curso: genética clínica semestre: octavo tema: variación...
TRANSCRIPT

Curso: Genética Clínica
Semestre: Octavo
Tema: Variación génica en los
individuos y las poblaciones
Instructor: Dra. Perla Niño

Conceptos a evaluar
Examinar la naturaleza de las diferencias determinadas
genéticamente entre los individuos.
Reconocer como algunas de las diferencias en la secuencia de DNA
tienen poco o ningún efecto sobre el fenotipo, mientras que otras son
responsables directas de enfermedades.
Evaluar como entre esos dos extremos, se sitúan las diferencias
responsables de la variabilidad fenotípica determinada genéticamente
en la anatomía, la fisiología, las intolerancias alimenticias, las
respuestas terapéuticas y las reacciones adversas a los
medicamentos, la susceptibilidad a las infecciones, la predisposición
al cáncer y, quizá, incluso la variabilidad en varios rasgos de la
personalidad, la aptitud atlética y el talento artístico.

Uno de los conceptos importantes de la genética humana y
de sus aspectos clínicos es que la enfermedad genética es solo
una de las manifestaciones mas evidentes y, a menudo, mas
notables de las diferencias genéticas.
Estas diferencias abarcan:
variantes raras que causan enfermedades.
variantes más comunes que pueden aumentar la predisposición
a las enfermedades.
variaciones más frecuentes en la población, sin relevancia
conocida con respecto a las enfermedades.
Variaciones génicas

Mutación: Se define como cualquier cambio heredado en la
información genética; los descendientes pueden ser células oindividuos.
Tipos de mutación
Mutaciones genómicas; Son las que afectan el numero decromosomas en la célula, se originan de errores en la segregaciónde los cromosomas durante la mitosis o la meiosis.
Mutaciones cromosómicas; Son las que alteran la estructurade un cromosoma en concreto. Implican solamente una parte de uncromosoma, como las duplicaciones o triplicaciones parciales, lasdeleciones, las inversiones o las traslocaciones que pueden ocurrirde manera espontanea o ser el resultado de una segregaciónanómala.
Mutaciones génicas; Son las que alteran genes concretos. Soncambios en la secuencia del DNA del núcleo o la mitocondria.

Mutaciones génicas
Por su mecanismo de producción:
En punto: Transiciones. Cambio de una base púrica o pirimídica por
otra púrica o pirimídica.
Transversiones. Cambio de una base púrica por una
pirimídica o viceversa.
Fuera de Fase: Adiciones. Adición de 1, 2 o más bases.
Deleciones. Supresión de 1, 2, o más bases

Fuera de faseAltera un solo codón
Cambia el marco de lectura y
puede alterar muchos codones

Mutaciones génicas.
Mutaciones por sustitución de una base por otra
distinta
Mutaciones por pérdida o inserción de bases
Mutaciones por variaciones de lugar de algunos
segmentos de ADN.

Por la naturaleza de las células afectadas
Germinales (Heredables). Se dan en organismos unicelulares o afectan a las células
reproductivas (asexuales o gametos) de los pluricelulares.
Somáticas. Afectan a cualquier célula somática de los seres pluricelulares. (No afectan a la
descendencia).
MUTACIÓN SOMÁTICA:
Origina una población de células mutantes
idénticas, todas ellas descendientes de la
célula en que ocurrió la mutación original
(clon).
Son importantes en el envejecimiento y el
cáncer
MUTACIÓN GERMINAL
Ocurre en un tejido que en última instancia
dará lugar a células sexuales.
Si estos gametos mutantes participan en la
fecundación, la mutación se transmitirá a la
siguiente generación.
Individuo normal de antepasados normales
puede ser portador de células germinales
mutantes no detectadas

Por la causa que produce el cambio Espontáneas. En cualquier momento en cualquier tipo de célula. Se pueden
producir por errores en la replicación, lesiones por azar en el ADN y
elementos genéticos transponibles.
Inducidas. Producidas por agentes mutágenos específicos

Sustituciones sinónimas (SILENCIOSAS)
El resultado de la sustitución es un nuevo codón para el mismo aminoácido.
Constituyen el cambio mas frecuente, se presentan sobretodo en la posición
de la tercera base.
Sin sentido
Es una sustitución no sinónima en la que un codón que codificaba para un
aminoácido es remplazado por un codón de detención. Es muy rara puesto que
una mutación de este tipo casi siempre genera disminución de la función
génica.
En sentido Erróneo
Sustituciones no sinónimas en las que el codón alterado codifica un aminoácido
distinto:
Sustituciones conservadoras; Remplazo de un aminoácido por otro
químicamente similar a él.
Sustituciones no conservadoras; Remplazo de un aminoácido por otro
químicamente distinto.
Efectos fenotípicos de las mutaciones

Mutaciones con pérdida de función. Producen la ausencia
completa o parcial de la función normal de la proteína, por alteración de
la estructura de la proteína o bien por mutaciones en las regiones que
regulan transcripción, traducción o corte y empalme. Suelen ser
recesivas y los individuos diploides deben ser homocigotos para
manifestar los efectos.
Mutaciones por ganancia de función. Existen mutaciones que
alteran el fenotipo bioquímico y potencian una o mas funciones
normales de una proteína. Producen un rasgo completamente nuevo o
causa la aparición de un rasgo en un tejido inapropiado o en un
momento inapropiado durante el desarrollo. Son generalmente de
expresión dominante.

AGENTES MUTÁGENOSUn agente mutágeno es todo factor capaz de aumentar la frecuencia de mutación natural.
Existen diversos factores, tanto físicos como químicos, capaces de actuar como agentes
mutágenos. En realidad, actuarán como agentes mutágenos todos aquellos agentes
capaces de alterar el material genético y en particular, aquellos que alteren la secuencia del
ADN.
Los principales agentes mutágenos son:
a) Agentes físicos:
Las radiaciones electromagnéticas como los rayos X y los rayos gamma.
Las radiaciones corpusculares como los rayos α, los rayos ß y los flujos de protones o
neutrones que generan los reactores nucleares u otras fuentes de radiactividad natural o
artificial.
Ciertos factores físicos como los ultrasonidos, los choque térmicos, la centrifugación,
etc.
b) Agentes químicos:
Los análogos de las bases nitrogenadas.
El ácido nitroso (HNO2), porque desamina ciertas bases nitrogenadas.
Los alcaloides como la cafeína, la nicotina, etc.
El gas mostaza, el agua oxigenada (H2O2), el ciclamato, etc.

