fedna anembe sobre alimentación del vacuno lechero · sobre alimentaciÓn de rumiantes madrid, 10...
TRANSCRIPT

Primera Jornada
FEDNA – ANEMBE
sobre alimentación del
vacuno lechero
ORGANIZAN:
PATROCINADORES:
BAYER – LUCTA S.A. – SETNA S.A.
Madrid, 10 de Noviembre de 2015


PROGRAMA
I Jornada FEDNA-ANEMBE 25 Aniversario
sobre Nutrición de Rumiantes Madrid, 10 de noviembre de 2015
Auditorio ETS Ingenieros Industriales, UPM
C/ José Gutiérrez Abascal 2, 28006 Madrid
10:00 h Inscripciones y entrega de documentación.
10:30 h Presentación de la jornada
Sesión 1: Nutrición. Moderador: A. González Garrido, ANEMBE.
10:45 h Importancia del laboratorio en la alimentación del vacuno lechero: valores de
referencia y nuevos parámetros. M. Hermida. Laboratorio Mouriscade.
11:45 h Descanso café.
12:15 h La importancia de los azúcares y sus tipos en la formulación de raciones para
los rumiantes. M. Fondevila.Universidad de Zaragoza.
13:15 h Últimos avances en nutrición vitamínico - mineral. Pedro Sayalero. Trouw.
14:15 h Comida.
15:45 h Proteínas, péptidos aminoácidos y otras fuentes nitrogenadas para la
formulación de raciones en rumiantes. M.D. Carro. UPM.
Sesión 2: Importancia del manejo en la eficiencia alimentaria. Moderador: A. González
Garrido, ANEMBE.
17:00 h Importancia del manejo alimentario sobre el cebo de terneros. M. Devant. IRTA.
18:00 Alimentación de rebaños lecheros con carro unifeed y robot de ordeño. Diego
Martínez del Olmo. Tolsa.
19:00 h Fin de la jornada.


CURRICULA PONENTES Dra. MARÍA HERMIDA FERRO
Licenciada en Farmacia por la Universidad de Santiago Compostela en 1988.. Tesis Doctoral “Aplicación de la espectrofotometría de infrarrojo cercano a la determinación de componentes mayoritarios en productos lácteos”. Universidad de Santiago Compostela 1996. Master en Prevención de riesgos Laborales, Especialidad Diputación de Pontevedra. Diploma de Directiva para empleados públicos de entidades locales de la provincia de Pontevedra . En la actualidad es Directora de Laboratorio, Funcionaría de carrera en el Laboratorio de Mouriscade (Diputación de Pontevedra)
---------------------------------------------------------------------------------------------------------- Dr. MANUEL FONDEVILA
Manuel Fondevila, Catedrático de Producción Animal en la Facultad de Veterinaria de la Universidad de Zaragoza, desde 2012. Ha trabajado fundamentalmente en valoración nutritiva y fisiología digestiva, así como en fermentación y actividad microbiana, tanto in vivo como in vitro, en rumiantes, porcino y conejos. Ha publicado más de 80 artículos científicos en revistas indexadas y 20 artículos de divulgación.
---------------------------------------------------------------------------------------------------------- D. PEDRO SAYALERO
Licenciado en Ciencias Biológicas, especialidad Biología Fundamental y en Veterinaria por la Universidad Complutense de Madrid. Hace gala de una trayectoria profesional extensa con más de 20 años como responsable del Departamento de Rumiantes de TROUW NUTRITION España. Cabe destacar que es miembro del ASC de Nutreco y responsable, actualmente, de la coordinación de alguno de los proyectos del RRC (Ruminant Research Center) de Nutreco.
---------------------------------------------------------------------------------------------------------- Dra. MARÍA DOLORES CARRO
Licenciada (1984) y Doctora (1989) en Veterinaria por la Universidad de León. Desde 1991 hasta 2012 fue profesora en la Facultad de Veterinaria de la Universidad de León y directora del grupo de investigación “Ecología Ruminal”. Desde finales de 2012 es profesora de la E.T,S.I. Agrónomos de la Universidad Politécnica de Madrid.

Ha desarrollado su actividad docente en el ámbito de la producción animalSu actividad investigadora se ha centrado en la nutrición de los rumiantes, valoración nutritiva y en la modificación de la fermentación ruminal para mejorar la eficiencia productiva y reducir su impacto medioambiental. Ha dirigido diversos proyectos de investigación y Tesis Doctorales, y es autora de numerosos artículos científicos. Ha sido editora de las revistas Animal Science, British Journal of Nutrition y Spanish Journal of Agricultural Research (2011- ). Actualmente es miembro del comité editorial de Frontiers in Microbiology. En los últimos diez años ha colaborado activamente con agencias estatales de evaluación científica (ANEP) y docente (ANECA).
---------------------------------------------------------------------------------------------------------- Dra. MARIA DEVANT
Licenciada en Veterinaria por la Universidad Autónoma de Barcelona, 1995; doctorado en Producción Animal por la misma Universidad en 2001. Actualmente es investigadora del Departamento de Rumiantes del IRTA. Posee amplia experiencia internacional en Investigación y Desarrollo en producción porcina y de rumiantes.
Con más de 15 años de experiencia en estudios para testar diferentes estrategias nutricionales y de manejo para mejorar la eficiencia productiva y la calidad de la carne en bovino. Sus principales líneas de investigación en cebo de terneros son la conducta animal y alimentaria, la fermentación ruminal y la calidad de la carne. Ha sido autora y coautora de más de 40 artículos en revistas internacionales indexadas y en más de 50 comunicaciones en congresos nacionales e internacionales. ---------------------------------------------------------------------------------------------------------- D. DIEGO MARTÍNEZ DEL OLMO
Licenciado en Veterinaria por la Universidad de León en 1999, Máster “Dirección estratégica y Management de empresas de Alimentación” por la Universidad Oberta de Cataluña, 2009, y Doctor en Veterinaria por la Universidad Complutense de Madrid, año 2012.
Actualmente Industrial Business Development Manager Animal Feed, en Tolsa, y con una amplia experiencia professional en Núter Feed S.A.U., y previamente en Grupo Leche Pascual, siempre ha mostrado una inquietud científica y docente, habiendo colaborado y trabajado en proyectos de investigación y en distintas Universidades y centros docentes. Posee diversas publicaciones en revistas especializadas dirigidas al sector productor y/o técnico y más de 100 ponencias en diferentes congresos técnicos españoles e internacionales.

I JORNADA FEDNA − ANEMBE 25 Aniversario
sobre ALIMENTACIÓN DE RUMIANTES
MADRID, 10 de Noviembre de 2015
INDICE
CAPÍTULO I
LA IMPORTANCIA DE LOS AZÚCARES EN LA ALIMENTACIÓN
DE LOS RUMIANTES
Manuel Fondevila
Instituto Agroalimentario de Aragón (IA2), Departamento de Producción Animal y
Ciencia de los Alimentos, Universidad de Zaragoza.
1.- Introducción ................................................................................................................... 1
2.- Los azúcares en los alimentos para los rumiantes ......................................................... 1
3.- Fermentación ruminal de los azúcares ........................................................................... 3
4.- Efecto de los azúcares solubles en dietas fibrosas ......................................................... 8
5.- Efecto de los azúcares solubles en dietas mixtas ......................................................... 10
6.- Referencias .................................................................................................................. 12
CAPÍTULO II
ÚLTIMOS AVANCES EN NUTRICIÓN VITAMÍNICO MINERAL
Pedro Sayalero
Trouw
1.- Introducción ................................................................................................................. 17
2.- Calcio ........................................................................................................................... 18
2.1.- Funciones ........................................................................................................ 19
2.2.- Modo de acción ............................................................................................... 19
2.3.- Absorción ........................................................................................................ 20
2.4.- Metabolismo .................................................................................................... 20
2.5.- Excreción ......................................................................................................... 21
3.- Hipocalcemia en vacas ................................................................................................ 21
4.- Uso de antagonistas del calcio para estimular la homeostasis ..................................... 22

I JORNADA FEDNA − ANEMBE 25 Aniversario
sobre ALIMENTACIÓN DE RUMIANTES
MADRID, 10 de Noviembre de 2015
5.- Metales traza ................................................................................................................ 25
5.1.- Funciones ........................................................................................................ 25
5.2.- Absorción de los metales traza ........................................................................ 27
5.3.- Regulación ....................................................................................................... 28
5.4.- Compuestos organicos de metales traza .......................................................... 29
6.- Vitaminas del grupo B ................................................................................................. 31
6.1.- Suplementacion ............................................................................................... 33
6.2.- Conclusiones sobre el uso de vitaminas del grupo B ...................................... 34
7.- Referencias .................................................................................................................. 34
CAPÍTULO III
PROTEÍNAS, PÉPTIDOS, AMINOÁCIDOS Y OTRAS FUENTES
NITROGENADAS PARA LA FORMULACIÓN DE RACIONES EN RUMIANTES
María Dolores Carro Travieso
Departamento de Producción Agraria de la Universidad Politécnica de Madrid
1.- Introducción ................................................................................................................. 39
2.- Degradación ruminal de las proteínas.......................................................................... 41
3.- Síntesis de proteína microbiana ................................................................................... 49
4.- Digestibilidad intestinal de la proteína ........................................................................ 55
5.- Utilización metabólica de los aminoácidos ................................................................. 56
6.- Utilización de aminoácidos protegidos ........................................................................ 60
7.- Comentarios finales ..................................................................................................... 64
8.- Referencias .................................................................................................................. 65
CAPÍTULO IV
LA IMPORTANCIA DEL MANEJO ALIMENTARIO
EN EL CEBO DE TERNEROS
M. Devant, A. Bach, A. Solé, B. Quintana, M. Verdú
Departamento Rumiantes IRTA
1.- Introducción ................................................................................................................. 71
2.- Forma presentación del concentrado ........................................................................... 74
2.1.- ¿Granulado o harina? ................................................................................................ 77

I JORNADA FEDNA − ANEMBE 25 Aniversario
sobre ALIMENTACIÓN DE RUMIANTES
MADRID, 10 de Noviembre de 2015
2.2.- ¿El tipo de cereal puede afectar los resultados? ....................................................... 77
2.3.- ¿La disponibilidad de paja puede afectar los resultados? ......................................... 77
2.4.- ¿Realmente la forma de presentación puede afectar al mermas? ¿en qué medida? . 78
2.5.- Calidad de granulación ............................................................................................. 79
2.6.- Migajas ..................................................................................................................... 80
3.- Tipo de comedero ........................................................................................................ 81
3.1.- Tipo de comedero y forma de presentación .............................................................. 82
4.- Conclusiones ................................................................................................................ 83
5.- Agradecimientos .......................................................................................................... 83
6.- Referencias .................................................................................................................. 84
CAPÍTULO V
ALIMENTACIÓN DE REBAÑOS LECHEROS CON CARRO UNIFEED Y
ROBOT DE ORDEÑO
Diego Martínez del Olmo
Tolsa S.A.
1.- ¿Realidad o ficción? .................................................................................................... 87
2.- Introducción; motivaciones, ventajas, desventajas y demandas .................................. 88
3.- Situación actual ............................................................................................................ 89
4.- Comportamiento nutricional en los sistemas automáticos de ordeño .......................... 90
5.- Estimular la frecuencia de ordeño a través de la formulación ..................................... 92
6.- Formular en función del tipo de tráfico en el AMS ..................................................... 95
7.- Otras consideraciones en el manejo con producciones basadas en AMS .................... 97
8.- Conclusiones ................................................................................................................ 98
9.- Referencias .................................................................................................................. 99


I JORNADA FEDNA − ANEMBE 25 Aniversario
sobre ALIMENTACIÓN DE RUMIANTES
MADRID, 10 de Noviembre de 2015
CAPÍTULO I
LA IMPORTANCIA DE LOS AZÚCARES EN LA
ALIMENTACIÓN DE LOS RUMIANTES
Manuel Fondevila
Instituto Agroalimentario de Aragón (IA2), Departamento de Producción Animal y
Ciencia de los Alimentos, Universidad de Zaragoza.
Miguel Servet 177, 50013, Zaragoza


LA IMPORTANCIA DE LOS AZÚCARES EN LA ALIMENTACIÓN DE LOS RUMIANTES 1
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
LA IMPORTANCIA DE LOS AZÚCARES EN LA ALIMENTACIÓN
DE LOS RUMIANTES
Manuel Fondevila
Instituto Agroalimentario de Aragón (IA2), Departamento de Producción Animal y Ciencia
de los Alimentos, Universidad de Zaragoza. Miguel Servet 177, 50013, Zaragoza
1.- INTRODUCCIÓN
Las dietas para los rumiantes incluyen entre un 5 y un 10% de azúcares, de los que
una fracción puede ser añadida a la dieta base para favorecer la utilización digestiva. El
concepto inicial de que la adición de azúcares solubles a la dieta basal se justifica
simplemente por favorecer la palatabilidad y por tanto la ingestión, ha ido matizándose:
por una parte, el sabor dulce es atractivo para el ganado vacuno, pero no para el ovino
(Ginane et al., 2011). Por otra parte, se barajan las hipótesis del efecto positivo de la
adición de azúcares sobre la fermentación ruminal, favoreciendo la sincronización entre la
utilización de la energía y el nitrógeno rápidamente degradable, y una producción de
ácidos grasos volátiles (AGV) favorable a una mayor síntesis de grasa láctea, aunque
también puede tener efectos negativos, como una mayor acidificación del rumen. Como
consecuencia, en la formulación de raciones para rumiantes se empieza a considerar el
nivel y tipo de azúcares incluidos en la dieta, aspectos que se discuten a continuación.
2.- LOS AZÚCARES EN LOS ALIMENTOS PARA LOS RUMIANTES
Los carbohidratos suponen más de un 70% del total de nutrientes aportados en la
dieta de los rumiantes. Entre ellos, las fracciones más estudiadas han sido tradicionalmente
los polisacáridos fibrosos, por su limitada y variable utilización, y el almidón como
polisacárido altamente fermentable en rumen. Sin embargo, los azúcares han adquirido un

2 M. FONDEVILA
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
interés creciente en la alimentación de estos animales. Se consideran como azúcares la
fracción de carbohidratos no estructurales solubles en agua o en etanol al 80% (Southgate,
1991). Entre ellos, se incluyen monosacáridos, fundamentalmente glucosa y fructosa,
disacáridos de origen vegetal como sacarosa (unidades de glucosa y fructosa) o animal
como lactosa (unidades de glucosa y galactosa) y, en menor medida, oligosacáridos de tres
y cuatro unidades, como rafinosa (galactosa, fructosa y glucosa), maltotriosa (tres unidades
de glucosa) y estaquiosa (dos unidades de galactosa unidas a glucosa y fructosa). El
contenido total de azúcares puede determinarse analíticamente de forma simple a partir de
su solubilidad en etanol al 80%, que extrae azúcares simples, disacáridos y oligosacáridos.
La extracción en agua puede incluir también algunos polisacáridos, como las fructanas o
fracciones de pectinas. No obstante, la identificación de los distintos monosacáridos
requiere análisis más complejos, como su determinación por métodos cromatográficos o
enzimáticos (Southgate, 1991).
El contenido en azúcares de los alimentos para el ganado varía considerablemente
en función de su naturaleza química (Tabla 1). Algunos subproductos, como la pulpa de
cítricos o las melazas de caña o remolacha tienden a tener contenidos considerablemente
altos de azúcares, mientras que la harina de soja, los guisantes y los forrajes, frescos o
conservados tienden a tener un contenido moderado, y los granos de cereales tienen un
contenido bajo. Los alimentos fermentados, como los ensilados y granos de destilería y
cervecería, también tienen un bajo contenido en azúcares, ya que éstos han sido
previamente fermentados por las enzimas microbianas. La variabilidad puede ser alta
incluso para un mismo tipo de alimento: Hall (2002) indica rangos entre 125 y 402 g/kg
materia seca (MS) para la pulpa de cítricos, y de 128 a 247 g/kg MS para la pulpa de
remolacha.
Tabla 1.- Contenido (g/kg MS) en azúcares y otros carbohidratos de distintos
alimentos (a partir de FEDNA 2010)
Azúcares1 Almidón Celulosa
2 Hemicelulosas
3 Pectinas
4
Cebada 18 566 58 119
Maíz 20 734 34 57
Harina de soja 80 6 77 64
Guisantes 40 482 76 53
Alfalfa deshidratada 36 6 244 128 80
Pulpa de cítricos 256 6 186 68 260
Pulpa de remolacha 67 0 236 221 210
Melaza de caña 624 0 0 0
Melaza de remolacha 612 0 0 0
1: Azúcares solubles, sin incluir oligosacáridos
2: Estimado por diferencia entre fibra ácido detergente y lignina sulfúrico
3: Estimada por diferencia entre fibra neutro detergente y fibra ácido detergente
4: Valores aproximados

LA IMPORTANCIA DE LOS AZÚCARES EN LA ALIMENTACIÓN DE LOS RUMIANTES 3
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
Así mismo, la composición de la fracción azucarada es diferente según el tipo o el
origen del alimento (Tabla 2). Los azúcares mayoritarios de las melazas de caña son la
sacarosa (65% del total), la fructosa (15%) y la glucosa (15%), mientras que las melazas de
remolacha apenas contienen fructosa, e incluyen un 65% de sacarosa y un 35% de glucosa
(Emanuele y Sniffen, 2014).
Tabla 2.- Composición (g/kg MS) de la fracción de azúcares de algunos alimentos
(Bach-Knudsen, 1997)
Azúcares
totales Monosacáridos Sacarosa Rafinosa Estaquiosa
Maíz 20 4 13 2 1
Cebada 21 4 12 5 1
Harina soja 137 7 70 10 47
Guisantes 881 9 30 5 23
Heno de hierba
(primer corte)
90 61 22 4 3
Harina de alfalfa 23 8 13 2 0
Pulpa remolacha 32 5 27 1 0
1: Incluye 22 g/kg verbascosa
3.- FERMENTACIÓN RUMINAL DE LOS AZÚCARES
Estudios in vitro con cepas puras de especies de microorganismos ruminales
muestran que la capacidad de fermentar diferentes azúcares solubles (monosacáridos y
disacáridos) es muy variable (Tabla 3). Por otra parte, a pesar de que los procesos de
fermentación de azúcares solubles y almidón a menudo se unifican por la similitud en el
ritmo de utilización de los sustratos, las especies microbianas sacarolíticas no
necesariamente tienen capacidad de fermentar almidón, por lo que tanto la población
especializada en la utilización de cada uno de estos nutrientes como las rutas metabólicas
implicadas, son diferentes.
La hidrólisis de disacáridos a azúcares simples es muy rápida, en parte porque su
alta solubilidad favorece una inmediata accesibilidad para los microorganismos. In vitro, el
ritmo de hidrólisis de la sacarosa varía entre 1200 y 1400% por hora, mientras el de la
lactosa oscila entre 200 y 250% por hora (Weisbjerg et al., 1998), variando la utilización
de glucosa y los otros monosacáridos liberados por dicha hidrólisis entre 300 y 700% por
hora. El Cornell Net Carbohydrate and Protein System asume un ritmo de fermentación
común para los azúcares de 300% por hora (Russell et al., 1992), lo que implica que serían
degradados en el rumen ya a los 20 minutos de su ingestión, y por tanto su utilización sería
prácticamente instantánea y completa. No obstante, la desaparición de un sustrato no
necesariamente implica su utilización, ya que asumiendo una tasa de tránsito de la fase

4 M. FONDEVILA
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
líquida ruminal de 0.14 por hora (Sutoh et al., 1996), un 5% de azúcares escaparían del
rumen sin ser fermentados (Oba, 2011). En condiciones in vivo con dietas altas en forraje,
Hristov et al. (2005) estimaron un ritmo de desaparición ruminal de azúcares del 124% por
hora, y un ritmo de fermentación de 115% por hora.
Tabla 3.- Capacidad de fermentación de carbohidratos solubles por microorganismos
ruminales (Stewart et al., 1997)
Glucosa Fructosa Galactosa Xilosa Sacarosa Almidón
Fibrobacter succinogenes
Prevotella ruminicola
Succinivibrio dextrinosolvens
Succinimonas amylolytica
Treponema bryantii
Selenomonas ruminantium
Lachnospira multiparus
Butyrivibrio fibrisolvens
Streptococcus bovis
Eubacterium ruminantium
Megasphaera elsdenii
Ruminobacter amylophilus
Ruminococcus bromii
+
+
+
+
+
+
+
+
+
+
+
-
d
-
+
d
-
-
+
+
+
+
+
+
-
+
-
+
+
+
+
d
+
+
-
-
-
-
-
-
d
d
-
+
+
d
d
-
d
-
-
-
+
d
d
-
+
d
+
d
+
d
d
-
-
-
d
-
+
-
+
d
d
+
+
+
+
Hongos ruminales + + - + + +
Dasytricha ruminantium
Isotricha spp.
Entodiniomorfos
+
+
+
+
+
-
+
+
-
-
-
+
+
+
+
d
d: variable en función de cepas/especies
Por otra parte, la fermentación ruminal de los monosacáridos depende también de
su estructura molecular, asumiéndose que un 84% de las hexosas se fermentan en el rumen,
mientras que la fermentación de las pentosas se considera inferior al 50% (Sniffen y
Tucker, 2011, citado por Emanuele y Sniffen, 2014). En condiciones in vitro, el ritmo de
utilización microbiana de arabinosa y xilosa es del 47 y 39%, respecto al ritmo medio de
utilización de las hexosas glucosa y fructosa y el disacárido sacarosa (Czerkawski y
Breckenridge, 1969). En el mismo sentido, la hidrólisis de oligosacáridos es más lenta, y a
menudo incompleta, y la rafinosa se utiliza 3 veces más lento que las mencionadas
hexosas. De hecho, el grado de fermentación de los distintos mono y disacáridos por las
especies bacterianas ruminales es variable (Russell y Baldwin, 1978, Tabla 4), por lo que
determinados azúcares favorecen el crecimiento de ciertas especies, afectando así el perfil
bacteriano del rumen. Las bacterias S. bovis y S. ruminantium, así como las bacterias
celulolíticas Ruminococcus albus y R. flavefaciens no utilizan las pentosas pero crecen
bien con celulosa, y R. amylophilus fermenta maltosa pero no glucosa (Stewart et al, 1997).

LA IMPORTANCIA DE LOS AZÚCARES EN LA ALIMENTACIÓN DE LOS RUMIANTES 5
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
Por todo ello, ritmos de fermentación entre 50 y 69% por hora (Henning et al., 1993) y
digestibilidades ruminales entre 45% para azúcares presentes en forrajes y 89% en
azúcares añadidos (Hoover et al., 2006) pueden ser más realistas. Además, el ritmo
fraccional de degradación de los azúcares solubles no es un valor fijo, sino que depende de
la actividad microbiana.
Tabla 4.- Ritmo de crecimiento específico (tiempo de duplicación de la biomasa, por
hora) de especies bacterianas sobre mono- y disacáridos como sustratos únicos
(Russell y Baldwin, 1978)
Glucosa Celobiosa Xilosa Sacarosa Maltosa
S. ruminantium 0,72 0,06 0,64 0,67 0,35
P. ruminicola 0,56 0,20 0,04 0,62 0,52
B. fibrisolvens 0,39 0,53 0,45 0,52 0,54
S. bovis 2,04 1,83 --- 2,10 1,85
M. elsdenii 0,45 --- --- 0,14 0,55
La composición de la dieta puede afectar a la fermentación de los azúcares. Así,
Weisbjerg et al. (1998) indican que, mientras la utilización microbiana de sacarosa no se
modifica por la adición de azúcares a la dieta basal, la de lactosa puede doblarse con la
adaptación de la población ruminal a la presencia de este disacárido. Kasperowicz et al.
(2014) no observan un efecto significativo de la inclusión de inulina como fuente de
fructosa, o sacarosa en la dieta sobre la actividad enzimática fructanolítica, sacarolítica o
amilolítica de la microbiota ruminal a las 4 horas de administración de la dieta.
La fermentación de los azúcares permite a la población ruminal la utilización de la
energía liberada para la síntesis de masa microbiana, liberando AGV, que son utilizados
como fuente de energía por el rumiante, así como CO2 y metano. En general, se considera
que la inclusión de azúcares en la dieta promueve una fermentación caracterizada por un
aumento de la proporción de butirato a expensas de acetato, con respuestas variables en
cuanto a la proporción de propionato, que puede aumentar (Lee et al., 2003; Hristov et al.,
2005) o permanecer constante (Heldt et al., 1999; Hoover et al., 2006). En algunos casos,
también la producción de ácidos grasos de cadena ramificada (isobutirato e isovalerato) se
ve reducida por la inclusión de azúcares solubles en sustitución de almidón (Vallimont et
al., 2004; Hristov et al., 2005). No obstante, el perfil de AGV puede variar en función del
tipo de azúcar: sin limitación en la disponibilidad de nitrógeno degradable, la proporción
de los AGV mayoritarios no difiere entre hexosas y sacarosa (Czerkawski y Breckenridge,
1969; Heldt et al., 1999), pero la proporción de acetato es mayor, y la de propionato
(Czerkawski y Breckenridge, 1969) o butirato (Sutton, 1968) menor, con pentosas.
La rápida fermentación de azúcares solubles también da lugar, en general, a
cantidades importantes de lactato (Strobel y Russell, 1986; Heldt et al., 1999), en mayor

6 M. FONDEVILA
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
medida a partir de hexosas y disacáridos que de pentosas (Cullen et al., 1986). No hay que
olvidar la importancia de posibles diferencias en las proporciones de estos catabolitos
microbianos sobre el metabolismo del rumiante, teniendo en cuenta el papel del butirato
como fuente energética de las células epiteliales ruminales e intestinales (Bergman, 1990),
o la capacidad acidificante del lactato en el rumen (Krause y Oetzel, 2006).
Como ya se ha comentado, el ritmo de fermentación microbiana en el rumen de los
azúcares es muy elevado, mayor que el de otros carbohidratos. Como consecuencia, el 83%
de la glucosa disponible es utilizado en las primeras 12 horas de incubación in vitro,
mientras que en ese tiempo sólo el 46% del almidón y el 60% de las pectinas son
fermentados (Figura 1). Estos resultados corresponden a unas condiciones de incubación in
vitro, a pH controlado y mantenido entre 6,2 y 6,7. No obstante, la fermentación
microbiana se ve afectada por el pH del medio, variando la producción de catabolitos en
función de éste. Así, a un pH de fermentación óptimo (6,7), la fermentación de azúcares
solubles después de 10 h de incubación rinde más acetato y butirato, mientras que
disminuye el lactato y no se afecta la proporción de propionato, respecto a un pH de 5,5.
Por otra parte, respecto a la fermentación de almidón, la sacarosa rinde más butírico a pH
6,7 y más lactato tanto a pH 6,7 como a 5,5 (Tabla 5, Strobel y Russel, 1986).
Figura 1.- Fermentación microbiana in vitro (ml de gas/g materia orgánica) de
carbohidratos puros, en comparación con dos forrajes (a partir de Fondevila et al.,
2002; Marinas et al., 2003)

LA IMPORTANCIA DE LOS AZÚCARES EN LA ALIMENTACIÓN DE LOS RUMIANTES 7
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
Tabla 5.- Fermentación bacteriana in vitro (10 h) de distintos carbohidratos puros, en
función del pH de incubación (Strobel y Russell, 1986)
Sustrato
pH final
%
fermentado
Acetato
mM
Propionato
mM
Butirato
mM
Lactato
mM
Sacarosa 6,7
5,5
92
81
4,7
1,7
2,1
1,1
1,1
0,7
3,7
8,3
Almidón 6,7
5,8
77
66
5,1
2,7
2,9
1,1
0,8
0,7
0,9
4,1
Xilano 6,7
5,9
54
42
3,6
2,9
1,6
1,2
0,2
0,3
N.D.
N.D.
Pectina 6,7
5,8
87
41
10,1
5,0
1,3
0,7
0,2
0,3
N.D.
N.D.
E.S.M. 0,30 0,16 0,03 0,23
N.D.: no determinado; E.S.M.: error estándar de la media
La rápida disponibilidad de energía que supone la fermentación de los azúcares
permite también la captación del nitrógeno rápidamente degradable disponible para la
síntesis microbiana. Una de las posibles ventajas de aportar una fuente de energía muy
rápidamente fermentable, como los azúcares, pudiera estar relacionada con su
sincronización con la utilización de nitrógeno, en dietas ricas en proteína rápidamente
degradable o nitrógeno no proteico. En teoría, la liberación en el rumen de energía
fermentable que coincida en el tiempo con la disponibilidad de una proporción importante
del nitrógeno de la dieta, evitaría el exceso de amoníaco en el rumen y su absorción a
través de la pared, con el consiguiente gasto energético para su excreción en forma de urea,
al mismo tiempo que favorecería la síntesis microbiana y por tanto el aporte de proteína
preformada al intestino (Chamberlain y Choung, 1995; Sinclair, 2008). A pesar del
atractivo de la teoría, y la clara reducción en la concentración de amoníaco ruminal en las
primeras horas postingestión (Obara et al., 1991; Chamberlain et al., 1993), los resultados
obtenidos a nivel experimental son contradictorios, y no reflejan una respuesta claramente
positiva en términos de síntesis microbiana, debido al rápido ritmo de reciclaje del
nitrógeno ruminal y, en parte, a la gran variabilidad de parámetros relacionados con la
síntesis microbiana, que dificulta las conclusiones a nivel experimental (Hall y Huntington,
2008).
Por otra parte, en situaciones de elevada disponibilidad de sustrato, Czerkawski y
Breckenridge (1969) observaron que, después de 4,5 h de incubación in vitro, un 39% de la
glucosa obtenida a partir de la hidrólisis de sacarosa se incorpora a la masa microbiana en
forma de glucógeno, como sustancia de reserva. Por el contrario, se ha justificado cierta
ineficiencia por desacoplamiento energético (energy spilling, Strobel y Russell, 1986;
Russell, 1998) debido al mayor ritmo de disponibilidad de energía en relación al de
nitrógeno para la síntesis microbiana. Este proceso tiene lugar en condiciones de acentuado

8 M. FONDEVILA
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
descenso de pH, en que una parte de la energía obtenida se deriva de procesos de
crecimiento a otros procesos, dado que el mantenimiento de un pH intracelular neutro
requiere un coste energético de expulsión de protones, disipándose finalmente en forma de
calor. Cuando la disponibilidad en el rumen de sustratos rápidamente fermentables, como
los azúcares solubles, es alta, bacterias como S. bovis y S. ruminantium cambian su patrón
de fermentación de la producción de acetato, propionato y formiato a la de lactato,
pudiendo provocar un descenso de pH. Si en dicha situación la disponibilidad de amoníaco
es alta, el cambio de patrón de fermentación ocurre a un ritmo más lento, pudiendo
compensar el aumento de la producción de lactato.
La presencia de azúcares en la dieta interactúa con otros componentes de la dieta,
como la proporción de otros carbohidratos rápidamente fermentables, de fibra o de
nitrógeno degradable, modulando su efecto tanto a nivel de rumen como en la respuesta
productiva del rumiante. Para facilitar la comprensión de la posible respuesta a la presencia
de azúcares solubles sobre la utilización digestiva y la respuesta productiva de los
animales, se van a agrupar a continuación en función del tipo de dieta, de alto contenido en
fibra o rica en concentrados.
4.- EFECTO DE LOS AZÚCARES SOLUBLES EN DIETAS FIBROSAS
Los forrajes frescos incluyen cantidades moderadamente altas de azúcares solubles
(entre el 10 y el 16%). Se ha observado que variedades de forrajes seleccionadas
genéticamente para un alto contenido en azúcares, hasta niveles que pueden superar el
20%, promueven un aumento de butirato y propionato a expensas de acetato, junto con una
reducción de la concentración ruminal de amoníaco y un aumento del flujo de nitrógeno no
amoniacal al duodeno (Lee et al., 2002). Cuando el contenido de azúcares en el forraje
incrementa a costa de la proporción de proteína o de una reducción conjunta de proteína y
fibra, la mejor utilización del nitrógeno se manifiesta en una menor excreción de nitrógeno
urinario (Ellis et al., 2011). Por otra parte, estudiando una simulación de la respuesta en
producción de metano a partir de resultados en la bibliografía, Ellis et al. (2012) muestran
que si el aumento de la proporción de azúcares solubles es a expensas del contenido en
fibra, el aumento de la producción de metano por unidad de sustrato fermentado se diluye
al expresarlo en base a la producción de leche, porque ésta aumenta en mayor medida. No
obstante, la respuesta en la producción de metano a la concentración de azúcares del
forraje parece variable, habiéndose observado en algunos casos reducciones (Lovett et al.,
2006) o ausencia de efecto (Staerfl et al., 2012) en la producción de metano. En cualquier
caso, la extrapolación de los resultados observados a un efecto positivo de una mayor
fracción de azúcares solubles debe realizarse con precaución, ya que el aumento de la
proporción de azúcares tiene lugar a costa de la disminución proporcional de la proporción
de pared celular (Oba, 2011).
Una situación diferente es la suplementación de un alimento fibroso con una fuente
de azúcares solubles. Kasperowicz et al. (2014) y Obara et al. (1991) no observaron efectos

