universidad de la repÚbilica facultad de agronomía
TRANSCRIPT

UNIVERSIDAD DE LA REPÚBILICA FACULTAD DE AGRONOMíA
CARACTERIZACIÓN DE LANAS FINAS PRODUCIDAS EN URUGUAY
por
María Magdalena PLATERO CLAVIER
TESIS presentada como uno de los requisitos para obtener el
título de Ingeniero Agrónomo
MONTEVIDEO URUGUAY
2013

II
Tesis aprobada por:
Director: __________________________________________________
Ing. Agr. Ignacio Abella
__________________________________________________
Ing. Agr. Jorge Urioste
__________________________________________________
Ing. Agr. José Rodríguez
Fecha: 10 de febrero de 2013
Autor: __________________________________________________
María Magdalena Platero Clavier

III
AGRADECIMIENTOS
Agradezco a todas las personas que me acompañaron a lo largo de la carrera. A mis tutores: Ignacio, Francisco y Jorge; al personal del laboratorio de lanas del SUL. A mis compañeros de trabajo, amigos y familia que supieron darme apoyo durante estos años; especialmente a mis padres y hermanos.

IV
TABLA DE CONTENIDO
Página PÁGINA DE APROBACIÓN……………………………………………. II AGRADECIMIENTOS…………………………………………………… III LISTA DE CUADROS E ILUSTRACIONES.…………………………. VI ABREVIATURAS………………………………………………………… IX 1. INTRODUCCIÓN……………………………………………………… 1 2. REVISIÓN BIBLIOGRÁFICA………………………………………… 3 2.1 CONTEXTO DE LA PRODUCCIÓN MUNDIAL DE LANAS… 3 2.1.1 Caracterización de la producción ovina en Uruguay…… 7 2.2 PRODUCCIÓN DE LANA Y FACTORES QUE LA AFECTAN. 9 2.2.1 Factores internos…………………………………………… 10 2.2.1.1 Sexo………………………………………………… 10 2.2.1.2 Edad………………………………………………… 10 2.2.1.3 Efecto materno……………………………………… 10 2.2.1.4 Estado fisiológico…………………………………… 11 2.2.2 Factores externos…………………………………………… 13 2.2.2.1 Sanidad……………………………………………… 13 2.2.2.2 Fotoperíodo…………………………………………. 13 2.2.2.3 Nutrición……………………………………………… 14 2.3 PRODUCCIÓN ESTACIONAL DE PASTURAS………………… 16 2.3.1 Región Basalto……………………………………………….. 17 2.3.1.1 Producción de forraje en campo natural…………… 17 2.4 RESISTENCIA A LA TRACCIÓN………………………………… 19 2.5 VARIACIÓN DEL DIÁMETRO Y RELACIÓN CON LA RESISTENCIA……………………………………………………... 22 2.5.1 Resistencia a la tracción…………………………………… 26 2.5.1.1 Relación entre diámetro máximo y diámetro mínimo (Dmáx y Dmín) con la resistencia………… 30 2.5.1.2 Relación de tasa de cambio de diámetro con la resistencia………………………………….…………. 31 2.5.1.3 Relación entre extensión de la mecha y resistencia……………………………………..……… 32 2.6 UTILIDAD PRÁCTICA DE PERFILES ………………………….. 32 2.7 HERRAMIENTAS Y MÉTODOS DE MEDIR PERFILES……… 34 3. MATERIALES Y MÉTODOS………………………………………… 36 4. RESULTADOS Y DISCUSIÓN……………………………………… 39 4.1 ANÁLISIS DESCRIPTIVO………………………………………. 39

V
4.2 ANÁLISIS DE VARIANZA………………………………………. 44 4.3 ANÁLISIS STEPWISE…………………………………………… 48 4.4 PERFILES DE DIÁMETRO……………………………………… 50 5. CONCLUSIONES……………………………………………………… 54 6. RESUMEN……………………………………………………………… 56 7. SUMMARY……………………………………………………………… 57 8. BIBLIOGRAFÍA………………………………………………………… 58

VI
LISTA DE CUADROS E ILUSTRACIONES Cuadro No. Página 1. Proporción de la participación en la producción total lana para los años 1990 y 2010…………………………... 4 2. Componentes de variación en diámetro…………………. 23 3. Media, desvío fenotípico (sp) y heredabilidades [h2(s.e)] para los caracteres medios de la muestra y a lo largo de esta…………………………………………… 27 4. Correlación fenotípica entre DMF, CVDMF y RT……….. 28 5. Características del perfil de diámetro de fibras en 2006: valores promedio y desvío estándar……………………... 39 6. Características del perfil de diámetro de fibras en 2007: valores promedio y desvío estándar…………………….. 40 7. Componentes de estimación de la variancia para RT en el primer año del trabajo (2006)...................................... 44 8. Componentes de estimación de la variancia para RT en el segundo año del trabajo (2007) ............................. 44 9. Componentes de estimación de la variancia para RT en el año 2007……………………………………………… 45 10. Resultados análisis stepwise……………………………… 48 Figura No. 1. Evolución de la producción mundial de lana sucia en toneladas (tt)………………………………………………... 3 2. Producción mundial de lana limpia según diámetro……. 5 3. Producción de lana en Australia………………………….. 6 4. Evolución de las exportaciones de lana en Uruguay

VII
según diámetro……………………………………………... 7 5. Distribución estacional de la producción de campo natural sobre Basalto………………………………………. 18
6. Producción anual de forraje (kgMS/ha) de los tres tipos de suelo de Basalto de la unidad Queguay chico, desde 1980 a 1994………………………………………… 18 7. Proporción de premios y descuentos en el valor del producto asociado a LM (mm) en lanas superfinas de Australia……………………………………………………… 20 8. Variación de premios y descuentos en el valor de lanas superfinas de Australia en función a la RT (N/ktex)……. 21 9. Influencia de los distintos componentes de calidad en el precio final de lana Merino en el rango de 16,6 a 19,5 micras en Australia para el período 2001/02 y las zafras 2004-2006…………………………………………… 22 10. Variación del diámetro en el cuerpo del animal…………. 25 11. Esquema de variación del diámetro entre fibras y a lo largo de la fibra……………………………………………... 25 12. Relación fenotípica entre RT y CVDMF en capones…… 29 13. Ubicación de los establecimientos……………………….. 36 14. Distribución de diámetro promedio de fibras (DPF) de animales dentro de cada majada…………………………. 41 15. Distribución de resistencia a la tracción promedio de animales dentro de cada majada…………………………. 42 16. Distribución del CVDMF de animales dentro de cada majada……………………………………………………….. 45 17. Distribución de diferencia entre DMáx y Dmín del perfil dentro de cada majada……………………………… 46

VIII
18. Distribución del diámetro en función a la distancia de la punta agrupado por productor………………………….. 51 19. Distribución del diámetro de los animales de las majadas que más se ajustaron a la media poblacional para el año 2006 y 2007…………………….. 52

IX
ABREVIATURAS SCMAU: Sociedad de criadores de Merino Australiano de Uruguay SUL: Secretariado uruguayo de la lana INIA: Instituto nacional de investigación agropecuaria CLU: Central lanera uruguaya PMF: Proyecto Merino fino AWEX: Australian wool exchange RT: Resistencia a la tracción LM: Largo de mecha DMF: Diámetro promedio/Diámetro medio de fibras ACVDF: Variación del diámetro a lo largo de la fibra CVDMF: Coeficiente de variación del diámetro medio de fibras Dmín: diámetro mínimo DMax: diámetro máximo RANGO/(DMax-Dmín): Diferencia entre el diámetro máximo y el mínimo RF: Resistencia de la fibra PDF: Perfil del diámetro de fibras CV 1-10: Coeficiente de variación del diámetro medido en 10 posiciones a lo largo de la mecha TASA: Diámetro máximo-diámetro mínimo/ distancia que las separa (Dmáx-Dmín/distancia). DF: diámetro de fibra (en lugar puntual) POB: Punto de ruptura de la mecha DS: Desvío estandar FC: Factor de confort (% de fibras menores a 30 micras) Hm: Hauter: altura media del top Noils: fibras cortas que resultan de la obtención del top

1
1. INTRODUCCIÓN
La producción de lanas finas en Uruguay ha incrementado su volumen en los últimos años, debido al desarrollo de diferentes proyectos en el país que impulsaron el incremento de la producción de fibras con 20 micras o menos (proyecto Merino fino y el club de Merino fino).
Este tipo de lanas son empleadas mayoritariamente para la confección de prendas de vestir, por lo que determinados estándares de calidad deben cumplirse. En Australia, principal país productor de este tipo de lanas, la resistencia a la tracción ha sido la segunda característica en importancia, luego del diámetro, al momento de determinar la variación en su precio de venta (Montossi et al., 2007).
En Uruguay la producción ovina se realiza principalmente sobre pasturas naturales en regiones arqueológicas con suelos con limitantes, dándose variaciones en la producción estacional de pasturas (en calidad y cantidad de pastura producida), afectando de esta forma la cantidad de nutrientes suministrados para la producción de la fibra. Diversos trabajos experimentales (Denny 1990, Schlink y Dollin 1995, Yamin et al. 1999, Adams et al. 2000, Brown et al. 2002, Ciappesoni et al. 2008) han mostrado una correlación fenotípica negativa entre la variación del diámetro a lo largo del año y la resistencia, sugiriendo que un aumento en la primera característica trae aparejado una reducción en la resistencia.
En consecuencia, el objetivo general del trabajo fue generar información objetiva acerca de la resistencia a la tracción de lanas finas producidas en Uruguay. Los objetivos específicos fueron:
1- generar información descriptiva de variables generadas a partir del análisis con OFDA, para caracterizar los perfiles de diámetro en estas majadas;
2- examinar la importancia relativa de la variación entre y dentro de majadas para las variables consideradas; y
3- determinar la existencia de asociaciones entre la resistencia a la tracción y las variables de la lana generadas en el análisis con el OFDA.

2
2. REVISIÓN BIBLIOGRÁFICA 2.1 CONTEXTO DE LA PRODUCCIÓN MUNDIAL DE LANAS
En el ámbito mundial y durante las últimas dos décadas se han
presentado cambios constantes en los precios y la producción e industrialización de lana, la cual repercutió negativamente en las economías de los productores ovejeros, particularmente en aquellos países con sistemas de alta especialización en la producción de lana (Australia, Nueva Zelanda, Uruguay, Sudáfrica y Argentina, Montossi et al., 2007).
1700000
1900000
2100000
2300000
2500000
2700000
2900000
3100000
3300000
3500000
1990 1995 2000 2005 2009 2010
Año
tt
Figura No. 1. Evolución de la producción mundial de lana sucia en toneladas (tt). Fuente: IWTO (2011).
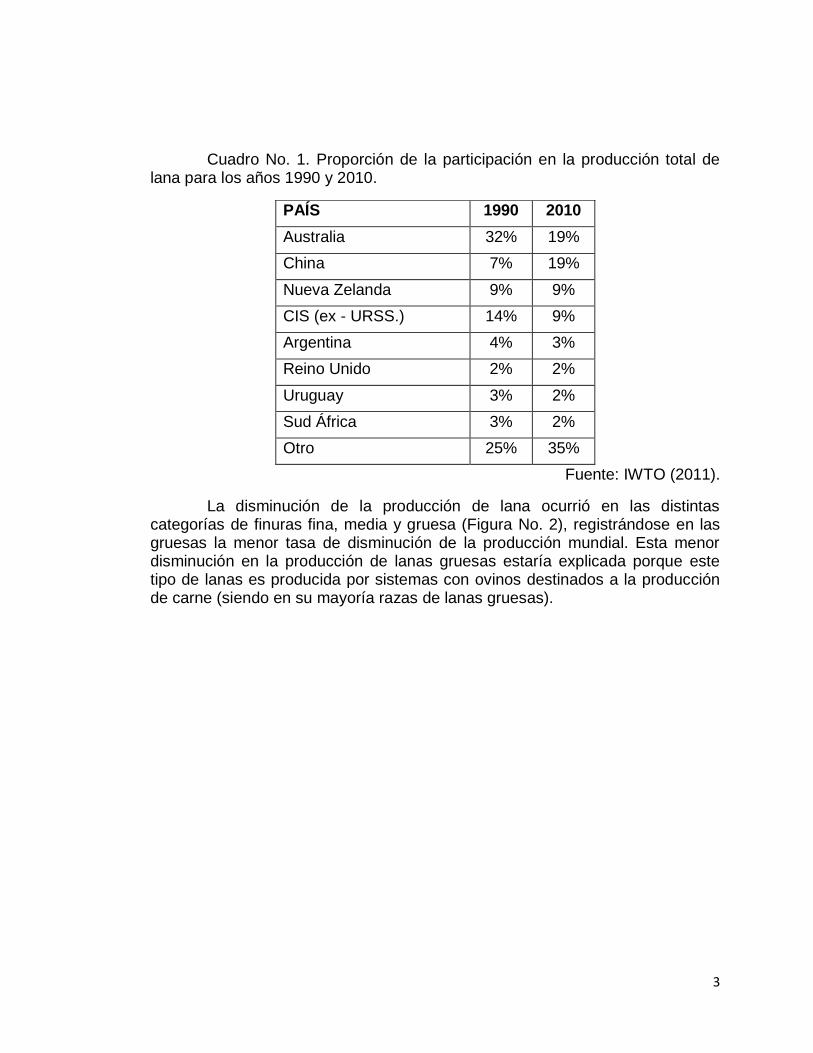
La disminución en la producción de lana (Figura No. 1) ocurrió a nivel de todos los principales países productores de lana y ocurrieron cambios a nivel de la incidencia relativa de ellos (Cuadro No. 1).
3
Cuadro No. 1. Proporción de la participación en la producción total de lana para los años 1990 y 2010.
PAÍS 1990 2010
Australia 32%
19%
China 7% 19%
Nueva Zelanda 9% 9%
CIS (ex - URSS.) 14% 9%
Argentina 4% 3%
Reino Unido 2% 2%
Uruguay 3% 2%
Sud África 3% 2%
Otro 25% 35%
Fuente: IWTO (2011).
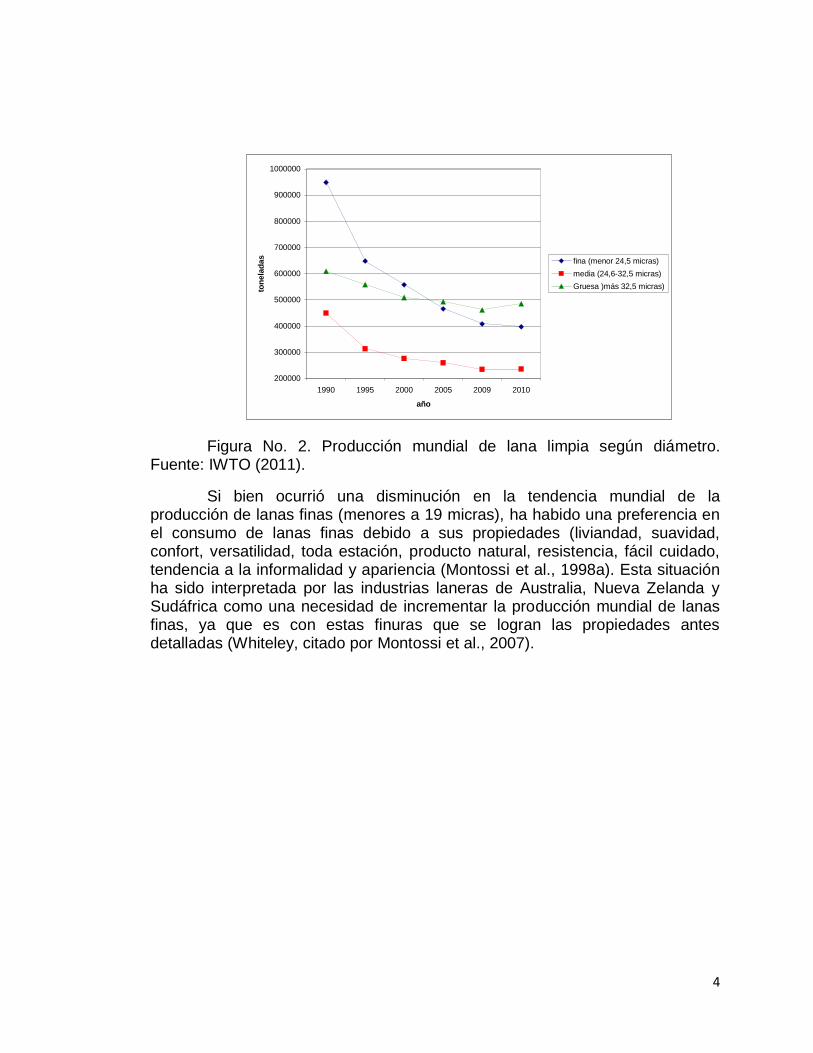
La disminución de la producción de lana ocurrió en las distintas categorías de finuras fina, media y gruesa (Figura No. 2), registrándose en las gruesas la menor tasa de disminución de la producción mundial. Esta menor disminución en la producción de lanas gruesas estaría explicada porque este tipo de lanas es producida por sistemas con ovinos destinados a la producción de carne (siendo en su mayoría razas de lanas gruesas).
4
200000
300000
400000
500000
600000
700000
800000
900000
1000000
1990 1995 2000 2005 2009 2010
año
ton
ela
das
fina (menor 24,5 micras)
media (24,6-32,5 micras)
Gruesa )más 32,5 micras)
Figura No. 2. Producción mundial de lana limpia según diámetro. Fuente: IWTO (2011).
Si bien ocurrió una disminución en la tendencia mundial de la producción de lanas finas (menores a 19 micras), ha habido una preferencia en el consumo de lanas finas debido a sus propiedades (liviandad, suavidad, confort, versatilidad, toda estación, producto natural, resistencia, fácil cuidado, tendencia a la informalidad y apariencia (Montossi et al., 1998a). Esta situación ha sido interpretada por las industrias laneras de Australia, Nueva Zelanda y Sudáfrica como una necesidad de incrementar la producción mundial de lanas finas, ya que es con estas finuras que se logran las propiedades antes detalladas (Whiteley, citado por Montossi et al., 2007).
5
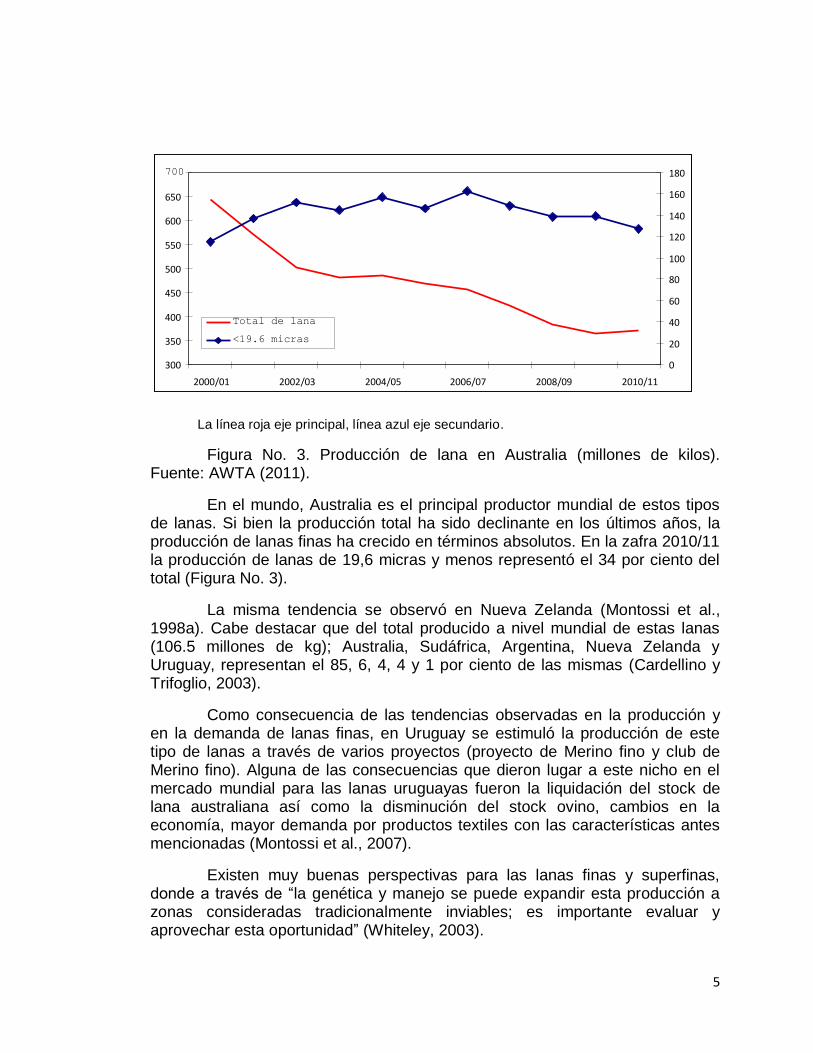
La línea roja eje principal, línea azul eje secundario.
Figura No. 3. Producción de lana en Australia (millones de kilos). Fuente: AWTA (2011).
En el mundo, Australia es el principal productor mundial de estos tipos de lanas. Si bien la producción total ha sido declinante en los últimos años, la producción de lanas finas ha crecido en términos absolutos. En la zafra 2010/11 la producción de lanas de 19,6 micras y menos representó el 34 por ciento del total (Figura No. 3).
La misma tendencia se observó en Nueva Zelanda (Montossi et al., 1998a). Cabe destacar que del total producido a nivel mundial de estas lanas (106.5 millones de kg); Australia, Sudáfrica, Argentina, Nueva Zelanda y Uruguay, representan el 85, 6, 4, 4 y 1 por ciento de las mismas (Cardellino y Trifoglio, 2003).
Como consecuencia de las tendencias observadas en la producción y en la demanda de lanas finas, en Uruguay se estimuló la producción de este tipo de lanas a través de varios proyectos (proyecto de Merino fino y club de Merino fino). Alguna de las consecuencias que dieron lugar a este nicho en el mercado mundial para las lanas uruguayas fueron la liquidación del stock de lana australiana así como la disminución del stock ovino, cambios en la economía, mayor demanda por productos textiles con las características antes mencionadas (Montossi et al., 2007).
Existen muy buenas perspectivas para las lanas finas y superfinas, donde a través de “la genética y manejo se puede expandir esta producción a zonas consideradas tradicionalmente inviables; es importante evaluar y aprovechar esta oportunidad” (Whiteley, 2003).
300
350
400
450
500
550
600
650
700
2000/01 2002/03 2004/05 2006/07 2008/09 2010/11 0
20
40
60
80
100
120
140
160
180
Total de lana
<19.6 micras
6
2.1.1 Caracterización de la producción ovina en Uruguay
Las lanas producidas en Uruguay se caracterizan por tener un alto rendimiento al lavado (superior al 70%) y bajo porcentaje de material vegetal (menor a 0,5%). Esto se debe a que la producción se realiza sobre pasturas naturales, presentando además resistencia a la tracción adecuada, lo que determina mayores rendimientos industriales (SUL, 2012).
0
5
10
15
20
25
<21 21.0-22.0 22,1-23 23,1-24,9 25-26.4 26.5-27.9 28-29.4 29.5-31.4 >31,5 Otros
Volu
men e
xp. (m
kg)
2009
2010
2011
2012
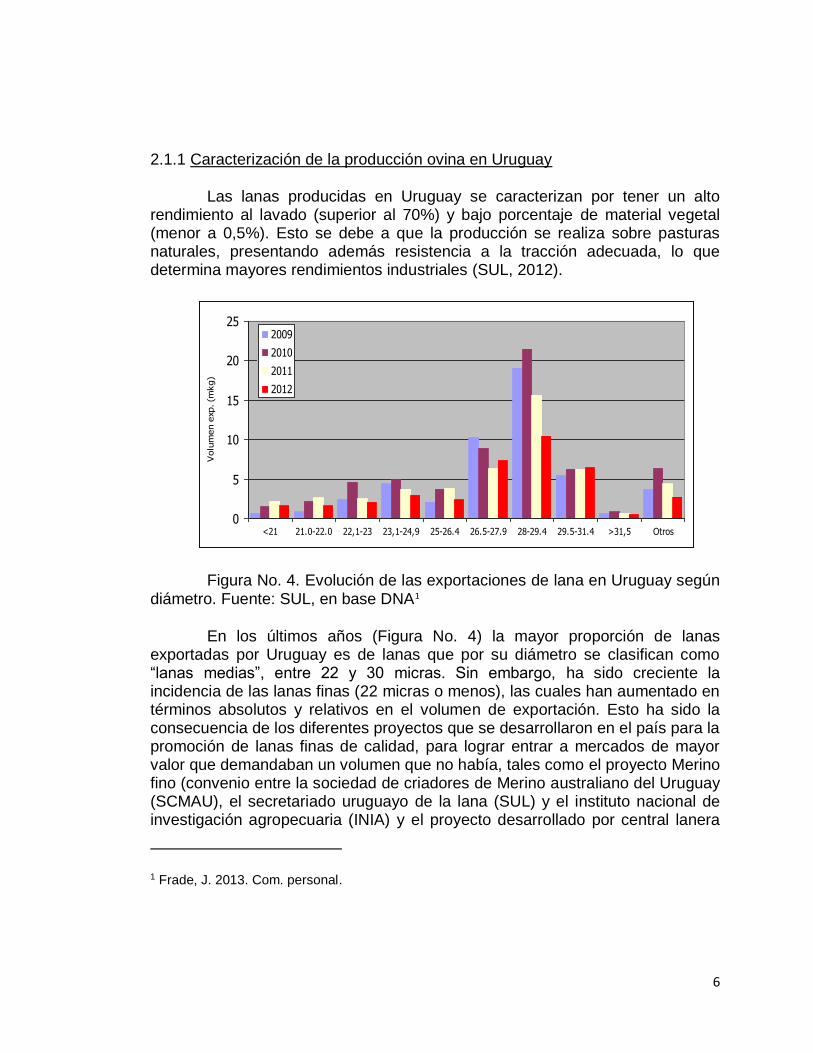
Figura No. 4. Evolución de las exportaciones de lana en Uruguay según diámetro. Fuente: SUL, en base DNA1
En los últimos años (Figura No. 4) la mayor proporción de lanas exportadas por Uruguay es de lanas que por su diámetro se clasifican como “lanas medias”, entre 22 y 30 micras. Sin embargo, ha sido creciente la incidencia de las lanas finas (22 micras o menos), las cuales han aumentado en términos absolutos y relativos en el volumen de exportación. Esto ha sido la consecuencia de los diferentes proyectos que se desarrollaron en el país para la promoción de lanas finas de calidad, para lograr entrar a mercados de mayor valor que demandaban un volumen que no había, tales como el proyecto Merino fino (convenio entre la sociedad de criadores de Merino australiano del Uruguay (SCMAU), el secretariado uruguayo de la lana (SUL) y el instituto nacional de investigación agropecuaria (INIA) y el proyecto desarrollado por central lanera
1 Frade, J. 2013. Com. personal.

