procesamiento mínimo de manzana: efecto de la radiación ... · tesis doctoral procesamiento...
TRANSCRIPT

Di r ecci ó n:Di r ecci ó n: Biblioteca Central Dr. Luis F. Leloir, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires. Intendente Güiraldes 2160 - C1428EGA - Tel. (++54 +11) 4789-9293
Co nta cto :Co nta cto : [email protected]
Tesis Doctoral
Procesamiento mínimo de manzana:Procesamiento mínimo de manzana:efecto de la radiación UV-C y la luzefecto de la radiación UV-C y la luzpulsada de alta intensidad sobre lapulsada de alta intensidad sobre la
calidadcalidad
Gómez, Paula Luisina
2010
Este documento forma parte de la colección de tesis doctorales y de maestría de la BibliotecaCentral Dr. Luis Federico Leloir, disponible en digital.bl.fcen.uba.ar. Su utilización debe seracompañada por la cita bibliográfica con reconocimiento de la fuente.
This document is part of the doctoral theses collection of the Central Library Dr. Luis FedericoLeloir, available in digital.bl.fcen.uba.ar. It should be used accompanied by the correspondingcitation acknowledging the source.
Cita tipo APA:
Gómez, Paula Luisina. (2010). Procesamiento mínimo de manzana: efecto de la radiación UV-Cy la luz pulsada de alta intensidad sobre la calidad. Facultad de Ciencias Exactas y Naturales.Universidad de Buenos Aires.
Cita tipo Chicago:
Gómez, Paula Luisina. "Procesamiento mínimo de manzana: efecto de la radiación UV-C y la luzpulsada de alta intensidad sobre la calidad". Facultad de Ciencias Exactas y Naturales.Universidad de Buenos Aires. 2010.

UNIVERSIDAD DE BUENOS AIRES Facultad de Ciencias Exactas y Naturales
Departamento de Industrias
PROCESAMIENTO MÍNIMO DE MANZANA: EFECTO DE LA RADIACIÓN UV'C Y LA LUZ PULSADA DE ALTA
INTENSIDAD SOBRE LA CALIDAD
Paula Luisina Gómez
Tesis presentada para optar al título de Doctora de la Universidad de Buenos Aires en el área Química Industrial
Directores de tesis: Dra. Stella Maris Alzamora
Dra. Daniela Marisol Salvatori
Buenos Aires, 2010.
����

II
������������ ������������������������������������������������������������������������������������������
�El objetivo general de esta tesis fue evaluar el efecto de la aplicación de radiación UV*C
y luz pulsada (LP) en combinación con otros factores de conservación microbianos y/o no
microbianos (pretratamientos de escaldado e inmersión en soluciones antipardeamiento),
sobre la calidad de rodajas de manzana (propiedades ópticas, características reológicas), con
miras al desarrollo de nuevas tecnologías de conservación de frutas mínimamente procesadas
que permitan conservar las características de calidad.
La irradiación de rodajas de manzana con luz UV*C o LP produjo un incremento
significativo del pardeamiento superficial, el cual fue función de la dosis aplicada y el tiempo
de almacenamiento a 4*5 ºC. Los cambios provocados sobre el comportamiento reológico
fueron menores y se asociaron con una leve pérdida de rigidez del tejido irradiado durante el
almacenamiento, reflejado fundamentalmente en una disminución de los módulos de
almacenamiento (G’) y de pérdida (G’’) y un aumento en las capacitancias J0, J1 y J2. Los
pretratamientos de escaldado e inmersión en solución antipardeamiento de ácido
ascórbico/cloruro de calcio fueron efectivos para inhibir el pardeamiento superficial
provocado por la luz UV*C en todas las dosis evaluadas, siendo la solución antipardeamiento
la que produjo mayor estabilidad en el color durante el almacenamiento y menores cambios
en las propiedades mecánicas y viscoelásticas del tejido. En las rodajas irradiadas con LP la
solución antipardeamiento fue menos efectiva para inhibir el pardeamiento en las muestras
irradiadas a las dosis más altas. Los cambios observados en el color y en el comportamiento
reológico de las muestras sometidas a los diferentes tratamientos pudieron ser correlacionados
en parte con las observaciones micro y ultraestructurales de los tejidos. Por otro lado, a pesar
de que la aplicación de la solución antipardeamiento redujo el efecto de la irradiación sobre la
inactivación de microorganismos, los recuentos de microorganismos inoculados y de flora
nativa al finalizar el almacenamiento fueron menores que en las rodajas no expuestas a la
irradiación UV*C.
Los resultados indican que tanto la radiación UV*C como la LP combinadas con un
pretratamiento antipardeamiento adecuado podrían ser utilizadas en la industria de frutas
minimamente procesadas para extender su vida útil/aumentar su inocuidad, como integrantes
de un sistema multifactorial de conservación.
Palabras claves: radiación UV*C, luz pulsada, frutas mínimamente procesadas, color,
reología, inactivación de microorganismos, manzana.

III
Minimal processing of apple: effect of UV'C radiation and high intensity pulsed light on quality
This thesis was aimed to evaluate the effect of UV*C radiation and pulsed light (PL) in
combination with other microbial and/or non*microbial preservation factors (blanching and/or
dipping into antibrowning solution pretreatments) on quality (mainly optical and rheological
properties) of apple slices, in order to develop new technologies for obtaining minimally
processed fruits that maintain quality characteristics.
Irradiation of apple slices with UV*C or PL caused a significant increase in surface
browning, which depended on the applied dose and time of refrigerated storage at 4*5 ºC.
Changes on rheological behavior were minor and were associated with a slight loss of
stiffness of irradiated tissue during storage, mainly reflected in lower values of storage (G´)
and loss (G'') modules and to an increase in J0, J1 and J2 capacitances. Blanching and dipping
in ascorbic acid / calcium chloride anti*browning solution were effective in inhibiting surface
browning caused by UV*C at all doses tested, although the anti*browning solution produced
higher color stability during storage and lesser changes in compression and viscoelastic
properties of tissue. In apple slices irradiated with PL, anti*browning solution was less
effective in inhibiting browning when fruit was irradiated at high doses. Changes in color and
rheological properties were correlated with microstructure and ultrastructure features.
Although the anti*browning solution reduced the effect of irradiation on the inactivation of
microorganisms, inoculated microorganisms and natural microflora counts after storage were
lower in UV*C irradiated that than in non*irradiated samples.
Results indicated that both UV*C and PL irradiation combined with a suitable anti*
browning pretreatment can be used in minimally processed fruit industry to extend shelf life.
Key words: UV*C radiation, pulsed light, minimally processed fruit, color, rheology,
microorganisms inactivation, apple.

IV
��������������
A Stella y Daniela por haberme dirigido durante estos años. Por el compromiso con el
que trabajan, por el apoyo y la dedicación que me brindaron. Muchas gracias.
A mis compañeros de trabajo Analía, Marcela, Silvina, Cielo, Silvia, Paola, Sebastián y
Joaquín por su amistad, compañerismo y por los gratos momentos compartidos. Quiero
agradecer especialmente a Analía por su ayuda con la estadística y a Marce por la ayuda con
la microbiología.
A Andrea y Sandra por su apoyo profesional incondicional.
A Leticia por estar siempre dispuesta a ayudarte en lo que necesites.
A la Agencia Nacional de Promoción Científica y Tecnológica, el CONICET y la
Universidad de Buenos Aires por el aporte financiero brindando para la realización de esta
tesis.
Al Sr. Alejandro Sebastiano de la empresa Sirex Médica por la buena predisposición
que ha tenido con nosotros para poder solucionar los inconvenientes técnicos ocurridos en el
equipo de luz pulsada.
A mi hermano Martín por haber sido una gran compañía en estos años.
A mis padres y mis hermanos Patri, Marian, Iva, Ale, Fer, Maru, Euge y Guille, por su
cariño y comprensión, y por estar siempre presentes a pesar de la distancia.
A los amigos que encontré en Buenos Aires, Emilia, Ricardo, Daniela y Blanca, por
todos los buenos momentos compartidos.
��
����������������������������������������������������������������������������������������������������������������������������������������������
�
�

1
�� !�"��
1. OBJETIVOS……………...………………………………………………………………………… 4
1.1. Objetivo general………………………….…………………………………………………………4
1.2. Objetivos específicos…………………………………………………………………………….....5
2. INTRODUCCIÓN…………………………………………………………………………………..6
2.1. Atributos de calidad de frutas………………………………………………………………………6
2.2. Procesamiento mínimo de frutas y vegetales……………………………………………………….9
2.2.1. Concepto y enfoques…………………………………………………………………….....…...9
2.2.2. Operaciones involucradas en los métodos de procesamiento míninimo estudiados en
esta tesis…………………………………………………………………………………………11
2.2.2.1. Luz ultravioleta de onda corta…………..……...…………………………………............11
2.2.2.2. Pulsos de luz de alta intensidad……………...…………………………….……………...19
2.2.2.3. Adición de calcio……………………………………….….……………………………...22
2.2.2.4. Tratamiento térmico*escaldado………………….………………………………………...24
2.2.2.5. Inmersión en soluciones antipardeamiento……………………….….…………………....25
2.3. Descripción del tejido vegetal…………………………………………………………………….27
2.3.1. Sistemas de tejido……………………….…………………………………………………….27
2.3.2. Estructura celular………………………….…………………………………………………..29
2.3.3. La manzana…………………….……………………………………………………………...34
2.3.3.1. Propiedades botánicas………………..……………………………………………….…...34
2.3.3.2. Taxonomía y morfología…………………………...……………………..……………....35
2.4. Color de tejidos vegetales: evaluación…………………………………………………………….35
2.5. Reología de tejidos vegetales: evaluación………………………………………………………...40
2.6. Relación entre la textura, las características reológicas y los cambios en la estructura
producidos por los diferentes tratamientos………………………………………………………..59
3. MATERIALES Y MÉTODOS……………………………………………………………………61
3.1. Materia prima……………………………………………………………………………………...61
3.2. Preparación de las muestras……………………………………………………………………….61
3.3. Tratamientos………………………………………………………………………………………62
3.3.1. Aplicación de luz ultravioleta de onda corta………………………………………………….62
3.3.1.1. Descripción del equipo……………………………………………………………………62

2
3.3.1.2. Determinación de la dosis de radiación UV*C……………………………………………62
3.3.1.3. Tratamientos de radiación UV*C………………………………………………………….64
3.3.2. Aplicación de pulsos de luz de alta intensidad…………………………………………..........65
3.3.2.1. Descripción del equipo……………………………………………………………………65
3.3.2.2. Medición de la temperatura……………………………………………………………….65
3.3.2.3. Tratamientos de irradiación con luz pulsada……………………………………………...66
3.4. Pretratamientos……………………………………………………………………………………67
3.4.1. Adición de calcio……………………………………………………………………………...67
3.4.2. Tratamiento térmico…………………………………………………………………………...67
3.4.3. Inmersión en soluciones antipardeamiento……………………………………………………68
3.5. Almacenamiento de las muestras………………………………………………………………….68
3.6. Medición instrumental del color…………………………………………………………………..69
3.7. Medición instrumental de las propiedades reológicas…………………………………………….71
3.7.1. Determinaciones a altas deformaciones……………………………………………………….71
3.7.2. Determinaciones a bajas deformaciones………………………………………………………71
3.7.2.1. Ensayos oscilatorios……………………………………………………………………….72
3.7.2.2. Ensayos rotatorios…………………………………………………………………………73
3.8. Técnicas microscópicas…………………………………………………………………………...74
3.8.1. Microscopía óptica (MO)…..………………………………………………………………….74
3.8.2. Microscopía electrónica de transmisión (MET)........................................................................75
3.8.3. Microscopía electrónica de barrido ambiental (MEBA)……………………………….…..…75
3.9. Determinación de calcio…………………………………………………………………………..75
3.10. Pérdida de peso…………………………………………………………………………………..76
3.11. Estudios microbiológicos………………………………………………………………………...76
3.11.1. Ensayos de inactivación con microorganismos inoculados…………………………….........76
3.11.2. Evolución de la flora nativa………………………………………………………………….78
3.12. Análisis estadístico……………………………………………………………………………....78
4. RESULTADOS Y DISCUSIÓN…………………………………………………………………..79
4.1. Aplicación de luz ultravioleta de onda corta……………………………………………………...79
4.1.1. Selección de la dosis de radiación UV*C………………… ……………………………......79
4.1.2. Determinación de la dosis de radiación UV*C…………………………………………….....81
4.1.3. Estudio de los cambios de color…………………………………………………...…………83
4.1.3.1. Tratamiento UV*C…………………………………………………...…………………...83
4.1.3.2. Aplicación de agentes antipardeamiento previo al tratamiento UV*C…………...……....88
4.1.3.3. Adición de calcio…………...…………………………………………………………….97

3
4.1.4. Caracterización de las propiedades reológicas de manzanas expuestas a la luz UV*C…...…118
4.1.4.1. Tratamiento UV*C……………………………………………………………………….118
4.1.4.2. Tratamiento UV*C con previo escaldado………………………………………………..131
4.1.4.3. Tratamiento UV*C con inmersión previa en la solución antipardeamiento……………..142
4.1.4.4. Tratamiento UV*C con inmersión previa en soluciones de sales de calcio..…………….154
4.1.5. Pérdida de peso………………………………………………………………………………165
4.1.6. Caracterización microestructural y ultraestructural de los tejidos de manzana
sometidos a los diferentes pretratamientos y/o a la radiación UV*C………………………..169
4.1.7. Estudios microbiológicos…………………………………………………………………….184
4.1.8. Integración de los resultados obtenidos a través de los ensayos instrumentales y de la
caracterización micro y ultraestructural de los tejidos de manzana sometidos a los
diferentes y/o la luz UV*C……………………………………………………...……………189
4.2. Aplicación de luz pulsada de alta intesidad……………………………………………………..195
4.2.1. Selección de la dosis………………………………………………………………………...195
4.2.2. Evolución de la temperatura…………..……………………………………………………195
4.2.3. Estudio de los cambios de color……………….……………….…………………………199
4.2.3.1. Irradiación con LP………………………………………………………………………199
4.2.3.2. Inmersión en solución de ácido ascórbico/cloruro de calcio previa a la
irradiación con LP………………..……………………………………………………...208
4.2.4. Caracterización de las propiedades reológicas de rodajas de manzana expuestas
a la luz pulsada……………..………………………………………………………………215
4.2.5. Pérdida de peso…………………………………………………………………………….232
4.2.6. Caracterización microestructural de los tejidos de manzana expuestos a la luz pulsada.....237
4.2.7. Estudios microbiológicos…………………………………………………………………..242
4.2.8. Integración de los resultados obtenidos a través de los ensayos instrumentales y de la
caracterización microestructural de los tejidos de manzana sometidos a la luz
pulsada con o sin tratamiento antipardeamiento…...………………………………………245
5. CONCLUSIONES………………………………………………………………………………..248
5.1. Aplicación de luz UV*C………………………………………………………………...……….248
5.2. Aplicación de pulsos de luz……………………………………………………………………..249
5.3. Conclusiones generales………………………………………………………………………….250
6. BIBLIOGRAFÍA…………………………………………………………………………………252
�

4
#$�%&'"(!�%)�
�
#$#$�%�*��+�����������
La inocuidad de los alimentos es una de las preocupaciones principales en relación a la
salud pública en todo el mundo. En particular, los materiales vegetales son vulnerables a la
contaminación, supervivencia y desarrollo de microorganismos y/o sus metabolitos que
pueden provocar ETA´s, cuyo riesgo se ve agravado por la tendencia al consumo de productos
minimamente procesados o por la ineficacia de los procesos de elaboración de subproductos
para destruir los contaminantes.
Además del enfoque en la prevención, es primordial remover o inactivar los
microorganismos patógenos y/o sus metabolitos. La presencia de una operación unitaria para
destruir los microorganismos en el procesamiento de los materiales vegetales es de
fundamental importancia para lograr la inocuidad y la estabilidad del producto.
En las últimas dos décadas se han propuesto y desarrollado técnicas alternativas “no
térmicas” físicas y químicas de destrucción microbiana (comúnmente denominadas “técnicas”
o “factores emergentes de conservación”) para lograr inocuidad manteniendo la calidad
sensorial, nutricional y/o funcional de productos de consumo directo o de materias primas y
productos intermedios antes de su industrialización. Entre ellas pueden mencionarse altas
presiones hidrostáticas, pulsos eléctricos de alto voltaje, ozono, ultrasonido, radiación
ultravioleta y luz pulsada (FDA, 2000). Si bien el avance en el conocimiento sobre dichas
tecnologías ha sido impresionante en los últimos años, muchas de las mismas se encuentran
todavía en estudio y su adopción industrial es en muchos casos incierta, ya que se requiere de
mayores investigaciones de naturaleza básica y tecnológica. Actualmente, no existen
demasiados estudios en la literatura que aborden sistemáticamente el efecto de agentes
emergentes aplicados en forma aislada o como factores no convencionales en tratamientos
multifactoriales, sobre todo en referencia a los atributos de calidad de los productos obtenidos.
El objetivo general de esta tesis fue evaluar el efecto de algunas tecnologías
emergentes (luz ultravioleta de onda corta, pulsos de luz de alta intensidad) en combinación
con otros factores de preservación, sobre la calidad de frutas mínimamente procesadas, con
miras al desarrollo de nuevas tecnologías de conservación.
�
�
�

5
#$,$�%�*��+�������� �����
Los objetivos específicos de esta investigación fueron: 1* Estudiar la aplicación de luz ultravioleta de onda corta y pulsos de luz de alta intensidad
sobre rodajas de manzana, tanto en forma individual como combinada con otros factores
de inhibición del deterioro microbiano y/o no microbiano (pretratamientos de escaldado y
de inmersión en una solución antipardeamiento).
2* Evaluar la respuesta de algunos parámetros de calidad (fundamentalmente color
superficial y propiedades reológicas) a la dosis de los factores emergentes seleccionados
y al tipo de pretratamiento aplicado, tanto al final del procesamiento como a lo largo del
almacenamiento
3* Evaluar el efecto de algunos tratamientos selectos en los cambios micro y
ultraestructurales producidos en el tejido de manzana para interpretar al menos
parcialmente las modificaciones observadas en el comportamiento reológico y en el color
del material.
4* Evaluar el efecto de la aplicación de los factores emergentes estudiados (como únicos
factores de estrés o en combinación con los pretratamientos seleccionados) en la
supervivencia de algunos microorganismos inoculados y de flora nativa a lo largo del
almacenamiento.
5* Integrar los resultados para diseñar tecnologías de conservación adecuadas para la
obtención de manzana mínimamente procesada que conserve al máximo sus
características de calidad.
6* Aplicar los tratamientos de conservación estudiados a manzanas previamente
enriquecidas con calcio mediante técnicas específicas para la incorporación de
componentes bioactivos en el interior de matrices porosas, con miras a diseñar
tecnologías de conservación de rodajas de manzana funcionales.
��

6
,$�!�(-% ���!.����
,$#$����������������������
La calidad de los productos de frutas es una combinación de atributos, propiedades o
características que determinan su valor para el consumidor. Los parámetros de calidad
incluyen apariencia, textura, “flavor” y valor nutritivo. La importancia relativa de cada
parámetro depende de cada producto y si va a ser consumido fresco o cocido (Kader, 2002).
Shewfelt (1999) sugirió que la combinación de las características propias del producto
debe definirse como “calidad”, mientras que la percepción del consumidor y la respuesta a
esas características se refieren a la “aceptabilidad”. Los consumidores usan todos sus sentidos
para evaluar la calidad (vista, olfato, gusto, tacto y oído) e integran todos los aportes
sensoriales en un juicio final de la aceptabilidad del producto (Abbott, 1999).
En frutas o vegetales enteros la calidad depende del cultivar, de las prácticas
precosecha, de las condiciones climáticas, de la madurez en la cosecha, de los métodos de
cosecha, de las condiciones de transporte y del almacenamiento poscosecha (Kader, 2002).
El deterioro natural en la calidad que presentan las frutas luego de la cosecha se debe a
que las células de sus tejidos constituyentes siguen vivas. El hecho de que los tejidos vivientes
respiren tiene consecuencias importantes desde el punto de vista de la calidad y la vida útil de
estos productos. Los factores que disminuyen la respiración pueden disminuir la senescencia;
sin embargo, algo de respiración debe continuar o los productos rápidamente senescen o
mueren. El enfriamiento del producto puede disminuir los cambios indeseables pero hay que
tener en cuenta que muchas frutas son intolerantes a las bajas temperaturas. El entendimiento
de la fisiología de cada fruta es fundamental para comprender su estabilidad y extender su
vida útil (Aked, 2002).
�
• ���������+�����
La apariencia es un factor de calidad muy importante que es tenido en cuenta por los
consumidores para adquirir un producto fresco. Los componentes principales de la calidad
visual incluyen el color, la uniformidad del color, la forma, el tamaño, el brillo, la ausencia de
defectos y de deterioro (Aked, 2002).
La aceptación del producto por parte del consumidor está determinada en primer lugar
por el color y en segundo lugar por otros atributos como la textura, “flavor”, etc. Muchos

7
protocolos de control de calidad usan al color como un atributo para medir el grado de calidad
de frutas, y de alimentos en general, y por consiguiente, determinar el valor comercial del
producto (Artéz y col., 2002)
El color de las frutas se debe fundamentalmente a la presencia de tres familias de
pigmentos: clorofilas, carotenoides y antocianinas. La estructura de estos pigmentos puede
verse afectada en cierto grado durante el desarrollo, la maduración y los tratamientos
poscosecha. Factores tales como la luz, el pH, la humedad relativa, la composición de gas y
sistemas enzimáticos estarían involucrados en el deterioro del color (Artéz y col, 2002).
Los otros aspectos de la apariencia que reducen la calidad son la pérdida de frescura,
como el marchitamiento, la pérdida de brillo en la superficie o arrugamiento de la piel, y el
desarrollo de defectos externos e internos, causados por la senescencia natural, desórdenes
fisiológicos y el desarrollo de organismos causantes de enfermedades (Aked, 2002)
En los productos de fruta destinados al procesamiento industrial, la importancia de la
apariencia va a depender de qué parte del producto va a ser usado y si la apariencia se puede
mejorar durante el procesamiento. En muchos productos, la piel se remueve; por lo tanto, las
imperfecciones en la piel son de poca importancia. En estos casos, el color interno es más
importante que la piel. Por otro lado, el tamaño y la forma son más importantes cuando el
procesamiento es automático en lugar de manual, aunque para algunos procesos estos
atributos son menos importantes, por ejemplo, para la extracción de jugo (Aked, 2002).
• (�/�����
�
La textura es otro de los atributos de calidad que determina la aceptabilidad de frutas. Si
bien el término textura es ampliamente utilizado, no es un atributo único, bien definido. Es un
término colectivo que involucra las propiedades mecánicas y estructurales de un alimento y su
percepción sensorial en la mano o en la boca (Abbott & Harper, 2004). La definición de la
norma ISO del año 1992 expresa que “la textura está dada por todos los atributos mecánicos,
geométricos y de superficie de un producto que se perciben por medio de los receptores
mecánicos, táctiles y, cuando corresponde, auditivos y visuales”. De esta definición se
desprende:
* la textura es una propiedad sensorial que es percibida y descripta por los seres humanos
* es un atributo multiparamétrico debido a que existen un gran número de términos que
son utilizados para describirla
* es detectada utilizando varios sentidos

8
Los atributos mecánicos incluyen parámetros básicos (dureza, cohesividad,
viscosidad, elasticidad y adhesividad) y parámetros secundarios (fragilidad, masticabilidad,
gomosidad, pegajosidad, crocancia). Los atributos geométricos están relacionados con el
tamaño, la forma y la orientación de las partículas (fibrosidad, granulosidad, aspereza),
mientras que los atributos de superficie se relacionan con el contenido de humedad y el
contenido de grasa (Szczesniak, 1963). Algunos de los términos usados para definir
sensorialmente la textura de frutas son duro, firme, blando, crujente, jugoso, arenoso.
Los atributos texturales están relacionados con las características bioquímicas,
fisiológicas y estructurales de las células que componen los tejidos de las frutas. Estas
características cambian a largo del tiempo y pueden ser alteradas durante el procesamiento
(Abbott & Harper, 2004).
Si bien muchas veces cierto grado de ablandamiento es requerido para una óptima
calidad de la fruta, el sobreablandamiento es indeseado y es un signo de senescencia o
deterioro interno (Aked, 2002).
�• 01��+��2��
�El “flavor” involucra la percepción de los sabores y aromas de muchos compuestos
presentes en las frutas. El “flavor” está determinado por el contenido de azúcares (dulzor),
ácidos orgánicos (acidez), compuestos fenólicos (astringencia) y compuestos volátiles
(aroma) (Kader, 2002).
El dulzor de algunas frutas puede incrementarse dramáticamente durante la maduración
debido a la conversión de almidón en azúcares, por ejemplo, en manzanas, bananas, mangos y
peras. Al mismo tiempo, los factores astringentes (taninos) desaparecen. Muchas veces los
niveles de azúcares son medidos para determinar si el producto ha llegado a la maduración
requerida para la comercialización. Los niveles de azúcares usualmente no disminuyen
durante el almacenamiento; sin embargo, los niveles de ácidos si decrecen. Si la relación
ácido/azúcar decrece demasiado, el producto puede llegar a ser insípido y perder la
aceptabilidad de consumo. Por otro lado, se pueden desarrollar componentes amargos en
varias frutas bajo ciertas condiciones de almacenamiento o cuando son infectadas con ciertos
patógenos. El perfil de aroma puede cambiar también dramáticamente durante la vida
poscosecha del producto fresco, particularmente en frutas climatéricas, en donde el volátil
dominante puede ser diferente en la fruta inmadura, madura y senescente (Aked, 2002).

9
• �����������+���
Las frutas y vegetales juegan un rol importante en la nutrición humana, especialmente
como fuentes de vitaminas (vitamina C, vitamina A, vitamina B6, tiamina, niacina), minerales
(Ca, Mg, Na, K, Fe, Cu), y fibras dietarias. Otros constituyentes importantes que pueden
reducir el riesgo de cáncer, enfermedades cardíacas, y otras enfermedades son los flavonoides,
carotenoides, polifenoles y otros fitonutrientes. Las pérdidas poscosecha en la calidad
nutricional pueden ser sustanciales e incrementarse por el daño físico, larga duración del
almacenamiento, altas temperaturas, baja humedad relativa y daño por frío en frutos sensibles
al frío (Kader, 2002).
�
,$,$������������� ���������������+���������
,$,$#$����������������3����
Las frutas y vegetales mínimamente procesados (MP) se definieron en un principio
como productos que mantienen sus atributos de calidad similares a la de los productos frescos.
Estos productos MP deben mantener las características de frescura y a su vez deben proveer
una vida útil conveniente, asegurando un nivel apropiado de inocuidad y de valor nutricional.
Por lo tanto las operaciones de manipulación, de procesamiento y almacenamiento deben ser
seleccionadas cuidadosamente (Palou y col., 2000).
La industria de vegetales y frutas mínimamente procesados fue inicialmente
desarrollada para abastecer hoteles, restaurantes, servicios de catering y otras instituciones,
pero con el tiempo se expandió para incluir a los minoristas de alimentos para el consumo
doméstico. El incremento en la popularidad de estos productos ha sido atribuido a los
beneficios para la salud que son asociados con el consumo de productos frescos, combinado
con la creciente tendencia de los consumidores de comer afuera y de ingerir alimentos “listos
para consumir” (Alzamora y col., 2000 a).
Uno de los principales problemas que presentan las frutas y vegetales mínimamente
procesados es su alto carácter perecedero. Durante las diferentes operaciones de preparación
(pelado, cortado, pulpeado, etc.) a las que es sometido el producto se favorecen las reacciones
de deterioro (respiración, transpiración, producción de etileno, deterioro enzimático) que
reducen la calidad sensorial y nutricional, y limitan la vida de anaquel. Por otro lado, el
aumento del tiempo y la distancia entre el procesamiento y el consumo pueden contribuir al
desarrollo de microorganismos patógenos. Algunos de los microorganismos patógenos

10
asociados con los productos frescos incluyen a Listeria monocytogenes, especies de
Salmonella, especies de Shigella, cepas enteropatogénicas de Escherichia coli, virus de la
hepatitis A, etc. Las posibles fuentes de contaminación en estos productos pueden provenir de
la contaminación presente en las frutas y verduras frescas, de los trabajadores de la planta, y
del ambiente del procesamiento (Alzamora y col., 2000 a).
Se han propuesto tres iniciativas principales para lograr productos con procesamiento
mínimo que mantengan una alta calidad (Alzamora & Salvatori, 2006):
* Optimización de los métodos de conservación tradicionales para mejorar la calidad
sensorial, nutricional y microbiológica de los alimentos, el rendimiento y la eficiencia
energética (extensión del concepto de procesamiento mínimo a las características de
excelencia de productos procesados).
* El desarrollo de procesos leves mediante la combinación de factores tradicionales de
preservación físicos y químicos, cada uno aplicado en baja intensidad, para conservar
los atributos de calidad del estado fresco o nativo de un dado alimento pero con una
mayor vida útil.
* El desarrollo de nuevas técnicas para obtener alimentos con atributos de calidad del
estado fresco utilizando combinaciones de factores de preservación emergentes o
combinaciones de factores emergentes con tradicionales, todos aplicados a bajas dosis.
A continuación, se presentan ejemplos de tecnologías de preservación seleccionadas
para cada uno de los enfoques mencionados anteriormente.
Tecnologías tradicionales mejoradas:
* empaquetamiento aséptico de alimentos procesados térmicamente
* calentamiento óhmico
* calentamiento con radiofrecuencias
* calentamiento infrarrojo
* microfiltración
* deshidratación osmótica
* deshidratación al vacío
* secado dieléctrico y por microondas
* enfriamiento criogénico
* enfriamiento en medio de dispersión dinámico

11
Nuevas tecnologías leves basadas en la combinación de factores de preservación
tradicionales:
* empaquetamiento en atmósferas controladas/modificadas
* técnicas de empaquetamiento activas
Tecnologías emergentes:
* Alta presión hidrostática
* Radiación ionizante
* Pulsos eléctricos
* Luz ultravioleta
* Pulsos de luz
* Antimicrobianos naturales
* Ultrasonido
* Biopreservación
�
,$,$,$� %���������� �+���������� ��� ���� 4������ ��� ������������ �������������������������� ,$,$,$#$�5��������+��������������������
• �������������������������
�La luz ultravioleta forma parte del espectro electromagnético y se encuentra ubicada
entre los rayos X y el espectro visible, abarcando un rango de longitudes de onda entre 10 y
400 nm (figura 2'1).
Esta radiación es frecuentemente referida como “no ionizante”; sin embargo, las
longitudes de onda ultravioleta más cortas pueden llegar a originar cierto grado de ionización.
La porción ultravioleta del espectro ha sido subdividida de forma más o menos arbitraria. El
término “ultravioleta de vacío” o “UV extremo” se reserva para las longitudes de onda
menores a 200 nm, debido a que esta región ultravioleta está atenuada fuertemente por el aire.
Por otro lado, es usual definir la región entre 200 y 300 nm como “ultravioleta lejano” y
aquella entre los 300 y 400 nm como “ultravioleta cercano”. Una subdivisión alternativa

12
Rayos
cósmicos
Rayos
gammaRayos X
aaaaaaaaaaaaaaa
aaaaaaaaaaaaaaa
aaaaaaaaaaaaaaa
aaaaaaaaaaaaaaa
aaaaaaaaaaaaaaa
aaaaaaaaaaaaaaa
aaaaaaaaaaaaaaa
aaaaaaaaaaaaaaa
Ultravioleta Visible Infrarrojo Ondas de radio
Rayos X
100 nm 200 nm 400 nm300 nm
VisibleGermicida
UV extremo UV lejano UV cercano
Rayos
cósmicos
Rayos
gammaRayos X
aaaaaaaaaaaaaaa
aaaaaaaaaaaaaaa
aaaaaaaaaaaaaaa
aaaaaaaaaaaaaaa
aaaaaaaaaaaaaaa
aaaaaaaaaaaaaaa
aaaaaaaaaaaaaaa
aaaaaaaaaaaaaaa
Ultravioleta Visible Infrarrojo Ondas de radio
Rayos X
100 nm 200 nm 400 nm300 nm
VisibleGermicida
UV extremo UV lejano UV cercano
Rayos
cósmicos
Rayos
gammaRayos X
aaaaaaaaaaaaaaa
aaaaaaaaaaaaaaa
aaaaaaaaaaaaaaa
aaaaaaaaaaaaaaa
aaaaaaaaaaaaaaa
aaaaaaaaaaaaaaa
aaaaaaaaaaaaaaa
aaaaaaaaaaaaaaa
Ultravioleta Visible Infrarrojo Ondas de radio
Rayos
cósmicos
Rayos
gammaRayos X
aaaaaaaaaaaaaaa
aaaaaaaaaaaaaaa
aaaaaaaaaaaaaaa
aaaaaaaaaaaaaaa
aaaaaaaaaaaaaaa
aaaaaaaaaaaaaaa
aaaaaaaaaaaaaaa
aaaaaaaaaaaaaaa
Ultravioleta Visible Infrarrojo Ondas de radio
Rayos X
100 nm 200 nm 400 nm300 nm
VisibleGermicida
UV extremo UV lejano UV cercano
Figura 2'1. Espectro electromagnético��Fuente:�Adaptación extraída de Guerrero*Beltrán & Barbosa*Cánovas, 2004.
�
frecuentemente utilizada en la literatura divide la región ultravioleta en tres regiones: (Shama,
2006):
* Radiación ultravioleta de onda corta (UV*C): entre 200*280 nm
* Radiación ultravioleta de onda media (UV*B): entre 280*320 nm
* Radiación ultravioleta de onda larga (UV*A): entre 320*400 nm
La radiación UV*A provoca cambios en la piel y conlleva al bronceado de la piel. La
radiación UV*B puede causar quemaduras en la piel y eventualmente cáncer. La radiación
UV*C es la más dañina debido a que es absorbida por proteínas, ARN y ADN y puede
provocar mutaciones en la célula, cáncer y/o muerte celular; también recibe el nombre de
“rango germicida” debido a que es muy efectiva en la inactivación de muchos tipos de
microorganismos tales como bacterias, virus, protozoos, hongos, levaduras y algas. (Bolton,
2001).
• "����������������������������
�
Las longitudes de onda ultravioleta más letales para los microorganismos son aquellas
que son absorbidas en mayor grado por las bases de ADN, y se encuentran en el rango entre
250*260nm. Como se observa en la figura2'2, el máximo efecto germicida se ubica alrededor
de los 254 nm y decae a valores cercanos a cero a 220 nm y 320 nm (Bintis y col., 2000).

13
Figura 2'2. Efecto germicida de las longitudes de onda del espectro ultravioleta. Fuente: Shama, 1999.
Si bien la luz UV*C puede provocar cambios en muchos componentes celulares, se ha
reportado que las reacciones más significativas que determinan la supervivencia de las células
ocurren por su interacción con los ácidos nucleicos. El daño provocado en el ADN se debe a
que induce la formación de “fotoproductos”. Entre estos los más importantes son los dímeros
de pirimidina (citosina y timina) que se forman entre dos bases de pirimidina adyacentes. Los
aductos de pirimidina son otro tipo de fotoproductos que pueden formarse entre bases
adyacentes de pirimidina, pero con velocidades de formación mucho menores que los
dímeros. Debido a la mutación de estas bases, la transcripción y la replicación del ADN es
bloqueada, comprometiendo las funciones celulares y conduciendo eventualmente a la muerte
celular. La irradiación a dosis altas también puede producir entrecruzamientos ADN*proteína
e incluso la ruptura de la cadena de ADN. El daño causado en el ADN es proporcional a la
cantidad de radiación UV a la que ha sido expuesto (Shama, 2006; Bintis y col., 2000;
Guerrero*Beltrán & Barbosa*Canovas, 2004).
Casi todas las células vivas poseen la capacidad de revertir el daño causado en su ADN
por la luz UV mediante el uso de uno o más mecanismos de reparación. Entre estos se
encuentra la reparación fotoenzimática, en donde los dímeros de pirimidina son
monomerizados enzimáticamente en presencia de luz. Otros procesos involucran la remoción
de las secciones de ADN dañado y su resíntesis utilizando como molde una hebra de ADN
Longitud de onda (nm)
Efe
ctiv
idad
ge
rmic
ida
rela
tiva

14
intacta (Shama, 2006). La capacidad de reparación de las células ha sido correlacionada con la
intensidad de la luz UV*C a la que son expuestas. Por eso es importante la aplicación de una
dosis adecuada de irradiación. Los procesos de fotoreactivación pueden ser inhibidos
almacenando el producto irradiado bajo refrigeración y/o en oscuridad (Guerrero*Beltrán &
Barbosa*Canovas, 2004).
• )��������������������������������������������
Con excepción de las bacterias fotosintéticas la mayoría de los microorganismos
muestran susceptibilidad al daño ultravioleta (López*Malo & Palou, 2005). Los distintos
microorganismos requieren de una dosis específica para ser inactivados, esta variación
dependerá de la estructura de la pared celular, del espesor y composición de la misma, de
proteínas que absorban la luz UV o de las diferencias en la estructura de loa ácidos nucleicos.
Diferencias entre especies y dentro de la misma (dependiendo de la cepa, el medio de
crecimiento y el estado del cultivo), la densidad de los microorganismos y otras características
como el tipo y la composición del alimento, pueden constituir factores importantes (Chang y
col, 1985). Los hongos y levaduras por ser microorganismos de mayor tamaño son más
resistentes que las bacterias. Los virus son más resistentes que las bacterias (Adams & Moss,
1995), mientras que los protozoos, que también son patógenos transmitidos por los alimentos,
presentan cierta resistencia al tratamiento con luz UV (Shama, 1999; Bolton, 2001; Sastry y
col., 2000). Las esporas bacterianas demuestran máxima resistencia y las dosis aplicadas
usualmente no llegan a destruirlas. El estado fisiológico de las células también es un factor
importante relacionado con la susceptibilidad de los microorganismos. Las células cuando se
encuentran en la fase estacionaria de crecimiento son más resistentes que las que se
encuentran en fase exponencial.
• 1�������������������
Las lámparas comúnmente utilizadas para la desinfección con luz UV*C consisten en
tubos de cuarzo que contienen un gas inerte, tal como el argón, y pequeñas cantidades de
mercurio. La diferencia que presentan con respecto a las lámparas fluorescentes es que no
están recubiertas, transmitiendo la radiación generada por el arco. Las fuentes de radiación
utilizadas para la desinfección UV*C pueden ser clasificadas en lámparas de arco de mercurio
de baja y mediana presión. Las lámparas de baja presión presentan una alta eficiencia de
15
energía (aproximadamente el 40% de la energía impuesta es convertida en radiación UV*C
con un pico máximo de emisión en 254 nm). Las lámparas de mediana presión producen más
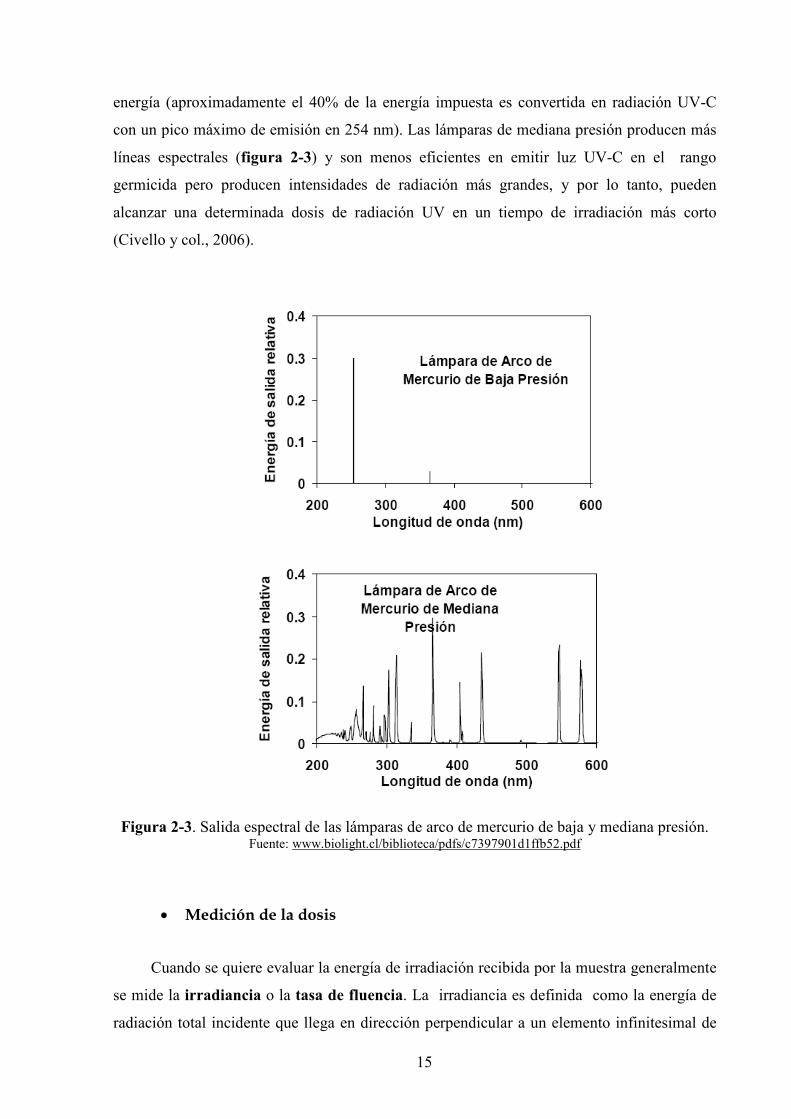
líneas espectrales (figura 2'3) y son menos eficientes en emitir luz UV*C en el rango
germicida pero producen intensidades de radiación más grandes, y por lo tanto, pueden
alcanzar una determinada dosis de radiación UV en un tiempo de irradiación más corto
(Civello y col., 2006).
Figura 2'3. Salida espectral de las lámparas de arco de mercurio de baja y mediana presión. Fuente: www.biolight.cl/biblioteca/pdfs/c7397901d1ffb52.pdf
• 6�����������������
�
Cuando se quiere evaluar la energía de irradiación recibida por la muestra generalmente
se mide la irradiancia o la tasa de fluencia. La irradiancia es definida como la energía de
radiación total incidente que llega en dirección perpendicular a un elemento infinitesimal de

16
superficie de área dA que contiene el punto en consideración dividido por dA. Por otro lado,
la tasa de fluencia se define como la energía de radiación total incidente que llega de todas
direcciones a una esfera pequeña infinitesimal con un área de sección dA, dividido por dA.
Este último término es más apropiado para la desinfección UV, ya que el microorganismo
puede recibir la irradiación desde cualquier dirección (Bolton y col, 2003). Las unidades de
irradiancia y de tasa de fluencia en el Sistema Internacional son W/m2.
El término dosis de UV es utilizado universalmente en la literatura y se obtiene
multiplicando la irradiancia o la tasa de fluencia, si se mantienen constantes, por el tiempo de
exposición a la radiación. Sin embargo, la dosis de UV es un término que en otros contextos
es usado para describir la energía total absorbida. En el caso de los microorganismos casi toda
la luz ultravioleta incidente pasa a través del organismo y sólo un porcentaje es absorbido.
Por lo tanto, el concepto de fluencia es apropiado ya que se relaciona con la energía UV
“incidente” en vez de la energía UV “absorbida” (Bolton y col, 2003). En este trabajo se
utilizará entonces el término más común, dosis de UV, para referirnos al producto de la
fluencia por el tiempo de exposición, expresada en J/m2.
La medición de la dosis de radiación UV puede llevarse a cabo por diferentes métodos.
Uno de los ellos se basa en la medición con un radiómetro, que es un aparato que consta de
un dispositivo selectivo el cual permite separar la parte del espectro de emisión que se desea
medir y un detector fotosensible que permite transformar la energía incidente en una medida
de voltaje o corriente (Shama, 2007). Otra de las alternativas es la medición por
actinometría. Esta técnica se basa en la medición de la luz UV incidente a través de una
reacción fotoquímica, para la cual la cantidad de moléculas de producto formado por fotones
absorbidos está bien establecida. Por lo tanto, un actinómetro es una reacción fotoquímica o
sistema químico donde la eficiencia cuántica (rendimiento cuántico) es conocida. La medición
del rendimiento químico después de la exposición a la luz permite conocer el flujo de fotones
incidentes (Bolton, 2001). Uno de los actinómetros químicos comúnmente utilizados es el par
ioduro*iodato (Rahn, 1997). La ventaja del uso del radiómetro es su fácil manejo y rápida
obtención de los datos, mientras que las ventajas que posee el método actinométrico en
relación a la radiometría es su bajo costo y además no requiere de calibraciones periódicas
con respecto a un estándar ya que la eficiencia cuántica de los actinómetros no varía para un
conjunto de condiciones (Bolton y col., 2003).
• ����������������������

17
La radiación ultravioleta ha sido ampliamente usada para la esterilización de superficies,
utensilios, aire y materiales de empaque. También se utiliza para la desinfección de grandes
cantidades de microorganismos en aguas potables y residuales (Gray, 1994; Shama, 1999;
Bintsis y col, 2000; Bolton, 2001) y en la piscicultura (Summerfelt, 2003). Sin embargo, con
los años ha crecido el interés en su aplicación como una tecnología de preservación de
alimentos.
En alimentos líquidos se ha evaluado su uso para la inactivación de microorganismos en
jugos de frutas y vegetales. El poder de penetración de la luz UV*C en líquidos depende del
tipo de líquido, de su absortividad y del contenido de sólidos y de materia suspendida. Un
incremento en la cantidad de sólidos puede provocar una disminución en la penetración de la
radiación, mientras que la presencia de grandes partículas suspendidas pueden bloquear la
incidencia de la luz sobre la población microbiana (Bintis y col., 2000; Shama, 2006). Por ello
es importante el uso de flujo turbulento durante el tratamiento de líquidos, fluidodinámica
que también posibilita la obtención de tiempos de residencia de los distintos elementos de
volumen más uniformes. En los EE.UU ha sido aprobado el uso de la radiación UV para el
tratamiento establecido de 5 ciclos logarítmicos de reducción del patógeno más resistente en
jugos frutales y vegetales, con el requerimiento adicional del flujo turbulento (FDA, 2000).
En frutas y vegetales, la aplicación de la luz UV*C ha sido estudiada con dos objetivos.
Uno de ellos es reducir la carga inicial de microorganismos en la superficie del producto, y el
otro, es inducir una respuesta al estrés. La aplicación de la luz UV*C con este último
propósito se denomina “hormesis”. La hormesis puede ser definida como una respuesta
beneficiosa de la planta que resulta de la aplicación de una dosis baja de un factor de estrés.
Existen una variedad de tratamientos físicos, incluyendo la radiación UV*C, que pueden
servir como factores de estrés. Las dosis óptimas para lograr efectos horméticos se encuentran
en el rango entre 0,12 a 9 KJ/m2. Este es un rango mucho más estrecho que el utilizado en la
irradiación de alimentos para obtener efectos germicidas (Shama & Anderson, 2005; Shama,
2006).
Dentro de los efectos horméticos que se han reportado se encuentran la inducción de la
síntesis de compuestos antifúngicos y el retraso de la maduración. En frutas cítricas se ha
reportado un aumento de la resistencia a hongos atribuido a la acumulación de las fitoalexinas
escaparona (Kim y col., 1991; D’hallewin y col., 1999) y escopoletina (D’hallewin y col.,
2000) luego del tratamiento UV*C. La inducción de la formación de fitoalexinas estaría
asociada con la estimulación de la producción de la enzima fenilalanina amonio*liasa (PAL).
En tomates se ha reportado un retraso de la maduración asociado con un incremento de la

18
concentración de poliaminas después de la aplicación de dosis bajas de radiación UV*C
(Dibble y col., 1988; Stevens y col., 1998; Maharaj y col., 1999). Las poliaminas son
policationes de bajo peso molecular que se acumulan en las plantas en respuesta al estrés
ambiental. Estos compuestos estarían implicados en las primeras etapas del desarrollo de las
frutas. La disminución de la concentración de poliaminas sería un factor desencadenante de la
senescencia (Shama & Anderson, 2005). Otros de los efectos benéficos reportados por
diversos autores en diferentes frutas y vegetales es el incremento de los niveles de compuestos
antioxidantes y fitoquímicos (Vicente y col., 2005; González*Aguilar y col., 2007; Lemoine y
col., 2007; Erkan y col., 2008).
En frutas y vegetales mínimamente procesados, la aplicación de la radiación UV*C se
ha evaluado fundamentalmente con el propósito de inactivar microorganismos presentes en la
superficie de estos productos. La irradiación UV*C fue efectiva para reducir la actividad
microbiológica en rodajas de zucchinis (Erkan y col., 2001), lechuga mínimamente procesada
(Allende & Artés, 2003) y cubos de sandía (Fonseca & Rushing, 2006). En melones cortados
que fueron procesados bajo la luz UV*C, la irradiación no sólo produjo una disminución de
microorganismos de deterioro tales como bacterias ácido lácticas y mesófilas, sino que
también indujo una respuesta de defensa hipersensible que condujo a una disminución de la
actividad respiratoria, un aumento de los niveles de peroxidasas (POD), una disminución de la
actividad esterasa y lipasa, y una mayor retención de la firmeza a lo largo del almacenamiento
(Lamikanra y col., 2005). Las reducciones decimales variaron entre las diferentes matrices y
las dosis aplicadas, variando en un rango entre 0,5 y 5,5 ciclos logarítmicos de reducción.
• �����*��������������
Las ventajas que presenta la irradiación con luz UV*C como tecnología de conservación
es que es un proceso simple y rápido, que no afecta significativamente la temperatura del
alimento, ni su contenido de humedad, y por lo tanto, no causa daño térmico al producto.
Otras ventajas son que tiene bajo costo de inversión, funcionamiento y mantenimiento y
puede ser aplicada en combinación con otras tecnologías de conservación. Cabe añadir
también que su uso está aprobado para la desinfección de superficies de alimentos (FDA,
2001).
La principal desventaja que presenta es su bajo poder de penetración. Otra de las
limitaciones, como se comentó anteriormente, es que en algunos casos los microorganismos
pueden revertir los efectos destructivos de la luz UV*C mediante mecanismos de
fotoreactivación.
19
,$,$,$,$��������������������������������
• �������
�
La radiación con luz pulsada (LP) es una nueva tecnología destinada a la
descontaminación de la superficie de alimentos mediante la aplicación de pulsos de corta
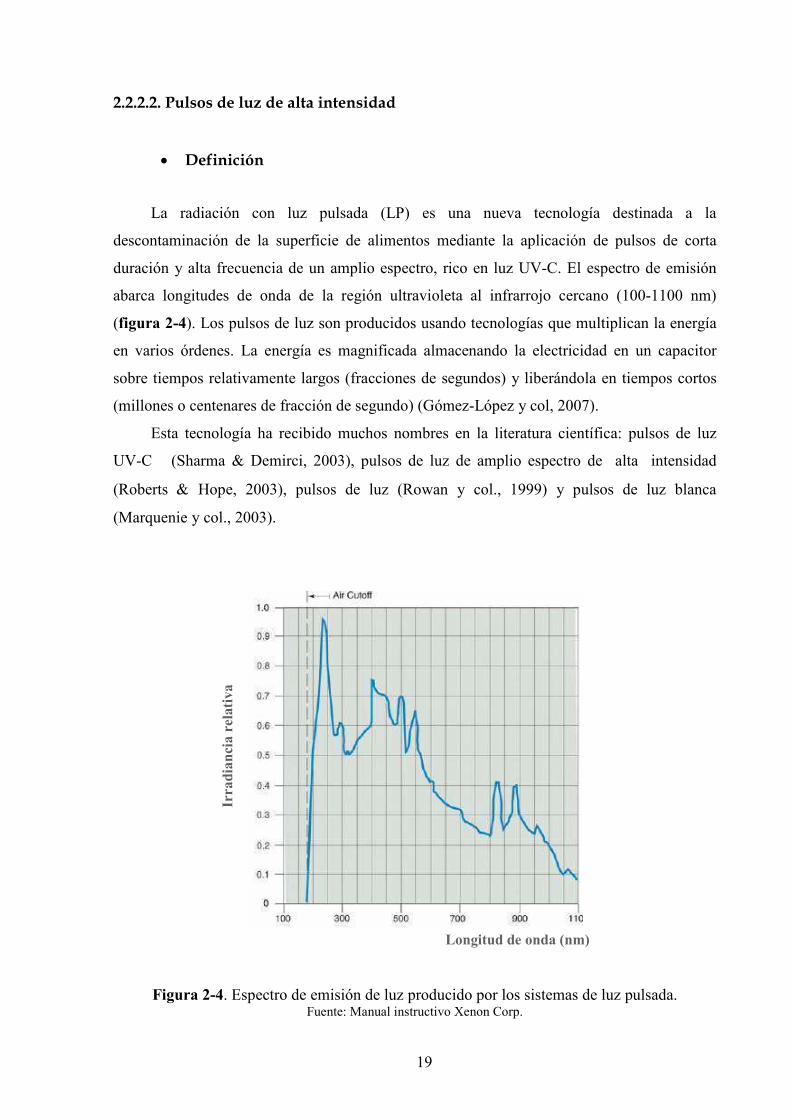
duración y alta frecuencia de un amplio espectro, rico en luz UV*C. El espectro de emisión
abarca longitudes de onda de la región ultravioleta al infrarrojo cercano (100*1100 nm)
(figura 2'4). Los pulsos de luz son producidos usando tecnologías que multiplican la energía
en varios órdenes. La energía es magnificada almacenando la electricidad en un capacitor
sobre tiempos relativamente largos (fracciones de segundos) y liberándola en tiempos cortos
(millones o centenares de fracción de segundo) (Gómez*López y col, 2007).
Esta tecnología ha recibido muchos nombres en la literatura científica: pulsos de luz
UV*C (Sharma & Demirci, 2003), pulsos de luz de amplio espectro de alta intensidad
(Roberts & Hope, 2003), pulsos de luz (Rowan y col., 1999) y pulsos de luz blanca
(Marquenie y col., 2003).
Figura 2'4. Espectro de emisión de luz producido por los sistemas de luz pulsada.
Fuente: Manual instructivo Xenon Corp.
Longitud de onda (nm)
Irra
dian
cia
rela
tiva

20
• )�����������������������
Las lámparas de luz pulsada contienen gas xenón y se encuentran conectadas
generalmente a un módulo controlador mediante un cable de alta tensión. Están recubiertas
externamente con un alambre muy fino que sirve de conductor cuando se aplica un disparo de
alto voltaje. Normalmente la lámpara se encuentra en un circuito abierto. Cuando se aplica el
disparo de voltaje, el gas xenón que contiene la lámpara se ioniza. Si el voltaje es lo
suficientemente alto, el gas xenón actúa como conductor y la descarga que se produce genera
la emisión de un pulso de luz en la región UV/visible. El espectro de emisión está
determinado por la densidad de corriente que existe dentro de la lámpara. Generalmente, altas
densidades de corriente conducen a la emisión de un espectro rico en la región UV*C. Para
disipar el calor generado en la lámpara como así también para evitar la acumulación de altos
niveles de ozono producidos por las longitudes de onda más cortas, generalmente las lámparas
se encuentran conectadas a un soplador, el cual genera un flujo de corriente de aire filtrado
que pasa continuamente por dentro de la caja que contiene la lámpara (Manual Xenon corp.).
�
• "����������������������������
�
El efecto letal de la luz pulsada sobre los microorganismos se debería
fundamentalmente a un mecanismo de acción fotoquímico y/o fototérmico. El mecanismo
fotoquímico involucraría el daño en el ADN provocado por la absorción de la luz de la región
UV*C, mientras que el mecanismo fototérmico estaría relacionado con el daño provocado por
el sobrecalentamiento generado por la emisión en la región visible*infrarroja. Es posible que
ambos mecanismos coexistan, y la importancia relativa de cada uno dependerá de la dosis y el
tipo de microorganismo (Gómez*López y col, 2007).
Muchos autores explican el mecanismo de inactivación microbiológica mediante la luz
pulsada en base a los estudios realizados en la irradiación con luz UV*C continua, en donde el
mecanismo es fotoquímico. Si bien pueden tener similitudes, podrían existir algunas
diferencias (Gómez*López y col., 2007). Wang y col. (2005) sostuvieron la hipótesis de que
el efecto fotoquímico es la principal causa de inactivación de microorganismos por la LP al
encontrar que la mayor eficiencia germicida se encontraba en el rango entre 230*300 nm, con
una máxima respuesta a 270 nm. Sin embargo, hay otros trabajos en donde se pone en
evidencia que la inactivación puede ocurrir por otros efectos. Wekhof (2000) propuso que
utilizando una dosis superior a 0,5 kJ/cm2 la desinfección se logra mediante la ruptura de la
bacteria durante el sobrecalentamiento momentáneo causado por la absorción de la luz

21
emitida por la lámpara. Fine & Gervais (2004) sugirieron que el sobrecalentamiento podría
causar vaporización y generar un pequeño flujo de vapor que causaría destrucción de
membranas. Takeshita y col. (2003) compararon la inactivación de S. cerevisiae mediante luz
UV*C continua y pulsada. Observaron que el daño en el ADN en las células de levadura era
esencialmente el mismo en los dos métodos. Sin embargo, las células expuestas a la luz
pulsada presentaron cambios estructurales (distorsión o daño de membranas, vacuolas
expandidas, forma circular) que no presentaban las células irradiadas con luz UV*C continua.
�
• )�������������������������������
�
Existen discrepancias en los estudios reportados sobre la susceptibilidad que presentan
los diferentes microorganismos a la luz pulsada. Anderson y col. (2000) y Rowan y col.
(1999) informaron que la susceptibilidad de los microorganismos a la luz pulsada decrecía en
el siguiente orden: bacterias Gram*negativas, bacterias Gram*positivas y esporas de hongos.
El color de las esporas puede jugar un rol significativo en la susceptibilidad de la espora del
hongo. Las esporas de Aspergillus niger son más resistentes que las esporas de Fusarium
culmorum, lo que podría deberse a que los pigmentos de A. níger absorben más en la región
UV*C que los de las esporas de F. culmorum, protegiendo a la espora contra la radiación UV*
C (Anderson y col., 2000). En contraste con esto, Gómez*López y col. (2005 a) no observaron
ningún patrón de sensibilidad entre los diferentes grupos de microorganismos, luego de
estudiar 27 bacterias, levaduras y hongos.
�
• ����������������������
�
Al tratarse de una tecnología más reciente que la luz UV*C continua, existe un menor
número de trabajos reportados en bibliografía que estudian la aplicación de la LP en
alimentos. Hoornstra y col. (2002) estudiaron la inactivación de microorganismos presentes
en la superficie de vegetales. Encontraron reducciones en el recuento aeróbico mayores a 1,6
log CFU/cm2 en los diferentes vegetales evaluados utilizando una dosis de 0,30 J/cm2. Gómez
y López (2005a) reportaron una reducción en el recuento de mesófilos aeróbicos entre 0,56 y
2,04 log CFU/cm2 en diferentes vegetales mínimamente procesados (espinaca, radicheta,
lechuga, zanahoria, pimientos y repollo blanco) y en brotes de soja luego del tratamiento con
más de 200 pulsos. Marquenie y col. (2003) reportaron una inactivación máxima de 3 a 4
unidades log de Botritis cinerea y Monilinia fructigena in vitro. Sin embargo, cuando se

22
inoculó B. cinerea en frutillas la LP no fue efectiva para lograr su inactivación. Ozer &
Demirci (2006) estudiaron la inactivación de E. coli O157:H7 y L. monocytogenes Scout A en
filetes de salmón fresco utilizando una lámpara que generaba 5,6 J/cm2 por pulso en la
superficie de la lámpara y 3 pulsos por segundo. Trataron dos superficies diferentes, el lado de
la piel y el del músculo. La reducción log máxima para E. coli O157:H7 fue de 1,09 sobre el
lado del músculo y de 0,86 sobre lado de la piel, mientras que para L. monocytogenes, la
máxima reducción log fue de 0,74 sobre el lado del músculo. Cuando los filetes de pescado
fueron colocados más cerca de la lámpara y expuestos a tiempos de irradiación más altos
presentaron un mayor sobrecalentamiento. �
• �����*��������������
La ventaja de la luz pulsada frente a la luz UV*C continua radicaría en que la alta
energía e intensidad del pulso amplificarían los mecanismos de destrucción de componentes
celulares asociados a longitudes de onda individuales y se producirían daños irreversibles en
DNA, proteínas y otras macromoléculas, impidiéndose los procesos de fotoreparación
asociados con la luz UV*C continua. Otra de las ventajas es el menor tiempo de proceso, ya
que permite obtener dosis de irradiación mayores en menores tiempos (Wang y col., 2005).
Por otro lado, las lámparas de xenón son más amigables con el medio ambiente que las
lámparas de luz UV*C continua ya que no contienen mercurio (Gómez*López y col., 2007).
Esta tecnología fue aprobada por la Federal Drug Administration para tratar alimentos (FDA,
1996) y se sugiere para esterilizar/pasteurizar superficies de empaques y alimentos, y
productos farmacéuticos transparentes, evitándose el uso de desinfectantes o conservadores.
Una de las principales desventajas que presenta la LP es el calentamiento de las
muestras irradiadas a dosis de exposición elevadas. Otras de las limitaciones de la LP, al igual
que en la luz UV*C continua, es su falta de penetración en muestras opacas y la atenuación de
la luz por los microorganismos a altos niveles de contaminación (efecto “sombra”) (Gómez y
col., 2007; Woodling y Moraru, 2005).
�,$,$,$7$�����������������
En los últimos años ha habido un creciente interés por el diseño de los llamados
alimentos funcionales, entre los cuales se encuentran los alimentos con componentes

23
fisiológicamente activos (CFA) agregados. En particular, los alimentos enriquecidos con
minerales han tenido una fuerte demanda comercial.
El calcio es un mineral esencial que interviene en el normal crecimiento y desarrollo.
Además de su función de mantener la integridad del esqueleto, el calcio regula la
excitabilidad nerviosa, la contracción muscular y la coagulación de la sangre. Diversos
estudios epidemiológicos han mostrado una relación inversa entre una baja ingesta de calcio
en la dieta y la incidencia de fracturas en la población anciana, con un incremento en la
morbilidad y mortalidad, especialmente en mujeres (Levenson and Bockman, 1994). En la
mayoría de las dietas el calcio proviene de productos lácteos; mientras que los granos,
vegetales verdes, frutas, bebidas y otros alimentos proveen menores cantidades. Se ha visto
que en Argentina existe una baja ingesta de calcio debido al bajo consumo de productos
lácteos, cuyas causas están asociadas, no a una situación económica, sino a los hábitos
alimenticios de la mayoría de la población (Zeni & Portela, 1988; Pacín y col., 1999).
Las frutas y vegetales debido a sus apreciadas propiedades funcionales, nutricionales y
sensoriales podrían ser alimentos ideales para ser enriquecidas con calcio. Algunos estudios
realizados han demostrado la viabilidad de las matrices vegetales para incorporar compuestos
bioactivos (Betoret y col., 2001; Fito y col., 2001; Gras y col., 2003; Ortiz y col., 2003; Tapia
y col., 2003; Anino y col., 2006).
La incorporación de CFA en los tejidos de frutas y vegetales puede realizarse mediante
procesos de impregnación a presión atmosférica (IA), impregnación a vacío (IV) o mediante
una combinación de impregnación a vacío seguido por largos periodos de impregnación a
presión atmosférica. La presión de operación es uno de los factores que afecta la composición
y las características estructurales del producto final.
Durante los procesos de impregnación atmosférica se produce una transferencia de CFA
desde la solución hacia la fruta debido al gradiente de potencial químico entre el medio y el
tejido celular vegetal. La impregnación se produce por lo tanto por un mecanismo de difusión
en donde la estructura celular de la planta actúa como una membrana semipermeable
(Torreggiani, 1993; Torregiani y Bertolo, 2002). En los procesos de impregnación al vacío la
solución de impregnación penetra en los poros (espacios intercelulares) de la matriz vegetal
por capilaridad y mediante gradientes de presión impuestos al sistema. Este fenómeno se
denomina mecanismo hidrodinámico y en algunos casos permite una mayor velocidad de
transferencia de masa, aunque la efectividad de la IV para la incorporación de un soluto
específico dentro del tejido esta limitada por su solubilidad y/o la porosidad de la matriz
vegetal. Si los procesos de IA y de IV se combinan con un tratamiento de escaldado previo se

24
pueden producir alteraciones estructurales en el tejido que pueden aumentar la velocidad de
transporte del soluto de interés hacia el interior del tejido, lográndose una mayor
concentración final del mismo.
,$,$,$8$�(����������4���������������
El escaldado se aplica usualmente antes de la congelación, la deshidratación y la
esterilización para destruir la actividad enzimática de frutas y verduras, causante de la
formación de aromas y sabores desagradables y cambios de color y textura. En las tecnologías
de procesamiento mínimo, no sólo se aplica para inactivar enzimas sino también para destruir
o dañar por calor los microorganismos presentes (entre otros hongos y levaduras), reduciendo
la carga microbiana inicial o sensibilizando a los sobrevivientes ante otros factores de estrés
(Alzamora y col, 1995). El escaldado además remueve el aire intercelular y provoca un
ablandamiento del tejido que en algunos vegetales resulta conveniente (Lewicki, 1998).
El tratamiento térmico a temperaturas mayores a las que el microorganismo puede
crecer produce, de acuerdo a su severidad y tiempo de aplicación, la inactivación o la lesión
subletal del mismo. Se han identificado cuatro blancos principales relacionados al daño
térmico letal y no letal: el ADN, el ARN y los ribosomas, las membranas citoplasmáticas y las
enzimas específicas (Ma y col., 1992).
Los dos métodos de escaldado comercialmente más empleados son: 1) escaldado en
atmósfera de vapor de agua saturado y 2) escaldado en un baño de agua caliente. El escaldado
en vapor es normalmente el método de elección ya que evita las pérdidas excesivas de
compuestos hidrosolubles como vitaminas, minerales y carbohidratos, aunque éste tenga
mayores gastos de inversión que el escaldado en agua caliente (Fellows, 1994).
Si el alimento no se escalda se producen normalmente durante el almacenamiento
cambios no deseados en su valor nutritivo y sus características organolépticas. Entre las
enzimas responsables de estos cambios se encuentran la lipooxigenasa (LOX), la peroxidasa
(POD), la polifenoloxidasa (PPO), la poligalacturonasa (PG) y la clorofilasa.
La adecuada inactivación de las enzimas requiere un calentamiento rápido hasta una
temperatura determinada, el mantenimiento de ésta durante un tiempo necesario y un
enfriamiento rápido hasta una temperatura próxima a la del ambiente. Los factores que
determinan el tiempo de escaldado son: (1) el tipo de fruta o vegetal, (2) su tamaño, (3) la
temperatura de escaldado, y (4) el sistema de calentamiento.
�

25
,$,$,$9$�!�����������������������������������
El pardeamiento ocurre durante el procesamiento de las frutas. Se conocen al menos
cinco causas de pardeamiento en las frutas y vegetales procesados y/o almacenados:
pardeamiento enzimático de fenoles, reacción de Maillard, oxidación de ácido ascórbico,
caramelización y formación de polímeros pardos mediante la oxidación de lípidos (Pizzocaro
y col. 1993).
En frutas y vegetales mínimamente procesados, el pardeamiento enzimático es una de
los factores más importantes que limitan la vida útil de estos productos. Durante las diferentes
etapas de preparación de las frutas y vegetales MP (pelado, corte, lavado), las células se
rompen, lo que provoca que las enzimas se descomportamentalicen y entren en contacto con
sus sustratos. El pardeamiento enzimático resulta de la acción de un grupo de enzimas
denominadas polifenoloxidasas (PPO), las cuales se encuentran en todas las plantas, y se
presentan en mayores cantidades en bananas, manzanas, peras, papas y duraznos. Estas
enzimas catalizan la reacción de oxidación de los compuestos fenólicos a o*quinonas. La
posterior oxidación y polimerización de las o*quinonas dan lugar a la formación de
compuestos pardos (Garcia & Barrett, 2002). Las peroxidasas también están involucradas en
reacciones de pardeamiento enzimático. Estas enzimas cuya principal función es oxidar
compuestos dadores de hidrógeno a expensas de los peróxidos, son altamente específicas para
el peróxido de hidrógeno pero pueden aceptar un amplio rango de dadores de hidrógeno
incluyendo a los polifenoles. Los productos principales de la oxidación de fenoles son
probablemente quinonas similares a aquellas obtenidas con las polifenoloxidasas. Aunque las
peroxidasas están distribuidas ampliamente, especialmente en las plantas, por lo general
parecen estar poco implicadas en el pardeamiento enzimático de frutas y hortalizas producido
después de un estrés mecánico (Nicolas y col., 1994).
Para que ocurran las reacciones de pardeamiento enzimático se requiere: la presencia de
PPO activa, oxígeno y sustratos fenólicos. La prevención del pardeamiento es posible, al
menos temporalmente, a través de la eliminación de los sustratos, productos y/o la inhibición
enzimática (Lamikanra, 2002).
Tradicionalmente, la prevención del pardeamiento enzimático se ha llevado a cabo
mediante la inactivación de la PPO por calor. Pero si bien la inactivación por calor es un
método efectivo para inhibir el pardeamiento, puede provocar cambios indeseables en algunos
atributos de calidad, como la textura y el “flavor”, y resultar en pérdidas nutricionales. En
lugar del (o en adición al) uso de calor, el control del pardeamiento puede lograrse mediante

26
el uso de diferentes tipos de compuestos químicos, generalmente denominados agentes
antipardeamiento. En general, estos agentes químicos son aplicados en soluciones,
frecuentemente como formulaciones que contienen uno o más compuestos (Lamikanra, 2002).
Los diferentes tipos de compuestos químicos que se utilizan pueden actuar directamente
como inhibidores de la PPO, generar un medio inadecuado para el desarrollo de la reacción de
pardeamiento, o reaccionar con los productos de la reacción de la PPO antes de que éstos
puedan conducir a la formación de compuestos pardos (Lamikanra, 2002).
Dentro de los compuestos que inhiben la actividad de la PPO, el dióxido de azufre es el
más efectivo y ha sido utilizado en la industria de alimentos por muchos años. Sin embargo,
los tratamientos con sulfitos pueden llegar a producir “flavors” indeseables como así también
el ablandamiento de las frutas. Además su uso en alimentos ha sido restringido por la FDA
debido a que puede presentar un riesgo de alergia para las personas asmáticas. Es por ello que
se han investigado alternativas al tratamiento con sulfitos (Zhu y col., 2007).
El uso de compuestos químicos que disminuyan el pH del producto, o acidulantes, es
una de las alternativas que se ha evaluado para el control del pardeamiento, ya que se ha visto
que la actividad de la PPO disminuye a pH menores a 4,5. El acidulante más común es el
ácido cítrico. Los acidulantes son frecuentemente utilizados en combinación con otros tipos
de agentes antipardeamiento, debido a que resulta difícil lograr una inhibición eficiente del
pardeamiento solamente a través del control del pH (Lamikanra, 2002).
Otros agentes antipardeamiento evaluados involucran compuestos reductores, quelantes
y formadores de complejos. Los agentes reductores inhiben el pardeamiento al reducir los o*
quinonas resultantes de la reacción de la PPO a o*difenoles, impidiendo la posterior oxidación
de las o*quinonas que da lugar a la formación de compuestos pardos. Dentro de los agentes
reductores se encuentran el ácido ascórbico y la cisteína. El ácido ascórbico no sólo tiene
propiedades reductoras sino que también se ha visto que tiene un efecto inhibidor directo
sobre la PPO (Whitaker, 1994). La cisteína presenta el problema que las cantidades requeridas
para inhibir el pardeamiento pueden afectar negativamente el sabor del producto (Richard*
Forget y col, 1992). Los agentes quelantes, como el EDTA, actúan formando quelatos con el
cobre presente en el sitio activo de la PPO. Los agentes formadores de complejos, como las
ciclodextrinas, atrapan los sustratos de la PPO o los productos de la reacción (Lamikanra,
2002).
Uno de los agentes antipardeamiento con mayor potencial en frutas cortadas es el 4*
hexilresorcinol, un compuesto químico que ha sido utilizado en medicamentos durante mucho
tiempo. Este compuesto actuaría como un eficaz inhibidor de la PPO.

27
Se ha reportado también que el cloruro de calcio es efectivo para inhibir el
pardeamiento (Pointing y col., 1972). Estos autores encontraron que la combinación de 1%
ácido ascórbico y 0,1 % de cloruro de calcio fue efectiva para inhibir el pardeamiento
superficial de manzanas cortadas y almacenadas en refrigeración. La acción inhibitoria del
cloruro de calcio sería causada por el anión cloruro que actuaría inhibiendo a la PPO.
�
,$7$� ���������������*���+�������
2.3.1. )�����������*���
Los frutos usualmente se consideran como el órgano reproductivo de las plantas,
conteniendo las semillas. Éstas generalmente tienen alto contenido en azúcar, acidez
relativamente elevada y perfume pronunciado. Los vegetales generalmente son clasificados
como las partes no reproductiva de las plantas, como las raíces, hojas, o tallos. De todas
maneras, la distinción entre frutas y vegetales no está del todo clara, y el uso común de los
términos a veces no coincide con la clasificación botánica estricta (Edwards, 1999).
La estructura de la planta a toda escala es en su mayoría anisotrópica, heterogénea y no*
continua, y por lo tanto exhibe una considerable variabilidad en su construcción (Jackman y
Stanley, 1995 a).
Cada una de las distintas partes de la planta está constituida de diferentes tipos de
tejidos. Cada uno de éstos tejidos está compuesto por un gran número de células de estructura
similar (Edwards, 1999), cada una de ellas dentro de su propia pared celular y unida con otras
células por medio de una sustancia intercelular cementante (Esau, 1982). Los tejidos del
cuerpo vegetal se clasifican según los tipos de células que los componen; según su función;
según el lugar y modo en que se originan y según su estado de desarrollo. Pueden dividirse
también en tejidos simples y complejos, según el número de tipos celulares que tengan. Los
principales tejidos de una planta se agrupan, según la continuidad topográfica, en tres sistemas
de tejido: 1) el sistema fundamental, que incluye los tejidos que forman el elemento básico
de la planta y al mismo tiempo, muestran varios grados de especialización; éstos son
parénquima, colénquima y esclerénquima; 2) el sistema vascular, que contiene dos tipos de
tejidos conductores, el xilema (conductor de agua) y el floema (conductor de alimento); y 3)
el sistema dérmico, que comprende la epidermis (Esau, 1982).
Las células del parénquima forman tejidos continuos en la corteza del tallo, de la raíz y
en el mesófilo de las hojas; aparecen también como cordones verticales y radios en los tejidos

28
vasculares. Las células parenquimáticas son típicamente células vivas, capaces de crecer y
dividirse. Tienen diversas formas, con frecuencia son poliédricas, pero pueden ser estrelladas
o muy alargadas. El parénquima está vinculado con la fotosíntesis, el almacenamiento de
diversos materiales, la cicatrización de heridas, la regeneración de tejidos y la formación de
nuevos vástagos y raíces adventicias. Las células parenquimáticas pueden especializarse como
estructuras secretoras o excretoras (Esau, 1982). La parte comestible de la mayoría de las
plantas está constituida por tejido parenquimático. Dependiendo del arreglo espacial y tamaño
relativo de las células, el tejido tiene cantidades significativas (1*25 %) de espacios
intercelulares llenos de aire que tienen un impacto considerable en las propiedades mecánicas
(Jackman y Stanley, 1995 a).
El colénquima y el esclerénquima son tejidos mecánicos de sostén de la planta (Fahn,
1985). Las células del colénquima aparecen en tallos, hojas, partes florales, frutos y raíces. Es
un tejido vivo estrechamente relacionado con el parénquima. La forma de las células varía
desde prismática corta a muy alargada; generalmente tienen paredes desigualmente
engrosadas (Esau, 1982). El colénquima es plástico y se deforma irreversiblemente cuando
crece el órgano en que se encuentra (Fahn, 1985). Las células del esclerénquima pueden
formar masas continuas o pueden aparecer en pequeños grupos, o individualmente en otras
células. Pueden desarrollar en cualquiera o en todas las partes de la planta. Tienen paredes
gruesas, secundarias, a menudo lignificadas, y pueden carecer de cloroplastos en la madurez.
Son células muertas y se distinguen dos tipos celulares: esclereidas y fibras. Las esclereidas
varían en forma desde poliédrica hasta alargada y pueden ser ramificadas. Las fibras
generalmente son células largas y delgadas (Esau, 1982). Las células del esclerénquima tienen
propiedades elásticas, al contrario que las del colénquima (Fahn, 1985).
El xilema es un tejido complejo que consta de varios tipos de células, de las cuales las
más importantes son los vasos o segmentos de tráquea, que son células muertas implicadas
fundamentalmente en el transporte de agua e iones disueltos, pero además desempeñan en
algún grado una función de sostén (Fahn, 1985). El floema también es un tejido complejo. Se
encuentra en el cuerpo de la planta junto al xilema. Está relacionado con la conducción y el
almacenamiento del alimento y con el sostén. (Esau, 1982). Las células fundamentales del
floema son los elementos cribosos, que conducen los productos de la fotosíntesis. Estos
elementos de tubo criboso están unidos por sus extremos (Fahn, 1985).
Las células epidérmicas forman una capa continua en la superficie del cuerpo de la
planta y muestran varias características especiales relacionadas con su posición superficial. La
epidermis proporciona protección mecánica. La presencia de material graso, cutina, en la

29
pared externa y sobre su superficie (cutícula) restringe la transpiración. Los estomas están
relacionados con el intercambio gaseoso (Esau, 1982).
2.3.2. "������������������
�
Las células eucarióticas son las unidades constitutivas del tejido parenquimático
vegetal, que como se mencionó anteriormente, constituye la parte comestible de la mayoría de
las plantas. Una representación esquemática se observa en la figura 2'5.�
Los componentes principales de la célula vegetal son la pared celular, la membrana
citoplasmática, el citoplasma y el núcleo. El citoplasma comprende el retículo
endoplasmático, el aparato de Golgi, mitocondrias, ribosomas, plástidos, vacuolas, sustancias
ergásticas, etc. (Fahn, 1985).
La matriz citoplasmática o citoplasma fundamental rodea al núcleo y forma parte del
protoplasto. Está compuesto por una sustancia semilíquida, la cual está en estado de sol o en
estado de gel, y cumple la función de proveer un medio para el desarrollo de los procesos
citoplasmáticos (Jackman y Stanley, 1995 a). El componente mayoritario (85*90 %) es el agua
����������
����� ��������
������� �����������
��������
����� � �����
��������������������
�����
�������
�� � ��� ��
����
!��������
"���������!��
�
Figura 2'5. Esquema de una célula eucariótica vegetal. Fuente: Curtis, 1985.