LA IMPORTANCIA DE LOS AZÚCARES EN LA ALIMENTACIÓN DE LOS RUMIANTES 9
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
de la inclusión de sacarosa a niveles de 8 y 15%, respectivamente, sobre la concentración
de AGV ni el pH, aunque sí un aumento de las proporciones de propionato y butirato en
ovino. En condiciones in vitro, Lee et al. (2003) observaron que niveles de 5 y 8% de
inulina y sacarosa como fuentes de azúcares solubles aumenta la concentración de AGV y
la proporción de propionato, acompañado de un descenso del pH ruminal y una reducción
de la digestibilidad de la fracción fibrosa. No obstante, el aumento de la disponibilidad de
energía favorece la utilización microbiana del nitrógeno, disminuyendo la concentración de
amoníaco y aumentando la síntesis microbiana (Tabla 6; Chamberlain et al., 1993). Este
efecto está ligado a menudo a la combinación entre la suplementación con azúcares y la
incorporación de una fuente de nitrógeno no proteico, debido a la dependencia de las
bacterias fibrolíticas del amoníaco como fuente de nitrógeno, observándose por tanto una
respuesta sinérgica al aporte de ambos sobre la síntesis microbiana (Chamberlain et al.,
1993; Heldt et al., 1999).
Independientemente de la contribución energética del azúcar añadido a la energía
disponible en el rumen, la respuesta a la suplementación con azúcares de dietas con un
elevado contenido en fibra se manifiesta fundamentalmente sobre la fermentación de los
polisacáridos estructurales. Este efecto puede depender tanto de la naturaleza del
carbohidrato añadido y del nivel de inclusión como de la interacción de ambos sobre las
condiciones ambientales, fundamentalmente el pH. Huhtanen y Khalili (1992) observaron
un descenso de la actividad fibrolítica celulasa y xilanasa al suplementar con un 16% de
sacarosa una dieta con un 70% de silo de maíz, desapareciendo este efecto con la adición
de bicarbonato sódico. Estos autores achacaron la respuesta a la reducción de la adhesión
bacteriana al sustrato, fundamental para el desarrollo de la actividad fibrolítica, por debajo
de pH 6,2, e incluso a la inhibición del crecimiento de las especies fibrolíticas a pH
inferiores. Por el contrario, Piwonka y Firkins (1993) indican que la inclusión de un 10%
de glucosa en el medio no afecta negativamente al ritmo de digestión in vitro de celulosa
purificada, pero sí cuando se incorpora más de un 23% de glucosa, siendo este efecto
independiente del pH si éste se mantiene por encima de 6,0. Tampoco Ortigues et al.
(1988) observaron variaciones en la digestibilidad de la fibra de un heno al suplementar
con 6,5% de melazas, ni tampoco sobre el pH ruminal ni la concentración total de AGV,
aunque la proporción de propionato aumentó con la inclusión de melazas. England y Gill
(1985) sugieren que niveles de sacarosa por debajo de 7,5% favorecen la degradabilidad
ruminal, mientras que el efecto contrario se observa a niveles superiores.
En cuanto al efecto de la naturaleza del sustrato fibroso, Barrios Urdaneta et al.
(2000) demostraron que la adhesión bacteriana a un sustrato de baja calidad (pared celular
de paja de cereal), y con ella su degradación, aumentan al suplementar con un 35% de
sacarosa, almidón o pectina, cuando se mantiene el pH del medio entre 6,1 y 6,3.
Tabla 6.- Efecto de la suplementación de un silo de hierba (4 kg/d) con 200 g/d de
distintos carbohidratos no estructurales sobre el pH, la concentración ruminal de

10 M. FONDEVILA
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
nitrógeno amoniacal (mg/L) y el aporte de nitrógeno microbiano al duodeno (g/d)
(Chamberlain et al., 1993)
Silo de
hierba + sacarosa + fructosa + almidón + lactosa
pH ruminal:
medio
mínimo
6,43
6,04
6,34
5,72
6,31
5,74
6,25
5,99
6,40
6,05
N amoniacal:
medio
máximo
255
354
157
240
164
233
213
288
158
233
Aporte N microbiano 10,2 14,8 13,7 11,9 14,3
5.- EFECTO DE LOS AZÚCARES SOLUBLES EN DIETAS MIXTAS
En dietas que incorporan un nivel medio o alto de concentrado, la problemática de
la incorporación de azúcares solubles se enfoca, además de al propio nivel de inclusión, a
la sustitución de almidón como carbohidrato altamente digestible en condiciones de pH por
debajo de 6,5. En este sentido, la sustitución de almidón por sacarosa no afecta (Broderick
et al., 2008) o incluso tiende a aumentar (Chamberlain et al., 1993; Heldt et al., 1999) el
pH ruminal. En parte, esto es debido a que la sacarosa aporta, por unidad de peso, menos
energía que el almidón, al tener una menor proporción de carbono (Broderick y Radloff,
2004). Por otro lado, si los azúcares aumentan el ritmo de tránsito (Sutoh et al., 1996) y la
síntesis microbiana (Chamberlain et al., 1993), la proporción de material disponible para su
fermentación en el rumen será menor que en dietas con una proporción equivalente de
almidón.
En un experimento in vitro con dietas a base de un 60% de forraje, la inclusión de
un 7,5% de sacarosa en sustitución de almidón promovió un aumento de la digestibilidad
de la fibra (Vallimont et al., 2004). Aunque tasas de inclusión de sacarosa más bajas no
afectaron a la concentración de amoníaco ni de AGV, se observó un aumento lineal de la
proporción de butirato. En un experimento paralelo in vivo (Broderick et al., 2000), se
observó un descenso en la relación acetato:propionato y un aumento en la digestibilidad de
la fibra, que presentó una tendencia cuadrática, alcanzando el máximo con un 5% de
sacarosa. Martel et al. (2011) observaron un aumento de la proporción de butirato, aunque
en este caso también un descenso de la de propionato, al sustituir un 5% del almidón por
melazas en la dieta de vacas lecheras. Por el contrario, tanto Broderick y Radloff (2004)
como Broderick et al. (2008), con dietas para vacas lecheras con niveles similares de
forraje, no observaron un efecto sobre los parámetros ruminales al sustituir maíz por una
fuente de azúcares (melazas o sacarosa, respectivamente), aunque detectaron mejoras en la
digestibilidad de la fibra.
LA IMPORTANCIA DE LOS AZÚCARES EN LA ALIMENTACIÓN DE LOS RUMIANTES 11
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
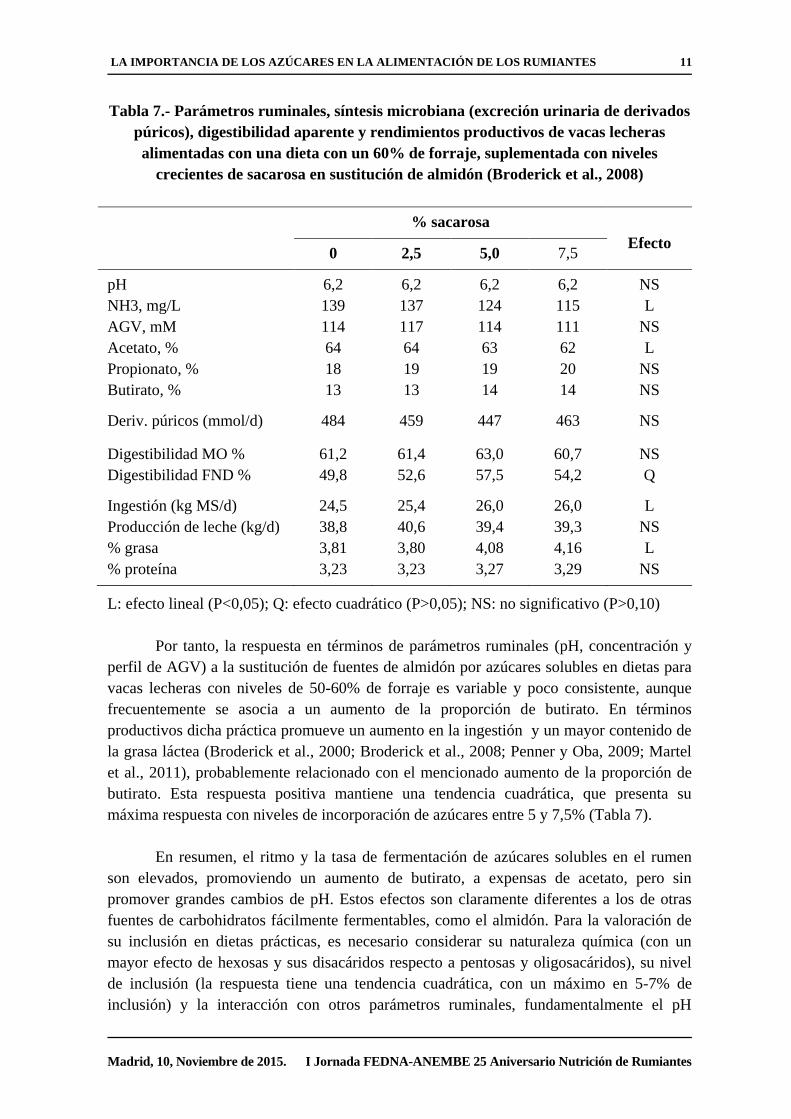
Tabla 7.- Parámetros ruminales, síntesis microbiana (excreción urinaria de derivados
púricos), digestibilidad aparente y rendimientos productivos de vacas lecheras
alimentadas con una dieta con un 60% de forraje, suplementada con niveles
crecientes de sacarosa en sustitución de almidón (Broderick et al., 2008)
% sacarosa
Efecto 0 2,5 5,0 7,5
pH
NH3, mg/L
AGV, mM
Acetato, %
Propionato, %
Butirato, %
6,2
139
114
64
18
13
6,2
137
117
64
19
13
6,2
124
114
63
19
14
6,2
115
111
62
20
14
NS
L
NS
L
NS
NS
Deriv. púricos (mmol/d) 484 459 447 463 NS
Digestibilidad MO %
Digestibilidad FND %
61,2
49,8
61,4
52,6
63,0
57,5
60,7
54,2
NS
Q
Ingestión (kg MS/d)
Producción de leche (kg/d)
% grasa
% proteína
24,5
38,8
3,81
3,23
25,4
40,6
3,80
3,23
26,0
39,4
4,08
3,27
26,0
39,3
4,16
3,29
L
NS
L
NS
L: efecto lineal (P<0,05); Q: efecto cuadrático (P>0,05); NS: no significativo (P>0,10)
Por tanto, la respuesta en términos de parámetros ruminales (pH, concentración y
perfil de AGV) a la sustitución de fuentes de almidón por azúcares solubles en dietas para
vacas lecheras con niveles de 50-60% de forraje es variable y poco consistente, aunque
frecuentemente se asocia a un aumento de la proporción de butirato. En términos
productivos dicha práctica promueve un aumento en la ingestión y un mayor contenido de
la grasa láctea (Broderick et al., 2000; Broderick et al., 2008; Penner y Oba, 2009; Martel
et al., 2011), probablemente relacionado con el mencionado aumento de la proporción de
butirato. Esta respuesta positiva mantiene una tendencia cuadrática, que presenta su
máxima respuesta con niveles de incorporación de azúcares entre 5 y 7,5% (Tabla 7).
En resumen, el ritmo y la tasa de fermentación de azúcares solubles en el rumen
son elevados, promoviendo un aumento de butirato, a expensas de acetato, pero sin
promover grandes cambios de pH. Estos efectos son claramente diferentes a los de otras
fuentes de carbohidratos fácilmente fermentables, como el almidón. Para la valoración de
su inclusión en dietas prácticas, es necesario considerar su naturaleza química (con un
mayor efecto de hexosas y sus disacáridos respecto a pentosas y oligosacáridos), su nivel
de inclusión (la respuesta tiene una tendencia cuadrática, con un máximo en 5-7% de
inclusión) y la interacción con otros parámetros ruminales, fundamentalmente el pH

12 M. FONDEVILA
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
ruminal esperable por la fermentación y la disponibilidad de nitrógeno no proteico, y
dietéticos, como el nivel y tipo de fibra.
6.- REFERENCIAS
BACH KNUDSEN K.E. (1997) Anim. Feed Sci. Technol. 67: 319-338.
BARRIOS URDANETA, A, FONDEVILA, M., BALCELLS, J., DAPOZA, C. y
CASTRILLO, C. (2000) Austr. J. Dairy Sci. 51: 393-400.
BERGMAN, E.N. (1990) J. Dairy Sci. 70: 567-590.
BRODERICK, G.A. y RADLOFF, W.J. (2004) J. Dairy Sci. 87: 2997-3009.
BRODERICK, G.A, LUCHINI, N.D., SMITH, W.J., REYNAL, S., VARGA, G.A. y
ISHLER, V.A. (2000) J. Dairy Sci. 83 (Suppl. 1): 248.
BRODERICK, G.A, LUCHINI, N.D., SMITH, W.J., REYNAL, S., VARGA, G.A. y
ISHLER, V.A. (2008) J. Dairy Sci. 91: 4801-4810.
CHAMBERLAIN, D.G. y CHOUNG, J.J. (1995) En: Recent Advances in Animal
Nutrition, P.C. Garnsworthy, D.J.A. Cole (eds.), Nottingham University Press,
Nottingham, pp: 3-27.
CHAMBERLAIN, D.G., ROBERTSON, S. y CHOUNG, J.J. (1993) J. Sci. Food Agric.
63: 189-194.
CULLEN, A.J., HARMON, D.L. y NAGARAJA, T.G. (1986) J. Dairy Sci. 69: 2616-2621.
CZERKAWSKI, J.W. y BRECKENRIDGE, G. (1969) Br. J. Nutr. 23: 925-937.
ELLIS, J.L., DIJKSTRA, J., BANNINK, A., PARSONS, A.J., RASMUSSEN, S.,
EDWARDS, G.R., KEBREAB, E. y FRANCE, J. (2011) J. Dairy Sci. 94: 3105-
3118.
ELLIS, J.L., DIJKSTRA, J., FRANCE, J., PARSONS, A.J., EDWARDS, G.R.,
RASMUSSEN, S., KEBREAB, E. y BANNINK, A. (2011) J. Dairy Sci. 95: 272-
285.
EMANUELE, S.M. y SNIFFEN, C.J. (2014) En: 2014 Penn State Dairy Cattle Nutrition
Workshop, pp: 99-107.
ENGLAND, P. y GILL, M. (1985) Anim. Prod. 40: 259-266.
FEDNA (2010) Tablas FEDNA de composición y valor nutritivo de alimentos para la
fabricación de piensos compuestos, 3ª ed, C. de Blas, G.G. Mateos y P. García
Rebollar (eds.), Fundación Española para el Desarrollo de la Nutrición Animal,
Madrid.
FONDEVILA, M., BARRIOS-URDANETA, A., BALCELLS, J. y CASTRILLO, C.
(2002) Anim. Feed Sci. Technol. 101: 1-15.
GINANE, C., BAUMONT, R. y FAVREAU-PEIGNÉ, A. (2011. Physiol. Behav. 104:
666-674.
HALL, M.B. 2002) En: Proceedings of the 13th Annual Florida Ruminant Nutrition
Symposium, Gainesville, FL, pp:146-158.
HALL, M.B. y HUNTINGTON, G.B. (2008) J. Anim. Sci. 86: E287-E292.
HELDT, J.S., COCHRAN, R.C., STOKKA, G.L., FARMER, C.G., MATHIS, C.P.,
TITGEMEYER, E.C. y NAGARAJA, T.G. (1999) J. Anim. Sci. 77: 2793-2802.

LA IMPORTANCIA DE LOS AZÚCARES EN LA ALIMENTACIÓN DE LOS RUMIANTES 13
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
HENNING, P.N., STEYN, D.G. y MEISSNER, H.H. (1993) J. Anim. Sci. 71: 2516-2528.
HOOVER, W.H., TUCKER, C., HARRIS, J., SNIFFEN, C.J. y DE ONDARZA, M.B.
(2006) Anim. Feed Sci. Technol. 128: 307-319.
HRISTOV, A.N., ROPP, J.K., GRANDEEN, K.L., ABERDI, S., ETTER, R.P., MELGAR,
A. y FOLEY, A.E. (2005) J. Anim. Sci. 83, 408-421.
HUHTANEN, P. y KHALILI, H. (1992) Br. J. Nutr. 67: 245–255.
KASPEROWICZ, A., STAN-GLASEK, K., KOWALIK, B., VANDZUROVA, A.,
PRISTAS, P., PAJAK, J., KWIATOWSKA, E. y MICHALOWSKI, T. (2014) Anim.
Feed Sci. Technol. 195: 38-46.
KRAUSE, K.M. y OETZEL, G.R. (2006) Anim. Feed Sci. Technol. 126: 215-236.
LEE, M.R.F., HARRIS, L.J., MOORBY, J.M., HUMPHREYS, M.O., THEODOROU,
M.K., MACRAE, J.C. y SCOLLAN, N.D. (2002) Anim. Sci. 74: 587-596.
LEE, M.R.F., MERRY, R.J., DAVIES, D.R., MOORBY, J.M., HUMPHREYS, M.O.,
THEODOROU, M.K., MACRAE, J.C. y SCOLLAN, N.D. (2003) Anim. Feed Sci.
Technol. 104: 59-70.
LOVETT, D.K., MCGILLOWAY, D., BORTOLOZZO, A., HAWKINS, M., CALLAN,
J., FLYNN, B. y O´MARA, F.P. (2006) Grass Forage Sci. 61: 9-21.
MARINAS, A., GARCÍA-GONZÁLEZ, R. y FONDEVILA, M. (2003) Anim. Sci. 76:
461-469.
MARTEL, C.A., TITGEMEYER, E.C., MAMEDOVA, L.K. y BRADFORD, B.J. (2011)
J. Dairy Sci. 94: 3995-4004.
OBA, M. (2011) Can. J. Anim. Sci. 91: 37-46.
OBARA, Y., DELLOW, D.W. y NOLAN, J.V. (1991) En: Physiological aspects of
digestion and metabolism in ruminants, T. Tsuda, Y Sasaki, R. Kawashima (eds.),
Academic Press, San Diego CA, pp: 515-539.
ORTIGUES, I., FONTENOT, J.P. y FERRY, J.G. (1988) J. Anim. Sci. 66: 975-985.
PENNER, G.B. y OBA, M. (2009) J. Dairy Sci. 92: 3341-3353.
PIWONKA, E.J. y FIRKINS, J.L. (1993) J. Dairy Sci. 79: 2196-2206.
RUSSELL, J.B. (1998) J. Anim. Sci. 76: 1955-1963.
RUSSELL, J.B. y BALDWIN (1978). Appl. Environ. Microbiol. 36: 319-329.
RUSSELL, J.B., O´CONNOR, J.D., FOX, D.G., VAN SOEST, P.J. y SNIFFEN, C.J.
(1992) J. Anim. Sci. 70: 3551-3561.
SINCLAIR, L.A. (2008) En: Gut efficiency: the key ingredient in ruminant production, S.
Andrieu, D. Wilde (eds.), Wageningen Academic Publishers, Wageningen, pp: 61-
78.
SOUTHGATE, D.A.T. (1991) Determination of food carbohydrates, 2nd
ed., Elsevier
Applied Science, Barking, UK.
STAERFL, S.M., AMELCHANKA, S.L:, KALBER, T., SOLIVA, C.S., KREUZER, M. y
ZEITZ, J.O. (2012) Livest. Sci. 150: 293-301.
STEWART, C.S., FLINT, H.J. y BRYANT, M.P. (1997) En: The rumen microbial
ecosystem, P.N. Hobson, C.S. Stewart (eds.), Blackie Academic & Professional,
London, pp: 10-72
STROBEL, H.J. y RUSSELL, J.B. (1986) J. Dairy Sci. 69: 2941-2947.

14 M. FONDEVILA
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
SUTOH, M., OBARA, Y. y MIYAMOTO, S. (1996. J. Agric. Sci. Camb. 126: 99-105.
SUTTON, J.D. (1968) Br. J. Nutr. 22: 689-712.
VALLIMONT, J.E., BARGO, F., CASSIDY, T.W., LUCHINI, N.D., BRODERICK, G.A.
y VARGA, G.A. (2004) J. Dairy Sci. 87: 4221-4229.
WEISBJERG, M.R., HVELPLUND, T. y BIBBY, B.M. (1998) Acta Agric. Scand., A, 48:
12-18.

I JORNADA FEDNA − ANEMBE 25 Aniversario
sobre ALIMENTACIÓN DE RUMIANTES
MADRID, 10 de Noviembre de 2015
CAPÍTULO II
ÚLTIMOS AVANCES EN NUTRICIÓN
VITAMÍNICO MINERAL
Pedro Sayalero
Trouw


ÚLTIMOS AVANCES EN NUTRICIÓN VITAMÍNICO MINERAL 17
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
ÚLTIMOS AVANCES EN NUTRICIÓN VITAMÍNICO MINERAL
Pedro Sayalero
Trouw
1.- INTRODUCCIÓN
La nutrición mineral del ganado vacuno de leche trata de equilibrar las necesidades
de los elementos minerales, según su estado fisiológico, nivel de producción… etc. y los
aportes que contiene la ración diaria. Las necesidades pueden determinarse usando un
método factorial, es decir, calculando las cantidades necesarias para reemplazar pérdidas
estimadas en orina, heces, leche, etc (Beaton, 1996) o con estimaciones de la dieta,
alimentando con diferentes concentraciones de un determinado mineral para ver sus
efectos (crecimiento, concentración en tejidos, producción…). Como no todo el mineral
suministrado se absorbe completamente se usan los coeficientes de absorción para las
diferentes fuentes minerales.
El NRC 2001 recomienda unos aportes minerales netos según el estado productivo
y las materias primas de la ración, lo que supone un avance respecto a las recomendaciones
como fracción de la materia seca ingerida, que es como se trabajaba anteriormente.
Después el sistema holandés (CVB, 2005) también recomienda unos aportes netos como
hace el sistema americano.
La nutrición mineral moderna va un poco más allá, no se detiene en aportar las
cantidades que marcan los distintos sistemas (NRC, 2001; INRA, 2003; CVB 2005) para
satisfacer unas necesidades concretas sino que busca el papel fisiológico de los minerales
sobre todo en situaciones que suponen un reto para los animales. Dentro de los minerales,
el calcio, por su directa implicación en problemas metabólicos tras el parto, sigue
acaparando la atención de los investigadores con el objetivo de establecer estrategias de
prevención de la hipocalcemia. La limitación del calcio disponible en la ración para que el
sistema de absorción intestinal responda rápidamente, vuelve a ser una alternativa a
considerar.

18 P. SAYALERO
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
En los oligoelementos, los metales de transición son quizás los que más
variabilidad presentan en su disponibilidad dada su gran predisposición a interaccionar con
otros minerales o por la presencia de antagonistas (Spears, 2003), por lo que merecen
atención no solo en cuanto a cantidades a aportar sino también a la forma de añadirlos en
la ración diaria, especialmente en momentos delicados para la vaca.
Por otro lado, las recomendaciones en cuanto a uso de vitaminas han estado sujetas
a modificaciones y cada vez es mayor el nivel propuesto para alguna de ellas e incluso ya
hay recomendaciones de vitaminas hidrosolubles (vitaminas del grupo B) asumiendo que
en determinadas condiciones la síntesis de éstas por los microorganismos del rumen puede
no ser suficiente (Santchi, 2005).
Hablaremos en este trabajo de los minerales que han tenido recientemente
novedades en cuanto a su uso en alimentación de vacas de leche, así como las novedades
en cuanto al uso de vitaminas hidrosolubles del grupo B.
2.- CALCIO
El Calcio es el mineral más abundante en el organismo. El 99% tiene función
estructural y se localiza en el esqueleto y el resto está en el líquido extracelular y en
distintos tejidos.
Calcio extracelular
Se encuentra en tres formas distintas:
1.- Unido a proteínas plasmáticas (albúminas y globulinas), su unión depende del pH
siendo menor si el pH desciende. Es el 40% del extracelular.
2.- Combinado con citrato y fosfato en el líquido extracelular. Puede difundir pero no está
ionizado. Es el 10% del extracelular.
3.- Ionizado. Capaz de difundir, lo que hace que sea la forma más activa. Es el 50% del Ca
plasmático total.
Calcio óseo
El hueso tiene una matriz orgánica compuesta por colágeno y proteoglicanos (35%)
y componentes inorgánicos (65%) que son calcio y fosfatos sobre todo. Este calcio óseo se
encuentra de dos maneras:
1.- Intercambiable o difusible, lo que le hace ser fácilmente depositado y movilizado. Son
cristales de fosfato monocálcico. Es sólo el 1% del calcio óseo.
2.- Calcio en depósito. Representa el 99% del total óseo. Son cristales de hidroxiapatito
con iones de sodio, magnesio, carbonatos y citratos.

ÚLTIMOS AVANCES EN NUTRICIÓN VITAMÍNICO MINERAL 19
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
2.1.- Funciones
1.- Estructural. En huesos y dientes. Representa el 99% del total corporal.
2.- Mensajero intracelular. Su radio iónico y la capacidad de formar enlaces de
coordinación (reversibles) con varios oxígenos (de los aminoácidos aspártico y glutámico)
le permite localizarse en los pliegues de proteínas ayudando a configurar la estructura de
las proteínas, facilitando así la funcionalidad de éstas. El calcio implicado sería calcio
ionizado (transmisor biológico más común que se conoce).
3.- Cofactor proteico. Activa distintos enzimas como la piruvato deshidrogenasa,
alfacetoglutarato deshidrogenasa, mieloproxidasa… y es también necesario en el proceso
de coagulación.
Algunas proteínas dependientes de calcio
Proteína Función
Calbindina Depósito y transporte de calcio
Caldesmón Contracción muscular
Troponina Contracción muscular
Calsecuestrinas Depósitos de calcio
Fosfolipasa A Síntesis de Ac. Araquidónico
Proteinquinasa C Fosforilación de proteínas
Calmodulina Regulación de Proteinquinasas
2.2.- Modo de acción
En condiciones de reposo en el interior de la célula, en el citosol, hay muy poco
calcio lo cual es lógico porque en caso contrario el calcio podría interaccionar con
proteínas dependientes de él, activándolas continuamente lo que conduciría a la muerte
celular, o bien interaccionaría con el fósforo citosólico precipitando y bloqueando así los
mecanismos energéticos de la célula que dependen de compuestos fosforilados (ATP). El
que la concentración en el citosol sea tan baja se debe a que la membrana celular limita la
permeabilidad del calcio y a que hay una eliminación desde el citosol al exterior por medio
de una ATPasa. Además se secuestra calcio hacia el retículo endoplásmico gracias a otra
ATPasa, fijándose a proteínas que lo retienen y almacenan para un uso posterior.
Cuando llega la señal a la célula (física, química o eléctrica) se abre un canal
específico para el calcio lo que permite su entrada. También se activa una fosfolipasa de la
membrana celular que hidroliza fosfatidilinositol produciendo inositoltrifosfato y
diacilglicerol. El primero provoca la liberación del calcio que estaba almacenado en el
retículo endoplásmico y el segundo, que tarda en actuar, lo que hace es sacar calcio al
exterior o provocar su entrada en el retículo endoplásmico de nuevo para volver a la
situación de reposo.

20 P. SAYALERO
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
2.3.- Absorción
La absorción del calcio se produce en el duodeno y yeyuno proximal y tiene lugar
de dos formas:
Paracelular. Sistema no saturable, se trata de difusión pasiva.
Transcelular. Es un mecanismo saturable muy bien regulado. El calcio entra por
canales específicos que se activan por mediación de la vitamina D3. Se han
identificado esos canales (TPRV6/5) así como la proteína transportadora que hay
en el interior del enterocito que, además, evita que el calcio interaccione con el
fósforo del citosol y con otras proteínas, es la Calbindina D9/28K.
Martín-Tereso y Verstegen. Nutrition Research Rerviews, 2011.
La vitamina D3 regula la formación tanto de los receptores intestinales (TPRV6) y
del túbulo proximal renal (TPRV5) como de la Calbindina D9/28K. La salida del calcio en
la cara basolateral es contra gradiente y requiere de una ATPasa que también depende de la
vitamina D3.
2.4.- Metabolismo
El calcio absorbido entra a formar parte del pool circulante extracelular y éste está
en equilibrio con el calcio óseo intercambiable.
La regulación homeostática del calcio plasmático es muy precisa y se establece un
nivel de 2 mmoles/l por debajo del cual habría hipocalcemia (equivalente a 8 mg/dl).
Cuando el nivel de calcio baja, la glándula paratiroides libera a sangre PTH y esta hormona
será responsable de:

ÚLTIMOS AVANCES EN NUTRICIÓN VITAMÍNICO MINERAL 21
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
1.- Frenar la salida de calcio por el riñón por aumento de la reabsorción en túbulo
proximal.
2.- Activación en el hígado de la hidroxilación del colecalciferol (vitamina D inactiva)
produciéndose 25OH colecalciferol.
3.- Activación en el riñón de una segunda hidroxilación del 25OH colecalciferol,
produciéndose 1-25 diOH colecalciferol, que es la hormona D, ya activa, que es
responsable de la absorción transcelular del calcio a nivel intestinal.
4.- Movilización del calcio óseo.
Es importante resaltar el papel del magnesio en la síntesis y secreción de PTH
(NRC, 2001), así como su papel en la cascada de reacciones enzimáticas que tienen lugar
en el hueso cuando la PTH estimula la liberación de calcio (Rude, 1998). La práctica
establece que con concentraciones de Mg en sangre por debajo de 1,6 mg/dl en vacas en
periparto, aumenta la incidencia de hipocalcemia (Van Braak, 1987).
En situaciones de hipercalcemia, otra hormona, sintetizada por la glándula tiroides, la
calcitonina, se encargará de hacer lo contrario de la PTH.
2.5.- Excreción
La excreción de calcio tiene lugar por las heces y la orina. En las heces está el
calcio que no ha sido absorbido, el de la descamación de células de la mucosa intestinal y
de secreciones de saliva, gástrica, intestinal y biliar. En el riñón se filtra el calcio ionizado
y normalmente se reabsorbe el 98%; dos tercios en el túbulo proximal y el tercio restante
en el Asa de Henle. El calcio no reabsorbido se excreta por orina.
3.- HIPOCALCEMIA EN VACAS
Cuando la vaca se encuentra produciendo leche, los dos mecanismos de absorción,
pasiva y activa, son necesarios ya que las necesidades de la vaca para este mineral son
altas. En el momento que secamos la vaca, ya no es necesario que el sistema activo siga
funcionando por lo que la vitamina D3, activa, es decir, el 1-25 diOH colecalciferol pasará
a 24-25 diOH colecalciferol y se eliminará por orina.
El problema aparece cuando las necesidades de Calcio se hacen, bruscamente,
importantes tras el parto. Existe un retraso en la activación del sistema y éste se debe a la
migración celular de los enterocitos desde el valle a la cresta de la microvellosidades
intestinales. El enterocito es sensible a la “orden” de generar el receptor de membrana
(TPRV6) en el valle pero la absorción se produce en la cresta, y la migración del valle a la
cresta dura unas 48 horas (Martín-Tereso and Verstegen, 2011).

