estudio de las relaciones ambientales, distribución y...
TRANSCRIPT

185
Estudio de las relaciones ambientales, distribución y composición de la fauna acompañante de merluza (Merluccius hubbsi) en primavera
de 2008.
Scarlato, Norberto A., P.Ibañez, L. Tringali
Estudio de las relaciones ambientales, distribución y composición de la fauna acompañante de merluza (Merluccius hubbsi) en primavera de 2008
Scarlato, Norberto A.1,2, P.Ibañez1, L. Tringali1
1.-Instituto Nacional de Investigación y Desarrollo Pesquero (INIDEP)2.-Facultad de Ciencias Exactas y Naturales, Departamento de Biología
Correo electrónico: [email protected]
RESUMEN: Se analizaron datos de una campa-ña global de evaluación de merluza de INIDEP de primavera de 2008 en la ZCP, con el objeto de caracterizar tanto el ambiente como los ensam-bles faunísticos. El área de estudio se dividió en cuatro secciones. La estructura de la comunidad se analizó mediante la biomasa (peso húmedo fijado en toneladas por milla náutica cuadrada). Se jerarquizaron las especies mediante sus valo-res de biomasa y sus frecuencias de ocurrencia porcentual. Se estimaron mediante métodos geoestadísticos los valores de temperatura y salinidad superficial y de fondo para lances sin mediciones directas. Luego se generó un CLUS-TER de las distancias Bray-Curtis de las bioma-sas logtransformadas, observándose separa-ción entre sectores. La estructura fue evaluada gráficamente mediante MDS y analíticamente mediante ANOSIM, encontrándose tanto buena separación como solapamiento entre zonas. Se identificaron las principales especies (merluza, congrio) contribuyentes a la similaridad de los grupos mediante SIMPER. Bassanago albescens fue la especie con mayor captura (43,9%) segui-do por la merluza (11,6%) y por Trachurus latha-mi (10,52%). Luego aparecen Zenopsis conchifer (5,61%) e Illex argentinus (4,99%). Se verifica que la conjunción de tres variables ambientales (sa-linidad de fondo y superficial, temperatura de fondo) se corresponde con el patrón encontra-do para el conjunto de especies considerado.
Palabras clave: Merluza común, Fauna acom-pañante, Composición de la comunidad, Estruc-tura de la Comunidad, Factores abióticos, Peces demersales
SUMMARY: Study of argentine hake (merluccius hubbsi) by-catch distribution, composition and relationships with environmental variables. Data from a global stock assessment research cruise carried out by INIDEP during springtime 2008 on Argentine hake in the Argentine-Uru-guayan Common Fishing Zone were analyzed with the aim of characterizing faunistic assem-blages and environmental patterns altogether. The studied area was split in four sections. Com-munity structure was analyzed using biomass (wet weight fixed in tons per square nautical mile). Species were arranged in order according to their biomass and mean percent frequency of occurrence. Missing surface and bottom tempe-rature and salinity values were geostatistically estimated for positions with no direct measu-rement. A CLUSTER analysis of Bray-Curtis dis-tances of log-transformed biomass values was carried out, thereby finding separation between sectors. The resulting structure was evaluated graphically by means of MDS and analytically through ANOSIM, finding both separation and overlap among zones. Species (Argentine hake, hairy conger) contributing to intergroup simi-larity were identified with SIMPER. Bassanago

186
FRENTE MARÍTIMO
Publicación de la Comisión Técnica Mixta del Frente Marítimo
albescens was the most abundant species in cat-ches (43.9 % of total catch), followed by Argenti-ne hake (11.6%) and Trachurus lathami (10.52%). Also included in this list was Zenopsis conchifer (5.61%) and Illex argentinus (4.99%). The combi-nation of three environmental variables (bottom
temperature and salinity, surface salinity) mat-ches the species pattern found in the area.
Key words: Argentine hake, By-catch, Commu-nity Composition, Community Structure, Abiotic Factors, Demersal Fish

187
Estudio de las relaciones ambientales, distribución y composición de la fauna acompañante de merluza (Merluccius hubbsi) en primavera
de 2008.
Scarlato, Norberto A., P.Ibañez, L. Tringali
Introducción
La operación con redes de arrastre es poco selectiva resultando en una diversidad de especies captu-radas. Se generan así modificaciones en el ecosistema marino por lo cual no sólo la especie objetivo se ve afectada sino que también aquéllas que de algún u otro modo están relacionadas en una de-terminada concentración íctica. De allí el interés suscitado en caracterizar la captura incidental de las pesquerías demersales de arrastre (Martínez et al., 1993; Tillman, 1993; Alverson et al., 1994; Kennelly, 1995; Crowder & Murawski 1998; Hall et al., 2000; Costa et al., 2001; Davis, 2002; Hall & Mainprize, 2005). Estos estudios se basan usualmente en datos provenientes de campañas de investigación (Gabriel & Tyler, 1980; Colvocoresses & Musick, 1984; Murawski & Finn, 1988; Kennelly, 1995; Ungaro et al., 1999; Mueter & Norcross, 2002) y permiten obtener índices estandarizados de abundancia relativa usualmen-te más precisos que los obtenidos a partir de datos de la flota comercial. Otro beneficio de este tipo de estudios es que permite anticipar en cierta forma la composición de la captura obtenida por los barcos comerciales que operan en la misma zona(Pennington & Strømme, 1998; Korsbrekke et al., 2001).
En aguas de Plataforma del Atlántico Sudoccidental se han efectuado diversos estudios destinados fundamentalmente a caracterizar la distribución, abundancia y áreas de concentración de las espe-cies demersales, tanto a partir de campañas de investigación(Arena et al., 1986; Rey et al., 1993; Dato, 1997; García de la Rosa et al., 1997; Renzi & Castrucci, 1998; Buratti, 2004), como a partir de datos de la flota comercial(Irusta 1990; Ehrlich 1998; Cañete et al., 2000; Irusta et al., 2001). No obstante, sólo recientemente se ha comenzado a definir límites temporales o geográficos prácticos orientados a la conservación o mantenimiento de los componentes de los ecosistemas, especialmente por encima del nivel específico(Jaureguizar et al., 2003, 2006a, 2006b, 2007; Jaureguizar, 2004; García et al., 2010). El propósito del presente trabajo es comenzar a examinar los cambios espaciales y temporales en la composición de las áreas de asociaciones de peces en el ecosistema de Plataforma y Talud del Atlán-tico Sudoccidental entre los 34°S y los 41°S, identificando para cada área propuesta las especies más comunes y las que permiten discriminar cada área de la adyacente. También se intentará identificar los parámetros físicos que controlan la estructuración de la comunidad íctica de Plataforma y Talud tanto espacial como temporalmente.
Materiales y Métodos
Para el presente análisis se tomaron los datos provenientes de una campaña de evaluación pesquera efectuada entre los 34°30´S y los 41°S, aproximadamente durante el año 2008. Este crucero cubrió la región de Plataforma y Talud del área, operando en un rango de profundidades de 46 a 334 m durante el mes de Octubre. Los 99 lances se dispusieron mediante un patrón estratificado al azar, de acuerdo a 27 estratos definidos (Figura 1). Se utilizó una red de arrastre de fondo tipo Engel de 35,5 m de relinga superior, 50,0 m de relinga inferior equipada con portones polivalentes de 1200 Kg. La luz de malla en el copo fue de 100 mm con un forro interno de 24 mm de luz de malla. La captura fue separada por especie, pesada y se determinó para cada una el número de ejemplares por kilo (D’Atri, 2008).

188
FRENTE MARÍTIMO
Publicación de la Comisión Técnica Mixta del Frente Marítimo
El área de estudio se dividirá en sectores tomando en cuenta la profundidad y la latitud, a saber:
· Somero Norte (SN): Involucra el sector entre 34º LS y 38º LS con profundidades menores o igua-les a 100m.
· Profundo Norte (PN): Entre 34º LS y 38º LS y profundidades mayores a 100m.· Somero Sur (SS): Desde 34º LS a 41º LS con profundidades menores o iguales a 100m.· Profundo Sur (PS): Entre 38º LS y 41º LS y profundidades mayores a 100m.
Figura 1 Posición de los lances de la Campaña EH-02/98.
Figura 1. Posición de los lances de la Campaña EH-02/98.
Figura 2 Sectores utilizados para diferenciar el área de estudio Figura 2. Sectores utilizados para diferenciar el área de estudio.

189
Estudio de las relaciones ambientales, distribución y composición de la fauna acompañante de merluza (Merluccius hubbsi) en primavera
de 2008.
Scarlato, Norberto A., P.Ibañez, L. Tringali
La estructura de la comunidad se analizará mediante la biomasa, expresada como el peso húmedo fijado en toneladas por milla náutica cuadrada. La jerarquización de las especies se efectuará me-diante la biomasa y la frecuencia de ocurrencia porcentual, aplicando la prueba de asociación no paramétrica de tipo Olmstead-Tukey denominada gráfico de cuadrantes (Sokal & Rohlf, 1981).
Con los valores de biomasa en peso húmedo, fijados en toneladas por milla náutica cuadrada se generará una matriz de datos, que será logtransformada (log x+1) previo al cálculo de las distancias Bray-Curtis. Esta transformación permitirá identificar patrones dentro de la ictiofauna, mezclando las contribuciones de las especies abundantes y raras (Clarke & Warwick, 2001). Es necesario aclarar que se opta por el uso de estas distancias debido a que cumplen con todos los siguientes criterios (Clarke & Warwick, 2001):
1.- Son iguales a 100 cuando dos muestras son idénticas, como pasa con otros coeficientes2.- Son iguales a 0 cuando dos muestras no tienen especies en común(la mayoría de los demás
coeficientes no lo hacen)3.- Los cambios en las unidades de medición no las afectan (como tampoco a otros coeficientes)4.- Sus valores no cambian por la inclusión o exclusión de una especie que no aparece en dos
muestras5.- La inclusión o exclusión de una tercer muestra C no afecta la similitud entre las muestras A y B
(la mayoría de los demás coeficientes no se comporta de esta manera, ya que dependen de la estandardización sobre cada especie)
6.- Tienen la flexibilidad necesaria para registrar diferencias en las abundancias totales en dos muestras como una similitud no tan perfecta aún cuando las abundancias relativas sean idénti-cas (otros coeficientes estandardizan automáticamente por los totales de las muestras, y de este modo no muestran estas similitudes/diferencias)
Las distancias Bray-Curtis entre los lances j y k (Sjk) se calcularán mediante la siguiente ecuación (Clarke & Warwick, 2001):
Donde: yij es la biomasa de la especie i en el lance j yik es la biomasa de la especie i en el lance k p es el número de especies
Con las distancias así calculadas se generará una matriz de similitudes, que se utilizará en los análisis subsiguientes.
A partir de la matriz de similitudes se generará un análisis de conglomerado (CLUSTER) de las dis-tancias Bray-Curtis de las biomasas logtransformadas. Este análisis es una representación particular-mente relevante en casos en los que se espera que las muestras se dividan en grupos bien definidos (Clarke & Warwick, 2001), con lo que resultará adecuado para probar la división de los conjuntos de lances propuesta para este estudio. De todas maneras, el cluster se genera en combinación con la rutina del perfil de similitudes (SIMPROF) Tipo II, que tiene como hipótesis nula que los perfiles de abundancia de las distintas especies son homogéneos y no tienen asociaciones (Clarke & Gor-

190
FRENTE MARÍTIMO
Publicación de la Comisión Técnica Mixta del Frente Marítimo
ley, 2001, 2005; Clarke et al., 2008). La estructura obtenida en el análisis anterior será evaluada grá-ficamente mediante escalamiento multidimensional (MDS). Esta técnica sirve para representar las muestras como puntos en un espacio, usualmente bidimensional, de manera tal que las distancias relativas entre los puntos se correspondan con las disimilitudes relativas de las muestras (calculadas mediante las distancias Bray-Curtis). La interpretación de este gráfico es directa: los puntos agrupa-dos representan muestras muy similares en su composición específica, y los puntos distantes entre sí corresponden a comunidades muy diferentes (Clarke & Warwick, 2001).
También se recurrirá al Análisis de Similitud no paramétrico de una vía (ANOSIM) para probar analí-ticamente las diferencias entre los grupos de muestras preestablecidos, operando con la hipótesis nula (H0) de la inexistencia de diferencias entre los conjuntos. Para examinar H0 se seguirán tres pasos(Clarke & Warwick, 2001):
1) Cálculo de un estadístico de prueba que refleje las diferencias observadas entre grupos, contras-tadas con las diferencias dentro del grupo
El estadístico propuesto tiene la forma:
Donde: (rB ) promedio de las similitudes de los pares de réplicas entre los distintos grupos, (rW ) promedio de las similitudes de los pares de réplicas dentro del grupo, n número total de muestras consideradas
2) Recálculo del estadístico permutando los nombres de los grupos un adecuado número de veces (999)
3) Cálculo del nivel de significación comparando el valor observado de R con la distribución de las permutaciones. Si H0 es verdadero, entonces el sesgo de mayor verosimilitud está dado por los arreglos al azar. Por tanto, si el valor real de R no parece provenir de esta distribución, se puede rechazar la hipótesis nula.
Se procederá a identificar qué especies generan las diferencias entre los grupos muestrales mediante un análisis de porcentajes de similitud (SIMPER)(Clarke & Gorley, 2001). Las especies discriminantes entre los distintos sectores se identificarán por sus altos valores de diferencia media y por los bajos niveles de desvío estándar de las diferencias. Así, el cociente entre estos valores será mayor para las especies discriminantes (Clarke & Warwick, 2001).
Posteriormente, se estimarán los valores de temperatura y salinidad superficial y de fondo para aque-llos lances en los que no hubo mediciones directas. Para esto, se considerarán los datos de estaciones de CTD efectuados durante la campaña para generar variogramas experimentales de temperaturas y salinidades superficiales y de fondo, a los que se ajustarán variogramas teóricos, que se utilizarán posteriormente para estimar los valores en posiciones sin mediciones(Bridges, 1985; Stein, 1999; Gringarten & Deutsch, 2001; Fischer & Getis, 2009). Se procurará contar con al menos 50 puntos para

