introduccion l - sociedad cientifica de estudiantes de ... · demostrado que los carbonos del...
TRANSCRIPT

1
INTRODUCCION
as micotoxinas son sustancias tóxicas producidas en el metabolismo secundario de los hongos, que aparecen como contaminantes naturales en los alimentos cuando las condiciones climáticas son propicias.
Se conocen cientos de micotoxinas, y una veintena aparecen como residuales en los alimentos. Ellas están distribuidas en diferentes familias tales como: aflatoxinas, ocratoxinas, tricotecenos, fumonisinas y alcaloides del ergot, entre otras. Conocer el efecto que provocan las micotoxinas en la salud humana y animal, así como el impacto económico que provocan las mismas, constituye una preocupación continua de todos los países del área. Sin embargo, los materiales de estudio en español que permiten adentrarse en el fascinante campo de las micotoxinas son pocos, lo que contrasta con lo que ocurre en países desarrollados donde la información acerca de la peligrosidad de estas sustancias tóxicas y en especial de las aflatoxinas, se ha realizado en ocasiones a través de la prensa, dándole gran difusión a esta problemática. En los últimos 30 años se ha puesto empeño en estudiar los orígenes, la naturaleza, la distribución y los posibles riesgos que para la salud humana y animal tienen las micotoxinas presentes en los alimentos y los piensos. Con la presente publicación por series para cada familia de micotoxinas se pretende ofrecer una información útil para los profesionales técnicos de la salud humana y animal, para los biólogos y para aquellos agentes que están responsabilizados con la producción o elaboración de alimentos en el campo de la industria. 1-Reseña histórica de las Aflatoxinas. En 1960, en el oeste de Inglaterra, ocurrió la muerte de más de 100000 pavos de una enfermedad desconocida (enfermedad X de los pavos). Aquí se vio que el hígado era el órgano principalmente afectado, presentaba necrosis de los hepatocitos, proliferación biliar y fibrosis, entre otros signos. Los síntomas clínicos de esta enfermedad ya habían sido descritos por Wanop (1960), quien observó en 1957, en el campo de Bedfordhere, la muerte de un gran número de pavos. Brotes adicionales ocurrieron en la primavera y verano de 1958 y 1959, respectivamente (Austwick, 1978). Producto de investigaciones realizadas, se concluyó que la causa de esta enfermedad de origen desconocido fue la ingestión por las aves de un lote de maní importado de Brasil, que estaba contaminado con una toxina producida por Aspergillus flavus. Este episodio fue el que llevó al hombre a la identificación de toxinas producidas por hongos, que fueron denominadas genéricamente micotoxinas. El descubrimiento de las propiedades hepatotóxicas y carcinogénicas, así como la elucidación de la estructura de las toxinas de A. flavus (aflatoxinas) modificó la estrategia de lucha en toda la
L

2
esfera de las micotoxinas y es a partir de este momento que se inicia un estudio sistemático de las toxinas producidas por hongos. Las enfermedades producidas por las micotoxinas en los hombres y los animales se conocen como micotoxicosis; sin embargo, los términos de micotoxinas y micotoxicosis no fueron establecidos hasta después de la década del 60 del presente siglo, época en la que estos metabolitos comenzaron a tener identidad propia. 2-Biosíntesis y producción de aflatoxinas. 2.1-Biosíntesis de las aflatoxinas. La biosíntesis de las aflatoxinas ocurre en el metabolismo secundario de los hongos. Este tipo de metabolismo, al no ser indispensable para los hongos es especie específico, explica la variabilidad en su producción, aun entre cepas de la misma especie y la gran variedad de metabolitos producidos por esta vía. La biosíntesis de las aflatoxinas ocurre por la vía metabólica de los policétidos, la más importante en el metabolismo secundario de los hongos. Este proceso involucra la condensación del acetil CoA con el malonil CoA en un proceso cíclico similar en algunos aspectos al de la biosíntesis de ácidos grasos de cadena larga (Ellis et al, 1991). Estudios metabólicos realizados con precursores (acetato) marcados isotópicamente, han demostrado que los carbonos del esqueleto carbonado de la aflatoxina provienen en su totalidad del acetato y el grupo metilo proviene del aminoácido metionina (Zaika y Buchanan, 1987). En la vía biosintética del acetato a la aflatoxina han sido reconocidos seis metabolitos intermediarios (figura 1). El decacétido lineal formado (policétido) sufre un proceso de ciclización a compuestos de la familia de las antronas hasta llegar a la aflatoxina. Este proceso biosintético ocurre en el citoplasma soluble (extramitocondrial) y utiliza los equivalentes reducidos (NADPH, H+) producidos en la vía de las pentosas.

3
2.2-Producción de aflatoxinas en el campo y almacenes. El origen de las micotoxinas está indisolublemente ligado al crecimiento fúngico. Las aflatoxinas, por ser un producto del metabolismo secundario de los hongos, como anteriormente se mencionó, no pueden aparecer o desarrollarse en los alimentos sin una previa contaminación de los mismos por hongos de las especies A. flavus o A. parasiticus. Sin embargo, la presencia de estos hongos en un producto no indica que automáticamente debe estar presente la aflatoxina. Especies de A. flavus y A. parasiticus han sido aisladas de una gran diversidad de productos vegetales como son: maní, semilla de algodón, arroz, maíz, higos, trigo, entre otros (Vedanayagan et al, 1987; Carballo y de Miguel, 1987; Elamin et al, 1988; Vesonder, 1991). Conjuntamente con las anteriores existen otras especies que producen aflatoxinas en condiciones in vitro, aunque en menores concentraciones (Cavalherio, 1981). En la tabla 1 se muestran las diferentes especies de hongos productores de aflatoxinas. Tabla 1. Especies productoras de aflatoxinas.
Especie
Aflatoxinas
B1 B2 G1 G2
A. flavus A. parasiticus A. oryzae A. niger A. wentii A. ruber P. paberulen P. variable P. frecuenutans P. citrinum Rhizopus sp
x x x x x x x x x x x
x x x
x x x
x
Cuatro factores están vinculados estrechamente a la contaminación fúngica de los alimentos en el campo, así como durante el almacenamiento. Estos son: humedad, temperatura, oxígeno y tiempo. Otros factores que influyen en el crecimiento de los hongos son: la calidad del grano, la composición del sustrato (Jones, 1977; González et al, 1989), la capacidad genética del hongo (Stubblefield et al, 1967; Dorner et al, 1984), la presencia de insectos (Barry et al, 1986; Albin y Bilbao, 1987) y el empleo de sustancias químicas (sales inorgánicas) durante el almacenamiento (Thanaboripat et al, 1992).

4
De los factores antes mencionados, la temperatura y la humedad están muy ligadas entre sí y tienen un efecto crucial sobre el crecimiento de los hongos y la formación de aflatoxinas en los alimentos. La especie A. flavus puede crecer en un intervalo de temperatura entre 7.5-40 °C; sin embargo, todos los resultados publicados coinciden en que el mayor crecimiento y posterior aumento en la producción de toxinas se encuentra entre los 25-30 °C. Algunos hongos pueden desarrollarse con más lentitud a 0 °C (A. flavus, Cladosporium spp y Fusarium spp) y por otra parte, hay especies que crecen a temperaturas superiores a los 55 °C como A. fumigatus . El crecimiento del hongo y la producción de toxinas se ven influenciados por la humedad relativa del ambiente, resulta el 85% la óptima, aunque puede oscilar entre 83-99% (Diener y Davies, 1968). Christensen (1978)(citado por Sánchez, 1987), informó que los hongos pueden crecer bajo diferentes intervalos de humedad relativa (65-100%) y distintos contenidos de agua (12-24 %) en los cereales y granos de soya. Sinha (1979) refiere como temperaturas límites para el crecimiento de hongos en granos post-cosecha entre 5 y 45°C, con un intervalo óptimo entre 25 y 30 °C, humedad relativa entre 65-90% y límites de pH de 2.0 a 10.2. En la tabla 2 se resumen los factores ya comentados y otros que influyen en la formación de micotoxinas en las diferentes etapas de la producción de los cereales. Tabla 2. Factores que influyen en la contaminación por hongos y producción de micotoxinas en los cereales.
Factores Campo Recolección Almace-namiento
Físicos Humedad Temperatura Daño mecánico Tiempo Mezclado del grano
x x x x
x x x x x
x x x x x
Químicos Oxígeno Dióxido de carbono Comp. del Sustrato Minerales Tto. Químico
x x
x x x x x
Biológicos Insectos Infección fúngica Capacidad Genética Dif. en las variedades de plantas
x x x x
x x x x

5
2.3-Producción de aflatoxinas en el laboratorio. Las cepas de A. flavus y A. parasiticus varían en su toxigenicidad. Algunas cepas no producen cantidades detectables de aflatoxinas, y entre las productoras puede haber mucha diferencia en la cantidad de toxina sintetizada. Los principales factores que afectan directamente la producción de aflatoxina in vitro son: potencial genético de la cepa, concentración del inóculo, sustrato, nutrientes, intensidad de la luz, pH, temperatura y. gases atmosféricos. 2.3.1-Potencial genético de la cepa. Las cepas de A.flavus producen, principalmente, las aflatoxinas del tipo B, mientras las cepas de A.parasiticus producen las cuatro aflatoxinas. Estas producciones se pueden afectar por la presencia de otros hongos en el cultivo. Mislivec et al (1988) reportaron que la capacidad genética de A. flavus no se vio afectada por la presencia de otras especies de Aspergillus (A.ochraceus y A.versicolor. Sin embargo, la presencia de Penicillium spp. disminuyó la producción de aflatoxina de 0.7 a 30%, en dependencia de la especie de Penicillium contaminante. 2.3.2-Concentración del inóculo. Existe un vínculo entre la producción de aflatoxinas y la ramificación micelial, donde a bajas concentraciones de esporas (103-104/mL) se aumenta dos veces la producción de aflatoxinas (Sharma et al, 1980). Bennett (1983) refirió que la fusión hifal, común en Deuteromyces, es más rápida cuando la concentración de esporas es mayor; por tanto el metabolismo secundario queda inhibido por los procesos de diferenciación y reproducción fúngica. Esto puede explicar los resultados de Batt (1983) y los de González et al (1989), donde se obtuvieron superiores concentraciones de aflatoxinas con los inóculos de baja concentración. Karanuratne y Bullerman (1990) informaron que la máxima producción de aflatoxina ocurre con una concentración de esporas de 103/mL, y menores o mayores concentraciones de esporas a esta, provocaban una disminución en la producción de aflatoxinas en la temperatura óptima de crecimiento. 2.3.3-Sustrato Pueden ser sintéticos o naturales, líquidos o sólidos y de diferente naturaleza química. Esta última puede desempeñar una función importante, ya que hay productos más susceptibles que otros a ser contaminados por un moho determinado; sin embargo, la mayor parte de los mohos pueden invadir numerosos sustratos pues, en general, tienen pocas exigencias nutricionales. Sus necesidades se limitan al carbono, nitrógeno y sales. Los carbohidratos y ácidos grasos, en los medios de cultivo, aumentan la producción de toxina. La producción de aflatoxinas es mínima en productos de origen animal (jamón) y en otros productos como el té y especias. Por el contrario, esta producción puede ser muy alta en cereales y semillas oleaginosas, y dentro de estass, la soya es un mal sustrato ya que se demostró que posee una cierta resistencia a la producción de aflatoxinas (Erlich y Ciegler, 1985).
6
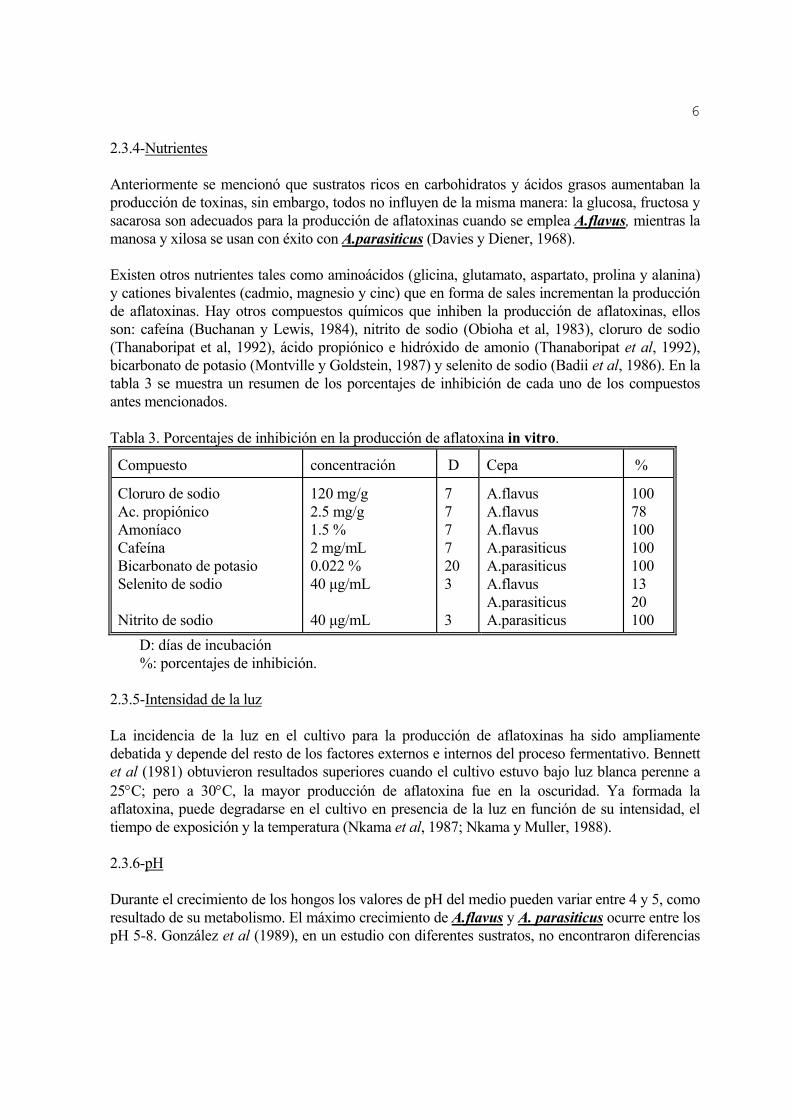
2.3.4-Nutrientes Anteriormente se mencionó que sustratos ricos en carbohidratos y ácidos grasos aumentaban la producción de toxinas, sin embargo, todos no influyen de la misma manera: la glucosa, fructosa y sacarosa son adecuados para la producción de aflatoxinas cuando se emplea A.flavus, mientras la manosa y xilosa se usan con éxito con A.parasiticus (Davies y Diener, 1968). Existen otros nutrientes tales como aminoácidos (glicina, glutamato, aspartato, prolina y alanina) y cationes bivalentes (cadmio, magnesio y cinc) que en forma de sales incrementan la producción de aflatoxinas. Hay otros compuestos químicos que inhiben la producción de aflatoxinas, ellos son: cafeína (Buchanan y Lewis, 1984), nitrito de sodio (Obioha et al, 1983), cloruro de sodio (Thanaboripat et al, 1992), ácido propiónico e hidróxido de amonio (Thanaboripat et al, 1992), bicarbonato de potasio (Montville y Goldstein, 1987) y selenito de sodio (Badii et al, 1986). En la tabla 3 se muestra un resumen de los porcentajes de inhibición de cada uno de los compuestos antes mencionados. Tabla 3. Porcentajes de inhibición en la producción de aflatoxina in vitro.
Compuesto concentración D Cepa %
Cloruro de sodio Ac. propiónico Amoníaco Cafeína Bicarbonato de potasio Selenito de sodio Nitrito de sodio
120 mg/g 2.5 mg/g 1.5 % 2 mg/mL 0.022 % 40 µg/mL 40 µg/mL
7 7 7 7 20 3 3
A.flavus A.flavus A.flavus A.parasiticus A.parasiticus A.flavus A.parasiticus A.parasiticus
100 78 100 100 100 13 20 100
D: días de incubación %: porcentajes de inhibición. 2.3.5-Intensidad de la luz La incidencia de la luz en el cultivo para la producción de aflatoxinas ha sido ampliamente debatida y depende del resto de los factores externos e internos del proceso fermentativo. Bennett et al (1981) obtuvieron resultados superiores cuando el cultivo estuvo bajo luz blanca perenne a 25°C; pero a 30°C, la mayor producción de aflatoxina fue en la oscuridad. Ya formada la aflatoxina, puede degradarse en el cultivo en presencia de la luz en función de su intensidad, el tiempo de exposición y la temperatura (Nkama et al, 1987; Nkama y Muller, 1988). 2.3.6-pH Durante el crecimiento de los hongos los valores de pH del medio pueden variar entre 4 y 5, como resultado de su metabolismo. El máximo crecimiento de A.flavus y A. parasiticus ocurre entre los pH 5-8. González et al (1989), en un estudio con diferentes sustratos, no encontraron diferencias

7
significativas en la producción de aflatoxinas cuando variaron los pH entre 4 y 7. Tsai et al (1984) hallaron que en medio YES los resultados diferían cuando el pH utilizado era de 6.5, en comparación con 4.5 y 5.5, de donde se puede inferir que el pH está muy relacionado con el sustrato que se esté utilizando al no existir un comportamiento homogéneo. 2.3.7-Temperatura Desde 1973, cuando West et al evaluaron varias temperaturas de incubación utilizando arroz, se evidenció que hay diferencias significativas en la producción de aflatoxinas en relación con este factor. Nandi y Haggblom (1984) informaron como óptima la temperatura de 30°C en sustratos naturales, sin embargo, Hill et al (1983) y Park y Bullerman (1983) obtuvieron resultados superiores con temperaturas inferiores. Por otra parte, Tuason y Madamba (1980) y Koehler et al (1985) lograron resultados similares de producción de aflatoxinas con temperaturas entre 20 y 21 °C, iguales a las obtenidas en nuestro medio (901 µg/kg)(González et al, 1989). 2.3.8-Gases atmosféricos La oxigenación constituye otro aspecto a tener en consideración en el crecimiento fúngico, puesto que los hongos micotoxigénicos son organismos aeróbicos. Se ha comprobado que bajas concentraciones de oxígeno y/o altas concentraciones de dióxido de carbono pueden deprimir el crecimiento fúngico y la consecuente formación de micotoxinas (Hesseltine, 1976). Sander et al (1968) obtuvieron altas concentraciones de aflatoxinas con un 21% de oxígeno y 0.033% de dióxido de carbono. Un estudio realizado en el almacenamiento de maní en condiciones de atmósfera controlada (CO2:N2 65:35) con diferentes clases de empacamiento (ASI, ASII y ASIII) mostró una reducción en la producción de aflatoxinas por A. flavus con concentraciones inferiores al máximo límite permisible (20 ng/g), sin embargo el efecto de las concentraciones de los gases fue dependiente del tipo de empacamiento y la temperatura de almacenamiento (Ellis et al, 1994 a,b). 3-Caracterización físico-química y efectos tóxicos de las aflatoxinas. 3.1-Propiedades físico-químicas. Se conocen 18 tipos de aflatoxinas, pero solo cuatro aparecen como contaminantes de los alimentos. Estas son: B1, B2, G1 y G2 (figura 2). El nombre genérico de aflatoxinas proviene de su principal especie productora (A.flavus) y la denominación B y G se debe al color de la fluorescencia que exhiben cuando son colocadas bajo la acción de la luz ultravioleta, donde B corresponde al azul (blue) y G al verde (green), mientras que los subíndices se refieren a la movilidad cromatográfica (Jones 1977). Existen otras aflatoxinas naturales que se producen en el metabolismo celular, como la M1, M2, B2a y G2a (figura 2) que también han sido aisladas de cultivos de A.flavus y A.parasiticus, en bajas concentraciones (Stubblefield et al, 1970; OPS-OMS, 1983; Dutton et al, 1985 ). Las aflatoxinas M1 y M2 pueden encontrarse en la leche y en la orina, así como la AFP1 y AFQ1 (figura 2). El parasiticol (AFB3) y el aflatoxicol (R0), compuestos que están estrechamente

8
relacionados con la biosíntesis de la aflatoxina B1, se han aislado de cultivos de A.flavus (Jones 1977). Algunas de la aflatoxinas antes mencionadas pueden ser obtenidas, además, mediante síntesis química. Ellas son: la aflatoxina M1, que ha sido sintetizada a partir de la oxidación de la aflatoxina B1 con el dióxido de selenio (Christou et al, 1985); las aflatoxinas B2a y G2a, que se sintetizan por la adición de una molécula de agua al doble enlace vinílico de las aflatoxinas B1 y G1 respectivamente, en un medio ácido (Pohland et al, 1968) y la AFB3, que puede obtenerse por hidrólisis de la aflatoxina G1 en la posición 5 del anillo lactónico seguido por una descarboxilación (Jones, 1977). Existen otras aflatoxinas no naturales, como la aflatoxina B1S que se produce por la acción del metabisulfito de sodio sobre el grupo vinilo de la molécula de la aflatoxina B1 y es muy poco tóxica. Otra no natural es la aflatoxina D1, producida por el empleo de amoníaco en su estado gaseoso o líquido, actuando sobre el anillo lactónico de la aflatoxina B1. Las aflatoxinas RB1 y RB2 son producidas por la reacción de las AFB1 y AFB2 con borohidruro de sodio como resultado de la ruptura del anillo de lactona, seguido de una reducción del grupo ácido de la ciclo pentanona (Jones, 1977).
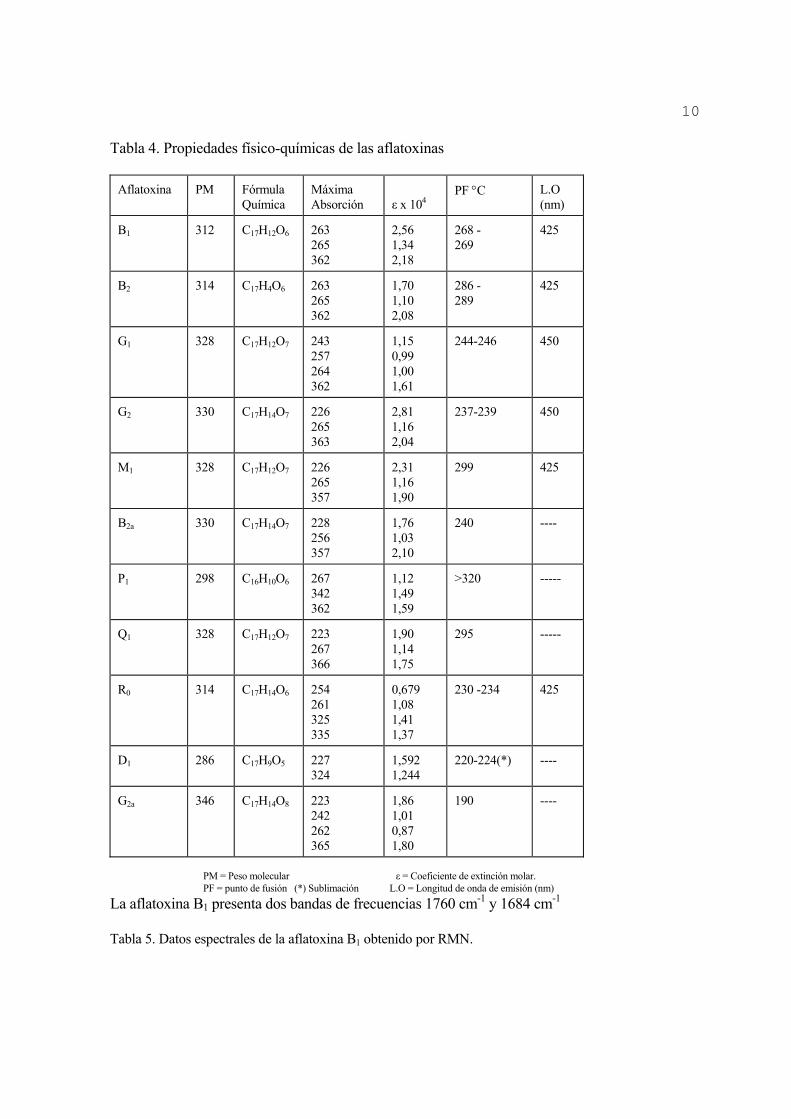
Las aflatoxinas son solubles en solventes orgánicos tales como: clorofomo, metanol, acetonitrilo, benceno, entre otros. Estos son usados para su extracción a partir de los medios de cultivo o de los alimentos contaminados. Las toxinas disueltas en cloroformo o en una mezcla de benceno:acetonitrilo (98:2 v/v) son estables durante años si se mantienen en lugares oscuros y fríos (Merck Index, 1983; AOAC, 1990); no obstante, cuando se mantienen en metanol su estabilidad disminuye con el tiempo. En la tabla 4 se muestran las principales propiedades físico-químicas de las aflatoxinas.