22 P. SAYALERO
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
4.- USO DE ANTAGONISTAS DEL CALCIO PARA ESTIMULAR LA
HOMEOSTASIS
La fiebre de la leche es uno de los problemas metabólicos más importantes en la
vaca de leche. La incidencia puede alcanzar el 44% (Jordan, 1993). Las vacas sufren un
periodo de unas 48 horas de hipocalcemia por el retraso antes descrito cuando las
necesidades han aumentado de forma rápida al inicio de la lactación (Horst, 1997). La
incidencia de problemas de hipocalcemia aumenta con la edad de los animales, un 9% en
cada lactación (Lean, 2006)
Reindhart et al, 2010. The Veterinary Journal
Las dietas deficientes en calcio durante el período de preparto, son altamente
eficaces para la prevención de la hipocalcemia (Thilsing-Hansen, 2002). Si se “entrena” la
absorción activa de calcio las semanas previas al parto, las vacas responden al aumento en
sus necesidades de forma rápida y eficaz, evitando así el problema (Goings, 1974). El
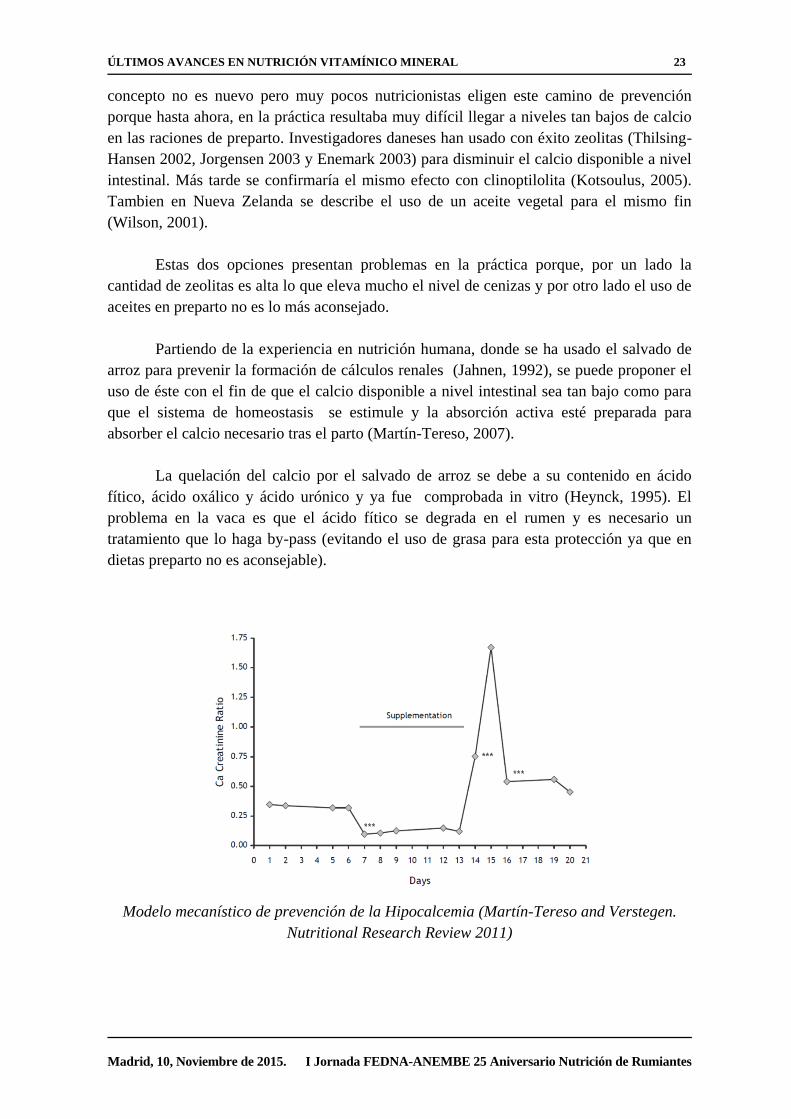
ÚLTIMOS AVANCES EN NUTRICIÓN VITAMÍNICO MINERAL 23
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
concepto no es nuevo pero muy pocos nutricionistas eligen este camino de prevención
porque hasta ahora, en la práctica resultaba muy difícil llegar a niveles tan bajos de calcio
en las raciones de preparto. Investigadores daneses han usado con éxito zeolitas (Thilsing-
Hansen 2002, Jorgensen 2003 y Enemark 2003) para disminuir el calcio disponible a nivel
intestinal. Más tarde se confirmaría el mismo efecto con clinoptilolita (Kotsoulus, 2005).
Tambien en Nueva Zelanda se describe el uso de un aceite vegetal para el mismo fin
(Wilson, 2001).
Estas dos opciones presentan problemas en la práctica porque, por un lado la
cantidad de zeolitas es alta lo que eleva mucho el nivel de cenizas y por otro lado el uso de
aceites en preparto no es lo más aconsejado.
Partiendo de la experiencia en nutrición humana, donde se ha usado el salvado de
arroz para prevenir la formación de cálculos renales (Jahnen, 1992), se puede proponer el
uso de éste con el fin de que el calcio disponible a nivel intestinal sea tan bajo como para
que el sistema de homeostasis se estimule y la absorción activa esté preparada para
absorber el calcio necesario tras el parto (Martín-Tereso, 2007).
La quelación del calcio por el salvado de arroz se debe a su contenido en ácido
fítico, ácido oxálico y ácido urónico y ya fue comprobada in vitro (Heynck, 1995). El
problema en la vaca es que el ácido fítico se degrada en el rumen y es necesario un
tratamiento que lo haga by-pass (evitando el uso de grasa para esta protección ya que en
dietas preparto no es aconsejable).
Modelo mecanístico de prevención de la Hipocalcemia (Martín-Tereso and Verstegen.
Nutritional Research Review 2011)
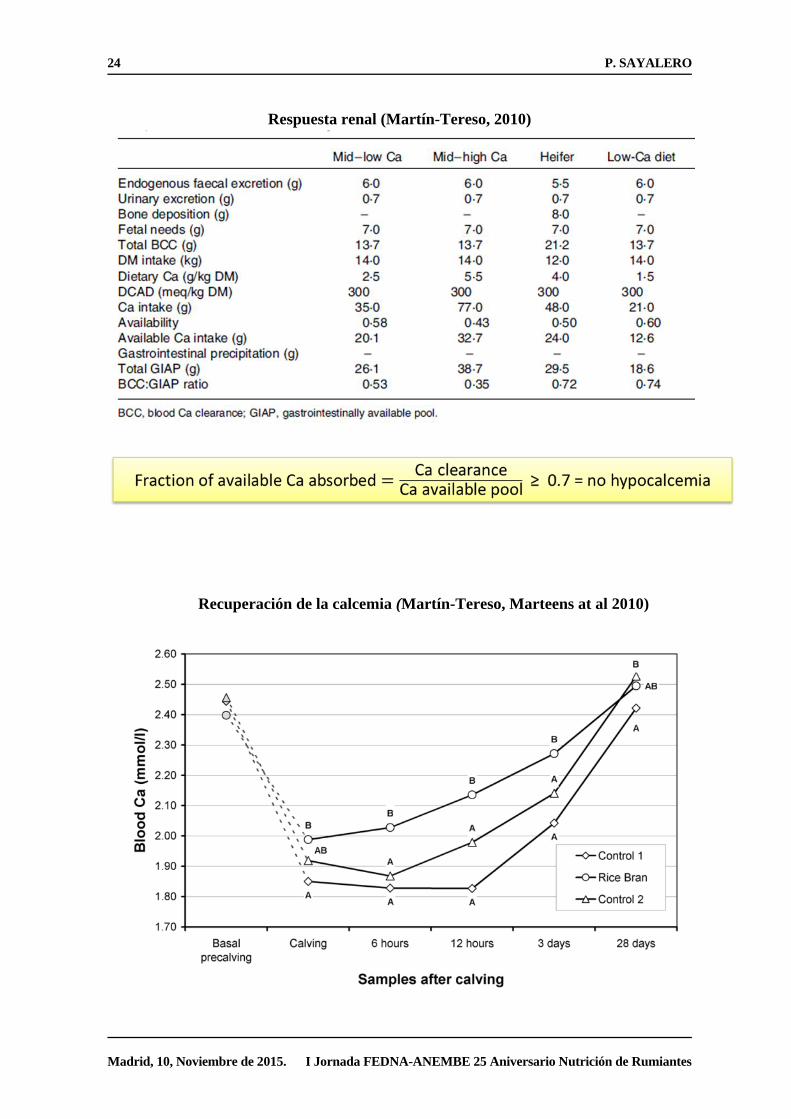
24 P. SAYALERO
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
Respuesta renal (Martín-Tereso, 2010)
Recuperación de la calcemia (Martín-Tereso, Marteens at al 2010)
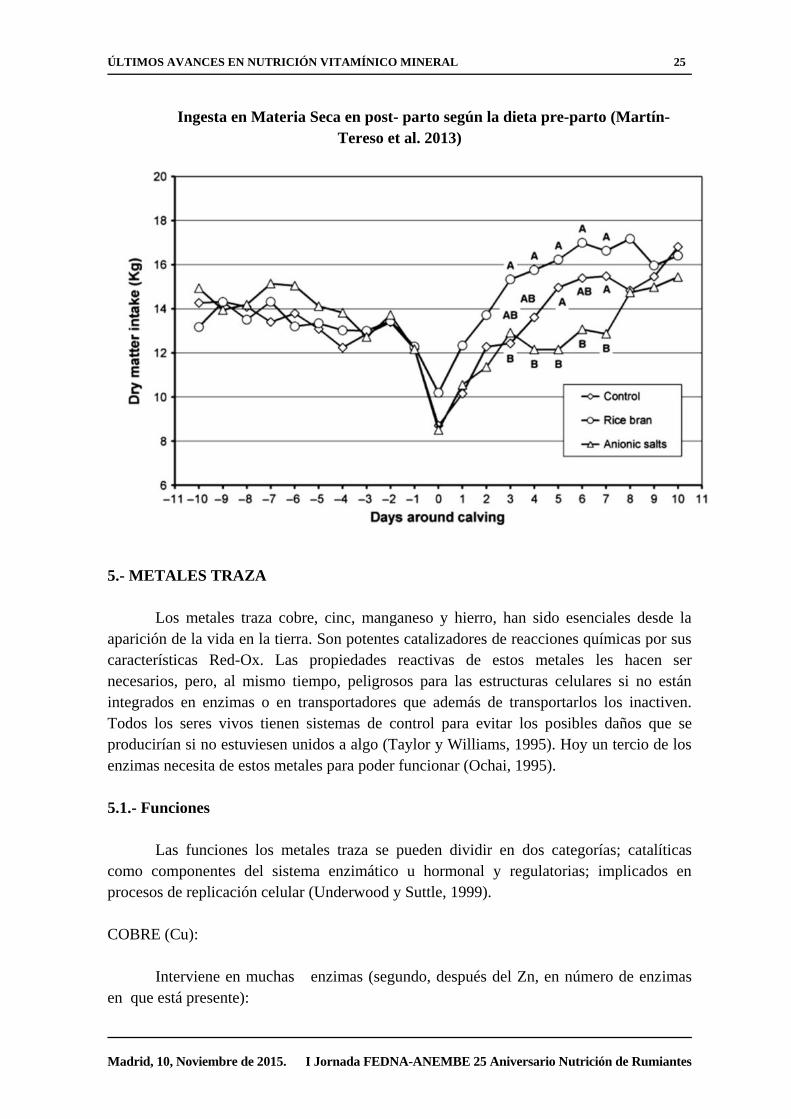
ÚLTIMOS AVANCES EN NUTRICIÓN VITAMÍNICO MINERAL 25
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
Ingesta en Materia Seca en post- parto según la dieta pre-parto (Martín-
Tereso et al. 2013)
5.- METALES TRAZA
Los metales traza cobre, cinc, manganeso y hierro, han sido esenciales desde la
aparición de la vida en la tierra. Son potentes catalizadores de reacciones químicas por sus
características Red-Ox. Las propiedades reactivas de estos metales les hacen ser
necesarios, pero, al mismo tiempo, peligrosos para las estructuras celulares si no están
integrados en enzimas o en transportadores que además de transportarlos los inactiven.
Todos los seres vivos tienen sistemas de control para evitar los posibles daños que se
producirían si no estuviesen unidos a algo (Taylor y Williams, 1995). Hoy un tercio de los
enzimas necesita de estos metales para poder funcionar (Ochai, 1995).
5.1.- Funciones
Las funciones los metales traza se pueden dividir en dos categorías; catalíticas
como componentes del sistema enzimático u hormonal y regulatorias; implicados en
procesos de replicación celular (Underwood y Suttle, 1999).
COBRE (Cu):
Interviene en muchas enzimas (segundo, después del Zn, en número de enzimas
en que está presente):

26 P. SAYALERO
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
Metabolismo del hierro; la ceruloplasmina que se sintetiza en el hígado y cuya
función es incorporar hierro a la hemoglobina necesita Cu.
Respiración celular; El Cu es componente esencial de la citocromo-oxidasa en el
transporte de electrones de la cadena respiratoria.
Formación del tejido conectivo; lysil-oxidasa también necesita Cu y sin su acción
no puede formarse la elastina.
Reproducción e inmunidad; La Cu-Zn superóxido dismutasa es un enzima que
protege frente al daño oxidativo que sufren las células reproductoras y del sistema
inmune, que se encuentra especialmente activa en el momento del periparto (Miller
et al, 1979; Bernabucci et al, 2005).
CINC (Zn)
Necesario en multitud de sistemas enzimáticos:
Expresión genética; Sería parte de los dedos de Zn del DNA (estructuras proteicas,
que coordinan Zn con cisteina e histidina y que estabilizan los pliegues del DNA).
El Zn interviene en la RNA polimerasa, DNA polimerasa, transcriptasa inversa
(Chester, 1997).
Crecimiento; ya que interviene de forma directa en la síntesis de proteína (Spear,
1999).
Reproducción e inmunidad; necesario en la Cu-Zn superóxido dismutasa como ya
se ha citado.
Metabolismo de la vitamina A. El Zn es necesario para que la vitamina A pueda
transportarse desde el hígado a los tejidos diana, ya que forma parte de la RBP
(retinol binding protein).
MANGANESO (Mn)
El manganeso interviene en muchos procesos ya mencionados para el cobre y cinc
y su principal función tiene que ver con la reproducción a través del enzima Mn
superóxido dismutasa, cuya función es similar a la descrita para la Cu-Zn superóxido
dismutasa, por reducir el riesgo de oxidación que se produce en las células reproductoras
especialmente en el periparto (Bernabucci, 2005).
HIERRO (Fe)
El hierro es el elemento traza más abundante en el organismo del rumiante. Se
encuentra unido a porfirina formando parte del grupo hemo en la hemoglobina, mioglobina
y enzimas como citocromos o bien en forma no hemo, formando parte de enzimas
necesarias en el metabolismo de hidratos de carbono (aconitasa, fosfoenolpiruvato-
carboxiquinasa..).

ÚLTIMOS AVANCES EN NUTRICIÓN VITAMÍNICO MINERAL 27
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
Sus funciones fundamentales son:
Transportar oxígeno.
Procesos Red-Ox en la transferencia de electrones en la cadena respiratoria
facilitando la fosforilación oxidativa que pasa el ADP a ATP.
5.2.- Absorción de los metales traza
La absorción de los metales traza tiene lugar en el intestino delgado, sobre todo en
el duodeno (Davies, 1980), y se produce de dos formas; pasiva, no saturable, y activa,
saturable. Para la entrada en la célula, de forma activa, el metal se une a una proteína que
actúa como proteína chaperona que facilita el paso del metal a través de la membrana o
bien pasar ambos, proteína y metal unidos, siendo entonces un co-transportador (Cousins,
1985).
Una vez que el metal ha entrado en la célula éste puede unirse a dos distintas
especies moleculares, una proteína transportadora, que en el caso del Zn se ha identificado:
CRIP (cystein rich intestinal protein) o bien a metalotioneina (MT) (Menard, 1981). La
concentración de MT determina la cantidad de metal que se unirá a su correspondiente
transportador, estando en razón inversa a la concentración de MT. En el caso del Zn,
cuanta más expresión de MT, menor unión a CRIP, lo que supone menor llegada a la cara
basolateral del enterocito, por lo tanto menos Zn llegará a sangre.
Este mecanismo conduce a distintos autores a decir que el intestino es el principal
órgano de regulación en el balance de los metales traza.
La metalotioneina (MT) se une a Fe, Cu y Zn por los grupos tiólicos de la cisteína
(tiene 20 Cys) y participa, como se ha descrito, en los procesos de regulación
homeostática. Es un bioindicador de toxicidad de metales y puede intercambiar estos
metales traza por metales pesados. La unión con metales tiene afinidades y fuerza de unión
distintas: de menos a más Fe<Zn<Pb<Bi<Cu<Ag<Hg<Cd (M.Santo & Tooyama, 2000).
Cuando la concentración de un metal traza en sangre es baja, la expresión de MT
disminuye para facilitar su entrada en sangre. El inconveniente es que la MT puede
expresarse por exceso de un metal y una vez producida en el enterocito unirse a otros los
metales dificultando su absorción, según la secuencia antes descrita.
Cuando el metal alcanza la membrana basolateral se une a otra proteína cuya
misión será sacarlo hacia la sangre, en un mecanismo también saturable, donde se unirá a
albúmina para entrar en la circulación y llegar al hígado y desde allí transportarse hacia
tejidos para su posterior uso.

28 P. SAYALERO
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
5.3.- Regulación
Las necesidades de los metales traza son variables según el estatus fisiológico de la
vaca, según varíe la necesidad de usar enzimas de cada metal y esto dependerá del
momento productivo, situación inmunitaria y reproductiva o grado de estrés.
La disponibilidad de los metales es muy variable. La absorción dependerá de tres
factores y por orden de importancia serían: el estatus nutricional, la presencia de
antagonistas y la forma química en la que se aportan.
Los metales traza sufren cambios químicos a medida que avanzan en el tracto
digestivo. Las formas químicas en las que se añaden a la dieta pueden variar en rumen y
ser hidrolizadas en abomaso. Es en duodeno donde las formas ionizadas se van a absorber
mejor ya que a medida que aumenta el pH digestivo la solubilidad va disminuyendo. La
presencia de metales y compuestos activos en las sales biliares complica aún más la
absorción por las interacciones.
Los dos factores que más influyen en la regulación de la absorción son la
concentración y la solubilidad. Un exceso de un metal soluble, provoca una regulación
negativa de la absorción. Si la concentración de un metal soluble es baja, ya por un aporte
escaso o bien por precipitación, se produce regulación positiva de la absorción. Esta
regulación positiva es muy eficaz y muy precisa, el enterocito pone en marcha los
transportadores y éstos al tener gran afinidad por los metales, los retiran de los complejos y
quelatos solubles para absorberlos e incorporarlos a las rutas fisológicas (Cousing, 2006).
En condiciones de absorción activa, aunque haya mecanismos compartidos (como en el
caso del Cu y Zn), si los metales se absorben activamente las interacciones se minimizan.
ÚLTIMOS AVANCES EN NUTRICIÓN VITAMÍNICO MINERAL 29
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
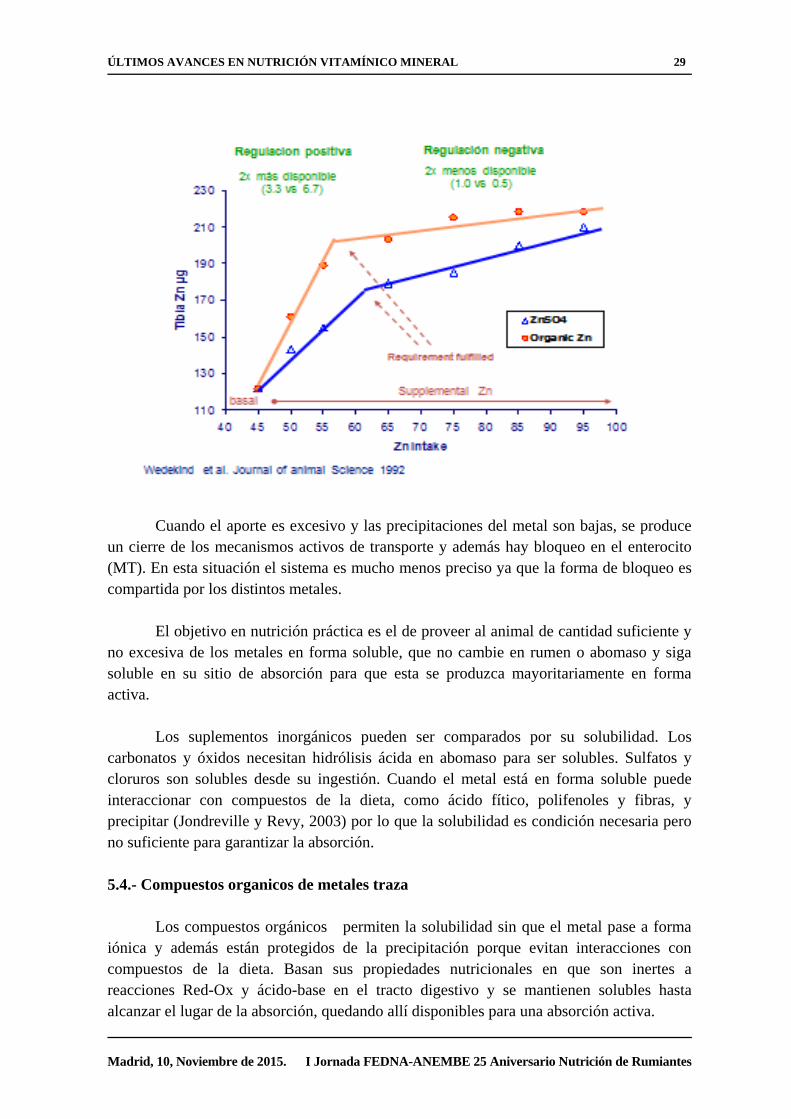
Cuando el aporte es excesivo y las precipitaciones del metal son bajas, se produce
un cierre de los mecanismos activos de transporte y además hay bloqueo en el enterocito
(MT). En esta situación el sistema es mucho menos preciso ya que la forma de bloqueo es
compartida por los distintos metales.
El objetivo en nutrición práctica es el de proveer al animal de cantidad suficiente y
no excesiva de los metales en forma soluble, que no cambie en rumen o abomaso y siga
soluble en su sitio de absorción para que esta se produzca mayoritariamente en forma
activa.
Los suplementos inorgánicos pueden ser comparados por su solubilidad. Los
carbonatos y óxidos necesitan hidrólisis ácida en abomaso para ser solubles. Sulfatos y
cloruros son solubles desde su ingestión. Cuando el metal está en forma soluble puede
interaccionar con compuestos de la dieta, como ácido fítico, polifenoles y fibras, y
precipitar (Jondreville y Revy, 2003) por lo que la solubilidad es condición necesaria pero
no suficiente para garantizar la absorción.
5.4.- Compuestos organicos de metales traza
Los compuestos orgánicos permiten la solubilidad sin que el metal pase a forma
iónica y además están protegidos de la precipitación porque evitan interacciones con
compuestos de la dieta. Basan sus propiedades nutricionales en que son inertes a
reacciones Red-Ox y ácido-base en el tracto digestivo y se mantienen solubles hasta
alcanzar el lugar de la absorción, quedando allí disponibles para una absorción activa.

30 P. SAYALERO
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
Habría que diferenciar entre Complejos, unión del metal a un aminoácido por un
único sitio y Quelatos, metal unido a dos o más sitios distintos dentro del mismo ligando
ya que el comportamiento en cuanto a solubilidad y sensibilidad a la hidrólisis ácida
varían. Un complejo simple es soluble a un amplio rango de pH pero sensible a la
hidrólisis ácida mientras que un quelato además de ser soluble a un amplio rango de pH, es
bastante resistente a la hidrólisis ácida.
Zn en forma iónica Zn unido a Metionina Quelato de Zn
Si comparamos con los aportes de metales traza en forma inorgánica, a nivel
práctico, sólo se verán diferencias si existe algún reto fisiológico o nutricional ya que la
regulación homeostática compensará las diferencias para garantizar un estatus adecuado.
Uno de los momentos más delicados para la vaca de leche es el período de
transición. En el momento del periparto la glándula mamaria retira de la sangre muchos
nutrientes para la producción del calostro, además el útero ha sufrido las agresiones
traumáticas del parto, hay sangrado del endometrio y éste debe ser reparado. A nivel
metabólico la actividad se multiplica y hay un uso importante de enzimas que necesitan ser
activadas por metales. Todo esto lleva consigo un drenaje importante de Metales Traza y si
las reservas del animal o los aportes a través de la nutrición no son suficientes habrá un
retraso en la producción y en la reproducción. Es en esta situación donde el uso de metales
traza en forma orgánica está justificado.
Los minerales en forma orgánica son más disponibles en condiciones de regulación
positiva y menos agresivos cuando la regulación es negativa de ahí la estrategia de aportar
una parte de éstos en forma orgánica.

ÚLTIMOS AVANCES EN NUTRICIÓN VITAMÍNICO MINERAL 31
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
6.- VITAMINAS DEL GRUPO B
Las vitaminas del grupo B son esenciales para la vida. Son cofactores necesarios
para activar enzimas cuya acción es fundamental para el metabolismo de los Principios
Inmediatos. Los microorganismos del rumen son capaces de sintetizar estas vitaminas, por
lo que los síntomas clínicos de sus deficiencias son prácticamente desconocidos en
rumiantes. El NRC 2001 indica que no es necesario suplementar para prevenir deficiencias
clínicas. Las conclusiones del NRC se basan en estudios de finales de los ochenta y
principio de los noventa y son extrapolaciones de datos obtenidos en cerdas.
Vitaminas B y algunos procesos fisiológicos en los que participan
Vitamina Nombre Funciones
B1 Tiamina Generación de energía a partir de carbohidratos.
Función nerviosa
Producción de ADN y ARN
B2 Riboflavina Catabolismo de ácidos grasos
Transporte de electrones
B3 Niacina Transferencia de energía
Metabolismo de glucosa y grasas
B5 Acido Pantoténico Oxidación de ácidos grasos y carbohidratos
Síntesis de ácidos grasos, cetonas y aminoácidos
Complejo B6 Piridoxina
Piridoxal
Piridoxamina
Metabolismo de aminoácidos y lípidos
Producción de vitamina B3
Gluconeogénesis
B7 Biotina Metabolismo de lípidos, proteínas y carbohidratos
Síntesis de ácidos grasos a partir de acetato
Gluconeogénesis
B9 Acido Fólico División celular, rápida, en gestación
Producción de glóbulos rojos
B12 Cianocobalamina Metabolismo de carbohidratos, lípidos y proteínas
Producción de glóbulos rojos

32 P. SAYALERO
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
Las vitaminas del grupo B que provienen del alimento, son degradadas en distinta
medida en el rumen,
Niveles by-pass de vitaminas B del alimento.
% By-pass NRC 2001 % By-pass Santchi 2005
Tiamina 52 32
Riboflavina 1 1
Niacina 6 1
Acido Pantoténico 22 2
Complejo B6 100 59
Biotina 100 55
Acido Fólico 3 3
Cianocobalamina 10 37
La absorción de estas vitaminas a través de la pared del rumen es casi nula. Los
microorganismos del rumen producen, utilizan y acumulan estas vitaminas. A medida que
estos microbios del rumen abandonan éste ceden estas vitaminas siendo una fuente natural
para proveer al animal.
Sintesis ruminal aparente, mg/día, de vitaminas B en el rumen
NRC 2001 Schwab et al., 2005 Santschi et al., 2005
Tiamina 127 41-61 26
Riboflavina 232 206-254 267
Niacina 1603 446-1547 2.213
Acido Pantoténico 38 - -
Complejo B6 85 14-30 -14
Biotina 12 -3 a -16 -1
Acido Fólico 6 13-20 21
Cianocobalamina 62 60-102 73
La síntesis ruminal aparente es la cantidad que llega al duodeno menos la ingerida.
No considera la degradación ruminal ni la posible absorción en el rumen.
La diferencias observadas son probablemente dependientes de la composición de la ración
(Schwab et al, 2005). Como tanto la síntesis como la degradación dependen de la ración,
es difícil hacer una estimación de cuanto deberíamos suministrar para unos óptimos
rendimientos.

ÚLTIMOS AVANCES EN NUTRICIÓN VITAMÍNICO MINERAL 33
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
6.1.- Suplementacion
TIAMINA (B1)
Cuando se suplementaba con 150 mg de tiamina/día en una ración basada en alfalfa
se veía una respuesta de producción positiva. Al hacer el ensayo con niveles de tiamina
superiores (300 mg/día) se veía que ésta podía deprimir el contenido de grasa en la leche lo
que llevó a la conclusión de que la tiamina era útil en dietas con forraje limitado (Shaver,
Ball, 2000).
Efecto de la adición de tiamina (150-300 mg/día) en dietas a base de alfalfa
Prop. F/C Variación IMS Leche (kg) Grasa (kg) Proteina (kg)
150 mg/día 55:45 0,9 2,7* 0,13* 0,10*
300 mg/día 50:50 -0,1 0,7 0,02 0,04
300 mg/día 60:40 -0,8 -0,2 -0,18* -0,03
*Efecto positivo
NIACINA (B3)
Ha sido bastante estudiada en las últimas décadas y se ha usado para prevenir el
hígado graso dada su acción antilipolítica al inicio de la lactación, sin embargo pocos
estudios que han medido el nivel de NEFAS y BHBA en vacas suplementadas con niacina
reportan resultados positivos (Niehoff et al, 2008). Probablemente esto es debido a que la
niacina usada no estaba protegida y ésta es muy sensible a la degradación en rumen
(Santschi, 2005). Cuando la niacina estaba protegida el efecto era significativamente
superior (Morey et al, 2011).
ACIDO PANTOTENICO (B5)
Está presente en muchos de los alimentos que componen la ración de las vacas de
leche aunque su contenido es variable. Se degrada mucho en rumen y la síntesis aparente
no es suficiente para llegar a las necesidades. La suplementación con formas desprotegidas
solo fue efectiva a dosis muy altas (Zinn et al, 1997). La forma protegida de esta vitamina
tiene efectos positivos en la producción de leche, grasa y proteína (Bonomi, 2000).
PIRIDOXINA (Complejo B6)
Hay pocos datos científicos, el aporte final de piridoxina al intestino depende de la
ración (Seck, 2010) y puede no ser suficiente.

34 P. SAYALERO
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
BIOTINA (B7)
Se empezó a usar por la mejora en las pezuñas y se vio efecto en la producción de
leche cuando se añadía 20 mg/día a la ración (Girard y Matte, 2006). Los efectos pueden
ser superiores si se usa biotina protegida ya que la degradación en rumen es mayor de lo
que se pensaba (Santschi, 2005).
ACIDO FOLICO (B9)
Juega un papel importante en la replicación del ADN y en el metabolismo de la
Metionina (Regaller, 2008). La molécula se degrada en rumen, pero al mismo tiempo los
microbios la generan. La suplementación con ácido fólico no afecta a la ingesta y los
efectos en producción son variables, mayores en vacas multíparas que en primíparas
(Regaller, 2009)
CIANOCOBALAMINA (B12)
Las bacterias del rumen necesitan cobalto para para producir B12. La efectividad
de los folatos (B9) puede verse disminuida si no hay suficiente B12 (Girard, 2010). La
degradación en rumen de la cianocobalamina es muy alta y su absorción intestinal baja
(Girard, 2009) por lo que si se añade debería ser en forma protegida para al menos
garantizar el paso del rumen.
6.2.- Conclusiones sobre el uso de vitaminas del grupo B
A pesar de que el NRC no haga recomendaciones concretas sobre vitaminas del
grupo B, parece haber claros beneficios cuando se suplementa al inicio de la lactación. Aún
es pronto para definir unas necesidades concretas porque tanto la destrucción como la
síntesis a nivel ruminal dependen de la ración. Parece claro que el momento más
interesante para suplementar con vitaminas del grupo B sería el inicio de la lactación ya
que la función ruminal puede ser subóptima y a la vez la demanda de éstas, máxima.
7.- REFERENCIAS
CAO, J. et al (2000) J. Anim. Sci. 78: 2039-2054.
COUSINS, R.J. (1985) Physiol. Rev. 65: 238-309.
ENEMARK, J.M.D., KIRKETERP-MOLLER, C.N. y JORGENSEN, R.J. (2003) Acta
Veterinaria Scandinavica Supplement 97: 119-136.
GIRARD, C.L, SANTSCHI, D.E., STABLER, S.P. y ALLEN, R.H. (2009) J. Dairy Sci.
GOINGS, R.L., JACOBSON, N.L., BEITZ, D.C., LITTLEDIKE, E.T. y WIGGERS, K.D.
(1974) J. Dairy Sci. 57: 1184-8.
HEYNCK, H., KRAMPITZ, G. y HESSE, A. (1995) European Journal of Clinical
Nutrition, Supplement 3. 49: 250-252.

ÚLTIMOS AVANCES EN NUTRICIÓN VITAMÍNICO MINERAL 35
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
HOLWERDA, R.A., ALBIN, R.C. y MADSEN, F.C. (1995) Feedstuffs 67: 12-13.
HORST, R.L., GOFF, J.P., REINHARDT, T.A. y BUXTON, D.R. (1997) J. Dairy Sci. 80:
1269-1280.
MARTÍN-TERESO et al. (2008) J. Anim. Physiol. Anim. Nutr.
MARTIN-TERESO et al. (2009) Effect of rumen-protected rice bran on mineral status of
non lactating dairy heifers. Anim. Feed Sci. Technol. 152: 286-297.
MARTÍN-TERESO J., TER WIJLEN, H., VAN LAAR, H. y VERSTEGEN, M.W.A.
(2010) J. Anim. Physiol. Anim. Nutr.
RAGALLER et al, 2009. Br. J. Nutr. 101: 153-164.
SANTSCHI et al. (2005) J. Dairy Sci. 88: 2043-2054.
SCHWAB et al. (2006) J. Dairy Sci. 89: 174-187.
TERRY, E. y ENGLE. (2003) Trace Mineral Metabolism in Ruminants. Department of
Animal Sciences, Colorado State University, USA.


I JORNADA FEDNA − ANEMBE 25 Aniversario
sobre ALIMENTACIÓN DE RUMIANTES
MADRID, 10 de Noviembre de 2015
CAPÍTULO III
PROTEÍNAS, PÉPTIDOS, AMINOÁCIDOS Y OTRAS
FUENTES NITROGENADAS PARA LA FORMULACIÓN
DE RACIONES EN RUMIANTES
María Dolores Carro Travieso
Departamento de Producción Agraria. Universidad Politécnica de Madrid


PROTEÍNAS, PÉPTIDOS, AMINOÁCIDOS Y OTRAS FUENTES NITROGENADAS 39
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
PROTEÍNAS, PÉPTIDOS, AMINOÁCIDOS Y OTRAS FUENTES
NITROGENADAS PARA LA FORMULACIÓN DE RACIONES EN RUMIANTES
María Dolores Carro Travieso
Departamento de Producción Agraria de la Universidad Politécnica de Madrid
1.- INTRODUCCIÓN
La cantidad y calidad de la proteína incluida en las raciones de los animales
rumiantes repercuten de forma importante en los costes de alimentación y condicionan en
gran medida el rendimiento productivo de estos animales, pero también afectan a la
cantidad de nitrógeno (N) excretado al medio ambiente y a la función reproductiva. Para
ilustrar estos aspectos en la Tabla 1 se muestran los resultados obtenidos en una prueba
práctica realizada en el AgriSearch de Irlanda (AgriSearch, 2010) en la que 90 vacas
Holstein-Friesian se dividieron en 3 grupos homogéneos que recibieron durante los
primeros 150 días de lactación una ración unifeed compuesta por 27% de ensilado de
hierba, 18% de ensilado de maíz y 55% de concentrado con diferente contenido en proteína
para aportar en la ración un total de 11,5, 14,5 ó 17,5% de proteína bruta. A medida que
aumentó el contenido en proteína en la ración se observó un aumento de la ingestión diaria
de materia seca y de la producción de leche, sin que se observaran diferencias en el
contenido en proteína de la leche, aunque la concentración en grasa fue mayor para el nivel
más bajo de proteína (11,5%). Como era de esperar, el aumento en la concentración de
proteína produjo una mayor excreción de N en el estiércol y resultó en una menor
eficiencia de utilización del N para la producción de leche, aunque también en una menor
producción de metano por litro de leche producida. El contenido en proteína también
afectó a la actividad reproductora, ya que el porcentaje de vacas gestantes a los 150 días de
lactación fue menor en los animales que recibían el nivel más alto de proteína. Como
consecuencia de todos estos efectos, la cantidad de proteína en la ración afectó al beneficio
económico de la explotación ganadera, aunque en este caso no pueden hacerse
generalizaciones, ya que el resultado final se ve afectado por diversos factores, entre los
que destacan el precio de los alimentos y de la leche.

40 M. D. CARRO
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
Tabla 1.- Efecto del contenido en proteína bruta en la ración en la productividad
y excreciones de 90 vacas lecheras en los primeros 150 días de lactación (adaptado de
AgriSearch, 2010)
Contenido en proteína bruta (% materia seca)
11,5% 14,5% 17,5%
Ingestión de MS (kg/d) 16,5 18,0 18,6
Producción de leche (l/d) 24,7 30,9 34,4
Grasa en leche (%) 4,20 3,83 3,81
Proteína en leche (%) 3,14 3,23 3,24
N ingerido (g/d) 322 445 562
N en leche (g/d) 100 132 144
N en estiércol (g/d) 227 300 380
% del N ingerido en la leche 31,1 29,7 25,6
Producción de metano (l/l
leche)
25,7 20,9 20,7
Vacas gestantes (%) 100 93 87
Beneficio (£/vaca) 335 457 514
El avance en el conocimiento de los procesos digestivos y metabólicos que sufre la
proteína ingerida por los rumiantes ha determinado que se haya pasado de formular las
raciones en términos de proteína bruta a la utilización de la proteína metabolizable (MP),
en un intento de ajustar de forma más precisa los aportes de aminoácidos (AA) en la ración
a las necesidades reales de los animales. La MP es definida como la mezcla de AA
verdaderamente absorbidos en el intestino, que provienen de tres orígenes diferentes: la
proteína del alimento no degradada en el rumen, la proteína microbiana sintetizada y, en
una pequeña proporción, las secreciones endógenas. Esta mezcla de AA que forma la MP
es utilizada para cubrir las necesidades proteicas de mantenimiento, reproducción y
producción de los animales, pero los AA también pueden ser catabolizados para obtener
energía o utilizados para sintetizar glucosa (AA glucogénicos) y otros AA no esenciales
(Figura 1). A pesar de los indudables progresos realizados en las últimas décadas en el
conocimiento de la nutrición proteica de los rumiantes, la eficiencia de utilización del N en
estos animales continúa siendo baja en comparación con la presentada por los animales no
rumiantes, por lo que su aumento es un objetivo importante en el ámbito de la nutrición de
rumiantes, especialmente en un mercado globalizado como el actual. Adicionalmente, la
creciente preocupación por la contaminación medioambiental provocada por la producción
animal ha contribuido a que exista un renovado interés en reducir la excreción de N sin
mermar la producción de los animales. El objetivo de este trabajo es repasar los principales
procesos que determinan la digestión de la proteína y la utilización metabólica de los AA
en los animales rumiantes, así como analizar los principales factores que afectan a estos
procesos y que pueden contribuir a estimaciones erróneas a la hora de formular raciones.

