centro de investigaciones marinas (cima)...
TRANSCRIPT

CENTRO DE INVESTIGACIONES MARINAS
(CIMA)
CONSELLERÍA DEL MAR Memoria 2016
m
m
m

2
CENTRO DE
INVESTIGACIONES MARINAS (CIMA)
CONSELLERÍA DEL MAR Memoria 2016
m
m

CIMA Memoria 2016 6 3
1 Presentación
EL CENTRO
2 Estructura y funciones
3 Personal
4 Servicios
INVESTIGACIÓN
5 Acuicultura
- Cultivo intensivo de especies de interés marisquero abordando la
producción rentable de semilla.
- Exploring the biological and socio-economic potential of new/emerging
candidate fish species for the expansion of the European aquaculture
industry.
- Inducción a la vitelogénesis, maduración ovocitaria, ovulación y
espermiogénesis en cherna (Polyprion americanus).
- Inducción de la fijación y metamorfosis en almeja fina, babosa y
japonesa.
6 Patología
- Estudio epidemiológico de la almeja rubia (Venerupis rhomboides) en
Galicia afectada por colonias procariotas intracelulares (CPI).
Identificación de las CPI. Seguimiento de la estructura de tallas de
poblaciones de almeja rubia de 4 bancos de la ría de Vigo.
- Estudio de la eliminación de virus entéricos patógenos en ostra
mediante procesos de reinstalación y depuración.
- Poblaciones bacterianas en criadero de bivalvos. Procesos de
patogénesis.
- Epidemiología de la enfermedad causada por Marteilia cochillia en las
poblaciones de berberecho Cerastoderma edule de Galicia.
- Evaluación del incremento de supervivencia en el engorde de ostra
rizada Crassostrea gigas por el uso de semilla descendiente de
supervivientes a la exposición prolongada al virus herpes de ostreidos
tipo 1. Identificación de marcadores moleculares de resistencia para
su aplicación en un programa de selección de estirpes resistentes al
virus.
7 Procesos oceanográficos costeros
- Desarrollo y optimización de métodos de determinación de toxinas de
tipo paralizante (PSP) y lipofílicas, en moluscos bivalvos para
maximizar el flujo de trabajo.
- Efecto de las biotransformaciones de toxinas de tipo diarreico (DSP)
en organismos del intermareal sobre las diferencias de toxicidad
estimada por LC-MS/MS y bioensayo de ratón y sus repercusiones
para los cierres de explotación y la salud humana.
- Identificación de toxinas DSP no detectadas por LC_MS/MS en
mejillón en fresco y en escabeche, biodisponibilidad de estos
compuestos, acumulación y distribución anatómica en el mejillón.

4
8 Recursos marinos
- Puesta a punto de las técnicas de cultivo en minicriadero de las
especies del género Cerastoderma de Galicia.
- Evaluación y seguimiento de las reservas marinas de interés pesquero
de Galicia.
- Determinación de las causas de la disminución de los bancos de
coquina. Condicionantes naturales y antropogénicos.
DIVULGACIÓN
9 Artículos en revistas SCI
10 Libros
11 Ponencias y paneles
12 Edición de publicaciones
ASESORAMIENTO
13 Informes
14 Reuniones
15 Otros
15.1. Asesoramiento. Otros
15.2. Revisión de artículos en publicaciones científicas
FORMACIÓN
16 Tesis doctorales
16.1. Dirección/tutorización de tesis
16.2. Participación en tribunales de tesis
16.3. Tesis presentadas
17 Becarios FP 2º grado
18 Cursos
18.1. Cursos impartidos
18.2. Asistencia a cursos
19 Estancias
19.1. Estancias del personal del CIMA en otros centros
19.2. Estudiantes en prácticas
19.3. Prácticas de máster
CUENTAS
20 Gastos
21 Fuentes de financiación

CIMA Memoria 2016 6 5
1 PRESENTACIÓN
El Centro de Investigaciones Marinas (CIMA) depende de la Consellería del Mar, está constituido por el Centro de
Investigaciones Marinas de Corón, en Vilanova de Arousa (Pontevedra) y el Centro de Cultivos Marinos de Ribadeo
(Lugo). La función básica de estos centros es el desarrollo de investigaciones encaminadas a conseguir una gestión
racional y eficiente de los recursos marinos renovables en el ámbito geográfico de competencia de la Administración
autonómica de Galicia.
El CIMA actúa como organismo asesor de la Administración pesquera gallega en las cuestiones de su competencia
que demandan los diferentes sectores productivos a través de las direcciones generales y oficinas territoriales de la
Consellería del Mar. También, mediante bolsas, se ocupa de la formación de nuevos investigadores (con tutorías y
dirección de tesis de doctoramiento) y de técnicos de laboratorio. La difusión de la investigación se realiza a través de
la asistencia y participación en reuniones científicas de diverso carácter (congresos, seminarios, foros, workshops, etc.)
y en numerosas publicaciones científicas de diferentes ámbitos.
Durante 2016 el Centro de Investigaciones Marinas (CIMA) continuó su labor de investigación científica y tecnológica
en temas marinos, orientada principalmente a la acuicultura de moluscos y peces, patología de moluscos bivalvos,
bioacumulación y eliminación de toxinas fitoplanctónicas y a la dinámica de poblaciones de invertebrados marinos con
interés comercial. Un trabajo cuyos resultados se muestran resumidos en la presente memoria anual de actividades
(número 18 de la serie iniciada en 1999).

6
EL CENTRO
CENTRO DE INVESTIGACIONES MARINAS
Dirección postal:
Pedras de Corón s/n
Apartado de Correos 13
36620 Vilanova de Arousa
Pontevedra
Teléfonos:
Teléfono: +34 886 20 63 64
Fax: +34 886 20 63 72
Correo electrónico:
CENTRO DE CULTIVOS MARIÑOS
Dirección postal:
Muelle de Porcillán s/n
27700 Ribadeo
Lugo
Teléfonos:
Teléfono: +34 982 88 94 03
Fax: +34 982 88 94 19
Correo electrónico:

CIMA Memoria 2016 6 7
2 ESTRUCTURA Y FUNCIONES
La actividad científica del CIMA se reparte en cuatro áreas:
Área de Acuicultura
Su objetivo es desarrollar y mejorar los procedimientos de cultivo de las especies
marinas consideradas de interés comercial. El ámbito de investigación abarca dos
grandes líneas: cultivo de moluscos y cultivo de peces.
Área de Patología
El objetivo es estudiar las alteraciones patológicas que afectan a los moluscos bivalvos
con interés comercial, para establecer estrategias eficaces de lucha que permitan
minimizar los efectos patogénicos.
Área de Procesos Oceanográficos Costeros
El objetivo del área es el conocimiento de los aspectos de la oceanografía relacionados
con la producción de los recursos pesqueros y marisqueros de Galicia. Las líneas de
investigación se relacionan principalmente con los episodios de toxicidad originados por
el fitoplancton.
Área de Recursos Marinos
El objetivo es conocer la biología, ecología y dinámica de población de las especies
marinas con interés comercial para mejorar la gestión sostenible de los recursos
marinos vivos.

8
3 PERSONAL
Director: (vacante)
Gerente: Dña. Lorenzo Losada, María Jesús (hasta 08.08.2016)
D. Villanueva Domínguez, Rafael (desde 04.10.2016)
Centro de Investigaciones Marinas de Vilanova de Arousa
Investigadores/as
Dr. Blanco Pérez, Juan Carlos
Dra. Cao Hermida, Asunción
asunció[email protected]
Dra. Carballal Durán, Mª Jesús
D. de Coo Martín, Alberto
D. Cuiñas Olmedo, Pedro
D. Febrero Mayor, Fernando
Dr. Fuentes González, José Miguel
D. García Fernández, Antonio
Dr. Iglesias Estepa, David
Dña. Linares Cuerpo, Fátima

CIMA Memoria 2016 9
Dra. López Gómez, Mª del Carmen
Dr. Montes Pérez, Jaime
D. No Couto, Edgar
Dña. Pérez Acosta, Carmen
Dr. Villalba García, Antonio
Analistas de laboratorio
Dña. Fernández Besada, Mercedes
D. Giráldez Rivero, Ramón
Dña. González Fontela, Ana Isabel
Dña. Gregorio Chenlo, Mª Victoria
Dña. Mariño Cadarso, Mª Carmen
Dña. Martín Sánchez, Helena
Dña. Martínez Verde, Gemma
Dña. Méndez Pérez, Belén
Dña. Pazos Sieira, Gema
Auxiliares de laboratorio
Dña. Brianes Beiras, Mª José
Dña. Campaña Ferro, Emilia
Dña. Cores González, Mª José
Dña. Meléndez Ramos, Isabel
Dña. Penas Pampín, Elena
Becarios/as
Dña. Acuña López, Iria
Dña. Alonso Costas, Juana Beatriz
D. Araujo Sendín, Alexis
Dña. Fernández González, Jenifer
Dña. Fuentes Villar, Noelia
D. Grande Ocampo, Oscar
D. Méndez Montenegro, José Carlos
Dña. Pena Thomas, Guadalupe
Dña. Riomayor Cernadas, Alicia
Dña. Ulla Refojo, Laura
Dña. Viturro García, Raquel
Encargado de mantenimiento
D. López Ares, Hermes
Jefe/a de negociado administrativo
D. Álvarez Araujo, Pedro Pablo (hasta 03.10.2016)
Dña. Fernández Sanmartín, Inmaculada C. (desde
04.10.2016)
Sistemas de información
D. Carballal Fortes, Félix Manuel
Biblioteca
Dña. Álvarez Francisco, Mª Esther
Auxiliares administrativos
D. Lago Torrado, José
Dña. Vidal Paz, Mª Xosefa (hasta 31.05.2016)
D. Martínez Paz, Ángel Rubén (desde 04.10.2016)
Subalternos-ordenanzas
D. Agra Carregal, Luis Manuel
D. Cubiella Granda, Rubén
D. Martínez Crespo, José Manuel
Limpiador
D. García Gamallo, Luis

10
Centro de Cultivos Marinos de Ribadeo
Responsable de planta
Dra. Martínez Patiño, Dorotea
Investigadoras
Dña. Fernández Álvarez, Aurora
Dra. Nóvoa Vázquez, Susana
Dra. Ojea Martínez, Justa
Analista de laboratorio
Dña. Bouzamayor Yánez, Mª Victoria
Auxiliares de laboratorio
Dña. Cotarelo Jardón, Mª José
D. Páez Pérez, Francisco Javier
Dña. Ruíz Misioné, Mercedes
Becarios/as
D. Aragunde Vila, R. (desde 11.01.2016 hasta
15.05.2016)
Dña. Castilla del Río, Sonia (desde 01.09.2016)
Dña. Guiance Pérez, Martiña (hasta 15.05.2016)
D. Matesanz Salgado, Diego (desde 07.10.2016)
Dña. Suárez Costa, I. (hasta 12.02.2016)
Oficial 1º de mantenimiento
D. Fernández Escudero, Antonio
Oficial administrativa
Dña. Neira Páez, Concepción
Administrativa
Dña. Cruzado Estévez, Ana
Subalterna
Dña. Loureiro Rodríguez, Mercedes (jubilación el
08.09.2016)
Dña. Díaz Rivas, Mª Antonia (contrato sustitución
hasta 08.09.2016)
Limpiadora
Dña. Fernández Goás, Carmen
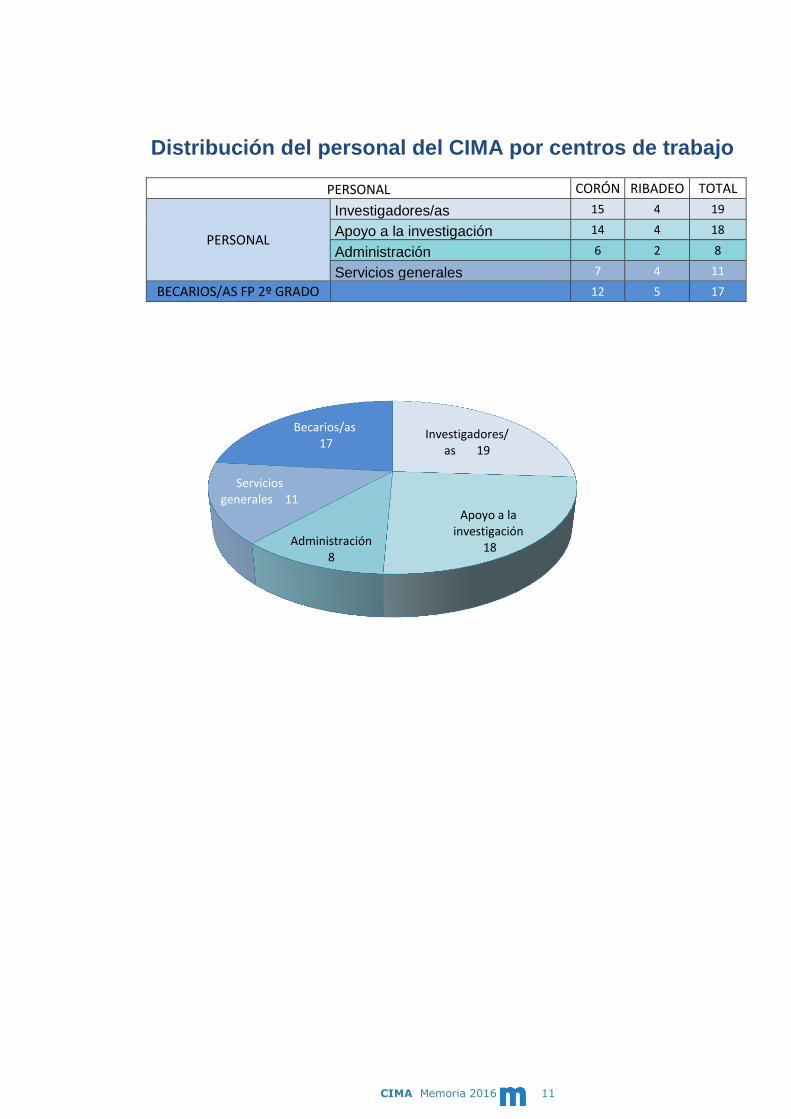
CIMA Memoria 2016 11
Distribución del personal del CIMA por centros de trabajo
Investigadores/ as 19
Apoyo a la investigación
18 Administración 8
Servicios generales 11
Becarios/as 17
PERSONAL CORÓN RIBADEO TOTAL
PERSONAL
Investigadores/as 15 4 19
Apoyo a la investigación 14 4 18
Administración 6 2 8
Servicios generales 7 4 11
BECARIOS/AS FP 2º GRADO
12 5 17

12
4 SERVICIOS
Biblioteca
La Biblioteca del CIMA es una biblioteca especializada en información
científica sobre recursos marinos, oceanografía, acuicultura,… que atiende las
necesidades de información del personal del Centro de Investigaciones
marinas de Corón (CIMA) y del Centro de Cultivos Mariños (CECUMAR) de
Ribadeo, así como consultas puntuales de otro personal de la Consellería. El
acceso para otros usuarios es público pero de carácter restringido (previa
autorización).
Instalaciones:
En el Centro de Investigaciones marinas de Corón, la Biblioteca se encuentra ubicada en la segunda planta, en una
sala única que cumple la triple función de despacho, depósito y espacio de consulta/lectura. La biblioteca pone a
disposición de los usuarios un par de mesas de trabajo individual/en grupo y un ordenador de consulta.
En el Centro de Cultivos Mariños de Ribadeo, se encuentra ubicada en la tercera planta, en una sala dedicada a
depósito, consulta y lectura.
Horario: de lunes a viernes de 9 a 14 horas.
Contacto:
CIMA-Corón: Telf.: 886 20 63 60 Correo electrónico: [email protected]
CECUMAR-Ribadeo Telf.: 982 88 94 02
Colección:
- Fondo bibliográfico: está compuesto por aproximadamente 1.500 volúmenes en Corón y 700 en Ribadeo.
Actualmente la Biblioteca ha iniciado un proceso de catalogación de estos fondos, con el objetivo de poner a
disposición de los posibles interesados un catálogo de consulta abierta accesible online que posibilite:
- La consulta conjunta de los fondos existentes en ambas localizaciones y disponibilidad (condiciones de
consulta, prestado hasta…).
- La localización e información sobre acceso a los fondos de interés (enlazado e información sobre derechos).
- La gestión en línea de las interacciones habituales entre cualquier biblioteca y sus usuarios: desideratas,
préstamos, renovaciones, reservas,…
- La interacción de los usuarios con su bibliografía de interés: listas personalizadas, descarga de registros al
gestor bibliográfico, puntuación y comentarios/reseñas en relación a los documentos,…
- Publicaciones periódicas: en 2015 el CIMA y el Consorcio de Bibliotecas Universitarias de Galicia (CBUG), firman un
convenio de colaboración que posibilita el acceso consorciado a tres colecciones de grandes editores (Elsevier,

CIMA Memoria 2016 13
Springer y Wiley), sumando un total que ronda los 5.000 títulos de revistas científicas accesibles. El CIMA mantiene a
mayores suscripción directa a un reducido número de títulos especializados que no forman parte de los paquetes
consorciados.
- Bases de Datos: en la actualidad, el CIMA cuenta con acceso a la Web of Science (WoS) de la empresa Thomson
Scientific, gracias a la licencia contratada por el Ministerio de Educación y Ciencia y gestionada por la FECyT, con el
fin de dar acceso a estos recursos de información científica a las instituciones públicas de investigación.
A mayores de los recursos disponibles mediante adquisición/contratación, desde la biblioteca se hace una labor de
localización y clasificación de fuentes de información en acceso abierto, tratando de visibilizar sus posibilidades de uso
al tiempo que se complementa la colección de base.
Servicios
Sala de lectura.
Información sobre los recursos disponibles y acceso a los mismos http://www.netvibes.com/bibliotecacima.
Préstamo interno.
Servicio de obtención de documentos.
Asesoramiento en planteamiento de búsquedas y consulta de fuentes de información secundarias.
Orientación en el uso de herramientas de gestión de información.
Formación en el uso de gestores bibliográficos (EndNote Desktop / Web, Mendeley, Zotero).
Atención a usuarios / sesiones de formación mediante videollamadas (Skype / Hangouts).
Salón de conferencias
El Centro de Investigaciones marinas de Corón, cuenta con un salón de
conferencias con capacidad para 84 personas.
El acceso es público pero de carácter restringido (presta servicio a usuarios
internos y a usuarios externos autorizados).
Sala de juntas
El CIMA-Corón cuenta con dos salas de juntas con capacidad para 10
personas y para 15 personas respectivamente.
El Centro de Cultivos Mariños de Ribadeo dispone de una sala con capacidad
para 15 personas.
El acceso es público pero de carácter restringido (prestan servicio a usuarios
internos y a usuarios externos autorizados).
14
INVESTIGACIÓN
CIMA Memoria 2016 15
5 ACUICULTURA

16
RESUMEN
La acción tiene como objetivo marcar los procedimientos
y protocolos para el cultivo de varias especies de
moluscos bivalvos de interés marisquero, desde la
obtención de semilla en criadero, el preengorde en
sistemas colocados en batea y su traslado a los bancos
naturales y la transferencia de tecnología a los
minicriaderos del IGAFA donde se forma el personal con
el fin de transmitir al sector los avances logrados en el
cultivo intensivo de bivalvos.
Consta de tres subproyectos:
Subproyecto I: Procedimientos y protocolos de
cultivo intensivo de las especies marisqueras.
El objetivo era marcar los procedimientos y protocolos
del cultivo intensivo de las especies de bivalvos de
interés marisquero, se llevó a cabo en el CECUMAR de
Ribadeo que cuenta con las instalaciones de la planta de
cultivo de moluscos, el minicriadero de O Vicedo y una
plataforma flotante tipo batea, que se colocó en la ría de
Ribadeo.
En este Subproyecto se trabajó con reproductores de las
siguientes especies de bivalvos: almeja fina (Ruditapes
decussatus), almeja babosa (Venerupis corrugata),
almeja japonesa (Ruditapes philippinarum), almeja
bicuda (Venerupis aurea), coquina (Donax trunculus) y
navaja (Ensis arcuatus).
A partir del seguimiento de cultivos, en los que se
consiguieron buenos resultados, se pudo establecer un
protocolo para cada especie.
Almeja fina (Ruditapes decussatus)
Los progenitores. Con esta especie se consiguen
desoves viables, espontáneos o inducidos, en los meses
en los que su gónada se encuentra en estado
gametogénico de madurez (abril-agosto) pero, también
es factible su acondicionamiento cuando se trabaja con
ejemplares en estado de no madurez. Puede ser
acondicionada en los meses de invierno y obtener
desoves a principios de primavera. Para ello es
estabulada en el criadero a temperaturas de 18-20ºC y la
ración de alimento, basada en una mezcla de
microalgas, es del 6% de peso seco de microalgas, en
relación al peso seco de los reproductores.
La inducción al desove se consigue sometiendo a los
reproductores a períodos en los que permanecen en
seco (aproximadamente 12 horas) y después a períodos
con agua fría (±10ºC, 30 minutos) y agua caliente (24-
26ºC, 60-90 minutos).
ACCIÓN DE INVESTIGACIÓN EQUIPO
TÍTULO:
Cultivo intensivo de especies de interés marisquero
abordando la producción rentable de semilla.
INICIO: junio, 2014
FINALIZACIÓN: diciembre, 2016
ENTIDAD FINANCIADORA:
Consellería do Medio Rural e do Mar. Xunta de
Galicia.
Código: PGIDIT-CIMA 14/06
INVESTIGADORA PRINCIPAL:
Martínez Patiño, D.
INVESTIGADORES/AS:
CIMA: Montes Pérez, J.; Nóvoa Vázquez, S.; Ojea Martínez,
J.
IGAFA: Andrés Rivas, M.C.; Lastres Couto, M.A.
USC: Barja Pérez, J.L.; Prado Plana, S.
APOYO A LA INVESTIGACIÓN:
Bouzamayor Yáñez, V.; Cotarelo Jardón, M.J.; Meléndez
Ramos, I.; Páez Pérez, F.J.; Penas Pampín, E.; Ruíz
Misioné, M.
BECARIOS/AS:
Aragunde Vila, R.; Castilla del Río, S.; Guiance Pérez, M.;
Matesanz Salgado, D.; Suárez Costa, I.

CIMA Memoria 2016 17
El desarrollo embrionario ocurre unas horas después
de haberse fertilizado el huevo, la aparición de la larva
es a las 26 horas. Cuando se inicia el cultivo larvario,
su duración desde larva D hasta pediveliger es de 14-17
días, produciéndose la fijación y la metamorfosis.
Llegados a este punto los individuos quedan retenidos
en tamiz de malla de 170 µm y necesitan un sustrato por
lo que se utilizan cilindros con fondo de malla de 150 µm,
consiguiendo porcentajes de supervivencia, desde larva
D, de un 70%. Pasados 34-36 días las postlarvas, con
todas sus estructuras de adultos formadas, quedan
retenidas en tamices de 300 µm y siguen su cultivo en el
sistema abierto del criadero. En este sistema llegan a
poder retenerse en tamices de 750 µm a los 64 días
desde el desove y con supervivencias del 34% desde
larva D que, en ocasiones, pueden llegar a superar el
70%. A los 72 días pasa la primera semilla a las
instalaciones del invernadero, para ello debe quedar
retenidas en tamices de malla de 1500 µm y durante dos
meses, se van pasando según alcanzan dicho tamaño.
Almeja babosa (Venerupis corrugata)
Los progenitores de esta especie presentan gametos
maduros durante todo el año, no existe un período de
reposo, por lo tanto, puede cultivarse durante todo el
año. Una buena propuesta de calendario, para poder
cultivar otras especies a la vez en el mismo criadero, es
la de ser recogidas y trasladadas al criadero en invierno
y de esta forma, iniciar los desoves de babosa a
primeros de año.
La inducción al desove se consigue mediante la
estimulación con choques térmicos, con una temperatura
mínima de 12ºC y máxima de 20ºC.
Después de la fecundación el desarrollo embrionario
ocurre en unas horas, la aparición de la larva trocófora
es a las 13-14 horas y de la larva D es a las 30-32 horas.
El cultivo larvario hasta la fijación tiene una duración
de 14-17 días después del desove. Se consiguen
supervivencias de un 87% y su cultivo continúa en los
mismos tanques pero, en cilindros con fondo de malla de
150 µm. Las postlarvas quedan retenidas en tamiz de
300 µm a los 21-35 días desde el inicio del cultivo, con
una supervivencia de un 58%, y esta etapa continúa
hasta que alcanzan la fase de semilla en el sistema
abierto del criadero. Quedan retenidas en tamiz de 750 a
los 51 días y cuando lo hacen en el de 1500 µm (a los 58
días son las primeras), el cultivo de semilla se realiza en
el sistema del invernadero.
Navaja (Ensis arcuatus)
Los progenitores son recogidos en el medio natural y
estabulados en el criadero en los meses de enero a abril,
meses, en los que su gónada está madura. A diferencia
de otras especies necesita que en los tanques exista un
sustrato, arena, en el que permanecen enterrados. La
ración de alimento suministrado suele ser del 6% en
peso seco de microalgas y las temperaturas a las que
permanecen en el criadero, oscilan entre 13-16ºC.
Para los desoves inducidos el estímulo más efectivo es
el cambio de niveles de agua, con breves períodos de
desecación.
El desarrollo embrionario dura unas horas observando
la larva trocófora a las 19 horas y la larva D a las 30
horas, desde la fecundación. La duración del cultivo
desde larva D a pediveliger es de 19-20 días quedando
retenidos en tamiz de 200 µm y la fijación, se realiza en
los tanques de 500 litros, en cilindros con malla de 150
µm.
En la etapa postlarvaria, cuando quedan retenidas en
tamices de 300 µm a los 34-40 días, se utiliza el sistema
abierto y se consiguen supervivencias, desde larva D,
del 43%. Aquí permanecen hasta los 60-100 días desde
el inicio del cultivo alcanzando los 5-6 mm, momento en
el que se inicia su preengorde en tres sistemas
diferentes según van creciendo.
Y cuando su tamaño es de 15-20 mm (a los 180-192
días de cultivo), puede continuar en el criadero,
enterrada en tamices con arena colocados en el fondo
de un tanque de 1.000 L.
Cuando la semilla tiene una talla de unos 30 mm (con
230-242 días), se inicia su engorde en el medio natural.

