mÓdulo de neurociencias aplicada a la fonoaudiologÍa
DESCRIPTION
Un documento que me mando a imprimir la Silvia D: me tire una resma casi completa T_TTRANSCRIPT

MÓDULO DE NEUROCIENCIAS APLICADA A LA FONOAUDIOLOGÍA
PRÁCTICAS DE LABORATORIO Y APUNTES PARA LAS CLASES
INSTITUTO DE BIOLOGÍA VEGETAL Y BIOTECNOLOGÍA
UNIVERSIDAD DE TALCA.
2010
PROFESOR HÉCTOR GANZ CONTRERAS

INDICE 1..Syllabus 21. Actividades Prácticas 21. El agua 22. Tensión superficial y vida 25. Propiedades del agua 26. Viscosidad del agua 29. Microscopía, uso microscopio Observación- histología 36. Observación de células procariontes y eucariontes 41. Identificación de carbohidratos y lípidos 44. Identificación de enlace peptídico-Proteínas 46. Bioenergética 48. Acción enzimática 51. Permeabilidad diferencial de la membrana 53. Fermentación alcohólica de la glucosa 53. Observación de mitocondrias 55. Actividad de lisosomas APOYO A CLASES TEÓRICAS 58. Elementos químicos 63. Química de la vida 62. Electronegatividad y polaridad de enlace 63. Puentes de Hidrógeno 64. Interacciones no covalente 66. Carbohidratos 67. Clasificación de Monosacáridos 70. Disacáridos 71. Polisacáridos 73. Amilosa, amilopectina, glucógeno, celulosa, quitina 75. Estudio con azúcares reductores, hidrólisis de sacarosa 77. Lípidos, estructuras y funciones 81. Ácidos grasos 83. Actividad en horas no presenciales 85. Ácidos grasos esenciales 86. Carencia de ácidos grasos 88. Síntesis de Ácidos grasos 91. Formación de prostaglandinas 95. Ácidos grasos y sistema nervioso 99. Esteroides 101. Colesterol 104. Proteínas 105. Estructura de aminoácidos 110. Síntesis de serotonina 111. Unión de los aminoácidos- enlace peptídico 112. Ácidos y Bases débiles 114. Estructura primaria 117. Estructura secundaria 120. Colágeno 121. Proteínas globulares- estructura terciaria 123. Efectos hidrófobo 124. Formación de enlaces disulfuro Estructura cuaternaria de las proteínas 125. Ácidos nucleicos, estructura y funciones 130. Gen 131.Cromosomas 132. Cromatina 133. Cariotipo humano 135. Genética y enfermedades Aberraciones cromosómicas, Aneuploidias, Monosomía , Trisomías 138. Célula Nerviosa- La neurona 142. Glucólisis 147. Cerebro y glucosa: Tema para discusión 148. Renovación de células neuronales

149. Citoesqueleto y Matriz citoplasmática 150. Citoesqueleto Microtúbulos 151. Centro organizador de microtúbulos 152. Proteínas MAPs, proteínas Tau 154. Proteínas motoras 155. Filamentos intermedios Membranas biológicas de células en general y Neuronas 161. Hacia el modelo de mosaico fluido Proteínas de membrana Bicapa lipídica Fluidez 167. Asimetría de la membrana, glicolípidos 169. Proteínas de membrana-funciones 170. Paso de Materiales a través de la membrana 171. Difusión 172. Diálisis 173. Osmosis 175. Transporte mediado de moléculas pequeñas 176. Difusión facilitada 177. Transporte activo . Sistemas de cotransporte 181. Transporte de grandes moléculas a través de las membranas 182. Endocitosis mediada por receptores de membrana 184. Intercambiador de bicarbonato cloro Membranas celulares y contacto 185. Uniones comunicantes tipo nexo 188. Clasificación de zonas de contacto entre las células 189. Uniones Adherente 191. Uniones oclusivas 191. Barrera hematoencefálica Células epiteliales intestinales Absorción en los epitelios- uniones adherentes 196. Estructura y función en la neurona, Movimiento de proteínas en la neurona, Vías de señalización, Péptido señal 203. Teoría neuronal. Principios de organización nerviosa 205. Redes neuronales Convergencia y divergencia neuronal Principio de polarización dinámica Potenciales, ley todo o nada Células neurogliales Producción de mielina, células, astrocitos Comunicación celular, microglias 218. Mecanismos fisiológicos de potenciales de membrana, potencial de reposo, potencial De acción, estímulo, canales de reposo, bomba de sodio y potasio 234. Mantención de las neuronas-factores de crecimiento- neurotrofinas 236. Comunicación celular Comunicación hormonal (endocrina) 239. Receptores, clasificación, neurotransmisores, acción moduladora, mecanismos de Transducción. 254 253. APOPTOSIS 260. Aprendizaje y memoria, Neuronas y comportamiento, Memoria y moléculas, Sistema límbico, Circunvoluciones, cerebro y aprendizaje . Mecanismos celulares y moleculares del aprendizaje y memoria Aprendizaje no asociativo: Sensibilización y habituación Potenciación a largo plazo. Clasificación de las neuronas, Aprendizaje perceptivo, Estudio con neuroimagenes funcionales, Condicionamiento clásico, Aprendizaje nervioso gustativo, interoceptivo, Condicionamiento Instrumental Aprendizaje motor, Amnesia, Aprendizaje espacial, Genética de la memoria y el aprendizaje Aprendizaje, memoria y circuitos neuronales, Regulación transcripcional de la expresión génica


Centro de Innovación y Calidad de la Docencia CICAD
Syllabus 2010
Descripción formal-
a) Nombre módulo Neurociencia Aplicada a la Fonoaudiología
b) Nº créditos (horas)
4 ECTS: Distribuidos: 2 horas cronológicas de asistencia a cátedra; 2 horas cronológicas de laboratorio- taller y 2 horas cronológicas de trabajo personal.
Total horas semestre: 108
c) Nivel
I
d) Requisitos
Ingreso
e) Responsables de la construcción del syllabus
Héctor Ganz Contreras
Magister en Educación de las Ciencias
Master en Psicobiología y Neurociencias Cognitivas
Profesor de Estado En Biología y Ciencias
Químico

f) Competencia(s) propósito del módulo
- Comprender los aspectos biológicos, tanto estructurales como funcionales, involucrados en la conducta humana y sus alteraciones.
g) Competencias y capacidades de egreso a la que contribuye el módulo:
Competencias:
- Evaluar y diagnosticar fonoaudiológicamente el lenguaje usando instrumentos de evaluación y clasificaciones diagnósticas pertinentes, considerando las características biopsicosociales del cliente y su entorno.
- Evaluar y diagnosticar fonoaudiológicamente la voz usando instrumentos de evaluación y clasificaciones diagnósticas pertinentes, y considerando las características biopsicosociales del cliente.
- Evaluar y diagnosticar fonoaudiológicamente el habla y la deglución usando instrumentos de evaluación y clasificaciones diagnósticas pertinentes, y considerando las características biopsicosociales del cliente.
- Evaluar y diagnosticar fonoaudiológicamente la audición usando instrumentos de evaluación y clasificaciones diagnósticas pertinentes, y considerando las características biopsicosociales del cliente.
Capacidades:
-Planifica el proceso de evaluación del lenguaje.
-Planifica el proceso de evaluación de la voz.
-Planifica el proceso de evaluación de la audición.
-Planifica el proceso de evaluación del habla.
-Planifica el proceso de evaluación de la deglución.
h) Capacidades secuenciadas del módulo
Asociar estructura y función de los componentes celulares en el proceso de autopoiesis.
Explicar las funciones de los diferentes componentes de la ultraestructura celular especialmente referido a la neurona.
Describir en términos bioquímicos o citológicos las características normales y anormales en el ejercicio de la función de proteínas en relación con metabolismo, organelos celulares y sistemas membranosos que inciden en la producción de patologías caracterizadas en el ámbito de la medicina y fonoaudiología en particular.
Comprender los mecanismos involucrados en la Herencia Biológica y de cómo estos influyen el d ontogénico del individuo.
Explicar los mecanismos de aprendizaje, memoria y comunicación, y asociarlas con la estructura neuronal y circuitos neuronales.


Planificación curricular (proceso de aprendizaje-enseñanza).
Propósito de aprendizaje del módulo.
Al finalizar el módulo el estudiante comprenderá los fundamentos biológicos de la conducta humana y los factores (genéticos, otros) involucrados en los distintos síndromes de naturaleza biológica (deficiencia mental, otros) que incluyen alteraciones de la comunicación. Para ello, el alumno aprenderá a describir los componentes de la célula, su función y sus interrelaciones, con especial referencia a la neurona; a aplicar procedimientos simples de análisis genético, tales como genética mendeliana, análisis de pedigree, herencia autosómica dominante y recesiva, epistasis; a asociar factores biológicos que subyacen a la conducta humana normal y a los distintos síndromes que incluyen alteraciones de la comunicación. Las actividades de aprendizaje se realizarán en el marco de sesiones presenciales, de talleres y laboratorio, y de trabajo personal del alumno.
Producto(s) esperado(s) del módulo
1) Interpreta en esquemas gráficos e imágenes la emergencia de problemas biológicos que ocurren entre la célula y su desarrollo e interrelación con el ambiente.
2) Resuelve situaciones problemáticas y de reconocimiento cualitativo y aplicación de nomenclatura en actividades de laboratorio, los diferentes componentes moleculares de la estructura celular, y asocia dichos componentes a funciones de transporte, obtención de energía y rol en el metabolismo del ser vivo.
3) Resuelve en forma teórica problemas documentados concernientes al análisis genético de tipo clásico y establece árboles genealógicos con análisis de pedigrí en genética humana relacionados con herencia autosómica dominante, recesiva y ligada al sexo. Estas últimas en relación con trastornos de la comunicación y del aprendizaje.
4) Distingue y describe en esquemas gráficos y planteamiento de problemas prácticos y teóricos la estructura y función de la neurona. Asocia dicha estructura y funcionalidad a los procesos de aprendizaje, memoria y comunicación celular.
Procedimiento evaluativo (dispositivo de evaluación). Componentes y criterios con que se hará la evaluación del logro de cada uno de los productos esperados
En pruebas de tipo objetivo se requiere un 60% de respuestas correctas para obtener la nota mínima de aprobación 4.0. El número de respuestas correctas se obtiene sumando puntaje ideal con el puntaje real. El puntaje promedio de esta suma se entiende como la nota máxima 7.0, y constituye la base para la obtención del puntaje correspondiente a 60%.
En pruebas de tipo ensayo se requiere exactitud y precisión en el desarrollo de las respuestas.
En ambos tipos de prueba se considera el uso de pauta de corrección.
En caso de trabajos se contempla además otorgar puntajes por respeto de normas establecidas para la confección de informes. Puntaje por creatividad, puntualidad y uso de bibliografía pertinente con los temas. Se empleará pauta de corrección.
Reiteradamente se realizará controles semanales para control de lectura analítica y descriptiva de los tópicos tratados en clases teóricas.

La evaluación será en cuatro unidades con un valor de 25% cada una. Cada unidad contempla la evaluación de trabajos de investigación, exposiciones, pruebillas de control de estudios en horas no presenciales, (Estos llámese: control de estudios en horas no presenciales-TCHP)
El desglose de cada unidad: 80% prueba y 20% TCHP). Cada minicontrol tiene un valor unitario y los trabajos son equivalentes a dos de ellos.
La asistencia a Laboratorio es obligatoria. La cátedra es de asistencia libre. Quedando este último criterio a resolución del consejo de escuela. Al no asistir al laboratorio, el alumno obtiene automáticamente la nota mínima correspondiente al minicontrol. Pudiendo recuperarla en caso de justificar la inasistencia.
Los demás trabajos deberán ser entregados en la fecha solicitada. El no cumplimiento de esta norma automáticamente contempla la calificación mínima. No obstante se podrá justificar mediante certificación médica o por motivos de fuerza mayor como son los siniestros, de tal manera deberá justificar ante el Director de Escuela.
Unidades de aprendizaje con sus productos esperados
Tiempo de trabajo del estudiante para cada unidad.
Desglose/descripción de capacidades, considerando en ella sus componentes: Cognitivo, Procedimental, Interpersonal
Estrategias y procedimientos metodológicos
Procedimientos evaluativos
Instalaciones, medios, fuentes de información
Unidad 1: Estructura y propiedades de moléculas y macromoléculas: importancia biológica.
Producto N°1:
Interpreta fenómenos en situaciones problemáticas planteadas en texto escrito y con base en torno a las propiedades que surgen como consecuencias de las estructuras moleculares de la célula, tales como: agua,
Capacidad 1:Asociar estructura y función de los componentes celulares en el proceso de autopoiesis.
Componente interpersonal: la actividad requiere que los alumnos se organicen para realizar trabajo grupal: Ello implica trabajo en ambiente ordenado y disciplinado, respetando las opiniones de los compañeros y monitoreando su propio quehacer y el de los compañeros en cuanto a las
Todo el módulo
- Asistir a Clases expositivas con la participación activa del alumno en el proceso de realizar preguntas y satisfacer inquietudes relacionadas con la disciplina.
- Participación de actividades en laboratorio virtual de macromoléculas.
- Asistencia y participación activa en laboratorio experimental de
-- Pruebas de tipo selección múltiples.
-Pauta de corrección
Con los siguientes componentes:
1.-Estructura y función de las moléculas de agua, hidratos de carbono, lípidos proteínas y ácidos nucleicos.
2.-Intercambio de sustancias entre células y medio
.- Laboratorio experimental de biología celular.
.- Laboratorio de computación.
.- CD Biología Celular.
- Educandus Utalca.
- Biblioteca
Bibliografía recomendada:
Bibliografía Fundamental:

hidratos de carbono, lípidos, proteínas y ácidos nucleicos, y de las relaciones entre estas al interior de la célula y con su entorno.
Tiempo estimado : 24 horas
Evaluación producto: 21 de Abril de 2006.
eficiencias de cada uno.
1.1. Reconocer las estructuras moleculares de la célula tales como: agua, hidratos de carbono, lípidos, proteínas y ácidos nucleicos y las relaciones entre ellos que se producen al interior de la célula y con su entorno.
1.2. Describir observaciones experimentales de comportamiento del agua ante la variación de factores ambientales.
1.3 Explicar difusión del agua y de solutos a través de una membrana semipermeable.y permeable.
Inferir las consecuencias que tiene para la célula la presencia de una barrera permeable e impermeable para una u otra sustancia.
Interpretar datos de variables dependientes e independientes recogidos del estudio de manipulación de factores que afectan la difusión de una sustancia.
Interpretar hechos derivados de observación de las proteínas, lípidos e hidratos de carbono en la interacción con
biología celular.
- Control semanal de estudios por desarrollo de minipruebas en laboratorio experimental de biología celular.
- Preparación de informes de laboratorio con participación grupal.
- Construcción de miniensayos con temas relevantes y relacionados con el desarrollo de capacidades.
ambiente
3.- Reconocimiento y control de variables en el estudio realizado acerca de funciones asociadas a las estructuras.
4.- Inferencias y deducciones acerca de la relación entre estructura y función de las moléculas y macromoléculas al interior de una célula.
.-Alberts Bray y otros Biología Molecular de la Célula. Ed. Omega 1994.
.- De Robertis. Biología Celular y Molecular Ed. El ateneo 2000
.- Junqueira Carneiro. Biología Celular y Molecular. Ed. McGraw-Hill Interamericana 1998.
Jiménez Luis Felipe y Merchant Horacio. Biología Celular y Molecular. Ed. Prentice May (educación Pearson)2003.
-Paniagua Ricardo, Biología Celular, Ed. McGraw-Hill. Interamericana. 2000.

el agua y con factores relacionados con acidez y temperatura.
Analizar datos experimentales de tipo cualitativo en reacciones de tipo enzima-sustrato.
Interpretar e inferir acerca de la reacción enzima sustrato a partir de datos experimentales.
Predecir el comportamiento de las enzimas a partir de las propiedades de las proteínas y relaciones de afinidad con el sustrato.
Unidad 2: Organización y estructura de la neurona.
Producto N°2:
Demostrar comprensión en el desarrollo de informes y pruebas de selección múltiple acerca de las relaciones entre elementos de estructura y función de la neurona en el modelo celular expresado como aspectos teóricos y prácticos acerca del fenómeno de la comunicación celular y su problemática en el desarrollo de partologías, de manera que se encuentre habilitado para
Capacidad 2
Explicar las funciones de los diferentes componentes de la ultraestructura celular especialmente referido a la neurona.
Componente interpersonal: la actividad requiere que los alumnos se organicen para realizar trabajo grupal: Ello implica trabajo en ambiente ordenado y disciplinado, respetando las opiniones de los compañeros y monitoreando su propio quehacer y el de los compañeros en cuanto a las eficiencias de cada uno.
- Asistir a Clases expositivas con la participación activa del alumno en el proceso de realizar preguntas y satisfacer inquietudes relacionadas con la disciplina.
- Asistencia y participación activa en laboratorio experimental de biología celular.
- Preparación de informes de laboratorio con participación grupal.
- Construcción de miniensayos con
Pauta de corrección con los siguientes componentes:
.- Estructura y función de organelos.
. Fundamentos de las anormalidades que conducen a las patologías y funcionamiento inadecuado del sistema.
.- Comunicación celular y señalización celular y relación con el sistema organismo.
.- Laboratorio experimental de biología celular.
.- Laboratorio de computación.
.- CD Biología Celular.
- Educandus Utalca.
- Biblioteca
Bibliografía recomendada:
Bibliografía Fundamental:
.-Alberts Bray y otros Biología Molecular de la Célula. Ed. Omega 1994.

adquirir conocimiento acerca de los fenómenos relacionados con patologías asociadas al ámbito de la fonoaudiología.
Tiempo estimado: 24 horas
Fecha evaluación producto:
19 de Mayo de 2006
Explicar las funciones de los diferentes componentes de la ultraestructura celular especialmente referido a la neurona.
2.1 Describe la estructura microscópica de organización de la neurona en términos citológicos estructurales y bioquímicos.
Explica los mecanismos de obtención de energía y principios de metabolismo energético celular.
Comprende los mecanismos principales que emergen de anormalidades en las estructura mitocondrial y que conduce al desarrollo de enfermedades relacionadas con la fonoaudiología.
Relaciona la estructura y función de la membrana en cuanto a los procesos de comunicación celular en los circuitos neuronales.
Describe el fenómeno de la sinapsis como una acción de comunicación intercelular en el seno de un organismo y cuyo objeto es la
temas relevantes y relacionados con el desarrollo de capacidades.
.- De Robertis. Biología Celular y Molecular Ed. El ateneo 2000
.- Junqueira Carneiro. Biología Celular y Molecular. Ed. McGraw-Hill Interamericana 1998.
Jiménez Luis Felipe y Merchant Horacio. Biología Celular y Molecular. Ed. Prentice May (educación Pearson)2003.
-Paniagua Ricardo, Biología Celular, Ed. McGraw-Hill. Interamericana. 2000.

supervivencia, adaptación y establecimiento de la homeostasis del organismo.
Describe los fenómenos de tipo sináptico en relación con la actividad de neurotransmisores, polarización y despolarización de la membrana celular de la neurona.
Explica el origen del potencial de reposo y fundamentar en términos de actividad molecular definida en el proceso de comunicación celular.
Capacidad 2.A
Describir en términos bioquímicos o citológicos las características normales y anormales en el ejercicio de la función de proteínas en relación con metabolismo organelos celulares y sistemas membranosos que inciden en la producción de patologías

Producto 2.A. unidad N° 3. Síntesis, tráfico y exportación de proteínas.
caracterizadas en el ámbito de la medicina y fonoaudiología en particular
2.A.1 Describir los procesos de Síntesis, tráfico y exportación de proteínas.
2A.2 Explicar Mecanismos mediante los cuales se producen anomalías en la síntesis, tráfico y exportación de proteínas.
2.A.3 Explicar los mecanismos de transporte a nivel de la célula en relación con la estructura intracelular y matriz extracelular y los procesos que conducen al desarrollo de enfermedades y patologías diversas en el ser humano.
Capacidad: 3
Comprender los mecanismos involucrados en la Herencia Biológica y de cómo estos
- Asistir a Clases expositivas con la participación activa del alumno en el proceso de realizar preguntas y satisfacer inquietudes relacionadas con la disciplina.
- Participación de actividades en laboratorio virtual de macromoléculas.
- Asistencia y participación activa en laboratorio experimental de biología celular.
- Control semanal de estudios por desarrollo de minipruebas en laboratorio
Producción de material de comunicación en Medio informático-Power Point y presentación oral con público.
Componentes:
Síntesis, tráfico y exportación de proteínas.
.- Laboratorio experimental de biología celular.
.- Laboratorio de computación.
.- CD Biología Celular.
- Educandus Utalca.
- Biblioteca
Bibliografía recomendada:
Bibliografía Fundamental:
.-Alberts Bray y otros Biología Molecular de la Célula. Ed. Omega 1994.
.- De Robertis. Biología Celular y Molecular Ed. El ateneo 2000
.- Junqueira Carneiro. Biología Celular y Molecular. Ed. McGraw-Hill Interamericana 1998.
Jiménez Luis Felipe y Merchant Horacio. Biología Celular y Molecular. Ed. Prentice May (educación
Muestra comprensión en el desarrollo de informes, investigaciones bibliográficas y resolución y planteamiento de problemas en producción medios de comunicación, power point y presentación oral publica acerca de las relaciones de función y estructura de las proteínas, membranas y organelos de manera que le permita alcanzar comprensión de los fundamentos biológicos de las patologías relacionadas con la fonoaudiología.
Tiempo estimado 12 horas.
Fecha evaluación
determinan el desarrollo ontogénico del individuo.
3.1. Explicar los mecanismos que permiten la perpetuación y sobrevivencia de las especies como son la mitosis y la meiosis.
3.2. Calcular porcentajes de fenotipos y genotipos de acuerdo a herencia de tipo mendeliano, herencia del sexo y ligada al sexo, alelos múltiples y fenómenos de epistasis.
3.3.Explicar anormalidades genéticas que pueden conducir a trastornos de la comunicación desde la perspectiva que se basa en el número de cromosomas o variación en el número de ellos, mutaciones, translocaciones y delecciones relacionados con la producción de dopamina y alteraciones de la conducta locomotora y la motivación.
3.4. Explicar las relaciones entre genes y conducta, mutaciones de genes de codificación de la leptina y relacion con la conducta alimentaria,
experimental de biología celular.
- Preparación de informes de laboratorio con participación grupal.
- Construcción de miniensayos con temas relevantes y relacionados con el desarrollo de capacidades.
.- Utilización de recursos de laboratorio de informática Internet para recopilar antecedentes, artículos, revistas, imágenes, etc
Trasporte en relación a sistema de microtúbulos.
Problemas asociados al transporte de sustancias u organelos y procesos asociados a la estructura de membrana y matriz extracelular.
.- De la presentación misma: Pauta de evaluación para presentación en público.
Pearson)2003.
-Paniagua Ricardo, Biología Celular, Ed. McGraw-Hill. Interamericana. 2000.
Laboratorio de microscopía para histología y biología celular.
Educandus sitio
Bilblioteca
CD especial para el desarrollo de cada una
producto:
2 de Junio de 2006.
Unidad 3: Principios generales de genética y Genética de la conducta.
receptores serotoninérgicos. Intensificación de la conducta impulsiva.
- Asistir a Clases expositivas con la participación activa del alumno en el proceso de realizar preguntas y satisfacer inquietudes relacionadas con la disciplina.
- Participación de actividades en laboratorio virtual de macromoléculas.
- Asistencia y participación activa en laboratorio experimental de biología celular.
- Control semanal de estudios por desarrollo de minipruebas en laboratorio experimental de biología celular.
- Preparación de informes de laboratorio con participación grupal.
- Construcción de miniensayos con temas relevantes y relacionados con el desarrollo de capacidades.
.Pauta de corrección
Componentes:
Ciclo celular y mecanismos de control del ciclo.
Mitosis y Meiosis su importancia biológica.
Cálculo de relaciones fenotípicas y genotípica.
Construcción y análisis de Pedigree.
Variaciones del número cromosómico, delecciones, mutaciones y translocaciones y sus consecuencias en el desarrollo de fenotipos.
de las unidades.
Libros sugerida para el estudio de los temas:
1.- Pinel John P.J. Biopsicología 4ª edición 2001. Prentice Hall
2.- Kandel Eric R., Schwartz James H., Jessel Thomas M. Principios de Neurociencia. 4ª edición. McGraw-Hill-Interamericana. 2000
3.- Griffith, JF Miller, DT Suzuki, RC Lewontin ... Genetica 7ª edición. AJF MacGraw-Hill. 2002

Producto N°3.
Demostrar habilidades en los procesos de construcción, cálculo de proporciones fenotípicas y genotípicas, así como las de construcción gráfica de pedigree, en pruebas de tipo desarrollo, de tal manera que pueda realizar estudios preliminares de genética en la población humana.
Demostrar comprensión acerca de las consecuencias para un ser vivo, especialmente el ser humano, las variaciones del número y delecciones de cromosomas, y mutaciones del ADN. Ello le permitirá da solidez a sus conocimientos básicos para la evaluación y criterios diagnósticos en la fonoaudiología.

Tiempo estimado:

24 horas
Fecha evaluación de Producto: 30 de Junio de 2006
Unidad 4: Neurociencia de la conducta y del aprendizaje.
Producto 4
Distingue y describe en esquemas gráficos y planteamiento de problemas prácticos y teóricos la estructura y función de la neurona. Asocia dicha estructura y funcionalidad a los procesos de aprendizaje, memoria y comunicación celular.
Describe en mapas topográficos del sistema nervioso central las estructuras relacionadas con el aprendizaje, la conducta y la memoria.
Tiempo estimado
Capacidad N° 4
Explicar los mecanismos de aprendizaje, memoria y comunicación, y asociarlas con la estructura neuronal y circuitos neuronales.
4.1. Explicar los mecanismos de condicionamiento clásico, sensibilización y habituación.
4.2. Explicar los mecanismos del condicionamiento instrumental.
4.3. Explicar los fundamentos que permiten el aprendizaje asociativo, inferencia causal, potenciación a largo plazo y aprendizaje hipocámpico.
4.4. Describir las
- Asistir a Clases expositivas con la participación activa del alumno en el proceso de realizar preguntas y satisfacer inquietudes relacionadas con la disciplina.
- Participación de actividades en laboratorio y aprendizaje.
- Asistencia y participación activa en laboratorio experimental de biología celular.
- Control semanal de estudios por desarrollo de minipruebas en laboratorio experimental de biología celular.
- Preparación de informes de laboratorio con participación grupal.
Pauta de corrección.
Componentes:
Circuitos neuronales en relación con conductas innatas.
Componentes del condicionamiento clásico de tipo pavloviano.
Componentes del condicionamiento instrumental.
Mecanismos de aprendizaje y modelos de formación de memoria a corto y largo plazo.
Mecanismos de inferencia causal, aprendizaje hipocámpico, potenciación y recuperación de memorias.
Laboratorio de microscopía para histología y biología celular.
Educandus sitio
Biblioteca
CD especial para el desarrollo de cada una de las unidades.
Libros sugeridos para el estudio de los temas:
1.- Anderson John R. Aprendizaje y Memoria- un enfoque integral.
McGRAW-HILL. 2001
2.- Pinel John P.J. Biopsicología 4ª edición 2001. Prentice Hall

24 horas
Fecha Evaluación producto: 28 de Julio.
características de adquisición y formación y recuperación de memorias.
Describir las estructuras del sistema nervioso central relacionadas con el aprendizaje, la conducta y la memoria.
Describir metodologías de estudio para observación de cerebro humano.
3.- Kandel Eric R., Schwartz James H., Jessel Thomas M. Principios de Neurociencia. 4ª edición. McGraw-Hill-Interamericana. 2000
4.- Skinner B.F. Aprendizaje y Comportamiento Una antología.
Ed. Martinez Roca. 1985 Barcelona
5.- Carlson Neil Fisiología de la Conducta. Editorial Pearson, 2005
6.- Apuntes entregados en clases. Guía de laboratorio.

21
ACTIVIDADES PRÁCTICAS
PRÁCTICO N° 1: “EL AGUA”
Durante el desarrollo de esta guía de laboratorio se espera que usted comience a comprender el método científico y sus formas de pensamiento racional y lógico. EL método científico requiere una forma de pensar ordenada, por lo tanto, al operar con instrumentos y realizar mediciones usted deberá registrar datos que sean claros, clasificados, agrupados, etc., en tablas. Se requiere que usted comience a manipular variables e identifique aquellas que son independientes y dependientes del fenómeno en estudio. En esta guía trataremos sobre las propiedades del agua y de cómo estas pueden influenciar o determinar la vida en la célula.
Tensión superficial:
En un fluido cada molécula interacciona con las que le rodean. El radio de acción de las fuerzas moleculares es relativamente pequeño, abarca a las moléculas vecinas más cercanas. En la figura se puede observar como las moléculas en el seno o interior de un líquido ejerce su acción en todas direcciones. No ocurre así con las moléculas que se encuentran en la superficie del líquido, aquellas que están en contacto con el aire. De manera que tenemos dos tipos de fuerzas: a) De interacción entre aire y líquido y b) las de interacción líquido-líquido. Así, la energía potencial de la molécula en la interfase(aire-líquido) es mayor que la de las moléculas en el interior del líquido.
Existe la tendencia de todo sistema a evolucionar hacia un estado de equilibrio estable, con una energía potencial mínima, por esto el fluido adquiere un estado en que el área de la interfase de separación con otros medios sea lo más pequeña posible.
Así, las moléculas en el interior del líquido sobre las cuales se están ejerciendo fuerzas intermoleculares de cohesión, se establece una simetría en que las fuerzas se ejecutan en todos sentidos y direcciones, y en consecuencia la resultante es nula.
El fenómeno se puede observar en la figura de enfrente. Las moléculas de superficie se encuentran rodeadas en parte por moléculas de aire. En el caso de este gas la cantidad de moléculas es muy pequeña si se le compara con el líquido. Por ello, debemos despreciar la interacción entre moléculas de ambos medios.

22
La fuerza de cohesión molecular entonces impide que las moléculas de la superficie del líquido se escapen. De una manera más elegante, se produce una fuerza tangencial neta en la superficie del líquido, dirigida hacia su interior y por lo tanto, oponiéndose a las fuerzas impulsoras de escape de las moléculas de superficie.
La fuerza superficial generada posee una energía equivalente al trabajo necesario para que una molécula salga despedida de la superficie. Esta energía superficial es proporcional al área de la superficie libre del líquido.
γ= Tensión superficial
Tensión superficial y vida:
El estudio de la tensión superficial y la vida resulta interesante desde el momento que observamos a los insectos posarse sobre la superficie del agua. Desde el punto de vista evolutivo de los seres vivos, observando el nivel celular se ha razonado lo siguiente: un ser unicelular es una estructura donde se realizan todas las funciones vitales básicas relacionadas con la nutrición y reproducción. Estos seres unicelulares habitan en ambientes acuáticos, allí se originaron los seres vivos. Al operar la selección natural no es de extrañar que las presiones selectivas, entre otras, tuvieran que ver con la adquisición de alimentos y las fuerzas de tensión superficial. El organismo unicelular seleccionado es aquel que puede aislarse del medio por medio de una membrana o pared que le permita adquirir alimentos y que a su vez soporte las fuerzas de tensión superficial del medio en que se desarrolla. En su lucha por la existencia el organismo va adquiriendo funciones nuevas como por ejemplo, el poder formar conglomerados celulares y en tal circunstancia surgen modos de comunicación celular para poder establecer relaciones. Conforme avanza desarrolla organelos hasta un cierto límite, pues, la presión del medio impide que la célula pueda adquirir o desarrollar tamaños y volumen como los que conocemos hoy. Comienza la evolución a pluricelulares, las células se especializan en funciones varias, avanzan hasta formar tejidos, órganos y sistemas de órganos.
La historia evolutiva de los seres vivos es interesante, pero debemos preocuparnos de entender la tensión superficial y su relación con los seres vivos. Hace algunos años atrás, Danielli y Davson observaron que al añadir triglicéridos sobre agua, estos se disponían con las cabezas polares hacia fuera. Sin embargo, los triglicéridos formaban gotitas y la tensión superficial era mucho más alta que la de las células. Más, al añadir proteínas al medio, la tensión superficial bajaba en forma notable. En razón de estos resultados los investigadores propusieron un modelo de membrana celular. Dicho modelo perduró en la mente de los científicos hasta que surgió el modelo de Mosaico Fluido. De todas formas es interesante que las propiedades de los líquidos y en especial del agua se relacionan con la tendencia de estos a formar esferas o gotas con el fin de reducir al máximo la superficie y de ahí la definición de tensión superficial que dice de la fuerza que trata de mantener el área superficial lo más pequeña posible. Esta fuerza tiende a generar burbujas, gotas esféricas; y
ε = γ ● S

23
es capaz de soportar objetos livianos. La lucha de los seres vivos contra la tensión superficial consiste en fabricar una membrana que disminuyera la tensión superficial, por cuanto se da el aporte de proteínas hacia la membrana. La forma de posicionamiento de los lípidos en la membrana consiste en el establecimiento de monocapas, permitiendo el intercambio de sustancias entre la célula y su medio ambiente. La adición de proteínas permite la disminución de la tensión superficial y por ende de la disminución de la resistencia a la penetración.
ACTIVIDAD 1:
Hipótesis: En un líquido, las moléculas experimentan atracciones entre sí casi simétricas en su seno y asimétricas en su superficie, en consecuencia, las moléculas de superficie presentan mayor atracción hacia el interior del líquido generando superficies esféricas resistentes a la inmersión de objetos livianos.
1.- Experimente con diversos objetos tratando de que floten en el agua. Utilice vasos de pp, monedas de $1 y 10$, hilo, bote de papel, etc.
2.- Agregue solución de detergente si es que alguno de los objetos flota. Anote observaciones y discuta con sus compañeros si la hipótesis se mantiene.
3.- Ensaye con bombillas de plástico (para bebida) y detergentes. Determine condiciones bajo las cuales algunas de las características de la tensión superficial se demuestran. En este caso lo que usted se encuentra manipulando se conoce con el nombre de variables. ¿Cuáles son las variables? Registre observaciones y realice esquemas y dibujos que ayuden a explicar.
4.- Sobre una moneda de $100 coloque gotas de agua con un gotario, cuente gotas hasta el límite en que no cae el agua de la moneda. Registre observaciones. Una o dos gotas antes de llegar al límite, ensaye con una gota de detergente. Ensaye con glicerina y soluciones salinas.
5.- Realice experiencia con un barquito de papel confeccionado con dicho material. Coloque el barco en el agua y luego agregue unas gotas de detergente por debajo del barco y en los alrededores. Observe y explique.
6.- Utilice el siguiente lenguaje técnico que deberá comprender e investigar acerca de su significado antes de realizar los experimentos: Tensión, cohesión, adhesión, absorción, adsorción, tensión superficial, tensión interfacial, dispersión, atracción molecular, menisco, menisco cóncavo, convexo y tenso activo.
7.- Investigue antes de la experiencia de laboratorio acerca del agente surfactante en los alvéolos y su importancia para prevenir el colapso respiratorio. Explique qué importancia médica posee este agente ¿Cómo se produce este agente? ¿Cuáles son las causas de que no se produzca este agente? ¿Cuál es su composición química? ¿Qué importancia alcanza la dipalmitoilfosfatidil colina (DPPC)? ¿Se encuentra fosfatidil colina en la membrana celular, y si es así, cuál es su conclusión?

24
8.- Investigue acerca de las relaciones entre tensión superficial y características de los adhesivos utilizados en la práctica odontológica en caso de ser estudiante de odontología.
9.- Ensaye las observaciones de la Tensión Superficial en agua dura y agua blanda.
10.- Distinga variables dependiente independiente en el siguiente párrafo informativo. Construya una tabla con la respuesta.
La tensión superficial depende de la naturaleza del líquido, del medio que le rodea y de la temperatura. En general, la tensión superficial disminuye con la temperatura, ya que las fuerzas de cohesión disminuyen al aumentar la agitación térmica. La influencia del medio exterior se comprende ya que las moléculas del medio ejercen acciones atractivas sobre las moléculas situadas en la superficie del líquido, contrarrestando las acciones de las moléculas del líquido.
Tensión superficial de los líquidos a 20ºC
Líquido (10-3 N/m)
Aceite de oliva 33.06
Agua 72.8
Alcohol etílico 22.8
Benceno 29.0
Glicerina 59.4
Petróleo 26.0
11.- Repita los ensayos realizados antes, pero empleando aceite, alcohol etílico y glicerina. Registre sus observaciones. Explique el fenómeno observado utilizando terminología científica de tal modo que se establezca la relación entre causa y efecto.
12.- En la medicina podemos encontrar tratamiento para evitar el resecamiento de los ojos. Para ello el médico receta lágrimas artificiales. ¿Qué características químicas de contenido deben tener esas lágrimas artificiales? ¿Por qué? ¿Cuál sería la explicación teórica que satisfaga los avances dados en la introducción? Los productos tales como “lagrimas artificiales son muy utilizados en medicina para aliviar el resecamiento de la conjuntiva en ojos, hecho que ocurre debido a la paralización de los músculos relacionados con el parpadeo y por tanto muy frecuente en parálisis facial.

25
13.- Realice un esquema de los alvéolos pulmonares de manera que se explicite el fenómeno de actividad de intercambio gaseoso, y de actividad de neumocitos tipo II.
14.- ¿Qué características químicas debe presentar los jabones o detergentes para transformarse en agentes surfactantes para disminuir la tensión superficial?
ACTIVIDA 2 : “EL AGUA”
En el práctico anterior acerca de la Tensión superficial ya hemos estudiado algunas características del agua y continuaremos en este práctico con otras propiedades. Sin embargo, aunque las tratemos por separado. No olvidemos que todas las propiedades se encuentran ligadas unas a otras y son causa solamente de la estructura química del agua. Las fuerzas de cohesión atómica- moleculares se pueden mostrar en el fenómeno de capilaridad ilustrado por la figura siguiente: Cuando usted realice un experimento similar al de la figura. Puede hacerlo introduciendo un tubo delgado en un recipiente con agua. Usted deberá corresponder con la observación de que el agua asciende por el tubo. La explicación es que este fenómeno ocurre gracias a las fuerzas de cohesión de las moléculas de agua, o sea que, las moléculas de agua deben presentar atracción entre ellas como para deslizarse juntas y ascender por el tubo. Tenemos otra explicación complementaria y es que deben existir fuerzas adhesivas entre las moléculas del tubo y el agua. Resuelva:
1) En la superficie del agua al interior del tubo se observa una curvatura ¿a qué se debe esta curvatura?
2) ¿Qué ocurrirá con la tensión superficial a medida que el agua asciende por el capilar? 3) ¿Qué caracterizará al fenómeno de capilaridad en la detención del movimiento
ascendente? 4) ¿Qué mecanismo ocurre con los árboles que deben subir el agua hasta su copa? 5) ¿Qué influencia tienen el diámetro del tubo? 6) Demuestre que el ascenso es inversamente proporcional al radio del tubo.
H2O

26
Viscosidad del agua
Un parámetro físico del agua que determina la movilidad del agua. Cuando la temperatura aumenta, la viscosidad disminuye; esto significa que el agua será más móvil a mayores temperaturas.
Es una medida de la resistencia a fluir que presentan los líquidos. Esta definición significa
que a mayor viscosidad un líquido escurre más lentamente. La viscosidad de un líquido disminuye con el aumento de temperatura, porque la viscosidad esta relacionada directamente con las fuerzas de atracción entre las moléculas del líquido. El agua tiene mayor viscosidad que otros líquidos porque sus fuerzas intermoleculares se deben a los enlaces de hidrógeno.
Hipótesis de trabajo: Si aumenta la viscosidad entonces la velocidad de movimiento debe
disminuir. Diseñe un experimento para realizar en laboratorio. El experimento debe ser factible y
pertinente con la hipótesis anterior.
Hipótesis trabajo: Si aumenta la temperatura del fluido entonces disminuye la viscosidad y en consecuencia el movimiento de partículas en el interior del fluido será mayor.
Experimento
Para realizar experimento:
• Probetas de igual medida
• Un cronómetro de precisión que mida en 0,1 ó 0,01 de segundos
• Bolitas de piedra o de cristal, todas del mismo tamaño y forma, selecciona usando balanza
• Un termómetro
• Aceite o jarabe
• Miel líquida
• Temperatura • Tiempo ensayo N°1
• (Promedio de tres)
• Agua
• Tiempo ensayo N°2
• (Promedio de tres)
• Aceite
• Tiempo EnsayoN°3
• (Promedio de tres)
• Clara de huevo
• Tiempo EnsayoN°3
• (Promedio de tres) Miel líquida
• 10° C • • • •
• 25°C • • • •

27
• 35°C • • • •
• 55° C • • • •
Resuelva:
1.- Distinga variables dependientes y variables independientes de los experimentos. Grafique los datos
2.- Realice observaciones a la siguiente definición de viscosidad: la resistencia de un líquido al movimiento o flujo. ¿Qué se puede decir con respecto a las moléculas del fluido?
RELACIONE LOS EXPERIMENTOS Y LUEGO REGISTRE EN EL INFORME DE LABORATORIO:
El siguiente experimento muestra que un líquido puede subir verticalmente por los poros de ciertos materiales (por ejemplo papel absorbente) debido al fenómeno de capilaridad. Usted observará que la velocidad de ascenso depende de la viscosidad.
Esquema de experimento:
NOTAS:OBSERVACIONES
Regla
Papel
Fluido
Experimente con agua destilada, agua potable, almíbar, aceite para motores bencineros o petroleros. Mida la velocidad de desplazamiento y confeccione tablas.

28
ACTIVIDAD 3 : MICROSCOPÍA MICROSCOPIO, TECNICAS PARA HACER PREPARACIONES
MICROSCOPICAS E INFORMES DE LABORATORIO I. INTRODUCCION.
La biología es una ciencia netamente experimental, la que ha avanzado gracias a la implementación tecnológica, el éxito de esta disciplina depende de varios factores tales como la observación, el empleo de instrumentos y técnicas especiales; para lo cual es necesario apoyar las explicaciones teóricas con prácticas de laboratorio; en donde cada persona puede realizar sus propias observaciones e investigaciones y de esta forma llegar a lograr las competencias relacionadas con el método científico; adquirir una forma de pensar con la lógica y razonamiento del método en sí, ello le permitirá en estos primeros pasos lograr un desarrollo básico para la acción de investigar en cualquier disciplina relacionada con las ciencias. En segundo lugar y no menos importante es la adquisición de conocimiento básico para construir aspectos básicos y fundamentales en el ejercicio de aprendizaje de las disciplinas que sustentan la práctica médica y por ende el logro de perfiles que orientan al profesional.
De la Observación en Biología Los seres vivos presentan variedad en tamaño, forma, estructura, función, etc.
Con relación al tamaño éste varía, el ojo humano logra identificar objetos hasta de 200 micras, por lo que se hace necesario recurrir a instrumentos ópticos que faciliten la observación de objetos de menor tamaño, este instrumento es el microscopio, que consiste en conjuntos de lentes que agrandan la imagen.
Al principio, el microscopio surgió como una lente que agranda la imagen (lupa),
luego se desarrolló el microscopio óptico compuesto, que consiste en varios lentes (parte óptica), sostenida por estructuras metálicas (parte mecánica).
Para observar los objetos a través del microscopio óptico compuesto, es necesario
colocarlos sobre una lamina de vidrio (Porta objetos) y generalmente, si es necesario, se incorpora una lamina de vidrio más delgada que cubre la preparación (Cubre objetos); en esta forma se obtiene la preparación microscópica; la que puede durar poco tiempo (preparación temporal), o puede ser guardada (preparación permanente).
En esta práctico se darán los lineamientos generales para el uso del microscopio, así
como para hacer preparaciones microscópicas y efectuar el informe de laboratorio.

29
II. El alumno deberá desarrollar las siguientes capacidades o habilidades
1. Identificar las partes ópticas y mecánicas del microscopio.
2. Realiza preparaciones microscópicas temporales.
3. Elabora adecuadamente sus informes de laboratorio.
4. Conoce el uso adecuado de cada una de las partes del microscopio óptico compuesto.
III. MATERIALES:
• Microscopio óptico compuesto
• Frascos gotero con agua destilada
• Porta Objetos y Cubre Objetos
• Papel impreso (periódico) con letra pequeña
• Tijeras y hoja de afeitar nueva
• Papel y lápiz
• Corcho
• Papel limpialentes
• Paño amarillo de limpieza
• Solvente para limpieza
• Xilol
• Pipeta Pasteur IV. PROCEDIMIENTO. 1. MICROSCOPIO PARTES Y FUNCION: El alumno identificará las partes ópticas y mecánicas del microscopio y Microscopio Optico Compuesto PARTES OPTICAS: -Oculares -Objetivos -Condensador -Filtros de luz -Prismas PARTES MECANICAS: -Pie, base o soporte -Columna o brazo -Platina -Carro y tornillos del carro -Tornillo macrométrico -Tornillo micrométrico -Mecanismo de revolver -Tubo del ocular -Diafragma

30
2. ENFOQUE DEL MICROSCOPIO
1. Baje completamente la platina del microscopio.
2. Haciendo uso del mecanismo del revólver, coloque frente a la preparación el objetivo de menor aumento (seco débil 10 X).
3. Coloque la preparación microscópica sobre la platina sujetándola con las pinzas del carro y centre la preparación haciendo uso de los tornillos del carro.

31
4. Regule la luz mediante el uso del mecanismo del diafragma.
5. Haciendo uso del tornillo macrométrico suba al máximo la preparación hasta que encuentre tope sin observar por los oculares.
6. Observando a través del ocular, accione el tornillo macrométrico hasta que visualice la imagen en el campo microscópico.
7. Una vez enfocada la imagen, afine el enfoque microscópico, haciendo uso del tornillo micrométrico.
8. Si desea mayor detalle, cambie de aumento rotando el mecanismo del revolver a un objetivo que le proporcione el aumento deseado, corrigiendo el enfoque de imagen moviendo el tornillo micrométrico, y graduando la intensidad de luz con el diafragma.
9. Después de haber realizado la observación, se limpian las lentes con papel limpia lentes o siguiendo instrucciones del profesor, se coloca el objetivo de menor aumento en posición de enfoque, se baja la platina, se desconecta y se tapa. El microscopio queda listo para la próxima observación.
2.1 PARA OBSERVAR EN UN MICROSCOPIO Correctamente, se debe poner la mano derecha en los tornillos que mueven la platina y la mano izquierda en los tornillos macro y/o micromètricos se deben graduar los oculares a nuestros ojos si el microscopio es binocular, si es monocular la observación se debe hacer con los dos ojos abiertos.
2.2 DESCRIPCION DEL MICROSCOPIO.
PARTE MECANICA. Está compuesta por el pie, la platina y el tubo. EL PIE. Es una pieza maciza y pesada, para asegurar la estabilidad del aparato y servir de soporte a sus demás partes. LA PLATINA: Es una pieza metálica donde se colocan las preparaciones; tiene en el centro una abertura circular por la que pasarán los rayos luminosos procedentes del sistema de iluminación. En los microscopios corrientes puede ser fija o estar endosada a un carro con dos tornillos y cremallera que permitan dos movimientos de translación, para centrarla, y también, si los tornillos están graduados, para medir sus desplazamientos. La preparación se sujeta, en las platinas fijas, con dos sujetadores móviles, y en las platinas de carro, por un reborde, en forma de escuadra y pestillo, que le impide cualquier movimiento imprevisto.
El pie se prolonga por encima de la platina, en arco más o menos curvo. La parte superior
de este arco es la que sostiene el tubo y su mecanismo de traslación vertical. Esta tiene suma importancia, pues permite enfocar el objetivo mediante dos movimientos uno rápido, gracias a una cremallera, y otro lento, con un tornillo micrométrico.
EL TUBO: En él está instalado el sistema óptico. Está constituido por dos tubos. Uno de ellos, externo, en el que se encuentran la cremallera y el ocular,
y otro, interno, adosado al anterior, donde está el objetivo. En la parte superior hay una división milimétrica que permite modificar la distancia entre objetivo y ocular. Son corrientes en los aparatos modernos los binoculares, que facilitan la visión con los dos ojos, y

32
los revólveres portaobjetivos, con los cuales se pueden cambiar los objetivos instantáneamente, sin desenfocar la preparación. PARTE OPTICA. OBJETIVOS. Están formados por la unión de varios lentes para corregir las aberraciones. Deben tratarse con mucho cuidado, pues cualquier golpe puede variar la posición de las lentes y averiarlas. Se atornillan a la parte inferior del tubo o al revólver portaobjetivos. La lente inferior del objetivo se denomina lente frontal. De ella depende principalmente la mayor o menor ampliación. Es siempre plano-convexa, de foco muy corto y de diámetro tanto menor cuanto mayor sea el aumento. Detrás de esta lente hay otras, que son las que corrigen las aberraciones cromáticas y esféricas. OBJETIVOS EN SECO: Son los que se emplean más corrientemente. Entre la lente frontal y el cubreobjetos sólo hay aire. A este grupo pertenecen los objetivos de menores aumentos. Las lentes frontales tienen de 3 a 10 milímetros de diámetro. Poseen gran profundidad de foco, lo cual permite observar diferentes planos paralelos del objeto. OBJETIVOS DE INMERSION: Se llama así a aquellos en los cuales, para la observación, deben interponerse entre la lente frontal y la preparación un líquido que, por su índice de refracción apropiado, permita una mayor luminosidad. Este líquido puede ser agua, aceite, monobromuro de naftaleno, etc. Son, estos objetivos, de gran aumento y de gran poder definidor. Se emplean en Bacteriología y Parasitología. Requieren gran luminosidad y empleo de condensador. OBJETIVOS APOCROMATICOS: Todos los objetivos, secos o de inmersiòn, están acromatizados sólo para dos rayos del espectro, el rojo y el azul. Son los llamados cromáticos, pero si conseguimos corregir este defecto para tres rayos, se elimina casi completamente el llamado espectro secundario, y tenemos los objetivos apocromáticos. Estos lentes corrigen además la esfericidad. Para una visita virtual a los laboratorios productores de lente tiene la siguiente dirección: http://www.zeiss.de/C12567A8003B58B9/allBySubject/34D946E306AF131DC1256A2A00436472 Para conseguir datos históricos se encuentra la siguiente dirección: http://www.ko-be.com/articuloMicroscopio.htm http://iessuel.org/salud/micro.htm http://teleformacion.edu.aytolacoruna.es/FISICA/document/fisicaInteractiva/OptGeometrica/Instrumentos/Microscopio/Hist_microscopio.htm
CUALIDADES DE LOS OBJETIVOS: Los objetivos deben poseer tres cualidades: poder definidor, poder penetrante y poder resolvente. El primero consiste en la propiedad de presentar con limpieza y corrección los contornos de la imagen. El poder penetrante es la propiedad de presentar, sin variar el enfoque, perfectamente detallados varios planos del

33
espesor de una preparación. El poder resolvente permite apreciar delicados detalles de estructura.

34
SISTEMA DE ILUMINACION. Se encuentra situado bajo la platina y tiene la misión de iluminar los objetivos por medio de luz transmitida, a causa de que la mayoría de las observaciones se realizan por transparencia. Consta de un espejo y de un diafragma. El espejo, redondo y adaptable a las más variadas posiciones, tiene una superficie plana y otra cóncava, que pueden intercambiarse a voluntad. Es espejo plano, para objetivos de escaso aumento, y el cóncavo, para grandes aumentos. La fuente luminosa puede ser natural o artificial. Esta última es idónea cuando proviene de una lámpara especial para estos aparatos. El diafragma va montado bajo la platina. Es de sistema iris, y permite, por medio de una palanca, obtener a voluntad conos luminosos de distinto tamaño y, mediante condensadores, conos luminosos muy grandes. Los condensadores constan de un sistema de lentes de gran abertura sujetos a una montura y colocados entre la platina y el espejo, pueden subirse y bajarse a voluntad y tienen un diafragma unido al conjunto. REGLAS GENERALES DE OBSERVACION Es regla general enfocar la preparación de abajo arriba: se aproxima el objetivo a la preparación de modo que la distancia entre ambos sea menor que la distancia focal. Se mira por el ocular y se hace retroceder el tubo lentamente mediante el tornillo rápido, hasta conseguir ver la preparación más o menos enfocada. Seguidamente, practique el enfoque exacto con el tornillo micrométrico. El objetivo de inmersión requiere más cuidado: Toda vez que es depositada la gota de líquido sobre la preparación, bájese el objetivo hasta que la toque. Para enfocar se utiliza el tornillo micrométrico, pero sin perder contacto con la gota y sin tocar la preparación. LIMPIEZA. Una vez terminada la observaciòn debe limpiarse el objetivo con papel limpia lentes, empapado de xilol o de tolueno. 2.3 CUIDADO DEL MICROSCOPIO. El microscopio es un instrumento costoso. Debemos darle el mejor cuidado posible. Siga siempre estas instrucciones generales cuando lo utilice.
• Transporte el microscopio sujetándolo con las dos manos: una por debajo de la base y la otra en el brazo.
• Colóquelo alejado del borde de la mesa. Si hay una lámpara unida al microscopio, tenga cuidado con los cables. Cuando trabaje con el microscopio quite de la mesa de laboratorio aquellas cosas que no sean absolutamente necesarias.
• Las lentes del microscopio cuestan casi tanto como todas las demás partes juntas. NUNCA LAS LIMPIE CON OTRA COSA QUE NO SEA PAPEL LIMPIALENTES).
• Cuando termine su trabajo guarde el microscopio, NO SIN DESMONTAR LAS PREPARACIONES MICROSCOPICAS QUE HA OBSERVADO Y COLOQUE EL OBJETIVO DE MENOR AUMENTO EN LA POSICION DE ENFOQUE, CON LA PLATINA EN SU POSICIÓN MAS BAJA. Realice la devolución de material al ayudante.

35
3. PREPARACIONES MICROSCOPICAS TEMPORALES. El profesor explicará la técnica correcta de hacer las preparaciones microscópicas temporales, con los materiales que el estudiante aporte utilizando agua destilada como medio de montaje. 1. PREPARACIÓN DE CORCHO:
Realice varios cortes delgados de corcho utilizando la hoja de afeitar nueva y escoja el más fino. En un porta objetos coloque una gota de agua destilada y coloque en él el pedazo de corcho seleccionado, y coloque un cubre objetos, procurando que no lo quede burbujas de aire. Proceda a su observación microscópica. Haga su esquema correspondiente utilizando los objetivos de aumento menor y aumento mayor.
2. LETRA IMPRESA: Busque en el periódico la letra “e” más pequeña que encuentre, córtela y colóquela en un porta objetos que tenga una gota de agua destilada, cúbrala con el cubre objetos y obsérvela en el microscopio usando los objetivos de aumento mayor y menor.
3.Solicite a su profesor preparaciones histológicas diversas Tejidos: Cortes transversales de vena, arteria, cartílago, hueso, páncreas, hígado, estómago, piel, cerebro, cerebelo, corteza, médula, hipocampo. En cada una de ellas observe y esquematice, dibuje, vea las distintas formas de Células y relaciónelas con el tejido y función de este. ¿A qué se debe la forma celular?, ¿Cuáles son los rangos de tamaño celular?, dibuje distintas células, ¿Cuál es la forma de una célula epitelial de intestino? ¿Por qué tendrá esa forma? Recuerde que las respuestas a todas estas preguntas deben encontrarse en su informe, y además deben formar parte de su conocimiento. 4. INFORME DE LABORATORIO. Al hacer observaciones de cualquier índole, es necesario describir lo observado con la mayor cantidad de detalles posibles, esto, es lo que se conoce como informe de laboratorio. Para los informes de los experimentos de laboratorio, debe proceder de la manera siguiente (según hoja adjunta):
• Datos generales
• Nombre de la Universidad, Facultad, Área, número de laboratorio, nombre completo del estudiante, día, número de carnet, nombre del profesor
• Número de práctico, título de la misma
• Propósito del práctico- Introducción
• Reporte de la parte experimental: resultados obtenidos
• Breve descripción de lo observado de acuerdo a sus esquemas
• Análisis y conclusiones
• Bibliografía utilizada- solicite al ayudante es estilo de citar bibliografías
• Utilizar Letra Arial N° 12, espacio simple. No olvide utilizar lápices de colores en práctico y en informe.

36
Es optativo utilizar cámaras digitales para obtener fotos, pero deben ser indicadas las zonas en observación mediante programas o software de dibujo.
• El tamaño del esquema debe ser proporcional al objeto observado
• Aumentos microscópicos del ocular, del objetivo y aumentos totales
• Hacer una pequeña descripción de lo que representa, señalando detalles sobresalientes de la observación.
ANALISIS Y CONCLUSIONES:

37
ACTIVIDAD 4
OBSERVACION MICROSCOPICA DE CELULA
PROCARIONTE Y EUCARIONTE I. INTRODUCCION. Los seres vivos presentan diversidad en forma, dimensión y estructura,
Características que dependen de las células que lo integran, de la actividad que realizan y del entorno que les rodea.
La célula como unidad morfológica lleva un patrón general de estructura, pero
existen variantes que establecen diferencia entre ellas, como son, ausencia o presencia de material genético rodeado por una membrana, lo cual permite la clasificación en dos grandes grupos: células procariontes (presentan nucleoide) y células eucariontes (presentan núcleo verdadero).
Es importante caracterizar que aún dentro de las células eucariontes existen
diferencias estructurales que permiten dividirlas en célula animal y vegetal. La célula
vegetal presenta pared celular, vacuolas prominentes y plastidios y la célula animal
carece de ellos, presentando centriolos y lisosomas.
II. El alumno debe lograr el desarrollo de las siguientes capacidades
a) Diferenciar por microscopía de luz, células procariontes de eucariontes y células animales de vegetales.
b) Utilizar correctamente las partes del microscopio. III. MATERIALES.
• Cebolla
• Palillos de dientes de punta roma
• Frasco gotero con lugol
• Frasco gotero con agua destilada
• Microscopio compuesto
• Porta y Cubre objetos (una caja)
• Hoja de afeitar nueva
• Papel limpia lentes
• Pinzas
• Frote bacteriano coloreado con Gram
• Guantes desechables

38
IV. PROCEDIMIENTO
A) OBSERVACION DE CELULAS PROCARIONTES (OBSERVACION DE FROTES BACTERIANOS
CON TECNICA DE GRAM): El alumno en trabajo previo de investigación bibliográfica deberá explicar que es un frote coloreado con técnica de GRAM, y cuál es su utilidad en la medicina antes de iniciar su observación microscópica. TINCIÓN DE GRAM: - Aplique una capa delgada del espécimen en el portaobjetos, déjelo secar ó fíjelo
con calor utilizando un mechero. - Cúbralo con cristal violeta durante 1 minuto, lávelo con agua destilada - Aplique solución yodada durante 1 minuto, lávelo con agua destilada - Cuidadosamente decolore la laminilla con alcohol-acetona hasta que el color azul
se torne en celeste claro. - Aplique solución de safranina por un minuto, lávelo con agua, secando la laminilla
con papel filtro.
• Las bacterias gram positivas se tiñen de color azul violeta.
• Las bacterias gram negativas se tiñen de color rosado.
Coloque el frote en la platina del microscopio, enfoque primero con aumento menor y luego con aumento mayor. ¿Qué otros detalles ve con seco mayor? Agregue una gota de aceite de inmersión en el frote y observe ahora con el objetivo de inmersión. El aceite forma un medio continuo entre el vidrio y el sistema de lentes del objetivo, lo cual permite un mayor aumento de la imagen del objeto observado. Haga esquemas y complete su informe de laboratorio.
B) OBSERVACION DE CELULAS EUCARIONTES:
1. CELULA VEGETAL.
1. Tome la cebolla y con la hoja de afeitar cubierta por uno de los constados con cinta adhesiva, proceda a cortar la cebolla separando la parte radical y la caulinar.
2. La parte que queda del bulbo, divídala en dos, haciendo un corte ecuatorial de tal forma que obtenga dos partes iguales.
3. De los segmentos obtenidos, haga cortes radiales para obtener trozos pequeños, levante cuidadosamente la epidermis de la parte cóncava del trocito del catáfilo.
4. En una gota de lugol sobre la parte central del portaobjetos, coloque la epidermis de la cebolla, cúbrala con el cubre objetos y coloque la preparación en la platina del microscopio. Enfoque y observe núcleo, pared celular y nucléolos. Haga esquemas de sus observaciones con objetivo mayor. 2. OBSERVACION DE CELULA ANIMAL.
A) Con un palillo de dientes de punta roma, raspe cuidadosamente la parte interna de la mejilla y coloque la muestra sobre el portaobjetos en el que previamente ha agregado una gota de lugol, cubriéndola con un cubreobjetos, colóquele en la platina y proceda a enfocar y observar. Haga esquemas de sus observaciones con objetivo mayor.

39
B) Utilizando el mismo material: Células de descamación de la mucosa oral.
Material de laboratorio:
• Abatelenguas estéril.
• Portaobjetos.
• Cubreobjetos
Reactivos:
• Solución de aceto-orceína al 2%.
• Aceite de inmersión.
Desarrollo de la práctica: 1. Enjuagarse la boca repetidas veces con agua y raspar la mucosa interna de la mejilla con un abatelenguas. Eliminar el primer raspado con un algodón y repetir la operación. 2. Hacer un frotis del segundo raspado sobre un portaobjetos. Añadir una o dos gotas de la solución de aceto-orceína. Colocar un cubreobjetos. Eliminar el exceso de colorante, use una piseta con agua destilada 3. Observar a seco débil hasta encontrar un campo en donde se localicen células. Observar después a inmersión para identificar el corpúsculo de Barr el cual se aprecia como una mancha obscura adherida a la membrana nuclear y que no desaparece al mover el tornillo micrométrico del microscopio. NOTA: EN MUJERES NORMALES EL NÚMERO PROMEDIO DE CÉLULAS EN FROTIS BUCALES CON CUERPO DE BARR ES DE 18-60% .

40
ACTIVIDAD 5 IDENTIFICACION DE CARBOHIDRATOS Y LIPIDOS POR REACCIONES
QUIMICAS, DE COLOR , Y MICROSCOPIA DE LUZ
I. INTRODUCCION. Los compuestos químicos de la célula se clasifican en dos grandes grupos:
substancias inorgánicas y substancias orgánicas, las primeras se caracterizan por la ausencia de uniones carbono-carbono en su estructura química; pudiéndose mencionar entre este grupo: agua, gases y sales disueltas, las segundas se caracterizan por presentar uniones carbono-carbono y carbono-hidrógeno y átomos de oxígeno en su estructura química, además de estos elementos, pueden existir átomos de nitrógeno, fósforo, azufre y algunos metales.
Las principales substancias orgánicas responsables de las características estructurales
y funcionales del protoplasma son: carbohidratos, lípidos, proteínas y ácidos nucleicos, dichas substancias pueden ser identificadas por una gran variedad de pruebas químicas y físicas. En esta práctica se efectuarán algunas de estas pruebas químicas para identificar carbohidratos y lípidos, en forma cualitativa.
II. CAPACIDAD A DESARROLLAR 1. Identificar carbohidratos y lípidos a través de reacciones químicas y microscopía de luz. 2. Utilizar correctamente el microscopio óptico compuesto.
III. EQUIPO Y MATERIALES NECESARIOS.
• Microscopio compuesto
• Porta y cubre-objetos
• 8 tubos de ensayo, resistentes al calor
• 8 pipetas de 2 a 5 ml o goteros
• Gradilla para tubos de ensayo
• Una hoja de afeitar nueva
• Solución de lugol (3 frascos)
• Solución de Sudán III ó IV (3 frascos)
• Solución de Benedict u otra para reconocimiento de H de C
• Solución de Glucosa al 1%
• Solución de Sacarosa al 1 %
• Solución de Almidón al 1%
• Una papa
• Un trozo pequeño de grasa animal
• Aceite de cocina
• Agua destilada
• Pinzas

41
• Tripode, anillo lámina de asbestos
• Vasos de precipitados
• Baño María
• Mechero
• 8 Agitadores de vidrio
• Lámpara de alcohol
• Cinta adhesiva IV. PROCEDIMIENTO. IDENTIFICACION DE CARBOHIDRATOS: El almidón se identifica con lugol (solución de I2/KI) dando un azul o morado intenso y los azúcares reductores se identifican con la solución de Benedict, los cuales después de calentarse en baño maría durante 3 minutos, dan un precipitado color rojo ladrillo. OBSERVACION MACROSCOPICA. 1. Numere 6 tubos de ensayo de 1 a 6, a los tubos 1 y 2 agregue 1 ml de almidón. 2. A los tubos 3 y 4 agregue 1 ml de sacarosa 3. A los tubos 5 y 6 agregue 1 ml de glucosa 4. A los tubos 1,3 y 5 agrégueles una gota de solución de lugol, observe y anote resultados
en el cuadro 5. A los tubos 2,4 y 6 agregue aproximadamente 10 gotas de solución de Benedict, y 6. Colóquelos en baño maría por 3 minutos o usando e mechero de alcohol llévelo a
ebullición, evite quemarse), observe y anote resultados en la tabla correspondiente, en el cuadro que se adjunta.
TUBO PRUEBA DE BENEDICT
PRUEBA DE
LUGOL
Color Inicial Color Final Color Inicial Color Final
1. Almidón
2. Almidón
3. Sacarosa
4. Sacarosa
5. Glucosa
6. Glucosa
IDENTIFICACION DE LIPIDOS. Los lípidos son identificados usando el reactivo Sudán III ó IV, los cuales al mezclarse dan un color rojo brillante. Coloque en un tubo de ensayo 1 ml. de aceite de cocina y 1 ml. de agua destilada; agréguele unas gotas de Sudán; agítelo, observe inmediatamente y anote los resultados; deje reposar el tubo y al cabo de 5 minutos obsérvelo y describa los resultados

42
OBSERVACION MICROSCOPICA. Gránulos de Almidón. 1. Por medio de la hoja de afeitar haga un corte muy fino del tubérculo de papa y colóquelo
sobre un porta objetos, haga una preparación utilizando lugol diluido como medio de montaje y obsérvela al microscopio usando el objetivo 40X para hacer el esquema de lo observado.
2. Observación de tejido adiposo. Usando una hoja de afeitar obtenga un corte delgado de la grasa animal.
Móntela en el portaobjetos y agréguele una gota de Sudán IV cúbrala con el cubreobjetos y presione suavemente, coloque la preparación sobre la platina del microscopio enfoque y observe con el objetivo de aumento menor (10X), y luego con el objetivo de aumento mayor (40X); construya un esquema de lo observado y descríbalo

43
ACTIVIDAD 6 PROTEÍNAS
IDENTIFICACION DEL ENLACE PEPTIDICO POR REACCIONES QUIMICAS I. INTRODUCCION. Las proteínas son compuestos cuaternarios formados por (C, H, 0, N) que tienen como
unidades básicas aminoácidos los cuales se unen por enlaces peptídico. Ellas cumplen en los seres vivos gran variedad de funciones, entre las cuales están: ser catalizadores de reacciones biológicas (enzimas), defender al organismo (anticuerpos) y formar estructuras.
Las proteínas pueden ser afectadas por cambios en la temperatura, ó concentración
de iones H+ (pH) y pueden ser identificadas por reacciones de color como Xantoproteica, Millón y Biuret; así como por cromatografía en capa fina o de gases. La identificación podemos hacerla en forma cualitativa o cuantitativa.
CAPACIDAD A DESARROLLAR: Al finalizar la práctica cada estudiante será capaz de identificar el enlace peptídico
presente en proteínas a través de reacciones químicas. III. MATERIALES. -3 Tubos de ensayo -3 Pipetas de 2 a 5 ml o goteros -Gradilla para tubos de ensayo -Solución de clara de huevo -Solución de hidróxido de sodio al 10% (3 frascos) -Solución de sulfato de cobre al 10% (3 frascos) -Orina -Leche -3 Agitadores de vidrio -Guantes descartables

44
IV. PROCEDIMIENTO.
A) IDENTIFICACION DEL ENLACE PEPTIDICO DE PROTEINAS. Reacción de Biuret (Identificación del enlace peptídico). En un tubo de ensayo ponga 2 ml. De solución de clara de huevo al 10%; luego
agregue 10 gotas de hidróxido de sodio al 10%, agréguelo cuidadosamente gota a gota y agite después de cada adición. Luego agregue solución de sulfato de cobre II al 10% hasta que se produzca una coloración violeta o púrpura, color que indica que la prueba es positiva.
Repita el mismo procedimiento con la leche y la orina. Anote sus resultados en el
cuadro, y explique a qué se deben los cambios observados.
CUADRO: RESULTADOS CON REACTIVO DE BIURET
MUESTRA CAMBIOS OBSERVADOS
LECHE
CLARA DE HUEVO
ORINA
ANALISIS DE LOS RESULTADOS: CONCLUSIONES:

45
ACTIVIDAD 7
BIOENERGETICA
I. INTRODUCCION. La oxidación celular, es un proceso por medio del cual se libera parcial o totalmente la
energía almacenada en los combustibles orgánicos; este proceso se verifica por etapas y en eucariontes se puede llevar a cabo en el citoplasma (oxidación anaeróbica); y parte en la mitocondria (respiración u oxidación aeróbica).
La mayoría de los organismos dependen del suministro de O2, principalmente los Eucariontes superiores, para oxidar totalmente la molécula orgánica (carbohidratos), verificándose una transformación energética que se lleva a cabo por medio de reacciones químicas controladas enzimáticamente, durante estas reacciones se forma ATP, como molécula que almacena la energía, además de CO2 y agua. Durante las transformaciones energéticas las células ponen de manifiesto las leyes de la termodinámica, buscando nuevos equilibrios químicos. En esta práctica Ud. Pondrá de manifiesto las leyes de la termodinámica y podrá concluir que mecanismos puede emplear un ser vivo para compensar los cambios durante la transformación de energía.
II. CAPACIDAD A DESARROLLAR Interpretar los datos obtenidos en la experiencia de laboratorio considerando los aspectos teóricos del fenómeno en estudio acerca de transformación energética en seres vivos. Explicar en términos de causas y efectos los fenómenos aludidos en la experiencia de laboratorio III. MATERIALES.
• Reloj ó cronómetro
• Fenoftaleína
• Pajillas o pajuelas para bebida
• Solución de Na0H al 2.5%
• Agua destilada
• Vasos de precipitado de 250 ml.
• Papel pH NOTA: Antes de asistir a la práctica el estudiante debe investigar: 1. Qué es pH? 2. ¿Qué importancia tiene el pH en el conocimiento del cuerpo humano

46
3. ¿Qué es un indicador de pH? 4. Rangos de pH en que se utiliza la fenoftaleína para los análisis de laboratorio 5 Qué reacción ocurre cuando se mezcla C02 y H20? (Escriba la ecuación). 7. ¿Cuáles son las relaciones que se producen entre respiración celular y mitocondrias?
8. Describa los procedimientos que ocurren en su cuerpo para obtener energía. IV. PROCEDIMIENTO. 1. En el vaso de precipitado coloque 100 ml de agua destilada y agregue 3 gotas de
Fenoftaleína, si el agua no se colorea de rosado agregue solución de Na0H gota a gota, agite suavemente al agregar cada gota hasta que la solución permanezca rosada más de 15 seg. Registre en la tabla el número de gotas que utilizó y anote los cambios observados que la servirán de control. Medir pH antes y después de agregar fenoftaleína.
2. A través de una pajilla expulse aire dentro de la solución por 40 segundos medir pH. 3. Después de haber borboteado agregue Na0H gota a gota a la solución. Cuente el
número de gotas necesarias para que la solución cambie de coloración y anótelo en la tabla. ¿Qué indica esto? Medir pH nuevamente.
4. Haga 20 abdominales o flexiones, sin descansar, inmediatamente después exhale por la
pajilla en la solución durante 40 s Con la frecuencia respiratoria producida por el ejercicio. Medir pH.
5. Repita el paso 3. Anote los cambios observados. ¿Qué otros factores pueden influir
en la cantidad de C02 que exhalamos?
6. Explique por qué aumenta la frecuencia respiratoria durante el ejercicio. Qué efecto tiene el ejercicio sobre el metabolismo? Qué relación hay entre consumo de 02 y producción de C02?
No. DE GOTAS DE Na0H CAMBIO OBSERVADO
Control (sin soplar)
Normal (sin ejercicio)
Después del ejercicio

47
ACTIVIDAD 8
ACCION ENZIMATICA I. INTRODUCCION. Cuando se realizan reacciones químicas a veces es necesario agregar sustancias que
aceleren o faciliten la reacción, a estas sustancias se les llama catalizadores. Las enzimas son proteínas, también llamadas catalizadores biológicos, porque son de naturaleza orgánica, producidas por la célula para acelerar la velocidad de las reacciones químicas, quedando estas intactas al terminar estas reacciones.
Las enzimas actúan sobre sustancias llamadas sustratos, que es la molécula que van a transformar, son altamente específicas, variando su actividad con la modificación del ph, temperatura, concentraciones de enzima y sustrato; estas pueden ser inhibidas por sustancias llamadas venenos enzimáticos. Durante esta práctica se tratará de poner de estudiar algunos de los factores que afectan la actividad enzimática.
II. CAPACIDAD A DESARROLLAR Interpretar los datos obtenido de experiencias de laboratorio que muestran la
acción enzimática sobre sustratos específicos y la influencia de factores. Explicar en términos de causas y efectos y fundamentando en aspectos teóricos la
experiencia recogida en laboratorio de actividad enzimática. III. MATERIAL.
• 3 Pinzas para sujetar tubos de ensayo
• Solución de almidón al 1%
• Saliva
• Lugol enfrasco gotero
• Reactivo de Benedict
• 1 Docena de tubos de ensayo, resistentes al calor
• Lámpara de alcohol
• 4 Varillas de vidrio
• Pastillas de cuajo (renina)
• HCl al 10%
• 30 cc. de leche
• 4 goteros
• 4 pipetas de 5 ml.
• Guantes desechables IV PROCEDIMIENTO. Acción de la Amilasa sobre el almidón

48
a. Rotule dos tubos con las letras A y B. b. Agregue a cada tubo dos ml de saliva. c. el tubo B caliéntelo hasta ebullición. d. Rotule 4 tubos del número 1 al 4, agregándoles 4 ml de solución de almidón al 1 %. e. Del tubo A extraiga una muestra, y agregue 2 gotas de el al tubo número 1, y 10
gotas al tubo número 2. f. Del tubo B extraiga una muestra, y agregue 2 gotas al tubo número 3, y 10 gotas al
tubo número 4. g. Divida el contenido de los tubos 1 Y 4 a la mitad. h. Después de 5 minutos, realice pruebas de lugol y benedict, a los tubos del número 1
al número 4. i. Utilice una varilla diferente para cada tubo, al mezclar. j. Cada vez que use goteros y pipetas lávelos con agua. k. Observe, anote y analice sus resultados en el cuadro de registro.
Acción de la Renina sobre la Caseína de la Leche. 1. Disuelva ¼ de pastilla de cuajo (renina) en 25 ml de agua destilada. 2. Numere 2 tubos de ensayo (1 y 2). 3. En el tubo # 1 coloque 5 ml de leche. 4. En el tubo # 2 coloque 5 ml de leche más 2 gotas de HCl al 10%. 5. Agregue 10 gotas de solución de renina a cada tubo numerado. 6. Observe anote y analice sus resultados.
ACCION DE LA AMILASA SOBRE EL ALMIDON
PRUEBA A LOS CINCO MINUTOS
NO. DE TUBO Calidad de enzima LUGOL BENEDICT
1 ACTIVA
2
3 INACTIVA
4
ACCION DE LA RENINA SOBRE LA CASEINA DE LA LECHE
TUBO No. DESCRIPCION DE OBSERVACIONES
1 (Leche + Renina)

49
2 (Leche + Renina + HCl)
En base a sus observaciones qué puede concluir acerca de cómo afecta a la actividad enzimática el tiempo, la concentración y la temperatura? Explique los procesos ocurridos que llevaron a los resultados obtenidos.

