anÁlisis molecular de la microbiota ... resumen el caracol pala (strombus gigas) posee una...
TRANSCRIPT

I
ANÁLISIS MOLECULAR DE LA MICROBIOTA BACTERIANA PRESENTE EN
MUESTRAS ASOCIADAS AL CARACOL PALA (Strombus gigas) Y
DETERMINACIÓN DE LAS ACTIVIDADES ANTIBACTERIANAS Y
ENZIMÁTICAS DE BACTERIAS AISLADAS
OLGA MARÍA PÉREZ CARRASCAL
Universidad Nacional de Colombia
Sede Medellín
Facultad de Ciencias
2012

II
ANÁLISIS MOLECULAR DE LA MICROBIOTA BACTERIANA PRESENTE EN
MUESTRAS ASOCIADAS AL CARACOL PALA (Strombus gigas) Y
DETERMINACIÓN DE LAS ACTIVIDADES ANTIBACTERIANAS Y
ENZIMÁTICAS DE BACTERIAS AISLADAS
OLGA MARÍA PÉREZ CARRASCAL
Tesis o trabajo de investigación presentada como requisito parcial para optar al título
de:
Magister en Ciencias Biotecnología
Directora:
Claudia Moreno (Bacterióloga, Ph.D)
Codirectora:
Magally Romero (Bióloga, Ph.D)
Asesora:
Gloria E. Cadavid (Bacterióloga, M.Sc. Ph.D)
Universidad Nacional de Colombia
Sede Medellín
Facultad de Ciencias
2012

III
DEDICATORIA
A Dios, a mis padres y hermanos por su confianza y apoyo incondicional, y a todos los
que me han ayudado a construir un propósito.
Si de todos los organismos creados por Dios los más pequeños y aparentemente menos
útiles fueran suprimidos, la vida se tornaría imposible, ya que el regreso a la atmósfera y
al reino mineral de todo lo que dejó de vivir sería bruscamente suprimido.
LOUIS PASTEUR

IV
AGRADECIMIENTOS
Esta investigación no hubiese sido posible sin el apoyo de la Universidad Nacional sede
Medellín a través del proyecto DIME bicentenario 20101007734. Mis más sinceros
agradecimientos a la Doctora Magally Romero por permitirme participar en este
proyecto y por su valiosa asesoría científica, a mi tutora la Doctora Claudia Moreno por
todos los conocimientos compartidos, su asesoría constante y confianza depositada en
mí, a la Doctora Gloria Cadavid por su asesoría y valioso apoyo, al Ingeniero biológico
Moisés Posada de quien logré aprender mucho y a quien agradezco su valiosa amistad y
cooperación en este proyecto, a mis compañeros de laboratorio y del grupo de
investigación Microbiop por sus buenos consejos y amistad. Muchas gracias a Mónica y
Tatiana por escucharme y motivarme cuando fue necesario. Muchas gracias a los
profesores de la Facultad de Ciencias y al Posgrado en Ciencias Biotecnología, quienes
de una u otra forma contribuyeron en mi formación como estudiante e investigadora a
nivel personal y profesional y en la solución de los problemas planteados a través de
este proyecto.
A las personas más importantes en mi vida y a quienes más quiero, mi familia, mis
padres Hugo y Carmen, mis hermanos Hugo, Francisco y Carlos a ellos siempre gracias
por permitirme crecer y avanzar, por su apoyo, amor y motivación en todo momento y
porque a pesar de la distancia siempre han estado allí. Infinitas gracias al más especial
de todos, Dios padre celestial con quien siempre estaré en deuda.

V
RESUMEN
El caracol pala (Strombus gigas) posee una diversidad bacteriana poco estudiada que
puede influir en la nutrición, el metabolismo, el estado físico del hospedero, entre otros
aspectos, por esa razón su microbiota bacteriana representa un recurso de imperativo
interés ecológico y biotecnológico. El objetivo principal de este estudio fue determinar
la diversidad bacteriana asociada al caracol, sus afiliaciones filogenéticas y el potencial
bioactivo de algunos aislados bacterianos. Metodologías de microbiología convencional
y herramientas moleculares dependiente e independientes de cultivo como la
electroforesis por gradiente de desnaturalización (DGGE), fueron usadas para estudiar
la diversidad presente en la baba, el alimento y el intestino de caracoles silvestres y en
cautiverio de las Islas del Rosario, Colombia.
Los perfiles de bandeo resultantes del DGGE fueron evaluados usando el programa
GelCompar II (Applied Maths TX, USA) y los análisis filogenéticos de las
comunidades bacterianas estudiadas revelaron una diversidad moderada, la cual se
estableció por un número variable de bandas detectadas por DGGE. Se encontraron
además diferencias entre las estructuras de las comunidades bacterianas asociadas a los
caracoles en cautiverio con respecto a las encontradas en caracoles silvestres, sugiriendo
una estrecha relación de poblaciones con el hábitat de los caracoles. Las afiliaciones
filogenéticas fueron determinadas por secuenciación del gen rRNA 16S; las secuencias
fueron agrupadas en cuatro clases: β-proteobacterias, γ-proteobacterias, Firmicutes y
Actinobacterias; algunas de estas secuencias están relacionadas con microorganismos
importantes en procesos de defensa y descomposición orgánica. Adicionalmente, los
estudios sobre el potencial bioactivo en bacterias asociadas al caracol pala indican que
algunas poseen propiedades antibacterianas e hidrolíticas. Actividades amilasas,
celulasas, proteasas y lipasas fueron exhibidas por bacterias asociadas a la baba y el
intestino de caracoles. Los aislados de Exiguobacterium y Psychrobacter fueron los
géneros más versátiles en cuanto a producción de enzimas. En general las bacterias
encontradas con actividad podrían desempeñar un papel en la transformación de materia

VI
orgánica y nutrición en caracoles, funciones que aún no han sido evaluadas. En este
estudio, el análisis de las secuencias del rDNA 16S y de los perfiles obtenidos por
DGGE permitieron inferir sobre la presencia de comunidades bacterianas únicas,
diversas e igualmente complejas en el caracol, asimismo este trabajo es el primer
reporte sobre la evidencia de hidrolasas extracelulares en bacterias asociadas a S. gigas,
que pueden ser de importancia ecológica y un recurso de interés biotecnológico para
nuestro país.
Palabras claves: caracol pala, S. gigas, DGGE, rRNA 16S, diversidad, hidrolasas
extracelulares, antibacterianos.

VII
ABSTRACT
The Queen Conch (Strombus gigas) has a poorly studied bacterial diversity that may
influence on the animal metabolism, nutrition, physical condition and others aspects, for
that reason its bacterial microbiota represent a resource of ecological and
biotechnological interest. The aim of this study was to determinate the bacterial
diversity associated with the Queen Conch and bioactive potential of some bacterial
isolates. Conventional microbiological methods and culture-dependent and independent
molecular tools such as denaturing gradient electrophoresis (DGGE) were used to assess
the diversity presence in mucus, food and gut from wild and captive conchs from
Rosario Islands, Colombia.
DGGE profiles were analyzed using GelCompar II software package (Applied Maths
TX, USA). The phylogenetic analysis of bacterial communities showed a moderate
diversity established by a variable number of bands detected by DGGE. We also found
differences between bacterial communities structures associated with captive and wild
conch, suggesting that populations are more related to the habitat.
Phylogenetic affiliation of conchs-associated bacteria was assessed by 16S rRNA gene
sequencing; the sequences were grouped into four classes: β-proteobacteria, γ-
proteobacteria, Firmicutes, and Actinobacterias, some of these sequences are related to
microorganisms involved in defense processes and organic decomposition. Additional
studies about the bioactive potential in bacteria associated with conch suggest that there
are a significant proportion of bacteria that display antibacterial and hydrolytic
activities.
Bacteria associated with mucus and gut from conchs exhibited production of hydrolytic
enzymes as well as amylases, cellulases, proteases and lipases. Psychrobacter and
Exiguobacterium have been found to be the most versatile genus because of the
production of multiple enzymes activities. As a result, these bacteria would have an
important facultative role in the transformation of organic matter and conch’s nutrition,
which has not yet been discussed.

VIII
In this study, analysis of 16S rDNA sequence and DGGE profiles obtained allow us to
infer about the presence of unique, different and equally complex bacterial communities
in the Queen Conch. To date this is the first study about the characterization and
extracellular enzymatic activities of bacteria associated with the Queen Conch in
Colombia which will be of interest in an ecological and biotechnological context.
Keywords: Queen Conch, DGGE, 16S rRNA, diversity, extracellular hydrolases
antibacterial.

IX
CONTENIDO
RESUMEN ....................................................................................................................... V
LISTA DE TABLAS ..................................................................................................... XII
LISTA DE FIGURAS .................................................................................................. XIII
LISTA DE ANEXOS .................................................................................................... XV
LISTA DE SÍMBOLOS Y ABREVIATURAS .......................................................... XVI
1. INTRODUCCIÓN ........................................................................................................ 1
2. HIPÓTESIS .................................................................................................................. 4
3. ESTADO DEL ARTE .................................................................................................. 5
3.1. Strombus gigas .......................................................................................................... 5
3.1.1. Distribución y hábitats .................................................................................... 5
3.1.2. Microbiota de caracoles silvestres y en cautiverio .......................................... 6
3.2. ECOLOGÍA MICROBIANA .................................................................................... 7
3.2.1. Herramientas genómicas para el estudio de la diversidad microbiana............ 7
3.2.2. Estrategias usadas para la identificación de especies bacterianas en ecología
microbiana ................................................................................................................. 8
3.2.2.1. Caracterización de fenotipos ........................................................................ 8
3.2.2.2. Reacción en cadena de la polimerasa (PCR) ................................................ 8
3.2.2.3. Secuenciación del DNA ............................................................................... 8
3.2.2.4. Estudio de la diversidad y bioprospección en bacterias asociadas al caracol
pala ............................................................................................................................ 9
3.3. BÚSQUEDA DE NUEVOS ANTIBACTERIANOS EN LAS BACTERIAS
ASOCIADAS AL CARACOL PALA ........................................................................... 11
3.4. HIDROLASAS EXTRACELULARES EN BACTERIAS ..................................... 12
3.4.1. Amilasas ........................................................................................................ 15
3.4.2. Celulasas........................................................................................................ 16
3.4.3. Proteasas ........................................................................................................ 18
3.4.4. Lipasas ........................................................................................................... 19

X
3.5. EL PRESENTE ESTUDIO ..................................................................................... 21
4. OBJETIVOS ............................................................................................................... 22
4.1. OBJETIVO GENERAL .......................................................................................... 22
4.2. OBJETIVOS ESPECÍFICOS .................................................................................. 22
5. MATERIALES Y MÉTODOS ................................................................................... 23
5.1. CULTIVO, AISLAMIENTO Y CARACTERIZACIÓN FENOTÍPICA DE LOS
AISLADOS BACTERIANOS ....................................................................................... 23
5.2. CARACTERIZACIÓN DE LA DIVERSIDAD BACTERIANA
PERTENECIENTE A LA MICROBIOTA ASOCIADA AL CARACOL PALA POR
TÉCNICAS MOLECULARES (PCR-DGGE) .............................................................. 24
5.2.1. Amplificación por PCR ................................................................................. 25
5.2.2. Análisis de electroforesis en geles de poliacrilamida en gradiente denaturante
(DGGE) ................................................................................................................... 26
.5.2.3. Análisis de secuencias .................................................................................. 27
5.3. CARACTERIZACIÓN MOLECULAR DE LAS BACTERIAS USADAS PARA
DETERMINAR LAS ACTIVIDADES ANTIBACTERIANAS Y ENZIMÁTICAS ... 28
5.4. DETERMINACIÓN DE LAS ACTIVIDADES ANTIBACTERIANAS DE
AISLADOS BACTERIANOS ....................................................................................... 28
5.4.1. Extracción de metabolitos secundarios ......................................................... 28
5.4.2. Evaluación de actividad antibacteriana ......................................................... 29
5.5. DETERMINACIÓN DE LOS MICROORGANISMOS CAPACES DE
PRODUCIR HIDROLASAS EXTRACELULARES Y EVALUACIÓN DE LAS
ACTIVIDADES ENZIMÁTICAS ................................................................................. 29
5.5.1. Determinación de microorganismos productores de algunas hidrolasas
extracelulares ........................................................................................................... 29
5.5.1.1. Determinación de microorganismos productores de amilasas ................... 29
5.5.1.2. Determinación de microorganismos productores de celulasas (CMCasa) . 30
5.5.1.3. Determinación de microorganismos productores de proteasas .................. 30
5.5.1.4. Determinación de microorganismos productores de lipasas ...................... 30
5.5.2. Evaluación de la actividad de algunas hidrolasas extracelulares .................. 31
5.5.2.1. Cuantificación de actividad amilasas ......................................................... 31
5.5.2.2. Cuantificación de actividad celulasas ........................................................ 31

XI
5.5.2.3. Cuantificación de actividad proteasas ........................................................ 32
5.5.2.4. Cuantificación de actividad lipasas ............................................................ 33
5.5.2.5. Determinación de proteínas en los extractos crudos enzimáticos .............. 34
6. RESULTADOS .......................................................................................................... 35
6.1. ANÁLISIS DE LOS PERFILES DE DGGE ........................................................... 35
6.2. CARACTERIZACIÓN MOLECULAR DE AISLADOS BACTERIANOS
ASOCIADOS AL CARACOL PALA. .......................................................................... 42
6.2.1. Aislamiento de bacterias ............................................................................... 42
6.2.2. Filogenia de los aislados bacterianos ............................................................ 42
6.3. PERFIL ANTIBACTERIAL DE LOS AISLADOS ASOCIADOS AL CARACOL
PALA .............................................................................................................................. 45
6.4. ACTIVIDAD ENZIMÁTICA DE CEPAS BACTERIANAS ASOCIADAS AL
CARACOL ..................................................................................................................... 48
6.4.1. Análisis cualitativo de enzimas hidrolíticas .................................................. 48
6.4.2 Análisis cuantitativo de las hidrolasas en los extractos crudos enzimáticas .. 49
6.4.2.1. Amilasas ..................................................................................................... 49
6.4.2.2. Proteasas ..................................................................................................... 51
6.4.2.3. Lipasas ........................................................................................................ 53
7. DISCUSIÓN ............................................................................................................... 56
7.1. ANÁLISIS DE LOS PERFILES DE DGGE. .......................................................... 56
7.2. FILOGENIA DE LOS AISLADOS BACTERIANOS ........................................... 62
7.3. ACTIVIDAD ANTIBACTERIANA ....................................................................... 64
7.4. ACTIVIDAD ENZIMÁTICA ................................................................................. 66
8. CONCLUSIONES ...................................................................................................... 70
BIBLIOGRAFÍA ............................................................................................................ 74
ANEXOS ........................................................................................................................ 95

XII
LISTA DE TABLAS
Tabla 1. Nomenclatura de las muestras obtenidas de la baba, el intestino y el alimento
de caracoles silvestres y en cautiverio. DNA total de las muestras originales (O), DNA
de la fracción cultivable o barridos (B). ............................................................................... 25
Tabla 2. Secuencias de los cebadores usados en las reacciones de amplificación del gen
rRNA 16S ............................................................................................................................. 26
Tabla 3. Homología de las secuencias de las bandas obtenidas por DGGE con las
secuencias de genes rRNA 16S en la base de datos del GenBank ....................................... 40
Tabla 4. Identificación taxonómica y posición filogenética de los aislados desde
muestras asociadas al caracol pala........................................................................................ 44
Tabla 5. Patrones de actividad antibacteriana (a-g) de bacterias asociadas a Strombus
gigas. .................................................................................................................................... 45
Tabla 6. Afiliación de las bacterias asociadas a Strombus gigas a los grupos
taxonómicos (OTU) y patrones de actividad (a-g) + Actividad antibiótica, − no
actividad antibiótica. ............................................................................................................. 47
Tabla 7. Amilasas extracelulares de aislados bacterianos asociados al caracol pala ........... 50
Tabla 8. Proteasas extracelulares de aislados bacterianos asociados al caracol pala ........... 52
Tabla 9. Lipasas extracelulares de aislados bacterianos asociados al caracol pala .............. 53
Tabla 10. Resultados de las absorbancias para diluciones diferentes de α.- amilasa de
Bacillus amyloliquefaciens. Los azucares reductores fueron medidos usando el método
de DNS. ................................................................................................................................ 54
Tabla 11. Resultados de las absorbancias para diluciones diferentes de proteasas de
Bacillus amyloliquefaciens. Las proteasas fueron medidas por el método de Lowry. ......... 55

XIII
LISTA DE FIGURAS
Figura 1. Patrones de bandeo en geles de DGGE del producto amplificado del gen
rRNA 16S desde muestras originales (O) y la fracción cultivable o barridos (B) de
intestino (i), baba o mucus (b) y alimento (a) de caracoles silvestres (S) y en cautiverio
(C) ......................................................................................................................................... 36
Figura 2. Dendograma obtenido con el programa GelCompar II a partir de los patrones
de bandeo en geles de DGGE del producto amplificado del gen rRNA 16S desde el
DNA original (OS y OC) y el DNA de los barridos (BS y BC) de muestras de caracoles
en estado silvestre y en cautiverio ........................................................................................ 37
Figura 3. Árbol filogenético de las secuencias parciales del gen rRNA 16S
correspondientes a las bandas obtenidas por DGGE. ........................................................... 41
Figura 4. Árbol filogenético basado en secuencias de rDNA 16S obtenidas de bacterias
aisladas de la baba, el intestino y el alimento de S. gigas. ................................................... 43
Figura 5. Halo de inhibición del extracto metanólico crudo derivado de la cepa
bacteriana identificada como Uncultured Psychrobacter sp. (6CpOI15) frente a Bacillus
subtilis. .................................................................................................................................. 45
Figura 6. Halo de inhibición del extracto metanólico crudo derivado de una cepa
bacteriana identificada como Uncultured Psychrobacter sp. (25CpB1) frente a Bacillus
subtilis ................................................................................................................................... 46
Figura 7. Halo de inhibición del extracto metanólico crudo derivado de una cepa
bacteriana identificada como Micrococcus sp. (20CpOB13) frente a E. coli. ..................... 46
Figura 8. Halos de inhibición del extracto metanólico crudo derivado de la cepa
bacteriana identificada como Psychrobacter sp. (3CpBB10) a) Frente a a) Bacillus
subtilis b) Pseudomona aeruginosa y c) E. coli. .................................................................. 47
Figura 9. Cepas productoras de enzimas hidrolíticas, aisladas de la baba, el intestino y el
alimento de caracoles silvestre y en cautiverio .................................................................... 48
Figura 10. Número de aislados identificados con actividad de enzimas hidrolíticas. .......... 49

XIV
Figura 11. Curva de calibración de glucosa usada para determinar la cantidad de
azúcares reductores liberados por las amilasas extracelulares. ............................................ 50
Figura 12. Curva estándar de BSA usada para la medición del contenido de proteínas en
los extractos crudos enzimáticos. ......................................................................................... 51
Figura 13. Curvar estándar de L-tirosina usada para la determinar la actividad proteasa .... 52
Figura 14. Curva estándar de p-nitrofenol usada para la cuantificación de lipasas
extracelulares. ....................................................................................................................... 53

XV
LISTA DE ANEXOS
Anexo 1. Caracterización morfológica microscópica y macroscópica de los aislados
bacterianos asociados al caracol pala ................................................................................... 95
Anexo 2. Geles de poliacrilamida teñidos con nitrato de plata (Fragmentos del gen
rRNA 16S amplificados con los primers 341F y 907R) ..................................................... 100
Anexo 3. Matrices .............................................................................................................. 101
Anexo 4. Análisis de similitud ANOSIM usando el software PAST ................................. 103
Anexo 5. Trazas de secuencias de las bandas obtenidas a partir del DGGE ...................... 104
Anexo 6. Establecimiento de las afiliaciones filogenéticas a través del RDP y el Blastn
para las secuencias de nucleótidos de las bandas obtenidas por DGGE. ........................... 107
Anexo 7. Alineamiento múltiple con Clustal W de las secuencias obtenidas desde las
bandas escindidas en los geles de DGGE ........................................................................... 111
Anexo 8. Identificación taxonómica y posición filogenética de los aislados bacterianos
asociados al caracol pala..................................................................................................... 111
Anexo 9. Alineamiento múltiple con Clustal W de las secuencias correspondientes a los
aislados. .............................................................................................................................. 112
Anexo 10. Evaluación de producción de enzimas hidrolíticas de las cepas aisladas de
Strombus gigas. .................................................................................................................. 113
Anexo 11. Resultados colorimétricos para la curva de glucosa usada para la
determinación de azucares reductores. ............................................................................... 115
Anexo 12. Resultados de la determinación de proteínas usando el reactivo de Bradford .. 115
Anexo 13. Resultados colorimétricos para la curva de L- tirosina usada para la
determinación de proteasas. ................................................................................................ 116
Anexo 14. Resultados colorimétricos para la curva de p-NP usada para la determinación
de lipasas. ........................................................................................................................... 117
Anexo 15. Árbol filogenético de los aislados bacterianos usando el método de
Neighbor- joining y el estimador de distancias Tamura–Nei. ............................................ 118

XVI
LISTA DE SÍMBOLOS Y ABREVIATURAS
°C. Grados centígrados
a. Alimento
b. Baba o mucus
B. Barrido
Blast. Basic Local Alignment Search Tool (Herramienta básica para el alineamiento de
secuencias)
bp. Pares de bases
BSA. Albúmina de suero bovino
C. Cautiverio
CaCl2. Cloruro de calcio
CITES. Convención sobre el Comercio Internacional de Especies Amenazadas de Fauna
y Flora Silvestres.
CMC. Carboximetil celulosa
DGGE. Electroforesis por gradiente de desnaturalización
DNA. Ácido desoxirribonucleico
DNS. 3,5-ácido dinitrosalicílico
dNTPs. Desoxirribonucleótidos
EDTA. Ácido etilendiaminotetraacético
F. Forward
g. Gramos
GC. Guanina- Citosina
GenBank. Base de datos de secuencias genéticas
h. Horas
i. Intestino
I2. Yodo
K2HPO4. Difosfato de potasio
KI. Yoduro de potasio

XVII
KOH. Hidróxido de potasio
LB. Medio de cultivo Luria-Bertani
LUV. Luz ultravioleta
M. Molar
Mg. Miligramo
MgSO4. Sulfato de magnesio
min. Minutos
ml. Mililitros
mm. Milímetros
mM. Milimolar
NaCl. Cloruro de sodio
NCBI. Centro Nacional para la Información Biotecnológica
ng. Nanogramo
nm. Nanómetro
o. Original
P/V. Peso/Volumen
PCR. Polymerase Chain Reaction (Reacción en cadena de la polimerasa)
PCR-DGGE. Reacción en cadena de polimerasa y electroforesis en gel con gradiente
desnaturalizante
p-NP. Paranitrofenol
p-NPP. Paranitrofenil palmitato
R. Reverse
rDNA 16S. 16S del DNA ribosómico
RDP. Ribosomal Database Project (Base de datos Ribosomal)
RNA. Ácido ribonucleico
RNA. Ácido ribonucleico ribosómico
rpm. Revoluciones por minuto
rRNA 16S. Subunidad 16S del RNA ribosómico
s. Silvestre
TAE. Buffer tris-acetato-EDTA
Taq polimerasa. Thermus aquaticus DNA polimerasa
TCBS. Agar tiosulfato citrato bilis sacarosa

XVIII
Tris. Tris (hidroximetil) aminometano
U. Unidad de actividad enzimática
UFC. Unidad formadora de colonia
UPGMA. Unweighted Pairgroup Method with Arithmetic Averages (Análisis de
agrupamientos por ligamiento promedio)
V. Voltios
v. Volumen
μg. Microgramo
μl. Microlitro
μm. Micrometro
μM. Micromoles

1
1. INTRODUCCIÓN
Los microorganismos se encuentran dispersos en los océanos libres o formando
asociaciones complejas con organismos marinos (Ashen y Goff, 2000). El creciente
interés en el estudio de estas asociaciones, es consecuencia de los caracteres únicos que
los microorganismos aportan a su individuo hospedero. Identificar en detalle la
diversidad microbiana asociada y las características metabólicas, es fundamental para
empezar a entender la conexión entre huésped-microbio, las variaciones en las
poblaciones de microorganismos entre especies y de acuerdo al hábitat, y sí éstos
microbios son fuentes de moléculas bioactivas importantes para el hospedero con
aplicaciones adicionales en la industria y la biotecnología. El caracol pala, habita en las
aguas de las costas del mar Caribe (Brownell y Stevely, 1981), es un molusco de gran
importancia ecológica y socioeconómica en el área caribeña colombiana, una especie
bajo amenaza y representa uno más, en la amplia lista de hospederos de
microorganismos marinos con una microbiota poco estudiada.
Estudios previos sobre la diversidad en caracoles fueron llevados a cabo por métodos
clásicos en microbiología (Ducklow, 1979). Se sabe que las bacterias necesitan
requerimientos nutricionales más exigentes para su crecimiento, estimándose que el
99% de los microorganismos presentes en la naturaleza no pueden ser estudiados
usando métodos dependientes de cultivo (Schloss y Handelsman, 2005). El desarrollo
de técnicas basadas en la amplificación de genes ha permitido a los investigadores
acceder a las poblaciones no cultivables hasta el momento.
Aunque algunos estudios se han centrado en la diversidad microbiana, la obtención de
compuestos bioactivos y la búsqueda de nuevos genes biocatalizadores, (Jiang et al.,
2010; Nishimura et al., 2010), la diversidad marina permanece aún poco explorada. El
ambiente marino es realmente complejo, con condiciones especiales de luminosidad,
alta salinidad, altas presiones y bajas temperaturas, lo cual puede contribuir a

2
significativas diferencias entre las enzimas generadas por los microorganismos marinos
y enzimas análogas obtenidas de microorganismos terrestres (Zhang y Kim, 2010). Las
bacterias del suelo siguen siendo ampliamente estudiadas, sin embargo, hay una notable
disminución en el hallazgo de nuevos productos a partir de éstas, estimándose que más
del 90% de los cultivos microbianos productores de compuestos bioactivos descubiertos
producen agentes ya descritos (Fenical, 1993). Por ello, la exploración se ha dirigido
hacia otros ambientes, como los sedimentos de los ríos, lagos y océanos, así como a las
plantas y animales acuáticos, que brindan la posibilidad de encontrar cepas microbianas
nativas no descritas, capaces de producir nuevos metabolitos secundarios
biológicamente activos (Jensen y Fenical, 1994).
Muchos estudios relacionados con la diversidad marina se han enfocado en bacterias
extremófilas. Zhang et al., (2007) caracterizaron un gen de lipasa de Psychrobacter sp.,
adaptado al frio y aislado de sedimentos en las profundidades del mar. Otros estudios se
han orientado hacia bacterias mesófilas como los realizados por Shanmughapriya et al.,
(2010) en el que caracterizaron una enzima celulasa de una bacteria asociada a una
esponja la cual es estable a pH alcalinos. La actividad óptima de las enzimas marinas
ocurre en alta salinidad, haciendo que estas enzimas sean realmente útiles en muchos
procesos industriales que involucran condiciones similares. Adicionalmente, muchas de
esas enzimas son termotolerantes, permaneciendo estables a temperatura ambiente por
largos períodos de tiempo (Mohapatra et al., 2003).
Esta investigación pretendió hacer un análisis molecular de la diversidad de las
poblaciones presentes en materiales biológicos (baba o mucus, intestino y alimento)
asociados al caracol pala, utilizando técnicas dependientes (Cultivo, extracción de
DNA, PCR, y DGGE-cultivables) e independientes de cultivo (Extracción de DNA,
PCR y DGGE-no cultivables). El campo de la biología molecular ha avanzado con
tecnologías innovadoras que han permitido nuevos descubrimientos. La habilidad para
extraer y aislar el DNA, la amplificación de genes de grupos específicos de
microorganismos por PCR y la obtención de datos de las secuencias de genes ahora son
eventos rutinarios. Los genes ribosomales son ampliamente usados como marcadores

3
moleculares para identificar especies morfológicamente indistinguibles o para inferir
sus relaciones filogenéticas y elucidar la diversidad microbiana.
Este trabajo tuvo también como propósito adicional, la exploración de propiedades
bioactivas y enzimáticas de algunos de los aislados de la microbiota del caracol. En
estos aislados fue evaluada la actividad antibacteriana de los metabolitos secundarios
producidos y la presencia de enzimas como las amilasas, proteasas, celulasas y lipasas
en general en medios de cultivos y su posterior cuantificación. Las cepas bacterianas
fueron caracterizadas morfológica y genotípicamente.
En ese contexto a través de las metodologías desarrolladas en este estudio se da un
acercamiento para inferir sobre la diversidad bacteriana del caracol pala del Caribe
Colombiano, así como del potencial bioactivo de la microbiota asociada a caracoles
silvestres y en cautiverio.

4
2. HIPÓTESIS
La hipótesis general está fundamentada en que existen modificaciones en las
poblaciones microbianas de un hospedero específico de acuerdo a su hábitat; la
habilidad de los microorganismos marinos para establecer relaciones simbióticas con su
hospedero y producir moléculas asociadas a los procesos de defensa del huésped y
enzimas participantes en la degradación orgánica, digestión y conservación del hábitat
natural donde se encuentren.
¿Cómo varían las poblaciones microbianas entre hospederos? ¿Existe una relación entre
las poblaciones microbianas de los caracoles silvestres y en cautiverio? ¿Son estos
microorganismos importantes en el área biotecnológica? Explorando esta hipótesis en el
caracol pala, esperando que existan relaciones entre las poblaciones microbianas
asociadas a los caracoles silvestres y entre los caracoles en cautiverio, y que las
bacterias aisladas estén vinculadas a la producción de moléculas bioactivas
(antimicrobianas) e hidrolasas. Teniendo en cuenta lo anterior las hipótesis son las
siguientes:
Hipótesis 1: La diversidad microbiana entre caracoles en estado silvestre y entre
caracoles en cautiverio está relacionada, teniendo en cuenta que los microorganismos
colonizan hospederos específicos de acuerdo a su hábitat.
Hipótesis 2: Las poblaciones microbianas en el caracol están relacionadas con la
producción de antimicrobianos y enzimas hidrolíticas, considerando evidencias
reportadas sobre la producción de biocatalizadores y antimicrobianos en especies
bacterianas asociadas a otros huéspedes marinos eucariotas, en donde se ha sugerido la
participación de las hidrolasas en procesos de digestión y nutrición del huésped y la
presencia de metabolitos implicados en la respuesta y acción frente a patógenos
microbianos de interés para la industria.

5
3. ESTADO DEL ARTE
3.1. Strombus gigas
Es un meso gastrópodo prosobramquio de la familia Strombidae, que tienen una concha
gruesa y pesada con una aguja cónica ornamentada. A menudo tienen un labio
extendido en forma de ala y la coloración varía desde blanco a marrón claro, con
abertura y labio externo de color rosa intenso. Es el más grande de las siete especies de
Strombus en el Caribe (Randall, 1964).
Conocido como caracol pala, que habita el Caribe, el golfo de México, sur de Florida,
las Bahamas y Bermudas, su pesca fue alguna vez la segunda con más valor en la región
(Berg y Olsen, 1989) con una producción estimada en 30 millones de dólares en 1992
(Creswell, 1994). El continuo decline de esta especie provocó que fuese clasificada
como comercialmente amenazada por la Convención Internacional del Comercio de
Especies de fauna y flora silvestre en Peligro (CITES) en 1992. La disminución en la
abundancia del caracol persistió y la CITES lo ubicó en el apéndice II, lo cual requería
del tratado entre naciones para el manejo de las reservas desde cerca, y monitorear
cuidadosamente las exportaciones para prevenir la extinción de la especie. Durante
siglos el caracol ha sido cosechado como alimento, sin embargo, en las últimas décadas
la pesquería comercial a gran escala se desarrolló, principalmente como respuesta al
aumento de la demanda internacional de la carne. El precio mayorista de los
desembarcos anuales se ha estimado en 60 millones de dólares estadounidenses. Las
caracolas también se usan y se comercian como curiosidad y recuerdos turísticos, pero
son consideradas como subproducto del comercio de la carne (Glazer y Berg, 1992).
3.1.1. Distribución y hábitats
Se encuentra en el Océano Atlántico Occidental, está distribuido a lo largo de todo el
Atlántico tropical noroccidental incluyendo Bermudas, los Cayos de la Florida, las

6
Antillas Mayores y Menores y las costas del Caribe de América Central y América del
Sur hasta Brasil, encontrándose en el Golfo de México (Brownell y Stevely, 1981). El
caracol es conocido por varios nombres a lo largo de su área de distribución,
incluyendo: botuto o guarura (Venezuela); cambombia (Panamá); cambute (Costa
Rica); caracol abulon (Guatemala); caracol gigante (Honduras); caracol pala
(Colombia); caracol rosado (México); carrucho (Puerto Rico); cobo (Cuba); y lambi
(isla de La Española y Antillas Francesas). Su presencia ha sido reportada en arrecifes
de coral, fondos rocosos y arena, localizado en praderas de fanerógamas marinas
(Thalassia testudinum y Syringodium filiforme), donde se alimenta de epifitos, detritos
de pastos, macroalgas (Cladophora sp. Polysiphonia, Laurencia sp., Enteromorpha sp.
y otras especies de rodófitas y clorófitas), (Randall 1964; Alcolado 1976) y microalgas
En Colombia a comienzos de la década de los años setenta, las principales áreas de
pesca comercial del caracol pala del Caribe eran el Archipiélago de San Bernardo y las
Islas del Rosario. Sin embargo, en 1977 estas áreas fueron clausuradas después de que
tuvo lugar una sobrepesca sustancial y por consiguiente los pescadores se retiraron a
nuevas áreas, principalmente en los Archipiélagos de San Andrés y de Providencia
(Mora, 1994). La mayor parte de la pesquería opera en la Península de la Guajira y en
los Archipiélagos de San Andrés y de Providencia (González, 2002). A principios de la
década de los años noventa se llevaron a cabo varios estudios, incluyendo morfológicos
y muestreos de abundancia en las principales áreas de pesca de los Archipiélagos de San
Andrés y de Providencia (Ospina et al., 1996).
3.1.2. Microbiota de caracoles silvestres y en cautiverio
Actualmente los estudios sobre el caracol están orientados hacia la búsqueda de
alternativas para su preservación, desde la biología de estos invertebrados hasta la
comprensión de su microbiota. En la última década, investigadores han reunido
esfuerzos en procura de identificar los microorganismos (bacterias y parásitos)
asociados al caracol (Acosta et al., 2009; Aldana et al., 2011; Rodríguez et al., 2011).
¿Cómo cambian las poblaciones de parásitos de acuerdo a su nicho?, ¿el efecto de estos
microbios sobre el caracol? Son preguntas que están empezando a ser respondidas, sin
embargo, aún son pocos los estudios en esta área.

