membranas activas: motores rotatorios - ffn.ub.esisaacll/membranes.pdf · mos en el papel de los...
TRANSCRIPT

Membranas activas: motores rotatorios
Isaac Llopis
6 de septiembre de 2005
Resumen
En este trabajo hacemos un estudio de la estructura y funcionalidadde las membranas ası como la necesidad de considerar el hecho que estasmembranas se encuentran fuera del equilibrio termodinamico. Nos centra-mos en el papel de los centros activos, en concreto de los que actuan comomotores rotatorios, como es el caso de la ATP-asa. A traves del metodode simulacion de lattice-Boltzmann estudiamos el campo hidrodinamicoque generan dichos rotores y analizamos su comportamiento colectivo.
Indice
1. Introduccion: Membranas y motores rotatorios 21.1. Estructura y funcionalidad de la membrana . . . . . . . . . . . . 2
1.1.1. Estructura . . . . . . . . . . . . . . . . . . . . . . . . . . 21.1.2. Funcionalidad . . . . . . . . . . . . . . . . . . . . . . . . . 5
1.2. Fluidez de la membrana . . . . . . . . . . . . . . . . . . . . . . . 51.3. Procesos de transporte . . . . . . . . . . . . . . . . . . . . . . . . 6
1.3.1. La hidrolisis del ATP . . . . . . . . . . . . . . . . . . . . 61.4. Membranas activas . . . . . . . . . . . . . . . . . . . . . . . . . . 8
1.4.1. Bacteriorhodopsin . . . . . . . . . . . . . . . . . . . . . . 91.5. Ejemplos de motores rotatorios (rotores) . . . . . . . . . . . . . . 10
1.5.1. La ATP-asa . . . . . . . . . . . . . . . . . . . . . . . . . . 111.5.2. Los cilios . . . . . . . . . . . . . . . . . . . . . . . . . . . 13
2. Un modelo de motores rotatorios 142.1. Modelo teorico . . . . . . . . . . . . . . . . . . . . . . . . . . . . 142.2. Metodo de simulacion . . . . . . . . . . . . . . . . . . . . . . . . 15
2.2.1. Lattice gas (LG) . . . . . . . . . . . . . . . . . . . . . . . 152.2.2. Lattice-Boltzmann . . . . . . . . . . . . . . . . . . . . . . 16
2.3. Dinamica de partıculas en LB . . . . . . . . . . . . . . . . . . . . 172.4. Motores rotatorios en LB . . . . . . . . . . . . . . . . . . . . . . 19
2.4.1. Partıculas activas . . . . . . . . . . . . . . . . . . . . . . . 192.4.2. Mecanismo de rotacion . . . . . . . . . . . . . . . . . . . . 20
1

1. Introduccion: Membranas y motores rotatorios 2
3. Resultados 203.1. Analisis de 1 rotor . . . . . . . . . . . . . . . . . . . . . . . . . . 21
3.1.1. Campo hidrodinamico inducido por los rotores . . . . . . 223.2. Estudio de 2 rotores . . . . . . . . . . . . . . . . . . . . . . . . . 253.3. Cadena de rotores . . . . . . . . . . . . . . . . . . . . . . . . . . 283.4. Comportamiento colectivo: . . . . . . . . . . . . . . . . . . . . . 29
3.4.1. Movimiento inducido . . . . . . . . . . . . . . . . . . . . . 293.4.2. Distribucion de velocidades . . . . . . . . . . . . . . . . . 293.4.3. Tiempos largos . . . . . . . . . . . . . . . . . . . . . . . . 32
3.5. Cristal de Wigner triangular . . . . . . . . . . . . . . . . . . . . . 333.5.1. Difusion activa . . . . . . . . . . . . . . . . . . . . . . . . 36
3.6. Fluctuaciones: acoplamiento entre los rotores y las fluctuacionesde la membrana . . . . . . . . . . . . . . . . . . . . . . . . . . . . 37
4. Conclusiones 37
1. Introduccion: Membranas y motores rotato-rios
1.1. Estructura y funcionalidad de la membrana
El termino membrana biologica uno tiende a relacionarlo con la membranaplasmatica de la celula, que separa el citoplasma celular del exterior. Aun ası,en una celula eucariota no solo existe esta membrana, sino que muchos de losorganulos celulares (mitocondrios, retıcula endoplasmatico, aparato de Golgi,etc.) estan delimitados por una membrana del mismo tipo.
1.1.1. Estructura
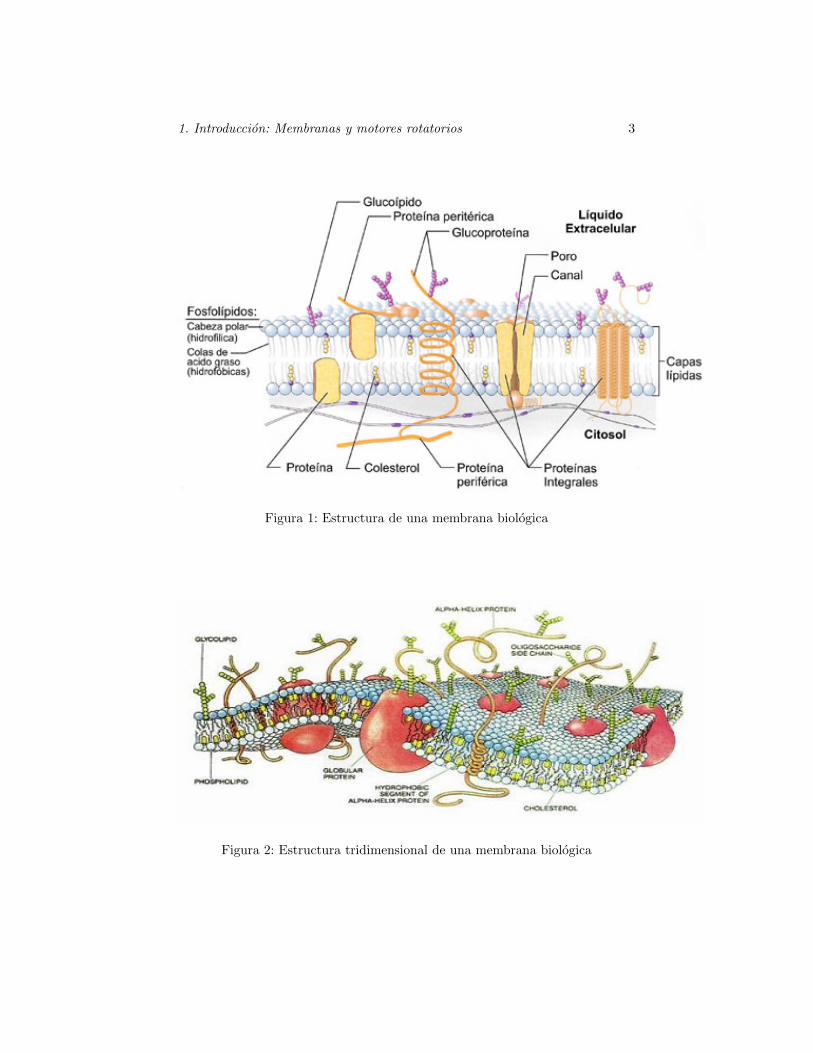
Una membrana biologica es una fina capa ( 5− 10 nm) compuesta por unamezcla compleja de moleculas lipıdicas y proteicas, y tambien puede contenerglucidos (Fig. 1) y (Fig. 2). Estas moleculas se mantienen unidas fundamental-mente por interacciones no covalentes.
Lıpidos: Los lıpidos son el componente principal de las membranas, concre-tamente la mitad de la masa de la membrana. Son moleculas amfipaticas, esdecir, tienen un extremo hidrofobico (no se asocia libremente con el agua) y elotro hidrofılico (sı se asocia libremente con el agua).
Existen tres clases principales de moleculas lipıdicas de membrana:
1. Fosfolıpidos: lo son la mayorıa de los lıpidos de membrana. Ver en Fig. 3su estructura detallada.
Parte hidrofılica: un grupo fosfato esterificado y un grupo alcohol.
1. Introduccion: Membranas y motores rotatorios 3
Figura 1: Estructura de una membrana biologica
Figura 2: Estructura tridimensional de una membrana biologica