50
ACTIVIDAD 9
PERMEABILIDAD DIFERENCIAL DE LA MEMBRANA
I. INTRODUCCION.
Las membranas que rodean a las células y a algunos organelos de ella, poseen permeabilidad diferencial, lo cual significa que la membrana celular deja pasar unas sustancias y otras no, sirviendo de barrera para el paso de una sustancia disuelta (soluto) dándose el fenómeno llamado DIALISIS, y la difusión del solvente (agua) dándose el fenómeno llamado OSMOSIS.
Las células responden en diferente forma según el medio que las rodea; si se encuentran ante una solución diluida, con una concentración de soluto menor al contenido interno de la célula se dice que la solución es HIPOTONICA o HIPOOSMOTICA, y habrá difusión neta de agua (solvente) a través de la membrana hacia el interior de la célula: ENDOSMOSIS; al entrar agua a la célula, esta aumentará de volumen hasta llegar a romperse, (si no posee pared celular): CITOLISIS; en caso que sea el eritrocito el que se rompe, se libera la hemoglobina y el fenómeno se denomina HEMOLISIS.
Por el contrario, si se coloca a una célula en una solución de mayor concentración de
soluto que la concentración interna de la célula, se dice que la solución es HIPERTONICA O HIPEROSMOTICA, y la difusión del solvente será en sentido contrario, al anterior, con la difusión del agua de adentro hacia afuera de la célula: EXOSMOSIS, la célula perderá líquido del citoplasma y se encogerá, el fenómeno se denomina CRENACION.
En las células vegetales, frente a soluciones hipertónicas o hiperosmóticas, puede verse
el efecto de la salida de líquido porque la membrana se contrae hasta poderla apreciar separada de la pared celular, a este fenómeno se le llama PLASMOLISIS. El caso contrario, la CITOLISIS, no se aprecia porque la pared celular protege a la membrana.
Si la célula es colocada en un medio ISOTONICO o ISOOSMOTICO, la célula no sufre
ningún cambio visible ya que la difusión del solvente a ambos lados está equilibrada. En esta práctica usted observará el comportamiento de células eucariontes ante
soluciones HIPO, ISO e HIPERTONICAS. II. CAPACIDADES A DESARROLLAR Interpretar los datos obtenidos de resultados de experimentación de los cambios
celulares por la permeabilidad de su membrana citoplasmática. Discutir las interpretaciones de datos obtenidos contemplando los aspectos teóricos y
empíricos acerca de la membrana celular.
1. Utilizar el microscopio óptico compuesto. III. MATERIALES. -Microscopio óptico compuesto -Portaobjetos y cubreobjetos (1 caja)

51
-Soluciones de cloruro de sodio (al 0.2% = hipotónica, al 0.9% = isotónica y al 5% = hipertónica)
-Agua destilada -Cajas de Petrí -Goteros -Jeringa hipodérmica desechable de 5 c.c. -Guantes desechables -Frasco con anticoagulante (Tarea de ayudante)
IV PROCEDIMIENTO.
a) Extraer 2 c.c. de sangre, y verterlo en el frasco que contiene el anticoagulante
b) Numerar tres tubos de ensayo
c) Al tubo número 1 colocar 3 ml. de solución de NaCl 0.2%
d) Al tubo número 2 colocar 3 ml de solución de NaCl 0.9%
e) Al tubo número 3 colocar 3 ml. de solución de NaCl 5%
f) Agregar 5 gotas de sangre a cada uno de los tubos numerados.
g) Mezcle el contenido de los tubos y describa las observaciones macroscópicas de cada uno de ellos, indique en que tubo hay hemólisis.
h) Haga una preparación microscópica del contenido de cada tubo, obsérvelos al microscopio con objetivo de aumento mayor y anote resultados respecto a la hemólisis, la crenación y las células que no se han modificado en su morfología.
i) Analice en qué casos se han dado los fenómenos de endosmosis y exosmosis.
j) Complete su reporte de laboratorio con análisis y conclusiones.

52
ACTIVIDAD 10
LA FERMENTACION ALCOHÓLICA DE LA GLUCOSA I. INTRODUCCION. La fermentación es la oxidación de sustratos energéticos en ausencia de oxígeno.
Los carbohidratos, metabolizados en este proceso, principalmente las Hexosas (Glucosa, Fructosa), tienen como productos finales entre otros Etanol más C02,
dependiendo del tipo de organismo que se trate. El hombre ha podido sacar provecho del conocimiento de los pasos metabólicos
que intervienen en la fermentación produciendo derivados alcohólicos, mediante el uso de hexosas (glucosa y fructosa) como sustratos sobre los cuales actúan las enzimas específicas producidas por microorganismo (levaduras, bacterias, etc).
II. CAPACIDAD A DESARROLLAR
Interpretar los datos obtenidos en forma experimental acerca de la fermentación alcohólica. Explicar en términos de causas y efectos los aspectos teóricos de la fermentación alcohólica fundamentando a partir de los datos obtenidos.
III. MATERIALES
• Termómetros
• Hornilla eléctrica
• 3 Tubos de ensayo
• Solución de glucosa al 1%
• Solución de fructosa al 1%
• Solución de maltosa al 1%
• Levadura seca o granulada
• 3 Tubos de hemólisis
• Agua destilada
• Gradillas
• 1 Vaso de precipitado de 500 ml.
• 3 pipetas de 2 a 5 ml.
• 3 goteros
• Jabón líquido
• Frasco con cloro, para descartar material
• IV. PROCEDIMIENTO.
a) Numere 3 tubos de ensayo, de 1 a 3 y proceda a identificar de la misma forma los tubos de hemólisis.
b) Añada a cada tubo de hemólisis 2 ml. de solución de carbohidratos así: N°1 Glucosa, N°2 Fructosa, N°3 Maltosa; agregue levadura en cantidad suficiente a cada uno de los tubos El equivalente a media cucharada de té.

53
c) Introducir los tubos de hemólisis invertidos dentro de los tubos de ensayo. (Campana de
Durham) d) Coloque un vaso con agua y caliente a 37°C, ponga los tubos dentro del vaso durante 30’,
teniendo cuidado de que la temperatura se mantenga a 37°C siempre. e) Anote el tiempo en qué inicia el procedimiento. f) Observe los cambios que se operan en los tubos, compare el tiempo y la intensidad en que
se van dando los cambios en cada uno de los 3 tubos. g) Anote los resultados al final de la observación, interprete los fenómenos ocurridos y
elabore sus conclusiones del proceso realizado. h) Aplicar el procedimiento de Normas de manejo de material biológico. i) De acuerdo a los resultados observados responda las siguientes preguntas: 1. Cuál es el rol de la levadura en el proceso de fermentación 2. Cuál es el sitio celular donde ocurrió el metabolismo de los carbohidratos 3. Haga un resumen de la secuencia enzimática qué participó en el proceso catabólico 4. Explique sí existe alguna(s) diferencia(s) entre fermentación y glucólisis, y porque

54
ACTIVIDAD 11
OBSERVACION DE MITOCONDRIAS I. INTRODUCCION. Al proceso de producción de energía en presencia de O2 a partir de un sustrato se
le llama respiración, la cual se realiza en las células en tres fases fundamentales: la primera fase concluye con la formación del ácido Pirúvico si el sustrato es glucosa; luego entra a una segunda fase donde éste es metabolizado en el ciclo de Krebs, dando como producto electrones o equivalentes reductores, los cuales pasan a la cadena respiratoria, llamada también cadena de transporte de electrones donde finalmente se produce el nucleótido adenosin-trifosfato o ATP.
Estas dos últimas fases se producen solo si el medio es aeróbico. En todo el proceso participa gran cantidad de enzimas y se verifican muchas reacciones de óxido reducción. El cíclo de Krebs y la fosforilación oxidativa se desarrollan en las Mitocondrias en eucariontes.
II. CAPACIDAD A DESARROLLAR Identificar mitocondria en células de levadura a través de tinción supravital y
microscopía de luz. Manejar el microscopio de luz. III. MATERIALES.
• Solución de glucosa al 5%
• Microscopio compuesto
• 24 horas antes de la práctica, agregar 10 granos de levadura a 5 cc de solución de glucosa al 5% en temperatura ambiente* dejarlo sin cubrir (ambiente aeróbico). Transportarla de igual manera.
• Porta y cubre objetos
• Verde de Janus al 0.001 P/V
• Aceite de inmersión
• Agua destilada
• Caja de Petrí (3 por laboratorio)

55
PROCEDIMIENTO.
A. Observación Microscópica de Levaduras:
Coloque sobre un portaobjetos bien limpio dos gotas de agua y agregue un granito de levadura. Agite suavemente la suspensión hasta que la levadura está bien repartida en toda el agua.
Observe al microscopio con lente de aumento menor primero, luego con aumento
mayor y finalmente con objetivo de inmersión. Construya esquemas de lo observado y complete su informe de laboratorio.
B. Observación de Mitocondrias: Colocar en una caja de Petri 5 gotas de levadura en incubación y agregarle una gota
de colorante verde de Janus, dejarlo actuar durante 2 minutos; luego de hacer una preparación microscópica y obsérvela con el objetivo de inmersión; selecciones de preferencia levaduras grandes donde pueda observar las mitocondrias y el núcleo de las células.
Haga esquemas de lo observado y complete su informe de laboratorio.

56
ACTIVIDAD 12
1. ACTIVIDAD DE LISOSOMAS I. INTRODUCCION.
Los lisosomas están presentes en casi todas las células eucarióticas, son organelos membranosos (vesículas membranosas).
Los lisosomas contienen algunas de las proteínas fabricadas en el retículo endoplamático rugoso que son enviadas al complejo de Golgi, éstas proteínas son enzimas (hidrolasas) digestivas intracelulares que pueden degradar sustratos orgánicos en las subunidades que los componen.
La principal función de los lisosomas es digerir las partículas alimenticias, que van desde proteínas individuales hasta microorganismos complejos.
Muchas células “comen” o digieren mediante FAGOCITOSIS, mecanismo que consiste en rodear con extensiones de la membrana a partículas extracelulares que pueden ser alimenticias, organelos defectuosos, sustancias dañinas o microorganismos infecciosos. Estas partículas ya rodeadas de membrana se mueven entonces dentro del citoplasma, dentro de sacos o vesículas membranosas que reciben el nombre de fagosoma, los lisosomas (primarios) reconocen estas vesículas y se fusionan con ellas (secundarios). El contenido de las dos vesículas de mezcla y las enzimas lisosómicas digieren los alimentos (o cualquier otra partícula extraña), convirtiéndolos en aminoácidos, monosacáridos, ácidos grasos y otras moléculas pequeñas. Las moléculas pequeñas o simples se difunden luego fuera de esta vesícula hacia el citosol para nutrir a la célula. Cuando los microorganismos o sustancias extrañas no pueden digerirse se forman los cuerpos residuales.
Los glóbulos blancos o leucocitos presentes en la sangre, son células, capaces de efectuar el proceso de fagocitosis. Los leucocitos tienen como función defender al organismo humano mediante el reconocimiento de microorganismos extraños, dañinos o infecciosos a los cuales fagocitan y digieren mediante las enzimas presentes en los lisosomas.
Si entre los microorganismos que ingieren los leucocitos se encuentran bacterias que todavía no han sido digeridas completamente, estas son observables en microscopía de campo claro, y con coloración de Gram; entre estas esta la Neisseria gonorrhoeae (bacteria infecciosa), la cual se observa como diplococos gram negativo dentro de los leucocitos que se obtienen a partir de muestras de secreciones de personas con infección.
II. OBJETIVOS. -Identificar diplococos intra y extra celulares, en leucocitos humanos. MATERIALES:

57
• Microscopio óptico compuesto
• Preparaciones microscópicas fijas de frotes de secreción coloreados con técnica de Gram. (Labor de ayudante)
• Aceite de inmersión
• Papel limpialentes IV. PROCEDIMIENTO. Observación de Preparaciones Microscopicas.
1. Tome el frote coloreado con coloración de Gram y colóquelo sobre la platina del microscopio.
2. Haga su enfoque inicial, utilizando el objetivo seco débil para localizar el área con más muestra.
3. Luego observe con el objetivo de 40 X y trate de localizar leucocitos.
4. Coloque una gota de aceite de inmersión sobre el área localizada y enfoque ahora utilizando el objetivo de inmersión.
5. Haga un esquema de lo observado con el objetivo de inmersión y complete su informe de laboratorio.
6. Cuando termine sus observaciones con el papel limpialentes, limpie al objetivo de inmersión y la preparación del frotis coloreado.
2. ACTIVIDAD DE CATALASA
I. INTRODUCCION.
El término peroxisoma fue introducido por De Duve para identificar cualquier microcuerpo que presente la secuencia flavín oxidasa-catalasa. Los peroxisomas son cuerpos delimitados por una membrana y contienen algunas enzimas. Son abundantes en las células hepáticas y en la de los túbulos renales, pero muy escasos o ausentes en otros tipos de células. Los peroxisomas contienen 3 enzimas que actúan catalizando reacciones en las que se forma peróxido de hidrógeno y una enzima que constituye alrededor de 40% del total de enzimas del peroxisoma, que degrada el peróxido de hidrógeno en agua y oxígeno gaseoso, recibiendo el nombre de CATALASA.
Flavín oxidasa
R H2 + 02 R + H2O2
catalasa 2H202 2H2O + O2 (gas)
II. CAPACIDAD A DESARROLLAR
Analizar e Interpretar los datos experimentales obtenidos en la experiencia de la actividad de la catalasa en tejido animal y vegetal.
III. MATERIALES:
• Vidrio de reloj

58
• Papa y rábano
• Agua oxigenada* (Peróxido de hidrógeno)
• Trozo de hígado y riñón de res
• Hojas de afeitar (dos)
IV. PROCEDIMIENTO: Para poder estimar la velocidad de reacción considere una escala entre 0 y 5 (O = no hay reacción, 1 = muy lento, 5 = muy rápido). Registre los datos en una tabla. A. Actividad de la Catalasa de Tejido Vegetal sobre Agua Oxigenada. 1. Haga un corte fino de rábano y uno de papa, cuyas dimensiones sean iguales, colóquelos
cada uno en diferente vidrio de reloj. Agregue tres gotas de H2O2. Qué observa?
B. Actividad de la Catalasa de tejidos animales sobre Agua Oxigenada.
1. Hacer un corte fino de tejido hepático y renal, cuyas dimensiones sean iguales, colóquelos sobre diferentes vidrios de reloj, agregue tres gotas de agua oxigenada.
COMPLETE
TEJIDO/ESCALA o 1 2 3 4 5
Papa
Rábano
Hígado
Riñón
INTERPRETACION DE RESULTADOS: ¿Cuál es el tejido con mayor actividad y cuál el que tiene menor actividad? ¿Qué tienen en común y en qué se diferencian? Mencione algunos procesos metabólicos de la célula que producen H2O2. El H2O2 se usa frecuentemente, como antiséptico. ¿Por qué? Cuando es agregado a heridas produce burbujas. ¿Qué indica esto?

59
ELEMENTOS QUÍMICOS
Se denominan elementos biogénicos o bioelementos a aquellos elementos químicos que forman parte de los seres vivos. Atendiendo a su abundancia (no importancia) se pueden agrupar en tres categorías:
Bioelementos primarios o principales: C, H, O, N
Son los elementos mayoritarios de la materia viva, constituyen el 95% de la masa total.
PROPIEDADES FÍSICO Y QUÍMICAS
1.-Forman entre ellos enlaces covalentes, compartiendo electrones. Tanto En la materia inanimada como en la inanimada existen los enlaces covalentes. Son importantes porque en los elementos que forman parte de los seres vivos tienen la capacidad de formar compuestos estables (difícil descomponer entre sus constituyentes).
Las moléculas son agrupaciones estables de átomos unidos por un tipo de enlace químico que se denomina enlace covalente. Además de este enlace entre átomos. Las moléculas se representan mediante fórmulas químicas y mediante modelos.
Cuando dos o más átomos iguales o diferentes se unen entre sí formando una agrupación estable, dan lugar a una molécula. Así, los gases hidrógeno (H2) y oxígeno (O2) están constituidos por moléculas diatómicas, en las cuales los dos átomos componentes son esencialmente iguales. Cuando dos átomos de hidrógeno y uno de oxígeno se unen forman el agua (H2O). Y cuando cuatro átomos de hidrógeno se enlazan con uno de carbono forman el gas metano (CH4).
CARBONO, NITRÓGENO Y OXÍGENO
1.-El carbono, nitrógeno y oxígeno, pueden compartir más de un par de electrones, formando enlaces dobles y triples, lo cual les dota de una gran versatilidad para el enlace químico
2.-Son los elementos más ligeros con capacidad de formar enlace covalente, por lo que dichos enlaces son muy estables.
3.- Las combinaciones del carbono con otros elementos, como el oxígeno, el hidrógeno, el nitrógeno, etc., permiten la aparición de una gran variedad de grupos funcionales que dan lugar a las diferentes familias de sustancias orgánicas . Estos presentan características físicas y químicas diferentes, y dan a las moléculas orgánicas propiedades específicas, lo que aumenta las posibilidades de creación de nuevas moléculas orgánicas por reacción entre los diferentes grupos.
4.-Los enlaces entre los átomos de carbono pueden ser simples (C - C), dobles (C = C) o triples.

60
Ello permite que puedan formarse cadenas más o menos largas, lineales, ramificadas y anillos.
Bioelementos secundarios S, P, Mg, Ca, Na, K, Cl
Los encontramos formando parte de todos los seres vivos, y en una proporción del 4,5%.
S Se encuentra en dos aminoácidos (cisteína y metionina) , presentes en todas las proteínas. También en algunas sustancias como la Coenzima A
P
Forma parte de los nucleótidos, compuestos que forman los ácidos nucleicos. Forman parte de coenzimas y otras moléculas como fosfolípidos, sustancias fundamentales de las membranas celulares. También forma parte de los fosfatos, sales minerales abundantes en los seres vivos.
Mg Forma parte de la molécula de clorofila, y en forma iónica actúa como catalizador, junto con las enzimas , en muchas reacciones químicas del organismo.
Ca Forma parte de los carbonatos de calcio de estructuras esqueléticas. En forma iónica interviene en la contracción muscular, coagulación sanguínea y transmisión del impulso
nervioso.
Na Catión abundante en el medio extracelular; necesario para la conducción nerviosa y la contracción muscular
K Catión más abundante en el interior de las células; necesario para la conducción nerviosa y la contracción muscular
Cl Anión más frecuente; necesario para mantener el balance de agua en la sangre y fluido intersticial
Oligoelementos

61
Se denominan así al conjunto de elementos químicos que están presentes en los organismos en forma vestigial, pero que son indispensables para el desarrollo armónico del organismo.
Se han aislado unos 60 oligoelementos en los seres vivos, pero solamente 14 de ellos pueden considerarse comunes para casi todos, y estos son: hierro, manganeso, cobre, zinc, flúor,
iodo, boro, silicio, vanadio, cromo, cobalto, selenio, molibdeno y estaño. Las funciones que desempeñan, quedan reflejadas en el siguiente cuadro:
Hierro Fundamental para la síntesis de clorofila, catalizador en reacciones químicas y formando parte de citocromos que intervienen en la respiración celular, y en la hemoglobina que interviene en el transporte de oxígeno.
Manganeso Interviene en la fotolisis del agua , durante el proceso de fotosíntesis en las plantas.
Yodo Necesario para la síntesis de la tiroxina, hormona que interviene en el metabolismo
Flúor Forma parte del esmalte dentario y de los huesos.
Cobalto Forma parte de la vitamina B12, necesaria para la síntesis de hemoglobina .
Silicio Proporciona resistencia al tejido conjuntivo, endurece tejidos vegetales como en las gramíneas.
Cromo Interviene junto a la insulina en la regulación de glucosa en sangre.
Zinc Actúa como catalizador en muchas reacciones del organismo.
Litio Actúa sobre neurotransmisores y la permeabilidad celular. En dosis adecuada puede prevenir estados de depresiones.
Molibdeno Forma parte de las enzimas vegetales que actúan en la reducción de los nitratos por parte de las plantas.

62
QUÍMICA DE LA VIDA
Necesariamente, para comprender la química de la vida, es estricto adquirir conocimientos acerca de los principios fundamentales para la química. El avance en el campo de la química comienza en el siglo XVI y hasta el siglo XVII, cuando los experimentos de laboratorio llevaron al desarrollo de las teorías cinéticas de reacción y de la termodinámica y al descubrimiento de la composición atómica de algunas moléculas. Macromoléculas y moléculas relacionadas con la vida La importancia del estudio de las macromoléculas que participan en la matriz estructural y funcional de la célula, radica en la estructura e interacciones que se dan entre los átomos para formar las moléculas. Entre las interacciones se encuentran los enlaces covalentes y las interacciones no covalentes que se producen entre iones, moléculas y partes de la molécula. Ocurre que, las moléculas presentes en la estructura celular poseen cierta arquitectura especial. Por ejemplo, el ADN, esta molécula posee una secuencia lineal de residuos que se mantiene estable mediante enlaces covalentes. Contribuye a esta estabilidad su conformación tridimensional y la interacción no covalente entre algunos de sus elementos. Las interacciones son de vital importancia para que surja el fenómeno de la vida. Relacionadas con estas interacciones se encuentra el fenómeno energético entre los constituyentes moleculares. Así al comparar las energías de enlace covalente y no covalente: Los enlaces covalentes, como por ejemplo, C-C y C-H tienen energía de enlace que oscilan entre 300 y 400 kJ/mol. Por otra parte, los enlaces no covalentes presentes en los seres vivos son de alrededor de 10 a 100 veces más débiles. Podemos llamarlos esenciales, puesto que la interacción débil permite que las moléculas se rompan y se formen con facilidad; permitiendo de esta forma las interacciones dinámicas que caracterizan la vida. Conceptos y definiciones para recordar Enlace químico: Fuerza que mantiene unidos los átomos en un compuesto o elemento. Los átomos al reaccionar pueden ceder, captar o compartir electrones, dando origen a distintos tipos de enlace. Enlace covalente: es aquella unión donde existe participación mutua de pares electrónicos entre los átomos enlazados. Ejemplo: Molécula de hidrógeno: Es un ejemplo de enlace covalente. Para que dos átomos de hidrógeno formen una molécula, los dos átomos deben aproximarse lo suficiente para que sus respectivos orbitales atómicos “1S” se superpongan formando un orbital molecular. De esta forma los dos electrones pueden ocupar cualquier espacio en este orbital, moviéndose sin perder la fuerza de atracción de al menos uno de los dos núcleos. Al formarse este enlace se desprende energía y la molécula gana estabilidad. H* + H* H2 + 436 kJ/mol-1

63
En la ecuación anterior podemos deducir que este enlace covalente es fuerte porque se requiere 436kJ para disociar un mol de moléculas de hidrógeno en sus átomos constituyentes. Esta energía recibe el nombre de energía de enlace La estabilidad alcanzada por la molécula puede explicarse porque alcanza la estructura de un gas noble para cada átomo al compartir dos electrones. Más aún, los dos núcleos cargados positivamente se mantienen juntos por la fuerza de atracción que ellos ejercen hacia los electrones. Se define energía de enlace como la energía que es necesario aplicar (energía positiva) para romper un mol de enlaces, o la energía que se libera (energía negativa) cuando se forma un mol de enlaces. En base a lo anterior es posible pensar que la energía de un enlace triple será mayor que la de uno doble y la de éste mayor que la de uno simple. Electronegatividad y polaridad de enlace La electronegatividad se define como la capacidad de un átomo en una molécula para atraer los electrones de enlace hacia su núcleo. 1.- Considerando una molécula de cloro, ambos átomos atraerán con igual fuerza el par electrónico de enlace, ya que ambos tienen la misma electronegatividad. En este caso, en que el par electrónico está igualmente compartido por ambos átomos, tenemos un enlace covalente o covalente puro. 2.- Si cambiamos un átomo de cloro por un átomo de hidrógeno, por ejemplo, el átomo de cloro atraerá con más fuerza el par electrónico de enlace, ya que es más electronegativo que el hidrógeno, y por lo tanto polariza el enlace. En este caso tenemos un enlace covalente polar, donde “δ de carga” corresponde a una separación de carga que es inferior a la carga de un electrón. 3.- Si ahora cambiamos el átomo de hidrógeno, por un átomo como el sodio, por ejemplo, que tenga una gran diferencia de electronegatividad con el átomo de cloro, la polarización del enlace se hace tan grande que ambos átomos se ionizan. El átomo de sodio cede un electrón al átomo de cloro, quedando cargado positivamente y el átomo de cloro, al captar el electrón, queda cargado negativamente. En este caso tenemos un enlace iónico y, en consecuencia, se trata de un compuesto iónico. Nota: Volviendo al caso N°2 anterior, la molécula de HCl presenta una separación de carga que da como resultado una polarización de enlace: como en este caso se trata de una molécula diatómica, si el enlace es polar, la molécula también es polar. Para conocer la magnitud de esta polaridad se determina experimentalmente el momento dipolar de la molécula. El momento dipolar es una magnitud vectorial (con dirección y sentido), que corresponde al producto de la carga por la distancia, que separa las cargas iguales y de signos opuestos. Interacciones del tipo dipolo-dipolo

64
Las moléculas covalentes polares presentan interacciones de tipo permanente dipolo-dipolo, debido a la atracción de la carga positiva: δ+ del átomo de una molécula con respecto a la carga δ- del átomo de otra molécula. Las energías de interacción promedio dipolo-dipolo son aproximadamente de 4 kJ por mol de enlaces, en contraste con valores promedio de energías típicas de enlaces iónicos y covalentes de aproximadamente 400 kJ por mol de enlaces. Todas las interacciones permanente dipolo-dipolo, incluyendo los puentes de hidrógeno, son algo direccionales. Un aumento de la temperatura produce un aumento de las energía traslacionales, rotacionales y vibracionales de las moléculas. Esto produce mayor aleatoriedad en la orientación de las moléculas. En consecuencia, las fuerzas de las interacciones dipolo-dipolo disminuyen a medida que la temperatura aumenta.
Puentes de Hidrógeno El puente de hidrógeno es un tipo de enlace que se establece entre moléculas capaces de generar cargas parciales. No son realmente enlaces químicos en el sentido estricto, son casos muy particulares de interacciones dipolo-dipolo. Se presentan entre moléculas covalentes polares que contienen hidrógeno y algún otro elemento pequeño altamente electronegativo, como el F, O, o N. Así como ocurre con las interacciones ordinarias dipolo-dipolo, los puentes de hidrógeno resultan de la atracción entre la carga δ+ en los átomos de una molécula, en este caso los átomos de hidrógeno, y la carga δ- de los átomos de la otra molécula. El ejemplo recurrente en este tipo de información es la molécula de agua donde los electrones que intervienen en sus enlaces, se encuentran más cercanos al átomo de oxígeno que a los átomos de hidrógeno. Por ello se forman dos cargas parciales positivas en donde es posible encontrar los átomos de hidrógeno y dos cargas parciales negativas en las cercanías del oxígeno. Estas cargas parciales en ambos lados lleva a describir entonces que la molécula de agua funcione como “imán”. En tal forma una molécula de agua puede unirse a otras 4 moléculas de agua por medio de 4 puentes de hidrógeno.
colineales
Fig. Encontrar más detalles de la figura en http://bq.unam.mx/~evazquez FUERZAS DE LONDON

65
También son conocidas como dipolo instantáneo-dipolo inducido: Son fuerzas atractivas intermoleculares, muy débiles que sólo son importantes a distancias muy cortas. También son conocidas como fuerzas de Van der Waals. Las fuerza de London resultan de la atracción de un átomo cuyo núcleo está cargado positivamente, por la nube electrónica de un átomo de las moléculas vecinas. Esto induce dipolos temporales en las moléculas, porque a medida que las nubes electrónicas se hacen más grandes y más difusas son menos atraídas por su propio núcleo cargado positivamente. Así, de algún modo, se ven distorsionadas o polarizadas por núcleos adyacentes. La polarización aumenta con el número de electrones y con el tamaño de la moléculas. Estas fuerzas de London existen en todas las substancias.
Interacciones no covalentes
Las moléculas y los iones pueden interactuar de forma no covalente. Fundamentalmente, las interacciones son de naturaleza electrostática, es decir, dependen de las fuerzas que las cargas eléctricas ejercen entre ellas. Interacciones carga-carga: Es la interacción electrostática entre un par de partículas cargadas. Muchas de las moléculas que se encuentran en las células, entre ellas las macromoléculas como el ADN y las proteínas, tienen una carga eléctrica neta. Además de estas moléculas, las célula contiene abundantes iones pequeños, tanto como cationes Na+ , K+ ,y Mg++ , y aniones como Cl- y HPO4
-2 . Todo este tipo de substancias con carga ejercen fuerza sobre las demás. Las fuerzas entre un par de cargas q1 y q2 , separadas en el vacío por una distancia r, viene determinada por la ley de Coulomb.
En la ecuación anterior k representa una constante cuyo valor depende de las unidades utilizadas. Si q1 y q2 tienen el mismo signo F es positiva, de modo que un valor positivo significa repulsión. Si una carga es + y la otra e -, F es negativa, lo que representa atracción. Debemos considerar, sin embargo, que el entorno biológico no es el vacío. En una célula, las cargas siempre están separadas por agua u otras moléculas. Se forma así un medio dieléctrico entre las cargas. Este fenómeno muestra el efecto de apantallar las cargas, de modo que la fuerza real es siempre inferior a los resultados que muestra la ecuación. Este efecto apantallador se expresa añadiendo a la ecuación un número sin dimensión llamado la constante dieléctrica ε. Toda substancia que actúe como un medio dieléctrico tiene un valor característico de ε. Cuanto más elevado es este valor, más débil es la fuerza entre las cargas separadas. La
r2
q1 q2 F=
K
r2 ε

66
constante dieléctrica del agua es elevada, aproximadamente 80, mientras que las sustancias orgánicas generalmente tienen valores muy inferiores, entre 1 y 10. La consecuencia es que las partículas cargadas interaccionan débilmente entre ellas en un medio acuoso a menos que estén muy cerca.

67
CARBOHIDRATOS Los carbohidratos son los azúcares simples, sus polímeros y otros derivados azúcar. Se encuentran ampliamente distribuidos en la naturaleza y en términos de masa son las moléculas orgánicas más abundantes. Provienen como resultado de la fotosíntesis- proceso mediante el cual los vegetales convierten el CO2 atmosférico a la forma de carbohidratos. Desempeñan variados roles dentro de la célula viva. En este rol se destaca el aspecto reducido de sus moléculas (del proceso reducción) a partir del cual, por medio de procesos oxidativos entregan la energía necesaria para el metabolismo. Por ello, se dice que son un reservorio de energía. A mayor complejidad estructural aumenta la diversidad de funciones, encontrándose en las paredes celulares y en recubrimientos protectores de muchos organismos. Cuando forman parte de las membranas celulares contribuyen a la identidad celular. También se pueden encontrar como derivados en un número indeterminado de moléculas biológicas, por ejemplo, ácidos nucleicos, antibióticos, coenzimas. Conocidos como sacáridos se pueden clasificar atendiendo al número de unidades monoméricas que contienen. Monosacáridos: son las unidades fundamentales de la estructura de los carbohidratos. Los oligosacáridos son polímeros de entre 2 y 20 residuos de monosacáridos. De estos tenemos los más conocidos, los disacáridos con dos monosacáridos enlazados y los polisacáridos que contienen por lo regular, más de 20 residuos de monosacáridos. En general, los monosacáridos y los disacáridos puros son sólidos cristalinos, blancos, solubles en agua y tienen un saber dulce. Los ejemplos, glucosa y la fructosa (monosacáridos) y la sacarosa y la lactosa (disacáridos).

68
CLASIFICACIÓN DE MONOSACÁRIDOS 1.- Se pueden clasificar como aldosas y cetosas: todos los monosacáridos contienen al menos tres átomos de carbono. Uno de éstos es un carbono carbonílico (es decir, el monosacárido es un aldehído o una cetona), y cada uno de los átomos de carbono restantes lleva un grupo hidroxilo (-OH). Las dos clases generales de monosacáridos son aldosas y cetosas. En las aldosas el carbono con mayor estado de oxidación, se designa como C1, es aldehídico. En las cetosas, el carbono más oxidado, usualmente es C2, es cetónico. 2.- Los monosacáridos más sencillos son triosas, o azúcares de tres carbonos. Como puede observarse en la figura el gliceraldehído es quiral- tiene un centro de simetría en su carbono central C2. En otras palabras, este átomo de carbono tiene 4 sustituyentes diferentes. En consecuencia el gliceraldehído tiene dos estereoisómeros del tipo llamado enantiómeros. La dihidroxiacetona es aquiral- no tiene centro de simetría. Todos los otros monosacáridos se pueden ver como versiones de cadena más larga de estos dos azúcares. Todos los otros monosacáridos son quirales. Los enantiómeros D y L, es decir, moléculas quirales, son activas ópticamente. Al igual que los aminoácidos, en los organismos vivos domina una forma enantiomérica de los monosacáridos. En las proteínas se encuentran los aminoácidos L; en los hidratos de carbono se trata de los monosacáridos D. El gliceraldehído y la dihidroxiacetona poseen la misma composición atómica. Son tautómeros (isómeros estructurales que difieren en la disposición de sus hidrógenos y dobles enlaces) y pueden interconvertirse.

69
ESTRUCTURAS DE ANILLO
Una característica notable respecto de las pentosas y hexosas. La presencia de cinco o seis carbonos en la cadena proporciona a estos compuestos la posibilidad de formar estructuras de anillo muy estables mediante la formación de un hemiacetal interno. En la formación de anillo hemiacetálico en una aldopentosa, como la D-ribosa. Existen dos alternativas. La reacción del oxígeno del C1 de la D- ribosa con el hidroxilo del carbono C-4 produce una estructura de anillo de 5 eslabones denominada furanosa, el nombre refleja su semejanza estructural con el compuesto heterocíclico furano. Otra posibilidad es la formación de un anillo de seis eslabones si la reacción se produce con el hidroxilo del C5. Este anillo de seis eslabones se denomina piranosa, semejante con el compuesto heterocíclico pirano.
Fig. Las pentosas formación de estructuras cíclicas
En condiciones fisiológicas- en disolución, los monosacáridos con cinco o más carbonos se encuentran habitualmente en más del 99% en las formas de anillo. La distribución entre las formas piranosa y furanosa depende de la estructura concreta del azúcar, del pH, de la composición del disolvente y de la temperatura. También influye en la forma cuando se establecen como polímeros. En polisacáridos se estabilizan formas específicas. Por ejemplo, el ARN contiene exclusivamente ribofuranosa, mientras que los polisacáridos de las paredes celulares de algunas plantas tienen pentosas únicamente en la forma piranosa.

70
Debemos atender que la ciclación ha creado un nuevo centro asimétrico en el carbono 1. Observe la figura anterior, en ella se ha dibujado dos estereoisómeros de la D- ribofuranosa,
a los que se denomina αααα-D-ribofuranosa y ββββ-D ribofuranosa, observe además el par correspondiente de ribopiranosas.
Así como ocurre con otros tipos de estereoisómeros, estas formas αααα y ββββ rotan el plano de la luz polarizada de forma diferente, y pueden diferenciarse de esta forma. Estos isómeros, que se diferencian en la configuración tan sólo del carbono 1, se denominan anómeros, y al carbono 1 se le denomina átomo de carbono anomérico. Los monosacáridos pueden
experimentar una interconversión entre las formas αααα y ββββ, utilizando como intermediario la estructura de cadena abierta. Este proceso se denomina mutarrotación Las estructuras cíclicas tal como hemos visto representadas se denominan proyecciones de Haworth. Con imaginación puede usted observar el anillo en perspectiva, los grupos unidos a los carbonos del anillo (H, OH, CH2OH) se representan situados por encima o por debajo del anillo. En todos los monosacáridos D el –CH2OH está por encima del anillo.
La relación entre ambas conformaciones es tal que lo que se representa a la derecha de la cadena de carbonos en proyección de Fischer, en la de Haworth aparece por debajo del anillo, y lo que en Fischer aparece a la izquierda, en la de Haworth aparece hacia arriba. Una excepción a esta regla es el sustituyente en el último carbono asimétrico. En este caso, el grupo -CH2OH queda por encima del anillo en proyección de Haworth. En el carbono
anomérico, el OH en la forma αααα se representa hacia abajo, y el OH en forma ββββ hacia arriba.

71
En este esquema puede apreciarse como se cierra la molécula de un monosacárido, en este caso una hexosa. El grupo carbonilo del C1 queda próximo al C5 y entre ellos reaccionan sus radicales en una reacción intramolecular entre un grupo aldehido (el del C1) y un grupo alcohol (el del C5), formándose un hemiacetal. Ambos carbonos quedarán unidos mediante un átomo de oxígeno. El C1 se denomina Carbono anomérico y posee un grupo -OH llamado hemiacetálico y según la posición de este grupo, se originan dos anómeros (alfa y beta).
DISACÁRIDOS
Los disacáridos están formados por la unión de dos monosacáridos, que se realiza de dos formas:
1. Mediante enlace monocarbonílico, entre el C1 anomérico de un monosacárido y un C no anomérico de otro monosacárido, como se ve en las fórmulas de la lactosa y maltosa. Estos disacáridos conservan el carácter reductor.
La enzima se denomina lactasa, β-D-galactosidasa, es sintetizada si al menos uno de los dos genes que la codifican están presentes. Sí la expresión de ambos genes está afectada se reduce la síntesis de la enzima lactasa, lo cual a su vez reduce la digestión de lactosa. La producción de la enzima es por acción de un gen autosómico dominante y por lo tanto la intolerancia fisiológica a la lactosa es una mutación autosómica recesiva.
La lactosa está formada por unión de una molécula de glucosa y otra de galactosa. Se trata de una ß-galactopiranosa y una ß-glucopiranosa unida por los carbonos 1 y 4 respectivamente. Al formarse el enlace entre los dos monosacáridos se desprende una molécula de agua. Además, este compuesto posee el hidroxilo hemiacetálico, por lo que da positivo la reacción de Benedict.
Intolerancia a la lactosa es una incapacidad de la mucosa intestinal para producir la enzima , la consecuencia es una imposibilidad de digerir la lactosa de la leche. La incapacidad puede ser congénita o adquirida.

72
La maltosa o azúcar de malta es un disacárido formado por dos glucosas unidas por un enlace glucosídico producido entre el oxigeno del primer carbón anomérico (proveniente de -OH) de una glucosa y el oxígeno perteneciente al cuarto carbón de la otra. Al establecerse el enlace se desprende una molécula de agua y ambas moléculas de glucosa quedan unidas mediante un oxígeno monocarbonílico que actúa como puente entre las dos moléculas.
La maltosa presenta en su estructura el OH hemiacetálico que le otorga la propiedad de comportase como azúcar reductor Resulta positivos a Fehling y reacción de Benedict. A la maltosa se le llama también azúcar de malta, ya que aparece en los granos de cebada germinada. Se puede obtener mediante la hidrólisis del almidón y glucógeno.
2. Mediante enlace dicarbonílico, si se establece entre los dos carbonos anomérico de los dos monosacáridos, con lo que el disacárido pierde su poder reductor, por ejemplo como ocurre en la sacarosa
Sacarosa: Es el azúcar común es un disacárido formado por alfa-glucopiranosa y beta-fructofuranosa.
El disacárido contiene 2 átomos de carbono anomérico libres y su unión mediante un enlace O-glucosídico, favorece a la sacarosa como un azúcar no reductor.
El enlace que une los dos monosacáridos es de tipo O-glucosídico. Además, dicho enlace es dicarbonílico ya que son los dos carbonos reductores de ambos monosacáridos los que forman el enlace.
La enzima que puede hidrolizar el enlace es la sacarasa, también conocida como invertasa ya que la sacarosa es llamada también azúcar invertido.
POLISACÁRIDOS:
Los polisacáridos están formados por la unión de muchos moléculas de monosacáridos (puede variar entre 11 y varios miles), mediante enlace O-glucosídico, similar al visto en disacáridos, con pérdida de una molécula de agua por cada enlace. Tienen pesos moleculares muy elevados, no poseen poder reductor y pueden desempeñar funciones de reserva energética o función estructural. Los polisacáridos que tienen función de reserva energética presentan enlace α-glucosídico y son:
1. Almidón: Es el polisacárido de reserva propio de los vegetales, y está integrado por
dos tipos de polímeros:

73
� Amilosa, formada por unidades de maltosa, unidas mediante enlaces α (1-4). Presenta estructura helicoidal.
� Amilopectina, formada también por unidades de maltosas unidas mediante enlaces α (1-4), con ramificaciones en posición α (1-6).
AMILOSA
AMILOPECTINA:
2. Glucógeno: Es el polisacárido propio de los animales. Se encuentra
abundantemente en el hígado y en los músculos. Molécula muy similar a la amilopectina; pero con mayor abundancia de ramificaciones. Como usted puede observar, se trata de un polímero ramificado con sólo un tipo de monómero, la glucosa. La mayoría de las unidades de azúcar de una molécula de glucógeno está unida con otra por medio de enlaces del tipo α (1 4) glucosídico. En los puntos de ramificación un azúcar se une a tres monómeros. El tercero se encuentra unido por enlace (1 6).
Esta macromolécula es un reservorio o almacén de energía química. Por ejemplo, el músculo esquelético humano en condiciones normales contiene glucógeno suficiente para uso combustible para alrededor de unos 30 minutos de actividad moderada. La gran molécula con un peso molecular de entre 1 a 4 millones de daltones, al almacenarse en las células se concentra y puede ser observado mediante

74
microscopía electrónica, las fotografías lo muestran como gránulos irregulares oscuros (Debido a la tinción).
La estructura del glucógeno es parecida a la estructura de la amilopectina, en cuanto a las ramificaciones, enlaces (1 4) y (1 6).
3. Celulosa: Posee una función principalmente estructural, de sostén, constituye la pared
celular de la célula vegetal. Esta pared constituye una cubierta en el que queda encerrada la célula, que persiste tras la muerte de ésta. Por ejemplo, el algodón y el lino son fibras de celulosa y constituyen el principal componente de las paredes celulares de los vegetales que originan estos productos. Las telas de algodón son muy durables por moléculas no ramificadas y largas que aparecen ordenadas formando cordones moleculares. Esta propiedad les permite ofrecer resistencia a fuerzas tensionales.
La celulosa está constituida por unidades de β-glucosa, y la peculiaridad del enlace β hace a la celulosa invulnerables al ataque por enzimas digestivas humanas, por ello, este polisacárido no tiene interés alimentario para el hombre. Las unidades de glucosa están unidas por enlaces β (1 4).

75
Celulosa
La estructura lineal o fibrosa, se establecen con puentes de hidrógeno entre los grupos hidroxilo de distintas cadenas de glucosa, ello conduce a que sean insolubles al agua.
Los animales no pueden utilizar la celulosa como fuente de energía, ya que no cuentan con la enzima necesaria para romper los enlaces β-1,4-glucosídicos, es decir, no es digerible por los animales; sin embargo, es importante incluirla en la dieta humana (fibra dietética) porque al mezclarse con las heces, facilita la digestión y defecación, así como previene los malos gases.
En el intestino de los rumiantes, de otros herbívoros y de termitas, poseen una enzima llamada celulasa que rompe el enlace β-1,4-glucosídico, con ello se rompe la estructura formando unidades de glucosa, forma disponible para la obtención de energía y el metabolismo en general.
4. QUITINA: Es un polímero no ramificado compuesto del azúcar N-acetilglucosamina, semejante a la glucosa, diferente debido a la presencia del grupo aminoacetilo que se une a una molécula de glucosa.

76
La quitina es una molécula que forma parte de la estructura de invertebrados, por ejemplo en el exosqueleto de los insectos, arañas, crustáceos. Poseen propiedades de resistencia, dureza y flexibilidad.
Los condroitinsulfatos son glicosaminoglicanos: Formados por restos
alternantes de ácido glucurónico y N-acetilgalactosamina. Ésta puede presentarse sulfatada en C4, dando lugar al condroitin-4-sulfato o bien en C6, lo que constituye el condroitin-6-sulfato. Son abundantes en la sustancia fundamental del cartílago. Otros glicosaminoglicanos son:
El Dermatan sulfato con una estructura análoga al condroitin-6-sulfato, excepto que contiene ácido L-idurónico en vez de ácido glucurónico; su disacárido constituyente es, por lo
tanto...L-idurónico ββββ-(1,3) N-acetilgalactosamina 6-sulfato ββββ-(1,4).
El Queratan sulfato, que es el único glicosaminoglicano que no contiene ácidos urónicos; su
estructura es D-Galactosa ββββ-(1,3) N-acetilgalactosamina 6-sulfato ββββ-(1,4). Todos ellos, en distintas proporciones, aparecen en la sustancia fundamental de tejidos de origen mesodérmico, constituyendo complejos con proteínas que reciben el nombre de Proteoglicanos.
ESTUDIO DE AZÚCARES REDUCTORES
Los monosacáridos y la mayoría de los disacáridos poseen poder reductor, que deben al grupo carbonilo que tienen en su molécula. Este carácter reductor puede ponerse de manifiesto por medio de una reacción redox llevada a cabo entre ellos y el sulfato de Cobre (II). Las soluciones de esta sal tienen color azul. Tras la reacción con la molécula reductora se forma óxido de Cobre (I) de color rojo. De este modo, el cambio de color indica que se ha producido la citada reacción y que, por lo tanto, el glúcido presente es reductor.
HIDRÓLISIS DE LA SACAROSA
La sacarosa es un disacárido que no posee carbonos anoméricos libres por lo que carece de poder reductor y la reacción con el licor de Fehling es negativa, tal y como ha quedado demostrado en el experimento 1. Sin embargo, en presencia de ClH y en caliente, la sacarosa se hidroliza, es decir, incorpora una molécula de agua y se descompone en los monosacáridos que la forman, glucosa y fructosa, que sí son reductores. La prueba de que se ha verificado la hidrólisis se realiza con el licor de Fehling y, si el resultado es positivo, aparecerá un precipitado rojo. Si el resultado es negativo, la hidrólisis no se ha realizado correctamente y si en el resultado final aparece una coloración verde en el tubo de ensayo se debe a una hidrólisis parcial de la sacarosa.

77
INVESTIGACIÓN DE POLISACÁRIDOS (ALMIDÓN)
El almidón es un polisacárido vegetal formado por dos componentes: la amilosa y la amilopectina. La primera se colorea de azul en presencia de yodo debido a la adsorción o fijación de yodo en la superficie de la molécula de amilosa, lo cual sólo ocurre en frío. Como reactivo se usa una solución denominada lugol que contiene yodo y yoduro potásico. Como los polisacáridos no tienen poder reductor, la reacción con Fehling como prueba resulta negativa.
En las actividades de laboratorio usted deberá ensayar métodos y técnicas de análisis de hidratos de carbono. Junto a su equipo o grupo, revise todo este material antes de ingresar al laboratorio. La revisión implica que debe aprender y tomar conocimiento de las estructuras moleculares, pues estás forman parte de las células. Al tomar conocimiento usted estará contribuyendo a su propia formación para la adquisición de capacidades y habilidades que le permitirán finalmente alcanzar las competencias que están dentro del perfil de su carrera.

78
LÍPIDOS ESTRUCTURA Y FUNCIONES Los lípidos poseen múltiples funciones. Las grasas, por ejemplo, se utilizan para almacenamiento de energía y aislamiento térmico. En general, los lípidos son ocupados para formar parte de las células, en especial, el sistema de membranas. La estructura especial de los lípidos le permite a la célula constituirse en organismo seleccionador de sustancias de intercambio entre el exterior e interior de la célula. La propiedad se reconoce como permeabilidad selectiva de la membrana. Las propiedades de los lípidos y sus constituyentes permiten contribuir a que en primer lugar la membrana celular establezca una diferencia entre lo exterior a la membrana y lo que se encuentra delimitado al interior de la célula. Las complejas interacciones lipídicas permiten las interacciones con diversas proteínas y glúcidos, que en segundo lugar permiten se establezca la interacción de la célula con el medio. Al contrario de las proteínas, ácidos nucleicos y polisacáridos, los lípidos no son polímeros, son moléculas pequeñas que presentan gran tendencia a asociarse mediante fuerzas no covalentes. Por lo tanto no debemos considerarlas como macromoléculas.
Los lípidos son moléculas orgánicas y como tal de origen biológico, compuestas principalmente por carbono e hidrógeno y oxígeno, son hidrofóbicas (aversión por el agua), insolubles en agua y solubles en disolventes orgánicos como la bencina, el alcohol, el benceno y el cloroformo. A los lípidos se les llama incorrectamente grasas, pero en realidad son un tipo de lípidos procedentes de animales. Los aceites son otro tipo de lípidos que se distinguen de los primeros por ser líquidos a temperatura ambiente. Los lípidos cumplen funciones diversas en los organismos vivientes, entre ellas la de reserva energética, estructural (formación de endomembranas) y funciones reguladoras (esteroides).
Estructuralmente los lípidos están formados por cadenas alifáticas saturadas o insaturadas, en general lineales, pero algunos tienen anillos (aromáticos).Las cadenas corresponden a ácidos grasos.
Fig: Ver más en http://sapiens.ya.com/bio_ic_barca/ai_LIPIDOS/ae_ac_gra_pleg.jpg

79
Los lípidos en su mayoría son de carácter polar, y una parte apolar hidrofóbica que ya se ha mencionado antes. Esto mismo significa que no es soluble en solventes polares como el agua.
La región polar del lípido o región hidrofílica tiene afinidad por el agua debido a la capacidad de formar puentes de hidrógeno). Cuando una molécula tiene una región hidrófoba y otra hidrófila es llamada anfipática. La región hidrófoba presenta átomos de carbono unidos a átomos de hidrógeno, como la larga "cola" alifática de los ácidos grasos. La zona hidrófila posee grupos polares o con cargas eléctricas, como el hidroxilo (–OH) del colesterol, el carboxilo (–COO–) de los ácidos grasos, también tenemos el grupo fosfato (PO4-) que forma los llamados fosfolípidos y que adquieren importancia en el funcionamiento de las membranas biológicas.
En la figura se observa la estructura de un fosfolípido con las siguientes partes: dos moléculas de ácido grasos, una molécula de glicerol y una de ácido fosfórico. La zonas señaladas como apolar y polar, son las zonas hidrófobas e hidrofílicas respectivamente.
Sin duda que para mucha personas el concepto de grasa se relaciona con la obesidad, pero esta creencia no es exacta, al menos para el especialista que será usted. “Grasa” alude a un tipo de clasificación de los lípidos, y el término corresponde a un tipo de lípidos que se encuentra en forma sólida a temperatura ambiente. Al respecto, se hará referencia en relación con los alimentos, de esta forma se contextualiza encontrándose desde ya significado al conocimiento de las moléculas de tipo biológico.

80
El conocimiento de estas moléculas, así como también de otras, atendidas en otros capítulos, nos ayudan a comprender lo que es un ser biológico. Los seres vivos estamos compuestos de elementos químicos, los que son identificables en estructuras por lo general de tipo molecular, luego moléculas de mayor tamaño conocidas como macromoléculas, y en un nivel de complejidad mayor, la organización de los seres vivos, se constituye como célula. Esto es lo que somos, desde aquí, desde este ser biológico, surge también lo que comprendemos como ser humano, y por tanto, desde este conocimiento biológico se alimenta la comprensión del ser humano a sí mismo, lo que es, lo que será y lo que fue.
Hasta no hace mucho las grasas habían quedado prohibidas, sobredimensionadamente en el País del norte, se señaló las cantidades óptimas de consumo de alimento dentro de un esquema dietario. Las principales recomendaciones era que deberíamos disminuir el consumo de grasas y aceites. Con esto se conseguiría derrotar la aterosclerosis. Posteriormente esta opinión cambió (Taubes Gary. Science 30 March 2001; 291:2536-2545). Lo cierto es que el consumo de lípidos es necesario para asegurar un aporte de ácidos grasos sobre todo esenciales, estos últimos son llamados así, porque el organismo no lo puede formar. También son esenciales para el transporte de las vitaminas liposolubles, A, D, E, y K, necesarias para las reacciones metabólicas de las células.
Por tanto, el déficit de lípidos puede tener efectos devastadores en el organismo. Piense por ejemplo, que el cerebro está constituido en un 70% de lípidos, que como toda célula de nuestro organismo, su membrana está formada por bicapas lipídicas, las neuronas y sus prolongaciones con cubierta de mielina, etc.
Figura Membrana Visitar http://docencia.izt.uam.mx/acbc/images/celula/membrana_celular.png
En nuestra dieta diaria, no podemos prescindir de los lípidos, cualquier alimento que provenga de algún ser vivo, tendrá muchos millones de moléculas de lípidos, pues, son parte constituyentes de los organismos vivos, de principio, se encuentran en los sistemas de membrana celular.

81
Aparte de su rol estructural, entre sus funciones se encuentra el aportar energía, calorías. Los lípidos poseen alto valor energético, aproximadamente 9 kilocalorías, los hidratos de carbono aportan unas 4 kilocalorías, en ambos casos, considerando como unidad el gramo. Los valores por supuesto son aproximados dependiendo del tipo de constituyentes de los lípidos.
Grasas son las mantequillas, manteca, que son en realidad el tipo de moléculas llamadas triglicéridos. Estas moléculas se forman mediante combinaciones de moléculas de glicerol que son alcoholes (en sentido estricto) formados por tres átomos de carbono, asociados a átomos de hidrógeno y oxígeno. Al combinarse se manifiesta la función química conocida como grupo carboxilo.
Visitar:http://www.virtual.unal.edu.co/cursos/ciencias/2000024/images/biomoleculas/glicerol.gif
Al ingerirse con los alimentos los triglicéridos, se hidrolizan en las primeras porciones del intestino delgado, liberándose los ácidos grasos que se absorben como tales por la pared intestinal para pasar al torrente sanguíneo.

82
Fig:Visitarhttp://images.google.cl/images?hl=es&source=hp&q=triglic%C3%A9ridos&um=1&ie=UTF-8&sa=N&tab=wi
Desde allí son transportados a los tejidos, donde se almacenan de nuevo como triglicéridos, constituyendo la principal reserva energética del organismo. Existen diferentes tipos de ácidos grasos, que se caracterizan ya sea por la longitud de su cadena, como por el grado de saturación de sus átomos de carbono. Así podemos clasificarlos como ácidos grasos de cadena corta, mediana o larga. Si la cadena está formada por 16 carbones o aún más, se denominan ácidos grasos de cadena larga. Si la cadena tiene entre 8 a 10 carbones, se llaman de cadena media y si tienen menos, se llaman de cadena corta.
Los lípidos en general pueden clasificarse como de cadenas saturadas e insaturadas. Las saturadas forman cadenas más rígidas, por el contrario, las insaturadas le confieren propiedades de mayor flexibilidad.
Para conocer sus propiedades en mayor profundidad se explicará de forma más sencilla comenzando por sus constituyentes básicos:
Ácidos grasos:
Los ácidos grasos son los lípidos más sencillos, y al mismo tiempo son los componentes estructurales de lípidos más complejos.

83
Los ácidos grasos constan de una cadena con un grupo carboxílico terminal, siendo la configuración más sencilla la de una cadena lineal completamente saturada con hidrógeno, de enlace simple C-C. En la figura se puede observar como la cadena lineal de carbono presente enlaces de tipo doble, lo cual indica que se trata de una molécula insaturada, nótese además que la curvatura de la cadena está indicando que hay mayor flexibilidad en ella.
La fórmula básica es CH3-(CH2)n-COOH.
La idea que condujo a los consejos de disminuir el consumo de las carnes rojas y aún de los productos lácteos era debida a la presencia de ácidos grasos saturados, se sostenía que estas contribuían a elevar el colesterol en la sangre, ello, posteriormente conducía a la formación de placas en las arterias y en consecuencia se produce un incremento de enfermedades cardiovasculares y coronarias. Sin embargo, esta secuencia de reacciones a juicio de los expertos no ha sido probada, menos aún, que la privación de grasas saturadas en la dieta conduzca a prolongar la vida de las personas( Scientific American Enero 2003- citado por revista CRECES.2008- Dependiente del INTA). Más aún, los “franceses tienen baja incidencia de enfermedades cardiacas a pesar del elevado consumo de grasas saturadas. Lo interesante es que al parecer el mayor consumo de grasas insaturadas facilitaría la permeabilidad de las membranas, puesto que estas la incorporan a su estructura.
Como el problema se relaciona con la salud humana, podemos sostener que se trata de consumir adecuadamente con ciertas proporciones de grasas mono y poliinsaturadas lo que permitiría reducir las enfermedades coronarias. Eso es lo que al parecer sucede en la Isla de Creta, con la denominada dieta mediterránea, esta contiene aceite de oliva, rico en aceites monosaturados y pescado ricos en aceites poli-insaturados. En la misma revista de Science citada antes (2001) y documentado también en la revista CRECES por especialistas en nutrición humana, los datos parecen indicar que las personas con altos niveles de colesterol mueren prematuramente, pero no se ha probado que la causa se deba a un elevado consumo de grasas saturadas, por otra parte las dietas bajas en grasas saturadas casi no influyen en el colesterol sanguíneo.
Hay sustancias lipídicas realmente nocivas. Por ejemplo, los ácidos grasos insaturados de tipo “trans”, que se producen por hidrogenación parcial de los aceites vegetales y de

84
pescado, el proceso muestra que los aceites líquidos pueden convertirse de esta forma a grasas sólidas. Con el proceso se produce variación desde la forma “cis” en los vegetales y pescado hacia la forma “trans”. El consumo de ácidos grasos “trans” eleva los triglicéridos y las LDL (Del “colesterol malo”) en la sangre, se ha demostrado que estas producen daño al cerebro. Los “Trans” se encuentran en margarinas y mantecas.
En la figura: Los ácidos grasos insaturados son moléculas de ácidos grasos que contienen al menos un doble enlace. Según la estructura que tengan los dobles enlaces dentro de la molécula, pueden clasificarse como “cis” (forma curva) o “trans” (forma recta).
Los ácidos grasos de importancia para los humanos tienen configuraciones relativamente sencillas, aparecen comúnmente insaturaciones con hasta seis dobles enlaces por cadena, siendo casi siempre los enlaces, los ácidos grasos más comunes de los sistemas biológicos que tienen un número par de átomos de carbono.
Dentro del estudio de la neurociencias es necesario relacionar el conocimiento de estas estructuras con conocimientos de sucesos que ocurren en el ser humano. Así por ejemplo, se sabe que existe relación entre el consumo de determinados ácidos grasos y las depresiones, defectos atencionales, cuadros clínicos cerebrales diversos, desarrollo cerebral.
Ciertos estudios indican que durante el desarrollo fetal hay un aumento de los ácidos grasos poliinsaturados de cadena larga de los tipos w6 y w3. EL aumento corre transporte placentario. Por su parte la leche humana contiene ácido araquidónico, docosaheanoenoico, dihomo y linolénico (Revista de nutrición Hospital materno infantil). Esto una vez más indica cuán importante es el consumo de leche materna por los lactantes.
ACTIVIDAD EN HORAS NO PRESENCIALES:
1.- Esquematice las estructuras de ácidos grasos, por lo menos con los primeros seis de la
primera serie de saturados. Coloque el nombre correspondiente al lado de cada estructura.
Luego esquematice para las estructuras de los insaturados.
2.- Desarrolle una lista con las propiedades de los ácidos grasos e indique la importancia de
cada una de ellas en relación con la célula y con los organismos en general.

85
3. ¿Por qué no es conveniente consumir leche descremada en personas que consumen dietas
balanceadas y equilibradas?
4.- ¿Cuáles son las funciones que cumplen los lípidos en un organismo cualquiera?
5.- ¿Cuáles son los beneficios a nivel celular del consumo de aceite de pescado? Fundamente
su respuesta.
6.- Para ejercitar como técnica de estudio, construya un mapa de trayectoria de los lípidos
hasta el consumo celular.
La mayor parte de los ácidos grasos del cuerpo humano tienen 16, 18 ó 20 átomos de carbono, si bien, hay varios con cadenas más largas que se encuentran principalmente en los lípidos del sistema nervioso. Si la cadena de carbonos tiene menos de 8 carbones, de 8 a 10 se llaman ácidos grasos de cadena media y si tienen más de 16 son llamados de cadena larga.
Si la cadena tiene la presencia de un doble enlace es un ácido graso monoinsaturado, si poseen doble enlace pasan a ser clasificados como poliinsaturados. Los monosaturados se encuentran en aceites de maní y la grasa de cerdo. Los poliinsaturados se encuentran en aceites vegetales y de pescado. Los ácidos grasos poliinsaturados son conocidos como PUFA del inglés “Poly Unsatured Fatty Acid”. Si poseen más de cinco dobles enlaces reciben el nombre de HUFA del inglés “Highly Unsatured Fatty” Acids”.
Hay estudios que indican que la dieta con PUFA y HUFA contribuyen a disminuir el colesterol sanguíneo. En los países desarrollados se ha producido en los últimos años una disminución de los accidentes vasculares de tipo cardiaco y se atribuye al aumento del consumo de los ácidos grasos poliinsaturados, oleico y linoleico.
Los dobles enlaces le otorgan a los ácidos grasos características especiales. Por ejemplo, la temperatura de fusión depende de la presencia de las dobles ligaduras. El ácido esteárico, formado por 18 carbonos sin doble enlace tiene una temperatura de fusión de 69ºC. El ácido oleico de 18 carbonos con un doble enlace tiene un punto de fusión a 10ºC. El ácido linoleico, con 18 carbonos, con dos dobles enlaces, se fusiona a -5ºC. El ácido linolénico, 18 átomos de carbono, con tres dobles enlaces, se fusiona a -11ºC.

86
La presencia de dobles enlaces tiene también consecuencias no deseables, algunas de ellas es que son vulnerables a experimentar reacciones de oxidación en presencia de oxígeno del aire, o a nivel celular por la presencia de oxígeno proveniente de la respiración. En estos procesos se producen radicales libres nocivos para la célula.
Ácidos grasos esenciales
Son aquellos que el organismo no puede sintetizar, son del tipo de los insaturados, se trata de los ácidos linoleico y araquidónico, que deben ser aportados por la dieta. Su ausencia produce enfermedades y su ausencia en los animales se debe a que se carece de las enzimas necesarias para colocar el doble enlace en una determinada posición.
Usando las reglas u nomenclatura: EL carbón de la cadena en que está el doble enlace, se cuenta desde izquierda a derecha, se coloca la letra w (Omega) junto al primer carbón del doble enlace.
El primer doble enlace caracteriza diferentes familias de ácidos grasos, así tenemos: Familia Omega-9, familia Omega-6 y familia Omega-3. El organismo puede sintetizar la familia Omega-9, pero no Omega-6 u Omega-3, entre ellos el linoleico, araquidónico y linoleico. Los vegetales pueden sintetizar Omega-6 y las algas marinas microscópicas pueden sintetizar Omega-3. Sumando, los Omega 6 y 3 provienen del reino vegetal, y al encontrarse en tejidos animales podemos deducir que estos han consumido vegetales que los tienen incorporados a sus tejidos. Por ejemplo, los peces acumulan Omega 6 y 3 mediante el consumo de plancton marino. Examine las fórmulas más adelante.
Una de las principales
funciones es formar las
estructuras en bicapas como
se da en las membranas
biológicas. En estas bicapas
se insertan proteínas con
diversas funciones, entre
ellas la selección de señales
o presencia de nutrientes. En
estos procesos adquiere
relevancia la presencia de
dobles enlaces en los ácidos
grasos, y de la actividad de
estos mismos compuestos en
el metabolismo celular.

87
Carencia de ácidos grasos:
De Omega 6: linoleico y araquidónico su deficiencia en ratas produce alteraciones de la piel, pérdida de pelo, aumento del metabolismo, freno del crecimiento, hasta la muerte del animal. Los síntomas se revierten si se agrega a la dieta el ácido linoleico.
Los ácidos grasos de la familia Omega-3 se encuentran en las células formando parte de otros compuestos, como la fosfatidil etanolamina y la fosfatidil serina. Dichos compuestos se encuentran en la retina, corteza cerebral, testículos, etc. Se cree que este ácido graso contribuye a la fluidez y como tal, determinadas funciones de la membrana. Por lo mismo, podría estar influenciando la acción de enzimas de membranas y receptores.
Datos de investigaciones en ratas muestran que estos ácidos grasos Omega-3, se encuentran en altas concentraciones en las vesículas sinápticas de la corteza.
Omega 3:
El ácido eicosapentanóico (EPA, o también ácido icosapentaenoico) es un ácido graso poliinsaturado esencial de la serie omega 3 (ω-3). Se utiliza en clínica como fármaco para el tratamiento de algunas formas de hiperlipidemias.

88
Es un tipo de la forma 20:5(n-3), por tener una cadena de 20 carbonos (un eicosanoides) con cinco enlaces dobles cis a partir del carbono número 3. Químicamente es un ácido carboxílico.
Es precursor de la prostaglandina-3 (un agregador plaquetario), del tromboxano-3 y el leucotrieno-5 (eicosanoides todos).
Se le puede consumir en dietas de pescados: aceite de hígado de bacalao, arenques, salmón, sardinas, etc. También se encuentra en la leche materna.
Un estudio publicado en la revista Journal of American Medical Association del 2003, y realizado sobre una población de 43.671 hombres, un grupo de la Harvard Medical School se propuso comprobar si era cierta la hipótesis de que los ácidos grasos omega-3 de cadena larga contenidos en el pescado graso de la dieta, reducen el riesgo del accidente vascular cerebral isquémico, aunque no así el del accidente vascular cerebral hemorrágico. Los estudios mostraron, que la población de raza esquimal del norte de Groenlandia, un elevado consumo de pescado graso en sus dietas reducía el riesgo de sufrir un accidente vascular cerebral isquémico, pero aumentaba el riesgo de padecer un accidente cerebral hemorrágico. Esta fue la hipótesis de trabajo del equipo.
La explicación dada para el fenómeno sería que los ácidos grasos omega-3 contenidos en el pescado, sobre todo graso (ácido eicosapentaenoico o EPA y ácido docosahexaenoico o DHA- ambos del tipo omega 3) disminuyen la agregación progresiva de las plaquetas de la sangre, mediante la inhibición de la síntesis del tromboxano A2, ello retarda el proceso de coagulación in situ disminuyendo la tasa de formación de trombos sobre las placas de ateromas depositadas en la cubierta interna o endotelio de los vasos produciendo la obstrucción de las arterias y de la isquemia del territorio que irrigan; si esta inhibición llegara a ser excesiva se produciría, por el contrario, un accidente cerebral vascular por hemorragia.
En el estudio, un total de 43.671 varones, entre 40 y 75 años de edad, libres de enfermedad cardiovascular en el año 1986, quienes completaron un cuestionario detallado y semi cuantitativo sobre sus dietas alimentarias.
En esta población, y a lo largo del seguimiento, que ha durado 12 años, se documentaron 608 accidentes vasculares cerebrales, de los cuales 377 fueron isquémicos (por interrupción brusca del riesgo sanguíneo en un área cerebral), 106 hemorrágicos (por ruptura de una arteria y consiguiente hemorragia cerebral) y 125 no pudieron ser clasificados. El riesgo de padecer un accidente vascular cerebral isquémico fue un 40% más bajo en los individuos que habían consumido pescado graso (como atún, salmón, arenques, sardinas, pez espada y otros) al menos una vez al mes o más, comparado con el riesgo de aquéllos que no habían consumido pescado.
Por otra parte, no se encontró asociación entre el consumo de pescado rico en ácidos grasos omega-3 y el riesgo de padecer un accidente vascular cerebral hemorrágico.(Texto modificado desde www.saludlandia.com/pescado-graso-y-riesgo-de-accidente-vascular-cerebral-isquemico-12659.html)). Mayor información se puede ver en el Journal mencionado antes.

89
Ácido docosahexaenoico
En la figura anterior el El ácido docosahexaenoico (DHA) es un ácido graso esencial, como usted puede observar, es un ácido graso poliinsaturado de la serie omega-3, (DHA) Es un ácido graso vital para el desarrollo y mantenimiento óptimo de la salud. Científicos de la Universidad de California han encontrado que el consumo de este ácido detiene el deterioro que causa la enfermedad de Alzheimer.
Importancia de la estructura:
Es conveniente dar una breve mirada a la estructura de estos ácidos grasos, debe observarse la presencia de enlaces simples y dobles, la ubicación precisa de los carbonos que acceden a doble ligadura con otro carbono. También es importante conocer la longitud de la cadena. Lo esencial que puede ser algún tipo de sustancias nos está indicando que nuestro organismo no puede sintetizarla. Sintetizarla quiere decir en términos de metabolismo, construcción de alguna sustancia, esto se logra por la presencia de enzimas, que son proteínas y estas últimas necesitan de genes que deben estar presentes en la molécula de ADN.
En la ocasión de presenciar la estructura de los ácidos grasos, podemos claramente deducir que si no se encuentra en nuestro organismo, alguna molécula de ácido graso, con un doble enlace en alguna determinada posición, se debe a la ausencia de la enzima capaz de catalizar la formación del enlace, por tanto, también es deducible que no exista el gen necesario para producir esta enzima. La importancia radica es que la presencia o ausencia hoy en día pueden determinar en parte la salud de las personas, y tiene que ver con todo tipo de afecciones, incluidas especialmente las de tipo cerebral, que han sido las menos investigadas hasta hoy, si se les compara con las enfermedades de tipo vascular.
Si aún persisten las dudas consulte con su profesor al respecto.
Síntesis de ácidos grasos:
Se requieren enzimas adicionales para un alargamiento posterior de la cadena y la desaturación. La síntesis de una gran variedad de ácidos grasos requieren un conjunto de enzimas (Por tanto, un conjunto de genes). Estas enzimas son retenidas en el Retículo Endoplásmico y mitocondrias. Las células animales contienen varias desaturasas que son las enzimas que rescatan átomos de hidrógeno, catalizando por tanto, la formación de dobles enlaces. Esta desaturación en las células animales sólo llega hasta el carbono 9 contando desde el extremo carboxílico. Entonces debe comprenderse porque un ácido graso es indispensable o esencial en la dieta de los animales, incluido el hombre. Véase el siguiente ejemplo, el ácido linoleico de 18 átomos de C, 18:2 (9,12). Ver en figura siguiente.

90
El ácido linoleico es una molécula precursora (Qué da origen a….) del ácido araquidónico, que es una molécula que se forma en una serie de reacciones de desaturación y alargamiento. En realidad todos estos ácidos deben llamarse según el estado fisiológico en que se encuentran. De manera que en lugar de ácido araquidónico se puede decir araquidonato.
El araquidonato o ácido araquidónico que deriva de la estructura de fosfolípidos de membrana es precursor de eicosanoides, grupo de sustancias importantes en la fisiología y fisiopatología humana que usted estudiará en profundidad en otros módulos.
Como se ha dicho antes, los ácidos linoleico (C18:2 w6) y α-linolénico (C18:3w3) son esenciales para el ser humano porque no puede insertar dobles enlaces por falta de enzimas (D12 y D15 desaturasas) que pueden introducir dobles enlaces en las posiciones w6 y w3.
A partir de los ácidos linoleico, α- linolénico y linoleico mediante elongación de la cadena y desaturación se puede sintetizar ácidos poliinsaturados de cadena más larga, de 20 átomos de carbono.

91
En la figura siguiente se puede observar el ácido araquidónico que resulta de la síntesis partiendo desde ácido linoleico, luego a partir del araquidonato se producen prostaglandinas.
Con estímulos principalmente hormonales, una fosfolipasa específica presente en la mayoría de las células de los mamíferos actúa sobre los fosfolípidos de membrana liberando araquidonato. Las enzimas del retículo endoplásmico liso (REL) transforman el araquidonato en prostaglandinas.

92
En humanos, el precursor más importante del las PGs es el ácido araquidónico (ácido 5,8,11,14-eicosatetraenoico) C20 con 4 insaturaciones no conjugadas. Esta molécula se sintetiza a partir del ácido linoleico por desaturación y elongación.
Secuencia de reacciones de formación de prostaglandinas
larga con funciones en regulación metabólica. Existen dos clases de moléculas eicosanoides. Una clase deriva de la ciclación del araquidonato, catalizada por ciclooxigenasa, los productos resultantes son prostaglandinas, prostaciclinas y tromboxano, llamados reguladores locales, puesto que su lugar de acción es en células vecinas, no viajan por la sangre, ni se producen en glándulas.
Por ejemplo, las plaquetas sanguíneas producen tromboxano A2, que es una sustancia que puede provocar la contracción de la musculatura lisa de las paredes arteriales, y por tanto, cambia el flujo de sangre local. El útero produce prostaglandinas que provocan contracción muscular durante el parto. Los eicosanoides median la sensibilidad al dolor, inflamación y la hinchazón.
Araquidonato
Prostaglandina G2 (PGG2)
Prostaglandina H2(PGH2)
Prostaciclina
(PG1)
Prostaglandina E2
(PGE2)
Prostaglandina F 2α
Tromboxano A2
Ciclooxígenasa Leucotrieno A4
En estas figuras se observan las vías de reacciones bioquímicas para la formación de eicosanoides, la primera de estas reacciones es catalizada por la enzima ciclooxigenasa que conduce a la formación de prostaglandinas, cada una de estas reacciones es catalizada por enzimas diversas. En la vía que conduce a leucotrieno A4, un precursor de otros leucotrienos, algunos de estos desencadenas reacciones alérgicas. El ácido araquidónico se encuentra en la capa interna de la membrana plasmática, esterificado al C2 del esqueleto del glicerol de los fosfolípidos. La liberación del araquidonato desde la membrana es catalizada por la fosfolipasa A2. El araquidonato puede ser utilizado como precursor de moléculas de eicosanoides, que son un grupo de ácidos grasos insaturados de cadena

93
La prostaglandina PGE2 participa en la regulación de la presión sanguínea, inducción al parto y la inducción a la coagulación sanguínea.
Es interesante conocer algo de los fármacos, la aspirina puede bloquear algunos de los efectos, de estas sustancias, el ingrediente activo de la aspirina es el ácido acetilsalicílico, capaz de inactivar en forma irreversible a la ciclooxigenasa y por tanto evita la formación de eicosanoides.
El segundo grupo de eicosanoides son aquellos productos catalizados por lipooxigenasas, conducentes a los leucotrienos que participan de la respuesta alérgica.
En la figura siguiente la síntesis de ácido graso de 20 átomos de carbono y deshidrogenación, que deja establecidos los dobles enlaces.
Los tres ácidos precursores, ácido oleico (C18:1w9), el ácido linoleico: C18:2 w6 (arriba en la figura) y ácido α-linolénico C18:3 w3, compiten por la D6 desaturasa, convirtiéndose en una enzima limitante. La mayor afinidad siempre la posee el ácido α-linolénico, seguido del linoleico y luego el oleico.

