uso de la aglutinación diferencial de eritrocitos como un ...148.206.53.84/tesiuami/uam3040.pdf ·...
TRANSCRIPT

UNIVERSIDAD AUTONOMA METROPOLITANA-IZTAPALAPA
DIVISION DE CIENCIAS BIOLOGICAS Y DE LA SALUD
Uso de la aglutinación diferencial de eritrocitos como un carácter para separar las subespecies del género
Oreochromis presentes en México
Tesis que presenta el Hidrobiólogo Sergio H. Alvarez Hernández para obtener
el grado de Maestro en Biologia Experimental

La Maestría en Biología Experimental de la División de Ciencias Biol6gicas y de la Salud se encuentra dentro del Padr6n de
Excelencia de CONACyT con número de registro 309-0.
Agradezco a CONACyT el apoyo recibido con la beca para estudios de Posgrado No. 88084.

lndice
111 . Materiales y Métodos ................................................................................. 111 . a Colecta de material ............................................................................ 111 . b Preparación de extracto algal crudo ................................................... 111 . c Solución de eritrocitos ........................................................................ 111 . d Ensayo de actividad hemaglutinante .................................................. 111 . e [nhibiclon por azúcares ...................................................................... 111 . f Obtención de proteínas ....................................................................... 111 g . Purificación de la lectina .....................................................................
111 . g 1 Cromatografía de afinidad ........................................................ 111 . g 2 Cromatografía de intercambio iónico ......................................... 111 . g 3 Métodos analltlcos .................................................................... 111 . g 4 Electroforesis en gel SDS-PAGE .............................................. 111 . g 5 Punto isoeléctrico ......................................................................
Ill . h Pruebas de aglutinación de eritrocitos de peces ................................ 111 . i Analisis estadístico de la aglutinación diferencial ................................
.,
..
V . Resultados y Discusión .............................................................................. V . a Pruebas de inhibición ......................................................................... V . b Cromatografía de afinidad .................................................................. V . c Cromatografía de intercambio iónico .................................................. V . d Electroforésis SDS-PAGE .................................................................. V . e Electroforésis en condiciones no desnaturalizantes ............................
V . g Eficiencia de aislamiento .................................................................... V . h Variabilidad específica por medio de la aglutinación de eritrocitos ......
V . f lsoelectroenfoque ...............................................................................
23 23 23 24 25 25 26 26 27 27 27 28 29 29 30
33 33 34 35 38 40 42 44 44
VI Conclusiones .............................................................................................. 50
VI1 Anexo 1 Las Tilapias en México ................................................................. 52
VIII . Literatura citada ....................................................................................... 59
~ ~ _ _ . - . ~ - .... . .

lndice de Tablas y Figuras .
Tablas
1 . Propiedades y usos de las lectinas ............................................................ 2 . Características que permiten separar los géneros de Tilapia y
3 . Títulos de aglutinación de eritrocitos de varias especies animales
4 . Características de las lectinas algales de la División Rhodophyta .............. 5 . Características de las lectinas algales de la División Phaeophyta .............. 6 . Características de las lectinas algales de la División Chlorophyta .............. 7 . Resultados de la prueba de inhibición por azúcares ................................... 8 . Rendimiento en la purificación de la lectina de Codium giraffa ................... 9 . Variabilidad específica por medio de la aglutinación con eritrocitos
Oreochromis presentes en México ..............................................................
por extractos de angiospermas ..................................................................
de Tilapias ................................................................................................
Figuras
1 . Morfología externa de las Tilapias ............................................................ 2 . Reacción del acid0 bincinconinico y las proteínas ..................................... 3 . Cromatograma de afinidad ........................................................................ 4 . Cromatograma obtenido por HPLC ........................................................... 5 . Perfil de elución de la lectina purificada .................................................... 6 . Gel SDS-PAGE ........................................................................................ 7 Gel en condiciones no desnaturalizantes ...................................................
9 . Gráfica del promedio de aglutinación de cada especie .............................. 1 O . Estados de la República Mexicana con mayor captura y producción
de Cíclidos ................................................................................................ I 1 . Oreochromis mossambicus ...................................................................... 12 . Oreochromis mossambicus (variedad roja) .............................................. 13 . Oreochromis aureus ................................................................................. 14 . Oreochromis aureus (variedad roja) ......................................................... 1 5 . Oreochromls nllotlcus ............................................................................... 16 . Oreochromis urolepis hornorum ............................................................... 17 . Tllapla rendalli ..........................................................................................
8 . Gel de Isoelectroenfoque ..........................................................................
. . .
. .
3
7
9 14 15 16 33 44
45
6 28 35 36 37 38 40 42 47
54 55 55 56 56 57 57 58
.......... . _Iup__ .... ..I

1
I. INTRODUCCION
La importancia de la clasificación de los seres vivos va más allá de la simple,
agrupación de los organismos jerárquicamente, o definición de especies. La
necesidad de reconocer a plantas y animales individualmente y a nivel de
población es imprescindible cuando se trata de especies de las cuales el ser
humano puede obtener beneficios inmediatos.
Las pruebas que ayudan al definir a los organismos a nivel de especie han
evolucionado, desde la sistemática fenética hasta el uso actual de herramientas
técnicas que han permitido evidenciar otras características que puedan ayudar al
taxónomo a resolver con mayor precisión su objeto de estudio. Los avances en
Embriología y Bioquímica, así como en técnicas de microscopía óptica y
electrónica posibilitan estudiar en detalle la estructura interna de las formas de vida
más pequeñas. Estas pruebas comprenden aspectos morfológicos en el ámbito de
ultraestructura, genéticos, isoenzimáticos y recientemente se ha explorado sobre
las características de los sitios antigénicos presentes en la superficie celular de
eritrocitos. Con el descubrimiento de las lectinas y su aplicación en estas pruebas,
se ha abierto una posibilidad más para el esclarecimiento de las identidades
específicas de los animales.
Las lectinas de origen vegetal han sido estudiadas principalmente debido a sus
propiedades químicas y bioquímicas únicas, que las hacen sustancias útiles como
pruebas para el estudio de las funciones biológicas en animales. Una característica
que comparten todas las lectinas es que pueden unir especificamente azúcares
situados sobre la superficie de la membrana y pared celular, incluyendo las de

2
linfocitos y eritrocitos, haciéndolas moléculas importantes en la caracterización de
la superficie de todas las células de mamíferos.
Las lectinas han sido utilizadas también como modelo para estudiar las
interacciones de tipo antígeno-anticuerpo en la repuesta inmunológica. El
mecanismo por el cual actúan es a través de la unión de moléculas de
carbohidratos de las glicoproteínas o glicolípidos de la superficie celular, a los sitios
activos de la molécula de la aglutinina (Sharon y Lis, 1972). Esta característica ha
favorecido el uso de estas moléculas en variadas aplicaciones. El potencial de
utilización de estas moléculas es aún desconocido, sin embargo, cada vez más
investigadores, en el mundo se interesan en encontrar aplicaciones que permitan
hacer más fácil el trabajo en todos los campos del conocimiento donde las lectinas
pueden ser herramientas importantes.
La mayoría de los estudios sobre lectinas han sido realizados con moléculas
provenientes de plantas fanerógamas, en particular, se han aislado estas
moléculas de las semillas de los integrantes de la familia Leguminosae. Aunque la
mayor concentración de lectinas se presenta en las semillas, se pueden encontrar
diferentes proporciones de lectinas en otras estructuras de la planta tales como
tallos, hojas y raíces. Algunas de las principales propiedades y usos que se les han
dado a las lectinas se muestran en la tabla 1.

3
Tabla l. Propiedades y usos de las lectinas
Especificidad por grupos Tipificación de grupos sanguíneos; estudios estructurales sanguíneos humanos de sustancias útiles en la tipificación; identificación de
nuevos grupos sanguíneos
Toxicidad en animales Estudios del valor nutrimental de los alimentos para animales
Inducción de mitosis en Estudios de la constitución cromos6mica de las Células Y linfocitos detección de anormalidades cromosómicas
Aglutinación de células Investigación de la arquitectura de las superficies malignas celulares y de su cambio debido a la transformación
Precipitación de Aislamiento, purificación y estudios estructurales de polisacáridos y polimeros que contienen carbohidratos; modelo de glucoproteínas reacciones antígeno-anticuerpo
Unión de azúcares Estudios de sitios de unión específica sobre proteínas
Fuente Sharon y Lis (1972)
Las lectinas de las macroalgas marinas han recibido escasa atención, no obstante,
actualmente se sabe que se les encuentra en una gran cantidad de representantes
de las Divisiones: Chlorophyta, Phaeophyta y Rhodophyta y que tienen
propiedades hemaglutinantes características semejantes a las de las aglutininas
reportadas en angiospermas (Hori et a/. , 1988b)

4
La identificación de aglutininas algales y su uso suponen una herramienta potencial
en el estudio de la superficie celular, sobre todo aquellas lectinas cuyos azúcares
inhibidores sean conocidos, pudiendo ser utilizadas en la diferenciación de razas
de peces, de acuerdo con la respuesta diferencial de aglutinación de los tipos
sanguíneos propios de cada subespecie o raza (Mufioz et a1.,1987c). Esta
posibilidad de servir como marcadores genéticos, capaces de mostrar la
variabilidad de los determinantes antigénicos presentes en la superficie celular,
hace que las lectinas puedan servir para la tipificación de las modificaciones en la
membrana de los eritrocitos de varias especies animales, no solamente de los
peces.
Considerando las características de las lectinas expuestas en el párrafo anterior,
parece atractiva la idea de poder aplicarlas en la identificación de variabilidad
debida a al aglutinación que pueda presentarse como consecuencia del manejo
genético que se ha dado a peces sometidos a cultivo. Esta aplicación ayudaría a
solucionar el problema del mejoramiento genético en las especies de tilapias
introducidas a México.
Arredondo-Figueroa et al. (1994) explican la problemática de la tilapias en nuestro
país afirmando que la producción actual se deriva de 75 ejemplares que fueron
traídos a México en diferentes etapas (Anexo 1) y además exponen que el manejo
que se ha dado a estas especies no ha sido el correcto.
Si consideramos que en su gran mayoría, estos peces han sido liberados en
presas y cuerpos de agua epicontinentales, sin un seguimiento adecuado de sus
condiciones reproductivas.. Como resultado de lo anteriormente expuesto, se

5
tienen problemas de hibridación no controlada y, aún más peligroso, un alto índice
de endogamía. Esta actividad reproductora sin control, provoca una reducción
critica en el intercambio genético de las especies presentes en los cuerpos de
agua y trae como consecuencia una mayor probabilidad de producir peces con
deformaciones y más propensos a enfermedades.
A continuación se describen las características de las especies de la familia
Cichlidae; los representantes de esta familia presentan coloración atractiva, sobre
todo las especies nativas de Africa, América Central y la región tropical de
Sudamérica. Es un carácter distintivo de los cíclidos la presencia de un sólo orificio
nasal a cada lado de la cabeza el cual posee la doble función de entrada y salida
de la cavidad nasal. El cuerpo es comprimido y generalmente discoidal, raramente
alargado. Se presenta en los machos de algunas especies un mayor crecimiento
en la cabeza con respecto a las hembras y se desarrollan en éstos tejidos grasos
en la región anterior y dorsal de la cabeza, conforme avanzan en edad.
La boca es protráctil, generalmente ancha y a menudo con labios gruesos.
Presentan dientes cónicos y en algunas ocasiones incisivos. Pueden o no
presentar freno en el maxilar inferior en la parte media debajo del labio. Las
membranas branquiales están unidas por 5 ó 6 radios branquióstegos y se
presentan branquiespinas en número variable según la especie. La parte anterior
de las aletas dorsal y anal siempre es corta y consta de varias espinas y la parte
terminal posee radios suaves los que en los machos están fuertemente
pigmentados. La aleta caudal está redondeada, trunca o raramente escotada. La

6
línea lateral está interrumpida presentándose en dos partes, la porción superior se
extiende desde el opérculo hasta los últimos radios de la aleta dorsal, mientras que
la porción inferior aparece varias escamas por debajo de donde termina la línea
lateral superior y se continúa hasta el final de la aleta caudal. Presentan escamas
de tipo cicloideo; el número de vértebras puede ser de 8 a 40 (Arredondo-Figueroa
y Guzman, 1986) (Figura 2)
4 Aleta dorsal
Radios dorsales Espinas dorsales
Aleta I I cau d
Boca
Aleta anal
Aleta p6lvica Aleta pectoral
Fig. 2. anatomía externa de los representantes de los géneros Tilapia y Oreochmmis.
A manera de resumen, se muestra en la siguiente tabla las principales
características que permiten diferenciar entre los géneros Tilapia y Oreochmrnis,
tomadas de Arredondo-Figueroa et a/. (1 994).