1. Introduccion: Membranas y motores rotatorios 4
Figura 3: Esquemas de una molecula de fosfolıpido
Parte hidrofobica: dos cadenas de acidos grasos, una saturada y otrainsaturada, lo cual genera curvatura en la membrana. Son dos cade-nas largas y voluminosas, y las interacciones entre ellas son fuertes,es por ello que se puede considerar que la parte hidrofobica tienegeometrıa cilındrica.
Debido a este comportamiento amfipatico y a que sus colas son muy largasy voluminosas, los fosfolıpidos en la membrana se disponen en una bicapacon sus colas hidrofobicas dirigidas hacia el interior, quedando de estamanera entre las cabezas hidrofılicas que delimitan la superficie externa einterna de la membrana [1].
2. Colesterol: es un lıpido esteroide, formado por una molecula de ciclopenta-noperhidrofenantreno (o esterano). Las membranas plasmaticas de celulaseucariotas contienen gran cantidad de colesterol, cuya funcion es reforzarla permeabilidad de la bicapa. Se encuentra en el area hidrofobica de lamisma, su presencia contribuye a la estabilidad de la membrana ya queinteracciona con las colas de la bicapa y disminuye su fluidez evitando quelas colas se empaqueten, vuelvan mas rıgida la membrana. Las membranasde las celulas vegetales no contienen colesterol, tampoco las de la mayorıade las celulas bacterianas.
3. Glucolıpidos: solamente en la monocapa externa. Se asocian entre ellospara formar microagregados. Son oligosacaridos unidos a los lıpidos, aun-que estos oligosacaridos tambien pueden unirse a las proteinas y formarglucoproteınas.
La bicapa lipıdica es una doble capa contınua de moleculas lipıdicas en laque estan inmersas varias proteınas de membrana. Esta bicapa es asimetrica, la

1. Introduccion: Membranas y motores rotatorios 5
composicion lipıdica y proteica de la monocapa externa es diferente a la de lainterna, lo que refleja las diferentes funciones de las dos caras de una membranacelular.
Estas bicapas tienden a cerrarse sobre sı mismas hasta formar comparti-mientos hermeticos, para minimizar el efecto de que colas hidrofobicas tengancontacto con agua.
Proteınas: Hay diferentes tipos de proteınas de membrana, cuyas funcionesdependen de la manera en que estas se asocian a la bicapa lipıdica:
Las proteınas transmembrana se extienden a traves de la bicapa, yestan diferenciadas en dominios hidrofılicos e hidrofobicos. Debido a estaestructura, pueden transportar moleculas a traves de la membrana, tien-den a ser canales de transporte.
Las proteınas perifericas no se extienden en el dominio hidrofobico, soloactuan en una de las monocapas.
Nosotros, en este trabajo, estamos interesados en las proteınas transmem-brana.
1.1.2. Funcionalidad
La funcion principal de una membrana es diferenciar el fluido interior delentorno. Aun ası, las membranas tambien participan en otras funciones celulares,normalmente por el papel de las proteınas:
Transporte de soluto via bombas o canales ionicos. De esta manera, pode-mos considerar una membrana como una barrera semipermeable, ya quemantiene la mayor parte de los productos producidos dentro de ella peropermite el paso de algunas moleculas.
Locomocion y adhesion celular.
Proteccion.
Permitir el reconocimiento celular.
1.2. Fluidez de la membrana
Los lıpidos y muchas proteınas de membrana estan en movimiento constante.Los lıpidos se difunden en una de las monocapas con un coeficiente de di-
fusion de 10−8cm2/s, que significa que un lıpido da una vuelta a la membranacelular en aproximadamente 1s. Este movimiento lateral es rapido, sin embar-go el movimiento de una molecula lipıdica entre las dos capas de la membrana(flip-flop) es muy lento, debido al efecto hidrofobico.
La difusion de las proteınas es mas lenta, un 1 − 10 % mas pequena que lade los lıpidos.

1. Introduccion: Membranas y motores rotatorios 6
La fluidez de una membrana aumenta con la temperatura. Los acidos grasospueden existir en un estado ordenado (gel) y en uno desordenado (lıquido), cuyatransicion de fase se da a una cierta temperatura Tm, que depende de la longitudde las cadenas de los acidos grasos y de su grado de insaturacion.
La fluidez tambien aumenta con la presencia de lıpidos insaturados y decadena corta.
Esta fluidez implica que los componentes en su mayorıa solo estan unidospor uniones no covalentes.
1.3. Procesos de transporte
La bicapa lipıdica es basicamente impermeable a los iones, independiente-mente de su tamano. Por eso el transporte de iones inorganicos y de pequenasmoleculas organicas hidrosolubles a traves de la bicapa lipıdica es medianteproteınas transmembrana, de las que hay 2 tipos:
1. Proteınas transportadoras: se unen al soluto que va a ser transportadoy hacen cambios conformacionales para que se transfiera dicho soluto atraves de la membrana.
2. Proteınas de canal: forman poros hidrofılicos que atraviesan la bicapalipıdica y, cuando los poros estan abiertos, permiten que diferentes solutospasen pasivamente a su traves y atraviesen la membrana.
El soluto tiene una carga neta, su transporte esta influido tanto por su gra-diente de concentracion como por el gradiente electrico a traves de la membrana.La combinacion es el gradiente electroquımico.
El gradiente electroquımico define 2 tipos basicos de transporte a traves dela membrana:
1. Transporte pasivo: si el gradiente electroquımico es favorable (normal-mente el potencial interior es mas negativo que el exterior) se favorece eltransporte de iones cargados del potencial positivo al negativo.
2. Transporte activo: algunas proteınas bombean activamente ciertos solu-tos en contra de su gradiente electroquımico, siempre mediante proteınastransportadoras.
La accion de las bombas es crucial para mantener las diferencias de con-centracion ionicas.
Una gran proporcion de toda la energıa que recibe el cuerpo humano esdedicada a las bombas ionicas.
1.3.1. La hidrolisis del ATP
El ATP es un nucleotido trifosfato derivado de la adenina. a pH fisiologicotiene una carga negativa en cada grupo fosfato. Los grupos fosfato se unen

1. Introduccion: Membranas y motores rotatorios 7
Figura 4: Estructura del ATP
mediante enlaces fosfoanhidrilo. Si en lugar de tres tiene dos fosfatos es ADP ysi tiene uno es AMP (Fig. 4).
El ATP es la mayor fuente de energıa quımica de las celulas.El transporte activo esta acoplado a una fuente de energıa metabolica como
la hidrolisis del ATP, un gradiente de concentracion ionica o un agente externo,donde el transporte mediante hidrolisis del ATP es el mas tıpico.
Consiste en la siguiente reaccion, considerando que el aceptor es el agua:
ATP +H20 → ADP + Pi(∆G = −31kJ/mol) (1)
Para que se produzca esta reaccion, se requieren encimas que la catalicen,son las llamadas ATP-sintasas o ATP-asas. En la siguiente seccion trataremosestas encimas con mas detalle.
Ejemplos:
Na+ −K+ −ATPasa: bombea Na+ de dentro a fuera de la celula y K+
de fuera a dentro.
Ca2+ − ATPasa: mantiene el gradiente de concentracion de Ca2+ entreel interior y el exterior celular.

1. Introduccion: Membranas y motores rotatorios 8
1.4. Membranas activas
Las membranas pasivas son membranas en equilibrio termodinamico, lasfluctuaciones de su forma son el ruido termico, que es el movimiento brownianode los elementos de la bicapa.
Sin embargo, desde un punto de vista estadıstico, una membrana biologicaesta muy lejos del equilibrio. Las membranas biologicas contienen proteınas (ca-nales ionicos, bombas ionicas o proteınas fotoactivas) que consumen la energıaquımica del ATP y la disipan en el medio, generan fuerzas que llevan a lamembrana fuera del equilibrio. A la que las proteınas difunden alrededor dela membrana, las fluctuaciones resultantes en esta fuerza producen ruido notermico para las fluctuaciones de forma de la membrana.
Este hecho se debe introducir a la hora de describir una membrana. Ası pues,es basico para la descripcion de una membrana suponerla activa.
Las membranas que contienen centros activos sufren fluctuaciones de noequilibro adicionales a las de equilibrio debido a la actividad de estos centrosactivos. El teorema de fluctuacion-disipacion no es valido y debemos tener encuenta las interacciones hidrodinamicas de largo alcance que estan mediadaspor el solvente. Debido a que no hay teorema de fluctuacion-disipacion hayuna amplificacion de las fluctuaciones, las fluctuaciones de no equilibrio son lasdominantes [5].
Estos procesos de no equilibrio tienen efecto incluso en los globulos rojos,que tradicionalmente han sido explicados a partir de fluctuaciones de forma deequilibrio termicas.
Las proteınas activas de membrana pueden ser modelizadas como discossuspendidos en un fluido bidimensional viscoso rodeado de fluido de viscosidadbaja [2].
Para determinar el efecto de las interacciones hidrodinamicas en la difusivi-dad de las proteınas en suspensiones no diluıdas, solucionamos numericamente laecuacion de Navier-Stokes para un sistema de discos en un fluido bidimensionalusando una expansion multipolar [3].
Consideramos las proteınas moviles, aunque tambien pueden haber algunasfijas.
Para estas proteınas moviles, podemos calcular autodifusiones a tiemposcortos tanto translacionales como rotacionales y el gradiente difusivo crece lo-garıtmicamente con el nombre de discos, que es la paradoja de Stokes.
El modelo del mosaico fluido (fluid mosaic model) [4] postula que la mem-brana es como un mosaico fluido en el que la bicapa lipıdica es una red donde sesituan las proteinas, que interaccionan unas con otras y con los lıpidos. Tantolas proteinas como los lıpidos pueden desplazarse lateralmente.
Por lo tanto, podemos considerar la membrana como proteınas suspendidasen un fluido (la bicapa lipıdica) que esta en movimiento constante. Las proteınastambien se mueven lateralmente en la bicapa, formando diferentes estructuras,que iran cambiando con el tiempo.