191
Estudio de las relaciones ambientales, distribución y composición de la fauna acompañante de merluza (Merluccius hubbsi) en primavera
de 2008.
Scarlato, Norberto A., P.Ibañez, L. Tringali
cada intervalo de distancia considerado en los variogramas experimentales, según lo recomendado (Journel & Huijbregts, 1978; Journel, 1989). La coherencia de los valores estimados respecto de los medidos in situ se constatará gráficamente mediante mapas de isolíneas.
Finalmente, los datos así estimados se estandarizarán para generar una matriz de similitudes de dis-tancias Euclideanas, debido a que las diferentes escalas de medición de las variables impiden el uso de las distancias Bray-Curtis, que asumen el uso de una escala común (Clarke & Gorley, 2001; Clarke & Warwick, 2001). Esta matriz se utilizará para calcular el grado de coincidencia con la matriz de simili-tudes biótica y para identificar las variables ambientales que maximizan el coeficiente de concordan-cia utilizando como método de correlación el de rangos de Spearman (Clarke & Gorley, 2001).
Resultados y Discusión
Los resultados de la aplicación de la prueba de asociación no paramétrica de tipo Olmstead-Tukey se pueden observar en la Figura 3. Mediante este análisis se categorizaron las especies como: 1) Do-minantes: Densidad media y frecuencia porcentual altas (Merluza común, Congrio de profundidad, especies que presentaron valores de densidad y frecuencia medias mayores al promedio), 2) Fre-cuentes: Densidad media baja y frecuencia porcentual alta (19 especies con valores de densidades medias menores al promedio y con frecuencias porcentuales mayores al promedio), 3) Ocasiona-les: Densidad media alta y frecuencia porcentual baja (7 especies con densidades medias mayores al promedio y con frecuencias porcentuales menores al promedio), y 4) Raras: Densidad media y frecuencia porcentual bajas (44 especies cuyo valor de densidad media y frecuencia porcentual es menor al promedio) (Tabla 1).
Figura 3. Diagrama de Olmstead y Tukey de las especies de peces e invertebrados colectados en la campaña
EH-02/08.
Diagrama de Olmstead-TukeyCampaña EH-02/08
Acabra AmbdoeAncmarAtlpla
Basalb
Batalb BatbraBatgri
BatmacBatmulBatscaBatsp. Brabra
Cheber
Coefas ConperCotgob DipchiDistscDiseleDulaur
Enganc
Galgal
Genbla
Heldac IllargIlufim
LolsanMacancMaccarMachol
MacmagMerhub
MicausMularg
Mussch
Myc
Mylgoo
Notfer Pariso ParpatPatram
PerbraPinbraPrinudPsaext Psalen PsanorPsarudPsarut
PsesemRajfla
RhasapSarsar Schbiv
Scojap
Sebocu SerporSpapag
Sphzyg
SquacaSqublaSqucubSquargSqusp.Strbra
SymbonThylep
Tralat
TrilepUmbcan
Zencon
-20,00% 0,00% 20,00% 40,00% 60,00% 80,00% 100,00%
Frecuencia Media (%)
-2
0
2
4
6
8
10
12
14
16
Den
sida
d M
edia
(tn/
m2 )
Acabra AmbdoeAncmarAtlpla
Basalb
Batalb BatbraBatgri
BatmacBatmulBatscaBatsp. Brabra
Cheber
Coefas ConperCotgob DipchiDistscDiseleDulaur
Enganc
Galgal
Genbla
Heldac IllargIlufim
LolsanMacancMaccarMachol
MacmagMerhub
MicausMularg
Mussch
Myc
Mylgoo
Notfer Pariso ParpatPatram
PerbraPinbraPrinudPsaext Psalen PsanorPsarudPsarut
PsesemRajfla
RhasapSarsar Schbiv
Scojap
Sebocu SerporSpapag
Sphzyg
SquacaSqublaSqucubSquargSqusp.Strbra
SymbonThylep
Tralat
TrilepUmbcan
Zencon
OCASIONAL DOMINANTE
FRECUENTERARO
Figura 3. Diagrama de Olmstead y Tukey de las especies de peces e invertebrados colectados en la campaña EH-02/08.

192
FRENTE MARÍTIMO
Publicación de la Comisión Técnica Mixta del Frente Marítimo
Tabla 1. Composición, Porcentaje de ocurrencia y Densidad (t/mn2) de las especies de peces e invertebrados colectados en la campaña EH-02/08.
CODIGO ESPECIE Porcentaje Densidad Categoría Ocurrencia (t/mn2)
Basalb Bassanago albescens 43,43% 15,375 DOMINANTEMerhub Merluccius hubbsi 91,92% 1,812 DOMINANTEBatalb Bathyraja albomaculata 19,19% 0,237 FRECUENTEBatbra Bathyraja brachyurops 31,31% 0,186 FRECUENTEBatmac Bathyraja macloviana 45,45% 0,156 FRECUENTECheber Nemadactylus bergi 24,24% 1,124 FRECUENTECoefas Caelorhynchus fasciatus 15,15% 0,184 FRECUENTEConper Congiopodus peruvianus 34,34% 0,042 FRECUENTEDipchi Dipturus chilensis 49,49% 0,197 FRECUENTEGenbla Genypterus blacodes 39,39% 0,156 FRECUENTEHeldac Helicolenus dactylopterus lahil. 26,26% 1,206 FRECUENTEIllarg Illex argentinus 74,75% 0,962 FRECUENTEIlufim Iluocoetes fimbriatus 30,30% 0,650 FRECUENTEParpat Paralichthys patagonicus 35,35% 0,080 FRECUENTEPatram Patagonotothen ramsayi 67,68% 0,492 FRECUENTEPsanor Psammobatis normani 45,45% 0,117 FRECUENTEPsarud Psammobatis rudis 37,37% 0,061 FRECUENTESchbiv Schroederichthys bivius 26,26% 0,078 FRECUENTESquaca Squalus acanthias 39,39% 0,262 FRECUENTEStrbra Stromateus brasiliensis 58,59% 0,686 FRECUENTEEnganc Engraulis anchoita 5,05% 5,412 OCASIONALGalgal Galeorhinus galeus 13,13% 1,366 OCASIONALMacmag Macruronus magellanicus 11,11% 1,371 OCASIONALMussch Mustelus schmitti 12,12% 1,762 OCASIONALSphzyg Sphyrna zygaena 1,01% 1,969 OCASIONALTralat Trachurus lathami 11,11% 13,831 OCASIONALZencon Zenopsis conchifer 10,10% 8,007 OCASIONALAcabra Acanthistius brasilianus 3,03% 0,101 RAROAmbdoe Amblyraja doellojuradoi 10,10% 0,078 RAROAncmar Anchoa marinii 1,01% 0,017 RAROAtlpla Atlantoraja platana 1,01% 0,149 RAROBatgri Bathyraja griseocauda 4,04% 0,594 RAROBatmul Bathyraja multispinis 3,03% 0,160 RAROBatsca Bathyraja scaphiops 4,04% 0,101 RARO

193
Estudio de las relaciones ambientales, distribución y composición de la fauna acompañante de merluza (Merluccius hubbsi) en primavera
de 2008.
Scarlato, Norberto A., P.Ibañez, L. Tringali
Batsp. Bathyraja sp. 2,02% 0,124 RAROBrabra Brama brama 8,08% 0,257 RAROCotgob Cottoperca gobio 2,02% 0,026 RARODistsc Discopyge tschudii 6,06% 0,058 RARODisele Dissostichus eleginoides 2,02% 0,053 RARODulaur Dules auriga 2,02% 0,004 RAROLolsan Loligo sanpaulensis 7,07% 0,379 RAROMacanc Macrodon ancylodon 2,02% 0,139 RAROMaccar Macrourus carinatus 1,01% 0,006 RAROMachol Macrourus holotrachys 3,03% 0,266 RAROMicaus Micromesistius australis 8,08% 0,006 RAROMularg Mullus argentinus 3,03% 0,005 RAROMyc Myctophidae 2,02% 0,084 RAROMylgoo Myliobatis goodei 2,02% 1,135 RARONotfer Notopogon fernandezianus 2,02% 0,003 RAROPariso Paralichthys isosceles 12,12% 0,043 RAROPerbra Percophis brasiliensis 3,03% 0,178 RAROPinbra Pinguipes brasilianus 1,01% 0,055 RAROPrinud Prionotus nudigula 5,05% 0,015 RAROPsaext Psammobatis extenta 1,01% 0,003 RAROPsalen Psammobatis lentiginosa 11,11% 0,038 RAROPsarut Psammobatis rutrum 1,01% 0,004 RAROPsesem Pseudopercis semifasciata 4,04% 1,266 RARORhasap Rhamdia sapo 3,03% 0,078 RAROSarsar Sarda sarda 1,01% 0,029 RAROScojap Scomber japonicus 5,05% 0,929 RAROSebocu Sebastes oculatus 1,01% 0,032 RAROSerpor Seriolella porosa 10,10% 0,212 RAROSpapag Sparus pagrus 5,05% 0,134 RAROSqubla Squalus blainvillei 12,12% 0,139 RAROSqucub Squalus cubensis 1,01% 0,020 RAROSquarg Squatina argentina 6,06% 0,188 RAROSqusp. Squatina sp. 5,05% 0,153 RAROSymbon Sympterygia bonapartei 10,10% 0,118 RAROThylep Thyrsitops lepidopodea 3,03% 0,010 RAROTrilep Trichiurus lepturus 8,08% 0,786 RAROUmbcan Umbrina canosai 2,02% 0,887 RARO

194
FRENTE MARÍTIMO
Publicación de la Comisión Técnica Mixta del Frente Marítimo
Abadejo
0
10
20
30
40
2 6 7 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 30 31 32 33
Estratos <= 100 m
Cap
tura
(kg)
0
2
4
6
8
10
Cap
(kg)
/lanc
es
Abadejo
0
10
20
30
40
50
60
2 8 11 12 15 16 19 20 23 24 28 33
Estratos > 100 m
Cap
tura
(kg)
0
5
10
15
20
25
30
Cap
(kg)
/lanc
es
Calamar
0
100
200
300
400
500
600
2 6 7 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 30 31 32 33
Estratos <= 100 m
Cap
tura
(kg)
0
40
80
120
160
200
Cap
(kg)
/lanc
es
Calamar
0
30
60
90
120
150
2 8 11 12 15 16 19 20 23 24 28 33
Estratos > 100 mC
aptu
ra (k
g)
0
10
20
30
40
50
Cap
(kg)
/lanc
es
Castañeta
0
50
100
150
200
2 6 7 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 30 31 32 33
Estratos <= 100 m
Cap
tura
(kg)
0
40
80
120
160
200
Cap
(kg)
/lanc
es
Castañeta
0
50
100
150
200
250
300
2 8 11 12 15 16 19 20 23 24 28 33
Estratos > 100 m
Cap
tura
(kg)
0
10
20
30
40
50
60
Cap
(kg)
/lanc
es
Congrio
0
50
100
150
200
250
300
2 6 7 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 30 31 32 33
Estratos <= 100 m
Cap
tura
(kg)
0
40
80
120
160
200
Cap
(kg)
/lanc
es
Congrio
0
500
1000
1500
2000
2500
2 8 11 12 15 16 19 20 23 24 28 33
Estratos > 100 m
Cap
tura
(kg)
0
200
400
600
800
Cap
(kg)
/lanc
es
Nototenia
0
50
100
150
200
2 6 7 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 30 31 32 33
Estratos <= 100 m
Cap
tura
(kg)
0
40
80
120
160
200
Cap
(kg)
/lanc
es
Nototenia
0102030405060708090
2 8 11 12 15 16 19 20 23 24 28 33
Estratos > 100 m
Cap
tura
(kg)
0
10
20
30
Cap
(kg)
/lanc
es
Rubio
0
5
10
15
20
2 6 7 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 30 31 32 33
Estratos <= 100 m
Cap
tura
(kg)
0
3
6
9
12
15
Cap
(kg)
/lanc
es
Rubio
0
100
200
300
400
2 8 11 12 15 16 19 20 23 24 28 33
Estratos > 100 m
Cap
tura
(kg)
0
50
100
150
200
Cap
(kg)
/lanc
es