PROTEÍNAS, PÉPTIDOS, AMINOÁCIDOS Y OTRAS FUENTES NITROGENADAS 41
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
Adicionalmente, se analizarán algunos de los resultados obtenidos recientemente al utilizar
AA protegidos en la ración. Debido a que algunos de los ejemplos numéricos que se
muestran están sacados de la publicación sobre las necesidades nutritivas del vacuno
lechero elaborada por el National Research Council (NRC, 2001), en este trabajo se
utilizarán las siglas usadas en dicha publicación.
Figura 1.- Origen y utilización de los aminoácidos en los animales rumiantes
2.- DEGRADACIÓN RUMINAL DE LAS PROTEÍNAS
Aunque normalmente se hace referencia al contenido en proteína bruta de los
alimentos, es más correcto referirse a los compuestos nitrogenados, ya que en la práctica el
contenido en proteína de un alimento se determina mediante un análisis de N y se
multiplica el resultado por 6,25, asumiendo así un contenido medio del 16% de N en estas
proteínas. Sin embargo, los alimentos contienen también compuestos nitrogenados de
naturaleza no proteica, como ácidos nucleicos, aminas, amidas, AA, péptidos y amoníaco.
Estos compuestos se engloban en la fracción de N no proteico (NPN) y pueden
cuantificarse como el N presente en el filtrado obtenido tras la precipitación de la proteína
verdadera con ácido túngstico o ácido tricloroacético (Licitra et al., 1996). El contenido en
NPN suele ser inferior al 12% del N total en la mayoría de los alimentos concentrados (no
forrajeros), pero alcanza valores más altos en los forrajes y dentro de ellos es
especialmente elevado en los forrajes conservados (henos y ensilados) debido a la
proteólisis producida durante los tratamientos de conservación (Van Soest, 1994).

42 M. D. CARRO
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
Las proteínas están formadas por cadenas lineales de AA (estructura lineal) que
giran y se pliegan al formarse uniones entre algunos AA para dar lugar a estructuras más
complejas (estructura secundaria, terciaria y cuaternaria), en función de la proteína de que
se trate. Las proteínas difieren en su secuencia de AA, pero también en su tamaño, forma,
función y solubilidad y algunas de estas características van a afectar a su degradación
ruminal. Cuando los alimentos ingeridos llegan al rumen, las proteínas son degradadas por
enzimas producidas por los microorganismos ruminales, que producen una gran variedad
de proteasas, peptidasas y desaminasas, lo que provoca la liberación de péptidos, AA y
finalmente amoníaco al medio ruminal. Sin embargo, los productos intermedios generados
son captados simultáneamente por los microorganismos para formar sus propias estructuras
en el proceso conocido como síntesis de proteína microbiana (MCP), de tal forma que las
concentraciones de péptidos y AA en el líquido ruminal suelen ser bajas. Algunos péptidos
y AA que no son degradados ni incorporados a la MCP pueden abandonar el rumen
mezclados con la digesta, pero su cantidad suele ser pequeña en animales que reciben
raciones convencionales. Por el contrario, en el caso de la administración de proteínas que
se degradan muy rápidamente (p.e. caseína) puede haber un flujo importante de péptidos
que abandone el rumen sin ser degradados (Broderick et al., 1991).
El ritmo y extensión de la degradación proteica dependerá del tipo y concentración
de enzimas producidas, así como del número de enlaces peptídicos susceptibles de ser
rotos. La estructura secundaria y terciaria de las proteínas determina la accesibilidad de las
proteasas microbianas a algunos de los enlaces peptídicos y por ello condiciona la
degradabilidad de la proteína. Las proteínas que contienen numerosos enlaces covalentes
(p.e. puentes disulfuro en albúminas e inmunoglobulinas) o aquellas en las que estos
enlaces se han formado como consecuencia de los tratamientos con ácido o calor son
menos accesibles para los microorganismos y por ello se degradan más lentamente. El
tratamiento térmico también puede coagular las proteínas y reducir su solubilidad, y el
enfriado posterior puede provocar la formación de nuevos enlaces que modifican las
características de la proteína y su degradación ruminal.
Debido a que existen un gran número de enlaces peptídicos diferentes dentro de una misma
proteína, es necesaria la acción sinérgica de diferentes proteasas para completar la
degradación proteica y por ello se requiere la participación de diferentes microorganismos
ruminales. Se calcula que entre el 30 y 50% de las bacterias ruminales tienen actividad
proteolítica sobre las proteínas extracelulares y entre ellas destacan por su gran actividad
Prevotella ruminicola, Ruminobacter amylophilus, Butirivibrio fibrisolvens y
Streptococcus bovis. En algunas bacterias la actividad proteolítica es extracelular (p.e. B.
fibrisolvens), pero en otras, como Prevotella ruminicola y Ruminobacter amylophilus, las
enzimas permanecen unidas a las membranas celulares y adsorben rápidamente las
proteínas presentes en el medio ruminal (Wallace, 1996). Como se ha comentado, la
conversión de las proteínas a amoníaco es un proceso secuencial (ver Figura 2), en el que
el punto crítico parece ser la ruptura de los oligopéptidos a dipéptidos, ya que existen pocas
especies bacterianas que puedan realizar esta actividad catabólica, destacando entre ellas
por su gran actividad Prevotella ruminicola, y con menor actividad S. bovis y R.

PROTEÍNAS, PÉPTIDOS, AMINOÁCIDOS Y OTRAS FUENTES NITROGENADAS 43
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
amylophylus (Wallace et al., 1997). Debido a que en este proceso están implicadas pocas
bacterias, se ha sugerido que la modificación de estas poblaciones podría ser un medio para
reducir el ritmo de degradación de los oligopétidos en el rumen (Calsamiglia et al., 2010).
Los péptidos se degradan finalmente a AA por la acción de diferentes peptidasas.
Estas peptidasas pueden ser extracelulares (asociadas a la pared celular) o intracelulares y
en este caso requieren la incorporación de los péptidos al interior de las bacterias. El
género Prevotella se ha identificado como el responsable de la mayoría de la actividad
peptidasa del rumen y su actividad dipeptidil peptidasa libera en mayor medida dipéptidos
que AA (Wallace et al., 1999). Numerosos estudios han demostrado que las bacterias
pueden captar tanto AA como péptidos de pequeño tamaño y que el ritmo de incorporación
y preferencia por unos u otros parece ser variable entre especies bacterianas (Armstead y
Ling, 1993). En el interior de las células los péptidos y AA pueden ser utilizados
directamente para la síntesis de MCP o ser degradados y desaminados hasta NH3-N y
esqueletos carbonados que pueden ser utilizados en el metabolismo bacteriano, pero la
gran mayoría del NH3-N producido es excretado al medio ruminal. El proceso de
desaminación es llevado a cabo por un pequeño grupo de bacterias que presentan una gran
actividad y por otras muchas bacterias con una actividad desaminativa baja, como
Prevotella spp. (Wallace et al., 1997). Debido a que la desaminación es llevada a cabo por
numerosas bacterias no parece que este proceso pueda ser modificado fácilmente mediante
la inhibición de determinadas poblaciones bacterianas, aunque algunos aceites esenciales
han mostrado cierta eficacia para reducir la desaminación y con ello las concentraciones
ruminales de NH3-N (Calsamiglia et al., 2007).
Figura 2.- Proceso de degradación ruminal de la proteína (adaptado de Wallace et al.,
1997)

44 M. D. CARRO
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
El pH óptimo para la actividad proteolítica ruminal oscila entre 5,5 y 7,0, aunque
algunas enzimas muestran una actividad óptima incluso a valores mucho más altos
(Wallace et al., 1997). Dado el elevado número de bacterias que producen enzimas
proteolíticos, la actividad proteolítica en el rumen se ve poco afectada por la ración,
aunque se ha demostrado que la degradabilidad proteica puede reducirse a bajos valores de
pH como los que se producen en animales que reciben raciones con alto contenido en
concentrados (Calsamiglia et al., 2008). Este hecho se ha atribuido a una reducción en el
acceso de los microorganismos a las proteínas de los alimentos (debido a la menor
degradación de la fibra) o a cambios en las poblaciones microbianas con una disminución
de los microorganismos con actividad proteolítica (Bach et al., 2005). Los protozoos
ruminales también muestran actividad proteolítica, pero en este caso se trata de una
actividad intracelular, ya que fagocitan las proteínas y partículas de alimento y no ejercen
una actividad importante sobre las proteínas solubles (Hino y Russell, 1987). Un aspecto
fundamental es la predación de bacterias por los protozoos, debido a que condiciona en
gran medida el turnover proteico en el rumen. En un estudio realizado por Wallace y
McPherson (1987) el turnover proteico bacteriano ausencia de protozoos osciló entre 0,3 y
2,7% h-1
, mientras que en presencia de protozoos ascendió a valores entre 2,4 y 3,7% h-1
.
Debido a ello el principal papel de los protozoos en relación a la proteína es el que ejercen
al metabolizar la proteína bacteriana, lo que tendrá una repercusión importante en la
eficiencia de síntesis de proteína microbiana, como se verá posteriormente. Otro hecho
diferencial de los protozoos respecto a las bacterias es su incapacidad para sintetizar AA
esenciales, por lo que han de obtenerlos de las bacterias o de las partículas de alimento que
fagocitan. En cuanto a los hongos algunos parecen tener cierta actividad proteolítica, pero
se cree que su contribución a la actividad proteolítica total del rumen es muy baja (Wallace
et al., 1997).
Los compuestos nitrogenados no proteicos (NPN) administrados en la ración, como
la urea y sus derivados o las sales de amonio, son degradados rápida y completamente en el
rumen a NH3-N. La actividad ureasa es fundamentalmente de origen bacteriano, ya que no
se ha encontrado esta actividad en algunos protozoos y hongos, aunque este aspecto no ha
sido investigado en profundidad (Wallace et al., 1997). En muchas de las bacterias
estrictamente anaerobias que se encuentran en las fases sólida y líquida del rumen se ha
encontrado una baja actividad ureasa, mientras que las bacterias que se encuentran en la
pared ruminal muestran una alta actividad, debido al reciclaje de la urea que se produce a
través de la pared. La actividad ureasa en el rumen es muy variable y se ha comprobado
que se reduce cuando existen altos niveles de reciclaje de urea o elevadas concentraciones
ruminales de NH3-N. La utilización de compuestos NPN en la ración debe ir acompañada
de la inclusión de una fuente de energía rápidamente disponible, ya que si ésta no existe se
produce un exceso de NH3-N que no puede ser utilizado por los microorganismos y debe
ser transformado en urea en el hígado, con el consiguiente gasto energético para el animal
hospedador. Cuando existen niveles muy elevados de NH3-N que superan la capacidad del
hígado para transformarlo en urea se puede producir una intoxicación del animal, por lo
que se recomienda que la administración de urea no supere el 1% de la materia seca de la
ración o el 25% del total del N en la misma (FEDNA; 2010).

PROTEÍNAS, PÉPTIDOS, AMINOÁCIDOS Y OTRAS FUENTES NITROGENADAS 45
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
La cuantificación de la degradabilidad de la proteína de los alimentos en el rumen
es un punto crítico en todos los sistemas de valoración nutritiva para los animales
rumiantes, ya que este valor condiciona la síntesis de MCP y determina la fracción de
proteína del alimento no degradada que abandona el rumen (RUP). Uno de los mayores
impedimentos para obtener información fiable sobre este aspecto es la falta de un método
simple y preciso para diferenciar la RUP, MCP y proteína endógena en el flujo proteico
duodenal. La cuantificación de la MCP se puede llevar a cabo únicamente mediante la
utilización de marcadores microbianos, pero ninguno de los utilizados habitualmente
parece idóneo (Carro, 2001). Además, es necesario el uso de animales provistos de cánulas
en el rumen y el duodeno y de marcadores de flujo para cuantificar la digesta duodenal, lo
que hace que la determinación de la degradabilidad de la proteína in vivo sea un proceso
complejo y laborioso que no es apto para las valoraciones rutinarias que se realizan en la
práctica. Por ello se han desarrollado técnicas alternativas, entre las que destaca la técnica
in situ, también denominada in sacco o de las bolsas de nailon. Esta técnica consiste en
colocar el alimento o ración a valorar en bolsas de material sintético y poroso que son
introducidas en el rumen y retiradas a diferentes tiempos de incubación para analizar
posteriormente la cantidad de proteína en el residuo no degradado. Los valores obtenidos
permiten elaborar curvas de degradación de la proteína como la que se muestra en la
Figura 3. El ajuste de los datos obtenidos a cada tiempo de incubación a modelos
matemáticos permite determinar la degradabilidad a un tiempo determinado de
permanencia en el rumen y también el ritmo de degradación. El modelo matemático más
utilizado es el descrito por Mehrez y Ørskov (1977): D = a + b (1 – e –Kd
* t
), en el que “a”
es la fracción soluble y rápidamente degradable, “b” es la fracción insoluble pero
potencialmente degradable, kd es el ritmo fraccional de degradación y “t” es el tiempo de
incubación. La suma “a+b” es la fracción degradable (RDP) y la diferencia hasta 100 es la
fracción no degradable (RUP) o proteína by-pass.
Figura 3.- Evolución de la degradación de la proteína con el tiempo de incubación

46 M. D. CARRO
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
Como puede observarse la degradación de la proteína varía con el tiempo de
incubación, por lo que puede calcularse degradabilidad efectiva (ED) para un tiempo
determinado de permanencia en el rumen mediante la siguiente fórmula propuesta por
Ørskov y McDonald (1979): ED = (a + b*kd) / (kd + kp), en la que a, b y kd son los
parámetros descritos anteriormente y kp es el ritmo fraccional de paso a través del rumen.
El valor de kp es el valor inverso del tiempo de retención de la proteína en el rumen; por
ejemplo, para un tiempo de retención en el rumen de 12,5 horas el valor de kp sería 0,080
(8% h-1
). Algunos sistemas como el NRC (2001) utilizan un único valor de kp para la
proteína, pero otros utilizan diferentes valores para cada fracción proteica como se
describirá posteriormente.
La técnica in situ es relativamente sencilla, pero es laboriosa y exige disponer de
animales canulados en el rumen. La validez de esta técnica se ha contrastado debido a la
estrecha correlación existente entre los valores de degradabilidad in situ de la proteína de
diferentes alimentos y los valores obtenidos in vivo en animales canulados (Madsen y
Hvelplund, 1985), por lo que diferentes sistemas de alimentación como el NRC (2001),
INRA (1988) y AFRC (1992) la han utilizado para elaborar sus tablas de valor proteico de
los alimentos. Sin embargo, la técnica también presenta diversas fuentes potenciales de
error. Algunos factores como el tamaño de poro del nailon, la relación muestra/superficie
(g/cm2), el lavado de las bolsas tras ser retiradas del rumen o la ración de los animales
pueden afectar en gran medida a los resultados obtenidos, por lo que se han elaborado
protocolos estandarizados para su utilización en los laboratorios (AFRC, 1992; Vanzant et
al., 1998).
Al utilizar la técnica in situ se asume que toda la proteína soluble (fracción a) es
rápidamente degradable, pero a través de los poros de las bolsas escapan partículas de
pequeño tamaño que pueden abandonar el rumen sin ser degradadas, provocando así una
sobreestimación de la degradabilidad. Además, se ha comprobado que parte del N soluble
puede escapar a la degradación ruminal y contribuir al aporte intestinal de AA (Volden et
al., 1998), por lo que algunos autores recomiendan utilizar modelos más complejos para la
evaluación proteica de alimentos con un alto contenido en N soluble, como los ensilados y
los forrajes verdes. Por otra parte, la fracción b está compuesta por proteínas que presentan
diferentes ritmos de degradación, pero el modelo descrito utiliza un único ritmo de
degradación (kd) para todas ellas. Si bien este modelo es el utilizado por el NRC (2001),
INRA (Sauvant et al., 2004) y AFRC (1992) en sus tablas de valoración, otros sistemas
proponen una evaluación más compleja de las fracciones proteicas.
El sistema de valoración energética y proteica de Cornell (CNCPS; Sniffen et al.,
1992) diferencia cinco fracciones en la proteína de los alimentos (A, B1, B2, B3, and C) y
le adjudica a cada una un ritmo de degradación diferente. La fracción A (NPN) es el
porcentaje de proteína que es solubilizada instantáneamente al llegar al rumen y tiene un kd
infinito, por lo que entraría directamente a formar parte del pool de NH3-N. La fracción C
es la proteína no degradable y contiene las proteínas asociadas con la lignina y taninos, así
como las dañadas por la acción del calor (p.e. productos de la reacción de Maillard). La

PROTEÍNAS, PÉPTIDOS, AMINOÁCIDOS Y OTRAS FUENTES NITROGENADAS 47
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
suma de las tres fracciones B es la proteína verdadera potencialmente degradable, pero las
tres fracciones se diferencian en su ritmo de degradación: B1 tiene un kd rápido (120–
400% h-1
) y es casi totalmente degradada en el rumen, B2 tiene un kd menor (3–16% h-1
) y
es degradada parcialmente, y B3 tiene un kd lento (0,06–0,55% h-1
) debido a su asociación
a la pared celular. Las fracciones RDP y RUP se calculan sumando la contribución de cada
fracción a las mismas, considerando que A solo contribuye a la RDP y C únicamente a
RUP. Para el cálculo de la ED se usa el mismo valor de kp para todas las fracciones de un
mismo alimento, el cual está en función del nivel de ingestión, porcentaje de forraje en la
ración y el contenido en NDF físicamente efectiva, y para el cálculo de kp se utilizan
ecuaciones diferentes según se trate de forrajes verdes, secos o concentrados.
La evaluación de las diferentes fracciones de la proteína de los alimentos en el
sistema de Cornell se lleva a cabo determinando su solubilidad en diferentes buffers
(Figura 4), de tal forma que la fracción A es la proporción soluble en buffer borato-fosfato
que no es precipitada en ácido tricloroacético (TCA) y la fracción C es el porcentaje de la
proteína total que se recupera con la fibra ácido detergente (ADICP). La fracción B1 es
soluble en buffer borato-fosfato y precipitada en TCA, la fracción B3 es la diferencia entre
la proteína recuperada en la fibra neutro detergente (NDICP) y la ADICP, mientras que la
fracción B2 constituye el resto de las proteínas y se calcula como diferencia entre la
proteína total y la suma de las fracciones A, B1, B3 y C. Este procedimiento no implica
técnicas analíticas complejas ni requiere el uso de animales canulados, pero se necesitan
varias determinaciones para cuantificar todas las fracciones proteicas, por lo que el cálculo
de las fracciones RDP y RUP es más complejo.
Figura 4.- Fraccionamiento de las proteínas de los alimentos en el sistema de
valoración energética y proteica de Cornell (CNCPS; Sniffen et al., 1992)
En el Reino Unido se ha desarrollado el sistema Feed into Milk (FiM; Thomas,
2004) para vacas lecheras que utiliza la técnica in situ pero tiene en cuenta la pérdida de
pequeñas partículas a través de los poros de las bolsas de nailon. Para ello determina el
componente soluble (“s”) de la proteína mediante la incubación de una muestra de

48 M. D. CARRO
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
alimento en agua durante 1 hora y filtrado posterior del líquido resultante a través de un
papel de filtro de 16 µm, siendo s la fracción no retenida en el papel. La cantidad de
pequeñas partículas se calcula como la diferencia entre la fracción “a” y “s” y se les
atribuyen las mismas características de degradación (valores de b y kd) que a las partículas
de mayor tamaño, pero se aplica el ritmo de paso de la fase líquida (kpliq) para calcular su
ED, mientras que a la fracción b se le aplica el ritmo de paso de la fase sólida, que tiene
diferentes valores para el forraje (kpfor) y el concentrado (kpcon) de la ración.
Un problema importante de la técnica in situ es la contaminación de los residuos no
degradados con N microbiano debido a la presencia de microorganismos ruminales
adheridos a las partículas no degradadas. Este hecho provoca una subestimación de la
degradabilidad proteica que puede llegar al 30-35% en alimentos fibrosos de degradación
lenta y con bajo contenido en proteína, como algunos forrajes y subproductos, pero
también puede ser considerable (9-10%) en forrajes de calidad media-alta y cereales de
degradación lenta como el maíz (Rodríguez y González, 2006; Arroyo y González, 2013).
Para evitar este problema se han propuesto tratamientos que permiten desligar los
microorganismos de los residuos de incubación, como la congelación, sonicación o el
tratamiento con un homogeneizador tipo Stomacher®. Sin embargo, estos procedimientos
complican la técnica, eliminan solo parcialmente los microorganismos (Ramos et al.,
2009a) y en algunos casos incluso pueden eliminar partículas que no han sido degradadas.
Alternativamente, puede medirse el grado de contaminación con N microbiano utilizando
un marcador microbiano (p.e. 15
N), pero este procedimiento es muy costoso económica y
laboralmente y rara vez se realiza. Como alternativa a la técnica in situ se han propuesto
métodos in vitro que utilizan proteasas comerciales (Krishnamoorthy et al., 1983; Licitra et
al., 1998) e incluso la utilización de la técnica NIRS, pero estos procedimientos todavía
requieren una mejor validación y estandarización antes de su uso rutinario en la práctica.
La degradabilidad de la proteína de la ración está determinada por sus propias
características, pero también por su tiempo de permanencia en el rumen. Debido a la
relación inversa existente entre el tiempo de permanencia de la digesta en el rumen y el
nivel de ingestión, el NRC (2001) muestra en sus tablas los valores de la RUP de la
proteína de diferentes alimentos calculados para una ingestión de materia seca equivalente
al 2 y al 4% del peso vivo. Como se observa en la Tabla 2, los valores de RDP se reducen
al aumentar el nivel de ingestión, y simultáneamente se produce un aumento en la fracción
RUP. Por ello, en aquellas condiciones en las que se prevea una reducción de la
degradabilidad ruminal de la proteína hay que tener en cuenta que estará aumentando
proporcionalmente la fracción by-pass. Sin embargo, también hay que considerar que si la
reducción se debe a tratamientos realizados para proteger la proteína (calor, ácidos, …) se
puede haber afectado negativamente la digestiblidad intestinal de la fracción by-pass
(Pereira et al., 1998). Otros sistemas también muestran diferentes valores de
degradabilidad de la proteína en función del tiempo de retención en el rumen. El AFRC
(1992) asume un valor de Kp de 0,02 h-1
para vacas y ovejas a niveles de ingestión bajos,
0,05 h-1
para animales alimentados hasta dos veces mantenimiento y 0,08 h-1
para vacas
lecheras con una producción de leche diaria superior a 15 kg, aunque también ofrece una

PROTEÍNAS, PÉPTIDOS, AMINOÁCIDOS Y OTRAS FUENTES NITROGENADAS 49
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
ecuación para poder calcular el valor de kp en función del nivel de alimentación real de los
animales. Por el contrario, el INRA (2007) no tiene en cuenta la influencia del tiempo de
retención en el rumen en la ED y asume un valor fijo de kp de 0,06 h-1
para su cálculo.
Tabla 2.- Efecto del nivel de ingestión de materia seca en el contenido en proteína
degradable (RDP) y no degradable (RUP) en el rumen de diferentes alimentos (% de
proteína bruta; adaptado del NRC, 2001)
RDP RUP
Ingestión de materia seca (% peso vivo) 2% 4% 2% 4%
Heno (gramíneas-leguminosas) 65,2 62,9 34,8 37,1
Ensilado de maíz 66,7 64,7 33,3 35,3
Harina de maíz 63,0 52,7 37,0 47,3
Harina de soja (45%) 42,0 31,0 58,0 69,0
Soja entera tratada 70,9 60,6 29,1 39,4
3.- SÍNTESIS DE PROTEÍNA MICROBIANA
La proteína microbiana representa la fracción mayoritaria de la proteína que llega al
duodeno de los rumiantes y aunque su proporción puede variar en función del nivel de
producción, en la mayoría de los casos representa entre el 50 y 80% de la MP (Storm and
Ørskov, 1983; AFRC, 1992). Las bacterias constituyen la mayoría de la MCP, ya que los
protozoos sufren un intenso reciclado y son retenidos selectivamente en el rumen, por lo
que su contribución al flujo de MCP es reducida, con valores que oscilan entre el 11 y 15%
del total del N que llega al duodeno (Shabi et al., 2000; Yáñez-Ruiz et al., 2006).
Una característica importante de la MCP es su buen perfil aminoacídico en
comparación con el de la carne y leche de los rumiantes. En las vacas lecheras la metionina
y la lisina han sido identificadas con frecuencia como los dos primeros AA limitantes
(NRC, 2001) y la histidina es considerado el tercero cuando se utilizan raciones basadas en
ensilados de hierba y con cebada y avena como cereales en lugar de maíz (Schwab, 2012).
Como puede observarse en la Figura 5, el contenido (% de CP) en metionina y lisina de las
bacterias ruminales es similar al de la leche y superior al de la carne de los animales, pero
el contenido en estos dos AA en los alimentos es inferior, con la excepción de la harina de
pescado, cuyo uso no está permitido en la UE en la actualidad. En cuanto a la histidina, su
contenido en las bacterias ruminales es menor que en la leche y la carne, pero en este caso
algunos alimentos como el maíz, soja, algodón y harina de pescado presentan
concentraciones superiores a las de la leche y carne. Las diferencias en el perfil
aminoacídico de la MCP y los alimentos reflejan la capacidad que tienen las bacterias
ruminales para sintetizar todos los AA, incluidos los esenciales.

50 M. D. CARRO
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
Figura 5.- Contenido medio (% de la proteína total) en metionina, lisina e histidina en
la leche y carne de rumiantes, las bacterias ruminales y algunos alimentos (adaptado
de NRC (2001) y Schwab (2012))
Los microorganismos ruminales necesitan energía y N para su mantenimiento y
crecimiento, por lo que la disponibilidad de ambos es el principal factor determinante de la
cantidad de MCP sintetizada. La energía se obtiene fundamentalmente de la fermentación
de los hidratos de carbono, ya que la grasa no aporta energía para la síntesis microbiana
(únicamente la obtenida en la fermentación del glicerol de los triacilglicéridos) y la energía
obtenida al fermentar la proteína es muy reducida. Por ello, para estimar la cantidad de
MCP que puede sintetizarse y calcular la cantidad necesaria de N degradable que debe
aportase en la ración se tiene en cuenta el contenido en materia orgánica fermentada en el
rumen (FOM) u otra medida del contenido en energía fermentable de la ración. El NRC
(2000; 2001) usa el total de nutrientes digestibles (TDN) corregido para el aporte de la
grasa (que no puede ser utilizada por los microorganismos como fuente energética) y da un
valor de 130 g MCP/kg TDN(corregido) para vacuno de cebo y vacas lecheras. El INRA
(2007) propone también un único valor de 145 g de MCP/FOM, calculando la FOM a
partir de la materia orgánica digestible corregida para el contenido en grasa, la proteína
indegradable y los productos de fermentación cuando se utilizan ensilados. El AFRC
(1993) da valores que oscilan entre 9 y 11 g de MCP/MJ de energía metabolizable
fermentada en el rumen (37,5 y 46 g de MCP/Mcal) según el nivel de ingestión (9 para
mantenimiento, 10 para crecimiento y 11 para lactación) y para el cálculo de la energía
fermentable también descuenta la contribución de la grasa y los productos de fermentación

PROTEÍNAS, PÉPTIDOS, AMINOÁCIDOS Y OTRAS FUENTES NITROGENADAS 51
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
en los ensilados. El sistema de Cornell (Russell et al., 1992) divide a los microorganismos
en dos grupos, según su capacidad para fermentar hidratos de carbono estructurales o no
estructurales y la eficiencia de síntesis se calcula para cada grupo por separado en función
de las características de los hidratos de carbono de la ración. Una vez calculada la cantidad
de MCP que puede sintetizarse a partir de la energía fermentable que aporta la ración,
puede determinarse la cantidad de RDP que se debe aportar. Para ello se tiene en cuenta la
eficiencia de captura del RDP de los microorganismos, que es del 85% según el NRC
(2001), 80% según el AFRC (1993) y 90% según el INRA (2007; 80% para el NPN).
Como se ha comentado, la mayoría de sistemas proponen un único valor para la
síntesis de MCP en función de la energía disponible, pero la eficiencia de la síntesis de
MCP es muy variable. En una revisión de diferentes estudios realizada por el ARC (1984)
los valores oscilaron entre 13,8 y 45,4 g N microbiano/kg FOM (86-284 g MCP/kg FOM)
en vacuno y entre 19,7 y 30,8 g N microbiano/kg FOM (123-193 g MCP/kg FOM) en
ovino. En ambas especies los valores más bajos se encontraron en animales que recibían
raciones con altas proporciones de concentrado o de ensilados de hierba. Si bien parte de la
variabilidad puede deberse a diferencias en los métodos de medida (marcadores de flujo,
marcadores microbianos, etc.), es indudable que el tipo de ración y otros factores afectan
de forma importante a la síntesis de MCP, por lo que a continuación se tratarán brevemente
algunos de ellos. El primer factor determinante de la cantidad de MCP sintetizada es la
energía obtenida a partir de la FOM, la cual está determinada por el tipo de hidratos de
carbono en la ración. Los hidratos de carbono rápidamente fermentables (almidón,
azúcares, pectinas) estimulan en mayor medida la síntesis de MCP en comparación con los
hidratos de carbono estructurales (celulosa, hemicelulosa), por lo que, en general, la
eficiencia de la síntesis de MCP es más alta con raciones mixtas que con raciones
exclusivamente forrajeras, pero el efecto de la relación forraje:concentrado sobre la
eficiencia es variable. Algunos estudios en ovejas han señalado que la eficiencia de síntesis
puede reducirse al sobrepasar un 70% de concentrados en la ración (Mathers y Miller,
1981), pero en otros se ha observado un aumento en la eficiencia hasta alcanzar el 70-80%
de concentrados (Carro et al., 2000; Ramos et al., 2009b). Por ello, el efecto global de la
relación forraje:concentrado dependerá de las características de ambos componentes. Por
ejemplo, cuando existe un exceso de hidratos de carbono rápidamente fermentables se
provoca un descenso acusado del pH y de la actividad fibrolítica, se reduce el ritmo de
paso de la digesta y disminuye la eficiencia de la síntesis de MCP (Russell and Wallace,
1997). En definitiva, es necesaria una combinación adecuada de hidratos de carbono rápida
y lentamente fermentables para alcanzar elevadas eficiencias de síntesis, ya que ello
proporciona a los microorganismos un aporte energético sostenido en el tiempo. En este
punto es importante resaltar que cualquier fallo en la estimación de la FOM de la ración
resultará en estimaciones erróneas de la cantidad de RDP que debe aportarse y por ello, de
la MCP sintetizada.
El segundo factor que determina el crecimiento microbiano es la disponibilidad de
N. Las bacterias pueden usar NH3-N, AA y péptidos para la síntesis de MCP, mientras que
los protozoos no pueden usar NH3-N (Jouany y Ushida, 1999). Si bien durante años se

52 M. D. CARRO
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
asumió que las bacterias que fermentan los hidratos de carbono estructurales únicamente
utilizaban NH3-N como fuente de N (Russell et al., 1992), diferentes estudios han mostrado
que la disponibilidad de N no amoniacal (AA, péptidos o proteína) provoca una
estimulación del crecimiento microbiano sobre sustratos fibrosos, así como un aumento de
su eficiencia (Griswold et al., 1996; Carro et al.,1999; Carro y Miller, 2001). Las distintas
especies bacterianas difieren en su preferencia por péptidos o AA para la síntesis de sus
estructuras, así como en el grado de captación de NH3-N. El grado de captación de NH3-N
por las bacterias ruminales depende de la disponibilidad de las diferentes fuentes de N y
puede oscilar desde el 26 hasta 100% cuando solo disponen NH3-N (Wallace, 1997). La
captación de NH3-N depende de numerosos factores y todavía no se conoce cuál es la
relación NH3-N / N proteico idónea para el crecimiento microbiano, aunque posiblemente
esta relación dependa del tipo de ración y de las poblaciones ruminales que se desarrollen
con la misma. Independientemente de la fuente de N utilizada con prioridad por los
microorganismos ruminales, éstos deben disponer de suficiente RDP para optimizar el uso
de la energía generada en la fermentación de la materia orgánica.
Además de aportar una adecuada cantidad de energía y N para los
microorganismos, es necesario que la disponibilidad de ambos componentes esté
sincronizada para lograr una mayor eficiencia de utilización. Aunque este punto parece
obvio, en muchos de los experimentos realizados para investigarlo no se han encontrado
diferencias y los resultados han sido inconsistentes. En una revisión de experimentos con
vacas lecheras realizada por Cabrita et al. (2006) se concluyó que la sincronización en el
aporte de energía y N en el rumen no tuvo un efecto consistente en las concentraciones de
NH3-N, la MCP sintetizada, la ingestión de materia seca o la producción de leche. Con
frecuencia los tratamientos experimentales de estos estudios se han diseñado modificando
los componentes de la ración (tipo y nivel de inclusión), por lo que es difícil determinar si
los resultados se deben a la sincronización en la liberación de energía y N o a la
modificación de los ingredientes (Dewhurst et al., 2000). Otra limitación es que en la
mayoría de estos estudios el ritmo de degradación de la energía y N se ha determinado in
situ, con las limitaciones que tiene la técnica. Por otra parte, las poblaciones microbianas
ruminales son complejas y es posible que la liberación de ambos componentes esté
sincronizada para una subpoblación y no lo esté para otra. Adicionalmente, el reciclado de
la urea en la saliva y directamente al rumen hace que exista un aporte de N que podría
compensar la falta de sincronización en la liberación de energía y N. En este sentido se ha
comprobado que la cantidad de N reciclada al rumen aumenta cuando se administran
raciones con un bajo contenido en proteína y que puede llegar a ser el total de la urea
generada en el hígado en animales que reciben raciones deficientes en N (Patra, 2015).
Este hecho se interpreta como un mecanismo que permite aportar N a los microorganismos
ruminales de una forma relativamente constante cuando el aporte de N alimenticio es
limitado y permite evitar a corto plazo los efectos negativos de una asincronía en la
liberación de energía y N. Un exceso de RDP en relación a la disponibilidad de energía
reduce la eficiencia de la síntesis de MCP y la proteína es degradada a NH3-N, que es
absorbido y debe ser metabolizado a urea en el hígado.