1. Introduccion: Membranas y motores rotatorios 9
Figura 5: Estructura de la Bacteriorhodopsin.
Pero la situacion real es que las proteınas asociadas con la superficie de lamembrana tienen que competir con varios obstaculos a la que experimentan elmovimiento Browniano.
Usando microscopia de fuerzas atomicas (AFM) en rotores suspendidos enuna membrana lipıdica, D.J. Muller et al. [6] mostraron, analizando las tra-yectorias individuales, que las proteınas podıan moverse segun difusion libreo difusion entorpecida, que tienen coeficientes de difusion considerablementediferentes. Lo hacıan en funcion de la distribucion de rotores a su alrededor.
El analisis con AFM mostraba que algunos rotores formaban clusters densos(difusion entorpecida) y otros, en cambio, no tenıan contacto con otros roto-res (difusion libre). Las trayectorias observadas sugieren que las proteınas demembrana experimentan transiciones entre estos dos tipos de movimiento.
El citoesqueleto, justo debajo de la membrana juega un papel fundamentalen el control de la mobilidad de las proteınas de membrana en una gran variedadde celulas, como es el caso de las nerviosas, las epiteliales o los globulos rojos.
1.4.1. Bacteriorhodopsin
La Bacteriorhodopsin (bR) es una proteına fotoactiva de la que se conoce suestructura a nivel atomico y con gran resolucion (Fig. 5), de hecho es la bombade iones mejor entendida [7] [8]. Cuando se ilumina con luz verde amarillenta se

1. Introduccion: Membranas y motores rotatorios 10
activa el bombeo de protones a traves de un fotociclo de unos 5ms.Su peculiaridad es que utiliza directamente la energıa que proviene de la
luz exterior en vez de requerir la hidrolisis de ATP o GTP para bombear io-nes. Cuando es iluminada con luz amarillenta-verdosa se activa el bombeo deprotones a traves de un fotociclo de τ ∼ 5ms.
Se han estudiado los cambios estructurales de la bR durante el fotociclo paraası saber la trayectoria de cada proton a traves de la proteına.
Experimentos con micropipetas en el estudio de vesıculas gigantes fluctuan-tes que contienen BR en la bicapa, mostraron que la actividad de bombeo deiones inducıa una amplificacion en las fluctuaciones de la forma de la membrana,lo cual se puede interpretar como un incremento de la temperatura efectiva dela membrana [9].
Con la tecnica de micropipetas se puede cuantificar el area acaparada en lasfluctuaciones de membrana. De hecho, la variacion de este exceso de area (∆α)se puede relacionar con la tension superficial (σ) de la membrana [9]:
∆α =KBT
8πκln
σ
σ0(2)
donde κ es la rigidez de la membrana y σ0 la tension superficial de equilibrio.En el caso de membranas pasivas T es la temperatura, pero en membranas
activas T es la temperatura efectiva, ya que estamos fuera del equilibrio.La pendiente de la recta determina el valor de la rigidez de la membrana.Se observo que cuando se emitıa luz sobre estas vesıculas gigantes y se variaba
la frecuencia de la luz emitida desde luz roja hasta luz amarillenta-verdosa, lapendiente de la recta, y en consecuencia, la rigidez disminuıa.
Esto indica que el exceso de area es mayor cuando se ilumina con luzamarillenta-verdosa, es decir, la actividad de la bR induce una amplificacionde las fluctuaciones de la membrana.
De esta seccion concluımos que los centros activos cambian considerablemen-te el estado de una membrana. Este experimento es muy claro, pues simplementevariando la frecuencia de la luz emitida sobre la membrana se observaban cam-bios importantes en el valor de la rigidez, lo que indicaba que los centros activosinducen una amplificacion importante de las fluctuaciones.
A partir de ahora, nos centraremos en otro tipo de proteınas, los motoresrotatorios, pero motivados por el hecho que con la bR hemos visto experimen-talmente un realzamiento de las flucutuaciones de membrana.
1.5. Ejemplos de motores rotatorios (rotores)
J. Prost et al. [10], [11] estudian una membrana activa como un conjunto demotores rotatorios en una superfıcie plana.
Este estudio esta motivado por sistemas biologicos en que procesos activosllevan a movimientos rotatorios en membranas y generan un flujo hidrodinamico.
Vamos a describir 2 posibles ejemplos de motores rotatorios: la ATP-sintasay los cilios. Ambos son posibles realizaciones de membranas activas ya que

1. Introduccion: Membranas y motores rotatorios 11
estan anclados en la membrana y cambian la dinamica del sistema debido a sumovimiento.
1.5.1. La ATP-asa
La ATP-asa es una encima universal que fabrica ATP a partir de ADP yfosfato, lo hace utilizando la energıa del transporte de protones a traves de lamembrana. Tambien se puede invertir ella misma e hidrolizar ATP para bombearprotones en contra del gradiente electroquımico. Los ciclos de sıntesis e hidrolisisllevan a la ATP-asa a un movimiento de rotacion [12] [13].
Cumple un papel importante en las celulas ya que proporciona la mayorıa dela energıa quımica que los organismos aerobicos y fotosinteticos necesitan paramantenerse vivos.
La ATP-asa es un complejo proteınico grande y asimetrico que esta com-puesto por distintas unidades. Basicamente, consiste en 2 multi-subunidadescomplejas distintas en estructura y funcion que estan enlazadas. La porcion hi-drofobica F0 esta anclada en la membrana y realiza el transporte de protones atraves de la membrana, mientras que la porcion F1 reside en la periferia acuosay cataliza la sıntesis/hidrolisis del ATP (Fig. 6).
Juntas acoplan el flujo de protones bajo el gradiente electroquımico con lasıntesis del ATP [14].
F0 y F1 funcionan como un par de motores rotatorios enlazados por un rotorcentral y un estator (que es el circuito fijo dentro del cual gira el rotor) periferico.
Un experimento que se hizo fue atar un filamento de actina fluorescenteen un final de la subunidad C12 de F0. A traves de microscopıa fluorescente,se vio que durante la hidrolisis del ATP el filamente experimentaba multiplesrotaciones en el sentido contrario al de las agujas del reloj. La rotacion de estasubunidad C12 ocurrıa durante la sıntesis del ATP [15].
Esta subunidad, y por consiguiente este filamento, esta rotando usando laenergıa de la hidrolisis del ATP.
El torque producido por esta subunidad es de 40pN ·nm, que es bastante pa-recido al generado por la subunidad γ del motor F1. Por lo tanto las subunidadesγ y C12 rotan juntas durante la sıntesis y la hidrolisis del ATP.
La direccion de los eventos quımicos acoplados (ATP sıntesis e hidrolisis) yel transporte de protones (bajo o en contra del gradiente electroquımico) puedeser revertida si invertimos la direccion de la rotacion de dicha subunidad [15].
Esta rotacion acoplada es esencial para el acoplamiento energetico entre eltransporte de protones a traves de F0 y la sıntesis e hidrolisis de ATP en F1.
Cuando los protones fluyen a traves de F0, se sintetiza ATP en F1. Estohace que la cola gire rapidamente, induciendo a la cabeza a fabricar ATP. Elmotor es reversible y un exceso de ATP provoca una rotacion en la direccionopuesta y un flujo reverso de protones. Durante la catalisis un complejo formadopor ciertas subunidades se mueve rotando en relacion al resto de la encima. LaATP-asa es el motor rotatorio mas pequeno nunca conocido (Fig. 7).
Esta encima es la fuente principal de ATP en la mayorıa de especies vivas dela tierra, incluidos los seres humanos. El cuerpo humano genera mas de 100 kg

1. Introduccion: Membranas y motores rotatorios 12
Figura 6: Esquema sencillo de la ATP-asa.
Figura 7: Diferentes perspectivas de la ATP-asa.