7
Tabla 2. Características que permiten identificar los géneros Tilapia y Oreochmmis, presentes en México. Propuestas por Trewavas (I 973) y modificadas por Lowe (1 959), Goldstein (1 970) y Fryer e Liies (1 979) [citados en Arredondo-Figueroa et a/. ,19941.
Preferentemente plantófagos Generalmente herbívoros Entre 14 y 15 branquiespinas en la parte ~
Entre 8 y 12 branquiespinas en la parte ...... . . ........... "" ............. .... ................. " . . - ._.-I -. ..... - . . . . . . . . " .................. I
.... " I"". inferior .... del "_ primer ~.~."__..__I arco branquial .......... c ~ . ". ...... inferior """ "_ del - ." .... primer - arco " ....... branquial - ... " " ....... i
.......... Pocos "" " huevos __.____I (menos .... ~. .......... de " ... .... . - .................... "" Muchos . ..~. .. ...... huevos ...... . - . . . .
Huevos con gran cantidad de vitelo de Huevos con poca cantidad de vitelo, de I color amarillo naranja (2.2 a 3 mm), no
presentan .. - -. -. .... - .... una "_ .. cubierta . - ..................................... adherente externa- El macho desarrolla una coloración muy
marcada en la época de reproducción y fija ......... su territorio, ~" .. en donde . establece su nido ..... .
. - . . Tiene _._ . . . . . . . . un periodo .~- prenupcial corto . . . . . . . . . . . . . - . . . . . . .
El macho es polígamo y usa el nido como un sitio temporal para el cortejo y la
fertilización de los huevos
Las hembras guardan los huevos y los alevines en la boca por espacio de 20 a 30
días
.e.
color amarillo (1.5 mm de diámetro) y una . . . . . . . ". .. capa " . exterior adhesiva j
Ambos sexos desarrollan una fuerte coloración en la época de reproducción y ; i
"~ fijan ....... su territorio ". "_ _I. I_ ......... para "" construir .............................. ahí su nido Tiene un periodo prenupcial largo i
.................................
...... --- -.I
............ ._ .............................. . ~,
Una pareja puede permanecer junta y desovar repetidas veces en una relación
monógama. Los huevos son cuidadosamente depositados usando el
ovipositor ..... . . . . . . . . . . . . . . . . .
Ambos padres cuidan los huevos y alevines; los huevos eclosionan a los 2 ó 3
días, son móviles y tienen una glándula especial para sujetarse. Los cuidan de 45 a
............ -. . . . . . . . . . " ... -.
- - " 50 días. ."- ........ . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . ............ . . . . . . . . .
Tienen gónadas pequeñas, pero los Tienen gónadas grandes con huevos '
...-I huevos " -. - . ". contienen ...... - mayor ... " ...... .~. cantidad - .... de - vitelo " .... pequeñosy ..I .... ". menor " cantidad - ...... " ..... de "._. vitelo - . - ....... ._,
. . . . . . . . . . . . . . . . . . . La sobrevivencia I - . ~. de crías es alta : estadios y menor cuidado parental
!
? --~- " I Una gran mortalidad en los diferentes
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
Los nidos tienen la forma de cráteres I Los nidos se presentan en forma de i circulares ligeramente mayores que la ' pequeños orificios (5-IO), colocados sobre
longitud de la hembra y se construyen en I el borde del estanque y cerca de la . . . . . . . . . . . . . . el fondo " de ... ." los ....................... estanques superficie del agua . . . . . . . . - . . . . . . . . . . . . . . . ~- . . . . . . . . . . . . . . . . . . . . ,
Tejeda (1987) concluye que en México se cuenta con un total de cinco especies de
diferentes orígenes, por tanto se estima conveniente realizar un análisis de las
poblaciones introducida en el país, con el fin de precisar con mayor exactitud la
pureza de las líneas.

8
I. a Hipótesis
Los eritrocitos de peces muestran aglutinación diferencial frente a las lectinas, por
tanto, el título de aglutinación de muestras de sangre de organismos del género
Oreochromis, permitirá segregar las diferentes razas de cíclidos de esta categoría
taxonómica presentes en México.
1. b Objefivos
Objetivo general
Utilizar la lectina del alga Codium girafia, Silva, para diferenciar las razas de
cíclidos del género Oreochromis introducidos en México.
Objetivos específicos
* Realizar el trabajo experimental que permita aislar y purificar la lectina presente
en el alga Codium girafia.
* Caracterizar parcialmente la proteína aislada del alga Codium giraffa.
* Montar pruebas de aglutinación de eritrocitos de cíclidos.
* Segregar las razas de cíclidos usando a la aglutinación diferencial obtenida.
9
II. ANTECEDENTES
Desde 1888 se descubrió que los extractos de la semilla de Ricinus communis
Linnaeus y, un poco más tarde, el extracto de la semilla de Abrus precaforius
Linnaeus (Leguminosae), provocaban la agrupación de eritrocitos humanos, a las
moléculas responsables de la aglutinación se les dio el nombres de ricina y abrina,
respectivamente (Sharon y Lis, 1981).
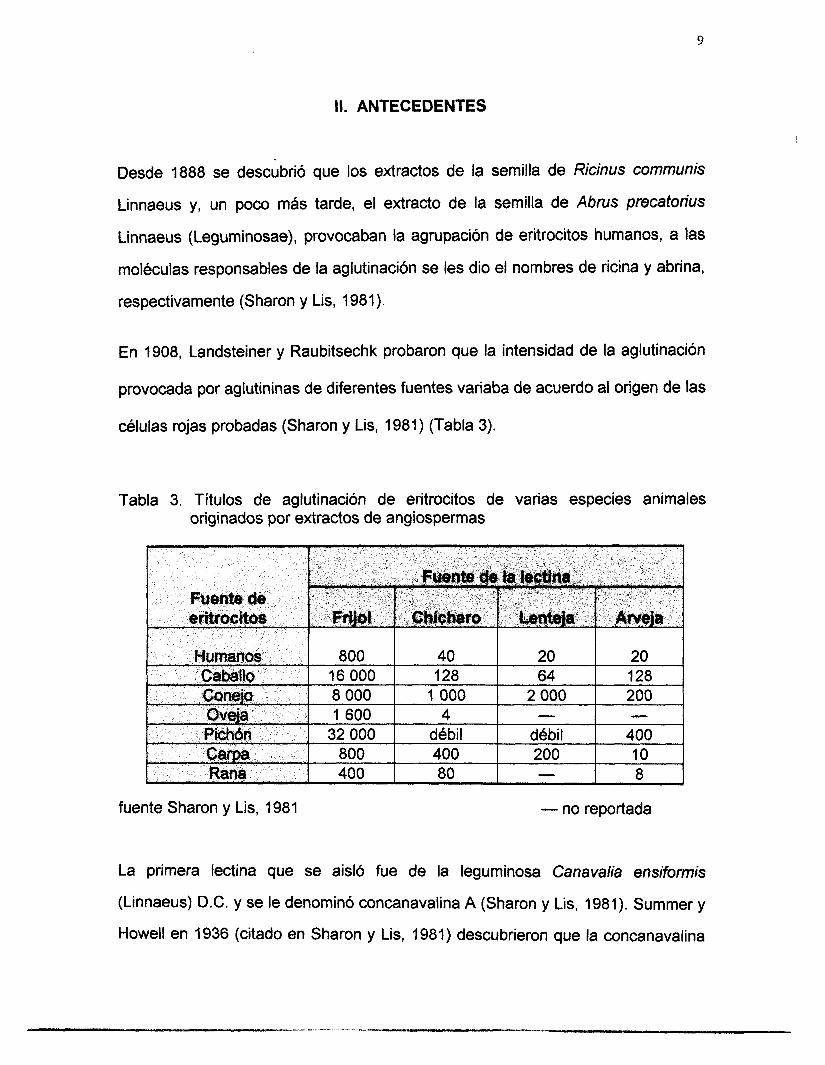
En 1908, Landsteiner y Raubitsechk probaron que la intensidad de la aglutinación
provocada por aglutininas de diferentes fuentes variaba de acuerdo al origen de las
células rojas probadas (Sharon y Lis, 1981) (Tabla 3).
Tabla 3. Títulos de aglutinación de eritrocitos de varias especies animales originados por extractos de angiospermas
fuente Sharon y Lis, 1981 - no reportada
La primera lectina que se aisló fue de la leguminosa Canavalia ensifomis
(Linnaeus) D.C. y se le denominó concanavalina A (Sharon y Lis, 1981). Summer y
Howell en 1936 (citado en Sharon y Lis, 1981) descubrieron que la concanavalina

10
A purificada precipitaba glucógeno en una solución de almidón y que la actividad
hemaglutinante se inhibía al agregar azúcar de caña, Esto sugirió que la actividad
de la concanavalina A podría ser una consecuencia de la reacción entre la proteína
con los carbohidratos presentes en la superficie de los eritrocitos (Sharon y Lis,
1972).
En 1945, Boyd y Reguera observaron que la lectina de Phaseolus limensis Macf.
(haba) aglutinaba especificamente los eritrocitos del tipo sanguíneo A, sin
reaccionar con los tipos O y B. Tres años después, Renkonen en 1948 destaca la
aglutinación exclusiva de células sanguíneas del tipo O positivo por el extracto de
la semilla de Lotus tetragonolobus Linnaeus.
El interés sobre las sustancias aglutinantes de células sanguíneas creció y la
investigación se centró en las provenientes de plantas fanerógamas,
particularmente las pertenecientes a la familia Leguminosae.
Uno de los descubrimiento más importantes sobre el uso de las lectinas y que abrió
un nuevo campo de la investigación biológica fue el evidenciar que la lectina de
Phaseolus vulgaris Linnaeus o fitohemaglutinina (PHA), estimulaba la
transformación de linfocitos, desde pequeñas células en reposo a grandes células
que sufren mitosis, con lo cual se facilita el examen visual de los cromosomas
humanos y de otros animales, promoviendo el crecimiento de la Citogenética y la
comprensión de las anormalidades cromosómicas (Nowell, 1960 citado en Sharon
y Lis, 1981).
La aglutinación diferencial de células malignas contra células sanas fue otro
descubrimiento importante (Aub et al., 1963; Burger y Goldberg, 1967, citados en

11
Sharon y Lis, 1981), esto permitió el uso de las lectinas para el estudio de los
cambios sufridos por las células en crecimiento, diferenciación y transformación a
células malignas.
Sin embargo, es hasta la década de los sesenta que se probaron por primera vez
extractos de 23 macroalgas y una Cianofita de Puerto Rico contra eritrocitos
humanos encontrando respuesta positiva en nueve especies, de las cuales
solamente se encontró especificidad en la aglutinación por el grupo sanguíneo A
en el extracto del alga Spyridia filamentosa (Wulfen) Harvey in Hooker, mientras
que los extractos acuosos de Dictyota bartaresii Lamouroux, D. cervicomis
Kützing, D. divaricata Linnaeus, Dicfyopteris delicatula Lamouroux, Sargassum
rigidulum Kützing, Padína vickersiae (Kützing) Sonder (= Padina gymnospora
(Kützing) Sonder) y la cianofita Lyngbya majuscula (Dillwyn) Harvey, mostraron la
extraña característica anti-(A+H) (Boyd et al., 1966).
En Inglaterra, se probaron 105 algas marinas de las cuales en 19 de ellas se
encontró actividad hemagtutinante inespecifica, solo el extracto del alga Ptilota
plumosa (Hudson) C. Agardh presentó especificidad por los eritrocitos del tipo B
(Blunden et al., 1975). Lo anterior fue corroborado más tarde al aislar y
parcialmente caracterizar el principio activo, el cual resultó ser una proteína con
dos subunidades, una con peso molecular de 65 y la otra con 170 kD (Rogers y
Blunden, 1980). Un trabajo previo realizado por Rogers (1977) reveló que de 96
especies de algas marinas, 36 aglutinaron eritrocitos humanos previamente
tratados con papaína o al añadir albúmina, lo que permitió desarrollar un método
más sensible para detección de actividad aglutinante.

