evaluación del potencial del cultivo de células en ... · máxima de biomasa de 12,11 g/l. se...
TRANSCRIPT

Evaluación del potencial del cultivo de células en suspensión de papa
(Solanum tuberosum subsp. andígena) como plataforma de
producción de proteínas recombinantes
Luisa Fernanda Granger Serrano
Universidad Nacional de Colombia
Facultad de Ciencias
Medellín, Colombia
2016


Evaluación del potencial del cultivo de células en suspensión de papa
(Solanum tuberosum subsp. andígena) como plataforma de
producción de proteínas recombinantes
Luisa Fernanda Granger Serrano
Tesis presentada como requisito parcial para optar al título de:
Magíster en Biotecnología
Director (a):
Ph.D. Rafael Arango Isaza
Codirector (a):
Ph.D. Mario Arias Zabala
Línea de Investigación:
Cultivo de células vegetales en suspensión para producción de proteínas recombinantes
Grupos de Investigación:
Biotecnología vegetal y Biotecnología industrial
Universidad Nacional de Colombia
Facultad de Ciencias
Medellín, Colombia
2016


A mis padres, familia y amigos, quienes creen
tanto en mí que me hacen mover mis propios
límites cuando yo he llegado a creer que mi
capacidad ha llegado a su tope.


Resumen VII
Resumen
La ingeniería genética ha permitido la producción de proteínas recombinantes en diversos
sistemas heterólogos, entre los cuales el cultivo de células vegetales ha ganado gran
importancia en la última década. En la búsqueda de plataformas altamente productivas,
se propone a Solanum tuberosum L. como una alternativa promisoria, por sus
características fisiológicas y genéticas. El objetivo de esta investigación fue establecer
cultivos de células en suspensión de la nueva variedad colombiana Pastusa Suprema
(Solanum tuberosum subsp. andígena), a partir de plantas transformadas con el gen
cry1Ac de Bacillus thurigiensis. Este sistema modelo permitió explorar el potencial de esta
variedad vegetal como alternativa a los sistemas tradicionales de producción de proteínas
heterólogas. Se determinó la composición hormonal del medio de cultivo que favorece la
inducción de callos (ANA 1,5 mg/L) y el establecimiento de suspensiones (ANA 0,5 mg/L)
en medio Murashigue y Skoog (MS). Se evaluó la cinética de crecimiento de los cultivos
en suspensión, obteniendo un tiempo de duplicación de 8,15 días y una concentración
máxima de biomasa de 12,11 g/L. Se detectó la expresión de la proteína recombinante
Cry1Ac en los cultivos en suspensión establecidos a partir de plantas transformadas
mediante el ensayo DAS-ELISA. Los bajos requerimientos hormonales, la alta tasa
específica de crecimiento y la expresión de la proteína Cry1Ac, sugieren el potencial de
Solanum tuberosum subsp. andígena var. Pastusa Suprema como plataforma de
producción de proteínas recombinantes de interés biotecnológico. El presente trabajo
constituye el primer reporte del cultivo de callos y células en suspensión de esta nueva
variedad vegetal.
Palabras clave: Cultivo de células vegetales en suspensión, proteínas
recombinantes, Solanum tuberosum L.

Contenido VIII
Contenido
Pág.
Resumen ................................................................................................................... VIIVII
Lista de figuras ............................................................................................................... X
Lista de tablas ............................................................................................................... XII
Introducción .................................................................................................................... 1
1. Estado del arte ......................................................................................................... 5 1.1 Proteínas terapéuticas ..................................................................................... 5 1.2 Sistemas de producción de proteínas terapéuticas ......................................... 7 1.3 Cultivo in vitro y producción de proteínas terapéuticas en papa ...................... 9 1.4 Antecedentes de Solanum tuberosum subsp. andígena variedad Pastusa Suprema .................................................................................................................. 11
2. Objetivos ................................................................................................................ 13 2.1 Objetivo general ............................................................................................ 13 2.2 Objetivos específicos..................................................................................... 13
3. Metodología ............................................................................................................ 14 3.1 Material vegetal ............................................................................................. 14 3.2 Inducción de callos ........................................................................................ 14 3.3 Establecimiento de suspensiones celulares .................................................. 16 3.4 Cinética de crecimiento de biomasa y consumo de sustrato.......................... 16 3.5 Establecimiento de suspensiones celulares a partir de plantas transformadas .......................................................................................................... 17 3.6 Métodos analíticos ........................................................................................ 18
3.6.1 Viabilidad celular .................................................................................. 18 3.6.2 Volumen de células sedimendatas ....................................................... 18 3.6.3 Concentración de biomasa en peso seco ............................................. 18 3.6.4 Concentración de azúcares residuales ................................................. 18 3.6.5 Detección de la expresión de proteína recombinante ........................... 19
4. Resultados y discusión .............................................. ¡Error! Marcador no definido.1 4.1 Inducción de callos ........................................................................................ 21 4.2 Establecimiento de suspensiones celulares .................................................. 27 4.3 Cinética de crecimiento de biomasa y consumo de sustrato.......................... 32 4.4 Establecimiento de suspensiones celulares a partir de plantas transformadas .......................................................................................................... 37

Contenido IX
5. Conclusiones y recomendaciones ........................................................................ 43 5.1 Conclusiones ................................................................................................. 43 5.2 Recomendaciones ......................................................................................... 43
Bibliografía .................................................................................................................... 45

Contenido X
Lista de figuras
Pág.
Figura 1-1: Mapa linear del cassette con los genes nptII y cry1Ac .......................... 12
Figura 4-1: Inducción de callos de Solanum tuberosum subsp. andígena var. Pastusa
suprema a partir de cortes de tallo en condiciones de oscuridad y fotoperiodo 16 h luz .. 21
Figura 4-2: Efecto del tipo de explante (nudo/entrenudo) sobre la respuesta a los
tratamientos de inducción in vitro de callos. .................................................................... 23
Figura 4-3: Inducción de callos a partir de entrenudos. ............................................ 26
Figura 4-4: Efecto de la concentración de ácido α-naftalenacético (ANA) en el medio
de cultivo sobre el crecimiento de las suspensiones celulares de Solanum tuberosum
subsp. andígena var. Pastusa suprema .......................................................................... 27
Figura 4-5: Morfología de los cultivos de células en suspensión de Solanum tuberosum
subsp. andígena variedad Pastusa Suprema en los primeros meses del establecimiento,
aumento total 200X ......................................................................................................... 28
Figura 4-6: Cultivo de células en suspensión de Solanum tuberosum subsp. andígena
variedad Pastusa Suprema tras 10 meses del establecimiento, aumento total 100X. ..... 29
Figura 4-7: Suspensiones de Solanum tuberosum subsp. andígena variedad Pastusa
Suprema de alta densidad celular obtenidas mediante inducción de callos en medio MS +
1,5 mg/l de ANA y establecimiento de suspensiones en medio MS + 0,5 mg/l de ANA ... 30
Figura 4-8: Suspensiones celulares que produjeron compuestos de coloración roja
que se observó en el medio extracelular y en la biomasa .............................................. 31
Figura 4-9: Cinética de crecimiento de biomasa del cultivo de células en suspensión
de Solanum tuberosum L. subsp. andígena var. Pastusa Suprema cultivadas in vitro. ... 33
Figura 4-10: Cinética de consumo de azúcares del cultivo de células en suspensión de
Solanum tuberosum L. subsp. andígena var. Pastusa Suprema cultivadas in vitro. ........ 34

Contenido XI
Figura 4-11: Ensayo DAS-ELISA de entrenudos de plantas de la línea PS 40.27 de
Solanum tuberosum subsp. andígena var. Pastusa Suprema para la detección de proteína
recombinante Cry1Ac. .................................................................................................... 37
Figura 4-12: Ensayo DAS-ELISA del cultivo de células en suspensión de Solanum
tuberosum subsp. andígena var. Pastusa Suprema para la detección de proteína
recombinante Cry1Ac ..................................................................................................... 38
Figura 4-13: Morfología y viabilidad del cultivo de células en suspensión de Solanum
tuberosum subsp. andígena var. Pastusa Suprema, línea PS 40.27 .............................. 40

Contenido XII
Lista de tablas
Pág.
Tabla 4-1: Porcentaje de formación de callos friables a partir de entrenudos de Solanum
tuberosum subsp. andígena var. Pastusa Suprema, en medio MS suplementado con
diferentes composiciones hormonales. ........................................................................... 24
Tabla 4-2: Comparación de la composición hormonal del medio de cultivo en diferentes
reportes de inducción de callos de Solanum tuberosum L., según la variedad y tipo de
explante……. .................................................................................................................. 25
Tabla 4-3: Comparación de la tasa específica máxima de crecimiento (µmáx) y el tiempo
de duplicación en la fase exponencial (td) del cultivo de células en suspensión de Solanum
tuberosum subsp. andígena var. Pastusa Suprema, con otros reportes de cultivo de células
vegetales …... ................................................................................................................. 35
Tabla 4-4: Resultados del ensayo DAS-ELISA para la detección de la expresión de la
proteína recombinante Cry1Ac en la biomasa de los cultivos de células en suspensión
establecidos a partir de plantas de la línea PS 40.27 de Solanum tuberosum L. subsp.
andígena variedad Pastusa Suprema. ............................................................................ 39

Introducción
La tecnología de ADN recombinante ha hecho posible la manipulación de genomas para
introducir y expresar genes heterólogos, herramienta que se constituye en la base de la
biotecnología actual. Hoy en día es posible utilizar las técnicas de ingeniería genética para
combinar características deseables de organismos distintos. Se han podido desarrollar,
por ejemplo, variedades vegetales que producen toxinas bacterianas en contra de insectos
patógenos o aprovechar la alta tasa de crecimiento de microorganismos para producir
grandes cantidades de enzimas con aplicación industrial a un costo global de producción
muy bajo.
Las proteínas producidas en sistemas heterólogos por técnicas de ingeniería genética son
muy diversas y tienen aplicaciones en la industria de fármacos, alimentos, detergentes,
textiles, cuero, papel, polímeros y plásticos (Demain & Vaishnav, 2009). En medicina,
además de ser útiles en las pruebas de diagnóstico, las proteínas recombinantes son
usadas como fármacos para el tratamiento de enfermedades, haciendo posible mejorar la
calidad y esperanza de vida de muchas personas con graves afecciones de salud. Las
proteínas terapéuticas han permitido desarrollar tratamientos de reemplazo enzimático,
vacunas y anticuerpos para enfermedades como la diabetes, la enfermedad de Gaucher,
la anemia, la hepatitis, la esclerosis múltiple y el cáncer (Leader, Baca, & Golan, 2008). En
comparación al aislamiento de tejidos animales, la obtención de estas proteínas por las
técnicas de ADN recombinante ofrece significativas ventajas como disponibilidad por
procedimientos más eficientes y menos costosos, actividad específica, reducido riesgo de
causar rechazo inmunológico y de transmitir enfermedades contagiosas causadas por
virus o priones que podrían estar presentes en las fuentes de aislamiento (Doran, 2000).
A pesar de que durante muchos años las células de mamíferos han sido el sistema de
preferencia para obtener biofármacos recombinantes, con un 45% del total producidos en
2009 (Huang & McDonald, 2009), en la última década se ha girado la atención hacia las
células vegetales debido al bajo costo global de producción y a la ausencia de riesgo de

2 Introducción
contaminación y transmisión de patógenos humanos como virus y priones. Sin embargo,
la baja productividad y la falta de autenticidad del producto aún son obstáculos por superar
(J. Xu, Ge, & Dolan, 2011), principalmente debido a que existen diferencias en el patrón
de glicosilación de las plantas y de los mamíferos, lo cual puede producir variaciones
significativas en la estabilidad y antigenicidad de las glicoproteínas recombinantes
humanas producidas en estos sistemas heterólogos.
La ventaja de los procesos fermentativos controlados, por su parte, es que se dispone de
múltiples enfoques para optimizar parámetros como la productividad y el costo global,
recurriendo a estrategias como la manipulación de la composición del medio y condiciones
de cultivo, modificando el diseño del biorreactor, el modo de operación, las técnicas de
ingeniería genética y metabólica, los métodos de separación y purificación, entre otras. De
esta manera se pueden encontrar casos exitosos que han logrado aumentar notablemente
la productividad de algunas proteínas heterólogas en sistemas vegetales (Huang &
McDonald, 2009; J. Xu et al., 2011) y a la bandera está la única proteína comercializada
que se produce en cultivo de células vegetales, la β-glucocerobrosidasa, producida en
células de zanahoria (Daucus carota L.) y comercializada en Israel, Estados Unidos, Brasil
y Uruguay por la empresa Protalix BioTherapeutics© (www.protalix.com).
Dado el relativamente reciente interés en las células vegetales como plataforma de
producción, la investigación ha estado centrada en unas pocas especies, principalmente
zanahoria (Daucus carota L.), tabaco (Nicotiana tabacum L.) y arroz (Oryza sativa L.). Cada
organismo cuenta con una maquinaria metabólica diferente, lo que conlleva a diferencias
en los niveles de expresión y productividad. Así, por ejemplo, la expresión y rendimiento
de interleuquina-12 fue mejorada cambiando únicamente la plataforma de producción de
células de tabaco a arroz (Shin, Chong, Han, Yang, & Kwon, 2010). De esta manera, se
hace necesario buscar otras alternativas que podrían resultar fuertes competidoras de las
plataformas tradicionales, teniendo en cuenta que no existe un único sistema capaz de
alcanzar el mayor rendimiento/productividad en la producción de todas las proteínas
heterólogas.
Se han destacado las siguientes características deseables en los sistemas vegetales que
pretendan explorarse como plataformas de producción de proteínas recombinantes
(Huang & McDonald, 2009): fácil transformación celular, robustez/vitalidad, baja

Introducción 3
sensibilidad a las condiciones de crecimiento, alta tasa específica de crecimiento, habilidad
de expresar alta cantidad de proteína, baja actividad proteolítica extracelular (proteínas
secretadas), baja producción de metabolitos secundarios (fenoles y/o quinonas),
capacidad de modificación post-transduccional, patrón correcto y uniforme de glicosilación
y estabilidad genética.
En este escenario el cultivo de células en suspensión de papa (Solanum tuberosum L.)
aparece como una alternativa promisoria para la producción de proteínas recombinantes.
La importancia agronómica de esta planta ha hecho que se haya acumulado gran cantidad
de información sobre sus características bioquímicas y fisiológicas y las técnicas de cultivo
in vitro y de manipulación genética. Se han establecido cultivos in vitro de células de papa
desde hace casi tres décadas para realizar estudios bioquímicos y genéticos de variedades
resistentes, y estudios de estabilidad genética y rutas metabólicas implicadas en
respuestas a condiciones de estrés o ataque por patógenos. Se ha reportado el crecimiento
de las células en suspensión en diferentes medios y con composiciones hormonales que
varían significativamente de una investigación a otra, evidenciando que esta especie
responde de manera robusta al cultivo in vitro. Además, debido a su importancia
agronómica, se han desarrollado numerosos estudios de transformación para
mejoramiento genético de variedades silvestres, por lo que se sabe que la especie es fácil
de transformar; se ha reportado, por ejemplo, 37% de eficiencia en la transformación de
papa Pastusa Suprema (Torres, Torres, Moreno, & Arango, 2012). La manipulación
genética también ha permitido desarrollar plantas transgénicas productoras de proteínas
terapéuticas con actividad biológica, como la interleuquina 2 (IL-2) (Park & Cheong, 2002),
revelando que este sistema tiene la capacidad de hacer las modificaciones post-
transduccionales correctas para que el biofármaco sea funcional.
Teniendo en cuenta las características mencionadas de Solanum tuberosum L. y que
además existen en el país grupos de investigación que han desarrollado importantes
fortalezas en las técnicas de manipulación genética de plantas de interés agronómico como
la papa, se plantea la siguiente pregunta de investigación: ¿Es el cultivo de células en
suspensión de Solanum tuberosum subsp. andígena variedad Pastusa Suprema una
promisoria plataforma de producción de proteínas recombinantes?