1. Introduccion: Membranas y motores rotatorios 13
de ATP, que es secuencialmente usado para proveer de energıa para diferentesreacciones bioquımicas, incluyendo la sıntesi proteica y de DNA, la contraccionde musculos, el tranporte de nutrientes y la actividad neuronal.
En plantas y bacterias fotosinteticas es esencial para convertir la energıasolar.
Es una encima membranosa. Esta situada en la membrana plasmatica, encloroplastos y en la membrana mitocondrial interna de celulas eucariotas.
El flujo hidrodinamico generado por estos motores puede cambiarlas propiedades de las membranas en las que estan situados y puederealzar la difusion de proteınas de membrana. Un ejemplo de ellos esen los globulos rojos, ya que las fluctuaciones de la forma de los globulos rojosdepende no solo de la viscosidad del medio sino de las concentraciones de ATP.
Las ATP-asas influencian a la hidrodinamica del sistema de 2 maneras:
1. Es un motor rotatorio que ejerce un torque dipolar.
2. Es una bomba de protones, la cual ejerce una fuerza cuadrupolar local.
Estos dos mecanismos estan desacoplados en el regimen lineal en el quenosotros estamos interesados.
Estos motores rotatorios actuan como pequenos microvortices e inducen unflujo en el fluido que rodea la membrana. Como estamos usando hidrodinamicalineal, el flujo inducido siempre sera paralelo al plano medio de la membrana.
Segun [11] una fuerza cuadrupolar es la distribucion de fuerza mas sencillaque es consistente con el movimiento rotacional del motor y que cumple elrequerimiento de no tener fuerzas externas presentes.
En la siguiente seccion presentaremos un modelo que simula estos objetos ynos centraremos en las consecuencias hidrodinamicas que podemos extraer.
1.5.2. Los cilios
Los cilios son estructuras largas y delgadas (0,25µm) parecidas a los ca-bellos, ver Fig. 8. Estan formados por grupos especializados de microtubulos(motores de los cilios) llamados cuerpos basales. Estos cilios se extienden desdela superficie de varios tipos de celulas eucariotas.
Las celulas pueden autopropulsarse y generar un flujo gracias a los cilios, yaque estos perturban el fluido que rodea la superficie y mueven la celula a travesdel fluido.
El movimiento cıclico de un cilio puede ser muy complicado, pero siempreexhibe un movimiento rotacional. Podemos decir que los cilios se comportan demanera efectiva como motores rotatorios.
Una celula tiene normalmente centenares de cilios, que se mueven de unamanera coordinada, con movimientos ondulatorios: ondas metacronales. Paradescribir este fenomeno es necesario tener en cuenta las interacciones hidro-dinamicas entre cilios vecinos [16].

2. Un modelo de motores rotatorios 14
Figura 8: Cilios respiratorios.
2. Un modelo de motores rotatorios
2.1. Modelo teorico
El modelo teorico que vamos a seguir es el propuesto por J. Prost et al. [10],[11], en el que, como hemos dicho antes, estudian una membrana activa comoun conjunto de motores rotatorios en una superfıcie plana.
En esta superficie, los rotores actuan como microvortices que inducen unflujo en el fluido que los rodea.
El hecho de no tener en cuenta la curvatura es solo una aproximacion de unamembrana, pero el flujo inducido por los rotores es paralelo al plano medio dela membrana, ası que la aproximacion de no tener en cuenta el bending no estan mala.
De hecho lo que estamos haciendo es ignorar el acoplamiento entre las bom-bas y la curvatura de la membrana.
Ademas, no tenemos en cuenta que una membrana no es una superfıcie,sino una bicapa. Estamos usando la representacion de Monge, en la que nuestramembrana se puede describir como una superficie, los puntos de la cual vienendados por la altura sobre el plano xy. Esta aproximacion es util en membranascasi planas y como para nosotros una membrana es plana, la representacion deMonge es muy apropiada para nuestro estudio.
Debemos recordar que estamos suponiendo la bicapa lipıdica como una flui-do superficial continuo de viscosidad cinematica ν (fluid mosaic model), las

2. Un modelo de motores rotatorios 15
proteınas estan suspendidas en este fluido.Es posible variar la concentracion de proteınas en el fluido hasta mas del
50 %, ya que es lo que podemos encontrar en membranas biologicas.
2.2. Metodo de simulacion
Para simular un fluido computacionalmente se pueden modelizar las molecu-las individuales que forman dicho fluido (Dinamica Molecular). Si las interac-ciones entre ellas estan correctamente modelizadas, el sistema que estamos si-mulando se comportara como un fluido, estaremos resolviendo apropiadamentelas ecuaciones de Navier-Stokes partiendo de un metodo puramente molecular.
Sin embargo, la gran desventaja de este metodo es el tiempo computacional.En cada paso de tiempo se deben identificar las parejas que pueden colisionaren el siguiente paso de tiempo y se deben aplicar los campos externos. Ademas,cada paso de tiempo de dinamica molecular es aproximadamente t ∼ 10−13s deltiempo real. Por lo tanto, se requiere mucho tiempo computacional para hacervariar la velocidad y la posicion de cada partıcula lo suficiente para tener unadinamica a tiempos largos.
Incluso en el caso de gases, donde el paso de tiempo es mas largo ya queel recorrido libre medio de cada molecula es mayor, el numero de moleculasque se deben considerar es demasiado grande para poder simular el sistemaeficientemente.
2.2.1. Lattice gas (LG)
En los anos 70 se desarrollo este metodo de computacion, que es cualitati-vamente diferente de la dinamica molecular.
En vez de considerar las moleculas individualmente, se consideran partıculasde fluido, donde cada una de estas es un grupo de moleculas (coarse-graining),que ocupan un cierto volumen, que es una fraccion aun muy pequena del volu-men del sistema, es decir, estas partıculas fluidas aun son muy pequenas.
Esto reduce drasticamente los datos que se necesitan computar.Este coarse-graining esta justificado en el hecho que las propiedades ma-
croscopicas no dependen directamente del comportamiento microscopico delfluido.
El modelo impone que estas partıculas de fluido estan obligadas a moverseen enlaces (links) de una red regular, y que este movimiento se da en pasos detiempo discretos. Las leyes de conservacion fısicas se incorporan en el momentodel update de las posiciones y velocidades, y se aplican a cada paso de tiempo.Ver en Fig. 9 un caso sencillo de aplicacion de este modelo.
En un paso de tiempo dejamos que las partıculas fluidas se propaguen ycolisionen con otras. Solo se pueden mover hacia nodos vecinos, es decir, soloexiste un conjunto finito de velocidades ~ci, donde i es el ındice que va de 1 aNv, siendo Nv el numero de vecinos. Tenemos ni(~r, t) partıculas situadas en elnodo ~r con velocidad ~ci.