Nomenclatura uniforme de las mutacionesPara nombrar de forma correcta una mutación, se seguirán las siguientes
indicaciones:
Para determinar el origen se utiliza el prefijo g, c, m o p, según sea DNA genómico, DNA
complementario, DNA mitocondrial o en una proteína.
Para indicar un cambio de nucleótido, se coloca primero el número de esa base, seguido
del nucleótido original, el símbolo > y el nuevo nucleótido en esa posición. En el DNAg
se escribe con mayúscula y en el DNAc con minúscula.
Si no se conoce la secuencia genómica entera como en caso de los intrónes, se utiliza
IVS seguido por el numero de intrón y los nucleótidos que varían se numeran +1, +2,
etc. donde +1 es la G invariable del sitio donante (GT) o -1, -2, etc. contando hacía
atrás desde la G invariable del sitio aceptor (AG).
Las pequeñas deleciones se indican por los números de los nucleótidos delecionados
separados por un guion bajo y seguidos por el termino del y finalmente los nucleótidos
que han sido delecionados.
Las pequeñas inserciones se designan por ins después de los dos nucleótidos en que
surgió la inserción, seguido s de los nucleótidos que han sido insertados.
En las proteínas una mutación se indica el aminoácido correcto, la posición de este,
seguido del aminoácido que ha remplazado al normal.
En el cDNA, la A del ATG de inicio se nombra como +1 y el nucleótido que le antecede
como -1. En la proteína la metionina amino-terminal se designa +1.

Problema 1
Nombre correctamente los siguientes ejemplos de mutaciones.
c.1444g>a
c.1524_1527del cgta
Glu6Val
c.1277_1278ins tatc
Gln39X
g.358T>G

Tasa de mutación
Definición: la tasa de mutación de un gen suele expresarse como el numero de
nuevas mutaciones por locus por generación.
Proporciona información acerca de la frecuencia con la que surge una mutación
y se ve afectada por:
Frecuencia con la cual se produce un cambio en el DNA
Probabilidad de que sea reparado
Probabilidad de reconocer y registrar una mutación
Las tasas de mutación varían entre los organismos y entre los genes y las
especies.
Procariontes 1-100 mutaciones por cada 10,000 millones de células
Eucariontes 1-10 mutaciones por cada millón de gametos
Se puede medir la incidencia de casos nuevos y esporádicos de enfermedades
genéticas autosómicas dominantes o Ligadas a la X, con penetrancia completa
y un fenotipo reconocible con claridad en el momento de nacimiento o un poco
después.

Calculo de la tasa de mutación
Ejemplo
En un estudio se detectaron 7 niños acondroplásicos en una serie de
242, 257 nacimientos consecutivos. Los padres presentaban estatura
normal, dado que la acondroplasia es considerada de penetrancia
completa, los siete casos se consideran mutaciones nuevas. En base a
estos datos calcule la tasa de mutación de acondroplasia en esta
población.
7 nuevos nacimientos / 2 (242,257) = 7/484514 = 1.44 x 10-5 mutaciones
por locus por generación.

Problemas
Calcular la tasa de mutación y diga que tipo de herencia tiene
esta enfermedad:
La Hemofilia A en México,
si se tienen 7 niños Hemofílicos en 1,242,257 nacimientos
consecutivos..
Distrofia muscular de Duchenne,
Si se tienen 5 mutaciones cada 3,100,000 nacimientos en la
población caucásica.
Retinoblastoma
Si se tiene1 mutación 10,000 nacimientos en la población
amerindia.
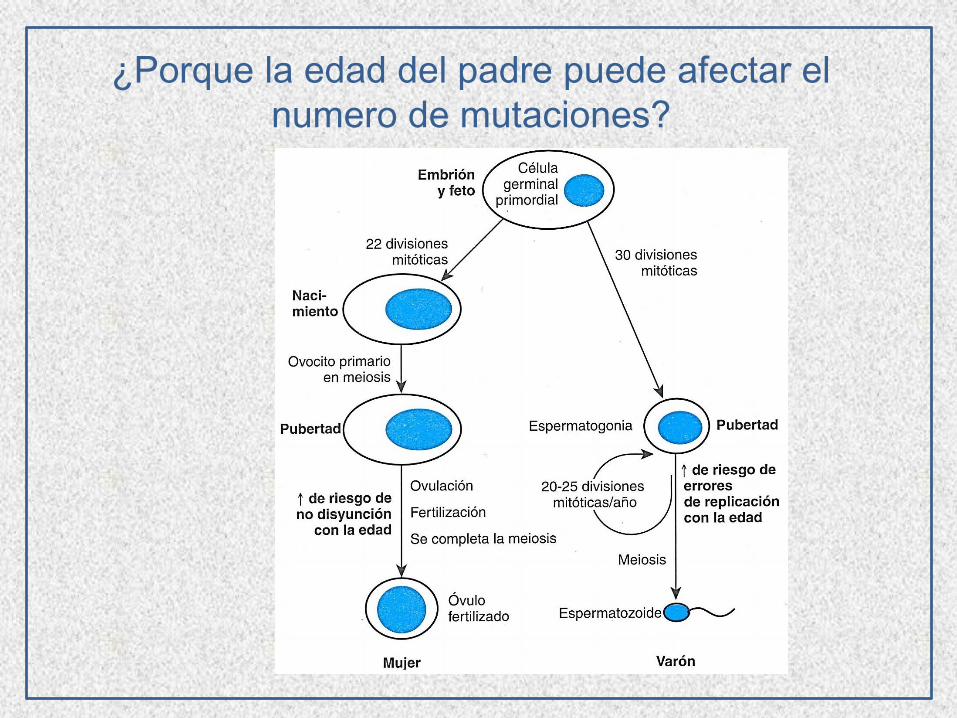
¿Porque la edad del padre puede afectar el
numero de mutaciones?

Mutaciones inducidas, causadas por sustancias químicas,
ambientales o radiación.
Problema 1Un codón que codifica el aminoácido Glicina sufre la sustitución de una
base, dando lugar a Ácido aspártico.
Enumere la secuencias de DNA posibles para el codón original y el
mutado.
La mutación es transición o transversión?
Glicina GGA GGC GGG GGT
Aspártico GAC GAT
Es una transición
TAREA

Problema 3Determine que tipo de mutación se presenta en las siguientes secuencias.
TTT AGT CAA CCG CAT GTT
Phe Ser Glu Pro His Val
Mutación 1
Phe Val Asn Arg Met
TTT AGTC AAC CGC ATG TT Existe una deleción de A
Mutación 2 dependiente de la primera mutación
Phe Val Lys Pro His Val
TTT GTC AAA CCG CAT GTT Existe una inserción de A

Conceptos básicos relacionados con la variación
genética.
Locus genético El locus se refiere a la posición o localización de un
gen en el genoma.
Los locus genéticos se definen por la localización
cromosómica, utilizando las bandas de los
cromosomas (banda G o banda R) o marcadores
moleculares microsatelites) como punto de referencia.
Ejemplo: Gen de determinación del sexo (SRY) se
localiza en el sitio p 11 del cromosoma Y.