9
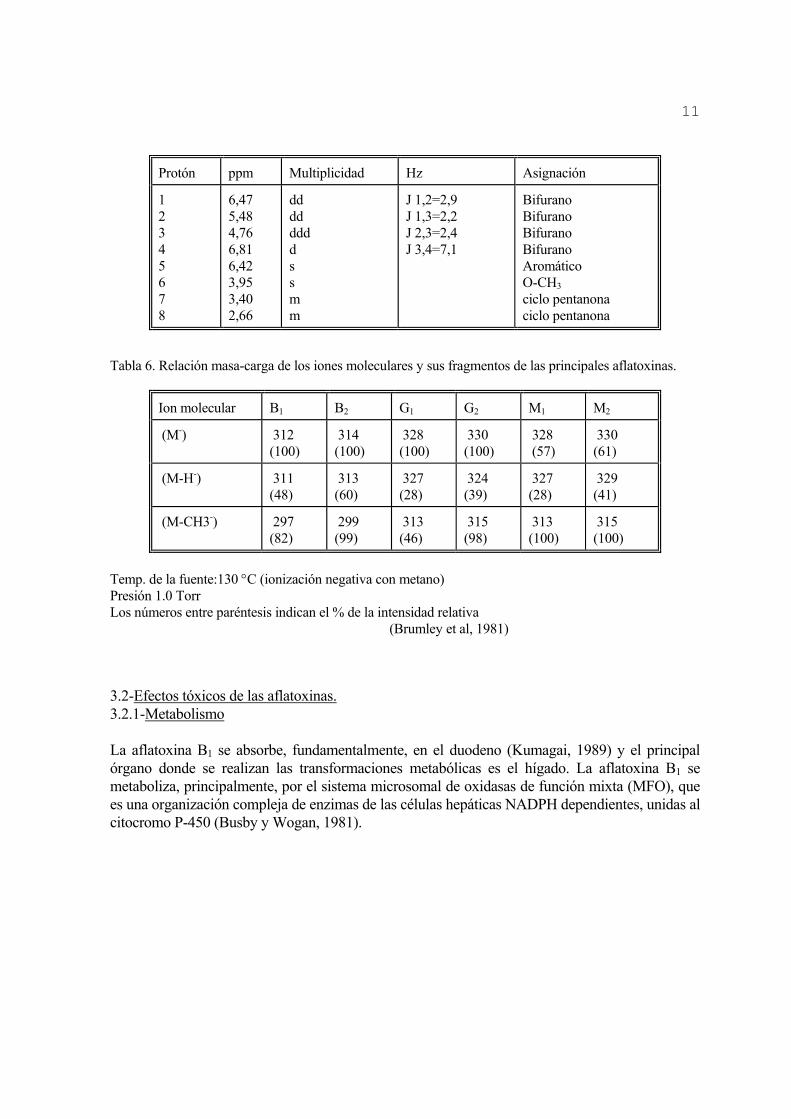
3.1.l-Espectroscopía Infrarroja (IR), Resonancia Magnético Nuclear (RMN) y Espectroscopía de Masa (EM). El análisis estructural de las aflatoxinas se ha basado, en parte, en sus espectros infrarrojos (IR). Aunque este es característico de la molécula entera, siempre ciertos grupos en ella dan lugar a bandas un determinado intervalo de frecuencia, independientemente de la naturaleza del resto de la molécula. La permanencia de estas bandas, características de grupos funcionales, permite una amplia utilización de la espectroscopía IR en la obtención de información estructural por simple inspección del espectro y el uso de tablas de frecuencia de grupos característicos. Existen otras bandas de menor frecuencia como son: 1632 cm-1 2850 cm-1 (-O-CH3) 1598 cm-1 1600 cm-1 (-HC=CH-) 1562 cm-1 1130 cm-1 (C-O-C) Estas bandas aparecen en la mayoría de las aflatoxinas y en el caso de los compuestos hidroxilados se encuentra otra en la región de 3400 cm-1, correspondiente al grupo OH (Asao et al, 1965; Jones, 1977). Un estudio estructural (IR) del aflatoxicol reveló que este no presenta las bandas a 1760 y 1684 cm-1 provenientes de la estructura cumarínica de las aflatoxinas, debido a que esta molécula no tiene el grupo carbonilo en el anillo de ciclo-pentanona. En su lugar aparecen dos bandas a 1070 y 1020 cm-1, indicativas de la presencia de un alcohol secundario y de un grupo carbonilo en el anillo saturado de la lactona (Detroy y Hesseltine, 1969). Conjuntamente, en la región de 3400 cm-1 presenta una banda de absorción ancha, correspondiente al grupo hidroxilo. La resonancia magnético-nuclear (RMN) juega un papel importante en la elucidación estructural de cualquier compuesto, conjuntamente con la espectroscopía infrarroja (IR). Esta se fundamenta en las propiedades magnéticas de los núcleos atómicos. La frecuencia de resonancia de un conjunto de protones (H+) que forman parte de un sistema molecular depende del entorno electrónico de los mismos. Estudios estructurales de las aflatoxinas por RMN han sido realizados por David et al (1987). En la tabla 5 se muestran algunos datos espectrales de la aflatoxina B1 obtenidos por RMN. La espectroscopía de masa permite identificar una muestra, inferir la estructura de un compuesto desconocido, estudiar la pureza de una muestra y analizar cualitativa y cuantitativamente una mezcla. Este procedimiento puede ser de ionización química positiva o ionización química negativa, y es específico para cada compuesto. Los fragmentos de aflatoxina que se han encontrado con mayor frecuencia relativa son el ión molecular (M-), la pérdida de un hidrógeno (M-H-) y la pérdida del grupo metoxi (M-CH3) (Brumley et al, 1981). En las tablas 6 y 7 se muestran los diferentes iones moleculares y sus fragmentos de las principales aflatoxinas.
10
Tabla 4. Propiedades físico-químicas de las aflatoxinas
Aflatoxina PM Fórmula Química
Máxima Absorción
ε x 104
PF °C L.O (nm)
B1 312 C17H12O6 263 265 362
2,56 1,34 2,18
268 - 269
425
B2 314 C17H4O6 263 265 362
1,70 1,10 2,08
286 - 289
425
G1 328 C17H12O7 243 257 264 362
1,15 0,99 1,00 1,61
244-246 450
G2 330 C17H14O7 226 265 363
2,81 1,16 2,04
237-239 450
M1 328 C17H12O7 226 265 357
2,31 1,16 1,90
299 425
B2a 330 C17H14O7 228 256 357
1,76 1,03 2,10
240 ----
P1 298 C16H10O6 267 342 362
1,12 1,49 1,59
>320 -----
Q1 328 C17H12O7 223 267 366
1,90 1,14 1,75
295 -----
R0 314 C17H14O6 254 261 325 335
0,679 1,08 1,41 1,37
230 -234 425
D1 286 C17H9O5 227 324
1,592 1,244
220-224(*) ----
G2a 346 C17H14O8 223 242 262 365
1,86 1,01 0,87 1,80
190 ----
PM = Peso molecular ε = Coeficiente de extinción molar. PF = punto de fusión (*) Sublimación L.O = Longitud de onda de emisión (nm)
La aflatoxina B1 presenta dos bandas de frecuencias 1760 cm-1 y 1684 cm-1 Tabla 5. Datos espectrales de la aflatoxina B1 obtenido por RMN.
11
Protón ppm Multiplicidad Hz Asignación
1 2 3 4 5 6 7 8
6,47 5,48 4,76 6,81 6,42 3,95 3,40 2,66
dd dd ddd d s s m m
J 1,2=2,9 J 1,3=2,2 J 2,3=2,4 J 3,4=7,1
Bifurano Bifurano Bifurano Bifurano Aromático O-CH3 ciclo pentanona ciclo pentanona
Tabla 6. Relación masa-carga de los iones moleculares y sus fragmentos de las principales aflatoxinas.
Ion molecular B1 B2 G1 G2 M1 M2
(M-) 312 (100)
314 (100)
328 (100)
330 (100)
328 (57)
330 (61)
(M-H-) 311 (48)
313 (60)
327 (28)
324 (39)
327 (28)
329 (41)
(M-CH3-) 297 (82)
299 (99)
313 (46)
315 (98)
313 (100)
315 (100)
Temp. de la fuente:130 °C (ionización negativa con metano) Presión 1.0 Torr Los números entre paréntesis indican el % de la intensidad relativa (Brumley et al, 1981) 3.2-Efectos tóxicos de las aflatoxinas. 3.2.1-Metabolismo La aflatoxina B1 se absorbe, fundamentalmente, en el duodeno (Kumagai, 1989) y el principal órgano donde se realizan las transformaciones metabólicas es el hígado. La aflatoxina B1 se metaboliza, principalmente, por el sistema microsomal de oxidasas de función mixta (MFO), que es una organización compleja de enzimas de las células hepáticas NADPH dependientes, unidas al citocromo P-450 (Busby y Wogan, 1981).

12
Tabla 7. Relación masa-carga de los iones moleculares y sus fragmentos de las principales aflatoxinas por ionización química positiva y negativa.
Ionización B1 B2 G1 G2 M1
Positiva MH+
313(100) 285(29) 270(6) 269(7) 257(5) 241(12) 214(4) 183(3)
315(100) 299(1) 297(6) 287(23) 272(6) 271(8) 259(30)
329(100) 311(18) 300(1) 283(16) 269(4) 257(3) 243(62) 225(3) 215(16)
331(100) 313(18) 303(3) 285(6) 275(5) 257(6) 245(11) 229(3) 217(4)
329(47) 301(11) 285(4) 283(5) 273(100) 259(49) 229(18) 216(5)
Negativa M-
312(1) 297(100) 269(4) 253(1) 209(1)
314(1) 299(100) 271(6) 243(8)
328(-) 313(33) 269(100) 241((16)
330(-) 315(28) 271(100) 243(11) 215(3)
328(1) 313(100) 270(1) 269(1)
En la figura 3 se resumen las principales vías metabólicas de la aflatoxina B1, y en la figura 4 se muestran sus posibles transformaciones estructurales. Como se observa, las hidroxilaciones en (e) y (c) dan las aflatoxinas M1 y Q1, respectivamente; la o-desmetilación (b), da la aflatoxina P1 y la 8,9 oxidación (2,3 oxidación, en el antiguo sistema de numeración) (a), llamada también reacción de activación metabólica, da el 8,9-óxido de aflatoxina B1. La formación de aflatoxicol es catalizada por una 17-OH-esteroide deshidrogenasa citoplasmática, siendo esta reacción reversible (d). El aflatoxicol, además, puede experimentar una reacción de oxidación en el sistema MFO que da por resultado el aflatoxicol H1. Este metabolito puede ser reducido en el citoplasma a AFQ1 (Wei y Hsieh, 1980). Como se observa, el metabolismo de la aflatoxina B1 produce tanto metabolitos destoxificados como activados. El predominio de una u otra vía metabólica depende, en parte, de la especie animal (Gorelick, 1990). En la tabla 8 se muestran los porcentajes de producción relativa de los diferentes metabolitos de aflatoxina B1 de forma comparativa entre especies. No solo los factores genéticos determinan la producción de los porcentajes relativos de los metabolitos de aflatoxina B1. Se ha demostrado que algunos productos químicos inducen una relativa mayor producción de algunos metabolitos, por ejemplo, los bifenilos policlorinados (PCB) o polibrominados (PBB) y el metil colantreno aumentan la producción de AFM1 y AFQ1 en el sistema MFO de ratas y hamsters, mientras que el fenobarbital deprime la producción de AFM1 y aumenta la de AFQ1 (Shepherd et al, 1984; Fukuhara et al, 1990).

13
En general, la destoxificación hepática de las aflatoxinas se realiza por el mecanismo de formación de productos más polares que puedan ser conjugados con aminoácidos, ácido
glucurónico, sulfatos o ácidos biliares que ayuden a su excreción, tal como se realiza para otros xenobióticos. El 8,9-óxido de AFB1 se excreta conjugado al glutatión (Swick, 1984; Cole et al, 1985). Tabla 8. Metabolismo in vitro de la afaltoxina B1 en diferentes especies. (Distribución porcentual de los
productos metabólicos).
Especie AFB1 AFQ1 AFP1 AFM1 Ro AFB2a
mono
hámster
pato
humano
ratón
rata
48.3
40.5
35.1
33.3
21.6
18.7
10.1
8.3
1.6
8.5
1.7
1.3
-
-
-
-
2.8
-
4.0
4.0
1.1
3.8
1.8
0.9
2.0
0.6
6.0
0.7
1.3
1.5
3.3
0.4
0.4
0.7
0.4
0.2
(Patterson,1977)
Las aflatoxinas P1 y Q1 son consideradas poco tóxicas, debido a que son rápidamente conjugadas con sustancias hidrosolubles (Swick, 1984), lo que les permite una rápida excreción. Como se ve (tabla 8), estas aflatoxinas son los metabolitos que predominan en el ratón y en el mono, especies resistente y poco sensible respectivamente, a los efectos carcinogénicos (Tulpule, 1981; Swick,

14
1984). La aflatoxina M1 (AFM1) mantiene una considerable toxicidad y sus propiedades carcinogénicas (OPS-OMS, 1983). Los primeros estudios que se realizaron de la toxicidad de la AFM1 revelaron que esta poseía similar toxicidad a la AFB1, pero actualmente se conoce que es menos tóxica (Ferrando et al, 1984) y menos hepatocarcinogénica que su antecesora metabólica (2-10 % de la AFB1) (Hsieh et al, 1986). La mayor importancia toxicológica de la AFM1 radica en que al poder encontrarse en la leche o productos lácteos y no ser un producto destoxificado, constituye un peligro para la salud pública. El aflatoxicol es también tóxico. Debido a la reversibilidad de la reacción de formación, se considera una reserva metabólica de aflatoxina B1. El 8,9 óxido de aflatoxina B1 es el metabolito activado. Su principal característica es la capacidad de unirse a las macromoléculas (proteínas, ADN y ARN) formando aductos (figura 5). Los primeros en ser descubiertos fueron los aductos AFB1-ADN. Posteriormente fue comprobado que después de una dosis única de AFB1, del 1 al 3% de esta toxina se une a la albúmina sérica formando aductos AFB1:albúmina, que pueden también ser observados en la exposición crónica. Existe una relación constante entre los aductos hepáticos (AFB1-ADN) y los plasmáticos AFB1-albúmina (Wild et al, 1986). 3.2.2-Efectos bioquímicos. Las acciones óxicas de la aflatoxina B1 a nivel bioquímico se ejercen en el metabolismo de las proteínas, lípidos y carbohidratos, en el transporte electrónico de la cadena respiratoria y en las membranas celulares y subcelulares (Patterson, 1977). Sin embargo, uno de sus efectos tóxicos más importantes es el relativo a su interacción con el ácido desoxirribonucleico (ADN), por los efectos mutagénicos y carcinogénicos que esta implica.

15
En una revisión realizada por Patterson (1977) se plantea que entre los principales efectos bioquímicos está la inhibición de la síntesis del ácido ribonucleico (ARN) ADN dependiente, en el núcleo de las células hepáticas. Ello provoca rápidamente una inhibición de la biosíntesis de las proteínas nucleares, mientras que posteriormente es inhibida la síntesis de proteínas en el sistema microsomal (proteínas de exportación). En cuanto a sus efectos en el metabolismo de los carbohidratos, se plantea que la AFB1 disminuye las actividades UDP glucosa-glucógeno transglucosilasa y glucosa 6-fosfatasa (G-6-Pasa), trastornos bioquímicos que se relacionan con la depleción de las reservas de glucógeno hepático y los cambios morfológicos observados en el retículo endoplasmático, respectivamente, teniendo en cuenta que la G-6-Pasa es específica del retículo endoplasmático. Los cambios grasos hepáticos observados en la intoxicación por aflatoxina B1 han sido explicados por los efectos que esta ejerce en el transporte de los lípidos del hígado. Se ha encontrado también que esta toxina inhibe la actividad tioquinasa, esencial para la activación del acetato en acetil CoA. Otra alteración bioquímica relacionada con el metabolismo de los lípidos es el trastorno del metabolismo de los carotenoides observado en las aves (Tyczkowski y Hamilton, 1987). Los efectos tóxicos metabólicos de la aflatoxina B1, a nivel de los organelos celulares, han sido descritos como un aumento en la permeabilidad de las membranas mitocondriales, que interrumpe el transporte de electrones de la cadena respiratoria y también el aumento en la permeabilidad de las membranas lisosomales con salida al exterior de las hidrolasas ácidas (OPS-OMS, 1983). Hay que destacar que las alteraciones bioquímicas descritas son las causantes de los efectos adversos que se observan en la intoxicación aflatoxínica en el hombre y en los animales, que en estos últimos provoca una disminución de sus capacidades productivas. 3.2.3-Carcinogenicidad.

16
La carcinogenicidad de la aflatoxina B1 fue probada por primera vez por Lancaster et al (1961), quienes produjeron hepatomas en ratas a las que se les suministró el mismo alimento de maní, implicado en la enfermedad X de los pavos, con anterioridad a descubrirse la verdadera identidad de esta toxina. Posteriormente, se probaron experimentalmente, sus efectos carcinogénicos en otras especies de animales, tales como patos, (Carnaghan, 1965), trucha arco iris (Salmo gardneri) (Halver, 1967), monos (Gopalan et al, 1972), entre otras. Boutibonnes et al (1984) realizaron un estudio de la actividad genotóxica de 33 micotoxinas utilizando la prueba de reparación del ADN o efecto "REC" y encontraron que la AFB1 presenta una actividad genotóxica potente. Martin y Garner (1977) postularon que la capacidad de producir mutaciones de la aflatoxina B1 podría deberse a la habilidad de un metabolito activado de unirse covalentemente a la guanina del ADN en la posición N-7 (aductos) (figura 5), además de que esta reacción podría ser importante en la iniciación del cáncer. Hoy está bien establecido que los carcinógenos químicos, como es el caso de la aflatoxina B1, se unen covalentemente al ADN formando aductos (Modali y Yang, 1986; Yu et al, 1990) y que el mecanismo de producción de la mutagénesis de AFB1 es el mismo que el de los agentes alquilantes, produciendo mutaciones puntuales (Modali y Yang, 1986) (figura 6). Estos mismos autores plantearon un posible mecanismo de la iniciación oncogénica en el cual también está involucrado el 8,9 óxido de AFB1 a través de la activación química de un proto-oncogen (figura 7).

17
3.2.4-Teratogenicidad Los efectos teratogénicos de la AFB1 se informaron, por primera vez, por Elis y DiPaolo (1967) en hamsters que recibieron una dosis de 4 mg AFB1/kg de peso el octavo día de gestación. Las principales lesiones encontradas estuvieron a nivel del sistema nervioso central. Otros autores (Ong, 1975) también han informado estos efectos. Por otra parte, Llewellyn et al (1977) encontraron que la AFB1 retarda el proceso de blastulación y gastrulación en Medaka Japonesa (Oryzias latipes) en forma proporcional a la dosis utilizada, así como produce efectos teratogénicos variados a nivel del sistema circulatorio, vejigas urinaria y natatoria y en el sistema óptico. Sin embargo, no pudo demostrarse la aparición de malformaciones groseras en embriones de pollo a los que se les administró dosis subletales de AFB1, que mostraron solo retardo en el crecimiento (Shibko et al, 1968). Otras acciones tóxicas de la AFB1 relacionadas con la teratogenicidad, son la relativa a la inducción de aberraciones cromosomales que esta toxina produce en células humanas (Promchainant et al, 1972) y de ratones (Krishnamurthy, 1986) y el efecto que ocasiona en el índice meiótico en curieles (Ranjan y Sinha, 1989). 3.2.5-Inmunodepresión En la década del 70 comenzaron los estudios relacionados con el efecto negativo de las aflatoxinas en el sistema inmunológico (Pier et al, 1977). Así, numerosos autores han informado que el consumo de bajas dosis de aflatoxinas afecta la resistencia natural o adquirida a enfermedades infecciosas en animales de experimentación (Wei y Hsieh, 1980) y en animales productivos (pollos) (Ilgaz, 1985), que la AFB1 afecta el sistema inmune humoral y celular en pollos (Giambrone, 1978), que las inmunoglobulinas séricas en terneros disminuyen por la acción tóxica de la AFB1 (Balaraman y Arora, 1987), que la actividad del complemento se ve reducida y que retarda la producción de interferón (Wei y Hsieh, 1980). Todos estos hechos, unidos a la también disminuida capacidad de los linfocitos y fagocitos producida por la ingestión de aflatoxinas, reducen las defensas contra los microorganismos (Wei y Hsieh, 1980; Slowick et al, 1985; Raisuddin et al, 1990; Pier, 1992).

18
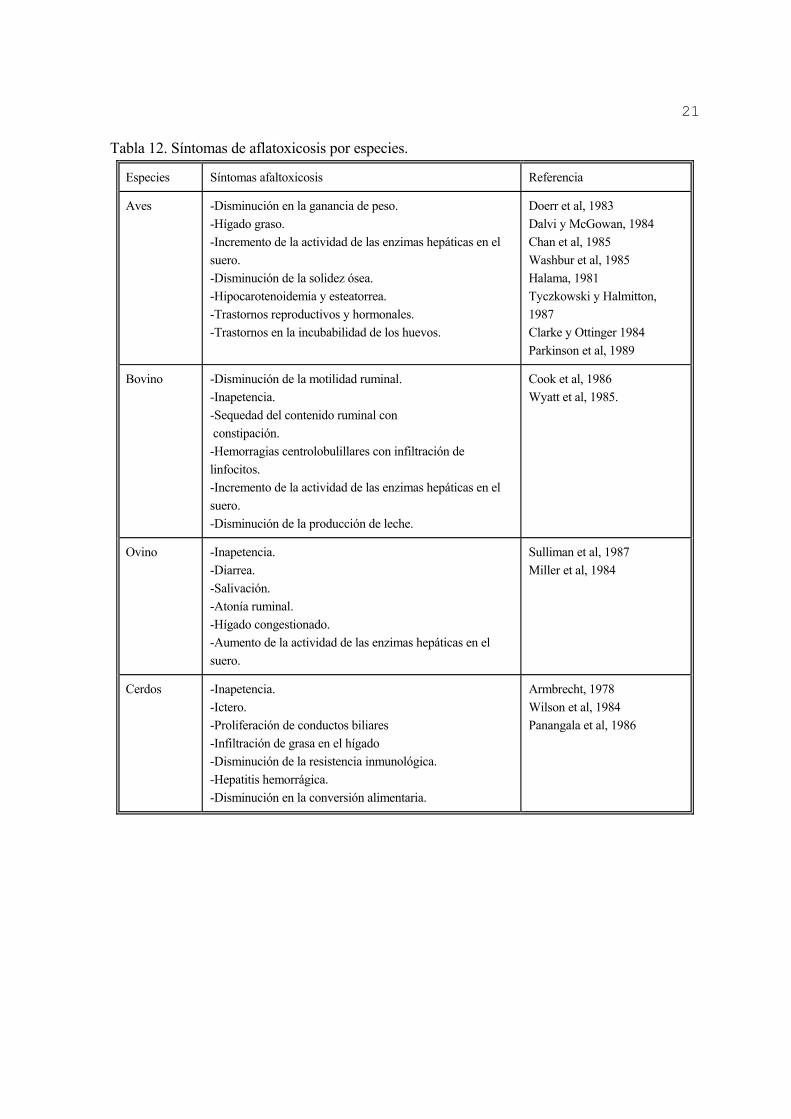
3.2.6-Efectos tóxicos en animales A pesar de las diferencias en susceptibilidad, prácticamente en todas las especies animales, se han informado los efectos tóxicos de las aflatoxinas. Entre estas, la de mayor toxicidad aguda es la aflatoxina B1, lo que puede apreciarse por las bajas DL50 que presentan (Tabla 9).
Tabla 9. Toxicidad aguda de la Aflatoxina B1 en diferentes especies.
Especie DL50 (mg/kg)
Especie DL50 (mg/kg)
patico conejo pavo gato cerdito perro curiel ovejas
0.3 - 0.6 0.3 - 0.5 0.5 - 1.0 0.55 0.62 1.0 1.2 - 2.0 2.0
cerdo caballo mono pollo rata ratón hámster trucha
2.0 2.0 2.0 2.0 - 6.3 5.5 - 18.0 9.0 10.0 0.8
Pequeñas variaciones en la estructura de las aflatoxinas implican también variaciones en su toxicidad. En la tabla 10 se muestra la dosis tóxica de varias aflatoxinas en paticos de un día de nacidos (Applebaum et al 1982). Tabla 10. Toxicidad relativa de las diferentes aflatoxinas en patos.
aflatoxina DL50 (mg/kg)
B1 B2 G1 G2 M1 M2
0.36 1.7 0.8 2.5 0.8 3.1
El principal efecto tóxico crónico de las aflatoxina B1 es la carcinogenicidad. La susceptibilidad a estos efectos entre especies está relacionada con la velocidad de su metabolismo en general y con el predominio de una u otra ruta metabólica. Tulpule (1981) plantea que las especies que metabolizan rápidamente la aflatoxina son más susceptibles a los efectos agudos y las que la metabolizan más lentamente, lo son más a los efectos carcinogénicos (tabla 11). Tabla 11. Velocidad metabólica comparativa del metabolismo de la aflatoxina B1 (in vitro).