12
En 1980, Rogers et a/. investigaron 49 macroalgas marinas, 7 líquenes
supralitorales, 2 especies de pastos marinos, 3 esponjas, 5 celenterados, 4
anélidos poliquetos, 6 artrópodos crustáceos, 1 sipuncúlido, 3 equinodermos, 2
holotúridos y 4 cordados (ascídias), sin obtener respuesta específica hacia ningún
tipo de células rojas, en este trabajo también se reportó la hemólisis provocada por
los extractos de algunos animales.
El primer trabajo donde se probó una lectina algal contra eritrocitos de otras
especies animales sin considerar los humanos fue el realizado por Shiomi et al.
( I 980), donde además aislaron y parcialmente caracterizaron una proteína con
25,000 daltones de peso molecular del extracto del alga Serraticardia maxima
(Yendo) Silva, siendo esta la responsable de la actividad aglutinante. En este
trabajo se reportó aglutinación para todos los eritrocitos de animales probados
(caballo, vaca, oveja, conejo, cobayo, ratón y pollo).
Shiomi et al. (1981) purificaron una aglutinina de la rodofícea Gracilaria vermcosa
(Hudson) Papenfuss presente en la primera fracción resultante de cromatografía
de exclusión molecular y que denominaron por esto GVA-1. Esta proteína fue
activa con células rojas de varios animales, las mismas especies probadas por
Shiomi et al. (1980), añadiendo a la lista de eritrocitos aglutinados por esta lectina
los de la carpa Carassius auratus Linnaeus.
Hori et al. (1981) probaron la presencia de aglutininas en algunas algas japonesas;
de 53 especies probadas, 14 fueron activas frente a eritrocitos de conejo y 1 O con
alguno o varios de los tipos de sangre humana A, B, O y AB positivos, sin mostrar
especificidad por ningún grupo.

13
Por su parte, Kamiya et al. (1 982) aislaron una glicoproteína del alga Palmaria
palrnata (Linnaeus) O. Kuntze, que aglutinó células rojas de conejo, caballo y
humanas de los grupos A, B y O, asimismo, aglutinó selectivamente las células de
leucemia de ratón (L5178Y). Los eritrocitos fueron tratados con una enzima
proteolítica (pronasa, tripsina y papaína).
Rogers y Toplis (1983) caracterizaron y purificaron una aglutinina del alga
rodofícea Soliera chordalis (C. Agardh) J. Agardh, que resultó poseer ácido siálico
y, que requirió la presencia de albúmina para obtener resultados aglutinantes en
altas diluciones.
En España se han buscado aglutininas en algas rojas (Fábregas et al., 1984;
Fábregas et al., 1985); algas verdes (Muñoz ef al., 1985) y algas pardas (Fábregas
et al., 1986) que tengan actividad sobre eritrocitos de animales (incluidos los
humanos), sin encontrar especificidad por algún tipo de sangre, excepción hecha
con el extracto de Giffordia granulosa (Sm.) Hamel, que sólo aglutinó las células
del grupo O positivo. Muñoz et al. (1 987a.), trabajaron con eritrocitos de algunas
especies animales, encontrando que las aglutininas algales son mas activas con
este tipo de células que con los eritrocitos humanos. Se ha investigado sobre la
posible aplicación práctica de las aglutininas de origen algal como reactivos para el
reconocimiento y caracterización de subpoblaciones de peces o como marcadores
genéticos en acuicultura (Muñoz et al., 1987b.; Muñoz et al., 1987~). También
como auxiliares en la investigación de procesos fisiológicos, anatomía y función
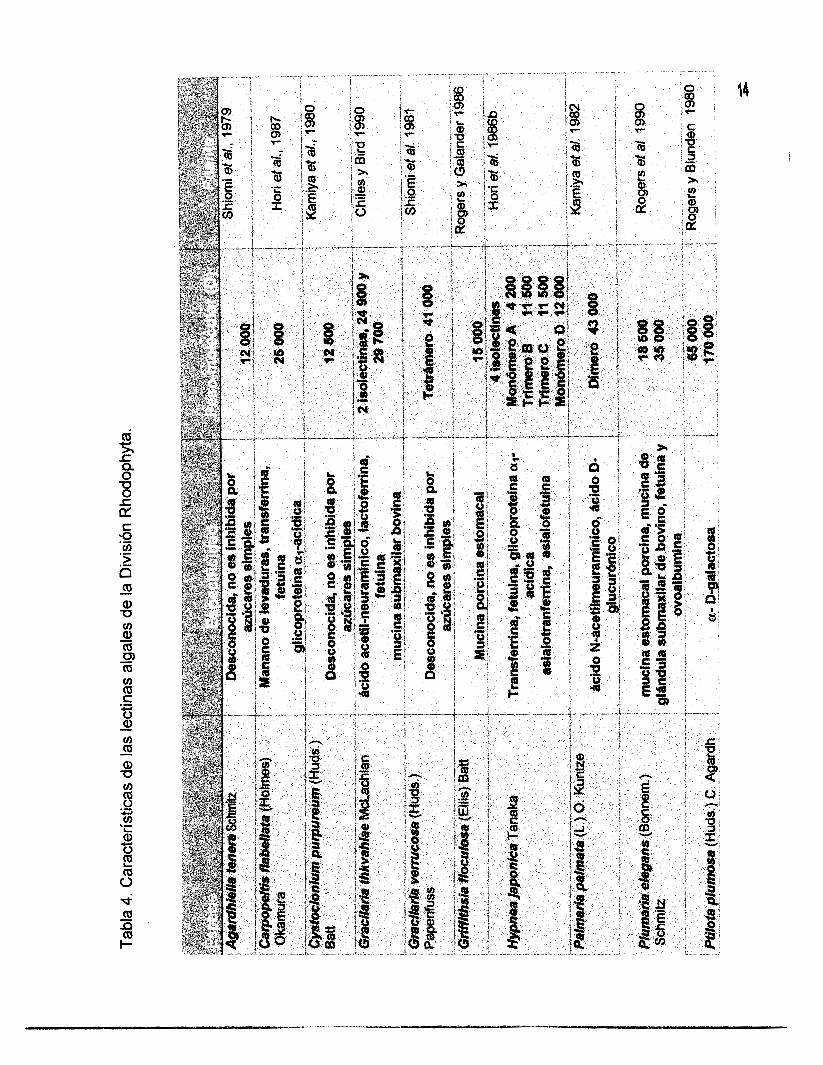
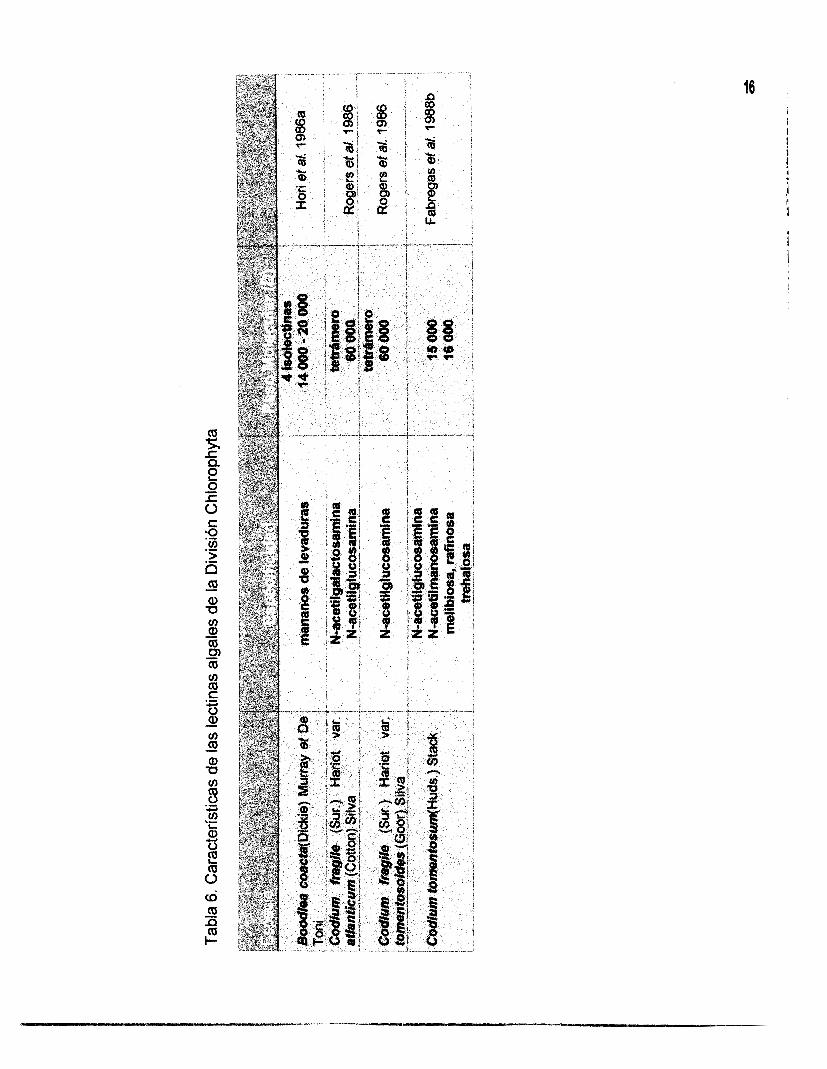
celular (Hori et al., 1987; Fábregas et al., 1988a y 1988~). En las Tablas 4, 5 y 6 se
muestran las lectinas aisladas de algas marinas y se describen algunas de sus
características más importantes.
c v)
> :o ii I-

c
M
8 + .z ([I
F
([I
W a,
z ([I
ctl
15
.._" "_ P "
16
” .

17
II. a Usos y aplicaciones de las lectinas algales
El mucopolisacárido con acción de lectina aislado de Fucus vesiculosus Linnaeus
ha mostrado tener efecto inmumodulatorio en ratones y también induce la
blastogénesis en linfocitos in vitro (Criado y Ferreiros, 1983a). La lectina de Fucus
vesiculosus también ha probado aglutinar selectivamente e inhibir el crecimiento de
Candida guillemondi y un débil efecto sobre C. kursei (Criado y Ferreiros, 1983b).
La lectina del alga roja Carpopeltis flabellata ha mostrado actividad mitogénica en
linfocitos T de bazo de ratón (Hori et al., 1987). Estas lectinas pueden ser útiles en
la investigación de los procesos que ocurren durante la activación de linfocitos.
Un uso popular de las lectinas de plantas terrestres ha sido en la agrupación de
tipos sanguíneos. Algunas lectinas de algas marinas poseen esta especificidad,
por ejemplo, la lectina de Ptdofa plumosa aglutina eritrocitos humanos que
expresan el antigen0 B (B, A1 B y A2 B) y no reacciona con los grupos A y O
(Rogers et al. 1977b).
Dos son los factores que han limitado el uso rutinario de las lectinas en las pruebas
de tipificación de sangre. El primero es la detección de los eritrocitos con el grupo
AIB que ha demostrado ser poco confiable y el segundo es que existen
anticuerpos mono y policlonales específicos que son m& fáciles de conseguir
(Fábregas et al. 1988a).