7
uruguaya (CLU) denominado el club de Merino fino (Central Lanera Uruguaya, 2010).
El Proyecto Merino fino (PMF) comenzó a desarrollarse en 1998 como consecuencia de la poca información nacional sobre mejoramiento genético, alimentación, sanidad, reproducción y manejo para la producción de lanas Merino finas y superfinas, tratando de buscar soluciones tecnológicas a los problemas de origen productivo y económico que tenían los productores laneros en la región de Basalto (Montossi et al., 2007).
A comienzos del PMF la proporción de la raza Merino dentro del stock nacional era un 10 por ciento (1.8 millones de cabezas), con una producción de 7 millones de kilos de lana en base sucia (Montossi et al., 1998a). URUGUAY. MGAP. DIEA (2002) muestra una proporción dentro del stock nacional de 18,8 por ciento (2.17 millones de cabezas), con una producción cercana a los 8 millones de kilos, siendo un 10 a 14 por ciento del volumen menor a 19,5 micras.
Además de cambios en el volumen de la fibra producida, también se dieron cambios en el diámetro. Según Montossi et al. (2007), la finura de la población Merino de Uruguay en 1998 era de 21,8 micras de promedio, teniendo alto rendimiento al lavado, aceptable largo y resistencia a la tracción, siendo el color y brillo de la fibra insatisfactoria. Aguerre y Coronel (2008) expusieron que la finura promedio para Uruguay era de 18.3 micras (en la base de datos de registros de producción flock-testing) para el período 2004-2006, disminuyendo 3.5 micras en un período de siete años. Esta lana corresponde al análisis de lana de borregos, que es el primer vellón producido por el animal en su vida y es el de menor diámetro.
El objetivo del proyecto consistió en desarrollar una alternativa de producción ovina que por medio de su difusión y posterior adopción permitiera mejorar la sustentabilidad socioeconómica de los productores de lana en las regiones de Basalto y Cristalino, considerando las demandas actuales y futuras de la cadena agroindustrial de lana del país y de los mercados consumidores.
Luego de observar los resultados de varios años de trabajo, una empresa lanera (Lanas Trinidad SA) estableció con la SCMAU un acuerdo comercial, teniendo como referencia los precios del mercado lanero australiano publicados por el SUL en base a la información suministrada por Australian Wool Exchange (AWEX), tomando como base el valor promedio de los precios de los remates realizados en Australia, con premios o descuentos en lanas con diámetro menor o igual a 20,2 micras de acuerdo a los parámetros logrados en

8
color (Y-Z menor o igual a 1), resistencia a la tracción (RT) (mayor o igual a 36 N/ktex) y largo de mecha (LM) (mayor o igual a 7,5 cm), de cumplirse en su totalidad podrían recibir un premio del 2 por ciento sobre el valor de Australia.
En un trabajo de caracterización de lanas Merino realizado por el SUL2 se midió en más de 70 lotes de lana de las zafras 1995, 1996 y 1997 las principales características de los mismos: diámetro promedio (DMF), rendimiento al lavado, RT, LM y grado de amarillamiento (Y), entre otras. Estos lotes provenientes de majadas comerciales tuvieron un valor de DMF de 21,8 µm (en un rango de 20,4 a 24,0 µm). En lo que respecta a la RT, el promedio de estos lotes fue de 32,2 N/ktex (con un mínimo de 23,5 N/ktex y un máximo de 35,0 N/ktex) teniendo el 80 por ciento de los lotes valores iguales o superiores a 30 N/ktex. A su vez, no hubo asociación entre los valores de DMF y RT (R2=0,05).
2.2 PRODUCCIÓN DE LANA Y FACTORES QUE LA AFECTAN
En el proceso de producción de lana diferentes factores genéticos como ambientales actúan independientemente e interactúan entre sí, determinando en definitiva la cantidad de lana por animal producida para determinadas condiciones (Rodríguez, citado por Gaggero et al., 2006). Entre los factores ambientales internos del animal que afectan la producción de lana se encuentran: sexo, edad, efecto materno y estado fisiológico. Con respecto a los factores ambientales externos al animal se pueden considerar: la nutrición, el clima (fotoperíodo) y la sanidad.
2.2.1 Factores internos
2.2.1.1 Sexo
La producción de lana de los carneros es mayor a la de capones, y éstos a vientres solteros. La mayor diferencia está dada por el tamaño corporal, asociado también a la capacidad de consumo (Corbett, 1979).
La mayor producción de los carneros es consecuencia de su mayor tamaño, pero también por efecto del estado hormonal, lo que puede aumentar la síntesis de proteína de la lana y la eficiencia de conversión del alimento en lana (Brown et al., citados por Corbett, 1979) estimaron que tres cuartos de la
2 Garín, M. 2009. Com. personal.

9
producción extra de lana limpia por carneros estaba atribuida principalmente a una mayor población folicular y a un mayor largo de mecha.
2.2.1.2 Edad
Para ovejas Merino, la producción de lana y varias características del vellón se alteran con la edad de la oveja. La tasa de producción de lana se incrementa hasta los cuatro años de vida y luego decrece en ovejas adultas, así como otras características de calidad del vellón (Corbett, 1979).
Esta disminución de la producción está explicada en un 20 a 30 por ciento por el menor número de folículos produciendo fibra por unidad de superficie, y un 70 a 80 por ciento se explica por una reducción en el volumen de la fibra (Gaggero et al., 2006).
2.2.1.3 Efecto materno
El efecto materno se observa en animales hijos de borregas, y los nacidos como mellizos. Estos producen como adultos entre 5 y 10 por ciento menos de lana por cabeza que los nacidos únicos como progenie de ovejas adultas. La diferencia se debe principalmente al menor número de folículos presentes en los hijos de borregas y en los nacidos mellizos; el total de folículos es menor por una deficiencia de folículos secundarios; no hay diferencias en cuanto al número de primarios (Turner, citado por Gaggero et al., 2006).
File y Whale encontraron que los corderos nacidos como mellizos son bastante más sensibles que los únicos a la alimentación deficitaria que puedan sufrir sus madres durante la gestación y lactancia, aunque con una buena alimentación de éstas existe poco efecto del tipo de nacimiento. Esta interacción entre el tipo de nacimiento y la nutrición en la producción de lana ya había sido concluido por Schinckel y Short, ambos trabajos citados por Gaggero et al. (2006). Estos autores encontraron que la maduración de los folículos secundarios en mellizos se veía retrasada debido a un aporte limitado de nutrientes al principio del período postnatal, que es cuando se espera el mayor incremento en la maduración de los mismos.
Gambetta et al. concluyeron que la población folicular no estuvo afectada en forma permanente por el nivel nutritivo pre-destete y tipo de nacimiento, determinando únicamente un retraso en la maduración (Gaggero et al., 2006). Con una buena nutrición luego del nacimiento se promueve la maduración post natal de los folículos secundarios (Corbett, 1979).

10
2.2.1.4 Estado fisiológico
Al comparar la producción de peso de vellón anual de una oveja vacía a una oveja gestando, la disminución de peso es del orden del 3 al 10 por ciento. Trabajos con ovejas de raza Merino señalan que la reducción en crecimiento de lana durante la preñez tardía fue de un 9-24 por ciento en relación a ovejas secas (Bianchi y Gambetta, 1991).
Según Corbett (1979), todo el ciclo reproductivo reduce la producción de lana 10 a 14 por ciento. Los mayores valores se dieron por lo general en ovejas criando mellizos. La preñez de mellizos reduce la producción de lana limpia y sucia en 7,6 y 5,9 por ciento comparado con una reducción de 5,4 y 1,8 por ciento en preñez de únicos (Ray y Sidwell, citados por Corbett, 1979).
Durante el fin de preñez y principio de lactancia en ambiente Mediterráneo, Stewart et al., citados por Masters (1992), registraron un 50 por ciento de reducción de producción de lana. Estos resultados coinciden con lo expuesto por Bianchi y Gambetta (1991) donde se plantean reducciones en la producción de lana limpia del orden del 51 al 56 por ciento en comparación a ovejas no encarneradas.
Al comparar ovejas Merino lactando y no lactando a 5 niveles diferentes de carga (entre 9,4 y 35,3/ovejas por hectárea), la producción de lana limpia en 12 semanas de lactación se redujo en promedio 29 por ciento, no habiendo efecto de la carga (Langlands, Ray y Sidwell, citados por Corbett, 1979) plantean que esta reducción en la producción depende del número de corderos lactando. Si bien la proporción de reducción de producción de lana fue menor, se mantiene la diferencia de producción de animales fallados en comparación a ovejas en lactancia (5 a 8 por ciento para únicos, y si son mellizos este valor se duplica).
La disminución en la producción es explicada por Brown et al., quienes atribuyen un tercio de la reducción del crecimiento en lana durante la preñez y lactancia a una disminución en el número de fibras y dos tercios por una disminución del volumen de la fibra. La disminución del número de fibras explica la aparición de acapachamiento (Lyne); la disminución del volumen es explicado en parte por menor largo de mecha y en parte por disminución del diámetro (Bosman, Slen y Whiting, Brown et al., Turner et al., citados por Corbett, 1979).
Estos cambios en la producción de la fibra afectan caracteres de calidad de la lana, como el aumento de rupturas, especialmente en ovejas con mellizos (Stevens y Wright), reducción en la resistencia a la tensión reportado por

11
Monteath, sobretodo en ovejas con subnutrición durante la preñez y cambio de color (Sanderson et al., citados por Corbett, 1979).
En vientres con vellones con resistencia menores a los 30 N/ktex (Schlink et al., 1992) observaron una mayor proporción de folículos que dejan de producir durante la preñez; esta discontinuidad en el crecimiento de la fibra contribuirá a la disminución de la RT. A su vez, en ovejas en pastoreo coincidió el punto más débil de la fibra con el fin de la gestación.
Luego de culminada la lactancia, dependiendo del estado en que se encuentren las ovejas comienzan a recuperar el nivel productivo, debido a una nueva funcionalidad de los folículos y a un mayor largo y diámetro de lana producida; no teniendo efectos permanentes aunque las ovejas que están con mejor nivel nutritivo, se recuperan más rápidamente (Corbett, Rodríguez, citados por Gaggero et al., 2006). Cambios en el crecimiento de lana durante la preñez serán el reflejo del cambio de la eficiencia de producción (Corbett, 1979).
2.2.2 Factores externos
Si bien el potencial de producción de lana de la oveja está determinado por características genéticas, variaciones estacionales y anuales ocurren en la cantidad y calidad de lana producida en ovinos en pastoreo, habiendo situaciones en las que el potencial es logrado (Robards, 1979). La variación en la producción de lana es un reflejo del estado nutricional del animal, combinado con los efectos del fotoperiodo, temperatura, estrés y enfermedades.
2.2.2.1 Sanidad
El efecto de parasitosis y enfermedades varía desde la producción de acapachamiento del vellón hasta una leve reducción en la producción de lana, siendo similares cualitativamente a los efectos causados por los cambios nutricionales, provocando cambios en el diámetro de fibra y en el largo dependiendo de la severidad y duración.
Mientras una enfermedad puntual o temporaria producirá una pronunciada reducción en el diámetro de fibra (suficiente para causar ruptura de fibras durante el procesamiento de ésta, pero no suficiente para producir pérdidas significativas en el peso de vellón en esquilas anuales), enfermedades crónicas o bajos grados de infección producirán menor largo de fibra y fibras más finas dando como resultado un vellón más liviano, pero que no rompe (Donald, 1979).

12
2.2.2.2 Fotoperiodo
No todas las razas ovinas ven afectada su producción de lana por el fotoperiodo. En algunas se observa una variación en la tasa de producción de lana, que está explicado por el ritmo fotoperiódico en la tasa de producción de crecimiento de lana (Nagorcka, 1979).
Los menores valores tanto de crecimiento de lana como de diámetro se alcanzan en los meses invernales (Laporte, 1975), cuando existen altos requerimientos relacionados con la termorregulación, y en etapas fisiológicas del ciclo reproductivo, como la gestación y la lactancia (Camejo et al., citados por Polanco de Vedia, 2005). Mientras tanto, los mayores valores se logran en el verano, donde el estado nutricional de los animales es generalmente superior, existiendo semejanza entre los valores de otoño y primavera.
En la raza Merino no se observa variación de la producción de lana explicado por el fotoperiodo. La variación en la producción observada (tasa de crecimiento y diámetro) está influenciada por la alimentación y los requerimientos animales en función al estado fisiológico. En las condiciones pastoriles en las que se desarrolla la cría ovina en Uruguay se observa una acentuada disminución en la calidad y cantidad del forraje durante el invierno (como se observa en la Figura No. 5 de distribución de la producción en los suelos donde se desarrolla la cría ovina en Uruguay), coincidiendo el fin del período invernal con los máximos requerimientos de los vientres ya que se encuentran en el último tercio de la gestación. La oferta de 400 a 700 kg MS/ha en los sistemas laneros tradicionales del Basalto, no permiten alcanzar consumos adecuados para cubrir el incremento de los requerimientos de mantenimiento para ovejas de cría en gestación avanzada (Montossi et al., 1998) Por lo tanto esta disminución de la tasa de crecimiento y/o del diámetro durante los meses de invierno se ve explicada por el plano nutritivo y estado fisiológico del animal, no viéndose afectado por el fotoperiodo.
2.2.2.3 Nutrición
La nutrición es el factor más importante que modifica el ritmo de crecimiento de la lana. El diámetro de fibra está directamente relacionado con la tasa de crecimiento de la lana: a mayor tasa de crecimiento, mayor diámetro de fibra (Doyle et al., 1991). Existe una desuniformidad del crecimiento y una diferencia significativa del diámetro promedio de la fibra entre las distintas estaciones del año (Laporte et al., citados por Polanco de Vedia, 2005).

