análisis arqueozoológico, tafonómico y de distribución ... · en cada periodo –...
TRANSCRIPT

Análisis arqueozoológico, tafonómico y de distribuciónespacial de la fauna de mamíferos
de la Cueva de la Fragua (Santoña, Cantabria)
Archaeozoological, taphonomical and spatial distribution analysis ofmammalian faunas from la Fragua Cave (Santoña, Cantabria)
PALABRAS CLAVE: Arqueozoología, Tafonomía, Distribución Espacial, Magdaleniense, Mesolítico.KEY WORDS: Archaeozoology, Taphonomy, Spatial Distribution, Magdalenian, Mesolithic.
Ana Belén MARÍN*
RESUMEN
Este trabajo analiza los restos de Mamíferos hallados en los niveles Magdaleniense, Aziliense, Mesolítico y Calcolítico de la Cueva de LaFragua (Santoña, Cantabria). En el nivel Magdaleniense existe una economía dual centralizada en la caza del ciervo y la cabra. A partir del nivelAziliense y sobre todo en el Mesolítico, a raíz del cambio climático, empieza a hacerse patente la diversificación de recursos con la relevanciaque adquieren taxones como el corzo y el jabalí, además de restos de malacofauna terrestre y marina.
Una zanja realizada en época Calcolítica (datada a partir de los restos cerámicos en ella recogidos) corta parcialmente parte de los estratoshasta ese momento estables, produciendo el revuelto de materiales en algunos de los cuadros excavados. Aparecen restos de animales do-mésticos. Esta zanja debió realizarse con fines sepulcrales a tenor de los restos humanos en ella encontrados.
Gran parte de los restos muestran trazas antrópicas, huellas de alteraciones naturales y actividades de carnívoros y roedores que se hanestudiado en un detallado análisis tafonómico.
Además, gracias a un Sistema de Información Geográfica (SIG) que se ha aplicado al análisis arqueozoológico de esta cueva se ha podidoobservar de una manera muy gráfica la distribución de cada uno de los restos en los diferentes niveles, lo cual ha permitido avanzar en el co-nocimiento de las unidades sociales de consumo y de los diversos pobladores que ocuparon esta cueva en las distintas etapas.
ABSTRACT
In this paper the Mammal remains found in the Magdalenian, Azilian, Mesolithic and Calcolithic levels, in La Fragua Cave (Santoña,Cantabria) are analyzed. There is evidence of a dual economy focused on red deer and goat during the Magdalenian. It is in the Azilien leveland, overall, in Mesolithic when faunal diversification begins, a result of change climatic, with the presence of boar and roe deer, and also seashells and land snails.
A trench dug in Calcolithic times (dated by the ceramic remains founded in it) cut down partly all the previous levels, which caused a mixon materials in some of the excavated squares. In this trench domestic animals have been found. Probably, the trench included a burial, ac-cording to the presence of some human bones.
Great part of the material shows anthropic fractures, natural alterations and marks of carnivores and rodents; both have been studied in adetailed taphonomic analysis.
In addition to the archaeozoological and taphonomical analysis, a Geographical Information System (GIS) has been applied to study thespatial distribution of the remains in each level. This analysis has produced relevant information in order to recognize the social and consump-tion areas in the different periods of occupation.
LABURPENA
Lan honek Cueva de la Fragua (Santoña, Kantabria) izeneko haitzuloko Madeleine, Azil, Mesolitiko eta Kalkolitiko aldietako mailetan aurki-turiko ugaztun aztarnak aztertzen ditu. Madeleine aldiko mailan, orein eta ahuntz ehizaren oinarrituriko ekonomia duala dago. Azil eta batik batMesolitiko mailetatik aurrera, eguraldi aldaketaren ondorioz, baliabide dibertsifikatzea nabari egiten hasten da, taxon batzuek garrantzia hartze-arekin batera, orkatza eta basurdea kasu, lehorreko eta itsasoko malakofaunaren aztarnez gain.
Kalkolitiko aldian eginiko lubaki batek (bertan aurkitutako zeramika hondarren arabera dataturik) partzialki mozten du ordura arte egonkor zi-ren geruzen parte bat eta, hola, hondeaturiko koadro batzuetan material nahasketa eragiten du. Etxe abereen hondarrak agertzen dira.
Hondarren parte handi batek aztarna antropikoak, aldaketa naturalen arrastoak eta haragijaleen eta karraskarien jarduerak erakusten dituz-te, azterketa tafonomiko batean zehaztasunez ikertu dituztenak.
Bestalde, haitzulo honen azterketa arkeozoologikoari aplikatu zaion Informazio Geografikoko Sistemak (SIG) mailetako aztarna bakoitzarenbanaketa oso era grafikoan ikusten lagundu du eta horrek haitzuloa hainbat garaitan bizitoki izan zutenen gizarte eta kontsumo unitateen eza-gutzan aurreratzeko bide eman du.
Recibido: 2004-02-17Aceptado: 2004-03-20
* Becaria Predoctoral del Gobierno Vasco. Instituto Internacional de Investigaciones Prehistóricas de Cantabria (unidad asociada al CSIC).e-mail: [email protected]
ISSN 1132-2217MUNIBE (Antropologia-Arkeologia) nº 56 19-44 SAN SEBASTIAN 2004

S. C. Aranzadi. Z. E. Donostia/San SebastiánMunibe (Antropologia-Arkeologia) 56, 2004
1. EL YACIMIENTO DE LA CUEVA DE LAFRAGUA
La cueva de La Fragua está situada en la lade-ra SE. del monte Buciero, en Santoña (Cantabria).La boca se orienta al SSE., en un entrante de labase del monte junto a la denominada Peña delFraile. El acceso a ella resulta difícil en la actuali-dad, pues es preciso descender desde el caminoque circunda el monte, a través de canchales yrampas de fuerte inclinación. Sus coordenadasUTM son VP46551050 y su altitud en torno a 125m.s.n.m. La cueva es de reducidas dimensiones -no más de unos ocho de metros de longitud portres escasos de ancho máximo- pero muy abriga-da y favorablemente orientada (Figura 1).
En 1990 se llevaron a cabo los primeros traba-jos arqueológicos en la cueva, bajo la dirección deM. R. GONZALEZ MORALES & Y. DIAZ CASADO, que secontinuaron en sucesivas campañas de corta dura-ción en 1991, 1993, 1994 y 1996.
En el cuadro A5 se observó esta secuencia es-tratigráfica (Figura 2), que sirve de referencia alconjunto del yacimiento:
• Superficie:- Nivel 0.1: Serie de hogares y capas de ceniza
y carbón alternando con capas de arcilla quemada(fuegos de pastores)
- Nivel 0.2: capa de limo arcilloso marrón, re-movido en parte
• Nivel 1: subdivido en • Nivel 1.0: superficie del nivel de conchero,
todavía con matriz terrosa del estrato superior• Nivel 1.1: nivel de conchero, muy rico en ce-
nizas• Nivel 2: Sedimento compacto amarillo con
éboulis finos• Nivel 3: Bolsadas de caracoles de tierra con
abundante carbón y pequeños fragmentos de hueso. • Nivel 4: Sedimentos pardos oscuros con
fragmentos de carbón y restos líticos y óseos, conalgunas conchas.
El yacimiento, en la zona excavada, no era ho-mogéneo: la parte central estaba cortada por unazanja compleja de dirección N-S, que incluía unacapa de tierras revueltas y otras de bloques y frag-mentos calizos, en cuya base aparecía una seriede huesos de animales. Más abajo, la zanja corta-ba profundamente la estratigrafía, y estaba rellenade material removido del conchero. La relativa an-tigüedad de la zanja quedaba certificada por el ni-vel de fuegos de pastores que la sellaba, pero sufecha posterior al conchero es indudable, toda vezque su relleno estaba formado por material de esenivel. Los hallazgos de la zanja incluyeron un con-junto de fragmentos de cerámica y materiales líti-cos de posible cronología calcolítica, que no apare-cen en absoluto en el resto del depósito del con-chero intacto.
Más detalles sobre el yacimiento y sus mate-riales se pueden ver en GONZALEZ MORALES, M. R.(1999 a y b) & GONZALEZ MORALES, M.R. & DIAZ
CASADO, Y. (1992).
2. OBJETIVOS DEL ANALISIS ARQUEOZOOLÓ-GICO
Los objetivos que se han tratado de alcanzar através del análisis del conjunto óseo de la Cuevade La Fragua no se limitan meramente al análisispaleontológico, sino que se ha pretendido conocerhasta qué punto el comportamiento de las socie-dades pasadas ha podido modificar la muestra fau-nística. Con dicho fin se ha tratado de precisar almáximo los siguientes aspectos:
– Representatividad de los diferentes taxonesen cada periodo
– Estacionalidad de las diferentes ocupacionesdel yacimiento
– Talla de las diferentes especies y su evolu-ción paleontológica
20 A. B. MARÍN
Fig. 1. Plano general de la zona excavada en la Cueva de La Fragua. Fig. 2. Corte estratigráfico de la Cueva de La Fragua.

Munibe (Antropologia-Arkeologia) 56, 2004 S. C. Aranzadi. Z. E. Donostia/San Sebastián
importancia de evaluar la naturaleza del registroreside en que, si se considera que el conjunto derestos de fauna encontrados en el yacimiento esresultado de la actividad antrópica, se debe confir-mar el posible grado de modificación que haya su-frido el registro, bien por causas antrópicas bienpor causas naturales recientes, como puede sertambién el mismo proceso de excavación. Hayque tener en cuenta que el conjunto de fauna quellega al presente es cuantitativamente más reduci-do y cualitativamente diferente de los conjuntosoriginales de los que derivan (MEADOW, 1976).
Se ha considerado como resto de fauna o ele-mento óseo cualquier fragmento de hueso (inclui-dos dientes y astas) o hueso completo recuperadodurante el proceso de excavación. Cada uno deestos restos puede englobarse en uno de los si-guientes grupos:
– Como material no identificable se han con-siderado los fragmentos óseos con partes no diag-nósticas o esquirlas de hueso (generalmente infe-riores a tres centímetros), material que ha resulta-do imposible identificar a nivel anatómico y taxo-nómico. Gran parte de este material procede delproceso de criba en seco. El análisis de este mate-rial consistió exclusivamente en cuantificar y pe-sar, para así poder observar posteriormente el gra-do de fragmentación existente y conocer los posi-bles procesos post-depósito que pudiese habersufrido el material en el yacimiento.
– Como material identificable se consideranlos huesos conservados enteros en su mayor par-te o fragmentos que presenten algún caráctermorfológico que permita identificarlos al menosanatómicamente.
3.1.1. Análisis anatómico y taxonómico
Como primer paso el material identificable seorganizó por partes anatómicas, sin hacer distin-ción de especies.
Se clasificó anatómicamente gracias a la ayu-da de los diversos atlas y publicaciones específi-cas (BARONE,1976; FORMIGO,1998; LAVOCAT,1966;PALES & LAMBERT,1972; SCHMID,1972) existentesen la Universidad de Cantabria y al préstamo provi-sional de la Colección Comparativa de Fauna delLaboratorio de Arqueozoología de la UniversidadAutónoma de Barcelona dirigido por DR. JORDI
ESTÉVEZ ESCALERA. Al mismo tiempo, se utilizó laColección Comparativa del Laboratorio deArqueozoología que se está formando en laUniversidad de Cantabria, y debemos agradecer alMuseo de Prehistoria y Arqueología de Cantabria
– Observación de las estrategias de gestión delos recursos animales (patrón de caza, procesadoy consumo). Actividad paleoeconómica.
– Procesos biológicos y geológicos acaecidosen el yacimiento
– Distribución espacio-temporal (concreción delas actividades y la funcionalidad de cada espacioocupado)
Mediante el análisis arqueozoológico se hapretendido reconstruir los patrones sociales deuso de recursos y del territorio por parte de los po-bladores del Bajo Asón en cada periodo, ya quehemos entendido por material arqueológico el re-flejo de la estructura social y económica de las co-munidades prehistóricas. Los restos arqueozooló-gicos también forman parte del complejo estructu-rado que es el registro arqueológico integral, encuya organización interna y espacial ha quedadoreflejada todo lo que podemos saber de las socie-dades pasadas. Este artículo recoge una parte deesas tareas, recogidas in extenso en un trabajomás amplio (MARIN, 2003).
3. METODOLOGIA
La metodología aplicada al análisis arqueofau-nísitico de este yacimiento tiene tres enfoquesfundamentales:
– El arqueozoológico, dirigido a extraer princi-palmente información anatómica, taxonómica, deedad y de sexo.
– El tafonómico, dirigido a conocer los posi-bles acontecimientos que hayan podido sesgar ymodificar la muestra ósea y por extensión la ar-queológica en general.
– La distribución espacial, destinada a conocerla organización espacial dentro del yacimiento enlos diferentes niveles de ocupación. Ésta es pro-ducto de la gestión social del espacio, por ello suestudio implica el análisis de los procesos de tra-bajo y de las actividades sociales representadas ysu jerarquización. (ESTÉVEZ, 1995b)
Se han considerado los restos de fauna comoproductos de trabajo, distribución y consumo es-pecíficos, con el fin de extraer de ellos informa-ción sobre cuales eran las estrategias de gestión yexplotación de los recursos, y, por tanto, cualeseran las relaciones establecidas entre la sociedady el medio natural.
3.1. Clasificación de los restos
Previo al análisis faunístico, se observó el pro-cedimiento de registro y recogida del material de-sarrollado durante el proceso de excavación. La
ANÁLISIS ARQUEOZOOLÓGICO, TAFONÓMICO Y DE DISTRIBUCIÓN ESPACIAL DE LA FAUNA DE MAMÍFEROS DE LA CUEVA DE LA FRAGUA (SANTOÑA, CANTABRIA) 21

