 38
Tabla 2. Lista de las especies identificadas en la Cuesta Santo Domingo que también se
encuentran en otras formaciones miocénicas: Navidad, Ranquil y Lacui. Tener en cuenta las
siguientes especificaciones: X₁, Perissodonta chilensis; X₂, Exilia alanbeui; X₃, géneros
de la familia Olividae; X₄, géneros de familia Turridae; X₅, genero Limopsis; X₆, Nucula
sp.; X₇, especies del orden Brachyura. ................................................................................. 41
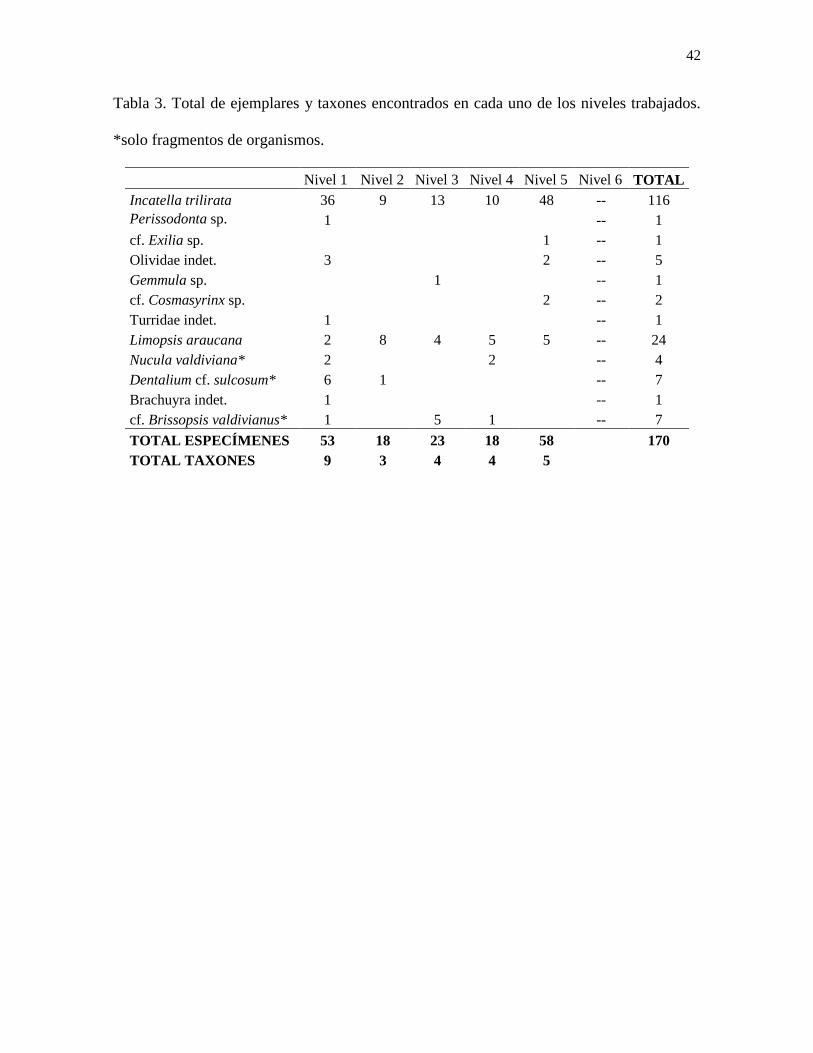
Tabla 3. Total de ejemplares encontrados en cada uno de los niveles trabajados. *solo
fragmentos de organismos. ................................................................................................... 42

IV
ÍNDICE DE LÁMINAS
Lámina 1. A-F, Gastrópodos fósiles encontrados en la Cuesta Santo Domingo. A, vista
dorsal de cf. Exilia sp. B, vista vental de cf. Exilia sp. C, vista vental Gemmula sp. D-F,
Incatella trilirata. La barra representa 5 mm de largo. ........................................................ 43
Lámina 2. A-E, Gastrópodos fósiles encontrados en la Cuesta Santo Domingo. A, vista
ventral Turridae indet. B-C, vistas ventrales y dorsales de dos ejemplares de Olividae indet.
D-E, vistas ventrales de los dos ejemplares de cf. Cosmasyrinx sp. La barra representa
5 mm de largo. ...................................................................................................................... 44
Lámina 3. A-E, Bivalvos fósiles encontrados en la Cuesta Santo Domingo. A, vista externa
e interna de valva izquierda de Limopsis araucana. B, vista interna de valva incompleta de
Limopsis araucana. D, valva parcial de Nucula valdiviana. La barra representa 5 mm de
largo. ..................................................................................................................................... 45
Lámina 4. A-F, Fósiles encontrados en la Cuesta Santo Domingo. A, vista de Perissodonta
sp. B, C, D, fragmentos de concha de Dentalium cf. sulcosum. E, ambulacros petaloídeos
de cf. Brissopsis valdivianus. F, fragmento de dáctilo de Brachyura indeterminada. Las
barras representan 1 mm de largo. ........................................................................................ 46

1
RESUMEN
La Formación Santo Domingo (Mioceno inferior) aflora en diferentes localidades
desde el área de Temuco (~38°S) hasta la zona norte de la Isla de Chiloé (~42°S),
presentando sucesiones compuestas principalmente por areniscas, limolitas y limolitas
arenosas con abundante material invertebrado fósil, siendo Gastropoda, Bivalvia, Crustacea
y Echinoidea los taxones más comúnmente encontrados en estos sedimentos. El presente
trabajo tiene como objetivo principal la identificación de marcoinvertebrados desde la
Cuesta Santo Domingo (39°56’S-73°6’O), ubicada a unos 20 km al sureste de la ciudad de
Valdivia. Se extrajo sedimento de una sección de 7 m de potencia, adyacente a la carretera
que une las ciudades de Valdivia y Paillaco, la cual fue dividida en seis niveles de muestreo.
Los sedimentos extraídos fueron disgregados de manera manual y posteriormente
tamizados con un tamiz de 2 mm de abertura, colectándose así un total de 170 ejemplares
de macroinvertebrados. Se identificaron cinco familias de gastrópodos: Turritellidae,
Struthiolariidae, Ptychatractidae, Olividae y Turridae; dos familias de bivalvos: Nuculidae
y Limopsidae; una familia de scaphopoda: Dentaliidae; una familia de echinoidea:
Brissidae; y el infraorden Brachuyra perteneciente a los decápodos. Las especies más
abundantes fueron Incatella trilirata y Limopsis araucana, ambas encontradas en todos los
niveles de muestreo.

2
SUMMARY
The Santo Domingo Formation (lower Miocene) crops out in many localities from
the Temuco area (~38°S) to the north of Chiloe Island (~42°S), presenting sequences
composed mainly of sandstones, siltstones and sandy siltstones with abundant invertebrate
fossil, being Gastropoda, Bivalvia, Echinoidea and Crustacea the taxa most commonly
found in these sediments. The present paper has as main objective the identification of
macroinvertebrates from Cuesta Santo Domingo (39°56’S-73°6’W), located 20 km
southeast of Valdivia. Sediments was extracted from a 7 m high section right next to the
road that link the cities of Valdivia y Paillaco, which was divided into six sampling levels.
The extracted sediments were dispersed by hand and then sieved with a 2 mm width sieve,
collecting a total of 170 fossil invertebrate. Five gastropod families: Turritellidae,
Struthiolariidae, Ptychatractidae, Olividae and Turridae; two bivalve families: Nuculidae
and Limopsidae; one scaphopoda family: Dentaliidae; one echinoidea family: Brissidae;
along with decapod infraorder Brachuyra were identified from the samplings collected. The
most abundant species were Incatella trilirata and Limopsis araucana, both found in all the
sampling levels.

