“descripciÓn de un nuevo gÉnero de algas rojas …biblio.uabcs.mx/tesis/te3148.pdf · rafael...
TRANSCRIPT

UNIVERSIDAD AUTÓNOMA DE BAJA CALIFORNIA SUR ÁREA DE CONOCIMIENTO DE CIENCIAS DEL MAR
DEPARTAMENTO ACADÉMICO DE BIOLOGÍA MARINA
TESIS
“DESCRIPCIÓN DE UN NUEVO GÉNERO DE ALGAS ROJAS
CORALINAS (CORALLINALES, RHODOPHYTA)”
QUE, COMO REQUISITO PARA OBTENER EL TÍTULO DE
BIÓLOGO MARINO
PRESENTA:
ALEJANDRA ANGELES PEREZ
DIRECTOR:
DR. RAFAEL RIOSMENA RODRIGUEZ
La Paz B.C.S. a Junio del 2014

1

2
DEDICATORIA
Dedico este esfuerzo a Elvira y Amelia por su amor, apoyo pero
sobre todo su ejemplo y fuerza las amo.
A Rafael y Pablo quienes hacen mí vida.
Para la familia Báez-Angeles por su amor y apoyo.
Para mis hermanos y hermanas quienes son incondicionales de la
vida.

3
AGRADECIMIENTOS
Esta tesis forma parte del proyecto “Determinación de amenazas y
posibles estrategias de mitigación en mantos de rodolitos como objetos de
conservación en el Caribe. Parte I: Quintana Roo.” que financio el Fondo
SAM, del proyecto “Monitoreo de la vulnerabilidad de mantos de rodolitos
respecto al cambio global en México: Analísis de las implicaciones
socioeconomicas y ecológicas con financiamiento del fondo sectorial
CONACYT-SEMARNAT 23425, y Scientific basis for conservation and
management of Rhodolith beds in the caribbean: a long term educational
program financiado por el Smthsonian Tropical Research Institute en
Panama al Dr. Rafael Riosmena Rodríguez. Agradezco el apoyo en el
trabajo de campo del Dr. Rafael Riosmena Rodríguez, Dr. Julio Espinoza
Avalos, Dra. Jazmín Hernández Kantún y M. en C. Marina Hirales Cota.
Los comentarios del Dr. Rafael Riosmena Rodriguez, Dr. Gustavo
Hernandez Carmona y la M. en C. María del Carmen Rodriguez Jaramillo
ayudarón a mejorar el presente documento.

4
ÍNDICE
Dedicatoria 2
Agradecimientos 3
Índice 4
Lista de tablas y figuras 5
Resumen 8
Abstract 9
Introducción 10
Antecedentes 19
Material y métodos 24
Resultados 29
Discusión 37
Referencias 40

5
LISTA DE TABLAS Y FIGURAS
Tabla 1. Organización sistemática actualmente aceptada para las algas
coralinas basado en Harvey et al. (2003).
Tabla 2. Características diagnosticas de familias en Corallinales.
Tabla 3. Características diagnosticas a nivel subfamilia de la familia
Corallinaceae y Hapalidiaceae.
.Tabla 4. Relación de los sitios de muestreo del proyecto y su referencia
geográfica (ND es No determinada).
Tabla 5. Análisis comparativo de los géneros conocidos para la subfamilia
Lithophylloideae basado en Woelkerling et al (2002) con relación al
nuevo género propuesto.

6
Tabla 6. Análisis comparativo respecto de las especies parasíticas y
eneolíticas en algas coralinas geniculados.
Figura.1. Mapa de distribución de las zonas de muestreo en la zona sur de
Quintana Roo.
Figura 2. Holotipo de Adelus harveyanum almacenado en FBCS 1400 que
esta como parasito de una planta de Neogoniolithon brassica-
florida, los ejemplares se observan como pequeñas ámpulas
(flecha) que sobresalen de la superficie del hospedero.
Figura 3. Corte longitudinal donde se muestra el aspecto general del
parasito donde se observa una región diferenciada entre el
huésped y el hospedero (flecha). En el recuadro se delimita la
(FBCS 1400).

7
Figura 4. Área de transición (línea blanca) entre el parasito (P) y el
huésped (H) en donde se muestra que el parasito tiene conexiones
pit secundarias entre los filamentos (flechas) mientras que el
hospedero tiene fusiones celulares (doble flecha) y tricocisto
(FBCS 1400).
Figura 5. Corte longitudinal donde se muestra la presencia de haustoria
(flecha) del parasito internándose en el hospedero. Barra de escala
1400).
Figura 6. Corte longitudinal del conceptáculo carpoesporofítico mostrando
que los filamentos gonimoblasticos están en posición superficial con
respecto al
14000X).

8
RESUMÉN
Los mantos de rodolitos son ambientes críticos para la conservación
debido a la biodiversidad de especies asociadas y a la intervención que
tienen estas comunidades en procesos ecológicos relevantes (como
reproducción, reclutamiento, crianza y como hábitat alternativo) en la zona
costera. El reconocimiento del valor de estos mantos en el Pacífico
Mexicano ha quedado claro, no se han hecho evaluaciones para la zona
del Atlántico, en particular para el arrecife Mesoamericano en donde se
desarrollaron muestreos intensivos entre 2006 a 2008 para caracterizar a
los mantos de rodolitos en la región. Como resultado de este análisis se
determinó la presencia de un género nuevo y, por lo tanto especie nueva,
dentro de la familia Lithophylloideae debido a que presenta conexiones pit
secundarias, no presenta fusiones celulares y tiene un conceptáculo
uniporado. Se propone el nombre de Adelus harveyanum para este nuevo
taxa. Con la descripción actual se incrementa el número de especies
parasíticas conocidas y es el primer registro de una especie parasítica
para zonas tropicales. Además, se demuestra el valor del conceptáculo
tetrasporangial como estructura taxonómica para diferenciar géneros en la
subfamilia Lithophylloideae lo que fortalece su utilización en la subfamilia
Mastophoroideae.

9
ABSTRACT
Maerl beds environments are critical to the conservation of biodiversity
because of associated species and intervention in this community relevant
ecological processes (e.g. reproduction, recruitment, and as an alternative
breeding habitat) in the coastal zone . Recognizing the value of these
garments in the Mexican Pacific is clear, no assessments have been made
in the Atlantic area, including the Mesoamerican reef where intensive
surveys between 2006 and 2008 were developed to characterize maerl
beds in the region . As a result of this analysis the presence of a new
genus was determined and , therefore new species within the family
Lithophylloideae because it has secondary pit connections, presenting no
cell fusions and has a uniporate conceptacle . Adelus harveyanum is the
name for this new taxa proposed . With the current description the number
of known parasitic species increases and is the first record of a parasitic
species in tropical areas. In addition , the value of tetrasporangial
conceptacle as taxonomic structure is shown to differentiate genera in
subfamily Lithophylloideae which strengthens Mastophoroideae use in the
subfamily. This paper shows how little known this coral fauna in other parts
of Mexico and the urgent need for detailed case studies.