18
Un uso más prometedor para estas lectinas es la diferenciación de los antígenos
de los grupos sanguíneos B genéticamente modificados de aquellos que los han
adquirido como un resultado de la modificaci6n por enzimas bacterianas o por la
adsorción pasiva de lipopolisacáridos bacterianos. Solamente los eritrocitos
determinados genéticamente son aglutinados por la lectina de ffilofa plumosa
(Rogers et al., 1979), mientras que los anticuerpos aglutinan indiferenciadamente
los antígenos del tipo B adquiridos o determinados genéticamente.
Se sabe muy poco sobre el posible papel que juegan las lectinas en las algas
marinas. El Único estudio que reporta una posible función de éstas moléculas en
las macroalgas es el de la lectina aislada de la superficie de las células
espermáticas de Fucus serratus Linnaeus (Bolwell et al., 1979 y 1980). Esta alga
normalmente no tiene fertilización cruzada con otros fucoides (Bolwell et al. I 1977)
y se cree que existe un reconocimiento en el gameto de tipo especie-específico,
debido a su asociación con los ligandos que contienen residuos fucosil y manosil
en la superficie de los óvulos con la lectina complementaria de la superficie de las
células espermáticas.
II. b Uso de las lectinas para definir variabilidad específica
Una de las aplicaciones de las lectinas algales que ha sido poco explorada es su
posible uso en pruebas serológicas con peces y en particular con especies

19
sometidas al cultivo, de las cuales se pone en tela de juicio la pureza de sus
características específicas.
Los antecedentes sobre la aplicación de las lectinas de angiospermas en el mundo
son realmente escasos al igual que el uso de lectinas algales con este propósito. A
continuación se describen los trabajos más importantes en este campo.
Sinderman (1962) y Utter et al. (1964) encontraron diferencias individuales en los
títulos de aglutinación entre especies de la familia Cuploideae utilizando lectinas de
plantas terrestres como la de Dolichos biflorus Linnaeus (= Vigna unguiculata
[Linnaeus] Walp. subsp. cylindrica [Linnaeus] Eselt.).
Utilizando el mismo modelo anteriormente expuesto, Utter et al. (1963), citado en
Ridway (1 969), descubren variabilidad antigénica en los eritrocitos de varias
especies de salmones del género Oncorhynchus.
Posteriormente, Vann y Cushing (1966) reportaron variaciones individuales en la
respuesta a la aglutinación de los eritrocítos del bonito de California (Sarda
chilensis Cuvier) con la lectina de Dolichos biflorus y la relacionaron con la clase de
edad de los peces, es decir, con el tamaño del pez, encontrando que a mayor
tamaño del animal, se presentó mayor título de aglutinación.

20
También, Gutiérrez (1967a y 1967b) estableció grupos sanguíneos en Thunnus
thynus Linnaeus Gibbs & Collete, usando lectinas de leguminosas.
Kuhns y Chuba.(l968) demuestran la presencia de diferencias intragenéricas en
los tipos de sangre de Ictalúridos. Utilizaron técnicas de isoinmunización, lectinas
de angiospermas y heteroaglutininas de otras especies de peces y humanas.
Sin el uso de lectinas y por métodos como isoinmunización o antisueros de otras
especies animales, destacan los trabajos de: Sanders y Wright (1962) que
inmunizaron conejos con eritrocitos de trucha arcoiris Salmo gairdned Walbaum y
de trucha café Salmo tnrtta Linnaeus, utilizando el antisuero obtenido en pruebas
de aglutinación con eritrocitos de la especie de trucha correspondiente. Sus
resultados mostraron dos tipos de antígenos que denominaron R-1 y R-2 para la
trucha arcoiris y otros dos B-1 y B-2 para la trucha café presentes en diferentes
proporciones.
Uno de los trabajos más recientes sobre la tipificación de sangre de peces lo
realizaron Tong y Wu (1993) los cuales inmunizaron conejos con sangre de
Carassius auratus (carpa común), obtuvieron el suero de los conejos y lo probaron
con los mismos peces. Sus resultados demostraron la presencia de dos antigenos
diferentes en las carpas.

21
El primer hallazgo de reacción de eritrocitos de peces dulceacuícolas con lectinas
de macroalgas marinas lo realizó Shiomi et a/. (1981) al descubrir actividad positiva
de las hypninas, aisladas de Hypnea japonica, con eritrocitos de Cyprinus carpio
Linnaeus.
Muñoz et al. (1987~) realiza un estudio en el cual, por primera vez utiliza lectinas
algales y eritrocitos de peces marinos, este trabajo pone de manifiesto que existen
receptores en la superficie de estas células para las lectinas; aquí se sometieron a
estudio 16 especies de peces marinos presentes en la costa norte española contra
extractos crudos de I 6 algas marinas de la División Phaeophyta.
Posteriormente, Muiioz et al. (1987b), proponen que la aglutinación diferencial de
eritrocitos de dos razas del pez marino Labrus bergylta Ascanius, puede servir
como un nuevo criterio auxiliar en la detección de diferencias intraespecíficas en
este organismo.
Fábregas et a/. (1992) aumenta el número de especies algales probadas a 29
pardas, 37 rojas y 4 verdes, contra los eritrocitos de las 16 especies de peces
marinos, encontrando respuesta positiva de los eritrocitos con todas las algas
probadas y estableciendo que estas pruebas serológicas pueden ser una
herramienta útil para reconocer el origen de las poblaciones de peces y que estas
diferencias serológicas permitirán distinguir entre peces cultivados y los
provenientes de poblaciones en la naturaleza.

22
Tomando en consideración los antecedentes expuestos en las páginas anteriores,
este trabajo aporta conocimiento en dos áreas aún no estudiadas en México. La
primera es la relativa a la caracterización de proteínas presentes en macroalgas
marinas y particularmente a moléculas tan útiles como son las lectinas. La segunda
área de incidencia de esta tesis, corresponde al campo inmunológico de peces,
que si bien, se ha investigado con algunas especies, debe hacerse hincapié que la
mayoría de los trabajos que comprueban la variación de los antígenos de la
superficie de los eritrocitos de peces, se han desarrollado en gran medida con
peces marinos y aunque sí existen antecedentes de trabajos realizados con
especies dulceacuícolas, los integrantes de la tribu Tilapinii no han sido sujetos a
experimentación con lectinas
Debe considerarse que el alga Clorofita Codium giraffa, reportada por Silva (1979)
permanece como una especie endémica de México. Este trabajo es importante, ya
que será el primero en nuestro país donde se identifica una lectina de origen algal.
Es también fundamental resaltar que estos estudios implicaron el desarrollo de
antisueros en otras especies animales como conejos y que la gran diferencia en la
aplicación de una lectina es que se utiliza directamente con la sangre de los peces
sin necesidad de desarrollar estos antisueros, convirtiéndola en una prueba fácili y
rápida para revisar las condiciones de las especies bajo cultivo. Por tanto, la
aplicación de esta lectina algal para diferenciar subespecies de peces, constituye
una herramienta de utilidad taxonómica barata y sencilla de realizar.

23
111. MATERIALES Y METODOS
111. a Colecta de material
El material algal se recolectó en la zona intermareal de dos localidades: Puerto
Vicente Guerrero (Puerto Escondido), municipio de Papanoa en la costa del estado
de Guerrero y en la costa Michoacana, en Chiquiapan, a 12 Km. de Puerto Lázaro
Cárdenas, los ejemplares se desprendieron de la roca con la ayuda de una
espátula. Las muestras se limpiaron in situ con agua de mar para evitar en lo
posible la contaminación por organismos epibiontes; se guardaron en bolsas de
plástico etiquetadas y se congelaron inmediatamente utilizando COZ sólido para su
transporte al laboratorio.
111. b Preparacidn de extracto algal crudo
Se descongeló el material algal a temperatura ambiente y se enjuagó con agua de
la llave y posteriormente con agua destilada para limpiarlo de arena y
contaminantes. Se limpió de epibiontes y epífitas manualmente bajo el microscopio
de disección, una parte del material se conservó en formo1 al 4 YO glicerinado, para
su posterior identificación y elaboración de ejemplares de herbario; el resto del
material se congeló a -29 "C en un Ultracongelador Forma Scientific. El alga se
molió en un homogenizador Ultra Turrax modelo TP 1812N usando regulador de
fosfatos 10-mM, pH 7.2 con O. 15 M de Na CI (PBS) utilizando pulsos de 30
segundos hasta completar 10 minutos, se agregó COZ sólido pulverizado para

24
congelar el material algal y así realizar una extracción más eficientemente. El
extracto obtenido se centrifugó en una centrífuga Beckman modelo J2-21 usando
un rotor JA-12 durante 20 min. a 18 O00 r.p.m. y el sobrenadante obtenido se filtró
usando filtros de papel Whatman del número 5, técnica modificada de Fábregas et
al. (1 98813).
111. c Solución de erifrocitos
Se usaron eritrocitos de conejo formalinizados (ECF). Se obtuvo la sangre de
conejo por decapitación, recogiéndola en recipientes plásticos con solución Alsever
con citrato de sodio al 3 % como anticoagulante, se transportó a 4-6 OC. En el
laboratorio se formalinizaron los eritrocitos de acuerdo con el método propuesto por
Nowak y Barondes (1975). Las células se lavaron tres veces en 20 volúmenes de
solución reguladora de fosfatos 100-mM, pH 7.2 (PB), centrifugando a 1 O00 x g
durante 5 min y manteniendo la temperatura a 4 OC. Se resuspendió el paquete
celular en PB hasta una concentración de 8 YO (46 vol / 1 vol), agregando el mismo
volumen de formalina al 3 % en el mismo regulador de fosfatos, ajustando el pH a
7.2 con Na OH 0.1 M. Esta mezcla se incubó a 37 O C por 16 hs con agitación
moderada en un baño de incubación y temperatura controlada marca Forma
Scientific. Pasado el periodo de incubación, se lavaron las células 4 veces con 5
volúmenes de PB por cada lavado para retirar el exceso de formalina. LOS
eritrocitos pueden conservarse a 4 OC como una solución al 10 % (45 vols. PBS
vol. de paquete celular).

25
111. d Ensayo de actividad hemaglutinante
La pruebas para comprobar la actividad hemaglutinante se realizaron utilizando
ECF que son los más sensibles para detectar la aglutinación (Hori et al., 1981;
Fábregas et al., 1988b); en placas de microtitulación de fondo en “U” en el
siguiente orden: a todos los pozos se les agregó 50 pl de PB, al segundo pozo se
le añadió 50 pi de extracto algal y a partir de este pozo se realizaron diluciones
dobles seriadas, el último volumen tomado de la dilución se desechó. Finalmente
se agregaron 50 pI de solución al 2 % de ECF. Se dejaron reposar las placas por 2
horas. La aglutinación se determinó macro y microscópicamente. El título se
reportó como la más alta dilución doble en la cual se presentó hemaglutinación.
111. e lnhibicibn por azúcares
Con el fin de identificar el sacárido o sacáridos que se unen al sitio activo de la
lectina, se realizaron pruebas de inhibición en placas de microtitulación de fondo
“U”, siguiendo el orden que se presenta: cada pozo de la placa se cargó con 25 pl
de PB, al segundo pozo de cada fila de la placa se le agregaron respectivamente,
25, 50, 75 y 100 pl de solución al 250-mM de los siguientes azúcares, Celulosa
(Merck Darmstadt), D-Dextrosa (Baker), D-Fructosa, D-Galactosa, D-Glucosa, D-
Lactosa, D-Maltosa, D-Raffmosa, D-Manosa, N-acetil-glucosamina, N-acetil-D-
manosamina, N-acetil-D-galactosamina, ácido N-acetil-neuraminic0 (SIGMA
Chemical Co.).

26
Se realizaron diluciones dobles seriadas de cada azúcar, posteriormente se
añadieron 25 1.11 de extracto algal y se dejaron reposar las placas por 10 minutos.
Transcurrido este tiempo, se agregaron 25 pl de solución al 1 % de ECF a todos
los pozos. Las placas se mantuvieron en reposo a temperatura ambiente y se
registraron los resultados de inhibición de la aglutinación después de 1 hr
(Fábregas ef al. , 1988c; Rogers y Topliss, 1983).
111 f. Obtencibn de proteínas
El extracto algal crudo se precipitó con sulfato de amonio sólido a 40, 60 y 85 % de
saturación manteniendo la temperatura a 4 OC, centrifugando cada vez a 15,000
r.p.m. durante 25 minutos. Las proteínas obtenidas de cada intervalo de
precipitación se dializaron exhaustivamente contra agua destilada y después contra
el amortiguador PB. Los intervalos de precipitación con sulfato de amonio que
resultaron positivos al aglutinar los eritrocitos de conejo fueron sometidos a
cromatografia de afinidad.
111. g Purificacidn de la Lectina
Para la purificación de la lectina de Codium girafla, se utilizaron dos métodos de
cromatografía.