PROTEÍNAS, PÉPTIDOS, AMINOÁCIDOS Y OTRAS FUENTES NITROGENADAS 53
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
Además del aporte de energía y N otro factor que ejerce una influencia importante
sobre la síntesis de MCP es el ritmo de dilución de las fases sólida y líquida del rumen. El
ritmo de dilución se ve afectado por varios factores, como el nivel de ingestión, las
características de la ración (p.e., la proporción de forraje, tamaño de partícula, …) y las
condiciones fisiológicas de los animales (Stern et al., 2006). En varios estudios in vitro se
ha observado un aumento en la síntesis de MCP al aumentar el ritmo de dilución de las
fases sólida y líquida del rumen, lo que se ha atribuido a una selección de microorganismos
con elevados ritmos de crecimiento, un menor gasto energético en el mantenimiento
bacteriano y a una reducción de la lisis bacteriana y la depredación por los protozoos. Sin
embargo, los resultados in vivo han sido contradictorios y se han observado aumentos,
reducciones e incluso una ausencia de efectos en la eficiencia de síntesis de MCP al
aumentar el ritmo de dilución (Bach et al., 2005). Estas inconsistencias en los resultados se
han atribuido a que al tratar de modificar el ritmo de dilución se provocan cambios en otras
variables que pueden afectar al crecimiento microbiano. A pesar de ello, como se ha
comentado anteriormente, el AFRC (1992) adjudica mayores eficiencias del crecimiento
microbiano a los niveles altos de ingestión, es decir, a altos ritmos de dilución. En los
estudios in vitro el ritmo de dilución suele incrementarse aumentando el flujo de saliva
artificial que se introduce en los fermentadores, por lo que se provoca un aumento
simultáneo del pH, que también puede afectar al crecimiento microbiano. Para diferenciar
estos efectos Martínez et al. (2009) realizaron un experimento en el que compararon dos
ritmos de dilución de la fase líquida y sólida manteniendo valores similares de pH
mediante modificaciones en la composición de la saliva artificial. El aumento del ritmo de
dilución de la fase líquida produjo un aumento de la digestibilidad de la ración, la
producción de ácidos grasos volátiles, pero no afectó significativamente al crecimiento
microbiano, aunque aumentó la actividad fibrolítica y proteolítica ruminal. El aumento del
ritmo de paso de la fase sólida provocó una reducción de la digestibilidad de la ración, de
la producción de ácidos grasos volátiles y del crecimiento microbiano y su eficiencia, pero
no modificó la degradabilidad de la proteína. La existencia de interacciones significativas
entre el ritmo de dilución de la fase líquida y de la fase sólida para numerosos parámetros
en este estudio pone de manifiesto la dificultad de interpretar los efectos de cada factor.
El pH ruminal es otro factor que afecta a la eficiencia de síntesis de MCP. En una
revisión realizada por St-Pierre (2001) con 187 datos de experimentos in vivo se observó
que la eficiencia de síntesis se mantiene relativamente constante en un amplio rango de pH
(5,5 – 7,0), pero la cantidad de MCP sintetizada aumentó al reducirse el pH ruminal,
debido a que las raciones con mayor aporte de FOM para el crecimiento microbiano
también provocan un menor pH ruminal. Sin embargo, los bajos valores de pH ruminal que
se observan en animales con acidosis pueden causar un efecto negativo sobre el
crecimiento microbiano, especialmente sobre el de los protozoos. Además, cuando el pH es
muy bajo las bacterias necesitan dedicar más energía a mantener su pH intracelular, lo que
reduce la energía disponible para su crecimiento (Russell y Dombrowski, 1980). Estos
hechos contribuyen a explicar las bajas eficiencias de síntesis de MCP observadas en
animales en cebo intensivo a base de concentrados. El sistema de Cornell tiene en cuenta la
influencia del pH en el crecimiento microbiano y cuando el contenido en fibra neutro

54 M. D. CARRO
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
detergente (NDF) que aporta el forraje de la ración es inferior al 20% de la materia seca
reduce la síntesis de MCP en un 2,5% por cada 1% de disminución en el contenido en NDF
(Russell et al., 1992).
La concentración de protozoos en el rumen es mucho menor que la de las bacterias,
pero debido a su mayor tamaño pueden representar entre un 10 y un 50% del total de la
biomasa ruminal (Jouany y Ushida, 1999). Sin embargo, su reciclado intraruminal es
también mayor que el de las bacterias y se calcula que sólo un 15% de los protozoos del
rumen pasan al duodeno, por lo que su contribución al flujo duodenal de N puede oscilar
entre el 5 y 30% del total (Jouany y Ushida, 1999), aunque los valores varían con el tipo de
ración administrada y sus efectos en el medio ambiente ruminal. Los protozoos ejercen una
actividad depredadora de bacterias importante, lo que aumenta el reciclado ruminal interno
del N bacteriano y tiene un elevado coste energético. Por ello, en numerosos estudios se ha
observado que la defaunación reduce las concentraciones de NH3-N en el rumen y aumenta
el crecimiento microbiano y su eficiencia. Además, los protozoos ejercen efectos
beneficiosos sobre la fermentación ruminal, ya que tienen actividad fibrolítica (contribuyen
a la degradación de la fibra), algunas especies metabolizan el ácido láctico (reducen el
riesgo de acidosis), engullen gránulos de almidón (regulan su ritmo de fermentación) y
algunos ejercen un papel importante en la degradación de compuestos tóxicos. Por ello, la
defaunación puede causar efectos negativos sobre la fermentación ruminal aunque aumente
el flujo duodenal de MCP. En cualquier caso, actualmente no existen compuestos que
puedan usarse en la práctica para provocar una defunación total o parcial de los animales.
La síntesis de MCP también requiere minerales y vitaminas, que suelen estar presentes en
las cantidades adecuadas en las raciones que reciben los rumiantes. Uno de los elementos
más importantes es el azufre (S), ya que es necesario para la síntesis de los AA azufrados.
Una ingestión limitada de S puede reducir la síntesis de MCP, por lo que se recomienda
que la relación N:S esté entre 10:1 y 12:1 (Van Soest, 1994). Este hecho es especialmente
importante cuando se administra NPN, ya que cuando las raciones contienen un nivel
adecuado de proteína verdadera la relación N:S suele encontrarse dentro de estos límites.
El fósforo es otro mineral necesario para la síntesis de MCP, ya que forma parte de los
ácidos nucleicos e interviene en el intercambio energético. Sin embargo, con las raciones
utilizadas normalmente en la práctica no se producen deficiencias que puedan afectar a la
síntesis microbiana.
Finalmente, hay que mencionar que no toda la MCP es proteína verdadera, ya que
un 20-25% del N de los microorganismos se encuentra en forma de ácidos nucleicos
(aproximadamente un 15% del total del N) y de otros compuestos no proteicos (Storm y
Ørskov, 1983), por lo que para el cálculo de la cantidad de AA que formarán parte de la
MP debe tenerse en cuenta este hecho, así como la digestibilidad intestinal de la MCP. El
NRC (2001) y el INRA (2007) consideran que la MCP contiene un 80% de proteína
verdadera con una digestibilidad intestinal del 80%, lo que implica que el 64% de la MCP
puede ser absorbida en el duodeno y contribuir a la mezcla de AA que forma la MP. El
AFRC (1993) considera que la MCP contiene un 75% de proteína verdadera con una

PROTEÍNAS, PÉPTIDOS, AMINOÁCIDOS Y OTRAS FUENTES NITROGENADAS 55
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
digestibilidad intestinal del 85%, llegando al mismo porcentaje de MCP absorbida (64%) y
el sistema de Cornell propone una cifra ligeramente inferior (60%).
4.- DIGESTIBILIDAD INTESTINAL DE LA PROTEÍNA
El flujo duodenal de proteína está compuesto por la fracción RUP del alimento, la
MCP sintetizada y la proteína endógena. La fracción endógena incluye fundamentalmente
mucoproteínas de la saliva, células epiteliales y restos celulares del tracto digestivo (boca,
esófago, retículo-rumen, omaso y abomaso) y secreciones del abomaso (NRC, 2001),
aunque parte de las mucoproteínas y células epiteliales son degradadas por los
microorganismos ruminales y no llegan al duodeno. La cuantificación del flujo duodenal
de proteína endógena es compleja, pero algunos estudios han mostrado que su cantidad
depende de la materia seca ingerida, por lo que el NRC (2001) propone una ecuación para
estimarla (N endógeno (g / d) = 1,9 x kg de materia seca ingerida (g/d)). Según estas
estimaciones, la cantidad de proteína endógena sería de 47,5 g de N/d (297 g de CP) en una
vaca de 600 kg que consumiese diariamente 25 kg de materia seca. Esta cifra concuerda
con los datos de algunos autores que indican que la proteína endógena supone entre un 9 y
12% del flujo proteico duodenal (Brandt et al., 1980), aunque otras estimaciones elevan
esta contribución hasta el 20% del total (Ouellet et al., 2002). A pesar de que existen pocos
datos sobre el contenido en proteína verdadera de la proteína endógena que llega al
duodeno, el NRC (2001) asume que es un 50% y que tiene una digestibilidad del 80%, por
lo que su contribución a la MP sería el 40% del total de proteína endógena. Los sistemas
INRA (2007) y AFRC (1993) no proponen un cálculo específico de la cantidad de proteína
endógena, sino que tienen en cuenta su contribución en algunos de los factores que
utilizan.
En cuanto a la fracción RUP del alimento, el NRC (2001), INRA (2007) y AFRC
(1993) asumen que el 100% es proteína verdadera. El AFRC (1993) estima la
digestibilidad verdadera de la fracción RUP asumiendo que el N ligado a la fibra ácido
detergente (ADIN) es completamente indigestible y el resto tiene una digestibilidad
verdadera del 90%. Para los sistemas NRC (2001) e INRA (2007) la digestibilidad
intestinal varía en función del alimento considerado, estando la mayoría de los valores
entre el 50 y 100%. El sistema de Cornell asume que la digestibilidad intestinal de los AA
que abandonan el rumen sin degradar es del 100% para las fracciones B1 y B2 y del 80%
para la fracción B3.
La determinación de la digestibilidad intestinal in vivo es un proceso complejo que
requiere el uso de animales fistulados y presenta problemas similares a los señalados
anteriormente para la determinación de la degradabilidad ruminal de la proteína. Una
alternativa es la técnica de las bolsas móviles, que consiste en incubar muestras del
alimento en cuestión, previamente incubadas en el rumen, en bolsas de nailon que se
introducen en el duodeno y se recuperan en el íleon o en las heces. Este método de
valoración es el utilizado por el NRC (2001) y el INRA (2007), aunque no está exento de

56 M. D. CARRO
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
aspectos críticos. Por una parte, el valor de digestibilidad intestinal está condicionado por
el tiempo que el alimento ha sido incubado en el rumen previamente, ya que normalmente
la digestibilidad intestinal se reduce al aumentar el tiempo de incubación ruminal
(González, 2006). Por ello, a veces se incuban los alimentos en el rumen durante un tiempo
fijo, pero ello no representa lo que realmente ocurre en la fisiología ruminal y puede
modificar la composición de la proteína by-pass. Como alternativa, González (2006)
propuso la generación de una muestra representativa del flujo post-ruminal del alimento
sobre la que se determinaría la digestibilidad intestinal. Esta muestra se obtiene mezclando
de forma ponderada los residuos de la incubación ruminal a diferentes tiempos, en función
del flujo de alimento, el cual se establece a partir del ritmo de degradación y paso del
alimento a través del rumen. Si bien este procedimiento refleja mejor la fisiología ruminal,
también es más costoso laboralmente. La utilización de la técnica de las bolsas de nailon
móviles también tiene otros aspectos críticos, ya que cuando las bolsas se recogen en las
heces las partículas no digeridas pueden estár contaminadas con la proteína de los
microorganismos del intestino grueso que las han colonizado.
Una alternativa a la técnica de las bolsas móviles pueden ser las técnicas in vitro, de
las cuales la más utilizada es la propuesta por Calsamiglia y Stern (1995). Este
procedimiento consiste en tres pasos, durante los cuales el residuo no degradado del
alimento tras 16 h de incubación en el rumen se incuba con pepsina y con pancreatina y
finalmente se precipita con ácido tricloroacético. Los valores obtenidos con este método
mostraron una correlación excelente con la digestibilidad intestinal de 34 muestras
determinada in vivo. Los valores de digestibilidad intestinal que utiliza el NRC (2001) han
sido obtenidos de 54 estudios realizados con las bolsas móviles y 6 estudios con la técnica
in vitro de Calsamiglia y Stern (1995) y en el caso de alimentos para los que no se disponía
de valores se utilizó el valor del INRA (Jarrige, 19897). Los valores del INRA (2007) han
sido también obtenidos con la técnica de las bolsas móviles y como ya se ha comentado el
AFRC (1993) utiliza el contenido en ADIN para estimar su digestibilidad intestinal.
5.- UTILIZACIÓN METABÓLICA DE LOS AMINOÁCIDOS
Los AA absorbidos en el intestino pueden tener diferentes destinos en los tejidos
corporales, siendo el primero de ellos la síntesis proteica. La síntesis proteica se lleva a
cabo en todos los tejidos del organismo, debido a que las proteínas tisulares experimentan
continuamente procesos de degradación y síntesis. Para la síntesis las células disponen de
los AA absorbidos, los AA no esenciales que pueden sintetizar y los AA movilizados de
los tejidos corporales. Otro posible destino de los AA absorbidos es la desaminación,
proceso en el que se genera un grupo amino y un esqueleto carbonado. El grupo amino
puede transferirse a un cetoácido y dar lugar a un nuevo AA (no esencial) o a otros
compuestos nitrogenados como los nucleótidos y aminas (ver Figura 6). Si el grupo amino
no es utilizado se transforma en urea en el hígado y posteriormente la urea se excreta en la
orina. En cuanto al esqueleto carbonado, puede servir para la síntesis de un nuevo AA o
transformarse en diferentes metabolitos (varían según el AA del que procedan) que entran

PROTEÍNAS, PÉPTIDOS, AMINOÁCIDOS Y OTRAS FUENTES NITROGENADAS 57
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
en el ciclo de Krebs y son oxidados para producir energía o dan lugar a la producción de
oxalacetato que se transforma en fosfoenolpiruvato y glucosa en el caso de los AA
glucogénicos.
Figura 6.- Representación esquemática del metabolismo de los aminoácidos
absorbidos
Las vacas lecheras necesitan AA para sintetizar la proteína láctea y su proteína
corporal, pero también para el crecimiento del feto cuando están gestantes y para formar
todas las proteínas necesarias para el mantenimiento de las funciones vitales (enzimas,
hormonas, etc.). Cada una de esas proteínas tiene una secuencia característica de AA, por
lo que las necesidades aminoacídicas del animal variarán en función de su estado de
lactación, crecimiento y gestación. Durante la lactación aumenta de forma muy marcada el
ritmo de síntesis proteica, especialmente en la glándula mamaria y en el hígado (Bell,
1995). La capacidad de síntesis proteica en la glándula mamaria es muy elevada y la
proteína láctea sintetizada en las vacas lecheras supone de media la mitad de la MP
disponible (Lapierre et al., 2012). Sin embargo, la transferencia de un AA absorbido a la
proteína de la leche no es un proceso lineal, ya que implica complejas interacciones entre
varios órganos corporales, aunque la glándula mamaria juega un papel central en el
proceso. Los tejidos esplácnicos (ntestino, grasa mesentérica, páncreas, bazo e hígado)
pueden alterar el perfil de los AA absorbidos, de tal forma que los AA que llegan a la
glándula mamaria no se corresponden exactamente con los AA absorbidos. A pesar de que
estos tejidos representan menos del 10% del peso corporal en la vaca lechera, contribuyen
casi al 50% del total de la síntesis de proteína corporal (Lapierre et al., 2006). El paso de
los AA a través de la pared intestinal va acompañado de la oxidación de algunos de ellos,
por lo que este es el primer punto en el que puede existir una pérdida de AA absorbidos.

58 M. D. CARRO
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
En vacas lecheras se ha observado una cierta desaparición de AA ramificados (leucina,
isoleucina y valina) y treonina, mientras que en ovejas se observó una reducción de la
cantidad de leucina y metionina, pero no hubo cambios en la lisina y fenilalanina (Lobley y
Lapierre, 2003).Posteriormente, el hígado capta AA para diferentes funciones, como la
síntesis de proteínas (que posteriormente son vertidas a la circulación sanguínea), la
síntesis de glucosa (uso preferente de AA no esenciales) y la retirada de un exceso de AA
para evitar una hiperaminoacidemia. Sin embargo, la captación hepática de AA es
selectiva, es decir, no todos los AA son captados en la misma proporción. Existe un grupo
de AA esenciales (Grupo I; histidina, metionina, fenilalanina, treonina y triptófano) que
son captados en gran medida por el hígado, con valores que oscilan entre el 36% para la
metionina y el 48% para la fenilalanina (Lapierre et al., 2005). Por el contrario, otro grupo
de AA (Grupo II; isoleucina, leucina, lisina, valina y arginina) prácticamente no son
captados por el hígado. Adicionalmente, se ha observado que el metabolismo de los AA en
el hígado se modifica al iniciarse la lactación. Doepel et al. (20009) observaron que en
vacas lecheras al inicio de la lactación aumentaba el flujo aminoacídico en la porta
respecto al observado antes del parto (debido al incremento en la ingestión), pero no se
producía un aumento de la captación de AA por el hígado, sino que incluso disminuyó para
algunos AA (histidina, metionina, fenilalanina, glicina y tirosina). Esta observación
confirma que la concentración de AA en la sangre portal no regula por sí sola la captación
hepática de AA, y que la prioridad metabólica al inicio de la lactación es destinar los AA a
la síntesis de proteína láctea en lugar de a otras funciones como la gluconeogénesis. Otros
estudios en vacas lecheras también han mostrado que en condiciones de bajo aporte
proteico el hígado reduce su captación de AA (Raggio et al., 2004).
Debido a los procesos que se producen en los tejidos esplácnicos, el perfil de AA
que llega a la glándula mamaria es diferente del absorbido, pero además la glándula
mamaria también realiza una captación selectiva de AA (Lapierre et al., 2012). Como
puede observarse en la Figura 7, la cantidad de AA del Grupo I (histidina, metionina,
fenilalanina y treonina) captada por la glándula mamaria es muy similar a la cantidad
secretada en la leche, es decir, la relación captación/secreción es cercana a 1. Sin embargo,
los AA del Grupo II (isoleucina, leucina, lisina, valina y arginina) son captados en mayor
cantidad de la que aparecen en la leche, es decir, la relación captación/secreción es superior
a 1. Esto implica que los AA del Grupo I son exclusivamente utilizados para la síntesis de
la proteína láctea, mientras que el exceso de AA del Grupo II se utiliza para sintetizar AA
no esenciales o se oxida para obtener energía.
Los AA que no son usados para las funciones metabólicas son retirados de la
circulación por los tejidos en los que se encuentran las enzimas responsables de su
catabolismo. Por ejemplo, los AA del Grupo I son retirados principalmente por el hígado,
mientras que los AA del Grupo II son retirados por la glándula mamaria, el intestino y los
tejidos periféricos y solo en muy baja proporción por el hígado (Lapierre et al., 2012).
Debido a que la retirada del exceso de AA no ocurre en el lugar principal de la síntesis
proteica implicada en el proceso productivo (es decir, la glándula mamaria) sino en
diversos tejidos, Lapierre et al. (2015) proponen la utilización de una eficiencia combinada

PROTEÍNAS, PÉPTIDOS, AMINOÁCIDOS Y OTRAS FUENTES NITROGENADAS 59
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
de mantenimiento y lactación en lugar de eficiencias separadas para cada uno de estos
procesos, y la última versión del sistema de Cornell (Van Amburgh et al., 2015) ya utiliza
una eficiencia combinada. Adicionalmente, la captación de los AA por los diferentes
tejidos corporales y la glándula mamaria depende en gran medida del aporte de AA
(Doepel et al., 2004; Raggio et al., 2004). A partir de un metaanálisis de estudios en los
que se realizaron infusiones de AA a vacas lecheras, Lapierre et al. (2007) calcularon una
eficiencia combinada de utilización de diferentes AA para mantenimiento y lactación en
diferentes situaciones de aporte de AA. Como puede observarse en la Tabla 3, la
eficiencia de utilización de los AA aumentó al reducirse el aporte de MP, pero se redujo
ante un exceso de MP. Los AA aportados en exceso deben ser catabolizados por la
glándula mamaria, el hígado y otros tejidos corporales, con el consiguiente gasto
energético. Por el contrario, en situaciones de escasez de AA se reduce su catabolismo en
la glándula mamaria, tejidos esplácnicos y teijdos periféricos, de tal forma que la eficiencia
de uso aumenta. Estos resultados contribuyen a explicar la falta de respuesta en la
producción de leche obtenida a veces en la práctica al administrar raciones ligeramente
deficientes o excedentes en MP.
Figura 7.- Relación entre la cantidad de aminoácidos captada por la glándula
mamaria y la cantidad secretada en la leche (adaptado de Lapierre et al., 2012)
A pesar de todas las evidencias existentes que muestran una eficiencia variable de
utilización de los AA en la vaca lechera, el NRC (2001) propone una eficiencia fija de
utilización de la MP de 67% para mantenimiento y lactación, mientras que las primeras
versiones del sistema de Cornell (O’Connor et al., 1993) proponían eficiencias diferentes
para cada AA, pero fijas dentro de cada proceso productivo (valores entre 66 y 85% para
mantenimiento y gestación y entre 42 y 100% para lactación). El AFRC (1993) y el INRA
(2007) también proponen eficiencias fijas de utilización de la MP para lactación, con
valores del 68% y 64%, respectivamente. En el caso del vacuno de cebo, el NRC (2000)
tiene en cuenta que la eficiencia de uso de los AA para la ganancia de peso se va
reduciendo a medida que aumenta el peso vivo, al igual que lo hacen las necesidades. Por
ello da una ecuación que permite estimar la eficiencia de utilización de los AA en función

60 M. D. CARRO
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
del peso vivo vacío para animales de menos de 300 kg (p.e. para un ternero de 150 kg de
peso la eficiencia sería 66,3%) y a partir de 300 kg de peso la eficiencia es solamente del
49,2%.
Tabla 3.- Eficiencia combinada (mantenimiento y lactación) de utilización de
aminoácidos y de la proteína metabolizable (MP) en relación a su aporte óptimo
(adaptado de Lapierre et al., 2015)
Aporte óptimo
50% 75% 100% 125%
Arginina 0,68 0,63 0,58 0,52
Histidina 0,96 0,85 0,76 0,68
Isoleucina 0,77 0,72 0,67 0,60
Leucina 0,74 0,67 0,61 0,55
Lisina 0,81 0,75 0,69 0,62
Metinonina 0,85 0,74 0,66 0,59
Fenilalanina 0,70 0,63 0,57 0,51
Treonina 0,68 0,68 0,66 0,60
Valina 0,79 0,72 0,66 0,59
MP 0,72 0,67 0,62 0,56
6.- UTILIZACIÓN DE AMINOÁCIDOS PROTEGIDOS
El principal objetivo de la suplementación de las raciones con AA protegidos es
lograr un perfil de AA en la MP que se ajuste a las necesidades de los animales. Las
recomendaciones del NRC (2001) en cuanto al contenido en lisina y metionina en la MP
para optimizar su uso para la producción de proteína láctea son 7,2 y 2,4%,
respectivamente, pero dado que normalmente no pueden conseguirse, las recomendaciones
“prácticas” son 6,5 y 2,2%, respectivamente, con una relación lisina/metionina cercana a
3,1. Como se ha visto anteriormente esto es muy difícil de conseguir debido al bajo
contenido en metionina y lisina de los alimentos, y más si se considera la prohibición
europea de usar harinas de origen animal. En definitiva, la utilización de AA protegidos
trata de lograr un equilibrio entre los aportes de AA en la MP y las necesidades del animal,
que permita la máxima eficiencia de utilización de los AA.
Existen numerosas evidencias de que la suplementación de las raciones de vacas
lecheras con AA esenciales puede aumentar la producción de leche y la cantidad de
proteína láctea producida, pero también se ha observado que no existe respuesta a la
suplementación con AA no esenciales (Doepel y Lapierre, 2010). Los esfuerzos
investigadores se han centrado en la metionina y la lisina por ser con mayor frecuencia los
primeros AA limitantes de la producción lechera. Sin embargo, la ventaja de utilizar estos
AA no es solo el aumento de la producción de leche o de proteína láctea, sino que permiten

PROTEÍNAS, PÉPTIDOS, AMINOÁCIDOS Y OTRAS FUENTES NITROGENADAS 61
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
reducir la excreción de N (al mejorar la eficiencia de utilización de los AA) y pueden
mejorar el balance energético del animal al disminuir el gasto energético que conlleva la
síntesis de urea para eliminar el NH3-N generado en la desaminación de AA no esenciales
que no pueden utilizarse al faltar un AA limitante. Por otra parte, el aumento en la
producción de proteína láctea observado en varios estudios se concentra en la caseína, lo
que beneficia especialmente a la industria de transformación quesera.
A pesar del gran interés que presenta la suplementación con AA protegidos, los
resultados obtenidos en la práctica han sido muy variables, ya que en numerosas ocasiones
se han observado aumentos significativos en la producción de proteína láctea y de leche,
pero en otras no se ha observado efecto o incluso se han producido descensos en la
producción (Robinson et al., 2000; Benefield et al., 2009). Existen varias razones que
pueden justificar estos resultados. En primer lugar, es fundamental utilizar una fuente de
AA con una buena protección, es decir, que no se degrade en el rumen, pero que se libere y
absorba en el intestino. En este sentido, algunos estudios han puesto de manifiesto
diferencias importantes en el aporte real de metionina logrado con distintos productos
comerciales (Südekum et al., 2004; Patton, 2010), por lo que sería aconsejable comparar
los resultados de estudios que hayan utilizado el mismo producto. Otro posible factor de
variación es la fase de la lactación en la que se ha realizado la prueba, ya que en general la
suplementación con AA protegidos es más efectiva en las fases iniciales de la lactación que
en fases más avanzadas, aunque este efecto se puede ver influenciado por otros factores
(Schwab et al., 1992). De forma similar, suelen ser mejores las respuestas productivas
obtenidas con raciones con niveles “normales” de proteína (14-18%) que con niveles
inferiores o superiores, por lo que este factor también influencia la comparación de
resultados.
Por otra parte, la suplementación con AA protegidos puede producir un retorno
económico positivo sin que haya un aumento de la producción de leche y/o de proteína
láctea, por lo que la producción no debería ser el único aspecto valorado. En este sentido,
se pueden utilizar AA protegidos para reducir el contenido en proteína de la ración sin
mermas en la producción y también para reducir la movilización de reservas en el
postparto, contribuyendo así a prevenir patologías típicas de este período (Patton, 2009).
Finalmente hay que señalar que aunque nos estamos refiriendo al uso de AA protegidos, se
podría lograr el mismo efecto si fuera posible el aporte de los dichos AA a través de
modificaciones en la RUP, es decir, su efecto fisiológico es la modificación del perfil
aminoacídico de la RUP.
Patton (2010) realizó un metaanálisis de los resultados de 35 experimentos en los
que se utilizó metionina protegida (entre 5 y 72 g/d) y observó que el principal efecto era
un aumento (0,07%) en el porcentaje de proteína verdadera en la leche, lo que supuso un
incremento medio de 27 g de proteína láctea. Este valor es similar al 0,09% de incremento
observado en varios estudios revisados por el NRC (2001) para estimar las concentraciones
idóneas de metionina en la MP para optimizar la síntesis de proteína láctea. En la revisión
de Patton (2010) los efectos de la suplementación con metionina protegida en la ingestión,

62 M. D. CARRO
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
producción de leche y grasa de la leche fueron mínimos. Un aspecto interesante es que la
respuesta obtenida no dependió de la cantidad de metionina suplementada, de la
deficiencia en metionina de las raciones control ni de su contenido en lisina (expresada en
g/d o en % de la MP), lo que se atribuyó a una reducción de la eficiencia de utilización de
la metionina al aumentar su disponibilidad o a la limitación de la síntesis de leche por la
falta de otros nutrientes. En cuanto a la influencia de la ración, se observó que las vacas
que recibían raciones con alfalfa tenían mayores aumentos en la producción de leche al
recibir la suplementación con metionina que las que recibían raciones con otros forrajes,
aunque la mayor respuesta en el aumento de la proteína láctea se observó con raciones
basadas en ensilado de maíz. El autor de este estudio sugiere que la metionina podría
ejercer algún papel regulador en el flujo sanguíneo a la glándula mamaria o en la síntesis
de proteína láctea. Sin embargo, no pudo predecirse la respuesta a la suplementación con
metionina a partir de las características de la ración o de la situación productiva de los
animales. Una conclusión importante de este estudio fue que la respuesta a la metionina no
depende necesariamente de la adición de lisina o de sus niveles en el flujo duodenal.
Robinson (2010) realizó otro metaanálisis de estudios en los que se había utilizado
metionina protegida (54 estudios), lisina protegida (12 estudios) o una combinación de
ambas (47 estudios). La suplementación con metionina (13 g/d de media) aumentó el
contenido en proteína y grasa de la leche y la eficiencia de utilización del N para la
producción de leche, mientras que la suplementación con lisina (24 g/d de media) redujo la
ingestión de materia seca y aumentó la eficiencia de la producción lácte (kg leche/kg
materia seca ingerida). La suplementación con ambos AA produjo efectos similares a los
observados con la metionina. En este estudio tampoco se observaron correlaciones
significativas entre las características de la ración y/o de los animales y la respuesta
obtenida a la suplementación con los AA protegidos. Un aspecto importante resaltado en
este estudio es que es que en muy pocos trabajos se ha medido el flujo real de AA al
duodenal o se ha tratado de estimar. Por ello, la discrepancia entre resultados también
puede deberse a fallos en la estimación del flujo duodenal de metionina y lisina en las
vacas que reciben la ración control, de tal forma que el nivel de suplementación diseñado
para el estudio no se correspondiera con el que debería ser para obtener una respuesta
productiva. En este sentido, una mejora en la precisión de la predicción del flujo
aminoacídico duodenal supondría un gran avance en el estudio de los factores que pueden
influenciar la respuesta a la suplementación con AA protegidos.
Otro metanálisis interesante es el realizado recientemente por DairyCo (2013) en el
que se analizaron los efectos de la suplementación con metionina y lisina protegidas
obtenidos en 12 estudios realizados vacas entre el inicio y mitad de la lactación que
recibían raciones con un contenido en proteína bruta igual o inferior al 15%. En la mayoría
de cada uno de los estudios se observaron aumentos numéricos de la producción de
proteína y de leche, pero las diferencias fueron significativas en muy pocos. Sin embargo,
en el metanálisis se observó un aumento significativo (P=0,002) en la producción de
proteína y de leche (P=0,05) al suplementar las raciones con ambos AA. Los autores de
este estudio señalan de nuevo la dificultad de predecir la respuesta productiva a la
suplementación con AA protegidos, pero sus resultados indican que la suplementación con

PROTEÍNAS, PÉPTIDOS, AMINOÁCIDOS Y OTRAS FUENTES NITROGENADAS 63
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
ambos AA produce mejores resultados que utilizados individualmente, especialmente en
las primeras fases de la lactación. La respuesta a la suplementación con estos AA
protegidos puede depender de la composición de la ración y características de los animales
(fase de la lactación, balance energético, producción de leche, etc.), pero en este estudio
tampoco pudo establecerse una relación clara entre estos factores y la respuesta observada.
Vyas y Erdman (2009) analizaron los resultados de varios estudios en los que se
suplementó con lisina (18 experimentos) o metionina (35 experimentos) la ración de vacas
lecheras o se infundieron a nivel postruminal. Los resultados mostraron que la eficiencia de
utilización de estos AA se reducía al aumentar su disponibilidad. El incremento en la
producción de proteína láctea pasó de 16 a 4 g de proteína/g de metionina metabolizable al
variar la ingestión de metionina suplementaria de 25 a 70 g/d. En el caso de la lisina, los
valores se redujeron de 5,0 a 3,2 g proteína láctea/g de lisina metabolizable al variar la
ingestión de lisina suplementaria de 80 a 203 g/d. Estos resultados indican que una
elevada suplementación reduce la respuesta obtenida, confirmando otros estudios previos
(Robinson et al., 2000).
Por último, hay que mencionar que además de los beneficios descritos en la
producción de proteína láctea y de leche, el aporte de metionina puede ejercer otros efectos
beneficiosos en el metabolismo del animal, aunque estos han sido menos investigados. Una
mayor disponibilidad de metionina podría contribuir a la formación de las lipoproteínas de
baja densidad necesarias para la exportación de los triacilglicéridos fuera del hígado,
debido a que la metionina es necesaria para la formación de la apoproteína B100 y actúa
como donadora de grupos metilo para la síntesis de fosfatidilcolina. Esta acción podría
reducir la infiltración grasa en el hígado, por lo que la suplementación con metionina
estaría especialmente indicada en las vacas en transición (Durand et al., 1992). Osorio et al.
(2013) observaron que la administración de metionina protegida (para alcanzar
concentraciones 2,12% de la MP) a vacas en transición producía una mejora del estado
inmune y reducía la incidencia de cetosis. Otro efecto posible de la metionina es su
participación en los mecanismos antioxidantes del organismo. Recientemente, Osorio et al.
(2014) han observado que la suplementación con metionina protegida en vacas en
transición produjo un aumento de la síntesis hepática de antioxidantes (glutatión y
carnitina) y redujo la concentración de interleucina 6, indicando una reducción en el estado
inflamatorio y una mejora del estado inmunometabólico tras el parto. Otras posibles
acciones de la metionina se enmarcan en el funcionamiento de la función reproductora.
Robert et al. (1997) observaron que en vacas que recibían una ración equilibrada en
metionina y lisina se producía una mejor involución uterina tras el parto (% de animales
con tamaño uterino normal) y se redujo el número de inseminaciones, aunque hay que
señalar que estos efectos no llegaron a ser significativos. Thiaucourt (1996) observó
también una reducción de 5 días del intervalo entre el parto y la primera inseminación en
vacas que recibían una ración con niveles adecuados de lisina y metionina en pruebas de
campo con 2000 vacas. La otra acción a través de la cual los AA protegidos pueden ejercer
una acción positiva en la función reproductora es a través de la reducción del contenido en
proteína de la ración, lo que resulta en menores niveles de urea en el organismo.