S. C. Aranzadi. Z. E. Donostia/San SebastiánMunibe (Antropologia-Arkeologia) 56, 2004
22 A. B. MARÍN
mación incompleta y sesgada. El NR no es igual ala importancia económica de las especies(UERPMANN, 1972). A veces esta unidad se inter-preta también de manera errónea, no diferencian-do entre la biomasa potencialmente suministraday la potencialmente consumida. La relación entrecarne disponible y carne consumida se ve afecta-da por un gran número de factores, y los huesosrecuperados representan únicamente parte de losanimales sacrificados.
3.3. Determinación de la edad y el sexo
Para la determinación de la edad se han utiliza-do el desgaste dental (HILLSON, 1990) y la fusiónde las epífisis. Se ha tenido en consideración quelas fechas de erupción de los dientes y la fusiónde las epífisis difiere muy significativamente entrelas diferentes razas que corresponden a una mis-ma especie, sobre todo en animales domésticos.
Para los grados de desgaste dentario se hanutilizado los procedimientos presentados porPAYNE (1973) & PÉREZ RIPOLL (1988), para los cápri-dos. Para los cérvidos ha servido de referencia elestudio de MARIEZKURRENA (1983) y para los suidosel de BULL & PAYNE (1982). El número de mandíbu-las y maxilares con la serie dentaria completa hasido muy escaso, lo que ha obligado a observar elgrado de desgaste de las piezas dentarias indivi-dualmente aparecidas.
El otro método para calcular la edad de las es-pecies identificadas ha sido la observación de lafusión de las epífisis de los diferentes elementosanatómicos aparecidos en el registro arqueológi-co. Se realizaron cinco clasificaciones generalesatendiendo al momento de fusión en que se en-cuentre las epífisis:
– NE: neonato o infantil– JU: juvenil– SA: subadulto– AD: adulto– SE: senil
La determinación de la edad en los animalesdomésticos se ha basado en las tablas de edad deSILVER (1980) que indican las fechas de fusión delas diferentes epífisis de los animales domésticos.Entre los individuos identificados la diferenciaciónsexual ha sido posible gracias a algunos elemen-tos aparecidos, los cuales han permitido diferen-ciar el animal macho de la hembra. Estos elemen-tos han sido un canino de jabalí macho y otro dehembra y algunos restos de asta de ciervo, aun-que estos últimos realmente no son significativos
el acceso a sus fondos para constituir una base dedicha colección.
Tras el análisis anatómico, el segundo pasofue el análisis taxonómico. Este análisis se reali-zó con todos los restos clasificados anatómica-mente como un mismo elemento para, de estemodo, poder realizar una primera observación dela presencia de las diferentes especies, el patrónde fracturación del mismo elemento en los diferen-tes estratos arqueológicos, comparación paleonto-lógica de las especies en los diferentes niveles yobservación de los procesos tafonómicos refleja-dos en el material.
3.2. Unidades de cuantificación
Se han utilizado tres tipos de unidades funda-mentales: el Número de Restos (NR), el NúmeroMínimo de Individuos representados (NMI), y elpeso (W) de los restos de cada taxón. La primeray última unidad son operaciones directas mientrasque el cálculo del Número Mínimo de Individuosse ha estimado tomando como base la parte es-quelética más representada de cada taxón y te-niendo en cuenta su lateralidad, sexo y talla delanimal respecto a los otros individuos de la mismaespecie aparecidos en el mismo estrato. El NMIse ha calculado de forma individualizada por nive-les y taxones.
Este cálculo nos indica la significación relativade los diferentes taxones que forman parte delconjunto de fauna (CASTEEL, 1977). No es una esti-mación directa, sino indirecta, calculada a partirdel número de restos. Es un cálculo que hay queutilizar con cuidado, ya que en los yacimientos enlos que aparecen los huesos muy fragmentadoslas especies representadas por un pequeño núme-ro de restos resultan sobrevaloradas respecto alas que han dado muchos restos.
La obtención del peso de los restos nos indicaa su vez el grado de fragmentación de cada espe-cie dividiendo el peso total de los restos de cadataxón entre el número de restos de la misma es-pecie. El peso de los restos es una medida indis-pensable porque ayuda a superar muchos de losinconvenientes que comportan el NR y el NMI,por lo que es fundamental en cualquier análisisfaunístico para una correcta interpretación tafonó-mica y valoración paleoeconómica del conjunto.
Se ha tratado de calcular la biomasa potencial-mente explotable, cuya estimación se hace nece-saria al considerar que el NR y el NMI tienen pocarelación con el alimento que cada ejemplar repre-senta, proporcionando en este aspecto una infor-

Munibe (Antropologia-Arkeologia) 56, 2004 S. C. Aranzadi. Z. E. Donostia/San Sebastián
de la caza de ciervos machos, ya que debido alcambio estacional de las astas éstas pueden ha-berse recogido después del desmogue sin necesi-dad de cazar al animal. La otra forma de diferen-ciar el sexo del animal es mediante la compara-ción osteométrica de un mismo hueso, aunqueeste estudio no se ha podido realizar debido a laescasez de restos mensurables en la muestraósea.
Aparte, con los restos de "ovicaprinos" encon-trados en el relleno de la Zanja se trató de realizaral máximo la identificación taxonómica utilizandoel trabajo de BOESSNECK e.a.(1980), PRUMMEL &FRISCH (1986) Y FORMIGO (1999) para diferenciar en-tre cabras y ovejas.
3.4. Biometría
Los datos biométricos se han tomado en to-dos los elementos posibles, pertenecientes tantoa animales juveniles como adultos, siempre ycuando no hayan sufrido alteración natural o antró-pica que haya podido modificar su dimensión origi-nal. Dicha actividad se ha realizado no solamentecon el objeto de comprobar la diferenciación se-xual entre especies, sino también con el fin de for-mar una base de datos que permita describir enun futuro la evolución biológica de las especies ysu homogeneidad o heterogeneidad en la zonacantábrica a nivel diacrónico. Sin embargo, sólo semuestran en este trabajo las medidas de animalesadultos.
Los criterios empleados han sido tomados devon den DRIESCH (1976). A continuación se mues-tran los códigos empleados en la toma de medi-das del material óseo.
A M3 Anchura de M3Ad Anchura máxima distalAmD Anchura mínima de la diáfisisAM Anchura máximaAmII Anchura mínima de la rama del ileonAp Anchura máxima proximalAS Anchura de la superficie articularCmIl Circunferencia mínima de la rama del ileonEl Espesor lateralEmO Espesor mínimo del olecranonEMP Espesor máximo del procesoEPA Espesor del proceso ancóneoLM1-M3 Longitud de la serie molarLM3 Longitud de M3LA Longitud del Acetabulum incluyendo el labio
ANÁLISIS ARQUEOZOOLÓGICO, TAFONÓMICO Y DE DISTRIBUCIÓN ESPACIAL DE LA FAUNA DE MAMÍFEROS DE LA CUEVA DE LA FRAGUA (SANTOÑA, CANTABRIA) 23
LAb Longitud del Acetabulum en el borde interior
Ldo Longitud de la superficie dorsal
LDS Longitud diagonal de la suela
Ll Longitud lateral
LM Longitud máxima
Lm Longitud medial
LMP Longitud máxima del proceso articular
LS Longitud de la superficie articular
Osteométricamente, los individuos adultos sir-ven de referencia para la observación del diformis-mo sexual, si bien los elementos disponibles a talefecto han resultado escasos y, por tanto, ha re-sultado imposible realizar dicha observación, comose dijo en el apartado 3.3.
Mediante la biometría se ha pretendido cono-cer:
– la evolución paleontológica de las especiesaparecidas en los diferentes niveles del yacimien-to
– la variabilidad interna de la talla animal (den-tro del mismo yacimiento)
– la variabilidad externa de las diferentes espe-cies respecto a otros yacimientos
– la identificación y separación taxonómica en-tre animales salvajes y domésticos (p.e. Sus do-
mesticus / Sus scrofa)
3.5. Análisis tafonómico
En 1940 EFREMOV propuso el concepto deTafonomía como "el estudio de la transición de losrestos orgánicos desde la biosfera a la litosfera".Actualmente, la Tafonomía se ocupa de estudiarlos procesos que sufre el material tras su depósitohasta el momento de su recogida en la excava-ción. En dicho proceso, debido a la naturaleza delos organismos o a la intervención de agentes ex-ternos, éstos han actuado a modo de filtros elimi-nando los restos orgánicos menos resistentes opreservables.
Se define la Tafonomía como un sistema con-ceptual de la Paleontología que aspira a explicarcómo han sido producidos y qué modificacioneshan experimentado los restos conservados en la li-tosfera, desde su producción biogénica o tafogéni-ca hasta la actualidad (ESTÉVEZ , 1984). LaTafonomía trata de explicar cómo se ha producidoy qué factores han modificado el registro fósil(FERNANDEZ-LOPEZ, 1999).

S. C. Aranzadi. Z. E. Donostia/San SebastiánMunibe (Antropologia-Arkeologia) 56, 2004
Toda muestra de fauna que llega hasta la ac-tualidad ha pasado por diferentes fases, en lascuales ha ido perdiendo información progresiva-mente. Este proceso es necesario tenerlo encuenta a la hora de realizar la interpretación delconjunto total. Dicho proceso pasa por cinco eta-pas según KLEIN & CRUZ-URIBE (1984): de un con-junto vivo se transforma en un conjunto muerto, asu vez en un conjunto depositado, y éste en unconjunto fósil, para encontrar finalmente en la ex-cavación una muestra ósea de lo que en su mo-mento fue el conjunto vivo. ESTÉVEZ (2000b) plan-tea el proceso arquetafonómico como algo dinámi-co y dialéctico en el cual al mismo tiempo que sepierde una información original se añade al conjun-to nueva información que ilustra esa historia.
En la Fragua, gracias a la recuperación exhaus-tiva de todo el material mediante cribas en secode 4 y 2 mm, se recogieron los fragmentos míni-mos de hueso que pueden indicar cómo fue elconjunto vivo y que transformaciones post-depósi-to pudo sufrir.
Si bien es bastante difícil deducir de la mues-tra recogida el conjunto muerto y por lo tanto, elconjunto vivo, el análisis debe remitirse solamentea la muestra recogida, lo cual presenta un obstá-culo para la investigación paleoecológica de la zo-na. Además, hay que tener en cuenta que todaslas partes de un elemento o todos los elementosde una asociación ósea procedentes de una enti-dad paleobiológica, bien antrópica o bien natural,no sufren variaciones en su estado de conserva-ción a menos que existan agentes que los destru-yan o modifiquen su estructura, composición e in-cluso su articulación espacial. Por ello el análisistafonómico de la Cueva de La Fragua, con el fin deconocer que agentes o proceso formó el conjuntoy qué agentes modificaron el mismo, se ha centra-do en la observación de dos tipos de alteraciones:
– Alteraciones antrópicas– Alteraciones naturales
Para poder observar este tipo de alteracionestodo el material se ha observado mediante una lu-pa binocular de entre 12X y 60X.
Se ha entendido por alteraciones antrópicas
todas aquellas actividades realizadas por agentessociales sobre los restos animales. El significadode las marcas encontradas en los restos óseos re-lacionan el proceso de carnicería realizado por lasgentes, que permiten la transformación del animalen un bien consumible (LYMAN, 1987). El estudiode las alteraciones antrópicas en los restos, desdeel punto de vista tafonómico, trata de conocer có-
mo los grupos humanos trataron e intervinieronsobre los restos óseos.
Existen diversos tipos de actuaciones huma-nas sobre los restos óseos. En el caso de LaFragua las marcas antrópicas más comunes hansido: incisiones o trazas de corte realizadas coninstrumentos líticos, rascados, corte con percu-sión, huellas de pisoteo y marcas de cremación(aunque dos últimas también pueden ser post-de-pósito). Con el estudio de todas estas marcas(PÉREZ RIPOLL, 1992) se ha pretendido llegar a co-nocer la fase del proceso de carnicería a la cual secorresponde con el fin de poder reconstruir los pa-sos de dicho proceso antrópico.
De igual modo, con la realización de este análi-sis se ha tratado de conocer si existe o no un pa-trón estandarizado del proceso de carnicería, com-probar si se sigue un mismo método en el proce-sado de las diferentes especies, y conocer si estepatrón varía a lo largo de los diferentes periodoscronológicos.
Desde el mismo momento en que se adquiereel animal empieza el proceso de carnicería. La for-ma de adquisición dependerá de la estrategia delos grupos cazadores o ganaderos. Esta adquisi-ción conlleva un estudio detallado de la frecuenciade las partes del esqueleto aparecidas de cada es-pecie y la articulación espacial de la muestra ósea,lo que nos puede indicar el patrón de caza y trans-porte de las especies del lugar de caza al yaci-miento. El "efecto schleep" (DALY, 1969:149) defi-ne que cuanto más grande sea el animal cazado ymás lejos sea el lugar de caza respecto al yaci-miento de ocupación, el transporte de las distintaspartes de la presa será realizará de un modo dife-rencial y más selectivo (BLASCO, 1992: 64).
Por otro lado, el procesamiento de los anima-les muertos abarca desde el despellejamiento has-ta su preparación para el consumo o posterior con-servación. Por eso cada traza ha sido anotada enla ficha arqueozoológica; tipo, posición, orientacióny cantidad. Todo ello con la intención de poder lle-gar a conocer el paso del proceso de carnicería alque corresponde. Estos procesos suelen producir-se en el siguiente orden: despellejado, eviscera-ción, descuartizado, desmembrado, descarnado,aprovechamiento medular y del cerebro (SAÑA,1992). El proceso de carnicería sigue siempreunos mismos pasos aunque estos no tienen porqué acontecer de la misma forma y en el mismoorden (WHITE, 1953).
La preparación para el consumo y el manteni-miento de los productos alimentarios también
24 A. B. MARÍN