1. Estado del arte
1.1 Proteínas terapéuticas
Las proteínas son quizás las biomoléculas más versátiles de la naturaleza, involucradas
en diversos procesos celulares como la duplicación del material genético, la transducción
de señales del ambiente, la catálisis de todas las reacciones del metabolismo y el
transporte de moléculas entre tejidos. Se ha estimado que el genoma humano contiene
entre 30.000 y 40.000 genes que transcriben a proteínas funcionales (Lander et al., 2001);
si a esto se le suma el efecto del splicing alternativo del ARNm y las modificaciones post-
transduccionales (como corte, fosforilación, acetilación y glicosilación), el número de
proteínas distintas es probablemente mucho mayor (Leader et al., 2008). La función que
cada una de ellas cumple es altamente específica y compleja, a partir de una relación
estrecha entre secuencia y estructura tridimensional que determina su actividad biológica.
Aunque el papel que cumplen las proteínas es vital, puede ser alterado por la fijación de
mutaciones, desencadenando enfermedades asociadas al mal funcionamiento, cantidades
anormales o ausencia de la proteína. Por esta razón, muchas investigaciones han hecho
esfuerzos para aprovechar los avances en técnicas de ingeniería genética e ingeniería de
procesos, para diseñar tratamientos más efectivos de enfermedades. Tal vez el ejemplo
más reconocido es el caso de la insulina, una hormona administrada a pacientes que
sufren de diabetes mellitus tipo I (DM-I) y tipo II (DM-II). Sin tratamiento, la DM-I es una
enfermedad que causa un debilitamiento severo del paciente e incluso la muerte, por
alteraciones en el metabolismo de la glucosa (Leader et al., 2008). Desde la purificación
de la insulina en 1922, a partir de páncreas bovino y porcino, esta proteína se convirtió en
el tratamiento diario que permitió mejorar la calidad y esperanza de vida de las personas
con diabetes. Sin embargo, existen múltiples desventajas en el uso de insulina de origen
animal, principalmente la disponibilidad, el costo de purificación y la inmunogenicidad.
Estos problemas fueron superados gracias a los avances de ingeniería genética, que

6 Evaluación del potencial del cultivo de células en suspensión de papa (Solanum
tuberosum subsp. andígena) como plataforma de producción de proteínas
recombinantes
permitieron la expresión del gen de la insulina humana en la bacteria Escherichia coli,
posibilitando la producción in vitro a gran escala.
Al igual que la insulina, muchas proteínas terapéuticas han revolucionado la medicina
moderna, ofreciendo un tratamiento para una larga lista de enfermedades, entre las que
se encuentran además de la diabetes, la enfermedad de Gaucher, la anemia, la hepatitis,
la esclerosis múltiple, la leucemia mieloide aguda y el cáncer (Xu et al., 2011). Entre ellas,
muchas son enfermedades huérfanas, que afectan a un número muy bajo de personas en
la población y, por tanto, muchas veces son olvidadas en las investigaciones y en las
políticas de salud pública gubernamentales.
Las proteínas terapéuticas obtenidas por técnicas de ADN recombinante ofrecen varias
ventajas. En primer lugar, la producción a gran escala permite tener mayor disponibilidad
por procedimientos más eficientes y menos costosos. Un claro ejemplo de esto se
encuentra en la enfermedad de Gaucher, un defecto congénito del metabolismo lipídico,
cuyo tratamiento es la enzima glucocerebrosidasa, y antes de la producción de la proteína
recombinante, era necesario procesar 50.000 placentas humanas para suministrar el
tratamiento anual de un paciente. En segundo lugar, gracias a que se utilizan genes
humanos, la actividad de la proteína recombinante es más específica y hay un reducido
riesgo de causar rechazo inmunológico, en comparación con las que provienen de fuentes
animales. Finalmente, la producción de proteínas terapéuticas por técnicas recombinantes
también elimina el riesgo de transmitir enfermedades contagiosas causadas por virus o
priones que podrían estar presentes en las fuentes de aislamiento.
Las proteínas terapéuticas tienen aplicación en muchos campos, como tratamiento de
reemplazo enzimático de muchas enfermedades, pero también son producidas con el
objetivo de usarlas como vacunas y anticuerpos para el tratamiento de cáncer. Pueden
clasificarse de acuerdo al mecanismo de acción en cuatro categorías (Leader et al., 2008):
Grupo I: Proteínas con actividad enzimática o regulatoria
Grupo II: Proteínas con actividad de reconocimiento específico
Grupo II: Vacunas

Capítulo 1 7
Grupo IV: Proteínas para diagnóstico
1.2 Sistemas de producción de proteínas terapéuticas
Tradicionalmente la mayoría de proteínas recombinantes, incluyendo las usadas como
biofármacos, son producidas en sistemas eucarióticos como células de mamífero (45%),
particularmente células de Ovario de Hámster Chino – CHO (35%), levaduras (15%) y
células de insecto (sistemas de baculovirus) o en sistemas bacterianos (40%) (Rader,
2008). Sin embargo, las células vegetales presentan muchas ventajas y en la última
década se han transformado en un sistema de producción muy promisorio.
En más de cinco décadas de trabajo con cultivos de células vegetales en suspensión para
la producción de metabolitos secundarios se han establecido decenas de líneas celulares
y se ha logrado acumular una gran cantidad de conocimiento del metabolismo de la célula
vegetal, de su comportamiento en cultivo y de la optimización del crecimiento de las
mismas. Las células vegetales son capaces de llevar a cabo casi todas las modificaciones
pos-traduccionales que ocurren en las células humanas, pueden llevar a cabo un
plegamiento correcto y ensamblar proteínas multiméricas (Wang & Ma, 2012). Esto
representa una ventaja significativa frente a los sistemas de producción procarióticos y de
levaduras.
Durante muchos años las células de mamífero fueron el sistema de preferencia para
obtener proteínas biofarmacéuticas en particular por la autenticidad del producto, a pesar
de esto presentan como desventajas el alto riesgo de contaminación y transmisión de
patógenos humanos (principalmente virus y priones) y los altos costos del cultivo, debido
a que se requieren medios de cultivo muy elaborados, ricos en vitaminas, factores de
crecimiento y otros componentes (Huang & McDonald, 2011; Wang & Ma, 2012). Estos
medios con alto contenido proteico también dificultan las operaciones de separación y
purificación haciendo que el procesamiento corriente abajo tenga también un costo
elevado (Huang & McDonald, 2009).
Las células vegetales se fortalecieron como plataforma para la producción de proteínas
recombinantes probablemente en 2009, luego de que la empresa biotecnológica Genzyme
tuvo que interrumpir la producción de la droga Cerezyme® para la terapia de la enfermedad

8 Evaluación del potencial del cultivo de células en suspensión de papa (Solanum
tuberosum subsp. andígena) como plataforma de producción de proteínas
recombinantes
de Gaucher, debido a la contaminación de la línea celular CHO con un calicivirus (Rybicki,
2010). Después de esto, Protalix Biotherapeutics reconoció y aprovechó el potencial de las
células vegetales y actualmente produce en cultivos de células de zanahoria la
glucocerebrosidasa, que es la primera proteína recombinante producida en células
vegetales que ha salido al mercado (Shaaltiel et al., 2007). Más aún, el perfil
farmacocinético de la enzima producida en células de zanahoria mostró que ésta tiene una
mayor vida media que su contraparte producida en células de mamífero (Aviezer et al.,
2009), demostrando el potencial de las plataformas vegetales para producir proteínas
terapéuticas de buena calidad.
Otro sistema de producción de proteínas recombinantes son las plantas transgénicas, con
muy bajos costos del proceso, comparable con los sistemas procarióticos. Sin embargo,
en este caso el tiempo de cultivo es muy prolongado y existe riesgo de contaminación por
micotoxinas, pesticidas, herbicidas y metabolitos secundarios endógenos (Franconi et al.,
2010; Shih & Doran, 2009). Adicionalmente, la producción en cultivos de células vegetales
permite un mayor control del ambiente, ofreciendo rendimientos reproducibles y mayor
consistencia en la homogeneidad y calidad el producto (Doran, 2013; Huang & McDonald,
2011; Xu et al., 2011). La producción en biorreactores permite, además, la optimización de
las condiciones del cultivo, del modo de operación, el uso de promotores inducibles para
separar las fases de crecimiento de la biomasa y de producción de la proteína (Huang &
McDonald, 2011; Wang & Ma, 2012). Todo lo anterior contribuye a que, mediante la
producción in vitro, sea más sencillo cumplir las normas de buenas prácticas de
manufactura que regulan la producción del biofármacos. Es importante considerar que un
sistema de producción no es necesariamente el mejor en todos los casos; se requiere el
análisis particular de cada proteína teniendo en cuenta el costo de producción, el volumen
del mercado, la eficacia, la seguridad, la estabilidad del producto y si la proteína
recombinante tiene propiedades bioquímicas y farmacológicas diferentes a la proteína
nativa (Doran, 2000). El cultivo de células vegetales en suspensión puede ser el sistema
de producción más adecuado cuando se requieren cantidades pequeñas a medianas de
una proteína especial, de alto valor comercial y con requerimientos de pureza muy
elevados, como es el caso de muchas aplicaciones terapéuticas (Doran, 2013).

Capítulo 1 9
1.3 Cultivo in vitro y producción de proteínas terapeúticas en papa
Muchas investigaciones han tenido como objeto de estudio las especies pertenecientes a
la familia Solanaceae debido a su importancia económica. En particular, los estudios
genéticos y genómicos se han enfocado principalmente en la papa y el tomate ya que, a
diferencia de otras especies modelo como Arabidopsis thaliana y Nicotiana tabacum,
producen órganos de interés comercial (el fruto en el caso del tomate y el tubérculo en la
papa) (Rigano et al., 2013). La papa (Solanum tuberosum L.) es la especie de la familia
Solanaceae más cultivada en todo el mundo (International Solanaceae Initiative, n.d.). Esta
planta ha sido modelo para el estudio de diversos procesos biológicos como la tuberización
(Bachem et al., 1996; Jackson, 1999; Müller et al., 1992; Taylor et al., 1992), la síntesis de
almidón (Riesmeier et al., 1994; Roessne et al., 2000; Visser et al., 1991) y la respuesta a
ataques de patógenos (Kombrink et al., 1988; Leister et al., 1996).
Hace más de tres décadas se han establecido cultivos in vitro de Solanum tuberosum L.,
en la mayoría de los casos con el objetivo último de mejorar características agronómicas.
Las suspensiones celulares han sido utilizadas como un modelo para estudiar rutas
metabólicas (Romano et al., 2003; Zacharius et al., 1985), epigenética (Law & Suttle,
2005), variación somaclonal (Munir et al., 2011), expresión génica (Ambrosone et al., 2012;
El-Banna et al., 2010; Johnson, 2001), entre otros aspectos genéticos, bioquímicos y
fisiológicos relevantes (Mullins et al., 2006). Sin embargo hasta el momento no se han
establecido bioprocesos a partir de cultivos de células en suspensión de papa por lo que
se desconocen reportes de parámetros que serían fundamentales como la cinética de
crecimiento (tasa específica de crecimiento, tiempo de duplicación), sensibilidad al estrés
hidrodinámico, demanda de oxígeno, capacidad de acumular proteína extracelular, entre
otros. Además, se ha reportado que la composición adecuada del medio de cultivo para
lograr un establecimiento de callos friables y de crecimiento rápido, depende del genotipo
(Shahab-ud-din et al., 2011; Turhan, 2004). Para establecer cultivos de células en
suspensión de una nueva variedad de papa es necesario entonces encontrar la
composición del medio que favorece el crecimiento del callo y la suspensión de este
genotipo.

10 Evaluación del potencial del cultivo de células en suspensión de papa (Solanum
tuberosum subsp. andígena) como plataforma de producción de proteínas
recombinantes
La comprensión de la genética también ha sido el objetivo de muchas investigaciones en
papa para ganar herramientas de manipulación y mejoramiento de las características
genéticas (Mullins et al., 2006). El genoma completo de la papa, que ya fue secuenciado,
tiene un tamaño relativamente pequeño (844 Mb), con alrededor de 39.000 genes que
codifican proteínas (Xu et al., 2011). Esta especie tiene además un tiempo de generación
corto, se adapta bien a las condiciones de cultivo in vitro y se han establecido numerosos
protocolos de transformación (Rigano et al., 2013). En Colombia, se han logrado manipular
genéticamente variedades de la subespecie andígena utilizando Agrobacterium
tumefaciens y obteniendo eficiencias de transformación superiores al 30% (López &
Chaparro, 2007; Torres et al., 2012). Todas estas características permiten el desarrollo
rápido y exitoso de herramientas genéticas y genómicas que contribuyen a hacer de esta
especie una plataforma vegetal promisoria para la producción de proteínas recombinantes.
La manipulación genética también ha permitido desarrollar plantas de papa productoras de
proteínas terapéuticas con actividad biológica. Tacket et al. (1998) produjeron una proteína
antigénica de E. coli en tubérculos de papa para ser administrada como vacuna oral. Los
ensayos clínicos mostraron la estimulación en la producción de inmunoglobulinas IgG e
IgA en los pacientes que ingirieron los tubérculos transformados. También se ha reportado
la expresión de vacunas orales de uso agropecuario, como es el caso del antígeno VP1
del virus de la fiebre aftosa (He et al., 2007).
La interleuquina-2 (IL-2) y la interleuquina-4 (IL-4) humanas se han producido
exitosamente en plantas de papa transgénicas (Ma et al., 2005; Park & Cheong, 2002).
Estas proteínas son citoquinas de importancia farmacológica y tienen gran potencial en el
tratamiento del cáncer, enfermedades virales y desórdenes autoinmunes por su papel en
la regulación del sistema inmunológico. El bioensayo reportado por Park & Cheong (2002)
reveló que la proteína humana expresada en los tubérculos estimuló la proliferación de
células dependientes de IL-2 (CTLL-2). En el estudio de Ma et al. (2005) también se reportó
proliferación de células T en los ensayos in vitro, mostrando que la proteína recombinante
producida en papa conservó su actividad biológica.