2. Un modelo de motores rotatorios 16
Figura 9: Sistema muy pequeno, de 11 partıculas en 15 nodos de fluido y 5 nodosde pared. Una simulacion tıpica usa 106 nodos y 2106 partıculas.
Otra ventaja de este modelo es que todas las colisiones entre partıculasocurren en el mismo tiempo, debido al hecho que los pasos de tiempos sondiscretos. Las reglas que gobiernan las colisiones son tales que el movimientomedio de las partıculas sea consistente con la ecuacion de Navier-Stokes.
Los calculos con LG requieren promedios para recuperar las ecuaciones ma-croscopicas. Con estos promedios a tiempos largos, esta tecnica esta restringidaa flujos que varıan lentamente [17].
2.2.2. Lattice-Boltzmann
LB usa distribuciones probabilısticas de partıculas, en vez de partıculas dis-cretas, es aplicar el metodo de punto flotante sobre LG.
Cada punto de la red es un elemento de fluido, y es representado por unafuncion distribucion de partıculas. Es decir, reemplazamos las variables de reddel Lattice gas ni(~r, t) por sus promedios en la colectividad fi(~r, t), dondefi(~r, t) =< ni(~r, t) >. Estas cantidades promediadas fi(~r, t) son ahora fun-ciones reales en el rango 0 ≤ fi ≤ 1, donde la masa y el momento promediosestan dados por
ρ(~r, t) =∑
i
fi(~r, t) (3)
ρ(~r, t)~u(~r, t) =∑
i
fi(~r, t)~ci (4)

2. Un modelo de motores rotatorios 17
La evolucion (translacion y colision) de fi esta dada por el promedio en lacolectividad de la funcion de colisiones (ecuacion de Boltzmann):
fi(~r + ~ci, t+ 1)− fi(~r, t) =< Ωi(n) > (5)
El termino de colisiones < Ωi(n) > puede aproximarse, segun la expresionde tiempo de relajacion:
< Ωi(n) >= −fi − feqi
τ(6)
Donde 1/τ es un tiempo relaxacional y feqi es la distribucion de equilibrio,
tıpicamente la distribucion de Maxwell-Boltzmann.Este termino de colisiones tan sencillo es suficiente para reproducir la dinami-
ca de la ecuacion de Navier-Stokes.No tenemos en cuenta las fluctuaciones termicas, nos queremos centrar en
los efectos hidrodinamicos.
El metodo lattice Boltzmann es una tecnica computacional potente paramodelizar muchas variedades de problemas sobre flujos de fluidos complejos,incluyendo diferentes fases y geometrıas complejas. Es un metodo computacionaldiscreto, basado en la ecuacion de Boltzmann.
Las simulaciones de este trabajo estan hechas con lattice-Boltzmann.
2.3. Dinamica de partıculas en LB
Queremos simular motores rotatorios suspendidos en un fluido, que esta des-crito por la red de nodos de lattice-Boltzmann. Para ello debemos modelizarpartıculas solidas. Por simplicidad, a partir de ahora siempre trataremos conpartıculas esfericas (3d) o discos (2d), lo que llamaremos coloides.
Podemos modelizar coloides suspendidos en dicho fluido como partıculas soli-das y esfericas en las que se impone la condicion de contorno de stick mediantelas reglas de bounce-back, es decir, imponer que la velocidad del fluido alrededorde la partıcula sea igual que la velocidad del centro de masas de la partıcula.Estas partıculas, a su vez, evolucionan siguiendo las reglas de la dinamica mo-lecular, es decir, las ecuaciones de Newton.
Por lo tanto estamos acoplando la hidrodinamica del solvente con este con-junto de partıculas, como es habitual en suspensiones coloidales.
Reglas de bounce-back: las partıculas solidas estan definidas por una su-perficie, que corta algunos enlaces entre nodos de la red. Una partıcula fluidamoviendose en uno de estos enlaces interactua con la superficie en nodos fron-teras situados a mitad de camino del enlace. Tenemos una descripcion discretade la superficie (Fig. 10).

2. Un modelo de motores rotatorios 18
Figura 10: Reglas de bounce-back
Cada nodo frontera tiene 2 funciones distribucion, que provienen de los 2extremos del enlace. Al colisionar con la superficie se transfiere la densidad deprobabilidad a traves del nodo frontera, cambiando la densidad de momentolocal tal que tenga en cuenta la velocidad del cuerpo rıgido, pero sin afectar lamasa ni la presion.
fi(~r + ~ci, t+ 1) = f∗i (~r + ~ci, t) +2aciρ~ub · ~ci
c2s
fi′(~r, t+ 1) = f∗i (~r, t)− 2aciρ~ub · ~cic2s
cs es la velocidad del sonido y el coeficiente aci del termino ~ub ·~ci esta deter-minado por el hecho que toda distribucion consistente con la velocidad de nodofrontera ~ub es estacionaria respecto a las interacciones con los nodos frontera[18].
Por lo tanto, se cambia momento localmente entre el fluido y la partıculasolida, pero el momento total del sistema se conserva.
El fluido llena todo el sistema, incluıdo el interior de las partıculas, pero elfluido interior relaja rapidamente a la velocidad del cuerpo rıgido.
Ademas, debido a la dinamica, las partıculas colisionan entre ellas. Cada

2. Un modelo de motores rotatorios 19
colision debe cumplir la condicion de conservar el momento total del sistema.
2.4. Motores rotatorios en LB
Queremos modelizar un motor rotatorio, encontrar un mecanismo con el quepodamos simular partıculas que tengan una cierta velocidad angular propia, seautopropulsen angularmente.
2.4.1. Partıculas activas
Antes de modelizar un motor rotatorio observemos la modelizacion de unmotor lineal, es decir una partıcula activa autopropulsada, ya que el fundamentodel mecanismo es el mismo.
He desarrollado un mecanismo simple que lleva a la autopropulsion de uncoloide [19]. Introducimos este mecanismo en vez de intentar simular un modelode propulsion mas realista. Tratamos esferas de radio R y seleccionamos unadireccion de movimiento y un conjunto de nodos frontera alrededor de estadireccion, que seran los que cumplen la condicion siguiente: ser vecinos de nodosinteriores a la partıcula y estar dentro de un cono de angulo α, ver Fig. 11.
En nuestras simulaciones siempre escogemos α = π, aunque tambien hemoscomprobado que los resultados no dependen de α.
La regla consiste en extraer una cierta cantidad de momento ∆~p equitativa-mente distribuida en todos los nodos que cumplen la condicion antes mencionaday esta misma cantidad de momento la anadimos al momento de la partıcula.
Esta partıcula estara, mediante este mecanismo, autopropulsada. Ademas,este mecanismo no viola la conservacion del momento lineal total.
El campo de velocidades inducido por este mecanismo es dipolar.Alternativamente, podemos tambien seleccionar los nodos del fluido que sean
frontera que sean interiores a un cono de angulo ψ0 con respecto a π en la mismadireccion que la direccion de movimiento seleccionada previamente. Ahora ∆~p esextraido equitativamente de todos los nodos seleccionados y anadido al momentode la partıcula, ver Fig. 11. De esta manera, la partıcula se propulsa generandoun campo de velocidades a su alrededor que sera asimptoticamente cuadrupolar.
Como estamos interesados en la autopropulsion de objetos que son pequenos,debemos escoger el parametro ∆~p de manera que el numero de Reynolds (Re =uL/ν) sea lo suficientemente pequeno.
Este modelo es muy simple y no tiene en cuenta la biologıa de estos mi-croorganismos, pero tiene en cuenta el acoplamiento hidrodinamico entre lasuspension de microorganismos y el solvente que los rodea, y recupera la ade-cuada interaccion entre biocoloides activos. De esta manera podemos saber elpapel de la hidrodinamica en suspensiones de objetos auto-propulsados.
Estas partıculas, partiendo del reposo, llegan a tener una velocidad estacio-naria debido unicamente al mecanismo y al hecho que estan sumergidas en unfluido viscoso. La velocidad asintotica viene dada por el balance entre la trans-ferencia de momento y la friccion. En el caso tridimensional, es la siguiente:

3. Resultados 20
δpα
δp2δp
Figura 11: Mecanismo introducido para simular partıculas activas.
~u =∆~p
6πρηR12
(7)
No hay una fuerza neta aplicada sobre esta partıcula, es por ello que la ve-locidad inducida por estas partıculas activas es menor que en partıculas pasivasbajo la accion de una fuerza externa aleatoria.
2.4.2. Mecanismo de rotacion
A partir del modelo de partıculas activas construyo un modelo de trans-ferencia de momento angular para tener un mecanismo de autopropulsionangular, que tambien conserve el momento lineal y angular total, ver Fig. 12.
Usando que ~L = 2|δ~p|Rx, donde el eje x es perpendicular al plano de lahoja o la pantalla donde esta inscrita, impongo que el sentido de rotacion sea elcontrario del de las agujas del reloj [20].
3. Resultados
A continuacion presento los resultados obtenidos a partir de simulacionesmediante el metodo lattice-Boltzmann antes explicado. Primeramente me cen-trare en el campo de velocidades creado por un motor rotatorio. A continuacion

3. Resultados 21
2δpδp α δp
L
Figura 12: Mecanismo propuesto para simular partıculas autorpropulsadas an-gularmente.
analizare el caso de 2 rotores y el de una cadena de rotores para comprobar si secorresponde con lo predicho por el modelo teorico. Finalmente me centrare en elcaso de un conjunto N de rotores y en el analisis del comportamiento colectivoasociado.
3.1. Analisis de 1 rotor
El mecanismo anteriormente presentado es suficiente para captar el hecho deque una partıcula inicialmente en reposo empezara a rotar cada vez mas rapidohasta alcanzar una velocidad angular constante estacionaria. Ver Fig. 13.
Debido a su orıgen hidrodinamico, la velocidad angular asintotica es alcan-zada algebraicamente. Siendo d la dimensionalidad del sistema:
ω(t)− ω∞ ∼ t−d/2 (8)
Esta expresion es equivalente a la obtenida en [19] para las partıculas activas.La escala de tiempo necesaria para alcanzar dicha velocidad angular esta con-
trolada por el tiempo en el que el flujo hidrodinamico se establece ya que elmomento se ha difundido: τr ∼ R2/ν, en el caso de una partıcula suspendidaen un fluido de viscosidad cinematica ν.

