modelado, análisis y control de un sistema biológico...
TRANSCRIPT

Modelado, análisis y control de un sistema biológico biestable
Enfoque Estocástico 81
Enfoque estocástico
A diferencia del enfoque determinista, donde se asocia una ecuación diferencial ordinaria a cada especie para describir su evolución, en el enfoque estocástico se asocia una probabilidad a cada interacción entre las moléculas, y mediante un algoritmo se simula el conjunto de proteínas, y sus interacciones de forma aleatoria, pero siguiendo una distribución conocida
.
En esta sección se tratará el modelado de los sistemas Biestable Simple y Biestable, mejorado a partir del enfoque mostrado por Gillespie, entre otros autores, y la resolución del sistema a partir del algoritmo que lleva su nombre.

82 Enfoque Estocástico

Modelado, análisis y control de un sistema biológico biestable
Enfoque Estocástico 83
1. La visión de Gillespie
El método determinista asocia una ecuación diferencial para la evolución de cada población molecular. Con el conjunto de ecuaciones se plantea el sistema a resolver. Éste método en ecuaciones diferenciales se basa en una serie de hipótesis falsas para nuestro supuesto:
• Las poblaciones son variables continuas: FALSO. Las poblaciones de la especies son entes enteros que varían su magnitud de forma discreta.
• Conocer las poblaciones en un instante de tiempo determinado es suficiente para predecir su evolución: FALSO. El choque entre moléculas es un proceso estocástico, ya que no podemos hacer un seguimiento de las posiciones, velocidad, aceleraciones, fuerzas intervinientes e incertidumbre cuántica; lo cual significa que no podemos predecir el estado final a través de unas condiciones iniciales generales.
• Los sistemas químicos no están mecánicamente aislados (la temperatura no es constante) y la bacteria en función del estado de madurez crea un ambiente diferente.
Estos son algunas por las que la solución aportada por este método es muy sesgada; debido a que la aproximación al comportamiento real es muy laxa.
Daniel T Gillespie comenzó a describir su método sobre sistemas químicos [10], lo cual terminó extrapolando a sistemas biológicos debido a las semejanzas que hay entre sistemas de este tipo.
La cinética química estocástica trata de describir la evolución temporal de un sistema químico homogéneo de una forma que tenga en cuenta la discreción y la aleatoriedad. Los fundamentos teóricos se exponen a continuación.
Fundamentos de la Cinética Química Estocástica
Sea un sistema homogéneo de 𝑁 especies químicas (𝑆1, … , 𝑆𝑁) que interaccionan a través de 𝑀 canales de reacción (𝑅1, … ,𝑅𝑀). El sistema se supone confinado en un volumen constante Ω y en equilibrio térmico a una temperatura constante. Sea 𝑋𝑖(𝑡) el número de moléculas de la especie 𝑆𝑖 en el sistema en el tiempo 𝑡, queremos estudiar la evolución del vector de estado 𝑋(𝑡) = (𝑋1(𝑡), . . . ,𝑋𝑁(𝑡)) dado, asumiendo que parte de un estado inicial 𝑋(𝑡0) = 𝑥0. Cada canal reactivo 𝑅𝑗 se supone elemental, i.e. describe un evento físico que ocurre instantáneamente. El canal reactivo 𝑅𝑗 se caracteriza por dos cantidades:

84 Enfoque Estocástico
• El vector de cambio de estados 𝑣𝑖𝑗 = (𝑣𝑖1, … , 𝑣𝑖𝑁), donde cada componente 𝜈𝑖𝑗 se define como el cambio de la población molecular de la especie 𝑆𝑖 causada por una reacción 𝑅𝑗. Si el sistema se encuentra en el estado 𝑥, y ocurre una 𝑅𝑗, el sistema cambia su estado a 𝑥 + 𝜈𝑖 . El vector 𝜈𝑖𝑗 se conoce como la matriz estequiométrica del sistema reactivo.
• La función de propensidad 𝑎𝑗 se define de forma que 𝑎𝑗(𝑥)𝑑𝑡 de la probabilidad de que, dado 𝑋(𝑡) = 𝑥, ocurra 𝑅𝑗 dentro de Ω en el siguiente instante infinitesimal [𝑡, 𝑡 + 𝑑𝑡).
Si 𝑅𝑗 es una reacción mono-molecula Si → producto/s r, la física que hay detrás de la mecánica cuántica implica la existencia de una constante 𝑐𝑗 tal que la función de propensidad asociada queda aj(x) = cj · xj
Si 𝑅𝑗 es una reacción bi-molecular 𝑆𝑖 + 𝑆𝑗 → 𝑝𝑟𝑜𝑑𝑢𝑐𝑡𝑜/𝑠, la física cuántica implica la existencia de una constante 𝑐𝑗 tal que la función de propensidad asociada queda:
aj(x) = �cj · xi · xi′ , i ≠ i′ cj · 1
2 · xi · (xi − 1), i = i′ en función de si las especias sean la misma o
diferentes.
Notar como 𝑐𝑗 para una reacción mono-.molecular es la constante cinética 𝑘𝑗 de la cinética química determinista convencional, mientras que 𝑐𝑗 para una reacción bi-molecular es 𝑘𝑗/𝛺 en el caso de reactivos diferentes, o 2 · 𝑘𝑗/𝛺 si son el mismo.
La Ecuación Química Maestra
Intentamos calcular la probabilidad 𝑃(𝑥, 𝑡|𝑥0, 𝑡0) de que 𝑋(𝑡) sea igual a 𝑥, dado 𝑋(𝑡0) = 𝑥0. Usando las leyes de la probabilidad:
𝑃(𝑥, 𝑡 + 𝑑𝑡|𝑥0, 𝑡0) = 𝑃(𝑥, 𝑡|𝑥0, 𝑡0) ∗ �1 −�𝑎𝑗(𝑥)𝑑𝑡𝑀
𝑗=1
� + �𝑃�𝑥 − 𝑣𝑗 , 𝑡�𝑥0, 𝑡0� ∗ (𝑎𝑗�𝑥 − 𝑣𝑗�𝑑𝑡𝑀
𝑗=1
El primer término de la derecha es la probabilidad de que, estando el sistema en el estado 𝑥 y en el instante de tiempo 𝑡, no ocurra ninguna reacción en el intervalo [𝑡, 𝑡 + 𝑑𝑡) y el segundo término, es la probabilidad de que el sistema cambie su estado actual 𝑥 en el instante de tiempo 𝑡, a través de una reacción 𝑅𝑗 en el instante consecutivo [𝑡, 𝑡 + 𝑑𝑡).