Capítulo 1 11
Otras proteínas terapéuticas expresadas en plantas de papa son la estafiloquinasa
(Gerszberg et al., 2012), que es un activador plasminógeno de origen bacteriano utilizado
en terapias trombolíticas, y el antígeno superficial del virus de la hepatitis B (Rukavtsova
et al., 2015).
1.4 Antecedentes de Solanum tuberosum subsp. andígena variedad Pastusa Suprema
Los sistemas vegetales en muchos casos son la mejor alternativa para la producción a
gran escala de proteínas recombinantes; sin embargo, se requieren largos períodos de
tiempo (entre meses y años) para el desarrollo y la optimización de la técnica de
transformación, la selección de un evento independiente de transformación y la
regeneración de plantas (Huang & McDonald, 2009; Twyman et al., 2003). Por esta razón,
para este estudio fue importante explorar el potencial como plataforma de producción de
proteínas recombinantes de una variedad de papa en la cual se han obtenido previamente
resultados de transformación genética y regeneración in vitro de plantas.
Un equipo de investigadores de la Universidad Nacional de Colombia Sede Bogotá
desarrolló tres nuevas variedades de papa de la subespecie andígena, entre las que se
destaca la variedad Pastusa Suprema obtenida mediante técnicas convencionales de
mejoramiento genético, por el cruce de un clon interespecífico (Solanum stoloniferum
230490 × Solanum phureja var. Yema de huevo) con Solanum tuberosum subsp. andígena
var. Parda pastusa (López & Chaparro, 2007).
En general hay muy pocos reportes de transformación genética en cultivares
suramericanos de la subespecie andígena, probablemente debido a la baja eficiencia de
regeneración de brotes en estos genotipos (Trujillo et al., 2001). Sin embargo, ya se ha
logrado introducir y expresar genes foráneos en la variedad androestéril Pastusa Suprema,
alcanzando incluso eficiencias de transformación superiores al 30% (López & Chaparro,
2007; Torres et al., 2012). López & Chaparro (2007) lograron la transformación y expresión
del gen reportero de la β-glucoronidasa, mientras que Torres et al. (2012) lo hicieron con
el gen cry1Ac de Bacillus thuringiensis, obteniendo líneas potencialmente resistentes a
Tecia solanivora.

12 Evaluación del potencial del cultivo de células en suspensión de papa (Solanum
tuberosum subsp. andígena) como plataforma de producción de proteínas
recombinantes
En el reporte de Torres et al. (2012) se utilizó el vector p1AcPRD con un promotor 2X
CaMV35S (Figura 1). Lograron obtener un 44% de regeneración de brotes transfectados
gracias la modificación del tiempo y medio de infección en el protocolo reportado
previamente por Trujillo et al. (2001) para variedades de la subespecie andígena. La
adición de acetosiringona que es un activador transcripcional de los genes vir (Stachel et
al., 1986) en el medio de infección fue probablemente uno de los factores más importantes.
La eficiencia de transformación obtenida fue de 22 y 37%. El porcentaje de la proteína
Cry1Ac estuvo entre 2,5 y 6,4% del total de proteína soluble, que corresponde a niveles
de expresión entre 88 y 639 ng/mg de tejido fresco.
Figura 1-1. Mapa linear del cassette con los genes nptII y cry1Ac. RB, borde derecho de
la region T-DNA; LB, borde izquierdo de la región T-DNA; CaMV35S-2X, promotor del virus
mosaico del coliflor 2X; P nos, promotor de la nopalin sintetasa; T nos, terminador de la
nopalin sintetasa; nptII, gen de la neomicin fosfotransferasa; Cry1Ac, gen cry1Ac B.
thuringiensis (Tomado de Torres et al., 2012).

2. Objetivos
2.1 Objetivo general
Establecer cultivos de células en suspensión de Solanum tuberosum subsp. andígena var.
Pastusa Suprema y explorar su potencial como plataforma de producción de proteínas
recombinantes utilizando Bt-Cry1Ac como proteína modelo.
2.2 Objetivos específicos
Evaluar el fotoperiodo, tipo de explante y composición hormonal que favorecen la
inducción de callos de Solanum tuberosum subsp. andígena var. Pastusa Suprema
en medio basal MS.
Evaluar la composición hormonal que favorece el crecimiento de suspensiones
celulares de Solanum tuberosum subsp. andígena var. Pastusa Suprema en medio
basal MS.
Caracterizar la cinética de crecimiento y consumo de sustrato del cultivo de células
en suspensión de Solanum tuberosum subsp. andígena var. Pastusa Suprema en
medio basal MS.
Establecer cultivos de células en suspensión a partir de plantas de Solanum
tuberosum subsp. andígena var. Pastusa Suprema transformadas con el gen
cry1Ac de Bacillus thuringiensis y evaluar si hay producción de la proteína
recombinante.


3. Metodología
3.1 Material vegetal
El material vegetal de partida fueron plantas libres de virus de Solanum tuberosum subsp.
andígena var. Pastusa Suprema proporcionadas por la Corporación para Investigaciones
Biológicas – CIB. La propagación in vitro se realizó en medio MS (Murashige & Skoog,
1962) sólido (agar 7 g/l), suplementado con 30 g/l de sacarosa y pH ajustado a 5.7 antes
de la esterilización en autoclave (121ºC, 15 psi, 15 min.). Los cultivos se incubaron a 20°C
y fotoperiodo 16 horas luz/ 8 horas oscuridad (Torres et al., 2012).
3.2 Inducción de callos
Se realizaron experimentos exploratorios para evaluar la condición de fotoperiodo y el tipo
de explante que favorecen la formación de callo friable. Se tomaron cortes de tallo como
explantes y se sembraron en dos frascos con medio de igual composición hormonal; en
total 25 parejas de cultivos correspondientes a los tratamientos hormonales descritos más
adelante. De cada pareja un cultivo se expuso al fotoperiodo 16 horas luz/8 horas
oscuridad y el otro se mantuvo en oscuridad permanente durante dos semanas.
Transcurrido este tiempo se evaluó cualitativamente el efecto de estas dos condiciones
lumínicas sobre la respuesta de los explantes al cultivo in vitro, en un análisis de muestras
pareadas de cada cultivo incubado en fotoperiodo frente a su contraparte en oscuridad.
Los cortes de tallo fueron tomados como explante para todos los experimentos por ser el
tejido de mayor abundancia y más rápido crecimiento en las plantas de la variedad de
interés cultivadas in vitro, en comparación con las hojas y los tubérculos que también han
sido reportados como explantes en otras variedades de S. tuberosum (Naik & Widholm,
1993; Zacharius et al., 1985). Se comparó la respuesta a la inducción de callos de nudos
y entrenudos, marcando una división en cada frasco de cultivo correspondiente a los

16 Evaluación del potencial del cultivo de células en suspensión de papa (Solanum
tuberosum subsp. andígena) como plataforma de producción de proteínas
recombinantes
tratamientos de composición hormonal y sembrando de un lado nudos y del otro,
entrenudos. Al igual que en el análisis del efecto del fotoperiodo, el análisis de muestras
pareadas hizo posible separar el efecto de la composición hormonal del medio y el tipo de
tejido. La respuesta fue evaluada cualitativamente en términos de organogénesis y
formación de callo friable.
Una vez encontradas el fotoperiodo y tipo de explante que favorecen la inducción de callos
se evaluó el efecto de la composición hormonal sobre la formación de callos friables. Se
sembraron cortes de entrenudos de 2 a 5 mm de longitud en medio MS suplementado con
30 g/l de sacarosa, 100 mg/l de ácido ascórbico y distintas composiciones hormonales de
una auxina y/o una citoquinina. Se determinó el efecto de la composición hormonal del
medio de cultivo sobre la inducción de callos friables en un total de 25 tratamientos, cada
uno con una combinación de una auxina y una citoquinina, en un diseño de bloques
completos al azar con dos repeticiones y cada unidad experimental de 10 explantes. Las
fitohormonas empleadas fueron ácido 2,4-diclorofenoxiacético (2,4-D), ácido α-
naftalenacético (ANA), 6-bencil aminopurina (BAP) y 6-furfurilaminopurina (quinetina), en
concentraciones de 0; 1,5 y 3,0 mg/l cada una. El pH del medio fue ajustado a 5,8 antes
de la esterilización en autoclave (121ºC, 15 psi, 15 min.). La inducción y cultivo de callos
se hizo a 20°C en oscuridad.
Tras dos semanas de cultivo, se determinó el porcentaje de formación de callos friables,
así:
𝑃𝑜𝑟𝑐𝑒𝑛𝑡𝑎𝑗𝑒 𝑑𝑒 𝑓𝑜𝑟𝑚𝑎𝑐𝑖ó𝑛 𝑑𝑒 𝑐𝑎𝑙𝑙𝑜 𝑓𝑟𝑖𝑎𝑏𝑙𝑒 =𝑛ú𝑚𝑒𝑟𝑜 𝑑𝑒 𝑒𝑥𝑝𝑙𝑎𝑛𝑡𝑒𝑠 𝑞𝑢𝑒 𝑓𝑜𝑟𝑚𝑎𝑟𝑜𝑛 𝑐𝑎𝑙𝑙𝑜 𝑓𝑟𝑖𝑎𝑏𝑙𝑒
𝑛ú𝑚𝑒𝑟𝑜 𝑑𝑒 𝑒𝑥𝑝𝑙𝑎𝑛𝑡𝑒𝑠 𝑡𝑜𝑡𝑎𝑙𝑒𝑠×
100 (1)
Los explantes fueron transferidos cada dos semanas a medio fresco. En el tercer
subcultivo, transcurridas seis semanas desde la inducción, se rasparon los callos para
separarlos del explante inicial. Los callos friables se transfirieron cada dos semanas a
medio fresco.

3. Metodología 17
En el análisis de resultados se realizó un análisis de varianza, previa verificación de los
supuestos de normalidad según la prueba de Shapiro-Wilk, y comparación de medias con
la prueba de Duncan para determinar si existía diferencia estadísticamente significativa
entre los tratamientos. En todos los casos se utilizó un nivel de significancia α= 0,05.
3.3 Establecimiento de suspensiones celulares
Los cultivos en suspensión se establecieron transfiriendo el callo friable a un matraz de
150 ml con 50 ml de medio MS líquido suplementado con 30 g/l de sacarosa, 100 mg/l de
ácido ascórbico y pH ajustado a 5.8 antes de la esterilización en autoclave (121ºC, 15 psi,
15 min.). Las suspensiones celulares fueron tapadas con papel aluminio, selladas con
vinipel y mantenidas en agitador rotatorio a 120 rpm, 20°C y oscuridad. Cada dos semanas
se cambió el medio gastado por medio fresco.
Se determinó el efecto de la composición hormonal sobre el crecimiento de las células en
suspensión, en un diseño completamente al azar con tres repeticiones y cuatro
tratamientos: ANA 0; 0,5; 1,5 y 3,0 mg/L. Estos tratamientos se escogieron de acuerdo con
los resultados obtenidos en la etapa de inducción de callos friables. Se utilizó un inóculo
de aproximadamente 1 g/l (peso seco), con un volumen final de 20 ml de medio de cultivo
en matraz Erlenmeyer de 100 ml de volumen útil. Luego de cuatro semanas se midió el
peso seco de la biomasa final, para determinar el tratamiento en el que hubo mayor
crecimiento.
3.4 Cinética de crecimiento de biomasa y consumo de sustrato
Se determinó la curva de crecimiento de biomasa y consumo de sustrato de los cultivos de
células en suspensión de Solanum tuberosum subsp. andígena var. Pastusa Suprema. Se
preparó el inóculo mezclando tres cultivos vigorosos y de alta viabilidad y realizando dos
lavados con medio MS fresco suplementado con 30 g/L de sacarosa y 0,5 g/L de ANA. La
concentración inicial de biomasa en cada cultivo fue aproximadamente de 2 g/L en peso
seco (5 ml de inóculo en 15 ml de medio fresco). El volumen de trabajo fue de 20 ml en
matraces Erlenmeyer de 100 ml de volumen útil. Los cultivos se mantuvieron a las mismas
condiciones descritas anteriormente. Se realizaron mediciones de concentración de
biomasa y de azúcares residuales por duplicado cada dos días durante 30 días.

18 Evaluación del potencial del cultivo de células en suspensión de papa (Solanum
tuberosum subsp. andígena) como plataforma de producción de proteínas
recombinantes
Los parámetros cinéticos en la etapa de crecimiento exponencial se calcularon así (Doran,
1995):
𝜇𝑚á𝑥 =𝐿𝑛 (𝑋𝑓 𝑋𝑖⁄ )
(𝑡𝑓 − 𝑡𝑖)⁄ (2)
𝑡𝑑 = 𝐿𝑛 2𝜇𝑚á𝑥
⁄ (3)
𝜇𝑚á𝑥: Tasa específica de crecimiento en la fase exponencial (d-1)
𝑋𝑓: Concentración final de biomasa en la fase exponencial (g/l)
𝑋𝑖: Concentración inicial de biomasa en la fase de crecimiento exponencial (g/l)
𝑡𝑓: Tiempo final de biomasa en la fase exponencial (d)
𝑡𝑖: Tiempo inicial de biomasa en la fase de crecimiento exponencial (d)
𝑡𝑑: Tiempo de duplicación en la fase de crecimiento exponencial (d)
3.5 Establecimiento de suspensiones celulares a partir de plantas transformadas
La línea PS 40.27 de plantas de Solanum tuberosum subsp. andígena var. Pastusa
Suprema transformadas con el gen Cry1Ac de Bacillus thuringiensis, fue reportada
previamente como la línea de más alta expresión de proteína recombinante (Torres et al.,
2012) por lo que se escogió como modelo de sistema transgénico para este trabajo.
Se realizó inducción de callos y establecimiento suspensiones utilizando el procedimiento
estandarizado para las plantas no transformadas (Solanum tuberosum subsp. andígena
var. Pastusa Suprema). Las plantas fueron provistas por la Unidad de Biotecnología
Vegetal de la Corporación para Investigaciones Biológicas – CIB.