3. Resultados 22
0 2 4 6 8 10t/τ
r
0
0.25
0.5
0.75
1
ω/ω
0
Figura 13: Velocidad angular de un rotor en funcion del tiempo, escalado porel tiempo de difusion del momento. En este caso τr = 2025 ya que R = 4,5y ν = 0,01. El vector velocidad angular es negativo porque es en el sentidocontrario de las agujas del reloj.
Debido al balance entre la transmision de momento, que en realidad generamomento angular, y a la friccion, esperamos una relacion de la velocidad angularcon los parametros del sistema del mismo tipo que encontramos con partıculasactivas [19]:
~ω =∆~p
6πρηR2
12
(9)
He analizado, a partir de simulaciones, el mecanismo de propulsion basicoy como depende la velocidad angular estacionaria de los parametros relevantesdel sistema: R, ν and ∆~p.
La dependencia en la viscosidad es la misma que en el caso de partıculasactivas: v ∼ 1/ν, ver Fig. 14. La dependencia en R es diferente, pero sim-plemente es porque ahora estamos calculando una velocidad angular en vez deuna velocidad lineal, ahora ω ∼ 1/R2, ver Fig. 15. El error en este grafico apequenos valores de R es debido a que en lattice Boltzmann la estimacion quese hace del radio es mala para valores pequenos del radio.
3.1.1. Campo hidrodinamico inducido por los rotores
Un motor rotatorio de radio R en una superfıcie de viscosidad despreciablesuspendido en un fluido newtoniano de viscosidad η para Re → 0 induce uncampo de velocidades en el fluido que es solucion de la ecuacion de Navier-Stokes.

3. Resultados 23
0 5 10 15 201/ν
0
4×10-3
8×10-3
ω(ν)
R=2.5R=3.5R=4.5R=5.5R=6.5R=7.5
Figura 14: Velocidad angular estacionaria del rotor en funcion de la viscosidaddel fluido (ν).
0 0.05 0.1 0.15 0.2
1/R2
0
2×10-3
4×10-3
6×10-3
8×10-3
ω(R)
ν=0.05ν=0.1ν=0.3ν=0.5ν=0.7ν=1
Figura 15: Velocidad angular estacionaria del rotor en funcion del radio de lapartıcula (R).

3. Resultados 24
x
z
yω
Figura 16: Motor rotatorio en el plano yz, su vector velocidad angular es paraleloal eje x.
ρ
(∂~v
∂t+ ~v · ~∇~v
)= −~∇p+ η~∇2~v (10)
Queremos saber el estado estacionario, por lo tanto no consideramos la de-rivada parcial temporal.
Nuestro problema tiene simetrıa rotacional (cilındrica), ver Fig. 16: ~ω = ωx,lo que significa que ~v = v(r)ϕ, donde ϕ es el angulo polar del plano yz. Estadependencia hace que el termino convectivo (el termino no-lineal) de Navier-Stokes se anule y, ademas, que el flujo sea incompresible: ~∇ · ~v = 0.
Esta simetrıa rotacional impone tambien que la presion sea constante, nohay nada en el sistema que pueda imponer gradientes de presion.
Por lo tanto, la ecuacion de Navier-Stokes (10) en esta situacion es simple-mente una ecuacion de Laplace para la velocidad del rotor:
~∇2~v = ~0 (11)
Al ser un fluido incompresible, podemos expresar ~v como el rotacional deotro vector.
Teniendo en cuenta las condiciones de contorno (~v(r = R) = ~ω ∧ ~r y v(r →∞) = 0) obtenemos el campo de velocidades inducido por el rotor:
~v(~r) =R3
r3~ω ∧ ~r (12)

3. Resultados 25
Figura 17: Campo de velocidadesoriginado por 1 rotor. t ∼ 0,5τr.
Figura 18: Campo de velocidadesoriginado por 1 rotor. t ∼ 5τr.
Una conclusion que podemos sacar de la ultima expresion es que el campohidrodinamico es independiente de la viscosidad.
En las Fig. 17 y 18 vemos la evolucion del campo de velocidades creado porun rotor de radio R = 4,5 suspendido en un fluido de viscosidad ν = 0,01 adiferentes tiempos. En ellas vemos que el campo inducido es cuadrupolar, comopredecıa el modelo teorico [11].
En la Fig. 19 vemos el caso en el que hay 9 rotores de este tipo distribuidosal azar en el mismo fluido.
3.2. Estudio de 2 rotores
Segun el modelo teorico de Prost et al. [11], en el caso de 2 rotores, estossimplemente rotan alrededor de su centro de masas y su interaccion no tienecomponente radial. Es necesario la presencia de rotores adicionales, que daranlugar a una interaccion radial que, generalmente, es repulsiva.
Para ver el efecto de las interacciones hidrodinamicas, estudiamos el sistemade dos rotores separados una distancia d y que giran con una cierta velocidadangular ω dentro de un fluido de viscosidad cinematica ν (Fig. 20).
Las interacciones dependen inversamente de la distancia al cuadrado, por lotanto la interaccion con las imagenes sera muy debil si el sistema es suficien-temente grande y los rotores estan suficientemente cerca. Es decir, imponemosque d L, donde L es el tamano del sistema.
Como hemos visto en la eq. 12, la velocidad lineal depende del producto

3. Resultados 26
Figura 19: Campo de velocidades del fluido originado por 9 rotores suspendidosen un fluido de ν = 0,01 a t ∼ 5τr.

3. Resultados 27
d
2R
ωω
Figura 20: Configuracion inicial dela pareja de rotores. Figura 21: 2 partıculas inicialmen-
te separadas en el eje de las abcisas,se moveran siguiendo una orbita res-pecto al centro de masas de ambaspartıculas, en este caso coincide conel centro geometrico debido a quesus masas son iguales.

3. Resultados 28
vectorial de la velocidad angular con el vector que une el rotor con el puntoque queremos evaluar. Debido a la anticommutatividad del producto vectorialdos partıculas rotando en el mismo sentido y separadas una distancia d se mo-veran inicialmente en direcciones perpendiculares a su orientacion relativa ycada una en un sentido diferente. Como la direccion relativa entre los motoresvariara debido al desplazamiento de cada uno de ellos, las direcciones de mo-vimiento tambien cambiaran. De hecho, el movimiento que seguiran sera el deorbitas elıpticas alrededor del centro de masas del sistema de 2 partıculas, comopredice el modelo teorico, ver Fig. 21.
Este sistema nos dice que cuando en una membrana tenemos a 2 rotoresmuy cercanos y los otros estan muy alejados, ellos rotaran alrededor del centrode masas del sistema de 2 partıculas, hasta que una tercera partıcula perturbeesta configuracion.
Para un R y d fijados, vamos variando el parametro ∆p y obtenemos unarelacion v = v(ω) (Fig. 22):
0 0.01 0.02 0.03 0.04 0.05ω
0
0.02
0.04
0.06
v(ω)
Figura 22: Velocidad de 2 rotores en orbita alrededor del centro de masas delsistema de 2 partıculas en funcion de la velocidad angular de ambas partıculas.
Se debe decir que este calculo esta hecho en 3d, la relacion es parecida para2d.
3.3. Cadena de rotores
Imponemos una distribucion inicial de rotores equiespaciados en una cadenaunidimensional recta, rotando todos ellos en el mismo sentido (Fig. 23).
La interaccion hidrodinamica sobre el rotor i, debido a que decrece como1/r2, sera basicamente la ejercida por los rotores i− 1 e i+ 1. Debido a que lasinteracciones entre rotores dependen de ~ω∧~r, estas dos fuerzas se compensaran.

