investigaciòn cientifica
TRANSCRIPT

El aumento de acoplamiento de Redes intrínsecos
en Remitido jóvenes con depresión predice la
rumiación y control cognitivo
x
Publicado: 27 Agosto 2014
http://dx.doi.org/10.1371/journal.pone.0104366
Artículo
autores
Métrica
comentarios
contenido relacionado
Abstracto
Introducción
métodos
resultados
Discusión
información de soporte
Expresiones de gratitud
Contribuciones de autor
referencias
Comentarios de los lectores (1)
Cobertura de los medios de comunicación (0)
figuras
Abstracto
Objetivo
La conectividad funcional de resonancia magnética (fcMRI) estudios de individuos
diagnosticados actualmente con los principales hyperconnectivities documento trastorno
depresivo mayor (TDM) dentro de la red en modo automático (DMN) y entre el DMN y
las redes de la prominencia (SN) con las regiones de la red de control cognitivo
(CCN). Se necesitan estudios de los individuos en el estado remitido a abordar si los
efectos se derivan de rasgo, y no indica o las características de la carga crónicas de
MDD.
Método
fcMRI datos de dos escáneres 3.0 Tesla GE se obtuvieron de 30 jóvenes sin medicación
(47% del medicamento ingenuo) (18-23 años de edad, episodios depresivos modales =
1, la edad media de inicio = 16,2, SD = 2,6) con TDM remitidas (rMDD; modal año así

= 4) y se compara con los datos de 23 controles sanos (HC) utilizando cuatro semillas
bilaterales en el DMN y SN (corteza cingulada posterior (PCC), subgenual cingulada
anterior (sgACC), y la amígdala), seguido de comparaciones basada en voxel de todo el
cerebro.
resultados
En comparación con los HC, rMDD jóvenes exhibió hyperconnectivities de ambas
semillas PCC y con SGACC lateral, parietal, y las regiones frontales del CCN, que se
extiende a la pared medial dorsal. Un análisis factorial redujo datos extraídos y un factor
de PCC se correlacionó inversamente con la rumia entre rMDD juventud. Dos factores
de los grupos hiperconectividad SGACC estaban relacionados con el rendimiento en el
control cognitivo en una tarea pasa / no pasa, una positiva y otra inversa.
conclusiones
Hallazgos documentan hyperconnectivities de la DMN y SN con el CCN (BA 8/10),
que se relaciona con la rumia y la atención sostenida. Teniendo en cuenta estos
marcadores cognitivos son predictores conocidos de la respuesta y la recaída,
hyperconnectivities pueden aumentar el riesgo de recaída o representar a los
mecanismos de compensación.
figuras



Cita: Jacobs RH, Jenkins LM, Gabriel LB, Barba A, Ryan KA, Weisenbach SL, et
al.(2014) Aumento del acoplamiento de Redes intrínsecos en Remitido jóvenes
deprimidos Predice rumiación y control cognitivo. PLoS ONE 9 (8): e104366. doi:
10.1371 / journal.pone.0104366
Editor: Daniel Margulies, Instituto Max Planck para Cognitiva Humana y Ciencias del
Cerebro, Alemania
Recibido: 12 de febrero de, 2014; aceptado: 11 de julio de, 2014; Fecha de
publicación: 27 de de agosto de, 2014
Copyright: © 2014 Jacobs et al. Este es un artículo de acceso abierto distribuido bajo
los términos de la licencia Creative Commons Atribución , que permite el uso ilimitado,
distribución y reproducción en cualquier medio, siempre que el autor original y la fuente
se acreditan.
Financiación: Este estudio fue financiado por el NIH MH 091.811 y NCATS
UL1TR000050. Los donantes no tenía papel en el diseño del estudio, la recogida y
análisis de datos, decisión a publicar, o la preparación del manuscrito.

Conflicto de intereses: Los autores han declarado que no existen intereses en
competencia.
Introducción
El estudio de individuos con antecedentes de trastorno depresivo mayor (TDM) que se
encuentran actualmente en el estado remitido permite un examen único de los posibles
mecanismos basados en el rasgo de la depresión y la depresión recaída (por
ejemplo, [1] ).Como tal, las expresiones fenotípicas evaluados durante la remisión
pueden representar marcadores fiables de la evolución de la enfermedad, ofreciendo
objetivos refinados para futuras investigaciones en las cohortes de alto riesgo. El estudio
de los mecanismos putativos temprano en el curso del trastorno depresivo mayor
(evitando la carga crónica de cicatrices enfermedad repetitiva), durante los (efectos del
estado evitando) Estado remitido, y hacia el final del desarrollo (evitando la variabilidad
del desarrollo en la adolescencia temprana) puede proporcionar una comprensión más
clara de mecanismos de recaída y recurrencia determinado riesgo de recaída de
depresión aumenta en función de los episodios anteriores [2] y puede dar lugar a
mayores insultos neurobiológicos (por ejemplo, [3] ). Es importante destacar que, los
mecanismos identificados a través de este enfoque puede informar sobre el desarrollo de
la detección temprana y programas de prevención primaria y secundaria.
Un método para la comprensión de marcadores a base de combinaciones genéticas para
el TDM consiste en el estudio de la función de red a través de mediciones de la
conectividad de red. Estado de reposo fMRI ha surgido como un enfoque para la
identificación de biomarcadores basados en el cerebro, en particular en la detección de
variaciones en la conectividad de la red se derivan de las características clínicas [4] . Por
otra parte, en estado de reposo fMRI se ha convertido en una técnica útil para el estudio
de poblaciones psiquiátricas debido a la buena relación señal-ruido, reducción de la
carga de los participantes, y se presta a la traducción clínica [5] . Conectividad de red
interrumpida se ha documentado entre individuos dentro de un episodio depresivo
mayor (MDE [6] , [7] ). En particular, las perturbaciones en un conjunto de regiones,
incluyendo la corteza cingulada posterior (PCC), corteza prefrontal (córtex prefrontal
medial), y la corteza parietal inferior (IPC) se han reportado y son la hipótesis de
contribuir a la depresión [8] , [9] . Estas regiones están incluidos en una red de modo de
tarea por defecto negativo (DMN), que abarca las regiones que muestran disminuciones
en la activación durante la realización de tareas de atención exigente y los
correspondientes incrementos en la activación durante el descanso, vagabundeo de la
mente, o durante pensamiento auto-reflexivo (para una reseña ver [10] ).
Por el contrario, una red positivo tarea incluye regiones que aumentan de activación
durante la atención a las tareas exigentes [11] . Tarea redes positivas y la tarea negativos
actúan en oposición, ya que se han demostrado que se anticorrelated durante ambas
tareas cognitivas y durante el estado de reposo. Dos redes de trabajo positivas disociable
incluyen la Red de Control Cognitivo (CCN) y la red de prominencia (SN; [12] ). El SN
soporta el procesamiento de las emociones y la regulación autonómica e incorpora
regiones como la corteza cingulada anterior dorsal (DACC) y el frontoinsula
orbital [12] .
Estado de reposo fMRI examina el funcionamiento de la red intrínseca mediante la
captura de las correlaciones temporales entre las regiones del cerebro en la señal de

