respuestas vocales en ranas neotropicales
TRANSCRIPT

8/3/2019 Respuestas Vocales en Ranas Neotropicales
http://slidepdf.com/reader/full/respuestas-vocales-en-ranas-neotropicales 1/7
Journalof Herpetology, Vol. 36 , No. 4, pp. 615420, 2002Copyright 2002 Society for the Study of Amphibians and Reptiles
Vocal Responses to Conspecific Call Virariation in the Neotropical Frog
Physalaemus enesefae
ZAIDA TARANO'
Postgrado en Cietncias Biol6gicas, Universidad Simn6n Bolfvar, Venezuela
ABSTRACT.-Female Physalaemus enesefae prefer long calls, low and average dominant frequency calls,
and calls repeated at short intercall intervals over short calls, high dominant frequency calls, and calls re-
peated at long intercall intervals. I tested the hypothesis that males modify their vocal behavior when faced
with potential competitors in ways that make them more attractive to females and that the more attractive
the competitor, the greater the modification. In playback experiments, I examined the responses of males tothe same stimuli used to test female responses: long and short calls, calls with low or high dominantfrequency, long and short intercall intervals, and average calls for all traits. Among the variables analyzed,
only the intercall interval was modified in the presence of the experimental stimuli. Males significantly
reduced the intercall interval (called at a faster rate) in response to all experimental calls, compared with
that when vocalizing alone, except in response to short calls and to long intercall interval calls. Males did
not respond differently to calls of average and above aver-age attractiveness. There was no difference in the
intercall interval in response to average and above average attractive stimuli. However, the intercall interval
in response to unattractive calls was significantly longer t:han that in response to calls of average attractive-
ness. The results partially support the hypothesis: males modify their spontaneous calling behavior when
faced with a competitor but do not show finely tuned graded responses in relation to the attractiveness ofthe competitor.
The advertisement calls of anurans are in-
volved in female choice and male-male compe-
tition (Greenfield, 1994; Schwartz, 1994). Male
responses to the calls of other males often in-
volve escalation: males increase intensity, rate,
duration, or complexity of their calls, especially
when confronted with males that are attractive
to females (Narins and Capranica, 1978;Schwartz and Wells, 1985; Ryan, 1985; Wagner,
1989; Bosch and Marquez, 1996). Competitive
escalation between males often parallels female
preferences based on acoustic traits. For in-
stance, female tungara frogs (Physalaemus pus-
tulosus) prefer more complex calls, and males
increase call complexitv in response to calls of
other males (reviewed in Ryan, 1985).Acoustic competition between males often in-
volves call duration, intensity, and rate; theseare call properties that tend to vary substantial-ly within individuals (Gerhardt, 1991). Male
Leptodactylus albilabris,Acris crepitans, Hyperolius
marmoratus, and Rana clamitans, however, lower
the dominant frequency of their calls, a propertythat tends not to vary dramatically within in-
dividuals in most anurans studied (summarized
in Howard and Young, 1998), when they hear
the calls of bigger or heavier males (L6pez et al.,
1988; Wagner, 1989; Grafe, 1995; Bee et al., 2000).
Phonotaxis experiments show that call dura-
tion, rate, intensity, and dominant frequency in-
fluience female choice in Physalaemus enesefae(TArano, 1999). Females prefer longer calls, more
intense calls (differences > 3 dB), calls repeated
at higher rates, and calls with lower dominantfrequency (Tarano, 1999). Among these charac-
te istics, only the dominant frequency of the callis negatively correlated with male mass, al-thaugh mass explains only a small fraction of
the variation of this trait (Tarano, 2001). Female
preferences based on the dominant frequency ofthe call may be related to the slight mating ad-
vantage observed among larger males. Obser-
vations of male behavior in the field showed that
males call from relatively stable positions. Fe-males approach them and initiate contact. Malesdo not search for females or attempt to dislodge
peired males (pers. obs.).
If females choose males based on their vocalcharacteristics, one might expect that males alter
their vocal output in relation to the attractive-
ness of their neighbors. Males could increase theattfractiveness of their calls when confronted
w th attractive competitors by reducing intercall
in terval (increasing call rate), increasing call du-ra :ion, or lowering the dominant frequency of
their calls. The more attractive the competitor,the greater the expected change in the male's

8/3/2019 Respuestas Vocales en Ranas Neotropicales
http://slidepdf.com/reader/full/respuestas-vocales-en-ranas-neotropicales 2/7
ZAIDA TARANO
cal behavior in relation to the attractiveness of
potential competitors to females, specifically,
that males modify their vocal output in ways
that increase their attractiveness to females.