30
y contiene proteínas, lípidos y otras sustancias solubles en agua, es decir, no ligadas a cuerpos
estructurados. Está separado de la pared celular por la membrana plasmática o plasmalema y
de las vacuolas por otra unidad de membrana que se denomina tonoplasto (Fahn, 1985).
La vacuola es un componente importante del protoplasto de la célula vegetal; una o
varias vacuolas ocupan más del 90 % del volumen de casi todas las células vegetales adultas.
Contiene agua y una variedad de sustancias orgánicas e inorgánicas, muchas de las cuales
están presentes en estado disuelto (azúcares, proteínas, ácidos orgánicos, fosfátidos, taninos,
flavonoides y oxalato cálcico). El tonoplasto que encierra a la vacuola tiene permeabilidad
diferencial y está por lo tanto relacionado con la regulación de los fenómenos osmóticos
asociados con las vacuolas, especialmente con el mantenimiento de la turgencia en la célula.
Con el crecimiento de las células, las vacuolas aumentan de tamaño y se fusionan,
constituyendo en muchas ocasiones una gran vacuola central que comprime al citoplasma
contra la pared celular. Las vacuolas funcionan como reguladoras del contenido de agua y de
sustancias disueltas en la célula en procesos tales como la regulación de la presión osmótica,
el almacenamiento de sustancias de reserva o la digestión. Hay evidencias de que las vacuolas
contienen enzimas digestivas capaces de disociar componentes citoplasmáticos y metabolitos
(Lehninger, 1988).
La membrana plasmática controla el transporte de agua y de solutos y es sede de
numerosos procesos bioquímicos. Para explicar su estructura, tiene actualmente gran
aceptación el modelo de mosaico fluido propuesto por Singer y Nicolson (1972), en el que la
membrana está constituida por una bicapa fosfolipídica fluida con moléculas de proteínas
globulares penetrando por cada lado de la membrana o extendiéndose enteramente por ella
(figura 2'6).
Por fuera del plasmalema se encuentra la pared celular. La pared celular es un
compartimento dinámico que cambia a lo largo de la vida de la célula. Las paredes sirven
como sostén mecánico a los órganos de las plantas, especialmente las paredes gruesas y
rígidas. Las paredes celulares actúan en actividades tan importantes de los tejidos vegetales
como son la absorción, la transpiración, el traslado y la secreción (Esau, 1982).
La pared celular primaria nace durante la división celular y rápidamente incrementa su
área superficial durante la expansión de la célula. La laminilla media forma la interfase entre
las paredes primarias de las células vecinas. Finalmente, puede existir una diferenciación:
algunas células elaboran dentro de la pared primaria una pared celular secundaria.
La pared primaria de las plantas es una estructura altamente organizada que contiene
aproximadamente 65 % de agua y 35 % de diferentes polisacáridos, proteínas y sustancias

31
Figura 2'6. Esquema del modelo de mosaico fluido propuesto por Singer y Nicolson. Fuente:
Madigan y col., 1997.
aromáticas (Tabla 2'1).
La composición y el arreglo molecular de los polímeros de la pared celular difiere entre
las especies, entre tejidos de la misma especie, entre células individuales y también entre
regiones de la pared que rodea a un solo protoplasto (Carpita & McCann, 2000).
El compuesto principal de las paredes de las células vegetales es la celulosa, polisacárido
formado por cadenas 1,4*β*D*glucosa. El marco celulósico es un sistema de fibrillas de
diferente magnitud; las más grandes son visibles con un microscopio óptico y se llaman
macrofribillas. Con un microscopio electrónico convencional estas fibrillas se resuelven en
microfibrillas de aproximadamente 100 Å. Con el aumento en el poder de resolución de los
microscopios electrónicos, se ven fibrillas más y más pequeñas que se describen como
subunidades de la microfibrilla (Esau, 1982).
La celulosa tiene propiedades cristalinas a causa de la disposición ordenada dentro de la
microfibrilla. Dicha disposición está restringida a las partes conocidas como micelas. Las
cadenas de glucosa menos regularmente dispuestas se encuentran entre las micelas y
constituyen las regiones paracristalinas de la microfibrilla. La estructura cristalina de la
celulosa hace que la pared de la célula sea anisótropa. Este hidrato de carbono forma el marco
interpenetrado por la matriz constituida por los hidratos de carbono no celulósicos.
Un modelo desarrollado para la pared celular primaria (Carpita y Gibeaut, 1993)

32
Tabla 2.1. Composición química de la pared celular
Polímero Solubilidad en agua a Composición centesimal b
Polisacáridos Celulosa Hemicelulosas xiloglucano xilano Pectinas homogalacturonano rhamnogalacturonano I rhamnogalacturonano II Glicoproteínas Arabinogalactanos proteínas Extensina
insoluble
soluble soluble
soluble soluble soluble
soluble soluble
30
25 5
15 15 5
variable ~5
a solubilidad después de la extracción del polímero de la pared. b expresada como porcentaje en base seca. La pared primaria tiene ~ 65 % p/p de agua. Fuente: Jackman y Stanley, 1995 a.
propone la existencia de tres dominios independientes que interactúan entre sí para formar la
estructura final (figura 2'7). Las hemicelulosas constituyen el principal componente de unión.
Poseen muchas ramificaciones de conformación lineal que conducen a la orientación de las
microfibrillas de celulosa ligándolas por uniones puente de hidrógeno. Esto resulta en un
dominio de hemicelulosa*celulosa, que representa el 50*60 % del peso seco de la pared y el
cual está embebido en un segundo dominio constituido por sustancias pécticas, que
constituyen el 30 % adicional de la masa de la pared. El mecanismo más frecuente de enlace
de las cadenas helicoidales de homogalacturonano de pectina de*esterificada es por medio de
puentes Ca2+ para formar las zonas de unión. Las macromoléculas de ramnogalacturomananos
I (RG I) representan una porción del polímero rica en arabinogalactanos a ambos lados de la
cadena e interrumpen las uniones con iones calcio. Por lo tanto la matriz péctica controla el
tamaño del poro de la pared celular. El tamaño y la frecuencia de las zonas de unión y el
tamaño y conformación de las cadenas laterales de RG pueden influir en la porosidad del gel
de pectina. Un tercer dominio estructural contiene unidades de extensina (glicoproteína rica
en hidroxiprolina), que forma enlaces cruzados covalentemente (Jackman y Stanley, 1995 a).
Conceptualmente existen tres planos de orientación de los dominios. El arreglo de
microfibrillas en el eje transversal, los xiloglucanos y pectinas en el eje longitudinal y la
extensina en el eje radial. La organización ultraestructural de los compuestos polisacáridos de

33
#������� �� $� ��� �� ��������%����
�� & � �� ��������� �
������������'(�����
�
Figura 2'7. Representación de la pared celular primaria. Fuente: Carpita y Gibeaut, 1993)
�
la pared primaria le confiere propiedades que le permiten soportar las tensiones de estrés y el
crecimiento simultáneamente.
Las sustancias pécticas se hallan concentradas en la laminilla media, que se encuentra
entre las paredes celulares de dos células adyacentes y actúa como sustancia cementante entre
ellas. Es lábil al calor y en su ausencia las células de las plantas se separan fácilmente
(Jackman y Stanley, 1995 a).
La resistencia mecánica de la pared celular de las plantas depende de la orientación de
las microfibrillas de celulosa, de las propiedades mecánicas, y de la unión entre las sustancias

34
pépticas y las fibrillas de celulosa. El efecto en la firmeza debido a las pectinas en los tejidos
involucran dos fenómenos separados: 1) en el tejido fresco, la formación de los grupos
carboxilos libres incrementa la posibilidad y la fuerza de la unión del calcio entre polímeros
de pectinas, y 2) en el tejido tratado térmicamente hay una combinación del incremento de
uniones con calcio y una disminución en la susceptibilidad de las pectinas a la
depolimerización por β*eliminación. En algunos tejidos como manzana y tomate, la
disminución normal en el grado de metoxilación no está acompañada por la firmeza durante el
almacenamiento. El ablandamiento durante el almacenamiento de frutas frescas está atribuido
a la degradación enzimática y a la solubilización de las protopectinas (Thakur, 1997).
,$7$7$�5�������� �,$7$7$#$���������������:������
En las frutas se manejan clasificaciones en función de su naturaleza, estado, composición
y características botánicas. El fruto es la parte de los vegetales que tiene como función
principal proteger las semillas y asegurar su dispersión. Es el resultado de la fecundación del
ovario, especialmente por el engrosamiento de las paredes de éste, aunque algunos frutos
tienen otro origen ya que pueden proceder del engrosamiento del receptáculo floral o de otro
lugar de la flor. Botánicamente, el primer paso en la clasificación de los frutos consiste en
diferenciar si provienen de una sola flor o de varias, dando lugar, en este último caso, a las
infrutescencias (granada, higo, piña, etc.). Los frutos que provienen de una sola flor se dividen
entre aquéllos en los que en su formación han entrado órganos o elementos ajenos al propio
ovario, lo que da lugar a frutos complejos, denominados pomo (manzana, pera, etc.) y
pepónide (melón, sandía, etc.), o los simples, formados únicamente a partir del ovario (kiwi,
plátano, etc.).
El fruto, desde afuera hacia adentro, consta de la piel o cáscara relativamente resistente
con ceras epicuticulares dispuestas en plaquetas superpuestas, epidemis externa con una
cutícula que incrementa su grosor durante el crecimiento del fruto y tejido subepidérmico
compacto de unas pocas capas cuyas células tienen las paredes algo engrosadas. La masa de
tejido extracarpelar (porción comestible) está constituida por parénquima con notables
espacios intercelulares y por los haces vasculares del tubo floral.
La parte central del fruto constituye el pericarpo, que consta de un parénquima donde se
localizan los haces vasculares dorsales y ventrales de los carpelos, y un tejido cartilagenoso

35
constituido por esclereida, que recubre interiormente los lóbulos y constituye el endocarpo.
Pomo: Es el fruto característico del grupo de las rosaceas a las que pertenece el
manzano, el peral, el níspero y el membrillo. El fruto se desarrolla a partir de un pistilo
compuesto formado por dos o más carpelos y en un ovario ínfero. La porción comestible, es
de origen apendicular (formada por el tubo floral o hipando). También se vuelve carnosa la
parte externa del pericarpo, mientras que el endocarpo se torna más o menos cartilaginoso
formando el corazón del fruto, que contiene varias o muchas semillas. Puesto que una parte
importante del fruto no procede del ovario, el pomo es un fruto accesorio.
�,$7$7$,$�(�/��� ����������� ��
Familia: Rosaceae. Subfamilia: Pomoideas.
Especie: Malus pumila
Planta: Fuerte y con un masa foliar abundante. Sus flores son blancas y perfumadas, muy
visitadas por las abejas, que facilitan la polinización cruzada que necesitan la mayoría de las
variedades para producir más fruta.
Fruto: pomo globoso, con pedúnculo corto y numerosas semillas de color pardo brillante.
Granny Smith: Es la variedad más difundida en el mundo, luego de Red Delicious y Golden
Delicious. Nuestro país es uno de los productores más importantes junto con Chile, Brasil y
Sudáfrica, entre otros. La manzana es una fruta de buen tamaño, esférica y simétrica. Tiene
color verde intenso que se vuelve más claro en la madurez, con numerosas lenticelas de color
blanquecino. La pulpa es blanquecina, compacta y jugosa, de sabor acidulado. El fruto del
cultivar Granny Smith durante el almacenamiento tiene las siguientes propiedades: muy
firme, quebradizo y no es harinoso.
,$8$������������*����+�����������+��������
Como se mencionó anteriormente, el color es un atributo de calidad importante en
frutas y vegetales, cuya percepción suele constituir el primer paso en la evaluación de estos
productos por parte del consumidor y es condicionante de su elección.
La sensación del color es un fenómeno psicofísico, el cual es sólo parte de la
percepción visual total de la información detectada por el ojo e interpretada por el cerebro. La
percepción del color es producto de la interacción de tres factores fundamentales: las

36
propiedades espectrales de la fuente de luz que iluminan el objeto, la naturaleza del material
observado y del proceso visual humano (MacDougall, 2002).
El ojo humano es sensible a la radiación electromagnética en el intervalo de 380 a 770
nm de longitud de onda, la cual es comúnmente llamada “luz”. Asimismo, el sistema visual
tiene la capacidad de discriminar entre radiaciones de distinta longitud de onda determinando
su “color” a través de un complejo mecanismo trivariante. El color, es por lo tanto, una
característica tridimensional, cuyas tres dimensiones corresponden a un atributo de claridad,
percibido por células especializadas de la retina llamadas bastones, y dos atributos
cromáticos, percibidos por otras células especializadas llamadas conos. Hay tres conjuntos de
conos: uno sensible al rojo, el otro al verde y el otro al azul, y la respuesta total es la
combinación de las tres señales en el cerebro (Lozano, 1978).
Los objetos son coloreados porque interactúan de distinta forma con la luz, usualmente
blanca, incidente sobre ellos. Se clasifican respecto a su comportamiento frente a la radiación
en:
* Opacos: absorben y/o reflejan toda la luz.
* Transparentes: transmiten la mayor parte de la luz sin reflejarla ni difundirla
* Translúcidos: parte de la luz es reflejada y parte transmitida, dando lugar a una
importante componente difusa tanto transmitida como reflejada.
En los alimentos el factor más importante que influye en la luz reflejada es la difusión
debido a las características físicas de la superficie. De forma general puede afirmarse que la
reflexión ocular ocurre en un ángulo de 90º respecto a la luz incidente y es la responsable
principal del brillo del material, mientras que la reflexión difusa ocurre en un ángulo de 45º y
es la principal responsable del color. Estas particularidades son importantes al momento de
efectuar mediciones porque del ángulo de incidencia de la luz en la muestra y la posición de
ésta con respecto al sensor, dependerá el atributo medido (Manresa Gonzalez & Vicente,
2007).
Los principios fundamentales de la colorimetría están basados en las tres leyes de
Grassman, de las cuales, la más importante establece que cualquier color puede ser igualado
por la suma de tres colores primarios en cantidades convenientes. Estos colores primarios
deben ser elegidos de forma tal que sean independientes entre sí, es decir, ninguno puede ser
obtenido por combinación de los otros dos. En la práctica, los colores primarios a utilizar en
forma aditiva son tres: rojo, verde y azul. La suma de todos ellos produce la sensación de luz
blanca o acromática (Lozano, 1978).

37
El color puede ser definido por los siguientes atributos:
* La claridad es el atributo que hace corresponder a cada color una equivalencia con respecto a
una escala de grises. A la cualidad psicológica claridad le corresponde la magnitud psicofísica
luminosidad.
* El tono o matiz es el atributo que adjudica al color una cualidad que se define como rojo,
naranja, amarillo, verde, azul, púrpura o cualquier combinación de ellos. A la cualidad
psicológica tono le corresponde la magnitud psicofísica longitud de onda dominante.
* La saturación es el atributo que, fijado el tono, describe al color por su similitud con un color
espectral puro, cuanto más parecido a este, tanto más saturado. A la cualidad psicológica
saturación le corresponde la magnitud psicofísica pureza.
La medida objetiva del color de un material puede obtenerse del análisis de su espectro
visible, por transmisión o reflexión, obtenido con un espectrofotómetro. De los sistemas
propuestos para la especificación del color el más difundido universalmente es el de la CIE
(Comission Internationale de l’Eclairage), en el cual el color es indicado por tres variables X, Y,
Z, conocidas como los valores triestímulo y que representan a tres colores primarios imaginarios
(relacionados con el verde, el rojo y el azul).
El cálculo necesario para obtener los valores triestímulo CIE para un objeto se
esquematiza de la siguiente manera (figura 2'8). El valor del estímulo visual es la resultante de
una combinación del espectro de la luz incidente (E vs λ) con el espectro de la muestra (R vs λ)
y con las funciones de distribución de la sensibilidad del ojo para cada uno de los tres primarios
Figura 2'8. Suma de radiaciones espectrales que conforman el estímulo o flujo radiante que
percibe el observador.
DISTRIBUCIÓN DE LA POTENCIA
ESPECTRAL DEL ILUMINANTE
REFLECTANCIA ESPECTRAL DEL
OBJETO
�
SENSIBILIDAD ESPECTRAL DEL
OBSERVADOR
�
�ESTÍMULO
�
λλλλ�
������
λλλλ� λλλλ� λλλλ�

38
imaginarios (S vs λ). El área bajo la curva resultante se integra en el espacio visible para obtener
los valores numéricos de la contribución de cada uno de los tres primarios ideales (X, Y, Z) al
color dado.
El factor de transmitancia o reflectancia espectral se obtiene mediante un
espectrofotómetro, en el que se determina la luz transmitida ó reflejada para cada λ. La
distribución energética para varios iluminantes es conocida. Para estandarizar las comparaciones
la CIE ha establecido fuentes de luz estándar: el iluminante A tiene una distribución energética
similar a la luz emitida por una lámpara incandescente de tungsteno, el iluminante B corresponde
a la luz solar con componente de cielo a medio día, el iluminante C tiene una distribución
energética similar a la luz blanca solar (difusa, cielo cubierto), el iluminante D corresponde a la
irradiación de la luz solar directa. Otras de las variables a fijar es el ángulo del observador ó
campo visual. Cuando se miran objetos la información general proviene de ángulos de 10*20º,
pero cuando se observan detalles, el ángulo es de 2 º. En general, en colorimetría de alimentos se
emplea este último. En los espectrofotómetros actuales este cálculo se realiza automáticamente
por un sistema de computación acoplado al instrumento. De esta manera el color queda
determinado por un punto en un espacio tridimensional de coordenadas X, Y, Z. Sin embargo, la
forma habitual de representación es calcular las llamadas coordenadas cromáticas x, y definidas
por:
ZYX
Xx
++= (2.1)
ZYX
Yy
++= (2.2)
El color definido por (x, y) se representa en el diagrama cromático CIE (figura 2'9)
donde sólo tiene lugar la cromaticidad del color en cuestión. Para que la especificación del
color sea completa, eventualmente se utiliza la luminosidad Y como tercera coordenada, en un
plano perpendicular que representa la propiedad de reflejar más ó menos luz. El plano de las
coordenadas cromáticas está delimitado por una línea recta que une los extremos del espectro
visible, conocida como línea de púrpuras (Kelly, 1943).
El espacio CIE 1931 es muy sencillo de manejar y es excelente para representar mezclas
aditivas, pero no es homogéneo (distancias geométricas iguales no suelen representar diferencias
de color iguales en la percepción por el ojo humano). Para subsanar el problema de su no
uniformidad se han desarrollado una serie de transformaciones del espacio CIE que permiten

39
Figura 2'9. Diagrama Cromático CIE.
obtener mejores resultados en la evaluación de la diferencia de color. Así surgen las
transformaciones CIELab (1974) y CIELuv (1976). El CIELuv es una transformación lineal del
espacio cromático CIE. En cambio, líneas rectas en el espacio CIE corresponden a líneas
curvas en el espacio CIELab. El espacio CIElab es un espacio de color rectangular de 3
dimensiones basado en la teoría de colores opuestos. Hay dos escalas de color populares de este
espacio en uso hoy en día, Hunter Lab y CIE L*a*b*. Aunque son similares en organización, un
mismo color tendrá valores diferentes en estos dos espacios. Ninguna de las dos escalas es
visualmente uniforme. Hunter Lab se concentra en la región azul del espacio de color y
CIEL*a*b* se expande en la región amarilla. La recomendación actual de la CIE es usar la
escala CIEL*a*b*.
Las ecuaciones de transformación del espacio CIE a CIE L*a*b* son las siguientes:
L*ab = 116.(Y/Yn)1/3 – 16 (2.3)
a* = 500.[ (X/Xn)1/3 * (Y/Yn)
1/3 ] (2.4)
b* = 200.[ (Y/Yn)1/3 * (Z/Zn)
1/3 ] (2.5)
donde L*ab es el parámetro luminosidad que toma valores entre 0 (negro) y 100
(blanco); a* es la coordenada cromática en el eje de las abscisas ( rojo*verde); b* es la

40
coordenada cromática en el eje de las ordenadas (amarillo*azul); X,Y,Z son los valores
triestímulo; Xn, Yn, Z n son los valores correspondientes al iluminante.
Dentro de este espacio es posible definir otras funciones que pueden ser indicativas de
determinados atributos de color, como la saturación y el tono. Estas funciones son la
cromaticidad o croma métrica (C*) y el ángulo de tono (h*). También puede calcularse la
“diferencia de color global” (aE*ab).
2.5. -����� �������*����+�����������+��������
La reología se define como el estudio de la deformación y el flujo de la materia. Estudia
la forma en que los materiales responden a la aplicación de un esfuerzo o de una deformación.
La reología ha sido extensamente aplicada en frutas y vegetales con el objeto de
comprender la relación entre la estructura, la textura, y los cambios inducidos por los distintos
procesos (Edwards, 1999).
Los alimentos son materiales complejos cuyas características y propiedades
especialmente las mecánicas, dependen principalmente de su estructura, esto es, del arreglo
espacial de los elementos estructurales a niveles micro y submicro, que está determinada por
la composición química y las fuerzas físicas. Existe una relación directa entre la estructura y
la textura (Stanley & Tung, 1975).
El análisis y la cuantificación del comportamiento reológico de las frutas y vegetales y
la investigación de las causas químicas y estructurales que lo determinan es un aspecto de
gran interés en la ciencia de los alimentos. Su importancia radica fundamentalmente en tres
razones. La primera es que existe en la naturaleza un gran número y variedad de estos
productos que son económicamente importantes y que además son imprescindibles en la dieta
humana contemporánea. La segunda razón consiste en que las propiedades mecánicas del
tejido de las frutas y vegetales son dependientes del tiempo, haciendo esencial cualquier
estudio reológico. La tercera razón es que las propiedades mecánicas son relevantes para
muchos aspectos del estudio de estos materiales, incluyendo las causas y la magnitud del daño
durante la cosecha, el transporte, el procesamiento y el almacenamiento, la percepción
humana sobre la calidad del producto y los cambios fisiológicos que tienen lugar en el
producto durante el crecimiento, la maduración, y el almacenamiento luego de la cosecha. En
conclusión, las propiedades reológicas de frutas y vegetales son importantes para el estudio de
la fisiología de las plantas, la horticultura, la ingeniería y la ciencia de los alimentos (Pitt,
1992).

41
• ���������������������������������������������
La denominación de sólidos elásticos se aplica a aquellos materiales, que tienen una
forma definida y se deforman bajo la aplicación de fuerzas externas entrando en una nueva
forma de equilibrio. Cuando estas fuerzas externas se remueven, la forma vuelve exactamente
al estado original. El sólido almacena toda la energía que obtiene a través del trabajo hecho
por las fuerzas externas durante la deformación. Esta energía luego está disponible para
restablecer el cuerpo a la forma original cuando se remueven las fuerzas aplicadas. El
principio básico de la reología de los sólidos elásticos es la ley de Hooke, que define el sólido
ideal como aquel que se deforma instantánea y proporcionalmente a la magnitud de la fuerza
aplicada y se recupera también instantáneamente al retirar la fuerza:
τ = E γ (2.6)
donde: τ es la fuerza por unidad de superficie, E el módulo de elasticidad y γ la
deformación.
Se denominan líquidos a aquellos materiales que no tienen una forma definida
(adoptan la del recipiente que los contienen) y fluyen irreversiblemente bajo la acción de
fuerzas externas. La energía de deformación de los fluidos se disipa en forma de calor. La
ecuación de Newton es la que describe a un líquido viscoso ideal bajo la acción de un
esfuerzo.
.•
γη=τ (2.7)
donde: τ es la fuerza por unidad de superficie, η es el coeficiente de viscosidad y .•
γ la
velocidad de deformación.
Los materiales alimenticios tienen propiedades que son intermedias entre un sólido
elástico y un líquido viscoso. Esta combinación de la conducta de los materiales es
generalmente llamada conducta viscoelástica. (Ward, 1990).
Los materiales viscoelásticos se clasifican en:
* Viscoelástico lineal: las propiedades sólo dependen del tiempo
* Viscoelástico no lineal: las propiedades dependen del tiempo y de la magnitud de la
fuerza o la deformación aplicada.
Cuando la relación entre la tensión aplicada y la deformación resultante es sólo función

42
del tiempo se considera que el material es viscoelástico lineal. En muchos materiales reales se
verifica este comportamiento si las deformaciones son pequeñas (Rao, 1992; Costell y col.,
1997). Si la magnitud de la tensión es tal que la deformación resultante no se recupera
totalmente cuando se libera el esfuerzo, la relación entre la tensión y la deformación no sólo
es función del tiempo sino también función de la magnitud del esfuerzo aplicado y el
comportamiento viscoelástico es no lineal. No existe una teoría general sencilla que
caracterice el comportamiento viscoelástico no lineal, y por lo tanto, para estudiar la reología
de los alimentos se aplican condiciones experimentales que permitan la aplicación de la teoría
de viscoelasticidad lineal
• �����������:�����
Fuerza: Cuando una fuerza actúa externamente sobre un cuerpo se pueden distinguir
diferentes casos: tensión, compresión y cizalla. El pandeo envuelve tensión y compresión, el
torque incluye cizalla, y la compresión hidrostática involucra a los tres tipos de fuerza. Todos
los demás casos involucran a estos tres casos individuales o combinación de estos (deMan,
1975).
Esfuerzo ó tensión: Respuesta o reacción interna de un material a la fuerza aplicada:
Esfuerzo [ ][ ] [ ]Pam
N
A
F
Área
Fuerza2
==== (2.8)
Esfuerzo normal (σσσσ): Perpendicular al área.
Esfuerzo de cizalla (ττττ): Paralelo al área.
Deformación: Cambio relativo en las dimensiones o forma de un cuerpo expuesto a un
esfuerzo. Es una cantidad adimensional.
Deformación normal (εεεε): Cambia una dimensión. El cambio por unidad de longitud es
debido a una fuerza (tensión o compresión) en una dimensión lineal original.
Deformación de cizalla (γγγγ): Deslizamiento. La tangente del ángulo cambia, debido a una
fuerza (cizalla), entre dos líneas originalmente perpendiculares una a otra a través de un punto
en un cuerpo (Mohsenin, 1978).
Módulo de elasticidad (E): También llamado Módulo de Young. Se define como la relación
entre el esfuerzo normal aplicado (σ) y la deformación normal (ε) que se obtiene.

43
��
������
�
[ ][ ]
[ ]Pa1
PaE ==
ε
σ= (2.9)
[ ][ ]m
m
L
L=
�=ε (2.10)
donde: σ es el esfuerzo, ε es la deformación normal, �L es el cambio en la longitud L es la
longitud original del espécimen.
Módulo de cizalla (G): También llamado Módulo de Rigidez. Se obtiene como la relación
entre el esfuerzo de cizalla aplicado (τ) y la deformación de cizalla (γ) que se obtiene.
������
��
�
[ ] [ ]PaPa
G ===1γ
τ (2.11)
[ ][ ]
( )θγ tan==�
=m
m
h
L (2.12)
donde: τ es el esfuerzo de cizalla, γ es la deformación de cizalla, �L es el cambio en la
longitud, h es el espesor del espécimen.
Velocidad de deformación (•
γ ):
[ ]
[ ]1ss
1
t−
•
==∂γ∂
=γ (2.13)
donde: γ es la deformación de cizalla y t es el tiempo.
Viscosidad (ηηηη): Fricción interna del fluido que provoca la resistencia a fluir.
[ ][ ] [ ]s.Pas
Pa1
==γ
τ=η
−• (2.14)
�
θ

44
• 6���������������������������������������������
�
�"�������������������������������������
�
Los ensayos reológicos realizados a altas de deformaciones más comúnmente utilizados
son: ensayo de compresión uniaxial, ensayo de penetración, ensayo de extrusión y análisis de
perfil de textura.
A continuación se describirá el ensayo de compresión, el cual fue utilizado en este
trabajo.
Tal como su nombre lo indica, la compresión es un ensayo de la resistencia del alimento a
la compresión. En él se comprime una muestra, de dimensión prefijada, hasta su ruptura o
hasta una determinada deformación y se observa el comportamiento del alimento. Los
ensayos que se aplican con más frecuencia son los que estudian la relación esfuerzo–
deformación con fuerzas de compresión uniaxial. Estas fuerzas son normales, es decir, se
aplican de modo perpendicular a la superficie sobre la que actúan. Para la realización de las
experiencias, en la mayoría de los casos la muestra de alimento se presenta en forma de
cilindro o de cubo y se somete a la acción de un émbolo que se desplaza verticalmente a una
velocidad prefijada (Costell y col., 1997).
Los equipos universales denominados “texturómetros” son los instrumentos más
utilizados para la realización de los ensayos de compresión. En este tipo de equipo se
comprime el espécimen con velocidad de desplazamiento constante dentro de un intervalo
continuo y amplio de valores y se registra la resistencia que aquél opone a la deformación
impuesta (Costell y col., 1997).
Debido a la naturaleza viscoelástica de los alimentos, la magnitud desarrollada no es sólo
función de la deformación sino también de la velocidad impuesta. La aplicación de distintas
velocidades afecta significativamente la respuesta mecánica del material. La naturaleza de
esta variación es característica de cada material. En general, cuanto mayor es la velocidad de
deformación, la tensión originada es mayor. (Costell y col., 1997).
En un ensayo ideal de deformación uniaxial la relación tensión*tiempo es independiente
de las dimensiones del espécimen por definición, pero en la práctica esto no sucede así y
varias pueden ser las causas. En algunos materiales reales existen estructuras orientadas (por
ejemplo, alimentos fibrosos) que pueden influir en la respuesta del material si alcanzan
dimensiones comparables a la del espécimen. La existencia de fuerzas de fricción entre la
muestra y las superficies del equipo, hace que el material se comporte con mayor resistencia

45
aparente, lo que probablemente modifique su pauta de deformabilidad dependiendo de sus
dimensiones. Por otro lado, muchos materiales biológicos y alimentos contienen líquidos; en
ellos, parte de la tensión alcanzada puede deberse al desarrollo de presiones hidrostáticas
durante la compresión. La disipación de estas tensiones dependerá de la naturaleza porosa, la
densidad o la microestructura de la matriz del material. En estos casos, especímenes con
mayores diámetros desarrollarán mayores tensiones aparentes (Costell y col., 1997).
La primera información que se puede obtener de un ensayo de compresión es a partir de
la forma de la curva. Las curvas que se registran al comprimir alimentos duros o rígidos, que
suelen romperse durante el ensayo, muestran un pico abrupto que marca el momento de la
ruptura o fractura. En algunos casos antes de la ruptura pueden presentar un pequeño cambio
en la pendiente, correspondiente al punto de “bioyield”, que está relacionado con una fractura
en la microestructura del material asociada con una disrupción inicial de la estructura celular
(Steffe, 1996). En cambio, las curvas que se registran para alimentos blandos, que se
comprimen sin ruptura aparente, presentan una forma ascendente continua hasta que finaliza
el ensayo (figura 2'10).
Figura 2'10��Curvas típicas obtenidas al comprimir alimentos duros y alimentos blandos.
Alimento duro
Alimento blando “bioyield”
Porcentaje de compresión (%)
Fuerza
Fuerza de ruptura

46
Para poder independizarse del área de aplicación de la fuerza, las curvas de fuerza vs
deformación pueden ser transformadas en curvas de esfuerzo vs deformación. Pero debido a
que estos ensayos se realizan en la región de altas deformaciones, el área de exposición a la
fuerza se expande considerablemente durante la compresión y por lo tanto no puede ser
aproximada al área original (Calzada & Peleg, 1978).
Teniendo en cuenta el cambio del área a lo largo del ensayo, el esfuerzo normal puede
ser expresado como el esfuerzo real (σR) según:
σR = )(
)(
tA
tF (2.15)
donde A (t) es el valor del área del espécimen deformado a un tiempo (t) de iniciado el
ensayo. Si se supone que el valor del volumen permanece constante, la ecuación (2.15) puede
expresarse como:
σR = [ ]
00
0 )()(
HA
tHHtF �− (2.16)
donde H0 es la altura inicial del espécimen, cH es la disminución absoluta de la altura
inicial del espécimen en la dirección de la fuerza aplicada y A0 es el área inicial de la muestra.
La deformación normal (ε) puede ser expresada como la deformación real definida por
Hencky (εR) según:
εR =
�− HH
H
0
0 ln (2.17)
Como se mencionó anteriormente, el módulo de elasticidad (E) es el esfuerzo dividido
la deformación en la región elástica o lineal. Pero debido a que los alimentos son básicamente
materiales viscoelásticos no lineales, en los ensayos de compresión, en general, no presentan
una región lineal bien definida. Existen excepciones; por ejemplo, las manzanas usualmente
tienen una región lineal definida en la compresión (Rebouillat & Peleg, 1988).
En alimentos, el término módulo de deformabilidad (Ed) es usado en vez de módulo de
elasticidad, y se obtiene de la pendiente de la curva de esfuerzo vs deformación en un rango
de deformación definido donde la deformación se aproxima a cero (Monhsenin & Mittal,
1977) o de la región lineal de la curva si es que existe (Rebouillat & Peleg, 1988).

47
Entre los parámetros que se pueden obtener de las curvas de compresión figuran el
esfuerzo de ruptura (σR), la deformación de ruptura (εR) y el módulo de deformabilidad (Ed).
Estos parámetros resultan muy útiles para evaluar la rigidez de los alimentos. En el caso de
alimentos blandos que se comprimen sin ruptura aparente, la información que se obtiene es el
esfuerzo a una dada deformación y el Ed.
"���������������������*���������������
En estos ensayos las deformaciones aplicadas al material son pequeñas. Dentro de los
mismos se encuentran los ensayos de cizallamiento rotatorio y oscilatorio que pueden
realizarse con un reómetro dinámico. Un ensayo rotatorio como el de fluencia permite
predecir separadamente las características elásticas, viscoelásticas y de flujo viscoso (Jackman
& Stanley, 1992), mientras que los ensayos oscilatorios, otorgan datos de viscosidad y
elasticidad relacionados con el tiempo de respuesta (Schramm, 1994).
Los ensayos oscilatorios, los ensayos de relajación y los ensayos de fluencia –
recuperación son normalmente utilizados para determinar las propiedades del material en la
región viscoelástica lineal, permitiendo caracterizar la microestructura sin disrupción de la
misma durante el proceso (Khan y col., 1997). En materiales frutihortícolas se utiliza la teoría
de la viscoelasticidad lineal con esfuerzos y deformaciones pequeñas. De esta manera es
posible obtener modelos matemáticos simples (combinación de elementos elásticos y
viscosos), que permiten estimar un gran número de parámetros, los cuales pueden ser
relacionados con la estructura del material (De Baerdemaeker & Segerlind, 1976; Ferry, 1980;
Jackman y Stanley, 1992).
������������ ������ ���� ����
El clásico ensayo de fluencia – recuperación se divide en dos etapas; en la primera etapa
se aplica un esfuerzo de cizalla constante hasta un tiempo t1, luego del cual se remueve el
esfuerzo. Durante el ensayo se registra la deformación que sufre la muestra en función del
tiempo (figura 2'11).
Cuando las curvas de fluencia (capacitancia vs. tiempo) para diferentes valores de
esfuerzo de cizalla coinciden, puede afirmarse que la conducta es viscoelástica lineal, es decir,
la magnitud de la respuesta es independiente de la magnitud del estímulo. En muchos casos,

48
0,00
0,04
0,08
0 20 40 60 80 100 120Tiempo (s)
Deformación(%)
Esfuerzo (Pa)
50
Fase de Fluencia Fase de Recuperación
Figura 2'11. Deformación de distintas muestras en un ensayo de fluencia / recuperación (Respuesta viscoelástica (▬); Esfuerzo aplicado (▬); Respuesta sólido ideal (▬); Respuesta
líquido Newtoniano (▬)).
solamente con deformaciones muy pequeñas la respuesta es viscoelástica lineal y por lo tanto
el ensayo puede aplicarse sólo en un rango muy estrecho de esfuerzos de cizalla.
Un material viscoelástico sometido a un esfuerzo de cizalla instantáneo y constante, por
un tiempo suficiente de manera tal de prevenir que el material muestre una elasticidad pura,
pero sin superar el rango viscoelástico lineal, exhibe una conducta de un sólido elástico en las
primeras etapas del cizallamiento seguido de una conducta de un fluido viscoso “en el sentido
tal que el trabajo de la deformación de cizalla no es conservado totalmente, como en un
sólido, ni es disipado completamente como en un fluido” (Sherman, 1970).
En la mayoría de los casos, la deformación total γ de un material viscoelástico es la
suma de tres partes separadas γ0, γR y γN (figura 2'12). γ0 y γR corresponden a la deformación
elástica instantánea y a la deformación elástica de retardo, respectivamente; mientras que γN
es la deformación debido al flujo Newtoniano. Debido a que el material muestra una conducta
lineal, la magnitud de las deformaciones γ0, γR y γN son exactamente proporcionales a la
magnitud del esfuerzo aplicado. Se define a la capacitancia como (Ward, 1990):
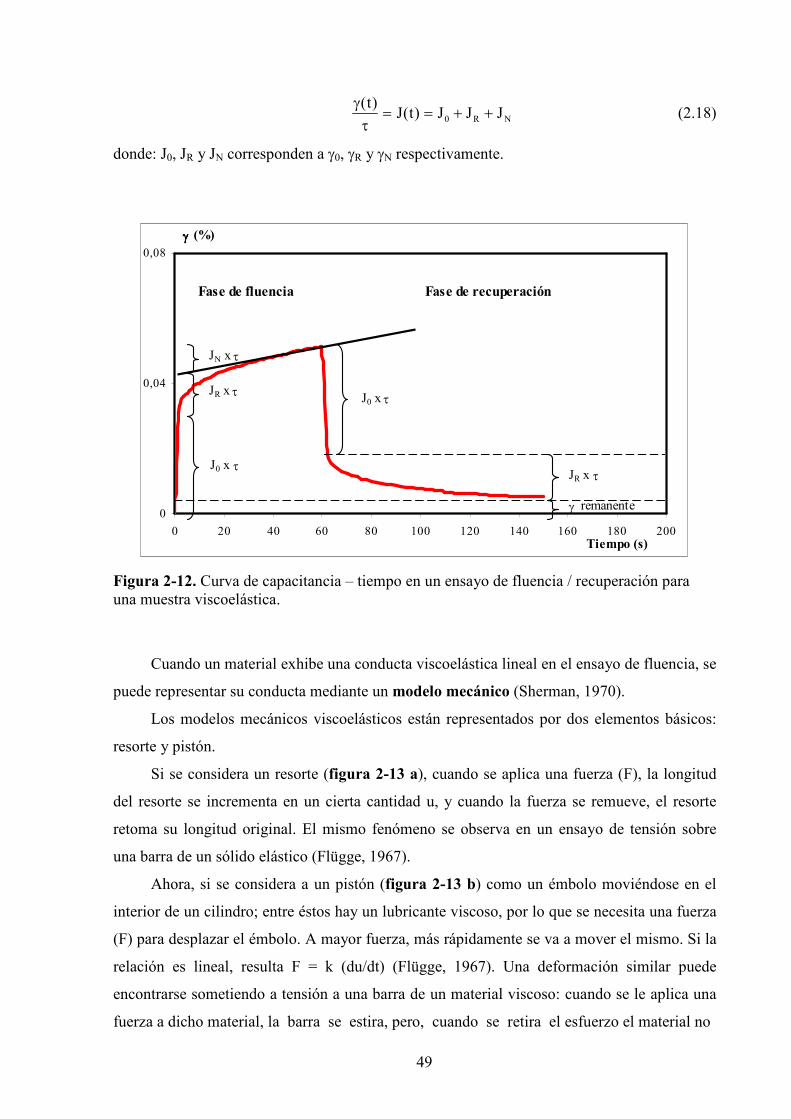
49
NR0 JJJ)t(J)t(
++==τ
γ (2.18)
donde: J0, JR y JN corresponden a γ0, γR y γN respectivamente.
0
0,04
0,08
0 20 40 60 80 100 120 140 160 180 200Tiempo (s)
γ γ γ γ (%)
J0 x τ
JR x τ
JN x τ
JR x τ
γ remanente
J0 x τ
Fase de fluencia Fase de recuperación
Figura 2'12. Curva de capacitancia – tiempo en un ensayo de fluencia / recuperación para una muestra viscoelástica.
Cuando un material exhibe una conducta viscoelástica lineal en el ensayo de fluencia, se
puede representar su conducta mediante un modelo mecánico (Sherman, 1970).
Los modelos mecánicos viscoelásticos están representados por dos elementos básicos:
resorte y pistón.
Si se considera un resorte (figura 2'13 a), cuando se aplica una fuerza (F), la longitud
del resorte se incrementa en un cierta cantidad u, y cuando la fuerza se remueve, el resorte
retoma su longitud original. El mismo fenómeno se observa en un ensayo de tensión sobre
una barra de un sólido elástico (Flügge, 1967).
Ahora, si se considera a un pistón (figura 2'13 b) como un émbolo moviéndose en el
interior de un cilindro; entre éstos hay un lubricante viscoso, por lo que se necesita una fuerza
(F) para desplazar el émbolo. A mayor fuerza, más rápidamente se va a mover el mismo. Si la
relación es lineal, resulta F = k (du/dt) (Flügge, 1967). Una deformación similar puede
encontrarse sometiendo a tensión a una barra de un material viscoso: cuando se le aplica una
fuerza a dicho material, la barra se estira, pero, cuando se retira el esfuerzo el material no

50
Figura 2'13. Representación gráfica de los elementos básicos utilizados para los modelos mecánicos. (a) Resorte, (b) Pistón.
retoma su longitud original. Por lo tanto, la tensión aplicada es proporcional a la velocidad de
la deformación.
Como se mencionó, un cuerpo hookeano puede ser representado por un resorte cuyo
comportamiento queda definido por su constante elástica E y un cuerpo viscoso o fluido
Newtoniano por un pistón cuyo comportamiento queda definido por su viscosidad η. Los
materiales reales son muy complejos y se necesita una combinación de resortes y pistones
para describir el comportamiento viscoelástico en los ensayos de fluencia y recuperación. El
modelo más utilizado para representar los materiales viscoelásticos en los ensayos de fluencia
es el modelo de Kelvin – Voigt generalizado. (Schramm, 1994). Este modelo consiste en
nelementos de Kelvin en serie (cada elemento consiste en un resorte y pistón dispuestos en
paralelo) con un resorte inicial y un elemento viscoso al final (figura 2'14).
η*
η�
'*
'�
'�
η+ '+
λ η* * *, - '
λ η+ + +, - '
λ η� � �, - '
Figura 2'14. Representación del Modelo de Kelvin – Voigt Generalizado.
�
.
(a)
�
(b)
.

51
La ecuación del modelo generalizado de Kelvin – Voigt es la siguiente:
γ(t) = τ0 (1/E0 + 1/E1 (1 * e*t/λ1) + 1/E2 (1 * e –t/λ2) + .....+ 1/En (1 * e*t/λn) + t/ηN) (2.19)
donde: τ0 es el esfuerzo aplicado, E0…y En son los módulos de elasticidad, λ1,λ2,....y λn
son los tiempos de retardo y ηN es el coeficiente de viscosidad asociado con el flujo
Newtoniano.
A continuación, por razones de simplicidad, se analiza el comportamiento de un
material que sigue el modelo de Burger de 4 elementos (consiste en 1 elemento de Kelvin con
un resorte inicial y un elemento viscoso al final) para describir las curvas de fluencia y
recuperación (figura 2'15).
Figura 2'15. Representación del Modelo de Burger)
Una curva típica de fluencia (capacitancia vs. tiempo) (etapa de fluencia), como ya se ha
mencionado, puede dividirse en tres regiones principales y según el modelo de Burger se
observa (figura 2'12) (Schramm, 1994):
1) La muestra reacciona ante el esfuerzo aplicado con una deformación espontánea γ0,
representada por el resorte R1. En esta región, las uniones entre la estructura primaria
son extendidas elásticamente.
2) Todos los elementos trabajan para la respuesta de la curva: el resorte R2 y el pistón P2.
La constante de tiempo de este movimiento se llama tiempo medio de retardo. Esta
región es elástica de retardo dependiente del tiempo con una capacitancia (JR). Acá las
uniones se rompen y reforman. Todas las uniones no se rompen y reforman a la misma
velocidad. Las uniones más débiles se rompen a menores valores de tiempo que las
η* '*
'�
η�
P2 R2
P1
R1

52
uniones más fuertes. Esta región está representado por:
JR = J1 (1–e t/λ) (2.20)
3) Ambos resortes están completamente extendidos; el constante incremento en la
deformación es causado por el pistón P1. Esta es la región lineal de la capacitancia
Newtoniana (JN). Se prosigue con la ruptura de algunas de las uniones (Sherman, 1970).
Esta región está representado por:
JN = t / ηN (2.21)
Cuando se remueve el esfuerzo aplicado, el comportamiento de la etapa de recuperación
es similar al de la etapa de fluencia si se desprecia la parte del fluido Newtoniano (γN). Y la
recuperación instantánea elástica es seguida por la recuperación elástica de retardo. Esto es
consecuencia de la conducta viscoelástica lineal (Ward, 1990).
La etapa de recuperación del ensayo también puede dividirse en 3 regiones:
1) La muestra reacciona con una espontánea re*formación γ0, causada por el resorte R1.
2) El resorte, que aún se encuentra activado, comienza a recuperar su forma; el pistón P2
vuelve a su estado original.
3) Todos los procesos de re*formación finalizan y la deformación remanente es debida
al pistón P1.
Los parámetros característicos de la etapa de fluencia son:
J0: Capacitancia instantánea
Ji: Máxima capacitancia viscoelástica
λi: Tiempo medio de retardo
ηN: Coeficiente de viscosidad asociado con el flujo Newtoniano
Jmáx: Capacitancia máxima de la curva de fluencia
JN: capacitancia Newtoniana
Para los ensayos de fluencia son válidas las siguientes ecuaciones:
J(t) = γ (t) / τ0

53
Etapa de fluencia:
J(t) = J0 + JR(t) + JN(t) τ0 = constante > 0
Si se utiliza el modelo de Kelvin Voigt Generalizado para modelar la curva J(t)
(Sherman, 1970):
J (t) = J0 + J1 (1 * e*t/λ1) + J2 (1 * e –t/λ2) + .....+ Jn (1 * e*t/λn) + t/ηN (2.22)
���������� ����������
En los ensayos dinámicos oscilatorios, las muestras se someten a un esfuerzo o a una
deformación de cizalla que varía en forma armónica (generalmente sinusoidal) (Ferry, 1980).
Por lo tanto existen dos posibilidades:
• Se impone un esfuerzo pequeño a una muestra y se mide la deformación,
• Se impone una deformación fija y se mide el esfuerzo desarrollado por la muestra.
Si se aplica una deformación sinusoidal (< 1%) sobre la muestra se obtiene:
γ = γ0 sen(ωt) (2.23)
donde: γ0 es la amplitud de la deformación y ω es la frecuencia de oscilación.
Al realizar un ensayo oscilatorio con un reómetro dinámico el rotor, tanto del plato o del
cono superior, se mueve alternativamente con un pequeño ángulo de deflexión (ϕ) hacia la
izquierda y hacia la derecha, con una dependencia sinusoidal del tiempo (figura 2'16).
La muestra sufre la máxima deformación en los puntos donde R (radio de la muestra) es
igual a r (diámetro del sensor), por lo tanto:
h
R
h
1
2R2R0
ϕ=
π
ϕπ=γ=γ (2.24)
donde: R es el diámetro de la muestra, h es la altura de muestra y ϕ el ángulo de deflexión.

54
Figura 2'16. Ensayo dinámico: Aplicación de una deformación o un esfuerzo sobre la muestra. Fuente: Schramm, 1994.
Respuesta de los diferentes tipos de materiales durante los ensayos dinámicos
a)� Sólido elástico:
Como se ha mencionado, un sólido elástico ante un esfuerzo de cizalla responde a la Ley
de Hooke y es representado mecánicamente a través de un resorte.
Aplicando una deformación sinusoidal durante el ensayo dinámico:
γ = γ0 sen (ωt) (2.25)
Según la Ley de Hooke :
τ = G γ (2.26)
donde el G es el módulo de cizalla ó módulo de rigidez. Reemplazando γ en la ecuación
anterior se obtiene:
τ = G (γ0 sen(ωt)) (2.27)
Por lo tanto se observa que para un sólido elástico la deformación y el esfuerzo están en fase.
−ϕ
+ϕ
����� ��� � ������� �� ���������
���� ���� ������� �����
��������
�+
�*
�
*
+
�*/�0 �/12 +/12 1/12
���� ��������������������������

55
�+
�*
�
*
+
�*/�0 */�0 1/0* �����
����� ���
'�3��4�
53�����%�
�������
+
*
�
�*
�+
+0�6
�6
7�6
*8�6
��
Figura 2'17. Ensayo dinámico: Respuesta del esfuerzo ante una deformación sinusoidal
aplicada a un sólido elástico. Fuente: Schramm, 1994.
b) Líquido viscoso ideal
Un líquido viscoso ideal, ante un esfuerzo de cizalla, responde a la Ley de Newton y es
representado mecánicamente a través de un pistón.
Según la Ley de Newton:
.•
γη=τ = η dγ/dt (2.28)
Derivando la deformación respecto del tiempo:
dγ/dt = ωγ0 cos(ωt) (2.29)
Reemplazando en la ecuación 2.28.

56
τ = η ω γ0 cos(ωt) = η ω γ0 sen(ωt + δ) (2.30)
donde: δ = 90º es el ángulo de desfasaje
Por lo tanto, se observa que la deformación y el esfuerzo están desfasados 90º (figura 2'
18).
Figura 2'18. Ensayo dinámico: Respuesta del esfuerzo ante una deformación sinusoidal aplicada a un líquido Newtoniano. Fuente: Schramm, 1994.
c) Materiales viscoelásticos
Los materiales viscoelásticos poseen propiedades que son intermedias entre un sólido
elástico y un líquido viscoso, por lo que el ángulo de desfasaje toma valores entre 0º y 90º
(figura 2'19).
Por lo tanto, se consideran las siguientes ecuaciones:
+0�6
�6
7�6
*8�6
��
� +
� *
�
*
+
� * / � 0 * / �0 1 / 0* � � � � �
� � �� � � � �
'�3 � � 4 � 5 3 � �� � �% �
������� +
*
�
�*
�+

57
γ = γ0 sen(ωt)
τ = τ 0 cos(ωt)
ω (1/s ó rad/s) = 2πf
f = frecuencia (ciclos/s)
En el caso de los materiales viscoelásticos se define el módulo complejo, G*, como la
relación entre la amplitud del esfuerzo y la amplitud de la deformación. G* representa a la
resistencia total del material a la deformación aplicada. Tanto el módulo complejo como el
ángulo de desfasaje son función de la frecuencia. Entonces se tiene que (Schramm, 1994):
( ) ( )[ ] ´´iG´G´´G´G*G2/122 +=+= (2.31)
G´= G* cosδ = τ0/γ0 cosδ = Módulo elástico ó de almacenamiento
G´´= G* senδ = τ0/γ0 senδ = Módulo viscoso ó de pérdida
�+
�*
�
*
+
�*/�0 �/�� */�0 2/*1 1/0* �����
����� ���
�+
�*
�
*
+
�*/�0 �/�� */�0 2/*1 1/0* �����
�������
��9���� �:������;%���� '������� �����������
+0�6
�6
7�6
*8�6 ��
Figura 2'19. Ensayo dinámico: Respuesta de un material viscoelástico ante la aplicación de
una deformación sinusoidal. Fuente: Schramm, 1994.

58
El módulo de almacenamiento (componente en fase) está asociado con el
almacenamiento y liberación de energía durante la aplicación periódica de la deformación;
por el contrario el módulo de pérdida (componente desfasado) está asociado a la disipación de
energía como calor.
En la mayoría de los casos, para alimentos sólidos y semisólidos, G´´ es mucho menor
que G´. Por consiguiente, G* es aproximadamente igual a G´ (Ward, 1990).
La relación entre la parte viscosa y la parte elástica del material, G´´ / G´, es el valor de
la tangente del ángulo de desfasaje (G´´ / G´ = tan δ). A mayor valor de tan δ, el material es
relativamente más viscoso y menos elástico, y por lo tanto la mayor parte de la energía
utilizada para deformar el material se disipa viscosamente, y el resto se almacena. Por el
contrario, a menor valor de tan δ, el material tiene un comportamiento más parecido a un
sólido, y por lo tanto las deformaciones serían esencialmente elásticas o recuperables (Rao,
1992).
• Si una sustancia es puramente viscosa, entonces: δ = 90º, G´= 0 y G´´ = G*
• Si una sustancia es puramente elástica, entonces: δ = 0º, G´= G* y G´´ = 0
Para poder utilizar las ecuaciones descriptas anteriormente y para realizar un ensayo no
destructivo se debe trabajar dentro del RVL. Para ello, antes de cualquier ensayo dinámico
para caracterizar un material, se realiza la determinación del RVL del mismo. Si se llevara a
cabo el ensayo oscilatorio fuera del RVL, no se dispondría de ecuaciones diferenciales
lineales y la resolución sería compleja. Por otro lado, la muestra se deformaría de tal forma
que las uniones moleculares temporarias o los agregados se destruirían, y la mayor parte de la
energía adquirida se perdería irreversiblemente como calor. El límite entre el rango lineal y el
no lineal se obtiene barriendo al módulo complejo o el módulo elástico (en alimentos sólidos
o semisólidos) en función de la amplitud de deformación ó de esfuerzo; cuando el valor de G*
o G´ deja de ser constante, la respuesta es no lineal (figura 2'20).
La representación gráfica de ambos módulos (G´y G´´) en función de la frecuencia de
oscilación (barrido de frecuencia) determina el espectro mecánico dinámico del material. Este
gráfico es muy útil ya que representa el comportamiento de la microestructura del material
“huella digital del material”.
A través de los ensayos dinámicos se obtienen medidas viscoelásticas diferentes en
comparación con las derivadas del ensayo de fluencia – recuperación. Por tal motivo, ambos
ensayos se complementan para describir de manera más amplia los aspectos viscoelásticos de
los materiales (Schramm, 1994).
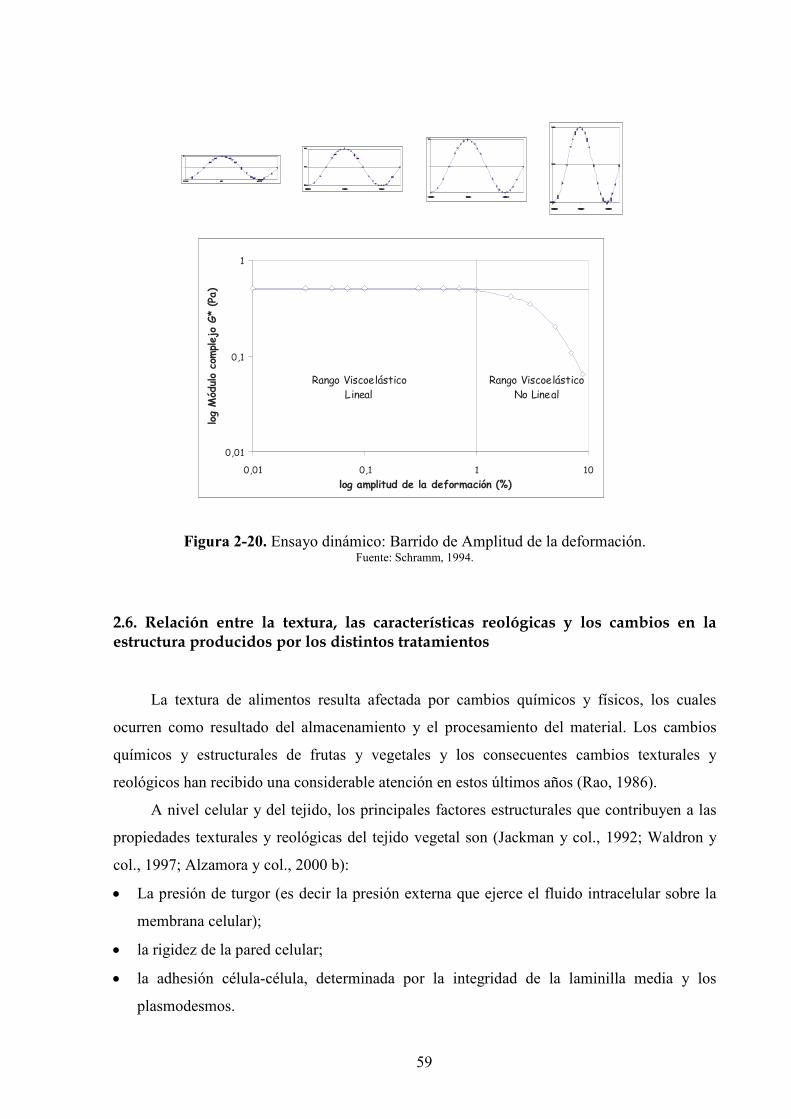
59
�/�*
�/*
*
�/�* �/* * *���� �������� �� �� ������ ��� �!�
���"����� �����#�$%�&��
���� ����������� ����
���� ����������� �� ����
Figura 2'20. Ensayo dinámico: Barrido de Amplitud de la deformación.
Fuente: Schramm, 1994.
�,$;$� -������� ������ ��� ��/����<� ���� �������� ������ ���������� �� ���� ������ ��� �������������������������������������������������� �
La textura de alimentos resulta afectada por cambios químicos y físicos, los cuales
ocurren como resultado del almacenamiento y el procesamiento del material. Los cambios
químicos y estructurales de frutas y vegetales y los consecuentes cambios texturales y
reológicos han recibido una considerable atención en estos últimos años (Rao, 1986).
A nivel celular y del tejido, los principales factores estructurales que contribuyen a las
propiedades texturales y reológicas del tejido vegetal son (Jackman y col., 1992; Waldron y
col., 1997; Alzamora y col., 2000 b):
• La presión de turgor (es decir la presión externa que ejerce el fluido intracelular sobre la
membrana celular);
• la rigidez de la pared celular;
• la adhesión célula*célula, determinada por la integridad de la laminilla media y los
plasmodesmos.

60
En adición a estos elementos estructurales principales, son también factores importantes
el porcentaje relativo de los diferentes tejidos, el tamaño y la forma de las células, la relación
entre el citoplasma y las vacuolas, el volumen de los espacios intercelulares (que pueden
contener fluidos o aire intersticial), el tipo de solutos presentes y la presencia de almidón y el
estado en que se encuentra.
Por lo tanto, el ablandamiento del tejido vegetal durante el procesamiento se atribuye
principalmente a (Bourne, 1976; Lin & Pitt, 1986; Jackman y col., 1992; Sajnin y col., 1999;
Alzamora y col., 2000):
• Cambios en los componentes de la pared celular, así como cambios en el espesor de la
pared celular;
• cambios en las propiedades viscoelásticas de la pared celular, la laminilla media y la
permeabilidad hidráulica del plasmalema;
• tamaño y forma de las células y espacios intercelulares;
• pérdida de turgor.

61
7$�6�("-!�5")�=�6>(% %)�
�
7$#$�6����������
�
Se utilizaron manzanas (Malus pumila), variedad Granny Smith (aw = 0,98 ± 0,01, ºBrix
= 12 ± 1, pH = 3,5 ± 0,3) adquiridas en un comercio local. En todas las experiencias las frutas
fueron seleccionadas teniendo en cuenta el tamaño, la forma y el contenido de sólidos
solubles (ºBrix) para trabajar con un lote lo más homogéneo posible. �
7$,$���������������������������
�
Antes de ser procesadas, las frutas enteras fueron lavadas en agua, inmersas en una
solución de hipoclorito de sodio (100 ppm) durante 3 min y enjuagadas nuevamente en agua.
Luego fueron peladas y cortadas en rodajas (en la mayoría de los casos de 3 cm de diámetro x
0,6 cm de espesor, salvo aclaración contraria) utilizando una cortadora manual y un
sacabocados cilíndrico de acero inoxidable (figura 3'1). Inmediatamente después del corte,
las muestras fueron sumergidas en agua destilada (4*5ºC) durante 1 min para eliminar los
fluidos celulares desprendidos y secadas con papel absorbente. Todos los elementos de corte y
los recipientes utilizados fueron sanitizados previamente con la solución de hipoclorito de
sodio mencionada anteriormente.
�
�/< ��
2 ��
�/< ��
2 ��
�/< ��
2 ��
�/< ��
2 ��
Figura 3'1. Esquema de pasos para el corte de la muestra.

62
Las muestras tratadas térmicamente (sección 3.4.2) presentaron un ligero encogimiento
(aprox. 3 cm de diámetro y 0,5 cm de espesor) debido al tratamiento, por lo que los controles
de tejido fresco con los que eran comparadas fueron cortados de esas dimensiones. Por otra
parte, las muestras utilizadas en los ensayos de impregnación con soluciones de calcio
(sección 3.4.1) se cortaron en cuadrados (4 cm de lado x 0,6 cm de espesor), y luego de la
impregnación se cortaron en rodajas de 3 cm de diámetro. En este caso, el espesor de las
muestras impregnadas no se vió alterado por el tratamiento.
7$7$�(����������
�
7$7$#$����������������������+��������������������
�
7$7$#$#$� ��������������3����
�
La cabina de exposición a la luz UV*C estaba provista por dos lámparas germicidas
(máxima emisión a 253,7 nm, TUV* 15W G13 T8 55V, Philips, Holanda) colocadas en
posición horizontal dentro de una caja de madera herméticamente cerrada (40 cm de alto x
100 cm de largo x 30 cm de ancho). Las muestras a ser irradiadas se ubicaban sobre un estante
forrado con papel aluminio colocado a 10 cm de distancia por debajo de las lámparas. En una
esquina de la cabina se instaló un sistema de ventilación (ventilador Ecoclima, Argentina)
para evitar el incremento de temperatura producido por la radiación UV*C. La temperatura
promedio alcanzada durante los tratamientos fue de 29 ± 2 °C. Antes de su uso las lámparas
se dejaban estabilizar prendidas durante 15 min (ver figura 3'2)
7$7$#$,$�� ��������������������������������������
• ���� �������� ��������
�
Se utilizó el par actinométrico ioduro/iodato de potasio para determinar las dosis de
radiación UV*C (Rhan, 1997). Se preparó una solución de ioduro de potasio (0,6 M) y iodato
de potasio (0,1 M) en buffer borato (0,01 M), pH 9,25. Todos los reactivos utilizados eran de
grado analítico (Anedra, Argentina). De esta solución se tomaron alícuotas de 20 ml que
fueron colocadas en placas de petri, las cuales tenían recubiertas las caras laterales para evitar

63
ventilador Lámparas
Estante
d = 10 cm
Figura 3'2. Cabina de radiación UV*C.
la incidencia de la luz UV*C por los costados. Se ubicaron dentro de la cabina y se dejaron
expuestas a la radiación UV*C a diferentes tiempos (1*5 min). La reacción producida bajo la
exposición a la luz UV*C se detalla a continuación:
8 KI + KIO3 + 3 H2O + hν → 3 I3* + 6 OH* + 9 K+
La cantidad de triioduro formado se determinó espectrofotométricamente
(espectrofotómetro UV/visible Hewlett Packard 8453, Hanover, Alemania) a una longitud de
onda de 352 nm (ε352 = 26400 M*1cm*1).
Conocidos los moles de triioduros formados, se calculó el número de einsteins de
fotones absorbidos por la muestra dividiendo el número de moles de producto por la
eficiencia cuántica, la cual a su vez es función de la temperatura y de la concentración inicial
de ioduro de potasio según la siguiente ecuación:
Φ = 0,75 [1 + 0,02 (T – 20,7)][1 + 0,23 (C – 0,577)] (3.1)
donde: Φ es la eficiencia cuántica, C es la concentración molar inicial de ioduro de potasio, y
T es la temperatura (°C) a la cual se llevó a cabo la reacción.
La concentración inicial de ioduro de potasio se determinó midiendo la absorbancia de
la solución sin irradiar a 300 nm (ε300 = 1,061 M*1cm*1).
Para convertir el número de fotones incidentes en energía incidente (joule), se
multiplicó el número de fotones por la energía de cada fotón. Asumiendo que todos los

64
fotones absorbidos tenían una longitud de onda de 254 nm, este factor era 4,72 x 105
J/einstein de fotones. Finalmente la dosis de UV*C se expresó en J/m2, dividiendo la energía
expresada en joules por el área expuesta a la fuente de irradiación. Para cada tiempo de
exposición las mediciones se realizaron por cuadriplicado.
La formación de triioduro se incrementó linealmente con el tiempo de exposición a la
radiación UV*C. A tiempos mayores, la misma se vió limitada por el consumo de los sustratos
de la reacción; por lo tanto, en ese caso las dosis se calcularon utilizando como factor de
proporcionalidad la pendiente de la curva dosis vs tiempo de exposición obtenida a partir de
los tiempos medidos.
Estudios realizados por otro integrante del mismo grupo de investigación (Schenk y
col., 2006) demostraron que la dosis recibida de radiación UV*C variaba según la posición en
el estante de la cabina. Por lo tanto, se tomó la precaución de ubicar siempre las muestras en
un área definida alrededor del punto central del estante con el fin de que recibieran una dosis
uniforme de radiación. Las dosis reportadas en este trabajo corresponden a las obtenidas en la
posición central.
• ���� ��� ����������
Las dosis obtenidas a través de la técnica actinométrica se compararon con mediciones
realizadas con un radiómetro (Broadband Power/energy Meter 13 PEM 001, Melles Griot,
EEUU), utilizando un sensor de 0,5 cm de radio (área: 0,78 cm2). Las mediciones se
realizaron por triplicado en el punto central del estante.
7$7$#$7$�(��������������������������
�
Los tiempos de exposición a la radiación UV*C utilizados en los ensayos fueron
seleccionados teniendo en cuenta los estudios de inactivación microbiológica realizados en la
matriz de fruta estudiada, de manera que las dosis utilizadas para evaluar los efectos del
tratamiento sobre los atributos de calidad estuvieran dentro de un rango adecuado para lograr
la reducción de la carga microbiana.
Los tiempos de irradiación seleccionados estuvieron en un rango entre 2*25 min. La
irradiación se realizó en rodajas de manzana fresca y en muestras sometidas previamente a
alguno de los tratamientos descriptos posteriormente en las secciones 3.4.1, 3.4.2 y 3.4.3. Las
muestras sujetas solo a la radiación UV*C fueron irradiadas durante 10, 15 y 25 min, mientras

65
que en las muestras con pretratamientos los tiempos de exposición evaluados fueron 2, 8, 14 y
20 min. Las muestras fueron irradiadas sobre una de sus caras, salvo aclaración contraria, y
las mediciones instrumentales se realizaron sobre esa parte expuesta.
��7$7$,$��������������������������������������������
�
7$7$,$#$� ��������������3�����
�
Se utilizó un sistema de esterilización Steripulse*XL modelo RS*3000B (Xenon
Corporation, Woburn, MA, EEUU), compuesto por un módulo controlador RC747 y una
lámpara de xenón (no tóxica, libre de mercurio) que emitía pulsos de luz de alta energía en un
rango de longitud de onda entre 200 y 1100 nm. La lámpara (41 cm de largo) estaba montada
dentro de una caja metálica (19 cm de alto x 76,2 cm de largo x 17,9 cm de ancho), la cual
contenía una ventana de cuarzo en uno de sus lados para dejar pasar la luz emitida, un sistema
de enfriamiento para evitar el sobrecalentamiento de la lámpara durante su uso y un sistema
eléctrico de conexión de la lámpara al módulo controlador. El sistema de enfriamiento
consistía en un ventilador que suministraba aire filtrado a través de un conducto metálico
conectado a la caja de la lámpara y dos filtros, ubicados en la parte superior de la misma, por
donde se removía el aire suministrado sin permitir el paso de luz. A su vez, la caja estaba
dispuesta en forma horizontal sobre una cabina de acero inoxidable (40 cm de alto x 76,2 cm
de largo x 20 cm de ancho) herméticamente cerrada, que disponía de estantes ubicados a 5, 10
y 15 cm de la lámpara (ver figura 3'3). El sistema liberaba pulsos de luz UV*Visible a una
velocidad fija de 3 pulsos por segundo, siendo la energía de cada pulso de 1,27 J/cm2 a 1,92
cm de la lámpara (datos provistos por el fabricante). El ancho de pulso (duración de cada
pulso) era de 360 �s.
�
7$7$,$,$�6�����������������������
Se analizó la evolución de la temperatura en diferentes posiciones dentro de la cabina
durante la emisión de pulsos en un lapso de tiempo de 100 segundos. Los perfiles de
temperatura se registraron en la superficie de la muestra expuesta a la irradiación, en la parte

66
Figura 3'3. Equipo de pulsos de luz.
inferior no expuesta y en el aire. Para las determinaciones se utilizaron termocuplas tipo T
(Cu*constantán) y un registrador de temperatura de 12 canales (Digi*Sense modelo 69202*30,
Barnant Company Division, EEUU). Las mediciones se realizaron a diferentes distancias de
la lámpara (5, 10 y 15 cm) y en distintas posiciones sobre el estante (1*7) (figura 3'4).
Figura 3'4. Esquema que representa los puntos de medición de la temperatura en el estante de
la cabina del equipo de pulsos de luz. �
�
7$7$,$7$�(��������������������������������������
Las rodajas de manzana (fresca o con pretratamientos) fueron colocadas sobre el estante
ubicado por debajo de la lámpara e irradiadas a distintas dosis. Las diferentes dosis se
�������� �����������������������
��������
�� ��� ��
����
���
7
�����
�����
Controlador
Lámpara
Sistema de enfriamiento

67
obtuvieron variando la distancia del estante con respecto a la lámpara (5 y 10 cm) y el tiempo
de exposición a la irradiación (2, 20, 60 y 100 segundos). Como la luz no incidía de manera
homogénea sobre todo el estante, para asegurarse que la irradiación de las muestras en los
tratamientos fuera uniforme, éstas fueron ubicadas siempre en fila debajo de la lámpara y a
una distancia no mayor de 10 cm del punto central de la lámpara.�
�
7$8$�����������������7$8$#$����������������
La adición de calcio se realizó mediante el proceso de impregnación a presión
atmosférica (IA), utilizando soluciones formuladas adecuadamente con el objeto de introducir
calcio en la estructura. La solución de impregnación estaba compuesta por monohidrato de
glucosa (12,6 % (p/p), grado alimentario, refinerías de Maiz S.A., Argentina), lactato de
calcio (1,1% (p/p), calidad farmacopea, Saporiti S.A., Argentina) y gluconato de calcio (4,4 %
(p/p), calidad farmacopea, Saporiti S.A., Argentina). Con el propósito de inhibir y/o retardar
el desarrollo microbiano y evitar los posibles cambios en el pH del tejido, se adicionó a la
solución sorbato de potasio (1500 ppm) y se ajustó el pH a 3,5 (pH natural de la fruta) con
ácido cítrico (calidad farmacopea, Saporiti S.A., Argentina). La cantidad de monohidrato de
glucosa fue calculada de manera que la solución final fuera isotónica con respecto a la
concentración de azúcares presentes en la matriz vegetal (10*13%) y de esta forma evitar
mecanismos de transferencia de agua entre la fruta y la solución.
Las impregnaciones se realizaron en un recipiente con agitación forzada a temperatura
ambiente (24*26ºC) durante 4,5 hs. Luego de impregnadas, las muestras eran escurridas y
secadas en forma estandarizada.
Las muestras impregnadas sometidas a un pretratamiento de escaldado fueron expuestas
a vapor saturado a presión atmosférica durante 2 minutos e inmediatamente enfriadas en agua
(4*5ºC) durante cinco minutos.
Las condiciones experimentales utilizadas fueron seleccionadas teniendo en cuenta
trabajos previos realizados en el grupo de trabajo sobre enriquecimiento de matrices de
manzana con sales de calcio (Salvatori y col., 2007).
�7$8$,$�(����������4����

68
El tratamiento térmico se realizó previo a los tratamientos de irradiación (luz UV*C)
con el objeto de inactivar las enzimas involucradas en el pardeamiento enzimático. Las
muestras fueron inmersas en agua en convección forzada a 80 ºC durante 5,5 min.
Inmediatamente después del escaldado se sumergieron en agua destilada a 4 ºC durante 5 min.
El tiempo de escaldado (t) se seleccionó para lograr la inactivación de la enzima
polifenoloxidasa (PPO) y se calculó según la siguiente ecuación:
t = tc + 2 D80ºC (3.2)
donde: tc es el tiempo en el cual la temperatura en el centro de la muestra alcanza 80
ºC y D80ºC = 1,78 min es el tiempo de reducción decimal de la PPO de manzana a 80 ºC)
(Weemaes y col., 1998).
El tiempo que tarda el centro de la muestra en alcanzar la temperatura de 80ºC se
determinó experimentalmente utilizando termocuplas tipo T (Cu*constantán) y el registrador
de temperatura antes descripto (ver sección 3.3.2.3 ).
�
7$8$7$�!����������������������������������
�
Una inmersión de las rodajas en una solución con agentes “antipardeamiento”
constituida por cloruro de calcio/ácido ascórbico se realizó como pretratamiento alternativo al
tratamiento térmico para inhibir el pardeamiento superficial de las muestras.
Las soluciones se prepararon con ácido ascórbico (1 % (p/v), grado alimentario,
Química Oeste S.A, Argentina) y cloruro de calcio (0,1% (p/v), grado alimentario, Saporiti
S.A, Argentina) a pH 3,5. La inmersión se realizó en las muestras frescas, antes de los
tratamientos de irradiación, durante 5 min a 4*5 ºC (Ponting y col., 1972). En el caso de las
muestras impregnadas con calcio, la inmersión en la solución antipardeamiento se realizó
antes y después del proceso de impregnación.
�
7$9$����������������������������
Las muestras sometidas a los tratamientos descriptos anteriormente y las muestras
control (sin tratamiento), fueron almacenadas en bandejas plásticas de polietileno (PET) de
alta densidad con tapa permeables al aire a 4*5ºC durante una semana, y analizadas a
diferentes tiempos a lo largo del almacenamiento desde el punto de vista de diferentes