27
111. g 1 Cromafografia de Afinidad
Se utilizó cromatografía de baja presión con una minicolumna empacada con gel
de N-acetilglucosamina-agarosa (Sigma Chemical Co.) con dimensiones de 1.4 x
6.5, volumen de cama de 10.0 ml y unión de 17 mg por mi de lectina de Triticum
vulgare Vill. Se equilibró con PB y se recolectó la fracción sin afinidad por el gel en
un solo tubo. Se eluyó la lectina usando el mismo amortiguador de fosfatos con N-
acetilglucosamina 0.2 M. Se probó la actividad aglutinante las dos fracciones
obtenidas después de haber sido dializadas contra agua destilada contra ECF.
111. g 2 Cromatografia de Intercambio iónico
Se utilizó HPLC de intercambio iónico (aniónico) con una columna de DEAE-
Celulosa, con un equipo Beckman SystemGold, usando amortiguador de fosfatos
0.05 M con gradiente de O a 1 M de Na CI, se colectaron fracciones de 1 ml y se
probó su actividad aglutinante.
111 g 3 Métodos Analíticos
Para la continua medición de la concentración de proteínas se utilizó el método de
absorción de luz en el ultravioleta. Las proteínas absorben con dos máximos,a 280
y 200 nanómetros (Stoscheck, 1990). Se midió la absorbancia a 280 nm de las
fracciones obtenidas en un espectrofotómetro Ultrospec I I 4050 UV/visible de LKB
Pharmacia.

28
El contenido de proteinas se determinó por el método colorimétrico del ácido
bincinconinico (Smith et al., 1985, citado en Stoscheck, 1990) [Figura 21. Se
efectuaron lectura de las absobancias a 532 nm., comparándolas posteriormente
en una curva de calibración con albúmina bovina (Pierce) los resultados se
expresan en microgramos de proteína por mililitro.
Proteínas 2 B C A
+ + cu 1
O H + H O 2
Figura 2. Reacción del ácido bicinconinico y las proteínas para formar un complejo colorido.
111. g 4 Nectrofor4sis en gel de SDS-PAGE
Las fracciones que resultaron positivas al aglutinar eritrocitos de conejo se
sometieron a electroforésis en gel. Se utilizó un equipo Phast system (Pharmacia),
con un gel homogéneo de 12.5 % de acrilamida. Las fracciones fueron preparadas
agregando Tris-HCI 20 mM pH 8.0 con EDTA 2-mM, SDS al 5% y p-

29
mercaptoetanol al 5 YO. Se hirvieron las muestras a 80 OC durante 3 minutos,
posteriormente se aplicaron al gel. Se reveló la corrida electroforética usando azul
de Comassie. Los siguientes marcadores de peso molecular fueron usados: a-
lactalbúmina (14,400), inhibidor de la tripsina (20, IOO), anhidrasa carbónica
(30,000), ovoalbúmina (43,000), albúmina (67,000) y fosforilasa b (94,000).
111. g 5 Punto Isoelécfrico
Para determinar el punto isoeléctrico de la lectina pura se usó un gel de acrilamida
con el mismo equipo, utilizando los marcadores de punto isoeléctrico siguientes:
aminoglucosidasa (3.50), inhibidor de la tripsina de soya (4.59, p-Iactoglobulina A
(5.20), anhidrasa carbónica bovina B (5.85), anhidrasa carbónica humana B (6.55),
mioglobina de caballo acídica (6.85), mioglobina de caballo básica (7.35), lectina
de lenteja acídica (8.15), lectina de lenteja media (8.45), lectina de lenteja básica
(8.63) y tripsinógeno (9.3).
111. h Pruebas de aglufinación de eritrocifos de peces
Las pruebas se realizaron con eritrocitos de las especies: Oreochrornis aureus
(variedad roja), Oreochromis aureus Stirling y la variedad Rocky Mountain que es
un híbrido producto de la cruza de O. aureus y O. niloficus, ejemplares mantenidos
en Planta Experimental de Producción Acuícola (PExPA) de la Universidad
Autónoma Metropolitana, plantel iztapalapa,

30
Los ejemplares que se encuentran en la PExPA proceden de diferentes lugares:
O. aureus (variedad roja) fue obtenido del Centro de Investigación y Estudios
Tropicales, El Clarín. Este centro pertenece a la Facultad de Medicina Veterinaria y
Zootecnia de la UNAM y se encuentra en Martinez de la Torre, Veracruz. La
variedad Rocky mountain fue donada por el Centro Acuícola de Zacatepec,
Morelos y O. aureus (Stirling) la donó el Centro Acuicola de Temazcal, Oaxaca.
La sangre de los peces se obtuvo utilizando punción cardiaca, según la técnica
sugerida por Llovo et al. (1987) para Salmo gairdniefl Richardson. El ángulo de
entrada sugerido de 80-90 grados se modificó a 70-80 grados con respecto al
plano dorsal, con punto de entrada en el primer cuarto resultante de dividir en 4 la
distancia entre la V-opercular y el comienzo de las aletas ventrales. También se
ajustó la penetración de la aguja hasta 1-1.5 cm aproximadamente, esto último
tomado en consideración el tamaño del pez. La sangre fue recogida directamente
en una jeringa desechable que contenía como antimagutante, 1 m1 de &rato de
sodio al 3 % en PB IOO-mM pH 7.2.
Para los ensayos de aglutinación, se preparó una solución de eritrocitos al 2 % de
la misma manera como se hizo con la sangre de conejo. Se usaron eritrocitos
nativos y con ellos se realizaron las pruebas de aglutinación con la lectina del alga
Codium girafla, siguiendo la metodología antes descrita en este texto para la
detección de aglutinación con eritrocitos de conejo formalinizados (Muñoz et al.,
I 987c).

31
111. i. Analisis Estadístico de la aglutinacidn diferencia NI 225’721.
Como variable aleatoria se midió la aglutinación provocada en eritrocitos de peces,
por la acción de la lectina aislada del alga Codium girafa (Chlorophyta). El número
de muestra fue de 30, diez ejemplares de cada especie o variedad de pez y se
realizaron 3 repeticiones de cada ensayo de aglutinación. La unidad experimental
estuvo representada por cada uno de los pozos de la placa de microtitulación.
Como grupo testigo se utilizó PB y eritrocitos de pez sin añadir lectina.
Para comprobar que los datos obtenidos pertenecían a una población con
distribución normal, se realizó una análisis de homogeneidad de varianzas de
Bartlett, consistente en una modificación de la prueba de Neyman-Pearson (Steel y
Torrie, 1988). Esta prueba parte de la base de que los datos son normales y
comprueba la distribución gausiana, mas que la heterogeneidad de los mismos
(Sokal y Rohlf, 1969).
Se planteó la siguiente prueba de hipótesis:
Ho : x’ = x‘ los datos poseen distribución normal
Ha ; x’ # x2 los datos no poseen distribución normal

32
Para estudiar la variabilidad en la aglutinación media, se realizó un Análisis de
varianza (ANOVA), donde la fuente de variación fue la máxima dilución que
presentó aglutinación de la sangre. Los niveles estuvieron representados por las
especies o variedades de peces.
Se plantearon las siguientes Hipótesis:
Ho = pz= j.~3 =,...,= pn
Ha: AI menos una de las medias es distinta, es decir,
existe diferencia en las medias de aglutinación
para al menos una subespecie de pez.
Una vez realizado el ANOVA se aplicó la prueba de Newman para obtener la
agrupación de medias con respuesta similares y aquellas que formaron grupos
distintivos.

33
V. RESULTADOS Y DISCUSION
V. a Pruebas de inhibición con azúcares
Las pruebas de inhibición con azúcares y derivados permitieron definir el grupo
funcional al cual se unió eficientemente la lectina de Codiurn girafa. La lectina fue
inhibida por una concentración tan baja como 8-mM de N-acetil-D-glucosamina,
por lo que se decidió utilizar este derivado unido a una matriz de agarosa para
aislar la proteína.
También se observó afinidad por N-acetil-D-galactosamina aunque con menor
efectividad 16-mM y por raffinosa con una concentración inhibitoria de 250-mM
(Tabla 7).
Tabla 7. Resultados de la prueba de inhibición de la actividad aglutinante con azúcares y derivados.

34
La lectina de C. giraffa (girafina) es capaz de unir N-acetil-D-glucosamina y N-
acetil-D-galactosamina, asimismo es inhibida por alta concentración de rafinosa.
Estas características la hacen diferente de las lectinas del mismo género aisladas
hasta la fecha. Las lectinas de las dos variedades de C. fragile son inhibidas
únicamente por N-acetilgalactosamina y el derivado N-acetilglucosamina solo las
afecta levemente (Rogers et al., 1986). La lectina tomentina aislada de C.
tomentossurn (Fábregas et a/., 1988b) inhibe la actividad con N-acetilmanosamina
y no reacciona ante N-acetilgalactosamina como la girafina. No obstante,
coinciden estas dos últimas lectinas en que son inhibidas por rafinosa.
V. b Cromatografía de Afinidad
La figura 3 muestra el perfil de elución (cromatografía de afinidad) de los
precipitados al 40 y 60 %. Principalmente se observó una mayor concentración de
lectina en el corte al 60 %; lo anterior se comprobó al dializar intensivamente las
soluciones obtenidas contra agua destilada y montando ensayos de
hemaglutinación con eritrocitos de conejo. Se observa marcada con (SA) la
fracción sin afinidad por el gel y con (CA) la fracción que se retuvo en la columna y
se desprendió con N-acetil-glucosamina. La corrida cromatográfica se llevó a cabo
en un periodo de dos horas promedio, para los dos precipitados.
El método utilizado para aislar a lectinas en una primera etapa ha sido
recomendado por Fábregas et al. (1988b) siempre y cuando se conozca et azúcar
inhibitorio de la actividad de la lectina en cuestión.

35
La fracción protéica con actividad aglutinante fue dializada contra agua destilada y
contra amortiguador de fosfatos con el propósito de someterlo a HPLC de
intercambio iónico.
o2 4
0.1 i
O -
SA CA
L O
Figura 3. Cromatograma de Afinidad, las primera curvas corresponden a la fraccidn sin afinidad por el gel (SA), la segunda curva (CA) fue obtenida después de agregar una solución de N-acetilglucosamina 0.2 M en PB. ( --------- ) PB 0.1 M. (I I m ) N- acetilglucosamina 0.2 M
V. c Cromatografia de alta resolución de intercambio iónico
La Figura 4 muestra el perfil de elución obtenido al utilizar HPLC de intercambio
iónico de la fracción con afinidad por el gel de N-acetilglucosamina-agarosa.
Marcada con una flecha se encuentra la fracción que aglutinó ECF y que tuvo un
tiempo de retención de 18 minutos a un porcentaje de salinidad de 60 Yo. Cabe
destacar, que los perfiles de elución tanto de la cromatografía de afinidad corno 10s

36
obtenidos por HPLC fueron idénticos, por lo que solamente se muestra uno de
cada uno.
i
i / I I / 1
/ , l- I I 1 l / l / / I I I I I I
0.0 10.0 20.0 30.0 35.0 tiempo (minutos)
A0
Figura 4 Cromatograma obtenido por HPLC de intercambio iónico a partir de la fracción con afinidad por el gel de N-acetilglucosamina-agamsa, se muestra con una flecha la fracción que aglutinó eritrocitos de conejo formalinizados.
Nea o

37
Esta fracción fue nuevamente separada por HPLC de intercambio iónico,
obteniéndose el perfil que se muestra en la Figura 5
1 I
0.0
Perfil de elución de la fracción que aglutinó eritrocitos de conejo formalinizados.
Con el fin de comprobar que la proteína se encontraba totalmente purificada, se
realizó electrofor6sis en gel de poliacrilamida en dos formas, nativa y
desnaturalizada. Para esto se utilizó la fracción marcada en la figura 5.