MATERIALS AN]) MET1IODS
I conducted the study at Hato Masaguaral
(8°34'N, 67°35'W), a cattle ranch located 50 km
south of the town of Calabozo at Guarico state,
in the central llanos of Venezuela, from June to
August in 1997 and 1998. 1 collected males from
choruses from 1900 to 2100 h nightly and per-
formed playback experiments from 2200 h to
0100 h, in a dimly lit room. I placed individual
males, each in a plastic bag with water, on a
table 50 cm apart from each other. Males were
able to call from the small volume of water at
the bottom of the plastic bags. I placed a speak-
er (JBL M5 or SONY) 60-80 cm in front of themales in the bags and stimulated them to callby broadcasting a recording of a natural chorus.
When a male called in response to the chorus
for 5 min or more, I carefully removed the other
males from the table; thus, during the experi-
mental session only one male was on the table.I positioned the speaker in front of the calling
male and placed a microphone (Sennheiser
ME62/K6) on the table, at mid distance between
the speaker and the male, oriented to the male.
I connected the microphone to an analog tape
recorded (Marantz PMD 201) and used metaltapes to record male responses during the ex-
periments. I adjusted the stimulus intensity atthe male to 82 dB SP L (re 20 p.Pa) using a sound
pressure level meter (Radio Shack, fast response,
flat weighting). This corresponds to the average
intensity of male calls at 50 cm from the male
(Tarano, 2001). Rvan and Rand (1998) used a
similar approach (broadcast and recorded callsthrough plastic bags) and found that acoustical
distortion was minimal.
I tested males singly and usually only one
male each night. The experimental session con-sisted of an initial control stimulus followed by
several experimental stimuli and a final control.
I presented to the test male the control for 2 min
and recorded his responses during the follow-
ing 2 min. I repeated the same procedure with
all the experimental stimuli. Time interval be-
tween subsequent stimuli was that necessary toretrieve the file containing it, and usually lasted
less than 1 min. Thus, there were short silent
periods between trials. Usually males briefly in-
terrupted calling activity during this silent pe-
riod or immediately after a new stimulus wasbroadcast. This interruption suggests that males
detect changes in their vocal environment and
TABLE 1. Characteristics of the experimental stim-
uli.
Dominant IntercallDuration frequency interval
Experimiienital call (ins) (Hz) (s)
Average call (Control) 720) 900 3.6Long call 860 900 3.6
Short call 440 900 3.6
Low dominant frequency 720 630 3.6
High dominant frequency 720 1000 3.6
Short intercall interval 720 900 2.3
Long intercall interval 720 900 6
I tested each male with several successive
stimuli so that the experimental session of one
male that responded to all the sounds presented
(controls and experimental) lasted around 40min. Trial sequence was randomized fo r eachmale, to control for any order effect. Calls, either
control or experimental, were broadcast while
recording; thus, both the stimulus and the male
response were recorded. I also recorded males
that vocalized spontaneously without stimula-
tion and used these recordings to estimate male
vocal activitv without competitors. Males that
stopped responding during or after a test were
stimulated to resume calling by playing the cho-rus stimulus. I finished the session when the
male did not resume calling after 10 min ofstimulation.
Stiiiili.-Stimuli consisted of typical natural
calls recorded in the field during 1996 and 1997,so that they were within the natural range ofvariation for the population under study (Tara-
no, 2001). 1prepared and broadcast the stimulus
with a PowerBook 1400 cs using SoundEdit 16
software. Control stimuli consisted of a typical
nattural call 720 ms in duration, 900 Hz in dom-
inant frequency and repeated with a period of3.6 sec. Controls represent average attractive
calls for females. Ihe experimental stimuli con-sisted of natural calls, average in all traits but
one. Thus, I chose natural calls from the popu-
lation that were close to the population mean in
two of the three variables analyzed and more
extreme in one of the others. Characteristics ofthe experimental stimuli are summarized in Ta-
ble 1. Experimental stimuli represent above and
below average attractive calls for females.