13
Esta desuniformidad en el crecimiento es explicada por Kempton (1979) quien argumentó que el crecimiento de la lana está afectado por la disponibilidad de nutrientes, asociado a la cantidad de alimento consumido y la composición del mismo. La mayor limitante en la producción de lana se debe a la cantidad y composición de los aminoácidos en los folículos de lana.
La oferta de forraje influye sobre el estado nutricional de los animales, por ende, constituye un factor fundamental que regula el crecimiento de la lana y el diámetro de fibra (Laporte, citado por Polanco de Vedia, 2005).
De acuerdo a Formoso (2005), existe una marcada estacionalidad en las condiciones de desarrollo de la cría ovina en Uruguay, lo cual determina fluctuaciones en calidad y cantidad del forraje disponible para los ovinos en pastoreo. En consecuencia, la tasa de crecimiento de la lana tendrá una importante variación a lo largo del año, lo cual es también señalado por Doyle y Thompson (1993).
Los requerimientos proteicos de los rumiantes son influenciados por factores del animal como: tasa potencial de crecimiento de tejidos, crecimiento fetal y producción de lana. Los aminoácidos (provenientes de la proteína ingerida en el alimento) están disponibles para el animal a través de la digestión y absorción por los microorganismos y la proteína no degradada que va al duodeno (Kempton, 1979). El suministro de proteína degradable en el duodeno tiene mayor influencia en la tasa de crecimiento de lana. Muchas veces se confunde la relación entre ingestión de proteína y crecimiento de lana, sobre todo cuando la proteína es degradada en el rumen.
En estudios realizados en animales con libre acceso a dietas de bajo contenido proteico (Kempton, 1979), se observó que el crecimiento de lana depende primeramente del suministro y absorción de aminoácidos (provenientes de la dieta o de los microorganismos), mientras el crecimiento de tejidos depende del suministro de aminoácidos y de la disponibilidad de energía. La disponibilidad de aminoácidos es el principal factor limitante en la tasa de crecimiento de lana y todos los factores que reducen en especial el suministro de aminoácidos (reducirán la relación fósforo/energía metabolizable) tendrán un efecto negativo en la producción de lana.
En el norte de New South Wales (Australia), la tasa de crecimiento de las pasturas en el invierno es muy baja, en contraste con los requerimientos de ovejas de cría que aumentan a medida que avanza la preñez Con la suplementación de proteína bypass en ovinos en crecimiento y preñados sobre

14
pasturas se observaron buenas respuestas (Beattie et al., Leng, citados por Abella, 2005).
En Western Australia se plantea que utilizando mejores pasturas, se podría ayudar a reducir el estrés nutricional de verano y otoño (Hansford y Geenty, 2004), las pasturas en ambiente mediterráneo tienen en primavera buena calidad y cantidad, pasando a senescer hacia fin de verano y temprano en otoño, siendo baja la digestibilidad (40-50 por ciento) y poca la proteína; posteriormente germinan hacia fin del otoño-invierno. Algunos valores de consumo de alimento y eficiencia de conversión del alimento a lana son expuestos por Langlands, citado por Corbett (1979), donde ovejas Merino en pastoreo con un cordero lactando consumían 80 por ciento más materia orgánica durante los 105 días de lactación que ovejas sin cordero en igual período. La tasa de producción de lana limpia fue de 1 gramo cada 365 gramos de materia orgánica para las lactantes y de 1 gramo de lana limpia cada 137 gramos de materia orgánica consumida para las no lactantes.
Dicha información coincide con lo citado por Corbett (1979) quien manifiesta que ovejas solteras producen de 60 a 150 por ciento más lana por unidad de alimento consumido que ovejas lactando.
La eficiencia de conversión de alimento en lana para ovinos variará si el ovino es bien alimentado, subnutrido o si se le suministró suficiente alimento para realizar crecimiento compensatorio (McCance y Widdowson). Subnutrición en vientres jóvenes, sin ser severa, aparenta tener efecto en la habilidad de producción de lana, o en la eficiencia de producción ya que la evidencia disponible indica que la tasa de crecimiento de la lana esta directamente relacionada al consumo (Corbett, 1979). En cuanto a los corderos, la subnutrición postnatal afecta la segunda ola de maduración de folículos secundarios (Short, citado por Corbett, 1979).

15
2.3 PRODUCCIÓN ESTACIONAL DE PASTURAS
Las pasturas naturales comprenden el 71 por ciento del territorio nacional (URUGUAY. MGAP. DIEA, citado por Formoso, 2005) y son la fuente principal de alimento de los herbívoros domésticos (ovinos y bovinos).
Las pasturas naturales son definidas como un tipo de cobertura vegetal formada por gramíneas y plantas herbáceas o subarbustivas, donde los árboles son raros (Berreta y do Nascimento, citados por Formoso, 2005).
Los productores de la raza Merino se encuentran principalmente localizados en los suelos más marginales de la región de Basalto, y otras regiones de similar problemática, donde esta raza representa al menos el 33% de las cabezas ovinas manejadas por los productores laneros de la región (Montossi et al., 2007).
2.3.1 Región Basalto
Según su grado de desarrollo a los suelos de esta región se los puede agrupar en suelos superficiales y profundos. La profundidad de los mismos varía desde la roca desnuda hasta aproximadamente un metro.
En los Litosoles (suelos superficiales), su uso es pastoril con énfasis en la cría vacuna y ovina. Los dos tipos más importantes de Litosoles que se encuentran en la región son los Litosoles negros y Litosoles pardo rojizos, llamados así por su color.
Los dos principales tipos de suelos medianamente profundos y profundos que se encuentran en la región basáltica, asociados a los suelos superficiales en proporciones variables, son Brunosoles y Vertisoles.
Son suelos con perfil desarrollado, de color pardo oscuro o negro, alta fertilidad natural y una profundidad que puede ser mayor a un metro.
2.3.1.1 Producción de forraje en campo natural
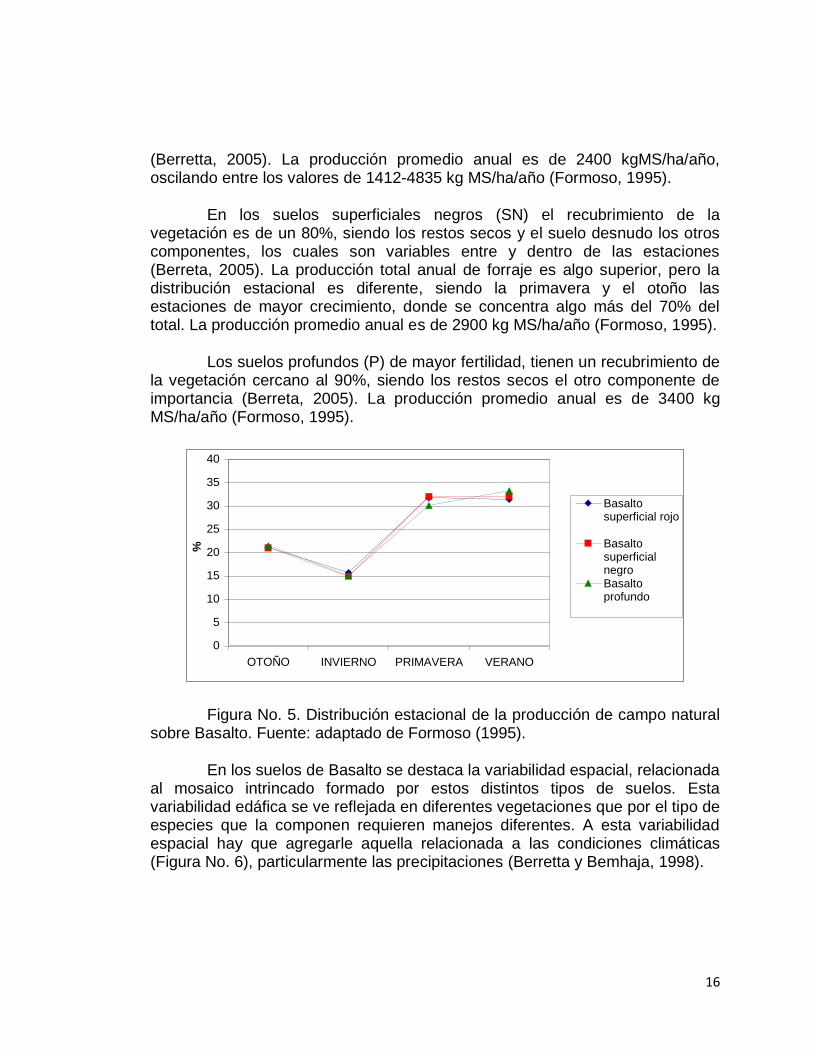
En los suelos superficiales pardos rojizos (SPR) la tasa de crecimiento (TCD), expresada en kgMS/ha/día es variable según las estaciones y también entre años (Figura No. 5). La mayor proporción de la producción anual de forraje se registra en primavera y verano, siendo esta estación la que presenta mayor variabilidad debido al alto riesgo de sequía de este tipo de suelo
16
(Berretta, 2005). La producción promedio anual es de 2400 kgMS/ha/año, oscilando entre los valores de 1412-4835 kg MS/ha/año (Formoso, 1995).
En los suelos superficiales negros (SN) el recubrimiento de la vegetación es de un 80%, siendo los restos secos y el suelo desnudo los otros componentes, los cuales son variables entre y dentro de las estaciones (Berreta, 2005). La producción total anual de forraje es algo superior, pero la distribución estacional es diferente, siendo la primavera y el otoño las estaciones de mayor crecimiento, donde se concentra algo más del 70% del total. La producción promedio anual es de 2900 kg MS/ha/año (Formoso, 1995).
Los suelos profundos (P) de mayor fertilidad, tienen un recubrimiento de la vegetación cercano al 90%, siendo los restos secos el otro componente de importancia (Berreta, 2005). La producción promedio anual es de 3400 kg MS/ha/año (Formoso, 1995).
0
5
10
15
20
25
30
35
40
OTOÑO INVIERNO PRIMAVERA VERANO
%
Basaltosuperficial rojo
BasaltosuperficialnegroBasaltoprofundo
Figura No. 5. Distribución estacional de la producción de campo natural sobre Basalto. Fuente: adaptado de Formoso (1995).
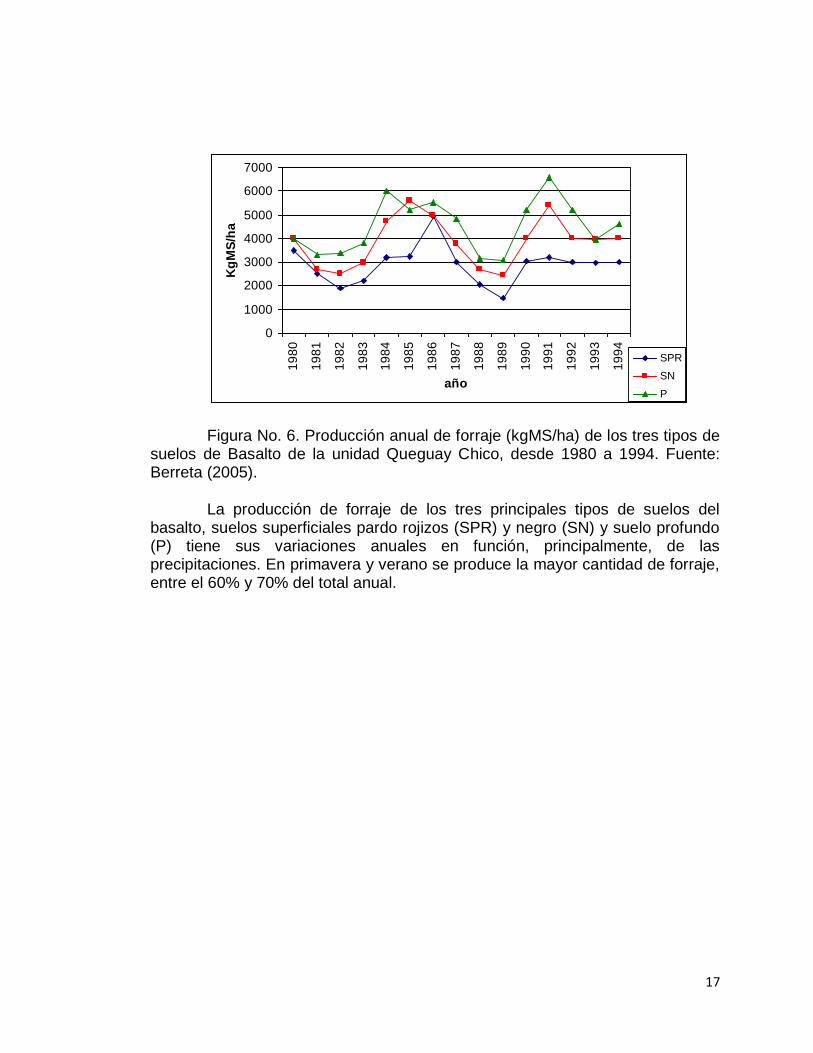
En los suelos de Basalto se destaca la variabilidad espacial, relacionada al mosaico intrincado formado por estos distintos tipos de suelos. Esta variabilidad edáfica se ve reflejada en diferentes vegetaciones que por el tipo de especies que la componen requieren manejos diferentes. A esta variabilidad espacial hay que agregarle aquella relacionada a las condiciones climáticas (Figura No. 6), particularmente las precipitaciones (Berretta y Bemhaja, 1998).
17
0
1000
2000
3000
4000
5000
6000
7000
19
80
19
81
19
82
19
83
19
84
19
85
19
86
19
87
19
88
19
89
19
90
19
91
19
92
19
93
19
94
año
Kg
MS
/ha
SPR
SN
P
Figura No. 6. Producción anual de forraje (kgMS/ha) de los tres tipos de suelos de Basalto de la unidad Queguay Chico, desde 1980 a 1994. Fuente: Berreta (2005).
La producción de forraje de los tres principales tipos de suelos del basalto, suelos superficiales pardo rojizos (SPR) y negro (SN) y suelo profundo (P) tiene sus variaciones anuales en función, principalmente, de las precipitaciones. En primavera y verano se produce la mayor cantidad de forraje, entre el 60% y 70% del total anual.

18
2.4 RESISTENCIA A LA TRACCIÓN
La resistencia a la tracción (RT) es una característica importante de la lana, ya que afecta su capacidad de procesamiento textil. Su medición objetiva permite identificar lanas con diferente potencial industrial al procesarlas, dada la importancia que tiene en el hauteur y noils de tops (Hansford y Kennedy, 1990).
La unidad de medida de RT es el Newton por ktex, donde la fuerza se mide en Newton (1 kg de peso = 9.81 N) y tex es la densidad lineal de la mecha (peso limpio/largo).
La RT es la fuerza necesaria aplicada a la mecha de lana para lograr la rotura de la misma (Franz, 2008). Es una característica compleja, que involucra la fuerza necesaria para romper la muestra y la densidad linear de la misma.
RT (N/Ktex)= Fuerza (Newton)/ Densidad linear (g/m)
Densidad linear (g/m) = lana limpia (g)/largo de mecha(m)
En Australia, fueron desarrolladas fórmulas de predicción (Williams, 1998) para estimar el largo de fibras en lana peinada o tops –conocido como hauteur- La altura media del top es el parámetro que tipifica la materia prima para la hilandería y junto con el diámetro define el precio final de la lana peinada (Polanco, 2005). Los ensayos de evaluación de mediciones adicionales (TEAM, en inglés) incorporaron el valor de resistencia a la tracción para predecir el hauteur en tops, junto con otras características como el largo de mecha, el diámetro promedio de fibras y el contendido vegetal (Jackson et al., 2004).
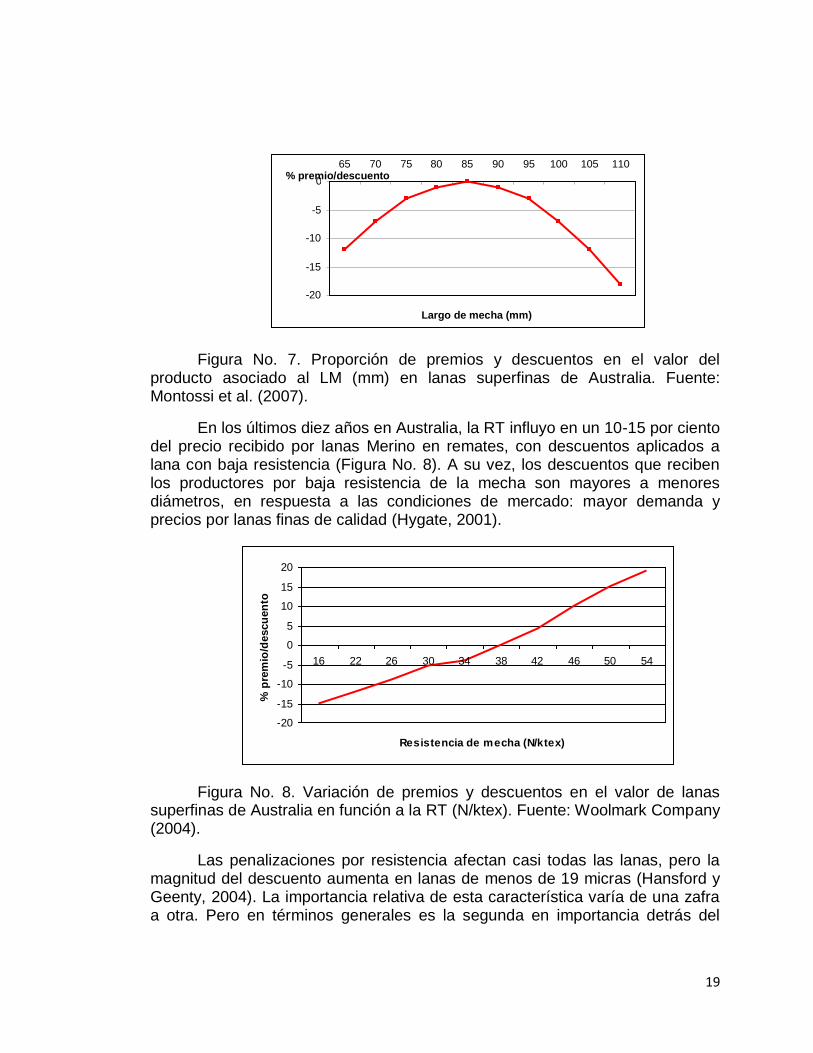
En el sistema de venta de lana por remates en Australia casi todos los vellones vendidos en la zafra 2007/2008 fueron ofertados con mediciones objetivas de sus principales características (A.W.E.). Los productores de lana reciben premios y/o castigos de acuerdo al largo de mecha logrado al momento de vender su lote, esto se debe a que las cardas de las industrias laneras se encuentran calibradas para trabajar con longitudes de 85 mm y de no lograrse determinado Hm ocurren penalizaciones (tanto por excesos como por déficit de largo óptimo para el cardado Figura No. 7). Lanas con resistencia mayor a 30 N/ktex se clasifican como resistentes, entre 30 y 15 N/ktex débiles y menores de 15 N/ktex quebradizas (Montossi et al., 2007).
19
Figura No. 7. Proporción de premios y descuentos en el valor del producto asociado al LM (mm) en lanas superfinas de Australia. Fuente: Montossi et al. (2007).
En los últimos diez años en Australia, la RT influyo en un 10-15 por ciento del precio recibido por lanas Merino en remates, con descuentos aplicados a lana con baja resistencia (Figura No. 8). A su vez, los descuentos que reciben los productores por baja resistencia de la mecha son mayores a menores diámetros, en respuesta a las condiciones de mercado: mayor demanda y precios por lanas finas de calidad (Hygate, 2001).
-20
-15
-10
-5
0
5
10
15
20
16 22 26 30 34 38 42 46 50 54
Resistencia de mecha (N/ktex)
% p
rem
io/d
escu
en
to
Figura No. 8. Variación de premios y descuentos en el valor de lanas superfinas de Australia en función a la RT (N/ktex). Fuente: Woolmark Company (2004).
Las penalizaciones por resistencia afectan casi todas las lanas, pero la magnitud del descuento aumenta en lanas de menos de 19 micras (Hansford y Geenty, 2004). La importancia relativa de esta característica varía de una zafra a otra. Pero en términos generales es la segunda en importancia detrás del
-20
-15
-10
-5
0 65 70 75 80 85 90 95 100 105 110
Largo de mecha (mm)
% premio/descuento

20
diámetro. En el rango de 16,6 a 19,5 micras, la RT explicó el 22 por ciento de la variación en el precio en la zafra 2001/2002 y alcanzó el 29 por ciento en el promedio de las tres zafras desde 2004 al 2006 (Figura No. 9).
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
2001/02 2004-2006Período
%
Mediciones adicionales
Factores de mercado *
Estilo
Color
Material Vegetal
Largo
Punto de ruptura
Resistencia
Diámetro
Figura No. 9. Influencia de los distintos componentes de calidad en el precio final de lana Merino en el rango de 16.6 a 19,5 micras en Australia para el período 2001/02 y las zafras 2004-2006. Fuente: Montossi et al. (2007).
Según Hynd y Schlink (1992) la densidad linear no es solo una medición del material en el punto de ruptura sino el promedio de todas las mediciones en la muestra. Los cambios en el valor de la RT reflejan los cambios en el material testado y de su fuerza intrínseca. La medición de la RT asume que la densidad linear es constante a lo largo de la muestra, por lo que cualquier factor que reduzca el diámetro de fibra a lo largo de la muestra disminuirá la RT. Reducciones del diámetro de fibra son producidos por disminución en el consumo o cambios en el balance de nutrientes absorbidos (aminoácidos), enfermedades y parasitismo, preñez y lactación (consumo, partición de nutrientes), época del año (asociado a la producción de pasturas, Hynd y Schlink, citados por Abella, 2005). Esto coincide con lo expuesto por Polanco (2005), quien plantea que es una propiedad que está fuertemente condicionada al medio ambiente productivo y a las condiciones climáticas del año.