Un alelo es la “versión” de un gen que
está presente en un locus dado.
Estas diferencias alélicas se
relacionan con las alteraciones en la
secuencia de nucleótidos de un gen.
Ejemplo: gen β hemoglobina:
eritrocito normal (alelo de tipo silvestre) .............GAG..... (ácido glutámico) hemo A
eritrocito forma de hoz (alelo mutado) .................GTG..... (valina) hemo S
Conceptos básicos relacionados con la variación
genética.
Alelo

Polimorfismos; describe múltiples y posibles estados de unaúnica propiedad. Son los múltiples alelos de un gen entre unapoblación, normalmente expresados como diferentes fenotipos(p.ej. el color de la piel es un polimorfismo).
Diversidad genética humana

Polimorfismo. Cuando una variante es tan común que se encuentra en más
del 1% de los cromosomas en la población en general.
Polimorfismos de nucleótido único, Son los más sencillos y frecuentes , en
general tienen sólo dos alelos que corresponden a dos bases distintas que
ocupan un determinado sitio en el genoma.
Polimorfismos de inserción-deleción: Microsatélites y minisatélites
Microsatélites, consisten en unidades de dos, tres o cuatro nucleótidos,
como TGTG……TG, CAACAA……CAA, o AAATAAAT…….AAAT. Que se
repiten entre una y pocas docenas de veces.
Minisatélites, Provienen de la inserción en tándem de un numero variable
(en general, de centenares o miles) de copias de una secuencia de DNA de
10 a 100 pares de bases de longitud.
Diversidad genética humana

Polimorfismos de nucleótido único
El cambio de un nucleótido aislado. En la mayor parte de los SNP´s (single
nucleotide polymorphism) se sustituye un nucleótido por otro diferente, pero el
termino incluye también insersiones o deleciones. Por lo general sólo tienen 2
alelos. El DNA codificante solo constituye el 1.5% del genoma humano y casi
todos los SNP´s se encuentran en regiones no codificantes como secuencias
dentro de intrones e intergénicas.
Polimorfismos simples de numero variable de repetición
tándemSuele describir alelos en locus que contienen tiras repetidas en forma tándem
de una secuencia simple:
Microsatélites (polimorfismo RSS repetición de secuencia simple) la secuencia
simple es de uno a varios nucleótidos de largo y la longitud del arreglo varía de
menos de 10 a mas de 100 nucleótidos.
Minisatélite, que incluye arreglos que con frecuencia abarcan cientos de
nucleótidos y consisten de repeticiones tándem de una secuencia entre nueve
y varias decenas de nucleótidos de largo.
Suelen tener múltiples alelos y se encuentran muy rara vez en DNA
codificante.

La mayoría de los polimorfismos no tienen efecto sobre el fenotipo (caen en
regiones no codificantes).
Algunos pocos afectan nuestro fenotipo (Estatura: alta/baja; Cabello:
claro/oscuro; Color de ojos)
Un numero muy pequeño de polimorfismos son responsables de
enfermedades genéticas
(Ej: 1/20 Habitantes de Europa del Norte portan el gene de la Fibrosis Quística)
Diversidad genética humana
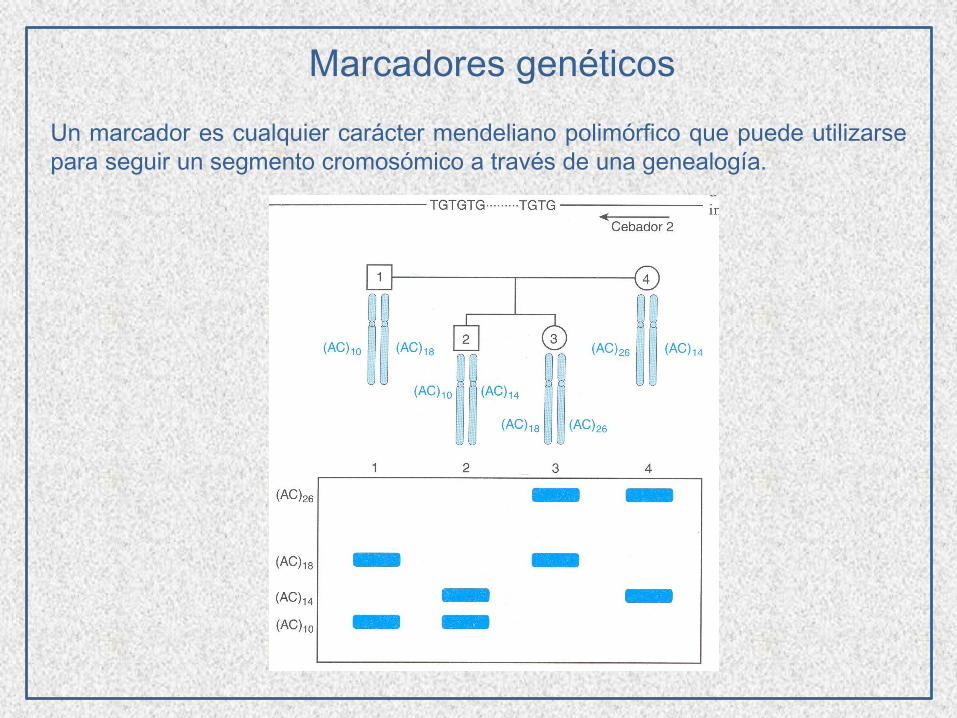
Marcadores genéticos
Un marcador es cualquier carácter mendeliano polimórfico que puede utilizarse
para seguir un segmento cromosómico a través de una genealogía.








Herencia codominante

Huella digital genética


Variación heredada yPolimorfismos en las proteínas
Aunque los polimorfismos son, el resultado de diferencias en la secuenciadel DNA, se han estudiado algunos loci polimórficos mediante el examende las variaciones en las proteínas codificadas por los respectivos alelos, envez de examinar las diferencias en las secuencias de DNA de los propiosalelos.
En la especie humana existe un notable grado de individualidadbioquímica en la fabricación de enzimas y otros productos génicos, es decirque posee una conformación química única y genéticamente determinaday, por tanto, responde de una manera única a las influencias ambientales,dietéticas y farmacológicas.

Polimorfismos de los grupos sanguíneos

Complejo mayor de histocompatibilidad

Complejo mayor de histocompatibilidad:Moléculas HLA

Complejo mayor de histocompatibilidad:Moléculas HLA

Complejo mayor de histocompatibilidad:
Moléculas HLA

Genotipos y fenotipos en poblaciones
Variación genética en poblaciones,
La genética de poblaciones es el estudio cuantitativo de la distribución de las
variaciones genéticas en las poblaciones y de la manera en que la frecuencia de
los genes y los genotipos se mantienen o cambian.
Se ocupa de los factores genéticos, como las mutaciones y la reproducción, y de
los factores ambientales y sociales, como la selección y la migración, que
determinan conjuntamente la frecuencia y la distribución de los alelos y los
genotipos en las familias y las comunidades.
En la práctica de la genética médica, la genética poblacional proporciona los
conocimientos sobre varios genes causantes de enfermedades que son frecuentes
en distintas poblaciones. Esta información es necesaria para el diagnóstico clínico y
el consejo genético, e incluye la determinación de las frecuencias alélicas
necesarias para el cálculo del riesgo.

Genética de poblaciones1.Concepto de población:
Una población es un conjunto de individuos de la misma especie que
viven en un lugar geográfico determinado (nicho ecológico) y que real o
potencialmente son capaces de cruzarse entre sí, compartiendo un acervo
común de genes.
2. Genética de Poblaciones estudia:
- la constitución genética de los individuos que componen las poblaciones
(frecuencias génicas y alélicas).
- la transmisión de los genes de una generación a la siguiente
(gametos=nexos de unión entre una generación y la siguiente).
- utilizando modelos matemáticos sencillos, cuando se considera 1 sólo
locus y una sola fuerza actuando sobre la población, diseñados para
individuos diploides con reproducción sexual.