19
Especies Tiempo* Velocidad
conejo
patico
mono (Rhesus)
cobayo
humano
39.6 seg
49.8 seg
6.4 min
11.8 min
13.3 min
rápidos
pollo
ratón
cerdo
carnero
32-97 min
1.57 h
2.50 h
4.26 h
intermedios
rata 0.8-2.6 días lentos (Tulpule 1981) * Tiempo para metabolizar una DL50 Los síntomas de la intoxicación por aflatoxinas son inespecíficos y son similares para todas las especies de animales: pérdida de apetito y disminución de sus capacidades productivas en general, según la especie; disminución en la ganancia en peso y disminución en la conversión alimentaria (Dieckman y Green, 1992), como disminución en la puesta de huevos en las gallinas (Iqbal et al, 1983); disminución en la producción de leche en las vacas (Key, 1978); trastornos reproductivos y hormonales (Guthrie, 1979; Clarke y Ottinger, 1984) y disminución de la resistencia inmunológica (Pier et al, 1977). Los signos clínicos más frecuentes son los relacionados a la disfunción hepática: incremento en la actividad sérica de enzimas hepáticas y trastornos en la coagulación. En todas la especies el hígado se encuentra congestionado, con infiltración de grasa y al examen microscópico pueden observarse, hemorragias centrolobulillares y proliferación de los conductos biliares que pueden llevar a la atrofia del órgano. En la tabla 12 se muestra un resumen de los principales síntomas de aflatoxicosis en animales de importancia productiva. 3.2.7-Relación de las aflatoxinas con algunas enfermedades en el hombre. Entre las enfermedades que han sido relacionadas con la ingestión de alimentos contaminados con aflatoxinas están en primer lugar, el carcinoma hepatocelular (CHC), el cáncer de pulmón y de colon, el síndrome de Reye, el kwashiorkor y la hepatitis tóxica. Para los estudios de los efectos cancerígenos de la aflatoxina B1 en el hombre, ha sido utilizado, principalmente, el método estadístico que relaciona el nivel de contaminación encontrado en los alimentos con los porcentajes de aparición de carcinoma hepatocelular y otros tumores en ciertas poblaciones. También se ha dosificado la aflatoxina B1 con sus metabolitos (Fukal y Reisnerova, 1990) y sus aductos (Wild et al, 1990) en fluidos biológicos (orina, sangre, leche) de pacientes, en población normal, y en los órganos internos (hígado) de fallecidos en enfermedades oncológicas u

20
otras en las que fue sospechada su relación con la aflatoxicosis (Dvorackova et al, 1977, Harrison et al 1993). Los informes iniciales de la relación de la ingestión de aflatoxina B1 y la aparición de CHC fueron realizados en Uganda, en Swazilandia, en Tailandia, Kenya y en Mozambique durante la década del 70 (OPS-OMS, 1983). Un estudio realizado en Filipinas por Bulatao-Jayme et al, (1982) demostró una fuerte asociación entre la ingestión de cantidades crecientes de aflatoxina y el riesgo de desarrollar CHC, habiéndose encontrado también que el consumo de alcohol y el tabaquismo son factores de riesgo asociados. Más modernamente se conoce que el virus de la hepatitis B es otro factor de riesgo al CHC (Austin et al, 1986; Wild et al, 1993).
21
Tabla 12. Síntomas de aflatoxicosis por especies.
Especies Síntomas afaltoxicosis Referencia
Aves -Disminución en la ganancia de peso. -Hígado graso. -Incremento de la actividad de las enzimas hepáticas en el suero. -Disminución de la solidez ósea. -Hipocarotenoidemia y esteatorrea. -Trastornos reproductivos y hormonales. -Trastornos en la incubabilidad de los huevos.
Doerr et al, 1983 Dalvi y McGowan, 1984 Chan et al, 1985 Washbur et al, 1985 Halama, 1981 Tyczkowski y Halmitton, 1987 Clarke y Ottinger 1984 Parkinson et al, 1989
Bovino -Disminución de la motilidad ruminal. -Inapetencia. -Sequedad del contenido ruminal con constipación. -Hemorragias centrolobulillares con infiltración de linfocitos. -Incremento de la actividad de las enzimas hepáticas en el suero. -Disminución de la producción de leche.
Cook et al, 1986 Wyatt et al, 1985.
Ovino -Inapetencia. -Diarrea. -Salivación. -Atonía ruminal. -Hígado congestionado. -Aumento de la actividad de las enzimas hepáticas en el suero.
Sulliman et al, 1987 Miller et al, 1984
Cerdos -Inapetencia. -Ictero. -Proliferación de conductos biliares -Infiltración de grasa en el hígado -Disminución de la resistencia inmunológica. -Hepatitis hemorrágica. -Disminución en la conversión alimentaria.
Armbrecht, 1978 Wilson et al, 1984 Panangala et al, 1986

22
Los estudios de la relación entre la ingestión de alimentos contaminados con aflatoxinas y la presentación del carcinoma hepatocelular siguen realizándose en Zimbabwe (Nyathi et al, 1987), en Swaziland (Peer et al, 1987), Africa del Sur (Lotter y Krohn, 1988), en norteamérica (Groopman et al, 1988), Tailandia (Wild et al, 1989) y China (Groopman et al 1993). Recientemente se ha planteado que entre los consumidores de heroína existe una alta probabilidad de adquisición de aflatoxinas por vía endovenosa provenientes de la contaminación de la planta, lo que provoca que estos sean más susceptibles a contraer SIDA y hepatitis a virus B por estar inmunodeprimidos. De hecho, la aflatoxina B1 se encontró en nueve muestras de orina procedentes de trece heroinómanos callejeros (9616 pg/g) y en el 20% de 113 adictos de Inglaterra y Holanda (Hendrickse et al, 1989). En el monitoreo del riesgo a CHC por aflatoxinas y en investigaciones de sus causas, se recomienda utilizar la medición de los aductos AFB1-ADN en el hígado de pacientes fallecidos de CHC (Garner et al, 1988 a) y en la orina (Groopman et al, 1985), la de los aductos AFB1-albúmina en el suero (Wild et al, 1990), los cuales reflejan la exposición a aflatoxinas en los cuatro a seis días anteriores al estudio (Xavier y Muñoz, 1987). Las técnicas que más ampliamente se utilizan para medir exposición a aflatoxinas son los procedimientos inmunológicos RIA (Sizaret et al, 1982; Fukal y Reisnerova, 1990) o ELISA (Denning et al, 1988; Garner et al, 1988 b). Otra forma de determinar exposición a las aflatoxinas es medir la aflatoxina M1 en leche materna. En un estudio realizado en Zimbabwe, en mujeres durante el período de lactación se encontró la presencia de aflatoxina M1 en muestras de leche materna (Wild et al, 1987), lo que constituye un riesgo enorme para los bebés. Los individuos que trabajan en procesadoras de alimentos también están expuestos a una contaminación por aflatoxinas, debido a sus propiedades electrostáticas. Autrup et al (1993) encontraron la presencia de aductos de aflatoxina B1-albúmina en el suero de 7 trabajadores de un total de 45 muestreados que trabajaban en compañías procesadoras de alimentos para el consumo animal en Dinamarca. En cuanto a las otras enfermedades que han sido relacionadas a la aflatoxina B1, existen evidencias al respecto, tales como las de Dvorackova et al (1981), quienes encontraron aflatoxina B1 en pulmones de dos pacientes fallecidos de cáncer del pulmón; las de Stora et al (1982) relacionadas con el cáncer de pulmón con aspiración de polvo contaminado con A. flavus, y las de Deger (1976), quien encontró relación entre el cáncer de colon y la ingestión de aflatoxinas. En cuanto al síndrome de Reye, Shank et al (1971), Dvorackova et al (1977) y Stora et al (1983) han notificado la presencia de aflatoxinas en los órganos de niños fallecidos de esta enfermedad. Por otra parte, la relación aflatoxinas-kwashiorkor fue señalada por Hendrickse et al (1983), Apeagyei et al (1986) y Hendrickse (1991), quienes también encontraron aflatoxinas en los hígados de niños con esta enfermedad. Otras enfermedades hepáticas han sido asociadas a las aflatoxinas: la cirrosis infantil de la India (Yadgan et al, 1970) y la hepatitis tóxica, ocurrida también en la India (Krishmamachari et al, 1975).

23
En Cuba un estudio realizado por Alvarez et al (1991) informaron la presencia de aflatoxina B1 en orina de pacientes pediátricos, donde el 56% de los positivos correspondieron a pacientes diagnosticados con hepatitis crónica y portadores de antígeno de superficie del virus de la hepatitis B, mientras que en personas sanas se encontró solo un 7,5% de positividad. También en 1988 se había reportado un 58% (140 muestras analizadas) de positividad a aflatoxina B en la orina de pacientes pediátricos con enfermedades hepáticas provenientes del municipio de San Cristóbal en Pinar del Río (Perguero, 1988). 4-Métodos de análisis. El éxito en la determinación de un residual en los alimentos, o en materiales biológicos, se debe a un conjunto de procedimientos que hacen posible una determinación más precisa y exacta independientemente del método analítico. Entre las etapas a considerar para llevar a cabo la detección de aflatoxinas en los alimentos están: muestreo, extracción, purificación o limpieza y detección. 4.1-Muestreo. El muestreo implica una selección de una muestra representativa de un lote (la cantidad de producto o granos entregados o recibidos, en un determinado momento que tiene propiedades comunes o características uniformes) que posibilita un análisis más preciso de la sustancia objeto de estudio. La distribución de micotoxinas en los productos tiene un carácter discreto o heterogéneo, y es posible que solo un pequeño porcentaje de partículas de un lote esté contaminado con niveles extremadamente altos de toxina. Un estudio realizado por Whitaker et al (1979) de un lote de maní con una concentración de aflatoxina B1 de 1 mg/kg, evidenció variaciones en el contenido de esta en granos individuales de 106 mg/kg. En los resúmenes realizados por Campbell et al (1986) y Jewers et al (1989) sobre el muestreo, se hace referencia a la variabilidad que existe en la concentración de aflatoxina B1 en diferentes muestras de un lote único (tabla 13). Estos resultados nos indican que si la muestra de prueba no es representativa, el resultado obtenido en el laboratorio caracteriza solamente a la muestra enviada y no al lote completo, por lo que el muestreo constituye la principal fuente de error en la determinación de micotoxinas.

24
Tabla 13. Concentración de aflatoxina en diferentes muestras colectadas a partir de un lote único de varios productos.
Cosecha total de muestras
<5-50
µg/kg
50-100
µg/kg
100-200
µg/kg
200-300
µg/kg
>300
µg/kg
semillas de algodón
semillas
de palmas
maíz
harina de maní
torta de
maní
264
96
96
130
165
214
76
7
7
-
21
9
33
75
9
11
11
38
47
50
8
-
11
1
6
10
-
7
-
-
Tejada, 1992 Whitaker et al (1979) plantearon que el error total en la determinación de aflatoxinas en alimentos era la suma de todos los errores durante el muestreo y el análisis (figura 8). Ellos obtuvieron coeficientes de variación de 21, 8, 11, y 26% para la muestra (4.55 kg), submuestra gruesa y fina (1 kg) y la muestra de análisis, respectivamente, con una concentración promedio de 20 µg/kg. En un estudio similar realizado en un lote de maní con una concentración de 20 µg/kg se obtuvieron coeficientes de variación de 55, 19 y 16 % para una muestra de 21,8 kg, submuestra de 1.1 kg y el análisis de dos porciones de la muestra, respectivamente, con un error total del 80%. Otro factor que influye en la determinación de micotoxinas es el tamaño del lote y este varía en dependencia del producto (tabla 14). Para disminuir los errores durante la preparación de la muestra se han desarrollado muestreadores que operan continuamente durante la pulverización y homogeneización, lo que permite un proceso más eficiente, esos son: el mezclador separador cortante vertical Hobbart y los molinos Dickens-Satterwhite y Rommer.
Figura 8. Pasos típicos empleados para estimar la concentración de aflatoxinas en granos asociados a la fuente de variación.
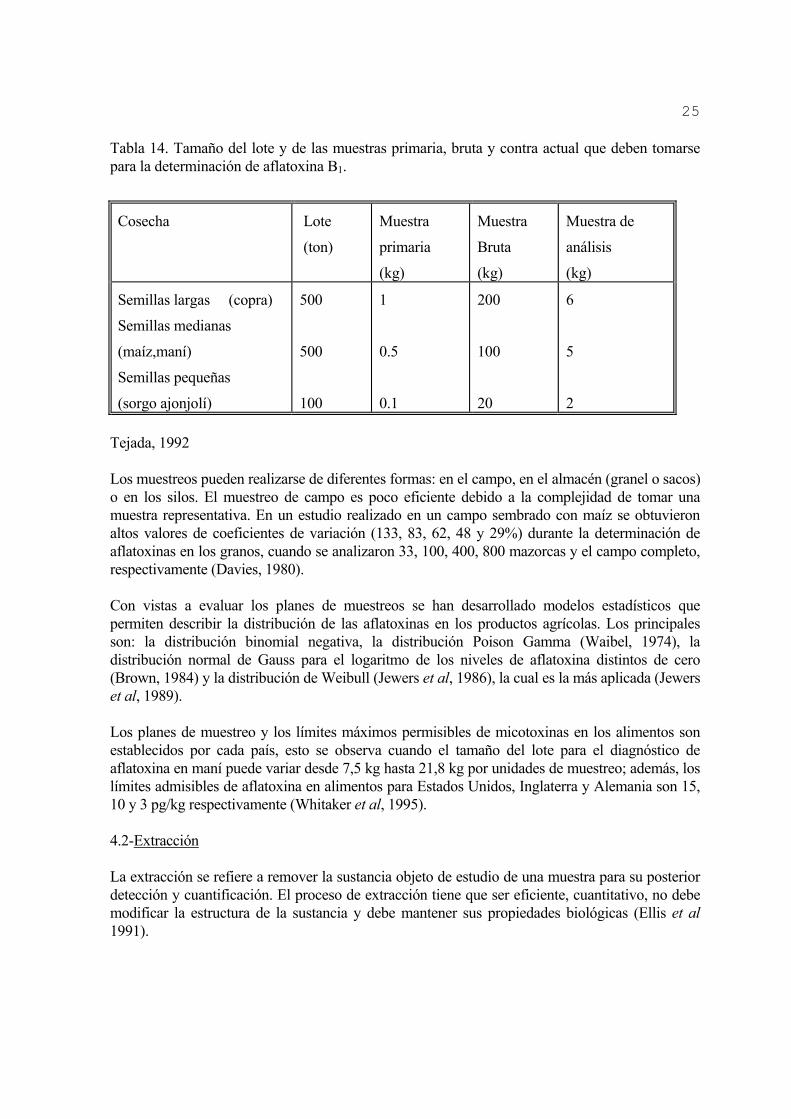
25
Tabla 14. Tamaño del lote y de las muestras primaria, bruta y contra actual que deben tomarse para la determinación de aflatoxina B1.
Cosecha Lote
(ton)
Muestra
primaria
(kg)
Muestra
Bruta
(kg)
Muestra de
análisis
(kg)
Semillas largas (copra)
Semillas medianas
(maíz,maní)
Semillas pequeñas
(sorgo ajonjolí)
500
500
100
1
0.5
0.1
200
100
20
6
5
2 Tejada, 1992 Los muestreos pueden realizarse de diferentes formas: en el campo, en el almacén (granel o sacos) o en los silos. El muestreo de campo es poco eficiente debido a la complejidad de tomar una muestra representativa. En un estudio realizado en un campo sembrado con maíz se obtuvieron altos valores de coeficientes de variación (133, 83, 62, 48 y 29%) durante la determinación de aflatoxinas en los granos, cuando se analizaron 33, 100, 400, 800 mazorcas y el campo completo, respectivamente (Davies, 1980). Con vistas a evaluar los planes de muestreos se han desarrollado modelos estadísticos que permiten describir la distribución de las aflatoxinas en los productos agrícolas. Los principales son: la distribución binomial negativa, la distribución Poison Gamma (Waibel, 1974), la distribución normal de Gauss para el logaritmo de los niveles de aflatoxina distintos de cero (Brown, 1984) y la distribución de Weibull (Jewers et al, 1986), la cual es la más aplicada (Jewers et al, 1989). Los planes de muestreo y los límites máximos permisibles de micotoxinas en los alimentos son establecidos por cada país, esto se observa cuando el tamaño del lote para el diagnóstico de aflatoxina en maní puede variar desde 7,5 kg hasta 21,8 kg por unidades de muestreo; además, los límites admisibles de aflatoxina en alimentos para Estados Unidos, Inglaterra y Alemania son 15, 10 y 3 pg/kg respectivamente (Whitaker et al, 1995). 4.2-Extracción La extracción se refiere a remover la sustancia objeto de estudio de una muestra para su posterior detección y cuantificación. El proceso de extracción tiene que ser eficiente, cuantitativo, no debe modificar la estructura de la sustancia y debe mantener sus propiedades biológicas (Ellis et al 1991).

26
Las primeras extracciones de aflatoxinas de los alimentos se realizaron con una previa delipidación con n-hexano o éter de petróleo (Beljaars et al, 1972). Sin embargo, fue comprobado que la extracción no se afecta por la presencia de lípidos o pigmentos. Asimismo, si se considera que los lípidos son solubles en los mismos solventes que se emplean en la extracción de aflatoxinas, parte de ellas se arrastrarían con la delipidación y conllevaría pasos adicionales para realizar una extracción selectiva, lo que en lugar de reducir pasos en el análisis para simplificarlo, lo complica y lo hace menos eficiente (Chu 1992). Los procedimientos más empleados en la extracción de aflatoxinas de los alimentos y que han sido oficializados por la AOAC, son: Contaminates Bureau (CB) y Best Food (BF) que emplean cloroformo y metanol : agua (55:45 v/v) respectivamente. Existen otros que también se han aprobado como es acetona:agua (85:15 v/v) (AOAC, 1990; Richard et al, 1993). No todos los solventes presentan la misma eficiencia de extracción y esto varía en dependencia del sustrato y la relación del extrayente con la muestra (Cole y Doner, 1994; Bradburn et al, 1995). Cuando se analizan fluidos biológicos, la extracción de la aflatoxina se puede realizar de forma directa adicionando un solvente a una muestra acidificada o desproteinizada previamente (Shepherd et al, 1986; Castegnaro et al, 1990). Así se alcanza un recobrado de la toxina entre 60-80 %; sin embargo, cuando la extracción de la toxina se realiza a través de columnas de Sep-Pak (C10 o sílica) (Ferguson-Foos y Warren, 1984; Bijl y vanPeteghem, 1985) o de afinidad (Groopman y Donahue, 1988; Fremy y Chu 1989) los recobrados oscilan entre 90-100%. No existe un método de aplicación general a la extracción de aflatoxinas, ya que este varía en dependencia del producto (tabla 15). Los tipos de solventes más comúnmente empleados durante la extracción se muestran en la tabla 15. 4.3-Purificación. La purificación previa de las toxinas en el extracto del material analizado es necesaria cuando se emplean procedimientos físico-químicos en la detección, pero cuando se utilizan procedimientos inmunológicos, el paso de purificación se omite analizándose solamente el extracto diluido (Escobar et al, 1991b). Mientras más pasos se usen para la purificación, será menor el recobrado de las toxinas. La eficiencia en el recobrado de la toxina durante la purificación de la muestra aumenta con el empleo de columnas pre-empacadas, debido a que se eliminan pasos de la purificación como son: la rotoevaporación, partición con otros solventes para lograr una extracción selectiva y el empleo de grandes volúmenes de solventes, que dificultan su manipulación. Las columnas más empleadas para la purificación de la aflatoxinas en los alimentos son: Sep-Pak (Chu, 1991), Bond Elut (Scott y Lawrence, 1988), Ciano (Cohen y Lapointe, 1986), Fenil (Bradburn et al, 1990a), Chem Elut (Hutchins et al, 1989) y de afinidad (Kaveri et al, 1987). Con los procedimientos anteriores se logran los máximos recobrados de las aflatoxinas presentes en el medio con un grado de pureza mayor de 90%; sin embargo, con el empleo de columnas de inmunoafinidad se logran extracciones selectivas de las diferentes aflatoxinas, pero el recobrado
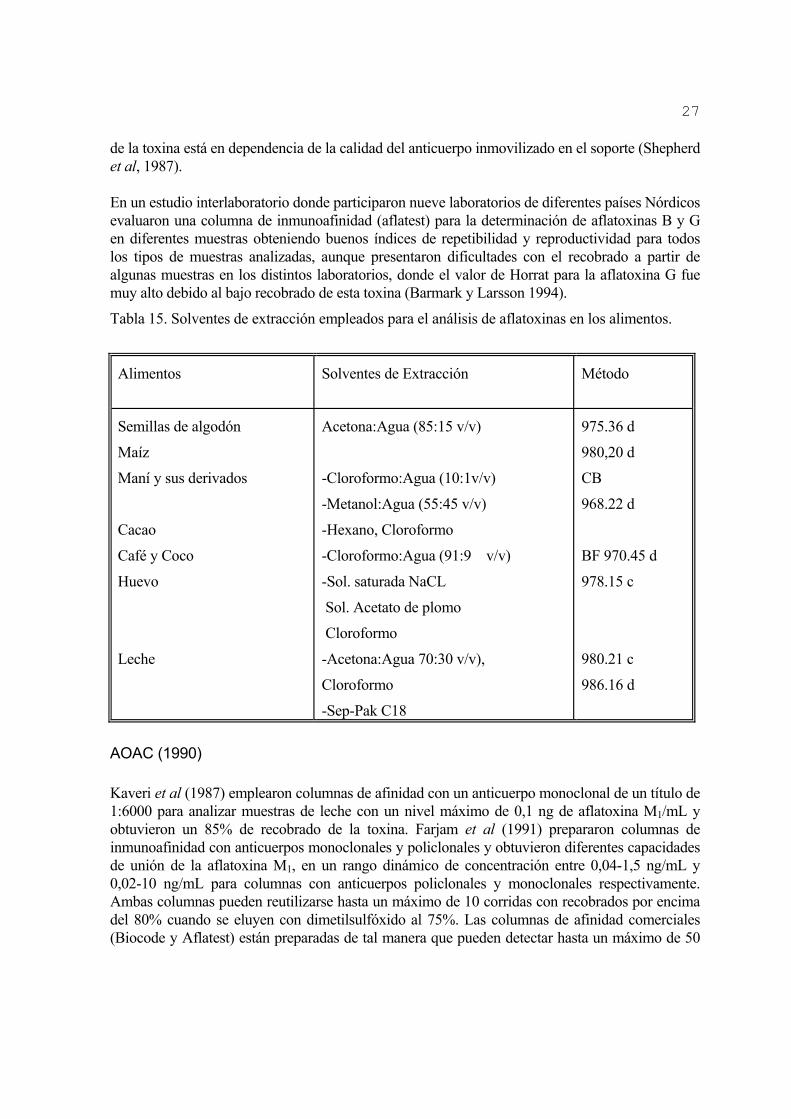
27
de la toxina está en dependencia de la calidad del anticuerpo inmovilizado en el soporte (Shepherd et al, 1987). En un estudio interlaboratorio donde participaron nueve laboratorios de diferentes países Nórdicos evaluaron una columna de inmunoafinidad (aflatest) para la determinación de aflatoxinas B y G en diferentes muestras obteniendo buenos índices de repetibilidad y reproductividad para todos los tipos de muestras analizadas, aunque presentaron dificultades con el recobrado a partir de algunas muestras en los distintos laboratorios, donde el valor de Horrat para la aflatoxina G fue muy alto debido al bajo recobrado de esta toxina (Barmark y Larsson 1994).
Tabla 15. Solventes de extracción empleados para el análisis de aflatoxinas en los alimentos.
Alimentos Solventes de Extracción Método
Semillas de algodón
Maíz
Maní y sus derivados
Cacao
Café y Coco
Huevo
Leche
Acetona:Agua (85:15 v/v)
-Cloroformo:Agua (10:1v/v)
-Metanol:Agua (55:45 v/v)
-Hexano, Cloroformo
-Cloroformo:Agua (91:9 v/v)
-Sol. saturada NaCL
Sol. Acetato de plomo
Cloroformo
-Acetona:Agua 70:30 v/v),
Cloroformo
-Sep-Pak C18
975.36 d
980,20 d
CB
968.22 d
BF 970.45 d
978.15 c
980.21 c
986.16 d
AOAC (1990) Kaveri et al (1987) emplearon columnas de afinidad con un anticuerpo monoclonal de un título de 1:6000 para analizar muestras de leche con un nivel máximo de 0,1 ng de aflatoxina M1/mL y obtuvieron un 85% de recobrado de la toxina. Farjam et al (1991) prepararon columnas de inmunoafinidad con anticuerpos monoclonales y policlonales y obtuvieron diferentes capacidades de unión de la aflatoxina M1, en un rango dinámico de concentración entre 0,04-1,5 ng/mL y 0,02-10 ng/mL para columnas con anticuerpos policlonales y monoclonales respectivamente. Ambas columnas pueden reutilizarse hasta un máximo de 10 corridas con recobrados por encima del 80% cuando se eluyen con dimetilsulfóxido al 75%. Las columnas de afinidad comerciales (Biocode y Aflatest) están preparadas de tal manera que pueden detectar hasta un máximo de 50