195
Estudio de las relaciones ambientales, distribución y composición de la fauna acompañante de merluza (Merluccius hubbsi) en primavera
de 2008.
Scarlato, Norberto A., P.Ibañez, L. Tringali
Figura 4. Captura (kg) y relación captura por lance por estrato de especies de fauna acompañante obtenidas en
la campaña EH 02/08 en estratos con profundidades menores o iguales a 100 m (izquierda) y
mayores de 100 m (derecha).
Figura 5 Estratos de la campaña EH-02/08.
El detalle de las capturas por estratos y la captura media por lance por estrato de especies de fauna
acompañante puede verse en la Figura 4. La ubicación de los estratos se detalla en la Figura 5. Por lo general,
Merluza
0
250
500
750
1000
1250
1500
2 6 7 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 30 31 32 33
Estratos <= 100 m
Cap
tura
(kg)
0
30
60
90
120
150
Cap
(kg)
/lanc
es
Merluza
0
50
100
150
200
2 8 11 12 15 16 19 20 23 24 28 33
Estratos > 100 m
Cap
tura
(kg)
0
0,25
0,5
0,75
1
Cap
(kg)
/lanc
es
Merluza de cola
0
20
40
60
80
100
2 8 11 12 15 16 19 20 23 24 28 33
Estratos > 100 m
Cap
tura
(kg)
0
0,25
0,5
0,75
1
Cap
(kg)
/lanc
es
El detalle de las capturas por estratos y la captura media por lance por estrato de especies de fauna acompañante puede verse en la Figura 4. La ubicación de los estratos se detalla en la Figura 5. Por lo general, las tendencias observadas para la captura total por estrato se mantienen para el caso de las capturas medias. El abadejo (Genypterus blacodes), apareció principalmente a profundidades superiores a 100 m, en el estrato 33, asociado al Talud, y también en el estrato 22, de Plataforma Intermedia. El calamar (Illex argentinus) se capturó preferentemente en los estratos 18 y 21, en pro-fundidades menores de 100 m, y en el estrato 20, en profundidades mayores a 100 m. La castañeta
Figura 4. Captura (kg) y relación captura por lance por estrato de especies de fauna acompañante obtenidas en la campaña EH 02/08 en estratos con profundidades menores o iguales a 100 m (izquierda) y mayores de 100 m (derecha).
Figura 5. Estratos de la campaña EH-02/08.
Figura 4. Captura (kg) y relación captura por lance por estrato de especies de fauna acompañante obtenidas en
la campaña EH 02/08 en estratos con profundidades menores o iguales a 100 m (izquierda) y
mayores de 100 m (derecha).
Figura 5 Estratos de la campaña EH-02/08.
El detalle de las capturas por estratos y la captura media por lance por estrato de especies de fauna
acompañante puede verse en la Figura 4. La ubicación de los estratos se detalla en la Figura 5. Por lo general,
Merluza
0
250
500
750
1000
1250
1500
2 6 7 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 30 31 32 33
Estratos <= 100 m
Cap
tura
(kg)
0
30
60
90
120
150
Cap
(kg)
/lanc
es
Merluza
0
50
100
150
200
2 8 11 12 15 16 19 20 23 24 28 33
Estratos > 100 m
Cap
tura
(kg)
0
0,25
0,5
0,75
1
Cap
(kg)
/lanc
es
Merluza de cola
0
20
40
60
80
100
2 8 11 12 15 16 19 20 23 24 28 33
Estratos > 100 m
Cap
tura
(kg)
0
0,25
0,5
0,75
1
Cap
(kg)
/lanc
es
196
FRENTE MARÍTIMO
Publicación de la Comisión Técnica Mixta del Frente Marítimo
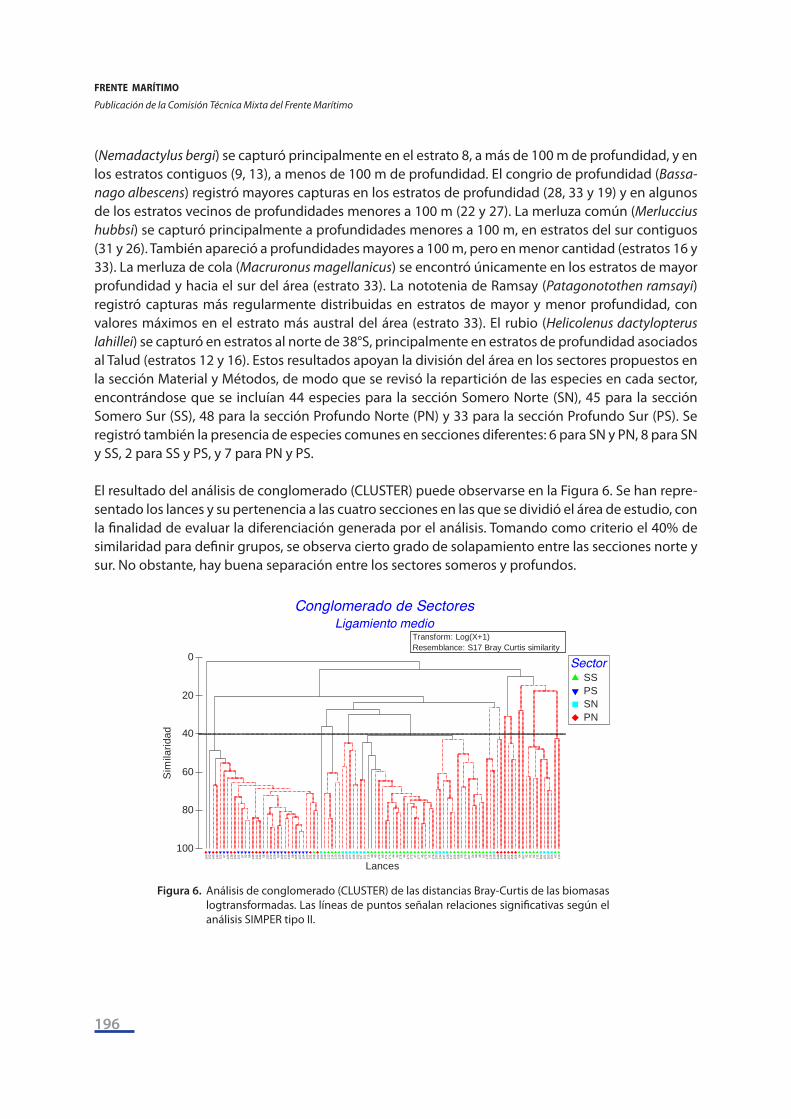
(Nemadactylus bergi) se capturó principalmente en el estrato 8, a más de 100 m de profundidad, y en los estratos contiguos (9, 13), a menos de 100 m de profundidad. El congrio de profundidad (Bassa-nago albescens) registró mayores capturas en los estratos de profundidad (28, 33 y 19) y en algunos de los estratos vecinos de profundidades menores a 100 m (22 y 27). La merluza común (Merluccius hubbsi) se capturó principalmente a profundidades menores a 100 m, en estratos del sur contiguos (31 y 26). También apareció a profundidades mayores a 100 m, pero en menor cantidad (estratos 16 y 33). La merluza de cola (Macruronus magellanicus) se encontró únicamente en los estratos de mayor profundidad y hacia el sur del área (estrato 33). La nototenia de Ramsay (Patagonotothen ramsayi) registró capturas más regularmente distribuidas en estratos de mayor y menor profundidad, con valores máximos en el estrato más austral del área (estrato 33). El rubio (Helicolenus dactylopterus lahillei) se capturó en estratos al norte de 38°S, principalmente en estratos de profundidad asociados al Talud (estratos 12 y 16). Estos resultados apoyan la división del área en los sectores propuestos en la sección Material y Métodos, de modo que se revisó la repartición de las especies en cada sector, encontrándose que se incluían 44 especies para la sección Somero Norte (SN), 45 para la sección Somero Sur (SS), 48 para la sección Profundo Norte (PN) y 33 para la sección Profundo Sur (PS). Se registró también la presencia de especies comunes en secciones diferentes: 6 para SN y PN, 8 para SN y SS, 2 para SS y PS, y 7 para PN y PS. El resultado del análisis de conglomerado (CLUSTER) puede observarse en la Figura 6. Se han repre-sentado los lances y su pertenencia a las cuatro secciones en las que se dividió el área de estudio, con la finalidad de evaluar la diferenciación generada por el análisis. Tomando como criterio el 40% de similaridad para definir grupos, se observa cierto grado de solapamiento entre las secciones norte y sur. No obstante, hay buena separación entre los sectores someros y profundos.
Figura 6. Análisis de conglomerado (CLUSTER) de las distancias Bray-Curtis de las biomasas logtransformadas. Las líneas de puntos señalan relaciones significativas según el análisis SIMPER tipo II.
Figura 6 Análisis de conglomerado (CLUSTER) de las distancias Bray-Curtis de las biomasas
logtransformadas. Las líneas de puntos señalan relaciones significativas según el análisis SIMPER tipo II.
Conglomerado de SectoresLigamiento medio
160
102
140
143
122 67 125
138
142
101 57 52 66 146
141
144 56 130
124
126 53 106
121
139 58 105
103
104
123
131
109
132
164
119
112
114
110
120
135
150
127
136
152
147
151
115 39 49 170 65 171
174 44 43 178 46 173
172 47 177 45 175 51 176
133
134
137
128
117
166
108 55 179
107 54 59 48 50 118
116
129
148
149
158
157
155
156 64 167 42 61 62 111
162
161
153
154 63 113
Lances
100
80
60
40
20
0
Sim
ilarid
ad
Transform: Log(X+1)Resemblance: S17 Bray Curtis similarity
SectorSSPSSNPN
197
Estudio de las relaciones ambientales, distribución y composición de la fauna acompañante de merluza (Merluccius hubbsi) en primavera
de 2008.
Scarlato, Norberto A., P.Ibañez, L. Tringali
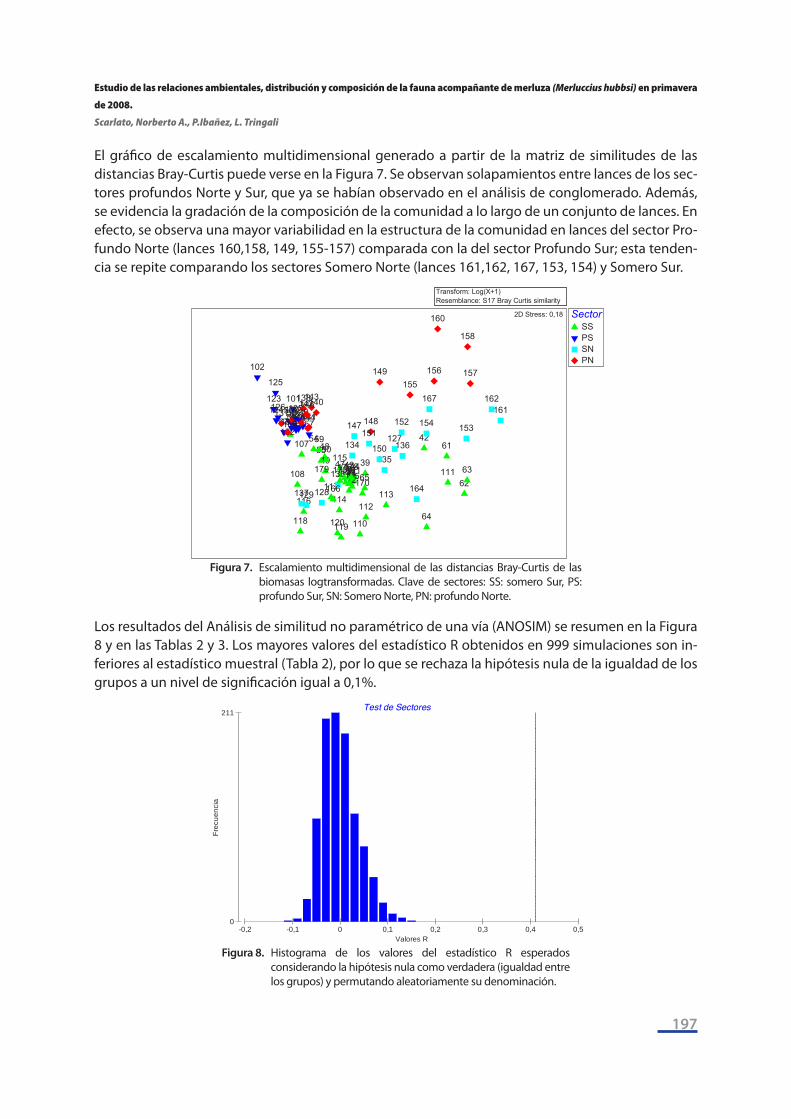
El gráfico de escalamiento multidimensional generado a partir de la matriz de similitudes de las distancias Bray-Curtis puede verse en la Figura 7. Se observan solapamientos entre lances de los sec-tores profundos Norte y Sur, que ya se habían observado en el análisis de conglomerado. Además, se evidencia la gradación de la composición de la comunidad a lo largo de un conjunto de lances. En efecto, se observa una mayor variabilidad en la estructura de la comunidad en lances del sector Pro-fundo Norte (lances 160,158, 149, 155-157) comparada con la del sector Profundo Sur; esta tenden-cia se repite comparando los sectores Somero Norte (lances 161,162, 167, 153, 154) y Somero Sur.
Figura 7 Escalamiento multidimensional de las distancias Bray-Curtis de las biomasas logtransformadas.
Clave de sectores: SS: somero Sur, PS: profundo Sur, SN: Somero Norte, PN: profundo Norte.
Figura 8 Histograma de los valores del estadístico R esperados considerando la hipótesis nula como
verdadera (igualdad entre los grupos) y permutando aleatoriamente su denominación.
Test de Sectores
-0,2 -0,1 0 0,1 0,2 0,3 0,4 0,5Valores R
0
211
Frec
uenc
ia
Figura 7 Escalamiento multidimensional de las distancias Bray-Curtis de las biomasas logtransformadas.
Clave de sectores: SS: somero Sur, PS: profundo Sur, SN: Somero Norte, PN: profundo Norte.
Figura 8 Histograma de los valores del estadístico R esperados considerando la hipótesis nula como
verdadera (igualdad entre los grupos) y permutando aleatoriamente su denominación.
Test de Sectores
-0,2 -0,1 0 0,1 0,2 0,3 0,4 0,5Valores R
0
211
Frec
uenc
ia
Figura 7. Escalamiento multidimensional de las distancias Bray-Curtis de las biomasas logtransformadas. Clave de sectores: SS: somero Sur, PS: profundo Sur, SN: Somero Norte, PN: profundo Norte.
Los resultados del Análisis de similitud no paramétrico de una vía (ANOSIM) se resumen en la Figura 8 y en las Tablas 2 y 3. Los mayores valores del estadístico R obtenidos en 999 simulaciones son in-feriores al estadístico muestral (Tabla 2), por lo que se rechaza la hipótesis nula de la igualdad de los grupos a un nivel de significación igual a 0,1%.
Figura 8. Histograma de los valores del estadístico R esperados considerando la hipótesis nula como verdadera (igualdad entre los grupos) y permutando aleatoriamente su denominación.
198
FRENTE MARÍTIMO
Publicación de la Comisión Técnica Mixta del Frente Marítimo
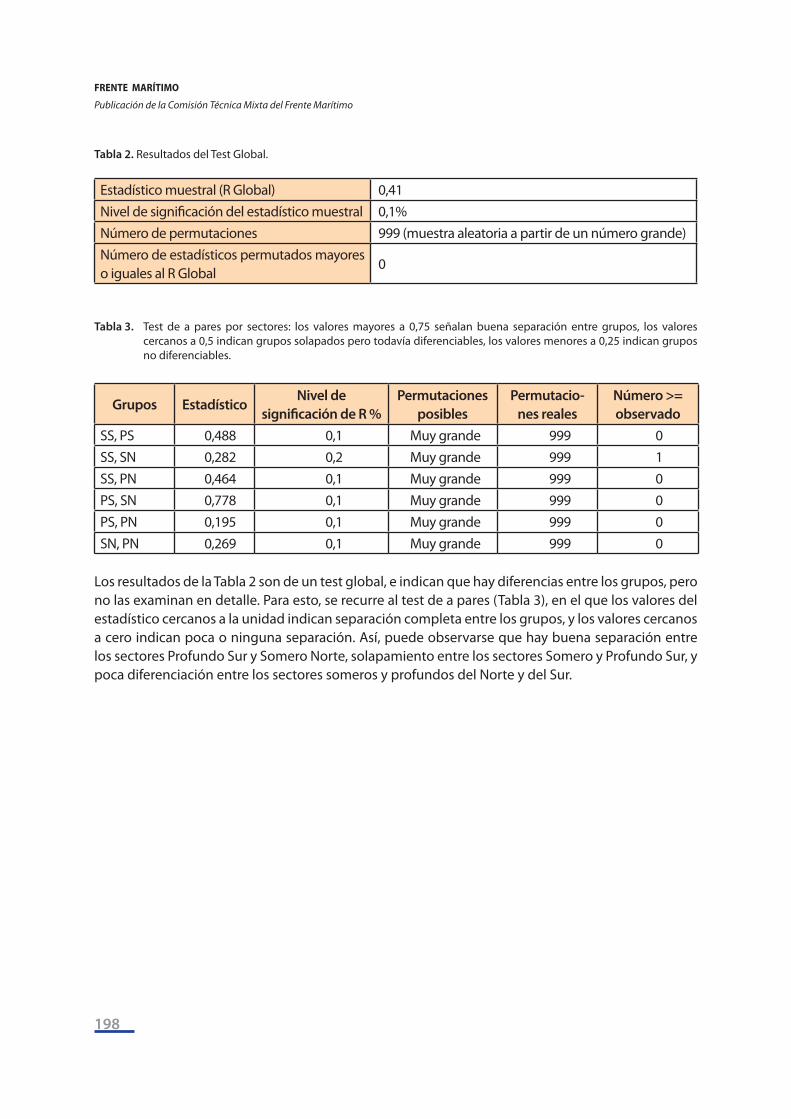
Tabla 2. Resultados del Test Global.
Estadístico muestral (R Global) 0,41Nivel de significación del estadístico muestral 0,1%Número de permutaciones 999 (muestra aleatoria a partir de un número grande)Número de estadísticos permutados mayores o iguales al R Global
0
Tabla 3. Test de a pares por sectores: los valores mayores a 0,75 señalan buena separación entre grupos, los valores
cercanos a 0,5 indican grupos solapados pero todavía diferenciables, los valores menores a 0,25 indican grupos no diferenciables.
Grupos EstadísticoNivel de
significación de R %Permutaciones
posibles Permutacio-
nes realesNúmero >= observado
SS, PS 0,488 0,1 Muy grande 999 0SS, SN 0,282 0,2 Muy grande 999 1SS, PN 0,464 0,1 Muy grande 999 0PS, SN 0,778 0,1 Muy grande 999 0PS, PN 0,195 0,1 Muy grande 999 0SN, PN 0,269 0,1 Muy grande 999 0
Los resultados de la Tabla 2 son de un test global, e indican que hay diferencias entre los grupos, pero no las examinan en detalle. Para esto, se recurre al test de a pares (Tabla 3), en el que los valores del estadístico cercanos a la unidad indican separación completa entre los grupos, y los valores cercanos a cero indican poca o ninguna separación. Así, puede observarse que hay buena separación entre los sectores Profundo Sur y Somero Norte, solapamiento entre los sectores Somero y Profundo Sur, y poca diferenciación entre los sectores someros y profundos del Norte y del Sur.

