ing. genética
TRANSCRIPT



1.7.1 Primeros estudios y evidencias de la existencia del ADN
1.7.2 Los genes y el dogma central de la Biología molecular
- Replicación
- Transcripción
- Traducción (síntesis de proteínas)


¿Cómo controlar la expresión génica? Mientras que las células procariotas transcriben
casi todos sus genes, las células eucariotas eligen qué genes transcribir. Cada tipo celular eucariota expresa sólo una fracción de los genes que tiene, alrededor del 20%. Es decir, sólo el 20% de los genes que contiene el ADN se transcribe en ARN y luego se traduce en proteínas.
De la misma manera en que en nuestras casas no abrimos todas las canillas de agua al mismo tiempo sino sólo las que necesitamos usar, las células sintetizan solamente aquellas proteínas que necesitan. Aún mas, pueden también regular la cantidad de proteínas que necesitan, produciendo 2, 3 o hasta 100 copias cada vez.
Las células regulan, entonces, la expresión de sus genes mediante diferentes procesos, pero la forma más eficiente y usual es por medio del control transcripcional, por el cual la célula puede aumentar y disminuir la cantidad de ARN transcripto

Procesos de control de la expresión génica:
Controles transcripcionales
Controles postranscripcionales
Controles postraduccionales

Controles transcripcionales
Por medio de la regulación del inicio de la transcripción se puede elegir qué genes “encender” (activar) o “apagar” (inhibir) en un momento determinado, así como también regular la cantidad de ARNm producido. Esto se logra a través de mecanismos muy diferentes. El primero implica mantener apagados ciertos genes: en determinados tipos celulares, hay regiones de ADN que están siempre apagadas y cuyos genes no se transcriben nunca.
El segundo involucra el uso de regiones regulatorias, ubicadas en la secuencia anterior al inicio de la transcripción. Estas secuencias permiten la unión de factores de transcripción que pueden facilitar o impedir la unión de la ARN polimerasa al promotor, regulando de esta manera la cantidad de mensajeros producidos. Para que la transcripción ocurra es necesario que la ARN polimerasa se una al promotor del gen. Esta unión puede regularse utilizando ciertas proteínas, denominadas factores de transcripción, que se unen a secuencias específicas cercanas al promotor y pueden facilitar o impedir la transcripción. Esta es una manera de regular la cantidad de moléculas de ARN que se transcriben

Controles postranscripcionales
Mecanismos de corte y empalme (del inglés, Splicing). Mediante el splicing, los intrones –secuencias no codificantes- son eliminados del pre-ARNm y los exones son unidos: el ARNm maduro está formado sólo por exones. En ciertos casos, pueden quedar entre las secuencias de exones algunos intrones que son utilizados como molde en la traducción. Esto se denomina splicing alternativo y en teoría puede generar proteínas diferentes, de manera proporcional al número de intrones que contenga el gen. Por medio de este mecanismo de regulación se “elige” qué intrones sacar de la secuencia que va a ser traducida. Si una proteína contiene tres intrones, se puede crear un ARNm con uno, dos o tres intrones entre la secuencia codificante. Como resultado obtenemos proteínas similares, ya que los exones son iguales, pero su diferencia está dada por la secuencia de los intrones

Edición del ARN. En algunos casos, la secuencia del ARNm es modificada luego de ser transcripta. Los cambios incluyen la inserción de nucleótidos en regiones específicas, lo que motiva un corrimiento en el marco de lectura y genera la expresión de proteínas muy diferentes.
Transporte del ARNm al citoplasma. Para poder ser transportado, el ARNm debe haber sido procesado correctamente; de lo contrario es degradado en el núcleo
Iniciación de la síntesis proteica. El ARNm contiene en sus extremos secuencias que no son traducidas y que sirven para regular la traducción (5’ UTR y 3’ UTR, del inglés untraslated regions). Si esas secuencias son bloqueadas, el ARN no es reconocido por el ribosoma y no se puede traducir el mensajero

Por otro lado, las secuencias que flanquean el codón inicial (AUG) son determinantes. Si el ribosoma se “saltea” el primer AUG, comenzará la traducción en el segundo codón produciendo una proteína con menos aminoácidos y/o secuencia diferente.
Estabilidad del ARNm. La vida media de un ARNm está determinada principalmente por la longitud de la cola poliA. Una vez en el citoplasma, la secuencia comienza a acortarse; cuando se hace demasiado corta el mensajero es degradado.
Eliminación de ARNm con errores. Los ribosomas –junto con otras proteínas- son capaces de detectar codones de terminación en lugares erróneos de la secuencia del mensajero (generados por algún error previo en el splicing o por mutaciones). ¿Cómo lo hacen? Reconociendo las secuencias de unión entre los exones. Si el mensajero es adecuado, todos los lugares de unión entre los exones deberían encontrarse antes del codón de terminación. Si esto no ocurre, el ARNm fallado es degradado