7
El proceso de recuperación de las poblaciones del caracol debe integrar los
conocimientos genético molecular, la variabilidad genética, el crecimiento, la
reproducción, el estudio de la ecología microbiana, con el fin de mejorar las estrategias
de repoblamiento y cultivo para la conservación de esta especie.
3.2. ECOLOGÍA MICROBIANA
Hace 3800 millones de años la vida procariota emergió, 2000 millones de años más
tarde la vida eucariota surgió. Se ha observado que las formas de vida procariota se han
encontrado en imaginables nichos ecológicos en la tierra (Xu, 2006). El estudio de los
microorganismos: archaeas, bacterias, protistas y hongos a través de la ecología
microbiana ha permitido conocer sus hábitats, esclarecer sus interacciones entre ellos y
otros grupos de organismos, y entender como individuos tan pequeños son capaces de
conservar su ambiente y su diversidad. En el pasado parte de la información sobre la
diversidad y actividad microbiana de hospederos marinos algas, corales, peces,
caracoles en general, era derivada de técnicas dependientes de cultivo, sólo una pequeña
fracción de los microorganismos era cultivado, así que nuestro conocimiento sobre la
densidad (Whitman et al., 1998) y diversidad microbiana hasta entonces era limitado e
incierto, sin embargo, desde mediados de la década de los 80, la perspectiva que se tenía
sobre la diversidad microbiana mejoró con la implementación de nuevas herramientas
moleculares para el análisis filogenético de organismos no cultivados (Head et al.,
1998). El estudio de la diversidad bacteriana (microbios benéficos y patógenos) en
vertebrados e invertebrados marinos ahora es más frecuente y sin dudas ha mejorado
(Radjasa et al., 2008; Kvennefors et al., 2010; Tanu et al., 2011).
3.2.1. Herramientas genómicas para el estudio de la diversidad microbiana
Genómica es un término utilizado para describir una disciplina en genética relacionada
con el mapeo, secuenciación y análisis funcional de genomas completos. El análisis
funcional incluye aspectos relacionados con el RNA transcrito (transcriptómica),
proteínas (proteómica), y metabolitos (metabolómica).La disciplina que usa métodos
genómicos para el análisis de las comunidades ecológicas naturales ha sido llamada

8
metagenómica, entre los métodos genómicos se destacan la secuenciación y análisis de
secuencias de DNA, microarreglos, electroforesis por gradiente de desnaturalización
(DGGE), entre otros.
3.2.2. Estrategias usadas para la identificación de especies bacterianas en ecología
microbiana
Son varias las estrategias utilizadas para la identificación y caracterización de
microorganismos, a continuación se describen algunas de ellas:
3.2.2.1. Caracterización de fenotipos
La utilización de sustratos y la producción de productos bacterianos proveen
importantes datos para el estudio de la diversidad funcional. La caracterización
fenotípica es útil para el estudio e identificación de genes que codifican proteínas, las
cuales pueden tener aplicaciones industriales. Una combinación de información del
genotipo y el fenotipo es requerida para la correcta descripción y clasificación de
bacterias (Rosselló et al., 2001).
3.2.2.2. Reacción en cadena de la polimerasa (PCR)
La exploración de la biodiversidad por PCR es el método más frecuentemente utilizado
en estudios de genes rRNA 16S; la información obtenida del proceso de amplificación
del DNA por PCR puede compararse con técnicas como ARDRA (Análisis de
Restricción de rDNA amplificado); DGGE (Electroforesis en Gel por Gradiente de
Desnaturalización); TTGE (Electroforesis en Gel por Gradiente de Temperatura); RFLP
(Polimorfismo de Longitud por Fragmentos de Restricción) y microarreglos a través del
análisis de patrones. Esas técnicas permiten la detección rápida y sensible de la
diversidad bacteriana independiente de cultivo (Dahllöf, 2002).
3.2.2.3. Secuenciación del DNA
Los principios básicos para la secuenciación del DNA fueron establecidos a mediados
de 1970, no obstante, hubo que esperar hasta 1990 para que un método eficiente de
secuenciación basado en el uso de dideoxiribonucleotidos fuera desarrollado. En el
presente un alto volumen de datos de secuencias de DNA es encontrado en instituciones

9
académicas y laboratorios de biología molecular. Las bases de datos de genomas listan
aproximadamente 1268 genomas completos, que en su gran mayoría son de microbios.
Hoy en día es necesario el uso de varios métodos para la identificación y caracterización
de especies bacterianas, para el estudio y determinación de la diversidad de sus
dominios (Bernardet et al., 2005; Dahllöf, 2002).
3.2.2.4. Estudio de la diversidad y bioprospección en bacterias asociadas al caracol
pala
El uso de nuevos enfoques dependientes e independientes de cultivo para el estudio y
análisis de la composición de las comunidades bacterianas en hábitats marinos y
terrestres cada vez es más frecuente. El análisis del gen rRNA 16S, con secuencias
altamente conservadas entre especies distantes ha llegado a ser un modelo útil para el
estudio de la evolución y las relaciones filogenéticas entre especies bacterianas
(McInerney et al., 2002) a pesar de que ofrece poca información sobre la evidencia de
genes funcionales (Hugenholtz et al., 1998; Schloss et al., 2004). La diversidad
microbiana asociada al caracol pala puede ser objeto de estudio mediante el uso de los
enfoques mencionados con anterioridad, en procura de descubrir la diversidad e
identificar los miembros vinculados a este huésped eucariota. Muchos microorganismos
son habitantes naturales de otros seres vivos, y en general se localizan dentro de sus
cavidades, sobre epitelios o el lumen de órganos. Normalmente estos microorganismos
desempeñan funciones benéficas para el hospedero, puesto que se encuentran en
algunos casos participando en procesos de degradación de los alimentos que de otra
manera serían imposibles de digerir por el organismo anfitrión. El caracol pala no es la
excepción, y se ha podido establecer que es hábitat de gran diversidad de
microorganismos, sin embargo, es escasa la información existente sobre la flora
asociada al caracol y su relación con su desarrollo. En ciertas especies de caracoles se
han descrito características de la baba, involucrando su acción bactericida por acción de
una glicoproteína (Kubota, 1985). La flora bacteriana en otros caracoles ha sido descrita
a través de caracterización fenotípica de las cepas aisladas en cultivo y ha sido
relacionada con el tamaño de los caracoles. Los géneros bacterianos encontrados
incluyeron Pseudomonas, Acinetobacter, Aeromonas, Vibrio y algunos pertenecientes a
la familia Enterobacteriaceae. Con un predominio de la presencia de Vibrio extorquens

10
y Pasteurella sp. Estas bacterias corresponden tanto a microbiota normal como a
bacterias presentes en la alimentación del caracol (Ducklow, 1979). El uso de técnicas
de microbiología molecular como la amplificación de las regiones espaciadoras entre
los genes del rRNA 16S-23S y el análisis de las secuencias del gen rRNA 16S en un
estudio previo en el caracol pala, ha conducido al descubrimiento de géneros no
descritos previamente en este caracol como Psychrobacter y Halomonas (Acosta et al.,
2009). Usualmente los microorganismos establecen con su hospedero una relación
simbionte en la que estos aportan ventajas a cambio de protección y alimento, teniendo
en cuenta además que las superficies y los espacios internos de los huéspedes eucariotas
son más ricos en nutrientes que el agua de mar y muchos sedimentos (Mohapatra et al.,
2003). Entre las ventajas aportadas por la microbiota natural de un organismo se pueden
citar protección frente a microorganismos patógenos invasores y aporte enzimático en la
degradación de sustratos utilizados como alimento; esta última es quizá una de las
ventajas más importantes debido a que el uso de bacterias productoras de
polisacaridasas y proteasas como probióticos en el cultivo de crustáceos con resultados
positivos sobre su crecimiento demuestran la importancia de éstos microbios y sus
enzimas en los procesos digestivos (Ahmadnia et al., 2012). La presencia de bacterias
productoras de moléculas biocatalizadoras en algas, caballitos de mar (Hippocampus
kuda) y peces marinos (Sugita et al., 1996) (Holmstrom y Kjelleberg, 1999) (Tanu et
al., 2011) indican que es posible encontrar bacterias en el caracol productoras de
enzimas con una notable actividad, las cuales posteriormente pueden ser caracterizadas
para ser utilizadas en diversos procesos industriales. Los microorganismos y
particularmente sus propiedades catabólicas, son utilizadas en una amplia gama de
procesos como la descontaminación de aguas residuales, transformación de desechos de
origen vegetal, producción de biopesticidas y metabolitos de interés farmacéutico,
industrial, alimenticio, agrícola, entre otros (Gupta et al., 2003). Estos organismos son
utilizados en países desarrollados como fuente de enzimas amilasas, celulasas,
xilanasas, proteasas, DNA polimerasas, lipasas, esterasas, etc.

11
3.3. BÚSQUEDA DE NUEVOS ANTIBACTERIANOS EN LAS BACTERIAS
ASOCIADAS AL CARACOL PALA
Actualmente se utilizan un gran número de antibióticos para proteger a los humanos y
animales contra infecciones, y en la producción de alimentos en el sector agrícola e
industrial. Por múltiples razones se ha reportado el aumento en el número de bacterias
resistentes a los antibióticos disponibles en el mercado, lo cual ha conllevado a la
necesidad y por tanto búsqueda de nuevas estructuras moleculares (Cabello et al.,
2004). Los microorganismos cuentan con mecanismos diferentes para resistir la acción
de los antibióticos tales como la síntesis de enzimas como las β-lactamasas, mutaciones
en su material genético y la presencia de plásmidos, entre otros (Fuchs et al., 1994).
En este momento la búsqueda y descubrimiento de nuevos antibióticos producidos por
colecciones de microorganismos aislados de forma natural, y/o de cepas modificadas
genéticamente es constante. La obtención de nuevos compuestos es un elemento
importante en la lucha contra un gran número de infecciones causadas por patógenos
resistentes a antibióticos (Nathan, 2004).
Hasta el momento, el mayor esfuerzo en la búsqueda de nuevos antibacterianos se ha
concentrado en ambientes terrestres, en comparación con la pequeña atención que se le
ha prestado a los metabolitos derivados de microbios marinos. Para finales del año
2008, tan sólo aproximadamente 3000 metabolitos microbianos de recursos marinos
habían sido reportados, mucho menos, que el número de compuestos aislados de otros
recursos naturales (Rahman, 2008). Una vieja creencia científica que argumentaba que
el mar contenía pocos microbios fue la razón principal para pasar por alto los
metabolitos secundarios de microorganismos marinos por mucho tiempo. El ambiente
marino con su microflora diversa se ha convertido en un recurso promisorio de nuevas
fábricas de antibióticos (Faulkner et al., 2004).
Los compuestos derivados de actinomicetos aislados del suelo, han proporcionado una
fuente rica en moléculas bioactivas por más de cinco décadas, produciendo más de
100000 compuestos, incluyendo el 70% de los antibióticos descubiertos a partir de
fuentes naturales (Berdy, 2005). La escasez de nuevos antibióticos emerge a finales de
1960 y el reto actual en la lucha contra enfermedades infecciosas resistentes a ellos,
resalta la importancia de los descubrimientos sobre la nueva diversidad química de

12
fuentes naturales. Las bacterias marinas podrían convertirse en una fuente importante de
compuestos útiles en el tratamiento de enfermedades nosocomiales y recalcitrantes,
como las causadas por el MRSA y VRE (Rahman et al., 2010).
La adaptación al ambiente marino supone un factor impulsor de la diversidad química,
considerando que el entorno competitivo de algunos nichos, como ocurre con las
bacterias epífitas asociadas con las superficies de las algas (Boyd et al., 1999; Mearns-
Spragg et al., 1998) puede fomentar el desarrollo de especies capaces de producir
metabolitos antibacterianos.
Existen reportes de bacterias aisladas en medios marinos con actividades enzimáticas,
producción de metabolitos secundarios y de nuevos antibióticos (Wiese et al., 2009),
que se han reportado para ser usados como probióticos (Das et al., 2008) entre otras
características. La existencia de comunidades microbianas únicas con propiedades
antimicrobianas han sido descrita previamente en un número significativo de
hospederos marinos: peces perciformes (Callionymus sp.), esponjas, briozoos, (Sugita et
al., 1998; Anand et al., 2006; Heindl, 2010) e incluso moluscos (Romanenko et al.,
2006). En ese orden de ideas podría sugerirse la presencia de microorganismos
productores de sustancias biológicamente activas en el caracol pala. El estudio de las
propiedades bioactivas de las bacterias asociadas al caracol constituye el principio hacia
el esclarecimiento del rol de estos microorganismos en el caracol, su participación en la
respuesta innata en la defensa frente a patógenos y el desarrollo de nuevos
antimicrobianos.
3.4. HIDROLASAS EXTRACELULARES EN BACTERIAS
En bioquímica una hidrolasa es una enzima que cataliza la hidrolisis de una unión
química.
Por ejemplo, una enzima que cataliza la siguiente reacción es una hidrolasa:
A-B + H2O → A–OH + B–H
El nombre sistemático de las hidrolasas está formado como hidrolasa sustrato, sin
embargo, el nombre común esta dado típicamente en la forma de sustrato, por ejemplo,

13
las esterasas incluyen lipasas, que rompen uniones éster (entre un ácido carboxílico y un
alcohol) en lípidos. Usualmente las hidrolasas dividen moléculas grandes en dos
moléculas pequeñas. Ejemplos de hidrolasas comunes incluyen esterasas, proteasas,
glicosidasas, nucleosidasas y lipasas.
El estudio en bacterias es de gran importancia, puesto que ellas pueden producir
compuestos de interés en la industria, como hidrolasas extracelulares que tienen
diversos usos en las ciencias biomédicas y la industria química (Margesin y Schinner,
2001).
Enzimas como las amilasas, celulasas y proteasas son ampliamente usadas en la
industria para la manufactura de drogas, alimentos, jarabes y dulces, o bien en el
procesamiento textil y del cuero (Wiseman, 1985). Las aplicaciones potenciales de
amilasas (Bailey et al., 1977) celulasas y proteasas (Outtrup et al., 1990) han sido
revisadas previamente. La mayoría de las enzimas usadas en la industria son de origen
microbiano debido a que son más estables que las enzimas derivadas de plantas y
animales (Bull et al., 2000). Con el reciente advenimiento de la biotecnología, se ha
generado un creciente interés y demanda por enzimas con nuevas propiedades.
Considerables esfuerzos han sido dedicados a la selección de microorganismos
mediante sofisticadas técnicas de selección y metodologías para la producción de
enzimas con nuevas propiedades fisiológicas y físicas, y tolerancia a condiciones
extremas usadas en procesos industriales (temperatura, sales y pH). Los ambientes
marinos proveen un rico recurso en diversidad biológica y química (Pomponi et al.,
1999). El potencial de organismos marinos incide en todas las áreas de la biotecnología.
Microorganismos marinos han irrumpido recientemente como un recurso valioso para el
aislamiento de enzimas industriales (Chandrasekaran et al., 1997). Adicionalmente
muchas enzimas derivadas de bacterias marinas son considerablemente termotolerantes,
permanecen estables a temperatura ambiente por largos periodos. Los microhábitats
para bacterias marinas están en el mar, en los sedimentos, superficies animadas e
inanimadas. Los organismos marinos sedentarios (plantas marinas y animales sésiles)
forman relaciones simbióticas altamente específicas con bacterias (Althoff et al., 1998)
que en casos extremos constituyen el 60% del volumen tisular. La superficie y los
espacios internos de esos organismos son ricos en nutrientes, al igual que el mar y
muchos sedimentos, así que ellos seguramente pueden ser un nicho único para el

14
aislamiento de diversas comunidades bacterianas, bacterias asociadas que son un
recurso potencial de compuestos biológicamente activos de bajo peso molecular
(Fenical et al., 1993).
Las enzimas producidas por los microorganismos marinos poseen propiedades
excepcionales como su adaptabilidad a ambientes salinos. La industria cada vez está
más interesada en las enzimas marinas y algunas de las razones para esto obedecen a la
necesidad de implementar metodologías que requieran menor consumo de energía y que
puedan ajustarse al resto de las exigencias de sus procesos. Básicamente se esperaría
que estas enzimas fueran mejores a sus homologas derivadas de especies bacterianas
habitantes del suelo, bajo ciertas condiciones físico-químicas: salinidad, temperatura,
presión y pH, todo depende del fin que se persiga y de las condiciones a las cuales se
llevara a cabo el proceso en donde se utilizarán los biocatalizadores, así por ejemplo, se
espera que aquellas enzimas que son muy buenas a temperaturas bajas no lo sean a
temperaturas superiores. Los problemas a los que se enfrentan la mayoría de los países
son similares: el crecimiento de la población, el consumo de recursos y la
contaminación, la solución de estos problemas depende de la forma en la cual son
optimizados los recursos. Si se requiere de enzimas para el tratamiento de aguas
residuales templadas (2-5 °C) una forma de optimizar el proceso es utilizar
microorganismos capaces de crecer y producir enzimas que sean activas bajo esas
condiciones en lugar de hacer un gasto energético innecesario para el ajuste de la
temperatura del agua a la temperatura a la cual son funcionales enzimas de especies
bacterianas como el Bacillus subtilis y el Bacillus amyloliquefaciens (35-55 °C)
ampliamente usadas en la industria.
Por otra parte las proteínas especialmente enzimas derivadas de bacterias asociadas a
esponjas y a algas marinas han sido descuidadas. No se sabe si estos microorganismos
son un recurso potencial o si las concentraciones de estas enzimas son de poca
importancia (Mohapatra et al., 2003). La detección de hidrolasas en bacterias asociadas
a organismos marinos ha sido correlacionada con procesos de nutrición (Tanu et al.,
2011) ciclos biogeoquímicos y transformación de materia orgánica (Mohapatra et al.,
2003; Dang et al., 2009).

15
3.4.1. Amilasas
El almidón es la mayor reserva de carbohidratos en plantas, consiste en un gran número
de unidades de glucosa unidas por enlaces glicosídicos. Todas las semillas de las plantas
y tubérculos contienen almidón, el cual está presente como amilosa y amilopectina. El
almidón puede ser hidrolizado en carbohidratos simples, por ácidos, enzimas
amilolíticas, o combinación de ambos. No obstante el interés se ha centrado en las
enzimas porque ellas poseen un alto nivel de selectividad, las condiciones de reacción
no son peligrosas, están fácilmente disponibles y son ambientalmente amigables,
(Utong et al., 2006). Las amilasas (E.C.3.2.1.1) son enzimas que hidrolizan moléculas
de almidón para obtener diversos productos, incluyendo dextrinas y pequeños polímeros
compuestos de unidades de glucosa (Chen et al., 2005). Las amilasas pueden derivarse
de plantas, animales, bacterias y hongos.
Por su corto periodo de crecimiento, la diversidad bioquímica y la facilidad con la cual
la concentración de las enzimas podría ser incrementada por el ambiente y la
manipulación genética, las enzimas microbianas generalmente son demandadas
industrialmente (De Oliveira et al., 2007). Muchas de las enzimas usadas al presente
son obtenidas de microorganismos mesófilos, estas enzimas son inestables a pH muy
alcalinos o muy ácidos, fuerza iónica elevada y temperaturas extremas (Utong et al.,
2006) a diferencia de las enzimas derivadas de bacterias halófilas y termófilas que
pueden ser usadas en muchos procesos industriales a altas concentraciones de sal y la
temperatura elevada (Saxena et al., 2007).
Las amilasas son producidas a gran escala para su uso en la industria de la alimentación
y de la producción de combustibles. Entre los usos más importantes, se pueden citar: la
hidrólisis parcial del almidón para generar dextrinas que son utilizadas como
espesantes; la obtención de jarabe de alta maltosa y alta glucosa utilizados en la
fabricación de la cerveza, panes, bebidas no alcohólicas, repostería, dulces y obtención
de fuentes de carbono alternativas para ser utilizadas en la producción de etanol con
diversos propósitos. Estas enzimas también están involucradas en la remoción final del
almidón utilizado en los procesos de fabricación de prendas (Olempska-Beer et al.,
2006).

16
3.4.2. Celulasas
Las celulasas son enzimas hidrolíticas que rompen enlaces glicosídicos ß-1,4 de los
polisacáridos de celulosa. Las enzimas celulasas pueden clasificarse de la siguiente
manera: las endoglucanasas o 1,4- ß-D-glucan-glucanhidrolasas (EC 3.2.1.4) que
rompen los enlaces internos de la molécula. Las exoglucanasas o 1,4-ß-D-
glucanhidrolasas también conocidas como celodextrinasas (EC 3.2.1.74) y 1,4- ß-D-
glucan celobiohidrolasas (EC 3.2.1.91) que remueven unidades de glucosa o celobiosa a
partir del extremo libre no reductor de la cadena de celulosa. Finalmente las ß-
glucosidasas o ß-glucosidasas glucohidrolasas (EC 3.2.1.21) hidrolizan la celobiosa
producida por las actividades anteriores, dando como producto final la glucosa (Teeri,
1997). Las celulasas juegan un papel muy importante en la industria textil, son
utilizadas para dar al azul índigo la apariencia de desteñido, lo que suplió el proceso de
tratamiento con piedra pómez, pues unos miligramos de enzima sustituyen kilos de
piedra necesaria. Igualmente son utilizadas en detergentes quita pelusas, la enzima
degrada las fibras que se separan restituyendo la superficie suave de la prenda. En la
industria alimenticia, las celulasas son utilizadas en la extracción y filtración de jugos de
frutas o verduras, mostos y aceites comestibles y en la en la hidrólisis parcial de
materiales lignocelulósicos mejorando la digestión de rumiantes. Igualmente se han
aprovechado los dominios de unión a celulosa sobre una matriz de celulosa para realizar
procesos de purificación de proteínas recombinantes (Demain et al., 2005).
Ambos hongos y bacterias han sido rigurosamente explotados por sus habilidades para
producir una amplia variedad de celulasas y hemicelulasas. Sin embargo, los hongos
han sido preferidos por producir grandes cantidades de celulasas y hemicelulasas las
cuales son secretadas al medio para una fácil extracción y purificación. En adición, las
enzimas fúngicas son a menudo menos complejas que las hidrolasas glicosidasas de las
bacterias y pueden por lo tanto ser rápidamente clonadas y producidas en forma
recombinante en bacterias de rápido crecimiento como la Escherichia coli. Pese a las
características favorables de las celulasas derivadas de hongos, el aislamiento y
caracterización de nuevas hidrolasas glicosidasas desde Eubacterias como el Bacillus
circulans y Estreptomyces sp. está adquiriendo importancia (Bao et al., 2011). Algunas
razones para este cambio, se debe primero a que las bacterias a menudo tienen una
mayor rata de crecimiento que los hongos, segundo las hidrolasas glicosidasas

17
(celulasas, hemicelulasas y pectinasas) bacterianas son a menudo expresadas
simultáneamente formando combinaciones de complejos enzimáticos que causan un
incremento en la función y la sinergia. Lo más importante, es que las bacterias habitan
en una amplia variedad de ambientes y nichos industriales siendo extremadamente
resistentes al estrés ambiental. Eso incluye cepas que son termófilas o psicrófilas,
alcalófilas o acidófilas, y cepas que son halófilas. Algunas de estas bacterias no pueden
sobrevivir a las condiciones severas encontradas en los procesos de bioconversión, pero
ellas a menudo producen enzimas que son estables bajo condiciones extremas las cuales
pueden estar presentes en éstos procesos incrementando la rata de hidrólisis enzimática,
fermentación y recuperación del producto. Ahora bien, la detección de actividad de
enzima celulasa en cepas microbianas, se realiza típicamente en carboximetilcelulosa
(CMC), en placas (Hankin et al., 1977). Recientemente, Kasana y sus colegas (2008)
encontraron que el yodo de Gram puede ser utilizado para inundar las placas de agar
CMC en lugar de bromuro de amonio o rojo Congo, dando un resultado más rápido y
altamente discernible. Sin embargo, los métodos de detección de placa utilizando
colorantes no son cuantitativos, o lo suficientemente sensibles como consecuencia de la
escasa correlación entre la actividad enzimática y el tamaño del halo. Estos métodos se
limitan principalmente a las bacterias cultivables que producen celulasas y por tanto el
potencial total de las celulasas (microorganismos cultivables y no cultivable), no está
siendo examinado a fondo. Los investigadores han centrado en la identificación y
explotación de los genes de las celulasas de microorganismos no cultivables presentes
en ambientes extremos con la esperanza de que las enzimas aisladas sean nuevas y
tengan aplicaciones específicas en la industria de la biorefinación debido a la mayor
resistencia a condiciones ambientales adversas.
Adicionalmente, las bacterias celulolíticas que son capaces de producir complejas
estructuras proteicas que soportan enzimas para la hidrólisis de la celulosa, como el
celulosoma, el xilosoma y enzimas bifuncionales o multifuncionales, actualmente han
ganado mucha atención.
Si estas enzimas pueden ser producidas de manera recombinante, ellas podrían tener un
gran potencial en la mejora de los costos de hidrólisis para la producción de
biocombustibles, reduciendo las necesidades para la producción de múltiples enzimas
para la hidrólisis eficiente. Por ejemplo, una endoglucanasa/ endoxilanasa bifuncional

18
fue aislada de Flavigena cellulomonas útil para el mejoramiento de diferentes procesos
industriales como la producción de biocombustibles (Pérez-Avalos et al., 2008). Del
mismo modo una enzima multifuncional resultó ser producida por Terendinibacter
turnerae, una bacteria simbionte aislada de los bivalvos marinos Pedicellatus lydrodus
perteneciente al filo molusca al igual que el caracol pala (Ekborg et al., 2007).
3.4.3. Proteasas
Las enzimas proteolíticas son enzimas degradadoras las cuales catalizan el clivaje de
uniones peptídicas en proteínas. En la actualidad, las proteasas están clasificadas sobre
la base de tres criterios mayores, tipo de reacción catalizada, naturaleza química del
sitio catalítico y relación evolucionaria con referencia a su estructura (Barett, 1994).
Las proteasas están subdivididas en forma ordinaria en dos grupos mayores: las
exopectidasas y las endopeptidasas, dependiendo de su sitio de acción. Las
exopeptidasas clivan las uniones peptídicas próximas al carboxilo o amino terminal del
sustrato, mientras que las endopeptidasas clivan uniones peptídicas distantes del amino
y carboxilo terminal del sustrato.
Las proteasas alcalinas son enzimas proteolíticas las cuales trabajan a un pH alcalino, la
gran diversidad de proteasas, en contraste a la especificidad de su acción, ha atraído la
atención mundial intentando explorar sus aplicaciones biotecnológicas y fisiológicas
(Fox et al., 1991; Gupta et al., 2002; Poldermans, 1990).
Por ejemplo en la industria de comidas y alimentos, síntesis de péptidos, industria del
cuero, manejo de residuos industriales y de los hogares, industria de la fotografía, usos
médicos, e industria de los detergentes.
Las proteasas representan uno de los tres mayores grupos de enzimas industriales y
constituyen el 60% de total de las enzimas vendidas en el mundo. Las proteasas son
fisiológicamente necesarias para los organismos, ellas son ubicuas, y se encuentra en
una amplia diversidad de recursos tales como plantas, animales y microorganismos. La
incapacidad de las proteasas de las plantas y animales para reunir la demanda actual en
el mundo ha conducido al creciente interés en proteasas microbianas. Los
microorganismos representan excelente recurso de estas enzimas por su amplia
diversidad bioquímica y su susceptibilidad a la manipulación genética. Muchas
proteasas comerciales, principalmente neutras y alcalinas son producidas por

19
organismos pertenecientes al género Bacillus como el Bacillus aquimaris. Las bacterias
productoras de proteasas pueden ser seleccionadas inicialmente en el laboratorio usando
medios de cultivo suplementados con caseína. La presencia de una zona clara alrededor
de las colonias confirmara la hidrolisis de la caseína y por ende la producción de
proteasas (Rohban et al., 2009; Zobell et al., 1941).
Las proteasas bacterianas neutras son activas a pH entre 5 y 8 y tienen una
termotolerancia relativamente baja. Debido a su rata de reacción intermedia, proteasas
neutras generan menos amargo en las proteínas hidrolizadas en alimentos que las
proteasas animales y por tanto son valiosas en la industria de los alimentos. Las
proteasas neutras se caracterizan por su alta afinidad por aminoácidos hidrofóbicos, su
baja termo tolerancia es ventajosa para el control de su reactividad durante la
producción de hidrolizados con bajo grado de hidrólisis. Algunas de las proteasas
neutras pertenecientes a las metaloproteasas requieren de iones divalentes para su
activación (Rao et al., 1998).
3.4.4. Lipasas
Las lipasas (EC 3.1.1.3) son enzimas ubicuas encontradas en animales, plantas y
microorganismos incluyendo hongos y bacterias. Las lipasas microbianas tienen un
considerable potencial industrial (Harwood, 1989; Jaeger et al., 1999).
Las reacciones lipolíticas ocurren en la interface lípido agua, donde los sustratos
lipolíticos usualmente forman un equilibrio entre los estados monomérico, micelar y
emulsificado. Hasta hace poco, dos criterios se habían utilizado para clasificar una
enzima lipolítica como una verdadera lipasa: primero ésta debía ser activada por la
presencia de una interface, es decir, su actividad debe aumentar considerablemente tan
pronto como los triglicéridos formen una emulsión. Este fenómeno se denominó
activación interfacial (Sarda et al., 1958). Segundo debía contener una tapa, que es una
superficie del bucle de la proteína que cubre el sitio activo de la enzima y evita el
contacto con la interface (Derewenda et al., 1992). Sin embargo, estos criterios no son
adecuados para la clasificación, debido principalmente a una serie de excepciones donde
se han descrito enzimas que tienen una tapa pero que no exhiben activación interfacial.
Por lo tanto las lipasas son definidas simplemente como carboxilesterasas que catalizan
la hidrólisis de largas cadenas de acilgliceroles. No existe una definición estricta

20
disponible para el término "cadena larga", pero los esteres de glicerol con una longitud
de la cadena de acilo mayor a 10 átomos de carbono puede considerarse como sustratos
para las lipasas, con trioleoilglicerol como sustrato estándar. La hidrólisis de esteres de
glicerol con una longitud de la cadena acilo menor de 10 átomos de carbono con
tributirina como el sustrato estándar usualmente indican la presencia de esterasas.
Los microbiólogos generalmente, desean utilizar un ensayo simple y fiable en placa que
permita la identificación de las bacterias productoras de lipasas. Los sustratos más
utilizados son tributirina y trioleína, que son emulsificados mecánicamente en varios
medios de cultivo vertidos en placas de petri. La producción de lipasas es indicada por
la formación de halos alrededor de las colonias crecidas en esas placas de agar (Atlas,
1996).
Algunos géneros bacterianos importantes en la producción de lipasas incluyen Bacillus,
Pseudomonas y Burkholderia. Las lipasas bacterianas son por lo general extracelulares,
comúnmente purificadas por cromatografía a través de interacciones hidrofóbicas.
Muchas lipasas pueden actuar en un rango amplio de pH y temperatura (Gupta et al.,
2004). Estas enzimas tienen aplicaciones en diversos campos, como agentes de
descontaminación en efluentes provenientes de plantas productoras de aceites,
mataderos y aguas residuales domesticas que arrojan diariamente un gran número de
grasas al medio ambiente de difícil degradación dada su baja solubilidad en el agua
(Matsumiya et al., 2007).
Actualmente existe un creciente interés por los microorganismos que pueden ser
utilizados en la solución de problemas ambientales, de hecho aquellos con gran
capacidad de producir lipasas, tienen un papel fundamental en la obtención de
elementos menos contaminantes y accesibles al metabolismo microbiano (Akoh et al.,
2006; Kaieda et al., 2001).
El uso comercial de las lipasas representa un billón de dólares y comprende una amplia
variedad de diferentes aplicaciones. En el área de los detergentes, cerca de 1000
toneladas de lipasas son vendidas cada año (Godfrey et al., 1996). Las lipasas juegan un
papel importante en la producción de ingredientes para alimentos (Jaeger, 1998). Otras
aplicaciones de interés creciente incluyen el uso de lipasas en la remoción de resinas de

21
la pulpa en la industria papelera (Farrell et al., 1997) y en la síntesis de compuestos
orgánicos (Boland et al., 1991; Drauz et al., 1995).
3.5. EL PRESENTE ESTUDIO
Esta investigación ha sido enmarcada dentro del proyecto DIME bicentenario
20101007734 de la Universidad Nacional de Colombia, sede Medellín titulado
Búsqueda y evaluación de la actividad antibiótica y enzimática de bacterias marinas
asociadas al caracol pala (Strombus gigas). Estudios iniciales sobre la caracterización de
la microbiota en caracoles pala mediante, el uso de secuencias espaciadoras intergénicas
(ITS) y el análisis de las secuencias del gen rRNA 16S de algunos aislados, permitieron
dar el primer paso hacia el esclarecimiento de su microbiodiversidad (Acosta et al.,
2009). Por lo tanto, la caracterización filogenética de las poblaciones microbianas
presentes en él, y la presencia de compuestos bioactivos de importancia biotecnológica
y ecológica producidos por las bacterias pertenecientes a la microbiota del caracol,
probablemente ayuden a mejorar la percepción sobre el caracol y los microbios
asociados. Este proyecto se ha enfocado en el estudio mediante técnicas dependientes e
independientes de cultivo de un recurso natural poco explorado como lo es la diversidad
bacteriana asociada a caracoles pala silvestres y en cautiverio. Se ha obtenido una
colección de bacterias caracterizadas fenotípica y genotípicamente, haciendo énfasis en
aquellas que presentaron características biológicas que permitirían a largo plazo su
utilización en procesos industriales y describir hipótesis sobre la función de estos
microbios en la ecología del caracol.

22
4. OBJETIVOS
4.1. OBJETIVO GENERAL
Evaluar la diversidad microbiana y las propiedades antibacterianas y enzimáticas de
bacterias asociadas a los caracoles pala “Strombus gigas” del Caribe Colombiano.
4.2. OBJETIVOS ESPECÍFICOS
Caracterizar la diversidad bacteriana de la microbiota asociada al caracol pala
por técnicas moleculares (PCR-DGGE)
Caracterizar molecularmente las bacterias con actividades antibacterianas y
enzimáticas.
Determinar la actividad antimicrobiana de aislados de la microbiota del caracol
frente a bacterias Gram positivas y Gram negativas.
Determinar la actividad enzimática de las cepas bacterianas asociadas al caracol.

23
5. MATERIALES Y MÉTODOS
5.1. CULTIVO, AISLAMIENTO Y CARACTERIZACIÓN FENOTÍPICA DE
LOS AISLADOS BACTERIANOS
Para realizar el presente proyecto se trabajó con muestras del cepario (aislados
bacterianos) y la genoteca (DNAs) de un estudio previo realizado por Acosta (2009).
Las muestras fueron colectadas en julio y diciembre del 2007. Las muestras de baba o
mucus (b), de intestino (i) y de alimento (Algas pertenecientes a los géneros Isochrysis
y Chaetoceros) (a) de caracoles silvestres (S) y en cautiverio (C) habían sido
preservadas a -80°C durante un año y posteriormente procesadas en el Laboratorio de
Biología Celular y Molecular de la Universidad Nacional de Colombia, Sede Medellín.
Las muestras de baba (b), intestino (i) y de alimento (a) de caracoles silvestres (S) y en
cautiverio (C) fueron sembradas en medios de cultivo agar marino y agar TCBS (Agar
tiosulfato citrato bilis sacarosa), después del crecimiento, se tomaron el total de las UFC
que crecieron en estos medios, se colectaron y se denominaron como barridos o fracción
cultivable (B) (Tabla 1).
Veinte cepas fueron seleccionadas directamente de la colección de bacterias aisladas.
Adicionalmente de los 23 barridos almacenados a -20°C en el laboratorio en el trabajo
de Acosta (2009), se aislaron 80 cepas. Las muestras para análisis dependientes de
cultivo, es decir, los barridos fueron procesados de acuerdo al protocolo de resucitación
de bacterias en posible estado viable pero no cultivable (VNC), descrito por Whitesides
y Oliver (1997) y Du et al., (2007). Después de la incubación de estos preinóculos por
48 horas a temperatura ambiente 100 µl de éstos fueron cultivados sobre agar TCBS y
agar marino. La incubación fue realizada a temperatura ambiente hasta la visualización
de crecimiento bacteriano. Las cepas fueron seleccionadas aleatoriamente y cultivadas
dos veces más para asegurar la pureza de los aislados. Los aislados fueron
caracterizados macro y microscópicamente, y almacenados a -20 °C hasta ulteriores
análisis (Anexo 1). Todos los medios de cultivo, tanto líquidos como sólidos, fueron

24
preparados utilizando agua de mar previamente filtrada (papel filtro de 0,45 µm) y
esterilizada por autoclave.
5.2. CARACTERIZACIÓN DE LA DIVERSIDAD BACTERIANA
PERTENECIENTE A LA MICROBIOTA ASOCIADA AL CARACOL PALA
POR TÉCNICAS MOLECULARES (PCR-DGGE)
Para estudiar la diversidad de las comunidades bacterianas dominantes, se usaron los
DNAs de los materiales biológicos correspondientes a la baba (b), el intestino (i) y el
alimento (Algas pertenecientes a los géneros Isochrysis y Chaetoceros) (a) de caracoles
silvestres (S) y en cautiverio (C) que fueron procesados en el Laboratorio de Biología
Celular y Molecular de la Universidad Nacional de Colombia, Sede Medellín. El DNA
fue obtenido según el método de Romero et al., 2002 y en nuestro estudio fue
denominado como DNAs originales (DNA O) para determinar las poblaciones
bacterianas presentes en la muestra inicial. Los barridos bacterianos o fracción
cultivable (B), es decir, el total de las UFC obtenidas en los medios de cultivo, también
fueron sometidos a extracción de DNA y denominados como DNA barridos (DNA B)
(Tabla 1).
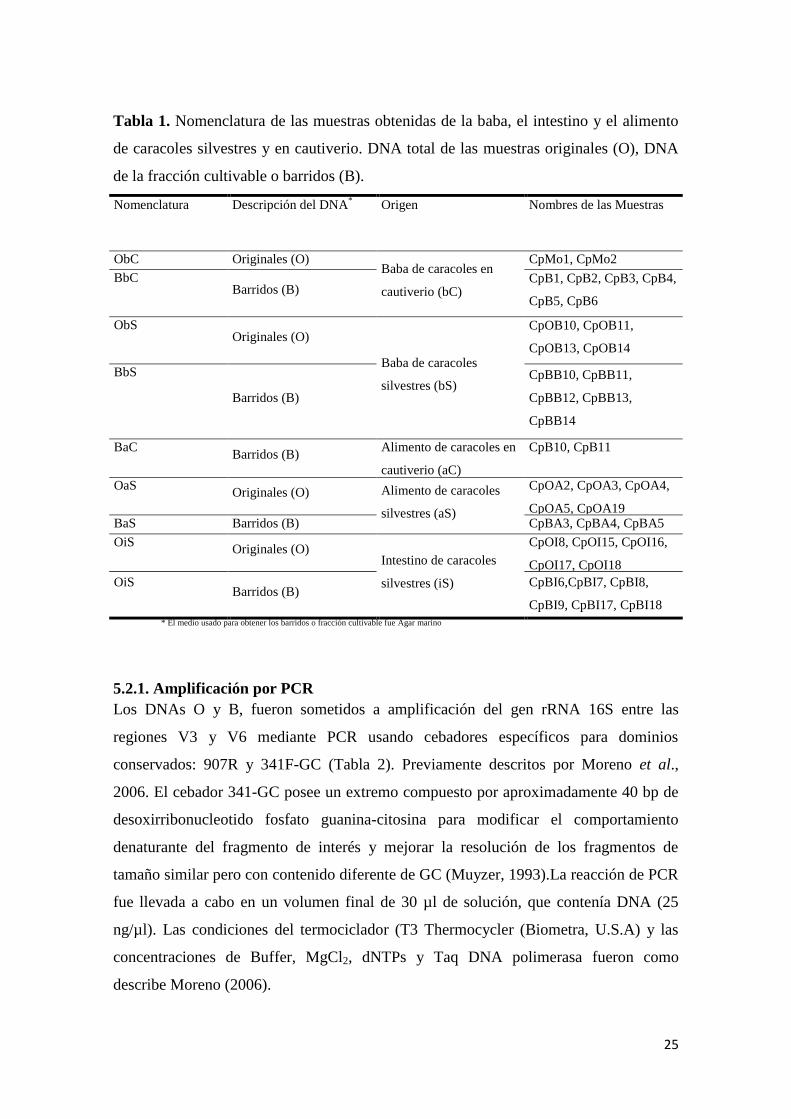
25
Tabla 1. Nomenclatura de las muestras obtenidas de la baba, el intestino y el alimento
de caracoles silvestres y en cautiverio. DNA total de las muestras originales (O), DNA
de la fracción cultivable o barridos (B).
Nomenclatura Descripción del DNA* Origen Nombres de las Muestras
ObC Originales (O) Baba de caracoles en
cautiverio (bC)
CpMo1, CpMo2
BbC Barridos (B)
CpB1, CpB2, CpB3, CpB4,
CpB5, CpB6
ObS Originales (O)
Baba de caracoles
silvestres (bS)
CpOB10, CpOB11,
CpOB13, CpOB14
BbS
Barridos (B)
CpBB10, CpBB11,
CpBB12, CpBB13,
CpBB14
BaC Barridos (B)
Alimento de caracoles en
cautiverio (aC)
CpB10, CpB11
OaS Originales (O) Alimento de caracoles
silvestres (aS)
CpOA2, CpOA3, CpOA4,
CpOA5, CpOA19 BaS Barridos (B) CpBA3, CpBA4, CpBA5
OiS Originales (O)
Intestino de caracoles
silvestres (iS)
CpOI8, CpOI15, CpOI16,
CpOI17, CpOI18
OiS Barridos (B)
CpBI6,CpBI7, CpBI8,
CpBI9, CpBI17, CpBI18
* El medio usado para obtener los barridos o fracción cultivable fue Agar marino
5.2.1. Amplificación por PCR
Los DNAs O y B, fueron sometidos a amplificación del gen rRNA 16S entre las
regiones V3 y V6 mediante PCR usando cebadores específicos para dominios
conservados: 907R y 341F-GC (Tabla 2). Previamente descritos por Moreno et al.,
2006. El cebador 341-GC posee un extremo compuesto por aproximadamente 40 bp de
desoxirribonucleotido fosfato guanina-citosina para modificar el comportamiento
denaturante del fragmento de interés y mejorar la resolución de los fragmentos de
tamaño similar pero con contenido diferente de GC (Muyzer, 1993).La reacción de PCR
fue llevada a cabo en un volumen final de 30 µl de solución, que contenía DNA (25
ng/µl). Las condiciones del termociclador (T3 Thermocycler (Biometra, U.S.A) y las
concentraciones de Buffer, MgCl2, dNTPs y Taq DNA polimerasa fueron como
describe Moreno (2006).
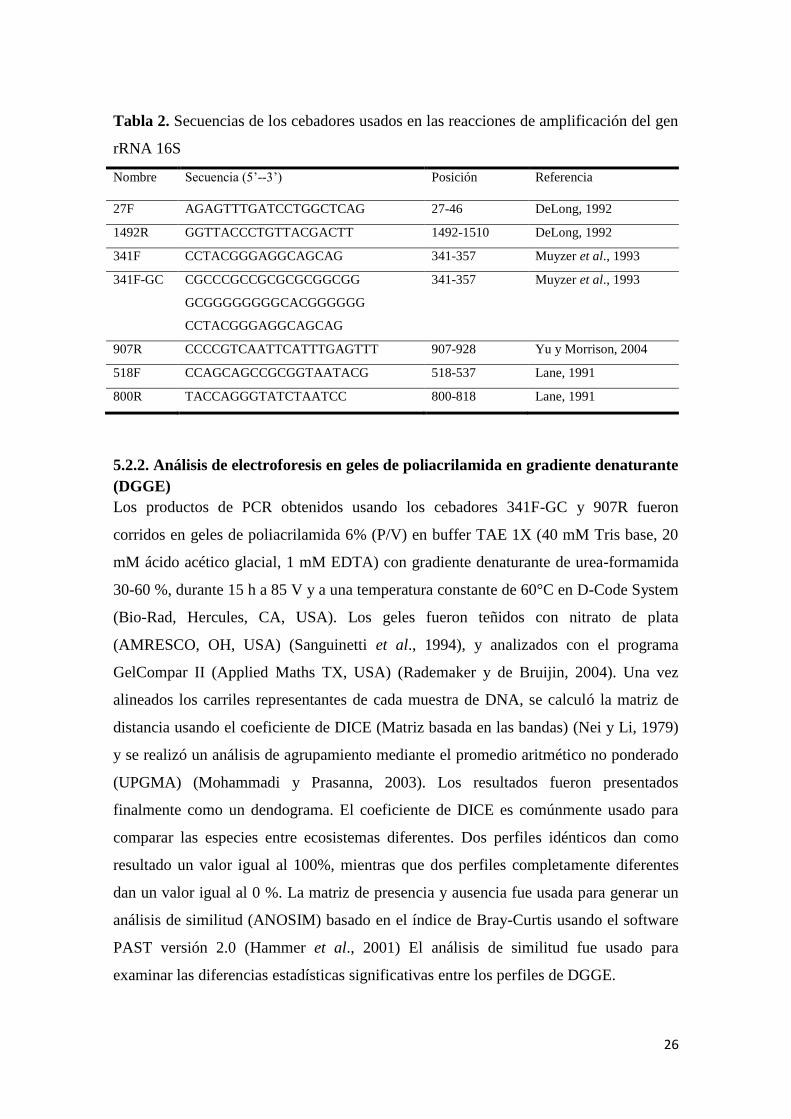
26
Tabla 2. Secuencias de los cebadores usados en las reacciones de amplificación del gen
rRNA 16S
Nombre Secuencia (5’--3’) Posición Referencia
27F AGAGTTTGATCCTGGCTCAG 27-46 DeLong, 1992
1492R GGTTACCCTGTTACGACTT 1492-1510 DeLong, 1992
341F CCTACGGGAGGCAGCAG 341-357 Muyzer et al., 1993
341F-GC CGCCCGCCGCGCGCGGCGG
GCGGGGGGGGCACGGGGGG
CCTACGGGAGGCAGCAG
341-357 Muyzer et al., 1993
907R CCCCGTCAATTCATTTGAGTTT 907-928 Yu y Morrison, 2004
518F CCAGCAGCCGCGGTAATACG 518-537 Lane, 1991
800R TACCAGGGTATCTAATCC 800-818 Lane, 1991
5.2.2. Análisis de electroforesis en geles de poliacrilamida en gradiente denaturante
(DGGE)
Los productos de PCR obtenidos usando los cebadores 341F-GC y 907R fueron
corridos en geles de poliacrilamida 6% (P/V) en buffer TAE 1X (40 mM Tris base, 20
mM ácido acético glacial, 1 mM EDTA) con gradiente denaturante de urea-formamida
30-60 %, durante 15 h a 85 V y a una temperatura constante de 60°C en D-Code System
(Bio-Rad, Hercules, CA, USA). Los geles fueron teñidos con nitrato de plata
(AMRESCO, OH, USA) (Sanguinetti et al., 1994), y analizados con el programa
GelCompar II (Applied Maths TX, USA) (Rademaker y de Bruijin, 2004). Una vez
alineados los carriles representantes de cada muestra de DNA, se calculó la matriz de
distancia usando el coeficiente de DICE (Matriz basada en las bandas) (Nei y Li, 1979)
y se realizó un análisis de agrupamiento mediante el promedio aritmético no ponderado
(UPGMA) (Mohammadi y Prasanna, 2003). Los resultados fueron presentados
finalmente como un dendograma. El coeficiente de DICE es comúnmente usado para
comparar las especies entre ecosistemas diferentes. Dos perfiles idénticos dan como
resultado un valor igual al 100%, mientras que dos perfiles completamente diferentes
dan un valor igual al 0 %. La matriz de presencia y ausencia fue usada para generar un
análisis de similitud (ANOSIM) basado en el índice de Bray-Curtis usando el software
PAST versión 2.0 (Hammer et al., 2001) El análisis de similitud fue usado para
examinar las diferencias estadísticas significativas entre los perfiles de DGGE.