Modelado, análisis y control de un sistema biológico biestable
Enfoque Estocástico 85
Sustrayendo el término 𝑃(𝑥, 𝑡|𝑥0, 𝑡0) de ambos lados, dividiendo por 𝑑𝑡 y tendiendo al límite 𝑑𝑡 → 0:
𝝏𝑷(𝒙, 𝒕|𝒙𝟎, 𝒕𝟎)𝝏𝒕
= ��𝒂𝒋�𝒙 − 𝒗𝒋, 𝒕�𝒙𝟎, 𝒕𝟎� − 𝒂𝒋(𝒙)𝑷(𝒙, 𝒕|𝒙𝟎, 𝒕𝟎)�𝑴
𝒋=𝟏
Ecuación Química Maestra
A pesar del aspecto que muestra, la resolución no puede hacerse de manera analítica excepto en casos muy simples. Si todas las reacciones fuesen mono-moleculares, todas las funciones de propensidad serían lineales con las variables de estado. Pero su alguna ecuación fuese de un orden mayor, el aspecto de la ecuación y su resolución se vuelven mucho más complejos.
El Algoritmo de Simulación Estocástico
La resolución de la Ecuación Quimifica Maestra no resulta útil en nuestros problemas, por eso buscamos un enfoque diferente. De nuevo Daniel T Gillespie nos enseña el camino por proceder [10]. Se ha comprobado que el siguiente método es muy efectivo: generar trayectorias numéricas para el vector de estado 𝑋(𝑡).
• Se define 𝑝(𝜏, 𝑗|𝑥, 𝑡)𝑑𝑡 como la probabilidad de que, dado 𝑋(𝑡) = 𝑥, la próxima reacción que ocurrirá en el sistema en el siguiente intervalo infinitesimal de tiempo [𝑡 + 𝜏, 𝑡 + 𝜏 + 𝑑𝜏) y que será la reacción 𝑅𝑗. Para derivar una expresión analítica, además,
• Se define 𝑃0(𝜏|𝑥, 𝑡) como la probabilidad de que, dado 𝑋(𝑡) = 𝑥, no ocurra ninguna reacción en el siguiente intervalo [𝑡, 𝑡 + 𝜏).
Por las leyes de probabilidad se obtienen estas ecuaciones:
𝑝(𝜏, 𝑗|𝑥, 𝑡) = 𝑃0(𝜏|𝑥, 𝑡) ∗ 𝑎𝑗(𝑥)𝑑𝜏
𝑃0(𝜏 + 𝑑𝜏|𝑥, 𝑡) = 𝑃0(𝜏|𝑥, 𝑡) ∗ �1− � 𝑎𝑗′(𝑥)𝑑𝜏𝑀
𝑗′=1
�
Si en la última ecuación aplicamos el límite de 𝑑𝜏 → 0 se encuentra que:
P0(τ|x, t) = exp (−a0(x)τ) , donde
𝑎0(𝑥) = � 𝑎𝑗′(𝑥)𝑀
𝑗′=1
Quedando finalmente la expresión cómo:
𝑝(𝜏, 𝑗|𝑥, 𝑡) = 𝑎𝑗(𝑥) · 𝑒𝑥𝑝 (−𝑎0(𝑥) · 𝜏)

86 Enfoque Estocástico
Haciendo uso del método de inversión de la teoría de Monte-Carlo concluimos el método general para el Algoritmo de Simulación Estocástico.
MÉTODO GENERAL PARA EL ALGORITMO DE SIMULACIÓN ESTOCÁSTICO
1. Inicialización del tiempo y del estado del sistema 𝑡 = 𝑡0 𝑥 = 𝑥0
2. Con el sistema actualizado en tiempo y estado, evaluamos las funciones de propensidad y su suma: 𝑎𝑗(𝑥)
𝑎0(𝑥) = � 𝑎𝑗′(𝑥)𝑀
𝑗′=1
3. Se generan los valores del paso temporal y de las reacciones a partir de dos
valores aleatorios (variable aleatoria entre 0 y 1) 𝑟1, 𝑟2 → 𝑉𝑎𝑙𝑜𝑟𝑒𝑠 𝑎𝑙𝑒𝑎𝑡𝑜𝑟𝑖𝑎𝑠
𝜏 =1
𝑎0(𝑥)ln �
1𝑟1�
Buscamos el valor de índice j’, para conocer la reacción asociada que se producirá en el siguiente paso temporal. Para ello, el índice debe cumplir la siguiente condición:
� 𝑎𝑗′(𝑥) ≤ 𝑟2𝑎0(𝑥) ≤ � 𝑎𝑗′(𝑥)𝑗
𝑗′=1
𝑗−1
𝑗′=1
4. Se aplicará el efecto que corresponda con el reemplazo del nuevo estado
temporal y del nuevo estado: 𝑡 ← 𝑡 + 𝜏 𝑥 ← 𝑥 + 𝑣𝑗
5. Se guardan los valores de (𝑥, 𝑡) y se vuelve al paso 2 o se termina la simulación si se ha alcanzado el tiempo total de simulación.