18
Coquina (Donax trunculus)
Desde marzo hasta julio se puede disponer de
progenitores en estado gametogénico de madurez y se
obtienen los primeros desoves espontáneos en marzo,
sin embargo, es una especie a la que se le puede
adelantar la puesta, con respecto al medio natural, ya
que se consiguen acondicionar cuando se mantienen a
17 ºC y con una ración de alimento del 6% en peso seco
de microalgas.
Se induce el desove permaneciendo los reproductores
en seco a temperatura ambiente (14º±1ºC) durante
aproximadamente 10 horas y posteriormente,
introduciéndolos en agua a una temperatura en torno a
los 16-18ºC.
En el transcurso de unas horas se produce su
desarrollo embrionario, observando la aparición de la
larva trocófora a las 15 horas y la larva D a las 41 horas.
El cultivo larvario dura unos 24-26 días, posteriormente
inician la fijación. Las larvas pediveliger quedan
retenidas en tamiz con malla de 180 µm, su
supervivencia desde larva D es del 24,5% aunque, se
pueden alcanzar porcentajes del 54% y se pasan a los
cilindros con fondo de malla de 150 µm.
Transcurridos 38 días desde el desove, se consiguen
postlarvas que quedan retenidas en tamiz de 300 µm y
que son cultivadas en el sistema abierto del criadero. Se
mantienen en los tanques de este sistema hasta que los
individuos quedan retenidos en tamices de 1500 µm (a
los 90 días de cultivo), momento en el que se pasan a
las instalaciones del invernadero, la supervivencia desde
la etapa de fijación hasta el preengorde puede variar
desde un 13% hasta un 55%.
En la actualidad, cuando la coquina queda retenida en
tamices de 2000-3000 µm, se está ensayando su
preengorde en la batea tratando de evitar los meses de
invierno dado que, las abundantes lluvias, provocan
bajadas importantes de salinidad, factor que afecta
bastante a esta especie aumentando su mortalidad.
Almeja japonesa (Ruditapes philippinarum)
Para iniciar el cultivo de esta especie el procedimiento a
seguir de estabulación y acondicionamiento de los
reproductores y de obtención de desoves, puede ser el
mismo que el de R. decussatus. La duración de su
estado de madurez en Galicia es un poco más larga que
esta especie y abarca, hasta el mes de octubre.
Al igual que la almeja fina, su desarrollo embrionario
dura unas horas y el cultivo larvario se realiza en circuito
cerrado, consiguiendo las primeras larvas pediveliger a
los 14-18 días desde el desove y con una supervivencia
del 62% desde larva D. Al llegar a esta etapa también
necesitan los cilindros con fondo de malla de 150 µm
para que puedan realizar la fijación y metamorfosis. Las
postlarvas, a los 37 días, quedan retenidas en tamiz de
300 µm con una supervivencia del 52% desde larva D.
En este momento se pasan al sistema abierto del
criadero, quedan retenidas en tamiz de 750 µm a los 60
días y al quedar en el de 1500 µm (a los 76 días), se
inicia el cultivo de semilla en las instalaciones del
invernadero.
Almeja bicuda (Venerupis aurea)
Se logra realizar su cultivo por primera vez en criadero.
Los desoves se consiguieron en el mes de mayo y a los
21 días se obtuvieron larvas pediveliger que se pusieron
en cilindros con fondo de malla para que realizaran la
fijación y metamorfosis. La semilla retenida en tamiz de
2000 µm pasó a las instalaciones del invernadero a los
115 días desde el desove (4 meses) y necesitó 4 meses
más (235 días desde el desove) para alcanzar tallas de
6-9 mm e iniciar el preengorde en la batea. Las
condiciones para el crecimiento en el invierno no son las
más adecuadas para ninguna especie por lo que, al
pasarlas a las instalaciones de la batea en enero,
necesitan más tiempo para alcanzar las tallas óptimas de
siembra en el medio natural. Este cultivo necesitó dos
meses más (295 días desde el desove) para conseguir
individuos de 7-10 mm, talla de siembra en el medio
natural.
Se está estudiando su ciclo gametogénico para optimizar
el momento de estabulación de progenitores así como la
obtención de desoves y para determinar cuándo y cómo
se debe de llevar a cabo su posible acondicionamiento.
Subproyecto II: Aplicación de las técnicas de cultivo
de semilla de moluscos bivalvos en instalaciones
tipo minicriadero.
Se llevó a cabo en los minicriaderos del IGAFA y abordó
la aplicación de las técnicas de cultivo de semilla de
moluscos bivalvos en instalaciones tipo minicriadero.
La actividad en este minicriadero empezó en octubre de
2014, con la contratación de 4 becarios, aunque la
producción de semilla no se comenzó hasta principios
del 2015, porque después de llevar un año sin actividad

CIMA Memoria 2016 19
fue necesario realizar tareas de puesta a punto de las
instalaciones:
- Limpieza integral del minicriadero.
- Comprobación del funcionamiento de maquinaria.
Reparar fugas, averías y modificaciones necesarias.
- Comprobación de bomba general y filtro de tambor.
- Revisión del listado de inventario existente y
actualización del mismo.
- Inicio del cultivo de cepas procedentes del IGAFA.
Posteriormente con estas cepas se inocularon botellones
y éstos se emplearon en bolsas en sistema discontinuo.
- Limpieza de tanques de larvario y repintado de los
que estaban en peor estado.
- Al recibir los primeros reproductores se puso en
marcha el sistema de fitoplancton continuo, con revisión
y mejoras del sistema de distribución del medio y de la
cosecha de fitoplancton.
- A la finalización de la beca, las instalaciones están
funcionando relativamente bien, con fallos o desajustes
puntuales, quedando pendiente la reparación del
automatismo de los pulverizadores de agua que regulan
la temperatura en la zona de producción de fitoplancton
en continuo y la instalación de la segunda bomba
principal.
Después de 24 meses de actividad en las instalaciones
del minicriadero, se puede decir que esta experiencia
aportó una serie de conocimientos y habilidades
necesarias para el desarrollo, puesta en marcha y
gestión integral de un criadero de moluscos:
- Aprendizaje completo del cultivo de diferentes
especies.
- Conocimiento en profundidad del funcionamiento de
un criadero.
- Trabajo y gestión en equipo de las instalaciones y el
cultivo.
- Solución de diferentes incidencias que se produjeron
en las instalaciones al inicio de la actividad y a lo largo
de la misma.
- Resultados satisfactorios en cuanto a producción de
semilla a pesar de las dificultades que genera la propia
instalación y a situaciones que se produjeron fuera de la
misma (temporales, robos de cuerdas con semilla,
depredadores, etc) escapándose a nuestro control.
Subproyecto III: Optimización de las técnicas de
producción industrial de moluscos bivalvos desde el
punto de vista patológico y microbiológico.
En este Subproyecto se aplicaron los conocimientos
previos del grupo desarrollados durante años en el
marco de diferentes proyectos de investigación, para
ayudar a la mejora de las técnicas y protocolos de
manejo y gestión de la producción.
Histopatología
Los controles histopatológicos, mediantes técnicas
rutinarias de histología, permitieron el análisis patológico
de los individuos utilizados como reproductores tanto en
el IGAFA como en el Centro de Cultivos Marinos de
Ribadeo.
Se analizaron un total de 12 lotes de progenitores de
diferentes especies de bivalvos con distintos orígenes: 3
lotes de almeja fina (Portugal), 7 lotes de almeja babosa
(ría de Arousa y ría de Ferrol), 1 lote de almeja japonesa
(ría de Arousa) y 2 lotes de almeja rubia (ría de Arousa y
ría de Vigo).
De los resultados obtenidos después de la observación
al microscopio de las preparaciones resultantes, hay que
destacar la presencia de Perkinsus sp. como principal
problema patológico detectado, llegando a alcanzar
prevalencias del 50% en algunos casos. Según la
prevalencia e intensidad, este protista puede ocasionar
mortalidades. También hay que mencionar otras
parasitologías presentes ocasionalmente, como
organismos tipo Rickettsia, hallados sobre todo en
almeja rubia o protozoos del género Nematopsis.
Además de los reproductores, se analizaron lotes de
semilla de ambos orígenes (IGAFA y Ribadeo), en el
momento de la siembra en los parques de cultivo. No se
detectaron parasitologías reseñables en ninguna de las
muestras evaluadas.
Microbiología
A lo largo de este proyecto, se inició la implantación de
protocolos de control microbiológico en las instalaciones
de los minicriaderos del IGAFA. Para ello, se partió de
los resultados obtenidos durante años de investigación
en colaboración entre la USC y el Centro de Cultivos
Mariños de Ribadeo. Estos trabajos permitieron
identificar los parámetros básicos a controlar en el
criadero y diseñar un protocolo adecuado, así como
poner en práctica su aplicación. Estos conocimientos
sirvieron de base para implementar un protocolo de

20
control adaptado a las instalaciones de los minicriaderos
del IGAFA.
En primer lugar, fue conocer en detalle el circuito de
agua, desde la toma exterior hasta los diferentes usos
que se le da en el criadero. Se detallaron no sólo los
distintos tratamientos a que se somete el agua según el
destino final, sino también los tramos de tuberías que se
utilizan para su distribución. En paralelo, se estudió el
sistema de alimentación. Al igual que con el agua, en
este apartado se tuvieron en cuenta los diferentes tipos
de cultivo de fitoplancton (en diferentes volúmenes y con
diferentes tratamientos del agua) y el circuito de
distribución.
Esta identificación detallada de los circuitos de agua y
alimentación, comunes a todas las especies de bivalvos
que se cultiven, fue la base para el diseño de un
protocolo preliminar de control adaptado, permitiendo
establecer los puntos de muestreo necesarios para
obtener la información que garantice la detección de
potenciales problemas microbiológicos, y asegurando su
accesibilidad.
Para poder llevar a cabo los trabajos propuestos, se
entrenó al personal en formación en la recogida de
muestras para análisis microbiológicos y en siembras
bacteriológicas. Los esfuerzos se orientaron sobre todo a
la detección de vibrios, los principales patógenos
bacterianos identificados en criaderos de moluscos,
responsables de numerosos episodios de mortalidad que
comprometen la regularidad de la producción. Por ello,
como medio bacteriológico para las siembras se utilizó el
medio selectivo para vibrios Tiosulfato-Citrato-Bilis-
Sacarosa (TCBS), cuya utilidad práctica se ha
demostrado en trabajos anteriores del grupo USC-CIMA.
Complementariamente, de forma ocasional empleó
también en medio para bacterias heterótrofas marinas
Agar Marino.
Dado que el protocolo a diseñar tenía una orientación
eminentemente práctica, y considerando la falta de
conocimientos básicos y experiencia del personal de la
instalación en aspectos microbiológicos, se optó por un
protocolo inicial sencillo que permitiera al mismo tiempo
su entrenamiento. El objetivo era identificar los posibles
problemas al final de los circuitos.
Los muestreos del circuito de agua se concentraron
principalmente en el agua utilizada para los cultivos
larvarios. Mediante muestreos semanales se determinó
la presencia/ausencia de vibrios, y además se evaluó la
eficiencia del tratamiento con radiación ultravioleta a que
se somete antes del llenado de los tanques de cultivo.
Una estrategia similar se aplicó a los muestreos del agua
utilizada para los cultivos de fitoplancton, previamente
tratada en un pasteurizador. En paralelo, para el control
de la presencia/ausencia de vibrios en los cultivos de
microalgas, se tomaron semanalmente muestras de los
tanques de mezcla de cultivo continuo de fitoplancton. La
presencia regular de vibrios llevó al control de diferentes
bolsas de cultivo de fitoplancton que se utilizan para la
mezcla en los tanques.
De entre los resultados obtenidos, destacó la buena
calidad microbiológica del agua de los cultivos larvarios,
junto con el correcto funcionamiento de la unidad de
radiación ultravioleta cuando se detectaron vibrios en el
agua de entrada. Similares resultados se obtuvieron del
agua utilizada para los cultivos microalgales.
En cuanto a los cultivos de fitoplancton, la presencia de
vibrios en los tanques de mezcla con más frecuencia de
la deseada constituyó un motivo de preocupación. Los
análisis de las bolsas de cultivos monoespecíficos
utilizados para la mezcla dieron resultados positivos para
la presencia de vibrios en un tercio de las bolsas
analizadas, por lo que el problema debería ser estudiado
a más largo plazo para identificar las causas, dado que,
aunque no se observen efectos directos sobre las
microalgas, el aporte continuo de vibrios a los cultivos de
bivalvos a través del alimento es un riesgo que no puede
ser asumido por una instalación de producción.
Hay que mencionar también que, además de los
controles bacteriológicos, se realizaron en la USC
análisis virológicos puntuales para la detección de
herpesvirus en cultivos larvarios de almeja babosa y
japonesa, con resultados negativos.
En conjunto, la aplicación del protocolo preliminar ha
permitido hacer pequeños cambios de manejo y limpieza
para la mejora de la producción, así como eliminar
aquellos cultivos de microalgas con vibrios. El desarrollo
de los trabajos ha puesto de manifiesto la necesidad de
un buen entrenamiento en técnicas básicas de
microbiología para la toma de muestras y siembra en
placa, indispensable para obtener resultados fiables. La
detección de potenciales riesgos a lo largo del período
de muestreos demostró que es imprescindible establecer
un protocolo de control rutinario con una aplicación
rigurosa, que permita la obtención de series temporales
de datos. Sólo esta información permitiría optimizar las
tareas de manejo y limpieza para garantizar la calidad

CIMA Memoria 2016 21
microbiológica general de la instalación. A partir de ahí,
se podría avanzar en la implementación de un protocolo
de mantenimiento, con el número mínimo de muestras, y
las pautas de ampliación adecuadas en caso de
detección de problemas.

22
RESUMEN
En este proyecto se aborda el estudio de los aspectos
clave de las 6 especies seleccionadas para su
desarrollo en la acuicultura europea: la corvina
(Argyrosomus regius) y el halibut (Hippoglossus
hippoglossus), cuyo estado de producción e
investigación es el más avanzado, seguidas por la perca
(Sanders lucioperca) y la seriola (Seriola dumerili) con
alguna producción e investigación, la cherna (Polyprion
americanus) de la que no existe producción y está en
fase de investigación y la lisa (Mugil cephalus) cuya
acuicultura se realiza a partir de alevines capturados del
medio natural.
La Consellería del Mar (CIMA e IGAFA) está involucrada
en el proyecto, conjuntamente con el IEO y el Acuario de
A Coruña (AF) en el desarrollo del cultivo de la cherna
(Polyprion americanus). Además, otros centros de
investigación españoles y extranjeros participan en el
desarrollo de esta especie: Hellenic Center for Marine
Research (HCMR), Universidad de las Palmas de Gran
Canaria (ULPGC), Institut de Recerca i Tecnología
Agroalimentaries (IRTA), Institut Francais de Recherche
Pour L'Exploitation de la Mer (IFREMER) y Universidad
de La Laguna (ULL). Asimismo se cuenta con la
colaboración del acuario de O Grove.
La cherna (Polyprion americanus) perteneciente a la
familia Serranidae es una especie muy atractiva para la
acuicultura comercial ya que tiene un crecimiento rápido,
madurez reproductiva tardía, alto precio en el mercado,
fácil manipulación en cautividad, etc. Su pesquería se ha
visto reducida drásticamente en los últimos años.
El control de la reproducción y el establecimiento de
protocolos de cultivo larvario se consideran los
principales retos para el desarrollo del cultivo de esta
especie. Además, se carece de estudios sobre
requerimientos nutricionales y de obtención de dietas
óptimas. Por ello las tareas a desarrollar se centran en
tres áreas de trabajo (WPs):
WP6. Reproducción de la cherna
WP12. Nutrición de la cherna
WP18. Cultivo larvario de la cherna
PROYECTO DE INVESTIGACIÓN EQUIPO
TÍTULO:
Exploring the biological and socio-economic potential of
new/emerging candidate fish species for the expansion of
the European aquaculture industry.
Acrónimo: DIVERSIFY
INICIO: diciembre, 2013
FINALIZACIÓN: diciembre, 2018
ENTIDAD FINANCIADORA:
7º Programa Marco. KBBE2013.1.2.09 Diversification of
fish species and products in European Aquaculture Area
2.1.2. Increased Sustainability of all Production Systems
(agriculture, forestry, fisheries and aquaculture).
INVESTIGADOR PRINCIPAL:
Coordinador del proyecto:
Constantinos C. Mylonas, HCMR. GREECE.
INVESTIGADORA PRINCIPAL (CIMA):
Linares Cuerpo, F.
INVESTIGADORES/AS:
IGAFA: Rodríguez Villanueva, J.L.
Investigadores pertenecientes a 20 centros de
investigación, 9 pequeñas y medianas empresas, 3
grandes empresas, 5 asociaciones profesionales y 1 de
consumidores de 12 países diferentes.
APOYO A LA INVESTIGACIÓN:
Cores González, M.J.; Méndez Pérez, B.; Pazos Sieira,
G. (hasta julio 2016)
BECARIOS/AS:
Acuña López, I.; Méndez Montenegro, J.C. (hasta mayo
2016)
Araujo Sendín, A.; Fuentes Villar, N. (desde septiembre
2016)

CIMA Memoria 2016 23
WP6. Reproducción de la cherna
Las actividades realizadas en la reproducción de la
cherna van encaminadas al cumplimiento de los
siguientes objetivos:
I. Incremento de los stocks de reproductores
existentes mediante la captura de ejemplares
salvajes.
II. Descripción del ciclo reproductivo en cautividad.
III. Desarrollo de protocolos de inducción al desove
en cautividad así como de fertilización artificial.
IV. Desarrollo de protocolos para análisis de
esperma: Computer Assisted Sperm Analysis (CASA)
y criopreservación de esperma.
V. Determinación de la influencia de la alimentación
de reproductores sobre la fecundidad y la calidad del
desove (WP12. Nutrition wreckfish).
Tarea 6.1. Captura de ejemplares salvajes para
establecer nuevos stocks de reproductores.
En relación con esta tarea se capturaron 5 ejemplares
entre 2014 y 2015 con pesos comprendidos entre 1 y 4
Kg que se mantuvieron en cuarentena en las
instalaciones del acuario de O Grove. Para muestrearlos
se anestesiaron con fenoxietanol (3.5 ml/litro de agua de
mar) y se realizaron mediciones de longitud total y
standard, perímetro y peso. También se recogieron
muestras de aletas para posterior análisis genético. Dos
de los ejemplares se mantuvieron controlados en las
instalaciones del IEO separados del resto del stock.
Todos los ejemplares fueron muestreados en el Acuario
de O Grove y en el IEO durante el año 2016
alcanzándose pesos medios en octubre de 2016 de
11,94 ±1,37Kg en el acuario de O Grove. En el IEO uno
de los ejemplares murió (Peso aprox. 2 Kg) y el otro
tiene un peso aproximado de 6 Kg.
Es de resaltar la dificultad existente para conseguir
ejemplares e incrementar el stock dada la brusca caída
de la pesquería de la cherna en los últimos años.
Tarea 6.2. Descripción del ciclo reproductivo.
Durante el año 2016 al igual que en años anteriores se
realizó un seguimiento exhaustivo de los stocks de
reproductores de cherna con la finalidad de describir el
ciclo reproductivo en cautividad.
Actualmente se dispone de los siguientes stocks de
reproductores (Tabla I):
Tabla I. Stocks de reproductores de cherna controlados en Diversify.
HCMR- Fotoperiodo y temperatura natural AF- Temperatura natural y fotoperiodo natural simulado IGAFA- Temperatura natural y fotoperiodo natural IEO- Temperatura y fotoperiodo natural
Cada stock se muestreó mensualmente durante el
período febrero-julio (estación reproductiva) y
bimensualmente durante el período agosto-enero
(período de reposo) en 2015 y 2016. Los pesos se
incrementan durante la maduración de los ejemplares.
Se extrajo sangre de parte de los ejemplares de cada
stock para estudiar niveles de esteroides (HCMR) y se
canuló a las hembras para extracción de ovocitos y
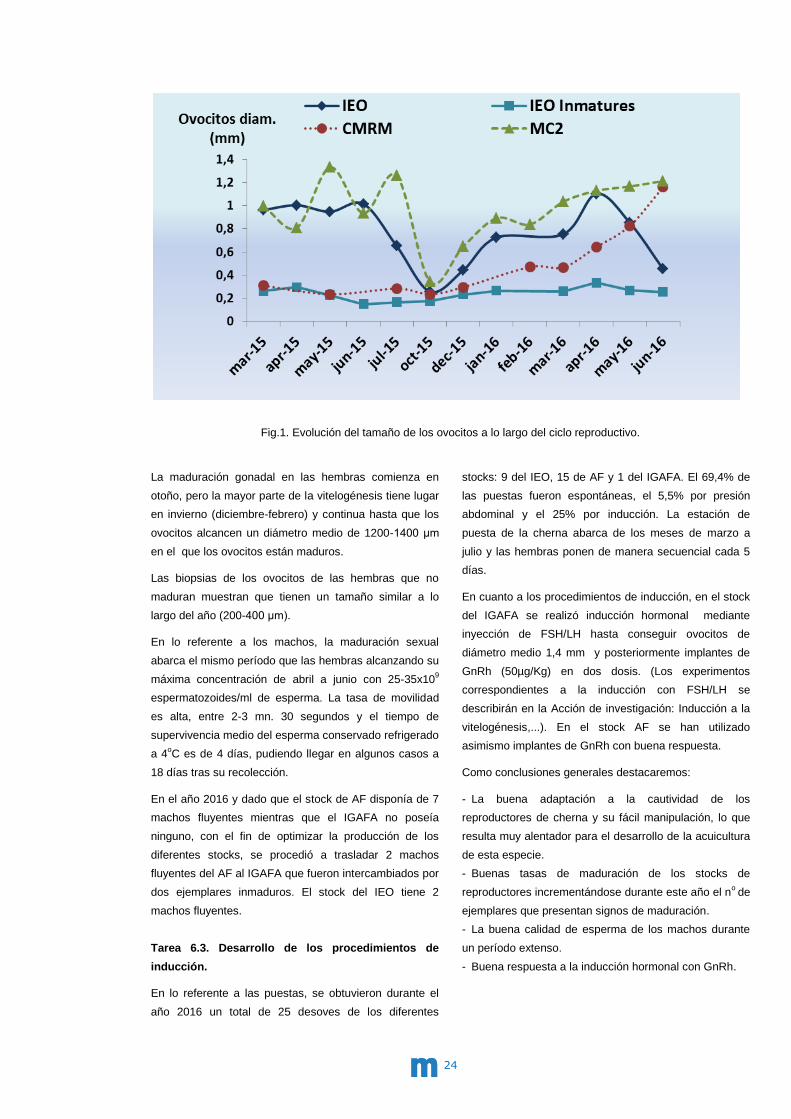
medición a lo largo del ciclo reproductivo (Fig.1). El
tamaño medio de ovocitos a lo largo del año sigue la
secuencia esperada en las hembras que han conseguido
completar el desarrollo reproductivo, obteniéndose
valores máximos en la época de maduración.
HCMR (Creta)
AF Aquarium Finisterrae.
MC2 (A Coruña)
IGAFA. Consellería del Mar (Illa de Arousa)
IEO. Centro Oceanográfico
de Vigo
TOTAL
MACHOS 1 6 4 3 14
HEMBRAS 1 10 5 8 24
INDETERMINADOS 3 2 3 8
TOTAL 2 19 11 14 46
MACHOS/HEMBRAS 1:1 0.6:1 0.8:1 0.4:1
24
Fig.1. Evolución del tamaño de los ovocitos a lo largo del ciclo reproductivo.
La maduración gonadal en las hembras comienza en
otoño, pero la mayor parte de la vitelogénesis tiene lugar
en invierno (diciembre-febrero) y continua hasta que los
ovocitos alcancen un diámetro medio de 1200-1400 μm
en el que los ovocitos están maduros.
Las biopsias de los ovocitos de las hembras que no
maduran muestran que tienen un tamaño similar a lo
largo del año (200-400 μm).
En lo referente a los machos, la maduración sexual
abarca el mismo período que las hembras alcanzando su
máxima concentración de abril a junio con 25-35x109
espermatozoides/ml de esperma. La tasa de movilidad
es alta, entre 2-3 mn. 30 segundos y el tiempo de
supervivencia medio del esperma conservado refrigerado
a 4oC es de 4 días, pudiendo llegar en algunos casos a
18 días tras su recolección.
En el año 2016 y dado que el stock de AF disponía de 7
machos fluyentes mientras que el IGAFA no poseía
ninguno, con el fin de optimizar la producción de los
diferentes stocks, se procedió a trasladar 2 machos
fluyentes del AF al IGAFA que fueron intercambiados por
dos ejemplares inmaduros. El stock del IEO tiene 2
machos fluyentes.
Tarea 6.3. Desarrollo de los procedimientos de
inducción.
En lo referente a las puestas, se obtuvieron durante el
año 2016 un total de 25 desoves de los diferentes
stocks: 9 del IEO, 15 de AF y 1 del IGAFA. El 69,4% de
las puestas fueron espontáneas, el 5,5% por presión
abdominal y el 25% por inducción. La estación de
puesta de la cherna abarca de los meses de marzo a
julio y las hembras ponen de manera secuencial cada 5
días.
En cuanto a los procedimientos de inducción, en el stock
del IGAFA se realizó inducción hormonal mediante
inyección de FSH/LH hasta conseguir ovocitos de
diámetro medio 1,4 mm y posteriormente implantes de
GnRh (50µg/Kg) en dos dosis. (Los experimentos
correspondientes a la inducción con FSH/LH se
describirán en la Acción de investigación: Inducción a la
vitelogénesis,...). En el stock AF se han utilizado
asimismo implantes de GnRh con buena respuesta.
Como conclusiones generales destacaremos:
- La buena adaptación a la cautividad de los
reproductores de cherna y su fácil manipulación, lo que
resulta muy alentador para el desarrollo de la acuicultura
de esta especie.
- Buenas tasas de maduración de los stocks de
reproductores incrementándose durante este año el no
de
ejemplares que presentan signos de maduración.
- La buena calidad de esperma de los machos durante
un período extenso.
- Buena respuesta a la inducción hormonal con GnRh.
CIMA Memoria 2016 25
WP12. Nutrición de la cherna.
Tarea 12.1. Nutrición larvaria. Influencia del
enriquecimiento de presas vivas y su eficacia en
larvas de peces.
Se enviaron muestras de huevos de cherna desde el
CIMA a la Universidad de las Palmas de Gran Canaria
(ULPGC) para realizar análisis bioquímicos para obtener
datos para el desarrollo de productos de enriquecimiento
de alimento vivo para larvas de cherna.
Se desarrolló un nuevo enriquecedor (ULPGC) basado
en harina de soja, fosfolípidos de krill, lecitina de soja,
aceite de pescado, microalgas y mezclas de vitaminas y
minerales con la siguiente composición: Proteína cruda
35%, lípidos totales 41%, humedad 8%, cenizas 11% y
carbohidratos 5%. El enriquecedor no pudo ser testado
ya que la cantidad de larvas obtenida no fue suficiente
para llevar a cabo los experimentos.
Aunque la mortalidad larvaria fue del 100% a día 26 de
vida se obtuvieron muestras de larvas a días 1, 3, 4, 18 y
26 de vida con la finalidad de completar los datos de
perfiles de ácidos grasos de la cherna iniciados en el año
2015 (hasta 10 días de vida).
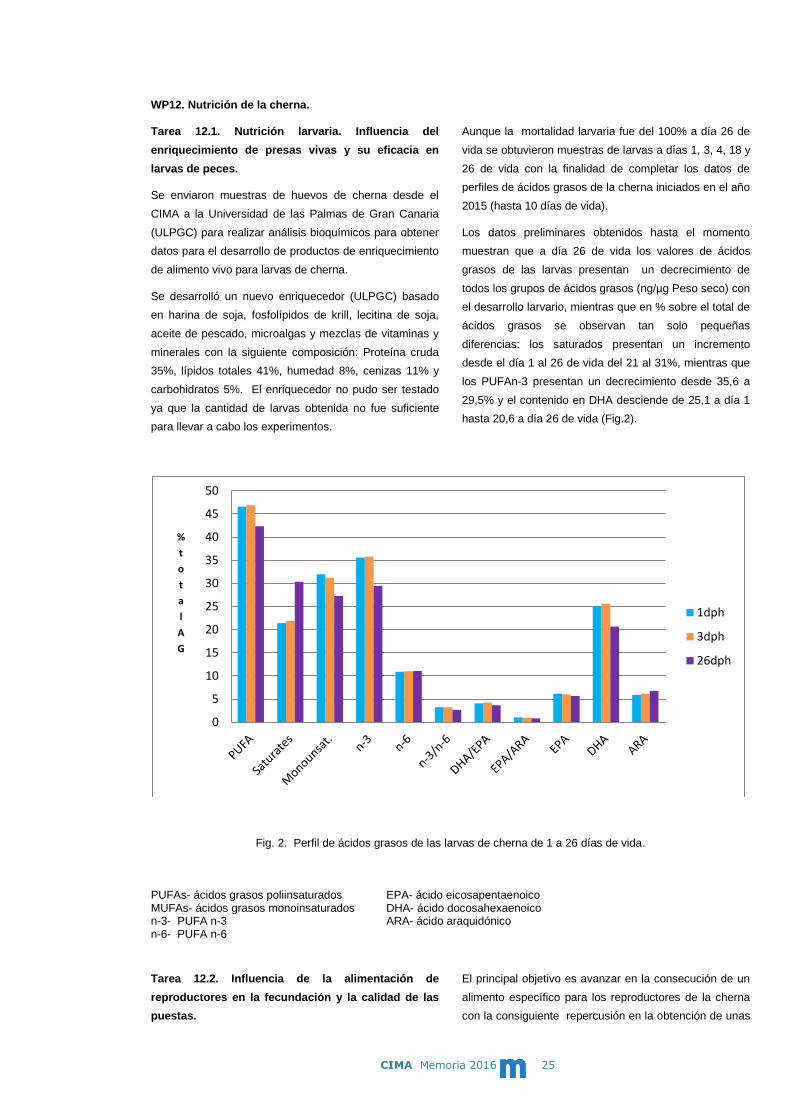
Los datos preliminares obtenidos hasta el momento
muestran que a día 26 de vida los valores de ácidos
grasos de las larvas presentan un decrecimiento de
todos los grupos de ácidos grasos (ng/µg Peso seco) con
el desarrollo larvario, mientras que en % sobre el total de
ácidos grasos se observan tan solo pequeñas
diferencias: los saturados presentan un incremento
desde el día 1 al 26 de vida del 21 al 31%, mientras que
los PUFAn-3 presentan un decrecimiento desde 35,6 a
29,5% y el contenido en DHA desciende de 25,1 a día 1
hasta 20,6 a día 26 de vida (Fig.2).
Fig. 2. Perfil de ácidos grasos de las larvas de cherna de 1 a 26 días de vida. PUFAs- ácidos grasos poliinsaturados EPA- ácido eicosapentaenoico MUFAs- ácidos grasos monoinsaturados DHA- ácido docosahexaenoico n-3- PUFA n-3 ARA- ácido araquidónico n-6- PUFA n-6
Tarea 12.2. Influencia de la alimentación de
reproductores en la fecundación y la calidad de las
puestas.
El principal objetivo es avanzar en la consecución de un
alimento específico para los reproductores de la cherna
con la consiguiente repercusión en la obtención de unas
0
5
10
15
20
25
30
35
40
45
50
%
t
o
t
a
l
A
G
1dph
3dph
26dph
26
buenas tasas de fecundación y en la buena calidad de
las puestas.
Relacionadas con esta tarea se han realizado las
siguientes actividades:
- Análisis de tejidos de ejemplares de cultivo y
comparación con los de ejemplares del medio natural
(datos 2014 y 2015).
- Experimentos de comparación de la alimentación de
reproductores con dieta semihúmeda y pienso seco.
- Perfil de ácidos grasos de ovocitos y larvas
procedentes de hembras alimentadas con dieta
semihúmeda y con pienso seco.
En lo referente a la composición de ejemplares salvajes
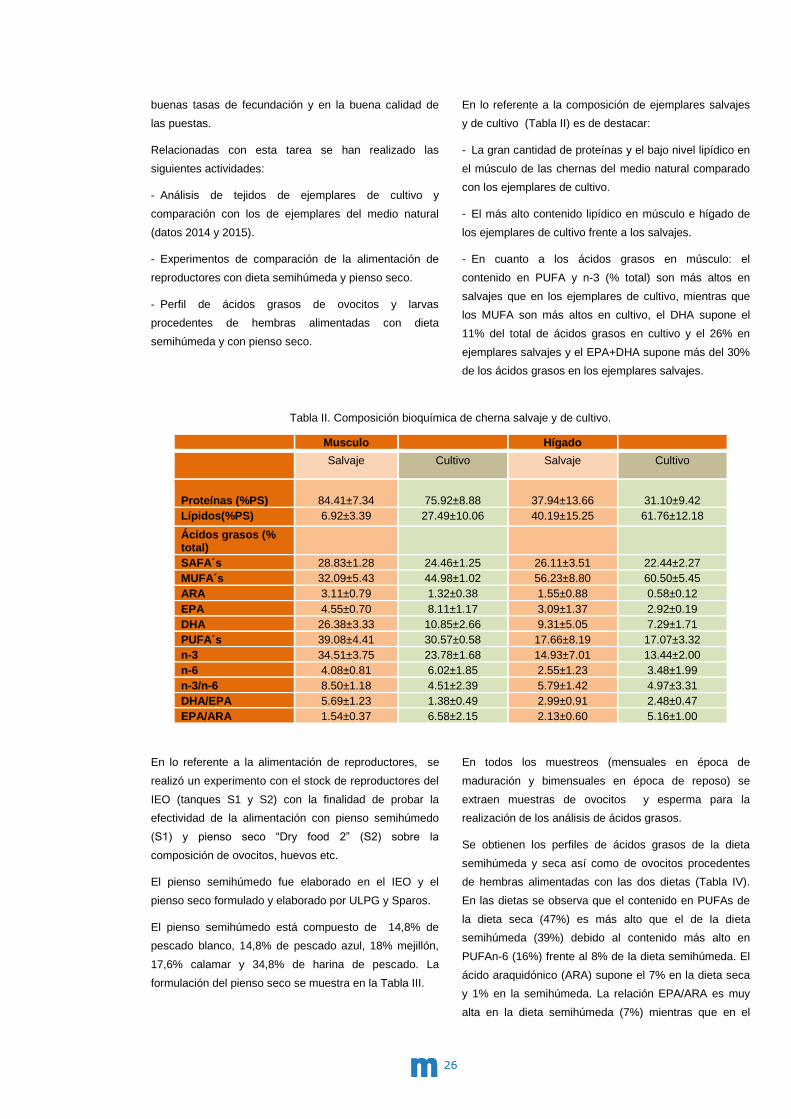
y de cultivo (Tabla II) es de destacar:
- La gran cantidad de proteínas y el bajo nivel lipídico en
el músculo de las chernas del medio natural comparado
con los ejemplares de cultivo.
- El más alto contenido lipídico en músculo e hígado de
los ejemplares de cultivo frente a los salvajes.
- En cuanto a los ácidos grasos en músculo: el
contenido en PUFA y n-3 (% total) son más altos en
salvajes que en los ejemplares de cultivo, mientras que
los MUFA son más altos en cultivo, el DHA supone el
11% del total de ácidos grasos en cultivo y el 26% en
ejemplares salvajes y el EPA+DHA supone más del 30%
de los ácidos grasos en los ejemplares salvajes.
Tabla II. Composición bioquímica de cherna salvaje y de cultivo.
En lo referente a la alimentación de reproductores, se
realizó un experimento con el stock de reproductores del
IEO (tanques S1 y S2) con la finalidad de probar la
efectividad de la alimentación con pienso semihúmedo
(S1) y pienso seco “Dry food 2” (S2) sobre la
composición de ovocitos, huevos etc.
El pienso semihúmedo fue elaborado en el IEO y el
pienso seco formulado y elaborado por ULPG y Sparos.
El pienso semihúmedo está compuesto de 14,8% de
pescado blanco, 14,8% de pescado azul, 18% mejillón,
17,6% calamar y 34,8% de harina de pescado. La
formulación del pienso seco se muestra en la Tabla III.
En todos los muestreos (mensuales en época de
maduración y bimensuales en época de reposo) se
extraen muestras de ovocitos y esperma para la
realización de los análisis de ácidos grasos.
Se obtienen los perfiles de ácidos grasos de la dieta
semihúmeda y seca así como de ovocitos procedentes
de hembras alimentadas con las dos dietas (Tabla IV).
En las dietas se observa que el contenido en PUFAs de
la dieta seca (47%) es más alto que el de la dieta
semihúmeda (39%) debido al contenido más alto en
PUFAn-6 (16%) frente al 8% de la dieta semihúmeda. El
ácido araquidónico (ARA) supone el 7% en la dieta seca
y 1% en la semihúmeda. La relación EPA/ARA es muy
alta en la dieta semihúmeda (7%) mientras que en el
Musculo Hígado
Salvaje Cultivo Salvaje Cultivo
Proteínas (%PS)
84.41±7.34
75.92±8.88
37.94±13.66
31.10±9.42
Lípidos(%PS) 6.92±3.39 27.49±10.06 40.19±15.25 61.76±12.18
Ácidos grasos (% total)
SAFA´s 28.83±1.28 24.46±1.25 26.11±3.51 22.44±2.27
MUFA´s 32.09±5.43 44.98±1.02 56.23±8.80 60.50±5.45
ARA 3.11±0.79 1.32±0.38 1.55±0.88 0.58±0.12
EPA 4.55±0.70 8.11±1.17 3.09±1.37 2.92±0.19
DHA 26.38±3.33 10.85±2.66 9.31±5.05 7.29±1.71
PUFA´s 39.08±4.41 30.57±0.58 17.66±8.19 17.07±3.32
n-3 34.51±3.75 23.78±1.68 14.93±7.01 13.44±2.00
n-6 4.08±0.81 6.02±1.85 2.55±1.23 3.48±1.99
n-3/n-6 8.50±1.18 4.51±2.39 5.79±1.42 4.97±3.31
DHA/EPA 5.69±1.23 1.38±0.49 2.99±0.91 2.48±0.47
EPA/ARA 1.54±0.37 6.58±2.15 2.13±0.60 5.16±1.00
CIMA Memoria 2016 27
pienso seco es de 1,2%, similar a la obtenida en
ejemplares salvajes.
Los perfiles de ácidos grasos de los ovocitos de hembras
alimentada con las dos dietas reflejan las mismas
tendencias que las observadas en los alimentos con un
mayor contenido en PUFAn-6 (12%) en los ovocitos
procedentes de hembras del tanque S2 alimentado con
pienso seco frente al 6% obtenido en los ovocitos
procedentes de hembras del tanque S1 alimentadas con
dieta semihúmeda. La misma tendencia se observa en el
ARA que representa el 7% en ovocitos S2 frente al 2%
en ovocitos S1.
Tabla III. Formulación del pienso seco (Dry food 2. 2016).
Ingredients Dry food 2 2016
%
Fishmeal 70 LT FF Skagen 25.000
CPSP 90 10.000
Squid meal 34.200
Krill meal (Aker Biomarine) 7.500
Wheat Gluten 7.000
Wheat meal 7.250
Tuna oil 1.000
Algatrium 70% DHA 0.200
Incromega DHA 500TG 1.000
VEVODAR 1.300
Vit & Min Premix PV01 2.000
Lutavit E50 0.050
Soy lecithin - Powder 1.500
Macroalgae mix 1.000
Antioxidant powder (Paramega) 0.200
Antioxidant liquid (Naturox) 0.200
SelPlex - Se yeast 0.020
Carophyll Pink 10% - astaxanthin 0.050
Nucleotides (Nucleoforce) 0.030
L-Taurine 0.500
Total 100.000
28
Tabla IV. Contenido en ácidos grasos de las dietas semihúmeda y seca y de los ovocitos procedentes de hembras alimentadas con dieta semihúmeda (S1) y dieta seca (S2).
Ácidos grasos (%total)
Dietas
Ovocitos
Semihúmeda Pienso seco
S1 S2
SAFA´s 29.96±1.19 27.1±0.21
23.45±0.80 21.35±1.62
MUFA´s 31.55±0.97 26.01±0.11
34.60±2.31 32.22±0.39
ARA 1.25±0.28 6.9±0.12 1.60±0.10 6.86±0.57
EPA 8.84±0.48 8.19±0.11 7.35±0.48 5.77±0.39
DPA 1.29±0.24 1.01±0.14 2.67±0.15 3.78±3.03
DHA 15.91±1.16 17.94±0.15
23.46±1.66 22.78±3.26
PUFA´s 38.49±1.65 46.89±0.1
41.94±2.07 46.43±1.72
Total n-3 29.40±1.57 30.31±0.14
35.52±2.08 34.10±1.26
Total n-6 8.28±0.56 16.25±0.06
6.11±0.10 12.11±1.20
n-3/n-6 3.56±0.23 1.87±0.01 5.82±0.40 2.84±0.30
DHA/EPA 1.43±0.66 2.19±0.02 3.19±0.04 3.98±0.69
EPA/ARA 7.45±1.86 1.19±0.04 4.58±0.08 0.85±0.10
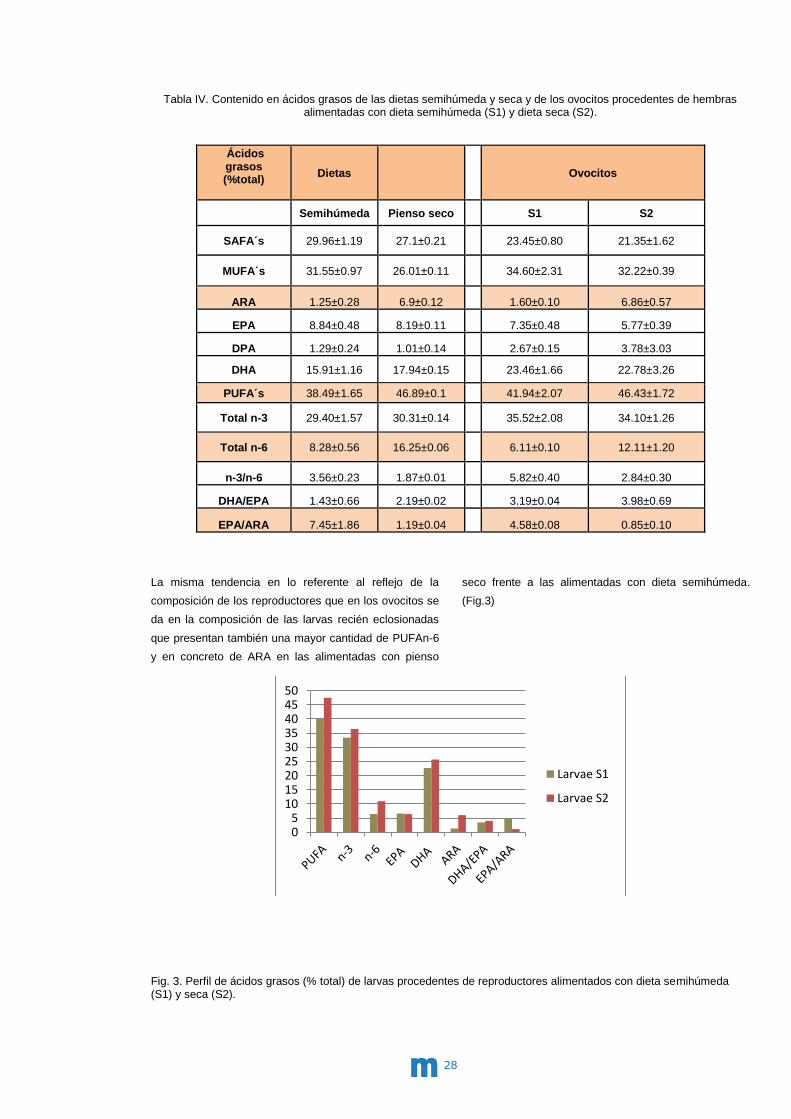
La misma tendencia en lo referente al reflejo de la
composición de los reproductores que en los ovocitos se
da en la composición de las larvas recién eclosionadas
que presentan también una mayor cantidad de PUFAn-6
y en concreto de ARA en las alimentadas con pienso
seco frente a las alimentadas con dieta semihúmeda.
(Fig.3)
Fig. 3. Perfil de ácidos grasos (% total) de larvas procedentes de reproductores alimentados con dieta semihúmeda (S1) y seca (S2).
05
101520253035404550
Larvae S1
Larvae S2