199
Estudio de las relaciones ambientales, distribución y composición de la fauna acompañante de merluza (Merluccius hubbsi) en primavera
de 2008.
Scarlato, Norberto A., P.Ibañez, L. Tringali
Los resultados del Análisis de Similitud de Porcentajes pueden observarse en las Tablas 4 a 9. En la Tabla 4 se comparan los sectores Somero Sur y Profundo Sur. Como se puede ver, la diferencia media es elevada (alrededor del 80), y una buena parte de ésta (alrededor de 39%) se debe a la presencia de Bassanago albescens, mostrando mayores valores de abundancia media en el sector Profundo sur. Por este motivo, y por su valor relativamente elevado del cociente Diferencia media/Desvío Standard(Dif/DS), se la considera como especie discriminante. También lo son por los mismos motivos Merluccius hubbsi e Illex argentinus (más abundantes en el sector somero), Iluocoetes fimbria-tus y Patagonotothen ramsayi (más abundantes en el sector profundo).
Tabla 4. Análisis SIMPER entre los sectores Somero Sur (SS) y Profundo Sur (PS). Diferencia media= 80,12%. Las especies discriminantes se señalan con un asterisco.
SS PS
EspecieBiomasa
mediaBiomasa
mediaDif.media Dif./DS Contrib% Acum.%
Bassanago albescens 0,26 3,06 31,53 2,96* 39,35 39,35Merluccius hubbsi 0,99 0,54 8,75 1,34* 10,93 50,27Illex argentinus 0,56 0,13 6,60 0,92* 8,24 58,51Iluocoetes fimbriatus 0,00 0,53 5,49 1,32* 6,86 65,37Patagonotothen ramsayi 0,22 0,37 3,59 1,29* 4,48 69,84Macruronus magellanicus 0,00 0,32 3,38 0,66 4,22 74,06Stromateus brasiliensis 0,34 0,02 3,38 0,64 4,21 78,27Trachurus lathami 0,27 0,00 2,06 0,31 2,58 80,85Genypterus blacodes 0,05 0,11 1,29 0,58 1,61 82,46Bathyraja brachyurops 0,05 0,09 1,22 0,85 1,52 83,98Bathyraja albomaculata 0,02 0,11 1,21 0,63 1,52 85,50Dipturus chilensis 0,10 0,04 1,21 0,92 1,51 87,01Engraulis anchoita 0,12 0,00 1,05 0,22 1,32 88,32Psammobatis normani 0,09 0,00 1,02 0,78 1,28 89,60Mustelus schmitti 0,13 0,00 0,99 0,28 1,23 90,83
200
FRENTE MARÍTIMO
Publicación de la Comisión Técnica Mixta del Frente Marítimo
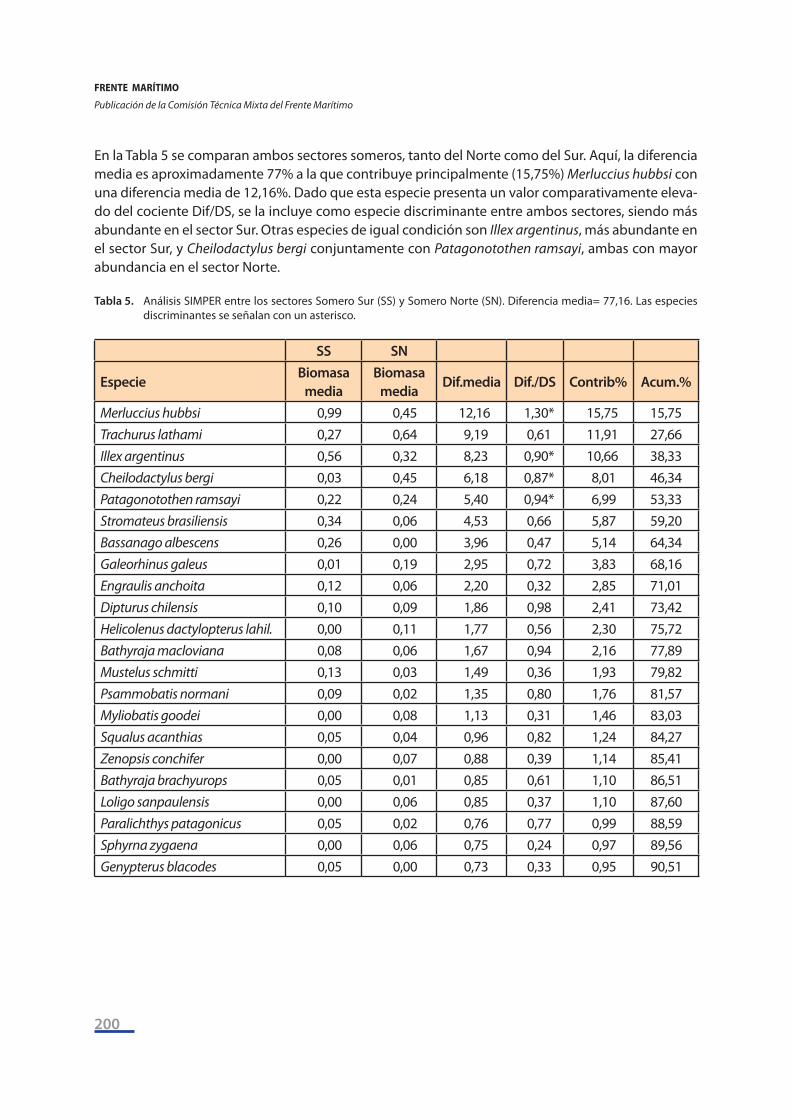
En la Tabla 5 se comparan ambos sectores someros, tanto del Norte como del Sur. Aquí, la diferencia media es aproximadamente 77% a la que contribuye principalmente (15,75%) Merluccius hubbsi con una diferencia media de 12,16%. Dado que esta especie presenta un valor comparativamente eleva-do del cociente Dif/DS, se la incluye como especie discriminante entre ambos sectores, siendo más abundante en el sector Sur. Otras especies de igual condición son Illex argentinus, más abundante en el sector Sur, y Cheilodactylus bergi conjuntamente con Patagonotothen ramsayi, ambas con mayor abundancia en el sector Norte.
Tabla 5. Análisis SIMPER entre los sectores Somero Sur (SS) y Somero Norte (SN). Diferencia media= 77,16. Las especies discriminantes se señalan con un asterisco.
SS SN
EspecieBiomasa
mediaBiomasa
mediaDif.media Dif./DS Contrib% Acum.%
Merluccius hubbsi 0,99 0,45 12,16 1,30* 15,75 15,75Trachurus lathami 0,27 0,64 9,19 0,61 11,91 27,66Illex argentinus 0,56 0,32 8,23 0,90* 10,66 38,33Cheilodactylus bergi 0,03 0,45 6,18 0,87* 8,01 46,34Patagonotothen ramsayi 0,22 0,24 5,40 0,94* 6,99 53,33Stromateus brasiliensis 0,34 0,06 4,53 0,66 5,87 59,20Bassanago albescens 0,26 0,00 3,96 0,47 5,14 64,34Galeorhinus galeus 0,01 0,19 2,95 0,72 3,83 68,16Engraulis anchoita 0,12 0,06 2,20 0,32 2,85 71,01Dipturus chilensis 0,10 0,09 1,86 0,98 2,41 73,42Helicolenus dactylopterus lahil. 0,00 0,11 1,77 0,56 2,30 75,72Bathyraja macloviana 0,08 0,06 1,67 0,94 2,16 77,89Mustelus schmitti 0,13 0,03 1,49 0,36 1,93 79,82Psammobatis normani 0,09 0,02 1,35 0,80 1,76 81,57Myliobatis goodei 0,00 0,08 1,13 0,31 1,46 83,03Squalus acanthias 0,05 0,04 0,96 0,82 1,24 84,27Zenopsis conchifer 0,00 0,07 0,88 0,39 1,14 85,41Bathyraja brachyurops 0,05 0,01 0,85 0,61 1,10 86,51Loligo sanpaulensis 0,00 0,06 0,85 0,37 1,10 87,60Paralichthys patagonicus 0,05 0,02 0,76 0,77 0,99 88,59Sphyrna zygaena 0,00 0,06 0,75 0,24 0,97 89,56Genypterus blacodes 0,05 0,00 0,73 0,33 0,95 90,51
201
Estudio de las relaciones ambientales, distribución y composición de la fauna acompañante de merluza (Merluccius hubbsi) en primavera
de 2008.
Scarlato, Norberto A., P.Ibañez, L. Tringali
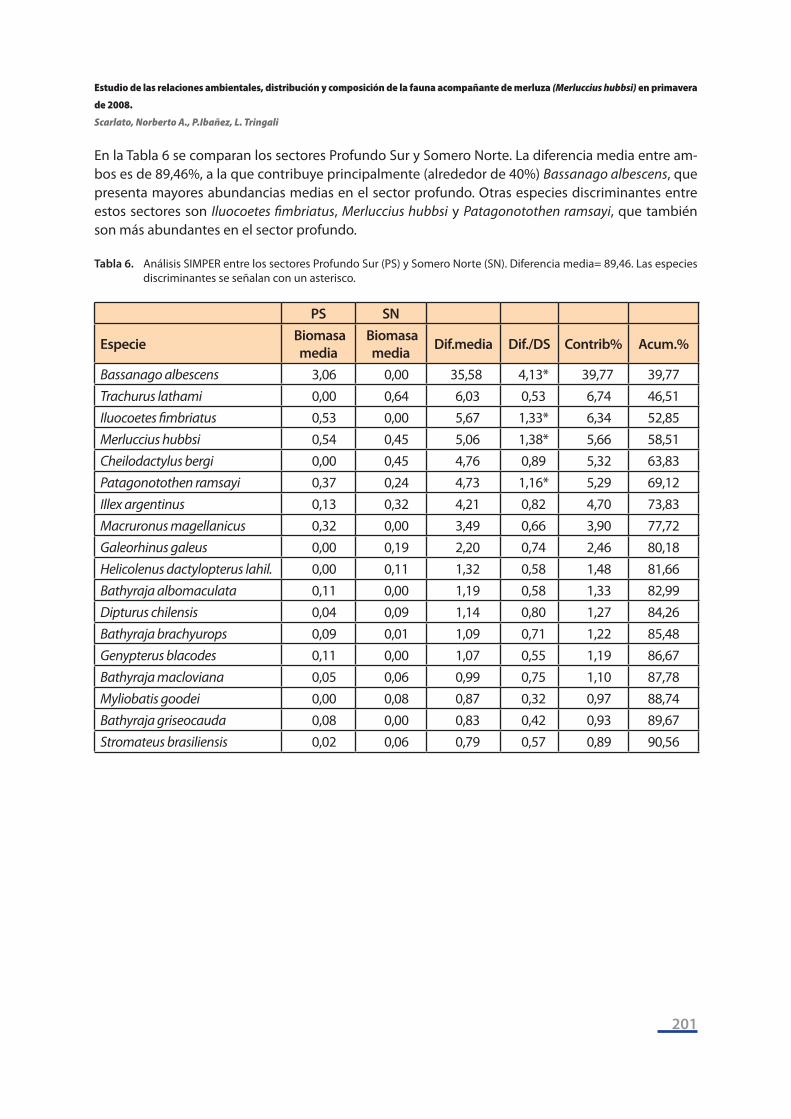
En la Tabla 6 se comparan los sectores Profundo Sur y Somero Norte. La diferencia media entre am-bos es de 89,46%, a la que contribuye principalmente (alrededor de 40%) Bassanago albescens, que presenta mayores abundancias medias en el sector profundo. Otras especies discriminantes entre estos sectores son Iluocoetes fimbriatus, Merluccius hubbsi y Patagonotothen ramsayi, que también son más abundantes en el sector profundo.
Tabla 6. Análisis SIMPER entre los sectores Profundo Sur (PS) y Somero Norte (SN). Diferencia media= 89,46. Las especies discriminantes se señalan con un asterisco.
PS SN
Especie Biomasa media
Biomasa media Dif.media Dif./DS Contrib% Acum.%
Bassanago albescens 3,06 0,00 35,58 4,13* 39,77 39,77Trachurus lathami 0,00 0,64 6,03 0,53 6,74 46,51Iluocoetes fimbriatus 0,53 0,00 5,67 1,33* 6,34 52,85Merluccius hubbsi 0,54 0,45 5,06 1,38* 5,66 58,51Cheilodactylus bergi 0,00 0,45 4,76 0,89 5,32 63,83Patagonotothen ramsayi 0,37 0,24 4,73 1,16* 5,29 69,12Illex argentinus 0,13 0,32 4,21 0,82 4,70 73,83Macruronus magellanicus 0,32 0,00 3,49 0,66 3,90 77,72Galeorhinus galeus 0,00 0,19 2,20 0,74 2,46 80,18Helicolenus dactylopterus lahil. 0,00 0,11 1,32 0,58 1,48 81,66Bathyraja albomaculata 0,11 0,00 1,19 0,58 1,33 82,99Dipturus chilensis 0,04 0,09 1,14 0,80 1,27 84,26Bathyraja brachyurops 0,09 0,01 1,09 0,71 1,22 85,48Genypterus blacodes 0,11 0,00 1,07 0,55 1,19 86,67Bathyraja macloviana 0,05 0,06 0,99 0,75 1,10 87,78Myliobatis goodei 0,00 0,08 0,87 0,32 0,97 88,74Bathyraja griseocauda 0,08 0,00 0,83 0,42 0,93 89,67Stromateus brasiliensis 0,02 0,06 0,79 0,57 0,89 90,56