3
1-. INTRODUCCIÓN
1.1-. MIOCENO MARINO DEL CENTRO Y SUR DE CHILE
A lo largo del territorio chileno, es posible encontrar numerosas localidades donde
se presentan estratos marinos del Neógeno (Cecioni, 1980; Encinas et al., 2008; Feldmann
et al., 2010; Encinas et al, 2012), los cuales fueron estudiados primeramente por Darwin
(1846), quien describió por primera vez la Formación Navidad, la cual es ahora la unidad
de referencia para el Neógeno marino en Chile (Martínez-Pardo, 1990; Encinas et al., 2008;
Feldmann et al., 2010; Encinas et al., 2012).
El origen de estos sedimentos marinos es explicado por Martinez-Pardo (1990),
quien reconoce dos eventos de transgresión marina en Chile y Perú denominados Neogene
South East Pacific Sequence I y II. El evento transgresivo NSEPS-I se extendió desde el
término del Mioceno temprano hasta la base del Mioceno tardío (19 - 10 Ma) con una
transgresión máxima en los 14 Ma; mientras que el NSEPS-II se situó dentro del Plioceno
tardío (3.5 - 1.6 Ma). La “Transgresión Navidad” corresponde en Chile al primero de esos
ciclos de transgresión marina, durante el cual se depositó la Formación Navidad en la zona
costera entre Valparaíso (~33°00´S) y Punta Topocalma (~34°30´S). Martínez-Pardo (1990)
se refiere a todos los depósitos marinos del Mioceno como Formación Navidad, incluyendo
bajo esa misma distinción a las Formaciones Ranquil en la Península de Arauco y Lacui en
el área de Chiloé (Finger, 2013), sin embargo, en el presente escrito se mantendrá la
distinción para diferenciar las unidades regionales.
La edad de la Formación Navidad y sus unidades correlativas Ranquil y Lacui, han
sido un foco de discusión por muchos años. Originalmente, Tavera (1979) asignó a la
Formación Navidad una edad Miocena inferior, en base a correlaciones con moluscos de la

4
Patagonia Argentina. La Formación Ranquil ha sido asignada al Mioceno en base a
especies de moluscos y foraminíferos, mientras que Lacui pertenecería al Mioceno medio
superior de acuerdo a análisis con foraminíferos (Encinas et al., 2008). Finger et al., (2007)
afirman que estas tres unidades geológicas de Chile central y sur son depósitos batiales del
Mioceno superior al Plioceno inferior, basándose en estudios con foraminíferos y
ostrácodos. Nielsen y Glodny (2009) midieron las razones de isótopos de estroncio en
diferentes taxones de gastrópodos, bivalvos y un cefalópodo provenientes nuevamente de
Navidad, Ranquil y Lacui, llegando a concluir que todas ellas tienen una edad Oligocena
tardía-Miocena temprana, mientras que Finger (2013) afirma que la edad debe ser acotada
solo al Mioceno temprano, de acuerdo a su estudio con foraminíferos planctónicos de las
tres formaciones geológicas.
Los estratos marinos del Neógeno son bastante frecuentes en las cercanías de la
ciudad de Valdivia (Chirino-Gálvez, 1985), aflorando en ambos flancos de la Cordillera de
la Costa (Covacevich et al., 1992) y con potencias variables entre 200 y 270 metros (Illies,
1960; Pino, 1990). En primera instancia, el origen de estos yacimientos sería el mismo que
originó las Formaciones de Navidad, Ranquil y Lacui. Martínez-Pardo y Pino (1979)
indican que los estratos de los alrededores de Valdivia son el resultado del proceso de
transgresión marina dentro del Cenozoico tardío, partiendo en el Mioceno medio (Chirino-
Gálvez, 1985; Pino, 1990) hasta la parte inferior del Mioceno tardío (Elgueta, 1990). Sin
embargo, los estudios de Elgueta et al. (2000) son mucho más precisos al momento de
hablar de las transgresiones marinas en la provincia, señalando que éstas se iniciaron en el
límite del Oligoceno tardío-Mioceno temprano, terminando en el Serravalliano tardío (15
Ma).

5
1.2-. FORMACIÓN SANTO DOMINGO
El primer registro del Terciario marino para la Región de Los Ríos es el entregado
por Philippi (1887), quien describió un afloramiento a 4 km al SE de Valdivia en la
localidad de Llancahue, encontrando estratos similares en Corral, Isla del Rey, Catamutún y
San Juan de la Costa (Mai, 1982), siendo Brüggen (1950) quien erróneamente nombró estos
estratos como parte de la Formación Navidad (Encinas et al., 2012).
La Formación Santo Domingo fue definida por Martínez-Pardo y Pino (1979),
caracterizándola en términos litológicos, micropalentológicos y sedimentológicos de
acuerdo a muestras obtenidas de la cuesta del mismo nombre, situada a 19 km al SE de la
ciudad de Valdivia. En cortes adyacentes a la carretera que une las ciudades de Valdivia y
Paillaco, se pudo observar dos unidades fácilmente reconocibles: un Basamento
Metamórfico denominado Formación Piedra Laja (Illies, 1970) y rocas sedimentarias
designadas en esa oportunidad y por primera vez como Formación Santo Domingo.
Los afloramientos de la formación se presentan en la zona costera, Cordillera de la
Costa y la Depresión Intermedia, comenzando desde el borde este del Valle Central en el
área de Temuco (~38°S) hasta el norte de la Isla de Chiloé (~42°S). Dentro de esta misma
zona existen distintas secciones de depósitos marinos con distintos nombres locales, como
la Formación Cholchol, cercana a Temuco, o la Formación Huilma en el área Valdivia-
Osorno. Todas ellas son ahora referidas como Formación Santo Domingo debido a sus
similitudes en cuanto a sus facies sedimentarias y contenido de microfósiles (Encinas et al.,
2012).

6
Originalmente, 1os estudios con foraminíferos de Martínez-Pardo y Pino (1979)
asignaron una edad Miocena media a la base de la formación, específicamente entre las
zonas N13 y N15 (Mai, 1982; Pino, 1990; Elgueta et al. 2000; Encinas et al. 2008; Fig. 4),
llegando a un total de 8 especies planctónicas descritas con un cierto grado de
incertidumbre al usar el indicador “cf.” al momento de identificarlas. Dentro de las especies
descritas por Martínez-Pardo y Pino (1979) se encuentran Globigerinoides cf. trilobus y
Globorotalia cf. continuosa las que tienen un rango geológico que va desde la zona N9
hasta N16 (Langhiano hasta Tortoniano; Mioceno medio a tardío). En el área de Catamutún,
Marchant (1990) registró la especie planctónica Globigerina pachyderma, dentro de
estratos marinos estratigráficamente equivalentes con la Formación Santo Domingo
(Encinas et. al., 2008), la cual tiene una primera aparición en la zona N16 (Tortoniano).
1.2.1-. Sedimentología
Según Martínez-Pardo y Pino (1979), Santo Domingo aflora entre los 30 y 140
metros sobre el nivel del mar, aunque Encinas et al. (2008) afirman que no se puede
obtener un rango estimado, debido a que no existe un lugar donde la sección pueda ser
medida completamente. Martínez-Pardo y Pino (1979), propusieron también la subdivisión
de la formación en dos miembros: uno inferior (74 m) caracterizado por la presencia de
limolitas, baja cementación y abundante presencia de caparazones pertenecientes a
organismos marinos, y un miembro superior (36 m) compuesto de grano más grueso pero
fácilmente erosionable con baja abundancia de especies fósiles.
En su estudio de la misma Cuesta Santo Domingo, Pino y Beltrán (1979)
identificaron areniscas y limolitas con bajo porcentaje de arenas (3% y 8%) para los
sedimentos de la parte inferior de la columna; mientras que para las capas superiores se

7
observaron areniscas con 28% de limo y 1% de arcillas, junto con limolitas arenosas con un
porcentaje de 43% de arena y limolitas con 29% de arena y 2% de arcilla. Si bien esta
sucesión es la más representativa y mejor estudiada, ella no incluye todas las litologías que
componen la formación, que presenta importantes cambios de facies laterales y verticales
(Elgueta et al., 2000; Fig. 1).
Cabe destacar que la Cuesta Santo Domingo se ha constituido en el área tipo del
estudio de depósitos terciarios cercanos a la ciudad de Valdivia (Covacevich et al., 1992),
lugar donde la formación alcanza los 110 m de espesor. Los sedimentos consisten en una
secuencia subhorizontal, de areniscas de grano fino a muy fino, limolitas y arcillolitas,
cuarzo-líticas, muy micáceas, con horizontes fosilíferos, de color gris claro (Elgueta et al.,
2000).
1.2.2-. Microfauna
Los microfósiles son particularmente abundantes, encontrándose aproximadamente
4000 individuos por 100 gr de sedimento (Martínez-Pardo y Pino, 1979; Chirino-Gálvez,
1985). Hasta el momento se han descrito aproximadamente 70 especies de foraminíferos
bentónicos y planctónicos para la formación (Martinez-Pardo y Pino, 1979; Chirino-Gálvez,
1985; Marchant, 1990; Encinas et al., 2008; Encinas et al., 2012; Finger, 2013), siendo las
formas bentónicas las que permiten determinar antecedentes de paleobatimetría, gracias a
su gran abundancia y diversidad en los mares actuales. Debido a que la mayoría del
desplazamiento del bentos es gracias a la acción de la gravedad, se considera que el límite
superior de distribución espacial para las especies foraminíferas es la profundidad mínima
de depositación (Encinas et al., 2012).