oxígeno de la sangre dependiente del nivel de (BOLD), que ofrece una
fiable [13] - [15] método para vincular las redes neuronales para aquellos rasgos que
pueden hacer que un individuo vulnerable a recaída. Estas fluctuaciones espontáneas de
baja frecuencia (<0,1 Hz) Rendimiento de mapas de los sistemas neuronales que
conforman conectoma funcional de un individuo [16] . La red positivo tarea (SN, CCN)
también puede ser estudiada durante el estado de reposo. Regiones de la SN, incluyendo
el subgenual corteza cingulada anterior (sgACC), se ha observado que se
hiperconectado en reposo entre los individuos con trastorno depresivo mayor [7] y
oscilaciones de baja frecuencia en estado de reposo redes de apoyo sesgos en el
procesamiento de la información [17] . A medida que estas redes interactúan e incluso
pueden competir entre sí para modular la atención tanto a los mundos externos e
internos [18] , y dado que la SN, en particular, puede iniciar el cambio entre CCN y
DMN; la comprensión del funcionamiento de la red durante el resto será avanzar en el
estudio del trastorno depresivo mayor. Por ejemplo, observado déficits en la activación
relacionada con las tareas entre las personas con trastornos psiquiátricos ha llevado a la
hipótesis de que la conectividad aberrante es una característica fundamental de la
enfermedad mental [19] , pero la falta de coherencia a través de experimentos de
resonancia magnética funcional basadas en tareas ha ralentizado el progreso hacia
integrada modelos de la disfunción de la red en la psicopatología.Funcionamiento de la
red aberrante es probable que la base de apoyo y síntomas clínicos observables como la
rumia (DMN) y la reactividad emocional (SN por ejemplo, [8] ), en consonancia con los
modelos de redes de psicopatología [19] .
El aumento de la conectividad dentro de y entre las redes es la hipótesis de contribuir a
la tendencia de los individuos deprimidos para asistir a los estímulos internos y para
volver de forma inadvertida a los pensamientos internos a expensas de tareas
externas [6] , [20] , [21] .Por otra parte, los síntomas depresivos, incluyendo los niveles
de mala adaptación de enfoque interno pensamiento se han asociado con una mayor
conectividad DMN [22] , mientras que la disfunción en el SN contribuye a sesgos en el
procesamiento de las emociones y la regulación autonómica ( [23] , [24] , para una
revisión ver [ 25] ). Adultos no medicados con TDM muestran una mayor conectividad
de la DMN con el SN [6] . Además, la porción lateral medial dorsal y dorsal del PFC
puede representar un nexo en la conectividad asociada depresión disfuncional entre las
tres redes [22] . Hasta la fecha; Sin embargo, la relación entre las anomalías
relacionados con la conectividad y las características clínicas y la evolución sigue
siendo relativamente inexplorada, con pocos resultados positivos (pero
véase [21] ). Además, sólo limitado trabajo ha explorado si existen estas diferencias en
los períodos de remisión [26]- [28] . No se han realizado estudios hasta la fecha con las
muestras-adolescente tardío o temprano adultos remitidos - esta época del desarrollo
puede servir como un período de evaluación y la intervención crítica del desarrollo
neuronal casi completa y la maduración de la red antes de la progresión de la
enfermedad crónica y secuelas. Por último, los investigadores han explorado enlaces
preliminares entre la disfunción de la red, rumiación y el control cognitivo [29] ; Sin
embargo, ningún estudio hasta la fecha vinculan directamente estos mecanismos.
Por lo tanto, una pregunta sin respuesta crítica es si conectividades neuronales
anormales representan marcadores de rasgo de vulnerabilidad a la enfermedad y la
recurrencia que no están relacionados con las cicatrices de enfermedades crónicas o
estatales. Para abordar esta cuestión, hemos examinado la conectividad funcional entre
los jóvenes en un estado remitido. Este diseño reduce varios factores de confusión

potenciales, incluyendo estado actual estado de ánimo, gravedad de la enfermedad, el
aumento de la cicatriz debido a episodios repetidos, y los efectos de la medicación
actual. La hipótesis de que hyperconnectivities se observarían en y entre las regiones de
la DMN y SN con las regiones del CCN entre los jóvenes no medicados con rMDD en
comparación con los controles sanos (HC), lo que refleja el potencial de riesgo basada
en el rasgo de recaída, o por el contrario la capacidad de recuperación. Por último, dada
la investigación preliminar de la correlación de la conectividad funcional DMN con
pensamientos negativos repetitivos como la rumia (por ejemplo, [21] ), así como
pruebas de que el sgACC contribuye a los procesos rumiativos [30] , se llevaron a cabo
análisis exploratorios comprobar que es auto-informe de la rumia se correlacionó con
conectividades entre aberrantes rMDD juventud. Por último, con el fin de investigar
específicamente los correlatos clínicos y de comportamiento de la hiperconectividad con
el CCN, se realizaron correlaciones de exploración de medidas de conectividad con el
rendimiento en el paramétrico Go / No-Go de tareas [31] , [32] a la luz de los trabajos
previos que sugieren inhibitoria el control predice curso de la enfermedad entre los
adultos con trastorno depresivo mayor [33] .
métodos
Participantes
El presente estudio fue aprobado por la Universidad de Michigan (UM) y la
Universidad de Illinois en Chicago (UIC) Juntas de Revisión Institucional y todos los
participantes firmaron el consentimiento. Los participantes fueron reclutados mediante
volantes y las múltiples formas de la publicación en internet. Todos los participantes
completaron un protocolo de evaluación idéntica, incluyendo la entrevista diagnóstica
para estudios genéticos (DIGS; [34] ), la Escala de Depresión de Hamilton (HAM-
D; [35] ), el paramétrico Go / Prueba de No-Go [31] , [ 32] , y la capacidad de respuesta
Ruminative Escala (RRS, [36] ). Los participantes fueron considerados remitidos desde
MDD si previamente se reunieron los criterios de al menos un MDE, pero en la
actualidad se calificó por debajo de un 7 en la HAM-D (administrado durante la pantalla
del teléfono y durante la entrevista diagnóstica inicial). HC no pudo cumplir con los
criterios actuales o pasados (Nunca Enfermedades Mentales, NMI) para TDM o
cualquier otro del Eje I o II trastorno psiquiátrico y no tenía parientes de primer grado
con antecedentes de enfermedad psiquiátrica. Además, se pidió a los participantes a ser
libre de la medicación por un período de 30 días previos a la exploración y aquellos con
abuso o dependencia de sustancias dentro de los últimos seis meses fueron excluidos. El
diagnóstico de trastorno depresivo mayor o pasado NMI se determinó con DIGS, y
confirmó usando una entrevista con la familia modificada para estudios genéticos
completado con un padre o tutor legal [34] . La muestra final fue de 30 rMDD (19 UM,
11 UIC) y 23 controles sanos (HC; 16 UM, 7 UIC) entre las edades de 18-23 años (66%
mujeres). Demográficas de los participantes y las características clínicas se presentan en
la Tabla 1 . Historia de tratamiento anterior dentro del grupo rMDD (datos disponibles
para n = 19) incluyen la medicación (n = 13) y la psicoterapia (n = 17).

Descargar:
PPT Diapositiva de PowerPoint
PNG Ampliar imagen (74KB)
PELEA imagen original (363KB)
Tabla 1. Datos demográficos y características clínicas de ejemplo.
http://dx.doi.org/10.1371/journal.pone.0104366.t001
Rumia
Autoinforme rumiación se recogió mediante la Escala de Respuesta Ruminative
(RRS; [36] ).Cuarenta individuos (23 rMDD, 17 HC) completaron el RRS. Los
individuos con puntuaciones superiores a dos desviaciones estándar de la media se les
dio una puntuación truncada [37] .Las correlaciones se calculan entre la puntuación total
de la rumia y los valores de ROI extraídos de las regiones que diferían
significativamente entre los grupos de conectividad basada en semillas.
Paramétrica Go / No-Go
Esta tarea (descrito anteriormente, [31] ) se administró a todos los sujetos durante las
pruebas cognitivas antes de la exploración para medir la exactitud atención sostenida (Ir
Precisión), la inhibición basada en el contexto (n-Go Precisión), y el tiempo la
velocidad de procesamiento (Reacción, [32] ), todos los aspectos de control
cognitivo. Dos valores atípicos (1 HC, 1 rMDD) se les dio una puntuación truncada
rendimiento para Go Precisión (dos desviaciones estándar, [37] ). Correlaciones
exploratorios se calcularon utilizando los factores de rendimiento de la inversión
extraídos con estas medidas de control cognitivo.
Adquisición fcMRI
En la UM una consulta del estado de reposo ojos-abiertos fue adquirido más de ocho
minutos en un escáner GE Signa 3.0 T (Milwaukee, WI) usando T2 * ponderadas solo
tiro secuencia inversa espiral con los siguientes parámetros: 90 grados flip, el campo de
visión 20, tamaño de la matriz = 64 × 64, rebanada de espesor = 4 mm, 30 ms tiempo de
eco, 29 rebanadas. Se recogieron los ojos abiertos, que descansan las exploraciones en
la UIC más de ocho minutos en un escáner 3.0 T GE Descubrimiento (Milwaukee, WI)
utilizando imágenes paralelo con ASSET y T2 * ecogradiente EPI axial con los
siguientes parámetros: 90 grados flip, campo de -view 22, tamaño de la matriz = 64 ×
64, rebanada de espesor = 3 mm, 22.2 ms tiempo de eco, 44 rebanadas. En ambos sitios,
se obtuvieron exploraciones anatómica T1 de alta resolución para la normalización