Modelado, análisis y control de un sistema biológico biestable
Enfoque Estocástico 87
Tau-Leaping
El tau-leaping es una estrategia de simulación acelerada. Se trata de una forma simple donde se requiere que τ sea pequeño para satisfacer la condición de salto, El cambio inducido por un salto debe ser lo suficientemente pequeño como para que los cambios en las funciones de propensidad sean lo suficientemente pequeños.
Si 𝑋(𝑡) = 𝑥, y si elegimos un tau lo suficientemente pequeño, las funciones de propensidad serán casi constantes, entonces, la reacción 𝑅𝑗 debe ocurrir aproximadamente en veces en el intervalo de tiempo [𝑡, 𝑡 + 𝜏).
𝑋(𝑡 + 𝜏) = 𝑥 + �𝑣𝑗 · 𝑃𝑗(𝑎𝑗(𝑥)𝑑𝜏)𝑀
𝑗=1
Fórmula básica del algoritmo tau-leaping
En ella tenemos que:
x = X(t) Pj(mj) Variable aleatoria independiente de Poisson con media (y varianza) 𝑚𝑗
𝑃(𝑎, 𝜏) se define como el número de eventos que ocurrirán en el tiempo 𝜏 dado que 𝑎 · 𝑑𝑡 es la probabilidad de que el evento ocurra en algún intervalo infinitesimal de tiempo dt, donde a es una constante real y positiva.
El método de integración de la ecuación química maestra de Gillespie, es muy lento ya que en cada paso de integración, supone que sólo se produce una reacción, mientras que en tau-leaping en cada paso, todas las reacciones se producen según una variable de Poisson
Aspectos de interés del método
• A partir de la expresión de la ecuación química maestra, el algoritmo de simulación estocástico permite la resolución numérica de sistemas químicos homogéneos. El método consiste en asignar pasos temporales entre eventos reactivos moleculares.
• El algoritmo de simulación estocástica como procedimiento de simulación numérica de sistemas químicos homogéneos, asigna pasos temporales entre eventos reactivos moleculares según las directrices de la ecuación química maestra.
• El algoritmo tiene en cuenta la naturaleza discreta y estocástica de los sistemas químicos reactivos. Por este motivo es más útil que el método basado en ecuaciones diferenciales ordinarias.

88 Enfoque Estocástico
• La simulación de cada reacción molecular hace que el método sea lento al aplicarlo en un sistema reactivo realista.
• El método de integración por tau-leaping es una mejora en la rapidez de la resolución. La suposición de cada función de propensidad como variable de Poisson permite ejecutar cada reacción en cada paso en integración.

Modelado, análisis y control de un sistema biológico biestable
Enfoque Estocástico 89
2. Scripts en Matlab
Algoritmos de simulación
A partir de lo explicado en el anterior, pasamos mostrar los programas que hemos realizado en matlab para la resolución y simulación de los modelos estudiados hasta el momento.
gillespie.m ...
%inicialización vector tiempo vtiempo(1)=0; %condiciones iniciales especies(:,1)=0.01*ones(nesp,1); %iniciación del algoritmo ind=2; while(vtiempo(ind-1)<tsim) %cálculo de las propensidades modelo %elección de la reacción cumprop=cumsum(prop); al1=rand(); al2=rand(); Rind=find(al1*sum(prop)<cumprop,1); %elección del salto temporal tau=log(1/al2)/(sum(prop)); %actualización del tiempo vtiempo(ind)=vtiempo(ind-1)+tau; %actualización de las poblaciones de las especies especies(:,ind)=especies(:,ind-1)+R(:,Rind); for i=1:nesp if(especies(i,ind)<0) especies(i,ind)=0; end end %visor de evolución del algoritmo vtiempo(ind)/tsim %actualización del índice ind=ind+1; end
...

90 Enfoque Estocástico
Además, hemos implementado el algoritmo tau-leaping para lograr simulaciones de los sistemas de forma más rápida.
tauleaping.m ...
%inicialización vector tiempo vtiempo(1)=0; %condiciones iniciales especies(:,1)=0.01*ones(nesp,1); %inicialización del algoritmo ind=2; tau=0.1; while vtiempo(ind-1)<tsim %cálculo de propensidades modelo %actualización del vector de estados for i=1:nesp Rn(i)=sum(random('poiss',prop(:)*tau).*R(i,:)'); especies(i,ind)=especies(i,ind-1)+Rn(i); if(especies(i,ind)<0) especies(i,ind)=0; end end %actualización vector tiempo vtiempo(ind)=vtiempo(ind-1)+tau; %actualziación del índice ind=ind+1; %visor de evolución del algoritmo vtiempo(ind-1)/tsim end
...

Modelado, análisis y control de un sistema biológico biestable
Enfoque Estocástico 91
Modelos de los sistemas
BIESTABLE SIMPLE
bsimple\Modelo.m ...
% 1 2 3 4 5 6 7 8 9 10 11 12 % Ol Oc Ml Mc L C L2 C2 L2Oc C2Ol RFP GFP % 1. Ol -> Ol + Ml % 2. Ml -> Ml + L + RFP % 3. L + L -> L2 % 4. L2 -> L + L % 5. L2 + Oc -> L2Oc % 6. L2Oc -> L2 + Oc % 7. Oc -> Oc + Mc % 8. Mc -> Mc + C + GFP % 9. C + C -> C2 % 10.C2 -> C + C % 11.C2 + Ol -> C2Ol % 12.C2Ol -> C2 + Ol % 13.Ml -> null % 14.Mc -> null % 15.L -> null % 16.C -> null % 17.L2 -> null % 18.C2 -> null % 19.RFP -> null % 20.GRP -> null %La dimerización se producirá en función del factor de cooperatividad a=1+boolean(random('poiss',alfa)-1); b=1+boolean(random('poiss',beta)-1); %Cálculo de las propensidades prop(1) = par(1) * especies(1,ind-1); prop(2) = par(2) * especies(3,ind-1); prop(3) = par(3) * especies(5,ind-1)^a; prop(4) = par(4) * especies(7,ind-1); prop(5) = par(5) * especies(7,ind-1) * especies(2,ind-1); prop(6) = par(6) * especies(9,ind-1); prop(7) = par(7) * especies(2,ind-1); prop(8) = par(8) * especies(4,ind-1); prop(9) = par(9) * especies(6,ind-1)^b; prop(10)= par(10)* especies(8,ind-1); prop(11)= par(11)* especies(8,ind-1) * especies(1,ind-1); prop(12)= par(12)* especies(10,ind-1); prop(13)= par(13)* especies(3,ind-1); prop(14)= par(14)* especies(4,ind-1); prop(15)= par(15)* especies(5,ind-1); prop(16)= par(16)* especies(6,ind-1); prop(17)= par(17)* especies(7,ind-1); prop(18)= par(18)* especies(8,ind-1); prop(19)= par(19)* especies(11,ind-1); prop(20)= par(20)* especies(12,ind-1);