queda patente en los restos óseos. Una de lasprincipales técnicas de preparación de alimentoses el cocinado de los mismos (cocción, hervido,asado...). El testimonio de este proceso son lasmarcas que deja el fuego directa o indirectamentesobre los restos. Dependiendo de la duración e in-tensidad del calor aplicado, los cambios efectua-dos sobre el hueso son diferentes. Se han identifi-cado cuatro grados de cremación de mayor a me-nor intensidad de calor: marrón, negro, gris y blan-co. Los restos totalmente quemados no implicannecesariamente un uso nutritivo, ya que puedenhaberse quemado accidentalmente o ser productode acciones de limpieza. En cambio, los huesosparcialmente quemados pueden indicar las partesque son expuestas al fuego.
Por otro lado, es necesario señalar los proce-sos post-depósito, es decir, las modificacionesposteriores al consumo de los restos producidasen alguna medida por los agentes sociales. Unavez depositados los restos, los humanos continúansiendo causa, de forma intencionada o no, de la al-teración y modificación de los restos. Estos proce-sos suelen modificar la posición espacial de losrestos deshaciendo su posición deposicional pri-maria. Normalmente, las actividades que alteran laposición original de los huesos son actividades delimpieza de los hogares o el pisoteo constanteproducido por los ocupantes del yacimiento.
El otro tipo de alteraciones presentes son lasalteraciones naturales, que nos indican los agen-tes que han contribuido a la integración en el de-pósito, alteración y modificación de los conjuntosde restos de fauna, tanto en su composición co-mo en su distribución espacial.
Este tipo de alteraciones naturales más fre-cuentes han sido: marcas de agua, concreción,deshidratación, hongos saprofitos, meteorizacióny raíces.
Las marcas de agua (brillos, concavidades irre-gulares, estrías finas) y las marcas de meteoriza-ción (escamas óseas que se producen en la super-ficie de los huesos debido a los fuertes contras-tes de temperatura) son debidas a fenómenos at-mosféricos y kársticos en función del tiempo quehan sido expuestos a la intemperie y se reflejancon mayor o menor intensidad en la muestraósea. También en este grupo de alteraciones natu-rales debidas a los fenómenos atmosféricos se in-cluyen los huesos deshidratados por una exhibi-ción prolongada al sol.
La hiedra era una planta que existía por toda lacueva en el momento de empezar la excavación,
por ello se cree que los ácidos de las raíces de es-tá planta han podido estar entre los agentes bióti-cos que han dejado improntas en los huesos.También se ha verificado la presencia de hongossaprofitos de color negruzco. Estos hongos se ali-mentan de materia orgánica en descomposición ysuelen aparecer adheridos a los restos óseos.Finalmente, las concreciones cálcicas aparecen enforma de costra adherida a algunos de los restosóseos, principalmente a los restos aparecidos enla Zanja.
Dentro del campo de las alteraciones observa-das en los restos aparecen las marcas de anima-les que muestra las alteraciones producidas porcarnívoros o roedores. Las alteraciones apreciadashan sido marcas de mordeduras principalmentepor carnívoros, huellas de roedores y huellas deácidos gástricos al digerir el hueso, sobre todo porcarnívoros.
La acción de los roedores ha dejado huella enalguno de los restos. Estos animales realizan consus dientes unas marcas en forma de finas estria-ciones paralelas y de fondo plano, con disposiciónregular, generalmente alrededor de las epífisis. Lapresencia en el yacimiento de restos con este tipode alteraciones ha sido muy escasa.
Otro tipo de marcas presentes en el yacimien-to son las de carniceras de carnívoros, afectando ala superficie de algunos huesos (la sección de lasmarcas dientes de carnívoro suele ser en U, conun fondo plano de bordes redondeados pero sinuna disposición regular como lo hacían los roedo-res) y también se ha documentado la acción de in-gerir y regurgitar los huesos.
3.6. Distribución espacio-temporal
Para facilitar el estudio de la dispersión anató-mica de los restos se han agrupado estos en cincograndes grupos anatómicos: craneal, axial, extre-midad anterior, extremidad posterior y extremidad(metápodos, falanges y sesamoideos).
En los planos de distribución espacial de losrestos, el etiquetado de cada parte anatómica seha realizado en un color diferente y se ha posicio-nado en una determinada zona de cada sector dela siguiente forma: Craneal (CR): extremo superiorizquierdo; Axial (AX): extremo superior derecho;Extremidad anterior (EA): extremo inferior izquier-do; Extremidad posterior (EP): extremo inferior de-recho; Extremidad (EX): mitad del sector.
ANÁLISIS ARQUEOZOOLÓGICO, TAFONÓMICO Y DE DISTRIBUCIÓN ESPACIAL DE LA FAUNA DE MAMÍFEROS DE LA CUEVA DE LA FRAGUA (SANTOÑA, CANTABRIA) 25
Munibe (Antropologia-Arkeologia) 56, 2004 S. C. Aranzadi. Z. E. Donostia/San Sebastián

Estas cinco clasificaciones anatómicas obser-vadas en el plano de la cueva facilitan la observa-ción de la presencia o ausencia de las diferentespartes anatómicas de las especies halladas en ca-da nivel. Lo que se ha pretendido con el estudiode la distribución espacial de los restos ha sido de-terminar la organización del procesado y consumodentro de los diferentes grupos humanos que ha-bitaron la Cueva de La Fragua e igualmente obser-var la dinámica espacial a lo largo de los sucesivosniveles. Por ello ha sido importante conocer lasposibles alteraciones antrópicas y naturales quehan podido modificar la articulación espacial de losrestos dentro del yacimiento (MARIN, 2003).
3.6.1. Remontajes
La técnica del remontaje es una aplicaciónbastante reciente en los análisis de fauna. Se hanconsiderado remontajes a todos los fragmentosde hueso con fractura antigua que han permitidoreconstruir el hueso originalmente fracturado. Estatécnica aporta no exclusivamente información ta-fonómica (cómo han actuado los agentes antrópi-cos y naturales en la formación del conjunto fau-nístico, o sobre la estabilidad estratigráfica del ya-cimiento) sino también información en el nivel so-cio-económico (ENLOE, 1995) y sobre la duraciónde la ocupación, la organización de las diferentescomunidades, diferentes unidades sociales o dife-rentes unidades de consumo. (ESTÉVEZ &MARTINEZ, 1995).
En total, entre todo el material óseo se han lo-calizado veintiún remontajes. Normalmente, losremontajes identificados constan de dos fragmen-tos, si bien existen algunos constituidos hasta porcinco fragmentos. Sin embargo, en ningún de loscasos se ha podido reconstruir el hueso completo.Debido a la apreciable cantidad de remontajes en-contrados en el yacimiento, ha sido posible obser-var el tipo de procesamiento y la dispersión espa-cial de los restos consumidos en los diferentesmomentos cronológicos (ESTÉVEZ, 1995b). Además,esta labor ha permitido estudiar el patrón de frac-turación de los elementos remontados, conocerhasta qué grado fueron aprovechados dichos ele-mentos (aprovechamiento medular, etc.) y discer-nir si el proceso de fractura es común a todas lasespecies o si por el contrario es exclusivo de cadauna de ellas.
De igual modo, el análisis de la distribución es-pacial de los remontajes posibilita el conocimiento
de la depositación primaria del resto, informandosobre los posibles procesos post-depósito o alte-raciones naturales que ha sufrido cada fragmentodesde el momento de su fractura hasta el mo-mento de su recogida en la excavación.
3.6.2. Rearticulaciones
Se ha denominado rearticulación a los elemen-tos que, habiendo sido recogidos por separado,forman parte de un mismo conjunto, conectandoanatómicamente entre sí para constituir un ele-mento complejo. (Por ejemplo: maxilar con sus co-rrespondientes piezas dentarias). En total, se hanlocalizado catorce rearticulaciones.
El estudio de la dispersión de las rearticulacio-nes permite conocer por dónde se han distribuidodichos elementos dentro del yacimiento y deducirsi su distribución corresponde a un proceso casualo si ha sido provocada por acciones antrópicas. Eneste sentido, algunos de estos elementos hanaparecido en un mismo lugar, lo que parece indi-car que se encontraban originalmente en procesode fusión o articulados.
3.7. Objetivos del estudio de la distribución es-pacial
Con la realización del estudio de la distribuciónde los restos encontrados en La Fragua medianteun Sistema de Información Geográfica (SIG) hapermitido llegar a conocer los siguiente puntos:
– Identificación de patrones de localización dedistintos tipos de restos para cada una de las es-pecies y su evolución temporal (unidades de con-sumo)
– Asignación de zonas de actividad dentro delyacimiento (unidades sociales)
– Análisis de la interrelación espacial del mate-rial óseo
– Observación de los posibles movimientospost-depósito a través de los remontajes y rearti-culaciones
Gracias a este sistema se ha podido disponerde la información procedente de una base de da-tos arqueozoológica típica de una manera más grá-fica, interactiva y directa, que ha permitido aunartanto la información propiamente dicha como sudistribución espacial, permitiendo la presentaciónde los resultados de una forma muy manejable(MARIN: en preparación).
26 A. B. MARÍN
Munibe (Antropologia-Arkeologia) 56, 2004 S. C. Aranzadi. Z. E. Donostia/San Sebastián