8
Según Encinas et al. (2008), Encinas et al. (2012) y Finger (2013), existen
asociaciones de foraminíferos bentónicos con distintas distribuciones espaciales, todas ellas
dentro de la zona batial, confirmándose así el desplazamiento descendente del bentos.
Con las características paleoecológicas de las especies Sphaeroidina bulliodes,
Gyroidinoides soldanii, Melonis soldanii, Bulimina mexicana y Pullenia bulloides,
Martínez-Pardo y Pino (1979) concluyen que la base de la Formación fue parte de una
biocenosis marina predominantemente bentónica desarrollada entre los 200 y 500 m, donde
las temperaturas de las aguas en el fondo eran de 10°C, mientras que en la superficie
llegaban hasta los 18°C.
1.2.3-. Macrofauna
Se conocen alrededor de 30 especies de macrofósiles en toda la Formación (Elgueta
et al., 2000), siendo d´Orbigny (1847) quien entregó el primer registro de
macroinvertebrados al describir el bivalvo Arcopagia valdiviana (Covacevich et al., 1992).
Sin embargo, fueron Philippi (1887), Illies (1970), Chirino-Gálvez (1985), Covacevich et
al., (1992), Kutscher et al., (2004), Nielsen (2005) y Feldmann et al., (2010) quienes
describieron escafópodos, nautiloídeos, corales, braquiópodos, crinoideos, crustáceos
equinodermos, bivalvos y gastrópodos, siendo las más comunes estos últimos cuatro
taxones (Tabla 1).
1.2.4-. Icnofósiles
La presencia del icnotaxón Chondrites isp., sumado a la baja icnodiversidad sugiere
que las aguas del fondo marino estaban pobremente oxigenadas (Encinas et al., 2008).
Sumado a esto, la abundancia de minerales lábiles sugiere una depositación con un mínimo

9
de transporte (Martinez-Pardo y Pino, 1979). Ambas afirmaciones concuerdan con Pino y
Beltrán (1979), quienes hablan de un ambiente reductor gracias a la alta cantidad de materia
orgánica y bajas concentraciones de oxígeno, debido muy probablemente a la ausencia de
corrientes (Chirino-Gálvez, 1985).
1.2.5-. Paleoflora
Existen escasos registros de improntas de flora fósil dentro de los estratos de la
Formación, siendo el más importante la descripción de la especie Lapageria rosea dentro
de la Cuesta Santo Domingo, concluyéndose que el clima en el momento de la depositación
era templado húmedo con una alta pluviosidad (Pino y Núñez, 1979). La existencia también
de improntas de Nothofagus obliqua y N. procera, nos hablan de una cercanía relativa con
el continente (Pino y Beltrán, 1979), así como también de un aporte de energía al
ecosistema para organismos depredadores (Chirino-Gálvez, 1985).
1.3-. HIPÓTESIS
Si el ambiente al momento de la depositación presentaba una baja energía cinética
con un mínimo de transporte, además de un déficit de oxígeno, entonces la macrofauna
debería estar representada en su mayoría por organismos detritívoros y depredadores.
1.4-. OBJETIVO
El objetivo del presente trabajo es identificar macroinvertebrados fósiles de la
Formación Santo Domingo, en la zona de la cuesta del mismo nombre, para así conocer las
características taxonómicas del ensamble faunístico.

10
1.4.1-. OBJETIVOS ESPECÍFICOS
Describir la sistemática paleontológica de macroinvertebrados presentes en la
Cuesta Santo Domingo.
2-. MATERIALES Y MÉTODOS
2.1-. ÁREA DE MUESTREO
La Cuesta Santo Domingo (39°56’S-73°6’O) se ubica entre las ciudades de Valdivia
y Paillaco, a unos 19 km al SE de la primera localidad (Pino y Nuñez, 1979). Comienza a
unos 300 metros al SE de la vía que lleva a la localidad de Corral, continuando por
alrededor de 2 km (Encinas et al., 2008; Fig. 2).
Las muestras fueron colectadas de una columna adyacente a la carretera que une
ambas ciudades (39°56’55’’S-73°6’35’’O), tomando en cuenta las capas portadoras de
fósiles identificadas con anterioridad por Pino y Beltrán (1979; ver Fig. 1). La columna
estudio se dividió en 6 niveles de muestreo, obteniendo de cada una alrededor de 3 kg de
sedimento. El nivel N°1 se encuentra a la misma altura que la carretera, mientras que el
nivel N°5 se posiciona a unos 6 m por sobre la misma. El nivel N°6 se encuentra sobre la
misma vertical pero desplazada en 4 m en dirección Este, debido a una pequeña planicie.
Este último nivel alcanza una altura de 7 m en relación a la carretera (Fig. 3).
2.2-. METODOLOGÍA
Posteriormente a su traslado al laboratorio, todas las muestras fueron hidratadas en
agua durante un tiempo aproximado de 14 horas, preparándolas así para su posterior
disgregación manual. Sólo se colectaron ejemplares de macroinvertebrados completos o

11
casi completos que poseyeran caracteres taxonómicos. Luego de la disgregación manual, se
procedió a un tamizado con tamiz de 2 mm para así obtener cualquier ejemplar no
colectado en el procedimiento anterior.
3.- RESULTADOS
3.1.- SISTEMÁTICA PALEONTOLÓGICA
Filo Mollusca Linnaeus, 1758
Clase Gastropoda Cuvier 1797
Subclase Caenogastropoda Cox, 1959
Orden Cerithiimorpha Golikov y Starobogatov, 1975
Familia Turritellidae Lovén, 1847
Género Incatella DeVries, 2007
Incatella trilirata (Philippi, 1887)
(Lámina 1, D-F)
Turritella trilirata Philippi, 1887, p. 71, pl. 9, fig. 8.
Turritella parvula Philippi, 1887, p. 73, pl. 57, fig. 4.
Incatella trilirata DeVries, 2007, p. 337, fig. 3.18, 3.19.
Diagnosis: Concha cónica alargada. Tres cordones espirales finamente nudosos.
Descripción: Concha turriforme con vueltas poco convexas, donde cada una posee tres
cordones espirales primarios nudosos continuos, presentando también cordones secundarios
delgados perpendiculares a la escultura axial. Suturas poco profundas. Abertura ovalada
cóncava. Labio exterior delgado y liso.

12
Rango geológico: Mioceno temprano (DeVries, 2007).
Observaciones: Covacevich et al. (1992) identificaron un solo ejemplar de la especie
Incatella cf. leptogramma (Philippi, 1887; =Turritella breantiana) en los estratos del Río
Futa, sin embargo, en la descripción de la misma afirman que en las vueltas se reconocen
tres cordones espirales finamente nudosos, con algunos hilos espirales intercalados, lo que
podría indicar que se trataría de la misma especie encontrada en esta oportunidad.
I. leptogramma difiere de I. trilirata al presentar un mayor número de cordones espirales
secundarios e hilos terciarios (DeVries, 2007).
Phillipi (1887) e Illies (1970) dieron cuenta del género Incatella dentro de la
Formación (Covacevich et al., 1992), siendo el primero de ellos quien da a conocer la gran
abundancia de ejemplares en la localidad de Catamutún. Chirino-Gálvez (1985) identificó
el género dentro de la Cuesta Santo Domingo sin llegar a una especie concreta.
Ejemplares de la especie I. trilirata presentes en los estratos de la Formación Pisco,
Mioceno medio-superior en el sur de Perú, llegan hasta los 50 mm de largo (DeVries, 2007);
talla muy por encima que la de todos los especímenes de la especies encontrados en la
presente investigación.
Material y dimensiones: Es el macroinvertebrado más abundante dentro de los estratos
estudiados en la cuesta, con un total de 116 ejemplares que van desde los 2 mm hasta los
15 mm de largo.