espacial y el movimiento se reducen al mínimo con almohadillas de espuma, una línea
de seguimiento visual (UIC solamente) y / o transversal (UIC y UM) en la pantalla, y
mediante la transmisión de la importancia de para permanecer inmóvil a los
participantes, con TR de 2.000 ms y 240 TR en total. Efectos de sitio de los parámetros
de adquisición y el escáner se evaluaron y se reportan en Material complementario
( Figura S2 ).
Preprocesamiento fc-RM
Se tomaron varias medidas para reducir las posibles fuentes de ruido y
artefactos. Rebanada de tiempo se completó con SPM8
( http://www.fil.ion.ucl.ac.uk/spm/doc/ ) y algoritmos de detección de movimiento se
aplicaron usando FSL ( http://fsl.fmrib.ox.ac.uk / FSL / fslwiki / ).Coregistration de
imágenes estructurales en imágenes funcionales, con normalización espacial de la
plantilla coregistradas T1-SPGR al Instituto Neurológico de Montreal (MNI). A
continuación, la matriz de normalización resultante se aplicó a la porción de tiempo
corregida, fisiológicamente corregida, los datos de series de tiempo. Estos T2 *
normalizado de datos de series de tiempo fueron suavizadas espacialmente con un
núcleo de Gauss 5 mm que resulta en imágenes T2 * con vóxeles isotrópicos, 2 mm de
lado.
Análisis de correlación cruzada
Series de tiempo se detrended y la media de centrado. Corrección fisiológica se realizó
mediante la regresión a cabo la sustancia blanca y señales de líquido
cefalorraquídeo [38] .Parámetros de movimiento se retrocedido a cabo [39] . Con base
en la literatura reciente [39] ,[40] , se identificaron los volúmenes de movimiento
basados en cualquier TR TR al movimiento superior a 0,5 mm y no difirió entre los
grupos ( Figura S1 ) [40] . Se evaluaron todas las diferencias significativas con respecto
al movimiento, y el movimiento no influyeron en las diferencias significativas
identificadas usando las semillas del PCC y SGACC. Sin embargo, conectividades con
la semilla de la amígdala izquierda se convirtió en no significativa después de covarying
medidas del movimiento. La señal global no fue una regresión debido a violaciónes de
colinealidad con la señal de la materia gris, problemáticas estimaciones erróneas de
anticorrelations [41] , y debido a que no afecta a las relaciones a distancia
Micromovimiento [39] . Finalmente series de tiempo fuera de banda de paso se filtró
sobre 0,01-0,10 Hz. Las semillas se obtuvieron en base a la literatura anterior examen
de conectividad estado de reposo de la amígdala [42] , [43] , PCC [44] , [45] , y
sgACC [46] , [47]. Se utilizaron las siguientes coordenadas: PCC (DMN, -5/5, -50, 36),
la amígdala (SN, -23/23, -5, -19), sgACC (SN, -4/4, 21, -8 ). Las regiones de interés
(ROI; 2,9 mm de radio, 19 voxels) se definieron en el espacio MNI y espacialmente
promediados datos del curso de tiempo se extrajeron de regiones de interés para cada
participante. Las semillas fueron superpuestos en la anatomía estructural deformado
promedio de la muestra actual y ajustarse cuando se requiera.
Los coeficientes de correlación se calcularon entre supuesto tiempo medio de regiones
de semillas y todos los otros vóxeles del cerebro, dando como resultado (imagen r) una
imagen coeficiente de correlación 3-dimensional en. Estas imágenes r se transformaron
en puntuaciones z utilizando una transformación de Fisher. Z resultantes imágenes se
utilizaron en la muestra 2-Student t pruebas aplicadas en SPM8. AlphaSim se utilizó

con 1000 simulaciones de Monte Carlo para determinar la corrección de todo el cerebro
con un umbral conjunta de altura y extensión ( p <0,005, medida racimo de 440 mm 3 )
para comparaciones de grupos con un corregida p valor de 0,05. Las imágenes se
muestran en una anatomía del cerebro promedio derivado de la muestra actual.
Es importante destacar que el movimiento se abordó mediante regresión de la señal de
la sustancia blanca como se recomienda en la literatura reciente [39] , [40] . Además,
llevamos a cabo análisis adicionales (véase la Tabla S3 en Archivo S1 y Figura S1 )
para explorar plenamente el potencial de (micro) confunde el movimiento. Todos los
grupos de PCC y SGACC reportados siendo significativas cuando los sujetos con
cualquier TR a TR movimientos superior a 0,5 mm (por lo general el desplazamiento Z)
fueron excluidos.
El análisis factorial exploratorio
El análisis factorial es una técnica estadística que se puede utilizar para descubrir qué
variables formar subconjuntos coherentes que son algo independiente. Los factores son
la hipótesis de reflejar los procesos subyacentes que resultan en las correlaciones entre
las variables. El análisis factorial se diferencia del análisis de componentes principales
en que el análisis factorial examina varianza compartida solamente y se hacen intentos
para estimar y eliminar la varianza que se deriva de error [37] . El análisis de factores
puede ser eficaz en la reducción de datos y también puede separar el ruido de señal de la
conectividad en fMRI [23] ;Por lo tanto, el análisis factorial exploratorio se llevó a cabo
en los valores z extraídos de cada grupo de diferencias significativas entre los grupos
por separado para cada región de la semilla. El número de factores retenidas se
determinó utilizando máxima verosimilitud como un método de extracción y un umbral
de valor propio> 1, seguido por la rotación oblicua.También se verificó que los factores
extraídos superaron el 50% de la varianza total.
resultados
conectividad PCC
En todos los grupos, la semilla PCC izquierda se correlacionó con las regiones que
abarcan el DMN incluyendo la corteza prefrontal medial (PFC), posterior y
circunvoluciones temporal superior, y el hipocampo bilateral. Figura 1 ilustra la DMN
entre los participantes de HC así como conectividades que eran mayores entre rMDD en
comparación con los participantes de HC. Tabla 2 detalles diferencias significativas
entre los grupos. En comparación con los HC, los jóvenes con rMDD demostró una
mayor conectividad de la semilla PCC izquierda en la ínsula derecha y circunvolución
frontal superior y media derecha (BA 9), así como los precuneus izquierda (BA 7) y
PFC dorsolateral izquierda (BA 8/10 ). Resultados para el PCC derecha se incluyen
como material complementario para la comparación (Tabla S1 en S1 Archivo ).

Descargar:
PPT Diapositiva de PowerPoint
PNG Ampliar imagen (1.45MB)
PELEA imagen original (1.65MB)
Figura 1. Conectividad de semillas cingulada posterior izquierda y diferencias entre los grupos.
Panel A: conectividades entre los jóvenes HC ilustran la red en modo automático.Panel
B: Los jóvenes con depresión remitidas demostró una mayor conectividad con la ínsula
derecha, circunvolución frontal superior y media, el putamen, giro angular, y
circunvolución frontal media izquierda.
http://dx.doi.org/10.1371/journal.pone.0104366.g001
Descargar:
PPT Diapositiva de PowerPoint
PNG Ampliar imagen (105KB)
PELEA imagen original (371KB)
Tabla 2. Diferencias entre controles sanos y remitido depresión mayor para las semillas cingulada posterior izquierda.
http://dx.doi.org/10.1371/journal.pone.0104366.t002
conectividad sgACC
En todos los grupos, la activación sgACC la izquierda se correlacionó
significativamente con áreas cercanas, incluyendo la corteza orbital frontal, tálamo, el
hipocampo y el PCC. La figura 2 ilustra el SN entre los participantes de HC y
conectividades que eran mayores entre rMDD jóvenes en comparación con los HC. La
Tabla 3 detalla esas diferencias significativas. rMDD juventud demostró una mayor
conectividad de la semilla sgACC izquierda a la corteza bilateral superior y frontal
medial (BA 8/10), el cerebelo y tálamo, así como la circunvolución temporal media
izquierda (BA 39/40), parahipocampo, y el derecho opérculo / ínsula anterior, en
comparación con HC. Resultados para la sgACC derecha se incluyen como material
complementario para la comparación (Tabla S2 en S1 Archivo ).