69
parámetros de calidad: color superficial, propiedades mecánicas y características
microestructurales. �
En la figura 3'5 se muestra un esquema que engloba todos los tratamientos evaluados
en este trabajo.
�
������������������������������������������������������(-�(�6!"�(%)��
�
����!�����������������������������������������������������������!��������������������������
����������������������������������������������������������������Sin pretratamiento ����������������������������������������������Sin pretratamiento������������(Tiempo de exposición: 10,15 ���������������������������������������������(Tiempo de exposición: 2, 20, 60 y���������������������
y 25 min) 100 seg.) ����������d =10 cm d = 5 y 10 cm�
����������Con pretratamiento ����������������������������������������������Con pretratamiento�����������(Tiempo de exposición: 2, 8, 14������������������������������������������(Tiempo de exposición: 2, 20, 60 y y 20 min) 100 seg.) ������������d = 10 cm d = 5 y 10 cm ��
� Escaldado �������������������������������������������������������������■��Inmersión en solución antipardeamiento � Inmersión en solución
antipardeamiento �
� Adición de calcio (con y sin escaldado previo)
��������
�
�Figura 3'5. Esquema de los tratamientos realizados.
�
�
7$;$�6��������������������������
�
El color en la superficie de las muestras fue medido con un espectrofotómetro
triestímulo de reflectancia con esfera integradora (modelo CM*508*d, Minolta Co, Japón)
usando una apertura de medida de 1,4 cm, iluminante C y 2º de ángulo de observador. Las
caras laterales de las muestras fueron recubiertas con una cartulina negra para evitar la

70
dispersión de la luz incidente por los laterales. Las mediciones se realizaron sobre un fondo
blanco.
Se registraron las coordenadas de color del espacio CIE (X, Y, Z) y los componentes
L*, a*, b* del espacio CIELAB, donde L* indica luminosidad, a* indica cromaticidad sobre
un eje que va del verde (*) al rojo (+), y b* cromaticidad sobre un eje que va del azul (*) al
amarillo (+). A partir de estos valores numéricos se calcularon distintas funciones de color
“diferencia de color global” (aE*ab), “índice de browning” (IB), “croma” (C) y “ángulo de
tono” (h) usando las siguientes ecuaciones:
∆E*ab = [(∆L*ab)2
+ (∆a*)2
+ (∆b*)2]
1/2 (3.3)
(las diferencias fueron calculadas teniendo en cuenta los valores de L*, a* y b* de las
muestras frescas antes del tratamiento)
IB = [100 (x – 0,31)] / 0,172 (3.4)
donde: x = X / (X + Y + Z) (3.5)
C = (a*2
+ b*2
)1/2 (3.6)
h = arctg (b*/a*) (3.7)
La diferencia de color global indica la magnitud del cambio de color luego del
tratamiento. h* representa un ángulo en una rueda de color de 360º y puede ser distribuido en
cuatro cuadrantes del plano a*b*. Los valores se definen según: rojo*púrpura (0º), amarillo
(90º), azul*verdoso (180º) y azul (270º). El valor de croma representa la intensidad o pureza
del valor de h* y toma un valor de cero en el centro y se incrementa de acuerdo a la distancia
con respecto al centro.
Las mediciones se realizaron al día 0 (sobre las rodajas frescas e inmediatamente
después de realizado el tratamiento), al tercer y al séptimo día del almacenamiento a 4*5 ºC.
Se utilizaron 10 replicados para cada condición, haciendo mediciones en al menos cuatro
posiciones diferentes de la superficie de cada muestra. Los resultados reportados se basaron
en el valor promedio de estas mediciones. Para minimizar la variabilidad biológica entre
frutas, los valores colorimétricos promedio obtenidos se expresaron como diferencias con

71
respecto al correspondiente valor promedio de la fruta fresca cortada inmediatamente antes
de su tratamiento.
��7$?$��6�����������������������������������������������7$?$#$� ���������������������������������
La evaluación de las propiedades mecánicas a altas deformaciones se realizó mediante
un test de compresión en una máquina de ensayos universales INSTRON Testing Machine
Model 1011 (EEUU).
Se obtuvieron las curvas de esfuerzo*deformación de 20 muestras para cada tratamiento
y tiempo de almacenamiento analizado (0, 3 y 7 días). Se utilizó una velocidad de cabezal de
10 mm/seg, un rango de carga de 500 N y un porcentaje de compresión de 80 %. Los datos de
fuerza*deformación fueron transformados en valores de esfuerzo real (σR) vs. deformación
real (εR), según las siguientes ecuaciones:
Ed = σR / εR (3.8)
Si el volumen permanece constante σR puede expresarse como:
σR = [ ]
00
0 )()(
HA
tHHtF �− (3.9)
εR =
�− HH
H
0
0 ln (3.10)
donde: σR es el esfuerzo real (N/m2), F(t) es la fuerza a tiempo t (N), H0 es la altura
inicial del espécimen (m), cH es la disminución absoluta de la altura inicial del espécimen en
la dirección de la fuerza aplicada (m), A0 es el área inicial de la muestra (m2) y εR es la
deformación real definida por Hencky (Peleg, 1984).
A partir de de las curvas de σR vs εR, se obtuvieron los valores de esfuerzo en el punto
del bioyield (σRB) y de ruptura (σR
R), y de módulo de deformabilidad (Ed), el cual se calculó
mediante regresión lineal de los datos de las curvas en el rango de 10*20 % de deformación.
7$?$,$� ����������������*���������������

72
Se determinaron las propiedades reológicas de las muestras a bajas deformaciones en el
rango viscoelástico lineal. La medición se realizó a 25 ºC en un Reómetro Dinámico Paar
Physica CR 300 (Antón Paar GMBH, Alemania). Para la medición se usó una geometría de
platos paralelos de 3 cm de diámetro (sensor PP30 con superficie rugosa).
Se realizaron dos tipos de ensayos:
* ensayos oscilatorios (barrido de amplitud y barrido de frecuencia)
* ensayos rotatorios (ensayo de fluencia – recuperación).
Se utilizaron 10 replicados para cada condición. Las muestras se analizaron al día 0 y
séptimo de almacenamiento.
Para proveer una mayor área de contacto entre la muestra y el sensor, y así minimizar el
deslizamiento de la muestra durante los ensayos, se aplicaba una fuerza normal de 1N durante
150 segundos antes de los ensayos (oscilatorio y rotatorio) y se mantenía dicha fuerza durante
el transcurso de los ensayos oscilatorios.
�
7$?$,$#$�"������������������
En primera instancia se realizó un barrido de amplitud con control de la deformación de
cizalla (BA CDC) para determinar el rango viscoelástico lineal (RVL). Se fijó una frecuencia
angular de 10 s*1, barriendo la amplitud de deformación entre 0,001 y 10 %. El valor del RVL
fue determinado utilizando el software del equipo (Paar Physica US 200). El software calcula
el RVL detectando mediante un punto de inflexión la caída de la pendiente de la curva
módulo elástico (G´) vs. deformación (γ). Determina el valor límite del RVL y un valor
inferior al mismo, como el recomendado para utilizarse en los ensayos de barrido de
frecuencia.
En una segunda etapa se realizó un barrido de frecuencia donde se obtuvieron los
valores experimentales de los módulos de almacenamiento (G´) y de pérdida (G´´), y del
factor de pérdida (tang δ = G´´ / G´). Se utilizó un rango de frecuencia angular entre 0,1 y 100
s*1, y un valor constante de amplitud de deformación de 0,01%, el cual se seleccionó para que
todas las muestras se encontraran dentro de los límites de linealidad establecidos en el BA
CDC.
En las tablas que acompañan a los espectros de frecuencia se reportaron los valores
promedios de G´ y tang δ tomando 4 valores de frecuencia angular a lo largo del barrido (0,1;

73
1; 10 y 100 s*1). Los valores del módulo elástico del barrido de frecuencia fueron modelados a
través de una regresión logarítmica:
log (G´) = n log ( ω ) + k (3.11)
donde: n es la pendiente de la regresión, y k es la ordenada al origen.
�
7$?$,$,$�"�����������������
Los ensayos de fluencia*recuperación fueron realizados aplicando un esfuerzo de cizalla
constante (τ) igual a 35 Pa durante 100 segundos, y un periodo de recuperación de 200
segundos sin aplicar esfuerzo. Se registró el porcentaje de deformación (γ %) de la muestra en
ambas etapas y a partir del mismo se calculó la capacitancia (J = γ / τ) en función del tiempo.
Para poder asegurar que durante el ensayo de fluencia se estuviera trabajando dentro del rango
viscoelástico lineal, se realizó previamente un barrido de amplitud con control de esfuerzo de
cizalla (CEC) en las muestras con los diferentes tratamientos para determinar el RVL. A partir
de estos ensayos, se decidió utilizar un valor de esfuerzo de 35 Pa para que todas las muestras
se encontraran dentro del RVL.
Previamente al ensayo de fluencia, las muestras fueron sujetas a repetidos ciclos de
esfuerzo y relajación para destruir la memoria lejana del material en estudio, remover
cualquier irregularidad y de esta forma obtener resultados reproducibles.
Las curvas de capacitancia obtenidas se representaron matemáticamente utilizando un
modelo de Kelvin Voigt generalizado de 6 elementos. El mismo consiste en un resorte
conectado en serie con dos elementos de Kelvin*Voigt (cada uno de los elementos de Kelvin*
Voigt tiene un resorte y un pistón conectados en paralelo) y con un pistón (Sherman, 1970).
En la figura 3'6 se muestra el esquema del modelo utilizado. La ecuación correspondiente es:
� �
( ) ( )N
t
i
iteJJtJ i
ητ λ +−+= −
=∑ /
2
10 1)()(, (3.12)
donde: J (t,� ττττ) es la capacitancia (= γ(t) / τ con γ (t) la deformación al tiempo t y τ el
esfuerzo constante aplicado); Jo es la capacitancia instantánea a t=0; Ji son las capacitancias
de retardo; λλλλi (= ηi x Ji ) son los tiempos de retardo y ηi son los coeficientes de viscosidad

74
asociados con los elementos de Kelvin*Voigt; y ηηηηN es el coeficiente de viscosidad asociado al
fluido Newtoniano inversamente proporcional a la fluidez del material en estado estacionario.
�
�
�
�
�
Figura 3'6. Esquema del modelo de Kelvin Voigt generalizado constituido por 6 elementos.
��
Las regresiones no lineales para el modelado de las curvas de fluencia (ec. 3.12) se
realizaron utilizando el programa Origin versión 7.0 (OriginLab Corporation, Northampton,
EEUU). Se utilizó el método de Marquardt para minimizar la suma de cuadrados residuales.
Los valores iniciales que deberían fijarse para que el programa comenzara la iteración fueron
obtenidos utilizando el software US200, el cual tiene incorporado el modelado de la curva de
fluencia para el modelo de Kelvin Voigt generalizado con un elemento de Kelvin*Voigt.
��7$@$�(4�����������������
�
7$@$#$�6������� ��������A6%B�
Las muestras de manzana fresca (control) y tratadas se cortaron con hoja de afeitar en
rectángulos (aprox. 2 mm x 5 mm de lado y 5 mm de espesor) identificando con una marca,
en el caso de las muestras irradiadas, la superficie expuesta al tratamiento. Estas muestras se
fijaron en una solución de glutaraldehído 3 % (v/v) preparada en buffer fosfato de potasio (20
mM), pH 7,4 a 4*5ºC durante 12 h. Luego fueron lavadas con solución buffer fosfato de
potasio (0,1 M), pH 7,4 y vueltas a fijar en solución acuosa de OsO4 1,5% (p/v) a 20°C
durante 2 h. Posteriormente fueron deshidratadas con una serie de soluciones sucesivas de
η2
η1
ηΝ
J1
J2
J0

75
concentración ascendente de acetona y embebidas en resina Spurr de baja viscosidad
(Sorrivas y Morales, 1983). Todos los reactivos utilizados eran de grado analítico (Merck
Química Argentina S.A, Argentina)
A partir de las muestras incluidas en resina Spurr, se obtuvieron secciones de 1 jm de
grosor que fueron teñidas con azul de toluidina y examinadas en un microscopio Zeiss
Axioskop 2 plus (Zeiss, Oberkochen, Alemania).
7$@$,$�6������� �������������������������A6"(B�
Para realizar las observaciones en el microscopio electrónico de transmisión, las
muestras embebidas en resina Spurr se cortaron en secciones ultrafinas con una cuchilla de
vidrio utilizando un ultramicrótomo (Sorvall MT2B Ultracut, USA) y se montaron en grillas
de cobre. Luego se colorearon con una solución de acetato de uranilo (5 % p/p) por 45
minutos y posteriormente con citrato de sodio y nitrato de plomo (solución de Reynolds) y
fueron examinadas con un microscopio JEOL modelo JEM*1200 ExII (Japón) utilizando una
aceleración de voltaje de 80 kv.
7$@$7$�6������� �������������������������������A6"&�B�
Se realizaron observaciones ultraestructurales de la superficie de las rodajas de manzana
fresca y tratada en un microscopio electrónico de barrido ambiental XL30 (Philips, Holanda).
Las muestras fueron cortadas en secciones de aprox. 7 mm x 3 mm de lado y 5 mm de
espesor, y colocadas directamente sobre la platina del microscopio. En todos los casos se usó
una aceleración de voltaje de 20,0 kV y condiciones de presión entre 1,1 * 3,5 Torr.
7$C$� �������������������
El contenido de calcio incorporado en los pretratamientos de impregnación (sección
3.4.1) se midió mediante la técnica de espectroscopia de absorción atómica. Después de
secarse en una estufa de vacío a 60 ºC hasta peso constante, las muestras fueron divididas en
dos mitades para analizar su contenido de calcio. Cada mitad fue digerida o mineralizada con
ácido nítrico (Merck, Alemania) en bombas Parr (Parr Instrument Company, EEUU) y en un
horno microondas. Luego 2*3 alicuotas de cada muestra digerida fueron medidas en un
espectrofotómetro de absorción atómica (espectrofotómetro Varian modelo SpectrAA*20,

76
Australia), utilizando LaCl3 (6500 �g/g, Merck, Alemania) como supresor de interferencia,
una llama de aire*acetileno, 0,5 nm de apertura y una longitud de onda de 422,7 nm (AOAC,
2000). Para verificar la precisión de los procedimientos analíticos se utilizó el material de
referencia RM 8435 (leche en polvo entera). Las determinaciones fueron realizadas por
duplicado. Para cada rodaja de manzana se determinó el valor medio de concentración de
calcio expresado como contenido total de la muestra en base húmeda.
�7$#D$��4�������������
La pérdida de peso de las rodajas de manzana fresca y tratadas se evalúo midiendo el
peso de las mismas al día 0, 3 y 7 del almacenamiento. El peso de las muestras fue registrado
en una balanza con una precisión de ± 0,0001 g. Los resultados fueron expresados como
porcentaje de variación de peso con respecto al de la muestra fresca sin tratamiento, según la
ecuación 3.13:
PP (%) = 100 x (pi – p0)/p0 (3.13)
donde: PP (porcentaje de pérdida de peso de la muestra), p0 (peso inicial de la muestra
fresca) y pi (peso de la muestra a un tiempo i).
Se utilizaron 10 replicados por cada tratamiento.
3.11$�"���������������������
�
7$##$#$�"��������������+�������������������������������
�
En los estudios de inactivación se utilizaron inóculos simples de Listeria innocua
ATCC 33090, Escherichia coli ATCC 11229 y Saccharomyces cerevisiae KE 162. Las
bacterias se sembraron en estrías de Agar Tripteína Soja (ATS*E) (Laboratorios Britania S.A.,
Argentina) y la levadura en Agar Sabouraud (Sab) (Biokar Diagnostics, Francia), y se
incubaron por 24 hs a 37°C y 27 ºC respectivamente, almacenando luego las estrías a 4 °C
para su posterior utilización.
Los inóculos se prepararon transfiriendo dos ansadas del cultivo en estría a un
erlenmeyer conteniendo 20 mL de Caldo Tripteína Soja (CTS*E) (Laboratorios Britania S.A.,
Argentina) (para bacterias) ó Caldo Sabouraud Dextrosa (CSD) (Biokar Diagnostics, Francia)

77
(para levadura) y se incubaron a 37 °C ó 27 ºC durante un periodo de tiempo adecuado para
que los microorganismos llegaran a la fase estacionaria (≈ 24 horas).
Las rodajas de manzana a ser inoculadas fueron preparadas como se describió en la
sección 3.2. (con o sin inmersión previa en la solución antipardeamiento de ácido
ascórbico/cloruro de calcio) trabajando dentro de una cabina de seguridad biológica (Nuaire,
Plymouth, MA) para prevenir una posible postcontaminación. Las rodajas se inocularon con
0,1 mL de inóculo, el cual fue dispersado sobre toda la superficie de la muestra con un
rastrillo de vidrio. Luego las rodajas inoculadas fueron irradiadas con luz UV*C durante 2*20
minutos o con luz pulsada durante 2*100 segundos (a una distancia de la lámpara de 5 o 10
cm). Las experiencias se realizaron por duplicado. Las muestras inoculadas no irradiadas (con
o sin solución antipardeamiento) fueron utilizadas como controles. Los recuentos iniciales de
microorganismos inoculados fueron de aproximadamente 4 x 10 4 – 8 x 106 UFC/cm2.
Para realizar la enumeración, las muestras se colocaron dentro de una bolsa estéril
(Whirl*Pack, Nasco, USA) con 20 mL de agua de peptona estéril y se homogeneizaron en un
destructor de muestra Stomacher (AES Laboratoire, Francia) durante 3 minutos. Con la
solución resultante se realizaron diluciones seriadas en agua de peptona 0,1 (p/v %) y luego se
se realizó el plaqueo en superficie (por duplicado) en medio ATS*E (para bacterias) o Sab
(para levadura), utilizando 0,1 mL de suspensión de la muestra. Las placas de Petri se
incubaron durante 48 hs a 37°C (± 1°C) en el caso de las bacterias, o 27 ºC (± 1°C) en el caso
de la levadura. El recuento de colonias se realizó con un contador automático (Comecta S.A.,
España). Las curvas de supervivencia se generaron a partir de la información experimental
para cada condición ensayada, por medio de la representación gráfica del logaritmo de (N/N0)
(donde N es el número de UFC/cm2 a un dado tiempo de exposición y N0 el número inicial de
UFC/cm2) versus el tiempo de irradiación.
La supervivencia de los microorganismos inoculados a lo largo del almacenamiento fue
analizada en las rodajas de manzana irradiadas con luz UV*C durante 20 minutos (con o sin
previa inmersión en solución antipardeamiento). Luego de la irradiación, las muestras se
colocaron en forma individual dentro de bolsas estériles, se almacenaron a 4*5 ºC en
oscuridad para evitar procesos de fotoreactivación y se analizaron al día 0, 3 y 7 del
almacenamiento. Las muestras irradiadas fueron comparadas con controles no irradiados. Las
experiencias se realizaron por duplicado. Los resultados se expresaron mediante la
representación gráfica del Log (N/N0) (donde N es el número de UFC/cm2 a un dado tiempo
del almacenamiento y N0 el número inicial de UFC/cm2) versus tiempo de almacenamiento.

78
7$##$,$�"+����������������������+��
Se evaluó el efecto de la radiación UV*C y del almacenamiento posterior sobre la flora
nativa (aerobios, hongos y levaduras) presente en rodajas de manzana. Las muestras fueron
preparadas como se describió en la sección 3.2. (con o sin inmersión en la solución
antipardeamiento). En estas experiencias el procesamiento de las muestras no se realizó en
ambiente estéril y los utensilios utilizados no fueron sanitizados previamente con agua
clorada. Las rodajas obtenidas fueron posteriormente irradiadas con luz UV*C durante 20
minutos por ambos lados. Luego se almacenaron en bandejas plásticas a 4*5 ºC en oscuridad y
se analizaron al día 0, 3 y 7 del almacenamiento. Se examinaron dos replicados para cada
condición. El procedimiento de enumeración fue similar al descripto en la sección 3.11.1. a
excepción de que en este caso los medios de cultivo utilizados fueron Agar de Recuento en
Placa (APC) (Merck, Alemania) para el recuento de microorganismos aerobios y Agar de
Papa Dextrosa (APD) (Laboratorios Britania S.A, Argentina) para el recuento de hongos y
levaduras. Las cajas de petri fueron incubadas durante 72 hs a 37 ºC ± 1 ºC (APC) o 27 ºC ±
1 °C (APD). Los resultados fueron expresados como N (donde N es el número de UFC/g) en
función del tiempo de almacenamiento.
7$#,$���:��������� �����
Los resultados fueron expresados como la media ± desviación estándar de la media y
sujetos al análisis estadístico utilizando el software Infostat v.2009 (Universidad Nacional de
Córdoba, Argentina). Los resultados de las mediciones instrumentales de las propiedades
reológicas fueron analizados mediante un ANOVA de dos factores (factores: tiempo de
almacenamiento y dosis o tratamiento). Para el análisis de los resultados de las mediciones de
color y de pérdida de peso, como las mediciones a lo largo del almacenamiento se realizaron
siguiendo la evolución de las mismas muestras, se utilizó un ANOVA de dos factores
(factores: tiempo de almacenamiento y dosis o tratamiento) con medidas repetidas en el factor
tiempo. En el caso de haber interacción significativa entre los factores, los resultados se
analizaron mediante efectos simples (efecto de un factor dejando fijo el otro factor). Cuando
no hubo interacción se analizaron los efectos principales. En todos los casos, las diferencias
significativas entre medias se establecieron aplicando el test de Tuckey con un nivel de
confianza del 95%.

79
8$�-")�5(� %)�=� !)��)!.��
8$#$���5!���!.�� "�5�E��5(��!%5"(�� "�%� ���%-(����8$#$#$�)��������������������������������������
El rango de dosis de radiación UV*C utilizado en este trabajo fue seleccionado a partir
de estudios microbiológicos preliminares realizados sobre la fruta estudiada.
En la figura 4'1 puede observarse el efecto de la aplicación de diferentes dosis de luz
UV*C en la supervivencia de algunas especies de microorganismos inoculados en rodajas de
manzana. Todas las curvas semilogarítmicas de supervivencia obtenidas fueron no lineales.
Luego de 20 minutos de irradiación (dosis ~ 11,2 kJ/m2), los ciclos logarítmicos de reducción
variaron en un rango entre 1 y 1,9 entre las diferentes especies analizadas. Para E. coli y S.
cerevisiae, la velocidad de inactivación disminuyó o fue casi nula luego de 2 minutos de
irradiación (dosis ~ 1,12 kJ/ m2), mientras que para L. innocua la inactivación fue en aumento
incluso hasta los 20 minutos de irradiación.
En la figura 4'2 se muestra con fines comparativos la cinética de destrucción de L.
innocua, Listeria monocytogenes, Zygosaccharomyces bailli y E. coli (experiencias
realizadas por otro integrante del grupo de investigación) en rodajas de pera irradiadas con
diferentes dosis de luz UV*C (Schenk y col., 2008). La reducción de las diferentes especies
varió entre 2,6 y 3,4 ciclos logarítmicos luego de irradiar las muestras durante 20 minutos.
Las diferencias en la inactivación observadas entre estas frutas utilizando una misma
dosis de irradiación posiblemente se deba, al menos parcialmente, a la mayor porosidad que
presenta la manzana (≈24%) en comparación con la pera (≈5%) (Salvatori, 1997; Salvatori y
col., 1998). Los microorganismos inoculados en manzana penetrarían hacia el interior de la
matriz con mayor facilidad y quedarían de esta forma protegidos de la acción de la luz UV*C,
la cual tiene baja penetración. La presencia de irregularidades en la superficie de la matriz, de
grietas superficiales e injurias en el tejido son otros factores que pueden favorecer la
penetración de los microorganismos y reducir la accesibilidad de la luz UV*C (Gómez*López
y col., 2008). También la composición de la matriz puede afectar la inactivación ya que
muchos componentes químicos de los alimentos pueden absorber la luz UV*C y actuar como
efectivos absorbedores químicos de la radiación incidente (Lagunas*Solar y col., 2006). Por lo
tanto, la efectividad de la radiación ultravioleta como método de descontaminación de frutas

80
2
1,6
1,2
0,8
0,4
0
0 5 10 15 20 25
����������
����� �
�
������
�������
��� � �����
Figura 4'1. Curvas semilogarítmicas de supervivencia de algunas especies de microorganismos en rodajas de manzana irradiadas con luz UV*C. N0 E. coli: 8 x 106 UFC/cm2); N0 L. innocua: 1,1 x 106 UFC/cm2
; N0 S. cerevisiae: 5 x 104 UFC/cm2.
4
3
2
1
0
0 5 10 15 20 25
����������
����� �
�
�������
����������� �
�������
���������
� Figura 4'2. Curvas semilogarítmicas de supervivencia de algunas especies de microorganismos en rodajas de pera irradiadas con luz UV*C. Valores experimentales (puntos) y ajuste mediante un modelo tipo Weibull (líneas). Fuente: Schenk y col., 2008.

81
dependería de las características de la matriz irradiada.
Teniendo en cuenta los patrones de inactivación observados, se decidió utilizar un rango
de dosis de 1 á 14 kJ/m2, correspondientes a tiempos de irradiación entre 2 y 25 minutos, para
evaluar los efectos de la irradiación UV*C sobre los atributos de calidad de las rodajas de
manzana mínimamente procesadas. Este rango de dosis no se aleja del utilizado en la mayoría
de los trabajos existentes en la bibliografía en donde se analiza la descontaminación de frutas
y vegetales mediante la radiación UV*C.
��8$#$,$� ���������������������������������������
En la figura 4'3 se muestran las dosis de radiación UV*C obtenidas a diferentes tiempos
de irradiación (0 – 5 min) mediante la técnica actinométrica, utilizando el par ioduro/iodato.
El valor de tasa de fluencia obtenido de la pendiente de la regresión lineal de la gráfica dosis
vs tiempo fue de 9,7 W/m2. Las dosis medidas a tiempos mayores dejaron de presentar un
comportamiento lineal con el tiempo, por lo tanto, se calcularon multiplicando el valor de tasa
de fluencia por el tiempo de irradiación (segundos).�
0
0,5
1
1,5
2
2,5
3
3,5
0 50 100 150 200 250 300 350
���������
�����������
�
Figura 4'3. Dosis de radiación UV*C determinadas mediante actinometría a diferentes tiempos de irradiación.
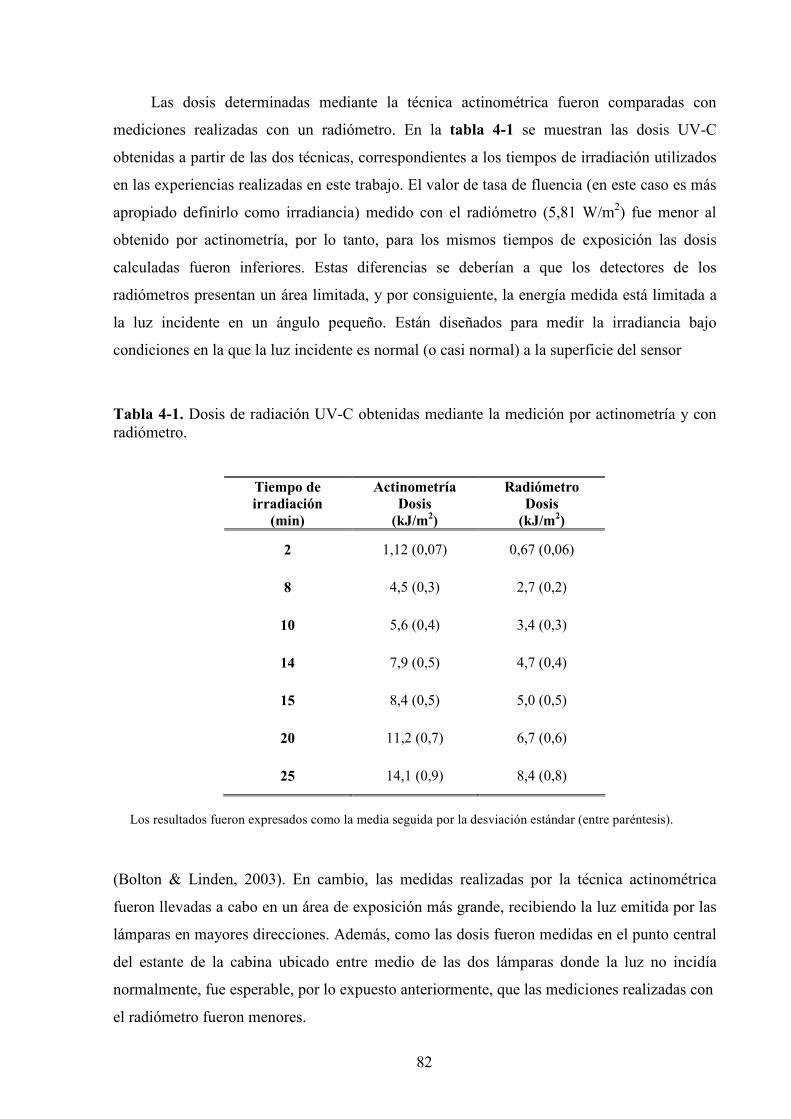
82
Las dosis determinadas mediante la técnica actinométrica fueron comparadas con
mediciones realizadas con un radiómetro. En la tabla 4'1 se muestran las dosis UV*C
obtenidas a partir de las dos técnicas, correspondientes a los tiempos de irradiación utilizados
en las experiencias realizadas en este trabajo. El valor de tasa de fluencia (en este caso es más
apropiado definirlo como irradiancia) medido con el radiómetro (5,81 W/m2) fue menor al
obtenido por actinometría, por lo tanto, para los mismos tiempos de exposición las dosis
calculadas fueron inferiores. Estas diferencias se deberían a que los detectores de los
radiómetros presentan un área limitada, y por consiguiente, la energía medida está limitada a
la luz incidente en un ángulo pequeño. Están diseñados para medir la irradiancia bajo
condiciones en la que la luz incidente es normal (o casi normal) a la superficie del sensor
Tabla 4'1. Dosis de radiación UV*C obtenidas mediante la medición por actinometría y con radiómetro. �
Tiempo de irradiación
(min)
Actinometría Dosis
(kJ/m2)
Radiómetro Dosis
(kJ/m2)
2 1,12 (0,07) 0,67 (0,06)
8 4,5 (0,3) 2,7 (0,2)
10 5,6 (0,4) 3,4 (0,3)
14 7,9 (0,5) 4,7 (0,4)
15 8,4 (0,5) 5,0 (0,5)
20 11,2 (0,7) 6,7 (0,6)
25 14,1 (0,9) 8,4 (0,8)
Los resultados fueron expresados como la media seguida por la desviación estándar (entre paréntesis).
(Bolton & Linden, 2003). En cambio, las medidas realizadas por la técnica actinométrica
fueron llevadas a cabo en un área de exposición más grande, recibiendo la luz emitida por las
lámparas en mayores direcciones. Además, como las dosis fueron medidas en el punto central
del estante de la cabina ubicado entre medio de las dos lámparas donde la luz no incidía
normalmente, fue esperable, por lo expuesto anteriormente, que las mediciones realizadas con
el radiómetro fueron menores.

83
Otra desventaja que presentan los radiómetros, y que puede llevar a mediciones
erróneas, es que son instrumentos que requieren ser calibrados periódicamente. En muchos
casos esta calibración se realiza utilizando actinómetros químicos (Bolton & Linden, 2003).
�8$#$7$�"�������������������������������8$#$7$#$�(����������������
Se analizó la evolución del color a lo largo del almacenamiento de rodajas de manzana
expuestas a la luz UV*C durante diferentes tiempos (10, 15 y 25 min). Las rodajas irradiadas
fueron comparadas con controles de muestras frescas ubicadas dentro de la cabina de
irradiación con las lámparas UV*C apagadas durante los mismos tiempos utilizados en los
tratamientos de irradiación.�
Las rodajas de manzana fresca tenían un color inicial blanco*crema, representado por
valores de L* entre 69,8 y 71,9, valores de a* entre *5,5 y *4,7 y valores de b* entre 16,6 y
18,8. En la figura 4'4 se presentan los valores promedio de los parámetros colorimétricos L*,
a* y b* obtenidos en las muestras irradiadas a las diferentes dosis (expresados como
diferencias con respecto a los parámetros obtenidos de la muestra fresca inmediatamente antes
del tratamiento de irradiación) a lo largo del almacenamiento. En todos los parámetros de
color analizados los resultados del ANOVA de 2 factores mostraron que había interacción
significativa entre la dosis aplicada y el tiempo de almacenamiento (p< 0,0001). Por lo tanto,
la evolución de los parámetros de color a lo largo del tiempo dependió de la dosis aplicada.
Los valores del parámetro L* de las muestras controles no presentaron variaciones
significativas a lo largo de los siete días de almacenamiento (figura 4'4 a). Al día 0 de
almacenamiento, se observó un decrecimiento del parámetro L* luego de la irradiación UV*C
solo en las muestras tratadas a la dosis más alta. Sin embargo, al tercer y séptimo día del
almacenamiento, en todas las dosis evaluadas, la irradiación produjo un decrecimiento leve
pero significativo (p < 0,0001) de los valores de L*. Esto indicaría un mayor oscurecimiento
(mayor aL*) de las muestras tratadas con respecto a los controles. Por otra parte, los
parámetros a* y b* se incrementaron significativamente luego de los 10, 15 y 25 min del
tratamiento UV*C (figura 4'4 b y c). En general, todas las muestras (tratadas y controles)
mostraron un fuerte incremento en el parámetro a* al día 3 y 7, pero este comportamiento fue
más pronunciado en las manzanas tratadas, especialmente a dosis altas. El parámetro b* de las
muestras irradiadas, presentó un aumento inicial inmediatamente después del tratamiento de
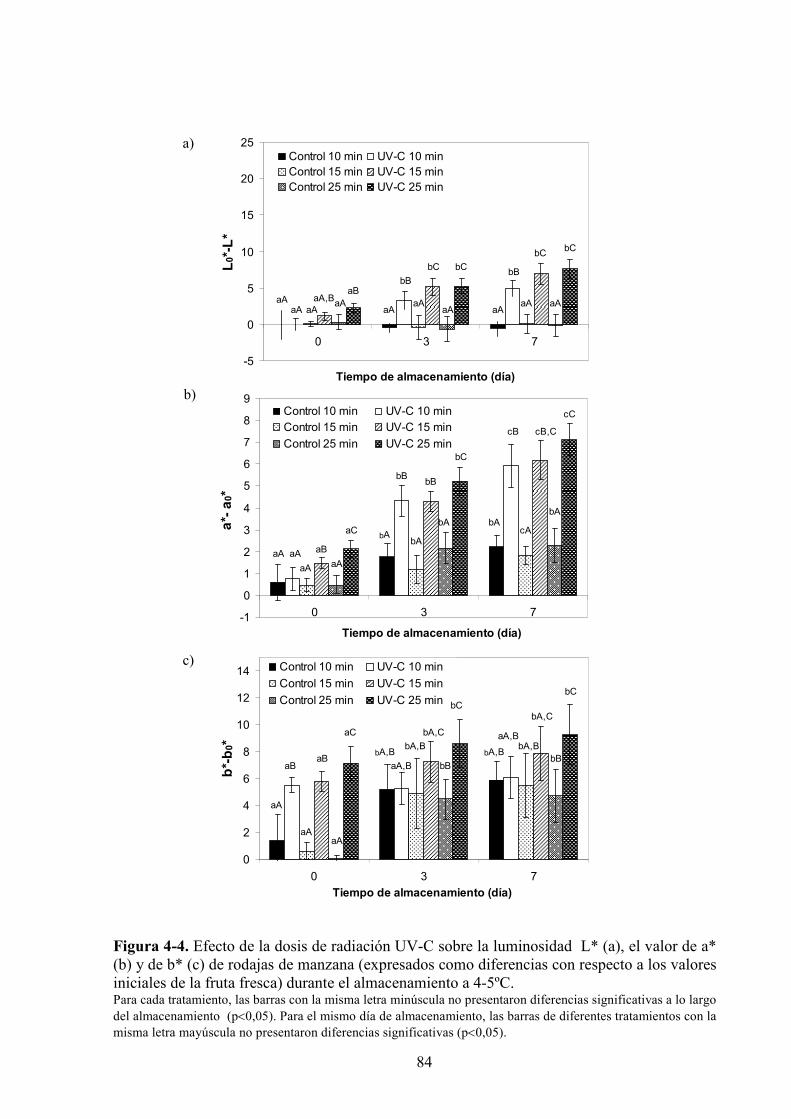
84
Figura 4'4. Efecto de la dosis de radiación UV*C sobre la luminosidad L* (a), el valor de a* (b) y de b* (c) de rodajas de manzana (expresados como diferencias con respecto a los valores iniciales de la fruta fresca) durante el almacenamiento a 4*5ºC. Para cada tratamiento, las barras con la misma letra minúscula no presentaron diferencias significativas a lo largo del almacenamiento (p<0,05). Para el mismo día de almacenamiento, las barras de diferentes tratamientos con la misma letra mayúscula no presentaron diferencias significativas (p<0,05).
5
0
5
10
15
20
25
0 3 7
���������������������������
������
Control 10 min UVC 10 min
Control 15 min UVC 15 min
Control 25 min UVC 25 min
aA aA aA
aA,B aA
aB
aA
bB
aA
bC
aA
bC
aA
bB
aA
bC
aA
bC
a)
1
0
1
2
3
4
5
6
7
8
9
0 3 7
���������������������������
�����
��
Control 10 min UVC 10 min
Control 15 min UVC 15 min
Control 25 min UVC 25 min
aA aA
aA
aB
aA
aC bA
bB
bA
bB
bA
bC
bA
cB
cA
cB,C
bA
cC
b)
0
2
4
6
8
10
12
14
0 3 7
���������������������������
������
Control 10 min UVC 10 min
Control 15 min UVC 15 min
Control 25 min UVC 25 min
aA
aB
aA
aB
aA
aC
bA,B
aA,B
bA,B
bA,C
bB
bC
bA,B
aA,B bA,B
bA,C
bB
bC
c)

85
irradiación, y luego se incrementó hasta el tercer día en las muestras irradiadas a las dosis más
altas. Por el contrario, los valores de b* para las muestras no irradiadas no presentaron
variaciones importantes al día 0 y se incrementaron al tercer día, permaneciendo luego sin
variación hasta el séptimo día de almacenamiento. Los valores al final del almacenamiento
fueron similares a aquellos exhibidos en las muestras irradiadas por 10 y 15 min.
Numerosos autores han reportado que un decrecimiento en el valor de L* y un
incremento en el valor de a* son indicativos de pardeamiento (Monsalve*González y col.,
1993; Goupy y col., 1995; Rocha & Morais, 2003). Sapers y Douglas (1987) sugirieron
también que el pardeamiento enzimático en la superficie de manzanas mínimamente
procesadas podría ser monitoreado midiendo los cambios en los valores de L* y a*, mientras
que el parámetro b* no estaría relacionado con la extensión del pardeamiento. Patrones
similares en la tasa de decrecimiento de la luminosidad a lo largo del almacenamiento han
sido también reportados por otros autores. Rocha & Morais (2003) encontraron un rápido
oscurecimiento de cubos de manzana mínimamente procesados durante los tres primeros días
de almacenamiento a 4 ºC, seguido por un decrecimiento en la luminosidad hasta los 10 días
de almacenamiento. Kim y col. (1993) reportaron un rápido decrecimiento en los valores de
L* en 12 cultivares de manzana. Soliva*Fortuny & Martín*Belloso (2003) indicaron que el
tratamiento con ácido peroxiacético sobre manzanas mínimamente procesadas resultó en un
aumento significativo de a* y una disminución significativa de L* en comparación con el
control. Atribuyeron esta tendencia al severo daño del tejido de manzana causado por el
tratamiento, provocando la liberación de la enzima polifenoloxidasa y promoviendo
reacciones de oxidación con los sustratos fenólicos de los tejidos de manzana.
Como se observa en la figura 4'4 los cambios simultáneos en ambos parámetros, L* y
a*, fueron un buen indicador del pardeamiento durante el almacenamiento, probablemente
debido al resultado de reacciones de pardeamiento oxidativo. Las superficies de las muestras
se tornaron más oscuras (menores valores de L*) y menos verdes (mayores valores de a*) en
comparación con las muestras frescas. Al final del almacenamiento, las muestras expuestas a
la radiación UV*C se presentaron más pardeadas que los controles y este efecto fue más
pronunciado con el incremento de la dosis de UV*C.
En la figura 4'5 se observa la evolución de las funciones de color ángulo de tono (h) y
croma (C) en las diferentes muestras. En estas funciones también se presentó interacción
significativa entre el tiempo de almacenamiento y la dosis de radiación aplicada (p<0,0001).
Los valores de h de las muestras frescas rondaron en 105*108º y decrecieron durante el
almacenamiento tanto en las muestras tratadas como en los controles, cuando las rodajas de

86
a)
�
0
5
10
15
20
25
30
35
0 3 7
���������������������������
����
Control 10 min UVC 10 min
Control 15 min UVC 15 min
Control 25 min UVC 25 min
aA aB
aA
aB,C
aA
aC
bA
bB
bA
bB,C
bA
bC
cA
cB
cA
cB,C
bA
cC
����
��
Figura 4'5. Valores promedio de las funciones de color ángulo de tono (h) (a) y croma (C) (b) (expresados como diferencias con respecto a los valores iniciales de la fruta fresca) obtenidos durante el almacenamiento (4*5 ºC) de rodajas de manzana expuestas a diferentes dosis de radiación UV*C. Para cada tratamiento, las barras con la misma letra minúscula no presentaron diferencias significativas a lo largo del almacenamiento (p<0,05). Para el mismo día de almacenamiento, las barras de diferentes tratamientos con la misma letra mayúscula no presentaron diferencias significativas (p<0,05).
0
2
4
6
8
10
12
14
0 3 7
���������������������������
���
�
Control 10 min UVC 10 min
Control 15 min UVC 15 min
Control 25 min UVC 25 min
aA
aB
aA
aB
aA
aB
bA,B aA,B
bA,B
a,bA,C
bB
a,bC
bA,B
aA,B bA,B
aA,C
bB
bC
b)

87
manzanas se tornaron pardas (figura 4'5 a). La disminución de h para los controles al final
del almacenamiento fue de alrededor de un 8%, y en las manzanas irradiadas, de alrededor de
un 16*20%. Los valores de croma (C) presentaron cambios significativos en las muestras
tratadas y los controles (figura 4'5 b). Los valores de C para los controles se incrementaron
gradualmente de 18 a 24 (incremento del 33%) a lo largo del almacenamiento, mientras que el
tratamiento UV*C provocó un aumento de 28% a un 36% en el valor de croma de acuerdo a la
dosis y se incrementó levemente al final del almacenamiento en las muestras tratadas a las
dosis más altas.
La figura 4'6 muestra los valores del índice de pardeamiento (IB) (expresado como
diferencia con los valores de IB iniciales de las rodajas frescas) de las muestras irradiadas a
las distintas dosis de radiación UV*C a los 0, 3 y 7 días del almacenamiento. Al inicio del
almacenamiento, no se presentaron diferencias significativas en los valores de �IB para las
muestras irradiadas por 10 y 15 min pero la irradiación durante 25 min provocó un incremento
significativamente mayor en el valor de �IB (p < 0,001). Estos valores se incrementaron
gradualmente al tercer y séptimo día, presentando el mismo patrón mencionado para el día 0
con el aumento de la dosis.
�
!�
!
��
�
"�
!� ! �� � "�
��������������������#������$����
�� ��%&
aA aA
bA
aB aB bB
a,bC
bC
aC
Figura 4'6. Efecto de la dosis de radiación UV*C sobre la diferencia del índice de pardeamiento (IB) de rodajas de manzana a diferentes tiempos del almacenamiento a 4*5ºC. ▲ día 0, ♦ día 3 ■ día 7. Las barras verticales representan las desviaciones estándar. Las medias con la misma letra minúscula no presentaron diferencias significativas (p<0,05) en los distintos tiempos de exposición a la radiación UV*C. Para un tiempo de exposición específico, las medias con la misma letra mayúscula no presentaron diferencias significativas (p<0,05).

88
Los cambios descriptos en el color de las rodajas de manzana irradiadas, pueden
observarse visualmente en la figura 4'7. Al final del almacenamiento, las rodajas irradiadas
presentaron claramente un mayor pardeamiento superficial incrementado con el aumento de la
dosis de radiación UV*C.
Otros autores reportaron también cambios en el color producidos por la aplicación de
radiación UV*C. Erkan y col. (2001) observaron la aparición de pardeamiento en la superficie
de rodajas de zucchinis expuestos a dosis UV*C mayores a 5 KJ/m2. Atribuyeron el
pardeamiento observado a una acumulación de compuestos fenólicos inducidos por la
radiación UV*C. Fonseca & Rushing (2006) reportaron que la aplicación de dosis superiores a
13 KJ/m2 en sandías mínimamente procesadas producía un efecto adverso en el color de la
pulpa al inducir la decoloración del tejido.
8$#$7$,$��������������������������������������+����������������������
Con el propósito de inhibir o retardar el pardeamiento superficial de las muestras
expuestas a la radiación UV*C, se evaluó la aplicación de dos pretratamientos diferentes:
* escaldado en agua
* inmersión en una solución antipardeamiento de ácido ascórbico/cloruro de calcio
El escaldado es un tratamiento térmico leve utilizado para la inactivación de enzimas
oxidativas de frutas y vegetales involucradas en reacciones de deterioro. En este trabajo, se
realizó con el propósito de lograr la inactivación térmica de la enzima polifenoloxidasa (PPO)
presente en manzanas.
El escaldado se realizó en convección forzada seleccionando una relación de tiempo y
temperatura (80ºC, 5,5 minutos) adecuada para lograr la inactivación de la PPO en toda la
muestra. El tiempo de escaldado utilizado se determinó mediante la ecuación 3.2, teniendo en
cuenta el tiempo de reducción decimal de la PPO a 80ºC y el tiempo en que se alcanzaba
dicha temperatura en el centro de la muestra (figura 4'8).
Para la selección de la solución antipardeamiento, se realizaron estudios preliminares
con tres soluciones reportadas en bibliografía como eficaces para inhibir el pardeamiento
enzimático en manzana. Estas soluciones eran: ácido ascórbico 1%(p/v) + ácido cítrico
0,2%(p/v), ácido ascórbico 1%(p/v) + cloruro de sodio 0,05%(p/v) (Pizzocaro y col., 1993) y
ácido ascórbico 1%(p/v) + cloruro de calcio 0,1%(p/v) (Ponting y col., 1972). Se seleccionó
la solución de ácido ascórbico/cloruro de calcio porque se comprobó sensorialmente que era
la que producía mayor inhibición del pardeamiento y menores cambios de “flavor” en las

89
�
���'��� ()���!���� ()���! ��� ()���� ������'��� ()���!���� ()���! ��� ()���� ������'��� ()���!���� ()���! ��� ()���� ���()���!���� ()���! ��� ()���� ���
Figura 4'7. Color superficial de rodajas de manzanas expuestas a diferentes dosis de radiación UV*C al séptimo día de almacenamiento a 4*5 ºC.
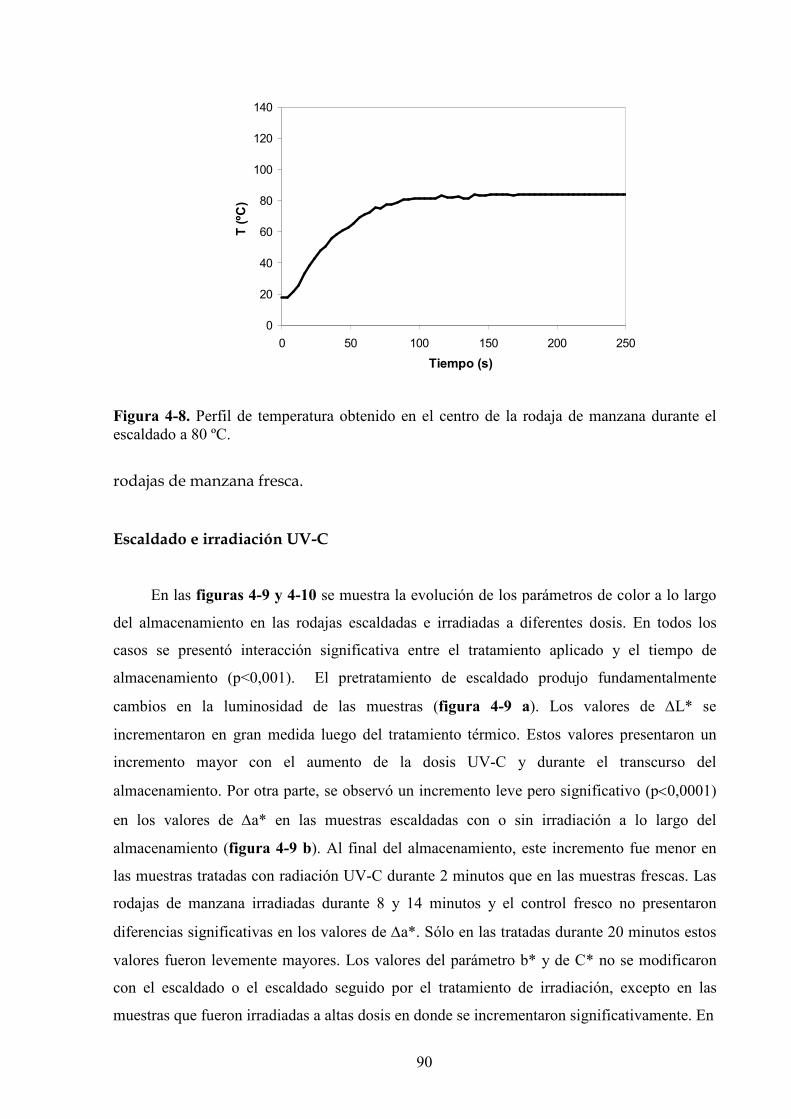
90
0
20
40
60
80
100
120
140
0 50 100 150 200 250
���������
���*�
Figura 4'8. Perfil de temperatura obtenido en el centro de la rodaja de manzana durante el escaldado a 80 ºC. �
����������� �� ����������
�
"�������������������������
En las figuras 4'9 y 4'10 se muestra la evolución de los parámetros de color a lo largo
del almacenamiento en las rodajas escaldadas e irradiadas a diferentes dosis. En todos los
casos se presentó interacción significativa entre el tratamiento aplicado y el tiempo de
almacenamiento (p<0,001). El pretratamiento de escaldado produjo fundamentalmente
cambios en la luminosidad de las muestras (figura 4'9 a). Los valores de �L* se
incrementaron en gran medida luego del tratamiento térmico. Estos valores presentaron un
incremento mayor con el aumento de la dosis UV*C y durante el transcurso del
almacenamiento. Por otra parte, se observó un incremento leve pero significativo (p<0,0001)
en los valores de �a* en las muestras escaldadas con o sin irradiación a lo largo del
almacenamiento (figura 4'9 b). Al final del almacenamiento, este incremento fue menor en
las muestras tratadas con radiación UV*C durante 2 minutos que en las muestras frescas. Las
rodajas de manzana irradiadas durante 8 y 14 minutos y el control fresco no presentaron
diferencias significativas en los valores de �a*. Sólo en las tratadas durante 20 minutos estos
valores fueron levemente mayores. Los valores del parámetro b* y de C* no se modificaron
con el escaldado o el escaldado seguido por el tratamiento de irradiación, excepto en las
muestras que fueron irradiadas a altas dosis en donde se incrementaron significativamente. En

91
0
5
10
15
20
25
0 3 7���������������������������
�������
Control EE + UVC 2 min E + UVC 8 minE +UVC 14 min E + UVC 20 min
aA
aA aA
bB bB aB a,bC
bB,C aC aC
aD bD
a,bC,D bE
cD
bC
cE
a)
0
1
2
3
4
5
6
7
8
9
0 3 7
���������������������������
�����
��
Control EE + UVC 2 min E + UVC 8 minE + UVC 14 min E + UVC 20 min
aA,C
bA
aA,B
bB
cB
aA bB
bB aB,C
bA
cA cA,C
bA,C
aC
bC
cC
aB,C
b)
3
1
1
3
5
7
9
11
13
15
0 3 7
���������������������'������
�����
��
Control EE + UVC 2 min E + UVC 8 minE + UVC 14 min E + UVC 20 min
aA
bA
aA
aB aB
aA
bB
bB,C aA
aB
aB,C
aA
bB a,bC
aB
bC
bD
c)
Figura 4'9. Efecto de la dosis de radiación UV*C sobre la luminosidad (L*) (a), el valor de (a*) (b) y de (b*) (c) de rodajas de manzana escaldadas (expresados como diferencias con respecto al valor inicial de la fruta fresca) durante el almacenamiento a 4*5 ºC. (E: escaldado). Para cada tratamiento, las barras con la misma letra minúscula no presentaron diferencias significativas a lo largo del almacenamiento (p<0,05). Para el mismo día de almacenamiento, las barras de diferentes tratamientos con la misma letra mayúscula no presentaron diferencias significativas (p<0,05).

92
todas las muestras tratadas, estos parámetros permanecieron en general sin variación o
decrecieron levemente con el tiempo de almacenamiento, presentando valores
significativamente menores (p<0,0001) que los exhibidos por el control (figura 4'9 c y figura
4'10 b). Los valores de h* de las muestras escaldadas e irradiadas a las dosis más altas
disminuyeron a lo largo del almacenamiento, siendo esta disminución mayor con el
incremento de la dosis (figura 4'10 a). Sin embargo, la disminución de h* de las muestras
tratadas térmicamente a lo largo del almacenamiento fue menor o similar a la que presentaron
las muestras control, excepto en las irradiadas durante 20 minutos que presentaron valores
levemente inferiores.
)�������������������������������������
�
En contraste con el comportamiento observado en las muestras irradiadas, escaldadas y
escaldadas e irradiadas, los valores de L* de las rodajas de manzana inmersas en la solución
antipardeamiento previo a la irradiación se incrementaron levemente, y en general, no hubo
un efecto significativo de la dosis UV*C aplicada (figura 4'11 a). Al día 3 y 7 de
almacenamiento se observó un incremento en los valores de a* pero en mucho menor grado
que en las muestras irradiadas con o sin escaldado previo (figura 4'11 b). En los valores del
parámetro b*, no se presentaron diferencias significativas ni por efecto de la dosis de
radiación UV*C ni por el tiempo de almacenamiento (figura 4'11 c). En general, los valores
de h* de las muestras tratadas disminuyeron levemente al final del almacenamiento mientras
que los valores de croma no presentaron variaciones significativas (figura 4'12).
El efecto inhibitorio de la solución de ácido ascórbico y cloruro de calcio sobre las
reacciones de pardeamiento fue evidenciado por los valores altos de L* y bajos valores de a*
en las muestras inmersas en esta solución e irradiadas, en comparación con las muestras
sometidas sólo al tratamiento de irradiación o en las muestras control. El incremento en la
luminosidad podría ser correlacionado con el blanqueamiento observado en las muestras.
Estos resultados estuvieron en concordancia con conclusiones previas de Fan y col. (2005).
Estos autores reportaron que el ascorbato de calcio podría inducir decoloración en los
pigmentos de la manzana (ej. blanqueo de tejidos), evidenciado por el blanqueamiento visual
observado en la superficie de esta fruta luego de la inmersión en la solución antipardeamiento.
La inhibición del pardeamiento en las rodajas de manzana irradiadas por la aplicación
de los pretratamientos de escaldado e inmersión en la solución antipardeamiento puede
observarse visualmente en la figura 4'13.

93
�
� Figura 4'10. Efecto de la dosis de radiación UV*C sobre las funciones de color ángulo de tono (h)(a) y croma (C)(b) de rodajas de manzana escaldadas (expresados como diferencias con respecto a los valores iniciales de la fruta fresca) obtenidas durante el almacenamiento (4*5 ºC). (E: escaldado) Para cada tratamiento, las barras con la misma letra minúscula no presentaron diferencias significativas a lo largo del almacenamiento (p<0,05). Para el mismo día de almacenamiento, las barras de diferentes tratamientos con la misma letra mayúscula no presentaron diferencias significativas (p<0,05).
4
2
0
2
4
6
8
10
12
14
1 2 3
���������������������������
�����
�
Control E
E + UVC 2min E + UVC 8 min
E + UVC 14 min E + UVC 20 min
aA
aA aA
aA
aB
aA
bB aB
bB
bC
a,bB
bA
bB bB aB
bB
bC
b)
0
5
10
15
20
25
30
35
0 3 7���������������������������
����
Control E
E + UV 2 min E +UVC 8 min
E + UVC 14 min E + UVC 20 min
bC
a,bB,CaA
cC bA,C
aA
bA
aB
aA,B
aB
bD
aB
aA cA,C
aC
bB
cD
a)

94
Figura 4'11. Efecto de la dosis de radiación UV*C sobre la luminosidad (L*) (a), el valor de (a*) (b) y de b* (b*) (c) de rodajas de manzana previamente inmersas en la solución antipardeamiento (expresados como diferencias con respecto a los valores iniciales de la fruta fresca) durante el almacenamiento a 4*5ºC. (SA: solución antipardeamiento) Para cada tratamiento, las barras con la misma letra minúscula no presentaron diferencias significativas a lo largo del almacenamiento (p<0,05). Para el mismo día de almacenamiento, las barras de diferentes tratamientos con la misma letra mayúscula no presentaron diferencias significativas (p<0,05).
3
2
7
12
17
22
0 3 7
���������������������������
�����
��
Control SASA + UVC 2 min SA + UVC 8 minSA + UVC 14 min SA + UVC 20 min
aA,B
aA,B aB aB
aA
bB bB bB
bB
bB
aA
bB bB bB
a,bB
bB
aA
a)
2
1
0
1
2
3
4
5
6
7
8
9
0 3 7
���������������������������
������
Control SASA + UVC 2 min SA + UVC 8 minSA + UVC 14 min SA + UVC 20 min
aA
bA
bA,C
cA,B
aA
bB
bA,B
aA
bB,C bB
aA
bA,B,C
cA,B
cA
aA
bA,B,C
aA
b)
2
0
2
4
6
8
10
12
14
0 3 7
���������������������������
�����
��
Control SA
SA + UVC 2 min SA + UVC 8 min
SA + UVC 14 min SA + UVC 20 min
aA aA
aA
aA
aA aA
aA aA
aA aA
aA aA
aA
aA
aA
aA
c)

95
Figura 4'12. Efecto de la dosis de radiación UV*C sobre las funciones de color ángulo de tono (h)(a) y croma (C)(b) de rodajas de manzana previamente inmersas en la solución antipardeamiento (expresados como diferencias con respecto a los valores iniciales de la fruta fresca) durante el almacenamiento a 4*5ºC. (SA: solución antipardeamiento) Para cada trratamiento, las barras con la misma letra minúscula no presentaron diferencias significativas a lo largo del almacenamiento (p<0,05). Para el mismo día de almacenamiento, las barras de diferentes tratamientos con la misma letra mayúscula no presentaron diferencias significativas (p<0,05).
0
5
10
15
20
25
30
0 3 7���������������������������
�������
Control SA
SA +UVC 2 min SA + UVC 8 min
SA + UVC 14 min SA + UVC 20 min
aA aA
aBbA
aA
a,bB bA
aAaB
bA
aA
aA,B
bA
aA aA,B
bA
a)
4
2
0
2
4
6
8
10
12
14
0 3 7
���������������������������
���
�
Control SA
SA + UVC 2 min SA + UVC 8 min
SA + UVC 14 min Sa + UVC 20 min
aA aA aA
aA
aA
aB
bB
aB aB
bB
aA
aA,B
aB
aB
aB
aA,B
b)

96
���'�� + +�,�()��
����
+�,�()��
-���
+�,�()��
�����
�
���'�� + +�,�()��
����
+�,�()��
-���
+�,�()��
�����
�
���'�� ./ ./�,�()���
����
./�,�()���
-���
./�,�()���
�����
./�,�()���
!0���
�
���'�� ./ ./�,�()���
����
./�,�()���
-���
./�,�()���
�����
./�,�()���
!0���
���'�� ./ ./�,�()���
����
./�,�()���
-���
./�,�()���
�����
./�,�()���
!0���
�
Figura 4'13. Color superficial de rodajas de manzanas escaldadas (a) e inmersas en solución de ácido ascórbico/cloruro de calcio (b) y expuestas a diferentes dosis de radiación UV*C, al séptimo día de almacenamiento a 4*5 ºC. E: escaldado; SA: solución antipardeamiento.

97
La figura 4'14 compara el efecto de la dosis de radiación UV*C sobre las funciones
diferencia de color total (�E*ab) y el índice de pardeamiento (IB) de las rodajas de manzana
irradiadas con o sin pretratamiento al día 7 del almacenamiento. En las rodajas de manzana
expuestas solo a la luz UV*C o en aquellas que tuvieron un escaldado previo, la diferencia de
color entre las muestras tratadas y el control fue bien marcada y se incrementó con el tiempo
de exposición a la radiación UV*C, debido fundamentalmente a los cambios en los parámetros
a* y L*. Por el contrario, los niveles de radiación UV*C aplicados no afectaron a esta función
de color en las muestras previamente inmersas en la solución antipardeamiento. Los valores
de IB rondaron en un rango entre 19 y 21 para las muestras frescas despúes del corte. Como
se muestra en la figura 4'14 b, al día 7, los valores de IB de las muestras irradiadas sin
pretratamiento se incrementaron significativamente, y este incremento fue mayor con el
aumento de la dosis UV*C. Tanto el pretratamiento de escaldado como la inmersión en la
solución antipardeamiento redujeron marcadamente los valores del IB de las muestras de
manzana irradiadas y no irradiadas, almacenadas durante una semana.
La evolución del color durante el almacenamiento de las rodajas de manzana, con o sin
pretratamientos, e irradiadas a las dosis más alta (20*25 min), puede ser apreciada en el
diagrama de cromaticidad CIE x*y mostrado en la figura 4'15. La superficie de las manzanas
irradiadas se tornó más pardeada con el aumento del tiempo de almacenamiento. Por el
contrario, se observaron cambios menores en el color de las muestras escaldadas e irradiadas
y el control. Se puede verificar en este diagrama que la mayor estabilidad en el color durante
el almacenamiento se observó en las muestras irradiadas que previamente fueron inmersas en
la solución de ácido ascórbico*cloruro de calcio.
8$#$7$7������������������
En el grupo de trabajo se han realizado numerosos estudios tendientes a formular
productos de fruta funcionales enriquecidos con calcio (Anino y col., 2006; Salvatori y col.,
2007; González*Fésler, 2008). En base a estudios realizados sobre biodisponibilidad de
calcio, se ha demostrado que este nutriente, cuando es incorporado en el tejido de manzana
puede ser rápidamente absorbido, y por lo tanto, estas frutas enriquecidas pueden ser un buen
vehículo para el aporte de calcio en la dieta (González*Fésler, 2008).
Si se piensa en desarrollar un producto funcional con características similares a las de la
fruta fresca, además de la conservación por refrigeración, se debe considerar la aplicación de
una tecnología adicional que asegure la inocuidad microbiológica durante el almacenamiento
sin producir efectos adversos sobre los atributos de calidad. Con este propósito, se quiso
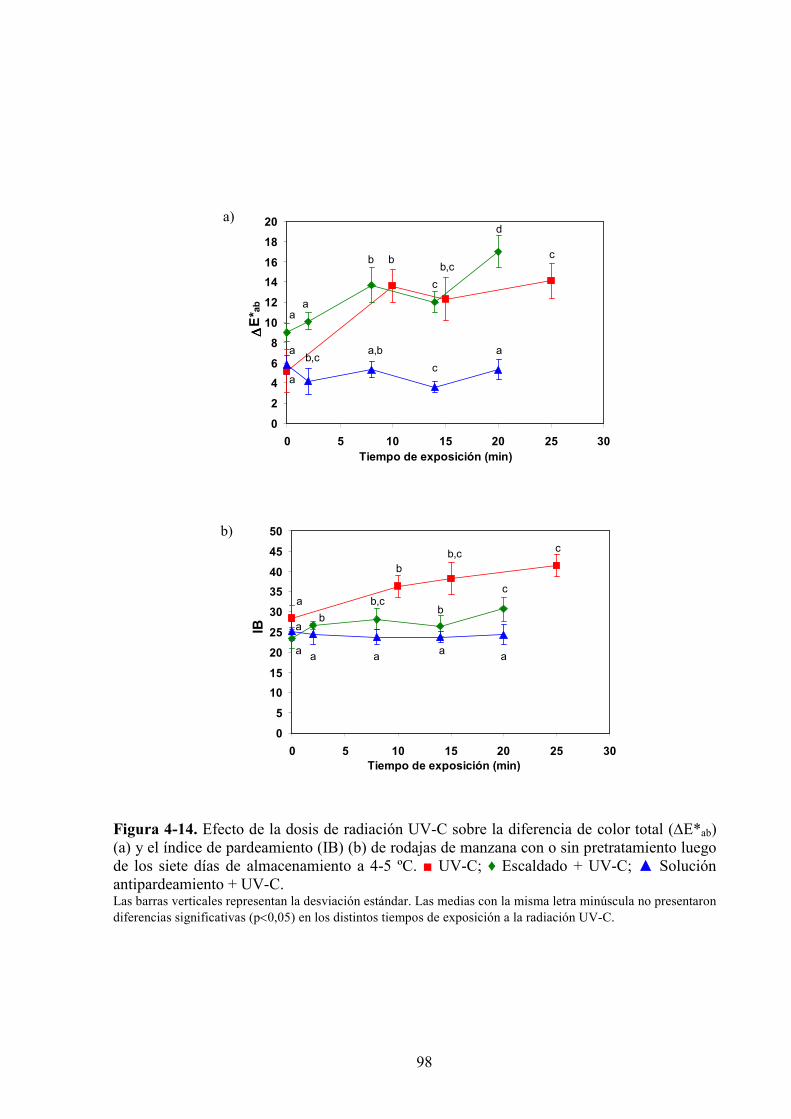
98
�
�
0
1
-
!�
!�
!0
!1
!-
��
� !� ! �� � "�
�����������#������$����
�� ��+� ��
a
b b,c
c
a a
b
c
d
a
b,c a,b
c
a
a)
�
!�
!
��
�
"�
"
0�
0
�
� !� ! �� � "�
�����������#������$����
%&
a
b
b,c c
a
a
b
b,c b
c
a a a a
b)
Figura 4'14. Efecto de la dosis de radiación UV*C sobre la diferencia de color total (�E*ab) (a) y el índice de pardeamiento (IB) (b) de rodajas de manzana con o sin pretratamiento luego de los siete días de almacenamiento a 4*5 ºC. ■ UV*C; ♦ Escaldado + UV*C; ▲ Solución antipardeamiento + UV*C. Las barras verticales representan la desviación estándar. Las medias con la misma letra minúscula no presentaron diferencias significativas (p<0,05) en los distintos tiempos de exposición a la radiación UV*C.

99
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
0 0,2 0,4 0,6 0,8
#
2
Figura 4'15. Evolución del color en la superficie de rodajas de manzana irradiadas con o sin pretratamientos en el diagrama de cromaticidades CIE x*y. (La flecha indica un aumento del tiempo de almacenamiento. Las manzanas fueron irradiadas utilizando la dosis más alta ensayada en cada tratamiento. Control (•), UV*C (•), escaldado + UV*C (•), solución antipardeamiento+UV*C(•).