92 Enfoque Estocástico
% 1 2 3 4 5 6 7 8 9 10 11 12 % Ol Oc Ml Mc L C L2 C2 L2Oc C2Ol RFP GFP %Matriz estequiométrica R=[ 0 0 1 0 0 0 0 0 0 0 0 0; %1 0 0 0 0 1 0 0 0 0 0 1 0; %2 0 0 0 0 -a 0 1 0 0 0 0 0; %3 0 0 0 0 a 0 -1 0 0 0 0 0; %4 0 -1 0 0 0 0 -1 0 1 0 0 0; %5 0 1 0 0 0 0 1 0 -1 0 0 0; %6 0 0 0 1 0 0 0 0 0 0 0 0; %7 0 0 0 0 0 1 0 0 0 0 0 1; %8 0 0 0 0 0 -b 0 1 0 0 0 0; %9 0 0 0 0 0 b 0 -1 0 0 0 0; %10 -1 0 0 0 0 0 0 -1 0 1 0 0; %11 1 0 0 0 0 0 0 1 0 -1 0 0; %12 0 0 -1 0 0 0 0 0 0 0 0 0; %13 0 0 0 -1 0 0 0 0 0 0 0 0; %14 0 0 0 0 -1 0 0 0 0 0 0 0; %15 0 0 0 0 0 -1 0 0 0 0 0 0; %16 0 0 0 0 0 0 -1 0 0 0 0 0; %17 0 0 0 0 0 0 0 -1 0 0 0 0; %18 0 0 0 0 0 0 0 0 0 0 -1 0; %19 0 0 0 0 0 0 0 0 0 0 0 -1]'; %20
BIESTABLE MEJORADO
bmejorado\Modelo.m
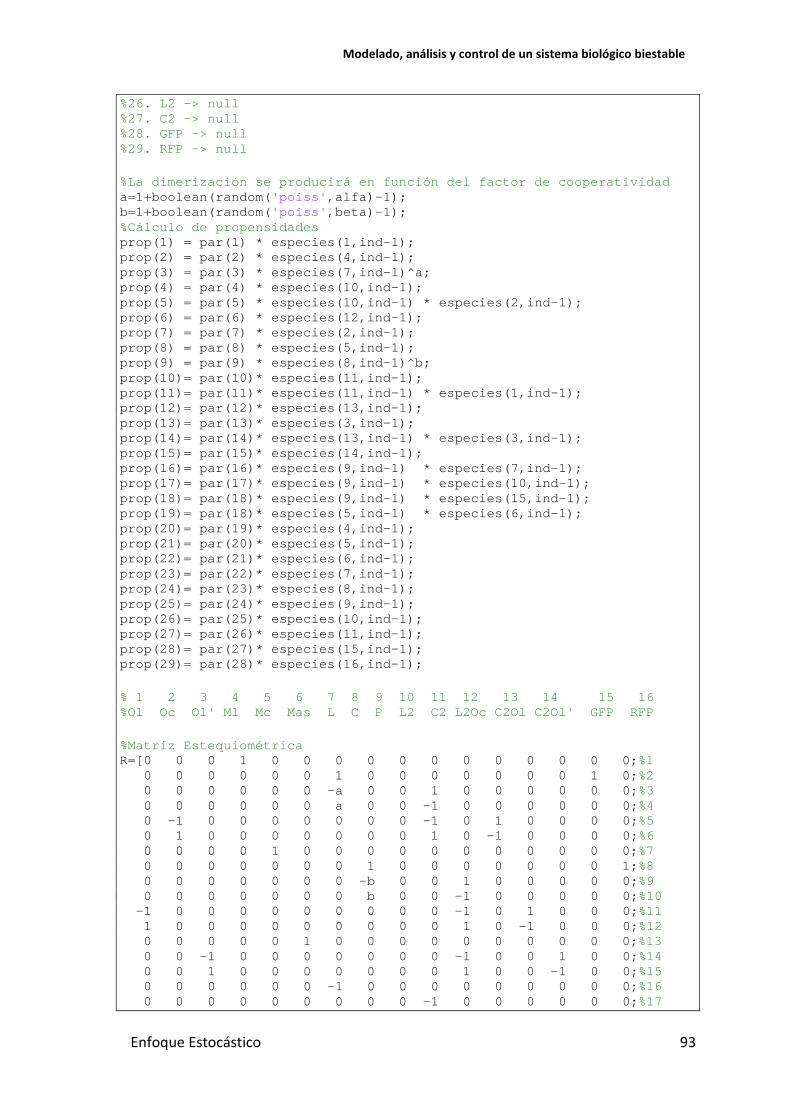
... % 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 %Ol Oc Ol' Ml Mc Mas L C P L2 C2 L2Oc C2Ol C2Ol' GFP RFP %1. Ol -> Ol + Ml %2. Ml -> Ml + L + GFP %3. L + L -> L2 %4. L2 -> L + L %5. L2 + Oc -> L2Oc %6. L2Oc -> L2 + Oc %7. Oc -> Oc + Mc %8. Mc -> Mc + C + P %9. C + C -> C2 %10. C2 -> C + C %11. C2 + Ol -> C2Ol %12. C2Ol -> C2 + Ol %13. Ol' -> Ol' + Mas %14. C2 + Ol' -> C2Ol' %15. C2Ol' -> C2 + Ol' %16. P + L -> P %17. P + L2 -> P %18. P + GFP -> P %19. Mas + Mc -> null %20 Ml -> null %21. Mc -> null %22. Mas -> null %23. L -> null %24. C -> null %25. P -> null
Modelado, análisis y control de un sistema biológico biestable
Enfoque Estocástico 93
%26. L2 -> null %27. C2 -> null %28. GFP -> null %29. RFP -> null %La dimerización se producirá en función del factor de cooperatividad a=1+boolean(random('poiss',alfa)-1); b=1+boolean(random('poiss',beta)-1); %Cálculo de propensidades prop(1) = par(1) * especies(1,ind-1); prop(2) = par(2) * especies(4,ind-1); prop(3) = par(3) * especies(7,ind-1)^a; prop(4) = par(4) * especies(10,ind-1); prop(5) = par(5) * especies(10,ind-1) * especies(2,ind-1); prop(6) = par(6) * especies(12,ind-1); prop(7) = par(7) * especies(2,ind-1); prop(8) = par(8) * especies(5,ind-1); prop(9) = par(9) * especies(8,ind-1)^b; prop(10)= par(10)* especies(11,ind-1); prop(11)= par(11)* especies(11,ind-1) * especies(1,ind-1); prop(12)= par(12)* especies(13,ind-1); prop(13)= par(13)* especies(3,ind-1); prop(14)= par(14)* especies(13,ind-1) * especies(3,ind-1); prop(15)= par(15)* especies(14,ind-1); prop(16)= par(16)* especies(9,ind-1) * especies(7,ind-1); prop(17)= par(17)* especies(9,ind-1) * especies(10,ind-1); prop(18)= par(18)* especies(9,ind-1) * especies(15,ind-1); prop(19)= par(18)* especies(5,ind-1) * especies(6,ind-1); prop(20)= par(19)* especies(4,ind-1); prop(21)= par(20)* especies(5,ind-1); prop(22)= par(21)* especies(6,ind-1); prop(23)= par(22)* especies(7,ind-1); prop(24)= par(23)* especies(8,ind-1); prop(25)= par(24)* especies(9,ind-1); prop(26)= par(25)* especies(10,ind-1); prop(27)= par(26)* especies(11,ind-1); prop(28)= par(27)* especies(15,ind-1); prop(29)= par(28)* especies(16,ind-1); % 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 %Ol Oc Ol' Ml Mc Mas L C P L2 C2 L2Oc C2Ol C2Ol' GFP RFP %Matriz Estequiométrica R=[0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0;%1 0 0 0 0 0 0 1 0 0 0 0 0 0 0 1 0;%2 0 0 0 0 0 0 -a 0 0 1 0 0 0 0 0 0;%3 0 0 0 0 0 0 a 0 0 -1 0 0 0 0 0 0;%4 0 -1 0 0 0 0 0 0 0 -1 0 1 0 0 0 0;%5 0 1 0 0 0 0 0 0 0 1 0 -1 0 0 0 0;%6 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0;%7 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 1;%8 0 0 0 0 0 0 0 -b 0 0 1 0 0 0 0 0;%9 0 0 0 0 0 0 0 b 0 0 -1 0 0 0 0 0;%10 -1 0 0 0 0 0 0 0 0 0 -1 0 1 0 0 0;%11 1 0 0 0 0 0 0 0 0 0 1 0 -1 0 0 0;%12 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0;%13 0 0 -1 0 0 0 0 0 0 0 -1 0 0 1 0 0;%14 0 0 1 0 0 0 0 0 0 0 1 0 0 -1 0 0;%15 0 0 0 0 0 0 -1 0 0 0 0 0 0 0 0 0;%16 0 0 0 0 0 0 0 0 0 -1 0 0 0 0 0 0;%17