94
Los ácidos grasos se encuentran principalmente en forma de ésteres del glicerol, como se muestra en la figura siguiente:
Los compuestos con un ácido graso esterificado (monoacilgliceroles) o dos (diacilgliceroles) se encuentran solamente en cantidades relativamente pequeñas y aparecen en gran medida como intermediarios metabólicos en la biosíntesis y degradación de lípidos que contienen glicerol. La mayor parte de los ácidos grasos del cuerpo humano existen en forma de triacilgliceroles en los que los tres grupos hidroxilo del glicerol están esterificados con un ácido graso. Históricamente a estos compuestos se les ha denominado grasas neutras o triglicéridos.
ACTIDAD NÚMERO 2.-
¿Cómo se forman estos compuestos?
2.1.- Rotule las estructuras e identifique las partes:
2.2.- Complete las ecuaciones a manera de ejercicio
2.3.- La ecuación que se observa ¿Qué está indicando?, ¿Dónde se produce esta reacción y para qué?

95
2.4.- Destalle como se forma el enlace entre las dos estructuras: Utilice la presentación power point respectiva dado en clases y que puede descargar de la intranet.
El enlace
Propiedades notables:
Una de las propiedades más prominentes y significativas de los ácidos grasos y de los triacilgliceroles es, ciertamente, su falta de afinidad por el agua.
Por ello se dice que es de carácter hidrofóbico resultando una propiedad esencial para la construcción de las complejas estructuras biológicas y para la separación del entorno acuoso.

96
También es esencial para el uso por moléculas biológicas de superficies activas, tal como sucede en el tracto intestinal.
La naturaleza hidrofóbica de los triacilgliceroles y su estado relativamente reducido los transforman en compuestos eficientes para el almacenamiento de energía:
La cantidad de ATP que se obtiene de la oxidación completa de los triacilgliceroles es dos veces y media superior a la correspondiente al glucógeno.
También los triacilgliceroles se pueden almacenar como lípido puro sin asociarse con el agua, mientras que el glucógeno es muy hidrofílico y cuando se almacena en los tejidos fija el doble de su peso de agua.
Una gran proporción de los ácidos grasos utilizados por el hombre se obtiene de la dieta. Diversos lípidos animales y vegetales son ingeridos, hidrolizados, al menos parcialmente por enzimas digestivos, y absorbidos a través de la pared intestinal para ser distribuidos a través del cuerpo, primero en el sistema linfático y después en el torrente circulatorio.
Los ácidos grasos son las unidades estructurales básicas de los lípidos. El organismo puede sintetizar ácidos grasos, pero, algunos deben ser obtenidos de la dieta y son denominados esenciales. Los dos ácidos grasos esenciales se denominan ácido linolénico (ácido graso Omega-3) y ácido linoleico (ácido graso Omega-6).
Los estudios actuales indican que si se incluye en la dieta más alimentos ricos en Omega -3 y reduciendo el consumo de carnes rojas, aceites hidrogenados y otras grasas saturadas contribuye a una buena salud. Se recomienda el consumo de pescado por su contenido en el ácido graso ante dicho.
Los ácidos grasos libres se almacenan como triglicéridos y estos son los lípidos que el organismo utiliza como fuente de energía. También son utilizados para funciones hormonales en regulación fisiológica y metabólica.
Ácidos Grasos y Sistema nervioso
Considérese el ácido linoléico de la familia omega-6 y el ácido linolénico de la familia Omega-3. Se sabe que su carencia en el organismo produce alteraciones metabólicas y en consecuencia se pueden desarrollar enfermedades. Estos ácidos grasos son importantes puesto que el linoleico da origen al ácido araquidónico (AA) y el linolénico al ácido docohexaenoico (DHA).
Los ácidos mencionados desarrollan importantes funciones metabólicas, para ello integran los fosfolípidos de la membrana celular. Dado que son insaturados se logra mayor fluidez de la membrana, debido a ello, las proteínas integrales de la membrana pueden lograr movilidad necesaria para ejercitar sus funciones de receptores de membrana, canales iónicos, inducción del desarrollo de formaciones vesiculares, etc.
En los procesos de formación del tejido nervioso, especialmente en el cerebro, la fluidez de las membranas adquiere gran relevancia en la formación de la estructura encefálica sobre todo en los últimos tres meses de embarazo y luego extendiéndose hasta los dos años

97
después del nacimiento. Los procesos morfogenéticos que han sido iniciados desde la cresta neural, se caracterizan por migración, neurogénesis, apoptosis, sinaptogénesis y procesos de mielinización. La morfogénesis requiere de concentraciones considerables de ácido araquidónico y docohexaenoico, ambos tipos de sustancias se acumulan en los conos de crecimiento axonal, en las vesículas sinápticas y parecen tener mucha relevancia en el movimiento de las vesículas acumuladoras de neurotransmisores.
Con respecto al DHA se le supone una tremenda importancia en el cerebro y su desarrollo dada su gran concentración. Las mediciones de las cantidades de lípidos en el cerebro muestran aproximadamente que un 65% del total de lípidos cerebrales son ácidos grasos poliinsaturados. Dentro de estos, el DHA y el AA que es un w6. El AA constituye el principal de la madre al feto, aunque este ácido es abundante en todos los tejidos, también no se le resta importancia en la construcción de células cerebrales.
El DHA se concentra principalmente en tejido nervioso, neuronas y glia, conos y bastones en la retina. Este ácido se concentra o acumula en la fosfatidilcolina, fosfatidilserina y fosfatidiletanolamina o permanece asociado a inositol, plasmalógenos (esfingolípidos). La etolamina, colina, serina e inositol se asocian al C1 del glicerol, El DHA se esterifica con el OH del C2, el C3 casi siempre se asocia con un ácido graso saturado.
El DHA, ácido de cadena larga adquiere una conformación semihelicoidal, los 6 dobles enlaces que otorgan fluidez con baja tendencia a formar estructuras cristalinas más densas. La mayor fluidez en la membrana permitiría facilitar el movimiento de moléculas tales como, receptores, proteínas G, canales iónicos, enzimas, factores de crecimiento neuronal (Sanhueza y colaboradores en Review, Revista Nutrición, Agosto 2004).
En estudios con sinaptosomas y mitocondrias neuronales muestran que presentan DHA en proporciones considerables. Los sinaptosomas son estructuras de los terminales presinápticos que se obtienen mediante fraccionamiento celular de tejido encefálicos.
Figura con fracción de sinaptosomas Revista de Neurologia 2001,32; 1031.1040
Las neuronas no tienen la capacidad de formar DHA a partir de su precursor el ácido alfa-linolénico. Se ha observado que las células gliales, en especial los astrocitos los que desaturan y elongan el alfa linolénico para convertirlo en DHA, el que posteriormente se

98
envía a las neuronas. La mayor fluidez de las membranas de la neurona se debería a la presencia de DHA en el tejido cerebral, esto tendría relación con el desarrollo de los conos de crecimiento del axón, el establecimiento de sinapsis, interacciones con dendritas. El DHA intervendría además en la inducción de genes para los procesos de apoptosis, neurogénesis. Se ha indicado por algunos investigadores (Véase la revisión de Sanhueza ya citada antes) que la suplementación con ácidos grasos omega-3, en especial con DHA produciría una estimulación de la termogénesis al desacoplar la oxidación mitocondrial y con la estimulación de oxidaciones peroxisomales de los ácidos grasos.
Diversos estudios manifiestan que la incorporación de DHA en el tejido cerebral (vía suplementos) produce mejores capacidades de aprendizaje y memorización en ratas, primates incluyendo el hombre. En los anteriores excepto el hombre hay correlación entre la incorporación de DHA en hipocampo y corteza frontal con mejor desempeño en pruebas de laberinto elevados con espacios ciegos, en laberinto de Morris o en caja de Skinner. En el hombre no ha sido posible aún dilucidar claramente el rol del DHA en relación con el aprendizaje y la memoria. Es más una recopilación de una treintena de estudios de observación de suplementación con DHA y AA, a lactantes de pretérmino y término, y en combinación con lactancia materna, no muestran resultados concluyentes con respecto al desempeño que requieren tareas de aprendizaje o evaluación de la inteligencia.
Propiedades en general
Los triacilglicéridos contienen una mezcla de ácidos grasos saturados e insaturados. Si las cadenas saturadas se colocan unas al lado de otras el resultado es una estructura semicristalina, más bien de tipo regular. La mezcla con cadenas insaturadas forma estructuras más flexibles que llegan a imponer los dobles enlaces de tipo cis.
Fuente: Mathews Christopher, Bioquímica

99
Figura arriba: Se observa una bicapa con moléculas de ácidos grasos saturados, el aspecto es ordenado, un estado semicristalino. Las estructuras formadas con este orden no son propias de los seres vivos.
Figura arriba: Se observa en esta bicapa lipídica la incorporación ácidos grasos de cadena con insaturaciones o dobles enlaces.
En la industria química se utiliza procesos de hidrogenación parcial de los aceites grasos insaturados como el aceite de maíz, la finalidad es que se transforman de esta manera en una grasa más sólida, luego pueden utilizarse como sustitutos de la mantequilla, por ejemplo, margarina.
En el laboratorio usted experimentará con ácidos grasos y triacilgliceroles. En el caso de los últimos, la esterificación de los ácidos grasos con glicerol reduce el carácter hidrófilo de los grupos de cabeza polares de los ácidos. Como consecuencia, los triacilgliceroles se convierten en formas insolubles en agua y no forman micelas en forma eficaz, por lo tanto, las grasas que se acumulan en las células forman pequeñas gotas aceitosas en el citoplasma. En los adipocitos, células que almacenan grasas, casi todo el volumen de la célula lo constituye una gota de grasa. Estas células adipocitos constituyen el tejido adiposo de los animales.

100
En la figura se puede observar las células del tejido adiposo humano.
En la industria química las grasas se transforman en jabones y detergentes. Esto se realiza mediante hidrólisis con álcalis como el NaOH o el KOH. El proceso se denomina saponificación. Los ácidos se liberan debido a la hidrólisis y forman sales sódicas o potásicas ionizadas. Por estas circunstancias, los ácidos precipitan en presencia con iones de Calcio o Magnesio, fáciles de encontrar en las aguas duras.
Esteroides
Los esteroides son un grupo de lípidos con estructura de lípidos diferentes a las que se ha visto antes. Todos los esteroides poseen cuatro anillos de carbono unido entre ellos, los que pueden presentar oxhidrilos o radicales. Entre los esteroides se encuentran:
Se encuentran en animales: ecdisteroides, andrógenos, estrógenos, progestágenos, corticosteroides, esteroides anabólicos, colesterol), en plantas: (fitosteroles, brasinosteroides) y hongos (ergosteroles). Los esteroides son lípidos insaponificables, su estructura química básica es la molécula formado con anillos del ciclopentano-perhidrofenantreno, que corresponde a cuatro anillos, tres de estos contienen seis átomos de carbono; y el cuarto sólo cinco.

101
La longitud, estructura y localización de las cadenas sustituyentes y de los dobles enlaces, define el tipo de esteroide. Entre ellos se pueden citar los esteroles, las sales biliares, las hormonas sexuales y otras hormonas como los mineralocorticoides y glucorticoides. Esas hormonas, en los crustáceos, insectos y vertebrados, están implicadas en procesos de regulación metabólica.
Entre las hormonas de la corteza suprarrenal, son importante los glucocorticoides (cortisol), por cuanto median las respuestas fisiológicas al estrés, favoreciendo el uso de ácidos grasos y proteínas para la obtención de energía por parte de todas las células, excepto las musculares y nerviosas que se beneficiarán de la energía obtenida directamente de las fuentes de glucosa (glucógeno), y mineralocorticoides (aldosterona).

102
Clasificación de los lípidos
Lípidos
Lípidos complejos o saponificables
Derivados de los ácidos grasos
Glicéridos
Acilglicéridos AG + Glicerina
Fosfoglicéridos AG + Glicerina + Fosfato + Sustancia polar
Glucoglicéridos AG + Glicerina + Monosacárido
Esfingolípidos
Esfingomielinas AG + Esfingosina +Fosfato + Sustancia polar
Glucoesfingolípidos AG + Esfingosina +Uno o varios monosacáridos
Céridos AG + Sustancia apolar
Lípidos simples
Terpenos
Esteroides
Prostaglandinas
Tabla: Clasificación de los lípidos
Colesterol
Es un componente especial que forma parte de las membranas de las células animales. Como observó en las figuras anteriores, esta sustancia pertenece al grupo de los esteroides. Además es la estructura básica desde la cual se sintetizan hormonas y entre ellas las hormonas sexuales y sales biliares. La esterificación de los ácidos grasos al grupo hidroxilo del C3 del colesterol forman los ésteres de colesterilo, los cuáles son más hidrofóbicos que el colesterol solo. Estos ésteres de colesterilo se encuentran en las lipoproteínas de la sangre y se forman cuando el colesterol va a ser almacenado dentro de las células.
En la Figura más abajo: Al estudiar las propiedades de este compuesto se puede describir como una molécula débilmente anfipática, debido al grupo hidroxilo en el carbono Nº 3. Por esta misma situación, el colesterol puede adquirir un carácter más hidrófobo, situación que surge a causa de la esterificación del grupo hidroxilo con un ácido graso.
El colesterol dada su estructura tiene un gran volumen y además es muy rígida. La molécula de colesterol al formar parte de la membrana tiende a alterar la estructura regular

103
de esta. Ello tiene como consecuencias, la alteración de la rigidez y permeabilidad de la membrana.
El colesterol y los triacilgliceroles de tipo no polar pueden ser transportados en la sangre solo si forman complejos con fosfolípidos y proteínas de tipo anfipáticas denominadas apoproteínas, con ello forman las partículas denominadas lipoproteínas. En el plasma humano se han identificado varias apoproteínas. Estas presentan centros hidrofóbicos y superficies hidrofílicas. El centro dada sus características contiene triacilgliceroles y ésteres de colesterilo, la superficie son moléculas anfipáticas, colesterol, fosfolípidos y apoproteínas.
Las lipoproteínas se clasifican de acuerdo a sus densidades relativas. Ya que los lípidos son menos densos que las proteínas, mientras mayor sea el contenido de lípidos en las lipoproteínas menor será la densidad.
En humanos las proteínas son los quilomicrones, que transportan triacilgliceroles y colesterol desde el intestino delgado a los tejidos, las lipoproteínas de muy baja densidad (VLDL), que transportan triacilgliceroles endógenos, colesterol y ésteres de colesterilo del hígado (órgano principal de síntesis de colesterol) a los tejidos; las lipoproteínas de baja densidad (LDL) que se forman durante la degradación de las VLDL, luego capturan más colesterol y ésteres de colesterilo; Por último tenemos las lipoproteínas de alta densidad (HDL), transportan colesterol endógeno y ésteres de colesterilo de vuelta al hígado.
Las partículas de LDL circulantes en el plasma sanguíneo llegan a los tejidos, excepto los de tipo hepático, luego allí, las células utilizan el colesterol en la síntesis de la membrana celular. Las LDL son capaces de fijarse a los receptores específicos presentes en la membrana plasmática de la célula. Luego de la fijación sobreviene un proceso de endocitosis, la membrana se invagina, cerrándose con el material LDL en su interior. La estructura formada recibe el nombre de vesícula, la cual además se cubre con clatrina, una proteína que forma una estructura similar a una red o clatrato. Una vez en el interior de la célula, la vesícula se encuentra con endosomas y luego con lisosomas. En el interior de estos últimos elementos, la apoproteína de las LDL se hidroliza dejando libre el colesterol el que se incorpora a la membrana celular.
Se ha realizado el largo camino de los párrafos anteriores que permiten establecer la comprensión de la importancia del colesterol en relación con la genética y que esta circunstancia permite explicar en parte, los orígenes de la aterosclerosis. Sin duda, el colesterol es indispensable para la vida de los animales, sin embargo, por arriba de ciertas concentraciones plasmáticas puede originar las formación de placas en arterias. Dichas placas contienen ésteres de colesterol, muy insolubles. El resultado es la aterosclerosis, en esta situación los vasos sanguíneos se deterioran y estrechan, disminuyendo por tanto el flujo sanguíneo, en otros términos, aumenta la resistencia al paso de sangre con sus nutrientes a los tejidos y órganos del cuerpo. Las consecuencias son aumentos de la presión arterial, enfermedades vasculares que suelen terminar en ataques cardiacos e infartos cerebrales. De estos el último tiene importancia para su desarrollo profesional, pues seguramente le tocará intervenir en el tratamiento de estas patologías en el futuro próximo.
La hipercolesterolemia familiar es un desorden hereditario que se caracteriza por altos niveles plasmáticos de LDL-colesterol, esto provoca enfermedad cardiaca y la muerte antes de los 20 años de edad. La enfermedad se caracteriza por sobreproducción de colesterol endógeno, ello como consecuencia de funciones deficientes en los receptores de membrana para LDL. La explicación es la siguiente: la producción endógena de colesterol se encuentra

104
sujeta a un control por retroalimentación, si ingresa colesterol en forma normal a la célula, esta detendrá la síntesis de la sustancia. Puesto que el receptor se encuentra defectuoso no habrá retroalimentación y la célula sigue produciendo sustancia. La enzima principal del proceso es la HMG-reductasa que inicia la síntesis de colesterol.
La forma homocigótica de la hipercolesterolemia familiar es rara, la forma heterocigota es la más común, la deficiencia del receptor es parcial y en consecuencia sobrevienen las enfermedades de tipo vascular.

105
PROTEÍNAS Las proteínas y su esencia comenzaron a descubrirse en el siglo XIX, la primera utilizada para el estudio fue la albúmina del huevo y de la leche. Se encontró que estas sustancia al calentarlas se coagulaban, esto era un proceso irreversible y se conoce que el proceso es irreversible. Las proteínas contenían carbono, oxígeno, hidrógeno, nitrógeno y azufre. Funciones biológicas: 1) Proteínas que son enzimas, esto quiere decir que son catalizadores de reacciones químicas, como se trata de que intervienen en los seres vivos, entonces es más apropiada llamarle reacciones de tipo bioquímico. Toda reacción de tipo bioquímico que se produce en los seres vivos es catalizada por enzimas. 2) Pueden participar en almacenamiento y trasporte. Por ejemplo, la hemoglobina, una de las proteínas más estudiadas se fija y transporta al O2 y el CO2. Se encuentra en las células llamadas glóbulos rojos de la sangre. 3) Hay proteínas que son netamente estructurales, otorgan soporte y sostén mecánico a células y tejidos, por ejemplo, el colágeno. 4) Proteínas que realizan trabajo mecánico, por ejemplo, proteínas que permiten el movimiento de los cromosomas durante las etapas de meiosis y mitosis celular, la contracción muscular. 5) Proteínas que son hormonas y en consecuencia regulan las actividades de tipo fisiológico-bioquímico en las células o tejidos. 6) Proteínas que leen códigos de información celular que físicamente emanan desde las moléculas de ácido desoxirribonucleico, ADN y ARN. Por ejemplo, aquellas que forman parte de las estructuras llamadas ribosomas, estas son necesarias para los procesos de traducción de proteínas que no es otra cosa que seguir instrucciones para la construcción de las moléculas de proteínas. También hay proteínas que regulan la expresión de los genes, ya sea, permitiendo o inhibiendo la actividad genómica. 7) Hay proteínas que son inmunoglobulinas que participan en la defensa del organismo contra bacterias y ataques virales. Las proteínas son del grupo de las macromoléculas por su capacidad de formar polímeros, y formar grandes complejos de proteínas asociadas a glúcidos y lípidos. Los monómeros de estas macromoléculas son los aminoácidos, moléculas pequeñas que contienen un grupo carboxilo (COOH) y n grupo amino (NH2). Los dos grupos poseen propiedades ácidas y básicas débiles, respectivamente.
Químicamente, las proteínas son macromoléculas, polímeros de aminoácidos (más de 100) dispuestos en una secuencia lineal, sin ramificaciones. Una secuencia de menos de 100 aminoácidos se denomina péptido.
Una proteínas con 20 aminoácidos diferentes puedan ser ordenados en cualquier secuencia para formar polipéptidos de cientos de aminoácidos, de ello surge la capacidad potencial de producir una gran cantidad de variantes de polipéptidos, o proteínas. Esta

106
variedad faculta a las proteínas para realizar las funciones diversas que han sido mencionadas antes.
Diagrama de los Aminoácidos-Estructura y propiedades
En nuestra estructura biológica tenemos 20 diferentes tipos de aminoácidos, todos con la configuración α. En la figura siguiente usted puede observar el ácido glutámico.
Los aminoácidos (aa) son moléculas orgánicas pequeñas con un grupo amino (NH2) y un grupo carboxilo (COOH), le acompaña un grupo R lateral que puede ser un resto de cadena de hidrocarburo con alguna función especial.
Todos los aminoácidos tienen la misma fórmula general:
Los 20 aminoácidos comunes de denominan α-aminoácidos porque poseen un grupo amino y un grupo carboxilo ácido unidos al C-2, conocido también como el carbono α. La cadena lateral se representa con la letra R, esta es distinta para cada aminoácido. A pH neutro, el grupo amino está protonado (-NH3+), en tanto el grupo carboxilo está ionizado como –COO-. Casi todos los aminoácidos muestran un átomo de carbono α que es quiral o sea, asimétrico porque posee cuatro grupos diferentes unidos entre sí. La excepción a la regla lo representa la glicina:
Como usted puede observar el grupo R está representado por un átomo de H, por lo tanto el carbono α está unido a dos átomos de hidrógeno, y es obvio que estos dos enlaces del carbono están unidos a la misma clase de átomo, o “grupo”.

107
Los aminoácidos pueden existir como estereoisómeros, que son cuerpos con la misma fórmula molecular, pero son diferentes en estructura en cuanto a la configuración espacial de sus átomos. Si una molécula contiene un carbono asimétrico, podemos observar dos estereoisómeros, en tal caso una molécula es la imagen especular de la otra, o sea, no pueden superponerse, como en la figura a continuación.
Los aminoácidos pueden agruparse según las propiedades que otorgan las cadenas de tipo lateral. En esta cadena se encuentra una diversidad de grupos químicos, y por ello se observa variedad en las propiedades de las proteínas. La presencia de grupos químicos permite que los aminoácidos adquieran características hidrofóbicas, hidrofílicas, polares, no polares, presencia o ausencia de grupos ionizables.
Aminoácidos con cadenas laterales de tipo alifáticos
Observe desde la glicina, la alanina, valina, leucina e isoleucina, en estos, el grupo lateral se extiende y por ello se hace más hidrófobo. La isoleucina debido a esta notable característica se encuentra normalmente en el interior de las macromoléculas proteicas. En este grupo Se encuentra la prolina porque comparte muchas propiedades con los aminoácidos alifáticos, no obstante ser un aminoácido cíclico. A juicio de los expertos, esta característica cíclica le dificulta el plegado como residuo de prolina en las estructuras proteicas. Con respecto a la metionina una nota importante, su característica principal es que se trata de una molécula muy hidrófoba, por ello su inclusión dentro de esta forma de clasificación, pero en su estructura se destaca la presencia de azufre, motivo por el cual se le puede clasificar dentro del grupo de aminoácidos que contienen S. La metionina es un tipo de aminoácido esencial para los mamíferos, estos animales no la sintetizan. La metionina junto a la cisteína son los aminoácidos que contienen azufre.
Aminoácidos Aromáticos: Tres aminoácidos contienen cadenas laterales de tipo aromático, estos son la fenilalanina, la tirosina y triptófano. La fenilalanina es una molécula muy
Los enantiómeros pueden dar dos formas, los llamados formas D y L. Estos son considerados isómeros ópticos. Los aminoácidos que incorporan las proteínas de los seres vivos son de la forma L, pero hay excepciones a la regla, por ejemplo, el D-glutámico se encuentra en paredes celulares bacterianas.

108
hidrofóbica y podría agruparse por esta propiedad junto con moléculas ya nombradas y clasificadas en los párrafos anteriores, tales como, valina, leucina e isoleucina. La tirosina y el triptófano son menos hidrófobos, atenuado por los grupos polares de sus cadenas laterales.
Los aminoácidos aromáticos puesto que poseen anillos aromáticos conjugados como puede observarse en la figura anterior, se caracterizan por absorber luz en la región del espectro ultravioleta cercano. Esta característica es muy explotada en los laboratorios clínicos y de experimentación para la detección y cuantificación de proteínas, midiendo la absorción a 280 nm. Consulte con su profesor si desea conocer más información al respecto.
El triptófano es un aminoácido esencial en la nutrición humana .Se clasifica entre los aminoácidos apolares, o hidrofóbicos, es precursor de la serotonina, un neurotransmisor neurotransmisores más importantes de nuestro sistema nervioso, por ello habrá una extensión más acerca de sus propiedades.
El triptófano es hidroxilado por medio de una oxigenasa y tetrahidrobiopterina en el carbono 5, de esta reacción resulta el 5-hidroxitriptófano que precede a la formación de serotonina y de 5-hidroxitriptamina.
Existe evidencia de que la concentración de serotonina en el cerebro es producida directamente proporcional a la concentración del triptófano en el plasma y el cerebro. La ingesta dietética de triptófano influye directamente en la cantidad de serotonina en el plasma, el cerebro y los niveles en todo el cuerpo. Parece claro en los trabajos de investigación realizados por Marina Diaz-Marsá y colaboradores en la revisión de Psiquiatría del 2006, acerca de la relevancia del consumo de triptófano para evitar conductas relacionadas con trastornos de la conducta alimentaria.
El experimento para mostrar la relación fue el siguiente: se utilizó la técnica de depleción de triptófano para evaluar la función serotoninérgica cerebral. Se estudió la respuesta sintomática de 57 pacientes mujeres diagnosticadas con trastornos de la conducta alimentaria, TCA y de 20 mujeres control a una prueba de depleción de triptófano. Para ello se administró una solución con todos los aminoácidos esenciales, excepto el triptófano, luego esto fue precedido por una dieta pobre en triptófano de 2 días de duración.
A las 2 y 5 horas posteriores a la administración de la bebida se midieron las concentraciones de triptófano. A lo largo de las 5 horas siguientes a la ingesta de la bebida se midió también las respuestas alimentarias y emocionales. Se realizó un estudio de las características básicas de la personalidad y de los rasgos de impulsividad de los pacientes.

109
Los resultados indicaron que la prueba fue efectiva en la reducción de triptófano plasmático, con descenso del 76% en la muestra global. La prueba fue bien tolerada y no se produjeron efectos adversos graves. Dos pacientes abandonaron la prueba por intolerancia gástrica a la bebida. En conclusión, la prueba de depleción de triptófano es eficaz en la reducción del triptófano plasmático en los TCA y esto resulta válido para el estudio de la función serotoninérgica y de su relación con los síntomas alimentarios e impulsivos de los TCA.
de triptófano e inhibe el transporte a través de la barrera hematoencefálica, en consecuencia la síntesis de serotonina disminuye.
Existe evidencia de que la serotonina tiene un rol relevante en la patogenia de la depresión mayor en relación con las alteraciones que presentan los pacientes deprimidos en la actividad serotoninérgica cerebral. También se fundamenta más debido a la efectividad de los fármacos antidepresivos que aumentan la neurotransmisión serotoninérgica de forma selectiva. Sin embargo, no parece muy claro que la deficiencia en serotonina sea la causa de signos clínicos de trastorno depresivo mayor.
La bulimia nerviosa es un trastorno de conducta alimentaria frecuente en mujeres con peso normal. El trastorno se caracteriza por alteraciones en el apetito, atracones, conductas purgativas, alteraciones en la imagen corporal, ánimo disfórico, alteraciones neuroendocrinas, ánimo depresivo, obsesiones y ansiedad. Otros estudios indican que el descenso en neurotransmisión de serotonina podría contribuir a la patogenia de la bulimia y de la anorexia nerviosa.
La serotonina participa en la regulación de la ingesta alimentaria. La estimulación de los receptores de serotonina (5HT) postsinápticos parecen inhibir de forma específica el consumo de carbohidratos en ratas; una estimulación más intensa es capaz de reducir la ingesta total de carbohidratos. Se ha propuesto (hipótesis) que los atracones en la bulimia nerviosa están precipitados por una reducción de de la actividad serotoninérgica cerebral. Experimentalmente, además, la reducción de la transmisión serotoninérgica en el cerebro debido a fármacos estimula la ingesta alimentaria. Otros estudios sugieren que las pacientes bulímicas no recuperadas tienen una actividad serotoninérgica disminuida.
La serotonina es sintetizada en el cuerpo y el cerebro a partir del aminoácido triptófano en una reacción que involucra dos enzimas: triptófano hidroxilasa (TPH) y una L-aminoácido aromático decarboxilasa (DDC).
La TPH existe en dos formas: TPH1, encontrada en varios tejidos, y la TPH2, que es una isoforma cerebro-específica. Hay evidencia de polimorfismos genéticos en ambos tipos y al
La depleción aguda de triptófano reduce la función serotoninérgica cerebral, esto ha sido mostrado también utilizando la tomografía por emisión de positrones. La síntesis cerebral de serotonina depende de la disponibilidad del triptófano plasmático. El estudio es posible dada la posibilidad de producir una rápida disminución de la disponibilidad de triptófano sustituyendo una dieta normal por una solución (“sopa”) de aminoácidos, sin considerar el triptófano. Este hecho produce una disminución de las concentraciones plasmáticas

110
parecer pueden influenciar en el desarrollo de estados depresivos. Tal es la importancia del estudio de los aminoácidos como el triptófano que es conducente y necesario para la síntesis de la serotonina.
La serotonina ingerida por vía oral no pasa al sistema nervioso central porque ésta no cruza la barrera hematoencefálica. Sin embargo, el triptófano y sus metabolitos 5-hidroxitriptófano (5-HTP), con los cuales la serotonina es sintetizada, pueden cruzar la barrera hematoencefálica. Estos agentes están disponibles como suplementos dietarios y pueden ser agentes serotoninérgicos efectivos.
Ver figura en: http://www.choser.es/5-categorie-10757831.html Un grupo ecologista presentan pancartas para solicitar que las personas sean felices. El cortisol se relaciona con el estrés y las tensiones de la vida actual.

111
Figura: Síntesis de serotonina a partir de triptófano
Aminoácidos Básicos:
En este grupo se encuentra Histidina, lisina y arginina, llevan grupos básicos en sus cadenas laterales. Son muy polares y a menudo se encuentran en la superficie de las proteínas, en esta posición pueden hidratarse desde el medio acuoso que rodea las proteínas.

112
En la figura anterior se encuentra además los aminoácidos ácidos que llevan carga negativa a pH 7. Las amidas de estos aminoácidos ácido aspártico y glutámico, las conocidas como asparagina y la glutamina.
En resumen se muestra las fórmulas de aminoácidos constituyentes de las proteínas.
Unión de los aminoácidos-enlace peptídico Los aminoácidos pueden permanecer unidos mediante enlaces del tipo peptídicos. Este enlace peptídico es de tipo covalente y ocurre entre el grupo amino (–NH2) de un aminoácido y el grupo carboxilo (–COOH) de otro aminoácido. El enlace peptídico se produce con pérdida de una molécula de agua y la formación de un enlace covalente CO-NH. Es, en realidad, un enlace amida sustituido.

113
En la figura anterior usted puede observar un tripéptido, o sea, tres aminoácidos unidos por enlaces de tipo peptídico
El enlace peptídico es un enlace de tipo amida que se forma entre el grupo carboxilo de un aa con el grupo amino de otro, con eliminación de una molécula de agua. La longitud de la cadena polipeptídica es variable y depende del código genético. En esta cadena se encuentra siempre un extremo amino terminal y un extremo carboxilo terminal que permanecen intactos. Los productos de los enlaces peptídicos reciben nombre según la cantidad de aminoácidos constituyentes en la unión. Por ejemplo, la unión entre dos aminoácidos es un péptido llamado dipéptido. La reacción puede seguir añadiendo más aminoácidos a la cadena, se forma así un tetrapéptido, o con pocos residuos (entiéndase también, restos, o, aminoácidos) se pueden denominar oligopéptido. Si la cadena es muy larga se puede denominar polipéptido. La mayoría de los oligopéptidos y los polipéptidos que conservan un grupo amino terminal a la izquierda y el grupo carboxilo terminal hacia la derecha. La escritura de estos se realiza desde el extremo amino hacia el carboxilo terminal y se puede utilizar letras o abreviaturas según la tabla siguiente: Ácido y Bases débil: donadores y aceptores de protones Los ácidos son donadores de protones y las bases son aceptoras de protones. Un ácido fuerte tal como los ácidos minerales, ácido sulfúrico, clorhídrico, nítrico, etc. se disocian totalmente en agua: HCl H+ + Cl- y permanecen así mientras se encuentren en solución acuosa, o sea disuelto en agua. Estos compuestos corresponden a ácidos y los álcalis corresponden a las bases fuertes, el ejemplo más común de este grupo son el NaOH, que se disocia en Na+ y OH- Por otra parte, los ácidos orgánicos, presentes en los seres vivos, y ya visto en parte al estudiar los aminoácidos, son ácidos o bases débiles. En una solución acuosa un ácido débil existe un cierto equilibrio entre las partes constituyentes del ácido que se han disociado y el ácido débil: Ácido débil Base conjugada + H+ HCOOH HCOO- + H+ En el ejemplo tenemos el ácido orgánico conocido como ácido fórmico que se ha disociado en la solución, la reacción descrita indica que se ha formado la base conjugada (Así
Enlace peptídico

114
denominada) HCOO-, si usted observa esta base conjugada y especialmente el signo -, que está indicando que se disoció del protón que se encuentra más a la derecha y se escribe como H+. Piense en esto, si HCOOH se puede separar del protón H+, entonces normalmente la base conjugada HCOO- que resulta, puede volver a unirse con el H+. Esto es muy análogo a los matrimonios que se divorcian y vuelven a casarse una y otra vez. Valga la analogía. Piense en lo siguiente y con recuerdos vagos de las fórmulas de los aminoácidos, aquellas sustancias que poseen grupos ácidos como el grupo carboxilo, o grupos básicos como el grupo amino. Estas sustancias también tienen la facultad para establecer divorcios, o sea, sus grupos COOH y NH2, pueden perder protones (divorcio) y quedar como COO-, pueden ganar protones (casorio) y quedar como NH3. Esto es de la mayor importancia para que ocurra el fenómeno de existencia en los seres vivos. Las proteínas poseen cientos de aminoácidos, con cientos de grupos COOH y NH2, o sea, con cientos de posibilidades de casorio y divorcios, de capturar y ceder protones respectivamente. Los que poseen el potencial de capturar protones son los grupos aminos NH2, y los que poseen la capacidad de ceder protones son los grupos carboxilo, más correcto carboxilato, COO-. Se dice que las proteínas son amortiguadoras de los excesos de protones, o sea, amortiguadoras de los excesos de sustancias que son ácidas. Por ejemplo, suponga que usted consume alcachofas con vinagre (ácido acético o CH3- COOH) que al consumirlo se encuentra en solución, diluido con agua, y por tanto, se encuentra disociado en un estado COO- + H+. Estas sustancia ingresan a nuestro organismo, ¿Cómo puede usted no sufrir daño con el ácido acético (vinagre)? En el resto de alimentos que usted consume si hay proteínas no hay problemas, puesto que los protones del ácido acético encontraran iones carboxilato (COO-) que estarán dispuestos a recibir estos protones y en consecuencia no habrá daños. Cuidado todo esto tiene sus límites no experimente sino en el laboratorio con supervisión de sus profesores. Debido a hechos como este podemos llamar amortiguadoras a las proteínas. Otro ejemplo, si usted ha sentido acidez estomacal le han recomendado que tome un vaso de leche, o un poco de bicarbonato, o “sal de fruta”. Explique porque en cada caso se puede aliviar la acidez. Durante toda nuestra vida el cuerpo estará batallando por lograr equilibrios, por lograr homeostasis. Ligeros desequilibrios pueden acabar con la existencia en cosa de minutos. Problema: En ciertas enfermedades de los centros de control respiratorio del cerebro o como pérdida de función de algunos órganos del cuerpo, se produce cambio del pH de la sangre, la tendencia es al aumento de la acidez, por aumento de la concentración de protones en el plasma sanguíneo. En condiciones normales ¿Por qué no se produce acidificación del plasma sanguíneo?, ¿Qué evita la sobrepoblación de protones? Los seres vivos deben mantener el pH interno estable, manteniéndose dentro de estrechos márgenes, entre 6,5 y 8,0. Puesto que los aminoácidos poseen carga positiva o negativa, debido a la ionización de sus grupos carboxilo o amino. Es de esperar que cuando forman las estructuras proteicas, las proteínas resultantes también manifiesten una carga, positiva o negativa, esto por supuesto dependerá a su vez de las condiciones del medio. El medio ambiente de las células por lo

115
general tiene un pH cercano a 7,0, y esta es una de las condiciones para la arquitectura. Por ejemplo, Las proteínas ricas en arginina, lisina, que son aminoácidos básicos, constituyen gran parte de las proteínas básicas que son las histonas. Estas al ser positivas se pueden asociar con gran fuerza al ADN que tiene cargas negativas debido al ión fosfato presente en su estructura. Pero, esta asociación también depende del medio en el cual se encuentra la cromatina. Vemos entonces, con este ejemplo, como las estructuras biológicas alcanzan forma definida con un alto grado de compactación, que luego tendrá repercusiones en la diferenciación celular y especialmente en la expresión de los genes.
Estructura primaria La secuencia de aminoácidos de una proteína es determinada por el gen que la codifica. Esto es, al diferenciarse un gen de otro, también una proteína será diferente de otra. Las diferencias en los genes se deben por ejemplo, a las secuencias de nucleótidos presentes en el gen. De manera que hay correspondencia entre la secuencia de nucleótidos en el ADN y secuencia de aminoácidos en una proteína. La figura siguiente tomada del texto de bioquímica de Mathew representa la secuencia de aminoácidos de la mioglobina de cachalote, que han determinado la estructura compleja de la proteína. A continuación se presenta una tabla presentada por el mismo autor, donde se compara la secuencia de aminoácidos de la Mioglobina de cachalote con la Mioglobina humana. En los mamíferos y en los dos tipos de Mioglobina de ejemplo presentadas usted observará que son muy parecidas en secuencia, se encontrará algunas diferencias en la posición de algún aminoácido, o algún cambio de aminoácidos en una determinada posición según si se observa una u otra. Lo principal que podemos observar al ver estas estructuras es que somos muy parecidos al cachalote, por lo menos en la Mioglobina. Partiendo de la similitud entre las dos proteínas, cachalote y humano, podemos deducir que somos parientes. Según otros miles de criterios e investigaciones asociadas, los antepasados que dieron origen a cachalotes y humanos vivieron en la Tierra hace millones de años. La mioglobina muestra algunas diferencias entre las dos especies, al observar la tabla podemos ver, por ejemplo, en la posición 1 que el aminoácido correspondiente a humano es glicina y para cachalote es valina. La mayoría de los aminoácidos se conservan para ambas especies. La presencia de aminoácidos diferentes, al respecto, nos está indicando que en estas estructuras operó la evolución. Una de las formas como opera la evolución se da por cambios puntuales en la secuencia de nucleótidos en el ADN, en este caso, el gen para mioglobina. El cambio puntual debe ser suficiente para cambiar la secuencia de tres nucleótidos y producir un cambio efectivo de la señal para un determinado aminoácido.

116
Figura siguiente: Mioglobina de cachalote
La mioglobina de cachalote y humanos contienen 153 aminoácidos. Se encuentran entre las proteínas más pequeñas. Las dos tienen secuencias similares, pero no son idénticas. Esta gran similitud es suficiente para que ambas realicen la misma función. Las proteínas evolucionan mediante cambios de las secuencias de aminoácidos, que su vez derivan de cambios en la secuencia de nucleótidos en los genes que las codifican. En resumen, la estructura primaria de cada proteína está determinada por un gen concreto.

117
En la tabla siguiente se encuentra el nombre de los aminoácidos, su abreviatura y una letra para identificación.

118
adopta una estructura helicoidal, que se mantiene mediante puentes de hidrógeno, con los grupos R orientados hacia el exterior. Para establecer la estructura es necesario que el grupo carboxilo de un aminoácido se una mediante un puente de hidrógeno, al grupo amino de otro aminoácido. Las hélices de tipo α son de tipo dextrógiras, con pocas excepciones se encuentran algunas de tipo levógira. La estabilidad es notable por los cientos de puentes de hidrógeno intrahelicoidales. Las cadenas laterales de los aminoácidos apuntan hacia afuera del eje de la hélice.
La mayoría de las proteínas tienen una conformación espacial determinada por la orientación de sus moléculas constituyentes y estás a su vez, están determinadas por la orientación de sus átomos en el espacio. Las consecuencias son variadas y se relacionan con la función que debe cumplir una proteína. Existen dos niveles de plegamiento tridimensional de las cadenas polipeptídica. En el primero de ellos nos encontramos con el tipo de plegamiento que describe un tipo de enrollamiento por regiones de las proteínas y esta puede ser de tipo helicoidal, y se establece de tal forma que establece regularidades en la estructura molecular. También se encuentra otras formas de regularidad como son las denominadas formas de lámina plegada.
Estructura secundaria: Se forma una estructura en α-hélice cuando la cadena

119
Figura obtenida de Mathew. Texto de bioquímica. Conformaciones de la estructura secundaria.
La α-queratina,es el componente principal de la lana, el pelo, la piel y las uñas, la proteína se clasifica como fibrosa compuesta casi totalmente por α-hélice. La unidad básica de la α-queratina es el protofibrilo, que son 4 hélices α dextrógiras interrumpidas por regiones no helicoidales que se entretegen. Las hélices se aparean formando cables superenrollados. Estas protofibrillas forman estructuras mayores denominadas microfibrillas. Las protofibrillas y la microfibrillas se entrecruzan, acción posible gracias a la formación de puentes disulfuro. Las estructuras así formadas son más estables. Las queratinas de las uñas contienen muchos puentes disulfuro, esto les confiere dureza y poca flexibilidad. Las queratinas con pocos enlaces disulfuro se encuentran por ejemplo, en la lana, son flexibles y se estiran con facilidad, también loc cabellos pueden ser rizados o lisos si se les trata con una solución que reduzca los puentes disulfuro. Se puede

120
trata el cabello con sustancias oxidantes para oxidar los residuos de cisteína y que formen nuevos puentes disulfuro. El contenido α helicoidal es variable en proteínas de tipo globular, la mioglobina y la hemoglobina contienen más de un 75% de residuos en α-hélice. En otras proteínas la fracción de hélices alfa es más pequeña.
La figura anterior es la estructura propuesta para los filamentos intermedios de queratina.Primero, dos monómeros se aparean a través de ovillos enrollados paralelos para formar formar un dímero. Estos se asocian para formar un protofilamento de 4 cadenas y una protofibrilla de 8 cadenas. Figura tomada del texto de Bioquímica de Mathew.
Conformación β: La estructura beta, de hoja plegada es un plegamiento en zig-zag, formando láminasconplegamientos regulares. Las cadenas polipeptídicas quedan extendidas con diferente sentido de orientación, una en sentido paralelo al eje de la lámina. Las cadenas β se estabilizan por puentes de hidrógeno entre oxígenos carbonílicos del COO- e hidrógenos amídicos. Estos enlaces pueden unir a dos o más cadenas polipeptídicas. Los puentes de hidrógeno son perpendiculares a las cadenas. Las cadenas laterales apuntan de modo alternativo hacia arriba y debajo de la lámina.

121
Figura de la forma β-plegada. Tomada del texto de Bioquímica del autor Mathew.
Figura capullos de seda, ver en: http://www.crisaguil.com/Talleres/Talle%20de%20seda/tipos%20de%20seda/Seda_tipos.htm
La estructura secundaria de muchas proteínas globulares contiene regiones de conformación en lámina β.
Colágeno Es una proteína fibrosa con una estructura helicoidal diferente a las mencionadas. Se trata de un componente proteico que forma principalmente la mayor parte del tejido conectivo, muy abundante en los vertebrados, llegando al 35% de las proteínas totales de un

122
mamífero. Hay variedades del colágeno con formas y funciones diferentes. Por ejemplo, la estructura considerada como matriz, sobre la que precipitan los minerales del hueso contiene un polímero de colágeno y fosfato de calcio. El colágeno en la piel adquiere la forma de fibras tejidas poco apretadas. El colágeno de los tendones forma fibras rígidas, fuertes. El colágeno es un agregado que consiste en tres cadenas que poseen hélices levógiras que se enrollan y luego hasta formar superenrollados. Este último estado se estabiliza mediante puentes de hidrógeno intracatenarios. La unidad repetitiva de la fibra de colágeno es la molécula de tropocolágeno una hélice triple de tres cadenas polipeptídicas, cada una contiene cerca de 1000 aminoácidos, entre ellos algunos destacan, por ejemplo, la prolina y la hidroxiprolina, cuyos grupos OH- (hidroxilo) que al parecer participan dando estabilidad a la molécula. Esto en parte sustentado por la observación de que la enzima promotora de la hidroxilación de la prolina necesita vitamina C (ácido ascórbico), al bajar su nivel de concentración produce la carencia y en consecuencia la enfermedad conocida como escorbuto. Esto conduce al debilitamiento de las fibras de colágeno debido a la falta de hidroxilación de la prolina y lisina, luego aparecen lesiones en la piel y las encías, se debilitan los vasos sanguíneos.
Proteínas globulares-Estructura terciaria Una gran parte de las proteínas tienen funciones en el metabolismo, en síntesis, transporte, para esto requieren una estructura globular. Esta forma la adquieren mediante el plegamiento de las cadenas polipeptídicas formando estructuras compactas muy distintas de las estructuras filamentosas que se describieron antes. La mioglobina es una proteína globular, la cual puede volver a observar en las figuras anteriores, actualmente se pueden realizar simulaciones de la estructura usando programas informáticos.
La mioglobina es una proteína bastante estudiada y se conoce mucho acerca de su estructura, aproximadamente el 70% de su estructura es del tipo hélice α, esta se dobla y pliega para formar la molécula compacta. En su estructura permanece un espacio que contiene un grupo prostético, el correspondiente al grupo hemo, unido en forma no covalente y se caracteriza especialmente por contener el lugar de unión del oxígeno de la mioglobina.
La cadena polipeptídica va cambiando su conformación a medida que se avanza. Así, se logra una mezcla de conformación β, α, etc. Para que desarrolle la estructura compacta, las regiones deben plegarse y se puede observar como en la figura al frente donde se describe su aspecto tridimensional.

123
Existe una gran variedad de plegados de las cadenas proteica, pero son comunes a ellos ciertas regularidades en el plegado y disposición de las cadenas de aminoácidos, estos son llamados motivos o principios comunes. El primer principio es que las proteínas están formadas por más de un dominio, siendo este una región compacta, con plegamiento local. Diferentes proteínas pueden tener dominios múltiples, únicos, y desempeñan algún rol en la funcionalidad de la proteína. En ocasiones un determinado dominio se puede encontrar en diferentes proteínas. Al observar las proteínas globulares en forma tridimensional, invariablemente vemos que en el desarrollo de la estructura terciaria los residuos hidrofóbicos se encuentran en el interior, mientras que los residuos hidrófilos se encuentran en la superficie. Desde luego, se puede señalar la grandeza de la naturaleza en la construcción o arquitectura de cada una de estas proteínas. Obviamente, la información está contenida en el gen, o sea en su secuencia de nucleótidos, cada triplete de nucleótidos está indicando que un determinado aminoácido se incorpore en un cierto orden a la cadena polipeptídica. El orden de la secuencia la
En la figura al frente se muestra la estructura hemo, que es común a mioglobina y hemoglobina. La estructura tridimensional se observa en la figura abajo. Se observa el plegamiento de las cadenas polipeptídicas. Cada aminoácido está indicado por un círculo correspondiente al átomo de carbono α. Las cadenas laterales han sido omitidas.

124
codifica el gen, la secuencia determina la forma de la proteína, también su funcionalidad y el lugar donde realizará su trabajo o función. Cierta secuencia de aminoácidos le ha indicado el camino a seguir para su ubicación en la estructura donde debe acometer sus funciones. Toda la información del cómo y el cuándo está en el gen, en la secuencia de nucleótidos. Eso ya no es un problema para entender, lo problemático es entender la coordinación de la expresión del gen, o sea, el cómo y el cuándo de la expresión genómica. ¿Cómo sabe el gen cuando expresarse? Veremos parte de la respuesta en el momento que se estudie los desarrollos embrionarios, especialmente, el desarrollo del sistema nervioso. Las conformaciones β se encuentran generalmente enrolladas, o envueltas en estructuras de forma cilíndricas. El enrollamiento es hacia la izquierda. Es interesante conocer mediante observaciones experimentales del tratamiento de las proteínas con ácidos minerales, variando el pH de su medio con ácidos o álcalis, alcohol, urea. Con estas sustancias la naturaleza nativa de la proteína desaparece. El proceso se denomina desnaturalización, o sea, la pérdida de la estructura natural de la proteína y en consecuencia se ha perdido las propiedades naturales de ellas. El plegado se realiza con ayuda de otras proteínas, llegando a estabilizarse con el establecimiento de puentes salinos (denominada así en algunos textos) internos. Estos se forman bajo ciertas condiciones de pH en la cual surgen atracciones electrostáticas que en general permiten la formación de enlaces iónicos entre los grupos de cadenas laterales. Este tipo de interacciones que ayudan a la estabilidad estructural y funcional de la proteína, pueden perderse si la proteína es sometida a condiciones del medio con pH bajo o elevados con lo cual la carga de las cadenas laterales de los aminoácidos pierdan la carga. Tal es la explicación en parte, para la desnaturalización ácida o básica de las proteínas. Otra forma de ayudar a la estabilidad de las proteínas es con formación de puentes de hidrógeno. Esto ocurre por la existencia de cadenas laterales en los aminoácidos que son buenos aceptores o dadores de puentes de hidrógeno, como por ejemplo, los hidroxilos de la serina o de la treonina, los grupos amino y los oxígenos de los grupos carboxilo de la asparagina o la glutamina, y en general cualquier grupo carboxilo o amino que no participe en la formación de la estructura secundaria. Contribuyen también a dar estabilidad a la estructura terciara las interacciones de van der Waals, que son interacciones débiles entre grupos moleculares sin carga. El aporte de este tipo es mayor si el interior de la proteína se encuentra densamente empaquetado, lo que permite el máximo contacto entre los átomos de las cadenas laterales.
Efecto hidrófobo Este efecto contribuye a la estabilidad de la estructura terciaria de las proteínas globulares. La explicación es la presencia de aminoácidos hidrófobos, los cuales en contacto con el agua, permiten que esta molécula forme estructuras con forma de jaulas, un término más correcto es clatrato. El siguiente ejemplo puede conducirnos a mayor conocimiento y claridad de esta situación: suponga una proteína que en su secuencia de aminoácidos, contenga aminoácidos (residuos) con largas cadenas laterales de tipo hidrófoba, por ejemplo, leucina, isoleucina y fenilalanina. Cuando esta proteína se encuentra desplegada y estos aminoácidos entran en contacto con el agua, produciendo la ordenación de estas moléculas alrededor. Luego al plegarse la cadena formando la estructura globular,

125
los aminoácidos quedan al interior liberándose las moléculas de agua. En términos físicos se produce un aumento de la entropía, en otras palabras, se produce una contribución negativa a la energía libre del plegado y en consecuencia aumenta la estabilidad de la estructura proteica. (El tema aparece más ampliamente desarrollado en textos de física-Biológica).
Formación de enlaces disulfuro La formación de enlaces disulfuro entre residuos de cisteína contribuye a la estabilización de la estructura terciaria. Pero, la formación de tales estructuras limita el número de conformaciones posibles que pueda llegar a tener una proteína desplegada si se le compara con proteínas sin posibilidad de estableces puentes disulfuro. Muchas proteínas no poseen enlaces disulfuros, entonces, ¿porque unas pocas poseen estas ligaduras? La respuesta hipotética es que las proteínas que contienen este tipo de enlaces, son del tipo exportable, por ejemplo, la ribonucleasa, insulina, etc., ello sucedería así porque el ambiente interno de las células es reductor, configurando un ambiente para mantener a los grupos sulfhidrilo reducido, en cambio, al salir al ambiente externo, (como en el caso de estas proteínas) por lo general, de tipo oxidante, lo que mantiene el estado de enlaces disulfuro o puentes disulfuro.
Estructura cuaternaria de las proteínas Una gran variedad de proteínas dentro de la célula forman tipos de agregados de dos o más cadenas polipeptídicas plegadas, también llamadas subunidades. Si la asociación es entre cadenas polipeptídicas idénticas o casi idénticas se le denomina homotípicas; si la asociación es entre unidades con estructura diferente se les denomina heterotípicas. La formación de estas proteínas involucra la asociación entre las subunidades.

126
MOLECULAR STRUCTURE OF NUCLEIC ACIDS
A Structure for Deoxyribose Nucleic Acid
We wish to suggest a structure for the salt of deoxyribose nucleic acid (D.N.A.). This structure has novel features which are of considerable biological interest. A structure for nucleic acid has already been proposed by Pauling and Corey (1). They kindly made their manuscript available to us in advance of publication. Their model consists of three intertwined chains, with the phosphates near the fibre axis, and the bases on the outside. In our opinion, this structure is unsatisfactory for two reasons: (1) We believe that the material which gives the X-ray diagrams is the salt, not the free acid. Without the acidic hydrogen atoms it is not clear what forces would hold the structure together, especially as the negatively charged phosphates near the axis will repel each other. (2) Some of the van der Waals distances appear to be too small. Another three-chain structure has also been suggested by Fraser (in the press). In his model the phosphates are on the outside and the bases on the inside, linked together by hydrogen bonds. This structure as described
is rather ill-defined, and for this reason we shall not comment on it. We wish to put forward a radically different structure for the salt of deoxyribose nucleic acid. This structure has two helical chains each coiled round the same axis (see diagram). We have made the usual chemical assumptions, namely, that each chain consists of phosphate diester groups joining ß-D-deoxyribofuranose residues with 3',5' linkages. The two chains (but not their bases) are related by a dyad perpendicular to the fibre axis. Both chains follow right- handed helices, but owing to the dyad the sequences of the atoms in the two chains run in opposite directions. Each chain loosely resembles Furberg's2 model No. 1; that is, the bases are on the inside of the helix and the phosphates on the outside. The configuration of the sugar and the atoms near it is close to Furberg's 'standard configuration', the sugar being roughly perpendicular to the attached base. There is a residue on each every 3.4 A. in the z-direction. We have assumed an angle of 36° between adjacent residues in the same chain, so that the structure repeats after 10 residues on each chain, that is, after 34 A. The distance of a phosphorus atom from the fibre axis is 10 A. As the phosphates are on the outside, cations have easy access to them. The structure is an open one, and its water content is rather high. At lower water contents we would expect the bases to tilt so that the structure could become more compact. The novel feature of the structure is the manner in which the two chains are held together by the purine and pyrimidine bases. The planes of the bases are perpendicular to the fibre axis. The are joined together in pairs, a single base from the other chain, so that the two lie side by side with identical z-co-ordinates. One of the pair must be a purine and the other a pyrimidine for bonding to occur. The hydrogen bonds are made as follows : purine position 1 to pyrimidine position 1 ; purine position 6 to pyrimidine position 6. If it is assumed that the bases only occur in the structure in the most plausible tautomeric forms (that is, with the keto rather than the enol configurations) it is found that only specific pairs of bases can bond together. These pairs are: adenine (purine) with thymine (pyrimidine), and guanine (purine) with cytosine (pyrimidine).

127
In other words, if an adenine forms one member of a pair, on either chain, then on these assumptions the other member must be thymine ; similarly for guanine and cytosine. The sequence of bases on a single chain does not appear to be restricted in any way. However, if only specific pairs of bases can be formed, it follows that if the sequence of bases on one chain is given, then the sequence on the other chain is automatically determined. It has been found experimentally (3,4) that the ratio of the amounts of adenine to thymine, and the ration of guanine to cytosine, are always bery close to unity for deoxyribose nucleic acid. It is probably impossible to build this structure with a ribose sugar in place of the deoxyribose, as the extra oxygen atom would make too close a van der Waals contact. The previously published X-ray data (5,6) on deoxyribose nucleic acid are insufficient for a rigorous test of our structure. So far as we can tell, it is roughly compatible with the experimental data, but it must be regarded as unproved until it has been checked against more exact results. Some of these are given in the following communications. We were not aware of the details of the results presented there when we devised our structure, which rests mainly though not entirely on published experimental data and stereochemical arguments. It has not escaped our notice that the specific pairing we have postulated immediately suggests a possible copying mechanism for the genetic material. Full details of the structure, including the conditions assumed in building it, together with a set of co-ordinates for the atoms, will be published elsewhere. We are much indebted to Dr. Jerry Donohue for constant advice and criticism, especially on interatomic distances. We have also been stimulated by a knowledge of the general nature of the unpublished experimental results and ideas of Dr. M. H. F. Wilkins, Dr. R. E. Franklin and their co-workers at King's College, London. One of us (J. D. W.) has been aided by a fellowship from the National Foundation for Infantile Paralysis. J. D. WATSON F. H. C. CRICK Medical Research Council Unit for the Study of Molecular Structure of Biological Systems, Cavendish Laboratory, Cambridge. April 2.
1. Pauling, L., and Corey, R. B., Nature, 171, 346 (1953); Proc. U.S. Nat. Acad. Sci., 39, 84 (1953).
2. Furberg, S., Acta Chem. Scand., 6, 634 (1952). 3. Chargaff, E., for references see Zamenhof, S., Brawerman, G., and Chargaff, E., Biochim. et Biophys. Acta, 9, 402 (1952). 4. Wyatt, G. R., J. Gen. Physiol., 36, 201 (1952). 5. Astbury, W. T., Symp. Soc. Exp. Biol. 1, Nucleic Acid, 66 (Camb. Univ. Press, 1947). 6. Wilkins, M. H. F., and Randall, J. T., Biochim. et Biophys. Acta, 10, 192 (1953).
VOL 171, page737, 1953

128
Así comienza la reciente historia de la biología, en antes y después del descubrimiento de la estructura del ADN. Es encantadora la introducción de Gary Zweiger al libro titulado “El Genoma”. En cierto modo sus vivencias relatadas guardan relación con nuestras esperanzas y sueños de estudiantes del área biológica; recuerda: “Cuando era estudiante graduado en el departamento de Genética de la Escuela de Medicina de Stanford, a finales de la década de los ochenta, acudí a numerosas conferencias por científicos visitantes. Los asistentes escuchábamos de primera fuente los más recientes triunfos de la biología molecular, de las moléculas y su función en enfermedades humanas. Escuchábamos esfuerzos elegantes, ingeniosos e incluso heroicos para descifrar la arquitectura molecular celular y disecar las vías mediante las cuales las acciones moleculares conducían a la fisiología normal o algunas veces a enfermedad”. Los ácidos nucleicos son polímeros de nucleótidos. Cada nucleótido se encuentra estructurado por una molécula de ácido fosfórico, una de pentosa y una base púrica o pirimídica. Las bases púricas son adenina y guanina y las bases pirimídicas son la timina, la citosina y el uracilo. Las pentosas son desoxirribosa en el ADN y en el ARN se trata de una ribosa (ver diferencia en apartado de carbohidratos) Una pentosa y una base nitrogenada unidas constituyen lo que se ha denominado nucleósidos. Precisamente un nucleósido es un producto de la degradación de un nucleótido; el otro producto es el ácido fosfórico. Los tres compuestos moleculares, es decir, la base nitrogenada, el ácido fosfórico y la pentosa al unirse forman el nucleótido. Los ácidos nucleicos son moléculas de tipo “informáticas”, pues proveen información. Con esta información se controla procesos vitales y con ellos el concepto de autopoiesis parece enriquecerse. La información que proporciona el ADN es relevante para la síntesis de macromoléculas, para la diferenciación celular y más que ello, la trascendencia del ser vivo más allá de su existencia. El ADN es portador de la transmisión hereditaria de una célula a sus descendientes.

129
Estructuralmente existen dos tipos de Ácidos nucleicos: El ácido desoxirribonucleico y ácido ribonucleico; ADN y ARN respectivamente. En el ADN la pentosa es la desoxirribosa y en el ARN es la ribosa. Para el ADN las bases nitrogenadas son: Adenina, Guanina, Citosina y Timina. En el ARN, el Uracilo reemplaza a la Timina. El ADN se encuentra localizado en el núcleo de las células eucariotas, también es posible de ubicar dentro de algunos organelos como las mitocondrias y cloroplastos. En el núcleo de las células eucariotas se encuentra asociado con proteínas histonas y proteínas del tipo no histonas. La molécula de ADN consiste en un gran polímero. Dos cadenas de nucleótidos que se encadenan mediante enlaces covalentes y luego forman una hélice con un sentido de izquierda a derecha. La unión es del tipo diéster-fosfato 3’-5’. Ambas cadenas se orientan en sentido antiparalelo.
Fig. 2. Estructura de base nitrogenada

130
Las bases se ubican hacia el interior de la doble hélice, en planos paralelos entre sí y con orientación perpendicular a las cadenas azúcar fosfato. Ver figuras 3 y 4. La estructura alcanza una apariencia de escalera. Como usted puede observar, se dan ciertas regularidades en esta estructura, por ejemplo, que la base de una cadena siempre se aparea con la base complementaria de la otra cadena. Es decir, A con T y C con G. De tal manera que siempre se encuentra que el número de moléculas de A y T es siempre el mismo y que de igual manera ocurre con la G y C. Al formar la hélice se logra estabilidad de la forma toda vez que se forman puentes de hidrógeno entre las bases. Entre G y C y entre T y A, tres y dos puentes respectivamente. Como usted debe recordar los puentes de hidrógeno se pueden romper por la acción del calor, esto trae, por cierto, una desnaturalización del ADN; las hebras se separan. El proceso es reversible.

131
CLASE DE ÁCIDOS NUCLEICOS
En la figura se puede observar en parte la arquitectura de la molécula de ADN, se observa los surcos ancho y angosto. Esas cavidades dan lugar o permiten las interacciones con moléculas como proteínas.
Podemos aprovechar los esquemas de la figura anterior y aparte de renombrar las dimensiones de la molécula. Puede anunciarse que el carácter hidrofóbico de las bases permite que estas se ubiquen al interior. Externamente el ácido fosfórico a pH fisiológico se encuentra cargado negativamente. Por ello mismo es de carácter hidrofílico. Volviendo al interior de las bases es conveniente anunciar que se producen interacciones hidrofóbicas. El ácido fosfórico cargado negativamente se permite asociarse con proteínas histonas que poseen carga positiva. La distribución helicoidal de los desoxinucleótidos determina la formación de una superficie exterior con un surco mayor ancho y uno menor angosto. Las 10 bases de una vuelta de hélice quedan expuestas al surco mayor si se les observa desde un ángulo, y hacia el surco menor, cuando se les observa con otro ángulo. Utilice un programa para visualizar estas dimensiones- consulte con su profesor. Muchas de las proteínas al interactuar con ciertas secuencias de bases del ADN lo hacen a través del surco mayor, ya que en este sitio las bases se encuentran más expuestas. Téngase presente que estas interacciones son muy importantes para la regulación de la expresión genética.
Gene: El gen como unidad de herencia: Los genes son estructuras químicas retenidas a
través de la vida de un organismo y pasan a los descendientes en sucesivas generaciones. Estas estructuras químicas que son los factores hereditarios se encuentran formando parte de los cromosomas. Están formados por ADN. Del término gen deriva el concepto de genoma que contiene todos los genes necesarios para construir un organismo.

132
La genética comenzó a surgir en el siglo XIX con los trabajos realizados por Gregorio Mendel en el cultivo de plantas como la arveja. En su investigación realizó cruces con variedades de semilla que al germinar surgían plantas con rasgos diferentes. Mendel eligió 7 características tales como la altura de la planta, color de las flores, textura de las semillas, etc. En esto estudios Mendel llega a las conclusiones siguientes:
� Las características de las plantas dependían de factores o unidades hereditarias, posteriormente a estos se les llamó genes. Cada planta poseía dos alelos del factor que es responsable de la característica. Uno de los alelos podía comportarse en forma dominante con respecto al otro, este a su vez se le reconoce como el alelo recesivo. El concepto de dominancia indica que es el rasgo que aparece en la expresión fenotípica y que por lo mismo impide que el alelo recesivo ejerza alguna influencia.
� Cada célula germinal o gameto recibe solo una copia del gen correspondiente a la característica. Nunca puede ocurrir que el gameto transporte los dos alelos. Cada planta se origina por la unión de dos gametos, uno correspondiente al progenitor masculino y el otro al progenitor femenino. En la misma planta, los dos alelos permanecen junto, pero luego al originar los gametos, estos se separan o segregan. “Ley de segregación” de Mendel.
� La separación de los alelos con respecto a una característica o afecta la separación de otros alelos responsables de otra característica. Por ello, se dice, son independientes. Esta es la “ley de la permutación independiente” de Mendel.
Cromosomas Hacia 1903 Sutton destacó que los cromosomas eran los portadores específicos de los factores genéticos de Mendel. Observó que la formación de células en el espermatozoide de saltamontes tiene cromosomas grandes. En las células germinales de la gónada masculina ocurren dos tipos de divisiones: La mitótica, en la cual las espermatogonias producen más espermatogonias, y la meiótica, en la cual la espermatogonia produce espermatozoides. Sutton distinguió la individualidad de los cromosomas y como estos se aparean en la meiosis. Luego viene el reconocimiento de que los factores de la herencia propuestos por Mendel, coinciden con la distribución de los cromosomas en las divisiones celulares correspondiente a la meiosis, lo que sugería que los factores estaban relacionados con los cromosomas. En el ser humano, los cromosomas son 46, de ellos, en la célula se reconocen 22 pares de autosomas (para distinguirlos del par sexual) y un par de cromosomas sexuales. Ello por lo menos en la especie humana, luego en la meiosis, los gametos reciben la mitad de los cromosomas. La secuencia de nucleótidos del ADN corresponde con la secuencia de aminoácidos de una proteína. Un gen se define como la secuencia o información requerida para la fabricación de un ARN, y por medio de esta molécula la información es utilizada para la síntesis de proteínas. La unidad de información recibe el nombre de locus. Cromatina - Nucleosoma:
El nucleosoma está formado por dos vueltas completas de la fibra de ADN, alrededor de un núcleo de histonas que forman un octámero con proteínas H2A, H2B, H3 y h4, cada una de ellas multiplicadas por dos, la unidad total las une el ADN separador adyacente unido a la

133
histona H1, se forma así una estructura regular que constituye la unidad básica de enrollamiento cromatínico. El octámero alcanza un diámetro de unos 10 nm.
El ADN que recorre el octámero lo hace casí con dos veces, y cada vuelta equivale a 83 pares de nucleótidos, el segmento de ADN asociado al nucleosoma contiene un total de 146 pares de nucleótidos. Los nucleosomas se hallan separados entre sí por tramos de ADN “espaciadores”, cuya longitud es variable, entre 20 y 60 pares de nucleótidos. El aspecto de esta estructura aparece al microscopio electrónico como un collar. El tratamiento de la cromatina en este nivel puede tratarse con nucleasas o ADNasas que degradan el ADN, ello provoca cortes en los segmentos espaciadores.
Dos tipos de cromatina en el núcleo interfásico:
Una forma altamente condensada llamada heterocromatina (inactiva) y el resto que está menos condensada eucocromatina (más activa).
Los cromosomas mitóticos están formados por cromatina en su forma más condensada.
La mayor parte de los cromosomas en interfase están demasiados extendidos, son tan delgados y están tan entremezclados que no son detectables.
Los cromosomas de todas las células son perfectamente visibles en la Mitosis.

134
Para que la cromatina pueda ser contenida en un espacio tan reducido como el núcleo, debe procesar hacia grados más altos de enrollamiento. Los nucleosomas se enrollan entre sí y generan una estructura helicoidal que reciben el nombre de solenoide, estructura que posee un diámetro de unos 30 nm. Cada vuelta del solenoide contiene unos 6 nucleosomas. Luego la cromatina se compacta más formando lazos o bucles de diversas longitudes, y puede observarse que surgen de un eje constituido por proteínas no histónicas. Como ya se ha advertido antes, la cromatina puede ser eucromatica o heterocromática. En la interfase, la cromatina experimenta un alto nivel de compactación, se le distingue como heterocromatina y el ADN s encuentra inactivo desde el punto de vista transcripcional, lo contrario con el resto de cromatina, menos condensada y con mayor actividad transcripcional, o sea, con capacidad de sintetizar moléculas de ARN. El ADN comprometido en esta escena, se encuentra cuantificado en alrededor de un 10 % del genoma. La heterocromatina puede ser constitutiva, altamente condensada, constante en todos los tipos celulares del organismo, es estable en el genoma, no convertible en eucromatina. En ella se distingue la cromatina de los brazos cortos de los cromosomas acrocéntricos y de las regiones que poseen ADN satélite, como en los centrómeros, otro ejemplo surge en parte del brazo largo del cromosoma Y. La heterocromatina facultativa se observa en los cromosomas en regiones diversas y depende del tipo celular. Ello significa que aparecerá como eucromatina en algunos tipos celulares y como heterocromatina en otros. Es ejemplo, el cuerpo de Barr, que es cromosoma X compactado en las células de la mujer.
EL CARIOTIPO HUMANO
La dotación cromosómica normal de la especie humana es de 46,XX para las mujeres y de 46, XY para los varones.
En el cariotipo humano los cromosomas se ordenan de mayor a menor. Hay cromosomas grandes, medianos y pequeños. Al ordenar los cromosomas es posible reconocer 7 grupos categorizando por tamaño y forma de las parejas cromosómicas, dentro del cariotipo humano podemos encontrar cromosomas metacéntricos (tienen los dos brazos aproximadamente iguales en longitud), submetacéntricos (con un brazo más pequeño que otro) y acrocéntricos (con un brazo corto muy pequeño).

135
Un idiograma es la representación esquemática del tamaño, forma y patrón de bandas de todo el complemento cromosómico, los cromosomas se sitúan alineados por el centrómero, y con el brazo largo siempre hacia abajo. La técnica utilizada en este trabajo es por bandeado G
Los grupos que comprende el cariotipo humano son los siguientes:
Cromosomas grandes
Grupo A, (cromosomas 1, 2 y 3), meta y submetacéntricos
Grupo B, (cromosomas 4 y 5), submetacéntricos
Los metacéntricos poseen el centrómero casi en posición central. Los submetacéntricos muestran el centrómero alejado del punto central, las cromátidas poseen un brazo largo y un brazo corto. En los acrocéntricos el centrómero se halla cerca de uno de los extremos del cromosoma.
- Cromosomas medianos
Grupo C, (cromosomas 7, 8, 9, 10, 11, 12 y además los cromosomas X), submetacéntricos
Grupo D, (cromosomas 13, 14 y 15) acrocéntricos. Poseen pequeñas masas de cromatina, llamadas satélites (no confundir con ADN satélite), ubicadas en el extremo libre de los brazos cortos. Los satélites se unen a los brazos cortos por filamentos delgados de cromatina llamados constricciones secundarias (la constricción primaria es el centrómero).

136
- Cromosomas pequeños
Grupo E, (cromosomas 16, 17 y 18) submetacéntricos
Grupo F, (cromosomas 19 y 20) metacéntricos
Grupo G, (cromosomas 21 y 22) acrocéntricos
Por acuerdo los cromosomas sexuales X e Y se separan de sus grupos correspondientes y se ponen juntos aparte al final del cariotipo.
Las técnicas de bandeado son tinciones que permiten la visualización de bandas oscuras y claras intercaladas en los cromosomas. La distribución de las bandas es constante en cada tipo de cromosoma y facilita la identificación de los cromosomas en el análisis del cariotipo, ello permite además establecer guías para el diagnóstico de trastornos genéticos y que se expresan como translocaciones, delecciones, inversiones y duplicaciones cromosómicas.
Genética y enfermedades El estudio de la genética es fundamental para entender las patologías y enfermedades en el ser humano. Considérese que en la actualidad se conocen más de tres mil enfermedades descritas con trastornos genéticos o bien, de tipo congénito y dentro de esta clasificación aquellas que son estrictamente hereditarias y que se heredan de padres a hijos. Comenzando el desarrollo en este tipo de estudios, para usted es bueno que busque más información en los siguientes temas y que aquí, por razones de espacio sólo se dará un breve espacio. En genética se considera la variación del número de cromosoma como un tipo de mutación, denominada mutación cromosómica. Otra variedad de mutación cromosómica son las delecciones, inversiones, etc., que ya se ha mencionado antes.
Aberraciones cromosómicas: Estas se pueden observar por cambios en el fenotipo. Normalmente el material genético se mantiene constante en las células somáticas, las cuales poseen un número diploide de cromosomas, o sea, dos juegos haploides de 23 cromosomas que se analizan en el cariotipo. Accidentalmente pueden producirse cambios en el cariotipo y ello tiene diversas consecuencias. Por ejemplo, cambios en el número de cromosomas, o en la estructura. Esto recibe el nombre aberraciones cromosómicas numéricas y aberraciones cromosómicas estructurales.
Poliploidias: Las células contienen múltiplos del número haploide de cromosomas, diferentes del diploide. En la poliploidia, se describe la modificación en el número de conjuntos haploides, pero el cariotipo aparece equilibrado. En las células triploides existen tres conjuntos haploides similares, en las tetraploides cuatro, etc. En las células somáticas las Poliploidias pueden surgir debido a duplicación del número de cromosomas sin que se logre alcanzar la citocinesis, también en los gametos se produce por la no separación de los cromosomas en cualquiera de las divisiones meióticas.