21
2.5 VARIACIÓN DEL DIÁMETRO Y RELACIÓN CON LA RESISTENCIA
La variación en el diámetro de la fibra dentro de una mecha se debe a la variación en diámetro a lo largo de las mechas y entre fibras (Ritchie y Ralph, 1990). En trabajos con borregos Merino de lana fina, estos autores mostraron que entre el 70 y 80 por ciento de la variación en la RT puede ser explicada por el coeficiente de variación del diámetro de una mecha, irrespectivamente del diámetro promedio de la misma. A mayor valor del coeficiente de variación del diámetro, menor será la resistencia a la tracción.
También, dentro de una majada, son variadas las fuentes de variación del diámetro de la fibra, tal como se presenta en el Cuadro No. 2.
Cuadro No. 2. Componentes de variación en diámetro.
Fuente de variación Buena resistencia Baja resistencia
% del total de variación
% del total de variación
Entre fibras dentro de una mecha
64 43
Entre mechas dentro de un vellón
4 3
Entre vellones dentro de una majada
16 11
A lo largo de la mecha 16 43
Total 100 100
Fuente: Whiteley (1987).
Como es mostrado en el Cuadro No. 2, la variación en diámetro entre fibras dentro de una mecha es la mayor fuente de variación en el diámetro de fibra en lanas de buena resistencia. Sin embargo, su importancia relativa decrece en lanas de baja resistencia ya que la variación en diámetro a lo largo de la mecha adquiere mayor importancia. Esto brinda soporte a lo aseverado por Mitchell et al. (1992), de que la variación a lo largo de la fibra tiene influencia en la resistencia a la tracción.
Como un ejemplo de esto, en su trabajo con capones y ovejas Merino, Denney (1990) encontró correlaciones fenotípicas significativas y negativas (-0,30) entre la variación del diámetro a lo largo de la mecha (ACVDF) y la RT, sugiriendo que aumentos en la primera se asocian a una reducción en la RT. En el mismo sentido Mac Kinley et al. (1976) encontraron que la ACVDF tuvo la misma proporción de la variación que la variación entre fibras (34 por ciento del total, para ambos componentes) al medir la variación total del diámetro

22
promedio de fibras en lanas de baja resistencia de ovejas Merino Peppin que gestaron y lactaron.
Thompson y Hynd (1998) mostraron que el diámetro mínimo (Dmín.) era la característica con mayor asociación a la RT, representando el 66 por ciento de la variación total. Estos autores mostraron que en la medida que el Dmín. se incrementaba 1 micra, la RT aumentaba 5 N/ktex. Y a su vez, el Dmín. y la tasa de variación en diámetro de fibras (micras por milímetro) combinadas explicaron el 72 por ciento de la variación de RT.
Contrariamente, Hansford y Kennedy (1990) concluyeron que la mayor variación en la resistencia a la tracción fue explicada por la tasa de cambio en el diámetro y no por el DMF, el diámetro en la posición de rotura, el Dmín. o el rango de diámetro de fibras (RANGO). Sin embargo, no fueron explicitadas relaciones biológicas que explicaran estas asociaciones.
De todos estos estudios referidos al tema queda en evidencia que hay diversos factores controlando e influyendo en la RT de la lana. A su vez, que no es una característica simple.
El diámetro de fibras del animal no es constante a lo largo del cuerpo (Figura No. 10), éste aumenta (en sentido axial-caudal) desde la cabeza hacia el anca y de abajo hacia arriba en el tren delantero y de arriba hacia abajo en el tren posterior, considerándose un punto representativo del diámetro de fibras del animal el medio del costillar (Hygate, 2001). En el trabajo de Rottenbury et al. (1981) se demostró que este lugar en el animal es representativo del promedio del vellón. En dicho trabajo se analizó el diámetro de cinco majadas Merino de diferentes localidades (New South Wales y Western Australia) y edades (a partir de 3 años de edad). Dentro de cada majada se seleccionó aleatoriamente a las ovejas a muestrear. Del total de los vellones obtenidos se sacaron muestras en 25 vellones de las 9 zonas a estudiar (100 gramos del centro de cada zona) y de cada una de las muestras se tomaron diez mechas para analizar la resistencia a la tracción. De las correlaciones obtenidas entre el valor medio de resistencia para el vellón y el valor de resistencia para cada una de las 9 zonas analizadas, siempre se obtuvo mayor valor de correlación en las zonas 2 y 5 (correspondientes al medio del costillar derecho e izquierdo). El valor de correlación para estas zonas fue de 0,97 y el valor de correlación para el resto de las zonas oscilo para el promedio de las 5 majadas de 0,88 en la zona 9 (anca) a 0,96 en la zona 8 (lomo).

23
Figura No. 10. Variación del diámetro en el cuerpo del animal: el diámetro promedio en la mitad del costillar de este animal es de 22.1 micras. Fuente: Mahar (2004).
Las fuentes de variación son tanto a lo largo como entre fibras (Figura No. 11). Adams et al. (2000), indican que la variación a lo largo de la fibra tiene baja heredabilidad (0,17).
Figura No. 11. Esquema de variación del diámetro entre fibras y a lo largo de la fibra.
La variación del diámetro entre fibras, está explicado básicamente por factores genéticos, en cambio la variación del diámetro a lo largo de la fibra está explicada en una mayor proporción por aquellos factores que influyen en la producción de lana (factores ambientales externos e internos, Swan, 2010b).

24
Según Swan (2010b) la RT involucra varias propiedades de la fibra de lana a nivel individual y colectivo:
Largo promedio de las fibras
Variación del diámetro entre fibras (efecto genético)
Variación del diámetro a lo largo de las fibras (efecto de manejo)
Producción de fibras discontinuas (efecto genético)
Daño por radiación UV (efecto del manejo)
Fibras con el mismo rizo y largo estarán en su máxima extensión al mismo tiempo, si hay mucha variación en el largo de las fibras de la mecha, la fuerza aplicada por el ATLAS no será en todas las fibras (Schlink et al., 2001), dando como consecuencia una mecha con menor resultado de resistencia a la tracción.
Las distintas relaciones entre resistencia de la fibra (RF) y resistencia de la mecha son un resultado de la correlación entre RF y propiedades de la fibra que influyen en la RT (Peterson et al., 1998).
Habría que considerar que al momento de realizar recomendaciones por selección indirecta para la RT, se está analizando correlaciones fenotípicas y no genéticas. Yamin et al. (1999) indican que correlaciones fenotípicas pueden ser usadas como predictores de correlaciones genéticas.
2.5.1 Resistencia a la tracción
Se ha demostrado que ciertas características del perfil del diámetro de fibras (PDF) se encuentran relacionadas a la RT, como el CVDMF, LM, DMF (Hansford y Kennedy, Ritchi y Ralph, Lewer y Li, Swan, Greeff et al., Brown, citados por Brown et al., 1999). Dichas características se comportaron mejor como indicadores de la performance del procesamiento que de la RT (Smith, 2006).
Sin embargo la variación de los resultados de la RT se debe a que el PDF varía entre ambientes, diferentes sangres, grupo al que pertenecen y animales individuales (Brown y Crook, 2005).

25
En un ensayo realizado por Yamin et al. (1999) en ovejas Merino en la zona sur de Australia, se observaron resultados de heredabilidad elevada (Cuadro No. 3) para varias de las características del PDF como el DMF, CVDMF, DMáx y Dmín; Hill, citado por Yamin et al. (1999) presentaron valores muy similares de heredabilidad para DMF (h2= 0,68), CVDMF (h2= 0,69) y RT (h2= 0,32).
Cuadro No. 3. media, desvío fenotípico (p) y heredabilidades [h2(s.e)] para los caracteres medios de la muestra y a lo largo de esta.
CARÁCTER MEDIA p h2(s.e)]
Media de la muestra
DMF 24,91 2,14 0,60(0,16)
CVDMF 22,55 2,5 0,78(0,17)
RT 28,1 11,09 0,24(0,11)
A lo largo de la muestra
ACVFD 9,36 2,73 0,17(0,10)
Dmín. 21,49 2,53 0,47(0,15)
DMáx. 28,11 2,36 0,57(0,16)
Máx.-mín. 6,62 1,81 0,20(0,10)
CV 1-10 17,83 2,7 0,59(0,16)
Referencias: DMF (diámetro medio de fibra), CVDMF (coeficiente de variación del diámetro de fibra), RT (resistencia a la tracción), ACVDF (coeficiente de variación del diámetro a lo largo de la fibra), Dmín. (diámetro de fibra mínimo), DMáx. (diámetro de fibra máximo), Máx.-mín. (diferencia entre el diámetro máximo y el mínimo), CV 1-10 (coeficiente de variación del diámetro de fibra medido en 10 lugares).
Fuente: Yamin et al. (1999).
Valores similares a los mencionados se presentaron de lanas Merino de Uruguay en el trabajo de Ciappesoni et al. (2008) en animales del núcleo fundacional en la estación experimental Glencoe de INIA, siendo la heredabilidad para DMF, CVDMF y RT de 0,62, 0,26 y 0,19 respectivamente. En los trabajos de Safari et al. (2007) fueron similares para DMF (h2=0,57) pero más altos en el caso de la RT (h2=0,52).
En función de que se observan valores de heredabilidad altos para algunos caracteres del PDF como el DMF, CVDMF, DMáx., CV 1-10, es importante analizar si estos están correlacionados a los resultados de RT (ya

26
que este carácter presentó alta heredabilidad, pero su medición es más compleja).
Al analizar resultados de correlación fenotípica presentada por Yamin et al. (1999) para distintos caracteres del PDF, se observó correlación significativa y negativa entre el CVDMF y la RT y entre el ACVDF y la RT (Cuadro No. 4). También se observó una correlación alta y positiva entre el DMF y el Dmín. y DMáx.
Cuadro No. 4. Correlación fenotípica entre DMF, CVDMF y RT.
CARÁCTER DMF CVDMF RT
Media de la muestra
DMF 1,00 -0,13 0,32
CVDMF -0,24 1,00 -0,44
RT 0,32 -0,44 1,00
A lo largo de la muestra
ACVFD -0,28 0,48 -0,43
Dmín. 0,83 -0,3 0,47
DMáx. 0,84 0,00 0,19
Máx.-mín. 0,02 0,41 -0,39
CV 1-10 0,69 0,73 -0,30
Error estándar para la correlación fenotípica: 0,01 a 0,04. Referencias: DMF (diámetro de fibra),
CVDMF (coeficiente de variación del diámetro de fibra), RT (resistencia a la tracción), ACVDF (coeficiente de variación del diámetro a lo largo de la fibra), Dmín. (diámetro de fibra mínimo), DMáx. (diámetro de fibra máximo), Máx.-mín. (diferencia entre el diámetro máximo y el mínimo), CV 1-10 (coeficiente de variación del diámetro de fibra medido en 10 lugares).
Fuente: Yamin et al. (1999).
Brown et al. (2002) presentaron en su análisis valores de correlación significativa y negativa entre el CVDMF y la RT que oscilan en R= -0,20 a -0,51; coincidiendo dicho resultado con lo citado anteriormente y con lo reportado por otros autores (Bray et al. 1995, Greeff et al. 1995, Adams et al. 1998, Brown et al. 1999). En el trabajo realizado por Adams et al. (2000) se observó correlación fenotípica significativa en doce de las veinte majadas analizadas, oscilando entre los valores de -0,02 a -0,54.
27
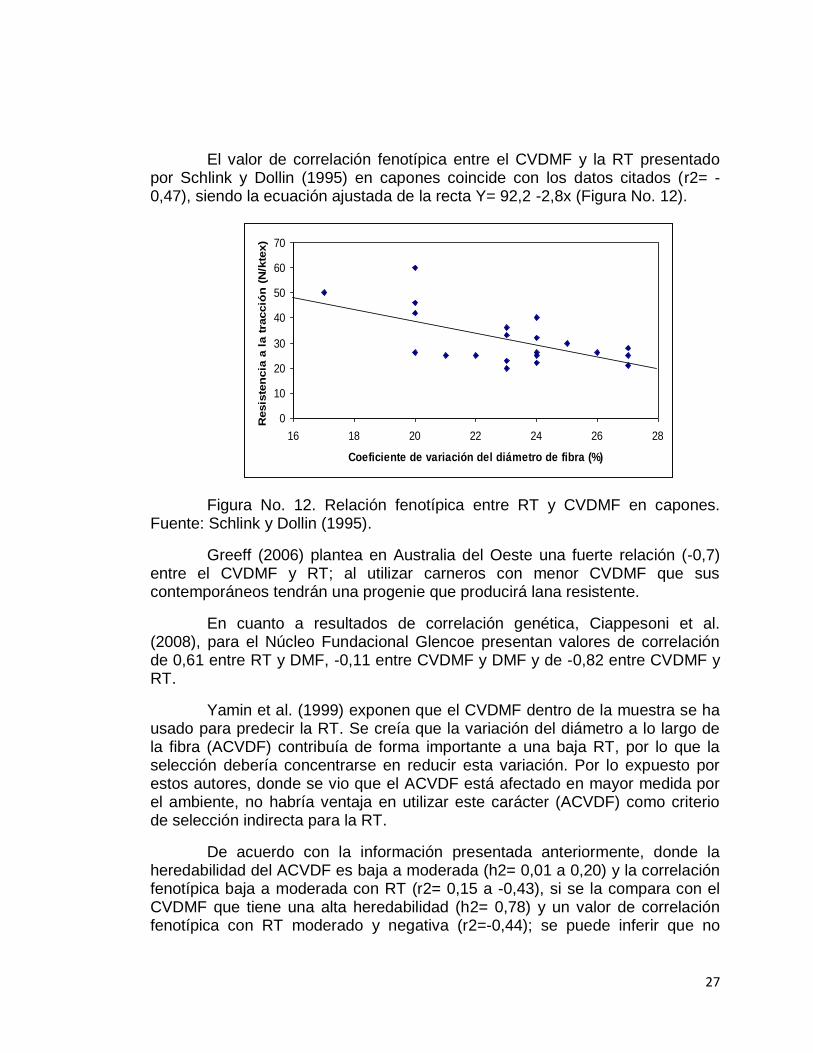
El valor de correlación fenotípica entre el CVDMF y la RT presentado por Schlink y Dollin (1995) en capones coincide con los datos citados (r2= -0,47), siendo la ecuación ajustada de la recta Y= 92,2 -2,8x (Figura No. 12).
0
10
20
30
40
50
60
70
16 18 20 22 24 26 28
Coeficiente de variación del diámetro de fibra (%)
Resis
ten
cia
a l
a t
racció
n (
N/k
tex)
Figura No. 12. Relación fenotípica entre RT y CVDMF en capones. Fuente: Schlink y Dollin (1995).
Greeff (2006) plantea en Australia del Oeste una fuerte relación (-0,7) entre el CVDMF y RT; al utilizar carneros con menor CVDMF que sus contemporáneos tendrán una progenie que producirá lana resistente.
En cuanto a resultados de correlación genética, Ciappesoni et al. (2008), para el Núcleo Fundacional Glencoe presentan valores de correlación de 0,61 entre RT y DMF, -0,11 entre CVDMF y DMF y de -0,82 entre CVDMF y RT.
Yamin et al. (1999) exponen que el CVDMF dentro de la muestra se ha usado para predecir la RT. Se creía que la variación del diámetro a lo largo de la fibra (ACVDF) contribuía de forma importante a una baja RT, por lo que la selección debería concentrarse en reducir esta variación. Por lo expuesto por estos autores, donde se vio que el ACVDF está afectado en mayor medida por el ambiente, no habría ventaja en utilizar este carácter (ACVDF) como criterio de selección indirecta para la RT.
De acuerdo con la información presentada anteriormente, donde la heredabilidad del ACVDF es baja a moderada (h2= 0,01 a 0,20) y la correlación fenotípica baja a moderada con RT (r2= 0,15 a -0,43), si se la compara con el CVDMF que tiene una alta heredabilidad (h2= 0,78) y un valor de correlación fenotípica con RT moderado y negativa (r2=-0,44); se puede inferir que no

28
habría ventajas en el uso del ACVDF como criterio de selección indirecto para la RT en comparación a la utilización del CVDMF como criterio de selección indirecto para la RT.
Otros caracteres del PDF como el diámetro medio de fibra y largo de mecha están fenotípica y genéticamente correlacionados a la RT, Brown et al. (2002) citan estudios que han observado fuertes correlaciones (r = 0,70 a 0,85) entre la RT y el DMF: Bigham et al., Fitzgerald et al., Orwin et al., Hansford y Kennedy, Bray et al., Peterson, Adams et al., Peterson et al., Thompson y Hynd, Brown et al., Yamin et al. Esta relación se mantiene para diferentes genotipos, sexo, estado fisiológico y condición nutricional.
2.5.1.1 Relación entre diámetro máximo y diámetro mínimo (DMáx. y Dmín.) con la resistencia
El DMáx. es aquella medición donde se da el máximo micronaje en el PDF; el Dmín. es la medición del menor micronaje en el PDF.
Dada la alta heredabilidad del DMF, es de esperar que los componentes del DMF como DMáx. y Dmín. sean de alta heredabilidad. Combinado con la alta variabilidad fenotípica, estas son características que por si solas producen cambios genéticos a través de la selección (Smith et al., 2006).
En diversos estudios se encontró que el Dmín. está correlacionado significativamente (r= 0,25 a 0,85) con RT (Peterson et al. 1998, Hansford y Kennedy 1998, Brown et al. 1999, Yamin et al. 1999, Brown et al. 2002). Estas relaciones se mantienen con animales de diferente genotipo, sexo, estado fisiológico y condición nutricional (Brown et al., 2002). En un estudio realizado por Sacchero y Muller (2007) las correlaciones encontradas entre Dmín. y RT fueron significativas pero bajas (r= 0,25).
Adams et al. (1998) sostienen que la RT responde al Dmín. a lo largo de la mecha, coincidiendo el Dmín. con el período en que el animal sufre algún estrés (Thompson y Hynd, 1998). La correlación positiva entre RT y Dmín. muestra que para mejorar la RT se necesitaría seleccionar por un mayor Dmín. Esto llevaría a lanas con características no deseables, por lo que pasaría a ser un criterio de selección indirecto no utilizado (Yamin et al., 1999).
2.5.1.2 Relación de tasa de cambio de diámetro con la resistencia
La tasa de cambio de diámetro de fibras (TASA) es la diferencia entre el máximo y mínimo diámetro sobre la distancia que las separa (Dmáx.-Dmín./distancia).

