estudio de la capacidad de degradación de fenol del
TRANSCRIPT

Estudio de la capacidad de degradación de fenol del consorcio microalga/bacteria y obtención de la cinética de crecimiento y consumo de sustrato. Maria V. Castro1, Vivianne A. Garzón1,2
1Departamento de Ingeniería Química, Universidad de los Andes, Bogotá, Colombia.
2Departamento de Microbiología, Universidad de los Andes, Bogotá, Colombia.
Objetivo General
Analizar y describir el comportamiento cinético del consorcio microbiano Pseudomonas
aeruginosa / Chlorella vulgaris cuando es empleado en la remoción de fenol un medio
preparado con el contaminante; estudiando en primera instancia la factibilidad de su
empleo.
Objetivos Específicos
1. Seleccionar la cepa de P. aeruginosa degradadora de fenol mediante selección en
agar mínimo de sales con fenol como única fuente de carbono.
2. Estudiar los parámetros de la cinética de degradación de fenol para el consorcio
microalga-bacteria con fenol como única fuente de carbono mediante
espectroscopía UV-VIS.
3. Determinar la relación de concentración de inóculo adecuada para el crecimiento
del consorcio microbiano Pseudomonas aeruginosa / Chlorella vulgaris y para la
biodegradación de fenol por parte del mismo.
4. Establecer si los dos microorganismos pertenecientes al consorcio presentan un
mejor desempeño en la remoción de fenol cuando crecen de forma conjunta o de
forma individual.

Estudio de la capacidad de degradación de fenol del consorcio
microalga/bacteria y obtención de la cinética de crecimiento y
consumo de sustrato.
Maria V. Castro1, Vivianne A. Garzón12
1Estudiante de Ingeniería Química, Facultad de Ingeniería, Universidad de los Andes, Bogotá, Colombia.
2Estudiante de Microbiología, Facultad de Ciencias Biológicas, Universidad de los Andes, Bogotá, Colombia.
Asesor:
Andrés González Barrios
Profesor Asociado, Departamento de Ingeniería Química, Universidad de los Andes, Bogotá, Colombia.
Co-asesora:
Martha J. Vives
Profesora Asociada, Departamento de Ciencias Biológicas, Universidad de los Andes, Bogotá, Colombia.
Resumen
El presente estudio pretende evaluar la capacidad de remoción de fenol por parte de un consorcio Chlorella vulgaris-
Pseudomonas aeroginosa estableciendo las condiciones favorables para el crecimiento del mismo. Se han reportado
casos exitosos de remoción de compuestos fenólicos con microorganismos como Pseudomonas spp., Acetobacter spp. y
Scenedesmus obliquus. Asimismo, estudios recientes han establecido que los consorcios alga-bacteria son una
alternativa viable para la descontaminación de aguas. Para determinar la capacidad de remoción se realizó una selección
entre 7 cepas bacterianas. Además, mediante el modelo cinético de Monod se determinaron los parámetros cinéticos de
crecimiento y consumo de sustrato en el medio Tris-Acetato-Fosfato modificado (TAPm) empleando diferentes
relaciones de pre-inóculo (1:1, 2:1 y 3:1 de bacteria y microalga, respectivamente). Se encontró que a una concentración
de 800 ppm de sustrato no fue posible recuperar las bacterias, mientras que a 400 ppm 5 cepas fueron tolerantes y se
determinó que la cepa M8A1 de Pseudomonas aeroginosa presentó una mayor remoción de fenol por lo que fue
seleccionada para este estudio. El porcentaje de remoción de fenol con la cepa M8A1 fue del 13% mientras que el
porcentaje de remoción de fenol en el consocio fue de 59.7%. Finalmente, se corroboró la capacidad de remoción de
fenol de la cepa de Pseudomonas aeruginosa seleccionada, y se concluyó que el desempeño de la misma mejoró cuando
se encontraba presente en consorcio con la microalga Chlorella vulgaris.
Palabras clave: Pseudomonas aeruginosa, Chlorella vulgaris, consorcio, fenol, remoción.
1. Introducción
La industria petrolera produce una variedad de materiales orgánicos tóxicos, que traen
consigo efectos de bioacumulación nocivos para el medio ambiente. Los compuestos fenólicos son
los representantes más comunes de contaminantes orgánicos tóxicos y pueden permanecer en el
ambiente por un largo tiempo debido a su estabilidad y capacidad de bioacumulación [1]. Este tipo

de compuestos es ampliamente utilizado en procesos industriales, por lo cual surge la necesidad
de implementar métodos seguros y eficaces para la eliminación de desechos fenólicos antes de su
disposición.
El fenol se puede eliminar mediante técnicas convencionales tales como extracción con
disolventes, adsorción, oxidación química, incineración y otros métodos de tratamiento no
biológicos. Algunas tecnologías no biológicas han presentado resultados satisfactorios en la
remediación de fuentes de aguas. En los últimos años, el proceso de oxidación fotocatalítica
heterogénea (HPO) empleando luz UV y TiO2 se ha convertido en una nueva ruta prometedora
para la degradación de contaminantes orgánicos persistentes, produciendo sustancias
biológicamente degradables y menos tóxicas para el medio ambiente [2]. Sin embargo, estos
métodos traen consigo serios inconvenientes, tales como altos costos y la formación de
subproductos peligrosos. La degradación biológica, es un método que implica menores costos y
trae consigo la posibilidad de mineralización completa de las moléculas de dióxido de carbono y
agua en condiciones aerobias [3]. La biodegradación de hidrocarburos por medio de poblaciones
naturales de microorganismos adaptados para metabolizar el contaminante es uno de los
principales mecanismos contemplados para eliminar el fenol del medio ambiente [4]. Sin embargo,
la incorporación de microorganismos libres en los sistemas de tratamiento tiene ciertas
limitaciones debido a las propiedades tóxicas de los contaminantes, y a la competencia entre las
poblaciones nativas y las exógenas [5]. A través del uso de microorganismos inmovilizados, es
posible evitar dichos problemas, puesto que la inmovilización brinda a las células una protección
contra el efecto tóxico de las sustancias presentes en el medio y a la depredación por parte de
otras poblaciones [6]. Este tipo de técnicas se encuentra en fase de desarrollo y únicamente se
han hecho pruebas a escala laboratorio.
Existen varios tipos de microorganismos capaces de degradar compuestos fenólicos. Se han
reportado casos exitosos con hongos, bacterias y micro-algas; Pseudomonas spp. y Acetobacter
spp., Chlorella spp., Scenedesmus obliquus [7, 8]. Sin embargo, cada tipo presenta un
comportamiento diferente en términos de su capacidad degradativa. Los organismos individuales
pueden metabolizar sólo una gama limitada de sustratos del contaminante [7], de modo que se
requiere la adaptación de poblaciones mixtas con capacidades enzimáticas altas capaces de
degradar mezclas complejas de contaminantes. Se ha visto que los consorcios alga-bacteria son
una alternativa viable para la descontaminación de aguas, puesto que se da un reciclaje de
nutrientes, las algas consumen el CO2 producido por las bacterias mientras estas consumen el O2
que producen las algas [9]. La interacción mutualista alga (fotótrofa)-bacteria (heterótrofa) podría
mejorar considerablemente la degradación de contaminantes ambientales. El estudio de la
naturaleza fisiológica y metabólica de las algas ha aumentado considerablemente dada su
eficiencia en procesos de biorremediación en aguas contaminadas. Entre las microalgas clorofitas,
existen 7 géneros específicos para el tratamiento de efluentes, dentro de los cuales se encuentra
Chlorella vulgaris debido a su tolerancia a elevadas concentraciones de nutrientes [10]. Sin
embargo, los cultivos de microalgas, especialmente a gran escala, presentan un alto
requerimiento de luz. Por lo tanto, es importante enfocarse en la búsqueda de sistemas más