64 M. D. CARRO
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
7.- COMENTARIOS FINALES
La complejidad de los procesos digestivos de los rumiantes y los cambios
fisiológicos que se producen durante la lactación dificultan claramente el racionamiento
para estos animales. Sin embargo, los avances en el conocimiento de su fisiología y
metabolismo realizados en los últimos años han permitido realizar avances en este campo,
los cuales están siendo o serán incorporados por los diferentes sistemas de alimentación
para ajustar de forma más precisa los aportes proteicos de la ración a las necesidades
aminoacídicas de los animales. En este sentido, es previsible una mejora si se consideran
eficiencias variables para la síntesis de proteína microbiana, en función de las
características de la ración y del nivel de ingestión, así como eficiencias variables de
utilización de cada AA en función del aporte proteico y del nivel productivo. Cualquier
mejora en la estimación de la degradabilidad de la proteína de la ración y de la
disponibilidad de energía fermentable para los microorganismos redundará en una
estimación más precisa de la síntesis de proteína microbiana. Adicionalmente, determinar
la contribución real de la proteína endógena al flujo duodenal y analizar los factores que la
afectan es otro punto en el que existen posibilidades de mejora. Aunque no se ha tratado en
esta revisión, una estimación más precisa de las necesidades aminoacídicas de los animales
también contribuiría en gran manera a la elaboración de raciones más adecuadas.
Desde un punto de vista práctico, es fundamental optimizar la síntesis de proteína
microbiana administrando forrajes y concentrados de buena calidad, así como equilibrar
los aportes de energía y RDP para los microorganismos. Un aporte deficiente de RDP
reduce la digestión ruminal, la ingestión de alimento, la producción de leche y la síntesis
de proteína microbiana, lo que empeorará casi siempre el perfil aminoacídico de la
proteína metabolizable. Un exceso de RDP también es perjudicial, ya que reduce la
eficiencia de la síntesis microbiana, supone un gasto energético para eliminar el exceso de
NH3-N, puede afectar negativamente a la reproducción y casi siempre supone un aumento
del coste de la ración. El aporte de RUP también debe ser adecuado para satisfacer las
necesidades de MP, pero no debe aportarse un exceso de RUP, ya que supone un mayor
coste económico y un gasto energético para el animal, “ocupa espacio” en la ración (que
podría ser reemplazado por hidratos de carbono que aportan energía) y reduce la eficiencia
de utilización de los aminoácidos. La utilización de AA protegidos en la ración es una
práctica en auge, que parece especialmente eficaz en las primeras fases de la lactación y
cuando se desea reducir el contenido en proteína de la ración y la excreción nitrogenada sin
reducir la producción, aunque también puede utilizarse para incrementar la producción de
proteína láctea. Sin embargo, las limitaciones actuales en la predicción del flujo duodenal
de aminoácidos y de la eficiencia de su utilización metabólica hacen que sea difícil
determinar las situaciones en las que son más eficaces y la respuesta productiva esperada.

PROTEÍNAS, PÉPTIDOS, AMINOÁCIDOS Y OTRAS FUENTES NITROGENADAS 65
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
8.- REFERENCIAS
AFRC (1992) Nutritive Requirements of Ruminant Animals: Protein. Technical Committee
on Responses to Nutrients. Report No. 9. Farnham Royal, Commonwealth
Agricultural Bureaux International, Wallingford, Reino Unido.
AFRC (1993) Energy and protein requirements for ruminants. An advisory manual
prepared by the AFRC Technical Committee on Responses to Nutrients.
Commonwealth Agricultural Bureaux International, Wallingford, Reino Unido.
ARC (1984) The nutrient requirements of ruminant livestock. Supplement nº 1.
Commonwealth Agricultural Bureaux, Reino Unido.
ARMSTEAD, I.P. y LING, J.R. (1993) Appl. Environ. Microbiol. 59: 3360–3366.
ARROYO, J.M. y GONZÁLEZ, J. (2013) J. Anim. Physiol. Anim. Nutr. 97: 109-118.
BACH, A., CALSAMIGLIA, S. y STERN, M.D. (2005). J. Dairy Sci. 88 (Suppl.): E1-29.
BELL, A.W. (1995) J. Anim. Sci. 73: 2804-2819.
BENEFIELD, B.C., PATTON, R.A., STEVENSON, M.J. y OVERTON, T.R. (2009) J.
Dairy Sci. 92: 4448-4455.
BRANDT, M., ROHR, K. y LEZIEN, P. (1980) J. Anim. Physiol. Anim. Nutr. 44: 26.
BRODERICK, G.A., WALLACE, R.J. y ØRSKOV, E.R. (1991) En: Physiological
Aspects of Digestion and Metabolism in Ruminants. (eds. T. Tsuda, Y. Sasaki y R.
Kawashima), Academic Press, New York, USA. pp: 541-594.
CABRITA, A.R.J., DEWHURST, R.J., ABREU, J.M.F. y FONSECA., A.J.M. (2006)
Anim. Res. 55: 1-24.
CALSAMIGLIA, S. y STERN, M.D. (1995) J. Anim. Sci. 73: 1459-1465.
CALSAMIGLIA, S., BUSQUET, M., CARDOZO, P.W., CASTILLEJOS, L. y FERRET,
A. (2007) J. Dairy Sci. 90: 2580-95.
CALSAMIGLIA, S., CARDOZO, P.W., FERRET, A. y BACH, A. (2008) J. Anim. Sci.
86: 702-711.
CALSAMIGLIA, S., FERRET, A., REYNOLDS, C.K., KRISTENSEN, N.B. y VAN
VUUREN, A.M. (2010) Animal 4: 1184-1196.
CARRO, M.D. y MILLER, E.L. (1999) Br. J. Nutr. 82: 149-157.
CARRO, M.D., LÓPEZ, S., VALDÉS, C. y GONZÁLEZ, J.S. (1999) J. Anim. Physiol.
Anim. Nutr. 81: 212-222.
CARRO, M.D., VALDÉS, C., RANILLA, M.J. y GONZÁLEZ, J.S. (2000) Anim. Sci. 70:
127-134.
CARRO, M.D. (2001) Inv. Agr.: Prod. San. Anim. 16: 5-27.
COLEMAN, G.S. (1985) J. Agric. Sci., Camb. 105: 39-43.
DAIRYCO (2013) The effect of reducing dietary crude protein on nitrogen utilisation,
milk production, health and fertility in dairy cows. Agriculture and Horticulture
Development Board. Warwickshire, Reino Unido.
DEWHURST, R.J., DAVIES, D.R. y MERRY, R.J. (2000) Anim. Feed Sci. Technol. 85:
11-21.
DOEPEL, L., LOBLEY, G.E., BERNIER, J.F., DUBREUIL, P. y LAPIERRE, H. (2009)
J. Dairy Sci. 92: 3233-3243.
DOEPEL, L. y LAPIERRE, H. (2010) J. Dairy Sci. 93: 3264-3274.
DUBREUIL, P. y LOBLEY, G.E. (2005) Anim. Sci. 80: 11-22.

66 M. D. CARRO
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
DURAND, D., CHILLIARD, Y. y BAUCHART, D. (1992) J. Dairy Sci. 75: 279.
FEDNA (2010). Tablas FEDNA de composición y valor nutritivo de alimentos para la
fabricación de piensos compuestos. (eds. C. de Blas, G.G. Mateos y P. García-
Rebollar), 3ª edición. Fundación Española para la Nutrición Animal. Madrid, España.
GONZÁLEZ, J. (2006) En: XXII Curso de Especialización FEDNA. Fundación Española
para la Nutrición Animal. Madrid, España. pp: 203-216.
GRISWOLD, K.E., HOOVER, W.H., MILLER, T.K. y THAYNE, W.V. (1996) J. Anim.
Sci. 74: 483-491.
HINO, T. y RUSSELL, J.B. (1985) Appl. Environ. Microbiol. 50: 1368-1374.
INRA (1988) Alimentation des bovins, ovins et caprins. R. Jarrige (ed.), INRA edition,
Paris, Francia.
INRA (2007) Alimentation des bovins, ovins et caprins: besoins des animaux, valeurs des
aliments. Quae, Versailles, Francia.
JARRIGE, R. (1989) Ruminant Nutrition: Recommended Allowances and Feed Tables.
Institut National de la Recherche Agronomique, Libbey, Eurotext, Paris, France.
JOUANY, J.P. y USHIDA, K. (1999) Asian Australas. J. Anim. Sci. 12: 113-128.
KRISHNAMOORTHY, U., SNIFFEN, C.J., STERN, M.D. y VAN SOEST, P.J. (1983)
Br. J. Nutr. 50: 555-568.
LAPIERRE, H., BERTHIAUME, R., RAGGIO, G., THIVIERGE, M.C., DOEPEL, L.,
PACHECO, D. DUBREUIL, P. y LOBLEY, G.E. (2005) Anim. Sci. 80 : 11-22.
LAPIERRE, H., RAGGIO, G., OUELLET, D.R., BERTHIAUME, R., DOEPEL, L. y
PACHECO, D. (2006) En: 21st Annual Southwest Nutrition & Management
Conference. Tempe, AZ, USA. pp: 15-27.
LAPIERRE, H., LOBLEY, G.E., OUELLET, D.R., DOEPEL, L. y Pacheco, D. (2007) En:
Proceedings of the Cornell Nutrition Conference for Feed Manufacturers. Cornell
University, NY, USA. pp: 39-59.
LAPIERRE, H., LOBLEY, G.E., DOEPEL, L., RAGGIO, G., RULQUIN, H. y
LEMOSQUET, S. (2012) J. Anim. Sci. 90: 1708–1721.
LAPIERRE, H., DOEPEL, L., PACHECO, D. y OUELLET, D.R. (2015) En: 26th Florida
Ruminant Nutrition Symposium. Gainesville, Florida, USA. pp: 166-178.
LICITRA, G., HERNANDEZ, T.M. y VAN SOEST, P.J. (1996) Anim. Feed Sci. Technol.
57: 347– 358.
LICITRA, G., LAURIA, F., CARPINO, S., SCHADT, I., SNIFFEN, C.J. y VAN SOEST,
P.J. (1998) Anim. Feed Sci. Technol. 72: 1-10.
LOBLEY, G.E. y LAPIERRE, H. (2003) En: Progress in research on energy and protein
metabolism. (eds. W.B. Souffrant y C.C. Metges). EAAP plublication No.109.
Wageningen Academic Publishers, Wageningen, Holanda. pp : 737-756.
MADSEN, J. y HVELPLUND, T. (1985) Acta Agric. Scand. Suppl. 25: 103-124.
MADSEN, J. y HVELPLUND, T. (1994) Livest. Prod. Sci. 39: 201-212.
MARTÍNEZ, M.E., RANILLA, M.J., RAMOS, S., TEJIDO, M.L. y CARRO, M.D. (2009)
J. Dairy Sci. 92: 3930-3938.
MATHERS, J.C. y MILLER, E.L. (1981) Br. J. Nutr. 45: 587-604.
MEHREZ, A.Z. y ØRSKOV, E.R. (1977) J. Agric. Sci. Camb. 88: 645-650.

PROTEÍNAS, PÉPTIDOS, AMINOÁCIDOS Y OTRAS FUENTES NITROGENADAS 67
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
NRC (2000) Nutrient Requirements of Beef Cattle. Seventh Revised Edition: Update 2000.
The National Academic Press. Washington D.C., USA.
NRC (2001) En: Nutrient Requirements of Dairy Cattle. Seventh revised edition. pp. 43-
104. The National Academic Press. Washington D.C., USA.
O'CONNOR, J.D., SNIFFEN, C.J. FOX, D.G. y CHALUPA, W. (1993) J. Anim. Sci. 71:
1298-1311.
ØRSKOV, E.R. y MCDONALD, I. (1979) J. Agri. Sci. Camb. 92: 499-503.
OSORIO, J.S., JI, P., DRACKEY, J.K., LUCHINI, D. y LOOR, J.J. (2013) J. Dairy Sci.
96: 6248-6263.
OSORIO, J.S., TREVISIM E., JI, P., DRACKEY, J.K., LUCHINI, D., BERTONI, G. y
LOOR, J.J. (2014) J. Dairy Sci. 97: 7437-7450.
OUELLET, D.R., DEMERS , M., ZUUR , G., LOBLEY, G.E., SEOANE, J.R., NOLAN,
J.V. y LAPIERRE, H. (2002). J. Dairy Sci. 85: 3013-3025.
PATRA, A. K. (2015) En: Rumen Microbiology: From Evolution to Revolution (eds.
D. N. Kamra). Springer, New Delhi, India. pp: 329-342.
PATTON, R.A. (2009) En: Proceedings of the 20th
Florida Ruminant Nutrition
Symposium. University of Florida, Gainesville, Florida. USA. pp: 39-51.
PATTON, R.A. (2010) J. Dairy Sci. 93: 2105-2118.
PEREIRA, J.C., CARRO, M.D., GONZÁLEZ, J., ALVIR, M. R. y RODRÍGUEZ, C.A.
(1998) Anim. Feed Sci. Technol. 74:107-121.
RAGGIO, G., PACHECO, D., BERTHIAUME, R., LOBLEY, G.E., PELLERIN, D.,
ALLARD, G., DUBREUIL, P. y. LAPIERRE, H. (2004) J. Dairy Sci. 87: 3461-
3472.
RAMOS, S., TEJIDO, M.L., RANILLA, M.J., MARTÍNEZ, M.E., SARO, C. y CARRO,
M.D. (2009a) J. Dairy Sci. 92: 5659-5668.
RAMOS, S., TEJIDO, M.L., MARTÍNEZ, M.E., RANILLA, M.J. y CARRO, M.D.
(2009b) J. Anim. Sci. 87: 2924-2934.
ROBERT, J.E., WILLIAMS, P.E.V. y BOUZA, B. (1997) J. Dairy Sci. 80 (Suppl. 1): 248.
ROBINSON, P.H., CHALUPA, W., SNIFFEN, C.J., JULIEN, W.E., SATO, H.,
FUJIEDA, T., UEDA, T. y SUZUKI, H. (2000) J. Anim. Sci. 78: 1067-1077.
ROBINSON, P.H. (2010) Livest. Sci. 127: 115-126.
RODRÍGUEZ, C.A. y GONZÁLEZ, J. (2006) Br. J. Nutr. 96: 316-325.
RUSSELL, J.B. y DOMBROWSKI, D.B. (1980) Appl. Environ. Microbiol. 39: 604–610.
RUSSELL, J.B., O’CONNOR, J.D., FOX, D.G., VAN SOEST, P.J. y SNIFFEN, C.J.
(1992) J. Anim. Sci. 70: 3551-3561.
RUSSELL, J.B. y WALLACE R.J. (1997) En: The Rumen Microbial Ecosystem. (eds. P.N.
Hobson y C.S. Stewart), Springer, The Netherlands. pp: 246-282.
SAUVANT, D., PEREZ, J. M. y TRAN, G. (2004) Tables INRA-AFZ de composition et de
valeur nutritive des matières premières destinées aux animaux d'élevage. 2ème
édition. Ed. INRA Editions. Versailles, Francia.
SCHWAB, C.G., BOZAK, C.K., WHITEHOUSE, N.L. y OLSON, V.M. (1992) J. Dairy
Sci. 75: 3503-3518.
SCHWAB, C.G (2012) En: Proc: of the 23rd
Annual Florida Ruminant Nutrition
Symposium. University of Florida, Gainesville, Florida, USA.

68 M. D. CARRO
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
SHABI, Z., TAGARI, H., MURPHY, M.R., BRUCKENTAL, I., MABJEESH, S.J.,
ZAMWEL, S., CELIK, K. y ARIELI, A. (2000) J. Dairy Sci. 83: 2326-2334.
SNIFFEN, C.J., O’CONNOR, J.D., VAN SOEST, P.J., FOX, D.G. y RUSSELL, J.B.
(1992) J. Anim. Sci. 70: 3562-3577.
STERN, M.D., BACH, A. y CALSAMIGLIA, S. (2006) En: Proceedings of the 21st
Annual Southwest Nutrition and Management Conference, University of Arizona,
Tempe, AZ, USA. pp: 45-66.
STORM, E. y ØRSKOV, E.R. (1983) Br. J. Nutr. 50: 463-470.
ST-PIERRE, N.R. (2001) J. Dairy Sci. 84: 741-755.
SÜDEKUM, K.H., WOLFFRAM, S., ADER, P. y ROBERT, J.C. (2004) Anim. Feed Sci.
Technol. 113: 17-25.
THIAUCOURT, L. (1996) L' opportunite de la methionine protegee en production laitiere.
Bulletin des GTV 2B.
THOMAS, C. (2004) Feed into Milk: A New Applied Feeding System for Dairy Cows,
Nottingham, Nottingham University Press. UK.
VAN AMBURGH, M.E., COLLAO-SAENZ, E.A., HIGGS, R.J., ROSS, D.A.,
RECKTENWALD, E.B., RAFFRENATO, E., CHASE, L.E., OVERTON, T.R.,
MILLS, J.K., y FOSKOLOS, A. (2015) J. Dairy Sci. 98: 6361-6380.
VAN SOEST, P. J. (1994) Nutritional Ecology of the Ruminant. 2nd
Ed. Cornell University
Press, Ithaca, NY.
VANZANT, E.S., COCHRAN, R.C. y TITGEMEYER, E.C. (1998) J. Anim. Sci. 76:
2717-2729.
VOLDEN, H., MYDLAND, L.T. y OLAISEN, V. (1998) J. Anim. Sci. 80: 2704-2716.
VYAS, D. y ERDMAN, R.A. (2009) J. Dairy Sci. 92: 5011-5018.
WALLACE., R. y MCPHERSON, C.A. (1987) Br. J. Nutr. 88: 313-323.
WALLACE R.J. (1996) Ann. Zootech. 45: 301-308.
WALLACE, R.J. (1996) J. Nutr. 126: 1326S-1334S.
WALLACE R.J., ONODERA R. y COTTA M.A. (1997) En: The Rumen Microbial
Ecosystem. (eds. P.N. Hobson y C.S. Stewart), Springer, The Netherlands. pp: 283-
328.
WALLACE, R.J. (1997) En: Rumen Microbes and Digestive Physiology in Ruminants (ed.
R. Onodera, H. Itabashi, K. Ushida, H. Yano y Y. Sasaki), Japan Sci. Soc. Press,
Tokyo, Japón. pp: 95-105.
WALLACE R.J., ATASOGLU, C. y NEWBOLD, C.J. (1999) Asian Australas. J. Anim.
Sci. 12: 139-147.
YAÑEZ-RUIZ, D.R., SCOLLAN, N.D. y NEWBOLD, C.J. (2006) Br. J. Nutr. 96: 861-
869.

I JORNADA FEDNA − ANEMBE 25 Aniversario
sobre ALIMENTACIÓN DE RUMIANTES
MADRID, 10 de Noviembre de 2015
CAPÍTULO IV
LA IMPORTANCIA DEL MANEJO ALIMENTARIO
EN EL CEBO DE TERNEROS
M. Devant, A. Bach, A. Solé, B. Quintana, M. Verdú
Departamento Rumiantes IRTA


LA IMPORTANCIA DEL MANEJO ALIMENTARIO EN EL CEBO DE TERNEROS 71
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
LA IMPORTANCIA DEL MANEJO ALIMENTARIO
EN EL CEBO DE TERNEROS
M. Devant, A. Bach, A. Solé, B. Quintana, M. Verdú
Departamento Rumiantes IRTA
1.- INTRODUCCIÓN
De todos es conocido que la nutrición tiene un importante impacto sobre los costes
y los resultados productivos en terneros cebados intensivamente (Santos, 2014). En
anteriores ponencias FEDNA (Bacha et al., 2005) se analizaron los efectos de la nutrición
sobre el crecimiento de los teneros; el presente resumen se centra en estudios que han sido
realizados en el manejo de la nutrición (alimentación) más allá de la propia fórmula, ¿es
importante en qué presentación ofrecemos el pienso? ¿Es importante el diseño del corral,
en concreto el comedero? Así pues, la presente comunicación pretende resumir los
resultados de una serie de estudios realizados en cebo intensivo de terneros bajo el
concepto “postfórmula”. Los estudios que se describirán están realizados en el cebo
intensivo basado en ofrecer al ternero pienso y paja ad libitum en comederos separados.
Parte de los resultados presentados forman parte de estudios realizados en el IRTA en
colaboración con empresas, algunos están publicados y otros no.
Esquema 1.- Resumen de los factores post-fórmula

72 M. DEVANT, A. BACH, A. SOLÉ, B. QUINTANA y M. VERDÚ
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
En el esquema 1 se resumen los principales factores post-fórmula. Las materias
primas se han incluido a pesar de ser un factor característico de la fórmula porque existen
interacciones entre las materias primas (principalmente cereales, grasas y fibra) y factores
postfórmula como el proceso de fabricación (molienda, granulación, etc.). Los principales
factores postfórmula están relacionados con la fabricación, con el sistema de distribución y
los comederos. El animal también es importante porque puede interaccionar con los
anteriores factores; por ejemplo, la edad del animal (Kelly et al., 2010; Verdú et al., 2015a)
y la raza (Schwartzkopf-Genswein et al., 2001; Robison y Oddy, 2004; Cummins et al.,
2007) pueden afectar a la conducta alimentaria y con ello a factores como los comederos.
De dicho esquema, también queda claro que la cronología es importante, es decir, lo que
ocurra en fábrica afectará a los pasos posteriores.
En la presente comunicación para valorar los diferentes factores postfórmula
utilizaremos los siguientes criterios:
productivos: el efecto será positivo si se mejora la eficiencia y/o aumenta el
crecimiento (o kg canal). Es decir, aunque el análisis económico depende de los
precios del pienso y de la canal, en general, si una estrategia mejora la eficiencia
pero no aumenta el crecimiento (menos kg canal), dicha estrategia suele reducir los
beneficios y se considerará negativa.
de salud ruminal: timpanismo, laminitis, caquexia, acidosis, etc.
Los factores post-fórmula que analizaremos (forma presentación, diseño de
comedero de pienso, etc) pueden afectar diferentes aspectos como:
la selección y mermas del alimento.
a la digestibilidad de los nutrientes, sobretodo, a la disponibilidad de los almidones,
que pueden condicionar su tipo de fermentación ruminal (repentinas o graduales,
las consecuencias de la cuáles serán distintas).
Las diferentes estrategias alimentarias pueden afectar a la vez diferentes aspectos
(selección y digestibilidad), dificultando la interpretación de sus efectos. Así pues, si la
estrategia reduce el mermas, evidenciándose en una disminución del consumo total, sin
afectar la GMD, el IC respecto al tratamiento control se reducirá (mejorará). En el supuesto
que dicha estrategia causará fermentaciones ruminales de almidón muy repentinas, el
consumo descendería (Stock et al., 1995) y, aunque no siempre, también descendería la
GMD (Owens et al., 1997; Schwartzkopf-Genswein et al., 2004); en este caso, el IC podría
no verse afectado o a veces incluso se reduciría (mejoraría). Esta es una de las estrategias
que puede causar más confusión cuando se evalúa si tiene efectos positivos o negativos
sobre los resultados productivos y de salud ruminal, ya que a pesar que el IC pueda ser más
bajo (mejorarse), los animales no crecen y suele aumentar la incidencia de patologías
digestivas. Finalmente, se puede mejorar la digestibilidad de los almidones de forma que
estos sean fermentados más gradualmente; con ello se suele mejorar la disponibilidad de
energía y, en consecuencia, el crecimiento con un consumo similar mejorando el IC

LA IMPORTANCIA DEL MANEJO ALIMENTARIO EN EL CEBO DE TERNEROS 73
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
(Huntington, 1997). Huntington (1997) describe acertadamente (perfectamente) la
complexidad de incrementar la digestibilidad ruminal del almidón. La digestión del
almidón en el rumen parece no estar limitada (enzimas microbianos, tiempo retención) a
diferencia del intestino delgado (enzimas, ritmo de paso), pero puede ocasionar
consecuencias negativas sobre la salud animal y ruminal (timpanismos, laminitis, bajos
consumos, desmedro, etc.) cuando la fermentación es muy rápida (súbita). Por ello, se
buscan estrategias para reducir la velocidad de fermentación y/o neutralizar los efectos
negativos de dicha fermentación (Sauvant et al., 1999; González et al., 2012). Aumentar la
fermentación ruminal de almidones de forma más gradual, además de estar correlacionado
positivamente con la digestibilidad total de almidones y la disponibilidad de energía, tiene
ventajas en la estimulación de la síntesis microbiana y el aumento de la absorción
postruminal de glucosa ya debido al aumento de la secreción de amilasa pancreática
(Theurer, 1986; Huntington, 1997). En el siguiente esquema (Esquema 2) se resumen los
efectos que tienen dichos aspectos sobre los datos productivos.
Esquema 2.- Posibles consecuencias de los factores postfórmula y sus efectos sobre los
índices productivos
Como veremos a lo largo de la comunicación, conocer la conducta alimentaria es
un aspecto crítico para entender el éxito o el fracaso de las diferentes estrategias
alimentarias.
Hay estrategias postfórmula que en la presente comunicación no se abordarán ya
sea porque no hay datos, o porque su aplicabilidad es difícil y los posibles beneficios no
compensan las reformas necesarias para su implementación. Este es el caso de la
alimentación restringida (Hick et al., 1990; Loerch et al., 1990, Murphy and Loerch, 1994),
dar varias veces de comer al día (Robles et al., 2007), dar unifeed seco o húmedo (Zea,
2010; Iraira et al., 2014), o el tipo de forraje/fuente de fibra (Iraira et al., 2013), e incluso,
interacciones entre la alimentación y otros factores como el alojamiento o el estado
sanitario. Sin embargo, cada granja es una situación única, hay granjas que por su

74 M. DEVANT, A. BACH, A. SOLÉ, B. QUINTANA y M. VERDÚ
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
ubicación geográfica, o por su personal pueden aplicar estrategias muy interesantes y de
futuro que no se discuten en la presente comunicación.
2.- FORMA PRESENTACIÓN DEL CONCENTRADO
Cuando hablamos de forma de presentación nos referimos principalmente al
proceso de fabricación del concentrado. El procesado de alimentos tiene diferentes
objetivos (Bacha y Villamide, 2010): aumentar el valor nutritivo (más digestibilidad,
menos factores antinutritivos), mejorar las características físicas (más palatabilidad),
facilitar el manejo y transporte (más densidad). En la literatura norteamericana hablaríamos
principalmente de “grain processing”, del maíz en USA (Theurer, 1986) o la cebada en
Canadá (Dehghan-Bandaky et al. 2007). En estas revisiones muy extensas no suele haber
referencias sobre granulación, extrusión o el uso de un expander.
Uno de los aspectos más importante y controvertido es el efecto que tiene el
procesado sobre la digestibilidad ruminal de los almidones. Svihus et al. (2005) resumen
que dicha digestibilidad depende de la ratio amilosa/amilopectina, del tamaño y
organización de los gránulos de almidón, de la digestión de la matriz proteica que rodea el
gránulo de almidón, y de otros factores como inhibidores de la alfa-amilasa.
El procesado más simple es la molturación. Este primer paso destruye las capas
externas del grano (pericarpo) y expone el endospermo (granos de almidón) a la digestión,
aumentado la disponibilidad de superficie para el ataque enzimático microbiano en el
rumen y mejorando la fermentación del sustrato. La velocidad de digestión del almidón en
el rumen es inversamente proporcional al tamaño de partícula (Galyean et al., 1981; Offner
et al., 2003), aunque también está muy relacionado con el tipo de cereal, siendo su efecto
superior en el caso del maíz comparado con la cebada (Offner et al., 2003). En el maíz la
unión entre la proteína y el almidón es muy fuerte, la molturación tiene un mayor impacto.
Hay muchas cuestiones por resolver:¿cuál es el tamaño de molturación óptimo para
maximizar el uso energético del concentrado? ¿cuál es el tamaño crítico para evitar
trastornos digestivos? ¿Qué tipo de molturación deberíamos aplicar-grinding or rolling?
¿Debe ser el mismo tamaño de molturación para un concentrado en forma de harina que en
granulado?¿En una granulometría , qué es más importante, el valor medio del tamaño de
partículas o la distribución del tamaño de partículas? No hay un método estandarizado para
valorar la granulometría, y el método (tiempo, velocidad, número y tipo tamices) afecta
mucho a los resultados. Además, en la escasa bibliografía existente hay datos obtenidos
con tamices de diferentes tamaños dificultando la comparativa entre estudios (Dehghan-
Bandaky et al. 2007).Estandarizar el método con el cual realizamos las granulometrías para
cebo, y dar unas guías prácticas es un importante reto de futuro.
Tras la molturación se pueden aplicar otros métodos para procesar el concentrado:
la granulación (30-40 g/kg de agua y 80ºC ), la expansión (80 g/kg agua a 100ºC), antes de
la granulación, y la extrusión (180 g/kg agua a 110 ºC y presión). De hecho, durante la
granulación, tras la molturación, el producto final es una aglomeración de partículas
pequeñas en un pellet de mayor tamaño obtenido mediante un proceso mecánico resultante

LA IMPORTANCIA DEL MANEJO ALIMENTARIO EN EL CEBO DE TERNEROS 75
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
de la mezcla de humedad, calor y presión. Temperaturas por encima de 80 ºC, con exceso
de agua, pueden causar un aumento de la susceptibilidad de la degradación amilolítica, ya
que se pierde la estructura cristalina del almidón (Svihus et al., 2005). En general la
granulación, y el expander, afectan ligeramente la gelatinización, pero dicho parámetro no
es el único que determina la digestibilidad ruminal del almidón. Por ejemplo, la proteólisis
de la matriz que rodea el almidón también es importante (Svihus et al., 2005). Además,
cuando hay elevados contenidos de agua (en la extrusión, antes que se seque y enfríe)
pueden darse procesos de retrogradación que reducen la digestibilidad del almidón. Existen
datos contradictorios respecto al efecto de la granulación sobre la digestibilidad ruminal
del almidón (Ljøkjel et al., 2003; Offner et al., 2003). En general, no se observa un efecto
positivo de la granulación en cereales como la cebada, ya que tiene una elevada
digestibilidad de los almidones sin ser procesada. En cambio, el maíz experimenta un
aumento sustancial de la digestibilidad ruminal de los almidones cuando es granulado, ya
que sin ser procesado tiene una degradación mucho más lenta. De hecho, dichos trabajos
(Ljøkjel et al., 2003; Offner et al., 2003) se basan en la técnica in situ de degradación
ruminal del almidón; por lo tanto, otros factores como la conducta alimentaria (velocidad
de ingestión) no se tienen en cuenta al estimar la digestibilidad ruminal del almidón. Ello
explicaría que en los pocos trabajos in vivo (Castillo et al..; 2006) se observen resultados
que indicarían que la digestibilidad de los almidones de los pellets es superior a la de la
harina; los animales alimentados con pellet (vs harina) tenían un balance ácido-base más
estable en el tiempo, pero los elevados niveles de L-lactato séricos podrían denotar que los
animales tenían un mayor riesgo de acidosis ruminal. Bertipaglia et al. (2010), en una serie
de trabajos in vitro, observaron que la forma de presentación del pienso (harina vs pellet)
afecta a la velocidad de formación de gases durante las primeras horas (superior en harina)
Dichos autores lo atribuyen parcialmente a que la desintegración del pellet determina la
superficie de ataque de los microorganimos, retardando el acceso de estos a los nutrientes y
la posterior fermentación; el pellet tardaba 45 min en desintegrarse en el agua. Así pues, no
es fácil entender los efectos relacionados con la forma de presentación que afectan a la
fermentación ruminal. Por un lado, la mayor velocidad de ingesta podría conllevar a
acidosis (menos salivación), pero una menor disponibilidad inmediata de almidones a
causa de una lenta desintegración del pellet podría ocasionar una fermentación ruminal de
almindón más gradual (lenta). Como ya se ha mencionado, todo ello no se contempla en
las técnicas in situ. Finalmente, es muy difícil comparar entre estudios, ya que las
condiciones de fabricación suelen diferir. Aunque intentemos hacer estudios repetibles, de
todos es conocido que el desgaste de los martillos, la velocidad de granulación, entre otros,
son factores importantes que “evolucionan” durante el proceso de fabricación (cambian sus
condiciones óptimas o iniciales). Además, queda por esclarecer si la granulación puede
mejorar la disponibilidad de almidones, ya que se suele reducir el tamaño de molturación
para mejorar la calidad de la granulación. Bertipaglia et al.(2010) observaron en
concentrados con un 30% de cebada y un 10% de maíz que entre una harina de 3mm y un
pellet (a 2,5 mm) de 3,5 mm no había diferencias en digestibilidad in vitro de MS, y en
ambos casos tras una re-molturación a 1 mm mejoraba de digestibilidad de la MS.
Finalmente, quedaría por esclarecer el efecto que tiene la reducción de molturación a 1 mm
sobre la salud animal. La reducción del tamaño de partículas suele tener efectos negativos