CIMA Memoria 2016 29
Como resumen del área de nutrición:
- Se han obtenido los datos preliminares de los perfiles
de ácidos grasos de las larvas de cherna hasta día 26 de
vida
- Se ha comenzado el desarrollo de nuevos
enriquecedores del alimento vivo (Rotífero y Artemia)
para las larvas de cherna.
- En relación a la alimentación de reproductores se ha
formulado un nuevo pienso seco y los resultados
obtenidos hasta el momento demuestran que el nivel de
grasa debe ser más bajo que el de la mayoría de los
piensos comerciales y deben contener una gran cantidad
de PUFAn-3 y la tasa EPA/ARA debe ser similar a la
obtenida en los ejemplares salvajes.
- Existe una clara relación entre el perfil de ácidos
grasos de las dietas de reproductores y el perfil de
ácidos grasos de los ovocitos y larvas, que es muy
evidente en el contenido en PUFAn-6.
WP18. Cultivo larvario de cherna
Durante el año 2016 se han realizado progresos en el
cultivo larvario de la cherna en lo que se refiere a la
optimización de los parámetros ambientales. Se
realizaron un total de 12 cultivos larvarios, 5 en el IEO (3
procedentes de puestas naturales y 2 de puestas
inducidas) y 7 en AF (3 de puestas naturales y 4
inducidas) con resultados variables en cuanto a
supervivencia larvaria, desde 3 hasta 27 días de vida de
las larvas.
Se han probado diferentes temperaturas de incubación
obteniéndose en los diferentes cultivos tasas de
fecundación entre el 49 y el 100% y de eclosión entre el
3 y el 65% y supervivencia larvaria hasta 27 días de vida.
Se demostró que la temperatura de incubación óptima es
de 16±0.8oC obteniéndose a esta temperatura los
mejores resultados en cuanto a desarrollo embrionario y
tasas de eclosión de los huevos que pueden llegar a
alcanzar el 65%.
De los cultivos larvarios realizados se puede extraer lo
siguiente:
La longitud total de las larvas fue de 4.70±0.27 mm a 1
día de vida.
- El saco vitelino se consume a día 11 de vida a
temperaturas de cultivo entre 14-17°C y a día 8 cuando
se cultiva a 17-20°C.
- La apertura de la boca ocurre a días 7 y 4 de vida a 14-
17°C y 17-20°C, respectivamente.
Durante el cultivo larvario se ha observado un alto grado
de malformaciones que oscilan desde el 10% en larvas
de 3 días hasta el 89% en larvas de 17 días. Aunque se
desconoce el origen de las malformaciones podría
deberse a la inadecuada nutrición ya que se ha
observado un alto porcentaje de larvas que no comen,
condiciones ambientales, stress oxidativo, manejo, etc.
Los resultados obtenidos en cultivo larvario nos indican
que:
- Se requiere intensificar el trabajo en el cultivo larvario
de la cherna para definir las condiciones óptimas de
cultivo.
- Aunque las deformidades en las larvas parecen
incrementarse a lo largo del cultivo sería importante
poder establecer cuando aparecen si en los inicios o al
final del cultivo.
- A pesar de que la mortalidad larvaria es del 100% a día
27 de las larvas, los resultados obtenidos en 2016
suponen un avance sobre los años anteriores y pueden
servir de punto de partida para nuevos experimentos que
definan las condiciones óptimas del cultivo larvario de
cherna.
- La mejora en la alimentación de reproductores y en los
métodos de inducción a la puesta, así como la obtención
de enriquecedores de presas vivas para la alimentación
larvaria pueden ser de gran importancia para obtener
éxito en el cultivo larvario de la cherna.
Información detallada de este proyecto en la web: www.diversifyfish.eu/

30
ACCIÓN DE INVESTIGACIÓN EQUIPO
TÍTULO:
Inducción a la vitelogénesis, maduración ovocitaria,
ovulación y espermiogénesis en cherna (Polyprion
americanus).
INICIO: marzo, 2016
FINALIZACIÓN: diciembre, 2018
ENTIDAD FINANCIADORA:
Consellería del Mar. Xunta de Galicia.
Código: CIMA 16/04
INVESTIGADORA PRINCIPAL:
Linares Cuerpo, F.
INVESTIGADORES/AS:
IGAFA: Rodriguez Villanueva, J.L.
IEO: Álvarez Blázquez, B.; Pérez Rial, E.
Rara Avis. Biotec S.L.: Giménez Nebot, I.
APOYO A LA INVESTIGACIÓN:
Cores González , M.J.; Méndez Pérez, B.
BECARIOS/AS:
Acuña López, I.; Méndez Montenegro, J.C. (hasta
mayo 2016)
Araujo Sendín, A.; Fuentes Villar, N. (desde
septiembre 2016)
RESUMEN
En el desarrollo de esta acción además del CIMA e
IGAFA participan la Planta de Cultivos del CO de Vigo y
la Empresa Rara Avis Biotec, S.L.
A pesar de los avances logrados mediante el desarrollo
del proyecto Diversify (2014-2018, UE), la ausencia de
puestas regulares en cautividad de las hembras de la
cherna (Polyprion americanus), dificulta el control de la
reproducción que es de gran importancia para el
desarrollo de esta especie en acuicultura. La inducción
hormonal de las fases finales del ciclo ovárico y testicular
se han llevado a cabo con éxito tanto en cherna como
en otras especies de peces (implantes de GnRh, HCG,
etc.) sin embargo, gran proporción de los ejemplares de
cherna conservados en cautividad, no logran llegar a un
estado de madurez ovárica suficiente para ser
susceptibles de tratamiento con los preparados
hormonales mencionados. El abordaje de este problema
mediante técnicas de reproducción asistida ofrece la
posibilidad de una solución que permitiría el desarrollo
de la acuicultura industrial de esta especie.
La estrategia de esta acción de investigación tiene como
fundamento fisiológico la maduración gonadal en cherna
mediante la administración de FSH y LH recombinantes.
Dado que estas hormonas actúan directamente sobre las
células gonadales induciendo su maduración, su
utilización permite obviar la intervención del eje
hipotálamo-hipofisario y, por tanto, todas las variables
ambientales que influyen sobre el ciclo reproductivo
espontáneo.
Esta metodología de inducción del ciclo es la que se
lleva a cabo regularmente en reproducción asistida
humana y en ganadería, y se ha demostrado eficaz en
aves y reptiles, y actualmente ya se está experimentando
con especies de peces que presentan disfunción en su
reproducción como el lenguado senegalés y la lubina.
En el caso de la cherna, el método de inducción del ciclo
con FSH y LH presenta gran interés para posibilitar el
completo desarrollo ovocitario en ejemplares que a pesar
de su tamaño no logran completarlo en cautividad.
Además, existe la posibilidad de obtener maduración
gonadal completa incluso en ejemplares que por su
edad y tamaño no entrarían en ciclo reproductivo de
forma espontánea.
Esta acción se presenta como un complemento de las
actuaciones que ya se están realizando en el ámbito de
la reproducción de la cherna en el marco del proyecto
Diversify.
El objetivo fundamental es definir un protocolo de la
maduración gonadal en Polyprion americanus

CIMA Memoria 2016 31
mediante la administración de FSH y LH
recombinantes con el objetivo final de la obtención
de puestas regulares en cautividad.
Objetivos específicos:
- Valoración de la viabilidad de la utilización de la
hormona FSH en ejemplares de cherna.
- Determinación de la dosis adecuada en cherna
µg/Kg (FSH) probando diferentes dosis utilizadas en
otras especies.
- Influencia de la administración de FSH en la
evolución del desarrollo reproductivo de las
hembras de cherna.
- Influencia de la administración de LH en la
evolución del desarrollo reproductivo de las hembras
de cherna.
Descripción de los experimentos:
1. Obtención de FSH y LH recombinantes
Esta tarea es realizada por la empresa “Rara Avis.Biotec
S.L.” que es la encargada de la producción de hormonas.
No existe información acerca de las secuencias
aminoacídicas específicas para FSH y LH del género
Polyprion. Sin embargo, dada la similitud de otras
secuencias sí conocidas (citocromo b, rodopsina, G
protein-coupled receptor 61, etc.) con perciformes como
Dicentrarchus labrax, se considera que la utilización de
FSH y LH recombinantes con las secuencias conocidas
de esta última especie será satisfactoria.
El proceso para la obtención de FSH y LH
recombinantes en cantidades suficientes para la
inducción de la maduración gonadal tiene una duración
aproximada de dos a tres meses y se realiza en cultivos
celulares CHO.
2. Inducción de maduración gonadal en Polyprion
Se utilizan las hormonas disponibles para trabajar en
cherna, que fueron usadas anteriormente en lubina
(Dicentrarchus labrax). Las hormonas FSH y LH que se
utilizan habitualmente en peces suelen proceder de
hormonas producidas en Pichia pastoris (hongo).
Los experimentos se llevaron a cabo desde el mes de
marzo (15/3/2016) hasta finales de julio de 2016 en el
IGAFA en un tanque de reproductores de cherna (tanque
T2) que contiene 8 ejemplares (4 hembras, tres machos
y un indeterminado). Estos ejemplares están marcados
individualmente con microchips. La alimentación de los
reproductores se realizó manualmente y consistió en
calamar suministrado dos veces por semana a saciedad
o “ad libitum”.
Diariamente se midieron los parámetros de temperatura,
pH, oxígeno, etc. Periódicamente se controló el
crecimiento de los ejemplares (talla, peso). Para el
control de los reproductores y previamente a la inyección
con FSH se procedió a su anestesia con 2-fenoxietanol
(0,4 mg/litro).
Antes de iniciar la inyección de la hormona (9-10µg/Kg)
se realizó una inspección de las hembras mediante
biopsia ovárica y se midieron los ovocitos. En base a
esta medición se eligieron 2 hembras del stock para la
inyección de FSH/LH: ♀26C8 (Peso medio 11,93 Kg) y la
♀7B19 (Peso medio 13,48 Kg) y dos controles: ♀4BDB
(Peso medio 16,9 Kg) y la ♀3FF2 (Peso medio 14,32 Kg)
a los que se les inyecta la misma dosis pero de suero
salino. La inyección se lleva a cabo 1 vez/semana, desde
el 15/3/2016 hasta el 14/6/2016, extrayéndose los
ovocitos para en función de sus tamaños controlar su
evolución. La medición de los ovocitos se realiza en el
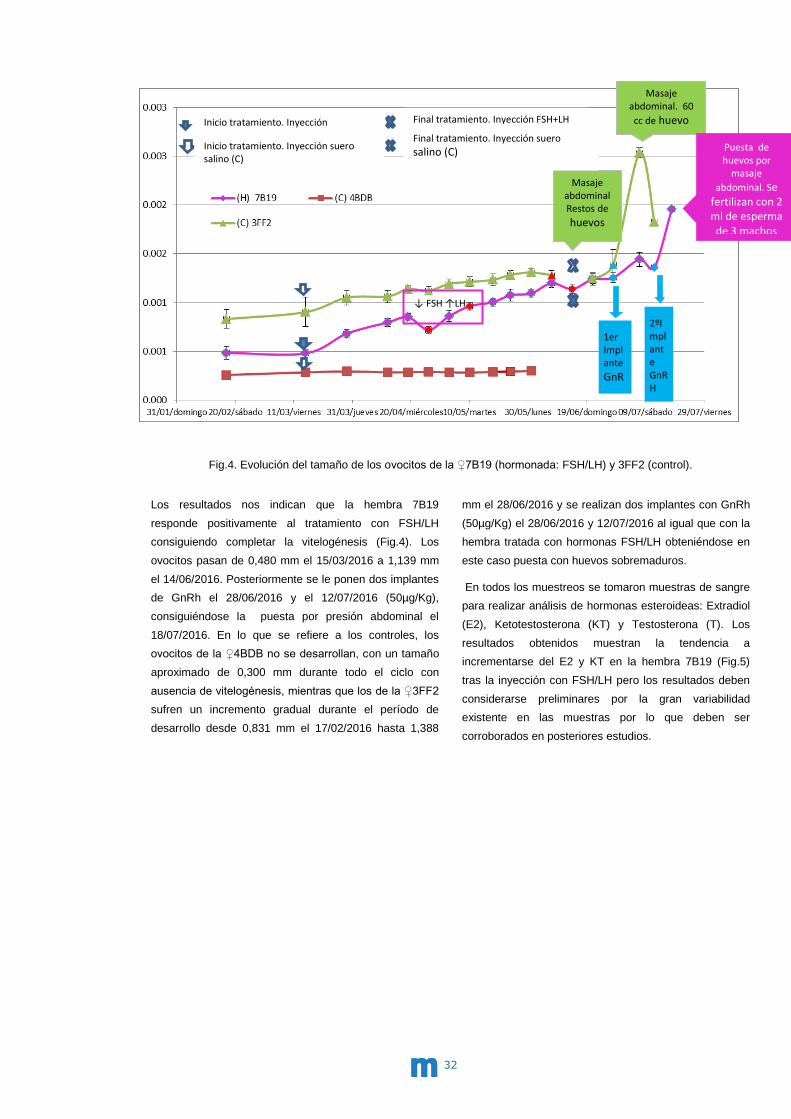
CO del IEO de Vigo. En la Fig.4 se presenta la evolución
de los tamaños de los ovocitos de las hembras 7B19
(hormonada) y de los controles 3FF2 y 4BDB. La 26C8
se desechó por no responder al tratamiento.
32
Fig.4. Evolución del tamaño de los ovocitos de la ♀7B19 (hormonada: FSH/LH) y 3FF2 (control).
Los resultados nos indican que la hembra 7B19
responde positivamente al tratamiento con FSH/LH
consiguiendo completar la vitelogénesis (Fig.4). Los
ovocitos pasan de 0,480 mm el 15/03/2016 a 1,139 mm
el 14/06/2016. Posteriormente se le ponen dos implantes
de GnRh el 28/06/2016 y el 12/07/2016 (50µg/Kg),
consiguiéndose la puesta por presión abdominal el
18/07/2016. En lo que se refiere a los controles, los
ovocitos de la ♀4BDB no se desarrollan, con un tamaño
aproximado de 0,300 mm durante todo el ciclo con
ausencia de vitelogénesis, mientras que los de la ♀3FF2
sufren un incremento gradual durante el período de
desarrollo desde 0,831 mm el 17/02/2016 hasta 1,388
mm el 28/06/2016 y se realizan dos implantes con GnRh
(50µg/Kg) el 28/06/2016 y 12/07/2016 al igual que con la
hembra tratada con hormonas FSH/LH obteniéndose en
este caso puesta con huevos sobremaduros.
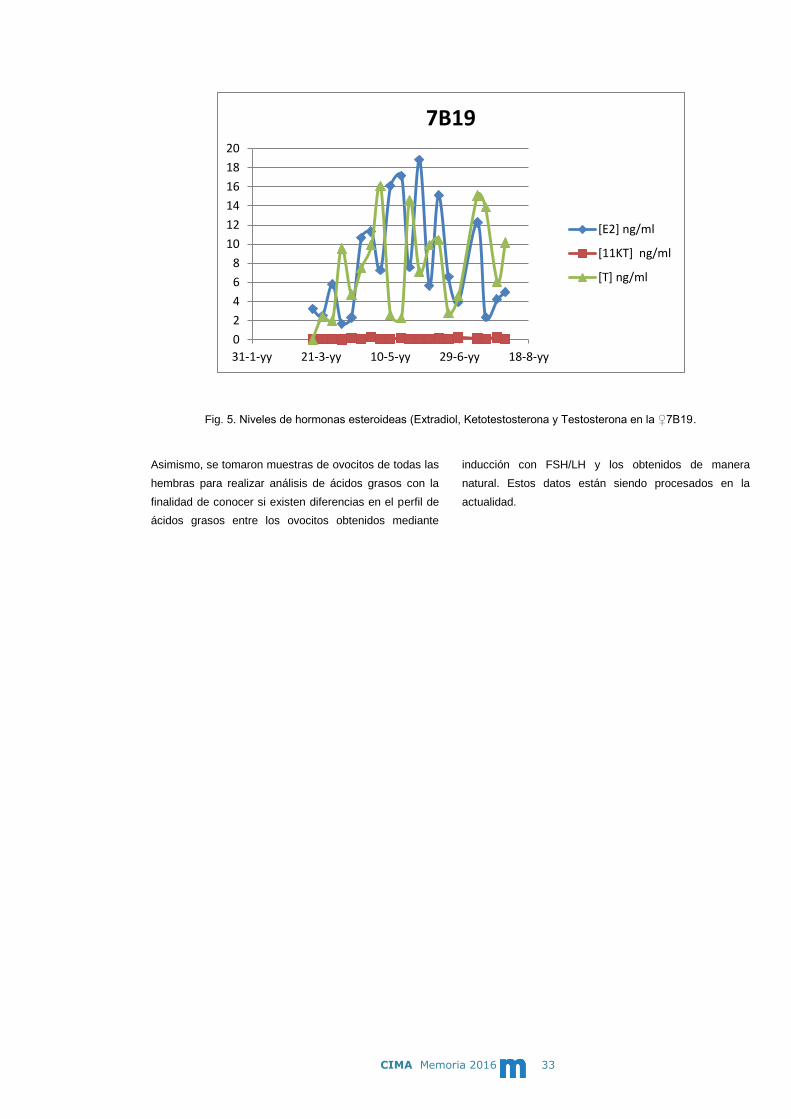
En todos los muestreos se tomaron muestras de sangre
para realizar análisis de hormonas esteroideas: Extradiol
(E2), Ketotestosterona (KT) y Testosterona (T). Los
resultados obtenidos muestran la tendencia a
incrementarse del E2 y KT en la hembra 7B19 (Fig.5)
tras la inyección con FSH/LH pero los resultados deben
considerarse preliminares por la gran variabilidad
existente en las muestras por lo que deben ser
corroborados en posteriores estudios.
↓ FSH ↑LH
1er Implante GnRH 50µg/Kg
2ºImplante GnRH 50µg/Kg
Masaje abdominal Restos de huevos
Masaje abdominal. 60
cc de huevo sobremaduro
Puesta de huevos por
masaje
abdominal. Se fertilizan con 2 ml de esperma de 3 machos
Inicio tratamiento. Inyección FSH+LH Inicio tratamiento. Inyección suero salino (C)
Final tratamiento. Inyección suero salino (C)
Final tratamiento. Inyección FSH+LH
CIMA Memoria 2016 33
Fig. 5. Niveles de hormonas esteroideas (Extradiol, Ketotestosterona y Testosterona en la ♀7B19.
Asimismo, se tomaron muestras de ovocitos de todas las
hembras para realizar análisis de ácidos grasos con la
finalidad de conocer si existen diferencias en el perfil de
ácidos grasos entre los ovocitos obtenidos mediante
inducción con FSH/LH y los obtenidos de manera
natural. Estos datos están siendo procesados en la
actualidad.
0
2
4
6
8
10
12
14
16
18
20
31-1-yy 21-3-yy 10-5-yy 29-6-yy 18-8-yy
7B19
[E2] ng/ml
[11KT] ng/ml
[T] ng/ml

34
RESUMEN
Galicia ocupa una posición privilegiada en la acuicultura
mundial de moluscos bivalvos. Las especies de almeja
predominantes en el cultivo son por orden de
abundancia: almeja japonesa (Ruditapes philippinarum,
Adams & Reeve, 1850), almeja babosa (Venerupis
corrugata, Gmelin, 1791) y almeja fina (Ruditapes
decussatus, Linnaeus, 1758). Hasta el momento la
capacidad de producción de semilla en criadero y la
captación natural están muy lejos de aproximarse a las
necesidades del sector. Es fácil justificar por tanto la
necesidad de criaderos propios que garanticen el
número y la calidad de semilla que puedan satisfacer las
demandas del mercado y el desarrollo de protocolos
robustos en las diferentes fases del cultivo en criadero.
Los protocolos obtenidos hasta ahora, tienen un punto
crítico que se corresponde, con el momento en el que los
individuos cultivados intentan superar la fase de fijación y
metamorfosis, puesto que muchos no lo consiguen y las
supervivencias disminuyen considerablemente. Se
piensa que la fijación y la metamorfosis no sólo están
condicionadas por características propias de la especie y
de la propia larva, sino que también están afectadas por
las características físicas del medio y por diferentes
inductores. Una gran variedad de sustancias naturales y
químicas se mostraron capaces de inducir estos
procesos en larvas de diferentes especies de
invertebrados marinos pero no hay aún resultados
consistentes que puedan ser trasladados al sector en el
caso de moluscos bivalvos.
Con este trabajo de investigación se pretende
profundizar en el estudio de la fijación y la metamorfosis
en las tres especies más importantes de almejas
cultivadas en Galicia. El objetivo es establecer un
método rutinario, efectivo y de bajo coste para la
inducción de la fijación y la metamorfosis de las larvas de
almeja. De esta forma se podría mejorar la producción
en criadero industrial durante todo el año, algo
imprescindible para el desarrollo y la subsistencia de
este sector productivo.
Los objetivos que se pretenden abordar en los tres años
que dura el proyecto son:
1. Efecto de iones y compuestos neuroactivos sobre la
fijación y metamorfosis de larvas de almeja.
2. Efecto de “biofilms” naturales sobre la fijación y
metamorfosis de larvas de almeja.
3. Efecto de diferentes substratos sobre la fijación y
metamorfosis de larvas de almeja.
En este primer año, solamente se dispuso de 6 meses
desde la aprobación del proyecto por lo que, se
ACCIÓN DE INVESTIGACIÓN EQUIPO
TÍTULO:
Inducción de la fijación y metamorfosis en almeja fina,
babosa y japonesa.
INICIO: marzo, 2016
FINALIZACIÓN: diciembre, 2018
ENTIDAD FINANCIADORA:
Consellería del Mar. Xunta de Galicia.
Código: CIMA 16/05
INVESTIGADORA PRINCIPAL:
Nóvoa Vázquez, S.
INVESTIGADORES/AS:
CIMA: Martínez Patiño, D.; Ojea Martínez, J.
USC: Pazos Castelos, A.J.; Pérez-Parallé Mera, M.L.;
Sánchez López, J.L.
APOYO A LA INVESTIGACIÓN:
Bouzamayor Yáñez, V.; Cotarelo Jardón, M.J.; Páez
Pérez, F.J.; Ruíz Misioné, M.
BECARIOS/AS:
Aragunde Vila, R.; Castilla del Río, S.; Guiance Pérez,
M.; Matesanz Salgado, D.; Suárez Costa, I.