3. Resultados 29
De esta manera, una cadena de este tipo sera estable. Lo hemos comprobadoen nuestro modelo, donde las velocidades de cada rotor son muy pequenas (∼10−7) y van oscilando alrededor de 0. De esta manera validamos la interaccionentre partıculas predicha por el modelo teorico.
3.4. Comportamiento colectivo:
3.4.1. Movimiento inducido
Un rotor, por si mismo, no tendra movimiento lateral (velocidad lineal), peroun rotor suspendido en un fluido con otros rotores sı se llegara a mover, y lohara debido a las interacciones hidrodinamicas con los otros rotores. Es decir,la propulsion de cada partıcula no es una cantidad propia sino que es debida alhecho que la partıcula se encuentra dentro del campo hidrodinamico generadopor otras partıculas.
Las interacciones entre partıculas cambiaran la velocidad de cada una, que-remos ver la dependencia de la velocidad con la fraccion volumica. Obviamente,el caso lımite de fracciones volumicas pequenas dara una velocidad nula. Estono es ası en un conjunto de partıculas autopropulsadas, debido a que allı lapropulsion es propia, las interacciones solamente cambian las velocidades. Aquı,ademas del cambio que originan, esta el hecho de que una partıcula no se mo-vera si no tiene otras partıculas cerca.
Es por esta razon que la velocidad de translacion no es monotona decrecientecomo funcion de la fraccion volumica, la velocidad angular sı es decreciente (Fig.24).
Hacemos el analisis a tiempos cortos y a Re pequeno. La velocidad de trans-lacion tiene un maximo (Fig. 25) debido a la competicion de los dos efectos: estemaximo φ∗ ∼ 0,13 es una caracterıstica del sistema y en principio es constante,no depende de la viscosidad ni de otros parametros del sistema.
3.4.2. Distribucion de velocidades
Hacemos un analisis a tiempos cortos, que consiste en hacer la simulacionsin dejar mover a las partıculas aunque dejando evolucionar las velocidades. Esdecir, es hacer el analisis cuando los rotores inducen un campo pero aun noha pasado el suficiente tiempo como para que ellos se hayan movido debido alcampo inducido por otros.
Analizamos las distribuciones de velocidades de los rotores y de los nodosdel fluido. Los siguientes graficos son de una suspension de 400 rotores en unfluido de ν = 0,01. La fraccion volumica es φ ∼ 0,3.
Para los rotores vemos que la distribucion no es gaussiana (Fig. 26). En vezde eso tiene un comportamiento algebraico (tipo 1/v) para velocidades pequenas,seguido de un decaimiento exponencial que puede ser gaussiano.
Para los nodos del fluido vemos que, a pesar de que en la escala lineal pareceuna distribucion gaussiana (Fig. 27), al usar la escala semilogarıtmica vemosdesviaciones importantes, el pendiente de la recta disminuye segun vamos avelocidades mayores (Fig. 28).

3. Resultados 30
d
2R
ωω ωω
Figura 23: Cadena de rotores equiespaciados
0 0.1 0.2 0.3 0.4 0.5 0.6φ
0
2×10-3
4×10-3
ω (φ)
Figura 24: Velocidad angular en funcion de la fraccion volumica.

3. Resultados 31
0 0.1 0.2 0.3 0.4 0.5 0.6φ
0
2×10-3
4×10-3
v(φ)
Figura 25: Velocidad lineal en funcion de la fraccion volumica.
0 5 10
(v/u)2
-4
-2
0
2
ln n
(v)
Figura 26: Distribucion de velocidades de los rotores.

3. Resultados 32
-4 -2 0 2 4v/u
0
0.2
0.4
0.6
0.8n(v
)
Figura 27: Distribucion de velocida-des del fluido en escala lineal, tieneun gran parecido con una gaussiana.
0 2 4 6 8
(v/u)2
-8
-6
-4
-2
0
ln n
(v)
Figura 28: Distribucion de velocida-des del fluido en la escala semilo-garıtmica, en ella vemos las desvia-ciones respecto a una gaussiana.
Sin embargo, debido a que la estadıstica necesaria para resolver bien losgraficos es enorme, no podemos decir nada concreto sobre los comportamientosde estas distribuciones, solo hacer la observacion de que tenemos desviacionesrespecto a las distribuciones gaussianas, lo que significa que la naturaleza deesta dinamica de tiempos cortos es claramente fuera del equilibrio.
3.4.3. Tiempos largos
Definimos como tiempo largo al tiempo para el cual las partıculas (los roto-res) se han desplazado una fraccion considerable del tamano del sistema.
Calculamos el desplazamiento cuadratico medio, definido como:
< ∆r2(t) >=1N
N∑i=1
(r2i (t+ τ)− r2i (τ)
)(13)
siendo N el numero de partıculas suspendidas en el fluido.Vemos (Fig. 29) que despues de un regimen transitorio (regimen I) en el
que las partıculas, partiendo del reposo, se aceleran el desplazamiento pasa a serbalıstico (regimen II), en el que las partıculas se desplazan como < ∆r2 >∼ τ2,cada uno de los rotores viaja independientemente de los demas.
Despues hay un crosover a un regimen superdifusivo (regimen III), en el que< ∆r2 >∼ τα, donde α ∼ 1,5. Este exponente no es muy lejano al 5/3 predichoen la teorıa RTP [21] [5].
Posteriormente un regimen difusivo (regimen IV), en el que el desplazamientocuadratico medio es lineal con el tiempo.

3. Resultados 33
4 6 8 10ln t
-2
0
2
4
6
8
ln <
∆r2
(t)>
I
II
III
IV
Figura 29: Desplazamiento cuadratico medio para una suspension de 400 rotoresen un fluido de ν = 0,01. La fraccion volumica es φ ∼ 0,3.
3.5. Cristal de Wigner triangular
Este sistema muestra, en contraste con el caso de membranas con canaleso bombas, un comportamiento colectivo no trivial debido a que el movimientorotatorio puede inducir interacciones hidrodinamicas repulsivas entre los rotores.
Las interacciones hidrodinamicas efectivas entre motores rotatorios son delargo alcance y repulsivas, decaen como 1/x. En ausencia de fluctuaciones termi-cas, el estado estacionario es un cristal de Wigner triangular, ver Fig. 30.
Cuando la velocidad angular estacionaria de las partıculas es alta, los moto-res pueden formar esta estructura cristalina triangular. Para velocidades meno-res forman una fase desordenada.
Esta estructura triangular es marginalmente estable. Solo puede ser estabi-lizada a traves de la fuerza inercial de Magnus FM o su analogo viscoso FV
M .Deberıamos obtener orden hexatico en la distribucion de partıculas en el
fluido. Se deberıa notar en el comportamiento del desplazamiento cuadraticomedio, que deberıa tener un plateau a tiempos largos.
Entonces se podrıa caracterizar la transicion de fase para un cierto valor dela velocidad angular ω∗ para la cual si ω > ω∗ los rotores se ordenan como uncristal de Wigner y para ω < ω∗ no se ordenan.

3. Resultados 34
Figura 30: Cristal de Wigner triangular
Parametro de orden hexatico Es una medida de si la distribucion de moto-res rotatorios tiende a ordenarse segun una red triangular definimos el parametrohexatico:
Ψ6 =1N
N∑i=1
ψ6(~xi) (14)
ψ6(~xi) =1Ni
Ni∑j=1
ei6θj(~xi) (15)
cogemos la parte real de este parametro de orden:
ψ6(~xi) =1Ni
Ni∑j=1
cos (6θj(~xi)) (16)
donde N es el numero total de partıculas, Ni es el numero de vecinos dela partıcula i y θj(~xi) es el angulo entre la horizontal y la recta que une laspartıculas i y j [22].
θj(~xi) = arctanzi − zj
yi − yj(17)
De hecho este parametro de orden no es mas que < cos(6θ) >, que geometri-camente tiene bastante sentido (Fig. 31).
Lo que falta es decidir el criterio de vecinidad, es decir, cuando dos partıculasson vecinas. La definicion teorica es: dos partıculas son vecinas si comparten unlımite de una celda de Voronoi. Pero nosotros haremos una aproximacion, debidoa que estamos en 2 dimensiones: una partıcula j es vecina de i si esta centradadentro de la circunferencia de radio R0 = 3R centrada en i.

