caracterización bioquímica del consumo de reservas vitelinas en
TRANSCRIPT

REDVET. Revista electrónica de Veterinaria 1695-7504 2011 Volumen 12 Número 3
Caracterización bioquímica del consumo de reservas vitelinas en peces teleósteos de ontogenia indirecta.- http://www.veterinaria.org/revistas/redvet/n030311/031102.pdf
1
REDVET Rev. electrón. vet. http://www.veterinaria.org/revistas/redvet Vol. 12, Nº 3 Marzo/2011– http://www.veterinaria.org/revistas/redvet/n030311.html
Caracterización bioquímica del consumo de reservas vitelinas en peces teleósteos de ontogenia indirecta (Cemical caracterization of vitelline reserves consumption in teleost fish of indirect ontogeny)
Zavala, Iram Dpto. Desarrollo de Tecnologías. Centro Interdisciplinario de Ciencias Marinas, Instituto Politécnico Nacional. A. P. 592. La Paz, B.C.S., México. C.P. 23000. (Nick: ziram28)
Contacto: [email protected] Resumen El presente trabajo es un ejercicio de revisión bibliográfica en el cual se describen los principales componentes bioquímicos del vitelo y glóbulo de aceite, su importancia, metabolismo, patrón de consumo y la influencia del contenido de las reservas vitelinas (saco de vitelo y glóbulo de aceite) sobre el desarrollo embrionario y la supervivencia larval de peces teleósteos de ontogenia indirecta. Como patrón general se puede destacar que, en este grupo de peces, el principal componente en términos de volumen fueron las proteínas seguido de los lípidos y después los carbohidratos. En términos de consumo, se consideran tres fases durante el desarrollo embrionario: pre-eclosión, post-eclosión y fase terminal. Durante la pre-eclosión son consumidos principalmente carbohidratos, durante la post-eclosión las proteínas y lípidos en diferentes proporciones, mientras que durante la fase terminal los lípidos. Desde un punto de vista práctico, mediante la caracterización bioquímica de las reservas vitelinas, es posible producir larvas de “mayor calidad”. Una vez que se conozca el contenido de sus componentes, utilización e importancia, es posible modificar la composición del vitelo en posteriores grupo de huevos mediante la adición del componente de interés en la dieta de los reproductores. Por ejemplo, se ha observado que en algunas especies la relación de ácidos grasos poliinsaturados (DHA/EPA) afecta la supervivencia larval. Después de conocer la relación DHA/EPA que permita lograr un porcentaje de supervivencia alto, se suministra en la dieta de los reproductores para ser incorporados al vitelo durante la vitelogénesis a los huevos. Palabras claves: Ontogenia indirecta | Teleósteos | Reservas vitelinas| Saco de vitelo| Glóbulo de aceite.

REDVET. Revista electrónica de Veterinaria 1695-7504 2011 Volumen 12 Número 3
Caracterización bioquímica del consumo de reservas vitelinas en peces teleósteos de ontogenia indirecta.- http://www.veterinaria.org/revistas/redvet/n030311/031102.pdf
2
Abstract This work is a bibliography review exercise in which are described the major biochemical’s compounds of yolk sac and oil droplet, his importance, metabolism, consumption pattern and the influence of the vitelline reserves content (yolk sac and oil droplet) on embryonic development and larval survival in teleost fishes of indirect ontogeny. How general pattern, in this group the major compound were proteins, the next lipids and carbohydrates later. With respect to food endogenous consumption three phases were considered during embryonic development: pre-hatching, post-hatching and terminal phase. In pre-hatching, carbohydrates were utilized principally. In post-hatching phase, the proteins and lipids were utilized at different proportion. While that in the final phase, the lipids were utilized. From a practical standpoint, by the biochemical characterization of the vitelline reserves, it is possible to produce larvae of "higher quality". Once you know the content of its components, use and importance, it is possible to alter the composition of the yolk of eggs in subsequent group by adding the interest component in the diet of the broodstock. For example, it appears that in some species the ratio of polyunsaturated fatty acids (DHA / EPA) affects larval survival. After knowing the ratio DHA / EPA permits to achieve a high survival rate, is supplied in the diet of the broodstock to be incorporated into the yolk during vitellogenesis to eggs. Keys words: Indirect ontogeny | Teleost | Vitelline reserves | Yolk sac| oil droplet. Introducción El grupo de los teleósteos comprende más de 21 000 especies que habitan una gran variedad de ambientes, tanto marinos como de agua dulce y profundidades abismales como superficiales. A lo largo de 200 millones de años de evolución este grupo se ha adaptado a diferentes hábitats y ha adoptado diversas estrategias reproductivas. No obstante, una característica reproductiva que se puede observar en todas las especies es la producción de vitelo durante el desarrollo del oocito (Arukwe & Goksøyr, 2003). La incorporación de vitelo al oocito se da mediante un proceso llamado vitelogénesis que ocurre tanto en peces como en invertebrados ovíparos. El principal elemento de reserva captado por el oocito es una proteína de origen hepático conocida como vitelogenina (VTG), la cual tras su incorporación por micropinocitosis al oocito, sufre proteolisis para incorporarse a los gránulos de vitelo que son la fuente de reserva de durante el periodo de alimentación endógena. El huevo de un pez puede ser considerado como un sistema semicerrado. Después de la fertilización, las membranas del huevo se

REDVET. Revista electrónica de Veterinaria 1695-7504 2011 Volumen 12 Número 3
Caracterización bioquímica del consumo de reservas vitelinas en peces teleósteos de ontogenia indirecta.- http://www.veterinaria.org/revistas/redvet/n030311/031102.pdf
3
endurecen y se vuelven relativamente impermeable a muchos solutos, pero permiten todavía el intercambio gaseoso. Como consecuencia, los embriones dependen de las reservas endógenas para obtener la energía destinada al desarrollo y crecimiento (Heming & Buddington, 1988). La fuente endógena de alimentación está constituida por el saco de vitelo y el (los) glóbulo(s) de aceite(s). La composición nutricional de ambos, es especie-específica y varía en función de la edad de la madre, el peso y la alimentación de esta (Kamler, 1976). Tanto la tasa de absorción como la eficiencia de utilización del vitelo son determinantes en el desarrollo inicial, crecimiento y supervivencia de las larvas de peces. Son afectados por un número de factores tales como la temperatura, la luz, la concentración de oxígeno y la salinidad (Heming & Buddington, 1988). Metodológicamente, pueden determinarse por el contenido calórico del vitelo, peso seco, peso húmedo y volumen. Sin embargo, existen otros métodos que brindan una mayor información, como los bioquímicos, que además permiten conocer de manera específica el consumo de los nutrientes (ej. carbohidratos, lípidos y proteínas) tanto en los huevos, como en embriones y larvas. Componentes bioquímicos del vitelo y su importancia La vitelogénesis es el principal evento responsable del crecimiento de los oocitos en muchos teleósteos. Consiste en la producción por el hígado de la vitelogenina la cual es un complejo glucolipofosfoproteico que se une al calcio y al hierro y es precursora de las proteínas del vitelo y otros nutrientes necesarios para el desarrollo del embrión (Brooks et al., 1997; Sarasquete et al., 2002). El proceso hormonal que rige la vitelogénesis consiste en la liberación del factor liberador de la hormona luteinizante (LHRH) por el hipotálamo. Esto estimula la síntesis y secreción de la hormona luteinizante (LH) y la hormona folículo estimulante (FSH) por parte de la hipófisis. En peces, tanto la LH como la FSH provocan la producción de estradiol-17β en las células foliculares a partir de la testosterona (Patiño y Sullivan, 2002). El estradiol-17β es liberado al sistema circulatorio para llegar al hígado donde estimula la síntesis y secreción de vitelogenina (Idler & Ng, 1983). La vitelogenina viaja por el sistema circulatorio hacia las gónadas para incorporarse al oocito. La incorporación ocurre por procesos de pinocitosis de las células foliculares y las microvellosidades conectadas al corion del oocito (Patiño & Sullivan, 2002). De manera general, las reservas vitelinas en los teleósteos pueden encontrarse como sacos de vitelo o glóbulos de aceite. El principal componente dentro del saco de vitelo son las proteínas, mientras que los glóbulos de aceite contienen lípidos, principalmente triglicéridos (Heming &
REDVET. Revista electrónica de Veterinaria 1695-7504 2011 Volumen 12 Número 3
Caracterización bioquímica del consumo de reservas vitelinas en peces teleósteos de ontogenia indirecta.- http://www.veterinaria.org/revistas/redvet/n030311/031102.pdf
4
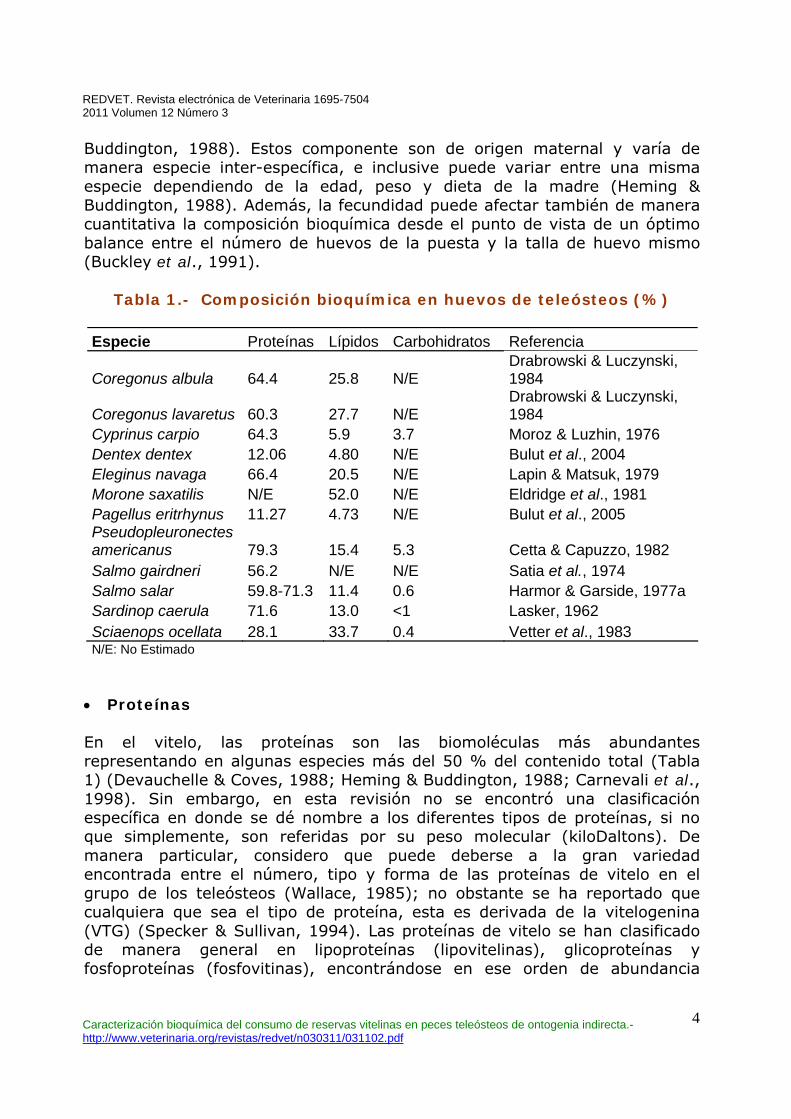
Buddington, 1988). Estos componente son de origen maternal y varía de manera especie inter-específica, e inclusive puede variar entre una misma especie dependiendo de la edad, peso y dieta de la madre (Heming & Buddington, 1988). Además, la fecundidad puede afectar también de manera cuantitativa la composición bioquímica desde el punto de vista de un óptimo balance entre el número de huevos de la puesta y la talla de huevo mismo (Buckley et al., 1991).
Tabla 1.- Composición bioquímica en huevos de teleósteos (%)
Especie Proteínas Lípidos Carbohidratos Referencia
Coregonus albula 64.4 25.8 N/E Drabrowski & Luczynski, 1984
Coregonus lavaretus 60.3 27.7 N/E Drabrowski & Luczynski, 1984
Cyprinus carpio 64.3 5.9 3.7 Moroz & Luzhin, 1976 Dentex dentex 12.06 4.80 N/E Bulut et al., 2004 Eleginus navaga 66.4 20.5 N/E Lapin & Matsuk, 1979 Morone saxatilis N/E 52.0 N/E Eldridge et al., 1981 Pagellus eritrhynus 11.27 4.73 N/E Bulut et al., 2005 Pseudopleuronectes americanus 79.3 15.4 5.3 Cetta & Capuzzo, 1982 Salmo gairdneri 56.2 N/E N/E Satia et al., 1974 Salmo salar 59.8-71.3 11.4 0.6 Harmor & Garside, 1977a Sardinop caerula 71.6 13.0 <1 Lasker, 1962 Sciaenops ocellata 28.1 33.7 0.4 Vetter et al., 1983 N/E: No Estimado
• Proteínas En el vitelo, las proteínas son las biomoléculas más abundantes representando en algunas especies más del 50 % del contenido total (Tabla 1) (Devauchelle & Coves, 1988; Heming & Buddington, 1988; Carnevali et al., 1998). Sin embargo, en esta revisión no se encontró una clasificación específica en donde se dé nombre a los diferentes tipos de proteínas, si no que simplemente, son referidas por su peso molecular (kiloDaltons). De manera particular, considero que puede deberse a la gran variedad encontrada entre el número, tipo y forma de las proteínas de vitelo en el grupo de los teleósteos (Wallace, 1985); no obstante se ha reportado que cualquiera que sea el tipo de proteína, esta es derivada de la vitelogenina (VTG) (Specker & Sullivan, 1994). Las proteínas de vitelo se han clasificado de manera general en lipoproteínas (lipovitelinas), glicoproteínas y fosfoproteínas (fosfovitinas), encontrándose en ese orden de abundancia