27
5.2.3. Análisis de secuencias
Las bandas de DNA con patrones únicos de migración con respecto al marcador
establecido y las bandas de mayor intensidad en cada gel de DGGE fueron escindidas
por duplicado en lo posible y colocadas en un vial estéril. El DNA de cada una de las
bandas fue recuperado por el método de elución de “Crush and Soak” (Sambrook y
Russel, 2001) sustituyendo 2 volúmenes de etanol a 4°C por 0,6 volúmenes de
isopropanol a 4°C durante la precipitación del DNA, obteniendo 10 µl del DNA eluido
de los cuales 6 µl fueron usados para llevar a cabo la re-amplificación usando los
cebadores 341F y 907R. Con el propósito de garantizar la pureza de las bandas después
de la amplificación por PCR usando los primers 341F y 907R, el DNA fue observado en
geles de poliacrilamida al 8% por tinción con nitrato de plata, usando controles
positivos y negativos (Anexo 2). Los productos fueron purificados usando UltraClean
PCR Clean-up (MoBio Laboratories, CA, USA). Los amplicones fueron enviados a
Macrogen (Korea) para ser secuenciados en un ABI PrimR 3100 Genetic Analyzer
(Applied Biosystems, CA, USA). Las secuencias obtenidas fueron editadas usando
BioEdit ® (Hall, 1999) y la presencia de secuencias quimeras se evaluó mediante el
software CHIMERA CHECK (http://www.bioinformatics-toolkit.org). Las secuencias
editadas fueron comparadas con secuencias conocidas en el GenBank usando BLAST
(Basic Local Alignment Search Tool) (Altschul et al., 1997) y el software Sequence
Match del sitio web Ribosomal Database Project (RDP). Las secuencias editadas fueron
alineadas usando Clustal W (Thompson et al., 1994). El software MEGA 5
(http://megasoftware.net/) se utilizó para construir un árbol filogenético por el método
de neighbor-joining (Saitou y .Nei, 1987) con un bootstrap de 1000 repeticiones. Las
distancias evolutivas fueron calculadas utilizando el método de Jukes-Cantor (Jukes y
Cantor, 1969).

28
5.3. CARACTERIZACIÓN MOLECULAR DE LAS BACTERIAS USADAS
PARA DETERMINAR LAS ACTIVIDADES ANTIBACTERIANAS Y
ENZIMÁTICAS
El DNA de las cepas aisladas fue obtenido desde cultivos líquidos por el método de
fenol-cloroformo y una precipitación con etanol, método descrito previamente
(Sambrook y Russel, 2001) Después de la extracción de DNA, éste fue analizado por
medio de un gel de agarosa al 1% (P/V), teñido con EZ-vision (Amresco, OH, USA),
según las recomendaciones del fabricante. Las bandas de DNA fueron visualizadas por
iluminación con LUV y fotografiadas por Sistem UV Transiluminator (Biometra). El
DNA extraído fue almacenado a -20°C.
Para identificar los aislados, se realizó una amplificación del gen rRNA 16S desde la
posición 28 a 1492 de acuerdo con Espejo et al., (1998) usando los cebadores
previamente descritos (DeLong, 1992) (Tabla 2). Los productos de la PCR fueron
analizados por electroforesis sobre un gel de agarosa al 1 % (P/V).
Los amplicones de 1465 bp aproximadamente fueron secuenciados por Macrogen
(Korea) en un ABI PrimR 3100 Genetic Analyzer (Applied Biosystems, CA, USA). Los
análisis posteriores se llevaron de acuerdo a los métodos y procedimientos descritos
anteriormente para las secuencias de las bandas obtenidas desde los geles de DGGE.
Las secuencias fueron depositadas en la base de datos del GenBank con los números de
acceso JN602212-JN602254.
5.4. DETERMINACIÓN DE LAS ACTIVIDADES ANTIBACTERIANAS DE
AISLADOS BACTERIANOS
5.4.1. Extracción de metabolitos secundarios
Se llevó a cabo siguiendo el método previamente descrito por Romero et al., 2006. Un
Erlenmeyer conteniendo 50 ml de agua de mar con caldo LB (Luria Bertani) al 2%
(P/V), resina amberlita XAD-16 al 2% (P/V) (Spyere, 2003), se inoculo con 0,5 ml de
un cultivo de cada cepa seleccionada crecido durante toda la noche. Después de siete
días de incubación a 30 ºC y 180 rpm, la resina fue separada del medio de cultivo por

29
decantación y lavada con agua destilada, los productos absorbidos fueron
posteriormente eluidos con 40 ml de metanol al 100% durante 30 minutos (Sangnoi et
al., 2009). Después de los cuales cada extracto fue concentrado a 1,5 ml en un
rotaevaporador a 40 °C (Heidolph Laborota 4001 Efficient Rotary Evaporator).
5.4.2. Evaluación de actividad antibacteriana
Se utilizó la prueba de difusión en agar (El-Masry et al, 2000) contra cepas bacterianas
Gram positivas (Bacillus subtilis) y Gram negativas (Escherichia coli y Pseudomona
aeruginosa). Discos de papel filtro Whatman N° 1 de 6- mm de diámetro estériles
fueron impregnados con 10, 20 y 50 µl de cada extracto. Los discos fueron puestos
sobre la superficie de cajas de Petri con agar Mueller-Hinton (Becton Dickison)
inoculado uniformemente con un cultivo líquido de las cepas blanco en una
concentración aproximada de 1.2 x 108 UFC/ml (Absorbancia 600 nm = 0,1). Las cajas
fueron incubadas a 37 °C por 18 horas, después de las cuales se midió el diámetro del
halo de inhibición del crecimiento alrededor de cada uno de los discos. El antibiótico
ampicilina (10 µg por disco) fue seleccionado como control positivo.
5.5. DETERMINACIÓN DE LOS MICROORGANISMOS CAPACES DE
PRODUCIR HIDROLASAS EXTRACELULARES Y EVALUACIÓN DE LAS
ACTIVIDADES ENZIMÁTICAS
5.5.1. Determinación de microorganismos productores de algunas hidrolasas
extracelulares
5.5.1.1. Determinación de microorganismos productores de amilasas
La presencia de actividad amilasa fue determinada sobre un medio de cultivo
constituido por agar 12 g/l, caldo nutritivo 3 g/l y almidón soluble 10 g/l preparado en
agua de mar, con un pH inicial de 7,2. Después de la incubación a temperatura ambiente
por 48 horas, las placas fueron reveladas con una solución de lugol (I2 -0,3% KI-0,6%).
Una zona clara alrededor del crecimiento indicó la presencia de cepas productoras de
amilasas (Rohban, et al., 2009).

30
5.5.1.2. Determinación de microorganismos productores de celulasas (CMCasa)
La actividad fue evaluada sobre un medio de cultivo constituido por extracto de
levadura 2 g/l, agar 20g/l y carboximetil celulosa (CMC) 10 g/l, preparado en agua de
mar, con un pH inicial de 7,2. Después de una incubación por 48 horas, las placas
fueron reveladas con una solución de lugol (I2 -0,3% KI-0,6%). Una zona clara
alrededor del crecimiento evidencio la presencia de CMCasa (Kasana et al., 2008;
Rohban et al., 2009)
5.5.1.3. Determinación de microorganismos productores de proteasas
La producción de proteasas fue detectada en un medio de cultivo constituido por agar 15
g/l, caldo nutritivo 5 g/l, leche descremada 100 g/l y NaCl 20 g/l, con un pH inicial de
7,2 preparado en agua destilada. Una zona clara alrededor del sitio de crecimiento
después de siete días a temperatura ambiente fue tomada como evidencia de actividad
proteolítica (Rohban et al., 2009)
La actividad proteasa usando gelatina como sustrato fue determinada usando medio de
cultivo constituido por 120 g/l de gelatina, caldo nutritivo 8 g/l y NaCl 20 g/l, preparado
en agua destilada (Lanyi,. 1988). La licuefacción de la gelatina después de siete días de
incubación a temperatura ambiente fue tomada como evidencia de actividad gelatinasa.
5.5.1.4. Determinación de microorganismos productores de lipasas
Para observar la producción de lipasas las cepas fueron cultivadas sobre placas de agar
lipasa con una solución de reactivo lipasas. El agar lipasa estaba compuesto por triptona
10 g/l, extracto de levadura 5 g/l, agar 20 g/l, azul de anilina 0,15 g/l; el reactivo de
lipasa contenía 100 ml de aceite de oliva, 1 ml de polisorbato 80 y 400 ml de agua
destilada. 30 ml del reactivo de lipasas fueron añadidos a un litro de agar lipasas con un
pH inicial de 6,8-7,0.Una zona clara alrededor del crecimiento después de 72 horas de
incubación a temperatura ambiente evidenció la presencia de cepas productoras de
lipasas (Frank et al., 1993).
Para los ensayos de actividad amilolítica, celulolítica y proteolítica fue usada como
control positivo una cepa de Bacillus subtilis, mientras que para la determinación de la
actividad lipolítica como control positivo se usó una cepa Staphylococcus aureus.

31
5.5.2. Evaluación de la actividad de algunas hidrolasas extracelulares
5.5.2.1. Cuantificación de actividad amilasas
Un volumen de 1 ml de pre-inoculo fue preparado en medio de cultivo para la
producción de amilasa e incubado durante toda la noche a 30 °C. De allí 0,1 ml fue
inoculado en un tubo de ensayo de 20 ml de volumen que contenía 4 ml de medio para
la producción de amilasas y fue incubado a 30 °C, 150 rpm por 48 h. Las células y el
material insoluble fueron removidos por centrifugación a 5000 rpm por 20 min a 4°C.
El sobrenadante libre de células fue usado como fuente de la enzima amilasa. Los
experimentos fueron llevados a cabo por triplicado. El medio para la producción de
amilasa contenía almidón soluble 20g/l, extracto de carne 10 g/l, extracto de levadura 2
g/l, K2HPO4 2 g/l, CaCl2 0,2 g/l y MgSO4.7H2O, 0,1 g/l. El pH fue ajustado a 7 (Babu y
Satyanarayana, 1993). Las proteínas, aminoácidos y otros constituyentes fueron
autoclavados por separado (a 121 °C por 20 min) y mezclados después del enfriamiento.
La actividad amilasa fue determinada de acuerdo con Bernfeld (1955) usando almidón
soluble como sustrato. La mezcla de la reacción contenía 125 µl del extracto crudo
enzimático y125 µl de almidón soluble (10 g/l) disuelto en buffer de fosfato 0,1 M a pH
7. La mezcla fue incubada por 30 minutos a 30°C. La reacción fue detenida adicionando
250 µl de solución de 3,5-ácido dinitrosalicílico (DNS), seguido del calentamiento en
baño de agua hirviendo por 5 min y la posterior incubación en un baño con hielo, la
absorbancia de cada solución fue medida a 540 nm en un fotómetro para micro placas
Multiskan EX (Miller, 1959). Se hizo una curva estándar de glucosa con diferentes
concentraciones (0,01 - 0,05 - 0,1 - 0,2- 0,4- 0,6 – 0,8 – 1,0 – 1,2 – 1,6 -1,8 g/l) que se
trataron como se describe previamente con DNS.
Una unidad de actividad enzimática fue definida como la cantidad de enzima necesaria
para liberar 1 μmol de azúcares reductores por minuto bajo las condiciones del ensayo.
5.5.2.2. Cuantificación de actividad celulasas
El pre-inoculo correspondiente de las cepas a evaluar fue preparado en un medio sólido
para la producción de celulasas, una asada (10 µl) del cultivo de las placas de agar
carboximetil celulosa fue inoculado en un tubo que contenía 5 ml del medio para la
producción sin agar, e incubado a 30 °C, a 150 rpm por 48 h. Los volúmenes de los

32
cultivos usados en este estudio fueron menores a los descritos en un estudio previo
(Gomare et al., 2011). Las células y el material insoluble fueron removidos por
centrifugación a 5000 g por 20 min a 4°C. El sobrenadante libre de células fue usado
como fuente de la enzima celulasa. Los ensayos fueron llevados a cabo por triplicado.
El medio para la producción de celulasa contenía carboximetil celulosa (CMC) 10 g/l,
extracto de levadura 2 g/l, agar 20 g/l y NaCl 20 g/l. El pH fue ajustado a 7. Las
proteínas, aminoácidos y otros constituyentes fueron autoclavados por separado (a 121
°C por 20 min).y mezclados después del enfriamiento.
La mezcla de la reacción contenía 125 µl del extracto enzimático más 125 del sustrato
(CMC 0,5% P/V) solubilizado en buffer de fosfato 100 mM, pH 7,0 (Baily et al., 1992).
Esta mezcla fue incubada por 30 min a 30°C. y la reacción fue parada por la adición de
250 µl DNS, seguido del calentamiento en baño de agua hirviendo por 5 min y la
posterior incubación en un baño con hielo; la absorbancia de cada solución fue medida a
540 nm en un fotómetro para micro placas (Multiskan ® EX, ThermolabSystem) en
lugar de un espectrofotómetro como describe Miller (1959). La curva estándar fue
realizada como se describe previamente para amilasas.
Una unidad de actividad enzimática fue definida como la cantidad de enzima necesaria
para liberar 1 μmol de azúcares reductores por minuto bajo las condiciones del ensayo.
5.5.2.3. Cuantificación de actividad proteasas
La producción de proteasa por los aislados fue llevada a cabo en un medio constituido
por extracto de levadura 5 g/l, peptona 10 g/l, glucosa 0,5 g/l, K2HPO4 1 g/l, MgSO4 0,2
g/l y NaCl 20 g/l (Kuddus y Ramteke, 2011)
Las proteínas, aminoácidos y otros constituyentes fueron autoclavados por separado
(121 °C por 20 min) y mezclados después del enfriamiento.
Un volumen de 200 µl de un cultivo crecido durante 18 horas sobre caldo marino fue
inoculado en un tubo de ensayo de 20 ml con 5 ml de medio para la producción de
proteasas e incubadas a 30 °C, 150 rpm por 72 h. Las células y el material insoluble
fueron removidos por centrifugación a 10000 rpm por 15 min a 4°C. El sobrenadante
libre de células fue usado como recurso de la enzima proteasa. Los ensayos fueron
llevados a cabo por triplicado.

33
La actividad proteolítica fue medida por el método de Lowry (1951). La mezcla de la
reacción estaba constituida por 50 µl del extracto crudo enzimático, 250 µl de caseína
0,65 % (P/V) disuelta en buffer de fosfato (50 mM) a pH 7. La cual fue incubada en
baño María a 30 °C por 30 min.
La reacción enzimática fue terminada por la adición de 250 µl de ácido tricloro acético
(TCA) 0,6 N e incubada por 20 minutos a 37°C.
La mezcla de la reacción fue centrifugada a 10000 g por 10 min at 4 °C. Después 250 µl
del sobrenadante más 625 µl de Na2CO3 (500 mM) y 125 µl del reactivo Folin fueron
mezclados e incubados a 37 °C por 30 minutos.
Finalmente la absorbancia fue medida contra un blanco (muestra no incubada) a 650
nm. Una unidad de actividad proteasa está definida como la cantidad de enzima
necesaria para liberar el equivalente de µmol de tirosina por minuto bajo las condiciones
definidas del ensayo.
Una solución stock de tirosina (1,1 µM) se prepara usando 0,2 mg/ml de L-tirosina en
agua destilada estéril calentando suavemente hasta disolver. A partir de esta solución se
prepararon las siguientes diluciones 0,000 - 0,022 - 0,044 - 0,110 - 0,220 - 0,440 - 0,660
µM para crear la curva estándar.
5.5.2.4. Cuantificación de actividad lipasas
Esta técnica está basada en el protocolo publicado por Hasanuzzaman (2004). Cinco
mililitros de medio para la producción de lipasas fueron inoculados con 0,1 ml de un
caldo de cultivo crecido durante toda la noche a 30°C, e incubado a 30°C, 150 rpm por
48 h. Las células y el material insoluble fueron removidos por centrifugación a 6000 g
por 30 min a 4°C. El sobrenadante libre de células fue usado como recurso de enzimas
lipasas. Los ensayos fueron llevados a cabo por triplicado.
El medio para la producción de lipasa contenía peptona 10 g/l, extracto de levadura 10
g/l, aceite de oliva 10 ml y glucosa 5 g/l. El pH fue ajustado a 7. Las proteínas,
aminoácidos y otros constituyentes fueron autoclavados por separado (a 121 °C por 20
min) y mezclados después del enfriamiento.
La lipasa extracelular fue evaluada por un método espectrofotométrico usando p-
nitrofenil palmitato (p-NPP) (Sigma-Aldrich) como sustrato en el que se mide la
hidrólisis de p-NPP por las lipasas, para liberar p-nitrofenol. 250 µl de buffer de fosfato

34
50 mM (pH 7,0) más 200 µl del extracto crudo enzimático fueron mezclados e
incubados a 30 °C por 2 minutos. 50 µl de 2,5 mM de p-NPP (disuelto en 2-propanol)
fue adicionado sobre la mezcla e incubado a 30°C por 30 minutos. Después de la
incubación la mezcla fue enfriada en agua con hielo, y la reacción fue terminada con la
adición de 100 µl de 0,5 M de KOH. La actividad de lipasa fue determinada en función
del incremento de la absorbancia medida a 405 nm.
Una unidad de lipasa está definida como los µmoles de p-NP liberado por minuto. Una
unidad de actividad es expresada como la cantidad de enzima necesaria para liberar 1
µmol de p-NP por minuto.
Una curva estándar fue preparada usando las siguientes concentraciones de p-NP: 2 -
4 – 10 – 20 – 40 – 60 – 80 - 100 micro molar.
5.5.2.5. Determinación de proteínas en los extractos crudos enzimáticos
La concentración de proteínas fue determinada de acuerdo al método de Bradford
(1976). 450 µl del reactivo de Bradford (Amresco) fueron añadidos a 50 µl de la
muestra y la absorbancia fue medida después de 5 minutos a 620 nm. Diferentes
concentraciones (0,03125 - 0,0625 - 0,125 - 0,25 - 0,5 - 0,75 - 1,0, mg/ml) de BSA
fueron usados para crear la curva estándar.

35
6. RESULTADOS
6.1. ANÁLISIS DE LOS PERFILES DE DGGE
La evaluación de la diversidad de las poblaciones bacterianas asociadas a muestras de
caracoles silvestres y en cautiverio en Islas del Rosario en el Caribe Colombiano se
llevó a cabo mediante la técnica DGGE, la cual permitió separar los fragmentos del gen
rRNA 16S de un tamaño equivalente a 585 bp.(Fig. 1). Los perfiles de bandeo fueron
evaluados a través del programa Gelcompar II (Applied Maths TX, USA) observando
diferencias entre las comunidades asociadas a caracoles silvestres y en cautiverio. De
acuerdo con la matriz binaria de datos sobre la presencia (1) y ausencia (0) (Anexo 3) de
las bandas, un total de 55 bandas diferentes fueron identificadas en los geles, en donde
las muestras asociadas a caracoles silvestres (n=25) y a caracoles en cautiverio (n=10)
aportaron 271 y 76 bandas respectivamente. La matriz de presencia y ausencia de
bandas resultantes del DGGE también fue usada para un análisis de similitud
(ANOSIM) usando el índice de Bray Curtis entre las muestras asociadas a caracoles
silvestres y en cautiverio. Este análisis fue llevado a cabo usando el software PAST
versión 2.0 (Hammer et al., 2001). Diferencias significativas fueron encontradas entre
las muestras asociadas a caracoles silvestres y en Cautiverio r = 0,26 (Bray Curtis) y
(Jaccard) r = 0,25, ambos con p < 0,0021. Mediante el análisis de similitud (SIMPER)
se determino la diferencia entre las comunidades bacterianas asociadas a caracoles en
Cautiverio y Silvestres, este análisis reveló un 75,45 % de disimilitud (Anexo 4).

36
b i i b i a b a b i
Figura 1. Patrones de bandeo en geles de DGGE del producto amplificado del gen
rRNA 16S desde muestras originales (O) y la fracción cultivable o barridos (B) de
intestino (i), baba o mucus (b) y alimento (a) de caracoles silvestres (S) y en cautiverio
(C) (Tabla 1). Líneas 1 a 3, 8, 10-12, 21-27, 34: OS; Líneas 4-7, 13-15, 19, 28, 29-30:
BS; Líneas 16-18, 31-33: BC; Líneas 9, 20: marcadores de referencia de 100 bp.
Los patrones de bandas obtenidos por DGGE fueron agrupados en tres clústeres por
medio del método UPGMA para la construcción de árboles filogenéticos moleculares
(Fig. 2).
El clúster 1 corresponde a patrones de bandeo de poblaciones bacterianas asociadas a la
baba de caracoles en cautiverio, el clúster 2 se asocian con muestras de alimento y el
clúster 3 se asocia con muestras de intestino y de la baba de caracoles silvestres.
Los perfiles de bandeo de muestras del alimento se relaciona más con las muestras de la
baba de caracoles en cautiverio que con las muestras de baba e intestino de caracoles
silvestres de acuerdo al dendograma. Sin embargo, a pesar de que el cluster 1 donde se
encuentran los perfiles de bandeo correspondientes a la baba de caracol en cautiverio
está más relacionado con el cluster 2 el análisis de similitud Anosim muestra que hay
diferencias significativas entre estos dos grupos (r=0,52; p<0,0003).

37
Figura 2. Dendograma obtenido con el programa GelCompar II a partir de los patrones
de bandeo en geles de DGGE del producto amplificado del gen rRNA 16S desde el
DNA original (OS y OC) y el DNA de los barridos (BS y BC) de muestras de caracoles
en estado silvestre y en cautiverio. Grupo 1: DNAs originales y de barridos de bC
(CpMo1, CpMo2, CpB2-CpB6). Grupo 2: DNAs originales y de barridos de aS (OA y
BA) y DNAs de barridos aC (CpB11), Grupo 3: DNAs originales y de barridos de bS
(OB y BB) e iS (OI y BI). El dendograma fue elaborado usando el coeficiente de Dice
(Nei y Li, 1979) y el promedio aritmético no ponderado (UPGMA) (Mohammadi y
Prasanna, 2003). Los valores sobre los nodos representan el porcentaje de similitud.
Muestra

38
Las bandas con patrones únicos de migración con respecto al marcador establecido (100
bp) y de mayor intensidad fueron escindidas por duplicado cuando fue posible. El DNA
obtenido como resultado de la extracción desde 47 bandas se utilizó para ser re-
amplificado y enviado a secuenciar. De las secuencias obtenidas solo 15 pudieron ser
editadas exitosamente utilizando el programa BioEdit debido a la presencia de
cromatógramas con ruidos excesivos y difíciles de alinear con una inapropiada
correspondencia con las secuencias registradas en el GenBank (Anexo 5). Los
resultados se muestran en la tabla 3.
Una vez editadas las secuencias de nucleótidos, éstas fueron sometidas tanto al
programa de búsqueda BLASTn (Altschul et al., 1990) como al RDP (Ribosomal
Database Project) (Cole et al., 2005) (Anexo 6) para establecer sus afiliaciones
filogenéticas. Con el propósito de probar la identidad de las secuencias se llevo a cabo
la construcción de árboles filogenéticos a través del método de Neighbor- joining y el
estimador de distancia Jukes-Cantor usando para esto alineamientos múltiples de las
secuencias en estudio más secuencias de referencia relacionas (Fig. 3) (Anexo 7).
En ese orden de ideas los resultados del análisis de las secuencias obtenidas de los re-
amplificados de la secuencia parcial gen rRNA 16S, revelaron una alta similitud con
secuencias de gammaproteobacterias (20%) y betaproteobacterias (60%) (Tabla 3). En
este estudio la secuencia de la banda L1 obtenida del DNA de BbS en el gel de
poliacrilamida del DGGE (Fig. 1), ha sido identificada como Pseudoalteromona y se
encontró tanto en los perfiles de los DNAs originales (DGGE no cultivable) como en
los DNAs de barridos (DGGE cultivable) de muestras obtenidas de la baba y el intestino
de caracoles silvestres.
Las secuencias de las bandas escindidas denominadas M1, G1, R1, A21, I3, J3, X1, K2,
H2, L31, G3 y O3 en los geles de poliacrilamida (Fig. 1) se agrupan en las afiliaciones
filogenética que se muestra en la Tabla 3 (Fig. 3).
La banda M1 se encontró en el DNA original y el DNA de los barridos de las muestras
de baba y del intestino de caracoles silvestres. Las bandas identificadas como R1, A21,
I3 y J3 estaban presentes en los DNAs de los barridos de baba de caracoles silvestres.
Las bandas X1, Y1 y K2 se encontraron en el DNA original de las muestras del
intestino de caracoles silvestres. B11 estaba presente en DNAs de lo barridos de

39
intestino de caracoles silvestres y H2 se observó en los DNAs de los barridos del
intestino de caracoles silvestre y de la baba de caracoles en cautiverio y en los DNAs
originales del intestino de caracoles silvestres. La banda L31 estaba presente en los
DNAs de los barridos y G3 en los DNAs originales obtenidos desde el alimento de
caracoles silvestre. O3 se encontró asociada al DNA de los barridos de baba de
caracoles en cautiverio. Las secuencias de las bandas X1, K2 y H2 asociadas al intestino
de caracoles pertenecen al mismo género. Los resultados se resumen en la Tabla 3.
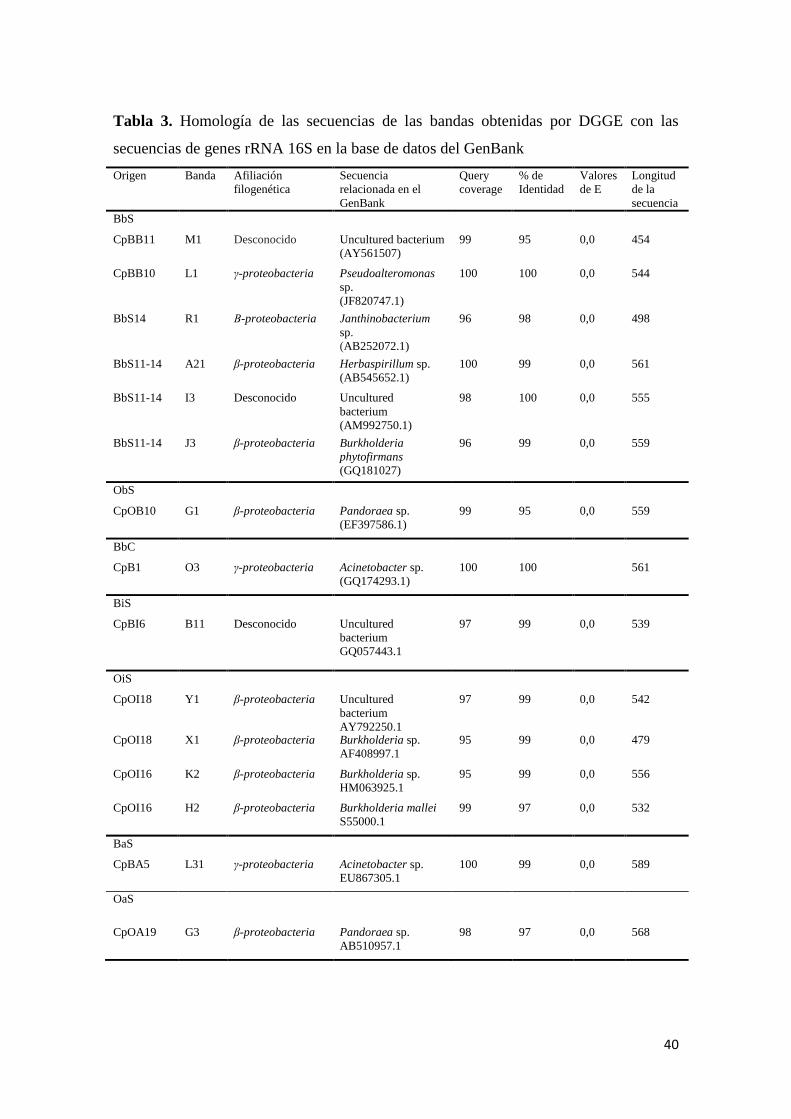
40
Tabla 3. Homología de las secuencias de las bandas obtenidas por DGGE con las
secuencias de genes rRNA 16S en la base de datos del GenBank
Origen Banda Afiliación
filogenética
Secuencia
relacionada en el
GenBank
Query
coverage
% de
Identidad
Valores
de E
Longitud
de la
secuencia
BbS
CpBB11 M1 Desconocido Uncultured bacterium
(AY561507)
99 95 0,0 454
CpBB10 L1 γ-proteobacteria Pseudoalteromonas
sp.
(JF820747.1)
100 100 0,0 544
BbS14 R1 Β-proteobacteria Janthinobacterium
sp.
(AB252072.1)
96 98 0,0 498
BbS11-14 A21 β-proteobacteria Herbaspirillum sp.
(AB545652.1)
100 99 0,0 561
BbS11-14 I3 Desconocido Uncultured
bacterium
(AM992750.1)
98 100 0,0 555
BbS11-14 J3 β-proteobacteria Burkholderia
phytofirmans
(GQ181027)
96 99 0,0 559
ObS
CpOB10 G1 β-proteobacteria Pandoraea sp.
(EF397586.1)
99 95 0,0 559
BbC
CpB1 O3 γ-proteobacteria Acinetobacter sp.
(GQ174293.1)
100 100 561
BiS
CpBI6 B11 Desconocido Uncultured
bacterium
GQ057443.1
97 99 0,0 539
OiS
CpOI18 Y1 β-proteobacteria Uncultured
bacterium
AY792250.1
97 99 0,0 542
CpOI18 X1 β-proteobacteria Burkholderia sp.
AF408997.1
95 99 0,0 479
CpOI16 K2 β-proteobacteria Burkholderia sp.
HM063925.1
95 99 0,0 556
CpOI16 H2 β-proteobacteria Burkholderia mallei
S55000.1
99 97 0,0 532
BaS
CpBA5 L31 γ-proteobacteria Acinetobacter sp.
EU867305.1
100 99 0,0 589
OaS
CpOA19 G3 β-proteobacteria Pandoraea sp.
AB510957.1
98 97 0,0 568
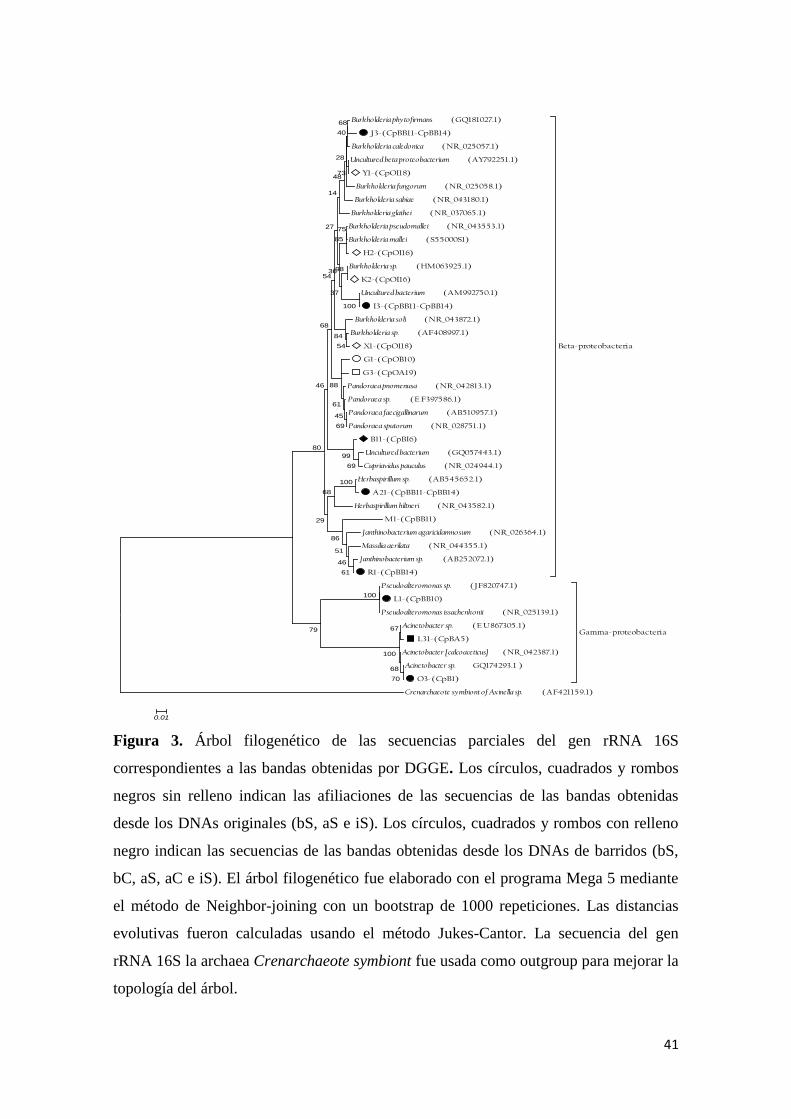
41
Figura 3. Árbol filogenético de las secuencias parciales del gen rRNA 16S
correspondientes a las bandas obtenidas por DGGE. Los círculos, cuadrados y rombos
negros sin relleno indican las afiliaciones de las secuencias de las bandas obtenidas
desde los DNAs originales (bS, aS e iS). Los círculos, cuadrados y rombos con relleno
negro indican las secuencias de las bandas obtenidas desde los DNAs de barridos (bS,
bC, aS, aC e iS). El árbol filogenético fue elaborado con el programa Mega 5 mediante
el método de Neighbor-joining con un bootstrap de 1000 repeticiones. Las distancias
evolutivas fueron calculadas usando el método Jukes-Cantor. La secuencia del gen
rRNA 16S la archaea Crenarchaeote symbiont fue usada como outgroup para mejorar la
topología del árbol.
(GQ181027.1) Burkholderia phytofirmans
J3-(CpBB11-CpBB14)
(NR_025057.1) Burkholderia caledonica
(AY792251.1) Uncultured beta proteobacterium
Y1-(CpOI18)
(NR_025058.1) Burkholderia fungorum
(NR_043180.1) Burkholderia sabiae
(NR_037065.1) Burkholderia glathei
(NR_043553.1) Burkholderia pseudomallei
(S55000S1) Burkholderia mallei
H2-(CpOI16)
(HM063925.1) Burkholderia sp.
K2-(CpOI16)
(AM992750.1) Uncultured bacterium
I3-(CpBB11-CpBB14)
(NR_043872.1) Burkholderia soli
(AF408997.1) Burkholderia sp.
X1-(CpOI18)
G1-(CpOB10)
G3-(CpOA19)
(NR_042813.1) Pandoraea pnomenusa
(E F397586.1) Pandoraea sp.
(AB510957.1) Pandoraea faecigallinarum
(NR_028751.1) Pandoraea sputorum
B11-(CpBI6)
(GQ057443.1) Uncultured bacterium
(NR_024944.1) Cupriavidus pauculus
(AB545652.1) Herbaspirillum sp.
A21-(CpBB11-CpBB14)
(NR_043582.1) Herbaspirillum hiltneri
M1-(CpBB11)
(NR_026364.1) Janthinobacterium agaricidamnosum
(NR_044355.1) Massilia aerilata
(AB252072.1) Janthinobacterium sp.
R1-(CpBB14)
Beta-proteobacteria
(JF820747.1) Pseudoalteromonas sp.
L1-(CpBB10)
(NR_025139.1) Pseudoalteromonas issachenkonii
(E U867305.1) Acinetobacter sp.
L31-(CpBA5)
(NR_042387.1) Acinetobacter [calcoaceticus]
GQ174293.1 ) Acinetobacter sp.
O3-(CpB1)
Gamma-proteobacteria
(AF421159.1) Crenarchaeote symbiont of Axinella sp.
100
100
100
69
99
98
69
45
61
88
61
46
51
86
75
85
54
84
73
68
68
40
28
48
37
36
14
27
54
68
46
29
80
79 67
100
68
70
0.01

42
6.2. CARACTERIZACIÓN MOLECULAR DE AISLADOS BACTERIANOS
ASOCIADOS AL CARACOL PALA.
6.2.1. Aislamiento de bacterias
Después del protocolo de resucitación, 15 de 23 muestras constituidas por varios tipos
de bacterias, mostraron crecimiento sobre medio de cultivo agar LB con agua de mar,
12 de los cuales crecieron sobre agar TCBS. El protocolo fue repetido con las 8
muestras restantes, incubando las placas de agar a temperatura ambiente y a 37 °C, pero
no se observó crecimiento alguno. 89 colonias fueron aisladas para evaluar la
producción de enzimas hidrolasas. En tanto solo un total de 57 cepas (37 aisladas en
este estudio por el protocolo de resucitación más 20 aisladas, las cuales habían sido
obtenidas en un estudio previo por Acosta et al., 2009) fueron seleccionadas para llevar
a cabo el test. Entre estas, 31 fueron aisladas de la baba, 15 de intestino, 9 de alimento y
2 de las paredes de los estanques donde los caracoles se encontraban en cautiverio.
6.2.2. Filogenia de los aislados bacterianos
De un total de 57 aislados bacterianos sólo 41 pudieron ser identificados
molecularmente mediante secuenciación del gen rRNA 16S desde la posición 27 a la
1492; todas las secuencias tuvieron una alta similitud (99 a 100 %) con las secuencias
más relacionadas reportadas en el GenBank, la mayoría de las cuales fueron
determinadas como bacterias no cultivables reportadas en trabajos realizados en
esponjas, algas, agua de mar, hidrotermales y otros ambientes marinos (Xu et al., 2005;
Nithyanand et al., 2011) (Tabla 5). Basándose en la filogenia de las secuencias más
relacionadas del BLAST y las secuencias de referencia obtenidas del RDP (Ribosomal
Database Project), tres grupos bacterianos a nivel de filo fueron identificados, entre
ellos Proteobacteria (89%), Firmicutes (9%) y Actinobacteria (2%), dentro de los
cuales encontramos cinco géneros diferentes: Psychrobacter 74% (30 aislados),
Halomonas 15% (6 aislados), Exiguobacterium 7% (3 aislados), Oceanobacillus 2% (1
aislado) y Micrococcus 2% (1 aislado) (Tabla 4, Fig. 4) (Anexo 8, Anexo 9).