76 M. DEVANT, A. BACH, A. SOLÉ, B. QUINTANA y M. VERDÚ
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
sobre la pared ruminal incrementando las fusiones de papilas, la queratinitzación y las
papilas con ramificaciones (Hironaka et al., 1979; Greenwood et al., 1997; Beharka et al,
1998).
No hay datos disponibles acerca de cuantos terneros cebados intensivamente se
alimentan con harina y granulado, aunque parece ser que hay zonas donde predomina más
un tipo de presentación que otro. Sin embargo, a nivel comercial hay un debate recurrente
sobre qué forma de presentación es mejor para cebar terneros: la harina o el granulado. En
la tabla 1 se resumen algunas ventajas e inconvenientes a tener en cuenta que van más allá
de los efectos que la forma de presentación tiene sobre los datos productivos.
Tabla 1.- Resumen de las ventajas e inconvenientes del pienso en harina o granulado
Harina Granulado
Disponibilidad almidón Aumenta en el caso del maíz
Higienización Mejora
Ingredientes Permite ir a niveles más
altos de grasa
Permite añadir ingredientes
que no permite la harina
Destrucción de vitaminas
y/o aditivos sensibles a altas
temperaturas
Aumenta
Coste de transporte y
almacenamiento
Más alto
Puntos críticos de calidad Control del tamaño de
partícula importante –
patologías digestivas
Control calidad: durabilidad
(calidad física del gránulo)
Manejo silos Se apelmaza más, más
mantenimiento
Selección Menor si la durabilidad es
buena y el porcentaje de
finos bajo
Mermas Alto, dependiente del tipo
de comedero
Por otro lado, la extrusión causa una rotura de la matriz proteica que rodea los
gránulos de almidón, así como una gelatinización de almidón (Solanas et al., 2005), y
puede causar una desnaturalización parcial de la proteína. Cuando la extrusión se aplica a
cereales, aumenta la degradabilidad proteica y la síntesis microbiana (Solanas et al. 2008);
pero cuando se aplica a fuentes proteicas solas o mezcladas con cereales desciende la
degradabilidad proteica (Solanas et al., 2007). Dichos efectos sobre los almidones y la
proteína se suelen traducir en menores consumos y mejores IC en animales alimentados
con extrusionado vs animales control (Solanas et al., 2005; 2008). Sin embargo no es un
proceso común en terneros de cebo por su elevado coste. De los pocos datos que hay sobre

LA IMPORTANCIA DEL MANEJO ALIMENTARIO EN EL CEBO DE TERNEROS 77
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
el efecto del expander, in situ se observó un descenso de la degradabilidad proteica en la
cebada y el trigo, pero un aumento de la digestibilidad de los almidones del maíz (Ljøkjel
et al., 2003).
Así pues, para la industria del ternero de cebo, optimizar el uso de los cereales y de
su procesado, manteniendo el funcionamiento ruminal y la salud animal, es uno de los
retos para mejorar la rentabilidad. De todas las posibles opciones, a continuación se
presentan algunos resultados de diferentes pruebas realizadas con diferentes empresas,
algunas están publicadas y otras no.
2.1.- ¿Granulado o harina?
Una de las discusiones más frecuente en la industria del cebo es que opción es
mejor, granulado o harina. No hay una respuesta única. En un primer estudio realizado con
machos frisones (220 kg de PV inicial), con 40% de maíz y 15% de trigo (1,03 UFC,
13,8% PB, 42% almidón), se observó un incremento de un 7% de la GMD, un descenso de
-2,5% del consumo, y un -8,8% del IC cuando los terneros se cebaron con pellet (3.5 mm)
vs harina (todo molturado a 2,75 mm excepto el maíz a 6mm) (Devant et al., 2010, no
publicado). Esta mejora en el crecimiento puede deberse a un mejor aprovechamiento del
almidón al contener la ración un 40% de maíz. Tal y como se ha descrito anteriormente, la
granulación mejora la digestibilidad ruminal del almidón del maíz. El consumo de harina
fue superior al de granulado, posiblemente debido a un mayor mermas (visualmente se
apreciaba). Castillo et al. (2006), en un estudio con un número más limitado de animales
(25% maíz), observó una mejora numérica de la GMD, un descenso del consumo y una
mejora del IC cuando los terneros consumían granulado respecto a harina.
2.2.- ¿El tipo de cereal puede afectar los resultados de la forma de presentación?
Se realizaron dos estudios con pasteros (240 kg de PV inicial); en el primero, el
pienso contenía un 40% de cebada y un 20% maíz, y en el segundo, un 40% de maíz y un
20% de cebada. En ambos estudios se contrastó una harina, con todos los ingredientes
molturados a 3 mm, y un granulado de 3.5 mm de diámetro. Cuando el pienso contenía un
40% de maíz, la GMD incrementó en un 12%, sin embargo cuando el pienso contenía un
40% de cebada no se observaron diferencias entre harina y pellet (Devant et al. 2014, 2015,
no publicado). Ello se podría explicar por el diferente efecto que tiene la granulación sobre
la digestibilidad de los almidones según el tipo de cereal. Mientras no parece aumentar la
digestibilidad de almidones en fuentes de cereales de alta digestibilidad (cebada), en
cereales de baja digestibilidad de almidones (maíz) la granulación tiene un efecto positivo
(Ljøkjel et al., 2003).
2.3.- ¿La disponibilidad de paja puede afectar los resultados de la forma de
presentación?

78 M. DEVANT, A. BACH, A. SOLÉ, B. QUINTANA y M. VERDÚ
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
Faleiro et al. (2011) realizaron dos estudios donde evaluaron el efecto de alimentar
terneros de cebo sin paja; en dichos estudios el concentrado contenía un 30% de cebada y
32% de maíz, y se presentaba en forma de harina molturada a 5 mm. En dichos estudios, a
pesar que algunos parámetros de comportamiento (rumia y estereotipias) fueron afectados,
no se observó un efecto sobre el consumo y el crecimiento. ¿En caso que el concentrado
fuera rico en maíz y en forma de granulado (mayor disponibilidad de almidones), tipo de
pienso predominante en Cataluña, los resultados serían los mismos? Se realizó un estudio
(Devant et al., 2015a) para evaluar el efecto de la forma de presentación del concentrado
(harina vs granulado) y la presencia o ausencia de paja sobre la conducta animal, la
morfología macroscópica y microscópica del rumen, y sobre la expresión de genes
relacionados con la conducta animal e inflamación. En este estudio, la harina estaba
molturada a 2,75 mm, excepto el maíz a 6mm, y el granulado molturado a 2,75 mm y con
3,5 mm de diámetro; los cereales eran 35% de maíz, 19% de cebada y 8% de trigo. Los
datos productivos no eran el principal objetivo de dicho estudio, pero se observó un efecto
opuesto sobre la GMD cuando no se aportaba paja según la forma de presentación. Cuando
no se suplementa paja, si la forma de presentación era granulado, la GMD se redujo en un
20%; por el contrario, aportando paja la GMD aumentó un 16%. El consumo de
concentrado, cuando el concentrado era granulado y no disponían paja, descendió un 13%
respecto a la forma de harina y sin paja. Las diferencias en GMD entre el granulado y la
harina con paja se observaron de nuevo en un estudio realizado con un mayor número de
animales (apartado 3.1). Dichos resultados ponen en evidencia que hay interacciones entre
factores postfórmula, cuando el concentrado es granulado, la presencia de paja parece ser
más crítica para evitar descensos de consumo y crecimiento; enmarcando los resultados
dentro de los tamaños de molturación y de granulado estudiados. Además, de este estudio
se desprendieron datos interesantes relacionados con el comportamiento y las lesiones
ruminales; independientemente de la forma de presentación, la ausencia de paja aumentó
las conductas orales no-nutritivas, descendió la rumia y aumentó la fusión de papilas
ruminales y la presencia de pelo en el rumen. Es decir, cuando no se aporta paja a los
terneros alimentados con concentrado hay una situación crítica de salud ruminal, los
animales pueden encontrarse en un mayor riesgo de padecer patologías digestivas que van
en contra del bienestar y, en un momento dado, contra el crecimiento.
2.4.- ¿Realmente la forma de presentación puede afectar al mermas? ¿en qué
medida?
Alrededor de los comederos se observa pienso que indica que hay mermas, reducir
dicho pienso supondría una mejora en la eficiencia sin perjudicar el crecimiento. Se realizó
un estudio (Devant et al., 2015b) para estimar el mermas y saber si este está afectado por la
forma de presentación. A priori, se hipotetizaba que los animales desperdiciarían más
harina que granulado. Otro factor que se evaluó fue el efecto de la experiencia alimentaria
previa. La experiencia alimentaria previa puede afectar a las preferencias de alimentos y al
comportamiento alimentario. En terneros lactantes se ha constatado que diferentes formas
de presentación afectan al comportamiento alimentario (Miller-Cushon et al., 2013). El
comportamiento alimentario (tiempo en comedero) está afectado por la forma de

LA IMPORTANCIA DEL MANEJO ALIMENTARIO EN EL CEBO DE TERNEROS 79
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
presentación (Ray y Drake, 1959). En dicho estudio se utilizaron 24 terneros Holstein (137
± 2,9 kg de PV y 117 ± 3,0 días de edad) alojados en corrales individuales cubiertos con
slat (1,2 m x 1,45 m). Cada corral estaba provisto de un comedero para paja y pienso.
Durante dos semanas 12 terneros recibieron el pienso en forma de harina (HA, tamaño
molturación 2,75 mm todos los ingredientes, excepto el maíz a 6 mm) y 12 terneros
recibieron granulado fabricado a partir de la misma harina (GRA, 3,5 mm de diámetro y 9-
10 mm de longitud). La fórmula del concentrado era la misma (cereales: 30,1% maíz,
18.08% cebada, 7,01% centeno). Tras estas dos primeras semanas experimentales
(semanas antes del cambio), y durante 3 semanas (semanas tras el cambio), a la mitad de
los terneros HA se les ofreció granulado y a la mitad de los terneros GRA se les ofreció
harina. Por lo tanto, durante estas 3 semanas los tratamientos tenían un diseño factorial 2
(forma de presentación previa: HA vs GRA) x 2 (forma de presentación actual: HA vs
GRA) quedando 4 tratamientos. En las dos primeras semanas se observó que el mermas
cuando los animales consumían harina era de un 5,9% vs un 2,8% del granulado .Tras el
cambio de presentación, los animales que consumían harina, y continuaban consumiendo
harina, mantenían un porcentaje de mermas elevado (6,8%), tal y como era de esperar. Sin
embargo, los animales que consumieron harina, y pasaron a granulado, redujeron el
porcentaje de mermas en un 2,3%, indicando que la experiencia previa no tenía ningún
efecto. Pero cuando la experiencia previa fue el consumo de harina, y pasaron a consumir
granulado, se observó un descenso del mermas del 2,5%, valor similar a los animales que
anteriormente consumían granulado y se mantuvieron consumiendo granulado (2,6%). Se
debería estudiar si dicho efecto se mantendría más allá de las 3 semanas estudiadas. Este es
un primer estudio que cuantifica el mermas, donde queda claro que la forma de
presentación es importante. No obstante, tal y como veremos posteriormente, el comedero
también es factor a tener en cuenta para minimizar el mermas de pienso. Dado los
resultados de mermas observados (6% en harina), hay que pensar en estrategias de manejo
para reducirlo, el mermas, y hay que replantearse cómo hacemos las cosas y cómo el
manejo en las primeras fases puede afectar a fases posteriores del cebo.
2.5.- Calidad de granulación
Otra cuestión muy debatida es si la calidad del granulado, definida por la
durabilidad, tiene un impacto sobre la eficiencia y el crecimiento de los terneros. En
concreto, la durabilidad depende de los ingredientes, las características de fabricación, y de
manipulación tras la fabricación (Thomas and Van der Poel, 1996; Thomas et al., 1997).
La durabilidad es uno de los parámetros que definen la calidad de la granulación; es el
concepto que expresa el comportamiento de los gránulos a distintas manipulaciones que
sufre desde la fabricación hasta el comedero. Para medir este parámetro puede recurrirse a
varios métodos, entre los que destacan la caja Pfost y el método Holmen, en los cuales se
mide el porcentaje de finos ocasionado tras someter los gránulos a una proceso de
abrasión.
Es difícil estudiar el efecto de la durabilidad, ya que para diseñar los tratamientos se
debería manipular los ingredientes o bien el proceso de granulación, y ambas estrategias
afectan la disponibilidad de energía (diferente composición y degradabilidad de

80 M. DEVANT, A. BACH, A. SOLÉ, B. QUINTANA y M. VERDÚ
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
almidones). Este planteamiento dificultaría discernir si el efecto es debido a la durabilidad,
o al método elegido para diseñar los tratamientos con diferentes durabilidades y su efecto
sobre la disponibilidad de los almidones.
En un primer estudio, se contrastó un pienso granulado con una durabilidad de un
95% vs a un 98%; la durabilidad se obtuvo a través de diferentes velocidades de
fabricación (18 tn/h vs 14 tn/h). La prueba se realizó durante el acabado (40% maíz, 20%
cebada, 12% trigo). El principal resultado obtenido fue una reducción del IC en un 8% con
una durabilidad superior (98%) respecto a una menor (95%).
2.6.- Migajas
¿Pero son esos finos en los comederos realmente importantes? Cuando observamos
los comederos, si la durabilidad es mala, la cantidad de finos será muy elevada.
Observaciones de vídeos nos muestran que los terneros buscan preferiblemente el gránulo
entero, y los finos se van acumulando y finalmente derramándose fuera del comedero; es
decir, el mermas aumentaba. Ello originó un estudio para contrastar un buen granulado vs
el mismo granulado tras un procesado de migajas (desmenuzadora de 3 mm). Al pasar el
granulado por unos rodillos se buscaba simular una mala calidad (mala durabilidad) del
gránulo sin cambiar los ingredientes ni el proceso de fabricación.
Se realizó un estudio (Verdú et al., 2015b) donde se contrastó un granulado (2,75
mm, 3,5 mm diámetro y 10 mm de longitud; 25% maíz, 25% cebada, 15% trigo; 98%
durabilidad); con un granulado en migajas; de cada partida de fabricación de granulado, la
mitad era desmenuzado a 3 mm (migajas). El diseño fue un cuadrado latino para que todos
los animales pasaran por todos los tratamientos con el fin de reducir la variación animal.
De dicho estudio se desprendieron diferentes resultados con implicaciones prácticas. La
calidad del gránulo (la durabilidad) empeoró, como era de esperar, de la fábrica al
comedero; sin embargo, empeoró mucho más cuando la calidad inicial (migajas) era mala.
Una mala durabilidad no sólo aumenta el porcentaje de finos en comedero, sino también la
densidad (y el coste de transporte y almacenaje). En dicho estudio, debido a que se
hipotetizaba que la durabilidad podría afectar al mermas de pienso, se colocaron unos
colectores en los comederos para recoger y pesar el mermas de pienso generado. Tal y
como se esperaba, los animales que consumían migajas desperdiciaban más pienso (1,6%
mermas) que los que comían granulado de buena calidad (0,8% mermas). Cuando la
durabilidad inicial era alta (buena calidad), los terneros crecieron un 6% más y
consumieron un 5% más de concentrado que los terneros que consumían migajas. Pero los
datos más interesantes fueron que los animales que consumían migajas lo hacían a menor
velocidad, dedicaban más minutos por visita, reduciendo el número de visitas, y,
finalmente, comían menores cantidades de pienso que los animales que consumían un
granulado de buena calidad. Es como si a los terneros les costara más comer migajas
(suposición), tardaran más tiempo en comer, se cansaran de comer, y al final acudieran
menos al comedero (aunque hay más tiempo de ocupación), y al final el consumo fuera
menor y, en consecuencia, el crecimiento también menor.

LA IMPORTANCIA DEL MANEJO ALIMENTARIO EN EL CEBO DE TERNEROS 81
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
Pero, ¿por qué comen más lentamente? Les cuesta deglutir y/o los animales
seleccionan favorablemente por un gránulo entero en lugar de uno disgregado tardando
más tiempo. Para responder esta última pregunta se realizó un estudio donde los terneros
podían elegir simultáneamente el granulado de alta durabilidad y las migajas. Los terneros
consumieron un 65% de granulado vs 35% de migajas; ello indicaría que un mal granulado
reduce la velocidad de ingestión (aumentando tiempo ocupación comederos y
maximizando el mermas) porque los animales prefieren el buen granulado dejando los
finos. Los datos del presente estudio coinciden con otros estudios donde los animales
prefirieron el granulado vs la harina (Ray y Drake, 1959; Arave et al., 1983; Spörndly and
Åsberg, 2006).
Así pues, la durabilidad, al afectar a la forma de presentación final en comederos,
puede tener un efecto sobre el consumo y el crecimiento independientemente de los
ingredientes y el proceso de fabricación.
3.- TIPO DE COMEDERO
El efecto del tipo de comedero (ratio animales: comedero) ha sido estudiado con
anterioridad por González et al. (2008). Los resultados de dicho estudio indicaron que al
aumentar la ratio animales:comedero aumentaba la variación en el PV final, descendía el
consumo, aumentaban los desplazamientos en los comederos y algunos parámetros
indicativos de acidosis ruminal. Por ello, al instaurar un comedero con una boca tipo túnel
por cada 20 terneros, en una granja comercial, para registrar los consumos individuales,
nos sorprendieron dos aspectos: no observábamos indicios de acidosis (laminitis, abscesos
hepáticos) y el consumo acumulado era inferior sin observar descensos en crecimiento
respecto a los datos históricos. Ello nos llevó a iniciar estudios para evaluar si el túnel
podía tener un efecto protector reduciendo las interacciones en comederos y, a su vez, al
verse reducidos los movimientos laterales, se podía reducir el mermas. Paralelamente, se
evaluó si el reducir la cantidad de pienso presente en comedero podía ser una buena
estrategia para reducir el consumo total acumulado como se ha observado en porcino
(Bergstrom et al., 2012; Myers et al., 2012).
En un primer estudio (Verdú et al., 2015a) se evaluó el efecto del tipo de comedero:
comedero corrido con 4 bocas, comedero corrido con 4 bocas y bajo nivel de concentrado
o uniboca con túnel
En dicho estudio se evaluó el efecto del tipo de comedero sobre los rendimientos, la
conducta animal y la salud ruminal. De dicho estudio destacan tres resultados. El primero
es que el consumo acumulado de pienso se redujo en un 4% al reducir el nivel de
concentrado, y en un 6% al instaurar el comedero uniboca con túnel respecto a un
comedero corrido con 4 bocas. Sin embargo (segundo resultado), durante la fase de
adaptación al comedero uniboca con túnel hubo un aumento sérico de la concentración de
AGNE, un bajo consumo y un bajo crecimiento que evidenciaron problemas de adaptación

82 M. DEVANT, A. BACH, A. SOLÉ, B. QUINTANA y M. VERDÚ
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
de los animales a dichos comederos. A pesar de estos problemas de adaptación,
posteriormente no se hallaron diferencias entre tipos de comederos; sin embargo, en
condiciones de manejo no óptimo o edades más precoces podría suponer un problema de
bienestar y productivo a considerar. Finalmente, otro resultado con implicaciones
prácticas, fue la evolución de la ratio animales:número de bocas en el comedero; cuando el
PV se encontraba alrededor de 130 kg, en los comederos de 4 bocas se observaron una
media 2 animales en el comedero (con 4 bocas disponibles); dicha ratio se redujo a 1,5
animales en el comedero a los 320 kg y a los 440 kg. Este resultado constatan que el
número de bocas o la ratio animales:bocas es un aspecto más crítico durante las primeras
fases de crecimiento.
¿Es importante aumentar el número de bocas al inicio? ¿El túnel puede ser una
buena estrategia durante el acabado?
En un segundo estudio (Devant et al., 2015, no publicado) se contrastó el túnel
uniboca con una tolva con 3 bocas con la posibilidad de regular el nivel de pienso (más
generoso al inicio, menos nivel al final). En dicho estudio no se observaron diferencias
entre tipo de tolvas en crecimiento, consumo acumulado de pienso, y kg canal. Ello
evidencia que cuando el nivel de pienso de una tolva es regulable, y se hace un buen uso de
esta, se pueden obtener resultados similares a una tolva uniboca en tolvas con 3 bocas. De
hecho, en los últimos meses, los ganaderos han realizado diferentes tipos de adaptaciones
en comederos para reducir el mermas.
3.1.- Tipo de comedero y forma de presentación
Todos los estudios realizados para contrastar el tipo de comedero fueron realizados
con concentrado. Durante la realización de dichos estudios se planteó si la forma de
presentación (harina vs granulado) podrían afectar a los resultados; ¿era posible que el
comedero uniboca tipo túnel pudiera tener un mayor efecto sobre el consumo total
acumulado cuando los animales consumieran harina? Por ello se realizó un estudio (Verdú
et al., 2015c) cuyo diseño experimental de los tratamientos fue un 2 x 2 factorial que
combinaba dos presentaciones de concentrado (harina vs granulado) con dos diseños de
comedero de pienso (tolva de 3 bocas con bajo nivel de pienso vs tolva de 1 boca tipo
túnel): harina/tolva (H3), granulado/tolva (G3), harina/túnel (H1), y granulado/túnel (G1).
Los primeros 28 días de estudio (fase de adaptación) solo se evaluó el efecto de la forma
de presentación del concentrado. El día 29 los túneles fueron bajados, coincidiendo con un
PV animal de 230 kg, iniciándose el estudio que evaluó el efecto de la forma de
presentación del pienso y el tipo de comedero durante 154 días de cebo (98 de crecimiento
y 54 de acabado). Se utilizaron dos formulaciones comerciales de pienso, una de
crecimiento (UFC: 1,02, PB: 13,8% sobre MF), y otra de acabado (UFC: 1,04, PB: 12,5%
sobre MF). Las fórmulas contenían 35% maíz, 19% cebada, 8% trigo, y la harina estaba
molturada a 2,75 mm excepto el maíz a 6mm, y el granulado tenía un diámetro de 3,5 mm.
Los resultados más destacados fueron los siguientes:

LA IMPORTANCIA DEL MANEJO ALIMENTARIO EN EL CEBO DE TERNEROS 83
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
− durante la fase adaptación (sin túnel), tal y como se había observado anteriormente,
los animales alimentados con granulado crecían más (6%) y tenían un menor IC
(5.2%) que los animales alimentados con harina.
− durante la fase de crecimiento se observaron efectos por separado de cada factor; el
granulado redujo (6.6%) el consumo respecto a la harina, y el túnel redujo el
crecimiento (7%) y el consumo (2.8%) respecto a la tolva 3 bocas con regulación.
− durante el acabado, el tipo de comedero no tuvo efecto, y el granulado redujo el
consumo (3.6%) y el IC (5.4%) respecto a la harina.
− en global, cuando los animales consumían harina la reducción del pienso
acumulado cuando la tolva era el túnel (1488 kg) era importante respecto a la tolva
3 bocas (1534 kg), no observándose diferencias entre comederos cuando el
concentrado era granulado.
4.- CONCLUSIONES
a) Los factores postfórmula (forma presentación y tipo de comedero) y sus
interacciones pueden ser una herramienta útil para mejorar la eficiencia y el
crecimiento.
b) Es importante observar el comportamiento alimentario pues permite detectar puntos
de mejora y puntos críticos donde poder intervenir y adecuar la mejor estrategia
según la edad y comportamiento alimentario de los animales. A nivel de campo, sin
embargo, la implementación de métodos de monitorización del comportamiento es
difícil.
5.- AGRADECIMIENTOS
Agropecuaria Montgai, Gepisa, Agropecuaria de Guissona S. Coop. Ltda., al
MAGRAMA (Proyecto 20130020000779, “Mejora de la eficiencia productiva en terneros
de cebo a través de la forma de presentación del pienso y del diseño de los comederos:
postfórmula”).
6.- REFERENCIAS
AARSETH (2004) Biosys. Eng. 89: 197.
BACHA, F. et al. 2005) Alimentación de terneros en ausencia de promotores de
crecimiento de tipo antibiótico: Control de timpanismo y acidosis. Publicaciones
FEDNA.
BACHA, F. y VILLAMIDE, M.J. (2010) Comparación y evaluación de los sistemas de
presentación de piensos. Rumiantes. Publicaciones FEDNA.
BEHARKA et al. (1998) J. Dairy Sci. 81: 1946.
BERTIPAGLIA et al. (2010) Anim. Feed Sci. Technol. 159: 88.
CASTILLO et al. (2006) Vet. Res. Communications 30: 823.
CUMMINS et al. (2007) Irish J. Agric. Food Res. 46: 149.

84 M. DEVANT, A. BACH, A. SOLÉ, B. QUINTANA y M. VERDÚ
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
DEHGHAN-BANADAKY et al. (2007) Anim. Feed Sci. Technol. 137: 1.
DEVANT et al. (2015a) J. Dairy Sci. 98 (Suppl.2): 829.
DEVANT et al. (2015b) J. Dairy Sci. 98 (Suppl.2): 442.
FALEIRO et al. (2011) Animal. 5: 294-303.
GALYEAN et al. (1981) J. Dairy Sci. 64: 1804.
GONZÁLEZ et al. (2008) J. Anim. Sci. 86: 419.
GONZÁLEZ et al. (2012) Anim. Feed Sci. Technol. 172: 66.
GREENWOOD et al. (1997) J. Dairy Sci. 80: 2534.
HICKS et al. (1990) J. Anim. Sci. 68: 233.
HIRONAKA et al. (1979) Can. J. Anim. Sci. 59: 395.
HUNTINGTON (1997) J. Anim. Sci. 75: 852.
IRAIRA et al. (2013) J. Anim. Sci. 91: 3827.
IRAIRA et al. (2014) Albeitar. http://albeitar.portalveterinaria.com/noticia/12703/articulos-
nutricion/pienso-y-paja-en-mezclas-unifeed-para-el-cebo-de-terneros.html.
KELLY et al. (2010) J. Anim. Sci. 88:109–123.
LJØKJEL et al. (2003) J. Anim. Feed. Sci. 12: 435.
LOERCH et al. (1990) J. Anim. Sci. 68: 3086.
MILLER-CUSHON et al. (2013) J. Dairy Sci. 96: 3257.
MURPHY y LOERCH (1994) J. Anim. Sci. 72: 2497.
OFFNER et al. (2003) Anim. Feed Sci. Technol. 106: 81.
OWENS et al. (1997) J. Anim. Sci. 75: 868.
RAY y DRAKE (1959) J. Anim. Sci. 18: 1333.
ROBISON y ODDY (2004) Lives. Prod. Sci. 90: 255.
ROBLES et al. (2007) J. Anim. Sci. 85: 2538.
SANTOS (2014) Albeitar http://albeitar.portalveterinaria.com/noticia/13495/articulos-
rumiantes/puntos-clave-en-la-gestion-de-cebaderos.html.
SAUVANT et al. (1999) INRA Prod. Anim. 12: 49.
SCHWARTZKOPF-GENSWEIN (2001)
http://www1.agric.gov.ab.ca/$department/deptdocs.nsf/all/beef4023.
SCHWARTZKOPF-GENSWEIN (2004) J. Anim. Sci. 82: 3357.
SOLANAS et al. (2005) Livest. Prod. Sci. 94: 225.
SOLANAS et al. (2007) J. Anim. Physiol. Anim. Nutri. 91: 269.
SOLANAS et al. (2008) Livest. Sci. 117: 203.
SPÖRNDLY y ÅSBERG (2006) J. Dairy Sci. 89: 2188.
STOCK et al. (1995) J. Anim. Sci. 73: 39.
SVIHUS et al. (2005) Anim. Feed Sci. Technol. 122: 303.
THOMAS y VAN DER POEL (1996) Anim. Feed Sci. Technol. 61: 89.
THOMAS et al. (1997) Anim. Feed Sci. Technol. 64: 173.
THEURER (1986) J. Anim. Sci. 63: 1649.
VERDÚ et al. (2015a) J. Anim. Sci. 93: 1.
VERDÚ et al. (2015b) J. Dairy Sci. 98 (Suppl.2): 444.
VERDÚ et al. (2015c) XVI Jornadas sobre Producción Animal, I: 239-241. AIDA
ZEA (2010) Albeitar. http://albeitar.portalveterinaria.com/noticia/5674/articulos-nutricion-
archivo/el-cebo-de-terneros-con-ensilados-de-forrajes.html

I JORNADA FEDNA − ANEMBE 25 Aniversario
sobre ALIMENTACIÓN DE RUMIANTES
MADRID, 10 de Noviembre de 2015
CAPÍTULO V
ALIMENTACIÓN DE REBAÑOS LECHEROS CON
CARRO UNIFEED Y ROBOT DE ORDEÑO
Diego Martínez del Olmo
Tolsa S.A.


ALIMENTACIÓN CON CARRO UNIFEED Y ROBOT DE ORDEÑO 87
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
ALIMENTACIÓN DE REBAÑOS LECHEROS CON CARRO UNIFEED Y
ROBOT DE ORDEÑO
Diego Martínez del Olmo
Tolsa S.A.
1.- ¿REALIDAD O FICCIÓN?
La adaptación del ser humano a las nuevas tecnologías es una realidad y el sector
agropecuario no iba ser una excepción. El automatic milking sistem (AMS), tal y como se
conoce en todo el mundo al ordeño automatizado a través de sistemas robotizados, ha ido
ganando adeptos principalmente en Europa pero también en el resto del mundo, y aunque
no haya cifras oficiales publicadas, el ritmo de nuevos equipos que se instalan, año tras
año, va creciendo a un ritmo nada desdeñable. Con las cifras publicadas por los propios
fabricantes de ordeño, el ritmo de nuevas incorporaciones al AMS por parte de productores
de todo el mundo se aproxima a las 3.000 unidades nuevas durante los últimos años,
terminando el año 2014 con más de 37.000 unidades vendidas y repartidas por todo el
mundo (DeLaval, 2015); con más de 10.000 granjas repartidas por todo el mundo, con 1,2
millones de vacas ordeñadas (Rodríguez, 2012) y 1,5 de robots por granja, siendo el Norte
de Europa, con Holanda, Alemania y Francia liderando este segmento (pensando que el 90
por ciento de los nuevos equipos instalados en Suiza y Finlandia son AMS, y más del 50%
en países como Alemania son cifras que indican que para granjas de tamaño pequeño y
mediano es una realidad este tipo de elección). Aunque el primer robot no se instaló hasta
el año 1992, no fue hasta el año 2000, cuando se instaló por primera vez en España.
La combinación de, ausencia de mano de obra cualificada, el buscar incrementar la
eficiencia, reducir el número de costes y la búsqueda por parte de los productores de estilos
de vida más flexibles hacen que la implantación de robot de ordeño aumente año tras año.
Otras zonas geográficas están buscando en estos momentos como adecuar su situación
particular a estos sistemas automáticos de ordeño, como son Australia y Nueva Zelanda,
donde hay muchas granjas que combinan el pastoreo con el ordeño automático. En
E.E.U.U., donde las granjas tienen sistemas de producción basados en economía de escala
y gran intensificación, empezó a venderse AMS en la mitad de los años noventa; y hoy en
día está desarrollándose en Canadá y en los países de América del Sur, principalmente en

88 D. MARTÍNEZ DEL OLMO
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
Argentina y Brasil (donde los costes laborales son mayores), con más de un 15% de la
producción destinada al AMS (Beekman y Bodde, 2015), aunque en general, debido a la
mano de obra más barata en estos países que en Europa y al tamaño de las explotaciones
mayor, es un sistema mucho menos popular todavía.
2.- INTRODUCCIÓN; MOTIVACIONES, VENTAJAS, DESVENTAJAS Y
DEMANDAS
Las motivaciones para que una granja invierta en un robot en vez de en una sala de
ordeño pueden ser diversas como ya hemos mencionado, pero las que coinciden para la
mayor parte de productores son (Hogeveen et al., 2004); menor coste laboral, mejorar la
flexibilidad en el horario laboral, la posibilidad de ordeñar más de dos veces al día,
eliminar mano de obra, y la necesidad de cambiar el sistema tradicional de ordeño.
Por encima de 3 robots, la eficiencia de la mano de obra por robot se mejora
(Rodríguez, 2012), como se puede observar en la siguiente tabla, aunque el número de
vacas por empleado mejora a partir del segundo robot.
Eficiencia por empleado con sistemas AMS
Nº
robots
Nº
de vacas
Nº
de empleados
Robots/empleados
Vacas por
empleado
1 60 1 1 60
2 120 1,5 1,3 80
4 240 2 2 120
8 480 3 2,7 160
20 1.200 5 4 240
El potencial beneficio del AMS vendría por un retorno de 100 $ por animal y año,
siempre y cuando se incremente la producción en un 5 % adicional (Rotz, et al., 2003); por
lo que es necesario e imprescindible ese incremento productivo para que sea viable la
inversión por parte del productor, y aquí la nutrición es actor fundamental, debido a que
independientemente del sistema elegido o del tipo de alimentación empleada el refuerzo de
las vacas para elegir libremente ir a ordeñarse es la alimentación, teniendo mayor éxito
cuanto mejor se elige el tipo de concentrado o los estímulos que lo hacen posible (Prescott,
et al., 1998).
Hay tres tipos de automatismos hoy en la producción lechera, que se pueden
definir, en unidades automatizadas individuales con un solo brazo mecánico por cada lote
(la más popularizada); unidades automatizadas que atienden a varios lotes de producción, y
el último avance que es la sala automática rotativa (AMR), donde emplea una plataforma
rotativa con múltiples brazos robóticos que pueden por si mismos preparar, ordeñar, y
realizar el cuidado post-ordeño. Un cuarto modelo de automatización que puede