13
Orden Strombimorpha Bandel, 1991
Superfamilia Stromboidea Rafinesque, 1815
Familia Struthiolariidae Fischer, 1884
Género Perissodonta Martens, 1878
Perissodonta sp.
(Lámina 4, A)
Diagnosis: Fragmento de ápice con gran cantidad de costillas axilares.
Descripción: Es posible reconocer la protoconcha y dos y media vueltas de la espira.
Surcos profundos y vueltas convexas. La primera vuelta se presenta lisa sin ornamentación,
pero a partir de la segunda es posible ver costillas axilares medianamente prominentes y
débiles costillas espirales.
Observaciones: Las únicas dos especies pertenecientes al género Perissodonta presentes
en Chile son P. ameghinoi Ihering, 1897, y P. chilensis Phillipi, 1887; ambas descritas para
la Formación Santo Domingo por Nielsen (2005a). P. chilensis (=Struthiolarella chilensis)
fue identificada en los estratos del Rio Futa por Covacevich et al. (1992), quienes dan a
notar la importancia biocronoestratigráfica de la especie, al ser ésta uno de los elementos
característicos del Miembro Inferior de la Formación Navidad.
Material y dimensiones: Solo un fragmento de concha de 3 mm de largo.

14
Orden Neogastropoda Thiele, 1929
Familia Ptychatractidae Stimpson, 1865
Genero Exilia Conrad, 1860
cf. Exilia sp.
(Lámina 1, A-B)
Diagnosis: Concha fusiforme con costillas prominentes.
Descripción: Ejemplar incompleto de forma fusiforme estrecho con espira alta y
moderadamente puntiaguda. Hombro poco prominente. Protoconcha desconocida. La
teleoconcha está compuesta por seis vueltas moderadamente convexas. Cada vuelta posee
costillas sobresalientes y finas costillas espirales. En la última vuelta es posible observar
alrededor de doce costillas axiales. Columela y el labio columelar son lisos sin ningún tipo
de dientes o estrías. Canal sifonal angosto.
Rango geológico: El género Exilia está presente desde el Cretácico superior hasta la
actualidad (Kantor et al., 2001).
Observaciones: El género está descrito en varios lugares alrededor del mundo, con las
excepciones de Sudamérica y África, lo cual podría ser el resultado de estudios
insuficientes en estas regiones (Kantor et al., 2001). Sin embargo, Nielsen (2005b)
identifica la especie E. alanbeui en la Formación Navidad y Ranquil, haciendo de este el
único y primer registro para Sudamérica hasta el momento. E. alanbeui difiere de la especie
encontrada principalmente por las notorias costillas espirales y a las vueltas a lo largo de la
escultura que se presentan más convexas.

15
Todos los especímenes hallados por Nielsen (2005b), poseen un rango de tallas de
entre 30 mm a 9 mm, muy por encima de la longitud total del ejemplar encontrado en la
Cuesta Santo Domingo.
Material y dimensiones: Solo un ejemplar de 7 mm de largo.
Superfamilia Olivoidea Latreille, 1825
Familia Olividae Latreille, 1825
Olividae indet.
(Lámina 2, D-E)
Diagnosis: Concha oval. Hombro poco prominente. Ápice achatado.
Descripción: Concha gruesa oval alargada con una superficie lisa y pulida. Espira corta
con las primeras vueltas angostas y aplanadas poco prominentes. Las suturas son poco
profundas y débiles. La vuelta del cuerpo representa un 70% de la altura total de la concha.
La abertura es corta y ovalada terminando en un canal sifonal corto y delgado. El labio
exterior se presenta grueso y liso sin ningún tipo de diente.
Observaciones: Lamprodomina dimidiata Sowerby, 1846, posee un ápice puntiagudo y
hombro breve en la vuelta del cuerpo, además del surco espiral en la última vuelta que
divide la concha en dos zonas (Frassinetti, 2006). L. dimidiata es común en sedimentos
miocénicos desde Navidad hasta el Archipiélago de Chonos, siendo muy abundante dentro
de los lugares donde se le ha identificado (Griffin y Nielsen, 2008). Los ejemplares
encontrados en este estudio tampoco guardan similitud con representantes del género

16
Olivancillaria, debido a que estos últimos poseen un fuerte surco espiral en la vuelta
corporal, el cual es inexistente en este caso.
Material y dimensiones: Tres ejemplares completos de hasta 6 mm de largo y dos conchas
parciales.
Infraorden Turrina Riedel, 2000
Superfamilia Conoidea Rafinesque, 1815
Familia Turridae H. y A. Adams, 1853
Turridae indet.
(Lámina 2, A)
Diagnosis: Concha incompleta con fuertes costillas espirales.
Descripción: Espira representa un poco menos de la mitad del largo de la concha. Cinco
vueltas en total más protoconcha bien conservada. Costillas axiales comienzan a aparecer
desde la segunda vuelta haciendose cada vez mas prominentes, llegando a la cantidad de
diez para la vuelta del cuerpo. Se presentan entre nueve y diez costillas espirales en las
vueltas, abarcando tambien la totalidad de la vuelta corporal. Zona anterior quebrada.
Observaciones: El único registro de la familia Turridae en Santo Domingo es el entregado
por Illies (1970) al describir el género Pleurotoma sin poder llegar a una especie en
concreto. Hasta hoy, el taxón Pleurotoma ha sido restringido en su uso y dividido en varios
nuevos géneros, todos ellos dentro de la familia Turridae.
Material y dimensiones: Concha incompleta de 6 mm de largo.

17
Genero Gemmula Weinkauff, 1875
Gemmula sp.
(Lámina 1, C)
Diagnosis: Concha fusiforme con protuberancias en el seno de las vueltas.
Descripción: Concha de forma fusiforme, de espira medianamente larga. Hombro poco
prominente. Protoconcha desconocida. Con un total de 6 vueltas fácilmente reconocibles
por la profundidad relativa de las suturas y la presencia de bandas subsuturales gruesas. Se
presentan protuberancias circulares y continuas en todas las crestas de cada vuelta que
también poseen costillas espirales débiles pero fácilmente reconocibles. Se llegan a
contabilizar entre 16 a 18 nudos en la corrida de la vuelta corporal. Columela con estrías
pero sin pliegues. Canal sifonal angosto. Labro y rostro quebrados.
Observaciones: Gemmula subaequalis Sowerby, 1846, fue descrita por Frassinetti (2000)
para el Plioceno tardío en la Isla Guafo al SE de Chiloé. La especie destaca por la presencia
de dos cordones espirales por debajo de la corrida de nudos principal en la vuelta del
cuerpo, los cuales son inexistentes en el único ejemplar encontrado en este estudio. Varios
especímenes de G. subaequalis estudiados en la Isla Guafo poseen la misma cantidad de
nudos en vuelta corporal que en el hallado en esta oportunidad, sin embargo, la
identificación a nivel de especie no puede ser solo concluida a partir del número de nudos
presentes en la última vuelta (Frassinetti, 2000). Gemmula (Ptychosyrinx) pseudodiscors
Sowerby, 1846, está presente dentro de los estratos de la Formación Navidad (Griffin y
Nielsen, 2008), sin embargo, ésta posee una escultura más robusta y un menor número de
nudos en todas las vueltas de la estructura, además de que estos mismos son

18
considerablemente menos prominentes en comparación con los del ejemplar hallado para
este escrito.
Material y dimensiones: Ejemplar de 13 mm de largo.
Género Cosmasyrinx Marwick, 1931
cf. Cosmasyrinx sp.
(Lámina 2, B-C)
Diagnosis: Concha fusiforme robusta con crestas protuberantes.
Descripción: Espira corta con protoconcha. Surcos profundos que permiten contar un total
de 6 vueltas, en donde cada una presenta protuberancias débilmente nudosas en las crestas
haciéndolas más convexas. Líneas de crecimiento paralelas al eje axial. Columela lisa.
Estrías muy débiles en la base de la vuelta corporal. Labio exterior presenta estrías delgadas.
La zona anterior se encuentra incompleta, pero permite apreciar un canal sifonal angosto.
Rango geológico: Todas las especies pertenecientes al género están dentro del Mioceno
(Beu y Maxwell, 1990; p. 250).
Observaciones: La forma maciza de la concha tiene mucha similitud con varias especies
del género presentes en Nueva Zelanda, siendo una de ellas Cosmasyrinx monilifera
Marwick, 1931, sin embargo, ésta presenta visibles estrías en todo lo alto de la vuelta
corporal, mientras que la misma zona en los ejemplares encontrados se presenta totalmente
lisa. Ejemplares de la especie C. monolifera llegan a medir hasta 15 mm de largo, teniendo
una talla mínima de 12 mm (Beu y Maxwell, 1990; p. 250).