43
Figura 4. Árbol filogenético basado en secuencias de rDNA 16S obtenidas de bacterias
aisladas de la baba, el intestino y el alimento de S. gigas. El árbol fue elaborado
mediante el método de Neighbor-joining con un bootstrap de 1000 repeticiones. Las
distancias evolutivas fueron calculadas usando el método Jukes-Cantor. La secuencia
del gen rRNA 16S la archaea Crenarchaeote symbiont fue usada como outgroup.
5CpBB11
2CpBB12
2CpBB11
21CpBI7
5CpBB13
25CpB1
19CpB3
18CpOB10
10CpBB10
4CpBB13
7CpBB10
8CpBB11
(FJ695576.1) Uncultured Psychrobacter sp.
1CpBB10
1CpBA4
3CpBA4
5CpBA3
3CpBB11
(HM566079.1) Psychrobacter sp.
2CpBB14
6CpOI15
(EF101550.1) Psychrobacter celer
28CpOI15
(EU371568.1) Uncultured Psychrobacter sp.
1CpBA3TCBS
(FJ695592.1) Uncultured Psychrobacter sp.
(FJ695589.1) Uncultured Psychrobacter sp.
1CpBB14
3CpBB10
(DQ399761.1) Psychrobacter sp.
24CpOI15
13CpOI15
8CpBB14
6CpBB14
(NR_028948.1) Psychrobacter nivimaris
(NR_025457.1) Psychrobacter submarinus
(NR_025531.1) Psychrobacter fozii
1CpB4
(GU570642.1) Psychrobacter sp.
3CpB4
10CpB4
Psychrobacter
(NR_027219.1) Halomonas axialensis
2CpOI6
(AB166968) Halomonas sp.
5CpOI8
(AM403725.1 ) Halomonas sp.
1CpB10
12CpOA5
26CpOI8
22CpB4
Halomonas
Gammaproteobacteria
20CpOB13
(GU213502.1) Micrococcus sp.Micrococcus
(NR_037113.1) Micrococcus luteus
Actinobacteria
29CpOB11
(DQ358670.1) Oceanobacillus sp.
(NR_028952.1) Oceanobacillus picturae
Oceanobacillus
(NR_024812.1) Exiguobacterium oxidotolerans
(DQ407720.1) 3CpBI9TCBS
Exiguobacterium sp.
5CpBI9
27CpOI17
FM203124.1 Exiguobacterium arabatum
Exiguobacterium
Firmicutes
(AF421159.1) Crenarchaeote symbiont of Axinella sp.
5255
54
100
100100
99
100
90100
87
66
62
98
100
64
100
100
64
64
14
58
38
69
62
58
36
43
99
43
100
52
0.01
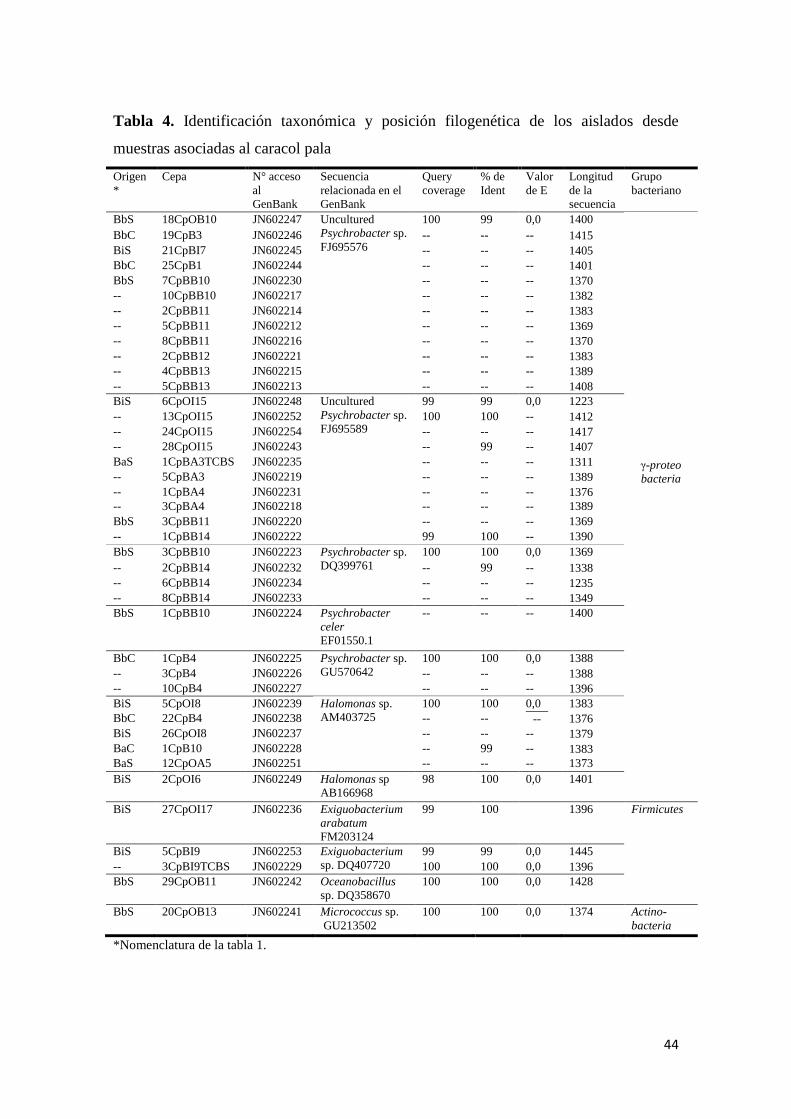
44
Tabla 4. Identificación taxonómica y posición filogenética de los aislados desde
muestras asociadas al caracol pala
Origen
*
Cepa N° acceso
al
GenBank
Secuencia
relacionada en el
GenBank
Query
coverage
% de
Ident
Valor
de E
Longitud
de la
secuencia
Grupo
bacteriano
BbS 18CpOB10 JN602247 Uncultured
Psychrobacter sp.
FJ695576
100 99 0,0 1400
γ-proteo
bacteria
BbC 19CpB3 JN602246 -- -- -- 1415
BiS 21CpBI7 JN602245 -- -- -- 1405
BbC 25CpB1 JN602244 -- -- -- 1401
BbS 7CpBB10 JN602230 -- -- -- 1370
-- 10CpBB10 JN602217 -- -- -- 1382
-- 2CpBB11 JN602214 -- -- -- 1383
-- 5CpBB11 JN602212 -- -- -- 1369
-- 8CpBB11 JN602216 -- -- -- 1370
-- 2CpBB12 JN602221 -- -- -- 1383
-- 4CpBB13 JN602215 -- -- -- 1389
-- 5CpBB13 JN602213 -- -- -- 1408
BiS 6CpOI15 JN602248 Uncultured
Psychrobacter sp.
FJ695589
99 99 0,0 1223
-- 13CpOI15 JN602252 100 100 -- 1412
-- 24CpOI15 JN602254 -- -- -- 1417
-- 28CpOI15 JN602243 -- 99 -- 1407
BaS 1CpBA3TCBS JN602235 -- -- -- 1311
-- 5CpBA3 JN602219 -- -- -- 1389
-- 1CpBA4 JN602231 -- -- -- 1376
-- 3CpBA4 JN602218 -- -- -- 1389
BbS 3CpBB11 JN602220 -- -- -- 1369
-- 1CpBB14 JN602222 99 100 -- 1390
BbS 3CpBB10 JN602223 Psychrobacter sp.
DQ399761
100 100 0,0 1369
-- 2CpBB14 JN602232 -- 99 -- 1338
-- 6CpBB14 JN602234 -- -- -- 1235
-- 8CpBB14 JN602233 -- -- -- 1349
BbS 1CpBB10 JN602224 Psychrobacter
celer
EF01550.1
EF101550
-- -- -- 1400
BbC 1CpB4 JN602225 Psychrobacter sp.
GU570642
100 100 0,0 1388
-- 3CpB4 JN602226 -- -- -- 1388
-- 10CpB4 JN602227 -- -- -- 1396
BiS 5CpOI8 JN602239 Halomonas sp.
AM403725
100 100 0,0 1383
BbC 22CpB4 JN602238 -- -- --
--
1376
BiS 26CpOI8 JN602237 -- -- -- 1379
BaC 1CpB10 JN602228 -- 99 -- 1383
BaS 12CpOA5 JN602251 -- -- -- 1373
BiS 2CpOI6 JN602249 Halomonas sp
AB166968
98 100 0,0 1401
BiS 27CpOI17 JN602236 Exiguobacterium
arabatum
FM203124
99 100 1396 Firmicutes
BiS 5CpBI9 JN602253 Exiguobacterium
sp. DQ407720
99 99 0,0 1445
-- 3CpBI9TCBS JN602229 100 100 0,0 1396
BbS 29CpOB11 JN602242 Oceanobacillus
sp. DQ358670
100 100 0,0 1428
BbS 20CpOB13 JN602241 Micrococcus sp.
GU213502
100 100 0,0 1374 Actino-
bacteria
*Nomenclatura de la tabla 1.

45
6.3. PERFIL ANTIBACTERIAL DE LOS AISLADOS ASOCIADOS AL
CARACOL PALA
Uno de los objetivos de este estudio fue caracterizar las comunidades bacterianas
cultivables que exhibieran actividades antimicrobianas. 23 aislados fueron agrupados en
tres unidades taxonómicas (Psychrobacter, Micrococcus y Halomonas), seis de estos
aislados (26%) mostraron actividad antibiótica contra al menos una de las cepas
evaluadas (Tabla 5). La inhibición de bacterias Gram negativas por los aislados
asociados al caracol pala fue más común que la inhibición de Gram positivos (Fig. 5,
Fig. 6, Fig. 7 y Fig. 6) (Tabla 6).
Tabla 5. Patrones de actividad antibacteriana (a-g) de bacterias asociadas a Strombus
gigas.
Inhibición de las cepas evaluadas Patrones de actividad Número de
aislados
Porcentajes (%)
de aislados
Sólo E. coli a 0 0
Sólo P. aeruginosa b 2 33,4
Sólo B. subtilis c 0 0
E. coli y P. aeruginosa d 0 0
E. coli y B. subtilis e 0 0
P. aeruginosa y B. subtilis f 1 16,7
E. coli, P. aeruginosa y B.subtilis g 3 50
Figura 5. Halo de inhibición del extracto metanólico crudo derivado de la cepa
bacteriana identificada como Uncultured Psychrobacter sp. (6CpOI15) frente a Bacillus
subtilis.
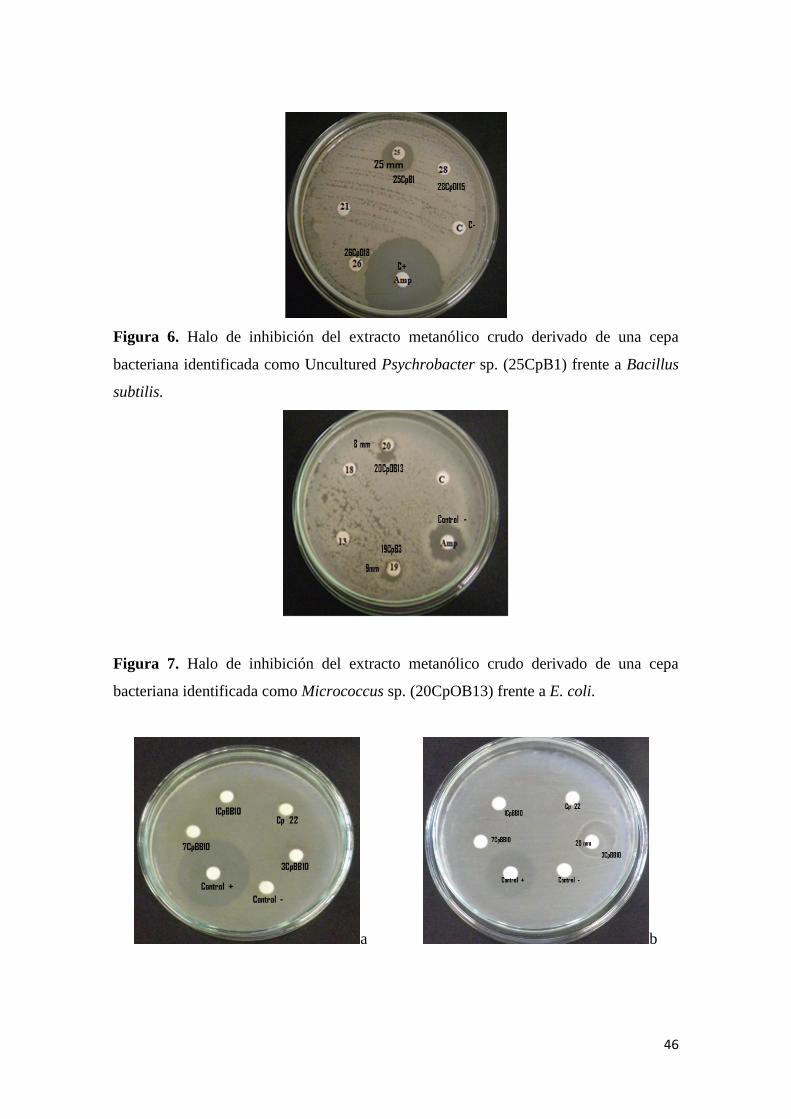
46
Figura 6. Halo de inhibición del extracto metanólico crudo derivado de una cepa
bacteriana identificada como Uncultured Psychrobacter sp. (25CpB1) frente a Bacillus
subtilis.
Figura 7. Halo de inhibición del extracto metanólico crudo derivado de una cepa
bacteriana identificada como Micrococcus sp. (20CpOB13) frente a E. coli.
a
b
25 mm
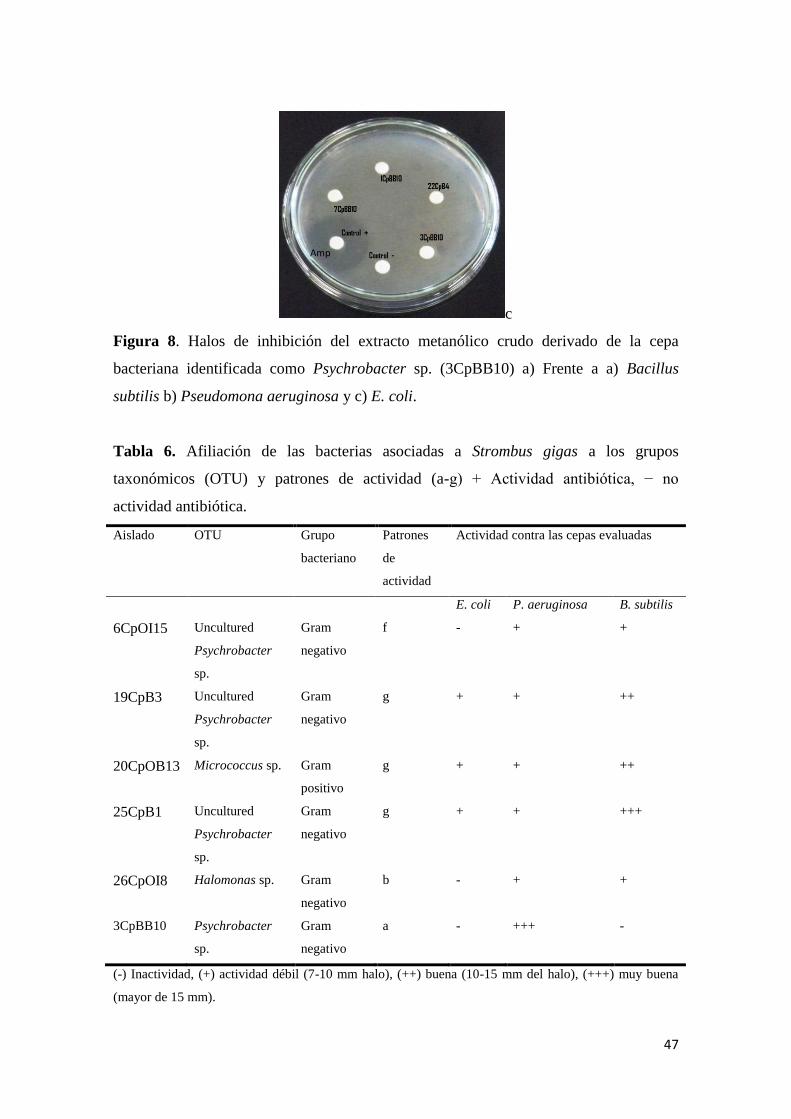
47
c
Figura 8. Halos de inhibición del extracto metanólico crudo derivado de la cepa
bacteriana identificada como Psychrobacter sp. (3CpBB10) a) Frente a a) Bacillus
subtilis b) Pseudomona aeruginosa y c) E. coli.
Tabla 6. Afiliación de las bacterias asociadas a Strombus gigas a los grupos
taxonómicos (OTU) y patrones de actividad (a-g) + Actividad antibiótica, − no
actividad antibiótica.
Aislado OTU Grupo
bacteriano
Patrones
de
actividad
Actividad contra las cepas evaluadas
E. coli P. aeruginosa B. subtilis
6CpOI15 Uncultured
Psychrobacter
sp.
Gram
negativo
f - + +
19CpB3 Uncultured
Psychrobacter
sp.
Gram
negativo
g + + ++
20CpOB13 Micrococcus sp. Gram
positivo
g + + ++
25CpB1 Uncultured
Psychrobacter
sp.
Gram
negativo
g + + +++
26CpOI8 Halomonas sp. Gram
negativo
b - + +
3CpBB10 Psychrobacter
sp.
Gram
negativo
a - +++ -
(-) Inactividad, (+) actividad débil (7-10 mm halo), (++) buena (10-15 mm del halo), (+++) muy buena
(mayor de 15 mm).
Amp
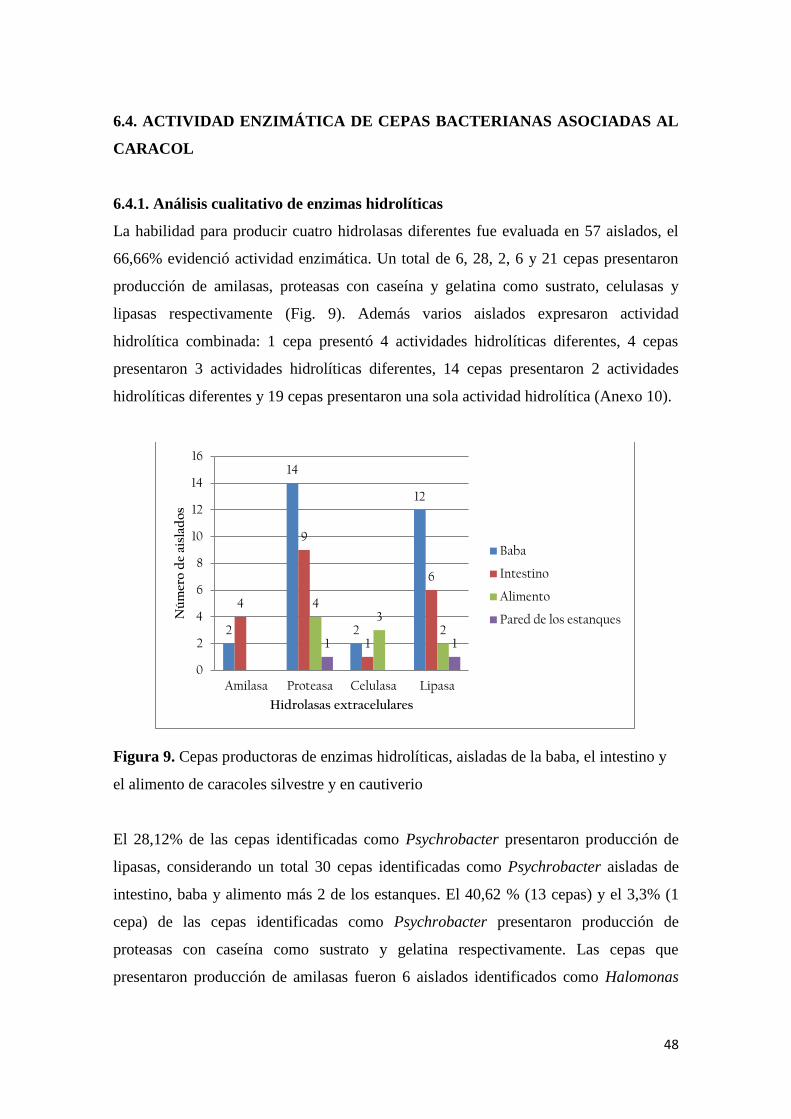
48
6.4. ACTIVIDAD ENZIMÁTICA DE CEPAS BACTERIANAS ASOCIADAS AL
CARACOL
6.4.1. Análisis cualitativo de enzimas hidrolíticas
La habilidad para producir cuatro hidrolasas diferentes fue evaluada en 57 aislados, el
66,66% evidenció actividad enzimática. Un total de 6, 28, 2, 6 y 21 cepas presentaron
producción de amilasas, proteasas con caseína y gelatina como sustrato, celulasas y
lipasas respectivamente (Fig. 9). Además varios aislados expresaron actividad
hidrolítica combinada: 1 cepa presentó 4 actividades hidrolíticas diferentes, 4 cepas
presentaron 3 actividades hidrolíticas diferentes, 14 cepas presentaron 2 actividades
hidrolíticas diferentes y 19 cepas presentaron una sola actividad hidrolítica (Anexo 10).
Figura 9. Cepas productoras de enzimas hidrolíticas, aisladas de la baba, el intestino y
el alimento de caracoles silvestre y en cautiverio
El 28,12% de las cepas identificadas como Psychrobacter presentaron producción de
lipasas, considerando un total 30 cepas identificadas como Psychrobacter aisladas de
intestino, baba y alimento más 2 de los estanques. El 40,62 % (13 cepas) y el 3,3% (1
cepa) de las cepas identificadas como Psychrobacter presentaron producción de
proteasas con caseína como sustrato y gelatina respectivamente. Las cepas que
presentaron producción de amilasas fueron 6 aislados identificados como Halomonas
2
14
2
12
4
9
1
6
4 3
2 1 1
0
2
4
6
8
10
12
14
16
Amilasa Proteasa Celulasa Lipasa
Nú
mer
o d
e ai
slad
os
Hidrolasas extracelulares
Baba
Intestino
Alimento
Pared de los estanques
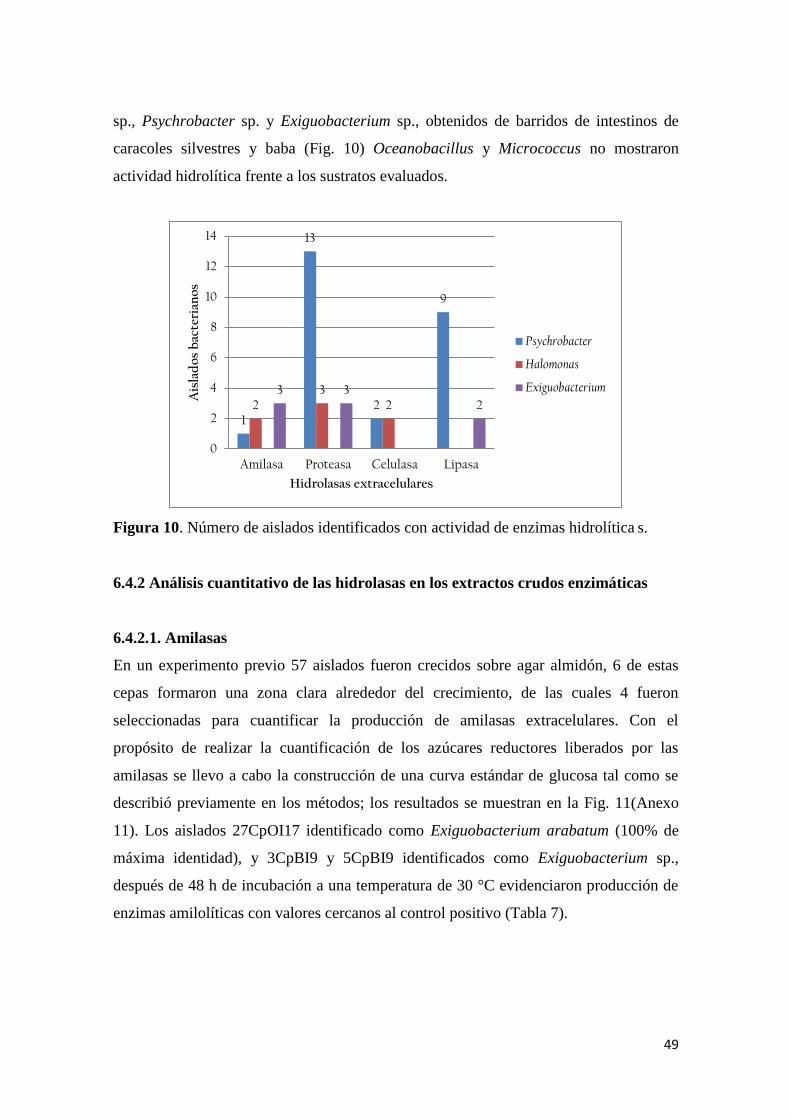
49
sp., Psychrobacter sp. y Exiguobacterium sp., obtenidos de barridos de intestinos de
caracoles silvestres y baba (Fig. 10) Oceanobacillus y Micrococcus no mostraron
actividad hidrolítica frente a los sustratos evaluados.
Figura 10. Número de aislados identificados con actividad de enzimas hidrolítica s.
6.4.2 Análisis cuantitativo de las hidrolasas en los extractos crudos enzimáticas
6.4.2.1. Amilasas
En un experimento previo 57 aislados fueron crecidos sobre agar almidón, 6 de estas
cepas formaron una zona clara alrededor del crecimiento, de las cuales 4 fueron
seleccionadas para cuantificar la producción de amilasas extracelulares. Con el
propósito de realizar la cuantificación de los azúcares reductores liberados por las
amilasas se llevo a cabo la construcción de una curva estándar de glucosa tal como se
describió previamente en los métodos; los resultados se muestran en la Fig. 11(Anexo
11). Los aislados 27CpOI17 identificado como Exiguobacterium arabatum (100% de
máxima identidad), y 3CpBI9 y 5CpBI9 identificados como Exiguobacterium sp.,
después de 48 h de incubación a una temperatura de 30 °C evidenciaron producción de
enzimas amilolíticas con valores cercanos al control positivo (Tabla 7).
1
13
2
9
2 3
2 3 3
2
0
2
4
6
8
10
12
14
Amilasa Proteasa Celulasa Lipasa
Ais
lad
os
bac
teri
ano
s
Hidrolasas extracelulares
Psychrobacter
Halomonas
Exiguobacterium
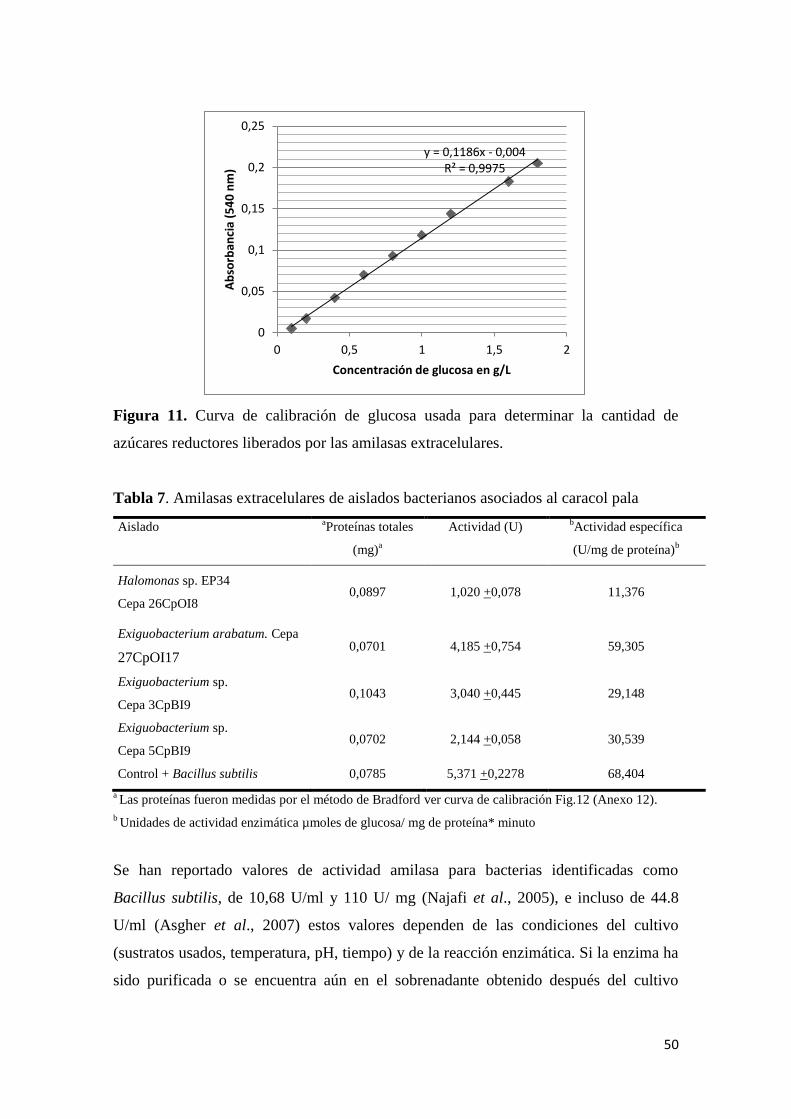
50
Figura 11. Curva de calibración de glucosa usada para determinar la cantidad de
azúcares reductores liberados por las amilasas extracelulares.
Tabla 7. Amilasas extracelulares de aislados bacterianos asociados al caracol pala
Aislado aProteínas totales
(mg)a
Actividad (U) bActividad específica
(U/mg de proteína)b
Halomonas sp. EP34
Cepa 26CpOI8 0,0897 1,020 +0,078 11,376
Exiguobacterium arabatum. Cepa
27CpOI17 0,0701 4,185 +0,754 59,305
Exiguobacterium sp.
Cepa 3CpBI9 0,1043 3,040 +0,445 29,148
Exiguobacterium sp.
Cepa 5CpBI9 0,0702 2,144 +0,058 30,539
Control + Bacillus subtilis 0,0785 5,371 +0,2278 68,404
a Las proteínas fueron medidas por el método de Bradford ver curva de calibración Fig.12 (Anexo 12).
b Unidades de actividad enzimática µmoles de glucosa/ mg de proteína* minuto
Se han reportado valores de actividad amilasa para bacterias identificadas como
Bacillus subtilis, de 10,68 U/ml y 110 U/ mg (Najafi et al., 2005), e incluso de 44.8
U/ml (Asgher et al., 2007) estos valores dependen de las condiciones del cultivo
(sustratos usados, temperatura, pH, tiempo) y de la reacción enzimática. Si la enzima ha
sido purificada o se encuentra aún en el sobrenadante obtenido después del cultivo
y = 0,1186x - 0,004 R² = 0,9975
0
0,05
0,1
0,15
0,2
0,25
0 0,5 1 1,5 2
Ab
sorb
anci
a (5
40
nm
)
Concentración de glucosa en g/L
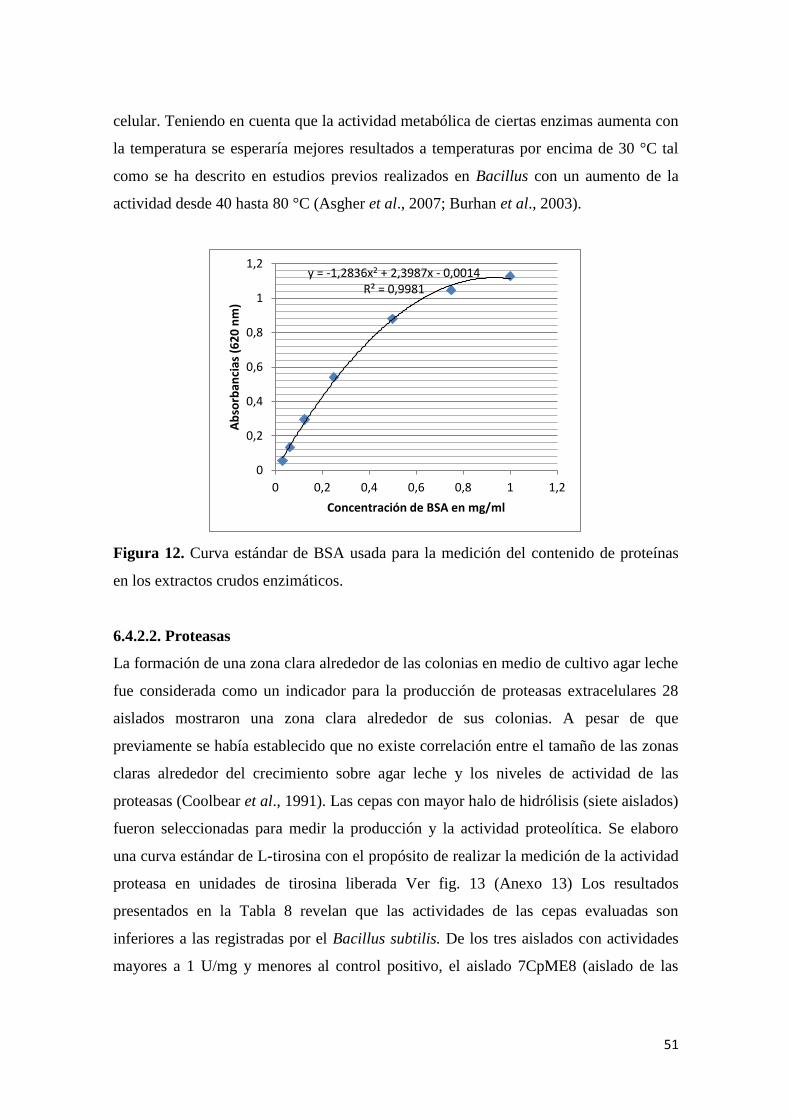
51
celular. Teniendo en cuenta que la actividad metabólica de ciertas enzimas aumenta con
la temperatura se esperaría mejores resultados a temperaturas por encima de 30 °C tal
como se ha descrito en estudios previos realizados en Bacillus con un aumento de la
actividad desde 40 hasta 80 °C (Asgher et al., 2007; Burhan et al., 2003).
Figura 12. Curva estándar de BSA usada para la medición del contenido de proteínas
en los extractos crudos enzimáticos.
6.4.2.2. Proteasas
La formación de una zona clara alrededor de las colonias en medio de cultivo agar leche
fue considerada como un indicador para la producción de proteasas extracelulares 28
aislados mostraron una zona clara alrededor de sus colonias. A pesar de que
previamente se había establecido que no existe correlación entre el tamaño de las zonas
claras alrededor del crecimiento sobre agar leche y los niveles de actividad de las
proteasas (Coolbear et al., 1991). Las cepas con mayor halo de hidrólisis (siete aislados)
fueron seleccionadas para medir la producción y la actividad proteolítica. Se elaboro
una curva estándar de L-tirosina con el propósito de realizar la medición de la actividad
proteasa en unidades de tirosina liberada Ver fig. 13 (Anexo 13) Los resultados
presentados en la Tabla 8 revelan que las actividades de las cepas evaluadas son
inferiores a las registradas por el Bacillus subtilis. De los tres aislados con actividades
mayores a 1 U/mg y menores al control positivo, el aislado 7CpME8 (aislado de las
y = -1,2836x2 + 2,3987x - 0,0014 R² = 0,9981
0
0,2
0,4
0,6
0,8
1
1,2
0 0,2 0,4 0,6 0,8 1 1,2
Ab
sorb
anci
as (
62
0 n
m)
Concentración de BSA en mg/ml
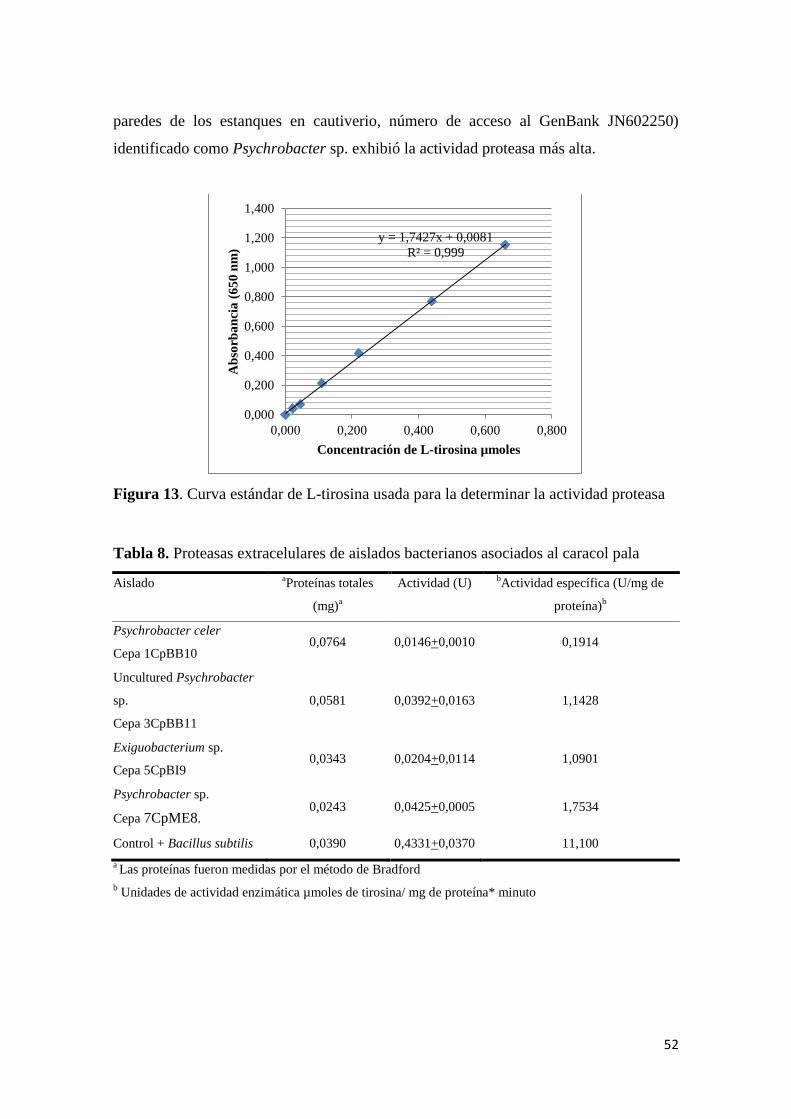
52
paredes de los estanques en cautiverio, número de acceso al GenBank JN602250)
identificado como Psychrobacter sp. exhibió la actividad proteasa más alta.
Figura 13. Curva estándar de L-tirosina usada para la determinar la actividad proteasa
Tabla 8. Proteasas extracelulares de aislados bacterianos asociados al caracol pala
Aislado aProteínas totales
(mg)a
Actividad (U) bActividad específica (U/mg de
proteína)b
Psychrobacter celer
Cepa 1CpBB10 0,0764 0,0146+0,0010 0,1914
Uncultured Psychrobacter
sp.
Cepa 3CpBB11
0,0581 0,0392+0,0163 1,1428
Exiguobacterium sp.
Cepa 5CpBI9 0,0343 0,0204+0,0114 1,0901
Psychrobacter sp.
Cepa 7CpME8. 0,0243 0,0425+0,0005 1,7534
Control + Bacillus subtilis 0,0390 0,4331+0,0370 11,100
a Las proteínas fueron medidas por el método de Bradford
b Unidades de actividad enzimática µmoles de tirosina/ mg de proteína* minuto
y = 1,7427x + 0,0081
R² = 0,999
0,000
0,200
0,400
0,600
0,800
1,000
1,200
1,400
0,000 0,200 0,400 0,600 0,800
Ab
sorb
an
cia
(6
50
nm
)
Concentración de L-tirosina µmoles
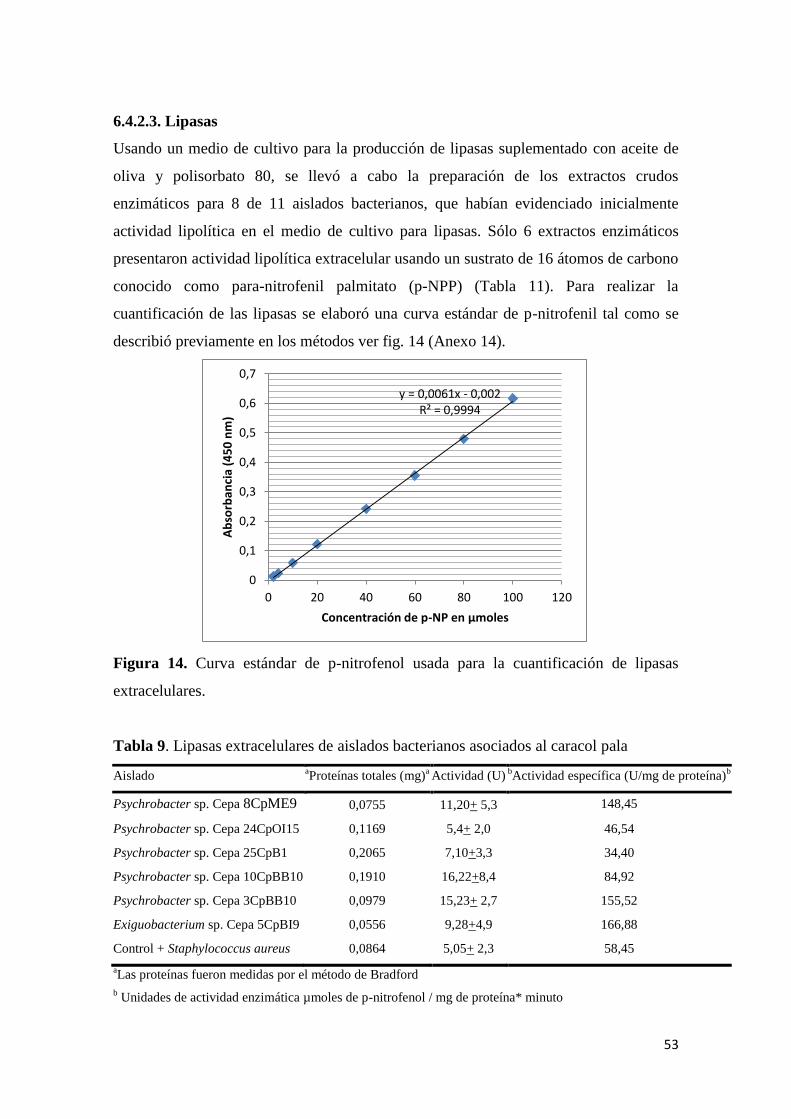
53
6.4.2.3. Lipasas
Usando un medio de cultivo para la producción de lipasas suplementado con aceite de
oliva y polisorbato 80, se llevó a cabo la preparación de los extractos crudos
enzimáticos para 8 de 11 aislados bacterianos, que habían evidenciado inicialmente
actividad lipolítica en el medio de cultivo para lipasas. Sólo 6 extractos enzimáticos
presentaron actividad lipolítica extracelular usando un sustrato de 16 átomos de carbono
conocido como para-nitrofenil palmitato (p-NPP) (Tabla 11). Para realizar la
cuantificación de las lipasas se elaboró una curva estándar de p-nitrofenil tal como se
describió previamente en los métodos ver fig. 14 (Anexo 14).
Figura 14. Curva estándar de p-nitrofenol usada para la cuantificación de lipasas
extracelulares.
Tabla 9. Lipasas extracelulares de aislados bacterianos asociados al caracol pala
Aislado aProteínas totales (mg)
a Actividad (U)
bActividad específica (U/mg de proteína)
b
Psychrobacter sp. Cepa 8CpME9 0,0755 11,20+ 5,3 148,45
Psychrobacter sp. Cepa 24CpOI15 0,1169 5,4+ 2,0 46,54
Psychrobacter sp. Cepa 25CpB1 0,2065 7,10+3,3 34,40
Psychrobacter sp. Cepa 10CpBB10 0,1910 16,22+8,4 84,92
Psychrobacter sp. Cepa 3CpBB10 0,0979 15,23+ 2,7 155,52
Exiguobacterium sp. Cepa 5CpBI9 0,0556 9,28+4,9 166,88
Control + Staphylococcus aureus 0,0864 5,05+ 2,3 58,45
aLas proteínas fueron medidas por el método de Bradford
b Unidades de actividad enzimática µmoles de p-nitrofenol / mg de proteína* minuto
y = 0,0061x - 0,002 R² = 0,9994
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0 20 40 60 80 100 120
Ab
sorb
anci
a (4
50
nm
)
Concentración de p-NP en µmoles