137
Aneuploidias: Este estado se describe como la ganancia o pérdida de uno o más cromosomas, de tal modo se llega a un desequilibrio, sin embargo, la información de los cromosomas permanece. La anomalía se produce por falla en la no disyunción de los cromosomas homólogos durante la anafase ya sea en mitosis o meiosis, esta última es la más informada. La meiosis no disyuntiva origina gametos aneuploides, que en la fusión con gametos normales puede dar origen a cigotos monosómicos o trisómicos. Una variedad importante son los casos en que se presenta nulisomía, condición que es letal. Los procesos moleculares que fallan en la no disyunción aún se desconocen, a pesar de los años que han pasado desde que el fenómeno se conoce. Pero, se ha observado en experimentos que la frecuencia de no disyunción puede incrementarse si se interfiere la acción de los microtúbulos. Las observaciones también indican que la no disyunción es más probable durante la meiosis I, puesto que la disyunción normal durante anafase I requiere que se mantenga el emparejamiento adecuado de los cromosomas de los cromosomas homólogos durante la profase I y la metafase I. Por su parte, la correcta disyunción durante la anafase II o durante la mitosis depende sólo de que el centrómero de cada cromosoma se divida de forma adecuada, por tanto, no requiere un proceso de mayor complejidad como ocurre en la profase y metafase. La no disyunción durante la meiosis I, se puede considerar como una falla en la formación o mantenimiento de la tétrada hasta anafase I. Algunas observaciones indican que los entrecruzamientos de las cromátidas, que permiten las asociaciones y la recombinación pueden ser esenciales para la disyunción. El fundamento de tal hipótesis es que en animales de experimentación, aquellas mutaciones que interfieren con la recombinación provocan incrementos sustanciales de no disyunción.
Monosomía-Ejemplo Síndrome de Turner: Fenotipo reconocible porque son mujeres estériles, de estatura baja, suelen tener repliegues membranosos entre el cuello y los hombros. Su inteligencia se acerca a lo normal, aunque están afectadas algunas funciones cognitivas. Se trata de la monosomía con respecto a los cromosomas sexuales, su nomenclatura es 44 X0, lo que significa 44 autosomas y un cromosoma X. La frecuencia en la población mundial es de 1 por cada 5000 mujeres. Los individuos monosómicos para cualquier autosoma mueren en el útero.
Trisomías- ejemplo Síndrome de Klinefelter: La trisomía produce un desequilibrio cromosómico y ocasiona anormalidades o la muerte del individuo. Sin embargo, se encuentran muchos casos de individuos trisómicos viables. Por ejemplo, en la trisomía XXY, que surge en 1 de cada 1000 nacimientos de varones. El cuerpo se presenta alto desgarbado, presentan retraso mental y son estériles. Otro ejemplo interesante para la discusión es el caso de la combinación XYY, que ocurren en cerca del 50% de los varones. Se han presentado controversia con respecto a la idea de de que esta combinación estaría asociada con conductas de violencia. Estos varones son fértiles.

138
El caso de mayor importancia en el ámbito de la psicología y por supuesto, de aquellas profesiones que se relacionan con el estudio del aprendizaje y la conducta, es el síndrome de Down. Es un tipo de aneuploidia viable, aparece con una frecuencia de 015% de todos los nacimientos, no obstante que el síndrome puede aparecer con una translocación, lo más frecuente es que se deba a una trisomía del cromosoma 21. Se conoce otros dos tipos de trisómicos para autosomas que sobreviven hasta el nacimiento; estos son: el síndrome de Patau, trisomía del cromosoma 13, síndrome de Edward, trisomía del cromosoma 18. En ambos casos los fenotipos indican anormalidades físicas y mentales. En la trisomía 13 se observa labio leporino, cabeza pequeña y deformada, pies arqueados y una esperanza de vida de 130 días. Con respecto a la trisomía del cromosoma 18 se observa posición baja de las orejas, mandíbula pequeña, pelvis estrecha y pies arqueados, mueren a las primeras semanas del nacimiento.

139
LA CÉLULA NERVIOSA- LA NEURONA
No nos excluyamos, nosotros los seres humanos pertenecemos a la clase de los mamíferos del reino animal. Al igual que los otros animales percibimos y nos comunicamos con el entorno por medio de los órganos de los sentidos, luego emitimos respuestas a los estímulos tales como, luz, color, frío, calor, sonidos, dolor, fragancias, sabores, etc. Estos estímulos producen respuestas en órganos especializados como el ojo, la piel, el oído, la nariz o la lengua. Estas respuestas, se producen por la transmisión de señales eléctricas que se propagan por medio de las células nerviosas desde diversas regiones del cerebro. Desde las regiones del cerebro las respuestas son elaboradas y procesadas. A veces las respuestas son emitidas desde la médula espinal o de regiones subcorticales sin que se registre el hecho en la conciencia. Funcionamos de la misma forma que funcionan los animales, las partes del organismo necesitan funcionar en armonía, como un todo. Para ello nos sirven las neuronas y todo lo que acontece en su interior, es fundamental conocerlo para dar sentido al conocimiento de si mismo.
A medida que usted avanza en la lectura, podrá vislumbrar los fenómenos celulares y por supuesto que todo ello involucra moléculas, iones, e interacción entre ellas darán lugar en primer lugar, fenómenos observables desde los aspectos microscópicos hasta los de tipo macroscópicos, entre ellos las asociaciones entre tejidos y las comunicaciones entre estos que llegan a constituir la función organismo. Las funciones del riñón, o del corazón, cuyas funciones macroscópicas las podemos percibir mediante la formación de la orina, en el caso del corazón, las podemos percibir con los latidos. La base de todo aquello que podemos percibir tiene su representación en el nivel de lo más pequeño, de aquello que es microscópico.
Con el comportamiento ocurre algo similar, cualquier comportamiento que usted muestre, podrá describirlo en términos de interacciones con el medio, con el mundo. Estas conductas tienen su explicación a nivel molecular, celular. En el establecimiento del conocimiento es necesario que se encuentren los correlatos, correlaciones entre conductas y funcionamiento o entre producciones de neurotransmisores por las neuronas y conductas. Las conductas vistas y observadas al nivel macro, en nuestras relaciones e interacciones con nosotros mismos y el resto del mundo deben estar acompañadas de incrementos o decrementos de señales en la neurona, estas a su vez habrán de movilizar organelos, sintetizar proteínas, activar genes, etc.
Una de las últimas propuestas para comprender la actividad nerviosa que denominamos consciencia es de Penrouse, indica que la consciencia se puede explicar por actividades de microtúbulos y de procesos que ocurren en este nivel, las actividades involucran teoría cuántica, tomada desde las ciencias físicas.
Evidentemente, el nombrar la conciencia puede conducir al borde entre ciencias y la filosofía. Pero los alumnos en más de alguna ocasión durante su desarrollo se preguntarán ¿qué es la conciencia? Posee algún sustrato biológico y si es así, ¿Dónde se ubica en nuestro estudiado cerebro. Es aún un misterio. Se recomienda la lectura de Damasio y de Koc…..
Kandel se ha referido a estos misterios de la conciencia como la última frontera de las ciencias biológicas, “su reto definitivo, es la comprensión del sustrato biológico de la conciencia y los procesos mentales por los que percibimos, actuamos, aprendemos y recordamos. Kandel me trae buenos recuerdos, en el caso de algunos experimentos y

140
mediciones del aprendizaje de las ciencias, ante hechos que deseaba abordar desde la perspectiva de la psicología del aprendizaje, terminé complementando, (muy bien), con aspectos de la biología celular y molecular, entre ellos citando algunos artículos del brillante investigador. En suma aconteció que la psicología y la biología celular y molecular se encontraron y complementaron en un estudio de cómo aprenden los seres humanos.
Estamos con la moda de hoy, en la biología se ha adquirido la capacidad de determinar la secuencia de genes, de aminoácidos en una proteína. Por ello, invito al alumno de que participe en el proceso unificador de la biología en el estudio de la conducta, gracias al conocimiento de la genética hoy en día los procesos mentales y su dinámica nos parece más comprensible.
CELULAS NERVIOSAS
Los estudios histológicos del siglo XIX de Cajal, de Golgi y de sus sucesores condujeron al consenso de que las células del sistema nervioso pueden ser divididas en dos clases: las neuronas (o células nerviosas propiamente dichas) y las células de sostén. Las primeras están especializadas en la transmisión de señales eléctricas a gran distancia, mientras que las segundas no tienen actividad. En el sistema nervioso central (encéfalo y médula espinal) estas células de sostén consisten principalmente en células neurogliales.
Desde el punto de vista morfológico las neuronas presentan gran variabilidad en su forma. Sin embargo, presentan características comunes que les diferentes de las células de los otros tejidos. Las funciones celulares están compartimentalizadas de tal manera que la disposición de sus componentes debe dar lugar a que ocurra en ella su principal función, que es la comunicación mediante señales eléctricas.
Las neuronas muestran polarización denotada por las siguientes estructuras fundamentales: cuerpo celular, dendritas, axones y terminales. Estas se encuentra separadas por distancias bastantes considerables y que ayuda al establecimiento de polaridad funcional.
Las células nerviosas se asemejan en la mayoría de los rasgos a las células normales. Se reconocen fácilmente, las siguientes estructuras:
• núcleo • retículo endoplásmico • aparato de Golgi • mitocondrias y orgánulos • membrana célular, etc
En la mayoría de las neuronas o célula nerviosa se presenta un cuerpo celular o soma con prolongaciones protoplasmáticas denominadas dendritas y axón, prolongación cilíndrica, a veces de gran longitud el soma contiene al núcleo y los organelos. La neurona, célula

141
altamente especializada se caracteriza por realizar funciones relacionadas con la génesis, la propagación, transmisión, integración de señales u unidades básicas de información.
El curso de la evolución ha conducido que las células alcancen estados maduros donde realizan sus funciones, luego este estado se caracteriza por permanecer estable, la célula alcanza así su estado final de maduración. La neurona no escapa al principio y una vez establecido su función y estado final de diferenciación pierde la capacidad de dividirse. La capacidad de regenerarse del sistema nervioso es my inferior a las de otros sistemas tales como el sistema sanguíneo.
Por esta razón cualquier injuria que recibe el sistema nervioso central, sobretodo, el organismo presenta déficit o pérdida, o deterioro de funciones tales como el habla, pérdida de memoria, alteraciones sensoriales, pérdida de capacidades motrices, etc.

142
Estructura y Ultraestructura de la neurona /Procesos de señalización intercelular/ Sinapsis y conducta. Desde el microcosmos hacia el macrocosmos.
Estructura y función /Obtención de Energía
Energía: La neurona como toda célula animal necesita de la obtención de nutrientes para la obtención de energía. La mayor parte de esta energía necesaria para realizar los trabajos celulares proviene del ATP, molécula que es producida en el citoplasma y en las mitocondrias de las células.
La energía es tomada de las moléculas de ATP, pues en esta estructura que ha sido descrita más arriba se encuentra depositada en las uniones químicas entre los fosfatos del ATP, estas son las uniones de alta energía. Cuando el ATP se rompe o más hablando en términos más técnicos, se hidroliza, se libera energía y se genera ADP (Adenosín Di fosfato) y una molécula de fosfato.
Las estructuras que principalmente generan ATP son las mitocondrias, por medio de procesos que consisten en la captura de energía que se encuentra depositada en los enlaces covalentes de las moléculas que en principio derivan desde los alimentos que en sucesivas transformaciones adquieren la forma necesaria que permite el proceso de capturar energía. El ADP resultante posterior a la entrega de energía ingresa a la mitocondria para la regeneración de ATP.
La energía de los alimentos proviene del sol. El flujo energético en la biósfera comienza cuando los vegetales utilizan el CO2 y el H20 y la energía luminosa para dar curso a una serie
ATP ( trifosfato de adenosina o adenosín trifosfato): es una molécula derivada de
los nucleótidos, es vital para la
obtención de energía en los seres vivos.
Lo componen una base nitrogenada
(adenina) unida al carbono 1 de un
azúcar de tipo pentosa, la ribosa, que en
su carbono 5 tiene enlazados tres grupos
fosfato. Se encuentra incorporada en los
ácidos nucleicos.
Las células necesitan energía para
sintetizar y degradar moléculas,
transporte de solutos en contra de
gradientes electroquímicos a través de
sus membranas (transporte activo),
incorporación de macromoléculas,
incorporar moléculas y partículas desde
el exterior (endocitosis) y secretar
macromoléculas al medio
extracelular, lo que se denomina
exocitosis, tráfico de sustancias,
organelos dentro de la célula.

143
de reacciones químicas. La energía lumínica queda transformada en energía química depositada en las uniones covalentes (fuertes y de alta energía) de las moléculas que conforman el cuerpo del vegetal. Esta energía es tomada por los animales herbívoros que a su vez constituyen la fuente de energía de los animales carnívoros.
Las sustancias alimenticias son los hidratos de carbono, lípidos (grasas y aceites), proteínas. No toda la energía depositada en este tipo de moléculas se transforma en ATP, parte de la energía es transformada en calor.
La energía que utilizan las neuronas son de utilidad principalmente para la mantención de flujos e intercambio de iones entre el citoplasma neuronal y su medio, de este intercambio se observa el mantenimiento de los equilibrios, o de homeostasis entre la célula y medio. Ello mismo facilitará las condiciones que permiten el flujo de la información que transita por las prolongaciones neuronales en forma de potenciales de acción.
Las neuronas al igual que las demás células extraen la energía mediante oxidaciones de las moléculas alimenticias, luego de las cuales el oxigeno se une al hidrógeno proveniente de las oxidaciones para formar agua.
Los productos alimenticios una vez que han pasado por los procesos de la digestión que involucran reacciones enzimáticas, se degradan a hidratos de carbono del tipo monosacáridos como la glucosa, los lípidos se transforman en ácidos grasos y glicerol y las proteínas a aminoácidos. Estas moléculas son absorbidas por el epitelio intestinal e ingresan a la sangre desde donde pasan al interior de las células. En el interior de la célula los monosacáridos como la glucosa pasa por dos procesos de liberación de energía. La Primera de ellas en el citoplasma es denominada glucólisis y la segunda ocurre en el interior de la mitocondria. La segunda provee de mayor cantidad de energía.
La mayor parte de los ácidos grasos son degradados en la mitocondria, puesto que ingresan a la célula y luego al interior de ellas. Los aminoácidos también pueden ser utilizados para generar energía, se pueden utilizar para otros fines, pero en general, su destino es formar estructuras proteicas.
Glucólisis: Este proceso es llevado a cabo en el citosol y son una serie de 10 reacciones
químicas mediadas por enzimas. Se inicia con una molécula de glucosa que posee 6 átomos

144
de carbono, que posteriormente finaliza en estas reacciones como dos moléculas de piruvato, cada una compuesta por 3 carbonos
Entonces, la glucólisis es el primer paso de la respiración, es una secuencia de reacciones que se realizan en el citoplasma de la célula y por el cual la molécula de glucosa sufre transformaciones llegando a formar dos moléculas de ácido pirúvico.
Es la vía metabólica más difundida en la naturaleza, también se le ha denominado vía de Embden-Meyerhof, se encuentra presente en una gran mayoría de seres vivos. La vía está promovida por una serie 10 enzimas que se encuentran en el citoplasma de la célula, pero no en las mitocondrias.
Este proceso puede continuar con la respiración celular (si existe oxígeno) o con la fermentación (en ausencia del oxígeno).
Al iniciarse el proceso de glucólisis, cada molécula que posee 6 átomos de C se oxida y degrada hasta dar origen a 2 moléculas de piruvato, cada una de estas dos moléculas resultantes aún poseen 3 átomos de carbono. En este proceso se invierte la energía de 2 moléculas de ATP y generan al final 4 moléculas de ATP. Por lo tanto la ganancia es energía que se almacena en 2 moléculas de ATP. Sin embargo, parte de la energía liberada durante la glucólisis no es transferida y dar forma a la molécula de ATP. En efecto, la reducción de dos moléculas de NAD+, una por cada molécula de piruvato obtenido, es la forma de energía transportada que luego se transfiere al ADP para formar ATP, en el interior de la mitocondria. Las moléculas resultantes piruvatos ingresan a la mitocondria. En este proceso parte de la energía liberada es transferida indirectamente a la célula mediante la reducción de dos moléculas de NAD+, uno por cada molécula de piruvato.
NAD+ es la abreviación de dinucleótido de nicotinamida adenina. El signo + indica la forma oxidada. LA forma NADH indica que la forma oxidada se ha reducido. En las reacciones bioquímicas desempeña el papel de coenzima. La función es captar electrones e hidrogeniones que posteriormente transportará a la mitocondria para obtención de más energía.
.
El NAD es un dinucleótido, ya que consta de dos nucleótidos unidos a
través de sus grupos fosfato con un nucleótido que contiene un anillo
adenosina y el otro que contiene nicotinamida.
En el metabolismo, el NAD+ participa en las reacciones redox
(oxidorreducción), llevando los electrones de una reacción a otra. El
NAD+, que es un agente oxidante, acepta electrones de otras moléculas
y pasa a ser reducido, formándose NADH, que puede ser utilizado
entonces como agente reductor para donar electrones. También es
utilizado en otros procesos celulares, en especial como sustrato de las
enzimas que añaden o eliminan grupos químicos de las proteínas, en
modificaciones post-traduccionales.

145
Este primer paso de la glucólisis es identificado como una de las fases del metabolismo anaeróbico. El término deriva de que los organismos anaerobios son seres vivos que viven en ambientes sin oxígeno y pueden obtener su energía metabólica por este proceso. Los organismos aeróbicos que dependen de ambientes con presencia de oxígeno, también pueden obtener energía mediante este proceso, pero la oxidación completa de la glucosa, para lo cual utiliza el oxígeno en el interior de la mitocondria, conduce a una mayor obtención de energía. De manera que la glucólisis es una ruta casi universal para la obtención de energía en los seres vivos, es una ruta central que posee intermediarios metabólicos para otras rutas o vías de tipo bioquímicos que en síntesis permiten se desarrolle la vida.
El cerebro utiliza glucosa como combustible, casi como única fuente de energía. Sin embargo, las células del resto de nuestro organismo pueden utilizar otros azúcares, y estos a su vez pueden convertirse en moléculas intermediarias de la glucólisis.

146
En la figura siguiente se puede observar la formación de ATP en el citoplasma, la formación de NADH que transporta los iones hidruro hacia la mitocondria, también se observa el piruvato resultante de la glucolisis que es descarboxilado, formando los grupos acetilo. La descarboxilación oxidativa se produce en la matriz mitocondrial.
Cada molécula de piruvato es
atacada por un complejo
multienzimático denominado
piruvato deshidrogenasa,
presente en la mitocondria. Se
ha descrito tres tipos de
piruvato deshidrogenasa y son:
1.-Piruvato deshidrogenasa (acetil transferidora) o PDH.
• Piruvato deshidrogenasa (NADP+).
• Piruvato deshidrogenasa (citocromo).
En los tres tipos, su función
es realizar la
descarboxilación oxidativa
del piruvato a acetato
desprendiéndose dióxido de
carbono.

147
La matriz mitocondrial contiene entre otras proteínas, aquellas enzimas que forman el complejo enzimático piruvato deshidrogenasa que lleva a cabo la descarboxilación oxidativa convirtiendo al piruvato en acetil CoA, se genera CO2, y energía para formar un NADH. El detalle de la reacción es el siguiente: Cada molécula de piruvato de 3 átomos de carbono se convierte en una molécula de 2 átomos de carbono que recibe el nombre de acetilo. Luego esta última se une a una coenzima, llamada coenzima A (CoA), con ella forma la llamada la acetilcoenzima A o acetil CoA. El carbono que ha sido removido del piruvato junto a dos átomos de oxígeno produce una molécula de CO2. El piruvato en esta reacción pierde
además un ión hidruro o H- (Un H+ y dos e-). Esta es la serie de reacciones conocidas como descarboxilación oxidativa.
EL ión hidruro debe transportarse hacia la matriz mitocondrial y lo hace mediante la unión con NAD+. El proceso de unión es en realidad un proceso de reducción del NAD+ que en realidad se convierte en una molécula diferente que el NADH. En terminología usada por los químicos la reacción es de oxidación del piruvato que en si es una parte de la oxidación total de la glucosa. Muchas de las reacciones que siguen a continuación en el ciclo de Krebs son reacciones de oxidación de compuestos que provienen de la glucosa, partiendo de la misma acetil CoA. Los iones hidruros que son protones y electrones, impulsarán la síntesis de ATP en la matriz mitocondrial.

148
Figura arriba: Ciclo de Krebs
Cerebro y glucosa: (TEMA PARA DISCUSIÓN). Neuronas
El cerebro posee elevado coeficiente metabólico, necesita de mucha azúcar, o glucosa principalmente para suplir las exigencias de energía necesarias para la actividad cerebral. El suministro de sustancias energéticas, depende del sistema circulatorio, específicamente de la sangre. La glucosa es captada en una proporción de 15 mmol/g de cerebro por hora. Una captación de oxígeno de 80-90 mmol/g de cerebro por hora, el dióxido de carbono liberado a la sangre venosa de valores similares a la de oxígeno. Ello permite inferir que la glucosa se oxida casi totalmente a CO2 y explica porque en la sangre venosa aparecen cantidades relativamente pequeñas (5-13%) de lactato y piruvato.
El metabolismo de la glucosa, conduce mediante series de reacciones químicas dióxido de carbono, agua, piruvato y lactato. Otras propiedades características del metabolismo de la glucosa cerebral son la formación de ácido g-aminobutírico (GABA) y la generación de grandes depósitos de glutamato y aspartato.
Existe una correlación consistente entre los niveles de actividad funcional y los niveles de captación de glucosa y de fosforilación en el sistema nervioso. Esto implica que, durante la respuesta funcional, se esta llevando a cabo una modulación rápida, prolongada y efectiva del metabolismo energético, que se acompaña de incrementos equivalentes en el flujo cerebral local. Esto viene a remarcar que los requerimientos energéticos varían constantemente en lo referente a aquellos componentes que utilizan energía del sistema nervioso: los relacionados con la producción de actividad eléctrica, con el tráfico de impulsos nerviosos y con la restauración de los potenciales de membrana neuronal. Esencialmente, este uso de la energía se centra sobre la actividad de la bomba de sodio, Na+K+-ATPasa, que es responsable del transporte interno acoplado del K+ y del transporte externo de Na+ para mantener los potenciales de membrana en reposo y restaurar sus gradientes iónicos

149
asociados tras la actividad eléctrica. La proporción de consumo de energía debido a este proceso clave es del 40%.
En años recientes Raichle en la revista Science: el cerebro humano consume un 20% de la energía del cuerpo vía alimentos consumidos, pero su peso representa sólo el 2%. Anteriormente ya se había realizado experimentos de medición de consumo de oxígeno en estudiantes mientras estos se encontraban trabajando en la resolución de problemas. Se mostró que el cerebro no consumía más oxígeno durante la actividad pensante. La cantidad de oxígeno consumida es similar al estado de descanso con los ojos cerrados. En realidad este órgano consume muy poca energía, según Raichle sus estudios muestran que el 60 – 80% de la energía del cerebro está dedicado a mantener la conexión entre las neuronas. Entre un 0,5-1 % está orientado a respuestas o demandas del medio externo. El cerebro recibe señales constantemente del medio, luego estas siguiendo procesamientos internos, sin embargo, las investigaciones muestran que las actividades de razonamiento gastan muy poca energía.
Las mitocondrias y los peroxisomas utilizan oxígeno molecular. Las mitocondrias son generadoras de ATP. Los peroxisomas participan en procesos de desintoxicación mediante reacciones de peroxidación y rompimiento de moléculas de peróxidos de hidrógeno.
Por lo general todos los organelos presentes en el citoplasma del cuerpo celular también se encuentran en las dendritas. Pero su concentración disminuye proporcionalmente a medida que la prolongación se encuentre más alejada del soma o cuerpo. En el axón la situación es un tanto diferente, por ejemplo, el retículo endoplasmático rugoso, los ribosomas el aparato de Golgi. En suma, aquellas estructuras relacionadas con la síntesis de proteínas de las neuronas no se encuentran en los axones. Asimismo se excluye también a los lisosomas.
En Neuronas:
La escasa renovación de las células neuronales una vez que degeneran o una vez que desaparecen miles de neuronas, no hay renovación de estas. Sin embargo, este fenómeno se está reconsiderando en la actualidad, hay investigaciones indicadoras de neurogénesis, por lo menos es seguro que muchas funciones desaparecidas debido a la muerte neuronal vuelven a surgir por la acción de otras neuronas que reemplazan en funciones a las células desaparecidas, Ese es el caso, por ejemplo, de aquellas personas que han sufrido infarto cerebral y que tiempo después han recuperado sus funciones motoras.
Pero, los axones son ricos en vesículas sinápticas y otros tipos vesiculares que participarán de los procesos de transmisión de señales. También en estas estructuras podemos encontrar mitocondrias y retículo endoplásmico liso cuya principal misión es guardar iones de calcio y participación en el metabolismo lipídico.

150
Citoesqueleto la matriz citoplasmática
La matriz citoplasmática o citoplasma fundamental de una célula eucariótica posee casi los mismos componentes que una célula de tipo procariótico, poseen por ejemplo, enzimas, proteínas globulares, enzimas, metabolitos diversos. Sin embargo, las células eucariotas y entre ellas, las neuronas, poseen estructuras más sofisticadas, de mayor complejidad. Algunos de estos nuevos componentes que surgieron dentro de los procesos evolutivos de los seres vivos y que acontecieron en millones de años, son los elementos del citoesqueleto y las membranas intracelulares del sistema endomembranoso o vacuolar. En el proceso evolutivo, surgieron también las mitocondrias, cloroplastos y peroxisomas, en resumen aparecieron diversos compartimientos y subcompartimientos.
En toda célula eucariótica en el citoplasma se puede reconocer dos regiones principales: La región que involucra e incluye a los organoides y estructuras membranosas y zona representada por la matriz citoplasmática o citosol. De esta última dependen las propiedades coloidales de la célula, por ejemplo, de las transformaciones de sol a gel, las modificaciones de la viscosidad. En esta matriz se producen numerosas reacciones enzimáticas, como las reacciones bioquímicas de la glucólisis, se encuentran los ribosomas, inclusiones o cuerpos de inclusión con depósitos de glucógeno, gotas lipídicas, cristales, que no están rodeadas por membranas. La matriz contiene agua, iones, metabolitos, proteínas enzimas, etc.
El glucógeno es la forma como almacenamos la glucosa en nuestras células animales. Al obtener este nutriente energético con los procesos de digestión y llegar a las células se acumula, entonces la enzima glucógeno sintetasa citosólica comienza a unir las moléculas de glucosa formando polímeros ramificados denominados glucógeno. Estas partículas son visibles con microscopía electrónica. Así se puede identificar a los glucosomas, cuerpos esféricos de un tamaño aproximado de 30 nm, con un centro de polisacárido rodeado por una envoltura proteica consistente en enzimas para la síntesis de la estructura, o bien para su disgregación. Los glucosomas forman acúmulos mayores denominadas partículas alfa.
Las células del hígado conocidas como hepatocitos, células musculares denominadas miocitos, las células de la sangre leucocitos neutrófilos, etc., almacenan glucógeno en grandes cantidades y a veces de tal magnitud que son visibles a microscopía óptica. En patologías como la diabetes o glucogenosis se acumulan a los niveles mencionados. Luego se hará referencia a estas enfermedades por la relevancia que poseen en el área de la medicina o de la neurobiología.
Los hepatocitos sólo acumulan glucógeno para luego destinarlo a los demás tejidos y órganos, previo envío a la sangre cuando la concentración de glucosa se reduce a valores bajo lo normal, aproximadamente un gramo por litro de sangre. La disminución de glucosa en la sangre se denomina hipoglucemia y se produce normalmente después de varias horas de ayuno.

151
Cuando se dan las condiciones de hipoglucemia se produce la glucogenólisis para liberar glucosa, esta acción la realizan enzimas que se encuentran en la superficie de los glucosomas. Estas proteínas enzimas, las glucógeno fosforilasas degradan el glucógeno con lo cual se libera la glucosa. Luego los hepatocitos inician el movimiento de la glucosa hacia la sangre.
Citoesqueleto
Determinante para la forma de la célula y de la neurona, célula que especialmente debemos estudiar para iniciar nuestros estudios de la neurobiología, es la expresión de genes que conducen a la síntesis de proteínas del citoesqueleto, este tipo de estructura es la responsable en cierta medida de la distribución asimétrica de los organelos en el citoplasma. Tres tipos principales de estructuras filamentosas: microtúbulos, neurofilamentos (filamentos intermedios en otras células) y los microfilamentos de actina. Todas estas estructuras son fácilmente visibles al microscopio electrónico.
Las proteínas que forman el citoesqueleto otorgan una organización fibrosa del citoplasma.
Microtúbulos
En general, los microtúbulos se encuentran en todos los tipos de células eucariotas y forman estructuras como el huso mitótico, los elementos que recorren la longitud de los axones en las neuronas, en los cilios, flagelos, etc. Físicamente aparecen como cilindros alargados de 20 a 30 nm de diámetro. La forma de visualizarlos al microscopio electrónico es usando la técnica que incluye glutaraldehido como fijador, también se utilizan técnicas inmunocitoquímicas.
La pared de cada microtúbulo está integrada por proteínas de estructura globular de un diámetro aproximado de unos nm, se disponen formando 13 columnas longitudinales, estos son los protofilamentos y dejan un centro sin estructuras.
Los microtúbulos están compuestos principalmente por la tubulina llegando a constituir el 85 % del total. El 15% restante se compone por proteínas microtubulares asociadas o MAPs.
Los microtúbulos forman armazones que se extienden por toda la célula. Uno de ellos puede alcanzar una longitud de 1 mm, están formados por dímeros de moléculas de tubulina, estas son de las formas alfa y beta. El microtúbulo crece por adición de dímeros α y β al extremo positivo del microtúbulo, en tanto, que su extremo negativo crece menos. Por medio de esta actividad esta estructura se mantiene constantemente en procesos de organización y reorganización. Estos, procesos involucran acortamiento y alargamiento de los microtúbulos o dicho de una forma más técnica, son procesos debido a un desequilibrio entre polimerización y despolimerización.
El citosol mantiene un pool de dímeros de tubulina, lo cual no necesita de síntesis de proteínas en forma constante. La polimerización requiere de iones Ca+2 y de las proteínas MAPs. Las unidades diméricas de las formas α y β, forman una unidad que se organizan

152
siempre “cabeza con cola”, esta polaridad indica que a la subunidad alfa, le sigue la subunidad beta.
Centro organizador de microtúbulos
Dentro de la célula existen centros organizadores de microtúbulos (COMTs) representados fundamentalmente por los centrosomas y por los cinetosomas o cuerpos basales de los cilios, a partir de ellos se produce el surgimiento de los polímeros. El comienzo de la formación de los microtúbulos se denomina “nucleación”
In vitro, es posible sintetizar o elongar microtúbulos con soluciones de microtúbulos en presencia de GTP y Mg+2 ,. Las observaciones en estos experimentos indican que los microtúbulos crecen más rápidos por uno de los extremos, por ello se le denomina extremo de crecimiento rápido (+), por oposición el otro extremo de crecimiento es más lento y se denomina extremo menos (-).
El centrosoma está formado por estructuras en forma de anillo que contiene otra forma de tubulina, la gama tubulina. Estos anillos actúan como centros de nucleación (crecimiento) de microtúbulos. Los dímeros de tubulina se añaden al anillos de gama tubulina con una orientación específica, siempre el "extremo -" queda dentro del centrosoma y el crecimiento se produce por el "extremo +".
Existe un comportamiento en la formación de los microtúbulos, se elongan a partir del centrosoma, luego detienen su crecimiento y se acortan hasta desaparecer o retomar el crecimiento. Todas estas variaciones se producen continuamente y principalmente polimerización y despolimerización, este fenómeno se ha denominado “inestabilidad dinámica de los microtúbulos y permite una rápida redistribución de los microtúbulos citoesqueléticos en el citoplasma en la división celular, los cambios de forma, distribución de organelos y vesículas en el citosol.
La elongación depende de la asociación de tubulina y GTP. Requiere en primer lugar de la presencia del dímero de tubulina unido a dos moléculas de GTP. Uno de los GTP permanece unido sin cambios a la tubulina α, la otra molécula de GTP se une a la tubulina β, y luego se hidroliza hasta GDP una vez que el dímero se incorpora al extremo (+).

153
Experimentalmente, se obtiene que los heterodímeros de tubulina poseen capacidad de unión a la colchicina o una derivado de este, el colcemid, estos evitan que se agreguen más dímeros a los extremos (+), ello produce una rápida desaparición de microtúbulos como los del huso mitótico. La propiedad también la poseen otros alcaloides diferentes, tales como la vinblastina y vincristina. Ambas se utilizan como drogas en el tratamiento de tumores por ser claramente antimitóticas. El taxol es de acción opuesta a la de la colchicina, se une a los microtúbulos e impide la despolimerización de la tubulina, por tal razón también, al igual que las anteriores sustancias es capaz de impedir la formación del huso y por ende de la separación de los cromosomas en la mitosis, por esto mismo se le puede utilizar en tratamientos antineoplásicos.
Las proteínas MAPs
Representan en promedio un 10% de las proteínas totales correspondientes a microtúbulos. Las proteínas MAPs estabilizan las relaciones estructurales de los microtúbulos, y de estos con diversos componentes estructurales de la célula. Algunas de estas notables proteínas actúan como proteínas motoras de movimientos o de transporte a lo largo de los microtúbulos.
En neuronas, las MAPs se encasillan en dos familias, cada una de ellas con varios participantes. Las de peso molecular alto, MAP 1 de axones y dendritas, las MAPs que se encuentran en la s dendritas, las de peso molecular bajo, conocidas como proteínas Tau que se encuentran en los axones. Una clase proteína MAP 1C conectan a los microtúbulos con vesículas, a las cuales luego transportan.
Proteínas Tau
La demencia aparece en individuos de edad avanzada. Por ejemplo un grupo de patologías denominado tauopatías. La tauopatía mejor conocida es la enfermedad de Alzheimer. Es una enfermedad de evolución lenta caracterizada por la pérdida progresiva de la memoria, la orientación, el juicio y el lenguaje. El examen postmortem de los cerebros de los individuos afectados muestra placas seniles, constituidas por proteínas amiloide, y los ovillos neurofibrilares, formados por la proteína tau. Ambas estructuras dan lugar a la degeneración del tejido cerebral y al funcionamiento deteriorado.

154
Existe una estrecha correlación entre la presencia de los filamentos helicoidales y el desarrollo de la demencia, al parecer no basta con la sola presencia del amiloide beta para producir la enfermedad. Normalmente, la proteína tau se une a la tubulina, contribuyendo a la estabilización de los microtúbulos, estos se extienden desde el cuerpo neuronal hasta el terminal nervioso, y como se ha dicho, participan en el transporte intracelular y el mantenimiento de la estructura de la célula. Hipotéticamente, la ruptura del sistema microtubular originaría defectos en el transporte axonal y probablemente degeneración celular, lo cual ha sido observado en las neuronas afectadas por la enfermedad de Alzheimer.
En el citoplasma, la proteína tau está normalmente fosforilada y este hecho regula la función de la proteína. Los filamentos helicoidales están compuestos por agregados de tau que, a diferencia de la proteína normal -que lleva unidos tres fosfatos- contiene un número elevado de estos, aproximadamente nueve, por lo cual se dice que tau se halla hiperfosforilada.
Las neuronas que muestran degeneración neurofibrilar mueren luego de un lento proceso, el espacio interneuronal que dejan es ocupado por ovillos "fantasmas" en. Es posible que la enfermedad se deba a una hipofunción de las que se encuentran en proceso degenerativo y luego con la exacerbación por muerte de las neuronas. Se descubrió que tau hiperfosforilada no tiene capacidad de promover la formación de microtúbulos ni de estabilizar a aquellos previamente formados. La proteína tau inhibe el ensamblado de la tubulina en los microtúbulos y puede desarmar aquellos formados con la proteína normal. Experimentalmente, si la proteína tau es defosforilada por tratamiento con fosfatasas, recupera su actividad biológica normal, lo que confirma la hipótesis en cuanto a que la fosforilación anormal de tau constituye para las neuronas una señal errónea, ya que en esas condiciones la proteína no cumple su función biológica y, además, destruye los microtúbulos que ya estaban formados.
Figura: Fuente: http://www.cienciahoy.org.ar/hoy41/protei1.htm
También algunas investigaciones dan cuenta de que las mutaciones en los genes para la codificación de la proteína Tau implica el desarrollo de demencias frontotemporales y ellas ocurren con deterioro del lenguaje. En estos hallazgo se asocia también la presencia de
Degeneración neurofibrilar en neuronas de corteza cerebral; obsérvese el gran ovillo neurofibrilar en el interior de una neurona. El tejido ha sido teñido con el colorante fluorescente Tioflavina S (Aumento: 400x)
Ejemplos de la forma en que se deposita el amiloideβ en el cerebro. A) Placas neuríticas que han invadido masivamente la corteza cerebral (Aumento: 40x). B) Localización extraneuronal de las placas neuríticas. Se observa la zona compacta central de sustancia amiloide en una placa neurítica (Aumento 200x)

155
ubicuitina por sobre lo normal en neuronas. La ubicuitina (proteína aparece asociada a protéinas que deben ser degradas en la célula.
Proteínas motoras
En este manual interesan especialmente dos tipos: 1) Las quinesinas y 2) Las dineínas citoplasmáticas.
Son necesarias para el transporte de macromoléculas y de organoides de una parte a otra de la célula. El transporte es guiado por elementos citoesqueléticos, en particular por los microtúbulos que se proyectan desde el centrosoma hacia la periferia celular. El movimiento se caracteriza por su rapidez. La quinesina se mueve hacia el extremo más, la dineína citoplasmática (MAP 1C) se mueve hacia el extremo menos.
Microfilamentos
Figura: Estructura de monómeros y polímeros de actina
La actina es la proteína constituyente de los microfilamentos y se encuentra en el citoplasma en dos formas, como forma de actina G, monomérica, de tipo globular y la actina de forma F, polímero de tipo helicoidal doble, de forma filamentosa, conformada por subunidades de actina G.
Estas estructuras de actina, en particular de la actina F tienen un diámetro mucho menor que los microtúbulos y que los filamentos intermedios, de ahí el nombre de microfilamentos. Su diámetro es de 6 -8 nm.
Los microfilamentos son estructuras polarizadas, tal como se observa en los microtúbulos, presentan un extremo de crecimiento rápido (+) y uno de crecimiento lento (-). Estas estructuras son fundamentales para dar lugar a los movimientos celulares, para determinar la forma celular, adhesividad entre células y matrices. La función principal es la participación en la construcción del esqueleto de membrana y para la mantención de estructuras permanentes relacionadas con la membrana. Tiene también su importancia en la formación de memoria de largo plazo en relación con la potenciación a largo plazo que resulta de

156
estimulaciones tetánicas de la neurona. La importancia en este caso radica en la participación de las espinas dendríticas que se relacionan con el aprendizaje y memoria.
Filamentos intermedios
Son estructuras muy estables, resistentes a cambios de temperatura, forman redes intracelulares y son parte de las uniones intercelulares. Están compuestos de proteínas fibrosas que se asocian como dímeros helicoidales y estos a su vez forman tetrámeros. Son apolares y las proteínas constituyen las llamadas láminas A, B y C que forman una capa fina en la superficie interna, en conjunto recibe el nombre de lámina fibrosa.
Otras protéinas forman filamentos intermedios y son la citoqueratina (de los epitelios), glioproteína fibrilar ácida (Astrocitos y células de Schawn), desmina (Músculo), neurofilamentos (Neurona), vimentina (Presente en derivados mesenquimáticos).
Los neurofilamentos poseen unos 10 nm de diámetro. Son los componentes fibrilares más abundantes en el axón, forman el citoesqueleto de las neuronas. Se distribuyen uniformemente con orientación paralela al eje longitudinal de las prolongaciones celulares.
Los microfilamentos de 3-5 nm de diámetro, son las fibras más finas del citoesqueleto, compuestos de moléculas de actina que forman polímeros cortos en el ectoplasma estableciendo redes con otras proteínas tales como espectrina, anquirina.

157
MEMBRANAS BIOLÓGICAS DE CÉLULAS
EN GENERAL Y NEURONAS
En las clases y cursos anteriores se ha planteado de forma reiterada que el establecimiento de la membrana plasmática en los seres vivos constituye un paso fundamental en la evolución de los seres vivos. La separación de la materia animada de la inanimada debe de necesitar, estrictamente hablando, de una barrera que pueda establecer la diferencia entre el adentro y el afuera de la membrana-de la célula. Lo que está dentro constituye aquello que definimos como vida y lo que está fuera es aquello que no tiene vida. Adentro el orden- fuera el desorden. Ciertamente la célula debe regular sus reacciones de metabolismo y por ello debe controlar asimismo la entrada de materia prima y por ende la salida de productos y sustancias desechables. La membrana en sí constituye el mecanismo de regulación de las actividades celulares que tiene que ver con el abastecimiento. No sólo ello se ve involucrada en otros aspectos y no menos primordiales. Moléculas para construir: Para construir membranas los lípidos constituyen sustancias adecuadas. En ellos debemos reconocer algunas propiedades fundamentales para los seres vivos. Algunas grasas se utilizan para el almacenamiento de energía y aislamiento térmico. Pero, en su mayor parte los lípidos son empleados para la construcción de membranas. Recordando acerca de las propiedades del agua: las moléculas con grupos capaces de formar enlaces de hidrógeno tienden a unirse con el agua mediante este tipo de enlace. De esta manera se disuelven fácilmente los compuestos que llevan grupos hidroxilo en su molécula, las que llevan grupos amina, ésteres, cetonas, etc. Por otra parte, las sustancias que son no polares, no iónicos y que no pueden formar puentes de hidrógeno, poseen limitada solubilidad en el agua. De hecho esta propiedad tiene relación con factores energéticos, y la descripción de esta propiedad escapa a los contenidos de competencia que se dan para el desarrollo de este módulo. Cuando se trata de disolver moléculas hidrófobas como los lípidos, no se forman capas de hidratación, a diferencia de las sustancias hidrófilas. En lugar de ello, el agua forma estructura de clatrato o jaulas, similares a las estructuras de hielo alrededor de moléculas no polares. Esta sorprendente disposición ordenada de las moléculas de agua se produce por una disminución de la entropía, o aleatoriedad de la mezcla. La disminución de la entropía favorece la baja solubilidad de las sustancias hidrófobas en agua. También explica la tendencia de las sustancias hidrófobas a formar agregados en agua. Puede usted realizar el experimento de mezclar el aceite con el vinagre. El aceite forma gotitas cuando se agita la mezcla.

158
Figura Nº 1. Clatrato o jaula. Tomada de Bioquímica Mathew.2002.
SUSTANCIAS ANFIPÁTICAS Ya hemos visto que los lípidos muestran propiedades antipáticas, siendo un poco más específico, recuerde usted la estructura de un ácido graso. Los detergentes también son antipáticos.

159
Fig. En la figura observamos un ácido graso, un detergente, y un fosfolípido. Los tres tipos se caracterizan por contener estructuras antipáticas. Cuando se intenta disolverlas en agua, las moléculas anfipáticas forman monocapas en la superficie del agua donde establecen enlaces con las cabezas polares. El resto de la cadena hidrocarbonada no establece contacto con el agua. Si la mezcla se agita se forma lo que ha sido denominado como micelas. Estas son estructuras formadas por una capa de moléculas. Las colas hidrocarbonadas se orientan de forma paralela, interactuando entre ellas mediante fuerzas de Van der Waals. Las cabezas permanecen hidratadas. Esta es la forma en que puede haberse originado la membrana plasmática. Si las moléculas incluyen a los fosfolípidos por lo menos podemos construir membranas sintéticas. Propiedades de ácidos grasos Los constituyentes básicos de las membranas son los ácidos grasos y las características de su estructura determinan que sean las más apropiadas para la función que deben llevar a cabo. Sabemos que la vida tiene lugar si existe el agua, pero para que algo tenga vida es necesario establecer la diferencia entre estar y no estar animado. De manera que, como ya se planteado, es necesario establecer una barrera entre ambos estado. La barrera la constituyen los lípidos.
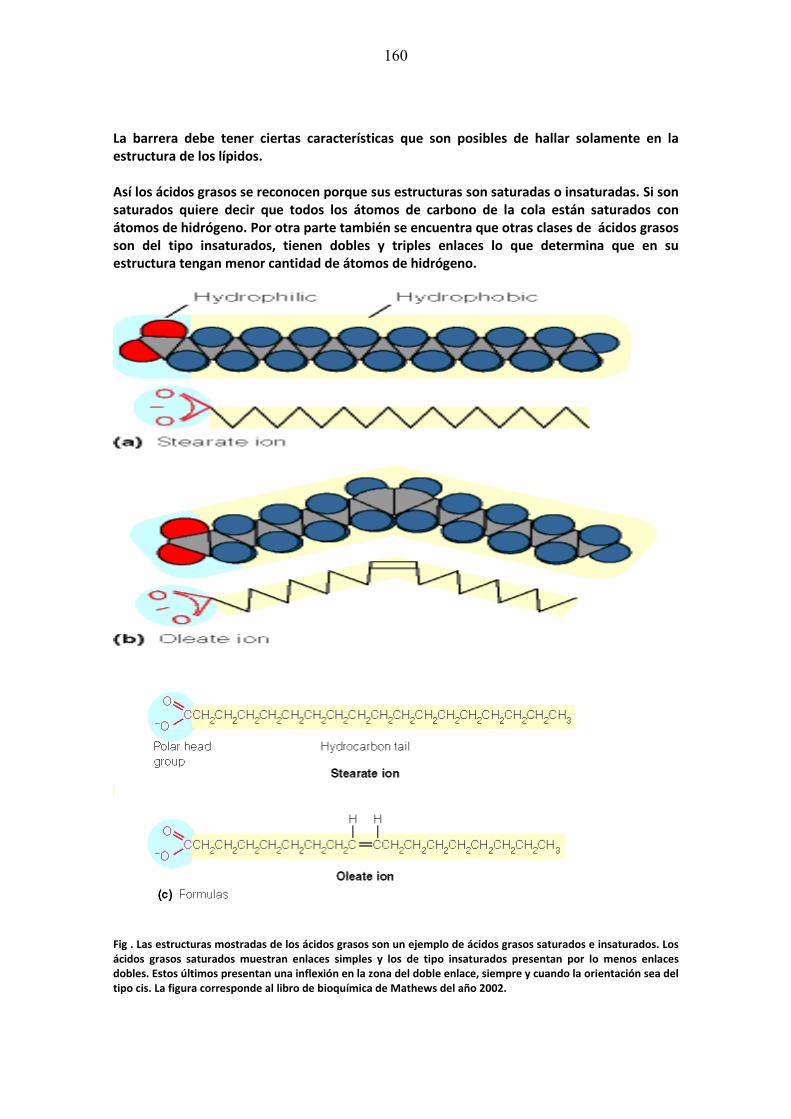
160
La barrera debe tener ciertas características que son posibles de hallar solamente en la estructura de los lípidos. Así los ácidos grasos se reconocen porque sus estructuras son saturadas o insaturadas. Si son saturados quiere decir que todos los átomos de carbono de la cola están saturados con átomos de hidrógeno. Por otra parte también se encuentra que otras clases de ácidos grasos son del tipo insaturados, tienen dobles y triples enlaces lo que determina que en su estructura tengan menor cantidad de átomos de hidrógeno.
Fig . Las estructuras mostradas de los ácidos grasos son un ejemplo de ácidos grasos saturados e insaturados. Los ácidos grasos saturados muestran enlaces simples y los de tipo insaturados presentan por lo menos enlaces dobles. Estos últimos presentan una inflexión en la zona del doble enlace, siempre y cuando la orientación sea del tipo cis. La figura corresponde al libro de bioquímica de Mathews del año 2002.

161
Los primeros estudios de la membrana, surgieron de la observación de células hinchadas cuando son sumergidas en soluciones hipotónicas, las células además se contraen y arrugan al ser sometidas a una solución hipertónica. En estos instantes de la historia (hacia el 1990), los biólogos las asemejaron a los osmómetros (sistema de dos cámaras separadas por una membrana semipermeable, que permite pasar moléculas de agua, pero no moléculas más grandes). Ello sugería que las células estaban rodeadas por una membrana semipermeable. La idea no fue bien aceptada en principio, puesto que nadie la había visto, considérese qué solo existía el microscopio electrónico. También fue puesto en duda cuando se le observó al microscopio electrónico, en época después de la segunda guerra mundial. La membrana en condiciones de observación para microscopía electrónica debe ser teñida con tetróxido de osmio y aparece como un par de líneas paralelas, pero de hecho, los científicos en aquella época no se convencieron del todo; la visión de un “artefacto” no era posible de descartar. A principios del siglo XX, Gortel y Grendel, tomaron glóbulos rojos, los vaciaron de hemoglobina, disolvieron lo que quedó de ellos (“fantasmas”) con solventes y esparcieron sobre el agua. La película que quedó sobre la superficie era la correspondiente a los lípidos de la membrana. La mancha tenía el espesor de dos veces el área de los glóbulos rojos, es decir, la mancha podía envolver dos veces al glóbulo rojo. Entonces se dedujo también que los lípidos escondían su porción hidrofóbica en el interior de la doble capa. Como puede usted haber pensado surgió un modelo de membrana celular y como modelo debe explicar satisfactoriamente algunos hechos. Hecho 1: En un tubo de ensayo con 50 cc de agua y 50 cc de aceite, se agrega una sustancia x, se agita y se deja pasar algunos minutos hasta que se separan las fases acuosa de la fase lipídica. Se puede encontrar que la sustancia x se disolvió más en el agua o en el aceite. Dicho de otra forma, encontramos que se repartió en cierta proporción- lo que se denomina- coeficiente de partición. La repetición de la prueba con una variedad de sustancias y que alguna de ella tienen mayor capacidad de disolverse en lípidos se encuentra que pueden atravesar más fácilmente la membrana, si es que esta se compone de lípidos. Este hecho apoya muy bien el modelo de la doble capa lipídica de la membrana celular. Hecho 2: Se encontró que moléculas de tamaño pequeño (ejemplo, urea) penetran en la célula con mayor facilidad de lo esperado de acuerdo a su solubilidad en lípidos. Esta situación no estaría de acuerdo con el modelo. Tratan de salvar el modelo postulando que deben existir poros a través de los cuales se puede atravesar la membrana. Deben hacerse nuevas pruebas para fortalecer la hipótesis de la presencia de poros. Se realizaron pruebas con distintas sustancias para medir su permeabilidad. La diferencia en la sustancias era el tamaño de ellas. Con lo cual se estimó hasta el tamaño de los poros. El problema se agudizo cuando dos moléculas diferentes, pero de igual tamaño, tenían permeabilidad diferente. Una de ellas considerablemente diferente, unas diez mil veces mayor. Un razonamiento viene a liquidar el modelo de membranas con poros: para que una molécula de glucosa pueda pasar desde el lado extracelular, luego se disuelva en los lípidos y entre al agua citoplasmática, necesita gran cantidad de energía para sacarle las moléculas

162
de agua unida a los grupos OH (oxhidrilos). Esta energía es tan alta que imposibilitan a la glucosa para entrar a la célula. Estos últimos problemas son conflictivos para el modelo, si es así, el modelo debe modificarse o sugerir sencillamente otro modelo.
Hacia el modelo de mosaico fluido Se trata de un modelo de la estructura de la membrana plasmática propuesto en 1972 por S. J. Singer y Garth Nicolson, se fundamentó en una gran variedad de experimentaciones y observaciones sobre todo permitido por el desarrollo de la microscopía electrónica, el estudio químico de interacciones hidrófilas, al estudio de enlaces no covalentes como puentes de hidrógeno y el desarrollo en biología celular de técnicas como la criofractura y el contraste negativo.
PROTEÍNAS DE MEMBRANA La conformación de las proteínas depende de los tipos de ambientes en que se encuentran, depende también de las condiciones de extracción a las que se ven sometidas, me refiero a la presencia de iones y detergentes presentes en la solución de extracción. Sin embargo el sencillo hecho de extraer las proteínas desde su entorno asegura un cambio de conformación y es seguro cambia las propiedades de ella y por ende, es muy difícil averiguar su función. La función puede ser de un receptor, una bomba o un canal iónico. Sin duda, una vez purificada la proteína no realizará ninguna de las funciones. Una vez más nos encontramos con los criterios de la autopoiesis. Todas las explicaciones que podamos otorgar respecto a la estructura no serán satisfactorias a menos que encontremos la relación entre los diversos componentes. Es decir, para una explicación válida necesitamos conocer a todos los componentes y estudiar cómo actúan en conjunto. Por ejemplo para los procesos de comunicación celular supongamos que poseemos un artefacto- una radio- podemos obtener sus parlantes y luego describirlos. Pero, los parlantes poco nos podrán decir acerca de cómo es la radio y mucho menos explicarnos su funcionamiento. Eso ocurre cuando aislamos una proteína de la membrana. Las proteínas de membrana suelen asociarse en dímeros, tetrámeros. Además las proteínas de membrana pueden unirse a proteínas del citoplasma con unión a proteínas del citoesqueleto tales como microfilamentos, filamentos intermedios, o pueden formar parte de una cadena transductora de señales. También tenemos proteínas que se unen a componentes de la matriz extracelular, o forman estructuras que les permiten asociarse a células vecinas, ese es el ejemplo, de los desmosomas, uniones oclusoras y uniones comunicantes.
Si la proteína es un receptor de señales, la señal debe ser enviada a la proteína G que tiene que decodificar esa señal y finalmente dentro de la célula, la señal debe ser amplificada y causar una variación en la función celular al interior de la célula. La señal regula funciones biológicas como división, diferenciación, movilidad o secreción celular.

163
Las proteínas también juegan un rol fundamental en las membranas biológicas, se trata de las funciones relacionadas con la permeabilidad. Pueden actuar como canales o como transportadores. También se puede encontrar algunas con características de enzimas. Cada clase de células posee su propio juego de proteínas, así encontramos diferencias entre una neurona, un eritrocito o un hepatocito. Clasificación de proteínas en las membranas: Se han clasificado de acuerdo a la posición dentro de la membrana; así, algunas fueron llamadas intrínsecas y periféricas o extrínsecas. Las proteínas integrales son en su mayoría transmembranosas, una porción de ella permanece en el interior de la bicapa, presenta dominios que alcanzan la superficie. En general, este tipo de proteínas se encuentra asociada a glúcidos formando glucoproteínas. Otro tipo de proteínas integrales no son transmembranosas y se encuentran unidas a lípidos en forma covalente. Este mismo tipo de enlaces requiere que su aislamiento sea drástico y con sustancias como detergentes que destruyen la membrana. Por acción de algunas fosfolipasas se puede romper enlaces entre lípidos y proteínas integrales no transmembranosas.
En el modelo de mosaico fluido la membrana plasmática, los lípidos se disponen formando una bicapa. Las proteínas se intercalan en esa bicapa de dependiendo de las interacciones con los lípidos. Existen dos tipos de proteínas según su disposición en la bicapa:
• Proteínas integrales (o intrínsecas): Son aquellas que atraviesan la bicapa de la membrana una o varias veces, asomando por una o ambos lados u hojas interna y externa, se les llama también proteínas transmembrana.
• Proteínas periféricas (o extrínsecas). A un lado u otro de la bicapa lipídica, pueden estar unidas débilmente por enlaces no covalentes. Al contrario de las anteriores son fácilmente separables de la bicapa mediante soluciones salinas, sin provocar su ruptura.
Este modelo fue desarrollado para demostrar la asimetría entre ambas capas, lo que explicaría porque no entran los mismos nutrientes que los que salen.

164
Debido a que la mayor parte de las proteínas tiene un diámetro mayor a 10 nm, siendo este el grosor de la membrana, este hecho constituía uno de los principales problemas para comprender la estructura básica de las membranas consistía en determinar la forma en que las moléculas se disponían en un espacio tan pequeño. Hacia 1972, S.T. Singer y G.L. Nicholson propusieron el modelo de estructura de membranas que sintetizaba las propiedades conocidas de las membranas biológicas.
Según este modelo del mosaico fluido, la bicapa lipídica constituye la estructura
básica de la membrana y actúa de barrera relativamente impermeable al paso de la mayoría de moléculas hidrosolubles.
Las moléculas proteicas, que normalmente se hallan distribuidas en la bicapa lipídica,
llevan a cabo una buena parte de las funciones de la membrana, por ejemplo transportando moléculas específicas a través de ella. Algunas actúan como eslabones estructurales que relacionan la membrana plasmática al citoesqueleto y/o con la matriz extracelular, con las células adyacentes, otras actúan como receptores que reciben y transmiten las señales procedentes del medio ambiente celular.
Bicapa lipídica
La bicapa lipídica ha sido establecida como la base universal de la estructura de la membrana celular. Es posible de observar en una micrografía electrónica utilizando microscopio electrónico con difracción de rayos X y técnicas de criofractura para revelar los detalles de su organización.
Los lípidos son insolubles en agua pero se disuelven fácilmente en disolventes
orgánicos. Constituyen aproximadamente el 50% de la masa de la mayoría de las membranas plasmáticas de las células animales, siendo casi todo el resto proteínas y estas en su forma nativa asociada a carbohidratos y lípidos.
Las propiedades físicas de los fosfolípidos, en particular la forma en que dichas
moléculas se asocian en el agua, son las que permiten la formación de capas dobles. Algunas propiedades de estos lípidos así como sus propiedades serán observadas por usted en las

165
actividades de laboratorio. Ya se ha mencionado que los fosfolípidos están formados por dos cadenas de ácidos grasos unidas a dos de los tres carbonos de la molécula del glicerol. Las dos cadenas de ácidos grasos de la molécula son hidrófobas (no afines al agua) y no polares. Pueden tener diferente longitud (habitualmente contienen de 14 a 24 átomos de carbono). Normalmente una de estas cadenas presenta uno o más dobles enlaces (es decir, es insaturada) de tipo cis mientras que la otra normalmente no tiene dobles enlaces (es saturada). Las diferencias en longitud y grado de instauración entre las colas hidrocarbonadas son relevantes porque afectan la capacidad de las moléculas de fosfolípidos para empaquetarse una contra otra y modifican su fluidez.
El tercer carbono del glicerol está unido a una molécula orgánica hidrofílica, de tal manera que la molécula muestra una región hidrofílica y una región hidrofóbica.
Por lo mismo, las moléculas del tipo de los fosfolípidos tienen un extremo que se
asocia libremente con el agua y otro que no lo hace, cuando se encuentran dispersas en agua adoptan por lo general una conformación de capa doble. Esta estructura permite que los grupos del extremo hidrofílico se asocien con el medio acuoso, y que las cadenas hidrófobas de ácidos grasos permanezcan en el interior de la estructura, lejos de las moléculas de agua.
No todos los lípidos son capaces de formar bicapas, algunos forman micelas con las
colas hidrocarbonadas hacia el interior como es el caso de los ácidos grasos libres. Los triglicéridos, por ejemplo, son hidrófobos, de manera que forman gotas de aceite dentro de la célula. Las características más importantes de los lípidos que forman bicapas son las siguientes: 1. Tener dos polos distintos, uno hidrófobo y otro hidrofílico (que juntos las hacen fuertemente anfipáticas).
2. Su forma les permite asociarse con el agua en forma de una estructura de doble capa. En consecuencia de su estructura química, estas bicapas lipídicas tienden a cerrarse sobre sí mismas formando compartimientos herméticos, eliminando así los bordes libres en los que las colas hidrofóbicas podrían estar en contacto con el agua. Por lo mismo, los compartimientos formados por bicapas lipídicas tienden a cerrarse de nuevo después de haber sido rotos.
La bicapa lipídica como un líquido bidimensional
Una característica importante de las bicapas de fosfolípidos es que en determinadas condiciones se comportan como cristales líquidos. Las bicapas tienen propiedades semejantes a los cristales, pero también tienen propiedades semejantes a los líquidos porque a pesar de la ordenada disposición de sus moléculas, los grupos hidrocarbonados están en movimiento constante. Por lo tanto, una molécula se desplaza rápidamente de un punto al otro en un mismo lado de la estructura. Este movimiento confiere a la bicapa la propiedad de un fluido bidimensional'. En condiciones normales, esto significa que una molécula de fosfolípido puede recorrer la superficie de una célula eucariota en pocos segundos.
Las propiedades de líquido de la bicapa lipídica también permiten el desplazamiento
de las moléculas que se encuentran insertas en ella sobre el plano de la membrana (siempre que no estén ancladas mediante algún otro mecanismo). David Frye y Michael Ediden mostraron lo anterior mediante experimentos en los cuales seguían el curso de las proteínas

166
de membrana en dos células adyacentes. Para ello prepararon anticuerpos contra proteínas de membrana de ratón y de hombre y unieron cada anticuerpo a un colorante fluorescente de diferente color (ver figura siguiente). Al fusionarse una célula de ratón con una humana se observó que las proteínas de ratón pasaban a la región humana de la membrana celular, y viceversa. Después de un tiempo pudo observarse la distribución al azar de ambos grupos de proteínas en la superficie celular. Estos experimentos confirmaron que las proteínas de membrana poseen una gran capacidad de movimiento.
La fluidez de las membranas celulares es relevante para algunos procesos de
transporte y actividades enzimáticas que pueden detenerse cuando la viscosidad de la membrana (parámetro inversamente relacionado con la fluidez). (Revise lo aprendido en el primer laboratorio) incrementa más allá de un nivel crítico (umbral de fluidez).
La fluidez de la bicapa depende tanto de su composición como de la temperatura.
Una menor longitud de las cadenas reduce la tendencia de las colas a interaccionar entre sí y los dobles enlaces cis producen pliegues en las cadenas hidrocarbonadas que dificultan su empaquetamiento, de forma que las membranas permanecen fluidas a temperaturas más bajas.
La mayor parte de las membranas biológicas se encuentran en un “estado cristalino
líquido”; sin embargo, a temperaturas bajas, las fuerzas de van der Waals entre las cadenas de hidrocarburos dispuestas una cerca de otra convierten las bicapas de fosfolípidos en un gel sólido. Para ejemplificar este proceso podemos mencionar lo que sucede con los triacilglicéridos. Algunos triacilglicéridos (grasas) son sólidos a temperatura ambiente, en tanto que otros son líquidos (aceites), y, ello depende de la presencia de las insaturaciones presentes en la molécula. El pliegue de los dobles enlaces en las cadenas de hidrocarburos del aceite de cocina evita que las moléculas se acerquen entre sí y formen interacciones de van der Waals; así, disminuyen la temperatura a la cual el aceite (o los lípidos de membrana) se cristalizará, lo mismo ocurre en una membrana biológica.

167
Bacterias, levaduras y otros organismos cuyas temperaturas varían con la de su medio ambiente, controlan la composición de ácidos grasos de sus lípidos de membrana para mantener una fluidez relativamente constante. Si la temperatura disminuye sintetizan ácidos grasos insaturados, de manera de evitar la pérdida de fluidez de sus membranas por efecto de la disminución de la temperatura.
Los principales fosfolípidos que se encuentran en las membranas son la
fosfatidilcolina, fosfatidiletanolamina, fosfatidilserina y esfingomielina. La razón por la cual hay tal variedad de fosfolípidos quizás se deba a que serían necesarios para los distintos tipos de proteínas que se hallan asociadas a la membrana, que únicamente podrían funcionar en presencia de grupos polares específicos, como los que brinda la cabeza polar de los distintos tipos de fosfolípidos.
Las membranas
plasmáticas bacterianas están compuestas por un único tipo de fosfolípido y no contienen colesterol. La composición de la membrana celular de la mayoría de las células eucarióticas es más variada, contiene grandes cantidades de colesterol a excepción de los vegetales que no presentan dicho compusto; también presentan una mezcla de diferentes fosfolípidos, además de colesterol y glicolípidos.
Algunas células animales contienen cantidades especialmente elevadas de colesterol hasta una proporción de más de una molécula de colesterol por cada molécula de fosfolípido.
Este diseño incluyendo al colesterol estaría siendo explicado en la siguiente forma:
Las moléculas de colesterol se acomodan entre los fosfolípidos y actúan como "reguladores de fluidez". A bajas temperaturas las moléculas de colesterol se interponen entre las cadenas de hidrocarburos, con lo cual evitan que se acerquen y formen interacciones de van der Waals, los cuales provocarían la cristalización de la membrana. Por otro lado, a temperaturas elevadas, las moléculas de colesterol restringen el movimiento excesivo de las cadenas de ácidos grasos disminuyendo la fluidez de la membrana.
Las bicapas tienen también otras propiedades biológicas importantes. Por ejemplo, tienden a resistir la formación de extremos libres; como resultado de esto, tienden a autosellarse y casi en cualquier circunstancia forman espontáneamente vesículas cerradas. Por último, en condiciones apropiadas, las bicapas son capaces de fusionarse con otras. Las consecuencias son vitales para la normal función de la célula, entre ellas el transporte intra y
cabeza polar del
colesterol
región más
rígida
región más
fluida

168
extracelular de sustancias. Este fenómeno ocurre con la fusión de membranas, hecho que requiere un estado fluido de la bicapa para producirse. Cuando una vesícula se fusiona con otra membrana, ambas bicapas y sus compartimientos forman una continuidad. Esto permite tanto la transferencia de material de un compartimiento a otro, o el movimiento de una vesícula secretora hacia afuera de la célula, mediante un proceso Ilamado exocitosis. De modo similar, aunque inverso, la endocitosis permite la incorporación de grandes moléculas del exterior mediante la formación de vesículas en alguna porción de la membrana.
Asimetría de la membrana
La bicapa lipídica es asimétrica, hecho que se explica mediante relaciones de tipo funcional, y ello debido a la diferente composición lipídica de cada una de sus monocapas. En los glóbulos rojos, la mayoría de las moléculas lipídicas que tienen colina en su grupo cabeza (fosfatidilcolina y esfingomielina) se encuentran en la mitad externa de la bicapa, mientras que la mayoría de las moléculas de fosfolípido que contienen un grupo amino (fosfatidiletanolamina y fosfatidilserina) se hallan en la mitad interna.
Algunos fosfolípidos, como el fosfatidilinositol, actúan como intermediarios en el proceso de señalización celular: ante estímulos extracelulares, el fosfatidilinositol ubicado en el interior de la membrana, durante los procesos de señalización es hidrolizado en trifosfato de inositol y diacilglicerol. Ambos moléculas resultantes actúan dentro de la célula como mensajeros solubles que permiten la difusión de la señal hacia el interior de la célula. El primero actúa sobre canales de calcio ubicados en el retículo endoplásmico: el calcio liberado es necesario para que actúe la proteína C quinasa (C porque es calcio dependiente), que es activada por el diacilglicerol. La proteína C quinasa incrementa la transcripción de genes específicos.
La síntesis de fosfolípidos ocurre en el retículo endoplásmico liso allí se genera la asimetría por traslocadores que trasladan específicamente moléculas de fosfolípidos de una capa a la otra. Los fosfolípidos son sintetizados en la cara externa (citosólica). Si bien se sintetizan tanto fosfatidilcolina como el resto de los fosfolípidos, sólo la fosfatidilcolina pasa a la cara interna de la membrana del retículo gracias a un traslocador fosfolipídico específico (una “flipasa”). Existen también enzimas que se encargan de mantener esta asimetría en la membrana plasmática, por ejemplo una enzima denominada “aminotraslocasa” retorna la fosfatidilamina y la fosfatidilserina a la monocapa interna. La pérdida de la asimetría de la membrana es una señal de muerte celular ya que la exposición de fosfatidilserina en la monocapa externa es un índice de apoptosis o muerte celular programada que favorece la fagocitosis de estas células por macrófagos.
Glicolípidos: Aparecen en las membranas plasmáticas de todas las células animales, constituyendo el 5% de las moléculas de lípido de la monocapa externa y son las moléculas que presentan una asimetría más marcada en cuanto a su distribución en las membranas celulares. Estas moléculas se encuentran exclusivamente en la cara externa de la membrana plasmática, de manera que los azúcares quedan al descubierto en la superficie de la célula, lo que sugiere que deben desempeñar una función en las interacciones de la célula con su entorno.
Puesto que los azúcares se añaden en la cara luminal del Golgi, al formarse la
vesícula de transporte, el residuo glicosídico queda hacia el interior de la misma, debido a

169
ello cuando se fusiona con la membrana plasmática, la porción glicosilada, que es hidrófila, queda hacia el exterior de la célula.
Hay variados tipos de glicolípidos, Los glucolípidos más complejos contienen
oligosacáridos con uno o más residuos de ácido siálico que les proporciona carga negativa. Estos lípidos son más abundantes en la membrana plasmática de células nerviosas. Hasta el momento se han encontrado más de 40 gangliósidos diferentes. La función de los glicolípidos puede ser variada: en las células epiteliales tapizan la cara que da al epitelio, donde las condiciones son extremas (bajos o altos valores de pH, enzimas degradativas), protegerían la integridad de las propias proteínas de membrana; también cumplen funciones aislantes, como ocurre en la membrana que rodea el axón de las células nerviosas, tapizada totalmente por glicolípidos en la cara externa. La presencia de carga eléctrica negativa en su molécula es responsable también de la concentración de iones, especialmente Ca+2 en la superficie externa. Además desempeñan una importante función en procesos de reconocimiento celular, ayudando a su vez a las células a unirse a la matriz extracelular y a otras células. Proteínas de membrana Aunque la estructura básica de las membranas biológicas es provista por los fosfolípidos, la mayoría de las funciones específicas de la membrana son llevadas a cabo por proteínas. De acuerdo con ello, las cantidades y tipos de proteínas de membrana son muy variables: en la membrana mielínica, que sirve de aislación eléctrica al axón de la neurona, menos del 25% en peso son proteínas, en tanto que en las membranas donde hay transducción energética (mitocondrias y cloroplastos) el porcentaje alcanza al 75%.
Las proteínas integrales de membrana poseen algunas regiones insertadas en las regiones hidrófobas de la bicapa lipídica. Algunas atraviesan toda la membrana, de manera que gran parte de ellas se encuentra en alguno de los lados de la membrana; estas proteínas integrales se llaman también proteínas transmembrana. Otras proteínas integrales poseen solo una pequeña porción dentro de la bicapa que sirve como ancla y el resto de la molécula en el citoplasma o hacia la superficie celular. Otras están casi por completo insertas en la región hidrófoba y poseen cadenas polipeptídicas que atraviesan una y otra vez (incluso hasta doce veces) la bicapa lipídica.
Las proteínas transmembranosas pueden estar unidas por una cadena de ácido graso a la cara interna de la membrana, pasar una sola vez a través de la misma o muchas veces, estar solubles en el citosol pero ancladas a la cara interna por un resto acilo o prenilo, o haber sido sintetizadas como proteínas transmembrana en el retículo endoplásmico rugoso y unidas a un resto de glicosilfosfatidilinositol, que las ancla a la cara externa de la membrana.

170
En las proteínas integrales, la parte que se halla dentro de la membrana usualmente
adopta una estructura en αααα-hélice, con predominio de aminoácidos hidrofóbico. Además este enrollamiento asegura que todas las uniones peptídicas (que son polares) estén disminuidas en su polaridad debido a la formación de puentes de hidrógeno. Sólo hacen
falta 20-30 aminoácidos para atravesar la membrana en forma de αααα-hélice y sólo unos diez
para hacerlo en forma de hoja ββββ-plegada. La mayoría de las proteínas integrales atraviesan
la membrana en forma de hélices αααα, pero algunas (como las porinas de bacterias y de
mitocondrias) formas láminas ββββ que se disponen en forma de barril (ββββ“-barrel”). Funciones de las proteínas de membrana
¿Por qué motivo la membrana plasmática requiere de tantas proteínas distintas? La diversidad de proteínas en una membrana refleja el número de funciones que se llevan a cabo en ella.
Habitualmente la manera en que una proteína se asocia a la bicapa lipídica es un indicativo de la función de la proteína. Así, solo las proteínas transmembrana pueden actuar a ambos lados de la bicapa o transportar moléculas a través de ellas. Por regla, las proteínas de la membrana plasmática se clasifican en diferentes grupos funcionales:
a) Proteínas de adhesión celular, unen firmemente las membranas de células adyacentes y actúan como puntos de anclaje con componentes del citoesqueleto.
b) Proteínas como canales proteicos entre dos células (uniones de hendidura, “gap junctions”) permiten el paso de moléculas pequeñas entre dos células vecinas.
c) Proteínas de transporte que permiten el transporte selectivo de moléculas esenciales, ya sea en forma pasiva o en forma activa, mediante procesos que requieren de energía.
d) Proteínas transductoras receptoras de señales son proteínas que se unen a moléculas portadoras de señales externas y que luego transfieren el mensaje al interior de la célula.
e) Proteínas como bombas, dependientes de ATP transportan activamente iones de un compartimiento a otro, constituyendo así un mecanismo de almacenamiento de energía.
f) Proteínas de membrana actúan como enzimas, con sitios activos localizados en la superficie de la membrana o en el interior de ella.