I used calls from five males to prepare the
stimuli to reduce the effect of uncontrolled var-
iables. Thus, I had five different controls, and
five different experimental stimuli for each var-iable analyzed. Each stimulus contained one
natural call from one male repeated at a fixed
616
8/3/2019 Respuestas Vocales en Ranas Neotropicales
http://slidepdf.com/reader/full/respuestas-vocales-en-ranas-neotropicales 3/7
VOCAL RESPONSES IN PHYSALAEMUS ENESEFAE
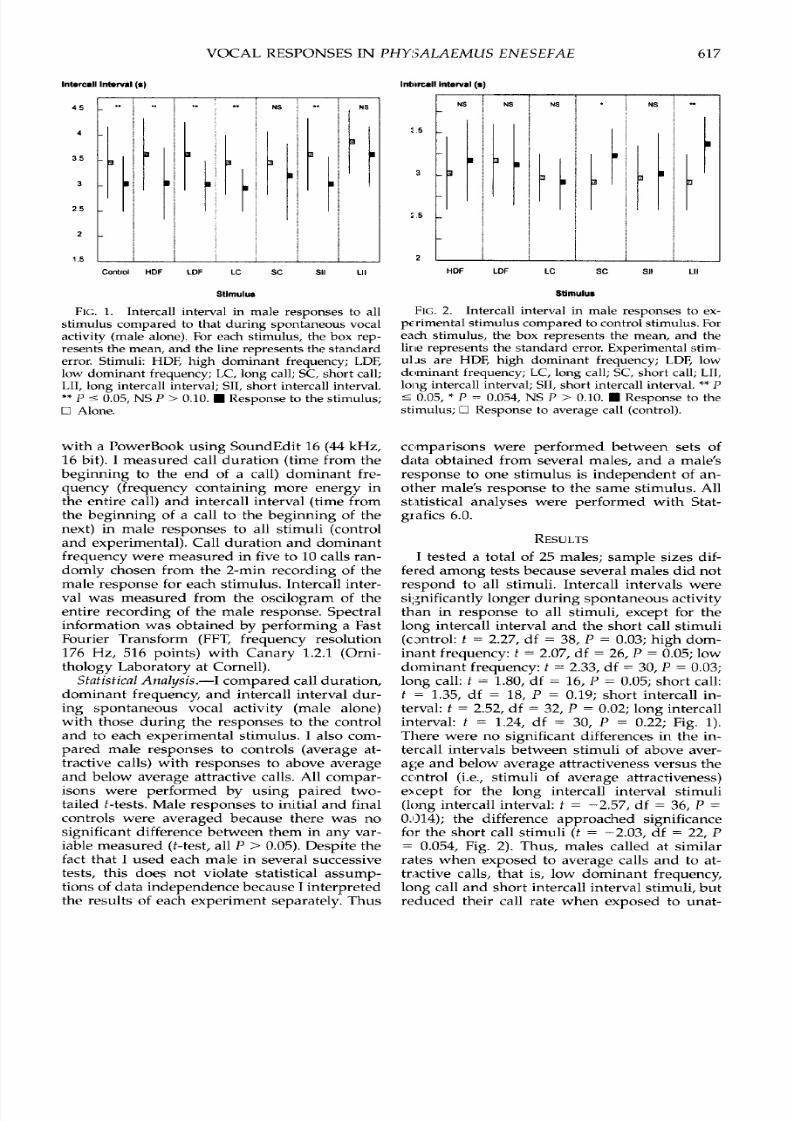
Intmrcall Interval (s)
45
4
3
25
2
1.5
Control HDF LDF LC SC SIl LIl
Stimulus
FIG. 1. Intercall interval in male responses to allstimulus compared to that during spontaneous vocal
activity (male alone). For each stimulus, the box rep-resents the mean, and the line represents the standard
error. Stimuli: HDF, high dominant frequency; LDF,low dominant frequency; LC, long call; SC, short call;
II, long intercall interval; SII, short intercall interval.**P - 0.05, NS P > 0.10. UResponse to the stimulus;E Alone.