202
FRENTE MARÍTIMO
Publicación de la Comisión Técnica Mixta del Frente Marítimo
En la Tabla 7 se comparan los sectores Somero Sur y Profundo Norte, que presentan una diferencia media de 84,01%, a la que contribuyen principalmente (alrededor de un 23%) Bassanago albescens y Merluccius hubbsi (alrededor de un 12%). La primer especie es más abundante en el sector profundo, y la segunda en el sector somero.
Tabla 7. Análisis SIMPER entre los sectores Somero Sur (SS) y Profundo Norte (PN). Diferencia media= 84,01. Las especies discriminantes se señalan con un asterisco.
SS PN Especie Biomasa
mediaBiomasa
mediaDif.media Dif./DS Contrib% Acum.%
Bassanago albescens 0,26 1,79 19,23 1,23* 22,89 22,89Merluccius hubbsi 0,99 0,50 10,45 1,11* 12,44 35,33Illex argentinus 0,56 0,06 6,94 0,75 8,27 43,60Helicolenus dactylopterus lahil. 0,00 0,52 4,90 0,79 5,84 49,43Zenopsis conchifer 0,00 0,45 4,76 0,39 5,67 55,10Stromateus brasiliensis 0,34 0,01 3,78 0,60 4,50 59,60Patagonotothen ramsayi 0,22 0,15 2,99 0,82 3,56 63,16Galeorhinus galeus 0,01 0,28 2,70 0,49 3,22 66,38Cheilodactylus bergi 0,03 0,23 2,45 0,48 2,92 69,29Squalus acanthias 0,05 0,23 2,28 0,69 2,71 72,00Trachurus lathami 0,27 0,00 2,20 0,30 2,62 74,63Dipturus chilensis 0,10 0,08 1,55 0,79 1,85 76,48Iluocoetes fimbriatus 0,00 0,15 1,49 0,80 1,77 78,25Trichiurus lepturus 0,00 0,13 1,40 0,31 1,67 79,91Bathyraja macloviana 0,08 0,04 1,23 0,79 1,47 81,38Mustelus schmitti 0,13 0,01 1,15 0,31 1,37 82,75Engraulis anchoita 0,12 0,00 1,15 0,20 1,37 84,13Psammobatis normani 0,09 0,01 1,13 0,70 1,35 85,47Loligo sanpaulensis 0,00 0,04 0,93 0,26 1,11 86,59Coelorhynchus fasciatus 0,00 0,11 0,92 0,50 1,10 87,69Bathyraja brachyurops 0,05 0,05 0,91 0,79 1,09 88,77Macruronus magellanicus 0,00 0,10 0,86 0,31 1,03 89,80Genypterus blacodes 0,05 0,03 0,71 0,37 0,84 90,64
203
Estudio de las relaciones ambientales, distribución y composición de la fauna acompañante de merluza (Merluccius hubbsi) en primavera
de 2008.
Scarlato, Norberto A., P.Ibañez, L. Tringali
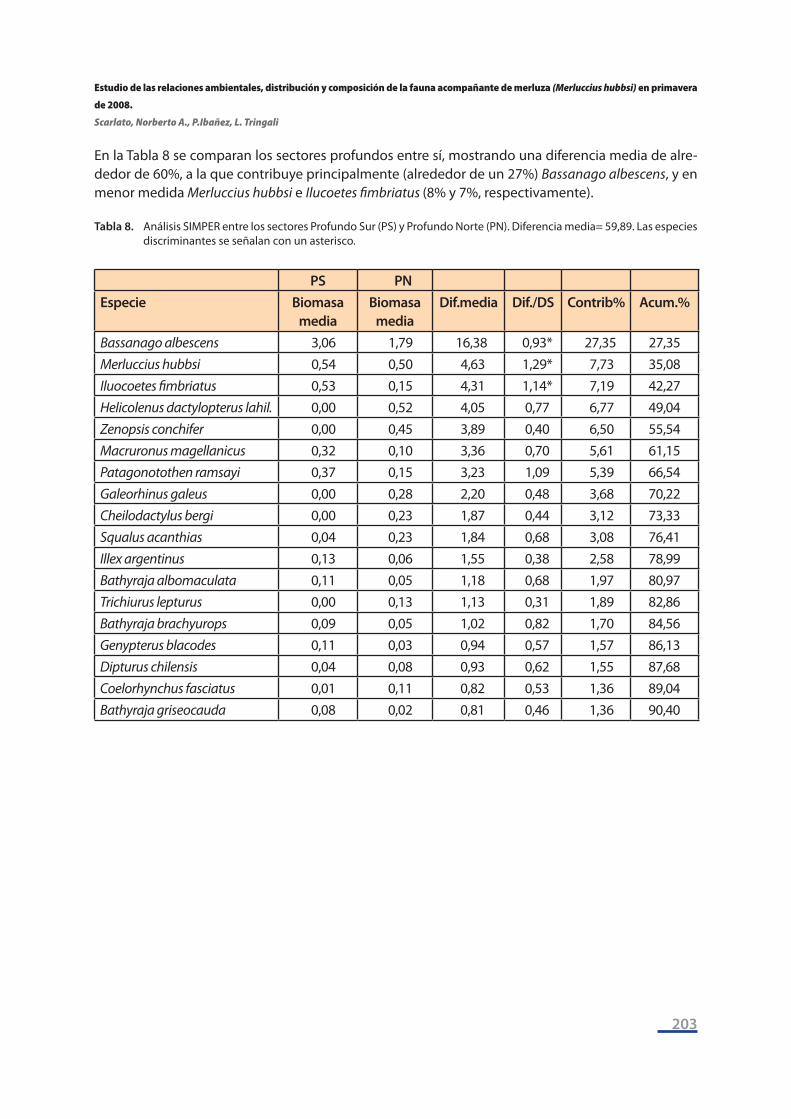
En la Tabla 8 se comparan los sectores profundos entre sí, mostrando una diferencia media de alre-dedor de 60%, a la que contribuye principalmente (alrededor de un 27%) Bassanago albescens, y en menor medida Merluccius hubbsi e Ilucoetes fimbriatus (8% y 7%, respectivamente).
Tabla 8. Análisis SIMPER entre los sectores Profundo Sur (PS) y Profundo Norte (PN). Diferencia media= 59,89. Las especies discriminantes se señalan con un asterisco.
PS PN Especie Biomasa
mediaBiomasa
mediaDif.media Dif./DS Contrib% Acum.%
Bassanago albescens 3,06 1,79 16,38 0,93* 27,35 27,35Merluccius hubbsi 0,54 0,50 4,63 1,29* 7,73 35,08Iluocoetes fimbriatus 0,53 0,15 4,31 1,14* 7,19 42,27Helicolenus dactylopterus lahil. 0,00 0,52 4,05 0,77 6,77 49,04Zenopsis conchifer 0,00 0,45 3,89 0,40 6,50 55,54Macruronus magellanicus 0,32 0,10 3,36 0,70 5,61 61,15Patagonotothen ramsayi 0,37 0,15 3,23 1,09 5,39 66,54Galeorhinus galeus 0,00 0,28 2,20 0,48 3,68 70,22Cheilodactylus bergi 0,00 0,23 1,87 0,44 3,12 73,33Squalus acanthias 0,04 0,23 1,84 0,68 3,08 76,41Illex argentinus 0,13 0,06 1,55 0,38 2,58 78,99Bathyraja albomaculata 0,11 0,05 1,18 0,68 1,97 80,97Trichiurus lepturus 0,00 0,13 1,13 0,31 1,89 82,86Bathyraja brachyurops 0,09 0,05 1,02 0,82 1,70 84,56Genypterus blacodes 0,11 0,03 0,94 0,57 1,57 86,13Dipturus chilensis 0,04 0,08 0,93 0,62 1,55 87,68Coelorhynchus fasciatus 0,01 0,11 0,82 0,53 1,36 89,04Bathyraja griseocauda 0,08 0,02 0,81 0,46 1,36 90,40