19
Material y dimensiones: Dos ejemplares casi completos de 8 y 6 mm de altura.
Clase Bivalvia Linnaeus, 1785
Subclase Protobranchia Pelseneer, 1889.
Orden Nuculida Dell, 1889
Familia Nuculidae Gray, 1824
Género Nucula Lamarck, 1799
Nucula valdiviana Philippi, 1887
(Lámina 3, C)
Nucula valdiviana Philippi, 1887, p. 188, pl. 41, fig. 2.
Diagnosis: Valva de forma triangular equilateral.
Descripción: Los moldes internos hallados permiten observar una forma triangular, casi
igual de alta como de ancha. Los restos de valvas encontrados en los moldes presentan
estrías radiales y concéntricas. Margen ventral ligeramente crenulado.
Rango geológico: Desde el Ordovícico hasta el presente (Whittle et al., 2012)
Observaciones: La especie ya fue descrita en la formación por Philippi (1887), Illies (1970)
y Chirino-Gálvez (1985). Destaca la presencia del género en otros sedimentos marinos
miocénicos del centro-sur como Nucula barrosi Philippi, 1887, en la Formación Navidad y
N. lebuensis Philippi, 1887, en la Formación Ranquil (Nielsen y Valdovinos, 2008), en el
Archipiélago de Chonos (Frassinetti, 2004) y en el Golfo Tres Montes (Frassinetti. 2006),
ambos de la región de Aysén.

20
Material y dimensiones: Cuatro moldes internos con algunos fragmentos de valvas que
van desde 3 mm hasta 5 mm de largo.
Subclase Pteriomorphia Beurlen, 1944
Orden Arcoida Stoliczka, 1871
Familia Limopsidae Dall, 1895
Género Limopsis Sassi, 1827
Limopsis araucana Philippi, 1887
(Lámina 3, A-B)
Limopsis araucana Philippi, 1887, p. 185, pl. 43, fig. 4.
Diagnosis: Concha redondeada en la zona dorsal, haciéndose más ancha hacia el área
ventral. Charnela delgada y larga con varios dientes.
Descripción: Concha redondeada casi equilateral con umbos pequeños y sobresalientes. La
zona dorsal de los ejemplares articulados encontrados muestra una equidad de las valvas.
La superficie interna se presenta lisa, mientras que la externa posee abundantes cordones
concéntricos bien definidos. En la cara interna de algunas valvas es posible apreciar la
cicatriz posterior del músculo retractor. Borde del manto grueso y liso. La charnela es
estrecha y ligeramente curva con hasta cinco dientes a cada lado de la foseta. Sobre la
charnela se aprecia un hoyuelo triangular para la inserción del ligamento.
Observaciones: Limopsis insolita Sowerby, 1846, es identificada en la Formación Guadal,
(Región de Aysén) por Frassinetti y Covacevich (1999). Fue una especie muy abundante
desde el Oligoceno superior hasta el Mioceno superior (Whittle et al., 2011), sin embargo

21
ésta posee un concha más ancha y larga en la zona ventral posterior, además de presentar
una ornamentación distinta en la superficie de la misma (Philippi, 1887). Difieren también
en tamaño, donde L. insolita llega a ser hasta cinco veces más alta que el espécimen de
mayor tamaño de este género en la Cuesta Santo Domingo. El único registro del género
Limopsis y de la especie L. araucana para la región, es el dado por Philippi (1887) al
describir esta especie en la localidad de Llancahue. Así también, existen dos especímenes
de este género encontrados en la Formación Navidad, pero hasta el momento no han sido
descritos (Kiel y Nielsen, 2010).
Material y dimensiones: Una concha articulada completa, trece valvas derechas, ocho
valvas izquierdas y dos valvas donde no es posible reconocer si son derechas o izquierdas,
haciendo un total de 24 ejemplares que van desde los 2 mm hasta los 5 mm de largo. Cabe
destacar que se observó material fragmentado de la especie en toda la columna estudiada.
Clase Scaphopoda Bronn, 1862
Orden Dentaliida Da Costa, 1776
Familia Dentaliidae Gray, 1847
Género Dentalium Linnaeus, 1758
Dentalium cf. sulcosum Sowerby, 1846.
(Lámina 4, B-D)
Dentalium sulcosum Sowerby, 1846, p. 263, pl. 2, fig. 2.
Dentalium sulcosum, Griffin y Nielsen, 2008, p. 310, pl. 24, fig. 17, 20, 21.

22
Diagnosis: Fragmentos de conchas tubulares alargadas.
Descripción: En algunos ejemplares se puede observar una ligera curvatura de la concha,
pero en todos ellos se aprecian entre 10 a 12 costillas longitudinales.
Observaciones: La familia Dentaliidae está bien representada en el Mioceno marino
chileno, sin embargo hasta el momento no se ha realizado una revisión del grupo (Nielsen y
Valdovinos, 2008). En la región existen tres especies descritas del género Dentalium
(Covacevich et al., 1992), siendo D. sulcosum la única de ellas que está presente en la
Cuesta Santo Domingo (Chirino-Gálvez, 1985).
Material y dimensiones: Siete fragmentos de concha que van desde los 3 mm hasta los 5
mm de largo.
Filo Arthropoda Latreille, 1829
Clase Malacostraca Letreille, 1802
Orden Decapoda Latreille, 1802
Infraorden Brachyura Latreille, 1802
Brachyura indet.
(Lámina 4, F)
Diagnosis: Dáctilo aserrado con denticulaciones.
Descripción: Fragmento de dáctilo recto con superficie rugosa y punta redonda. Línea de
denticulaciones paralelas al eje estructural. En todo el fragmento es posible reconocer
incisiones de salida de pilosidades o setas.

23
Rango Geológico: Desde el Cretácico superior hasta la actualidad (Varela, 2013).
Observaciones: Hasta el momento, se han registrado 7 géneros y 5 especies de Brachyura
para la formación (ver Tabla 1), gracias a los estudios de Illies (1970), Chirino-Gálvez
(1985) y Feldmann et al. (2010). La única especie identificada y que entrega cierta
información en cuanto al paleoambiente es Chaceon quadrata Feldmann et al. 2010, que
registra un rango de profundidad que va desde los 120 hasta los 2800 m (Feldmann et al.,
2010), lo cual concuerda con Chirino-Gálvez, quien identificó el género Geryon, cuyos
representantes son exclusivos habitantes de fondos sedimentarios profundales.
Material y dimensiones: Un solo fragmento encontrado de 4 mm de largo.
Filo Echinodermata Klein, 1734
Clase Echinoidea Leske, 1778
Orden Spatangoida L. Agassiz, 1840
Familia Brissidae Gray, 1855
Género Brissopsis L. Agassiz, 1840
cf. Brissopsis valdivianus (Philippi, 1887)
(Lámina 4, E)
Micraster valdivianus Philippi, 1887, p. 222, pl. 52, fig. 3.
Brissopsis valdivianus Kutscher, Neumann, Nielsen, Reich y Villier, 2004, p. 455, fig. 3.
Diagnosis: Fragmentos de ambulacros petaloídeos.