54
En un estudio relacionado con la caracterización de una lipasa extracelular obtenida de
una cepa de Staphylococcus aureus, la actividad específica medida en el sobrenadante
del cultivo usando tributirina como sustrato fue de 10 U/mg, menor a la reportada en
este estudio, (Horchani et al., 2009) otra lipasa aislada de un Staphylococcus xylosus
registro una actividad de 54.61 U/mg a 50 °C cuando se utilizó p-nitrofenil butirato
como sustrato; adicionalmente fueron medidos valores menores por debajo de esta
temperatura, lo cual relaciona en forma positiva el aumento de la actividad enzimática
con el aumento de la temperatura. La cinética de las enzimas depende de factores como
la temperatura, el pH u otros descritos previamente (Borges et al., 2011).
Para comparar en este estudio la actividad de las hidrolasas de los aislados con enzimas
comerciales se usaron las siguientes enzimas:
α.- amilasa de Bacillus amyloliquefaciens de la casa comercial Sigma-Aldrich
Proteasa de Bacillus amyloliquefaciens de la casa comercial Sigma-Aldrich
Los resultados encontrados muestran que la actividad de estas enzimas es notoriamente
superior a las de los extractos enzimáticos evaluados. Tal como se evidencia en la
siguiente tabla si se comparan la densidad óptica de 0,15 con el resultado obtenido por
el aislado 5CpBI9 (0,160) se espera que al aumentar las concentraciones en términos de
unidades de actividad, los niveles sean superiores a los registrados por el aislado.
Tabla 10. Resultados de las absorbancias para diluciones diferentes de α.- amilasa de
Bacillus amyloliquefaciens. Los azucares reductores fueron medidos usando el método
de DNS.
Unidades /gramo Absorbancia (650 nm)
0,001 0,031
0,002 0,082
0,003 0,115
0,004 0,135
0,005 0,143
0,006 0,15

55
La actividad de la enzima comercial evaluada sin diluir es equivalente a 250
unidades/gr. Cuando concentraciones superiores a 100 unidades/gramo fueron
evaluadas, los valores de las absorbancias permanecieron iguales lo cual se puede
explicar por el consumo completo de la cantidad de almidón en una solución al 1%
(P/V) y a que tan sólo una unidad de almidón de la enzima comercial puede degradar
5,26 gramos de almidón seco por hora.
Al comparar una enzima producida bajo condiciones que garantizan su máxima
eficiencia y pureza con un extracto crudo enzimático en términos de actividad, se ha
observado que la actividad enzimática de la hidrolasa en estado puro es mayor (Reddy
et al., 1998)
Cuando fue evaluada la presencia de proteasas en los extractos crudos se obtuvo para el
control positivo (Bacillus subtilis) una absorbancia promedio de 0,713 cercana a la
registrada para 0,1 unidades/gramo de la enzima comercial (Tabla 11), sin embargo, los
valores de las absorbancias registrados para el resto de los aislados se mantuvo por
debajo de 0,1 unidades.
Tabla 11. Resultados de las absorbancias para diluciones diferentes de proteasas de
Bacillus amyloliquefaciens. Las proteasas fueron medidas por el método de Lowry.
Unidades/gramo Absorbancia (650 nm)
0 0,206
0,1 0,708
0,2 0,768
0,3 0,916
0,4 0,989
0,5 1,052
La actividad de la enzima comercial sin diluir es de 0,8 unidades/gramo.

56
7. DISCUSIÓN
7.1. ANÁLISIS DE LOS PERFILES DE DGGE.
El caracol pala Strombus gigas pertenece a la clase Gastropoda habita en los fondos
marinos arenosos poco profundos y está distribuido a lo largo de todo el atlántico
tropical noroccidental (Warmke y Abbott, 1961; Randall 1964; Brownell y Stevely
1981, McCarthy, 2007). Sobre la microbiota del caracol pala y de los microorganismos
simbiontes que éste hospeda poco es conocido (Acosta et al., 2009). La microbiota
bacteriana ha sido estudiada en varios organismos marinos algas, corales (Kvennefors et
al., 2010), peces (Sun et al., 2009) y en caracoles (Rodríguez et al., 2011) con el
propósito de conocer y entender el efecto de las poblaciones bacterianas sobre la
ecología, la conservación, la salud y la protección frente a patógenos en los hospederos,
las variaciones en la diversidad microbiana de acuerdo al ecosistema habitado, entre
otros aspectos.
En este estudio se examinó la diversidad microbiana asociada a los caracoles de las Islas
de Rosario en Colombia, en hábitats silvestres y en cautiverio, usando aproximaciones
dependientes de cultivo (cultivo en placas de agar y PCR-DGGE) e independientes de
cultivo (PCR-DGGE y análisis de secuencias del gen rRNA 16S) encontrando
variaciones en las poblaciones bacterianas de acuerdo al material biológico de donde
éstas fueron obtenidas y al hábitat de los caracoles. Hasta hace muy poco la información
sobre la microbiota bacteriana en el caracol pala era escasa, sin embargo, gracias al uso
de estrategias dependientes e independientes de cultivo en el laboratorio, se comenzó a
describir la microbiota del caracol pala en términos de diversidad y clasificación de
bacterias Acosta (2009). Él presente trabajo, aporta información que puede llegar a ser
notable en la ecología, en la industria y en la biotecnología, información que será
discutida en detalle en los siguientes apartes. Los DNAs originales obtenidos por
extracción directa desde el material biológico (baba, intestino y alimento) y los DNAs

57
bacterianos extraídos desde los barridos crecidos sobre medios de cultivos (agar marino)
ambos asociados a muestras de caracoles en cautiverio y silvestres, fueron utilizados
para analizar la diversidad por DGGE (cultivables y totales (cultivables y no
cultivables).
Cuando los perfiles de DGGE de las secuencias parciales del gen rRNA 16S de los
barridos (UFC) fueron comparados con los perfiles de DGGE del DNA original
(obtenido directamente desde el material biológico), se observaron diferencias en el
número de bandas de acuerdo a la matriz de presencia y ausencia de datos con un
número mayor de bandas diferentes para las muestras originales. Al contrastar los
índices de diversidad (Shannon-Wiener) entre las muestras originales y las
correspondientes a los barridos (H’= 3,44) de caracoles silvestres las diferencias no
fueron notables (p>0,05), pese a que el índice de diversidad resulto ser más alto para las
muestras originales (H’=3,55). Cuando los perfiles de DGGE independiente (DNAs
originales-totales) y dependientes de cultivo (DNAs barridos- Cultivable) asociados a
caracoles silvestres fueron examinados se encontraron 52 bandas diferentes. El 13.5%
de las bandas fueron únicas a los perfiles de DGGE dependientes de cultivo (barridos),
el 17,3% a los perfiles de DGGE independientes de cultivo (originales) y el 69,2%
fueron encontradas en ambas comunidades. La presencia de bandas únicas tanto en los
DNAs de los barridos como en los DNAs originales y de bandas comunes en ambas
comunidades, demuestran que los perfiles de DGGE independientes de cultivo y
dependientes de cultivo pueden resolver comunidades bacterianas únicas, diversas e
igualmente complejas (Edenborn y Sexstone, 2007).
La matriz de presencia y ausencia de bandas resultantes del DGGE también fue usada
para un análisis de similitud (ANOSIM) entre las muestras asociadas a caracoles
silvestres y en cautiverio como se describe en los resultados previamente; encontrando
diferencias significativas entre las muestras asociadas a caracoles silvestres y en
cautiverio. El coeficiente de Bray-Curtis ha sido sugerido como un coeficiente ideal
para la construcción de matrices de similitud, es un método robusto (Faith et al., 1991).
El Sorensen or Dice también ha sido descrito en el análisis de comunidades microbianas
(Eichner et al., 1999). Es importante anotar que cuando se usan el coeficiente de Bray-
Curtis para analizar presencia y ausencia de datos, éste es idéntico al coeficiente de
Sorensen el cuál usa presencia y ausencia de datos por definición.

58
La diversidad microbiana en caracoles había sido estudiada usando ITS (Acosta el al.,
2009) con patrones que sugerían la existencia de comunidades diversas tanto en los
DNAs originales obtenidos por extracción directa desde muestra de baba, alimento e
intestino como en la fracción cultivable o barridos. Probablemente el almacenamiento
prolongado durante aproximadamente 2 años de las muestras originales y los barridos,
estuvo asociado al deterioro del DNA, y por consiguiente a una reducción en el número
de bandas que se esperaba observar por DGGE. Aunque el DGGE no es la opción más
apropiada para la caracterización completa de las comunidades bacterianas o para medir
la diversidad microbiana por si mismo, es aún ideal para el cribado inicial cuando se
comparan múltiples muestras (Green et al., 2009). La técnica del DGGE puede ser
afectada por bajas concentraciones de DNA para las diferentes especies o por altas
concentraciones de DNA que compiten durante las reacciones de PCR, (Ogier et al.,
2002). La abundancia relativa de los miembros de una comunidad puede afectar la
detección de ciertas especies por PCR y DGGE.
Una de las mayores ventajas de la técnica DGGE sobre otros métodos usados para el
análisis de comunidades microbianas es que permite hacer una comparación cualitativa
entre diferentes muestras con relativa facilidad por separación de los productos de PCR
de acuerdo al comportamiento de la temperatura de melting de los diferentes amplicones
(Muyzer et al., 1998). No obstante la técnica del DGGE no es un indicador
suficientemente preciso de la composición de las comunidades con una diversidad
desconocida (Kisand y Wikner, 2003). La representación visual de los perfiles de cada
una de las poblaciones analizadas, a través de patrones de bandeo permiten enfocarse en
las bandas de interés que pueden ser escindidas, en ese orden de ideas las bandas
podrían ser procesadas de dos formas: a) secuenciación de los productos de PCR o b)
clonación, selección de los clones por DGGE y secuenciación (Murray et al., 1996;
Sekiguchi et al., 2002). La opción b suministra una calidad superior en las secuencias,
pero la opción a proporciona información importante sobre la pureza de las bandas
escindidas (Green et al., 2009). La baja calidad en las secuencias obtenidas desde las
bandas escindidas en este estudio (68 %), es un indicador de los sesgos que pueden estar
ligados a la técnica de DGGE. Así mismo, reportes previos han demostrado que una
única banda puede contener rDNA 16S de más de dos especies bacterianas (Co-

59
migración de bandas con secuencias similares) (Sekiguchi et al., 2001) o que varias
bandas son generadas desde una sola especie lo cual puede ser dilucidado por la
presencia de varios cistrones presentes en un mismo genoma o polimorfismos del gen
rRNA 16S (Nubel et al., 1996; Moreno et al., 2002; Vásquez et al., 2005). Las causas
de una información filogenética imprecisa pueden ocurrir antes o después de los
procesos de separación de las bandas como fallas en el proceso de extracción del DNA,
amplificación preferencial de ciertas secuencias (Reysenbach et al., 1992), formación de
quimeras (Kopczynski et al., 1994; Wang y Wang, 1997; Speksnijder et al., 2001) y
hererodúplex (Muyzer y Smalla 1998; Ferris y Ward 1997). Otra fuente de error en
PCR-DGGE es que las comunidades muy diversas pueden estar representadas por smear
o manchas que pueden ser interpretadas erróneamente (Torsvik et al., 1998; Nikolausz
et al., 2005)
En el análisis de los clusters resultantes de los patrones de bandeo de las muestras de
DNA obtenidas del alimento de los caracoles en cautiverio y silvestres están más
relacionados con las muestras de DNA de la baba de caracoles en cautiverio (Fig. 2).
Posiblemente debido a la mayor concentración de alimento en los estanques donde los
caracoles eran cultivados y a la reducción del flujo del agua comparado con el hábitat
natural de los caracoles silvestres.
Es poco probable que las secuencias de rDNA 16S que representan menos del 1 % de
todos los DNAs molde en una comunidad bacteriana sean detectadas por la técnica de
DGGE (Muyzer et al., 1993; Muyzer y Smalla, 1998). Sin embargo, los análisis basados
en aproximaciones independientes de cultivo como el DGGE a partir de DNAs
extraídos directamente desde muestras ambientales (suelo, agua, tejidos biológicos)
podrían expresar mejor la diversidad y la riqueza de las comunidades microbianas
debido a que los cultivos destruyen las interacciones que ocurren entre los organismos
en sus hábitats naturales (Joint et al., 2010).
La estructura de las comunidades bacterianas no necesariamente refleja la
heterogeneidad fisiológica de las poblaciones ecológicamente importantes y pueden
ignorar las poblaciones que podrían desempeñar un papel fundamental en los
ecosistemas. En caracoles pala es muy poco conocido sobre la diversidad microbiana
con escasos estudios que puedan ser usados como referentes para comparar los
resultados encontrados. Los resultados de las secuencias de las bandas obtenidas por la

60
técnica DGGE (Barridos o fracción cultivable) fueron diferentes a los de las secuencias
de los aislados sobre los medios de cultivo que fueron usados para las pruebas
enzimáticas. La discrepancia puede presentarse por los medios de cultivos empleados y
el método de resucitación de bacterias que no aseguró la recuperación completa de las
cepas que crecieron inicialmente a partir del material biológico como las
Pseudoalteromonas. Los datos de las secuencias de los estudios dependiente e
independientes de cultivo están relacionados filogenéticamente a nivel de la clase
perteneciente a las Gammaproteobacteria, no obstante los géneros encontrados fueron
diferentes (Tablas 3 y 4), exceptuando las secuencias correspondientes al género
Pseudoalteromona, que fue detectada por DGGE (Banda L1) (Fig.1) y la secuencia del
aislado DI1Sg (GQ253505) reportado anteriormente en los barridos por Acosta et al.,
(2009).
Las secuencias del gen rRNA 16S de las bandas escindidas en los geles obtenidos por
DGGE han sido agrupadas en dos clases taxonómicas las Betaproteobacterias y las
Gammaproteobacterias, éstas últimas, reportadas como el grupo bacteriano dominante
en comunidades bacterianas asociadas a invertebrados marinos (Wegley et al., 2007;
Kvennefors et al., 2010). Las Betaproteobacterias también han sido descritas
previamente como habitantes comunes en ambientes marinos (Kirchman et al., 2005;
Zhang et al., 2006).
Las secuencias de las bandas asociadas a muestras del intestino de caracoles silvestres
(iS) H2, K2 y X1(Tabla 3) tuvieron un porcentaje de similitud en sus secuencias entre el
97 y 99% en el Blastn con Burkholderia sp., género bacteriano extremadamente
heterogéneo que ha sido descrito previamente como fijador de nitrógeno en el arroz
(Kennedy et al., 2004) y en los suelos salinos costeros (Barua et al., 2011; y como
patógeno oportunista de plantas y humanos (Onofre-Lemus et al., 2009). El papel de las
Burkholderia en organismos acuáticos aún es poco comprendido. En un estudio sobre la
microbiota intestinal de peces como Epinephelus coioides, Sun et al., (2009) se informó
que la presencia de una mayor densidad celular de Burkholderia cepacia estaba
asociada al crecimiento lento en éstos juveniles.
La secuencia del amplificado de la banda A21 (Fig.1) corresponde a Herbaspirillum un
género relacionado con la especie Aquaspirillum aislada en medios acuáticos (Ding y
Yokota, 2004) y asociado también como bacterias fijadoras de nitrógeno a través de las

61
secuencias más relacionadas reportadas en el GenBank, sin embargo, algunas de las
especies pertenecientes a este género son considerados como patógenos oportunistas. El
rol de las bacterias fijadoras puede ser de interés, pues se sabe que el nitrógeno no es un
nutriente limitante sólo para plantas, muchos insectos tienen una dieta pobre en
nitrógeno, así que la existencia de bacterias fijadoras de nitrógeno en el intestino de
termitas y moscas de fruta (Behar et al., 2005) permite cuestionar el impacto ecológico
extendido de la fijación biótica de nitrógeno asociativa en animales (Bothe et al., 2007).
La secuencia correspondiente a L1 (bS) de baba de caracoles silvestres con un
porcentaje de similitud del 100% en el Blastn está relacionada con el género
Pseudoalteromona frecuentemente aislado de ambientes marinos, corales, briozoos y
peces (Pukall et al.; 2002; Shnit-Orland y Kushmaro, 2009; Svanevik y Lunestad,
2011). Algunas especies de Pseudoalteromonas producen compuestos antibacterianos.
Chen et al., (2010) reportaron la presencia de una Pseudoalteromona aislada de los
corales Mantipora aequituberculata, con actividad contra algunos Staphylococcus
aureus resistentes a la meticilina, adicionalmente muchas de las especies de
Pseudoalteromonas han sido encontradas en asociación con huéspedes eucariotas en
ambientes marinos produciendo metabolitos contra un amplio rango de bacterias y
hongos (Gauthier et al., 1995; Holmstrom et al., 1999).
La secuencia de las bandas L31 (aS) y O3 (bC) guardan una similitud del 99 y 100%
con Acinetobacter, relacionado con procesos de comensalismo en el intestino de
algunos invertebrados marinos, además han sido reportados como perteneciente a la
micro-flora normal en otras especies de caracoles (Ducklow et al., 1979) esta secuencia
también ha sido relacionada en el GenBank con microorganismos asociados a las
semillas de leguminosas esto tal vez se deba a la amplia versatilidad de este género en
diferentes ambientes.
Las secuencias de las bandas G1 (bS) y G3 (aS) con porcentajes de similitud de 95 y
96% respectivamente para Pandoraea sp. género bacteriano que está relacionado con
mecanismos de oxidación de tiosulfato en el agua y en el suelo, y en la estimulación del
crecimiento de las plantas (Anandham et al., 2008), encontrándose en los ambientes
marinos en forma libre o asociada al plantón (Fera et al., 2007). Aunque algunos
invertebrados marinos que habitan en las profundidades del océano han sido reportados

62
en procesos de transformación del sulfuro de hidrogeno a tiosulfato el cual finalmente
puede utilizado por las bacterias endosimbiontes en la quimiosíntesis (Childress y
Girguis, 2010), no hay evidencias suficientes que permitan concluir que algunas de
estas bacterias pertenecen al género Pandoraea y también estén asociadas al caracol
pala, a pesar de que éste se ha encontrado en profundidades de hasta 100 metros.
La secuencia correspondiente a la banda R1 fue identificada como Janthinobacterium
sp. con un porcentaje de similitud del 99%. Este género ha sido recientemente aislado a
partir de tres especies de anfibios entre estos la Rana mucosa, e inhibe un hongo
patógeno el Batrachochytrium dendrobatidis por secreción de violaceína, un metabolito
antimicrobiano (Nakamura et al., 2002; Bosch y McFall-Ngai, 2011). En estas bacterias
también ha sido detectada la presencia de amilasas y quitinasas. Si se avanza hacia el
estudio de las interacciones que ocurren entre el caracol y su microbiota, es posible que
se pueda conocer más sobre la diversidad microbiana e importancia para su hospedero,
en procesos de defensa, descomposición de materia orgánica y los ciclos
biogeoquímicos, aislar microorganismos aún no cultivados es uno de los pasos que
ayudara a entender cómo ocurren los procesos relacionados con la síntesis de
antimicrobianos como la violaceína y si realmente ocurren en el caracol pala, a causa de
que la identificación de una única secuencia correspondiente a una banda extraída desde
el DGGE no nos aporta suficiente información sobre el microorganismo en cuestión.
7.2. FILOGENIA DE LOS AISLADOS BACTERIANOS
En este estudio el haber empleado métodos de cultivo como la reactivación de células
en estado viable no cultivable, dio como resultado el aislamiento de cepas
pertenecientes a grupos taxonómicos que no se habían podido cultivar después de ser
almacenadas como barridos; además permitió aislar géneros que no se habían descrito
anteriormente para caracol pala (Stevenson et al., 2004; Davis et al., 2005; Stott et al.
2008; Acosta et al., 2009)
Los aislados bacterianos identificados a través de las secuencias del gen rRNA 16S
como Psychrobacter sp. y Halomonas sp. en este estudio, habían sido previamente
descritos en el caracol pala (Acosta et al., 2009), exceptuando los géneros

63
Oceanobacillus, Micrococcus y Exiguobacterium. Bacterias pertenecientes al género
Psychrobacter han sido relacionadas en el GenBank con bacterias no cultivables (Tabla
4). Las diversas estrategias como el uso de agua de mar para la preparación de los
medios de cultivo, el protocolo de resucitación, así como la temperatura de incubación,
posiblemente propiciaron la recuperación de bacterias que hasta ahora habían sido
reportadas como no cultivables (Whitesides y Oliver, 1997).
El uso del método de resucitación de bacterias en estado viable no cultivable, aseguro la
recuperación de algunos grupos bacterianos previamente descritos, sin embargo, al
contrastar con el número de bandas observadas en los perfiles de DGGE de los barridos
(cultivables), se podría concluir que el proceso de resucitación no fue del todo exitoso.
La construcción del árbol filogenético usando el método de Neighbor-joining y el
estimador de distancias Jukes Cantor con secuencias de referencia relacionadas permitió
dilucidar la presencia de cinco agrupaciones taxonómicas a nivel de género en los
aislados como se describió previamente en los resultados (Fig. 4).
Para la construcción de árboles filogenéticos Neighbor-joining es el método más
utilizado frecuentemente debido a su velocidad y precisión. Todos los métodos para la
construcción de árboles filogenéticos tienen un mismo punto de origen el alineamiento
múltiple. Cuando un alineamiento múltiple es creado una hipótesis evolutiva es
generada automáticamente (Yang y Zhang, 2008).
Muchos de los algoritmos actuales son heurísticos, por ende conducen a una solución
aproximada. Los alineamientos múltiples son sensibles a los cambios en los parámetros
de puntuación, así que en muchos casos el alineamiento múltiple que refleja la
verdadera historia evolutiva es desconocido. Un estudio previo muestra que cuando se
usan secuencias con una longitud entre 500 y 1500 bp Jukes-Cantor y Kimura (Kimura,
1980), básicamente, tenían el mismo rendimiento en el agrupamiento al azar, mientras
que el rendimiento para Tamura-Nei fue más alto (Yang y Zhang, 2008). Cuando se
utilizo en este estudio el estimador de distancias Tamura–Nei (Anexo 15) los resultados
en la estructura de los árboles construidos usando Neighbor-joining fue similar a cuando
se usaron los modelos de Kimura y Jukes-Cantor. Estudios sobre el análisis filogenético
de comunidades microbianas muestran la aplicación de varios estimadores de distancias
(Tian et al., 2008; Kobayashi et al., 2012). Si el número de sustituciones de nucleótidos
por sitio para la mayoría de las secuencias divergentes es relativamente pequeño (por

64
ejemplo < 0,2), muchas de las medidas de distancia ofrecen básicamente la misma
estimación. En éste caso, es suficiente utilizar Jukes-Cantor o Kimura como estimadores
de distancias para la construcción de árboles filogenéticos usando Neighbor- joining
(Tateno et al., 1994)
El comportamiento de un método filogenético basado en un modelo puede depender del
ajuste del modelo a los datos. Para un grupo de secuencias largas con niveles bajos de
polimorfismo, el modelo puede tener pocos efectos sobre los resultados del análisis. Sin
embargo, cuando se trabajan con secuencias más divergentes, el uso de un modelo en
lugar de otro puede alterar los resultados del análisis e incluso conducir a la selección de
un árbol con una topología inapropiada (Bos y Posada, 2005).
7.3. ACTIVIDAD ANTIBACTERIANA
Durante este estudio 23 cepas fueron seleccionadas para evaluar su actividad
antibacteriana frente a bacterias Gram negativas (Escherichia coli y Pseudomona
aeruginosa) y Gram positivas (Bacillus subtilis). Los aislados fueron organizados en
tres agrupaciones taxonómicas (Psychrobacter, Micrococcus y Halomonas), sólo 6
aislados exhibieron actividad antibiótica. Sin embargo, no se encontró ninguna
correlación entre las distintas agrupaciones taxonómicas y la actividad antibacteriana
registrada. Hasta el momento se han reportado bacterias productoras de extractos con
actividades biológicas en varias especies de corales (Porites luta, Galaxea fascicularis u
otras) colectadas en la isla Panjang en Indonesia los cuales fueron probados contra
Escherichia coli, Bacillus subtilis y Micrococcus encontrando que éstas inhiben el
crecimiento de al menos una de las cepas evaluadas (Radjasa et al., 2008).
Algunas especies de Micrococcus asociadas a corales inhiben el crecimiento de Gram
negativos y Gram positivos (Tanaka et al., 2001). Los miembros del genero
Micrococcus son bacterias heterotróficas distribuidas en varios ambientes (Tanaka et
al., 2001), con un gran potencial para la producción de compuestos antimicrobianos
como las bacteriocinas las cuales inhiben el crecimiento de otras bacterias Gram
positivas, pero no Gram negativas.