171
PASO DE LOS MATERIALES A TRAVÉS DE LAS MEMBRANAS
Debido a su interior hidrofóbico, la bicapa lipídica de una célula constituye una barrera altamente impermeable a la mayoría de las moléculas polares o solubles en agua. Esta función de barrera es de especial importancia ya que permite a una célula mantener en su citosol ciertos solutos a concentraciones diferentes a las que están en el fluido extracelular y en cada uno de los compartimientos intracelulares. Sin embargo, para poder salvar la barrera cuando es conveniente, las células desarrollado sistemas para transportar específicamente moléculas hidrosolubles a través de la membrana y así poder ingerir nutrientes esenciales, excretar los productos de desecho del metabolismo y regular las concentraciones intracelulares de iones.
El transporte de iones inorgánicos y de pequeñas moléculas orgánicas hidrosolubles
a través de la bicapa lipídica se consigue mediante proteínas transmembrana especializadas, cada una de las cuales es responsable de la transferencia de una molécula o un ion específico o de un grupo de moléculas afines.
El hecho de que una membrana permita el paso de ciertas sustancias depende de la estructura de aquélla y del tamaño y carga eléctrica de las moléculas. Se dice que una membrana es permeable para alguna sustancia si permite que ésta la cruce e impermeable si no permite el paso de dicha sustancia. Una membrana
selectivamente permeable permite
el paso de algunas sustancias pero
no el de otras. Todas las membranas biológicas (aquellas que rodean a las células, núcleos, vacuolas, mitocondrias, cloroplastos y otros organelos celulares) son selectivamente permeables.
Al reaccionar a las condiciones ambientales cambiantes o a las diversas necesidades de la célula, la membrana puede constituir una barrera al paso de un compuesto determinado en cierto momento, mientras promueve activamente su
paso en otro momento. Mediante la regulación del tráfico químico de esa manera, la célula controla su propia composición interna de iones y moléculas que puede ser muy diferente a la del exterior. En el mundo abiótico, los materiales se mueven pasivamente por procesos físicos como la difusión. En los seres vivos, los materiales se mueven según la dirección señalada por la concentración de material, es decir, desde una región de mayor a menor

172
concentración, pero mueven materiales en forma activa, proceso que requiere energía, para ello, evidentemente son procesos fisiológicos y los ejemplos son, transporte activo, exocitosis y endocitosis.
Difusión
Algunas sustancias se desplazan hacia adentro y afuera de las células, y se mueven dentro de éstas por medio de un proceso llamado difusión simple, el cual se basa en el desplazamiento al azar. A temperaturas mayores al cero absoluto (0° Kelvin o -273°C, el punto en el cual se detiene el movimiento), todos los átomos y moléculas poseen energía cinética, o energía de movimiento.
Los tres estados de la materia (sólido, líquido y gaseoso) difieren con respecto a la
libertad de movimiento de las moléculas que los constituyen. Las moléculas del estado sólido se encuentran muy cerca una de otra, y las fuerzas de atracción entre ellas les permiten vibrar, pero no desplazarse. Las moléculas de un líquido se encuentran más separadas entre sí, respecto a las de un sólido; las fuerzas intermoleculares son más débiles y las moléculas se desplazan con relativa libertad; en el estado gaseoso, las moléculas están tan separadas que las fuerzas intermoleculares son mínimas y por tanto el desplazamiento de las moléculas sólo está limitado por las paredes del recipiente que las contiene. Esto significa que los átomos y moléculas de líquidos y gases se separan en una especie de desplazamiento al azar.
A este desplazamiento se atribuye el proceso de difusión, movimiento neto de
partículas (átomos, iones, moléculas, etc.) de una región de alta concentración a una de concentración más baja, de manera que las partículas adquieren una distribución uniforme.
Cuando se deja caer un terrón de azúcar en un vaso de precipitado lleno con agua,
las moléculas de aquél se disuelven y luego comienzan a difundirse hacia toda el agua del recipiente, como consecuencia de que las moléculas de azúcar individuales se desplazan al azar en todas direcciones. En última instancia, la difusión ocasiona una distribución uniforme de las moléculas de azúcar en toda el agua del vaso.
La difusión implica el movimiento de partículas en favor de una gradiente de
concentración (diferencia de concentración de una sustancia de un punto a otro). Esto no significa que las partículas no puedan desplazarse en sentido "equivocado" (en contra del gradiente). Sin embargo, si al inicio hay mayor cantidad de partículas en una región de concentración elevada, lógicamente más partículas se desplazarán desde la zona de alta concentración hacia la de baja concentración, que a la inversa. La proporción de difusión está en función del tamaño y forma de las moléculas, de sus cargas eléctricas y de la temperatura. Al aumentar la temperatura, las moléculas se mueven con mayor rapidez y aumenta la proporción de difusión.
El examen de una gota de agua al microscopio no revela el movimiento de sus
moléculas, ya que éstas son demasiado pequeñas para ser vistas; sin embargo, cuando se agrega al agua una gotita de tinta china, que contiene partículas de carbón muy finas, el movimiento de dichas partículas sí es visible al microscopio. Cada partícula de carbón es golpeada constantemente por las moléculas de agua, de manera que el rebote ocasionado por las colisiones hace que la partícula se mueva de un lugar a otro en forma zigzagueante y aleatoria. Este movimiento de las partículas pequeñas -llamado movimiento browniano, en honor a Robert Brown, el botánico inglés que lo observó por primera vez al examinar bajo el

173
microscopio granos de polen suspendidos en agua- constituye un modelo que explica el mecanismo de difusión de las moléculas.
Conforme se verifica la difusión, cada molécula individual se desplaza en línea recta
hasta que choca contra algo, que puede ser otra molécula o la pared del recipiente. Luego rebota y se desplaza en otra dirección. Cada molécula individual puede moverse a una velocidad de varios centenares de metros por segundo; sin embargo, su desplazamiento en el espacio es de sólo una fracción de nanómetro antes de chocar con otra molécula y rebotar. Por esta razón, el movimiento de avance de una molécula en línea recta es muy lento. Las moléculas siguen moviéndose, incluso a pesar de que se distribuyan uniformemente en un espacio dado; sin embargo, con la misma rapidez con que ciertas moléculas se mueven en un sentido, hay otras que se mueven en el sentido opuesto, de modo que el sistema, considerado como un todo, se conserva en equilibrio (es decir, que no se observa un cambio neto).
Las moléculas de cualquier número de sustancias diferentes se difunden
independientemente de las otras en la misma solución; al final, todas ellas tendrán una distribución uniforme Con mayor frecuencia, en los sistemas biológicos no se alcanza nunca una condición de equilibrio. Por ejemplo, el dióxido de carbono se forma en las células durante el metabolismo de los azúcares y de otras moléculas, y durante la respiración celular; se difunde rápidamente a través de la membrana plasmática, y luego se elimina rápidamente del torrente circulatorio. Esto limita la oportunidad de las moléculas para reentrar nuevamente en las células, de manera que siempre existe un gradiente de concentración de moléculas de dióxido de carbono a través de la membrana.
Diálisis La difusión de un soluto (una sustancia disuelta) a través de una membrana
diferencialmente permeable se llama diálisis. Para demostrar la diálisis se utiliza una bolsa de celofán, llena con una solución de azúcar, que luego se sumerge en un matraz que contiene agua pura. Si la membrana de celofán es permeable al azúcar y al agua, las moléculas de azúcar pasarán a través de ella hasta que la concentración de azúcar en el agua de los dos lados de la membrana sea exactamente igual. A partir de ese momento, las moléculas de soluto (así como también las moléculas de agua) seguirán pasando a través de la membrana, pero ya no habrá ningún cambio neto en las concentraciones, ya que la velocidad de movimiento será igual en ambos sentidos.

174
La diálisis renal es una aplicación práctica de este proceso; los productos de desecho, que se difunden a través de las membranas artificiales del aparato, pueden retirarse del organismo, pero los eritrocitos, proteínas sanguíneas y otras moléculas grandes, no se difunden a través de la membrana y por tanto se retendrán en el organismo.
El celofán con frecuencia se utiliza como "membrana artificial" para experimentos de laboratorio. Está compuesto por moléculas de polisacáridos y puede formar una lámina delgada que permite el paso de moléculas de agua. Estas membranas se fabrican con permeabilidad variable para diferentes solutos.
Aunque la difusión es muy rápida en las microdistancias, una molécula individual tarda mucho tiempo en recorrer distancias medidas en centímetros. Esta baja velocidad global de difusión tiene importantes implicaciones biológicas, ya que limita el número de moléculas de nutrientes y oxígeno que pueden llegar hasta un organismo por difusión simple. Con suficiente tiempo, prácticamente cualquier molécula que pueda atravesar la bicapa lipídica acabará difundiendo a través de ella. Sin embargo la velocidad a la que se produce esta difusión varía enormemente dependiendo en parte del tamaño de la molécula y principalmente de su solubilidad relativa en aceite. En general cuanto menor es la molécula y cuanto más soluble en aceite (es decir, mas hidrofóbica o no polar) más rápidamente difunde a través de la bicapa. Las moléculas pequeñas no polares (O2, CO2) se disuelven fácilmente en la bicapa lipídica y por lo tanto difunden con rapidez. Las moléculas polares no cargadas también difunden rápidamente a través de la bicapa lipídica si su tamaño es reducido. Por ejemplo agua, etanol y urea atraviesan rápidamente una bicapa, el glicerol lo hace con menor rapidez y la glucosa prácticamente no la atraviesa. Permeabilidad de la bicapa lipídica a diferentes sustancias
Tipo de molécula Ejemplo Permeabilidad
Hidrófoba N2, O2• hidrocarburos Permeabilidad libre
Polar pequeña H20, CO2, glicerol, urea Permeabilidad libre
Polar grande Glucosa y otros monosacáridos y disacáridos sin carga
No permeable
Iones y moléculas con carga Aminoácidos, H+, HCO-3, Ca+, Cl-
; Mg+ No permeable
Ósmosis
La ósmosis es una variedad especial de difusión que implica el movimiento de moléculas de solvente (en este caso, el agua) a través de una membrana de permeabilidad selectiva. Las moléculas de agua pasan libremente en cualquier dirección, pero al igual que en todos los procesos de difusión, el movimiento neto ocurre a partir de la región de mayor concentración a la de menor.
Los principios que intervienen en el proceso de ósmosis se ilustran mediante la
utilización de un aparato llamado tubo en U. El tubo en U se divide en dos secciones por una

175
membrana de permeabilidad selectiva que impide el paso de las moléculas de soluto (glucosa, sal y otras)
En una parte del tubo se coloca una solución de agua y solutos; en la otra se coloca
agua pura. La solución de agua y solutos contiene una concentración de agua menor a la del agua pura, porque las moléculas de soluto han "diluido" las moléculas de agua. Por tanto, hay un movimiento neto de moléculas de agua del lado del agua pura (con mayor concentración de moléculas de agua) hacia el lado del agua con soluto (que tiene menor concentración de moléculas de agua), como resultado de esto el nivel de líquido del lado del agua pura disminuye, mientras que se eleva el del lado del agua con soluto.
No obstante, aún existe una diferencia en la concentración de las moléculas de agua
entre ambos lados, debido a que las moléculas de soluto no pueden moverse a través de la membrana. El movimiento neto de agua continuará, y el nivel de líquido del lado del agua con soluto seguirá aumentando. En condiciones no sujetas a la gravedad, este proceso continuaría indefinidamente, pero en la tierra, el peso de la columna de líquido en aumento finalmente ejercerá una presión suficiente para detener el cambio en los niveles de líquido, aunque las moléculas de agua continuarán pasando a través de la membrana en ambas direcciones.
La presión osmótica de una solución está relacionada con la tendencia que presenta
el agua de moverse hacia dicha solución mediante ósmosis. En el ejemplo del tubo en U se podría medir la presión osmótica insertando un pistón del lado del agua con soluto y midiendo la presión necesaria que debe ejercer el pistón para evitar el aumento en el nivel del líquido de dicho lado del tubo. Una solución con una alta concentración de soluto tendrá una baja concentración de agua y una elevada presión osmótica; en cambio, una solución con una baja concentración de soluto tendrá una elevada concentración de agua y una baja presión osmótica.
Soluciones isotónicas, hipertónicas e hipotónicas
Con frecuencia deseamos comparar las presiones osmóticas de dos soluciones. En todo líquido de los compartimientos de las células vivas se encuentran disueltas sales, azúcares y otras sustancias que le confieren a dicho líquido una determinada presión osmótica.
Cuando una célula se coloca en una solución cuya presión osmótica es igual a la suya,
no hay movimiento neto de moléculas de agua, ni hacia afuera ni hacia adentro de ella; por

176
tanto, la célula no se hincha ni se encoge. Se dice que el líquido en el cual se colocó la célula es un líquido isotónico (es decir que tiene presión osmótica igual) con respecto al líquido del interior de la célula. Normalmente, el plasma de la sangre (componente líquido de ella) y los demás líquidos corporales son isotónicos con respecto al líquido intracelular; es decir, contienen una concentración de agua igual a la del líquido intracelular. Una solución de cloruro de sodio al 0,9% (suero salino) es isotónica respecto a las células humanas y a las células de otros mamíferos. Los eritrocitos humanos colocados en una solución de cloruro de sodio al 0.9% no se encogen ni se hinchan.
Concentración de solutos en la Solución A
Concentración de solutos en la Solución B
Tonicidad Dirección de movimiento del agua
Mayor Menor A es hipertónica respecto a B; B es hipotónica respecto a A
B hacia A
Mayor Mayor B es hipertónica respecto a A. A es hipotónica respecto a B
A hacia B
Igual Igual Isotónica No hay movimiento neto
Si el líquido circundante tiene una concentración de solutos mayor que la del líquido intracelular y, por tanto, una presión osmótica mayor que la de éste se dice que es una solución hipertónica; una célula colocada en una solución hipertónica pierde agua y por tanto, se encoge. Así, los eritrocitos humanos colocados en una solución de cloruro de sodio al 1,3% pierden agua y se encogen. Cuando una célula con pared celular se coloca en un medio hipertónico pierde agua, y entonces su contenido disminuye dentro de la pared celular; este proceso se llama plasmólisis. Dicho fenómeno se observa en las plantas cuando se depositan grandes cantidades de sales o fertilizantes en la tierra o agua que las rodea.
Si el líquido circundante posee una concentración de solutos menor que la del líquido intracelular, y por lo tanto tiene una presión osmótica menor que la de éste, se denomina solución hipotónica; en estas circunstancias, el líquido se desplazará hacia el interior de la célula, provocando que ésta se hinche. En una solución de cloruro de sodio al 0,6%, el agua entra hacia el interior de los eritrocitos, provocando el hinchamiento de éstos y su ruptura o lisis.
Transporte mediado de moléculas pequeñas
La membrana celular es relativamente impermeable a casi todas las grandes moléculas polares. Esto constituye una ventaja biológica para la célula, ya que casi todos los compuestos metabolizados en su interior son polares y la impermeabilidad de la membrana impide su pérdida por difusión. Para transportar nutrientes polares, como glucosa y aminoácidos, a través de la membrana lipídica hacia el interior de la célula, han aparecido por evolución sistemas de proteínas transportadoras que se unen a esas moléculas y las transfieren a través de la membrana. El paso de solutos a través de la membrana celular por el sistema de transporte se llama transporte mediado.

177
Comparación entre un transporte pasivo a favor del gradiente electroquímico y el transporte activo en contra del gradiente electroquímico(Albert)
Difusión facilitada
La célula utiliza la energía almacenada por el gradiente de concentración de una sustancia cuya concentración es mayor en el líquido extracelular que en el intracelular. En estas circunstancias, mientras la membrana sea permeable a dicha sustancia, ésta se desplazará hacia el interior de la célula. En el caso de iones, azúcares, aminoácidos, nucleótidos y muchos metabolitos, para que esto suceda se necesita de la presencia de proteínas especiales de membrana. Este tipo de transporte se llama difusión facilitada.
La difusión facilitada depende de la existencia de proteínas transportadoras, las
cuales se combinan temporalmente con la molécula de soluto para acelerar el paso de ésta a través de la membrana celular. Cada proteína está destinada al transporte de un tipo particular de molécula y con frecuencia de una cierta especie molecular de la clase por lo que presentan especificidad. La proteína transportadora no se modifica por esta acción; después de transportar la molécula de soluto, queda libre para unirse a una nueva molécula. Todas las proteínas de transporte estudiadas son proteínas transmembrana múltiples.
El transporte de las moléculas de glucosa en los eritrocitos es un buen ejemplo de
difusión facilitada por transportador. Las moléculas que transportan glucosa son glucoproteínas; éstas comprenden un 2% de las proteínas totales de la membrana. Las células mantienen una baja concentración interna de glucosa mediante la adición inmediata de un fosfato a las moléculas de glucosa que entran en ellas: de esta manera convierten dichas moléculas en fosfatos de glucosa con elevada carga eléctrica y, así, no pueden regresar al otro lado de la membrana.
Otra clase de proteínas transportadoras son las formadoras de canal; estas no se
unen al soluto sino que forman poros hidrofílicos que atraviesan la bicapa lipídica, que al estar abiertos permiten que determinados solutos (habitualmente iones inorgánicos de tamaño y carga apropiados) puedan pasar a su través y por lo tanto atravesar la membrana).

178
Transporte activo
Las moléculas se transportan a través de la membrana mediante el proceso de difusión; a otras las requiere la célula en concentraciones mayores a su concentración extracelular. Estas moléculas se incorporan mediante mecanismos de transporte activo. Este mecanismo exige una fuente de energía debido a que el transporte activo implica el "bombeo" de una molécula en contra de su gradiente de concentración (de una zona de baja concentración hacia una de concentración elevada). Lo extraordinario de este tipo de transporte es que se produce acumulación de sustancias, moléculas o iones a un lado de la membrana, en su entorno, es el caso de la bomba de sodio y especialmente en las neuronas. La neurona por medio de la proteína bomba de sodio, prepara su entorno para su actividad que es la producción de potenciales de acción.
Los sistemas de transporte activo utilizan energía generada por el metabolismo
celular en forma de trifosfato de adenosina (ATP) o bien utilizan algún otro tipo de energía almacenada, derivada de la hidrólisis del ATP.
Uno de los ejemplos más sorprendentes de los mecanismos de transporte activo es la
bomba de sodio y potasio que se observa en todas las células animales. Esta bomba consta de una proteína específica, localizada en la membrana plasmática, que utiliza ATP para intercambiar iones de sodio del interior de la célula por iones de potasio de su exterior. Esto provoca un desequilibrio en la concentración de iones de sodio y potasio en los lados opuestos de la membrana, de manera que en condiciones normales, la concentración de potasio sea de 10 a 15 veces mayor en el interior que en el exterior de la célula y a la inversa para el sodio.
Las células necesitan estos gradientes de concentración para generar un potencial
eléctrico (separación de cargas eléctricas) a través de la membrana, el cual constituye la base para la transmisión de los impulsos eléctricos necesarios para la transmisión de los impulsos nerviosos.
Los gradientes de concentración también almacenan energía, la cual puede
utilizarse para la conducción de otros mecanismos de transporte activo. El gradiente electroquímico producido por estas bombas es tan importante que de hecho algunas células (p. ej., las células nerviosas) utilizan el 70% de su energía en el funcionamiento de este sistema de transporte.

179
Ver figura en Albert – Biología celular y molecular
Debido al bombeo de 3 iones Na+ positivamente cargados hacia el exterior de la
célula cada dos que bombea hacia el interior, se dice que la bomba es electrogénica, es decir dirige una corriente neta a través de la membrana tendiendo a crear un potencial eléctrico,
con el interior negativo con relación al exterior. Por otra parte la bomba tiene un papel directo regulando el volumen celular a través de sus efectos osmóticos que pueden hacer que la célula se hinche o se retraiga. Por último, también se utiliza para dirigir el transporte de azúcares y aminoácidos hacia el interior de la célula.
Sistemas de cotransporte
Algunas proteínas de transporte actúan como transportadores acoplados, en los que la transferencia de un soluto depende de la transferencia simultánea o secuencial de un segundo soluto, ya sea en la misma dirección (transporte unidireccional) o en dirección opuesta (bidireccional).
Transporte activo secundario
Muchos sistemas de transporte activo no son impulsados directamente por la hidrólisis del ATP sino por la energía almacenada por los gradientes iónicos. El gradiente electroquímico generado por la bomba de sodio y potasio proporciona suficiente energía para propiciar el transporte activo de otras moléculas esenciales. En estas reacciones, el gradiente de concentración de Na+ cotransporta las moléculas requeridas, junto con los iones de Na+. La energía liberada durante el desplazamiento de un ion a favor de su gradiente electroquímico se utiliza como fuerza impulsora para bombear otros solutos en contra de su gradiente. Así, las proteínas transportadoras actúan como transportadores

180
acoplados. El Na+ que entra en la célula durante este transporte es bombeado hacia el exterior mediante la ATPasa Na+-K+. Sistemas de transporte múltiple integrado
La distribución asimétrica de las proteínas transportadoras en la membrana plasmática de una célula epitelial intestinal da lugar al transporte transcelular de glucosa a través del epitelio intestinal.
En algunas células se observa el funcionamiento de más de un sistema de transporte
para una sustancia determinada. Por ejemplo, el transporte de la glucosa del intestino hacia el torrente circulatorio se lleva a cabo a través de una delgada capa de células epiteliales que recubren la luz del intestino y que poseen regiones especializadas, o dominios, en su
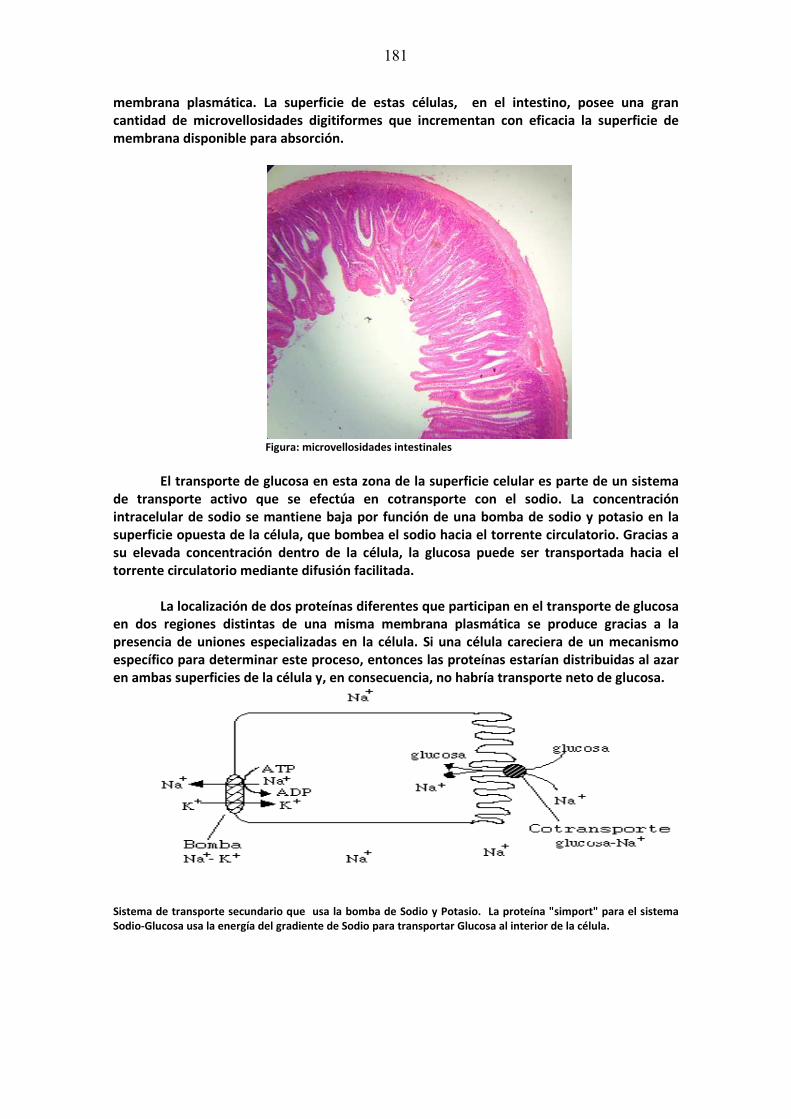
181
membrana plasmática. La superficie de estas células, en el intestino, posee una gran cantidad de microvellosidades digitiformes que incrementan con eficacia la superficie de membrana disponible para absorción.
Figura: microvellosidades intestinales
El transporte de glucosa en esta zona de la superficie celular es parte de un sistema
de transporte activo que se efectúa en cotransporte con el sodio. La concentración intracelular de sodio se mantiene baja por función de una bomba de sodio y potasio en la superficie opuesta de la célula, que bombea el sodio hacia el torrente circulatorio. Gracias a su elevada concentración dentro de la célula, la glucosa puede ser transportada hacia el torrente circulatorio mediante difusión facilitada.
La localización de dos proteínas diferentes que participan en el transporte de glucosa
en dos regiones distintas de una misma membrana plasmática se produce gracias a la presencia de uniones especializadas en la célula. Si una célula careciera de un mecanismo específico para determinar este proceso, entonces las proteínas estarían distribuidas al azar en ambas superficies de la célula y, en consecuencia, no habría transporte neto de glucosa.
Sistema de transporte secundario que usa la bomba de Sodio y Potasio. La proteína "simport" para el sistema Sodio-Glucosa usa la energía del gradiente de Sodio para transportar Glucosa al interior de la célula.

182
Transporte de grandes moléculas a través de las membranas
En la difusión simple, en la difusión facilitada y en el transporte activo las moléculas individuales y los iones pasan a través de la membrana celular. Sin embargo, en ocasiones también es necesario el desplazamiento de cantidades más grandes de material o de partículas de alimento o incluso de células completas, hacia afuera o adentro de una célula. Esto implica un gasto de energía por parte de la célula y en ocasiones necesita de la fusión de membranas.
Figura: Tomada de Albert: se ilustra Tres rutas de degradación en los lisosomas
Endocitosis: Mecanismo mediante el cual la célula incorpora materiales hacia su interior.
En los sistemas biológicos operan varios mecanismos endocitóticos. Por ejemplo, en la fagocitosis (literalmente "ingesta de células"), la célula ingiere partículas sólidas como bacterias o nutrientes. La fagocitosis es el mecanismo utilizado por protozoarios y leucocitos para ingerir partículas, incluso algunas tan grandes como una bacteria completa. Durante la ingestión, los pliegues de la membrana celular engloban a la partícula, que se ha unido a la superficie celular, y forman una vesícula alrededor de ella. Una vez que la membrana ha encerrado a la partícula en cuestión, se fusiona en el punto de contacto e ingresa en el citoplasma. Posteriormente la vesícula se fusiona con los lisosomas, donde el material es ingerido y degradado.
En otro tipo de endocitosis, llamada pinocitosis, la célula incorpora materiales disueltos. Algunos pliegues de la membrana plasmática engloban gotas de líquido, las cuales emergen en el citoplasma celular en forma de pequeñas vesículas. El contenido líquido de estas vesículas se libera lentamente en el citoplasma celular y las vesículas van disminuyendo poco a poco de tamaño, hasta el punto en que parecen desvanecerse.
En un tercer tipo de endocitosis, llamado endocitosis mediada por receptor, algunas
proteínas específicas de determinadas partículas se unen a proteínas receptoras, localizadas en la membrana plasmática de la célula. Luego, las moléculas ligadas al receptor emigran
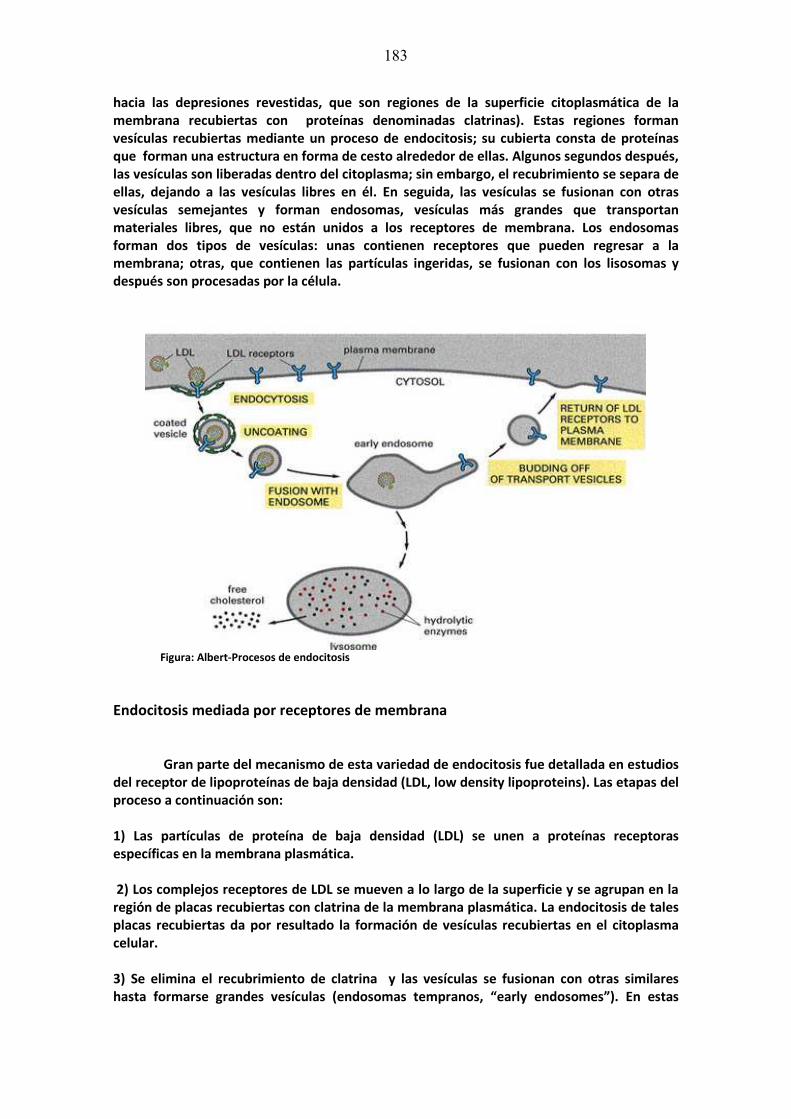
183
hacia las depresiones revestidas, que son regiones de la superficie citoplasmática de la membrana recubiertas con proteínas denominadas clatrinas). Estas regiones forman vesículas recubiertas mediante un proceso de endocitosis; su cubierta consta de proteínas que forman una estructura en forma de cesto alrededor de ellas. Algunos segundos después, las vesículas son liberadas dentro del citoplasma; sin embargo, el recubrimiento se separa de ellas, dejando a las vesículas libres en él. En seguida, las vesículas se fusionan con otras vesículas semejantes y forman endosomas, vesículas más grandes que transportan materiales libres, que no están unidos a los receptores de membrana. Los endosomas forman dos tipos de vesículas: unas contienen receptores que pueden regresar a la membrana; otras, que contienen las partículas ingeridas, se fusionan con los lisosomas y después son procesadas por la célula.
Figura: Albert-Procesos de endocitosis
Endocitosis mediada por receptores de membrana
Gran parte del mecanismo de esta variedad de endocitosis fue detallada en estudios
del receptor de lipoproteínas de baja densidad (LDL, low density lipoproteins). Las etapas del proceso a continuación son:
1) Las partículas de proteína de baja densidad (LDL) se unen a proteínas receptoras específicas en la membrana plasmática.
2) Los complejos receptores de LDL se mueven a lo largo de la superficie y se agrupan en la región de placas recubiertas con clatrina de la membrana plasmática. La endocitosis de tales placas recubiertas da por resultado la formación de vesículas recubiertas en el citoplasma celular.
3) Se elimina el recubrimiento de clatrina y las vesículas se fusionan con otras similares hasta formarse grandes vesículas (endosomas tempranos, “early endosomes”). En estas

184
estructuras, los receptores y las partículas de LDL se disocian, y dirigen hacia diferentes regiones de la vesícula. 4) A partir de los endosomas se forman nuevas vesículas. Las vesículas que contienen los receptores se desplazan hacia la superficie y se fusionan con la membrana plasmática, en la cual los receptores son reciclados. Las vesículas que contienen partículas de LDL se fusionan con los lisosomas. 5) El colesterol se libera por medio de diversas enzimas hidrolíticas: sólo entonces puede utilizarlo la célula. Las personas con incapacidad hereditaria para la producción de receptores de membrana para el LDL (puede faltar el gen o fabricar un receptor defectuoso, incapaz de unirse a la cubierta de clatrinas) son propensas a arterioesclerosis prematura y a sufrir ataques cardíacos.
El reciclaje del receptor de LDL a la membrana plasmática a través de la formación de vesículas ilustra un problema común a todas las células que emplean los mecanismos de endocitosis y exocitosis. En las células que secretan sustancias en forma continua, una porción equivalente de membrana debe incorporarse al interior de la célula por cada vesícula que se fusiona con la membrana plasmática; de no ocurrir así, la superficie de la célula estaría en expansión constante, aunque el crecimiento de la célula esté limitado. Existe una situación similar para las células que realizan endocitosis. Los macrófagos, por ejemplo, incorporan en forma de vesículas el equivalente del total de su volumen en cerca de 30 minutos, y requieren un reciclamiento similar para que las células mantengan su área de superficie y su volumen.
En la exocitosis una célula expulsa productos de desecho o productos específicos de secreción (como hormonas), mediante la fusión de una vesícula con la membrana plasmática de la célula. La exocitosis consiste en la fusión de la membrana de la vesícula secretora con la membrana plasmática. Este es también un mecanismo primario de crecimiento de la membrana plasmática.
Clasificación del transporte a través de la membrana
UNIPORT: Transportan sólo un tipo de sustancia a través de la membrana, como ejemplo de este tipo de transporte tenemos a Glut-4 de las células llamadas adipocitos y de las células musculares. Ellos captan la glucosa del torrente sanguíneo y la deposita en el citoplasma.

185
SIMPORT: Transportan dos sustancias en una misma dirección, ejemplo, el péptido/H que transporta hacia el lado intracelular de las células epiteliales intestinales protones y tripéptidos que se producen por la digestión de proteínas.
ANTIPORTE: Transportan dos sustancias en direcciones opuestas. Por eso mismo se les llama intercambiadores. Ejemplo; intercambiador Na+/ /Ca+2 , se encuentra en muchas células eucariontes. Toma tres iones de sodio del lado externo a la célula y los transporta hacia el interior, al mismo tiempo toma un ión calcio del citosol y lo saca de la célula. De esta manera mantiene bajas las concentraciones del ión en el citosol.
Ejemplo de transporte a través de la membrana:
INTERCAMBIADOR DE BICARBONATO/CLORO (RESPIRACIÓN)
Durante el proceso de respiración las células de los vertebrados toman y eliminan el CO2. Los glóbulos rojos transportan ambos gases por el torrente sanguíneo. Al avanzar por tejidos periféricos, el Co2 difunde hacia el interior, reacciona con una molécula de agua y produce el ión bicarbonato (HCO3
-). La reacción es catalizada por la anhidrasa carbónica. El intercambiador saca los iones HCO3
- de los glóbulos rojos y al mismo tiempo introduce iones cloruro (Cl-); se establece así un ambiente ácido en el citoplasma del glóbulo. Esto facilita la disociación del O2 de la hemoglobina.a continuación el oxígeno difunde hacia las células vecinas.
Los glóbulos rojos al llegar a los capilares pulmonares, donde la concentración de oxígeno es mayor se asocia a la hemoglobina. En este momento la anhidrasa carbónica trabaja al revés, capta ahora el bicarbonato y protones para producir CO2 y H2O. El intercambiador también trabaja al revés, saca Cl- e introduce bicarbonato.
El intercambiador es conocido como la proteína de la banda 3 de la membrana del glóbulo rojo y se une a proteínas del cortex, bajo la membrana del glóbulo. Las proteínas a las cuales se une son proteínas de la banda 4.1, la ankirina, la espectrina, la actina y la miosina.
Las membranas celulares establecen contacto con las células vecinas: La aparición de organismos pluricelulares o multicelulares plantea el desarrollo de nuevas estructuras preparadas a soportar las presiones de la selección natural, la nueva estructura debe adquirir una función. También puede ocurrir que estructuras incorporadas antiguamente en el organismo comienzan a mostrar nuevas funciones. Cada célula individual en un organismo debe informarse de cuando realizar sus actividades para asistir con su trabajo para el conjunto de células, tejidos y órganos que conforman el organismo. Por ello las células deban poseer un sistema de generación, transmisión, recepción y respuesta de miles de señales que permitan la comunicación e interrelación de ellas, el afán es responder a las exigencias que realiza el medio ambiente,

186
esto debe hacerlo en armonía, conservando su propia homeostasis. Las señales permiten que unas células influyan en el comportamiento de otras. Uniones comunicantes tipo nexo Hacia el interior de los territorios tisulares (tejidos), las comunicaciones celulares pueden efectuarse por medio de contactos directos entre las células. Estas son uniones de tipo comunicantes o nexos (bautizadas así). Este tipo de interacciones es esencial para la coordinación de las actividades, en especial para señales de control de crecimiento y diferenciación celular. Las uniones nexos son canales de unión y capaces de compartir concentraciones determinadas de metabolitos y iones, que pueden pasar libremente de una célula a otra. Cada célula, sin embargo, conserva su identidad, puesto que semejante tipo de organización no permite que las macromoléculas, incluyendo ADN o ARN pasen desde una célula a otra. Las interconexiones celulares- las uniones comunicantes, dejan espacio un espacio extracelular de unos 20 nm por ello se les da el nombre de gap junction o uniones con espacio.
Los gap junction son dos conexones que forma hemicanales,
estos se unen conectando a las dos células

187
Revel, J. P.; Karnovsky, M. J. (1967). "Hexagonal array of subunits in intercellular junctions of the mouse heart and liver". The Journal of cell biology 33 (3):
Las proteínas denominadas conexones comunican ambas células puesto que su estructura crea canales. El conexón individual es un tubo cilíndrico que atraviesa la capa de lípidos de cada una de las membranas celulares conectadas. Cada hemiconexón es un hexámero de conexina, polipéptido de unos 26 kDa, poseen un diámetro aproximado de 8 nm y dejan un orificio que es el canal de tipo hidrofílico de 1,5 nm. Existe un mecanismo de apertura y cierre del canal respecto del cual usted puede buscar más datos de estructura y funcionamiento en texto de Biología Celular de Robertis, entre otros. La regulación de apertura y cierre del poro es importante para el traslado de sustancias, tales como iones. Experimentalmente se ha observado que al agregar calcio+2 se produce el cierre del canal. En realidad el calcio+2 está muy concentrado en el exterior celular, varios miles de veces. Esta diferencia de concentración en ambos lados de la membrana, provoca el establecimiento de un gradiente entre el citoplasma y el exterior de la célula. Es útil saber que cuando una membrana celular muere o se desorganiza el calcio ingresa por gradiente, también lo hacen otros iones, con ello la células obviamente termina por sucumbir. Las observaciones indican que a través de estas uniones pueden pasar moléculas como péptidos con un máximo de tamaño de entre 1300 a 1900 Da. Una consecuencia de este tipo de interacciones celulares es que se produce un acoplamiento de tipo eléctrico en células adyacentes dentro del mismo tejido. Así las células están acopladas eléctricamente, dicho de otra forma, por las células fluye una corriente eléctrica transportada por iones. El acoplamiento así producido se denomina comunicación a través de uniones. Especialmente en los animales adultos presentan células que no presentan uniones comunicantes, por ejemplo, los leucocitos, la mayoría de las neuronas y las células musculares esqueléticas. El acoplamiento eléctrico que depende de las uniones con hendidura o nexos, reviste gran importancia en la contracción cardiaca, en esta, el potencial de acción se transmite de una célula a otra (cardiocitos) por acoplamiento eléctrico en uniones comunicantes los cuales forman parte de sus discos intercalares.

188
Figuras autorizadas creadas por Mariana Ruiz Villarreal En las neuronas respecto de este modelo de comunicación existen dos tipos de comunicación o sinapsis: las sinapsis eléctricas de mayor rapidez que las sinapsis de tipo químico. Las sinapsis de tipo eléctrico se presentan en los clásicos ejemplos de reacciones de huida de los insectos. Las comunicaciones de tipo nexo tienen otras funciones, por ejemplo, en los tejidos de músculo cardiaco y en las sinapsis eléctricas de algunas neuronas tienen como objeto la transmisión de señales de una célula a otra. Estas uniones comunicantes son abundantes en los períodos embrionarios, son muy funcionales en los procesos de diferenciación celular. El acoplamiento entre las células es también de tipo metabólico, puesto que también se permite el flujo de moléculas pequeñas tales como, nucleótidos, azúcares, aminoácidos, vitaminas, y otros metabolitos. Este tipo de interacción es lo que en biología celular se conoce con el nombre de Cooperación Metabólica intercelular.

189
Relación con cáncer: En las células cancerosas existe una ausencia de acoplamiento e incapacidad para comunicarse. CLASIFICACIÓN DE LAS ZONAS DE CONTACTO ENTRE LAS CÉLULAS Estas zonas de contacto o uniones intercelulares son especializaciones desarrolladas a partir de las membranas plasmáticas entre células vecinas. A través de ellas, las células de los distintos tejidos se mantienen unidas unas a otras formando estructuras complejas tales como la epidermis, impermeabilizando los epitelios del medio externo, como en el tubo digestivo, o posibilitando el trabajo coordinado de grandes masas celulares, como ocurre en el músculo cardiaco. Ya hemos visto que el trabajo de células epiteliales es por ejemplo el transporte de glucosa hacia la sangre. El mecanismo es que la glucosa y su paso debe ser regulado y esta regulación ocurre gracias a los mecanismos de regulación que posee la célula. Hasta donde se conoce el paso de glucosa se debe a la acción de un transportador de tipo simporte, que utiliza la energía del transporte de sodio a favor del gradiente (transporte secundario). Lo interesante es que las uniones estrechas establecidas entre las células no permiten que la glucosa ingrese por otras zonas.

190
JAM (JUNTIONAL ASSOCIATED MEMBRANE PROTEIN)
Esta proteína se expresa en células epiteliales y endoteliales. Posee una sola región transmembranal y en la zonas correspondientes al lado extracelular contiene dos dominio con enlaces disulfuro. Su localización es en las uniones estrechas. La transfección de la proteína conduce a la disminución de la permeabilidad de las monocapas al flujo paracelular de moléculas.
Algunos hechos observados como por ejemplo, la presencia de JAM en epitelios y endotelios sugieren que puede estar relacionada con el paso de leucocitos a través de la ruta paracelular.
PROTEÍNAS ZO.Z0-1, ZO-2 Y ZO-3
Z0-1 es una abreviatura de zonula occludens. Z0.1 es una proteína de las células epiteliales y endoteliales. También se encuentra presente en el citoplasma y los contactos puntuales de astrocitos, células de Schawnn, fibroblastos, glioma, sarcoma, etc. En las células epiteliales a asocia a las uniones adherentes en las primeras etapas del desarrollo embrionario. En la actualidad se realizan variadas investigaciones con respecto al rol de este tipo de proteínas en los procesos de diferenciación celular, cáncer, y el rol funcional- estructural en los procesos relacionados con absorción o permeabilidad de los túbulos proximales, distales y colectores del riñón. ZO-2 es una proteína de las uniones estrechas de las células epiteliales y endoteliales y se encuentra en las uniones adherentes de fibroblastos y células musculares cardiacas. Las proteínas ZO-1, ZO-2 y ZO-3 son miembros de la familia MAGUK, que en general se localizan en estructuras de unión celular especializadas. Este tipo de moléculas presentan dominios que han sido muy conservados en la evolución. La función es asociarse a otras proteínas y transducir señales. Un ejemplo de ello es la proteína PSD95 y CASK que se localizan en las sinapsis y se asocian a canales y proteínas transmembranales. Las proteínas ZO al parecer se establecen como puentes entre la actina (proteína del citosqueleto) y proteínas transmembranales de las uniones estrechas. En este caso participan la claudinas. UNIONES ADHERENTES: Los contactos entre las células en los tejidos se inicia con la formación de uniones adherente. Este tipo de uniones, como hemos visto en clases, no pueden distinguirse con una criofractura a nivel de la membrana plasmática lateral.

191
En este tipo de uniones también participan complejos multiproteicos. En el caso se ha estudiado con detalle la participación de la cadherina. Esta es una proteína transmembrana, que en la unión se asocia a proteínas citoplasmáticas tales como las cateninas y estas a su vez con la actina del citoesqueleto. Las cadherinas son un grupo perteneciente a las moléculas de adhesión, las cuales se dividen en cuatro grupos: integrinas, inmunoglobulinas, selectinas y cadherinas. Como se ha dicho, las cadherinas se localizan en las uniones adherentes y consisten químicamente en glicoproteínas dependientes de calcio para la adhesión celular.
En general, la clasificación de las uniones intercelulares ha sido confusa durante mucho tiempo, por tanto, es sugerente seguir la nomenclatura y ordenamiento dado en el texto de Biología Celular y Molecular de Robertis del año 2000, ello permitirá una mayor comprensión.
Tipo de Unión Sinónimo Filamentos asociados
Proteínas de Unión
A) Unión oclusiva Zonula occludens
Unión estrecha
Actina Ocludina
B)Uniones adherentes:
Desmosoma puntiforme
Hemidesmosoma (célula-lámina basal)
Banda de adhesión
Mácula adherens
Desmosoma
--
Zonula adherens
Unión intermedia
Desmosoma en banda
Desmosoma en cinturón
Intermedios
Intermedios
Actina
Desmogleínas Desmocolinas
Integrinas
Cadherinas tradicionales (E,P,N)
Punto adherente
Contacto focal (célula- matriz)
Punctum adherens
Placa de adhesión
Actina
Actina
Cadherinas
Integrinas
c) Unión comunicante Unión con -- conexina

192
espacio
Unión con hendidura (gap junction)
Nexo o nexus
Figura: Tabla tomada de texto de Robertis
Uniones oclusivas Uniones estrechas: Regiones diferenciadas para cerrar el espacio
intercelular y evitar el pasaje de sustancias a través de dicho espacio.
Estas uniones estrechas (tight junctions) son aquello contactos más íntimos conocidos entre células. Previenen la difusión de moléculas entre células adyacentes y la migración lateral de las proteínas y lípidos de membrana. En las células epiteliales del intestino contribuyen a la polarización celular diferenciando en la membrana dos dominios funcionalmente diferentes: el apical, que es aquel que contiene las microvellosidades y se especializa en la absorción de nutrientes desde el lumen del intestino, y el basolateral cuya función es transferir los nutrientes a la sangre. Las uniones estrechas se sitúan por debajo de la superficie apical de las microvellosidades.
Se denominó proteína ZO-1 al hallazgo de esta macromolécula en las uniones estrechas, pero hasta el día de hoy se ha descubierto más de 30 proteínas asociadas. Por ejemplo, la ocludina y proteínas de la familia de las claudinas interaccionan con interacciones homofílicas o heterofílicas con ocludinas y claudinas de células adyacentes y proteínas ZO-1, ZO2, ZO3, AF6 y cingulina, se acoplan a otras proteínas y con microfilamentos de actina.
Barrera hematoencefálica: (Un paréntesis)
En neurociencias debe destacarse la importancia de la barrera hematoencefálica para el sistema nervioso central. Los aportes nutritivos, sustratos energéticos, especialmente oxígeno y glucosa, son suministrados por red de capilares sanguíneos. Esta barrera aísla al sistema nervioso del resto de la circulación sanguínea. Los efectos de esta separación aparecen en concreto cuando tras la administración de un tratamiento farmacológico por vía intravascular, surge una ausencia de efecto terapeútico, ello se debe a que el fármaco no puede traspasar la barrera y llegar al cerebro. Los ejemplos son variados, en caso de algunos antibióticos, que aunque idóneos para eliminar bacterias patógenas como en el caso de la meningitis, no pueden utilizarse por la dificultad de atravesar la barrera, otro ejemplo, es con respecto al uso de citostáticos en la terapia contra los tumores primarios del sistema nervioso central.
La permeabilidad de la BHE en el endotelio vascular, es muy restringido al paso de solutos plasmáticos, se exceptúan el agua, gases como oxígeno y el bióxido de carbono, moléculas liposolubles pequeñas menores de 600 Da, las moléculas orgánicas no pueden atravesar libremente el endotelio, para hacerlo deben hacerlo a través de sistemas de transporte específicos y finamente regulados.
Las observaciones con microscopio electrónico muestran que las células endoteliales son fundamentales en la constitución de la barrera. Las células endoteliales de los vasos del

193
encéfalo y la médula espinal poseen características morfológicas y funcionales que son diferentes a las células del endotelio del resto de vasos sanguíneos.
Las células del endotelio son selladas con uniones estrechas (tigh junction). La barrera hematoencefálica está constituida por células endoteliales de la red capilar del sistema nervioso central. Además hay participación de los pericitos, la lámina basal abluminal, los astrocitos perivasculares y la microglía. El endotelio de los capilares cerebrales es notable porque sus bordes celulares están íntimamente unidos a la célula adyacente que hace impermeable a la pared interna del capilar.
Los capilares en el tejido neuronal están constituidos por una capa simple de células endoteliales, asociadas a una membrana basal, pericitos y una capa casi continua de astrocitos.
La relevancia en el estudio de esta barrera se deja ver por su participación en enfermedades neurológicas tales como infartos cerebrales, esclerosis, Alzheimer. Hay evidencias que la inflamación altera la expresión de proteínas de la unión estrecha. Por ejemplo, en la enfermedad de Alzheimer la proteína β amiloide induce la liberación de citoquinas y la migración de monocitos a través de la barrera, los depósitos de β amiloide inducen degeneración de la membrana basal de la microvasculatura.
Podemos entender la Barrera como una separación entre el SNC y el sistema circulatorio y que por tanto, regula el medio ambiente de las células cerebrales. Las uniones estrechas disminuyen la permeabilidad paracelular.
Células epiteliales intestinales
Al frente se compara la estructura
de un capilar cerebral con barrera
hematoencefálica y un capilar sin la
barrera.1) Uniones estrechas entre
dos células endoteliales,2) Célula
endotelial, 3) Pericito, 4) Pie
astrocitario pericapilar, 5) Lámina
basal subendotelial, 6) Hendidura
interendotelial, 7) Fenestración
intraendotelial. Tomada desde
Revista de Neurología, Pascual y
col, 2004.

194
Fig. Sistema de transporte y el aporte de tigh junctions a las funciones de absorción de nutrientes al organismo.
La unión estrecha se ubica en el límite entre las caras apical y basolateral y su función es actuar como barrera de difusión que regula el paso de iones y moléculas a través de la ruta paracelular (espacio lateral entre célula y célula). También es importante en la función de mantenimiento de la polaridad celular, puesto que impide el libre movimiento de los lípidos y de las proteínas a través de la membrana plasmática, esto se da por la diferenciación que provoca en el establecimiento de los dominios apical y basolateral.
En suma, constituye un sistema de sellado entre las células vecinas, en consecuencia moléculas hidrosolubles no pueden difundir entre las células. La criofractura revela que están compuestas por una red filiforme que rodea la zona apical. Estas estructuras son proteínas transmembranales específicas, situadas en cada una de las membranas plasmáticas, ellas concluyen con unión directa y ocluyen el espacio intercelular.
La unión de las proteínas que forman el sellado puede ser descrita en términos de un dominio extracelular adhesivo constituyente de proteínas transmembranales, éstas a su vez se encuentran con proyección en la regiones citoplasmáticas (dominio submembranal) estableciendo allí contacto con proteínas transductoras de señales o proteínas constituyentes del citoesqueleto.
Constitución Química
Ocludina: Es una proteína que forma parte de la Unión estrecha. Se trata de una proteína transmembranal. Se le observa claramente con técnicas de criofracturas. El análisis de su secuencia aminoacídica nos revela cuatro dominios transmembranales y dos dominios extracelulares. Estos dos dominios forman la región de contacto con la célula vecina. Los estudios con técnicas de biología molecular muestran que la ocludina mutada introducida a los sistemas de unión, permite que la unión se vuelva permeable al flujo paracelular de iones y moléculas.
Por otra parte, se ha observado que el ARNm de la ocludina se expresa intensamente en epitelios que presentan abundante uniones estrechas, no ocurre así en los fibroblastos que no poseen uniones estrechas. En las células endoteliales de cerebro, que tienen alta resistencia transepitelial, hay altos niveles de expresión de ocludina que establece contacto con la Z0-I en los sitios de unión célula. Lo contrario podemos encontrar en las células

195
endoteliales de la aorta, cuyas uniones estrechas muestran baja resistencia transepitelial, en este caso la ocludina se encuentra escasamente; en los túbulos renales la expresión de Ocludína aumenta a medida que crece la resistencia eléctrica transepitelial. Por ello, hay más ocludina en los segmentos colectores que en los proximales.
Bioquímicamente se ha sugerido que existe algún tipo de regulación de la ocludina en la formación de la unión estrecha. Ello sería posible pro procesos de fosforilación y defosforilación.
CLAUDINAS
Otro tipo de proteínas de la unión estrecha son las llamadas claudinas, también tienen como función oponerse al paso de moléculas por la vía paracelular. Al introducirlas a fibroblastos forman redes filamentosas. Hasta el momento se ha identificado una familia de claudina formadas por 18 miembros. La claudina número 4 se identificó como receptor de la enterotoxina de clostridium perfringens;, la claudina 11 de los oligodendrocitos, células de Sertoli y de cubiertas laminares de mielina. Otros descubrimientos importantes han conducido a la hipótesis de el tipo 16 de claudina participaría con función de canal selectivo para ión magnesio. La reabsorción del ión magnesio se diferencia de los iones Na, K, Ca, porque ocurre por la ruta paracelular. El hallazgo es importante por su concordancia con el modelo de función de la unión estrecha, que no es otro que el regular el flujo de iones y moléculas por la ruta paracelular.
La reabsorción de magnesio y calcio en el segmento grueso de la rama ascendente del asa de Henle debido a una mutación del gen PCLN1, que codifica una proteína, la paracelina-1, que interviene en la reabsorción de ambos cationes.
El intercambio de solutos y de agua a través de un epitelio se realiza por distinta vías. Como hemos visto en clases estas vías son: la transcelular y la paracelular. La transcelular se encuentra regulada por bombas y transportadores. El transporte paracelular en cambio es de tipo pasivo, estableciéndose intercambio por medio de gradientes electro-osmóticos generados por el transporte transcelular.
En los últimos años se ha producido algunos avances en la comprensión del transporte paracelular. Ya se sabe que está regulado por los “tight junction” (clásicamente conocidas como “zonula occludens”), en ella las membranas de las células epiteliales se encuentran en estrecho contacto.
Absorción en los epitelios
La absorción de monosacáridos como hexosas y pentosas ocurre en el intestino delgado. Las moléculas pasan de las mucosas a la sangre en los capilares que drenan hacia la vena porta. Un factor importante para el transporte de hexosas es la concentración de sodio en la mucosa, si esta es alta entonces se facilitará la entrada a las células epiteliales y una baja concentración permite el efecto opuesto. Por supuesto como se ha dicho antes, la glucosa y el sodio comparten un transportador. En el organismo hay otros diseños de transportadores de sodio y glucosa y han sido denominados en forma distinta SGLT1, SGLT2. También algunos de estos transportadores están encargados del transporte de glucosa hacia fuera de los túbulos renales.

196
Uniones adherentes
Las uniones adherentes incluyen a los desmosomas, cuya función es mantener unidas a las células del epitelio, en esta circunstancia de unión, se produce una asociación de filamentos intermedios, de células vecinas, con ello da lugar a una red transcelular con alta resistencia a la tracción mecánica. De esta forma, ha contribución a que las células mantengan su forma y que la lámina epitelial exista en forma estable. Las cadherinas desmosómicas, desmogleína y desmocolina, conectan con los filamentos intermedios a través de la plakoglobina, plakofilina y desmoplakina.
Los desmosomas en banda y desmosomas puntiformes, contribuyen a la adhesión intercelular mediante los puntos adherentes. Estas uniones se caracterizan por la composición molecular, filamentos asociados, etc.
En los desmosomas puntiformes existen miembros de las cadherinas, que son las desmogleínas y las desmocolinas.
La desmogleína 1 es un polipéptido de 160 kDa, tiene gran importancia médica, por cuanto en pacientes que sufren de pénfigo foliáceo se encuentran autoanticuerpos dirigidos contra las estas proteína.
La desmogleína 3 aparece en epitelios planos estratificados, también de importancia médica, los pacientes afectados en relación con esta proteína presentan síntomas caracterizados como pénfigo vulgar.
Figura: Más información en http://www.nlm.nih.gov/medlineplus/spanish/ency/esp_imagepages/2459.htm
Los hemidesmosomas son también un grupo de uniones que se clasifican dentro de las uniones adherentes, se encuentran en contacto con la lámina basal de las células epiteliales, contactan con filamentos intermedios. Asocian el citoesqueleto de citoqueratina con la matriz extracelular por medio de moléculas transmembrana, las llamadas integrinas.
Otra categoría de uniones adherentes se encuentra caracterizada por la forma de contactos puntuales o asociadas en línea o cintas alrededor de las células (adhesión belts).

197
Está presente en células adyacentes de la mayoría de los tejidos humanos. A través de estas uniones se produce el contacto de citoesqueletos de células vecinas, la función surge en la resistencia mecánica al tejido y proporciona puntos de anclaje para los movimientos del citoesqueleto.
Las proteínas transmembrana cadherinas forman contactos homotípicos en este tipo de uniones adherentes. La principal cadherina en el tejido epitelial es la E-cadherina que requiere interactuar con los filamentos de actina del citoesquelético. Estas proteínas están implicadas en la transducción de señales, adhesión celular y morfogénesis, señalización y diferenciación.
Estructura y función de la neurona. Generalidades
Por lo general, del cuerpo celular sale un solo axón. Este es una prolongación capaz de propagar los impulsos eléctricos a grandes distancias en dirección de los terminales sinápticos.
Una principal característica de las neuronas es que son excitables. Experimentan variaciones de potencial eléctrico, lo cual es posible gracias a la presencia de estructuras membranosas de tipo proteico, tal como la bomba de iones y los canales iónicos. La propiedad de excitabilidad no es exclusiva de las neuronas, también la poseen las células musculares, pero, en un grado menor de especialización. El proceso por el cual el axón transmite una señal se denomina potencial de acción, una onda eléctrica que se propaga desde el punto de iniciación (llamado cono axónico) hasta el extremo del axón.
Las terminaciones axónicas presentan regiones especializadas denominadas botones o terminaciones sinápticas. Las sinapsis son las uniones entre las células nerviosas pudiendo ser carácter eléctrico (si hay contacto físico entre las dos células) o químico (si la célula envía la señal en forma de transmisor químico o neurotransmisor). Los neurotransmisores se sintetizan en las terminaciones sinápticas y se almacenan en las vesículas sinápticas, a la espera de ser liberados en la hendidura sináptica a la llegada de una señal. Los neurotransmisores producidos por una neurona de tipo presináptico difunde en la hendidura sináptica fijándose a los receptores de la célula post-sináptica que emite respuesta o señal, el primer evento a ocurrir es la entrada de iones del medio extracelular (lo que genera de nuevo un potencial de acción), bien indirectamente a través de los segundos mensajeros.
La diversidad citológica de las neuronas que se obtiene a consecuencia de los procesos de diferenciación durante el desarrollo es muy evidente al nivel molecular. Cada neurona es el resultado de la expresión de genes, es decir, de las proteínas y enzimas que constituyen de fondo los fundamentos para todo tipo de acción metabólica de la célula.
Las neuronas se desarrollan a partir de células epiteliales y conservan la polaridad propia de dichas células. Poseen polos diferenciados, para el caso, desde la superficie basolateral surgen las dendritas, y desde la superficie apical surge el axón. La membrana citoplasmática recibe el nombre de plasmalema y representan una barrera de tipo hidrófoba e impermeable a muchos tipos de sustancias. Mayores detalles se pueden encontrar en los apuntes del primer semestre.

198
Movimiento de proteínas en la neurona:
Los procesos de tipo fisiológico de los organismos, si bien pueden ser observados a nivel microscópico mediante técnicas de laboratorio que evidencian reacciones moleculares o hasta nivel celular. En el nivel macroscópico encontramos evidencias del funcionamiento de órganos, sistemas de órganos, por ejemplo, con electroencefalogramas se evidencia que el cerebro humano posee actividad que verificamos en el instante por observaciones o interacciones con el paciente. Luego ese evento tiene sus correlatos con actividad celular y molecular. Para lograr claridad mayor, suponga que usted memoriza la letra de una canción de moda, lo escuchamos en el nivel de interacciones con nuestro medio ambiente y que podemos llamar el medio macroscópico. Luego podemos registrar su canto, al hacerlo verificamos que usted en realidad aprendió la letra, la melodía, podemos tener pruebas de ello, por ejemplo, mediante la grabación en un CD. Luego podemos preguntarnos que estaría sucediendo en el cerebro mientras usted memorizaba. A este nivel microscópico podemos observar con instrumentos, técnicas bioquímicas, análisis electroforético, podemos utilizar distinto tipo de microscopios, ópticos, electrónicos, etc. El objeto del investigador en este caso es buscar la correspondencia entre la conducta del sujeto y los eventos de tipo molecular que ocurren en las neuronas, entre neuronas o al interior de ella.
Como estudiante es bueno que usted se entere que cuando memoriza están ocurriendo cientos de reacciones bioquímicas, se activan determinados genes, se inactivan otros, se establecen cambios en el citoesqueleto, etc.
En la mayor parte de los procesos fisiológicos y sus alteraciones de tipo patológico se encontrará relaciones con la superficie celular. Los tratamientos farmacológicos por lo general involucran aspectos relacionados con la superficie celular. Por ello es importante conocer los mecanismos de generación y mantención de la organización celular, de la superficie celular, aspecto muy vital, porque se relaciona con los procesos de comunicación celular. De manera que se debe considerar aspectos relevantes de la superficie celular, en cuanto a su génesis, su recambio y la reestructuración continua.
Las funciones nombradas anteriormente serán más conocidas si se estudian los mecanismos de tráfico vesicular de proteínas al interior de la célula. Así por ejemplo, podemos conocer la forma en que llega a posicionarse una determinada proteína en la membrana celular, membrana mitocondrial o de cualquier organelo al interior de la célula.
El sistema de membranas y endomembranas que posee la neurona o cualquier otro tipo de célula eucarionte, es el lugar donde se realiza funciones determinadas según la información genómica que surge desde el núcleo y luego con esta información se realiza la síntesis proteica en los ribosomas asociados al retículo endoplasmático o de ribosomas libres en el citoplasma. Las proteínas una vez fabricadas deben dirigirse a los distintos organelos donde median las funciones que ocurren en el sistema de endomembranas.

199
Las proteínas fabricadas (sintetizadas) contienen señales de destino, y por tanto de direccionamiento del lugar en que realizarán su actividad, también puede tratarse de una señal que indique exocitosis, o sea, traspasar la membrana celular y luego realizar una determinada función en células distintas de aquellas que la fabricaron.
Recordemos que la membrana celular o plasmática es el límite de la unidad, separa los componentes externos e internos de la célula. La membrana tiene una estructura que adopta distinta función, según si las interacciones de ella son con los componentes extracelulares o intracelulares. Las interacciones con el ambiente externo pueden ser con la matriz extracelular, con nutrientes, iones, estímulos variados o con elementos patógenos. Estas interacciones son mediadas por proteínas que se han integrado a la membrana, logran atravesarla y mostrar parte de su estructura en la lámina externa de dicha cubierta. La

200
lámina interna de la membrana establece cientos de interacciones moleculares que regulan la forma, el movimiento celular, la secreción, el tráfico endocítico y en general cascadas de señales en respuesta a señales-estímulos provenientes del medio externo. Funciones tales como la secreción de enzimas, hormonas, citoquinas, interpretación de señales desde el medio externo, nutrición celular, mecanismos de defensa del sistema inmune, interacciones célula-patógeno dependen de las proteínas de membrana.
El soma de la neurona en general es el lugar principal relacionado con el origen de las señales. En otras células neuronales el soma no recibe señales sinápticas. El soma participa activamente en la síntesis de proteínas. Para ello requiere del uso de macromoléculas como el ADN que se encuentra en el núcleo, de los ARNs que abandonan el núcleo en dirección del citoplasma y se asocian con ribosomas para dar lugar a la síntesis de proteínas. Los ribosomas son una mezcla de proteínas con ARN ribosómico. Es de particular importancia el movimiento de las proteínas recién sintetizadas hacia los diferentes compartimientos de la célula neuronal y es lo que veremos a continuación.
El tráfico de proteínas en el citoplasma se realiza mediante dos rutas: las rutas exocítica y la endocítica. La primera surte con proteínas la membrana plasmática, su característica principal es la destinación de proteínas lisosomales y de secreción, en la segunda ruta la activación de los procesos endocíticos remueve proteínas de la membrana, luego estas son degradadas en los lisosomas, los aminoácidos resultantes de esta degradación serán útiles para la construcción de nuevas proteínas. El tráfico o el transporte de estas proteínas hacia su destino son realizados en pequeñas vesículas que surgen de sistemas de membranas y luego después del transporte se fusionan con otros compartimientos membranosos.
Las proteínas pueden ingresar a la ruta exocítica si es que poseen péptidos señales que consisten en pequeñas secuencias de aminoácidos. De esta manera, mientras transcurre la síntesis de estas proteínas, la señal permite un acoplamiento entre el polipéptido o cadena naciente de aminoácidos y ribosomas unidos a membranas del retículo endoplasmático.
Los ribosomas se unen con el retículo endoplasmático por la subunidad 60 S, el péptido señal comienza por atravesar la membrana mediante un complejo proteico denominado sec61, especie de canal que mediante una de sus subunidades se asocia a la cadena polipeptídica en síntesis.
Este tipo de Translocación Cotraduccional comienza con una fase de orientación durante el cual el ribosoma complejo y la cadena naciente se dirige a la membrana de la partícula de reconocimiento de señal (SRP) y su receptor de membrana (receptor de SRP) (para más información, véase Halic Beckmann & 2005, & Sinning Luirink 2004). Una vez que el ribosoma se une a la proteína-canal conductor, alargando la cadena polipeptídica que se mueve desde estos hacia el canal de membrana; la hidrólisis del GTP durante la traducción proporciona la energía para la translocación. Cuando el ribosoma sintetiza un dominio citosólico de la proteína de la membrana, la cadena de polipéptidos surge de la unión al ribosoma canal de lado en el citosol (Mothes et al. 1997). El péptido señal es eliminado, la cadena ingresa al lumen del RE, las proteínas solubles o de secreción resultantes son útiles en el mismo RE, o Aparato de Golgi, luego de pasar estos organelos experimentando procesos de arreglos o terminaciones finas de las proteínas, resultan en proteínas de

201
membrana plasmática, de secreción y lisosomales, podemos agregar proteínas de algunos virus que se ensamblan en los compartimientos de la ruta exocítica o en la membrana plasmática.
En el RE, las proteínas se pliegan estableciéndose una conformación compatible con su función. Las proteínas experimentan glicosilación y son controladas por un sistema de “control de calidad” mediado por chaperonas. El sistema devuelve las proteínas que tienen errores en el plegamiento (proceso de retro-translocación), luego en el citoplasma son degradadas por el proteosoma. El incorrecto funcionamiento de este complejo de reacciones, obviamente, conduce al desarrollo de patologías, especialmente, de enfermedades neurodegenerativas.
Las proteínas luego de haber pasado por el RE, ingresan al AG, por las cisternas de tipo Cis, de ahí a las cisternas mediales y por último alcanzan las cisternas trans. Habrá algunas que les corresponde permanecer allí y realizarán funciones propias de la estructura, otras pasarán en tránsito, experimentando modificaciones, remodelamiento de monosacáridos de oligosacáridos y cortes con eliminación de pequeñas secuencias de aminoácidos, finalmente pasan de región trans-Golgi a vesículas y túbulos transportadores con destino lisosomas, membrana plasmática.
Ubicadas o insertadas en la membrana celular algunas proteínas se integran de manera constitutiva a rutas de tipo endocítico y con ello, reciclan. Por ejemplo, receptores de lipoproteínas de baja densidad (LDRL) y de transferrina (TfR).
La familia de Receptores de Lipoproteínas de Baja Densidad (LDLRs) está formada en mamíferos por nueve proteínas transmembrana relacionadas estructural y funcionalmente. El LDLR, tiene como función principal la regulación de la homeostasis del colesterol. Se expresa principalmente en el hígado donde media la endocitosis de las partículas LDL, ricas en colesterol. Su alteración o disfunción provoca hipercolesterolemia y enfermedades cardiovasculares. Al igual que el LDLR, la principal actividad de los otros miembros de la familia es también mediar la endocitosis de múltiples ligandos, que abarcan desde las distintas lipoproteínas, proteasas, complejos proteasa-inhibidor de proteasa, complejos de vitaminas con sus proteínas trasportadoras, algunas toxinas como la exotoxina A de Pseudomonas, la Amiloid Precursor Protein etc.
Varios de estos LDLRs también participan en señalización intercelular. Dos ejemplos claros son ApoER2 y VLDLR que actúan de receptores redundantes para la proteína extracelular Reelin, responsable de la correcta migración neuronal durante el desarrollo del cerebro, y LRP5/LRP6 (Arrow en Drosophila) que son esenciales para la transducción de la señal Wnt.

202
Figura péptido señal
La vía de señalización Wnt está siendo conocida, para ver más consulte la cita de la figura mostrada antes.
La síntesis de proteínas es realizada en el cuerpo celular y en las dendritas. El destino final de una proteína se encuentra codificada en la secuencia de aminoácidos de la misma, el péptido señal.

203
Los procesos secretores en las neuronas son similares a las de otras células. Excepto que el lugar principal de secreción es la terminal axónica que está a considerable distancia del soma, y las dendritas donde son sintetizadas las proteínas de secreción. El movimiento de plasma del axón- axoplasma, es considerado como movimiento de dos tipos, lento y rápido.
Además de la velocidad desarrollada en el movimiento, también se pueden clasificar en dos tipos de acuerdo a la dirección. Movimiento hacia el axón se denomina movimiento anterógrado y desde el axón hacia el cuerpo se denomina movimiento retrógrado.
Transporte axónico rápido: supera los 400 mm/día.
Transporte lento: Las proteínas citoplasmáticas y del citoesqueleto se mueven sólo en dirección anterógrada y de forma lenta.
Los orgánulos membranosos son transportados por movimientos rápidos en cualquier dirección, hacia el axón o desde el axón. Los orgánulos son referidos a vesículas de la vía secretoria constitutiva, membranas precursoras de vesículas sinápticas, mitocondrias y elementos del retículo endoplásmico liso.
El transporte anterógrado del axón es dependiente de los microtúbulos que parecen pistas sobre las cuales los orgánulos se mueven mediante motores moleculares.
Las moléculas motoras del transporte anterógrado son la kinesina y proteínas relacionadas KIF, que son una familia de ATPasa, cada una de las cuales transporta diferentes cargas membranales.

204
En dirección retrógrada desde la terminación del axón hacia el soma, los órganulos son principalmente endosomas producidos por endocitosis en las terminales nerviosas.
Teoría neuronal-principios de organización nerviosa.
- Los potenciales de acción son señales que el cerebro recibe, analiza y transmite información. Las señales son estereotipadas aún cuando sean iniciadas por una amplia variedad de sucesos del entorno.
- El principio se extiende al segundo principio de que la información transmitida por un potencial de acción no está determinada por la forma se la señal sino por la vía que recorre en el cerebro.
- El cerebro analiza e interpreta los patrones de señales eléctricas que recibe, luego crea las sensaciones habituales de l a vista, el tacto, etc.
- El potencial de acción es unidireccional y ello implica también que las neuronas en su organización no muestran contacto de continuidad citoplasmática entre ellas, es decir, no forman un sincitio.
1º Principio de la polarización dinámica
· En una neurona las señales eléctricas sólo viajan en una dirección, desde los receptores de la neurona (dendritas y cuerpo celular) al axón (reacción desencadenante), y a partir de ahí, el potencial de acción se propaga unidireccionalmente a la Terminal presináptica.
2º Principio de la especificidad de conexión
· Las neuronas no se conectan indiscriminadamente entre sí para formar redes al azar. Al contrario, las neuronas establecen conexiones en determinados puntos y en determinadas células.
Forma neuronal:
Lo que diferencia una neurona de otra es la forma y número de prolongaciones.
Neuronas unipolares: Poseen una prolongación primaria que a menudo es el origen de otras ramas, axones o dendritas. Se encuentran en el sistema nervioso autónomo.

205
Neuronas bipolares: el soma presenta dos o más prolongaciones, dendritas que llevan información desde la periferia al cuerpo y un axón que lleva información hacia el SNC. Son ejemplo, las células sensoriales como las de la retina ocular, epitelio olfatorio en la nariz.
Neuronas pseudounipolares: al desarrollo de estas células se observa como sus prolongaciones bipolares se fusionan y se extienden desde el cuerpo celular como una única prolongación. Luego esta se divide en dos prolongaciones y ambas funcionan como axones. Una dirigida hacia la piel o músculos periféricos y el otro hacia la médula espinal.