10
INTRODUCCIÓN
Las algas coralinas pertenecen al Phyllum Rhodophyta y se distingue
por presentar en su pared celular carbonato de calcio en forma de calcita
(Silva & Johansen 1986). Estas algas tienen una amplia distribución a lo
largo/ancho del mundo, son abundantes en diversos hábitats marinos
(ambientes rocosos, praderas de pastos, arrecifes tropicales, templados y
formando mantos de rodolitos). Pueden crecer en un rango amplio de
temperaturas desde aguas frías hasta aguas tropicales, tienen un rango de
profundidad por debajo de la zona intermareal hasta el límite conocido
para macroalgas a 286 m de profundidad (Littler et al.1991).
Ecológicamente estas algas son ingenieros ecosistémicos ya que
intervienen en el proceso de reclutamiento, crianza, refugio y producción
de hábitat para animales y otras algas en ambientes marinos (Coleman &
William 2002). Las coralinas representan también, fuentes de alimento
para el ramoneo de numerosos peces e invertebrados. Uno de los
principales componentes de la dieta de diferentes invertebrados como
equinodermos, moluscos y algunos crustáceos son las algas coralinas
(James 2000). Existe un beneficio mutuo entre algas coralinas y cangrejos

11
herbívoros en las áreas de pastos marinos, las algas proporcionan refugio
a los cangrejos y estos a su vez limpian la superficie de las algas de
epifitos. Las coralinas son importantes componentes de los arrecifes
tropicales, por ejemplo cubren el 39% del fondo arrecifal de Waikiki Hawaii
(Littler 1973), al sureste de Japón el 33% de la cobertura total (Iryu &
Matsuda 1988). En el arrecife de la Gran Barrera en Australia el 20%
(Fabricius & Deáth 2001) de la cobertura total está representado por estas
macroalgas. Los arrecifes de coral dependen de las algas coralinas para
reforzar y mantener resistencia frente al embate de las olas, absorbiendo
su energía y ayudando a reducir la erosión del arrecife (Littler & Littler
1988).
Algas coralinas no geniculadas presentan formas de vida libre
conocidas como rodolitos o maerl, y que abarcan grandes extensiones
debido a los sedimentos carbonatados que generan (Boscence 1983). El
carbonato del cual están hechos los rodolitos es utilizado como un paleo
indicador del medio ambiente (Foster et al. 1997). Depósitos de rodolitos
han sido utilizados por el hombre como enriquecedores de suelos
europeos (Blunden, Farnham et al. 1977). Rodolitos individuales presentan
una gran variación morfológica que, aparentemente, responde a

12
variaciones físicas del ambiente (Steller & Foster 1995). Además los
mantos de rodolitos soportan una comunidad rica de diversas especies de
flora y fauna, (Steller et al. 2003). Se ha propuesto que los mantos de
rodolitos son buenos indicadores para el monitoreo de áreas naturales
protegidas (Avila & Riosmena 2010) y pueden ser utilizados para
comprender el calentamiento global. Las algas coralinas se ha utilizado en
estudios o investigaciones de tipo fisiológico, ecológicos o de
conservación, pero depende de la habilidad para identificar los
especímenes a nivel de especies la calidad de los datos a encontrar
(Harvey & Woelkerling 2007).
Las Taxonomía de las coralinas tiene una historia interesante ya que
Lennaeus (AÑO) consideró a las algas coralinas dentro de un mismo grupo
con los corales, lo que genero una gran controversia sobre la diferencia
entre animales y plantas. Esper (1830) usando las estructuras
reproductivas como carácter distintivo fundamento apropiadamente la
naturaleza vegetal de este grupo y apoyo la segregación entre animales y
plantas. Entre 1830 y 1980 las coralinas lograron distinguirse como una
familia de algas rojas, sin embargo las coralinas han sido aceptadas su
posición como orden Coralinales hasta hace poco (Silva & Johansen 1986)

13
y en estudios recientes de biología molecular, se ha sugerido considerarlas
como subclase (Yoon et al. 2006).
Desde su reconocimiento como familia Corallinaceae del reino
vegetal (Esper 1830) no se habían propuesto cambios a este nivel. En
términos generales se consideraba que todas las algas coralinas que
presentan calcita también tenían sus estructuras reproductivas en
conceptáculos. Sin embargo, el reconocimiento de la presencia de soros
en organismos que secretaban calcita justifico el reconocimiento de una
segunda familia a la que se le nombro como Familia Sporalithaceae
(Verheij 1993) y que además presentaba la característica de presentar
tanto conexiones pit secundarias como fusiones celulares entre filamentos,
un carácter poco valorado en el pasado pero con gran utilidad sistemática
(Woelkerling 1988). Además, estos organismos presentan células
epiteliales en forma de campana, otra característica diagnostica del grupo
(Verheij 1993).
Contemporáneo a esta propuesta también se había estado
evaluando el valor sistemático de varias características clave entre las más
relevantes hasta nuestra época son: 1) presentar células epiteliales en

14
forma plana o redonda contra presentar forma acampanada; 2) presentar
una célula subepitelial larga o corta; 3) presentar conexiones entre
filamentos compuestas por pit secundarios o fusiones celulares; 4)poseer
un conceptáculo tetrasporangial uniporados o multiporado, ya que los
conceptáculos gametangial en todos los grupos (incluido Sporalithaceae)
son uniporados; 4) presentar espermatia simples o dendroides y 5) la
organización de los filamentos gonimoblasticos de manera periférica o
superficial (Woelkerling 1987, 1988, 1996a, 1996b, 1996c, 1996d, 1996e,
1996; Woelkerling & Harvey 1996) para diferenciar subfamilias. Este
esfuerzo desemboco en la propuesta de reconocer una tercer familia
Hapalidiaceae (Harvey et al. 2003) basada en datos anatómicos, ultra
estructurales y ontogénicos muestran que las coralinas poseen placas
multiporada que distingue a la familia Hapalidiaceae, incluye a la
subfamilia Choreonematoideae, Melobesioideae y Austrolithoideae.
Mientras en la anterior Familia Corallinaceae se reconoce a las subfamilias
Lithophylloideae, Mastophoroideae y Metagoniolithoideae (Tabla 1).