65
El interés hacia la búsqueda de nuevos metabolitos desde recursos como las macro
algas, tunicados y octocorales ha disminuido, sin embargo, los metabolitos obtenidos
desde bacterias han aumentado considerablemente, puesto que el número de metabolitos
derivados de algas e invertebrados pueden ser producidos por microorganismos
asociados (Faulkner 2000).
Existen pocos reportes de metabolitos secundarios bioactivos relacionados con la
bacteria psicrófila Psychrobacter sp. no obstante, estudios previos han descrito 16
dipéptidos cíclicos aislados desde un Psychrobacter sp., obtenido de una esponja marina
Stelletta sp., los dipéptidos cíclicos poseen diversas actividades biológicas como
propiedades antibacterianas (Li et al., 2008).Otro género ampliamente reportado es
Halomonas de las cuales varias especies exhiben actividades antibacterianas frente a
bacterias Gram positivas con halos de inhibición entre 15 y 25 mm y actividad
citotóxica frente a líneas celulares tumorales (Bitzer et al., 2006)
A pesar de haber encontrado varios géneros bacterianos en el caracol con propiedades
antimicrobianas contra bacterias Gram positivas y negativas, que incluso han sido
reportados previamente como productores de sustancias inhibitorias, aún no es posible
deducir el verdadero potencial antimicrobiano de las comunidades bacterianas asociadas
a este molusco, las métodos de cultivo en placas en agar marino para el aislamiento de
bacterias asociadas al caracol son restrictivos, puesto que el total de los
microorganismos cultivados no representan la gran diversidad biológica de las
comunidades bacterianas. Si no se tiene en cuenta a todos los microorganismos es
posible subestimar el verdadero potencial de las especies microbianas. El uso de medios
de caldos de cultivo para la obtención de metabolitos secundarios podría afectar la
producción de estos, las células sometidas a condiciones diferentes a las acostumbradas
en su ambiente natural pueden dejar de producir moléculas que antes eran necesarias
para su supervivencia, Diggle et al., 2007 describieron que la ausencia de agentes que
estimulan la producción de proteasas en Pseudomona aeruginosa no afectan su
desarrollo cuando ésta es cultivada en caldo LB, debido a que los subproductos
resultantes de la acción de las proteasas no son necesarios para el crecimiento celular en
ese medio de cultivo. Los cambios en los medios de cultivos también pueden afectar la
producción de metabolitos secundarios con propiedades antimicrobianas (Yan, 2002;
Yan et al., 2002) Los procesos de comunicación célula-célula (quórum sensitivo) en la

66
producción de las moléculas de señalización son importantes en ambientes altamente
competitivos como los océanos. Es poco lo que conocemos sobre microorganismos
asociados al caracol, además aún no se ha comprendido cuál es la función que
desempeñan éstos microbios en él, la producción de sustancias por parte de bacterias
asociadas al caracol podrían tener un propósito y dadas las condiciones especiales en
ese hábitat ser producidas exclusivamente allí, teniendo en cuenta que se han descrito en
bacterias que coexisten con plantas u otras bacterias, la elaboración de moléculas de
señalización que estimulan la producción de compuestos necesarios para la defensa de
esos organismos (Diggle et al., 2007). La escasa información sobre el sistema
inmunológico en gastrópodos, no permite precisar sobre la función que su microbiota
desempeña en la respuesta innata a través de la producción de péptidos antimicrobianos.
Herramientas moleculares para el estudio meta genómico posiblemente ayuden a
elucidar y caracterizar los genes o familias de genes microbianos que codifican para
proteínas o péptidos antimicrobianos en ambientes naturales. Además la purificación de
proteínas a partir de los extractos antimicrobianos es indispensable para conocer con
exactitud cuáles son las moléculas metabólicamente activas. Estudios realizados en
caracoles pertenecientes a la familia planorbidae reportados en un artículo no publicado
describen cuan diversa puede ser la microbiota bacteriana (23 filos bacterianos) de estos
caracoles tan sólo en el intestino, sugiriendo la necesidad de estudios adicionales para
determinar el papel de estos microbios en el fisiología de los caracoles (Van Horn et al.,
en prensa). La presencia de bacterias asociadas al caracol pala con propiedades
antibacterianas reportadas en este estudio, hacen parte de la vasta diversidad que hasta
hace muy poco había sido ignorada en este molusco, no se sabe en realidad si estos
microbios son benéficos o patógenos para el caracol puesto que algunos se comportan
como patógenos oportunistas en humanos, sin embargo, éste es el momento para
empezar a describir las propiedades bioactivas de estas bacterias sobre otros procariotas.
7.4. ACTIVIDAD ENZIMÁTICA
Nithyanand et al., 2011 y Süβ et al., 2004 reportaron el aislamiento de γ-
Proteobacteria, Actinobacteria y Firmicutes, los mismos tres grupos bacterianos a nivel

67
de filo encontrados en este estudio. Los grupos bacterianos cultivables dominantes
fueron Psychrobacter (74%) y Halomonas (15%); resultados similares fueron obtenidos
en un estudio realizado en el sur de Okinawa en los sedimentos marinos (Dang et al.,
2009). Además tres aislados desde el intestino de los caracoles pala fueron identificados
como Exiguobacterium sp. cepas productoras de amilasas y proteasas.
Los microorganismos del intestino son muy importantes para la nutrición de muchos
invertebrados e incluso moluscos (Harris, 1993). Conociendo que el caracol pala se
alimenta de algas marinas, las cuales contienen grandes cantidades de carbohidratos
(Becker, 2007). Se ha descrito previamente que la microbiota depende de los hábitos
alimentarios de sus hospederos, las amilasas producidas por las especies de
Exiguobacterium podrían tener un papel importante en los procesos digestivos del
caracol pala. Hinsa-Leasure et al., 2010 estudiaron dos bacterias adaptadas al frio,
Exiguobacterium y Psychrobacter, que habían sido aisladas en suelos congelados.
Algunos miembros pertenecientes a estos dos géneros pueden crecer a temperaturas de
-5 y -10 °C. Todas las muestras evaluadas en este estudio fueron obtenidas en un
estudio previo (Acosta et al., 2009), las cuales fueron almacenadas por
aproximadamente un año a temperaturas entre -10 y -20 °C, esto probablemente sea una
razón que podría explicar el aislamiento de varias cepas identificadas como
Psychrobacter, y el aislamiento de tres cepas identificadas como Exiguobacterium,
trabajando con muestras obtenidas inicialmente desde aguas cálidas y poco profundas.
Las algas marinas son un buen recurso de proteínas, carbohidratos y lípidos. La
producción de hidrolasas extracelulares por las bacterias asociadas al caracol reina
podrían tener importantes implicaciones fisiológicas para el huésped. Los microbios del
intestino de moluscos como los abulones los cuales se alimentan de algas que contienen
complejos carbohidratos que resisten a la digestión, excretan enzimas capaces de
degradar esos compuestos cuando crecen fuera del tracto digestivo de los abulones
(Sawabe et al., 1995).
Las poblaciones bacterianas del caracol pueden variar entre ellas dependiendo del
hábitat al cuál pertenecen. Estudios previos demuestran que la microbiota intestinal de
algunos organismos varía de acuerdo a la dieta (Yamada et al., 2007).
El enfoque en los microorganismos no patógenos para los eucariotas cada vez toma más
importancia entre los zoólogos, la sinergia e interdependencia entre organismos

68
macroscópicos, bacterias, archaeas y hongos como consecuencia de milenios de
coevolución (Turnbaugh et al., 2007, Biagi el al., 2011).
Las enzimas extracelulares detectadas en este estudio podrían proveer un recurso para el
descubrimiento de nuevas enzimas sugiriendo por lo tanto continuar con la evaluación
de las poblaciones bacterianas relacionadas con el caracol pala.
A pesar de que no existen reportes sobre bacterias como Exiguobacterium y las
propiedades metabólicas de estos aislados asociados al caracol, estudios relacionados
con la producción de hidrolasas, llevados a cabo en sobre el género Exiguobacterium
han permitido concluir sobre la existencia de enzimas como las pululanasas
pertenecientes a la familia de las α-amilasas (Salleh et al., en prensa). Las pululanasas
participan en la hidrólisis del pululano un polisacárido formado por unidades de
maltriosa. Las células de Exiguobacterium son capaces de hidrolizar el almidón
(Kulshreshtha et al., 2010), la caseína y la gelatina, tal como se ha evidenciado en este
estudio. Bacterias psicrotolerantes aisladas desde el suelo identificadas como
Exiguobacterium sp. conocidas por sus habilidades extremofilas y la producción de
enzimas amilasas, 400 µ/ml después de nueve horas de incubación (Hosseini et al.,
2009) manifiestan que la producción de estas enzimas en Exiguobacterium aislados
desde el caracol podría mejorar. La optimización de las condiciones de crecimiento en
cultivo tales como temperatura, pH, tiempo y el uso de diferentes sustratos como fuente
de carbono y nitrógeno ayudarían a establecer el punto para la máxima producción de
estas exoenzimas.
Estudios relacionados con la producción de proteasas extracelulares por
Exiguobacterium en sobrenadantes de los cultivos muestran valores similares a los
obtenidos en esta investigación en unidades por miligramo de proteína, con una mayor
actividad cuando la enzima es purificada (Kasana y Yadav, 2007). Estudios adicionales
tales como el aislamiento de genes y la completa caracterización de las enzimas usando
sustratos específicos son sugeridos.
En un estudio previo relacionado con la purificación y caracterización de proteasas
producidas por Bacillus subtilis, la actividad enzimática medida para el extracto crudo
enzimático fue de 5 U/mg, esta actividad mejoró después de los procesos de
purificación, además la temperatura óptima para la actividad proteasa fue de 60°C (Kim

69
y Kim, 2005). Se cree que las proteasas de las bacterias asociadas al intestino de
crustáceos marinos juegan un papel importante en los procesos de nutrición de sus
huéspedes (Donachie y Zdanowski, 1998). Denner et al., 2001 reportaron una especie
de Psychrobacter capaz de hidrolizar la gelatina y la caseína aislada de un crustáceo.
La actividad lipolítica medida a partir de un extracto celular de un producto de la
clonación de un gen para lipasas obtenido desde un Psychrobacter, es similar a las
obtenidas para las especies de Psychrobacter que registraron actividad lipasa en este
estudio (Zhang et al., 2007).
Esterasas y lipasas derivadas de Exiguobacterium acetylicum han sido reportadas
previamente, con una alta actividad esterasa sobre pNPA (p-nitrofenil acetato) pero una
baja actividad lipasa sobre pNPP (p-nitrofenil palmitato) (Hwang et al., 2005), los
resultados obtenidos en este estudio no permiten concluir sobre la actividad esterasa
pero si sobre la actividad lipasa medida en el sobrenadante obtenido desde el cultivo de
la especie Exiguobacterium aislada desde el caracol pala, la cual es un poco mayor a la
registrada por la cepa de Exiguobacterium acetylicum.
Microorganismos pertenecientes al género Psychrobacter que han sido aislados en
lugares con bajas temperaturas han sido previamente reportados como productores de
enzimas lipolíticas. Además han logrado elucidar cuales son los genes implicados en el
metabolismo de lípidos, y expresar tales genes utilizando la maquinaria celular de otros
organismos procariotas. Estas hidrolasas son importantes dada las condiciones abióticas
(temperatura, pH y salinidad) bajo las cuales crecen los microorganismos a partir de los
cuales han sido aisladas. En el presente trabajo se ha llevado a cabo un estudio
preliminar de la microbiota vinculada a caracoles pala, se sabe que muchos de los
microorganismos asociados a invertebrados participan en procesos de defensa y
degradación de proteínas carbohidratos y ácidos grasos, gracias a la producción de
péptidos, enzimas.

70
8. CONCLUSIONES
Un metaorganismo está representado por más de un individuo, los procesos de
coevolución han permitido mantener unidos a cientos de individuos en comunidades
complejas en el tiempo y en el espacio. Los cambios ambientales o de lugar,
probablemente afecten este equilibrio, propiciando la formación de nuevas asociaciones
que de alguna u otra forma harán más susceptible o más exitosos a algunos. La muerte
de los corales debido a la pérdida de este equilibrio y la extinción de ciertas especies
como resultado de la influencia antropogénica sobre los ecosistemas son tan sólo un par
de ejemplos, para dimensionar el problema, es decir, el efecto de los cambios sobre un
organismo. Las relaciones entre múltiples genotipos (individuos de especies diferentes)
y entre fenotipos (proteínas y hormonas), son estudiadas ahora. Aún es poco lo
conocido sobre la diversidad microbiana del caracol comparado con otros invertebrados
marinos, no obstante el desarrollo de este estudio mediante la implementación de
técnicas dependientes e independientes de cultivo ha permitido empezar a hacer juicios
e hipótesis sobre la compleja estructura de sus comunidades bacterianas y el potencial
de las bacterias asociadas con propiedades metabólicas antibacterianas e hidrolíticas. Es
claro que el conocimiento sobre la diversidad genética del caracol pala es importante,
pero también lo es, el conocer la diversidad microbiana, para explicar en forma conjunta
los cambios que suceden entre poblaciones; ¿cuál es la dinámica de las poblaciones
microbianas en el caracol, de acuerdo al hábitat, en individuos sanos y enfermos, en
caracoles pertenecientes a poblaciones distantes y aisladas reproductivamente?
De acuerdo con el planteamiento anterior y los resultados encontrados en este estudio se
ha llegado a las siguientes conclusiones:
Se ha podido apreciar diferencias significativas entre las poblaciones bacterianas
asociadas a caracoles silvestres y en cautiverio, lo cual sugiere el cambio de las
poblaciones de acuerdo al hábitat donde se encontraban los caracoles, sin

71
embargo, se debe ser muy cuidadoso con los análisis realizados cuando se utiliza
solamente el DGGE.
En el análisis de los clusters los patrones de bandeo de las muestras de DNA
obtenidas del alimento de caracoles en cautiverio y silvestres estaban más
relacionados con las muestras de DNA de la baba de caracoles en cautiverio. El
cambio en las condiciones ambientales, posiblemente esté vinculado a estas
variaciones entre las poblaciones bacterianas naturales de los caracoles, con
predominio de nuevas especies sobre las especies nativas, o el florecimiento de
especies bacterianas con mejores respuestas adaptativas, las cuales estaban
presentes en el alimento de los caracoles. La restricciones en el flujo del agua y
una mayor densidad de alimento en los estanques comparado con el hábitat de
los caracoles silvestres, podría también explicar la similitud en los perfiles de
bandeo entre las muestras derivadas de la baba de caracoles en cautiverio y los
perfiles de las muestras de alimento.
Los grupos bacterianos identificados a partir de las secuencias de las bandas de
DGGE son diferentes a los identificados a partir de las secuencias de los
microorganismos cultivados usados en las pruebas enzimáticas. La mayoría de
los aislados identificados por secuenciación fueron relacionadas con bacterias
que no habían sido cultivadas y reportadas en trabajos llevados a cabo en
hospederos y hábitats marinos. Así, se cuenta con un nuevo grupo de bacterias
que no han sido estudiadas, y de acuerdo a los resultados preliminares, son
prometedoras en cuanto a producción de enzimas, además de tener otras posibles
aplicaciones, por ejemplo, la producción de péptidos antimicrobianos.
Adicionalmente algunas de las secuencias encontradas se relacionaron patógenos
oportunistas de plantas y animales, el papel de estos microbios en organismos
acuáticos es aún poco comprendido, otros de los microorganismos identificados
han sido reportado por haberse encontrado en asociación con otros eucariotas
secretando metabolitos con propiedades bioactivas contra varios organismos.
Los microorganismos identificados en este estudio no representan toda la

72
diversidad microbiana asociada al caracol pala, pero probablemente sean
importantes para el hospedero.
La actividad antibacteriana de los aislados asociados al caracol, no es específica
de un grupo bacteriano, la recuperación de más bacterias cultivables
pertenecientes a otras agrupaciones filogenéticas desde los caracoles palas, sería
de gran utilidad para complementar esta investigación e inferir sobre las
propiedades antibacterianas de cada grupo. Miembros del mismo género a los
identificados en este estudio Halomonas y Psychrobacter, han sido reportados
como productores de moléculas antimicrobianas (biocinas, dipéptidos) en
investigaciones anteriores. La elucidación de las estructuras moleculares de los
metabolitos con propiedades antibacterianas en los extractos evaluados debe ser
considerada en estudios posteriores.
Teniendo en cuenta los resultados obtenidos, las cepas que presentaron
producción de enzimas hidrolíticas podrían llegar a tener una aplicación
potencial a nivel industrial, en el caso de procesos fermentativos (amilasas y
lipasas), la fabricación de detergentes (proteasas) la industria de la leche
(lipasas), el tratamiento de residuos industriales (proteasas), la producción de
papel (celulasas), entre otros. Las enzimas halotolerantes especialmente aquellas
aisladas de organismos marinos son ampliamente estudiadas gracias a su amplia
versatilidad, la capacidad para adaptarse a diferentes pH, temperaturas, en
presencia de iones u otras moléculas. Evaluar todas estas condiciones en las
enzimas de los organismos descritos en este trabajo, permitiría conocer el mejor
comportamiento de estas moléculas e incluso localizar y caracterizar los genes
enzimáticos o mejorar un proceso industrial. Al comparar las extractos
enzimáticos con enzimas comerciales se pudo apreciar que la actividades
estaban por debajo de los valores de actividad registrados para enzimas
comerciales como las α-amilasas y las proteasas, por lo tanto los métodos y las
condiciones en las que ocurre el proceso deberán ser replanteados en términos de
la temperatura, pH, salinidad, sustrato, entre otros aspectos, en aras de conseguir
una mejor producción de las enzimas en los extractos que puedan ser

73
comparables con las actividades de las enzimas comerciales e importantes para
la industria en el país.
En líneas generales en este estudio el análisis de las secuencias del rDNA 16S y
de los perfiles obtenidos por DGGE mediante análisis de similitud, permiten
inferir sobre comunidades bacterianas únicas, diversas e igualmente complejas
en el caracol, adicionalmente es el primer informe sobre la presencia de cepas
productoras de hidrolasas extracelulares. La información obtenida en estudio
permitirá enfocarse en aquellas cepas que podrían llegar a ser importantes en
procesos biotecnológico tras una mejora en las condiciones en los procesos de
producción de biocatalizadores por parte de los aislados que han registrado
mejores actividades hidrolasas.

74
BIBLIOGRAFÍA
1 Acosta EA, Gómez E, Romero M, Cadavid G, Moreno CX. Identificación
molecular de poblaciones bacterianas asociadas al caracol pala (Strombus gigas) del
Caribe colombiano. Acta biol Colomb. 2009;14(2):69-84.
2 Ahmadnia HR, Farhangi M, Rafiee G, Noori F. Modulating gut microbiota and
digestive enzyme activities of Artemia urmiana by administration of different levels
of Bacillus subtilis; and Bacillus licheniformis. Aquacult Int. 2012:1-13.
3 Akoh C, Chang S, Lee G, Shaw J. A review on hydrolytic enzymes in the treatment
of wastewater with high oil and grease content. Bioresour Technol. 2006;97:2195-
2210.
4 Alcolado PM. Crecimiento, variaciones morfológicas de la Concha y algunos datos
biológicos del cobo Strombus gigas L. (Mollusca, Mesogastrópoda). Acad. Cien.
Cuba Ser. Oceanol. 1976 34-36.
5 Aldana D, Frenkiel L, Brulé T, Montero J, Baqueiro E. Occurrence of
Apicomplexa-like structures in the digestive gland of Strombus gigas throughout the
Caribbean. J Invertebr Pathol. 2011;106(2):174-178.
6 Althoff K, Schütt C, Steffen R, Batel R, Müller W. Evidence for a symbiosis
between bacteria of the genus Rhodobacter and the marine sponge Halichondria
panicea: harbour also for putatively toxic bacteria. Mar Biol. 1998;130:529-536.
7 Altschul SF, Madden TL, Schäffer AA, Zhang J, Zhang Z, Miller W. Gapped
BLAST and PSI-BLAST: a new generation of protein database search programs.
Nucleic Acids Res. 1997;25(3389-3402).
8 Anand TP, Bhat AW, Shouche YS, Roy U, Siddharth J, Sarma SP. Antimicrobial
activity of marine bacteria associated with sponges from the waters off the coast of
South East India. MicrobiolRes. 2006;161(3):252-262.
9 Anandham R, Gandhi P, Madhaiyan M, Sa T. Potential plant growth promoting
traits and bioacidulation of rock phosphate by thiosulfate oxidizing bacteria isolated
from crop plants. J Basic Microbio. 2008;48:439-447.

75
10 Asgher M, Javaid Asad M, Rahman SU, Legge RL. A thermostable α-amylase from
a moderately thermophilic Bacillus subtilis strain for starch processing. J Food Eng.
2007;79:950-955.
11 Ashen JB, Goff LJ. Molecular and ecological evidence for species specificity and
coevolution in a group of marine algal-bacterial symbioses. Appl Environ
Microbiol. 2000;66(7):3024-3030.
12 Atlas RM. Handbook of Microbiological Media. Segunda ed. Boca Raton, FL:
CRC 1996.
13 Babu KR, Satyanarayana T. Extracellular calcium inhibited α-amylase of Bacillus
coagulans B49. Enzyme Microb Technol. 1993;15:1066-1069.
14 Bailey JE, Ollis DF. In: Biochemical Engineering Fundamentals. Tokyo: McGraw-
Hill Ltd, Kogakusha; 1977. p. 155–220.
15 Baily MJ, Biely P, Pountanen K. Interlaboratory testing of methods for assay of
xylanase activity. J Biotechnol. 1992;23:257-270.
16 Bao L, Huang Q, Chang L, Zhou J, Lu H. Screening and characterization of a
cellulase with endocellulase and exocellulase activity from yak rumen metagenome.
J Mol Catal B Enzym. 2011;73:104-110.
17 Barett AJ. Proteolytic enzymes: serine and cysteine peptidases. Methods Enzymol.
1994;244:1-15.
18 Barua S, Tripathi S, Chakraborty A, Ghosh S, Chakrabarti K. Characterization and
crop production efficiency of diazotrophic bacterial isolates from coastal saline
soils. Microbiol Res. En prensa 2011.
19 Becker EW. Micro-algae as a source of protein. Biotechnol Adv. 2007;25:207-210.
20 Behar AYB, Jurkevitch E. Enterobacteria-mediated nitrogen fixation in natural
populations of the fruit fly Ceratitis capitata. Mol Ecol. 2005;14:2637-2643.
21 Berdy J. Bioactive microbial metabolites a personal view. J Antibiot. 2005;58:1-26.
22 Berg CJ, Olsen DA. Conservation and management of Queen Conch (Strombus
gigas) fisheries in the Caribbean. NY: JF Caddy (ed.) Marine invertebrate
fisheries:Their assessment and management. Wiley and Sons; 1989. p. 421-442.
23 Bernardet JF, Vancanneyt M, Matte-Taillieza O, Grisez L, Tailliez P, Bizet C,

76
Nowakowski M, Kerouault B, Swings J. Polyphasic study of Chryseobacterium
strains isolated from diseased aquatic animals. Syst Appl Microbiol. 2005;28:640–
660.
24 Bernfeld P. Amylase, alpha and beta. Methods Enzymol. 1955;1:149.
25 Biagi E, Candela M, Fairweather-Tait S, Franceschi C, Brigidi P. Aging of the
human metaorganism: the microbial counterpart. Age; 2011.
26 Bitzer J, Grosse T, Wang L, Lang S, Beil W, Zeeck A. New Aminophenoxazinones
from a Marine Halomonas sp: Fermentation, Stucture Elucidation, and Biological
Activity. J Antibiot. 2006;59(2):86-92.
27 Boland W, Frößl C, Lorenz M. Esterolytic and lipolytic enzymes in organic
synthesis. Synthesis. 1991 1049-1072.
28 Borges J, Razzera G, Vernal J, Cristiano F, Brod A, Maisonnave AC, et al.
Structural stability of Staphylococcus xylosus lipase is modulated by Zn2+
ions.
Biochim Biophys Acta. 2011;1814(9):1120-1126.
29 Bos D, Posada D. Using models of nucleotide evolution to build phylogenetic trees.
Dev. Comp Immunol. 2005;29:211-227.
30 Bosch T, Mcfall-Ngai MJ. Metaorganisms as the new frontier. Zoology.
2011;114(4):185-190.
31 Bothe H, Ferguson SJ, Newton WE. Biology of the Nitrogen Cycle. Amsterdam:
Elsevier; 2007. p. 180-192.
32 Boyd KG, Adams DR, Burgess JG. Antibacterial and repellent activities of marine
bacteria associated with algal surfaces. Biofouling. 1999;14:227-236.
33 Bradford MM. A rapid and sensitive method for the quantification of microgram
quantities of protein utilizing the principle of protein-dye binding. Anal Biochem.
1976;72:248-254.
34 Bräutigam A. CITES: A Conservation Tool. Washington, DC: Center for Marine
Conservation; 1992.
35 Brownell WN, Stevely JM. The biology, fisheries and management of the Queen
Conch,Strombus gigas. Mar Fish Rev. 1981;43(7):1-12.
36 Bull A, Ward A, Goodfellow M. Search and discovery strategies for biotechnology:

77
the paradigm shift. Microbiol Mol Biol Rev. 2000;64(3):573-606.
37 Burhan A, Nisa U, Gokhan C, Omer C, Ashabil A, Osman G. Enzymatic properties
of a novel thermophilic, alkaline and chelator resistant amylase from an alkalophilic
Bacillus sp. Isolate ANT-6. Process Biochem. 2003;38:1397-1403.
38 Cabello FC. Antibiotics and aquaculture in Chile: implications for human and
animal health. Rev Méd Chile. 2004;132:1001-1006.
39 Chandrasekaran M. Industrial enzymes from marine microorganisms: The Indian
Scenario. J Mar Biotechnol. 1997;5:86–89.
40 Chen WM, Chang J, Chiu CH, Chang SC, Chen WC, Jiang CM. Caldimonas
taiwanensis sp. nov., α-amylase producing bacterium isolated from a hot spring.
Syst Appl Microbiol. 2005;28:415-420.
41 Chen WM, Lin CY, Chen CA, Wang JT, Sheu SY. Involvement of an L-amino acid
oxidase in the activity of the marine bacterium Pseudoalteromonas flavipulchra
against methicillin-resistant Staphylococcus aureus. Enzyme Microb Technol.
2010;47(1-2):52-58.
42 Childress J, Girguis P. The metabolic demands of endosymbiotic chemoautotrophic
metabolism on host physiological capacities. J Exp Biol. 2010;214:312-325.
43 Coolbear T, Eames CW, Casey Y, Daniel RM, Morgan HW. Screening of strains
identified as extremely thermophilic bacilli for extracellular proteolytic activity and
general properties of the protease from two of the strains. J Appl Bacteriol.
1991;71:252-264.
44 Creswell R. An historical overview of queen conch mariculture. Caracas,
Venezuela: R.S. Appeldoorn and B. Rodríguez (Eds.): Queen Conch biology,
fisheries and mariculture, Fundación Científica Los Roques; 1994. p. 223-230.
45 Dahllöf I. Molecular community analysis of microbial diversity. Curr Opin
Biotechnol. 2002;13:213-217.
46 Dang H, Zhu H, Wang J, Li T. Extracellular hydrolytic enzyme screening of
culturable heterotrophic bacteria from deep-sea sediments of the Southern Okinawa
Trough. World J Microbiol Biotech. 2009;25:71-79.
47 Das S, Ward LR, Burke C. Prospects of using marine actinobacteria as probiotics

78
in aquaculture. Appl Microbiol Biotechnol. 2008;81:419-429.
48 Davis KE, Joseph SJ, Janssen PH. Effects of growth, inoculum size, and incubation
time on culturability and isolation of soil bacteria. Appl Environ Microbiol.
2005;71:826-834.
49 De Oliveira AN, De Oliveira ILA, Andrade JS, Chagas JAF. Rhizobia amylase
production using various starchy substances as carbon substrates. Braz J Microbiol.
2007;38:208-216.
50 Delong EF. Archaea in coastal marine environments. Proc Natl Acad Sci USA.
1992;89:5685-5689.
51 Demain AL, Newcomb M, Wu J. Cellulase, clostridia, and ethanol. Microbiol Mol
Biol Rev. 2005;69:124-154.
52 Denner EB, Mark B, Busse HJ, Turkiewicz M, Lubitz W. Psychrobacter
proteolyticus sp. nov., a psychrotrophic, halotolerant bacterium isolated from the
Antarctic krill Euphausia superba Dana, excreting a cold-adapted metalloprotease.
Syst Appl Microbiol. 2001;24:44-53.
53 Derewenda U, Brzozowski AM, Lawson DM, Derewenda ZS. Catalysis at the
interface: the anatomy of a conformational change in a triglyceride lipase.
Biochemistry. 1992;31:1532-1541.
54 Diggle S, Crusz S, Cámara M. Quorum sensing. Curr Biol. 2007;7:907-910.
55 Ding L, Yokota A. Proposals of Curvibacter gracilis gen. nov., sp. nov. and
Herbaspirillum putei sp. nov. for bacterial strains isolated from well water and
reclassification of [Pseudomonas] huttiensis, [Pseudomonas] lanceolata,
[Aquaspirillum] delicatum. Int J Syst Evol Microbiol. 2004;54:2223–2230.
56 Donachie SP, Zdanowski MK. Potential digestive function of bacteria in krill
Euphausia superba stomach. Aquat Microb Ecol. 1998;14(2):129-136.
57 Drauz K, Waldmann H. Enzyme Catalysis in Organic Synthesis: A Comprehensive
Handbook. New York: VCH Publ; 1995. p. 504-545.
58 Du M, Chenj, Zhang X, Li A, Li Y. Characterization and resuscitation of viable but
nonculturable Vibrio alginolyticus VIB 283. Arch Microbiol. 2007;188(3):283-288.
59 Ducklow HW, Boyle PJ, Maugel PW, Strong C, Mitchell R. Bacterial Flora of the

79
Schistosome Vector Snail Biomphalaria glabrata. Appl Environ Microbiol.
1979;38(4):667-672.
60 Ducklow HW, Clausen K, Mitchell R. Ecology of bacterial communities in the
schistosomiasis vector snail Biomphalaria glabrata. Microb Ecol. 1981;7:253-274.
61 Edenborn SL, Sexstone AJ. DGGE fingerprinting of culturable soil bacterial
communities complements culture-independent analyses. Soil Biol Biochem.
2007;39(7):1570–1579.
62 Eichner C, Erb RW, Timmis KT, Wagner-Döbler I. Thermal gradient gel
electrophoresis analysis of bioprotection from pollutant shocks in the activated
sludge microbial community. Appl Environ Microbiol. 1999;65:102–109.
63 Ekborg NA, Morrill W, Burgoyne AM, Li L, Distel DL. CelAB, a multifunctional
cellulase encoded by Teredinibacter turnerae T7902T, a culturable symbiont
isolated from the wood-boring marine bivalve Lyrodus pedicellatus. Appl Environ
Microbiol. 2007;73(23):7785-7788.
64 El-Masry HA, Fahmy HH, Abdelwahed ASH. Synthesis and antimicrobial activity
of some new benzimidazole derivatives. Molecules. 2000;5:1429-1438.
65 Espejo RT, Feijo CG, Romero J, Vasquez M. PAGE analysis of the heteroduplexes
formed between PCR-amplified 16S rRNA genes: estimation of sequence similarity
and rADN complexity. Microbiology. 1998;144:1611-1617.
66 Faith DP, Humphrey CL, Dostine PL. Statistical power and BACI designs in
biological monitoring: comparative evaluation of measures of community
dissimilarity based on benthic macroinvertebrate communities in Rockhole Mine
Creek, Northern Territory, Australia. Aust. J. Mar. Freshwat. 1991. Res. 42:589–
602.
67 Farrell RL, Hata K, Wall MB. Solving pitch problems in pulp and paper processes
by the use of enzymes or fungi. Adv Biochem Eng Biotechnol. 1997;57:197-212.
68 Faulkner DJ, Newman DJ, Cragg GM. Investigations of the marine flora and fauna
of the Islands of Palau. Nat Prod Rep. 2004;21(1):50-76.
69 Faulkner DJ. Marine pharmacology. Antonie Van Leeuwenhoek. 2000;77(2):135-
145.

80
70 Fenical W. Chemical studies of marine bacteria: Developing a new resource. Chem
Rev. 1993;93:1673-1683.
71 Fera MT, Maugeri T, Gugliandolo C, Bonanno D, La Camera E, Papasergi S, et al.
Occurrence of Burkholderia cepacia complex, Ralstonia and Pandoraea species
DNAs in the coastal environment of the Straits of Messina (Italy). Mar Pollut Bull.
2007;54(6):803-808.
72 Ferris MJ, Ward DM. Seasonal distributions of dominant 16S rRNA-defined
populations in a hot spring microbial mat examined by denaturing gradient gel
electrophoresis. Appl Environ Microbiol. 1997;63:1375-1381.
73 Fox JW, Shannon JD, Bjarnason JB. Proteinases and their inhibitors in
biotechnology. Enzymes in biomass conversion. ACS Symp Ser Am Chem Soc.
1991;460:62-79.
74 Frank JF, Christen GL, Bullerman LB. Marshall. Standard methods for the
examination of dairy products. 16th ed. American Public Health Association
Washington, D.C.; 1993.
75 Fuchs Y, Chihu L, Conde C, González VM, Noguez AH, Calderón E, et al.
Mecanismos Moleculares de Resistencia Bacteriana. Salud Pública de México.
1994;36:428-438.
76 Gauthier G, Gauthier M, Christen R. Phylogenetic analysis of the genera
Alteromonas, Shewanella, and Moritella using genes coding for small-subunit
rRNA sequences and division of the genus Alteromonas into two genera,
Alteromonas (emended) and Pseudoalteromonas gen. nov. Int J Syst Bacteriol.
1995;45(4):755-761.
77 Glazer RA, Berg CJ. Growth and mortality of the Queen Conch, Strombus gigas, in
Florida. Proc Gulf Carib Fish Inst. 1992;42:153-157.
78 Godfrey T, West S. Industrial Enzymology - Application of Enzymes in Industry.
2nd ed. Macmillan, London 1996.
79 Gomare S, Kim HA, Ha JH, Lee MW, Park JM. Isolation of the polysaccharidase
producing bacteria from the gut of sea snail, Batillus cornutus. Korean Journal of
Chemical Engineering. 2011;28(5):1252-1259.

81
80 González E. Catch and Effort in Queen Conch Strombus gigas (Mesogastropoda:
Strombidae) fishery in the Archipelago of San Andres, Providencia and Santa
Catalina, Colombia. Mexico 55th Proc. Gulf. Carib. Fish Inst. 2002.
81 Green SJ, Leigh MB, Neufeld JD. Denaturing gradient gel electrophoresis (DGGE)
for microbial community analysis.Microbiology of Hydrocarbons, Oils, Lipids, and
Derived Compounds (Timmis KN, ed), Springer, Heidelberg. 2009. p. 4137–4158.
82 Gupta R, Beg QK, Lorenz P. Bacterial alkaline proteases:molecular approaches and
industrial application. Appl Microbial Biotechnol. 2002;59(1):15-32.
83 Gupta R, Gupta N, Rathi P. Bacterial lipases: an overview of production,
purification and biochemical properties. Appl Microbiol Biotechnol.
2004;64(6):763-781.
84 Gupta R, Rathi P, Gupta N, Bradoo S. Lipase assays for conventional and
molecular screening. Biotechnol Appl Biochem. 2003;37:63-71.
85 Hall TA. BioEdit: a user-friendly biological sequence alignment editor and analysis
program for Windows 95/98/NT. Nucl Acids Symp Ser. 1999;41:95-98.
86 Hammer Ø, Harper DAT, Ryan PD. PAST: Paleontological statistics software
package for education and data analysis. Palaeontologia Electronica. 2001;4:1-9.
87 Hankin L, Anagnostakis S. Solid media containing carboxymethyl cellulose to
detect CM celllulase activity of microorganisms. J Gen Microbiol. 1977;98:109-
115.
88 Harris JM. The presence, nature, and role of gut microflora in aquatic invertebrates:
a synthesis. Microb Ecol. 1993;25(3):195-231.
89 Harwood J. The versatility of lipases for industrial uses. Trends Biochem Sci.
1989;14(4):125-126.
90 Hasanuzzaman M, Umadhay K, Zsiros S, Morita N, Nodasaka Y, Yumoto I, et al.
Isolation, identification, and characterization of a novel, oil-degrading bacterium,
Pseudomonas aeruginosa T1. Curr Microbiol. 2004;49(2):108-114.
91 Head IM, Saunders JR, Pickup RW. Microbial Evolution, Diversity, and Ecology:
A Decade of Ribosomal RNA Analysis of Uncultivated Microorganisms. Microb
Ecol. 1998;35(1):1-21.

82
92 Heindl H, Wiese J, Thiel V, Imhoff JF. Phylogenetic diversity and antimicrobial
activities of bryozoan-associated bacteria isolated from Mediterranean and Baltic
Sea habitats. Syst Appl Microbiol. 2010;33(2):94-104.
93 Hinsa-Leasure SM, Bhavaraju L, Rodrígues JL, Bakermans C, Gilichinsky DA,
Tiedje JM. Characterization of a bacterial community from a Northeast Siberian
seacoast permafrost sample. FEMS Microbiol Ecol. 2010;74(1):103-113.
94 Holmstrom C, Kjelleberg S. Marine Pseudoalteromonas species are associated with
higher organisms and produce biologically active extracellular agents. FEMS
Microbiol Lett. 1999;30(4):285-293.
95 Horchani H, Mosbah H, Salem NB, Gargouri Y, Sayari A. Biochemical and
molecular characterization of a thermoactive, alkaline and detergent-stable lipase
from a newly isolated Staphylococcus aureus strain. J Mol Catal B Enzym.
2009;56(4):237-245.
96 Hosseini F, Safari A, Rad SA, Khaniki GRB, Zahiri HS. Biochemical
characterization of an amylase produced by a psychrotolerant Exiguobacterium sp.
isolated from soils around Tehran. N Biotechnol. 2009;25:S64.
97 Hugenholtz P, Goebel B, Pace NR. Impact of culture independent studies on the
emerging phylogenetic view of bacterial diversity. J Bacteriol. 1998;180(18):4765–
4774.
98 Hwang BY, Kim JH, Kim J, Kim BG. Screening of Exiguobacterium acetylicum
from soil samples showing enantioselective and alkalotolerantesterase activity.
Biotechnol Bioprocess Eng. 2005;10(4):367–371.
99 Jaeger KE, Dijkstra BW, Reetz MT. Bacterial biocatalysts: molecular biology,
three-dimensional structures, and biotechnological applications of lipases. Annu
Rev Microbiol. 1999;53:315-351.
100 Jaeger KE, Reetz MT. Microbial lipases form versatile tools for biotechnology.
Trends Biotechnol. 1998;16(9):396-403.
101 Jensen PR, Fenical W. Strategies for the discovery of secondary metabolites from
marine bacteria: ecological perspectives. Annu Rev Microbiol. 1994;48:559-584.
102 Jiang C, Wu LL, Zhao GC, Shen PH, Jin K, Hao ZY, et al. Identification and

83
characterization of a novel fumarase gene by metagenome expression cloning from
marine microorganisms. Microb Cell Fact. 2010;9:91.
103 Joint I, Mühling M, Querellou J. Culturing marine bacteria an essential prerequisite
for biodiscovery. Microb Biotechnol. 2010;3(5):564-575.
104 Jukes TH, Cantor CR. Evolution of protein molecules. In Munro HN, Mammalian
Protein Metabolism. New York: Academic Press; 1969. p. 21-132.
105 Kaieda M, Samukawa T, Kondo A, Fukuda H. Effect of methanol and water
contents on production of biodiesel fuel from plant oil catalyzed by various lipases
in a solvent-free system. J Biosc Bioeng. 2001;91(1):12-15.
106 Kasana RC, Salwan R, Dhar H, Dutt S, Gulati A. A rapid and easy method for the
detection of microbial cellulases on agar plates using gram’s iodine. Curr
Microbiol. 2008;57(5): 503-507.
107 Kasana RC, Yadav SK. Isolation of a psychrotrophic Exiguobacterium sp. SKPB5
(MTCC 7803) and characterization of its alkaline protease. Curr Microbiol.
2007;54(3):224–229.
108 Kennedy IR, Choudhury AT, Kecskes ML. Non-symbiotic bacterial diazotrophs in
crop-farming systems: can their potential for plant growth promotion be better
exploited?. SoilBiolBiochem. 2004;36(8):1229–1244.
109 Kim WJ, Kim SM. Purification and characterization of Bacillus subtilis JM-3
protease from anchovy sauce. J Food Biochem. 2005;29:591-610.
110 Kimura M. A simple method for estimating evolutionary rate of base substitutions
through comparative studies of nucleotide sequences. J Mol Evol 1980;16:111–20.
111 Kirchman DL, Dittel AI, Malmstrom RR, Cottrell MT. Biogeography of major
bacterial groups in the Delaware estuary. Limnol Oceanogr. 2005; 50:1697–1706.
112 Kisand, Wikner J. Combining culture-dependent and - independent methodologies
for estimation of richness of estuarine bacterioplankton consuming riverine
dissolved organic matter. Appl Environ Microbiol. 2003;69(6):3607–3616.
113 Kobayashi H, Endo K, Sakata S, Mayumi D, Kawaguchi H, Ikarashi M, et al.
Phylogenetic diversity of microbial communities associated with the crude-oil,
large-insoluble-particle and formation-water components of the reservoir fluid from

84
a non-flooded high-temperature petroleum reservoir. J Biosci Bioeng.
2012;113(2):204-210.
114 Kopczynski ED, Bateson MM, Ward DM. Recognition of chimeric small-subunit
ribosomal DNAs composed of genes from uncultivated microorganisms. Appl
Environ Microbiol. 1994;60(2):746-748.
115 Kubota Y, Watanabe Y, Otsuka H, Tamiya T, Tsuchiya T, Matsumoto JJ.
Purification and characterization of an antibacterial factor from snail mucus. Comp
Biochem Physiol C. 1985;82(2):345-348.
116 Kuddus M, Ramteke PW. Production optimization of an extracellular cold-active
alkaline protease from Stenotrophomonas maltophilia MTCC 7528 and its
application in detergent industry. Afr J Microbiol Res. 2011;5(7):809-816.
117 Kulshreshtha NM, Kumar A, Dhall P, Gupta S, Bisht G, Pasha S, et al.
Neutralization of alkaline industrial wastewaters using Exiguobacterium sp. Int
Biodeterior Biodegradation. 2010;64(3):191-196.
118 Kvennefors ECE, Sampayo E, Ridgway T, Barnes AC, Hoegh-Guldberg O.
Bacterial communities of two ubiquitous Great Barrier Reef corals reveals both
site- and speciesspecificity of common bacterial associates. PLoS One.
2010;5:e10401.
119 Lanyi B. Classical and rapid identification methods for medically important
bacteria. Methods Microbiol. 1988;19:1-67.
120 Li H, Lee BC, Kim TS, Bae KS, Hong J, Choi SH, et al. Bioactive cyclic dipeptides
from a marine sponge-associated bacterium, Psychrobacter sp. Biomol Ther.
2008;16:356-363.
121 Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. Protein measurement with the
Folin phenol reagent. J Biol Chem. 1951;193(1):265–275.
122 Margesin R, Schinner F. Potential of halotolerant and halophilic microorganisms
for biotechnology. Extremophiles. 2001;5(2): 73-83.
123 Matsumiya Y, Wakita D, Kimura A, Sanpa S, Kubo M. Isolation and
Characterization of a Lipid-Degrading Bacterium and Its Application to Lipid-
Containing Wastewater Treatment. J Biosc Bioeng. 103(4):325-330.