204
FRENTE MARÍTIMO
Publicación de la Comisión Técnica Mixta del Frente Marítimo
En la Tabla 9 se comparan los sectores Somero y Profundo del Norte. Se encuentra una diferencia media de 85,89%, a la que contribuye principalmente (alrededor de un 23%) Bassanago albescens, seguida por Cheilodactylus bergi y Merluccius hubbsi (alrededor de un 7%). La primera y la última especie son más abundantes en el sector profundo, mientras que la segunda lo es en el sector so-mero.
Tabla 9. Análisis SIMPER entre los sectores Somero Norte (SN) y Profundo Norte (PN). Diferencia media= 85,89. Las especies discriminantes se señalan con un asterisco.
SN PN
Especie Biomasa media
Biomasa media
Dif.media Dif./DS Contrib% Acum.%
Bassanago albescens 0,00 1,79 19,98 1,16* 23,27 23,27
Trachurus lathami 0,64 0,00 6,60 0,51 7,68 30,95
Cheilodactylus bergi 0,45 0,23 6,27 0,90* 7,30 38,24
Merluccius hubbsi 0,45 0,50 5,93 1,17* 6,90 45,15
Zenopsis conchifer 0,07 0,45 5,40 0,44 6,29 51,44
Helicolenus dactylopterus lahil. 0,11 0,52 5,22 0,85 6,08 57,52
Galeorhinus galeus 0,19 0,28 4,47 0,81 5,21 62,73
Patagonotothen ramsayi 0,24 0,15 4,21 0,65 4,90 67,63
Illex argentinus 0,32 0,06 3,88 0,79 4,52 72,15
Squalus acanthias 0,04 0,23 2,36 0,69 2,74 74,89
Trichiurus lepturus 0,04 0,13 1,80 0,38 2,10 76,99
Loligo sanpaulensis 0,06 0,04 1,56 0,39 1,82 78,81
Iluocoetes fimbriatus 0,00 0,15 1,53 0,80 1,79 80,60
Dipturus chilensis 0,09 0,08 1,52 0,72 1,77 82,37
Bathyraja macloviana 0,06 0,04 1,21 0,58 1,41 83,78
Myliobatis goodei 0,08 0,00 0,98 0,30 1,14 84,92
Coelorhynchus fasciatus 0,00 0,11 0,95 0,50 1,11 86,03
Macruronus magellanicus 0,00 0,10 0,89 0,31 1,03 87,06
Squalus blainvillei 0,03 0,04 0,87 0,70 1,02 88,07
Brama brama 0,02 0,08 0,85 0,68 0,99 89,06
Stromateus brasiliensis 0,06 0,01 0,83 0,52 0,96 90,02
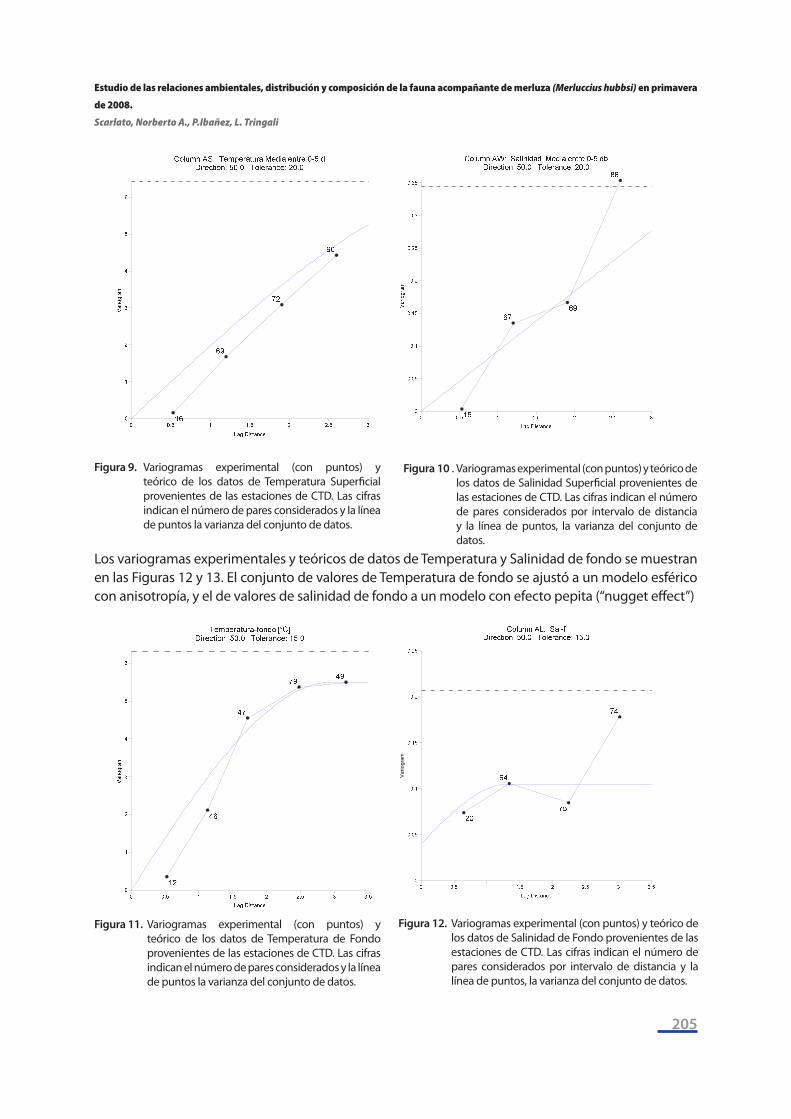
Los variogramas experimentales y teóricos de datos de Temperatura y Salinidad superficiales se muestran en las Figuras 10 y 11. El conjunto de valores de Temperatura superficial se ajustó a un modelo esférico con anisotropía, y el de valores de salinidad superficial a un modelo lineal con ani-sotropía.
205
Estudio de las relaciones ambientales, distribución y composición de la fauna acompañante de merluza (Merluccius hubbsi) en primavera
de 2008.
Scarlato, Norberto A., P.Ibañez, L. Tringali
Los variogramas experimentales y teóricos de datos de Temperatura y Salinidad de fondo se muestran en las Figuras 12 y 13. El conjunto de valores de Temperatura de fondo se ajustó a un modelo esférico con anisotropía, y el de valores de salinidad de fondo a un modelo con efecto pepita (“nugget effect”)
Figura 9 Variogramas experimental (con puntos) y
teórico de los datos de Temperatura Superficial
provenientes de las estaciones de CTD. Las cifras
indican el número de pares considerados y la línea
de puntos la varianza del conjunto de datos
Figura 10 Variogramas experimental (con puntos) y
teórico de los datos de Salinidad Superficial
provenientes de las estaciones de CTD. Las cifras
indican el número de pares considerados por
intervalo de distancia y la línea de puntos, la
varianza del conjunto de datos
Figura 9. Variogramas experimental (con puntos) y teórico de los datos de Temperatura Superficial provenientes de las estaciones de CTD. Las cifras indican el número de pares considerados y la línea de puntos la varianza del conjunto de datos.
Figura 10 . Variogramas experimental (con puntos) y teórico de los datos de Salinidad Superficial provenientes de las estaciones de CTD. Las cifras indican el número de pares considerados por intervalo de distancia y la línea de puntos, la varianza del conjunto de datos.
Figura 11. Variogramas experimental (con puntos) y
teórico de los datos de Temperatura de Fondo
provenientes de las estaciones de CTD. Las cifras
indican el número de pares considerados y la línea
de puntos la varianza del conjunto de datos
Figura 12. Variogramas experimental (con puntos) y
teórico de los datos de Salinidad de Fondo
provenientes de las estaciones de CTD. Las cifras
indican el número de pares considerados por
intervalo de distancia y la línea de puntos, la
varianza del conjunto de datos
Figura 11. Variogramas experimental (con puntos) y teórico de los datos de Temperatura de Fondo provenientes de las estaciones de CTD. Las cifras indican el número de pares considerados y la línea de puntos la varianza del conjunto de datos.
Figura 12. Variogramas experimental (con puntos) y teórico de los datos de Salinidad de Fondo provenientes de las estaciones de CTD. Las cifras indican el número de pares considerados por intervalo de distancia y la línea de puntos, la varianza del conjunto de datos.