CIMA Memoria 2016 35
desarrollaron las tareas T0 y T1 que estaban ya
marcadas en el cronograma.
T0. Acondicionamiento de progenitores y cultivos
larvarios
Esta tarea fue desarrollada en el CECUMAR de Ribadeo.
Los reproductores se recogen del medio natural en la
época que coincida son su madurez sexual para cada
una de las especies que va a ser estudiada y una vez
estabuladas en el criadero, en tanques de 150 litros, se
mantendrán polo menos 24 horas sin comida, con agua
de mar filtrada en circuito abierto y a temperatura similar
a la del medio natural para proceder a su depuración y
rebajar la carga bacteriana que traen del medio natural.
Posteriormente se irá subiendo la temperatura del agua
de manera progresiva hasta alcanzar la temperatura de
acondicionamiento (18-19ºC). Si en unos días no se
obtienen desoves de forma espontánea se procede a la
inducción a la puesta para la obtención de gametos.
Una vez conseguidas las larvas, se realiza su cultivo
utilizando tanques de fibra de vidrio de 500 litros. El agua
de cultivo se trata por filtros de arena, de cartucho y
ultravioleta. La densidad larvaria estuvo entre 5-10
larvas/ml. El protocolo de cultivo (Cerviño-Otero, et al.,
2010; da Costa et al., 2012; Aranda et al., 2014) será el
utilizado habitualmente en dichas instalaciones:
temperatura del agua entre 18-20ºC, renovación total del
agua cada dos días y alimentación de las larvas con una
mezcla de por lo menos cuatro especies microalgales
cultivadas en sistema de producción continua. En dichos
ensayos se utilizarán aquellas condiciones y sustancias
inductoras que dieran mejores resultados en el
laboratorio.
T1. Efecto de iones y compuestos neuroactivos
sobre la fijación de larvas de almeja
Dada la estación del año en la que se iniciaron los
trabajos correspondientes a la tarea 0 (T0), solamente
fue posible disponer de larvas competentes de almeja
japonesa (Ruditapes philippinarum, Adams & Reeve,
1850).
De inicio se utilizaron 3 compuestos neuroactivos
diferentes: epinefrina, L-3,4dihydroxyphenilalanine (L-
DOPA) y ácido gamma-amino-n-butírico (GABA).
Estos experimentos se desarrollaron a escala de
laboratorio siguiendo los protocolos previamente
descritos en García-Lavandeira, et al. (2005) y Mesías-
Gansbiller, et al. (2013). Por problemas derivados de una
avería larga e importante en las instalaciones del
Instituto de Acuicultura, de acuerdo con el grupo do
CECUMAR de Ribadeo, se decidió llevar a cabo estos
primeros ensayos en las instalaciones del CECUMAR de
Ribadeo para lo que en diferentes momentos miembros
del IA se desplazaron a Ribadeo para poner en común
aspectos metodológicos y desarrollar los trabajos
específicos. Los ensayos de laboratorio fueron
realizados por triplicado en placas Petri en un volumen
final de 25 mL de agua de mar filtrada (FSW) con una
malla de 10 µm y tratada con radiación ultravioleta. En
cada una de las placas se colocaron un total de 100
larvas pediveliger de almeja aproximadamente. Las
larvas fueron expuestas a varias concentraciones (10-4,
10-5 y 10-6 M) de los compuestos antes referidos.
GABA, epinefrina, L-DOPA. Los experimentos
transcurrieron en oscuridad a 18 ± 2ºC durante 24 h y 48
h. Cada ensayo tenía un control con FSW sin inductor
potencial añadido. La fijación fue monitorizada con un
microscopio Nikon SMZ-2T transcurridas 24 y 48 horas.
Los experimentos fueron completados y los resultados
están pendientes de análisis estadístico para su
evaluación, pero de forma cualitativa se puede adelantar
que el neurotransmisor GABA parece ser efectivo como
inductor de la metamorfosis en larvas de almeja
japonesa y que dentro de las concentraciones
estudiadas, 10-5 parece ser la más efectiva sin afectar a
la viabilidad larvaria por posibles efectos tóxicos. En
resumen podemos considerar que estos primeros
resultados fueron satisfactorios, la colaboración entre las
dos instituciones muy positiva y la previsión es que el
proyecto progresará perfectamente durante el próximo
año 2017.
García-Lavandeira, M.; Silva, A., Abad, M., Pazos, AJ.,
Sánchez, J.L. & Pérez-Parallé, M.L. 2005. Effects of
GABA and epinephrine on the settlement and
metamorphosis of the larvae of four species of bivalve
molluscs. J. Exp. Mar. Biol. Ecol. 316: 149-156
Mesías-Gansbiller, C., Silva, A., Maneiro, V., Pazos, A.l,
Sánchez, J.L. & Pérez-Parallé, M.L. 2013. Effeels of
chemical cues on larval settlement of the flat oyster
(Ostrea edulis L.): A hatchery approach. Aquaculture
376-379: 85-89.
36
6 PATOLOGÍA

CIMA Memoria 2016 37
ACCIÓN DE INVESTIGACIÓN EQUIPO
TÍTULO:
Estudio epidemiológico de la almeja rubia (Venerupis
rhomboides) en Galicia afectada por colonias
procariotas intracelulares (CPI). Identificación de las
CPI. Seguimiento de la estructura de tallas de
poblaciones de almeja rubia de 4 bancos de la ría de
Vigo.
INICIO: octubre, 2016
FINALIZACIÓN: octubre, 2017
ENTIDAD FINANCIADORA:
Consellería del Mar. Xunta de Galicia.
Código: CIMA 16/01
INVESTIGADORA PRINCIPAL:
López Gómez, C.
INVESTIGADORES/AS:
CIMA: Febrero Mayor, F.M.; No Couto, E.
Xefatura Territorial CM Vigo: Rodríguez González,
L.M.
INTECMAR: Darriba Couñago, S.
APOYO A LA INVESTIGACIÓN:
Brianes Veiras, M.J.; Meléndez Ramos, M.I.
BECARIO:
Grande Ocampo, O.
RESUMEN
En el año 2012 la Consellería do Medio Rural e do Mar
encargó, a investigadores del CIMA, una acción corta
para un “Análisis preliminar de la problemática de
disminución de la producción de almeja rubia (Venerupis
rhomboides) en la ría de Vigo”. Los resultados de este
estudio presentaron en conjunto abundancias de
población muy reducidas y, en muchos casos,
estructuras de tamaños desequilibrados, sugiriendo la
existencia de un déficit en la incorporación de juveniles a
la fracción comercial. El estudio histopatológico llevado a
cabo en estas poblaciones mostró la presencia de
Colonias Procariotas Intracelulares en la branquia (CPI-I)
con prevalencias muy altas e Intensidades de infección
moderadas y bajas, así como CPI hiperparasitadas por
un virus (CPI-II). Las CPI-I y II ya fueran detectadas en el
2010 y 2011 asociadas a mortalidades.
Como continuación de ese estudio preliminar, en el año
2013 se pone en marcha una acción de investigación
para profundizar en el estudio de la almeja rubia en la ría
de Vigo. En el marco de esta acción se muestrearon 4
bancos sublitorales, en los que trabajan los barcos del
plan de libre marisqueo a flote de la ría de Vigo: Cabo de
Mar, Tirán, Liméns e Islas Cíes. En el año 2016 se
solicitó una prórroga de la acción para continuar con el
estudio.
A continuación, se presenta un resumen de los
resultados obtenidos por objetivos.
Objetivo 1. Análisis de la población
Ría de Vigo.
En el muestreo de otoño de 2015 la estructura de
tamaños de la población parece equilibrada, con una alta
fijación. En el año 2016, el muestreo de primavera
muestra el crecimiento de la fijación del otoño de 2015 y
en otoño de 2016 la continuación del crecimiento y un
nuevo reclutamiento.
La conclusión de los muestreos (primavera y otoño)
desde el año 2013 hasta 2015 indicaba, en general, que
los bancos tenían una estructura de tallas de población
desequilibrada y presentaban una baja fracción juvenil,
incluyendo el muestreo de primavera de 2015. En el
muestreo de otoño de 2015 se encuentra un nuevo
reclutamiento que equilibra la estructura de tallas. En los
muestreos de primavera y otoño de 2016, se observa el
crecimiento (primavera y otoño) del reclutamiento de
otoño 2015 y un nuevo reclutamiento (aunque menor que
en 2015) en otoño de 2016. Las cohortes que
obtenemos, aplicando el método de Battacharya son las
siguientes: otoño 2015: 15,18; 41,86; 45,17 mm.
Primavera 2016: 18,77; 25,58; 40,37 mm. Otoño 2016:
14,59; 38,73; 43,84 mm.
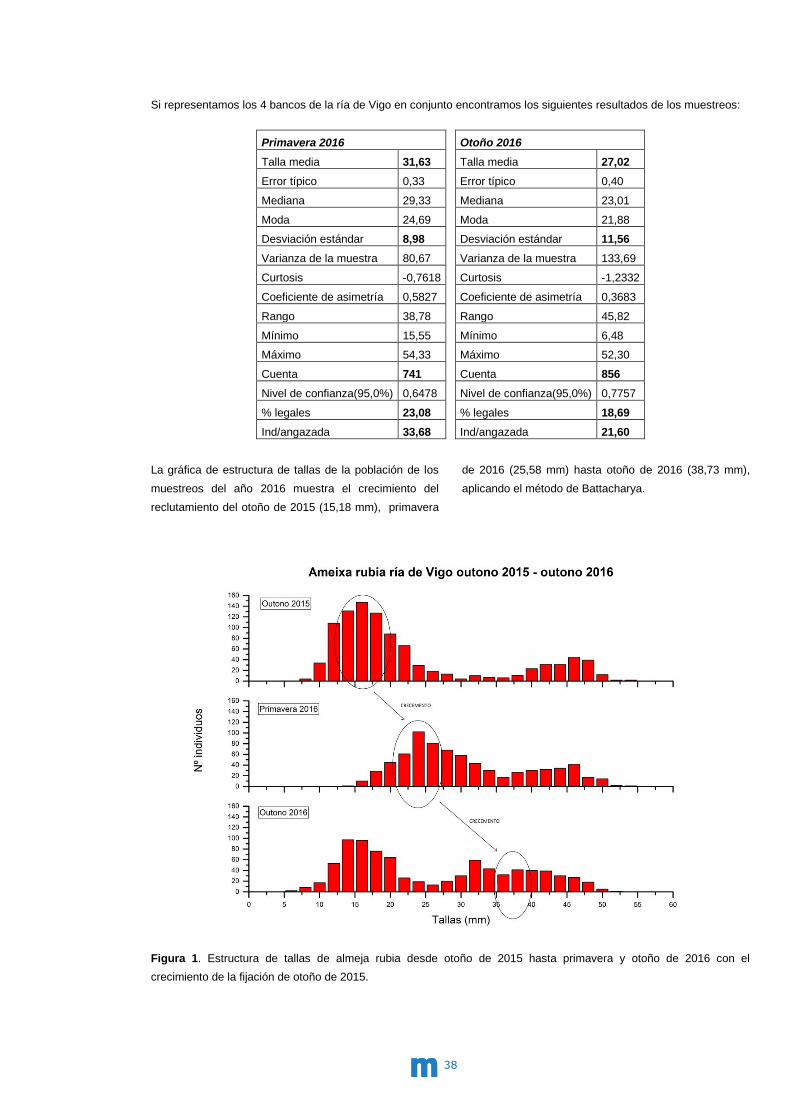
38
Si representamos los 4 bancos de la ría de Vigo en conjunto encontramos los siguientes resultados de los muestreos:
Primavera 2016
Otoño 2016
Talla media 31,63
Talla media 27,02
Error típico 0,33
Error típico 0,40
Mediana 29,33
Mediana 23,01
Moda 24,69
Moda 21,88
Desviación estándar 8,98
Desviación estándar 11,56
Varianza de la muestra 80,67
Varianza de la muestra 133,69
Curtosis -0,7618
Curtosis -1,2332
Coeficiente de asimetría 0,5827
Coeficiente de asimetría 0,3683
Rango 38,78
Rango 45,82
Mínimo 15,55
Mínimo 6,48
Máximo 54,33
Máximo 52,30
Cuenta 741
Cuenta 856
Nivel de confianza(95,0%) 0,6478
Nivel de confianza(95,0%) 0,7757
% legales 23,08
% legales 18,69
Ind/angazada 33,68
Ind/angazada 21,60
La gráfica de estructura de tallas de la población de los
muestreos del año 2016 muestra el crecimiento del
reclutamiento del otoño de 2015 (15,18 mm), primavera
de 2016 (25,58 mm) hasta otoño de 2016 (38,73 mm),
aplicando el método de Battacharya.
Figura 1. Estructura de tallas de almeja rubia desde otoño de 2015 hasta primavera y otoño de 2016 con el
crecimiento de la fijación de otoño de 2015.
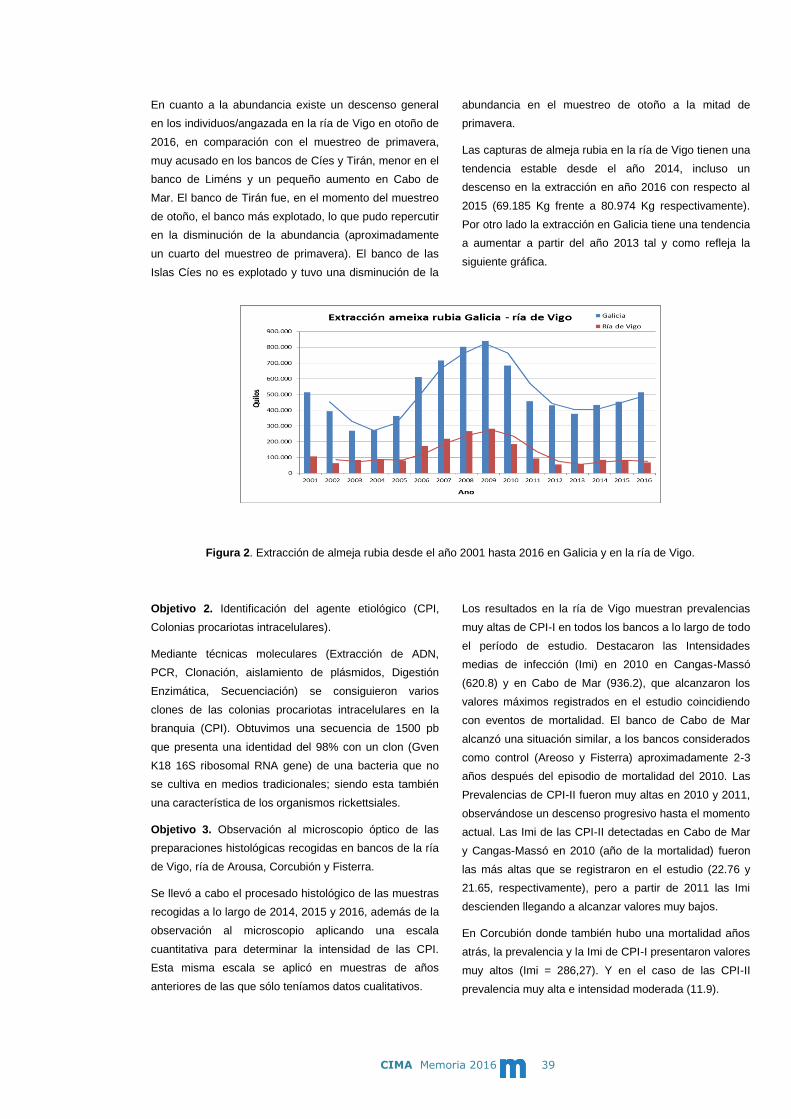
CIMA Memoria 2016 39
En cuanto a la abundancia existe un descenso general
en los individuos/angazada en la ría de Vigo en otoño de
2016, en comparación con el muestreo de primavera,
muy acusado en los bancos de Cíes y Tirán, menor en el
banco de Liméns y un pequeño aumento en Cabo de
Mar. El banco de Tirán fue, en el momento del muestreo
de otoño, el banco más explotado, lo que pudo repercutir
en la disminución de la abundancia (aproximadamente
un cuarto del muestreo de primavera). El banco de las
Islas Cíes no es explotado y tuvo una disminución de la
abundancia en el muestreo de otoño a la mitad de
primavera.
Las capturas de almeja rubia en la ría de Vigo tienen una
tendencia estable desde el año 2014, incluso un
descenso en la extracción en año 2016 con respecto al
2015 (69.185 Kg frente a 80.974 Kg respectivamente).
Por otro lado la extracción en Galicia tiene una tendencia
a aumentar a partir del año 2013 tal y como refleja la
siguiente gráfica.
Figura 2. Extracción de almeja rubia desde el año 2001 hasta 2016 en Galicia y en la ría de Vigo.
Objetivo 2. Identificación del agente etiológico (CPI,
Colonias procariotas intracelulares).
Mediante técnicas moleculares (Extracción de ADN,
PCR, Clonación, aislamiento de plásmidos, Digestión
Enzimática, Secuenciación) se consiguieron varios
clones de las colonias procariotas intracelulares en la
branquia (CPI). Obtuvimos una secuencia de 1500 pb
que presenta una identidad del 98% con un clon (Gven
K18 16S ribosomal RNA gene) de una bacteria que no
se cultiva en medios tradicionales; siendo esta también
una característica de los organismos rickettsiales.
Objetivo 3. Observación al microscopio óptico de las
preparaciones histológicas recogidas en bancos de la ría
de Vigo, ría de Arousa, Corcubión y Fisterra.
Se llevó a cabo el procesado histológico de las muestras
recogidas a lo largo de 2014, 2015 y 2016, además de la
observación al microscopio aplicando una escala
cuantitativa para determinar la intensidad de las CPI.
Esta misma escala se aplicó en muestras de años
anteriores de las que sólo teníamos datos cualitativos.
Los resultados en la ría de Vigo muestran prevalencias
muy altas de CPI-I en todos los bancos a lo largo de todo
el período de estudio. Destacaron las Intensidades
medias de infección (Imi) en 2010 en Cangas-Massó
(620.8) y en Cabo de Mar (936.2), que alcanzaron los
valores máximos registrados en el estudio coincidiendo
con eventos de mortalidad. El banco de Cabo de Mar
alcanzó una situación similar, a los bancos considerados
como control (Areoso y Fisterra) aproximadamente 2-3
años después del episodio de mortalidad del 2010. Las
Prevalencias de CPI-II fueron muy altas en 2010 y 2011,
observándose un descenso progresivo hasta el momento
actual. Las Imi de las CPI-II detectadas en Cabo de Mar
y Cangas-Massó en 2010 (año de la mortalidad) fueron
las más altas que se registraron en el estudio (22.76 y
21.65, respectivamente), pero a partir de 2011 las Imi
descienden llegando a alcanzar valores muy bajos.
En Corcubión donde también hubo una mortalidad años
atrás, la prevalencia y la Imi de CPI-I presentaron valores
muy altos (Imi = 286,27). Y en el caso de las CPI-II
prevalencia muy alta e intensidad moderada (11.9).

40
En la ría de Arousa, donde nunca se detectó mortalidad
las prevalencias de CPI-I fueron siempre muy altas. Sin
embargo la Imi fue variable alcanzando valores
moderados en muestreos puntuales sin seguir un patrón
estacional. Las prevalencias de CPI-II fueron variables y
la Imi siempre baja.
Las Imi fueron moderadas en Liméns 2014 y 2015, y en
Cíes 2012 siendo el resto de los años bajas o muy bajas.
En el caso de Liméns, Tirán e Islas Cíes las prevalencias
de CPI-II fueron variables y las Imi de bajas a muy bajas.
Objetivo 4. Estudio epidemiológico centrándose en los
agentes patógenos que fueron identificados como
posibles responsables de causar efectos deletéreos en la
almeja rubia.
Pendientes de analizar los datos para el estudio
epidemiológico. Hace falta resaltar que, entre otros
protozoos, destaca por su prevalencia e intensidad la
presencia de la gregarina Nematopsis sp, así como la
presencia con bajas prevalencias e intensidades
Coccidios, Perkinsus sp., Turbelarios y Trematodos.
Conclusiones:
- En los muestreos de primavera y otoño de 2016 la
estructura de tallas sigue equilibrada y con aumento de
la talla de reclutamiento de otoño de 2015 (15,18 →
25,58 → 38,73 mm).
- Existe un descenso de la abundancia desde el
muestreo de primavera al de otoño de 2016 (33,68 por
21,40 ind./angazada respectivamente).
- Descenso de las capturas en la ría de Vigo en 2016 con
respecto a 2015 al contrario que el resto de Galicia.
- La presencia de CPI-I y CPI-II es normal en almeja
rubia ya que se encuentran prevalencias altas
indistintamente en bancos afectados por mortalidad y en
bancos control.
- Es el aumento de la intensidad de infección del CPI-I
lo que desemboca en los episodios de mortalidad.
- Se registró un descenso de la intensidad de las CPI-I a
partir del año 2012 hasta el año 2015 (en Cabo de Mar),
coincidiendo con la mejoría detectada en el estudio de la
población.

CIMA Memoria 2016 41
ACCIÓN DE INVESTIGACIÓN EQUIPO
TÍTULO:
Estudio de la eliminación de virus entéricos patógenos
en ostra mediante procesos de reinstalación y
depuración.
INICIO: octubre, 2016
FINALIZACIÓN: octubre, 2018
ENTIDAD FINANCIADORA:
Consellería del Mar. Xunta de Galicia.
Código: CIMA 16/08
INVESTIGADORA PRINCIPAL:
López Gómez, C.
INVESTIGADORES/AS:
CIMA: Febrero Mayor, F.M.
INTECMAR: Álavares Álvarez, C.; Darriba Couñago,
S.
USC: López Romalde, J.
APOYO A LA INVESTIGACIÓN:
CIMA: Meléndez Ramos, M.I.
INTECMAR: Rodríguez Ruibal, L.
BECARIO:
CIMA: Grande Ocampo, O.
RESUMEN
El proyecto tiene como objetivos, por una parte, el
establecimiento de un plan integrado de vigilancia que
reduzca de forma significativa los efectos del riesgo de
contaminación microbiológica y viral durante la
producción primaria de las ostras comerciales producidas
en Galicia, enfocando el esfuerzo en el estudio de la
efectividad de la depuración en la eliminación de estos
virus.
Por otra parte, el proyecto pretende establecer la
capacidad infectiva de estos virus después del proceso
de depuración. Este parámetro nos llevará a establecer
el riesgo real de estos patógenos, ya que no todas las
partículas virales detectadas por las técnicas de
cuantificación actuales son infectivas.
El plan de trabajo establecido fue el siguiente:
1. Selección de las zonas de estudio
Se escogieron como zonas representativas de las zonas
de producción dos zonas que disponían de parques de
cultivo de bateas y de bancos de infaunales, y que
dispongan de ambas especies comerciales de ostra,
Ostrea edulis y Crasssotrea gigas.
Ostrea edulis: la zona elegida fue el polígono Cambados
D (ría de Arousa) (Figura 1).
Crasssotrea gigas: Se eligió la ría de O Barqueiro por ser
la zona donde hay abundancia de esta especie de ostra.
Como zona de reinstalación se escogió la ría de Aldán
(Figura 2).
2. Diseño del plan de muestreo
Se diseñaron 10 experiencias de
reinstalación/depuración con cada especie de ostra. La
frecuencia de muestreo durante la reinstalación quedó
fijada con el siguiente régimen: días 0 (antes de llevar a
la zona de reinstalación), 15, 22 y 30. El día 30,
dependiendo de los niveles de contaminación, las ostras
se trasladan a la depuradora experimental en el CIBUS
(Universidad de Santiago de Compostela) para su
depuración y se realizan análisis los días 32 (48 h de
depuración) y 37 (1 semana de depuración).
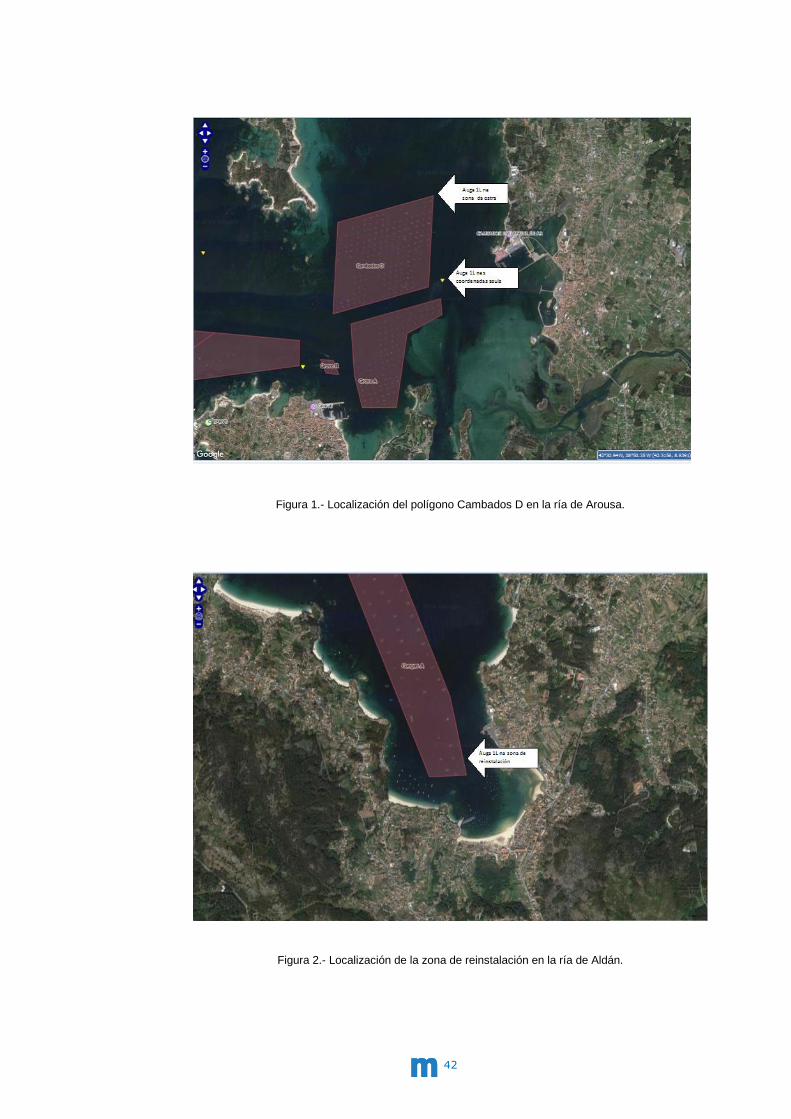
42
Figura 1.- Localización del polígono Cambados D en la ría de Arousa.
Figura 2.- Localización de la zona de reinstalación en la ría de Aldán.