29
Brown et al. (2002) demostraron que la TASA está negativamente correlacionada a la RT (r=-0,16 a -0,38). Correlaciones más altas fueron presentadas por Hansford y Kennedy (1988) –0,77 a –0,42, –0,64 por Peterson (1997), –0,67 por Thompson y Hynd (1998), y –0,21 a –0,63 por Brown et al. (1999).
Otra característica de la mecha que está afectando la TASA (y por lo tanto la RT) es el largo de mecha (LM). El LM es un determinante clave de la RT por su efecto sobre TASA. Lanas con altos valores de crecimiento en LM pueden tener cambios más atenuados en el diámetro (y menor TASA) que aquellas con bajos valores de crecimiento en LM (si la proporción de cambio es la misma en ambas). Thompson y Hynd (1998) observaron que cuanto más abrupto es el cambio en DF menor la RT, es decir, las lanas son más débiles cuando la reducción o aumento del diámetro se produce en una sección longitudinal menor de la fibra. Por ejemplo, es más grave si un cambio de 24 a 18 micras se produce en 2 milímetros que si se produce en 6 milímetros.
En el trabajo de Adams et al. (2000) se observó correlación significativa (5 por ciento) entre la RT y el LM en trece de 20 majadas estudiadas, siendo r= -0,46. Se observó también que la correlación era negativa, cuando la mecha rompe al medio de la muestra, si la ruptura se daba hacia las puntas la correlación era positiva. Hansford y Kennedy (1990) identificaron que estimar el cambio del diámetro de fibra más allá del POB (punto de ruptura) es clave para la RT.
2.5.1.3 Relación entre extensión de la mecha y resistencia
Cuando cada fibra se tensiona a la fuerza de POB de la mecha, esta se rompe. Cuanto más uniforme el largo de fibra, más fibras van a estar tensionadas al mismo momento llevando a una mayor fuerza para romper (Swan). Según el mismo autor un aumento (2,5 a 5 por ciento) en el CV a lo largo de la fibra (ACVDF) reduciría la RT un 40 por ciento. Pero en esta investigación el ACVDF fue de 4,6 a 20,4 por ciento y no afectó la resistencia (Peterson et al., 1998).

30
2.6 UTILIDAD PRÁCTICA DE PERFILES
El perfil de diámetro (PDF) de una mecha muestra el crecimiento de la lana, medido como diámetro, desde la punta hasta la base de la mecha y mide la respuesta fenotípica del ovino al ambiente y/o condiciones fisiológicas que lo afectan (Hansford, 1997).
El perfil de diámetro fue inicialmente descripto por Jackson y Downes (1979). Si una mecha es cortada en una serie de segmentos y el diámetro promedio de cada segmento es medido, se obtiene como resultado el promedio del cambio del diámetro de fibras a través de todo el crecimiento de la fibra en el período, representando el perfil de diámetro de fibra. Los PDF son generados para estudiar la forma en que el diámetro de fibra cambia en el período de crecimiento de la lana. Los PDF son el resultado de la medición total de la muestra en segmentos de 2 mm. El diámetro de fibra es medido en estos segmentos, produciendo un patrón de cambio de diámetro de fibra a lo largo de la muestra.
Otro método de medición de perfiles de mechas de lana fue descripto por Hansford et al. (1985), mechas con las puntas alineadas son cortadas en trozos consecutivos de 2 mm que son lavados con solventes y acondicionados, para ser medidos con el FFDA (Fibre Fineness Distribution Analyser). Al graficar cada uno de los valores de diámetro obtenidos y ser graficados con el largo de mecha (considerando la distancia desde la punta) se obtiene el perfil de diámetro. Estos autores coinciden que de esta forma se podría avanzar en la relación que tiene el diámetro de fibras con otras características de la lana como la resistencia a la tracción.
Los PDF pueden ser utilizados para observar las variaciones a lo largo del año (Brown et al., 2002). Las características del PDF son asociadas con la RT y explican (diez a veinte porciento) de mejor manera la variación en la resistencia que las características fácilmente medibles como el DMF, CVDMF y LM (Brown et al. 1999, Brown 2000a).
Relacionado a esta técnica, Brown et al. (2000b) mencionaron que al trabajar con perfiles de diámetro pueden ser descriptas características como el DMáx., el Dmín., DMáx.-Dmín., TASA y ACVDF.
Sin embargo, estas técnicas para medir perfiles demandan mucho tiempo, son de difícil automatización y antieconómicas (Brown et al., 2000). El uso del OFDA 2000 (Optical Fibre Diameter Analyser) permitió la construcción de PDF, siendo además bajo su costo de medición (Baxter, 2001).

31
Con el OFDA 2000 el perfil de diámetro estaría proporcionando una herramienta para mejorar la resistencia a la tracción (Brown et al., 2000) ya que las características del perfil de diámetro están asociadas a la RT (Bigham et al., Hansford y Kennedy, Denney, Hansford, Peterson, Adams et al., Thompson y Hynd, Brown et al., citados por Brown et al., 2002).
Diversos estudios muestran que algunas características del PDF pueden ser asociadas y potenciales indicadores de la RT y POB (Hansford y Kennedy 1988, Peterson et al. 1998, Brown et al. 2002), comportándose como mejores indicadores de la performance del procesamiento que de la RT (Smith et al., 2006).
Estudios de Greeff et al., Swan et al., citados por Smith et al. (2006), muestran que el CVDMF puede ser usado como un criterio indirecto de selección para mejorar la RT en programas de cría de Merino. Greeff (2006) concluye que el CVDMF se transformó en una herramienta importante para los criadores. Muestra la variabilidad relativa y se expresa como porcentaje. Matemáticamente, el CVDMF es la estimación del desvío estándar dividido sobre el promedio.
CVDMF (%)= (desvío estándar/diámetro medio)*100
Usando el CVDMF como un criterio de selección indirecto podría prevenirse el deterioro de la RT en el tiempo, podría manejarse menor susceptibilidad al fleece-rot y mejoraría la performance de procesamiento (Greeff, 2006).
La RT de la mecha es una característica heredable y puede ser mejorada mediante el manejo. Sin embargo, hay una correlación genética positiva entre la RT y el DMF. Esto significa que una reducción en el DMF de una majada Merino llevaría a una reducción de la RT. También hay una correlación genética negativa entre la RT y CVDMF (Greeff et al., Swan et al., citados por Hygate, 2001).
Los animales podrían ser seleccionados directamente en las mediciones de RT realizadas a las muestras tomadas. Un mínimo de 10 muestras se necesitarían medir para obtener resultados significativos (pero aunque sea una forma más directa de medir, es menos predecible que el diámetro) basado en mediciones (Swan, 2010b).
La heredabilidad media encontrada para la RT demuestra el potencial existente para seleccionar por esta característica. Asimismo, al presentar una alta correlación positiva (desfavorable económicamente) con el DMF, es de gran importancia incluir a esta característica dentro de los objetivos de selección de

32
la raza Merino. La alta correlación negativa encontrada entre la RT y el CVDMF, indica que esta última característica podría ser incluida como criterio de selección en la evaluación genética de Uruguay, puesto que todos los animales con registro de diámetro cuentan a su vez con información de CVDMF y su heredabilidad es superior a la de RT (Ciappesoni et al., 2008).

33
2.7 HERRAMIENTAS Y MÉTODOS DE MEDIR PERFILES
Con el uso del OFDA es posible medir la variación del diámetro a lo largo de la mecha de lana y es creciente el interés del uso de los perfiles de diámetro al poder medirlos de forma económica con este equipo (Baxter, 2001). Tiene la capacidad de medir el diámetro de las fibras a lo largo de la mecha con intervalos de 5 mm y es posible el uso de estos perfiles para monitorear cambios en las características de lana producida según el manejo recibido por la majada (Peterson y Gherardi, 2001).
Las mediciones hechas con el OFDA 2000 tienen diversas aplicaciones. Por ejemplo, según Baxter (2001) permiten hacer un ranking de animales por diámetro, refugar aquellos con lana más gruesa, preparar la lana para la venta y ajustar prácticas nutricionales para optimizar el diámetro de fibras y la resistencia a la tracción.
Más recientemente, el análisis de la información derivada de los perfiles de diámetro puede ser usada para predecir el Hauteur de un lote de lana al ser procesado a top (Peterson y Oldham, 2002). A su vez, en un taller realizado en Armidale (Hansford y Geenty, 2004), fue sugerido que los perfiles tienen el potencial de ser usados en la investigación, en las mediciones objetivas de lana a campo y como herramienta de predicción del desempeño industrial. Sin embargo, Atkins et al. (2004) presentaron un punto de vista escéptico, argumentando la dificultad de visualizar el valor extra que agrega el uso de perfiles al que ya brindan otras características como el diámetro promedio, su coeficiente de variación, el largo de mecha y la resistencia a la tracción.
Para evaluar las relaciones entre resistencia y otras características de la mecha, se realizó un trabajo durante dos años en cinco majadas comerciales de la raza Merino ubicadas en la zona del Basalto -representativa de la producción de lanas finas- donde se analizó el comportamiento de la fibra a lo largo del año (a través del análisis de perfiles de diámetro de fibra y de diferentes características que de ellos se obtiene).

34
Las hipótesis consideradas fueron:
Que existen diferencias en la resistencia a la tracción entre diferentes majadas y entre animales de una misma majada.
Que existen diferencias en los perfiles de diámetro de fibra a lo largo del tiempo entre diferentes majadas y años.
Que existe una asociación fenotípica entre la resistencia a la tracción y ciertas características del perfil del diámetro (diámetro promedio, coeficiente de variación del diámetro, diámetro mínimo, diámetro máximo y diferencia entre diámetro máximo y mínimo).

35
3. MATERIALES Y MÉTODOS
Participaron en este trabajo cinco majadas Merino ubicadas en campos de Basalto (Figura No. 13). Se considera que los animales de estas majadas son representativos de la producción de lanas finas del Uruguay. Las mismas correspondieron a los establecimientos “Los Manantiales” de Sylvia Jones de Pérez (Paysandú, 31º50’ S, 56º 58’ O), “Los Arrayanes” de Alfredo y Álvaro Fros (Salto, 31º46’ S, 56º59’ O), “Los Pampas” de Fernando Fillat (Durazno, 32º53’S, 56º45’ O), “El Rancho” de Fernando Dutra (Paysandú, 32º0’ S, 56º22’ O) y “Cerro Camoatí” de Eduardo Brito (Paysandú, 31º54’ S, 56º42’ O).
Figura No. 13. Ubicación de los establecimientos. Fuente: Google Earth (2012).
El trabajo se realizó durante dos años, ocupando el período productivo de invierno 2005 a invierno 2006 (esquila del 2006) y del invierno 2006 al invierno 2007 (esquila 2007). En todos los casos, se hizo esquila preparto, con

36
doce meses de crecimiento de lana. En cada uno de estos dos años, de cada una de estas cinco majadas fueron seleccionadas al azar 30 ovejas de cría adultas que se habían manejado conjuntamente. En la esquila, de cada vellón se tomaron muestras de la mitad del costillar. Las muestras de lana se midieron en el laboratorio del Secretariado Uruguayo de la Lana (SUL) con el OFDA 2000 (Baxter, 2001). Las mediciones se realizaron en mechas naturales (sin lavar) aplicándose un factor de corrección de 1.6.3
De cada animal se determinó el diámetro promedio de fibras (DPF), coeficiente de variación del diámetro promedio de fibras (CVDPF), diámetro mínimo (Dmín.), diámetro máximo (DMáx.) y la diferencia entre éstos (DMáx.-mín.). A su vez, de cada muestra individual se tomaron 10 mechas para medir la RT (resistencia a la tracción) con el equipo Agritest del laboratorio del SUL. Se obtuvo el valor promedio de RT (N/Ktex) a partir de estas mediciones bajo la norma IWTO 30-07 (IWTO, 2012). El Agritest es un extensiómetro semiautomático que mide la resistencia de la mecha en muestras de lana sucia. Para la medición de la densidad linear se toma una muestra de lana sucia del espesor de un lápiz que es colocada en el medio de placas que se cierran y miden la sección transversal del material. Luego de medir la densidad de la muestra se procede a medir el largo, mediante una cinta transportadora con un scanner para posteriormente realizar la medición de la resistencia. Para romper la mecha se coloca la misma en los clips que dispone el aparato (tomándose por la base y por la punta), una vez sujetos se acciona la separación de los mismos, extendiéndose y midiendo la fuerza necesaria para la ruptura de la muestra (Baxter, s.f.).
Se realizó la descripción cuantitativa de cada majada –y en cada uno de los años- de todas las variables estudiadas: diámetro (µm), desvío estándar (µm), coeficiente de variación (%), diámetro máximo (µm), diámetro mínimo (µm), diferencia entre Máx.-mín. (µm) y resistencia a la tracción (N/Ktex).
Además, de cada majada y en cada año, se estudió con una regresión múltiple si las variables estudiadas (DPF, CVDPF, Dmín., DMáx. y DMáx.-Mín.) fueron significativas en la explicación de la variabilidad observada en RT de acuerdo a la ecuación:
y = b0 + b1 x1 + b2 x2 + b3 x3 + b4 x4 + b5 x5
Donde y = RT, x1 = DPF, x2 = CVDPF, x3 = DMín., x4 = DMáx. y x5 = DMáx.-Mín.
3 De Paula, R. 2012. Com. personal.

37
El procedimiento consistió en seleccionar con el análisis Stepwise aquellas variables independientes de mayor capacidad de predicción con respecto a la variable dependiente (RT = resistencia a la tracción), tomando como criterio el AIC (Akaike's Information Criterion) más bajo como indicador del modelo que mejor ajuste logró. Para cada una de las ecuaciones se obtuvo el coeficiente de determinación (R2).
Como el OFDA 2000 mide el diámetro a lo largo de la fibra, fue posible determinar la variación que esta característica tuvo. Dicho equipo trabaja con mechas de lana sucia, realizando mediciones de diámetros sobre 100 a 150 fibras en secciones transversales de mechas de lana cada 5 mm. Con los datos correspondientes a cada mecha, se obtuvo una secuencia de mediciones que representa el período anual de crecimiento de la fibra, que se denomina “perfil de diámetro de fibras” (Sacchero et al., 2010). Con el objetivo de resolver los distintos largos de mecha, se ajustaron todas las muestras individuales a una base común del 100% del crecimiento anual. Se ajustaron curvas suavizadas denominadas “Splines” a los perfiles de diámetro promedio de cada una de las majadas, determinándose también el grado de ajuste correspondiente. El uso de estas líneas suavizadas permite obtener de buena manera la forma que adquiere la variable Y a lo largo de la variable X, teniendo los puntos de cada tramo una influencia mayor en la curva que se ajusta.
Finalmente, y con el objetivo de estudiar la variabilidad de la RT, se consideraron como variables aleatorias los establecimientos y los animales anidados en cada establecimiento de acuerdo al siguiente modelo:
Yij = µ + Ei + Aij
Donde µ es la media general, Ei el efecto aleatorio de establecimiento, Aij el efecto aleatorio de animal anidado dentro de establecimiento (variancia residual).
Los datos de cada año se analizaron por separado. De esta forma, en cada año se estimó la variación entre y dentro de cada establecimiento. El análisis estadístico y el ajuste de Splines fue realizado con el programa JMP.IN (2002).
38
4. RESULTADOS Y DISCUSIÓN
En el siguiente capítulo se presentan los resultados obtenidos al procesar las muestras de las majadas para los años 2006 y 2007 y los resultados de los análisis realizados.
4.1 ANÁLISIS DESCRIPTIVO
Los datos obtenidos de las mediciones de las ovejas pertenecientes a las cinco majadas analizadas los años 2006 y 2007 se resumen en los Cuadros No. 5 y 6.
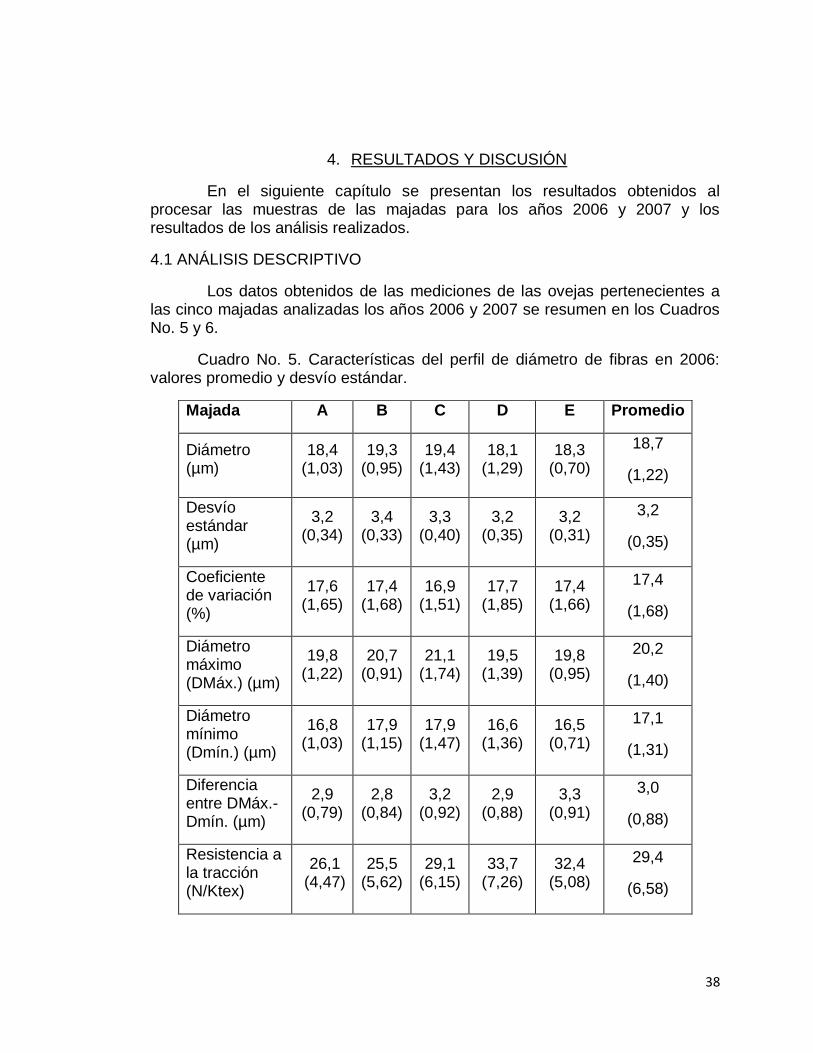
Cuadro No. 5. Características del perfil de diámetro de fibras en 2006: valores promedio y desvío estándar.
Majada A B C D E Promedio
Diámetro (µm)
18,4 (1,03)
19,3 (0,95)
19,4 (1,43)
18,1 (1,29)
18,3 (0,70)
18,7
(1,22)
Desvío estándar (µm)
3,2 (0,34)
3,4 (0,33)
3,3 (0,40)
3,2 (0,35)
3,2 (0,31)
3,2
(0,35)
Coeficiente de variación (%)
17,6 (1,65)
17,4 (1,68)
16,9 (1,51)
17,7 (1,85)
17,4 (1,66)
17,4
(1,68)
Diámetro máximo (DMáx.) (µm)
19,8 (1,22)
20,7 (0,91)
21,1 (1,74)
19,5 (1,39)
19,8 (0,95)
20,2
(1,40)
Diámetro mínimo (Dmín.) (µm)
16,8 (1,03)
17,9 (1,15)
17,9 (1,47)
16,6 (1,36)
16,5 (0,71)
17,1
(1,31)
Diferencia entre DMáx.-Dmín. (µm)
2,9 (0,79)
2,8 (0,84)
3,2 (0,92)
2,9 (0,88)
3,3 (0,91)
3,0
(0,88)
Resistencia a la tracción (N/Ktex)
26,1 (4,47)
25,5 (5,62)
29,1 (6,15)
33,7 (7,26)
32,4 (5,08)
29,4
(6,58)
39
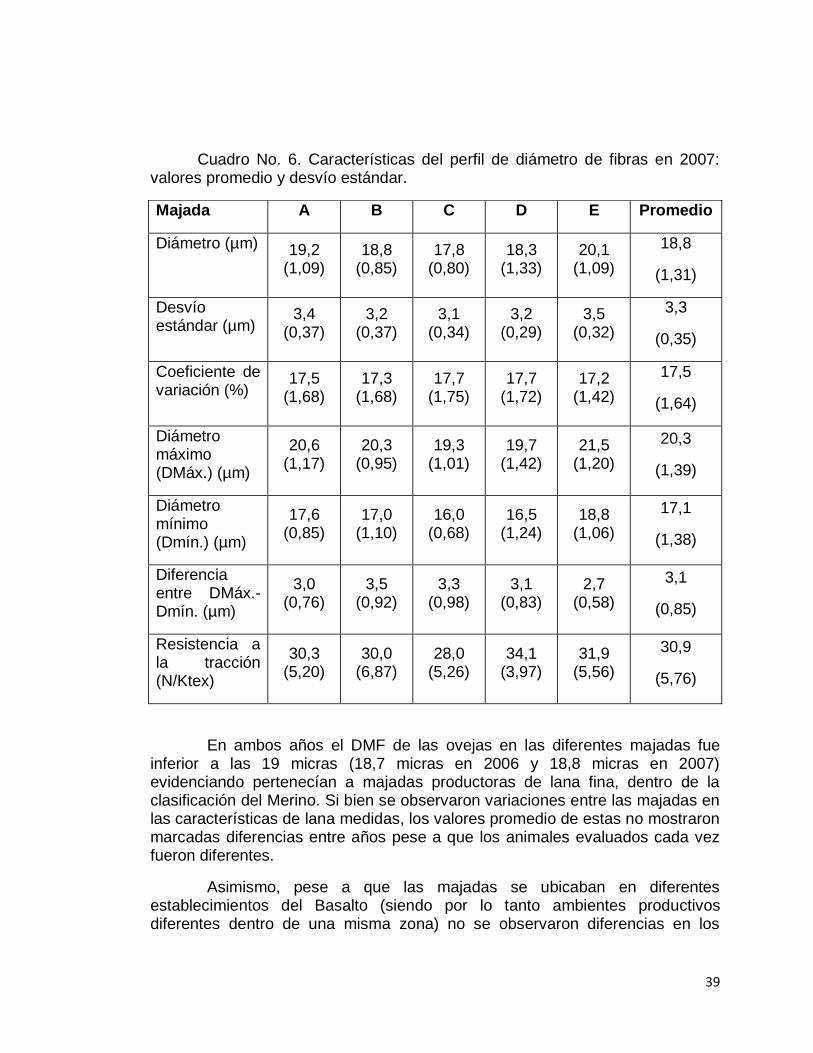
Cuadro No. 6. Características del perfil de diámetro de fibras en 2007: valores promedio y desvío estándar.
Majada A B C D E Promedio
Diámetro (µm) 19,2 (1,09)
18,8 (0,85)
17,8 (0,80)
18,3 (1,33)
20,1 (1,09)
18,8
(1,31)
Desvío estándar (µm)
3,4 (0,37)
3,2 (0,37)
3,1 (0,34)
3,2 (0,29)
3,5 (0,32)
3,3
(0,35)
Coeficiente de variación (%)
17,5 (1,68)
17,3 (1,68)
17,7 (1,75)
17,7 (1,72)
17,2 (1,42)
17,5
(1,64)
Diámetro máximo (DMáx.) (µm)
20,6 (1,17)
20,3 (0,95)
19,3 (1,01)
19,7 (1,42)
21,5 (1,20)
20,3
(1,39)
Diámetro mínimo (Dmín.) (µm)
17,6 (0,85)
17,0 (1,10)
16,0 (0,68)
16,5 (1,24)
18,8 (1,06)
17,1
(1,38)
Diferencia entre DMáx.-Dmín. (µm)
3,0 (0,76)
3,5 (0,92)
3,3 (0,98)
3,1 (0,83)
2,7 (0,58)
3,1
(0,85)
Resistencia a la tracción (N/Ktex)
30,3 (5,20)
30,0 (6,87)
28,0 (5,26)
34,1 (3,97)
31,9 (5,56)
30,9
(5,76)
En ambos años el DMF de las ovejas en las diferentes majadas fue inferior a las 19 micras (18,7 micras en 2006 y 18,8 micras en 2007) evidenciando pertenecían a majadas productoras de lana fina, dentro de la clasificación del Merino. Si bien se observaron variaciones entre las majadas en las características de lana medidas, los valores promedio de estas no mostraron marcadas diferencias entre años pese a que los animales evaluados cada vez fueron diferentes.
Asimismo, pese a que las majadas se ubicaban en diferentes establecimientos del Basalto (siendo por lo tanto ambientes productivos diferentes dentro de una misma zona) no se observaron diferencias en los