Descargar:
PPT Diapositiva de PowerPoint
PNG Ampliar imagen (1,4 MB)
PELEA imagen original (1,6 MB)
Figura 2. Conectividad de la semilla cingulada anterior subgenual izquierda y diferencias entre los grupos.
Panel A: conectividades entre los jóvenes HC ilustrar la relevancia de la red. Panel B:
Los jóvenes con depresión remitidas demostró una mayor conectividad de la semilla
cingulada anterior subgenual izquierda a la derecha anterior frontal inferior
circunvolución / ínsula, medial bilateral, corteza frontal superior y medio, el tálamo y el
giro temporal medial izquierdo, lóbulo parietal inferior, y del hipocampo
circunvolución.
http://dx.doi.org/10.1371/journal.pone.0104366.g002
Descargar:
PPT Diapositiva de PowerPoint
PNG Ampliar imagen (186KB)
PELEA imagen original (642KB)
Tabla 3. Diferencias entre controles sanos y remitido depresión mayor para las semillas cingulada anterior subgenual la izquierda.
http://dx.doi.org/10.1371/journal.pone.0104366.t003
hiperconectividad amígdala
En todos los grupos, la activación de la izquierda de la amígdala se correlacionó
significativamente con la que rodea la amígdala bilateral, hipocampo y regiones
Uncal. La figura 3 ilustra las regiones del SN conectada de forma significativa con la
semilla de la amígdala izquierda entre los HC y los que fueron significativamente
diferentes entre los participantes rMDD y HC. Tabla 4 detalles de estas diferencias. El
grupo rMDD exhibió una mayor conectividad entre la amígdala izquierda y la
circunvolución frontal medial derecho, lóbulo parietal medial, el CAC rostral, y la
circunvolución del hipocampo izquierdo (no todas las regiones se muestran en la Figura

3 ). No hubo diferencias significativas entre los grupos para la conectividad con la
amígdala derecha.
Descargar:
PPT Diapositiva de PowerPoint
PNG Ampliar imagen (1.39MB)
PELEA imagen original (1.54MB)
Figura 3. Conectividad de la semilla de la amígdala izquierda y diferencias entre los grupos.
Figura 3 Panel A: conectividades entre los jóvenes de HC con la semilla de la amígdala
izquierda. Panel B: Los jóvenes con depresión remitidas demostró hyperconnectivities
con la derecha circunvolución frontal medial, lóbulo parietal medial, la circunvolución
poscentral, y la corteza cingulada anterior.
http://dx.doi.org/10.1371/journal.pone.0104366.g003
Descargar:
PPT Diapositiva de PowerPoint
PNG Ampliar imagen (34KB)
PELEA imagen original (181KB)
Tabla 4. Las diferencias entre los controles sanos y remitido depresión mayor para las semillas de la amígdala izquierda.
http://dx.doi.org/10.1371/journal.pone.0104366.t004
Reducción de datos con análisis factorial exploratorio
Los resultados del análisis factorial exploratorio sugirieron una solución de dos factores
para que la semilla PCC y una solución de tres factores para que la semilla
sgACC. Factor 1 PCC tenía cargas más altas en seis de los ocho grupos, como se indica
en la Tabla 2 . PCC Factor 2 tenía una carga superior a la media derecha y medial giros
frontales y cargas comparables en un segundo grupo circunvolución frontal media
derecha. sgACC Factor 1 incluye 11 de los 17 grupos que fueron Hiperconectado entre
rMDD juventud, como se indica en la Tabla 3 . Cuatro grupos tenían cargas dominantes
en sgACC Factor 2, y cuatro grupos tenían cargas dominantes en sgACC Factor 3.
Como se detalla en las tablas 2 y 3 , algunos grupos cargados altamente en más de un
factor. El análisis factorial de los grupos de conectividad basada en la amígdala fallo de
convergencia.

Relación de Redes intrínsecos a la rumia y control cognitivo
Como era de esperar, rMDD jóvenes reportaron niveles más altos de la rumia de los HC
(t = -3,78, df = 38, p <.01). Para el paramétrica Go / No-Go, rMDD jóvenes mostraron
mayores problemas de control de impulsos (n-Go Precisión, t = 2,30, df = 49, p = 0,03),
pero no fue diferente en Ir Precisión (t = -0.59, df = 49, p = .99) o Go Tiempo de
respuesta (t = -0,70, df = 49, p = .47). Entre los jóvenes con rMDD, Ir Tiempo de
respuesta se correlacionó inversamente con la rumia (r = -.48, p <.05). También entre
los jóvenes rMDD, el PCC Factor 2 (conectividad circunvolución frontal superior
derecho y medio) se correlaciona inversamente con la rumia (r = -.49, p = 0,03; Figura
4 ). Entre los jóvenes rMDD solamente, el sgACC Factor 2 (frontal izquierdo media,
temporal inferior, frontal inferior derecha, y la conectividad circunvoluciones frontal
media) se asoció positivamente con Go Precisión (r = 0,40, p = 0,04;Figura 5 , panel 1)
y sgACC Factor 3 (circunvolución del hipocampo izquierdo, caudado, tálamo dorsal
medial bilateral, conectividad circunvoluciones poscentral derecha) se correlacionó
inversamente con n-Go Precisión (r = -.44, p = 0,02; Figura 5 , panel B).
Descargar:
PPT Diapositiva de PowerPoint
PNG Ampliar imagen (31KB)
PELEA imagen original (118KB)
Figura 4. Correlación de Factor basada en semillas con rumiación.
La correlación de la corteza cingulada posterior izquierda factor 2 con rumiación.
http://dx.doi.org/10.1371/journal.pone.0104366.g004

Descargar:
PPT Diapositiva de PowerPoint
PNG Ampliar imagen (56KB)
PELEA imagen original (281KB)
Figura 5. Correlación de los factores basada en semillas con rumiación y el control cognitivo.
A) Correlación de la subgenual Factor cingulada anterior izquierda con Go 2 Precisión y
B) correlación de subgenual Factor cingulada anterior izquierda 3 con n-Go exactitud
por ciento.
http://dx.doi.org/10.1371/journal.pone.0104366.g005
Los posibles factores de confusión
Para descartar posibles factores de confusión, se realizaron análisis exploratorios post-
hoc de las características clínicas y los desafíos tecnológicos y también examinó la
influencia del sexo de los participantes (véase Material complementario). Estos factores
no afectan de manera significativa entre los grupos para los resultados de las semillas de
PCC y SGACC.
Discusión
Los resultados actuales abordan un conjunto poco estudiada, sin embargo, muy
importante de preguntas con respecto a la disociación del estado, rasgo, compensatoria,
y las características de la cicatriz de MDD. Para empezar a desentrañar estas preguntas,
hemos obtenido los datos del estado de reposo de una muestra homogénea de cuidado
caracterizado juventud, sin medicación en un estado remitido, que nos permite estudiar
curso MDD en ausencia de funciones estatales y la carga crónica que puede haber
confundido los resultados anteriores. Este es el primer estudio de adultos jóvenes con
rMDD y que demuestran la hiperconectividad, tanto de la DMN y SN con porciones de
la derecha giros frontales superior y media - un nodo central en el CCN. Por otra parte,
hyperconnectivities observadas estaban relacionadas con dos predictores conocidos de
la recaída: la rumia (correlación inversa) y la atención sostenida (correlación positiva).
Autoinforme rumiación ha sido documentado anteriormente que se correlaciona con la
conectividad sgACC-PCC durante la tarea entre los adultos activamente con
depresión [29] .Encontramos una correlación inversa de la rumia, hyperconnectivities lo
que sugiere que este aumento de las correlaciones temporales pueden tener un efecto
protector, compensatoria, o incluso pueden reducir el riesgo de recaída. Por el contrario,
otra hipótesis es que más enriquecida con conexiones a regiones fuera del CCN pueden
ser perjudiciales. Por ejemplo, la hiperconectividad a un factor de la sgACC izquierda a