REDVET. Revista electrónica de Veterinaria 1695-7504 2011 Volumen 12 Número 3
Caracterización bioquímica del consumo de reservas vitelinas en peces teleósteos de ontogenia indirecta.- http://www.veterinaria.org/revistas/redvet/n030311/031102.pdf
5
(Nakagawa, 1970) con muy poca variación en la proporción encontrada durante el desarrollo (Nakagawa & Tsuchiya, 1972). Sin embargo, en algunas especies como Hucho perryi (Hiramatsu & Hara, 1996), Verasper moseri (Matsubara & Sawano, 1995) y Acipencer ruthenus (Hiramatsu et al., 2002) se ha reportado otro tipo de proteína denominada β-componente. Pese al orden de abundancia que se ha reportado de las proteínas, es de esperarse que durante el desarrollo de los organismos las propiedades físicas y químicas de estas moléculas cambien. Por ejemplo, las lipoproteínas incrementan su peso molecular, los grupos lipídicos prostéticos disminuyen y algunos aminoácidos son liberados (Nakagawa & Tsuchiya, 1974). En las últimas décadas, se ha observado que en los huevos pelágicos los compuestos nitrogenados más abundantes son los aminoácidos en su forma libre (AAL) (Rønnestad et al., 1998a). Los AAL pueden contribuir con el 20 al 50 % del contenido en huevos recién desovados (Rønnestad & Fyhn, 1993; Finn, 1994). El contenido de AAL es muy variable dentro del diverso grupo de los teleósteos (Tabla 2). Una de las especies que mayor cantidad de aminoácidos libres presenta es el lenguado del Atlántico Hippoglossus hippoglossus (2300 nmol/huevo, aproximadamente), en contraste, la macarela Scomber scombrus es una de las especies que presenta menor cantidad de aminoácidos libres (30 nmol/huevo, aproximadamente). Pese a la gran variación existente entre el volumen absoluto del contenido de los AAL, existe una gran relación entre el volumen de AAL y la talla del huevo (Rønnestad & Fyhn, 1993). Además, el contenido de AAL también está relacionado con el tipo de huevo. En especies con huevos pelágicos, el contenido de AAL (%) es mayor que en aquellos demersales en los cuales solo representan alrededor del 2-3 % de los aminoácidos totales (Rønnestad & Fyhn, 1993). Mientras que en especies de agua dulce como Coregonus albula y C. lavaretus constituyen el 4.7 y 4.5, respectivamente (Dabrowski et al., 1985). Dentro de los aminoácidos existen los esenciales que son aquellos que no pueden ser sintetizados por el organismo y se ingieran a través del alimento, y los no esenciales que el organismo sintetiza. Se ha observado que la proporción entre esenciales y no esenciales dentro de los aminoácidos libres es alrededor del 50% y que se consumen en igual proporción (Rønnestad et al., 1994; Rønnestad et al., 1998a). Los más abundantes son leucina, lisina, valina e isoleucina (esenciales), alanina y serina (no esenciales). Contribuyen hasta con el 60 % del volumen de aminoácidos libres en Dicentrarchus labrax (Rønnestad et al., 1998a) y el 50 % en Sparus aurata (Rønnestad et al., 1994).

REDVET. Revista electrónica de Veterinaria 1695-7504 2011 Volumen 12 Número 3
Caracterización bioquímica del consumo de reservas vitelinas en peces teleósteos de ontogenia indirecta.- http://www.veterinaria.org/revistas/redvet/n030311/031102.pdf
6
Tabla 2.- Cantidad de aminoácidos en huevos de teleósteos
Especies AAL (nmol/huevo)
AAL:AA (%) Referencia
Cyclopterus lumpus 66 N/E Thorsen & Fyhn, 1991 Dicentrarchus labrax 90 19 Ronnestad et al., 1993 Fundulus heteroclitus 141 3 Thorsen & Fyhn, 1991 Gadiculus thori 61 40 Thorsen & Fyhn, 1991 Gadus morhua 223 31 Thorsen & Fyhn, 1991 Hippoglossus hippoglossus 2300 23 Finn et al., 1991 Microstomus kitt 134 31 Ronnestad et al., 1992a Pleuronectes platessa 649 35 Thorsen & Fyhn, 1991 Pollachius pollachius 166 38 Thorsen & Fyhn, 1991 Scophthalmus maximus 61 21 Ronnestad et al., 1992b Sparus aurata 43 15 Ronnestad et al., 1993
AA: Aminoácidos totales AAL: Aminoácidos libres N/E: No Estimado
De manera general se ha observado que los AAL son utilizados mayormente durante el desarrollo del embrión después de iniciada la epibolia y hasta el agotamiento del vitelo (Rønnestad & Fyhn, 1993). Sin embargo, se les relaciona en la osmorregulación del proceso de hidratación final del oocito y la embriogénesis (Hølleland & Fyhn, 1986; Fyhn, 1993; Thorsen & Fyhn, 1996). Los AAL también actúan como combustible en el metabolismo energético de huevos y larvas de peces marinos (Fyhn et al., 1987; Fyhn, 1989; Finn et al., 1996). Hay evidencia que sugiere que la importancia cuantitativa de los aminoácidos libres, principalmente en el metabolismo energético durante el desarrollo embrionario en huevos pelágicos de peces marinos puede ser menor en especies que poseen glóbulos de aceite comparado con aquellos que no tienen (Rønnestad et al., 1992a; Rønnestad et al., 1994; Fyhn & Govoni, 1995; Finn et al., 1995a, b; 1996). Lo cual puede ser atribuido a que los huevos que contienen glóbulo de aceite, tienen un alto contenido en lípidos que al parecer son utilizados como energía durante el desarrollo (>15% Finn et al., 1996) por lo que son menos dependientes de los AAL. Desde hace un poco más de tres décadas, se han atribuido dos funciones principales a las proteínas, la formación de tejido y la generación de energía mediante procesos catabólicos (Heming & Buddington, 1988). Existe un grupo de proteínas conocida como enzimas, cuya principal función es la catálisis de diversos procesos metabólicos. Las más estudiadas son las catepsinas, que en primera instancia, degradan la VTG en proteínas, antes de llegar a la gónada, para su almacenamiento en el oocito. Después de la fertilización, estas

REDVET. Revista electrónica de Veterinaria 1695-7504 2011 Volumen 12 Número 3
Caracterización bioquímica del consumo de reservas vitelinas en peces teleósteos de ontogenia indirecta.- http://www.veterinaria.org/revistas/redvet/n030311/031102.pdf
7
enzimas median la degradación de las proteínas almacenadas, en aminoácidos libres para su uso en el desarrollo del embrión (Sire et al., 1994). Las proteínas que forman parte de la envoltura vitelina tienen otras funciones como protección contra daños mecánicos, desecación, cambios químicos bruscos en el ambiente y ejercen actividad bactericida y fungicida (Kudo, 1992). En cuanto a los aminoácidos libres, se ha reportado que su utilización o función pueden ser separada en tres: 1) hidratación o regulación osmótica, 2) síntesis de compuestos, y 3) catabolismo aeróbico. Respecto a la primera función, se ha observado que los AAL son incorporados durante la maduración final del oocito para iniciar un flujo osmótico de agua que permita al oocito hincharse previo al desove (Thorsen & Fyhn, 1991). Este proceso tiene lugar justo antes de la ovulación y resulta en un prominente incremento en el volumen del oocito de hasta 3 o 5 veces (Selman & Wallace, 1989). La segunda función de los AAL es la formación de proteínas durante el desarrollo del embrión. Tanto en Gadus morhua (Fyhn & Serigstad, 1987) como en Scophthalmus maximus (Rønnestad et al., 1992a), Microstomus kitt (Rønnestad et al., 1992b) y Hippoglossus hippoglossus (Rønnestad et al., 1993) se ha observado una correlación entre el incremento en el contenido de proteína de la larva con un decremento en el contenido de aminoácidos libres durante el periodo de alimentación endógena. Para Scophthalmus maximus y Microstomus kitt se ha determinado que aproximadamente el 20 y 30 % (respectivamente) del contenido total de aminoácidos libres es transformado en proteínas. No obstante, aunque la dinámica de la interacción entre los AAL y las proteínas estructurales no sea muy conocida, existe evidencia de que las proteínas estructurales también pueden fungir como almacenamiento temporal de aminoácidos provenientes de AAL durante el estadio de huevo que serán movilizados para el metabolismo energético en estadios posteriores (Rønnestad et al., 1992a; Fyhn & Govoni, 1995). Además, los AAL son empleados para la síntesis de otros compuestos de importancia como las hormonas, los neurotransmisores y cofactores aunque estos cuantitativamente menos importantes, lo cual se ha demostrado para Gadus morhua (Fyhn & Serigstad, 1987). La tercera función de los AAL consiste en proveer energía mediante el catabolismo aeróbico. Esta producción de energía se da cuando se remueve el grupo α-amino y se forma un grupo ácido α-ceto mediante diversas vías enzimáticas. El principal destino del ácido α-ceto es la oxidación mediante el ciclo del ácido tricarboxílico. La energía liberada es conservada a través de la síntesis de fosfatos de alta energía (ATP’s) (Rønnestad & Fyhn, 1993). El catabolismo aeróbico de los AAL con frecuencia es medido por la producción de amonio y el consumo de oxígeno (Rønnestad et al., 1992a). El cual será

REDVET. Revista electrónica de Veterinaria 1695-7504 2011 Volumen 12 Número 3
Caracterización bioquímica del consumo de reservas vitelinas en peces teleósteos de ontogenia indirecta.- http://www.veterinaria.org/revistas/redvet/n030311/031102.pdf
8
tratado de manera más detallada en el apartado siguiente (metabolismo de las macromoléculas). • Lípidos Seguido de las proteínas, los lípidos son el siguiente componente más abundante de las reservas de vitelo (saco de vitelo y glóbulo de aceite) en la mayoría de los huevos de peces (Tabla 1) (Heming & Buddington, 1988). El contenido lipídico en término de cantidad varía de manera interespecífica desde el 0.1 % del peso del huevo en Pleuronectes platessa hasta el 45 % en Labeotropheus sp. (Balon, 1977). Dicha variación ha sido atribuida a la adaptación a diferentes estrategias reproductivas (Balon, 1977) o a la duración del periodo de alimentación endógena (Kaitaranta & Ackman, 1981). Los lípidos del vitelo se encuentran principalmente en los glóbulos de aceite, que pueden variar en número y talla, desde múltiples glóbulos pequeños, hasta la presencia de uno solo de gran tamaño como en Coregonus alpinus (Heming & Buddington, 1988; Kunz, 2004). De acuerdo con análisis histoquímicos, los lípidos neutros o triglicéridos con sus constituyentes ácidos grasos (AG), son los más abundantes en el (los) glóbulo (s) de aceite, (Kaitaranta & Ackman, 1981; Planas et al., 1993; Rønnestad et al., 1998a; Kunz, 2004). Finn (1994) sugiere que la presencia de un solo glóbulo de aceite reflejará un contenido lipídico mayor al 15 %. Los lípidos se encuentran también en el saco de vitelo (Tabla 3) y son considerados como fracción de alta densidad (FAD). Los fosfolípidos son los lípidos mas abundantes en los huevos que no tiene glóbulo de aceite (Ohkubo et al., 2006). En especies que poseen un glóbulo de aceite se ha observado que es mayor la cantidad de ácidos grasos procedentes de lípidos neutros especialmente triglicéridos que aquellos procedentes de los fosfolípidos (Rønnestad et al., 1998a). En general, los triglicéridos son considerados como mejor fuente de energía disponible que los lípidos polares (Planas et al., 1993; Finn, 1994). No obstante, existe una diferencia en el tipo de lípido utilizado como principal fuente de energía entre peces de aguas cálidas y de aguas frías. En Sciaenops ocellata (especie cálida) son preferiblemente consumidos los lípidos neutros (Vetter et al., 1983), mientras que en especies como Gadus morhua y Pleuronectes platessa (especies de aguas frías) utilizan como principal reserva, lípidos polares como fosfatidilcolina y fostatidiletanolamina (Rainuzzo et al., 1992). Los principales ácidos grasos de lípidos neutros que son catabolizados durante el desarrollo son los n-3 (ácido decosahexanoico, DHA; ácido eicosapentanoico, EPA). Estos ácidos grasos son los más abundantes pese a su grado de insaturación. Los n-3, al igual que el ácido araquidónico (ARA, n-6) son considerados esenciales (Rønnestad et al., 1998a). En términos de síntesis de los ácidos grasos poliinsaturados (PUFAS, por sus siglas en inglés), se observa una clara división entre peces dulceacuícolas y

REDVET. Revista electrónica de Veterinaria 1695-7504 2011 Volumen 12 Número 3
Caracterización bioquímica del consumo de reservas vitelinas en peces teleósteos de ontogenia indirecta.- http://www.veterinaria.org/revistas/redvet/n030311/031102.pdf
9
marinos. Los peces de agua dulce satisfacen sus requerimientos de ácidos grasos esenciales (AGE) como lo son el EPA, DHA y ARA a partir del ácido linoleico (18:2n-6) y ácido linolénico (18:3n-3). Estos ácidos grasos son precursores del ARA 18:2n-6 y del EPA y DHA, y los peces de agua dulce pueden elongar las cadenas y disminuir saturaciones (reducir) de estos AG para formar el ácido araquidónico (ARA, 20:4n-6), el ácido eicosapentaenoico (EPA, 20:5n-3) y el ácido docosahexaenoico (DHA, 22:6n-3), para su incorporación a los fosfolípidos de las membranas celulares (Kanazawa et al., 1980; Sargent et al., 1995). El precursor es el 18:3n-3 (Sargent et al., 2003). Al contrario los peces marinos presentan una escasa capacidad de sintetizarlos el ARA, el EPA y el DHA. (Owen et al., 1975; Cowey et al., 1976; Yamada et al., 1980; Sargent et al., 1995). Se ha reportado que insuficientes niveles en las dietas de AA, EPA y DHA son la causa principal de crecimiento lento y altas mortalidades observadas en las larvas de peces marinos y se requiere la presencia de ellos en las concentraciones y proporciones adecuadas a cada especie para su desarrollo inicial (Fukusho et al., 1984; Kitajima et al., 1980a, b; Mourente et al., 1999; Sargent et al., 1999; Watanabe et al., 1983, Watanabe, 1993). Otro rol importante del DHA es que es indispensable para la formación de tejido nervioso y la formación de la retina (Kanazawa, 1993). En estudios realizados por Bell et al. (1995) se demostró una baja actividad depredadora de las larvas de Clupea harengus alimentadas con Artemia deficiente en DHA. La importancia del ARA radica en que es un precursor primario de eicosanoides y prostaglandinas cuya función esta relacionada con la actividad del sistema nervioso central y el sistema inmunológico (Castell, 1978; Bell & Sargent, 2003). Estudios comparativos de larvas alimentadas y no alimentadas, mostraron que la concentración de ARA, EPA y DHA no varía a expensas de otros ácidos grasos en las larvas no alimentadas. Lo cual sugiere que deben ser componentes esenciales para las larvas de Sparus aurata (Koven et al., 1989; Rodríguez, 1994), Pagrus major (Tandler et al., 1989), Scophthalmus maximus (Rianuzzo et al., 1994), Gadus morhua (Van der Meeren et al., 1991) y Perca fluviatilis (Abi-Ayad et al., 2000). • Carbohidratos De las biomoléculas presentes en el huevo, los carbohidratos son los menos abundantes (Tabla 1). Se localizan principalmente asociadas a la membrana del huevo aunque también los hay en forma libre en el saco de vitelo (Tabla 3) (Heming & Buddington, 1988). Los carbohidratos son los nutrientes mayormente empleados desde la fertilización hasta la eclosión y se le han atribuido roles nutricionales en el proceso inicial de la segmentación (Heming & Buddington, 1988; Moroz & Luzhin, 1976). Sin embargo, también se encuentran carbohidratos en el vitelo, ya sea de forma libre o asociados a