28
ng de aflatoxina por gramo de muestra para ser usadas en una sola ocasión, ya que se eluyen con metanol o acetonitrilo para obtener el máximo de recobrado. Una columna multifuncional para la purificación de aflatoxina a partir de diferentes muestras fue validada en un estudio interlaboratorio, la columna multifuncional está preparada con una mezcla de diferentes soportes (fase reversa, exclusión e intercambio iónico) con la finalidad de retener interferencia tales como: grasa, pigmentos, carbohidratos y proteínas, los recobrados obtenidos para todas las aflatoxinas fueron superiores al 95% (Trucksess et al, 1994). 4.4-Detección. Los métodos de detección pueden dividirse en biológicos y físico-químicos (Buckle y Sanders, 1990). Las varias divisiones y subdivisiones de los métodos de determinación de aflatoxinas en cada grupo se muestran en la figura 9. 4.4.1-Cromatografía de capa delgada. La cromatografía en capa delgada (CCD) es la técnica más utilizada en el análisis de aflatoxinas y otras micotoxinas. El éxito de su aplicación depende del tipo de adsorbente, del sistema de solventes y del método de detección. Los adsorbentes más comunes son: la sílica gel G60 y la sílica gel F 254, que pueden estar sobre soporte de cristal, plástico o metal (5x20 cm, 10x20 cm ó 20x20 cm) con un grosor que varía desde 0,1 hasta 1mm.
Para las aflatoxinas, el soporte más usado es el de sílica gel G 60 (Miguel y de Andrés, 1982) y los solventes más utilizados son: cloroformo, metanol, benceno, acetona y éter etílico, preparados en diferentes proporciones. En la tabla 16 se muestran algunos de los sistemas de solventes y los Rf de las principales aflatoxinas. El sistema de detección de las aflatoxinas se basa en la

29
fluorescencia que ellas emiten, aunque su confirmación se realiza por su reacción con una sustancia química (Nesheim y Brumley, 1981). La sensibilidad, exactitud y resolución del análisis de aflatoxinas por cromatografía de capa delgada se vieron mejoradas con la introducción de la cromatografía de capa delgada de alta resolución (HPTLC) (Bradburn et al, 1989, 1990 b), donde el soporte de los platos empleados pueden ser de fase normal (sílica) o de fase reversa (Abramson et al, 1989). Tabla 16. Solventes empleados en cromatografía de capa delgada y valores de Rf para las principales aflatoxinas.
Sistema de solvente B1 B2 G1 G2
Cloroformo:acetona (9:1v/v)
Benceno:metanol:ac. acético (90:5:5 v/v/v)
Cloroformo:acetona:hexano
(85:15:20)
Tolueno:etil-acetato: ac. fórmico (6:3:1 v/v/v).
Benceno:etanol (95:1 v/v)
44
26
48
35
25
37
23
43
28
23
34
20
36
25
23
29
19
30
15
20
Los valores numéricos corresponden a los Rf x 100 (Isaaq y Cutchin 1981) En un estudio comparativo de diferentes métodos, Bradburn et al (1990a) encontraron que el procedimiento de HPTLC bidimensional con una fase estacionaria modificada (fenil) presentó los mejores recobrados de aflatoxinas, con respecto a las técnicas de CB, BF y Rommer. En otro estudio comparativo de los procedimientos de HPTLC, HPLC y ELISA en la determinación de aflatoxinas en mantequilla de maní se encontró que la exactitud y la reproducibilidad por HPTLC y HPLC fueron similares, sin embargo, el ELISA presentó coeficientes de variación (CV) mucho mayores y menor reproducibilidad con respecto al HPTLC (Dell et al, 1990). Un procedimiento que combina las ventajas del HPTLC y la cromatografía líquida (HPLC) fue desarrollado para aumentar la resolución entre las muestras y el número de muestras a analizar. Este método se fundamenta en el desarrollo de una cromatoplaca en forma horizontal sometida a una sobrepresión y se conoce en inglés como OPLC (Overpressure Layer Chromatography). En un estudio donde se compararon diferentes procedimientos cromatográficos se encontró que la resolución entre las aflatoxinas G1/B2 aumentó de 0,82 a 1,25 cuando se analizaron por HPTLC y OPLC respectivamente. Lo mismo sucedió para las combinaciones B2/B1 (1,12 a 1,6) y G2/G1 (1,12 a 1,45), además el tiempo de desarrollo de la cromatoplaca disminuyó de 40 minutos a 12 minutos con HPTLC bidimensional y OPLC respectivamente (Váradi et al, 1986).

30
4.4.2-Minicolumnas. La determinación de aflatoxinas en alimentos por el método de minicolumnas (AOAC, 1990) conlleva menos gasto de reactivos y tiempo que la CCD. Para esto se utilizan tubos de cristal de 6 mm de diámetro y 190 mm de largo que se llenan con alúmina, florisil y sulfato de sodio. Las aflatoxinas se detectan por la aparición de una banda fluorescente entre las capas de alúmina y florisil que será mayor con mayores concentraciones de aflatoxina. Con este método pueden detectarse concentraciones de 5 µg/kg en almendras y 10 µg/kg en maní (AOAC, 1990). Juegos comerciales de la Oxoid (Koeltzow y Tanner, 1990) y Biocode (Bisson et al, 1994) emplean minicolumnas de florisil y alúmina para la determinación de la aflatoxina B1, en extractos purificados por afinidad. Otra firma comercial Terratek emplea minicolumnas con soportes de aluminosilicatos para la determinación de diferentes micotoxinas como son aflatoxinas, fumonisinas y vomitoxina con límites de detección de 20 µg/Kg, 5 mg/Kg y 1 mg/Kg respectivamente. 4.4.3-Cromatografía Líquida de alta resolución (CLAR o HPLC). La cromatografía líquida de alta resolución o alta presión (CLAR) constituye otro de los métodos de determinación de aflatoxinas en diferentes medios. Esta técnica presenta una serie de ventajas con respecto a la cromatografía de placa y minicolumnas, ellas son: aumento de la sensibilidad, exactitud, precisión y resolución, además que el sistema puede ser automatizado muy fácilmente. El éxito de este método radica en el tipo de columna, detector y sistema de solventes que se utilicen. En la determinación de aflatoxinas se utilizan las columnas de sílica gel y de fase reversa (Stubblefield y Shotwell, 1977; Gilbert y Shepherd, 1985; Beaver 1989; Wilson 1989) y entre los detectores, los más usados son el ultravioleta y el de fluorescencia (Seiber y Hsieh, 1973; Gilbert y Shepherd, 1985). Los detectores de fluorescencia tienen mayor sensibilidad, y puede ser mejorada por el uso de una celda empaquetada con sílica gel (Panalaks y Scott, 1977) y la incorporación de ácido fórmico a la fase móvil que evita el "quenching" en la fluorescencia de las aflatoxinas B1 y B2 (Manabe et al, 1978; Francis et al, 1982). La formación de los derivados hemiacetálicos de las aflatoxinas B1 y G1 (B2a y G2a) aumenta su fluorescencia y su polaridad y permite detectar concentraciones en el orden de los ng y pg (Tarter et al 1984; Shepherd et al, 1986; Yen y Bidasee, 1993). La derivación de la toxina con iodo o bromo, después de la separación por columna (Tuinstra y Haasnoot, 1983; Dorner y Cole, 1988) o el empleo de una precolumna de derivatización (Chu, 1991), también incrementan la fluorescencia de las aflatoxinas. Una de las desventajas de la CLAR es que requiere de una purificación previa de la muestra, lo que encarece la determinación. Shepherd et al (1986) evaluaron seis métodos de extracción y procedimientos de limpieza química para la determinación de aflatoxina M1 en leche y encontraron que el método que utiliza columna Sep-Pak fue el que presentó los mejores resultados en términos de velocidad, costo y limpieza del producto a analizar.

31
Se han desarrollado diferentes protocolos que automatizan los análisis en los alimentos. Ellos incluyen líneas de diálisis y concentración (Tuinstra et al, 1990) o cromatografía de inmunoafinidad (Carmen et al, 1989; Sharman y Gilbert, 1991), como paso de limpieza o purificación seguido de una separación y cuantificación por CLAR. 4.4.4-Cromatografía Gaseosa. La alta polaridad, baja volatilidad e inestabilidad térmica de las moléculas de aflatoxinas ha limitado el uso de la cromatografía gaseosa para la determinación de estas sustancias (Beaver 1986). Sin embargo, el empleo de columnas capilares de sílica fundida y el uso de detectores de espectrómetros de masa ha permitido detectar aflatoxina B1 en maíz y mantequilla de maní por este método (Rosen et al, 1989). Con el empleo de detectores de ionización de llama con columnas capilares de sílica fundida de diferentes tamaños, se han podido separar las cuatro aflatoxinas naturales (B1, B2, G1 y G2), sin embargo, cuando se emplearon columnas mayores la sensibilidad para las aflatoxinas del tipo G fue menor que para las del tipo B (Goto et al, 1988). 4.4.5-Otros métodos de detección. Los métodos instrumentales complementan a los procedimientos antes mencionados para aumentar el poder resolutivo y detección de las sustancias. Densitómetros (200-800 nm): Se aplican en la detección y cuantificación de aflatoxinas u otras micotoxinas una vez realizada la CCD y visualizada con anisaldehído o algún otro reactivo químico (Sydenham et al, 1990) o por la fluorescencia que emite la propia sustancia al hacer incidir la luz ultravioleta (Wu et al, 1990). Espectrómetro de masa: Puede utilizarse solo o en combinación con la cromatografía gaseosa (Plattner et al, 1984; Mirocha et al, 1989) o líquida (Kostiainen, 1991; Kostiainen y Kurone, 1991; Hurst et al, 1991; Capiello et al 1995). Los espectrómetros de masa pueden ser de ionización química positiva o negativa (Park et al, 1985; Kostiainen, 1988) y detectan iones moleculares. Además existen otros métodos de ionización como la desorción y bombardeo atómico rápido (Ackermann et al, 1987). Entre los espectrómetros de masas más usados, se encuentran los TANDEM, que son dos espectrómetros de masa acoplados entre sí. Su principio se fundamenta en una separación primaria de las diferentes sustancias y posteriormente la identificación de estas (Uyakual et al, 1989). 4.4.6-Métodos biológicos. Los bioensayos se emplean como métodos complementarios de los procedimientos físico-químicos para determinar la actividad biológica de las aflatoxinas (teratogenicidad y

32
mutagenicidad) y son técnicas simples, inespecíficas y poco sensibles. Los bioensayos incluyen procedimientos de: Cultivos de tejidos: Embrión de pollo, células de fibroblasto de ratón (Abbas et al, 1984) y células de riñón de hámster (Senter et al, 1991). Prelusky et al (1987) optimizaron el ensayo de embriones de pollo y encontraron la menor variación cuando se suministraba la toxina en los sacos aéreos del huevo. Este método ha sido oficializado por la Asociación Oficial de Químicos Analistas (AOAC) para la confirmación de la toxicidad de la aflatoxina B1 (AOAC, 1990). Animales de laboratorio: Larvas de camarón, larvas de pescado cebra, planaria carmelita (Lewelly, 1973), paticos de un día de nacidos y truchas, entre otros. La trucha es una de las especies más sensibles en exhibir los cambios patológicos que ocurren en el hígado durante el ensayo de la aflatoxina B1. Microorganismos: Bacillus megaterium, B.brevis, Escherichia coli, B. subtilis, B. mycoides y Brevibacterium spp. Discos impregnados con B. megaterium se han utilizado como método confirmatorio para el diagnóstico de las aflatoxinas, similar al empleado en la determinación de la sensibilidad de los antibióticos. 4.4.7-Métodos inmunológicos. El descubrimiento de la técnica inmunológica por Yalow y Berson (1959) y el desarrollo ulterior de los procedimientos inmunoenzimáticos por Engvall y Perlman (1971) y van Weemen y Schuurs (1971) permitieron que estos métodos sean de gran aceptación por diversos laboratorios para la determinación de sustancias tóxicas en los alimentos y fluidos biológicos. Las técnicas inmunológicas se fundamentan en la reacción antígeno anticuerpo y en dependencia del compuesto marcado se clasifican. Si es un radioisótopo se denominan Radioinmunoanálisis (RIA) y si es una enzima se le llaman Enzimoinmunoensayos (EIA). La reacción inmunológica se caracteriza por su alta especificidad y su capacidad de poder detectar concentraciones en el orden de los ng y pg por mililitro de muestra. 4.4.7.1-Antígeno o Hapteno. En las reacciones inmunológicas intervienen un grupo de moléculas que en dependencia de su peso molecular son antígenos o haptenos. Las sustancias de alto peso molecular (PM> 5 000 daltons) que son portadoras de signos de extrañeza genética que al ser introducidas en el organismo provocan el desarrollo de reacciones inmunológicas específicas (respuesta humoral o celular), reciben el nombre de antígenos. Estas sustancias deben presentar una estructura estable y la posibilidad de ser degradadas (Petrov, 1987; Tizard, 1989). Los haptenos son moléculas extrañas, pero de bajo peso molecular (PM< 5000 daltons) y no pueden por sí solas iniciar una repuesta inmunológica. Las aflatoxinas pertenecen a este último grupo, por lo que se hace necesario unirlas a una macromolécula (proteína) que les dé carácter antigénico. La proteína que

33
se acopla al hapteno para estos fines se denomina portadora o "carrier". Para llevar a cabo el acoplamiento entre el hapteno y la proteína es necesario que la estructura del hapteno presente grupos funcionales libres (carboxilos o aminos) capaces de reaccionar con los grupos carboxilos, aminos, tioles o fenoles presentes en la proteína para formar un enlace covalente. Como proteínas portadoras, las más usadas son: albúmina de suero bovino (BSA), ovoalbúmina (OVA), gamma globulina (BGG), key hole limpet hemocianina (KLH) y polímeros sintéticos como la polilisina. La conjugación entre el hapteno y la proteína puede realizarse de forma directa o indirecta, en dependencia de las características estructurales del hapteno. La aflatoxina B1, al no presentar un grupo carboxilo en su estructura, es necesario realizar una derivación química que permita obtener una aflatoxina más reactiva que pueda acoplarse de manera directa a la proteína. En la figura 10 se muestran diferentes estrategias para lograr un compuesto reactivo de aflatoxina B1 (Wilkinson et al, 1992). La más utilizada es la introducción de un carboxilo en el grupo ceto del anillo de la ciclopentanona a través de la reacción con hidrocloruro de carboximetiloxima (Chu et al, 1977). Los diferentes procedimentos que se emplean para la conjugación covalente entre la proteína y el hapteno modificado se relacionan a continuación. Anhídrido mezclado. La conjugación tiene lugar en dos etapas. En la primera se forma el anhídrido mezclado en condiciones anhidra y alcalina entre el grupo carboxilo del hapteno y el isobutilcloroformato. En

34
la siguiente etapa el anhídrido reacciona con los grupos aminos de los residuos de lisina de la proteína portadora disuelta en una mezcla de agua:dioxano con la formación de un enlace covalente entre la dos sustancias (Escobar et al, 1991a). Carbodiimida. Utiliza como principal agente acoplante el 1 etil propil amino carbodiimida (EDPC). Ocurre en dos etapas: primero se forma el anhídrido con el grupo carboxilo del hapteno y una urea disustituida a pH 5,5 ó 6, y después ocurre una reacción entre el éster de la isourea y los grupos aminos de la proteína formando un enlace amida (Chu et al, 1982). Método del éster N-hidroxisuccinimida. Es una variante del método de carbodiimida. El grupo carboxilo del hapteno y la n-hidroxisuccinimida (NSH) reaccionan con la diciclohexil carbodiimida para formar un éster activo que posteriormente se acopla a la proteína a través de un enlace amida, con la posterior liberación de la n-didroxisuccinimida. Una de las ventajas de este procedimiento es que permite obtener el derivado y conservarlo en forma sólida para su posterior utilización, y aumenta los rendimientos en la conjugación. Condensación de Manish . Se fundamenta en la formación de un ión fenolato por la apertura del anillo furánico de la aflatoxina B2a en condiciones alcalinas, que posteriormente se une al grupo amino de la proteína formando una base de Schiff en presencia de formaldehído. El enlace formado se estabiliza por reducción con borohidruro de sodio (Jackman, 1985). Los conjugados obtenidos se pueden clasificar en dependencia de la función que realizan. Conjugados inmunogénicos. Son aquellos que se emplean en la inmunización de los animales para obtener una repuesta específica al antígeno o hapteno objeto de análisis. Deben presentar una relación molar superior a los 20 moles de haptenos por mol de proteína (Morgan 1985). Conjugado de sensibilización o cubrimiento. Es aquel que se emplea para ser inmovilizado en una fase sólida para la detección de anticuerpos. Esta inmovilización puede ser pasiva o química. Conjugados enzimáticos. Se emplean como marcadores en los ensayos para poder detectar la sustancia objeto de análisis. Los conjugados pueden ser enzima-hapteno (Escobar et al, 1986) o enzima-inmunoglobulina

35
(Catty y Raykundalia, 1989) y deben mantener las propiedades inmunológicas y enzimáticas de sus componentes. 4.4.7.2-Anticuerpos. Los anticuerpos son proteínas que se refieren a una u otra clase de inmunoglobulinas producidas por las células plasmáticas como consecuencia de la interacción entre los linfocitos B sensibles y el antígeno. Las inmunoglobulinas presentan su máxima concentración en el suero sanguíneo donde constituyen cerca del 2,5% (residuo seco), es decir más de 1/3 de todas las proteínas. Los anticuerpos pueden ser policlonales o monoclonales en dependencia de la cantidad de clones celulares que lo produzcan. El anticuerpo policlonal es producido por distintos clones de linfocitos de un solo animal como el resultado de una inmunización con el antígeno, mientras el anticuerpo monoclonal es derivado a partir de un solo clon de linfocitos producidos por un hibridoma, como el resultado de la unión de linfocitos normales de animales inmunizados con células mielómicas cultivadas en un medio nutritivo (Gavilondo, 1987). La obtención de los anticuerpos policlonales con fines de diagnóstico se realiza en diferentes especies y de diversas formas, y no existe un esquema general para la obtención de los mismos. Compuestos de alto peso molecular pueden producir antisueros con altos títulos en conejos, mientras que para aquellos de bajo peso molecular se han obtenido las mejores respuestas en cabras y carneros (Morris, 1985). Recientemente se obtuvieron anticuerpos específicos para la micotoxinas a partir de la inmunización de gallinas ponedoras obteniendo las inmunoglobulinas específicas a partir de la yema del huevo (Clarke et al, 1993; Li et al, 1994), este procedimiento presenta la ventaja con respecto a los sistemas convencionales de obtención de anticuerpos que no requiere de una extracción de sangre, se alcanzan altos rendimientos de las inmunoglobulinas y las mismas son altamente específicas al antígeno estudiado. Otros aspectos que influyen en la producción y rendimientos de los anticuerpos es el empleo de sustancias que permitan la liberación lenta del antígeno. Estas sustancias se conocen con el nombre de adyuvantes. Los adyuvantes de Freund (completo e incompleto) son los más empleados, aunque existen otros como la saponina (Quil-A) y los geles de aluminio, que se utilizan muy poco en inmunógenos de haptenos. La inmunización puede llevarse a cabo por diferentes vías que incluyen, la intraperitoneal, intramuscular, intradérmica, subcutánea, o la combinación entre ellas. La frecuencia de inmunización depende de la repuesta del animal y de la experiencia práctica obtenida en cada laboratorio. El esquema más empleado son las inmunizaciones intradérmicas a los tiempos 0, 28, 38 días, con una extracción de sangre a los 10 días después de la última inmunización (Wilkinson et al, 1992). 4.4.7.3-Reacción antígeno anticuerpo.

36
Como ya se mencionó, los métodos inmunológicos se basan en la reacción antígeno (Ag) anticuerpo (Ac). Esta reacción puede ser dividida en reacciones primarias y secundarias. (Buttler, 1987). Reacciones primarias: ocurren rápidamente (milisegundos), son macroscópicamente invisibles y la molécula de Ac presenta sitios combinantes. Reacciones secundarias: ocurren lentamente debido a la multivalencia del Ag, además son microscópicas o parcialmente visibles al humano a través de una reacción de precipitación. La reacción Ag:Ac es reversible, obedece a la ley de Acción de las masas y se puede expresar de la siguiente manera: Ag + Ac ===== Ag : Ac [Ag:Ac] K = --------------- [Ag][Ac] donde: K es la constante de equilibrio. [Ag:Ac], [Ag] y [Ac] representa la concentración del complejo y de los reactivos libres en la mezcla final del equilibrio. Las fuerzas que unen al Ag con el Ac son interacciones no covalentes. Estas proporcionan, en los sistemas biológicos, una manera rápida y reversible de formar complejos que permiten reutilizar las moléculas de anticuerpos de una forma que no sería posible en las uniones covalentes. Las uniones no covalentes se forman con distancias intermoleculares relativamente pequeñas, por eso, la unión más fuerte entre Ag y Ac se produce cuando las formas del epítope (característica de un antígeno que define el patrón de reconocimiento de un anticuerpo) y del lugar de combinación del Ac se adaptan de manera muy estrecha, como sistema de llave/cerradura. Las fuerzas que contribuyen a la interacción Ag-Ac son los enlaces electrostáticos, los enlances de hidrógeno, los enlaces hidrófobos y las fuerzas de van der Waals. Las más importantes son las uniones hidrófobas. Las fuerzas de la unión entre un epítope y una molécula de inmunoglobulina (Ac) recibe el nombre de afinidad y se calcula por la Ley de Acción de las Masas a través de la curva de Scatchard Plot y se refiere al antígeno. Otro término, que es frecuentemente usado como sinónimo de la afinidad, es la avidez; pero en este caso se refiere al anticuerpo. En ambos términos reflejan la energía con la cual se combinan los sitios de unión del anticuerpo con el antígeno específico. 4.4.8-Clasificación de los inmunoensayos. 4.4.8.1-Radioinmunoensayos o radioinmunoanálisis (RIA).