15
Tabla 1. Organización sitemática actualmente aceptada para las algas
coralinas basado en Harvey et al. (2003).
Familia Familia Familia
Corallinaceae Hapalidiaceae Sporolithaceae
Subfamilia Subfamilia
Corallinoideae Austrolithoideae
Lithophylloideae Choreonematoideae
Metagoniolithoideae Melobesioideae
Mastophoroideae
Las 3 familias se pueden reconocer claramente usando 4
características diagnosticas (Tabla 2): 1) arreglo de las esporas en el
tetraesporangio; 2) Tetra-biesporangial con tapones apicales; 3) Tetra-
biesporangial con techos multiporados y 4) Tetra-biesporangial nacido en
conceptáculos o compartimientos calcificados. Estas características se han
utilizado ampliamente en la literatura (revisado por Harvey et al 2005) y
representan caracteres consistentes en todos los estudios realizados hasta
la fecha.

16
Tabla 2. Características diagnosticas de familias en Corallinales.
Carácter Corallinaceae Sporolithaceae Hapalidiaceae
Arreglo de las esporas en el tetraesporangio
Zonado Cruciado Zonado
Tetra-biesporangial con tapones apicales
No Si Si
Tetra-biesporangial con techos multiporados
No No Si
Tetra-biesporangial nacido en conceptáculos o compartimientos calcificados.
Conceptáculos Compartimientos calcificados
conceptáculos
Sin embargo, el reconocimiento de las subfamilias todavía
representa un reto a resolver es la posición sistemática de las subfamilias
dentro de las familias reconocidas actualmente (Tabla 3) ya que no se
tiene suficiente información para determinar si cada una de estas
subfamilias representan una familia potencialmente a reconocer.

17
Tabla 3. Características diagnosticas a nivel subfamilia de la familia
Corallinaceae y Hapalidiaceae.
. Familia Subfamilia Caracteres diagnósticos
Corallinaceae Corallinoideae Células con filamentos vegetativos pegados células fusionadas, conexiones pit secundarias desconocidas, Genícula compuesta de una hilera de células.
Metagoniolithoideae Células con filamentos vegetativos ligados a células fusionadas, conexiones pit secundarias no conocidas, ge nicula compuesta de una hilera filamentos multicelulares.
Mastophoroideae Células con filamentos vegetativos contiguos ligados principalmente o exclusivamente por células fusionadas, conexiones pit secundarias conocidas solamente en un genero (Metamastophora), ge nicula ausente.
Lithophylloideae Células con filamentos vegetativos contiguos ligados principalmente o exclusivamente
Hapalidiaceae Austolithoideae Células con filamentos vegetativos no ligados a células fusionadas o conexiones pit secundarios, genícula ausente, placas multiporadas compuestas de células maduras
Choreonematoideae Células con filamentos vegetativos contiguos no ligadas a células fusionadas o conexiones pit secundarias , genícula ausente, placa multiporada madura, compuesta su matriz solamente de carbonato de calcio.
Melobesioideae Células con filamentos vegetativos contiguos ligados a células fusionadas, no se conocoe conexiones pit secundarios, genicula ausente, placa multiporada compuesta de células maduras.
Otro de la grandes problemas que tienen cada una de las subfamilias
es el incompleto inventario de los géneros y especies debido a la gran
cantidad de formas que presentan algunas especies pero que al analizar la

18
anatomía encontramos que solo representa a una especie, a la poca
atención que ha tenido este grupo en algunas regiones del mundo o los
pobres conceptos en que están basa la taxonomía en regiones estudiadas
con mucha anticipación (Woelkerling et al. 1993, Riosmena- Rodríguez et
al. 1999). Sin embargo algunas estructuras anatómicas son cruciales para
la identificación taxonómica. Para obtener estas estructuras es necesario
cortes histológicos y análisis en microscopio electrónico. Los procesos
histológicos incluyen parafina (Riosmena-Rodríguez et al 1999) o resina
blanca (Woelkerling 1988). Una buena lámina requiere tener una buena
observación de la anatomía vegetativa, donde se observe la región epitelial
si son células redondeadas o aplanadas.
Una segunda estructura es la observación clara de conexiones pit
secundarias o células fusionadas. Un grupo de cuidado son los parásitos
ya que no tienen ninguna de las características anteriores que ayuden a
diferenciar, sin embargo el tamaño de las células nos podrá proporcionar
ayuda para su diferenciación. Los poros de las estructuras reproductivas
es muy importante como es proporcionarnos información si son
conceptáculos uniporados bi – tetrasporangial o conceptáculos
multiporados bi - tetrasporangial, formando Soros. En algunos

19
conceptáculos uniporados es importante determinar la posición de los
filamentos gonimoblasticos o en caso de conceptáculos machos la
presencia de simple conceptáculos espermatangial o un conceptáculo
dendroide espermatangial.
Choreonematoideae.- presenta conexiones entre los filamentos
vegetativos adjuntos y tienen una placa multiporada que es acellular al
tiempo de madurez, con una matriz solamente de carbonato de calcio.
ANTECEDENTES
Rodolito se le ha denominado a los individuos pertenecientes a las
algas rojas calcáreas no geniculadas (Orden Corallinales), que no están
sujetas a un sustrato (Woelkerling 1988). La denominación que recibe en
Europa es “maerl”, pudiéndose ambos utilizar indistintamente. El origen de
estos rodolitos es el recubrimiento de un fragmento de concha, una roca o
por el asentamiento libre de esporas. Estos individuos al crecer adquieren
una forma cercana a la esférica, en la mayoría de los casos, debido a
procesos de bioturbación y oleaje (Foster 2001). Los rodolitos pueden
formar agregaciones muy densas (mantos) en la zona costera submareal,

20
que puede variar en extensión desde unos cuantos metros a varios
kilómetros (Hetzinger et al. 2006).
Debido a que recubren amplias extensiones de fondo, y a que están
en continuo crecimiento, estos organismos son productores natos de
sedimento biogénico, el cual puede llegar a ser muy relevante en los
procesos sedimentarios costeros o en función de la biodiversidad presente
(Birkett et al. 1998). Estos mantos se presentan en una amplia gama de
estratos geológicos, desde el Paleoceno (hace 60 millones de años) hasta
el Pleistoceno (hace 5 millones de años), siendo uno de los pocos registros
continuos en yacimientos fósiles para macroalgas marinas que pueden ser
comparados en los cinco continentes.
Los estudios en biodiversidad han determinado que existe una alta
riqueza y abundancia de organismos asociados, principalmente de
macroalgas, invertebrados y peces. Los mantos de rodolitos constituyen un
hábitat alternativo para especies, tanto de hábitats rocosos como arenosos
(Steller et al. 2003). En estos mantos se ha llevado a cabo el reclutamiento
y desarrollo de especies de importancia ecológica. Debido a su amplia
distribución vertical (profundidad) y horizontal (geográfica), se puede