Genotipo. Es el conjunto de genes que contiene un organismo heredado
de sus progenitores. En organismos diploides, la mitad de los genes se
heredan del padre y la otra mitad de la madre.
Fenotipo. Es la manifestación externa del genotipo, es decir, la suma de
los caracteres observables en un individuo. El fenotipo es el resultado de
la interacción entre el genotipo y el ambiente. El ambiente de un gen lo
constituyen los otros genes, el citoplasma celular y el medio externo
donde se desarrolla el individuo.
Gen. Unidad hereditaria que controla cada carácter en los seres vivos. A
nivel molecular corresponde a una sección de ADN, que contiene
información para la Síntesis de una cadena proteínica.
Para ilustrar los principios básicos que determinan las frecuencias
alélicas y genotípicas en las poblaciones podemos utilizar un ejemplo
importante correspondiente a un rasgo autosómico común, determinado
por un solo par de alelos.

Frecuencias fenotípicas, genotípicas y alélicas.
Locus A: alelos A1 y A2
Genotipos: A1A1
A1A2
A2A2
En codominancia existen 3 genotipos = 3 fenotipos
En dominancia completa existen 3 genotipos = 2 fenotipos
Frecuencias fenotípicas de una población son las proporciones o porcentajes
de individuos de cada fenotipo que están presentes en la población.
Número de individuos de un fenotipo
Número total de individuos
Frecuencias genotípicas de una población son la proporciones o porcentajes de
individuos de cada genotipo que están presentes en la población
Número de individuos de un genotipo
Número total de individuos
La suma de las frecuencias genotípicas será 1

Las frecuencias alélicas. son las proporciones de los diferentes alelos
en cada locus, presentes en la población.
Total de alelos A1/A2
Total de aleos A1 + A2
La suma de las frecuencias alélicas será 1
En codominancia: frecuencias fenotípicas = frecuencias genotípicas.
En dominancia: frecuencias fenotípicas ≠ frecuencias genotípicas
Genotipos Nº de individuos Frecuencias genotípicas
A1A1 n1 n1/N
A1A2 n2 n2/N
A2A2 n3 n3/N
Total N 1 →100%
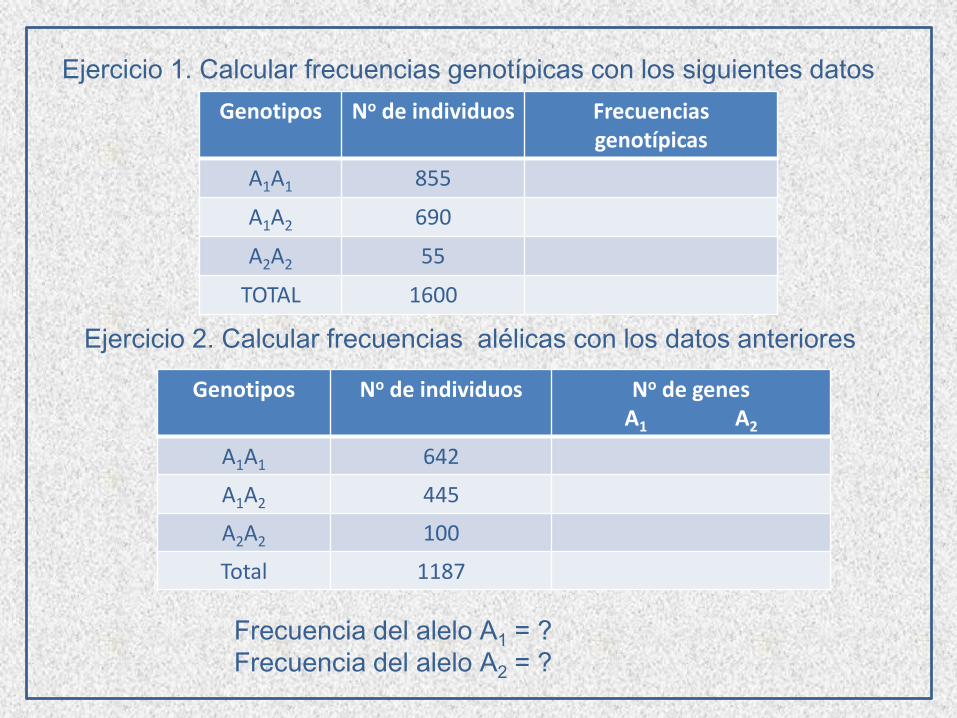
Genotipos No de individuos Frecuencias genotípicas
A1A1 855
A1A2 690
A2A2 55
TOTAL 1600
Genotipos No de individuos No de genesA1 A2
A1A1 642
A1A2 445
A2A2 100
Total 1187
Frecuencia del alelo A1 = ?
Frecuencia del alelo A2 = ?
Ejercicio 1. Calcular frecuencias genotípicas con los siguientes datos
Ejercicio 2. Calcular frecuencias alélicas con los datos anteriores

Ejemplo: Factores genéticos en la resistencia al
virus de la inmunodeficiencia humana.
El gen CCR5
ΔCCR5 32 pares de bases
Ejercicio 4:
Determinar frecuencias genotípicas del gen CCR5 y ΔCCR5 observadas
de una población europea .
788 individuos europeos
647 CCR5/CCR5
134 CCR5/ΔCCR5
007 ΔCCR5/ΔCCR5
Donde CCR5 es A1, ΔCCR5 es A2 y CCR5/ΔCCR5 es A1A2

Ejercicio 5. Cálculo de la frecuencia alélica de esa población:788 individuos europeos647 CCR5/CCR5134 CCR5/ΔCCR5007 ΔCCR5/ΔCCR5
RESULTADOS:
La sumatoria de los alelos debe ser 1

La genética poblacional se ocupa de la composición genética de una
población y de cómo esta cambia con el tiempo. El conjunto génico de
una población puede describirse a partir de las frecuencias de los
genotipos y los alelos en la población.

Ejercicio
Los antígenos MN del grupo sanguíneo humano se determinan por dos
alelos codominantes LM y LN. Se estudiaron 398 finlandeses de Karjala y
se obtuvieron los siguientes datos.
Fenotipo Genotipo Número
MM LM LM 182
MN LM LN 172
NN LN LN 44
Calcule las frecuencias alélicas y genotípicas en el locus MN para la
población de Karjala.