eficientes para su crecimiento. Por otro lado, la bacteria Pseudomonas aeruginosa ha sido utilizada
con éxito en la degradación de diversos productos como la gasolina, el queroseno, aceite diesel y
petróleo crudo [7].
Generalmente los compuestos fenólicos son biodegradables en condiciones aerobias o
anaerobias bajo las cuales la recuperación del medio contaminado se da eficazmente [9].
Actualmente puede considerarse a la degradación anaerobia como una tecnología desarrollada
que presenta ventajas significativas respecto a otras alternativas [11]. Teniendo en cuenta que el
principal problema de la degradación de compuestos aromáticos es el rompimiento del anillo, los
microorganismos utilizados deben tener la capacidad de activarlo mediante el oxígeno y formar
derivados dihidroxilados (catecol) en los que se facilita la ruptura del anillo [12]. La degradación
aeróbica de los hidrocarburos es considerablemente más rápida que el proceso anaeróbico, de
modo que la oxigenación será necesaria para mantener las condiciones aeróbicas para una rápida
degradación [13].
Las condiciones de operación del sistema son fundamentales en la determinación del
comportamiento del mismo y afectan considerablemente las condiciones de degradación del
sustrato y las limitaciones de transferencia de masa que se puedan presentar a causa de estas.
Dentro de las variables relevantes se encuentra la concentración inicial de fenol y la concentración
de biomasa. La biodegradación de fenol en bajas concentraciones es proceso natural que se lleva a
cabo bajo condiciones normales y la velocidad del mismo dependerá del tipo de microorganismos
presentes. Los microorganismos empleados en este estudio se caracterizan por vivir en
concentraciones medias de fenol. Mediante un acondicionamiento previo, fue posible adaptar el
consorcio a concentraciones mayores de fenol.
Este estudio pretende evaluar si la capacidad de remoción de fenol por parte de dos
microorganismos distintos aumenta cuando éstos interactúan de forma conjunta. Para ello, se
determinará un modelo cinético para la predicción de la velocidad de reacción en respuesta a
diferentes concentraciones de sustrato. Para determinar el crecimiento microbiano es esencial
establecer la interacción entre los dos organismos del cultivo mixto puesto que los modelos que
describen el crecimiento se basan en estas interacciones; las interacciones más frecuentes son la
de competencia, el mutualismo y la depredación [14]. Para este caso particular, donde el objetivo
es la degradación del contaminante, los estudios cinéticos permiten establecer cuán bien el
sistema microbiano está llevando a cabo su función. Además, permitirían la detección y la
determinación de posibles interferencias entre los diversos componentes de los medios de cultivo.
La obtención de un consorcio que cumpla con los objetivos planteados en este estudio, permitirá
futuras aplicaciones a nivel industrial. Actualmente existen medios de almacenamiento para algas
como tanques tubulares de CO2 [15], los cuales podrían ser dimensionados de acuerdo a las
cinéticas de degradación obtenidas en este estudio.

2. Modelo matemático
La degradación biológica se lleva a cabo a través del rompimiento del anillo de fenol utilizando
la enzima presente en el microorganismo. En el caso de las bacterias se presenta un
comportamiento diferente cuando se exponen a diferentes concentraciones iniciales de fenol,
entre otras condiciones experimentales. Por lo tanto, se realizó un acondicionamiento para
evaluar el comportamiento de estos organismos en diferentes concentraciones de sustrato. Dichos
resultados pueden ser útiles en el diseño y optimización de reactores biológicos para el
tratamiento de residuos fenólicos. La velocidad de degradación es un factor esencial para la
determinación de la viabilidad del estudio.
La determinación de la cinética de crecimiento de los microorganismos es esencial para
establecer la capacidad de degradación. El modelo de Monod indica que la tasa de crecimiento
microbiano está determinada por la concentración real de un metabolito en particular [16]. Existe
una relación entre el consumo del fenol y el crecimiento del consorcio. El término S(t) denota la
concentración (ppm) del fenol en el momento t. El parámetro es la máxima tasa de
crecimiento de biomasa y es la constante de saturación. En consecuencia, la dinámica de la
concentración de células X (t) está descrita por ecuación 1:
( ) ( )
El modelo de Monod establece que la tasa de crecimiento aumenta con la concentración del sustrato. El crecimiento y el consumo del sustrato están relacionados linealmente a través del coeficiente de rendimiento. De esta forma, la relación entre el crecimiento celular y el consumo de sustrato está dada por:
( )
[ ]
[ ]
[ ]
[ ]
[ ]
[ ]
[ ]
[ ]
[ ]
[ ]

Donde,
( )
Para este estudio el modelo se modificó teniendo en cuenta la interacción entre el alga y la
bacteria con el fenol. Teniendo en cuenta que hay un punto en el que la velocidad de crecimiento
de los microorganismos a lo largo del tiempo se mantiene constante, se asumió que el medio
estaba saturado de fenol a excepción de las últimas horas de crecimiento donde la concentración
de sustrato era menor. Asimismo, teniendo en cuenta que hay muerte celular del alga y la
bacteria, se introduce un término que represente la tasa de defunción de los mismos. Debido a
que la concentración de sustrato con respecto al tiempo no se ve afectada por la muerte de
microorganismos sino por el crecimiento de los mismos, la ecuación 6 no involucra el término de
tasa de defunción. Las ecuaciones 1 y 2 quedan de la siguiente forma:
( )
( )
( )
Cabe mencionar que el modelo de Monod aplica cuando el sustrato no inhibe el
crecimiento de la biomasa. En este caso se realizó un análisis de sensibilidad con el parámetro de
inhibición y no se presenciaron cambios significativos. Por lo tanto, es el modelo correcto para la
determinación de los parámetros cinéticos de este estudio. Teniendo en cuenta que las
concentraciones de fenol estudiadas no fueron lo suficientemente altas para inhibir el
crecimiento, la ecuación cinética no inhibidora de Monod queda como se indica a continuación:
3. Materiales y métodos
3.1. Microorganismos y medios de cultivo
Se seleccionaron 5 cepas de Pseudomonas aeruginosa nativas colombinas y 2 de referencia,
PA14 y PA01 suministradas por el Centro de Investigaciones Microbiológicas (CIMIC) de la
Universidad de los Andes (Tabla 1). Las cepas 1 y 6 son cepas clínicas, las cepas 2 y 3 fueron
aisladas de aguas contaminadas con hidrocarburos de Caño Limón, Arauca; la cepa 4 fue obtenida