38
V. d Electrofor4sis SDS-PAGE
En este gel en condiciones desnaturalizantes se puede observar en el carril 2 el
extracto algal crudo, carril 3 la lectina purificada, carril 4 los marcadores de peso
molecular; en los carriles 5 ,6 y 7 se colocó el precipitado al 40 % (Fig. 6). El peso
molecular calculado por este método fue de 13 KD.
Figura 6. Gel SDS-PAGE de la lectina de Codium giraffa.
Esta técnica se utiliza rutinariamente para determinar la composición de las
proteínas, es decir, si éstas presentan subunidades, así como establecer los

39
pesos moleculares de las mismas. En general, el fraccionamiento por
electroforésis en gel de poliacrilamida, está basado en los tamaños,
conformaciones y cargas netas de las macromoléculas. Los sistemas para
proteínas en conformación nativa no distinguen entre los efectos del tamaño, perfil
o forma y carga de la molécula en la movilidad electroforética. Como una
consecuencia, proteínas con diferente peso molecular pueden tener la misma
movilidad en el sistema empleado, por tanto no son recomendables como criterio
de pureza (Garfin, 1990).
AI ser agregado a la muestra de proteína el detergente sodio dodecil sulfato
caliente, éste se une debido a su carga aniónica, impartiendo carga negativa a los
complejos resultantes. La interacción con SDS, rompe todas las uniones de
proteína no covalentes causando que las macromoléculas se desdoblen. Si
además, se trata la muestra con un agente reductor de puentes disulfuro, como el
2-mecapto-etanol o ditiotreitol, la proteína se desnaturaliza desdoblándose a sus
subunidades constituyentes. Una sola banda en estos geles es criterio de pureza
(Garfin, 1990).
Como puede observarse en este gel y de acuerdo a las consideraciones vertidas
anteriormente, pudo considerarse que la proteína se encontraba purificada, debido
a la aparición de una única banda en el carril 3, además se observa que no se
presentan subunidades, por tanto se trata de una proteína monomérica. El criterio
de pureza se corroboró realizando el gel de isoelectroenfoque.
Las lectinas aisladas de algas del mismo género: Codium fragile vars. atlanticum y
tomentosoides y C. tomentossurn en que las primeras dos son tetraméricas
(Rogers et al., 1986) y la segunda es dimérica (Fábregas et al., 1988b), la lectina

40
de C. girafla es un monómero y posee un peso molecular menor que las
anteriores.
V. e €lectrofor&is en condiciones no desnaturalizantes
De izquierda a derecha se puede observar, en el carril 2 el extracto algal, carril 3 la
fracción considerada como pura, carril 4 los marcadores de peso molecular antes
descritos, carril 5 el precipitado al 40 %, carril 6 el precipitado al 60 % y en el carril
7 se colocó una fracción impura al 40 Oh (Fig. 7). El peso molecular obtenido fue
de 16.8 KD.
1 2 3 4 5 6 7 8
Figura 7. Gel en condiciones no desnaturalizantes
El peso molecular estimado por este método se considera confiable, debido a que
las proteínas migran de acuerdo a su conformación y peso molecular real. Las
diferencias con otras proteínas reportadas, son evidentes en función de que las

+ 225721 41
lectinas aisladas de especies relacionadas, poseen masas relativas por arriba de
los 30,000 Daltones C. fragile (op. cit.) con sus dos variedades tienen pesos
moleculares de 60 000, C. tornentossurn (op. cit.) 31 O00 y Boodlea coacta con Mr
de 34 kD (Hori et al., 1986a). La lectina de C. giraffa posee un peso molecular
mucho menor, solamente comparables con las lectinas aisladas de algunas
rodofitas como: Agardhiella tenera con 12 kD (Shiomi et al., 1979), Cysfoclonium
purpureurn de 12.5 kD (Kamiya et al., 1980) y las cuatro isolectinas del alga
Hypnea japonica con 4.2, 11.5, 11.5 y 12 kD (Hori et al., 1986b). El Único caso
extraño debido a su pequeña masa relativa de 2 O00 que puede comparase con
un péptido está representado por la única lectina aislada de un alga caf6 Fucus
vesiculosus (Ferreiros y Criado, 1983). No obstante, se debe mencionar que esto
no es indicativo de la conformación y sitios activos de la lectina, porque las
proteínas de las algas rojas son inhibidas por carbohidratos complejos, mientras
que las lectinas de la División Chlorophyta suspenden su actividad por unión con
azúcares simples y no reaccionan con carbohidratos de cadena larga.
Si consideramos que las lectinas de leguminosas poseen en general mayor masa
molecular, por ejemplo, la lectina del germen de trigo con 36 kD, la del frijol de
lima 265 kD, el peso de la lectina Concanavalina A de 51 kD a pH 6 hasta 102 a
pH fisiológico, la del frijol de soya con 120 kD y por último, la lectina de cacahuate
con 11 O kD (Lis y Sharon, 1981), es por tanto, evidente que en general las lectinas
de macroalgas marinas, poseen pesos moleculares menores que las mismas
moléculas aisladas de Leguminosas e igualan a éstas últimas en funciones y
actividad características.

42
V. f lsolectroenfoque
En este gel se destaca en el carril 4 y 6 la proteína purificada cuyo pl estimado fue
de 6.36, en el carril 5 se muestran los marcadores de punto isoeléctrico descritos
en materiales y métodos (Figura 8).
Figura 8 Gel de isoelectroenfoque.
La técnica de isoelectroenfoque se considera de alta resolución, debido a que la
proteínas que difieren en sus valores de PI por solamente 0.02 unidades de pH, o
menos, y que parecen homogéneas al ser corrida electroforéticamente por otros
métodos, al ser probadas por esta técnica pueden ser separadas a menudo en
varios componentes. Tal microheterogeneidad puede ser indicativa de diferencias

43
en la estructura primaria, isómeros conformacionales, diferencias en el tipo y
número de grupos prostéticos o desnaturalización (Garfin, 1990).
La lectina de C.giraffa, mostró homogeneidad, y no presentó isoformas
moleculares, asimismo se observó una estabilidad conformacional en las
condiciones bajo las que se asiló que permitieron que no se desnaturalizara. Con
este método se confirmó que se trata de una proteína monomérica con punto
isoeléctrico de 6.36. Las lectinas de las variedades de C. fragile citadas (op.cit.),
poseen un rango de PI de 3.8 a 3.9, mientras que la tomentina aislada de C.
tomentossum (op. cit.) con PI de 4.0 y 4.64 son considerada proteínas ácidas,
mientras que la giraffma está dentro del rango de las proteínas neutras.
Es importante resaltar que la mayoría de las lectinas aisladas, tanto de
Leguminosas como de macroalgas marinas, poseen isoformas moleculares,
resulta interesante que la giraffina es una proteína única que tiene una alta
actividad específica, a deiferencia de la tomentina de C. tomentossurn que posee
9 isoformas, dos de ellas pudieron ser separadas y fueron denominadas TOM I y
TOM I I . Cuando se probó su actividad, TOM II mostro menores títulos de
aglutinación que TOM I (Fábregas et al., 1988b). Debe considerarse también que
la actividad de las lectinas con isoformas, se debe a la sinergia de las isoformas
en conjunto, debido a que es difícil separalas por sus puntos isoeléctricos tan
cercanos. La importancia de que la giraffina sea una proteína única permite que se
aun más fácil su aislamiento y purificación.

44
V. g Eficiencia de aislamiento
La eficiencia en el aislamiento de la lectina de Codiurn giraffa por medio de
cromatografía de afinidad, duplicó la obtenida usando HPLC de intercambio iónico,
lo cual se evidencia en la tabla que se presenta, se pudo recuperar mayor
cantidad de proteína por el primer método, lo que se reflejó en una mayor
actividad específica. Recomiendo este método para aislar lectinas algales (Tabla
8.)
Tabla 8. Rendimiento en la Purificación de la lectina de Codiurn giraffa (giraffine)
V. i Variabilidad específica por medio de la aglutinacibn con eritrocitos
En la tabla 8 se observan las medias de los resultados de las pruebas de
aglutinación de eritrocitos de las especies del genero Oreochrornis probadas
contra la lectina del alga Codiurn giraffa. Los resultados se muestran en dos tipos
diferentes de unidades: como la media de tres repeticiones de la dilución que
aglutinó positivamente a los eritrocitos de los peces y como la media del recíproco

45
de la más alta dilución triple que los aglutinó. No se presentaron diferencias en los
títulos de aglutinación en ninguna de las 3 repeticiones que se efectuaron por lo
cual solamente se presenta la aglutinación media.
Tabla 9. Medias de los resultados de las pruebas de aglutinacibn con eritrocitos de tilapias. Los resultados se presentan en forma de titulo (izquierda) y en la dilucibn correspondiente (derecha).
1
2’ 512 212 4096 2’ 512 3
2* 256 2” 2048 2’ 512 2
2’ 256 212 4096 2’ 512
4 2’ 512 212 4096 2’ 512
5 2’ 512 2” 2048 2” 1024
6 2’ 512 212 4096 2” 1024
7 28 256 212 4096 2’ 512
I 8 I 2’ 512 1 2‘2 4096 I 2’ 256
I 9 I 2’ 512 I 212 4096 I 28 256
I lo 1 2’ 512 1 Z1* 4096 1 28 256

46
Con estos resultados se procedió a realizar el análisis de homogeneidad de
varianzas, se escogió la prueba de Bartlet, por ser la más confiable según Sokal y
Rohlf (1969). Se planteó la siguiente prueba de hipótesis:
Ho : x2 = x2 los datos poseen distribución normal
Ha : x* z x2 los datos no poseen distribución normal
El resultado del estadístico ajustado de Bartlett X' = 0.86713 se comparó con el
valor obtenido de tablas de x* o.5[9~ = 16.919, quedando el valor en la región de
aceptación, por tanto se aceptó la hipótesis nula y se concluyó que los datos
poseen distribución normal. Esta prueba permitió decidir si los datos eran
susceptibles de un análisis estadística paramétrico o si se tenía la necesidad de
transformarlos para realizar este tipo de análisis.
Obtenidos los resultados de la prueba de Bartlett, se procedió con el análisis de
varianza (ANOVA) para una vía, las medias de aglutinación representadas por la
dilución que aglutinó a los eritrocitos mostró diferencias significativas (p = 0.000).
En la siguiente figura se muestra una gráfica de agrupación de medias, obtenidas
con la prueba de Newman donde se destacan los grupos formados con medias
similares y los que resultaron diferentes. Gráficamente se observa que el híbrido
Rocky mountain se separó totalmente de las variedades de O. niloficus (roja y
Stirling).