REDVET. Revista electrónica de Veterinaria 1695-7504 2011 Volumen 12 Número 3
Caracterización bioquímica del consumo de reservas vitelinas en peces teleósteos de ontogenia indirecta.- http://www.veterinaria.org/revistas/redvet/n030311/031102.pdf
10
proteínas como glicoproteínas. Las glucoproteínas alcanzan hasta un 10 % del total de las proteínas encontradas en Salmo gairdneri (Nakagawa, 1970). En Salmo salar, se ha reportado que después de la membrana del huevo, el líquido perivitelino contiene la mayor cantidad de carbohidratos alcanzando 48 y 46 %, respectivamente, mientras que en el vitelo se reporta solo el 2 % en base al peso total (Hamor & Garside, 1977a). El glucógeno presente en el vitelo es el principal carbohidrato. Se utiliza como una fuente de energía relacionada con etapas embrionarias tempranas (Vetter et al., 1983). En Sciaenops ocellata, el glucógeno disminuye un 63% entre la fertilización y la eclosión (Vetter et al., 1983). Lahnsteiner & Patarnello (2003) mencionan que esta especie es la única en la que se ha estudiado a detalle el metabolismo de los carbohidratos y dado que encontraron que el glucógeno disminuye de manera considerable durante el desarrollo del huevo, se ha asociado con la principal fuente de energía en esa etapa. Sin embargo, en Sparus aurata, la cantidad de carbohidratos (monosacáridos: glucosa, fructosa y galactosa) no decreció, lo cual indica que estos no juegan un rol como fuente energética durante el desarrollo (Lahnsteiner & Patarnello, 2003; 2004). En ese sentido, se ha considerado que los monosacáridos tienen otro tipo de funciones como por ejemplo la síntesis de ácidos nucléicos (ARN, ADN) (Rawn, 1983). Carnevali y colaboradores (2001) y Lahnsteiner & Patarnello (2003) reportaron un constante incremento en la síntesis de ácidos nucléicos durante la embriogénesis. Los monosacáridos pueden también contribuir a la hidratación de los oocitos. La reacción cortical y la subsecuente entrada de agua entre el ambiente y el espacio perivitelino son procesos regulados osmóticamente, los cuales ocurren después de que el huevo ha sido liberado (Alderdice, 1988). Las vesículas corticales contienen principalmente carbohidratos (Alderdice, 1988). Inmediatamente después de que los huevos han sido liberados, el agua puede fluir dentro de ellos dado que la permeabilidad del oolema aun es alta (Lahnsteiner & Patarnello, 2003). Tabla 3.- Localización de los componentes nutrimentales en huevos de Salmo salar. Los valores están dados en porcentaje de peso total. Tomado de Hamor & Garside (1977a)
Componentes Membrana del huevo
Líquido perivitelino Vitelo
Agua 5.2 23.3 71.8 Masa seca 0.8 16.9 82.1 Proteínas 0.3 22.7 76.9 Lípidos 0.1 7.7 92.2 Carbohidratos 48.0 46.0 2.0 Cenizas 7.1 7.1 85.7

REDVET. Revista electrónica de Veterinaria 1695-7504 2011 Volumen 12 Número 3
Caracterización bioquímica del consumo de reservas vitelinas en peces teleósteos de ontogenia indirecta.- http://www.veterinaria.org/revistas/redvet/n030311/031102.pdf
11
• Otros nutrientes Pese a que las proteínas y los lípidos constituyen la mayor cantidad de las reservas para el desarrollo de los embriones, existen otras moléculas que pueden ser derivadas de fuentes externas al oocito como las vitaminas y metales traza (Brooks et al., 1997). Aunque estos componentes se encuentran en muy pequeñas cantidades, son igualmente necesarios para promover huevos viables y subsecuentemente larvas viables. Las vitaminas y minerales son requeridos como co-factores (Brown & Lynam, 1981). La función principal atribuida a las vitaminas es la síntesis de proteínas y por ende el crecimiento (Woodward, 1994; Mæland et al., 2003) o bien propiedades antioxidantes como es el caso del ascorbato y α-tocoferol (Yamamoto et al., 2001). Se han observado diferentes complejos vitamínicos en algunos teleósteos como la vitamina A (trans-retinal; trans-retinol, y 13-cis-retinol) (Rønnestad et al., 1998b), vitamina B (Rønnestad et al., 1997; Mæland et al., 2003), vitamina C (ascorbato) (Rønnestad et al., 1999) y vitamina E (α-tocoferol) (Rønnestad et al., 1999). En Hippoglossus hippoglossus, se observó que la principal fuente de vitamina A son los trans-retinal alcanzando hasta un 90 % del contenido total de este complejo vitamínico (Rønnestad et al., 1998b). La vitamina A ha sido implicada en diversos procesos biológicos. Durante la embriogénesis, se relaciona con la proliferación y diferenciación celular (Audouin-Chevallier et al., 1993). Durante el desarrollo larval se le asocia con la pigmentación, en donde un déficit de esta vitamina puede ocasionar un pseudoalbinismo (Miki et al., 1990; Kanazawa, 1993; Takeuchi et al., 1995). Además, es de vital importancia para el funcionamiento de la retina al momento de la primera alimentación. En la misma especie, se reporta una disminución de vitamina B (ácido fólico) desde la eclosión hasta la primera alimentación de aproximadamente el 50 % de su contenido total (Mæand et al., 2003), mientras Rønnestad et al. (1997) reportan en esta misma especie una disminución en vitamina B6 del 25 % a partir del día 10 después de la eclosión y hasta la primera alimentación. Así mismo, una gran variedad de hormonas, factores de crecimiento y ARNs depositados en el vitelo que codifican para estos componentes están involucrados en la embriogénesis (Brooks et al., 1997). Los pigmentos, específicamente los carotenoides, son otro tipo de componente bioquímico del vitelo. Entre los carotenoides identificados dentro de los ovarios o huevos de peces están las astaxantinas, tunaxantinas, luteínas, zeaxantinas e idoxantinas y otras más encontradas en cantidades trazas (Lubzens et al., 2003). El pigmento más representado en los huevos de peces es la astaxantina, constituyendo hasta un 80 % de pigmentos totales y son

REDVET. Revista electrónica de Veterinaria 1695-7504 2011 Volumen 12 Número 3
Caracterización bioquímica del consumo de reservas vitelinas en peces teleósteos de ontogenia indirecta.- http://www.veterinaria.org/revistas/redvet/n030311/031102.pdf
12
transportados del vitelo, principalmente a la dermis, aunque también pueden ser utilizados por otros órganos (Kitahara, 1984). Metabolismo de las macromoléculas La acumulación de macromoléculas en los oocitos provee de material de reserva que es utilizado en diferentes actividades biosintéticas esenciales durante la embriogénesis como fuente de energía (Boulekbache, 1981). Las principales fuentes energéticas de reserva en teleósteos son glucógeno, lípidos (triglicéridos, lípidos neutros y ácidos grasos) proteínas de vitelo (lipoproteínas y fosfoproteínas). Estas fuentes de reserva son metabolizadas por distintos metabolitos o enzimas (Boulekbache, 1981). Los estudios del metabolismo se llevan a cabo evaluando la concentración de diversos componentes. Para el metabolismo de las proteínas y aminoácidos, se cuantifican las proteínas totales, aspartato-proteasa o catepsina D, cisteina-proteasa o catepsina L y catepsina B, aminoácidos libres, aspartato-aminotransferasa, alanina-aminotransferasa, glutamato-deshidrogenasa (Kestemont et al., 1999; Lahnsteiner et al., 1999; Lahnsteiner & Partenello, 2003; 2004; Britton & Muray, 2004); Para los lípidos: colesterol, ácidos grasos esterificados y no-esterificados, glicerol, fosfatidil-colina, fosfatidil-etanolamina, fostatidil-inositol, triglicéridos, acetilcarnitina transferasa, lipasa (Lahnsteiner et al., 1999; Zhu et al., 2001; Salze et al., 2005); y para los carbohidratos: fructosa, galactosa, glucosa, glucógeno, lactato, transaldolasa, glucosa-6-fosfatasa, fosfofructoquinasa, piruvatoquinasa, lactatodeshidrogenasa (Terner, 1979; Boulekbache, 1981; Mommsen & Walsh, 1988; Lahnsteiner et al., 1999; Lahnsteiner & Partenello, 2003; 2004). El metabolismo de los carbohidratos es alto durante el desarrollo de huevos, como sucede en Sparus aurata (Lahnsteiner & Patarnello, 2003). En esta especie, los niveles de monosacáridos libres y totales, así como la actividad piruvato carboxilasa se incrementan durante el desarrollo. La piruvato carboxilasa cataliza la conversión intra-mitocondrial de piruvato a oxalacetato. Posteriormente, el oxalacetato es transaminado en aspartato por la aspartato aminotransferasa y transportado fuera de la mitocondria; donde es retransaminado a oxalacetato, para llegar a tomar parte en reacciones durante la glucogénesis (Rawn, 1983). Así, los monosacáridos no tienen que ser almacenados, si no que son reutilizados y sirven para la síntesis de ácidos nucléicos. En cuanto a las proteínas, durante el desarrollo se ha reportado un nivel constante para algunos teleósteos (Fyhn & Govoni, 1995; Rønnestad et al., 1998a; Carnevali et al., 2001; Lahnsteiner & Patarnello, 2004). La actividad proteolítica, sobre todo de la catepsina L, localizada en el sincitio, se incrementa inmediatamente después de la fertilización (Sire et al., 1994; Kestesmont et al., 1999; Carnevali et al., 2001), lo mismo sucede con la

REDVET. Revista electrónica de Veterinaria 1695-7504 2011 Volumen 12 Número 3
Caracterización bioquímica del consumo de reservas vitelinas en peces teleósteos de ontogenia indirecta.- http://www.veterinaria.org/revistas/redvet/n030311/031102.pdf
13
catepsina B (Gerhartz et al., 1997). Mientras que la catepsina D solo actúa durante la endocitosis de la VTG y disminuye significativamente después. (Kestesmont et al., 1999), no obstante, no es una generalidad en teleósteos, ya que se ha observado que algunas proteasas como las catepsinas B no son detectadas durante la vitelogénesis ni en el desarrollo embrionario de Oncorhynchus mykiss (Sire et al., 1994). En términos de aminoácidos, se observa de manera general un descenso en el contenido de AAL. La principal diferencia entre especies es el momento en que ocurre. Por ejemplo, en Hippoglussus hippoglossus (Fyhn, 1989; Finn et al., 1991; Rønnestad et al., 1993) ocurre principalmente después de la eclosión, es decir, en la larva vitelina, estadio al que Balon (1975) llama eleuteroembrión; término poco utilizado en la actualidad (Kamler, 2002). En Scophthalmus maximus, (Rønnestad et al., 1992a), Sparus aurata (Rønnestad et al., 1993) y Dicentrarchus labrax (Gatesoupe, 1986), el descenso se observa en estadios embrionarios y poco después de la eclosión. Finalmente, se ha observado que la disminución de los AAL en Microstomus kitt (Rønnestad et al., 1992b) y Gadus morhua (Fyhn & Serigstad, 1987), es a través de todo el periodo de alimentación endógena. El metabolismo o consumo de los AAL es similar entre AAL esenciales y no-esenciales (Fyhn & Govoni, 1995). No obstante, existen excepciones en dos aminos ácidos no esenciales (Rønnestad et al., 1998a). La cantidad de taurina permanece constante a través del desarrollo embrionario (Fhyn, 1989; Fyhn & Govoni, 1995; Rønnestad et al., 1998a). La función de la taurina en los primeros estadios de vida aun no es clara, sin embargo, en peces adultos esta ligada a procesos de regulación del volumen celular, inhibición de la transmisión de impulsos nerviosos, regulación de la excitación de células sensoriales; por lo que, probablemente, alguna de esas funciones realice en el desarrollo temprano (Rønnestad et al., 1998a). La fosfoserina también permanece en cantidades constantes a través del desarrollo (Rønnestad et al., 1994) lo cual puede ser ocasionado por la degradación de la fosvitina, la cual contiene un 12 % de fosforo y 45 % de serina (Murakami et al., 1990). De manera general, los AAL han sido asociados con la producción de energía durante el desarrollo embrionario (Rønnestad et al., 1994; 1998a) y con la osmoregulación para ayudar a la flotabilidad del huevo (Thorsen & Fhyn, 1996). En Sparus aurata, el catabolismo de los aminoácidos fue demostrado por la presencia de actividad aspartato-aminotransferasa y glutamato deshidrogenasa. Un agudo incremento en la actividad de la aspartato-aminotransferasa en estadios embrionarios puede ser indicador de un alto catabolismo de aminoácidos, lo cual está relacionado con la gluconeogénesis (Rønnestad et al., 1998a). Durante el metabolismo de los lípidos, se ha observado que en el desarrollo embrionario la composición lipídica permanece sin cambios significativos