100
evaluar la factibilidad de la aplicación de la radiación UV*C para la conservación de rodajas
de manzana enriquecidas con calcio, analizando la evolución de algunos de los atributos de
calidad durante el almacenamiento refrigerado de las muestras tratadas.
La adición de calcio se realizó mediante el proceso de impregnación a presión
atmosférica (IA). Este proceso consiste en la inmersión de las muestras durante un cierto
tiempo en soluciones formuladas adecuadamente de forma de lograr la incorporación de uno o
varios solutos de interés. En este caso el medio de impregnación fue preparado de forma de
poder introducir el calcio en la estructura, evitando o minimizando al mismo tiempo la
deshidratación del tejido (Alzamora y col., 2005; Anino y col., 2006).
La impregnación se realizó en rodajas de manzana con o sin escaldado previo. Otros
estudios demostraron que el escaldado previo a la impregnación permite incrementar la
incorporación de calcio dentro de matrices vegetales muy diversas como manzana, papa,
champiñón (Salvatori y col., 2007; Ortiz y col., 2003; Indaco y col., 2005). Estos tratamientos
térmicos producen alteraciones estructurales (hinchamiento de paredes celulares, rotura de
membranas, desintegración de paredes celulares, etc), afectando los fenómenos de transporte
de masa y dando como resultado una mayor incorporación de solutos en el interior del tejido
(Alzamora y col. 2000 b; Salvatori & Alzamora, 2000).
La cantidad de calcio incorporado luego de la aplicación de los diferentes tratamientos
se muestra en la tabla 4'2. El contenido de calcio aumentó considerablemente en las rodajas
de manzana impregnadas en comparación con el contenido presente en la fruta fresca
(control), alcanzando en aquellas muestras que habían recibido un escaldado previo a la
impregnación, cantidades mucho mayores (≈ 10 veces ) que en las sólo impregnadas.
Tabla 4'2. Contenido de calcio de rodajas de manzana fresca (control), impregnadas a presión atmosférica (IA) y escaldadas e impregnadas a presión atmosférica (E + IA).
Tratamiento Contenido de calcio (����g/g)
Control 7,4
IA 721
E + IA 2734
�

101
A continuación se describen los cambios en el color a lo largo del almacenamiento
refrigerado de las rodajas de manzanas enriquecidas con calcio e irradiadas con luz UV*C.
�
!���������������4��������������������
En la figura 4'16 y 4'17 se muestran los valores promedio de los parámetros de color
L*, a*, b*, h y C obtenidos a lo largo del almacenamiento de las muestras impregnadas con
calcio y posteriormente expuestas a la luz UV*C durante diferentes tiempos (2, 8, 14 y 20
minutos). Estos valores se expresaron como diferencias con respecto a los obtenidos
inicialmente en las rodajas frescas sin tratamiento. En todos los parámetros analizados se
presentó interacción significativa entre la dosis aplicada y el tiempo de almacenamiento (p <
0,0001).
Al día 0 del almacenamiento, las rodajas de manzana tratadas con IA (con o sin
irradiación) no presentaron variaciones importantes en los valores de los parámetros de color
con respecto a los valores iniciales de las muestras frescas que no recibieron tratamientos. Sin
embargo, después de los tres días del almacenamiento, los cambios en todos los parámetros de
color de estas muestras fueron marcados y mucho mayores a los que presentaron las muestras
control. El oscurecimiento (mayor aL*) de estas muestras fue notorio al tercer día del
almacenamiento, siendo levemente mayor en las muestras que fueron irradiadas a dosis altas
(14 y 20 min). Luego de los siete días de almacenamiento, el decrecimiento en la luminosidad
de las muestras con IA e irradiadas fue similar al de las muestras impregnadas que no fueron
irradiadas (figura 4'16 a). Por otra parte, los valores de a* se incrementaron
significativamente en todas las muestras tratadas al tercer y séptimo día del almacenamiento,
presentando un cambio levemente mayor las muestras irradiadas a dosis altas (figura 4'16 b).
El parámetro b* presentó para las diferentes muestras una evolución similar al parámetro a*.
(figura 4'16 c). Como se mencionó anteriormente, un decrecimiento de L* y un aumento de
a* simultáneos se relacionarían con un incremento del pardeamiento superficial.
En concordancia con las variaciones que presentaron a* y b*, las funciones ángulo de
tono y croma disminuyeron y aumentaron, respectivamente, durante el almacenamiento de
las muestras tratadas. Al día 7 de almacenamiento, las muestras impregnadas e irradiadas
mostraron un decrecimiento levemente mayor de h* que las no irradiadas, no encontrándose
diferencias significativas entre las diferentes dosis aplicadas (figura 4'17).
De acuerdo a lo analizado, el tratamiento de IA indujo cambios importantes en el
color de las muestras durante el almacenamiento, asociados con un incremento del

102
5
0
5
10
15
20
25
0 3 7
���������������������������
������
Control IAIA + UVC 2 min IA + UVC 8 minIA + UVC 14 min IA + UVC 20 min
aA
aAaA
aA aA
bB
bB,C,D bB,C
bC,D bD
cBbB cB
bB
bB
aAaA
0
2
4
6
8
10
12
14
16
18
20
0 3 7���������������������������
�
��
Control IAIA + UVC 2 min IA + UVC 8 minIA + UVC 14 min IA + UVC 20 min
aA aA aA
aA aA
bB
bB,CbB
bC bC
aA
cBbB cB bB
cC
0
5
10
15
20
25
0 3 7���������������������������
������
Control IAIA + UVC 2 min IA + UVC 8 minIA + UVC 14 min IA + UVC 20 min
aA
aAaA
aAaA
bB,CbB bB
bC
bB,C
aA
cB
cB,C
AcC
bB,C
cB,C
aA
Figura 4'16. Efecto de la dosis de radiación UV*C sobre la luminosidad (Lo** L*) (a), el valor de (a**ao*) (b) y de b* (b**bo*) (c) de rodajas de manzana impregnadas a presión atmosférica (IA) y almacenadas a 4*5ºC. Para cada tratamiento, las barras con la misma letra minúscula no presentaron diferencias significativas a lo largo del almacenamiento (p<0,05). Para el mismo día de almacenamiento, las barras de diferentes tratamientos con la misma letra mayúscula no presentaron diferencias significativas (p<0,05).
a)
b)
c)
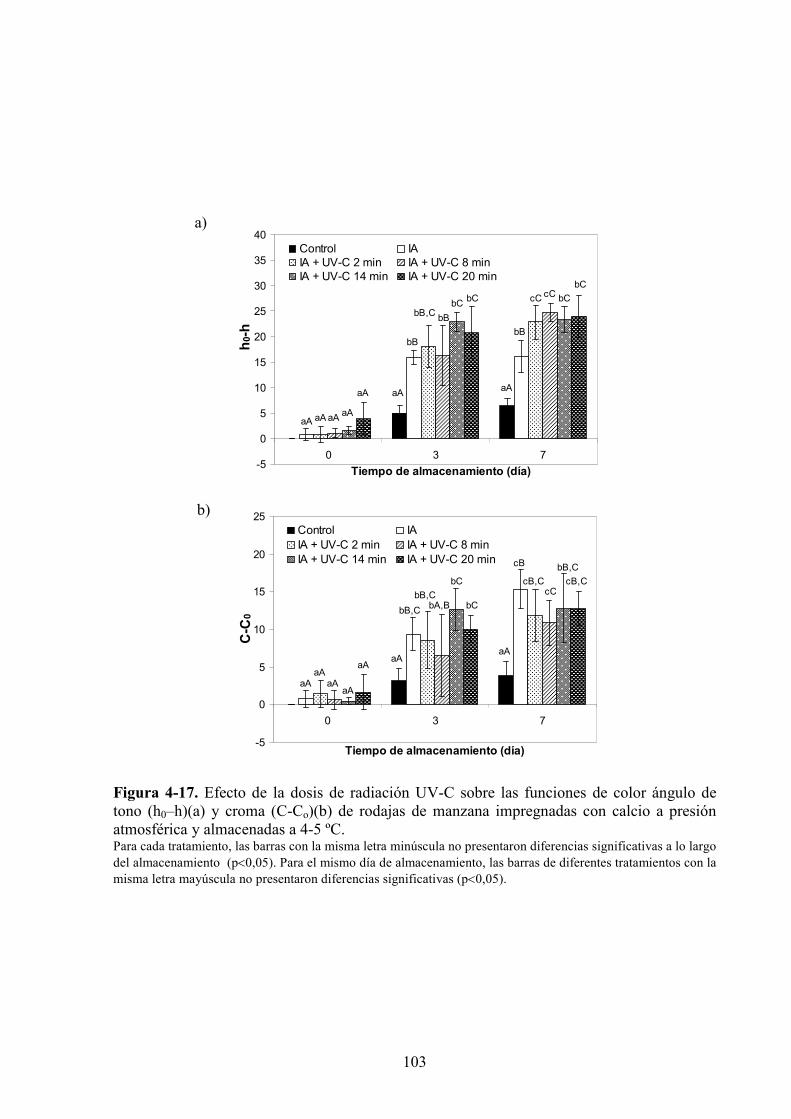
103
�
5
0
5
10
15
20
25
30
35
40
0 3 7
���������������������������
����
Control IA
IA + UVC 2 min IA + UVC 8 min
IA + UVC 14 min IA + UVC 20 min
aA aAaA
aA aA
bB
bB,C bB
bC bC
aA
bB
cCcC
bC
bC
aA
5
0
5
10
15
20
25
0 3 7
���������������������������
���
�
Control IA
IA + UVC 2 min IA + UVC 8 min
IA + UVC 14 min IA + UVC 20 min
aAaA
aA
aA
bB,C
bB,C bA,B
bC
bC
aA
cB
cB,C cC
bB,C
cB,C
aA
aA
Figura 4'17. Efecto de la dosis de radiación UV*C sobre las funciones de color ángulo de tono (h0–h)(a) y croma (C*Co)(b) de rodajas de manzana impregnadas con calcio a presión atmosférica y almacenadas a 4*5 ºC. Para cada tratamiento, las barras con la misma letra minúscula no presentaron diferencias significativas a lo largo del almacenamiento (p<0,05). Para el mismo día de almacenamiento, las barras de diferentes tratamientos con la misma letra mayúscula no presentaron diferencias significativas (p<0,05).
a)
b)

104
pardeamiento superficial, siendo levemente afectado por la radiación a dosis altas. En la
figura 4'18 se puede observar a través de imágenes fotográficas el color que presentaron las
diferentes muestras al final del almacenamiento. En algunas muestras tratadas el color
desarrollado no fue uniforme en toda la superficie, lo que se evidencia en las elevadas
desviaciones estándares obtenidas en los parámetros de color medidos.
Para intentar reducir el pardeamiento desarrollado en estas muestras se aplicó un
tratamiento previo con solución antipardeamiento de ácido ascórbico/cloruro de calcio, la
cual, como se ha demostrado en los resultados previos, resultó altamente efectiva para inhibir
el pardeamiento de rodajas de manzana expuestas a la radiación UV*C.
La evolución de los parámetros de color a lo largo del almacenamiento de las muestras
impregnadas e irradiadas que fueron inmersas en la solución antipardeamiento se muestran en
las figuras 4'19 y 4'20. En todos los parámetros se presentó interacción significativa (p <
0,01) entre el tiempo de almacenamiento y los tratamientos aplicados.
Como puede observarse en la figura 4'19 la aplicación de la solución antipardeamiento
de ácido ascórbico/cloruro de calcio redujo la variación de la luminosidad a lo largo del
almacenamiento de las muestras impregnadas que fueron o no expuestas a la luz UV*C. Al día
3 y 7 del almacenamiento, si bien las rodajas de manzana tratadas presentaron valores de L*
menores que el control (mayor cL*), está disminución fue mucho menor a las que mostraron
las muestras a las que no se les había aplicado la solución antipardeamiento. Por otro lado, los
valores de a* no presentaron variaciones significativas (p>0,05) entre las diferentes muestras
al día 0 y 3 del almacenamiento, mientras que al séptimo día solo hubo un leve incremento de
a* en las muestras con IA sin irradiar (figura 4'19 b). Los valores de b* disminuyeron
levemente en las rodajas impregnadas sin irradiar al tercer día, mientras que en las rodajas con
IA expuestas a la irradiación durante 20 minutos no presentaron variaciones significativas (p
> 0,05) a lo largo del almacenamiento (figura 4'19 c).
En los valores de croma y de ángulo de tono de las muestras tratadas no se observaron
variaciones significativas (p > 0,05) durante el almacenamiento, en concordancia con la leve
o nula variación que presentaron los parámetros a* y b* en estas muestras (figura 4'20).
La figura 4'21 compara el efecto de la aplicación de la solución antipardeamiento a las
rodajas de manzana impregnadas con calcio (con o sin irradiación) sobre las funciones de
color diferencia de color total (�E*) e índice de pardeamiento (cIB). Los valores de �E* de
las muestras impregnadas (con o sin irradiación) se redujeron notablemente cuando fueron
inmersas en la solución antipardeamiento, presentando variaciones similares al control a lo
largo del almacenamiento (figura 4'21 a). Por otro lado, los valores del índice de

105
� �
������
��Figura 4'18. Color superficial de rodajas de manzanas impregnadas con calcio a presión atmosférica e irradiadas a diferentes dosis de radiación UV*C, al séptimo día del almacenamiento a 4*5 ºC. IA: Impregnación a presión atmosférica.
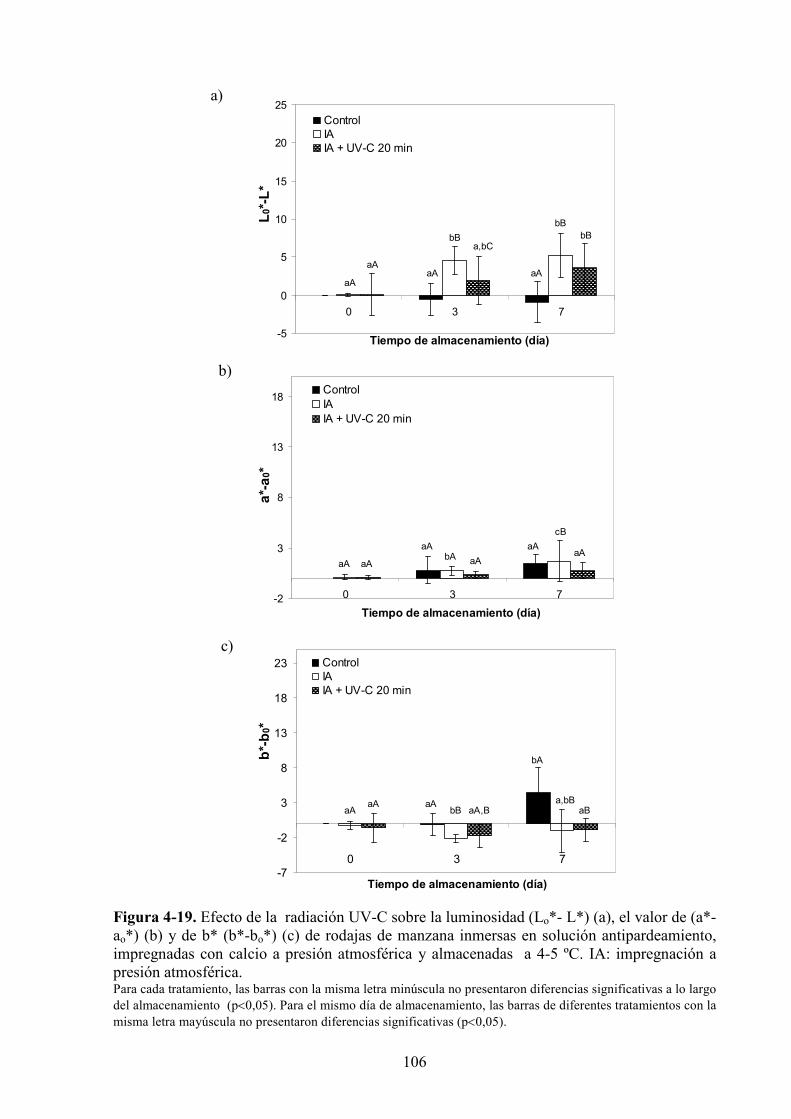
106
5
0
5
10
15
20
25
0 3 7
���������������������������
������
Control
IA
IA + UVC 20 min
aA
aAaA
bBa,bC
aA
bB
bB
2
3
8
13
18
0 3 7
���������������������������
������
Control
IA
IA + UVC 20 min
aA aA
aAbA
aA
aA
cB
aA
7
2
3
8
13
18
23
0 3 7
���������������������������
������
Control
IA
IA + UVC 20 min
aAaA aA
bB aA,B
bA
a,bBaB
Figura 4'19. Efecto de la radiación UV*C sobre la luminosidad (Lo** L*) (a), el valor de (a**ao*) (b) y de b* (b**bo*) (c) de rodajas de manzana inmersas en solución antipardeamiento, impregnadas con calcio a presión atmosférica y almacenadas a 4*5 ºC. IA: impregnación a presión atmosférica. Para cada tratamiento, las barras con la misma letra minúscula no presentaron diferencias significativas a lo largo del almacenamiento (p<0,05). Para el mismo día de almacenamiento, las barras de diferentes tratamientos con la misma letra mayúscula no presentaron diferencias significativas (p<0,05).
a)
b)
c)
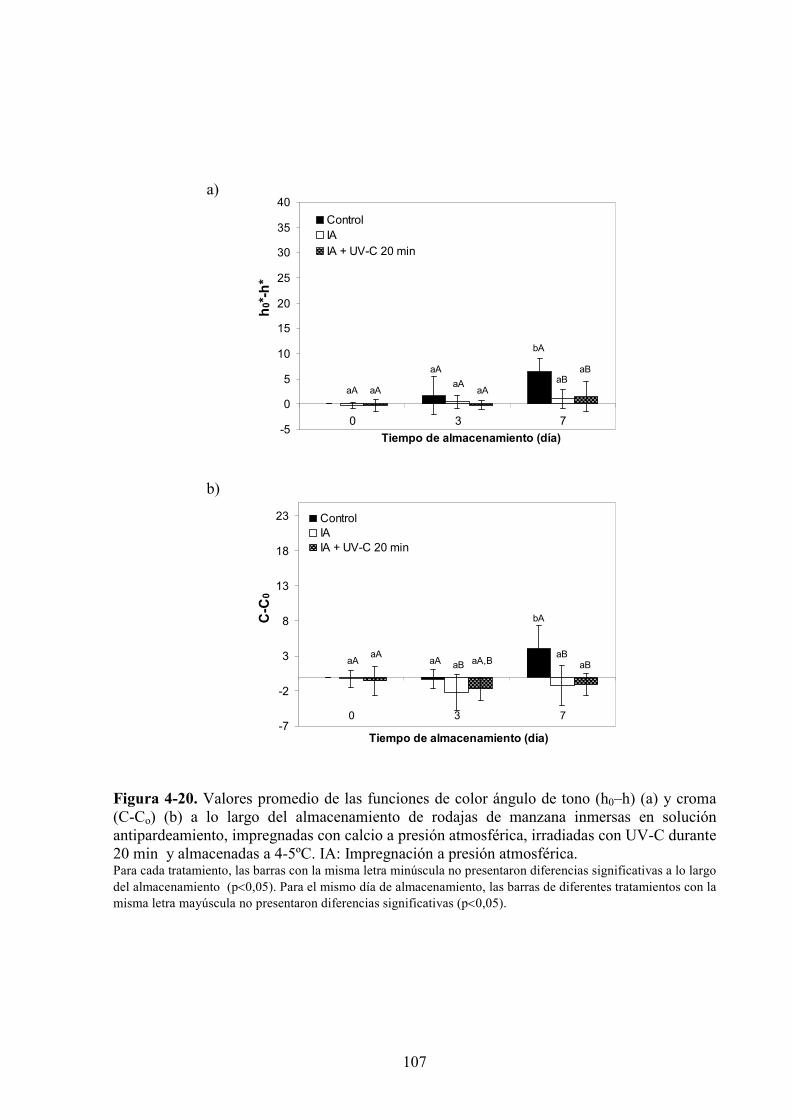
107
5
0
5
10
15
20
25
30
35
40
0 3 7
���������������������������
������
Control
IA
IA + UVC 20 min
aA aA
aA
aAaA
bA
aBaB
7
2
3
8
13
18
23
0 3 7
���������������������������
���
�
Control
IA
IA + UVC 20 min
aAaA aA aB aA,B
bA
aBaB
Figura 4'20. Valores promedio de las funciones de color ángulo de tono (h0–h) (a) y croma (C*Co) (b) a lo largo del almacenamiento de rodajas de manzana inmersas en solución antipardeamiento, impregnadas con calcio a presión atmosférica, irradiadas con UV*C durante 20 min y almacenadas a 4*5ºC. IA: Impregnación a presión atmosférica. Para cada tratamiento, las barras con la misma letra minúscula no presentaron diferencias significativas a lo largo del almacenamiento (p<0,05). Para el mismo día de almacenamiento, las barras de diferentes tratamientos con la misma letra mayúscula no presentaron diferencias significativas (p<0,05).
a)
b)
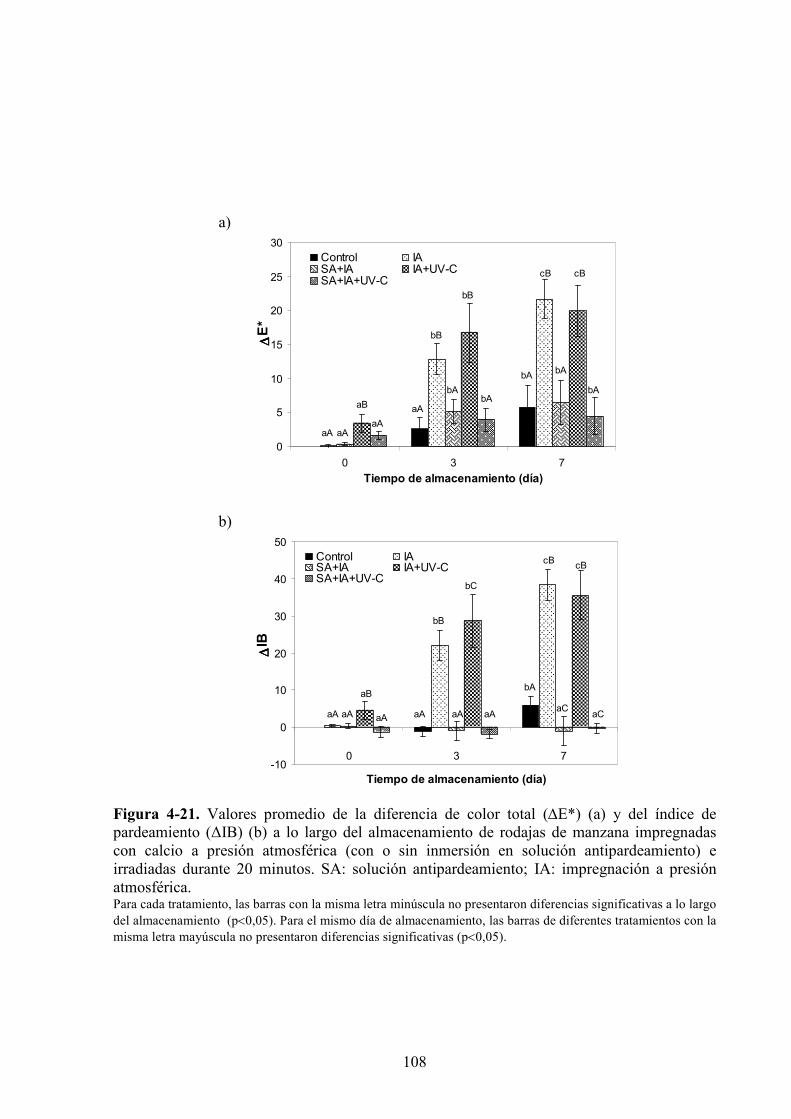
108
0
5
10
15
20
25
30
0 3 7
���������������������������
�� ��+�
Control IASA+IA IA+UVCSA+IA+UVC
aA aA
aB
aA
aA
bB
bA
bB
bA
bA
cB
bA
cB
bA
10
0
10
20
30
40
50
0 3 7
���������������������������
�� ��%&
Control IASA+IA IA+UVCSA+IA+UVC
aA aA
aB
aA aA
bB
bC
aA aA
bA
cB
aC
cB
aC
Figura 4'21. Valores promedio de la diferencia de color total (�E*) (a) y del índice de pardeamiento (cIB) (b) a lo largo del almacenamiento de rodajas de manzana impregnadas con calcio a presión atmosférica (con o sin inmersión en solución antipardeamiento) e irradiadas durante 20 minutos. SA: solución antipardeamiento; IA: impregnación a presión atmosférica. Para cada tratamiento, las barras con la misma letra minúscula no presentaron diferencias significativas a lo largo del almacenamiento (p<0,05). Para el mismo día de almacenamiento, las barras de diferentes tratamientos con la misma letra mayúscula no presentaron diferencias significativas (p<0,05).
a)
b)

109
pardeamiento también se redujeron marcadamente por efecto de la aplicación de la solución
de ácido ascórbico/cloruro de calcio en comparación con las muestras tratadas que no fueron
inmersas en esta solución (figura 4'21 b).
La efectividad de la solución ácido ascórbico/cloruro de calcio para inhibir el
pardeamiento superficial de las muestras impregnadas con calcio (con o sin irradiación) puede
observarse visualmente en la figura 4'22.
"��������<����������������4��������������������
�
La evolución de los parámetros de color durante el almacenamiento de las rodajas de
manzana impregnadas con calcio e irradiadas (2, 8, 14 y 20 minutos) que recibieron un
escaldado previo a la impregnación se muestran en las figuras 4'23 y 4'24. En todos los
parámetros evaluados la interacción entre los factores analizados (tiempo de almacenamiento
y tratamiento) fue significativa (p<0,05).
El escaldado previo a la impregnación produjo una disminución en la luminosidad de
las muestras al día 0, permaneciendo en general sin variaciones en las diferentes muestras
tratadas hasta el final del almacenamiento (figura 4'23 a). Por otro lado, las muestras
impregnadas con calcio (con o sin irradiación) que fueron escaldadas presentaron variaciones
menores en los parámetros a* y b* que las muestras no escaldadas. Los valores de a* sólo
presentaron un leve incremento a lo largo del almacenamiento en las muestras tratadas que
fueron irradiadas a las dosis UV*C más altas (8, 14 y 20 minutos) (figura 4'23 b). En cuanto
a los valores de b*, las muestras tratadas no presentaron cambios significativos, excepto en
aquellas que fueron irradiadas durante 20 minutos, que presentaron una disminución de b* al
día 7 (figura 4'23 c).
El ángulo de tono disminuyó al tercer día en las muestras escaldadas e impregnadas en
mayor proporción que el control, y en general, no se presentaron variaciones significativas
por efecto de las dosis de irradiación aplicadas. Al séptimo día, los valores de h* sólo
aumentaron levemente en las rodajas que fueron irradiadas a la dosis más alta (figura 4'24a).
La evolución de la función croma en las muestras analizadas fue similar a la que presentó el
parámetro b* (figura 4'24 b).
De acuerdo a lo analizado y como puede observarse en la figura 4'25, la aplicación del
escaldado previo a la impregnación no solo favoreció la incorporación de calcio en el interior
de la estructura sino que también tuvo un efecto inhibidor del pardeamiento superficial de las
muestras. Sin embargo, como se observa en la figura 4'25, la reducción del pardeamiento de

110
���'���������� ./�,�%/������ ./�,�%/�,�
()���������
���'���������� ./�,�%/������ ./�,�%/�,�
()���������
Figura 4'22. Color superficial al séptimo día del almacenamiento de rodajas de manzanas impregnadas con calcio a presión atmosférica (IA) con previa inmersión en solución antipardeamiento (SA) e irradiadas con luz UV*C durante 20 minutos. IA: Impregnación a presión atmosférica.

111
�
��
�
��
�
� �
� �������������� ���������
���
��
Control E+IA
E+IA+UVC 2 min E+IA+UVC 8 min
E+IA+UVC 14 min E+IA+UVC 20 min
aAaA
aA aA
aA
aBaB,C
a,bB,C aB,C
aC
aA
aBaB,C
bD aC,D
aB,C,D
aA
�
�
�
�
�
��
��
��
��
��
��
0 3 7
� �������������� ���������
���
��
������� ����
�������������� ��������������
��������������� ���������������
aA aA aA aAaA,B
aA
aA,B
bB a,bA,B
bA,B aA,B
aA aA
bBbA,B
cC
aA
�
�
��
�
��
�
� �
� �������������� ���������
���� �
��
Control E+IA
E+IA+UVC 2 min E+IA+UVC 8 min
E+IA+UVC 14 min E+IA+UVC 20 min
aA aA aA aA
aA
aBaB
aB
aB
bC
aA
aB aB
aB aB
cCaA
Figura 4'23. Efecto de la dosis de radiación UV*C sobre la luminosidad (Lo** L*) (a), el valor de (a**ao*) (b) y de b* (b**bo*) (c) de rodajas de manzana escaldadas, impregnadas con calcio a presión atmosférica y almacenadas a 4*5ºC. E: escaldado; IA: impregnación atmosférica. Para cada tratamiento, las barras con la misma letra minúscula no presentaron diferencias significativas a lo largo del almacenamiento (p<0,05). Para el mismo día de almacenamiento, las barras de diferentes tratamientos con la misma letra mayúscula no presentaron diferencias significativas (p<0,05).
a)
b)
c)

112
�
��
�
��
�
��
�
��
� �
� �������������� ���������
���
Control E+IA
E+IA+UVC 2 min E+IA+UVC 8 min
E+IA+UVC 14 min E+IA+UVC 20 min
aAaA aA
aA
aA
bB
bB
bB
bA
bB
aA
bB
bB
bA
a,bA
cC
aA
�
��
�
��
�
� � �
� �������������� ���������
��
�
Control E+IA
E+IA+UVC 2 min E+IA+UVC 8 minE+IA+UVC 14 min E+IA+UVC 20 min
aAaA
aAaA
aA
aB
aA,B
aB
aA,B
bA,B
aA
aB aB
aB
aB cBaA
Figura 4'24. Efecto de la dosis de radiación UV*C sobre las funciones de color ángulo de tono (h0–h)(a) y croma (C*Co)(b) de rodajas de manzana escaldadas, impregnadas con calcio a presión atmosférica y almacenadas a 4*5ºC. Para cada tratamiento, las barras con la misma letra minúscula no presentaron diferencias significativas a lo largo del almacenamiento (p<0,05). Para el mismo día de almacenamiento, las barras de diferentes tratamientos con la misma letra mayúscula no presentaron diferencias significativas (p<0,05).
�
a)
b)
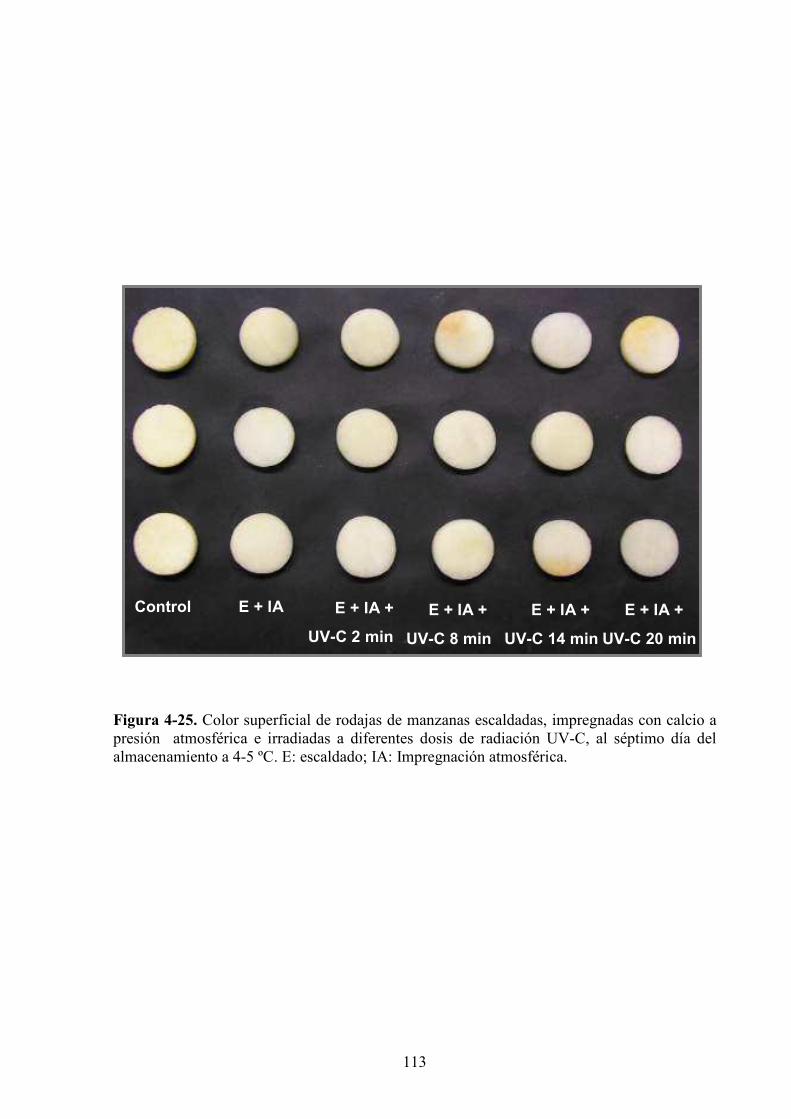
113
���'���������� +�,�%/������ +�,�%/�,
()�������
+�,�%/�,
()���-���
+�,�%/�,
()���!0���
+�,�%/�,
()��������
���'���������� +�,�%/������ +�,�%/�,
()�������
+�,�%/�,
()���-���
+�,�%/�,
()���!0���
+�,�%/�,
()��������
Figura 4'25. Color superficial de rodajas de manzanas escaldadas, impregnadas con calcio a presión atmosférica e irradiadas a diferentes dosis de radiación UV*C, al séptimo día del almacenamiento a 4*5 ºC. E: escaldado; IA: Impregnación atmosférica.

114
las muestras irradiadas a las dosis más altas fue menos efectiva que en aquellas muestras a las
que se les aplicó la solución antipardeamiento. Por otra parte, en estas muestras, al igual que
lo observado en las rodajas impregnadas e irradiadas sin escaldado, el pardeamiento
desarrollado no fue uniforme en toda la superficie de la muestra.
Para intentar reducir el pardeamiento observado al final del almacenamiento en las
muestras escaldadas, impregnadas e irradiadas a dosis altas, se evaluó también la posibilidad
de aplicar una etapa adicional de inmersión en solución antipardeamiento en estas muestras.
Los parámetros de color obtenidos se muestran en la figura 4'26. En todos los parámetros
medidos se presentó interacción significativa entre el tiempo de almacenamiento y el
tratamiento aplicado (p<0,05).
La luminosidad de las muestras escaldadas e impregnadas con calcio (con o sin
irradiación) que fueron inmersas en la solución antipardeamiento disminuyó marcadamente al
día 0 del almacenamiento, siendo la disminución mayor en comparación con las rodajas que
no fueron tratadas con esta solución. Al día 3, L* disminuyó en las muestras tratadas, siendo
el decrecimiento levemente mayor en las rodajas irradiadas. Sin embargo, al final del
almacenamiento los valores de L* disminuyeron en las rodajas sin irradiar y no presentaron
diferencias significativas con respecto a las muestras irradiadas (figura 4'26 a). Por otra
parte, no se obtuvieron variaciones significativas (p<0,01) a lo largo del almacenamiento en
los valores de a* de las muestras tratadas (figura 4'26 b). El parámetro b* disminuyó
levemente en las rodajas irradiadas al día 3 del almacenamiento, mientras que al séptimo día,
aumentó en las rodajas irradiadas y no irradiadas, siendo el incremento similar al que
presentaron las muestras control (figura 4'26 c). En cuanto a los valores de h* y C*, las
muestras tratadas no presentaron variaciones en la tonalidad a lo largo del almacenamiento
pero si en la cromaticidad. Al día 7, las muestras tratadas mostraron un aumento de C* similar
al control (figura 4'27). Este aumento de C* se relacionaría con el incremento en el
parámetro b*, lo que indicaría que las muestras presentaron una tonalidad amarillenta de
mayor grado de saturación.
En la figura 4'28 pueden observarse a través de imágenes fotográficas los cambios en el
color de las diferentes muestras al final del almacenamiento. Si bien las muestras escaldadas e
impregnadas que fueron inmersas en la solución antipardeamiento presentaron una mayor
translucencia en comparación con las rodajas que no fueron tratadas con esta solución, el
color desarrollado fue más uniforme en toda la superficie, lo que hizo que presentaran una
mejor apariencia visual.
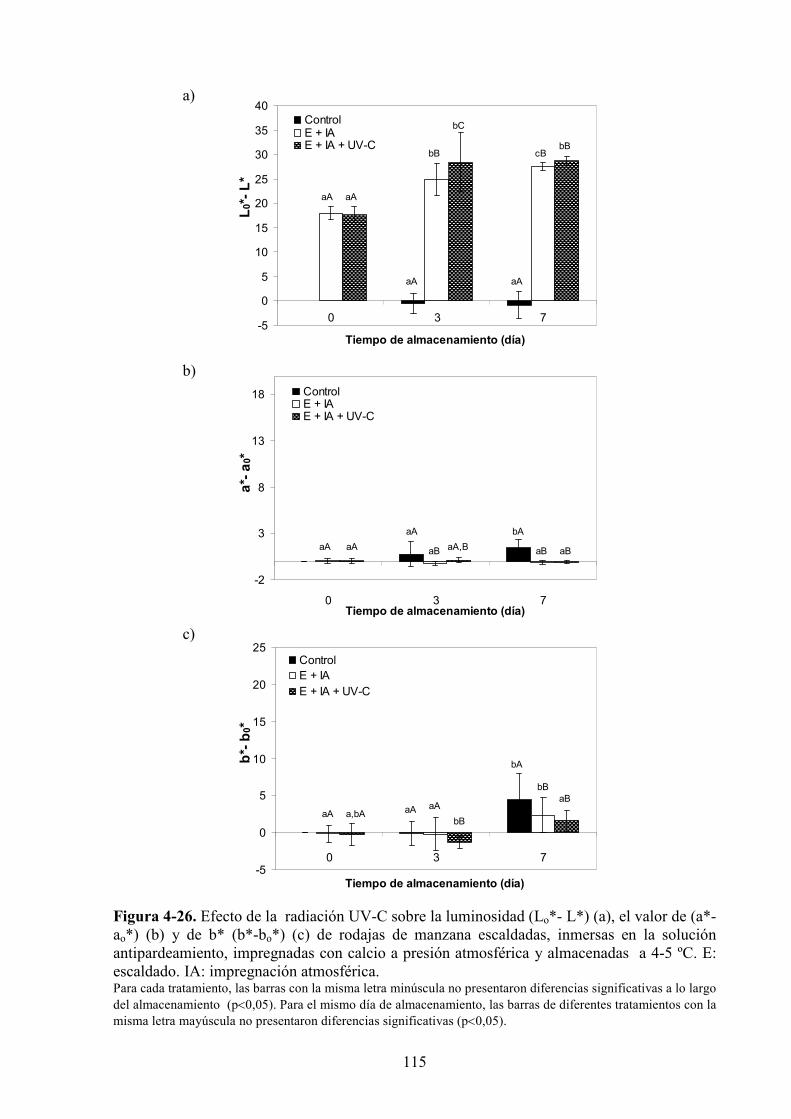
115
5
0
5
10
15
20
25
30
35
40
0 3 7
���������������������������
�������
ControlE + IAE + IA + UVC
aA aA
aA
bB
bC
aA
cBbB
2
3
8
13
18
0 3 7���������������������������
�����
��
ControlE + IAE + IA + UVC
aA aA
aA
aB aA,B
bA
aB aB
5
0
5
10
15
20
25
0 3 7
���������������������������
�����
��
Control
E + IA
E + IA + UVC
aA a,bA aA aA
bB
bA
bB
aB
Figura 4'26. Efecto de la radiación UV*C sobre la luminosidad (Lo** L*) (a), el valor de (a**ao*) (b) y de b* (b**bo*) (c) de rodajas de manzana escaldadas, inmersas en la solución antipardeamiento, impregnadas con calcio a presión atmosférica y almacenadas a 4*5 ºC. E: escaldado. IA: impregnación atmosférica. Para cada tratamiento, las barras con la misma letra minúscula no presentaron diferencias significativas a lo largo del almacenamiento (p<0,05). Para el mismo día de almacenamiento, las barras de diferentes tratamientos con la misma letra mayúscula no presentaron diferencias significativas (p<0,05).
a)
b)
c)

116
a)
5
0
5
10
15
20
25
30
35
40
0 3 7
���������������������������
������
Control
E + IA
E + IA + UVC
aAaA
aA
aAaAaBaB
bA
b)
5
0
5
10
15
20
25
0 3 7
���������������������������
�����
�
Control
E + IA
E + IA + UVC
bA
bB
bA,B
aAaA
aAaA aA
Figura 4'27. Efecto de la dosis de radiación UV*C sobre las funciones de color ángulo de tono (h0–h)(a) y croma (C*Co)(b) de rodajas de manzana escaldadas, inmersas en solución antipardeamiento e impregnadas con calcio a presión atmosférica y almacenadas a 4*5ºC. Para cada tratamiento, las barras con la misma letra minúscula no presentaron diferencias significativas a lo largo del almacenamiento (p<0,05). Para el mismo día de almacenamiento, las barras de diferentes tratamientos con la misma letra mayúscula no presentaron diferencias significativas (p<0,05).

117
���'���������� +�,�./�,�%/������+�,�./�,�%/�,
()�����������'���������� +�,�./�,�%/������
+�,�./�,�%/�,
()��������
Figura 4'28. Color superficial al séptimo día del almacenamiento de rodajas de manzanas escaldadas, inmersas en solución antipardeamiento, impregnadas con calcio a presión atmosférica e irradiadas con luz UV*C durante 20 minutos. E: escaldado. SA: solución antipardeamiento. IA: Impregnación atmosférica.

118
8$#$8$�������������������������������������������������������/������������������������������������8$#$8$#$�(�����������������
Las propiedades reológicas a altas deformaciones de las rodajas de manzana irradiadas
fueron evaluadas mediante ensayos de compresión. Las propiedades reológicas a bajas
deformaciones se determinaron a partir de ensayos oscilatorios y de fluencia*recuperación.
�"���������������������
En la figura 4'29 se muestra, a modo de ejemplo, las curvas típicas de fuerza*distancia
obtenidas en los ensayos de compresión al día 0 y séptimo del almacenamiento para las
muestras control e irradiadas con UV*C durante 25 min. Los perfiles de compresión del tejido
fresco (control) y de las muestras irradiadas presentaron un pico de ruptura marcado,
característico de alimentos duros, tanto al día 0 como al final del almacenamiento. En todas
las curvas también se presentó antes de la ruptura un pico más pequeño correspondiente al
denominado “bioyield”. Este pico se produce cuando la fuerza aplicada provoca la fractura
inicial del tejido o el movimiento de células con respecto a las células vecinas, produciendo
un notable decrecimiento en la pendiente de la curva sin llegar a la fractura visible del
material (Abbott & Harker, 2004).
Los datos obtenidos en los ensayos de compresión fueron transformados en valores de
esfuerzo o tensión real (σR) vs deformación real o de Henky (εR) según las ecuaciones 3.9 y
3.10, asumiendo que el volumen permanece constante. La respuesta de los materiales a la
compresión depende de las dimensiones del espécimen (Chu & Peleg, 1985; Canet &
Sherman, 1988). Por ello, es más ventajoso definir el valor de “esfuerzo” en lugar de “fuerza”
porque el esfuerzo caracteriza la habilidad que tiene la superficie del material para responder
a fuerzas externas, independientemente de la forma y del tamaño de la muestra (Varela y col.,
2007).
En la figura 4'30 se muestran las curvas obtenidas de σR vs εR correspondientes a
muestras control e irradiadas al inicio y al tiempo final del almacenamiento. En todas las
curvas, se observó inicialmente una relación lineal de σR vs εR, seguido por una relación no
lineal en donde la tensión siguió aumentando. Luego, al producirse el “bioyield”, la tensión
disminuyó ligeramente volviendo luego a incrementarse hasta alcanzar un valor
máximo(correspondiente a la ruptura del espécimen), a partir del cual comenzó a decaer.
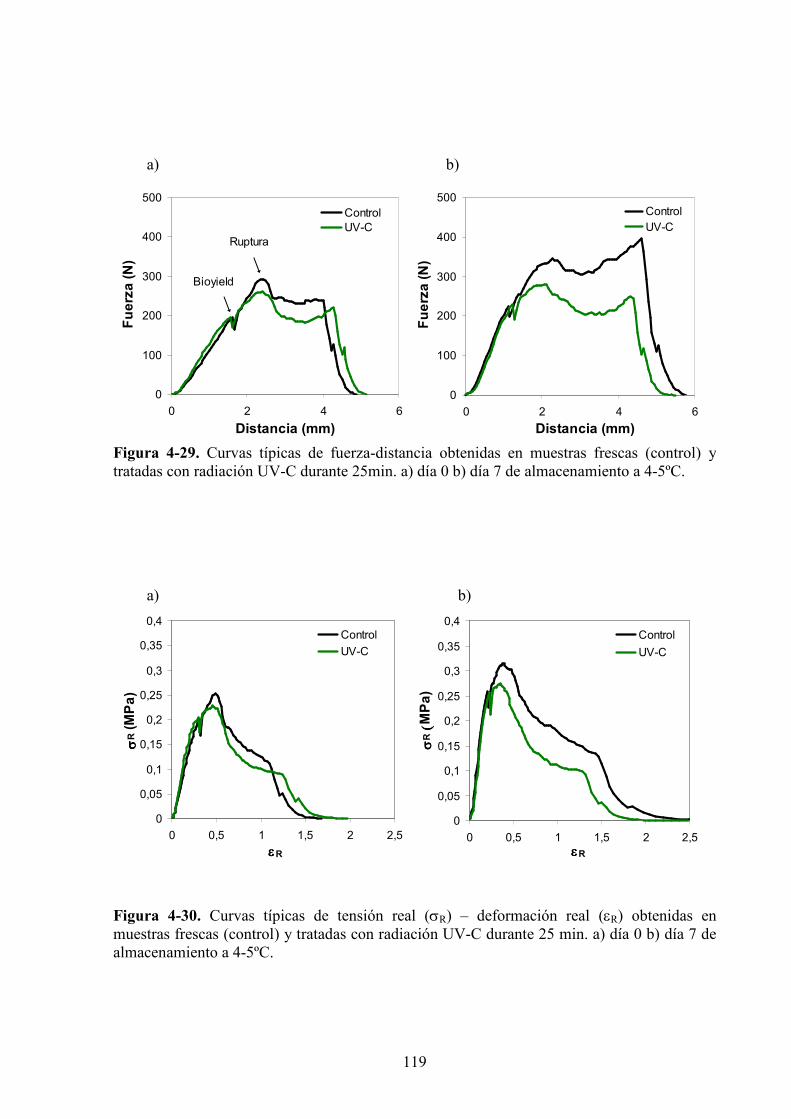
119
0
100
200
300
400
500
0 2 4 6
������������
34�'5���
Control
UVC
Bioyield
Ruptura
0
100
200
300
400
500
0 2 4 6
������������34�'5���
Control
UVC
Figura 4'29. Curvas típicas de fuerza*distancia obtenidas en muestras frescas (control) y tratadas con radiación UV*C durante 25min. a) día 0 b) día 7 de almacenamiento a 4*5ºC. �
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
0,4
0 0,5 1 1,5 2 2,5
�εεεε6
σσ σσ6��78�
Control
UVC
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
0,4
0 0,5 1 1,5 2 2,5
εεεε6�
��σσ σσ6 ( ( ( (78�
Control
UVC
a) b)
a) b)
Figura 4'30. Curvas típicas de tensión real (σR) – deformación real (εR) obtenidas en muestras frescas (control) y tratadas con radiación UV*C durante 25 min. a) día 0 b) día 7 de almacenamiento a 4*5ºC.

120
A partir de las curvas de σR vs εR se obtuvieron los valores de esfuerzo correspondientes
al “bioyield” (σRB) (Tabla 4'3) y al pico de ruptura (σR
R) (Tabla 4'4), y los valores del
módulo de deformabilidad (Ed) (Tabla 4'5). El ANOVA de 2 factores aplicado a los valores
obtenidos de estos parámetros mostró que en todos los casos se presentó interacción entre la
dosis aplicada de radiación UV*C y el tiempo de almacenamiento (p < 0,0001). Las muestras
control presentaron un incremento significativo (p < 0,0001) en los valores de σRB y σR
R a lo
largo del almacenamiento. Los valores de σRB de las rodajas de manzana irradiadas, en
general, no presentaron diferencias significativas a lo largo del tiempo analizado en las dosis
evaluadas. Por otro lado, los valores de σRR aumentaron al tercer día del almacenamiento en
las muestras irradiadas por 10 min, permaneciendo luego sin cambios hasta el final del
almacenamiento. Las muestras irradiadas a tiempos mayores (15 y 25 min) no presentaron
diferencias significativas en los valores de tensión de ruptura con el almacenamiento. El
aumento de la tensión de ruptura que presentaron las muestras control e irradiadas por 10 min
reflejarían un incremento de la dureza durante el almacenamiento.
El incremento temporario de σRR al día 3 del almacenamiento en las muestras irradiadas
durante 10 minutos podría estar asociado a una respuesta hipersensible provocada por la
irradiación UV*C. Lamikanra y col. (2005) asociaron la retención de la firmeza observada
durante el almacenamiento de melones mínimamente procesados irradiados con dosis bajas de
luz UV*C con un aumento en la concentración de poliaminas. Las poliaminas son policationes
de bajo peso molecular que están implicadas en el desarrollo de las frutas y pueden
acumularse en respuesta al estrés ambiental (Shama & Alderson, 2005). Estos compuestos
actuarían reforzando las paredes celulares mediante un mecanismo similar al del calcio,
formando uniones con las pectinas y otros polisacáridos presentes en la pared (Maharaj y col.,
1999). Por otro lado, el incremento de σRR que presentaron las rodajas de manzanas frescas
durante el almacenamiento también fue reportado por otros autores en manzanas y en tomates
cortados almacenados en refrigeración (Lana y col., 2005; Costelló Gómez, 2007).
El módulo de deformabilidad fue calculado como la pendiente en la región lineal inicial
de la curva esfuerzo real*deformación real, hasta un 10% de la deformación real. Este
parámetro es un indicador de la rigidez del tejido cuando se aplican esfuerzos pequeños
(Mohsenin & Mittal, 1977). Los valores de Ed de las muestras expuestas a la radiación UV*C
disminuyeron levemente al séptimo día de almacenamiento en las rodajas irradiadas por 10 y
15 min, y al tercer día en aquellas irradiadas durante 25 min. A diferencia de las muestras
irradiadas, las manzanas no irradiadas (control) no presentaron diferencias significativas en el
valor de Ed a lo largo del almacenamiento (Tabla 4'5). Estos resultados indicarían una leve

121
Tabla 4'3. Valores promedio de esfuerzo real en el punto del bioyield (σRB) (MPa) a lo largo
del almacenamiento a 4*5ºC de muestras de manzanas fresca (control) e irradiadas a diferentes dosis de radiación UV*C. C: control �
Tiempo de irradiación
(min)
Tiempo de almacenamiento (día)
0 3 7
0 (C) 0,20(0,01) a A 0,23(0,02) b A 0,25(0,02) c A
10 0,22(0,02) a B 0,25(0,02) b B 0,22(0,01) a B
15 0,21(0,01) a,b A,B 0,22(0,02) b A 0,20(0,02) a C
25 0,21(0,04) a A,B 0,22(0,02) a A 0,22(0,02) a B
Los resultados fueron expresados como la media seguida de la desviación estándar (entre paréntesis). Para cada tiempo de irradiación, las medias seguidas por la misma letra minúscula no presentaron diferencias significativas a lo largo del almacenamiento (p<0,05). Para el mismo día de almacenamiento, las medias de los diferentes tratamientos seguidas por la misma letra mayúscula no presentaron diferencias significativas (p<0,05).
�
Tabla 4'4. Valores promedio de esfuerzo real en el pico de ruptura (σR
R) (MPa) a lo largo del almacenamiento a 4*5ºC de muestras de manzanas fresca (control) e irradiadas a diferentes dosis de radiación UV*C. C: control
Tiempo de irradiación
(min)
Tiempo de almacenamiento (día) 0 3 7
0 (C) 0,22 (0,03) a A 0,25 (0,03) b A 0,32 (0,05) c A
10 0,24 (0,02) a A 0,29 (0,03) b B 0,28 (0,04) b B
15 0,23 (0,03) a A 0,24 (0,03) a A 0,23 (0,04) a C
25 0,24 (0,05) a A 0,25 (0,02) a A 0,27 (0,05) a B
�
Los resultados fueron expresados como la media seguida de la desviación estándar (entre paréntesis). Para cada tiempo de irradiación, las medias seguidas por la misma letra minúscula no presentaron diferencias significativas a lo largo del almacenamiento (p<0,05). Para el mismo día de almacenamiento, las medias de los diferentes tratamientos seguidas por la misma letra mayúscula no presentaron diferencias significativas (p<0,05).�

122
Tabla 4'5. Valores promedio de módulo de deformabilidad (Ed) (MPa) a lo largo del almacenamiento a 4*5ºC en muestras de manzanas fresca (control) e irradiadas a diferentes dosis de radiación UV*C. C: control �
Tiempo de irradiación
(min)
Tiempo de almacenamiento (día)
0 3 7
0 (C) 1,4 (0,3) a A 1,4 (0,2) a A 1,4 (0,3) a A
10 1,5 (0,2) a A 1,6 (0,3) a B 1,1 (0,2) b B
15 1,5 (0,3) a A 1,4 (0,2) a A 0,9 (0,2) b C
25 1,5 (0,4) a A 1,2 (0,1) b A 1,1 (0,2) b B
�Los resultados fueron expresados como la media seguida de la desviación estándar (entre paréntesis). Para cada tiempo de irradiación, las medias seguidas por la misma letra minúscula no presentaron diferencias significativas a lo largo del almacenamiento (p<0,05). Para el mismo día de almacenamiento, las medias de los diferentes tratamientos seguidas por la misma letra mayúscula no presentaron diferencias significativas (p<0,05). �
pérdida de rigidez del tejido de manzana irradiado.
Probablemente las diferencias pequeñas o no significativas encontradas en los
parámetros de compresión del tejido irradiado comparado con el tejido fresco se deban a
que, a grandes deformaciones, la conducta mecánica estaría siendo determinada por las
características del tejido en toda la rodaja. La irradiación UV*C tiene un efecto sólo
superficial en el tejido, que podría verse más reflejado en los ensayos en donde el estímulo
sea tangencial a la superficie del material, como ocurre durante ensayos oscilatorios o
rotacionales.
�
"������������������
�
En primera instancia, para determinar el rango viscoelástico lineal (RVL) de las
muestras se realizó un barrido de amplitud con control de la deformación de cizalla (BA
CDC). El barrido se llevó a cabo para deformaciones entre 0,001 % y 100 %, utilizando un
valor de frecuencia constante de 10 ϖ s*1. Este valor de frecuencia fue seleccionado teniendo
en cuenta estudios previos realizados en el grupo de investigación en donde se observó que
trabajando con valores de frecuencia más bajos existía mucha dispersión en los datos de
medición (Martínez, 2005). Como se observa en la figura 4'31, las muestras control al día 0 y
7 del almacenamiento y las irradiadas al día 0 presentaron un RVL similar (límite RVL

123
recomendado: γ = 0,01 %), mientras que las muestras irradiadas y almacenadas durante 7 días
exhibieron un RVL mayor (límite RVL recomendado: γ = 0,1%). Estos resultados coinciden
con lo reportado por Martínez (2005), quien encontró que los tejidos de frutas sometidos a
distintos tratamientos (térmico, osmótico, etc) presentaban un RVL mayor al de las muestras
frescas, y al ir aumentando la severidad del tratamiento, este RVL se iba haciendo cada vez
mayor. En los ensayos de barrido de frecuencia, para asegurarse que todas las muestras
evaluadas estuvieran dentro de los límites de linealidad, se utilizó el menor valor
recomendado de γ = 0,01 %, obtenido para las muestras frescas y las irradiadas al día 0.
El ensayo de barrido de frecuencia se llevó a cabo para valores de frecuencia entre 100
s*1 hasta 0,1 s*1, utilizando entonces la deformación definida en el barrido de amplitud de
0,01%. En la figura 4'32 se muestra el espectro mécanico de las muestras control e irradiadas
durante 20 minutos, obtenido al día 0 y séptimo de almacenamiento. En todas las muestras,
los valores del módulo de almacenamiento (G’) excedieron a los valores del módulo de
pérdida (G’’) a lo largo de todo el rango de frecuencia, indicando un predominio del
������
�����
�����
�����
�����
����
�����
����
����� ���� ��� � �� ���
γγγγ ����
� �
�!
�
Figura 4'31. Barrido de amplitud con control de la deformación de cizalla para las muestras de manzanas fresca y expuestas a la radiación UV*C durante 20 min, al día 0 y 7 del almacenamiento en refrigeración. Control día 0 (■), Control día 7 (□), irradiada 20 min día 0 (▲), irradiada 20 min día 7 ( ). Valor límite recomendado de RVL (■).

124
a)
������
�����
�����
�����
�����
����
�����
��� � �� ���ϖϖϖϖ ��"
��
� �
�!
�
b)
������
������
������
������
������
�����
� � �� ���ϖϖϖϖ ��"
��
�#�
�!
�
Figura 4'32. Barrido de frecuencia a γ = 0,01 % en función de la frecuencia angular (ϖ = 100 s*1 – 0,1 s*1) de manzanas fresca e irradiada y almacenadas en refrigeración a 4*5 ºC. a) Módulo de almacenamiento (G’); B) módulo de pérdida (G”). Control día 0 (■), Control día 7 (□), irradiada 20 min día 0 (▲), irradiada 20 min día 7 ( ).

125
comportamiento sólido. Por otro lado, las muestras tratadas presentaron valores de G’ y G’’
menores que el control fresco tanto al día 0 como al final del almacenamiento.
En la tabla 4'6 se muestran los valores de G’ obtenidos a diferentes frecuencias
angulares. El análisis estadístico aplicado mostró que no había interacción entre el tiempo de
almacenamiento y el tiempo de irradiación (p > 0,05); por lo tanto, los resultados se evaluaron
por efectos principales. En todas las muestras, los valores de G’ aumentaron con el
incremento de la frecuencia angular, siendo la pendiente (n) de las muestras irradiadas
ligeramente mayor al de las muestras frescas sólo al día 0 de almacenamiento. Para el resto de
las muestras la variación de G’ con la frecuencia fue similar. Las rodajas expuestas a la
radiación UV*C mostraron una disminución inicial de G’ (con respecto a las muestras frescas)
de un 30%, mientras que al final del almacenamiento esta disminución fue de un 75%. Las
muestras control también presentaron una disminución de G’ durante el almacenamiento, pero
el porcentaje de decrecimiento (alrededor de un 40%) fue mucho menor al de las muestras
irradiadas. La mayor disminución de G’ en las muestras irradiadas, acentuada durante el
almacenamiento, indicaría una pérdida del carácter elástico por efecto de la irradiación.
La dependencia de G” con la frecuencia angular fue diferente a la de G’. En todas la
muestras, G” disminuyó con el aumento de la frecuencia angular a bajas frecuencias (0,1*
1%), permaneció sin variación a frecuencias entre 1*10 %, y luego presentó un aumento
Tabla 4'6. Valores promedio del módulo G´ (kPa) a distintas frecuencias angulares de muestras de manzanas frescas e irradiadas y almacenadas en refrigeración. Parámetros obtenidos de la regresión lineal de Log (G’) vs Log�ϖ). �
Tiempo de almace'
namiento (día)
Tiempo de
irradiación (min)
ϖϖϖϖ�= 0,1 s'1� ϖϖϖϖ�= 1 s'1� ϖϖϖϖ�= 10 s'1� ϖϖϖϖ�= 100 s'1�
� n R2
0
367,6 (87,3)
399,1 (89,9)
449,1 (96,8)
509,4 (10,9)
0,05 0,98 (0,009) 0
20
249,6 (59,3)
282 (67)
318,4 (79,1)
382,6 (95,7)
0,06 0,98 (0,007)
0
220,8 (86,9)
245,7 (86,7)
270,5 (95,2)
308,9 (10,9)
0,05 0,99 (0,007)
7
20
90,8 (21,6)
98,3
(25,5)
112,1 (30,3)
129,4 (35,6)
0,05 0,97 (0,005)
Los resultados fueron expresados como la media seguida por la desviación estándar (entre paréntesis).

126
gradual a frecuencias más altas (10*100%). Al igual que G’, el módulo de pérdida disminuyó
en las muestras irradiadas y además, tanto en la muestra control como en la muestra tratada,
decreció con el tiempo de almacenamiento.
En general, el factor de pérdida (tan δ (G”/G’)) no fue un parámetro muy sensible para
distinguir diferencias en las propiedades viscoelásticas de las diferentes muestras (figura 4'33
y tabla 4'7). A frecuencias entre 1*100 s*1 este parámetro presentó interacción significativa
entre el tiempo de irradiación y el tiempo de almacenamiento (p<0,05), mientras que a ϖ=0,1
s*1 la interacción no fue significativa (p=0,47). Se observó un aumento leve pero significativo
(p < 0,001) en los valores de tan δ por efecto de la irradiación a frecuencias angulares entre
10*100 s*1 al inicio del almacenamiento. Al final del almacenamiento, los valores de tan δ de
las muestras irradiadas no presentaron cambios, mientras que los de las muestras control
disminuyeron levemente a frecuencias entre 1*100 s*1. Esto reflejaría un leve aumento de la
contribución elástica en las muestras control al final del almacenamiento, y una disminución
de la misma en las muestras irradiadas inmediatamente después del tratamiento y al cabo del
almacenamiento.
�
���
���
���
���
���
��� � �� ���ϖϖϖϖ ��"
��
��
��δδ δδ
�
�
Figura 4'33. Factor de pérdida (tan� δ ) para las muestras de manzana fresca e irradiadas durante 20 minutos �� almacenadas en refrigeración. Control día 0 (■), Control día 7 (□), irradiada 20 min día 0 (▲), irradiada 20 min día 7 ( ).

127
Tabla 4'7. Valores promedio del factor de pérdida (tan�δ)�a distintas frecuencias angulares de muestras de manzanas frescas e irradiadas y almacenadas en refrigeración.
Tiempo de almacenamiento
(día)
Tiempo de irradiación
(min) ϖϖϖϖ�= 0,1 s'1� ϖϖϖϖ�= 1 s'1� ϖϖϖϖ�= 10 s'1� ϖϖϖϖ�= 100 s'1�
0 0,14 (0,04) 0,11(0,02)aA 0,10 (0,01)aA 0,12 (0,01)aA
0 20 0,16 (0,02) 0,12 (0,01)aA 0,12 (0,01)aB 0,14 (0,01)aB
0 0,14 (0,02) 0,090(0,005)bA 0,090(0,004)bA 0,110(0,005)bA
7 20 0,14 (0,01) 0,12 (0,01)aB 0,12 (0,01)aB 0,14 (0,01)aB
Los resultados fueron expresados como la media seguida por la desviación estándar (entre paréntesis). En caso de interacción significativa tiempo x tratamiento, para un dado tratamiento, las medias seguidas por la misma letra minúscula no presentaron diferencias significativas durante el almacenamiento (p<0,05). Para el mismo día de almacenamiento, las medias de los diferentes tratamientos seguidas por la misma letra mayúscula no presentaron diferencias significativas (p<0,05).
Ensayos de fluencia' recuperación
�
Para determinar el esfuerzo a aplicar en los ensayos de fluencia*recuperación, se realizó
un barrido de amplitud con control del esfuerzo de cizalla (BA CEC) para determinar el rango
viscoelástico lineal. Para todas las muestras evaluadas, el límite del RVL recomendado por el
software del equipo fue de τ = 50 Pa (figura 4'34). Teniendo en cuenta estos resultados, para
realizar los ensayos rotatorios se decidió utilizar un esfuerzo de 35 Pa, valor en el cual todas
las muestras analizadas se encontraban dentro de los límites de linealidad.
En la figura 4'35 se observan las curvas promedio de deformación vs tiempo para las
muestras control e irradiadas durante 20 min al día 0 y séptimo de almacenamiento. Las
muestras irradiadas no presentaron cambios importantes en la forma de las curvas al inicio del
almacenamiento con respecto a las muestras control. Sin embargo, al final del
almacenamiento las diferencias fueron notorias tanto en la etapa de fluencia como en la de
recuperación. Las curvas de capacitancia correspondientes se muestran en la figura 4'36 para
la etapa de fluencia. Dichas curvas experimentales se ajustaron mediante el modelo de Kelvin
Voigt generalizado (constituido por un resorte en serie con dos elementos de Kelvin y un
amortiguador) con un coeficiente de correlación > 0,99 de acuerdo a la ecuación 3.12. de la
sección Materiales y Métodos.
Los parámetros del modelo obtenidos mediante el ajuste (J0, J1, J2, λ1, λ2 y ηN) se
muestran en la tabla 4'8. Los valores de capacitancia instantánea ó elástica (J0) y de las
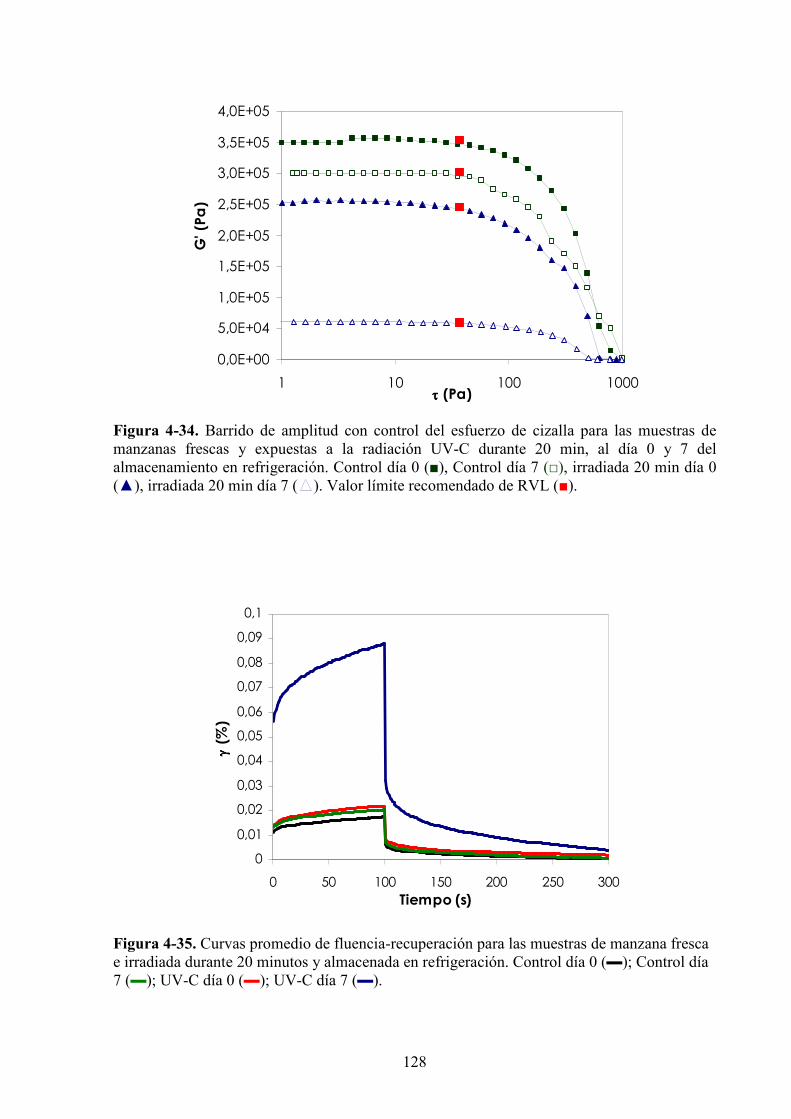
128
�������
������
������
�����
������
�����
������
�����
������
� �� ��� ����ττττ��!�
� �
�!
�
Figura 4'34. Barrido de amplitud con control del esfuerzo de cizalla para las muestras de manzanas frescas y expuestas a la radiación UV*C durante 20 min, al día 0 y 7 del almacenamiento en refrigeración. Control día 0 (■), Control día 7 (□), irradiada 20 min día 0 (▲), irradiada 20 min día 7 ( ). Valor límite recomendado de RVL (■).
�
����
����
����
����
���
����
���
����
����
���
� � ��� �� ��� �� ���
� ������"�
γ γ γ γ ��
�
Figura 4'35. Curvas promedio de fluencia*recuperación para las muestras de manzana fresca e irradiada durante 20 minutos y almacenada en refrigeración. Control día 0 (▬); Control día 7 (▬); UV*C día 0 (▬); UV*C día 7 (▬).
�
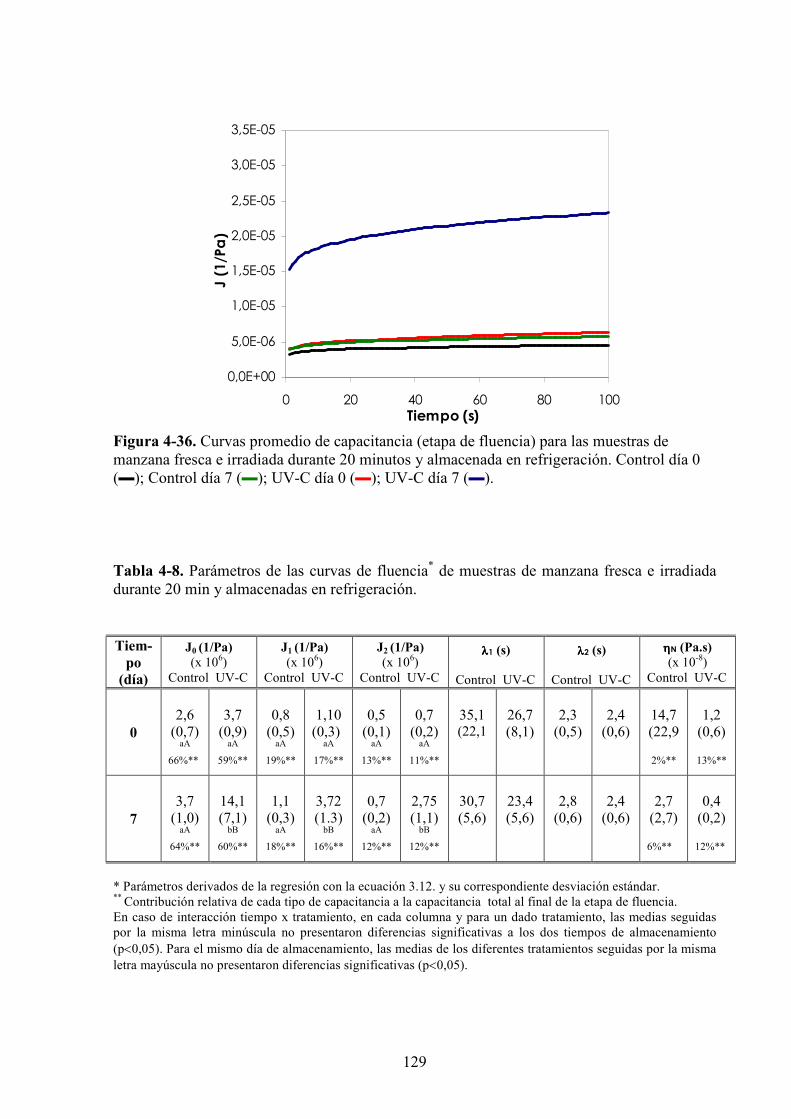
129
�
�������
�����
�����
����
�����
����
�����
����
� �� �� �� �� ���
� ������"�
$���
%!
�
�Figura 4'36. Curvas promedio de capacitancia (etapa de fluencia) para las muestras de manzana fresca e irradiada durante 20 minutos y almacenada en refrigeración. Control día 0 (▬); Control día 7 (▬); UV*C día 0 (▬); UV*C día 7 (▬). � Tabla 4'8. Parámetros de las curvas de fluencia* de muestras de manzana fresca e irradiada durante 20 min y almacenadas en refrigeración.
Tiem' po
(día)
J0 (1/Pa) (x 106)
Control UV*C
J1 (1/Pa) (x 106)
Control UV*C
J2 (1/Pa) (x 106)
Control UV*C
λλλλ��(s)��
Control UV*C
λλλλ��(s)��
Control UV*C
ηηηη&�(Pa.s)�(x 10*8)
Control UV*C�
0
2,6
(0,7) aA
66%**
3,7
(0,9) aA
59%**
0,8
(0,5) aA
19%**
1,10
(0,3) aA
17%**
0,5
(0,1) aA
13%**
0,7
(0,2) aA
11%**
35,1 (22,1
26,7 (8,1)
2,3
(0,5)
2,4
(0,6)
14,7 (22,9
2%**
1,2
(0,6)
13%**
7
3,7
(1,0) aA
64%**
14,1 (7,1)
bB
60%**
1,1
(0,3) aA
18%**
3,72 (1.3)
bB
16%**
0,7
(0,2) aA
12%**
2,75 (1,1)
bB
12%**
30,7 (5,6)
23,4 (5,6)
2,8
(0,6)
2,4
(0,6)
2,7
(2,7)
6%**
0,4
(0,2)
12%**
* Parámetros derivados de la regresión con la ecuación 3.12. y su correspondiente desviación estándar. ** Contribución relativa de cada tipo de capacitancia a la capacitancia total al final de la etapa de fluencia. En caso de interacción tiempo x tratamiento, en cada columna y para un dado tratamiento, las medias seguidas por la misma letra minúscula no presentaron diferencias significativas a los dos tiempos de almacenamiento (p<0,05). Para el mismo día de almacenamiento, las medias de los diferentes tratamientos seguidas por la misma letra mayúscula no presentaron diferencias significativas (p<0,05).

130
capacitancias elásticas de retardo ó viscoelásticas (J1 y J2) presentaron interacción
significativa entre el tratamiento y el tiempo del almacenamiento (p < 0,05), mientras que en
los valores de los tiempos de retardo (λ1 y λ2) y de los coeficientes de viscosidad (ηN)
asociados al flujo newtoniano la interacción no fue significativa (p > 0,1)
A pesar de que los ensayos se realizaron con un número considerable de replicados, los
valores medios de los parámetros de las curvas de capacitancia presentaron valores de
desviación estándar grandes, especialmente en el caso de las muestras frescas. Estos errores
no se alejan de los que normalmente se obtienen en las mediciones de propiedades de fluencia
de tejidos frutihortícolas. Esta gran variabilidad puede atribuirse a varias causas, entre otras
anisotropía de los tejidos vegetales, heterogeneidad en las propiedades reológicas en células
aparentemente homogéneas de un mismo tejido, edad de las células, aspectos climáticos y
prácticas agronómicas y madurez en la cosecha (Mittal y Mohsenin, 1987; Pitt, 1992;
Alzamora y col., 2008).
Las capacitancias J0, J1 Y J2 de la muestra control no variaron significativamente con el
almacenamiento. Por el contrario, la capacitancia instantánea de las rodajas irradiadas se
incrementó significativamente (p < 0,0001) durante el almacenamiento. Este incremento se
relacionaría con una disminución en el módulo elástico (E0=1/J0). Las capacitancias elásticas
de retardo ó viscoelásticas (J1 y J2) de dichas muestras también sufrieron un incremento al
final del almacenamiento.
El aumento de las capacitancias instantánea y de retardo en las muestras irradiadas
almacenadas sería simultáneo con una disminución en el coeficiente de viscosidad (ηN) del
flujo estacionario, lo que denotaría un aumento en la fluidez del tejido (tabla 4'8). En el caso
de las muestras control los elevados errores asociados no permiten concluir acerca del
comportamiento de este parámetro a lo largo del almacenamiento.
Los tiempos de retardo de los dos elementos de Voigt (λ1 y λ2) no presentaron
diferencias significativas entre las muestras control y las irradiadas, lo que indicaría que la
velocidad de la respuesta de los elementos estructurales asociados no se vió afectada ni por
efecto de la irradiación ni por el almacenamiento (tabla 4'8). Además, los tiempos de retardo
de los dos elementos de Voigt difirieron aproximadamente en un orden de magnitud en todas
las muestras. Como estos parámetros representan el tiempo requerido para que la deformación
de los elementos estructurales asociados con el comportamiento viscoelástico alcancen el 63
% de la máxima deformación, el menor tiempo de retardo perteneciente al segundo elemento
de Kelvin refleja la conducta viscoelástica de los tejidos a tiempos relativamente cortos,
siendo el responsable de una mayor estabilidad estructural. (Jackman & Stanley, 1995 b). Por

131
lo tanto, para ambos tejidos, tratados y frescos, los elementos estructurales asociados con la
segunda unidad de Kelvin alcanzarían el equilibrio más rápidamente que aquellos elementos
pertenecientes a la primera unidad de Kelvin.
Las muestras expuestas a la luz UV*C presentaron un incremento de la capacitancia
total al final de la fase de fluencia, con respecto a las muestras frescas, de alrededor de un
60% inmediatamente después del tratamiento (día 0) y de un 500% al final del
almacenamiento.
La contribución relativa de cada capacitancia a la capacitancia total para las diferentes
muestras se encontró dentro de los siguientes rangos: 59*66 % para la capacitancia elástica
instantánea (J0), 16*19 % para la capacitancia viscoelástica que respondió a menor velocidad
(J1), 11*13 % para la capacitancia viscoelástica que respondió a mayor velocidad (J2) y 2*12
% para la inversa del flujo viscoso en estado estacionario (1/ηN). Esta última contribución,
que se denomina plasticidad (% P), es la relación entre la deformación no recuperable o
permamente y la deformación total (J) calculada a los 100 s. Se puede concluir que, en todos
los casos la mayor contribución estuvo dada por la capacitancia J0 (tabla 4'8).
Los ensayos realizados a bajas deformaciones, a diferencia de los ensayos de
compresión, fueron más sensibles para detectar diferencias en las propiedades reológicas de
las muestras irradiadas. Sin embargo, como la radiación UV*C tiene baja penetración, los
cambios encontrados en la viscoelasticidad de las muestras irradiadas se darían
fundamentalmente en las regiones superficiales de las muestras.
8$#$8$,$�(�������������������������������+��
�"��������������������
El comportamiento de compresión para las rodajas de manzana que fueron sometidas a
un escaldado previo a la irradiación puede observarse en la figura 4'37. A diferencia de las
muestras frescas control, los perfiles de las curvas fuerza*distancia para las rodajas escaldadas
con o sin irradiación no presentaron “bioyield” y el incremento de la fuerza fue más gradual
hasta llegar a la fractura del material. Este comportamiento estaría indicando una pérdida de
dureza en la fruta, necesitándose una menor fuerza para obtener la misma deformación.
En la figura 4'38 se muestran las curvas típicas de esfuerzo real vs deformación real
obtenidas a partir de las curvas de fuerza*distancia de las muestras control, escaldadas y
escaldadas e irradiadas durante 20 minutos para el día 0 y 7 de almacenamiento. Como puede
observarse en esta figura, las curvas de las rodajas escaldadas con o sin irradiación
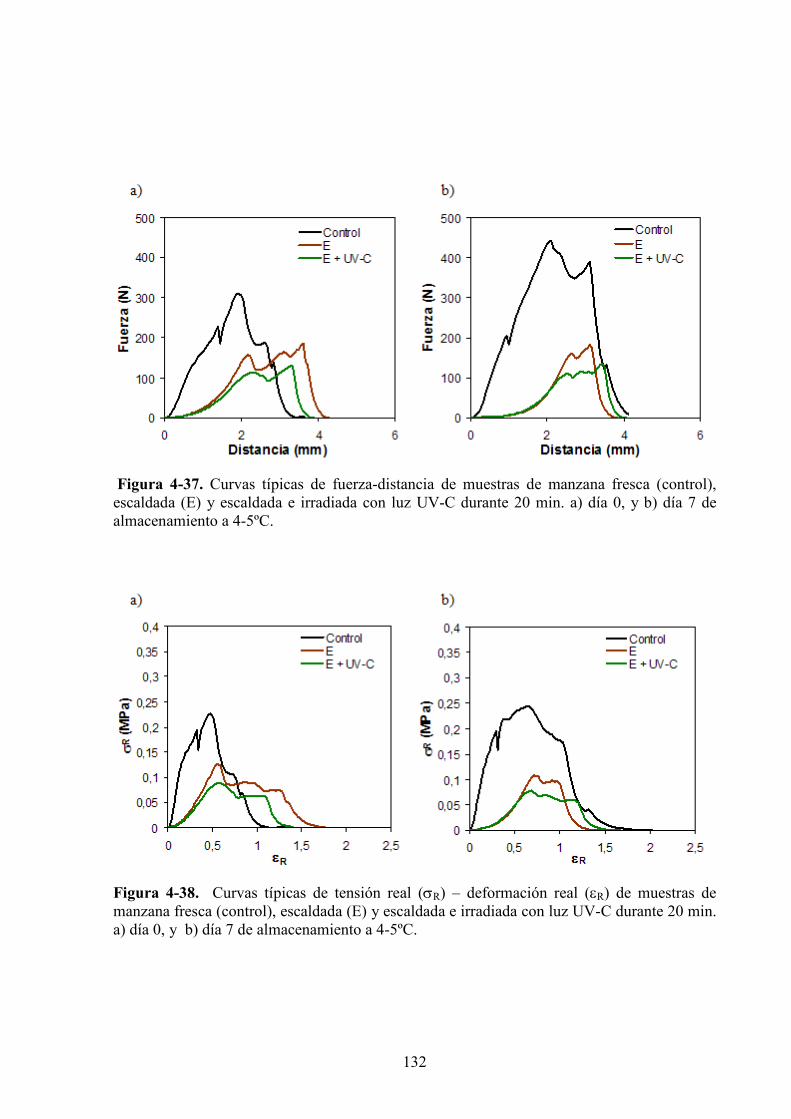
132
Figura 4'37. Curvas típicas de fuerza*distancia de muestras de manzana fresca (control), escaldada (E) y escaldada e irradiada con luz UV*C durante 20 min. a) día 0, y b) día 7 de almacenamiento a 4*5ºC.
Figura 4'38. Curvas típicas de tensión real (σR) – deformación real (εR) de muestras de manzana fresca (control), escaldada (E) y escaldada e irradiada con luz UV*C durante 20 min. a) día 0, y b) día 7 de almacenamiento a 4*5ºC. �

133
presentaron una pendiente inicial y esfuerzos de ruptura marcadamente menores que el
control. Al final del almacenamiento, los perfiles de estas muestras mostraron picos de ruptura
menos marcados y una leve disminución en el valor de la pendiente inicial. Esto estaría
indicando que las matrices se vuelven más deformables, no sólo luego de los tratamientos,
sino a lo largo del almacenamiento.
En la tabla 4'9 y 4'10 se muestran los valores de esfuerzo de ruptura y del módulo de
deformabilidad respectivamente, obtenidos de las curvas de σR vs εR de las muestras control,
escaldadas y escaldadas e irradiadas a diferentes dosis UV*C. El análisis estadístico aplicado a
estos parámetros mostró interacción entre el tratamiento aplicado y el tiempo de
almacenamiento (p< 0,0001). Los valores de σRR de las muestras control aumentaron a lo
largo del almacenamiento, mientras que en las muestras escaldadas estos valores
disminuyeron y fueron significativamente menores (p < 0,0001) que los del control. Las
muestras escaldadas e irradiadas, si bien no exhibieron diferencias importantes a lo largo del
almacenamiento, presentaron valores de esfuerzo de ruptura menores que las escaldadas sin
irradiar, especialmente cuando se aplicaron dosis mayores de radiación UV*C (tabla 4'9).
En lo que respecta al módulo de deformabilidad, las muestras previamente escaldadas,
con o sin irradiación posterior, presentaron valores promedio de Ed marcadamente menores
que el control fresco, y estos valores disminuyeron con el almacenamiento (tabla 4'10). En
general no se observó un efecto importante de la irradiación, en las dosis empleadas, en el
valor del módulo de deformabilidad de las muestras tratadas térmicamente. Esto reflejaría una
importante pérdida de rigidez de la fruta provocada por el tratamiento térmico.
"������������������
�
Se realizó un barrido de amplitud con control de la deformación de cizalla para
determinar el rango viscoelástico lineal de las rodajas de manzana frescas y las sometidas a
los tratamientos de escaldado e irradiación. El RVL de las muestras se incrementó por efecto
de los tratamientos y del almacenamiento. En la figura 4'39 se muestra, a modo de ejemplo,
el barrido de amplitud obtenido para las muestras frescas al día 0 y para las escaldadas e
irradiadas al día 7 del almacenamiento. Estas muestras fueron las que presentaron el menor y
mayor RVL respectivamente. En los ensayos de frecuencia, para que todas las muestras se
encontraran dentro de los límites de linealidad, se utilizó el límite de RVL recomendado para
las muestras frescas (γ = 0,01%).
El espectro mecánico de las muestras control, escaldadas y escaldadas e irradiadas

134
Tabla 4'9. Valores promedio de esfuerzo real en el pico de ruptura (σRR) (MPa) a lo largo del
almacenamiento a 4*5 ºC de muestras de manzana fresca (control), escaldada y escaldada e irradiada a diferentes dosis de radiación UV*C. C: control, E: escaldado. �
Tiempo de irradiación
(min)
Tiempo de almacenamiento (día)
0 3 7
0 (C) 0,25 (0,04)a A 0,32 (0,03)b A 0,33 (0,03)b A
0 (E) 0,13 (0,01)a B 0,11 (0,01)b B 0,10 (0,02)b B
2 0,11 (0,01)a C 0,11 (0,01)a B 0,09 (0,02)b B,C
8 0,09 (0,01)a D 0,10 (0,01)a B,C 0,09 (0,01)a B,C
14 0,09 (0,01)a D 0,09 (0,01)a C,D 0,09 (0,01)a B,C
20 0,09 (0,01)a D 0,08 (0,01)a D 0,08(0,01)a C
Los resultados fueron expresados como la media seguida de la desviación estándar (entre paréntesis). Para cada tiempo de irradiación, las medias seguidas por la misma letra minúscula no presentaron diferencias significativas a lo largo del almacenamiento (p<0,05). Para el mismo día de almacenamiento, las medias de los diferentes tratamientos seguidas por la misma letra mayúscula no presentaron diferencias significativas (p<0,05).
Tabla 4'10. Valores promedio de módulo de deformabilidad (Ed) a lo largo del almacenamiento a 4*5 ºC de muestras de manzana fresca (control), escaldada y escaldada e irradiada a diferentes dosis de radiación UV*C. C: control, E: escaldado. �
Tiempo de irradiación
(min)
Tiempo de almacenamiento (día)
0 3 7
0 (C) 1,1 (0,2)a A 1,2 (0,2)b A 0,9 (0,2)c A
0 (E) 0,09 (0,02)a B 0,04 (0,01)b B 0,020 (0,006)c B
2 0,05 (0,01)a C 0,04 (0,01)b B 0,03 (0,01)c C
8 0,06 (0,01)a C,D 0,04 (0,01)b B 0,028 (0,005)c C
14 0,07 (0,02)a D 0,04 (0,01)b B 0,03 (0,01)c C
20 0,07 (0,02)a D 0,034 (0,007)b B 0,018 (0,003)c B
Los resultados fueron expresados como la media seguida de la desviación estándar (entre paréntesis). Para cada tiempo de irradiación, las medias seguidas por la misma letra minúscula no presentaron diferencias significativas a lo largo del almacenamiento (p<0,05). Para el mismo día de almacenamiento, las medias de los diferentes tratamientos seguidas por la misma letra mayúscula no presentaron diferencias significativas (p<0,05).