3. Metodología 19
3.6 Métodos analíticos
3.6.1 Viabilidad celular
La viabilidad celular se estimó mediante el método de exclusión de colorante vital con Azul
de Evans, mezclando en volúmenes iguales la suspensión celular y una solución 0,05%
(p/v) del colorante. Las células coloreadas fueron consideradas no viables y las hialinas
viables (Bhojwani & Razdan, 1996).
3.6.2 Volumen de células sedimentadas
El volumen de células sedimentadas (VCS) se determinó transfiriendo por lo menos 20ml
de la suspensión celular a una probeta (incertidumbre ±1 ml). Luego de 20 minutos de
sedimentación, se midió el volumen de la biomasa y el volumen total. El volumen de células
sedimentadas es expresado así (Mustafa et al., 2011):
𝑉𝐶𝑆 (%) = (𝑣𝑜𝑙𝑢𝑚𝑒𝑛 𝑑𝑒 𝑏𝑖𝑜𝑚𝑎𝑠𝑎 𝑣𝑜𝑙𝑢𝑚𝑒𝑛 𝑡𝑜𝑡𝑎𝑙)⁄ × 100 (4)
3.6.3 Concentración de biomasa en peso seco
Se separó la biomasa del medio extracelular utilizando filtración al vacío y papel filtro
cualitativo (gramaje 84 g/m2, tamaño de poro 12-15 µm). La concentración de biomasa en
peso seco fue determinada secando la biomasa fresca filtrada en horno a 60ºC durante 24
horas.
𝑋 = 𝑝𝑒𝑠𝑜 𝑠𝑒𝑐𝑜 𝑑𝑒 𝑙𝑎 𝑏𝑖𝑜𝑚𝑎𝑠𝑎 𝑣𝑜𝑙𝑢𝑚𝑒𝑛 𝑑𝑒𝑙 𝑐𝑢𝑙𝑡𝑖𝑣𝑜⁄ (5)
3.6.4 Concentración de azúcares residuales
La medición de azúcares residuales se hizo por HPLC acoplado a un detector de índice de
refacción (rid-10ª) usando una columna amino, fase móvil de acetonitrilo/agua (80:20), flujo
de 2 ml/min a 30ºC, resolviendo picos de fructosa (tR = 7.013 min) y glucosa (tR = 8.037
min). El análisis de las muestras se realizó junto con estándares de fructosa y glucosa a
20 g/L cada uno.

20 Evaluación del potencial del cultivo de células en suspensión de papa (Solanum
tuberosum subsp. andígena) como plataforma de producción de proteínas
recombinantes
3.6.5 Detección de la expresión de proteína recombinante
Se evaluó la expresión de la proteína recombinante Cry1Ac en el entrenudos de las
plantas, callos y suspensiones, por medio del ensayo DAS-ELISA según el procedimiento
descrito por Torres et al. (2012). Se utilizó el kit Bt-Cry1Ab/1Ac (PSP 06200, Agdia Inc.,
Elkhart, IN, USA), siguiendo el procedimiento recomendado por el fabricante con algunas
modificaciones. Las muestras fueron preparadas macerando 100 mg de material vegetal
(tejido o biomasa fresca) en 1 ml de buffer de sales fosfato con Tween-20 (PBST). Se
sirvieron 100 µl del sobrenadante resultante del macerado en el plato de ELISA, seguidos
de 100 µl del conjugado de la enzima. Se incubó en cámara húmeda durante 2 horas, tras
lo cual se realizaron lavados con buffer PBST, se sirvieron 100 µl de sustrato TMB por
pozo y después de 20 minutos de incubación a temperatura ambiente se determinó la
absorbancia a 660 nm utilizando un lector de microplacas (Model 680, Bio Rad
Laboratories, Inc., Hercules, CA, USA). Visualmente se podía pudo determinar el resultado
cualitativo de la prueba mediante la aparición del color azul como indicativo de la presencia
del antígeno de Bt-Cry1Ac en la muestra. Para el ensayo se utilizó el control positivo
Cry1Ab/Cry1Ac del kit y como controles negativos, el buffer PBST y una suspensión no
transformada. Se determinó el valor de corte para reportar un resultado positivo de
expresión de proteína Bt-Cry1Ac con una significancia α= 0,05 así:
𝑉𝑎𝑙𝑜𝑟 𝑑𝑒 𝑐𝑜𝑟𝑡𝑒 = 𝑚𝑒𝑑𝑖𝑎 + 3 ∗ 𝐷𝐸 (6)
Media: promedio de los valores de absorbancia de los controles negativos
DE: desviación estándar de los controles negativos.

4. Resultados y discusión
4.1 Inducción de callos
La respuesta de los tejidos vegetales al cultivo in vitro obedece a mecanismos celulares
complejos que dependen de un gran número de variables. Además de la especie vegetal
y el genotipo, algunas de las más importantes son la cantidad y relación auxina/citoquinina,
el fotoperiodo y el tipo de explante (Bhojwani & Razdan, 1996). Estas variables fueron
evaluadas para estandarizar un protocolo eficiente de obtención de callos friables a partir
de plantas de S. tuberosum subsp. andígena variedad Pastusa Suprema.
La evaluación del efecto de dos condiciones de exposición a la luz, oscuridad y fotoperiodo
16 horas luz/8 horas oscuridad, mostró que en todos los tratamientos de diferente
composición hormonal donde hubo formación de callos, el crecimiento de los explantes
incubados en oscuridad fue mayor que aquellos incubados en fotoperiodo 16 h luz (Figura
4-1).
Figura 4-1. Inducción de callos de Solanum tuberosum subsp. andígena var. Pastusa
Suprema a partir de cortes de tallo en condiciones de oscuridad (izquierda) y fotoperiodo
16 h luz (derecha). Se observó consistentemente un mayor crecimiento en los cultivos
mantenidos en condiciones de oscuridad permanente.

22 Evaluación del potencial del cultivo de células en suspensión de papa (Solanum
tuberosum subsp. andígena) como plataforma de producción de proteínas
recombinantes
Esta respuesta diferenciada ocurre debido a que la luz regula casi todos los procesos
fisiológicos y bioquímicos en las plantas (Kreslavski et al., 2009); la longitud de onda, la
intensidad y los ciclos de luz/oscuridad inciden en las diversas respuestas (Neff et al.,
2000), que además varía entre genotipos y tejidos de una misma planta. En cultivos de S.
tuberosum L. se ha reportado ampliamente el efecto del tipo de luz y el fotoperiodo en el
crecimiento, morfogénesis y tuberización durante el cultivo in vitro de plantas (Seabrook,
2005). El cultivo de callos friables se ha establecido en condiciones de oscuridad para
algunas variedades (Hawkins & Lips, 1997; Naik & Widholm, 1993) y en ciclos de
luz/oscuridad para otras (Lu et al., 2006; Queirós et al., 2007). Sin embargo, en la variedad
Pastusa Suprema no se encuentran reportes previos de la evaluación del efecto de
diferentes condiciones de exposición a la luz sobre la formación de callos friables. Dados
los resultados obtenidos se decidió fijar la condición de oscuridad en el protocolo de
inducción de callos.
En la evaluación del efecto del tipo de explante se encontró que en general los nudos
presentaron mayor organogénesis en comparación con los entrenudos (Figura 4-2), por lo
cual en adelante se utilizaron los entrenudos como tejido de explante para la inducción de
callos. El estado fisiológico del tejido meristemático de los nudos le confiere una mayor
capacidad de respuesta regenerativa ante los estímulos hormonales porque este en la
planta, tiene precisamente la función de formar nuevos tejidos (Taiz & Zeiger, 2002). Al
igual que en otras especies, se ha evidenciado que los requerimientos hormonales para el
cultivo in vitro de S. tuberosum L. son dependientes del tipo de explante (Anjum & Ali,
2004). Se ha reportado la obtención de callos friables a partir de diferentes tejidos como
hojas (Gottstein & Schliemann, 1994; Naik & Widholm, 1993; Queirós et al., 2007; Torabi
et al., 2008), tubérculos (Hawkins & Lips, 1997; Zacharius et al., 1985) y tallos (Lu et al.,
2006; Vargas et al., 2005; Vargas et al., 2008). Hay un reporte de inducción de callos en
la variedad Pastusa Suprema en el que se utilizan entrenudos como explantes (Jiménez
et al., 2009); sin embargo no se conocen reportes previos de la evaluación del tipo de
explante en la callogénesis de esta variedad.

4. Resultados y discusión 23
Figura 4-2. Efecto del tipo de explante (nudo/entrenudo) sobre la respuesta a los
tratamientos de inducción in vitro de callos. En MS suplementado con diferentes
composiciones hormonales se observó siempre mayor tendencia hacia la organogénesis
(formación de tallos y raíces adventicias) en los nudos (N) en comparación con los
entrenudos (EN).
Fijando la condición de oscuridad permanente y los entrenudos como explante en el
protocolo de inducción de callos, se procedió a evaluar la suplementación del medio. Se
encontró que la respuesta varió significativamente según la composición hormonal (Tabla
4-1); en general la formación de callos friables predominó en los tratamientos de medio
suplementado únicamente con una auxina. Luego de dos semanas de cultivo en los
tratamientos ANA 1,5 mg/l y 3 mg/l se logró obtener abundante formación de callo friable
en el 90 y 95% de los explantes cultivados, respectivamente (Tabla 4-1). No hubo
diferencia significativa en el porcentaje de inducción obtenido en estos dos tratamientos
(α=0,05), por lo que se escogió como medio de inducción el medio MS suplementado con
la menor concentración de ANA, es decir 1,5 mg/l. En los tratamientos de medio
suplementado únicamente con 2,4-D (1,5 y 3,0 mg/L) también se encontró formación de
callos fiables pero con un crecimiento más lento, ya que sólo fue evidente luego de cuatro
semanas de cultivo (datos no mostrados). La presencia de la auxina ANA se ha reportado
previamente que favorece la formación de callos a partir de entrenudos en la variedad
Pastusa Suprema (Jiménez et al., 2009).
N
EN
N
EN
(a) (b)

24 Evaluación del potencial del cultivo de células en suspensión de papa (Solanum
tuberosum subsp. andígena) como plataforma de producción de proteínas
recombinantes
Tabla 4-1. Porcentaje de formación de callos friables a partir de entrenudos de Solanum
tuberosum subsp. andígena var. Pastusa Suprema, en medio MS suplementado con
diferentes composiciones hormonales.
* Los porcentajes de inducción con la misma letra no tienen diferencia significativa con
α=0,05.
Auxina Concentración
auxina
(mg/L)
Citoquinina Concentración
citoquinina
(mg/L)
Porcentaje de
formación de callos*
(%)
Ninguna -- Ninguna -- 0d
BAP 1,5 0d
3,0 0d
Quinetina 1,5 0d
3,0 0d
ANA 1,5 Ninguna -- 90a
BAP 1,5 0d
3,0 0d
Quinetina 1,5 0d
3,0 0d
3 Ninguna -- 95a
BAP 1,5 0d
3,0 0d
Quinetina 1,5 0d
3,0 0d
2,4-D 1,5 Ninguna -- 0d
BAP 1,5 0d
3,0 0d
Quinetina 1,5 5d
3,0 0d
3 Ninguna -- 45b
BAP 1,5 0d
3,0 25c
Quinetina 1,5 0d
3,0 0d

4. Resultados y discusión 25
En la Tabla 4-2 se compara la composición hormonal del medio de cultivo para la inducción
de callos de diferentes variedades de Solanum tuberosum L. Debido al efecto del genotipo
sobre la respuesta al cultivo in vitro, la composición hormonal para la formación de callos
es dependiente de la variedad vegetal. Entre las variedades de S. tuberosum comparadas
en la Tabla 4-2, se evidencia que la variedad Pastusa Suprema requiere de forma neta
menor cantidad de hormonas para el crecimiento de callos, lo cual en un bioproceso
significa una reducción del costo total de producción. Más aún, queda abierta la posibilidad
de evaluar en experimentos posteriores si se forman callos friables de crecimiento rápido
y vigoroso a concentraciones inferiores a 1,5 mg/l de ANA, de tal manera que se logre
disminuir en mayor medida los requerimientos de suplementación del medio de cultivo.
Tabla 4-2. Comparación de la composición hormonal del medio de cultivo en diferentes
reportes de inducción de callos de Solanum tuberosum L., según la variedad y tipo de
explante.
Referencia Variedad Explante Auxina Citoquinina
(Zacharius et al.,
1985)
Kennebec y
Merrimack
Tubérculo
2,4-D 2 mg/L
ANA 6 mg/L
Quinetina
0,21 mg/L
(Naik &
Widholm, 1993)
Kennebec,
Norchip, Red
Pontiac, Russet
Burbank, Russet
Norkotah,
Superior
Raquis de
hoja
ANA 5 mg/L --
(Lu et al., 2006) Chieftain Tallo 2,4-D 0,5mg/L BAP 2 mg/L
(Torabi et al.,
2008)
White Desiree Tallo 2,4-D 2 mg/L
ANA 2 mg/L
Quinetina
2 mg/L
Granger (2016)
(Este trabajo)
Pastusa Suprema Tallo
(entrenudos)
ANA 1,5 mg/L --

26 Evaluación del potencial del cultivo de células en suspensión de papa (Solanum
tuberosum subsp. andígena) como plataforma de producción de proteínas
recombinantes
Las condiciones evaluadas de tipo de explante, fotoperiodo y composición hormonal del
medio permitieron llegar a un protocolo eficiente de obtención de callos friables. Las
plantas propagadas a partir de esquejes rápidamente crecen hasta alcanzar alrededor de
8 cm en sólo cuatro semanas, formando tallos jóvenes con entrenudos largos (Figura 4-
3a). De estas plantas se logran sacar entre uno y dos cultivos para iniciar callos,
sembrando 10 explantes por frasco (Figura 4-3b). En el medio MS suplementado con 1,5
mg/l de ANA y en oscuridad, luego de dos semanas se comienza a evidenciar la formación
de callo friable y entre la semana tres y cuatro es lo suficientemente abundante para hacer
el raspado (Figura 4-3c). Se evidenció que el raspado es un paso crítico en estos cultivos
porque a partir de ahí los callos se oxidan y, aunque continúan creciendo, esto puede
reducir la viabilidad de las células y disminuir la eficiencia en la obtención de suspensiones.
Este fenómeno se podría controlar utilizando aditivos como el carbón activado (Bhojwani
& Razdan, 1996) o ensayando un protocolo alternativo para establecer suspensiones en
el que se evite el paso de raspado de los callos.
Figura 4-3. (a) Plantas de Solanum tuberosum subsp. andígena var. Pastusa Suprema
cultivadas in vitro. (b) Entrenudos utilizados como explantes para la inducción de callos en
medio Murashige & Skoog (MS). (c) Callos friables obtenidos luego de tres semanas de
cultivo en medio MS suplementado con ácido naftalenacético (ANA) 1,5 mg/l.
(b)
(c)
(a)