de tierra de jardín de Villeta, Cundinamarca; la cepa 5 fue aislada de estiércol de vaca de Puerto
Boyacá, Boyacá. De todas las cepas se conservaron 4 mL en crioviales con caldo BHI y glicerina a -
80°C.
Tabla 1. Numeración de las cepas de P. aeruginosa usadas
Cepa Número de identificación
1 PA14
2 M8A1
3 M8A4
4 M6A1
5 M28A1
6 PAO1
7 PB18
Por otra parte, la microalga Chlorella vulgaris fue proporcionada por el laboratorio de
biotecnología del departamento de Ingeniería Química de la Universidad de los Andes. El cultivo
de algas se encuentra en medio TAP a 27°C en condiciones autótrofas, con CO2 como fuente de
carbono y luz artificial como fuente de energía.
3.2. Medio de selección de la cepa de P.aeruginosa
Se inocularon 100 µl de cada cepa en 10 mL de caldo BHI y se incubó por 12 horas, en seguida
fueron usadas para tener un cultivo masivo en el medio de selección. El medio usado para la
selección fue un agar de medio mínimo de sales minerales (MMS) que contiene (en g/l) 0.5
KH2PO4, 1.0 NH4CI, 2.0 Na2SO4, 2.0 KNO3, 0.001 CaCl2, 1.0 MgSO4.7H2O, 0.0004 FeSO4.7H2O, a un
pH de 7.09 y con concentraciones de 400 ppm y 800 ppm de fenol como única fuente de carbono.
La selección se realizó paralelamente con dos metodologías diferentes: la primera de ellas fue la
técnica Réplica Platina que consistió en adaptar una caja de petri con cuadrícula. Se sembraron las
bacterias aisladas en diferentes cajas de Petri y en el centro de la caja se introduce un recipiente
con el contaminante a una concentración determinada. Este se volatilizó en la caja y únicamente
permitió el crecimiento de las bacterias que resistían dicha concentración del fenol. Las bacterias
que sobrevivieron al paso anterior se sembraron en una nueva caja de petri. De nuevo se introdujo
el contaminante en el centro pero en una concentración más alta. Se repitió el proceso realizando
2 réplicas y al final se seleccionan los microorganismos que resistieron mayores concentraciones
del contaminante conformando un pool. La segunda metodología, consistió en diluir el fenol en el
agar y se realizó el mismo proceso de la primera metodología (Figura 1).

Figura 1. Metodologías de adición del fenol como única fuente de carbono
3.3. Determinación del fenol
Las bacterias tolerantes al fenol en el medio de selección fueron cultivadas en medio líquido
LB y posteriormente ajustadas a un patrón de McFarland de 0.5 [17]. Luego, se extrajeron 6 mL de
dicho cultivo y fueron agregados a un Erlenmeyer con 54 mL de LB líquido con 230 ppm de fenol
como concentración inicial; este procedimiento se repitió 8 veces con el fin de tomar dos muestras
por Erlemeyer (Figura 2) y así garantizar que el medio conservara la concentración de nutrientes y
de fenol. De estos Erlenmeyer se tomaron las muestras para realizar la cuantificación de fenol y la
curva de crecimiento bacteriano que determinó las unidades formadoras de colonia que degradan
el compuesto.
Figura 2. Metodología, toma de muestras, curva de crecimiento bacteriano y cuantificación de fenol.

La curva de crecimiento de las bacterias se realizó midiendo la absorbancia de las muestras
en diferentes tiempos con una longitud de onda de 600nm [18], con la absorbancia obtenida se
determinaron las diluciones que fueron ser sembradas para cuantificar así las unidades
formadoras de colonias degradantes de fenol. Asimismo, la curva de crecimiento de la microalga
se realizó con un conteo celular mediante la cámara de Neubauer y se midió la absorbancia de las
muestras en diferentes tiempos con una longitud de onda de 750nm.
Por otra parte, para determinar el fenol degradado las muestras fueron centrifugadas por 15
minutos a 4500 rpm y posteriormente fueron sometidas a cuantificación de fenol por medio del
método colorimétrico con 4-aminoantipirina [19].
3.4. Formación del consorcio
Para determinar la condición a la cual los microorganismos no se encontraran estresados por
la presencia del otro, se realizó el estudio del consorcio a diferentes relaciones de inóculo: 1:1, 2:1
y 3:1 de bacteria y microalga, respectivamente.
El consorcio fue cultivado en medio líquido TAP que contiene (en g/l) 5.4 KH2PO4, 14.4
K2HPO4, 0.375 NH4CI, 0.1 MgSO4.7H2O, 0.05 CaCl2.2H2O, 2.0 Tris. Con una concentración inicial de
fenol de 230 ppm y se añadió 1 mL de solución de trazas de Hunter [18]. Para el estudio del
consorcio se realizó la misma metodología explicada anteriormente en la Figura 2.
3.5. Obtención de cinética
La concentración inicial de biomasa para las bacterias fue de 1000 células/mL. Ésta densidad
de población se reprodujo durante 3 días y alcanzó una concentración de bacterias/mL.
Se obtuvieron datos experimentales a partir de la interacción individual y conjunta de dos
microorganismos que utilizaban el mismo sustrato como fuente de carbono. Posteriormente se
definieron los parámetros del modelo y se obtuvo una aproximación de la cinética de crecimiento
del consorcio y consumo del sustrato.
Para obtención de la cinética de crecimiento de la bacteria y el alga, se midió la densidad
óptica de las muestras y adicionalmente se realizó un conteo celular. Asimismo, para determinar la
degradación de fenol se llevó a cabo el método colorimétrico de 4-aminoantipirina [19].
El sistema de ecuaciones del modelo propuesto es no lineal, en donde la fuente de no
linealidad es la ecuación de Monod. Los parámetros del modelo fueron determinados mediante
una optimización en MATLAB® utilizando el método de punto interior en donde se busca
minimizar el error cuadrado medio entre la solución de las ecuaciones diferenciales y los datos
experimentales. La resolución del sistema de ecuaciones diferenciales ordinarias se llevó a cabo
mediante un método de tipo Runge Kutta de cuarto orden.