37
El principio de este ensayo se fundamenta en la competencia del hapteno marcado (135I, 3H o 14C) y el hapteno no marcado por los sitios de unión del anticuerpo. Posteriormente se separan el hapteno libre del unido por una técnica apropiada(tabla 17) y la radioactividad de estas fracciones se determina por contadores gamma o beta, en dependencia del isótopo empleado como marcador. La concentración del hapteno de la muestra desconocida se determina a través de una curva de calibración, la cual se establece por la relación de la fracción unida y la libre contra el logaritmo en base 10 de la concentración del hapteno no marcado (patrón de referencia). El primer informe de un análisis de aflatoxina B1 por RIA fue hecho por Langone y Vunakis (1976) quienes emplearon la aflatoxina B1 marcada con H3 y anticuerpos policlonales obtenidos en conejos por la inmunización con un conjugado de polilisina-aflatoxina B1. Resultados similares fueron obtenidos por Chu y Ueno (1977), con el uso de un inmunógeno de albúmina bovina-aflatoxina B1. La sensibilidad de ambos antisueros osciló entre 0.2-2 ng/mL y se concluyó que esta podía ser mejorada aumentando la actividad específica del compuesto marcado (Chu y Ueno 1977). Tabla 17. Métodos empleados para la separación de fases por RIA
Principio Técnicas
Cromatografía
Precipitación química
Absorción
Inmunología
Fase sólida
-Electroforesis
-Filtración en gel
-Capa delgada de papel osílica
-Sulfato de sodio o amonio
-Etanol
-Polietilenglicol
-Carbón activado
-Spherosil
-Talco
-Proteína A o G
-Doble anticuerpo
-Inmuno Adsorbente (tubos)
-Partículas magnéticas En la tabla 18 se muestran algunas aplicaciones del radioinmunoensayo para la determinación de aflatoxina B1 en alimentos y fluidos biológicos, con un límite de detección de 0.1 a 5 µg/kg o µg/L.

38
Tabla 18. Aplicaciones del radioinmunoensayo en el diagnóstico de aflatoxinas en alimentos y fluidos biológicos.
Aflatoxina Aplicación Referencia
Aflatoxina B1
Aflatoxina B1
Aflatoxina B1 y metabolitos.
Aductos de aflatoxina B1
Maní
Tejido de hígado
Orina
Suero
Fukal et al 1987
Stora y Dvorackova 1987
Dragsted et al 1988
Gan et al 1988 Los procedimientos de RIA presentan la desventaja que los juegos de reactivos tienen una vida limitada en dependencia del isótopo, y además, el costo del equipamiento es alto y se requiere de regulaciones especiales para el personal que trabaja con isótopos radioactivos. 4.4.8.2-Enzimoinmunoensayos (EIA). En los enzimoinmunoensayos, el compuesto marcador es una enzima que debe reunir un grupo de requisitos (Sauer et al, 1985): 1- Debe ser económica y poder obtenerse con un alto grado de pureza. 2- Elevada actividad específica. 3- Estable en condiciones de almacenamiento y prueba. 4- Soluble. 5- Sencilla, sensible y medible con gran rapidez. 6- No debe estar presente en el medio analizado. 7- Los sustratos inhibidores y factores de interferencia procedentes del material a examinar no deben interferir en el ensayo. 8- No debe perder totalmente la actividad cuando se conjuga. 9- La actividad debe conservarse en las condiciones del ensayo. Las enzimas que se emplean con mayor frecuencia para los enzimoinmunoensayos son: la fosfatasa alcalina, la peroxidasa y la ß galactosidasa provenientes de la mucosa de ternero, de rábano y de Escherichia coli, respectivamente (Catty y Raykundalia, 1989). Los enzimoinmunoensayos pueden clasificarse en homogéneos y heterogéneos (Sauer et al, 1985. 4.4.8.2.1-Enzimoinmunoensayo homogéneo. El enzimoinmunoensayo homogéneo, conocido en inglés como EMIT (enzyme mediated-immunoassay-technique), debe su nombre a que no hay separación de fases. Su fundamento es la competencia del hapteno marcado y el no marcado presentes en la muestra o patrón por el anticuerpo, donde la actividad enzimática se inhibe por la interacción específica con el anticuerpo

39
(Bastiani et al, 1973). El ensayo homogéneo solo puede ser usado en la determinación de sustancias de bajo peso molecuar (haptenos). Es muy rápido y puede ser automatizado, pero es poco sensible debido a que la inhibición de la actividad enzimática es tan escasa que tan solo pueden medirse pequeñas diferencias de extinción (100-200 ME) entre una reacción inhibida y otra no inhibida. Esta dificultad puede ser superada en parte usando heterología de puente en los conjugados haptenos-proteína (BSA) (Mella et al, 1988). Existen otras variantes del método homogéneo (Ngo y Lenhoff, 1985). En lugar de usar una enzima como marcador pueden emplearse otros tipos de marcadores tales como: -Un sustrato fluorescente o precursor de sustratos (SLFIA). -Un inhibidor enzimático (EMMIA). -Un grupo prostético (ARIS). -Un liposoma que contenga enzima. 4.4.8.2.2-Enzimoinmunoensayo heterogéneo. El enzimoinmunoensayo heterogéneo, conocido en inglés como ELISA (enzyme linked immunosorbent assay), se diferencia del procedimiento anterior en que siempre se realiza una separación de complejo antígeno-anticuerpo, no fijado después de una incubación inicial. Para lograr esta separación es necesario haber inmovilizado con anterioridad los anticuerpos o los antígenos a la fase sólida o emplear algún método de centrifugación, sedimentación o precipitación con segundos anticuerpos. En el primer caso las fases sólidas más empleadas son: las placas de 96 huecos de poliestireno o polivinilo o tiras reactivas de nitrato de celulosa. El ensayo heterogéneo se clasifica en ensayos directos e indirectos, directo cuando se marcan los reactivos primarios (antígeno o anticuerpo) e indirecto cuando se utilizan segundos anticuerpos (antiinmunoglobulina) marcados. En ambos procedimientos hay dos variantes competitivo y no competitivo. A continuación se muestra como pueden ser clasificados estos ensayos en dependencia de qué y cómo se determina: -Competivo directo: Determinación de antígenos utilizando un antígeno marcado o un anticuerpo marcado con una enzima. -No competitivo indirecto: Determinación de anticuerpos empleando una antiinmunoglobulina marcada con una enzima. -Competitivo indirecto: Determinación de antígenos utilizando una antiinmunoglobulina marcada con una enzima. -Doble anticuerpo o "two site": Determinación de antígeno, donde uno de los anticuerpos está marcado y los otros pueden ser de diferentes especies. -USERIA (Ultrasensitive-Enzymatic-Radioimmunoassay): Determinación de antígenos

40
empleando dos anticuerpos Recientemente ha comenzado a utilizarse la vinculación de procedimientos inmunológicos con procedimientos químicos, tales como la cromatografía de capa delgada, surgiendo así el denominado ELISAGRAM (Pestka, 1991). Este se fundamenta en el principio de transferencia donde se vincula la sensibilidad y especificidad del ELISA con el poder resolutivo de la cromatografía. Otra técnica cromatográfica que se ha vinculado al inmunoanálisis es la CLAR que aumenta su potencialidad porque se emplea un anticuerpo genérico que reconoce diferentes haptenos de un mismo grupo (Chu y Lee 1989). 5-Validación de un ensayo. Antes que un ensayo pueda ser aplicado a un análisis de micotoxinas en alimentos o fluidos biológicos, debe reunir las siguientes características (Barnes, 1992; Ward et al, 1993): - El título de los anticuerpos policlonales y monoclonales a utilizar debe ser mayor de 1:10 000 y 1:100, respectivamente. - El límite de detección debe ser menor de 10 pg/ensayo. - Especificidad adecuada para el ensayo según la toxina. - Extracción y efecto matriz. - Recobrado. - Reproducibilidad. - Correlación con otros métodos. Además de las características anteriores que debe reunir un juego de reactivos, la AOAC ha establecido, como requisito fundamental para oficializar un método de análisis su validación a través de estudios interlaboratorios donde se incluye los siguientes acápites (Chu, 1990). a) Sensibilidad (rango de desplazamiento de la curva patrón; máximo color de patrón cero;
cambio de color por unidad de concentración y concentración al 50% de inhibición). b) Exactitud (reproducibilidad intra e inter ensayo, efecto matriz, especificidad del antisuero y
comparación con otros procedimientos). c) Simplicidad (fácil manipulación, pocos pasos de ejecución y rapidez) d) Estabilidad de los reactivos e) Costos 5.1-Titulación de los antisueros. La dilución óptima de un antisuero debe ser determinada en el tipo de formato que se emplee durante el análisis, ya que este depende de la concentración de los inmunoreactivos, el tiempo de incubación y la temperatura. La dilución óptima del antisuero se determina cuando existe una diferencia en densidad óptica de 1.5 entre la señal y el ruido, y se calcula a través de experimentos cruzados con diferentes

41
variables (Wilkinson et al, 1992 y Ward et al, 1993). 5.2-Sensibilidad. Diferentes son los criterios que se emplean para determinar la sensibilidad de un ensayo. El más utilizado es el que la determina a través de la curva de calibración donde existe una relación entre la medida y la concentración (C) de un analítico (x=fc). Este define la sensibilidad como el más pequeño cambio de concentración que puede ser medido por un procedimiento analítico, es decir como la pendiente de la curva (m=dx/dc) (FAO 1990). La sensibilidad de un ensayo también se ha expresado como la concentración que se detecta al 50% de la curva de calibración o al 95% (Feldman y Rodbar, 1971). La sensibilidad de un ensayo tiende a confundirse y se informa a veces como el límite de detección, que es la concentración más pequeña de sustancia que puede ser detectada con un alto grado de confianza por un método analítico. En la práctica este valor se calcula como la media del primer punto (patrón cero) menos dos o tres desviaciones estándar de la media. Este valor puede coincidir con la sensibilidad del ensayo. La sensibilidad depende de las concentraciones de los inmunoreactivos, en particular del antisuero. Una mayor constante de afinidad del antisuero favorece que el ensayo sea más sensible. 5.3-Especificidad. La especificidad de un antisuero (reacción cruzada), no es más que la habilidad que tienen algunas sustancias de enlazarse al anticuerpo con mayor o igual facilidad que el antígeno que le dio origen. Tales sustancias pueden ser precursores naturales, productos degradados del antígeno o sustancias de estructuras similares. La reacción cruzada de un antisuero policlonal varía en dependencia de la respuesta inmune del animal. Diferentes laboratorios, que emplean el mismo inmunógeno (aflatoxina B1:CMO-BSA), han obtenido antisueros con especifidades diferentes para las distintas aflatoxinas (Chu, 1984). Incluso cuando se obtienen anticuerpos monoclonales, estos pueden reconocer con gran afinidad a otros metabolitos. Dixon-Holland et al (1988) obtuvieron un anticuerpo monoclonal que presentó mayor afinidad por la aflatoxina B1, mientras otro anticuerpo monoclonal contra la aflatoxina B1 reconocía de igual manera a la aflatoxina G1 (Lee et al, 1992). 5.4-Efecto matriz. Para poder realizar el ensayo es necesario que el extracto de la muestra no interfiera en el análisis. Esta determinación del efecto matriz se realiza a través de dos curvas de calibración, donde el patrón se encuentra disuelto en solución tampón fosfato salina (PBS) y otra en el extracto de una muestra negativa o blanco. No existe interferencia cuando las dos curvas se superponen, o las pendientes de ambas curvas no difieren entre sí.

42
Si existe interferencia entre la curva de calibración blanco y la de extracto, entonces se requiere de purificar la muestra o buscar un sistema de extracción selectivo que permita igualar las dos curvas. Si las curvas son paralelas, pueden igualarse por la adición de pequeñas concentraciones del solvente de la matriz a la curva patrón con PBS (Mills et al, 1988) o desarrollar la curva en un extracto control negativo (Wilkinson et al, 1988a; Escobar et al, 1991b). La linealidad es otro indicador que se emplea para determinar si existe interferencia en la reacción antígeno-anticuerpo. Se obtiene a partir de las concentraciones del analítico en diferentes volúmenes de un extracto positivo. No existe interferencia cuando no se produce desviación de la linealidad y se obtiene una correlación alta (>0.90). 5.5-Exactitud y Precisión. La exactitud es la fidelidad de la concordancia entre el valor real del compuesto analizado y la media de los resultados obtenidos al aplicar el procedimiento. La exactitud de un ensayo se determina por el recobrado de la toxina de una muestra artificialmente contaminada (Wilkinson et al, 1992; van Egmond et al, 1994) o a partir de una muestra certificada por un laboratorio de referencia como: International Agency for Research on Cancer (IARC), Community Bureau of Reference Brussels (BCR), FAPAS y otros. Los recobrados se consideran adecuados cuando sus valores se aproximan al 100 % (± 5 %). Si el mismo se desvía bastante del valor real, entonces es necesario corregir el cálculo final (van Egmond y Wagstaffe, 1990). En las técnicas inmunológicas, que no requieren de una limpieza total de la muestra, se obtienen recobrados cercanos al 100 % cuando se emplean sistemas de solvente metanol:agua 65:35 v/v (Escobar et al, 1991b) o acetonitrilo:agua 55:45 v/v (Morgan et al, 1986a). Una vez establecido el recobrado de la toxina a partir de la muestra, se procede a determinar la precisión, que es la fidelidad de concordancia entre los resultados obtenidos por un método estándar, sobre un material idéntico y bajo determinadas condiciones. La precisión se determina a través de la repetibilidad y reproducibilidad (Thielmann, 1973). Los términos de intraensayo, interensayo e interlaboratorio también son utilizados como medida de precisión y pueden ser de 15, 25 y 50 %, respectivamente (Heitzman, 1984). Es recomendable que los estudios de precisión se realicen con más de una muestra contaminada. 5.6-Estudios de correlación. La correlación con otros métodos nos indica la exactitud de la técnica investigada en lo que respecta a verdaderos positivos y verdaderos negativos de un conjunto de muestras analizadas paralelamente con un método de referencia. A partir de los datos obtenidos por el procedimiento de referencia y el método propuesto pueden determinarse los indicadores de calidad (sensibilidad, especificidad, prevalencia, eficacia y valor predictivo positivo o negativo).

43
La validación de procedimientos inmunológicos en la detección de aflatoxinas en alimentos y fluidos biológicos se ha realizado con distintos métodos convencionales, en la tabla 19 se muestran algunas de estas correlaciones.
44
Tabla 19. Estudios de correlación entre los procedimientos inmunológicos y los métodos convencionales.
Aflatoxina Muestra Método investigado
Método de referencia
correlación Referencia
M1 Leche ELISA HPLC ---- Fremy y Chu, 1989
B1 Mantequilla de maní
ELISA (Biokits)
HPTLC r>0,90 Dell et al, 1990
B1 Materia primas
ELISA HPLC r=0.90 Adachi et al, 1991
B1 Suero RIA HPLC ---- Tsuboi et al, 1984
B1 Higo seco ELISA TLC r=0,97 Reichert et al, 1988
B1 Maní ELISA HPLC 98,6% Cole et al, 1988
B1 Maní EZ-Screen Afla cup
HPLC 82,5% 81,5%
Dorner y Cole, 1989
B1 Maíz Afla cup Afla test Agri Screen EZ-Screen Oxoid Sam-A
TLC Minicolumna
r=0,91 r=0,75 r=0,98 r=0,80 r=0,56 r=0,97
Koeltzow y Tanner, 1990
Conjuntamente con los estudios de correlación, se ejecutan experimentos interlaboratorios para el pesquisaje y la determinación cuantitativa de aflatoxina B1 en alimentos. Este último aspecto es indispensable para que un método pueda ser reconocido internacionalmente por la Asociación Oficial de Químicos Analistas y debe realizarse por las orientaciones generales establecidas en la AOAC. Se ha recomendado adoptar el ELISA (Agri-Screen y Afla cup) como métodos oficiales de análisis para el pesquisaje de aflatoxina B1 en semillas de algodón, maíz, maní y productos derivados del maní cuando la concentración de aflatoxina es superior a los 15, 20 y 30 µg/kg respectivamente (Park et al, 1989 a y b; Trucksess et al, 1989; AOAC, 1990; Trucksess, 1995). Un ensayo cuantitativo para el diagnóstico de aflatoxina B1 por ELISA (Biokits Aflatoxin Kit(Cortecs)) fue sometido a un experimento internacional y aprobado por la AOAC como primera acción oficial para el diagnóstico cuantitativo de aflatoxina B1 en mantequilla de maní en un intervalo de concentración de 9-90 µg/kg (Patey et al, 1992).
La aplicación de los inmunoensayos para el análisis de aflatoxinas en alimentos y fluidos
45
biológicos se resume en la tabla 20. Se observa que en un futuro muy cercano los procedimientos inmunológicos se realizarán como rutina para el diagnóstico cuantitativo de aflatoxinas u otras micotoxinas.
Tabla 20. Aplicación de los inmunoensayos de aflatoxinas en alimentos y fluidos biológicos.
Aflatoxina Tipo de muestra Tipo de ensayo referencias
M1 M1 M1 M1 M1 B1 B1 B1 Aflas B1 Aductos Aductos Aductos Aductos B1 y M1 aflas B1 B1 B1 B1,B2 y G1
Leche Leche Leche Leche Leche Leche Maíz y productos de maní Mantequilla Alimentos Maíz Suero Fluidos biol. Hígado Suero Alimento y orina Orina y suero Suero Alimentos Maiz Maiz
ELISA CA ELISA ELISA ELISA ELISA ELISA ELISA ELISA ELISA ELISA ELISA y CA ELISA y CA ELISA y CA ELISA ELISA ELISA IMMUNOBLOT ELISA ELISA-CUP
Wild et al, 1987 Mortimer et al, 1987 Pestka et al, 1981 Li y Chu, 1982 Fremy y Chu, 1984 Margolles et al,1990b Park et al, 1988 Morgan et al, 1986a Mortimer et al, 1988 Patey et al, 1992 Truksess et al, 1989 Wilcke et al, 1990 Okoye y Neal, 1988 Wild et al, 1988 Garner et al, 1988a Groopman y Donahue, 1988. Yu et al, 1989 Autrup, 1989 Denning et al, 1988 Wilkinson et al,1988a Wilkinson et al,1988b Wilkinson et al, 1989 Abouzied y Pestka, 1994 Hirano et al, 1991 Trucskess y Stack, 1994

46
En la actualidad existe una gran oferta de juegos de reactivos para la determinación de aflatoxina B1 en alimentos por parte de diversas casas comerciales (tabla 21), que facilitan los planes de prevención y control establecidos para el diagnóstico de esta micotoxina en los alimentos. En la tabla 22 se muestra un resumen de las características generales de los métodos analíticos discutidos anteriormente. Tabla 21. Casas comerciales que ofertan juegos de reactivos para el diagnóstico de aflatoxinas.
Casa comercial Afaltoxina Principio Aplicación Chemopolt Ltd (Checolovaquia)
B1 M1/B1
RIA Cuantitativo (cuant.)
Cortecs Diagnostic (UK) B/G ELISA Cuant. Cualitativo
Environmental Diagnostic (USA)
B/G M1
Card Cualitativo
Genego SPa (Italia) B1 M1
ELISA Cuant.
Idexx (USA) B1 M1
Cup Cualitativo
International diagnostic (USA)
B/G Cup Cualitativo
Neogen Corp (USA) B/G M1
ELISA Cualitativo
Noak (Austria) B/G M1
ELISA Cuant. Cualitativo
Oxoid Ltd (UK) B/G M1
Afinidad Cualitativo
Penicillins Assays Inc (USA)
B/G M1/M2
RIA Cuant.
Rhône-Poulenc Diagnostic (USA)
B1 B/G
ELISA Afinidad
Cuant. Cualitativo
Riedel-de Haën (Alemania) B1 M1
ELISA Cuant.
Transia Ltd (Francia) B1 M1 B/G
ELISA CARD
Cuant. cualitativo

47
Tabla 22 Resumen de las características generales de los métodos de análisis.
Ensayo Rango de aplicación
Precisión Límite de detección
Costo de equipamiento
Posible automatización
Bioensayo Minicolumna TLC HPLC RIA ELISA
limitado limitado amplio amplio limitado limitado
baja moderada alta alta alta alta
mg/kg µg/kg a mg/kg ng/kg a µg/kg ng/kg a µg/kg pg/kg a µg/kg pg/kg a µg/kg
bajo bajo bajo alto alto moderado
no no no parcial si si
6. Descontaminación de aflatoxinas Después de conocerse la potencia tóxica de las aflatoxinas y en especial de la aflatoxina B1, la forma de eliminarla de los alimentos una vez contaminados, eliminar su producción por los hongos e incluso eliminar o aminorar sus efectos tóxicos in vivo con el uso de sustancias adsorbentes, ha sido un objetivo humano. Las aflatoxinas en estado seco son químicamente muy estables, debido a su alto punto de fusión (268-269°C), pero en presencia de humedad y altas temperaturas ocurre la destrucción parcial de estas sustancias (Jones, 1977), así por ejemplo ha sido informada su destrucción parcial en el tostado del maní, en la fabricación del pan y durante el tratamiento alcalino del maíz para hacer “tortillas” (OPS-OMS, 1983). La aflatoxina M1 es también difícil de ser descontaminada, por ser termorresistente (Destro y Gelosa, 1986). Munch y Stein (1985) plantean que los principales métodos de descontaminación de aflatoxinas en piensos u otros productos alimenticios son los físicos y químicos, mencionado también los microbiológicos. Los métodos físicos incluyen la separación mecánica, los procesos térmicos, las irradiaciones y los pulsos eléctricos, mientras que los químicos se fundamentan en la reacción entre la molécula de aflatoxina y una sustancia química que puede ser un agente oxidante o reductor, un ácido o una base fuerte. Cuando se utilizan procesos físicos para destruir las aflatoxinas en productos alimenticios suele ocurrir simultáneamente la destrucción de otras sustancias nutritivas. Al utilizar las radiaciones ultravioletas para descontaminar piensos u otros alimentos sólidos, además de nos ser muy efectivas por el poco poder de penetración que tienen, se produce el deterioro del valor nutritivo del alimento. En la descontaminación de aflatoxina M1 en leche pasteurizada y sin pasteurizar por radiaciones UV, se redujeron la cantidad de aflatoxina hasta un 56,2% y 53,9% respectivamente (Yousef y Marth, 1986). La luz visible descontamina el arroz hasta un 10% de la contaminación inicial con una dosis de 112 mw/cm2 (Nkama et al, 1987). Las radiaciones gamma, a pesar de tener un poder de penetración muy grande, descontaminan la aflatoxina B1 en soluciones y en alimentos solo a altas dosis. Escobar y Fernández-Miranda (1989) obtuvieron solo un 50% de descontaminación de un pienso usando dosis de 60 kgrey y Patel et al (1989) requirieron usar 400 kgrey + 5% H2O2 para la total descontaminación de 100 µg de AFB1; sin embargo, estas radiaciones producen un efecto beneficioso en la inhibición de la producción, tal como fue probado por Rodríguez y Rodríguez (1983) y Sharma et al (1990).