Loci con alelos múltiplesPara calcular las frecuencias alélicas a partir de los números de genotipos
se sigue el mismo principio.
EjemploPara un locus con tres alelos (A1, A2, A3) y seis genotipos (A1A1, A1A2, A1A3, A2A2,
A2A3, A3A3) las frecuencias de los alelos son:
F(A1) = 2nA1A1 + nA1A2 + nA1A3
2N
F(A2) = 2nA2A2 + nA2A1 + nA2A3
2N
F(A3) = 2nA3A3 + nA3A2 + nA3A1
2N
También se pueden calcular de la siguiente manera:
F(A1) = f(A1A1) + ½ f(A1A2) + ½ f(A1A3)
F(A2) = f(A2A2) + ½ f(A1A2) + ½ f(A2A3)
F(A3) = f(A3A3) + ½ f(A1A3) + ½ f(A2A3)

Loci ligado al XSe sigue el mismo principio, sin embargo, debe recordarse que una mujer posee
dos cromosomas X, y por consiguiente dos alelos ligados al X, mientras que un
varón tiene un solo cromosoma X y por lo tanto un solo alelo ligado al X.
Ejemplo
Tenmos dos alelos ligados al X, que son XA y Xa, Las mujeres pueden ser
homocigotos (XAXA, XaXa) o heterocigotos (XAXa). Todos los varones son
hemicigotos (XAY o XaY)
Para determinar la frecuencia del alelo XA
F(XA) = 2n XAXA + n XAXa + n XAY
2nmujeres + nvarones
F(Xa) = 2n XaXa + n XAXa + n XaY
2nmujeres + nvarones
Pueden calcularse también a partir de las frecuencias genotipicas
F(XA) = f(XAXA) + f(XAXa) + f(XAY)
F(Xa) = f(XaXa) + f(XAXa) + f(XaY)

El color del pelaje anaranjado en los gatos se debe a un alelo ligado al X
(X0) que es codominante con el alelo para el negro (X+) . Se determinaron
los genotipos del locus anaranjado de los gatos en Minneapolis y St.
Paul, Minnesota y se obtuvieron los siguientes datos:
X0X0 hembras 11
X0X+ hembras 70
X+X+ hembras 94
X0Y machos 36
X+Y machos 112
Calcule las frecuencias de los alelos X0 y X+ para esta población

El objetivo principal de la genética poblacional es comprender los
procesos que forman el conjunto génico de una población.
Ley de Hardy- WeinbergDescribe como la reproducción y los principios mendelianos afectan las
frecuencias alélicas y genotípicas de una población.
Formulada en 1908 de forma independiente por Godfrey H. Hardy y Wilhelm
Weinberg, se trata realmente de un modelo matematico.
Para un locus autosómico con dos alelos, la ley de Hardy-Weinberg puede ser
establecida de la siguiente manera:
Supuesto Si una población es grande, apareada al azar y no afectada por
mutación, migración o selección natural, entonces:
Predicción 1 las frecuencias alélicas de una población no cambian
Predicción 2 las frecuencias genotípicas se estabilizan (no cambiarán) después
de una generación en las proporciones p2, 2pq y q2, donde p es igual a la
frecuencia del alelo A y q es igual a la frecuencia del alelo a.
Cuando los genotipos están en las proporciones esperadas (p2, 2pq y q2), se dice
que la población se encuentra en equilibrio de Hardy-Weinberg

La ley de Hardy-Weinberg tiene dos componentes fundamentales. El primero es
que, en determinadas condiciones ideales, existe una relación simple entre las
frecuencias alélicas y las genotípicas en una población.
alelo A = p
alelo a = q
Los emparejamientos en la población ocurren completamente al azar con
respecto a los genotipos en ese locus.
p2 = AA
q2 = aa
2pq = Aa
La ley de Hardy-Weinberg postula que la frecuencia de los tres genotipos, AA, Aa y aa viene dada por los términos de la fórmula binomial (p + q)2 = p2 + 2pq + q2.
Un segundo componente de la ley es que si las frecuencias alélicas no cambian
de generación en generación, la proporción relativa de los genotipos tampoco
cambiará, es decir, las frecuencias de los genotipos permanecerán constantes, en equilibrio, de generación en generación, si las frecuencias alélicas p y q permanecen constantes.Entonces:
p2 = p x p
q2 = q x q
2pq = (p x q) + (p x q)

Si aplicamos esto a nuestro ejemplo de 788 sujetos europeos:
p2 = p x p = 0.906 x 0.906 = 0.821
q2 = q x q = 0.094 x 0.094 =0.009
2pq = (p x q) + (p x q) = (0.906 x 0.094) + (0.094 x 0.906) = 0.170
Por lo tanto
788 x 0.821 = 647
788 x 0.009 = 7
788 x 0.170 = 134
0.821: 0.170: 0.009

Genotipos No de individuos Frecuencias genotípicas
A1A1 0.1384180
A1A2 0.6666666
A2A2 0.1949152
TOTAL 1770
Genotipos No de individuos Frecuencias genotípicas
A1A1
A1A2
A2A2
Total 100
Frecuencia del alelo A1 = 0.6
Frecuencia del alelo A2 = 0.4
Ejercicio 1. Aplicar el principio de Hardy-Weinberg a los siguientes datos para
encontrar el numero de individuos que poseen ese genotipo
Ejercicio 2. Calcular frecuencias genotípicas y numero de individuos
aplicando la ley de Hardy-Weinberg

Así pues, la distribución de Hardy-Weinberg de genotipos en las poblaciones es
simplemente una distribución binomial
(p + q)n,
donde p + q = 1 y n = 2.
Para mas alelos (p1 + p2 + . . . . .pn)2.


Análisis de los supuestos de la Ley de Hardy-
Weinberg
La población es grande, el supuesto de Hardy-Weinberg habla de
poblaciones infinitas, sin embargo las desviaciones importantes solo
ocurren cuando la población es pequeña.
Los individuos se aparean al azar, es decir que cada genotipo se aparea
en proporción a su frecuencia.
Las frecuencia alélicas no son afectadas por la migración, selección
natural o selección.
Los supuestos de la ley de Hardy-Weinberg se aplican a un locus
individual. Ninguna población real se aparea al azar para todos los
rasgos, ni hay una población completamente libre de selección natural
para todos los rasgos.
La Ley de Hardy-Weinberg requiere estas condiciones solo para el locus
en consideración . Una población puede hallarse en equilibrio de Hardy-
Weinberg para un locus pero no para otros.

Consecuencias de la Ley de Hardy-Weinberg
-Una población no puede evolucionar
- Las frecuencias genotípicas son determinadas por las frecuencias alélicas

El propósito de la Ley de Hardy-Weinberg consiste en examinar solo el efecto
de la reproducción en el conjunto génico

Extensiones de la Ley de Hardy-Weinberg
Extensiones de la Ley de Hardy-Weinberg
Situación Frecuencias alélicas Frecuencias genotípicas
Tres alelos f(A1) = p f(A1 x A1) = p2
f(A2) = q f(A1 x A2) = 2pq
f(A3) = r f(A2 x A2) = q2
f(A2 x A3) = 2qr
f(A3 x A3) = r2
Alelos ligados al X f(X1) = p f(X1 X1 hembra) = p2
f(X2) = q f(X1 X2 hembra) = 2pq
f(X2 X2 hembra) = q2
f(X1 Y varón) = p
f(X2 Y varón) = q

Alelos Múltiples
• En el caso en que un gen en particular se encuentra en tres o más formas alélicas
en una población.
• Para los genes con múltiples alelos las proporciones de la Ley H-W se expanden:
(p+q+r)2= p2+q2+r2+2pq+2qr+2pr

Genes ligados a X (ejemplo)
• 4% de los hombres tienen daltonismo (Xc) y 96% son no daltónicos (Xc+), por lo
tanto: p(Xc+)=0.96 y q(Xc)=0.04. El genotipo y fenotipo esperado en mujeres
puede ser calculado:
q2(XcXc)= (0.04)2 =0.0016 ----------------------- 0.0016 daltónicas
2pq(Xc+ Xc)=2(0.96)(0.04)=0.0768
p2(Xc+ Xc+)=(0.96)2=0.9216
Cuando el número de mujeres afectadas es mucho menor que el de
hombres afectados indica que está envuelto un gen ligado a X.
0.9984 normales

La frecuencia de un rasgo recesivo ligado al X entre los varones es q,
mientras que la frecuencia entre las mujeres es q2. Cuando un alelo
ligado al X es infrecuente, el rasgo sera en consecuencia mucho mas
frecuente en los varones que en las mujeres.
Ejemplo
Considere la Hemofilia A , con una frecuencia de 1 en 10,000 en el
equilibrio de Hardy-Weinberg ¿cual será la frecuencia de la enfermedad
en las mujeres?
En los varones la frecuencia de la enfermedad será igual a 1 en 10,000
pero en las mujeres será (1\10000)2, que es solo 1 en 10 millones.
La hemofilia es 1000 veces mas frecuente en los varones que en las
mujeres.