ARN de interferencia. Cuando una célula humana es infectada por un virus que fabrica una doble cadena de ARN, ciertas enzimas lo reconocen y lo degradan. Esto mecanismo permite la eliminación de ciertos virus y puede ser utilizado también para regular la traducción de un mensajero: si se introduce (o se transcribe en la misma célula) una cadena de ARN complementaria a un mensajero, este no podrá ser traducido y además será detectado como foráneo y degradado

Controles postraduccionales Una vez sintetizadas, las proteínas pueden
ser modificadas mediante la unión de distintas moléculas (grupos fosfato, adenilatos, azúcares, etcétera). Estos agregados permiten regular la acción proteica de manera muy rápida porque no dependen del proceso de síntesis. Existen además ciertas proteínas que contienen segmentos que, bajo ciertas condiciones, se activan y se separan de la molécula, que puede cambiar su actividad.
Otro mecanismo de regulación es la degradación de proteínas. La vida media de la proteína puede ser un parámetro a regular que también actúa de manera rápida. El sistema ubiquitina-proteosoma lleva a cabo la degradación.


Los organismos multicelulares complejos están compuestos de diferentes tejidos cuyas características individuales dependen de las proteínas específicas expresadas por sus tipos celulares. La diferenciación, el desarrollo y la funcionalidad de los tejidos específicos dependen del conjunto de proteínas selectivamente expresadas por cada célula. Estas proteínas expresadas en forma diferencial pueden funcionar como componentes estructurales de las células, enzimas reguladoras del metabolismo, factores de transcripción, receptores celulares, componentes intracelulares de señalización, etc.
La expresión incorrecta de tales proteínas, su expresión en lugares equivocados, a destiempo, o la producción en cantidades anormales de proteínas específicas o de proteínas de función anómala subyace a toda patología celular de base genética.

Por consiguiente el conocimiento de los mecanismos de regulación de la expresión proteica en eucariontes contribuirá al conocimiento de las bases moleculares de diversas patologías.
En las células eucariotas, la capacidad de expresar proteínas biológicamente activas resulta de diferentes niveles regulatorios.


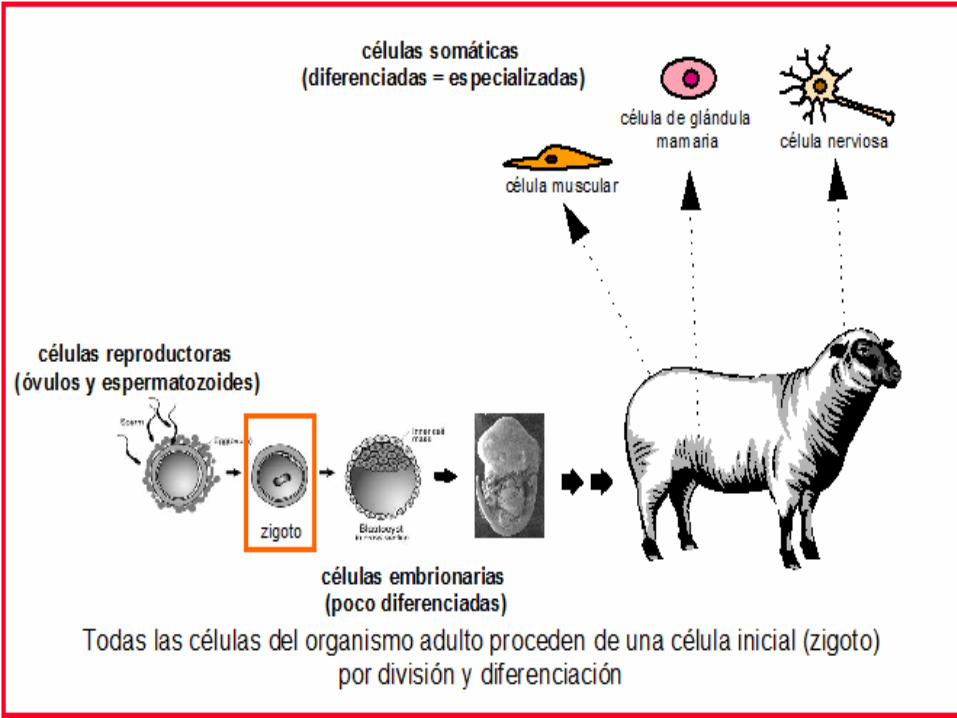
La clonación puede definirse como el proceso por el que se consiguen copias idénticas de un organismo ya desarrollado, de forma asexual. Estas dos características son importantes