4. Resultados y discusión
Se monitorearon las condiciones de crecimiento de las 7 cepas de Pseudomonas aeruginosa
bajo las dos metodologías propuestas para la utilización de fenol como única fuente de carbono.
Posteriormente se seleccionaron las cepas que presentaron mayor crecimiento de las bacterias.
Las muestras fueron tratadas con fenol para garantizar que las cepas seleccionadas fueran
resistentes a este compuesto.
4.1. Aislamiento y selección de cepas bacteriana tolerantes al fenol mediante presión de
selección
Se realizaron dos réplicas del experimento: en la primera se hicieron 4 pases y en la segunda 3
pases a partir de la siembra inicial para propiciar el crecimiento adecuado de las bacterias.
Después del tratamiento, de acuerdo a una apreciación cualitativa de cuatro niveles acorde al
grado de crecimiento dentro del área estipulada por la cuadrícula (figuras A1 y A2). Se determinó
que a 400 ppm de fenol 6 cepas (1, 2, 3, 5, 6, 7) presentaron mayor tolerancia al sustrato, mientras
que a 800 ppm solo 5 cepas (1, 2, 3, 5, 7) que se identificaron como tolerantes a esa concentración
de fenol. Según la Tabla 1, las cepas 3 y 5 alcanzaron un alto grado de crecimiento en el medio
sólido. Sin embargo, al introducir estas cepas en un medio de LB líquido la cepa 2 presentó mayor
tolerancia durante el proceso de adaptación a las dos concentraciones de fenol. Por lo tanto, esta
cepa fue la seleccionada para continuar con las fases siguientes de este estudio.
Tabla 2. Nivel de crecimiento de las cepas de P. aeroginosa
Replica 1 Replica 2
400 ppm 800 ppm 400 ppm 800 ppm
Cepa 1 Alto Nulo Bajo Nulo
Cepa 2 Alto Bajo Bajo Nulo
Cepa 3 Alto Alto Bajo Nulo
Cepa 4 Bajo Bajo Medio Nulo
Cepa 5 Medio Bajo Nulo Bajo
Cepa 6 Nulo Nulo Nulo Nulo
Cepa 7 Bajo Nulo Bajo Nulo
4.2. Selección de inóculo para el crecimiento del consorcio
Al estudiar las tres proporciones de la concentración de bacteria descritas en la sección 3.4
fue posible obtener la relación de inóculo favorable para la degradación de fenol. En la Figura 3 se
presenta la degradación de fenol por parte de la cepa 2. Las tres proporciones alcanzaron un grado
similar de degradación de sustrato. Sin embargo, cuando la concentración de bacteria era el doble
de la del alga se obtuvo una degradación en menor tiempo.
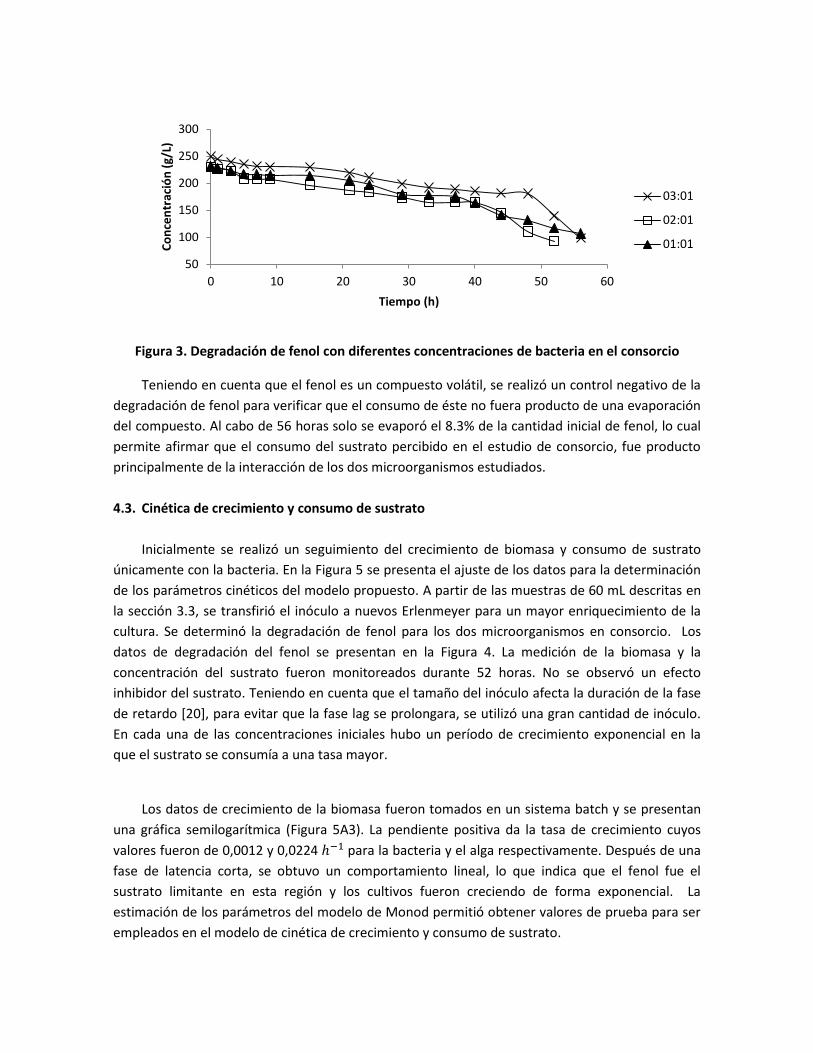
Figura 3. Degradación de fenol con diferentes concentraciones de bacteria en el consorcio
Teniendo en cuenta que el fenol es un compuesto volátil, se realizó un control negativo de la
degradación de fenol para verificar que el consumo de éste no fuera producto de una evaporación
del compuesto. Al cabo de 56 horas solo se evaporó el 8.3% de la cantidad inicial de fenol, lo cual
permite afirmar que el consumo del sustrato percibido en el estudio de consorcio, fue producto
principalmente de la interacción de los dos microorganismos estudiados.
4.3. Cinética de crecimiento y consumo de sustrato
Inicialmente se realizó un seguimiento del crecimiento de biomasa y consumo de sustrato
únicamente con la bacteria. En la Figura 5 se presenta el ajuste de los datos para la determinación
de los parámetros cinéticos del modelo propuesto. A partir de las muestras de 60 mL descritas en
la sección 3.3, se transfirió el inóculo a nuevos Erlenmeyer para un mayor enriquecimiento de la
cultura. Se determinó la degradación de fenol para los dos microorganismos en consorcio. Los
datos de degradación del fenol se presentan en la Figura 4. La medición de la biomasa y la
concentración del sustrato fueron monitoreados durante 52 horas. No se observó un efecto
inhibidor del sustrato. Teniendo en cuenta que el tamaño del inóculo afecta la duración de la fase
de retardo [20], para evitar que la fase lag se prolongara, se utilizó una gran cantidad de inóculo.
En cada una de las concentraciones iniciales hubo un período de crecimiento exponencial en la
que el sustrato se consumía a una tasa mayor.
Los datos de crecimiento de la biomasa fueron tomados en un sistema batch y se presentan
una gráfica semilogarítmica (Figura 5A3). La pendiente positiva da la tasa de crecimiento cuyos
valores fueron de 0,0012 y 0,0224 para la bacteria y el alga respectivamente. Después de una
fase de latencia corta, se obtuvo un comportamiento lineal, lo que indica que el fenol fue el
sustrato limitante en esta región y los cultivos fueron creciendo de forma exponencial. La
estimación de los parámetros del modelo de Monod permitió obtener valores de prueba para ser
empleados en el modelo de cinética de crecimiento y consumo de sustrato.
50
100
150
200
250
300
0 10 20 30 40 50 60
Co
nce
ntr
ació
n (
g/L)
Tiempo (h)
03:01
02:01
01:01
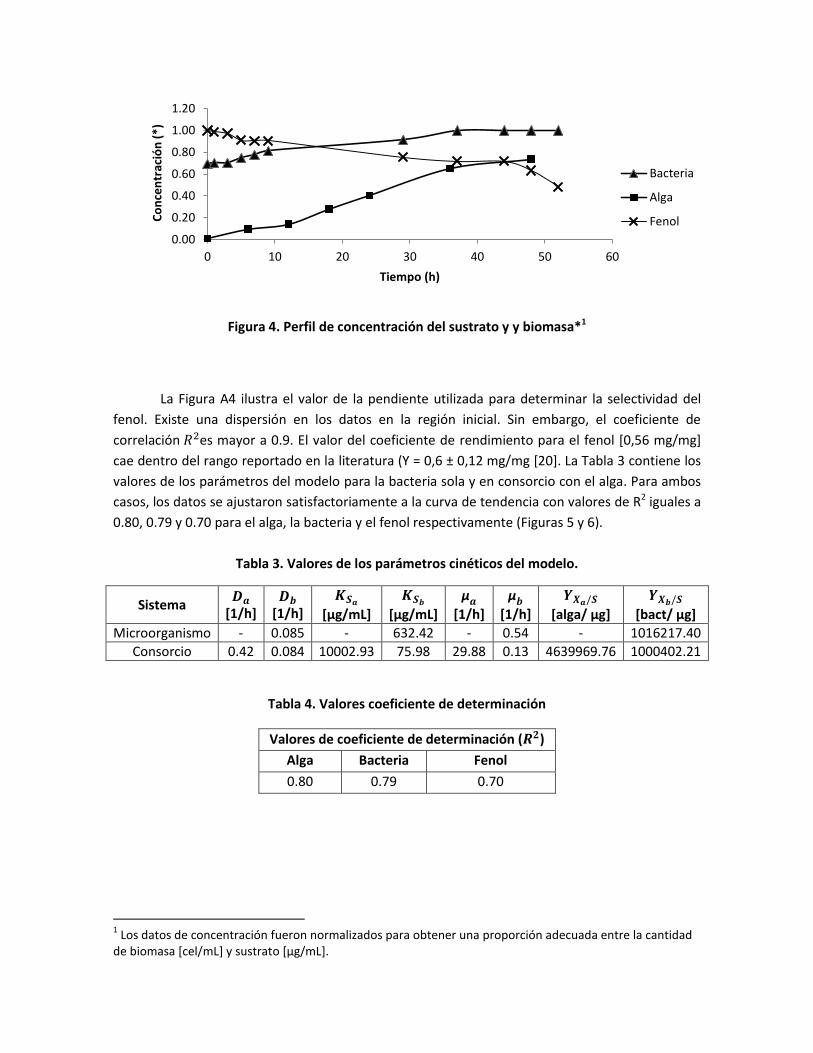
Figura 4. Perfil de concentración del sustrato y y biomasa*1
La Figura A4 ilustra el valor de la pendiente utilizada para determinar la selectividad del
fenol. Existe una dispersión en los datos en la región inicial. Sin embargo, el coeficiente de
correlación es mayor a 0.9. El valor del coeficiente de rendimiento para el fenol [0,56 mg/mg]
cae dentro del rango reportado en la literatura (Y = 0,6 ± 0,12 mg/mg [20]. La Tabla 3 contiene los
valores de los parámetros del modelo para la bacteria sola y en consorcio con el alga. Para ambos
casos, los datos se ajustaron satisfactoriamente a la curva de tendencia con valores de R2 iguales a
0.80, 0.79 y 0.70 para el alga, la bacteria y el fenol respectivamente (Figuras 5 y 6).
Tabla 3. Valores de los parámetros cinéticos del modelo.
Sistema
[1/h]
[1/h]
[µg/mL]
[µg/mL]
[1/h]
[1/h]
[alga/ µg]
[bact/ µg]
Microorganismo - 0.085 - 632.42 - 0.54 - 1016217.40
Consorcio 0.42 0.084 10002.93 75.98 29.88 0.13 4639969.76 1000402.21
Tabla 4. Valores coeficiente de determinación
Valores de coeficiente de determinación ( )
Alga Bacteria Fenol
0.80 0.79 0.70
1 Los datos de concentración fueron normalizados para obtener una proporción adecuada entre la cantidad
de biomasa [cel/mL] y sustrato [µg/mL].
0.00
0.20
0.40
0.60
0.80
1.00
1.20
0 10 20 30 40 50 60
Co
nce
ntr
ació
n (
*)
Tiempo (h)
Bacteria
Alga
Fenol
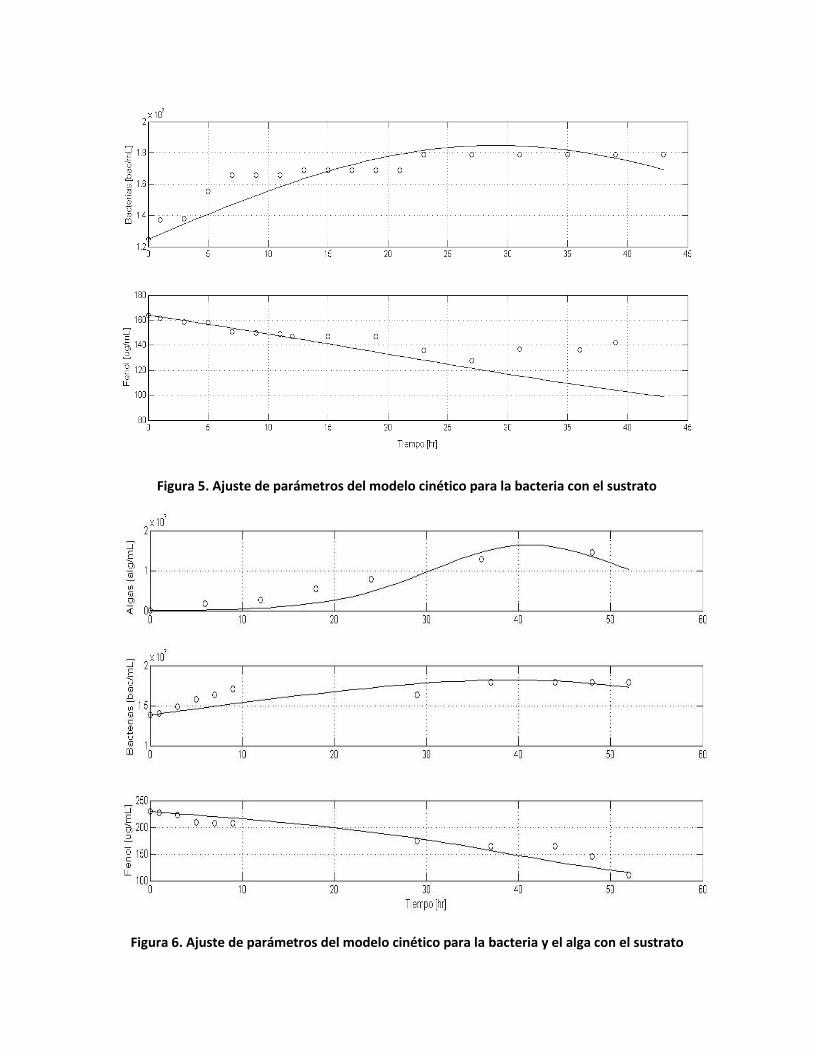
Figura 5. Ajuste de parámetros del modelo cinético para la bacteria con el sustrato
Figura 6. Ajuste de parámetros del modelo cinético para la bacteria y el alga con el sustrato