REDVET. Revista electrónica de Veterinaria 1695-7504 2011 Volumen 12 Número 3
Caracterización bioquímica del consumo de reservas vitelinas en peces teleósteos de ontogenia indirecta.- http://www.veterinaria.org/revistas/redvet/n030311/031102.pdf
14
(Vetter et al., 1983; Zhu et al., 2001; Salze et al., 2005) lo que indica que antes de la eclosión no hay una preferencia por el consumo de algún tipo o clase de lípido. Después de la eclosión, los fosfolípidos del vitelo son preferentemente consumidos (Heming & Buddington, 1988). En Dicentrarchus labrax, se observa que después de la eclosión, los ácidos grasos de los lípidos neutros derivados del glóbulo de aceite son mayormente empleados (Rønnestad et al., 1998a). Recientemente, en Hippoglossus hippoglossus se observa un incremento en triglicéridos y fosfolípidos (fosfatidilcolina y fosfatidieletanolmina) después de la eclosión (Zhu et al., 2001). Zhu y colaboradores (2001) concluyen que el hecho de que se observe un aumento en cantidad de todas las clases lipídicas es debido a que los aminoácidos libres son la fuente de carbono para la síntesis de lípidos en la eclosión. Al momento de la primera alimentación, en Gadus morhua (Finn et al., 1995a) y Pseudopleuronectes americanus (Cetta & Capuzzo, 1982) se observa que la cantidad de lípidos decrece. Caracterización del consumo del vitelo La absorción del vitelo en peces se da mediante una estructura especializada llamada sincitio, que desaparece una vez que el vitelo haya sido agotado (Yamamoto, 1982). El presuntivo sincitio o periblasto, puede ser reconocido en los huevos de teleósteos desde la fecundación (Yamamoto, 1982). Conforme la segmentación avanza, van apareciendo nuevos núcleos libres en el periblasto, convirtiendo esta capa en un sincitio más diferenciado. La formación del sincitio en teleósteos se da por la sobreposición del mesodermo y ectodermo sobre el saco de vitelo encerrándolo por completo. Lo que hace que la absorción del vitelo ocurra en ausencia de células del endodermo por lo cual el vitelo no es absorbido por el intestino que es de origen endodérmico (Bachop & Schwartz, 1974). La circulación del material de reserva de la masa vitelina hacia las células somáticas, se da mediante un sistema de vasos sanguíneos desarrollados entre las paredes del saco vitelino. Algunas regiones de estos capilares están desprovistos de pared interna lo que permite el contacto entre el saco vitelino y el sincitio (Shimizu & Yamada, 1980). Los componentes del vitelo entran en el sincitio por endocitosis. Se presenta luego una digestión intrasincitial y posterior síntesis de metabolitos los cuales son liberados a la circulación vitelina (Heming & Buddington, 1988). En el sincitio pueden diferenciarse dos regiones principales de actividad. La primera región se encuentra a través de todo el sincitio y se caracteriza por la presencia de retículo endoplásmico liso y un gran número de mitocondrias. La segunda región se extiende en parches a través del sincitio formando una estructura estratificada. Se caracteriza por la presencia de retículo endoplástico rugoso y aparato de Golgi. Las dos regiones del sincitio tienen funciones diferentes, se asocia la primera con el metabolismos de carbohidratos y lípidos mientras que

REDVET. Revista electrónica de Veterinaria 1695-7504 2011 Volumen 12 Número 3
Caracterización bioquímica del consumo de reservas vitelinas en peces teleósteos de ontogenia indirecta.- http://www.veterinaria.org/revistas/redvet/n030311/031102.pdf
15
la segunda región tiene como funciones la síntesis y transporte de sustancias proteicas (Shimizu & Yamada, 1980). De manera general, el consumo de vitelo se ha caracterizado en muchos teleósteos por presentar tres fases de absorción. La primera fase, también llamada fase de pre-eclosión, es caracterizada por un suave pero constante incremento en la tasa de absorción de vitelo, donde el saco de vitelo y el glóbulo de aceite son consumidos en aproximadamente la misma proporción (Nakagawa & Tsuchiya, 1972). Justo un poco antes y al momento de la eclosión, la tasa de absorción de vitelo se incrementa rápidamente, probablemente en respuesta al aumento de la superficie de absorción debido a los cambios en la forma del saco vitelino o bien al aumento en la actividad metabólica del sincitio. Esto marca el inicio de la segunda fase o fase post-eclosión, y es caracterizada por una relativamente alta y constante tasa de absorción. Durante esta fase, el saco de vitelo es preferentemente consumido (May, 1974; Eldridge et al., 1982; Li & Mathias, 1982; Quantz, 1985). Dado que se aproxima el agotamiento del saco de vitelo, su tasa de absorción disminuye, probablemente en respuesta a la disminución de la superficie del área de absorción por el encogimiento del saco de vitelo o al cambio en la composición del vitelo. De esta forma da inicio de la tercera fase o fase terminal de absorción, en la cual, el glóbulo de aceite es preferentemente consumido (Fig. 1) (Heming & Buddington, 1988). Este patrón de consumo de reservas vitelinas se ha reportado para diferentes especies como Siganus guttatus (Avila & Juario, 1987), Microstomus kitt (Rønnestad, et al., 1992a), Sparus aurata (Rønnestad et al., 1994), Brevooria tyranus y Leiostumus xanthurus (Fyhn & Govoni, 1995) Lutjanus campechanus (Williams et al., 2004), Pagrus pagrus (Aristizabal, 2006), Lutjanus peru (Zavala, 2007) entre otros.
Figura 1.- Patrón de consumo de reservas vitelinas en teleósteos. La línea punteada indica el 50 % de eclosión. (Modificado de Heming & Buddington, 1988).

REDVET. Revista electrónica de Veterinaria 1695-7504 2011 Volumen 12 Número 3
Caracterización bioquímica del consumo de reservas vitelinas en peces teleósteos de ontogenia indirecta.- http://www.veterinaria.org/revistas/redvet/n030311/031102.pdf
16
Se ha observado que el consumo es mayor durante las primeras horas después de la eclosión, coincidiendo con un incremento en la talla de la larva (May et al., 1974; Heming & Buddington, 1988; Zavala, 2007). En términos bioquímicos, se ha esquematizado un patrón de consumo de macromoléculas para la producción de energía. Antes de la eclosión, se consumen principalmente carbohidratos, lípidos y proteínas en ese orden. Después de la eclosión, lípidos y proteínas. No obstante, no se puede precisar una secuencia del consumo para todos los peces, ya que la composición del vitelo es variable (Heming & Buddington, 1988). Actualmente se tiene evidencia de que los carbohidratos son el principal sustrato para la energía en un periodo corto de divisiones celulares en diversas especies dulceacuícolas y marinas (Finn et al., 1995b). No obstante, en algunas especies como Gadus morhua (Finn et al., 1995b), y Scophthalmus maximus (Finn et al, 1996) se ha sugerido que los aminoácidos libres son la principal fuente de energía durante la embriogénesis. En contraste, después de la eclosión de Scophthalmus maximus, los lípidos son la principal fuente de energía (Rønnestad et al., 1992b). Finn & Govoni (1995) reportaron en esta misma especie que durante el periodo de alimentación endógena, la contribución de los aminoácidos totales (AAL + ligados a proteínas) contribuye con el 40 % de la energía, mientras que los lípidos el 60 %. En Sparus aurata se observó que los aminoácidos libres son la fuente principal de energía durante el desarrollo embrionario (60-70 % de energía), mientras que después de la eclosión y durante el periodo de alimentación endógena, los lípidos neutros derivados del glóbulo de aceite son el combustible dominante (Rønnestad et al., 1994). En Dicentrarchus labrax, la principal fuente de energía para el catabolismo durante la embriogénesis e inicio del periodo de alimentación endógena son los aminoácidos libres (Rønnestad et al., 1994). En Theragra chalcogramma la secuencia en la utilización durante la embriogénesis es similar, sin embargo después de la eclosión y durante el periodo de alimentación endógena, la secuencia de utilización es primero las proteínas (lipovitelina) y luego, los lípidos (fosfolipidos y triglicéridos) (Ohkubo et al., 2006). La tasa de absorción o consumo de vitelo puede ser determinado por diferentes metodologías: cambio en las calorías del vitelo a través de un tiempo dado, el peso seco, el peso húmedo, el volumen o el área como referencia entre el tiempo inicial (eclosión) y otro momento durante la alimentación endógena. No obstante, cada uno de estos métodos tiene sus limitantes, las cuales deben ser consideradas. Por ejemplo, el área y volumen del vitelo generalmente son estimados por la medición de dos dimensiones, lo

REDVET. Revista electrónica de Veterinaria 1695-7504 2011 Volumen 12 Número 3
Caracterización bioquímica del consumo de reservas vitelinas en peces teleósteos de ontogenia indirecta.- http://www.veterinaria.org/revistas/redvet/n030311/031102.pdf
17
cual requiere que la masa vitelina sea visible. La evaluación del volumen, área y peso húmedo pueden ser influenciados por factores ambientales como la salinidad (Alderdice et al., 1979) y la preservación de los organismos (Heming & Preston, 1981). El peso húmedo puede ser influenciado por la cantidad relativa de agua del vitelo, la cual puede variar durante el desarrollo (Heming & Buddington, 1988). Desde el punto de vista energético, la determinación del contenido de calorías totales del vitelo es la mejor aproximación. Por otro lado, la tasa de consumo de las reservas vitelinas está en función del área de la superficie de la capa absortiva (sincitio) y la actividad metabólica de dicha capa. En teleósteos, la superficie del sincitio es aproximadamente igual al área del vitelo y conforme cambia la superficie del saco de vitelo se reduce también el sincitio. En su revisión, Heming y Buddington (1988) muestran algunas fórmulas matemáticas para calcular el área absorbente y el volumen de la masa de vitelo de diversas formas.
1. Masa esférica Área: A = �D2 Volumen: V = 0.1667�D3, donde D es le diámetro del vitelo
2. Masa cónica y piriforme (forma de pera) Área: A = 0.250�[H1
2 + H22 + (H1 + H2) + [(H2-H1)2 + 4L2]0.5]
Volumen: V = 0.0833�L(H12 + H2
2 + H1H2) Donde H1 es la altura del extremo más pequeño (H1 = 0 en una masa de forma cónica), H2 es la altura del extremo más grande, y L es la longitud de la masa vitelina.
3. Masa cilíndrica Área: A = 0.50�(H2 + 2HL) Volumen: V = 0.250�H2L
4. Masa elipsoidal Área: A = 0.50�H2 + �HL2 (L2-H2)-0.5 senh-1 [(L2-H2)/L] Volumen: V = 0.1667�LH2
Los factores que afecten la actividad metabólica del sincitio afectan de manera directa la tasa de absorción del vitelo. La tasa de absorción es reducida por una baja concentración de oxígeno (Harmor & Garside, 1977b), por un sub o supraoptimo nivel de salinidad (May, 1974), altas concentraciones de amonio (Fedorov & Smirnova, 1978), concentraciones subletales de xenobióticos tóxicos (Crawford & Guarino, 1985). En contra parte, la tasa de absorción se incrementa por el efecto de la temperatura fuera de un rango óptimo (Heming & Buddington, 1988; Polo et al., 1991; Jobling 1997; Jordaan, 2002; Zavala, 2007), y se ha reportado que, en Morone saxatilis se incrementa la tasa de absorción del glóbulo de aceite al

REDVET. Revista electrónica de Veterinaria 1695-7504 2011 Volumen 12 Número 3
Caracterización bioquímica del consumo de reservas vitelinas en peces teleósteos de ontogenia indirecta.- http://www.veterinaria.org/revistas/redvet/n030311/031102.pdf
18
momento de la co-alimentación (traslape de la alimentación endógena y exógena) (Rogers & Westin, 1981; Eldridge et al., 1982). Sin embargo, no existe un patrón definido en las especies que presentan co-alimentación, en Oncorhynchus tshawytscha no se observa un efecto en la tasa de consumo (Heming, 1982). La eficiencia de utilización del vitelo es un aspecto muy importante para la supervivencia larval. Se mide por el crecimiento de la larva durante el consumo de vitelo. Sin embargo, también puede ser calculada como la proporción entre el consumo de vitelo (metodologías antes mencionadas para calcularlo) y formación de tejido en peso seco o calorías (Blaxter, 1969). Es de esperarse que la eficiencia con que son consumidas las reservas vitelinas sea menor al 100 %, debido principalmente a la pérdida de energía por el metabolismo de mantenimiento. La tasa de crecimiento embrionario está en función de la composición del vitelo, degradación del vitelo por medio del sincitio, la absorción y transporte de los nutrientes del vitelo, así como de síntesis de tejido somático y de la demanda metabólica de mantenimiento y actividad. Los factores que afectan la eficiencia de utilización del vitelo pueden clasificarse como bióticos y abióticos. Dentro de los bióticos destacan la calidad y composición del huevo, así como la talla del huevo (Heming & Buddington, 1988). Mientras que los factores abióticos que se han reportado son niveles extremos de pH (Nelson, 1982), temperaturas fuera del rango óptimo (Blaxter & Hempel, 1966), salinidad (May, 1974), bajas concentraciones de oxígeno disuelto (Hamor & Garside, 1977a), exposición y calidad de la luz (Hamor & Garside, 1975) y xenobióticos tóxicos (Tilseth & Ellertsen, 1984). Pese al número de factores abióticos que se han identificado, sin lugar a dudas la temperatura es el más estudiado. Después de un gran número de investigaciones realizadas del efecto de la temperatura sobre la eficiencia en la utilización del vitelo se puede decir que no existe un patrón definido dado que los resultados pueden ir en un sentido como en otro, es decir, la temperatura tiene un efecto especie-específico (Pittman, et al., 1989; Blaxter, 1992; Gracia-López et al. 2004). Por citar algunos ejemplos, en la cabrilla sardinera Mycteroperca rosacea (Gracia-López et al., 2004), Anarhichas minor (Sund & Falk-Petersen, 2005), Lutjanus peru (Zavala, 2007) las larvas presentan una mayor longitud a la temperatura más baja del rango probado, mientas que otras como Hippoglossoides elassondon (Alderdice & Forrester, 1974) la longitud es mayor a la temperatura más alta.