4. Resultados y discusión 27
4.2 Establecimiento de suspensiones celulares
A partir de los callos friables se establecieron suspensiones celulares en medio MS líquido.
Teniendo en cuenta los resultados obtenidos en la inducción de callos, se evaluó el efecto
de la concentración de ANA sobre el crecimiento de los cultivos de células en suspensión.
La mayor concentración de biomasa final fue 6,71 ± 1,65 g/l y se obtuvo en el medio
suplementado con ANA 0,5 mg/l. Esto quiere decir que en el medio con la menor
concentración evaluada se logró aumentar más de seis veces la concentración inicial de
biomasa tras cuatro semanas de cultivo. En los tratamientos se observó una tendencia a
aumentar el crecimiento de la biomasa a medida que la concentración de hormona
disminuye (Figura 4-4); por esta razón se recomienda evaluar el comportamiento de los
cultivos con concentraciones de ANA menores a 0,5 mg/l, ya que de obtenerse una mayor
producción de biomasa se favorece aún más el costo total del bioproceso.
Figura 4-4. Efecto de la concentración de ácido α-naftalenacético (ANA) en el medio de
cultivo sobre el crecimiento de las suspensiones celulares de Solanum tuberosum subsp.
andígena var. Pastusa Suprema.
28 Evaluación del potencial del cultivo de células en suspensión de papa (Solanum
tuberosum subsp. andígena) como plataforma de producción de proteínas
recombinantes
El protocolo de inducción de callos en medio MS suplementado con ANA 1,5 mg/l y
establecimiento de suspensiones en medio MS suplementado con ANA 0,5 mg/l se utilizó
durante 10 meses para hacer un seguimiento a los cultivos obtenidos, en cuanto a la
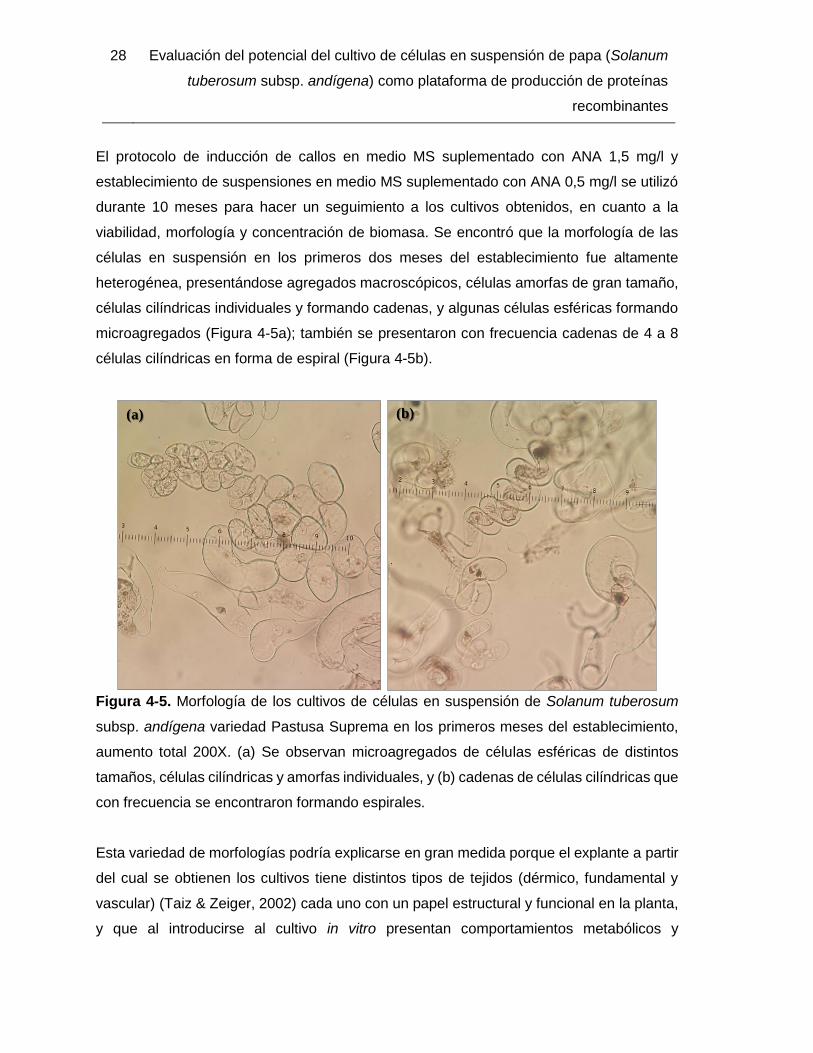
viabilidad, morfología y concentración de biomasa. Se encontró que la morfología de las
células en suspensión en los primeros dos meses del establecimiento fue altamente
heterogénea, presentándose agregados macroscópicos, células amorfas de gran tamaño,
células cilíndricas individuales y formando cadenas, y algunas células esféricas formando
microagregados (Figura 4-5a); también se presentaron con frecuencia cadenas de 4 a 8
células cilíndricas en forma de espiral (Figura 4-5b).
Figura 4-5. Morfología de los cultivos de células en suspensión de Solanum tuberosum
subsp. andígena variedad Pastusa Suprema en los primeros meses del establecimiento,
aumento total 200X. (a) Se observan microagregados de células esféricas de distintos
tamaños, células cilíndricas y amorfas individuales, y (b) cadenas de células cilíndricas que
con frecuencia se encontraron formando espirales.
Esta variedad de morfologías podría explicarse en gran medida porque el explante a partir
del cual se obtienen los cultivos tiene distintos tipos de tejidos (dérmico, fundamental y
vascular) (Taiz & Zeiger, 2002) cada uno con un papel estructural y funcional en la planta,
y que al introducirse al cultivo in vitro presentan comportamientos metabólicos y
(a) (b)

4. Resultados y discusión 29
morfológicos diferentes (Rodríguez & Galindo, 2003). La morfología de las células es
además una función de la especie vegetal, el método de preparación del inóculo, la
composición del medio, las condiciones del cultivo, la etapa de crecimiento y el tipo de
biorreactor; su caracterización es importante porque tiene efectos sobre los procesos de
transferencia de masa, energía y cantidad de movimiento (Huang & McDonald, 2009;
Kieran et al., 1997).
Figura 4-6. Cultivo de células en suspensión de Solanum tuberosum subsp. andígena
variedad Pastusa Suprema tras 10 meses del establecimiento, aumento total 100X. Tinción
con azul de Evans, las células no viables están coloreadas mientras que las viables
permanecen hialinas.
En los cultivos en suspensión de Solanum tuberosum subsp. andígena var. Pastusa
Suprema de más de diez meses aún se observó mucha heterogeneidad en la morfología
de las células, aunque con cierta predominancia de los microagregados tanto de células
cilíndricas como de células esféricas (Figura 4-6). Estos microagregados son comunes en
los cultivos de células vegetales en suspensión y se han reportado previamente en
diferentes especies vegetales como Solanum chrysotrichum (Trejo et al., 2001), Thevetia
peruviana (Arias, 2013), Oryza sativa, Nicotiana benthamiana y Trichosanthes kirilowii
(McDonald et al., 2001).

30 Evaluación del potencial del cultivo de células en suspensión de papa (Solanum
tuberosum subsp. andígena) como plataforma de producción de proteínas
recombinantes
Mediante el protocolo estandarizado para el establecimiento de cultivos de células en
suspensión, y luego de alrededor de cuatro meses de adaptación a las condiciones del
medio líquido, se logró obtener suspensiones celulares de muy alta densidad celular y
crecimiento vigoroso (Figura 4-7).
Figura 4-7. Suspensiones de Solanum tuberosum subsp. andígena variedad Pastusa
Suprema de alta densidad celular obtenidas mediante inducción de callos en medio MS +
1,5 mg/l de ANA y establecimiento de suspensiones en medio MS + 0,5 mg/l de ANA. (a)
Cultivo en matraz de 100 ml con volumen de células sedimentadas del 100%. (b)
Suspensiones en matraces de 250 ml con alta cantidad de biomasa.
(a)
(b)

4. Resultados y discusión 31
Los cultivos alcanzaron concentraciones de alrededor de 12 g/l de biomasa (peso seco)
con un volumen de células sedimentadas del 95 al 100%. Dada la alta viscosidad
observada en el medio de cultivo de las suspensiones con alta densidad celular, se
presume además que las células podrían estar produciendo polisacáridos extracelulares,
que no sólo mantendrían las interacciones intercelulares favoreciendo la formación de
agregados, sino que también aumentan la viscosidad del medio, disminuyendo el estrés
hidrodinámico (Kieran et al., 1997).
Durante el establecimiento de suspensiones se observó la eventual producción de
compuestos de coloración roja, que se presumen son de naturaleza fenólica, tanto en el
medio extracelular como en la biomasa de algunos de los cultivos (Figura 4-8). No fue
posible encontrar las causas que desencadenan la producción de estos compuestos e
incluso luego de la división de una suspensión celular en dos, bajo las mismas condiciones,
ocurrió que una de ellas se tornaba roja y la otra no. Este proceso ya ha sido reportado
previamente en cultivos de Taxus igualmente como un fenómeno aleatorio (Wickremesinhe
& Arteca, 1994). Una vez se producía este fenómeno no fue posible recuperar las
suspensiones mediante lavados y remplazo del medio, y como se observó que siempre se
frenaba el crecimiento celular se optó por descartar los cultivos una vez aparecía esta
coloración.
Figura 4-8. Suspensiones celulares que produjeron compuestos de coloración roja que se
observó (a) en el medio extracelular y (b) en la biomasa.
(a) (b)

32 Evaluación del potencial del cultivo de células en suspensión de papa (Solanum
tuberosum subsp. andígena) como plataforma de producción de proteínas
recombinantes
4.3 Cinética de crecimiento de biomasa y consumo de sustrato
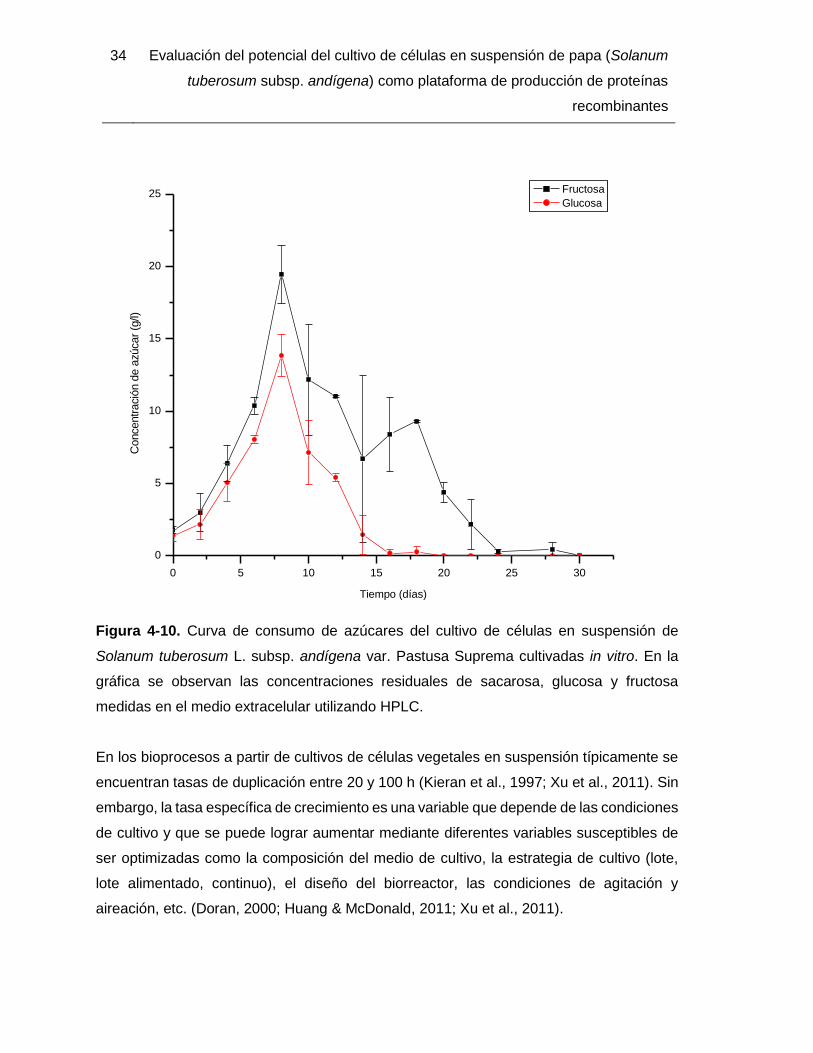
En la evaluación de la cinética de crecimiento de la biomasa se encontró una fase de
aceleración del crecimiento hasta alcanzar la fase exponencial, probablemente por
adaptación a las condiciones de cultivo durante los primeros cuatro días (Figura 4-9). Esta
primera fase se relaciona con el aumento de fructosa y glucosa en cantidades casi
equivalentes que se evidencian en la cinética de consumo de sustrato (Figura 4-10), lo que
bien podría explicarse como un proceso de hidrólisis de sacarosa mediado por la secreción
de enzimas extracelulares. Esta etapa finaliza en el día ocho cuando la glucosa y la
fructosa alcanzan su concentración máxima. A partir del día seis se evidencia el
crecimiento exponencial de la biomasa hasta el día 24, con una tasa específica de
crecimiento máxima (µmáx) de 0,085 d-1, correspondiente a un tiempo de duplicación (td) de
8,15 días y una concentración máxima de biomasa (Xmáx) de 12,11 g/l. Así mismo, en la
curva de consumo de sustrato se evidencia un consumo neto de los azúcares desde el día
ocho al día 30 de cultivo, más rápidamente de la glucosa en comparación con la fructosa.
El día 22 el cultivo entra en una fase estacionaria corta de cuatro días y finalmente en fase
de muerte entre el día 22 y 30, lo cual se relaciona con el agotamiento de la fuente de
carbono y energía que el día 24 llega a cero.