4. MATERIAL
El conjunto de restos de mamíferos (sin teneren cuenta los siete restos humanos) está com-puesto por 5.862 restos óseos, de los cuales4.323 son restos no identificables (NI), mientrasque 597 han podido ser asignados a 16 especiesdiferentes de Mamíferos, siendo 574 restos deUngulados.
Al mismo tiempo, se ha identificado un dientede Insectívoro, un hueso de Mustélido y tres ele-mentos más pertenecientes a la familia de lasPinnípedos, mientras que los 934 elementos res-tantes han sido englobados dentro de las catego-rías generales de Mamíferos (grande, medio y pe-queño) y Caprinos (en los Niveles 1, 2, 3 y 4) yOvicaprinos (en la Zanja).
Los restos de gran bóvido aparecidos en losNiveles 1, 2, 3 y 4, debido a su elevada fragmen-tación, ha impedido la observación de morfologías
discriminantes entre Bos primigenius y Bison pris-
cus. Por lo tanto, estos restos se han englobadodentro de la categoría general de Bovini. En cam-bio, los restos de bovinos encontrados en la Zanjase han atribuido a Bos taurus principalmente porsu tamaño. La presencia de ejemplares de vacunodoméstico parece clara y es probable su relacióncon la cerámica encontrada dentro de la Zanja.
Los restos de cabra encontrados en la Zanjahan sido englobados en la categoría Capra sp yaque, por una parte, la Zanja está formada en partepor el revuelto de los niveles 1, 2, 3 y parte del 4y, en segundo lugar, esta Zanja fue realizada enépoca Calcolítica, por lo tanto los restos de cabrapodrían pertenecer a Capra hircus. La diferencia-ción entre Capra pyrenaica y Capra hircus no seha podido comprobar osteométricamente entretodos los restos aparecidos en la Zanja debido aque sólamente uno de esos restos ha podido sermedido.
ANÁLISIS ARQUEOZOOLÓGICO, TAFONÓMICO Y DE DISTRIBUCIÓN ESPACIAL DE LA FAUNA DE MAMÍFEROS DE LA CUEVA DE LA FRAGUA (SANTOÑA, CANTABRIA) 27
Munibe (Antropologia-Arkeologia) 56, 2004 S. C. Aranzadi. Z. E. Donostia/San Sebastián
NIVEL 4 % NIVEL 3 % NIVEL 2 % NIVEL 1 % ZANJA % SUPUNGULADOSBos/Bison 5 0,33 1 0,16 1 0,43 4 0,73 - - -Bos taurus - - - - - - - - 29 1,01 -Cervus elaphus 79 5,14 22 3,62 3 1,30 18 3,28 45 1,57 1Capreolus capreolus 13 0,85 6 0,99 2 0,87 8 1,46 9 0,31 -Capra pyrenaica 114 7,42 4 0,66 2 0,87 6 1,09 - - -Capra sp. - - - - - - - - 41 1,43 -Rupicapra rupicapra 1 0,07 - - - - - - 6 0,21 -Ovis aries - - - - - - - - 1 0,03 -Sus scrofa 7 0,46 - - 5 2,16 19 3,47 6 0,21 -Sus sp. - - - - - - - - 46 1,60 1Total Ungulados 219 14,27 33 5,43 13 5,63 55 10,03 183 6,37 2
CARNÍVOROSCanis sp. - - - - - - 3 0,55 - - -Canis familiaris - - - - - - - - 6 0,21 -Meles meles - - - - - - 1 0,18 - - -Vulpes vulpes 5 0,33 - - 1 0,43 1 0,18 2 0,07 -Felis silvestris - - - - - - - 1 0,03 -INSECTÍVOROSErinaceus europeus - - - - - - - - 1 0,03 -PINNÍPEDOSPhoca sp. - - - - - - - - 1 0,03 1HUMANOHomo sapiens - - - - - - - - 4 0,14 1
Caprinos/ Ovicaprinos 18 1,17 4 0,66 - - 8 1,46 39 1,36 -Mamífero grande - - 1 0,16 - - 2 0,36 1 0,03 -Mamífero medio 379 24,67 126 20,72 7 3,03 48 8,76 275 9,58 -Mamífero pequeño 15 0,98 3 0,49 6 2,60 1 0,18 7 0,24 -
No identificable 900 58,59 441 72,53 204 88,31 429 78,28 2351 81,86 -TOTAL 1536 608 231 548 2872 4
Tabla 1. Número de restos (NR) y porcentajes de las diversas especies en los distintos niveles.
28 A. B. MARÍN
Munibe (Antropologia-Arkeologia) 56, 2004 S. C. Aranzadi. Z. E. Donostia/San Sebastián
NIVEL 4 % NIVEL 3 % NIVEL 2 % NIVEL 1 % ZANJA % SUP %UNGULADOSBos/Bison 39,4 1,80 2,7 0,53 2,5 1,79 91,9 12,79 - - - -Bos taurus - - - - - - - - 532,8 22,39 - -Cervus elaphus 746 34,12 253,9 49,59 5,3 3,79 138,3 19,24 313,7 13,18 0,2 0,46Capreolus capreolus 13,3 0,61 8 1,56 4 2,86 8,7 1,21 6,9 0,29 - -Capra pyrenaica 518,3 23,71 15,2 2,97 5,1 3,64 22 3,06 - - - -Capra sp. - - - - - - - - 122,3 5,14 - -Rupicapra rupicapra 0,7 0,03 - - - - - - 14,7 0,62 - -Ovis aries - - - - - - - - 1,5 0,06 - -Sus scrofa 8,9 0,41 - - 6,7 4,79 220,6 30,69 75,8 3,18 - -Sus sp. - - - - - - - - 133,8 5,62 0,6 1,39Total Ungulados 1326,6 60,68 279,8 54,65 23,6 16,87 481,5 66,99 1201,5 50,48 0,8 1,85
CARNÍVOROSCanis sp. - - - - - - 2,3 0,32 - - - -Canis familiaris - - - - - - - - 42,9 1,80 - -Meles meles - - - - - - 1,9 0,26 - - - -Vulpes vulpes 7,2 0,33 - - 0,9 0,64 1,3 0,18 1,4 0,06 - -Felis silvestris - - - - - - - 1,9 0,08 - -INSECTÍVOROSErinaceus europeus - - - - - - - - 1 0,04 - -PINNÍPEDOSPhoca sp. - - - - - - - 22,1 0,93 40,6 93,76HUMANOHomo sapiens - - - - - - - - 4,2 0,18 1,9 4,39
Caprinos/Ovicaprinos 61 2,79 3,4 0,66 - - 35,5 4,94 116,6 4,90 - -Mamífero grande - - 16,3 3,18 - - 16,2 2,25 2,2 0,09 - -Mamífero medio 293,3 13,42 89,1 17,40 43 30,71 45,5 6,33 195,1 8,20 - -Mamífero pequeño 3,8 0,17 1,3 0,25 0,5 0,36 0,6 0,08 3,1 0,13 - -
No identificable 494,2 22,61 122,1 23,85 72 51,43 133,9 18,63 775,7 33,11 - -TOTAL 2186,1 512 140 718,7 2380,1 43,3
Tabla 2. Peso de los restos (W) y porcentajes de las diversas especies en los distintos niveles.
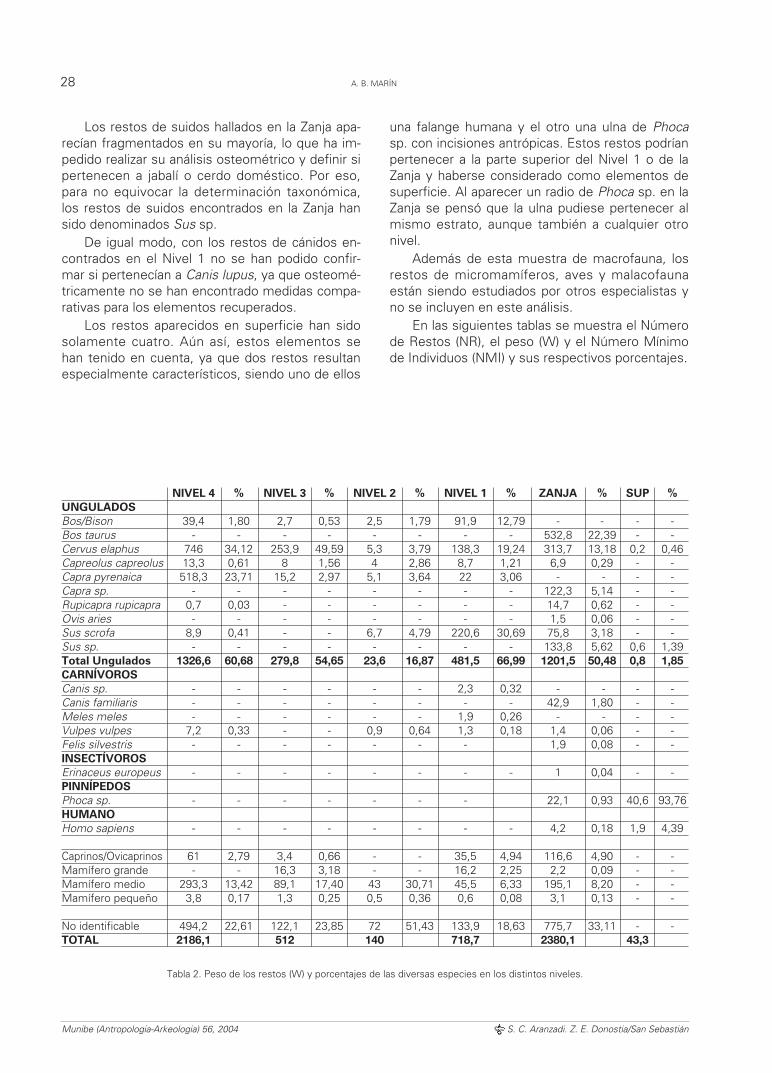
Los restos de suidos hallados en la Zanja apa-recían fragmentados en su mayoría, lo que ha im-pedido realizar su análisis osteométrico y definir sipertenecen a jabalí o cerdo doméstico. Por eso,para no equivocar la determinación taxonómica,los restos de suidos encontrados en la Zanja hansido denominados Sus sp.
De igual modo, con los restos de cánidos en-contrados en el Nivel 1 no se han podido confir-mar si pertenecían a Canis lupus, ya que osteomé-tricamente no se han encontrado medidas compa-rativas para los elementos recuperados.
Los restos aparecidos en superficie han sidosolamente cuatro. Aún así, estos elementos sehan tenido en cuenta, ya que dos restos resultanespecialmente característicos, siendo uno de ellos
una falange humana y el otro una ulna de Phoca
sp. con incisiones antrópicas. Estos restos podríanpertenecer a la parte superior del Nivel 1 o de laZanja y haberse considerado como elementos desuperficie. Al aparecer un radio de Phoca sp. en laZanja se pensó que la ulna pudiese pertenecer almismo estrato, aunque también a cualquier otronivel.
Además de esta muestra de macrofauna, losrestos de micromamíferos, aves y malacofaunaestán siendo estudiados por otros especialistas yno se incluyen en este análisis.
En las siguientes tablas se muestra el Númerode Restos (NR), el peso (W) y el Número Mínimode Individuos (NMI) y sus respectivos porcentajes.

5. ANALISIS POR NIVELES DE LOS RESTOS DEMACROMAMIFEROS
5.1. Nivel 4
Varios huesos de este nivel fueron fechadosproporcionando una datación de 12.960 ± 50 BP,lo que señala un momento de ocupación del yaci-miento durante el Magdaleniense Superior Final.
El número de restos aparecidos en este nivelasciende a 1.535 con un peso total de 2.186 gra-mos. La media del fragmento óseo en este niveles de 1,42 gramos. Los restos no identificables re-presentan un 58.63% del total. Entre los Unguladosmás destacados el espectro faunístico aparecemarcado por la elevada cantidad de restos de ca-bra montés y de ciervo. Únicamente aparecen res-tos de un carnívoro, Vulpes vulpes. A continuaciónse muestra en la Figura 3 el perfil de edades delos distintos individuos identificados (neonato, ju-venil y adulto).
A pesar de ser la cabra el taxón que mayor nú-mero de restos ha presentado fue el ciervo el ani-mal que mayor tanatomasa aportó a los gruposhumanos. Respecto al traslado de las piezas caza-das al yacimiento, se ha podido apreciar un trans-porte antrópico diferencial en los taxones más re-
ANÁLISIS ARQUEOZOOLÓGICO, TAFONÓMICO Y DE DISTRIBUCIÓN ESPACIAL DE LA FAUNA DE MAMÍFEROS DE LA CUEVA DE LA FRAGUA (SANTOÑA, CANTABRIA) 29
Munibe (Antropologia-Arkeologia) 56, 2004 S. C. Aranzadi. Z. E. Donostia/San Sebastián
NIVEL 4 NIVEL 3 NIVEL 2 NIVEL 1 ZANJAUNGULADOS Bos/Bison 2 1 1 2Bos taurus 4Cervus elaphus 8 2 1 2 4Capreolus capreolus 3 2 2 2 2Capra pyrenaica 5 1 1 3Capra sp 4Rupicapra rupicapra 1 3Sus sp 7Sus scrofa 3 1 4 1Ovis aries 1Total Ungulados 22 6 6 13 26
CARNÍVOROSCanis familiaris 1Canis species 1Felis silvestris 1Meles meles 1Vulpes vulpes 1 1 1 1INSECTÍVOROSErinaceus europeus 1PINNÍPEDOSPhoca sp. 1TOTAL 23 6 7 16 31
Tabla 3. Número Mínimo de Individuos (NMI) de las diversas especies en los distintos niveles.
presentativos en este nivel, la cabra y el ciervo. Elprimero, al ser un animal de hábitat de roquedopróximo a la cueva, tras su caza era trasladado en-tero al yacimiento, mientras que el ciervo, al mo-verse en zonas más alejadas, implicaba a los caza-dores al menos una hora de desplazamiento des-de la cueva (MARIN, 2003), y para un mayor rendi-miento transportaban únicamente las extremida-des y la cabeza abandonando en el lugar de cazael esqueleto axial (Figura 4).
Fig. 3. Distribución por edades del NMI de Ungulados y Carnívorosdel Nivel 4.