21
encontrar un gama de especies asociadas, lo que ha llevado a considerar
su valor como refugios espaciales (Riosmena-Rodríguez & Medina-López
2010). Esto ha propiciado que existan especies típicas de ambientes
rocosos en combinación de especies típicas de zonas arenosas (Foster
2001). Los mantos representan lugares donde viven poblaciones
completas de organismos expatriados de sus poblaciones originales por la
distancia de su población original. Además, existen especies exclusivas de
este hábitat. Se considera que son refugios tanto de especies protegidas
(por su baja densidad) como de especies conocidas (por su gran
abundancia), y se les ha incluido como un elemento crítico para la
conservación para el diseño de ANP y su monitoreo (Riosmena-Rodríguez
et al. 2010; Avila & Riosmena 2010).
Tradicionalmente se han estudiado los mantos de rodolitos en
Europa, donde se explotan comercialmente como fuente de fertilizante
(Blunden et al. 1971), para terrenos agrícolas que sufren de acidificación.
Adicionalmente, estos sitios se han reconocido por ser zonas donde la
actividad pesquera es alta y donde las actividades de la pesca de arrastre
se han practicado con mayor continuidad (Bordehore et al. 2003). Las
principales amenazas que han detectado para estos hábitats son la

22
extracción de los mantos, la presencia de especies invasoras, la
eutrofización, la maricultura y la pesca (Grall & Hall-Spencer 2003). En
esas condiciones se ha encontrado que una intensa actividad produce el
cambio de tipo de fondo de arenas hacia limos y el deterioro del
ecosistema con la pérdida de su biodiversidad y productividad económica
(Boscence & Wilson 2003). Una de las principales amenazas es la falta de
conocimiento de su existencia y, por lo tanto, subvaloración en la
implementación de políticas apropiadas de conservación.
Dentro del Atlántico oeste se ha reconocido la presencia y valor de
los mantos de rodolitos en aguas de Estados Unidos, desde Alabama
hasta el Golfo de México (Prager& Ginsbur 1989), incluyendo Bermuda
(Littler et al. 1991) y Puerto Rico (Balentine et al. 2000). Para el Sistema
Arrecifal Mesoamericano, los reportes sobre la presencia y distribución de
estos mantos de rodolitos están restringidos a Cozumel y Panamá
(Muckelbauer 1990 &). La escasez evidente de conocimiento de estos
mantos en el SAM se refleja claramente en que no han sido consideradas
como parte de ningún Plan de Manejo de las Áreas Naturales Protegidas
declaradas, al menos en el Estado de Quintana Roo. Aunque en las listas
de especies presentadas para la justificación existan algunas especies que

23
se han considerado en algún momento formadoras de rodolitos. Sin
embargo, las posibles amenazas, algunas ya identificadas como la
eutrofización y presión por la actividad turística (CONABIO 1996),
encaminan a una posible degradación o pérdida inadvertida de estos
hábitats. En consecuencia, no hay una descripción de la situación actual
tanto de la distribución como del conocimiento de la fauna/flora asociada y
de las posibles amenazas a los que están expuestos, por lo que el objetivo
de conservar los recursos naturales de esta región queda lejos de
cumplirse en su totalidad. Riosmena-Rodríguez (2008) determinó que los
mantos de rodolitos se encontraron compuestos por al menos 3 especies:
Neogoniolithon brassicaflorida, Amphiroa fragilissima y Amphiroa tribulus
con evidencia que bien podrían existir otras 3 especies menos abundantes
pero se requiere de hacer un estudio taxonómico formal de estas especies
formadoras de rodolitos. Como parte de estos esfuerzos se encontró un
ejemplar de alga parasita que no concuerda con ninguna de las especies o
géneros descritos en la actualidad y representa el primer registro de un
alga coralina no geniculada parasítica para el Océano Atlántico.
OBJETIVO

24
Describir un nuevo género y especie de alga coralina no geniculada
parasita de mantos de rodolitos para la reserva de Xcalak.
MATERIAL & MÉTODOS
TRABAJO DE CAMPO
Se determinó que dentro del estado de Quintana Roo los mantos de
rodolitos se encuentran ampliamente en al menos 18 sitios visitados (Tabla
4) en dos regiones principales Cozumel y Xcalak (Fig. 1). Se tomo la
determinación de desarrollar el trabajo de campo extenso en la zona de
Xcalak debido a su cercanía con proyectos turísticos y en donde se
consideró que bien podría existir un mayor riesgo con respecto a la
actividad turística. Se muestreo donde existen poblaciones importantes
(100% cobertura) de rodolitos asociados a pastos en Xcayal, Punta
Gavilanes, Santa Julia y Sur deXcalak. Batimétricamente se encontraron
mantos de forma discontinua desde los 10 cm de profundad (zona de
Mahaual y Xcalac) hasta los 5 m, después parches entre los 10 a los 20 m
de profundidad en algunas zonas de Cozumel. Considero que existe un
gradiente en función de la profundidad que se ve reflejado en la presencia
de especies de Amphiroa y Neogoniolithon en zonas someras (hasta 1 m

25
de profundidad) y la presencia de Neogoniolithon en zonas profundas
como al especie a los 20 m de profundidad máximo.
Tabla 4. Relación de los sitios de muestreo del proyecto y su referencia
geográfica (ND es No determinada).
Sitio de muestreo Latitud Longitud
Playa Bonita 20°20’ 46.9’’N 86°54’12.9’’W
Cerca del faro 20° 16.241’N 86°59.457’W
Chunchakaab 20° 17.175’N 87° 00.153’W
Colombia (parte sur)
20° 18.047’N 87° 01.197’W
Colombia bajo 20° 18.070’N 87° 00.832’W
Palancar 20°19.643’N 87°01.346’W
Yucab 20° 25.592’N 87°00.590’W
San Francisco 20° 20.643’ N 87°07.374’ W
Playa Mia 20° 23.892N 87°01.172W
Mahahual Sitio 1 18°42.98’ N 87° 42.44´W
Canal 87°15.832´ W
Cerca del arrecife 18°42.974N 87°47.366W
Sitio 4 18°42.9871N 87°42.309W
Cerca del faro 18° 43.357 N 87° 42.133W
Xcalak 18° 17.742’N, 87° 49.905’W
Sitio 2 18° 17.755’N, 87° 49.903’W
Frente al arrecife ND ND
Santa Julia 18° 21.021’ N 87° 48.86’ W