with a PowerBook using SoundEdit 16 (44 kHz,
16 bit). I measured call duration (time from the
beginning to the end of a call) dominant fre-
quency (frequency containing more energy in
the entire call) and intercall interval (time fromthe beginning of a call to the beginning of the
next) in male responses to all stimuli (control
and experimental). Call duration and dominant
frequency were measured in five to 10 calls ran-
domly chosen from the 2-min recording of the
male response for each stimulus. Intercall inter-
val was measured from the oscilogram of the
entire recording of the male response. Spectral
information was obtained by performing a FastFourier Transform (FFT, frequency resolution
176 Hz, 51 6 points) with Canary 1.2.1 (Orni-
thology Laboratory at Cornell).StatisticalAnalysis.-I compared call duration,
dominant frequency, and intercall interval dur-
ing spontaneous vocal activity (male alone)
with those during the responses to the control
and to each experimental stimulus. I also com-
pared male responses to controls (average at-tractive calls) with responses to above average
and below average attractive calls. All compar-
isons were performed by using paired two-
tailed t-tests. Male responses to initial and final
controls were averaged because there was no
significant difference between them in any var-iable measured (t-test, all P > 0.05). Despite the
fact that I used each male in several successive
..5
1.5
2HDF LDF LC SC Sil LIl
Stimulus
FIG. 2. Intercall interval in male responses to ex-pcrimental stimulus compared to control stimulus. For
each stimulus, the box represents the mean, and thelire represents the standard error. Experimental stim-ulas are HDF, high dominant frequency; LDF, lowdominant frequency; LC, long call; SC, short call; LII,long intercall interval; SII, short intercall interval. **F• 0.05, *P = 0.054, NS P > 0.10. UResponse to the
stimulus; P Response to average call (control).
comparisons were performed between sets ofdata obtained from several males, and a male's
response to one stimulus is independent of an-
other male's response to the same stimulus. All
statistical analyses were performed with Stat-giafics 6.0.
RESULTS
I tested a total of 25 males; sample sizes dif-
fe red among tests because several males did not
respond to all stimuli. Intercall intervals were
significantly longer during spontaneous activity
than in response to all stimuli, except for the
long intercall interval and the short call stimuli
(cDntrol: t = 2.27, df = 38, P = 0.03; high dom-
inant frequency: t = 2.07, df = 26 , P = 0.05; low
dominant frequency: t = 2.33, df = 30, P = 0.03;long call: t = 1.80, df = 16, P = 0.05; short call:t = 1.35, df = 18, P = 0.19; short intercall in-
terval: t = 2.52, df = 32, P = 0.02; long intercall
interval: t = 1.24, df = 30, P = 0.22; Fig. 1).There were no significant differences in the in-
tercall intervals between stimuli of above aver-
age and below average attractiveness versus the
control (i.e., stimuli of average attractiveness)
except for the long intercall interval stimuli
(long intercall interval: t = -2.57, df = 36, P =
0.014); the difference approached significance
for the short call stimuli (t = -2.03, df = 22 , P= 0.054, Fig. 2). Thus, males called at similar
rates when exposed to average calls and to at-
Intercall Interval (s)
-I l _______NN S NBS N
FKFF FK617

8/3/2019 Respuestas Vocales en Ranas Neotropicales
http://slidepdf.com/reader/full/respuestas-vocales-en-ranas-neotropicales 4/7
ZAIDA TARANO
tractive calls, that is , long intercall interval and
short call stimuli, with the exception of high
dominant frequency stimuli.
I did not find significant differences in call
duration nor in the dominant frequency of the
call in male responses between the spontaneous
vocal activity and the experimental stimuli
(paired t-test, NS for all comparisons), nor be-
tween the control and the experimental stimulus
(paired t-test, NS for all comparisons).
DISCUSSION
The responses of P enesefae males to variation
in conspecific calls supported the hypothesis
that males would alter their vocal activity when
exposed to potential conspecific competitors.
Males reduced their spontaneous intercall inter-
val in the presence of stimuli of average or high
attractiveness to females but did not modify
their spontaneous intercall interval in the pres-
ence of unattractive stimuli. The results, how-
ever, do not support the prediction that males
would adjust their vocal responses to the attrac-
tiveness of the stimuli because their responses
were similar for stimuli of average and above
average attractiveness.