94 Enfoque Estocástico
0 0 0 0 0 0 0 0 0 0 0 0 0 0 -1 0;%18 0 0 0 0 -1 -1 0 0 0 0 0 0 0 0 0 0;%19 0 0 0 -1 0 0 0 0 0 0 0 0 0 0 0 0;%20 0 0 0 0 -1 0 0 0 0 0 0 0 0 0 0 0;%21 0 0 0 0 0 -1 0 0 0 0 0 0 0 0 0 0;%22 0 0 0 0 0 0 -1 0 0 0 0 0 0 0 0 0;%23 0 0 0 0 0 0 0 -1 0 0 0 0 0 0 0 0;%24 0 0 0 0 0 0 0 0 -1 0 0 0 0 0 0 0;%25 0 0 0 0 0 0 0 0 0 -1 0 0 0 0 0 0;%26 0 0 0 0 0 0 0 0 0 0 -1 0 0 0 0 0;%27 0 0 0 0 0 0 0 0 0 0 0 0 0 0 -1 0;%28 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 -1]';%29
...

Modelado, análisis y control de un sistema biológico biestable
Enfoque Estocástico 95
3. Selección de parámetros
En los modelos mostrados, aparecían los parámetros como un vector cuyas componentes guardan los valores para cada una de las reacciones. Veamos estos scripts:
bsimple\parametros.m bmejorado\parametros.m
... parbs par(1) = TRN; par(2) = TRD; par(3) = DIM; par(4) = NDIM; par(5) = REP; par(6) = NREP; par(7) = TRN; par(8) = TRD; par(9) = DIM; par(10)= NDIM; par(11)= REP; par(12)= NREP; par(13)= DGM; par(14)= DGM; par(15)= DGP; par(16)= DGP; par(17)= DGP; par(18)= DGP; par(19)= DGR; par(20)= DGG; alfa=coop; beta=coop; nesp=12;
... parbs par(1) = TRN; par(2) = TRD; par(3) = DIM; par(4) = NDIM; par(5) = REP; par(6) = NREP; par(7) = TRN; par(8) = TRD; par(9) = DIM; par(10)= NDIM; par(11)= REP; par(12)= NREP; par(13)= TRN; par(14)= REP; par(15)= NREP; par(16)= DIGP; par(17)= DIGP; par(18)= DIGP; par(19)= INH; par(20)= DGM; par(21)= DGM; par(22)= DGM; par(23)= DGP; par(24)= DGP; par(25)= DGP; par(26)= DGP; par(27)= DGP; par(28)= DGG; par(29)= DGR; alfa=coop; beta=coop; nesp=16;