Cualquier célula del organismo adulto (células somáticas, no reproductoras) puede servir teóricamente para obtener un nuevo ser vivo de las mismas características, ya que tiene en su ADN la información de cómo es y como se desarrolla ese determinado organismo. Se trataría de tomar una célula cualquiera, exceptuando las células reproductoras que tienen una dotación incompleta, y conseguir que esa información se exprese, se ponga en funcionamiento y

nos produzca otro ser. Clonar consistiría por tanto en reprogramar una célula somática para que empiece el programa embrionario. Una vez comenzado su desarrollo se implantaría en un útero, ya que de momento no es posible que los embriones lleguen a término fuera de un útero.
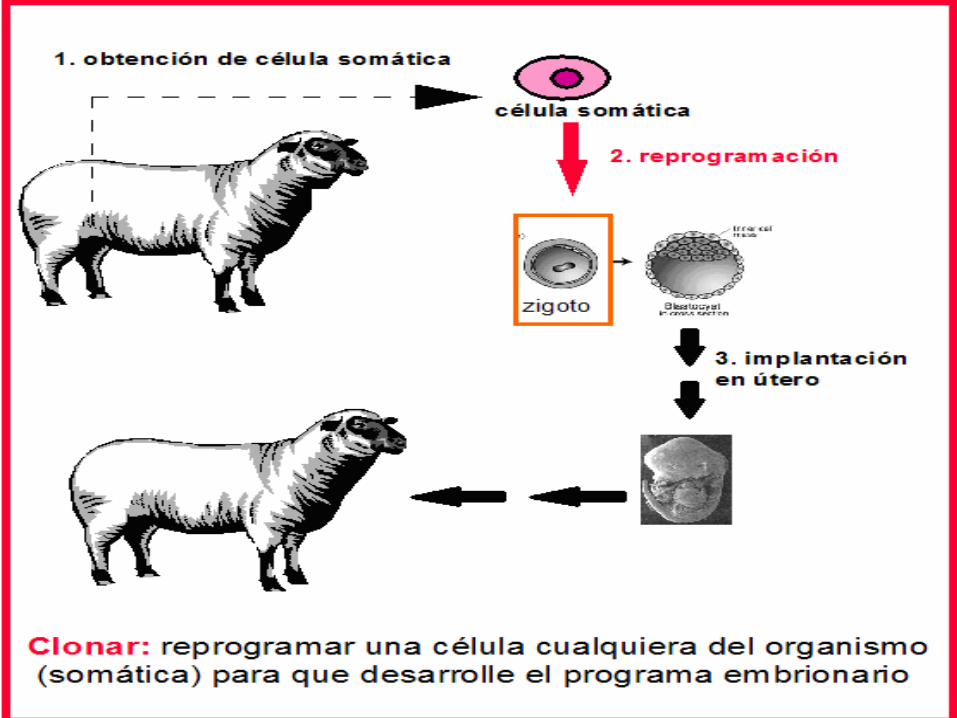


El proceso de obtención de Dolly fue muy costoso, y en la actualidad no se ha mejorado mucho. Dolly fue el único resultado positivo de 277 intentos, a partir de los cuales se consiguieron 29 embriones, muchos de estos no llegaron a desarrollarse y otros murieron al poco de nacer.
Con todo, Dolly fue un logro científico muy importante. Demostró que hay más de un modo de obtener nuevos animales. Por un lado tendríamos la reproducción natural, que es sexual y que produce diversidad; y, por otro, la clonación: una reproducción artificial, asexual, y que da lugar a individuos idénticos.

Por un lado tendríamos la reproducción natural, que es sexual y que produce diversidad; y, por otro, la clonación: una reproducción artificial, asexual, y que da lugar a individuos idénticos.




- copias idénticas de animales que nos interesan por diversos motivos:producción de leche, salud, longevidad...
- ampliar las posibilidades de manipulación genética
- investigación

el número de óvulos necesarios: hicieron falta más de 400 para conseguir a Dolly
El clonado carecería de las relaciones elementales de familia: no tendría en absoluto padre, ni propiamente hablando madre: tendría un hermando gemelo mayor, una madre ovular (¿citoplásmica?) y una madre de alquiler

El clonado sería generado con un fin: emular a alguien cuyas características interesan por algún motivo: un hijo fallecido al que se pretende sustituir, un genio cuyas habilidades interesa mantener, etc. Las consecuencias psicológicas de esa presión serían imprevisibles.
La posibilidad de curar enfermedades llevando a cabo transplantes no con órganos completos, sino con células, mediante la llamada terapia celular

La posibilidad de obtener células madre embrionarias. En el año 1998 dos grupos de Estados Unidos publicaron la obtención de células madre embrionarias a partir de embriones humanos que procedían de la fecundación in vitro
Nadie tiene derecho a la salud a cualquier precio, y menos si el precio es otra vida humana.

Existen alternativas a la clonación humana con fines terapéuticos que no presentan objeciones éticas tan serias. La más interesante es la posibilidad de conseguir células madre de origen no embrionario
También pueden obtenerse células madre del cordón umbilical y de la placenta del recién nacido, ya que proceden del embrión y sus células tampoco provocarían rechazo.

En definitiva: hay muchas vías terapéuticas que van haciéndose posibles por el desarrollo de la ciencia y que no vulneran el respeto debido a la vida humana en todas las fases de su desarrollo. Es deber de todos defender la vida humana y fomentar que se canalicen los esfuerzos de la investigación hacia lo que son verdaderos avances