206
Neuronas multipolares: Poseen un axón y muchas dendritas. Ejemplo neuronas motoras espinales que inervan fibras musculares esqueléticas. Las células piramidales con cuerpo aproximadamente triangular, sus dendritas surgen de la parte apical o basal. Se encuentran en el hipocampo y en la corteza cerebral. Tenemos también las células de Purkinje del cerebelo las dendritas forman ramas que semejan arboles. Ello sugiere que estas células forman gran cantidad de sinapsis.
Una neurona motora espinal puede recibir unos 10.000 contactos sobre el cuerpo celular y dendritas. En una célula de Purkinje puede establecer uno 150.000 contactos.
Las neuronas se clasifican también en tres grupos de acuerdo a su posición funcional. Estas son: sensitivas, motoras e interneuronas.
Neuronas sensitivas: transmiten información desde la periferia del cuerpo al sistema nervioso central.
Neuronas motoras: transmiten información órdenes del encéfalo o la médula espinal a los músculos y las glándulas.
Neuronas interneuronas: No son sensitivas ni motoras. Se dividen en dos clases:
Interneuronas de transmisión o proyección con largos axones y transmisión a largas distancias.
Interneuronas locales con axones cortos, trabajan en circuitos locales.
REDES NEURONALES:
Las manifestaciones conductuales del cerebro son expresiones de la información sensitiva, programación de respuestas motoras y emocionales, almacenamiento de información (memoria). Las neuronas correspondientes a tales actividades forman conjuntos de neuronas conectadas cuya acción conjunta puede pasar a constituir una conducta.
Ejemplo: Reflejo simple, rotuliano, miotático

207
El tendón del cuádricep femoral, músculo extensor que mueve la pierna, se encuentra unido a la tibia por el tendón rotuliano. Un golpe en el tendón (estímulo) con dirección bajo la rótula, estira los cuádriceps femorales. En este tipo de acción no hay participación de la voluntad.
Es el golpe bajo la rótula que inicia la serie de eventos que llevarán a la contracción refleja del cuádriceps femoral.
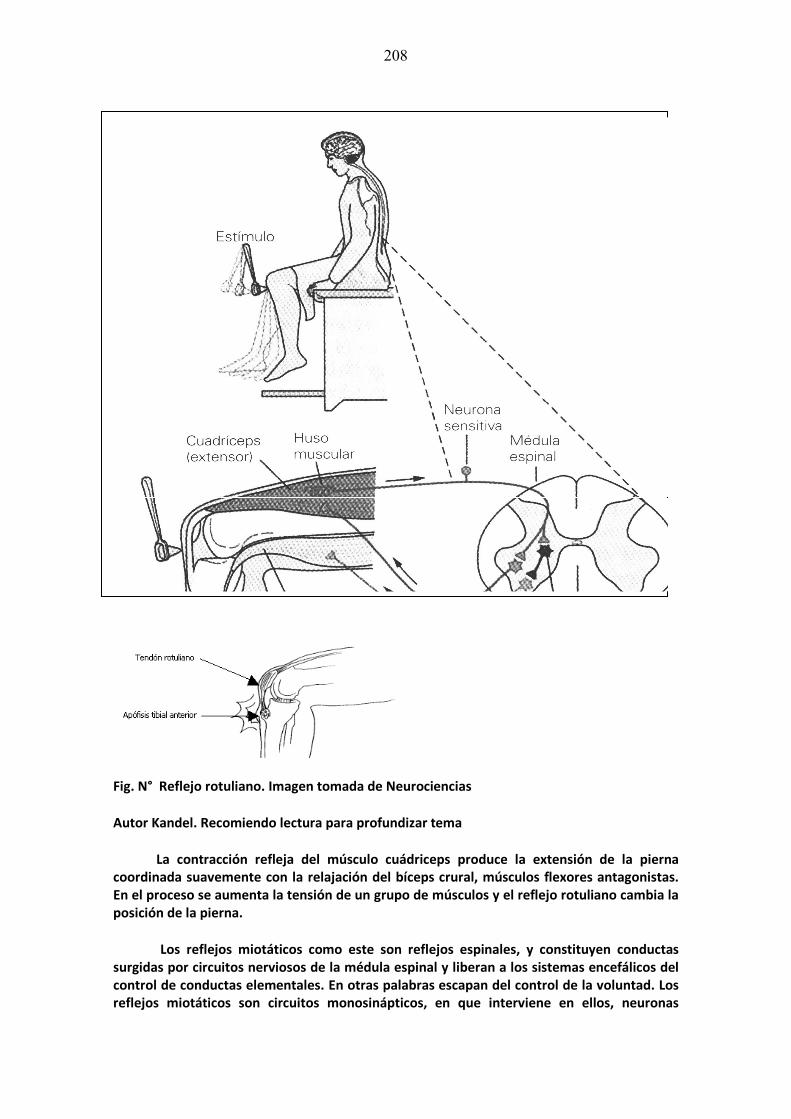
208
Fig. N° Reflejo rotuliano. Imagen tomada de Neurociencias
Autor Kandel. Recomiendo lectura para profundizar tema
La contracción refleja del músculo cuádriceps produce la extensión de la pierna coordinada suavemente con la relajación del bíceps crural, músculos flexores antagonistas. En el proceso se aumenta la tensión de un grupo de músculos y el reflejo rotuliano cambia la posición de la pierna.
Los reflejos miotáticos como este son reflejos espinales, y constituyen conductas surgidas por circuitos nerviosos de la médula espinal y liberan a los sistemas encefálicos del control de conductas elementales. En otras palabras escapan del control de la voluntad. Los reflejos miotáticos son circuitos monosinápticos, en que interviene en ellos, neuronas

209
sensitivas y neuronas motoras conectadas entre sí, sin la mediación de interneuronas. La mayoría de los reflejos espinales, sin embargo, están construidos con circuitos polisinápticos y pueden ser controlados por el encéfalo.
Las neuronas sensitivas de tipo mecanorreceptoras para el caso dado, se encuentran en los ganglios cercanos de la raíz dorsal de la médula. Son células pseudounipolares que proyectan una de sus ramas axónicas hacia el músculo cuádriceps y la otra se proyecta hacia la médula. La rama que establece contacto con el músculo media con receptores sensibles al estiramiento denominado husos musculares y es capaz de excitarse cuando el músculo es estirado. Por su parte, la rama que va hacia la médula establece conexiones excitadoras con las neuronas motoras que inervan al cuádriceps y controla su acción. Además, entra en contacto con interneuronas locales que inhiben las neuronas motoras que controlan los músculos flexores antagonistas. Es necesario aclarar que las interneuronas no participan en el reflejo miotático mismo, sin embargo, al controlar la acción motora, aumenta la estabilidad de la respuesta refleja.
Análisis: el circuito produce cuatro tipos de información:
1.- Transmisión de información sensitiva al sistema nervioso central (la médula espinal) desde la superficie del cuerpo.
2.- Envío de órdenes motoras desde el sistema nervioso central a los músculos ligados al reflejo rotuliano.
3.- Emisión de órdenes complementarias, de tipo inhibidoras, a las neuronas motoras que inervan los músculos antagonistas. Se permite así la coordinación de la acción muscular.
4.- Transmisión de información acerca del reflejo rotuliano a los centros superiores del sistema nervioso central. Permitiendo que el cerebro coordine las órdenes conductuales.
Divergencia neuronal:
La distensión del cuádricep produce la activación de cientos de neuronas sensitivas, cada una de ellas establece contacto con 100 a 150 neuronas motoras. En otras palabras, una neurona activa muchas células efectoras. El fenómeno se denomina divergencia neuronal.

210
Convergencia neuronal:
Existe 5 a 10 veces más neuronas sensitivas que neuronas motoras, por lo tanto es de esperar que una neurona motora reciba aferencias de muchas neuronas sensitivas. Este fenómeno es llamado convergencia. De esta manera la neurona motora es capaz de integrar información diversa de distinta procedencia.
Inhibición por alimentación anterógrada:
Hemos visto antes las neuronas sensitivas realizan conexiones excitatorias (sinapsis excitatorias) y realizan conexiones con interneuronas inhibitorias. Las conexiones excitadoras con los músculos extensores de la pierna determinan que ellos se contraigan, no así con las conexiones de neuronas inhibitorias, estas impiden que los músculos antagonistas se contraigan. Esto es lo que caracteriza al circuito, la inhibición por alimentación anterógrada permite que las vías flexoras y extensoras se inhiban mutuamente, por ello se dice que es recíproca. En otras palabras, se permite que sólo se recluten los músculos apropiados para el movimiento y no los que se oponen a él.
Inhibición por retroacción:
Este caso se refiere a una neurona activa que puede tener conexiones excitadoras con una célula efectora y con una interneurona inhibidora que tiene su propia conexión por retroacción con la neurona activa. Así, las señales de la neurona activa excitan la neurona diana y la interneurona inhibidora, que a su vez es capaz de limitar la capacidad de la neurona activa de excitar a la neurona diana.
Para generar una conducta cada neurona sensitiva y motora produce secuencialmente cuatro señales diferentes en lugares distintos de la célula:
a) Señal de entrada (componente receptivo) b) Señal desencadenante (componente de suma o integrador) c) Señal de conducción (de larga distancia-señalización) d) Señal de salida (componente secretor)
Neurona
sensitiva
Neuronas
motoras

211
Este modelo representa fisiológicamente el principio de polarización dinámica de Ramón y Cajal. El principio dice que la información entra por las dendritas y el soma, y sale por el axón. Principio que posee excepciones. De manera que planteamos y plantean algunos, algo así: “la polarización dinámica es generalmente válida en casi todas las neuronas”. Ciertamente no se trata de un principio absoluto. La intuición de Cajal es buena, y se acepta en la actualidad que la neurona posee regiones con funciones determinadas y que se relacionan con la clasificación dada en el párrafo anterior. Por lo tanto, el principio de polarización dinámica se reemplaza por el de “especialización regional de las funciones dentro de la célula neuronal”.
Las señales se encuentran determinadas fundamentalmente por las propiedades eléctricas de la membrana celular. Brevemente se hará alusión, por ahora.
El potencial de reposo es igual en toda la célula. En las neuronas sensitivas el flujo de corriente se inicia como consecuencia de un estímulo sensitivo, que activa proteínas receptoras especializadas en la superficie receptora de la célula. En el caso del reflejo rotuliano el estiramiento del músculo activa proteínas específicas y sensibles al estiramiento de la neurona sensitiva. La proteína receptora especializada forma canales iónicos en la membrana y a través de ellos fluye iones sodio y potasio. Estos canales se abren cuando la célula se estira. El potencial de reposo se transforma en potencia receptor. La amplitud y la duración del potencial receptor dependen de la intensidad del estiramiento muscular. A mayor estiramiento o mayor duración mayor y más duradero será el potencial receptor. La mayoría de los potenciales receptores son despolarizantes, es decir, excitadores. Existen potenciales receptores hiperpolarizantes, o sea, inhibidores en la retina ocular.
El potencial de reposo se propaga pasivamente a través del axón. De manera que su amplitud disminuye con la distancia y no puede transmitirse mucho más allá de 2mm. Al recorrer dicha distancia la amplitud disminuye más de dos tercios de la original. Luego para ser transportada al sistema nervioso central, la señal debe ser amplificada, es decir, generar un potencial de acción. En el reflejo rotuliano el potencial receptor de la neurona sensitiva se propaga hasta el primer anillo de Ranvier del axón, donde puede generar el potencial de acción que se propaga hasta la terminal axónica en la médula espinal. A consecuencia, luego, la sinapsis con la neurona motora promueve la señal de entrada a la neurona motora.
El potencial de acción de la neurona sensitiva libera una señal química- un neurotransmisor, a la hendidura sináptica. El neurotransmisor se une a las proteínas receptoras de la neurona motora, y la consecuencia es la transducción de energía química potencial del transmisor en energía eléctrica. El potencial de membrana de la célula motora se altera y la variación de este potencial se denomina potencial sináptico.
Potencial sináptico:
La amplitud del potencial sináptico depende de la cantidad de neurotransmisor que se libera y la duración de actividad del mismo.
Por otra parte, el potencial sináptico puede ser despolarizante o hiperpolarizante dependiendo del tipo de receptor que se activa. Los potenciales sinápticos son como los potenciales receptores por cuanto se comportan como alteraciones locales del potencial de membrana que se propaga en forma pasiva a lo largo de la neurona y luego puede generar un potencial de acción.

212
Los potenciales de acción se producen por flujos de iones de sodio hacia el interior de la neurona. El flujo avanza por canales sensible al voltaje (más adelante se profundiza en estos aspectos) existente en la membrana. Cuando una señal de entrada, ya sea un potencial sináptico o un potencial receptor, despolariza la membrana celular, la variación del voltaje abre los canales iónicos del sodio. Estos canales de sodio están concentrados en el primer segmento- primer nudo de Ranvier del axón, tal y como se observa en las neuronas sensitivas. En la interneuronas y las neuronas motoras la mayor densidad de canales de sodio surge en el montículo del axón ubicado en el cuerpo celular.
La mayor cantidad de canales en estas zonas tiene sus consecuencias, por una parte poseen un umbral más bajo, es decir, que cuesta menos alcanzar un estado de potencial de acción que en cualquier otro lugar de la célula. Estas zonas se conocen como zonas de iniciación de impulsos o zonas desencadenante. La suma de las actividades de todos los potenciales receptores o sinápticos ocurre en esta zona.
Concepto de ley de todo o nada:
Se refiere a que los estímulos subumbrales no producen señal. Para que se produzca la señal debe alcanzar un nivel umbral. Todos los estímulos que superen dicho umbral ya sea, variando en intensidad o duración, la respuesta será que la amplitud y la duración de cada potencial de acción son prácticamente iguales.
Diferencia con potencial receptor y sináptico:
Estos potenciales se propagan pasivamente y disminuyen su amplitud. El potencial de acción no se reduce y viaja por todo el axón hasta la terminal axónica.
Los potenciales de acción son estereotipados: varían muy poco en su forma de onda. Para entender esto se debe considerar dos características de la señal de conducción que transmiten información:
a) El número de potenciales de acción. b) Intervalos de tiempo entre ellos.
Por lo tanto, lo que determina la intensidad de sensación es la frecuencia. Claramente no es la magnitud o la duración de cada potencial de acción.
Problema: Si las señales son estereotipadas y no reflejan las propiedades del estímulo ¿Cómo transmiten las neuronas la información específica?
Un poco más claro: ¿Cómo hacemos la distinción entre un mensaje de tipo auditivo y uno de picada de insecto?
El mensaje se interpreta de acuerdo a la vía que lo transporta.
Componente de salida de las neuronas (componente secretor)
Al llegar el potencial de acción a la terminal, se produce liberación de transmisores químicos. Estos neurotransmisores se encuentran almacenados previamente a la señal. El almacenaje ocurre en vesículas sinápticas. Para la descarga del neurotransmisor la vesícula

213
se desplaza hacia la membrana de la neurona y se fusiona con ella; luego por exocitosis se libera el neurotransmisor hacia el espacio sináptico.
Característica: La cantidad de neurotransmisor liberado depende del número y frecuencia del potencial de acción.
Desde el espacio sináptico el neurotransmisor difunde hacia los receptores de membrana de la neurona postsináptica. La unión del neurotransmisor al receptor es la causa del nacimiento de un potencial sináptico en la neurona postsinática. El potencial sináptico puede tener un efecto excitador o inhibidor. Ello queda anunciado por el tipo de receptor de la neurona postsináptica y no por el tipo de neurotransmisor. De hecho, el mismo tipo de neurotransmisor puede tener efectos diferentes en receptores distintos.
Mayor comprensión: El potencial de reposo se altera cuando procesos químicos en la sinapsis producen un cambio en la permeabilidad a una molécula, o ión, por lo general, el ión Na+. A diferencia del K+, el Na+ está más concentrado fuera de la célula. Ello es causado por un mecanismo activo que bombea iones sodio hacia el exterior. Entre sus características es que el bombeo es gastador de energía en las neuronas.
La transmisión sináptica puede dar como resultado un aumento transitorio de la permeabilidad al Na+, este sigue un movimiento que depende de su gradiente de concentración e ingresa a la neurona. Puesto que el sodio es un ión positivo en consecuencia el movimiento del potencial de membrana en dirección positiva desde unos -70mV a una cifra más cercana a cero. Este gradiente de carga positiva se propaga de manera pasiva y casi simultánea, como la electricidad, reduciendo la polarización de la membrana a una cierta distancia desde la dendrita hacia el soma. A medida que esta despolarización se extiende y se difunde por la superficie de la membrana, comienza a disminuir su magnitud. Esta es la conducción de tipo electrotónica, rápida y decreciente, al llegar al soma experimenta una despolarización menor. En este caso, la magnitud del potencial depende críticamente de la distancia que debe recorrer, por ello, las sinapsis en ramificaciones dendríticas alejadas son menos eficaces que aquellas cercanas al soma de la neurona.
La alteración de la membrana celular de la neurona es espectacular y se aprecia con normalidad en los axones. En el terminal postsináptico de las dendritas y de la mayor parte del soma, la membrana no modifica en forma sustancial su permeabilidad. Así, los potenciales desarrollados en la sinapsis son propagados por conducción electrotónica, con rapidez, pero luego disminuye a lo largo de la dendrita. Por ello la conducción pasiva electrotónica está lejos de preservar la comunicación de señales en los axones. Como se ha dicho antes, a partir de la región del cono, en los inicios de la estructura axonal se produce el cambio de permeabilidad de la membrana de forma espectacular. Esta es la región donde se alcanza el umbral y donde los canales son controlados por el voltaje.
Células neurogliales
Las células neurogliales o sencillamente células de la glía no participan directamente en la transmisión de las señales eléctricas aunque ayudan al metabolismo de los neurotransmisores y en otras funciones de ayuda a las neuronas. Las células de la glía son más abundantes que las neuronas en la proporción de 3:1. Las funciones de la glía más o menos aceptadas son:

214
• mantenimiento del medio iónico de las neuronas • modulación de la velocidad de propagación de la señal • modulación de la sinapsis al captar parte de los neurotransmisores • recuperación de las lesiones nerviosas
Existen tres tipos de células neurogliales:
Producción de Mielina
Los oligodendrocitos son células gliales que envuelven a las neuronas en el SNC. La vaina formada por su actividad tiene la característica de servir de sustancia aislante. La presencia de esta sustancia aislante acelera la transmisión de impulsos nerviosos. En la enfermedad llamada esclerosis múltiple se produce un deterioro de zonas mielinizadas y son reemplazadas por tejido cicatrizal. Las lesiones producidas interfieren con la conducción de impulsos neurales, el paciente experimenta pérdida de coordinación, temblor y parálisis parcial o completa de zonas corporales.
Las células de Schawnn, otro tipo de células gliales, se ubican fuera del sistema nervioso central (sistema nervioso periférico) y forman vainas de mielina en neuronas. Las células de schawnn, ubicadas a lo largo de un axón forman segmentos de mielina ya vaina de mielina adopta su forma cuando la célula de schawnn da varias vueltas en torno al axón, lo envuelve en capas concéntricas de membrana. Los intervalos entre los segmentos producidos se denominan nódulos o anillos de Ranvier.
En suma, la producción de mielina corre por cuenta de los oligodendrocitos y las células de Schawnn. Cada oligodendrocito en el sistema nervioso central envuelve en promedio 15 espacios internodales axónicos. Las células de schawnn en cambio, en el sistema nervioso periférico, sólo pueden envolver un único espacio internodal de un axón por célula.
Debe quedar claro que los oligodendrocitos en la sustancia blanca proporcionan la mielina, y en la sustancia gris rodean y sostienen los cuerpos celulares de las neuronas, en este caso se trata de oligodendrocitos perineurales.
Existen varios trabajos sobre enfermedades asociadas a mielina, estas muestran dificultades en la transmisión del impulso nervioso, por ello, a manera de ejemplo, se altera las funciones motoras y sensitivas. Algunas proteínas básicas de la mielina, se conocen alrededor de siete de ellas (revisión de kandel), se encuentran relacionadas con enfermedades desmielinizantes, estas se caracterizan porque los potenciales de acción son más lentos en axones y dendritas. En ratones mutantes (transgénicos) que presentan temblores y convulsiones se ha observado delecciones de regiones de ADN del gen de la proteína básica del cromosoma 18. La enfermedad se puede transmitir hacia la descendencia como un tipo de herencia autosómica recesiva.
Se conoce otras proteínas asociadas con la mielina como las proteínas MAG (glucoproteínas asociadas a mielina) y se sospecha que es una molécula de adherencia que se expresa en las células de Schawnn en los inicios de los procesos de mielinización.
Los axones conductores de señales de las neuronas sensibles y motoras están envainados en mielina en toda su trayectoria. La mielina actúa como aislante, además acelera la transmisión a lo largo de las fibras, hechos importantes para los reflejos rápidos.

215
La capa de mielina se dispone en forma de vainas concéntricas. La composición de la mielina es semejante a la composición de la membrana celular, 70% de lípidos y 30% de proteínas.
En el desarrollo del sistema nervioso periférico, los axones ubicados a lo largo del nervio en depresiones formadas por las células de Schawnn a intervalos regulares, cada intervalos entre una y otra célula dará lugar posteriormente a los nudos de Ranvier. La membrana celular de cada célula de Schawnn rodea a un axón y forma una estructura de doble membrana llamada mesoaxón, que se enrolla en torno al axón en capas concéntricas.
Astrocitos: Son células con forma de estrellas con numerosas prolongaciones citoplasmáticas ramificadas que se extienden en todas direcciones. Sus núcleos son ovoides o esféricos, de color pálido, con pocos gránulos finos de cromatina que se distribuyen hacia la periferia en estrecho contacto con la membrana nuclear. Los nucléolos no son muy notorios. Se distingue dos tipos de astrocitos.
Poseen varias funciones:
.- Función de guía y soporte en la migración neuronal durante el desarrollo.
.- Participan en la mantención del medio ambiente neuronal-por ejemplo, mantener un medio químico en cuanto a concentraciones iónicas y de glucosa. Captar y metabolizar los neurotransmisores que han cumplido su función. En la transmisión sináptica, los astrocitos por medio de sus prolongaciones envuelven las regiones sinápticas, y algunos trabajos de investigación sugieren que pueden ejercer una influencia a través de la modulación del volumen, composición, y las concentraciones de iones, neurotransmisores, neuromoduladores, y fuentes de energía dentro del espacio extracelular y en particular en la hendidura sináptica. Poseen activa participación en la formación de la barrera hematoencefálica.Las investigaciones indican que de alguna forma intervienen en la formación, maduración y conexiones sinápticas entre neuronas, lo que se denomina en términos más amplios, sinaptogénesis. Otras investigaciones indican que entre sus funciones se encuentra el de incrementar la sinaptogénesis. En ciertos experimentos se ha observado que los astrocitos producen colesterol, este compuesto a su vez participa en la inducción de la sinaptogénesis. Los mecanismos de acción se encuentran sometidos a estudios.
Otros estudios indican que los astrocitos activan la maduración de las células troncales nerviosas, es decir, de la maduración de las células que dan origen a las neuronas.
Los astrocitos producen factores de crecimiento útiles para regeneración de tejidos cerebrales o espinales dañados por traumas o enfermedades.
Los astrocitos forman una red en neuropilo llamado sincitio astrocítico. Las moléculas difunden en el sincitio por las hendiduras entre los astrocitos
El calcio se propaga en oleadas través del sincitio, siendo estas oleadas inducidas por estimulación mecánica y por el glutamato. De esta manera, los astrocitos modulan la composición y concentración de moléculas en el espacio extracelular si el flujo de calcio a través de los mismos produce entrada o salida de iones y neuromoduladores Ca-dependientes. Así, el sincitio permite un medio de comunicación no sináptica en el cerebro. Los astrocitos pueden recibir e integrar información de varias sinapsis simultáneamente y recibir información de la composición del líquido extracelular y dentro de los capilares sanguíneos.

216
Una de las formas mediante las cuales los astrocitos están implicados en la función sináptica es facilitando metabolitos para la actividad neuronal. La energía es suministrada por la glucosa que entra en SNC a través de los astrocitos que están en contacto con los vasos sanguíneos. Parece ser que la glucosa es parcialmente metabolizada en los astrocitos siendo empleados algunos intermedios metabólicos para el uso neuronal. En efecto, cuando se bloquea el metabolismo de los astrocitos del hipocampo con flurocitrato (un inhibidor del ciclo de Krebs), la transmisión sináptica es abolida. Esto indica que las neuronas necesitan del combustible que reciben de los astrocitos.
Astrocitos fibrosos. (Con forma de araña) se encuentran principalmente en la sustancia blanca. Sus prolongaciones pasan entre las fibras nerviosas. Tienen prolongaciones largas, delgadas, lisas y no muy ramificadas. Contienen muchos filamentos en su citoplasma.
Astrocitos protoplasmáticos (citoplasmáticos) se encuentran en las sustancia gris del cerebro y médula espinal, sus prolongaciones pasan también entre los cuerpos de las células nerviosas, son anchas, cortas, gruesas y ramificadas, con un contenido menor en filamentos intermediarios (También se les reconoce como células satélite). Los filamentos intermedios están compuestos en su gran mayoría por una proteína denominada proteína fibrilar ácida
de la glía (GFAP). Ambos, los fibrosos y los protoplasmáticos, proporcionan un marco de sostén, son aislantes eléctricos, limitan la diseminación de los neurotransmisores, captan iones de K+, almacenan glucógeno y tienen función fagocítica, ocupando el lugar de las neuronas muertas (gliosis de reemplazo). Ambos tipos de astrocitos envían prolongaciones que envuelven la sinapsis y la superficie de las células nerviosas. El potencial de membrana de estas células varía con la concentración de K+ extracelular. Los efectos sobre las células neuronales son varios entre ellos, ayudan a mantener la concentración apropiada de iones y neurotransmisores por medio de la captura de ión K+ y neurotransmisores como glutamato, gama-aminobutirato(GABA).
Fig. Interacción entre astrocitos y neuronas glutaminérgicas. Los astrocitos captan el glutamato liberado y lo convierten en glutamina, la cual regresa a las neuronas convirtiéndose en glutamato y liberándose como neurotransmisor. El esquema es conocido como ciclo glutamato-glutamina. Según el esquema el glutamato que se encuentra en el espacio sináptico es captado por un transportador de glutamato que es dependiente de Na+. Otro aspecto interesante es que desde los capilares sanguíneos se transfiere la glucosa a los astrocitos y las neuronas. En los astrocitos la glucosa se metaboliza a lactato con ello

217
(Recuerde, el metabolismo de la glucosa) se obtiene dos moléculas de ATP. Una molécula de ATP se utiliza para transformar el glutamato en glutamina y la molécula de ATP restante la utiliza la ATPasa de Na+ y K+ para bombear tres átomos de sodio fuera de la célula y con dos átomos de potasio que ingresan a la célula (Ganong W. 2004)
Podemos resumir aquellos aspectos relevantes acerca de los astrocitos y luego considerar algunos aspectos que se investigan en la actualidad. Tenemos planteado ya que los astrocitos desempeñan varias funciones en relación con el desarrollo, metabolismo, patologías. Considérese la importancia de estas células en el soporte trófico y metabolismo neuronal, la supervivencia, diferenciación y guía neuronal, el crecimiento neurístico y la sinaptogénesis y muy especialmente en la regulación de la homeostasis cerebral toda vez que regula la concentración local de sustancias e iones.
Existen evidencias de la existencia de comunicación bidireccional entre astrocitos y neuronas de tal manera entonces que puede plantearse la hipótesis de que los astrocitos participan activamente en los procesos de información del sistema nervioso.
Evidencias: Con el desarrollo de técnicas de imagen y de marcadores fluorescentes que son sensible a iones se ha podido estimar la concentración de iones intracelulares en las células vivas, con esto se ha mostrado que las células vivas astrocitos son excitables, y esta excitabilidad es causada por las variaciones de la concentración de Ca++ intracelular. Clásicamente se ha excluido a los astrocitos de la fisiología del sistema nervioso y se aceptaba especialmente lo resumido en los párrafos anteriores. Pero vemos que la ciencia cambia con el tiempo y las evidencias aportadas principalmente por Cornell a partir de los años 90 y desde entonces algunos otros investigadores, han llevado a una pequeña revolución si nos referimos a las sinapsis.
La señal intracelular de calcio puede alcanzar a otros astrocitos vecinos. En estudios con astrocitos en cultivo se ha demostrado que los aumentos de calcio pueden propagarse sin disminución a las células vecinas. Así, de esta manera se propaga la onda de Calcio. Se ha calculado la velocidad y distancia de la propagación de este tipo de ondas.
Una hipótesis mantiene que las ondas se propagan debido a la difusión de inositol trifosfato (recuerde las clases de señales y comunicación celular), IP3, a través de uniones intercelulares comunicantes, o gap junctions donde se encuentran las proteínas conexinas, que son las que forman las uniones comunicantes.
Una segunda hipótesis propone que las ondas de Calcio son mediadas por señales químicas que serían captadas por receptores de los astrocitos, luego a consecuencia aumentaría el calcio intracelular y con ello, la propagación. En apoyo de esta hipótesis se encuentra el hecho de que la señal extracelular es el ATP. Podemos considerar que las dos hipótesis son complementarias puesto que las conexinas podría mediar la liberación de ATP.
Por lo menos, los astrocitos son capaces de comunicarse entre sí, mediante la transferencia de la señal intracelular de calcio++, lo que representa una nueva forma de comunicación intercelular ene. SNC.
En ambos tipos de astrocitos se observa que una o más prolongaciones citoplasmáticas terminan en un acúmulo aplanado situado en la adventicia de un vaso

218
sanguíneo. Son los llamados “pies perivasculares”, que se presentan en gran número por ello a veces se dice que forman una vaina externa o perivascular del vaso sanguíneo. Los elementos gliales, las prolongaciones de los astrocitos, más los cuerpos neuronales y sus axones y dendritas constituyen lo que se ha denominado neurópilo. El neurópilo es muy denso en la superficie del sistema nervioso central, justo por debajo de la piamadre y por abajo del epéndimo. Debido a esta arquitectura el tejido nervioso se encuentra separado de las estructuras de origen mesodérmico tales como, membranas y vasos sanguíneos. La separación la impone las células gliales y sus prolongaciones.
Microglías
Son células, derivadas de las células madre hematopoyéticas con muchas de las propiedades de los macrófagos tisulares. Por lo tanto son células representantes del sistema inmunológico que se establecen en tejidos correspondientes al sistema nervioso central. La microglía pasa de un estado inactivo a un estado activo. Prolifera después de una lesión del sistema nervioso, ayudando a reparar el daño neural. De modo que representan un sistema de reparación.
Las microglias tienen su origen en monocitos sanguíneos y alcanzan el cerebro en estados de desarrollo tempranos. Luego en etapas postnatal experimentan cambios morfológicos y permanecen sin actividad hasta reconocer señales- estímulos que conducen a la activación. El proceso indica proliferación, aumento en la expresión de moléculas propias correspondientes a macrófagos, como por ejemplo, complejos de histocompatibilidad. Adquieren propiedades fagocíticas y migratorias.
Estas células se activan por lesiones de tipo isquémico, inflamatorias, químicas y mecánicas. Enfermedades crónicas: esclerosis múltiple, encefalopatías, enfermedad de Alzheimer, de Parkinson, etc.
Las células microglias son especialmente notables en los procesos de daño tisular menor y cuando no hay destrucción de parénquima. Si los daños abarcan mayor espacio (o es masivo), las microglias son ayudadas por monocitos sanguíneos.

219
MECANISMOS FISIOLÓGICOS DE POTENCIALES DE MEMBRANA
A ambos lados de la membrana celular, se da una diferencia en la carga eléctrica, se establece de esta forma un potencial eléctrico. Se origina por las diferencias en la concentración iónica en ambos lados de la membrana. Normalmente la concentración de iones K+ en el citoplasma es superior a su concentración en el exterior y por el contrario, la concentración de iones Na+ es mayor en el exterior que en el interior celular.
En general la difusión iónica a ambos lados de la membrana está gobernada por tres factores o fuerzas:
A) Atracción y repulsión de cargas, cargas positivas y cargas negativas B) Difusión de partículas siguiendo las leyes de la difusión o gradientes de
concentración. C) Propiedades de la membrana
La membrana debido a su constitución química es impermeable a los iones orgánicos de gran tamaño como el Na+, y es algo más permeable a los iones K+. Más adelante se verá en mayor profundidad estos aspectos. Pero el asunto de permeabilidad para estos iones es superada por mecanismos tales como el bombeo de iones realizados por la bomba de sodio y potasio.
Las fuerzas que constantemente actúan sobre los iones K+ son responsables del potencial eléctrico de la membrana en reposo. Gracias al establecimiento del gradiente de concentración, estos iones tienden a moverse al lado extracelular, desarrollándose un exceso de carga negativa en el interior celular y por tanto, se impide que una mayor cantidad de iones se desplacen hacia fuera. El punto es que se alcanza un equilibrio en el cual no existe movimiento neto de iones K+ a través de la membrana. En este equilibrio, sin embargo, existe un exceso ligero de carga negativa en el interior de la célula, y al potencial resultante se conoce como potencial de reposo.
Todas las células poseen un potencial de membrana, con valores diferentes, pero las células excitables como las neuronas o las de tipo muscular pueden modificar su potencial de membrana es respuesta a estímulos. Dependiendo de las características y de la intensidad de los estímulos aplicados, el potencial de membrana pasa a ser potencial graduado o electrotónicos.
Los potenciales electrotónicos no tienen umbral ni período refractario (ver más adelante), pueden ser despolarizantes o hiperpolarizantes y pueden ser iniciados mediante canales iónicos dependientes de receptor, ya sea por estímulos mecánicos, ambientales y por neurotransmisores.
La sinapsis es un lugar especializado de contacto célula, llegando a ser el punto de comunicación y de interacción entre las células nerviosas, o entre células nerviosas y otras células como las musculares o glandulares. La comunicación se logra mediante señales eléctricas o químicas. En las sinapsis eléctricas, la codificación de la información no sufre ninguna transformación cuando pasa de una célula a la siguiente y son el tipo de sinapsis

220
menos numeroso. Las sinapsis químicas son más numerosas y presentan otras propiedades como las del fenómeno de la plasticidad, son lugares de acción para gran cantidad de sustancias de tipo farmacológicas que influyen la actividad del sistema nervioso.
POTENCIAL DE REPOSO:
El concepto se refiere a la diferencia de carga eléctrica en ambos lados de la membrana citoplasmática de una célula, me refiero a la neurona. Una neurona en reposo no transmite impulsos porque se encuentra polarizada, es decir, en ambos lados de la membrana- hacia el lado citosólico y hacia el lado del líquido intersticial que rodea a la célula, existe la diferencia de carga, ello significa que el interior de la membrana posee carga negativa si se le compara con el exterior. La separación de cargas por la membrana lleva al establecimiento de una diferencia de potencial eléctrico entre ambos lados. A este se le denomina potencial de membrana o potencial de reposo.
El potencial de reposo suele expresarse en milivolts (mV), que es una medida de potencial eléctrico. Dicho potencial de una neurona corresponde a unos 70 mV. Por convención se expresa como -70mV, puesto que la superficie interna de la membrana se encuentra más cargada negativamente al comparársele con la superficie externa. El potencial puede medirse colocando un electrodo dentro de la célula y un segundo electrodo en la superficie externa. Los dos electrodos se conectan mediante un instrumento- un galvanómetro, el que mide la corriente eléctrica. Experimentalmente si colocamos los dos electrodos en la superficie externa a la célula no encontraremos diferencia de potencial. Entonces, de esta manera, podemos considerar la neurona como una especie de batería.
El origen del potencial de reposo en la neurona se debe por una parte, a la presencia de las bombas de sodio y potasio que trabajan contra gradientes de concentración y por lo tanto requieren para su trabajo de la energía química del ATP. Recuerde usted la clase de transporte. Por otra parte se debe al flujo de iones por difusión facilitada por proteínas de membrana que forman canales para iones en particular. En este proceso el movimiento de iones se produce de zonas de alta concentración a zonas de menor concentración.
Para poder separar cargas de distinto signo es necesario una energía, para ser más precisos, se trata de energía eléctrica puesto que se trata de cargas eléctricas. Las cargas separadas de signo opuesto tienen una energía potencial. Esta energía para separar las cargas es denominada voltaje. El voltaje, en concreto, es la energía eléctrica que hace que los electrones se muevan, se llama también fuerza electromotriz. El voltaje depende de la diferencia entre las cargas de dos puntos y la distancia de separación entre ellos.
La corriente eléctrica que se produce entre ambos lados de la membrana se debe al flujo de cargas y este fenómeno se debe a la apertura y cierre de canales iónicos. Convencionalmente el flujo de corriente se define como el movimiento neto de carga positiva. De ahí en una solución de tipo iónico los cationes se mueven en la dirección de la corriente eléctrica y los aniones en dirección contraria.
Cada vez que hay un flujo de iones, ya sea de cationes o aniones hacia dentro o fuera de la membrana, se altera la carga en ambos lados de la célula en reposo y ello se dice que se altera la polarización.

221
En consecuencia, una reducción en la separación de carga, (cuando entra ion sódicol) conduce a un potencial de membrana menos negativo y recibe el nombre de despolarización. Si aumentamos la separación de carga, (si entra cloruro) conduce a un potencial de membrana más negativo y se denomina hiperpolarización.
Aquellos cambios en el potencial de membrana que membrana que no conducen la apertura de los canales iónicos reciben el nombre de potenciales electrotónicos. Al parecer son respuestas pasivas de la membrana, al igual que las respuestas hiperpolarizantes o las despolarizaciones pequeñas.
Cuando la despolarización se aproxima a un nivel crítico llamado umbral, la célula responde con la apertura de los canales sensibles a voltaje.
Como se llega al potencial de acción
Los iones de K+ salen de la membrana siguiendo su concentración hasta que las cargas positivas fuera de la membrana llegan a una concentración tal que los iones son repelidos por la presencia de cargas positivas fuera de la membrana. Mayores detalles se verán con el apartado de canales iónicos en adelante.
Lo interesante es que se alcanza una estabilidad cuando la salida de potasio iguala a la entrada de iones sodio. Es entonces cuando se alcanza la diferencia de potencial de -70mV de reposo de la membrana.
Otros contribuyentes al potencial de membrana lo constituyen proteínas y fosfatos orgánicos con carga negativa ubicados al interior de la neurona. Esto son moléculas grandes con dificultades para poder difundir.
PROFUNDIZANDO
¿Por qué están polarizadas las neuronas cuando están en reposo?
Como todas las sales que se encuentran en solución, las sales del tejido neuronal como es de esperarse, se separan en partículas con cargas positivas y negativas. Del mismo modo cuando usted agrega sal a la sopa, ocurre que la sal se disocia, esto es pasa de la forma molecular NaCl a la forma disociada Na+ y Cl-.
Las formas disociadas son átomos de sodio y cloro que tienen carga + y – respectivamente y se conocen como iones. Las carga positivas además tienen la capacidad de repelerse entre si y lo mismo ocurre con las cargas negativas. Por el contrario las cargas positivas con las negativas se atraen.
El potencial de reposo entonces tiene que ver con las cargas, estas cargas se concentran en ambos lados de la membrana celular. En el lado interno predominan las cargas negativas cuando la célula se encuentra en el llamado estado de reposo. Avanzando con el concepto, la célula se encuentra en estado de reposo cuando se alcanza el potencial de membrana de -70 mV. El estado de reposo es un concepto que debe entenderse como aquel estado distinto de aquel en que la célula se encuentra enviando impulsos.
El potencial de reposo ocurre porque la cantidad de cargas negativas al interior de las células es mayor. Esto se debe a cuatro factores: dos fuerzas cuya acción es distribuir los

222
iones uniformemente en los líquidos intracelular y extracelular del sistema nervioso, y otros dos factores que compensan las fuerzas que conducen a la uniformidad.
A) La primera fuerza es el movimiento aleatorio y la tendencia natural del movimiento al azar de las partículas es la distribución regular de las partículas. Por ello es muy probable que por estas fuerzas no se produzcan gradientes de concentración de sustancias, al contrario, dichos gradientes tienden a la reducción. En otras palabras, las sustancias se mueven desde una región donde se encuentran en alta concentración hacia una de baja concentración.
B) La segunda fuerza que promueve la distribución uniforme de iones es la presión electrostática. Ello significa que al acumularse cargas en una zona, estas mismas tienden a dispersarse por la repulsión de las cargas del mismo signo. El fenómeno aumenta debido a la atracción de las cargas de signo opuesto. Ambos fenómenos constituyen la presión electrostática.
Contra todo efecto de homogenización o de movimiento al azar de los iones, estos terminan distribuyéndose desigualmente a ambos lados de la membrana. Existen cuatro iones de los cuales depende el potencial de reposo: iones de Na+, K+, Cl- y proteínas con carga negativa.
En la mantención del potencial de reposo uno podría preguntarse porqué las concentraciones extracelulares de sodio o de potasio no siguen hacia el equilibrio de concentración en ambos lados de la membrana (recuerde las leyes de difusión-primer semestre). Es decir, por que el sodio no atraviesa la membrana hacia el lado donde se encuentra en menor concentración.
Podríamos preguntarnos o expresarnos de alguna otra forma: De qué manera los gradientes iónicos aportan al potencial de reposo.
Dos investigadores iniciaron esta investigación en la década de los años cincuenta. Ya se sabía que el potencial de reposo era de -70 mV, habría que calcular la presión electrostática para cada ión y compararla con el potencial de reposo.
Lo que habría que calcular entonces es la carga electrostática para cada ión y que necesita para contrarrestar la presión que les empujaba a moverse hacia un gradiente de concentración menor.
Para el caso de los iones de cloruro (Cl-) la carga electrostática era de -70 mV, igual a la del potencial de reposo. En conclusión cuando las neuronas están en reposo, la distribución desigual de iones de Cl- entre los lados de la membrana se mantenía estable debido al equilibrio entre la fuerza de 70 mV que tiende a impulsar a los iones de Cloruro hacia el gradiente de concentración más bajo del interior de la neurona y la presión electrostática de 70 mV que tiende a sacarlos hacia fuera de la membrana.
Con los iones de Potasio se calculó una presión electrostática de 90 mV para impedir que estos iones se muevan hacia un gradiente de concentración menor fuera de la neurona. Para el caso del sodio es en tanto diferente, porque el sodio tendería a moverse en virtud de dos factores que actúan en la misma dirección de flujo. Al estar la neurona en reposo, la concentración de sodio en el exterior posee una presión de 50 mV para que los iones se muevan hacia el lado de concentración menor y los 70 mV que les
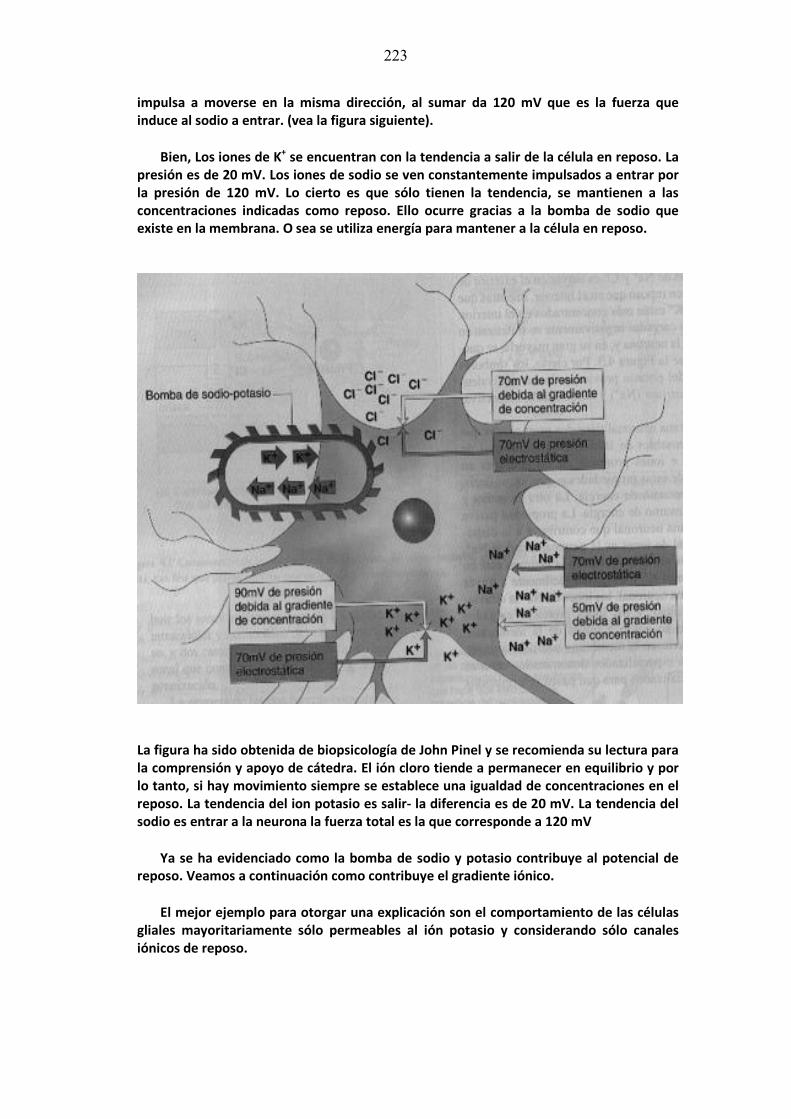
223
impulsa a moverse en la misma dirección, al sumar da 120 mV que es la fuerza que induce al sodio a entrar. (vea la figura siguiente).
Bien, Los iones de K+ se encuentran con la tendencia a salir de la célula en reposo. La presión es de 20 mV. Los iones de sodio se ven constantemente impulsados a entrar por la presión de 120 mV. Lo cierto es que sólo tienen la tendencia, se mantienen a las concentraciones indicadas como reposo. Ello ocurre gracias a la bomba de sodio que existe en la membrana. O sea se utiliza energía para mantener a la célula en reposo.
La figura ha sido obtenida de biopsicología de John Pinel y se recomienda su lectura para la comprensión y apoyo de cátedra. El ión cloro tiende a permanecer en equilibrio y por lo tanto, si hay movimiento siempre se establece una igualdad de concentraciones en el reposo. La tendencia del ion potasio es salir- la diferencia es de 20 mV. La tendencia del sodio es entrar a la neurona la fuerza total es la que corresponde a 120 mV
Ya se ha evidenciado como la bomba de sodio y potasio contribuye al potencial de reposo. Veamos a continuación como contribuye el gradiente iónico.
El mejor ejemplo para otorgar una explicación son el comportamiento de las células gliales mayoritariamente sólo permeables al ión potasio y considerando sólo canales iónicos de reposo.

224
La célula glial posee elevada concentración de K+ y aniones orgánicos situados en la parte interna de la célula y en la zona externa se puede encontrar gran cantidad de iones Na+ y Cl-.
Dado que el ion potasio se encuentra en gran concentración dentro de la célula existe la tendencia natural a que este ion fluya hacia fuera. Como resultado el exterior se torna positivo y el interior negativo. Sin embargo, este proceso tiene su limitación. La separación de cargas que produce el movimiento de potasio y que origina una diferencia de potencial eléctrico nos lleva a seguir razonando de la siguiente manera: En la fase sucesivas a este fenómeno, el ion potasio se irá acumulando fuera de la célula, generando por una parte, mayor diferencia de potencial; pero, esta misma diferencia de potencial se irá oponiendo al flujo de ion potasio hacia fuera y se desarrolla una fuerza que lo arrastra hacia dentro. Ello se explica por las fuerza de repulsión- el exceso de cargas positivas fuera reprime que sigan llegando al lugar cargas positivas.
De forma que los iones y sus movimientos se encuentran dependiendo de dos factores que actúan a través de la membrana:
a) Flujo que depende del gradiente de concentración a través de la membrana.( también se le llama fuerza química)
b) Flujo que depende de la diferencia de potencial eléctrico (Depende de las cargas).
El potencial de equilibrio surge de la oposición de las dos fuerzas. Para el caso dado tenemos el potencial de equilibrio del potasio. En una célula permeable sólo al potasio determina el potencial de membrana en reposo.
El potencial de equilibrio para cualquier ion puede calcularse con la ecuación de Nernst.
R= constante de los gases, T= temperatura en grados Kelvin, z=valencia del ion, F= constante de Faraday, y [x] son las concentraciones del ion fuera y dentro de la célula.
Nota: El flujo de potasio una vez llegado a cierto punto en que la fuerza son repelidos por los iones de fuera e ahí el equilibrio y que determina el potencial de reposo.
Los canales de reposo:
Como se ha planteado antes las neuronas son permeables a los iones de sodio, cloro y potasio. Los otros iones de proteínas y aminoácidos con carga negativa no pueden atravesar la membrana. ¿Cómo interactúan los tres gradientes, correspondientes a los tres iones para determinar el potencial de membrana o de reposo?
Coloquemos por caso una neurona con canales sólo para ion potasio y con gradientes de concentración para K+, Na+, Cl- y A-. En estas condiciones, el potencial de

225
membrana en reposo está determinado por el gradiente de concentración de ion potasio que es igual -75 mV.
Con unos pocos canales de sodio la situación es distinta. En este caso actúan dos fuerzas para el ión sodio para arrastre hacia el interior de la neurona; una de ellas por el gradiente de concentración químico y por la diferencia de potencial eléctrico.
El flujo hacia dentro del sodio despolariza la célula, pero sólo hasta el potencial de equilibrio de K+ de -75 mV. El nuevo potencial de sodio es distinto, un poco por encima del potencial de equilibrio para el potasio (el potencial de equilibrio de sodio es de +55 mV, y esto se debe a que hay más canales de potasio que de sodio en la membrana.
En la medida que el potencial de membrana se despolariza el flujo de potasio cambia. Como está aumentando la carga positiva dentro de la célula se reduce el movimiento de iones potasio hacia el interior y es encauzado hacia el exterior. Se produce un cierto equilibrio entre las cargas de potasio que salen y las de sodio que ingresan. Luego el potencial de membrana se establece en alrededor de -60 mV.
Para determinar el punto de equilibrio se necesita de la magnitud del flujo de un ion a través de una membrana celular es:
Flujo = (F de arrastre eléctrica + F de arrastre química ) x conductancia Membrana
La célula posee pocos canales de reposo de sodio, por lo cual la conductancia para el ion es muy baja, a pesar de que se encuentre sometido a grandes fuerzas químicas y eléctricas que lo inducen a ingresar al interior de la célula. Vale decir, el ion sodio ingresa muy poco porque existen pocos canales de reposo para el sodio. En el caso de los iones de potasio, hay mucho más canales para ellos. La conductancia para el potasio es muy alta.
El problema es entonces el cómo una célula puede tener un potencial de membrana en reposo de forma constante. En otras palabras, la separación de cargas debe mantenerse constante, el flujo de cargas hacia el interior como hacia el exterior debe estar equilibrado. De lo contrario el potencial variará. Pero, el movimiento pasivo de los iones de potasio y sodio se encuentra equilibrado, aunque los gradientes disminuyen a medida que pasa el tiempo, esto sin embargo no sucede.
La pérdida de los gradientes se evita gracias a la bomba de sodio y potasio. La bomba, como ya se ha explicado mueve los iones contra sus gradientes. Este proceso requiere energía del ATP.
Ion Cloruro (Cl-)
En las células con potencial de reposo determinado por los flujos de sodio y potasio y cuyas concentraciones intracelulares se deben a mecanismo de transporte activo conocido como bomba de sodio y potasio, la concentración de cloruro dentro de la célula es afectada sólo por fuerzas pasivas como los potenciales eléctricos y gradiente de concentración. Por ello, el movimiento de cloruro tiende hacia el equilibrio a través de la membrana. El potencial de equilibrio del cloruro es igual al potencial de reposo.

226
GENERANDO EL POTENCIAL DE ACCIÓN
La célula en reposo ya se ha visto en forma reiterada desde distintos ángulos que se trata de un flujo constante de sodio hacia dentro de la célula que se encuentra con un flujo constante de ion potasio hacia fuera. Este equilibrio cambia cuando la membrana se despolariza alcanzando un umbral y sobre este se logra un potencial de acción.
Las neuronas forman circuitos eléctricos que representan las formas que transmiten información. Ello lo realizan por medio de conexiones entre dendritas y prolongaciones axónicas de las neuronas, las sinapsis y las prolongaciones de las células gliales constituyen una densa red dentro de un tejido denominado neuropilo. Las células nerviosas que transmiten información desde la periferia hacia la médula espinal o el cerebro se llaman neuronas aferentes, mientras que las que transmiten información desde el encéfalo o la médula hacia las extremidades se denominan neuronas eferentes. Las neuronas locales que sólo participan en los aspectos locales de un circuito de llaman interneuronas.
Los circuitos de procesamiento suelen estar asociados en sistemas que desempeñan funciones muy amplias. Existen numerosos sistemas como el visual, el auditivo, el sistema nervioso autónomo, etc., cada uno de ellos especializado en controlar y dirigir las funciones de una serie de órganos.
Para poder establecer el sistema de comunicación las neuronas deben poseer propiedades tal como desarrollar la capacidad de ser altamente excitables. Esto es, de responder a estímulos, y convertir estos estímulos en impulsos neurales. Los estímulos eléctricos, de tipo químico o de tipo mecánico puede modificar el estado de reposo de las células. Esto lo hacen por medio del incremento de la permeabilidad de la membrana celular al ión sodio. Ingresa sodio a la célula.
Los mecanismos de bomba de sodio y potasio y los canales iónicos ya mencionados, la membrana plasmática del axón y el cuerpo celular contienen canales iónicos activados por voltaje, son de tipo específico y se abren cuando detectan un cambio de potencial de membrana. Si el voltaje alcanza el punto crítico denominado nivel umbral, la proteína sufre un cambio conformacional que le lleva a abrirse. El cierre y apertura de los canales permite la regulación de la concentración de los iones.
La membrana de la neurona puede experimentar despolarización en unos 15 mV, es decir, un potencial de reposo de unos -55 mV (rango entre -70 y -55 mV), sin experimentar un impulso nervioso. Pero, si la despolarización es mayor se alcanza el nivel umbral. En este preciso momento, los canales iónicos se abren y entra sodio a la célula. El movimiento de sodio sigue la dirección del gradiente, de zona de alta a baja concentración. También se abren canales de potasio cuando se alcanza el nivel umbral.
Cuando se produce el potencial de acción, la membrana de la neurona alcanza rápidamente el potencial cero y luego excede hasta cerca de los +35 mV; de este modo ocurre una inversión de la polaridad. La despolarización de una zona en particular hace que los canales regulados por voltaje que se encuentran adyacentes al punto de despolarización se abran, permitiendo que en esa zona entre el sodio; el proceso se repite hasta que se alcanza el extremo del axón. Este impulso neural se transmite entonces como una onda de despolarización que viaja por la neurona.

227
Cuando el potencial de acción ha recorrido unos pocos mm en el axón, sobre la membrana por la cual ha pasado comienza la repolarización. Las compuertas de sodio se cierran y la membrana se hace impermeable al ión. Las compuertas de potasio en la membrana se abren en ese tiempo y este ión es liberado. Estos iones de potasio al salir devuelven al interior de la membrana su carga negativa.
Factores que influencian al Potencial de Acción:
En la figura se puede observar como algunos factores influencian al estímulo. Tenemos entre estos factores a la intensidad del estímulo en la variable dependiente (eje Y) y el tiempo como factor independiente (eje X). La intensidad se mide en volts. La frecuencia es el número de veces que el estímulo es aplicado en determinada unidad de tiempo. En la gráfica se observa la aplicación de un estímulo con una duración de 3,5 segundos aproximadamente.

228
Los estímulos que producen respuesta son llamados estímulos umbrales. Aquellos que no logran una respuesta son llamados subumbrales. Los estímulos supraumbrales son aquellos estímulos cuya intensidad es superior y más que suficiente para lograr una respuesta.
Con todo, la respuesta frente a un estímulo umbral o supraumbral es cuantitativamente igual, ya que como hemos visto, la fibra nerviosa responde de acuerdo a la ley todo o nada.
Un estímulo subumbral puede convertirse en umbral considerando:
1.- Por aumento de su duración con la intensidad constante
Tiempo
2.- Aumentando su intensidad, manteniendo su duración constante, en el ejemplo siguiente son dos segundos.
Intensidad

229
Como ya se ha planteado, la respuesta frente a un estímulo umbral o supraumbral es cuantitativamente igual, puesto que la fibra nerviosa responde de acuerdo con la ley de todo o nada. La ley es válida tanto para una fibra en particular como para un nervio. Este último es un paquete de fibras nerviosas.
Dicho de otra forma, al aplicar sobre una fibra nerviosa o (un nervio) un estímulo umbral o subumbral se produce una inversión de la polaridad de reposo de su membrana que da origen al potencial de acción.
La inversión de polaridad se propaga, es decir, se invierte la polaridad de las áreas vecinas y el potencial de acción avanza a lo largo de la fibra. La velocidad de propagación es menor en las fibras amielínicas que en las mielínicas. En general, la velocidad de propagación es tanto mayor cuanto más gruesa es la fibra, es decir, es directamente proporcional a su diámetro. La velocidad es mayor si aumenta la distancia que existe entre los nudos de Ranvier. Recuerde de las clases que el modo de conducción del impulso es saltatoria, lo que quiere decir, que el impulso salta de un nudo a otro aumentando con ello la velocidad.
Características del impulso:
1.- Todo o Nada (ya hemos visto el significado)
2.- La excitabilidad de la fibra disminuye una vez que el potencial de acción ha pasado por cierta zona. El potencial de acción deja tras su paso una zona cuya excitabilidad es cero y no responde inmediatamente a ningún estímulo, lo que se denomina período refractario absoluto. La duración del período refractario absoluto es de fracciones de milisegundos, luego se recupera lentamente lo que se llama período refractario relativo.

230
3.- El potencial de acción se autopropaga, una vez que se genera se automantiene.
Potencial postsináptico excitatorio
Cuando una neurona se dispara, de los botones terminales del axón se liberan sustancias químicas denominadas neurotransmisores, que difunden hacia el espacio sináptico y luego interactúa con moléculas receptoras de las membranas receptoras de las neuronas.
Las despolarizaciones postsinápticas se denominan potenciales postsinápticos excitatorios.
Potencial postsináptico inhibitorio

231

232
CANALES IÓNICOS:
Las señalizaciones neuronales depende de las rápidas variaciones en la diferencia de potencial a través de la membrana neuronal. Cada célula puede dar origen a variaciones del potencial de membrana como respuesta a estímulos como la luz, sustancias odoríferas o sonoras, etc.
Las variaciones rápidas del potencial de membrana están mediadas por canales iónicos que son proteínas transmembranales y que se encuentran en todas las células del organismo.
Los canales iónicos poseen tres propiedades importantes:
1.- Conducen iones
2.-Reconocen y seleccionan iones específicos
3.- Se abren y cierran en respuestas a señales eléctricas, mecánicas o químicas.
Además la velocidad de conducción iónica es alta, pueden pasar hasta 100 millones de iones por segundo.
El potencial de membrana de las neuronas en reposo está determinado en gran parte por canales selectivamente permeables al K+. Lo normal es que estos canales son bastante más permeables al K+ que al Na+. Lo extraordinario es que al presentarse el potencial de acción se activan canales iónicos más permeables al ión sodio que al ión potasio.
Muchos canales están regulados, se abren o cierran como respuesta a diversos estímulos:
a) Canales regulados por voltaje b) Canales regulados por ligando c) Canales regulados por estímulos mecánicos- presión o estiramiento.
Existen canales no regulados, están normalmente abiertos con la célula en reposo. Contribuyen al potencial de reposo.
Es útil también explicar algunas características de los iones en relación con los canales. Los iones en solución están rodeados de una nube de moléculas de agua- se trata de iones hidratados (recordar materias del primer semestre)-las moléculas de agua son atraídas por la carga neta del ión. El conocimiento que debemos agregar ahora es que el ión de Na+ es menor en diámetro que el ión de K+, al encontrarse en solución estos iones adquieren un diámetro efectivo de solución y en este caso el del ión sodio es mayor, eso debido a su fuerza local de campo. Por ello, un canal puede seleccionar K+ en vez de Na+. Se puede excluir iones hidratados con diámetro mayor que el del poro.
Con respecto al canal de sodio, estos tienen un filtro de selectividad en algún sitio al que se unen débilmente los iones de Na+. Este ion de sodio se une transitoriamente en un sitio activo a medida que se mueve a través del filtro. En el lugar de unión la carga positiva del ion se estabiliza por un residuo (una molécula) de carga negativa ubicado en la pared del canal.(Kandel, pag.107).

233
Debemos entender que el paso de los iones por los poros no depende sólo del tamaño de ion, sino que también de la capa de solvatación (capa de agua) que se encuentra en su periferia.
Mientras más pequeño es el ión mayor potencia tiene su campo eléctrico. Comparando el sodio con el potasio, el de sodio tiene efectivamente un campo mayor y en consecuencia su atracción por el agua es de mayor potencia. De hecho, el sodio parece de mayor tamaño.
Que el canal sea selectivo existen hipótesis que explican el hecho de que pasan los iones al atravesar un filtro de selectividad, en este filtro el ion pierde su agua de hidratación y puede interactuar para formar enlaces químicos con aminoácidos de las paredes del canal. El canal puede apoyar en mayor o menor medida este hecho si son unas cuantas las moléculas de agua en su interior, en la medida que el poro esté más hidratado menos pérdida del agua de solvatación del ion podría esperarse. El problema no parece del todo explicado, pero ya se vislumbra el camino que conduce a la luz.
Características de los canales:
*El flujo de iones a través de los canales es de carácter pasivo, ello indica que no necesita energía.
*La dirección, el flujo y equilibrio de flujo se encuentra determinado por las fuerzas de difusión existente en las membranas.
*Los canales seleccionan los tipos de iones que pueden atravesar la membrana, pero no es del todo claro el proceso de selección de iones por parte del canal.
*Algunos tipos de canales no discriminan entre cationes y permiten el paso de iones de sodio, potasio, calcio y magnesio. No obstante, la mayoría de los canales si se permiten la discriminación de iones para darle libre paso, en otras palabras, son muy selectivos.
Modelos de regulación de los canales:
La función de los canales iónicos en las neuronas es generar señales eléctricas transitorias. Se ha postulado tres modelos de mecanismos de regulación para controlar el tiempo que un canal permanece abierto y activo. Los modelos son:
a) Canales regulados por ligandos químicos: Unión directa al canal, en el lado extracelular por transmisores.
b) Canales regulados por variaciones en el potencial de membrana. c) Canales regulados por tracción mecánica de la membrana.
En cualquiera de los canales anteriores se puede encontrar en los siguientes estados funcionales:
a) Cerrados y activables (en reposo) b) Abiertos (activos) c) No activables (refractarios)
En canales regulados por voltaje, la regulación es sensible a los niveles intracelulares de ATP, este es un ejemplo.

234
SEÑALES ELECTRICAS EN LAS CÉLULAS NERVIOSAS
Las sinapsis eléctricas permiten el paso de corrientes eléctricas (en forma de un flujo de iones) a través de unos canales que conectan las dos células, estando las células unidas una con la otra por un contacto intercelular denominado unión en hendidura ("gap junction"). Los poros de los canales que interconectan dos células son bastante grandes, de modo que se pueden intercambiar, además de iones, moléculas más grandes como el ATP y otros metabolitos importantes. La corriente eléctrica fluye pasivamente cuando se genera un potencial de acción. La célula fuente de corriente se denomina presináptica, mientras que la que recibe la corriente se denomina post-sináptica. Este tipo de sinapsis permite el paso de corriente bidireccionalmente y tiene la característica de ser extraordinariamente rápida (< 0.1 mseg).
Los poros que conectan las dos células con complejos proteicos hexaméricos denominados conexones, siendo cada una de las 6 subunidades una proteína transmembrana denominada conexina que, a su vez consta de cuatro regiones. El tamaño de estos poros permite el paso del ATP y de los segundos mensajeros, lo que permite que las sinapsis eléctricas coordinen el señalamiento intercelular.
Las sinapsis químicas, por el contrario, no presentan ninguna continuidad estructural, sino que las neuronas contiguas están separadas por la hendidura sináptica mucho más ancha que el espacio intercelular adyacente (30 a 40 nm). Como resultado de ello, la transmisión de la señal depende de la liberación de neurotransmisores.

235
Mantención de las neuronas- Factores de Crecimiento. Levi- Montalcini y Hamburger enla década del 40 del siglo pasado aislaron una molécula con relación a la supervivencia y el crecimiento de las neuronas. La molécula fue denominada Factor de crecimiento nervioso (NGF- nervous growth factor). En sus observaciones fue evidente que la supervivencia de neuronas en los procesos embrionarios, de aquellas relacionadas con funciones de tipo sensorial y simpáticas, dependía de la presencia del factor. En ausencia de NGF, o de su receptor las neuronas mueren. Los hallazgos se confirmaban en experimentos de tratamiento con NGF de tipo exógeno a neuronas en vías de eliminación que normalmente morirían, en lugar de ello, estas neuronas se mantenían vivas. Las neurotrofinas son factores neurotróficos, es decir, moléculas que modulan el proceso de diferenciación, maduración, regeneración, diferenciación, y muerte neuronal. Las observaciones condujeron a los científicos a plantear la hipótesis neurotrófica de acuerdo a ella las neuronas inmaduras compiten por células diana proveedoras de factores tróficos. Las neuronas que logran éxito en establecer las conexiones sinápticas para la obtención del factor trófico. Las neuronas que no obtienen el éxito desaparecen (mueren). Las neuronas más aptas son las que sobreviven. La proposición de Edelman es que se trata de “Darwinismo neuronal”, proceso en que ocurre una selección de grupos neuronalesbasados en el establecimiento de nuevas sinapsis y el uso de las conexiones establecidas entre grupos neuronales. Según esta hipótesis las conexiones crean redes neuronales que originan la emergencia de mapas cognitivos para la interacción con el ambiente. Edelman va aún más lejos con su propuesta, las redes neuronales que son arreglos neuronales complejos dan origen a la conciencia como una función emergente. Se advierte, sin embargo, que la competencia tal y como se conoce en la teoría de Darwin, se da entre seres vivos que obviamente pueden tener alguna deficiencia en aspectos corporales, de inteligencia, estrategias de caza, huida, etc., muchas de estas deficiencias tienen que ver con eliminación de seres que portan genes que no son lo suficientemente expresivos para lograr la sobrevivencia a través de la adaptación. Actividad del equipo: Discuta con su equipo la teoría de Edelman acerca de la competencia neuronal en los procesos de desarrollo, ¿será esta un acierto, porque? Problema: Si sabemos que hasta cierto punto existe un determinismo genético en el destino de las neuronas, hasta en los límites de la formación de sinapsis, ¿Cómo puede haber competencia?, en esta lógica habría una suerte de competencia entre nuestros mismos genes para la supervivencia, lógicamente la expresión de estos genes se logra en el desarrollo de la neurona misma y de las sinapsis que logra establecer. Resuelva usted esta problemática planteada. Se conocen cuatro tipos de neurotrofinas: el factor de crecimiento nervioso (NGF), el factor neurotrófico derivado del cerebro (BDNF), neurotrofinas 3 (NT3), neurotrofina 4 (NT4), NT5, NT6 y NT7. En el sistema nervioso central adulto, las neurotrofinas participan en acontecimientos sinápticos y su eficiencia en la adquisición de nueva información en la interacción del sujeto con su ambiente. Hay evidencia de que las neurotrofinas se relacionan con el sueño, en particular con procesos de inducción del sueño, así se ha visto que aumenta su expresión cuando se ha

236
privado experimentalmente el sueño en ratas. Se ha propuesto que estas moléculas jugarían algún rol en procesos de restauración neuronal, los cuales se activarían durante el sueño. Como usted puede ver, en esta lectura se va desde el aspecto microscópico las evidencias y descubrimiento de las moléculas y de sus interacciones en los procesos de comunicación celular y con el interior de la célula misma. A partir de esta información vemos como repercute este conocimiento en los aspectos macroscópicos, que son en su mayoría nuestras vivencias y nuestra vida. Fruto del conocimiento desde sus aspectos más mínimos, reductivos, se logra construir el conocimiento de las terapias biomédicas, de los aspectos de la cognición, la memoria, la inteligencia, etc. La medicina funciona con elevados grados de empirismo, y también a partir de estos se ha llegado al descubrimiento de mecanismos celulares y moleculares que desencadenan la curación. Es así, por ejemplo, el tratamiento de las úlceras oculares, con dosis y tratamiento tópico con factor de crecimiento nervioso (NGF). No se conoce cuales son los mecanismos celulares y moleculares que conducen a la curación de las úlceras. Los investigadores sugieren vías posibles tales como: 1) Activación de la proliferación y diferenciación del epitelio corneal, 2)Inducción indirecta de inflamación neurogénica, 3) Recuperación de la inervación corneal o proliferación de terminales nerviosos, 4) Estimulación de neuropéptidos u otros factores neurotróficos que estimulen la curación. Factor neurotrófico en los procesos de comunicación celular El sistema nervioso almacena información mediante eventos moleculares que se expresan en las sinapsis, así las señales tróficas son útiles para propiciar la comunicación intercelular. La presencia de neurotrofinas o la incorporación de las mismas impide la denervación o pérdida de contacto entre neuronas con lesiones recientes, estas moléculas parecen promover la reinervación entre neuronas que pierden sus contactos sinápticos. Las neuronas reciben señales por intermedio de sus sinapsis. La mantención de estas gracias a la acción de las neurotrofinas, entre las más estudiadas se encuentra el BDNF o factor trófico derivado del cerebro y la neurotrofina 3 que ejercen efectos sobre las motoneuronas lesionadas. Cuando usted realiza un movimiento, por ejemplo de las piernas, la orden de hacerlo proviene desde la corteza, esta luego se comunica con neuronas de niveles inferiores hasta que llegan a la última neurona que establece contacto con el músculo responsable del movimiento. Eso es lo que se puede llamar un flujo de información descendente. EL flujo ascendente es lo contrario, en estos existe participación de neurotrofinas. Cuando se corta un axón, con un accidente si se añade neurotrofinas inmediatamente, los contactos sinápticos comienzan a recuperarse. Las neurotrofinas impiden la denervación de neuronas recién lesionadas y promueven la reinervación de neuronas que ya han perdido sus contactos sinápticos. Las neurotrofinas realizan su actividad gracias a la presencia de receptores Trk, con actividad de tirosinoquinasa. La unión de las neurotrofinas a los Trk dan lugar a la cascada de reacciones químicas intracelulares, que finalizan con la expresión de los genes que responden a la neurotrofina.

237
Se ha identificado el gen del receptor Trk en el anfioxo, organismo invertebrado parecido a un pez, pero sin vertebras ni sistema nervioso complejo. Las indicaciones que derivan de estas investigaciones es que el origen de los receptores está ligado a la evolución de los vertebrados hace unos 500 millones de años atrás. El gen de las proteínas receptoras de Trk, el denominado AmphiTrk presenta alta homología a nivel de secuencia genética y funcional con tres tipos de Trk de vertebrados, en otras palabras, este es un indicio más de que el sistema nervioso comenzó a surgir hace más de 200 millones de años, ya estaba presente en los amphiosos. Luego en los vertebrados los receptores Trk aparecen implicados en los incrementos de complejidades del sistema nervioso, tales como aparición de funciones neuronales como la memoria, la inteligencia o la agresividad en los linajes de los vertebrados. Con el uso del conocimiento acerca de las neurotrofinas, como usted habrá pensado al leer estas páginas, se encuentra en etapa de experimentación el uso de terapias en enfermedades neurodegenerativas, como en la esclerosis lateral amiotróficas. Esto hecho otorga esperanzas a miles de personas damnificadas con este mal.
COMUNICACIÓN CELULAR Cuando usted escucha acerca de las comunicaciones seguramente asocia inmediatamente el término con los medios de comunicación que a diario vemos, la televisión, el periódico, la revista, etc. Me parece que ahora debe asociarlo también a las distintas señales que se envían las células. Un aspecto demasiado interesante de los seres vivos y que ha permitido el desarrollo de la vida en el tiempo evolutivo del universo, es que precisamente las células pueden comunicarse. Esta capacidad de comunicarse entre ellas les confiere adaptabilidad o más bien encajar con un medio ambiente siempre cambiante. Es decir, mediante las comunicaciones celulares podemos ajustarnos a las condiciones ambientales. Si no nos ajustamos perecemos. La capacidad de ajustarnos se denomina plasticidad, ¿cómo es esto?, bien, supongamos que capturamos un ser vivo del desierto y lo trasladamos a la zona de los mares australes.

238
Ciertamente el medio en este caso es diferente y ello exige respuesta del ser vivo, a objeto se seguir con vida. Se da una exigencia de ajustarse al medio. En la medida que se ajusta podemos decir que este ser posee plasticidad. Todo cambia en los momentos de ajuste, habrá nuevos y desconocidos alimentos, otra temperatura, el paisaje será diferente. ¿Cómo se ajusta? Precisamente porque hay comunicación, ello implica estímulo y respuesta. En toda persona que se ve expuesta al peligro le suceden fenómenos, por caso tenemos, temblor que denota intranquilidad, sudores de manos, transpiración, aumento de azúcar en la sangre, aumento del número de latidos cardiacos por minuto. ¿ cómo se produce estos cambios, tan de súbito? En realidad lo que sucede es que todas las células de este organismo se informaron y respondieron considerando la red de comunicación celular. Esta red de tipo sistémico nos permite separarla en dos componentes principales: el sistema nervioso y el sistema endocrino. La forma de operar en ambos casos es por medio de moléculas que constituyen en si, mensajeros químicos. Considerando ambos, podemos plantearlo como sistema neuroendocrino. Como sistema es capaz de captar los cambios en el medio externo y ajusta el medio interno por medio de la acción coordinada de las células del organismo. OPERATIVIDAD DE LOS SISTEMAS DE COMUNICACIÓN *COMUNICACIÓN HORMONAL (ENDOCRINA)
- Glándulas de secreción interna (hipófisis, suprarrenales, tiroides, ovarios, testículos, etc), vierten sus mensajeros hormonales al torrente sanguíneo con lo cual pueden tomar contacto con cualquier célula del organismo, pero solo realiza el verdadero contacto con las células blanco. Por lo tanto, podemos decir que la molécula mensajera es selectiva, es decir, selecciona a las células.
- Neuronas. Estas pueden comunicarse mediante el proceso de neurotransmisión. El proceso básico es que una neurona presináptica, establece contacto con una neurona postsináptica. Las membranas de ambas células establecen el contacto que se denomina sinapsis. Sin embargo, el contacto no es estrecho, existe un pequeño espacio entre las células. Ese es el espacio sináptico. El sentido del proceso es unidireccional desde la neurona presináptica a la neurona postsináptica. La molécula que interviene en el proceso se denomina neurotransmisor y es la molécula que lleva el mensaje de la neurona presináptica a la postsinática.
- Neurosecreción: Es una mezcla de ambas anteriores. En el caso, alguna célula del tejido nervioso envía su mensaje a distancia y el viaje se realiza por el torrente sanguíneo. Continua hasta encontrar a la célula blanco.

239
- Comunicación paracrina: Se produce entre células cercanas, de variado tipo de células. Se produce por medio de hormonas locales o mediadores locales, o autacoides.
- Comunicación yuxtacrina: Este fenómeno ocurre entre células adyacentes. En este caso el mensajero no difunde en el medio, permanece anclado a la superficie celular y así es capaz de establecer contacto con los receptores de la membrana de la célula adyacente. Ejemplo de este el facto de crecimiento y transformación alfa (TGF-α).
- Comunicación autocrina: Se trata de un monólogo. La misma célula es capaz de autocomunicarse. Por ejemplo. Una neurona emite un mensajero- un neurotransmisor que tras su cometido de comunicar a una célula postsináptica, es capaz de retroalimentar a la célula presináptica para que no siga produciendo más neurotransmisor. Otro caso es que algunas células producen factores de crecimiento y proliferación que actúan sobre la misma célula. En algunos tipos de cáncer ocurre que las células producen estos factores de forma incontrolada.
*CARÁCTERÍSTICAS DE LOS MENSAJEROS - Un mensajero puede ser útil para una variedad de receptores. Por ejemplo, la adrenalina (epinefrina) es producida por la glándula suprarrenal (sistema endocrino), sin embargo, también se le reconoce como neurotransmisor (sistema nervioso). - Estructura química: A) Estructura lipídica: esteroides y prostaglandinas: complementando la clase: los esteroides son de estructura semejante al colesterol.