CIMA Memoria 2016 43
Se están recogiendo de modo paralelo dos muestras de
agua en la zona de origen de la ostra, en un punto
próximo al emisario de la EDAR más influyente en la
zona y en el punto de recogida de la muestra, así como
una muestra de agua en la zona de reinstalación.
3. Transporte y recogida de las muestras
La reinstalación y recogida de las muestras, coordinadas
por el CETMAR, es realizada por personal cualificado,
así como por personal contratado específicamente para
este trabajo.
El traslado del producto se realiza siguiendo el método
Coleman de refrigeración según se realiza en la Unidad
de Microbiología y Virología del Intecmar. Las muestras
de agua se recogerán en botes estériles y se
conservarán también en refrigeración hasta el momento
del ensayo.
4. Realización de ensayos
Hasta el momento se llevan 3 experiencias con ostra
plana. Debido a algunos condicionantes externos al
proyecto, como la no existencia de ostras en cultivo o el
cierre en el verano de las zonas de reinstalación, las
experiencias comenzaron en el mes de octubre.
A. Carga bacteriana de E. coli / 100 g en el molusco
Cada fecha de muestreo se lleva al INTECMAR una
parte de la muestra para el cálculo de carga bacteriana
mediante la técnica del número más probable (NMP).
Los resultados obtenidos hasta el momento son
coherentes con los históricos tanto de la zona de
producción como de la zona de reinstalación, y no
indican eventos de contaminación extraordinarios en
dichas zonas.
B. Presencia de virus entéricos (Norovirus y hepatitis A
HAV)
Para la detección de virus entéricos se utiliza la técnica
de reacción en cadena de la polimerasa en tiempo real
(RT-PCR) con sondas TaqMan siguiendo las
recomendaciones de la tecina ISO/TS 15216-1:2013.
Este análisis se está realizando en paralelo en los
laboratorios de la Universidad de Santiago de
Compostela y del INTECMAR, como prueba de ensayo
interlaboratorio.
Hasta el momento no se detectaron los virus incluidos en
el proyecto en ninguna de las muestras correspondientes
a las tres experiencias en ninguno de los laboratorios, ni
de agua ni de ostra. Los controles tanto positivos como
negativos incluidos en cada uno de los análisis indican
que estos resultados no se deben a una falta de la
técnica, sino a la no presencia de los virus en las
muestras. Cabe destacar lo atípico de la meteorología
durante los meses de otoño y comienzo de invierno con
pocas lluvias lo que podría ser un factor importante para
la no presencia de los virus en las muestras de ostra y
de agua.
C. Estudio histológico e inmunohistoquímico
Así mismo, se recogieron muestras para llevar a cabo el
estudio histológico e inmunohistoquímico mediante
técnicas clásicas en tejidos de ostra en origen, después
de la reinstalación y después de la depuración, con el fin
de determinar la presencia de partículas virales en los
tejidos. La negatividad de las muestras por otros
métodos hizo innecesario el análisis de las muestras
aunque estas fueron procesadas para el estudio
histológico.
Plan de futuras acciones:
Se va a continuar con las experiencias de ostra plana y
además, está contemplada la realización, durante este
año de las primeras experiencias con ostra rizada. Con
el cambio en la meteorología es de esperar que los
niveles de contaminación con virus entéricos vuelvan a la
media de los datos históricos y comiencen a aparecer
muestras positivas para virus que permitan el análisis de
la eliminación por reinstalación y depuración.

44
PROYECTO DE INVESTIGACIÓN EQUIPO
TÍTULO:
Poblaciones bacterianas en criadero de bivalvos.
Procesos de patogénesis.
INICIO: junio, 2015
FINALIZACIÓN: diciembre, 2017
ENTIDAD FINANCIADORA:
Ministerio de Economía y Competitividad
INVESTIGADORA PRINCIPAL:
Barja Pérez, J.L. (USC)
INVESTIGADORES/AS:
CIMA: Montes Pérez, J.; Nóvoa Vázquez, S.; Ojea
Martínez, J.
USC: Prado Plana, S.; Dubert Pérez, J.
APOYO A LA INVESTIGACIÓN:
CIMA: Bouzamayor Yáñez, M.V.; Brianes Beiras,
M.J.; Cotarelo Jardón, M.J.; Meléndez Ramos, I.;
Páez Pérez, F.J.; Penas Pampín; E.; Ruíz Misioné, M.
USC: Núñez García, S.
BECARIOS/AS:
Castilla del Río, S.; Matesanz Salgado, D.; Ulla
Refojo, L.
RESUMEN
El proyecto “Poblaciones bacterianas en criaderos de
bivalvos. Procesos de patogénesis” alcanzó ya
importantes resultados, algunos de los cuales se
resumen a continuación.
En el campo de estudio de los mecanismos de infección
bacteriana en cultivos larvarios de moluscos bivalvos, se
demostró el modo de infección y secuencia espacio-
temporal. Este importante avance fue el resultado de
ensayo de inoculación in vivo en laboratorio de larvas de
bivalvos con vibrios patógenos marcados con “Green
fluorescent protein” (GFP). Se pudieron diferenciar tres
fases en el proceso infectivo, que comienza con la
filtración de las bacterias a través del velo, su entrada en
el sistema digestivo y el inicio de la proliferación. En la
segunda etapa (6-8 horas), los vibrios patógenos se
expanden por los diferentes órganos. La fase final (14-24
horas) implica la colonización completa de la larva. Hay
que destacar que se demostró en el transcurso de los
experimentos que en las fases iniciales del proceso la
larva es asintomática, manifestando los primeros signos
cuando la infección está muy avanzada, lo que explica la
falta de efectividad de los tratamientos.
En cuanto a la dinámica de los patógenos bacterianos en
el criadero, se demostró, mediante técnicas clásicas y
moleculares, la transmisión vertical de un Vibrio
patógeno naturalmente presente en un desove de almeja
fina (Ruditapes decussatus), que causó graves
mortalidades desde el principio del cultivo. El patógeno
se detectó ya en los huevos y, a partir de ahí, se pudo
observar cómo se convertía en el tipo dominante en las
muestras en medio selectivo para vibrios Tiosulfato-
Citrato-Bilis-Sacarosa (TCBS). Con esto se puso de
manifiesto el grave riesgo que supone la presencia de
potenciales patógenos en los huevos, aún en números
bajos, ya que las condiciones de cultivo favorecen su
proliferación ya en las etapas iniciales hasta niveles que
resultan limitantes de forma irreversible. Por otro lado, la
transmisión vertical desde los reproductores quedó
demostrada, ya que los otros posibles orígenes (agua,
fitoplancton) se descartaron mediante los análisis
necesarios.
Otro trabajo a comentar fue la evaluación del uso de
líneas celulares de cultivo continuo como alternativa al
método tradicional de inoculaciones in vivo de larvas
para establecer la patogenicidad de aislados
bacterianos. Primero se evaluó la citotoxicidad de los
productos extracelulares (ECPs) de patógenos
conocidos de cultivos larvarios de bivalvos, del género
Vibrio, en líneas celulares de animales homeotermo,
confirmando su capacidad de destrucción del tapiz

CIMA Memoria 2016 45
celular. En la segunda fase se trabajó con aislados
bacterianos del mismo género asociados a baja
supervivencia larvaria pero cuya patogenicidad se
desconocía, comparando la patogenicidad in vivo con la
citotoxicidad de sus ECPs. Los datos obtenidos
demostraron que los ensayos con líneas celulares son
útiles para detectar aislados patógenos con elevada
secreción de proteínas, pero no diferencian entre
distintos grados de virulencia. En conjunto, los resultados
constituyen una base sólida para la implementación de
una metodología alternativa para los ensayos de
virulencia de patógenos larvarios de bivalvos.
Por último, apuntar que también en el marco del proyecto
se secuenciaron los genomas completos de vibrios
patógenos de cultivos larvarios de bivalvos, Vibrio
europaeus PP-638 (antes Vibrio tubiashii subsp.
Europaeus) y Vibrio bivalvicida PP2-605, que permitirán
abordar análisis mediante herramientas bioinformáticas
para la búsqueda de genes candidatos relacionados con
la patogenicidad.
Esta será una de las tareas a desarrollar, dentro del
objetivo de estudio de mecanismos de infección, junto
con otras del apartado de conocimiento de la dinámica
de los patógenos en criadero, como aquellos
relacionados con las interacciones con el fitoplancton o
la competencia entre especies bacterianas. Asimismo, se
continuará con los trabajos que se vienen realizando de
análisis histopatológico de los reproductores de
diferentes especies de bivalvos utilizados en el criadero,
como complemento para el estudio de las vías de
entrada y transmisión de patógenos.

46
ACCIÓN DE INVESTIGACIÓN EQUIPO
TÍTULO:
Epidemiología de la enfermedad causada por Marteilia
cochillia en las poblaciones de berberecho
Cerastoderma edule de Galicia.
INICIO: marzo, 2015
FINALIZACIÓN: diciembre, 2017
ENTIDAD FINANCIADORA:
Consellería del Mar. Xunta de Galicia.
Código: PGIDIT-CIMA 15/02
INVESTIGADORA PRINCIPAL:
Carballal Durán , M.J.
INVESTIGADORES/AS:
CIMA: Cao Hermida, A.; Iglesias Estepa, D.; No Couto,
E.; Villalba García, A.
INTECMAR: Darriba Couñago, S.; Montero Vila, P.
IIM: González González, A.; Pascual del Hierro, S.
CETMAR: Abollo Rodríguez, E.
Cofradía de Pescadores “San Antonio” de Cambados:
Mariño Balsa, J.C.
Delegación de Carril de la Consellería del Mar:
Fernández Barreiro, J.
APOYO A LA INVESTIGACIÓN:
Brianes Beiras, M.J.; González Fontela, A.I.; Martínez
Verde, G.; Meléndez Ramos, I.; Penas Pampín, E.
BECARIA:
Fernández González, J.; Riomayor Cernadas, A.
RESUMEN
El parásito Marteilia cochillia afecta a la glándula
digestiva de Cerastoderma edule impidiendo la absorción
de alimento cuando la infección es intensa, lo que
provoca inanición y muerte del berberecho. Marteilia
cochillia se detectó por primera vez en Galicia en febrero
de 2012 en berberechos de Lombos do Ulla, causando
una mortalidad masiva de los mismos. En los meses
siguientes, la enfermedad se detectó en berberechos de
los otros bancos de la ría de Arousa; las intensas
mortalidades causadas por este parásito llevaron a la
práctica desaparición de este recurso en esta ría. En los
años 2013-2014 desarrollamos un proyecto de
investigación en el que se estableció un programa de
vigilancia de la enfermedad en las rías limítrofes a la ya
afectada, resultando que los berberechos de la ría de
Pontevedra y los berberechos de algunos bancos de la
ría de Vigo estaban afectados por esta parasitosis.
En marzo del año 2015 se inicia este nuevo proyecto de
investigación sobre M. cochillia, en el que se realizan
estudios epidemiológicos y ambientales, y se busca
avanzar en el conocimiento del ciclo de vida del parásito.
Uno de los objetivos del estudio es el desarrollo de un
programa de vigilancia de la enfermedad en las
principales poblaciones de berberecho de Galicia, con el
fin de conocer el estatus sanitario de las mismas. Con
esta intención, durante el año 2015, se recogieron
muestras de 30 individuos de berberecho C. edule de las
poblaciones seleccionadas en dos ocasiones: primavera
y otoño. Los bancos seleccionados fueron los que
figuran en la red de muestreo del programa oficial de
control patológico de moluscos bivalvos, que desarrolla
la Unidad de Patología del INTECMAR. Debido a las
mortalidades causadas por la marteiliosis, en el
muestreo de primavera (junio) en los bancos de
Redondela, Vilaboa, Lombos do Ulla, Cabo de Cruz y
Combarro no fue posible obtener un número suficiente
de individuos de C. edule y, como alternativa, se
analizaron berberechos de la especie C. glaucum. En el
muestreo de otoño (octubre-noviembre) se muestrearon
únicamente individuos de la especie C. edule; en
algunos bancos los individuos recolectados pertenecían
a la nueva cohorte debido a que no existían individuos
mayores a causa de las mortalidades originadas por la
marteiliosis. Los resultados de ambos muestreos

CIMA Memoria 2016 47
mostraron que los berberechos C. edule de Noia, Muros
y Anllóns no estaban afectados por la enfermedad. En el
mes de junio, los berberechos de Cambados (O Sarrido)
y Moaña estaban afectados por M. cochillia, y los
berberechos C. glaucum estaban libres de este parásito.
En octubre-noviembre, los berberechos de Lombos do
Ulla, Combarro y A Pobra estaban afectados por la
marteliosis. En el caso de la ría de Noia, el programa de
vigilancia se desarrolló inicialmente con una frecuencia
de recolección de muestras y análisis mensual, debido a
que presenta la mayor producción de berberecho de
Galicia. Los resultados de este estudio señalan que los
berberechos de Noia no están afectados por este
parásito y se ha pasado a una frecuencia de muestreo
bimestral.
Otro de los objetivos contemplados en el proyecto es el
estudio de la dinámica temporal de la marteiliosis, ya que
conocer si existe un patrón predecible en el desarrollo de
la enfermedad permitirá una mejor gestión del recurso
dirigida a minimizar las pérdidas originadas por la misma.
Para ello, durante los años 2015 y 2016 se recogieron
mensualmente muestras de berberecho en dos
importantes bancos de la ría de Arousa: Lombos do Ulla,
banco mayoritariamente submareal situado en la zona
más interna de la ría y muy influenciado por el río Ulla, y
O Sarrido, banco intermareal situado en el interior de la
ensenada de O Grove en la desembocadura del río
Umia. El tamaño de muestra fue de 30 berberechos y de
cada individuo se realizaron cortes histológicos que
fueron analizados al microscopio óptico con el fin de
diagnosticar la presencia de Marteilia cochillia. Se
calculó la prevalencia mensual del parásito en cada
banco como el porcentaje de berberechos afectados por
Marteilia cochillia del total de analizados. Se evaluó,
asimismo, la mortalidad contabilizando, cada mes, el
número de berberechos supervivientes del total de
individuos que previamente se habían dispuesto en cajas
de control que se situaban enterradas en el substrato, en
el caso del banco intermareal de O Sarrido, y en una
estructura dispuesta sobre el fondo, que se recuperaba
con ayuda de una embarcación dotada de grúa, en el
caso del banco submareal de Lombos do Ulla. Los
resultados obtenidos hasta el momento indican una
dinámica de la enfermedad diferente en ambos bancos,
en lo que respecta al inicio de la infección y evolución de
la enfermedad. Así la enfermedad se detecta siempre
antes en el banco de Lombos do Ulla que en O Sarrido.
Además, la progresión de la prevalencia y la mortalidad
es también más rápida en Os Lombos ya que, tras la
detección de los primeros individuos infectados, el
porcentaje de berberechos enfermos se incrementa de
forma exponencial conduciendo al agotamiento del
recurso apenas unos meses después. En O Sarrido la
progresión suele ser más lenta y transcurre más tiempo
hasta la desaparición del recurso. Así, en octubre de
2015, se detectó un nuevo brote de M. cochillia en el
banco de Lombos do Ulla, que siguió un modelo de
desarrollo rápido al igual que los brotes de años
anteriores, de manera que la prevalencia y mortalidad
aumentaron progresivamente hasta provocar la extinción
de la cohorte de berberecho en abril de 2016. En Sarrido
la enfermedad no se detectó hasta febrero de 2016 y se
pudieron recoger muestras de individuos de esta cohorte
hasta junio de 2016, momento en el que quedó
prácticamente agotada. En julio de 2016 se detecta
fijación de berberecho en ambos bancos y se recogieron
muestras para monitorizar el estatus sanitario de esta
nueva cohorte de 2016. A principios de noviembre de
2016 se detectaron los primeros casos de infección por
M. cochillia en berberechos de Lombos do Ulla, y en
diciembre en O Sarrido. Durante el año 2017 se
continuará con la monitorización de ambas poblaciones.
La posible transmisión de M. cochillia entre berberechos,
Cerastoderma edule, fue otro de los objetivos del
proyecto. Se realizaron experimentos de transmisión de
la enfermedad por cohabitación entre individuos sanos y
enfermos en vasos de vidrio y en circuito abierto en el
laboratorio. Se dispusieron 20 berberechos del Sarrido
(banco afectado por marteliosis) y 20 berberechos de
Noia (banco libre de la enfermedad) en cada uno de 3
vasos. Al realizarse el experimento en circuito abierto la
enfermedad podía transmitirse a través del agua, y por
ello se dispusieron 40 berberechos de Noia en cada uno
de los 3 vasos que se usaron como control. Los
berberechos que se morían se eliminaban, y se hacían
cortes histológicos para evaluar la presencia del parásito.
El experimento finalizó a los 71 días de su inicio, y la
enfermedad no se transmitió entre los berberechos. En
los berberechos de los vasos control tampoco se
observó marteiliosis.
En la actualidad no se conoce el ciclo de vida completo
de las especies de Marteilia que afectan a los moluscos
bivalvos. Varios estudios postulan la existencia de un
ciclo de vida complejo en el que participarían
hospedadores intermediarios. Otro de los objetivos del
proyecto es la identificación de potenciales
hospedadores intermediarios en el ciclo biológico de M.
cochillia. Para este estudio, se realizaron arrastres

48
pelágicos empleando una red de Bongo de dos mangas
con malla de 200 µm de luz, para obtener muestras de
zooplancton en la columna de agua de 3 zonas de la ría
de Arousa: O Sarrido, Lombos do Ulla y Corón. En cada
transepto, la muestra procedente de una de las mangas
de la red de Bongo fue conservada en alcohol de 96%
para extracción de ADN, y la procedente de la otra
manga fue fijada en 10% de formaldehído en agua de
mar para el estudio histológico. Se equipó la red de
Bongo con un medidor de flujo de corriente para estimar
abundancias (nº individuos/m3) de la comunidad
mesozooplanctónica a partir del volumen de agua de mar
filtrado en cada transepto. Para la identificación de
especies se pipeteó un volumen concreto en una cámara
de recuento de zooplancton. Cuando resultó posible
cada individuo fue identificado hasta nivel de especie. En
el mes de febrero se pudo observar una clara
disminución de la abundancia de individuos en todas las
zonas de estudio. Por el contrario, en los meses de abril
y mayo de 2016 se observó un incremento bastante
elevado de la abundancia de individuos respecto al resto
de los meses. Este resultado es el esperado por entrada
de la primavera, época de reproducción de la mayoría de
las especies. Los taxones identificados se reagruparon
en dos categorías, holoplancton (organismos que
durante todo su ciclo vital forman parte de la comunidad
planctónica) y meroplancton (que está constituido por
seres que están en esta comunidad sólo durante una
parte de su vida, como pueden ser fases larvarias de
moluscos, crustáceos o peces). En las 3 zonas de
estudio, en la mayoría de los meses la abundancia de
holoplancton superó al meroplancton, y esto se debe en
mayor medida al grupo de copépodos que suele ser
dominante en todo el periodo. En el periodo de análisis,
comprendido desde noviembre de 2015 hasta octubre de
2016, se identificaron un total de 35 taxones. Las
especies de zooplancton separadas en cada muestreo
fueron analizadas para determinar la presencia de
Marteilia cochillia mediante pruebas moleculares. Se han
identificado varias especies del holoplancton como
posibles hospedadores intermediarios. Se realizarán
pruebas de hibridación “in situ” para confirmar estos
resultados.

CIMA Memoria 2016 49
ACCIÓN DE INVESTIGACIÓN EQUIPO
TÍTULO:
Evaluación del incremento de supervivencia en el
engorde de ostra rizada Crassostrea gigas por el uso
de semilla descendiente de supervivientes a la
exposición prolongada al virus herpes de ostreidos tipo
1. Identificación de marcadores moleculares de
resistencia para su aplicación en un programa de
selección de estirpes resistentes al virus.
INICIO: marzo, 2016
FINALIZACIÓN: diciembre, 2017
ENTIDAD FINANCIADORA:
Consellería del Mar. Xunta de Galicia.
Código: CIMA 16/02
INVESTIGADOR PRINCIPAL:
Villalba García, A.
INVESTIGADORES/AS:
Cao Hermida, M.A; Fuentes González, J.M.
APOYO A LA INVESTIGACIÓN:
Brianes Beiras, M.J.; Giráldez Rivero, R.; González
Fontela, A.I.; Gregorio Chenlo, M.V.; Meléndez Ramos,
M.I.; Penas Pampín, E.
BECARIOS/AS:
Alonso Costas, J.B.; Campañó Piñeiro, M.S.
RESUMEN
El engorde de la ostra rizada, Crassostrea gigas, en la
ría de Arousa se ve afectado desde hace al menos 6
años por una mortalidad muy alta de la semilla, causada
por la infección de la microvariante del virus herpes 1 de
ostreidos (OsHV1-µvar). Se trata de una enfermedad
detectada inicialmente en Francia, en el verano del 2008,
que se ha expandido a otros países europeos así como a
Australia, Nueva Zelanda y Corea del Sur. Nuestro grupo
se ha implicado en el estudio de esta enfermedad, en
particular en la búsqueda de procedimientos que
permitan aumentar la supervivencia de la ostra rizada en
el engorde. Otros autores han demostrado que la
temperatura del agua y la edad y el tamaño de la ostra
influyen en la dinámica de la enfermedad; por debajo de
16 ºC no hay mortalidad relevante y por encima de 24 ºC
la mortalidad se reduce significativamente; la
susceptibilidad de la ostra a la infección y mortalidad se
reduce notablemente en los individuos mayores, con
peso superior a 20 g. En un proyecto previo evaluamos
la eficacia de procedimientos alternativos de engorde
cuyo diseño se había basado en la información sobre la
influencia de la temperatura y la edad y tamaño de las
ostras en la mortalidad pero los resultados no fueron
satisfactorios pues la supervivencia fue baja. Teniendo
en cuenta que en Francia se habían obtenido resultados
muy esperanzadores en la lucha contra la infección por
OsHV1-µvar mediante programas de mejora genética
para aumentar la resistencia a la enfermedad, basados
en utilizar como reproductores ostras supervivientes en
zonas afectadas por la enfermedad, repitiéndose el
proceso en generaciones sucesivas, decidimos
desarrollar un nuevo proyecto de investigación para
evaluar si, también en la ría de Arousa, la utilización de
supervivientes de una zona afectada como reproductores
conduciría a un aumento de la supervivencia de sus
descendientes durante el engorde y, en tal caso, sentar
las bases para desarrollar un programa de selección
genética. Además, se planteó la búsqueda de
marcadores moleculares de resistencia a la enfermedad
que sirvan de apoyo a un ulterior programa de mejora
genética, mediante dos enfoques, uno proteómico,
desarrollado por nuestro grupo, y otro genómico, a
desarrollar por el equipo de la Dra. María Prado Álvarez,
investigadora del Instituto de Investigaciones Marinas
(CSIC, Vigo).
La primera etapa del proyecto implicó la producción de
varias familias de semilla de ostra rizada de dos orígenes
diferentes, (1) una población nunca afectada por el virus
y (2) supervivientes tras la exposición al virus en un área
afectada. Un banco naturalizado de ostra rizada en la ría
de O Barqueiro se utilizó como fuente de reproductores
adultos de una población nunca afectada, lo que se ha
ido confirmando por muestreos y análisis sucesivos
desde 2012 y ausencia de mortalidad anormal. Como
“supervivientes”, se utilizaron ostras de un lote producido
en un criadero industrial a partir de reproductores
recogidos en ese mismo banco, que se transfirió para su
engorde a una batea de la ría de Arousa muy afectada

50
por el virus, en diciembre de 2014, y que sufrió una
mortalidad muy alta en la primavera de 2015. En la
primavera de 2016, reproductores de ambos orígenes se
repartieron en dos criaderos, el de la Estación de
Ciencias Marinas de Toralla (ECIMAT-Universidad de
Vigo) y el del CIMA en Vilanova de Arousa, donde se
procedió a su acondicionamiento para acelerar la
gametogénesis, se obtuvieron gametos maduros
mediante disección, se posibilitó la fecundación y se
cultivaron las larvas resultantes. En el criadero de la
ECIMAT, el equipo de Damián Costas produjo familias
de semilla descendientes de 6 parejas distintas de ostras
recogidas en la ría de O Barqueiro y de 3 parejas
distintas de supervivientes de la ría de Arousa; las larvas
de cada cruzamiento se cultivaron por separado en todo
momento. Las limitaciones del criadero del CIMA
impedían mantener por separado las larvas
descendientes de cada cruzamiento por lo que en el
caso de los reproductores de la ría de O Barqueiro se
mezclaron espermatozoides de una ostra con ovocitos
de ocho ostras y, en el caso de los reproductores
supervivientes de la ría de Arousa, se mezclaron
espermatozoides de 4 ostras con los ovocitos de 13
individuos; las larvas resultantes de cada origen se
cultivaron por separado.
En septiembre de 2016, semillas de las 6 familias de
origen ría de O Barqueiro y las 3 familias descendientes
de supervivientes de la ría de Arousa producidas en
ECIMAT y de los lotes de cada uno de los dos orígenes
producidas en el criadero del CIMA se transfirieron a una
batea de la ría de Arousa para su engorde;
mensualmente se está evaluando la mortalidad de cada
familia y lote; el crecimiento se evalúa con muestreos
más distanciados. Además se han realizado análisis para
determinar si hay infección por OsHV1-µvar en
momentos diferentes, mediante diagnóstico molecular
(PCR) incluido en las directrices de la UE para la
vigilancia de esta enfermedad.
Con respecto a los resultados obtenidos en el engorde
de la semilla producida en el criadero de la ECIMAT, en
el momento de la transferencia a la batea (01/09/2016)
todos los individuos analizados resultaron negativos para
el diagnóstico de infección por OsHV1-µvar, mientras
que en el muestreo de finales de septiembre de 2016 se
detectaron casos positivos en todas las familias. En el
siguiente muestreo (31/10/2016), se detectaron tasas de
mortalidad elevadas. En el muestreo de diciembre de
2016 se detectaron diferencias significativas (test de χ2)
en la mortalidad acumulada entre los dos orígenes de
reproductores, siendo mayor en las familias derivadas de
reproductores del banco de la ría de O Barqueiro, nunca
afectado por el virus, que en las descendientes de
supervivientes a un episodio de mortalidad masiva por la
infección vírica en la ría de Arousa (Tabla 1). En el caso
del engorde de los lotes de semilla producidos en el
criadero del CIMA, no se detectó ningún caso de
diagnóstico positivo para OsHV1-µvar en el momento de
la transferencia a la batea (22/09/2016) mientras que en
el muestreo siguiente (31/10/2017) detectaron casos
positivos en ambos lotes coincidiendo con mortalidad
alta; la mortalidad acumulada registrada en el muestreo
de diciembre de 2016 fue significativamente mayor en el
lote de semilla derivado de reproductores recogidos en la
ría de O Barqueiro que en el de semilla descendiente de
supervivientes de la ría de Arousa (Tabla 2). El
seguimiento del engorde continuará a lo largo de 2017
hasta que las ostras alcancen tamaño comercial.
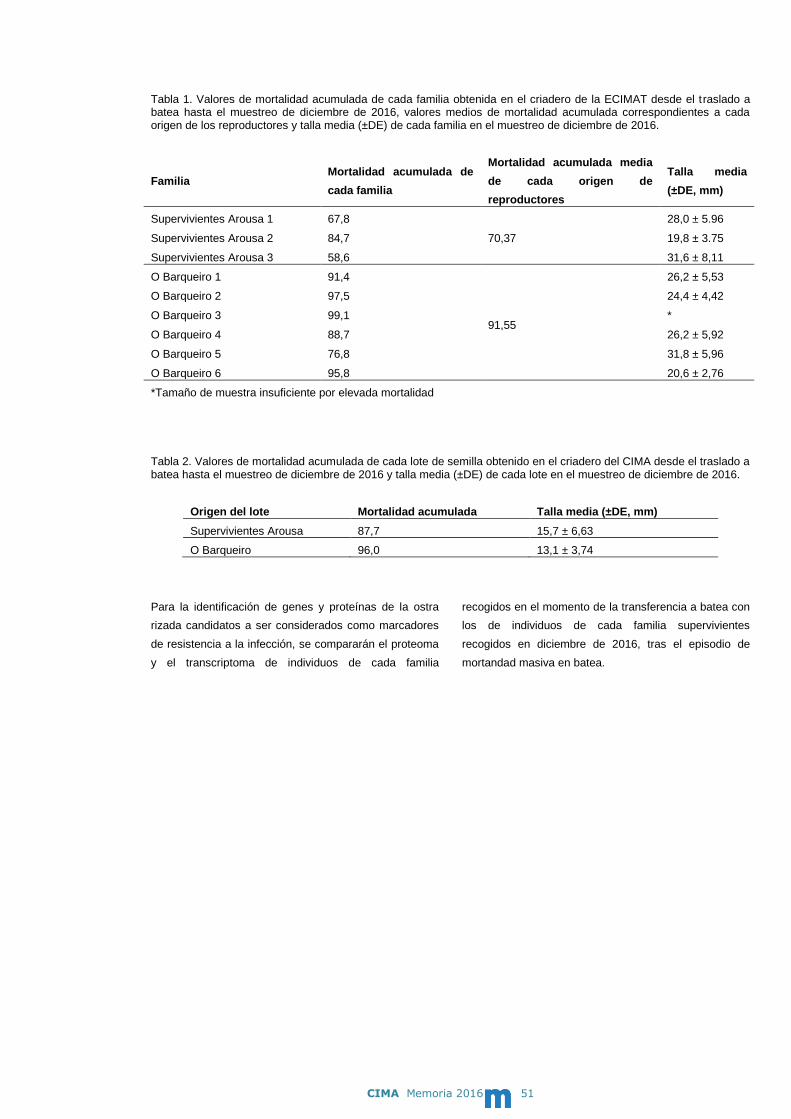
CIMA Memoria 2016 51
Tabla 1. Valores de mortalidad acumulada de cada familia obtenida en el criadero de la ECIMAT desde el traslado a batea hasta el muestreo de diciembre de 2016, valores medios de mortalidad acumulada correspondientes a cada origen de los reproductores y talla media (±DE) de cada familia en el muestreo de diciembre de 2016.
Familia Mortalidad acumulada de
cada familia
Mortalidad acumulada media
de cada origen de
reproductores
Talla media
(±DE, mm)
Supervivientes Arousa 1 67,8
70,37
28,0 ± 5.96
Supervivientes Arousa 2 84,7 19,8 ± 3.75
Supervivientes Arousa 3 58,6 31,6 ± 8,11
O Barqueiro 1 91,4
91,55
26,2 ± 5,53
O Barqueiro 2 97,5 24,4 ± 4,42
O Barqueiro 3 99,1 *
O Barqueiro 4 88,7 26,2 ± 5,92
O Barqueiro 5 76,8 31,8 ± 5,96
O Barqueiro 6 95,8 20,6 ± 2,76
*Tamaño de muestra insuficiente por elevada mortalidad
Tabla 2. Valores de mortalidad acumulada de cada lote de semilla obtenido en el criadero del CIMA desde el traslado a batea hasta el muestreo de diciembre de 2016 y talla media (±DE) de cada lote en el muestreo de diciembre de 2016.
Origen del lote Mortalidad acumulada Talla media (±DE, mm)
Supervivientes Arousa 87,7 15,7 ± 6,63
O Barqueiro 96,0 13,1 ± 3,74
Para la identificación de genes y proteínas de la ostra
rizada candidatos a ser considerados como marcadores
de resistencia a la infección, se compararán el proteoma
y el transcriptoma de individuos de cada familia
recogidos en el momento de la transferencia a batea con
los de individuos de cada familia supervivientes
recogidos en diciembre de 2016, tras el episodio de
mortandad masiva en batea.