En este nivel la mayor parte de los huesos hansufrido una intensa fracturación, fruto de la utiliza-ción de la médula como un aporte alimenticiomás. Esa exhaustiva fracturación del material seajusta a la tónica general de procesamiento de losrestos de fauna que tiene lugar en el Magdale-niense Medio pero sobre todo en el Superior.
Mediante el estudio tafonómico se ha distin-guido una elevada cantidad de trazas de corte, ras-cados y huellas de percusión como resultado delproceso de carnicería. Únicamente en la cabra y elciervo se ha podido reconstruir parcialmente elproceso antrópico de carnicería realizado en el pe-riodo magdaleniense.
El proceso primario o Primary Butchering
Activity es similar en ambos taxones, ya que esprobable que despellejasen los animales a partirde las primeras falanges y que abriesen al animalpor la parte interna de las costillas (hecho consta-tado únicamente con la cabra). Normalmente laextremidad anterior era desarticulada cortando porel cuello de la escápula (Figura 5) o por el cuellodel húmero, el radio-ulna solía desarticularse delhúmero por el proceso anconeo de la ulna y final-mente, el metacarpo se desligaba por los huesoscarpales del radio-ulna. De igual modo, en la extre-midad posterior el fémur solía desarticularse de lapelvis por la cabeza de éste, la tibia del fémur cor-tando los ligamentos y tendones que lo unen al ély por último, se cortaba por el talus y calcáneo pa-ra desarticular el metatarso de la tibia.
30 A. B. MARÍN
Munibe (Antropologia-Arkeologia) 56, 2004 S. C. Aranzadi. Z. E. Donostia/San Sebastián
Fig. 4. Distribución anatómica de los restos de Mamíferos del Nivel 4.
Fig. 5. Escápulas de Cervus elaphus con un mismo patrón de frac-turación e incisiones antrópicas.
FRB51/4.4
FRA52/4.3 NR 27

Se entiende por proceso secundario oSecondary Butchering Activity según BINFORD
(1981) la fracturación de los huesos una vez desar-ticulados. Se ha podido observar un mismo patrónde fracturación en huesos como el radio, húmeroo tibia en mamíferos de tamaño medio. La mandí-bula, después de su despellejamiento, era fractu-rada para su obtener la médula (Figura 6).
En referencia a los carnívoros, los escasos res-tos de zorro hallados han aparecido muy concen-trados a la entrada del yacimiento, en el cuadroA6, lo que refuerza la idea que pertenezcan a unmismo individuo. La presencia de marcas de corteen algunos de sus restos indica que este animalfue manipulado antrópicamente, probablementepara el uso de su piel y posiblemente para suaprovechamiento cárnico.
El Nivel 4 fue el menos afectado por la excava-ción de la Zanja, de ahí que haya sido en este niveldonde, exclusivamente, se han podido conocer ladistribución espacial de los restos de consumo delas unidades sociales que ocuparon la Cueva de LaFragua. Así, a partir del estudio de la distribuciónespacial de los restos óseos, la conclusión másnotable obtenida es la localización de una zona deactividad antrópica, de procesado y consumo ali-mentario, principalmente de cabra y ciervo, en lossectores 1 y 4 del cuadro B5. Además, en el casode los restos del ciervo, estos aparecen dispersospor los seis cuadros excavados, al igual que losrestos de bovino, mientras que los restos de cabraaparecen más concentrados en los cuadros A5,A6, B5 y B6. Por el contrario los restos de corzo,rebeco y jabalí aparecen muy próximos a la entra-da de la cueva.
La elevada concentración de restos de cabraen los sectores 1 y 4 del cuadro B5, junto a losseis remontajes y cuatro rearticulaciones que sehan podido realizar con sus restos, nos indicanque varias cabras fueron consumidas inmediata-mente antes de abandonar el yacimiento, ya quesi la ocupación hubiese continuado la dispersiónde dichos restos hubiese sido mucho mayor, co-mo así ha ocurrido con los restos de ciervo. Cincode los remontajes de cabra han correspondido arestos de metacarpos con un patrón de fractura-ción muy similar al que muestra el Remontaje-5(Foto 1). Este remontaje muestra una fractura lon-gitudinal en la metáfisis proximal, al haber sidofracturada la epífisis proximal previamente.
En la mayor parte de remontajes, siete en to-tal en este nivel, los fragmentos que los compo-nen se localizan en sectores inmediatos, y única-mente el Remontaje 10 (Figura 8), de ciervo, ha
ANÁLISIS ARQUEOZOOLÓGICO, TAFONÓMICO Y DE DISTRIBUCIÓN ESPACIAL DE LA FAUNA DE MAMÍFEROS DE LA CUEVA DE LA FRAGUA (SANTOÑA, CANTABRIA) 31
Munibe (Antropologia-Arkeologia) 56, 2004 S. C. Aranzadi. Z. E. Donostia/San Sebastián
Fig. 6. Mandíbulas de Capra pyrenaica con similar patrón de fractu-ración y con marcas de alteraciones antrópicas y naturales.
Fig. 7. Distribución espacial de los restos de Capra pyrenaica delNivel 4.
FRB54/4.4
FRB67/RZ NR 115
FRA64/4.3 NR 40
FRB43/RZ NR67
Fig. 8. Distribución espacial de los remontajes del Nivel 4.

permitido observar los movimientos que tuvieronlugar antes del enterramiento de los huesos (pro-cesos bioestratinómicos) del interior hacia el exte-rior de la cueva. Este tipo de movimientos post-depósito suelen producirse cuando el material,tras su fracturación y posterior abandono pero an-tes de su incorporación al cuerpo de la sedimenta-ción, sufre desplazamientos dentro del yacimientopor la circulación humana, animal o escorrentíasde agua.
Para finalizar, se ha podido conocer la estacio-nalidad de la ocupación a partir del estudio de va-rias piezas dentales de cabra y de ciervo.
En el caso de las cabras, las piezas dentalessitúan la época de caza en el otoño, cuando tienelugar el periodo de celo, que es cuando se con-centran los machos y las hembras para iniciar el ci-clo biológico. Se le considera un momento idóneopara la caza por su bajo coste en tiempo y enenergía. El molar de ciervo indica que la época deabatimiento del animal fue el otoño, si bien a partirde dos ulnas de dos individuos neonatos se haatestiguado la ocupación del yacimiento tambiénen temporada estival.
Por lo tanto, las ocupaciones antrópicas de LaFragua debieron producirse en otoño y también enverano, pudiendo corresponderse a asentamientostemporales de grupos del interior que se desplaza-ban a la costa temporalmente en busca de algúntipo de recursos existente cerca del yacimiento.
5.2. Nivel 3
Este nivel se definió como una serie de bolsa-das de caracoles de tierra y restos carbonosos dis-tribuidas de modo discontinuo en distintas áreasde la excavación. Varios de esos restos de carbónproporcionaron una datación de 9.600 ± 140 BP,que sirvió como principal criterio de su atribucióntentativa al Aziliense dada la pobreza de la indus-tria en dicho estrato.
El número de restos encontrados en este nivelha sido de 608, con un peso total de 512 gramos,por lo que la media del fragmento óseo de este ni-vel es 0,84 gramos. El 72.53 % de los restos halla-dos en el Nivel 3 corresponden a huesos no identi-ficables perteneciendo únicamente 33 a restosUngulados.
En la Cueva de La Fragua, durante este perio-do la especie dominante es el ciervo (66,7 %); noobstante también aparece corzo (27,3 %), cabra(18,18 %), y bovino (3,03 %). El predominio delciervo existe, en porcentajes similares, en los ni-veles de los yacimientos cantábricos de Urtiaga C,Ekain III y Aizbitarte IV (ALTUNA, 1995).
La caza en este periodo parece centrarse prin-cipalmente en zona de llanuras y bosques dondehabitan ciervos y corzos, relegando a un segundoplano los recursos de las zonas de roquedo, a pe-sar de que si tenemos en cuenta la subida del ni-vel de mar en este periodo, el porcentaje del áreamontañosa accesible aumenta respecto al nivelmagdaleniense.
La explicación de este fenómeno puede radi-car en el cambio climático que sobrevino con elinicio del Holoceno, lo que provocó un atempera-miento de las condiciones ambientales y, por tan-to, favoreció el desarrollo de especies propias dellanura y bosque como el corzo o el jabalí gracias ala expansión rápida y generalizada de bosquesmixtos y caducifolios. En este sentido, la diversifi-cación de los recursos de llanura y bosque juntocon la existencia de preferencias nutricionales ha-cia el ciervo provocó una disminución del consu-mo de animales de montaña. Este descenso po-dría deberse a que debido a la expansión de losbosques probablemente se produjo una migraciónaltitudinal de las cabras hasta cotas más altas ypor consiguiente, se produjo el alejamiento de lasáreas de caza más habituales.
El número mínimo de individuos encontradosha sido de seis individuos. Cuatro de ellos corres-ponden a ejemplares adultos y dos a juveniles. En
32 A. B. MARÍN
Munibe (Antropologia-Arkeologia) 56, 2004 S. C. Aranzadi. Z. E. Donostia/San Sebastián
Foto 1. Metacarpo de Capra Pyrenaica (Remontaje-5) del Nivel 4.

la siguiente figura se ilustra la distribución por eda-des de cada individuo identificado (juveniles yadultos).
El ciervo es el animal del que más aprovecha-miento cárnico y medular se obtuvo en este perio-do. Aún así, el número mínimo de individuos esti-mado de esta especie es de solamente dos, loque parece señalar una ocupación bastante cortade la cueva.
El brusco descenso de Capra pyrenaica en LaFragua no se observa en los Niveles 2a y 2b delAbrigo de la Peña del Perro (yacimiento situado enlas proximidades de la Cueva de La Fragua) dondesu consumo no experimenta un descenso tan acu-sado. Esta diferencia bien podría deberse a quelos estratos azilienses de Abrigo de la Peña delPerro, datados en el 10.160 BP, son inmediata-mente anteriores al inicio del Holoceno, la diversi-ficación de los recursos no era posiblemente tannotoria y, por tanto, la economía seguía aún patro-nes magdalenienses. En este sentido hay que re-saltar el vacío de dataciones de unos quinientosaños en la cornisa cantábrica, que separa precisa-mente en sus dos extremos a ese nivel de 10.160de Peña del Perro del nivel de 9.600 BP de LaFragua 3, y que coincide con el abrupto final de lafase del Dryas reciente (ESTÉVEZ, com. pers).
Respecto al transporte de los animales al yaci-miento, al igual que ocurría en el Nivel 4, única-mente la cabeza y extremidades de ciervo fuerontrasladadas a la cueva. Este transporte selectivo o"efecto schlepp" (PERKINS & DALY, 1968) hace pen-sar en que probablemente la caza de animales demayor peso y de hábitat a mayor distancia propi-ciase su transporte diferencial. Sin embargo, du-rante el Aziliense en el yacimiento de El Perro seapreció un traslado completo de las piezas de cier-vo (MORALES, ms.), constituyendo otro rasgo dife-rencial entre ambos asentamientos.
A pesar de la escasez de restos dentales, apartir de las piezas dentarias de ciervo se puedesugerir un periodo de ocupación invernal del yaci-miento.
En el análisis tafonómico se ha identificado unpatrón de fracturación antrópica en los huesos deciervo similar al nivel magdaleniense; sin embar-go, no ha sido posible conocer el proceso de carni-cería dada la escasez de marcas de corte registra-das. Únicamente, siguiendo con la tónica del Nivel4, se han advertido incisiones o rascados efectua-dos en las diáfisis de los huesos, producidos nor-malmente durante el proceso secundario de trata-
miento del animal. De igual modo que en el Nivel4, los restos óseos aparecen muy fracturados, evi-denciando también su aprovechamiento medular.
Entre las alteraciones naturales más frecuen-tes, las raíces y los hongos saprofitos destacansobre las demás. No se han apreciado marcas decarnívoros o roedores.
La Zanja ha cortado la mitad del total del áreaexcavada, lo que supone que sólo se hayan con-servado restos en la zona que quedó intacta. Laejecución de la Zanja provocó la mezcla y el des-plazamiento de parte del nivel, lo que ha provoca-do que una porción de los restos en él deposita-dos haya ido a parar al relleno de la Zanja. Estoqueda constatado en un metacarpo de ciervo -Remontaje 9 – (Foto 2) formado por dos restos, delos que uno fue localizado en la zanja. Ambos res-tos muestran un proceso tafónomico muy diferen-te.
ANÁLISIS ARQUEOZOOLÓGICO, TAFONÓMICO Y DE DISTRIBUCIÓN ESPACIAL DE LA FAUNA DE MAMÍFEROS DE LA CUEVA DE LA FRAGUA (SANTOÑA, CANTABRIA) 33
Munibe (Antropologia-Arkeologia) 56, 2004 S. C. Aranzadi. Z. E. Donostia/San Sebastián
Fig. 9. Distribución por edades del NMI de Ungulados del Nivel 3.
Fig. 10. Distribución anatómica de los restos de Ungulados delNivel 3.

5.3. Nivel 2
El Nivel 2 de La Cueva de La Fragua corres-pondía a un estrato de sedimento compacto ama-rillo con éboulis finos. Este tipo de estratos sueleformarse, normalmente, en épocas de desocupa-ción.
En general, el material de macrofauna encon-trado en esta capa ha sido muy escaso. El númerode restos encontrados en este nivel ha sido de294 con un peso total de 140 gramos. El peso delfragmento medio es de 0.48 gramos. Casi un 70% de los restos de este nivel corresponde a res-tos no identificables.
Se han identificado 13 restos de los siguientesUngulados: Bos/Bison, Cervus elaphus, Capreolus
capreolus, Capra pyrenaica y Sus scrofa y un únicoresto de carnívoro: Vulpes vulpes.
A partir del análisis arqueozoológico y tafonó-mico realizado con el material de este nivel y te-niendo en cuenta la existencia de la Zanja, esteestrato no puede ser considerado en si como unnivel de ocupación humana, sino que más bien pu-diera corresponder a un momento de desocupa-ción temporal entre el 9.600 y el 7.530 BP.
Además, el estrato no aparece de forma conti-nua en todo el yacimiento, sino que existen zonasdonde el Nivel 1 y el Nivel 3 incluso están en con-tacto directo, lo que viene a confirmar el origennatural del mismo, sin que pueda constatarse nin-gún tipo de presencia humana.
En el transcurso del análisis individualizado porespecies, se ha apreciado una total desconexiónentre los restos aparecidos, por lo que se piensaque los restos de Ungulados registrados comopertenecientes a este estrato pueden provenir enrealidad de estratos adyacentes, como sendosrestos de bovino y corzo. Incluso los restos deMamífero Medio no constituyen una cantidad muyrelevante y podrían pertenecer a una mezcla demateriales de diversos niveles como consecuen-cia de la ejecución de la Zanja.
Tafonómicamente, en el caso de los ungula-dos, no se han podido catalogar los restos comoelementos del procesado, consumo y deshechode grupos humanos, ya que no se han apreciadoevidencias de actividad antrópica en los mismos.
En resumen, este nivel de limos debió consti-tuirse de forma natural entre las ocupaciones delnivel 3 y las mesolíticas posteriores al 7.530 BP ,no habiendo existido ocupación humana, hechoque queda confirmado a la luz de los resultadosdel análisis faunístico y remarca el hiato recurrenteque se produce nuevamente hacia el 8.000BP entoda la zona cantábrica.
5.4. Nivel 1
El Nivel 1 corresponde a un potente concheromesolítico en el que abundaban las ostras y losmejillones, pero donde las conchas de Monodonta
y las lapas estaban también presentes de manerasignificativa, siendo Patella el género mas frecuen-te en conjunto, así como gran cantidad de restoscarbonosos y una casi total ausencia de industrialítica.
Se realizaron tres fechaciones de carbones en-contrados en la parte superior, media e inferior delnivel, proporcionando unas fechas de 6.650 ±120BP, 6.860±60 BP y 7.530±70 BP, respectivamen-te, lo que responde a varios momentos de ocupa-ción humana a lo largo del Mesolítico.
Entrado el Holoceno, esta fase cronológicacoincide con el Atlántico, periodo climático tem-plado en el que el paisaje se diversifica dando pa-so a bosques caducifolios con amplias zonas dellanura. El nivel del mar se sitúa a menos de 20metros por debajo del nivel actual y en progresivoascenso, disponiendo así el entorno de la Cuevade La Fragua de unas características orográficasmuy variadas durante el Mesolítico, adecuadas pa-ra la explotación de muy diferentes recursos, des-de mar abierto y zonas de estuario hasta las zonasde roquedo y de amplias llanuras.
34 A. B. MARÍN
Munibe (Antropologia-Arkeologia) 56, 2004 S. C. Aranzadi. Z. E. Donostia/San Sebastián
Foto 2. Metacarpo proximal de ciervo (Remontaje-9) del Nivel 3.