240
Entre los esteroides más importantes tenemos: a) Hormonas sexuales. b) Cortisol y cortisona (corteza glándula suprarrenales) regulan el metabolismo de la glucosa. B) Estructura peptídicos (polipéptidos): son ejemplos la insulina, glucagon, hormona antidiurética, oxitocina, angiotensina, factores de liberación de hormonas hipofisiarias, endorfinas, factores de crecimiento y transformación, etc. c) Estructura aminas.: las aminas son compuestos que contienen grupos aminos, por ejemplo para este caso de mensajeros; el ácido glutámico, el ácido aspártico y la glicina. Hormonas tiroideas, adrenalina, serotonina, histamina y dopamina y acetilcolina. Como hemos visto los mensajeros químicos son moléculas muy diversas, tenemos algunos péptidos, proteínas, complejos proteicos, moléculas pequeñas derivadas de aminoácidos, compuestos lipídicos derivados del ácido araquidónico y compuestos esteroideos derivados del colesterol. Estas moléculas las podemos clasificar según su solubilidad y asociación con receptores en:
a) Moléculas lipofílicas: pueden difundir a través d la membrana plasmática e interaccionar con receptores del citosol o del núcleo. Son ejemplos, las hormonas esteroideas, la tiroxina y los derivados del ácido retinoico. Al interactuar con receptores intracelulares forman complejos que interaccionan con el núcleo y pueden en consecuencia aumentar o disminuir la actividad de transcripción de genes. Estos mensajeros producen sus efectos después de horas o días posteriores a la interacción con el receptor. Por lo general se relacionan con el crecimiento y diferenciación de tejidos.
b) Moléculas hidrofílicas: no pueden difundir a través de la membrana de manera que interactúan con receptores de la membrana celular. Son ejemplos: péptidos como insulina, proteínas como hormonas del crecimiento, acetilcolina y derivados de aminoácidos como la epinefrina, histamina, serotonina y dopamina.
c) Moléculas lipofílicas con receptores de superficie: son ejemplos, las prostaglandinas que derivan del ácido araquidónico. Son responsables de la agregación plaquetaria.
RECEPTORES
Un receptor es una proteína capaz de recibir al mensajero y de transmitir el mensaje para que se produzca la respuesta de la célula. Es decir,el receptor puede : 1) reconocer al mensajero para interactuar con él y 2) activar la secuencia de eventos que conducen a la respuesta celular.
Las proteínas, como toda molécula poseen una estructura tridimensional, es decir, una forma determinada en el espacio. Esa estructura tridimensional es motivo de investigación actual para comprender como las superficies del mensajero y del receptor se adaptan perfectamente entre sí, como un guante y una mano. Esta perfecta adaptación de superficies es la base de la alta selectividad de los receptores para una hormona específica. Imaginemos al receptor no como una estructura rígida, sino como una estructura con cierta flexibilidad,

241
capaz de experimentar ciertos cambios en su forma. El sitio de reconocimiento también tiene cierta flexibilidad, lo cual nos lleva al concepto de afinidad.
La afinidad puede definirse como una medida de la facilidad de interacción entre dos sustancias, en este caso entre el receptor y el mensajero. Esto es similar a lo que ocurre entre los seres humanos; dos individuos que tienen que interactuar para la realización de un trabajo lo harán con facilidad si existe afinidad entre ellos y lo efectuarán con extrema dificultad si no son afines.
Los receptores reconocen al mensajero y luego transducen el mensaje al ambiente intracelular. La respuesta celular a una molécula mensajera que en general se le designa como ligando. Las interacciones entre el ligando y su receptor son de alta afinidad, no covalentes y producen un cambio conformacional en el receptor, el que inicia cambios en la actividad celular. Por otra parte, el ligando no se metaboliza en algún tipo de productos, tampoco es algún tipo de mediador en alguna reacción química. La única función del ligando es afectar al receptor. Se ha observado en algunas células que la degradación del ligando termina con la respuesta. Un efecto observable con la unión del ligando a los receptores es el aumento en la concentración de segundos mensajeros intracelulares. Ejemplo de estos segundos mensajeros son: 3’,5’- AMPcíclico, 3’,5’-GMPcíclico, 1,2 diacilglicerol, inositol 1,4,5-trifosfato y Ca+2 . Clasificación de receptores : Transducción de la señal en medio intracelular: Una forma de clasificación. Receptores tipo I: o ionotrópicos, son canales iónicos con compuerta, gatillados por un ligando. Ver la figura siguiente de Viguera Neurociencia y Biología del comportamiento.
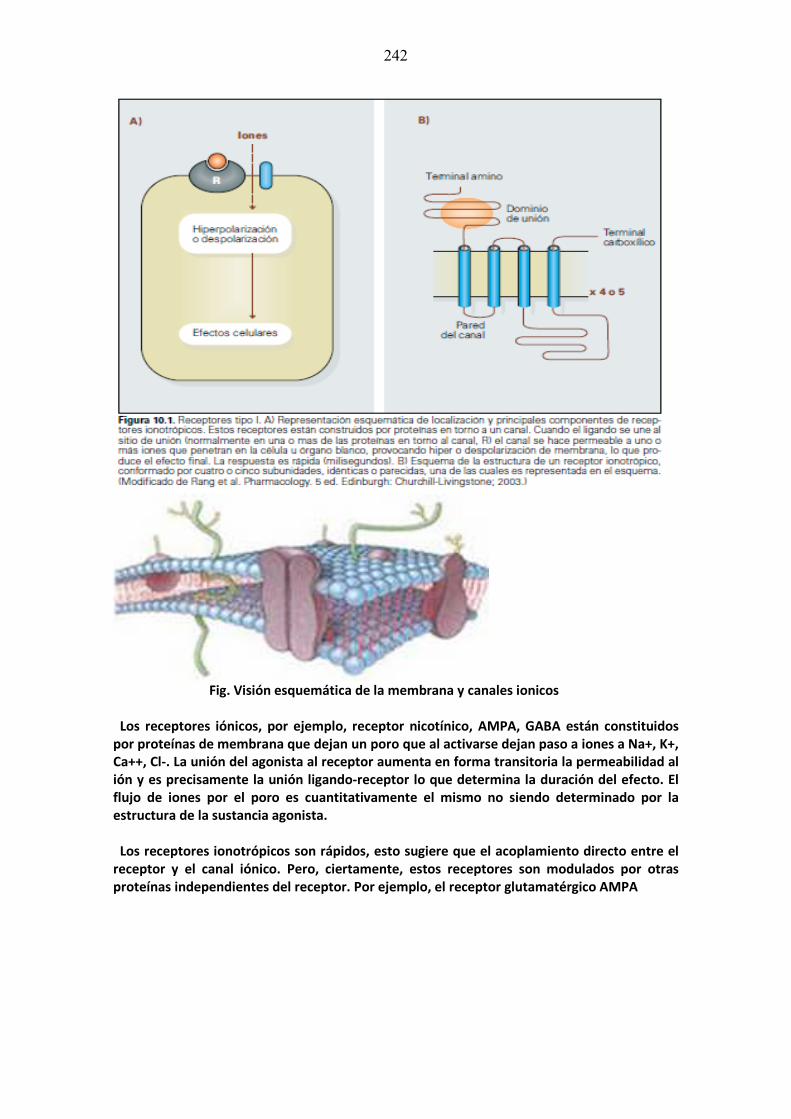
242
Fig. Visión esquemática de la membrana y canales ionicos Los receptores iónicos, por ejemplo, receptor nicotínico, AMPA, GABA están constituidos por proteínas de membrana que dejan un poro que al activarse dejan paso a iones a Na+, K+, Ca++, Cl-. La unión del agonista al receptor aumenta en forma transitoria la permeabilidad al ión y es precisamente la unión ligando-receptor lo que determina la duración del efecto. El flujo de iones por el poro es cuantitativamente el mismo no siendo determinado por la estructura de la sustancia agonista. Los receptores ionotrópicos son rápidos, esto sugiere que el acoplamiento directo entre el receptor y el canal iónico. Pero, ciertamente, estos receptores son modulados por otras proteínas independientes del receptor. Por ejemplo, el receptor glutamatérgico AMPA

243
y su corriente desencadenada por la activación es inhibida o aumentada por proteínas quinasas y proteínas quinasas II dependientes de calmodulina Ca+2. Otro ejemplo, es que los receptores AMPA que luego de ser sintetizadas en el retículo endoplasmático son acompañados por proteínas NSF, PICK1 hasta las dendritas en donde se acoplan a proteínas asociadas a la PDZ (postsynaptic density zone). Receptores Tipo II: o metabotrópicos son canales acoplados a proteínas G, responsables del acoplamiento de los receptores a los sistemas enzimáticos que son efectores en la transducción de señales intracelulares.

244
Fig. Ver en la página web: www.genomasur.com/lecturas/Guia07.htm

245
Esta figura ha sido obtenida del texto de bioquímica de Mathew. Describa el fenómeno de la transducción intracelular.
Estas son las proteínas de 7 dominios transmembranales, unidos por residuos hidrofóbicos. La región amino-terminal(N) está en el dominio extracelular, por su parte el extremo carboxílico se encuentra en el citosol.
Figura: Ver http://cuentospintados.com/portfolio/geodon/centro_enf/art3a.htm

246
Figura: Ver en http://cuentospintados.com/portfolio/geodon/centro_enf/art3a.htm
La unión del ligando permite el acoplamiento a una o varias proteínas G, muscarínicos, adrenoreceptores, dopaminérgicos. Las proteínas compuestas por subunidades alfa, beta y gama, catalizan por medio de la subunidad alfa, se produce el reemplazo de una molécula de GDP por una molécula de GTP, esto lleva a que se produzca la liberación de la subunidad alfa que difunde en la membrana y es capaz de asociarse a varias enzimas como la adenilato ciclasa, la fosfolipasa C, y/o canales iónicos, en este acto, activa o desactiva procesos de transducción que implican la formación de las moléculas llamadas segundos mensajeros, acoplados al AMPc, 1,4,5 Esta clasificación se realiza sobre la base similitudes de sus secuencias aminoacídicas, en la predicción de su estructura secundaria y terciaria:
- Receptores con actividad enzimática.
- Receptores acoplados a proteínas G.
- Receptores sin actividad enzimática, pero asociados a proteínas citosólicas.
- Receptores accesorios. RECEPTORES CON ACTIVIDAD ENZIMÁTICA Se clasifican en tres grandes grupos:
a) Receptores fosforiladores b) Receptores con actividad de proteína fosfatasa c) Receptores con actividad de guanilil ciclasa.

247
Receptores fosforiladores: Poseen actividad cinasa, esto es, enzimas que fosforilan transfiriendo grupos fosfatos a sus sustratos. En la actualidad gozan de tremenda importancia, puesto que son mediadores de respuestas de sustancias como la insulina y factores de crecimiento celular como el EGF (factor de crecimiento epidérmico) y el TGF-β (factor de crecimiento y transformación- beta). Las cinasas de proteínas se clasifican de acuerdo a su especificidad por el sustrato:
- Fosforilación de residuos de tirosina.
- Fosforilación de residuos de serina y treonina. Los receptores con actividad de cinasa de tirosina poseen un dominio de unión al ligando separado por la membrana del dominio con actividad de cinasa. Cuando el ligando está ausente estos receptores se encuentran al estado monomérico, y al momento de la unión al ligando se induce el estado dimérico. La dimerización permite la transmisión de un cambio conformacional del dominio extracelular al dominio citoplasmático activándose la acción catalítica de autotransfosforilación de los receptores. Este fenómeno es el inicio de la transducción y amplificación de la señal y tiene como objeto a blancos intracelulares. Ejemplo: receptor de insulina formado por dos cadenas beta y dos subunidades alfa localizadas en el lado extracelular; las cuatro cadenas están unidas por puentes disulfuro. Las subunidades beta del receptor de insulina son las que poseen la actividad cinasa (o quinasa). La unión de la insulina provoca un cambio conformacional que termina en la autofosforilación de un residuo de serina. Una vez fosforilado es innecesaria la presencia de insulina y la actividad quinasa puede ser inactivada sólo por acción de fosfatasas. La respuesta celular al final del proceso es el incremento en la entrada de glucosa. Como efecto también de la insulina puede activar la transcripción de genes, inicio de división celular. En general, la función de las cadenas fosforiladas de los receptores es ser de utilidad como puntos de unión de proteínas efectoras tales como fosfolipasa C gamma, o la cinasa del fosfatidilinositol -4,5,bifosfato, las cuales a su vez pueden ser fosforiladas por la cinasa del receptor y así propagar intracelularmente la señal. Otra proteína que se activa por la formación de estos complejos es la MAP cinasa (Mitogen Activated Protein Kinase), esta viaja al núcleo para activar la transcripción de genes para respuesta rápida. RECEPTORES CON ACTIVIDAD DE PROTEÍNA FOSFATASA Estas enzimas extraen el fosfato que colocan cinasas en las proteínas. RECEPTORES CON ACTIVIDAD DE GUANILIL CICLASA A) FORMAS MEMBRANALES: El corazón además de bomba se comporta como una
glándula, pues en la aurícula se producen y secretan péptidos conocidos como natriuréticos auriculares. Se llaman así porque favorecen la eliminación urinaria de sodio. Agreguemos que son vasodilatadores, es decir, que relajan las capas musculares de los vasos. Experimentaciones con activación de las células musculares con estos natriuréticos se produce un aumento de los niveles intracelulares de GMPcíclico. Los receptores para estos péptidos poseen actividad guanilil ciclasa en el dominio citosólico.

248
B) Guanilil ciclasas solubles: Algunos compuestos con grupos nitro, tales como la nitroglicerina son vasodilatadores- inducen la relajación de vasos sanguíneos en la capa endotelial. Son útiles para relajar las arterias coronarias y mejorar la circulación cardiaca. Este tipo de sustancias activan la guanilil ciclasa citoplasmática.
El óxido nítrico (NO)(segundo mensajero), puede actuar sobre proteínas citoplasmáticas. Puesto que pueden difundir libremente a través de la membrana celular. En el proceso un mensajero, la acetilcolina, actúa sobre las células endoteliales de los vasos sanguíneos y las induce a liberar óxido nítrico, este a su vez provoca la relajación de las fibras del músculo liso de dichos vasos sanguíneos. Allí activa la guanilil ciclasa, produciéndose GMPc.
Concepto de transducción celular y neuromodulación en Neurona Los neuromoduladores son sustancias químicas de tipo endógeno, productos del metabolismo celular que actúan a nivel presinápticamente, modulando la síntesis o la liberación de un neurotransmisor.
Los neuromoduladores son sustancias secretadas de manera natural que actúa de manera similar a un neurotransmisor, pero no queda restringido al espacio sináptico por el contrario difunde por el fluido extracelular. Un neuromodulador es un producto liberado en una sinapsis que influye directamente en la actividad postsináptica de la neurotransmisión, son ejemplo de esto las sustancias tales como, el ATP, la adenosina, el GTP, la feniletilamina, etc.
Los neurotransmisores producen dos efectos sobre las membranas postsinápticas, uno de ellos es la despolarización, con potenciales excitatatorios postsinápticos PEP, el otro tipo es la hiperpolarización con potenciales inhibitorios postsinápticos. De manera que se podría clasificar a los neurotransmisores como inhibitorios o excitatorios. Sin embargo, hay muchas otras sustancias que se relacionan con esta actividad de excitar o inhibir pero de una manera diferente. Por ejemplo, en el encéfalo, la mayor parte de la comunicación sináptica por medio de neurotransmisores se debe a los neurotransmisores, uno de ellos con efectos excitatorios denominado glutamato, la glicina también puede ser considerada dentro de esta modalidad, pero a nivel de la médula o tronco encefálico. Otro tipo de neurotransmisor de acción inhibitoria es el GABA.
El GABA se sintetiza en la terminación del axón, luego es almacenado en vesículas sinápticas. La vía de síntesis utiliza una vía alternativa del ciclo del ácido cítrico. La glucosa capturada

249
una vez en el proceso glucolítico produce ácido pirúvico. Este es a su vez ingresa al ciclo de ácido cítrico. En este se produce ácido oxalacético y alfacetoglutárico, que por transaminación produce ácido glutámico que sale del ciclo por una vía colateral. Luego por acción de la enzima glutámico descarboxilasa, el ácido glutámico se transforma en ácido gamma amino butírico.
El GABA almacenado en las vesículas sinápticas se libera al espacio intersináptico por exocitosis. Luego interacciona con receptores específicos de la membrana neuronal postsináptica. Posteriormente las moléculas de GABA son recaptados en las terminales gabaérgicas y en células de tipo glial. Finalmente, la neurona puede volver a utilizarlo, previa incorporación a las vesículas, o degradarlos hasta ácido succínico.
El GABA en el espacio sináptico interactúa con los receptores de tipo específico, puede ser con Receptor GABA-A o el receptor GABA-B. GABA-C, el primero es predominante a nivel supraespinal. Al activar a este receptor se producen efectos inhibitorios por hiperpolarización, el receptor opera mediante canales de cloro y aumento de la conductancia a este ión. El canal es de tipo ionotrópico, en tanto el de tipo B y C son de carácter metabotrópico.
Los receptores GABA, al abrir canales de cloro, se transforman en inhibidores de la conducción del impulso nervioso. Los receptores GABA-B permiten que aumente la permeabilidad al K+, transmiten la señal por segundo mensajero. Estos receptores se encuentran asociados a proteínas G. En cualquiera de los dos casos se obtiene el mismo resultado, la célula se hiperpolariza y se inhibe la producción de potenciales de acción.
Los agentes que bloquean la transmisión gabaérgica generan convulsiones mientras que agentes que aumentan la inhibición tienen efectos sedativos, anticonvulsivantes y ansiolíticos”.
EL GABA se encuentra en todo el cerebro, sin embargo, alcanza mayor concentración en el cerebelo. Es posible que las neuronas cerebelosas inhibitorias del cerebelo efectúen su neurotransmisión con este este tipo de moléculas, entre las neuronas que participan se encuentran las de Purkinje, las células en canasta, las estrelladas y las de Golgi.
Glutamato
El glutamato es el principal neurotransmisor excitador en el cerebro, se asocia con receptores ionotrópicos y metabotrópicos, estos últimos constituyen una familia de receptores acoplados a proteína G que unen el GTP a diferentes moléculas de señalización intracelular.
El glutamato interactúa con receptores acoplados a canales iónicos (ionotrópicos) con receptores acoplados a proteínas G (metabotrópicos). La activación de estos receptores es la responsable de la transmisión sináptica excitadora como por ejemplo con los fenómenos de la memoria y del aprendizaje.

250
Receptores ionotrópicos
Los receptores ionotrópicos de glutamato son complejos formados por 4 o 5 subunidades y se dividen en grupos según su comportamiento farmacólogico:
• receptores para AMPA (acido a-amino-3-hidroxi-5-metil-4-isoxazolpropiónico) • receptores para NMDA (N-metil-D-aspartato) • receptores para Kainato (ácido 2-carboxi-3-carboximetil-4-isopropenilpirrolidina) • receptores para Quisqualato (ácido a-amino-3,5-dioxo-1,2,4-oxadiazolidina-2-
propanoico)
En el encéfalo, la mayoría de las comunicaciones se realizan con GABA inhibitorio o con glutamato excitador. Estos dos neutrotransmisores deben encontrarse en cierto equilibrio en los circuitos neuronales.
El resto de los neurotransmisores se distinguen por que más bien poseen un efecto modulador antes que efecto de transmisión de información. Tienden a inhibir o activar circuitos neuronales. A modo de ejemplo, la acetilcolina es capaz de activar la corteza cerebral, pero la información que se recuerda y aprende se transmite por glutamato y GABA. Otro ejemplo similar es el de la noradrenalina que aumenta el nivel de vigilancia. La serotonina inhibe conductas impulsivas. La dopamina a nivel de la sustancia negra activa movimientos voluntarios, pero no es específica para el tipo de movimiento a realizar. En otras regiones del cerebro la dopamina actúa como refuerzo de conductas que se están desarrollando, aunque este último ejemplo, es tema de investigación aún.
Concepto de modulador: Descripción/tipo de acción:
1)Los neuromoduladores son sustancias de tipo endógeno, surgen del propio metabolismo
celular.
2) Influencian la síntesis y liberación de un neurotransmisor
3) En postsinápsis modifican la unión del ligando a su receptor
4) Influyen en los mecanismos de transducción de receptores
Serotonina acción moduladora:
Se presenta a continuación un tipo de acción de la serotonina como neuromodulador en
situaciones de memoria y aprendizaje. ¿Cuáles son los mecanismos moleculares en la

251
memoria a corto plazo?. Las investigaciones que se han desarrollado especialmente con el
famosos investigador Kandel ha permitido descubrir que la serotonina actúa como
modulador para incrementar la liberación de glutamato en la terminal presináptica de la
neurona sensorial. Para ello la serotonina, como primer mensajero, actúa sobre un receptor
de serotonina transmembrana que activa la enzima Adenil Ciclasa (AC), la cual desencadena
la formación del AMP cíclico. El AMPc es un segundo mensajero, que difunde la señal
proveniente de la membrana celular hacia el interior de la célula, activa a la Proteína
Quinasa A (PKA) que aumenta la liberación de glutamato en la terminal del axón mediante
una combinación de mecanismos de fosforilación (de los canales iónicos y de la exocitosis).
El glutamato en este momento puede actuar en las sinapsis.
El aprendizaje genera la memoria de corto plazo mediante cambios en la tenacidad de las conexiones sinápticas entre las neuronas sensoriales y las motoras. El AMPc y la PKA regulan la liberación del neurotransmisor glutamato que controla la fuerza sináptica, da lugar a la plasticidad sináptica que es el elemento clave para la formación de la memoria de corto plazo.
También actúan a nivel postsináptico, influyendo, modificando la unión del ligando con el receptor.
La Serotonina (5-hidroxitriptamina, o 5-HT), es
una monoamina sintetizada en las neuronas
serotoninérgicas en el Sistema Nervioso Central
(SNC) y las células enterocromafines (células de
Kulchitsky) en el tracto gastrointestinal de los
animales y del ser humano.

252
Estos moduladores influyen en los mecanismos de transducción de los receptores. Se denomina neurotransmisor a sustancias químicas sintetizadas por una neurona con objeto de ser utilizadas como señales en la comunicación con otras neuronas, otras células, como por ejemplo, vasos sanguíneos, fibras musculares, glándulas, etc. Podemos considerar que una sustancia es neurotransmisora cuando se cumplen ciertos criterios: 1.- El neurotransmisor debe ser sintetizado en la neurona y actuará como señal nerviosa. 2.- El neurotransmisor puede ser almacenado en vesículas sinápticas cerca de la zona de liberación, o zona activa. 3.- La liberación del neurotransmisor debe ser controlada por la actividad neuronal, lo que implica despolarización. La consecuencia es que debe liberarse un quantum de su contenido al espacio sináptico( Espacio extracelular) por un mecanismo de exocitosis dependiente del calcio. 4.. La señal liberada debe alcanzar concentración suficiente para la unión competitiva a receptores de membrana postsinápticas y presinápticas. 5.- La unión del neurotransmisor al receptor debe establecer un complejo agonista-receptor capaz en último término producir la actividad nerviosa. 6.- Debe existir un mecanismo de inactivación de la sustancia liberada, de manera de garantizar especificidad y oportunidad de su efecto. Cumplen los requisitos arriba mencionados los neurotransmisores tales como acetilcolina, noradrenalina, dopamina, serotonina, glutamato y gama aminobutírico (GABA), un gran número de neuropéptidos, como las encefalinas y neurocininas. Existen además sustancias endógenas, que no alcanzan a cumplir los criterios empleados que satisfagan el concepto de neurotransmisor, estas pueden modular los efectos neuronales o actividad nerviosa. Por ejemplo, el ATP, se sigue clasificando como neuromodulador por que hasta la fecha no se ha demostrado un mecanismo de almacenamiento presináptico, ni se ha demostrado que se libere de manera dependiente de la actividad neuronal. Sin embargo, la adenosina tiene gran relevancia en la psicofarmacología y formulación de teorías, puesto que un gran número de drogas tienen efecto agonista o antagonista en receptores adenosinérgicos.

253
Apoptosis
Hace más de 30 años aproximadamente se ha establecido que perdida espontánea de células es un fenómeno normal e importante en la homeostasis de tejidos normales (proliferacion celular y recambio versus muerte celular. Las células deben eliminarse continuamente en muchos tejidos normales para equilibrar la producción dada la división celular; sino aconteciera este fenómeno el balance en número de células conduciría a posibles expansiones tumorales.
Los términos apoptosis, muerte celular programada, muerte celular activa o suicidio celular, se utilizan para definir un modo particular de muerte celular, caracterizado por un modelo específico de cambios que involucran al núcleo y el citoplasma.
Las características morfológicas asociadas a este caso de transformación células, que alcanza al estado apoptótico, sugiere que es un fenómeno activo y programado, asociado o inhibido por una variedad de estímulos ambientales, relacionados con estados fisiológicos y patológicos.
La apoptosis se observa por ejemplo, con la caída de las hojas en otoño, la ovulación, el ciclo menstrual que es la expresión de un suicidio celular colectivo del epitelio uterino cuya misión era servir de nido a un óvulo fecundado por un espermatozoide. De la misma forma, es un proceso apoptótico el desarrollo y modelación del embrión en el mundo animal, la renovación de los epitelios durante la vida del animal. Las células no mueren por vejez o simplemente por agresiones, solamente. Por el contrario, la gran mayoría de células muere en cualquier etapa de su vida, particularmente en el embrión.
El desarrollo del embrión está caracterizado por fenómenos de crecimiento celulares , migraciones celulares en masa, modelación de órganos en formación a partir del crecimiento y desarrollo del cigoto. Las células que mueren en estos estadios del desarrollo, no lo hacen por procesos infecciosos o envejecimiento. Mueren por que existe un programa dado en el genoma, las muertes así ocurren en los mismos estadios y en los mismos tiempos de ocurrencia. Una metáfora interesante nos ilustra como un proceso de esculpido, de remodelado de estructuras semejantes a branquias, cola de renacuajo, que la rana adulta no presenta, sólo se ven en estado embriológico.
El concepto de muerte celular programada define el fenómeno para las células destinadas a desaparecer en relación al diseño básico de organización durante el período embrionario y en relación a los tejidos en el estado adulto.
NECROSIS
A diferencia de las células que mueren naturalmente, están aquellas que mueren por causa de infecciones, traumas, isquemia, exposición a sustancias tóxicas. Estos son casos que corresponden a muerte por necrosis.
La necrosis a diferencia de la apoptosis, involucra inflamación, es un proceso violento, destructivo, catastrófico. Las células invadidas por gérmenes, atacadas por sustancias tóxicas, o privadas de nutrientes, de oxígeno, se hinchan, la membrana celular se rompe y las enzimas intracelulares se liberan, las quimioquinas secretadas por células adyacente o por las mismas células en necrosis. El entorno extracelular así formado atraen células de tipo

254
inflamatorias. El proceso finaliza dando lugar a neoformaciones tisulares, es decir, una cicatrización con deformación del diseño original, o culmina con la muerte del organismo, si es que el agente agresor no ha sido combatido.
Cambios intracelulares: Son los cambios intracelulares los que permiten ver las mayores diferencias entre procesos necróticos y apoptóticos. De hecho, la apoptosis debe entenderse como un fenómeno contrario a la proliferación y neoplasia. En condiciones normales existe equilibrio entre la reproducción celular y la apoptosis con objeto de mantener la población celular adecuada hasta llegado el instante de alcanzar el estado adulto de desarrollo. En este recorrido de existencia se observa que hay equilibrio entre apoptosis y mitosis. En los próximos párrafos veremos el paso desde el nivel reductivo como son las interacciones moleculares, al interior de la células y sus repercusiones que alcanzan el estado macroscópico en la supervivencia del organismo, un nivel con una visión más holística.
En relación con el desarrollo nervioso, las neuronas que se generan a partir de células precursoras que una vez diferenciadas no se dividen más. Antes de emitir dendritas y axones, las neuronas inmaduras o precursoras emigran desde el lugar de nacimiento en busca de la localización definitiva utilizando la senda glial como camino u orientación. La migración tiene además como objetivo lograr las conexiones interneuronales correctas. Si los procesos no se concreta una causa probable puede ser debido a la ausencia de acción por parte de factores de crecimiento secretados por la célula receptora a que pertenece el axón conectado y la célula que hizo la conexión equivocada muere por apoptosis. O sea, es el mecanismo de apoptosis el que contribuye de manera directa al desarrollo del tejido nervioso, a su constitución y establecimiento de las redes neuronales, que son en definitiva el medio de comunicación y de operatividad de todas la células, tejido, órganos y organismos tales como nosotros.
La apoptosis también se observa en procesos tales como la isquemia e hipoxia. En las zonas de infartos lacunares, o infartos cerebrales se ha observado células en apoptosis con fragmentación de cromatina y sobreexpresión de caspasas. Como usted ya ha incorporado estas materias a su mente, es útil que se tenga siempre en cuenta, de que estos fenómenos a menudo observados con scanner o imagenología a nivel macro, que tienen su relación directa con el daño isquémico, es un fenómeno que la percepción humana es capaz de internalizar en el trato mismo con los pacientes, o sea, nos damos cuenta si interaccionamos con el paciente en los momentos que se produce el infarto, o después que este ha tenido lugar. Luego, los análisis que se pueden realizar a nivel histológico, a nivel celular, indican los detalles de químicos, bioquímicos, y celulares que debieron ocurrir y que posiblemente causaron el infarto.
Acontece también que las enfermedades neurodegenerativas, tales como las de Alzheimer, Hungtington y Parkinson. Las células apoptóticas aparecen especialmente en Alzheimer en neuronas del hipocampo. Varias hipótesis se han planteado, una de ellas es el descontrol genético de la apoptosis.
Características morfológicas de la apoptosis
Las células reducen su volumen, pierden contacto con las células vecinas y pierden elementos de superficie especializados como las microvellosidades y uniones célula-célula. Estos cambios ocurren rápidamente. Se observa condensación del núcleo y del citoplasma, probablemente por salida del líquido celular. Se forman pequeños fragmentos celulares que permanecen unidos o cercanos a la superficie celular. Estos remanentes celulares son los

255
llamados cuerpos apoptóticos se caracterizan porque están bien conservados y contienen organelos, los cuales pueden estar condensados pero están aparentemente intactos. Se observan además, vacuolas y masas densas de material nuclear en algunos cuerpos. Estos cuerpos no muestran evidencia de degeneración y es probable que en esa etapa ellos sean capaces de presentar actividad metabólica.
El retículo endoplasmico se dilata y aparece una serie de cavidades semejantes a cráteres, de tal manera que las cisternas dilatadas se fusionan con la superficie celular. Inicialmente y a diferencia de la necrosis, las mitocondrias son normales.
La cromatina se condensa en capsulas granulares densas bajo la membrana nuclear. Adyacente a eso, los poros nucleares desaparecen. El nucleolo se disocia y da origen a partículas estrechamente asociadas a la periferia de la cromatina condensada. Todos esos cambios nucleares llevan a la ruptura de la cromatina, de tal manera que el ADN de las células apoptóticas se reducen a una serie de fragmentos del tamaño de los nucleosomas.
Las células apoptóticas no inducen reacciones inflamatorias. Sin embargo, ellas son fagocitadas inmediatamente, por macrófagos presentes o por otras células adyacentes. Los cuerpos apoptóticos ingeridos por otras células, sufren un proceso dentro de los fagosomas, similar a la autolisis in vitro de células integras. Las enzimas lisosomales degradan los cuerpos fagocitados. De esta manera son rápidamente reducidos a cuerpos lisosomales.
La apoptosis determina la talla y forma del sistema nervioso de los vertebrados.
La visión predominante hasta ahora es que la muerte celular se lleva a cabo principalmente para igualar el tamaño de la población neuronal al de sus dianas, células con las cuales hacen contactos sinápticos, de esta forma elimina las neuronas con proyecciones erróneas o inadecuadas, en este sentido parece un tipo de selección natural de tipo como la teoría enunciada por Darwin en el origen de las especies.
Por otro lado, la posibilidad de que la muerte celular sea un mecanismo morfogénetico en el sistema nervioso ha sido poco investigada. Recientemente, estudios realizados en ratones knock out* para genes implicados en el proceso de apoptosis, así como el análisis genético de la apoptosis en el nematodo Caenorhabditis elegans, han llevado a considerar la muerte celular como un mecanismo para emparejar a la población neuronal con la población celular donde va a mandar sus proyecciones, y como mecanismo de ajuste del tamaño poblacional de células neuronales iniciales, necesario para una morfogénesis correcta del sistema nervioso.
Christine Pozniak y colaboradores en Science (2000 (289): 304-306), del Centro de Supervivencia Neuronal han mostrado tanto in vitro como in vivo, la función antiapoptótica de la proteína p73 (perteneciente a una familia de proteínas con función contraria, p53 y p63) en la muerte celular durante el desarrollo neuronal. Describen la expresión de una isoforma de p73 en neuronas en desarrollo y su dramática disminución cuando estas neuronas entran en apoptosis después de retirarles el suministro de NGF (esencial para su supervivencia). Además en los ratones knock out para p73 (p73-/-), la apoptosis en neuronas del sistema nervioso simpático se ve incrementada. Estos resultados llevan a los autores a postular una función esencialmente antiapoptótica de p73 en neuronas, permitiendo así contrarrestar la función proapoptótica realizada por p53.

256
La proteína* p53 es el miembro fundador de una familia de factores de transcripción (proteínas que se unen al DNA y controlan así la transcripción génica), que contiene dos otros miembros con secuencias de aminoácidos muy parecidas, p63 y p73. Pero este hecho no significa que estas distintas proteínas realicen una misma función. La proteína p53 activa la expresión de genes diana implicados en la detención del crecimiento celular o en la muerte celular en respuesta a un daño del DNA. Es decir, es una proteína implicada en un servicio de alarma, cuando detecta lesiones en el DNA activa una cascada de genes que llevarán a esa célula a su muerte para evitar que se reproduzcan esos defectos. Así encontramos mutaciones de esta proteína en tumores humanos, lo que llevó a denominarla proteína supresora de tumores. Su función de apoptosis es particularmente importante en el sistema nervioso sobre todo después de una lesión (isquemia, epilepsia), en enfermedades neurodegenerativas crónicas y en algunos tipos de muerte neuronal selectiva durante el proceso de desarrollo.
La elevada identidad de aminoácidos* entre p53, p63 y p73, llevó a pensar que todas ellas tenían una función parecida. Una serie de estudios recientes (entre ellos el de Pozniak et al.) ha demostrado claramente que, a pesar de estar evolutivamente muy conservados, los miembros de la familia p53 tienen distintas actividades biológicas, que llegan incluso a ser antagonistas.
Aunque p53 y p73 presentan algunas características comunes en ensayos in vitro, difieren en muchas otras. En contraste con los ratones knock-out para p53, los animales p73-/- no presentan un incremento de la potencialidad para formar tumores. Además los ratones p53-/- se desarrollan normalmente, mientras que los p73-/- presentan defectos importantes en el desarrollo cerebral. Por lo que p53 y p73 parecen estar implicadas en diferentes aspectos del desarrollo del sistema nervioso.
Para determinar qué hace p73 durante el desarrollo del ratón, los autores caracterizaron la expresión de las distintas variantes de p73 en el cerebro neonatal de ratón. Observaron que la variante predominantemente expresada es una forma truncada de p73 (aquella que carece del dominio capaz de activar directamente genes diana pertenecientes a la cascada de p53). Cuando se realizaron estudios de neuronas en cultivo con NGF, observaron sorprendentemente que, en la ausencia de éste, las neuronas presentaban niveles mucho más bajos de la forma truncada de p73 y entraban en un proceso de muerte celular dependiente de p53. Estos resultados junto con los obtenidos con la sobreexpresión de la proteína (a partir de adenovirus) evitando este proceso de apoptosis, sugieren que esta variante de p73 en neuronas es una proteína con efectos antiapoptóticos (contrariamente al efecto de su hermana p53).
En resumen, la variante de p73 en neuronas es una proteína con efectos antiapoptóticos. Lo que será interesante ahora es determinar si esta proteína está regulada positivamente en neuronas del sistema nervioso central en respuesta a una lesión o enfermedad. Si se llega a demostrar que p73 truncada inhibe la muerte neuronal en respuesta a diferentes lesiones, esta proteína sería la panacea terapéutica para rescatar y "curar" neuronas dañadas, a parte de las importantes implicaciones en otros tipos celulares, ya que es la forma predominante en la mayoría de tejidos adultos y embrionarios.

257
ASPECTOS BIOQUIMICO-MOLECULARES DE LA APOPTOSIS
La apoptosis se considera como un proceso activo de suicidio celular. Una característica común de la apoptosis es la participación activa de la célula en su propia aniquilación, pues esta moviliza una cascada de eventos que conduce a este fin. Existe considerable variación en la señal inicial y los eventos metabólicos celulares necesarios para inducir apoptosis, que dependen en gran medida del tipo celular y de la naturaleza del evento externo el cual dispara la apoptosis. Las señales externas que conducen a este tipo de muerte son tan variadas como aquellas que conducen a la proliferación y diferenciación celular, y pueden incluir la supresión de señales extracelulares así como su aparición.
La apoptosis es un proceso dirigido por genes y puede ser visto como uno más de los procesos dirigidos por genes, como parte del repertorio con que dispone la célula para responder a estímulos internos y externos.
Influencia de genes reguladores
Algunos oncogenes y genes oncosupresores han sido ya identificados y se conoce que regulan la susceptibilidad celular para entrar en apoptosis. Hasta el presente hay datos convincentes relacionando a p53, c-myc, bcl-2, ced-4 y ras (Williams, 1991; Alions y Sarraj, 1992).
El gen supresor tumoral p53 y el proto-oncogen c-myc han mostrado tener efectos sobre la apoptosis en algunas células. En timocitos, y otros tipos celulares, p53 es un componente esencial de la vía que conduce desde el daño al ADN hasta la apoptosis, aunque en otras células este puede conducir a la detención del crecimiento y la reparación del ADN.
El gen c-myc, al igual que el p53, es más conocido como un elemento importante en el control de la proliferación celular, su presencia bajo condiciones de detención del crecimiento, puede indicar la presencia de apoptosis. El gen c-myc puede, inducir ya sea proliferación como apoptosis y la decisión celular entre esas dos respuestas será determinada por otras señales, como pueden ser la presencia de factores de crecimiento u otros estímulos de sobrevivencia.
Segundos mensajeros
La apoptosis es inducida cuando, un receptor de membrana o un mediador citoplasmático, se une a un ligando, el cual provoca la generación del segundo mensajero. Varios sistemas de segundos mensajeros han sido asociados con la inducción de apoptosis y la respuesta final varia con el estado metabólico de la célula y el tipo de célula en particular.
Hay investigaciones que señalan que el AMPc desempeña un determinado rol en la apoptosis de timocitos y otros tipos de células linfoides. El efecto del AMPc es mediado por la proteína cinasa A (PKA) e involucra la activación de AMPc-dependiente de proteína cinasa 1 (PK1).

258
En la apoptosis posterior al establecimiento de los cambios en la cromatina, el calcio libre citosólico se eleva y se sostiene, al menos en algunos tipos celulares, lo cual parece ser importante para el resto del proceso. No se conoce si la fuente de calcio es externa a la célula o interna (por ejemplo, de mitocondria)(McConkey et al., 1989). La importancia de este incremento en los niveles de calcio en la muerte celular es indicado por el hecho de que la inhibición del incremento de calcio (por agentes farmacológicos) previenen la muerte celular. Se desconocen los cambios específicos inducidos por la elevación de los niveles de calcio que causan la muerte celular. Es probable que el calcio active directamente a una enzima nucleasa, sensible al calcio-magnesio endógeno (endonucleasa) (Nelipovich et al., 1988; Compton y Lowsky 1992). Al parece el calcio actúa directamente sobre esta enzima y que produce la ruptura de la cromatina en nucleosomas, de tal manera que el ADN de la célula en apoptosis se reduce a una serie de fragmentos.
La muerte celular programada (apoptosis) es un mecanismo importante que determina la talla y forma del sistema nervioso de los vertebrados. Tradicionalmente, el estudio de la muerte neuronal durante el proceso de desarrollo se ha basado en la investigación de la función de factores de supervivencia neuronal como el Neural Growth Factor (NGF) y otras neurotrofinas relacionadas. La visión predominante hasta ahora es que la muerte celular se lleva a cabo principalmente para igualar el tamaño de la población neuronal al de sus dianas y así eliminar las neuronas con proyecciones erróneas o inadecuadas. Por otro lado, la posibilidad de que la muerte celular sea un mecanismo morfogénetico en el sistema nervioso ha sido relativamente inexplorada. Recientemente, estudios realizados en ratones knock
out* para genes implicados en el proceso de apoptosis, así como el análisis genético de la apoptosis en el nematodo Caenorhabditis elegans, han llevado a considerar la muerte celular no sólo como un mero mecanismo para emparejar a la población neuronal con la población celular donde va a mandar sus proyecciones, sino como un mecanismo de ajuste del pool de progenitores neuronales iniciales, necesario para una morfogénesis correcta del sistema nervioso.
Mecanismos moleculares:
La serie de reacciones bioquímicas que ocurren al interior de la célula que ingresa al mecanismo de apoptosis es una activación de proteasas que desmantelan las células, lo hacen de forma rápida y controlada y no causan daños en el resto de células o en el entorno celular. Las proteasas (caspasas), son sintetizadas a partir de proenzimas inactivas, denominados procaspasas, estas se activan por proteólisis. Existen de varios tipos y al respecto han sido denominadas con un número. Por ejemplo, las caspasas iniciadoras 8 y 9 se activan a señales apoptóticas. Es interesante el hecho de que los ratones mutantes nulos para la caspasa-3 mueren al nacer y el análisis indica que poseen un número superior de células en el cerebro.
Se encuentran en este proceso de muerte celular las proteínas intracelulares reguladoras de la muerte celular programada, relacionadas con la proteínas Bcl-2, homóloga estructural y funcionalmente a la proteína CED-9 esencial para la prevención de la muerte celular en C.
elegans. Las proteínas Bcl-2, Bcl-Xl, y Bax participan en la muerte celular en sistema nervioso. Todos estos tipos son reguladores intracelulares.
Reguladores extracelulares: Las proteínas de la familia Bcl-2 y las procaspasas son reguladas por señales extracelulares de supervivencia o de muerte. Estas son familias de neurotrofinas y factores de crecimiento.

259
Otra vía de inducción de muerte es la existencia de receptores de muerte, entre ellos se encuentran los Fas y receptor de TNF (Factor de necrosis tumoral) a los que se unen los ligandos de Fas y TNF y que participan en la apoptosis del sistema inmune. En el desarrollo del sistema nervioso un fenómeno es la activación por NGF del receptor de neurotrofinas p75. Este receptor contiene una secuencia con homología de dominio de muerte, participa en la apoptosis inducida por NGF en retina de pollo, es esencial para la muerte celular de neuronas simpáticas. La unión de NGF puede producir liberación de ceramida.

260
Aprendizaje- memoria
Neuronas y comportamiento
Aprendizaje y memoria son dos procesos muy ligados, mediante los cuales pueden originar cambios en la conducta. Lo que aprendemos se retiene o almacena en la memoria. El siguiente aforismo resulta cierto cada vez que pensamos en la formulación de ambos conceptos, se trata de que memoria y aprendizaje y memoria son como las dos caras de la misma moneda. Ambos se encuentran presentes en una infinidad de procesos cerebrales, tales como percepción sensorial, las emociones, o el lenguaje. Se puede categorizar los sistemas de memoria y de aprendizaje atendiendo a las características de cada uno de estos procesos. Memoria implícita y explicita: Reconocemos la memoria porque esta se expresa mediante los recuerdos acerca de lo aprendido. Estos recuerdos pueden clasificarse según si son recuerdos implícitos o explícitos. La memoria implícita son en efecto recuerdos de tipo inconscientes, entre los que se encuentran nuestros hábitos perceptivos y motores. La expresión de estos recuerdos se caracteriza por aparecer en forma automática y difícil de verbalizar, son memorias fieles y rígidas, duraderas, posibles de perfeccionar con la práctica. Se relacionan con formas básicas de aprendizaje como la habituación, la sensibilización, aprendizaje perceptivo, condicionamiento clásico e instrumental y aprendizaje motor. La mayoría de estos aprendizajes son filogenéticamente muy antiguos, lo que significa que han permanecido con la aparición de las especies durante millones de años en los procesos de la evolución, son vitales para la adaptación y supervivencia de las especies. Algunos ejemplos para ejemplificar brevemente estos aprendizaje y memoria de tipo implícitos, en el caso de la habituación es el caso cuando nos acostumbramos a los golpes de martillo del “maestro” cuando realiza una tarea en su casa, o cuando nos percatamos de la música de la casa vecina, una vez que nos acostumbramos a ella y sentimos que no nos molesta, y que terminamos ignorando el ruido, decimos entonces que estamos habituados, es la llamada habituación. Cada vez que salivamos ante la presencia de un alimento estamos en presencia de una ejecución de un aprendizaje que ha surgido por condicionamiento, en este caso, se trata del condicionamiento clásico Pavloviano. Hemos aprendido a comportarnos de algún modo determinado en nuestras relaciones sociales, y ello porque ha operado un comportamiento que se ha producido como resultado de un proceso de aprendizaje conocido como condicionamiento instrumental. El reconocimiento de las personas conocidas en forma inmediata es porque ha ocurrido el proceso de aprendizaje perceptivo. Subirse y desplazarse sobre una bicicleta es un aprendizaje de tipo motor. Todos estos aprendizajes son del tipo de memoria implícita. Los recuerdos deliberados y conscientes corresponden a la categoría de memoria explícita, en esta tenemos el conocimiento del mundo, los recuerdo de las experiencias personales. Aquellos que pertenecen a esta memoria explícita son los recuerdos que pueden categorizarse como memoria semántica, por ejemplo, donde viven tus amigos, la declaración

261
de la renta que obtienes con tu trabajo de Fonoaudiólogo, Psicólogo, en que lóbulo del cerebro se encuentra la formación hipocampal, etc. Otro tipo de recuerdos en la categoría explícita son la memoria episódica, como ejemplo, lo que hoy te ocurrió, la hora que fuiste a cenar, el gol de la selección en el mundial de fútbol, etc. Esta memoria corresponde también al aprendizaje relacional, forma más compleja que permite utilizar operaciones del pensamiento tales como el comparar, utilizar información sobre hechos, circunstancias, utilizando para ello más de una modalidad sensorial. Es la que utilizamos muy a menudo en el aprendizaje de materias relacionadas con enfermedades, etc.
Muchos tipos de aprendizaje y memoria de tipo explícito comienzan siendo de tipo consciente y luego con la práctica se convierten en una conducta de memoria implícita. Por ejemplo, cuando escribimos en el computador, o cuando conducimos el automóvil, con la práctica se automatiza y se convierte en rutina inconsciente. Así, hay tareas que pueden aprenderse tanto de modo explícito como de modo implícito, ello requiere de regiones cerebrales distintas que originan memorias diferentes, pero en general ambos tipos de memorias interactúan entre sí.
El sistema nervioso en principio era conocido como una estructura casi inmutable, los estudios indicaban que el sistema no cambiaba. En este siglo la visión es diferente, mucha evidencia científica existe y da cuenta de la capacidad del sistema nervioso para modificar su actividad en virtud de algún cambio morfofuncional en las estructuras que se relacionan con la actividad sináptica. Es un hecho de de que el sistema nervioso puede cambiar, ello se presiente cuando observamos el desarrollo de los niños y no es tan evidente en los adultos. Sin embargo, los adultos, según las observaciones, también modifican sus estructuras nerviosas, ello es lo que se ha denominado plasticidad. Donald Hebb en su libro la “Organización del Comportamiento”, postula que las sinapsis podían modificarse y que existían mecanismos para fortalecer estas conexiones. De esta forma nace la teoría acerca de la plasticidad cerebral. Los cambios sinápticos han sido observados como de mayor o de menor duración. El cambio se mide en el incremento de la eficiencia en la transmisión del impulso nervioso. Así se ha llegado a determinar que existe una forma de corto plazo de transmisión de impulsos (potenciación= STP = Short-term Potentiation), en la cual las modificaciones de las sinapsis duran minutos. La segunda forma es la potenciación de largo plazo que puede durar días, meses o incluso años. Si la memoria es almacenada en el evento sináptico, entonces lo que tenemos es una memoria a largo y corto plazo. La lógica y la coherencia que encontramos a nivel macroscópico debemos también encontrarla en el nivel microscópico. De Tal manera que desde ambos puntos de vistas, al ser coincidentes en el establecimiento de este fenómeno de la memoria y del aprendizaje podremos establecer una mayor certeza en las explicaciones. Se ha encontrado a nivel molecular, que los principios bajo los cuales funciona la memoria son idénticos para todos los animales. En el establecimiento de la memoria a corto plazo, los recuerdos duran un corto período de tiempo, y no parece haber cambios estructurales notables en la sinapsis. En cambio, en el largo plazo al cabo de algunas horas habrá activación de genes que en última instancia conduce a la fabricación de proteínas y estas si

262
que afectan la estructura sináptica y los mecanismos funcionales de la célula integrados para la función de transmitir información. En experimento de laboratorio utilizando ratones se ha visto que aplicando una corta serie de estímulos eléctricos a frecuencias de 1 Hz sobre las fibras perforantes, musgosas o los axones de la vía colateral de Schaffer en el hipocampo, se produce el fenómeno de potenciación a corto plazo. Al aumentar la frecuencia de estimulación hasta 50-100 estímulos a 100 HZ, aparecen cambios sinápticos característicos de la potenciación a largo plazo, que duran varios días o meses. Lo interesante es que los patrones eléctricos de estimulación simulan las condiciones bajo las cuales se produce aprendizaje en los circuitos neuronales intactos. (Más adelante encontrará experimentos en Aplysia que lo ilustrará a usted con más detalle). Las modificaciones de la actividad neural se dan por cambios en la fuerza de las sinapsis existentes, estas pueden modificarse por períodos cortos o transitorios, o por períodos prolongados, estos pueden llevar a cambios en la expresión genética. Por otra parte, cada neurona tiene la capacidad de modificar el patrón de sus conexiones con otras neuronas, puede reemplazar el patrón de conexiones mediante eliminación y creación. Las conexiones neuronales se inician tempranamente en el desarrollo del ser humano, siendo determinadas genéticamente, las que se podrían llamar conexiones innatas. Por ejemplo, la secreción de hormonas al inicio de la pubertad es producto de modificaciones en las neuronas conectadas a la glándula neurohipofisiaria. Estos cambios son independientes de las fluctuaciones ambientales. No importa que tan distinto sea el ambiente en el Asia o en América, los adolescentes siempre tienen y tendrán hormonas a las cuales responderán con modificaciones de la estructura corporal y de la conducta en general. Es motivo de discusión y controversia es que si el ambiente (social) es capaz de provocar el desarrollo o modificación de conexiones sinápticas entre neuronas. Lo cierto es que si puede hacerlo, eso si, aceptando algunas investigaciones que han realizado científicos en roedores u otros mamíferos. Sin embargo, no debemos olvidar el sustrato de investigación, que siempre es biológico. Se puede modificar el ambiente y lograr el desarrollo o modificación de conexiones neuronales, pero estas, se encuentra prevenido en la naturaleza que ocurra. La plasticidad ocurre porque existe la posibilidad dada la estructura genética presente en el organismo, así puede acontecer. La posibilidad se encuentra en la estructura genética que posibilita la estructura y diseño del organismo, y este es modificable en función de las posibilidades que da el rango de expresión en interacción con el medio ambiente. Recientemente se ha descubierto proteínas que participan en la regulación del crecimiento y en la activación de las conexiones nerviosas del cerebro.

263
La proteína Dasm1 (llamada arborización de dendritas y maduración de sinapsis) tiene su
actividad en el cerebro en desarrollo, controla la proyección de nuevas conexiones y estimula conexiones entre neuronas jóvenes. Se cree que funciona en el cerebro maduro.
Las espinas dendríticas se extienden desde las dendritas. La dendrita y sus espinas son
lugar de sinapsis, sitio de suceso en que las dendritas reciben señales químicas como los neurotransmisores. El crecimiento de las dendritas nuevas puede incrementar la conexión entre neuronas.
Utilizando anticuerpos marcadores se ha observado la presencia de la proteína en las
dendritas y en mucha menor cantidad en los axones. La observación fue realizada en hipocampo de ratones. La proteína codificada por un gen homologo se ha observado en las dendritas de células neuronales de la mosca.
Cuando los investigadores bloquearon la actividad del gen en ratas se encontró que la
arborización de las dendritas se reducía. Por otra parte también en estos trabajos se investigó los efectos de la Dasm1 en la
maduración de las conexiones neuronales. Se sabe que las conexiones recién formadas o inmaduras no contienen receptores (“silenciosas”) AMPA, pero si poseen receptores NMDA, que se sabe están asociados con cambios a largo plazo en la fuerza de señalización neuronal: Durante la maduración, las dendritas adquieren receptores AMPA activos.
Experimentalmente, al interferir con la función de Dasm1 se reduce la actividad y por lo
tanto, se produce una disfunción de en la maduración sináptica. Por ello, se ha planteado que Dasm1 es una molécula de control de la maduración y de la arborización de la dendrita.
El interés en este tipo de hechos es mayor puesto que se da una razón entre los receptores
AMPA y NMDA, y esta aumenta durante el desarrollo y en la potenciación de largo plazo. Los hechos han llevado a los investigadores a levantar la hipótesis de que Dasm1 sea un
tipo de molécula receptor dado el hecho de sus características estructurales. El ejercicio mejora el aprendizaje
Fig1 En la foto se observa sección de cerebro de ratón adulto en las neuronas el ADN se ha teñido de rojo y la proteína Dasm1, que controla el desarrollo de dendritas en mamíferos, se ha teñido de verde. Dasm1 surge abundantemente en las dendritas de las neuronas, pero esto no ocurre en los axones.
Fuente: Yuh Nung Jan y col . controlof dendrite arborization by an Ig family member, dendrite arborization and synapse maturation 1 (Dasm1)2204 PNAS- September 7.2004

264
Investigadores Del Instituto Médico Howard Hughes (HHMI) han realizado investigaciones que permiten establecer la hipótesis de que el ejercicio físico estimula el crecimiento de nuevas células nerviosas y mejora el aprendizaje y la memoria. Pero, esto sólo se ha visto en ratones adultos que son los animales sometidos a ensayos por T. Sejonowski. El trabajo referido en si es “Running enhances neurogenesis, learning and long- term potentiation in mice” publicado en el año 1999.
Según los investigadores la actividad de correr incrementa la neurogenésis en el giro
dentado del hipocampo de cerebro de ratones. Algunas investigaciones de otros autores daban cuenta de la capacidad para el surgimiento de nueva neuronas en el cerebro adulto En la experimentación se midió la capacidad para memorizar de un grupo de ratones sedentarios comparados con ratones que realizaron ejercicios durante un mes. Los ejercicios se llevaron a cabo en una rueda para correr, los ratones corredores recorrían una distancia de 4,78 kilómetros diarios. Ambos grupos de ratones fueron entrenados para localizar una plataforma camuflada y sumergida que se encontraba debajo de una superficie de agua turbia. Su llegada a la plataforma se realizaba a través de un laberinto. Los ratones deben memorizar el camino a la plataforma, que es una forma de evitar la natación continua. Después de un entrenamiento pos seis días se encontró que el grupo de ratones sedentarios tomó caminos más largos y demoró más en encontrar la plataforma. Por lo tanto, el grupo de ratones buenos para ejercitarse debían poseer mejor memoria. Los cerebros estos ratones tenían cerca de 2,5 veces más neuronas nuevas. Sejnowski midió la potenciación a largo plazo examinando el cerebro en los dos grupos. Resultando el grupo de los sedentarios los que presentaban unas dos veces menos de potenciación a largo plazo que los del grupo de los buenos corredores. El incremento de la LTP (potenciación a largo plazo) sólo se dio en la circunvolución dentada. Las observaciones apoyan la idea de Sejnowski de que el ejercicio mejora la formación y supervivencia de células nerviosas nuevas, las conexiones entre las neuronas, las que a su vez mejoran la memoria a largo plazo. Los resultados experimentales de Sejnowki no parecen concordar con aquellos obtenidos de Bhardwaj, Pasco Raskic, etc. En el año 2006 ( Neocortical neurogenesis in human is restricted to development) el primero ha mostrado que la neurogénesis experimenta diversos grados en diferentes animales y entre ellos los roedores (animal de experimentación de Sejnowski). Bhardwaj lleva pasos adelantados al determinar en neocortex de cerebros humano que no habría procesos de neurogénesis hacia la zona de la mencionada estructura cuando se llega a la edad adulta. Pero si habría procesos relacionados con la plasticidad neocortical en el sentido de la modulación de las conexiones cerebrales. SISTEMA LÍMBICO: (Recuerde que el concepto de sistema límbico se encuentra en revisión constante) El conocimiento de la anatomía nos puede llevar a conocer mejor el sustrato, por ejemplo, en el cerebro- zona sistema límbico, acontecen la memoria, el aprendizaje, las emociones, etc. En el siglo XIX, Pierre Paul Broca, famoso Neurólogo francés, señaló que las estructuras cerebrales que tenían forma de C en la superficie cerebral medial debían poseer algún rol en las emociones. Esta afortunada mención del notable personaje encontró eco mucho tiempo después. La estructura en forma de C incluye además la

265
formación hipocampal y la amígdala, se encuentra además formada por las circunvoluciones del cíngulo y la parahipocampal. Estas estructuras rodean al diencéfalo y bordean al cortex. Básicamente el sistema límbico está formado por la formación hipocampal y la amígdala, estás estructura contienen parte de los circuitos nerviosos con funciones de participación en los procesos de memoria y aprendizaje. Dichos circuitos se conectan con distintas zonas del encéfalo. Los circuitos hipocampales están formados por estructuras telencefálicas y diencefálicas. Los circuitos de la amígdala intervienen en procesos más relacionados con la emoción, como ejemplo, notable se encuentra muy activa en los hechos que denotan estados de furia. No obstante, también participa de procesos de memoria y aprendizaje. El sistema limbico se desarrolla a partir del prosencéfalo. Los componentes telencefálicos de este sistema son: a) La corteza de asociación límbica, b) la formación hipocampal con su vía eferente, el fórnix, y c) parte de la amígdala y la vía de output llamada estría Terminal. La corteza de asociación límbica se ubica en la superficie medial de los lóbulos frontal, parietal y temporal:
Recibe información de áreas sensoriales de orden superior y de las áreas corticales de asociación: área parieto- temporo-occipital y área cortical de asociación prefrontal.
La información sensorial integrada es recibida por la formación hipocampal. Esta información se relaciona con el aprendizaje del entorno o entorno asociados a situaciones relacionadas con movimiento o de peligro. LA relación con la amígdala queda entonces de manifiesto. Circunvoluciones del sistema límbico: 1) Circunvolución del cíngulo 2) Circunvolución parahipocampal 3) Circunvoluciones orbitales medianas 4) Circunvoluciones del polo temporal. Formación hipocampal y fórnix: Forman un anillo concéntrico dentro de la corteza de asociación límbica rodeando el diencéfalo y el mesencéfalo, ubicado en la superficie cerebral medial del lóbulo. El fórnix constituye la vía eferente de la formación. La formación hipocampal es un estructura subcortical formada por arquicorteza primitiva. Formada por tres estructuras: el hipocampo, el subículo y la circunvolución dentada. Las tres estructuras parecen bandas constituidas en sentido rostrocaudal dentro del lóbulo temporal. Las tres forman una especie de cilindro. La formación hipocampal recibe su principal input desde la corteza entorrinal que pertenece a la corteza de asociación límbica. Las células neuronales output son de tipo piramidal ubicándose en el hipocampo y el subículo y sus axones colaterales contactan la superficie de la formación hipocampal formando el haz de fibras llamado Fórnix, este se proyecta a estructuras subcorticales, telencefálicas y diencefálicas. Existen dos sistemas de output, uno desde el hipocampo y el del subículo. Los dos se relacionan con cognición, aprendizaje y memoria. El output desde el subículo (Axones) sinaptan en los cuerpos mamilares del hipotálamo y este por medio del tracto mamilotalámico se proyecta a los

266
núcleos anteriores del tálamo, estos, a su vez se proyectan a la circunvolución del cíngulo. Se forma así una especie de circuito anatómico. Esta circunvolución como se ha dichos antes, forma parte de la corteza de asociación límbica que envía información a la corteza entorrinal. Esta informaciones constituye un input de la formación hipocampal.
MECANISMOS CELULARES Y MOLECULARES DEL APRENDIZAJE Y MEMORIA
No se trata de ver los aspectos reduccionistas o enfocar desde esa perspectiva. Al
contrario, todo estudioso en la actualidad, debe considerar en observar desde su posición hacia todos los campos que se encuentran relacionados con sus materias de estudio. La mirada desde este apartado significa reconocer que existen hechos psicológicos que se correlacionan con la conducta y los aspectos moleculares. Ellos son innegables, no pueden dejar de ser considerados a la hora del estudio y abordaje desde una perspectiva científica.
Los mecanismos celulares y moleculares que subyacen al aprendizaje y memoria en aplysia son parte de muy buenos modelos explicativos del fenómeno que nos concierne en la unidad temática. Son tan buenos que su principal investigador ha sido galardonado con el premio Nobel del año 2000.
¿De qué forma se adquiere la nueva información y, una vez adquirida, cómo se retiene?
La memoria (lo que resulta del aprendizaje y este, se demuestra por la memoria) implícita- no declarativa es una memoria de tipo inconsciente de habilidades cognitivas y motoras. Lo contrario parece ser la memoria de tipo declarativo- explícita que consiste en poder recordar personas, lugares, etc., como actividad consciente. Los dos tipos de memoria se pueden abordar desde la biología mediante estudios en invertebrados e invertebrados. Desde el punto de vista evolutivo a nivel molecular las formas aprendizaje son mecanismos moleculares que han sido muy conservados, es decir, hay muy poca variación: De hecho los mecanismos en las formas vivientes más complejas utilizan elementos básicos para dar forma al aprendizaje y a la memoria. La relación básica se encuentra en las variaciones de la eficacia de transmisión sináptica, de ello depende el almacenamiento de la memoria a corto plazo. Inicialmente se puede plantear el aprendizaje en sus formas más simples para comenzar nuestro estudio. En este tipo de aprendizajes ocurren modificaciones de la actividad sináptica en sus expresiones molecular y celular. Habituación: Es una forma de aprendizaje simple de tipo implícito: Se presenta al animal un estímulo inocuo nuevo, que no le trae beneficios ni daños. Después de la exposición repetida el animal aprende a ignorarlo. Trabajando con gatos Sherrington observó una disminución de la intensidad de ciertos reflejos. En el reflejo de retirada de una extremidad, en respuesta a la estimulación repetida. La respuesta refleja tenía la capacidad de retornar después de algunos minutos de descanso. La disminución de la respuesta refleja, en estas condiciones la llamó habituación y sugirió, a manera de hipótesis, que la disminución de la respuesta tendría como causa la disminución de la eficacia sináptica en las vías de las neuronas motoras que han sido activadas en forma repetida.

267
Mucho más tarde el problema fue investigado a nivel celular por Spencer y Thomsom. Ellos demostraron por medio de registros intracelulares en las neuronas motoras de gatos que la habituación disminuye la fuerza de la conexión sináptica entre neuronas excitadoras y las neuronas motoras.
Más tarde, los estudios de habituación se realizaron con Aplysia califórnica, con un
sistema nervioso más simple y debido a lo mismo, es más fácil abordarlo desde la perspectiva que requieren los estudios celulares y moleculares. El animal presenta reflejos similares a los de retirada de la pata estudiado por Spencer. Un ligero estímulo táctil en el sifón tiene como respuesta la retracción del sifón y la branquia. La estimulación repetida produce la habituación.
Fig. 2Tomada de Neurociencias pag 1248 vesículas de neurotransmisor liberadas del los terminales axónicos presinápticos de las
neuronas sensitivas. Estas neuronas sensitivas emplean el glutamato como neurotransmisor.
En respuesta al estímulo táctil las neuronas sensitivas que inervan el sifón generan potenciales sinápticos excitadores en interneuronas y células motoras. Los potenciales de ambas células se suman llegando a provocar la descarga repetida de las células motoras hincando la retracción refleja de la branquia. Si el estímulo se repite, lo potenciales van disminuyendo y con ello la respuesta refleja. La disminución de la fuerza sináptica es la consecuencia del descenso en el número de

268
Fig. N° 3 Tomado de Kandel con fines didácticos ocupa un lugar de privilegio, sino que se distribuye ampliamente por todo el circuito. El
lugar de la depresión puede situarse en cualquiera de los lugares de sinapsis. Por ejemplo, en el reflejo de flexión de la médula espinal la depresión se produce en una sinapsis distinta de la que se produce entre una neurona sensitiva con interneurona.
Los avances en el estudio molecular y celular nos permiten ampliar la visión acerca de los
fenómenos de aprendizaje. Los hechos que involucran aprendizaje deben ser explicados desde la perspectiva que el fenómeno implica variación de la fuerza sináptica, y la duración del almacenamiento de la memoria debe ser reflejada por la duración del cambio sináptico.
En los experimentos realizados en Aplysia se ha observado que una sesión de 10 estímulos
táctiles al sifón conduce a la memoria de habituación del reflejo de retracción branquial de corto plazo que dura unos pocos minutos. Se puede desarrollar la memoria de largo plazo que puede durar hasta tres semanas. Ello se consigue con cuatro sesiones de 10 estímulos separadas por períodos de horas, hasta un día. Las mediciones y experimentos realizados con Aplysia sin experiencia previa muestran que estos últimos establecen conexiones en un 90% entre neuronas sensitivas y neuronas motoras de la branquia. Como resultado de la habituación el número de conexiones se reduce a un 30%. Posteriormente este fenómeno se revierte en forma paulatina hasta completarse totalmente.
Un hecho es claro con el resultado anterior y es que se ha dado una modificación de la
conexión neuronal. De los resultados podemos trazar líneas hacia la complejidad en los aspectos biológicos
que tienen que ver con los mecanismos de adaptación. Por cierto, el mecanismo de depresión sináptica constituye un acto de adaptación al ambiente. No olvide que el individuo puede adaptarse al cambio ambiental si es que posee el potencial para poder cambiar. El ambiente selecciona aquellos individuos que tienen el potencial para cambiar, estos individuos ya se encuentran entre nosotros. Eso por una parte.
Se sabe que el glutamato interacciona con dos
tipos de receptores en las células receptoras:
uno de ellos similar al tipo NMDA de los
vertebrados y el segundo tipo no NMDA Los
informes señalan que no existe variación de la
sensibilidad de estos receptores con la
habituación. La reducción de la eficacia sináptica
puede prolongarse durante varios minutos.
Este hecho constituye en sí, una explicación de
cómo se produce la memoria a corto plazo para
la habituación. Este fenómeno de variación de la
eficacia sináptica, que es una variación plástica
se producen en cualquier lugar del circuito
reflejo, por ello, se plantea de que la memoria no

269
Por otro lado, tenemos el aspecto genético, el resultado sugiere que indudablemente las conexiones sinápticas, si bien sufren modificación, se deprimen, estas vuelven a reconstituirse luego de un determinado tiempo en que ha desaparecido el estímulo. Vuelven a aparecer las conexiones porque debe haber un programa genético que establece cuando y como establecer conexiones. Cuando y como producir vesículas sinápticas productivas. Todo ello requiere de genes activos y organizados para realizar el trabajo que se encuentra desde luego, prestablecido. Los genes preestablecen, y sin embargo, se puede modificar su actividad al menos transitoriamente que es lo que los experimentos en forma indirecta nos muestran. De todas manera necesariamente no todos los genes tienen que estar activos, muchos deben encontrarse en un estado silencioso, bastaría con recordar los procesos de diferenciación celular, desde el embrión, hasta el final de la vida de un organismo.
Aprendizaje no asociativo: Sensibilización y habituación: Se entiende que estas formas de aprendizaje son formas elementales de plasticidad
conductual y no requieren algún tipo de asociación entre estímulos o entre estímulos y respuestas del entorno del organismo. Estas formas son la habituación y la sensibilización. Una descripción simple de la habituación es que se trata de procesos mediante los cuales una respuesta refleja disminuye, o pierde intensidad ante la presentación repetida de un estímulo inocuo, es decir, que no produce daño, ni atenta contra la integridad física del animal. La habituación es ignorar estímulos irrelevantes como los dados a conocer antes, súmese los ejemplos de, olvidarse del ruido del tráfico de la cuidad. En general la respuesta de habituación más estudiada ha sido la disminución de la intensidad del reflejo de sobrecogimiento, o sea, el susto ante la emisión de un tono sonoro alto en la rata. A medida que se aumenta la repetición de la emisión del tono, la respuesta de susto en la rata cada vez es menor. Los experimentos han señalado que la respuesta de habituación puede disminuirse o eliminarse a la vez, o sea se puede producir la deshabituación. Esto puede ocurrir cuando el tono que es el estímulo que conduce a la habituación, es precedido por luz. El resultado es la deshabituación.
La sensibilización es una forma de aprendizaje que consiste en la intensificación de una
respuesta refleja ante estímulos moderados y que son precedidos por estímulos más intensos o nocivos. Es una respuesta anormalmente intensa ante una situación o estímulo habitual cuando previamente se tiene una experiencia negativa, dolorosa, o desagradable que sensibiliza al animal. Por ejemplo, en el mismo experimento con ratas, cuando esta emite una respuesta de sobrecogimiento ante un tono, esta puede intensificarse si previo a la emisión del tono se da al animal un pinchazo en cualquier parte de su cuerpo. Las respuestas caracterizadas como de sensibilización son un tipo de respuestas defensivas, con objetos de adaptación. Sin embargo, a diferencia de la habituación en que la respuesta es para un estímulo específico, la respuesta de sensibilización afecta a todo el organismo y se sensibilizará ante una diversidad de estímulos, por ejemplo, de tipo táctil o luminoso.
Con un estímulo nocivo el animal puede responder en forma más enérgica.
En el caso de Aplysia los reflejos defensivos de retracción y escape adquieren más fuerza. Como hemos visto en la figura N°3, el reflejo de habituación se relaciona con la
estimulación a una vía y que por medio de esta se produce el reflejo. En el caso de la sensibilización tenemos dos vías que interaccionan. Al aplicar un estímulo a una de ellas se produce efecto en la otra vía que es la producción de un reflejo con mayor intensidad. De igual modo, la sensibilización tiene una forma de memoria a largo y corto plazo. Una descarga en la cola produce sensibilización a corto plazo y dura algunos minutos. Cinco descargas o más pueden durar días o semanas.

270
Experimentalmente se aplica un estímulo nocivo en la cola ello facilita la transmisión
sináptica en el circuito del reflejo, las conexiones entre neuronas sensitivas con neuronas motoras e interneuronas. Lo interesante es que se trata de las mismas vías que se involucran en la habituación. De tal manera, se infiere, que una sinapsis puede participar en más de un tipo de aprendizaje y memoria, con la diferencia que la sensibilización requiere de un proceso de facilitación del establecimiento de la fuerza sináptica, cuya acción corre por cuenta de neuronas denominadas interneuronas facilitadoras.
Fig. N° 4 Tomada de neurociencias Autor Kandel Aparece ilustrado los procesos
conducentes a la sensibilización: Se aplica un estímulo nocivo a una zona distinta del sifón como es la cola. Ello estimula a las neuronas sensitivas de la zona correspondiente, estas neuronas forman sinapsis con las interneuronas que a su vez sinaptan sobre las terminales de neuronas sensitivas que inervan el sifón. Las sinapsis axoaxónicas, las interneuronas facilitadoras promueven la liberación del neurotransmisor desde las neuronas sensitivas, esto es identificado como facilitación presináptica.
En el caso de las interneuronas serotoninérgicas al liberar serotonina se observa que estas
se unen a receptores que entonces en el segundo paso activan a proteínas G trimérica que actúa bajo el influjo de reacciones de GTP:
La facilitación presináptica está mediada, a lo menos por tres vías de tipo bioquímico. La interneurona presináptica libera el neurotransmisor denominado serotonina (5-HT, 5-hidroxitriptamina). Se une a dos receptores. Uno de ellos establece interacción con una proteína G, la que a su vez aumenta la actividad de la adenilciclasa, esta enzima convierte el ATP en AMPc. Este compuesto activa la quinasa A que se une a la subunidad reguladora inhibidora. De este modo libera la subunidad catalítica quedando en condiciones para actuar siguiendo con la cascada bioquímica.

271
Figura tomada de Bioquímica Mathews. El esquema muestra una vía o sucesos de pasos bioquímicos en el
señalamiento celular. De esta forma se propone que sería una de las vías relacionadas con el Aprendizaje y la Memoria. El esquema se ajusta a lo que ocurre en la neurona sensitiva.
Figura N°6 Tomada de Kandel para la clase de Neurociencia y Psicobiología
5-HT hidroxitriptamina( Serotonina)

272
La subunidad catalítica de PKa actúa siguiendo tres vías: 1) Fosforila los canales de potasio, lo cual disminuye la corriente de potasio. La situación
prolonga el potencial de acción y aumenta la entrada de ión calcio, se sigue con un aumento en la liberación del neurotransmisor.
2) Las vesículas que contienen neurotransmisor se acerca a la membrana y la liberación de neurotransmisores.
3) Se abren los canales de ión calcio de tipo L. Para ello, la serotonina actúa a través de un segundo receptor activa la proteína G0, esta activa a su vez a la fosfolipasa C(PLC), y esta a la cinasa C (PKC) por mediación del diacilglicerol.
Actividad sináptica en mamíferos. Uniones neuromusculares En la actividad sináptica surgen cambios agudos como resultado de la actividad neuronal. La facilitación se caracteriza por cambios transitorios en la fuerza sináptica. Se desarrolla cuando dos o más potenciales de acción llegan a la terminación sináptica, ambos muy cercanos en el tiempo. El resultado es mayor liberación de neurotransmisor con cada potencial de acción. En consecuencia, el cambio de voltaje generado en la célula postsináptica aumenta en forma progresiva. Desde la década de los años cincuenta ya se descubre que parte de los mecanismos involucrados en la facilitación en la unión neuromuscular podían ser explicados desde la perspectiva que considera las concentraciones del ión calcio y su elevado nivel en la terminación sináptica. Cuando dos potenciales de acción llegan juntos, se eleva la concentración del ión y se libera más neurotransmisor. Luego de la repetición del suceso, la transmisión sináptica disminuye hasta causar la depresión sináptica. La liberación del neurotransmisor en cantidad sobrecarga el sistema de recaptación y ulterior recarga en las vesículas, como consecuencia la actividad del sistema se detiene hasta que sobreviene la recuperación. La misma actividad de sobresaturación de los niveles del ión calcio activa mecanismos que posibilitan disponibilidad mayor de vesículas con neurotransmisor y cuando surge nuevamente un potencial de acción puede conducir nuevamente al aumento de la liberación del neurotransmisor. Esto se conoce como potenciación postetánica que dura algunos minutos después de una descarga de alta frecuencia de potenciales de acción. Los tres tipos de fenómenos, facilitación, depresión y potenciación postetánica modifican la transmisión sináptica. Al parecer no se relacionan con los procesos de memoria y aprendizaje que estamos viendo en estas clases. Potenciación a largo plazo en el hipocampo Estos son los estudios realizados en mamíferos donde la fuerza sináptica y su modificación prolongada al parecer produce cambios en la memoria y el aprendizaje. Los estudios son realizados en el hipocampo.
La estimulación eléctrica durante algunos segundos con alta frecuencia de una vía de fibras aumenta la transmisión sináptica entre los axones estimulados y las células postsinápticas. El cambio dura semanas, por lo que se ha sido denominado potenciación a largo plazo. En cortes de hipocampo los somas de las neuronas se distribuyen en una capa empaquetada que a su vez se divide en varias regiones distintas, las principales son CA1 y CA3. Las dendritas de las células piramidales en la región CA1 reciben información desde las colaterales de Schaffer, que son aferencias originadas desde las células piramidales de CA3.