40
valores de los caracteres y en el comportamiento de la fibra a lo largo del año, como se ve en las Figuras No. 14 y 15 de perfiles.
Si bien se presenta en la Figura No. 14 cierta dispersión del DMF a nivel de animales individuales dentro de cada majada, más del 50% de las observaciones de cada majada (rectángulo rojo), tuvo menos de una micra de diferencia con el diámetro promedio de las cinco majadas (línea horizontal gris).
Figura No. 14. Distribución de diámetro promedio de fibras (DPF) de animales dentro de cada majada (2006 a la izquierda y 2007 a la derecha).
Respecto a la RT los resultados que se obtuvieron fueron muy similares en ambos años. Se presenta en la Figura No. 15, la RT promedio para cada año (línea horizontal gris) fue en el entorno de los 30 N/ktex (29,4 N/ktex en 2006 y 30,9 N/ktex en 2007).
15
16
17
18
19
20
21
22
Diá
metr
o
A B C D E
Productor
16
17
18
19
20
21
22
Diá
metr
o
A B C D E
Productor

41
Figura No. 15. Distribución de resistencia a la tracción promedio de animales dentro de cada majada (2006 a la izquierda y 2007 a la derecha).
Estos valores de RT fueron levemente inferiores a los determinados en el trabajo de caracterización de lanas realizado por el SUL siendo que correspondieron a lanas con casi 3 micras menos. Estos lotes provenientes de majadas comerciales tuvieron un valor de DMF de 21,8 µm (en un rango de 20,4 a 24,0 µm). En lo que respecta a la RT el promedio de estos lotes fue de 32,2 N/ktex (con un mínimo de 23,5 N/ktex y un máximo de 35,0 N/ktex) teniendo el 80% de los lotes valores iguales o superiores a 30 N/ktex. No hubo asociación entre los valores de diámetro y resistencia (R2=0,05).
Al comparar estos valores con los valores de lana fina provenientes de Australia -con un diámetro promedio de 20,5 micras- comercializadas en el sistema de remates de la zafra 2009/2010, tuvieron valores de RT de 33,8 N/ktex (AWEX, 2010). A su vez, en los centros de venta del norte (Sydney y Newcastle) los valores fueron incluso mayores: 36,2 N/tex. Los valores promedio encontrados en este trabajo de Uruguay se asemejaron a los encontrados en las lanas vendidas en el oeste de Australia (Fremantle), cuyo valor promedio fue 30,1 N/ktex en la zafra considerada.
Fue considerable el rango de valores observado: el rango de valores de RT en las majadas fue de 18,4 a 30,3 N/ktex en 2006 (siendo los extremos las majadas E y D) para el año 2006. A nivel individual, el registro mínimo fue de 8,2 N/ktex en una oveja de la majada C y el valor máximo fue de 42,4 N/ktex en la majada E. En el año 2007 el rango de RT medido fue de 16,6 a 32,1 N/ktex (majadas D y B), registrándose el menor valor individual en la majada B (7,3 N/ktex) y el mayor valor una oveja en la majada C con 40,5 N/ktex. Estos valores ejemplifican la marcada variación individual que existió dentro de cada majada a nivel de esta característica.
10
15
20
25
30
35
40
45
50
Resis
tencia
A B C D E
Productor
5
10
15
20
25
30
35
40
Resis
tencia
A B C D E
Productor

42
Al considerar el valor de 30 N/ktex como un valor absoluto por encima del cual las lanas pueden ser consideradas con buena RT, en el primer año el porcentaje de animales con valor igual o mayor a 30 N/ktex para las majadas A, B, C, D y E fue de 20, 20, 57, 73 y 70% respectivamente. Para el segundo año del trabajo el porcentaje de ovejas fue de 53, 67, 30, 87 y 70% para las majadas A, B, C, D y E respectivamente. Estos valores reafirman la marcada variación que existió a nivel de majadas y años.
La RT es un carácter de importancia económica en la lana, ya que afecta los procesos de cardado, las pérdidas en el peinado y las rupturas en el hilado. Al tener una heredabilidad de moderada a alta existe potencial para la selección y es factible el progreso genético como consecuencia de un correcto criterio de selección (0,23-0,51, Greeff, Hill y Ponzoni, citados por Yamin et al., 1999).
Estimaciones nacionales de parámetros genéticos fueron inferiores (h2 = 0,19) a lo expuesto en trabajos australianos (h2 = 0,52, Safari et al., citados por Ciappesoni et al., 2008).
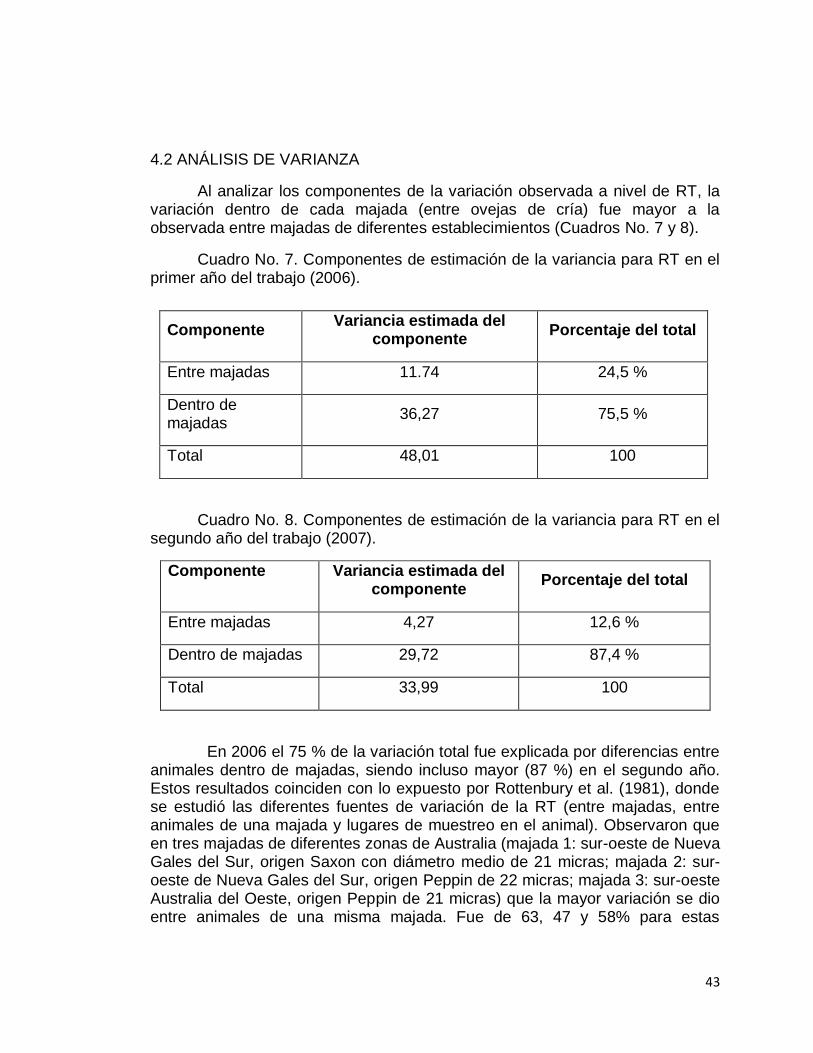
43
4.2 ANÁLISIS DE VARIANZA
Al analizar los componentes de la variación observada a nivel de RT, la variación dentro de cada majada (entre ovejas de cría) fue mayor a la observada entre majadas de diferentes establecimientos (Cuadros No. 7 y 8).
Cuadro No. 7. Componentes de estimación de la variancia para RT en el primer año del trabajo (2006).
Cuadro No. 8. Componentes de estimación de la variancia para RT en el segundo año del trabajo (2007).
Componente Variancia estimada del componente
Porcentaje del total
Entre majadas 4,27 12,6 %
Dentro de majadas 29,72 87,4 %
Total 33,99 100
En 2006 el 75 % de la variación total fue explicada por diferencias entre animales dentro de majadas, siendo incluso mayor (87 %) en el segundo año. Estos resultados coinciden con lo expuesto por Rottenbury et al. (1981), donde se estudió las diferentes fuentes de variación de la RT (entre majadas, entre animales de una majada y lugares de muestreo en el animal). Observaron que en tres majadas de diferentes zonas de Australia (majada 1: sur-oeste de Nueva Gales del Sur, origen Saxon con diámetro medio de 21 micras; majada 2: sur-oeste de Nueva Gales del Sur, origen Peppin de 22 micras; majada 3: sur-oeste Australia del Oeste, origen Peppin de 21 micras) que la mayor variación se dio entre animales de una misma majada. Fue de 63, 47 y 58% para estas
Componente Variancia estimada del
componente Porcentaje del total
Entre majadas 11.74 24,5 %
Dentro de majadas
36,27 75,5 %
Total 48,01 100
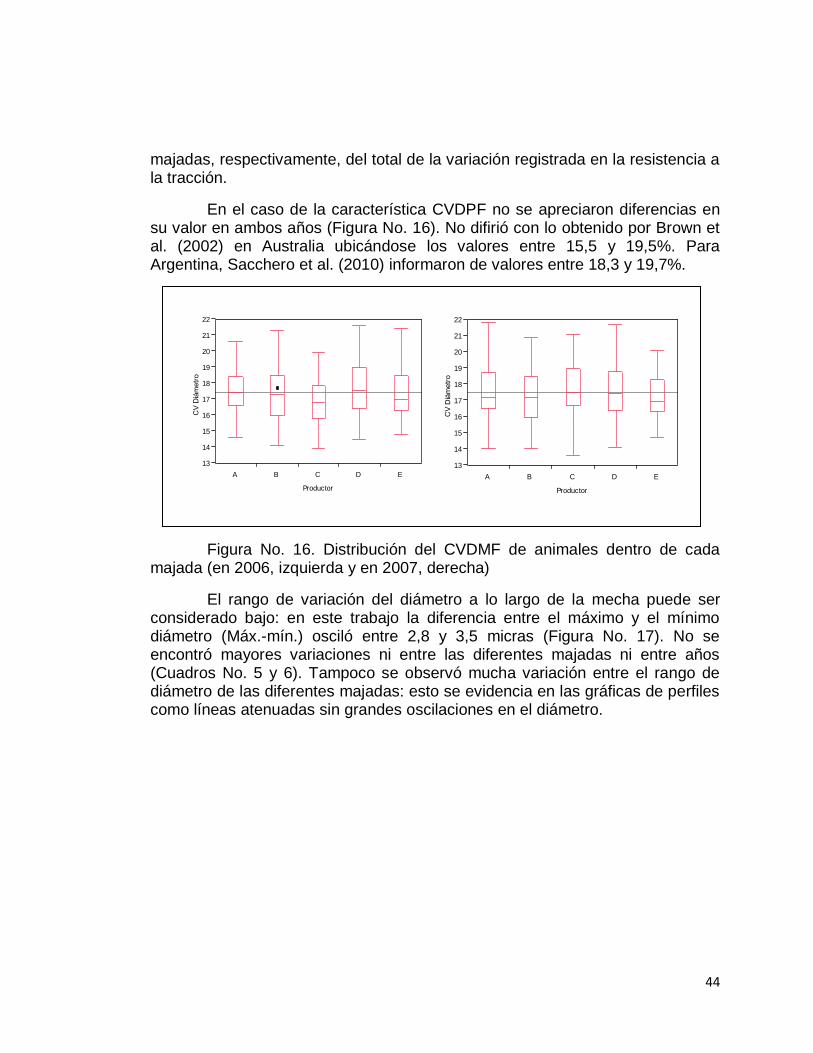
44
majadas, respectivamente, del total de la variación registrada en la resistencia a la tracción.
En el caso de la característica CVDPF no se apreciaron diferencias en su valor en ambos años (Figura No. 16). No difirió con lo obtenido por Brown et al. (2002) en Australia ubicándose los valores entre 15,5 y 19,5%. Para Argentina, Sacchero et al. (2010) informaron de valores entre 18,3 y 19,7%.
Figura No. 16. Distribución del CVDMF de animales dentro de cada majada (en 2006, izquierda y en 2007, derecha)
El rango de variación del diámetro a lo largo de la mecha puede ser considerado bajo: en este trabajo la diferencia entre el máximo y el mínimo diámetro (Máx.-mín.) osciló entre 2,8 y 3,5 micras (Figura No. 17). No se encontró mayores variaciones ni entre las diferentes majadas ni entre años (Cuadros No. 5 y 6). Tampoco se observó mucha variación entre el rango de diámetro de las diferentes majadas: esto se evidencia en las gráficas de perfiles como líneas atenuadas sin grandes oscilaciones en el diámetro.
13
14
15
16
17
18
19
20
21
22
CV
Diá
metr
o
A B C D E
Productor
13
14
15
16
17
18
19
20
21
22
CV
Diá
metr
o
A B C D E
Productor

45
Figura No. 17. Distribución de diferencia entre DMáx. y Dmín. del perfil dentro de cada majada (2006 derecha y 2007 izquierda).
Estos valores observados fueron similares a los resultados encontrados en trabajos australianos realizados en zonas típicamente productoras de lanas finas; se encontraron diferencias entre el máximo y mínimo diámetro del perfil de diámetro en el rango de 2,0 a 4,5 micras. En el trabajo hecho con ovejas Merino del proyecto “lana fina” de CSIRO en Armidale, Brown (2000) encontró valores de 4,5 micras para esta característica, en lana que tuvo 43,8 N/Ktex de resistencia a la tracción. También en Armidale, pero en trabajos con capones de dos líneas genéticas, los valores fueron de 2,9 y 3,1 micras. Otro trabajo hecho con ovejas (Brown et al., 2002) las diferencias entre diámetro máximo y mínimo del perfil de lana fueron de 3,3 y 3,2 micras. Por su parte, ovejas Merino del establecimiento Mount William en la localidad de Guyra (Nueva Inglaterra) tuvieron valores de aproximadamente 2 micras (Abella, 2005).
Puestos en perspectiva, los valores de diferencia entre valores de diámetro máximo y mínimo en el perfil citados no resultan elevados si se tiene en consideración que correspondieron a lana producida en condiciones de pastoreo. De forma comparativa, en producción de lana de animales en confinamiento y con dieta controlada -denominada Sharlea- se obtuvo un rango de 2-3 micras en esta característica (Smith et.al., 2006).
En ambiente templados como el de Northern Tablelands en Nueva Gales del Sur (Australia) la fluctuación estacional del diámetro es menor que la ocurrida en ambientes mediterráneos. Por ejemplo, Schlink et al. (1996) reportaron una diferencia de 8 micras a lo largo de la mecha en lana producida por capones Merino en pasturas, con el diámetro máximo ocurriendo en
1
2
3
4
5
6M
ax-M
in
A B C D E
Productor
1
1,5
2
2,5
3
3,5
4
4,5
5
5,5
Max-M
in
A B C D E
Productor