Ley de Hardy-Weinberg
• Para sacar la frecuencia del alelo A
– p(A)= [p2+½(2pq)] / (p2 + 2pq + q2)
• Para sacar la frecuencia del alelo a
– q(a)= [q2+½(2pq)] / (p2 + 2pq + q2)

Pruebas para las proporciones de Hardy-Weinberg
Para determinar si los genotipos de población están en equilibrio, las
proporciones genotípicas esperadas deben compararse con las
frecuencias genotípicas observadas.
Se comparan con una prueba de bondad de ajuste de X2, en donde la Ho
es que los genotipos se encuentren en equilibrio de Hardy-Weinberg.
p>0.05 se acepta Ho
Los gl para una prueba de X2 del equilibrio de Hardy-Weinberg igualan el
numero de clases genotípicas esperadas menos el numero de alelos
asociados.
Ejemplo
Unos investigadores encontraron tres genotipos (R2R2, R2R3 y R3R3)
en un locus que codifica la enzima peroxidasa en los arboles pino
ponderosa que crecen en el Lago Glacial, Colorado. Los números
observados de estos genotipos fueron
genotipos numero observado
R2R2 135
R2R3 44
R3R3 11

¿Están los arboles pino ponderosa del Lago Glacial, Colorado en
equilibrio de Hardy-Weinberg en el locus peroxidasa?
R2 = p
R3 = q
p = f(R2) = 2(135) + 44 = 0.826
2(190)
q = f(R3) = 2(11) + 44 = 0.174
2 (190)
Las frecuencias esperadas se calculan
R2R2 = p2 = (0.826)2 = 0.683
R2R3 = 2pq = 2(0.826) (0.174) = 0.287
R3R3 = q2 = (0.174)2 = 0.03
Los valores esperados para cada genotipo se calculan
R2R2 = 0.683 x 190 = 129.8 =130
R2R3 = 0.287 x 190 = 54.5 = 55
R3R3 = 0.03 x 190 = 5.7 = 6

Prueba de X2 se calcula
X2 = ∑(observado – esperado)2
esperado
X2 = (135 – 130)2 + (44 – 55)2 + (11 – 6)2
130 55 6
X2 = 0.21 + 2.02 + 4.39 = 7.16
gl = 3-2 = 1
p entre 0.01 y 0.001 por lo tanto no están en equilibrio.

Estimación de la frecuencias alélicas con la Ley de
Hardy-Weinberg
Un uso practico de la Ley de Hardy-Weinberg es el que permite calcular las
frecuencias alelicas cuando hay presencia de dominancia.
Si suponemos que una poblacion se encuentra en equilibrio de Hardy-Weinberg
con respecto a ese locus, entonces la frecuencia del genotipo recesivo (aa) será
q2 y la frecuencia alelica será la raíz cuadrada de la frecuencia genotípica
q = √f(aa)
Ejemplo
La frecuencia de la fibrosis quística en los norteamericanos blancos es de 1 en
cada 2000. ¿Cual es la probabilidad de portadores para el alelo de la fibrosis
quistica?
q = √(1\ 2000) = 0.02
p = 1 – q = 1 – 0.02 = 0.98
p2 = (0.98) (0.98) = 0.960
2pq = 2 (0.98 x 0.02) = 0.0392 = 0.04 x 100 = 4%

Postulado de Hardy-Weinberg
• Si la población se aparea al azar y existe equilibrio, entonces:
p² + 2pq + q² = 1
p + q = 1
p² = Proporción de personas con los dos alelos dominantes.
q² = Proporción de personas con dos alelos recesivos.
2pq = Proporción de personas con uno de cada alelo.

Población en equilibrio:
• No puede haber mutaciones
• No puede haber migraciones
• La población debe ser grande
• El apareamiento debe ser al azar
• No debe existir selección natural

Factores que alteran el equilibrio de Hardy-
Weinberg
La ley de Hardy-Weinberg supone que se cumplan diversos
supuestos.
El primero es que la población sea grande y que los
emparejamientos entre sus miembros se produzcan al azar:
Emparejamiento dirigido
Consanguinidad y endogamia
El segundo es que las frecuencias alélicas no cambian con el tiempo:
migración
Selección
Nuevas mutaciones

Apareamiento no aleatorioEl apareamiento no aleatorio afecta la manera por la cual los alelos se combinan
para formar los genotipos y altera la frecuencia genotípica de una población.
Apareamiento clasificado positivamente: Se refiere a una tendencia a
aparearse con individuos similares.
Apareamiento negativo: Se refiere a una tendencia para aparearse
preferencialmente entre individuos diferentes.
El apareamiento clasificado es para un rasgo particular y afectará sólo a
genes que codifican para el rasgo.
La endogamia, es una forma de apareamiento no aleatorio entre individuos
relacionados difiere del apareamiento clasificado positivamente porque afecta
todos los genes. Causa desviación del equilibrio de Hardy-Weinberg, al conducir
a un aumento en la proporción de homocigotos y una disminución en la
proporción de heterocigotos en una población.
El exocruzamiento es la prevención de apareamientos entre individuos
relacionados.

En un organismo diploide, cada individuo homocigoto puede tener dos copias de
un mismo gen. Estas dos copias pueden tener el mismo estado, es decir que los
dos alelos son similares en estructura y función pero no tienen un origen común.
La endogamia es una medida de la probabilidad de que dos alelos sean idénticos
por ascendencia.

La endogamia suele medirse
mediante el coeficiente de
endogamia, designado F, que es una
medida de la probabilidad de que dos
alelos sean idénticos por
ascendencia.
Los coeficientes de endogamia
pueden variar de 0 a 1. Un valor de 0
indica que el apareamiento de una
población grande es al azar; un valor
de 1 indica que todos los valores son
idénticos por ascendencia.
Con la endogamia, la proporción de
heterocigotos disminuye en 2Fpq, y la
mitad de ese valor (Fpq) se agrega a
la proporción de cada homocigoto en
cada generación.
f(AA) = p2 + Fpq
f(Aa) = 2pq- Fpq
f(aa) = q2 + Fpq

En la mayoría de las especies con exocruzamiento, la endogamia es perjudicial
porque aumenta la proporción de homocigotos y por eso estimula que la
probabilidad de que los alelos recesivos deletéreos y letales se combinen para
producir homocigotos con un rasgo perjudicial.