273
Estas conexiones sinápticas entre CA3 y CA1 son zonas de potenciación a largo plazo de gran calidad y la región CA1 Cobra importancia por los efectos sobresalientes sobre la memoria humana.
La estimulación eléctrica de las colaterales de Schaffer produce potenciales postsinápticos excitatorios (PPSE) en las células CA1 postsinápticas. Si se estimulan las colaterales de Schaffer dos o tres veces por minuto, el PPSE en las CA1 se mantiene constante. Pero, con un tren de alta frecuencia breve de estímulos en los colaterales se produce un incremento duradero en la amplitud del PPSE. Esto significa que la transmisión sináptica en el hipocampo se ve aumentada cuando la sinapsis es activada en forma reiterada.
Fig obtenida de www.down21.org/.../neurobiologia/g_hipocampo.htm
El hipocampo es una estructura importante para la formación de la memoria a largo plazo. Tiene el tamaño de un dedo pulgar de niño, y se encuentra localizado en la profundidad de la porción media del lóbulo temporal. La información fluye hacia y a lo largo

274
del hipocampo por medio de tres vías principales que están indicadas en la imagen ampliada que se ve a la izquierda de la figura: 1. la vía perforante que circula desde la corteza entorrinal a las células granulares del giro dentado. 2. La vía de las fibras musgosas que va desde las células granulares del giro dentado a las piramidales de la región CA3 del hipocampo. 3. la vía colateral de Schaffer que proyecta desde las células de región CA3 a las de la región CA1. (Figura modificada de: Memory:From Mind to Molecules. L.S.Squire y E.R. Kandel. Scientific American Library, Nueva York 2000). Hoy en día nadie puede dudar acerca de lo fundamental de conocer la estructura anatómica de la memoria. Nadie puede dudar asimismo que la actividad de memorizar sea un hecho que mantiene ocupado a todo el cerebro. Sin embargo, algunas zonas neuronales son más especializadas en diferentes tipos de memoria y cada una de ellas se caracteriza por estructuras y circuitos neuronales. En 1953 el médico William Scoville, con objeto de tratar un tipo de epilepsia realizó una remoción bilateral del lóbulo temporal medial y esto involucró la pérdida del hipocampo. El paciente manifestó pérdida selectiva de la memoria con severa dificultad para consolidar la memoria a corto plazo en largo plazo. En otras palabras, perdió la capacidad para almacenar información. El paciente olvidaba rápidamente los acontecimientos o episodios ocurridos en lo inmediato. Este tipo de olvido es caracterizado como amnesia anterógrada.
El aprendizaje y la memoria son mecanismos de adaptación o ajuste a las complejas
condiciones ambientales. Como consecuencia de los procesos de aprendizaje y memoria, los animales modifican su
conducta. La conducta es el resultado de la interacción entre los genes y el medio. El aprendizaje es el proceso por el que adquirimos el conocimiento sobre el mundo,
mientras que la memoria es el proceso por el que el conocimiento es codificado, almacenado y posteriormente recuperado.
Muchas conductas importantes son aprendidas. Somos lo que somos en gran parte por lo que aprendemos y lo que recordamos.
Aprendemos las capacidades motoras que nos permiten dominar nuestro medio, y aprendemos lenguajes que nos permiten comunicar lo que hemos aprendido, y de esta manera transmitir culturas que se pueden mantener durante generaciones.
No todo lo que aprendemos es beneficioso. El aprendizaje produce también alteraciones de
la conducta, y estas conductas pueden constituir trastornos psicológicos. El estudio del aprendizaje es, por consiguiente, fundamental para la comprensión de los trastornos del comportamiento, así como la conducta normal, puesto que lo que se aprende a menudo se puede olvidar. Cuando la psicoterapia tiene éxito en el tratamiento de los trastornos del comportamiento, lo hace a menudo creando un medio en el que las personas pueden aprender a cambiar sus patrones de conducta.
MEMORIAS:

275
La memoria no es una función diferenciada e independiente de otras funciones cognitivas como el lenguaje, la percepción o el movimiento. De hecho no posee una localización precisa en el cerebro, es más, el almacenamiento de la memoria afecta a muchas zonas cerebrales, cada una de aquellas puede almacenar diferentes tipos de información.
Las primeras pruebas de que los procesos de memoria podían localizarse en regiones específicas del cerebro humano fueron aportadas por Wilder Penfiel que fue discípulo de Charles Sherrington investigador quien aplicó electrodos a la corteza cerebral y registrando la actividad de los nervios motores pudo trazar un mapa de representación motora en la corteza cerebral. Hacia 1940 Penfiel aplicó electrodos a la corteza cerebral de pacientes sometido a cirugía para tratamiento de epilepsia, logró así la localización de regiones cerebrales relacionadas con funciones motoras, sensitivas y del lenguaje.
En algunas estimulaciones de los lóbulos temporales los pacientes recordaban alguna
experiencia.. Del estudio de pacientes que habían experimentado una extirpación bilateral del hipocampo y de las regiones vecinas, se llegó a la conclusión de la importancia de los lóbulos temporales para la memoria.
La memoria se puede clasificar como implícita o explícita basándose en cómo se almacena
y se recuerda la información. La memoria explícita nos permite aprender que es el mundo, adquirir conocimientos sobre personas, lugares y acontecimientos. Los recuerdo explícitos pueden tener carácter semántico, o sea, recuerdo de hechos o conocimientos en general, y episódicos formados por circunstancias pasadas del individuo. La memoria episódica nos permite recordar, por ejemplo, la persona que me acompaño a jugar al fútbol y la semántica nos permite recordar en que lugar se encuentran las casas de Pablo Neruda que obtuvo el premio Nóbel. También es semántica: “Las relaciones entre el volumen y la masa de una sustancia”. Es episódica: “ayer fui a pescar”
Fundamentalmente hay dos tipos de almacenamiento de la información: ellas son definidas como memoria declarativa y la memoria de procedimiento. La memoria declarativa se define como el almacenamiento y recuperación de información disponible para la mente consciente. Esta es codificada en base a símbolos y se expresa mediante el lenguaje, por ello, es declarativa. Ejemplos: recordar números de teléfono, cumpleaños, listas de compras, etc. La memoria de procedimiento se caracteriza por mostrar habilidades y asociaciones, adquiridas y recuperadas en un nivel inconsciente. Son ejemplos: Recordar como se toca un instrumento, andar en patines, o ejecutar asociaciones desarrolladas por condicionamiento clásico. Criterio que considera el tiempo de ejecución de memorias: A) Memoria inmediata: dura sólo algunos segundos, se trata del registro de algunas experiencias visuales, verbales, táctiles. B) Memoria de corto plazo: la información se mantiene en la mente durante períodos de segundos, minutos. Un tipo de memoria de corto plazo es la memoria de trabajo, que es la capacidad para mantener en la mente las cosas o acciones el tiempo suficiente y necesario para llevar a cabo acciones secuenciales. C) Memoria de largo plazo: es la retención de la información durante días, semanas, o años. Debemos aprender otro concepto relacionado: Engrama: Se refiere al sustrato físico para la memoria en cuanto a la maquinaria neural. Esta forma más permanente de la memoria depende de los cambios en las sinapsis ya sea por durabilidad o reordenamientos. ¿Cuál es el papel del hipocampo en la memoria? Los datos se obtienen del estudio de pacientes con amnesia anterógrada y retrógrada. La amnesia retrógrada es una pérdida de la

276
memoria para los eventos o acontecimientos que preceden a una lesión, que en general, puede ser de tipo encefálico, como enfermedades neurodegenerativas, o por traumatismos. El individuo no recuerda los sucesos acontecidos hasta antes del accidente. La amnesia anterógrada es la incapacidad para establecer nueva información en la memoria, en otras palabras, sólo afecta la adquisición de nuevos hechos y eventos, eso es, la memoria declarativa. Es decir, se relaciona con los hechos que registra y consolida la memoria de corto plazo. Estos dos hechos muestran que existe un sustrato anatómico diferente para cada tipo de amnesia. La amnesia anterógrada y pérdida de la memoria a largo plazo desde el accidente en adelante permite inferir que son sustratos diferentes para los dos tipos de memoria, aquella de corto plazo y la de largo plazo. La memoria declarativa se almacena en otro sustrato distinto al hipocampo. El caso es claro en daño al hipocampo, la cita clásica es el ejemplo del paciente H.M. del Dr. William Scoville y de la Dra Brenda Milner, colaboradora de Penfield, cuando queda de manifiesto el hecho de que el paciente olvida los episodios diarios y en forma rápida. Sin embargo, estos pacientes no tenían problemas para establecer información (o recuperarla) con respecto a la memoria de procedimiento. El hecho sugiere otra zona anatómica diferente de la memoria declarativa. La memoria declarativa es “la habilidad para recordar los detalles de los eventos, e incluye tiempo, lugar, circunstancia, y los conceptos.
El paciente H.M. joven de 27 años fue atropellado por alguien que circulaba en bicicleta, y, debido a las lesiones causadas sufrió durante 10 años crisis convulsivas bilaterales del lóbulo temporal. La enfermedad lo incapacitó para la vida laboral o para cualquier circunstancia de la vida normal. Fue sometido a una intervención quirúrgica en la se extirpó la formación del hipocampo, el núcleo amigdalino y partes del área de asociación multimodal de la corteza temporal de los dos hemisferios. Se controló las crisis convulsivas, pero, la extirpación de los lóbulos temporales mediales tuvo efectos indeseables por pérdida de memorias.
H.M. aún mostraba memoria a corto plazo normal, que duraba segundos o minutos y conservaba memoria de largo plazo de circunstancias o acontecimientos ocurridos hasta antes de la intervención quirúrgica. Por ejemplo, recordaba su nombre, profesión, aspectos de la infancia. Manifestó algunas evidencias de amnesia retrógrada, lo que es una incapacidad para recordar eventos ocurridos con anterioridad al surgimiento de la amnesia. Mostraba dominio del lenguaje, vocabulario y pruebas de C.I. con puntaje normal-alto.
H.M., en consecuencia, no poseía la capacidad de trasformar la memoria de corto plazo en
memoria de largo plazo. No retenía información sobre las personas, los lugares u objetos por más de un minuto. En pruebas que se le pedía recordar un número como 2389617 podía repetirlo durante cortos períodos de minutos. Poseía buena memoria de corto plazo (Kandel), al distraérsele brevemente olvidaba el número. Este paciente no podía reconocer a las personas a las que conoció después de la intervención quirúrgica. Además poseía dificultades con la orientación espacial. No aprendía el camino a casa.
Sin embargo, este tipo de pacientes pueden aprender algunas tareas y son capaces de
conservar el recuerdo. Brenda se dio cuenta de esto cuando observó que H.M. podía aprender nuevas capacidades motoras a un ritmo normal. Aprendió a dibujar el contorno de una estrella mientras miraba su mano y la estrella en un espejo, en este trabajo, no tuvo diferencias al compararse con sujetos normales.

277
Los estudios posteriores en H.M. y otros pacientes con afecciones al lóbulo temporal, permitieron establecer que además del aprendizaje de habilidades motoras podían aprender reflejos simples, habituación, sensibilización, condicionamiento clásico y condicionamiento operante. Por otra parte, podían mejorar tareas de percepción, por ejemplo, el recuerdo de las palabras o de los objetos mejora debido a la exposición previa a dichas palabras. Esta prueba consiste en mostrar las primeras letras de palabras que se han estudiado con anterioridad. Esta prueba corresponde a modalidad de memoria denominada memoria de sensibilización.
La memoria implícita en el caso anterior se denota por la conservación de la memoria de
aquellas tareas de naturaleza refleja relacionada con hábitos, capacidades motrices o perceptivas, no requiere de conocimiento consciente o de procesamiento cognitivo complejo, por ejemplo, comparación y evaluación. También la memoria implícita se conoce como memoria no declarativa, lo cual equivale a decir, que es un tipo de memoria que se recuerda de manera inconsciente.
Por otra parte, la memoria explícita o declarativa se relaciona con el acto de recordar el
conocimiento objetivo de las cosas, las personas, lugares. El recuerdo es consciente y deliberado. Es de característica flexible y afecta a la asociación de fragmentos de información. Por el contra, la memoria implícita es más rígida y obviamente conectada a las condiciones de los estímulos originales bajo las cuales se produjo el aprendizaje.
La memoria explícita puede clasificarse en episódica que es una memoria para los
acontecimientos y la experiencia personal o semántica que es para los hechos como los ya expuestos en esta guía.
Sin embargo, el intervalo de tiempo comprometido en el acto amnésico es corto. Esto
evidentemente, sugiere que el rol del hipocampo en la memoria de largo plazo es limitado a ciertos períodos. El paciente, por cierto, debe recordar hechos anteriores al intervalo de tiempo comprometido en la amnesia (Squire y Zola, citados por Passig, 1994).
Datos y observaciones acerca de la condición de H.M.:
• Su coeficiente intelectual permaneció inalterado (CI: de 104 a 118).
• No almacenaba nueva información semántica en su cerebro. Durante las tres noches que estuvo ingresado en el Centro hospitalario, preguntó a la enfermera dónde estaba y porqué, ya que no recordaba que había pasado aunque podía identificar que estaba en un hospital.
• No tenía problemas ni de la atención, ni del habla, ni de la expresión verbal compleja, y mantenía intacta la comprensión del lenguaje.
• Desarrolló problemas de olfacción ya que la operación había afectado también a las áreas cerebrales diana de los tractos olfatorios.
• Las baterías de tests incluyeron fotografías de gente, material verbal, secuencias de dígitos, diseños geométricos complejos o patrones sin sentido aparente, y todas confirmaron amnesia anterógrada y parcialmente retrógrada: recordando con certeza hasta los 16 años pero los otros 11 años (de los 16 a los 27) eran una nebulosa y nunca pudo recordarlos con claridad.

278
• Leía las mismas revistas una y otra vez sorprendiéndose cada vez de las mismas noticias.
• Cada día repetía los mismos rompecabezas sin parecer haber aprendido con la práctica.
• No podía recordar a nadie que hubiera conocido después de la operación, ni caras ni nombres (aunque le hubieran visitado continuamente).
• Podía recordar series de números en breve tiempo pero no en largos periodos de tiempo. No podía recordar dónde vivía o quien cuidaba de él, o que era lo último que había comido.
• Respecto a la memoria espacial, conseguía aprender lentamente: así, al tener que cambiarse de casa, le costó más de un año aprender el camino.
• Respecto a su situación emocional: era capaz de tener reacciones emocionales intensas de pocos minutos (se puede argüir que esta emoción duraba mientras permanecía la Información en la memoria). Por ejemplo, cada vez que se le informaba del fallecimiento de un pariente, tenía la misma reacción emocional pasajera intensa que era repetitiva una y otra vez. Incluso posteriormente preguntaba por sus pariente y manifestaba deseos de volver a verla.
• Vivía con recuerdos a corto plazo y con aquellos que mantenía de antes de la operación. Sus experiencias de cada día eran como las de las personas que comienzan a darse cuenta de lo que les rodea, sin comprensión de la información porque no podía recordar lo que acababa de ocurrir. Relataba su situación como
“todos los días son únicos en sí mismos, igual que cualquier alegría o cualquier
tristeza”. Hacía descripciones de su propio estado diciendo que era como “pasear por un sueño”. Como despertarse de un sueño del que no se puede recordar nada …
• Se atribuía a si mismo una edad inferior a la que realmente tenía, y no era capaz de reconocerse a si mismo en el espejo: en 1982 fue incapaz de reconocerse en una foto de su 40 cumpleaños tomada en 1966.
• Llegó a poder trabajar en una tarea manual automática (empaquetar encendedores en las cajas de cartulina) lo que mostró su capacidad de aprender tareas motoras con el mismo ritmo que una persona normal. Entre los estudios realizados por la Dra. Milner se encontraba el test de remarcar las líneas del contorno de una estrella o dibujar la estrella mirando a un espejo. El paciente H.M. tuvo un resultado igual que cualquier persona normal, mejorando con la práctica
Conclusiones
1) Queda establecido que el hipocampo, si bien es importante para almacenar la memoria, no es decisivo para todas las clases de memoria.
2) había una primera diferencia entre memoria de corto y largo plazo (el hipocampo estaría implicado en esta última y no en la de corto plazo).
3) Se identificó las capacidades cognitivas de H.M y sus déficit. Aun cuando no podía recordar ningún evento reciente, si que era capaz de aprender nuevas tareas

279
motoras (como las de la estrella en el espejo). Ello conduce a proponer que, existen dos sistemas de memoria bien diferenciados: episódico y procedural, que involucran diferentes áreas cerebrales.
Los estudios con el paciente H.M. han permitido establecer áreas especializadas según el tipo de aprendizaje: el hipocampo está implicado en la memoria episódica de largo plazo, luego son transferidas a áreas dónde se almacene de forma permanente. Así, H.M. podría aprender tenis, esquiar, baloncesto…. pero no recordaría que ha aprendido a realizar bien esas tareas… para eso necesitaría su hipocampo “desaparecido”.
CLASIFICACIÓN DE LAS MEMORIAS: La clasificación de las memorias es variada, así por ejemplo, tenemos la memoria espacial y su relación con el hipocampo (Sherry y cols; 1992, citado por Passig 1995). Lo que no parece dejar dudas, es la participación del hipocampo en la memoria de largo plazo declarativa. También tiene que ver con el comportamiento espacial y el recuerdo de este tipo de información. Lesiones hipocampales afectan la memoria de espacial, discriminaciones olfatorias, de tiempo, de configuraciones auditivo-visuales. El asunto es que independiente del tipo de memoria o de su contenido, tenemos una memoria de largo plazo que tiene un contenido que puede estar relacionado con lo espacial, o bien puede ser del tipo declarativa. En concreto, de acuerdo al tiempo de duración existen al menos dos tipos de memoria: memoria de corto y largo plazo. La clasificación la propuso Hermann Ebbinhaus en 1885, posteriormente, William James distinguió entre un tipo de memoria que duraba algunas horas y otra que duraba días, semanas, años. La memoria de corto plazo trata de la información reciente es frágil y muy fácil de borrarse. Sin embargo, la memoria de corto plazo se puede consolidar en memoria de largo plazo. El término “consolidación” fue aplicado al fenómeno por Müller en el año 1900. Hipótesis de los cambios neuronales en relación con proceso de memorización: La hipótesis de que la información puede almacenarse por medio de cambios en la comunicación sináptica es antigua. Santiago Ramón y Cajal propuso hace más de un siglo que la información se almacenaba en el cerebro y su expresión era cambios anatómicos entre las conexiones neuronales. Sherrington propuso el término plasticidad sináptica para describir los cambios en las propiedades sinápticas como resultado de su actividad. Hace más de 60 años Donald Hebb postuló que la interacción recurrente (repetida) entre dos neuronas conduce a procesos de crecimiento y cambios metabólicos. En consecuencia, aumenta la eficacia de la sinapsis y por ende se fortalece la conexión sináptica. De tal manera,- postula Hebbs- que es necesario el desarrollo de cambios estructurales para el mantenimiento de la memoria formada como consecuencia del aprendizaje. Entonces, el almacenamiento de la información ocurre por medio de cambios en la eficiencia de la comunicación o transmisión sináptica. Debemos conocer otro concepto, que es de la potenciación a largo plazo (LTP), es decir incremento sostenido en la fuerza de la comunicación sináptica debido a la activación repetida de la vía. La plasticidad sináptica es consecuencia de la LTP. Para inducir LTP se puede aplicar a una vía excitatoria aferente, estímulos cortos y repetidos de alta frecuencia. Esto tiene en consecuencia un incremento duradero y estable en la magnitud de la respuesta sináptica, puede durar días. Los efectos de la LTP se han investigado en el hipocampo y la neocorteza. Por cierto, esas son zonas claras de aprendizaje y registro de memoria.

280
La plasticidad incluye cambios en la estructura, distribución y el número de sinapsis ¿están estos cambios relacionados con la memoria? Los experimentos son buenos en cuanto responden a hipótesis que se puedan refutar. Analicemos el experimento de Rosenzweig aproximadamente en 1964. Los experimentos mostraban cambios morfológicos en la corteza cerebral en virtud de la experiencia. Se utilizó como sustrato a una especie animal de laboratorio, se utilizó un ambiente enriquecido. En este ambiente, una caja para 10 animales, con ambiente de diversos objetos que los animales podían explorar. Los objetos se cambiaban a diario. Los análisis histológicos (se utiliza cortes de tejido y se observa al microscopio) que los animales en el ambiente enriquecido mostraban un incremento del peso y grosor de la corteza cerebral al compararse con animales con depresión de estímulos, con cajas pequeñas y sin objetos que pudieran explorar. El experimento resulta interesante, sobre todo porque parece refutarse con el grupo control deprivado de estímulos. Las conclusiones debieran ser: cerebro más estimulado mayor desarrollo de corteza. El problema que tiene este tipo de experimentos es que tanto los animales control como aquellos en que opera la experimentación debieran poseer la misma línea de base, lo que ocurre es que a los animales deprivados es como si a ellos se les estuviera dirigiendo su desarrollo justo a lo contrario, no desarrollar su cerebro normalmente. Por ejemplo, si usted mantiene los animales en una caja oscura, con escaso movimiento, no es raro que logren escaso desarrollo, de hecho, este fenómeno ha sido sobretodo observado en el desarrollo de la visión en gatos, monos, ratas, etc.
En esta como en otras ocasiones durante las clases se recomienda lecturas a los alumnos, y se les recomienda no porque la actividad sugerida le haga a usted más inteligente, sino porque es necesario para el desarrollo profesional. Lea porque es necesario que lea, así podrá entender y comprender mejor al mundo y su profesión. El libro que se recomienda ahora es de muy fácil lectura, el título es El pasado de la mente de Michael Gazzaniga, tiene mucha fuerza cada uno de sus capítulos.
En el capítulo número dos nos cuenta que hacia 1997, época en que los Clinton hacían noticias por otras cosas, se dedicaban a dar patrocinios de algunas conferencias acerca de los bebés. Con Clinton podría pasar cualquier cosa, hace poco vino a Chile, y la prensa comentaba que su mensaje costó algo así como 100.000 o 200.000 dólares. Dicen que la culpa no la tiene el chancho……Bueno, este mensaje le costará a usted la módica suma de cero pesos por que podrá encontrarla en el sitio Web. El caso es que los Clinton- el Presidente y la primera Dama- algunos psicólogos y neurocientistas, exponen del maravilloso cuadro del desarrollo cerebral, el mensaje era que leer cuentos a los bebés es altamente beneficioso. Muchos seguramente encontraron todo esto muy razonable, a tal punto que el New York Times editorializó proclamando, que la neurociencia aseveraba que durante su desarrollo el cerebro debía ser modelado y un muy buen instrumento era la lectura. Desde ya, supongo que a usted también le parece razonable, pero, ¿Qué hay de aquellos mortales que poco o nada han leído?, hay que pensar en los millones de sujetos que dieron vida al desarrollo del mundo en, por ejemplo, el siglo XVI, muchos millones jamás conocieron las letras. A propósito conozco a algunos prósperos propietarios de parcelas, aquí mismo, en la región del Maule. De ellos hay muchos que jamás leyeron y es porque no tuvieron oportunidad para aprender o ir a la escuela, y sin embargo, son personas exitosas, prósperas y no tiene que ver con la lectura. De más está decir que el autor de este texto se pronuncia a favor de la postura de Gazzaniga, porque está convencido de que la construcción cerebral se encuentra sostenida

281
genéticamente. El cerebro puede realizar conexiones neuronales dentro del amplio rango que permite la genética. Dentro de este mismo rango se encuentra lo que se denomina plasticidad, que en el fondo se trata del reordenamiento de la circuitería cerebral mediante la experiencia que se adquiere en la interacción con el medio. En otras palabras, tanto el ambiente como los genes son necesarios para la expresión de la conducta, pero, no se puede ir más allá de la propia naturaleza. Sin duda, algunos entusiastas creen que pueden ir más allá, no es así. Son buenas las preguntas del Dr. Michael: ¿Así que el cerebro reacciona de manera diferente ante un padre que lee cuentos y ante los Rolling Stones que se escucha en la radio? ¿Dónde están las pruebas? ¿Podríamos tratar con los chanchos en piedra? En uno de los párrafos siguientes Michael me recuerda alguna clase en que escuché del maestro acerca de sus observaciones y estudios, que los hijos de profesionales tenían más seguro acceder a la universidad. Por supuesto, no nombraré al maestro, sería mala educación decirle que estaba equivocado, en general los alumnos no contradicen a los maestros, es una herencia de nuestro pasado que debemos romper. Si Gazzaniga y otros nos alertan de que no es tanto así. No podemos plantear que la corteza es más espesa y de mayor peso, porque el individuo escucha una mayor cantidad de palabras, y en consecuencia aumente la densidad neuronal o mayor desarrollo de conexiones neuronales-eso es ridículo- me huele a un sentido Lamarckiano. Las jirafas no lograron un cuello más largo por tratar de lograr las ramas más altas de las acacias. Tampoco podemos continuar con la piel morena todo el año si acudimos en verano a la playa. Las conexiones neuronales se realizarán en la medida que si la programación genética lo permita. Los hijos de profesionales disponen de mayores recursos tanto materiales como humanos, y también de mayor calidad. Claro es cierto, a veces la estadística da para cualquier cosa. El caso era el siguiente: Staso, Psicólogo y el periodista del New York Times apoyan la sentencia de que…..el volumen de información verbal que recibe el bebé es crucial para su éxito en la vida”. Los hijos de padres profesionales escuchan un promedio de dos mil cien palabras por hora, aquellos de los empleados y obreros, unas mil doscientos, y los de hogares sociales unas 600. Hasta aquí la correlación se ve positiva y claramente. ¿Cómo- dice el New York Times- no van a ser más exitosos los hijos de profesionales? Ciertamente, si bien puede haber correlación, puede no existir una relación causa-efecto. La correlación según Gazzaniga es con los genes, es decir, serán más exitosos aquellos que son hijos de padres exitosos. Da un ejemplo, la elocuencia-habilidad verbal es necesaria para permanecer en las clases altas, por el contrario, la torpeza verbal vaticina descender en la escala. Los hijos y padres seguros comparten los mismos genes relacionados con hablar y escribir. Por último vea y observe a nuestros políticos, vea que ocurre con sus descendencias, uno el presidente, el otro, ministro. Claro que puede ser parte del cuoteo, en todo caso se puede estudiar por la amigología. En todo caso, es muy creíble la posibilidad que el fenómeno tenga las causas aludidas por Gazzaniga. Sin embargo, también es posible tenga que ver con los procesos educativos que ocurren mayoritariamente al interior de un hogar de clase media, en general hay más comunicación, mayores deseos de progresar en el sentido que la sociedad promueve, y que por lo general es la promoción o el logro en el desarrollo de una profesión y que esta sea rentable. Se aplaude cada éxito escolar del retoño. Lo contrario ocurre más que a menudo en las clases bajas.

282
De genes y rendimiento escolar: en realidad es difícil creer que los genes se estratifican según las clases sociales, eso sería locura. Propuesta, si seguimos genes de la inteligencia (cuando se tenga certeza de que son lo que se dice de los genes de la inteligencia) o para el aprendizaje, encontraremos que en estos tiempos estos se distribuyen más al azar en las poblaciones. De manera que encontramos a los que tienen mayores capacidades de aprendizajes distribuidos normalmente en los diferentes estratos. Por eso mismo, es menos creíble la hipótesis de Gazzaniga y que tiene que ver con la elocuencia. ¿Donde se desarrollan mejor los niños de acuerdo a las buenas aspiraciones de la sociedad? Evidentemente donde se le entregan estímulos que permitan desarrollar respuestas en la dirección indicada. Precisamente, esos estímulos no se encuentran en las condiciones paupérrimas de una parte de la población. El sujeto que logra desarrollarse, “salir adelante”, a pesar de la falta de estímulos adecuados, podemos asumir que es genial. En los sectores de pobreza extrema lo que falta son los estímulos adecuados- no es que haya carencia estímulos- es posible que se encuentren sobre-estimulados para sobrevivir en el ambiente que es adverso a los requerimientos sociales deseables.
No es suficiente con disponer de estímulos adecuados en los colegios. Primero
debemos estimular a la gente en sus condiciones de trabajo, seguridad social y vivienda. La lectura no tiene que ver con el desarrollo del cerebro, la lectura nos informa de lo que ocurre en el medio, de las fantasías, de la recreación, de lo entretenido y muy deseable es que todos lean bastante. Pero no se puede decir que si lees aumentas tus conexiones neuronales. Si así fuera los invidentes de nacimiento estarían en serias dificultades, y me parece que no. Si usted le otorga la facilidad para acceder al conocimiento para que se desenvuelva créame que lo logrará, es raro encontrar una persona más perseverante que ellos. Volviendo a lo estructural celular y molecular: Estudios posteriores a los de Rozenzweig mostraron que tras la exposición de ratas por varios días a tareas de acrobacia ocurre sinaptogénesis, es decir, de la formación de sinapsis en la corteza motora y en el cerebelo. Las pruebas eran acerca del aprendizaje de nuevas habilidades. Otros estudios indican que la gran mayoría de las sinapsis excitatorias axodendríticas en los mamíferos ocurren en las espinas dendríticas. Por otro lado, estudios en animales respecto al entrenamiento en ambientes complejos, estimulante de las conductas exploratoria tiene como consecuencia el incremento de la densidad de las espinas dendríticas en la zona CA1 del hipocampo. Estos animales, posteriormente eran más eficientes que animales que no fueron sometidos a entrenamiento en ambientes complejos. Hoy en día se puede estudiar in vivo la dinámica de las espinas dendríticas utilizando microscopía láser de dos fotones. Svoboda, citado por Balberas, 2004, ha encontrado que durante la experiencia sensorial las espinas dendríticas son muy móviles en escala de tiempo de minutos. Se ha sugerido que la estimulación sensorial puede incrementar la cantidad de contactos sinápticos, pero, es necesario establecer la duración de estas sinapsis, el mismo Svoboda no lo tiene claro, se da cuenta de una estabilidad en la corteza sensorial que no todos los neurocientistas comparten en opinión. El 50% de las espinas tienen una duración de un mes, las demás un día o menos. Los valores se sustentan en que la aparición y retracción de espinas están asociadas a la eliminación de sinapsis y desde luego, esto puede estar relacionado con la remodelación de los circuitos neuronales asociados a la experiencia. Otros autores indican que en otras zonas de la corteza hay más estabilidad en la formación de las espinas dendríticas.

283
El hipocampo experimenta cambios sinaptogénicos durante los procesos de aprendizaje y memoria. De hecho, hay incremento de los contactos sinápticos de las fibras musgosas después de la manipulación e inducción d epilepsia, y de potenciación de largo plazo en vivo. Esto sugiere que los cambios morfológicos de las terminales de las fibras musgosas han ocurrido gracias a la estimulación repetida de alta frecuencia de 100 Hz, lo que es suficiente para inducir LTP, sin llegar a estados epilépticos. El hipocampo tiene regiones especializadas, existen neuronas que se activan cuando el animal está localizado en algún lugar de su entorno. Estas neuronas son células piramidales de la región CA1 y CA3. Las células que tienen que ver con la posición se les ha llamado “células de lugar” y al parecer formarían parte de un mapa cognitivo en el hipocampo que representa un mapa de coordinadas en el ambiente. Baldera y colaboradores han encontrado que se puede producir sinaptogénesis en las fibras musgosas del hipocampo en el stratum oriens después de someter a ratas adultas a sesiones repetidas de aprendizaje espacial en la situación experimental conocida como laberinto acuático de Morris. Este experimento de memoria espacial somete a las ratas a una piscina en la que se coloca animales y deben aprender a llegar a una plataforma de escape. El examen histológico muestra que los animales poseen incremento significativo de las terminales de la fibras musgosas en la CA3 en la región stratum orines (SO) después de 10 ensayos por día, durante varios días, esto comparado con ratas que entrenaron uno o dos días. Algunos animales con incremento de las terminales recordaban hasta un mes después de realizada la experiencia. ¿Tiene relación con la memoria de largo plazo?
Aprendizaje perceptivo Es un proceso que permite el reconocimiento inmediato de estímulos anteriormente presentados. En este caso, se da en los hechos la facultad de identificar personas, objetos y situaciones familiares. Esta forma de aprendizaje tiene lugar en los sistemas perceptivos de nuestro cerebro, en específico, algunas regiones corticales. Como ejemplo clásico, son las cortezas de tipo asociativas, de las cuales Wilhem Penfield, estimuló quirúrgicamente el cerebro de pacientes conscientes. Esta situación pudo realizarse con ocasión de intervención quirúrgica para el tratamiento de la epilepsia. La estimulación de regiones concretas de la superficie de la corteza temporal provocó percepciones alucinatorias de tipo visual y auditivo. Al estimular la corteza de asociación auditiva los pacientes según su relato, escuchaban melodías.

284
Figura tomada del libro de Kandel, “En busca de la memoria”
Estudios con neuroimágen funcional en primates muestran que las áreas corticales implicadas en los aspectos discriminativos (corriente ventral o inferotemporal) o espaciales (corriente dorsal o parietal) de la percepción visual almacenan memorias sobre los atributos del objeto percibido, como el color y movimiento, a ellos se accede durante la evocación o recuerdo de éste. Los monos presentan problemas de discriminación cuando tienen lesiones en la corteza
inferotemporal, por ejemplo, para distinguir estímulos visuales diferentes, por ejemplo, objetos comestibles y no comestibles. Los pacientes humanos con daño cerebral en regiones equivalentes tienen problemas para nombrar o reconocer ciertas palabras, por lo general verbos, o categorías de objetos como frutas o animales que se presentan a su vista. En la corteza inferotemporal de monos conscientes se ha encontrado neuronas que responden e forma selectiva a imágenes familiares de una o varias caras.
En estudios de “emparejamiento retardado con la muestra se ha experimentado y registrado la actividad de neuronas individuales para el aprendizaje perceptivo visual. En el experimento cada animal tenía que recordar durante algunos segundos el color de un objeto previamente presentado y luego seleccionar a continuación de entre cuatro objetos (botones) aquel del mismo color que la muestra previa. Si la respuesta era correcta el mono recibía jugo de fruta como refuerzo. Los resultados muestran que neuronas de la corteza inferotemporal respondían al color de la muestra, incluyendo períodos de demora en el ensayo, cuando se había retirado la muestra. Los resultados indicaron que la memoria se había consolidado. El resultado indica la evidencia de que la corteza inferotemporal posee circuitos nerviosos para el recuerdo de estímulos visuales conocidos.

285
Por otra parte, lesiones de la corteza temporal superior, específicamente, corteza de
asociación auditiva, en monos dan origen a la no formación de memorias auditiva en las mismas condiciones experimentales con emparejamiento retardado, en este caso, las memorias visuales no se ven afectadas. En seres humanos, las lesiones afectan la memoria lingüística, dando lugar a déficit selectivo para categorías gramaticales de palabras, en una gran cantidad de casos. Por ejemplo, los pacientes pueden tener dificultad para recordar verbos pero no nombres o palabras funcionales como artículos, preposiciones, etc.
Condicionamiento clásico En el condicionamiento clásico, observado y descrito por Iván Pavlov, tenemos un tipo de aprendizaje que tiene su fundamento en una asociación entre dos estímulos que desencadena una respuesta fisiológica mensurable. La presentación de alimento al animal, desencadena en este la salivación. La presentación del mismo alimento con otro estímulo que sea neutro, por ejemplo, el sonido de una campana produce la asociación. Luego la respuesta surge en cuando se produce el sonido, incluso en ausencia de alimento. El animal ha aprendido a asociar un estímulo incondicional (EI), que es la presentación de alimento, que desencadena una respuesta (la salivación) sin condicionamiento, con un estímulo condicional (EC) que es sonido de la campana, que entonces puede desencadenar una respuesta aprendida, un condicionamiento o simplemente, aprendizaje. La secuencia de presentación del EI y del EC es importante para que surja el condicionamiento. El EC debe preceder siempre unos segundos al EI. Necesariamente, el sonido debe preceder al alimento y la contigüidad en el tiempo es un elemento a considerar como necesario, puesto que si el EC (sonido) precede por mucho tiempo la aparición del EI (alimento) no surgirá el condicionamiento. Por ejemplo, con 30 segundos de intervalo no habrá aprendizaje. Se dice entonces, tiene que haber coincidencia temporal entre estos dos estímulos para que el condicionamiento se establezca. Una vez que se ha establecido el condicionamiento, la presentación repetida de tan sólo el estímulo condicionado (sonido) tiene como efecto la desaparición gradual de la respuesta, este fenómeno se denomina extinción de la respuesta condicionada. Los investigadores Rescoria y Wagner han mostrado que para que el condicionamiento clásico tenga lugar debe existir una verdadera relación de contingencia entre estímulo condicionado y estímulo incondicionado. Esto significa que no basta con presentar estímulos en contigüidad temporal. La explicación es que debe haber una posibilidad de ocurrencia inmediata del estímulo incondicionado, de esta manera, el animal determina una correlació predictiva entre esos estímulos. Si así no ocurriera, probablemente el cerebro se repletaría con información y asociaciones irrelevantes. El comportamiento del animal entonces acontece como el aprender a predecir acontecimientos que verdaderamente pueden surgir en el medio ambiente. El condicionamiento clásico, requiere además de las áreas del cerebro relacionadas con la percepción de los estímulos, de conexiones neuronales que podrían radicar en regiones como la amígdala, el tálamo, el cerebelo o tronco del encéfalo, ello ha sido avanzado en

286
distintas investigaciones de condicionamiento humano y animales. En los mamíferos, el condicionamiento clásico adquiere importancia porque se relaciona con la conducta emocional.
En la llamada “supresión condicionada”, aspecto en que se mide la respuesta emocional condicionada indirectamente, por su capacidad para suprimir otra conducta establecida en forma previa. El procedimiento experimental consiste en presentar un tono, que es un estímulo neutro, inmediatamente antes que una ligera descarga eléctrica en las patas del animal, esto es el estímulo incondicionado y la respuesta son los evidentes signos de ansiedad en la rata. Para la evaluación del condicionamiento clásico y la intensidad de esa respuesta y por tanto, de la asociación establecida, se presenta el tono justo cuando el animal está presionando una palanca para obtener comida. “A mayor fuerza emocional del tono”, más se inhibirá la conducta. Este es un ejemplo, de cómo se puede estudiar el condicionamiento de las emociones en los animales. Ledoux y otros investigadores han señalado que el condicionamiento clásico podría ocupar regiones subcorticales, tales como los núcleos geniculados medial e intralaminar posterior del tálamo, por una parte, y la amígdala basolateral, por otra. En estas regiones confluyen las actividades neurales correspondientes a los estímulos condicionados (tono) e incondicionado (descarga eléctrica), por lo tanto, es en estas zonas donde podría ocurrir la asociación. La hipótesis adquiere mayor fuerza toda vez que se ha observado la lesión de estas estructuras y/o de la amígdala, altera la respuestas emocionales condicionadas de las ratas, y más aún porque mediante la estimulación eléctrica, mediante electrodos intracraneales de los núcleos talámicos antes nombrados, constituyen un estímulo incondicionado, tal como la descarga eléctrica en las patas del animal. Hasta el momento presente, aún pese al alto cúmulo de investigaciones no ha sido posible determinar con algún grado de exactitud el lugar de almacenamiento permanente de la memoria para el condicionamiento clásico. Ciertas hipótesis dan la posibilidad, por ejemplo, en el condicionamiento que ocurre con participación de la amígdala, estos aprendizajes se trasladarían posteriormente a la corteza cerebral. Sin embargo, en trabajos muy recientes que contemplan lesiones de NMDA en la amígdala basolateral, se ha observado que las ratas pierden la mayor parte de la respuesta de miedo(La respuesta es inmovilización) no sólo para los condicionamientos de una 24 horas, sino también para los que se han formado 16 meses antes. Esto, evidentemente, es una fuerte indicación que la formación de la memoria permanente, de largo plazo, o remota como dicen los autores, radica en la amígdala basolateral. Pero la amígdala basolateral no es el único lugar, hay otras regiones involucradas, pues otros condicionamientos, tales como la evitación inhibitoria, el condicionamiento de miedo al contexto, no necesitan de una amígdala intacta y funcionando. La amígdala parece muy implicada en la memoria emocional de los seres humanos. Por ejemplo, en las observaciones de James McGaugh, acerca del paciente B.P. con síndrome de Urbach-Whiete, que presenta una enfermedad caracterizada por lesiones bilaterales cerebrales confinadas en el complejo amigdaloide, con conducta normal, con funciones cognitivas tales como, la atención, la memoria a corto plazo, la inteligencia general, pero notable en no presentar incremento de memoria (Como ocurre en personas normales) para asociaciones de estímulos emocionales, por ejemplo, imágenes de accidentes de tráfico. B.P., olvidaba este tipo de acontecimientos son extraordinaria facilidad.

287
En los seres humanos el estímulo condicionado aversivo, tal como la imagen de una cara
desagradable, puede provocar una respuesta de miedo, incluso cuando la presentación es breve e inmediatamente enmascarada por la imagen de una cara neutral, (un tipo de experimento con imágenes subliminares) de forma que el sujeto en experimentación no alcanza a percibir conscientemente. En estos experimentos se observa la activación de la amígdala derecha, en cambio se activa la amígdala izquierda cuando los estímulos aversivos son percibidos conscientemente.
Aprendizaje aversivo gustativo, “toxifobia”, Aprendizaje Interoceptivo Es un tipo particular de condicionamiento clásico que se da en mamíferos. Es conocido
que cuando una persona ingiere comida en mal estado, se puede generar indigestión con mareos, vómitos, malestar general gastrointestinal, ocurre que los estímulos asociados a esa comida, ya sea, gusto, sabor, generan aversión y rechazo. Experimentalmente puede observarse haciendo que las ratas reciban junto con la comida una solución de cloruro de Litio, con ello se producen las sensaciones de malestar descritas antes y luego la comida es rechazada.
La experiencia es suficiente con un ensayo para que se produzca el condicionamiento. Se
relaciona con el instinto de supervivencia, para prevenir la ingesta de venenos o comidas en mal estado. En el tratamiento del cáncer en seres humanos se da con bastante frecuencia el rechazo a las comidas, los pacientes asocian la dieta o hasta el contexto ambiental, con el malestar visceral que produce la quimioterapia.
A veces se plantea la discusión si el aprendizaje de tipo interoceptivo es un verdadero
condicionamiento clásico. La verdad es que lo que lo hace diferente es la contigüidad en el tiempo. Pues no parece necesario que la ingestión de comida y el malestar consecuente se sigan en el tiempo. La asociación se produce incluso después de minutos.

288
Condicionamiento instrumental- Condicionamiento operante Este tipo de aprendizaje implica la asociación entre una conducta y un refuerzo. El refuerzo es un estímulo agradable, placentero, capaz de aumentar la probabilidad de ocurrencia de la conducta o respuesta al estímulo. Lo que el animal aprende es que en determinada situación, una expresión conductual tiene consecuencias positivas. Los investigadores principales e históricos en estos tipos de condicionamiento son Thorndike y Skinner. La experiencia es que ratas hambrientas presionan una palanca desde el interior de una jaula para obtener alimentos. La acción de presionar la palanca corresponde a lo que se ha denominado respuesta instrumental, y los alimentos constituyen el refuerzo.
refuerzo. La asociación se produce luego de ensayos sucesivos. La retirada del refuerzo origina la disminución progresiva y la extinción de la conducta instrumental. En el último tiempo son cuantiosos los experimentos que se realizan para detectar los mecanismos neurales del condicionamiento instrumental, en tales experiencias los animales son capaces de presionar una palanca que conecta a un dispositivo electrónico que al activarse estimula eléctricamente regiones del cerebro, el objeto del animal es recibir el refuerzo. La conducta se ha denominado “autoestimulación eléctrica intracraneal”, ha sido muy útil para descubrir las estructuras y circuitos neuronales que son necesarios para el refuerzo y el placer. Por ejemplo, los estímulos reforzadores naturales capaces de condicionamiento instrumental, la comida, agua, sexo, etc., activan el sistema mesolímbico dopaminérgico, liberando dopamina en el núcleo accumbens septi región paleoestriada. Al mismo tiempo, las células del núcleo accumbens proyectan mediante vías directas o indirectas (vía talamodorsomedial) a regiones motoras como el pálido, los núcleos mesencefálicos y pontinos, y la corteza frontal. También se ha descubierto que las lesiones de los ganglios basales y de la corteza prefrontal pueden impedir el aprendizaje de respuestas complejas instrumentalmente condicionadas. Mediante la tecnología actual de neuroimágen, se ha establecido además que las mismas regiones mencionadas incrementan su metabolismo cuando se da este tipo de aprendizaje condicionado instrumental. Hasta el momento, las investigaciones indican que los ganglios basales y la corteza prefrontal son las regiones donde podrían confluir las señales de actividad neuronal correspondientes a los sistemas perceptivos y las respuestas motoras que corresponden al aprendizaje denominado como condicionamiento instrumental. Esas regiones podrían ser potenciadas por influencias desde el núcleo accumbens y desde otras regiones. En apoyo de esta hipótesis se ha observado que en experimentación de evitación activa, la autoestimulación eléctrica intracraneal postentrenamiento acelera la consolidación de la memoria, lográndose que aquellos animales que aprenden menos alcancen los niveles de memoria que logran los animales que aprenden más. Tal vez, los mejores rendimientos de
Existe otro tipo de condicionamiento que consiste en la evitación activa, en esta, los animales aprenden a moverse y cambiar de lugar para evitar una descarga eléctrica en las patas. (Ver en figura más adelante). Al igual que en el condicionamiento clásico instrumental se requiere contingencia entre la conducta y el

289
los estudiantes en el futuro, se logren en laboratorios especializados paralelos al salón de clases.
Tal vez las dos formas de aprendizaje, como el condicionamiento instrumental y el clásico son lo mismo, o por lo menos no son formas de aprendizaje radicalmente diferentes. Sin embargo hay una diferencia notoria, en el condicionamiento clásico el estímulo condicionado provoca una respuesta del organismo, una respuesta se podría decir, automática, aspecto que es diferente en el condicionamiento instrumental, en este, el animal emite una respuesta para obtener un refuerzo y ello puede considerarse una conducta voluntaria. Pese a la diferencia, muchos investigadores creen que el proceso básico de ambos tipos de aprendizaje debe ser el mismo para ambos. Los dos tipos son semejantes porque implican contingencias ambientales y su aprendizaje asociativo, es decir, la asociación debe ser contingente, segundo, el tiempo entre los estímulos y las respuestas asociadas debe ser breve, condición necesaria para el condicionamiento. Ambos procesos muestran gradualidad en las fases de adquisición y extinción de la respuesta aprendida.
Aprendizaje motor Las actividades motoras, andar, comer, conducir un vehículo, etc, se adquieren y se perfeccionan con la práctica, al inicio los movimientos suelen ser voluntarios y funcionan con mucha imprecisión. Luego estas conductas se automatizan, se tornan inconscientes y son muy precisas. Este tipo de aprendizaje es de tipo asociativo entre estímulo y respuesta. Hay aprendizajes más complejos, como la conducción de un automóvil, que se puede adquirir por condicionamiento clásico o instrumental o por combinación de ambos y puede participar el proceso de memoria explícita. Se acepta que las acciones voluntarias complejas están representadas en la corteza cerebral y las acciones más automáticas y simples se encontrarían representadas en estructuras subcorticales tales como, el cerebelo, el tronco del encéfalo y la médula espinal.

290
La hipótesis que soluciona el caso es que muchas memorias podrían elaborarse inicialmente en la corteza y luego emigrar a regiones subcorticales. La corteza parece contener programas y representaciones motoras novedosas, amplias y flexibles, capaces de guiar con versatilidad los movimientos necesarios para alcanzar un objetivo. Las estructuras subcorticales parecen preocuparse y contener programas motores más ensayados, hábitos motores bien aprendidos y ejecutables en forma más rígida. Estos aspectos han sido confirmados por las observaciones realizadas en monos y seres humanos que presentaban lesiones en áreas motoras corticales. Los sujetos lesionados pierden capacidad para realizar movimientos automáticos bien establecidos. Por otra parte, los análisis por tomografía de emisión de positrones (que permiten la obtención imágenes funcionales) indican que durante la adquisición de una habilidad motora, al avanzar con la práctica aumenta la actividad subcortical en regiones como el cerebelo y núcleos estriados, simultáneo al avance de la práctica la actividad cortical disminuye. La práctica que debe realizar el fonoaudiólogo lleva a profundizar más en estos aspectos del aprendizaje. EL desarrollo de un programa motor complejo puede incluir una variedad de movimientos que deben ser llevados a cabo en una secuencia temporal determinada, de manera que es importante la sintaxis de la acción, o lo que es lo mismo, la secuencia misma y la temporalidad precisa de esos movimientos. Al respecto, se ha propuesto (hipótesis) como lugar para esta memoria la corteza prefrontal, las lesiones en esta área alteran la planificación de la acción y la iniciación de nuevas secuencias de habla o conducta motora. Sobre todo en humanos, la conducta motora al parecer, implica una interacción entre representaciones de conducta automática y voluntaria, o, entre patrones innatos o adquiridos. En otras palabras, los hábitos y programas motores instintivos pueden ser modificados por estrategias y planificaciones de los individuos, lo que por cierto no ocurre en otras especies. Esto estaría indicando que las memorias o representaciones neurales de las conductas motoras pueden ubicarse en diferentes niveles y por lo tanto, tendrían la capacidad de distribuirse con amplitud en el sistema nervioso.
Los resultados de investigaciones de condicionamiento de respuestas motoras han mostrado que el cerebelo podría tener una importante función en el condicionamiento de las respuestas.
Aprendizaje relacional y memoria explícita Este tipo es una forma de adaptación al medio, operativamente se alcanza mediante evaluaciones, comparaciones, inferencias conscientes entre informaciones diversas. Requiere interacciones complejas entre la neocorteza y el lóbulo temporal medial que incluye el sistema hipocampal (hipocampo, circunvolución dentada, subículo y corteza entorrinal), y las cortezas parahipocampal y perirrinal adyacentes. El aprendizaje relacional tiene como característica la expresión de memoria explícita o declarativa, deliberada y consciente, aspectos que han quedado de manifiesto a partir del análisis de la amnesia que presentan pacientes con daños cerebrales. En los animales, el aprendizaje relacional origina una memoria similar que no puede ser declarada. Los conceptos y expresiones tales como memoria explícita, memoria declarativa y memoria relacional son equivalentes Amnesia y memoria declarativa

291
Este caso conceptualmente se analizó con el paciente H.M., el paciente olvidaba los acontecimientos diarios tan rápidamente como ocurrían. La invalidez desarrollada se debía a la pérdida de capacidad de aprender y retener nuevas informaciones, el déficit se denomina “amnesia anterógrada”. H.M. poseía ausencia de la memoria a largo plazo, pero no estaba afectada su memoria de corto plazo, por ello, era capaz de seguir una conversación a partir de su propio pensamiento. La cirugía le produjo incapacidad de recordar numerosos eventos vividos antes de la cirugía, o sea, no podía recordar memorias bien establecidas cuando el cerebro estaba intacto.
Figura: Estructuras del sistema hipocampal relacionadas con la memoria explícita
como la torre de Hanoi, con eficacia creciente, pero son incapaces de recordar y reconocer los materiales utilizados para el aprendizaje, no recuerdan explícitamente las experiencias anteriores con el rompecabezas. Cada ensayo es, por lo tanto, una novedad, pero, sin duda, el sujeto aprende y recuerda implícitamente lo aprendido. Las observaciones indican que los amnésicos conservan aquellas capacidades motoras y cognitivas que no requieren memoria consciente y que se expresan como conducta automática o rutinaria. La deficiencia en estos pacientes es la capacidad de adquirir información o memorias conscientes o explícitas, o sea. recuerdos deliberados que pueden declararse verbalmente o por escrito.
La incapacidad de formar nuevas memorias, es causada por la lesión bilateral de los lóbulos temporales del cerebro, por ejemplo, en los enfermos de Alzheimer la región más atrofiada es la corteza entorrinal, que es la estructura que conecta el hipocampo con la corteza cerebral. Los pacientes amnésicos con daño o lesión en los lóbulos temporales, conservan intacta su capacidad para la mayor parte de los aprendizajes que implican la formación de memorias implícitas. Poseen capacidad para el aprendizaje perceptivo, para el condicionamiento clásico e instrumental y aprendizaje motor. Pueden aprender y recordar con facilidad, utilizando memoria a largo plazo tareas del tipo perceptivo motoras complejas como por ejemplo, dibujar el perfil de objetos visualizados en un espejo, son capaces de reconocer objetos con solo indicios o partes de estos, esta conducta se denomina “priming”, pueden realizar rompecabezas y ejercicios algorítmicos. Los pacientes son capaces de realizar un rompecabezas manual

292
Figura: Rompecabezas “torre de Hanoi. La tarea es pasar todos los discos de un eje a otro, de uno en uno, sin colocar un disco mayor sobre uno menor.
La memoria declarativa en los seres humanos puede estudiarse mediante pruebas de asociación verbal. Los sujetos estudian pares de palabras asociadas y después se comprueba su memoria. Para ello, se le presenta la primera palabra de cada par y se espera la respuesta del sujeto para verificar el recuerdo de la otra.
Aprendizaje espacial Para el estudio de este tipo de aprendizaje se ha utilizado el denominado test de Morris. El diseño contempla una piscina circular de un metro de diámetro, se llena con agua y se tiñe con leche u otra sustancia inocua. Debajo de la superficie se coloca una plataforma, esta permanece invisible. En un experimento el animal se ubica en el borde y debe aprender a nadar hacia la plataforma, le sirven de guía algunas señales visuales localizadas en el entorno exterior de la piscina. De esa forma escapa del agua llegando a la plataforma y evita tener que nadar en forma permanente.
Los animales sin deficiencia, normales nadan de forma decidida hacia la plataforma, independiente del lugar de la piscina en que han sido ubicados. Las ratas con lesiones del hipocampo muestran desorientación y alcanzan la plataforma después de varios intentos azarosos. En estos experimentos se ha observado que si las ratas se sitúan siempre en la misma posición de partida, aprenden a encontrar la plataforma en ensayos posteriores, esto ocurre aún cuando su hipocampo se encuentre lesionado. La conclusión es que la estructura hipocámpica no es necesaria para estos aprendizaje de tipo espacial que están ocurriendo de forma rutinaria con la repetición de los ensayos. En ratones transgénicos a los cuales se les ha inducido la sobreexpresión de una subunidad del receptor NMDA y por lo tanto; con producción de potenciales excitadores más duraderos que los normales, en el hipocampo y en diversas regiones del encéfalo, se les facilitó el aprendizaje en el test de Morris. En el hipocampo de estos ratones se observó una potenciación de largo plazo (PLP) mayor que lo normal. Entonces es evidente la plasticidad del hipocampo para el aprendizaje relacional y la memoria explícita. Dos investigadores, Eichenbaum y Cohen, el primero de ellos autor de “The Cognitive Neuroscience of Memory”, basados en resultados de cientos de experimentos en animales y humanos, han propuesto la hipótesis de que el sistema hipocampal constituye la base neural de un sistema de memoria que se caracteriza por su capacidad de comparar, y contrastar información, realizar inferencias y generalizaciones a partir de datos procedentes de múltiples fuentes de procesamiento cerebral. Este sería un tipo de memoria flexible, que permite la asociación de diversas informaciones desde distintas áreas. El sistema propuesto

293
permitiría el recuerdo en contextos variados, diferentes a las situaciones originales. Ello permite el encaje con la memoria explícita o declarativa. La hipótesis ha sido fortalecida con los resultados obtenidos por medio de neuroimagenes funcionales en humanos y con registros electrofisiológicos en neuronas individuales de animales. Larry Squire ha observado mediante tomografías por emisión de positrones, incrementos específicos en la actividad de hipocampo de un sujeto mientras trataba de recordar palabras de una lista a partir de secuencias de letras que se le presentaban para estimular su recuerdo (memoria explícita). El incremento no ocurría si al sujeto en tareas de “priming”, o sea, si el sujeto recibía la instrucción previa de, más que tratar de recordar, mencionara la primera palabra que se le viniera a la mente, cuando veía la misma secuencia de letras, esto indicaba por lo tanto, que no había incremento de la actividad para memoria implícita. Por otra parte, registros electrofisiológicos en ratas dan cuenta que el hipocampo posee células cuya actividad parece representar relaciones entre estímulos relevantes percibidos durante el desarrollo de una tarea. Estas células fueron descubiertas por John O`Keefe, al observar que ratas explorando campos abiertos, muchas neuronas del hipocampo se activaban mientras el animal estaba en un lugar particular. A estas neuronas las denominó “células de lugar”. Según la teoría de O`Keefe, el hipocampo de la rata contiene un mapa cognitivo del entorno espacial, lo constituiría las células de lugar y su función sería señalar al animal para conducirse en ese entorno particular. Pero, las lesiones del hipocampo impiden
Figura arriba: Dispositivo de registro de células individuales del hipocampo de Ratas en movimiento.
Que otros tipos de aprendizajes, distintos de los espaciales, como aquellos de la discriminación de olores. También las investigaciones indican que el lugar donde se encuentra el animal no es el único factor capaz de activar las llamadas células de lugar. Estas también responden con la presentación de un olor particular en un lugar en un momento concreto; pero no responden ante el olor en cualquier lugar o momento. Estas células al parecer, codifican relaciones espaciales o temporales que son significativos para la supervivencia del animal. El hipocampo es una estructura que ha mostrado evidencia suficiente de que media el aprendizaje relacional. Su rol es crítico para la adquisición inicial de la información, pero, no es necesario para el recuerdo de las viejas memoria, que podrían radicar en áreas de asociación corticales, tal como indica el análisis de déficit en pacientes amnésicos.

294
El hipocampo al parecer se relaciona más con el procesamiento de nueva información que con aquella que podría parecer más familiar. Sin embargo, investigaciones más recientes por tomografía por emisión de positrones indican que el lóbulo temporal medial se activa durante el recuerdo explícito.
Genética de la memoria y el aprendizaje Desde que se instaló la genética a principios del siglo XX, se ha investigado acerca del rol de la herencia en el comportamiento humano. Los estudios con gemelos idénticos intentaban establecer la importancia de lo innato y de lo adquirido. Desde entonces, para los genéticos el concepto de que el comportamiento es moldeado por la interrelación compleja que se produce entre la información genética que porta el individuo y el medio ambiente que lo rodea. Establecido está en que el comportamiento en sí no es heredado, lo que se hereda es la información genética que se encuentra en los cromosomas localizados en el núcleo de las células sexuales. Es el ADN cromosomal lo que se transmite a la progenie. Las instrucciones vienen codificadas en el ADN, los genes dirigen el desarrollo de los organismos desde la formación del cigoto, la formación de tejidos, órganos y de sistemas. No hace mucho, en 1993 salió a luz pública una investigación que daba cuenta de una familia europea que tenía la mitad de los varones con una serie de rasgos particulares de cognición o conducta, por ejemplo, eran impulsivos, agresivos, y presentaban niveles bajos de inteligencia. El análisis molecular de los genes indicaba que había una mutación en el gen que codifica la enzima monooxidasa de aminas tipo A. La enzima es esencial para el metabolismo de varios tipos de neurotransmisores fundamentales para la función normal del cerebro y las actividades mentales. Posteriormente se encontró que mutaciones en el promotor (región del ADN que regula la expresión genética) del gen codificante de una proteína que posee una función de transportador sináptico de la serotonina, origina altos niveles de ansiedad y comportamiento neurótico en los seres humanos. Luego se descubrió que muchos trastornos del comportamiento son causados por diversas mutaciones. Los cromosomas como ya ha visto antes, se encuentran constituidos por ADN y proteínas asociadas, por ejemplo, las histonas. La información codificada en el ADN en forma de genes no se expresa directamente, sino que requiere un procesamiento que origina la producción de otro tipo de ácido nucleico conocido como ARN. La síntesis o producción de ARN a partir de ADN se denomina transcripción. Esta molécula sirve como mensajero en el interior de las células y para poder transmitir y llegar a la expresión del gen, deberá ser sometido a otro tipo de procesamiento que es conocido como traducción o síntesis de proteínas. De manera que ya tenemos un hecho fácil de comprender y asimilar, las neuronas que son células con organización similar a cualquier otra célula, presentan la maquinaria que permite la expresión de genes, los productos de esta expresión son las que determinan el comportamiento. En otras palabras, para que haya un comportamiento se necesita que algún tipo de proteína se haya sintetizado, estas moléculas permiten el desarrollo, el establecimiento, mantención y regulación de los circuitos neurales. No es que esto este determinado, o que la conducta esté determinada, se trata simplemente de que los seres humanos (Como cualquier otro ser vivo) al nacer somos un proyecto que puede ser consolidado en dependencias de las interrelaciones con el medio ambiente. No puede haber comportamiento sin la causa material que lo permite.

295
En el mismo sentido, los circuitos neurales subyacen todos los aspectos del comportamiento y se componen de varias neuronas, cada una de las cuales es capaz de expresar un arreglo particular de genes, así cada una de ellas es la contribución al circuito el que surge por la producción y función de proteínas específicas. Muchas características de comportamientos complejos, que se reconocen como normales o que se agrupan como de trastornos mentales son el resultado de la interacción e interrelación de varios genes. Estas interrelaciones entre genes, proteínas y circuitos neurales han sido informadas por los investigadores en forma muy cuantiosa; se ha observado en toda clase de organismo desde los invertebrados hasta los organismos vertebrados los que presentan mayor complejidad.
En la figura de arriba, se muestra en general, la regulación génica: El diagrama informa sobre el dogma central de la biología molecular. Los cromosomas, compuestos de proteínas y ADN, almacenan la información genética. Cada gen, determinado por una secuencia específica de nucleótidos, contiene una región reguladora de su expresión (Promotor) y porciones que irán a señalar la secuencia de aminoácidos de la proteína en particular, se puede distinguir en esta secuencia de nucleótidos de ADN, la presencia de exones e intrones. Durante el proceso de transcripción se produce el ARNm, el que representa una copia de información codificada por un gen en particular. El proceso en si es promovido por proteínas nucleares que funcionan como factores o activadores de transcripción. El ARNm primario debe transitar por varias etapas en un nuevo procesamiento que incluye empalme y corte, en esto se remueven las secuencias intrónicas, ocurre otras fases como la poliadenilación. El ARNm maduro llega al citosol, en donde la información genética que porta se traduce para la síntesis de proteínas indicada, esta luego a través de nuevos procesos quedará activa y funcionando, es el momento que se le puede considerar como etapa final de la expresión del gen.

296
Relación conceptual entre aprendizaje, memoria y circuitos neurales
Puede definirse el aprendizaje como un cambio en el comportamiento basado en experiencias.
La memoria, puede definirse como la facultad mental de retener y recordar experiencias del pasado; es el acto de recordar o evocar nuestras experiencias.
La memorización involucra que acontezcan varias fases complejas, comenzando con la adquisición de información por medio de mecanismos sensoriales y continuando con la consolidación o almacenamiento de información, el cual se da de una forma distribuida en el cerebro. Es útil recordar la diferencia conceptual entre los términos: La adquisición es la capacidad de codificar o aprender nuevos eventos, mientras que la consolidación es el almacenamiento de esta nueva información.
Tras la consolidación inicial de memorias que implica consolidación bioquímica y celular, llas memorias pueden reactivarse mediante su recuperación a niveles conscientes (el acto de recordar eventos del pasado) y tal reactivación puede promover un nuevo proceso de mantenimiento de memoria, mejor conocido como reconsolidación.
Las nuevas memorias son frágiles inicialmente, pero pueden reforzarse a través del tiempo.
Las estructuras subcorticales, por ejemplo el hipocampo y la amígdala, están involucradas en la adquisición y almacenamiento temporal de nueva información. El aprendizaje surge con la activación de circuitos neurales existentes en dichas estructuras, los cuales se van fortaleciendo con el tiempo. Son estos circuitos neurales distribuidos los que se consideran como bancos de información de experiencia.
Como ya se dijo antes, los trazos de memoria almacenados en áreas subcorticales se trasladan a áreas específicas de la corteza, luego mediante la expansión y distribución de circuitos, pasan a constituir vías de comunicación neural con gran especificidad.
A este proceso de traslado de información a la corteza a partir de estructuras subcorticales que consolidan la información se le puede llamar consolidación neural. La corteza se considera, por tanto, una estructura de almacenamiento permanente de memorias, en forma de circuitos neurales. De manera que el establecimiento y fortalecimiento de vías, redes o circuitos neuronales retransmite memorias mediante mecanismos celulares y moleculares que regulan la conectividad neuronal y la plasticidad sináptica en el cerebro. En general, la formación de memoria depende de mecanismos celulares y moleculares que de forma eficiente sirven para precisar las conexiones sinápticas entre neuronas.
Se ha planteado que las memorias se fijan y almacenan a través de redes neuronales específicas. El término de red o circuito neural se refiere a neuronas interconectadas que integran y envían mensajes entre ellas. La red es la conexión entre neuronas que forman circuitos mediante sinapsis específicas.
Las sinapsis se componen de terminales presinápticos que interactúan directa o indirectamente con dendritas postsinápticas. La estimulación de estas neuronas postsinápticas genera una señal de salida que puede inducir la activación de otras neuronas, causando un efecto dominó extraordinario. La activación de redes específicas de neuronas es el mayor ejemplo del dinamismo del cerebro que ocurre durante los procesos cognitivos. Eventualmente,

297
las redes neurales llegan a un equilibrio a través del tiempo, reforzando la conectividad neuronal.
En la figura arriba se representa un esquema de los circuitos del hipocampo, con las tres vías principales de tipo excitadoras del hipocampo: la vía perforante (VP), la vía de fibras musgosas (FM) y la vía de colaterales de Schaffer (CS). La circunvolución dentada (CD) y las neuronas piramidales de la región CA3 reciben señales de las capas II/IV y II/V de la corteza entorrina (CE) a través de las vías perforantes (VP). Células granulares de la CD conectan con las células piramidales de la región por CA3 a través del circuito de las FM. Las células piramidales de la región CA3 hacen sinapsis con las células piramidales de CA1 desde el hemisferio ipsilateral o contralateral a través de la vía de las CS o mediante la vía de fibras comisurales FC asociativas, respectivamente. Las células piramidales de la región CA1 inervan el subículo Sb.
En este capítulo ya se ha mencionado y descrito en parte el modelo de formación de memorias y aprendizaje, basado en el hipocampo y lóbulos temporales en general. Ahora se continuara con mayores detalles pero involucrando la acción de los genes. Anatómicamente, el hipocampo posee tres circuitos fundamentales para los estudios que nos interesan, estos van a través de la capa de células granulares de la circunvolución dentada (CD) y las capas piramidales CA3 y CA1. Las vías aferentes son: la vía perforante (VP), la vía de fibras musgosas (FM) y la vía de colaterales de Schaffer (CS). La vía perforante son axones provenientes de la corteza entorrinal (CE) y establecen sinapsis con dendritas de las neuronas granulares de la circunvolución dentada. Desde las células granulares se proyectan axones (Vía de fibras musgosas) hacia las células piramidales de la región CA3. Las fibras identificadas como colaterales de Schaffer (CS) son axones que se proyectan desde las neuronas piramidales de la región CA3 y establecen sinapsis con neuronas piramidales en la región CA1.
Es destacable que los circuitos del hipocampo es que siguen las reglas hebbianas, por cuanto se establece que cuando una neurona estimula a otra en forma repetida, las conexiones se refuerza y alcanzan mayor eficiencia. Esto se debe a los cambios químicos ocasionados por cambios de expresión genética. Las vías exhiben lo que se denomina potenciación a largo plazo y son parte del modelo explicativo para comprender los procesos de aprendizaje y memoria. Los procesos genéticos moleculares en relación con estos procesos de plasticidad sináptica, aprendizaje y memoria, son acoplamientos de señales mediadas por neurotransmisores, procesos de transcripción y traducción como respuesta a las señales. Parece claro que la memoria de corta duración requiere de modificaciones de proteínas sinápticas preexistentes para estabilizar las señales recibidas y modificar el estado de excitabilidad de las neuronas en el circuito que opera. El proceso no requiere de síntesis de proteínas y puede durar minutos, horas. La memoria de largo plazo, o durable, necesita de la

298
síntesis de ARNm, transcripción y traducción y por lo tanto, de proteínas. La duración puede ser de horas, meses, años. Recientes investigaciones indican que la memoria de largo plazo requiere de procesos de rearreglo genómico o recombinación de ADN. La información genética como sabemos está
contenida en un código de tipo lineal, en ocasiones estas secuencias de nucleótidos pueden experimentar rearreglos de distintos tipos. Por ejemplo, es factible que una región de la cadena se salga, a lo cual se le llama deleción; o un fragmento de la misma puede duplicarse y amplificarse; o bien, invertirse o ir a otro lugar del genoma. Estos fenómenos frecuentes son muy conocidos, por ejemplo, cuando la doble hélice del ADN se replica, en algún momento para dar lugar a células hijas con la misma información genética, a veces ocurre que la maquinaria de replicación (conjuntos de enzimas) comete errores, en consecuencia actúan otros grupos de enzimas que reparan los errores, pero no siempre hay reparación. Las enzimas que de forma natural reparan estos fragmentos encuentran porciones genéticas iguales unas a otras, y las juntan, pero lo hacen en el sitio incorrecto. Se trata de la llamada recombinación homóloga de ADN no alélica. La explicación es que los genomas no son estructuras estáticas; por un lado, mutan, es decir, se presentan transformaciones en las bases que los conforman y, por el otro pueden presentar rearreglos, estos mismos hechos permiten la evolución de los organismos.

299
En la figura arriba se puede observar un esquema general de los mecanismos moleculares relacionados con procesos de aprendizajes y memoria. Luego de una experiencia se produce una activación sináptica origina una serie de eventos intracelulares, que incluyen la activación de cascadas de segundos mensajeros, como por ejemplo, Ca +
2 y AMPc. La activación de las cascadas ocurre gracias a la acción de neurotransmisores como
glutamato que actúa sobre receptores metabotrópicos (GLuRm) o ionotrópicos (NMDA- que es un receptor de N-metil-D-aspartato o GluRi). Los receptores ionotrópicos funcionan como poros que se abren en la membrana cuando su neurotransmisor se liga a ellos. Los receptores metabotrópicos se acoplan a proteínas G y activan enzimas como la adenilciclasa (AC). Estos cambios causan la activación de una variedad de proteincinasas (enzimas) como la PKA, algunas de las cuales llegan al núcleo y activan la función de factores de transcripción como los denominados CREB. La transcripción y, finalmente, la síntesis de proteínas proveen de proteínas que adquieren relevancia en la producción de cambios en la eficiencia sináptica, que pueden o no asociarse a cambios estructurales en las conexiones neuronas-neurona. Se cree que algunos genes importantes para procesos de memoria se regulan a nivel de traducción local de ARNm latentes, y otros, por medio de procesos epigenéticos tales como la alteración de la conformación de la cromatina o la metilación de ADN. Otro grupo de genes podría estar regulado por mecanismos de diversificación genómica similares a los que se utilizan en el sistema inmune para producir proteínas diversas.
Regulación transcripcional de la expresión génica Con la activación de las neuronas se originan cambios celulares que originan la fosforilación de proteínas proteincinasas, incluyendo la PKA nombrada anteriormente, esta es dependiente de adenosina monofosfato cíclico (AMPc). Este tipo de proteincinasas regulan la función de factores de transcripción durante los procesos de aprendizaje y memoria. Los factores de transcripción son proteínas que regulan la síntesis de ARNm. Sólo para ilustrar este aspecto en el establecimiento de prueba, se ha demostrado que la síntesis de ARNm para la formación a largo plazo se puede detener mediante sustancias inhibidoras como la actinomicina D. Se ha descubierto varios factores de transcripción para los procesos de aprendizaje y memoria, el factor más estudiado por los biólogos moleculares es el CREB que se denomina proteína ligando del elemento responsivo a AMPc.

300
Los estudios realizados con CREB en la memoria son de gran cuantía y muy probatorios de la fenomenología descrita en este capítulo. Se ha mostrado diversos estudios y continúan realizándose en insectos, vertebrados e invertebrados diversos, pero especialmente en mamíferos y entre ellos, las ratas. Una fuerte evidencia del rol crítico de la transcripción mediada por CREB en la consolidación de la memoria a largo plazo se obtuvo de experimentos de silenciamiento traduccional, realizados por el grupo de investigadores dirigidos por Guzowski. Ellos inyectaron en ratas una microinyección en la región dorsal del hipocampo con ADN complementario o antisentido al ARNm codificante de CREB. EL resultado fue la inactivación específica del gen CREB. Los animales mostraron impedimento en la formación de memorias de largo plazo, ello no ocurrió en la expresión de memorias de corto plazo relacionadas con aprendizaje en el laberinto de agua de Morris, el cual depende de la función del hipocampo dorsal para la consolidación. De la misma forma, el grupo de investigación dirigidos por Yadin Dudai en Israel, mostraron que con microinyecciones de ADN complementario al ARN de CREB en la amígdala antes del desarrollo de aversión gustativa condicionada provocó impedimento de la memoria de largo plazo, no se observó efectos en la expresión de memoria a corto plazo. Otros informes de investigaciones realizadas muy recientemente acerca de microinfusiones bilaterales de ADN complementario al ARNm de CREB en la región CA3 del hipocampo provoca, al igual que en los casos anteriores, el silenciamiento o inactivación de genes, el impedimento de formación de memorias de larga duración, también realizados en laberintos de Morris. En este tipo de aprendizaje se requiere la función intacta del hipocampo para la consolidación. Cientos de estudios en nuestros días, realizados con ratones mutantes de la función CREBS, con ratones transgénicos, con mutaciones dirigidas, indican coincidencias con los resultados mostrados en los párrafos anteriores. De mayor importancia en este curso, es reconocer la importancia de la proteína CREB y esta misma de reclutar a su proteína ligando CBP, durante la transcripción. Ratones con mutaciones en CBP de los dominios para la interacción con los factores de transcripción como el CREB muestran impedimentos en la formación de memorias de largo plazo en tareas dependientes del hipocampo. Las mutaciones del gen que codifica CBP han mostrado correlación con el síndrome de Rubinstein- Taby en los seres humanos, esto es un trastorno de transmisión genética dominante que se caracteriza por retraso mental. De todas maneras falta por confirmar esta hipótesis.
Bibliografía Recomendada (Además de la entregada en clases y en Syllabus):
Referencias:
• Bayley PJ, Hopkins RO, Squire LR. The fate of old memories after Medial Temporal Lobe damage. J. Neurosci 26:13311-13317. 2006.
• Raskic Pasko No more cortical Neurons for you. 18 August 2006 Vol 313 Science
• Sejnowski Terrence y col. Running enhances neurogenesis,learning, and long-term potentiation in mice. PNAS November 9, 1999, Vol 96, N° 23, pp 13427-13431

301