206
FRENTE MARÍTIMO
Publicación de la Comisión Técnica Mixta del Frente Marítimo
Las temperaturas y salinidades superficiales estimadas mediante krigeado que incorporan los vario-gramas teóricos de las figuras 10 y 11 se incorporaron a mapas que incluyen los valores medidos y las isolíneas resultantes, con la finalidad de comprobar su coherencia y validez (Figura 13 y Figura 14), no detectándose errores ni incoherencias.
Figura 13. Temperaturas superficiales medidas
Figura 15. Temperaturas de fondo medidas (círculos
. Temperaturas superficiales medidas Figura 14. Salinidades superficiales medidas
. Temperaturas de fondo medidas (círculos Figura 16. Salinidades de fondo medidas (círculos
. Salinidades superficiales medidas
. Salinidades de fondo medidas (círculos
Figura 13. Temperaturas superficiales medidas
Figura 15. Temperaturas de fondo medidas (círculos
. Temperaturas superficiales medidas Figura 14. Salinidades superficiales medidas
. Temperaturas de fondo medidas (círculos Figura 16. Salinidades de fondo medidas (círculos
. Salinidades superficiales medidas
. Salinidades de fondo medidas (círculos
Figura 13. Temperaturas superficiales medidas (círculos negros) y estimadas (cruces grises)mediante krigeado que incorpora el variograma teórico de la Figura 9.
Figura 14. Salinidades superficiales medidas (círculos negros) y estimadas (cruces grises) mediante krigeado que incorpora el variograma teórico de la Figura 10.
Las temperaturas y salinidades de fondo estimadas mediante krigeado que incorporan los variogra-mas teóricos (Figura 11 y Figura 12) se incorporaron a mapas que incluyen los valores medidos y las isolíneas resultantes, de acuerdo a la metodología detallada en el párrafo anterior. Los valores esti-mados se ajustan favorablemente, no detectándose errores ni incoherencias (Figura 15 y Figura 16).
Figura 15. Temperaturas de fondo medidas (círculos negros) y estimadas (cruces grises)mediante krigeado que incorpora el variograma teórico de la Figura 11.
Figura 16. Salinidades de fondo medidas (círculos negros) y estimadas (cruces grises) mediante krigeado que incorpora el variograma teórico de la Figura 12.
207
Estudio de las relaciones ambientales, distribución y composición de la fauna acompañante de merluza (Merluccius hubbsi) en primavera
de 2008.
Scarlato, Norberto A., P.Ibañez, L. Tringali
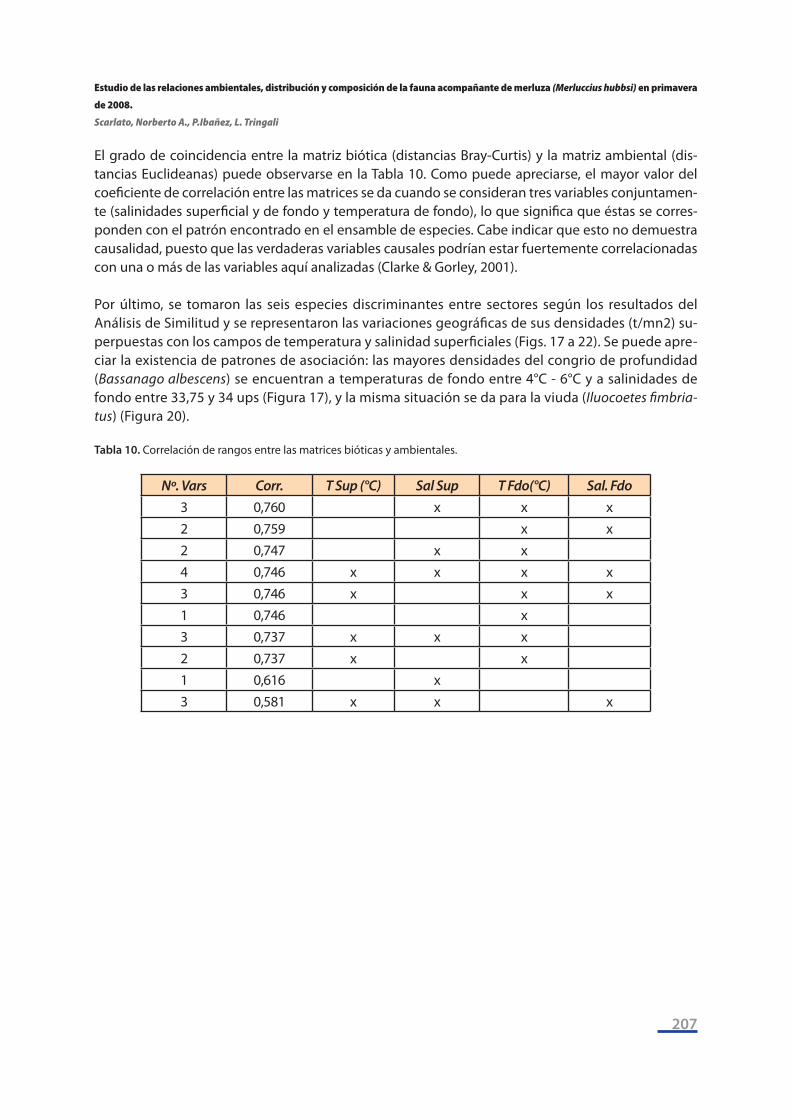
El grado de coincidencia entre la matriz biótica (distancias Bray-Curtis) y la matriz ambiental (dis-tancias Euclideanas) puede observarse en la Tabla 10. Como puede apreciarse, el mayor valor del coeficiente de correlación entre las matrices se da cuando se consideran tres variables conjuntamen-te (salinidades superficial y de fondo y temperatura de fondo), lo que significa que éstas se corres-ponden con el patrón encontrado en el ensamble de especies. Cabe indicar que esto no demuestra causalidad, puesto que las verdaderas variables causales podrían estar fuertemente correlacionadas con una o más de las variables aquí analizadas (Clarke & Gorley, 2001).
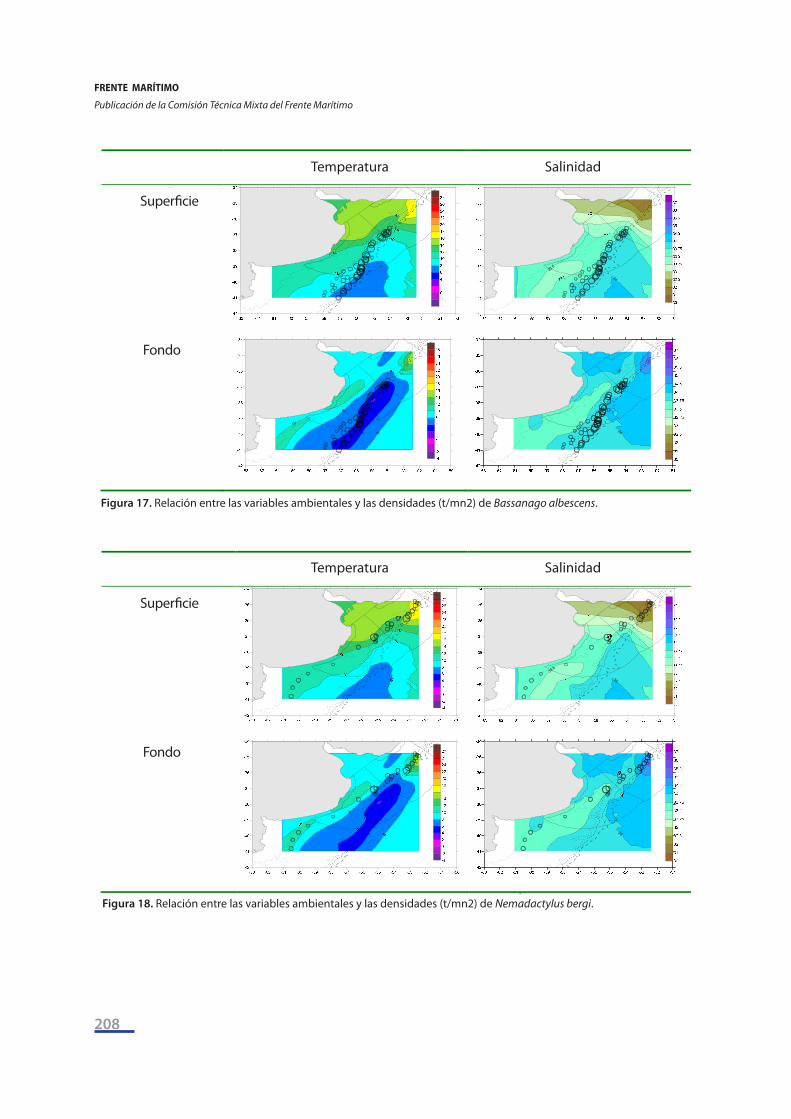
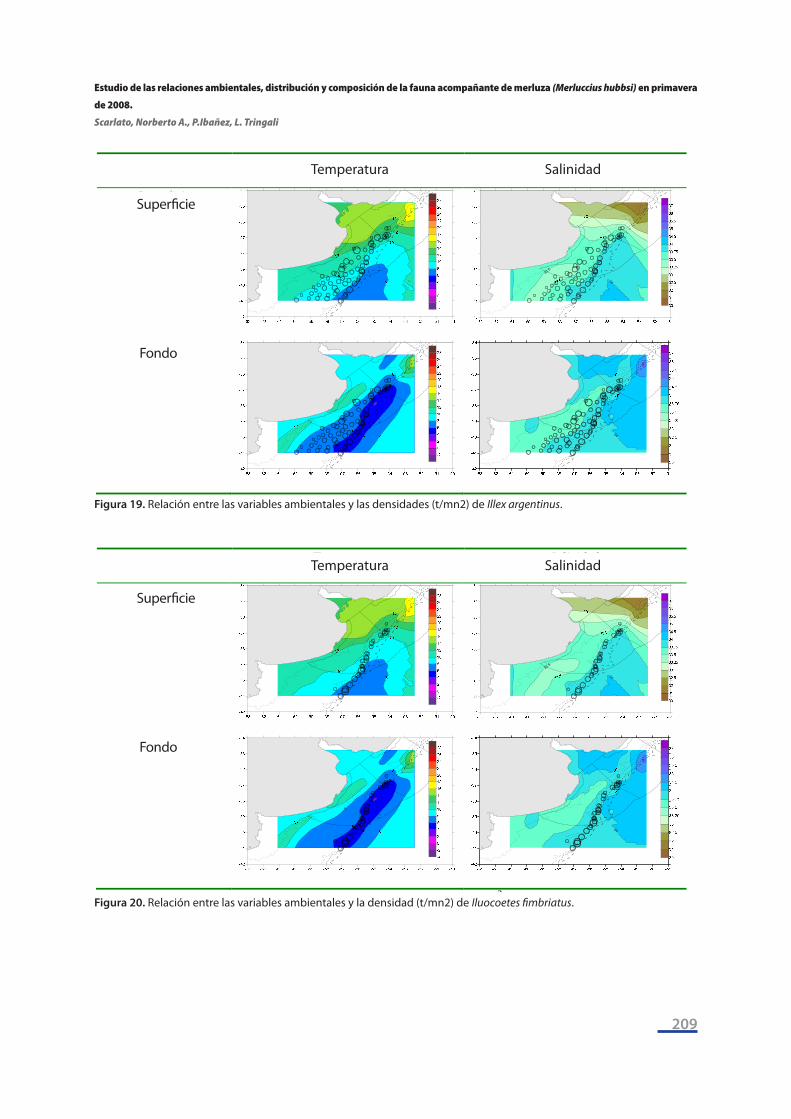
Por último, se tomaron las seis especies discriminantes entre sectores según los resultados del Análisis de Similitud y se representaron las variaciones geográficas de sus densidades (t/mn2) su-perpuestas con los campos de temperatura y salinidad superficiales (Figs. 17 a 22). Se puede apre-ciar la existencia de patrones de asociación: las mayores densidades del congrio de profundidad (Bassanago albescens) se encuentran a temperaturas de fondo entre 4°C - 6°C y a salinidades de fondo entre 33,75 y 34 ups (Figura 17), y la misma situación se da para la viuda (Iluocoetes fimbria-tus) (Figura 20).
Tabla 10. Correlación de rangos entre las matrices bióticas y ambientales.
Nº. Vars Corr. T Sup (°C) Sal Sup T Fdo(°C) Sal. Fdo3 0,760 x x x2 0,759 x x2 0,747 x x4 0,746 x x x x3 0,746 x x x1 0,746 x3 0,737 x x x2 0,737 x x1 0,616 x3 0,581 x x x
208
FRENTE MARÍTIMO
Publicación de la Comisión Técnica Mixta del Frente Marítimo
Temperatura Salinidad
Superficie
Fondo
Figura 17 Relación entre las variables ambientales y las densidades (t/mn2) de Bassanago albescens.
Temperatura Salinidad
Superficie
Fondo
Figura 18 Relación entre las variables ambientales y las densidades (t/mn2) de Nemadactylus bergi.
Temperatura Salinidad
Superficie
Fondo
Figura 17 Relación entre las variables ambientales y las densidades (t/mn2) de Bassanago albescens.
Temperatura Salinidad
Superficie
Fondo
Figura 18 Relación entre las variables ambientales y las densidades (t/mn2) de Nemadactylus bergi.
Figura 17. Relación entre las variables ambientales y las densidades (t/mn2) de Bassanago albescens.
Figura 18. Relación entre las variables ambientales y las densidades (t/mn2) de Nemadactylus bergi.
Temperatura Salinidad
Temperatura Salinidad
Superficie
Fondo
Superficie
Fondo
209
Estudio de las relaciones ambientales, distribución y composición de la fauna acompañante de merluza (Merluccius hubbsi) en primavera
de 2008.
Scarlato, Norberto A., P.Ibañez, L. Tringali
Temperatura Salinidad
Superficie
Fondo
Figura 19 Relación entre las variables ambientales y las densidades (t/mn2) de Illex argentinus.
Temperatura Salinidad
Superficie
Fondo
Figura 20 Relación entre las variables ambientales y la densidad (t/mn2) de Iluocoetes fimbriatus.
Temperatura Salinidad
Superficie
Fondo
Figura 19 Relación entre las variables ambientales y las densidades (t/mn2) de Illex argentinus.
Temperatura Salinidad
Superficie
Fondo
Figura 20 Relación entre las variables ambientales y la densidad (t/mn2) de Iluocoetes fimbriatus.
Temperatura Salinidad
Temperatura Salinidad
Superficie
Fondo
Superficie
Fondo
Figura 19. Relación entre las variables ambientales y las densidades (t/mn2) de Illex argentinus.
Figura 20. Relación entre las variables ambientales y la densidad (t/mn2) de Iluocoetes fimbriatus.