96 Enfoque Estocástico
Y estos variables toman los siguientes valores:
bmejorado\parbs.m ... TRN=0.5; TRD=4; REP=1; NREP=2.4; DIM=2; NDIM=0.001; INH=1.5; DIGP=1.5; DGM=0.462; DGP=0.2; DGR=0.01; DGG=0.001; coop=2;
Esta elección se ha basado en múltiples documentos y bibliografías:
- Documento de investigación de Michail Stamatakis y Nikolaos V. Mantzaris llamado Comparison of Deterministic and Stochastic Models of the lac Operon Genetic Network y publicado en Febrero de 2009 en la revista Biophysical Journal [8]. En este paper, tratan de comprender como un estudio estocástico del operón Lac puede aportar un conocer más amplio del comportamiento de esta red reguladora genética. Así, crean un modelo del operón Lac de reacciones simples partiendo del modelo determinista. A través de simulaciones de ambos sistemas alcanzan un estudio comparativo de las bondades de cada enfoque. Es en este desarrollo donde asocian valores a los parámetros utilizados. Dado que nuestro modelo se basa en gran parte en el operón lac, y las reacciones que se ha detallado en este proyecto no difieren en gran medida del desarrollado por estos investigadores, tomamos estos valores como referencia.
- Registro de partes de iGEM (http://parts.igem.org/Main_Page). Todas las estructuras genéticas creadas por cada equipo en el concurso iGEM, pasan a formar parte de una base de datos. Además de detallar la secuencia genética, algunos de ellos, llevan incluidos un pequeño estudio y caracterización del ente diseñado. Concretamente, nuestro grupo utilizó la proteína GFP (Green Fluorescent Protein) como indicadors del estado de las bacterias. Gracias a esta base de datos, tenemos valores reales de su caracterización. GFP: http://parts.igem.org/wiki/index.php?title=Part:BBa_E0040
- Base de datos binumbers de Harvard [7]: http://bionumbers.hms.harvard.edu/ Esta página compendia valores de propiedades biológicas extraidos de documentos de investigación.

Modelado, análisis y control de un sistema biológico biestable
Enfoque Estocástico 97
Ha resultado muy útil su consulta, ya que al no disponer de valores reales, hemos podido tomar decisiones en cuanto a qué órdenes de magnitud manejábamos para todo tipo de reacciones.
Antes de continuar con los resultados de matlab, cabe destacar el valor asignados a los indicadores RFP y, especialmente, al GFP. En la bibliografía encontramos que:
𝑑𝑅𝐹𝑃 ≈ 0.01 𝑚𝑖𝑛−1
𝑑𝐺𝐹𝑃 ≈ 0.00035 𝑚𝑖𝑛−1 a pesar de que en las simulaciones se toma: 0.001 𝑚𝑖𝑛−1
Resulta muy llamativa la elección de estas proteínas como indicadores por dos motivos principales.
- La degradación del indicador verde resulta hasta mil veces más lenta que las de las proteínas que pretende indicar. Esto implica que en las observaciones gran parte de las dinámicas quedan ocultas.
- Las tasas de degradación de ambas son muy dispares. La proteína roja es más de 10 veces más rápida.
Lo expuesto ha sido de vital importancia al intentar comprender los experimentos realizados en laboratorio por el equipo.
Podemos comprobar que con estos valores, se están por encima de los valores críticos definidos en el estudio de bifurcaciones llevado a cabo con los modelos deterministas.