85
124 Mccarthy K. Queen conch (Strombus gigas) standardized catch rates from the
Puerto Rico and US Virgin Island commercial fisheries. SEDAR 14-AW-06.
Sustainable Fisheries Division Contribution. 2007: 1-20.
125 Mcinerney JO, Wernecke M, Mullarkey M, Powell R. Bacteria and Archaea:
molecular techniques reveal astonishing novel diversity. Biodiversity. 2002;3(3):3-
10.
126 Mearns-Spragg A, Bregu M, Boyd KG, Burgess JG. Cross-species induction and
enhancement of antibiotic production by epiphytic bacteria from marine algae and
invertebrates, after exposure to terrestrial bacteria. Lett Appl Microbio.
1998;27:142-146.
127 Miller GL. Use of dinitrosalicylic acid reagent for determination of reducing sugar.
Anal Chem. 1959;31:426-428.
128 Mohammadi SA, Prasanna BM. Analysis of genetic diversity in crop plants -
Salient statistical tools and considerations. Crop Sci. 2003;43:1235-1248.
129 Mohapatra BR, Bapuji M, Sree A. Production of Industrial Enzymes (Amylase,
Carboxymethylcellulase and Protease) by Bacteria Isolated from Marine Sedentary
Organisms. Acta biotechnology. 2003;23(1):75-84.
130 Mora O. Análisis de la pesquería del caracol pala (Strombus gigas L.) en Colombia.
In: Appeldoorn RS, Rodríguez B, editors. Queen Conch biology, fisheries and
mariculture, Fundación Científica Los Roques, Caracas, Venezuela; 1994. p. 137-
144.
131 Moreno CX, Moy F, Daniels TJ, Godfrey HP, Cabello FC. Molecular analysis of
microbial communities identified in different developmental stages of Ixodes
scapularis ticks from Westchester and Dutchess Counties, New York.
Environmental Microbiology. 2006;8(5):761-772.
132 Moreno C, Romero J, Espejo RT. Polymorphism in repeated 16S rRNA genes is a
common property of type strains and environmental isolates of the genus Vibrio.
Microbiology. 2002;148:1233-1239.
133 Murray AE, Hollibaugh JT, Orrego C. Phylogenetic compositions of
bacterioplankton from two California estuaries compared by denaturing gradient gel
electrophoresis of 16S rDNA fragments. Appl Environ Microbiol.

86
1996;62(7):2676-2680.
134 Muyzer G, Smalla K. Application of denaturing gradient gel electrophoresis
(DGGE) and temperature radient gel electrophoresis (TGGE) in microbial ecology.
Antonie Van Leeuwenhoek. 1998;73:127-141.
135 Muyzer G, De Waal EC, Uitterlinden AG. Profiling of complex microbial
populations by denaturing gradient gel electrophoresis analysis of polymerase chain
reaction amplified genes coding for 16S rRNA. Appl Environ Microbiol.
1993;59(3):695-700.
136 Najafi MF, Deobagkar D, Deobagkar D. Purification and characterization of an
extracellular α-amylase from Bacillus subtilis AX20. Protein Expr Purif.
2005;41(2):349-354.
137 Nakamura Y, Sawada T, Morita Y, Tamiya E. Isolation of a psychrotrophic
bacterium from the organicresidue of water tank keeping rainbow trout
andantibacterial effect of violet pigment produced from thestrain. Biochem Eng J.
2002;12:79-86.
138 Nathan C. Antibiotics at the crossroads. Nature. 2004;431:899–902.
139 Nei M, Li WH. Mathematical models for studying genetic variation in terms of
restriction endonucleases. Proc Natl Acad Sci USA. 1979;76(10):5269-5273.
140 Nikolausz M, Sipos R, Révész S, Székely A, Márialigeti K. Observation of bias
associated with re-amplification of DNA isolated from denaturing gradient gels.
FEMS Microbiol Lett. 2005;244(2):385-390.
141 Nishimura H, Kitano Y, Inoue T, Nomura K, Sako Y. Purification and
characterization of membrane-associated hydrogenase from the deep-sea
epsilonproteobacterium Hydrogenimonas thermophila. Biosci Biotechnol Biochem.
2010;74(8):1624-1630.
142 Nithyanand P, Manju S, Karutha Pandian S. Phylogenetic characterization of
culturable actinomycetes associated with the mucus of the coral Acropora
digitiferafrom Gulf of Mannar. FEMS Microbiol Lett. 2011;314(112-118):2.
143 Nubel U, Engelen B, Felske A, Snaidr J, Wieshuber A, Amann R, et al. Sequence
heterogeneities of genes encoding 16s rRNAs in Paenibacillus polymyxa detected

87
by temperature gradient gel electrophoresis. J Bacteriol. 1996;178(19):5636-5643.
144 Ogier JC, Son O, Gruss A, Tailliez P, Delacroix-Buchet A. Identification of the
bacterial microflora in dairy products by temporal temperature gradient gel
electrophoresis. Appl Environ Microbiol. 2002;68(8):3691-3701.
145 Olempska-Beer ZS, Merker RI, Ditto MD, Dinovi MJ. Fda: Food-processing
enzymes from recombinant microorganisms- a review. Regul Toxicol Pharmacol.
2006;45(2):144-158.
146 Onofre-Lemus J, Hernández-Lucas I, Girard L, Caballero-Mellado J. ACC. (1-
aminocyclopropane-1-carboxylate) deaminase activity, a widespread trait in
Burkholderia species, and its growth promoting effect on tomato plants. Appl
Environ Microbiol. 2009;75(20):6581-6590.
147 Ospina JF, Chiquillo E, Gallo J. Evaluación de captura y esfuerzo del caracol pala
Strombus gigas (Linnaeus, 1758) en el Departamento Archipiélago de San Andrés,
Providencia y Santa Catalina (Caribe Colombiano).; Boletín Científico No 4. Santa
Fe de Bogotá. 1996. p. 125-139.
148 Outtrup H, Boyce COL. Microbial proteinases and biotechnology. In: Microbial
Enzymes and Biotechnology (Fogarty W,KCT,E, editor. London and New York.
Science, Elsevier Applied; 1990. p. 227-254.
149 Pérez-Avalos O, Sánchez-Herrera LM, Salgado LM, Ponce-Noyola T. A
Bifunctional endoglucanase/endoxylanase from Cellulomonas flavigena with
potential use in industrial processes at different pH. Curr Microbiol. 2008;57(1):39-
44.
150 Poldermans B. Proteolytic Enzymes. In: Proteolytic Enzymes inIndustry:
Production and Applications. Gerhartz W, editor. VCH Publishers, Weinheim,
Germany; 1990. p. 108-123.
151 Pomponi SA. The bioprocess-technological potential of the sea. J Biotechnol.
1999;35:5–13.
152 Pukall R, Kramer I, Rohde M, Stackebrandt E. Microbial diversity of cultivatable
bacteria associated with the North Sea bryozoan Flustra foliacea. Syst Appl
Microbiol. 2002;24:623-633.

88
153 Rademaker JLW, De Bruijn FJ. Computer-assisted analysis of molecular
fingerprint profiles and database construction. In: Kowalchuk GA, De Bruijn FJ,
Head IM, Akkermans ADL, Van Elsas JD, editors. Molecular Microbial Ecology
Manual, 2nd ed. Springer, Dordrecht; 2008. p. 1397-1446.
154 Radjasa OK, Wiese J, Sabdono A, Imhoff J. Corals as source of bacteria with
antimicrobial activity. Journal of Coastal Development. 2008;11(3):121-130.
155 Rahman H, Austin B, Mitchell WJ, Morris PC, Jamieson DJ, Adams DR, et al.
Novel anti-infective compounds from marine bacteria. Mar Drugs. 2010;8(3):498-
518.
156 Rahman H. Unusual Sesquiterpenes: Gorgonenes and Further Bioactive Secondary
Metabolites Derived from Marine and Terrestrial Bacteria. PhD Thesis. German:
Universität Göttingen; 2008.
157 Randall JE. Contributions to the biology of the queen conch Strombus gigas. Bull
Mar Sci Gulf Carib. 1964;14:246-295.
158 Rao MB, Tanksale AM, Ghatge MS, Deshpande VV. Molecular and
biotechnological aspects of microbial proteases. Microbiol Mol Biol Rev.
1998;62:597-635.
159 Reddy PRM, Swamy MV, Seenayya G. Puriification and characterization of
thermostable β-amylase and pullulanase from high-yielding Clostridium
thermosulfurogenes SV2. World J. Microbial Biotechnol. 1998;14:89-94.
160 Reysenbach AL, Giver LJ, Wickham GS, Pace NR. Differential amplification of
rRNA genes by polymerase chain reaction. Appl Environ Microbiol.
1992;58:3417–3418.
161 Rodríguez AI, Hariharan HA, Nimrod ST. Occurrence and Antimicrobial Drug
Resistance of Potential Bacterial Pathogens from Shellfish, Including Queen
Conchs (Strombus gigas) and Whelks (Cittarium pica) in Grenada. WebmedCentral
Microbiology. 2011;2-11.
162 Rohban R, Amoozegar MA, Ventosa A. Screening and isolation of halophilic
bacteria producing extracellular hydrolyses from Howz Soltan Lake, Iran. J Ind
Microbiol Biotechnol. 2009;36(3):333–340.

89
163 Romanenko LA, Uchino M, Kalinovskaya NI, Mikhailov VV. Isolation,
phylogenetic analysis and screening of marine mollusc-associated bacteria for
antimicrobial, hemolytic and surface activities. Microbiol Res. 2008;163(6):633-
644.
164 Romero J, García-Varela M, Laclette JP, Espejo RT. Bacterial 16S rRNA gene
analysis revealed that bacteria related to Arcobacter spp. constitute an abundant and
common component of the oyster microbiota (Tiostrea chilensis). Microb Ecol.
2002;44:365–371.
165 Romero-Tabarez M, Jansen R, Sylla M, Lünsdorf H, Häussler S, Santosa DA, et al.
7-O-malonyl macrolactin A, a new macrolactin antibiotic from Bacillus subtilis
active against methicillin-resistant Staphylococcus aureus, vancomycin-resistant
enterococci, and a small-colony variant of Burkholderia cepacia. Antimicrob
Agents Chemother. 2006;50(5):1701-1709.
166 Rosselló-Mora R, Amann R. The species concept for prokaryotes. FEMS Microbiol
Rev. 2001;25(1):39–67.
167 Saitou N, Nei M. The Neighbor-Joining Method: A New Method for
Reconstructing Phylogenetic Trees. Mol Biol Evol. 1987;4:406-425.
168 Salleh, Hassan, Illias, Shahab. Cloning and expression if pullulanase gene from
locally isolated bacillus. Project Report. Faculty of Science, Skudai, Johor. En
prensa. 2006.
169 Sambrook J, Russell DW. Molecular Cloning: a Laboratory Manual. 3rd ed. Cold
Spring Harbor: NY: Cold Spring Harbor Laboratory; 2001.
170 Sangnoi Y, Srisukchayakul P, Arunpairojana V, Kanjana-Opas A. Diversity of
marine gliding bacteria in Thailand and their cytotoxicity. Electron J Biotechnol.
2009;12(3):1-8.
171 Sanguinetti CJ, Dias Neto E, Simpson AJG. Rapid silver staining and recovery of
PCR products separated on polyacrylamide gels. Biotechniques. 1994;17:915-919.
172 Sarda L, Desnuelle P. Action de la lipase pancreatique sur les esters en emulsion.
Biochim Biophys Acta. 1958;30:513–521.
173 Sawabe T, Oda Y, Shiomi Y, Ezura Y. Alginate degradation by bacteria isolated

90
from the gut of sea urchins and abalones. Microb Ecol. 1995;30:193-202.
174 Saxena R, Dutt K, Agarwal L, Nayyar P. A highly thermostable and alkaline
amylase from a Bacillus sp. PN5. Bioresour Technol. 2007;98(2):260-265.
175 Schloss PD, Handelsman J. Metagenomics for studying unculturable
microorganisms: Cutting the Gordian Knot. Genome Biol. 2005;6:229.1-229.4.
176 Schloss PD, Handelsman J. Status of the microbial census. Microbiol Mol Biol
Rev. 2004;68(4):686-691.
177 Sekiguchi H, Watanabe M, Nakahara T, Xu B, Uchiyama H. Succession of
Bacterial Community Structure along the Changjiang River Determined by
Denaturing Gradient Gel Electrophoresis and Clone Library Analysis. Appl
Environ Microbiol. 2002;68(10):5142-5150.
178 Sekiguchi Y, Takahashi H, Kamagata Y, Ohashi A, Harada H. In situ detection,
isolation, and physiological properties of a thin filamentous microorganism
abundant in methanogenic granular sludges: a novel isolates affiliated with a clone
cluster, the green non-sulfur bacteria, subdivision I. Appl Environ Microbiol.
2001;67(12):5740–5749.
179 Shanmughapriya S, Kiran GS, Selvin J, Thomas TA, C R. Optimization,
Purification, and Characterization of Extracellular Mesophilic Alkaline Cellulase
from Sponge-Associated Marinobacter sp. MSI032. Appl Biochem Biotechnol.
2010;162(3):625-640.
180 Shnit-Orland M, Kushmaro A. Coral mucus-associated bacteria: a possible first line
of defense. FEMS Microbiol Ecol. 2009;67:371-380.
181 Speksnijder A, Kowalchuk GA, De Jong S, Kline E, Stephen JR, Laanbroek HJ.
Microvariation artifacts introduced by PCR and cloning of closely related 16S
rRNA gene sequences. Appl Environ Microbiol. 2001;67(1):469–472.
182 Spyere A, Rowley D, Jensen PR, Fenical W. New neoverrucosane diterpenoids
produced by the marine gliding bacterium Saprospira grandis. J Nat Prod.
2003;66(6):818-822.
183 Stevenson BS, Eichorst SA, Wertz JT, Schmidt TM, Breznak JA. New strategies
for cultivation and detection of previously uncultured microbes. Appl Environ

91
Microbiol. 2004;70(8):4748–4755.
184 Stott MB, Crowe MA, Mountain BW, Smirnova AV, Hou S, Alam M, et al.
Isolation of novel bacteria, including a candidate division, from geothermal soils in
New Zealand. Environ Microbiol. 2008;10(8):2030–2041.
185 Sugita H, Hinrose Y, Matsuo N, Deguchi Y. Production of the antibacterial
substance by Bacillus sp. strain NM 12, an intestinal bacterium of Japanese coastal
fish. Aquaculture. 1998;165:269-280.
186 Sun Y, Yang H, Ling Z, Chang J, Ye J. Gut microbiota of fast and slow growing
grouper Epinephelus coioides. Afr J Microbiol Res. 2009;3(11):713-720.
187 Süβ J, Engelen B, Cypionka H, Sass H. Quantitative analysis of bacterial
communities from Mediterranean sapropels based on cultivation-dependent
methods. FEMS Microbiol Ecol. 2004;51(1):109-121.
188 Svanevik CS, Lunestad BT. Characterisation of the microbiota of Atlantic mackerel
(Scomber scombrus). Int J Food Microbiol. 2011;151:164–170.
189 Tamura K, Nei M. Estimation of the number of nucleotide substitutions in the
control region of mitochondrial- DNA in humans and chimpanzees. Mol Biol Evol.
1993;10: 512–526.
190 Tanaka T, Burgess JG, Wright PC. High-pressure adaptation by salt stress in a
moderately halophilic bacterium obtained from open seawater. Appl Microbiol
Biotechnol. 2001;57(1-2):200–204.
191 Tanu, Deobagkar DD, Khandeparker R, Sreepada RA, Sanaye SV, Pawar HB. A
study on bacteria associated with the intestinal tract of farmed yellow seahorse,
Hippocampus kuda (Bleeker, 1852): characterization and extracellular enzymes.
Aquaculture Research. 2011;42:1-9.
192 Tateno Y, Takezaki N, Nei M.Relative efficiencies of the maximumlikelihood,
neighbor-joining, and maximum- parsimony methods when substitution rate varies
with site. Mol Biol Evol. 1994;11:261–277.
193 Teeri T. Crystalline cellulose degradation: new insight into the function of
cellobiohydrolases. Trends Biotechnol. 1997;15(5):160-167.
194 Thompson JD, Higgins DG, Gibson TJ. Clustal W: Improving the Sensitivity of

92
Progressive Multiple Sequence Alignment through Sequence Weighting, Position-
Specific Gap Penalties and Weight Matrix Choice. Nucleic Acids Res.
1994;22(22):4673–4680.
195 Tian F, Yu Y, Chen B, Li H, Yoa Y-F, Guo XK. Bacterial, archaeal and eukaryotic
diversity in Arctic sediment as revealed by 16S rRNA and 18S rRNA gene clone
libraries analysis. Polar Biol. 2008;32:93–103.
196 Torsvik V, Daae FL, Sandaa RA, Ovreas L. Novel techniques for analysing
microbial diversity in natural and perturbed environments. J Biotechnol.
1998;64(1):53-62.
197 Turnbaugh PJ, Ley RE, Hamady M, Fraser-Liggett CM, Knight R, Gordon JI. The
human microbiome project. Nature. 2007;449:804-810.
198 Utong JF, Al-Quagan F, Akel H. Efeect of varios growth conditions on production
of extracellular amylase from thermotolerant Bacillus species isolated from hot
springs in Jordan. J Biol Sci. 2006;6:621-625.
199 Van Horn DJ, Hall JR, Loker ES, et al. Gut bacterial community composition in
three planorbid snail species. En prensa.
200 Vásquez A, Molin G, Pettersson B, Antonssona M, Ahrné S. DNA-based
classification and sequence heterogeneities in the 16S rRNA genes of Lactobacillus
casei/paracasei and related species. Syst Appl Microbiol. 2005;28(5):430–441.
201 Wang GC, Wang Y. Frequency of formation of chimeric molecules as a
consequence of PCR coamplification of 16S rRNA genes from mixed bacterial
genomes. Appl Environ Microbiol. 1997;63(13):4645–4650.
202 Warmke GL, Abbott RT. Caribbean shells. Narbeth Penn, Livingstone; 1961. p.
346.
203 Wegley L, Edwards R, Rodríguez-Brito B, Liu H, Rohwer F. Metagenomic analysis
of the microbial community associated with the coral Porites astreoides. Environ
Microbiol. 2007;9:2707-2719.
204 Whitesides MD, Oliver JD. Resuscitation of Vibrio vulnificus from the Viable but
Nonculturable State. Appl Environ Microbiol. 1997;63(3):1002-1005.
205 Whitman WB, Coleman DC, Wiebe WJ. Prokaryotes: the unseen majority. Proc

93
Natl Acad Sci USA. 1998;95(12):6578-6583.
206 Wiese J, Thiel V, Nagel K, Staufenberger T, Imhoff JF. Diversity of Antibiotic-
Active Bacteria Associated with the Brown Alga Laminaria saccharina from the
Baltic Sea. Mar Biotechnol (NY). 2009;11(2):287-300.
207 Wiseman A. Handbook of Enzyme Biotechnology. New York: Ellis Horwood Ltd;
1985. p. 274–379.
208 Xu J. Microbial ecology in the age of genomics and metagenomics: concepts, tools,
and recent advances. Mol Ecol. 2006;15(7):1713-1731.
209 Xu MX, Wang P, Wang FP, Xiao X. Microbial diversity in a deep-sea Station of
Pacific Nodule Province, Studied via 16S rRNA Gene Sequences and Extremophile
Cultivation. Biodiv and Conserv. 2005;14(14):3363–3380.
210 Yamada A, Inoue T, Noda S, Hongoh Y, Ohkuma M. Evolutionary trend of
phylogenetic diversity of nitrogen fixation genes in the gut community of
woodfeeding termite. Mol Ecol. 2007;16(18):3768-3777.
211 Yan L. Production of antimicrobial compounds by marine epiphytic bacteria under
conditions which mimic their natural environment. Ph.D. thesis. Edinburgh, United
Kingdom: Heriot-Watt University; 2002.
212 Yan L, Boyd KG, Grant Burgess J. Surface attachment induced production of
antimicrobial compounds by marine epiphytic bacteria using modified roller bottle
cultivation. Mar Biotechnol. 2002;4(4):356–366.
213 Yang K. and Zhang, L. Performance comparison between k-tuple distance and four
model-based distances in phylogenetic tree reconstruction. Nucleic Acids Res.
2008:36, e33.
214 Yu ZT, Morrison M. Comparisons of different hypervariable regions of rrs genes
for use in fingerprinting of microbial communities by PCR-denaturing gradient gel
electrophoresis. Appl Environ Microbiol. 2004;70(8):4800-4806.
215 Zhang C, Kim SK. Research and application of marine microbial enzymes: status
and prospects. Mar Drugs. 2010;8(6):1920-1934.
216 Zhang J, Lin S, Zeng R. Cloning, expression, and characterization of a cold-adapted
lipase gene from an antartic deep-sea psychrotrophic bacterium, Psychrobacter sp

94
7195. J Microbiol Biotechnol. 2007;17(4):604-610.
217 Zhang Y, Jiao NZ, Cottrell MT, Kirchman DL. Contribution of major bacterial
groups to bacterial biomass production along a salinity gradient in the South China
Sea. Aquat Microb Ecol. 2006;43(3):233-241.
218 Zobell CE. Studies on marine bacteria. I. The culture requirements of heterotrophic
aerobes. J Mar Res. 1941;4:42.
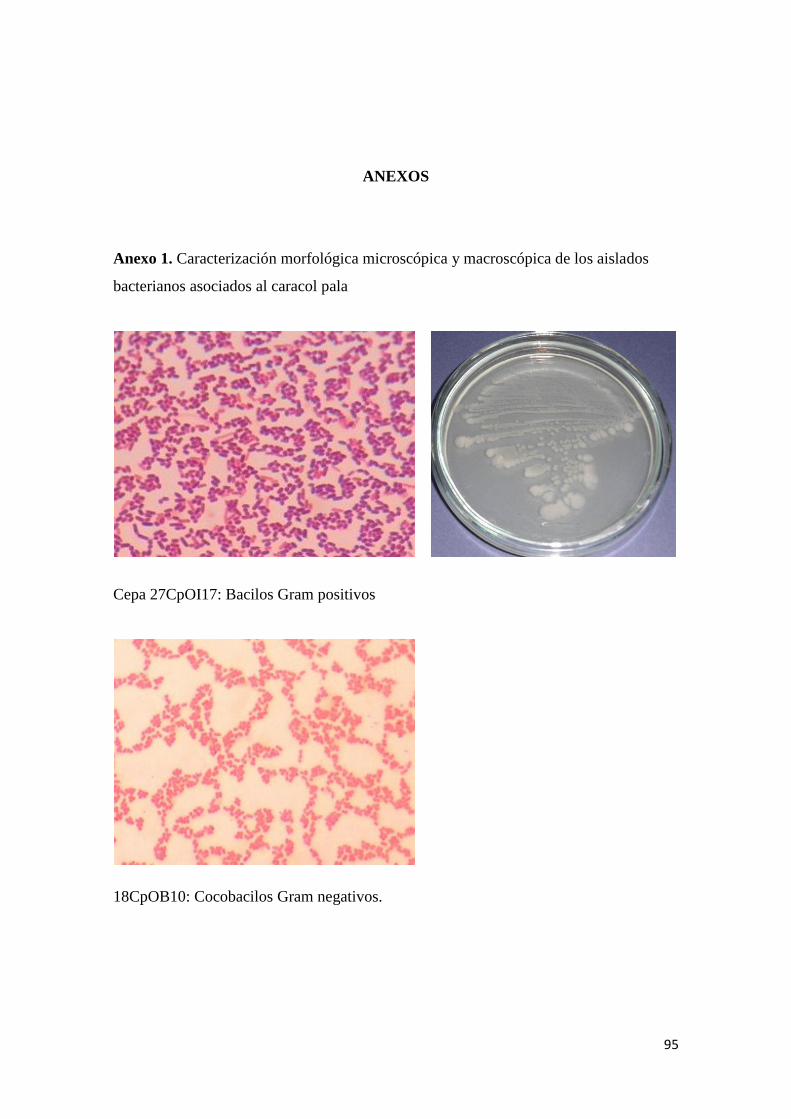
95
ANEXOS
Anexo 1. Caracterización morfológica microscópica y macroscópica de los aislados
bacterianos asociados al caracol pala
Cepa 27CpOI17: Bacilos Gram positivos
18CpOB10: Cocobacilos Gram negativos.
96
20CpOB13: Cocos Gram positivos
29CpOB11: Bacilos Gram positivos
Cepa 19CpB3: Cocobacilos Gram negativos
97
12CpOA5: Bacilos Gram negativos
Caracterización fenotípica de los aislados obtenidos desde las muestras (OiS, BiS, OiC,
BiC, ObS, BbS, ObC, BbC, OaS, BaS, OaC, BaC) asociadas al caracol pala.
Aislado Morfología
microscópica
Tinción de Gram Morfología de la colonia
18CpOB10
Cocos bacilos
Negativos
Forma circular, margen débilmente ondulado, superficie
lisa, elevación plana, color crema, tamaño mediano.
19CpB3
Cocobacilos
Negativos
Forma circular, margen débilmente ondulado, superficie
lisa, elevación convexa, color crema, tamaño mediano.
21CpBI7
Cocos
Negativos
Forma circular, margen débilmente ondulado, superficie
lisa, elevación plana, color crema, tamaño mediano.
25CpB1 Cocos Negativos Forma circular, margen débilmente ondulado, superficie
lisa, elevación plana, color crema, tamaño mediano.
7CpBB10 Bacilos Negativos Forma circular, margen débilmente ondulado, superficie
lisa, elevación plana, color crema, tamaño mediano.
10CpBB10 Cocos Negativos Forma circular, margen débilmente ondulado, superficie
lisa, elevación ligeramente convexa, color blanco, tamaño
pequeño.
2CpBB11 Cocos Negativos Forma circular, margen débilmente ondulado, superficie
lisa, elevación plana, color blanco, tamaño grande.
5CpBB11 Cocobacilos Negativos Forma circular, margen débilmente ondulado, superficie
lisa, elevación plana, color blanco, tamaño mediano.
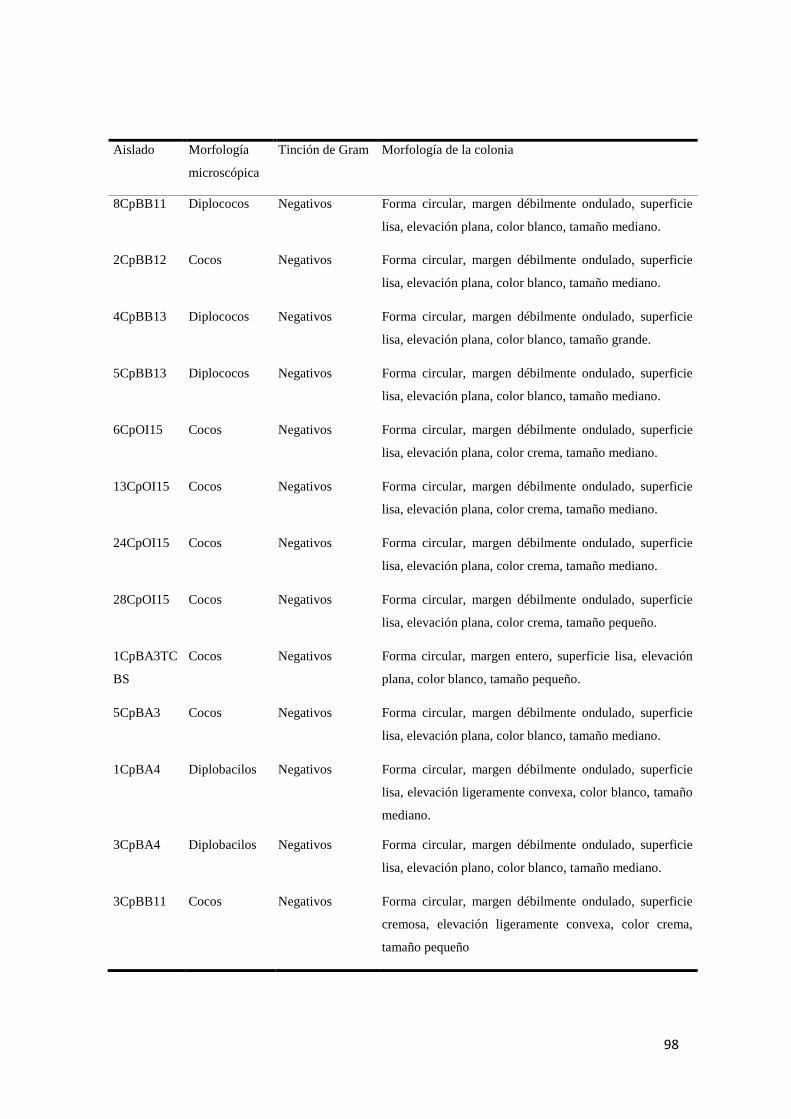
98
Aislado Morfología
microscópica
Tinción de Gram Morfología de la colonia
8CpBB11 Diplococos Negativos Forma circular, margen débilmente ondulado, superficie
lisa, elevación plana, color blanco, tamaño mediano.
2CpBB12 Cocos Negativos Forma circular, margen débilmente ondulado, superficie
lisa, elevación plana, color blanco, tamaño mediano.
4CpBB13 Diplococos Negativos Forma circular, margen débilmente ondulado, superficie
lisa, elevación plana, color blanco, tamaño grande.
5CpBB13 Diplococos Negativos Forma circular, margen débilmente ondulado, superficie
lisa, elevación plana, color blanco, tamaño mediano.
6CpOI15 Cocos Negativos Forma circular, margen débilmente ondulado, superficie
lisa, elevación plana, color crema, tamaño mediano.
13CpOI15 Cocos Negativos Forma circular, margen débilmente ondulado, superficie
lisa, elevación plana, color crema, tamaño mediano.
24CpOI15 Cocos Negativos Forma circular, margen débilmente ondulado, superficie
lisa, elevación plana, color crema, tamaño mediano.
28CpOI15 Cocos Negativos Forma circular, margen débilmente ondulado, superficie
lisa, elevación plana, color crema, tamaño pequeño.
1CpBA3TC
BS
Cocos Negativos Forma circular, margen entero, superficie lisa, elevación
plana, color blanco, tamaño pequeño.
5CpBA3 Cocos Negativos Forma circular, margen débilmente ondulado, superficie
lisa, elevación plana, color blanco, tamaño mediano.
1CpBA4 Diplobacilos Negativos Forma circular, margen débilmente ondulado, superficie
lisa, elevación ligeramente convexa, color blanco, tamaño
mediano.
3CpBA4 Diplobacilos Negativos Forma circular, margen débilmente ondulado, superficie
lisa, elevación plano, color blanco, tamaño mediano.
3CpBB11 Cocos Negativos Forma circular, margen débilmente ondulado, superficie
cremosa, elevación ligeramente convexa, color crema,
tamaño pequeño
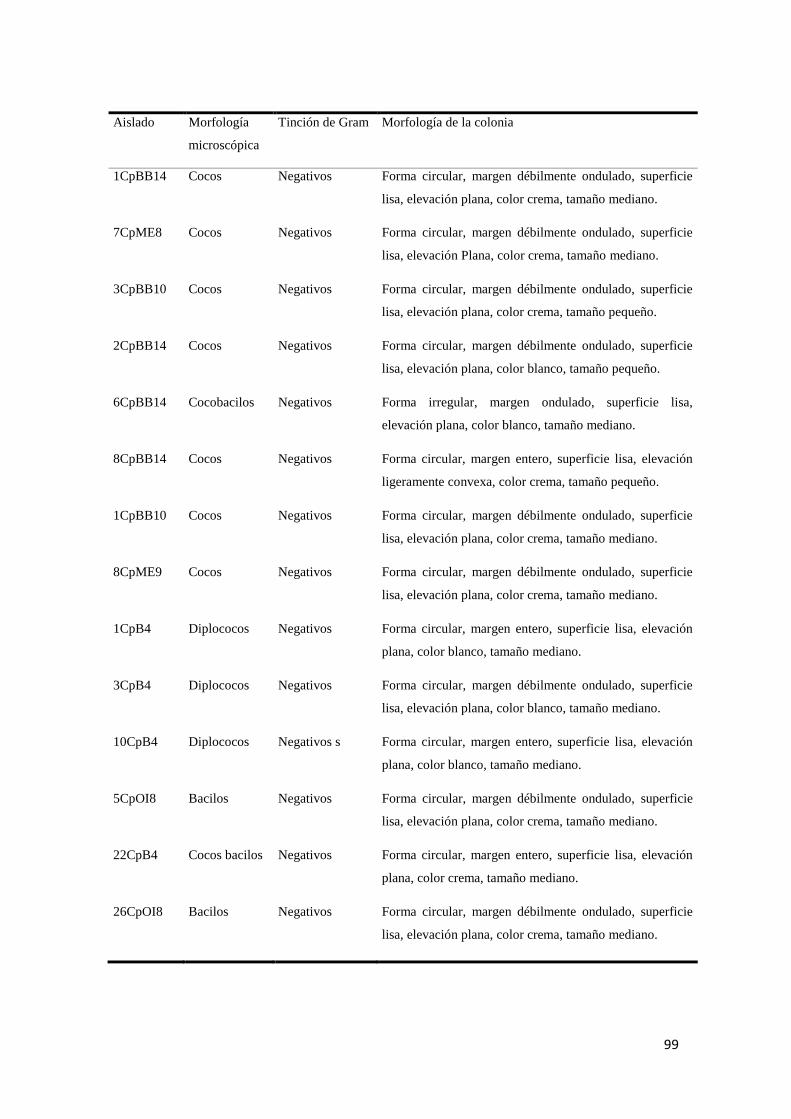
99
Aislado Morfología
microscópica
Tinción de Gram Morfología de la colonia
1CpBB14 Cocos
Negativos Forma circular, margen débilmente ondulado, superficie
lisa, elevación plana, color crema, tamaño mediano.
7CpME8 Cocos Negativos Forma circular, margen débilmente ondulado, superficie
lisa, elevación Plana, color crema, tamaño mediano.
3CpBB10 Cocos Negativos Forma circular, margen débilmente ondulado, superficie
lisa, elevación plana, color crema, tamaño pequeño.
2CpBB14 Cocos Negativos Forma circular, margen débilmente ondulado, superficie
lisa, elevación plana, color blanco, tamaño pequeño.
6CpBB14 Cocobacilos Negativos Forma irregular, margen ondulado, superficie lisa,
elevación plana, color blanco, tamaño mediano.
8CpBB14 Cocos Negativos Forma circular, margen entero, superficie lisa, elevación
ligeramente convexa, color crema, tamaño pequeño.
1CpBB10 Cocos Negativos Forma circular, margen débilmente ondulado, superficie
lisa, elevación plana, color crema, tamaño mediano.
8CpME9 Cocos Negativos Forma circular, margen débilmente ondulado, superficie
lisa, elevación plana, color crema, tamaño mediano.
1CpB4 Diplococos Negativos Forma circular, margen entero, superficie lisa, elevación
plana, color blanco, tamaño mediano.
3CpB4 Diplococos Negativos Forma circular, margen débilmente ondulado, superficie
lisa, elevación plana, color blanco, tamaño mediano.
10CpB4 Diplococos Negativos s Forma circular, margen entero, superficie lisa, elevación
plana, color blanco, tamaño mediano.
5CpOI8 Bacilos Negativos Forma circular, margen débilmente ondulado, superficie
lisa, elevación plana, color crema, tamaño mediano.
22CpB4 Cocos bacilos Negativos Forma circular, margen entero, superficie lisa, elevación
plana, color crema, tamaño mediano.
26CpOI8 Bacilos Negativos Forma circular, margen débilmente ondulado, superficie
lisa, elevación plana, color crema, tamaño mediano.
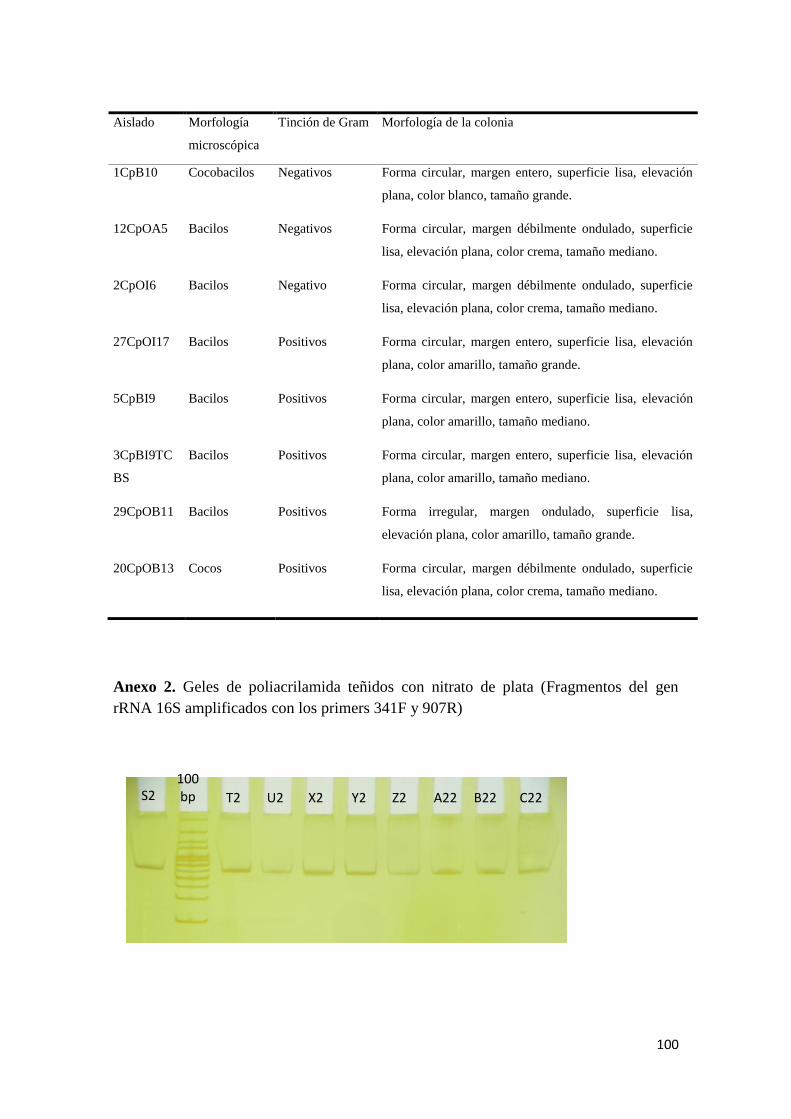
100
Aislado Morfología
microscópica
Tinción de Gram Morfología de la colonia
1CpB10 Cocobacilos Negativos Forma circular, margen entero, superficie lisa, elevación
plana, color blanco, tamaño grande.
12CpOA5 Bacilos Negativos Forma circular, margen débilmente ondulado, superficie
lisa, elevación plana, color crema, tamaño mediano.
2CpOI6 Bacilos Negativo Forma circular, margen débilmente ondulado, superficie
lisa, elevación plana, color crema, tamaño mediano.
27CpOI17 Bacilos Positivos Forma circular, margen entero, superficie lisa, elevación
plana, color amarillo, tamaño grande.
5CpBI9 Bacilos Positivos Forma circular, margen entero, superficie lisa, elevación
plana, color amarillo, tamaño mediano.
3CpBI9TC
BS
Bacilos Positivos Forma circular, margen entero, superficie lisa, elevación
plana, color amarillo, tamaño mediano.
29CpOB11
Bacilos Positivos Forma irregular, margen ondulado, superficie lisa,
elevación plana, color amarillo, tamaño grande.
20CpOB13
Cocos Positivos Forma circular, margen débilmente ondulado, superficie
lisa, elevación plana, color crema, tamaño mediano.
Anexo 2. Geles de poliacrilamida teñidos con nitrato de plata (Fragmentos del gen
rRNA 16S amplificados con los primers 341F y 907R)
S2 T2 U2 X2 Y2 Z2 A22 B22 C22
100 bp