26
Figura.1. Mapa de distribución de las zonas de muestreo en la zona sur de
Quintana Roo.
TRABAJO DE LABORATORIO
Se seleccionaron la mayor cantidad de talos con la única condición
de que éstos fuesen reproductivos y, con ayuda de un estereoscopio, se
localizaron los conceptáculos. Las estructuras seleccionadas fueron

27
cortadas con ayuda de unas pinzas y pasaron por un lavado de agua
corriente para eliminar el formol y después fueron puestas a descalcificar
en ácido nítrico al 0.6 M, realizando recambios de este cada 2 horas
aproximadamente.
Una vez descalcificadas, todas las estructuras fueron sometidas a un
proceso de tinción (KMnO4 por 25 min.), deshidratación, en una secuencia
de alcoholes de 30, 60, 90 y 100% de etanol, etanol-butanol (50 y 50%) y
butanol al 100%, cada uno por intervalos de 30 min. y, por último, un
proceso de inclusión en parafina de 2 pasos; 30 min. en parafina-butanol
(50 y 50%) y parafina al 100%, por 12 hrs, de acuerdo con los
procedimientos sugeridos en Riosmena-Rodríguez et al. (1999). Después
de dicho proceso las muestras fueron embebidas en parafina, con los
cuales se obtuvieron cortes histológicos de los tejidos de un grosor de .6
m.
Una vez hechos los cortes, éstos fueron puestos en un baño maría
para su estiramiento y montados en un portaobjetos. Después de su
observación al microscopio óptico y dependiendo de la calidad de los
cortes, éstos fueron desparafinados con Xylol al 100% y montados como

28
laminillas permanentes, añadiendo en la parte del corte Cytoseal™ como
resina y colocando encima un cubreobjetos. Evaluando los contenidos de
las laminillas, se realizó la medición y conteo de diferentes estructuras
para obtener una descripción del género nuevo y especie nueva de
acuerdo a la propuesta de Chamberlain (1983). Todas las recolectas que
se utilizaron para este trabajo, se le asigno un número de catalogo en el
Herbario Ficológico de la Universidad Autónoma de Baja California Sur
(FBCS).
TRABAJO DE ESCRITORIO
Con la información derivada de las laminas histológicas y las
mediciones de las características estudiadas se realizo una comparación
para evaluar a que familia, subfamilia podría ser asignado este nuevo
género/especie con referencia a las especies y géneros conocidos hasta
este momento. Se designo el material tipo e isotipos y se determinó tanto
la distribución geográfica como ecológica de los organismos estudiados.
La terminología utilizada para describir la anatomía siguió lo descrito en
Woelkerling (1988) y para las formas de crecimiento (morfología) se siguió
lo descrito en Woelkerling et al. (1993).

29
RESULTADOS
DISPOSICIÓN GENÉRICA DEL MATERIAL
En el material analizado, se presentó una construcción monomérica
(Figs. 2A y B) en donde solo un grupo de filamentos es posible observar,
así como la presencia de fusiones celulares (Fig. 3C) uniendo células de
filamentos adyacentes. En el caso de las células epiteliales se observaron
redondeadas o planas pero no acampanadas (Figs. 3C, D y E). Además,
consistentemente se observaron células subepiteliales más grandes que
las subsecuentes (Fig. 3D y E).

30
Tabla 5. Análisis comparativo de los géneros conocidos para la subfamilia
Lithophylloideae basado en Woelkerling et al (2002) con relación al
nuevo género propuesto.
Característica/género Adelus Amphiroa Ezo Lithophyllum/ Titanoderma
Lithothrix Paulsilviella Tenarea
Parasito P A P A A A A
Parte central organizada
A A A A
Genícula A P A A P A A
Crecimiento dímero a partir de la genícula
A A A A P A A
Organización isobilateral
A A A A A A P
Dimero/monómero (D/M)
M D/M M D/M D/M M A
Exclusivamente costra
A A A A P A A
Posición de filamentos gonimoblasticos en función del disco de fusión: superficial (S) o periférico (P)
S P P P P P P
DESCRIPCIÓN DEL NUEVO GÉNERO Y ESPECIE
Adelus harveyanum Ángeles-Pérez and Riosmena-Rodríguez
Figs. (2 - 6)
LOCALIDAD TIPO: Punta Gavilanes, Quintana Roo.

31
HOLOTYPE: El material tipo Es la collection 16 (Fig. 1) e isotipos están
depositados en el Herbario Ficológico de la Universidad Autónoma
de Baja California Sur con el número 14000 (holotipo) 14001
(isotipo).
Etimologia: Esta combinación específica se utiliza para reconocer el
trabajo de la Dra. Adela Harvey en el campo de la taxonomía de las
algas coralinas por su trabajo de más de 20 años dedicados a esta
labor.
Talos no geniculados parasíticos (Figs. 2 -3), forma de crecimiento
costroso, (Figs. 3); pseudoparenquimatoso, organización interna dorso-
ventral en la zonas incrustantes del talo, y construcción monomérica (Figs.
3); filamentos compuestos por células de 5 - 8µm de alto por 7-12µm de
largo; terminando en la parte superficial con células epiteliales de paredes
distantes redondeadas o planas, pero nunca acampanadas de 5-8µm de
largo por 2µm de alto. Células de los mismos filamentos unidas por
conexiones pit primarias; células de filamentos adyacentes unidas por

32
conexiones pit secundarias (Fig. 4); haustoria presente (Fig. 5); tricocistos
ausentes.
Figura 2. Holotipo de Adelus harveyanum almacenado en FBCS 14000
que esta como parasito de una planta de Neogoniolithon
brassica-florida, los ejemplares se observan como pequeñas
ámpulas (flecha) que sobresalen de la superficie del hospedero.

33
Figura 3. Corte longitudinal donde se muestra el aspecto general del
parasito donde se observa una región diferenciada entre el
huésped y el hospedero (flecha). En el recuadro se delimita la
zona qu
(FBCS 14000).

34
Figura 4. Área de transición (línea blanca) entre el parasito (P) y el
huésped (H) en donde se muestra que el parasito tiene conexiones
pit secundarias entre los filamentos (flechas) mientras que el
hospedero tiene fusiones celulares (doble flecha) y tricocisto
presente (flecha con cabeza cuadrada).
(FBCS 14000).