Se llevó a cabo el cultivo de cepas tolerantes de fenol por medio del enriquecimiento
progresivo del medio con fenol en MMS, donde fenol era la única fuente de carbono. Inicialmente
se estableció que las cepas que pudieran sobrevivir en este medio (cepas tolerantes al compuesto)
serían las que podrían degradar fenol. Durante el proceso, el número de cepas tolerantes al fenol
quedó reducido en las etapas subsiguientes. Después de someter las cepas a condiciones diversas
de concentración de sustrato el número de cepas con capacidad degradativa disminuyó. Al evaluar
las 3 relaciones de inóculo para el consorcio, se obtuvo un mejor desempeño cuando la
concentración de la bacteria era el doble de la concentración del alga. Bajo esa condición, la
presencia de la bacteria no inhibió el crecimiento del alga y se obtuvo un menor tiempo de
degradación. Sin embargo, las diferencias entre las 3 proporciones no fueron altamente
significativas.
Inicialmente se realizó una evaluación del crecimiento de los dos microorganismos de forma
individual y de forma conjunta. A partir de los datos de las figuras Figura 7 y Figura 8 es posible
afirmar que tanto la bacteria como el alga presentan un crecimiento más rápido cuando están en
el consorcio. Ambos microorganismos llegan a la fase exponencial en menor tiempo cuando
conviven en el mismo medio. Asimismo, se evaluó la degradación de fenol en ausencia del alga y
una vez más el desempeño del microorganismo se vio favorecido en el consorcio. La Figura 9
muestra que la tendencia de la línea de consumo de sustrato tiene un mayor pendiente cuando la
bacteria interactúa con el alga.
Figura 7. Crecimiento de la bacteria durante la degradación del sustrato.
0.5
0.6
0.7
0.8
0.9
1
1.1
0 10 20 30 40 50 60
De
nsi
dad
óp
tica
(6
00
nm
)
Tiempo (h)
Bacteria sola