3. Resultados 35
π/6
Figura 31: Definicion del angulo que define el parametro hexatico
Estabilidad del cristal En hidrodinamica lineal (ecuacion de Stokes) la redtriangular es marginalmente estable. La hidrodinamica no lineal tiene en cuentala correccion inercial de 1er orden a la ecuacion de Navier-Stokes, donde estacorreccion genera una fuerza de Magnus que actua sobre el rotor:
~FM = −2πρlR3rωeφ ∧ ~ω (18)
donde, debido al analisi lineal:
ω = 3ω∞∑
m=1
R3
m3a3= 3ξ(3)ω
R3
a3(19)
Esta fuerza de Magnus devuelve a su posicion inicial a los rotores que sedesplazan respecto a la estructura de red, estabiliza la red hexagonal. En nues-tro codigo tenemos la posibilidad de introducir esta correccion inercial en eloperador de colisiones de la ecuacion de Boltzmann.
Tiempo de relajacion a la estructura hexatica: balance entre la fuerza deMagnus y la friccion de Stokes:
6πηRdr
dt= −2πρlωr(t)ωR3 (20)
De esta manera:1tr∼ ρlω
2R5
ηa3(21)
donde a es la distancia interatomica, en nuestro caso la distancia carac-terıstica de la red. De esta manera, el tiempo de relajacion al cristal de Wigneres:
tr ∼ν
v2θφ
(22)

3. Resultados 36
siendo φ la fraccion volumica. Queremos que este tiempo sea menor queel tiempo para el que los rotores se han difundido y estan en equilibrio. Parafracciones volumicas mayores y viscosidades menores llegaremos antes a esteregimen en que la distribucion de rotores en el fluido esta en la configuracion deequilibrio.
Importancia de la geometrıa Comparacion entre discos y esferas.Hasta ahora hemos suspendido discos en una superficie plana que consi-
derabamos la membrana y que a parte era todo el fluido que tenıamos en cuentaen la simulacion.
Nos dimos cuenta que estos discos, en realidad cilindros infinitos, no tienenvorticidad, de manera que no existe una fuerza que estabilice una red hexagonal.
De esta manera decidimos suponer un fluido tridimensional en el que unaseccion del mismo es la membrana y es en esta superficie donde los rotores, eneste caso esferas, tienen libertad de movimiento. Imponemos la prohibicion deexplorar el fluido no contenido en la superficie de membrana.
Resultado negativo Hemos usado los parametros indicados para obtenerla red triangular en el caso tridimensional pero nos ha sido imposible.
De hecho hemos generado una condicion inicial consistente en N partıculasdistribuıdas segun una red hexagonal y hemos comprobado que esta red esinestable. Se producen dislocaciones que se propagan por todo el fluido y elparametro hexatico decrece de 1 a 0 en un tiempo relaxacional caracterıstico.
3.5.1. Difusion activa
Ya hemos dicho que el flujo hidrodinamico generado por motores rotatoriospuede cambiar las propiedades de la membrana. De hecho, la difusion de otraspartıculas suspendidas en el fluido aumenta debido a la interaccion hidrodinami-ca con estos rotores.
Cambio del coeficiente de difusion. Integral de Green-Kubo (autocorrelacionde las velocidades angulares).
δD =12
∫ ∞
0
dt < ~v(~x, t) · ~v(~x, 0) > (23)
donde Dm es la difusion de equilibrio, y segun [11]:
δD
Dm∼ 9πρd2
16(τ
kBT)2 (24)
Las fluctuaciones de la densidad local de motores rotatorios induce fluctua-ciones locales del campo de velocidades.

4. Conclusiones 37
3.6. Fluctuaciones: acoplamiento entre los rotores y lasfluctuaciones de la membrana
Introducir la temperatura en el modelo. Un ruido de no-equilibrio aumentalas fluctutaciones de la forma de la membrana.
Asumiendo que los motores permanecen con el eje de giro perpendicular ala membrana y que el flujo creado por el motor puede ser caracterizado porlos torques ejercidos sobre las 2 partes del motor en la direccion normal a lamembrana.
4. Conclusiones
En este trabajo he querido hacer una introduccion a la estructura y funcio-nalidad de las membranas, haciendo hincapie en la necesidad de considerar loscentros activos a la hora de describirlas.
He querido analizar el papel de los motores rotarios como es el caso de laATP-asa y los cilios, y para ello me he basado en el trabajo teorico realizadopor J. Prost et al..
A traves de simulaciones numericas mesoscopicas he realizado un modelosencillo de dichos motores rotatorios suspendidos en un fluido en el que las inte-racciones hidrodinamicas estan incluıdas. He querido reproducir los resultadosteoricos de Prost y, de esta manera, testar el modelo, el resultado ha sido positivoen lo que se refiere a los comportamientos individuales o de 2 partıculas.
Estudiando los movimientos colectivos, podemos llegar a entender fenomenoscooperativos que se dan lugar en las membranas biologicas.
Queremos indicar brevemente cual sera el trabajo futuro:
Estudio sistematico de las distribuciones de velocidades para esferas rota-torias y para el fluido.
Estudio de los diferentes regımenes dinamicos, caracterizacion de posiblestransiciones con la fraccion volumica. Estudio de la estructuracion de losrotores en el fluido.
Elaborar la misma teorıa con mezclas binarias. Hacer que la membranasepare 2 fluidos diferentes, caracterizados por distintas viscosidades (ν1 yν2).
Usar motores dipolares en vez de motores monopolares y ver que diferen-cias fundamentales hay en las interacciones entre ellos.
Una molecula de ATP hace una rotacion de 120o y luego esta paradadurante aproximadamente 30s, tiempo en el que se produce la catalisis.Hay modelos de dos estados, en el que los centros activos se encienden y seapagan, donde las transiciones se producen aleatoriamente para ası captarla naturaleza estocastica de estas bombas [21].

REFERENCIAS 38
El estudio de la difusion entorpecida [6] de las proteınas de membranapuede ser una prueba para saber la naturaleza de las interacciones entreproteınas y los constituyentes de la membrana donde residen. El conoci-miento de estas interacciones es la clave para conocer el funcionamientoglobal de las celulas.
Referencias
[1] B. Alberts et al., The Molecular Biology of the Cell, 4th edition (Garland,New York, 2002).
[2] P.G. Saffman and M. Delbruck, Proc. Natl. Acad. Sci. USA 72, 3111(1975).
[3] T.L. Dodd, D.A. Hammer, A.S. Sangani and D.L. Koch, J. Fluid. Mech.293, 147 (1995).
[4] S.J. Singer and G.L. Nicolson, Science 175, 720 (1972).
[5] S. Ramaswamy, J. Toner, J. Prost, Phys. Rev. Lett. 84, 3494 (2000).
[6] D.J. Muller, A. Engel, U. Matthey, T. Meier, P.Dimroth and K.Suda, J.Mol. Biol. 327, 925 (2003).
[7] H.Luecke et al., Science 286, 255 (1999).
[8] S.Subramamiam and R.Henderson, Nature 406, 653 (2000).
[9] J-B. Manneville, P.Bassereau, D. Levy and J.Prost, Phys.Rev. Lett. 82,4356 (1999).
[10] P. Lenz, J-F. Joanny, F. Julicher and J. Prost, Phys. Rev. Lett. 91, 108104(2003).
[11] P. Lenz, J-F. Joanny, F. Julicher and J. Prost, Eur. Phys. J. E 13, 379(2004).
[12] H.Wang and G.Oster, Nature 396, 279 (1998).
[13] Y. Sombongi, Y. Iko, M. Tanabe et al., Science 286, 1722 (1999).
[14] W. Junge, Proc. Natl. Acad. Sci. USA 96, 4735 (1999).
[15] R.L.Cross, Nature 427, 407 (2004).
[16] M. Cosentino Lagomarsino, B.Bassetti and P.Jona, Eur. Phys. J. B 26,81 (2003).
[17] A.J.C. Ladd and R.Verberg, J. Stat. Phys. 104, 516 (2001).
[18] C.P. Lowe, A. Masters and D. Frenkel, J. Chem. Phys. 103, 4 (1995).

REFERENCIAS 39
[19] I. Llopis and I. Pagonabarraga, preprint.
[20] Adachi K, Itoh H, Nishizaka T et al., Bioph. J. 80, 655.53 (2001).
[21] D. Lacoste and A.W.C. Lau, Eur. Phys. Lett. 10, 1209 (2005).
[22] S. Pronk and D. Frenkel, Phys. Rev. E 69, 066123 (2004).