35
Figura 5. Corte longitudinal donde se muestra la presencia de haustoria
(flecha) del parasito internándose en el hospedero. Barra de escala
14000).
Conceptáculos tetraesporangiales, biesporangiales, espermatangial y
femeninos no observados. Conceptáculos carpoesporangial maduros
ligeramente elevados sobre la superficie del talo (Fig. 6). Carposesporofito
desarrollándose junto con el conceptáculo femenino después de una
posible Cariogamia, consistiendo éste de una forma irregular y
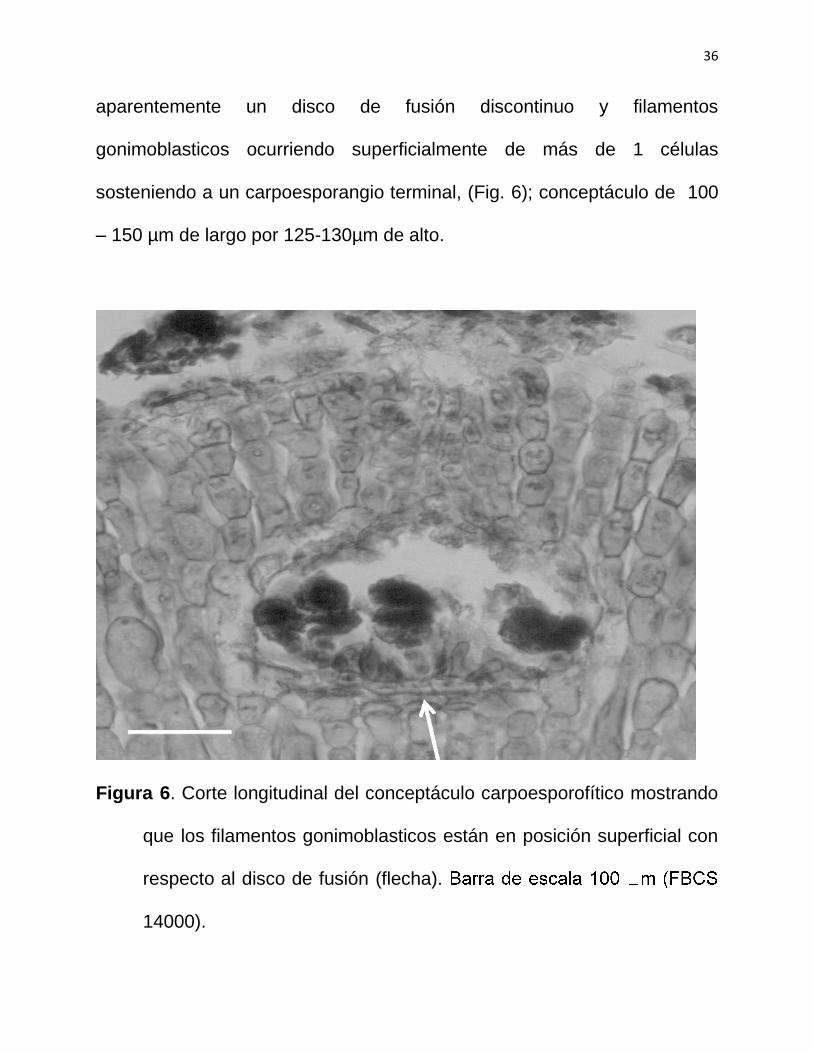
36
aparentemente un disco de fusión discontinuo y filamentos
gonimoblasticos ocurriendo superficialmente de más de 1 células
sosteniendo a un carpoesporangio terminal, (Fig. 6); conceptáculo de 100
– 150 µm de largo por 125-130µm de alto.
Figura 6. Corte longitudinal del conceptáculo carpoesporofítico mostrando
que los filamentos gonimoblasticos están en posición superficial con
respecto al disco de fusión (flecha).
14000).

37
Hábitat: Los talos colectados se encontraron a una profundidad de 0.30 m
hasta los 4 a 5 m, el parasito estaba sobre Neogoniolithon brassica
florida y sobre una especie indeterminada del género Lithophyllum.
Distribución geográfica: solo se encontraron ejemplares en Área Natural
Protegida de Xcalak.
Material Examinado: Punta Gavilanes, Hernandez-Kantun & Hirales-Cota,
Fecha FBCS14000.
Discusión
En los años recientes la sistemática de las algas coralinas ha sufrido
cambios relevantes y profundos (Bittner et al 2011; Aki et al 211) utilizando
técnicas de biologia molecular. Esto ha propuesto la resurrección de
géneros que se consideraban anteriormente como sinónmos de otros
grupos (caso Porolithon). Sin embargo, se ha propuesto una gran
diversidad de líneas nuevas a explorar por lo que estudios como el

38
presente son necesarios para comprender mejor la sistematica del grupo y
sus relaciones filogenéticas usando tanto la morfología como datos
moleculares.
Dentro de las algas coralinas no geniculadas se conoce 2 especies
endofiticas Choreonema thuretii (Bornet) Schmitz que pertenece a la
Familia Hapalidaceae; y Ezo epiyoense Adey, Maski et Akioka que
pertence a la Familia Corallinaceae, subfamilia Lithophylloideae. Mientras
que se conocen 2 consideradas parasíticas (por la presencia de un estilete
que penetra al hospedero sin que hasta el momento se haya demostrado
su funcionalidad): Kvaleya epilaeve Adey & Serapani que pertenece a la
Familia Hapalidaceae, y Lesueuria minderiana Woelkerling & Ducker que
se encuentra en la subfamilia Mastophoroideae (Tabla 6). Con la
descripción actual se incrementa el número de especies parasíticas
conocidas y es el primer registro de una especie parasítica para zonas
tropicales o del Caribe.
Tabla 6. Análisis comparativo respecto de las especies parasíticas y
eneolíticas en algas coralinas geniculadas.