98 Enfoque Estocástico
4. Simulaciones estocásticas del biestable simple
A continuación expondremos algunas simulaciones realizadas del modelo estocástico a través del algoritmo basado en las teorías de Daniel Gillespie [11].
Las condiciones iniciales para las variables de estado del sistema que se han utilizado suponen una pequeña población de cada una de las especies.
Población de represor LacI Población de represor cI
Excepto en alguna simulación, en general, el sistema no llega a alcanzar un estado de equilibrio dominante. Esto es lo lógico al haber impuesto unos parámetros iguales para cada reacción. Sin embargo, cabría esperar que, en cuanto uno de los represores alcanzara un estado de nivel alto, el contrario no tendría la capacidad de alcanzar la dominancia del sistema.
Modelado, análisis y control de un sistema biológico biestable
Enfoque Estocástico 99
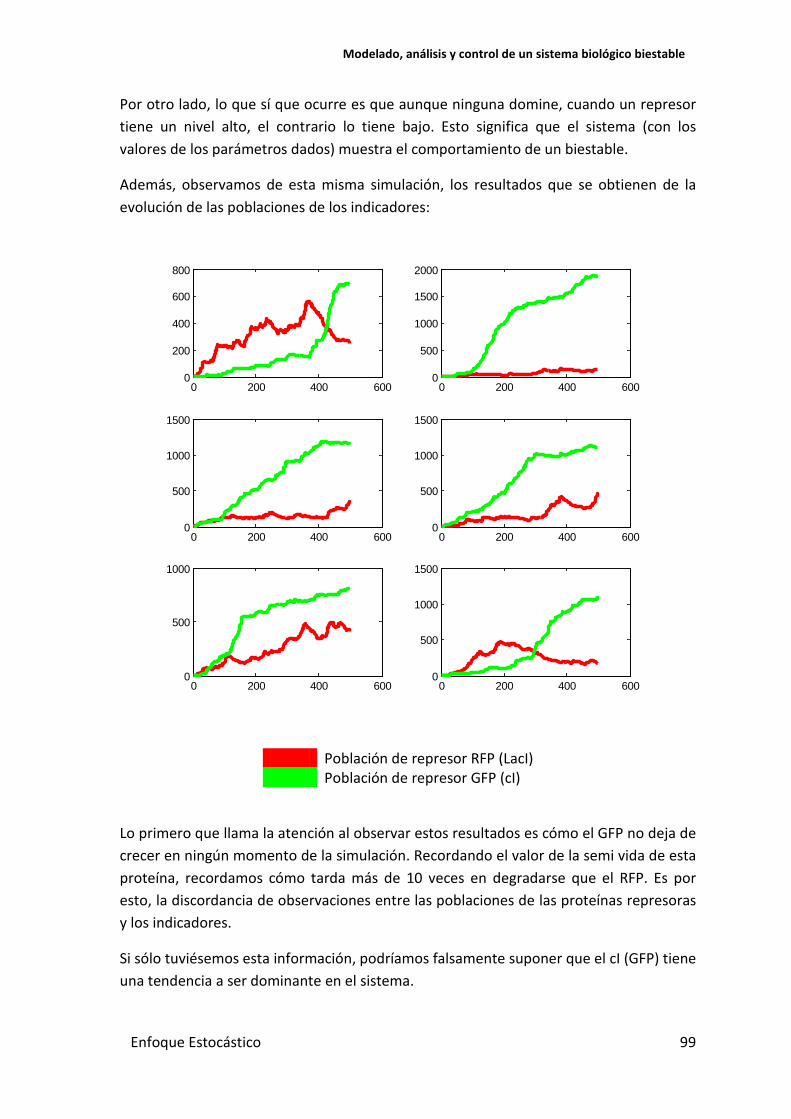
Por otro lado, lo que sí que ocurre es que aunque ninguna domine, cuando un represor tiene un nivel alto, el contrario lo tiene bajo. Esto significa que el sistema (con los valores de los parámetros dados) muestra el comportamiento de un biestable.
Además, observamos de esta misma simulación, los resultados que se obtienen de la evolución de las poblaciones de los indicadores:
Población de represor RFP (LacI) Población de represor GFP (cI)
Lo primero que llama la atención al observar estos resultados es cómo el GFP no deja de crecer en ningún momento de la simulación. Recordando el valor de la semi vida de esta proteína, recordamos cómo tarda más de 10 veces en degradarse que el RFP. Es por esto, la discordancia de observaciones entre las poblaciones de las proteínas represoras y los indicadores.
Si sólo tuviésemos esta información, podríamos falsamente suponer que el cI (GFP) tiene una tendencia a ser dominante en el sistema.
0 200 400 6000
200
400
600
800
0 200 400 6000
500
1000
1500
2000
0 200 400 6000
500
1000
1500
0 200 400 6000
500
1000
1500
0 200 400 6000
500
1000
0 200 400 6000
500
1000
1500
100 Enfoque Estocástico
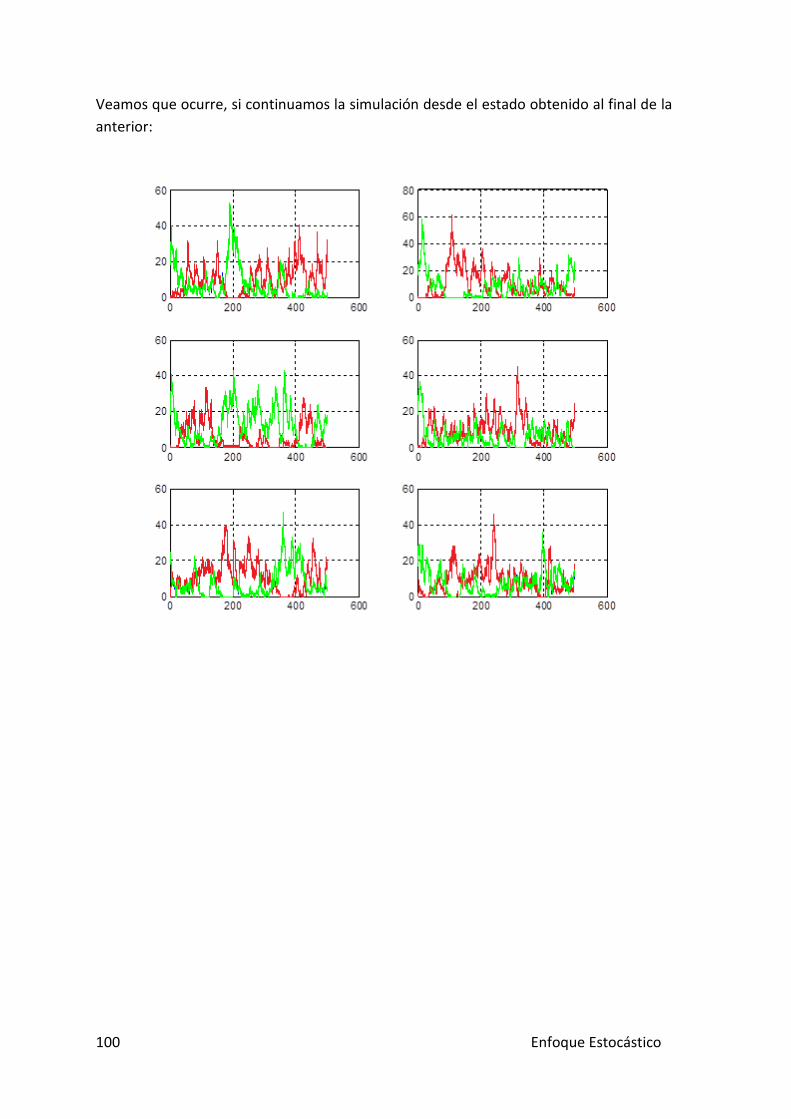
Veamos que ocurre, si continuamos la simulación desde el estado obtenido al final de la anterior:

Modelado, análisis y control de un sistema biológico biestable
Enfoque Estocástico 101
Las poblaciones de los represores mantienen esa lucha constante en la que nadie alcanza el poder; mientras que para los indicadores, se observa como hay un ganar aparente en cada caso, el GFP.
0 200 400 6000
500
1000
1500
2000
0 200 400 6000
500
1000
1500
2000
0 200 400 6000
1000
2000
3000
0 200 400 6000
500
1000
1500
0 200 400 6000
500
1000
1500
2000
0 200 400 6000
500
1000
1500
2000