210
FRENTE MARÍTIMO
Publicación de la Comisión Técnica Mixta del Frente Marítimo
Temperatura Salinidad
Superficie
Fondo
Figura 21 Relación entre las variables ambientales y la densidad (t/mn2) de Merluccius hubbsi.
Temperatura Salinidad
Superficie
Fondo
Figura 22 Relación entre las variables ambientales y la densidad (t/mn2) de Patagonotothen ramsayi.
Temperatura Salinidad
Superficie
Fondo
Figura 21 Relación entre las variables ambientales y la densidad (t/mn2) de Merluccius hubbsi.
Temperatura Salinidad
Superficie
Fondo
Figura 22 Relación entre las variables ambientales y la densidad (t/mn2) de Patagonotothen ramsayi.
Temperatura Salinidad
Temperatura Salinidad
Superficie
Fondo
Superficie
Fondo
Figura 21. Relación entre las variables ambientales y la densidad (t/mn2) de Merluccius hubbsi.
Figura 22. Relación entre las variables ambientales y la densidad (t/mn2) de Patagonotothen ramsayi.

211
Estudio de las relaciones ambientales, distribución y composición de la fauna acompañante de merluza (Merluccius hubbsi) en primavera
de 2008.
Scarlato, Norberto A., P.Ibañez, L. Tringali
Conclusiones
En el área de estudio se encontró un conjunto de especies que pueden caracterizarse en cuatro gru-pos, de acuerdo a sus valores de densidad media y frecuencia media de aparición. Sólo dos (2,78%) (Bassanago albescens y Merluccius hubbsi) se consideran dominantes, diecinueve (26,4%) frecuentes, siete (9,72%) ocasionales y 44 (61,11%) raras. La división en cuatro sectores atendiendo a límites ba-timétricos y latitudinales pareció ajustarse a las tendencias evidenciadas por las capturas de distintas especies discriminadas por estrato. Sin embargo, el Análisis de Conglomerados permite observar solapamiento entre distintos sectores (principalmente Norte y Sur), que se vuelven a detectar en el Escalamiento Multidimensional. Con esta técnica se evidencia una mayor variabilidad en la estructu-ra de la comunidad en regiones de los sectores del Norte (profundo y somero) comparada con las del Sur. El Análisis de Similitudes de una vía permite afirmar que los sectores son diferentes entre sí en cuanto a la estructuración de las comunidades, pero refuerza la evidencia de la escasa diferenciación entre los sectores limitados por la latitud, aunque registra una fuerte separación entre los sectores Somero Norte y Profundo Sur. Se identifica un total de cinco especies como discriminantes entre sectores mediante el análisis de Similitud de Porcentajes (no para todos los casos) y se evidencia que la conjunción de tres variables ambientales (salinidad de fondo y superficial, temperatura de fondo) se corresponde con el patrón encontrado para el conjunto de especies considerado. Sería recomen-dable plantear una diferenciación en sectores más coherente en análisis ulteriores, con la finalidad de lograr identificar cambios espaciotemporales a lo largo de series de tiempo.

212
FRENTE MARÍTIMO
Publicación de la Comisión Técnica Mixta del Frente Marítimo
Bibliografía
ALVERSON, D. L., FREEBERG, M., MURAWSKI, S. A. & POPE, J. G. 1994. A global assessment of fisheries bycatch and discards. FAO Fish. Tech. Paper 339: 1–233. Food & Agriculture Org.
ARENA, G., UBAL, W., GRUNWALDT, P. & A, F. 1986. Distribución latitudinal y batimétrica de la merluza (Merluccius hubbsi) y otros organis-mos demersales de su fauna acompañante, dentro de la Zona Común de Pesca Argentino-Uruguaya. Publ. Com.Téc.Mixta Fr.Mar. (Argen-tina-Uruguay) 1: 253–437.
BRIDGES, N. J. 1985. KRIGING - an interactive pro-gram to determine the best linear unbiased estimation. U.S. Geological Survey.
BURATTI, C. C. 2004. Fauna acompañante de la merluza (Merluccius hubbsi) en la zona común de pesca Argentino-Uruguaya y Mar Argenti-no hasta 41°S, en 1994, 1999 y 2001. Informe Técnico Interno INIDEP 10: 1–90.
CAÑETE, G., DATO, C. & VILLARINO, M. F. 2000. Characterization of hake Merluccius hubbsi discard process by freezers and factory fleets. Preliminary results stemming from data ga-thered by INIDEP observers during six trips ca-rried out in August-December 1995. In: INIDEP, Mar del Plata (Argentina). Vol. 32, 18 pp. 2000., pp. 1–18. INIDEP, Mar del Plata (Argentina).
CLARKE, K. & GORLEY, R. 2005. PRIMER: Getting started with v6. PRIMER-E Ltd: Plymouth, UK.
CLARKE, K. R. & GORLEY, R. N. 2001. Primer v5: User Manual/Tutorial. Plymouth Marine Labo-ratory.
CLARKE, K. R., SOMERFIELD, P. J. & GORLEY, R. N. 2008. Testing of null hypotheses in explora-tory community analyses: similarity profiles and biota-environment linkage Journal of Ex-perimental Marine Biology and Ecology 366: 56 – 69.
CLARKE, K. R. & WARWICK, R. M. 2001. Change in marine communities: an approach to statisti-cal analysis and interpretation. In: pp. 1–176. PRIMER-E.
COLVOCORESSES, J. & MUSICK, J. 1984. Species associations and community composition of middle Atlantic Bight continental-shelf demersal fishes. Fishery Bulletin 82: 295–313. NATL MARINE FISHERIES SERVICE SCIENTIFIC PUBL OFFICE 7600 SAND POINT WAY NE BIN C15700, SEATTLE, WA 98115.
COSTA, M. E., OLIM, S., PARIS, C. & BORGES, T. C. 2001. The importance of By-catch from Com-mercial Trawl Fisheries off the South Coast of Portugal. In: Deep-sea Fisheries Symposium, pp. 1–6.
CROWDER, L. B. & MURAWSKI, S. A. 1998. Fishe-ries bycatch: implications for management. Fisheries 23: 8–17. Taylor \& Francis.
D’ATRI, L. L. 2008. Campaña global de evaluación de merluza. Área norte de 41°S hasta 34°30’ S. INIDEP Inf. Campaña: 1–13.
DATO, C. V. 1997. Distribución e importancia de la fauna acompañante en la pesquería de merlu-za (Merluccius hubbsi) al sur de 41°S. Informe Técnico Interno INIDEP: 1–22.

213
Estudio de las relaciones ambientales, distribución y composición de la fauna acompañante de merluza (Merluccius hubbsi) en primavera
de 2008.
Scarlato, Norberto A., P.Ibañez, L. Tringali
DAVIS, M. W. 2002. Key principles for understan-ding fish bycatch discard mortality. Canadian Journal of Fisheries and Aquatic Sciences 59: 1834–1843. NRC Research Press.
EHRLICH, M. D. 1998. Composición y abundancia de la fauna acompañante de merluza ( Merluc-cius hubbsi) a través de datos de la flota comer-cial. Inf. Téc. DNI 125.
FISCHER, M. M. & GETIS, A. 2009. Handbook of Applied Spatial Analysis: Software Tools, Methods and Applications. Springer Berlin Heidelberg.
GABRIEL, W. L. & TYLER, A. 1980. Preliminary analysis of Pacific coast demersal fish assem-blages. Mar. Fish. Rev 42: 83–88.
GARCÍA DE LA ROSA, S. B., GIUSSI, A. R. & SÁN-CHEZ, F. 1997. Fauna acompañante de la mer-luza común (Merluccius hubbsi) en la patago-nia austral (45o - 55o S). Inf. Téc. DNI 135.
GARCÍA, M. L., JAUREGUIZAR, A. J. & PROTOGI-NO, L. C. 2010. From fresh water to the slope: fish community ecology in the Río de la Plata and the sea beyond. Latin American Journal of Aquatic Research 38: 81–94. SciELO Chile.
GRINGARTEN, E. & DEUTSCH, C. V. 2001. Teacher’s aide variogram interpretation and modeling. Mathematical Geology 33: 507–534. Springer.
HALL, M. A., ALVERSON, D. L. & METUZALS, K. I. 2000. By-catch: problems and solutions. Mari-ne Pollution Bulletin 41: 204–219. Elsevier.
HALL, S. J. & MAINPRIZE, B. M. 2005. Managing by-catch and discards: how much progress are we making and how can we do better? Fish and Fisheries 6: 134–155. Wiley Online Library.
IRUSTA, C. 1990. Relaciones espacio-temporales entre la merluza común (Merluccius hubbsi) y otros organismos de su fauna acompañante.
IRUSTA, C. G., CASTRUCCI, R. & BLANCO, G. 2001. Análisis espacio-temporal de los rendimientos de merluza ( Merluccius hubbsi) y de otros or-ganismos de su fauna acompañante. Inf. Téc. Int. INIDEP: 1–17.
JAUREGUIZAR, A. J. 2004. Patrón espacial y temporal de las áreas de asociaciones ícticas demersales costeras (34° S-41° S) y su relación con los factores ambientales.
JAUREGUIZAR, A. J., MENNI, R., BREMEC, C., MIAN-ZAN, H. & LASTA, C. 2003. Fish assemblage and environmental patterns in the Río de la Plata estuary. Est. Coast. Shelf Sci. 56: 921–933.
JAUREGUIZAR, A. J., MENNI, R., LASTA, C. & GUE-RRERO, R. 2006a. Fish assemblages of the northern Argentine coastal system: spatial patterns and their temporal variations. Fish. Oceanogr. 15: 326–344.
JAUREGUIZAR, A. J., RUARTE, C. & GUERRERO, R. A. 2006b. Distribution of age-classes of striped weakfish (Cynoscion guatucupa) along an es-tuarine-marine gradient: Correlations with the environmental parameters. Estuarine, Coastal and Shelf Science 67: 82–92. Elsevier.
JAUREGUIZAR, A., WAESSLE, J. & GUERRERO, R. 2007. Spatio-temporal distribution of Atlantic searobins (Prionotus spp.) in relation to estua-rine dynamics (Rio de la Plata, Southwestern Atlantic Coastal System). Estuarine, Coastal and Shelf Science 73: 30–42. Elsevier.
JOURNEL, A. G. 1989. Fundamentals of geostatis-tics in five lessons. Am.Geophys. Union.
JOURNEL, A. G. & HUIJBREGTS, C. J. 1978. Mining geostatistics. Academic Press.
KENNELLY, S. J. 1995. The issue of bycatch in Australia’s demersal trawl fisheries. Reviews in Fish Biology and Fisheries 5: 213–234. Springer.

214
FRENTE MARÍTIMO
Publicación de la Comisión Técnica Mixta del Frente Marítimo
KORSBREKKE, K., MEHL, S., NAKKEN, O. & PEN-NINGTON, M. 2001. A survey-based assessment of the Northeast Arctic cod stock. ICES Journal of Marine Science: Journal du Conseil 58: 763–769. Oxford University Press.
MARTINEZ, E. X., NANCE, J. M., COMMISSION, T. W. & OTHERS. 1993. Trawling bycatch in the Galveston Bay system. Galveston Bay National Estuary Program.
MUETER, F. & NORCROSS, B. 2002. Spatial and temporal patterns in the demersal fish com-munity on the shelf and upper slope regions of the Gulf of Alaska. Fishery Bulletin 100: 559–581. NATL MARINE FISHERIES SERVICE SCIEN-TIFIC PUBL OFFICE 7600 SAND POINT WAY NE BIN C15700, SEATTLE, WA 98115 USA.
MURAWSKI, S. & FINN, J. 1988. Biological bases for mixed-species fisheries: species co-distri-bution in relation to environmental and bio-tic variables. Canadian Journal of Fisheries and Aquatic Sciences 45: 1720–1735. NRC Re-search Press.
PENNINGTON, M. & STRØMME, T. 1998. Surveys as a research tool for managing dynamic stocks. Fisheries Research 37: 97–106. Elsevier.
RENZI, M. A. & CASTRUCCI, R. 1998. Análisis cuali-cuantitativo de la fauna acompañante en la pesquería de merluza (Merluccius hubbsi) al norte de 41°S. Informe Técnico Interno INIDEP: 1–20.
REY, M., CASCUDO, J. & GRUNWALDT, P. 1993. Análisis de algunas poblaciones de la fauna acompañante de la merluza. Rangos de distri-bución, concentración y rendimientos. Perío-do otoño 1986-verano 1987. CTMFM- Circular 4: 1–27.
SOKAL, R. & ROHLF, F. J. 1981. Biometry. In: pp. 1–259 (2nd edition). Freeman.
STEIN, M. L. 1999. Interpolation of Spatial Data: Some Theory for Kriging. Springer New York.
TILLMAN, R. F. 1993. By-catch- the issue of the 90’s. In: (R P Jones (ed))pp. 13–18. Southeas-tern Fisheries Association.
UNGARO, N., MARANO, C. A., MARSAN, R., MAR-TINO, M., MARZANO, M. C., STRIPPOLI, G. & VLORA, A. 1999. Analysis of demersal species assemblages from trawl surveys in the South Adriatic Sea. Aquatic Living Resources 12: 177–185. Cambridge Univ Press.