REDVET. Revista electrónica de Veterinaria 1695-7504 2011 Volumen 12 Número 3
Caracterización bioquímica del consumo de reservas vitelinas en peces teleósteos de ontogenia indirecta.- http://www.veterinaria.org/revistas/redvet/n030311/031102.pdf
19
Influencia del contenido bioquímico en el desarrollo y supervivencia Las reservas necesarias para el desarrollo de embriones y larvas de teleósteos son sintetizadas durante la vitelogénesis, acumulándose en el vitelo del oocito. Mediante estudios bioquímicos se ha encontrado una relación entre la dieta de los reproductores y la composición del huevo y de las reservas vitelinas de las larvas (Mazorra et al., 2003). En ese sentido, es posible evaluar mediante métodos bioquímicos la relación entre composición y la calidad del oocito. Los principales indicadores bioquímicos de la calidad de huevos y larvas son determinaciones lipídicas: lípidos totales, lípidos neutros, lípidos polares y ácidos grasos (Sargent, 1995; Bell & Sargent, 2003;), aminoácidos (Rønnestad & Fyhn, 1993) y vitaminas (Rønnestad et al., 1997, 1999; Mæland et al., 2003). Recientemente, algunas enzimas involucradas en el metabolismo energético han sido identificadas como buenos marcadores para definir la calidad (Lahnsteiner & Patarnello, 2004). Estos indicadores generalmente son relacionados o contrastados con la tasa de eclosión, supervivencia a la eclosión, malformaciones, tiempo en que tarda la larva en alcanzar la muerte por inanición, supervivencia a la metamorfosis (Lahnsteiner et al., 1999; Giménez et al., 2006). Sin embargo hoy en día las determinaciones bioquímicas de ácidos grasos son la herramienta más empleada para evaluar la calidad. Los ácidos grasos mayormente empleados para tales fines son los poliinsaturados (AGPI) como el ácido araquidónico (ARA, 20:4n-6), el ácido eicosapentaenoico (EPA, 20:5n-3) y el ácido docosahexaenoico (DHA, 22:6n-3). Se ha observado que el uso en reproductores de dietas con diferentes niveles de los ácidos grasos antes mencionados tiene un efecto sobre la tasa de fertilización, tasa de eclosión y supervivencia en Sparus aurata (Fernández-Palacios et al., 1995) y Dicentrarchus labrax (Bruce et al., 1999). Se ha reportado también que la proporción DHA/EPA y el contenido de ARA afecta la morfología de los blastómeros (Picknova et al., 1997) y la supervivencia larval (Furuita et al., 2000). Salze et al., (2005) reportan que en Gadus morhua, altos niveles de fosfatidil-inositol, ARA y carotenoides ayudan a alcanzar altos porcentajes de fertilización y eclosión, mejor simetría de los blastómeros durante la segmentación y supervivencia a la eclosión. Mientras que la proporción DHA/EPA no presentó diferencia en estos parámetros ni en otros. Pese a estos resultados, las investigaciones sobre la cantidad o nivel requerido de ácidos grasos deben hacerse para cada especie. En Gadus morhua (Zheng et al., 1996), Paralichthys olivaceus (Furuita et al., 1998), Seriola sp. (Ishizaki et al., 1998) se ha reportado que el crecimiento y supervivencia larval es menor con altos niveles de ARA en la dieta de los reproductores. Otro efecto negativo de un alto nivel de ARA es el albinismo que ha sido demostrado en Scophtalmus maximus (Estévez et al., 1999; Sargent et al., 1999). Uno de los trabajos más antiguos en el cual se evaluó

REDVET. Revista electrónica de Veterinaria 1695-7504 2011 Volumen 12 Número 3
Caracterización bioquímica del consumo de reservas vitelinas en peces teleósteos de ontogenia indirecta.- http://www.veterinaria.org/revistas/redvet/n030311/031102.pdf
20
la calidad del huevo a través de indicadores bioquímicos es el de Hirao et al. (1954) en el cual encontró una correlación entre el hierro (Fe) y el porcentaje de eclosión. Craik & Harvey (1984) observan una correlación entre la tasa de eclosión y los niveles de hierro, la composición proteica y lipídica del huevo. Agradecimientos A la Dra. Silvie Dumas y al Dr. Renato Peña su valiosa ayuda para la realización de esta revisión bibliográfica. Al Dr. Iván Murillo, Dr. José De la Cruz y a la Dra. Laura Sánchez por sus acertados comentarios y revisiones del presente trabajo. Al Centro Interdisciplinario de Ciencias Marinas (CICIMAR-IPN) por el apoyo otorgado. A CONACYT y PIFI por las becas otorgadas. Literatura citada • Abi-Ayad, S.M.E.A., P. Kestemont & C. Melard. DYNAMICS OF TOTAL
LIPIDS AND FATTY ACIDS DURING EMBRYOGENESIS AND LARVAL DEVELOPMENT OF EURASIAN PERCH (Perca fluviatilis). Fish Physiol. Biochem., 2000; 23: 233-243.
• Alderdice, D.F. 1988. OSMOTIC AND IONIC REGULATION IN TELEOST EGGS AND LARVAE. En: Hoar, W.S. & D.J. Randall (Eds). The physiology a development fish: Part A. Eggs and larvae. Fish Physiology, Vol. 11. Academis Press, London. 1988; 163-242 p.
• Alderdice, D.F. & C.R. Forrester. EARLY DEVELOPMENT AND DISTRIBUTION OF THE FLATHEAD SOLE (Hippoglossoides elassodon). J. Fish. Res. Board Can., 1974; 31:1899–1918.
• Alderdice, D.F., H. Rosenthal & F.P. Velsen. INFLUENCE OF SALINITY AND CADMIUM OF THE VOLUME OF PACIFIC HERRING EGGS. Helgol. Wiss. Meeresunters, 1979; 32: 163-178.
• Aristizabal, O.E. CONSUMO DE OXÍGENO, CRECIMIENTO Y UTILIZACIÓN DEL ALIMENTO DURANTE EL DESARROLLO LARVAL DEL BESUGO PAGRUS PAGRUS (Linné 1758) (Pisces: Sparidae). Rev. Biol. Mar. Ocean., 2006; 41 (2): 209-222.
• Arukwe, A. & A. Goksøyr. EGGSHELL AND EGG YOLK PROTEINS IN FISH: HEPATIC PROTEINS FOR THE NEXT GENERATION: OOGENETIC, POPULATION, AND EVOLUTIONARY IMPLICATIONS OF ENDOCRINE DISRUPTION. Comparative hepatology, 2003; 3: 1-21.
• Audouin-Chevallier, I., P. Higueret, V. Pallet, D. Higueret & H. Garcin. DIETARY VITAMIN A MODULATES THE PROPERTIES OF RETINOIC ACID AND GLUCOCORTICOID RECEPTORS IN RAT LIVER. J. Nutr., 1993; 123: 1195–1202.
• Ávila, E.M. & J.V. Juario. YOLK AND OIL GLOBULE UTILIZATION AND DEVELOPMENTAL MORPHOLOGY OF THE DIGESTIVE TRACT EPITHELIUM IN

REDVET. Revista electrónica de Veterinaria 1695-7504 2011 Volumen 12 Número 3
Caracterización bioquímica del consumo de reservas vitelinas en peces teleósteos de ontogenia indirecta.- http://www.veterinaria.org/revistas/redvet/n030311/031102.pdf
21
LARVAL RABBITFISH, Siganus guttatus (Bloch). Aquaculture, 1987; 65: 319-331.
• Bachop, W.E. & F.J. Schward. QUANTITATIVE NUCLEIC ACID HISTOCHEMESTRY OF THE YOLK SAC SYNCYTIUM OF OVIPAROUS TELEOSTS: IMPLECATIONS FOR HYPOTHESIS OF YOLK UTILIZATION. En: Blaxter, J.H.S (Ed). The early life history of fish. Springer- Verlag, Berlin. 1974; 345-353 p.
• Balon, E.K. TERMINOLOGY OF INTERVALS IN FISH DEVELOPMENT. J. Fish. Res. Bd. Can., 1975; 32: 1663-1670.
• Balon, E.K. EARLY ONTOGENY OF Labeotropheus Ahl, 1927 (Mbuna, Cichidae, Lake Malawi), WHIT A DISCUSSION ON ADVANCED PROTECTIVE STYLES IN FISH REPRODUCTION AND DEVELOPMENT. Eviron. Biol. Fishes, 1977; 2: 177-176.
• Bell, J.G. & J.R. Sargent. ARACHIDONIC ACID IN AQUACULTURE FEEDS: CURRENT STATUS AND FUTURE OPPORTUNITIES. Aquaculture, 2003; 218: 491-499.
• Bell, M.V., R.S. Batty, J.R. Dick, K. Fretwell, J.C. Navarro & J.R. Sargent. DIETARY DEFICIENCY OF DOCOSAHEXANOIC ACID IMPAIRS VISION AT LOW INTENSITIES IN JUVENILE HERRING (Clupea harengus L.). Lipids, 1995; 30: 443-449.
• Blaxter, J.H.S. Development: eggs and larvae. En Hoar, W.S & D.J.Randall (Eds). Fish Physiology, Vol. 3. Academi Press, New York. 1969; 177-252 p.
• Blaxter, J. H. S. THE EFFECTS OF TEMPERATURE ON LARVAL FISHES. Neth. J. Zool., 1992; 42 (2-3): 336-357.
• Blaxter, J. H. S. & G. Hempel. UTILIZATION OF YOLK BY HERRING LARVAE. J. Mar. Biol. Assoc. U. K., 1966; 46: 219-234.
• Boulekbache, H. ENERGY METABOLISM IN FISH DEVELOPMENT. Amer. Zool., 1981; 21:377-389.
• Britton, C. & L. Murray. CATHEPSIN L PROTEASE (CPL-1) IS ESSENTIAL FOR YOLK PROCESSING DURING EMBRYOGENESIS IN Caenorhabditis elegans. J. Cell Sci., 2004; 117 (21): 5133-5143.
• Brooks, S., C.R. Tyler & J.P. Sumpter. EGG QUALITY IN FISH: WHAT MAKES A GOOD EGG?. Reviews in Fish Biology and Fisheries, 1997; 7: 387-416.
• Brown, D.J.A. & S. Lynam. THE EFFECT OF SODIUM AND CALCIUM CONCENTRATIONS ON THE HATCHING OF EGGS AND THE SURVIVAL OF THE YOLK-SAC FRY OF BROWN TROUT, Salmo trutta L. AT LOW PH. J. Fish Biol., 1981; 19: 205-211.
• Bruce, M., F. Oyen, G. Bell, J.F. Asturiano, B. Farndale, M. Carrillo, S. Zanuy, J. Ramos & N. Bromage. DEVELOPMENT OF BROODSTOCK DIETS FOR THE EUROPEAN SEA BASS (DICENTRARCHUS LABRAX) WITH SPECIAL EMPHASIS ON THE IMPORTANCE OF N-3 AND N-6 HIGHLY UNSATURATED FATTY ACID TO REPRODUCTIVE PERFORMANCE. Aquaculture, 1999; 177: 85-97.

REDVET. Revista electrónica de Veterinaria 1695-7504 2011 Volumen 12 Número 3
Caracterización bioquímica del consumo de reservas vitelinas en peces teleósteos de ontogenia indirecta.- http://www.veterinaria.org/revistas/redvet/n030311/031102.pdf
22
• Buckley U., A.S. Smigielslu, T.A. Halavik, E.M. Caldarone, B.R. Burns & G.C. Laurence. WINTER FLOUNDER PSEUDOPLEURONECTES AMERICANUS REPRODUCTIVE SUCCESS. II. EFFECTS OF SPAWNING TIME AND FEMALE SIZE ON SIZE, COMPOSITION AND VIABILITY OF EGGS AND LARVAE. Mar. Ecol. Prog. Ser., 1991; 74: 125-135.
• Bulut, M. O. Ozden, S. Saka & K. Firat. BIOCHEMICAL COMPOSITION OF FERTILIZED DENTEX (Dentex dentex) EGGS. Pak. J. Biol. Sci., 2004; 7 (7): 1096-1098.
• Bulut, M., I. Celik & S. Billen. BIOCHEMICAL COMPOSITION OF FERTILIZED STRIPED SEABREAM (Pagellus erytrhinus Linnaeus, 1758) EGGS. Pak. J. Biol. Sci., 2005; 8 (10): 1342-1345.
• Carnevali, O., G. Mosconi, F. Centonze, J. Navas, S. Zanuy, M. Carrillo & N.R. BROMAGE. INFLUENCE OF DIETARY LIPID COMPOSITION ON YOLK PROTEIN COMPONENTS IN SEA BASS, Dicentrarchus labrax. Sci. Mar., 1998; 62 (4): 311-318
• Carnevali, O., G. Mosconi, M. Cardinali, I. Meiri & A. Polzonetti-Magni. MOLECULAR COMPONENTS RELATED TO EGG VIABILITY IN THE GILTHEAD SEA BREAM, Saparus aurata. Mol Reprod. Dev., 2001; 58: 330-335.
• Castell, J.D. REVIEW OF LIPID REQUIREMENTS OF FISH. En: EIFAC/FAO Symposium on fish nutrition and feed technology. Hamburgo, Germany. 1978; 40 p.
• Cetta, C.M. & J.M. Capuzzo. PHYSIOLOGICAL AND BIOCHEMICAL ASPECTS OF EMBRYONIC AND LARVAE DEVELOPMENT OF THE WINTER FLOUNDER Pseudopleuronectes americanus. Mar. Biol., 1982; 71: 327-337.
• Civera-Cerecedo, R., C. A., Álvarez-González & F. J., Moyano-López. NUTRICIÓN Y ALIMENTACIÓN DE LARVAS DE PECES. En: Cruz-Suárez, L., D., Ricquie Marie, M. G., Nieto López, D., Villareal, U., Scholz & M., González (Eds.). Avances en nutrición acuícola VII. Memorias del VII simposium internacional de nutrición acuícola, Sonora, México. 2004; 8-94 p.
• Cowey, C.B., J.M. Owen, J.W. Adron & C. Middleton. STUDIES ON NUTRITION OF MARINE FLATFISH: THE EFFECT OF DIFFERENT DIETARY FATTY ACIDS ON THE GROWTH AND FATTY ACID COMPOSITION OF TURBOT, Scophthalmus maximus. Br. J. Nutr., 1976; 36: 479-486.
• Craik, J.C.A. & S.M. Harvey. EGG QUALITY IN RAINBOW TROUT. THE RELATION BETWEEN EGG VIABILITY, SELECTED ASPECTS OF EGG COMPOSITION, AND TIME OF STRIPPING. Aquaculture, 1984; 40: 115-134.
• Crawford, R.B. & A.M. Guarino. EFFECTS OF ENVIRONMENTAL TOXICANTS ON DEVELOPMENT OF A TELEOST EMBRYO. J. Environ. Pathol. Toxicol., 1985; 6: 185-194.
• Dabrowski, K., M. Luczynski. UTILIZATION OF BODY STORES IN EMBRYONATED OVA AND LARVAE IN TWO COROGONID SPECIES