39
Huésped Hospedero Hábito Distribución
Familia Hpalidaceae Subfamilia Choeronematoideae
Choreonema thuretii 1 Cheilosporeum, Corallina, Jania, Haliptylon
Parasito Mundial
Kvaleya epilaeve 2 Leptophytum laeve Atlántico NE, NO
Familia Corallinaceae
Subfamilia Mastophoroideae
Lesueuria minderiana 3 Metagoniolithon chara
Endofitico Pacifico Sur Australia Sur y
Este
Subfamilia Lithophylloideae
Adelus harveyanus 4 Parasito Atlántico tropical Arrecife
Mesoamericano Ezo epiyoense 5 Lithophyllum
yessoense Endofitico Pacifico NO
Japo Atlantico NE
Inglaterra
1. Woelkerling 1988; 2. Adey & Serapani 1971; 3. Woelkerling & Ducker
; 4. Este estudio. 5. Adey et al 1974, Chamberlain 1999.
Tradicionalmente se había utilizado la posición de los filamentos
gonimoblasticos (periféricos vs superficiales) para diferenciar entre
Neogoniolithon y Spongites dentro de la subfamilia Mastophoroideae
(Woelkerling 1996 d), esta característica no se había utilizado con

40
anterioridad en otras subfamilias. Sin embargo, para diferenciar entre Ezo
y Adelus se ha encontrado que la posición de los filamentos
gonimoblasticos es muy similar y se demuestra el valor del conceptáculo
tetrasporangial como estructura taxonómica para diferenciar géneros en la
subfamilia Lithophylloideae lo que fortalece su utilización en la sistemática
del orden Corallinales.
El presente trabajo muestra lo poco conocida que esta la flora
coralina en otras regiones de México y la urgente necesidad de hacer
estudios monográficos detallados usando criterios modernos.
Bibliografía
Adey W.H. and Sperapani C.P. 1971. The biology of Kvaleya epilaeve, a
new parasitic genus and species of Corallinaceae. Phycologia 10: 29-
42.
Adey W.H., Masaki T. and Akioka H. 1974. 1974. Ezo epiyessoense, a new
parasitic genus and species of Corallinaceae (Rhodophyta,
Cryptonemiales). Phycologia 13: 329-344.

41
Ávila E, Riosmena-Rodríguez R. 2010. Rhodolith beds as critical habitat for
monitoring in Marine Protected Areas in the Gulf of California. In:
Columbus F, (Ed.). National Parks: Vegetation, Wildlife and Threats.
Nova Science Publishers, Inc., New York.
Ballantine, D.L., A. Bowden-Kerby and N. E. Aponte 2000. Cruoriella
rhodoliths from shallow-water back reef environments in La Parguera,
Puerto Rico (Caribbean Sea). Coral Reefs 19:75-81.
Birkett, D., C. Maggs, and M. Dring. 1998. Maerl (volume V). An overview
of dynamic and sensitivity characteristics for conservation
management of marine SACs. Scottish Association for Marine
Science (UK Marine SACs Project).
Bittner, L. et al. 2011. Evolutionary history of the Corallinales
(Corallinophycidae, Rhodophyta) inferred from nuclear, plastidial and
mitochondrial genomes. Molecular phylogenetics and evolution, 61:
697-713

42
Blunden, G., W. Farnham, N. Jephson, C. Barwell, R. Fenn, and B.
Plunkett. 1981. The composition of maerl beds of economic interest
in Northern Brittany, Cornwall and Ireland. International Seaweed
Symposium 10:651-656.
Blunden, G., W. Farnham, N. Jephson, R. Fenn, and B. Plunkett. 1977.
The composition of maerl from the Glenan Islands of Southern
Brittany. Botanica Marina 20:121-125.
Bordehore, C., A.A. Ramos-esplá and R. Riosmena-Rodríguez. 2003.
Comparative study of two maerl beds with different otter trawling
history, southeast Iberian Peninsula. Aquatic Conservation Marine
Freshwater Ecosystems 13:S43-S54.
Bosence, D. W. J. 1983. The occurrence and ecology of recent rhodoliths -
a review. Pages 225-242 in T. M. Peryt, editor. Coated Grains.
Springer-Verlag, Berlin.

43
Boscence, D. and J. Wilson 2003. Maerl growth, carbonate production
rates and accumulation rates in the northeast Atlantic. Aquatic
Conservation Marine Freshwater Ecosystems 13: S21-S31
Chamberlain YM 1983. Studies in the Corallinaceae with special reference
to Fosliella and Pneophyllum in the British Isles. Bulletin of the British
Museum (Natural History), Botany 11, 291-463.
Chamberlain Y.M.1The occurrence of Ezo epiyessoense Adey, Masaki &
Akioka (Rhodophyta, Corallinaceae) in England with a summary of
parasitism and endophytism in nongeniculate Corallinaceae.
Cryptogamie Algologie 20: 155-165
Coleman, FC. & William S.L. 2002. Overexploiting marine ecosystem
engineers: potential consequences for biodiversity. TREE 17:40-44.
CONABIO 1996. Regiones marinas prioritarias para la conservación en
México. CONABIO 123 pp.

44
Esper E.G.C. 1830. Die Pflanzenthiere. Vol. 3 Part17, pp. 285-363. Raspe,
Nurnberg.
Fabricius K. & De´ath G. 2001 Environmental factors associated with the
spatial distribution of crustose coralline algae on the Great Barrier
Reef Coral Reefs 19: 303-309.
Foster, M. S. 2001. Rhodoliths: Between rocks and soft places. Journal of
Phycology 37: 659-667.
Foster, M.S., Riosmena-Rodriguez, R., Steller, D.S. and Woelkerling, Wm
J. 1997. Living Rhodolith Beds in the Gulf of California and their
Implications for Interpreting Paleoenvironments. Geological Scoiety of
America Special Paper: 318: 127-139.
Grall J & Hall-Spencer JM (2003) Problems facing maerl conservation in
Brittany. Aquatic Conservation, Marine and Freshwater Ecosystems
13, 55-64.

45
Harvey, A.S. & Woelkerling Wm. J. 2007 . A guide to nongeniculate
coralline red algal (Corallinales, Rhodophyta) rhodolith identification.
Ciencias Marinas 33: 411-426.
Harvey, S. Broadwater. W. J. Woelkerling, and P. Mitrovski. 2003.
Choreonema (Corallinales, Rhodophyta): 18S rDNA phylogeny and
resurrection of the Hapalidiaceae for the subffamilies
Choreonematoidea, Austrolithoideae and Melobesioidae. Journal of
Phycology 39: 988-998.
Harvey, A., W. Woelkerling, T. Farr, K. Neill, and W. Nelson. 2005.
Coralline algae of central New Zealand: An identification guide to
common “crustose” species:. National Institute of Water &
Atmospheric Research Information Series No. 57, Wellington, 145 p.
Hetzinger, J, Halfar, J., Riegl, B., Godinez-Orta, L. 2006 “Sedimentology y
acoustic mapping of modern rhodolith beds on a non-tropical
carbonate shelf (Gulf of California, Mexico)”. Journal of Sedimentary
Research.