101
Anexo 3. Matrices
a) Matriz de similitud (Coeficiente de Dice)
a)
46.2
66.7
58.3
71.4
52.7
41.5
72.7
58.6
49.3
35.2
53.3
45.0
87.0
72.3
96.6
71.3
57.6
38.2
32.2
60.9
82.4
87.5
66.6
58.4
70.0
54.5
52.3
84.2
66.6
48.6
87.5
35.0
31.0
17.9
DGGE 15 h
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
CpOB13
CpMo2
CpB3
CpBI17
CpBI18
CpB4
CpB5
CpMo1
CpB2
CpOB11
CpB6
OA19
CpB11
OA2
OA4
OA5
OA2-4
CpBA3
CpBA5
OA3
CpBI7
CpBI8
CpOI15
CpOI18
CpOB10
CpOB14
CpBI6
CpB10
CpOI8
CpBB10
CpBB11
CpBB14
CpOI16
CpOI17
CpB1
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
100
46.2 100
44.4 46.2 100
41.7 42.1 66.7 100
50.0 31.6 50.0 66.7 100
28.6 44.4 42.9 40.0 50.0 100
55.6 30.8 66.7 50.0 66.7 71.4 100
37.5 54.5 62.5 54.5 45.5 33.3 37.5 100
15.4 25.0 61.5 31.6 21.1 0.0 15.4 72.7 100
33.3 57.1 33.3 33.3 11.1 25.0 16.7 60.0 57.1 100
31.6 42.9 52.6 48.0 32.0 40.0 31.6 58.8 42.9 46.2 100
37.0 27.3 29.6 30.3 48.5 26.1 29.6 32.0 18.2 19.1 28.6 100
38.1 37.5 38.1 29.6 37.0 35.3 38.1 42.1 12.5 26.7 18.2 53.3 100
43.5 44.4 34.8 41.4 34.5 21.1 26.1 38.1 22.2 23.5 25.0 43.8 46.2 100
30.0 13.3 30.0 30.8 23.1 25.0 20.0 44.4 26.7 14.3 38.1 41.4 43.5 48.0 100
38.1 25.0 38.1 29.6 37.0 23.5 19.1 52.6 37.5 26.7 36.4 46.7 58.3 46.2 87.0 100
47.6 25.0 57.1 37.0 44.4 23.5 47.6 52.6 37.5 26.7 36.4 26.7 33.3 38.5 69.6 75.0 100
52.2 11.1 43.5 41.4 34.5 31.6 52.2 38.1 22.2 11.8 33.3 37.5 38.5 35.7 72.0 69.2 69.2 100
58.3 21.1 50.0 40.0 40.0 20.0 50.0 45.5 31.6 22.2 40.0 36.4 44.4 41.4 69.2 74.1 74.1 96.6 100
23.5 16.7 35.3 17.4 26.1 15.4 11.8 40.0 33.3 18.2 33.3 23.1 30.0 18.2 52.6 60.0 60.0 54.5 60.9 100
55.6 30.8 44.4 50.0 33.3 14.3 33.3 50.0 46.2 33.3 21.1 14.8 38.1 34.8 30.0 38.1 47.6 34.8 41.7 35.3 100
34.8 11.1 34.8 41.4 34.5 10.5 26.1 38.1 33.3 23.5 25.0 31.3 38.5 35.7 24.0 30.8 38.5 28.6 34.5 18.2 60.9 100
35.3 33.3 23.5 43.5 26.1 0.0 11.8 40.0 33.3 36.4 22.2 15.4 30.0 36.4 21.1 30.0 30.0 27.3 43.5 12.5 58.8 63.6 100
33.3 30.8 33.3 41.7 16.7 0.0 11.1 37.5 46.2 33.3 31.6 22.2 38.1 43.5 20.0 28.6 28.6 34.8 41.7 11.8 55.6 52.2 82.4 100
37.5 18.2 37.5 36.4 18.2 0.0 25.0 28.6 36.4 40.0 23.5 24.0 42.1 28.6 33.3 31.6 42.1 28.6 27.3 13.3 50.0 57.1 66.7 62.5 100
55.6 30.8 33.3 41.7 25.0 0.0 11.1 37.5 15.4 33.3 21.1 22.2 38.1 26.1 30.0 28.6 28.6 26.1 33.3 11.8 77.8 52.2 70.6 66.7 87.5 100
52.6 28.6 31.6 40.0 32.0 13.3 21.1 35.3 42.9 30.8 20.0 28.6 36.4 33.3 28.6 45.5 27.3 33.3 40.0 11.1 63.2 50.0 66.7 63.2 58.8 73.7 100
52.6 14.3 52.6 32.0 40.0 26.7 31.6 35.3 28.6 46.2 30.0 35.7 54.5 41.7 38.1 54.5 36.4 41.7 48.0 22.2 42.1 50.0 44.4 42.1 47.1 52.6 70.0 100
30.0 13.3 20.0 38.5 38.5 0.0 10.0 22.2 26.7 14.3 9.5 20.7 34.8 48.0 27.3 34.8 17.4 32.0 38.5 10.5 40.0 56.0 63.2 60.0 44.4 50.0 57.1 47.6 100
44.4 15.4 66.7 41.7 58.3 14.3 66.7 50.0 30.8 33.3 21.1 14.8 38.1 17.4 20.0 28.6 47.6 43.5 50.0 35.3 55.6 52.2 47.1 44.4 50.0 44.4 42.1 52.6 30.0 100
52.6 28.6 52.6 48.0 56.0 26.7 63.2 47.1 28.6 30.8 30.0 21.4 36.4 16.7 19.1 36.4 45.5 41.7 48.0 33.3 63.2 50.0 55.6 42.1 58.8 52.6 60.0 50.0 28.6 84.2 100
36.4 23.5 45.5 50.0 42.9 0.0 36.4 40.0 11.8 25.0 17.4 25.8 40.0 37.0 25.0 32.0 32.0 29.6 35.7 38.1 54.5 51.9 57.1 45.5 40.0 45.5 52.2 43.5 41.7 63.6 69.6 100
44.4 30.8 33.3 58.3 41.7 14.3 33.3 37.5 15.4 33.3 21.1 29.6 28.6 34.8 20.0 28.6 28.6 26.1 25.0 11.8 44.4 52.2 58.8 33.3 25.0 44.4 42.1 42.1 30.0 33.3 42.1 54.5 100
37.5 36.4 25.0 54.5 27.3 16.7 37.5 14.3 0.0 20.0 11.8 24.0 21.1 38.1 22.2 10.5 21.1 28.6 27.3 13.3 25.0 28.6 40.0 25.0 14.3 25.0 23.5 23.5 33.3 25.0 23.5 50.0 87.5 100
26.7 20.0 13.3 0.0 28.6 18.2 13.3 30.8 0.0 0.0 0.0 25.0 22.2 20.0 23.5 22.2 33.3 30.0 28.6 14.3 26.7 30.0 0.0 0.0 15.4 13.3 25.0 25.0 11.8 26.7 25.0 10.5 13.3 15.4 100
V1 W1 X1 Y1 Z1
100 bp

102
b) Matriz de datos binarios (Presencia (1) y ausencia de bandas (0))

103
Matriz binaria de datos
Anexo 4. Análisis de similitud ANOSIM usando el software PAST
Jaccard Bray-Curtis
Análisis de disimilitud (SIMPER)
DGGE 15 h
18.2
%
20.7
%
22.5
%
32.1
%
33.9
%
34.9
%
37.1
%
38.2
%
43.2
%
47.8
%
49.3
%
50.3
%
51.4
%
52.6
%
53.1
%
54.5
%
55.1
%
56.3
%
56.9
%
57.5
%
57.9
%
59.7
%
61.3
%
62.8
%
63.4
%
64.2
%
64.8
%
65.6
%
66.9
%
67.7
%
68.8
%
69.9
%
70.8
%
71.9
%
73.0
%
73.8
%
75.1
%
76.9
%
77.7
%
79.0
%
80.2
%
81.2
%
82.8
%
84.1
%
84.8
%
86.4
%
87.9
%
89.3
%
90.7
%
92.1
%
93.0
%
94.5
%
95.6
%
DGGE and matrix
DG
GE
15 h
:18.
2%
DG
GE
15 h
:20.
7%
DG
GE
15 h
:22.
5%
DG
GE
15 h
:32.
1%
DG
GE
15 h
:33.
9%
DG
GE
15 h
:34.
9%
DG
GE
15 h
:37.
1%
DG
GE
15 h
:38.
2%
DG
GE
15 h
:43.
2%
DG
GE
15 h
:47.
8%
DG
GE
15 h
:49.
3%
DG
GE
15 h
:50.
3%
DG
GE
15 h
:51.
4%
DG
GE
15 h
:52.
6%
DG
GE
15 h
:53.
1%
DG
GE
15 h
:54.
5%
DG
GE
15 h
:55.
1%
DG
GE
15 h
:56.
3%
DG
GE
15 h
:56.
9%
DG
GE
15 h
:57.
5%
DG
GE
15 h
:57.
9%
DG
GE
15 h
:59.
7%
DG
GE
15 h
:61.
3%
DG
GE
15 h
:62.
8%
DG
GE
15 h
:63.
4%
DG
GE
15 h
:64.
2%
DG
GE
15 h
:64.
8%
DG
GE
15 h
:65.
6%
DG
GE
15 h
:66.
9%
DG
GE
15 h
:67.
7%
DG
GE
15 h
:68.
8%
DG
GE
15 h
:69.
9%
DG
GE
15 h
:70.
8%
DG
GE
15 h
:71.
9%
DG
GE
15 h
:73.
0%
DG
GE
15 h
:73.
8%
DG
GE
15 h
:75.
1%
DG
GE
15 h
:76.
9%
DG
GE
15 h
:77.
7%
DG
GE
15 h
:79.
0%
DG
GE
15 h
:80.
2%
DG
GE
15 h
:81.
2%
DG
GE
15 h
:82.
8%
DG
GE
15 h
:84.
1%
DG
GE
15 h
:84.
8%
DG
GE
15 h
:86.
4%
DG
GE
15 h
:87.
9%
DG
GE
15 h
:89.
3%
DG
GE
15 h
:90.
7%
DG
GE
15 h
:92.
1%
DG
GE
15 h
:93.
0%
DG
GE
15 h
:94.
5%
DG
GE
15 h
:95.
6%
DG
GE
15 h
:97.
1%
DG
GE
15 h
:98.
9%
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
CpOB13
CpMo2
CpB3
CpBI17
CpBI18
CpB4
CpB5
CpMo1
CpB2
CpOB11
CpB6
OA19
CpB11
OA2
OA4
OA5
OA2-4
CpBA3
CpBA5
OA3
CpBI7
CpBI8
CpOI15
CpOI18
CpOB10
CpOB14
CpBI6
CpB10
CpOI8
CpBB10
CpBB11
CpBB14
CpOI16
CpOI17
CpB1
104
Anexo 5. Trazas de secuencias de las bandas obtenidas a partir del DGGE
Cap Contig assembly program (L1)
L1- 341F

105
L1-907R
Cap Contig assembly program (G3)

106
Cap Contig assembly program (H2)
Trazas de secuencias que no pudieron ser editadas
Cap Contig assembly program (O1)
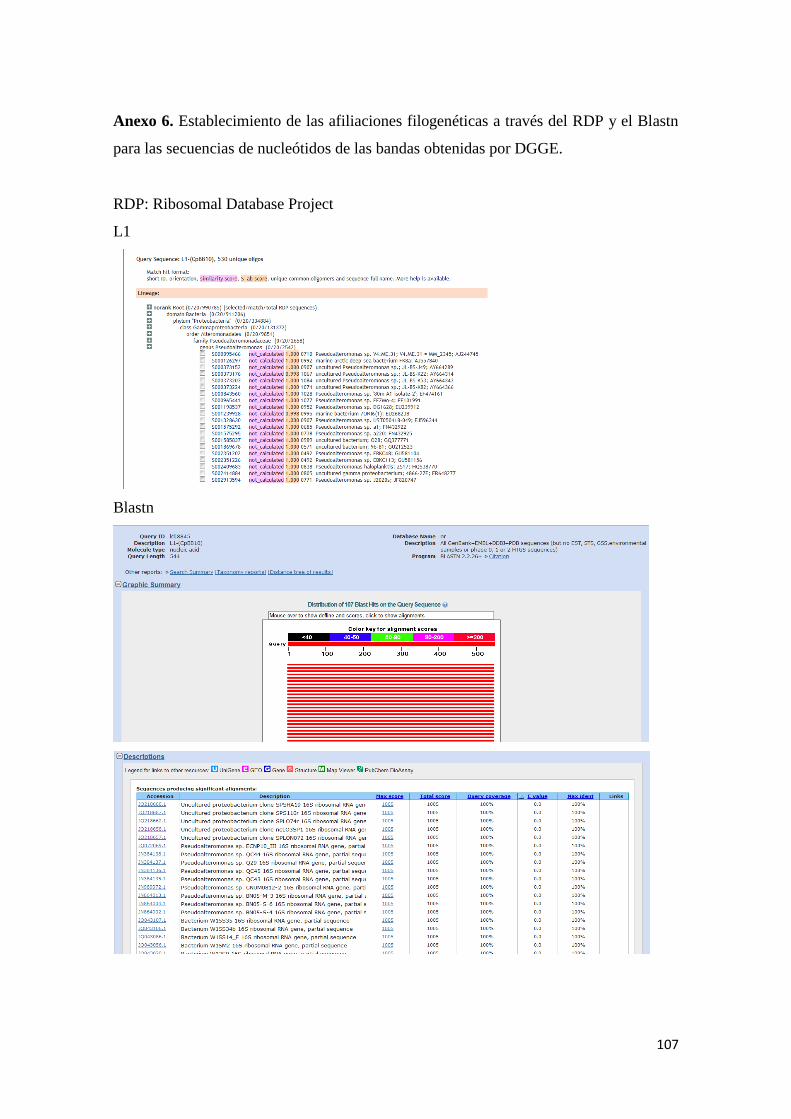
107
Anexo 6. Establecimiento de las afiliaciones filogenéticas a través del RDP y el Blastn
para las secuencias de nucleótidos de las bandas obtenidas por DGGE.
RDP: Ribosomal Database Project
L1
Blastn
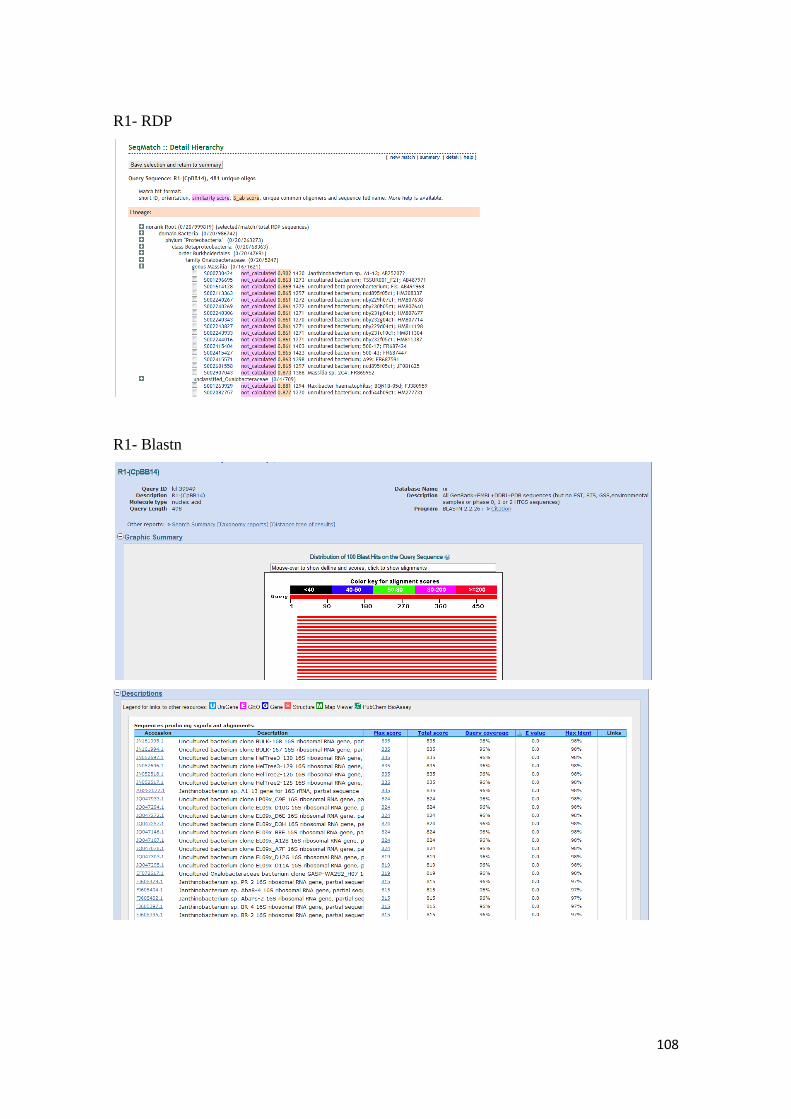
108
R1- RDP
R1- Blastn
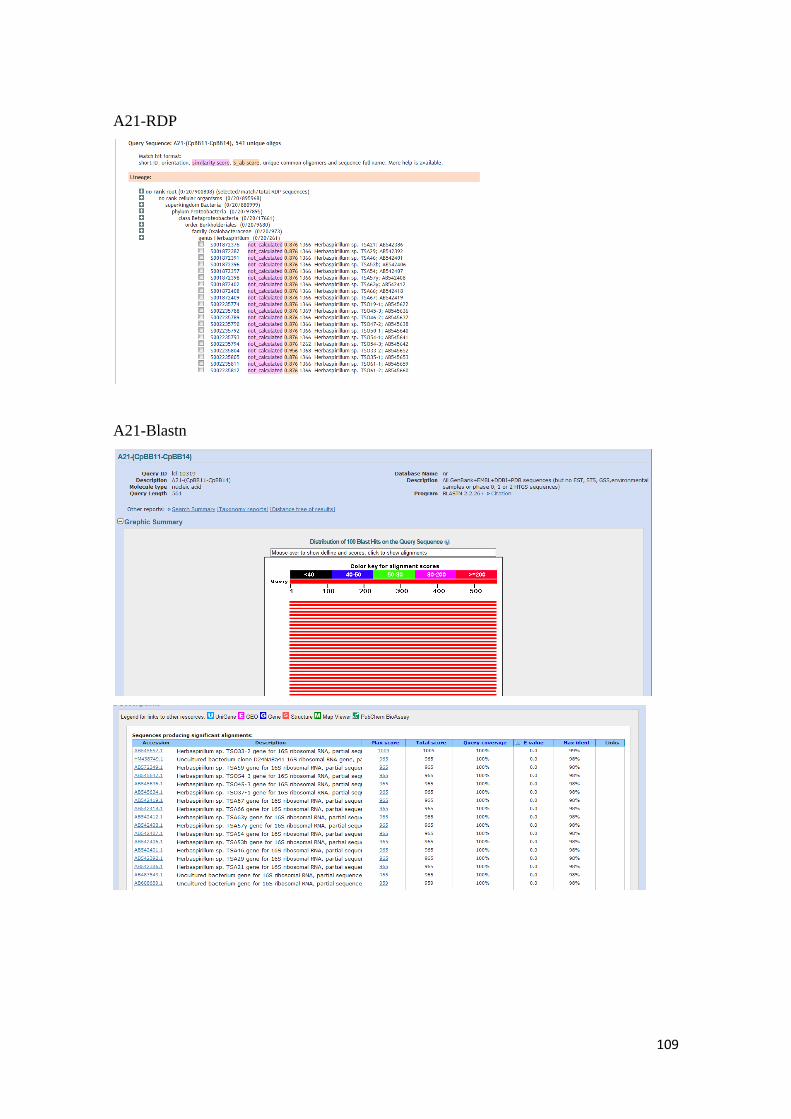
109
A21-RDP
A21-Blastn
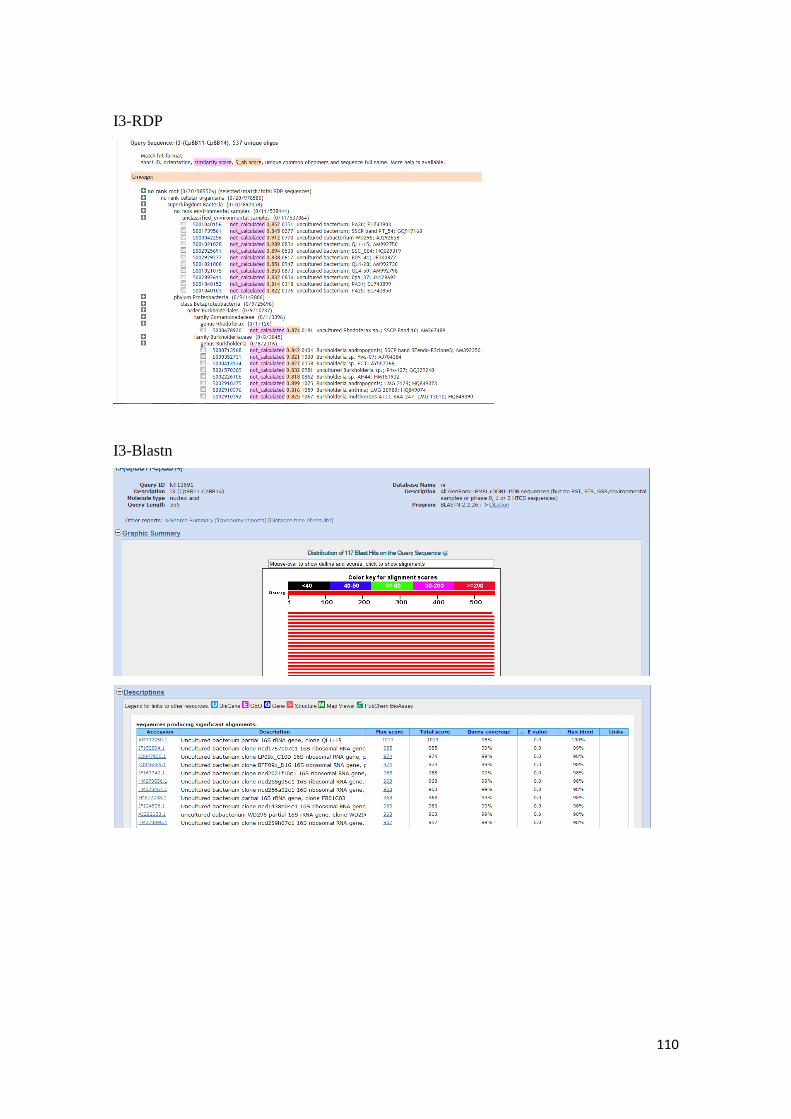
110
I3-RDP
I3-Blastn

111
Anexo 7. Alineamiento múltiple con Clustal W de las secuencias obtenidas desde las
bandas escindidas en los geles de DGGE
Anexo 8. Identificación taxonómica y posición filogenética de los aislados bacterianos
asociados al caracol pala
10CpBB10

112
Anexo 9. Alineamiento múltiple con Clustal W de las secuencias correspondientes a los
aislados.
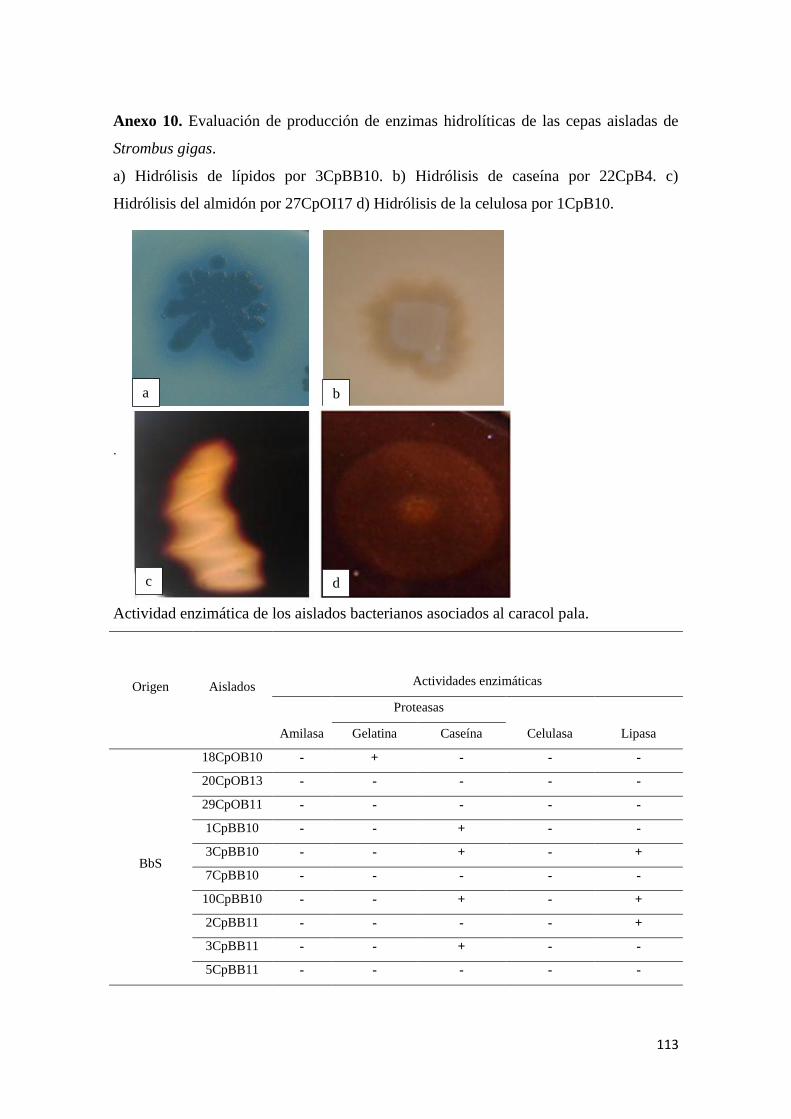
113
Anexo 10. Evaluación de producción de enzimas hidrolíticas de las cepas aisladas de
Strombus gigas.
a) Hidrólisis de lípidos por 3CpBB10. b) Hidrólisis de caseína por 22CpB4. c)
Hidrólisis del almidón por 27CpOI17 d) Hidrólisis de la celulosa por 1CpB10.
.
Actividad enzimática de los aislados bacterianos asociados al caracol pala.
Origen Aislados Actividades enzimáticas
Amilasa
Proteasas
Celulasa Lipasa Gelatina Caseína
BbS
18CpOB10 - + - - -
20CpOB13 - - - - -
29CpOB11 - - - - -
1CpBB10 - - + - -
3CpBB10 - - + - +
7CpBB10 - - - - -
10CpBB10 - - + - +
2CpBB11 - - - - +
3CpBB11 - - + - -
5CpBB11 - - - - -
a b
d c
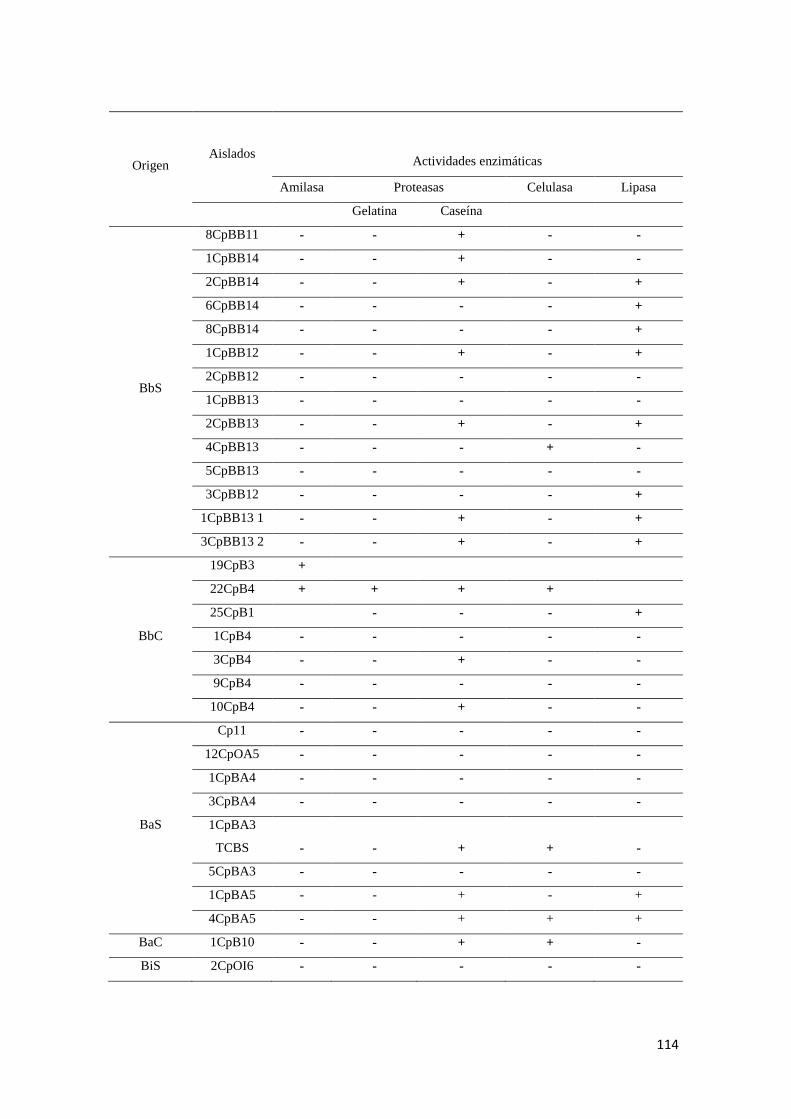
114
Origen Aislados
Actividades enzimáticas
Amilasa Proteasas Celulasa Lipasa
Gelatina Caseína
BbS
8CpBB11 - - + - -
1CpBB14 - - + - -
2CpBB14 - - + - +
6CpBB14 - - - - +
8CpBB14 - - - - +
1CpBB12 - - + - +
2CpBB12 - - - - -
1CpBB13 - - - - -
2CpBB13 - - + - +
4CpBB13 - - - + -
5CpBB13 - - - - -
3CpBB12 - - - - +
1CpBB13 1 - - + - +
3CpBB13 2 - - + - +
BbC
19CpB3 +
22CpB4 + + + +
25CpB1 - - - +
1CpB4 - - - - -
3CpB4 - - + - -
9CpB4 - - - - -
10CpB4 - - + - -
BaS
Cp11 - - - - -
12CpOA5 - - - - -
1CpBA4 - - - - -
3CpBA4 - - - - -
1CpBA3
TCBS - - + + -
5CpBA3 - - - - -
1CpBA5 - - + - +
4CpBA5 - - + + +
BaC 1CpB10 - - + + -
BiS 2CpOI6 - - - - -

115
Origen
Aislados Actividades enzimáticas
Amilasa Proteasas Celulasa Lipasa
Gelatina Caseína
BiS
5CpOI8 - - + - -
6CpOI15 - - - - -
13CpOI15 - - - - -
21CpBI7 - - - - -
24CpOI15 - - + - +
26CpOI8 + - - - -
27CpOI17 + - + - -
28CpOI15 - - + - -
2CpBI9
TCBS - - + + +
3CpBI9
TCBS + - + - +
5CpBI9 + - + - +
2CpBI7 - - + - +
5CpBI8 - - + - +
BaS Cp10 - - - - -
Estanque 8CpME9 - - - - +
Cp7 - - + - -
Anexo 11. Resultados colorimétricos para la curva de glucosa usada para la
determinación de azucares reductores.
Anexo 12. Resultados de la determinación de proteínas usando el reactivo de Bradford

116
Anexo 13. Resultados colorimétricos para la curva de L- tirosina usada para la
determinación de proteasas.

117
Anexo 14. Resultados colorimétricos para la curva de p-NP usada para la determinación
de lipasas.
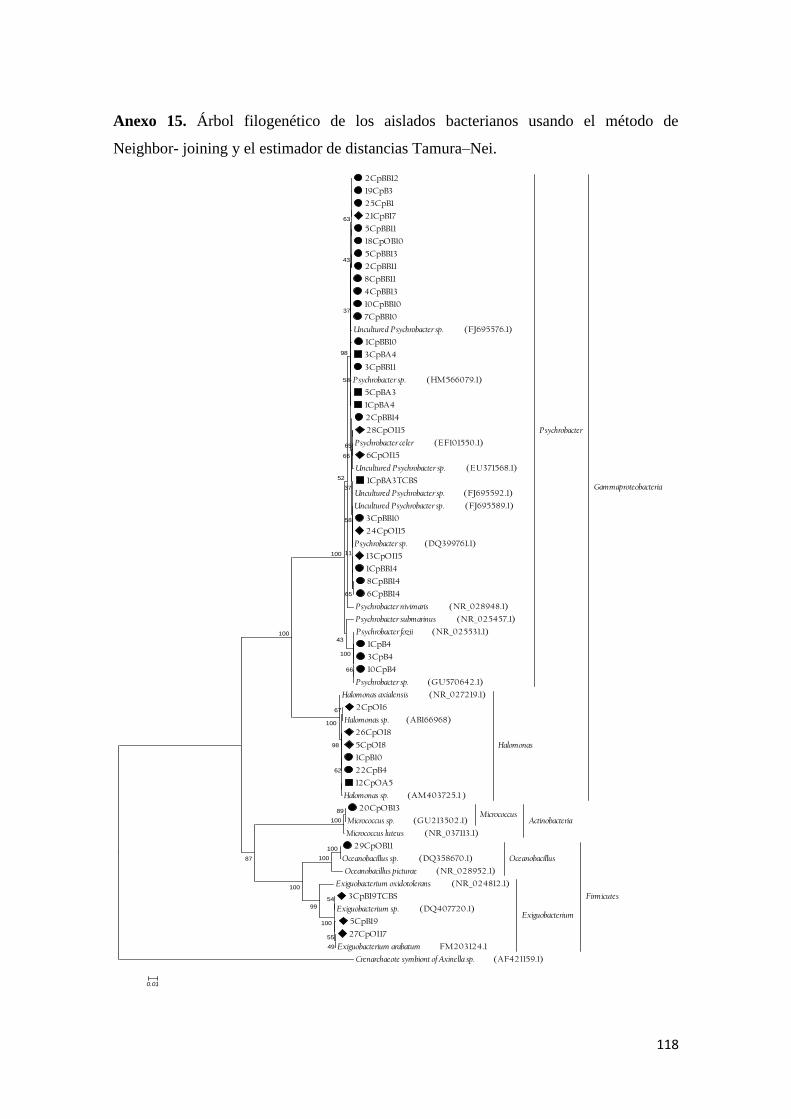
118
Anexo 15. Árbol filogenético de los aislados bacterianos usando el método de
Neighbor- joining y el estimador de distancias Tamura–Nei.
2CpBB12
19CpB3
25CpB1
21CpBI7
5CpBB11
18CpOB10
5CpBB13
2CpBB11
8CpBB11
4CpBB13
10CpBB10
7CpBB10
(FJ695576.1) Uncultured Psychrobacter sp.
1CpBB10
3CpBA4
3CpBB11
(HM566079.1) Psychrobacter sp.
5CpBA3
1CpBA4
2CpBB14
28CpOI15
(EF101550.1) Psychrobacter celer
6CpOI15
(EU371568.1) Uncultured Psychrobacter sp.
1CpBA3TCBS
(FJ695592.1) Uncultured Psychrobacter sp.
(FJ695589.1) Uncultured Psychrobacter sp.
3CpBB10
24CpOI15
(DQ399761.1) Psychrobacter sp.
13CpOI15
1CpBB14
8CpBB14
6CpBB14
(NR_028948.1) Psychrobacter nivimaris
(NR_025457.1) Psychrobacter submarinus
(NR_025531.1) Psychrobacter fozii
1CpB4
3CpB4
10CpB4
(GU570642.1) Psychrobacter sp.
Psychrobacter
(NR_027219.1) Halomonas axialensis
2CpOI6
(AB166968) Halomonas sp.
26CpOI8
5CpOI8
1CpB10
22CpB4
12CpOA5
(AM403725.1 ) Halomonas sp.
Halomonas
Gammaproteobacteria
20CpOB13
(GU213502.1) Micrococcus sp. Micrococcus
(NR_037113.1) Micrococcus luteus
Actinobacteria
29CpOB11
(DQ358670.1) Oceanobacillus sp.
(NR_028952.1) Oceanobacillus picturae
Oceanobacillus
(NR_024812.1) Exiguobacterium oxidotolerans
3CpBI9TCBS
(DQ407720.1) Exiguobacterium sp.
5CpBI9
27CpOI17
FM203124.1 Exiguobacterium arabatum
Exiguobacterium
Firmicutes
(AF421159.1) Crenarchaeote symbiont of Axinella sp.
49
55
54
100
100
100
99
100
89
100
87
66
100
67
62
98
100
100
65
65
11
58
37
66
63
58
37
43
98
43
100
52
0.01