46
primavera y el mínimo en otoño. Esto significó una amplitud estacional del 47% (Schlink et al., 1999). Esta diferencia puede ser incluso de hasta 12 micras, como fue informado por Hansford, citado por Naylor y Hansford (1999). En sus trabajos realizados en zonas con clima mediterráneo, estudiaron la forma de manipular los perfiles de diámetro de la lana para incrementar la resistencia a la tracción, encontrando que los mayores valores para esta característica fue en ovejas con menor variación en el diámetro (tanto entre fibras y a lo largo de las fibras), mayor valor de diámetro mínimo y mayor valor de diámetro máximo.
Con los resultados obtenidos hasta ahora de similares DMF, CVDMF moderados y bajos valores de rango (dentro y entre majadas) se podría inferir que los animales que presentan baja resistencia, no es a consecuencia de factores ambientales que ocasionen máximos o mínimos diámetros muy marcados generando una mayor variación a lo largo de la fibra (ya que dicha situación no se dio en el tipo de ambiente en el que estuvieron estas majadas).
Considerando que el CVDMF está determinado por la variación del diámetro a lo largo de la fibra y entre fibras de una misma mecha; y dado que la variación a lo largo de la fibra está afectada por condiciones ambientales (siendo muy bajo el valor registrado en las majadas estudiadas), podemos inferir que los bajos valores de RT están explicados por una mayor variación del diámetro entre fibras de una misma mecha.
Considerando lo citado por Hygate (2001), Swan (2010b) que la heredabilidad de la variación del diámetro entre fibras es alta, y viendo el efecto de esta sobre la RT, es importante la selección de animales con bajos valores de CVDMF, de origen genético principalmente.
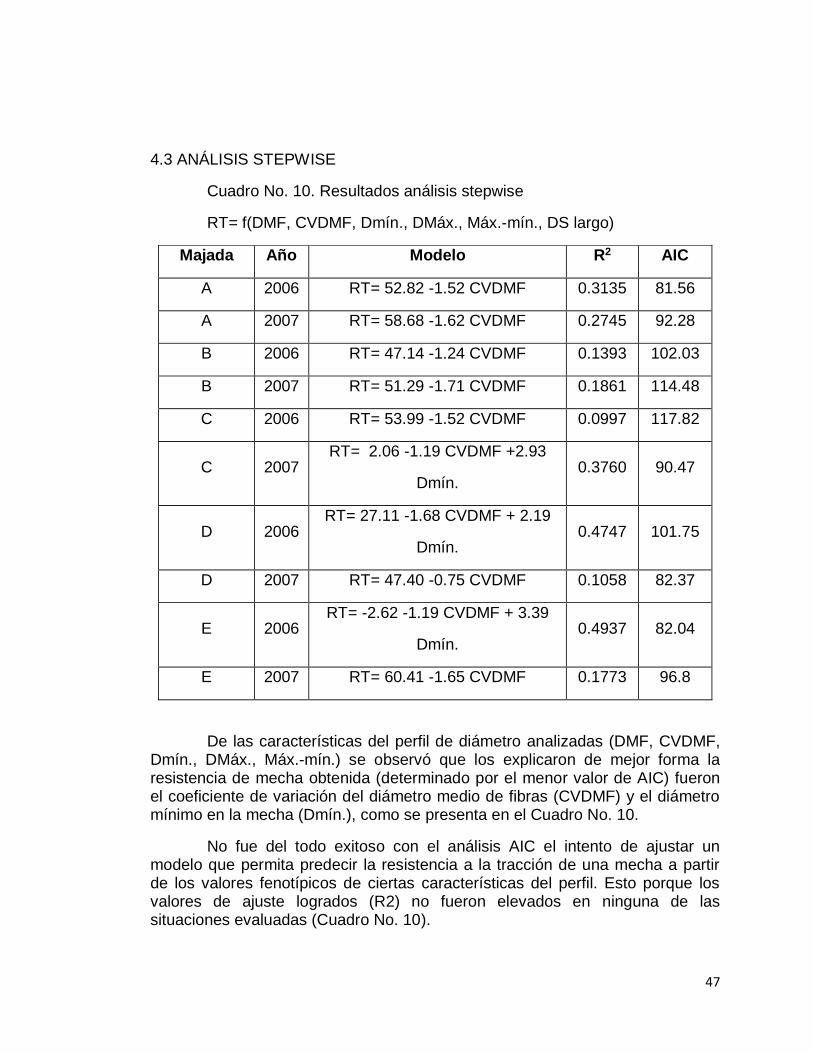
47
4.3 ANÁLISIS STEPWISE
Cuadro No. 10. Resultados análisis stepwise
RT= f(DMF, CVDMF, Dmín., DMáx., Máx.-mín., DS largo)
Majada Año Modelo R2 AIC
A 2006 RT= 52.82 -1.52 CVDMF 0.3135 81.56
A 2007 RT= 58.68 -1.62 CVDMF 0.2745 92.28
B 2006 RT= 47.14 -1.24 CVDMF 0.1393 102.03
B 2007 RT= 51.29 -1.71 CVDMF 0.1861 114.48
C 2006 RT= 53.99 -1.52 CVDMF 0.0997 117.82
C 2007 RT= 2.06 -1.19 CVDMF +2.93
Dmín. 0.3760 90.47
D 2006 RT= 27.11 -1.68 CVDMF + 2.19
Dmín. 0.4747 101.75
D 2007 RT= 47.40 -0.75 CVDMF 0.1058 82.37
E 2006 RT= -2.62 -1.19 CVDMF + 3.39
Dmín. 0.4937 82.04
E 2007 RT= 60.41 -1.65 CVDMF 0.1773 96.8
De las características del perfil de diámetro analizadas (DMF, CVDMF, Dmín., DMáx., Máx.-mín.) se observó que los explicaron de mejor forma la resistencia de mecha obtenida (determinado por el menor valor de AIC) fueron el coeficiente de variación del diámetro medio de fibras (CVDMF) y el diámetro mínimo en la mecha (Dmín.), como se presenta en el Cuadro No. 10.
No fue del todo exitoso con el análisis AIC el intento de ajustar un modelo que permita predecir la resistencia a la tracción de una mecha a partir de los valores fenotípicos de ciertas características del perfil. Esto porque los valores de ajuste logrados (R2) no fueron elevados en ninguna de las situaciones evaluadas (Cuadro No. 10).

48
En todas las situaciones en que el CVDMF formó parte del modelo, fue negativa la relación con la resistencia: a mayor valor de coeficiente, menor el valor de resistencia (en rango de -0,75 a -1,71). Por su parte, en aquellas situaciones en que el diámetro mínimo fue incluido en el modelo la relación fue positiva en los tres casos: a mayor valor de esta característica, mayor resistencia.
El coeficiente de determinación (R2) de los diferentes modelos osciló entre 0,09 y 0,49. Los mayores valores ocurrieron en los tres casos que el Dmín. fue también incorporado, junto con el CVDMF.
En trabajo de Ciappesoni et al. (2008) se recomendó la inclusión de la RT en los objetivos de selección tanto del NFG como de las cabañas comerciales, en estas se podrá usar como criterio de selección al CVDMF. La correlación genética entre RM y CVD fue -0,82 ± 0,08, demostrándose así el potencial de incluir el CVD como criterio para la selección por RM.
En ninguna de las situaciones analizadas la RT presentó relación significativa con el DMF, DMáx., Máx.-mín. por lo tanto se descarta parte de una de las hipótesis del trabajo que plantea que existe correlación fenotípica entre la resistencia y dichos caracteres.
Los valores de determinación obtenidos coinciden con lo citado en la bibliografía, donde Brown et al. (2002) presentaron valores de r= -0,2 a -0,51 coincidiendo con el rango de valores citados por Yamin et al. (1999), Adams et al. (2000). En su estudio con perfiles de diámetro, Brown (2000a) sugirió que la variación del diámetro a lo largo de la mecha brindó mayores beneficios para predecir la resistencia a la tracción cuando la variación a lo largo del perfil no fue pronunciada. En situaciones de mayor variación a lo largo de la mecha, el diámetro mínimo en el perfil adquirió mayor importancia.
En ambiente mediterráneo, Adams et al. (1998) encontraron que la importancia relativa de las diferentes características de la mecha diferían entre líneas de Merino: el diámetro mínimo en la mecha y el diámetro promedio fueron las más importantes para la tradicional genética fina de la zona de Nueva Inglaterra (Nueva Gales del Sur). Los coeficientes de correlación entre ambas características y la resistencia a la tracción fueron de 0,59 y 0,49, respectivamente.

49
4.4 PERFILES DE DIÁMETRO
Los perfiles de diámetro de fibra son reflejo de la forma en que los animales responden al ambiente de producción donde se encuentran (Brown et al., 1999). Las oscilaciones observadas en el diámetro en ambos años de estudio fueron similares para las cinco majadas, con una tendencia de incremento en el diámetro hacia la base de la mecha (Figura No. 18).
Dada la alta variación observada entre animales de una misma majada es que las curvas de las cinco majadas no tuvieron un alto porcentaje de ajustes entre ellas. De todas formas se pudo observar ciertas tendencias y comportamientos en todas las majadas. Estos resultados coinciden con trabajos australianos de Smith et al. (2006) quienes observaron un amplio rango de formas en los perfiles ajustados, como consecuencia de la mayor variación existente a nivel de animales individuales. Pese a esto, un formato general pudo ser ajustado a ese grupo de animales. A su vez, encontraron que ese perfil promedio de cada majada se repetía a través de los años pese a la importante variación individual existente.
En 2006, la tendencia de las majadas fue a incrementar el diámetro hacia la base de la mecha, es decir hacia el último tercio de la gestación. Sin embargo, en 2007 el comportamiento a lo largo del año fue diferente observándose un incremento del diámetro en el medio de la mecha y disminución hacia la base.
Como era esperable en animales de la raza Merino –sin incidencia del fotoperiodo- la estacionalidad en la producción de lana coincidió con la disponibilidad forrajera (Butler y Head, 1992).

50
17
18
19
20
21
0 10 20 30 40 50 60 70 80 90 100
Distancia de la punta (%)
Productor=="A" Productor=="B" Productor=="C" Productor=="D"
Productor=="E"
Productor A Productor B Productor C Productor D Productor E
Figura No. 18. Distribución del diámetro en función a la distancia de la punta agrupado por majada (izquierda 2006, derecha 2007). El grado de ajuste para el año 2006 oscilo entre 0,11-0,32 y para el año 2007 entre 0,11-0,33.
En concordancia, Hansford (1997) remarcó que es variada la respuesta individual a las condiciones ambientales, pese a que existe un patrón de comportamiento de esa majada. También remarcó que esa variación dentro de una majada es el resultado del potencial genético de cada animal para producir lana, ocurriendo como consecuencia amplias diferencias en resistencia a la tracción medidas en cada majada.
En las majadas E y C, fue en las que se observó mayor grado de ajuste cada año (R2: 0,32 y 0,33) para 2006 y 2007 respectivamente (Figura No. 19). Estos valores de ajuste relativamente bajos son consecuencia de la gran variación individual que se observó dentro de cada majada.
16
17
18
19
20
21
0 10 20 30 40 50 60 70 80 90 100
Distancia de la punta (%)
Diámetro
()
Diámetro ()

51
15
16
17
18
19
20
21
22D
iám
etr
o
0 10 20 30 40 50 60 70 80 90 100
DFT100
14
15
16
17
18
19
20
21
Diá
metr
o
0 10 20 30 40 50 60 70 80 90 100
DFT100
(DFT 100: distancia desde la punta en %).
Figura No. 19. Distribución del diámetro de los animales de las majadas que más se ajustaron a la media poblacional para el año 2006 y 2007 respectivamente (izquierda, majada E línea tendencia verde año 2006; derecha, majada C línea de tendencia azul año 2007).
En ambos años de estudio se observó una tendencia similar en el comportamiento del diámetro de las majadas, dándose en el último tercio de la mecha cierta estabilidad en cuanto al incremento del diámetro. Todas las ovejas Merino que participaron en este trabajo fueron encarneradas en otoño y con esquila preparto a fines del invierno.
La estabilidad en el diámetro se asocia al último tercio de la gestación (mayores requerimientos nutricionales) y fin del invierno, coincidiendo con el incremento de los requerimientos del animal para mantener la producción de la fibra y mayor desarrollo del feto.
Los mínimos registros de diámetro (punta de la mecha) coinciden con la época del año de bajo aporte forrajero (invierno) y con animales incrementando sus requerimientos debido a su estado fisiológico (último tercio de la gestación-lactación). Este comportamiento del DMF coincide con el trabajo de Polanco (2005) donde Laporte, plantea que los menores valores tanto de crecimiento de lana como de diámetro se alcanzan en los meses invernales.
Independientemente del estado fisiológico y alimento ofrecido, se observó (en la Figura No. 18 de perfiles) que el ambiente afectó de manera similar a las cinco majadas cada año.
Como se observa en la Figura No. 18 y de acuerdo con el trabajo de Hansford y Geenty (2004), el PDF obtenido por el OFDA muestra cuál fue el

52
comportamiento a lo largo del año del diámetro de una majada; coincidiendo con dicho trabajo los perfiles son una herramienta que se podrían utilizar para realizar un análisis de cuál fue el comportamiento del diámetro en las majadas ambos años asociado a los manejos retrospectivos que se le realizaron a las mismas.
Los perfiles por sí solos no son una herramienta que prediga el comportamiento o brinde un resultado específico, habría que asociarlo con algún parámetro del perfil que sea utilizado en los programas de mejoramiento (como el CVDF) y relativizarlo a una majada específica.

53
5. CONCLUSIONES
Este trabajo realizado en majadas Merino productoras de lanas finas en cinco establecimientos ubicados en diferentes zonas de suelos de Basalto presentó resultados originales en lo referente a la característica resistencia a la tracción de lanas finas producidas en Uruguay.
La lana de ovejas –con diámetro promedio de 18,7 y 18,8 micras- tuvo valores promedio de resistencia de 29,4 y 30,9 N/ktex en los dos años del trabajo, respectivamente. Estos valores fueron levemente inferiores (13%) al promedio de lanas finas australianas.
Dentro de cada majada se observó una marcada variación de resistencia a la tracción entre animales, con diferencias de hasta 32.1 N/ktex entre ovejas que compartieron el mismo ambiente de producción. Esta variación fue cuantificada, encontrándose que en el primer año el 73% de la variación total fue explicada por diferencias entre animales y el restante 27% entre diferentes establecimientos un año y 87% y 13%, respectivamente en el segundo.
La diferencia entre el valor promedio de diámetro máximo y mínimo en el perfil (DMáx.-Dmín.) fue de 3,0 y 3,1 micras en ambos años, respectivamente. Esto evidenció una no pronunciada variación de diámetro a lo largo del año en las condiciones productivas del Uruguay. Estos valores fueron similares a los logrados en zonas típicamente productoras de lanas finas de Australia.
La segunda hipótesis de este trabajo no fue descartada; del análisis “Stepwise” de las características del perfil de diámetro asociadas a la resistencia a la tracción se observó que el coeficiente de variación del diámetro (CVD) estuvo negativamente relacionado a la misma: a mayores valores de CVD, menores valores de resistencia. Esta relación negativa ocurrió en las cinco majadas en ambos años de estudio; sin embargo, el valor del coeficiente de determinación encontrado no fue alto en ninguna de las situaciones (R2 osciló entre 0,09 y 0,49). Otra característica que presentó una relación significativa –únicamente en tres situaciones- fue el valor de diámetro mínimo (Dmín.) en el perfil; a mayores valores de Dmín., mayor resistencia.
Fue posible ajustar perfiles para analizar en los animales de una majada la variación del diámetro a lo largo del año. Por las variadas respuestas individuales a las condiciones ambientales, el grado de ajuste no fue alto. Pese a esto, se pudo observar un patrón de comportamiento común dentro de cada majada con una tendencia general a incrementos del diámetro hacia el medio y una estabilización del diámetro en el último tercio de la mecha de las ovejas.

54
Los perfiles de diámetro podrían ser una herramienta para analizar retrospectivamente el comportamiento del diámetro de una majada a lo largo de un año (o el período que involucre el largo de la mecha). Sin embargo, no permitiría predecir el comportamiento de determinada fibra, ya que una vez obtenido el perfil no es posible realizar medidas correctivas que alteren la fibra de lana ya producida.

55
6. RESUMEN
En cinco majadas Merino de Uruguay (con diámetro medio de 18,7 - 18,8 micras) ubicadas en campos con suelos de Basalto se midió la resistencia a la tracción de lana producida por ovejas de cría durante dos años: produjeron lanas con valores promedio de resistencia a la tracción para cada año de evaluación de 29,4 y 30,9 N/ktex. Fue pronunciado el rango de valores de RT dentro de las majadas: de hasta 30,3 N/ktex en 2006 y hasta 32,1 N/ktex en 2007. Esto ejemplifica la marcada variación individual que existió dentro de cada majada a nivel de esta característica. De la variación observada en los valores de resistencia, la mayor parte se debió a diferencias entre animales de una misma majada (73 y 83 % del total para el año 2006 y 2007 respectivamente) y la restante a diferencias entre establecimientos. La variación del diámetro a lo largo de la mecha no fue pronunciada, con diferencias promedio de 3,0 y 3,1 micras entre el diámetro máximo y mínimo de cada animal. Se analizaron distintos caracteres del perfil de diámetro de fibra generados por el OFDA 2000 para determinar si hubo relación con la resistencia a la tracción. En el análisis “Stepwise” de características del perfil de diámetro, el coeficiente de variación mostró una correlación negativa con la resistencia aunque con un bajo coeficiente de determinación (0,09-0,49); el diámetro mínimo presentó relación significativa y positiva en tres situaciones. Al ajustar los perfiles de diámetro para las diferentes majadas -pese a que el grado de ajuste no fue alto- se pudo observar cierto patrón de comportamiento, con un incremento de diámetro hacia la mitad de la mecha y estabilización del mismo hacia el último tercio de la mecha. El perfil de diámetro puede ser una herramienta para observar retrospectivamente el comportamiento del diámetro y otros caracteres de determinado animal a lo largo de la mecha pero la medición de estas características no aportaría mayormente para mejorar indirectamente la resistencia de lanas finas.
Palabras clave: Diámetro medio de fibras; Resistencia a la tracción; Coeficiente de variación del diámetro; Perfil de diámetro de fibras; OFDA.

56
7. SUMMARY
On five Merino flocks in Uruguay (with 18,7-18,8 micron fibre diameter average) localised on Basalto soil, staple strength were mesaured two years on rearing ewes. The staple strength for each year was 29,4 and 30,9 N/ktex. Difference of 30,3 N/ktex were observed within flocks in 2006 and 32,1 N/ktex in 2007. This shows the high variability that exist within each flock. Most of staple strength variance was about difference between animals on a same flock (73 and 83% of the total for 2006 and 2007 respectively) the remaining difference was explain about difference between flocks. Fibre diameter variation along staple wasn´t high, with difference of 3,0 and 3,1 microns between maximun and minimum diameter of each animal. Different characteristics of fibre diameter profile generated by OFDA 2000 were analized to determine their relation with staple strength. In the “Stepwise” analysis, coefficient of variation showed negative correlation with staple strength with a low determination coefficient (0,09 to 0,49), minimum diameter showed significant and positive correlation in three situations. Trying to set profile diameter to the different flocks (adjustment degree was not high), it was a diameter increased to mid staple and stabilization toward last third of staple. Fibre diameter profile could be a tool to see what happend with diameter and other characteristics along staple but not to improve staple strength on fine wool.
Keywords: Medium fibre diameter; Staple strength; Coefficient of variation of diameter; Fibre diameter profile; OFDA (optical fibre diameter analyser).