24
Descripción: Los restos de testa encontrados poseen partes de ambulacros petaloídeos,
conformados por secciones de formas hexagonales.
Observaciones: La especie fue identificada por Philippi (1887) en los sedimentos terciarios
de Llancahue y Catamutún. Representantes del género Brissopsis son encontrados
generalmente en sedimentos arenosos del submareal (Kutscher et al., 2004).
Material y dimensiones: Siete fragmentos de testas, que van desde los 7 mm hasta los
14 mm de largo.
4-. DISCUSIÓN
Dentro de todos los autores que han identificado macroinvertebrados fósiles desde
la Formación Santo Domingo (Tabla 1), el único que lo ha hecho en la Cuesta Santo
Domingo ha sido Chirino-Galvez (1985), esperándose así una similitud entre el ensamble
faunístico hallado por este autor y el del presente trabajo. Sin embargo, las diferencias entre
ambos son significativas: solo el género Incatella y las especies Nucula valdiviana,
Dentalium sulcosum y Brissopsis valdivianus están presentes en ambos ensambles. Chirino-
Gálvez (1985) dio a conocer también una abundante paleofauna de Crustáceos y
Pelecypodos, la cual no pudo ser encontrada en este estudio.
Así también, se puede desglosar desde la Tabla 1 los escasos taxones que han sido
identificados por más de un autor para la Formación, lo que podría significar importantes
diferencias del ambiente donde estos fueron depositados, con localidades donde las
condiciones fueron más favorables para desarrollar la vida de solo algunos taxones de
macroinvertebrados.

25
Casi la totalidad de los géneros y especies descritos en esta oportunidad para la
Formación Santo Domingo, se encuentran presentes también dentro de la Formación
Navidad (Tabla 2) sugiriendo una importante correlación bioestratigráfica entre ambas.
Finger (2013) afirma que las Formaciones Navidad, Ranquil, Santo Domingo y
Lacui no pueden ser distinguidas entre ellas debido a las similitudes en cuanto a edad
geológica, depositación y composición de fauna foraminífera, otorgándole al grupo
completo una edad Miocena temprana, basado principalmente en la presencia de
foraminíferos planctónicos en todas las localidades antes señaladas.
La baja talla de todos los especímenes encontrados es particularmente llamativa. Tal
y como se explica en las observaciones de la sistemática paleontológica, los tamaños de
Incatella trilirata, cf. Exilia sp., Gemmula sp. y cf. Cosmasyrinx sp. son bastante reducidos
en comparación con especímenes de otras formaciones geológicas. No se puede descartar
que las tallas del resto de los taxones registrados sean consideradas pequeñas también,
debido a que no existe suficiente respaldo bibliográfico para aseverarlo. Dentro de los
estratos de la Cuesta Santo Domingo, es posible observar capas de limolitas entremezcladas
con capas de arena, lo cual demostraría un grado de inestabilidad sedimentológica (Pino,
com. pers. 2014). Este ambiente estresante o inestable podría haber ocasionado que las
especies que habitaban en él, desplegaran la estrategia de supervivencia R, optando por
gastar sus recursos energéticos en asegurar la descendencia produciendo gran cantidad de
crías, que en aumentar el tamaño corporal (Reznick et al., 2008).
Las diferencias de abundancia y diversidad entre los niveles analizados (Tabla 3) es
otro dato que resulta igualmente llamativo. Los Niveles 1 y 3 de muestreo, son los que

26
presentan una mayor cantidad de especímenes y diversidad, con 53 y 58 ejemplares
divididos entre 9 y 5 taxones identificados respectivamente. Por otro lado, el Nivel 2 es el
que presenta una menor cantidad de macrofauna total, hallándose solo 18 especímenes de 3
taxones distintos. Los niveles que presentan una mayor cantidad y diversidad de
macroinvertebrados posiblemente poseían condiciones ambientales más favorables para que
estos se desarrollaran, mientras que el ambiente de depositación de los niveles menos
fértiles probablemente fue más inhóspito o estresante, influyendo así en la baja diversidad y
abundancia.
Las características paleoecológicas de las especies Perissodonta sp. y Limopsis
araucana nos permiten conocer las temperaturas relativas del ambiente donde se
desarrollaron: la fauna stromboídea miocénica para el centro de Chile, está constituida por
una mezcla de especies de aguas frías y cálidas; sin embargo, los representantes del género
Perissodonta son habitantes exclusivos de aguas frías (Nielsen, 2005a). Dicha aseveración
concuerda con uno de los postulados de Martínez-Pardo y Pino (1979) en cuanto a la
paleoecología de la Formación Santo Domingo, afirmando que las temperaturas de las
aguas del fondo de la cuenca no sobrepasaban los 10°C, basándose en la presencia de
ciertas especies de foraminíferos bentónicos. Especies actuales del género Limopsis están
adaptadas a hábitats de aguas frías también, lo que podría haber sido crucial en el aumento
de representantes del taxón en las altas latitudes del hemisferio sur durante el Mioceno
temprano (Whittle et al., 2011).
Por otro lado, el género cf. Exilia sp. y el orden Brachyura nos permiten conocer un
poco de la paleobatimetría del ambiente: los únicos registros del género Exilia para nuestro
país son los encontrados en las formaciones Navidad y Ranquil, localizándose en limolitas

27
emplazadas en ambientes de profundidades batiales (Nielsen, 2005b). En cuanto a la
presencia del dáctilo de Brachyura, Feldmann et al. (2010) indican que el género Chaceon
(identificado por ellos en Santo Domingo) habita en fondos marinos profundos, que van
desde 120 a 2800 m de profundidad. Chirino-Gálvez (1985) afirma también que todos los
geriónidos son habitantes de fondos marinos profundales.
5-. CONCLUSIONES
En total, se identificaron doce taxones de macroinvertebrados dentro de la Cuesta
Santo Domingo: siete gastrópodos, dos bivalvos, un scafópodo, un braquiuro y un
equinodermo, llegando a un total de 170 ejemplares fósiles y contabilizándose 11
macroinvertebrados por kilogramo de sedimento extraído. Incatella trilirata es la especie
más abundante dentro de la columna estudiada, presentándose en todos los niveles de donde
se extrajo sedimento, a excepción del Nivel 6, donde no se pudo encontrar ningún registro
de macroinvertebrados fósiles (Tabla 3). Limopsis araucana también está presente en todos
los niveles, pero su abundancia es mucho menor en comparación con I. trilirata.
De los 12 taxones identificados en el punto de muestreo en la Cuesta Santo
Domingo, ocho de ellos corresponden a macroinvertebrados detritívoros o depredadores.
Sin embargo, la totalidad de especímenes de estos taxones es de sólo 25 ejemplares, los
cuales representan un 15% del total de invertebrados encontrados, por lo tanto la hipótesis
del presente trabajo debe ser rechazada.
Los gastrópodos Incatella trilirata, cf. Exilia sp., Gemmula sp. y cf. Cosmosyrinx sp.
nunca antes habían sido descritas para la Formación Santo Domingo, siendo éste el primer
registro de ellas en la Formación. Dentro de este nuevo registro, sólo I. trilirata pudo ser

28
identificada a nivel de especie, mientras Gemmula sp. sólo pudo ser identificada a nivel de
género. Si bien, los caracteres morfológicos de cf. Exilia sp. la posicionan dentro del género
Exilia, este último tiene a E. alanbeui como único representante del género para nuestro
país, el cual difiere en ciertas características como ya fue observado en los resultados de
este estudio. Se sugiere hacer una revisión de la especie cf. Cosmosyrinx sp., debido a que
con sus caracteres morfológicos no fue posible reconocerlo dentro de ninguna Formación
del centro-sur de nuestro país.
También se origina el primer registro de representantes de la familia Olividae en
Santo Domingo, aun cuando no fue posible una identificación a nivel de género ni mucho
menos a nivel de especie.
6-. BIBLIOGRAFIA
BEU, A.G. y MAXWELL, P.A., 1990 Cenozoic Mollusca of New Zealand. New Zealand
Geological Survey Paleontological Bulletin 58: 518 pp.
BRUGGEN, J., 1950. Fundamentos de la Geología de Chile. Instituto Geográfico Militar,
Santiago, Chile: 374 pp.
CECIONI, G., 1980. Darwin´s Navidad embayment, Santiago region, Chile, as a model of
the southeastern Pacific shelf. Journal of Petroleum Geology, 2: 309-321.
CHIRINO-GÁLVEZ, L., 1985. Paleoecología del bentos del Mioceno marino en Valdivia.
Actas 4to
Congreso Geológico Chileno, Antofagasta, 1: 133-151.