48
Dentro de las sustancias químicas, el agente destoxificante más empleado ha sido el amoníaco en su estado gaseoso y acuoso, que forma la aflatoxina D1, la cual es 130 veces menos tóxica que su antecesora (Pival et al, 1981). Este tratamiento ocasiona la destrucción de sustancias nutritivas como la de algunos aminoácidos. Otro agente químico que se ha utilizado en la descontaminación de aflatoxinas es el metabisulfito de sodio (Hagler et al, 1982). Echevarría y Escobar (1989) realizaron un estudio comparativo del poder de destoxificación de la aflatoxina B1 entre el amoníaco y el metabilsulfito, utilizando el método inmunoenzimático para la determinación de la residualidad de la aflatoxina B1 y encontraron que el primero tiene mayor poder de destoxificación que el segundo y además caracterizaron el metabolito que se produce (aflatoxina B1S). Otra desventaja de la destoxificación con agentes químicos es la pérdida de las propiedades organolépticas del producto destoxificado. Mucho mejores resultados en este sentido se han obtenido con la inhibición de la producción de las aflatoxinas con ozono (González et al, 1990), quienes probaron que se mantenían las propiedades organolépticas y el poder nutritivo de los alimentos ozonizados. Con el objetivo de prevenir el crecimiento de los hongos y por tanto la producción de aflatoxinas en los alimentos, se han empleado agentes químicos como son: ortofenilfenato de sodio, el dióxido de sulfuro, la sal de amonio del ácido propiónico y el thiabendazole. A partir de los estudios realizados por Fonseca et al (1994) con las sustancias anteriormente mencionadas se encontró que el tratamiento con la sal de amonio del ácido propiónico (5000 mg/Kg) fueron los más eficientes, donde las concentraciones de aflatoxina no sobrepasaron los 30 µg/kg, mientras en los otros tratamientos, incluyendo los controles, las concentraciones de aflatoxinas variaron desde de 150 hasta 108,333 µg/kg. También han sido utilizados agentes adsorbentes para destoxificar los alimentos de las aflatoxinas, cuyos resultados son comprobados in vivo. Ademoyero y Dalvi (1983) adicionaron carbón activado en alimento contaminado para pollos y más recientemente, han comenzado a emplearse con éxito los alúmino-silicatos hidratados de sodio y calcio con estos mismos fines en los alimentos destinados a: pollos (Phillips et al, 1988), cerdos (Colvin et al, 1989; Beaver et al, 1990; Schell et al, 1993 a,b) y bovino (Veldman, 1992). En Cuba, se ha ensayado con algunas rocas zeolíticas y se encontró que estass tienen poder adsortivo a la aflatoxina B1 in vitro, donde dos tipos de bentonitas (cálcica y sódica) y las zeolitas del tipo modernítico presentaron un poder de adsorción a las aflatoxinas por encima del 80% superior al producto comercial usado como referencia (Margolles et al, 1993). El empleo de alúmino-silicatos en piensos con la finalidad de disminuir los efectos tóxicos de las aflatoxinas fueron evaluados de forma directa en el pienso a través de un ELISA (Margolles et al, 1994) y con animales in vivo (Margolles et al, 1992). El estudio directo en pienso corroboró la alta efectividad de las zeolitas del tipo mordenítico y las bentonitas para adsorver aflatoxinas (>77%); sin embargo las zeolitas modificadas mostraron una menor efectividad (19%), la zeolita de referencia mostró un 39% de adsorción a las

49
aflatoxinas (Margolles et al, 1994), mientras el estudio en ocas comprobó el efecto protector de los alúmino-silicatos, cuando el grupo que consumió alimento contaminado con aflatoxina + zeolita, presentó un comportamiento similar a los grupos controles (pienso; pienso + zeolita), no así el grupo que se alimentaba con el pienso contaminado (Margolles et al, 1992).

50
REFERENCIAS
1) Abbas, H. K.; W. T. Shier y C. T. Mirocha (1984): Sensitivity of cultured human and mouse fibroblasts to trichothecene. J. Assoc. Off. Anal. Chem. 67:607-610.
2) Abouzied, M. M. y J. J. Pestka (1984): Simultaneous screening of fumonisin B1, aflatoxin B1 and zearalenone by line ammunoblot: A computer assisted multianalyte assay system. J. AOAC Int. 77:495-501
3) Abramson, D.; T. Thorsteinson y D. Forest (1989): Chromatography of mycotoxins on precoated reversed phase thin layer plates. Arch. Environ. Contam. Toxicol. 18:327.
4) Ackermann, B. L.; J. F. Holland y J. T. Watson (1987): Comparison of thermally assisted fast atom bobardmente (TA-FAB) with conventional FAB and EI spectrometry for the analysis of the Helminthosproium carbonum mycotoxins. Biomed. Environ. Mass Spectrom. 14:501.
5) Adachi, Y.; M. Hara; N. H. Kumasawua; K. Hirano; I, Ueno y K. Egawa (1991): Detection of aflatoxin B1 in imported foods products into Japan by enzyme linked immunoabsorbent assay and high performance liquid chromatography. J. Vet. Med. Sci. 53:49-52.
6) Ademoyero, A. A. y R. R. Dalvi 1983): Efficacy of activated charcoal and other agents in the reduction of hepatotoxic effects of a simple dose of aflatoxin B1. Toxicol. Letters 16:153-157.
7) Albin, Lutgarda y Tania Bilbao (1987): Ecología de los hongos toxigénicos. Curso internacional sobre contaminantes químicos con especial énfasis en micotoxinas. INHA - Cuba.
8) Alvarez, M. T.; C. Castañeda; A. Escobar y T. Fragoso (1991): Detección de aflatoxina en la orina en enfermedades hepáticas en la infancia. Rev. SOC. Venezolana de Gastroenterología 45:205-209.
9) AOAC (1990): Official methods of analysis of the Association of Official Analytical Chemist. Ed. K. Helrich. Fifteenth edition. Arlington V. A. E.U.A.
10) Apeagyei, F.; S. M. Lamplugh; R. G. Hendrickse; K. Afframy y S. Lucas (1986): Aflatoxins in the livers of children with Kwashiorkor in Ghana. Tropical & Geographical Medicine. 38:273-276.
11) Appleabaum, R. S.; R. E. Brackett; D. W. Wisemar y E. H. Marth (1982): Aflatoxin toxicity in dairy cattle and occurrence in milk and milk products. A review. J. Food. Protect. 45:752-777.
12) Armbrecht, B. H. (1978): Mycotoxic fungi Mycotoxins, Mycotoxicoses. An Encyclopedic Handbook. Vol II Eds. T. D Wyllie y L. G Morehouse. New York y Basel. pp 227-236.
13) Asao, T.; B. Buchi; M. A, Kader; S. B Chang; W. L Wick y G. N. Wogan (1965): The structures of aflatoxins B1 and G1. J.Amer.Chem.Soc. 87:882-886.

51
14) Austin, H.; E. Delzell; S. Grofforman; R. Levine; A. S. Morrison, P. D. Stollley y P. Cole (1986): A case control study of hepatocellular carcinoma and the hepatitis B virus, cigarette smoking and alcohol consumption. Cancer Res. 46:962-965.
15) Austwick, P. K. C. (1978): Mycotoxicoses in Poultry. En Mycotoxin fungi Mycotoxins, Mycotoxicoses. Encyclopedic Handbook. Vol II Eds. T. D Wyllie y L. G Morehouse. New York y Basel. pp 279-301.
16) Autrup, J. L.; J Schmidt y H. Autrup (1993) Exposure to aflatoxin B1 in animal feed production plant workers. Environ. Health Perspectives. 99: 195-197.
17) Autrup. H. (1989): Exposure to aflatoxins in Denmark. J. Toxicol. Toxin. Rev 8:53-67.
18) Badii, F.; M. O. Moss y K. Wilson (1986): The effect of sodium biselenite on the growth and aflatoxin production of Aspergillus parasiticus and the growth of the other Aspergillus. Lett. Appl. Microbiol. 2:61-64.
19) Balaraman, N. y S. P. Arora (1987): Effects of aflatoxin on immunoglobulin status in blood serum of noeonatal calves. Ind. J. Anim. Sci. 57:61-63.
20) Barmak Anna-Lena y K. Larsson (1994): Immunoaffinity column cleanup/liquid chromatographic determination of aflatoxins: an interlaboratory study. J. of AOAC Int. 77:46-53
21) Barnes, J. C. (1992): Importance of laboratory and accurate descriptions of analytical procedures for drug residues in foods. En: Analysis of antibiotic drug residues in food products of animal origin. Ed. V.K. Agarwal. Plenum Press New York. pp 1-4.
22) Barry, D.; E. B. Lillehoj; N. W. Widstrom; W,W, McMillan; M. S. Zuber; W. F. K. Wolek y W. D. Guthrie (1986): Effect of husk tightness and insect (Lepidotera). Infestation on aflatoxin contamination of preharvest maize. Environ. Entomol. 15:1116-1118.
23) Bastiaini, R. J.; R. C. Philips; R. S. Schneider y E. Ulman (1973): Homogeneous Immunochemical drug assays. Am. J. Med. Technol. 39:211-214.
24) Batt, C. (1983): Relationship of asexual development to aflatoxin production in Aspergillus parasiticus. J. Food Safety 5:31-40.
25) Beaver, R. W. (1986): Gas chromatography in mycotoxins analysis. En: Modern methods in the analysis and structural elucidation of mycotoxins. Ed R. J. Cole. Academic, Press New York pp 289.
26) Beaver, R, W, (1989): Determination of aflatoxins in corn and using high performance liquid chromatography. Arch. Environ. Contam. Toxicol. 18:315.
27) Beaver, R. W.; D. M. Wilson y M. W. Trucksess (1990): Comparison of postcolumn derivatization liquid chromatography with thin layer chromatography for determination of aflatoxins in naturally contaminated corn. J. Assoc. Off. Anal. Chem 73:579-581.
28) Beljaars, P. R.; F. H. M. Fabry; M. M. A. Pickott y M. J. Peeters (1972): Quantitative fluorodensitometric measurement of aflatoxin B1 with flying-spot densitometer. II

52
Comparative study of Aflatoxin B1 measurements in spiked and naturally contaminated peanuts products. J. AOAC 55:1310-1315.
29) Bennett. J. W.; J. J. Dunn y C. I. Goldsman (1981): Influence of white ligh on production os aflatoxins and anthraquinones in Aspergillus parasiticus. Experientia 42:848-851.
30) Bennett J. W. (1983): Secondary metabolism and differentiation in fungi. En: Differentiation and secondary metabolism in mycelia fungi. Eds, J.W. Bennett y A. Cigler. Marcel Dekker Inc New York y Basel.
31) Bijl, J. y C. vanPeteghem (1985): Rapid extraction and sample clean-up for fluorescence densitometric determination of aflatoxin M1 in milk and milk powder. Anal. Chemical. Acta 170:149-152.
32) Bisson, E.; S. Dring y M. Rosenbaum (1994): Análisis de aflatoxinas en maíz, maní y alimentos de gluten con columnas de inmunoafinidad, utilizando HPLC, TLC y columnas analíticas Easi-Extract. Libro Resumen. I Congreso Latino-Americano de Micotoxicología. Brasil. Rio de Janerio. 26-30 de septiembre.
33) Boutibonnes, P.; Y. Auffray; C. Malherbe; W. Kogboy y C. Morris (1984): Proprietes antibacteriennes et genotoxiques de 33 mycotoxins. Mycopathologia 87:43-49.
34) Bradburn, N.; K. Jewers, B. D. Jones y K. J Tomlins (1989): Rapid method for the clean-up of extracts of cottonseed prior to aflatoxin determination by bi-directional HPTLC. Chromatographia 28:541-544.
35) Bradburn, N.; R. D. Coke y K. Jewers (1990a): A comparative study of pheny bonded phase CB and Romer clean-up procedures for determining aflatoxin levels in maize by bi-directional HPTLC. Chromatographia 29:177-181.
36) Bradburn, N.; K. Jewers, B. D. Jones y K. J Tomlins (1990b): Evaluation of the ability of different concentrations of aqueous acetone, aqueous methanol and aqueous acetone:metanol (1:1) to extract aflatoxin from naturally contaminated maize. Chromatographia 29:435-440.
37) Bradburn, N.; R. D. Coke y G. Blunden (1995): A comparative study of solvent extraction efficiency and the performance of immunoaffinity and solid phase columns on the determination of aflatoxin B1. Food Chemistry 52:159-185.
38) Brown, G. H. (1984): The distribution of total aflatoxin levels in composited samples of peanuts. Food Techn. Australia 36:128-130.
39) Brumley, W. C.; S. Nesheim; M. W. Truksess; E. W. Trucksess; P. A. Dreyfus; J. A. Roach; D. Andrzejewski; R. M Eppley, A. E. Pohland, C. W. Thorpe y J. A Spho (1981): Negative ion chemical ionization mass spectrometry of aflatoxins and related mycotoxins. Anal Chem 53:2003-2006.
40) Buckle, A. E. y M. F. Sanders (1990): An appraisal of bioassay methods for detection of mycotoxins. Review. Lett. Appl. Microbiol. 10:155-160.
41) Buchanan, R. L. y D. F. Lewis (1984): Caffeine inhibition of aflatoxin synthesis. Probable

53
site of action. Appl Environ. Microbiol. 47:1216-1220.
42) Bulatao-Jayme, J.; E. M. Almero; Ma. Cecilia; A. Castro; Ma. T. Jardeleza y L. A. Salamat (1982): A case control dietary study of primary liver cancer risk from aflatoxin exposure. Int. J. Epidemiol. 11:112-119
43) Busby, W. F. y G. N. Wogan (1981): Aflatoxins. En Mycotoxins in foodstuffs. Proceedings of MIT Symposium. Ed. G. H. Wogan. March 18-19, MIT Press Cambridge M.A. pp 175-186.
44) Butler, J. E. (1987): Antibody-antigen and antibody-hapten reactions. En Enzyme-Immunoassay. Ed. E. T Maggio. CRC Press Inc Boca Raton Florida pp: 5-52.
45) Campbell, A. D.; T. B. Whitaker; A. E. Pohland; J. W. Dickens y D. L Park (1986): Sampling, sample preparation and sampling plans for foodstuffs for mycotoxins analysis. Pure Appl. Chem 58:305-314.
46) Capiello, A.; G. Famiglene y B. Trillini (1995): Determination of aflatoxins in peanut meal by LC_MS with a particle beam interface. Chromatographia 40:411-416.
47) Carballo, M. y J.A. de Miguel (1987): Rapid detection of aflatoxin producing strains of the Aspergillus flavus group isolated from mixed feed and cereal grains in Spain. J. Sci. Food Agric. 40:11-15.
48) Carmen, A. S.; S. S. Kuan; G. W. Ware; O. J. Francis y K. V. miller (1989): Automated assay of aflatoxina M1 in fluid milk using robotics. Presented at the 10 erd AOAC Annu Int. Mtg. Sept 25-28. St Louis M.O (abst).
49) Carnaghan, R. (1965): Hepatic tumors in ducks fed a low level of toxic groundnut meal. Nature (London)208:308.
50) Castegnaro, M.; V. Maru; G. Maru; y M. D. R. Lopéz (1990): High performance liquid chromatographic determination of ochratoxin A and its 4r-4hydroxy metabolite in human urine. Analyst 115:129-131.
51) Catty, D. y C. Raykundalia (1989): ELISA and related enzyme immunoassay. En Antibodies vol II. A practical Approach. Ed. D. Catty. IRL Press Oxford, New York, Tokio. pp:97-54.
52) Cavalheiro A, C. L. (1981): Aflatoxin and aflatoxicosis. A review. W. P. S. A. J. 37:34-38.
53) Clarke, R. N.; R. R. Marquardt; A. Dosterveld; A. A. Frohlich; F. J. Madrid y M. Dawood (1993): Development of a quantitative and sensitive Enzyme Linked Inmunoabsorbent Assay for Ochratoxin A using antibodies from the yolk of the layin hen. J. Agric. Food Chem. 41:1784-1789.
54) Clarke, R. N. y M. A. Ottinger (1984): Further investigation of reproductive hormonal changes associated with aflatoxicosis in the male White Leghorn. Poultry Sci. 63 (suppl 1):80.
55) Cohen, H. y M. LaPointe (1986): Determination of ochratoxin A in animal feed and ceral

54
grains by liquid chromatography with fluorescence. J. Assoc. Off. Anal. Chem. 69:957.
56) Cole, B.; D. J. Meyer; B. Kettere; C. Stanto y R. C Garner (1985): Studies on the detoxication of microsomally activated aflatoxin B by glutathione and glutathione transferases in vitro. Carcinogenesis 6: 693-698.
57) Cole, R. J.; J. Dorner; J. W. Kirksey y F. E, Dowell (1988): Comparison of visual, enzyme linked immunosorbent assay screening and HPLC methods in detecting aflatoxin in farmers stock peanut grade samples. Peanut Sci. 15:61-83.
58) Cole, R. J.; J. W. Dorner (1994): extraction of aflatoxins from naturally contaminated peanuts with different solvents and solvent/peanut ratios. J. of AOAC Int. 77:1509-1511.
59) Colvin, B. M.; L. T. Sangster; K. D. Haydon; R. W. Beaver y D. M. Wilson (1989): effect of high affinity alumino-silicate sorbente on prevention of aflatoxicosis in growing pig. Vet. & Human Toxicol 31:46-48
60) Cook, W. O.; J. L. Richard; G. D. Osweiler y D. W. Trampel (1986): Clinical and pathologic changes in acute bovine aflatoxicosis: rumen motility and tissue and fluid concentrations of aflatoxins B1 and M1. Am. J. Vet. Rev. 47:1817-1825.
61) Chan, C.; A. M. Pearson; T. H. Coleman; J. J. Gray y A. Wolzak (1985): Broiler aflatoxicosis with recovery after replacement of the contaminated diet. Br. Pultry Sci. 26:56-71.
62) Christou, P.; K. D. Miller y A. Poau (1985): Chemical convertion of aflatoxin B1 to M1. Phytochemistry 24:933-935.
63) Chu, F. S.; M. T. S. Hsia y P. S. Sun (1977): Preparation and characterization of aflatoxina B1-o carboxymethy oxime. J. AOAC 60:791-794
64) Chu, F. S. y I. Ueno (1977): Production of antibody against aflatoxin B1. Appl. Environ. Microbiol. 33:1125-1128.
65) Chu, F. S.; H. P. Lau; T. S. Fan y G. S. Zhanh (1982): Ethylendiamine modified bovine serum albumin as protein carrier in the production of antibody against mycotoxins. J. Immunol. Methods 55:73-78
66) Chu, F. S. (1984): Immunoassay for analysis of mycotoxins. J. Food Prot. 47:562-569.
67) Chu, F, S. y R. C. Lee (1989): Immunochromatography of group A trichothecene mycotoxyns. Food Agric. Immunol. 1:127-136.
68) Chu, F. S. (1990): Immunoassays for mycotoxins: Current state of the art commercial and epidemiological applications. Vet. Hum. Toxicol. 32 (supplement):42-50.
69) Chu, F. S. (1991): Development and use of immunoassays in detection of the ecologycally importante mycotoxins. En: Handbook of Applied Micology Vol V Mycotoxins. Eds. D, Bhatnagar; E. B. Lillihoj y D. K. Arora. Marcel, Dekker, New York.
70) Chu, F. S. (1992): Recent progress on analytical techniques for mycotoxins in feedstuffs.

55
J. Anim. Sci. 70:3950-3963.
71) Dalvi, R. R. y C. McGowan (1984): Experimental induction of chronic aflatoxicosis in chickens by purified aflatoxin B1 and charcoal, phenobarbital and reduced glutathione. Pultry Sci. 63:485-491.
72) David, L. A.; D. L. Orti y R. H. Hill (1987): Proton nuclear magnetic resonance evidence for two configurations of the hemiacetals of aflatoxin B1 and sterigmatocystin. J. Agri. Food Chem. 35:782-785
73) Davies, N. D. y U. L. Diener (1968): Growth and aflatoxin production by Aspergillus parasiticus from various carbon sources. Appl. Microbiol. 16:158
74) Davies, N. D. (1980): Protocols for surveys sampling post collection, handling and analysis of grain samples in mycotoxins problem. J. Assoc Off. Anal. Chem. 63:95-102.
75) Deger, G. E. (1976): Aflatoxin: human colon carcinogenenesis. Ann. Int. Med 85:204-208.
76) Dell, M. P. K.; S. J. Haswell; O. G. Roch; R. D. Coker; V. F. P. Medlock y K. Tomlins (1990): Analytical methodology for the determination of aflatoxins in peanut butter: comparison of high performance thin layer chromatography, enzyme linked immunosorbent assay and high performance liquid chromatographic methods. Analyst 115:1435-1440.
77) Denning, D. W.; J. K. Onwubalili; A. P. Wilkinson y M. R. A. Morgan (1988): Measurement of aflatoxin in Nigerian sera by ELISA. Trans Royal Soc. Trop. Med. Hygiene 82:169-171.
78) Destro, O. y L.Gelosa (1986): Thermoresistenza della aflatossina M1 nel latle. Industrie alimentarie 25:979-980
79) Detroy R. W. y C. W. Hesseltine (1969): Aflatoxicol: structure of a new transformation product of aflatoxin B1. Can J. Biochem. 48:830-832.
80) Dieckman, M. A. y M . L. Green (1992): Mycotoxin and reproduction in domestic livestock. J. Anim. Sci. 70:1615-1627.
81) Diener, U. L. y N. D. Davies (1968): Effect of environment on aflatoxin production in freshly dug peanuts. Trop. Sci. 10:22-28.
82) Dixon-Holland, D. E.; J. J. Pestka; B. A. Bidigare; W. L. Casale; R, L Warner; B. P. Ram y L. P Hart (1988): Production of sensitive monoclonal antibodies to aflatoxin B1 and aflatoxin M1 and their application to ELISA of naturally contaminated food. J. Food Prot. 51:201-204.
83) Doerr, J. A.; W. E. Huff; C. J. Wabeck; G. W. Chaloupka; J. D. May y J. W. Merkeley (1983): Effects of low level chronic aflatoxicosis in broiler chickens. Poultry Sci. 62:1971-1977.
84) Dorner, J. W.; R. J. Cole; y U. L Diener (1984): The relationship of Aspergillus flavus y Aspergillus parasiticus with reference to production of aflatoxins and cyclopiazonic acid. Mycopathologia 87:13-15.

56
85) Dorner, J. W. y R. J. Cole (1988): Rapid determination of aflatoxins in raw peanuts by liquid chromatography with post column iodination and modified minicolumn clean-up. J. Assoc. Off. Anal. Chem. 71:43-47.
86) Dorner, J. W. y R. J. Cole (1989): Comparison of two ELISA screening tests with liquid chromatography for determination of aflatoxins in raw peanuts. J. Assoc. Off. Anal. Chem. 72:962-964.
87) Dragsted, L. O.; I. Bu;; y H. Autrup (1988): Substances with affinity to a monoclonal aflatoxin B1 antibody in Danish urine samples. Fd. Chem. Toxic. 26:233-242.
88) Dutton, M. F.; W, Ehrlich y J. W. Bennett (1985): Biosynthetic relationships among aflatoxins B1, B2, M1 and M2. App. Environ. Microbiol. 49:1392-1395.
89) Dvorackova, I.; V. Kusak; D. Vesely; J. Vesela y P. Nesnidal (1977): Aflatoxin and encephalopathy with fatty degeneration of viscera (Reye). Ann. Nutr. Alim. 31:977-980.
90) Dvorackova, I.; C. Stora y N. Ayraud (1981): Evidence for aflatoxin B1 in two cases of lung cancer in man. J. Cancer Res. Clin. Oncol. 100:221-224.
91) Echevarría , Ileana Y Arturo Escobar (tutor) 1989: Validación del enzimoinmunoensayo de aflatoxina B1 en pienso descontaminado por métodos químicos. Tesis de Diploma. Facultad de Biología Universidad de la Habana. Cuba.
92) Elamin, N. H. H.; A. M. A. Rahim y A. E. Khalid (1988): Aflatoxin contamination of groundnuts in Sudan. Mycopathologia 104:25-31.
93) Elis, J. y J. A . DiPaolo (1967): Aflatoxin B1 induction of malformations. Arch. Path. 83:53-57.
94) Ellis, W. O.; J. P. Smith y B. K. Simpson (1991): Aflatoxins in foods: Occurrence, biosynthesis, effects on organism, detection and methods of control. Critical Reviews in Food Sci. Nut. 30:403-439.
95) Ellis, W. O.; J. P. Smith y B. K. Simpson, H Ramaswany y G. Doyon (1994 a ): Growth of and aflatoxin production by aspergillus flavus in peanut stored under modified atmosphere packaging (MAP) conditions. International J. of Food Microbiol. 22: 173-187.
96) Ellis, W. O.; J. P. Smith y B. K. Simpson, H Ramaswany y G. Doyon (1994 b ): Effect of gas barrier characteristic of film on aflatoxin production by aspergillus flavus in peanuts packaged under modified atmosphere packaging (MAP) conditions. Food Research International 27: 505-512.
97) Engvall, E. y P. Perlman (1971): Enzyme linked immunosorbent assay (ELISA). Quantitative assay of immunoglobulin G. Immunochemistry 8:871-874.
98) Erlich, K. y E. Ciegler (1985): Effect of phytate on aflatoxina formation by Aspergillus parasiticus grown on different grains. Mycopathologia 92:36.
99) Escobar, A.; C. Mella y O, Fernández (1986): Síntesis y caracterización de conjugados

57
afla B-oxima-albúmina bovina y afla B oxima peroxidasa. Rev. Salud Anim, 8:169-175.
100) Escobar, A.; Estrella. Margolles y A. Acosta (1991a): Synthesis and characterization of three aflatoxin B1-protein conjugates. Food Agric. Immunol. 13:57-61.
101) Escobar, A.; Estrella. Margolles y A. Acosta (1991b): Methodological aspects of aflatoxin B1 determination in food and feeds, and biological materials by enzyme linked immunosorbent assay (ELISA) Rev. Salud Anim 13:246-251.
102) Escobar, A.; J. A. AgÜero; María. C. Muñoz, E. Mendosa y M. Ramos (1995): Primer reporte en ocas de una intoxicación por tricotecenos en Cuba. Rev. Salud Anim. 17:311-315
103) FAO (1990): Manuals of food quality control 10. Training in mycotoxin analysis. FAO Food and Nutrition Paper 14/10 pp:39-46.
104) Farjam, A.; R. De Vries; H. Lingeman y U. A. Brinkman (1991): Immunoprecolumns for selective on-line sample pretreatment of aflatoxins in milk prior to column liquid chromatography. Int. J. Environ. Anal. Chem 44:175-184.
105) Feldman, H. y D. Rodbard (1971): Mathematical theory of radioimmunoassay. En Principles of competitive protein-binding assays. Eds. W. P O'Dell y W. H. Daughadug. Lippincott Co. Philadelphia. pp:158-203
106) Ferguson-Foos, J. y J. D. Warren (1984): Improved clean-up for liquid chromatographic analysis and fluorescence detection of aflatoxins M1 and M2 in fluid milk products. J. Assoc. Off. Anal. Chem. 67:1111-1114.
107) Ferrando, R.; M. Palisse-Roussel y L. Jacquot (1984): Toxicite de relais d l`aflatoxine M1 de la poudre de lait. Etude a moyen terme sur le caneton. C. R. Acad. Sc. Paris T.298 Serie III:355-358
108) Fonseca H.; (1994); Program for prevention of peanut contamination with aflatoxin Presented at the 2nd Asian Conference on Food Safety. Bangkok 18-23 September
109) Francis, O. J.; L. J. Lipinski; J. A. Gaul y A. Campbell (1982):High pressure liquid chromatographic determination of aflatoxins in peanut butter using a silica gel packed flow cell for fluorescence detection. J. Assoc. Off. Anal. Chem. 65:672-676
110) Fremy, J. M. y F. S Chu (1984):Direct linked immunosorbent assay for determining aflatoxin M1 at picogram level in dairy products. J. Assoc. Off, Anal. Chem. 67:1098-1101.
111) Fremy, J. M. y F. S. Chu (1989): Immunochemical method of analysis for aflatoxin M1. En Mycotoxins in Dairy products. Ed H.P. vanEgmond. Elsevier, Science, London pp97-126.
112) Fukal, L.; J. Prosek y Z. Sova (1987): The occurrence of aflatoxins in peanuts imported into Czechoslovakia for human consumption. Food Addit. Contam. 4:285-289
113) Fukal, L. Y H Reisnerova (1990): Monitoring of aflatoxins and ochratoxins A in Czechoslovak human sera by immunoassay. Bull Environ. Contam. Toxicol. 44:345-349.