4. Resultados y discusión 33
Figura 4-9. Curva de crecimiento de biomasa del cultivo de células en suspensión de
Solanum tuberosum L. subsp. andígena var. Pastusa Suprema cultivadas in vitro. La
concentración de biomasa se obtuvo por medición de peso seco.
0 5 10 15 20 25 30
2
4
6
8
10
12
14
Concentr
ació
n d
e b
iom
asa (
g/l)
Tiempo (días)
34 Evaluación del potencial del cultivo de células en suspensión de papa (Solanum
tuberosum subsp. andígena) como plataforma de producción de proteínas
recombinantes
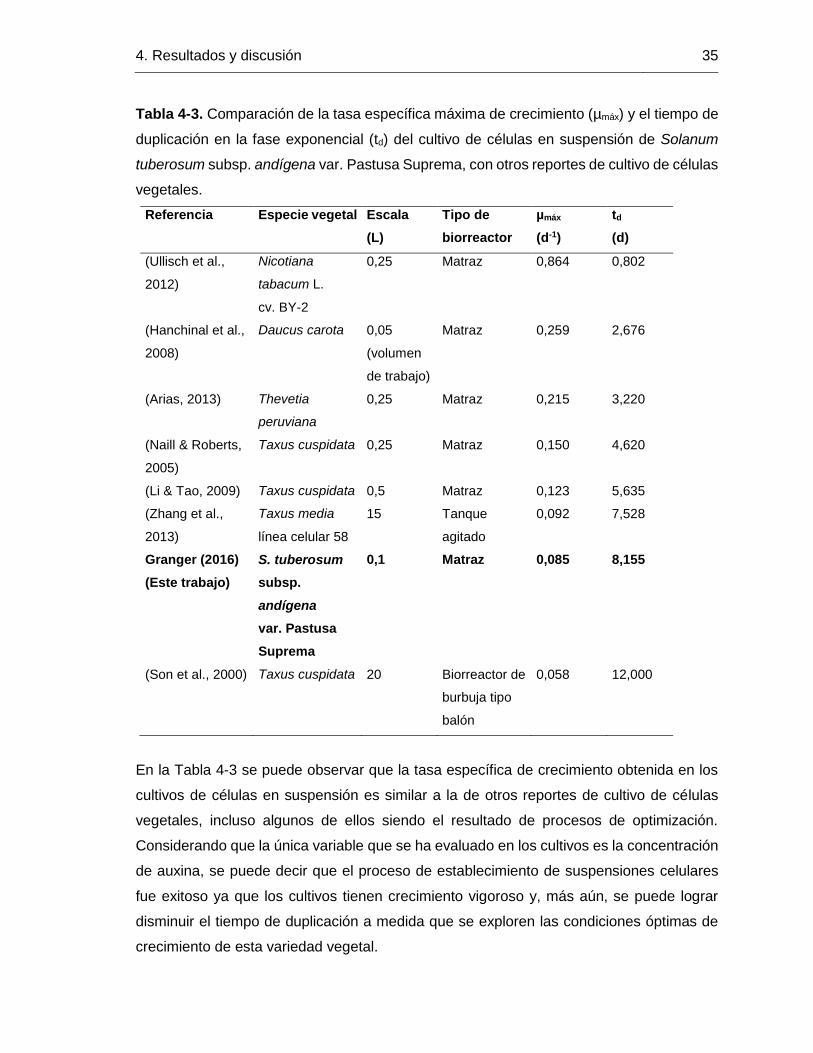
Figura 4-10. Curva de consumo de azúcares del cultivo de células en suspensión de
Solanum tuberosum L. subsp. andígena var. Pastusa Suprema cultivadas in vitro. En la
gráfica se observan las concentraciones residuales de sacarosa, glucosa y fructosa
medidas en el medio extracelular utilizando HPLC.
En los bioprocesos a partir de cultivos de células vegetales en suspensión típicamente se
encuentran tasas de duplicación entre 20 y 100 h (Kieran et al., 1997; Xu et al., 2011). Sin
embargo, la tasa específica de crecimiento es una variable que depende de las condiciones
de cultivo y que se puede lograr aumentar mediante diferentes variables susceptibles de
ser optimizadas como la composición del medio de cultivo, la estrategia de cultivo (lote,
lote alimentado, continuo), el diseño del biorreactor, las condiciones de agitación y
aireación, etc. (Doran, 2000; Huang & McDonald, 2011; Xu et al., 2011).
0 5 10 15 20 25 30
0
5
10
15
20
25
Concentr
ació
n d
e a
zúcar
(g/l)
Tiempo (días)
Fructosa
Glucosa
4. Resultados y discusión 35
Tabla 4-3. Comparación de la tasa específica máxima de crecimiento (µmáx) y el tiempo de
duplicación en la fase exponencial (td) del cultivo de células en suspensión de Solanum
tuberosum subsp. andígena var. Pastusa Suprema, con otros reportes de cultivo de células
vegetales.
Referencia Especie vegetal Escala
(L)
Tipo de
biorreactor
µmáx
(d-1)
td
(d)
(Ullisch et al.,
2012)
Nicotiana
tabacum L.
cv. BY-2
0,25 Matraz 0,864 0,802
(Hanchinal et al.,
2008)
Daucus carota 0,05
(volumen
de trabajo)
Matraz 0,259 2,676
(Arias, 2013) Thevetia
peruviana
0,25 Matraz 0,215 3,220
(Naill & Roberts,
2005)
Taxus cuspidata 0,25 Matraz 0,150 4,620
(Li & Tao, 2009) Taxus cuspidata 0,5 Matraz 0,123 5,635
(Zhang et al.,
2013)
Taxus media
línea celular 58
15 Tanque
agitado
0,092 7,528
Granger (2016)
(Este trabajo)
S. tuberosum
subsp.
andígena
var. Pastusa
Suprema
0,1 Matraz 0,085 8,155
(Son et al., 2000) Taxus cuspidata 20 Biorreactor de
burbuja tipo
balón
0,058 12,000
En la Tabla 4-3 se puede observar que la tasa específica de crecimiento obtenida en los
cultivos de células en suspensión es similar a la de otros reportes de cultivo de células
vegetales, incluso algunos de ellos siendo el resultado de procesos de optimización.
Considerando que la única variable que se ha evaluado en los cultivos es la concentración
de auxina, se puede decir que el proceso de establecimiento de suspensiones celulares
fue exitoso ya que los cultivos tienen crecimiento vigoroso y, más aún, se puede lograr
disminuir el tiempo de duplicación a medida que se exploren las condiciones óptimas de
crecimiento de esta variedad vegetal.

36 Evaluación del potencial del cultivo de células en suspensión de papa (Solanum
tuberosum subsp. andígena) como plataforma de producción de proteínas
recombinantes
Una muestra del alcance que se puede tener mediante procesos de optimización se puede
ver en el cultivo de células vegetales del género Taxus, que es muy reconocido como
bioproceso para la producción de paclitaxel y por lo tanto un punto de referencia en las
plataformas vegetales de producción. Se han reportado, por ejemplo, tasas específicas de
crecimiento entre 0,10 y 0,13 d-1 en el cultivo de suspensiones de Taxus chinensis a escala
de matraz agitado, que variaron dependiendo de la concentración inicial de sustrato y el
tipo de alimentación (lote y lote alimentado) (Wang et al., 1999). El mismo grupo de
investigación logró duplicar la tasa específica de crecimiento al cambiar el biorreactor de
un tanque agitado a una columna de burbujeo (Pan et al, 2000). Otra aproximación ha sido
elaborar modelos del crecimiento de la biomasa para utilizarlos en la optimización del
bioproceso. Se ha reportado el modelo no estructurado del crecimiento de Taxus media y
la optimización del bioproceso de producción de taxol, obteniendo como parámetros del
modelo Xmáx 9,7233 g/l y µmáx 0,1779 d-1 (Zhang et al., 2013).
Se considera que S. tuberosum subsp. andígena variedad Pastusa Suprema es una
plataforma promisoria al haber obtenido una respuesta robusta al cultivo in vitro, bajos
requerimientos hormonales para la inducción de callos y crecimiento de suspensiones y
parámetros cinéticos de crecimiento comparables a los que se han obtenido en otras
plataformas vegetales después de procesos de optimización. Más aún, múltiples enfoques
pueden ser utilizados en adelante para aumentar el crecimiento de la productividad de las
proteínas recombinantes de interés, desde la manipulación de la composición del medio y
de las condiciones del ambiente de cultivo (agitación, aireación, etc.), hasta el diseño y
modo de operación del biorreactor en el escalado. Sólo manipulando la concentración de
componentes del medio basal (nitrógeno, fósforo y micronutrientes) y la fuente de carbono
(sacarosa, glucosa) se puede incrementar entre 3 y 20 veces la productividad (Doran,
2000; Holland et al., 2010; Xu et al., 2011).

4. Resultados y discusión 37
4.4 Establecimiento de suspensiones celulares a partir de plantas transformadas y evaluación de la producción de la proteína Bt-Cry1Ac
Los entrenudos de las plantas de la línea transformadas PS 40.27 dieron positivo para la
expresión del gen Cry1Ac (Figura 4-11), al igual que los callos obtenidos a partir de estos
explantes (datos no mostrados). Este resultado permitió descartar pérdida de la expresión
del gen recombinante por diferentes causas como eventos de variación somaclonal,
pérdida del gen y silenciamiento postranscripcional que han sido reportados previamente
en plantas cultivadas in vitro (Huang & McDonald, 2011; Shih & Doran, 2009; Wang & Ma,
2012) o cambios de ploidía celular durante la formación de callos, que ya ha sido
previamente reportado en papa (Vargas et al., 2008).
Figura 4-11. Ensayo DAS-ELISA de entrenudos de plantas de la línea PS 40.27 de
Solanum tuberosum subsp. andígena var. Pastusa Suprema para la detección de proteína
recombinante Cry1Ac. Los pozos 1 y 2 corresponden a los controles negativos de buffer
PBST, 3 y 4 al control positivo de proteína Cry1Ab/1Ac del kit DAS-ELISA, 5 y 6 a plantas
no transformadas y 7 y 8 a plantas transformadas. Se evidencia cualitativamente el
resultado positivo para la expresión (coloración azul).
La inestabilidad genética de las células desdiferenciadas es un factor potencialmente
limitante en el desarrollo de cultivos para producción de proteínas recombinantes (Reiss,
2003). Se han reportado reducciones significativas en el rendimiento de producto
atribuidas a inestabilidad genética, pérdida del transgen, cambios en la estructura
cromosómica, variación de la tasa de crecimiento y otros cambios genéticos no deseados
en cultivos de células vegetales mantenidas en subcultivo constante (Doran, 2013; Gao et
al., 1991; van der Maas et al., 1994). La expresión de la proteína recombinante en estas
plantas que tienen más de cuatro años de haber sido establecidas in vitro, siendo repicadas

38 Evaluación del potencial del cultivo de células en suspensión de papa (Solanum
tuberosum subsp. andígena) como plataforma de producción de proteínas
recombinantes
cada cuatro a seis meses, refleja tanto la estabilidad genética intrínseca del genotipo como
la del método de transformación mediada por Agrobacterium tumefasciens.
El protocolo estandarizado en las plantas silvestres para establecer cultivos de células en
suspensión se utilizó en la línea PS 40.27 de plantas transformadas para obtener cultivos
en medio líquido. Al igual que en los cultivos de callos y suspensiones obtenidos a partir
de las plantas silvestres, se obtuvo oxidación de las células luego del raspado de los callos
y la consecuente obtención de cultivos con pardeamiento cuando éstos se transfirieron a
medio líquido. Se esperaba entonces que el crecimiento vigoroso de las suspensiones se
observaría luego de alrededor de 10 meses desde el establecimiento, que fue lo que tomó
a los cultivos no transformados adaptarse a las condiciones de medio líquido y aumentar
progresivamente la población de células viables. Sin embargo, se decidió evaluar la
expresión de la proteína recombinante en etapas muy tempranas. Mediante el ensayo de
DAS-ELISA se detectó la expresión de la proteína recombinante Bt-Cry1Ac en todas las
suspensiones evaluadas (Figura 4-12).
Figura 4-12. Ensayo DAS-ELISA del cultivo de células en suspensión de la línea PS 40.27
de Solanum tuberosum subsp. andígena var. Pastusa Suprema para la detección de
proteína recombinante Cry1Ac. Las fila 1 y 2 son duplicados de las muestras de ocho
suspensiones; las filas 3 y 4 son duplicados, donde los pozos A y B corresponden a los
controles negativos y del C al G son concentraciones crecientes del control positivo. Se
evidencia cualitativamente el resultado positivo para la expresión (coloración azul).
1
2
3
4
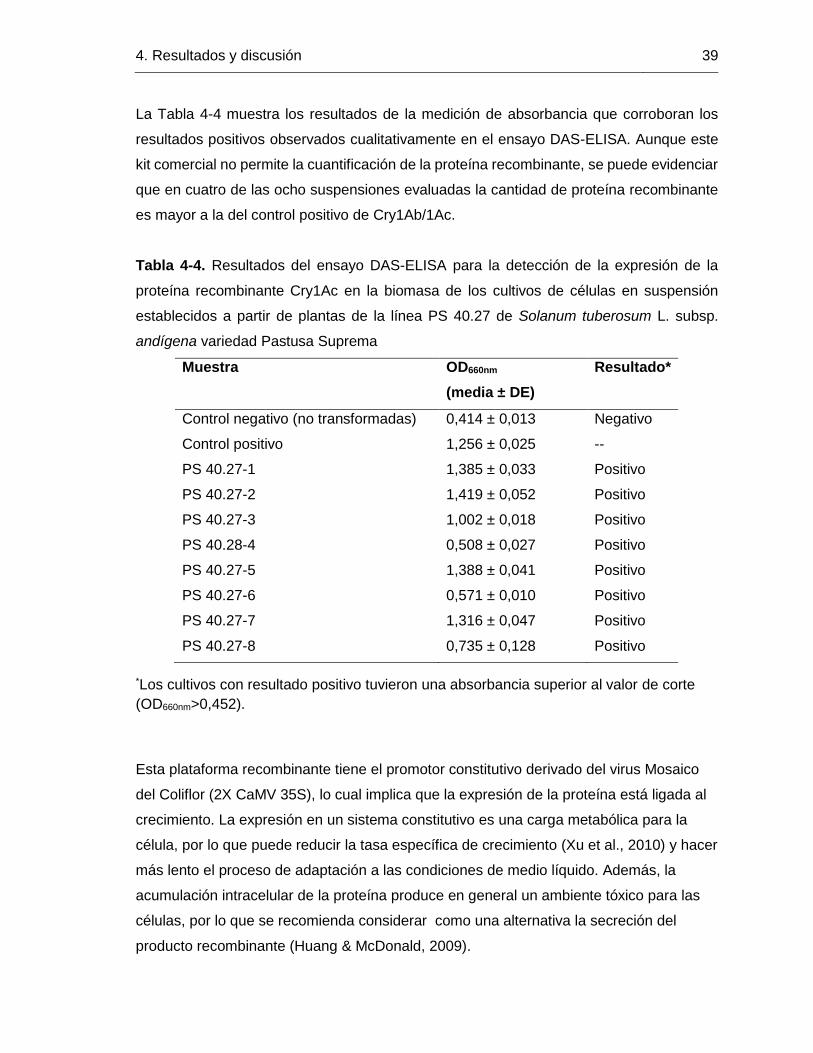
4. Resultados y discusión 39
La Tabla 4-4 muestra los resultados de la medición de absorbancia que corroboran los
resultados positivos observados cualitativamente en el ensayo DAS-ELISA. Aunque este
kit comercial no permite la cuantificación de la proteína recombinante, se puede evidenciar
que en cuatro de las ocho suspensiones evaluadas la cantidad de proteína recombinante
es mayor a la del control positivo de Cry1Ab/1Ac.
Tabla 4-4. Resultados del ensayo DAS-ELISA para la detección de la expresión de la
proteína recombinante Cry1Ac en la biomasa de los cultivos de células en suspensión
establecidos a partir de plantas de la línea PS 40.27 de Solanum tuberosum L. subsp.
andígena variedad Pastusa Suprema
*Los cultivos con resultado positivo tuvieron una absorbancia superior al valor de corte
(OD660nm>0,452).
Esta plataforma recombinante tiene el promotor constitutivo derivado del virus Mosaico
del Coliflor (2X CaMV 35S), lo cual implica que la expresión de la proteína está ligada al
crecimiento. La expresión en un sistema constitutivo es una carga metabólica para la
célula, por lo que puede reducir la tasa específica de crecimiento (Xu et al., 2010) y hacer
más lento el proceso de adaptación a las condiciones de medio líquido. Además, la
acumulación intracelular de la proteína produce en general un ambiente tóxico para las
células, por lo que se recomienda considerar como una alternativa la secreción del
producto recombinante (Huang & McDonald, 2009).
Muestra OD660nm
(media ± DE)
Resultado*
Control negativo (no transformadas) 0,414 ± 0,013 Negativo
Control positivo 1,256 ± 0,025 --
PS 40.27-1 1,385 ± 0,033 Positivo
PS 40.27-2 1,419 ± 0,052 Positivo
PS 40.27-3 1,002 ± 0,018 Positivo
PS 40.28-4 0,508 ± 0,027 Positivo
PS 40.27-5 1,388 ± 0,041 Positivo
PS 40.27-6 0,571 ± 0,010 Positivo
PS 40.27-7 1,316 ± 0,047 Positivo
PS 40.27-8 0,735 ± 0,128 Positivo