Una vez superado el hiato del nivel 3, el con-sumo de animales de roquedo comienza a remon-tar en la medida que aumenta la proporción delárea de montaña en la zona del Bajo Asón. Sinembargo, este incremento es bastante reducido yno llega a alcanzar los valores magdalenienses, yaque continúa la diversificación de especies de lla-nura y monte, como sucede en este nivel con eljabalí. Podría decirse que hay un mayor ajuste a laoferta del medio inmediato, que en definitiva talvez se podría relacionar con una reducción del ran-go de movilidad. Pero esta hipótesis también ha-brá que tratarla con el análisis del resto de la evi-dencia arqueológica (aprovisionamiento de molus-cos, materias primas minerales, etc.)
El número total de restos encontrados en estenivel ha sido de 548, con un peso de 718.7 gra-mos. El peso medio del fragmento es de 1.31 gra-mos. El 78.28 % del material encontrado en estenivel corresponde a restos no identificables.
En este nivel se han identificado cinco espe-cies de ungulados, dos de carnívoros y una demustélidos. No se han encontrado indicios de do-mesticación animal.
A partir de los restos de los diferentes taxo-nes identificados se ha calculado un mínimo de 16individuos (neonato, juvenil, adulto y senil).
El taxón más representativo de este nivel es elSus scrofa, tanto en número de restos como enpeso. Es una especie que empieza a ser represen-tativa a partir del Holoceno. En algunos yacimien-tos mesolíticos se repite como la especie más re-levante, tal y como ocurre en Zatoya Ib (MARIEZ-KURRENA Y ALTUNA, 1989). Consecutivamente, elciervo y el bovino han sido los taxones que mayoraporte cárnico han proporcionado después del ja-balí.
Se ha podido constatar el traslado exclusivode las extremidades y la cabeza de los jabalíes; encambio; en el ciervo su patrón de consumo pareceque varía según corresponda a animales juvenileso adultos. De los animales juveniles de Cervus
elaphus se encuentran restos de todas las partesdel esqueleto mientras que de los individuos adul-tos no se han registrado restos del esqueletoaxial.
A partir de los resultados del análisis faunísti-co, los restos de bovino parecen correspondersecon restos de uro. Esto concuerda con la corrientegeneral que sitúa la emigración del bisonte y suconsiguiente desaparición del Cantábrico a partirdel Holoceno (ALTUNA, 1992), ya que hasta el mo-mento no se ha podido constatar la presencia de
restos de Bison bonasus en la Cornisa Cantábricaen un momento tan avanzado del Holoceno.
El corzo ha sido el tercer ungulado con mayornumero de restos; en cambio, su aporte cárnicofue inferior al de la cabra.
La cabra ha sido la cuarta especie que mayoraporte cárnico proporcionó en este periodo, identi-ficándose tres individuos de edad juvenil y adulta.Tal y como sucedía en el Nivel 4, todo indica queeste animal se transportaba íntegro al yacimientoindependientemente de su edad. Además, a partirde las trazas antrópicas identificadas en sus restosse aprecia un proceso de carnicería similar al atri-buido a la cabra durante el periodo magdalenien-se.
Sin embargo, ha resultado muy difícil indicarque papel desempeñaron los cánidos en el depósi-to mesolítico, ya que según los restos identifica-dos ni siquiera se ha podido conocer si correspon-dían a lobo. Sin embargo, la presencia de la espe-cie doméstica está documentada en Europa desde
ANÁLISIS ARQUEOZOOLÓGICO, TAFONÓMICO Y DE DISTRIBUCIÓN ESPACIAL DE LA FAUNA DE MAMÍFEROS DE LA CUEVA DE LA FRAGUA (SANTOÑA, CANTABRIA) 35
Munibe (Antropologia-Arkeologia) 56, 2004 S. C. Aranzadi. Z. E. Donostia/San Sebastián
Fig. 11. Distribución por edades del NMI de Mamíferos del Nivel 1.
Fig. 12. Distribución anatómica de los restos de Unguladosdel Nivel 1.

el final del Paleolítico superior. Los yacimientosepipaleolíticos postazilienses cantábricos con res-tos de cánidos (ALTUNA, 1994) han sido: Marizulo(ALTUNA,1980), Arenaza (ALTUNA,1979) y Santi-mamiñe (CASTAÑOS, 1984).
Las especies identificadas en este nivel refle-jan el atemperamiento climático durante el perio-do Atlántico, con la expansión de los bosques ca-ducifolios que desencadena el incremento delconsumo de especies como el jabalí y el corzo, sibien continua existiendo el aprovechamiento des-tacado de especies como el ciervo.
Debido al corte de la Zanja la distribución es-pacial de los restos en este nivel, al igual que su-cedió en el Nivel 3, no ha permitido reconocer unazona de actividad y consumo que pudo haber exis-tido durante el periodo mesolítico. No obstante, sehan encontrado cinco remontajes, de los cualestres se encuentran dispersos por varios sectoresde la cueva, como se ilustra en la Figura 13. En elcaso de un canino de jabalí - Remontaje 1 - (Foto3) varios de sus fragmentos se han recuperado eneste nivel y un resto correspondiente a la zonamandibular se recogió en la Zanja. Una diáfisiscaudal de radio de ciervo - Remontaje 20 - (Foto 4)compuesta por dos restos, éstos se han localizadoen sectores distanciados más de un metro. Enambos remontajes, debido a que han estado de-positados en distintos lugares, se observa un pro-ceso tafonómico bastante diferenciado.
A tenor del análisis faunístico, parece que losgrupos humanos mesolíticos realizaron una inten-so aprovechamiento cinégetico, al menos en cier-tas épocas el año, de suidos de diferentes edadesy también de animales carnívoros, ya que las inci-
siones aparecidas en el resto de zorro bien podrí-an atribuirse a un aprovechamiento cárnico de di-cho animal. Para finalizar, el grado de fracturaciónde los restos óseos observados en este nivel hasido bastante elevado.
En conclusión, todas estos testimonios pareceseñalar que en determinadas épocas de ocupaciónmesolítica de esta cueva debió producirse unaprovechamiento de Ungulados de varios tipos dehábitat (predominando los de llanuras), cazando in-cluso animales seniles y consumiendo algunosanimales carnívoros.
36 A. B. MARÍN
Munibe (Antropologia-Arkeologia) 56, 2004 S. C. Aranzadi. Z. E. Donostia/San Sebastián
Fig. 13. Distribución espacial de los remontajes del Nivel 1.
Foto 4. Díafisis caudal de radio de Cervus elaphus (Remontaje-20)del Nivel 1.
Foto 3. Colmillo inferior de Sus scrofa (Remontaje-1) del Nivel 1.

5.5. Nivel "Relleno Zanja"
A partir del tipo de cerámica y de la industria lí-tica encontrada en el interior de la Zanja, ésta fueatribuida a un periodo de cronología calcolítica.Hasta ahora no se dispone de fechaciones de lamisma.
Es de suponer, a partir de los restos humanosencontrados, que la ejecución de esta Zanja tuvocomo finalidad el enterramiento humano de al me-nos un individuo, aunque estos restos han sufridouna enorme dispersión y una deficiente conserva-ción.
La realización de esta Zanja en una cueva tanpequeña tuvo como consecuencia que parte delos niveles arqueológicos hasta el momento esta-bles (concretamente hasta el Nivel 4, aunque a es-te último apenas llegó a afectarlo) fuesen altera-dos parcialmente. Por ello, las conclusiones extraí-das de este paquete no deben ser consideradasde total fiabilidad puesto que, aunque se ha inten-tado verificar la pertenencia de los restos a dichomomento calcolítico a través del aspecto de fosili-zación y distinguirlos de huesos removidos comoparte del sedimento movilizado procedente de losniveles inferiores, ello no ha sido posible con totalseguridad. Algunos restos que se han podido re-montar nos han indicado que el proceso de fosil-diagénesis ha seguido modificando los restos delos niveles inferiores desplazados hacia la Zanja.
Existen muy pocas referencias faunísticas deniveles calcolíticos en Cantabria, y la mayor partede ellas proviene de yacimientos en cuevas, quesuelen haber sufrido remociones por diversos ti-pos de ocupaciones, lo que proporciona datos po-co fiables en cuanto a cronologías. La mayoría deestos yacimientos corresponden a cuevas sepul-crales (RUIZ COBO, 1993).
El número total de restos encontrados en estaZanja ha sido de 2.872, con un peso total de 2.380gramos. El peso medio de cada fragmento ha sidode 0,83 gramos. Esta elevada fragmentación delos restos de la Zanja es, en gran parte, debida alrevuelto de material de varios estratos.
Un 81.86 % del material corresponde a restosno identificables. Del total del material determina-do un 70 % de los restos pertenecen a animalesdomésticos y un 30 % a salvajes.
Dentro de la cabaña ganadera dominan, en pri-mer lugar, los ovicaprinos (35,9 %), seguidos delos suidos (20,72 %) y el bovino (13,1 %). Todosestos porcentajes se refieren al total de restos deungulados, domésticos y salvajes. En cuanto a ta-natomasa se refiere ha sido el bovino el que ma-
yor aporte cárnico ha proporcionado, seguido delos suidos y los ovicaprinos. Y entre los unguladossalvajes, ha sido el ciervo el que mayor tanatoma-sa ha aportado.
En el caso de los suidos, en La Fragua ha re-sultado complicado diferenciar entre cerdo salvajey doméstico debido a la fragmentación de los res-tos y la imposibilidad de obtener medidas osteo-métricas, y por ello se han incluido todos los res-tos en la categoría de Sus sp.
Como restos de ovicaprinos se han contabili-zando tanto los restos de Capra sp., Ovis aries ylos restos de ovicaprinos en sentido genérico.Esta elevada representación de ovicaprinos se re-pite en yacimientos sepulcrales como Aer oPajucas. En Los Husos oscilan entre un 38-45 %,y en Arenaza entre un 46-66 % (RUIZ COBO, 1993).No se ha podido diferenciar exactamente entreCapra pyrenaica y Capra hircus debido a la esca-sez de restos mensurables.
Bos taurus es la especie a la que se han atri-buido los restos de bovino de la Zanja, debido prin-cipalmente a su talla y escasa robustez de los res-tos. En los yacimientos del País Vasco queda ates-tiguado en Arenaza en el nivel IC con un 33 %(APELLANIZ & ALTUNA, 1975) y en el nivel IIB3-1C delos Husos con un 46 %.
En cuanto a los ungulados salvajes encontra-dos en la Zanja, el ciervo (20,27 %) ha sido el ta-xón más representativo, seguido de lejos por elcorzo (4,1 %), el rebeco (2,7 %) y el jabalí (2,7 %).Todos estos porcentajes se refieren, igualmente,al total de restos de ungulados, domésticos y sal-vajes.
Asimismo, se han identificado restos de trescarnívoros (siendo uno de ellos doméstico), restosde un insectívoro y un Pinnípedo. La presencia delos carnívoros en la Zanja no ha sido muy notable.Se ha identificado un individuo de perro, uno dezorro y otro de gato silvestre. El zorro suele seruna especie paradójicamente rara en niveles holo-cenos de la Península Ibérica según comentaCASTAÑOS (1987), al contrario que ocurre con el ga-to silvestre, que es más frecuente en el Holoceno.
Entre todas las especies animales encontra-das en la Zanja se ha calculado un número mínimode 32 individuos: 26 son de ungulados salvajes ydomésticos, cuatro de carnívoros también salvajesy domésticos, uno de insectívoros y uno de pinní-pedos.
En la figura 14 se muestra la distribución poredades (neonato, juvenil y adulto) de los NMI detodas las especies halladas en la Zanja.
ANÁLISIS ARQUEOZOOLÓGICO, TAFONÓMICO Y DE DISTRIBUCIÓN ESPACIAL DE LA FAUNA DE MAMÍFEROS DE LA CUEVA DE LA FRAGUA (SANTOÑA, CANTABRIA) 37
Munibe (Antropologia-Arkeologia) 56, 2004 S. C. Aranzadi. Z. E. Donostia/San Sebastián