EjemploEl alelo recesivo a causa una enfermedad genética con una frecuencia q
de 0.01. Si la población se aparea al azar (F = 0). ¿Cual será la
frecuencia de individuos afectados por la enfermedad?
q2 = (aa) = (0.01)2 = 0.0001
por consiguiente solo 1 de cada 10,000 individuos tendrá la enfermedad.
Sin embargo, si existe apareamiento entre hermanos ¿Cual será la
frecuencia de individuos afectados por la enfermedad?
F = 0.25
q2 – Fpq
p = 1 - q = 1-0.01 = 0.99
q2 – Fpq = (0.01)2 – [0.25 (0.01) (0.99)] = 0.0026
Por consiguiente, la enfermedad genética es 26 veces mas frecuente en
este nivel de endogamia.

Los efectos perjudiciales se han reconocido en seres humanos, así como
en cultivos y animales domésticos, durante siglos.
La endogamia se utiliza a menudo para producir plantas y animales
domésticos que tengan rasgos deseables.

Efecto de las fuerzas evolutivas en las frecuencias
alélicasLos procesos que provocan cambios en la frecuencia alélica incluyen la mutación,
la migración, la deriva génica (efectos aleatorios debido al tamaño pequeño de la
población) y la selección natural.
MutaciónSi bien, pueden originarse combinaciones nuevas de genes existentes por
recombinación durante la meiosis, todas las variantes genéticas, en ultima
instancia, se generan por mutación.
La mutación puede influir en la tasa a la cual aumenta una variante genética a
expensas de otra.

Si hay 45 copias de G1 y 5 de G2 entonces p = 0.9 y q = 0.1
La mutación pasa un alelo G1 a G2 ahora hay 44 G1 y 6 G2, entonces q = 0.12

Si continua mutando copias de G1 a G2, la frecuencia de G2 aumentará y la
frecuencias de G1 disminuirá. La cantidad de G2 que cambiará (Δq) como
resultado de la mutación depende de:
1. La tasa de mutación de G1 a G2 (μ) y
2. La frecuencia de de G1 en la población (p)
El cambio en G2 como resultado de la mutación iguala la tasa de mutación por la
frecuencia alélica.
Δq = μp
Cuando la frecuencia de p disminuye como resultado de la mutación, el cambio
en la frecuencia debida a la mutación será cada vez menor.
También se producen mutaciones inversas a una tasa ν que es diferente a μ
Siempre que se produce una mutación inversa la frecuencia de G2 disminuye y
aumenta G1

La tasa de cambio debido a las mutaciones inversas es igual a la tasa de
mutación inversa por la frecuencia alélica de G2 (Δq = νq). El cambio
global en la frecuencia alélica es un equilibrio entre las fuerzas opuestas
de la mutación directa y la mutación inversa.
Δq = μp – νq
Ejemplo
Si las tasas de mutación directa e inversa en un locus de ratón
son 1 x 10 -5 y 0.3 x 10-5 por generación respectivamente y las
frecuencias alélicas son p = 0.9 y q = 0.1. Calcular el cambio neto en la
frecuencia alélica por generación debido a la mutación.
Δq = μp – νq
Δq = (1 x 10 -5 x 0.9) – (0.3 x 10-5 x 0.1)
Δq = 8.7 x 10 -6 = 0.000008.7
Por consiguiente, el cambio debido a la mutación en una sola generación
es sumamente pequeña.

Alcance del equilibrio de las frecuencias
alélicas
Se refiere que al final, el numero de genes que sufre mutación directa se
contrarrestara con el numero de genes que sufre mutación inversa. En
este punto el aumento de q debido a la mutación directa será igual a la
disminución de q debido a la mutación inversa, y no habrá ningún cambio
neto en la frecuencia alélica (Δq = 0).
El punto en el cual no hay cambios en la frecuencia alélica de una
población se denomina EQUILIBRIO, en equilibrio la frecuencia de G2
(ˆq) será:
ˆq = __μ___
μ + ν
Esta ecuación final nos dice que la frecuencia alélica en equilibrio es
determinada sólo por las tasas de mutación directa e inversa.

Ejemplo
¿Cuál será la frecuencia en el equilibrio de un alelo si sus tasas de
mutación directa e inversa son 0.6 x 10 -6 y 0.2 x 10-6, respectivamente ?
Suponga que no hay presente ninguna otra fuerza evolutiva.
ˆq = __μ___
μ + ν
ˆq = ____ 0.6 x 10 -6 ______
0.6 x 10 -6 + 0.2 x 10-6
ˆq = 0.75

Resumen de los efectos
El efecto de las tasas de mutación típica en el Equilibrio de Hardy-Weinberg
es insignificante y se requieren muchas generaciones para que una población
alcance el equilibrio mutacional. No obstante. Si la mutación es la única fuerza
que actúa en una población durante periodos prolongados, las tasas de
mutación determinaran las frecuencias alélicas.

MigraciónOtro proceso que puede provocar un cambio en las frecuencias alélicas
es el ingreso de genes provenientes de otras poblaciones, denominado
con frecuencia migración o flujo génico.
El efecto global de la migración es doble:
1. Previene la divergencia genética entre las poblaciones y
2. Aumenta la variación genética dentro de las poblaciones.
Efecto de la migración en las frecuencias alélicas

Después de la migración la
frecuencia del alelo a en la
población II combinada (q’II) es.
q’II = qI (m) + qII (1-m)
Donde qI (m) es la contribución
a q hecha por las copias del
alelo a en los inmigrantes y qII
(1-m) es la contribución a q de
las copias del alelo a de los
residentes.
El cambio en las frecuencias
alélicas debido a la migración
(Δq) será.
Δq = m (qI - qII)

Deriva genéticaLa deriva genética es el cambio de las frecuencias alélicas debida a factores
aleatorios. Cuanto más pequeña es la muestra de gametos mayor es la
posibilidad de que su composición se desvíe del conjunto génico completo. Esta
clase de desvición es una proporción esperada debido al tamaño limitado de la
muestra y se denomina error de muestreo.
Magnitud de la deriva genética
La magnitud del cambio de la frecuencia alélica debido a la deriva genética esta
relacionada de manera inversa con el tamaño efectivo de la población.
La magnitud de la deriva genética puede determinarse al examinar los cambios
en la frecuencia alélica dentro de una misma población o al examinar la
magnitud de las diferencias genéticas que se acumulan entre poblaciones.
La cantidad de deriva genética puede estimarse a partir de la varianza en la
frecuencia alélica..
Sp2 = pq/ 2N
Donde p y q son las frecuencias alélicas y N el tamaño de la población, La deriva
genética será máxima cuando p y q sean iguales (0.5 cada uno).

Ejemplo 1
En una población de 50 individuos cuyas frecuencias alelicas son
iguales (p = q = 0.5). ¿Cuál será la varianza en la frecuencia alélica?
Sp2 = (0.5 x 0.5) / 2 (50) = 0.0025
Y cuando p = 0.9 y q = 0.1
Sp2 = (0.9 x 0.1) / 2 (50) = 0.0009
La deriva genética será mas alta cuando el tamaño de la población sea
pequeño
Ejemplo 2
En una población de 10 individuos cuyas frecuencias alelicas son
iguales (p = q = 0.5). ¿Cuál será la varianza en la frecuencia alélica?
Sp2 = (0.5 x 0.5) / 2 (10) = 0.0125
Lo que es 5 veces mayor que cuando la población era de 50 individuos.