58
114) Fukuhara, M.; K. Mizokami; M. Sakaguchi; Y. Niimura; K. Kato; S. Induye y A. Takanaka (1990): Aflatoxin B1 specific cytochrome. P450 isozyme (P450 a+b) inducible by 3 methyl colanthrene in golden hamsters. Biochem. Pharm 399:463-469.
115) Gan, L.; P. L. Skipper; X. Peng; J. D. Groopman; J. Chen; G. N. Wogan y S. R. Tannebaum (1988): Serum albumin adducts in the molecular epidemiology of aflatoxin carcinogenisis. Correlation with aflatoxin B1 intake and urinary excretion of aflatoxin M1. Carcinogenesis 9:1323-1325.
116) Garner, R. C. (1988): Monitoring aflatoxin exposure at a macromolecular level in human with immunological methods. Abst.7th International Symposium of Mycotoxins and Phycotoxins. Aug. 16-19. Tokyo. Japan.
117) Garner, R. C.; I. Dvorackova y F. Tursi (1988a):Immunoassay procedures to detect exposure to aflatoxin B1 and benzo (a) pyrene in animals and man at the DNA level. Int. Arch. Ocup. Environ. Heath. 60:145-150.
118) Garner, R. C.; R. Ryder y R. Montesano (1988b): Monitoring aflatoxins in human an body fluids and applications to field studies. Cancer Res 45:922-928.
119) Gavilondo, J. (1987): Aspectos básicos y avances recientes en la tecnología de producción de anticuerpos monoclonales. Interferon y Biotecnologia 4:1-16.
120) Giambrone, J. J. (1978): Effect of aflatoxin on the humoral and cell mediated immune system of the chicken. Am. J. Vet. Res. 39:305-309.
121) Gilbert, J. y M. J. Shepherd (1985):A survey of aflatoxins in peanut butter, nuts and not confectionery products by HPLC with fluorescence detection. Food Addit. Contam. 2:171-183.
122) González, Marisol.; Elba, Alvarez y A. Acosta (1989): Evaluación y optimización de la producción de aflatoxina B1 en sustratos naturales. Rev. Salud Anim. 11:101-108.
123) Gopalan, C.; P. G. Tupule y D. Krishnamurthy (1972): Induction of hepatic carcinoma with aflatoxin in the rhesus monkey. Food Cosmet. Toxicol 10:519-521.
124) Gorelick, N. J. (1990): Risk assessment for aflatoxin metabolism of aflatoxin B1 by different species. Risk Analysis 10:539-559.
125) Goto, T.; M. Matsui y T. Kitsuwa (1988): Determination of aflatoxin by capillary column gas chromatography. J. Chromatog 447:410.
126) Groopman, J. D.; P. R. Donhaus; J. Zhu; J. Chen y G. N. Wogan (1985): Aflatoxin metabolism in human detection of metabolism and nucleic acid adducts in urine by affinity chromatography. Proc Natl Acad. Sci. (USA) 82:6492-6496.
127) Groopman, J. D.; P. R. Donahue; J. Zhu; J.Chen y C. N. Wogan (1987): Temperal patterns of aflatoxin metabolites in urine of people living in Guangxi province P.R.C. Proc. Am. Assoc. Cancer Res. 28:36.
128) Groopman, J. D. y K. F. Donahue (1988): Aflatoxin a human carcinogen determination in

59
food and biological samples by monoclonal antibody affinity chromatography. J. Assoc. Off. Anal. Chem. 71:861-867.
129) Groopman, J. D.; T. W. Kensler y L. G Cain (1988):Aflatoxin exposure to human populations measurements and relationship to cancer. CRC Council Rev. in Toxicol 19:113-115.
130) Groopman, J. D.; C. P. Wild. J. Hasler; C. Junshi; G. N. Wogan y T. W. Kensler (1993): Molecular epidemioloy of aflatoxin exposures: validation of aflatoxin N7 guanine levels in urine as a biomarker in experimental rat models and humans. Environ. Health Perspectives. 99:107-113.
131) Guthrie, L. D. (1979):Effects on aflatoxin in corn on production and reproduction in dairy cattle. J. Dairy Sci. 62:134.
132) Hagler , W. M.; J. E. Hutchins y P. B. Hamilton (1982): Destruction of aflatoxin B1 with sodium metabisulfite. Isolation of the mayor product aflatoxin B1S. J. Food Protect. 46:295-300.
133) Halama, A. K. (1981): Disorders of bone mineralization caused by mycotoxins in fattening chickens. Wienwe Tierarz Monasch 68:196-200.
134) Halver, J. E. (1967): Crystaline aflatoxin and other vector for trout hepatoma .Trout Hepatoma Research Conference Papers. Res Rept 70. Bureau of Sports Fisheries and Wild Life. Washington pp:78-102 .
135) Harrison, J. C.; M. Carvajal y R. C. Garner (1993): Does aflatoxin exposure in the United Kingdom constitute a cancer risk. Environ Health Perspective 99:99-105.
136) Heiztman, R. (1984): Immunoassay techniques for measuring veterinary drug residues in farm animals meat products. En Analysis of food contaminants. Ed. J. Gilbert. Elsevier. Applied Science Publishers. London y New York. pp:73-116.
137) Hendrickse, R. G.; J. B. S. Coulter; S. M. Camplugh; S. B. J. McFarlane; F. E. Williams; M. I. A. Omer; G.I. Sulliman y G. A. Elzorgani (1983): Aflatoxins and Kwashiorkor. Bull Soc. Path. 76:559-566.
138) Hendrickse, R. G.; S. M. Maxwell; R. Young; D. M. Yourtee; H. G Raj; H. R. Prasanna y H. Autrup (1989): Aflatoxins and heroin. Proc of the Int. Symp. on Agric. and Biol. aspects of aflatoxins related health hazards. J. Toxicol. Toxin Review pp:89-94.
139) Hendrickse, R. G. (1991): Clinical implications of food contaminated by aflatoxins. Ann. Acd. Med 20:84-90.
140) Hesseltine, C. V. (1976):. Conditions leading to mycotoxin contamination of food and feed. En: Mycotoxins and other fungi related food problems. Eds. J.V. Rodricks. Adv in Chemistry. Series 159 Am. Chem. Soc. Washington DC pp:1-22.
141) Hill, R. A.; P. D. BlanKeship; R. J. Cole y T. H. Sanders (1983): Effects of soil moisture, and temperature on preharvest invasion of peanuts by the Aspergillus flavus group and subsequent aflatoxin development. Appl. Environ. Microbiol. 45:629-633.

60
142) Hirano, K.; Y. Adachi; M. Hara; S. Ishibashi; M. Sueyoshi; I. Ueno; K. Egawa y N. H. Kumazawa (1991):Detection of aflatoxin B1 imported maize kernels used as feed by enzyme linked immunosorbent assay. J. Vet. Med. Sci 53:767-768..
143) Hsieh, D. P. H.; L. M. Beltran; M. Y. Fukuyama y D. W. Rice (1986): Production and isolation of aflatoxin M1 for toxicological studies. J. Assoc. Anal. Chem. 69:510-512.
144) Hurst, W. J.; R. A. Martin y C. H. Vestal (1991): Use of HPLC thermospray MS for the confirmation of aflatoxins in peanuts. J. Liquid Chromatographic 14:2541-2550
145) Hutchins, J. E.; Y. J. Lee; K. Tyczkowska y W. M. Hagler (1989):Evaluation of silica cartridge purification and hemiacetal formation for liquid chromatographic determination of aflatoxins in corn. Arch. Environ. Contam. Toxicol. 18:319-326.
146) Ilgaz, A. (1985): Immunosupressive effects of aflatoxicosis in chicks vaccinated against Newcastle disease. Vet. Fak Dergisi Istanbul Universitesi 9:62-72.
147) Iqbal, Q.; K. P. V. Rao y S. J. Reddy (1983): Dose response relationship of experimentally induced aflatoxicosis. Indian J. Anim. Sci 53:1277-1280.
148) Isaaq. H. J. y W. Cutchin (1981): A guide to thin layer chromatographic system for the separation of aflatoxins B1, B2, G1 y G2. J. Liquid Chrom. 4:1087-1096.
149) Jackman, R. (1985): Determination of aflatoxin by enzyme linked immunosorbent assay with special reference to aflatoxin M1 in raw milk. J. Sci. Food Agric. 36:685-698.
150) Jewers, K.; R. D. Coker; B. D. jones; J. Cornelius; M. J. Nagler; N. Bradburn; K. Tomlins; V. Medlock; P. Dell; G. Blunder; O.G. Roch y A. J. Sharkey (1989): Methological developments in the sampling of food and feed for mycotoxins analysis. J. Appl. Bact. Symposium Suppl pp:105s-116s.
151) Jewers, K.; R. D. Coker; G. Blun der; M. J. Jazwinski y A. J. Sharkey (1986):Problems involved in the determination of aflatoxin levels in bulk commodities. Int. Biodeterioration 22 (Suppl) pp:83-88.
152) Jones, B. D. (1977):Chemistry of mycotoxins En: Mycotoxic Fungi Mycotoxins, Mycotoxicoses. An Encyclopedic Handbook .vol 1 Eds. T.D Wyllie y L. G. Morehouse. New York y Basel.
153) Karanuranet, A. y L. B. Bullerman (1990): Interactive effects of spore load and temperature on aflatoxin production. J. Food Prot. 53.227.
154) Kaveri, S. V.; J. M. Fremy; C. Lapeyre y A. D. Strosberg (1987): Immunodetection and immunopurificatión of aflatoxins using a high affinity monoclonal antibody to aflatoxin B1. Lett. Appl. Microbiol. 4:71-75.
155) Key, A. C. (1978): Mycotoxicosis in cattle. En Mycotoxic Fungi Mycotoxins, Mycotoxicoses. An Encyclopedic Handbook vol 1 Eds. T.D Wyllie y L. G. Morehouse. New York y Basel. pp:9-28.
156) Koehler, P. E.; L. R. Beuchat y M. S. Chibinnan (1985):Influence of temperature and

61
water activity on aflatoxin production by Aspergillus flavus in cowpea (vigna unguicoulata) seed and meal. J. Food Prot. 48:1042-1043.
157) Koeltzow, D. E. y S. N. Tanner (1990): Comparative evaluation of commercially available aflatoxin test methods. J. Assoc. Off. Anal. Chem. 73:584-589.
158) Kok, W. T (1994): Derivatization reactions for the determination of aflatoxins by liquid chromatography with fluorescence detection. J. Chromatogr. B. Biomed. Appl. 659:127-137.
159) Kostiainen, R. (1988): Characterization of trichothecene by Tandem mass spectrometry using reactive collision with ammonia. Biomed. Environ. Mass Spectrom. 16:197
160) Kostiainen, R. (1991): Identification of trichothecenes by thermospray, plasmaspray and dynamic atom bombardment liquid chromatography-mass spectrometry. J. Chromatography. 562:555
161) Kostiainen, R. y K. Kuronen (1991): Use of 1-p(2-3 dihydroxypropoxy)phenyl 1-alkanones as retention index standard in the identification of trichothecenes by liquid chromatography thermospray and dynamic atom bombardment mass spectrometry. J. Chromatography. 543:39.
162) Krihmamachari, K. A. V. R.; R. V. Bhat; V. Nagarajan y T. B. G. Tilak (1975): Hepatitis due to aflatoxicosis. An outbreak in Western India. Lancet 1:1061-1063.
163) Krishnamurthy, P. B. (1986): Effect of dietary fat on aflatoxin B1 induced chromosomal aberrations in mice. Toxicol. Lett. 31:229-234.
164) Kumagai, S. (1989): Intestinal adsorption and excretion of aflatoxin in rats. Toxicol. Appl. Pharmacol. 97:88-97.
165) Lancaster, M. C.; F. P. Jekins y J. Mc Philp (1961): Toxicity associated with certain samples of groundnuts. Nature (London) 192: 1095-1096.
166) Langone, J. L. y H. vanVunakis (1976): Antibodies and their use in radioimmunoassay. J. Natl. Cancer. Inst. 56:591-595.
167) Lee, H. A.; A. P. Wilkinson; C. M. Ward; E, N. C. mills y M. R. A. Morgan (1992): Immunoassay of mycotoxins in food and human serum. En Food Quality of Safety Assurance Applications of Immunoassays Systems. Eds. M. R. A. Morgan; C. J. Smith y P. A. Williams. Elseviers, Applied Science (London, New york) pp:85-92.
168) Lewelly G. C. (1973): Evaluation of the response of Dugesia tiguna (Brown planaria) to aflatoxin B1. J. Assoc. Off. Anal. Chem 56:1119.
169) Li S.; R. Marquardt; A.A. Frohlich; H. Xiao y J. Clarke (1994). The development of a quantitative ELISA for aflatoxin B1 in ground peanut using antibodies isolated from the yolk of laying hen. J. Food Protection 57:1022-1024.
170) Li, Y. K. y F. S. Chu (1982): Kinetics of transformation of aflatoxin B1 into aflatoxin M1 in lactating mouse an ELISA analysis. Experientia 30:942-943.

62
171) Lotter, L. H. y H. J. Krohn (1988): Occurrence of aflatoxins in human foodstuffs in South Africa. Bull. Environ. Contam. Toxicol. 40:240-243.
172) Llewelyn, G. C.; G. A. Stephenson y J. Woffman (1977):Aflatoxin B1 toxicity and teratogenicity in Japanese Medaka eggs (oryzias latipes). Toxicon 15:582-587.
173) Manabe, M. V.; T. Goto y S. Matsura (1978): High performance liquid chromatography of aflatoxins with fluorescent detection. Agric. Biol. Chem. 42:2003-2007.
174) Margolles, Estrella; A. Escobar y A. Acosta (1990): Determinación de la residualidad de aflatoxina B1 directamente en leche de vaca utilizando una técnica inmunoenzimática. Rev. Salud Anim. 12:35-38.
175) Margolles, Estrella, Victoria Zaldivar; Ma Cristina Muñoz; Esmeralda Lon-Wo; Niurka Gomez y A. Escobar (1992): Descontaminación de aflatoxinas en alimentos para animales con el uso de zeolitas naturales Cubanas. Rev. Salud Anim. 14:171-188.
176) Margolles, Estrella, Victoria Zaldivar y A Escobar (1993): Evaluación de Bentonitas Cubanas como adsorbentes de aflatoxina B1 . Rev. Salud Anim. 15:335-337.
177) Margolles, Estrella; A Escobar y Victoria Zaldivar (1994): Evaluación de la descontaminación de aflatoxina B1 en piensos con aluminosilicatos Cubanos utilizando la técnica ELISA. . Rev. Salud Anim. 16:69-72
178) Martin, C. N. y R. C. Garner (1977): Aflatoxin B1-oxide generated by chemical or enzymic oxidation of aflatoxin B1 causes guanine substitution in nucleid acids. Nature (London) 267:863-865.
179) Mella, C. M.; A. Escobar y A. Acosta (1988): Ensayo inmunoenzimático homógeneo de aflatoxina B1. Rev. Salud Anim. 10:303-310.
180) Miguel, J. A. y V de Andrés (1982): Método simple de determinación simultánea de aflatoxinas, ocratoxina A y cearalenona en productos agrícolas. Serie Ganadería No 15 pp:101-110.
181) Miller, d. M.; J. D. Clark; R, C, Hatch y A. V. Jain (1984):Caprine aflatoxicosis serum electrophoresis and pathology changes. Am. J. Vet. Res. 45:1136-1141.
182) Mills, E. N. C.; J. M. Jhonston; H. A. Kemp y M. R. A. Morgan (1988):An enzyme linked immunosorbent assay for diacetoxyscirpenol applied to the analysis of wheat. J. Sci Food Agric. 42:225-233.
183) Mirocha, C. J.; R. J. Pawlosky y H. K Abbas (1989):Analysis of T2 toxin in biological matrix using multiple reaction monitoring. Arch. Environ. Contam. Toxicol. 18:349.
184) Mislivec, P. B., M. W. Trucksess y L. Stoloff (1988): Effect of other toxigenic mold species on aflatoxin production by Aspergillus flavus in sterile broth shake culture. J. Food Prot. 51:449-451.
185) Modali, R. y S. S. Yang (1986): Specificity of aflatoxin B1 binding on human proto-oncogene nucleotide sequence. En: Monitoring of Occupational genotoxicants. Ed. Alan

63
R. Liss. Inc (Copyright) pp:147-158.
186) Montville, T. J. y P. K. Goldstein (1987): Sodium bicarbonate reduced viabillity and alters aflatoxin distribution of aspergillus parasiticus in czapeks agar. Appl. Environ. Microbiol. 53:2303-2307.
187) Morgan, M.R. A. (1985): Newer techniques in food analysis immunoassays and their application to small molecules. J. Assoc. Pub. Analysts 23:59-63.
188) Morgan, M. R. A.; A. S. Kang y H. W. S. Chan (1986): Aflatoxin determination in peanut butter by enzyme-linked immunosorbent assay. J.Food Sci. Agric. 37:908-914.
189) Morris, B. A. (1985): Principles of immunoassays. En Immunoassays in Food Analysis. Eds. B.A. Morris y M. N. Clifford. Elsevier Applied Science Publishers pp:53-73.
190) Mortimer, D. N.; M. J. Shepherd; J. Gilbert y C. Clark (1988): Enzyme linked immunosorbent (ELISA) determination of aflatoxin B1 in peanut butter: collaborative trial. Food Addit. Contam 5:601-608.
191) Mortimer, D. N.; M. J. Shepherd; J. Gilbert y M. R. A. Morgan (1987): A survey of the occurrence of aflatoxin B1 in peanut butters by enzyme linked immunosorbent assay. Food Addit. Contam. 5:127-132.
192) Münch, E. W. y W. Stein (1985): Aflatoxin removal from feed and food. A review a presentation of a new procedure. 41st. DGF Meeting in Gottinger (sept. 11) 2-18 pp
193) Nandi, B. y P. Haggblom (1984): Production of aflatoxin in rough rice under different storage conditions. Acta. Agric. Scand. 34:128-132.
194) Nesheim, S. Y W. C. Brumley (1981): Confirmation of identity of aflatoxins. J. Assoc. Off. Chem, Soc. 58:945a-949a.
195) Ngo, T. T. y H. M. Lenhoff (1985):Separation free (homogeneous) enzyme mediated immunoassay. En: Enzyme mediated immunoassay Eds. T.T. Ngo y H.M. Lenhoff. Plenum Press New York and London. pp:33-202.
196) Nkama, J.; J. H. Nobbs y H. G. Muller (1987): Destruction of aflatoxins B1 in rice exposed to light. J. Cereal. Sci 5:167.
197) Nkama, Y y H. G Muller (1988): Effects of initial moisture content and light intensity on the photo degradation of aflatoxin B1 in contaminated rice. J. Cereal. Sci 8:269-274.
198) Nyathi, C. B.; C. F. Nutiro; J. A. Hasler y C. J. Chetsanga (1987): A survey of urinary aflatoxin in Zimbabwe. Int. J. Epidemiol. 16:516-519.
199) Obioha, W. I.; H. M. Stahr y A. A. Kraft (1983): Effect of sodium nitrite on aflatoxin production in pork sausage at different temperatures and the effect on the nitrite on growth of Aspergillus parasiticus in culture. J. Agric. Food Chem. 31:1039-1042.
200) Okoye, Z. S. y G. E. Neal (1988): Enhanced ethanol induced changes in disposition and toxic response to dietary aflatoxin B1 due to sacoglottis gabonensis bark extract a

64
Nigerian alcoholic beverage additive. Food Chem. Toxicol. 26:679-689.
201) Ong, T. M (1975): Aflatoxin Mutagenesis. Mutat. Res. 32:35-53.
202) OPS-OMS (1983): Criterios de salud ambiental 11. Micotoxinas Publicación científica No. 453.
203) Oyelami, O. A.; S. M. Maxwell; T. A. Aladekame y K. A. Adelusa (1995): Two unsual cae of kawashiorkor can protein deficiency explain the mystery. Ann. Trop. Paediatr. 15:217-219
204) Panalaks, T. y P. M Scott (1977): Sensitive silica gel packed flowcell of aflatoxins by high pressure chromatography. J. Assoc. Off. Anal. Chem. 60:583-584.
205) Panangala, V. S.; J. J. Giambrone; U. L. Diener; N. D. Davis; F. J. Hoerr; A. Mitra; R.D. Schultz y G. R. Wiltz (1986): Effects of aflatoxin on the growth performance and immune responses of weahling swine. Am. J. Vet. Res. 47:2062-2067.
206) Park, D. L; V. Diprossimo; E. Abdel-malek; M. W. Trucksess; S. Nesheim; W. C. Brumley; J. A. Sphon; T. L Barry y G. Petzinger (1985): Negative ion chemical ionization mass spectrometric method for confirmation of identity of aflatoxin B1: collaborative study. J.Assoc. Off. Anal. Chem. 68:636-640.
207) Park, D. L.; M. W. Trucksess; R. C. Lee y F. S. Chu (1988): Evaluation by enzyme linked immunosorbent assay of clean-up for thin layer chromatography of aflatoxin B1 in corn peanut and peanut butter. J. Assoc. Off. Anal. Chem. 71:953-956.
208) Park, D. L.; B. M. Miller; P. Hart; G. Yang; J. McVey; S. W. Page; J. Pestka y L. H. Brown (1989a): Enzyme linked immunosorbent assay for screening aflatoxin in cottonseed products and mixed feed.: Collaborative study. J. Assoc. Off. Anal. Chem. 72:326-332.
209) Park, D. L.; B. M. Miller; S. Nesheim; M. W. Trucksess; A. Vekich; B. Bidigare; J.L. McVey y L. H Brown (1989b): Visual and semiquantitative spectrophotometric screening method for aflatoxin B1 in corn and peanut products: follow-up study. J. Assoc. Off. Anal. Chem. 72:638-643..
210) Park, K. Y. y L.B. Bullerman (1983): Effect of cycling temperatures on aflatoxin production by A.parasiticus and A. Flavus in rice and cheddar cheese. J. Food Sci. 48:889-895.
211) Parkinson, J. F.; E. E. Steele; E. A. Sauter y C. F. Petersen (1989): Effect of low levels of aflatoxin B1 on hatchability of chicken eggs. Poultry Sci. 63:(suppl 1) pp:161-166.
212) Patel, U. D.; P. Govindarajan y P. J. Dave (1989): Inactivation of aflatoxin B1 by using the synergistic effect of hydrogen peroxide and gamma radiation. Appl. Environ. Microbiol. 55:465-467.
213) Patey, A. L.; M. Sharman y J. Gilbert (1992):Determination of total aflatoxin levels in peanut butter by enzyme linked immunosorbent assay: collaborative study. J. Assoc. Off. Ana. Chem. international. 75:693-697.
214) Patterson, D. S. P. (1977): Aflatoxins and related compounds. En Mycotoxic fungi,