REDVET. Revista electrónica de Veterinaria 1695-7504 2011 Volumen 12 Número 3
Caracterización bioquímica del consumo de reservas vitelinas en peces teleósteos de ontogenia indirecta.- http://www.veterinaria.org/revistas/redvet/n030311/031102.pdf
23
(Coregonus lavaretus L. AND C. albula L.). Comp. Biochem. Physiol., 1984; 79A: 329-334.
• Dabrowski, K., M. Luczynski & M. Rusiecki. FREE AMINO ACID IN THE LATE EMBRYOGENESIS AND PRE-HATCHING STAGE IN TWO COREGONID FISHES. Biochem. Syst. Ecol., 1985; 13: 349-356.
• Devauchelle N. & D. Coves. THE CHARACTERISTICS OF SEA BASS (Dicentrarchus labrax) EGGS: DESCRIPTION, BIOCHEMICAL COMPOSITION AND HATCHING PERFORMANCES. Aquat. Living Resour., 1998; 1: 223-230.
• Eldridge, M.B., J. Wipple & D. Eng. ENDOGENOUS ENERGY SOURCES AS FACTORS AFFECTING MORTALITY AND DEVELOPMENT IN STRIPED BASS (Morone saxatilis) EGGS AND LARVAE. Rapp. P.-V. Reun., Const. Int. Explor. Mer., 1981; 178: 568-570.
• Eldridge, M. B., J. A. Whipple, & M. J. Bowers. BIOENERGETICS AND GROWTH OF STRIPPED BASS, Morone saxatilis EMBRYOS AND LARVAE. Fish. Bull., 1982; 80: 61-474.
• Estévez, A., L.A. McEvoy, J.C. Bell & J.R. Sargent. GROWTH, SURVIVAL, LIPID COMPOSITION AND PIGMENTATION OF TURBOT (Scophthalmus maximus) LARVAE FED LIVE-PREY ENRICHED IN ARACHIDONIC AND EICOSAPENTAENOIC ACIDS. Aquaculture, 1999; 180: 321–343.
• Federov K.Y. & Z.V. Smirnova. DYNAMICS ON AMMONIA ACCUMULATION AND ITS EFFECT OF THE DEVELOPMENT OF THE PINK SALMO Onchorynchus gorbuscha, IN CLOSED CIRCUIT INCUBATION SYSTEMS. J. Ichthyol., 1978; 18: 288-295.
• Fernández-Palacios, H., M. Izquierdo, L. Robaina, A. Valencia, M. Salhi y J. Vergara. EFFECT OF HUFA N-3 LEVEL IN BROODSTOCK DIETS ON EGG QUALITY OF GILTHEAD SEABREAM (Sparus aurata). Aquaculture, 1995; 132: 325-337.
• Finn, R.N. PHYSIOLOGICAL ENERGETIC OF DEVELOPING FISH EMBRYOS AND LARVAE. Dr. Thesis. University Bergen. Bergen, Norway. 1994; 128p.
• Finn, R.N., H.J. Fyhn & M.S. Evjen. RESPIRATION AND NITROGEN METABOLISM OF ATLANTIC HALIBUT EGGS (Hippoglossus hippoglossus). Mar. Biol., 1991; 108: 11-19.
• Finn, R.N., H.J. Fyhn & M.S. Evjen. PHYSIOLOGICAL ENERGETIC OF DEVELOPMENT EMBRYOS AND YOLK-SAC LARVAE OF ATLANTIC COD (Gadus morhua L.) I. RESPIRATION AND NITROGEN METABOLISM. Mar. Biol., 1995a; 124: 355-369.
• Finn, R.N., I. Rønnestad & H.J. Fyhn. RESPIRATION, NITROGEN AND ENERGY METABOLISM OF DEVELOPING YOLK-SAC LARVAE OF ATLANTIC HALIBUT (Hippoglossus hippoglussus). Comp. Biochem. Physiol., 1995b; 111A: 647-671.
• Finn R.N., J. Widdows & H.J. Fyhn. CALORESPIROMETRY OF THE DEVELOPING EMBRYO AND YOLK-SAC LARVAE OF TURBOT (Scophthalmus maximus). Mar. Biol., 1996; 122: 157-163.

REDVET. Revista electrónica de Veterinaria 1695-7504 2011 Volumen 12 Número 3
Caracterización bioquímica del consumo de reservas vitelinas en peces teleósteos de ontogenia indirecta.- http://www.veterinaria.org/revistas/redvet/n030311/031102.pdf
24
• Fukusho, K., M. Okauchi, S. Nuraini, A. Tsujigado & T. Watanabe. FOOD VALUE OF ROTIFER, BRACHIONUS PLICATILIS CULTURED WHITH Tetraselmis tetrathele FOR LARVAL RED SEABREAM, Pagrus major. Bull. Jpn. Soc. Sci. Fish., 1984; 50: 1439- 1444.
• Furuita, H., T. Takeuchi & K. Uematsu. EFFECT OF EICOSAPENTAENOIC AND DOCOSAHEXAENOIC ACIDS ON GROWTH, SURVIVAL AND BRAIN DEVELOPMENT OF LARVAL JAPANESE FLOUNDER Paralichthys olivaceus. Aquaculture, 1998; 161: 269– 279.
• Furuita, H., H. Tanaka, T. Yamamoto, M. Shiraishi & T. Takeuchi. EFFECTS OF n-3 HUFA LEVELS IN BROODSTOCK DIET ON THE REPRODUCTIVE PERFORMANCE AND EGG AND LARVAL QUALITY OF THE JAPANESE FLOUNDER, Paralichthys olivaceus. Aquaculture, 2000; 187: 387–398.
• Fyhn, H.J. FIRST FEEDING OF MARINE FISH LARVAE: ARE FREE AMINO ACIDS THE SOURCE OF ENERGY?. Aquaculture, 1989; 80: 111-120.
• Fyhn, H.J. MULTIPLE FUNCTIONS OF FREE AMINO ACIDS DURING EMBRYOGENESIS IN MARINE FISH. En: Walther, B.T. & H.J. Fyhn (Eds.). Physiology and biochemistry of fish larval development. University Bergen, Bergen. 1993; 229-308 p.
• Fyhn, H.J. & B. Serigstad. FREE AMINO-ACIDS AS ENERGY SUBSTRATE IN DEVELOPING EGGS AND LARVAE OF THE COD Gadus morhua. Mar. Biol., 1987; 96: 335-341.
• Fyhn, H.J. & J.J. Govoni. ENDOGENOUS NUTRIENT MOBILIZATION DURING EGG AND LARVAE DEVELOPMENT IN TWO MARINE FISHES-ATLANTIC MENHADEN AND SPOT. ICES Mar. Sci. Symp., 1995; 201: 64-69.
• Fyhn, H.J., B. Serigstad & A. Mangor-Jensen. FREE AMINO ACIDS IN DEVELOPING EGGS AND YOLK-SAC LARVAE OF THE COD, Gadus morhua L. Sarsia, 1987; 72: 363-365.
• Gatesoupe, F.J. THE EFFECT OF STARVATION AND FEEDING ON THE FREE AMINO ACIDS COMPOSITION OF SEA BASS LARVAE (Dicentrarchus labrax). Océanis, 1986; 12: 207-222.
• Gerhartz, B., E.A. Auerswald, R. Mentele, H. Fritz, W. Machleidt, H.J. Kolb & J. Wittmann. PROTEOLYTIC ENZYMES IN YOLK-SAC MEMBRANE OF QUAIL EGG. PURIFICATION AND ENZYMATIC CHARACTERISATION. Comp. Biochem. Physiol., 1997; 118B: 159–166.
• Giménez G., A. Estévez, F. Lahnsteiner, B. Zecevic, J.G. Bell, J. Henderson, J.A. Pinñera & J.A. Sànchez-Prado. Egg QUALITY CRITERIA IN COMMON DENTEX (Dentex dentex). Aquaculture, 2006; 260: 232-243.
• Gracia-López, V., M. Kiewek-Martínez & M. Maldonado-García. EFFECTS OF TEMPERATURE AND SALINITY ON ARTIFICIALLY REPRODUCED EGGS AND LARVAE OF THE LEOPARD GROUPER Mycteroperca rosacea. Aquaculture, 2004; 237: 485-498
• Harmor, T. & E.T. Garside. REGULATION OF OXYGEN CONSUMPTION BY INCIDENT ILLUMINATION IN EMBRYONATED OVA OF ATLANTIC SALMO Salmo salar L. Comp. Biochem. Physiol., 1975; 52A: 277-280.

REDVET. Revista electrónica de Veterinaria 1695-7504 2011 Volumen 12 Número 3
Caracterización bioquímica del consumo de reservas vitelinas en peces teleósteos de ontogenia indirecta.- http://www.veterinaria.org/revistas/redvet/n030311/031102.pdf
25
• Harmor, T. & E.T. Garside. QUANTITATIVE COMPOSITION OF THE FERTILIZED OVUM AND CONSTITUENT PARTS IN ATLANTIC SALMO, Salmo salar L. Can. J. Zool., 1977a; 55: 1650-1655.
• Harmor, T. & E.T. Garside. SIZE RELATION AND YOLK UTILIZATION IN EMBRYONATED OVA AND ALEVIN OF ATLANTIC SALMON Salmo salar L. IN VARIOUS COMBINATIONS OF TEMPERATURE AND DISSOLVED OXYGEN. Can. J. Zool., 1977b; 55: 1892-1898.
• Heming, T.A. EFFECTS OF TEMPERATURE ON UTILIZATION OF YOLK BY CHINOOOK SALMON (Onchorhyncus tshawytscha) EGGS AND ALEVINS. Can. J. Fish. Aquat. Sci., 1982; 39: 184-190.
• Heming, T.A., & R.K. Buddington. YOLK ABSORPTION IN EMBRYONIC AND LARVAL FISHES. En: Hoar, W.S. & D.J. Randall (Eds.). Fish Physiology, Vol. 11 A. Academic Press Inc., London. 1988; 407-446 p.
• Hirao, S., J. Yamada & R Kikuchi. RELATION BETWEEN BIOCHEMICAL CONSTITUENTS OF RAINBOW TROUT EGGS AND HATCHING RATE. Bull. Jpn. Soc. Sci. Fish., 1954; 21: 240-243.
• Hiramatsu, N. & A. Hara. RELATIONSHIP BETWEEN VITELLOGENIN AND ITS RELATED EGG YOLK PROTEINS IN SAKHALIN TAIMEN (Hucho perryi). Comp. Biochem. Physiol., 1996; 115A: 243–251.
• Hiramatsu N, T. Matsubara, G.M. Weber, C.V. Sullivan & A. Hara. VITELLOGENESIS IN AQUATIC ANIMALS. Fish. Sci., 2002; 68 (1): 694–699.
• Hølleland, T. & H.J. Fyhn. OSMOTIC PROPERTIES OF EGGS OF THE HERRING Clupea harengus. Mar. Biol., 1986; 91: 377-383.
• Idler, D & T. Ng. YOLK FORMATION AND DIFFERENTIATION IN TELEOST FISH. En: Hoar, W., D. Randall & E. Donaldson (Eds). Fish Physiology, Vol. 9 A. Academic Press, New York. 1983; 373-397 p.
• Ishizaki, Y., T. Takeuchi, T, Watanabe, M., Arimoto & K., Shimizu. A PRELIMINARY EXPERIMENT OF THE EFFECT OF Artemia ENRICHED WITH ARACHIDONIC ACID ON SURVIVAL AND GROWTH OF YELLOWTAIL. Fish. Sci., 1998; 64: 295–299.
• Jobling, M. TEMPERATURE AND GROWTH: MODULATION OF GROWTH RATE VIA TEMPERATURE CHANGE. En: Wood C. M., & D. G. McDonald (Eds). Global warming: implications for freshwater and marine fish, Soc. Exp. Biol. Sem. Ser. Vol. 61. Cambridge University Press, Cambridge, UK. 1997; 225-253 p.
• Jordaan, A. THE EFFECT OF THE TEMPERATURE ON THE DEVELOPMENT, GROWTH AND SURVIVAL OF ATLANTIC COD (Gadus morhua) DURING EARLY LIFE-HISTORIES. Master thesis, University of Maine. 2002; 87p.
• Kaitaranta, J.K. & R.G. Ackman. TOTAL LIPIDS AND LIPID CLASSES OF FISH ROE. Comp. Biochem. Physiol., 1981; 69B: 725-725.
• Kamler, E. VARIABILITY OF RESPIRATION AND BODY COMPOSITION DURING EARLY DEVELOPMENTAL STAGES OF CARP. Pol. Arch. Hydrobiol., 1976; 23: 431-485.