The results of this study show that, among
the variables analyzed, only the intercall inter-
val is dependent on the social context in P ene-
sefae. The selective importance of the modifica-
tion of the intercall interval is straightforward:females show strong and unanimous preferenc-
es for calls repeated at short intervals (TArano,
1999).Despite the fact that males significantly re-
duced the intercall interval in the presence of a
stimulus, compared to their spontaneous vocal
output, a response observed in many anurans
(Wells, 1988), they did not call at relatively
shorter intervals in response to the most attrac-
tive calls to females. This does not seem to re-sult from a trade-off between matching the
stimulus rate and avoiding call overlap, because,albeit there is no correlation between response
rate and stimulus rate, call overlap in response
to stimuli repeated at 3.6-sec or 2.3-sec intervals
do not depart from random in either case (Ta-
rano, 1999). In contrast, the proportion of over-
lapping calls in response to stimuli repeated at
2.3-sec intervals is significantly greater than that
in response to stimuli repeated at 3.6-sec inter-
vals, as expected if males respond randomlv to
the stimulus. Because silence intervals between
successive calls in P enesefae are long, in relation
to call duration (3-4 times longer), randomprobability of placing a call in these intervals,
and thus avoid call overlap, is relatively high:
tween the stimulus calls and male responses re-
mains to be determined, these results indicate
that males do not shift the timing of their calls
in ways that avoid overlap with those of the
stimulus. Call matching has been described inseveraL anurans (Wells, 1988; Schwartz, 2001).
Male Philautus leucorh inus alternate calls with
those of neighbors with few instances of call
overlap, despite considerable variation in call
duration and intercall intervals during each
bout of alternation (Arak, 1983). However, play-
backs do not elicit an increase in call rate insome anurans. Hyla versicolor males always de-
creased calling rate and increase call duration,
regardless of the stimulus presented (Wells and
Taigen, 1986).The lack of correspondence between stimulus
attractiveness and male responses, in P enesefae,
suggests several interpretations. First, males donot perceive the differences between stimuli ofaverage and above average attractiveness. The
percent difference between calls of average and
below average attractiveness was slightly higher
than that between calls of average and above
average attractiveness, 36% and 30%o, respec-
tively. This explanation would be more likely if
there were a profound asymmetry in the re-
sponsiveness of the inner ear, so that differences
between certain pairs of stimuli need to be larg-
er than between others to be detected. Second,
males do perceive the differences between stim-uli of average and above average attractiveness,
but the differences lack biological meaning.
Neurophysiological studies are needed to assess
this hypotheses properly. Third, male responses
to all stimuli, except to unattractive ones, rep-
resent typical responses to a potential compet-
itor. When the social context changes from being
alone to being with another male, the male re-
sponds by increasing its call rate, regardless ofthe characteristics of the potential competitor.
Fixed responses to potential competitors might
result from a trade-off between competition fo rfemales and energy conservation. Calling is en-
ergetically expensive in anurans, and energy ex -penditure is related to call rate (Pough et al.,
1992; Prestwich; 1994). Wells and Taigen (1989)found that metabolic rate in Hyla microcephala
increased with increasing calling rate. If females
choose males based on their calling rate, males
would make a better use of their energy re-
serves if they call at their maximum call rate
only when females are in their vicinity, much asPhysalaennis pustulosus only adds chucks, which
increase attraction to both females and frog-eat-ing bats, only in response to vocal competition
with other males (Ryan, 1985). Males would call
618

8/3/2019 Respuestas Vocales en Ranas Neotropicales
http://slidepdf.com/reader/full/respuestas-vocales-en-ranas-neotropicales 5/7
VOCAL RESPONSES IN PHYSALAEMUS ENESEFAE
their maximum rate only when females ap-
proach them. Field observations support this hy-
pothesis: a chorusing male dramatically increas-
es his call rate, although for short periods, when
a female approaches him (pers. obs.). Similarly,
Wells (1988) found that males of three Neotrop-
ical treefrogs (Hyla ebraccata,H. phlebodes, H. mi-
crocephala) alter their vocal behavior when they
detect a female nearby. Thus the equally strong
response to the controls (average calls) and to
the experimental stimuli (with the exception oflong intercall interval calls and short calls) may
simply be a fixed response to any average or
above average attractive call but not the maxi-
mum response a male is able to produce.
It is also possible that apparent indifference
of males to variation in experimental stimuli ofaverage and above average attractiveness could
be caused by sampling error and not a popu-lation characteristic. Sample size was small for
several tests (short and long calls), and analyses
indicate that power of those tests was below
0.10. Thus, an effect might exist but could not
be detected. In addition, if male vocal respon-
siveness varies with age, population means
would not reflect individual behavior. Green
(1990) found that large male P pustulosus in-
creased the complexity of their calls more than
small males, in response to a novel calling
neighbor.