regiones, incluyendo la circunvolución del hipocampo izquierdo, caudado, tálamo
dorsal medial bilateral y circunvoluciones poscentral derecha se asoció con el control
inhibitorio más pobre. Cuando se interpreta a la luz de los puntos fuertes de la muestra
actual, nuestros resultados ofrecen nueva evidencia de que las diferencias de
conectividad pueden representar ambos factores de riesgo y de protección basados en
los rasgos de la recaída en el curso de MDD.
El sgACC largo ha sido implicado como un nodo disfuncional en MDD aguda y se ha
correlacionado con estados de ánimo negativos, la respuesta al tratamiento, y
"refractariedad tratamiento" [48] - [50] . Nuestra conclusión de que este nodo SN está
hiperconectado tanto con el DMN y EN rMDD entre los jóvenes es digno de
mención. Funcionamiento sgACC aberrante y la conectividad han sido conceptualizado
como una consecuencia de la enfermedad depresiva y la tristeza, en oposición a un
riesgo potencial o factor de protección para la futura MDEs. De hecho, una reciente
investigación de los adolescentes activamente deprimidos, no medicados encontró que
los niveles más altos de rumiación se asociaron conuna reducción conectividad entre el
sgACC y la circunvolución frontal medial [7] . Nuestros hallazgos sugieren que el
aumento de la conectividad entre estas mismas regiones puede fortalecer la capacidad
del adolescente para resistir la rumia y apoyar control de la atención, promoviendo con
ello y, potencialmente, el mantenimiento de la remisión. Remitido jóvenes cuya CCN es
excesivamente comprometida-puede regular a la baja de manera más eficaz vagabundeo
de la mente, el pensamiento auto-referencial con el apoyo de la DMN.
Nuestros resultados ampliar y contextualizar los resultados de un proyecto de anormal
'dorsal nexo' en el TDM [22] por lo que sugiere que la hiperconectividad de la DMN y
SN con el CCN es un fuera observable marcador curso temprano de la MDEs, similar a
un estudio de la función DMN entre remitido niños en edad preescolar [28] . La
circunvolución frontal superior y media puede representar una extensión del CCN que
modula la DMN y SN entre aquellos con antecedentes de trastorno depresivo mayor, lo
que puede funcionar para mejorar las tendencias rumiativos basados en los rasgos que a
menudo permanecen en períodos de bienestar [51] , [52 ] .
También vale la pena señalar, sin embargo, que los factores primarios (por tanto el PCC
como semillas SGACC) representaban la mayor parte de la varianza de los factores
entre redes no estaban relacionadas con características clínicas de la enfermedad o el
funcionamiento en el estudio actual - nuestros resultados pueden reflejar rasgo o
biomarcadores cicatriz. Un biomarcador rasgo representaría una característica
observable antes de la enfermedad y en un principio independiente de la enfermedad,
mientras que un biomarcador de la cicatriz es el resultado de la enfermedad y representa
un fracaso para lograr la recuperación completa entre los episodios. Los factores
segundo y tercero, que representa menos varianza, y más probabilidades de ser
específicas de muestra, se relacionaron con los marcadores cognitivos de la
enfermedad. Hyperconnectivities de la DMN y SN con partes de la CCN pueden causar
un aumento de los recursos neuronales dirigidas a la regulación emocional entre
individuos vulnerables. hyperconnectivities del desarrollo pueden tener un efecto
protector en cierta medida, pero pueden no ser siempre suficiente para el montaje de las
respuestas de adaptación a la mayor adversidad y los factores de estrés experimentado
por esta población en riesgo de múltiples MDEs y las comorbilidades. Actualmente
estamos siguiendo esta muestra longitudinal y seremos capaces de identificar
predictores de recaída y tanto la capacidad de recuperación.

Observamos varias limitaciones de nuestro estudio. En primer lugar, los datos actuales
no pueden discriminar entre las anomalías de la red que hacen que un individuo
vulnerable a la primera aparición de MDD frente consecuencias de la cicatriz de la
enfermedad, los procesos de maduración normal, o mecanismos de compensación
incluso de protección. En este contexto, los estudios prospectivos de cohortes de alto
riesgo representan una dirección importante para la investigación futura. En segundo
lugar, a pesar de la fuerza de la contratación de una muestra libre de la medicación, no
pudimos examinar la forma anterior el uso de medicación o la terapia de exposición
pueden haber afectado a los resultados actuales.Como un punto relacionado, porque
nuestros participantes han tenido relativamente menos MDEs y han estado bien durante
períodos relativamente prolongados de tiempo, pueden representar un nivel de gravedad
más leve o curso de la enfermedad. Para capturar las trayectorias de desarrollo que
contribuyen a la resiliencia y el riesgo en el TDM, futura investigación longitudinal
puede examinar si el exceso de acoplamiento de las redes intrínsecas predice primera
aparición de la depresión o la recaída como transición adolescentes en la edad adulta
temprana. Además, la especificidad anatómica es motivo de preocupación cuando se
realizan análisis basados en semillas, en particular en regiones como el
PCC [53] , [54] . Tampoco se examinó específicamente la conectividad de la red CCN
usando una semilla CCN. Actualmente estamos analizando el CCN durante el descanso
y de trabajo para examinar más a fondo la función CCN entre los individuos
remitidos. La investigación con futuras muestras más grandes también examinarán
específicamente lateralidad. Por último, el examen de la función de red tanto en reposo
como en respuesta a la tarea proporciona información complementaria y aditivo con
respecto a cómo funcionan las redes de apoyo a la salud o la enfermedad. Orientaciones
futuras podrían incluir la inducción de la rumia durante una tarea de resonancia
magnética funcional y el examen de la forma en la rumia impide el rendimiento en las
tareas de control cognitivo para examinar cómo funcionan estas redes entre sí, tanto en
reposo y en respuesta a tareas relacionadas con la etiología de la depresión. A pesar de
estas limitaciones, creemos que el examen de los mecanismos durante una fase
relativamente temprana de la enfermedad proporciona un nivel de protección frente a
factores de confusión potenciales, incluyendo historias complejas de tratamiento y
cicatrización de los nervios como resultado de décadas de la enfermedad, por lo que el
estudio actual innovadora e importante.
En resumen, este estudio proporciona evidencia de rasgos basados en el cerebro
asociados con el curso MDD que se pueden observar fuera de un MDE, temprano en el
curso de la enfermedad. Creemos que la transición de los adolescentes la edad adulta
temprana-tardía representa una ventana única para la observación de los mecanismos de
trastorno depresivo mayor, como la estabilidad de las redes se ha establecido su
desarrollo, pero siguen existiendo oportunidades para la prevención secundaria antes de
la iniciación de la enfermedad crónica. Por lo tanto, la comprensión de la continuidad de
la conectividad funcional de los patrones disfuncionales en el cerebro humano es
fundamental en el esclarecimiento de la psicopatología del desarrollo del trastorno
depresivo mayor.acoplamiento entre redes aumentado en nuestro curso temprano, la
muestra remitida sugiere que hyperconnectivities pueden ser evaluados como rasgo o
capacidad de recuperación factores de desarrollo, como dianas para el tratamiento, o
como los posibles resultados de la enfermedad temprana.
información de soporte
Figure_S1.tif
Fig cuota 1 / 3
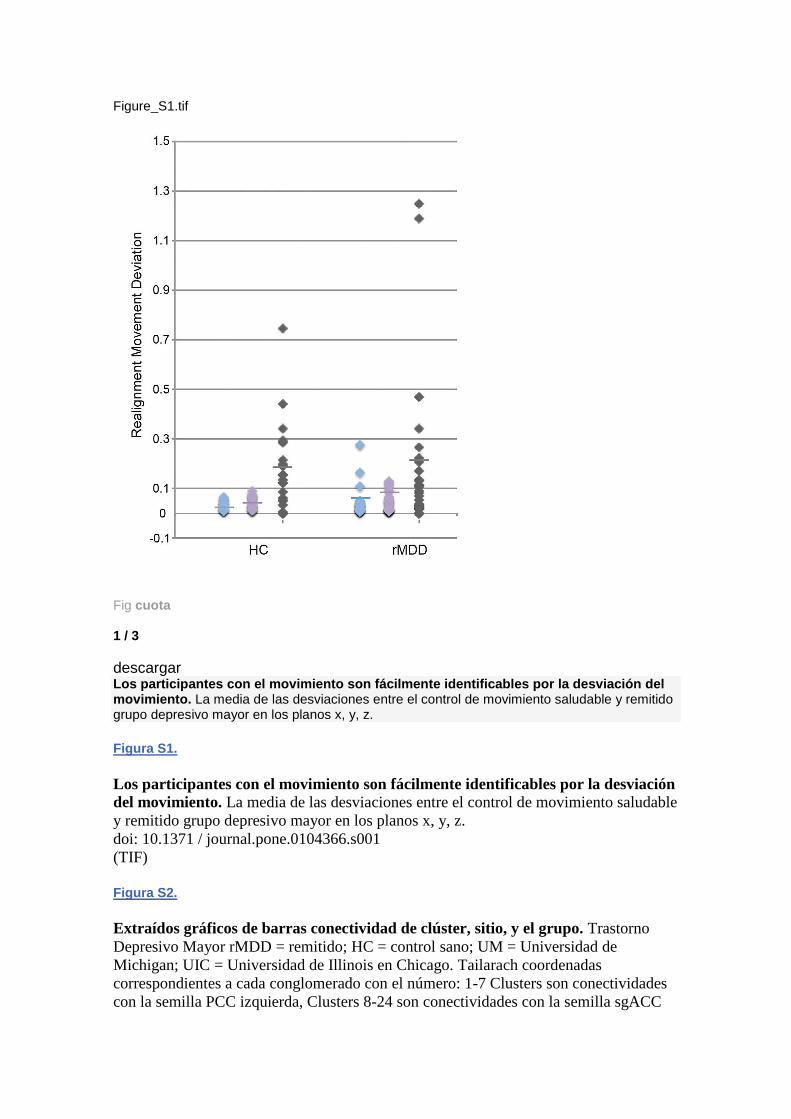
descargar Los participantes con el movimiento son fácilmente identificables por la desviación del movimiento. La media de las desviaciones entre el control de movimiento saludable y remitido grupo depresivo mayor en los planos x, y, z.
Figura S1.
Los participantes con el movimiento son fácilmente identificables por la desviación
del movimiento. La media de las desviaciones entre el control de movimiento saludable
y remitido grupo depresivo mayor en los planos x, y, z.
doi: 10.1371 / journal.pone.0104366.s001
(TIF)
Figura S2.
Extraídos gráficos de barras conectividad de clúster, sitio, y el grupo. Trastorno
Depresivo Mayor rMDD = remitido; HC = control sano; UM = Universidad de
Michigan; UIC = Universidad de Illinois en Chicago. Tailarach coordenadas
correspondientes a cada conglomerado con el número: 1-7 Clusters son conectividades
con la semilla PCC izquierda, Clusters 8-24 son conectividades con la semilla sgACC