40 Evaluación del potencial del cultivo de células en suspensión de papa (Solanum
tuberosum subsp. andígena) como plataforma de producción de proteínas
recombinantes
Se considera que el proceso de establecimiento de suspensiones celulares fue exitoso
porque se conservó la expresión de la proteína recombinante. Más aún, el potencial del
cultivo de células en suspensión transformadas es mucho mayor si se toma en
consideración que al momento de realizar el ensayo DAS-ELISA las suspensiones aún
estaban en etapas tempranas del establecimiento, adaptándose a las condiciones de
cultivo en medio líquido y en consecuencia con baja viabilidad celular (Figura 4-13). Una
vez estas suspensiones alcancen la vigorosidad y altas densidades de biomasa logradas
en los cultivos de células no transformadas, se esperaría superar ampliamente los
niveles de expresión.
Figura 4-13. Morfología y viabilidad del cultivo de células en suspensión de Solanum
tuberosum subsp. andígena var. Pastusa Suprema, línea PS 40.27. Tinción con el
colorante azul de Evans, las células no viables están teñidas por el colorante, mientras las
células viables permanecen hialinas. El porcentaje de viabilidad del cultivo se estimó entre
el 20 y el 30%.
Si bien los resultados obtenidos en la línea PS 40.27 de Solanum tuberosum subsp.
andígena constituyen únicamente un modelo de producción de proteínas recombinantes,
sí permiten evidenciar el potencial de esta plataforma vegetal para la producción de

4. Resultados y discusión 41
proteínas terapéuticas. En esta primera fase se logró con éxito establecer un protocolo
efectivo de obtención de cultivos en suspensión transformadas con expresión del gen
Cry1Ac. En una siguiente fase se evaluarán condiciones que permitan optimizar la
producción de biomasa y de proteína heteróloga.


5. Conclusiones y recomendaciones
5.1 Conclusiones
Este es el primer reporte de establecimiento de cultivos de células en suspensión de la
nueva variedad vegetal Pastusa Suprema. Las condiciones evaluadas de tipo de explante,
fotoperiodo y composición hormonal del medio permitieron llegar a un protocolo eficiente
de obtención de células en suspensión, utilizando entrenudos como explante, en medio
MS suplementado con ANA y en oscuridad permanente.
Se logró obtener suspensiones celulares de muy alta densidad celular y crecimiento
vigoroso. La cinética de crecimiento mostró una tasa específica de crecimiento máxima
(µmáx) de 0,085 d-1, correspondiente a un tiempo de duplicación (td) de 8,15 días y una
concentración máxima de biomasa (Xmáx) de 12,11 g/l. Estos parámetros son comparables
a los que se han obtenido en otras plataformas vegetales, incluso en algunos casos
después de procesos de optimización.
Los bajos requerimientos hormonales, la respuesta robusta al cultivo in vitro, la alta tasa
específica de crecimiento y la estabilidad genética evidenciada en la expresión de la
proteína Bt-Cry1Ac en las suspensiones celulares, ponen en evidencia el potencial de
Solanum tuberosum subsp. andígena var. Pastusa Suprema como plataforma de
producción de proteínas recombinantes de interés biotecnológico.
5.2 Recomendaciones
Se detectó que el raspado de los callos es un paso muy crítico en el que se produce
oxidación de la biomasa y se piensa que está relacionado con la baja viabilidad celular en
el establecimiento de las suspensiones. Es probable que, si se encuentran mecanismos
para reducir el pardeamiento de los callos, bien sea mediante aditivos en el medio sólido
o eliminando el paso de raspado, se obtengan cultivos en mejores condiciones fisiológicas

44 Evaluación del potencial del cultivo de células en suspensión de papa
(Solanum tuberosum subsp. andígena) como plataforma de producción de
proteínas recombinantes
y por tanto se disminuya el tiempo de adaptación y la eficiencia en el establecimiento de
suspensiones celulares. En el establecimiento de suspensiones se observó una tendencia
a aumentar el crecimiento de la biomasa a menor concentración de auxina; por esta razón
se recomienda evaluar el comportamiento de los cultivos con concentraciones de ANA
menores a 1,5 mg/l en callos y menores a 0,5 mg/l en suspensiones, con miras a reducir
los costos globables del bioproceso.

Bibliografía
Ambrosone, A., Di Giacomo, M., Leone, A., Grillo, M. S., & Costa, A. (2012). Identification of early induced genes upon water deficit in potato cell cultures by cDNA-AFLP. Journal of Plant Research. doi:10.1007/s10265-012-0505-7
Anjum, M. A., & Ali, H. (2004). Effect of Culture Medium on Shoot Initiation from Calluses of Different Origin in Potato (Solanum tuberosum L.). Biotechnology, 3(2), 194–199.
Arias, J. P. (2013). Optimización de parámetros de cultivo en suspensión de células de la especie vegetal Thevetia peruviana y su efecto en la producción de metabolitos secundarios. Universidad Nacional de Colombia Sede Medellín.
Aviezer, D., Brill-almon, E., Shaaltiel, Y., Hashmueli, S., Bartfeld, D., Liberman, Y., Galun, E. (2009). A Plant-Derived Recombinant Human Glucocerebrosidase Enzyme — A Preclinical and Phase I Investigation. PLoS ONE, 4(3), 1–6. doi:10.1371/journal.pone.0004792
Bachem, C. W. B., Hoeven, R. S. Van Der, Bruijn, S. M. De, Vreugdenhil, D., Zabeau, M., Visser, R. G. F., Visser, R. G. F. (1996). Visualization of differential gene expression using a novel method of RNA fingerprinting based on AFLP: analysis of gene expression during potato tuber development. The Plant Journal, 9(5), 745–753. doi:10.1046/j.1365-313X.1996.9050745.x
Bhojwani, S. S., & Razdan, M. K. (1996). Plant Tissue Culture: Theory and Practice, a Revised Edition. Amsterdam, The Netherlands: Elsevier Science B.V.
Demain, A. L., & Vaishnav, P. (2009). Production of recombinant proteins by microbes and higher organisms. Biotechnology Advances, 27, 297–306. doi:10.1016/j.biotechadv.2009.01.008
Doran, P. M. (1995). Bioprocess engineering principles. Elsevier Science & Technology Books.
Doran, P. M. (2000). Foreign protein production in plant tissue cultures. Current Opinion in Biotechnology, 11(2), 199–204. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/10753768
Doran, P. M. (2013). Therapeutically important proteins from in vitro plant tissue culture systems. Current Medicinal Chemistry, 20, 1047 – 1055. Retrieved from http://www.sinab.unal.edu.co:2109/login.aspx?direct=true&db=mnh&AN=23210789&lang=es&site=ehost-live
El-Banna, A., Hajirezaei, M.-R., Wissing, J., Ali, Z., Vaas, L., Heine-Dobbernack, E., Kiesecker, H. (2010). Over-expression of PR-10a leads to increased salt and osmotic tolerance in potato cell cultures. Journal of Biotechnology, 150, 277–87. doi:10.1016/j.jbiotec.2010.09.934

46 Título de la tesis o trabajo de investigación
Franconi, R., Demurtas, O. C., & Massa, S. (2010). Plant-derived vaccines and other therapeutics produced in contained systems. Expert Review of Vaccines, 9(8), 877–892. doi:10.1586/erv.10.91
Gao, J., Lee, J. M., & An, G. (1991). The stability of foreign protein production in genetically modified plant cells. Plant Cell Reports, 10(10), 533–536. doi:10.1007/BF00234589
Gerszberg, A., Wiktorek-Smagur, A., Hnatuszko-Konka, K., Łuchniak, P., & Kononowicz, A. K. (2012). Expression of recombinant staphylokinase, a fibrin-specific plasminogen activator of bacterial origin, in potato (Solanum tuberosum L.) plants. World Journal of Microbiology and Biotechnology, 28(3), 1115–1123. doi:10.1007/s11274-011-0912-2
Gottstein, D., & Schliemann, W. (1994). Purine glucosylating activity in cell suspension cultures of Solanum tuberosum L. Plant Cell Reports, 36, 265–267.
Hanchinal, V. M., Survase, S. A., Sawant, S. K., & Annapure, U. S. (2008). Response surface methodology in media optimization for production of β-carotene from Daucus carota. Plant Cell, Tissue and Organ Culture, 93, 123–132. doi:10.1007/s11240-008-9350-8
Hawkins, H. J., & Lips, S. H. (1997). Cell Suspension Cultures of Solanum tuberosum L . as a Model System for N and Salinity Response Effect of Salinity on N0 3 - Uptake and PM-ATPase Activity zrc. Journal of Plant Physiology, 150, 103–109. doi:10.1016/S0176-1617(97)80188-5
He, D.-M., Qian, K.-X., Shen, G.-F., Li, Y.-N., Zhang, Z.-F., Su, Z.-L., & Shao, H.-B. (2007). Stable expression of foot-and-mouth disease virus protein VP1 fused with cholera toxin B subunit in the potato (Solanum tuberosum). Colloids and Surfaces B: Biointerfaces, 55, 159–163. doi:http://dx.doi.org/10.1016/j.colsurfb.2006.11.043
Holland, T., Sack, M., Rademacher, T., Schmale, K., Altmann, F., Stadlmann, J., Hellwig, S. (2010). Optimal nitrogen supply as a key to increased and sustained production of a monoclonal full-size antibody in BY-2 suspension culture. Biotechnology and Bioengineering, 107(2), 278–289. doi:10.1002/bit.22800
Huang, T. K., & McDonald, K. a. (2011). Bioreactor systems for in vitro production of foreign proteins using plant cell cultures. Biotechnology Advances, 30(2), 398–409. doi:10.1016/j.biotechadv.2011.07.016
Huang, T. K., & McDonald, K. A. (2009). Bioreactor engineering for recombinant protein production in plant cell suspension cultures. Biochemical Engineering Journal, 45(3), 168–184. doi:10.1016/j.bej.2009.02.008
International Solanaceae Initiative. (n.d.). Sol Genomics Network. Retrieved June 30, 2016, from https://solgenomics.net/
Jackson, S. D. (1999). Multiple Signaling Pathways Control Tuber Induction in Potato. Plant Physiology, 119, 1–8. doi:10.1104/pp.119.1.1
Jiménez Barreto, J. P., Chaparro-Giraldo, A., & Blanco, J. (2009). Evaluación de diferentes combinaciones fitohormonales en la regeneración de Solanum tuberosum (Solanaceae) Var. Pastusa Suprema a partir de explantes internodales. Revista Colombiana de Biotecnología, XI(2), 66–74.
Johnson, A. A. T. (2001). Effect of ploidy elevation, copy number and parent-of-origin on

Bibliografía 47
transgener expression in potato. Virginia Polytechnic Institute and State University.
Kieran, P. M., MacLoughlin, P. F., & Malone, D. M. (1997). Plant cell suspension cultures: Some engineering considerations. Journal of Biotechnology, 59, 39–52. doi:10.1016/S0168-1656(97)00163-6
Kombrink, E., Schröder, M., & Hahlbrock, K. (1988). Several “pathogenesis-related” proteins in potato are 1,3-beta-glucanases and chitinases. Proceedings of the National Academy of Sciences of the United States of America, 85(3), 782–786. doi:10.1073/pnas.85.3.782
Kreslavski, V. D., Carpentier, R., Klimov, V. V., & Allakhverdiev, S. I. (2009). Transduction mechanisms of photoreceptor signals in plant cells. Journal of Photochemistry and Photobiology C: Photochemistry Reviews, 10, 63–80. doi:10.1016/j.jphotochemrev.2009.04.001
Lander, E. S., Linton, L. M., Birren, B., Nusbaum, C., Zody, M. C., Baldwin, J., … Szustakowki, J. (2001). Initial sequencing and analysis of the human genome. Nature, 409(February), 860–921. doi:10.1038/35057062
Law, R. D., & Suttle, J. C. (2005). Chromatin remodeling in plant cell culture: patterns of DNA methylation and histone H3 and H4 acetylation vary during growth of asynchronous potato cell suspensions. Plant Physiology and Biochemistry, 43, 527–34. doi:10.1016/j.plaphy.2005.03.014
Leader, B., Baca, Q. J., & Golan, D. E. (2008). Protein therapeutics: a summary and pharmacological classification. Nature Reviews. Drug Discovery, 7(1), 21–39. doi:10.1038/nrd2399
Leister, D., Ballvora, A., Salamini, F., & Gebhardt, C. (1996). A PCR-based approach for isolating pathogen resistance genes from potato with potential for wide application in plants. Nature Genetics, 14, 421–429. doi:10.1038/ng1296-421
Li, Y.C., & Tao, W.Y. (2009). Effects of paclitaxel-producing fungal endophytes on growth and paclitaxel formation of Taxus cuspidata cells. Plant Growth Regulation, 58, 97–105. doi:10.1007/s10725-008-9355-7
López, A., & Chaparro, A. (2007). Propuesta de un sistema de transformación de plantas de papa ( Solanum tuberosum sp . andigena var . Pastusa Suprema ) mediado por Agrobacterium tumefaciens. Agronomía Colombiana, 25(1), 16–25.
Lu, Q., Yang, Q., & Zou, H. (2006). Effects of Cerium on Accumulation of Anthocyanins and Expression of Anthocyanin Biosynthetic Genes in Potato Cell Tissue Cultures. Journal of Rare Earths, 24(4), 479–484. doi:10.1016/S1002-0721(06)60147-6
Ma, S., Huang, Y., Davis, A., Yin, Z., Mi, Q., Menassa, R., Jevnikar, A. M. (2005). Production of biologically active human interleukin-4 in transgenic tobacco and potato. Plant Biotechnology Journal, 3, 309–318. doi:10.1111/j.1467-7652.2005.00125.x
McDonald, K. a., Jackman, A., & Hurst, S. (2001). Characterization of plant suspension cultures using the focused beam reflectance technique. Biotechnology Letters, 23(4), 317–324.
Müller-Röber, B., Sonnewald, U., & Willmitzer, L. (1992). Inhibition of the ADP-glucose