REDVET. Revista electrónica de Veterinaria 1695-7504 2011 Volumen 12 Número 3
Caracterización bioquímica del consumo de reservas vitelinas en peces teleósteos de ontogenia indirecta.- http://www.veterinaria.org/revistas/redvet/n030311/031102.pdf
26
• Kamler, E. ONTOGENY OF YOLK-FEEDING FISH: AN ECOLOGICAL PERSPECTIVE. Rev. Fish Biol. Fish., 2002; 12: 79-103.
• Kanazawa A. NUTRITIONAL MECHANISMS INVOLVED IN THE OCCURRENCE OF ABNORMAL PIGMENTATION IN HATCHERY-REARED FLATFISH. J. World Aquac. Soc., 1993; 24: 162-166.
• Kanazawa A., S. Teshima, M. Sakamoto & A. Awal Mo. REQUERIMENT OF Tilapia zilli FOR ESSENTIAL FATTY ACIDS. Bull. Jpn. Soc. Sci. Fish., 1980; 46: 1353-1356.
• Kestemont, P, J. Cooremans, A. Abi-Ayad & C. Mélard. CATHEPSIN L IN EGGS AND LARVAE OF PERCH Perca fluviatilis: VARIATIONS WITH DEVELOPMENTAL STAGE AND SPAWNING PERIOD. Fish Physiology and Biochemistry, 1999; 21: 59–64.
• Kitajima, C., T. Arakawa, G. Oowa, S. Fujutaet, O. Imada & T. Watanabe. DIETARY VALUE FOR RED SEABREAM OF ROTIFER, Brachionus plicatilis, CULTURED WITH NEW TYPE OF YEAST. Bull. Jpn. Soc. Sci. Fish., 1980a; 46: 43-46.
• Kitajima, C., M. Yoshida & T. Watanabe. DIETARY VALUE FOR AYU, Plecoglossus altivelis OF ROTIFER, Brachionus plicatilis, CULTURED WITH BEAKER´S YEAST, Sacharomyces cerevisiae, SUPPLEMENTED WITH CUTTLEFISH LIVER OIL. Bull. Jpn. Soc. Sci. Fish., 1980b; 46: 47-50.
• Kitahara,T., BEHAVIOR OF CAROTENOIDS IN THE CHUM SALMON Onchorhynchus keta DURING DEVELOPMENT. Bull. Jpn. Soc. Sci. Fish., 1984; 50: 531–536.
• Koven, W.M., G.W. Kissil & A. Tandler. LIPID AND ω-3 REQUIREMENTS OF Sparus aurata LARVAE DURING STARVATION AND FEEDING. Aquaculture, 1989; 79: 185-191.
• Kudo, S. ENZYMATIC BASIS FOR PROTECTION OF FISH EMBRYOS BY THE FERTILIZATION ENVELOPE. Experientia, 1992; 4B: 277-281.
• Kunz, I.W. DEVELOPMENTAL BIOLOGY OF TELEOST FISHES. Springer. Netherlands. 2004; 636 p.
• Lahnsteiner, F. & P. Patarnello. INVESTIGATIONS ON THE METABOLISM OF VIABLE AND NONVIABLE GILTHEAD SEA BREAM (Sparus aurata) EGGS. Aquaculture, 2003; 223: 159-174.
• Lahnsteiner, F. & P. Patarnello. EGG QUALITY DETERMINATION IN THE GILTHEAD SEA BREAM, Sparus aurata, WITH BIOCHEMICAL PARAMETERS. Aquaculture, 2004; 237: 443-459.
• Lahnsteiner, F., T. Weismann & A. Patzner. PHYSIOLOGICAL AND BIOCHEMICAL PARAMETERS OF EGG QUALITY DETERMINATION IN LAKE TROUT, Salmo trutta lacustris. Fish physiol. Biochem., 1999; 20: 375-388.
• Lapin, V.I. & V. Matsuk. UTILIZATION OF YOLK AND CHANGE IN THE BIOCHEMICAL COMPOSITION OF THE EGGS OF THE NAVAGA, Eleginus navaga, DURING EMBRYONIC DEVELOPMENT. J. Ichthyol., 1979; 19:131-136.

REDVET. Revista electrónica de Veterinaria 1695-7504 2011 Volumen 12 Número 3
Caracterización bioquímica del consumo de reservas vitelinas en peces teleósteos de ontogenia indirecta.- http://www.veterinaria.org/revistas/redvet/n030311/031102.pdf
27
• Lasker, R. EFFICIENCY AND RATE OF YOLK UTILIZATION BYE DEVELOPING EMBRYOS AND LARVAE OF THE PACIFIC SARDINE, Sardinops caerulae (Girard). J. Fish. Board Can., 1962; 19: 867-875.
• Li, S. & J. A. Mathias. CAUSES OF HIGH MORTALITY AMONG OF CULTURED LARVAL WALLEYE. Trans. Am. Fish. Soc., 1982 ; 111: 710-721.
• Lubzens, E., L. Lissauer, B. Levavi-Sivan, J.C. Avarre & M. Sammar. CAROTENOID AND RETINOID TRANSPORT TO FISH OOCYTES AND EGGS: WHAT IS THE ROLE OF RETINOL BINDING PROTEIN?. Molecular Aspects of Medicine, 2003; 24: 441–457.
• Matsubara, T., & K. Sawano. PROTEOLYTIC CLEAVAGE OF VITELLOGENIN AND YOLK PROTEINS DURING VITELLOGENIN UPTAKE AND OOCYTE MATURATION IN BARFIN FLOUNDER (Verasper moseri). J. Exp. Zool., 1995; 272: 34–45.
• May, R. C. LARVAL MORTALITY IN MARINE FISHES AND THE CRITICAL PERIOD CONCEPT. En: Blaxter, J.H.S. (Ed.). The Early Life History of Fish. Springer Verlag, Berlin. 1974; 3-19 p.
• May, R. C., D. Popper, & J.P. McVey. REARING AND LARVAL DEVELOPMENT OF Siganus canaliculatus (Park) (PISCES: SIGANIDAE). Micronesica, 1974; 10: 285-298.
• Mazorra, C., M. Bruce, J.G. Bell, A. Davie, E. Alorend, N. Jordan, J. Rees, N. Papanikos, M. Porter & N. Bromage. DIETARY LIPID ENHANCEMENT OF BROODSTOCK REPRODUCTIVE PERFORMANCE AND EGG AND LARVAL QUALITY IN ATLANTIC HALIBUT (Hippoglossus hippoglussus). Aquaculture, 2003; 227:21-33.
• Mæland, A., I. Rønnestad & R. Waagbø. FOLATE IN EGGS AND DEVELOPING LARVAE OF ATLANTIC HALIBUT, Hippoglossus hipoglossus L. Aquacult. Nut., 2003; 9: 185-188.
• Miki, N. T. Taniguchi, H. Hamakawa, Y. Yamada, & J. Sakurai. REDUCTION OF ALBINISM IN HATCHERY-REARED FLOUNDER “HIRME” Paralichthys olivaceus BY FEEDING ON ROTIFER ENRICHED WITH VITAMIN A. Suisanzoshoku, 1990; 38: 147–155.
• Mommsen, T.P. & P.J. Walsh. VITELLOGENESIS AND OOCYTE ASSEMBLY. En: Hoar, W.S. & D.J. Randall. (Eds.). The Physiology of Developing Fish: Part A. Eggs and Larvae. Fish Physiology, Vol. 11. Academic Press, London. 1988; 347-406 p.
• Moroz, I. Y. & B. P. Luzhin. DYNAMICS OF METABOLISM IN THE EMBRYONIC AND EARLY POST-EMBRYONIC DEVELOPMENT OF THE CARP Cyprinus carpio. J. Ichthyol., 1976; 16: 964-970.
• Mourente, G., A. Rodríguez, A. Grau & E. Pastor. UTILIZATION OF LIPIDS BY Dentex dentex L. (OSTEICHTHYES, SPARIDAE) LARVAE DURING LECITOTROPHIA AND SUBSEQUENT STARVATION. Fish Physiol. Biochem., 1999; 21: 45-58.

REDVET. Revista electrónica de Veterinaria 1695-7504 2011 Volumen 12 Número 3
Caracterización bioquímica del consumo de reservas vitelinas en peces teleósteos de ontogenia indirecta.- http://www.veterinaria.org/revistas/redvet/n030311/031102.pdf
28
• Murakami, M., I. Iuchi & K. Yamagami. YOLK PHOSPHOPROTEIN METABOLISM DURING EARLY DEVELOPMENT OF THE FISH, Orizyas latipes. Dev. Growth Differentiation, 1990; 32: 619-627
• Nakagawa, H. STUDIES ON RAINBOW TROUT EGG (Salmo gairdnerii irideus). II. CARBOHYDRATE IN THE EGG PROTEIN. J. Fac. Fish. Anim. Husb., Hiroshima Univ., 1970; 9: 57-63.
• Nakagwa, H. & Y. Tsuchiya. STUDIES ON RAINBOW TROUT EGG (Salmo gairdnerii irideus). IV. CHANCES OF YOLK CONTENT DURING EMBRYOGENESIS CARBOHYDRATE IN THE EGG PROTEIN. J. Fac. Fish. Anim. Husb., Hiroshima Univ., 1972; 11: 111-118.
• Nakagwa, H. & Y. Tsuchiya. STUDIES ON RAINBOW TROUT EGG (Salmo gairdnerii irideus). V. FURTHER STUDIES ON THE YOLK PROTEIN DURING EMBRYOGENESIS. J. Fac. Fish. Anim. Husb., Hiroshima Univ., 1974; 13: 15-27.
• Nelson, J.A. PHYSIOLOGICAL OBSERVATIONS ON DEVELOPING RAINBOW TROUT, Salmo gairdneri (Richardson), EXPOSED TO LOW PH AND VARIED CALCIUM ION CONCENTRATIONS. J. Fish Biol., 1982; 20: 359-372.
• Ohkubo, N., S. Sawaguchi, T. Hamatsu & T. Matsubara. UTILIZATION OF FREE AMINO ACIDS, YOLK PROTEINS AND LIPIDS IN DEVELOPING EGGS AND YOLK-SAC LARVAE OF WALLEYE POLLOCK Theragra chalcogramma. Fish. Sci., 2006; 72: 620-630.
• Owen, J.M., J.W. Adron, C. Middleton & C.B. Cowey. ELONGATION AND DESATURATION OF DIETARY FATTY ACIDS IN TURBOT Scophthalmus maximus, AND RAINBOW TROUT Salmo gairdneri. Lipids, 1975; 10: 528-531.
• Patiño, R. & C. Sullivan. OVARIAN FOLLICLE GROWTH, MADURATION, AND OVULATION IN TELEOST FISH. Fish Physiol. Biochem., 2002; 26: 57-70
• Pickova, J., P.C. Dutta, P. Larsson & A. Kiessling. EARLY EMBRYONIC CLEAVAGE PATTERN, HATCHING SUCCESS, AND EGG-LIPID FATTY ACID COMPOSITION: COMPARATION BETWEEN TOW COD (Gadus morhua) STOCK. Can. J. Fish. Aquat. Sci., 1997; 54: 2410-2416.
• Pittman, K., A. Berit, & T. Harboe. EFFECT OF TEMPERATURE ON GROWTH RATES AND ORGANOGENESIS IN THE LARVAE OF HALIBUT (Hippoglossus hippoglossus L.). Rapp. P.v. Réun. Cons. Int. Explor. Mer., 1989; 191: 421-430.
• Planas, M., U. Labarta, M.J. Fernández-Reiriz, M.J. Ferreiro, R. Munilla & J.L. Garrido. CHEMICAL CHANGES DURING DEVELOPMENT IN TURBOT (Scophthalmus maximus) EGGS AND LARVAE. En: Walther, B.T. & H.J. Fyhn (Eds).physiology and biochemistry of fish larval development. University Bergen, Bergen. 1993; 267-278 p.
• Polo, A., M. Yúfera, & E. Pascual. EFFECTS OF TEMPERATURE ON EGGS AND LARVAL DEVELOPMENT OF Sparus aurata L. Aquaculture, 1991; 92: 367-375.