A pesar de la actividad ganadera, en la cuevade La Fragua la caza pudo seguir teniendo un pa-pel relevante en la dieta de estos grupos calcolíti-cos, sobre todo la caza del ciervo y secundaria-mente la de corzo, rebeco y jabalí.
A partir de la fusión de las epífisis y del des-gaste dental (GRIGSON, 1982) en los animales do-mésticos se ha estimado una fecha aproximadade sacrificio en los bovinos alrededor de los tresaños y medio, y para el caso de los suidos alrede-dor del año de vida, si bien se han encontrado res-tos de un individuo de más de tres años y medio.
Por otra parte, las alteraciones antrópicas quese han observado en los restos responden a las al-teraciones apreciadas en los niveles anteriores,por lo que se intuye una continuidad del patrón deprocesado y consumo de los restos en algunos delos taxones, como es el caso de la cabra. Del mis-mo modo, el tipo de fracturación de algunos res-tos señala el aprovechamiento no exclusivamentecárnico sino también medular, aunque bien pudie-sen pertenecer estos elementos a cualquiera delos niveles que la Zanja removió. En cuanto a lasalteraciones naturales más frecuentes, se hanidentificado costras de concreciones cálcicas y lasimprontas, principalmente, de las raíces de la hie-dra que crecía por toda la superficie de la cuevahasta el momento de la excavación.
A pesar de no haberse realizado el estudio dela distribución espacial de la Zanja, sí se ha tenidoen cuenta la distribución de las rearticulaciones yremontajes, cuyas conclusiones muestran que,por lo general, los movimientos postdepósito sehan desarrollado dentro del mismo cuadro, por lo
que la distancia máxima de desplazamiento de es-tos huesos es inferior a un metro. No obstante,muchas de las fracturas observadas en los remon-tajes han sido producidas por los movimientos delestrato en el que se encontraban depositados,que ha producido una fracturación postdeposicio-nal de los mismos.
En conclusión, esta zanja parece ser que ade-más de servir de recinto sepulcral también fue uti-lizada como lugar de habitación, ya que el materialde fauna analizado responde a restos de procesa-do y consumo humano. Se desconoce cuántasocupaciones hubo y qué duración tuvieron desdela realización de la Zanja. Habrá que esperar a quese obtengan fechaciones radiocarbónicas para co-nocer la cronología de las mismas.
5.6. Nivel de Superficie
A pesar de ser únicamente cuatro los restosrecogidos en superficie, ha sido ineludible reseñardos de ellos por ser una falange humana y una ul-na de foca, ambos pertenecientes a individuosadultos. Es muy probable que dichos restos proce-dan de la Zanja y que hayan sido removidos porocupaciones modernas de la cueva, saliendo al ni-vel superficial.
Sería interesante fechar los restos de foca ha-llados en este yacimiento para conocer la épocacorrecta en que fue cazada, ya que existen muypocos testimonios de presencia de pinnipedos enla Cornisa Cantábrica durante el PaleolíticoSuperior (SERANGELI, 2000). Hasta el momento en
38 A. B. MARÍN
Munibe (Antropologia-Arkeologia) 56, 2004 S. C. Aranzadi. Z. E. Donostia/San Sebastián
Fig. 14. Distribución por edades del NMI de los Mamíferos de la Zanja.

el norte de la Península solamente se han encon-trado restos de foca en Altamira (BREUIL &OBERMAIER 1935; ALTUNA & STRAUS 1976), dondese recogió un canino actualmente extraviado, enun nivel solutrense, y una falange primera dePhoca vitulina.
En La Riera se encontraron tres restos en ni-veles pertenecientes al Solutrense superior y en elnivel Aziliense se identificaron tres restos de focagris (PÉREZ RIPOLL & RAGA, 1998).
Y en Tito Bustillo se encontraron dos astrága-los y un húmero juvenil (ALTUNA, 1976; PÉREZ
RIPOLL & RAGA 1998). Un astrágalo podría corres-ponder a Phoca hispida y el otro a Phoca vitulina,aunque no está confirmado, ya que en el últimoresto no se podía excluir que pudiera pertenecer aHalichoerus grypus.
6. VISION GENERAL DE LA COMPOSICIONFAUNISTICA. CONCLUSIONES. (fig. 15)
La importancia de la Cueva de La Fragua en lavaloración de la composición faunística de los yaci-mientos del Cantábrico es vital, pues su secuenciaestratigráfica, que abarca el tránsito delPleistoceno al Holoceno, es muy adecuada paraestudiar el comportamiento y las estrategias desubsistencia adoptadas por los cazadores recolec-tores, así como los cambios ambientales en elCantábrico Oriental.
Al final de Pleistoceno empiezan a producirsecambios que se aceleran tras el comienzo delHoloceno. Las consecuencias principales se cen-tran en cambios en el territorio regional, en su co-bertera vegetal (empiezan a producirse procesosde reforestación) y en la simplificación de faunaspresentes, con llamativos procesos de desapari-ción definitiva, como es el caso del bisonte, el re-no y todas las especies frías (ALTUNA, 1990).
Una primera visión general nos indica que enel nivel Magdaleniense Superior-Final existía unaeconomía cazadora basada en dos especies: la ca-bra montés (52 % de los Ungulados) y en el ciervo(36 % de los Ungulados) seguidos a gran distanciadel corzo (5,9 %). Está economía dual se debeprincipalmente a que en las proximidades del yaci-miento existe un doble radio de acción al existirlos biotopos de ambas especies: zona de llanuraen el caso del ciervo y zonas montañosas para lacabra. En Urtiaga D (ALTUNA, 1972), La Riera(STRAUS; CLARK, 1986) o Ekain III (ALTUNA et al.
1984) sucede lo mismo que en La Fragua.
Sin embargo, en el Nivel 3 se produce un no-table decrecimiento de la cabra montés (12,1 %)en favor del corzo (18,1 %). Esta última especieaumenta en concordancia con la expansión de losbosques caducifolios, hecho que queda constata-do a través del polen y de los restos de maderaquemada. Se produce una extensión de las masasforestales, que van colonizando nuevos terrenoscon especies vegetales relativamente adaptadas aclimas templados y, por tanto, la fauna de paisajesmás abiertos conoce un impulso de crecimiento.Además, esta extensión vegetal favorece la explo-tación de otros recursos como son los moluscosterrestres comestibles encontrados en este mis-mo nivel al igual que sucede en la Cueva del Valle(FERNANDEZ-TRESGUERRES, 1980).
Según GONZALEZ SAINZ (1989), durante elMagdaleniense Superior-Final y sobre todo en elAziliense los grupos cantábricos enfocaron la in-tensificación del aprovechamiento mediante unadiversificación de recursos cada vez más decidida.De esta forma, la fauna del Nivel 3 marca el tránsi-to entre el Pleistoceno y el Holoceno, donde losgrupos cazadores-recolectores parecen realizaruna caza en función de sus preferencias alimenti-cias, gasto energético, dificultad de la caza y capa-cidades tecnológicas. La recolección de moluscosterrestres parece corroborar ese problema deltránsito y no es un hecho aislado en la vertientecantábrica, existiendo en Francia yacimientos con-temporáneos especializados en esa actividad co-mo Poëymau.
Por su parte, en el Mesolítico es el jabalí (34,5%) el taxón más representativo seguido del ciervo(32,7 %) y del corzo (10,9 %). Estos tres animalesson el reflejo de la subida de humedad y tempera-
ANÁLISIS ARQUEOZOOLÓGICO, TAFONÓMICO Y DE DISTRIBUCIÓN ESPACIAL DE LA FAUNA DE MAMÍFEROS DE LA CUEVA DE LA FRAGUA (SANTOÑA, CANTABRIA) 39
Munibe (Antropologia-Arkeologia) 56, 2004 S. C. Aranzadi. Z. E. Donostia/San Sebastián
Fig. 15. Porcentajes de los Ungulados según el NR en los distintosniveles del yacimiento.

tura del Óptimo Climático, a partir del Boreal y so-bre todo en el Atlántico; el ciervo y el corzo prefie-ren los llanos y los valles con ondulaciones y el ja-balí gusta de terrenos con matorrales, marismas,bosque mediterráneo o regiones cubiertas deabundante vegetación.
Es sobre todo en el Mesolítico cuando la diver-sificación como eje económico aparece más clara-mente definida. En La Fragua se ha podido apre-ciar dicha diversificación a través de los restos ma-rinos y de estuario recogidos. Así, el nivel mesolíti-co aparece como un potente conchero en el quese han encontrado gran cantidad de lapas de ta-maños reducidos. Este intenso aprovechamientoparece señalar una época de carestía de recursos.En la fauna, esta escasez parece estar representa-da por el consumo incluso de animales carnívoros(de baja calidad nutritiva y elevado coste de proce-sado), como lo demuestran las trazas antrópicasencontradas en los restos de zorro y de perro, nosolamente propias de la extracción de la piel sinotambién del consumo de su carne.
Por otro lado, en la Zanja, en cuanto a númerode restos, destacan en primer lugar los suidos
(28,5 %) seguidos de cerca por la cabra (22,5 %),el ciervo (24,7 %) y el bovino doméstico. Sin em-bargo, el corzo (4,9 %) decrece respecto a los an-teriores niveles analizados.
Aunque se ha constatado la presencia de ani-males domésticos en la Zanja (bovinos), el restode especies susceptibles de corresponder a la for-ma doméstica (Capra y Sus) no ha podido sercomprobado osteométricamente debido a la frac-turación que presentaban sus restos y la escasezde restos mensurables. En este sentido, convienedestacar que la presencia de bovinos en el yaci-miento es especialmente relevante en la Zanja,mientras que en los otros niveles apenas es repre-sentativo.
Además, es de reseñar la exigua presencia delrebeco en todos los niveles, limitada en el Nivel 4a una pieza dental, y en la Zanja a un porcentajerespecto al resto de ungulados de un 3,3 %, ex-clusivamente con seis restos. Esta escasa mues-tra coincide con la casi total desaparición de estetaxón en los yacimientos cantábricos durante elMagdaleniense a favor de la cabra (GONZALEZ
SAINZ, 1992).
40 A. B. MARÍN
Munibe (Antropologia-Arkeologia) 56, 2004 S. C. Aranzadi. Z. E. Donostia/San Sebastián
Mandíbula Metacarpo
Capra pyrenaica
L M1-M3 L M3 A M349,26 21,77 8,3
Nivel 4
Ap Ad27,53 -
- 33,6429,38 -
Nivel 4
ASp21,58Nivel 4
Ad31,88
29Nivel 4
LM AM72,1 20,671 20,2
Nivel 4
Em Lm LI Ad El21,05 36,83 37,95 25,57 20,9
Talus
Nivel 4
Ap AmD Ad LM16,04 12,09 11,82 31,9715,7 11,52 11,46 26,5
Falange II
Nivel 4
Ulna Tibia
Calcáneo
AM29,19Nivel 4
Centrotarsal
Ap LM16,51 43,3115,84 42,19
Nivel 4
Falange I
LDS Ldo34,24 26,1737,81 29,9537,89 29,1136,16 29,36
Nivel 4
Falange III
MEDIDAS OSTEOMÉTRICAS

Cervus elaphus
ANÁLISIS ARQUEOZOOLÓGICO, TAFONÓMICO Y DE DISTRIBUCIÓN ESPACIAL DE LA FAUNA DE MAMÍFEROS DE LA CUEVA DE LA FRAGUA (SANTOÑA, CANTABRIA) 41
Munibe (Antropologia-Arkeologia) 56, 2004 S. C. Aranzadi. Z. E. Donostia/San Sebastián
Em Lm LI Ad El32,36 54,83 59,28 36,12 31,83
Ap61.8
Nivel 3
Talus Húmero
Nivel 4
Ap45.72Nivel 3
Metacarpo
Ap16,6
19,38Nivel 1
Falange I
Ad39,57
Nivel 1
Metacarpo
Ad5,85
Nivel 1
Falange II
Sus scrofa
Sus sp.
Bos taurus
Rupicapra rupicapra Canis familiaris
LA LAb AmII CmII42,17 33,74 14,04 25,2
Pelvis
Nivel 4
Ap Ad LM14,62 12,79 21
Falange II
Zanja
Ap LM AmD Ad LM17,68 24,2 14,1 15,8 26,54
Falange II
Nivel 4
Falange III
LDS Ldo32,59 33,04
Nivel 4
M3
L M3 A M334,63 14,5
Zanja
Falange III
LDS Ldo31,2 28,32
31,89 29,75Zanja
L P2-P452,73Zanja
Maxilar
Ap32,86Zanja
Falange I
Ad AmD Ap LM21,21 21,08 26,67 36,9
Falange II
Zanja
Radio
Ap ASp26,8 24,45
Zanja
Vulpes vulpes
ASp EPA EmO10,71 18,53 15,23
Ulna
Nivel 4
EMP AS LS18,17 11,19 13,46
Escápula
Nivel 1
Ap7,84
Nivel 4
Metacarpo V
LM1-M334,31Zanja
Mandíbula
LM1-M3 LP1-P4 LP1-M317,81 49,3 65,72
Maxilar
Zanja