135
������
�����
�����
�����
�����
�����
�����
�����
�����
����� ���� ��� � �� ���
γ γ γ γ ���
� �
�!
�
Figura 4'39. Barrido de amplitud con control de la deformación de cizalla obtenido para las muestras de manzana frescas al día 0 (■) y escaldadas e irradiadas 20 minutos al día 7 del almacenamiento (▲). Valor límite recomendado de RVL: (■) (0,01% muestras frescas y 0,1% muestras escaldadas e irradiadas).
durante 20 minutos, al inicio y final del almacenamiento, se muestra en la figura 4'40. Todas
las muestras analizadas presentaron un predominio del comportamiento sólido, evidenciado
por valores de G’ mayores a los de G”. Por otra parte, los módulos de almacenamiento y de
pérdida de las muestras tratadas térmicamente, con o sin exposición a la radiación UV*C,
fueron menores a los del control, tanto al día 0 como al séptimo día de almacenamiento.
Los valores de G’ obtenidos a diferentes frecuencias angulares (0,1; 1; 10 y 100 s*1) y
los parámetros de la regresión lineal del log G’ vs log ϖ se observan en la tabla 4'11. Todos
los parámetros analizados presentaron interacción significativa entre el tratamiento aplicado y
el tiempo de almacenamiento (p< 0,0001). Todas las muestras presentaron un pequeño
incremento del modulo elástico G´ con la frecuencia angular, siendo la pendiente n
ligeramente mayor en las muestras tratadas. El control mostró un descenso en G’ de alrededor
de un 30% después de los siete días de almacenamiento. En las rodajas escaldadas (con o sin
irradiación) el módulo de pérdida disminuyó marcadamente (alrededor de un 90% en
comparación con las muestras frescas) al inicio del almacenamiento, y permaneció luego sin
cambios hasta el final del almacenamiento. No se presentaron variaciones significativas entre
las muestras escaldadas por efecto de la irradiación. Estos resultados reflejarían una
disminución importante en la rigidez del tejido provocada por el tratamiento térmico previo a
la exposición a luz UV*C.

136
a)
b)
0,E+00
1,E+05
2,E+05
3,E+05
4,E+05
5,E+05
6,E+05
7,E+05
0,1 1 10 100ϖϖϖϖ ��"�
�
� �
�!
�
������
������
������
������
������
�����
�����
�����
��� � �� ���ϖϖϖϖ ��"
��
�#�
�!
�
Figura 4'40. Barrido de frecuencia a γ = 0,01 % en función de la frecuencia angular (ϖ = 100 s*1 – 0,1 s*1), de muestras de manzana fresca, escaldada (E), y escaldada e irradiada 20 min, y almacenadas en refrigeración a 4*5 ºC. a) Módulo de almacenamiento (G’); b) módulo de pérdida (G”). Control día 0 (■), Control día 7 (□), E día 0 (■), E día 7 (□), E + UV*C día 0 (▲), E + UV*C día 7 ( ).

137
Tabla 4'11. Valores promedio del módulo G´ (kPa) a distintas frecuencias angulares de las muestras de manzana control (C), escaldada (E) y escaldada e irradiada durante 20 min y almacenadas en refrigeración. Parámetros obtenidos de la regresión lineal del Log (G’) vs Log (ϖ). �
Tiempo (día)
Tratamiento ϖϖϖϖ�= 0,1 s'1� ϖϖϖϖ�= 1 s'1� ϖϖϖϖ�= 10 s'1� ϖϖϖϖ�= 100 s'1��
n R2
C
341(57) aA
411(65)
aA
448(71)
aA
510(82)
aA
0,052 aA 0,99 (0,005)
E
31,5(7,3) aB
38,6(6,8)
aB
43,2(7,5)
aB
50,2(8,9)
aB
0,060 aB 0,99 (0,004)
0
E + UV'C
26,56(11,5) aB
28,1(2,7)
aB
29(3,1)
aB
32,6(3,5)
aB
0,044 aC 0,99 (0,002)
C
203(69) bA
278(9,3)
bA
309(100)
bA
350(119)
bA
0,054 aA 0,99 (0,005)
E
25,7(6,7) aB
30,3(7,9)
aB
34,2(8,8)
aB
38,7(9,9)
aB
0,059 aB 0,99 (0,003)
7
E + UV'C
17,8(5,4) aB
20,7(6,3)
aB
23,5 (7,4)
aB
27,3(8,8)
aB
0,064 bC 0,99 (0,002)
Los resultados fueron expresados como la media seguida de la desviación estándar (entre paréntesis). En cada columna y para un dado tratamiento, las medias seguidas por la misma letra minúscula no presentaron diferencias significativas a los dos tiempos de almacenamiento (p<0,05). En cada columna y para un mismo día de almacenamiento, las medias de los diferentes tratamientos seguidas por la misma letra mayúscula no presentaron diferencias significativas (p<0,05).
La dependencia del módulo G´´ con la frecuencia angular para las muestras de manzana
fresca fue similar a la descripta en los ensayos de irradiación sin pretratamiento, mostrando
valores relativamente constantes a frecuencias entre 1 s*1 y 10 s*1, pendientes negativas a
frecuencias entre 0,1*1 s*1 y positivas a frecuencias entre 10*100 s*1.
En la figura 4'41 y tabla 4'12 se muestran los valores promedio del factor de pérdida a
diferentes frecuencias angulares. A frecuencias angulares de 1, 10 y 100 s*1, estos valores
mostraron una interacción significativa entre el tratamiento y el tiempo de almacenamiento
(p< 0,01), mientras que a frecuencias bajas de 0,1 s*1 la interacción no fue significativa
(p>0,05). Las diferencias en la tan δ entre las muestras tratadas y el control fueron leves y no
presentaron la misma tendencia a lo largo del rango de frecuencia. A frecuencias de 0,1 s*1,
los valores de tan δ de las muestras tratadas y el control no presentaron diferencias
significativas en los tiempos del almacenamiento analizados. A frecuencias altas (10 y 100 s*
1), el parámetro de las rodajas escaldadas sin irradiar mostró un aumento leve al día 0 con
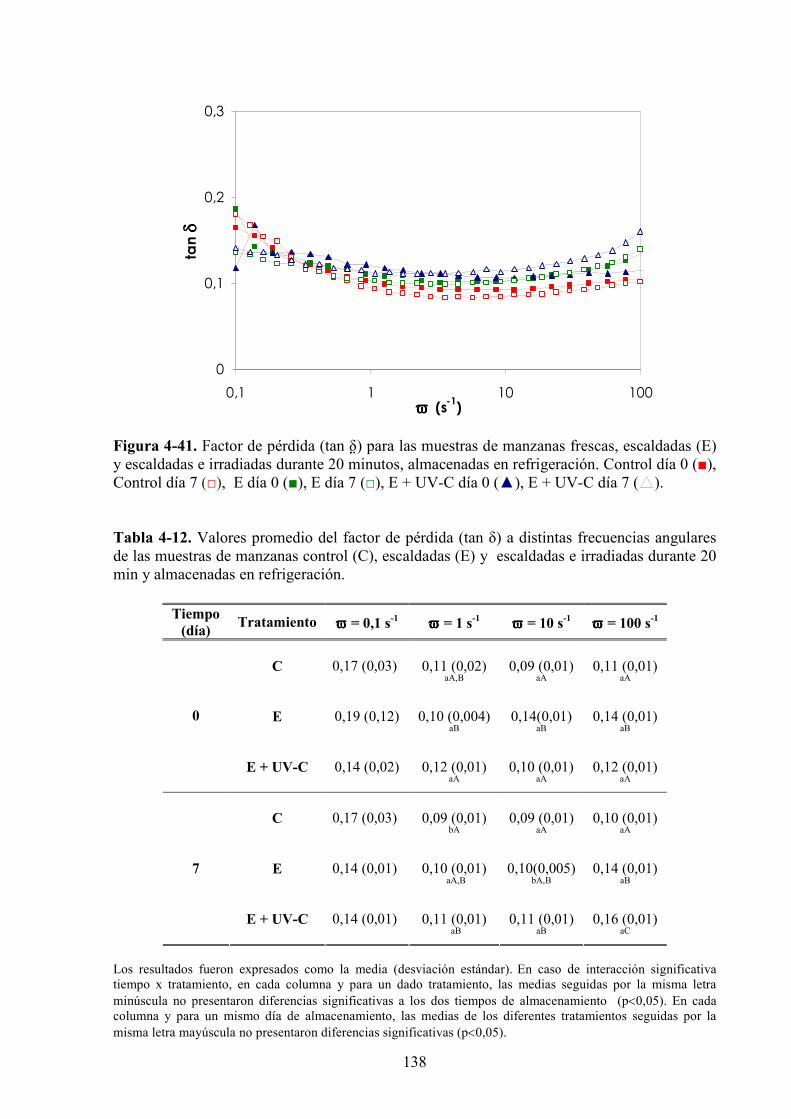
138
�
���
���
���
��� � �� ���ϖϖϖϖ ��"
��
��� δδ δδ
�
Figura 4'41. Factor de pérdida (tan δ) para las muestras de manzanas frescas, escaldadas (E) y escaldadas e irradiadas durante 20 minutos, almacenadas en refrigeración. Control día 0 (■), Control día 7 (□), E día 0 (■), E día 7 (□), E + UV*C día 0 (▲), E + UV*C día 7 ( ). Tabla 4'12. Valores promedio del factor de pérdida (tan δ) a distintas frecuencias angulares de las muestras de manzanas control (C), escaldadas (E) y escaldadas e irradiadas durante 20 min y almacenadas en refrigeración. �
Tiempo (día)
Tratamiento ϖϖϖϖ�= 0,1 s'1� ϖϖϖϖ�= 1 s'1� ϖϖϖϖ�= 10 s'1� ϖϖϖϖ�= 100 s'1�
C 0,17 (0,03)
0,11 (0,02) aA,B
0,09 (0,01)
aA
0,11 (0,01)
aA
E
0,19 (0,12)
0,10 (0,004)
aB
0,14(0,01)
aB
0,14 (0,01)
aB 0
E + UV'C
0,14 (0,02)
0,12 (0,01)
aA
0,10 (0,01)
aA
0,12 (0,01)
aA
C 0,17 (0,03)
0,09 (0,01) bA
0,09 (0,01)
aA
0,10 (0,01)
aA
E 0,14 (0,01)
0,10 (0,01) aA,B
0,10(0,005)
bA,B
0,14 (0,01)
aB 7
E + UV'C 0,14 (0,01)
0,11 (0,01) aB
0,11 (0,01)
aB
0,16 (0,01)
aC
Los resultados fueron expresados como la media (desviación estándar). En caso de interacción significativa tiempo x tratamiento, en cada columna y para un dado tratamiento, las medias seguidas por la misma letra minúscula no presentaron diferencias significativas a los dos tiempos de almacenamiento (p<0,05). En cada columna y para un mismo día de almacenamiento, las medias de los diferentes tratamientos seguidas por la misma letra mayúscula no presentaron diferencias significativas (p<0,05).

139
respecto al control, mientras que al día 7 el aumento fue gradual en las muestras escaldadas y
escaldadas e irradiadas respectivamente. Este aumento reflejaría una disminución de la
contribución de la componente elástica.
Ensayos de fluencia'recuperación
��
Al contrario de lo ocurrido en el BA CDC, cuando se hizo el barrido de amplitud con
control del esfuerzo de cizalla (BA CEC), las muestras tratadas presentaron un RVL menor
que las muestras frescas (figura 4'42). Las rodajas que presentaron menor RVL fueron las
escaldadas e irradiadas al séptimo día del almacenamiento, siendo el valor de esfuerzo límite
de τ = 50 Pa. Se decidió utilizar el mismo valor de esfuerzo (τ = 35 Pa) elegido en los ensayos
previos para las muestras irradiadas sin pretratamiento.
�
�������
������
������
�����
������
�����
������
�����
������
� �� ��� ����ττττ��!�
� �
�!
�
�Figura 4'42. Barrido de amplitud con control del esfuerzo de cizalla obtenido para las muestras de manzanas frescas al día 0 (■) y escaldadas e irradiadas 20 minutos al día 7 de almacenamiento (▲). Valor límite recomendado de RVL (■) (100 Pa muestras frescas y 50 Pa muestras escaldadas e irradiadas).
En las figura 4'43 y figura 4'44 se presentan las curvas experimentales promedio de
deformación y de capacitancia de los ensayos de fluencia*recuperación, respectivamente. El
ajuste de los datos experimentales mediante el modelo de Kelvin Voigt generalizado con 2
elementos de Kelvin resultó en coeficientes de correlación > 0,99. Los parámetros obtenidos

140
�
���
���
���
���
��
� � ��� �� ��� �� ���
� ������"�
γγ γγ���
�
Figura 4'43. Curvas promedio de fluencia*recuperación para las muestras de manzanas frescas, escaldadas (E) y escaldadas e irradiadas durante 20 minutos, almacenadas en refrigeración. Control día 0 (▬); Control día 7 (▬); E día 0 (▬); E día 7 (▬); E + UV*C día 0 (▬); E + UV*C día 7 (▬).
������
����
����
����
����
�����
�����
� �� �� �� �� ���
� ������"�
$���
%!
�
Figura 4'44. Curvas promedio de capacitancia (etapa de fluencia) para las muestras de manzanas frescas, escaldadas (E) y escaldadas e irradiadas durante 20 minutos, almacenadas en refrigeración. Control día 0 (▬); Control día 7 (▬); E día 0 (▬); E día 7 (▬); E + UV*C día 0 (▬); E + UV*C día 7 (▬).

141
se muestran en la tabla 4'13. Las capacitancias J0 y J2 presentaron interacción significativa
(p=0,027 y p=0,03 respectivamente) entre el tratamiento aplicado y el tiempo de
almacenamiento, mientras que en la capacitancia J1, los tiempos de retardo (λ1 y λ2) y ηN no
hubo interacción (p>0,05).
Tabla 4'13. Parámetros de las curvas de fluencia* para las rodajas de manzana fresca, escaldada y escaldada e irradiada 20 min y almacenadas en refrigeración.
Tiempo (día)
Tratamiento J0 (1/Pa) (x 106)
J1 (1/Pa) (x 106)
J2 (1/Pa) (x 106)�
λλλλ��(s) �
λλλλ��(s)��
ηηηη&�(Pa.s) (x 10'8)
Control
2,2 (0,3)
aA
49%
1,1(0,6)
24%**
0,5(0,09)
aA
11%**
31,7(25,7)
2,8(1,9)
1,4(0,6)
16%**
E
25,6(5,3)
aB
58%**
7,8 (1,5)
18%**
4,3(2,4)
aB
10%**
20,5(0,8)
2,4(0,1)
0,15(0,04)
15%**
0
E + UV'C
41,1(11,7)
aC
55%**
13,3(4,8)
18%**
7,7(3,1)
aC
10%**
20,6(1,4)
2,7(0,7)
0,08(0,03)
16%**
Control
3,4(0,9)
aA
61%**
0,8(0,4)
14%**
0,5(0,2)
aA
10%**
22,9(5,2)
2,4(0,5)
1,1(0,4)
15%**
E
34,0(9,8)
aB
57%**
10,2(5,9)
17%**
7,5(2,7)
bB
12%**
20,9(0,9)
2,4(0,1)
0,12(0,04)
14%**
7
E + UV'C
60,3(21,3)
bC
61%**
16,1(6,9)
16%**
12,7(6,1)
bC
13%**
22,1(3,4)
2,5(0,3)
0,10(0,01)
10%**
* Parámetros derivados de la regresión con la ecuación 3.12. y su correspondiente desviación estándar entre paréntesis. ** Contribución relativa de cada tipo de capacitancia a la capacitancia total al final de la etapa de fluencia. En caso de interacción significativa tiempo x tratamiento, en cada columna y para un dado tratamiento, las medias seguidas por la misma letra minúscula no presentaron diferencias significativas a los dos tiempos de almacenamiento (p<0,05). En cada columna y para un mismo día de almacenamiento, las medias de los diferentes tratamientos seguidas por la misma letra mayúscula no presentaron diferencias significativas (p<0,05).
A diferencia de los espectros mecánicos, los ensayos de fluencia mostraron diferencias
en las propiedades viscoelásticas de las rodajas de manzana escaldadas expuestas o no a la luz
UV*C. Las capacitancias instantánea (J0) y viscoelásticas (J1 y J2) de las muestras escaldadas
se incrementaron marcadamente (principalmente J0) al día 0 del almacenamiento en

142
comparación con las rodajas frescas, siendo mayor el aumento en las muestras irradiadas. La
capacitancia viscoelástica J1 no sufrió cambios durante el almacenamiento de la muestras. La
capacitancia viscoelástica J2 se incrementó levemente en las muestras escaldadas expuestas o
no a la irradiación, mientras que J0 se incrementó sólo en las muestras escaldadas e irradiadas.
Los tiempos de retardo λ1 y λ2 no presentaron diferencias significativas (p>0,3) ni por
efecto del tratamiento ni por el almacenamiento. Al igual que en las muestras analizadas en
los ensayos de irradiación sin pretratamiento, los valores de λ1 y λ2 difirieron
aproximadamente en un orden de magnitud.
Las rodajas de manzana escaldadas (con o sin irradiación), en comparación con las
muestras frescas, presentaron una disminución significativa al día 0 de almacenamiento en los
valores de la viscosidad en estado estacionario (ηN), reflejando un aumento en la fluidez del
tejido. No se observaron diferencias significativas (p>0,05) entre los valores de ηN para las
muestras escaldadas y sometidas a la radiación UV*C y las que no fueron expuestas a la
radiación. Tanto las muestras control como las tratadas no presentaron cambios significativos
en ηN después de transcurridos los siete días de almacenamiento.
El incremento de la capacitancia total de las muestras tratadas al final de la fase de
fluencia con respecto al tejido fresco fue muy marcado, no sólo inmediatamente después de
los tratamientos, sino fundamentalmente al final del almacenamiento, siendo para el día 0 de
aproximadamente 900 % para las muestras escaldadas y de 1500 % para las escaldadas e
irradiadas, y para el día 7, de 1200% y de 2000%, respectivamente. Las contribuciones
relativas de cada capacitancia a la capacitancia total fueron en general similares en todas las
muestras y estuvieron dentro de los siguientes rangos: J0 (49*61%), J1 (14*24%), J2 (10*13%)
y 1/ηN (10*16%) (tabla 4'13).
De acuerdo a lo expuesto, el pretratamiento de escaldado indujo cambios notables en las
propiedades de compresión y viscoelásticas del tejido de manzana, que se vieron
incrementados en menor proporción por la posterior exposición a la luz UV*C.
8$#$8$7$�(����������������������������+��������������������������������"�������������������
Las curvas típicas de fuerza*distancia y de esfuerzo real* deformación real que
presentaron las rodajas de manzana irradiadas previamente inmersas en la solución
antipardeamiento de ácido ascórbico/cloruro de calcio se muestran en las figura 4'45 y 4'46

143
0
100
200
300
400
500
0 2 4 6
������������
34�'5���
Control
SA
SA + UVC
0
100
200
300
400
500
0 2 4 6
������������34�'5���
Control
SA
SA + UVC
Figura 4'45. Curvas típicas de fuerza*distancia obtenidas en muestras de manzana frescas (control), inmersas en solución antipardeamiento (SA) e inmersas en solución antipardeamiento e irradiadas con luz UV*C durante 20 min. a) día 0, y b) día 7 de almacenamiento a 4*5 ºC.
a) b)
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
0,4
0 0,5 1 1,5 2 2,5εεεε6
σσ σσ6��7
8�
ControlSASA + UVC
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
0 0,5 1 1,5 2 2,5εεεε6
σσ σσ6��7
8�
ControlSASA + UVC
Figura 4'46. Curvas típicas de tensión real (σR) – deformación real (εR) de muestras de manzanas frescas (control), inmersas en solución antipardeamiento (SA e inmersas en solución antipardeamiento e irradiadas con luz UV*C durante 20 min. a) día 0, y b) día 7 de almacenamiento a 4*5 ºC.
a) b)

144
respectivamente. Las muestras control y las inmersas en la solución antipardeamiento con o
sin irradiación posterior presentaron perfiles similares con un pico pequeño correspondiente al
“bioyield” y picos de ruptura marcados característicos de alimentos duros, tanto al día 0 como
al día séptimo de almacenamiento.
Los valores de σRB del control aumentaron levemente a lo largo del almacenamiento,
mientras que en las rodajas inmersas en la solución antipardeamiento con o sin irradiación,
no cambiaron o disminuyeron ligeramente en las muestras irradiadas a altas dosis (tabla 4'
14). En cuanto a los valores de esfuerzo en el pico de ruptura, las muestras inmersas en la
solución de ácido ascórbico/cloruro de calcio con o sin irradiación presentaron valores
levemente inferiores al de las muestras control sólo al final del tiempo de almacenamiento
(tabla 4'15).
Como puede observarse en la tabla 4'16, en general, no se presentaron diferencias
apreciables en los valores de Ed entre las muestras control y las inmersas en la solución
antipardeamiento sin irradiar. Sin embargo, los valores de Ed de las muestras inmersas en la
solución antipardeamiento y expuestas a la luz UV*C durante 8 y 20 min fueron menores que
los de las muestras no irradiadas y los del control. En general esta tendencia se presentó a lo
largo de todo el almacenamiento.
Tabla 4'14. Valores promedio de esfuerzo real en el punto de “bioyield” (σR
B) (MPa) a lo largo del almacenamiento a 4*5 ºC de muestras de manzanas frescas (control), inmersas en solución antipardeamiento, inmersas en solución antipardeamiento e irradiadas a diferentes dosis de radiación UV*C. C: control, SA: solución antipardeamiento �
Tiempo de irradiación
(min)
Tiempo de almacenamiento (día)
0 3 7
0 (C) 0,20(0,01)a A 0,21(0,01)b A 0,21(0,01)bA
0 (SA) 0,20(0,01)a A 0,20(0,01)a B 0,20(0,01)a B
8 0,20(0,01)a A 0,20(0,01)a B 0,20(0,01)a B
20 0,20(0,01)a A 0,20(0,01)a B 0,19(0,01)bC
Los resultados fueron expresados como la media seguida de la desviación estándar (entre paréntesis). Para cada tiempo de irradiación, las medias seguidas por la misma letra minúscula no presentaron diferencias significativas a lo largo del almacenamiento (p<0,05). Para el mismo día de almacenamiento, las medias de los diferentes tratamientos seguidas por la misma letra mayúscula no presentaron diferencias significativas (p<0,05). �

145
Tabla 4'15. Valores promedio de esfuerzo real en el pico de ruptura (σRR) (MPa) a lo largo
del almacenamiento a 4*5 ºC de muestras de manzanas frescas (control), inmersas en solución antipardeamiento, e inmersas en solución antipardeamiento e irradiadas a diferentes dosis de radiación UV*C. C: control, SA: solución antipardeamiento. �
Tiempo de irradiación
(min)
Tiempo de almacenamiento (día)
0 3 7
0 (C) 0,30(0,03)a A 0,31(0,02)a A 0,32(0,05)a A
0 (SA) 0,30(0,04)a A 0,29(0,03)a,b A 0,27(0,03)b B
8 0,28(0,03)a A 0,29(0,03)a A 0,27(0,04)a B
20 0,28(0,03)a,b A 0,29(0,03)a A 0,26(0,03)b B
Los resultados fueron expresados como la media seguida de la desviación estándar (entre paréntesis). Para cada tiempo de irradiación, las medias seguidas por la misma letra minúscula no presentaron diferencias significativas a lo largo del almacenamiento (p<0,05). Para el mismo día de almacenamiento, las medias de los diferentes tratamientos seguidas por la misma letra mayúscula no presentaron diferencias significativas (p<0,05). �
Tabla 4'16. Valores promedio de módulo de deformabilidad (Ed) (MPa) a lo largo del almacenamiento a 4*5 ºC de muestras de manzanas frescas (control), inmersas en solución antipardeamiento, inmersas en solución antipardeamiento e irradiadas a diferentes dosis de radiación UV*C. C: control, SA: solución antipardeamiento. �
Tiempo de irradiación
(min)
Tiempo de almacenamiento (día)
0 3 7
0 (C) 1,8 (0,2)a A 1,9 (0,2)a A 1,6 (0,3)b A
0 (SA) 1,7 (0,3)a A 1,7 (0,2)a B 1,5 (0,3)a A
8 1,5 (0,2)a B 1,6 (0,2)a B 1,2 (0,2)b B
20 1,4 (0,2)a B 1,1 (0,3)b C 1,2 (0,2)b B
Los resultados fueron expresados como la media seguida de la desviación estándar (entre paréntesis). Para cada tiempo de irradiación, las medias seguidas por la misma letra minúscula no presentaron diferencias significativas a lo largo del almacenamiento (p<0,05). Para el mismo día de almacenamiento, las medias de los diferentes tratamientos seguidas por la misma letra mayúscula no presentaron diferencias significativas (p<0,05).

146
Zhu y col. (2007) reportaron que la inmersión de cubos de manzana en una solución de
ácido ascórbico 1% (p/v) y cloruro de calcio 1% (p/v) durante 5 minutos provocó una
disminución de la firmeza en el tejido. A diferencia de estos autores, en este trabajo no se
encontraron cambios importantes en las propiedades reológicas a altas deformaciones por la
aplicación de la solución de ácido ascórbico/cloruro de calcio.
Ensayos oscilatorios
�
En coincidencia con lo encontrado en los ensayos para las muestras irradiadas y
escaldadas e irradiadas, cuando se realizó el barrido de amplitud con control de la
deformación, las muestras de manzana fresca al día 0 fueron las que presentaron menor RVL.
En la figura 4'47 se muestra, a modo de ejemplo, el barrido de amplitud para las muestras
frescas al día 0 y las inmersas en la solución antipardeamiento e irradiadas durante 20 min al
día 7 del almacenamiento. El límite del rango viscoelástico lineal recomendado por el
software del equipo fue de 0,01% y 0,1%, respectivamente. Se utilizó en los ensayos
posteriores un valor de γ=0,01% para que todas las muestras estuvieran dentro del RVL.
������
�����
�����
�����
�����
����
�����
����
����� ���� ��� � �� ���
γγγγ �����
� �
�!
�
Figura 4'47. Barrido de amplitud con control de la deformación de cizalla obtenido para las muestras de manzanas frescas al día 0 (■) y tratadas con solución antipardeamiento e irradiadas 20 minutos al día 7 del almacenamiento (▲). Valor límite recomendado de RVL (■) (0,01% muestras frescas y 0,1% muestras tratadas con solución antipardeamiento e irradiadas)

147
En la figura 4'48 se observa el espectro mecánico para las muestras frescas y las
inmersas en la solución de ácido ascórbico/cloruro de calcio con o sin exposición a la luz UV*
C, al día 0 y séptimo de almacenamiento. Todas las muestras tuvieron una conducta de sólido
elástico con G´ dominando la respuesta viscoelástica en todo el rango de frecuencia.
Los valores de G’ a diferentes frecuencias angulares y los parámetros de la regresión
lineal del Log G’ vs Log ω pueden observarse en la tabla 4'17. Los valores de G’ no
presentaron interacción significativa entre el tratamiento y el tiempo de almacenamiento
(p>0,1), mientras que para la pendiente obtenida de la regresión lineal se observó interacción
entre ambos factores (p<0,001). En todas los muestras, el módulo de almacenamiento
aumentó con la frecuencia angular, obteniéndose por lo tanto pendientes positivas. No se
observaron diferencias significativas (p >0,05) entre las pendientes de las muestras tratadas y
el control al inicio del almacenamiento. Después de los siete días de almacenamiento, las
pendientes de las rodajas tratadas no tuvieron variaciones significativas mientras que la
correspondiente al control decreció levemente. Inmediatamente después de aplicados los
tratamientos (día 0), las rodajas de manzana tratadas no presentaron diferencias
significativas en los valores de G’ con respecto al control. Al final del almacenamiento, las
rodajas que habían recibido sólo el pretratamiento antipardeamiento tuvieron una disminución
de G’ similar al control (alrededor de un 40%), mientras que la disminución de las muestras
irradiadas fue de un 83%.
Los valores del factor de pérdida a distintas frecuencias angulares presentaron
interacción significativa entre el tratamiento y el tiempo de almacenamiento (p<0,05). A
frecuencias bajas, los valores de tan δ decrecieron en todas las muestras por efecto del
almacenamiento, no presentándose diferencias significativas entre las mismas. Esta
disminución reflejaría un decrecimiento mayor de G’’ en comparación con la disminución de
G’ durante el almacenamiento. A frecuencias más altas, las rodajas tratadas tendieron a
presentar una disminución levemente menor que el control (figura 4'49 y tabla 4'18).
�"�������������������������������
Al realizarse el ensayo de barrido de amplitud con control del esfuerzo de cizalla
(CEC), al igual que en ensayos anteriores, las muestras tratadas presentaron un límite de RVL
menor que las muestras frescas. Esto puede observarse en la figura 4'50, en donde se muestra
el BA CEC para las rodajas frescas al día 0 e inmersas en la solución antipardeamiento e
irradiadas durante 20 minutos al séptimo día de almacenamiento. En todas las muestras, el
valor de esfuerzo límite fue mayor a 35 Pa, por lo tanto, se decidió utilizar este valor de
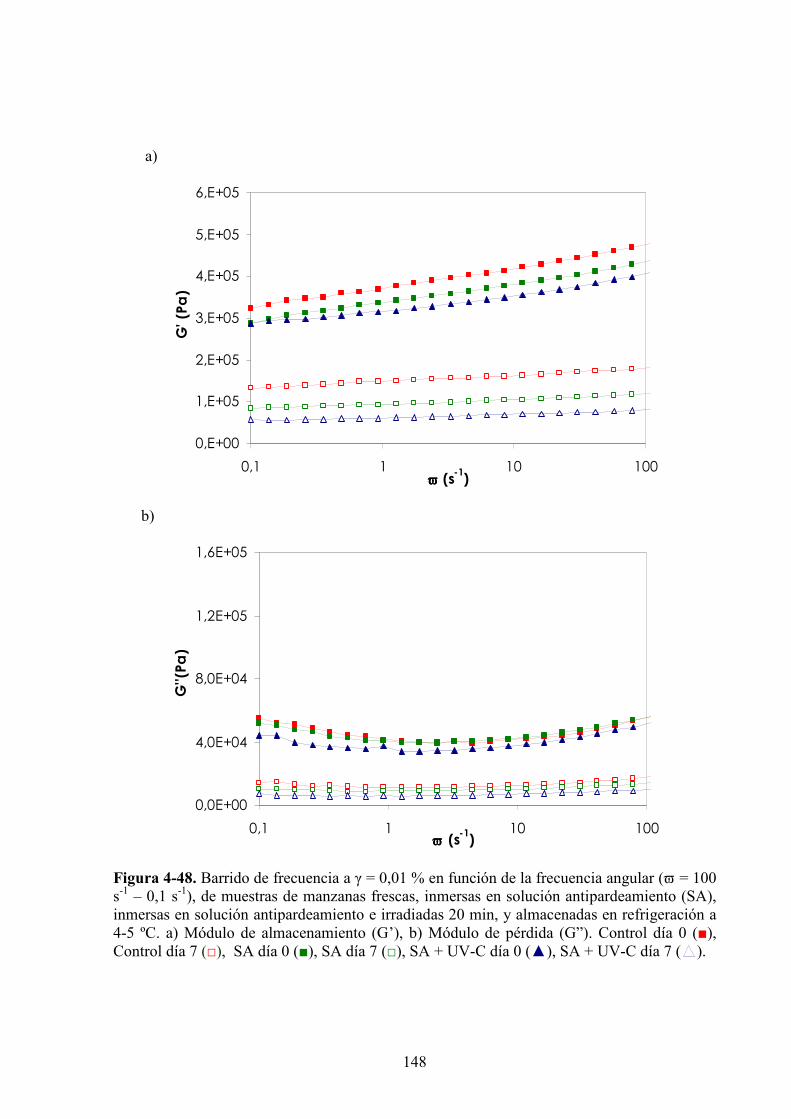
148
�
�
a)
������
�����
�����
�����
�����
����
�����
��� � �� ���ϖϖϖϖ ��"
��
� �
�!
�
b)
�������
�������
�������
������
������
��� � �� ���ϖ ϖ ϖ ϖ �"
��
� �
!
�
Figura 4'48. Barrido de frecuencia a γ = 0,01 % en función de la frecuencia angular (ϖ = 100 s*1 – 0,1 s*1), de muestras de manzanas frescas, inmersas en solución antipardeamiento (SA), inmersas en solución antipardeamiento e irradiadas 20 min, y almacenadas en refrigeración a 4*5 ºC. a) Módulo de almacenamiento (G’), b) Módulo de pérdida (G”). Control día 0 (■), Control día 7 (□), SA día 0 (■), SA día 7 (□), SA + UV*C día 0 (▲), SA + UV*C día 7 ( ).

149
Tabla 4'17. Valores promedio del módulo G´ (kPa) a distintas frecuencias angulares de las muestras de manzana control (C), inmersas en solución antipardeamiento (SA), inmersas en solución antipardeamiento e irradiadas durante 20 min, y almacenadas en refrigeración. Parámetros obtenidos de la regresión lineal del Log (G’) vs Log (ϖ). �
Tiempo (día)
Tratamiento ϖϖϖϖ�= 0,1 s'1� ϖϖϖϖ�= 1 s'1� ϖϖϖϖ�= 10 s'1� ϖϖϖϖ�= 100 s'1� ���n R2
C
324 (111)
370 (119) 415 (135) 479 (160) 0,057 aA 0,99 (0,01)
SA
288 (46)
342 (61) 377 (68) 437 (80) 0,056 aA 0,99
(0,003) 0
SA + UV'C
286 (63)
314 (72) 350(80) 409 (94) 0,053 aA 0,99 (0,003)
C
132 (46)
150 (50) 162 (54) 182 (61) 0,045 bA 0,99 (0,004)
SA
83,9 (21,8)
92,5 (29,5) 104 (25) 121(29) 0,053 aB 0,99 (0,004)
7
SA + UV'C
56,9 (9,1)
62 (14,7) 68,9 (16,2) 80,9 (19,3) 0,057 aB 0,99 (0,003)
Los resultados fueron expresados como la media seguida de la desviación estándar entre paréntesis. En caso de interacción significativa tiempo x tratamiento, en cada columna y para un dado tratamiento, las medias seguidas por la misma letra minúscula no presentaron diferencias significativas a los dos tiempos de almacenamiento (p<0,05). En cada columna y para un mismo día de almacenamiento, las medias de los diferentes tratamientos seguidas por la misma letra mayúscula no presentaron diferencias significativas (p<0,05).
�

150
�
���
���
���
���
���
���
��� � �� ���ϖϖϖϖ ��"
��
��� δδ δδ
Figura 4'49. Factor de pérdida (tan δ) de muestras de manzanas frescas, inmersas en solución antipardeamiento (SA), inmersas en solución antipardeamiento e irradiadas 20 min, y almacenadas en refrigeración a 4*5 ºC. Control día 0 (■), Control día 7 (□), SA día 0 (■), SA día 7 (□), SA + UV*C día 0 (▲), SA + UV*C día 7 ( ). Tabla 4'18. Valores promedio del factor de pérdida (tan δ) a distintas frecuencias angulares de las muestras de manzanas control (C), inmersas en solución antipardeamiento (SA), inmersas en solución antipardeamiento e irradiadas durante 20 min, y almacenadas en refrigeración. �
�
Tiempo (día)
Tratamiento ϖϖϖϖ�= 0,1 s'1� ϖϖϖϖ�= 1 s'1� ϖϖϖϖ�= 10 s'1� ϖϖϖϖ�= 100 s'1�
C 0,18(0,05)
aA 0,11(0,02)
aA 0,10(0,01)
aA 0,12(0,02)
aA
SA 0,18(0,02)
aA 0,12(0,01)
aA 0,11(0,01)
aB 0,13(0,01)
aA 0
SA + UV'C 0,15(0,03)
aA 0,11(0,01)
aA 0,11(0,01)
aB 0,13(0,01)
aA
C 0,11(0,03)
bA 0,08(0,01)
bA 0,08(0,01)
bA 0,10(0,01)
bA
SA 0,12(0,02)
bA 0,09(0,004)
bA,B 0,10(0,01)
bB 0,12(0,01)
bB 7
SA + UV'C 0,13(0,01)
bA 0,10(0,01)
aB 0,10(0,005)
bB 0,12(0,01)
bB
Los resultados fueron expresados como la media seguida de la desviación estándar. En cada columna y para un dado tratamiento, las medias seguidas por la misma letra minúscula no presentaron diferencias significativas a lo largo del almacenamiento (p<0,05). En cada columna y para el mismo día de almacenamiento, las medias de los diferentes tiempos de irradiación seguidas por la misma letra mayúscula no presentaron diferencias significativas (p<0,05)

151
Figura 4'50. Barrido de amplitud con control del esfuerzo de cizalla obtenido para las muestras de manzanas frescas al día 0 (■) y tratadas con solución antipardeamiento e irradiadas 20 minutos al día 7 de almacenamiento (▲). Valor límite recomendado de RVL (■) (100 Pa muestras frescas y 50 Pa muestras tratadas con solución antipardeamiento e irradiadas).
esfuerzo en los ensayos de fluencia*recuperación
En las figura 4'51 y figura 4'52 se representan las curvas promedio de deformación y
capacitancia respectivamente, para las muestras de manzana fresca y tratadas con solución
antipardeamiento con o sin exposición a la luz UV*C. Las curvas de capacitancia fueron
ajustadas con el modelo de Kelvin Voigt generalizado con un resorte en serie con 2 elementos
de Kelvin y un pistón obteniéndose coeficientes de correlación > 0,99. Las muestras
analizadas presentaron curvas promedio similares al inicio del almacenamiento, mientras que
después de los siete días, las rodajas tratadas presentaron perfiles diferentes al control.
En la tabla 4'19 se muestran los parámetros viscoelásticos obtenidos a partir del ajuste
de las curvas de capacitancia. Los parámetros J0, J1, J2, λ2 y ηN presentaron interacción entre el
tratamiento y el tiempo de almacenamiento (p<0,01), mientras que en el parámetro λ1 no hubo
interacción (p>0,3).
Las capacitancias instantánea y viscoelásticas no presentaron diferencias significativas
en ninguna de las muestras analizadas al inicio del almacenamiento. Después del séptimo día,
los valores de J0 y J2 aumentaron en todas las muestras, siendo levemente mayores en las
rodajas con el tratamiento de ácido ascórbico/cloruro de calcio que fueron o no expuestas a la
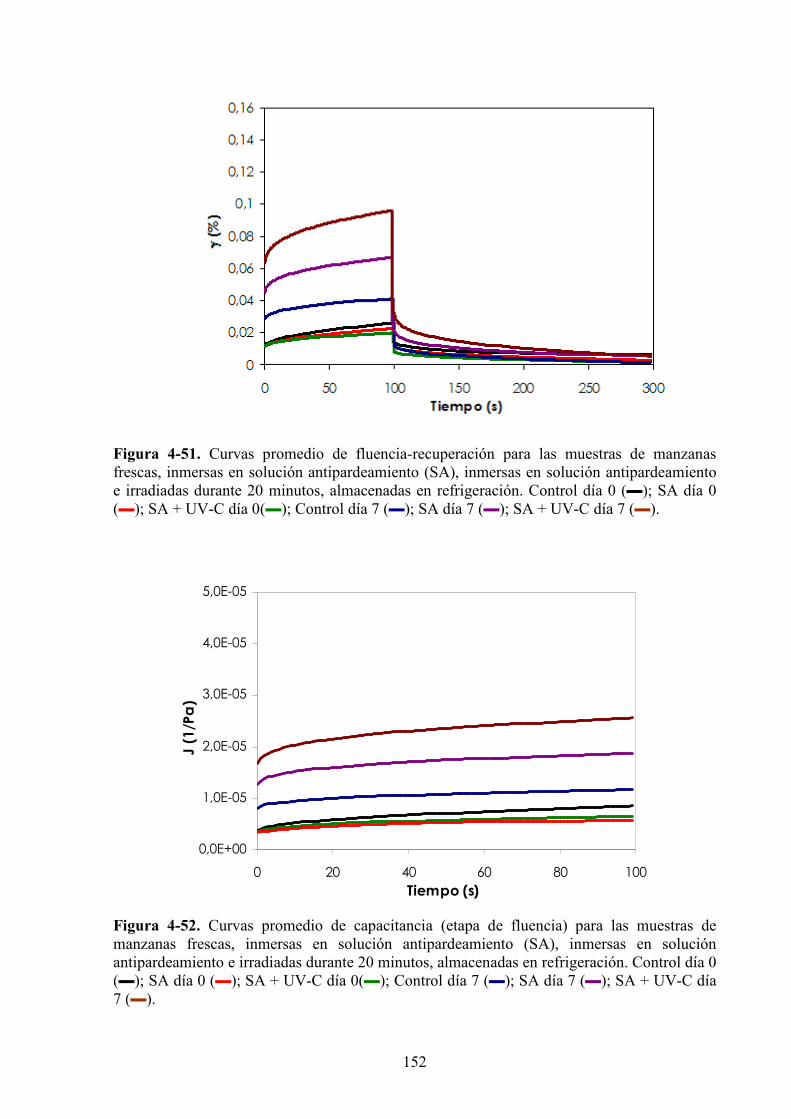
152
Figura 4'51. Curvas promedio de fluencia*recuperación para las muestras de manzanas frescas, inmersas en solución antipardeamiento (SA), inmersas en solución antipardeamiento e irradiadas durante 20 minutos, almacenadas en refrigeración. Control día 0 (▬); SA día 0 (▬); SA + UV*C día 0(▬); Control día 7 (▬); SA día 7 (▬); SA + UV*C día 7 (▬).
�������
�����
�����
�����
�����
����
� �� �� �� �� ���
� ������"�
$���
%!
�
Figura 4'52. Curvas promedio de capacitancia (etapa de fluencia) para las muestras de manzanas frescas, inmersas en solución antipardeamiento (SA), inmersas en solución antipardeamiento e irradiadas durante 20 minutos, almacenadas en refrigeración. Control día 0 (▬); SA día 0 (▬); SA + UV*C día 0(▬); Control día 7 (▬); SA día 7 (▬); SA + UV*C día 7 (▬).

153
Tabla 4'19. Parámetros de las curvas de fluencia* obtenidos durante el almacenamiento de rodajas de manzana fresca, inmersas en solución antipardeamiento y tratadas con solución antipardeamiento e irradiadas 20 min.
Tiempo (día)
Tratamiento J0 (1/Pa) (x 106)
J1 (1/Pa) (x 106)
J2 (1/Pa) (x 106)
λλλλ��(s)��
λλλλ��(s)��
ηηηη&�(Pa.s) (x 10'8)�
Control
3,4 (1,2)
aA
40%**
1,8(1,2)
aA
21%**
0,9(0,4)
aA
11%
27,1(6,1)
3,0(1,1) aA
0,4(0,3)
aA
28%
SA
2,9(0,7)
aA
44%**
1,5 (0,5)
aA
23%**
0,9(0,8)
aA
15%**
21,6(4,5)
1,4(0,6)
aB
0,8(0,5)
aA
18%**
0
SA + UV'C
3,0(0,4)
aA
51%**
1,6(1,4)
aA
27%**
0,7(0,1)
aA
12%**
29,4(16,1)
2,5(0,8)
aA
1,7(0,7)
aB
10%**
Control
7,5(2,1)
bA
64%**
1,7(0,5)
aA
14%**
1,2(0,5)
aA
11%**
22,8(6,5)
1,9(0,8)
aA
0,8(0,5)
aA
11%**
SA
11,8(4,4)
bB
63%**
2,6(0,8)
aA,B
14%**
2,1(0,6)
bB
11%**
21,2(2,8)
2,2(0,2) bA
0,4(0,2)
aA
12%**
7
SA + UV'C
15,6(4,9)
bB
61%**
3,7(0,5)
aB
14%**
2,9(0,4)
bB
11%**
21,9(2,8)
2,3(0,4) aA
0,3(0,06)
bA,B
13%**
* Parámetros derivados de la regresión de la ecuación 3.12. ** Contribución relativa de cada tipo de capacitancia a la capacitancia total al final de la etapa de fluencia. Los resultados fueron expresados como la media con la correspondiente desviación estándar (entre paréntesis).
En caso de interacción significativa tiempo x tratamiento, en cada columna y para un dado tratamiento, las medias seguidas por la misma letra minúscula no presentaron diferencias significativas a los dos tiempos de almacenamiento (p<0,05). En cada columna y para el mismo día de almacenamiento, las medias de los diferentes tiempos de irradiación seguidas por la misma letra mayúscula no presentaron diferencias significativas (p<0,05).
radiación UV*C. La capacitancia J1 se incrementó levemente sólo en las rodajas irradiadas.
Por lo tanto, los tratamientos a los que fueron sometidas las muestras indujeron una leve
pérdida de elasticidad y rigidez del tejido al cabo del almacenamiento.
Los tiempos de retardo (λ1 y λ2), al igual que en los otros tratamientos analizados, no
presentaron diferencias significativas entre las muestras, excepto los valores de λ2 en las
rodajas tratadas con solución antipardeamiento sin irradiar que fueron levemente inferiores al
resto de las muestras al día 0.

154
El coeficiente de viscosidad en el estado estacionario aumentó al inicio del
almacenamiento en las muestras tratadas con la solución de ácido ascórbico y cloruro de
calcio e irradiadas, y luego disminuyó durante el almacenamiento, presentando valores
levemente inferiores que el control.
Al final del almacenamiento, el incremento de la capacitancia total al final de la fase de
fluencia de las muestras tratadas comparadas con el tejido fresco fue de aproximadamente
120% en las rodajas tratadas con solución antipardeamiento sin irradiar, y de 200% en las
muestras irradiadas. Estos cambios fueron mucho menores a los encontrados en las muestras
irradiadas sometidas previamente al tratamiento térmico.
Los contribución relativa de cada capacitancia a la capacitancia total fue la siguiente: J0
(40*64 %), J1 (14*27%), J2 (11*15%) y 1/ηN (10*28%). En general, las contribuciones relativas
fueron relativamente similares en todas las muestras, excepto en la capacitancia 1/ ηN que fue
considerablemente superior a la de las muestras tratadas al inicio del almacenamiento. Por
otra parte, en todas las muestras, J0 fue la que presentó mayor porcentaje de contribución a la
deformación total, viéndose incrementada la misma al final del almacenamiento (tabla 4'19).
De acuerdo a los resultados obtenidos, las propiedades viscoelásticas de las rodajas de
manzana irradiadas no se vieron afectadas notoriamente por efecto de la aplicación previa de
la solución antipardeamiento de ácido ascórbico/cloruro de calcio.
8$#$8$8$�(�������������������������������+����������������������������������
Se analizaron las propiedades reológicas a bajas deformaciones de las rodajas de
manzana impregnadas con calcio (con o sin escaldado previo � a las que se les aplicó la
solución de ácido ascórbico/cloruro de calcio para inhibir el pardeamiento, y que fueron
irradiadas con luz UV*C durante 20 minutos.
�"������������������
�
Las rodajas de manzana fresca presentaron un rango viscoelástico lineal menor que las
muestras tratadas; por lo tanto, para determinar los espectros mecánicos, al igual que en los
ensayos anteriores, se utilizó el límite de RVL recomendado para las muestras frescas (γ =
0,01%).
Las figuras 4'53 y 4'54 muestran el espectro mecánico obtenido para las rodajas de
manzanas frescas y las muestras con los diferentes tratamientos, al día 0 y 7 de

155
a)
0,E+00
1,E+05
2,E+05
3,E+05
4,E+05
5,E+05
6,E+05
0,1 1 10 100ωωωω���
�!
9:��8�
b)
0,E+00
1,E+05
2,E+05
3,E+05
4,E+05
5,E+05
6,E+05
0,1 1 10 100ωωωω���
�!
9:��8�
Figura 4'53. Valores promedio del módulo de almacenamiento (G’) en función de la frecuencia angular (ϖ = 100 s*1 – 0,1 s*1) obtenidos a γ = 0,01 %, en muestras de manzanas frescas, impregnadas con calcio (con o sin escaldado), impregnadas con calcio (con o sin escaldado) e irradiadas durante 20 min, y almacenadas en refrigeración a 4*5 ºC. a) Día 0; b) Día 7. Control (■), IA (■), IA + UV*C (□), E + IA (■), E + IA + UV*C (□). IA: impregnación atmosférica; E: escaldado.

156
a)
0,E+00
1,E+04
2,E+04
3,E+04
4,E+04
5,E+04
6,E+04
7,E+04
8,E+04
0,1 1 10 100ωωωω���
�!
9::��8�
b)
0,E+00
1,E+04
2,E+04
3,E+04
4,E+04
5,E+04
6,E+04
7,E+04
8,E+04
0,1 1 10 100ωωωω���
�!
9::��8�
Figura 4'54. Valores promedio del módulo de pérdida (G”) en función de la frecuencia angular (ϖ = 100 s*1 – 0,1 s*1) obtenidos a γ = 0,01 %, en muestras de manzanas frescas, impregnadas con calcio (con o sin escaldado), impregnadas con calcio (con o sin escaldado) e irradiadas durante 20 min., y almacenadas en refrigeración a 4*5 º C. a) Día 0; b) Día 7. Control (■), IA (■), IA + UV*C (□), E + IA (■), E + IA + UV*C (□). IA: impregnación atmosférica; E: escaldado.

157
almacenamiento. Todas las muestras presentaron un predominio del comportamiento sólido,
con valores de G’ mayores a los de G’’ en todo el rango de frecuencia. Por otro lado, las
muestras tratadas presentaron valores de G’ y G’’ menores que el control fresco, tanto al día 0
como al final del almacenamiento.
En la tabla 4'20 se presentan los valores de G’ de las muestras analizadas a diferentes
frecuencias angulares y la pendiente de la regresión lineal Log G’ vs Log ω. En todos los
casos se presentó interacción significativa (p<0,05) entre los factores tiempo de
almacenamiento y tratamiento. Los valores de G’ de las muestras aumentaron con la
frecuencia angular, presentando, por lo tanto, pendientes positivas. Los valores de n no
mostraron variaciones significativas entre las muestras, tanto frescas como tratadas, al día 0.
Al día 7 de almacenamiento, las muestras control y las muestras escaldadas e impregnadas
con calcio (con o sin irradiación) mostraron una disminución y un incremento leve de la
pendiente, respectivamente, respecto del día 0. Hubo diferencias significativas entre los
valores de n de las muestras frescas, las muestras enriquecidas con calcio y las muestras
enriquecidas con calcio con tratamiento térmico previo al final del almacenamiento, los que se
incrementaron ligeramente en ese orden.
Después de los diferentes tratamientos, las muestras tratadas presentaron valores de G’
significativamente menores (p<0,0001) que el control fresco en todo el rango de frecuencia,
siendo la diferencia mayor en las rodajas de manzana impregnadas con calcio que habían
recibido un tratamiento de escaldado previo. El porcentaje de disminución con respecto al
control fue de alrededor de un 90% y 76% en las rodajas con o sin escaldado,
respectivamente. Estos resultados indicarían una pérdida de rigidez del tejido, incrementada
por la combinación del tratamiento de impregnación con un escaldado previo. La posterior
exposición de las muestras a la radiación UV*C produjo una disminución adicional leve en los
valores de G’ (alrededor de 84 % con respecto al control) de las rodajas impregnadas sin
escaldado, mientras que en las rodajas escaldadas no se presentaron diferencias significativas
(p>0,05). Estos cambios en el comportamiento mecánico aumentaron con el almacenamiento
de las muestras, sin influencia de la aplicación de la radiación UV*C. Después de los siete
días de almacenamiento, los porcentajes de disminución en los valores de G’ con respecto a
las muestras frescas al día 0 fueron de 62%, 88% y 94% para las muestras control,
impregnadas con calcio, escaldadas e impregnadas con calcio, respectivamente. No se
presentaron variaciones significativas entre las rodajas que fueron o no irradiadas.
Los valores promedio del factor de pérdida a las diferentes frecuencias angulares se
observan en la figura 4'55 y tabla 4'21. Para todas las frecuencias angulares se presentó
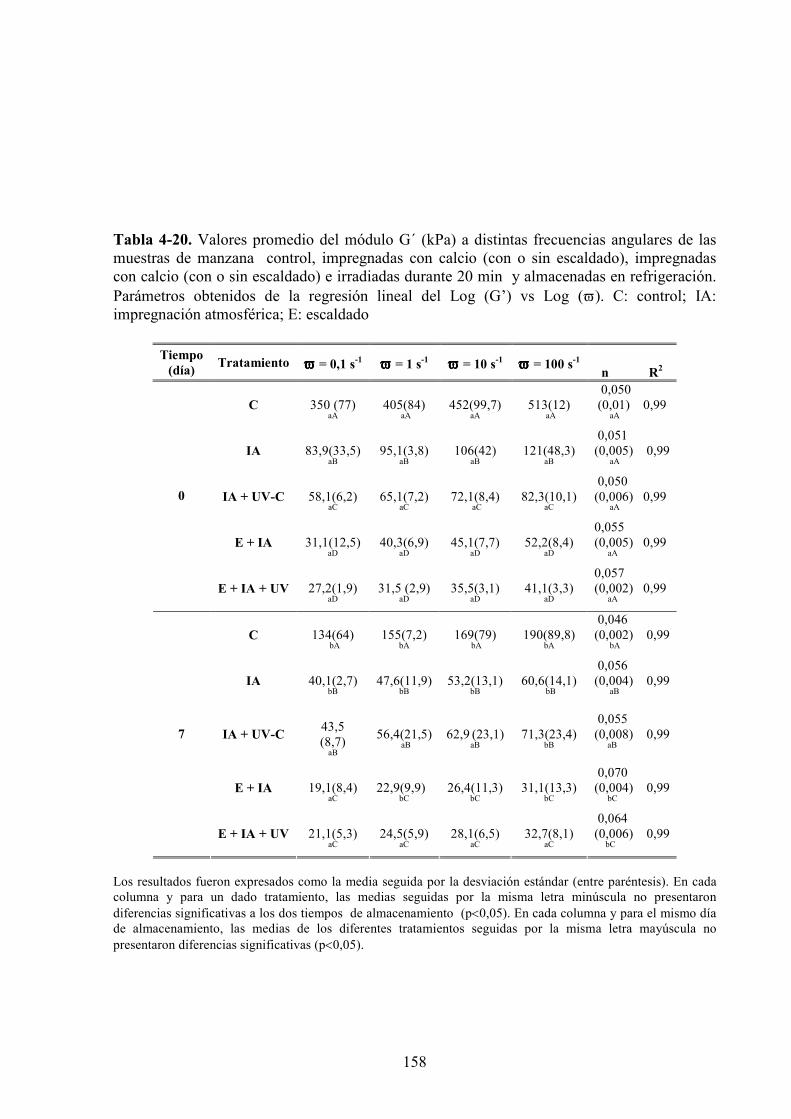
158
Tabla 4'20. Valores promedio del módulo G´ (kPa) a distintas frecuencias angulares de las muestras de manzana control, impregnadas con calcio (con o sin escaldado), impregnadas con calcio (con o sin escaldado) e irradiadas durante 20 min y almacenadas en refrigeración. Parámetros obtenidos de la regresión lineal del Log (G’) vs Log (ϖ). C: control; IA: impregnación atmosférica; E: escaldado��
Tiempo (día)
Tratamiento ϖϖϖϖ�= 0,1 s'1� ϖϖϖϖ�= 1 s'1� ϖϖϖϖ�= 10 s'1� ϖϖϖϖ�= 100 s'1� �n R2
C
350 (77) aA
405(84)
aA
452(99,7)
aA
513(12)
aA
0,050 (0,01) 0,99 aA
IA
83,9(33,5) aB
95,1(3,8)
aB
106(42)
aB
121(48,3)
aB
0,051 (0,005) 0,99 aA
IA + UV'C
58,1(6,2) aC
65,1(7,2)
aC
72,1(8,4)
aC
82,3(10,1)
aC
0,050 (0,006) 0,99 aA
E + IA
31,1(12,5) aD
40,3(6,9)
aD
45,1(7,7)
aD
52,2(8,4)
aD
0,055 (0,005) 0,99 aA
0
E + IA + UV
27,2(1,9) aD
31,5 (2,9)
aD
35,5(3,1)
aD
41,1(3,3)
aD
0,057 (0,002) 0,99 aA
C
134(64) bA
155(7,2)
bA
169(79)
bA
190(89,8)
bA
0,046 (0,002) 0,99 bA
IA
40,1(2,7) bB
47,6(11,9)
bB
53,2(13,1)
bB
60,6(14,1)
bB
0,056 (0,004) 0,99 aB
IA + UV'C
43,5 (8,7)
aB
56,4(21,5)
aB
62,9 (23,1)
aB
71,3(23,4)
bB
0,055 (0,008) 0,99 aB
E + IA
19,1(8,4) aC
22,9(9,9)
bC
26,4(11,3)
bC
31,1(13,3)
bC
0,070 (0,004) 0,99 bC
7
E + IA + UV
21,1(5,3) aC
24,5(5,9)
aC
28,1(6,5)
aC
32,7(8,1)
aC
0,064 (0,006) 0,99 bC
Los resultados fueron expresados como la media seguida por la desviación estándar (entre paréntesis). En cada columna y para un dado tratamiento, las medias seguidas por la misma letra minúscula no presentaron diferencias significativas a los dos tiempos de almacenamiento (p<0,05). En cada columna y para el mismo día de almacenamiento, las medias de los diferentes tratamientos seguidas por la misma letra mayúscula no presentaron diferencias significativas (p<0,05).
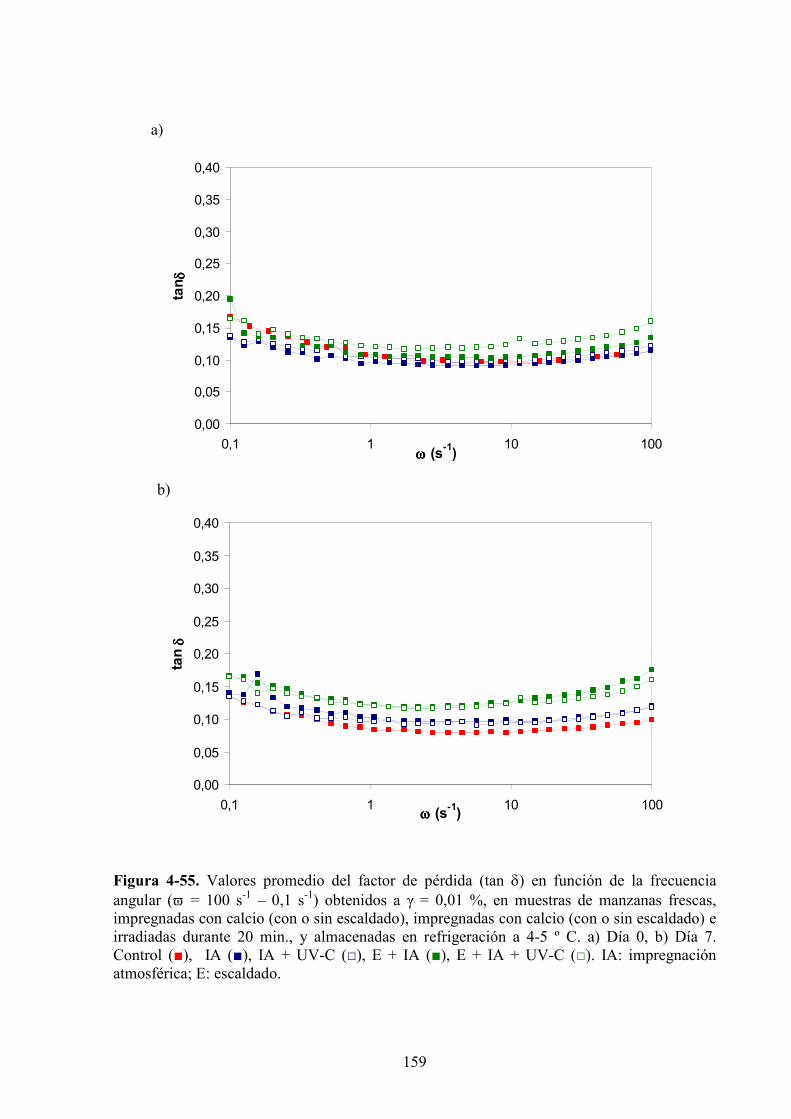
159
a)
0,00
0,05
0,10
0,15
0,20
0,25
0,30
0,35
0,40
0,1 1 10 100ωωωω���
�!
��
δδ δδ
b)
0,00
0,05
0,10
0,15
0,20
0,25
0,30
0,35
0,40
0,1 1 10 100ωωωω���
�!
��� δδ δδ
�
�
Figura 4'55. Valores promedio del factor de pérdida (tan δ) en función de la frecuencia angular (ϖ = 100 s*1 – 0,1 s*1) obtenidos a γ = 0,01 %, en muestras de manzanas frescas, impregnadas con calcio (con o sin escaldado), impregnadas con calcio (con o sin escaldado) e irradiadas durante 20 min., y almacenadas en refrigeración a 4*5 º C. a) Día 0, b) Día 7. Control (■), IA (■), IA + UV*C (□), E + IA (■), E + IA + UV*C (□). IA: impregnación atmosférica; E: escaldado.

160
Tabla 4'21. Valores promedio del factor de pérdida (tan δ) a distintas frecuencias angulares de las muestras de manzanas frescas, impregnadas con calcio (con o sin escaldado), impregnadas con calcio (con o sin escaldado) e irradiadas durante 20 min y almacenadas en refrigeración. C: control; IA: impregnación atmosférica; E: escaldado.
Tiempo (día)
Tratamiento ϖϖϖϖ�= 0,1 s'1� ϖϖϖϖ�= 1 s'1� ϖϖϖϖ�= 10 s'1� ϖϖϖϖ�= 100 s'1�
C
0,17(0,04) aA
0,11(0,02)
aA
0,097(0,009)
aA,B
0,11(0,01)
aA
IA
0,14(0,01) aB
0,096(0,008)
aA
0,091(0,003)
aB
0,114(0,003)
aB
IA + UV'C
0,14(0,01) aB
0,103(0,005)
aA
0,100(0,005)
aA,B
0,122(0,003)
aB,C
E + IA
0,16(0,03) aA,B
0,108(0,008)
aA
0,104(0,006)
aA
0,134(0,006)
aC,D
0
E + IA + UV
0,14(0,01) aB
0,108(0,008)
aA
0,106(0,005)
aA
0,141(0,005)
aD
C
0,13(0,02) bA
0,08(0,01)
bA
0,081(0,006)
bA
0,099(0,005)
bA
IA
0,14(0,01) aA
0,102(0,008)
aB
0,098(0,008)
bB
0,120(0,005)
aB
IA + UV'C
0,134(0,009) aA
0,09(0,02)
aA,B
0,09(0,01)
aB
0,12(0,01)
aB
E + IA
0,16(0,01) aB
0,122(0,009)
bC
0,12(0,01)
bC
0,17(0,03)
bC
7
E + IA + UV
0,16(0,02) bB
0,120(0,004)
bC
0,123(0,009)
bC
0,16(0,01)
bC
Los resultados fueron expresados como la media seguida por la desviación estándar (entre paréntesis). En cada columna y para un dado tratamiento, las medias seguidas por la misma letra minúscula no presentaron diferencias significativas a los dos tiempos de almacenamiento (p<0,05). En cada columna y para el mismo día de almacenamiento, las medias de los diferentes tratamientos seguidas por la misma letra mayúscula no presentaron diferencias significativas (p<0,05).
�

161
interacción significativa entre el tiempo de almacenamiento y el tratamiento (p<0,001). Las
diferencias en la tan δ entre las muestras tratadas y el control fueron leves y no siguieron la
misma tendencia a lo largo del rango de frecuencia analizado. Al día 0 del almacenamiento,
los valores de tan δ de las rodajas tratadas fueron menores que el control a frecuencias de 0,1
s*1 y mayores a frecuencias altas (10 s*1). Al final del almacenamiento, estos valores
disminuyeron en todas las frecuencias angulares en las muestras control y aumentaron en las
rodajas escaldadas e impregnadas, sin presentar un efecto adicional aquellas muestras que
fueron irradiadas.
Ensayos de fluencia'recuperación
En las figura 4'56 y figura 4'57 se presentan las curvas experimentales promedio de
deformación y de capacitancia de los ensayos de fluencia*recuperación, respectivamente. El
ajuste de los datos experimentales de capacitancia mediante el modelo de Kelvin Voigt
generalizado con un resorte en serie con 2 elementos de Kelvin en paralelo y un pistón
generó coeficientes de correlación > 0,99. Los parámetros obtenidos se muestran en la tabla
4'22. En todos los parámetros analizados se presentó interacción significativa entre el tiempo
de almacenamiento y el tratamiento, excepto en los valores de viscosidad en el estado
estacionario (ηN).
Luego de la aplicación de los tratamientos, pudo observarse un aumento significativo de
la capacitancia instantánea (J0), tanto en las rodajas de manzana impregnadas con calcio que
fueron irradiadas como en las impregnadas previamente escaldadas (con o sin irradiación).
Este incremento fue mayor en las muestras escaldadas, con poca influencia de la aplicación
posterior de irradiación. Las capacitancias viscoelásticas de retardo J1 y J2 aumentaron
significativamente sólo en las muestras escaldadas e impregnadas (con o sin irradiación). El
efecto de los tratamientos sobre las capacitancias J0 y J1 fue significativamente mayor luego
de los siete días del almacenamiento en las muestras con IA sin irradiar y en mayor medida en
las previamente escaldadas con o sin irradiación. Los valores de estas capacitancias fueron
similares entre las rodajas tratadas térmicamente que fueron o no expuestas a la irradiación.
Por otra parte, el parámetro J2 se incrementó sólo en las muestras impregnadas que fueron
escaldadas (con o sin irradiación).
La capacitancia viscosa en el estado estacionario (1/ηN) fue mayor en las muestras que
fueron impregnadas e irradiadas y en las rodajas previamente escaldadas (con o sin UV*C),
presentando valores mayores en estas últimas muestras. Al final del almacenamiento, ηN

162
a)
0
0,02
0,04
0,06
0,08
0,1
0,12
0 50 100 150 200 250 300
���������
γγ γγ��;
b)
0
0,02
0,04
0,06
0,08
0,1
0,12
0 50 100 150 200 250 300
���������
γγ γγ��;
Figura 4'56. Curvas promedio de fluencia*recuperación de muestras de manzanas frescas, impregnadas con calcio (con o sin escaldado), impregnadas con calcio (con o sin escaldado) e irradiadas durante 20 min, y almacenadas a 4*5 ºC. a) Día 0; b) Día 7. Control (▬); IA (▬); IA + UV*C (▬); E + IA (▬); E + IA + UV*C (▬). IA: impregnación atmosférica; E: escaldado.
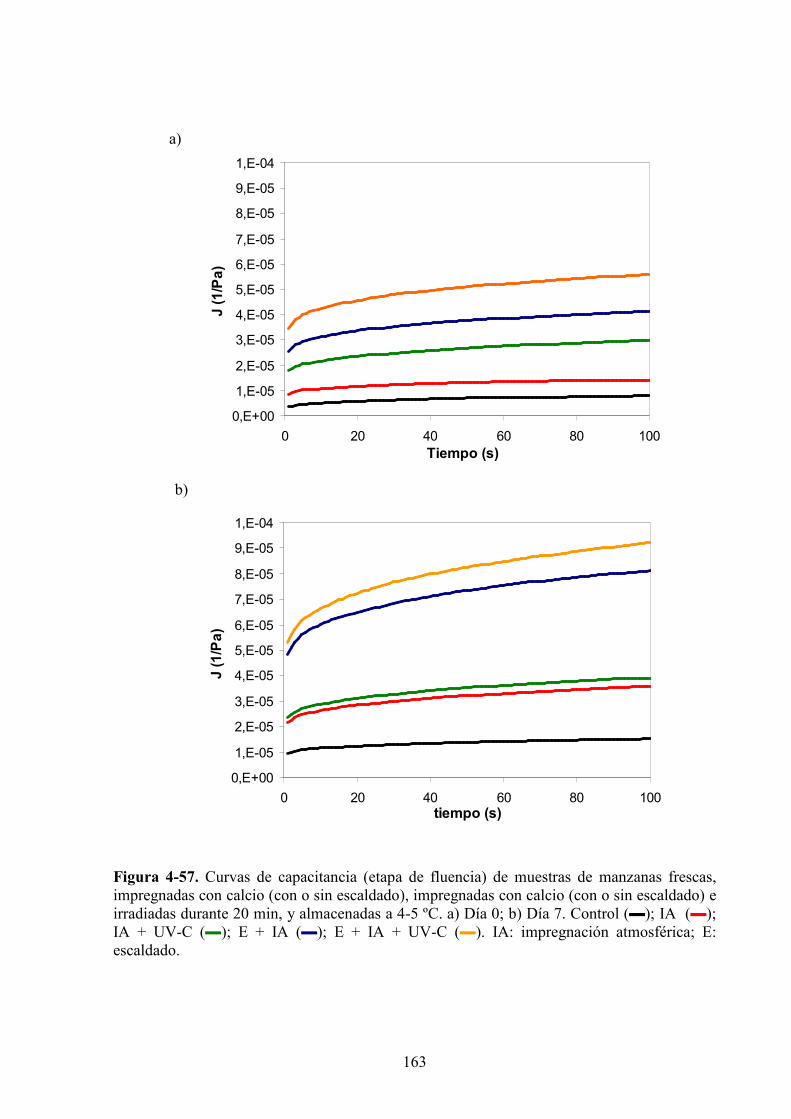
163
a)
0,E+00
1,E05
2,E05
3,E05
4,E05
5,E05
6,E05
7,E05
8,E05
9,E05
1,E04
0 20 40 60 80 100
���������
���!�8�
b)
0,E+00
1,E05
2,E05
3,E05
4,E05
5,E05
6,E05
7,E05
8,E05
9,E05
1,E04
0 20 40 60 80 100���������
���!�8�
Figura 4'57. Curvas de capacitancia (etapa de fluencia) de muestras de manzanas frescas, impregnadas con calcio (con o sin escaldado), impregnadas con calcio (con o sin escaldado) e irradiadas durante 20 min, y almacenadas a 4*5 ºC. a) Día 0; b) Día 7. Control (▬); IA (▬); IA + UV*C (▬); E + IA (▬); E + IA + UV*C (▬). IA: impregnación atmosférica; E: escaldado.
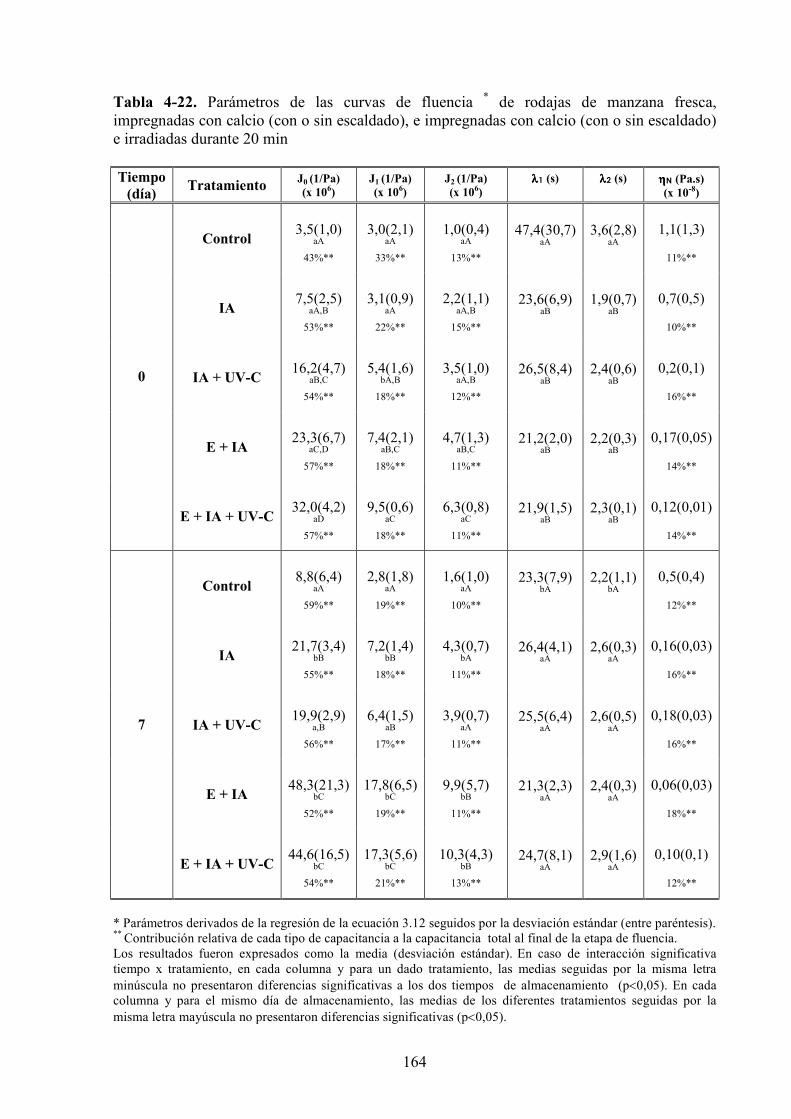
164
Tabla 4'22. Parámetros de las curvas de fluencia * de rodajas de manzana fresca, impregnadas con calcio (con o sin escaldado), e impregnadas con calcio (con o sin escaldado) e irradiadas durante 20 min Tiempo
(día) Tratamiento J0 (1/Pa)
(x 106) J1 (1/Pa) (x 106)
J2 (1/Pa) (x 106)
λλλλ��(s) �
λλλλ��(s)��
ηηηη&�(Pa.s) (x 10'8)�
Control
3,5(1,0)
aA
43%**
3,0(2,1)
aA
33%**
1,0(0,4)
aA
13%**
47,4(30,7) aA
3,6(2,8) aA
1,1(1,3)
11%**
IA
7,5(2,5)
aA,B
53%**
3,1(0,9)
aA
22%**
2,2(1,1)
aA,B
15%**
23,6(6,9)
aB
1,9(0,7)
aB
0,7(0,5)
10%**
IA + UV'C
16,2(4,7)
aB,C
54%**
5,4(1,6)
bA,B
18%**
3,5(1,0)
aA,B
12%**
26,5(8,4) aB
2,4(0,6) aB
0,2(0,1)
16%**
E + IA
23,3(6,7)
aC,D
57%**
7,4(2,1)
aB,C
18%**
4,7(1,3)
aB,C
11%**
21,2(2,0)
aB
2,2(0,3) aB
0,17(0,05)
14%**
0
E + IA + UV'C
32,0(4,2)
aD
57%**
9,5(0,6)
aC
18%**
6,3(0,8)
aC
11%**
21,9(1,5) aB
2,3(0,1) aB
0,12(0,01)
14%**
Control
8,8(6,4)
aA
59%**
2,8(1,8)
aA
19%**
1,6(1,0)
aA
10%**
23,3(7,9) bA
2,2(1,1) bA
0,5(0,4)
12%**
IA
21,7(3,4)
bB
55%**
7,2(1,4)
bB
18%**
4,3(0,7)
bA
11%**
26,4(4,1) aA
2,6(0,3) aA
0,16(0,03)
16%**
IA + UV'C
19,9(2,9)
a,B
56%**
6,4(1,5)
aB
17%**
3,9(0,7)
aA
11%**
25,5(6,4) aA
2,6(0,5) aA
0,18(0,03)
16%**
E + IA
48,3(21,3)
bC
52%**
17,8(6,5)
bC
19%**
9,9(5,7)
bB
11%**
21,3(2,3) aA
2,4(0,3) aA
0,06(0,03)
18%**
7
E + IA + UV'C
44,6(16,5)
bC
54%**
17,3(5,6)
bC
21%**
10,3(4,3)
bB
13%**
24,7(8,1) aA
2,9(1,6) aA
0,10(0,1)
12%**
* Parámetros derivados de la regresión de la ecuación 3.12 seguidos por la desviación estándar (entre paréntesis). ** Contribución relativa de cada tipo de capacitancia a la capacitancia total al final de la etapa de fluencia. Los resultados fueron expresados como la media (desviación estándar). En caso de interacción significativa tiempo x tratamiento, en cada columna y para un dado tratamiento, las medias seguidas por la misma letra minúscula no presentaron diferencias significativas a los dos tiempos de almacenamiento (p<0,05). En cada columna y para el mismo día de almacenamiento, las medias de los diferentes tratamientos seguidas por la misma letra mayúscula no presentaron diferencias significativas (p<0,05).