izquierda, Clusters 25-28 son conectividades con la amígdala izquierda: 1 = -41, 35,
12; 2 = -6, -63, 49; 3 = 12, 45, 14; 4 = 17, -9, 3; 5 = 26, 32, 42; 6 = 27, 24, 20; 7 = 43, -
36, 30; 8 = -22, -38, 0; 9 = -24, -30, 16; 9 = -24, 43, 13; 10 = -24, 43, 13; 11 = -31, -83,
-33; 12 = -36, 16, 45; 13 = -43, -65, 35; 14 = -54, -54, -1; 15 = -57, -15, -19; 16 = -1, -
15, 6; 17 = 8, 32, 49; 18 = 10, 63, 19; 19 = 15, -69, -39; 20 = 29, -71, -39; 21 = 31, -36,
49; 22 = 36, 24, 41; 23 = 43, 14, -3; 24 = 4, 45, 39; 25 = -18, -9, -27; 26 = 19, 12, 21; 27
= 19, 24, 19; 28 = 48, -19, 39.
doi: 10.1371 / journal.pone.0104366.s002
(TIF)
Presentar S1.
material complementario que contiene tablas de ayuda. doi: 10.1371 / journal.pone.0104366.s003
(DOCX)
Expresiones de gratitud
Los autores desean reconocer las contribuciones del Centro para el Programa de
Investigación de Resonancia Magnética 3T en la UIC y gracias al Dr. F. Xavier
Castellanos por sus útiles comentarios sobre el manuscrito, así como Rachel Ringrose y
Gloria Harrington por sus esfuerzos de recolección de datos. Estos datos fueron
presentados en parte en la reunión anual de 2013 del Colegio Americano de
Neuropsicofarmacología.
Contribuciones de autor
Concebido y diseñado los experimentos: SAL JKZ RCW. Realizado los experimentos:
LBG AMB KAR SLW. Analizados los datos: RHJ SAL LMJ AB AV LBG
NAC. Contribuido reactivos / materiales / herramientas de análisis: RCW. Escribió el
documento: RHJ SAL LMJ KLP IHG ATP AV SLW.
referencias
1. 1.Marchetti I, Koster EH, Sonuga-Barke EJ, De Raedt R (2012) La red en modo
automático y depresión recurrente: un modelo neurobiológico de factores de
riesgo cognitivos. Opiniones Neuropsicología 22: 229-251. doi: 10.1007 /
s11065-012-9199-9
o Ver artículo
PubMed / Revista
Google Académico
2. 2.Keller MB (2003) Pasado, presente y futuras direcciones para definir el
resultado óptimo del tratamiento de la depresión: la remisión y más allá. JAMA
289: 3152 a 3160. doi: 10.1001 / jama.289.23.3152
Ver artículo
PubMed / Revista
Google Académico
3. 3.De Raedt R, Koster EH (2010) Comprensión de la vulnerabilidad para la
depresión desde una perspectiva de la neurociencia cognitiva: Una reevaluación

de los factores atencionales y un nuevo marco conceptual. Cogn afecta Behav
Neurosci 10: 50-70.doi: 10.3758 / cabn.10.1.50
Ver artículo
PubMed / Revista
Google Académico
4. 4.Castellanos FX, Di Martino A, Craddock RC, Mehta AD, Milham MP (2013)
Aplicaciones clínicas de la conectoma funcional. Neuroimagen 80: 527-540. doi:
10.1016 / j.neuroimage.2013.04.083
Ver artículo
PubMed / Revista
Google Académico
5. 5.Fox MD, Greicius M (2010) Aplicaciones clínicas de la conectividad
funcional en reposo estado. Frente Syst Neurosci 4: 19. doi: 10.3389 /
fnsys.2010.00019
Ver artículo
PubMed / Revista
Google Académico
6. 6.Greicius MD, Flores BH, Menon V, Glover GH, Solvason HB, et
al. conectividad funcional (2007) en estado de reposo en la depresión mayor:
anormalmente aumentada contribuciones de corteza cingulada subgenual y el
tálamo. Biol Psychiatry 62: 429-437. doi: 10.1016 / j.biopsych.2006.09.020
Ver artículo
PubMed / Revista
Google Académico
7. 7.Connolly CG, Wu J, Ho TC, Hoeft M, Wolkowitz O, et al. La conectividad
funcional (2013) en estado de reposo de subgenual corteza cingulada anterior en
adolescentes deprimidos. Biol Psychiatry 12: 898-907. doi: 10.1016 /
j.biopsych.2013.05.036
Ver artículo
PubMed / Revista
Google Académico
8. 8.Hamilton JP, Etkin A, Furman DJ, Lemus MG, Johnson RF, et al. (2012) La
neuroimagen funcional del trastorno depresivo mayor: un meta-análisis y la
nueva integración de la activación de la línea de base y datos de respuesta
neural. American Journal of Psychiatry 169: 693-703. doi: 10.1176 /
appi.ajp.2012.11071105
Ver artículo
PubMed / Revista
Google Académico
9. 9.Pizzagalli DA (2011) Frontocingulate disfunción en la depresión: hacia
biomarcadores de la respuesta al tratamiento. Neuropsychopharmacology 36:
183-206. doi: 10.1038 / npp.2010.166
Ver artículo
PubMed / Revista
Google Académico
10. 10.Whitfield-Gabrieli S, Ford JM actividad de la red (2012) El modo por defecto
y la conectividad en la psicopatología. Annu Rev Clin Psychol 8: 49-76. doi:
10.1146 / annurev-clinpsy-032.511-143.049
Ver artículo
PubMed / Revista