47
4500
4000
3500
3000
2500 :o 2 2000
1500
1 O00
o .- n
500
O
Figura 9. Grafica del promedio de aglutinación de cada grupo F(2,18)=117.75; p<.OOOO
J O. niloticus O. niloticus x O. aureus O. niloticus
(variedad roja) (Stirling) Media de la aglutinación
En la prueba de Newman se reportan diferencias significativas entre O. niloticus
(variedad roja) y Rocky mountain, así como, entre esta última y O. niloticus
(Stirling)
La lectina giraffina aglutinó diferencialmente a los eritrocitos de las especies de
Oreochromis, al igual que lo reportó Fábregas et al. (1 992) donde encontró
diferencialidad en los títulos de aglutinación de los eritrocitos de las 16 especies
de peces marinos probados contra extractos de 70 macroalgas, en este trabajo se
destacó la actividad de C. tomentossurn, debido a que su lectina fue capaz de

48
aglutinar los glóbulos rojos de 13 de las 16 especies probadas con diferencias de
hasta dos unidades de título, la giraffina mostró este mismo comportamiento.
Afirmo, al igual que ellos lo hicieron apoyando la hipótesis de Sinderman (1 962),
que estás moléculas pueden formar parte de un amplio espectro de herramientas
serológicas aplicables en la investigación de pesquerías con el propósito de
identificar subpoblaciones o razas de peces.
Los resultados de aglutinación diferencial permitieron separar evidentemente, al
híbrido Rocky mountain de las variedades de Oreochromis aureus, aunque la
separación debida al título de aglutinación que mostraron las variedades no pudo
separarlas adecuadamente, esto puede significar que estas variedades están
relacionadas estrechamente en cuanto a sus características genéticas que les
permita compartir similares sitios antigénicos por los cual no se separaron con se
había previsto.
Tong y Wu (1993) dicen que el uso de los sueros heteroinmunes y las lectinas ha
permitido evidenciar la presencia de grupos sanguíneos en los peces. Sin
embargo en algunos casos estos reactivos son incapaces de reaccionan con la
sangre de estos vertebrados, por su parte, lngram en 1985 y posteriormente
Muñoz et al (1987) dicen que las diferencias en el contenido aglutinante de las
algas puede deberse a los estadios de maduración de los órganos reproductivos o
a las diferentes fases del ciclo vital del alga. En los resultados expuestos, la lectina
del alga C. giraffa fue capaz de aglutinar a los eritrocitos en el 100 % de los casos,

49
lo cual al convierte en una molécula con propiedades de aglutinación constantes y
le permite ser un reactivo idóneo para la investigación de los tipos sanguíneos
presentes en las tilapias, la detección de las modificaciones de los sitios
antigénicos debida al estado de madurez o condiciones del ambiente en el que se
desarrollan y fundamentalmente en la agrupación de organismos que compartan
características similares en la respuesta de la sangre frente a la lectina.
Aún persiste la duda sobre la posibilidad de separar a las variedades de O.
niloticus sometidas al examen antigénico, seguramente por la estrecha relación
que pudiera existir entre los progenitores de los organismos presentes
actualmente y de la progenie en sí misma, es decir, cabe la posibilidad de que los
peces de las variedades de 0.niloticus compartan una carga genética mayor entre
ellos que con el híbrido Rocky mountain.
Es también indispensable realizar esta experiencia con grandes poblaciones de
peces sometidos a cultivo en los Centro Acuícolas de nuestro país con el propósito
de validar aún más los resultados obtenidos y poder así generar conclusiones más
robustas sobre la utilidad de la prueba
Si los resultados se corroboran con altas densidades de peces, la prueba
demostraría ser fácil de realizar, eficiente y de bajo costo. Lo que puede resaltarse
de la misma es que al efectuarla no hay mortalidad, en comparación con otros
métodos en los que necesariamente se debe sacrificar al animal, perdiendo de

50
esta manera, el seguimiento del organismo en cuestión y lo más importante, de su
progenie.
VI. CONCLUSIONES
1. Los métodos cromatográficos utilizados en el aislamiento y purificación de la
lectina del alga Codium giraffa (giraffma) reportaron un elevado índice de
eficiencia lo que se reflejó en el aumento de la actividad específica.
2. Fue fácil purificar la lectina girafina, solamente se requirieron 3 pasos, una
corrida en cromatografía de afinidad y dos eorridas cromatográficas de alta
resolución de intercambio iónico.
3. El porcentaje de rendimiento de purificación (7.16) puede considerarse
incipiente, debido principalmente a que la lectina no es una molécula esencial
para el alga.
4. Se sugiere el uso de estos dos métodos como idóneos para aislar lectinas de
algas de la División Chlorophyta.
5. La giraffma mostró diferencias importantes con respecto a las lectinas aisladas
en especies del mismo género en Inglaterra y España y con especies de la
misma División Chlorophyta.

5 1
6. El peso molecular es inferior a la mitad del peso reportado en la lectinas
aisladas de especies del mismo género.
la giraffina son diméricas y tetraméricas.
8. Su punto isoeléctrico se encuentra en el rango de la neutralidad, mientras que
las lectinas del mismo grupo taxonómico poseen características ácidas.
9. Los datos obtenidos en las pruebas de aglutinación con eritrocitos de Cíclidos
permitieron separar confiablemente dos grupos fundamentales: el híbrido
Rocky mountain pudo ser diferenciado fácilmente de las variedades de
Oreochromis niloticus a las que se les realizó la prueba.
I O . Queda aún la incertidumbre si las variedades de Oreochromis aureus pueden
ser separadas de otras especies del género Tilapia.
1 1 . Las pruebas de aglutinación con la lectina giraffina probó ser fácil de realizar,
aunque se recomienda validar los resultados obtenidos realizando pruebas con
muestras mayores y con otras especies de la Familia Cichlidae.

52
VIL ANEXO I
Las Tilapias en México
Morales-Diaz en 1974 reporta que el 10 de junio de 1964 se introdujeron a México
varias especies de Tilapias procedentes de Estados Unidos de Norteamérica desde
la Universidad de Auburn, Alabama, las que fueron recibidas en el Centro Acuícola
de Temascal, Oaxaca. Estas especies fueron: Tilapia melanopleura, Tilapia nilotica
y Tilapia mossambica. Posteriormente, Arredondo-Figueroa y Guzmbn en 1986,
con los organismos obtenidos de un muestre0 realizado en 1976, aclaran la
composición específica de los organismos presentes en la Presa Miguel Alemán en
el estado de Oaxaca, llegando a la conclusión que los organismos introducidos
pertenecían a las especies Tilapia melanopleura, Oreochromis mossambicus y
Oreochromis niloticus; para el caso de Tilapia niloticus, se hace la corrección de
que el organismo iniciador de la población en realidad se trató de Oreochromis
aureus.
Los primeros ejemplares de O. niloficus fueron importados desde Panamá en 1978
y se llevaron al Centro de Acuicultura de Tezontepec de Aldama en Hidalgo,
México desde donde se trasladaron a Temascal, Oaxaca (Arredondo et al., 1994).
Es en el aiio de 1981 que la Secretaría de Pesca importa desde Palmeto, Florida
E.U.A., dos especies de ciclidos con el propósito de obtención de híbridos 100 YO
machos; Oreochromis urolepis homorum y Oreochromis mossambicus (de la linea
roja), estas especies fueron recibidas en el Centro Acuícola del Rodeo en Morelos,
México desde donde fueron distribuidas a todo el país (Arredondo et al., 1994).

53
En el mes de julio de 1986 se recibe por donación que realizó la Universidad de
Stirling, Escocia, nuevos ejemplares de Oreochromis niloticus de color rojo que
fueron depositados en el CINVESTAV, en la unidad Mérida en Yucatán. La
Secretaría de Pesca fue la encargada de distribuir estos ejemplares a los centros
acuícolas del país tales como Temascal, Oaxaca; Varejonal, Sinaloa y Zacatepec,
Morelos (Arredondo et al., 1994).
En 1987 se recibe la donación del Gobierno de Costa Rica de un lote de peces con
15 individuos de cada una de las siguientes especies: Oreochromis mossambicus.
Oreochromis urolepis homorum y Tilapia rendalli, lote que incluía 15 híbridos de la
cruza de las dos últimas especies depositándolas en Temascal, Oaxaca
(Arredondo et al., 1994).
Debe destacarse también que han habido introducciones de diversas especies y
variedades de estos organismos realizadas por instituciones oficiales y privadas sin
un registro adecuado se supone que los sitios de donde se han importado especies
de Cíclidos podrían ser paises como Cuba, Puerto Rico o Estados Unidos de
Norteamérica. Solamente se tiene la seguridad de las especies que han sido
traídas desde Egipto, Panamá y Costa Rica e introducidas a los Centros Acuícolas
que maneja la Secretaria de Pesca (Arredondo et. al., 1994).
La Tilapia ha encontrado condiciones ecológicas idheas para su adaptación en el
territorio mexicano, principalmente en los estados de Oaxaca, Tabasco, Chiapas,
Michoacán, Veracruz y Sinaloa (Fig. IO), reproduciéndose con éxito, lo cual se

54
refleja en las capturas anuales que registra la Secretaría de Pesca. En estos
estados se registran capturas anuales de 79,154 toneladas de biomasa
aproximadamente (Anónimo, 1996), lo cual permite un desarrollo importante de
fuentes de trabajo y una fuente de proteína animal que es útil para las poblaciones
propias de la región donde se explotan y también una posibilidad de abasto en la
República Mexicana.
Morales-Díat (1 991) propone una clave para identificar a las especies introducidas
en México. Se ilustran las especies y sus variedades con fotografías y dibujos.

55
1. De 1 O a 28 branquiespinas en el arco inferior del primer arco branquial, 30 a 31 escamas en una serie longitudinal, incubadoras orales ............................. ..(2)
De 7 a 16 branquiespinas en el arco inferior del primer arco branquial, desovan en el substrato .............................................................................................. (4)
2. De 13 a 19 branquiespinas en el arco inferior del primer arco branquial, D.XV/XVII, 10-12; A,III, 9-10; de 29 a 32 escamas en una serie longitudinal ....... .............................................. Oreochromis mossambicus Peters, (Figs. 1 1 y 12).
Figura 1 l. Oreochromis mossambicus
"
Figura 12. Oreochromis mossambicus (variedad roja)

56
De 18 a 22 branquiespinas en el arco inferior del primer arco branquial, D.XV.,
12-15; A.111, de 3 a 5 hileras de dientes delgados, de 30 a 33 escamas en una
serie longitudinal ..................... O~ochromis aureus Steidachner, (Figs. 13 y 14).
Figura 13. Oreochmmis aureus
Figura 14. Oreochromis aueus (variedad roja)
2, De 19 a 22 branquiespinas en el arco inferior del primer arco branquial; D.XVI,
XVII, 12-13; AM, 10-1 1; de 2 a 4 hileras de dientes robustos y gruesos, de 31 a
33 escamas en una serie longitudinal .................................................................
............................................................ Oreochmmis niloticus Linnaeus, (Fig. 15).

57
Figura 15. Oreochromis niloticus
De 19 a 27 branquiespinas en el arco inferior del primer arco branquial,
D.XIII/XVIII, 10-13; A-Ill, 9-1 1 de 29 a 30 vértebras .........................................
.............................................................. Oreochmmis urolepis homorum (Fig. 16).
Figura 16. Oreochromis urolepis homorum
3. De 8 a 12 branquiespinas en el arco inferior del primer arco branquial.
D.XV/XVI, 12-13; A.III, 9-1 I; de 29 a 32 escamas en una serie longitudinal ........

................... I ..
58
Tilapia melanopleura Dumeril (= Tilapia rendalli Boulenger) (Fig. 17)
Figura 17. Tilapia rendalli

59
VIL LITERATURA CITADA
ANONIMO, 1996. Anuario Estadístico de Pesca. SEMARNAP. (Fuente: Internet, httD://w.semarnap.gob.mx).
ARREDONDO-FIGUEROA J.L. y GUZMAN A.M., 1986. Actual situación taxon6mica de las especies de la tribu Tilapiini (Pisces:Cichilidae) introducidas en México. An. Inst. Biol. UNAM, Ser. Zool., 56(2): 555-572.
ARREDONDO-FIGUEROA J.L., FLORES-MUÑOZ V.F., GONZALEZ-TOVAR F., GARDUÑO-ARGUETA H. y CAMPOS-VERDUZCO R., 1994. Desarrollo Científico y Tecnolbgico del Banco de Genoma de Tilapia. Convenio SEPESCAIUAM-I. México, 89 p.
BLUNDEN G.D., ROGERS J. y FRANHAM W.F., 1975. Survey of British seaweeds for haemagglutinins. Lloydia, 38: 162-1 68.
BOLWELL G.P., CALLOW J.A., CALLOW M.E. y EVANS L.V., 1977 Cross- fertilization in fucoid seaweeds. Nature (London), 268: 626-627.
BOLWELL G.P., CALLOW J.A., CALLOW M.E. y EVANS L.V., 1979 Fertilization in brown algae. I I . Evidence of lectin-sensitive complementary receptors involved in gamete recognition in Fucus serratus. J. Cell, Sci., 36: 19-30.
BOLWELL G.P., CALLOW J.A. y EVANS L.V., 1980 Fertilization in brown algae. 111. Preliminary characterization of putative gamete receptors from eggs and sperm of Fucus serratus. J. Cell. Sci., 43: 209-224.
BOYD W.C. y REGUERA R.M., 1945. Haemagglutinating substances for human cells in various plants. J. Immunol., 62: 333-339.
BOYD W.C., ALMODOVAR L.R. y BOYD L.C., 1966. Agglutinins in marine algae for human erythrocytes. Transfusion, 66: 82-83.
CHILES T.C. y BIRD KT., 1990. Gracilaria tikvahiae agglutinin. Partial purification and preliminary characterization of its carbohydrate specificity. Carbohyd. Res., 207: 319-326.
CRIADO M.T. y FERREIROS C.M., 1983a. lmmunomodulatory effect produced in mice by complex-carbohydrate specific lectin-like mucopolisaccharide from Fucus vesiculosus. IRCS Med. Sci., 11: 286-287.
CRIADO M.T. y FERREIROS C.M., 1983b. Selective interaction of a Fucus vesiculosus lectin-like mucolopysaccharide with several Candida species. Ann. Microbiol. (Inst. Pasteur) 134A: 149-154.