165
disminuyó en todas las muestras analizadas siguiendo la misma tendencia que al día 0. Esto
estaría indicando que los distintos tratamientos aplicados provocarían una mayor fluidez de
los tejidos con una mayor deformación permamente.
Los tiempos de retardo (λ1 y λ2) de las muestras tratadas fueron menores al control al
día 0 del almacenamiento, mientras que al séptimo día, disminuyeron sólo en las muestras
control, no presentando diferencias significativas con respecto a los valores de las muestras
tratadas. Ello pondría de manifiesto que los elementos viscoelásticos de los tejidos tratados
exhibieron mayor velocidad para alcanzar un dado nivel de deformación que el tejido fresco
después de los tratamientos.
El incremento de la capacitancia total al final de la fase de fluencia de las muestras
tratadas comparadas con el tejido fresco fue para el día 0 de aproximadamente 76% para las
muestras impregnadas, 270% para las impregnadas e irradiadas, y de 400% y 600% para las
escaldadas e impregnadas sin y con exposición a la irradiación, mientras que al final del
almacenamiento, los incrementos respectivos fueron de 400% para las rodajas impregnadas
(con o sin UV*C) y de 1000% para las muestras previamente escaldadas (con o sin UV*C).
La contribución de J0 a la capacitancia total fue mayor en las rodajas tratadas que en el
control al tiempo 0, pero al final del almacenamiento, las contribuciones relativas de cada
capacitancia a la capacitancia total fueron similares en todas las muestras. Las contribuciones
relativas entre las diferentes muestras estuvieron dentro de los siguientes rangos: J0 (43*59
%), J1 (17*33 %), J2 (11*15 %) y 1/ηN (10*18 %).
De acuerdo a lo analizado, las rodajas de manzana enriquecidas con calcio y expuestas a
la radiación UV*C sufrieron cambios en su comportamiento viscoelástico (ensayos
oscilatorios y de fluencia/recuperación), producidos principalmente por el tratamiento de
impregnación con calcio y por el escaldado, y en menor medida, por la exposición a la
radiación UV*C. Dichos cambios en general se incrementaron con el almacenamiento, no
observándose para el día 7 diferencias significativas debido a la aplicación de luz UV*C.
�8$#$9$��4����������������
El agua es el componente más abundante de los tejidos de fruta (80 a 90%) y pequeños
cambios en su contenido pueden producir efectos adversos sobre la calidad. La pérdida de
agua en frutas está determinada por muchos factores, siendo probablemente el más importante
la resistencia que ofrece la epidermis exterior o cutícula al movimiento de vapor de agua por
transpiración. En las frutas mínimamente procesadas, la remoción de la piel y el corte

166
provocan un incremento de la pérdida de líquidos propios del tejido (líquido nativo). Esto
puede generar un impacto negativo no sólo en la apariencia del producto (menor brillo,
marchitamiento) sino también en las características texturales percibida (Toivonen & Deell,
2002; Garcia & Barret, 2005).
En la figura 4'58 y la tabla 4'23 se muestra el porcentaje de pérdida de peso
(relacionado con la pérdida de líquido nativo) durante el almacenamiento de rodajas de
manzana fresca (control) y expuestas a la luz UV*C con o sin la aplicación de los
pretratamientos de escaldado e inmersión en solución antipardeamiento. En todos los casos
analizados se presentó interacción significativa entre el tiempo de almacenamiento y el
tratamiento (p<0,0001). Todas las muestras (tratadas y control) mostraron una disminución
del peso a lo largo el almacenamiento. La exposición de las muestras a la irradiación UV*C
durante 8 minutos no produjo variaciones significativas (p<0,0001) en los porcentajes de
pérdida de peso en comparación con el control en ninguno de los tiempos analizados. Sin
embargo, las rodajas de manzana irradiadas durante 20 minutos presentaron una pérdida de
peso mayor que el control tanto al día 3 como al día 7. Los porcentajes de pérdida de peso al
final del almacenamiento fueron de alrededor de un 8 % para el control y las rodajas
irradiadas 8 minutos, y de un 14 % para las muestras irradiadas durante 20 minutos. Por lo
tanto, la exposición a dosis altas de irradiación provocaría un daño adicional al tejido de
manzana que el generado por las operaciones de pelado y corte, lo que se traduciría en un
aumento de la tasas de respiración y de transpiración de la fruta cortada.
Las rodajas que fueron inmersas en la solución antipardeamiento de ácido
ascórbico/cloruro de calcio antes de la irradiación presentaron un patrón similar en la
evolución del peso a la de las muestras irradiadas sin dicho pretratamiento. Por otro lado, en
las muestras previamente escaldadas, el peso disminuyó después del tratamiento (día 0 de
almacenamiento) en alrededor de 4*6%, debido probablemente a la disrupción general del
tejido (ruptura de membranas celulares y pérdida de integridad de paredes celulares)
provocada luego de la aplicación del tratamiento, con la consiguiente pérdida de fluidos
celulares. Al tercer y séptimo día del almacenamiento, el porcentaje de pérdida de peso se
incrementó en estas muestras y fue mayor que en el control, pero no se presentaron
diferencias significativas (p>0,05) entre las rodajas escaldadas que fueron o no expuestas a la
irradiación. La pérdida de peso luego de los siete días de almacenamiento fue de un 15*16%,
similar a la que presentaron las muestras irradiadas durante 20 minutos sin escaldado.

167
0
2
4
6
8
10
12
14
16
18
20
0 1 2 3 4 5 6 7 8�����������
88��;
Control
UVC 8 min
UVC 20 min
0
2
4
6
8
10
12
14
16
18
20
0 1 2 3 4 5 6 7 8
�����������
88��;
Control E
E + UVC 8 min E + UVC 20 min
0
2
4
6
8
10
12
14
16
18
20
0 1 2 3 4 5 6 7 8�����������
88�;
Control SA
SA + UVC 8 min SA + UVC 20 min
Figura 4'58. Pérdida de peso (PP) (%) durante el almacenamiento refrigerado de rodajas de manzana fresca (control) y tratadas. a) UV*C; b) Escaldado (E) + UV*C; c) Solución antipardeamiento (SA) + UV*C.
a)
b)
c)
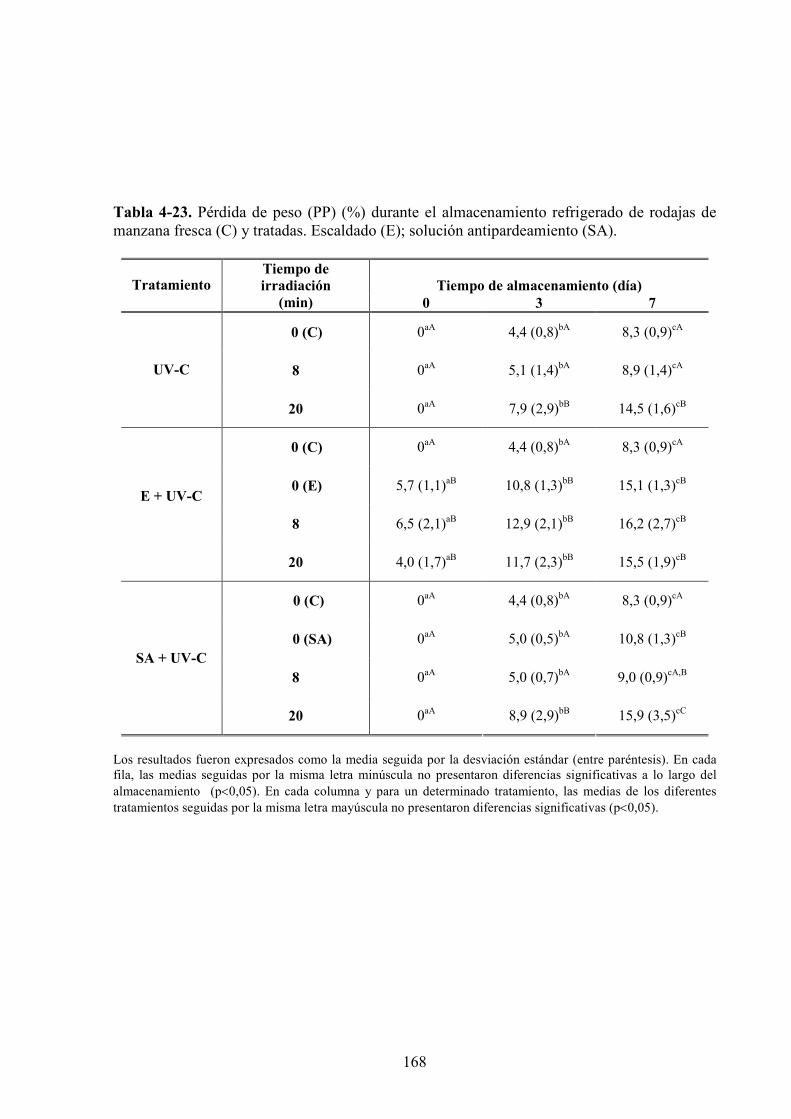
168
Tabla 4'23. Pérdida de peso (PP) (%) durante el almacenamiento refrigerado de rodajas de manzana fresca (C) y tratadas. Escaldado (E); solución antipardeamiento (SA).
Tiempo de almacenamiento (día) Tratamiento
Tiempo de irradiación
(min) 0 3 7
0 (C) 0aA 4,4 (0,8)bA 8,3 (0,9)cA
8 0aA 5,1 (1,4)bA 8,9 (1,4)cA UV'C
20 0aA 7,9 (2,9)bB 14,5 (1,6)cB
0 (C) 0aA 4,4 (0,8)bA 8,3 (0,9)cA
0 (E) 5,7 (1,1)aB 10,8 (1,3)bB 15,1 (1,3)cB
8 6,5 (2,1)aB 12,9 (2,1)bB 16,2 (2,7)cB
E + UV'C
20 4,0 (1,7)aB 11,7 (2,3)bB 15,5 (1,9)cB
0 (C) 0aA 4,4 (0,8)bA 8,3 (0,9)cA
0 (SA) 0aA 5,0 (0,5)bA 10,8 (1,3)cB
8 0aA 5,0 (0,7)bA 9,0 (0,9)cA,B SA + UV'C
20 0aA 8,9 (2,9)bB 15,9 (3,5)cC
Los resultados fueron expresados como la media seguida por la desviación estándar (entre paréntesis). En cada fila, las medias seguidas por la misma letra minúscula no presentaron diferencias significativas a lo largo del almacenamiento (p<0,05). En cada columna y para un determinado tratamiento, las medias de los diferentes tratamientos seguidas por la misma letra mayúscula no presentaron diferencias significativas (p<0,05).

169
8$#$;$����������������������������������������������������������*�����������������������������������������������������F��������������������
Los cambios microestructurales de las muestras con los diferentes tratamientos fueron
evaluados mediante microscopía óptica (MO) y los ultraestructurales mediante microscopia
electrónica de barrido ambiental (MEBA) y microscopía electrónica de transmisión (MET).
Las observaciones se realizaron sobre la parte superficial de las muestras, y en el caso de las
muestras irradiadas, sobre la cara superficial expuesta a la irradiación.
En las figuras 4'59, 4'60, 4'61 y 4'62 se muestran las microfotografías obtenidas con
MO y MEBA del tejido de manzana fresco (control) e irradiado con luz UV*C durante 20
minutos, con o sin aplicación previa de los tratamientos de escaldado e inmersión en la
solución de ácido ascórbico/cloruro de calcio, obtenidas al día 0 y 7 de almacenamiento. En
las figuras 4'63, 4'64, 4'65 y 4'66 se visualizan las imágenes obtenidas con MET del tejido
fresco e irradiado (sin pretratamiento), al inicio y final del almacenamiento.
En el tejido de manzana fresco, las células y los espacios intercelulares se observaron
distribuidos en un arreglo homogéneo y anisotrópico. Los espacios celulares exhibieron varias
formas y tamaños. Las células, más o menos regulares en forma, aparecieron túrgidas con una
estructura celular aparentemente bien definida. El volumen celular se mostró ocupado por una
gran vacuola central, y el protoplasto, rodeado por el plasmalema y el tonoplasto, se presentó
como una fina capa revistiendo la superficie celular (figura 4'59 A). En las observaciones
ultraestructurales con MEBA realizadas sobre el tejido fresco también se observaron células
túrgidas con paredes celulares consistentes, en algunas zonas muy reforzadas (figura 4'61 A).
En las observaciones con MET se visualizaron paredes celulares con buena densidad
electrónica, algunas con una laminilla media no muy bien definida ocupando una banda
central (figura 4'63 B, C y D). El citoplasma de las células se presentó unido a la pared
celular (figura 4'63 A, B y C) y en algunas regiones se observó la presencia de vesículas
(figuras 4'63 D).
La exposición del tejido a la luz UV*C provocó una mayor disrupción de membranas
celulares (plasmalema y tonoplasto) (figura 4'59 B). Las células se presentaron más
redondeadas y las paredes más lisas (figuras 4'59 B y 4'61 B). En las observaciones con
MET las paredes celulares aparecieron con buena densidad electrónica pero sin definición de
la laminilla media (figura 4'64 B, C, D) o bien con una laminilla media angosta y poco teñida
(figura 4'64 A). El citoplasma de las células se visualizó levemente separado de la pared
celular (plasmólisis incipiente) (figuras 4'64 A y C), observándose claramente la ruptura de
membranas (figuras 4'64 A, B, C y D).

170
El tejido escaldado presentó células con ruptura de membranas y pérdida consiguiente
de turgencia. Las paredes celulares aparecieron colapsadas y con plegamientos por la pérdida
de presión de turgor pero también se observaron más hinchadas y muy debilitadas en algunas
zonas. También se observó contracción del tejido (mayor cantidad de células por unidad de
área) y células más irregulares en cuanto a su forma (figura 4'59 C). En las observaciones
ultraestructurales se pudo observar una leve separación de las paredes celulares atribuida a la
degradación de pectinas provocada por el tratamiento térmico (figura 4'61 C). Las células del
tejido escaldado e irradiado presentaron un arreglo similar al del tejido irradiado sin
escaldado, aunque las paredes aparecieron menos teñidas (figura 4'59 D). En la
microfotografía de la figura 4'61 D se notó también una mayor separación entre células
debido al efecto del tratamiento térmico en la pared.
La inmersión en la solución ácido ascórbico/cloruro de calcio no modificó en gran
medida el tejido de manzana comparado al control. En general, no hubo disrupción de
membranas y el citoplasma fue parietal. Sólo en algunas células se observó ruptura de
membranas probablemente debido al efecto del corte. Las paredes celulares aparecieron
densamente teñidas pero con algunas interrupciones (figura 4'59 E). En la microfotografía de
la figura 4'61 E puede observarse mejor la disrupción de las paredes celulares. La irradiación
UV*C causó ruptura de membranas en las células de manzana tratadas previamente con la
solución antipardeamiento (figura 4'59 F). También se observaron células con paredes
celulares bien definidas y otras con paredes rotas (figuras 4'59 F y 4'61 F).
Es de destacar que todos los tejidos irradiados, con o sin pretratamientos, mostraron
después de la irradiación (día 0 de almacenamiento) células mucho más redondeadas que
aquellos no irradiados (figuras 4'59 B, D, F y 4'61 B, D, F)
Luego de siete días de almacenamiento, tanto los tejidos de las muestras tratadas como
del control presentaron colapso y plegamiento de paredes celulares. Sin embargo, estos
cambios fueron más notorios en el tejido de las muestras irradiadas (con o sin
pretratamientos) (figuras 4'60 y 4'62). El tejido escaldado e irradiado fue el que presentó
mayor daño, observándose paredes celulares menos teñidas (figura 4'60 D). Por otro lado,
llamativamente las paredes del control almacenado se visualizaron más teñidas que al día 0
(figura 4'60 A). En las observaciones de MET realizadas sobre el control también se
observaron células con una mejor definición de la laminilla media, que aparecía como una
línea fina muy densamente teñida, en comparación con lo observado en el tejido fresco al día
0 (figura 4'65 B, C, D). El citoplasma de estas células apareció levemente plasmolizado
(figura 4'65 A) o con disrupciones de membranas (figura 4'65 E), y en algunas regiones las

171
paredes celulares presentaron ensanchamientos (figura 4'65 F). En concordancia con lo
observado en las imágenes de MO y MEBA, en las observaciones de MET del tejido irradiado
las paredes celulares se visualizaron con buena densidad electrónica pero con plegamientos y
ondulaciones pronunciados al final del almacenamiento (figuras 4'66 A, B y C). Al igual que
lo observado en el control, se visualizaron células que presentaron ensanchamientos de la
pared celular en algunas zonas (figura 4'66 D), separación del citoplasma de la pared celular
(figura 4'66 D), ruptura de membranas (figura 4'66 A, B y C) y formación de vesículas
(figura 4'66 B). Pero a diferencia del control, no se observó en general en los tejidos
irradiados una buena definición de la laminilla media.
En las figuras 4'67 ' 4'70 se muestran las microfotografías correspondientes a los
tejidos que fueron impregnados con calcio (con o sin posterior irradiación), al día 0 y 7 de
almacenamiento. El tratamiento de impregnación atmosférica con calcio provocó plasmólisis
en la mayoría de las células, con algunos episodios de ruptura de membranas. Las células se
presentaron con formas más irregulares que el control (figuras 4'67 B y 4'68 B). La
exposición a la luz UV*C del tejido impregnado provocó una mayor ruptura de membranas
(figuras 4'67 C y 4'68 C). Los tejidos previamente escaldados e impregnados presentaron
contracción del tejido pero con paredes celulares con buena densidad óptica, bastante lisas,
con escasos plegamientos y pocas disrupciones (figuras 4' 67 D y 4'68 D). Esto indicaría
que la penetración de calcio pudo haber mejorado la estructura de la pared celular de los
tejidos escaldados. Este fenómeno también fue observado en trabajos previos realizados en el
grupo de trabajo sobre la misma matriz y bajo condiciones de impregnación similares
(González*Fésler y col., 2008). La posterior irradiación de estos tejidos produjo disrupción del
plasmalema y tonoplasto en la mayoría de las células (figuras 4'67 E y 4'68 E) y también un
mayor encogimiento (mayor número de células por unidad de área).
Luego del séptimo día de almacenamiento, los tejidos impregnados con calcio (con o
sin escaldado, con o sin irradiación) presentaron paredes celulares más plegadas que al día 0
pero en menor medida que el control (figuras 4'69 y 4'70). Las diferencias estructurales entre
las muestras irradiadas y no irradiadas disminuyeron con el almacenamiento en las muestras
impregnadas sin escaldado (figuras 4'69 B, C y 4'70 A, B). Por el contrario, si bien el arreglo
celular del tejido escaldado e impregnado sometido a irradiación fue similar al del tejido
escaldado e impregnado sin irradiar, el primero presentó paredes celulares ligeramente más
hinchadas y menos teñidas en el centro, con mayores interrupciones que el no irradiado
(figuras 4'69 D, E y 4'70 D, E).

172
100 m �
A B
C D
E F
100 m �
100 m � 100 m �
100 m � 100 m �
��
��
Figura 4'59. Imágenes de microscopía óptica (MO) de la superficie del tejido de manzana irradiado con luz UV*C con o sin pretratamientos, obtenidas al día 0 de almacenamiento. A: muestra fresca (control); B: irradiada 20 min; C: escaldada; D: escaldada e irradiada 20 min; E: inmersa en solución antipardeamiento; F: inmersa en solución antipardeamiento e irradiada 20 min. ei: espacio intercelular; mc: membrana celular.
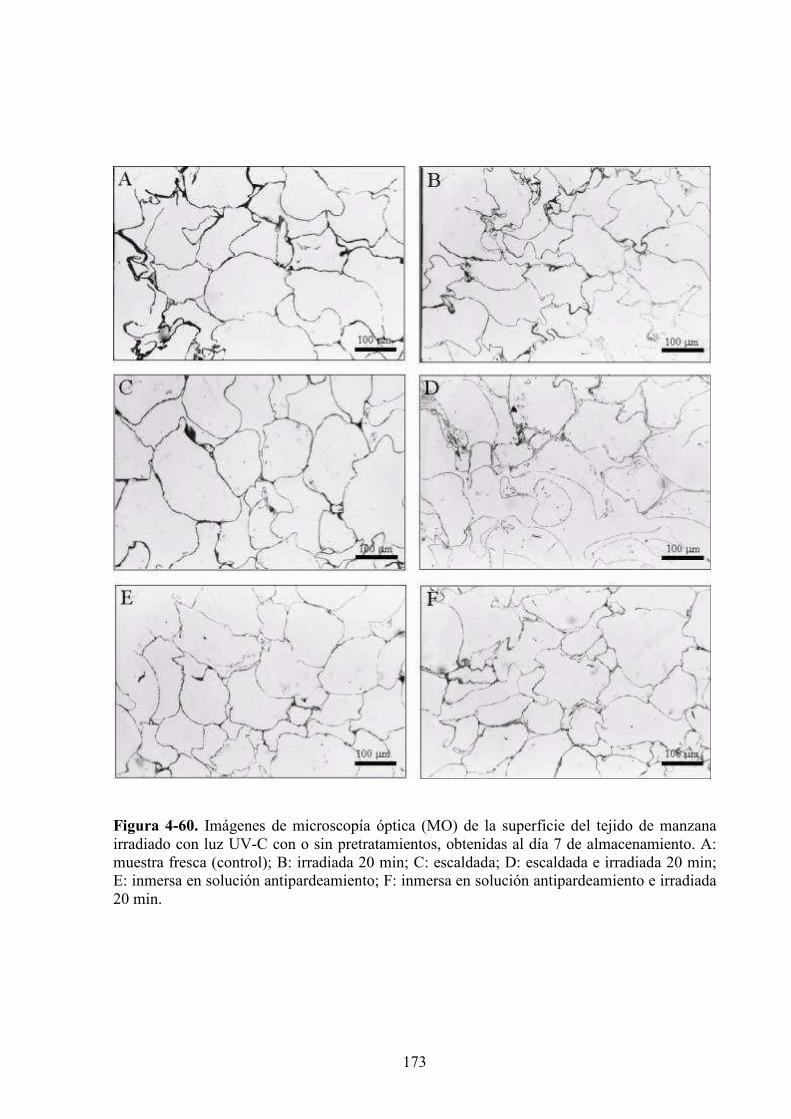
173
Figura 4'60. Imágenes de microscopía óptica (MO) de la superficie del tejido de manzana irradiado con luz UV*C con o sin pretratamientos, obtenidas al día 7 de almacenamiento. A: muestra fresca (control); B: irradiada 20 min; C: escaldada; D: escaldada e irradiada 20 min; E: inmersa en solución antipardeamiento; F: inmersa en solución antipardeamiento e irradiada 20 min.

174
A B
D
E F
C
Figura 4'61. Imágenes de microscopía electrónica de barrido ambiental (MEBA) de la superficie del tejido de manzana irradiado con luz UV*C con o sin pretratamientos, obtenidas al día 0 del almacenamiento. A: muestra fresca (control); B: irradiada 20 min; C: escaldada; D: escaldada e irradiada 20 min; E: inmersa en solución antipardeamiento; F: inmersa en solución antipardeamiento e irradiada 20 min.

175
A B
C
E
D
F
Figura 4'62. Imágenes de microscopía electrónica de barrido ambiental (MEBA) de la superficie del tejido de manzana irradiado con luz UV*C con o sin pretratamientos, obtenidas al día 7 de almacenamiento. A: muestra fresca (control); B: irradiada 20 min; C: escaldada; D: escaldada e irradiada 20 min; E: inmersa en solución antipardeamiento; F: inmersa en solución antipardeamiento e irradiada 20 min.

176
���� ����
���� ����
��
<
<�
A B
C D
��
��
�
Figura 4'63. Imágenes de microscopía electrónica de transmisión (MET) de la superficie del tejido de manzana fresco (control), obtenidas al día 0 del almacenamiento (A*D). pc: pared celular; v: vacuola; lm: laminilla media; c: citoplasma; ve: vesículas.
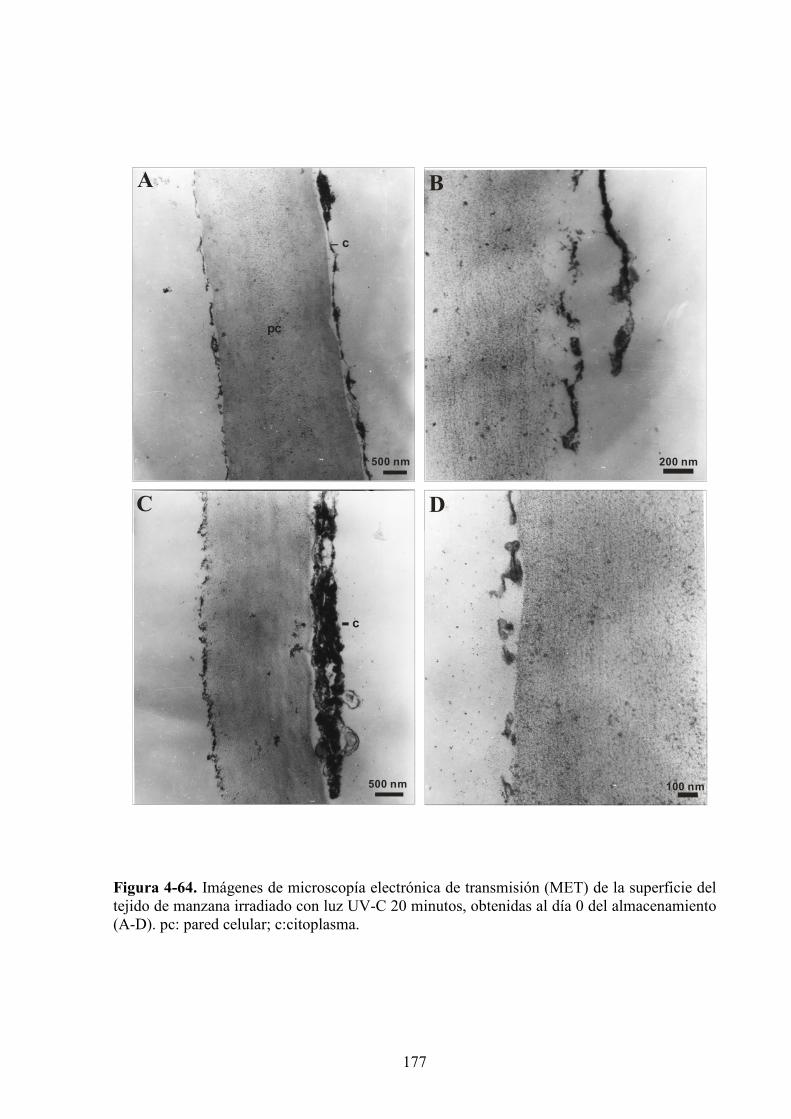
177
�����
����
����
A B
C
!����
D
��
�
�
Figura 4'64. Imágenes de microscopía electrónica de transmisión (MET) de la superficie del tejido de manzana irradiado con luz UV*C 20 minutos, obtenidas al día 0 del almacenamiento (A*D). pc: pared celular; c:citoplasma.

178
���� !����
! �� �����
���� ����
A B
C D
E F
��
��
��
�
��
�
Figura 4'65. Imágenes de microscopía electrónica de transmisión (MET) de la superficie del tejido de manzana fresco (control), obtenidas al día 7 del almacenamiento (A*F). pc: pared celular; c: citoplasma; lm: laminilla media.
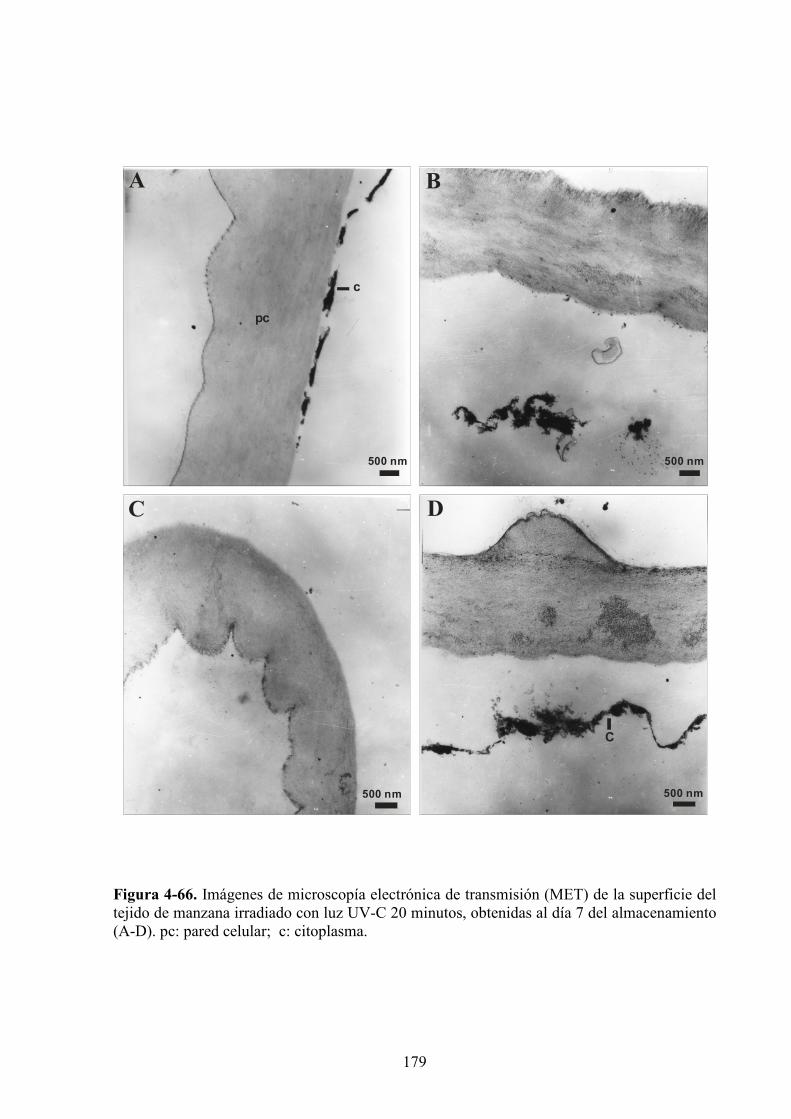
179
���� ����
���� ����
A B
C D
��
�
�
Figura 4'66. Imágenes de microscopía electrónica de transmisión (MET) de la superficie del tejido de manzana irradiado con luz UV*C 20 minutos, obtenidas al día 7 del almacenamiento (A*D). pc: pared celular; c: citoplasma.
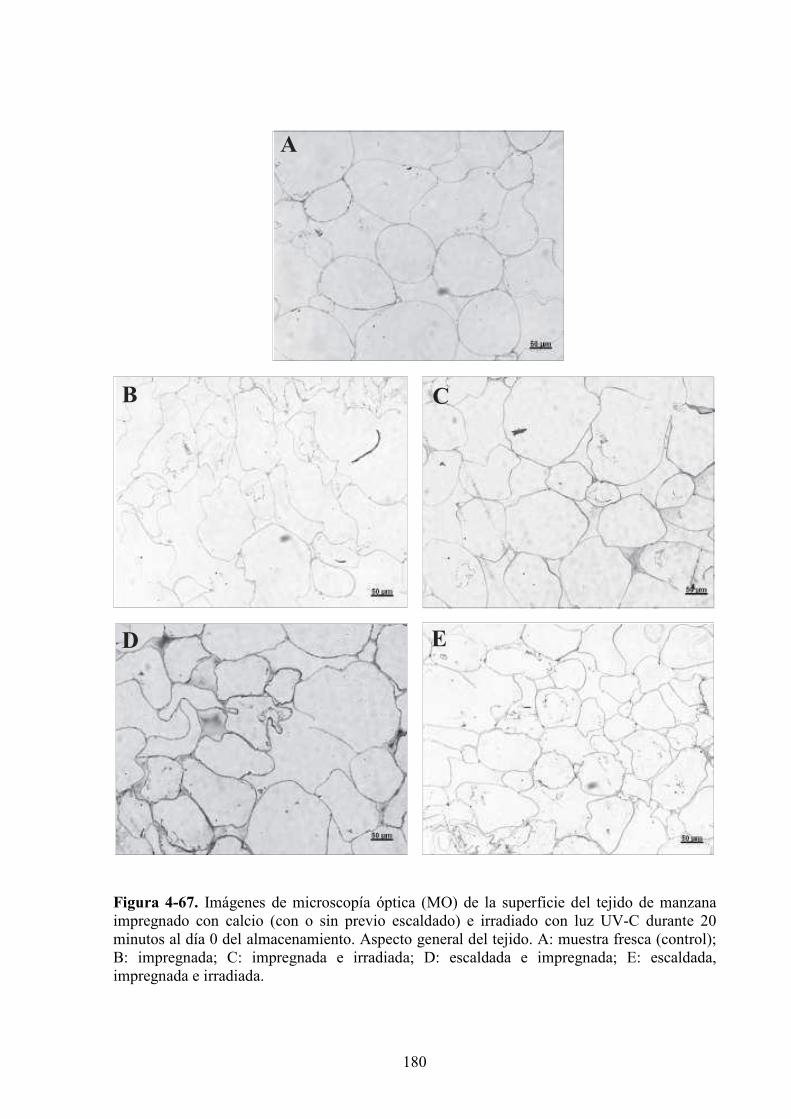
180
A
B C
D E
Figura 4'67. Imágenes de microscopía óptica (MO) de la superficie del tejido de manzana impregnado con calcio (con o sin previo escaldado) e irradiado con luz UV*C durante 20 minutos al día 0 del almacenamiento. Aspecto general del tejido. A: muestra fresca (control); B: impregnada; C: impregnada e irradiada; D: escaldada e impregnada; E: escaldada, impregnada e irradiada.

181
A
B C
D E
Figura 4'68. Imágenes de microscopía óptica (MO) de la superficie del tejido de manzana impregnado con calcio (con o sin previo escaldado) e irradiado con luz UV*C durante 20 minutos al día 0 del almacenamiento. Detalle. A: muestra fresca (control); B: impregnada; C: impregnada e irradiada; D: escaldada e impregnada; E: escaldada, impregnada e irradiada.
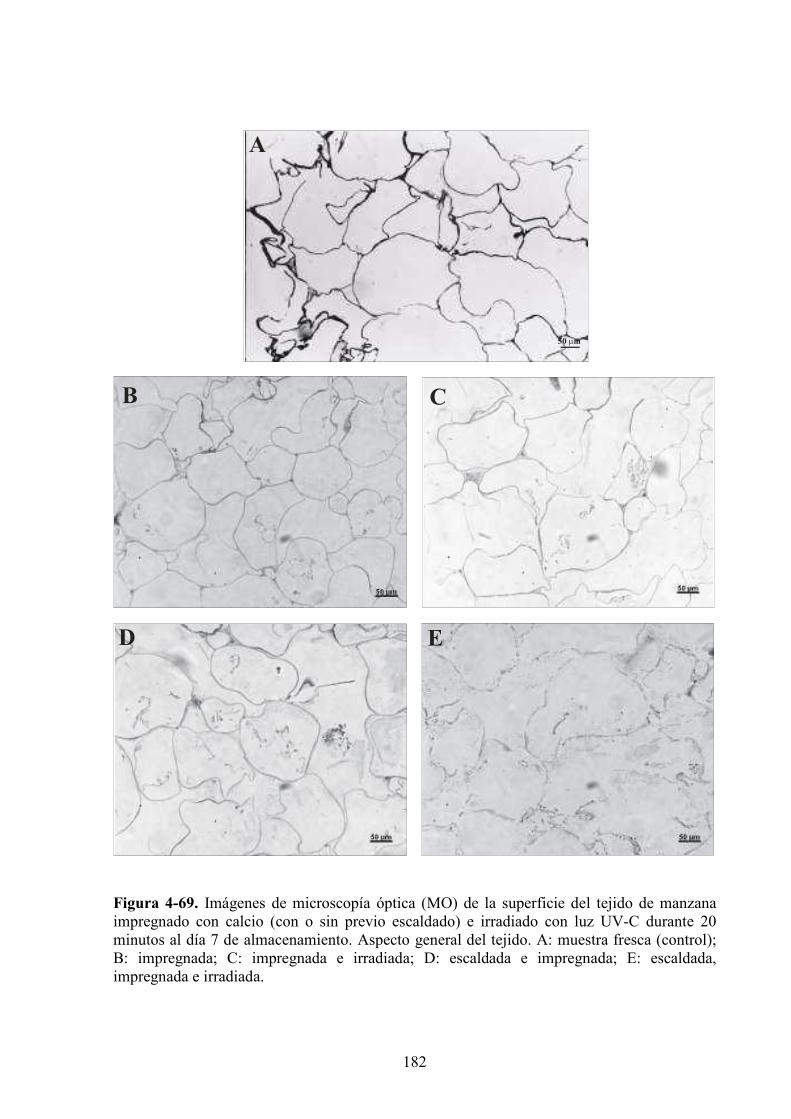
182
B C
50 m��
A
D E
Figura 4'69. Imágenes de microscopía óptica (MO) de la superficie del tejido de manzana impregnado con calcio (con o sin previo escaldado) e irradiado con luz UV*C durante 20 minutos al día 7 de almacenamiento. Aspecto general del tejido. A: muestra fresca (control); B: impregnada; C: impregnada e irradiada; D: escaldada e impregnada; E: escaldada, impregnada e irradiada.

183
A B
C D
Figura 4'70. Imágenes de microscopía óptica (MO) de la superficie del tejido de manzana impregnado con calcio (con o sin previo escaldado) e irradiado con luz UV*C durante 20 minutos al día 7 de almacenamiento. Detalle. A: muestra impregnada; B: impregnada e irradiada; C: escaldada e impregnada; D: escaldada, impregnada e irradiada.

184
8$#$?$�"���������������������
Se realizaron algunos estudios microbiológicos complementarios para analizar la
evolución de la población microbiana (ya sea microorganismos inoculados o flora nativa)
durante el almacenamiento de las rodajas de manzana irradiadas con luz UV*C, como así
también para evaluar si la aplicación de los pretratamientos antipardamiento propuestos
modificaban el patrón de inactivación observado en las muestras irradiadas. Con este último
fin, se analizó en particular el efecto de la aplicación de la solución de ácido ascórbico/cloruro
de calcio ya que fue el pretratamiento que mejor permitió conservar el color y las propiedades
mecánicas de las muestras durante el almacenamiento.
La figura 4'71 muestra la cinética de destrucción de L. innocua, E. coli y S. cerevisiae
en las rodajas de manzana pretratadas con la solución de ácido ascórbico/cloruro de calcio
(SA) y posteriormente irradiadas con luz UV*C. El tratamiento antipardeamiento modificó en
gran medida los perfiles de las curvas de inactivación en relación a los obtenidos en las
muestras irradiadas sin dicho pretratamiento (ver figura 4'1). Por ejemplo, en las muestras
inmersas en la SA e irradiadas durante 20 min (dosis ~11,2 kJ/m2) los ciclos de reducción
logarítmica de las diferentes especies analizadas variaron entre 0,2 y 0,7, mientras que en las
rodajas irradiadas sin pretratamiento las reducciones fueron mayores, variando en un rango
entre 1 y 1,9.
1,5
1,3
1,1
0,9
0,7
0,5
0,3
0,1
0 5 10 15 20 25����������
����� �
�
�������
�������
���� � �����
Figura 4'71. Curvas semilogarítmicas de supervivencia de algunas especies de microorganismos en rodajas de manzanas inmersas en solución antipardeamiento e irradiadas con luz UV*C. N0 E. coli: 1,5 x 106 UFC/cm2; N0 L. innocua: 1,5 x 106 UFC/cm2
; N0 S.
cerevisiae: 4,4 x 104 UFC/cm2.

185
La disminución en la inactivación observada al tratar las muestras con la solución de
ácido ascórbico/cloruro de calcio previamente a la irradiación probablemente se deba a la
capacidad antioxidante del ácido ascórbico, que pudo actuar revirtiendo el efecto germinicida
de luz UV*C sobre los microorganismos. Si bien no se han encontrado trabajos reportados que
estudien el efecto de la presencia de antioxidantes como el ácido ascórbico en el daño causado
por la luz UV*C sobre los microorganismos, en células epidérmicas de humanos se ha visto
que la aplicación conjunta de ácido ascórbico y D*α*tocoferol es efectiva para proteger contra
el daño causado por la luz UV (UV*B) en el ADN de estas células, el que puede conducir al
cáncer de piel (Placzek y col., 2005). Utilizando anticuerpos monoclonales contra los dímeros
de timina formados luego de la exposición a la luz UV, estos autores observaron que la
administración continua de estos antioxidantes reducía significativamente la formación de
dichos dímeros.
La evolución de los microorganismos inoculados durante el almacenamiento en las
rodajas de manzana irradiadas 20 minutos (con o sin previa inmersión en la SA) y en los
controles no irradiados se muestra en la figura 4'72. En el caso de E .coli, como era de
esperar, debido a la baja temperatura de almacenamiento y el bajo pH de las muestras, no se
produjo un incremento de la población durante el almacenamiento tanto en las muestras
controles como en las tratadas (figura 4'72 a). La población de L. innocua aumentó
levemente en los controles y en las rodajas inmersas en SA sin irradiar (≈ 0,24 ciclos
logarítmicos) luego de 7 días de almacenamiento. En las rodajas irradiadas (con o sin SA), los
recuentos permanecieron relativamente constantes durante el almacenamiento (entre 0,8 y 1
ciclo log de reducción) (figura 4'72 b). Por otro lado, la población inicial de S. cerevisiae se
incrementó durante el almacenamiento en las rodajas no irradiadas (0,5 y 0,6 ciclos
logarítmicos en el control y en las muestras con SA, respectivamente) y también en las
rodajas irradiadas sin pretratamiento. En este último caso, si bien inicialmente después de la
irradiación se logró una reducción de ≈ 1,3 ciclos logarítmicos, al final del almacenamiento
los ciclos log de reducción fueron de ≈ 0,5. Por el contrario, en las rodajas irradiadas inmersas
en la solución antipardeamiento los niveles iniciales de inactivación (≈ 0,2 ciclos log)
permanecieron constantes durante el almacenamiento (figura 4'72 c).
En la figura 4'73 puede observarse la respuesta de la flora nativa (aerobios mesófilos y
hongos y levaduras) luego de la aplicación de los tratamientos analizados y durante el
almacenamiento refrigerado. Los recuentos iniciales de microorganismos aerobios mesófilos
en las muestras control y tratada con SA fueron de 57*66 UFC/g, y los recuentos de hongos y

186
a)
2
1,5
1
0,5
0
0,5
1
0 3 6 9
���������������������������
����� �
�
Control SA
UVC SA+UVC
b)
2
1,5
1
0,5
0
0,5
1
0 2 4 6 8
���������������������������
����� �
�
Control SA
UVC SA+UVC
c)
2
1,5
1
0,5
0
0,5
1
0 2 4 6 8
���������������������������
����� �
�
Control SA
UVC SA+UVC
Figura 4'72. Supervivencia de microorganismos inoculados en rodajas de manzana irradiadas con luz UV*C durante 20 minutos (con o sin solución antipardeamiento (SA)) a lo largo del almacenamiento a 4*5 ºC en oscuridad. a) E. coli, N0: 1,5 x 106 UFC/cm2; b) L. innocua, N0: 9,2 x 105 UFC/cm2; c) S. cerevisiae, N0: 4,3 x 104 UFC/cm2.
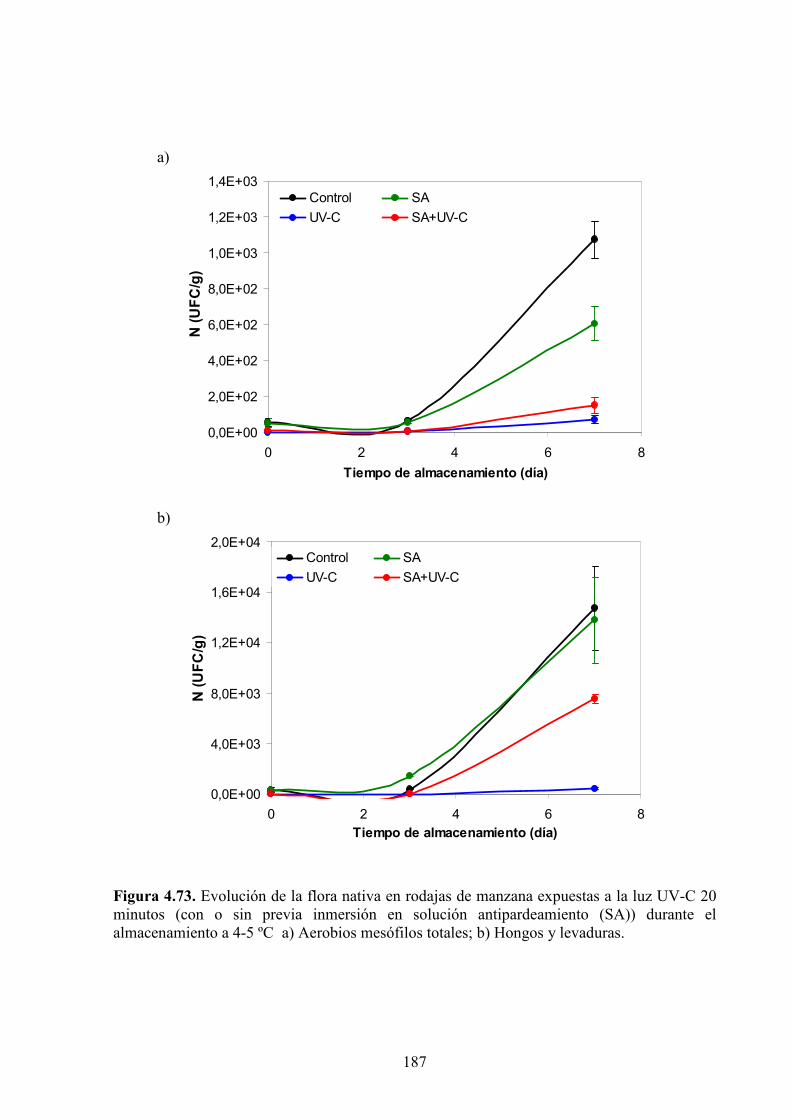
187
a)
0,0E+00
2,0E+02
4,0E+02
6,0E+02
8,0E+02
1,0E+03
1,2E+03
1,4E+03
0 2 4 6 8
���������������������������
��(3���
Control SA
UVC SA+UVC
b)
0,0E+00
4,0E+03
8,0E+03
1,2E+04
1,6E+04
2,0E+04
0 2 4 6 8
���������������������������
��(3���
Control SA
UVC SA+UVC
Figura 4.73. Evolución de la flora nativa en rodajas de manzana expuestas a la luz UV*C 20 minutos (con o sin previa inmersión en solución antipardeamiento (SA)) durante el almacenamiento a 4*5 ºC a) Aerobios mesófilos totales; b) Hongos y levaduras.

188
levaduras de 314*391 UFC/g. La aplicación de luz UV*C (dosis ~11,2 kJ/m2) produjo una
reducción de la flora nativa de aproximadamente 1,8 ciclos logarítmicos, mientras que en las
rodajas irradiadas previamente inmersas en SA la reducción fue de 0,8 ciclos logarítmicos
para aerobios mesófilos y de 1,2 ciclos logarítmicos para hongos y levaduras.
A pesar que durante el almacenamiento se observó crecimiento en todas las muestras,
los recuentos fueron siempre mayores en las rodajas no irradiadas que en las muestras tratadas
con luz UV*C. En todos los casos el mayor crecimiento se observó después del tercer día del
almacenamiento. Al día 7, el recuento de aerobios mesófilos en el control (1x103 UFC/g) fue
levemente mayor al de las rodajas con SA (6x102 UFC/g). En las rodajas irradiadas los
recuentos respectivos para las muestras con o sin SA fueron de 1,4x102 UFC/g y 72 UFC/g, lo
que equivaldría a una disminución con respecto al control de 0,8 y 1,2 ciclos logarítmicos. En
cuanto al recuento final de hongos y levaduras, en las muestras no irradiadas fue de 1,4x104 *
1,5x104 UFC/g, mientras que en las rodajas expuestas a la radiación UV*C la población se
incrementó a 4x102 UFC/g en las muestras sin tratamiento antipardeamiento y a 7,5 x103 en
las muestras que recibieron el pretratamiento con SA. Estas diferencias equivalen a una
reducción con respecto al control de 1,5 y 0,3 ciclos logarítmicos respectivamente.
Los resultados de trabajos publicados en bibliografía que analizan el efecto de la
radiación UV*C C sobre la flora nativa de diferentes frutas y vegetales mínimamente
procesados son diversos. Ello indica que la efectividad de la luz UV*C como tecnología
descontaminante depende de la matriz a la que se aplica, reafirmando lo mencionado en la.
Sección 4.1.1. Por ejemplo, Lamikanra y col. (2005) encontraron una disminución en el
crecimiento de la flora nativa de melones mínimamente procesados expuestos a la radiación
UV*C durante 4 minutos (dosis~ 11,8 J/m2) y almacenados a 10 ºC. Después de seis días de
almacenamiento, los melones irradiados presentaron una reducción con respecto al control no
irradiado de 3*3,5 ciclos logarítmicos en el recuento de microorganismos aerobios mesófilos y
de hongos y levaduras. Observaron además que se podían obtener mayores reducciones si la
irradiación se aplicaba durante el procesamiento de las muestras (≈5,5 ciclos de reducción en
el recuento de aerobios mesófilos) y no al final del mismo. Por otro lado, Erkan y col. (2001)
analizaron el efecto de la aplicación de luz UV*C sobre la población microbiana de rodajas de
zucchinis. Luego de almacenar las muestras durante 14 días a 10 ºC, observaron una
disminución en los recuentos de bacterias mesófilas y de hongos y levaduras en relación al
control de 2*3 ciclos logarítmicos en las muestras irradiadas con una dosis de 5 kJ/m2. La
aplicación de dosis mayores de irradiación no provocó una mayor inactivación de los
microorganismos. Por el contrario, López*Rubira y col. (2005), al irradiar granadas

189
mínimamente procesadas con dosis entre 0,6 – 14 kJ/m2, no encontraron una disminución
apreciable en el recuento de bacterias mesófilas, psicrotróficas, ácido lácticas, enterobacterias
y hongos y levaduras en relación al crecimiento observado en las muestras no irradiadas a lo
largo del almacenamiento a 5 ºC.
A partir de los resultados obtenidos en este trabajo, es posible concluir que el
tratamiento con luz UV*C con o sin pretratamiento antipardeamiento permitió reducir la carga
microbiana y disminuir o inhibir su crecimiento durante el almacenamiento. Por lo tanto,
desde el punto de vista microbiológico los tratamientos aplicados permitirían prolongar la
vida útil de las rodajas de manzana. Por otro lado, a pesar de que la solución de ácido
ascórbico/cloruro de calcio inhibió en cierta medida los efectos de la luz UV*C sobre los
microorganismos, la aplicación de un pretratamiento antipardeamiento sería necesaria debido
a que el color desarrollado durante el almacenamiento de las muestras sólo irradiadas sería
inaceptable desde el punto de vista del consumidor.
8$#$@$� !���������� ��� ���� ����������� ��������� �� ���+4�� ��� ���� ��������������������� �� ��� ��� ������������������ �� ����������������� ��� ���� ��*���� �������������������������������������������������F�����������������
Los estudios que se encuentran en bibliografía sobre luz ultravioleta aplicada a células
vivas suelen explicar el mecanismo de acción de la misma principalmente a través de los
cambios producidos en el ADN. Sin embargo, otros componentes de los tejidos pueden
absorber también la luz UV*C afectando en este caso la calidad del producto.
El pardeamiento enzimático, una de las principales reacciones de deterioro en vegetales
cortados, se produce principalmente por la oxidación de compuestos fenólicos catalizada por
la enzima polifenoloxida (PPO). En manzanas, se ha reportado que la PPO estaría localizada
tanto en organelas (cloroplastos y mitocondria), donde puede estar unida a la membrana,
como en la fracción soluble de la célula (Nicolas y col., 1994), mientras que los compuestos
fenólicos están localizados fundamentalmente en la vacuola. Las observaciones
microscópicas, realizadas en este trabajo (figuras 4'59 B, 4'60 B, 4'64 A'D y 4'66 A'C),
indicaron claramente que los cambios en el color de las rodajas de manzana irradiadas con luz
UV*C pueden atribuirse, al menos parcialmente, a la ruptura de las membranas celulares, lo
que ocasionaría la pérdida de compartimentalización celular. Esto aumentaría el contacto
enzima*sustrato con el consecuente incremento del pardeamiento enzimático del tejido. Más
aún, se ha reportado también que la irradiación de tejidos de plantas con luz UV*C estimula el

190
metabolismo fenilpropanoide estimulando el pool de sustratos para la reacción de
pardeamiento (Civello, 2006). Según estos autores, la síntesis de novo de los compuestos
fenólicos es inducida para mantener bajo control las especies reactivas de oxígeno generada
por la radiación UV*C, tales como peróxido de hidrógeno, radical superóxido e hidroxilo.
En las rodajas de manzana irradiadas que recibieron un pretratamiento previo de
escaldado o inmersión en solución de ácido ascórbico/cloruro de calcio, a pesar de la ruptura
de membranas (figura 4'59 C'F y 4'60 C'F), el pardeamiento fue controlado mediante la
inactivación de las enzimas, o afectando los sustratos y productos de la reacción de
pardeamiento. Cabe señalar también que ambos pretratamientos pudieron decrecer o eliminar
el fenómeno hormético en los productos. Por un lado el escaldado es un tratamiento térmico
que provoca la muerte celular en los tejidos y los tratamientos con ácido ascórbico antes de la
irradiación pudieron reducir los efectos de la luz UV*C en la expresión de muchos genes
(Civello y col., 2006).
A pesar del efecto mejorador del color que se obtuvo mediante la aplicación de una
inmersión previa en solución de ácido ascórbico/cloruro de calcio, la inactivación de
microorganismos no se vio igualmente afectada. Si bien la aplicación de luz UV*C con o sin
el pretratamiento redujo la carga inicial de microrganismos del producto y el crecimiento se
vio disminuido o inhibido durante el almacenamiento, esta reducción fue menor en el caso de
las manzanas que habían recibido una inmersión previa en la solución antipardeamiento.
Probablemente esto debería a la capacidad antioxidante del ácido ascórbico, que pudo actuar
revirtiendo el efecto germinicida de luz UV*C sobre los microorganismos.
En otra matriz vegetal, pera, y utilizando dosis similares de irradiación, la efectividad de
la aplicación de luz UV*C con respecto a la inactivación de microorganismos fue mayor
(Schenk y col., 2008). Las diferencias observadas en el efecto germicida logrado en ambas
matrices pueden atribuirse, al menos parcialmente, a las diferentes porosidades de los tejidos,
lo que permitiría en mayor o menor grado la penetración de los microorganismos inoculados
hacia el interior del tejido, disminuyendo así el efecto de la irradiación
El pardeamiento desarrollado en las rodajas de manzana enriquecidas con calcio puede
ser también explicado por la ruptura de membranas celulares en toda la rodaja provocada por
el tratamiento de impregnación con calcio a presión atmosférica (figura 4'67 B y 4'68 B). La
exposición posterior de estos tejidos a la luz UV*C indujo un daño adicional (figura 4'67 C y
4'68 C), que se vió reflejado en un leve incremento del pardeamiento a las dosis de
irradiación más altas. Al igual que en las rodajas de manzana fresca irradiadas, el escaldado y

191
la inmersión en la solución de ácido ascórbico/cloruro de calcio previos a los tratamientos de
IA e irradiación contribuyeron a controlar el desarrollo del pardeamiento.
El comportamiento mecánico de los tejidos de manzana sometidos a los diferentes
tratamientos también puede ser correlacionado con las observaciones microestructurales y
ultraestructurales. A nivel celular y de tejido, los tres principales factores estructurales que
contribuyen al comportamiento mecánico de frutas y vegetales son la presión de turgor
(fuerza ejercida sobre la pared celular por el fluido intracelular), la rigidez de la pared celular,
y la adhesión célula*célula, determinada por la integridad de la laminilla media y los
plasmodesmos (Jackman & Stanley, 1995; Waldron y col., 1997; Alzamora y col., 2000). La
presión de turgor conduce a la rigidez de las células y tejidos vegetales, y junto con la pared
celular, proveen el soporte mecánico para mantener la forma de las células y tejidos
(Alzamora y col., 2000).
La elasticidad inducida por la presión de turgor y el aire en los espacios intracelulares
en las células y tejidos podrían estar relacionados con los valores del módulo elástico (G’)
(Alzamora y col., 2008). Diversos autores han reportaron una relación lineal entre la presión
de turgor y el módulo elástico en diferentes frutas y vegetales (Lin & Pitt, 1986; Jackman &
Stanley, 1992; Rojas y col., 2001).
Las rodajas de manzana expuestas a la luz UV*C presentaron una disminución de los
módulos G’ y G’’ en relación con las muestras frescas (control), tanto al día 0 como al final
del almacenamiento. Estas diferencias en el espectro mecánico podrían relacionarse entonces
con una disminución de la turgencia del tejido irradiado como consecuencia de la ruptura de
membranas celulares provocada por la radiación UV*C. Dicha pérdida se incrementó con la
mayor pérdida de agua atribuible a la ruptura del tejido superficial y/o la mayor velocidad de
transpiración que presentaron los tejidos irradiados durante el almacenamiento.
Los ensayos de fluencia*recuperación, al igual que los ensayos oscilatorios, fueron
sensibles para detectar los cambios estructurales en el tejido irradiado. Estos resultados
pueden ser analizados teniendo en cuenta la propuesta estructural de Alzamora y col. (2008).
Estos autores adaptaron las interpretaciones realizadas por Jackman y Stanley (1995) para
poder llegar a relacionar los cambios producidos en los parámetros de fluencia en tejidos de
manzana y de melón con los cambios en la estructura de la pared celular y en la turgencia de
las células debido a distintos tratamientos. De este análisis surge que la capacitancia elástica
instantánea J0 podría relacionarse con la presión de turgor y la fuerza de la pared celular
primaria dada a través de la celulosa. Las modificaciones en las capacitancias viscoelásticas J1
y J2 podrían atribuirse a cambios dependientes del tiempo de las pectinas y las hemicelulosas

192
respectivamente. La viscosidad en estado estacionario podría relacionarse con el incremento
en la fluidez de la pared celular provocado por la solubilización y degradación de polímeros,
menor capacidad de unión de éstos con el agua y exoosmosis.
Las capacitancias J0, J1 y J2 de las manzanas irradiadas se incrementaron
significativamente durante el almacenamiento, mientras que en las muestras control no se
observaron cambios significativos. Estos cambios en J0 podrían ser relacionados en parte con
la pérdida de turgor observada en el tejido irradiado, cuyas paredes aparecieron mucho más
colapsadas a los 7 días de almacenamiento. Las observaciones realizadas con MET (figura. 4'
66 A'C) denotan paredes celulares plegadas y onduladas con poca definición de la laminilla
media, lo que justificaría el aumento observado en las capacitancias.
A pesar de que en los ensayos reológicos realizados a bajas deformaciones se
encontraron cambios en las propiedades viscoelásticas del tejido de manzana por efecto de la
irradiación UV*C, estas diferencias, en general, no fueron apreciadas en los ensayos de
compresión. Esto se debería a que la irradiación con luz UV*C es un tratamiento superficial
que tiene baja penetración en el tejido. Por lo tanto, los cambios estructurales observados se
darían en la superficie de las muestras. Como en los ensayos de compresión la conducta
mecánica se analiza a grandes deformaciones abarcando gran parte del tejido de la muestra,
estas diferencias no pudieron ser apreciadas. Sólo se evidenció una leve pero significativa
disminución del módulo de deformabilidad al final del almacenamiento, especialmente a las
mayores dosis de irradiación. Ello indicaría una ligeramente mayor habilidad del tejido para
resistir a la ruptura (el tejido fue más deformable), lo que coincidiría con el mayor colapso y
plegamiento de las paredes del tejido irradiado observado a los 7 días de almacenamiento.
El tratamiento de escaldado previo a la irradiación UV*C, a diferencia del
pretratamiento de inmersión con la solución antipardeamiento, indujo cambios notorios en las
propiedades reológicas del tejido de manzana, asociado con cambios estructurales profundos.
Las muestras escaldadas presentaron una disminución marcada en los valores del
esfuerzo de ruptura (σR) y del módulo de deformabilidad (Ed), reflejando una pérdida de
rigidez del tejido, una mayor habilidad para resistir la ruptura y una menor resistencia a la
ruptura. Estos cambios en las propiedades mécanicas estarían asociados con alteraciones
observadas en la estructura del tejido tales como la ruptura de membranas celulares, la pérdida
de presión de turgor y el daño y disociación de las paredes celulares, puestos de manifiesto en
la micrografía por zonas alternadas de mucha menor densidad de tinción (figura 4'59 C). La
marcada disminución de los valores de G’ luego del tratamiento térmico, indicativa también
de una pérdida de rigidez del tejido, estaría relacionada fundamentalmente con la pérdida de

193
la presión de turgor y la remoción del aire en los espacios intracelulares. Por otro lado, el
incremento de las capacitancias J0, J1, J2 y 1/ηN en las muestras escaldadas estaría asociado
también a la pérdida de turgor, a la disrupción de la red de microfibrillas y de la red de
sustancias pécticas y al aumento de la fluidez en el tejido provocadas por el tratamiento
térmico.
Si bien en algunos ensayos se detectaron cambios en las propiedades viscoelásticas de
las rodajas escaldadas por efecto de la irradiación UV*C, éstos fueron mucho menores a los
provocados por el tratamiento térmico. Este fenómeno respondería a que las alteraciones
estructurales provocadas por el escaldado fueron más profundas que las producidas por la
radiación UV*C.
El tratamiento de impregnación con sales de calcio produjo una disminución en el
módulo elástico G’ del tejido de manzana impregnado en comparación con el tejido fresco.
Sin embargo, en los parámetros del modelado de la curva de fluencia para el día 0 del
almacenamiento, si bien las capacitancias promedio J0, J1 y J2 fueron mayores que las del
control, los incrementos debido a la incorporación de calcio no fueron significativos. La
disminución de G’ estaría asociada con la pérdida de turgencia de los tejidos impregnados
provocada fundamentalmente por la plasmólisis de membranas celulares (figura 4'68 B). La
posterior irradiación de los tejidos impregnados provocó un incremento en la capacitancia
elástica J0, lo que podría relacionarse con la mayor disrupción de membranas celulares que se
observaron en estos tejidos. Al cabo del almacenamiento, los valores de G’ disminuyeron y
las capacitancias J0 y J1 se incrementaron en los tejidos impregnados, no presentándose
diferencias por efecto de la exposición a la luz UV*C. Estos cambios podrían estar asociados
con la pérdida de agua de los tejidos por transpiración y por la ruptura de membranas y
modificaciones producidas en las paredes celulares del tejido durante el almacenamiento.
El escaldado previo a la impregnación produjo un incremento de la concentración de
calcio en el tejido, lo que se vió reflejado en las observaciones de MO en paredes celulares
con buena densidad óptica (figura 4'68 D). Sin embargo, el posible reforzamiento de las
paredes celulares provocado por la interacción del calcio con los componentes de la pared no
se vio reflejado en la conducta viscoelástica de estos tejidos. Los valores de G’ y de las
capacitancias obtenidas a partir del modelado de la curva de fluencia (J0, J1, J2 y1/ηN)
disminuyeron y aumentaron respectivamente en las rodajas escaldadas e impregnadas, en
mayor proporción que en los tejidos impregnados no escaldados, tanto al día 0 como al final
del almacenamiento. La posterior irradiación de estos tejidos no produjo cambios en las
propiedades reológicas medidas, a pesar de que en las observaciones microscópicas se

194
observaron células con mayor disrupción de membranas y paredes celulares más debilitadas al
final del almacenamiento (figuras 4'69 E y 4'70 E).
En estudios previos realizados en el grupo de trabajo, donde se impregnaron rodajas de
manzana en soluciones de glucosa con o sin sales de calcio, a pesar de que las células de los
tejidos impregnados con calcio exhibieron paredes celulares más teñidas, y en algunos casos
con una laminilla media también más reforzada, tampoco se encontró un aumento en la
rigidez del tejido por efecto de la incorporación de calcio. Esto fue atribuido a que el
tratamiento de impregnación con calcio provocó también otros cambios estructurales, como
disrupción de células, plasmólisis y plegamiento de paredes celulares, provocando una
pérdida de la resistencia mecánica (Salvatori y col., 2010).
De acuerdo a la cantidad de calcio incorporada después de 4,5 h de tratamiento de
impregnación (IA), la cantidad presente en 200 g de fruta (ración diaria) satisfacerla alrededor
del 14 % de la Ingesta Adecuada (1000 mg / día). Con escaldado previo se consigue mejorar
significativamente la capacidad de impregnación de la matriz, ya que es posible alcanzar un
contenido de 2734 ppm de calcio frente a 721 ppm obtenidas en muestras no impregnadas.
Ello implicaría con una ración diaria de este producto cubrir el 55 % de la Ingesta Adecuada
para adultos de este macronutriente. Cabe aclarar que de acuerdo a los estudios de absorción
relativa en ratas, la manzana impregnada con lactato y gluconato de calcio es un vehículo para
proveer calcio fácilmente absorbible. Cuando las matrices son escaldadas e impregnadas a
presión atmosférica por largos periodos de tiempo se obtiene una mayor absorción de este
nutriente en el organismo (Salvatori y col., 2007).

195
8$,$���5!���!.�� "���5)%)� "�5�E� "��5(��!�("�)! � �� 8$,$#$�)���������������������
En la bibliografía existente sobre la aplicación de pulsos de luz (LP) en alimentos,
muchos autores coinciden en que el rango de dosis utilizado para lograr la inactivación de
microorganismos estaría condicionado por el aumento de temperatura generado en las
muestras a dosis crecientes de irradiación (Jun y col., 2003; Gómez*López y col., 2005 a y b;
Krishnamurthy y col. 2007). Por lo tanto, es importante analizar la tolerancia de cada alimento
a la irradiación sin que el incremento de temperatura y la luz absorbida lleguen a producir
efectos adversos sobre la calidad.
En este trabajo se decidió evaluar la tolerancia de las rodajas de manzana a la
irradiación con luz pulsada en un rango de dosis amplio, teniendo en consideración los
estudios de inactivación microbiológica reportados en bibliografía. Las diferentes dosis se
obtuvieron variando la distancia de las muestras a la lámpara (5 y 10 cm) y el tiempo
exposición a la irradiación (2*100 segundos). Por otro lado, teniendo en cuenta que en los
tratamientos de irradiación con luz UV*C continua el atributo de calidad que se veía más
afectado con el incremento de la dosis era el color, se decidió evaluar en primera instancia el
efecto de la dosis de LP sobre este parámetro de calidad.
A pesar de que la determinación de la dosis es un factor importante para caracterizar los
tratamientos de irradiación, las dosis que fueron utilizadas en las experiencias no pudieron ser
determinadas porque hasta el momento no se cuenta con un radiómetro adecuado para realizar
la medición ni tampoco fue posible utilizar algún otro método alternativo de medición. El
único dato de dosis disponible es el provisto por el fabricante del equipo de LP, que reporta
que la energía emitida por cada pulso a una distancia de 1,92 cm de la lámpara es de 12,7
kJ/cm2.
�8$,$,$�"+�������������������������
Se analizó la evolución de la temperatura durante la irradiación con luz pulsada en la
superficie de las muestras y en la cara inferior no expuesta a la irradiación. Las mediciones se
realizaron ubicando las muestras a diferentes distancias de la lámpara (5, 10 y 15 cm) y en
distintas posiciones sobre el estante (1*7) (ver figura 3'4 de la sección Materiales y Métodos).
La temperatura se registró en un intervalo de tiempo de 100 segundos equivalentes a la
emisión de 300 pulsos.

196
En la figura 4'67 se pueden observar los perfiles de temperatura obtenidos en la
superficie y en la cara inferior de las muestras ubicadas en el punto central del estante, a
diferentes distancias por debajo de la lámpara. Como se observa en esta figura, la temperatura
se incrementó con el aumento del número de pulsos emitidos, tanto en la superficie como en
la cara inferior de la muestras. Este incremento fue mayor a medida que disminuyó la
distancia a la cual se ubicaron las muestras con respecto a la lámpara. Por otra parte, en todas
las distancias evaluadas, el aumento de la temperatura en la parte inferior de la muestra fue
levemente menor al alcanzado en la superficie. En la tabla 4'25 se muestran las temperaturas
registradas en la parte superficial a diferentes tiempos. La temperatura inicial de las muestras
fue de aproximadamente 19*20 ºC, y se incrementó luego de 100 segundos de irradiación a
86, 60 y 44 ºC en las muestras ubicadas a 5, 10 y 15 cm, respectivamente. Estos resultados
permiten confirmar que tanto el tiempo de exposición a la irradiación como la proximidad de
las muestras a la lámpara serán condiciones operativas fundamentales para la elección de un
proceso efectivo desde el punto de vista microbiológico, sin deterioro de la calidad.
Se determinaron también los perfiles de temperatura obtenidos en la superficie de las
muestras colocadas en diferentes posiciones sobre el estante (figura 4'68). El incremento de
temperatura varió según la posición de las muestras en el estante, siendo más notoria está
variación cuando el estante fue ubicado a menores distancias de la lámpara (5 y 10 cm).
Cuando el estante fue colocado a 5 cm de la lámpara, los incrementos de la temperatura
en la posición central del estante (posición 3) y en los puntos ubicados debajo de la lámpara a
10 cm del centro (posiciones 2 y 4) fueron similares, y mayores a los registrados en las
posiciones 1 y 5 (debajo de los extremos de la lámpara) y posiciones 6 y 7 (puntos laterales
ubicados a 8 cm del centro) (figuras 4'68 a). Las variaciones en los perfiles de temperatura
entre las diferentes ubicaciones disminuyeron a medida que el estante fue colocado más
alejado de la lámpara (figuras 4'68 b y c). Esto indicaría que a mayores distancias de la
lámpara la incidencia de la luz sobre la superficie del estante fue más homogénea.
Gómez*López y col. (2005 a) analizaron la influencia en la eficiencia de la
decontaminación con LP de la posición relativa entre la muestra y la lámpara. Observaron
que cuanto mayor era la distancia vertical entre la muestra y la lámpara, menor era la letalidad
del proceso. Por otro lado, en las muestras ubicadas más próximas a la lámpara, la
decontaminación era mayor cuando se ubicaban directamente debajo de la lámpara, mientras
que en los costados la decontaminación era casi nula. Cuando se colocaban más alejadas de la
lámpara, la inactivación debajo de la lámpara era menor, sin embargo en los bordes
aumentaba.
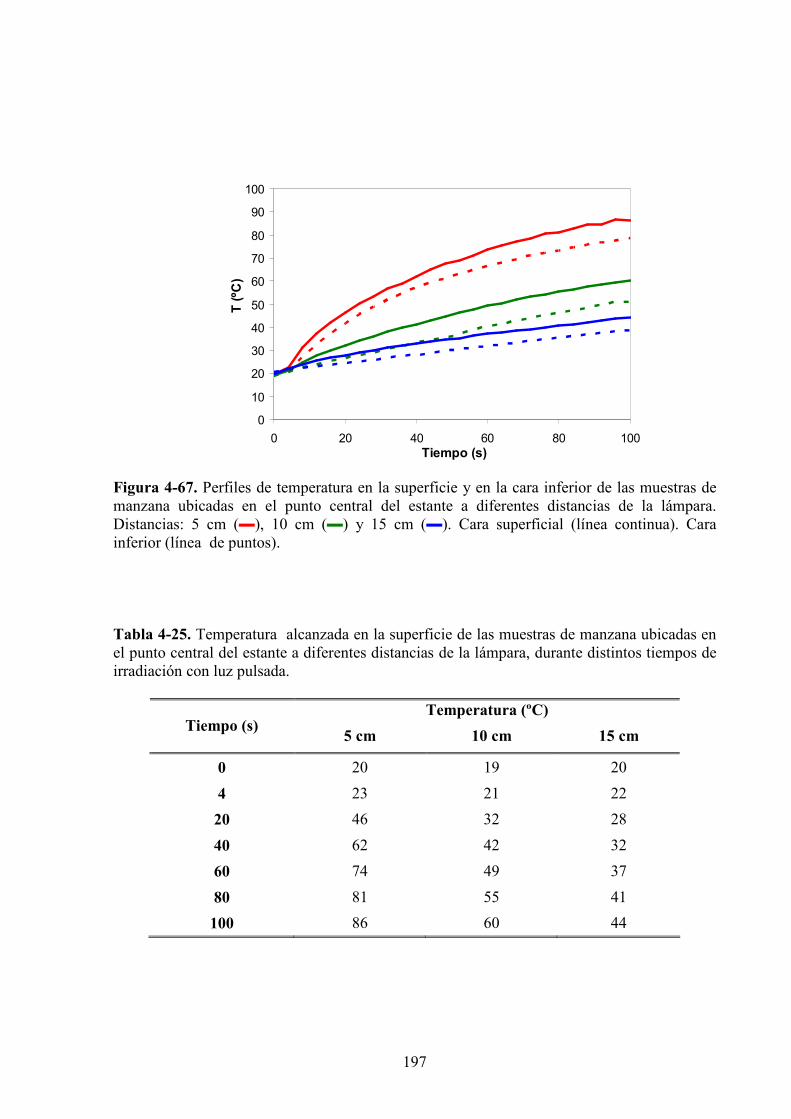
197
0
10
20
30
40
50
60
70
80
90
100
0 20 40 60 80 100
���������
���*�
Figura 4'67. Perfiles de temperatura en la superficie y en la cara inferior de las muestras de manzana ubicadas en el punto central del estante a diferentes distancias de la lámpara. Distancias: 5 cm (▬), 10 cm (▬) y 15 cm (▬). Cara superficial (línea continua). Cara inferior (línea de puntos). Tabla 4'25. Temperatura alcanzada en la superficie de las muestras de manzana ubicadas en el punto central del estante a diferentes distancias de la lámpara, durante distintos tiempos de irradiación con luz pulsada.
Temperatura (ºC) Tiempo (s)
5 cm 10 cm 15 cm
0 20 19 20
4 23 21 22
20 46 32 28
40 62 42 32
60 74 49 37
80 81 55 41
100 86 60 44
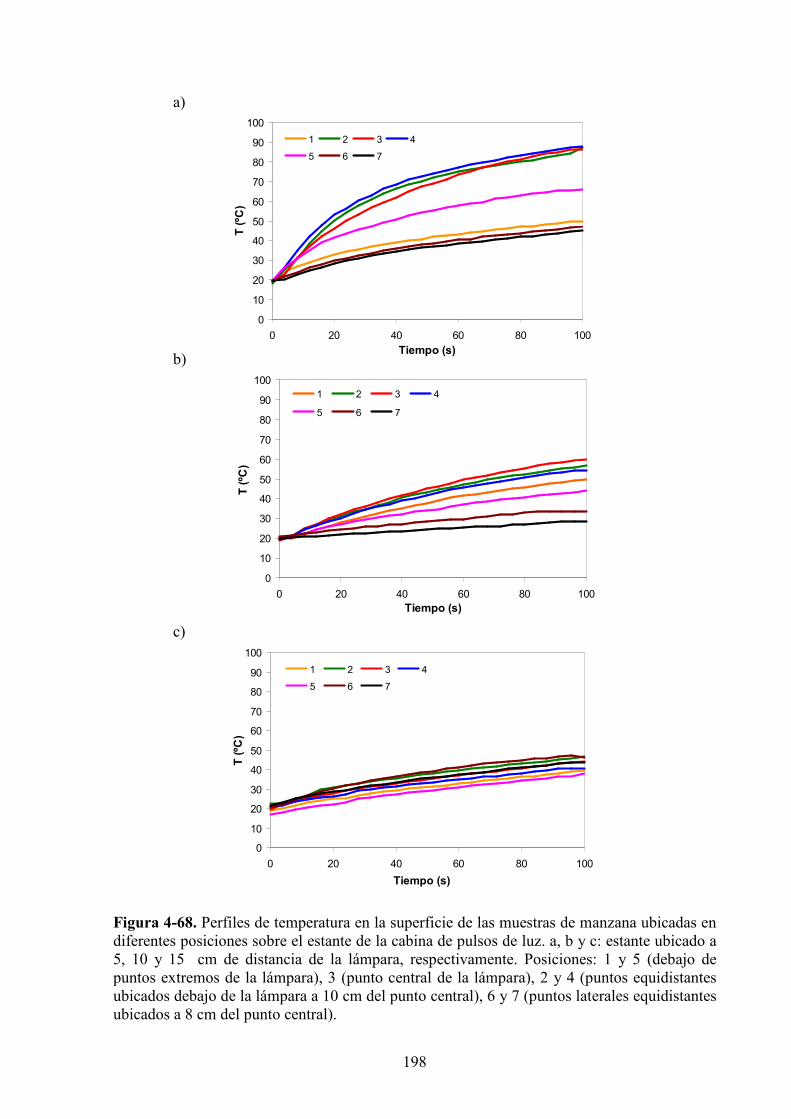
198
a)
b)
0
10
20
30
40
50
60
70
80
90
100
0 20 40 60 80 100
���������
���*�
1 2 3 4
5 6 7
0
10
20
30
40
50
60
70
80
90
100
0 20 40 60 80 100
���������
���*�
1 2 3 4
5 6 7
c)
0
10
20
30
40
50
60
70
80
90
100
0 20 40 60 80 100
���������
���*�
1 2 3 4
5 6 7
Figura 4'68. Perfiles de temperatura en la superficie de las muestras de manzana ubicadas en diferentes posiciones sobre el estante de la cabina de pulsos de luz. a, b y c: estante ubicado a 5, 10 y 15 cm de distancia de la lámpara, respectivamente. Posiciones: 1 y 5 (debajo de puntos extremos de la lámpara), 3 (punto central de la lámpara), 2 y 4 (puntos equidistantes ubicados debajo de la lámpara a 10 cm del punto central), 6 y 7 (puntos laterales equidistantes ubicados a 8 cm del punto central).