Google Académico
11. 11.Fox MD, AZ Snyder, Vincent JL, M Corbetta, Van Essen DC, et al. (2005)
El cerebro humano está organizado en redes funcionales intrínsecamente
dinámicos, anticorrelated. Actas de la Academia Nacional de Ciencias de los
Estados Unidos de América 102: desde 9673 hasta 9678. doi: 10.1073 /
pnas.0504136102
Ver artículo
PubMed / Revista
Google Académico
12. 12.Seeley WW, Menon V, Schatzberg AF, Keller J, Glover GH, et al. (2007)
disociables redes de conectividad intrínsecas para el procesamiento de la
prominencia y el control ejecutivo. J Neurosci 27: 2349-2356. doi: 10.1523 /
jneurosci.5587-06.2007
Ver artículo
PubMed / Revista
Google Académico
13. 13.Shehzad Z, Kelly AMC, Reiss PT, Gee DG, Gotimer K, et al. (2009) The
Resting cerebro: sin restricciones pero fiable. Corteza cerebral 19: 2209-
2229. doi: 10.1093 / cercor / bhn256
Ver artículo
PubMed / Revista
Google Académico
14. 14.Zuo XN, Kelly C, Adelstein JS, Klein DF, Castellanos FX, et al. (2010)
Reliable redes de conectividad intrínsecas: Test de evaluación del uso de ICA-
retest y el enfoque de regresión dual. Neuroimagen 49: 2163-2177. doi: 10.1016
/ j.neuroimage.2009.10.080
Ver artículo
PubMed / Revista
Google Académico
15. 15.Zuo XN, Di Martino A, C Kelly, Shehzad ZE, Gee DG, et al. (2010) El
cerebro oscilante: complejo y fiable. Neuroimagen 49: 1432-1445. doi: 10.1016
/ j.neuroimage.2009.09.037
Ver artículo
PubMed / Revista
Google Académico
16. dieciséis.Biswal BB, Mennes M, Zuo XN, S Gohel, Kelly C, et al. (2010) Hacia
la ciencia descubrimiento de la función del cerebro humano. Actas de la
Academia Nacional de Ciencias de los Estados Unidos de América 107: 4734
hasta 4739.
Ver artículo
PubMed / Revista
Google Académico
17. 17.Sadaghiani S, R Scheeringa, Lehongre K, B Morillon, Giraud AL, et al. redes
(2010) intrínsecas de conectividad, oscilaciones alfa, y el estado de alerta tónica:
un estudio de la electroencefalografía simultánea / funcional por resonancia
magnética. J Neurosci 30: desde 10.243 hasta 10.250. doi: 10.1523 /
jneurosci.1004-10.2010
Ver artículo
PubMed / Revista
Google Académico

18. 18.Bressler SL, Menon V (2010) las redes cerebrales a gran escala en la
cognición: métodos y principios emergentes. Tendencias Cogn Sci 14: 277-
290. doi: 10.1016 / j.tics.2010.04.004
Ver artículo
PubMed / Revista
Google Académico
19. 19.Menon V (2011) las redes cerebrales a gran escala y la psicopatología: un
modelo de red de triple unificador. Tendencias Cogn Sci 15: 483-506. doi:
10.1016 / j.tics.2011.08.003
Ver artículo
PubMed / Revista
Google Académico
20. 20.Broyd SJ, Demanuele C, Debener S, ayuda a SK, James CJ, et al. disfunción
cerebral (2009)-El modo por defecto en los trastornos mentales: una revisión
sistemática.Neurociencia y Bioconductual opiniones 33: 279-296. doi: 10.1016 /
j.neubiorev.2008.09.002
Ver artículo
PubMed / Revista
Google Académico
21. 21.Hamilton JP, Furman DJ, Chang C, Thomason ME, Dennis E, et al. (2011)
predeterminado-modo y la actividad de la red de trabajo positivo en el trastorno
depresivo mayor: implicaciones para la rumia de adaptación y de mala
adaptación. Biol Psychiatry 70: 327-333. doi: 10.1016 / j.biopsych.2011.02.003
Ver artículo
PubMed / Revista
Google Académico
22. 22.Sheline YI, Precio JL, Yan Z, Mintun MA (2010) Resonancia magnética
funcional en estado de reposo en la depresión desenmascara una mayor
conectividad entre las redes a través del nexo dorsal. Actas de la Academia
Nacional de Ciencias de los Estados Unidos de América 107: 11020-11025. doi:
10.1073 / pnas.1000446107
Ver artículo
PubMed / Revista
Google Académico
23. 23.Briceño EM, Weisenbach SL, Rapport LJ, Hazlett KE, Bieliauskas LA, et
al. (2013) desplazado lateralidad frontal inferior en las mujeres con trastorno
depresivo mayor se relaciona con déficits de procesamiento de la
emoción. Psychol Med 43: 1433-1445.doi: 10.1017 / s0033291712002176
Ver artículo
PubMed / Revista
Google Académico
24. 24.Drevets WC, Precio JL, Furey ML (2008) Brain anormalidades estructurales
y funcionales en los trastornos del humor: implicaciones para los modelos
neurocircuito de depresión. Estructura y función del cerebro 213: 93-118. doi:
10.1007 / s00429-008-0189-x
Ver artículo
PubMed / Revista
Google Académico
25. 25.Precio JL, Drevets WC (2010) neurocircuito de los trastornos del estado de
ánimo.Neuropsychopharmacology 35: 192-216. doi: 10.1038 / npp.2009.104

Ver artículo
PubMed / Revista
Google Académico
26. 26.Wang Z, Yuan Y, Bai F, J, Li L, et al. (2012) anormal de red en modo
predeterminado en la conversión de la angiotensina enzima portadores del alelo
D con la depresión geriátrica remitido. Behav Brain Res 230: 325-332. doi:
10.1016 / j.bbr.2012.02.011
Ver artículo
PubMed / Revista
Google Académico
27. 27.Yuan Y, Zhang Z, Bai F, Yu H, Shi Y, et al. (2008) la actividad neuronal
anormal en los pacientes con depresión geriátrica remitido: un estado de reposo
estudio de imágenes por resonancia magnética funcional. J afecta Disorders,
111: 145-152. doi: 10.1016 / j.jad.2008.02.016
Ver artículo
PubMed / Revista
Google Académico
28. 28.Gaffrey MS, Luby JL, Botteron K, G Repovs, Barch DM conectividad de red
de modo (2012) por defecto en los niños con antecedentes de depresión de inicio
preescolar. J Psychol Psiquiatría Infantil 53: 964-972. doi: 10.1111 / j.1469-
7610.2012.02552.x
Ver artículo
PubMed / Revista
Google Académico
29. 29.Berman MG, Peltier S, Nee DE, Kross E, Deldin PJ, et al. (2010) La
depresión, la rumia y la red por defecto. Soc Cogn afecta Neurosci doi: 10.1093
/ exploración / nsq080
Ver artículo
PubMed / Revista
Google Académico
30. 30.Cooney RE, Joormann J, M Eugene, Dennis EL, Gotlib IH (2010) neuronal
se correlaciona de la rumia en la depresión. Cognitiva y afectiva Behavioral
Neuroscience 10: 470-478. doi: 10.3758 / cabn.10.4.470
Ver artículo
PubMed / Revista
Google Académico
31. 31.Langenecker SA, Zubieta JK, EA joven, Akil H, Nielson KA (2007) Una
tarea de manipular la carga atencional, la configuración de cambio, y el control
inhibitorio: validez convergente y fiabilidad test-retest de la paramétrico Go /
No-Go prueba. J Clin Exp Neuropsychol 29: 842-853. doi: 10.1080 /
13803390601147611
Ver artículo
PubMed / Revista
Google Académico
32. 32.Votruba KL, Langenecker SA (2013) Estructura de los factores, la validez de
constructo, y los datos normativos por edad y basadas en la educación para la
paramétrico Go / No-Go prueba. J Clin Exp Neuropsychol 35: 132-146. doi:
10.1080 / 13803395.2012.758239
Ver artículo
PubMed / Revista