Ejemplo 3
Se establecieron 107 poblaciones de replicación, con moscas de la
fruta, para examinar la frecuencia de dos alelos (bw* y bw) que afectan
el color de los ojos de las moscas, cada generación compuesta por 8
machos y 8 hembras. La frecuencia promedio de bw* = 0.53125
¿Cuál será la varianza esperada en la frecuencia alélica debida a la
deriva genética?
Sp2 = pq/ 2N
bw = 1- bw* = 1 – 0.53125 = 0.46875
N = 8 + 8 = 16
Sp2 = (0.53125 x 0.46875)/ 2(16)
Sp2 = 0.0156

La deriva genética es el resultado de un tamaño de población pequeño
sostenido, del efecto fundador (establecimiento de una población por
parte de unos pocos fundadores) y del efecto de cuello de botella
(reducción de la población).
La deriva genética produce un cambio en las frecuencias alélicas dentro
de la población, una pérdida de variación genética mediante la fijación de
alelos y la divergencia genética entre poblaciones.

Selección natural
La selección natural sucede cuando los individuos con rasgos
adaptativos producen una cantidad de descendencia mayor que la
producida por otros en la población. Por tanto, un rasgo que proporciona
una ventaja reproductiva se incrementa con el tiempo y permite a las
ppoblaciones adecuarse mejor a sus ambientes

La selección natural se mide como aptitud, que es el éxito reproductivo
de un genotipo en comparación con otro genotipo dentro de una
población.
La aptitud (W) varia de 0 a 1. Para calcular la aptitud para cada genotipo
tomamos el numero promedio de descendencia producido por un
genotipo y lo dividimos por la media del numero de la descendencia
producida por el genotipo más prolífico.
Ejemplo
Genotipos A1A1 A1A2 A2A2
Media del numero de 10 5 2descendencia producida
W11 = (A1A1) = 10 / 10 = 1
W12 = (A1A2) = 5 /10 = 0.5
W22 = (A2A2) = 2 / 10 = 0.2

El coeficiente de selección (s) es una variable relacionada que mide la
intensidad relativa de selección contra un genotipo. Es igual a 1-W
s = 1 – W
s11 = (A1A1) = 1 - 1 = 0
s12 = (A1A2) = 1 - 0.5 = 0.5
s22 = (A2A2) = 1 – 0.2 = 0,8
Modelo de selección generalEl cambio en la frecuencia alélica debido a la selección puede
determinarse para cualquier tipo de rasgo genético con el uso del
modelo de selección general.

También es posible desarrollar formulas para determinar el cambio en la
frecuencia alélica cuando la selección es contra rasgos recesivos,
dominantes y codominantes, así como rasgos en los que el heterocigoto
tiene la aptitud mas alta

EjemploEl alcohol es una sustancia común en la putrefacción de la fruta, en la
que crecen y se desarrollan las larvas de la mosca de la fruta; las larvas
utilizan la enzima alcohol deshidrogenasa (Adh) para detoxificar los
efectos de este alcohol. En algunas poblacionesde moscas de la fruta
existen dos alelos presentes en el locus que codifica la Adh: Adh*, que
codifica una forma de la enzima que migra con rapidez en la
electroforesis en gel y la Adh, que codifica una forma de la enzima que
migra con lentitud. Las hembras de la mosca de la fruta con diferentes
genotipos Adh producen los siguientes numeros de descendientes en
presencia de alcohol:
Genotipo Media del numero de descendencia
Adh*/Adh* 120
Adh*/Adh 60
Adh/Adh 30
a) Calcule la aptitud relativa de las hembras que presentan estos
genotipos.
b) Si una población de moscas de la fruta tienen una frecuencia inicial a
0.2, ¿Cuál será la frecuencia en la generación siguiente cuando el
alcohol este presente?

a) Calcule la aptitud relativa de las hembras que presentan estos
genotipos.
Genotipo Numero medio de Aptitud
descendencia
Adh*/Adh* 120 W** = 120/120 =1
Adh*/Adh 60 W* = 60/120 = 0.5
Adh/Adh 30 W = 30/120 = 0.25
b) Si una población de moscas de la fruta tienen una frecuencia inicial a
0.2, ¿Cuál será la frecuencia en la generación siguiente cuando el
alcohol este presente?

Resultados de la selección
La selección natural modifica las frecuencias alélicas; la dirección y la
magnitud del cambio dependen de la intensidad de la selección, de las
relaciones de dominancia de los alelos y de las frecuencias alélicas.

La selección direccional favorece a un alelo sobre otro y, con el tiempo, lleva a la
fijación del alelo favorecido.
La sobredominancia lleva a un equilibrio estable con mantenimiento de los dos
alelos en la población
ˆq = f(A2) = s11____
s11 + s22
Donde s11 representa el coeficiente de selección del homocigoto A1A1 y s22
representa el coeficiente de selección del homocigoto A2A2.
La subdominancia produce un equilibrio inestable porque el heterocigoto tienen
una aptitud menor que los dos homocigotos.

La tasa con la cual la selección cambia las frecuencia s alélicas depende
de la frecuencia alélica en sí. Si un alelo A2 es letal y recesivo, W11 =
W12 = 1, mientras que W22 = 0. La frecuencia del alelo A2 disminuirá
con el tiempo, y la tasa de disminución será proporcional a la frecuencia
del alelo recesivo.
Cuando la frecuencia del alelo es alta, el cambio en cada generación es
relativamente grande, pero a medida que la frecuencia del alelo se
reduce, una mayor proporción de los alelos se encuentra en los
genotipos heterocigotos , donde son inmunes a la acción de la selección
natural.

Mutación y selección natural
La mutación y la selección natural actúan como fuerzas opuestas en los
alelos perjudiciales: las mutaciones tienden a aumentar y la selección
natural tiende a disminuirla, por lo que se produce, en última instancia,
un equilibrio.
La frecuencia de un alelo equilibrado ( q) es igual a la raíz cuadrada de
la tasa de mutación dividida por el coeficiente de selección:
q = õ\s
Cuando la selección actua en un alelo dominante se puede mostrar que
su frecuencia en equilibrio es:
q = µ\s
EjemploLa acondroplasia es un tipo común de enanismo humano, que es resultado de
un gen dominante. Las personas con esta afección son fértiles, aunque solo
producen alrededor del 74% de los niños en comparación con los individuos sin
acondroplasia.
Si asumimos que la tasa de mutación para la acondroplasia es alrededor de 3 x
10-5, ¿Cuál es la frecuencia de equilibrio para el alelo de la acondroplasia?

Aptitud
W = 74/100 = 0.74
Coeficiente de selección
s = 1 –W = 1 - 0.74 = 0.26
Frecuencia de equilibrio
q = µ\s = 3 x 10-5 / 0.26
q = 0.0001153

Efectos generales de las fuerzas evolutivas