60
FABREGAS J., MUÑOZ A., LLOVO J. y ABALDE J., 1984. Agglutinins in marine red algae. ICRS Med. Sci., 12: 298-299.
FABREGAS J., LLOVO J. y MUÑOZ A., 1985. Hernagglutinins in red seaweeds. Bot. Mar.. 28: 517-520.
FABREGAS J., LOPEZ A., LLOVO J. y MUÑOZ A., 1992. A comparative study of seafish erythrocytes and agglutinins from seaweeds. Comp. Biochem. Physiol. 103a: 307-31 3.
FABREGAS J., MUÑOZ A. y LLOVO J., 1986. Hemagglutinins in brown seaweeds. J. Exp. Mar. Biol. Ecol., 97: 21 3-21 9.
FABREGAS J., MUÑOZ A., LLOVO J. y CARRACEDO A., 1988a. Tomentine: a lectin for the detection of a glycoprotein polimorphysms. Med. Sci. Res., 16: 819-820.
FABREGAS J., MUÑOZ A., LLOVO J. y CARRACEDO A., 1988b. Purification and partial characterization of tomentine. An N-acetylglucosamine-specific lectin from the green alga Codiurn tomentosum (Huds,) Stackh. J. Exp. Mar. Biol. Ecol., 124: 21-30.
FABREGAS J., LLOVO J. y MUÑOZ A., 1988c. Agglutination activity of algal extracts against spermatozoa of the fish Diplofus sargus L. Nippon Suisan Gakkaishi, 54: 2121-2126.
FERREIROS C.H. y CRIADO M.T., 1983. Purification and partial characterization of a Fucus vesiculosus agglutinin. Rev. Esp. Fisíol., 39: 51-60.
GARFIN D.E., 1990. One dimesional gel electrophoresis. En: Deutscher M.P. (Ed.) Methods in Enzymology Vol. 182. Guide to protein purification. Academic Press, Inc., New York, USA, pp. 425-441.
GUTIERREZ M., 1967a. Estudios hematológicos en el atún Thunnus thynnus (L.) de la Costa Sudatlántica de España. Inves. Pesq., 31: 53-90.
GUTIERREZ M., 1967b. Estudios hematológicos del atún Thunnus thynnus (L.) de la Costa Sudatlántica de España. Inves. Pesq., 31: 137-143.
HORl K., IKEGAMI S., MIYAZAWA K. e IT0 K., 1988a. Mitogenic and antineoplastic isoagglutinins from the red alga Soliera robusta. Phytochemistry 27: 2063-2067.
HORl K., MIYAZAWA K. e IT0 K., 1981. Hemagglutinins in marine algae. Bull. Japan. Soc. Sci. Fish., 47: 793-798.

225721 61
HORl K., MIYAZAWA K. e IT0 K., 1986a. Isolation and characterization of glycoconjugate-specific isoagglutinins from marine green alga Boodlea coacta (Dickie) Murray et De Toni. Bot. Mar., 29: 323-328.
HORl K., MIYAZAWA K., FUSETANI N., HASHIMOTO K. e IT0 K., 1986b. Hypnins, low-molecular weight agglutinins isolated from a marine alga, Hypnea japonica. Biochim. Biophys. Acta., 873: 228-236.
HORl K., MATSUDA H., MIYAZAWA K. e IT0 K., 1987. A Mitogenic agglutinin from the red alga Carpopeltis flabellata. Phytochemistry, 26: 1335-1 338.
HORl K., OlWA C., MIYAZAWA K. e IT0 K., 1988b. Evidence of wide distribution of agglutinins in marine algae. Bot. Mar., 31: 133-138.
KAMIYA H., OGATA K. y HORl K., 1982. Isolation and characterization of a new agglutinin in the red alga Palmaria palmafa (L.) O. Kuntze. Bot. Mar., 25: 537-540.
INGRAM g. A., 1985. Lectins and lectin.like molecules in lower plants. I. Marine alge (review). Devel. Comp. Immunol., 9: 1-1 1.
KAMIYA H., SHlOMl K. y SHlMlZU Y., 1980. Marine biopolymers with cell especificity. 111. Agglutinins in the red alga Cystoclonium purpureum: isolation and characterization. J. Nat. Prod., 43: 136-139.
KUHNS W.J. y CHUBA J., 1968. lntrageneric blood group differences between lctalurids (fresh water catfishes). Fed. Proc., 27: 491.
LIS H. y SHARON N., 1981. Lectins in higher plants. En: Marcus A. (Ed.) The Biochemistry of plants. A comprehensive treatise Vol. 6. Academic Press, New York, E.U.A., pp, 372-425.
LLOVO J., MUÑOZ A., ROMARIS M. y FABREGAS J., 1987. Obtención de sangre de peces mediante punción cardíaca. Cuad. Marisq. Publ. Téc., 12: 261- 266.
MORALES-DIM A., 1974. El cultivo de la Tilapia en México. Datos Biológicos. Instituto Nacional de Pesca. INP., 25 p.
MORALES-DIM A., 1991. La Tilapia en México. Biología, cultivo y pesquerias. AGT editores, S.A. México, 190 p.
MUÑOZ A., LLOVO J. y FABREGAS J., 1985. Hemaglutininas de algas verdes. ACCCAW, 22: 873-878.

62
MUÑOZ A., LLOVO J. y FABREGAS J., 1987a. Different agglutinic activity of red marine algae against erythrocytes from several animal species. Thalassas, 5: 87-89.
MUROZ A., LLOVO J., ROMARIS M. y FABREGAS J., 1987b. Utilización de Algas Marinas como un nuevo criterio de diferenciación de las libreas de Labrus bergylta Ascanius, Cuad. Mariq. Publ. Tec., 12: 251-256.
MUÑOZ A., LLOVO J., ROMARIS M. y FABREGAS J., 1987c. Presencia de receptores para aglutininas algales sobre la superficie de eritrocitos de peces. Cuad. Marisq. Publ. Tec., 12: 245-250.
NOWAK T.P. y BARONDES S.H., 1975. Agglutinin from Limulus polyphemus. Purification with formalinized adsorbent. Biochim. Biophys. Acta, 393: 1 15-1 23.
RENKONEN K.O., 1948. Studies on hemagglutinins present in seeds of some representatives of the family of Leguminosae Ann. Med. Exp. Biol. Fenn., 26: 66.
RIDWAY G.J., 1969. Blood groups in salmonid fishes. En: Fish in Research. Neuhaus O.W. y Halver J.E. (Eds.). Academic Press. New York, EUA, pp. 185-1 94.
ROGERS D.J., 1977a. Antibody-like substances in marine organisms. En: Faulkner, D.J. y Fenical, W.H. (Eds.). Marine Natural Products Chemistry. Plenum Press, New York, pp 31 1-327
ROGERS D.J., BLUNDEN G. y EVANS P.R., 1977b. Ptilota plumosa, a new source of a blood-group B specific lectin. Med. Lab. Sci., 34: 193-200.
ROGERS D.J., TOPLISS J.A. Y GUIRY M.D., 1980. A survey of some marine organisms for haemagglutinins. Bot. Mar., 23: 569-577.
ROGERS D.J. y BLUNDEN G., 1980. Structural properties of the Anti-B lectin from the red alga Ptjlofa plumosa (Huds.)C. Ag. Bot. Mar., 23: 459-462.
ROGERS D. J. y TOPLISS J.A., 1983. Purification and characterisation of an Anti- sialic acid agglutinin from the red alga Soliera Chordalis (C. Ag.) J. Ag. Bot. Mar., 24: 301-305.

63
ROGERS D. J., FISH B.C. y BARWELL C.J., 1990. Isolation and properties of lectins from two red marine algae: Plumaria elegans and Ptilota serrata. En: Kocourek, J. y D.L.J. Freed (Eds.). Lectíns: Biology, Biochemistry, Clinical Biochemistry. Sigma Chemical Company, St. Luois MO, USA. Vol. 7, pp 49-52.
ROGERS D.J., LOVELESS R.W. y BALDING P., 1986. Isolation and characterization of the lectins from subspecies of Codium fragile. En: Bag- Hansen, T.C. y Van Driessche, E. (Eds.). Lectins: Biology, Biochemistry, Clinical Biochemistry. Walter de Gruyter, Berlin. Vol. 5, PP. 155-1 60.
ROGERS D.J., TOPLISS J.A. y BLUNDEN G., 1979. Reactions of Anti-B lectins with erythrocytes sesitized with lipopolysaccharide from Escherichia coli. Med. Lab. Sci., 36: 79-84.
SANDERS B.G. y WRIGHT J.E., 1962. Immunogenetic studies in two trout epecies of the genus Salmo. Ann. N. Y. Acad. Sci., 97: 116-130.
SHARON N. y LIS H., 1972. Lectins: Cell agglutinanting and sugar-specific proteins, Science, 177: 949-958.
SHARON N. y LIS H., 1981. Lectins in higer plants. Cap. 10. En: Marcus, A. (Ed.) The Biochemistry of Plants. Academic Press. Londres, 371-447.
SHlOMl K., KAMIYA H. y SHlMlZU Y., 1979. Purification and characterization of an agglutinin in the red alga Agardhiella tenera. Biochim. Biophys. Acta. 576: 1 18-1 27.
SHlOMl K., YAMANAKA H. y KlKUCHl T., 1980. Biochemical properties of hemagglutinins in the red alga Serraticardia maxima. Bull. Japan. SOC. SCi. Fish., 46: 1369-1373.
SHlOMl K., YAMANAKA H. y KlKUCHl T., 1981, Purification and physicochemical properties of a hemagglutinin (GVA-1) in the red alga Gracilaria verucosa. Bull. Japan. Soc. Sci. Fish., 47: 1077-1084.
SINDERMAN C., 1962. Use of plant hemagglutinins in serological studies of clupeoids fishes. Fish. Bull. 63: 137-141.
SOKAL R.R. y ROHLF F.J., 1969. Biometry. The principles and practice of statistics in biological research. W.H. Freeman & Co., San Francisco, USA, 776 p.
STEEL R.D. y TORRIE J.H., 1988. Bíoestadística: principios y procedimientos, segunda edición. McGrw-Hill/lnteramericana, México, 622 p.

64
STOSCHECK C.M., 1990. Quantitation of proteins. En: Deutscher M.P. (Ed.) Guide to protein purification Vol. 182. Academic Press, London, U.K., PP. 50-67.
TEJEDA S.M., 1987. Contribución al conocimiento de la sistemática de las especies de la tribu Tilapiini (Pisces:Cochlidae), presentes en México. Tesis de Licenciatura, ENEP-lztacala, UNAM, 75 p.
TONG J. y C. WU, 1993. Study on the boold type factors in red crucian carp (Carassius auratur var). Aquacul., 11 I: 129-1 38.
UTTER F.M., RIDWAY G.J. y HODGINNS H.O., 1964. Use of plant extracts in serological studies of fish. U. S. Fish and Wildlife Service, Special Scientific report, Fisheries No. 472.
VANN D.C. y CUSHlNG D.E., 1966. Reactions of the lectin from Dolichos bifloms with wrythrocytes from California bonito, Sarda chilensis. Fed. Proc., 25: 437.

El jurado designado por la División de Ciencias Biológicas y de la Salud de la Universidad Authoma Metropolitana, Unidad Iztapalapa, I
aprobó la presente tesis el día 1 I del mes de diciembre de 1997.