Google Académico
33. 33.Langenecker SA, Kennedy SE, Guidotti LM, Briceño EM, LS propios, et
al. (2007) la activación frontal y límbico durante el control inhibitorio predice la
respuesta al tratamiento en el trastorno depresivo mayor. Biol Psychiatry 62:
1272-1280. doi: 10.1016 / j.biopsych.2007.02.019
Ver artículo
PubMed / Revista
Google Académico
34. 34.Nurnberger JI Jr, Blehar MC, CA Kaufmann, York-C más frío, Simpson SG,
et al.(1994) Entrevista diagnóstica para estudios genéticos. Fundamento,
características únicas, y la formación. Iniciativa Genética NIMH. Arco General
Psychiatry 51: 849-859. doi: 10.1001 / archpsyc.1994.03950110009002
Ver artículo
PubMed / Revista
Google Académico
35. 35.Hamilton M (1960) Una escala de calificación para la depresión. J Neurol
Neurosurg Psiquiatría 23: 56-62. doi: 10.1136 / jnnp.23.1.56
Ver artículo
PubMed / Revista
Google Académico
36. 36.Treynor W, González R, Nolen-Hoeksema S (2003) rumiación
reconsiderada: Un análisis psicométrico. La terapia cognitiva e Investigación 27:
247-259.
Ver artículo
PubMed / Revista
Google Académico
37. 37.Tabachnick BG, Fidell LS (2007) Uso de la estadística multivariante. Boston,
MA: Pearson Education.
38. 38.Behzadi Y, Restom K, J Liau, Liu TT (2007) Un método de corrección
acústica provocada por el componente (CompCor) para fMRI BOLD y la
perfusión basada.Neuroimagen 37: 90-101. doi: 10.1016 /
j.neuroimage.2007.04.042
Ver artículo
PubMed / Revista
Google Académico
39. 39.Jo HJ, Gotts SJ, Reynolds RC, Bandettini PA, Martín A, et al. (2013) Los
procedimientos de preprocesamiento eficaz prácticamente eliminan los
artefactos por movimiento depende de la distancia en estado de reposo FMRI. J
Appl Matemáticas 2013: 1-9. doi: 10.1155 / 2013/935154
Ver artículo
PubMed / Revista
Google Académico
40. 40.JD Power, Barnes KA, AZ Snyder, Schlaggar BL, Petersen SE (2012)
correlaciones espurias pero sistemáticas en las redes de resonancia magnética
funcional de conectividad surgen de movimiento del sujeto. Neuroimagen 59:
2142-2154. doi: 10.1016 / j.neuroimage.2011.10.018
Ver artículo
PubMed / Revista
Google Académico

41. 41.Fox MD, Zhang D, AZ Snyder, Raichle ME (2009) La señal observada
global y anticorrelated descansando redes cerebrales estatales. J Neurophysiol
101: 3270-3283. doi: 10.1152 / jn.90777.2008
Ver artículo
PubMed / Revista
Google Académico
42. 42.Pannekoek JN, Veer IM, van Tol MJ, van der Werff SJ, Demenescu LR, et
al. (2013) aberrante conectividad límbico y la red de prominencia en estado de
reposo funcional en el trastorno de pánico sin comorbilidad. J afecta Disorders,
145: 29-35. doi: 10.1016 / j.jad.2012.07.006
Ver artículo
PubMed / Revista
Google Académico
43. 43.McCabe C, Mishor Z (2011) Los medicamentos antidepresivos reducen la
conectividad funcional en estado de reposo subcortical-cortical en voluntarios
sanos. Neuroimagen 57: 1317-1323. doi: 10.1016 / j.neuroimage.2011.05.051
Ver artículo
PubMed / Revista
Google Académico
44. 44.Bluhm RL, Clark CR, McFarlane AC, Moores KA, Shaw ME, et
al. conectividad de red (2011) por defecto durante una tarea de memoria de
trabajo. Hum cerebro Mapp 32: 1029-1035. doi: 10.1002 / hbm.21090
Ver artículo
PubMed / Revista
Google Académico
45. 45.Alexopoulos GS, Hoptman MJ, Kanellopoulos D, Murphy CF, Lim KO, et
al. (2012) La conectividad funcional en la red de control cognitivo y la red en
modo automático en la depresión tardía. J afecta Disorders, 139: 56-65. doi:
10.1016 / j.jad.2011.12.002
Ver artículo
PubMed / Revista
Google Académico
46. 46.Kelly AM, Di Martino A, Uddin LQ, Shehzad Z, Gee DG, et al. (2009)
Desarrollo de la conectividad funcional cingulada anterior desde finales de la
infancia a la edad adulta temprana. Cereb corteza 19: 640-657. doi: 10.1093 /
cercor / bhn117
Ver artículo
PubMed / Revista
Google Académico
47. 47.Margulies DS, Kelly AM, Uddin LQ, Biswal BB, Castellanos FX, et
al. (2007) Mapeo de la conectividad funcional de la corteza cingulada
anterior. Neuroimagen 37: 579-588.doi: 10.1016 / j.neuroimage.2007.05.019
Ver artículo
PubMed / Revista
Google Académico
48. 48.Konarski JZ, Kennedy SH, Segal ZV, Lau MA, Bieling PJ, et al. (2009) Los
predictores de la falta de respuesta a la terapia de comportamiento cognitivo o
venlafaxina utilizando metabolismo de la glucosa en el trastorno depresivo
mayor. J Psiquiatría Neurosci 34: 175-180.
Ver artículo

PubMed / Revista
Google Académico
49. 49.Siegle GJ, Carter CS, Thase ME (2006) El uso de fMRI para predecir la
recuperación de la depresión unipolar con la terapia de comportamiento
cognitivo. Am J Psiquiatría 163: 735-738. doi: 10.1176 / appi.ajp.163.4.735
Ver artículo
PubMed / Revista
Google Académico
50. 50.Keedwell P, Drapier D, Surguladze S, Giampietro V, Brammer M, et
al. (2009) Los marcadores neurales de la mejoría de los síntomas durante la
terapia con antidepresivos en la depresión severa: cingulada subgenual y las
respuestas corticales visuales para triste, pero no es feliz, estímulos faciales se
correlacionan con los cambios en la puntuación de los síntomas. J
Psychopharmacol 23: 775-788. doi: 10.1177 / 0269881108093589
Ver artículo
PubMed / Revista
Google Académico
51. 51.Nolen-Hoeksema S, J Morrow (1991) realizó un estudio prospectivo de los
síntomas de depresión y de estrés postraumático después de un desastre natural:
1989 terremoto de Loma Prieta. Journal of Personality and Social Psychology
61: 115-121. doi: 10.1037 // 0022-3514.61.1.115
Ver artículo
PubMed / Revista
Google Académico
52. 52.Riso LP, Newman CF (2003) La terapia cognitiva para la depresión crónica. J
Clin Psychol 59: 817-831. doi: 10.1002 / jclp.10175
Ver artículo
PubMed / Revista
Google Académico
53. 53.Margulies DS, Vincent JL, Kelly C, G Lohmann, Uddin LQ, et
al. Arquitectura funcional (2009) acciones precuneus intrínseca en los seres
humanos y los monos. Proc Natl Acad Sci EE.UU. 106: 20069 hasta 20074. doi:
10.1073 / pnas.0905314106
Ver artículo
PubMed / Revista
Google Académico
54. 54.Lynch CJ, Uddin LQ, Supekar K, Khouzam A, Phillips J, et al. modo de red
(2013) por defecto en el autismo infantil: la heterogeneidad corteza
posteromedial y la relación con los déficits sociales. Biol Psychiatry 74: 212-
219. doi: 10.1016 / j.biopsych.2012.12.013
Ver artículo
PubMed / Revista
Google Académico

Chilpancingo de los bravo a 28 de septiembre del 2016.
Nota de aprendizaje:
En esta clase vimos otro medio para compartir en nuestro blog una investigación
científica por medio de otra herramienta llamada “slideshare”.
De la cual, da una mejor calidad para compartirla en nuestra página personal,
donde continuamente estaré subiendo información relevante a las tic,s.
El aumento de acoplamiento de Redes intrínsecos en Remitido jóvenes con
depresión predice la rumiación y control cognitivo.
Me pareció una investigación científica eficiente y llamativa para el lector.
Agradecimientos por esta rigurosa investigación a:
Rachel H. Jacobs,
Lisanne M. Jenkins,
Laura B. Gabriel,
Alyssa Barba,
Kelly A. Ryan,
Sara L. Weisenbach,
Alvaro Verges,
Amanda M. Baker,
Amy T. Peters,
Natania A. Crane,
Ian H. Gotlib,
Jon-Kar Zubieta,
K. Luan Phan,
Scott A. Langenecker ,
Robert C. Welsh