29
COVACEVICH, V., FRASSINETII, D. y ALFARO, G., 1992. Paleontología y condiciones
de depositación del mioceno marino en las nacientes del rio Futa, Valdivia, Chile. Boletín
Museo Nacional de Historia Natural, 43: 143-154.
DARWIN, C. 1846. Geological observations on South America being the third part of the
Geology of the Voyage of the Beagle, under the command of Capt. Fitzroy, R.N. during the
years 1832 to 1836. Smith, Elder and Co., London: 279 pp.
D´ORBIGNY, A., 1847. Atlas de Géologie. Voyage au pole sud et dans l´Océanie sur les
corbettes l´Astrolabe et la Zeleé pendant les années 1837-1840 (Dumont-d´Urville, M.J.).
Géologie, Paleontologie. París.
DeVRIES, T.J., 2007. Cenozoic Turritellidae (Gastropoda) from southern Peru. Journal of
Paleontology, 81 (2): 331-351.
ELGUETA, S., 1990. Sedimentación marina y paleogeografía del Terciario superior en la
Cuenca de Temuco, Chile. Actas 2do
Simposio sobre el Terciario de Chile, 1: 85-96.
ELGUETA, S., Mc DONOUGH, M., Le ROUX, J., URQUETA, E. y DUHART, P., 2000.
Estratigrafía y sedimentología de las cuencas terciarias de la Región de Los Lagos (39°-
41°30´S). Boletín Sernageomin, 57: 50 pp.
ENCINAS, A., FINGER, K.L., BUATOIS, L.A. y PETERSON, D.E., 2012. Major forearc
subsidence and deep-marine Miocene sedimentation in the present Coastal Cordillera and
Longitudinal Depression of south-central Chile (38°30´S-41°45´S). Geological Society of
America Bulletin, 124: 1262-1277.
ENCINAS, A., FINGER, K.L., NIELSEN, S.N., LAVENU, A., BUATOIS, L.A,
PETERSON, D.E. y LE ROUX, J.P, 2008. Rapid and mayor coastal subsidence during the

30
late Miocene in south-central Chile. Journal of South American Earth Sciencies, 25: 157-
175.
FELDMANN, R., SCHWEITZER, C. y ENCINAS, A., 2010. Neogene decapod Crustacea
from southern Chile. Annals of Carnegie Museum, 78 (4): 337-366.
FINGER, K.L., NIELSEN, S.N., DeVRIES, T.J., ENCINAS, A. y PETERSON, D.E., 2007.
Paleontology evidence for sedimentary displacement in Neogene forearc basins of central
Chile. Palaios, 22: 3-16.
FRASSINETTI, D., 2000. Moluscos del Plioceno superior marino de Isla Guafo, Sur de
Chile. Parte II. Gastropoda. Boletín del Museo Nacional de Historia Natura, Chile, 49: 131-
161.
FRASSINETTI, D., 2004. Moluscos fósiles del Mioceno marino de Isla Ipún, Sur de Chile.
Boletín del Museo Nacional de Historia Natural, Chile, 53: 71-83.
FRASSINETTI, D., 2006. Moluscos fósiles del Mioceno marino de las Islas Crosslet y
Hereford (Golfo Tres Montes, Aisén). Boletín del Museo Nacional de Historia Natural,
Chile, 55: 61-74.
FRASSINETTI, D. y COVACEVICH, V., 1999. Invertebrados fósiles marinos de la
Formación Guadal (Oligoceno superior-Mioceno inferior) en Pampa Castillo, Región de
Aisén, Chile. Servicio Nacional de Geología y Minería, 1: 96 pp.
GRIFFIN, M. y NIELSEN, S.N., 2008. A revision of the type specimens of Tertiary
molluscs from Chile and Argentina described by d´Orbigny (1842), Sowerby (1846) and
Hupé (1854). Journal of Systematic Palaeontology, 6 (3): 251-316.

31
ILLIES, H., 1970. Geología de los alrededores de Valdivia y volcanismo y tectónica en
márgenes del Pacífico en Chile meridional. Instituto de Geología y Geografía, Universidad
Austral: 64 pp.
KANTOR, Y.I., BOUCHET, P. y OLEINIK, A., 2001. A revision of the recent species of
Exilia, formerly Benthovoluta (Gastropoda: Turbinellidae). Ruthenica, 11 (2): 81-136.
KIEL, S y NIELSEN, S.N., 2010. Quaternary origin of the inverse latitudinal diversity
gradient among southern Chilean mollusks. Geological Society of America, 38 (10):
955-958.
KUTSCHER, M., NEUMANN, C., NIELSEN, S.N., REICH, M. y VILLIER, L., 2004.
Echinoderms from the Miocene of Chile. En T. Heinzeller y J.H. Nebelsick, Echinoderms:
München, Londres: 453-456.
MAI, H., 1982. Nuevas observaciones sobre el terciario de la Cuesta Santo Domingo
(Coccolithoforidos), provincia de Valdivia, Chile. Actas 3er
Congreso Geológico Chileno,
Concepción, A: 147-158.
MARCHANT, M., 1990. Foraminiferos miocénicos de los estratos de Pupunahue
(Provincia de Valdivia: X Región): determinación de la edad probable y paleoambiente.
Actas 2do
Simposio sobre el Terciario de Chile, 1: 177-188.
MARTINEZ-PARDO, R., 1990. Major Neogene events of the southeastern Pacific: The
Chilean and Peruvian record. Palaeogegraphy, Palaeoclimatology, Palaeoecology, 77:
263-278.

32
MARTINEZ-PARDO, R. y PINO, M., 1979. Edad, paleoecología y sedimentología del
mioceno marino de la Cuesta Santo Domingo, provincia de Valdivia, X región. Actas 2do
Congreso Geológico Chileno, Arica, H: 103-124.
NIELSEN, S.N., 2005a. Cenozoic Strombidae, Aporrhaidae, and Struthiolariidae
(Gastropoda, Stromboidea) from Chile: Their significance to the interpretation of south-east
Pacific biogeography and climate: Journal of Paleontology, 79: 1120-1130.
NIELSEN, S.N., 2005b. Exilia alanbeui, a new species from the Neogene of central Chile:
the first record of Exilia (Gastropoda: Ptychatractidae) from South America. The Nautilus,
119 (4): 153-156.
NIELSEN, S.N. Y GLODNY, J., 2009. Early Miocene subtropical water temperatures in
the southeast Pacific. Palaeogeography, Palaeoclimatology, Palaeoecology, 208: 480-488.
NIELSEN, S.N. y VALDOVINOS, C., 2008. Early Pleistocene mollusks of the Tubul
Formation, South-Central Chile. The Nautilus, 122 (4): 201-216.
PHILIPPI, R.A., 1887. Los fósiles terciarios i cuartarios de Chile. F.A. Brockhaus, Leipzig:
256 pp.
PINO, M., 1990. Interpretación a través de análisis litoestratigráficos y granulométricos de
los ambientes de depositación en una cuenca miocénica, Valdivia, Chile. Actas 2do
Simposio sobre el Terciario de Chile, Concepción: 245-258.
PINO, M. y BELTRAN, C., 1979. Sedimentología del Mioceno Marino de la Cuesta de
Santo Domingo, Provincia de Valdivia. Revista Medio Ambiente, 4 (1): 51-61.