The response of male P enesefae to calls re-
peated at various intervals is similar to that ob-
served in other species. Male Eleutherodactylus
coqui increase their calling rate in response to
neighbor males, although there is no correlation
between stimulus call rate and male response
call rate (Benedix and Narins, 1999). Males ofthe grasshopper Ligurotettix planum responding
to a stimulus reduce their call rate when ex -posed to unattractive males but not below their
spontaneous call rate (Greenfield, 1994).In this study, I analyzed male P enesefae re-
sponses to calls of fixed levels of attractiveness,high, mid, or low attractiveness, for the study
population. But male responses could be affect-
ed by the relative attractiveness of the compet-
itor in relation to the focal male. Bosch et al.(2000) found that P pustulosus males responddifferently to "fixed frequency" conspecific calls
(calls of high, mid, and low dominant frequency
for the study population) than to "relative fre-
quency" calls (calls of high or low dominant fre-
quency in relation to those of the focal male).
Male responses did not vary among fixed fre-
quency tests. However, males responded withmore whines and chucks to calls of similar orhigher frequency than to calls of lower frequen-
Male P enesefae did not modify either the
dominant frequency or the duration of their
calls in the presence of potential competitors.
T -iis result is not unexpected because only a fewrrale anurans are able to modify the dominant
frequency of their calls, and even in such spe-cies, the change is small, about 2-5% (L6pez et
a]. 1988; Wagner, 1989; Grafe, 1995; Bee et al.,
20)00). The relative stability of the dominant fre-
quency is attributed to morphological restric-
tions (Martin, 1972; Ryan and Drewes, 1990).Morphological correlates of call duration in an-
u 2ans have not been determined but are prob-ably related to limits on air exchange between
tlhe lungs and vocal sac.
Acknowledgments.-This study was part of my
doctoral dissertation submitted at UniversidadSim6n Bolivar. I am grateful to E. A Herrera, M.J. Ryan, C. Bosque, and L. E. Levin for their sug-
gestions throughout the study. M.J. Ryan also
commented on this manuscript. I thank T
B-.ohm for his hospitality while conducting the
fieldwork at his ranch. H. Mirabal and J. I. Del-
gado gifted me their friendship during the long
and hot days at the field. I was supported by a
doctoral fellowship from Consejo Nacional de
Irivestigaciones Cientificas y Tecnol6gicas
(CONICIT) and bv Decanato de Estudios de
Postgrado of Universidad Sim6n Bolivar.
LITERATURE CITED
ARAK, A. 1983. Male-male competition and matechoice in anuran amphibians. In P. Bateson (ed.),Mate Choice, pp. 181-210. Cambridge UniversityPress, Cambridge.
BEE, M. A., S. A. PERRILL, AND P. C. OWENS. 2000.Male green frogs lower the pitch of acoustic signalsin defense of territories: a possible dishonest signalof size? Behavioral Ecology 11:169-177.
BENEDIX, J. H., AND P.M. NARINS. 1999. Competitivecalling behavior by male treefrogs, Eleutherodacty-lu s coqui (Anura: Leptodactylidae). Copeia 1999:1118-1122.
BoSCH, J., AND R. MARQUEZ. 1996. Acoustic compe-tition in male midwife toads Alytes obstetricansandAlytes cisternasii: response to neighbor size andcalling rate. Implications for female choice. Ethol-ogy 102:841-855.
BoscH, J., A. S. RAND, AND M. J. RYAN. 2000. Acousticcompetition in Phnysalaemus pustulosus, a differen-tial response to calls of relative frequency. Ethol-ogy 106:865-871.
G:RHARDr, H. C. 1991. Female mate choice in treefrogs: static and dynamic acoustic criteria. AnimalBehaviour 42:615-635.• SAFE, T. U. 1995. Graded aggressive calls in the Af-
619

8/3/2019 Respuestas Vocales en Ranas Neotropicales
http://slidepdf.com/reader/full/respuestas-vocales-en-ranas-neotropicales 6/7
ZAIDA TARANO
tition on advertisement calling in the tuingara frog
Physalaemus pustulosus (Leptodactylidae). Animal
Behaviour 38:620-638.
GREENFIELD, M. D. 1994. Synchronous and alternating
choruses in insects and anurans: common mecha-
nisms and diverse functions. American Zoology
34:605-615.HOWARD, R. D., AND J. R. YOUNG. 1998. Individual
variation in male vocal traits and female mating
preferences in Bufo ainericanus. Animal Behaviour
55:1165-1179.