REDVET. Revista electrónica de Veterinaria 1695-7504 2011 Volumen 12 Número 3
Caracterización bioquímica del consumo de reservas vitelinas en peces teleósteos de ontogenia indirecta.- http://www.veterinaria.org/revistas/redvet/n030311/031102.pdf
29
• Quantz, G. USE OF ENDOGENOUS ENERGY SOURCES BY LARVAL TURBOT Scophthalmus maximus. Trans. Am. Fish. Soc., 1985; 114: 558-563.
• Rainuzzo, J.R., K.I Retain & L. Jørgensen. COMPARATIVE STUDY ON THE FATTY ACID AND LIPID COMPOSITION OF FOUR MARINE FISH LARVAE. Comp. Biochem. Physiol., 1992; 103B: 21-26.
• Rainuzzo, J.R., K.I. Reitan, L. Jørgesen & Y. Olsen. LIPID COMPOSITION IN TURBOT LARVAE FEED LIVE FEED CULTURE BY EMULTIONS OF DIFFERENT LIPID CLASSES. Comp. Biochem. Physiol., 1994; 107: 699-710.
• Rawn, J.D. Biochemistry. Harper and Row, Publishers, New York. 1983. • Ribeiro, L., J.L. Zambonino-Infante, C. Cahu & M.T. Dinis. DEVELOPMENT
OF DIGESTIVE ENZYMES IN LARVAE OF Solea senegalensis, Kaup 1858. Aquaculture, 1999; 179: 465-473.
• Rodríguez, C. ESTUDIOS DE LOS REQUERIMIENTOS DE LOS ÁCIDOS GRASOS ESENCIALES DE LA DORADA EUROPEA (Sparus aurata, L.) DURANTE LAS DOS PRIMERAS SEMANAS DE ALIMENTACIÓN. Tesis Doctoral. Universidad de la Laguna, España. 1994; 284 p.
• Rogers, B.A. & D.T. Westin. LABORATORY STUDIES ON EFFECT ON THE TEMPERATURE AND DELAYED INITIAL FEEDING ON DEVELOPMENT IN STRIPED BASS LARVAE. Trans. Am. Fish. Soc., 1981; 110: 100-110.
• Rønnestad, I., & H.J. Fyhn. METABOLIC ASPECTS OF FREE AMINO ACIDS IN DEVELOPING MARINE FISH EGGS AND LARVAE. Rev. Fish. Sci., 1993; 1: 239-259.
• Rønnestad, I. H.J. Fyhn & K. Gravningen. THE IMPORTANCE OF FREE AMINO ACIDS TO THE ENERGY METABOLISM OF EGGS AND LARVAE TURBOT (Scophthalmus maximus). Mar. Biol., 1992a; 114: 517-525.
• Rønnestad, I., R.N. Finn, E.P. Groot & H.J. Fyhn. UTILIZATION OF FREE AMINO ACIDS RELATED TO ENERGY METABOLISM OF DEVELOPING EGGS AND LARVAE OF LEMON SOLE Microstomus kitt REARED IN THE LABORATORY. Mar. Ecol. Progr. Ser., 1992b; 88: 195-205.
• Rønnestad, I., E.P. Groot & H.J. Fyhn. COMPARTMENTAL DISTRIBUTION ON FREE AMINO ACIDS AND PROTEIN IN DEVELOPING YOLK-SAC LARVAE OF ATLANTIC HALIBUT (Hippoglossus hippoglussus). Mar. Biol., 1993; 116: 349-354.
• Rønnestad, I., W.M. Koven, A. Tandler, M. Harel & H.J. Fyhn. ENERGY METABOLISM DURING DEVELOPMENT OF EGGS AND LARVAE OF GILTHEAD SEA BREAM (Sparus aurata). Mar. Biol., 1994; 120: 187-196.
• Rønnestad, I., O. Lie & R. Waagbø. VITAMIN B6 IN ATLANTIC HALIBUT, Hippoglossus hippoglossus – ENDOGENOUS UTILIZATION AND RETENTION IN LARVAE FED NATURAL ZOOPLANKTON. Aquaculture, 1997; 157: 337-345.
• Rønnestad, I., W.M. Koven, A. Tandler, M. Harel & H.J. Fyhn. UTILIZATION OF YOLK FUELS IN DEVELOPING EGGS AND LARVAE OF EUROPEAN SEA BASS (Dincentrarchus labrax). Aquaculture, 1998a; 162: 157-170.

REDVET. Revista electrónica de Veterinaria 1695-7504 2011 Volumen 12 Número 3
Caracterización bioquímica del consumo de reservas vitelinas en peces teleósteos de ontogenia indirecta.- http://www.veterinaria.org/revistas/redvet/n030311/031102.pdf
30
• Rønnestad, I. K. Helland & O. Lie. FEEDING Artemia TO LARVAE OF ATLANTIC HALIBUT (Hippoglossus hippoglossus L) RESULTS IN LOWER LARVAL VITAMIN A CONTENT COMPARED WITH FEEDING COPEPODS. Aquaculture, 1998b; 165: 159-164.
• Rønnestad, I. K. Hamre, O. Lie & R. Waagbø. ASCORBIC ACID AND α-TOCOPHEROL IN LARVAE OF ATLANTIC HALIBUT BEFORE AND AFTER EXOGENOUS FEEDING. J. Fish Biol., 1999; 55: 720-731.
• Salze, G., D.R. Tocher, W.J. Roy & D.A. Robertson. EGG QUALITY DETERMINANTS IN COD (Gadus morhua L.): EGG PERFORMANCE AND LIPIDS IN EGGS FROM FARMED AND WILD BROODSTOCK. Aquacult. Res., 2005; 36: 1488-1499.
• Sarasquete, C., M.L. Gonzales de Canales, C. Piñuela, J.A. Muñoz, C. Rendon, E. Mañanós, F.M. Rodríguez & E. Pascual. HISTOCHEMICAL CHARACTERISTICS OF THE VITELLOGENIC OOCYTES OF THE BLUEFIN TUNA, Thunnus thynnus L. Ciencias Marinas, 2002; 28: 419-431.
• Sargent, J.R., J.G. Bell, M.V. Bell, J. Handerson & D.R. Tocher. REQUIREMENT CRITERIA FOR ESSENTIAL FATTY ACIDS. J. Appl. Ichthyol., 1995; 11: 183-198.
• Sargent, J., L. Mc. Evoy, A. Estevez, G. Bell, M. Bell, J. Henderson & D. Tocher. LIPID NUTRITION OF MARINE FISH DURING EARLY DEVELOPMENT: CURRENT STATUS AND FUTURE DIRECTIONS. Aquaculture, 1999; 179: 217-229.
• Sargent, J.R., D.R. Tocher & J.G. Bell. THE LIPIDS. En: Halver, J. (Ed.). Fish Nutrition. Academic Press, New York, U.S.A. 2003; 181-257 p.
• Satia, B.P., L.R. Donaldson, L.S. Smith & J.N. Nightingale. COMPOSITION ON OVARIAN FLUID AND EGGS OF THE UNIVERSITY OF WASHINGTON STRAIN OF RAINBOW TROUT (Salmo gardneri). J. Fish. Res. Boar. Can., 1974; 31: 1796-1799.
• Selman, K. & R.A. Wallace. CELLULAR ASPECTS OF OOCYTE GROWTH IN TELEOSTS. Zool. Sci., 1989; 6: 211-231.
• Shimizu, M. & J. Yamada. ULTRASTRUCTURAL ASPECT OF YOLK ABSORPTION IN THE VITELLINE SYNCYTIUM OF THE EMBRYONIC ROCKFISH, Sebastes schlegeli. J. Ichthyol., 1980; 27: 56-63.
• Sire, M.F., P.J. Babin & J.M. Vernier. INVOLVEMENT OF THE LYSOSOMAL SYSTEM IN YOLK PROTEIN DEPOSIT AND DEGRADATION DURING VITELLOGENESIS AND EMBRYONIC DEVELOPMENT IN TROUT. J. Exper. Zool., 1994; 269: 69-83.
• Specker, J.L. & C.V. Sullivan. VITELLOGENESIS IN FISHES: STATUS AND PERSPECTIVES. En: Davey, K.G., R.E. Peter, & S.S. Tobe. (Eds). Perspectives in Comparative Endocrinology. Ottawa, Canada: National Research Council of Canada, 1994; 304-315 p.
• Sund, T., & I. B. Falk-Petersen. EFFECTS OF INCUBATION TEMPERATURE ON DEVELOPMENT AND YOLK SAC CONVERSION EFFICIENCIES OF

REDVET. Revista electrónica de Veterinaria 1695-7504 2011 Volumen 12 Número 3
Caracterización bioquímica del consumo de reservas vitelinas en peces teleósteos de ontogenia indirecta.- http://www.veterinaria.org/revistas/redvet/n030311/031102.pdf
31
SPOTTED WOLFFISH (Anarhichas minor Olafsen) EMBRYOS UNTIL HATCH. Aquacult. Res., 2005; 36: 1133-1143.
• Takeuchi, T. J. Dedi, C. Ebisawa, T. Watanabe, T. Seikai, K. Hosoya & J.I. Nakazoe. THE EFFECT OF β-CAROTENE AND VITA A ENRICHED Artemia nauplii ON THE MALFORMATION AND COLOR ABNORMALITY OF LARVAL JAPANESE FLOUNDER. Fish. Sci., 1995; 61:141-148.
• Tandler A., T. Watanabe, S. Satoho & K. Fukusho. THE EFFECT OF FOOD DEPRIVATION ON THE FATTY ACID AND LIPID PROFILE OF RED SEABREAM LARVAE Pagrus major. Br. J. Nutr., 1989; 62: 349-361.
• Terner, C. METABOLISM AND ENERGY CONVERSION DURING EARLY DEVELOPMENT. En: Hoar, W.S. & D.J. Randall (Eds.). The Physiology of Developing Fish: Part A. Eggs and Larvae. Fish Physiology, Vol. 8. Academic Press, London. 1979; 261-278 p.
• Thorsen A. & H.J. Fyhn. OSMOTIC EFFECTORS DURING PREOVULATORY SWELLING IN MARINE FISH. En: Scott, A.P., J.P. Sumpter, D.E. Kime & M.S. Rolfe (Eds.), Proceedings 4th international symposium on the reproductive physiology of fish. Fish Symp., 91, University of Shiffield. 1991.
• Thorsen A. & H.J. Fyhn. FINAL OOCYTE MADURATION IN VIVO AND IN VITRO IN MARINE FISH WITH PELAGIC EGGS; YOLK PROTEIN HYDROLYSIS AND FREE AMINO ACIDS CONTENT. J. Fish Biol., 1996; 48: 1195-1209.
• Tilseth, S. & B. Ellertsen. FOOD CONSUMPTION RATE AND GUT EVACUATION PROCESSES OF FIRST FEEDING COD LARVAE (Gadus morhua L). En: Dahl, E.; D. S. Danelssen, E. Moksness & P. Solendal (Eds.). The propagation of cod Gadus morhua L. part I. An international symposium. Institute of Marine Research, Arendal, Norway. 1984; 167-182 p.
• Van der Meeren, T., J. Klungosoyr, S. Wilhelmsen & P.G. Kvensennth. FATTY ACID COMPOSITION OF UNFED AND GROWING COD LARVAE, Gadus morhua L. FEEDING ON NATURAL PLANKTON IN LARGE ENCLOSURES. En: B.T. Walther y H.J. Fyhn (Eds.). Physiological and Biochemical aspects of fish development. University Bergen, Norway. 1991; 34-35 p.
• Vetter, R.D., R.E. Hodson & C. Arnold. ENERGY METABOLISM IN A RAPIDLY DEVELOPING MARINE FISH EGG, THE RED DRUM (Sciaenops ocellata). Can. J. Fish. Aquat. Sci., 1983; 40, 627-634.
• Wallace, R.A. VITELLOGENESIS AND OOCYTE GROWTH IN NON-MAMMALIAN VERTEBRATES. En: Browder, L.W. (Ed). Developmental Biology, Vol. 1: Oogenesis. New York, Plenum Press. 1985; 127-177 p.
• Watanabe, T. IMPORTANCE OF DOCOSAHEXAENOIC ACID IN MARINE FISH LARVAE. J. World Aquacult. Soc., 1993; 24: 152-161.
• Watanabe, T., C. Kitajima & S. Fujita. NUTRITIONAL VALUE OF LIVE ORGANISMS USED IN JAPAN FOR MASS PROPAGATION IN FISH: A REVIEW. Aquaculture, 1983; 34: 115-143

REDVET. Revista electrónica de Veterinaria 1695-7504 2011 Volumen 12 Número 3
Caracterización bioquímica del consumo de reservas vitelinas en peces teleósteos de ontogenia indirecta.- http://www.veterinaria.org/revistas/redvet/n030311/031102.pdf
32
• Williams, K., N. Papanikos, R. Phelps, & J. Shardo. DEVELOPMENT, GROWTH AND YOLK UTILIZATION OF HATCHERY-REARED RED SNAPPER Lutjanus campechanus LARVAE. Mar. Ecol. Prog. Ser., 2004; 275: 231-239.
• Woodward, B. DIETARY VITAMIN REQUIREMENTS OF CULTURED YOUNG FISH, WITH EMPHASIS ON QUANTITATIVE ESTIMATES FOR SALMONIDS. Aquaculture, 1994; 124: 133-168.
• Yamada, K., K. Kobayashi & Y. Yone. CONVERSION OF LINOLENIC TO ω-3 HIGHLY UNSATURATED FATTY ACID IN MARINE FISHES AND RAINBOW TROUT. Bull. Jpn. Soc. Sci. Fish., 1980; 46: 1231-1233.
• Yamamoto, K. PERIBLAST IN THE EGG OF THE EEL, Anguilla japonica. Jpn. J. Ichthyol., 1982; 28: 423-430.
• Yamamoto, Y., A. Fujisawa, A. Hara & W.C. Dunlap. AN UNUSUAL VITAMIN E CONSTITUENT (α-TOCOMONOENOL) PROVIDES ENHANCED ANTIOXIDANT PROTECTION IN MARINE ORGANISMS ADAPTED TO COLD-WATER ENVIRONMENTS. PNAS, 2001; 98 (23): 13144–13148.
• Zavala, I. EFECTO DE LA TEMPERATURA, INTENSIDAD DE LUZ, TIPO Y DENSIDAD DE PRESA EN LA EFICIENCIA ALIMENTICIA DURANTE LA ONTOGENIA INICIAL DEL HUACHINANGO DEL PACÍFICO (Lutjanus peru). Tesis de maestría. CICIMAR-IPN. 2007; 52p.
• Zheng, F., T. Takeuchi, K. Yoseda, M. Kobayashi, J. Hirokawa & T. Watanabe. REQUIREMENT OF LARVAL COD FOR ARACHIDONIC ACID, EICOSAPENTAENOIC ACID AND DOCOSAHEXAENOIC ACID USING BY THEIR ENRICHED Artemia NAUPLII. Nippon Suisan Gakkaishi, 1996; 62: 669– 676.
• Zhu, P., C. Parrish & J. Brown. LIPID AND AMINO ACID METABOLISM DURING EARLY DEVELOPMENT OF ATLANTIC HALIBUT (Hippoglossus hippoglossus). Aquacult. Int., 2001; 11: 43–52.
REDVET: 2011, Vol. 12 Nº 3
Recibido 20.02.2010 / Ref. prov. FEB1021_REDVET / Revisado 20.05.2010 / Aceptado 10.01.2011 / Ref. def.
031102_REDVET / Publicado 01/03/2011
Este artículo está disponible en http://www.veterinaria.org/revistas/redvet/n030311.html concretamente en http://www.veterinaria.org/revistas/redvet/n030311/031102.pdf
REDVET® Revista Electrónica de Veterinaria está editada por Veterinaria Organización®.
Se autoriza la difusión y reenvío siempre que enlace con Veterinaria.org® http://www.veterinaria.org y con REDVET® - http://www.veterinaria.org/revistas/redvet