65
Mycotoxins, Mycotoxicoses. An Encyclopedic Handbook. Vol 1 Eds. T.D Wyllie y L. G. Morehouse. New York y Basel. pp 136-189.
215) Peer, F.; X. Bosch; J.Kaldor; A. Linsell y M. Pluumaen (1987): Aflatoxin exposure hepatitis B virus infection, and liver cancer in Swaziland. Int. J. Cancer. 39:545-553.
216) Perguero Martha (1988) Estudio de la hepatitis aguda y excreción urinaria de aflatoxina en una población pediátrica. Tesis presentada en opción al grado de especialista de Primer grado de Pediatria . Hospital Docente Comadante Pinares. Pinar de Río. Cuba
217) Pestka, J. J.; Y. Li; N. O. Harden y F. S, Chu (1981): Comparison of RIA and ELISA for determining aflatoxin M1 in milk. J. Assoc. Off. Anal. Chem. 64:294-301.
218) Pestka, J. J. (1991): High performance thin layer chromatography ELISAGRAM. Application of a multi-hapten immunoassay to analysis of the zearalenone and aflatoxin mycotoxins families. J. Immunol. Meth. 136:177-183.
219) Petrov, R. V. (1987): Anticuerpos y anticuerpogénesis. En Inmunología. Editorial Mir Moscu. pp:37-71.
220) Phillips, T. D.; L. F. Kubena; R. B. Harvey; D. R. Taylor y N. D. Heidelbaugh (1988): Hydrated sodium calcium aluminosilicate a high affinity sorbent for aflatoxin. Poultry Sci. 67:343-347
221) Pier, A. C.; R. E. Fitchner y S. J Cysewski (1977): Effects of aflatoxin on the cellular immune system. Ann. Nutr. Alim. 34:781-788.
222) Pier, A. C. (1992): Major biological consequences of aflatoxicosis in animal production. J. Anim. Sci. 70:3964-3967.
223) Pival, G.; A. Pietri; E. Santi y L Fiorentini (1981): inattivazione dell afatossina B1 con trattamenti chimica. Rev. Ital. Sostanze Grase 58:289-295.
224) Plattner, R. D.; G. A. Bennett y R. D. Stubblefield (1984): Identification of aflatoxins by quadrupule mass spectrometry/mass spectrometry. J. Assoc. Anal. Chem. 67:734-738.
225) Pohland, A. E.; M. E. Cushmac y P. J Andrellos (1968): Aflatoxin Hemiacetal. J. Am. Off. Anal. Chem. 51:907-910.
226) Prelusky, D. B.; R. M. Hamilton; B. C. Foster; H. L. Trenholm y B. K. Thompson (1987): Optimization of chicken embryotoxicity bioassay for testing toxicity potential of fungal metabolites. J. Assoc. Off. Anal. Chem. 70:1049-1055.
227) Promchainant, C.; V. Baimai y A. Nondasuta (1972): The Cytogenic effects of aflatoxin and gamma rays on human leukocytes in vitro. Mut. Res 16:373-380.
228) Raisuddin, R.; K. P. Singh; S. I. A. Zaidi; A. K. Saxena y P. K. Ray (1990):Effects of aflatoxin on lymphoid cell of weanling rats. J. Appl. Toxicol. 10:245-250.
229) Ranjan, K. S. y A. D. Sinha (1989): Effect of aflatoxin B1 on meiotic index in guinea pigs. Nat. Acad. Sci. Lett. (India) 12:301-302.

66
230) Reichert, N.; S. Steinmeyer y R. Weber (1988): Determination of aflatoxin B1 in dried figs by visual screening thin layer chromatography and ELISA. Z. Lebensm Unters Forsch 186:505-508.
231) Richard, J. L.; G.A. Bennett; P. F. Ross y P. E. Nelson (1993): Analysis of naturally occurring mycotoxins in feedstuffs and food. J. Anim. Sci. 71:2563-2574
232) Robert, J. F. y J. L. Richard (1992): Aflatoxins in animal and human health. Reviews of Environ. Contam. Toxicol. 127:69-94.
233) Rodríguez, Mayra y A. Rodríguez (1983): Reducción de la producción de aflatoxina en maní por radiaciones gamma. Ciencia y Técnica Agric. (vet.-Cuba) 5:103-110
234) Rosen, S.; N. O. Patei; A. O. Scott y M. F. Stringer (1989): Immunoassay kits for the detection of toxins associated with foodborne illness. Soc. Appl. Bact. Tech. Ser. pp:265-281.
235) Sánchez, Olga. (1987): Factores que afectan la formación de micotoxinas en los alimentos. Curso Internacional sobre contaminantes químicos con especial énfasis en las micotoxinas. Inst. de Nutrición e Higiene de los Alimentos (INHA). Ciudad de la Habana, Cuba.
236) Sanders, T. H.; N. D. Davis y V. L.Diener (1968): Effect of dioxide, temperature and relative humidity on production of aflatoxin in peanut. J. Am. Oil Chem. 45:683-685.
237) Sauer, M. J.; J. A. Foulkes y B. A. Morris (1985): Principles of enzymoimmunoassay. En Immunoassays in Food Analysis. Eds. B.A. Morris y M. N. Clifford. Elsevier Applied Science Publishers pp:21-53.
238) Scott P. M. y G. A. Lawrencen (1988): Liquid chromatographic determination of zearalenone and b zearalenone in milk. J.Assoc. Off. Anal. Chem. 71:1176.
239) Seiber, J. N. y D. P. Hsieh (1973): Application of high speed liquid chromatography to the analysis of aflatoxns. J. Assoc. Off. Anal. Chem. 56:827-830.
240) Senter, L. H.; D.R. Sanso; D.G. Corley; M. S. Tempseta; A. A. Rottinghaus y G. E. Rottinghaus (1991): Cytoxicity of trichothecene mycotoxins isolated from Fusarium sporotrichoides (MC 72083) and Fusarium sambucinum in baby hamster kidney (BHK-21) cells. Mycopathologia 113:127.
241) Shank, R. C.;C. H. Burgeois; N. Keschamtas y P. Chandavimol (1971): Aflatoxins in autopsy specimens from Thai children with an acute disease of unknown a ethiology. Food Cosmet. Toxicol. 9:501-507.
242) Sharma, A.; A. G. Beheve; S. R. Padwal-Desai y G. B Nadkarne (1980): Influence of inoculum size Aspergillus parasiticus spores on aflatoxin production. Appl. Environ. Microbiol. 40:989-993.
243) Sharman, M. y J. Gilbert (1991): Automated aflatoxin analysis of food and animal feed using immunoaffinity column clean-up and high performance liquid chromatographic. J. Chromatogr. 543:220-225

67
244) Schell, T. C.; M. D. Lindemann; E. T. Kornegay; D. J. Blodgett y J. A. Doerr(1993 a): Effectiveness of different types of clay for reducing the detrimental effects of aflatoxins contaminated diets on performance and serum profiles of weanling pig. J. Anim. Sci. 71:1226-1231.
245) Schell, T. C.; M. D. Lindemann; E. T. Kornegay y D. J. Blodgett (1993b): Effects of feeding aflatoxin contaminated diets with and without clay to weanling and growing pig on performance, liver function and mineral metabolism. J. Anim. Sci. 71:1209-1218
246) Shepherd, E. C.; T. D. Phillips y T. R. Irwin (1984): aflatoxin B1 metabolism in the rat polyhalogenated biphenyl enhanced conversion to aflatoxin M1. Xenobiotica 14:741-750.
247) Shepherd, M. J. (1986): High performance liquid chromatography and its applications to the analysis of mycotoxins. In R.J Cole (Ed) Modern Methods in the Analysis and Structural Elucidation of Mycotoxins. Academic Press New York pp 293-333.
248) Shepherd. M. J.; M. Holmes y J. Glibert (1986): Comparison and critical evaluation of six published extraction and cleanup procedures for aflatoxin M1 in liquid milk. J. Chromatogr. 359:305-315.
249) Shepherd. M. J.; D. N. Mortimer y J. Glibert (1987): A review of approaches to the rapid analysis of aflatoxins in foods. J.Assoc. Pub. Analyst 25:129-142.
250) Shibko, S. I.; D. L Arnold, J. Mornigstar y L. Friedman (1968): Studies on the effect of aflatoxin B1 on the development of the chick embryom. Proc. Soc. Exp. Biol. Med. 127:835-839.
251) Sinha, R. N. (1979): Ecology of mycloflora in stored grain. Ann. Techn. Agric. 28:191-209.
252) Sizaret, P.; C. Malaveille; R. Montesano y C. Frayssinet (1982):. Detection of aflatoxins and related metabolites by RIA. J. N. C. I. 69:1375-1381.
253) Slowick, J.; S. Graczyc y J. A. Madej (1985): The effect of a single dose of aflatoxin B1 on the value of nucleolar index of blood lymphocytes and on histological changes in the liver, bursa fabricii, suprarrenal gland, and spleen in ducklings. Folia Histochemical et Cytobiological 23:71-80.
254) Stora, C.; I. Dvorackova y N. Ayraud (1982): L' inhalation de poussiere polluce par l'aspergillus flavus une voice de contamination por l'homme. Med. Biol. Environ. Sep-Dec:9-12.
255) Stora, C.; I. Dvorackova y N. Ayraud (1983): Aflatoxins and Reyes syndrome. J.of Medicine 14:47-54.
256) Stora, C. y I. Dvorackova (1987): Aflatoxin, viral hepatitis, and primary liver cancer. J. Med (Clin. Exp. Theor) 18:23-41.
257) Stubblefield, R. D.; O. L. Shotwell; C. W. Hesseltine; M. L. Smith y H. H. Hall (1967): Production of aflatoxin on wheat and oats. Measurement with a recording densitometer. Appl. Microbiol. 15:186-190.

68
258) Stubblefield, R. D.; G. M. Shannon y O. L. Shotwell (1970): Aflatoxins M1 and M2 preparation and purification. J. Am Oil Chem. Soc. 47:389-390.
259) Stubblefield, R. D. (1980): Stability and molar adsortivity of aflatoxin M1 in acetonitrile-benzene (1+9) J. Assoc. Off. Anal. Chem. 63:634-636.
260) Sulliman, H. B.; A. F. Mohamed; N. A. Awadelsield y A. M. Shommein (1987): Acute mycotoxicoses in sheep field cases. Vet. Human. Toxicol. 29:241-243.
261) Swick, R. A. (1984). Hepatic metabolism and bioctivation of mycotoxins and plant toxins. J. Anim. Sci. 58:1017-1028.
262) Sydenham, E. W.; W. C. A. Gelderblom; P. G. Thiel y W. F. O. Marasas (1990): Evidence for the natural occurrence of fumonisin B1 a mycotoxin produced by Fusarium moniliforme in corn. J. Agri. Food Chem. 38:285-290.
263) Tarter, E. J.; J, P. Hanchay y P. M. Scott (1984): Improved liquid chromatographic method for determination of aflatoxins in peanut butter. J. Assoc. Off. Anal. Chem. 67:597-600.
264) Tejada, Irma. (1992): Consideraciones generales sobre muestreo y preparación de muestras. En Control de calidad y análisis de alimentos para animales. Editorial Secretaría de Educación Pública. México. pp:7-14
265) Thanaboripat, D.; W. Ramunsri; M. Apintanapong y U. Chusanatasana (1992): Effects of sodium chloride, propionic acid and ammonium hydroxide on growth of Aspergillus flavus on corn and aflatoxin production. ASEAN Food J. 7: 24-29.
266) Tizard, I. (1989): Anticuerpos. En: Inmunología Veterinaria. Tercera edición. Interamericana Mc Graw-Hill. pp:33-47.
267) Trucksess, M. W.; M. E. Stock; S. Nesheim; D. L. Park y A. E. Pohland (1989): Enzyme linked immunosorbent assay of aflatoxins B1, B2, G1 and G2 in corn cottonseed, peanut, peanut butter and poultry feed: collaborative study. J. Assoc. Off. Anal. Chem. 72:957-962.
268) Trucksess, M. W.; M. E. Stack; S. Nesheim; R. H. Albert; J. y T Romer (1994): Multifuntional column coupled with liquid chromatography for determination of aflatoxin B1, B2,G1 and G2 in corn, almond, Brasil nuts, peanuts and pistachio nuts; collaborative study . J. AOAC.Int. 77:1512-1520.
269) Trucksess, M. W. y M. E. Stack (1994): Enzyme linked immunosorbent assay of total aflatoxin B1 aflatoxin B2 and aflatoxin G1 in corn follow-up collaborative study. J.AOAC int. 77:655-659.
270) Trucksess, M. W. (1995): Mycotoxins. General refere reports. J. AOAC international 78:135-141
271) Tsai, W. Y. J.; J. D. Lambert y L. B Bullerman (1984): Simplified method for microscale production and quantification of aflatoxin in broth. J.Food Prot. 47:526-529.

69
272) Tsuboi, S.; T. Nakagawa, M. Tomita; T. Seo; H. Ono; K. Kawamura y N. Iwamura (1984): Detection of aflatoxin B1 in serum samples of male Japanese subjects by radioimmunoassay and high performance liquid chromatography. Cancer Res. 44:1231-1234.
273) Tuason, M. A. y L. S. F. Madamba (1980): Aflatoxin production in Copra Phil. Agric. 63:189-195.
274) Tuekam, T. D.; R. D. Miles G. D. Butcher (1994):Performance and cell mediated and humural immune response in broilers fed and aflatoxin supplemented diet. J.Appl. Anim. Rev. 6:27-35.
275) Tuinstra, L. G. M. y W. Hassnoot (1983): Rapid determination of aflatoxin B1 in dutch feeding stuffs by high performance liquid chromatography and post column derivatization. J. Chromatogr. 282:457-462.
276) Tuinstra, L. G. M.; P. M. G. Kienhuis y P. Dols (1990): Automated liquid chromatographic determination of aflatoxin M1 in milk using on line dialysis for sample preparation. J. Assoc. Off. Anal. Chem 73:969-973.
277) Tulpule, P. G. (1981): Aflatoxins experimental studies. J. Cancer Res. Cli. Oncol. 99:137-142.
278) Tyczckowski, J. K. y P. B. Hamilton (1987): Altered metabolism of carotenoids during aflatoxicosis in young chickens. Poultry Sci 66:1184-1188.
279) Uyakual, D.; M. Isube y T. Goto (1989): Mycotoxin analysis by fast atom bombardment tandem mass spectrometry. J. Assoc. Off. Anal. Chem. 72:491.
280) van Egmond, H. P. y P. J. Wagstaffe (1990): Aflatoxin B1 in compound feed reference materials an intercomparison of methods. Food Addit. Contam. 7:239-251.
281) van Egmond, H. P.; S. Patel; W. E. Paulsch; E. A. Sizoo; L. G. M. Th Tuinstra; G. Wood; A. Boenke B. Schure y P. J. Wagstaffe (1994): The development of five animal feed reference materials certified for their aflatoxin B1 content. Food Add. Contam. 11:449-477.
282) van Weemen, B. K. y A. H. W. Schuurs (1971): Immunoassay using antigen enzyme conjugates. FEBS- Lett. 15:232-236.
283) Varadi, M. (1986): Analysis of aflatoxins in different food and fodder by overpressure layer chromatography. Hugarian Scientific Instrument. 61:23-28.
284) Vedanayagam, H. S.; A. S. Indulkar y S. R Rao (1986): The occurrence of Aspergillus flavus and aflatoxins in Indian Cottonseed A survey. J.Food Sci. Techn. 23:73-76.
285) Veldam, A. (1992): Effect of sorbetia on carry over of aflatoxin from cow feed to milk. Milchwissenschaft 12:777-780.
286) Vesonder, R.; J. Haliburton; R. D. Stubblefield; W. Gilmore y S. Peterson (1991): Aspergillus flavus and aflatoxins B1, B2, and M1 corn associated with equine death.

70
Arch. Environ. Contam. Toxicol. 20:151-153.
287) Waibel, J. (1977): Stick probengrosse fur die bestimmong von aflatoxin in erdnussen. Deutsche Lebensmittel Rundschau. 73:353-357.
288) Wanop, C. C. (1960): Disease of turkey. Poultry Vet. Res. 72:671-672.
289) Ward, C. M.; A. P. Wilkinson y M. R. A. Morgan (1993): Emerging techniques: enzyme linked immunosorbent assay (ELISA) as alternatives to chromatographic methods. En J. Chromatography Library Vol 54. Chromatography of Mycotoxins Techniques and Application. Ed. V. Betina. Elservier-London-New York-Tokyo.
290) Washbur, K. W.; R. D. Wyatt; P. L. Potts y G. M. Lanza (1985): Effects and mechanism of aflatoxin on variation in shell strength. Poultry Sci. 64:1302-1305.
291) Wei, C. y D. P. H. Hsieh (1980). Aflatoxin in human and animal health. Proc. of the 84th Annual Meeting of the US Animal Health Ass. Louisville Kentucky USA pp:283-297.
292) West, S.; R. D. Wyatt y P. B. Hamilton (1973): Improved yield of aflatoxin by incremental increase of temperature. Appl. Microbiol. 25:1018-1019
293) Whitaker, T. B.; J. W. Dickens y R. J. Monroe (1979): Variability associated with testing corn for aflatoxin. J. Am. Oil Chem. Soc. 56:798-794.
294) Whitaker, T.B.; J. Springer; P. R. Defize; W. J. Dekoe y R. Coker (1995): Evaluation of sampling plans used in the Unite States, United Kingdom and the Netherlands to test raw shelled peanuts for aflatoxins. J. AOAC int. 78:1010-1018.
295) Wilcke, W. F.; M. R. Ehrinch y K. W. Ko (1990): Using ELISA kits to test corn for aflatoxin. Appl. Agric. Res. 5:32-36.
296) Wild, C. P.; R. C. Garner; R. Montesano y F. Tursi (1986): Aflatoxin B1 binding to plasma albumin and liver DNA upon chronic administration to rats. Carcinogenesis 7:853-858.
297) Wild, C. P.; F. A. Pionneau; R. Montesano; C. F Mutro y C. J. Chetsanga (1987): Aflatoxin detected in human breast milk by immunoassay. Int. J. Cancer 40:328-333.
298) Wild, C. P.; B. Chapot; E, Scherer; L. Den Engelse y R. Montesano (1988): Application of antibody methods to the detection of aflatoxin in human fluids. En Methods for detecting DNA damaging agent in humans. Applications in Cancer Epidemiology and Prevention. Eds. H. Sartsch; K. Hemminki y I.K.O'Neill. IARC 89:67-74.
299) Wild, C. P.; Z. Jiang; R. Montesano; M. Pakin; M. Khlat y P. Srivatanakul (1989): Correlation study of aflatoxin exposure and liver cancer incidence in five geographical regions of Thailand. Proc. Am. Assoc. Cancer Res. 30:317.
300) Wild, C. P.; Y. Z. Jiang; G. Sabbioni; B. Chaot y R .Montesano. (1990): Evaluation of methods for quantification of aflatoxin-albumin adducts and their application to human exposure assessment. Cancer Res. 50:245-251.
301) Wild, C. P.; L. A. M. Jansen; L. Cova y R. Montesano (1993): Molecular dosimetry of

71
aflatoxin exposure contribution to understanding the multifactorial etiopathogenesis of primary hepatocellular carcinoma with particular reference to hepatitis B virus. Environ. Health Perspect. 99:115-122.
302) Wilkinson, A. P.; C. M. Ward y M. R. A. Morgan (1992): Immunological analysis of mycotoxins. En Modern Methods of Plant Analysis. New Series Vol 13 Plant Toxin Analysis. Eds. H. F. Linskens y J. F. Jackson. Springer-Verlag-Berlin-Heidelberg-New York-London-Paris-Tokyo-Hong Kong-Barcelona-Budapest pp:185-223.
303) Wilkinson, A. P.; D. W. Denning y M. R. A. Morgan (1988a): Analysis of sera for aflatoxin by enzyme linked immunosorbent assay. Human Toxicol. 7:353-356.
304) Wilkinson, A. P.; D. W. Denning y M. R. A. Morgan (1988b): An ELISA method for the rapid and simple determination of aflatoxin in human serum. Food Add. Contam. 5:609-619.
305) Wilkinson, A. P.; D. W. Denning y M. R. A. Morgan (1989): Immunoassay o aflatoxin in food and human tissues. J. Toxicol. Toxin Reviews 8:69-79.
306) Wilson, D. M. (1989): Analytical method for aflatoxins in corn and peanut. Arch. Environ. Contam. Toxicol. 18:308-314.
307) Wilson, D. M.; L. T. Sangster y D. M. Bedell (1984): Recognizing the signs of porcine aflatoxicosis. Vet. Med. July:974-977.
308) Wyatt, R. D.; M. N. Neathery; W. H. Moods; W. J. Miller; R. P. Gentry y G. O. Ware (1985):Effects of dietary aflatoxin and Zn on enzymes and other blood constituents in dairy calves. J.Dairy Sci, 68:437-442.
309) Xavier, F. y N. Muñoz (1987): Methods for detecting DNA damaging agents in humans applications in cancer epidemiology and prevention. IARC Scientific Publ. No, 89 Eds. H. Sartsch; K. Hemminki y I.K.O'Neill.
310) Yadgan, B.; V. Reddy; P. G. Tulpule; S. G. Srikantia y C. Gopalan (1970): Aflatoxin and Indian Chilhood chirrosis. Am. J. Clin. Nutr. 23:94-98.
311) Yalow, R. S. y S. A. Berson (1959): Assay of plasma insulin in human subjects by immunological methods. Nature 184:1648-1649.
312) Yen I. C. y K. R. Bidasee (1993): Liquid chromatographic determination of aflatoxin in animal feeds and feed components. J. AOAC Int. 76:366-370.
313) Yousef, A. E y E. H. Marth (1986): Use of ultraviolet energy to degrade AFM1 in raw or heated milk with and without added perxide. J. Dairy Sci. 69:2243-2247.
314) Yu, F. L.; W. Render y I. H. Geronimo (1990): Base and sequence specificities of Aflatoxin B1 binding to single stranded and double stranded DNAs. Carcinogenesis 11:475-478.
315) Yu, S. Z.; Z. Cheng; Y. Liu; Z. Huang y Y Zhao (1989): The aflatoxins and contaminated water in the etiological study of primary liver cancer. En: Bioactive molecules Vol 10.

72
Mycotoxins and Phycotoxins 88 7th Int. IUPAC Symp.Tokyo. Eds. S.Natori; K.Hashimoto y I.Ueno. Elsevier-London pp:37-44.
316) Zaika, L. L. y R. L Buchanan (1987): Review of compounds affecting the biosynthesis or bioregulations of aflatoxins. J.Food Prot. 50:691-708.
Autor: Arturo Escobar Dr.C.
Departamento de Inmunoquímica Centro Nacional de Sanidad Agropecuaria (CENSA),
Apartado 10, San José de las Lajas, La Habana, CP 32700, Cuba
FAX: 53-64-63897,Telef. 53-7-63145 E-mail: [email protected]