LOPEZ, P. T., P. M. NARINS, E. R LEWIS, AND S. W.
MOORE. 1988. Acoustically induced call modifi-
cation in the white-lipped frog Leptodactylus albi-
labris. Animal Behaviour 36:1295-1308.
MARTIN, W F 1972. Evolution of vocalizations in the
genus Bufo, pp. 279-309. In W E Blair (ed.), Evo-lution in the Genus Bufo, pp. University of Texas
Press, Austin.
NARINs, P. M., AN D R. R. CAI'RANICA. 1978. Com-
municative significance of the two-note call of thetreefrog Eleutherodactyluscoqui. Journal of Compar-
ative Physiology 127:1-9.
PouCH, F H., W E. MAGNLSSON, M. J. RYAN, K. D.
WELLS, AND T. L. TAIGEN. 1992. Behavioral ener-
getics. In M. E. Feder and W. W. Burggren (eds.),
Environmental Physiology of the Amphibians, pp.
395-436. University of Chicago Press, Chicago.
PRESIWICH, K. N. 1994. The energetics of acoustic sig-
naling in anurans and insects. American Zoology
34:625-643.
RYAN, M. J. 1985. The Tungara Frog. A Study in Sex-
ual Selection and Communication. University of
Chicago Press, Chicago.RYAN, M. J., AND R. C. DREWES. 1990. Vocal mor-
phology of the P7tysalaemnus pustulosus species
group (Leptodactylidae): morphological response
to sexual selection for complex calls. Biological
Journal of the Linnean Society of London 4(I:37-52.RYAN, M. J., AND A. S. RAND. 1998. Evoked vocal
response in male tungara frogs: pre-existing biases
in male responses? Animal Behaviour 56:1509-
1516.SClHWARTZ, J. J. 1994. Male advertisement and female
choice in frogs: recent findings and new approach-
es to the study of communication in a dvnamic
acoustic environment. American Zoology 34:616-
624.-_____ 001. Call monitoring and interactive play-
back svstems in the study of acoustic interactions
among male anurans. In M. J. Ryan (ed.), Anuran
Communication, pp. 183-204. Smithsonian Insti-
tution Press, Washington, DC.
SCHWARTZ, J. J., AN D K. D. WELLS. 1985. Intra- and
interspecific vocal behaviour of the Neotropical
treefrog EN/la microcephala. Copeia 1985:27-38.
TARANO, Z. 1999. Reconocimiento coespecffico y se-
leccibn sexual en la rana Pkysalaemnus enesefae. Un-
publ. Ph.D. diss., Universidad Simon Bolfvar, Ca-
racas, Venezuela.
- . 2001. Variation in male advertisement calls in
the Neotropical frog Physalaemus enesefae. Copeia2001:1064-1072.
WAGNER JR., W. E. 1989. Graded aggressive signals
in Blanchard's cricket frog: vocal responses to op-
ponent proximity and size. Animal Behaviour 38:
1025-1038.
WELLS, K. D. 1988. The effects of social interactions
on anuran vocal behavior. In B. Fritszch, W Wil-
czynski, M. J. Ryan, I Etherington, and W. Wal-
kowiak (eds.), The Evolution of the Amphibian
Auditory System, pp. 433-454. John Wiley, New
York.WELLS, K. D., AND T. L. TAIGEN. 1986. The effects of
social interactions on calling energetics in the graytreefrog (Hyla zrrsicolor). Behavioral Ecology and
Sociobiology 19:9-18.
-_____ 989. Calling energetics of a Neotropical tree-
frog, Hyla microcephala. Behavioral Ecology and So -
ciobiology 25:13-22.
Accepted: 27 February 2002.
620

8/3/2019 Respuestas Vocales en Ranas Neotropicales
http://slidepdf.com/reader/full/respuestas-vocales-en-ranas-neotropicales 7/7
COPYRIGHT INFORMATION
TITLE: Vocal Responses to Conspecific Call Variation in the
Neotropical Frog Physalaemus enesefae
SOURCE: J Herpetol 36 no4 D 2002
WN: 0233505199010
The magazine publisher is the copyright holder of this article and it
is reproduced with permission. Further reproduction of this article in
violation of the copyright is prohibited.
Copyright 1982-2003 The H.W. Wilson Company. All rights reserved.