Figura 8. Crecimiento del alga durante la degradación del sustrato.
Figura 9. Comparación de la tasa de degradación de fenol con la bacteria
Sin embargo, las figuras Figura 7 a Figura 9 solo permiten obtener un comportamiento
aproximado de la interacción del los microorganismos con el fenol. Es por ello que los parámetros
cinéticos obtenidos son de vital importancia a la hora de llegar a cualquier conclusión en este
estudio. Evidentemente, el crecimiento celular está relacionado con la concentración de sustrato,
puesto que a mayor concentración de fenol, el crecimiento de la bacteria y el alga aumentó. La
tasa de crecimiento máximo ( ) de la bacteria aumentó cuando hubo interacción con el alga. En
un estudio similar realizado para la biodegradación de fenol y catecol, en el cual también se siguió
el modelo de Monod, se obtuvo una tasa de crecimiento de 0.46 [21] para la bacteria,
mientras que para una cultura mixta se obtuvo una tasa de 0.26 [22].
El valor de la constante de saturación del alga ( ) es mucho mayor que el de la bacteria
( ), lo cual indica que la bacteria tiene una afinidad más fuerte al fenol y requiere una menor
cantidad de sustrato para crecer. Sin embargo, al estudiar la degradación por parte del consorcio,
este duró un mayor tiempo en la fase exponencial y por lo tanto la degradación de fenol se generó
en un menor tiempo. Ahora bien, entre mayor sea el término de selectividad ( ), se requiere
0
0.2
0.4
0.6
0.8
1
0 50 100 150 200
De
nsi
dad
óp
tica
(6
00
nm
)
Tiempo (h)
Alga sola
Alga enconsorcio
50
100
150
200
250
0 10 20 30 40 50 60
Co
nce
ntr
acio
n (
pp
m)
Tiempo (h)
Consorcio
Bacteriasola

más biomasa para consumir el fenol. Es decir que entre menor sea el término de selectividad,
mayor es el grado de degradación de sustrato. En este caso particular, la constante de selectividad
indica que cuando los dos microorganismos coexisten en un mismo medio, la degradación de fenol
es mayor. En el caso de la bacteria, ésta se saturó más rápido que el alga. Los valores calculados de
máxima tasa de crecimiento, constante de saturación y tasa de muerte se encuentran en el rango
adecuado para el sistema de degradación de fenol por parte de Pseudomonas aeroginosa
(
[ ]
[ ]
[ ] ).
5. Conclusiones
La aclimatación de los microorganismos superó los problemas de inhibición de sustrato que
normalmente se producen en la biodegradación de fenol en alta concentración según [18]. Sin
embargo, en concentraciones suficientemente altas, se produjo un efecto inhibidor leve sobre
algunas bacterias. Por lo tanto la evaluación de la inhibición de sustrato se convierte en una
consideración importante que se deberá evaluar en las etapas subsecuentes de este estudio.
Se obtuvieron los parámetros para una cinética de tipo Monod mediante una optimización por
punto interior que permitió ajustar los datos experimentales al modelo planteado. Al comparar los
valores de los parámetros hallados con los encontrados en la literatura, es posible afirmar que se
encuentran dentro del orden de magnitud adecuado. Adicionalmente, éstos permitieron confirmar
la hipótesis principal que consistía en determinar la factibilidad del consorcio para la
biodegradación de fenol. Ambos microorganismos presentaron un mejor desempeño cuando
coexistían en un mismo medio. Bajo las condiciones establecidas para la adaptación del consorcio
P. aeroginosa – Chlorella vulgaris, fue posible obtener 57% de remoción de fenol a partir de una
concentración inicial de 230 ppm de solución y un tiempo de contacto de 52 horas. Sin embargo,
debido a que no se determinaron los parámetros cinéticos para el alga sola, es aconsejable que
para estudios posteriores se determinen dichos valores. De esta forma se podrá observar el efecto
que tuvo la bacteria sobre el alga en el consorcio.
Al adaptar el consorcio se determinó que la concentración factible de bacteria debía ser el
doble de la del alga. Aun así se recomienda evaluar otros factores como el pH, la temperatura y la
concentración de CO2 y O2 del medio para determinar el efecto de los mismos en la degradación
del sustrato y el crecimiento de la biomasa. Es necesario establecer las condiciones bajo las cuales
se desea que los morfotipos trabajen, para así poder optimizar la biorremediación, cambiando los
posibles parámetros físicos o químicos que puedan ir en contra de la actividad de los dos
microorganismos estudiados.
El ajuste directo de la velocidad de consumo de sustrato y crecimiento celular estuvo
garantizada sólo en ciertas condiciones. En el proceso de degradación, el rendimiento de la masa
de células varió en función del tamaño del inóculo. La disminución del crecimiento de
microorganismos en las concentraciones de sustrato estuvo relacionada a la disminución de la tasa
de consumo de fenol.