33
PINO, M. y NUÑEZ, L., 1979. Presencia de la especie Lapageria rosea, Ruiz et Pavos
“Copihue” en el Mioceno marino de la provincia de Valdivia, X región, Chile. Actas 2do
Congreso Geológico Chileno, Arica: 151-158.
REZNICK, D., BRYANT, M.J. y BASHEY, F., 2002. R- and K- selection revised: the role
of population regulation in life-history evolution. Ecology 83 (6): 1509-1520.
TAVERA, J., 1979. Estratigrafía y paleontología de la Formación Navidad, Provincia de
Colchahua, Chile (Lat. 30° 50´ - 34° S). Boletín Museo Nacional de Historia Natural, Chile,
36: 176 pp.
VARELA, C., 2013. Nuevos datos sobre los crustáceos fósiles (Decapoda: Brachuyra) de
Cuba. Solenodon 11: 5 pp.
WHITTLE, R.J., LINSE, K. y GRIFFITHS, H.J., 2011. The fossil record of Limopsis
(Bivalvia: Limopsidae) in Antarctica and the southern high latitudes. Paleontology, 54 (4):
935-952.
WHITTLE, R.J., QUAGLIO, F., CRAME, J.A. Y LINSE, K., 2012. Nuculidae (Bivalvia)
in the Cape Melville Formation, King George Island, Antarctica, with an overview of the
bivalve fauna. Antarctic Science, 24 (6): 625-633.

34
7-. ANEXOS
7.1-. FIGURAS
Figura 1. Columna estratigráfica definida por Pino y Beltrán (1979) en un área cercana al
sitio del presente estudio dentro de la Cuesta Santo Domingo. En azul, se muestra la
posición los niveles desde los cuales se extrajo sedimento para su análisis.
35
Figura 2. Ubicación del área de estudio. El punto marca la ubicación de Valdivia, mientras
que la estrella señala el lugar de la Cuesta Santo Domingo.

36
Figura 3. Posición de los 6 niveles de muestro en la columna de estudio.

37
Figura 4. Período Neógeno dividido en época y edades.
38
7.2-. TABLAS
Tabla 1. Lista de macroinvertebrados descritos para la Formación Santo Domingo de acuerdo con d´Orbigny (1847), Phillipi (1887),
Illies (1970), Chirino-Gálvez (1985), Covacevich et al. (1992), Kutscher et al. (2004), Nielsen (2005a) y Feldmann et al. (2010). Se
mantienen las denominaciones originales de estos autores, a excepción de los taxones encontrados en este trabajo. *especies
encontradas en la Cuesta Santo Domingo por estos autores.
d´Orbigny, Philippi, Illies,
Chirino-
Gálvez,
Covacevich
et al.,
Kutscher
et al., Nielsen,
Feldmann
et al.,
1847 1887 1970 1985 1992 2004 2005a 2010
Anthozoa
Lithomyces aequalis
X
Lithomyces costellanus
X
Bryozoa indet.
X
Braquiopoda
Terebratula sp.
X
Terebratula araucana
X
Terebratula foncki
X
Bivalvia
Amathusia cf. orbicularis
X
Arca valdiviana
X
Chlamys sp.
X
Cucullaea fricki
X
Limopsis araucana
X
Lithophaga (?) sp.
X
Lutraria vetula
X X
Malletia volckmanni*
X
39
d´Orbigny, Philippi, Illies,
Chirino-
Gálvez,
Covacevich
et al.,
Kutscher
et al., Nielsen,
Feldmann
et al.,
1847 1887 1970 1985 1992 2004 2005a 2010
Nucula valdiviana*
X X X
Ostrea (Crassostrea) sp.
X
Pinna costata
X
Tellinidae indet. X
Tellina fricki X
Tellina (Arcopagia) valdiviana X X
Venus volckmanni
X
Gastropoda
Borsonia sp.* X
Bittium (?) sp.
X
Cassis monilifera
X
Clio pyramidata
X
Crepidula sp
X
Naticidae indet.
X
Natica ovoidea
X
Opalia sp.
X
Pleurotoma sp.
X
Sinum sp.
X
Perissodonta ameghinoi
X
Perissodonta chilensis
X
X
Trochus fricki*
X X X
Turritella sp.*
X X X
Turritella cf. breantiana
X
Vermetidae indet.
X
Scaphopoda Dentalium lebuense
X
Dentalium majus?
X
Dentalium sulcosum*
X
40
d´Orbigny, Philippi, Illies,
Chirino-
Gálvez,
Covacevich
et al.,
Kutscher
et al., Nielsen,
Feldmann
et al.,
1847 1887 1970 1985 1992 2004 2005a 2010
Crustacea
Archaeogeryon sp.*
X
Axianassa? chilensis*
X
Bathynomus? sp.* X
Cancer araucanus X
Chaceon quadrata X
Crustacea indet. X
Ctenocheles notialis
X
Geryon sp.*
X
Lithodes? sp.* X
Megabalanus sp. psittacus X
Pilumnus cucaoensis X
Proterocarcinus navidad X
Trichopeltarion levis
X
Echinoidea Echinoidea indet.
X
Loxechinus cf. albus
X
Micraster valdivianus*
X
X
X
Cronoidea Pentacrinus sp.
X
X

41
Tabla 2. Lista de las especies identificadas en la Cuesta Santo Domingo que también se
encuentran en otras formaciones miocénicas: Navidad, Ranquil y Lacui. Tener en cuenta las
siguientes especificaciones: X₁, Perissodonta chilensis; X₂, Exilia alanbeui; X₃, géneros
de la familia Olividae; X₄, géneros de familia Turridae; X₅, genero Limopsis; X₆, Nucula
sp.; X₇, especies del orden Brachyura.
Formación Navidad Formación Ranquil Formación Lacui
Incatella trilirata X X
Perissodonta sp. X₁ cf. Exilia sp. X₂ X₂
Olividae indet. X₃ X₃ X₃
Gemmula sp. X
X
cf. Cosmasyrinx sp.
Turridae indet. X₄ X₄ X₄
Limopsis araucana X₅ Nucula valdiviana X₆
X₆
Dentalium cf. sulcosum X
Brachuyra indet. X₇ X₇ X₇
cf. Brissopsis valdivianus
42
Tabla 3. Total de ejemplares y taxones encontrados en cada uno de los niveles trabajados.
*solo fragmentos de organismos.
Nivel 1 Nivel 2 Nivel 3 Nivel 4 Nivel 5 Nivel 6 TOTAL
Incatella trilirata 36 9 13 10 48 -- 116
Perissodonta sp. 1
-- 1
cf. Exilia sp.
1 -- 1
Olividae indet. 3
2 -- 5
Gemmula sp.
1
-- 1
cf. Cosmasyrinx sp.
2 -- 2
Turridae indet. 1
-- 1
Limopsis araucana 2 8 4 5 5 -- 24
Nucula valdiviana* 2
2
-- 4
Dentalium cf. sulcosum* 6 1
-- 7
Brachuyra indet. 1
-- 1
cf. Brissopsis valdivianus* 1
5 1
-- 7
TOTAL ESPECÍMENES 53 18 23 18 58
170
TOTAL TAXONES 9 3 4 4 5

43
7.3-. LÁMINAS
Lámina 1. Gastrópodos fósiles registrados en la Cuesta Santo Domingo. A, vista dorsal de
cf. Exilia sp. B, vista vental de cf. Exilia sp. C, vista vental Gemmula sp. D-F, Incatella
trilirata. La barra representa 5 mm de largo.

44
Lámina 2. Gastrópodos fósiles registrados en la Cuesta Santo Domingo. A, vista ventral
Turridae indet. B-C, vistas ventrales de los dos ejemplares de cf. Cosmasyrinx sp. D-E,
vistas ventrales y dorsales de dos ejemplares de Olividae indet. La barra representa 5 mm
de largo.

45
Lámina 3. Bivalvos fósiles registrados en la Cuesta Santo Domingo. A, vista externa e
interna de valva izquierda de Limopsis araucana. B, vista interna de valva incompleta de
Limopsis araucana. D, valva parcial de Nucula valdiviana. La barra representa 5 mm de
largo.

46
Lámina 4. Fósiles registrados en la Cuesta Santo Domingo. A, vista de Perissodonta sp. B,
C, D, fragmentos de concha de Dentalium cf. sulcosum. E, ambulacros petaloídeos de cf.
Brissopsis valdivianus. F, fragmento de dáctilo de Brachyura indeterminada. Las barras
representan 1 mm de largo.