102 Enfoque Estocástico
5. Simulaciones estocásticas del biestable mejorado
Las condiciones iniciales para las variables de estado del sistema que se han utilizado suponen una pequeña población de cada una de las especies.
Tras realizar una gran cantidad de simulaciones del sistema, nos damos cuenta de cómo los parámetros cinéticos asociados las nuevas reacciones son capaces de modificar los resultados finales de forma apreciable. Ya que no disponemos de valores orientativos, y a la vista de los resultados mostrados, decidimos tomar finalmente los siguientes:
𝐼𝑁𝐻 = 0.1; 𝐷𝐼𝐺𝑃 = 2;
A continuación mostraremos algunas de las simulaciones por las que hemos elegido éstas, sin embargo, podemos hacer un desarrollo teórico:
Las nuevas interacciones incluidas, a diferencia del biestable simple, no son parejas en el sentido de acciones iguales y contrarias. Por un lado, la inhibición ARN antisentido actúa directamente sobre el ARN mensajero de una de las ramas (concretamente del ARNm cI); mientras que la proteasa digiere el represor, sus multímeros y su reporter.
Esto significa que cada una de las acciones se aplica en diferentes etapas de la expresión de los genes, afectando de manera diversa a la dinámica global del sistema.
Además, La población de ARNm es bastante menor que la de represor por varios motivos:
• La cinética asociada a la transcripción es bastante menor que la asociada a la traducción.
• La cinética asociada a la degradación del ARNm es mayor que la asociada a la degradación del represor.
𝑑𝐶𝑑𝑡
= 𝑘𝑡𝑑 · 𝑀𝐶 − 𝑑 · 𝐶
En el régimen permanente encontramos una relación entre la cantidad de ARNm y el represor asociado:
𝑘𝑡𝑑 · 𝑀𝐶 − 𝑑 · 𝐶 = 0;
𝛿𝐶 = �𝑘𝑡𝑑𝑑� · 𝛿𝑀𝐶
Valiendo esta proporción (con los valores utilizados en los modelos) 𝑘𝑡𝑑𝑑
= 20.
Significando que un pequeña variación en el ARN del represor afectará mucho a la

Modelado, análisis y control de un sistema biológico biestable
Enfoque Estocástico 103
población final de la proteína. Es por este motivo por el que se ha decido equiparar el efecto que puedan aplicar las reacciones, dándole un valor de un orden de magnitud menor a la inhibición por ARN antisentido frente a la digestión por proteólisis.
Población de represor cI Población de represor LacI
Estos resultados no muestran una estabilidad mayor que las del biestable simple. Vemos cómo en las tres primeras imágenes, el sistema tiende a oscilar entre ambos estados sin que ninguno de ellos logre dominar el sistema. En la quinta simulación, encontramos esa situación ideal, en la que un estado se mantiene por encima del otro y con un nivel de expresión suficientemente alto, ocurriendo un cambio de estados casi ideal.
Veamos los resultados para los indicadores:

104 Enfoque Estocástico
Población de represor RFP Población de represor GFP
En este caso, el GFP no tiende a crecer indefinidamente a pesar de poseer la misma semivida que en el sistema simple. El motivo, es que la proteólisis también se aplica a este indicador. Por tanto a medida que existe más RFP (Y por tanto cI), habrá más proteasa, y el GFP irá disminuyendo su población.
Ya que los resultados de las poblaciones de las proteínas, mostraban cierto potencial a mantener un estado expresión alto, vamos a realizar un conjunto de simulaciones a partir de un estado inicial marcado como dominancia del GFP.
0 200 400 6000
500
1000
1500
2000
0 200 400 6000
1000
2000
3000
0 200 400 6000
500
1000
1500
2000
0 200 400 6000
500
1000
1500
2000
0 200 400 6000
1000
2000
3000
0 200 400 6000
500
1000
1500
2000

Modelado, análisis y control de un sistema biológico biestable
Enfoque Estocástico 105
Población de represor cI Población de represor LacI
Partiendo desde el punto final de las 6 simulación anterior, el sistema se comporta de la manera que mostramos en esta imagen. Es interesante constatar que una vez el sistema puede lograr un punto de equilibrio, no lo abandona en ninguno de las 6 experimentos. Hemos encontrado por tanto un sistema que con una pequeña inducción de uno de los estados, parece mantenerlo indefinidamente. Efectivamente, la probabilidad nos asegura que hay posibilidades de que el sistema cambie aleatoriamente su estado. Sin embargo, podemos asegurar que este sistema muestra un comportamiento más estable que el anterior.
Si vemos ahora la imagen con los resultados para los indicadores:

106 Enfoque Estocástico
Población de represor RFP Población de represor GFP
Constatamos que el GFP sigue comportándose de la misma forma ya pesar de que el represor al que está asociado ha alcanzado el régimen permanente, ésta proteína sigue creciendo hasta alcanzar su régimen permanente.
Si partimos desde un estado contrario al anterior, los resultados muestran el mismo comportamiento para la otra rama del biestable. Logrando incluso mayor estabilidad, la proteasa digiere casi completamente al represor contrario como al indicador.
0 50 100 150 200 2500
5000
10000
0 50 100 150 200 2500
5000
10000
0 50 100 150 200 2500
5000
10000
0 50 100 150 200 2500
2000
4000
6000
8000
0 50 100 150 200 2500
5000
10000
0 50 100 150 200 2500
2000
4000
6000
8000

Modelado, análisis y control de un sistema biológico biestable
Enfoque Estocástico 107
Población de represor cI Población de represor LacI
Y los resultados para los indicadores:

108 Enfoque Estocástico
Población de represor RFP Población de represor GFP
0 50 100 150 200 2500
5000
10000
0 50 100 150 200 2500
5000
10000
15000
0 50 100 150 200 2500
5000
10000
15000
0 50 100 150 200 2500
5000
10000
0 50 100 150 200 2500
5000
10000
0 50 100 150 200 2500
5000
10000