57
8. BIBLIOGRAFÍA
1. ABELLA, I. 2005. Grazing management, supplementation and wool production of Merino ewes. Armidale, Australia, The University of New England School of Rural Science and Agriculture. 90 p.
2. ADAMS, N. R.; BRIEGEL, J. R.; RITCHIE, A. J. M. 1998. Relationship of
staple strength to variation in fibre diameter in sheep selected for or against staple strength. Animal Production in Australia. 22: 423.
3. ________.; HEWETT, L. J.; SCHLINK, A. C.; BRIEGEL, J. R. 2000.
Phenotypic correlates of staple strength. Asian Australian Journal Animal Science. 13: 289-292.
4. AGUERRE, J.; CORONEL, F. 2008. Lanas Merino finas y superfinas;
producción y perspectivas. In: Seminario Internacional (1º., 2003, Salto). Trabajos presentados. Salto, Uruguay, INIA/SUL/SCMAU/CLU. p. irr.
5. ATKINS, K. D.; POPE, C. E.; SEMPLE, S. J. 2004. Maximising the
benefits from on-farm fibre measurement. Animal Production in Australia. 1: 9–12.
6. AWTA. 2011. Micron profile data for the the australian wool clip. (en
línea). Kensington. s.p. Consultado 21 mar. 2012. Disponible en http://www.awta.com.au/index.php/en/
7. BAXTER, P. s.f. Description and performance of the Agritest Staple
Breaker. (en línea). Wellington, New Zealand. SGS Wool Testing Services. Consultado set. 2011. Disponible en http://www.sgs.com/~/med.ia/Global/Documents/Third%20Party%20Documents/Third%20Party%20Technical%20and%20Research%20Papers/TP_L4_Staple%20Length%20and%20Strength.
8. ________. 2001a. On-farm classing of animals and fleeces with the
OFDA2000. Wool Technology and Sheep Breeding. 49 (2): 133-155. 9. ________. 2001b. Precision of measurement of diameter, and diameter-
length profile, of greasy wool staples on-farm, using the OFDA2000 instrument. Wool Technology and Sheep Breeding. 49 (1): 42-52.

58
10. BERRETTA, E.; BEMHAJA, M. 1998. Producción estacional de comunidades naturales sobre suelos de basalto de la unidad Queguay chico. In: Seminario de Actualización en Tecnologías para Basalto (1998, Tacuarembó). Trabajos presentados. Montevideo, INIA. pp. 17-27 (Serie Técnica no. 102).
11. ________. 2005. Producción y manejo de la defoliación en campos
naturales de Basalto. In: Seminario de Actualización Técnica en Manejo de Campo Natural (2005, Treinta y Tres). Trabajos presentados. Montevideo, INIA. pp. 61-73 (Serie Técnica no. 151).
12. BIANCHI, G.; GAMBETTA, A. 1991. Efecto de la raza, del nivel nutritivo y del estado fisiológico en la producción de lanas en ovejas Merino y Corriedale. Montevideo, UdelaR. Facultad de Agronomía. 122 p.
13. BROWN, D. J.; CROOK, B. J.; PURVIS, I. W. 1999. Genotype and
environmental differences in fibre diameter profile characteristics and their relationship with staple strength in Merino sheep. (en línea). In: Meeting of the Association for the Advancement of Animal Breeding and Genetics (13th., 1999, Manduroh, WA). Proceedings. Armidale, CSIRO. pp. 274-277. Consultado set. 2011. Disponible en http://www.aaabg.org/livestocklibrary/1999/AB99062.pdf
14. ________. 2000a. The estimation and utilisation of variation in fibre
diameter profile characteristics between sheep. Post-Graduate Doctoral Thesis. Armidale, Australia. The University of New England. 251 p.
15. ________.; CROOK, B. J.; PURVIS, I. W. 2000b. The estimation of fibre
diameter profile characteristics using reduced profiling techniques. Wool Technology and Sheep Breeding. 48: 1-14.
16. ________.; ________.; ________. 2002a. Differences in fibre diameter
profile characteristics in wool staples from Merino sheep and their relationship with staple strength between years, environments, and bloodlines. Australian Journal of Agricultural Research. 53. 481-491.
17. ________.; ________.; ________. 2002b. Variation in fibre diameter
profile characteristics between wool staples in Merino sheep. Wool Technology and Sheep Breeding. 48: 86-93.

59
18. ________.; ________. 2005. Environment responsiveness in fibre diameter in grazing fine wool Merino sheep. Australian Journal of Agricultural Research. 56: 673-684.
19. BUTLER, L. G.; HEAD, G. M. 1992. Photoperiodic rhythm of wool growth
and its contribution to seasonal wool production by the Merino, Polwarth and their reciprocal crosses in southern Australia. Australian Journal of Agriculture. 34: 311-317.
20. CARDELLINO, R.; TRIFOGLIO, J. 2003. El mercado de lanas finas y
superfinas. Lanas Merino finas y superfinas; producción y perspectivas. In: Seminario Internacional de Lanas Finas y Superfinas (1º., 2003, Salto). Trabajos presentados. Salto, Uruguay, INIA/SUL/SCMAU/CLU. pp. 7-15.
21. CENTRAL LANERA URUGUAYA. 2010. Club de Merino fino. (en línea)
Montevideo. s.p. Consultado may. 2011. Disponible en http://www.central-lanera.com.uy/sitio/html/proyectos
22. CIAPPESONI, G.; MONTOSSI, F.; DE BARBIERI, I.; GIMENO, D.;
AGUILAR, I. 2008. Impacto de un núcleo de selección en la mejora genética de la raza Merino en Uruguay. In: Congreso Latinoamericano de Genética (13º.), Congreso Peruano de Genética (4º., 2008, Lima). Trabajos presentados. Lima, Perú, s.e. p. irr.
23. CORBETT, J. L. 1979. Physiological and enviromental limitations to wool
growth. Variation in wool growth with physiological state. Sydney, University of New England. 405 p.
24. DENNEY, G. D. 1990. Phenotypic variance of fibre diameter along wool
staples and its relationship with other raw wool characters. Australian Journal of Experimental Agriculture. 30: 463-467.
25. DONALD, A. D. 1979. Physiological and enviromental limitations to wool
growth. Effects of parasites and disease on wool growth. Sydney, University of New England. 405 p.
26. DOYLE, P.; THOMPSON A. 1993. Grazing management for fine and
sound wool. In: National Workshop on Management for Wool Quality in Mediterranean Environments (1992, Perth). Proceedings. Perth, Western Australia. Departament of Agriculture. pp. 115-124.

60
27. FRANZ, N. 2008. Una alternativa para la producción ovina; las lanas finas. Corrientes, INTA. pp. 1-3. (Noticias y comentarios no. 439).
28. FORMOSO, D. 2005. La investigación en utilización de pasturas
naturales sobre cristalino desarrollada por el secretariado uruguayo de la lana. In: Seminario de Actualización Técnica en Manejo de Campo Natural (2005, Treinta y Tres). Trabajos presentados. Montevideo, INIA. pp. 51-59 (Serie Técnica no. 151).
29. GAGGERO, J.; MUTTONI, M.; RODRÍGUEZ, B. 2006. Correlaciones
fenotípicas entre características de la piel y calidad de la lana en cuatro cabañas pertenecientes al núcleo Merino fino. Tesis Ing. Agr. Montevideo, Uruguay. Facultad de Agronomía. 75 p.
30. GREEFF, J. C.; LEWER, R.; PONZONI, R. W.; PURVIS, I. W. 1995.
Staple strength: Progress towards elucidating its place in Merino breeding. Proceedings of the Australian Association of Animal Breeding and Genetics 11: 595-601.
31. ________. 2006. Coefficient of variation of wool fibre diameter in Merino
breeding programs. (en línea). Katanning, Government of Western Australia. Department of Agriculture. pp. 1-4 (Farmnote 46/98). Consultado nov. 2011. Disponible en http://www.agric.wa.gov.au/objtwr/imported_assets/content/aap/sl/wool/f04698.pdf
32. GOOGLE. 2012. Google earth (en línea). s.l. s.p. Consultado mar. 2012.
Disponible en http://www.google.com/earth/ 33. HANSFORD, K.; EMERY, G. B.; TEASDALE, D. 1985. A method to
measure the diameter profile of wool staples. Wool Technology and Sheep Breeding. 33: 25-27.
34. ________.; KENNEDY, J. P. 1988. Relationship between the rate of
change in fibre diameter and staple strength. Proceedings of the Australian Society of Animal Production. 17: 415-419.
35. ________.; ________. 1990. The relationship between variation in fibre
diameter along staples and staple strength. In: International Wool Textile Research Conference (8th., 1990, Chirstchurch). Proceedings. Christchurch, New Zealand, s.e. pp. 590-598.

61
36. ________. 1997. Wool strength and topmaking. Wool Technology and Sheep Breeding. 45 (4): 309-320.
37. ________.; GEENTY, K. G. 2004. High rainfall areas. In: Fibre Diameter
Profiles Workshop (2004, Armidale, Australia). Proceedings. Armidale, Australian Sheep Industry CRC. pp. 7-9.
38. HYGATE, L. 2001. The genetics of staple strength in Merino sheep.
Proceedings of the Australian Sheep Veterinary Society. 11: 103-105.
39. HYND, P. I.; SCHLINK, A. C. 1992. Factors responsible for variation in
the strength of wool fibres. In: National Workshop on Management for Wool Quality in Mediterranean Environments (1992, Perth, Western Australia). Proceedings. South Perth, Western Australia, Woolwise. pp. 92-98.
40. INTERNATIONAL WOOL TEXTIL ORGANIZATION (IWTO). 2011.
Statistics for the global woll production and textile industry; market information. (en línea). Brussels. 66 p. Consultado abr. 2012. Disponible en http://www.iwto.org/uploaded/publications/Order_Forms/IWTO_Market_Information
41. ________. 2012. IWTO specifications "Red Book". Brussels. 13 p. 42. JACKSON, M.; ASHMAN, I.; MARLER, J.; LINDSAY, A.; MAHAR, T.;
COUCHMAN, B. 2004. Trial evaluating additional measurement: 2001-2004 (TEAM-3); final report. Wool Technology and Sheep Breeding. 52 (4): 260-277.
43. JACKSON, N.; DOWNES, A. 1979. The fibre diameter profile of wool
staples from individual sheep. Australian Journal of Agricultural Research. 30: 163-171.
44. JMP. IN. 2002. JMP start statistics version 5. (en línea). Cary, NC, SAS
Institute. s.p. Consultado nov. 2011. Disponible en http:// www.jmp.com
45. KEMPTON, T. J. 1979. Physiological and environmental limitations for
wool growth; protein to energy ratio of absorbed nutrients in relation

62
to wool growth. Armidale, The University of New England Publishing Unit. pp. 209-222.
46. LAPORTE, O. 1975. Variaciones del crecimiento y el diámetro de la fibra
de lana en la Patagonia desértica. Trelew, Argentina, INTA: 15 p. 47. LEWER, R.; LI, Y. 1994. Some aspects of selection for staple strength.
Wool Technology and Sheep Breeding. 42 (2): 103-111. 48. MAC KINLEY, A. H.; IRVINE, P. A.; ROBERTS, A. M.; ANDREWS, M. W.
1976. The direct partitioning of variation in fibre diameter in tender wool. Proceedings of the Australian Society of Animal Production. 11: 181-184.
49. MAHAR, T. 2004. Wool profile metrology. In: Fibre Diameter Profiles
Workshop (2004, Armidale, Australia). Proceedings. Armidale, Australian Sheep Industry CRC. pp. 59-61
50. MASTERS, D. G. 1992. The influence of reproduction on wool growth.
Proceedings of the Australian Society of Animal Production. 19: 211-212.
51. MITCHELL, W. J.; COTTLE, D. J.; ROBERTS, E. M. 1992. The
evaluation of different sampling methods for the measurement of mean and distribution of fibre diameter. Proceedings of the Association for the Advancement of Animal Breeding and Genetics. 10: 444-447.
52. MONTOSSI, F.; SAN JULIÁN, R.; DE MATTOS, D.; BERRETA, E.J.;
RÏOS, M.; ZAMIT, W.; LEVRATTO, J. 1998a. Alimentación y manejo de la oveja de cría durante el último tercio de la gestación en la región de basalto. In: Seminario de Actualización en Tecnologías para Basalto (1998, Tacuarembó). Trabajos presentados. Montevideo, INIA. pp. 195-208 (Serie Técnica no. 102).
53. ________.; ________.; ________.; ________.; ZAMIT, W.; LEVRATTO,
J.; RÍOS, M. 1998b. Producción de lana fina: una alternativa de valorización de la producción ovina sobre suelos superficiales del Uruguay con escasas posibilidades de diversificación. In: Seminario de Actualización en Tecnologías para Basalto (1998, Tacuarembó). Trabajos presentados. Montevideo, INIA. pp. 307-315 (Serie Técnica no. 102).

63
54. ________.; DE BARBIERI, I.; CIAPPESONI, G.; DE MATTOS, D.;
MEDEROS, A.; LUZARDO, S.; SOARES DE LIMA, J.; DE LOS CAMPOS, G.; NOLLA, M.; SAN JULIÁN, R.; GRATTAROLA, M.; PÉREZ JONES, J.; DONAGARAY, F.; FROS, A. 2007. Los productos logrados en los primeros 8 años (1998-2006) de existencia del Proyecto Merino Fino del Uruguay. In: Montossi, F.; De Barbieri, I. eds. Proyecto Merino Fino del Uruguay; una visión con perspectiva histórica. Montevideo, INIA. pp. 17-36 (Boletín de Divulgación no. 90).
55. NAGORCKA, B. N. 1979. The effect of photoperiod on wool growth. In:
Black, J. L.; Reis, P. J. eds. Physiological and environmental limitations to wool growth. Sydney, The University of New England. pp. 127-137.
56. NAYLOR, G. R. S.; HANSFORD, K. A. 1999. Fibre diameter properties in
processed top relative to the staple for wool grown in a Mediterranean climate and shorn in different seasons. Wool Technology and Sheep Breeding. 47 (2): 107-117.
57. PETERSON, A. D. 1997. The components of staple strength.
Postgraduate Masters thesis. Perth, Australia. The University of Western Australia. 92 p.
58. ________.; GHERARDI, S. G.; DOYLE, P. T. 1998. Components of
staple strength in fine and broad wool Merino hoggets run together in a Mediterranean environment. Australian Journal of Agricultural Research. 49: 1181-1186.
59. ________.; ________. 2001. The ability of the OFDA 2000 to measure
fleeces and sale lots on-farm. Wool Technology and Sheep Breeding. 49 (2): 110-132.
60. ________.; OLDHAM, C. M. 2002. Predicting hauteur from the fibre-
diameter profiles of the OFDA2000. Wool Technology and Sheep Breeding. 50 (4): 847-852.
61. POLANCO DE VEDIA, V. 2005. Efecto de la fecha de esquila sobre
características de interés comercial en lanas finas. Tesis Ing. Agr. Buenos Aires, Argentina. Universidad de Buenos Aires (UBA). Facultad de Agronomía. 49 p.

64
62. PURVIS, I.; SMITH, J. 2004. Overview of past work and uses of profiles.
(en línea). In: Fibre Diameter Profiles Workshop (2004, Armidale, Australia). Proceedings. Armidale, Australian Sheep Industry CRC. pp. 7-9. Consultado abr. 2012 Disponible en
http://www.sheepcrc. org.au/articles.php?rc=183 63. RITCHIE, A. J. M.; RALPH, I. G. 1990. Relationship between total fibre
diameter variation and staple strength. (en línea). Proceedings of the Australian Society of Animal Production. 18: 543. Consultado nov. 2011. Disponible en http://www.asap.asn.au/livestocklibrary/1990/Ritchie90.PDF
64. ROBARDS, G. E. 1979. Regional and seasonal variation in wool growth
throughout Australia. In: Black. J. L.; Reis, P. J. eds. Physiological and environmental limitations to wool growth. Sydney, The University of New England. pp. 1-42.
65. ROTTENBURY, R. A.; BOW, M. R.; KAVANAGH, W. J.; CAFFIN, R. N.
1981. Staple strength variation in Merino flocks. Wool Technology and Sheep Breeding. 29 (4): 143-148.
66. SACCHERO, D.; MULLER, J. 2007. Diferencias en el perfil de diámetro
de fibras, largo de mecha y resistencia a la tracción de la lana, en ovejas de una majada Merino seleccionada y otra no seleccionada. RIA. 36 (2): 49-61.
67. ________.; WILLEMS, P.; MULLER J.P. 2010. Perfiles de diámetro de
fibras en lanas preparto de ovejas Merino. 1 Estudio comparativo de líneas genéticas. Revista Argentina de Producción Animal. 30 (1): 31-42.
68. SAFARI, E.; FOGARTY, N.; GILMOUR, A.; ATKINS, K.; MORTIMER, S.;
SWAN, A.; BRIEN, F.; GREEFF, J. 2007. Across population genetic parameters for wool growth and reproduction traits in Australian Merino sheep. 2. Estimates of heritability and variance components. Australian Journal of Agricultural Research. 58: 177-184.
69. SCHLINK, A. C.; MASTERS, D. G.; DOLLIN, A. 1992. Effects of amino
acid on fibre sheeding in reproducing ewes. In: Annual Scientific Meeting Nutrition Society of Australia (17th, 1992, Perth).

65
Proceedings. Proceedings of the Nutrition Society of Australia. 17: 80.
70. ________.; DOLLIN, A. 1995. Abnormal shedding contributes to the
reduced staple strength of tender wool in Western Australian Merinos. Wool Technology and Sheep Breeding. 43: 268-284.
71. ________.; LEA, J. M.; RITCHIE, A. J. M.; SANDERS, M. 1996. Impact of
a Mediterranean environment on wool follicles ans fibre growth in high and low staple strength Merino wethers. Wool Technology and Sheep Breeding. 44 (2): 81-82.
72. ________.; MATA, G.; LEA, J. M.; RITCHIE, A. J. M. 1999. Seasonal
variation in fibre diameter and length in wool of grazing Merino sheep with low or high staple strength. Australian Journal of Experimental Agriculture. 39 (5): 507-517.
73. ________.; BROWN, D. J.; LONGREE, M. 2001. Role of fibre length
variation in staple strength of Merino wool. Wool Technology and Sheep Breeding. 49 (3):202-211.
74. SMITH, J.; PURVIS, I.; LEE, G. 2006. Fibre diameter profiles—potential
applications for improving fine-wool quality. International Journal of Sheep and Wool Science. 54 (2): 54-61.
75. SUL. 2012. Lanas del Uruguay. (en línea). Montevideo. s.p. Consultado
mar. 2012. Disponible en http://www.sul.org.uy/lana_produccion_ovina.asp
76. SWAN, P. 2010a. The future of wool as an apparel fibre. In: Cottle, D. J. ed. International Sheep and Wool Handbook. Nottingham, UK, Nottingham University Press. pp. 647- 660.
77. _________. 2010b. Managing wool staple strength in intensive Merino
systems. Principles of straple strength management for fine Merino producers. Denistone East, NSW, Paul G Swan & Associates Pty. 16 p.
78. THOMPSON, A. N.; HYND, P. I. 1998. Wool growth and fibre diameter
changes in young Merino sheep genetically different in staple strength and fed different levels of nutrition. Australian Journal of Agricultural Research. 49: 889-898.

66
79. URUGUAY. MINISTERIO DE GANADERÍA AGRICULTURA Y PESCA.
DIRECCIÓN DE INVESTIGACIONES ESTADÍSTICAS AGROPECUARIAS. 2002. Encuesta ganadera año 2001; boletín informativo. Montevideo. 57 p. (Serie Encuestas no. 211).
80. WHITELEY, K. J. 1987. Some observations on the classing of fleeces for
fineness. Wool Technology and Sheep Breeding. 35 (1): 51-52. 81. ________. 2003. Características de importancia en lanas finas y
superfinas. Lanas Merino finas y superfinas; producción y perspectivas. In: Seminario Internacional (1º., 2003, Salto). Trabajos presentados. Salto, Uruguay, INIA/SUL/SCMAU/CLU. pp.17-22.
82. WILLIAMS, V. A. 1998. From fleece to fabric, 1948-1998; 50 years of
wool textile research. Victoria, Australia. CSIRO Wool Technology Geelong. 198 p.
83. YAMIN, M. H.; P. I.; PONZONI, R. W.; HILL, J. A.; PITCHFORD, W. S.;
HANSFORD, K. A. 1999. Is fibre diameter variation along the staple a good indirect selection criterion for staple strength? Wool Technology and Sheep Breeding. 47 (3): 151-158.