199
Teniendo en cuenta las variaciones de la temperatura en las diferentes posiciones sobre
el estante, y para poder asegurar que la irradiación de las muestras en los tratamientos fuera
uniforme, en los estudios posteriores éstas fueron ubicadas siempre en fila debajo de la
lámpara y a una distancia no mayor de 10 cm del centro de la misma.
�
8$,$7$�"����������������������������
�
8$,$7$#$�!�������������5��
�
El efecto de la irradiación con luz pulsada sobre el color de las rodajas de manzana se
evaluó ubicando las muestras a una distancia de la lámpara de 5 y 10 cm e irradiándolas
durante 2, 20, 60 y 100 segundos.
En todos los parámetros de color analizados se presentó interacción significativa
(p<0,0001) entre el tiempo de almacenamiento y el tiempo de irradiación.
El color inicial de las rodajas de manzana frescas se caracterizó por presentar valores de
L* ≈ 70, a* ≈ 6,3, b* ≈ 22, h ≈106 y C ≈ 22. En las figuras 4'69 y 4'70 se muestran las
variaciones en los parámetros y en las funciones de color L*, a*, b*, h y C (con respecto a los
valores iniciales de la muestra fresca) a lo largo del almacenamiento de las rodajas irradiadas
a una distancia de 5 cm de la lámpara. Las muestras que fueron irradiadas durante 20, 60 y
100 segundos presentaron una disminución de L* y un aumento de a* mayor que el control a
lo largo de todo el almacenamiento, mientras que las irradiadas durante 2 segundos no
mostraron variaciones significativas con respecto al control. Al día 0, la disminución de la
luminosidad en las rodajas irradiadas 60 y 100 segundos fue similar, y mayor a la que
presentaron las muestras irradiadas 20 segundos, mientras que los valores de a* se
incrementaron gradualmente con el aumento del tiempo de irradiación. Después de
transcurridos siete días, la disminución de la luminosidad de las muestras control e irradiadas
2 s fue de aproximadamente 7%, y el aumento de a* de 60*67%, mientras que en las rodajas
irradiadas a tiempos mayores la disminución de L* fue de alrededor de 15*20%, y el
incremento de a* de 120*150%. (figura 4'69 a y b). Por otro lado, los valores del parámetro
b* se incrementaron en las muestras recién irradiadas, siendo el incremento mayor en las
muestras tratadas durante 60 y 100 segundos, pero al tercer y séptimo día, las variaciones de
b* de las muestras irradiadas no presentaron diferencias significativas en relación con el
control (figura 4'69 c).

200
Figura 4'69. Efecto de la dosis de radiación con pulsos de luz sobre la luminosidad (L*), el valor de a* y de b* (expresados como diferencias con respecto a los valores iniciales de la muestra fresca) de rodajas de manzana durante el almacenamiento a 4*5ºC.(d= 5 cm). Para cada tratamiento, las barras con la misma letra minúscula no presentaron diferencias significativas a lo largo del almacenamiento (p<0,05). Para el mismo día de almacenamiento, las barras de diferentes tratamientos con la misma letra mayúscula no presentaron diferencias significativas (p<0,05).
a)
0
5
10
15
20
25
0 3 7���������������������������
�������
Control 2 s LP
20 s LP 60 s LP
100 s LP
aA
cB
aB aB
aA
bB,C
bB
bA aAcA
bA
bC
cBcB
0
1
2
3
4
5
6
7
8
9
10
0 3 7
���������������������������
�����
��
Control 2 s LP20 s LP 60 s LP100 s LP bB
bB
bAaA
aC
aB
aA
aA
cAbA
bB bB
bB
cB
0
3
6
9
12
15
0 3 7
���������������������������
�����
��
Control 2 s LP
20 s LP 60 s LP
100 s LP
aA
bA
bA,B aA,BaB
aB
aA
aA
bA,B bA,B
bB bB
aA
cA
b)
c)

201
Figura 4'70. Valores promedio de las funciones de color ángulo de tono (h) (a) y croma (C) (b) (expresados como diferencias con respecto a los valores iniciales de la muestra fresca) durante el almacenamiento (4*5 ºC) de rodajas de manzana expuestas a diferentes dosis de radiación con luz pulsada.(d = 5 cm) Para cada tratamiento, las barras con la misma letra minúscula no presentaron diferencias significativas a lo largo del almacenamiento (p<0,05). Para el mismo día de almacenamiento, las barras de diferentes tratamientos con la misma letra mayúscula no presentaron diferencias significativas (p<0,05).
0
5
10
15
20
25
30
35
0 3 7
���������������������������
������
Control 2 s LP
20 s LP 60 s LP
100 s LP
bC cB,C
cB
cAbA
bCbB,C
bB
bAaA
aC
aB
aA
aA
a)
0
2
4
6
8
10
12
14
0 3 7
���������������������������
�����
�
Control 2 s LP
20 s LP 60 s LP
100 s LP
bA,B
bA
aA
bA
bAaAaB
aB
aA
aA
bB
aA,B
cA
bA,B
b)

202
El ángulo de tono disminuyó durante el almacenamiento, tanto en las muestras tratadas
como en el control, pero las rodajas irradiadas durante 20, 60 y 100 segundos presentaron
variaciones de h* mayores que el control (figura 4'70 a). Luego de siete días de
almacenamiento, las muestras irradiadas entre 20*100 segundos presentaron valores de h
cercanos a 84*89º, y las muestras control e irradiadas por 2 segundos, valores entre 94*95º.
Por otro lado, los valores de C* aumentaron al inicio del almacenamiento en las rodajas
irradiadas, siendo mayor el incremento en las muestras expuestas durante tiempos de
irradiación más prolongados. Sin embargo, al tercer y séptimo día del almacenamiento, las
variaciones en los valores de C* entre las muestras irradiadas y el control no fueron
significativas (figura 4'70 b). Los valores de C* de las diferentes muestras al final del
almacenamiento rondaron entre 26*29.
Las rodajas ubicadas a 10 cm de distancia de la lámpara también presentaron una
disminución de L* y un incremento de a* mayor que el control cuando fueron irradiadas
durante 20, 60 y 100 segundos (figura 4'71 a y b). El oscurecimiento de las muestras a lo
largo del almacenamiento (mayor cL*) fue más notorio en las irradiadas a las dosis más altas,
mientras que los incrementos en el parámetro a* fueron similares. Las rodajas expuestas a la
irradiación durante 2 segundos no presentaron diferencias significativas en las variaciones de
L* y a* con respecto al control. Al día 7, la disminución de L* de las muestras control e
irradiadas 2 s fue de 7 % y el aumento de a* de 50*60 %, mientras que en las rodajas
irradiadas entre 20*100 s los porcentajes de variación respectivos fueron de 13*18 % para L*
y de 110*120 % para a*. En cuanto al parámetro b*, en las muestras irradiadas durante 60 y
100 segundos el incremento de b* durante el almacenamiento fue mayor que en el control, y
menor en las irradiadas 20 segundos. Las muestras irradiadas 2 segundos no presentaron
variaciones significativas de b* durante los tiempos analizados (figura 4'71 c).
Los valores de h* disminuyeron en todas las muestras (tratadas y control) durante el
almacenamiento. Inmediatamente después de la irradiación (día 0), las variaciones del ángulo
de tono en las muestras irradiadas 60 y 100 segundos fueron mayores que en las irradiadas a
menores tiempos. Al finalizar el almacenamiento, no se presentaron diferencias significativas
en los valores de ch* de las rodajas irradiadas 2 segundos y el control, pero para las
irradiadas durante tiempos más altos las diferencias fueron significativas. Los valores de h* al
final del almacenamiento fueron de 95*99º en las muestras control e irradiadas 2 segundos y
de 87*91º en las rodajas irradiadas a tiempos mayores (figura 4'72 a). Por otra lado, los
valores de croma aumentaron en todas las muestras a lo largo del almacenamiento, excepto en
las irradiadas 2 segundos que no presentaron variaciones significativas en ninguno

203
�
�
�
�
�
�
�
�
�
�Figura 4'71. Efecto de la dosis de radiación con pulsos de luz sobre la luminosidad (L*), el valor de a*y de b* (expresados como diferencias con respecto a los valores iniciales de las muestras frescas) de rodajas de manzana durante el almacenamiento a 4*5ºC.(d = 10 cm). Para cada tratamiento, las barras con la misma letra minúscula no presentaron diferencias significativas a lo largo del almacenamiento (p<0,05). Para el mismo día de almacenamiento, las barras de diferentes tratamientos con la misma letra mayúscula no presentaron diferencias significativas (p<0,05).
0
5
10
15
20
25
0 3 7
���������������������������
�������
Control 2 s LP
20 s LP 60 s LP
100 s LP
aA aA
aB
aB
aA
bA,B bB
bC bC
bA
cA
cB
cB,C
cC
0
1
2
3
4
5
6
7
8
9
10
0 3 7���������������������������
�����
��
Control 2 s LP
20 s LP 60 s LP
100 s LP
aA
cB
cB
cB
bA
bA bB
bB
bA aA
aB
aA,B aA,B
bB
3
0
3
6
9
12
15
0 3 7
���������������������������
�����
��
Control 2 s LP
20 s LP 60 s LP
100 s LP
bCcC
bB
aB
aA aCbC
bB
aB
aA aB
aB
aA
aA
a)
b)
c)
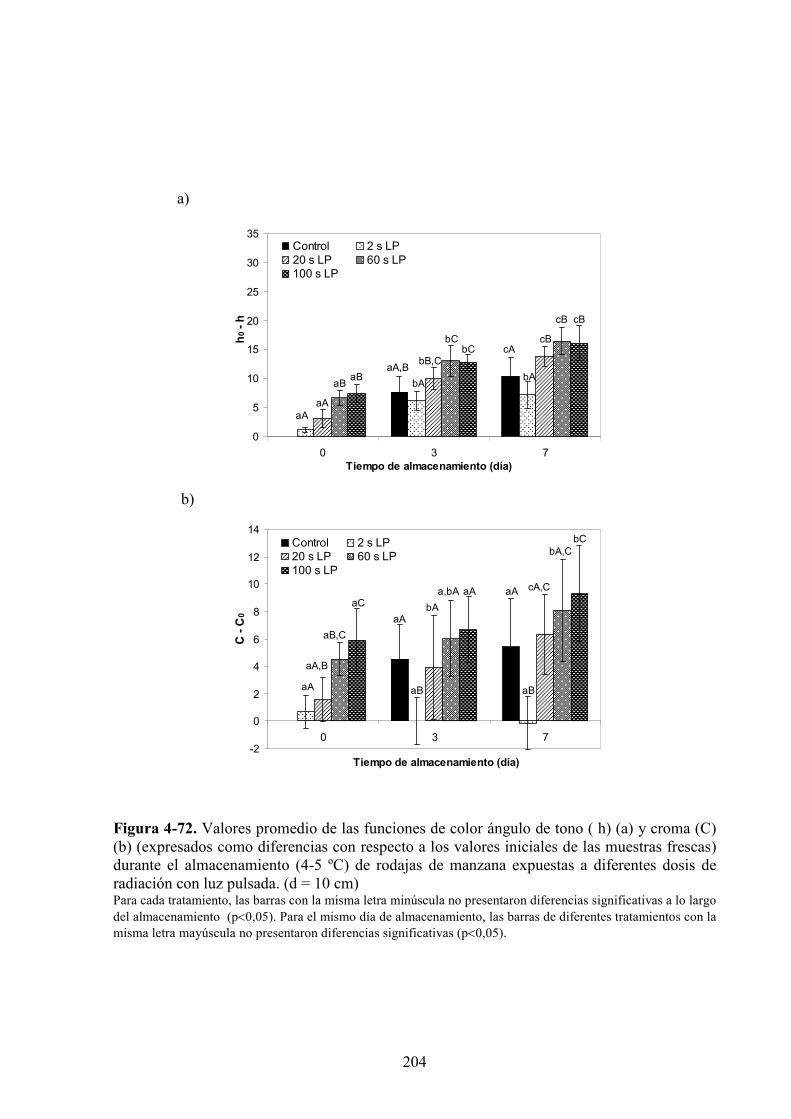
204
a)
0
5
10
15
20
25
30
35
0 3 7
���������������������������
������
Control 2 s LP
20 s LP 60 s LP
100 s LP
cB cB
cB
bA
cAbCbC
bB,C
bA
aA,BaB
aB
aA
aA
b)
2
0
2
4
6
8
10
12
14
0 3 7
���������������������������
�����
�
Control 2 s LP
20 s LP 60 s LP
100 s LP
aA
bC
bA,C
cA,C
aB
aA aA a,bA
bA
aB
aA
aC
aB,C
aA,B
�
�
Figura 4'72. Valores promedio de las funciones de color ángulo de tono ( h) (a) y croma (C) (b) (expresados como diferencias con respecto a los valores iniciales de las muestras frescas) durante el almacenamiento (4*5 ºC) de rodajas de manzana expuestas a diferentes dosis de radiación con luz pulsada. (d = 10 cm) Para cada tratamiento, las barras con la misma letra minúscula no presentaron diferencias significativas a lo largo del almacenamiento (p<0,05). Para el mismo día de almacenamiento, las barras de diferentes tratamientos con la misma letra mayúscula no presentaron diferencias significativas (p<0,05).

205
de los tiempos analizados. Al día 7, sólo las rodajas irradiadas 100 segundos presentaron un
aumento de croma mayor que el control. Los valores de C* al final del almacenamiento de las
rodajas irradiadas 2 segundos rondaron entre 21*22, y en las muestras control e irradiadas a
tiempos mayores entre 29*31 (figura 4'72 b).
En base a lo analizado, la irradiación con LP durante 20, 60 y 100 segundos, a 5 y 10
cm de distancia de la lámpara, produjo un incremento del pardeamiento de las muestras,
reflejado fundamentalmente en una mayor disminución de la luminosidad y un aumento del
parámetro a*.
En la figura 4'73 se muestra en forma comparativa la evolución del índice de
pardeamiento (expresado como diferencia con los valores de IB de las rodajas frescas) en
función del tiempo de irradiación, en las rodajas ubicadas a 5 y 10 cm de la lámpara, al día
0, 3 y 7 del almacenamiento. Al inicio del almacenamiento, las rodajas de manzana expuestas
a la luz pulsada mostraron un incremento gradual del IB con el aumento del tiempo de
irradiación. En las muestras irradiadas durante 60 y 100 segundos, el pardeamiento fue mayor
cuando fueron ubicadas a menor distancia de la lámpara, mientras que para tiempos de
irradiación menores no se presentaron diferencias significativas (p>0,05) (figura 4'73 a). Al
tercer y séptimo día, las variaciones del IB de las rodajas irradiadas a 5 cm de la lámpara
durante 20, 60 y 100 segundos fueron similares, y mayores a las que presentaron las muestras
irradiadas por 2 segundos y el control.
En las rodajas tratadas a 10 cm, las mayores variaciones del IB a lo largo del
almacenamiento las tuvieron las muestras irradiadas durante 60 y 100 segundos, y al final del
mismo, los valores de cIB de estas muestras no presentaron diferencias significativas con
respecto a las muestras irradiadas durante los mismos tiempos pero ubicadas a menor
distancia. Por otro lado, las muestras expuestas a la LP durante 2 segundos presentaron
valores menores de cIB que el control, lo que reflejaría una inhibición del pardeamiento por
efecto de la irradiación (figura 4'73 b y c). Dunn y col. (1989) observaron también una
inhibición del pardeamiento enzimático en rodajas de papa irradiadas con luz pulsada durante
tiempos cortos. Encontraron que la disminución del pardeamiento se relacionaba con una
disminución de la actividad de la enzima polifenoloxidasa.
La evolución del color durante el almacenamiento de las rodajas de manzana irradiadas
con luz pulsada durante un tiempo de irradiación intermedio (60 s), a las distancias de 5 y 10
cm, puede ser apreciada en el diagrama de cromaticidad CIE x*y que se muestra en la figura
4'74. Las superficies de las manzanas irradiadas presentaron un mayor pardeamiento con el
aumento del tiempo de almacenamiento, mientras que los cambios observados en el color del

206
0
5
10
15
20
25
30
35
40
0 20 40 60 80 100 120
�����������''������$���
%&���%&�
a a,bA
aA
dB
dA
cB
cA
bA
bA
0
5
10
15
20
25
30
35
40
0 20 40 60 80 100 120
�����������''������$���
%&���%&�
a
dA bA
c,dB
bA
a,cB
bA
bB
aA
0
5
10
15
20
25
30
35
40
0 20 40 60 80 100 120
�����������''������$���
%&���%&�
a
cA
bA cA
bA bA
aB
aA
bB
Figura 4'73. Valores promedio del índice de pardamiento (IB) (expresados como diferencias con respecto al valor inicial de la muestra fresca) de rodajas de manzana irradiadas a diferentes dosis de luz pulsada. a) día 0, b) día 3, c) día 7. Muestras irradiadas a d = 5 cm (■); muestras irradiadas a d = 10 cm (▲). Para una determinada distancia de la lámpara, los valores de (IB*IB0) de manzanas con diferentes tiempos de irradiación con la misma letra minúscula no presentaron diferencias significativas (p>0,05). Para un mismo tiempo de irradiación, los valores de (IB*IB0) de muestras ubicadas a diferentes distancias de la lámpara con la misma letra mayúscula no presentaron diferencias significativas (p>0,05).
a)
b)
c)
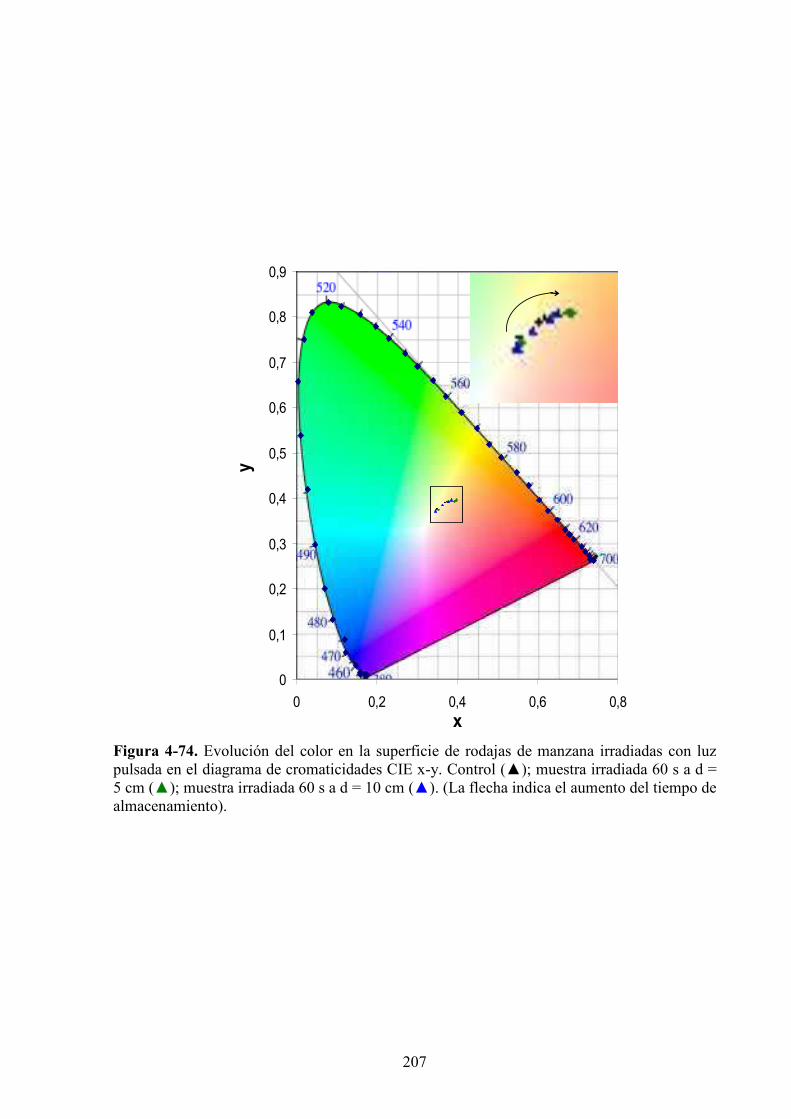
207
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
0 0,2 0,4 0,6 0,8
#
2
Figura 4'74. Evolución del color en la superficie de rodajas de manzana irradiadas con luz pulsada en el diagrama de cromaticidades CIE x*y. Control (▲); muestra irradiada 60 s a d = 5 cm (▲); muestra irradiada 60 s a d = 10 cm (▲). (La flecha indica el aumento del tiempo de almacenamiento).

208
control fueron menores.
Los cambios descriptos en el color de las rodajas de manzana irradiadas a diferentes
dosis de LP al final del almacenamiento pueden observarse en las imágenes de la figura 4'75.
Si bien a bajas dosis la irradiación con LP produjo una inhibición del pardeamiento, la
exposición a tiempos de irradiación más prolongados pudo provocar un mayor daño al tejido
(ruptura generalizada de membranas en toda la estructura), lo que posiblemente favoreció el
desarrollo de reacciones de pardeamiento.
��8$,$7$,$� !������� ��� �������� ��� :���� ��������F�������� ��� ������ ���+�� �� ����������������5���
Para tratar de inhibir el pardeamiento superficial provocado por la irradiación con LP, se
evaluó la aplicación de la misma solución antipardeamiento de ácido ascórbico/cloruro de
calcio (SA) utilizada previamente en las rodajas de manzana expuestas a la radiación UV*C.
Las figuras 4'76 y 4'77 muestran la evolución de los parámetros L*, a* y b* en función
de la dosis aplicada y el tiempo de almacenamiento, para las muestras ubicadas a 5 y 10 cm de
la lámpara respectivamente. La interacción entre los factores analizados fue significativa (p<
0,05).
Las rodajas de manzana frescas presentaron valores promedios iniciales de L*, a* y b*
de 70, *7,5 y 21 respectivamente. Las muestras pretratadas con solución antipardeamiento e
irradiadas 2 s a una distancia de 5 cm de la lámpara, a diferencia de las muestras irradiadas sin
inmersión en SA, no tuvieron una disminución apreciable en la luminosidad durante el
almacenamiento. En las muestras irradiadas 20 y 60 s se observó una disminución de L* (4%)
al día 7 del almacenamiento similar al control, mientras que en las expuestas a la LP durante
100 s el oscurecimiento fue mayor (10%) y se produjo inmediatamente después de la
irradiación (día 0), acentuándose luego hasta el tercer día del almacenamiento (figura 4'76 a).
En cuanto a las variaciones de a*, si bien todas las muestras tratadas y control mostraron un
incremento de este parámetro durante el almacenamiento, las irradiadas durante 20, 60 y 100 s
presentaron al final del mismo un aumento mayor (40*80%) al de las muestras con SA que no
fueron irradiadas (27%). Los mayores cambios se observaron en las rodajas irradiadas 100 s
(figura 4'76 b). Por otro lado, las variaciones en el parámetro b* de las muestras tratadas no
fueron significativas, observándose solo una leve disminución durante el almacenamiento en
las rodajas irradiadas a la dosis más alta (figura 4'76 c).
En las rodajas de manzana irradiadas a 10 cm de distancia, las muestras expuestas a la

209
���'�� �����8 ������8 1�����8 !������8
�
���'�� �����8 ������8 1�����8 !������8
�
���'�� �����8 ������8 1�����8 !������8
�
���'�� �����8 ������8 1�����8 !������8
�
���'�� �����8 ������8 1�����8 !������8
�
���'�� �����8 ������8 1�����8 !������8
�
Figura 4'75. Color superficial de rodajas de manzanas expuestas a diferentes dosis de radiación con luz pulsada, al séptimo día de almacenamiento a 4*5 ºC. a) Muestras irradiadas a d = 5 cm. b) Muestras irradiadas a d = 10 cm.

210
5
0
5
10
15
20
25
0 3 7
���������������������������
�������
Control SA
SA+LP 2s SA+LP 20sSA+LP 60s SA+LP 100 s
aA,B
aBaA
aA,B
aB aA,B
a,bA
bC
aA,B bA,B
bB bB
bC
a,bA
aAaA
2
0
2
4
6
8
10
0 3 7
���������������������������
��������
Control SA
SA+LP 2s SA+LP 20s
SA+LP 60s SA+LP 100 s
aA
aA
aB
aC
aA
bA,B
bA
bB,C
bC
bD
bA,C
bB cA,B
cC
cC
bD
3
1
1
3
5
7
9
11
13
15
0 3 7
���������������������������
�����
��
Control SA
SA+LP 2s SA+LP 20s
SA+LP 60s SA+LP 100 s
aA
aA
aA
aA
aA aB
aB
aB aB
bB
bA
aA,B
aB,C
aA
aB,C
bC
bA
Figura 4'76. Valores promedio de la luminosidad (L*), el valor de a* y de b* (expresados como diferencias con respecto a los valores iniciales de las muestras frescas) durante el almacenamiento (4*5ºC) de rodajas de manzana inmersas en solución antipardeamiento e irradiadas a diferentes dosis de luz pulsada a d = 5 cm. Para cada tratamiento, las barras con la misma letra minúscula no presentaron difer2encias significativas a lo largo del almacenamiento (p<0,05). Para el mismo día de almacenamiento, las barras de diferentes tratamientos con la misma letra mayúscula no presentaron diferencias significativas (p<0,05).
a)
b)
c)

211
LP durante 60 s y 100 s mostraron una leve disminución de la luminosidad al final del
almacenamiento (alrededor de 3*5%), si bien las diferencias con el control no fueron
significativas (figura 4'77 a). Las variaciones del parámetro a* entre las muestras irradiadas
20 s y las inmersas en la SA sin irradiar no fueron significativas (p<0,05), mientras que para
dosis mayores (60 s y 100 s) el incremento de a* durante el almacenamiento fue mayor al de
estas muestras, siendo al final del mismo, similar al de las muestras control (49*55%) (figura
4'77 b). Las variaciones en el parámetro b* de las muestras tratadas a lo largo del
almacenamiento en general no fueron significativas (figura 4'77 c). Cabe señalar, que en esta
distancia no se evaluó la aplicación de la solución antipardeamiento en las rodajas irradiadas 2
segundos porque, como se comentó anteriormente, el pardeamiento en estas muestras fue
inhibido por la irradiación.
En la figura 4'78 puede apreciarse la evolución del índice de pardeamiento con el
aumento de la dosis aplicada y el tiempo de almacenamiento. Para todas las dosis evaluadas,
la inmersión previa de las muestras en la SA redujo el incremento del pardeamiento
provocado por la irradiación a lo largo del almacenamiento. En las rodajas irradiadas a una
distancia de 10 cm, solo las muestras irradiadas a la dosis más alta presentaron un incremento
en el IB por efecto de la irradiación. En las muestras irradiadas a menor distancia, el IB se
incrementó con el aumento del tiempo de irradiación durante los primeros días del
almacenamiento (día 0 y 3), pero al final del mismo, el incremento fue similar en las muestras
irradiadas durante 20, 60 y 100 segundos. Por otro lado, al día 7 no se presentaron diferencias
significativas (p<0,05) en la variación del IB de las rodajas colocadas a diferentes distancias
de la lámpara e irradiadas 100 segundos.
Los cambios analizados en el color de las muestras al día 7 del almacenamiento pueden
visualizarse en la figura 4'79. Si bien la aplicación de la solución de ácido ascórbico/cloruro
de calcio redujo el pardeamiento desarrollado en las muestras por efecto de la irradiación, en
las rodajas irradiadas durante tiempos de irradiación más prolongados y a menor distancia de
la lámpara la inhibición fue menos efectiva.
Si bien la solución de ácido ascórbico/cloruro de calcio pudo haber inhibido las
reacciones de pardeamiento enzimático en muestras irradiadas a dosis altas, es probable que
las altas temperaturas que alcanzaron estas muestras, entre otros, hayan favorecido reacciones
de pardeamiento no enzimático (PNE).
Dentro del grupo de reacciones de PNE se encuentran reacción de Maillard,
caramelización, oxidación química de polifenoles y de ácido ascórbico. La reacción de
Maillard incluye las reacciones de aldehídos, cetonas y azúcares reductores con aminas,

212
5
0
5
10
15
20
25
0 3 7
���������������������������
�������
Control SA
SA+LP 20s SA+LP 60s
SA+LP 100 s
aA aA aA aA aA,B,C
aA aA aA aA aA,B
aA
bC bB,C
2
0
2
4
6
8
10
0 3 7
���������������������������
�����
��
Control SASA+LP 20s SA+LP 60sSA+LP 100 s
aA
aBaA,B
aA
aA
aA
aB aB
bA
aB bB
bA
bA
5
3
1
1
3
5
7
9
11
13
15
0 3 7
���������������������������
�����
��
Control SA
SA+LP 20s SA+LP 60s
SA+LP 100 s
aA a,bA
aA
aA,B
bA
aB aB
aA,B bA aA,B
aB,C bC
aA,C
Figura 4'77. Valores promedio de la luminosidad (L*), el valor de a* y de b* (expresados como diferencias con respecto a los valores iniciales de las muestras frescas) durante el almacenamiento (4*5ºC) de rodajas de manzana inmersas en solución antipardeamiento e irradiadas a diferentes dosis de luz pulsada a d = 10 cm. Para cada tratamiento, las barras con la misma letra minúscula no presentaron diferencias significativas a lo largo del almacenamiento (p<0,05). Para el mismo día de almacenamiento, las barras de diferentes tratamientos con la misma letra mayúscula no presentaron diferencias significativas (p<0,05).
a)
b)
c)
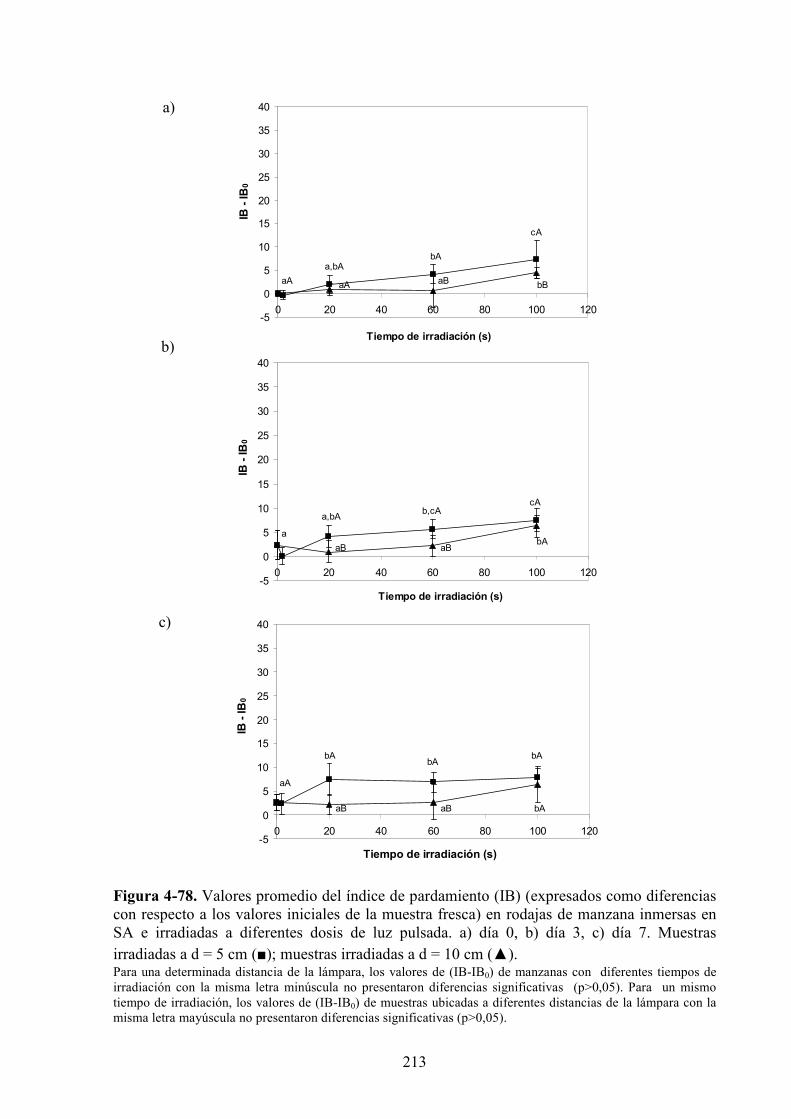
213
5
0
5
10
15
20
25
30
35
40
0 20 40 60 80 100 120
�����������''������$���
%&���%&�
bA
bB
cA
aA aB
a,bA
aA
5
0
5
10
15
20
25
30
35
40
0 20 40 60 80 100 120
�����������''������$���
%&���%&�
a,bA
a
aB aB
b,cA
bA
cA
5
0
5
10
15
20
25
30
35
40
0 20 40 60 80 100 120
�����������''������$���
%&���%&�
bA bA
bA
aB
aA
bA aB
Figura 4'78. Valores promedio del índice de pardamiento (IB) (expresados como diferencias con respecto a los valores iniciales de la muestra fresca) en rodajas de manzana inmersas en SA e irradiadas a diferentes dosis de luz pulsada. a) día 0, b) día 3, c) día 7. Muestras irradiadas a d = 5 cm (■); muestras irradiadas a d = 10 cm (▲). Para una determinada distancia de la lámpara, los valores de (IB*IB0) de manzanas con diferentes tiempos de irradiación con la misma letra minúscula no presentaron diferencias significativas (p>0,05). Para un mismo tiempo de irradiación, los valores de (IB*IB0) de muestras ubicadas a diferentes distancias de la lámpara con la misma letra mayúscula no presentaron diferencias significativas (p>0,05).
a)
b)
c)

214
���'�� ./ ./,�8�
��
./,�8�
���
./,�8�
1��
./,�8�
!���
�
���'�� ./ ./,�8�
��
./,�8�
���
./,�8�
1��
./,�8�
!���
�
���'�� ./ ./,�8�
��
./,�8�
���
./,�8�
1��
./,�8�
!���
�
���'�� ./ ./,�8�
���
./,�8�
1��
./,�8�
!���
�
���'�� ./ ./,�8�
���
./,�8�
1��
./,�8�
!���
�
���'�� ./ ./,�8�
���
./,�8�
1��
./,�8�
!���
�
Figura 4'79. Color superficial de rodajas de manzanas inmersas en solución antipardeamiento y expuestas a diferentes dosis de radiación con luz pulsada, al séptimo día de almacenamiento a 4*5 ºC. a) Muestras irradiadas a d = 5 cm. b) Muestras irradiadas a d = 10 cm.

215
aminoácidos, péptidos y proteínas. La caramelización ocurre cuando compuestos
polihidroxicarbonilicos (azúcares, ácidos polihidroxicarboxílicos) son calentados a
temperaturas altas en ausencia de compuestos aminos. En las reacciones de oxidación química
de polifenoles y ácido ascórbico, estos compuestos son convertidos a compuestos di o
policarbonílicos (Hodge, 1953, Monzocco y col., 2001). Estas reacciones están influenciadas
por numerosos factores tales como la naturaleza, concentración y proporción de los
reactantes, actividad de agua, temperatura y tiempo de calentamiento, pH, presencia de
oxígeno, luz o iones metálicos (Ames, 1992).
A pesar de que las temperaturas alcanzadas por las muestras irradiadas a dosis altas son
adecuadas para que puedan llegar a producirse reacciones de PNE (especialmente en las
rodajas irradiadas a 5 cm de distancia de la lámpara durante 60 s y 100 s que presentaron
incrementos de temperatura mayores a los 70 ºC), dado los numerosos factores que influyen
en la cinética de estas reacciones, y sobre todo debido al corto tiempo en que las muestras
fueron sometidas a esas temperaturas, la posibilidad de que realmente se hayan producido este
tipo de reacciones debería ser estudiada con mayor profundidad.
���8$,$8$� �������������� ��� ���� ����������� ���������� ��� ����*��� ��� ��������/���������������������������"���������������������
Se analizó la resistencia a la compresión de las rodajas de manzana expuestas a la luz
pulsada (LP) durante 2, 20, 60 y 100 segundos, a 5 y 10 cm de distancia de la lámpara.
Las curvas típicas de fuerza vs distancia y de esfuerzo real vs deformación real que se
obtuvieron en las rodajas de manzana sin irradiar (control) y en las rodajas irradiadas durante
100 segundos a 5 y 10 cm, al día 0 y 7 del almacenamiento, se muestran a modo de ejemplo
en las figuras 4'80 y 4'81$� �Como puede observarse, todas las curvas presentaron un pico
pequeño correspondiente al “bioyield” y picos de ruptura bien marcados. Al cabo del
almacenamiento, la ruptura en las muestras irradiadas se produjo a deformaciones ligeramente
menores que en el control.
Los valores promedio de esfuerzo en el punto de “bioyield” (σRB), de esfuerzo de
ruptura (σRR) y de módulo de deformabilidad (Ed) de las diferentes muestras analizadas se
presentan en las figuras 4'82 y 4'83 y tablas 4'26 y 4'27. En todos los parámetros evaluados

216
��������a) b)
0
50
100
150
200
250
300
350
400
450
0 1 2 3 4 5 6
������������
34�'5���
Control
100 s LP; d = 5 cm
100 s LP; d = 10 cm
0
50
100
150
200
250
300
350
400
450
0 1 2 3 4 5 6
������������34�'5���
Control
100 s LP; d = 5 cm
100 s LP; d = 10 cm
Figura 4'80.� Curvas típicas de fuerza*distancia� obtenidas en rodajas de manzana frescas (control) y expuestas a la luz pulsada durante 100 s a d = 5 cm y d =10 cm. a) día 0 b) día 7 del almacenamiento a 4*5 ºC.������ a) b)
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
0 0,5 1 1,5 2 2,5
εεεε6
σσ σσ6��78�
Control
100 s LP; d = 5 cm
100 s LP; d = 10 cm
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
0 0,5 1 1,5 2 2,5εεεε6�
σσ σσ6��78�
Control
100 s LP; d = 5 cm
100 s LP; d = 10 cm
�Figura 4'81.�Curvas típicas de tensión real (σR) – deformación real (εR) obtenidas en rodajas de manzana frescas (control) y expuestas a la luz pulsada durante 100 s a d = 5 cm y d =10 cm. a) día 0 b) día 7 del almacenamiento a 4*5 ºC.����
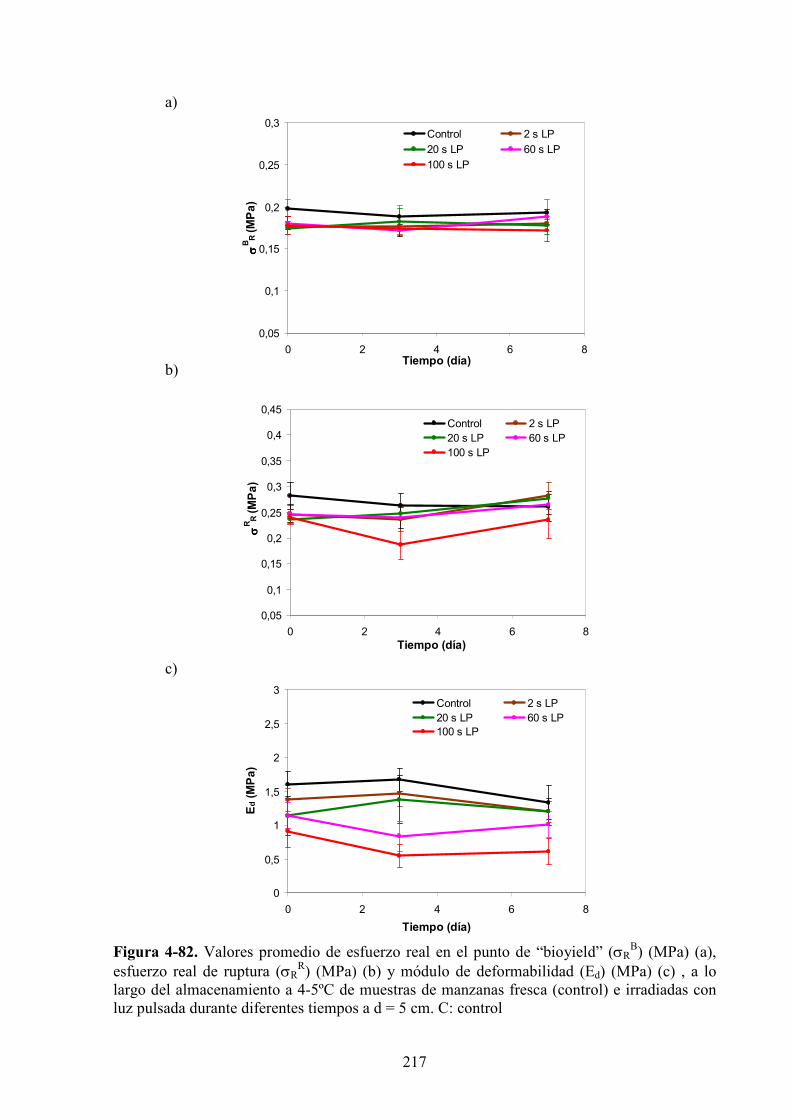
217
a)
b)
0,05
0,1
0,15
0,2
0,25
0,3
0 2 4 6 8�����������
σσ σσ&6��78�
Control 2 s LP
20 s LP 60 s LP
100 s LP
0,05
0,1
0,15
0,2
0,25
0,3
0,35
0,4
0,45
0 2 4 6 8
�����������
σσ σσ66��78�
Control 2 s LP
20 s LP 60 s LP
100 s LP
c)
0
0,5
1
1,5
2
2,5
3
0 2 4 6 8
�����������
+���78�
Control 2 s LP
20 s LP 60 s LP
100 s LP
�Figura 4'82. Valores promedio de esfuerzo real en el punto de “bioyield” (σR
B) (MPa) (a), esfuerzo real de ruptura (σR
R) (MPa) (b) y módulo de deformabilidad (Ed) (MPa) (c) , a lo largo del almacenamiento a 4*5ºC de muestras de manzanas fresca (control) e irradiadas con luz pulsada durante diferentes tiempos a d = 5 cm. C: control �
218
Tabla 4'26. Valores promedio de esfuerzo real en el punto de “bioyield” (σRB) (MPa) (a),
esfuerzo real de ruptura (σRR) (MPa) (b) y módulo de deformabilidad (Ed)(MPa) (c), a lo largo
del almacenamiento a 4*5ºC de muestras de manzanas fresca (control) e irradiadas con luz pulsada durante diferentes tiempos a d = 5 cm. C: control. �a)
Tiempo de irradiación
(s)
Tiempo de almacenamiento (día)
0 3 7
0 (C) 0,20 (0,01)aA 0,19 (0,01)bA 0,19 (0,02)a,bA
2 0,177 (0,006)aB 0,18 (0,09)aB 0,18 (0,01)aA,B
20 0,175 (0,007)aB 0,18 (0,02)aA,B 0,18 (0,01)aA,B
60 0,181 (0,007)aB 0,172 (0,007)bB 0,188 (0,009)aA
100 0,18 (0,01)aB 0,175 (0,007)aB 0,17 (0,01)aB
b)
Tiempo de irradiación
(s)
Tiempo de almacenamiento (día)
0 3 7
0 (C) 0,29 (0,03)aA 0,26 (0,02)bA 0,26 (0,03)bA
2 0,25 (0,02)aB 0,24 (0,03)aB 0,28 (0,03)bA
20 0,24 (0,02)aB 0,25 (0,03)aB 0,28 (0,02)bA
60 0,25 (0,02)aB 0,24 (0,02)aB 0,27 (0,02)bA
100 0,24 (0,02)aB 0,19 (0,03)bC 0,24 (0,04)aB
c)
Tiempo de irradiación
(s)
Tiempo de almacenamiento (día)
0 3 7
0 (C) 1,6 (0,2)aA 1,7 (0,2)aA 1,3 (0,2)bA
2 1,3 (0,3)a,bB 1,5 (0,2)bA,B 1,2 (0,1)aA
20 1,1 (0,3)aB 1,4 (0,4)aB 1,2 (0,2)aA
60 1,1 (0,2)aB 0,8 (0,2)bC 1,0 (0,2)aB
100 0,9 (0,2)aC 0,5 (0,2)bD 0,6 (0,2)bC
�Los resultados fueron expresados como la media seguida de la desviación estándar (entre paréntesis). Para cada tiempo de irradiación, las medias seguidas por la misma letra minúscula no presentaron diferencias significativas a lo largo del almacenamiento (p<0,05). Para el mismo día de almacenamiento, las medias de los diferentes tratamientos seguidas por la misma letra mayúscula no presentaron diferencias significativas (p<0,05).�
�

219
a)
0,05
0,1
0,15
0,2
0,25
0,3
0 2 4 6 8
�����������
σσ σσ&6��78�
Control 2 s LP
20 s LP 60 s LP
100 s LP
b)
0,05
0,1
0,15
0,2
0,25
0,3
0,35
0,4
0,45
0 2 4 6 8
�����������
σσ σσ66��78�
Control 2 s LP
20 s LP 60 s LP
100 s LP
c)
0
0,5
1
1,5
2
2,5
3
0 2 4 6 8
�����������
+���78�
Control 2 s LP
20 s LP 60 s LP
100 s LP
Figura 4'83��Valores promedio de esfuerzo real en el punto del “bioyield” (σRB) (MPa) (a),
esfuerzo real de ruptura (σRR) (MPa) (b) y módulo de deformabilidad (Ed)(MPa) (c), a lo largo
del almacenamiento a 4*5ºC de muestras de manzanas fresca (control) e irradiadas con luz pulsada durante diferentes tiempos a d = 10 cm. C: control.

220
Tabla 4'27. Valores promedio de esfuerzo real en el punto del “bioyield” (σRB) (MPa) (a),
esfuerzo real de ruptura (σRR) (MPa) (b) y módulo de deformabilidad (Ed)(MPa) (c), a lo largo
del almacenamiento a 4*5ºC de muestras de manzanas fresca (control) e irradiadas con luz pulsada durante diferentes tiempos a d = 10 cm. C: control. �a)
Tiempo de irradiación
(s)
Tiempo de almacenamiento (día)
0 3 7
0 (C) 0,20 (0,01)aA 0,19 (0,01)bA 0,19 (0,02)a,bA
2 0,18 (0,01)aB,C 0,18 (0,01)aB 0,20 (0,01)bA
20 0,18 (0,01)aB 0,19 (0,01)aA,B 0,20 (0,01)bA
60 0,170 (0,007)aC 0,18 (0,01)aB,C 0,195 (0,008)bA
100 0,174 (0,008)aB,C 0,169 (0,008)aC 0,19 (0,01)bA
b)
Tiempo de irradiación
(s)
Tiempo de almacenamiento (día)
0 3 7
0 (C) 0,29 (0,03)aA 0,26 (0,02)bA 0,26 (0,03)bA
2 0,25 (0,03)aB 0,25 (0,03)aA 0,28 (0,02)bA
20 0,24 (0,03)aB 0,26 (0,03)aA 0,28 (0,02)bA
60 0,25 (0,02)aB 0,25 (0,02)aA 0,28 (0,02)bA
100 0,25 (0,02)aB 0,25 (0,02)aA 0,30 (0,02)bB
c)
Tiempo de irradiación
(s)
Tiempo de almacenamiento (día)
0 3 7
0 (C) 1,6 (0,2)aA 1,7 (0,2)aA 1,3 (0,2)bA
2 1,3 (0,2)aB,C 1,6 (0,3)bA 1,3 (0,3)aA
20 1,4 (0,2)aB 1,7 (0,2)bA 1,2 (0,2)cA
60 1,1 (0,2)a,bC 1,0 (0,3)aB 1,2 (0,2)bA
100 1,2 (0,2)aC 0,93 (0,02)bB 1,2 (0,2)aA
�Los resultados fueron expresados como la media seguida de la desviación estándar (entre paréntesis). Para cada tiempo de irradiación, las medias seguidas por la misma letra minúscula no presentaron diferencias significativas a lo largo del almacenamiento (p<0,05). Para el mismo día de almacenamiento, las medias de los diferentes tratamientos seguidas por la misma letra mayúscula no presentaron diferencias significativas (p<0,05).�

221
la interacción entre el tiempo de irradiación y tiempo de almacenamiento fue significativa ( p
< 0,01)
Las rodajas de manzana sin irradiar (control) presentaron durante el almacenamiento
una ligera disminución de σRB, una disminución de σR
R de ≈ 10 % y de Ed de ≈ 20 %. En las
manzanas irradiadas a 5 cm de distancia de la lámpara, los valores de σRB disminuyeron en
relación a las muestras frescas ≈ 10 % luego de la irradiación y permanecieron constantes o
con variaciones leves durante el almacenamiento (figura 4'82 a y tabla 4'26 a). Los valores
de σRR también disminuyeron en estas muestras entre 14*17 % después de la irradiación, pero
al final del almacenamiento, estos valores se incrementaron levemente en las rodajas
irradiadas durante 2*60 segundos, no presentando diferencias significativas con respecto al
control. En las rodajas expuestas a la LP durante 100 segundos, los valores de σRR al día 7 del
almacenamiento siguieron siendo menores que los del control (figura 4'82 b y tabla 4'26 b).
Por otro lado, el módulo de deformabilidad se redujo inicialmente en las muestras irradiadas
entre 19*44%, siendo mayor la disminución en las rodajas irradiadas durante 100 segundos.
Al tercer día del almacenamiento, los valores de Ed aumentaron levemente en las rodajas
irradiadas durante cortos tiempos (2 s) y disminuyeron en aquellas irradiadas a tiempos más
prolongados (60 y 100 segundos). Al finalizar el almacenamiento, sólo las muestras irradiadas
durante 60 s y 100 s presentaron una reducción significativa de Ed con respecto al control. En
el caso de las rodajas irradiadas 100 segundos, la disminución en relación al valor inicial de
Ed de las muestras frescas fue de ≈ 70 % (figura 4'82 c y tabla 4'27 c).
Las manzanas irradiadas a mayor distancia de la lámpara presentaron una disminución
inicial en los valores de σRB y σR
R similar a la observada en las muestras irradiadas a menor
distancia. Sin embargo, durante el almacenamiento ambos parámetros aumentaron en todas
las muestras irradiadas, y al cabo del almacenamiento, no se observaron diferencias
significativas con respecto al control, excepto en las rodajas irradiadas 100 segundos en donde
los valores de σRR fueron levemente mayores (figura 4'83 a y b, tabla 4'27 a y b). El
módulo de deformabilidad disminuyó también inicialmente en estas muestras (entre 19*25%).
Pero si bien al tercer día del almacenamiento los valores de Ed aumentaron en las muestras
irradiadas entre 2 * 20 s y disminuyeron en las rodajas irradiadas 100 s, al séptimo día del
almacenamiento las diferencias entre las diferentes muestras irradiadas y el control no fueron
significativas (figura 4'83 c y tabla 4'27 c).
En la figura 4'84 se comparan los valores de σRR y del Ed obtenidos en las rodajas de
manzana irradiadas con LP durante 60 s (d = 5 cm ó d =10 cm) que fueron o no previamente
inmersas en la solución de ácido ascórbico/cloruro de calcio. En general, no se observaron

222
�
a) b)
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
0,4
0,45
0,5
0 7�����������
σσ σσ66��78�
LP
SA+LP
a a a a
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
0,4
0,45
0,5
0 7�����������
σσ σσ66��78�
LP
SA+LP
a a
a a
c) d)
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6
1,8
2
0 7
�����������
+���78�
LP
SA+LP
a a
a
b
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6
1,8
2
0 7
�����������
+� �78�
LP
SA+LP
a
a a
a
Figura 4'84. Valores promedio de esfuerzo real de ruptura (σRR) (MPa) y módulo de
deformabilidad (Ed) (MPa) obtenidos en rodajas de manzana irradiadas con luz pulsada durante 60 s (con o sin inmersión previa en solución antipardeamiento (SA)), al día 0 y 7 del almacenamiento. a) y c) muestras irradiadas a d= 5 cm; b) y d) muestras irradiadas a d= 10 cm. Para cada tiempo de almacenamiento, las barras seguidas por la misma letra minúscula no presentaron diferencias significativas (p<0,05).

223
variaciones significativas en los parámetros analizados por la aplicación de la solución
antipardeamiento (SA). Sólo se observó una leve reducción en los valores de Ed al día 7 del
almacenamiento en las rodajas con SA irradiadas a 5 cm de la lámpara.
La disminución inicial de σRR y de Ed en todas las rodajas de manzanas irradiadas
estaría reflejando una pérdida de rigidez de estos tejidos en relación al tejido fresco luego de
la exposición a la irradiación. La evolución que siguieron los parámetros durante el
almacenamiento, según lo expuesto anteriormente, fue diferente dependiendo del tiempo de
irradiación y la distancia a la que fueron irradiadas las muestras. Al final del almacenamiento,
sólo las muestras que fueron irradiadas durante tiempos de irradiación más prolongados y a
menores distancias de la lámpara presentaron una pérdida de rigidez del tejido mayor al
control.
El efecto de la aplicación de luz pulsada sobre las propiedades viscoelásticas de las
rodajas de manzana a bajas deformaciones (ensayos oscilatorios y de fluencia* recuperación),
se analizaron en las muestras expuestas a la irradiación durante 60 segundos a una distancia
de la lámpara de 10 cm (con o sin previa inmersión en la solución de ácido ascórbico/cloruro
de calcio).
Ensayos oscilatorios
En los ensayos de barrido de amplitud con control de la deformación de cizalla (BA
CDC) realizados, el comportamiento fue similar al que tuvo lugar en los tratamientos
evaluados anteriormente. Las rodajas de manzana fresca sin almacenar fueron las que
presentaron el menor rango viscoelástico lineal, por lo tanto, el valor límite de RVL
recomendado para estas muestras (γ = 0,01%) fue el que se utilizó para realizar el barrido de
frecuencia.
En la figura 4'85 se muestra el espectro mecánico obtenido para las diferentes muestras
analizadas, al día 0 y 7 de almacenamiento. Todas las muestras presentaron un predominio del
comportamiento sólido, con valores de módulo elástico (G’) mayores a los de módulo de
almacenamiento (G”) en todo el rango de frecuencia. Por otro lado, en general en las muestras
tratadas los valores de G’ y G” fueron siempre menores que los del control.
Los valores promedios de G’ a diferentes frecuencias angulares y los parámetros de la
regresión lineal Log G’ vs Log ω se presentan en la tabla 4'29, mientras que los valores
correspondientes a l factor de pérdida (tan δ) se muestran en la tabla 4'30. Para todos los

224
a)
0,E+00
1,E+05
2,E+05
3,E+05
4,E+05
5,E+05
6,E+05
0,1 1 10 100ϖϖϖϖ ���
�!
9:��8�
b)
0,E+00
2,E+04
4,E+04
6,E+04
8,E+04
1,E+05
0,1 1 10 100ϖϖϖϖ ���
�!
9=��8�
��Figura 4'85. Barrido de frecuencia a γ = 0,01 % en función de la frecuencia angular (ϖ = 100 s*1 – 0,1 s*1) de muestras de manzanas frescas e irradiadas con luz pulsada (con o sin aplicación previa de solución antipardeamiento (SA)), almacenadas a 4*5 ºC. a) Módulo de almacenamiento (G’), b) Módulo de pérdida (G”). Control día 0 (■); Control día 7 (□); SA día 0 (■); SA día 7 (□); LP día 0 (■); LP día 7 (□): SA + LP día 0 (■); SA + LP día 7 (□). ��
225
Tabla 4'29. Valores promedio del módulo G´ (kPa) a distintas frecuencias angulares de las muestras de manzana control (C), inmersas en solución antipardeamiento (SA), irradiadas con luz pulsada (LP), inmersas en solución antipardeamiento (SA) e irradiadas con LP, almacenadas en refrigeración. Parámetros obtenidos de la regresión lineal del Log (G’) vs Log (ϖ). �Tiempo
(día) Tratamiento ϖϖϖϖ�= 0,1 s'1� ϖϖϖϖ�= 1 s'1� ϖϖϖϖ�= 10 s'1� ϖϖϖϖ�= 100 s'1� n R2
C 302 (70)aA 371 (77)aA 419 (90)aA 488 (105)aA 0,06aA 0,96
SA 289 (24)aA 294 (33)aB 316 (36)aB 352 (38)aB 0,03aB 0,97
LP 158 (66)aB 170 (65)aC 184 (71)aC 210 (81)aC 0,04aC 0,93
0
SA + LP 236 (54)aA 250 (54)aB 271 (61)aB 312 (72)aB 0,04aC 0,96
C 232 (83)bA 262 (92)bA 285 (100)bA 320 (114)bA 0,04bA 0,99
SA 159 (48)bB 185 (56)bB 203 (60)bB 230 (69)bB 0,05bB 0,99
LP 132 (26)aB 142 (23)aB 156 (25)aB 180 (29)aB 0,05bB 0,97
7
SA + LP 77 (17)bC 77 (19)bC 87 (22)bC 101 (25)bC 0,06bC 0,99
�Los resultados fueron expresados como la media seguida de la desviación estándar (entre paréntesis). En cada columna y para un dado tratamiento, las medias seguidas por la misma letra minúscula no presentaron diferencias significativas en los dos tiempos de almacenamiento (p<0,05). En cada columna y para un mismo día de almacenamiento, las medias de los diferentes tratamientos seguidas por la misma letra mayúscula no presentaron diferencias significativas (p<0,05).
�����������

226
parámetros la interacción entre los factores tiempo de almacenamiento y tratamiento fue
significativa (p < 0,001).
En todas las muestras los valores de G’ aumentaron ligeramente con la frecuencia
angular, observándose una dependencia ligeramente mayor (> pendiente representada por el
parámetro n) al día 0 en las muestras control y al día 7 en las muestras tratadas. La irradiación
de las rodajas de manzana con luz pulsada produjo una disminución inicial de G’ (con
respecto al valor de las muestras frescas) entre 48*57 %, dependiendo de la frecuencia
angular. Esto reflejaría una pérdida de rigidez del tejido de manzana por efecto de la
irradiación. Después de los siete días de almacenamiento, los valores de G’ no variaron
significativamente en las muestras irradiadas, y si bien en el control se produjo una reducción
de 23*34 %, esta disminución fue menor a la observada inicialmente en las rodajas irradiadas.
Por otra lado, en las muestras irradiadas previamente inmersas en la solución
antipardeamiento (SA) la reducción de G’ con respecto al control fue menor (entre 32*36%)
para frecuencias angulares entre 1* 100 s*1 y no fue significativa para frecuencias más bajas.
Las diferencias iniciales de estas muestras con respecto a las muestras con SA sin irradiar no
fueron significativas. Sin embargo, al cabo del almacenamiento la disminución de G’ en las
rodajas con SA e irradiadas fue de74*79%, lo que estaría reflejando una marcada pérdida de
elasticidad del tejido irradiado cuando la aplicación de luz pulsada se combinó con el
tratamiento antipardeamiento (Tabla 4'29).
De manera similar a lo ocurrido anteriormente en los tratamientos con radiación UV*C,
el factor de pérdida (tan δ (G”/G’)) no fue un parámetro muy sensible para distinguir
diferencias entre tratamientos. En general, para todas las frecuencias angulares, se observó al
inicio del almacenamiento una ligera disminución de tan δ en las rodajas inmersas en SA con
o sin irradiación. Sin embargo, al cabo de 7 días, las diferencias entre las muestras tratadas y
no tratadas fueron leves o no significativas (figura 4'86 y tabla 4'30), lo que implicaría que
independientemente de los tratamientos aplicados, la contribución relativa de las componentes
viscosa y elástica sería similar.
��"��������������������������������
En la figura 4'87 se encuentran las curvas promedio de deformación vs tiempo
obtenidas al día 0 y 7 para las muestras control, irradiadas con LP e inmersas en SA con o sin
posterior irradiación. Las rodajas tratadas no presentaron cambios notorios en la forma de las �

227
Tabla 4'30. Valores promedio del factor de pérdida (tan δ) a distintas frecuencias angulares de las muestras de manzanas control (C), inmersas en solución antipardeamiento (SA), irradiadas con luz pulsada (LP), inmersas en solución antipardeamiento (SA) e irradiadas con LP, almacenadas en refrigeración.
Tiempo (día)
Tratamiento ϖϖϖϖ�= 0,1 s'1� ϖϖϖϖ�= 1 s'1� ϖϖϖϖ�= 10 s'1� ϖϖϖϖ�= 100 s'1�
C 0,19(0,05)aA 0,12(0,01)aA 0,11(0,01)aA 0,12 (0,01)aA
SA 0,15(0,03)aB 0,10(0,01)aB 0,09(0,01)aB 0,11(0,01)aB
LP 0,15(0,01)aB 0,10(0,01)aB 0,10(0,01)aC 0,13(0,01)aC
0
SA + LP 0,15(0,01)aB 0,11(0,01)aC 0,10(0,005)aC 0,12(0,01)aA
C 0,15(0,01)bA 0,11(0,01)bA 0,10(0,005)bA 0,12 (0,01)aA
SA 0,16(0,04)aA 0,10(0,01)aA 0,10(0,004)bA 0,11(0,004)aB
LP 0,13(0,02)bA 0,10(0,005)aA 0,10(0,004)aA 0,13(0,01)aC
7
SA + LP 0,14(0,01)aA 0,11(0,004)aA 0,11(0,003)bB 0,13(0,003)bC
�Los resultados fueron expresados como la media seguida de la desviación estándar (entre paréntesis). En cada columna y para un dado tratamiento, las medias seguidas por la misma letra minúscula no presentaron diferencias significativas en los dos tiempos de almacenamiento (p<0,05). En cada columna y para un mismo día de almacenamiento, las medias de los diferentes tratamientos seguidas por la misma letra mayúscula no presentaron diferencias significativas (p<0,05).

228
0,00
0,05
0,10
0,15
0,20
0,25
0,30
0,1 1 10 100ϖϖϖϖ ���
�!
��� δδ δδ
Figura 4'86. Factor de pérdida (tan δ) de muestras de manzanas frescas e irradiadas con luz pulsada (con o sin aplicación previa de solución antipardeamiento (SA)), almacenadas a 4*5 ºC. a) Módulo de almacenamiento (G’), b) Módulo de pérdida (G”). Control día 0 (■); Control día 7 (□); SA día 0 (■); SA día 7 (□); LP día 0 (■); LP día 7 (□): SA + LP día 0 (■); SA + LP día 7 (□).
0
0,01
0,02
0,03
0,04
0,05
0,06
0,07
0,08
0,09
0,1
0 50 100 150 200 250 300
���������
γγ γγ��;
1�ura 4'87. Curvas promedio de fluencia*recuperación para las muestras de manzana fresca, tratadas con solución antipardeamiento (SA) e irradiadas con luz pulsada (con o sin previa inmersión en SA). Control día 0 (▬); Control día7 (▬); SA día 0 (▬); SA día7 (▬); LP día 0 (▬); LP día 7 (▬); SA + LP día 0 (▬); SA + LP día 7 (▬). �
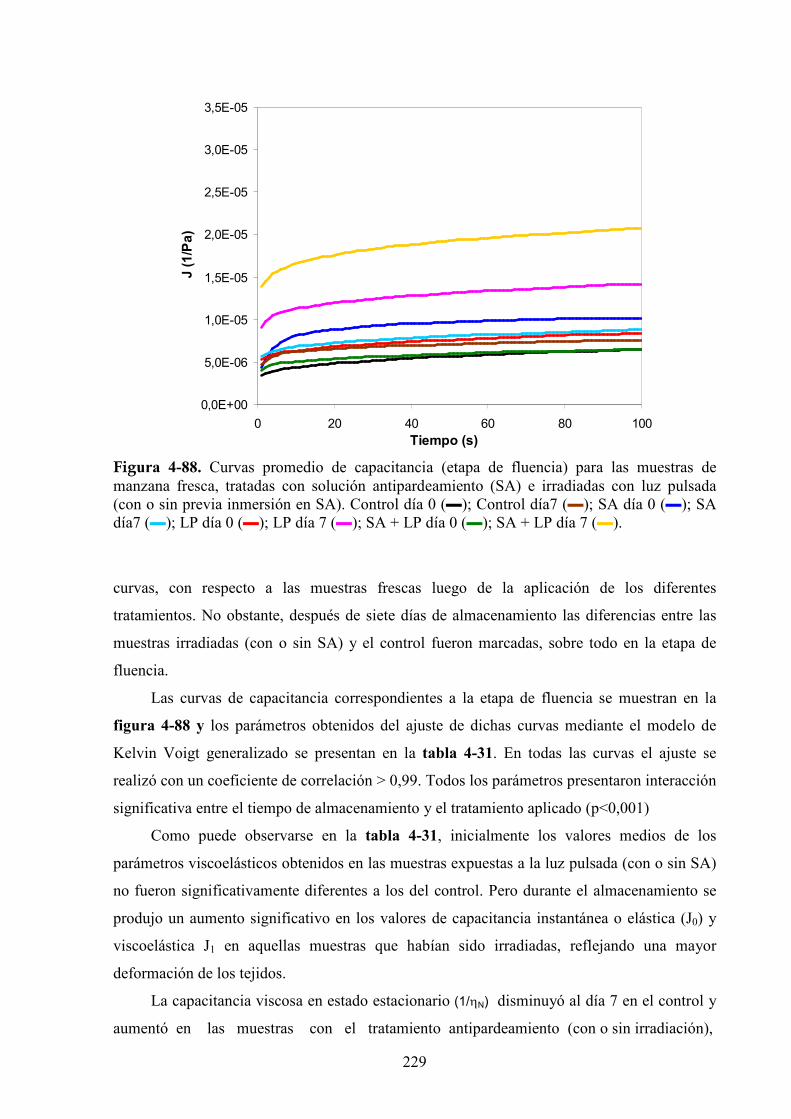
229
0,0E+00
5,0E06
1,0E05
1,5E05
2,0E05
2,5E05
3,0E05
3,5E05
0 20 40 60 80 100
���������
���!�8�
�1�ura 4'88. Curvas promedio de capacitancia (etapa de fluencia) para las muestras de manzana fresca, tratadas con solución antipardeamiento (SA) e irradiadas con luz pulsada (con o sin previa inmersión en SA). Control día 0 (▬); Control día7 (▬); SA día 0 (▬); SA día7 (▬); LP día 0 (▬); LP día 7 (▬); SA + LP día 0 (▬); SA + LP día 7 (▬). �
curvas, con respecto a las muestras frescas luego de la aplicación de los diferentes
tratamientos. No obstante, después de siete días de almacenamiento las diferencias entre las
muestras irradiadas (con o sin SA) y el control fueron marcadas, sobre todo en la etapa de
fluencia.
Las curvas de capacitancia correspondientes a la etapa de fluencia se muestran en la
figura 4'88 y los parámetros obtenidos del ajuste de dichas curvas mediante el modelo de
Kelvin Voigt generalizado se presentan en la tabla 4'31. En todas las curvas el ajuste se
realizó con un coeficiente de correlación > 0,99. Todos los parámetros presentaron interacción
significativa entre el tiempo de almacenamiento y el tratamiento aplicado (p<0,001)
Como puede observarse en la tabla 4'31, inicialmente los valores medios de los
parámetros viscoelásticos obtenidos en las muestras expuestas a la luz pulsada (con o sin SA)
no fueron significativamente diferentes a los del control. Pero durante el almacenamiento se
produjo un aumento significativo en los valores de capacitancia instantánea o elástica (J0) y
viscoelástica J1 en aquellas muestras que habían sido irradiadas, reflejando una mayor
deformación de los tejidos.
La capacitancia viscosa en estado estacionario (1/ηN) disminuyó al día 7 en el control y
aumentó en las muestras con el tratamiento antipardeamiento (con o sin irradiación),

230
Tabla 4'31. Parámetros de las curvas de fluencia*de rodajas de manzana fresca (control), inmersas en solución antipardeamiento (SA), irradiadas con luz pulsada (LP), inmersas en solución antipardeamiento (SA) e irradiadas con LP, almacenadas en refrigeración.
�
Tiempo (día)
Tratamiento J0 (1/Pa) (x 106)
J1 (1/Pa) (x 106)
J2 (1/Pa) (x 106)
λλλλ1 (s)
λλλλ2 (s)
ηηηηN (Pa.s) (x 10'8)
Control
3,1(0,9)
aA
48%**
1,4(0,7)
aA
21%**
0,7(0,3)
aA
10%**
26,4(3,9) aA
2,6(0,5) aA,B
0,7(0,5)
aA
21%**
SA
3,1(0,8)
aA
37%**
4,3(3,2)
aB
41 %**
0,5(0,2)
aA
10%**
32,9(12,2) aA
3,1(1,1) aA
1,9(1,2)
aB
12%**
LP
4,9(1,1)
aA
58%**
1,4(0,3)
aA
17%**
0,9(0,2)
aA
10%**
26,2(8,1) aA
2,4(0,6) aA,B
0,8(0,3)
aA
15%**
0
SA + LP
3,5(0,5)
aA
59%**
1,2 (0,5)
aA
19%**
0,6(0,1)
aA
11%**
35,0(19,1) aA
2,1(0,6) aB
1,5(0,8)
aA,B
11%**
Control
4,1(0,9)
aA
53%**
1,1 (0,4)
aA
14%**
1,9(1,8)
aA
25%**
30,3(13,4)
aA
2,6(0,6) aA
1,7(0,8)
bA
8%**
SA
5,2(1,4)
aA
59%**
1,4 (0,7)
bA
16%**
0,9(0,3)
aA
10%**
19,1(5,7) bA
1,9(0,7) bA
0,8(0,6)
bB
15%**
LP
8,2(4,4)
bB
58%**
2 (1) aA,B
14%**
2(1)
bA
15%
22,8(3,8) aA
2,1(0,6) aA
0,6(0,2)
aB
13%**
7
SA + LP
13(5,2)
bC
62%**
3,1 (1,8)
bB
15%**
2,2 (0,9)
bA
11%**
21,3(3,9) bA
2,6(0,9) aA
0,4(0,2)
bB
12%**
* Parámetros derivados de la regresión de la ecuación 3.12 y su correspondiente desviación estándar **Contribución relativa de cada tipo de capacitancia a la capacitancia total al final de la etapa de fluencia. Los resultados fueron expresados como la media seguida de la desviación estándar (entre paréntesis). En cada columna y para un dado tratamiento, las medias seguidas por la misma letra minúscula no presentaron diferencias significativas en los dos tiempos de almacenamiento (p<0,05). En cada columna y para un mismo día de almacenamiento, las medias de los diferentes tratamientos seguidas por la misma letra mayúscula no presentaron diferencias significativas (p<0,05).
�

231
denotando una reducción y un incremento de la fluidez respectivamente. En las rodajas sólo
irradiadas este parámetro no se modificó, siendo no significativas las diferencias en relación a
las otras muestras tratadas (tabla 4'31). De todas maneras, debe destacarse al considerar estas
conclusiones que las diferencias observadas fueron pequeñas y los valores de fluidez
exhibieron grandes errores asociados.
En general, los tiempos de retardo de los dos elementos de Voigt (λ1 y λ2) no variaron
significativamente entre las diferentes muestras, por lo tanto, la velocidad de la respuesta de
los elementos estructurales asociados no se vió afectada ni por los tratamientos ni por el
almacenamiento (tabla 4'31). En el caso de las muestras tratadas con SA (con o sin
irradiación) existieron algunas diferencias significativas pero de poca magnitud. Por otro lado,
al igual que lo observado en los ensayos analizados anteriormente (sección 4.1.4), los tiempos
de retardo de los dos elementos de Voigt difirieron aproximadamente en un orden de
magnitud en todas las muestras.
Los incrementos en la capacitancia total al final de la fase de fluencia al día 7 en las muestras
tratadas en relación al tejido fresco (control día 0) fueron de ≈ 134 % para las rodajas
inmersas en SA sin irradiar, de ≈ 220 % para las expuestas a la LP y de ≈ 310 % para las
rodajas expuestas a la LP previamente tratadas con SA.
La contribución relativa de cada capacitancia a la capacitancia total en las diferentes
muestras se encontró dentro de los siguientes rangos: 37*63 % para la capacitancia elástica
instantánea (J0), 14*41 % para la capacitancia viscoelástica que respondió a menor velocidad
(J1), 10*25 % para la capacitancia viscoelástica que respondió a mayor velocidad (J2) y 8*21
% para la inversa del flujo viscoso en estado estacionario (1/ηN) (tabla 4'31). En general, en
todas las muestras la mayor contribución estuvo dada por la capacitancia J0.
De lo analizado en los ensayos oscilatorios y de fluencia*recuperación en general se
puede concluir que la irradiación con luz pulsada, en la dosis evaluada, produjo cambios en la
conducta viscoelástica del tejido de manzana que condujeron tanto a una pérdida de rigidez
de los tejidos como a mayores valores de capacitancia. Dichos cambios se vieron
incrementados por la aplicación previa de la solución antipardeamiento y durante el
almacenamiento de las muestras. Es probable que debido a la baja penetración que puede
tener la irradiación con LP, al igual que la radiación UV*C, los cambios encontrados se den
fundamentalmente en las regiones superficiales de las muestras. Aunque para dosis altas el
daño en el tejido por efecto térmico no solo sería superficial.
�

232
8$,$9$��4�������������
�
En la figura 4'89 y en la tabla 4'32 se muestran los porcentajes de pérdida de peso
(PP), debido a la pérdida de fase líquida, que presentaron las rodajas de manzana fresca
(control) y las expuestas a diferentes dosis de luz pulsada (LP) a lo largo del almacenamiento.
Inmediatamente después de la exposición a la LP, las manzanas irradiadas entre 20 y 100
segundos presentaron una disminución de su peso inicial entre 2 y 12 %, que se vio
incrementada con el aumento del tiempo de irradiación y en las muestras irradiadas a menor
distancia de la lámpara. Esta reducción inicial sería causada fundamentalmente por la
evaporación del agua de los tejidos como consecuencia del incremento de la temperatura en la
superficie de las muestras. Es de esperar por esto que la reducción haya sido mayor en las
rodajas irradiadas más cercanas a la lámpara y durante tiempos de irradiación más
prolongados en donde los incrementos de temperatura fueron mayores. Durante el
almacenamiento de las muestras se observó una disminución del peso tanto en las muestras
control como en las rodajas irradiadas, pero la velocidad de disminución fue mayor en las
muestras no irradiadas. Al séptimo día del almacenamiento, el porcentaje de PP que presentó
el control (≈ 12 %) no fue significativamente diferente al de las muestras irradiadas, con
excepción de las rodajas irradiadas 2 segundos a 5 cm de la lámpara que presentaron una
pérdida de peso menor (≈ 6 %). Para poder explicar esta conducta se necesitarían mayores
investigaciones.
En las muestras irradiadas que fueron previamente inmersas en la solución
antipardeamiento (SA), si bien inicialmente la reducción del peso luego de la irradiación fue
similar al de las rodajas irradiadas sin SA, durante el almacenamiento presentaron mayores
variaciones en el peso. Al final del almacenamiento, el porcentaje de PP de las rodajas
irradiadas 60 y 100 s a d = 5 cm (17*22 %) fue mayor al control, pero en las demás muestras
tratadas las diferencias no fueron significativas (figura 4'90 y tabla 4'33). En estas muestras,
la aplicación previa de la solución antipardeamiento pudo haber provocado un mayor daño al
tejido que el generado solo por el tratamiento de irradiación, lo que pudo haber favorecido un
aumento de la tasa de transpiración.
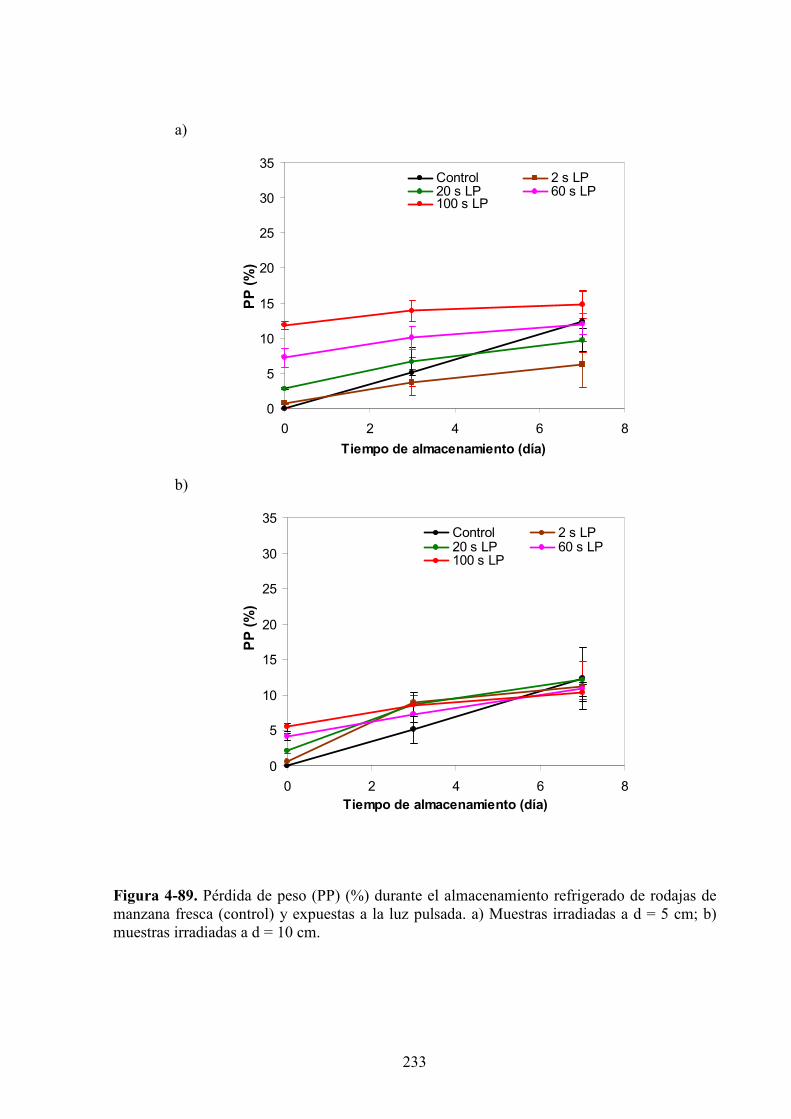
233
a)
0
5
10
15
20
25
30
35
0 2 4 6 8
���������������������������
88��;
Control 2 s LP20 s LP 60 s LP100 s LP
b)
0
5
10
15
20
25
30
35
0 2 4 6 8
���������������������������
88��;
Control 2 s LP20 s LP 60 s LP100 s LP
Figura 4'89. Pérdida de peso (PP) (%) durante el almacenamiento refrigerado de rodajas de manzana fresca (control) y expuestas a la luz pulsada. a) Muestras irradiadas a d = 5 cm; b) muestras irradiadas a d = 10 cm.