ALIMENTACIÓN CON CARRO UNIFEED Y ROBOT DE ORDEÑO 89
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
combinarse con cualquiera de los sistemas anteriores (incluso con sistemas de ordeño no
automático) es el alimentador dinámico y preciso de concentrado (Bach, 2014), en
llegando a un grado de precisión mucho mayor en la nutrición aportada, retroalimentando
diariamente con información on line las necesidades y aportes nutritivas.
Las ventajas de estos sistemas automáticos son la mejora de la calidad de vida,
mejorar la eficiencia laboral por trabajador, mejorando además la cualificación y, que pese
a que es necesario un adecuado entrenamiento inicial por parte de los animales, luego dado
la repetitividad del proceso es un proceso fácil de manejar, y dado a un flujo continuo y
permanente de animales yendo voluntariamente a ordeñarse, aumenta la eficiencia por
animal. La individualización de la alimentación, detectando necesidades individuales, y
personalizando en función de las necesidades, los sistemas de alerta creados que nos
permite detectar desviaciones que previamente hemos solicitado, y la diferenciación en el
manejo a la salida del AMS, pudiendo separar y actuar sobre ese animal, facilitando el
manejo final del productor son ventajas claras también de este tipo de sistemas.
Por otro lado el alto coste de la inversión, unido a los grandes cambios de la
producción lechera en Europa, y al desconocimiento inicial por parte del resto de sectores
técnicos de este tipo de necesidades y requerimientos especiales, necesitando un soporte
mucho más especializado, son los retos a los que se enfrentan este tipo de automatismos.
Demandas tradicionales del sector para este tipo de sistemas son la integración de
otras tecnologías en la práctica diaria con el AMS, de forma que de herramientas que
permita manejar el rebaño de forma global, además de una adecuada automatización.
3.- SITUACIÓN ACTUAL
¿Cuáles son las circunstancias actuales en este segmento? Tiene buenos factores de
productividad y eficiencia para granjas entre las 60 y las 240 vacas (aunque hay granjas
con más de 20 AMS y 1.500 animales), y especialmente dónde la mano de obra representa
un coste por litro elevado, o hay dificultades para contratar nueva (Salfer y Endres, 2011).
Hay dos tipos de sistemas de trabajo genéricos para este tipo de instalaciones, qué
consisten en sistemas de libre flujo, que permiten la libre circulación de los animales entre
los parques de alimentación, descanso y ordeño, y, en flujos guiados, con puertas de
selección que dirigen a los animales, o bien pasando antes del ordeño por la zona de
alimentación, o bien ordeñándose antes de poder acceder a la comida.
Los rangos medios por unidad de ordeño automatizada fluctúan entre los 140 a 190
ordeños cada periodos de 24 horas, y entre 2,4 y 3,0 ordeños por día, dependiendo de las
condiciones de la granja y del manejo (Salfer y Endres, 2011). Una de las ventajas técnicas
es el soporte que este tipo de sistemas aportan en la detección precoz de animales en celo,
actividades de rumia, recuentos de células somáticas, variaciones de peso vivo en el

90 D. MARTÍNEZ DEL OLMO
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
animal, producción de leche individualizada…todo este tipo de mejoras técnicas en los
software instalados tienen que ser valorados en términos de retorno de la inversión, aunque
todo este tipo de aplicaciones a mayores de las clásicas empleadas, permiten a los técnicos
y ganaderos ampliar la información disponible para la toma correcta de decisiones. Lo que
está demostrado es que este tipo de instalaciones puede tener éxito en todo tipo de
condiciones, desde pastoreo hasta sistemas de cubículos con todo tipo de cama, y por
supuesto cama caliente, siempre y cuando se den las condiciones de manejo idóneas.
El gasto por consumo de agua y el uso de productos químicos tiende a ser menor
que en las salas de ordeño tradicionales, pero el consumo de electricidad aumenta
(Rodenburg, 2002). Y sin diferencias descritas en cuanto a recuentos celulares somáticos,
salud del rebaño y reproducción con las adecuadas condiciones de manejo (Salfer, 2011).
Las consideraciones para lograr incrementar la eficiencia descrita del 3-5% (Salfer,
2011, Rodenburg, 2007) son aquellas que; logran unas condiciones de las instalaciones
adecuadas, manejando además de forma adecuada el estrés térmico (principalmente en las
zonas sensibles de entrada y salida de la estación), consiguen que los ajustes de jerarquía
entre animales sean rápidos y sin movimientos bruscos, con diseños de las salidas y
entradas a los AMS adecuadas para que no haya problemas de jerarquía (Katelaar, 1995), y
por supuesto aquellas condiciones que permitan a los animales un número de ordeños alto,
de tal forma que se reduzcan los tiempos de espera delante del robot de ordeño, y que esa
merma de visitas por animal al AMS, no disminuya la eficiencia del mismo (Katelaar,
1995).
Por último y no por ello menos importante, para poder tener éxito con los sistemas
automáticos de ordeño, la gestión de la nutrición y formulación de la ración de volumen
(unifeed) y del concentrado del robot, deben permitir al animal, expresar todo su potencial
genético, incrementando el número de ordeños, y por lo tanto su eficiencia productiva, para
que el periodo de amortización del AMS se reduzca lo más posible.
4.- COMPORTAMIENTO NUTRICIONAL EN LOS SISTEMAS AUTOMÁTICOS
DE ORDEÑO
La introducción del sistema de ordeño robotizado, no sólo trae cambios en el modo
en que se ordeñan los animales sino también, en el modo en el que se alimentan. Trabajos
antiguos hablaban de, cómo los animales alimentados con sistemas automáticos, se
distribuía, el tiempo dedicado por ellos a este fin, homogéneamente en a lo largo del día;
pero se ha demostrado que las veces en que las vacas van a alimentarse al comedero,
dependen de los mecanismos de saciedad que los animales alcancen (Tolkamp et al.,
1998); debido a estos mecanismos, el tiempo que pasa hasta la siguiente vez que el animal
iniciará la actividad para alimentarse, comienza desde la última comida que ha tenido.

ALIMENTACIÓN CON CARRO UNIFEED Y ROBOT DE ORDEÑO 91
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
Los patrones de alimentación y bebida por parte de los animales ordeñados en el
AMS desarrollan conductas individuales de comportamiento características que no tienen
que ver con los patrones desarrollados con otros sistemas de ordeño, ni con el resto del
grupo de animales dentro de ese mismo lote (Melin et al., 2005), por lo que es una
oportunidad clara que, estos sistemas permitan desarrollar información a partir de los datos
recogidos diariamente e individualmente en los AMS, desarrollen sistemas de alimentación
precisos y dinámicos.
El comportamiento alimentario de las vacas ordeñadas en un robot difiere del
comportamiento alimentario de las vacas ordeñadas en sala (Wagner, 2003). Mientras que
la actividad alimentaria se incrementa después del ordeño en las vacas ordeñadas en sala,
en el robot, se incrementa con el comienzo de la actividad humana, teniendo el pico de
ingesta en la ración de volumen tres horas más tarde de este inicio de actividad, por lo que
esto es importante para tener en cuenta cuando debemos de empezar a suministrar la ración
diaria de volumen o arrimar la comida al pesebre para estimular la tan ansiada ingesta de
materia seca.
Aunque inicialmente se creía que los ordeños en el AMS se distribuían
uniformemente a lo largo del día, se observa que los animales se ordeñan menos veces
durante la noche y a primera hora de la mañana, teniendo durante el resto del día más
actividad alrededor del AMS, por lo que hay menor actividad en total en la zona del
comedero que las vacas ordeñadas en sala.
La incidencia del estrés térmico en la ingesta y en el número de ordeños es muy
elevada en los AMS, por lo que es muy importante poner las condiciones nutricionales y de
manejo que minimicen estos problemas, ya que en condiciones de ITH (índice temperatura
humedad) elevado, se reduce la ingesta de materia seca, y por lo tanto la asistencia
voluntaria al AMS, por lo que la disminución de la producción de leche podría ser mayor
que incluso en sala de ordeño en situaciones de temperatura y humedad extremas (Wagner,
2003).
Las vacas en sistemas de ordeño tradicionales invierten 300 minutos por día en
comer, con 11 comidas de media de 5,7 lbs. a 3,7 lbs. dependiendo del nivel productivo
(Dado y Allen, 1994), mientras que en los sistemas de AMS el tiempo se reduce a 4,4
comidas al día con una duración media de 52,5 minutos y 8,8 kg. (Tolle et al., 2002).
Una ración bien balaceada totalmente mezclada (TMR) puede satisfacer a un
animal dentro de un mismo rebaño (independientemente de que se elijan bien las
necesidades modelo o animal tipo del rebaño), pero puede ser no apropiada para otro
dentro del mismo rebaño, y ocurre lo mismo para ese mismo animal en función de en qué
estado de lactación se encuentre, en lactación temprana o tardía; por lo que una de las
posibles ventajas que permiten este tipo de sistemas es una alimentación más encaminada a
satisfacer los nutrientes que se necesitan, por lo que, tanto en cantidad (en todos los AMS)
como en composición (si lo permite el sistema de alimentación del AMS, mediante

92 D. MARTÍNEZ DEL OLMO
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
alimentación dinámica o suministro de más de un concentrad), pueden sacar ventaja los
animales al satisfacer de una forma más precisa sus necesidades.
Otro de los problemas tradicionales de las raciones TMR (y donde este tipo de
sistemas con AMS que alimentan en función de las necesidades puede sacar ventaja), es la
selección que por parte de los animales se obtiene a favor del azúcar y del almidón de la
dieta (Leonardi et al., 2005), incluso en aquellas situaciones en las que la preparación del
carro unifeed se hace en las mejores condiciones, lo que hace todavía más impreciso el
diseño de raciones TMR, y que por lo tanto aspectos como la fertilidad sea clave a la hora
de acertar con los requerimientos ofertados a los necesitados, ya que una gran desviación
típica en nuestro rebaño con la media que hemos utilizado para el cálculo de las
necesidades, hace que el sistema fracase, sub o sobre-alimentando animales, en tanto
mayor proporción, cuanto mayor es la desviación típica.
Por lo tanto con el AMS podemos trabajar con la cantidad de concentrado ofertado,
disminuyendo el riesgo y la incertidumbre, trabajando con la cantidad de leche producida,
pero uno de los retos que tenemos nutricionistas e ingenieros de sistemas es diseñar los
aportes también en función de la cantidad de grasa y proteína producida diariamente y
tener en cuenta factores como la pérdida de peso, e incluso algunos indicadores que se
podrían monitorizar diariamente durante los primeros 21 días como es el
betahidroxibutirato plasmático (BHBA). En el caso de que nuestro AMS nos permitiera
poder aportar diferentes tipos de concentrados deberíamos de poder aprovecharlo, ya que
las necesidades de energía y proteína varían en función de la producción real producida por
el animal (INRA 2008).
También minimiza este tipo de manejo los cambios de lotes entre animales respecto
a los sistemas TMR y sistemas tradicionales de ordeño, para suministrar diferentes
raciones, y donde la disminución del tiempo de reposo puede llegar a ser de hasta un 12 %
menor (Guasch, 2009), disminuyendo por lo tanto la producción de leche.
Por otro lado existen también AMS que nos permiten la oportunidad de analizar la
actividad ruminal, y este factor combinado con el resto de indicadores de la granja es
válido para prevenir e identificación temprana de problemas metabólicos en rumiantes
(Ungar et al. 2005).
5.- ESTIMULAR LA FRECUENCIA DE ORDEÑO A TRAVÉS DE LA
FORMULACIÓN
Estos sistemas por lo tanto requieren que parte del concentrado se suministre en la
estación por lo que comúnmente se denominan raciones parcialmente mezcladas al resto de
ingredientes que se suministran en el comedero, incluido la otra parte del concentrado
(PMR), y como principales funciones se encuentra como acabamos de decir que la vaca se
sienta atraída para ir a la estación y mantenerla ocupada mientras se está ordeñando.

ALIMENTACIÓN CON CARRO UNIFEED Y ROBOT DE ORDEÑO 93
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
La frecuencia de ordeño en un AMS es influida por; el nivel productivo con una
relación positiva y proporcional (Caja et al., 2000), por el número de animales por grupo
de AMS (densidad), por la palatabilidad y cantidad del concentrado suministrado, por el
estrés social, por el miedo a entrar a la unidad (malos diseños) y por los problemas que
inhiban la locomoción (Klaas et al., 2003).
Por lo tanto, uno de los más importantes factores en conseguir tener éxito con
nuestro AMS es tener una ración balanceada y definido un protocolo de gestión nutricional
claro en la explotación, que permitan estimular la entrada al AMS, que incremente la
producción de leche, por lo tanto la necesidad del animal de ser ordeñado, y que evite
problemas relacionados con el síndrome de bajo pH ruminal, para evitar problemas entre
otros, de locomoción. La menor asistencia voluntaria al robot desencadenaría finalmente
en más problemas de de mamitis y laminitis, además de la consecuente pérdida de leche y
beneficio para el productor.
Está definido que las vacas entren al robot para consumir el concentrado que éste
les dispensa, no para ordeñarse, por lo que una de las principales circunstancias que tiene
que cumplir el concentrado del AMS es tener una buena palatabilidad. También es muy
importante una adecuada complementariedad del concentrado específico para el robot con
la ración de volumen, y donde una adecuada proporción de forraje es muy importante para
garantizar un número de visitas apropiado al robot con las propias instalaciones de las que
se disponga (de Jong et al., 2003). La cantidad de concentrado suministrado de media en
estudios hechos sobre el terreno muestran que el rango óptimo varía entre los 2 kg/día y los
9 kg/día (Salfer, 2011). Aportando siempre una cantidad mínima de concentrado cuando
las vacas asistan al AMS incluso en las producciones más desviadas a la baja, y aportando
siempre un máximo establecido que evite minimizar los problemas de acidosis para
aquellos animales más productores.
El concentrado es suministrado al AMS a través de diferentes sistemas, pero
siempre es muy importante controlar la velocidad de suministro de ese pienso, para
permitir que se ordeñe el animal sin problema, pero por otro lado, que nos permita el
sistema suministrar esa máxima cantidad de concentrado para los animales más
productores (La velocidad de suministro del concentrado debe de ser entre 0.3-0.4
kg/minuto). Hay que tener en cuenta la velocidad de suministro del concentrado pensando
que los animales pasan entre 6 y 8 minutos de media ordeñándose en el robot.
Es muy importante controlar la salud podal de nuestros animales, ya que este tipo
de problemas además de afectar a la salud en general reduce la movilidad de los animales,
y la motilidad restringida es un problema muy importante para las granjas con AMS, y por
otro lado, la relación existente con laminitis (Klaaset al., 2003), de la lactación temprana y
de la alta producción, hace que sea un factor que se debe de tener especial cuidado cuando
se diseñan las raciones PMR, así como las circunstancias de manejo nutricional para
minimizar el riesgo de problemas podales, ya que además de reducir el número de visitas
voluntarias diarias al AMS reduciría la eficiencia nutricional conseguida.

94 D. MARTÍNEZ DEL OLMO
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
Para conseguir reducir los problemas podales y la laminitis es importante que la
ración de volumen tenga una estructura física mínima que estimule la asistencia del animal
al AMS. Al final, debemos de tener en cuenta que las variaciones de pH ruminal descritas a
lo largo del día para raciones TMR (sorting) con los sistemas PMR podemos disminuir
esas variaciones, mejorando la salud ruminal, aunque el tamaño de esa bajada de pH sea
mayor. El resultado final es que el rumen, pese a lo que se creía hace unos años está mejor
adaptado con este tipo de sistemas.
Cuando hablamos de incrementar la frecuencia de ordeños en sistemas AMS,
debemos de conocer la capacidad de ordeño del AMS. La capacidad de un AMS puede ser
expresada en términos de ratio de ocupación, definido como el porcentaje de horas que el
AMS ordeña por día. Tenemos que tener en cuenta que el intervalo aceptado entre ordeños
para minimizar los problemas y maximizar los beneficios es el siguiente (André et al..,
2010):
Intervalos recomendados máximos entre ordeños con AMS
Producción primíparas
(kg/d)
Producción multíparas
(kg/d)
Intervalo admitido
(d)
< 20 <25 0,40
<20-37> <25-33> 0,29
<27-30> <33-40> 0,25
>30 >40 0,22
Incrementar la frecuencia de ordeño e incrementar el tamaño del rebaño en el AMS,
mejora los beneficios obtenidos de nuestra granja, pero elegir cuál de las dos estrategias es
la más óptima, dependerá de los costes de producir esa leche extra esperada (André y col.,
2010). Por lo tanto la relación entre coste del concentrado suministrado en el AMS y la
eficiencia de su utilización son los factores claves para buscar esa optimización anhelada.
Relación entre incremento de la frecuencia de ordeño y beneficio (André et al., 2010)
Intervalo entre ordeños
(d)
Producción leche por
AMS (kg/d)
Beneficio obtenido
(€/d)
O,334 1,939 513
0,295 1,963 519
0,264 1,982 525
0,238 1,997 529
La importancia de la palatabilidad del pienso en el AMS ha sido demostrado en la
bibliografía (Rodemburg y Wheeler, 2002), donde las visitas voluntarias al AMS se logran

ALIMENTACIÓN CON CARRO UNIFEED Y ROBOT DE ORDEÑO 95
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
incrementar en +0,64 y el número de ordeños en +0,34, a través de estrategias que la
optimicen. Para ello es muy importante conocer los ingredientes más palatables y
apetecibles para el animal, además de combinarlo con aditivos que favorezcan la asistencia
o bien que puedan camuflar determinados olores o sabores de materias primas que por
razones nutricionales nos interese conocer (Futcher, 2011).
En general los datos de más alta palatabilidad se asignan a derivados de la industria
cervecera, DDGs, melaza y pulpa de remolacha. En el rango medio de este tipo de
características es para la harina de soja, y cereales, y en el rango más bajo de apetecibilidad
está la harina de colza y habas de soja enteras. Ingredientes como el gluten meal, derivados
de sangre, carne y harinas de pescado, así como grasa bypass, manteca, minerales, y
buffers son muy poco apetecibles para el ganado vacuno lechero. Madsen et al., 2010,
comparando diferentes ingredientes en diferentes formulaciones llegan a la conclusión que
los ingredientes más apetecibles para vacuno de leche son la combinación de cebada y
avena seguido del trigo. El maíz y derivados fueron menos elegidos por parte de los
animales, y la adicción de grasa resultó en el menos palatable.
Respecto a la literatura para la textura ideal del concentrado hay claras evidencias
que la mejor forma de aportar el concentrado a la ración es en forma de pellet (Rodenburg
et al., 2004), garantizando el menor número de finos posibles, y la menor rotura de pellet
posible. Para lograr la correcta granulación, importante para favorecer la asistencia al
robot, la palatabilidad, reducir pérdidas económicas y evitar problemas metabólicos, la
adicción de aditivos muy puros en sepiolita (Exal®) al 2% en la fórmula del concentrado,
demuestra ser una de las elecciones más eficaces desde hace años (Angulo et al., 1995).
El número de asistencias al robot aumentó cuando la cantidad de concentrado
disminuyó en la ración de volumen un 30% (Hauspie, 2008), y se aumentó un 12% en la
estación, por lo que el resultado final, fue más producción de leche de acuerdo a un número
mayor de asistencias al robot de ordeño. Bach y col., 2007 también demostraron que
trabajar con altas densidades energéticas en el pesebre, resultan en un menor número de
visitas. La asistencia al robot se verá afectada cuando la densidad energética de la ración
exceda los 1,66 Mcal /Kg, y la materia seca forrajera sea inferior al 52% (Rodenburg y
Wheeler, 2002).
6.- FORMULAR EN FUNCIÓN DEL TIPO DE TRÁFICO EN EL AMS
Las diferencias entre los diferentes sistemas que regulan el flujo de los animales
hacia el robot de ordeño, sistema libre o guiado, tiene un impacto significativo en el
programa de alimentación; en el sistema libre, el nivel energético de la ración sobre el
comedero, tiene que ser lo suficientemente bajo como para permitir que los animales no
encuentren satisfechos sus requerimientos en ella, y que tengan que ir a ordeñarse para
completar sus necesidades. Este aspecto es el que principalmente diferencia este tipo de

96 D. MARTÍNEZ DEL OLMO
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
raciones PMR de las TMR, junto con el aporte de parte de los nutrientes fuera del
comedero.
Típicamente en los sistemas de libre circulación la PMR es balanceada con las
necesidades energéticas estimadas de los animales menos las correspondientes a los 7-8
últimos litros medios, de acuerdo a que esos requerimientos serán aportados con el pienso
en el robot, por lo que en la práctica esto significa que parte del concentrado es aportado en
la ración de volumen y la otra parte en el robot (Gardner, 2013).
Por otro lado, en los sistemas guiados donde se ordeña en primer lugar antes de
acceder a la zona de alimentación, los animales no pueden conseguir la energía necesaria
hasta que no son primeramente ordeñadas, por lo tanto el nivel energético en el PMR
dentro de la ración de volumen debería de ser lo suficientemente alto para asegurarnos que
van a visitar la ración un número suficiente de veces y con regularidad.
El tráfico dirigido donde se alimentan primero antes de ordeñarse, se usa con
raciones de volumen muy bajas en energía y por lo tanto la mayoría del concentrado se
utiliza o bien en la estación o en una estación suministradora de concentrado separada del
robot.
Al final, sea cual sea el sistema de tráfico elegido para nuestra instalación, es muy
importante que el concentrado usado sea muy palatable, usando la mayoría de veces
piensos granulados, aunque hay ocasiones en las que se emplean harinas u otro tipo de
texturas groseras, pero es muy importante que el concentrado en el AMS sea lo
suficientemente duro provocando el menor número de finos posible para conseguir la
mayor ingesta de materia seca como ya se ha comentado, ya que para los animales más
productores el tiempo de consumo de alimento puede ser muy limitado.
Por lo tanto para esos animales muy productores se programa el consumo de
alimento en varias tomas al día, habiendo trabajos que recomiendan que ese concentrado
sea alto en fibra soluble, para evitar acidosis y garantizar la durabilidad del pellet (Gardner,
2013). Lo cierto es que todos los sistemas de tráfico con los AMS tienen sus ventajas y
desventajas, pero parece establecido que los sistemas de libre flujo son más adecuados para
aquellos rebaños donde buscamos más producción, y que el trabajar con niveles
energéticos más bajos en la ración voluminosa puede llegar a ser un problema en los
sistemas forzados, creando “colas de trabajo” con animales que no irían a ordeñarse por sí
mismos. Al final el objetivo del sistema de libre circulación debería ser que no haya
muchos animales que satisfagan sus necesidades en el sistema PMR, para que tengan que ir
a alimentarse al AMS y por lo tanto ordeñarse. Los sistemas guiados permiten al productor
utilizar mayor cantidad de concentrado, almidón en el PMR, aún a costa de su producción.
También debemos de ser conscientes que el sistema de libre tráfico implica ocupar a más
mano de obra que los sistemas con puerta de selección o guiados (hasta un 3% más de
vacas a recoger; Rodríguez, 2012).

ALIMENTACIÓN CON CARRO UNIFEED Y ROBOT DE ORDEÑO 97
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
El número de comidas por animal y día es mayor con sistema de libre circulación,
que con sistemas de guiado forzado (con o sin preselección), aunque el número de ordeños
en estos sistemas con puertas de selección es mayor que en los modelos de libre
circulación (Rodenburg, 2011). Al final las vacas en sistemas forzados pasan más tiempo
esperando ser ordeñadas, y menos tiempo descansando. Ambos sistemas pueden funcionar
perfectamente bajo condiciones idóneas de manejo, pero cuando no es así, en sistemas de
libre circulación el productor sufre las consecuencias de tener que “empujar” vacas al
AMS, y en los sistemas forzados, son las vacas las que sufren una posible disminución de
la ingesta de materia seca diaria y por lo tanto productiva.
7.- OTRAS CONSIDERACIONES EN EL MANEJO NUTRICIONAL CON
PRODUCCIONES BASADAS EN AMS
Es importante recordar que debemos de tener en cuenta que es muy importante
calibrar el concentrado suministrado al AMS entre cada entrega por parte de la industria,
así como periódicamente, en función de nuestro protocolo en granja. Esto es así, por lo que
la entrada al AMS puede verse afectada por los ingredientes aportados en la formulación
del concentrado, la durabilidad, y la cantidad de concentrado.
Hablar de transición en vacuno lechero es hablar del periodo crítico en el que
preparamos al animal para el comienzo de una nueva lactación, y en el que el productor y
nutrólogo se juegan gran parte del éxito en la lactación futura, pero, añadido a este
concepto cuando en una explotación con AMS, hablamos de transición es también hablar
de adaptación de los animales al ordeño robotizado, cuando pasan de un ordeño en sala
convencional, o bien la adaptación de las primíparas a esta situación. En inicio de
lactación, la ingesta de materia seca se suele ver disminuida, por lo que para que no haya
desequilibrios con el aporte de concentrado en el AMS, se suele limitar la cantidad al parto
e irla aumentando progresivamente, hasta llegar al límite que nos propongamos en nuestra
tabla alimentaria. Esto es importante ya que pequeños desequilibrios en la ingesta de PMR
por los animales tras el parto provocaría problemas por el bajo consumo de fibra
procedente del forraje (si mantenemos alta la dispensación de concentrado en el AMS) en
relación con el aporte de concentrado total (PMR + AMS). Las vacas con desequilibrios en
el aporte de concentrado respecto al forraje pasan menos tiempo comiendo y rumiando y
en cambio invierten más tiempo descansando (Robinson, 1997), yendo por lo tanto menos
veces al comedero, por lo que se reduce el número de visitas.
Posteriormente a esto se suele ofertar en el AMS el máximo del concentrado
calculado para la ración hasta que lleguen a los días a los que alcanzan el pico de
producción los animales (dato que se puede obtener fácilmente de la información que me
aporta el software del AMS), pero que tradicionalmente se establece entre los 35 y los 50
días para las multíparas, y entre los 75 y los 100 días para las primíparas. A partir de ese
momento en el que los animales han alcanzado la máxima producción, la cantidad de
concentrado aportado por el AMS debería de ir corregida en función de los datos

98 D. MARTÍNEZ DEL OLMO
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
productivos obtenidos por la estación, y cuanto más información corrija, más precisa serán
los aportes dado por nuestros animales. De hecho en los sistemas dinámicos, cuanto más
precisa es la información, más eficiente será el aporte nutricional, por lo que el precio de la
ración se podría reducir (Wessenlink, 2011).
Unido a lograr un adecuado estado ruminal que permita aumentar la eficiencia del
aporte nitrogenal y energético que se le está dando, es necesario aportar un forraje de una
muy alta calidad y mejorar las condiciones de las instalaciones en las que se instala el
AMS, prestando especial atención al confort térmico, como se ha comentado con
anterioridad.
8.- CONCLUSIONES
El coste de un AMS varía pero en general es más elevado para ese número de
animales que una sala de ordeño convencional. Debemos de entender que una estación
puede mantener a 55 y 65 vacas con ahorros de mano de obra entre 4 y 5 horas por vaca,
pero para lograr la máxima eficiencia que permita justificar la inversión sólo lo
conseguiremos con una adecuada gestión nutricional que permita maximizar la eficiencia
de utilización de los nutrientes, minimizando los riesgos ruminales y buscando un
adecuado número de visitas que permita incrementar la frecuencia de ordeños (hasta que la
tecnología y las instalaciones haga posible que los robots busquen y vayan al animal.
Lo que parece claro que la tecnología en este aspecto ha venido a quedarse, y que
los nutricionistas necesitamos formular raciones en las que el coste sea importante pero
también ir de la mano de ésta tecnología para que permita al animal ir a ordeñarse muchas
veces sin provocarle problemas metabólicos. En todas las áreas productivas es importante
el entendimiento con el resto de aspectos técnicos, pero con los sistemas AMS, para
favorecer la máxima eficiencia, es importante aunar los conocimientos y disponibilidad de
alimento para elegir el mejor sistema, y una perfecta coordinación en los aspectos de salud
podal, salud de ubre y reproducción para conseguir obtener el máximo beneficio a estos
sistemas. Sin perder de vista las cuestiones etológicas donde deberíamos estudiar, aprender
y así sacar más rédito de las circunstancias que rodean a este tipo de animales en este tipo
de ambiente, ya que no debemos de olvidar que son animales gregarios, por lo que este tipo
de automatismos rompen esa rutina establecida, y debemos de aprender más acerca de
cómo se altera el comportamiento de los animales en este tipo de sistemas para conseguir
minimizar los problemas y maximizar el rendimiento económico de la explotación.
El siguiente reto para los nutricionistas debería de ser que la alimentación en los
AMS pasara de ser una necesidad a ser una estrategia nutricional que permita exprimir al
máximo todas las oportunidades que la tecnología brinda al productor en este tipo de
sistemas, ya que el ritmo demográfico actual y las necesidades de alimento no parece que
vayan a permitir la relajación de precios de los componentes típicos de una ración para
vacuno lechero. Por lo tanto ganar eficiencia en los nutrientes consumidos por parte de los

ALIMENTACIÓN CON CARRO UNIFEED Y ROBOT DE ORDEÑO 99
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
animales, es un reto permanente de la producción lechera, y en este sentido deberíamos de
sacar provecho de las ventajas que la tecnología nos brinda.
Por lo tanto la tecnología lleva implícita trabajar con sistemas dinámicos de
alimentación (donde también es medido y calculado lo que el animal debería de haber
comido y no lo ha hecho), por lo que nuestros sistemas de diseño de concentrados y de
raciones debería de tender a aprovechar todos esos datos que nos aporta este tipo de
sistemas para adaptarlos a nuestra base de información y calcular la mejor ración posible.
Todas las raciones que ahora estamos diseñando se basan en predicciones,
estimaciones, pero con el AMS tenemos la oportunidad de alimentar a los animales
basados en hechos, lo que nos hace que uno de los objetivos claros de todo buen nutrólogo
para este tipo de automatismos sea el conseguir que haya de media más de un rechazo por
día (es decir vacas que quieren ser ordeñadas-alimentarse en el AMS, pero que no las deja
porque es demasiado pronto). Porque la nutrición es como la digestión ruminal, es un
proceso continuo y dinámico, que hace que el diseño de los aportes sea preciso, por lo que
a nuestras ecuaciones de predicción de aportes debemos de incorporar los datos
(información) que el AMS nos da.
9.- REFERENCIAS
ANDRÉ, G., BERENTSEN, P., ENGEL, B., KONING, C. y OUDE, A. (2010) J. Dairy
Sci. 93: 942-953.
ANGULO, E., BRUFAU, J. y ESTEVE-GARCÍA, E. (1995) Anim. Feed Sci. Tech. 53:
233-241.
BACH, A., IGLESIAS, C., CALSAMIGLIA, S. y DEVANT, M. (2007) J. Dairy Sci. 90-
11:5049-5055.
BACH, A. 2014. WCDS Advances in Dairy Technology 26: 177-189.
BEEKMAN, J. y BODDE, R. (2015) Milking automation is gaining popularity. Published
in press, dairy global net articles.
CAJA, G., AYADI., M., CONILL, C., M´RAD, M., ALBANELL, E. y SUCH, X. (2000)
En: Robotic Milking. pp: 177-178.
DADO, R. y ALLEN, M. (1994) J. Dairy Sci. 77: 132.
DE JONG, W., FINNEMA, A. y REINEMANN, D.J. (2003) ASAE An Int Mtg Tech Paper
No. 033017.
FUCTHER, M. (2011) Personal communication. Delaval.
GARDNER, E. (2013) Nutritional Principles for Automated Milking Systems. Penn State
Dairy Cattle Nutrition Workshop.
GUASCH, I., y BACH, A. (2009) J. Dairy Sci. 92: 6.
HAUSPI, M. (2008) Reported in the Lely Monthly Management Magazine. Lely Industries.
HOGEVEEN H., HEEMSKERK, K. y MATHIJS, E. (2004) En: A better Understanding of
Automatic Milking. pp: 56-61.

100 D. MARTÍNEZ DEL OLMO
Madrid, 10, Noviembre de 2015. I Jornada FEDNA-ANEMBE 25 Aniversario Nutrición de Rumiantes
KETELAAR-DE LAUWERE, C.C., DEVIR, S. y METZ, J.H.M. (1995) Applied Animal
Behavior Science 49: 199-211.
KLAAS, I., ROUSING, T., FOSSING, C., HINDHEDE, J. y SORENSEN, T.J. (2003)
Anim. Welfare. 12: 599-603.
MADSEN, J., WEISBJERG, M. y HVELPLUND, T. (2010) Livest. Sci. 127: 45-50.
MELIN, M., WILKTORSSON, H. y NORELL, L. (2005) J. Dairy Sci. 88: 71-85
PRESCOTT, N., MOTTRAM, T. y WEBSTER, A. (1998) Anim. Behav. Sci. 57: 23-33.
RODENBURG, J. (2002) Robotic Milkers: En: Proc Ohio Dairy Management Conference.
pp: 1-18
RODENBURG, J. y WHEELER, B. (2002) En: Proceedings of the first North American
Conference on Robotic Milking, Toronto, Canada. Pp: III 18 –III 32.
RODENBURG, J., FOCKER, E. y HAND, K. (2004) En: Proceedings of the International
Conference on automatic milking. Lalystad, pp: 511.
RODENBURG, J, y HOUSE, H.K. (2007) En: Proc of Sixth International Dairy Housing
Conf. ASABE Pub No 701P0507e.
RODDENBURG, J. (2011) Designing Feeding Systems for Robotic Milking. Conference
Tri-State Nutrition. Indiana.
RODRIGUEZ, F. (2012) The realities of robotic milking technology today published in
Progressive Dairyman. U.S.
ROTZ, C., COINER, C.U. y SODER, K. (2003) J. Dairy Sci. 86: 4167-4177.
SALFER, J. y ENDRES, M. (2011) Robotic Milking: What producers have learned. 4-
State Dairy Management Conference, Dubuque, IA, 2011.
TOLKAMP, B., ALLCROFT, D., AUSTIN, E., NIELSEN, B. y KYRIAZAKIS, I. (1998)
J. Theor. Biol. 194: 235-250.
TOLLE, K., JOULAUD, G. y KRIETER, G. (2002) Zuchtungskunde 74(5): 330-40.
UNGAR, E. y RUTTLEr, S. (2005) Anim.. Behav. Sci. 98: 11-27.
Wagner, A. y Palmer, R. (2003) J. Dairy Sci. 86:494-1502.
WESSENLINK, W. (2011) Dynamic feeding in Western Dairy Farmer magazine, Jn, Fb.
pp: 20-26.