Agradecimientos
A nuestros padres por apoyarnos incondicionalmente día a día en el transcurso de nuestra
carrera universitaria. Queremos expresar nuestro más profundo y sincero agradecimiento a todas
aquellas personas que con su ayuda han colaborado en la realización del presente trabajo, en
especial a los profesores Martha Vives y Andrés González, asesores de esta investigación, por la
orientación, el seguimiento y la supervisión continua de la misma.
Gracias por el interés mostrado y las sugerencias brindadas por nuestro amigo Ángel
Galvis, quien nos brindó apoyo incondicional durante todo el proceso. También agradecemos al
personal de laboratorio por la ayuda y la excelente atención recibida durante todo el semestre.
Queremos hacer extensiva nuestra gratitud a nuestro compañero Daniel Bejarano, con
quien realizamos este trabajo. Su colaboración hizo posible la satisfactoria culminación de esta
investigación.
Referencias
[1] C. Lakshmi y V. Sridevi, «Effect of pH and inoculum size on phenol degradation by Pseudomonas aeruginosa (NCIM2074),» International Journal Science , vol. 4, nº 7, pp. 2246-2252, 2009. [2] Garcýa, A., Amat, A.M., Arques, A., Vicente, R., Lopez, M.F., Oller, I., Maldonado,M.I. and Gernjak,W. Increased biodegradability of Ultracide in aqueous solutions with solar TiO2. [3] S. Tiong-Lee, B. Yan-Pui y H. Jiang, «Rapid cultivation of stable aerobic phenol-degrading granules using acetate-fed granules as microbial seed,» Journal of Biotechnology, vol. 115, pp. 387-395, 2005. [4] J. Leahy y R. Colwell, «Microbial Degradation of Hydrocarbons in the Environment,» Microbiological Reviews, vol. 53, nº 3, pp. 305-315, 1990. [5] Fantroussi, S.E. y Agathos, S.N. (2005). Is bioaugmentation a feasible strategy for pollutant removal and site remediation? Current Opinion in Microbiology 8, 268-275. [6] Mishra, S., Jyot, J., Kuhad, R. y Lal, B. (2001). Evaluation of inoculum addition to stimulate it in situ bioremediation of oily-sludge-contaminated soil. Applied and Environmental Microbiology 67, 1675-16. [7] V. Santos y V. R. Linardi, «Biodegradation of phenol by a filamentous fungi isolated from industrial effluents - identification and degradation potential,» Process Biochemistry, vol. 39, nº 8, pp. 1001-1006, 2004. [8] Chetty, R. Ramanjaneyulu y G. Srinikethan, «Biological phenol removal using immobilized cells in a pulsed plate bioreactor. Effect of dilution rate and influent phenol cocnentration.,» Journal of Hazardousmaterials, nº 149, pp. 452-459, 2007. [9] S. Subashchandrabose, B. Ramakrishnan, M. Megharaj, K. Venkateswarlu y R. Naidu, «Consortia of cyanobacteria/microalgae and bacteria: Biotechnological potential,» Biotechnology Advances, nº 29, pp. 896 - 907, 2011. [10] M. Salazar, V. Bernal y E. Martín, «Sociedad mexicana de biotecnología de bioingeniería (Chlorella vulgaris y Sphaerocystis sp),» 18 Septiembre 2005. [En línea]. Available: http://www.smbb.com.mx/congresos%20smbb/merida05/TRABAJOS/AREA_IV/CIV-47.pdf. [Último acceso: 11 Septiembre 2013]. [11] Universidad Autónoma de Hidalgo, «Evaluación y modelado de la cinética de depuración anaerobia de vinazas de la inductria alcoholera,» Pachuca, 2007.

[12] Gernjak,W. Increased biodegradability of Ultracide in aqueous solutions with solar TiO2photocatalysis. Chemosphere 68, 2007, 293–300 [13] Science technica, “Alternativa de tratamiento para tierras fuller contaminadas con aceite ieléctrico,” Universidad tecnológica de Pereira, vol. 4, no. 32, pp. 419-424, 2006. [14] M. L. Shuler y F. Kargi, Bioprocess Engineering: Basic Concepts, New Jersey: Pentice Hall, Inc., 1993. [15] M. Candelaria, “Algas para producir combustible,” 14 Noviembre 2012. [Online]. Available: http://magazineoceano.com/algas-para-producir-combustible/#.UjZW8NJWySo. [Accessed 14 Septiembre 2013]. [16] S. Carcano, “A model for cell growth in batch bioreactors”, Politecnico de Milano, 2010. [17] C. Malbrán, “Manual de procedimientos para la determinación de la sensibilidad a los antimicrobianos en bacterias aisladas de humanos”, Instituto nacional de enfermedades infecciosas, Buenos Aires, Argentina,2001 [19] Centro de Investigaciones Microbiológicas (CIMIC), «Biorremediación: degradación de fenol,» Bogotá, 2010. [20] A.G. Livingston, H.A. Chase, Modeling phenol degradation in a fluidized bed bioreactor, AIChE J. 35 (1989) 1980–1992. [21] P. Kumaran, Y.L. Paruchuri, Kinetics of phenol biotransformation, Water Res. 31 (1997) 11–22. [22] U. Pawlowsky, J.A. Howell, Mixed culture biooxidation of phenol I. Determination of kinetic parameters, Biotechnol. Bioeng. 15 (1973) 889–896. [23] A. Tanyolac, H. Beyenal, Prediction of substrate consumption rate, average biofilm density and active thickness for a thin spherical biofilm at pseudo-steady state, Biochem Eng. J. 2 (1998) 207–216 [24] Á. M. Quiroga, A. González, J. Husserl y F. Vischi, «Evaluación del potencial de degradación de fenol y tolueno a partir del consorcio microalga-bacteria,» Universidad de los Andes, Bogotá, 2012. [25] A. Kumar y S. Kumar, «Biodegradation kinetics of phenol and catechol using Pseudomonas putida MTCC 1194,» Biochemical Engineering Journal, vol. XXII, pp. 151-159, 2005. [26] M. Chandana Lakshmi y V. Sridevi, «Effect of pH and inoculum size on phenol degradation by Pseudomonas aeruginosa (NCIM 2074),» International Journal of chemistri, vol. 7, nº 4, pp. 2246-2252, 2009. [27] A. Leonard y N. Lindly, «Growth of Ralstoniaeutropha on inhibitory concentration of phenol,» Enzyme and microbial technology, vol. 65, nº 4, pp. 407-414, 1999. [28] C. Lob y P. Tar, «Effect of additional carbon sources on biodegradation of phenol,» Buletin of Environmental Contamination and Toxicology, vol. 64, pp. 756-767, 2000. [29] D. Valera, “Obtención de un consorcio microbiano granular para la biodegradación de fenol en un reactor discontinuo de alimentación secuenciada,” Pachuca, 2006. [30] A. M. Cantillo Carrero, “Optimización de consorcio alga-bacteria para degradación de fenol,” Universidad de los Andes, Bogotá, 2013. [31] S. Wilson y K. Jones, «Bioremediation of soil contaminated with polynuclear aromatic hydrocarbons (PAHs):
a review,» Environmental pollution, vol. 81, nº 3, pp. 229-249, 1993.

Anexo 1: Medio de crecimiento para las cepas de P. aeroginosa
Figura A1. Crecimiento de las cepas a 400 ppm de fenol
Figura A2. Crecimiento de las cepas a 800 ppm de fenol

Anexo 2: Regresiones para obtener valores de inicialización de parámetros
cinéticos
Figura A3. Gráfica semilogarítmica para determinar valores iniciales de la tasa de crecimiento
máxima.
Figura A4. Regresión lineal para determinar valores iniciales del coeficiente de selectividad.
y = -1,7915x + 225,71 R² = 0,934
0
50
100
150
200
250
0 10 20 30 40 50 60
Co
nce
ntr
ació
n (
pp
m)
Tiempo (h)

Anexo 2: Determinación de parámetros de cinética de crecimiento y consumo
de sustrato mediante un ajuste de datos minimizando el error cuadrado con
un método de punto interior.
%% Funcion objetivo de minimos cuadrados para el ajuste de parametros a %% partir de datos experimentales function rta=LeastSq(P) %% Condiciones Iniciales de las ecuaciones diferenciales Ao=42529; %Concentracion inicial de algas [alg/mL] Bo=13842633.08; %Concentracion inicial de bacterias [bac/mL] So=229.7777778; %Concentracion inicial de fenol [ug/mL] %% Integracion de las ODEs tf=53; %Tiempo de simulacion [hr] [t,x]=ode45(@(t,x)ODEs(t,x,P),[0 tf],[Ao Bo So]); %% Captura de variables A=x(:,1); %Concentracion de algas [alg/mL] B=x(:,2); %Concentracion de bacterias [bac/mL] S=x(:,3); %Concentracion de fenol [ug/mL] %% Experimentales AlgExp=[0 42529 6 1742861 12 2671849 18 5394529 24 7930529 36 12842529 48 14582189]; texpAlg=AlgExp(:,1); Aexp=AlgExp(:,2); BacExp=[0 13842633.08 1 14054162.16 3 14831617.86 5 15763477.2 7 16353363.71 9 17112512.83 29 16374538.59 37 17888888.89 44 17888888.89 48 17888888.89 52 17888888.89]; texpBac=BacExp(:,1); Bexp=BacExp(:,2); FenExp=[0 229.7777778 1 226.5185185 3 222.8888889 5 208.7407407 7 207.6296296 9 207.6296296 29 173.6666667 37 164.8518519 44 164.4074074 48 145.0740741 52 110.6296296]; texpFen=FenExp(:,1); Sexp=FenExp(:,2); %% Funcion Objetivo suma1=0; suma2=0; suma3=0;