52
7 PROCESOS OCEANOGRÁFICOS COSTEROS

CIMA Memoria 2016 53
ACCIÓN DE INVESTIGACIÓN EQUIPO
TÍTULO:
Desarrollo y optimización de métodos de determinación
de toxinas de tipo paralizante (PSP) y lipofílicas, en
moluscos bivalvos para maximizar el flujo de trabajo.
Acrónimo: DeMeTox
INICIO: marzo, 2016
FINALIZACIÓN: marzo, 2019
ENTIDAD FINANCIADORA:
Consellería del Mar. Xunta de Galicia.
INVESTIGADOR PRINCIPAL:
Blanco Pérez, J.C.
INVESTIGADORES/AS:
INTECMAR: Arévalo Bartolomé, F.; Moroño Mariño,
A.; Correa Jorge, J.
APOYO A LA INVESTIGACIÓN:
CIMA: Mariño Cadarso, C.; Martín Sánchez, H.
INTECMAR: Muñiz Romero, S.
RESUMEN
Los controles de la toxicidad de tipo paralizante (PSP) y
de tipo lipofílico se llevan a cabo en la actualidad por
medio de bioensayo con ratón y LC-MS/MS,
respectivamente. Con todo, es inminente la adopción de
un método de cromatografía líquida de alta eficacia
(HPLC), con oxidación precolumna y detección de
fluorescencia, como referencia en la UE para la
determinación de la toxicidad PSP, substituyendo al
bioensayo de ratón, lo cual hace necesaria su adopción
en el sistema de control de la Comunidad Autónoma de
Galicia.
Ambas metodologías analíticas (HPLC y LC-MS/MS)
presentan problemas cuando se implementan en
sistemas de control con una gran carga de trabajo, como
es el caso del Intecmar. Dado que requieren tiempos de
preparación y análisis muy largos, en el primer caso, el
mantenimiento muy frecuente de los equipos, en el
segundo, resulta necesario modificar los métodos
empleados para que su utilización en el sistema de
control sea factible y ágil. En la acción se pretenden
desarrollar las modificaciones necesarias para optimizar
la respuesta del Intecmar.
Tal como estaba previsto, la ejecución del proyecto de
investigación DEMETOX comenzó en el mes de mayo
del año en curso, teniendo realizadas 5 de sus tareas
con resultados parciales de considerable interés.
La primera de las actividades desarrolladas consistió en
evaluar diferentes columnas cromatográficas, para
separar los compuestos a analizar y seleccionar las
mejores para acelerar al máximo el proceso analítico,
con objeto de encontrar una columna cromatográfica que
permita el análisis rápido de las toxinas PSP tras su
oxidación; que pueda ser usada para el control rutinario
de la toxicidad en los moluscos bivalvos; y que tenga una
química igual o similar a la de la columna de referencia
del método estándar, de manera que su utilización pueda
considerarse una optimización del mismo y no un nuevo
método.
Como resumen del trabajo desarrollado, se concluyó que
las columnas de mayor interés son la Atlantis T3 y más la
Poroshell EC18 4µm. Con cualquiera de las dos podrían
analizarse todos los derivados de las toxinas en
cromatogramas de 6 minutos o menos, no siendo
esperable que las columnas tengan poca duración, ya
que el tamaño de partícula es igual o superior a 4µm.
Con la misma química que la Atlantis T3 pero con
partículas de núcleo sólido, se consiguieron excelentes
resultados con una columna Cortecs T3, reduciéndose
la duración de los cromatogramas a 2,5 minutos, aunque
queda por evaluar la duración de este tipo de columnas
en el sistema de control.
El intento de reducción de la complejidad del conjunto de
toxinas a analizar, por medio de la reducción de las
toxinas N-OH con ácido ascórbico en presencia de

54
hemina, fue la segunda de las actividades realizadas en
el segundo semestre de 2016 y, tras los trabajos hechos,
se consideró que esta aproximación no es viable para
su uso en el sistema de control, ya que la presencia de
ácido ascórbico afecta a la necesaria oxidación de las
toxinas y su eliminación completa resulta laboriosa.
La tercera actividad abordada fue la evaluación de la
limpieza por extracción en fase sólida, con diferentes
cartuchos de fases C18 y poliméricas, llegándose a la
conclusión que, en contra de lo comentado en los pocos
trabajos que lo tratan, la mayor parte de los cartuchos
con fases C18 fueron más eficaces para la limpieza, que
los que contenían fases poliméricas, por lo que es
preferible no substituir los cartuchos originales.
La actividad desarrollada en cuarto lugar consistió en la
evaluación de la posibilidad de substituir la limpieza por
extracción en fase sólida con cartuchos, por una
equivalente (con las mismas fases), pero en modo
dispersivo. El método dio malos resultados, siendo
similares para todas las fases probadas.
Se concluyó, por tanto, que la SPE dispersiva no puede
ser utilizada para substituir a la SPE en cartucho, para la
limpieza de los extractos de toxinas PSP.
La quinta de las actividades realizadas se centró en
buscar un método para reducir el número de inyecciones
requeridas para el control de las principales toxinas
lipofílicas. En la experiencia, se comprobó la viabilidad
del método, modificando las condiciones en las que se
inyecta la muestra (volumen y dilución con agua),
encontrándose que, tanto con inyecciones por debajo de
3 µL, como diluyendo las muestras al 50% con agua, es
posible concentrar en un cartucho por lo menos 10
muestras, sin una pérdida apreciable de toxina. El
método puede emplearse, por tanto, para hacer un
cribado de muestras en las que es de esperar que la
concentración de ácido okadaico sea muy baja, sin tener
que inyectar cada muestra individualmente.

CIMA Memoria 2016 55
ACCIÓN DE INVESTIGACIÓN EQUIPO
TÍTULO:
Efecto de las biotransformaciones de toxinas de tipo
diarreico (DSP) en organismos del intermareal sobre
las diferencias de toxicidad estimada por LC-MS/MS y
bioensayo de ratón y sus repercusiones para los
cierres de explotación y la salud humana.
Acrónimo: ESTINFA
INICIO: mayo, 2013
FINALIZACIÓN: mayo, 2016
ENTIDAD FINANCIADORA:
Consellería del Mar. Xunta de Galicia.
INVESTIGADOR PRINCIPAL:
Blanco Pérez, J.C.
INVESTIGADORES/AS:
INTECMAR: Arévalo Bartolomé, F.; Moroño Mariño,
A.; Correa Jorge, J.
APOYO A LA INVESTIGACIÓN:
CIMA: Mariño Cadarso, C.; Martín Sánchez, H.
INTECMAR: Muñiz Romero, S.
RESUMEN
Esta acción de investigación surge de la aparición de
algunas discrepancias, especialmente en fases tardías
de los episodios de toxicidad DSP, entre la estimación de
la toxicidad por bioensayo de ratón y por HPLC-MS/MS,
en el sentido de que la última de estas técnicas estima
valores superiores. Para clarificar esta situación y
evaluar su impacto, se plantean los siguientes objetivos:
1.- Evaluación de la extensión del problema
identificando en qué especies se presenta.
2.- Identificación de los compuestos
responsables.
3.- Evaluación de la liberación de toxinas de los
compuestos responsables durante un proceso
simulado de digestión humana.
Se obtuvieron muestras de moluscos del intermareal (y
adicionalmente volandeira) expuestos a toxinas DSP, en
la medida de lo posible en varios momentos del ciclo de
intoxicación/desintoxicación. Se obtuvieron más de 60
muestras que comprenden diversas almejas (fina,
babosa, japonesa, rubia), navajas (navaja, longueirón),
berberecho y volandeira. En todas ellas se estimó la
toxicidad DSP por medio de bioensayo de ratón y por
HPLC MS-MS. En 8 de las muestras analizadas las
estimaciones obtenidas por LC-MS/MS fueron superiores
a las correspondientes por bioensayo de ratón (estimado
de forma semicuantitativa ya que el bioensayo de ratón
no permite una cuantificación precisa). Los resultados
fueron equivalentes para las dos técnicas en volandeira
pero no para algunas muestras de berberecho, almeja
(varias especies) y navaja, sin que las diferencias se
localizasen en una especie concreta.
El análisis de los extractos discrepantes por medio de
LC-MS/MS en modo “parent scan” mostró la presencia
de un compuesto que podría ser un derivado del ácido
okadaico y un azúcar, pero cuya identidad no pudo
confirmarse.
El fraccionamiento de algunos extractos por tamaños
moleculares por medio de cromatografía de exclusión
molecular con Sephadex LH20, mostró que existen
compuestos de mayor masa molecular que las toxinas
libres y que los acil-derivados que por hidrólisis alcalina
liberan las toxinas libres. Probablemente estos
compuestos sean los responsables de las discrepancias,
pero su identificación inequívoca no fue posible hasta el
momento.

56
ACCIÓN DE INVESTIGACIÓN EQUIPO
TÍTULO:
Identificación de toxinas DSP no detectadas por
LC_MS/MS en mejillón en fresco y en escabeche,
biodisponibilidad de estos compuestos, acumulación y
distribución anatómica en el mejillón.
Acrónimo: TOXNODE
INICIO: enero, 2015
FINALIZACIÓN: diciembre, 2016
ENTIDAD FINANCIADORA:
Consellería del Mar. Xunta de Galicia.
INVESTIGADOR PRINCIPAL:
Blanco Pérez, J.C.
INVESTIGADORES/AS:
INTECMAR: Arévalo Bartolomé, F.; Moroño Mariño,
A.; Correa Jorge, J.
APOYO A LA INVESTIGACIÓN:
CIMA: Mariño Cadarso, C.; Martín Sánchez, H.
INTECMAR: Muñiz Romero, S.
RESUMEN
Un estudio centrado en las discrepancias entre técnicas
de cuantificación de la toxicidad DSP, durante el
procesado industrial del mejillón expuesto durante
períodos prolongados a toxinas de este tipo, por un lado,
mostró que se libera ácido okadaico y DTX2 a partir de
compuestos no identificados con el proceso de cocción
industrial y, por otro, sugirió que en el proceso de
enlatado/esterilización en escabeche, se generan
derivados de compuestos con hidroxilos que son
esterificados por el grupo carboxilo de las toxinas.
Tanto la presencia de los compuestos comentados como
la distribución anatómica de las toxinas pueden tener
importantes repercusiones tanto para la industria como
para la salud pública ya que distintas técnicas de
detección de la toxicidad DSP responden de forma
diferente a la habitual en situaciones en las que los
compuestos citados están presentes (o tienen una
contribución importante a la cantidad total de toxina) y/o
en las que un porcentaje elevado de la toxina se halla
fuera de la glándula digestiva.
En el estudio comentado no fue posible identificar los
compuestos implicados, cuya presencia, parece
relacionada con el tiempo de exposición de los bivalvos a
las toxinas. Los compuestos que parecen dar poca
respuesta en el bioensayo en ratón, y que se encuentran
en los procesados en escabeche, parecen, sin embargo
relacionados con el proceso de enlatado/esterilización.
Dado que la presencia de los compuestos comentados
genera inconsistencias en las determinaciones de
toxicidad y que tanto su identidad como su
biodisponibilidad se desconocen, se pretende con este
estudio obtener información sobre esos aspectos y
desarrollar técnicas que permitan analizar el contenido
total de toxinas de forma fiable, independientemente de
que estos compuestos estén presentes o no, y que
permitan además el análisis de los compuestos en
cuestión.
Hasta el momento se han realizado diversos
experimentos y se han recogido muestras para su
análisis posterior.
Las digestiones enzimáticas con amilasa, alfa - y beta -
glucosidasa y glucuronidasa, proteinasa K, proteasa,
esterasa y lipasa, no dieron resultados consistentes en
ningún caso y no se constató una liberación significativa
de ácido okadaico con estos tratamientos.
Las extracciones, tanto con solventes más apolares,
como con asistencia por microondas no mejoraron
significativamente la extracción del ácido okadaico.
El estudio se vio adicionalmente complicado porque,
aparentemente, la extracción de los compuestos de
interés después del tratamiento industrial (no
reproducible en nuestro laboratorio) es mucho mejor que
después de un proceso de cocción más lento (hervido o
autoclavado), con lo que resulta difícil identificar nuevo
material que presente el problema estudiado.

CIMA Memoria 2016 57
8 RECURSOS MARINOS

58
ACCIÓN DE INVESTIGACIÓN EQUIPO
TÍTULO:
Puesta a punto de las técnicas de cultivo en
minicriadero de las especies del género Cerastoderma
de Galicia.
INICIO: junio, 2014
FINALIZACIÓN: diciembre, 2016
ENTIDAD FINANCIADORA:
Consellería del Mar. Xunta de Galicia.
Código: PGIDIT-CIMA 14/03
INVESTIGADOR PRINCIPAL:
Fuentes González, J.M.
INVESTIGADOR:
Villalba García, A.
INVESTIGADORES COLABORADORES/AS:
Iglesias Estepa, D.: Mariño Balsa, C.
APOYO A LA INVESTIGACIÓN:
Giráldez Rivero, R.; Gregorio Chenlo, M.V.
RESUMEN
En Galicia coexisten dos especies de berberecho:
Cerastoderma edule y Cerastoderma glaucum. Las
poblaciones naturales de C. edule están tradicionalmente
afectadas por episodios recurrentes de mortalidad
(Villalba et al., 2001), de los cuales siempre se
recuperaron con rapidez. Sin embargo, a partir del año
2012, gran parte de las poblaciones de C. edule de las
rías gallegas están siendo diezmadas por el parásito
Marteilia cochillia, (Villalba et al., 2014), sin que las
poblaciones de C. glaucum se vean afectadas. La lucha
contra este tipo de enfermedades en el medio natural
usando agentes terapéuticos es compleja. Una forma
diferente de lucha es la obtención de estirpes tolerantes
a la enfermedad mediante programas de selección
genética (da Silva et al., 2005; Fuentes y Villalba, 2007).
Para la obtención de estas estirpes, es necesario
dominar las técnicas de cultivo en criadero, técnicas que
todavía están poco desarrolladas para estas especies de
berberecho (Pronker et al., 2013).
Una vez realizada la puesta a punto de las técnicas
básicas de cultivo en minicriadero (acondicionamiento y
cultivo larvario) desde finales de 2014 a inicios del 2015,
obtuvimos durante el primer semestre del año 2015
varios cultivos de Cerastoderma edule (CE) que
alcanzaron la fase de semilla. Estos cultivos fueron pre-
engordados en condiciones de cautividad para más tarde
ser engordados en batea. El objetivo de este engorde fue
comprobar si es posible acelerar el crecimiento de los
berberechos en cultivo suspendido y obtener así
individuos de talla comercial antes de que la aparición
otoñal de la Marteiliosis diezme las poblaciones
cultivadas.
En la Figura 1 mostramos la evolución de la longitud
media (L, máxima distancia antero-posterior, símbolos
rojos) de los individuos de varios lotes de berberecho
Cerastoderma edule (CE) producidos en criadero y
engordados en batea desde el verano del año 2015
hasta el invierno de 2016. Como puede observarse, en
aquellos lotes cuyo engorde se inició más temprano
(02/07/2015, CE-S-F1 y CE-N-F1), casi el 100% de los
individuos superaron la talla mínima comercial (25 mm;
línea de trazos rojos de la gráfica) en el mes de octubre
del mismo año (20/10/2015), antes de que las
mortalidades más intensas debido a la Marteiliosis
tuvieran lugar durante el otoño del 2015 y el invierno de
2016 (símbolos negros en Figura 2). Sin embargo,
cuando el engorde se inició algo más tarde (18/08/2015,
CE-STC-F1 en Figura 1), apenas la mitad de los
individuos llegaron a alcanzar la talla mínima comercial
(25 mm) y lo hicieron varios meses más tarde
(19/01/2016) cuando las mortalidades acumuladas
superaron ya el 40% (CE-STC-F1, triángulos negros en
Figura 2).
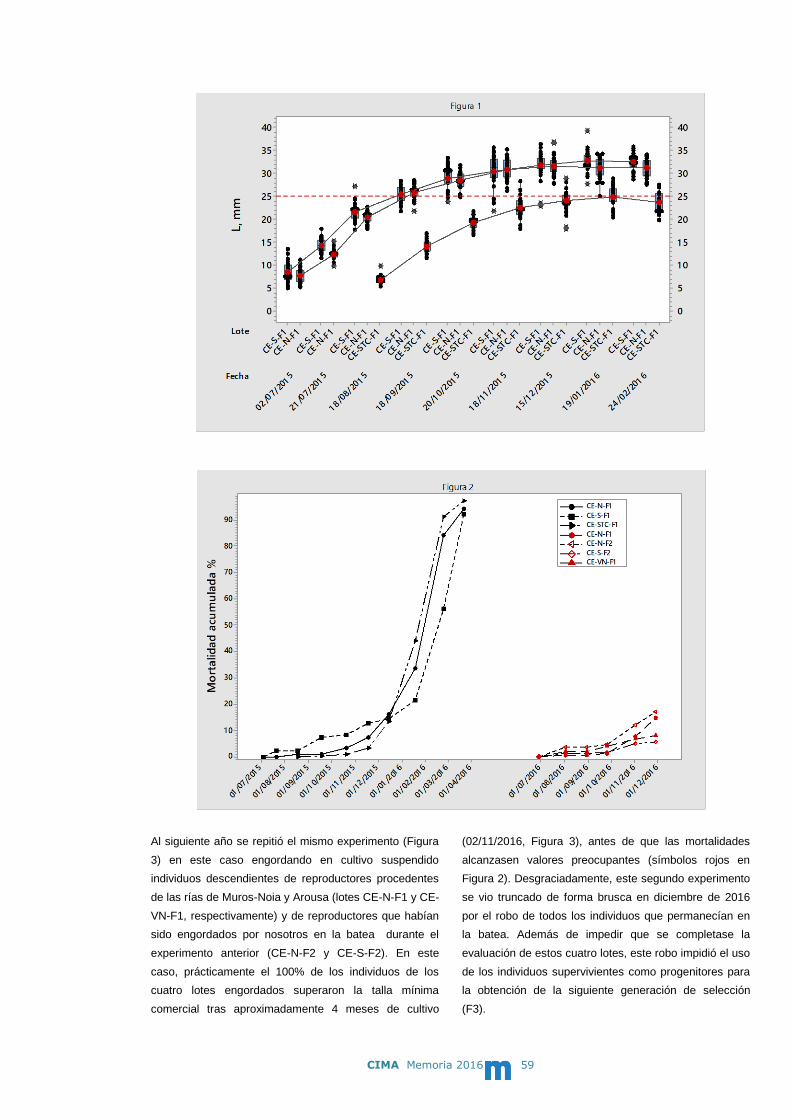
CIMA Memoria 2016 59
Al siguiente año se repitió el mismo experimento (Figura
3) en este caso engordando en cultivo suspendido
individuos descendientes de reproductores procedentes
de las rías de Muros-Noia y Arousa (lotes CE-N-F1 y CE-
VN-F1, respectivamente) y de reproductores que habían
sido engordados por nosotros en la batea durante el
experimento anterior (CE-N-F2 y CE-S-F2). En este
caso, prácticamente el 100% de los individuos de los
cuatro lotes engordados superaron la talla mínima
comercial tras aproximadamente 4 meses de cultivo
(02/11/2016, Figura 3), antes de que las mortalidades
alcanzasen valores preocupantes (símbolos rojos en
Figura 2). Desgraciadamente, este segundo experimento
se vio truncado de forma brusca en diciembre de 2016
por el robo de todos los individuos que permanecían en
la batea. Además de impedir que se completase la
evaluación de estos cuatro lotes, este robo impidió el uso
de los individuos supervivientes como progenitores para
la obtención de la siguiente generación de selección
(F3).
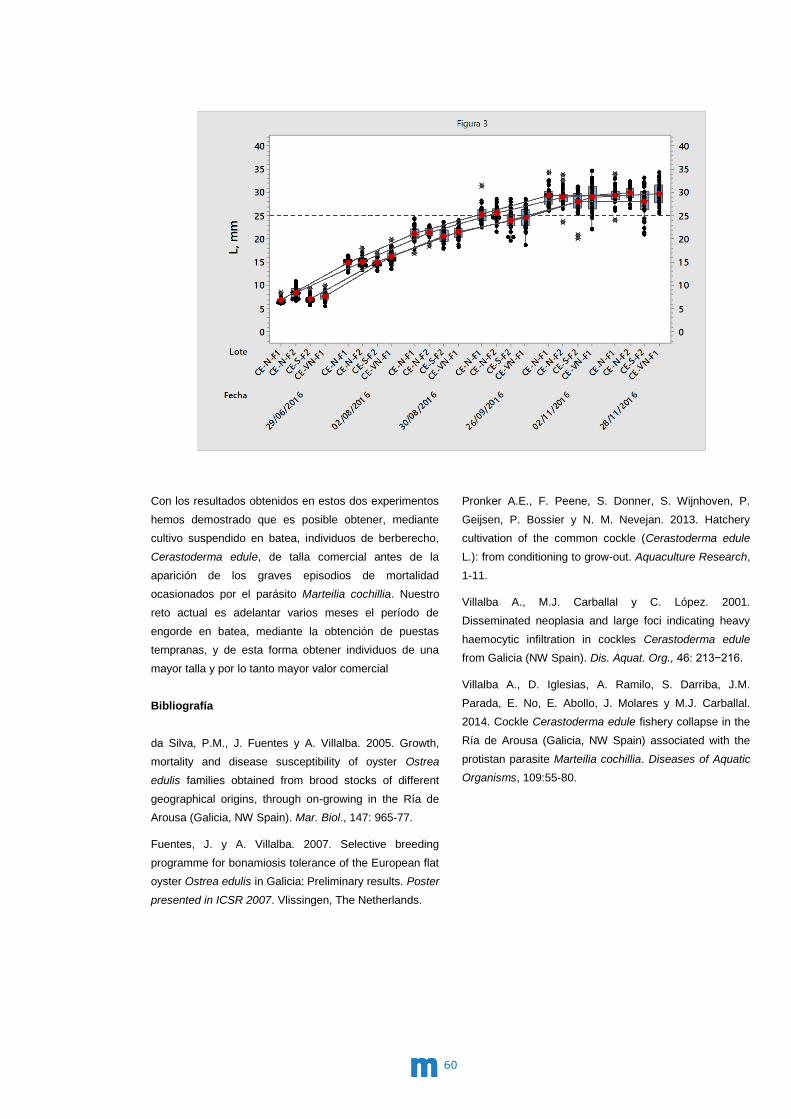
60
Con los resultados obtenidos en estos dos experimentos
hemos demostrado que es posible obtener, mediante
cultivo suspendido en batea, individuos de berberecho,
Cerastoderma edule, de talla comercial antes de la
aparición de los graves episodios de mortalidad
ocasionados por el parásito Marteilia cochillia. Nuestro
reto actual es adelantar varios meses el período de
engorde en batea, mediante la obtención de puestas
tempranas, y de esta forma obtener individuos de una
mayor talla y por lo tanto mayor valor comercial
Bibliografía
da Silva, P.M., J. Fuentes y A. Villalba. 2005. Growth,
mortality and disease susceptibility of oyster Ostrea
edulis families obtained from brood stocks of different
geographical origins, through on-growing in the Ría de
Arousa (Galicia, NW Spain). Mar. Biol., 147: 965-77.
Fuentes, J. y A. Villalba. 2007. Selective breeding
programme for bonamiosis tolerance of the European flat
oyster Ostrea edulis in Galicia: Preliminary results. Poster
presented in ICSR 2007. Vlissingen, The Netherlands.
Pronker A.E., F. Peene, S. Donner, S. Wijnhoven, P.
Geijsen, P. Bossier y N. M. Nevejan. 2013. Hatchery
cultivation of the common cockle (Cerastoderma edule
L.): from conditioning to grow-out. Aquaculture Research,
1-11.
Villalba A., M.J. Carballal y C. López. 2001.
Disseminated neoplasia and large foci indicating heavy
haemocytic infiltration in cockles Cerastoderma edule
from Galicia (NW Spain). Dis. Aquat. Org., 46: 213−216.
Villalba A., D. Iglesias, A. Ramilo, S. Darriba, J.M.
Parada, E. No, E. Abollo, J. Molares y M.J. Carballal.
2014. Cockle Cerastoderma edule fishery collapse in the
Ría de Arousa (Galicia, NW Spain) associated with the
protistan parasite Marteilia cochillia. Diseases of Aquatic
Organisms, 109:55-80.

CIMA Memoria 2016 61
ACCIÓN DE INVESTIGACIÓN EQUIPO
TÍTULO:
Evaluación y seguimiento de las Reservas Marinas de
Interés Pesquero de Galicia.
Acrónimo: ESCAMPE 2
INICIO: septiembre, 2014
FINALIZACIÓN: septiembre, 2017
ENTIDAD FINANCIADORA:
Consellería del Mar. Xunta de Galicia.
Código: PGIDIT-CIMA 14/05
INVESTIGADOR PRINCIPAL:
Cuiñas Olmedo, P.
INVESTIGADOR:
Febrero Mayor, F.
RESUMEN
Las Reservas Marinas de Interés Pesquero (Rmip) son
figuras administrativas creadas para asegurar la
conservación, la protección y la gestión sostenible de
espacios naturales en los cuales se lleva a cabo la
explotación pesquera y marisquera de recursos naturales
marinos.
En la actualidad en la Comunidad Autónoma de Galicia
existen dos Reservas Marinas de Interés Pesquero
(Rmip): “Os Miñarzos”, creada en el año 2007 y “Ría de
Cedeira” creada en el año 2009. Cada una de estas
Rmip cuenta con un “Órgano de Gestión, Seguimiento y
Control” (Oxesco) constituído por representantes de la
Administración Pesquera de la Xunta de Galicia y
representantes de las Organizaciones Sectoriales de
Pesca.
Esta acción pretende cubrir las necesidades de apoyo
científico-técnico que se requieren en la gestión de las
dos RMIP existentes en Galicia.
Dentro de esta Acción de Investigación, durante el año
2016, se llevaron a cabo actividades de actualización de
la información recogida el año anterior:
Actualización bibliográfica sobre la temática del
seguimiento científico-técnico de áreas marinas
protegidas (AMPs).
Selección de los indicadores útiles para la evaluación de
la evolución de la actividad pesquera, mensual (para
cada campaña) y anual (para comparar campañas).
Entre ellos, con los datos disponibles actualmente,
hemos seleccionado los siguientes:
- Esfuerzo pesquero: presenta la evolución del número
de embarcaciones/día explotando los recursos mediante
las distintas artes.
- Evolución temporal del volumen de capturas
acumuladas (kg) por especie y arte.
- Capturas por unidad de esfuerzo (CPUE) tomando
como unidad de esfuerzo la embarcación, aunque a
partir del año 2012 puede utilizarse también el tripulante
como unidad de esfuerzo.
- Evolución de los precios de primera venta.
- Número medio de especies/día capturadas por las
artes menos selectivas (enmalle y anzuelo).
- Número total de especies que son capturadas por las
distintas artes así como su importancia relativa en cuanto
a capturas.
Dentro de la actividad de apoyo Científico-Técnico la
Gestión de las RMIPs y como respuesta a la
preocupación del Órgano de Gestión, Seguimiento y
Control de la RMIP “Os Miñarzos” por el estado de
conservación de las poblaciones de erizo, que son
explotadas mediante marisqueo a flote dentro de la
reserva, se llevó a cabo la evaluación de las poblaciones
submareales de erizo existentes en la RMIP “Os
Miñarzos” de Lira, con el objetivo de obtener la
información en la que basar la adopción de las medidas
de gestión necesarias para asegurar la explotación
sostenible del recurso.