48 Título de la tesis o trabajo de investigación
pyrophosphorylase in transgenic potatoes leads to sugar-storing tubers and influences tuber formation and expression of tuber storage protein genes. The EMBO Journal, 11(4), 1229–1238. Retrieved from http://www.ncbi.nlm.nih.gov/pmc/articles/PMC556571/\nhttp://www.ncbi.nlm.nih.gov/pmc/articles/PMC556571/pdf/emboj00089-0015.pdf
Mullins, E., Milbourne, D., Petti, C., Doyle-Prestwich, B. M., & Meade, C. (2006). Potato in the age of biotechnology. Trends in Plant Science, 11(5), 254–260. doi:10.1016/j.tplants.2006.03.002
Munir, F., Naqvi, S. M. S., & Tariq, M. (2011). In vitro culturing and assessment of somaclonal variation of Solanum tuberosum var . desiree. Turkish Journal of Biochemistry, 36(4), 296–302.
Muñoz-Cruz, W., Vanegas-Monterrosa, O. A., Guzmán-Rosas, A. A., Capataz-Tafur, J., Hoyos-Sánchez, R. A., & Orozco-Sánchez, F. (2006). Estimación de variables de operación de un biorreactor con células de Azadirachta indica A. Juss. Revista de La Facultad Nacional de Agronomía, 59(2), 3467–3478.
Murashige, T., & Skoog, F. (1962). A revised medium for rapid growth and bioassays with tobacco cultures. Physiologia Plantarum, 15, 473–497.
Mustafa, N. R., de Winter, W., van Iren, F., & Verpoorte, R. (2011). Initiation, growth and cryopreservation of plant cell suspension cultures. Nature Protocols, 6(6), 715–42. doi:10.1038/nprot.2010.144
Naik, P. S., & Widholm, J. M. (1993). Comparison of tissue culture and whole plant responses to salinity in potato. Plant Cell, Tissue and Organ Culture, 33(3), 273–280. doi:10.1007/BF02319012
Naill, M. C., & Roberts, S. C. (2005). Cell cycle analysis of Taxus suspension cultures at the single cell level as an indicator of culture heterogeneity. Biotechnology and Bioengineering, 20(4), 491–500.
Neff, M. M., Fankhauser, C., & Chory, J. (2000). Light: an indicator of time and place. Genes & Development, 14, 257–271. doi:10.1101/gad.14.3.257
Pan, Z. W., Wang, H. Q., & Zhong, J. J. (2000). Scale-up study on suspension cultures of Taxus chinensis cells for production of taxane diterpene. Enzyme and Microbial Technology, 27, 714–723. doi:10.1016/S0141-0229(00)00276-3
Park, Y., & Cheong, H. (2002). Expression and production of recombinant human interleukin-2 in potato plants. Protein Expression and Purification, 25, 160–165. doi:10.1006/prep.2002.1622
Queirós, F., Fidalgo, F., Santos, I., & Salema, R. (2007). In vitro selection of salt tolerant cell lines in Solanum tuberosum L. Biologia Plantarum, 51(4), 728–734.
Reiss, B. (2003). Homologous Recombination and Gene Targeting in Plant Cells. International Review of Cytology, 228, 85–139. doi:10.1016/S0074-7696(03)28003-7
Riesmeier, J. W., Willmitzer, L., & Frommer, W. B. (1994). Evidence for an essential role of the sucrose transporter in phloem loading and assimilate partitioning. The EMBO Journal, 13(1), 1–7. Retrieved from http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=394773&tool=pmcentrez&

Bibliografía 49
rendertype=abstract
Rigano, M. M., Guzman, G. De, Walmsley, A. M., & Frusciante, L. (2013). Production of Pharmaceutical Proteins in Solanaceae Food Crops. International Journal of Molecular Sciences, 14, 2753–2773. doi:10.3390/ijms14022753
Rodríguez-Monroy, M., & Galindo, E. (2003). Las células vegetales, ¿frágiles para crecer en biorreactores? Biotecnología, 8(2), 6–17. Retrieved from http://scholar.google.com/scholar?hl=en&btnG=Search&q=intitle:Las+c?lulas+vegetales,+?fr?giles+para+crecer+en+biorreactores?#0
Roessner, U., Wagner, C., Kopka, J., Trethewey, R. N., & Willmitzer, L. (2000). Technical advance: simultaneous analysis of metabolites in potato tuber by gas chromatography-mass spectrometry. The Plant Journal, 23(1), 131–42. doi:10.1046/j.1365-313x.2000.00774.x
Romano, A., Vreugdenhil, D., Jamar, D., van der Plas, L. H. W., de Roo, G., Witholt, B., Mooibroek, H. (2003). Evidence of medium-chain-length polyhydroxyoctanoate accumulation in transgenic potato lines expressing the Pseudomonas oleovorans Pha-C1 polymerase in the cytoplasm. Biochemical Engineering Journal, 16, 135–143. doi:10.1016/S1369-703X(03)00030-5
Rukavtsova, E. B., Rudenko, N. V., Puchko, E. N., Zakharchenko, N. S., & Buryanov, Y. I. (2015). Study of the immunogenicity of hepatitis B surface antigen synthesized in transgenic potato plants with increased biosafety. Journal of Biotechnology, 203, 84–88. doi:10.1016/j.jbiotec.2015.03.019
Rybicki, E. (2010). Plant-made vaccines for humans and animals. Plant Biotecnology Journal, 8(5), 620–637.
Seabrook, J. E. A. (2005). Light effects on the growth and morphogenesis of potato (Solanum tuberosum) in vitro: a review. American Journal of Potato Research, 82(5), 353–367.
Shaaltiel, Y., Bartfeld, D., Hashmueli, S., Baum, G., Brill-almon, E., Galili, G., Aviezer, D. (2007). Production of glucocerebrosidase with terminal mannose glycans for enzyme replacement therapy of Gaucher’s disease using a plant cell system. Plant Biotechnology Journal, 5, 579–590. doi:10.1111/j.1467-7652.2007.00263.x
Shahab-ud-din, Sultan, I. N., Kakar, M. A., Yousafzai, A., Sattar, F. A., Ahmmad, F., Arif, B. (2011). The Effects of Different Concentrations and Combinations of Growth Regulators on the Callus Formation of Potato (Solanum tubrosum) Explants. Current Research Journal of Biological Sciences, 3(5), 499–503.
Shih, S. M.-H., & Doran, P. M. (2009). Foreign protein production using plant cell and organ cultures: Advantages and limitations. Biotechnology Advances, 27(6), 1036–42. doi:10.1016/j.biotechadv.2009.05.009
Shin, Y.-J., Chong, Y.-J., Han, K.-B., Yang, M.-S., & Kwon, T.-H. (2010). N-linked glycan analysis of glycoproteins secreted from rice cell suspension cultures under sugar starvation. Enzyme and Microbial Technology, 47, 189–193. doi:10.1016/j.enzmictec.2010.06.007
Son, S. H., Choi, S. M., Lee, Y. H., Choi, K. B., Yun, S. R., Kim, J. K., Park, Y. G. (2000).

50 Título de la tesis o trabajo de investigación
Large-scale growth and taxane production in cell cultures of Taxus cuspidata (Japanese yew) using a novel bioreactor. Plant Cell Reports, 19(6), 628–633.
Stachel, S. E., Nester, E. W., & Zambryski, P. C. (1986). A plant cell factor induces Agrobacterium tumefaciens vir gene expression. Proceedings of the National Academy of Sciences of the United States of America, 83, 379–383.
Tacket, C., Mason, H., Losonsky, G., Clements, J., Levine, M., & Arntzen, C. (1998). Immunogenicity in humans of a recombinant bacterial antigen delivered in a transgenic potato. Nature Medicine, 4(5), 607–609. doi:http://dx.doi.org/10.1038/nm0598-607
Taiz, L., & Zeiger, E. (2002). Plant physiology (3 edition.). Sinauer Associates.
Taylor, M. A., Arif, S. A., Kumar, A., Davies, H. V, Scobie, L. A., Pearce, S. R., & Flavell, A. J. (1992). Expression and sequence analysis of cDNAs induced during the early stages of tuberisation in different organs of the potato plant (Solanum tuberosum L.). Plant Molecular Biology, 20(4), 641–51.
Torabi, F., Majad, A., & Ehsanpour, A. A. (2008). Plant regeneration from cell suspension culture of potato (Solanum tuberosum L.). Pakistan Journal of Biological Sciences, 11(5), 778–782.
Torres, E. S., Torres, J., Moreno, C., & Arango, R. (2012). Development of transgenic lines from a male-sterile potato variety , with potential resistance to Tecia solanivora Povolny Desarrollo de líneas transgénicas de una variedad androestéril de papa , potencialmente resistentes a Tecia solanivora Povolny. Agronomía Colombiana, 30(2), 163–171. Retrieved from http://www.revistas.unal.edu.co/index.php/agrocol/article/view/19395
Trejo-Tapia, G., Jiménez-Aparicio, A., Villarreal, L., & Rodríguez-Monroy, M. (2001). Broth rheology and morphological analysis of solanum chrysotrichum cultivated in a stirred tank. Biotechnology Letters, 23, 1943–1946. doi:10.1023/A:1013789918463
Trujillo, C., Rodriguez-Arango, E., Jaramillo, S., Hoyos, R., Orduz, S., & Arango, R. (2001). One-step transformation of two Andean potato cultivars (Solanum tuberosum L . subsp . andigena). Plant Cell Reports, 20, 637–641. doi:10.1007/s002990100381
Turhan, H. (2004). Callus induction and growth in transgenic potato genotypes. African Journal of Biotechnology, 3(8), 375–378.
Twyman, R. M., Stoger, E., Schillberg, S., Christou, P., & Fischer, R. (2003). Molecular farming in plants: Host systems and expression technology. Trends in Biotechnology, 21(12), 570–578. doi:10.1016/j.tibtech.2003.10.002
Ullisch, D. A., Müller, C. A., Maibaum, S., Kirchhoff, J., Schiermeyer, A., Schillberg, S., Büchs, J. (2012). Comprehensive characterization of two different Nicotiana tabacum cell lines leads to doubled GFP and HA protein production by media optimization. Journal of Bioscience and Bioengineering, 113(2), 242–248. doi:10.1016/j.jbiosc.2011.09.022
van der Maas, H. M., de Jong, E. R., Rueb, S., Hensgens, L. a, & Krens, F. a. (1994). Stable transformation and long-term expression of the gusA reporter gene in callus lines of perennial ryegrass (Lolium perenne L.). Plant Molecular Biology, 24, 401–5. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/8111042

Bibliografía 51
Vargas, T. E., De García, E., & Oropeza, M. (2005). Somatic embryogenesis in Solanum tuberosum from cell suspension cultures: histological analysis and extracellular protein patterns. Journal of Plant Physiology, 162, 449–456. doi:10.1016/j.jplph.2004.07.001
Vargas, T. E., Xena, N., Vidal, M. del C., Oropeza, M., & de García, E. (2008). Genetic stability of Solanum tuberosum l. cv. désirée Plantlets obteined from embryogenic cell suspension cultures. Interciencia, 33(3), 213–218.
Visser, R. G. F., Somhorst, I., Kuipers, G. J., Ruys, N. J., Feenstra, W. J., & Jacobsen, E. (1991). Inhibition of the expression of the gene for granule-bound starch synthase in potato by antisense constructs. Molecular & General Genetics, 225(2), 289–296. doi:10.1007/BF00269861
Wang, A., & Ma, S. (Eds.). (2012). Molecular farming in plants: Recent advances and future prospects. Molecular Farming in Plants: Recent Advances and Future Prospects (Vol. 9789400722). Springer. doi:10.1007/978-94-007-2217-0
Wang, H. Q., Yu, J. T., & Zhong, J. J. (1999). Significant improvement of taxane production in suspension cultures of Taxus chinensis by sucrose feeding strategy. Process Biochemistry, 35, 479–483.
Wickremesinhe, E. R. M., & Arteca, R. N. (1994). Taxus Cell Suspension Cultures: Optimizing Growth and Production of Taxol. Journal of Plant Physiology, 144, 183–188. doi:10.1016/S0176-1617(11)80541-9
Xu, J., Ge, X., & Dolan, M. C. (2011). Towards high-yield production of pharmaceutical proteins with plant cell suspension cultures. Biotechnology Advances, 29(3), 278–99. doi:10.1016/j.biotechadv.2011.01.002
Xu, J., Okada, S., Tan, L., Goodrum, K. J., Kopchick, J. J., & Kieliszewski, M. J. (2010). Human growth hormone expressed in tobacco cells as an arabinogalactan-protein fusion glycoprotein has a prolonged serum life. Transgenic Research, 19, 849–867. doi:10.1007/s11248-010-9367-8
Xu, X., Pan, S., Cheng, S., Zhang, B., Mu, D., Ni, P., Visser, R. G. F. (2011). Genome sequence and analysis of the tuber crop potato. Nature, 475, 189–195. doi:10.1038/nature10158
Zacharius, R. M., Kalan, E. B., & Kimoto, W. I. (1985). Biotransformation of potato stress metabolites rishitin, lubimin, and 15-dihydro lubimin by potato and soybean cell cultures. Plan Cell Reports, 4, 1–3.
Zhang, C.-Y., Dong, Y.-S., Li, Y.-L., Fu, C.-H., Zhao, C.-F., & Yu, L.-J. (2013). Unstructured models for suspension cultures of Taxus media cells in a bioreactor under substrate-sufficient conditions. Biochemical Engineering Journal, 71, 62–71. doi:10.1016/j.bej.2012.11.014