234
Tabla 4'32. Pérdida de peso (PP) durante el almacenamiento refrigerado de rodajas de manzana fresca (C) y expuestas a la luz pulsada. a) Muestras irradiadas a d = 5 cm; b) muestras irradiadas a d = 10 cm. a)
Tiempo de almacenamiento (día)
Tiempo de irradiación
(s) 0 3 7
0 (C) 0aA 5,2(2,1)bA,B 12,3(4,4)cA,C
2 0,75(0,06)aA 3,7(1,8)bA 6,2(3,3)c B
20 2,8(0,1)aB 6,6(1,9)bB 9,7(1,7)cB,C
60 7,2(1,4)aC 10,1(1,6)bC 12,1(1,5)cA
100 11,8(0,6)aD 13,9(1,4)bD 14,8(2,1)bA
b)
Tiempo de almacenamiento (día)
Tiempo de irradiación
(s) 0 3 7
0 (C) 0aA 5,2(2,1)bA 12,3(4,4)cA
2 0,57 (0,05)aA 9,1(1,2)bB 11,2(1,8)cA
20 2,1(0,3)aB 8,7(1,4)bB 12,1(2,7)cA
60 4,1(0,4)aC 7,3(1,2)bA 10,9(0,9)cA
100 5,5(0,6)aD 8,7(1,7)bB 10,3(1,2)cA
Los resultados fueron expresados como la media seguida por la desviación estándar (entre paréntesis). En cada fila las medias seguidas por la misma letra minúscula no presentaron diferencias significativas a lo largo del almacenamiento (p<0,05). En cada columna, las medias de los diferentes tratamientos seguidas por la misma letra mayúscula no presentaron diferencias significativas (p<0,05).

235
a)
0
5
10
15
20
25
30
35
0 2 4 6 8
���������������������������
88��;
Control SASA+2 s LP SA+20 s LPSA+60 s LP SA+100 s LP
b)
0
5
10
15
20
25
30
35
0 2 4 6 8
���������������������������
88��;
Control SASA+2 s LP SA+20 s LPSA+60 s LP SA+100 s LP
Figura 4'90. Pérdida de peso (PP) (%) durante el almacenamiento refrigerado de rodajas de manzana fresca (control) y expuestas a la luz pulsada con inmersión previa en solución antipardeamiento (SA). a) Muestras irradiadas a d = 5 cm; b) muestras irradiadas a d = 10 cm.

236
Tabla 4'33. Pérdida de peso (PP) durante el almacenamiento refrigerado de rodajas de manzana fresca (control) y expuestas a la luz pulsada con inmersión previa en solución antipardeamiento (SA). a) Muestras irradiadas a d = 5 cm; b) muestras irradiadas a d = 10 cm.
a)
Tiempo de almacenamiento (día)
Tiempo de irradiación
(s) 0 3 7
0 (C) 0aA 4,5(0,9)bA 12,4(3,5)cA
0 (SA) 0aA 5,2(1,3)bA 12,5(2,5)cA
2 1,5(0,1)aA,B 9,9(0,3)bB 14,8(2,3)cA,B
20 3,9(2,5)aB 11,2(3,1)bB 13,8(3,6)bA,B
60 7,3(2,5)aC 10,5(2,2)bB 16,9(4,2)cB,C
100 11,8(1,4)aD 16,3(2,7)bC 21,7(6,9)cC
b)
Tiempo de almacenamiento (día)
Tiempo de irradiación
(s) 0 3 7
0 (C) 0aA 4,5(0,9)bA 12,4(3,5)cA,B
0 (SA) 0aA 5,2(1,3)bA,B 12,5(2,5)cA,B
2 1,5(2,1)aA,B 5,9(0,5)bA,B 11,4(3,2)cA
20 2,1(0,5)aB 7,7(0,5)bB,C 11,8(3,1)cA
60 3,7(0,4)aB,C 10,1(3,2)bC 15,8(3,9)cB
100 5,8(0,3)aC 9,6(1,4)bC 15,3(4,6)cB
Los resultados fueron expresados como la media seguida por la desviación estándar (entre paréntesis). En cada fila, las medias seguidas por la misma letra minúscula no presentaron diferencias significativas a lo largo del almacenamiento (p<0,05). En cada columna, las medias de los diferentes tratamientos seguidas por la misma letra mayúscula no presentaron diferencias significativas (p<0,05).

237
8$,$;$���������������������������������������*���������������/�������������������������
� Los cambios microestructurales en la superficie de las rodajas de manzana irradiadas
con luz pulsada fueron evaluados mediante microscopía óptica.
En las figuras 4'80, 4'81, 4'82 y 4'83 se muestran las microfotografías obtenidas a los
días 0 y 7 del almacenamiento del tejido de manzana fresco (control), del tratado con la
solución antipardeamiento (SA), y del tejido irradiado con LP durante 60 segundos a 10 cm
de distancia de la lámpara (con o sin inmersión previa en la SA).
Las células del tejido fresco y del tejido tratado con la solución antipardeamiento (sin
irradiar) se observaron turgentes y con paredes celulares bien definidas. Algunas células
presentaron ruptura de membranas celulares probablemente como consecuencia de las
operaciones de corte (figuras 4'80 y 4'81 A y C). También se observó en alguna de ellas
plasmólisis incipiente. En el tejido irradiado las células exhibieron disrupción de membranas
celulares y paredes más debilitadas (figuras 4'80 B y 4'81 B). En los tejidos irradiados
previamente inmersos en la SA se observó además paredes que presentaron ruptura en algunas
zonas (figuras 4'80 D y 4'81 D).
Los tejidos de las muestras control almacenados en refrigeración durante una semana
presentaron paredes celulares con cierto grado de plegamiento pero con una mayor densidad
óptica que la observada en el tejido fresco al día 0 (figuras 4'82 A y 4'83 A). Este
comportamiento también fue visualizado previamente en los controles realizados al analizar
los tratamientos de irradiación con luz UV*C (sección 4.1.6). Este mismo efecto del
almacenamiento fue observado también en los tejidos correspondientes a las muestras que
fueron inmersas en la solución antipardeamiento (figuras 4'82 C y 4'83 C).
Los tejidos irradiados (con o sin pretratamiento antipardeamiento) se visualizaron luego
de los siete días de almacenamiento con una disrupción generalizada. Las células aparecieron
muy colapsadas, con ruptura de membranas celulares y paredes con disrupciones y con
plegamientos pronunciados, en parte atribuibles a la pérdida de presión de turgor (figuras 4'
82 (B y D) y 4'83 (B y D)). También fue muy notoria la menor tinción de las paredes
celulares, tanto en la zona de unión de dos células como en las zonas de adhesión de tres
células. Ello indicaría un efecto del tratamiento a nivel de las sustancias pécticas de la pared
celular, incrementado a lo largo del almacenamiento. Las paredes de las células inmersas en la
SA e irradiadas se mostraron además más hinchadas (figura 4'83 D).
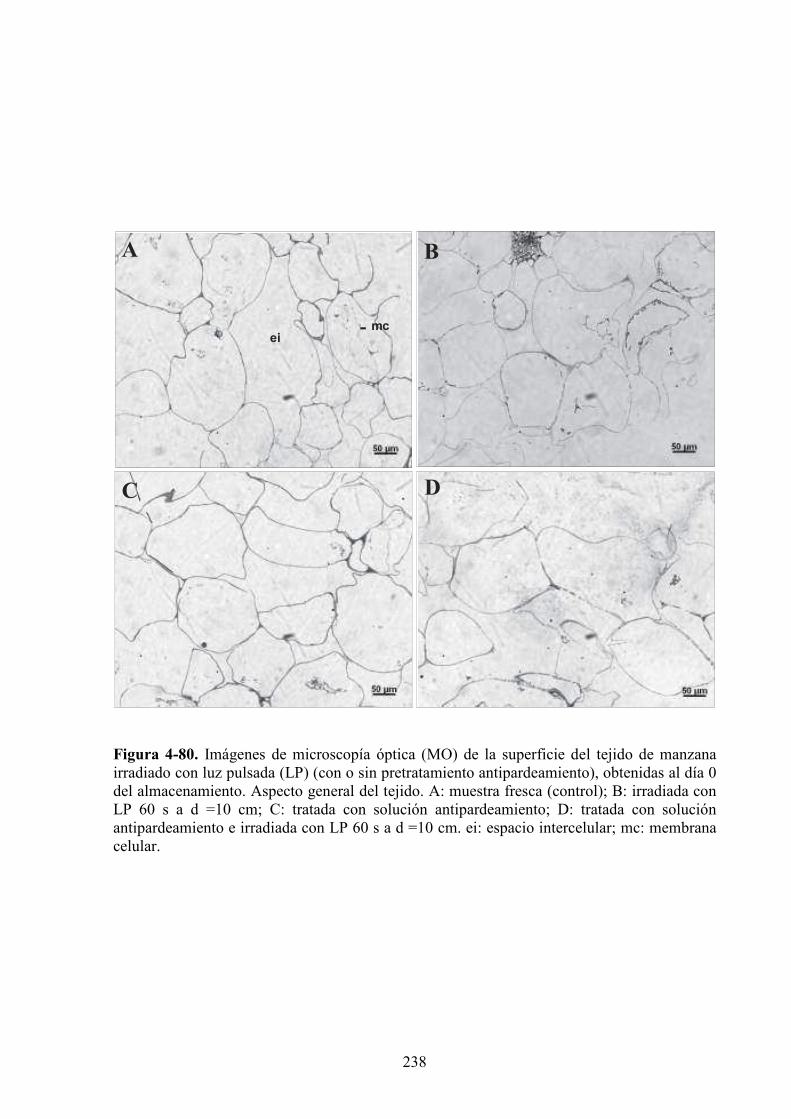
238
A B
C D
����
Figura 4'80. Imágenes de microscopía óptica (MO) de la superficie del tejido de manzana irradiado con luz pulsada (LP) (con o sin pretratamiento antipardeamiento), obtenidas al día 0 del almacenamiento. Aspecto general del tejido. A: muestra fresca (control); B: irradiada con LP 60 s a d =10 cm; C: tratada con solución antipardeamiento; D: tratada con solución antipardeamiento e irradiada con LP 60 s a d =10 cm. ei: espacio intercelular; mc: membrana celular.

239
A B
C D
��
Figura 4'81. Imágenes de microscopía óptica (MO) de la superficie del tejido de manzana irradiado con luz pulsada (LP) (con o sin pretratamiento antipardeamiento), obtenidas al día 0 del almacenamiento. Detalle. A: muestra fresca (control); B: irradiada con LP 60 s a d =10 cm; C: tratada con solución antipardeamiento; D: tratada con solución antipardeamiento e irradiada con LP 60 s a d =10 cm. lm: laminilla media.

240
A B
C D
Figura 4'82. Imágenes de microscopía óptica (MO) de la superficie del tejido de manzana irradiado con luz pulsada (LP) (con o sin pretratamiento antipardeamiento), obtenidas al día 7 del almacenamiento. Aspecto general del tejido. A: muestra fresca (control); B: irradiada con LP 60 s a d =10 cm; C: tratada con solución antipardeamiento; D: tratada con solución antipardeamiento e irradiada con LP 60 s a d =10 cm.
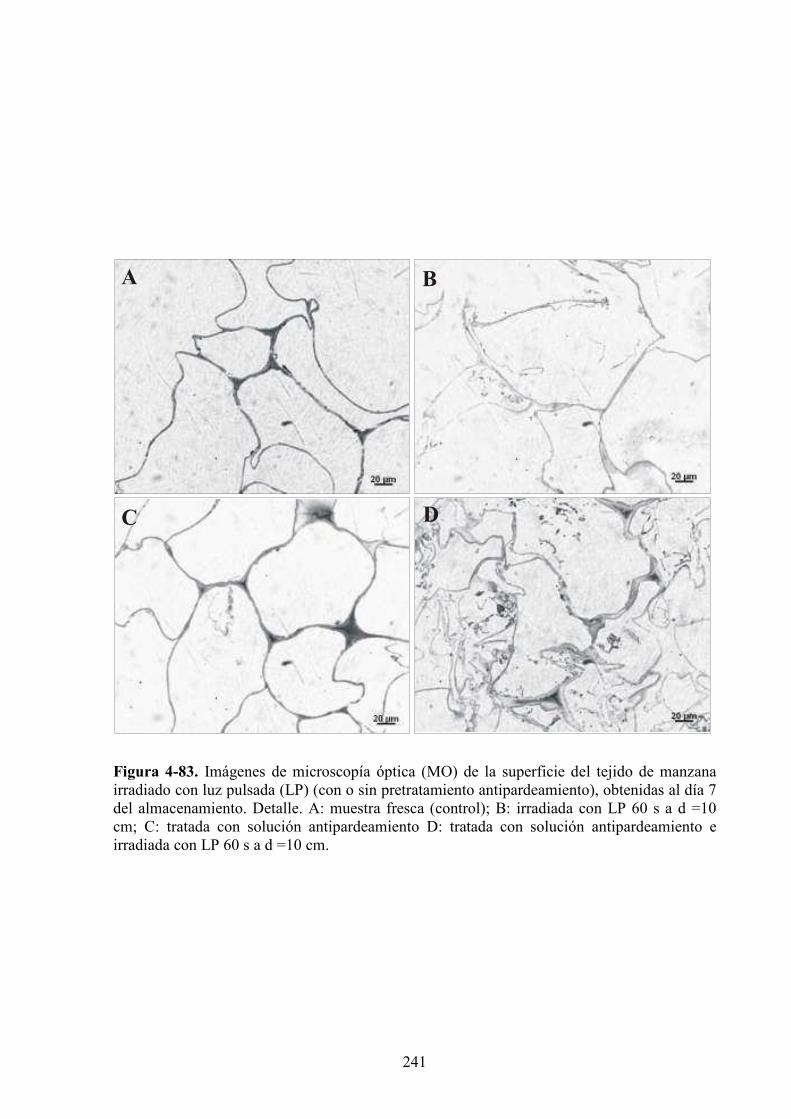
241
A B
C D
Figura 4'83. Imágenes de microscopía óptica (MO) de la superficie del tejido de manzana irradiado con luz pulsada (LP) (con o sin pretratamiento antipardeamiento), obtenidas al día 7 del almacenamiento. Detalle. A: muestra fresca (control); B: irradiada con LP 60 s a d =10 cm; C: tratada con solución antipardeamiento D: tratada con solución antipardeamiento e irradiada con LP 60 s a d =10 cm.

242
8$,$?$�"��������������������
Se realizaron algunos estudios microbiológicos preliminares en donde se evaluó la
supervivencia de E. coli y L. innocua inoculados en rodajas de manzana luego de la
exposición a diferentes dosis de luz pulsada. Los tiempos de irradiación utilizados fueron los
mismos seleccionados para realizar los estudios de calidad, como así también la distancia de
la lámpara a la que fueron ubicadas las muestras.
Como puede observarse en la figura 4'95 las curvas semilogarítmicas de supervivencia
fueron fuertemente no lineales. En las curvas de L. innocua se observó inicialmente una
abrupta reducción de ≈ 0,5 ciclos logaritmos y luego el recuento permaneció relativamente
constante, decayendo luego de los 40 segundos de irradiación, mucho más abruptamente en
las rodajas irradiadas a menor distancia (figura 4'95 a). En el caso de E. coli, la disminución
en la población siguió un patrón similar, si bien fue un poco más gradual con el aumento del
tiempo de irradiación a 10 cm de la lámpara. La curva de las muestras irradiadas a menor
distancia presentó algunas fluctuaciones (figura 4'95 b). Luego de 100 segundos de
irradiación, la reducción de L. innocua en las dos distancias evaluadas fue de ≈ 2 ciclos
logarítmicos, mientras que en E. coli la reducción fue de ≈ 2 ciclos logarítmicos a d = 5 cm y
de ≈ 1,2 ciclos logarítmicos a d = 10 cm.
Las diferencias encontradas entre estos microorganismos por un lado y la diferente
respuesta del mismo microorganismo según la distancia a la lámpara por otro, no sólo pueden
deberse a diferencias en la susceptibilidad al daño causado por la región UV*C del espectro de
irradiación y/o la diferente intensidad de la radiación según la ubicación, sino también al
calentamiento de las muestras y a las diferencias en la resistencia térmica entre ambas
especies cuando se aplican tiempos de irradiación más prolongados, sobre todo a la distancia
de 5 cm.
Los estudios reportados en bibliografía sobre el efecto de la luz pulsada en la
inactivación de microorganismos en superficies inorgánicas y de frutas y vegetales
mínimamente procesados son muy escasos. Uno de los trabajos existentes es el reportado por
Uesugi y col. (2007), quienes, utilizando un equipo de irradiación similar al de este trabajo,
estudiaron la cinética de inactivación de L. innocua en superficies de acero inoxidable para
simular un sustrato sólido, irradiando las muestras a una distancia de 5 cm de la lámpara y
durante tiempos de irradiación de hasta 15 segundos (dosis ≈ 170 kJ/m2). Las curvas de
supervivencia semilogarítmicas que se obtuvieron fueron no lineales y con un pronunciado
efecto “cola”, lográndose reducciones de ≈ 3,5 ciclos logarítmicos. Este “plateau” también fue

243
a)
2,5
2
1,5
1
0,5
0
0 20 40 60 80 100 120���������
����� �
�
d = 5 cm
d = 10 cm
b)
2,5
2
1,5
1
0,5
0
0 20 40 60 80 100 120
���������
����� �
�
d= 5 cm
d= 10 cm
Figura 4'95. Curvas semilogarítmicas de supervivencia de L. innocua (a) y E. coli (b) en rodajas de manzana irradiadas con luz pulsada a una distancia de la lámpara de 5 cm y 10 cm. N0 L. innocua: 6 x 106 UFC/cm2; N0 E. coli: 4 x 106 UFC/cm2

244
observado en las experiencias de este trabajo a dosis similares. En estas experiencias los
tiempos de irradiación utilizados fueron bajos, y por lo tanto, el mecanismo de inactivación
estaría dado sólo por efecto de la irradiación en sí y no participaría la componente térmica.
Bialka y col. (2008) estudiaron la cinética de inactivación de E. coli O157:H7 y de
Salmonella enterica inoculadas en frambuesas y frutillas utilizando también el mismo equipo
de irradiación. Las frambuesas fueron irradiadas a una distancia de la lámpara de 3 cm
durante tiempos de irradiación entre 5 y 60 s (rango de dosis: 60 kJ/m2* 720 kJ/m2) y las
frutillas durante los mismos tiempos de irradiación pero a una distancia de 13 cm (rango de
dosis: 54 kJ/m2*644 kJ/m2). Las curvas de supervivencia semilogarítmicas que se obtuvieron
fueron en todos los estudios no lineales, pero en el caso de los microorganismos inoculados en
frutilla las curvas presentaron un efecto cola que no fue observado cuando fueron inoculados
en frambuesas. Si bien los autores explican estas diferencias en base a diferencias en la
composición de la matriz, los mayores incrementos de temperatura que sufrieron las muestras
de frambuesa al ser irradiadas a menor distancia de la lámpara pudieron también afectar el
patrón de inactivación. Luego de 60 segundos de irradiación, la reducción de E. coli y
Salmonella en frutillas fue de ≈ 2,5 ciclos logarítmicos, mientras que en frambuesas la
reducción fue de ≈ 4*4,5 ciclos logarítmicos.
Gómez*López y col. (2005 b) estudiaron la descontaminación de diferentes vegetales
mínimamente procesados (espinaca, radicheta, lechuga, zanahoria, pimiento y repollo blanco)
y brotes de soja. Obtuvieron reducciones logarítmicas en los recuentos de los
microorganismos aerobios mesófilos entre 0,21 * 1,67 cuando los vegetales fueron irradiados
durante 45 segundos (en cada lado) a 12,8 cm de distancia de la lámpara, y entre 0,56 *2,04
cuando se irradiaron durante 180 segundos. La irradiación durante tiempos más prolongados
(180 s) no permitió obtener reducciones mucho mayores, y el deterioro en la calidad
provocado por el mayor calentamiento de las muestras fue notorio haciendo inaceptable la
aplicación de esas dosis de irradiación. Al realizar estudios de vida útil en repollo y lechuga, a
pesar de la reducción inicial en los recuentos, no observaron un incremento de la vida útil de
estos vegetales por la aplicación de LP en las condiciones ensayadas.
El efecto de la luz pulsada sobre la flora nativa de las rodajas de manzana, así como
también la evolución de los microorganismos irradiados durante el almacenamiento, no
fueron evaluados y serán abordados en estudios posteriores. También se analizará si la
aplicación previa de la solución antipardeamiento modifica el patrón de inactivación, tal como
fue observado en los tratamientos de irradiación con luz UV*C.

245
8$,$@$� !���������� ��� ���� ����������� ��������� �� ���+4�� ��� ���� ��������������������� �� ��� ��� �������������� ��������������� ��� ���� ��*���� �������������������������������������������������������������������
El efecto letal de la aplicación luz pulsada sobre los microorganismos puede ser
atribuido a su amplio espectro rico en la región UV*C y la emisión de pulsos de corta
duración y alta energía. Debido a que el espectro de emisión también abarca la región visible*
infrarroja, los incrementos de temperatura durante tiempos prolongados de irradiación
pueden ser importantes, y en estos casos, el mecanismo de inactivación podría estar dado no
solo por un efecto fotoquímico (como ocurre con la radiación UV*C continua) sino también
fototérmico.
Durante los tiempos de irradiación evaluados en este trabajo (2*100 s), la temperatura
superficial de las rodajas de manzana aumentó con el incremento de la dosis de irradiación
(mayor tiempo de irradiación y menor distancia a la lámpara), llegándose a temperaturas
cercanas a los 90º C en las muestras ubicadas a 5 cm de la lámpara e irradiadas 100 segundos.
Este incremento de temperatura no solo se produjo en la superficie irradiada ya que en la cara
inferior de la muestra no expuesta a la irradiación los incrementos fueron levemente menores.
Por otro lado, el aumento de temperatura varió según la posición de las muestras en el estante,
siendo más notoria esta variación cuando el estante fue ubicado a la menor distancia a la
lámpara. Si bien las dosis de irradiación no pudieron ser medidas, estos resultados permiten
inferir que la intensidad de irradiación que reciben las muestras varía de acuerdo a la posición.
Esta observación es importante, ya que considerando una potencial adopción de la tecnología
a nivel industrial, la posición y la orientación de las lámparas respecto a las muestras es un
aspecto a tener en cuenta para poder asegurar que la eficiencia de la descontaminación sea la
misma en todos los productos. A estas mismas conclusiones llegaron Gómez*López y col.
(2005 a) cuando analizaron la influencia en la eficiencia de la descontaminación con LP de la
posición relativa entre la muestra y la lámpara.
El incremento del pardeamiento superficial que presentaron las muestras irradiadas con
LP a dosis crecientes de irradiación puede atribuirse, en parte, al mayor grado de ruptura de
membranas celulares que fue observada en los tejidos irradiados y la consecuente
descompartimentalización de enzimas y sustratos (figuras 4'80 B, 4'81 B, 4'82 B y 4'83 B),
de igual manera que lo observado en las rodajas irradiadas con luz UV*C. Pero en este caso
no está claro si el pardeamiento fue solo enzimático, ya que las altas temperaturas registradas
en las rodajas irradiadas durante tiempos prolongados pudieron haber favorecido también
reacciones de pardeamiento no enzimático. Sería interesante además poder analizar la

246
actividad remanente de la enzima polifenoloxidasa, principal responsable del pardeamiento
enzimático, luego de la aplicación de distintas dosis de irradiación para poder explicar
también la inhibición del pardeamiento que fue observada cuando se aplicó una dosis baja de
irradiación.
La aplicación del tratamiento antipardeamiento con la solución de ácido
ascórbico/cloruro de calcio previamente a la irradiación, si bien redujo el desarrollo del
pardeamiento a todas las dosis evaluadas, en las rodajas irradiadas durante tiempos más
prolongados (100 s a d= 10 cm y 60 s y 100 s a d = 5 cm) la inhibición fue menos efectiva, y
el color desarrollado al final del almacenamiento en estas muestras podría no ser aceptable
para el consumidor.
La irradiación también provocó cambios en la conducta viscoelástica del tejido de
manzana. Los valores del módulo elástico (G’) y del módulo de almacenamiento (G”)
disminuyeron después de que las muestras fueran expuestas a la irradiación. Este
comportamiento podría relacionarse con una disminución de la turgencia del tejido irradiado
como consecuencia no sólo de la ruptura de membranas celulares durante la irradiación, sino
también, de la pérdida de agua por evaporación que sufrieron las muestras durante la
irradiación al incrementarse la temperatura. También contribuiría el debilitamiento de las
paredes celulares, observadas en la micrografía con una densidad óptica menor a la del control
(figuras 4'80 B y 4'81 B). El almacenamiento no causó cambios posteriores en G’ si bien en
las observaciones histológicas al día 7 las células se mostrarían ligeramente más colapsadas
(figura 4'82 B).
Los parámetros viscoelásticos derivados de las curvas de fluencia no revelaron cambios
al inicio del almacenamiento en las características viscoelásticas del tejido de manzana
irradiado en relación al tejido fresco (control). Sin embargo, en los tejidos irradiados
almacenados las capacitancias instantánea o elástica (J0) y viscoelástica J1 presentaron un
aumento significativo, reflejando la mayor deformabilidad de estos tejidos. A nivel estructural
estos cambios estarían asociados con la ruptura de membranas celulares y con el
debilitamiento de las paredes celulares, visualizadas con baja densidad en los tejidos
irradiados al día 7 (figuras 4'82 B y 4'83 B).
Cuando se analizaron las propiedades viscoelásticas a altas deformaciones, el módulo de
deformabilidad (Ed) fue el parámetro que reflejó mayores diferencias entre las muestras
irradiadas y el control. Esto era de esperarse, ya que este parámetro refleja el comportamiento
a bajas deformaciones, y como la LP al igual que la radiación UV*C tiene baja penetración en
el tejido, el mayor daño se produciría en las regiones superficiales. El Ed disminuyó en todas

247
las muestras irradiadas luego de la exposición a la irradiación, siendo mayor la disminución
cuando las muestras fueron ubicadas a menor distancia de la lámpara e irradiadas durante
tiempos más prolongados. Esto podría relacionarse con la mayor pérdida de agua observada
inicialmente en las muestras a dosis crecientes de irradiación y con un mayor daño en el tejido
que pudieron provocar la pérdida de turgencia en las células con la consecuente disminución
de la rigidez del mismo. A pesar de esta reducción inicial en el Ed que presentaron todas las
muestras irradiadas, al finalizar el almacenamiento sólo las muestras irradiadas durante 60 s y
100 s a una distancia de 5 cm presentaron una disminución significativa de Ed con respecto al
control.
Cuando las muestras irradiadas fueron pretratadas con la solución antipardeamiento, si
bien al inicio del almacenamiento no se observaron en general cambios en la respuesta a los
ensayos oscilatorios y de fluencia en relación a los cambios que presentaron las muestras sólo
irradiadas, luego de ser almacenadas durante siete días, la pérdida de rigidez y elasticidad y el
aumento de la deformación de estos tejidos fue mayor, y se vió reflejada en una mayor
disminución de Ed y G’ y en un aumento en las capacitancias J0 y 1/ηN. Estas diferencias
podrían asociarse, en parte, con la mayor pérdida de agua que presentaron las rodajas tratadas
con SA e irradiadas durante el almacenamiento en relación a las muestras sólo irradiadas. A
nivel estructural se observaron también diferencias entre estas muestras, ya que las
alteraciones producidas en la pared por el tratamiento antipardeamiento (figuras 4'80 D y 4'
81 D) se intensificaron durante el almacenamiento (paredes muy plegadas, con importantes
disrupciones) y las paredes aparecieron también más hinchadas (figuras 4'82 D y 4'83 D),
indicando una exosmosis importante que aumentaría el valor de l/ηN.
Si bien la aplicación de LP permitió obtener mayores reducciones decimales de los
microorganismos inoculados cuando se irradiaron las muestras durante tiempos más
prolongados y a menor distancia de la lámpara, si se tienen en cuenta los estudios de calidad,
la aplicación de tiempos de irradiación mayores a 20 segundos a una distancia de 5 cm y
mayores a 60 segundos a una distancia de 10 cm serían inaceptables, fundamentalmente por
los cambios provocados en el color superficial de las manzanas durante el almacenamiento,
que no pudieron ser totalmente controlados con la aplicación de la solución antipardeamiento.

248
9$��%��5�)!%�")���9$#$�����������������������
� La rodajas de manzana irradiadas con luz UV*C en un rango de dosis entre 5,6 y 14,1
KJ/m2, presentaron un aumento del pardeamiento superficial con el aumento de la
dosis aplicada, que se vio incrementado durante el almacenamiento de las mismas.
� La radiación UV*C produjo cambios menores en el comportamiento reológico de las
rodajas de manzana, asociados con una pérdida de rigidez del tejido. Los ensayos de
barrido de frecuencia y de fluencia*recuperación fueron más sensibles para detectar
estos cambios que los ensayos de compresión. Por otra parte, debido a la baja
penetración de la radiación UV*C, los cambios encontrados se darían
fundamentalmente en las regiones superficiales de las muestras expuestas a la
irradiación.
� Las rodajas irradiadas a las dosis más altas presentaron una pérdida de peso mayor
durante el almacenamiento que las rodajas no irradiadas. Esto estaría relacionado con
un incremento en la tasa de transpiración o en la pérdida de agua debido a la
eliminación o disminución de la resistencia ofrecida por las membranas.
� El incremento del pardeamiento superficial y de la pérdida de peso, como así también
los cambios en las propiedades viscoelásticas que presentaron las rodajas irradiadas
durante el almacenamiento, fueron relacionados con la mayor disrupción de las
membranas celulares y la pérdida de turgencia que fueron observadas en las células de
estos tejidos.
� Los tratamientos de escaldado e inmersión en la solución de ácido ascórbico/cloruro
de calcio aplicados a las muestras antes de la irradiación fueron efectivos para inhibir
el pardeamiento superficial provocado por la luz UV*C en todas las dosis evaluadas.
La mayor estabilidad en el color durante el almacenamiento se observó en las muestras
irradiadas que previamente fueron inmersas en la solución antipardeamiento.
� El pretratamiento de escaldado indujo cambios notables en las propiedades mecánicas
y viscoelásticas del tejido de manzana, que se vieron incrementados en menor
proporción por la posterior exposición a la luz UV*C. Estas modificaciones fueron
asociadas con la ruptura de membranas celulares, el daño en las paredes celulares y la
pérdida de la presión de turgor que presentaron estos tejidos. Por otro lado, las

249
propiedades reológicas de las rodajas de manzana irradiadas no se vieron afectadas
notoriamente por efecto de la aplicación previa de la solución antipardeamiento de
ácido ascórbico/cloruro de calcio.
� Los tratamientos de impregnación con calcio mediante el tratamiento de impregnación
atmosférica condujeron a muestras altamente enriquecidas ya que de acuerdo a los
requerimientos diarios para adultos de este nutriente se logra satisfacer entre el 14 y el
55 % de la Ingesta Adecuada (1000 mg/día) con la cantidad presente en 200 gramos de
fruta.
� Las rodajas de manzana enriquecidas con calcio presentaron cambios importantes en
el color durante el almacenamiento. Como estos cambios fueron marcados, sólo se
observó un leve incremento adicional del pardeamiento por efecto de la irradiación a
las dosis más altas.
� La aplicación del tratamiento de escaldado previo a la impregnación no solo favoreció
una mayor incorporación de calcio en la matriz sino que también disminuyó en gran
medida el desarrollo del pardeamiento superficial. Sin embargo, la inmersión de las
muestras en la solución de ácido ascórbico/cloruro de calcio fue más efectiva para
inhibir el pardeamiento.
� Las rodajas de manzana enriquecidas con calcio y expuestas a la radiación UV*C que
fueron impregnadas a presión atmosférica con o sin un escaldado previo, sufrieron
cambios en su comportamiento viscoelástico producidos principalmente por el
tratamiento de impregnación con calcio y por el escaldado, y en menor medida, por la
exposición a la radiación UV*C. Dichos cambios en general se incrementaron durante
el almacenamiento
� Los cambios en el color y en la conducta viscoelástica que presentaron las rodajas
enriquecidas con calcio e irradiadas con luz UV*C se relacionaron con modificaciones
estructurales provocadas por los tratamientos aplicados, tales como ruptura de
membranas, alteración de paredes celulares y pérdida de turgencia.
� A pesar de que la aplicación de la solución antipardeamiento redujo el efecto de la
irradiación sobre la inactivación de microorganismos, los recuentos de
microorganismos inoculados y de flora nativa fueron menores que en las rodajas no
expuestas a la irradiación.
9$,$���������������������������
�

250
� La irradiación de rodajas de manzana con luz pulsada produjo un incremento del
pardeamiento que fue mayor con el aumento de la dosis aplicada (mayor tiempo de
irradiación y menor distancia a la lámpara) y a medida que transcurría el
almacenamiento. Al igual que en los tratamientos con luz UV*C, el desarrollo del
pardeamiento fue relacionado en parte con la mayor disrupción de membranas
celulares observadas en los tejidos irradiados.
� La aplicación previa de la solución de ácido ascórbico/cloruro de calcio (SA) a la
irradiación con LP redujo el desarrollo del pardeamiento pero fue menos efectiva en
las muestras irradiadas a las dosis más altas.
� La exposición a la luz pulsada (con o sin SA) también indujo modificaciones a lo
largo del almacenamiento en la conducta viscoelástica del tejido de manzana asociadas
a una disminución en la rigidez, sobre todo en las regiones más superficiales de las
muestras. Dichas modificaciones se acentuaron cuando las mismas fueron irradiadas
más próximas a la lámpara y durante tiempos más prolongados, y fueron relacionadas
con la mayor disrupción y pérdida de agua que presentaron los tejidos irradiados como
consecuencia no solo de la irradiación sino también del incremento de temperatura.
� La utilización de tiempos de irradiación muy prolongados, sobre todo cuando las
muestras se ubicaron muy próximas a la lámpara, no fueron adecuados desde el punto
de vista del deterioro en la calidad (principalmente color) de la fruta, a pesar de que
resultaron en una mayor inactivación de los microorganismos.
�
9$7$�����������������������
�
� Al estudiar la aplicación de la radiación UV*C y de la luz pulsada en frutas y vegetales
mínimamente procesados o en cualquier otro alimento es necesario evaluar
conjuntamente el efecto de la dosis tanto sobre la inactivación microbiológica (flora
nativa e inoculada) como así también sobre los parámetros de calidad del alimento,
para luego poder seleccionar una dosis óptima para aplicar estos factores de
conservación sin producir un gran deterioro en la calidad. El conocimiento de dicha
dosis permitirá un mejor diseño de tecnologías combinadas de conservación, donde se
utilicen otros factores de estrés microbiano simultáneos y/o secuenciales para lograr
estabilidad microbiológica. �
� Los atributos de calidad y el comportamiento microbiano deben analizarse no sólo
después de aplicar los tratamientos sino también a lo largo del tiempo de

251
almacenamiento. En esta tesis se ha visto, por un lado, que los mayores cambios
adversos en el color y en las propiedades reológicas de las manzanas tratadas con luz
UV*C o LP se produjeron durante esta etapa. Por otro lado, el estudio de la flora
durante el almacenamiento de manzanas tratadas con UV*C permitió comprobar la
acción del factor de estrés en la injuria y supervivencia microbianas, y determinar el
potencial efecto benéfico de esta tecnología, en este caso, desde el punto de vista
microbiano. �
� Es importante al estudiar la aplicación de pretratamientos que permitan mejorar la
calidad del alimento irradiado evaluar también el efecto de su aplicación combinada
sobre la inactivación y supervivencia de los microorganismos.�
�
�

252
;$�&!&5!%G-�1���
Abbott, J.A. (1999). Quality measurement of fruits and vegetables. Postharvest Biology and
Technology, 15, 207*225. Abbott, J. A. & F. R. Harper. (2004). Texture. En: The commercial storage of fruits,
vegetables and florist and nursery stocks. USDA. ARS. Agriculture Handbook, 66. Adams, M.R. & Moss, M.O. (1995). Food Microbiology. The Royal Society of Chemistry, U.K. Aked, J. (2002). Maintaining the post*harvest quality of fruits and vegetables. En: Jongen, W. (ed.), Fruit and vegetable processing. Improving quality, Woodhead Publishing Limited and CRC Press, England. Pág. 119*149 Allende, A., & Artes, F. (2003). UV*C radiation as a novel technique for keeping quality of fresh processed “Lollo Rosso” lettuce. Food Research International, 36, 739*746. Alzamora, S.M., Castro, M.A., Nieto, A.B., Vidales, S.L & Salvatori, D.M. (2000 b). The role of tissue microstructure in the textural characteristics of minimally processed fruits. En: Alzamora S.M., Tapia M.S. & López*Malo A. (eds), Minimally Processed Fruits and
Vegetables. Maryland: Aspen Publishers Inc. Pág. 153*171. Alzamora, S.M.; Cerrutti, P.; Guerrero, S. & López*Malo, A. (1995). Minimally processed fruits by combined methods. En: Barbosa*Cánovas GV, Welti*Chanes J, (eds.). Food
preservation by moisture control: fundamentals and applications (International Symposium on the Properties of Water*ISOPOW Practicum II, June 19 24, 1994, México). Lancaster, PA: Technomics Publishing Co. Pág. 463*492. Alzamora, S.M., López*Malo, A. & Tapia, M.S. (2000 a). Overview. En: Alzamora S.M., Tapia M.S. & López*Malo A. (eds), Minimally Processed Fruits and Vegetables, Maryland Aspen Publishers Inc. Pág. 1*9. Alzamora, S.M. & Salvatori, D. M. (2006). Minimal processing: fundamental and applications En: Y.H. Hui (ed.), Handbook of Food Science, Technology and Engineering,
CRC Press / Taylor & Francis, Boca Raton, USA. Pág. 118*1 a 118*16. Alzamora, S.M., Salvatori, D.M., Tapia, M.S., López*Malo, A., Welti*Chanes, J. & Fito, P. (2005). Journal of Food Engineering, 67, 205*214. Alzamora, S.M., Viollaz, P.E., Martinez, V. Nieto, A.B. & Salvatori, D.M. (2008). Exploring the linear viscoelastic properties structure relationship in processed fruit tissues .En: G.E. Gutiérrez*López, G.V. Barbosa*Cánovas, J. Welti*Chanes & E. Parada*Arias (eds), Food
Engineering: Integrated Approaches, Springer, New York, USA. Cap. 9, Pág. 143*167. Ames, J.M. (1992). The maillard reaction. En: Hudson, B.J.F. (ed.), Biochemistry of food
proteins, Elsevier Applied Science Publishers, London.

253
Anderson, J. G., Rowan, N.J., MacGregor, S.J., Fouracre, R.A. & Farish, O. (2000). Inactivation of food*bourne enteropathogenic bacteria and spoilage fungi using pulsed*light. IEEE transactions on Plasma Science, 28, 83*88. Anino, S.V., Salvatori, D.M. & Alzamora, S.M. (2006). Changes in calcium level and mechanical properties of apple tissue due to impregnation with calcium salts. Food Research
International, 39, 154*164. Artés, F., Mínguez, M. I. & Hornero, D. (2002). Analysing changes in fruits pigments. En: Mac Dougall, D.B. (ed.), Colour in food. Improving quality, Woodhead Publishing Limited and CRC Press, England. Cap. 10. Pág. 248*282. Betoret, N., Fito, P., Martinez*Monzó, J., Gras, M.L. & Chiralt, A. (2001). Viability of vegetable matrices as support of physiolically active compounds. En: Welti*Chanes, J., Barbosa*cánovas, G.V. y Aguilera, J.M. (eds)., Proceedings del Congreso Internacional de Ingeniería y alimentos (ICEF 8), vol.2, 1366*1371,Lancaster, Technomic Publishing Co, Inc. Bialka, K.L., Demirci, A. & Puri, V. (2008). Modeling the inactivation of Escherichia coli O157:H7 and Salmonella enterica on raspberries and strawberries resulting from exposure to ozone or pulsed UV*light. Journal of Food Engineering, 85, 444*449. Bintsis, T., Litopoulou*Tzanetaki, E., & Robinson, R. (2000). Existing and potential applications of ultraviolet light in the food industry. A critical review. Journal of the Science
of Food and Agriculture, 80, 1*9. Bolton, J.R. (2001). Ultraviolet Application Handbook. J.R. Bolton (ed.) Photosciences Inc, Ayr, Ontario, Canada. Bolton, J.R & Linden, K.G. (2003). Standardization of methods for fluence (UV dose) determination in bench*scale UV experiments. Journal of environmental Engineering, 129, 209*215. Bourne M.C. (1976). Texture of fruits and vegetables. En: deMan JM, Voisly PW, Rasperr VF, Stabley DW. Westport (eds.), Rheology and Texture in Food Quality., CT: AVI Publishing Co.. Calzada, J.F., Peleg, M., (1978). Mechanical interpretation of compressive stress–strain relationships of solid foods. Journal of Food Science, 43, 1087–1092. Canet, W. & Sherman (1988). Influence of friction, sample dimensions and deformation rate on the uniaxial compression of raw potato flesh. Journal of Texture Studies, 19, 275*287. Carpita, N. & McCann, M. (2000). The cell wall. En: Buchanan, B.B.; Gruissem, W.; Jones, R.L. (eds), Biochemistry & Molecular Biology of Plants, American Society of Plant Physiologists. Rockville, Maryland. Pág. 52*108. Carpita, N.C. & Gibeaut, D.M. (1993). Structural models of primary cell walls in flowering plants: consistency of molecular structure with the physical properties of the walls during growth. The Plant Journal, 3, 1*30.

254
Chang, J.C.H., Ossoff, S.F., Lobe, D.C., Dorfman, M.H., Dumais, C.M., Dumais, C.M., Qualls, R.G. & Johnson, J.D. (1985). UV inactivation of pathogenic and indicator microorganisms. Applied and Environmental Microbiology, 49(6), 1361*1365. Chu, C.F. & Peleg, M. (1985). The compressive behaviour of solid food specimens with small height to diameter ratios. Journal of Texture Studies. 16, 451*464. Civello, P.M., Vicente, A.R., Martínez, G.A. (2006). UV*C technology to control postharvest diseases of fruits and vegetables. En: Troncoso*Rojas, R., Tiznado* Hernández, M.E. & Gónzalez*León, A (eds), Recent Advances in Alternative Postharvest Technologies to Control
Fungal Diseases in Fruits and Vegetable,, Transworld research Network, Kerala, India. Pág. 31*102. Costell, E.; Fiszman, S.M.; and Durán, L. (1997). Propiedades físicas I. Reología de sólidos y textura. En: Aguilera, J.M. (ed.), Temas en Tecnología de Alimentos, CYTED. Programa Iberoamericano de Ciencia Y Tecnología para el desarrollo, Instituto Politécnico Nacional, México. Pág. 215*260. Costelló Gómez, M. (2007). Efecto de las condiciones de operación en los cambios fisicoquímicos y fisiológicos de frutas mínimamente procesadas por deshidratación osmótica. Tesis doctoral, Universidad Politécnica de Valencia, España. Curtis, H. (1985). Biología. Editorial Médica Panamericana SAIC. Buenos Aires, Argentina. De Baerdemaeker, J.G. & Segerlind, L.J. (1976). Determination of the viscoelastic properties of apple flesh. Transactions of the ASAE, 346*353. DeMan, J.M. (1975). Mechanical properties of foods. En: De Man, J.M.; Voisey, P.W.; Rasper, V.F.; and Stanley, D.W.. Westport (eds), Rheology and Texture in Food Quality, Connecticut. The Avi Publishing Company, Inc. Pág. 507 D’hallewin, G., Schirra, M., Manueddu, E., Piga, A. & Ben*Yehoshua, S. (1999). Scoparone and scopaletin accumulation and ultraviolet*C induced resistance to posharvest decay oranges as influenced by harvest date. Journal of the American Society of Horticultural Science, 124, 702*707. D’hallewin, G., Schirra, M., Pala, M. & Ben*Yehoshua, S. (2000). Ultraviolet C irradiation at 0.5 KJ/m2 reduces decay without causing damage or affecting posharvest quality of star ruby grapefruit (C. paradisi Macf.). Journal of Agricultural and Food Chemistry, 48, 4571*4575. Dibble, A.R.G., Davies, P.J. & Mutschler, M.A. (1988). Polyamine content of long keeping alcobaca tomato fruit. Plant Physiology, 86, 338*340. Dunn, J.E., Clark, R.W., Asmus, J. F. Pearlman, J.S., Boyerr, K. & Painchaud, F. (1989). Methods for preservation of foodstuffs. US Patent, 4.871.559. Edwards (1999). Vegetables and fruit. En: Rosenthal A.J.. (ed.), Food Texture. Measurement
and Perception, An Aspen Publication. Pág. 259*268
Esau, K. (1982). Anatomía de las Plantas con Semilla. Editorial Hemisferio Sur SA, Buenos Aires, Argentina.

255
Erkan, M., Wang, C.Y. & Krizek, D.T. (2001). UV*C irradiation reduces microbial populations and deterioration in Cucurbita pepo fruit tissue. Environmental and Experimental
Botany, 45, 1*9. Erkan, M., Wang, S.& Wang, C.Y. (2008). Effect of UV treatment on antioxidant capacity, antioxidant enzyme activity and decay in strawberry fruit. Postharvest Biology and Technology, 48, 163*171.
Fahn A. (1985). Anatomía Vegetal. Ediciones Pirámide SA, Madrid, España.
Fan, X., Niemera, B.A., Mettheis, J.P., Zhuang, H.,& Olson, D.W. (2005). Quality of fresh cut apples slices as affected by low*dose ionizing radiation and calcium ascorbate treatment. Journal of Food Science, 70, 143*148. FDA (1996). Code of Federal Regulations. 21CFR179.41. FDA (2001). Food and Drug Administration, Center for Food Safety and Applied Nutrition Analytical Bacteriological Manual. FDA editions. Fellows, Peter (1994). Escaldado. En: Procesado de los Alimentos. Teoría y Práctica. Editorial Zaragosa Acribia. Ferry, J.D. (1980). Viscoelastic Properties of Polymers (3ed.). Wiley, New York. Fine, F. & Gervais, P. (2004). Efficiency of pulsed UV light for microbial decontamination of food powders. Journal of Food Protection, 67,787*792. Fito, P., Chiralt, A., Betoret, N., Gras M., Cháfer M., Martínez*Monzó, J.A., Andrés, A., Vidal, D. (2001). Vacuum impregnation and osmotic dehydratation in matriz engineering. Application in functional fresh food development. Journal of Food Enginering, 49, 175*183. Flügge, W. (1967). Viscoelasticity. Blaisdell Publishing Company. A division of Ginn and Company. Waltham, Massachusetts * Toronto – London. Fonseca, M.J. & Rushing, J.W (2006). Effect of Ultraviolet*C light on quality and microbial population of fresh*cut watermelon. Postharvest Biology and Technology, 40, 256*261. Garcia, E. & Barrett, D.M. (2002). Preservative treatments for fresh*cut fruits and vegetables. En: O. Lamikanra (ed.), FreshLcut Fruits and Vegetables. Science, Technology and Market. Boca Raton, Florida: CRC Press LLC. Pág. 267*303. Garcia, E. & Barrett, D.M. (2005). Fresh*cut fruits. En: Barrett, D.M, Somogyi, L. & Ramaswamy, H. (eds.), Processing Fruits. Science and Technology (2ed.). CRC press, EEUU. Pág. 53*72 Gómez*López, V.M., Devlieghere, F., Bonduelle, V. & Debevere, J. (2005 a). Factor affecting the inactivation of micro*organisms by intense light pulses. Journal of Applied
Microbiology, 99, 460*470.

256
Gómez*López, V.M., Devlieghere, F., Bonduelle, V. & Debevere, J. (2005 b). Intense light pulses decontamination of minimally processed vegetables and their shelf*life. International
Journal of Food Microbiology, 103, 79*89. Gómez*López, V.M., Ragaert, P., Debevere, J. & Devlieghere, V.M. (2007). Pulsed light for food decontamination: a review. Trends in Food Science & Technology, 18, 464*473. Gómez*López, V.M., Ragaert, P., Debevere, J. & Devlieghere, F. (2008). Decontamination methods to prolong the shelf*life of minimally processed vegetables, state*of*the*art. Critical
Reviews in Food Science and Nutrition, 48, 487*495. González*Aguilar, G.A., Zavaleta*Gatica, R. & Tiznado*Hernández, M.E. (2007). Improving postharvest quality of mango “Haden” by UV*C treatment. Postharvest Biology and
Technology, 45, 108*116. Gónzalez*Fésler, M., Salvatori, D., Gómez, P. & Alzamora, S.M. (2008). Convective air drying of apples as affected by blanching and calcium impregnation. Journal of Food
Engineering, 87, 323*332. Goupy, P., Amiot, M.J., Richard*Forget, F., Duprat, F., Aubert, S., & Nicolas, J. (1995). Enzymatic browning of model solutions and apple phenolic substrates by apple polyphenoloxidase. Journal of Food Science, 60(3), 497. Gras, M.L, Vidal, D., Btoret, N., Chiralt, A. & Fito, P. (2003). Calcium fortification of vegetables by vacuum impregnation interactions with cellular matrix. Journal of Food
Engineering, 56, 279*284. Gray, N.F. (1994). Drinking Water Quality. Problems y Solutions. Ed. N.F. Gray. Wiley, Chichester, USA. Guerrero*Beltrán, J.A., & Barbosa*Cánovas, G.V. (2004). Review: Advantages and limitations on processing foods by UV light. Food Science and Technology International, 10, 137*147. Hodge, J.E. (1953). Dehydrated Foods. Chemistry of browning reactions in model systems. Agricultural and Food Chemistry, 1, 928*943. Hoornstra, E., De Jong, G. & Notermans, S. (2002). Preservation of vegetables by light. En: Society for Applied Microbiology (Ed.), Frontiers in microbial fermentation and
preservation, Wageningen, The Netherlands. Pág. 75*77 Indaco, M., Martínez, V., Salvatori, D. y Alzamora, S.M. (2005). Rheological behaviour of potato tissue as affected by calcium impregnation at high temperature. En: Proceedings of the 4
rd MERCOSUR Congress on Process Systems Engineering, 2
st MERCOSUR Congress on
Chemical Engineering (ENPROMER 2005). ISBN 85*7650*043*4. Río de Janeiro, Brasil. Jackman, R.L.; and Stanley, D.W. (1992). Failure mechanisms of tomato pericarp tissue suggested by large and small deformation test. Journal of Texture Studies, 23, 475*489. Jackman, R.L.; and Stanley, D.W. (1995a). Perspectives in the textural evaluation of plant foods. Trends in Food Science & Technology, 6, 187*194

257
Jackman, R.L.; and Stanley, D.W. (1995b). Creep behaviour of tomato pericarp tissue as influenced by ambient temperature ripening and chilled storage. Journal of Texture Studies, 26, 537*552. Jun, S., Irudayaraj, J., Demirci, A. & Geiser, D. (2003). Pulsed UV*light treatment of corn meal for inactivation of Aspergillus niger spores. International Journal of Food Science and
Technology, 38, 883*888. Kader, A.A. (2002). Quality parameters of fresh*cut fruit and vegetable products. En: Lamikanra, O. (ed.), Fresh cut fruits and vegetables. Science, technology and market. CRC Press, LLC, Boca Raton, Florida. Pág. 11*20 Kelly, K.L.(1943). Color designations for lights. Journal of the Optical Society of America, 33, 627*31. Khan, S., Royer, J. & Srinivasa, R. (1997). Rheology: Tools and methods. Aviation Fuels with Improved Fire Safety: A Proceedings, 6, 31*46 Kim, D. M., Smith, N. L., & Lee, C. Y. (1993). Quality of minimally processed apple slices from selected cultivars. Journal of Food Science, 58, 1115*1117. Kim, J.J., Ben*Yehoshua, S., Shapiro, B., Henis, Y. & Camely, S. (1991). Accumulation of scaparone in heat*treated lemon fruit inoculated with Penicillium digitatum Sacc. Plant
Physiology, 97, 880*885. Krishnamurthy, K., Demirci, A. & Irudayaraj, J.M. (2007). Inactivation of Staphylococcus
aureus in milk using flow*through pulsed UV*light treatment system. Food Microbiology and
safety, 72, 233*239. Lagunas*Solar, M.C., Piña, C., MacDonald, J.D.& Bolkan, L. (2006). Development of pulsed UV light processes for surface fungal desinfection of fresh fruits, Journal of Food Protection, 69, 376*384. Lamikanra, O. (2002). Enzymatic effects on flavor and texture of fresh*cut fruits and vegetables. En: O. Lamikanra, FreshLcut Fruits and Vegetables. Science, Technology and
Market (pp.). Boca Raton, Florida: CRC Press LLC. Pág. 125*186 Lamikanra, O., Kueneman, D., Ukuku, D., & Bett*Garber, K.L. (2005). Effect of processing under ultraviolet light on the shelf life of fresh*cut cantaloupe melon. Journal of Food
Science, 70, C534*C539. Lana, M.M., Tijskens, L.M.M. & Van Kooten, O., 2005. Effects of storage temperatura and fruit riping on firmess of fresh cut tomatoes. Postharvest Biology and Technology, 35, 87–95. Lehninger, A. (1988). Bioquímica. Ediciones Omega S.A.; Barcelona, España Lemoine, M.L., Civello, P.M., Martinez, G.A. & Chaves, A. R. (2007). Influence of postharvest UV*C treatment on refrigerated storage of minimally processed brócoli (Brassica oleracea var. Italica). Journal of the Science of Food and Agriculture, 87, 1132*1139.

258
Levenson, D.I, Bockman, R.S. (1994). A review of calcium preparations. Nutrition Reviews, 51, 221*232. Lewicki, P.P. (1998). Effect of pre*drying and rehydration on plant tissue properties: a review. International Journal of Food Properties, 1, 1*22. Lin, T*T & Pitt R.E. (1986). Rheology of apple and potato tissue as affected by cell turgor pressure. Journal of Texture Studies, 17, 291*313 López Malo, A. & Palou, E. (2005). Ultraviolet light and food preservation. En: G. Barbosa Cánovas, M.D. Tapia & M.P Cano (eds.), Novel Food Processing Technologies, CRC Press, London, England. Cap. 18. Pág. 405*421. López*Rubira, V., Conesa, A.; Allende, A. & Artés, F. (2005). Shelf life and overall quality of minimally processed pomegranate arils modified atmosphere packaged and treated with UV*C. Postharvest Biology and Technology, 37, 174*185. Lozano, R. (1978). El color y su medición. America Lee, Buenos Aires, Argentina. Ma, S.; Silva, J.; Hearnsberger, J; and Garner, J. (1992). Prevention of enzymatic darkening in frozen sweet potatoes (Ipomoea batatas (L.) Lam.) by water blanching: Relationship among darkening, phenols, and polyphenol oxidase activity. Journal of Agricultural and Food
Chemistry, 40, 864*867. MacDougall, D.B. (2002). Colour measurment of food. En: MacDougall, D.B. (ed.), Colour
in food. Improving quality, CRC Press LLC, Boca Raton, USA. Madigan, M.T.; Martinko, J.M. & Parker, J. (1997). Brock Biology of Microorganisms. Eighth Edition. Prentice Hall Upper Saddle River. Manresa González, A. & Vicente, I. (2007). El color en la industria de los alimentos, Ed. Universitaria, La Habana, Cuba. Maharaj, R., Arul, J. & Nadeau, P. (1999). Fffect of photochemical treatment in the preservation of fresh tomato (Lycopersicon sculentum cv. Capello) by delaying senescence. Postharvest Biology and Technology, 15, 13*23. Marquenie, D., Geeraerd, A.H., Lammertyn, J., Soontjens, C., Van Impe, J.F. & Michielis , C.W (2003). Combination of pulsed white light and UV*C or mild heat treatment to inactivated conidia of Botrytis cinerea and Monilia fructigena. International Journal of Food
Microbiology, 85, 185*196. Martinez, V.Y. (2005). Alteraciones microestructurales y ultraestructurales de tejidos vegetales mínimamente procesados. Impacto en las características mecánicas. Tesis doctoral. Universidad de Buenos Aires, Argentina. Mittal, J.P.; and Mohsenin, N.N. (1987). Rheological characterization of apple cortex. Journal of Texture Studie,. 18, 65*93.

259
Mohsenin, N. N. (1978). Physical Properties of Plant and Animal Materials. Structure, Physical Characteristics and Mechanical Properties. Gordon and Breach Science Publishers. New York * London – Paris. Mohsenin,N.N. & Mittal, J.P.(1977). Use of rheological terms and correlation of compatible measurements in food texture research. Journal of Texture Studies, 8, 395*408. Monsalve*González, A., Barbosa*Cánovas, G.V., Cavalieri, R.P., McEvily, A., & Iyengar, R. (1993). Control of browning during storage of apple slices preserved by combined methods, 4*hexyl*resorcinol as antibrowning agent. Journal of Food Science, 58, 797*800. Monzocco, L., Calligaris, S., Mastrocola, D., Nicoli, M.C. & Lerici. C.R. (2001). Review of non*enzymatic browning and antioxidant capacity in processed foods. Trends in Food Science
& Technology, 11, 340*346. Nicolas, J.J., Richard*Forget, F.C., Goupy, P.M., Amiot, M.J. & Aubert, S.Y. (1994). Enzymatic browning reactions in apple and apple products. Critical Reviews in Food Science
and Nutrition 34, 109–157. Ortiz, C., Salvatori, D. M. & Alzamora, S.M. (2003) Mushroom tissue as matrix for calcium incorporation by vacuum impregnation. Latin American Applied Research, 33, 281*287. Ozer, N.P. & Demirci, A. (2006). Inactivation of Escherichia coli O157:H7 and Listeria monocytogenes inoculated on raw salmon fillets by pulsed UV*light treatment. International
Journal of Food Science and Technology, 41, 354*360. Pacín, A., Martínez, E., Pita Mertín de Portela, M.L, & Neira, M.S. (1999). Consumo de alimentos e ingesta de algunos nutrientes en la población de la Universidad Nacional de Luján, Argentina. Archivos Latinoamericanos de Nutrición, 49, 31*39. Palou, E., López*Malo, A., Barbosa*Cánovas, G.V., & Welti*Chanes J. (2000). High hydrostatic pressure and minimal processing. En: Alzamora, S.;Tapia, M and López*Malo, A. (eds.) Minimally Processed Fruits and Vegetables. Fundamental Aspects and Applications, Aspen Publication, Inc. Gasithersburg, Maryland. Pág. 205*222. Placzek, M., Gaube, S., Kerkmann, U., Gilbertz, K.P., Herzinger, T., Haen, E. & Przybilla, B. (2005). Ultraviolet B*induced DNA damage in human epidermis is modified by the antioxidants ascorbic acid and D*α* tocopherol. Journal of Investigative Dermatology, 124, 304*307. Ponting, J.D., Jackson, R. & Watters,G. (1972). Refrigerated apple slices: preservative effects of ascorbic acid, calcium and sulfites. Journal of Food Science, 37, 434*436. Pizzocaro, F., Torreggiani, D. & Gilardi, G. (1993). Inhibition of apple polyfhenoloxidase (PPO) by ascorbic acid, citric acid and sodium chloride. Journal of Food Processing and
Preservation, 17, 21*30. Pitt, R.E. (1992). Viscoelastic properties of fruits and vegetables. En: Rao, M.A.& Steffe,J.F.(eds.), Viscoelastic Properties of Foods, Elsevier Applied Science. London and New York.

260
Rahn, R.O. (1997). Potassium iodide as a chemical actinometer for 254 nm radiation: use of iodate as an electron scavenger. Photochemistry and Photobiology, 66, 450–455. Rao, M.A. (1986). Kinetics of thermal softening of foods * A review. Journal of Food
Processing and Preservation, 10, 311*329. Rao, V.N.M. (1992). Classification, description and measurement of viscoelastic properties of solid foods. En: Rao, M.A.; and Steffe, J.F.. (eds.), Viscoelastic Properties of Foods, Elsevier Applied Science. London and New York. Pág. 12*25 Rebouillat, S. & Peleg, M. (1988). Selected physical and mechanical properties of commercial apple cultivars. Journal of Textures Studies, 19, 217*230. Richard*Forget, F.C., Goupy, P.M., Nicolas, J.J. (1992). Cysteine as an inhibitor of enzymatic browning. Kinetic studies. Journal of Agricultural and Food Chemistry, 40, 2108*2113. Roberts, P. & Hope, A. (2003). Virus inactivation by hight intensity broad spectrum pulsed light. Journal of Virological Methods, 110, 61*65. Rocha, A. M. C. N., & Morais, A. M. M.B. (2003). Shelf life of minimally processed apple (cv. Jonagored) determined by colour changes. Food Control, 14, 13*20. Rojas, A.M.; Gerschenson, L.N.; Marangoni, A.G. (2001). Contributions of cellular components to the rheological behaviour of kiwifruit. Food Research International, 34, 189*195. Rowan, N., MacGregor, S.J., Anderson, J.G., Fouracre, R.A., McIIvaney, L. & Farish, O. (1999). Pulsed*light inactivation of food*related microorganisms. Applied and Environmental
Microbiology, 65, 1312*1315. Sajnin, C.; Gerschenson, L.N. & Rojas, A.M. (1999). Turgor pressure in vegetable tissues: comparison of the performance of incipient plasmolysis technique using mannitol and polyethylenglycol. Food Research International, 32, 531*537. Salvatori, D.M. (1997). Deshidratación osmótica de frutas: cambios composicionales estructurales a temperaturas moderadas. Tesis doctoral, Universidad Politécnica de Valencia, España. Salvatori, D.M., Andrés, A., Chiralt, A. & Fito, P. (1998). The response of some properties to vacuum impregnation. Journal of Food Process Engineering, 21, 59*73
Salvatori, D.M. & Alzamora, S.M. (2000). Structural changes and mass transfer during glucose infusion of apples as affected by blanching and process variables. Drying technology, 18, 21*48. Salvatori, D., Doctorovich, R. y Alzamora, S.M. (2010). Impact of calcium on viscoelastic properties of fortified apple tissue. Journal of Food Processing and Preservation. DOI: 10.1111/j.1745*4530.2009.00550.x.

261
Salvatori, D.M, González*Fesler, M., Weisstaub, A., Portela, M.L. & Alzamora, S.M. (2007). Uptake kinetics and absorption of calcium in apple matrices. Food Science and Technology
International, 13, 333*340. Sapers, G. M., & Douglas, F. W. (1987). Measurement of enzymatic browning at cut surfaces and in juice of raw apple and pear fruit. Journal of Food Science, 52, 1258*1262. Sastry, S.K, Datta, A.K. & Worobo, R.W. (2000). Ultraviolet light. Journal of Food Science,
Supplement, 65 (12), 90*92. Schenk, M., Guerrero, S. & Alzamora, S.M. (2008). Response of some microorganisms to ultraviolet treatment on fresh*cut pear. Food Bioprocess Technology, 1, 384*392. Schenk, M., Guerrero, S., Gómez, P., & Alzamora, S.M. (2006). En: Proceedings XIV
Seminario Latinoamericano y del Caribe de Ciencia y tecnología de los alimentos, La Habana, Cuba, 22*24 de octubre 2006. Schramm Gebhard (1994). A Practical Approach to Rheology and Rheometry (Haake
Rheometers). Gebrueder HAAKE Gmbh, Karlsruhe, Federal Republic of Germany. Shama, G. (1999). Ultraviolet light. En: Robinson RK, Batt, C., Patel, P. (eds.), Encyclopedia
of Food Microbiology, Academic Press, London, 2208*2214. Shama, G. (2007). Process challenges in applying low doses of ultraviolet light to fresh produce for eliciting beneficial hormetic responses. Postharvest Biology and Technology, 44, 1–8. Shama, G. (2006). Ultraviolet light. En: Hui, Y.H., Handbook of Food Science, Technology
and Engineering, Vol. 3 (pp 122*1 – 122*14). Boca Raton, Florida:.CRC / Taylor & Francis. Shama, G., & Alderson, P. (2005). UV hormesis in fruits: a concept ripe for commercialisation. Trends in Food Science and Technology, 16, 128*136. Sharma, R.R. & Demirci, A. (2003). Inactivation of Escherichia coli O157:H7 on inoculated alfalfa seeds with pulsed ultraviolet light and response surface modeling. Journal of Food
Science, 68, 1448*1453. Sherman, P (1970). Industrial Rheology. Academic Press. Shewfelt, R.L. (1999). What is quality?. Postharvest Biology and Technology, 15, 197*200. Singer, S.J & Nicolson, G.L. (1972). The fluid mosaic model of cell membranes. Science, 175, 720*731. Soliva*Fortuny, R. C. & Martín*Belloso, O. (2003). Microbiological and biochemical changes in minimally processed fresh*cut “Conference” pears. European Food Research and
Technology, 217, 4*9. Stanley, D.W. & Tung, M.A. (1975). Microstructure of food and its relation to texture. En: De Man, J.M.; Voisey, P.W.; Rasper, V.F.; Stanley, D.W.(eds.), Rheology and Texture in Food
Quality, Westport, Connecticut. The Avi Publishing Company, Inc. Pág. 28*78.

262
Steffe, J.F. (1996). Rheological methods in food process engineering (2ed.). Freeman Press, USA. Stevens, C., Khan, V. A., Lu, J.Y., Wilson, C.L., Pusy, P.L. & Kawake, M.K. (1998). The germicidal and hormetic effects of UV*C light on reducing brown rot disease and yeast microflora of peaches. Crop Protection, 17, 75*84. Summerfelt, S.T. (2003). Ozonation and UV irradiation. An introduction and examples of current applications. Aquacultural Engineering, 28, 21*36. Szczesniak, A.S. (1963). Classification of textural characteristic. Journal of Food Science, 28, 385*389. Szczesniak, A. S. (1963). Classification of textural characteristics. Journal of Food Science, 28, 385–389. Takeshita, K., Sibato, J., Sameshima, T., Fukunaga, S., Isobe, S., Arihara, K. & Itoh, M. (2003). DAmage of yeast cells induced by pulsed light irradiation. International Journal of
Food Microbiology, 85, 151*158. Tapia, M.S., Schulz, E., Gómez, V.,Lopz*Malo, A., Welti*Chanes, J. (2003). A new approach to vacuum impregnation and functional foods: melon impregnated with calcium and zinc. En: IFT (Institute of Food Technologist) Annual Meeting, Chicago, 60D*2. Thakur, B.R.; Singh, R.K. & Handa, A.K. (1997). Chemistry and uses of pectin * a review. Food Science and Nutrition. 37, 47*73. Toivonen, P.M.A. & Deell, J.R. (2002). Phisiology of fresh*cut fruits and vegetables. En G. Lamikanra (ed.) FreshLcut Fruits and Vegetables. Science, Technology and Market. Boca Raton, Florida: CRC Press LLC. Pág. 91*123 Torreggiani, D. (1993). Osmotic dehydration in fruit and vegetablesprocessing. Food
Research International. 26, 59*62. Torregiani, D. & Bertolo, G. (2002). The role of an osmotic step: combined processes to improve quality and control functional properties in fruit and vegetables. En Welti*Chanes, J., Barbosa Cánovas, G.V. & Aguilera, J.M.0. (Eds.),. Engineering and food for the 21
st century .
Florida, USA. Pág. 651*669 Uesugi, A.R., Woodling, S.E. & Moraru, C.I. (2007). Inactivation kinetics and factors of variability in the pulsed light treatment of Listeria innocua cells. Journal of Food Protection, 70, 2518*2525. Varela, P., Salvador, A. & Fiszman, S. (2007). Changes in apple tissue with storage time: Rheological, textural and microstructural analyses. Journal of Food Engineering, 78, 622*629. Vicente, A.R., Pineda, C., Lemoine, L., Civello, P.M., Martinez, G.A.& Chaves, A.R. (2005). UV*C treatments reduce decay, retain quality and alleviate chilling injury in pepper. Postharvest Biology and Technology, 35, 69*78.

263
Waldron, K.W.; Smith, A.C.; Parr, A.j.; Ng, A.; and Parker M.L. (1997). New approaches to understanding and controlling and controlling cell separation in relation to fruit and vegetable texture. Trends in Food Science & Technology, 8, 213*221. Wang, T., MAcGregor, S.J., Anderson, J.G., & Woolsey, G.A. (2005). Pulsed ultra*violet inactivation spectrum of Escherichia coli. Water Research, 39, 2921*2925. Ward, I.M. (1990). Mechanical Properties of Solid Polymers (2ed.) John Wiley & Sons. Chichester * New York * Brisbane * Toronto – Singapore. Weemaes, C.A. Ludikhuyze, L.R., Van den Broeck, I.,Hendrickx, M.E. & Tobback, P.P. (1998). Activity, Electrophoretic Characteristics and Heat Inactivation of Polyphenoloxidases from Apples, Avocados, Grapes, Pears and Plums. Lebensmittel�Wissenschaft undL
Technologie, 31, 44*49. Wekhof, A. (2000). Desinfection with flash lamps. PDA Journal of Pharmaceutical Science
and Technology, 54, 264*276. Whitaker, J.R. (1994). Principles of Enzymology for the Food Sciences (2ed.), Marcel Dekker, Nw YorK. Woodling S. E. & Moraru, C.I. (2005). Influence of surface topography on effectiveness of pulsed light treatment for the inactivation of Listeria innocua on stainless*steel surfaces. Journal of Food Science, 70,240*244. Zeni, S.M. & Portela, M.L. (1988). Estado nutricional respecto del calcio en Argentina. Archivos Latinoamericanos de Nutrición, 38, 209*218. Zhu, Y., Pan, Z. & Mchugh, T. (2007). Effect of dipping treatments on color stabilization and texture of apple cubes for infrared dry*blanching process. Journal of Food Processing and
Preservation, 31, 632*648. Páginas web consultadas FDA (2000). FDA approves the use of ultraviolet radiation for juice. Food Safety Initiative. Center for Food Safety and Applied Nutrition. (http://www.foodsafety.gov/~dms/fsiupd22.html) http://www.biolight.cl/biblioteca/pdfs/c7397901d1ffb52.pdf http://www.infoagro.com/frutas/frutas_tradicionales http://www.biologia.edu.ar/botanica/tema6/6_1fruto.htm http://www.elpais.com.uy/Suple/Agropecuario/03/04/30/agrope_39131.asp http://frutas.consumer.es/documentos/conozcamos/imprimir.php