62
El estudio del estado de la pesquería de marisqueo a
flote de erizo (Paracentrotus lividus, Lamarck 1816)
dentro de la reserva se concentró en la elaboración de
un informe que se resume a continuación:
1.- Evolución histórica de las capturas
comercializadas a través de la lonja de Lira entre los
años 2001 a 2016.
Se observa una tendencia a la disminución de las ventas
en la lonja de Lira, principalmente desde el año 2008
hasta hoy (datos de la plataforma tecnológica
Pescadegalicia).
2.- Delimitación del área a evaluar (Figura 1).
Corresponde a la zona costera submareal de la RMIP
“Os Miñarzos” que es la considerada por la OXESCO de
la RMIP, como zona a vedar temporalmente, debido a la
información suministrada por los mariscadores de
recursos específicos de Lira sobre la escasez de recurso
de talla comercial.
Figura 1. Área del muestreo.
3.- Metodología del muestreo de las poblaciones de
erizo.
Para la evaluación del stock de erizo presente en la
zona, se realizó un muestreo estratificado, con dos
estratos muy diferenciados, uno de ellos que presenta,
en general, baja densidad de individuos, por lo que la
denominaremos “ZBD” (Zona de Baja Densidad), el otro
estrato, que presenta relativamente altas densidades que
denominamos “ZA” (Zona Agregados). El muestreo
consistió, por una parte en un censo visual, llevado a
cabo mediante la realización de 25 transeptos de 100
metros de longitud, distribuidos por toda el área de
estudio, y por otra, en un muestreo extractivo en las dos
zonas diferenciadas por su densidad de población, para
el análisis de la estructura de tallas.
4.- Resultados.
4.1 Distribución espacial de la población de erizo
Los valores de densidades de erizo encontradas en esos
tramos de 5 metros evidencian que la clase de 0
individuos/m2 es muy mayoritaria (61% de los tramos) y
que la densidad de individuos, en general para toda el
área, es baja presentándose los máximos en la “ZA”
(Estrato de Agregados) con un valor máximo de 9,7
individuos/m2 y con sólo 6 observaciones por encima de
los 5 individuos/m2 que representan el 1% del total de
casos en toda el área de estudio.
4.2 Estructura de población (tallas)
La distribución de tallas es bastante similar en los dos
estratos contemplados.
Los valores medios de las tallas registradas fueron de
52,1 mm para la zona de “baja densidad” (con un 32%
de individuos de talla comercial) y 50,7 mm para la zona
de “Agregados” (con un 29% de individuos de talla
comercial).
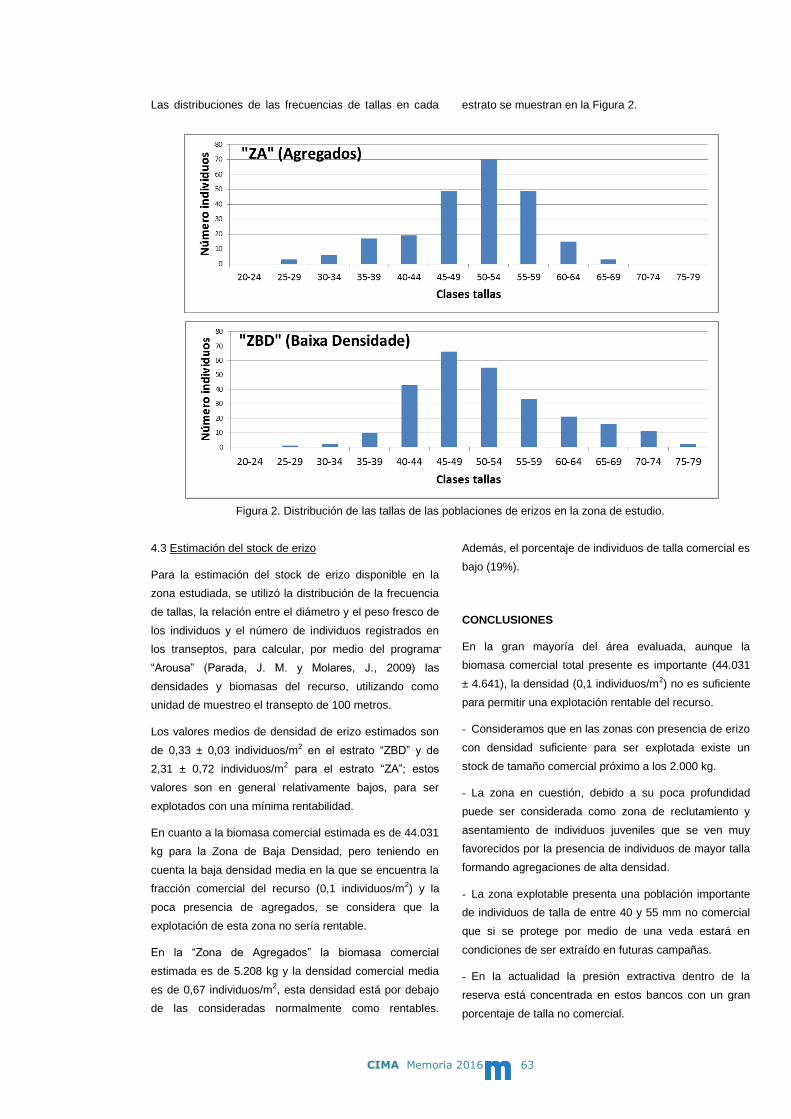
CIMA Memoria 2016 63
Las distribuciones de las frecuencias de tallas en cada estrato se muestran en la Figura 2.
Figura 2. Distribución de las tallas de las poblaciones de erizos en la zona de estudio.
4.3 Estimación del stock de erizo
Para la estimación del stock de erizo disponible en la
zona estudiada, se utilizó la distribución de la frecuencia
de tallas, la relación entre el diámetro y el peso fresco de
los individuos y el número de individuos registrados en
los transeptos, para calcular, por medio del programa
“Arousa” (Parada, J. M. y Molares, J., 2009) las
densidades y biomasas del recurso, utilizando como
unidad de muestreo el transepto de 100 metros.
Los valores medios de densidad de erizo estimados son
de 0,33 ± 0,03 individuos/m2 en el estrato “ZBD” y de
2,31 ± 0,72 individuos/m2 para el estrato “ZA”; estos
valores son en general relativamente bajos, para ser
explotados con una mínima rentabilidad.
En cuanto a la biomasa comercial estimada es de 44.031
kg para la Zona de Baja Densidad, pero teniendo en
cuenta la baja densidad media en la que se encuentra la
fracción comercial del recurso (0,1 individuos/m2) y la
poca presencia de agregados, se considera que la
explotación de esta zona no sería rentable.
En la “Zona de Agregados” la biomasa comercial
estimada es de 5.208 kg y la densidad comercial media
es de 0,67 individuos/m2, esta densidad está por debajo
de las consideradas normalmente como rentables.
Además, el porcentaje de individuos de talla comercial es
bajo (19%).
CONCLUSIONES
- En la gran mayoría del área evaluada, aunque la
biomasa comercial total presente es importante (44.031
± 4.641), la densidad (0,1 individuos/m2) no es suficiente
para permitir una explotación rentable del recurso.
- Consideramos que en las zonas con presencia de erizo
con densidad suficiente para ser explotada existe un
stock de tamaño comercial próximo a los 2.000 kg.
- La zona en cuestión, debido a su poca profundidad
puede ser considerada como zona de reclutamiento y
asentamiento de individuos juveniles que se ven muy
favorecidos por la presencia de individuos de mayor talla
formando agregaciones de alta densidad.
- La zona explotable presenta una población importante
de individuos de talla de entre 40 y 55 mm no comercial
que si se protege por medio de una veda estará en
condiciones de ser extraído en futuras campañas.
- En la actualidad la presión extractiva dentro de la
reserva está concentrada en estos bancos con un gran
porcentaje de talla no comercial.

64
RESUMEN
Esta Acción es la continuación de los estudios realizados
por diversos equipos de investigación que, con el
objetivo principal de recuperación de la coquina (Donax
trunculus Linnaeus, 1758) en los bancos naturales de
Galicia, unieron sus investigaciones en una Acción que
comenzó en el año 2013: "Estudio de los bancos
naturales de coquina: problemática de su desaparición
en Galicia".
En esta nueva Acción se determinarán las causas
naturales que están afectando a los bancos de Donax
sp. y los efectos que sobre ellos tiene la actividad
humana de diferentes orígenes (industrial, de la
agricultura, minería, construcción, urbanización o
deforestación entre otros). Las causas naturales pueden
estar vinculadas a la naturaleza de los sedimentos, a la
topografía de las playas y a sus cambios por efectos
ambientales y antropológicos, a diferentes toxicidades, a
ACCIÓN DE INVESTIGACIÓN EQUIPO
TÍTULO:
Determinación de las causas de la disminución de los
bancos de coquina. Condicionantes naturales y
antropogénicos.
INICIO: junio, 2016
FINALIZACIÓN: diciembre, 2018
ENTIDAD FINANCIADORA:
Consellería del Mar. Xunta de Galicia.
Código: CIMA 16/06
COORDINADORA:
Martínez Patiño, D.
INVESTIGADORES/AS:
CIMA: Febrero Mayor, F.; Montes Pérez, J.; Nóvoa
Vázquez, S.; Ojea Martínez, J.
UNI VIGO: Alejo Flores, I.; Diz Ferreiro, P.; Nombela
Castaño, M.A.
CSIC: Álvarez Salgado, X.A.; Otero Villar, J.
Xefatura Territorial de Celeiro. Consellería del Mar:
Cerdeira Arias, D.; Teijeiro Golpe, O.
Xefatura Territorial de Vigo. Consellería del Mar:
Rodríguez González, L.M.
Xefatura Comarcal de Ferrol. Consellería del Mar:
Brezmes Comesaña, C.; Liñán Gil, I.M.
Xefatura Territorial da Coruña. Consellería del Mar:
Catoira Gómez, J.L.
USC: Barja Pérez, J.L.; Olveira Hermida, J.G.; Prado
Plana, S.
ASISTENCIAS TÉCNICAS:
Cofradía de Vicedo: Gómez Brandariz, D.
Cofradía do Barqueiro: del Río Rocha, G.
Cofradía de A Coruña: Seoane Sánchez, P.
APOYO A LA INVESTIGACIÓN:
CIMA: Bouzamayor Yáñez, V.; Cotarelo Jardón, M.J.;
Meléndez Ramos, I.; Páez Pérez, J.; Penas Pampín,
E.; Ruíz Misioné, M.
Xefatura Territorial de Celeiro. Consellería del Mar:
Martín Salas, F.
BECARIOS/AS:
Aragunde Vila, R.; Castilla del Río, S.; Guiance Pérez,
M.; Matesanz Salgado, D.; Suárez Costa, I

CIMA Memoria 2016 65
cambios por causas hidrológicas o climáticas o a
diversas patologías que ya afecten a los individuos en
una talla pequeña. De la misma forma que se pretende
abordar el problema conociendo sus causas, se propone
comenzar a estudiar la posible recuperación futura de los
bancos, haciendo pequeños ensayos piloto de siembras
con semilla obtenida y marcada en el minicriadero de
Vicedo.
Los datos obtenidos por el grupo coordinado del
proyecto anterior y que se pretende continuar, sugieren
confirmar la disminución en todas las playas estudiadas,
causada por la escasez de individuos adultos (> 28mm).
Además en Vilarrube, la única playa que está siendo
explotada en la actualidad, comienzan a escasear los
individuos de talla comercial (> 35mm).
De los resultados obtenidos se llegó a la conclusión de
que la escasez de los individuos, en las principales
playas estudiadas (O Lombo das Navallas, América y
Vilarrube), no se debe a la contaminación química en los
sedimentos ni en el agua, ni a la reducción de la
diversidad genética, ni a mortalidades por patologías.
Pero de todas ellas, se deben abordar nuevos estudios
en esta Acción para de esta forma, llegar a extraer
conclusiones definitivas y firmes. Parece ser que el
tamaño del grano del sedimento y su grado de selección,
son los factores que pueden determinar la supervivencia
de la coquina, puesto que el sedimento existente en la
actualidad en la playa de Vilarrube, explicaría el
mantenimiento de su población.
Después del esfuerzo que supuso llevar a cabo todos
estos estudios coordinados, por un grupo de
investigación amplio, multidisciplinar y con numerosos
puntos de vista sobre la problemática existente con esta
especie en Galicia, se consideró necesaria la
continuación de las investigaciones en un punto donde
ya se pueden descartar muchas de las causas
analizadas hasta ahora para continuar con las definitivas
y al mismo tiempo, dar las soluciones más adecuadas.
El objetivo principal es: “Estudio de los factores naturales
y antropogénicos que afectan a la escasez de adultos en
las poblaciones de Donax en la costa gallega”.
Como objetivos parciales a realizar por los distintos
equipos de trabajo integrantes de la Acción serían:
1. Sedimentos y topografía intermareal
1.1. Control temporal de la variación de las texturas del
sedimento en cada una de las estaciones de muestreo
de poblaciones.
1.2. Caracterización y control de la variabilidad temporal
de los cambios en la topografía intermareal del banco de
Vilarrube.
2. Influencia de las condiciones hidrológicas y
meteorológicas
2.1.Estudiar la variabilidad interanual a lo largo del
tiempo en la densidad y estructura de tamaños
poblacionales, así como en la productividad de los
bancos de coquina de Playa América, Lombo das
Navallas y Vilarrube en función de los cambios
meteorológicos que afectan a sus condiciones
hidrográficas.
3. Estudios poblacionales
3.1.Evaluación de las densidades poblacionales en los
bancos objeto de estudio.
3.2.Estudio de los sedimentos de los diferentes bancos
en relación con la densidad y talla de las poblaciones.
4. Pruebas en criadero
4.1.Seguir estudiando la respuesta de la coquina ante
factores relacionados con las características del
sedimento (granulometría y compactación), haciendo
pruebas puntuales en el laboratorio, con adultos y con
semilla. Profundizar en los estudios de la fecundidad de
la coquina determinando su talla de madurez sexual y de
desoves.
4.2.Obtenidos los desoves, se transferirán los
conocimientos de cultivo de esta especie del criadero de
Ribadeo a las instalaciones del minicriadero de Vicedo.
En estas instalaciones se producirá una cantidad
suficiente de semilla que, servirá para hacer los primeros
ensayos piloto de puesta a punto de futuras
repoblaciones en las tres playas principales.
5. Histopatología
5.1.Situación actual de las patologías que afectan a las
poblaciones de coquina mediante análisis histológicos.
5.2.Evaluación de la prevalencia e intensidad de los
parásitos encontrados.
6. Microbiología
6.1.Continuar el estudio de la presencia/ausencia de
herpesvirus en los bancos de coquina.
Esta acción fue aprobada el 22 de junio de 2016, y se
comenzaron a realizar los trabajos correspondientes a
partir de la primera reunión de coordinación que tuvo
lugar el 12 de julio.

66
Hasta la fecha se hicieron 2 muestreos, uno el 20 de
septiembre y otro el 15 de diciembre de 2016. En la
Playa de Vilarrube. Para ello se estableció el criterio de
intermareal inferior, próximo a la línea de la bajamar;
intermareal superior, que corresponde a la zona que
tarda más en inundarse con la pleamar e intermareal
medio, situado entre las dos zonas anteriores.
De los resultados granulométricos preliminares, se
puede resaltar que el sedimento es bastante homogéneo
en los dos muestreos. A simple vista se observa que en
diciembre presenta más minerales pesados pero queda
todavía el análisis granulométrico para tener el estudio
completo.
Hasta la fecha se hicieron algunos análisis
granulométricos y en general, se observan arenas de
color ocre con presencia de pequeños fragmentos de
conchas y minerales pesados. Más del 75% de la
muestra queda retenida entre los tamices de 0.180,
0.125 y 0.090 mm.
De los trabajos correspondientes al apartado topográfico
llevados a cabo durante las dos campañas realizadas
(septiembre y diciembre), se pueden dar algunos
resultados preliminares.
La morfología general de la zona de estudio se
caracteriza por la presencia de barras arenosas con
surcos, o canales, entre ellas más o menos
desarrollados en función de la dinámica del oleaje. Las
Figuras 1 y 2 muestran imágenes panorámicas durante
las campañas de septiembre (Figura 1) y diciembre
(Figura 2) de 2016.
Figura 1.- Vista panorámica de la zona de trabajo durante la campaña de septiembre 2016.
Figura 2.- Vista panorámica de la zona de trabajo durante la campaña de diciembre 2016.
A partir de los datos topográficos realizados a lo largo de los
transeptos recorridos durante los muestreos se han
elaborado modelos digitales del terreno para ambas
campañas, utilizando el programa informático surfer y el
método de interpolación de triangulación lineal. Los
resultados preliminares se muestran en la Figura 3, y en la
que también se indican las estaciones muestreadas.
Se puede observar, que la morfología de la zona en ambas
campañas es muy similar, si bien lo más destacable es que
en el mes de diciembre la zona presenta una mayor
pendiente, es decir, unas condiciones más disipativas en
total concordancia con la mayor energía del oleaje en esta
época, junto con una marcada erosión de la playa y frente
dunar. Durante la campaña de septiembre se observa una
mayor acumulación de sedimentos hacia la parte alta de la
playa, característico de condiciones de bonanza energética.
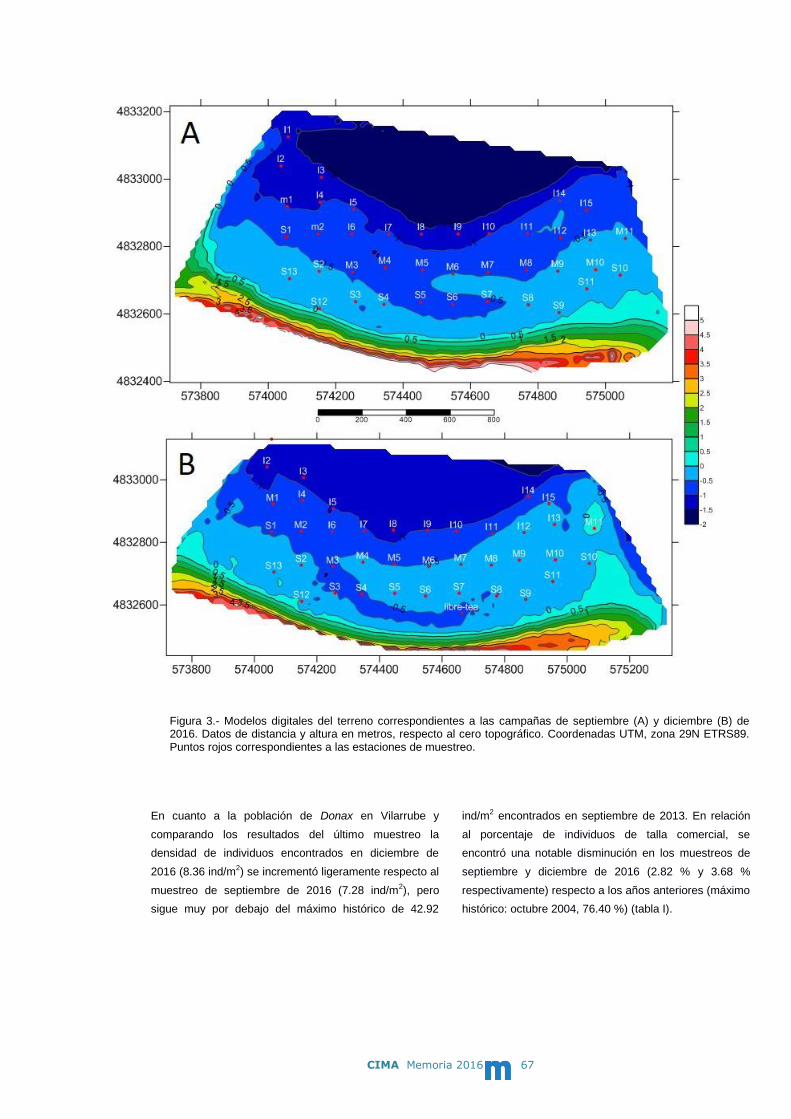
CIMA Memoria 2016 67
Figura 3.- Modelos digitales del terreno correspondientes a las campañas de septiembre (A) y diciembre (B) de 2016. Datos de distancia y altura en metros, respecto al cero topográfico. Coordenadas UTM, zona 29N ETRS89. Puntos rojos correspondientes a las estaciones de muestreo.
En cuanto a la población de Donax en Vilarrube y
comparando los resultados del último muestreo la
densidad de individuos encontrados en diciembre de
2016 (8.36 ind/m2) se incrementó ligeramente respecto al
muestreo de septiembre de 2016 (7.28 ind/m2), pero
sigue muy por debajo del máximo histórico de 42.92
ind/m2 encontrados en septiembre de 2013. En relación
al porcentaje de individuos de talla comercial, se
encontró una notable disminución en los muestreos de
septiembre y diciembre de 2016 (2.82 % y 3.68 %
respectivamente) respecto a los años anteriores (máximo
histórico: octubre 2004, 76.40 %) (tabla I).
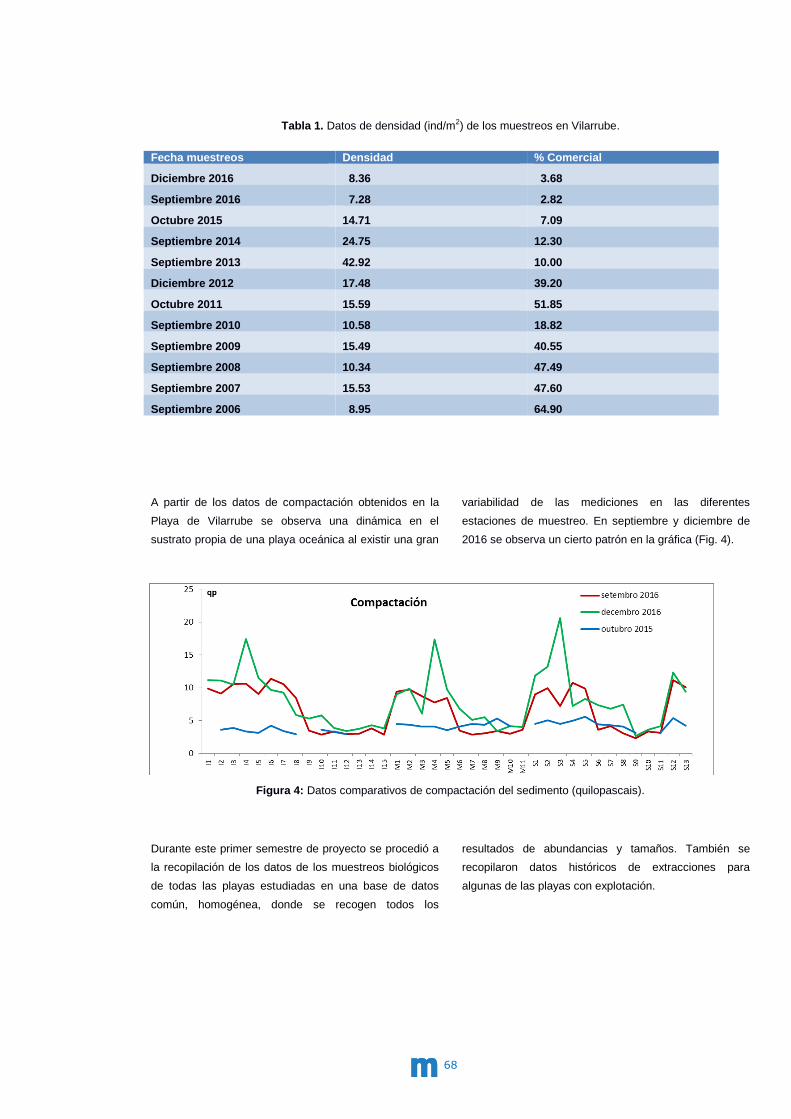
68
Tabla 1. Datos de densidad (ind/m2) de los muestreos en Vilarrube.
Fecha muestreos Densidad % Comercial
Diciembre 2016 8.36 3.68
Septiembre 2016 7.28 2.82
Octubre 2015 14.71 7.09
Septiembre 2014 24.75 12.30
Septiembre 2013 42.92 10.00
Diciembre 2012 17.48 39.20
Octubre 2011 15.59 51.85
Septiembre 2010 10.58 18.82
Septiembre 2009 15.49 40.55
Septiembre 2008 10.34 47.49
Septiembre 2007 15.53 47.60
Septiembre 2006 8.95 64.90
A partir de los datos de compactación obtenidos en la
Playa de Vilarrube se observa una dinámica en el
sustrato propia de una playa oceánica al existir una gran
variabilidad de las mediciones en las diferentes
estaciones de muestreo. En septiembre y diciembre de
2016 se observa un cierto patrón en la gráfica (Fig. 4).
Figura 4: Datos comparativos de compactación del sedimento (quilopascais).
Durante este primer semestre de proyecto se procedió a
la recopilación de los datos de los muestreos biológicos
de todas las playas estudiadas en una base de datos
común, homogénea, donde se recogen todos los
resultados de abundancias y tamaños. También se
recopilaron datos históricos de extracciones para
algunas de las playas con explotación.

9 ARTÍCULOS EN REVISTAS SCI
Álvarez, G., Uribe, E., Regueiro, J., Blanco, J., Fraga, S. Gonyaulax taylorii, a new yessotoxins-producer dinoflagellate
species from Chilean waters. Harmful Algae, 58: 8–15.
Amaro, R., Bouza, C., Gómez, B., Castro, J., San Miguel, E., Villalba, A., Lois, S., Outeiro, A., Ondina, P. Hyperspora
aquatica n.gn., n.sp. (Microsporidia), hyperparasitic in Marteilia cochillia (Paramyxida) is closely related to crustacean-
infecting microsporidian taxa. Zoological Journal of the Linnean Society. En prensa.
Blanco, J., Arevalo, F., Correa, J., Porro, M.C., Cabado, A. G., Vieites, J.M., et al. Effect of the industrial canning on
the toxicity of mussels contaminated with diarrhetic shellfish poisoning (DSP) toxins. Toxicon, 112:1–7.
Calvo-Iglesias, J., Pérez-Estévez, D., Lorenzo-Abalde, S., Sánchez-Correa, B., Quiroga, M.I., Fuentes, J.M.,
González-Fernández, A. Characterization of a monoclonal antibody directed against Mytilus spp larvae reveals an
antigen involved in shell biomineralization. PLOS ONE, Research Article,
http://dx.doi.org/10.1371/journal.pone.0152210.
Carballal, M.J., Iglesias, D., Darriba, S., Cao, A., Mariño, J.C., Ramilo, A., No, E., Villalba, A. Parasites and
pathological conditions and resistence to Marteilia cochillia in lagoon cockle Cerastoderma glaucum from Galicia (NW
Spain). Diseases of Aquatic Organisms, 122: 137-152.
Díaz, S., Iglesias, D., Villalba, A., Carballal, M.J. Long-term epidemiological study of disseminated neoplasia of cockles
in Galicia (NW Spain): temporal patterns at individual and population levels, influence of environmental and cockle-
based factors and lethality. Journal of Fish Diseases, 39: 1027-1042.
Díaz, S., Villalba, A., Carballal, M.J. Assessment of transmissibility of the disseminated neoplasia affecting cockles
Cerastoderma edule in Galicia (NW Spain). Bulletin of the European Association of Fish Pathologists. En prensa.
Dubert, J., da Costa, F., Aranda-Burgos, J.A., Martínez-Patiño, D., Prado, S., Barja, J.L. Beneficial effects of carpet
shell clam (Ruditapes decussatus) depuration during short periods of conditioning in shellfish hatchery: Role of the
temperature and phytoplankton on reduction and diversity of vibrios. Aquaculture, 459: 65-72.
Fernández-Boo, S., Villalba, A., Cao, A. Protein expression profile in haemocytes and plasma of the Manila clam
Ruditapes philippinarum in response to infection with Perkinsus olseni. Journal of Invertebrate Pathology, 39: 1369-
1385.
Fernández-Pardo, A., da Costa, F., Rial, D., Nóvoa, S., Martínez-Patiño, D., Vázquez, J.A. Use of response surface
methodology to determine optimum diets for Venerupis corrugata larvae: effects of ration and microalgal assemblages.
Aquaculture, 452: 283-290.
Hasanuzzaman, A.F.M., Robledo, D., Gómez-Tato, A., Alvarez Dios, J.A., Harrison, P.W., Cao, A., Fernández-Boo, S.,
Villalba, A., Pardo, B.G., Martínez, P. De novo transcriptome assembly of Perkinsus olseni trophozoite stimulated in
vitro with Manila clam (Ruditapes philippinarum) plasma. Journal of Invertebrate Pathology, 125: 22-33.
Hasanuzzaman, A.F.M., Robledo, D., Gómez-Tato, A., Alvarez-Dios, J.A., Harrison, P.W., Cao, A., Fernández-Boo,
S., Villalba, A., Martínez, P. Transcriptomic profile of Manila clam (Ruditapes philippinarum) haemocytes in response
to Perkinsus olseni infection. Aquaculture. En prensa.