for i=2:length(texpAlg) for j=1:length(t) if texpAlg(i)>t(j) && texpAlg(i)<=t(j+1) Amod=(A(j+1)-A(j))/(t(j+1)-t(j))*(texpAlg(i)-t(j))+A(j); end end suma1=suma1+(Aexp(i)-Amod)^2; end for i=2:length(texpBac) for j=1:length(t) if texpBac(i)>t(j) && texpBac(i)<=t(j+1) Bmod=(B(j+1)-B(j))/(t(j+1)-t(j))*(texpBac(i)-t(j))+B(j); end end suma2=suma2+(Bexp(i)-Bmod)^2; end for i=2:length(texpFen) for j=1:length(t) if texpFen(i)>t(j) && texpFen(i)<=t(j+1) Smod=(S(j+1)-S(j))/(t(j+1)-t(j))*(texpFen(i)-t(j))+S(j); end end suma3=suma3+(Sexp(i)-Smod)^2; end rta=suma1+suma2+suma3; end
%% Sistema de ecuaciones diferenciales del modelo function rta=ODEs(t,x,P) %% Captura de variables A=x(1); %Concentracion de algas [alg/mL] B=x(2); %Concentracion de bacterias [bac/mL] S=x(3); %Concentracion de fenol [ug/mL] %% Parametros del modelo uamx=P(1); %Velocidad de crecimiento exponencial de algas [1/hr] ubmx=P(2); %Velocidad de crecimiento exponencial de bacterias [1/hr] Ka=P(3); %Constante de saturacion del crecimiento de algas [ug/mL] Kb=P(4); %Constante de saturacion del crecimiento de bacterias [ug/mL] da=P(5); %Tasa de defuncion de algas [1/hr] db=P(6); %Tasa de defuncion de bacterias [1/hr] Yas=P(7); %Selectividad del crecimiento de algas al consumo de fenol [alg/ug] Ybs=P(8); %Selectividad del crecimiento de bacterias al consumo de fenol [bac/ug] %% Cineticas de Monod ua=uamx*S/(Ka+S); %Velocidad de crecimiento de algas en ausencia de saturacion
[1/hr] ub=ubmx*S/(Kb+S); %Velocidad de crecimiendo de bacterias en ausencia de
saturacion [1/hr] %% Ecuaciones Diferenciales dAdt=(ua-da)*A; dBdt=(ub-db)*B; dSdt=-(1/Yas*ua*A+1/Ybs*ub*B); %% Retorno rta=[dAdt dBdt dSdt]'; end
%% Codigo que minimiza una funcion objetivo de minimos cuadrados para %% ajustar los parametros del modelo function ParameterEstimation %% Valores estimados de los parametros para la inicializacion uamx=29.89943; %Velocidad de crecimiento exponencial de algas [1/hr]

ubmx=0.123901; %Velocidad de crecimiento exponencial de bacterias [1/hr] Ka=10002.928835; %Constante de saturacion del crecimiento de algas [ug/mL] Kb=75.974151; %Constante de saturacion del crecimiento de bacterias [ug/mL] da=0.428684; %Tasa de defuncion de algas [1/hr] db=0.082244; %Tasa de defuncion de bacterias [1/hr] Yas=4639969.763565; %Selectividad del crecimiento de algas al consumo de fenol
[alg/ug] Ybs=1000402.215155; %Selectividad del crecimiento de bacterias al consumo de
fenol [bac/ug] Po=[uamx ubmx Ka Kb da db Yas Ybs]; %Vector de parametros inicial %% Limites lb=[0 0 0 0 0 0 0 0]; ub=[30 20 12000 5000 10 10 Inf Inf]; %% Minimizacion options=optimset('Algorithm','interior-
point','MaxIter',500000,'Display','iter','MaxFunEvals',1000000); P=fmincon(@LeastSq,Po,[],[],[],[],lb,ub,[],options); %%%P=Po; %% Retorno de Parametros display('===========Valores de los parametros=========='); fprintf('Velocidad de crecimiento exponencial de algas [1/hr]: %f\n',P(1)); fprintf('Velocidad de crecimiento exponencial de bacterias [1/hr]: %f\n',P(2)); fprintf('Constante de saturacion del crecimiento de algas [ug/mL]: %f\n',P(3)); fprintf('Constante de saturacion del crecimiento de bacterias [ug/mL]:
%f\n',P(4)); fprintf('Tasa de defuncion de algas [1/hr]: %f\n',P(5)); fprintf('Tasa de defuncion de bacterias [1/hr]: %f\n',P(6)); fprintf('Selectividad del crecimiento de algas al consumo de fenol [alg/ug]:
%f\n',P(7)); fprintf('Selectividad del crecimiento de bacterias al consumo de fenol [bac/ug]:
%f\n',P(8)); %% Graficas Simulacion(P) end
%% Codigo que simula el sistema de crecimiento en competicion por sustrato %% (fenol) entre Bacterias y Algas. function Simulacion(P) %% Condiciones Iniciales Ao=42529; %Concentracion inicial de algas [alg/mL] Bo=13842633.08; %Concentracion inicial de bacterias [bac/mL] So=229.7777778; %Concentracion inicial de fenol [ug/mL] %% Integracion de las ODEs tf=52; %Tiempo de simulacion [hr] [t,x]=ode45(@(t,x)ODEs(t,x,P),[0 tf],[Ao Bo So]); %% Captura de variables A=x(:,1); %Concentracion de algas [alg/mL] B=x(:,2); %Concentracion de bacterias [bac/mL] S=x(:,3); %Concentracion de fenol [ug/mL] %% Experimentales AlgExp=[0 42529 6 1742861 12 2671849 18 5394529 24 7930529 36 12842529 48 14582189]; BacExp=[0 13842633.08 1 14054162.16 3 14831617.86 5 15763477.2 7 16353363.71

9 17112512.83 29 16374538.59 37 17888888.89 44 17888888.89 48 17888888.89 52 17888888.89]; FenExp=[0 229.7777778 1 226.5185185 3 222.8888889 5 208.7407407 7 207.6296296 9 207.6296296 29 173.6666667 37 164.8518519 44 164.4074074 48 145.0740741 52 110.6296296]; %% Graficas subplot(3,1,1) plot(t,A,'b',AlgExp(:,1),AlgExp(:,2),'or') ylabel('Algas [alg/mL]') grid on subplot(3,1,2) plot(t,B,'b',BacExp(:,1),BacExp(:,2),'or') ylabel('Bacterias [bac/mL]') grid on subplot(3,1,3) plot(t,S,'b',FenExp(:,1),FenExp(:,2),'or') ylabel('Fenol [ug/mL]') xlabel('Tiempo [hr]') grid on end