46
Iryu Y. and Matsuda S. 1988. Depth distribution, abundance and species
assemblages of nonarticulated coralline algae in the Ryukyu Islands,
southwestern Japan. Proceedings of the Sisth International Coral Reef
Symposium (Townsville) 3: 101-106.
James, D. W. 2000. Diet, movement, and covering behavior of the sea
urchin Toxopneustes roseus in rhodolith beds in the gulf of California,
México. Marine Biology 137, (5/6): 913-923.
Littler M.M. 1973. The distribution, abundance and communities of
deepwater Hawaiian crustose Corallinaceae (Rhodophyta,
Cryptonemiales). Pacific Science 27: 281-289.
Kato, A., Baba, M. and Suda, S. 2011. Revision of the mastophoroideae
(corallinales, rhodophyta) and polyphyly in nongeniculate species
widely distributed on pacific coral reefs. Journal of Phycology,
47: 662–672.

47
Littler M.M. and Littler D.S. 1988. Structure and role of algae in tropical reef
communities. In: Algae and Human Affairs (Ed. by C. A. Lembi and J.
R. Waaland), pp. 29-56. Cambridge University Press, Cambridge,
UK.
Littler M.M., Littler D.S. and Hanisak M.D. 1991. Deep-water rhodolith
distribution, productivity, and growth history at sites of formation and
subsequwent degradation. Journal of Experimental Marine Biology
and Ecology 150: 163-182.
Muckelbauer G.E. 1990. The Shelf of Cozumel, Mexico: Topography and
Organisms. Facies 23: 185 – 240.
Prager E.J. and Ginsburg R, N. 1989. Carbonate nodule growth on
Florida's Outer Shelp and its implications for fossil interpretations.
Palaios 4: 310-317.
Riosmena-Rodríguez R. 2008. Determinación de amenazas y posibles
estrategias de mitigación en mantos de rodolitos como objetos de

48
conservación en el Caribe. Parte I: Quintana Roo.SAM Informe final
MF-06-002.22 paginas
Riosmena-Rodríguez R. y Medina-López M.A. 2010. The role of rhodolith
beds in the recruitment of invertebrate species from the
southwestern Gulf of California, México In: Israel A., Einav R. &
Seckbach J. (eds.) Role of seaweeds in future globally changing
environments. Wiley NY
Riosmena-Rodríguez, R., W. J. Woelkerling, and M. S. Foster. 1999.
Taxonomic reassessment of rhodolith-forming species of Lithophyllum
(Corallinales, Rhodophyta) in the gulf of California, México. Phycologia
38, (5) (Sep): 401-417.
Riosmena-Rodriguez R., Steller D.L., Hinojosa-Arango G. y Foster M.S.
2010. Reefs that Rock and Roll: Biology and Conservation of
Rhodolith beds in the Gulf of California. En: Biodiversity and
Conservation of the Sea of Cortez (R.C. Bursca, ed.). University of
Arizona and Arizona-Sonora Desert Museum Press.

49
Silva, P. C. y H. W. Johansen. 1986. A reappraisal of the order
Corallinales (Rhodophyceae). Br. Phycol. J. 21: 1179.
Steller, D. L., and M. S. Foster. 1995. Environmental factors influencing
distribution and morphology of rhodoliths in Bahía Concepción,
B.C.S., México. Journal of experimental marine biology and ecology
194, (2): 201-212
Steller, D. L., R. Riosmena-Rodriguez, M. S. Foster, and C. A. Roberts.
2003. Rhodolith bed diversity in the Gulf of California: The importance
of rhodolith structure and consequences of disturbance. Aquatic
Conservation: Marine and Freshwater Ecosystems (S5-S20. Jan-Feb):
Vol. 13, suppl. 1, p.
Verheij, E. 1993. The genus Sporolithon (Sporolithaceae fam. nov.,
Corallinales,Rhodophyta) from Spermonde Archipelago, Indonesia.
Phycologia 32 (3): 184-196.

50
Woelkerling, Wm. J. 1987. The genus Choreonema in southern Australia
and its subfamilial classification within the Corallinaceae
(Rhodophyta). Phycologia 26: 111-127.
Woelkerling, Wm. J. 1988. The Coralline Red Algae: An Analysis of the
Genera and Subfamilies of Nongeniculate Corallinaceae
(Rhodophyta). British Museum (Natural History), London, and Oxford
University Press, Oxford. xi + 268 pp.
Woelkerling, Wm. J. 1996a. Sporolithaceae. In: The Marine Benthic Flora
of Southern Australia. Part IIIB. Gracilariales, Rhodymeniales,
Corallinales and Bonnemaisoniales. (Ed. by H.B.S. Womersley), pp.
153-158. Australian Biological Resources Study, Canberra.
Woelkerling, Wm. J. 1996b. Melobesioideae. In: The Marine Benthic Flora
of Southern Australia. Part IIIB. Gracilariales, Rhodymeniales,
Corallinales and Bonnemaisoniales. (Ed. by H.B.S. Womersley), pp.
164-210. Australian Biological Resources Study, Canberra.

51
Woelkerling, Wm. J. 1996c. Choreonematoideae. In: The Marine Benthic
Flora of Southern Australia. Part IIIB. Gracilariales, Rhodymeniales,
Corallinales and Bonnemaisoniales. (Ed. by H.B.S. Womersley), pp.
210-214. Australian Biological Resources Study, Canberra.
Woelkerling, Wm. J. 1996d. Lithophylloideae. In: The Marine Benthic Flora
of Southern Australia. Part IIIB. Gracilariales, Rhodymeniales,
Corallinales and Bonnemaisoniales. (Ed. by H.B.S. Womersley), pp.
214-237. Australian Biological Resources Study, Canberra.
Woelkerling, Wm. J. 1996e. Mastophoroideae (excluding Hydrolithon,
Neogoniolithon, Pneophyllum, SpongitesI). In: The Marine Benthic
Flora of Southern Australia. Part IIIB. Gracilariales, Rhodymeniales,
Corallinales and Bonnemaisoniales. (Ed. by H.B.S. Womersley), pp.
237-255. Australian Biological Resources Study, Canberra.
Woelkerling, Wm. J. & Harvey, A.S. 1996. Austrolithoideae. In: The Marine
Benthic Flora of Southern Australia. Part IIIB. Gracilariales,

52
Rhodymeniales, Corallinales and Bonnemaisoniales. (Ed. by H.B.S.
Womersley), pp. 160-163. Australian Biological Resources Study,
Canberra.
Woelkerling, W.J., Irvine L.M. & Harvey A.S. 1993. Growth-forms in non-
geniculate coralline red algae (Corallinales, Rhodophyta). Australian
Systematic Botany 6: 277-293.
Yoon H.W., Müller K. M., Sheath R. G., Ott F. D. & Bhattacharya D. 2006.
Defining the major lineages of red algae (Rhodophyta). Journal of
Phycology 42: 482–492.