70
Metzger, M.J., Villalba, A., Carballal, M. J., Iglesias, D., Sherry, J., Reinisch, C., Muttray, A. F., Baldwin, S. A., Goff, S.
P. Widespread transmission of independent cancer cells lineages within multiple bivalve species. Nature, 534 (7609):
705-709.
Pardo, B.G., Alvarez-Dios, J.A., Cao, A., Ramilo, A., Gómez-Tato, A., Planas, J.V., Villalba, A., Martínez, P.
Construction of an Ostrea edulis database from genomic and expressed sequence tags (ESTs) obtained from
Bonamia ostreae infected haemocytes: development of an immune-enriched oligo-microarray. Fish and Shellfish
Immunology, 59: 331-344.
Quijano-Scheggia, S., Barajas-Gonzalez, M., Lim, H. C., Leaw, C. P., Olivos-Ortiz, A., Gaviño-Rodríguez, J., Blanco
Pérez, J., Bates, S. The inhibitory effect of a non-yessotoxin-producing dinoflagellate, Lingulodinium polyedrum (Stein)
Dodge, towards Vibrio vulnificus and Staphylococcus aureus. Rev. Biol. Trop, 64(2): 805–816.
Ramilo, A., Pintado, J., Villalba, A., Abollo, E. Perkinsus olseni and P. chesapeaki detected in a survey of perkinsosis
of various clam species in Galicia (NW Spain) using PCR-DGGE as a screening tool. Journal of Invertebrate
Pathology, 133: 50-58.
Ruíz, M., López, C., Lee, R.S., Rodríguez, R., Darriba, S. A novel paramyxean parasite, Marteilia octospora n.sp.
(cercozoa) infecting the grooved razor shell clam Solen marginatus from Galicia (NW Spain). Journal of Invertebrate
Pathology, 135: 34-42.
Stentiford, G.D., Ramilo, A., Abollo, E., Kerr, R., Bateman, K.S., Feist, S.W., Bass, D., Villalba, A. Hyperspora aquatica
n.gn., n.sp. (Microsporidia), hyperparasitic in Marteilia cochillia (Paramyxida) is closely related to crustacean-infecting
microsporidian taxa. Parasitology. En prensa.
Tenorio, C., Uribe, E., Gil-Kodaka, P., Blanco, J., Álvarez, G. Morphological and toxicological studies of Pseudo-
nitzschia species from the central coast of Peru. Diatom Research, 31(4): 331–338.
Vera, M., Carlsson, J., Carlsson, J.E.L., Cross, T., Lynch, S., Kamermans, P., Villalba, A., Culloty, S., Martínez, P.
Current genetic status, temporal stability and structure of the remnant wild European flat oyster populations:
conservation and restoring implications. Marine Biology, 163: 239.
Villalba, A. What’s new in bivalve mollusc pathology? – Overview of articles published in the last two years. Bulletin of
the European Association of Fish Pathologists, 36: 24-37.

CIMA Memoria 2016 71
10 LIBROS
Oysters and clams: cultivation, habitat, threats and ecological impact.
Editor: J.L. Romalde. Nova Sci. Publ. NY.
BARJA, J.L., DUBERT, J., MONTES, J., OLVEIRA, J.G., PRADO, S., MARTÍNEZ-PATIÑO, D. Microbilogical
approaches to understand the threats for the wedge-shell clam (Donax trunculus) in natural beds. Pp. 67-77.
LOUZÁN, A., NÓVOA, S., OJEA, J., MARTÍNEZ PATIÑO, D. Culture of the wedge-shell clam Donax trunculus: new
developments. Pp. 15-28.

72
11 PONENCIAS Y PANELES
PONENCIAS
108th Annual Meeting National Shellfisheries Association (Las Vegas)
VILLALBA, A., METZGER, M.J., IGLESIAS, D., CARBALLAL, M.J., GOFF, S.P. At least two contagious cancer cell
lineages can transmit disseminated neoplasia in cockles Cerastoderma edule.
I International Symposium “Advances in Marine Mussel Research”, Vigo
VILLALBA, A., IGLESIAS, D., CARBALLAL, M.J. Transmission of independent cancer lineages within bivalve molluscs
in Galicia.
PANELES
XIX Foro dos Recursos Mariños e da Acuicultura das Rías Galegas
LASTRES, M., CARRAGAL, R., GONZÁLEZ, P., LÓPEZ, A., POTEL, I., FIGUEIRA, I., CID, E., ANDRÉS, C., CASAIS,
E, DA COSTA, F., NÓVOA, S., MARTÍNEZ-PATIÑO, D. Comparativa de luz L.E.D. versus luz fluorescente en el
cultivo de microalgas. Evaluación del coste energético por célula producida.
NÓVOA, S., OJEA, J., MARTÍNEZ, D. Protocolo de cultivo de la navaja Ensis magnus (Schumacher, 1817) en
criadero.
4th International Symposium on Genomics in Aquaculture, Athens, Greece
HASANUZZAMAN, A.F.MD.,GÓMEZ-TATO, A., ALVAREZ-DIOS, J.A., RUBIOLO, J.A., ROBLEDO, D., CAO, A.,
FERNÁNDEZ-BOO, S., VILLALBA, A., PARDO, B.G., MARTÍNEZ, P. Gene Expression in Ruditapes philippinarum
Haemocyte in Response to Natural Infection with Perkinsus olseni.
Aquaculture Europe 2016
ÁLVAREZ-BLÁZQUEZ, B., VILAR, A., PAPANDROULAKIS, N., LINARES, F., CAL, R., RODRÍGUEZ, J.L.,
MARTÍNEZ, J.M., LLUCH, N., GÓMEZ, C. AND PELETEIRO, J.B. First experiences of wreckfish (Polyprion
americanus) larval husbandry in NW Atlantic and East Mediterranean.
LINARES, F., RODRÍGUEZ, J.L., PELETEIRO, J.B., CAL, R., MARTÍNEZ, J.M., PAZOS, G., ÁLVAREZ-BLÁZQUEZ,
B. Influence of broodstock nutrition of wreckfish (Polyprion americanus) on the oocytes fatty acid composition.
MARTÍNEZ-VÁZQUEZ, J.M., PÉREZ-RIAL, E., PELETEIRO, J.B., LINARES, F., RODRÍGUEZ, J.L., VILAR, A., CAL,
R., ÁLVAREZ-BLÁZQUEZ, B. Description of the wreckfish (Polyprion americanus) reproductive cycle in captivity.

CIMA Memoria 2016 73
12 EDICIÓN DE PUBLICACIONES
BLANCO PÉREZ, J.C.
Revista Galega dos Recursos Mariños. Artigos e informes técnicos.
Revista Galega dos Recursos Mariños. Monografías.
VILLALBA GARCÍA, A.
Diseases of Aquatic Organisms.
Journal of Shellfish Research.

74
13 INFORMES
TÍTULO: Informe sobre la población de erizo (Paracentrotus lividus, Lamarck 1816) en la zona costera submareal de
la RMIP “Os Miñarzos” (Lira).
AUTORES: Cuiñas Olmedo, P., Febrero Mayor, F.
SOLICITANTE: Subdirección Xeral de Investigación e Apoio Científico-Técnico. Consellería del Mar. Xunta de Galicia.
TÍTULO: Informe sobre el Plan de Gestión Integral (PXI) de la Reserva Marina de interés Pesquero “Ría de Cedeira”
para el año 2017.
AUTORES: Cuiñas Olmedo, P., Febrero Mayor, F.
SOLICITANTE: Subdirección Xeral de Investigación e Apoio Científico-Técnico. Consellería del Mar. Xunta de Galicia.
TITULO: Análisis histopatológico de una muestra del molusco gasterópodo Nassarius reticulatus de la ría de Arousa
(Cabío).
AUTORES: Carballal Durán, M. J., Iglesias Estepa, D.
SOLICITANTE: Subdirección Xeral de Investigación e Apoio Científico-Técnico. Consellería del Mar. Xunta de Galicia.
TITULO: Análisis histopatológico de una muestra del molusco gasterópodo Nassarius reticulatus de la ría de Arousa
(Rianxo).
AUTORES: Carballal Durán, M. J., Iglesias Estepa, D.
SOLICITANTE: Subdirección Xeral de Investigación e Apoio Científico-Técnico. Consellería del Mar. Xunta de Galicia.
TITULO: Informe del año 2015 correspondiente a las “Experiencias para el cultivo del besugo realizadas en las
Jaulas Experimentales” de las que es titular el Centro de Investigaciones marinas (Consellería del Mar) ubicadas en
Bueu.
AUTORA: Linares Cuerpo, F.
SOLICITANTE: Subdirección Xeral de Acuicultura. Consellería del Mar. Xunta de Galicia.
TITULO: Informe para la solicitud de inclusión en el REGA de dos plataformas de cultivo ubicadas en el distrito de
Bueu.
AUTORA: Linares Cuerpo, F.
SOLICITANTE: Servizo de Inspección e Control de Recursos. Subdirección de Gardacostas. Consellería del Mar.
Xunta de Galicia.
TITULO: Informe y Exposición de resultados en la reunión de coordinación del Proyecto Diversify de las actividades
relacionadas con el desarrollo de la “Nutrición de la cherna ((Polyprion americanus) WP.12)”.
AUTORA: Linares Cuerpo, F.
SOLICITANTE: Coordinador WP Nutrición Proyecto Diversify.
TITULO: 2ndReport. Resumen de actividades WP12. Nutrición de la cherna. Proyecto Diversify.
AUTORA: Linares Cuerpo, F.
SOLICITANTE: Coordinador Proyecto Diversify.

CIMA Memoria 2016 75
14 REUNIONES
REUNIÓN: Órgano de Gestión Seguimiento y Control (Oxesco) de la Reserva Marina de interés pesquero Os
Miñarzos. (17 de marzo 2016).
ASISTENTES: Presidente, Secretaria y Vocales del OXESCO y personal asesor.
REUNIÓN: Órgano de Gestión Seguimiento y Control (Oxesco) de la Reserva Marina de interés pesquero ría de
Cedeira. (04 de abril 2016).
ASISTENTES: Presidente, Secretaria y Vocales del OXESCO y personal asesor.
REUNIÓN: Órgano de Gestión Seguimiento y Control (Oxesco) de la Reserva Marina de interés pesquero ría de
Cedeira. (27 de octubre 2016).
ASISTENTES: Presidente, Secretaria y Vocales del OXESCO y personal asesor.
REUNIÓN: Órgano de Gestión Seguimiento y Control (Oxesco) de la Reserva Marina de interés pesquero Os
Miñarzos. (09 de noviembre 2016).
ASISTENTES: Presidente, Secretaria y Vocales del OXESCO y personal asesor.
REUNIÓN: Reuniones de planificación y coordinación de actividades con los participantes de Galicia en el proyecto
“Exploring the biological and socio-economic potential of new/emerging candidate fish species for the expansión of the
European aquaculture industry”. “Diversify”.
ASISTENTES: Linares Cuerpo, F. (CIMA), Rodríguez Villanueva, J.L. (IGAFA) e investigadores del IEO de Vigo y del
Acuario de A Coruña.
REUNIÓN: Reunión anual de coordinación del proyecto “Exploring the biological and socio-economic potential of
new/emerging candidate fish species for the expansión of the European aquaculture industry”. “Diversify”.
ASISTENTES: Linares Cuerpo, F. e Investigadores Principales de todos los centros de investigación participantes en
el proyecto, representantes de empresas y Asociaciones.
REUNIÓN: Seminario “INTERREG Espacio Atlántico: Primera convocatoria”, organizado por la Dirección General de
Fondos Comunitarios (Ministerio de Hacienda y Administraciones Públicas).
ASISTENTE: Villalba García, A.
REUNIÓN: Presentación del inSeminario “INTERREG Espacio Atlántico: Primera convocatoria”, organizado por la
Dirección General de Fondos Comunitarios (Ministerio de Hacienda y Administraciones Públicas).
ASISTENTE: Villalba García, A.
REUNIÓN: Reunión de coordinación para la elaboración de la propuesta del proyecto de investigación “Co-Operation
for Restoring Cockle Shellfiseries and its Ecosystem Services in the Atlantic Area (COCKLES)”, a presentar en la
convocatoria de INTERREG Espacio Atlántico de la UE.
ASISTENTE: Villalba García, A.

76
15 OTROS
15.1. ASESORAMIENTO. OTROS
LINARES CUERPO, F.
Convenio de colaboración entre la Xunta de Galicia a través de la Consellería del Mar y la Asociación de Acuicultores
“Illa do Santo” de Bueu (Pontevedra), para el cultivo de peces en jaulas.
15.2. REVISIÓN DE ARTÍCULOS EN PUBLICACIONES CIENTÍFICAS
BLANCO PÉREZ, J.C.
Toxicon (3)
Toxins
Marine Drugs
Phycologia
Aquatic Toxicology
VILLALBA GARCÍA, A.
Diseases of Aquatic Organisms

CIMA Memoria 2016 77
16 TESIS DOCTORALES
16.1. DIRECCION/TUTORIZACIÓN DE TESIS
TEMA: Estudio proteómico del sistema inmunitario de la ostra Ostrea edulis y de su respuesta frente a la bonamiosis.
Estudio proteómico del sistema inmune de la ostra plana europea (Ostrea edulis) y su interacción con parásitos
Bonamia spp.
DOCTORANDO/A: Rodríguez de la Ballina, N.
TITULACIÓN: Licenciada en Biología
TUTOR/A Y DIRECTOR/A DE LA TESIS: Villalba García, A. (tutor e codirector), Cao Hermida, A. (codirectora)
TEMA: Estudio patológico de los solénidos comerciales en Galicia.
DOCTORANDO/A: Ruiz Pérez, M.
TITULACIÓN: Licenciada en Ciencias del Mar
TUTOR/A Y DIRECTOR/A DE LA TESIS: López Romalde, J.A. (USC, tutor), López Gómez, M.C. (directora) y Darriba
Couñago, S. (INTECMAR, directora).
TEMA: Consideracións biolóxicas, ecolóxicas e produtivas relativas ao cultivo de ostra rizada Crassostrea gigas
(Thunberg, 1793) nas rías galegas.
DOCTORANDO/A: Rodríguez González, L.
TITULACIÓN: Licenciado en Biología
TUTOR/A Y DIRECTOR/A DE LA TESIS: Montes Pérez, J.; Iglesias Estepa, D.
TEMA: Alimentación en una planta de cultivos. Calidad del alimento vivo y alternativas a su sustitución o
suplementación.
DOCTORANDO/A: Fernández Pardo, A.
TITULACIÓN: Licenciada en Biología
TUTOR/A Y DIRECTOR/A DE LA TESIS: Martínez Patiño, D.; Nóvoa Vázquez, S.

78
TEMA: Optimización del cultivo intensivo de la almeja fina Ruditapes decussatus en criadero.
DOCTORANDO/A: Aranda Burgos, J.A.
TITULACIÓN: Licenciado en Biología
TUTOR/A Y DIRECTOR/A DE LA TESIS: Martínez Patiño, D. (Tutora y Directora); da Costa González, F.
(Codirector).
TEMA: Desarrollo del cultivo integral de la coquina Donax trunculus (Linnaeus, 1758). Ciclo reproductivo, cultivo en
criadero y en el medio natural.
DOCTORANDO/A: Louzán Pérez, A.
TITULACIÓN: Licenciada en Biología
TUTOR/A Y DIRECTOR/A DE LA TESIS: Martínez Patiño, D; Nóvoa Vázquez, S.
16.2. PARTICIPACIÓN EN TRIBUNALES DE TESIS
MARTINEZ PATIÑO, D.
Título: Mejora de la supervivencia larvaria en cultivos de moluscos mediante la mejora de las técnicas de cultivo de
microalgas y la introducción de nuevas especies de microalgas.
Doctorando/a: Pedro Nuno Alexandre Nascimento da Fonseca
Universidad: Universidad de Santiago de Compostela
MONTES PÉREZ, J.
Título: Desarrollo del cultivo integral de la coquina Donax trunculus (Linnaeus, 1758). Ciclo reproductivo, cultivo en
criadero y en el medio natural.
Doctorando/a: Louzán Pérez, A.
Universidad: Universidad de A Coruña
VILLALBA GARCÍA, A.
Título: Modulation of gene expression in Manila Clam (Ruditapes philippinarum) and in molluscan parasite (Perkinsus
olseni) through host-parasite in vivo and in vitro interaction.
Doctorando/a: Abul Farah Md. Hasanuzzaman
Universidad: Universidad de Santiago de Compostela
Título: Caracterización molecular de las ostras del litoral atlántico andaluz y sus patologías.
Doctorando/a: López Sanmartín, M.
Universidad: Universidad de Granada

CIMA Memoria 2016 79
16.3. TESIS PRESENTADAS
Título: Desarrollo del cultivo integral de la coquina Donax trunculus (Linnaeus, 1758). Ciclo reproductivo, cultivo en
criadero y en el medio natural.
Doctorando/a: Louzán Pérez, A.
Universidad: A Coruña
Fecha de presentación: 28/04/2016
RESUMEN
Donax trunculus (Linnaeus, 1758), denominada comúnmente coquina y cadelucha en Galicia, es una especie de
molusco bivalvo de distribución atlántico–mediterránea, cuyos bancos naturales en la actualidad se encuentran en
regresión. Se trata de una especie con un elevado interés comercial y supone un recurso económico importante para
el sector marisquero de Galicia. El desarrollo del cultivo en criadero de la coquina, como estrategia para la obtención
de semilla, con fines de repoblación para aumentar la producción, supone un claro beneficio para el sector.
La población de D. trunculus estudiada, presenta un ciclo anual de reproducción con un corto período de reposo y un
amplio período de madurez y puesta que va del mes de marzo en adelante, finalizando hacia el mes de julio.
Macroscópicamente, a partir del estadío de gametogénesis avanzada el período en el que la gónada aparece
ocupada en su mayor parte por tejido folicular, se pueden definir los sexos de la coquina, presentando las hembras la
gónada de color violeta y los machos, de color blanco. La marcada estacionalidad a lo largo de la evolución del ciclo
gametogénico, determina cambios en la composición bioquímica mayoritaria de los individuos. Los carbohidratos
mantienen una relación inversa con el ciclo gametogénico, disminuyen conforme este avanza. A medida que los
carbohidratos disminuyen, los lípidos aumentan, mientras que las proteínas mantienen un patrón de evolución estable
y no relacionado con el ciclo.
El conocimiento de los diferentes estadíos embrionarios y tiempos de transformación, constituye una parte importante
en el proceso de optimización del cultivo de este bivalvo. El estudio de la variación en composición bioquímica
durante el desarrollo larvario de la coquina así como la determinación de los requerimientos nutricionales y
movimiento de reservas energéticas durante esta etapa, contribuyen a la optimización y establecimiento de un
protocolo de cultivo en criadero de D. trunculus. Con la optimización y estandarización de cada una de las fases de
cultivo en criadero y en el medio natural de la especie, se puede afirmar que existe la posibilidad de obtener semilla
de D. trunculus en criadero para su posterior siembra en bancos naturales hasta llegar a su talla comercial.

80
17 BECARIOS FP 2º GRADO
TUTOR/A: Carballal Durán, M. J.
BECARIO/A: Fernández González, J.
ESPECIALIDAD: Técnico especialista en FP 2º grado, Rama Sanitaria, especialidad de Anatomía, Patología y
Citología
TUTOR/A: Carballal Durán, M. J.
BECARIO/A: Riomayor Cernadas, A.
ESPECIALIDAD: Técnico especialista en FP 2º grado, Rama Sanitaria, especialidad de Anatomía, Patología y
Citología
TUTOR/A: Cao Hermida, M.A.
BECARIO/A: Alonso Costas, J.B.
ESPECIALIDAD: Técnica Superior en Análisis y Control de Calidad
TUTOR/A: Villalba García, A.
BECARIO/A: Alonso Costas, J.
ESPECIALIDAD: Técnica Superior en Análisis y Control de Calidad
TUTOR/A: Cao Hermida, M.A.
BECARIO/A: Campañó Piñeiro, M.S.
ESPECIALIDAD: Técnica Superior en Análisis y Control de Calidad
TUTOR/A: Cao Hermida, M.A.
BECARIO/A: Viturro García, R.
ESPECIALIDAD: Técnica Superior en Análisis y Control de Calidad
TUTOR/A: Cao Hermida, M.A.
BECARIO/A: Pena Thomas, G.
ESPECIALIDAD: Técnica Superior en Análisis y Control de Calidad
TUTOR/A: Linares Cuerpo, F.
BECARIO/A: Acuña López, I.
ESPECIALIDAD: Técnica Superior en Producción Acuícola
TUTOR/A: Linares Cuerpo, F.
BECARIO/A: Méndez Montenegro, J.C.
ESPECIALIDAD: Técnico Superior en Producción Acuícola

CIMA Memoria 2016 81
TUTOR/A: Linares Cuerpo, F.
BECARIO/A: Araujo Sendín, A.
ESPECIALIDAD: Técnico Superior en Producción Acuícola
TUTOR/A: Linares Cuerpo, F.
BECARIO/A: Fuentes Villar, N.
ESPECIALIDAD: Técnica Superior en Producción Acuícola
TUTOR/A: López Gómez, M.C.
BECARIO/A: Grande Ocampo, O.
ESPECIALIDAD: Técnico Superior Anatomía Patológica y Técnico Superior en Acuicultura
TUTOR/A: Montes Pérez, J.
BECARIO/A: Ulla Refojo, L.
ESPECIALIDAD: Técnico Superior Anatomía Patológica
TUTOR/A: Nóvoa Vázquez, S.
BECARIO/A: Castilla del Río, S. (desde 01.09.2016)
ESPECIALIDAD: Técnica Superior en Producción Acuícola
TUTOR/A: Ojea Martínez, J.
BECARIO/A: Matesanz Salgado, D. (desde 07.10.2016)
ESPECIALIDAD: Técnico Superior en Producción Acuícola

82
18 CURSOS
18.1. CURSOS IMPARTIDOS
Máster Interuniversitario de Acuicultura (UDC/USC/UVIGO)
BLANCO PÉREZ, J.C. (junto con Gago, A.-UVIGO)
Mareas Tóxicas.
MARTÍNEZ PATIÑO, D., NÓVOA VÁZQUEZ, S., OJEA MARTÍNEZ, J.
Impartición de prácticas sobre cultivo de moluscos bivalvos.
MARTÍNEZ PATIÑO, D., NÓVOA VÁZQUEZ, S.
Tutorización de Trabajo Fin de Máster.
VILLALBA GARCÍA, A.
Enfermedades en invertebrados:
- Enfermedades neoplásicas.
- Estrategias de lucha frente a enfermedades en invertebrados.
- Sesión de prácticas.
10th International Postgraduate Course" "Histology and Histopathology of Aquatic Animals" (Universidad del
País Vasco), correspondiente al programa de Master "European Master of Science In Marine Environment and
Resources.
VILLALBA GARCÍA, A.
General Histopathology of mollusks.
Neoplastic lesions in mollusks.
Practical session 2: Microscopical observations on general molluscan histopathology.
Practical session 3: Microscopical observations on general molluscan histopathology (cont.) and identification of
neoplastic lesions.
Líneas actuales en investigación en Biología Animal (Universidad de Alcalá), correspondiente al “Máster
Universitario en Investigación en Ciencias, Especialidad en Biología Animal”.
VILLALBA GARCÍA, A.
Epidemias de cáncer en moluscos bivalvos.
Estrategias de lucha frente a enfermedades de moluscos bivalvos.
Ciclo de Seminarios en la sede central del INIA
VILLALBA GARCÍA, A.
Epidemias de cáncer en moluscos bivalvos.

CIMA Memoria 2016 83
18.2. ASISTENCIA A CURSOS
Fundación Ramón Domínguez. Hospital Clínico Universitario de Santiago de Compostela.
Proteómica traslacional
Brianes Beiras, M.J.; Gregorio Chenlo, M.V.
Escola Galega de Administración Pública. EGAP. Santiago de Compostela.
Curso de Inglés B2
Linares Cuerpo, F.

84
19 ESTANCIAS
19.1. ESTANCIAS DEL PERSONAL DEL CIMA EN OTROS CENTROS
MARTÍNEZ PATIÑO, D.
NÓVOA VÁZQUEZ, S.
Hatchery de la empresa Azura Group en Dakhla, Marruecos.
Duración: 7 días.
Tema: Intercambio de conocimientos como complemento del desarrollo de los distintos trabajos relacionados con el
CECUMAR de Ribadeo y para abrir futuras vías de cooperación en proyectos de investigación.
19.2. ESTUDIANTES EN PRÁCTICAS
Alumno/a: Prieto Sierra, D.
Procedencia: Facultad de Ciencias. Universidad de A Coruña
Duración: 265 horas
Tutor/a: Ojea Martínez, J.
Alumno/a: Loor Mera, A.G.
Procedencia: Master of Science in Aquaculture. University of Gent
Duración: 114 horas
Tutor/a: Martínez Patiño, D.
19.3. PRÁCTICAS DE MÁSTER
Alumno/a: Caeiro Aguado, M.
Máster: Máster Interuniversitario en Acuicultura (UDC/USC/UVIGO)
Duración: 11/10/2016 a 26/01/2017
Tutoras: Martínez Patiño, D.; Nóvoa Vázquez, S.

CIMA Memoria 2016 85
CUENTAS
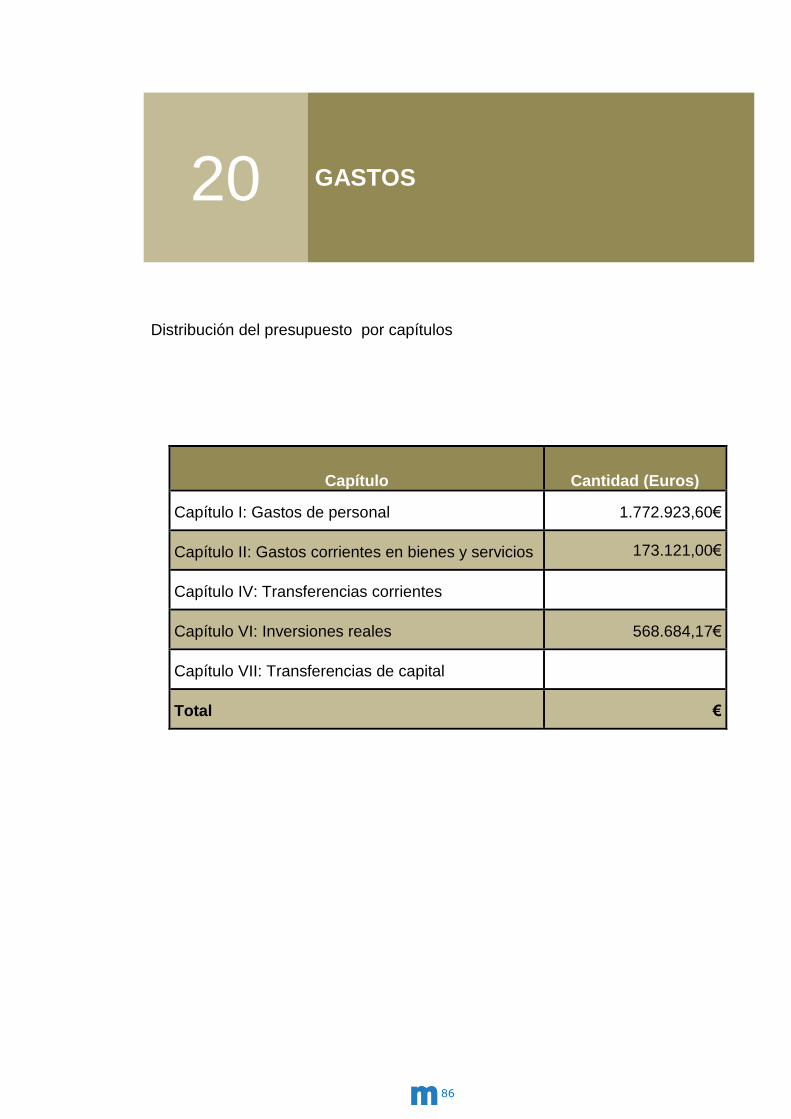
86
20 GASTOS
Distribución del presupuesto por capítulos
Capítulo
Cantidad (Euros)
Capítulo I: Gastos de personal 1.772.923,60€
Capítulo II: Gastos corrientes en bienes y servicios 173.121,00€
Capítulo IV: Transferencias corrientes
Capítulo VI: Inversiones reales 568.684,17€
Capítulo VII: Transferencias de capital
Total €

CIMA Memoria 2016 87
21 FUENTES DE FINANCIACIÓN
ORIGEN DE LOS FONDOS
CANTIDAD (€)
Propia: Dirección Xeral de Desenvolvemento Pesqueiro.
Consellería del Mar. Xunta de Galicia
a) Acciones de Investigación que desarrolla el CIMA
b) Convenios con entidades externas
368.749,32€
I+D de la Xunta de Galicia
Administración General del Estado €
Unión Europea (INTERREG, VI Programa Marco,...) 18.683,00€
Total €

88
m