42 A. B. MARÍN
Munibe (Antropologia-Arkeologia) 56, 2004 S. C. Aranzadi. Z. E. Donostia/San Sebastián
BIBLIOGRAFÍA
ALTUNA, J.
1972. "Fauna de mamíferos de los yacimientos prehistóricos deGuipúzcoa", Munibe 24: 1-464.
1976 "Los Mamíferos del yacimiento prehistórico de TitoBustillo (Asturias)". En: MOURE ROMANILLO, J. A. & CANO
HERRENA M. Excavaciones en la cueva de Tito Bustillo(Asturias). Instituto de Estudios Asturianos, 149-154.Oviedo
1979 "La faune des Ongulés du Tardiglaciaire en Pays Basqueet dans le reste de la Région Cantabrique". Colloq.Internat. CNRS 271: La fin des Temps Glaciaires en
Europe (1977), 85-96. Paris.
1980 "Historia de la domesticación animal en el País Vasco des-de sus orígenes hasta la Romanización". Munibe 32, 1-163.
1990 " La caza de herbívoros durante el Paleolítico y Mesolíticodel País Vasco", Munibe (Antropologia-Arkeologia) 42,
229-240.
1992 "El medio ambiente durante el Pleistoceno Superior en laregión Cantábrica con referencia especial a sus faunas demamíferos". Munibe (Antropologia-Arkeologia) 44, 13-29
1994 "El perro en los yacimientos arqueológicos del norte de laPenínsula Ibérica", Museo y Centro de Investigación de
Altamira, Monografía, 17, 159-162.
1995 "Fauna de mamíferos y cambios ambientales durante eltardiglaciar cantábrico", EN: MOURE ROMANILLO, A.;GONZALEZ SAINZ, C. El Final del Paleolítico Superior cantá-
brico. Universidad de Cantabria, 77-117.
ALTUNA, J., STRAUS, L. G.
1976 "The Solutrean of Altamira: the Artifactual and faunal evi-dence", Zephyrus 26, 175-182. Munibe 32, 1-163.
ALTUNA, J; MERINO, J.M.
1984 El yacimiento prehistórico de la Cueva de Ekain (Deba,
Guipúzcoa). Eusko Ikaskuntza. Sociedad de EstudiosVascos.
APELLÁNIZ, J. M. & ALTUNA, J.
1975 "Memoria de la 2ª campaña de excavación arqueológicade la Cueva de Arenaza I (San Pedro de Galdames,Vizcaya). Noticiario Arqueológico Hispano. Prehistoria 4.
BARONE, R.
1976 Anatomie comparée des mamiferes domestiques. VigotFréres Editeurs. París.
BINFORD, L. R
1981 Bones.Ancient men and modern myths. Academic Press.New York.
BLASCO M.F.
1992 Tafonomía y Prehistoria. Métodos y procedimientos de in-
vestigación. Universidad de Zaragoza. Zaragoza.
BOESSNECK, J.
1980 "Diferencias osteológicas entre las ovejas (Ovis aries
Linne) y cabras (Capra hircus Linne)". In: BROTHWELL, D. &HIGGS, E. (Ed.). Ciencias en Arqueología. Fondo de CulturaEconómico. Madrid.
BREUIL,H., OBERMAIER,H.
1935 The cave of Altamira at Santillana del Mar. Madrid, Spain.
BULL, G.; PAYNE, S.
1982 " Tooth eruption and epiphysial fusion in pigs and wild bo-ar", In: WILSON, B.; GRISON, C.; PAYNE, S. (eds). Ageing and
Sexing Animal Bones from Archaeological Sites. BAR,
British Series 109, 55-71.
CASTAÑOS, P.
1984 "Estudio de los Macromamíferos de la Cueva deSantimamiñe (Vizcaya)". Kobie 14, 235-318.
1987 "Los carnívoros prehistóricos de Vizcaya". Kobie XVI, 7-76.
CASTEEL, R.W.
1976-77 "A consideration of the behaviour of the minimum numberof individual index: A problems in faunal characterisation",Ossa 3/4, 141-153.
DALY, P.
1969 "Approaches to faunal analysis in archaeology. American
Antiquity 34, 146-153.
DRIESCH, A. van den.
1976 A guide to the measurement of animal bones from archae-
ological sites, (Peabody Museum Buletin,1). Cambridge.Peabody Museum. Harvard University.
EFREMOV, I.A.
1940 "Taphonomy: a new branch of Paleontolgy". Pan-American
Geologist 74 (2), 8-93.
ENLOE, J. G.
1995 "Remontatge en Zooarqueología: tafonomía, economía isocietat". Cota Zero 11, 31-37.
ESTÉVEZ, J.
1984 "Paleontología/Arqueología versus Paleoeconomía/Ar-queología", Primeras Jornadas de Metodología de Investi-
gación Prehistórica, 293-298. Soria.
1995a "Zooarqueología y Antracología: Estrategias de aprovecha-miento de los recursos animales y vegetales en TunelVII", Encuentros en los conchales fueguinos. Treballs d'et-noarqueologia 1. CSIC, 143-238.
1995b "Una historia inacabada: l`estudi de restes animals arque-ològiques davant un gran repte". Cota Zero 11, 13-24.
2000a "Experimentación y muestreo en Arqueozoología,"enMAMELI, L. & PIJOAN,J. (eds) Reunión de Experimentación
en Arqueología. Treballs d’Arqueologia (nº especial).Universitat Autònoma de Barcelona. Edición en CD ISSN:1134-9263
2000b "Aproximación dialéctica a la Arqueotafonomía" Revista
Atlántica-Mediterránea de Prehistoria y Arqueología Social
3, 7-27
ESTÉVEZ, J. & MARTÍNEZ, J.
1995 "Reflexiones metodológicas a partir del análisis de losrestos de pinnípedos de Túnel VII", en ESTÉVEZ, J. & VILA,A. (Ed), Encuentros en los concheros fueguinos (Treballsd`Etnoarqueologia). Barcelona. UAB-CSIC.

ANÁLISIS ARQUEOZOOLÓGICO, TAFONÓMICO Y DE DISTRIBUCIÓN ESPACIAL DE LA FAUNA DE MAMÍFEROS DE LA CUEVA DE LA FRAGUA (SANTOÑA, CANTABRIA) 43
Munibe (Antropologia-Arkeologia) 56, 2004 S. C. Aranzadi. Z. E. Donostia/San Sebastián
FERNÁNDEZ-LÓPEZ, S.
1999 "Tafonomía y fosilización", en Tratado de Paleontología 1.(Ed. B. MELÉNDEZ, 1998). CSIC, Madrid: 51-107, 438-441.
FREEMAN, L. G.
1973 "The siginificance of mammalian faunas from Paleolithicocupations in Cantabrian-Spain", American Antiquity 38,
3.41.
FORMIGO, C.
1998 Atlas digital osteológico. Diferencias osteológicas entre
cabras, ovejas y rebecos. UAB.
GONZÁLEZ MORALES, M. R.
1999a "La Prehistoria en Santoña", Monte Buciero 2, 17-28.
1999b "Varilla decorada del Magdaleniense de la Cueva de laFragua (Santoña, Cantabria), Sautuola VI, 177-183.Santander.
GONZÁLEZ MORALES, M.R. & DÍAZ CASADO, Y.
1992 "Excavaciones en los Abrigos de la Peña del Perro(Santoña, Cantabria). Estratigrafía, cronología y comenta-rio preliminar de sus industrias", Veleia 8-9, 43-64.
GONZÁLEZ SAINZ, C.
1989 El Magdaleniense Superior-Final de la región Cantábrica.
Tesis doctoral. Universidad de Cantabria. Santander.
1992 "Aproximación al aprovechamiento económico de las po-blaciones cantábricas", en: MOURE ROMANILLO et al.
Elefantes, ciervos y ovicaprinos. Universidad de Cantabria,129-145.
GRIGSON, C.
1982 "Sex and Age determination of some bones and teeth ofdomestic cattle: A review of the literature". In: WILSON, B.;GRIGSON, C.; PAYNE, S. (eds). Ageing and Sexing Animal
Bones from Archaeological Sites. BAR. British Series 109,55-71.
HILLSON, S.
1990 Teeth. Cambridge Manuals in Archaeology. Cambridge.
KLEIN, R. G. & CRUZ-URIBE, K.
1984 The Analysis of Animal Bones from Archaeological Sites,
Chicago. The University of Chicago Press.
LAVOCAT, R.
1966 Faunes et flores préhistoriques, París. Boubée.
LYMAN, L.R.
1987 "Archaeofaunas and butchery studies: a TaphonomicPerspective", In: SCHIFFER, M. (ed). Advances in:
Archaeological Method and Theory 10, 249-337.
MARIN ARROYO, A.B.
2003 "Análisis arqueozoológico, tafonómico y de distribución
espacial de la fauna de Mamíferos de la Cueva de La
Fragua (Santoña – Cantabria)". Trabajo de Investigación.Universidad de Cantabria. Inédito.
MARIN ARROYO, A.B.
Estudio Arqueozoológico de Distribución Espacial median-te un Sistema de Información Geográfica (SIG): aplicacióna la Cueva de La Fragua (Santoña – Cantabria). (en prepa-ración).
MARIEZKURRENA, K.
1983 "Contribución al conocimiento del desarrollo de la denti-ción y el esqueleto postcraneal de Cervus elaphus",Munibe 35, 149-202.
MARIEZKURRENA, K. & ALTUNA, J.
1989 "Análisis arqueológico de los macromamíferos del yaci-miento de Zatoya". Trabajos de Arqueología de Navarra 8,
237-266.
MEADOW, R. H.
1976 "Methodological Concerns in Zoo-Archaeology". X
Congres UISSP, 110-123. Nice, UISSP.
MORALES, A. (ms.).
El Abrigo de la Peña del Perro: el estudio óseo. Informeinédito.
PALES, L. & LAMBERT, C.
1972 Atlas ostéologiques des mammifères. París. CNRS.
PAYNE, S.
1973 "Kill-off Patterns in Sheep and Goats: The mandibles fromAsvan Kale". Anatolian Studies XXXIII, 281-303.
PÉREZ RIPOLL, M.
1988 "Estudio de la secuencia del desgaste de los molares dela "Capra pyrenaica" de yacimientos prehistóricos". Archivo
de prehistoria Levantina XVIII, 83-125.
1992 Marcas de carnicería, fracturas intencionadas y mordedu-
ras de carnívoros en huesos prehistóricos del Mediterrá-
neo español. Alicante.
PÉREZ RIPOLL, M., RAGA, J. A.
1998 "Los mamíferos marinos en la vida y en el arte de laprehistoria de la cueva de Nerja", En: SANCHIDRIAN TORI, J.L., SIMON VALLEJO, M.D. (coord.), Las culturas del pleisto-
ceno superior en Andalucia, 251-275
PERKINS, D & DALY, P.
1968 "A hunter´s vil lage in Neolithic Turkey". Scientific
American 219(5), 96-106
PRUMMEL, W. & FRISCH, H.J.
1986 "A guide for the distinction of species, sex and body sidein bone of sheep and goat". Journal of Archaeological
Science 13, 567-577.
RUIZ COBO, J.
1993 "El registro faunístico de las series de la Prehistoria concerámica de Cantabria". Cota Cero 3, 39-55.
SAÑA, M.
1999 Arqueología de la domesticación animal. Treballsd'Arqueologia del Pròxim Orient 1. Universitat Autònomade Barcelona.

44 A. B. MARÍN
Munibe (Antropologia-Arkeologia) 56, 2004 S. C. Aranzadi. Z. E. Donostia/San Sebastián
SCHMID, E.
1972 Atlas of Animal Bones. Amsterdam, Elsevier.
SERANGELI, J.
2001 La zona de Costa en Europa durante la última glaciación.Consideraciones al análisis de restos y representacionesde focas, cetáceos y alcas gigantes. Cypsela 13, 125-138.
SILVER, I. A.
1980 "La determinación de la edad en los animales domésti-cos". En: BROTHWELL, D. Y HIGGS, E. (Ed.) Ciencia en
Arqueología 289-309. Madrid. Fondo de CulturaEconómica.
STRAUS, L. G. & CLARK, G. A.
1986 La Riera Cave. Stone Age Hunter-Gatherner Adaptations
in Northern Spain. Anthropological Research Papers 36.Arizona State University.
FERNÁNDEZ-TRESGUERRES VELASCO, J.A.
1980 El Aziliense en las provincias de Asturias y Santander.
Centro de Investigación y Museo de Altamira. Monogra-fías 2. Santander.
UERPMANN, H.P.
1972 "Animal bone finds and economic archaeology: a criticalstudy of osteo-archaeological method". World
Archaeology 4, 307-322.
WHITE, T. E.
1953 "Observations on the butchering techniques of some abo-riginal peoples 2". American Antiquity 19 (2), 160-164.