aula08bioqii metabolismo de lipídeos
TRANSCRIPT

Aula de Bioquímica II
Tema:
Metabolismo de Lipídeos
Prof. Dr. Júlio César BorgesDepto. de Química e Física Molecular – DQFM
Instituto de Química de São Carlos – IQSCUniversidade de São Paulo – USP
E-mail: [email protected]

Lipídeos
Biomoléculas insolúveis em água;Quarto principal grupo de biomoléculas;Grande diversidade química e estrutural;
Não são poliméricos, mas podem formar agregados.Várias funções 3 principais:
Papel estrutural: principal constituinte membranas biológicas
Armazenamento de energia: metabolismo energéticoFornece 80% da energia para o coração e fígado
Sinalização inter- e intracelular
Vitaminas e hormônios

Lipídeos Substâncias de origem biológica (do grego lipos, gordura);
Não poliméricos, mas podem se agregar;
Importância no Metabolismo energético ~ 2 x mais energia do que o mesmo peso seco de
glicose ou aminoácidos. Principal reserva energética metabólica a longo
prazo. Inércia química e baixa osmolaridade
Balanço energético da célula Demanda de energia Degradação -
Catabolismo Excesso de energia Síntese - Anabolismo
Envolvem reações de oxido-redução Diferem na direção e enzimas, localização
celular, doador/aceptor de elétrons
Adipócitos: especializados na síntese e armazenamento de triacilgliceróis;
Tecido adiposo: Abundante na camada subcutânea e cavidade
abdominal; Pode suprir a necessidade energética por alguns meses;
Isolamento térmico

Estratégia geral do catabolismo de ácidos graxos Degradação por remoção sequencial de unidades de 2 carbonos
Experimento de Franz Knoop (1904)- Marcação de ácidos graxos no último carbono com ácido benzoico
Hipótese da β-oxidação Clivagem entre o carbono
α e β
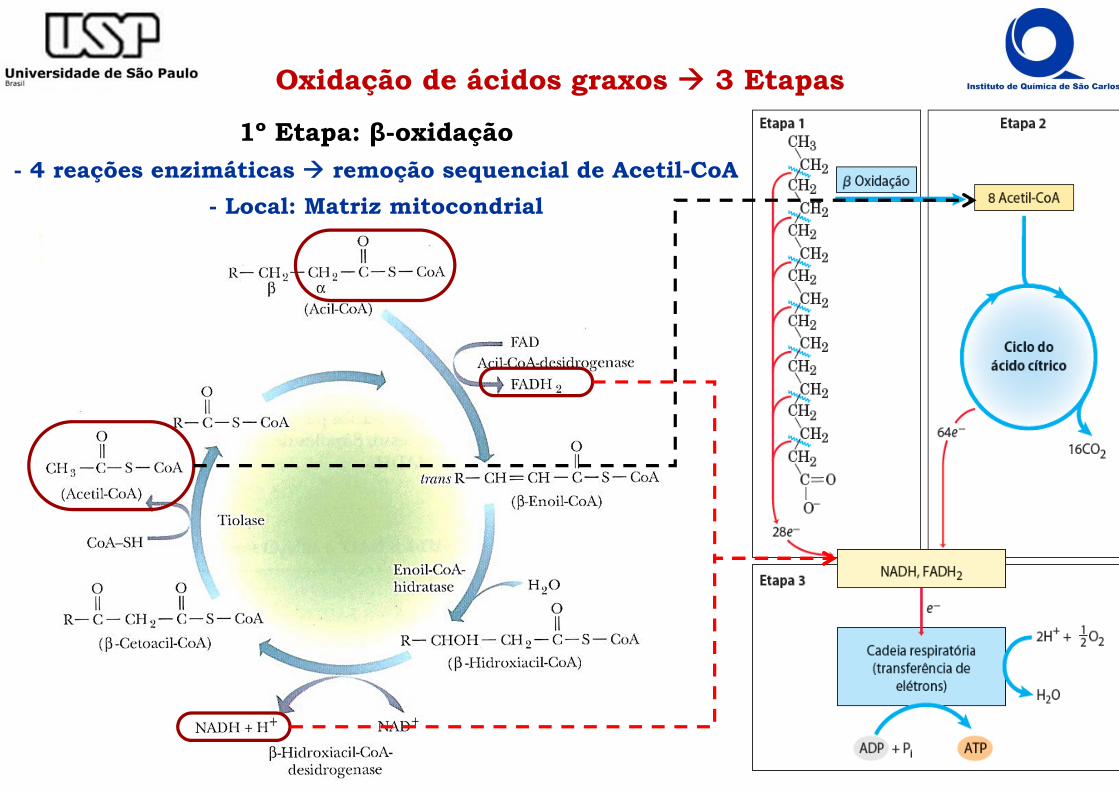
1º Etapa: β-oxidação- 4 reações enzimáticas remoção sequencial de Acetil-CoA
- Local: Matriz mitocondrial
Oxidação de ácidos graxos 3 Etapas

A mobilização de ácidos Graxos- Ação hormonal mobilizam ácidos graxos dos ADIPÓCITOS para o catabolismo
Adrenalina ou GlucagonFormação de FFA“Free Fatty acids”
1) Resposta a hormônios2) Via cAMP PKA
3) PKA fosforila e ativa a lipase sensível a hormônios (HSL)4) Fosforilação da Perilipina5) Liberação da CGI, que se
associa e ativa a lipase ATGL (adipose triacilglicerol lipase)
7) A Pi-HSL interage com a gotícula via Pi-perilipina
6) e 8) Ação conjunta da ATGLe Pi-HSL libera ácidos graxos
no citoplasma9) Os FFA atingem a corrente sanguínea e interage com a albumina sérica na razão de
10:1

Catabolismo de Triacilgliceróis Hidrólise da ligação éster entre o ácido graxo e glicerol
Triacilglicerol
LipasesGlicerol
Ácidos graxos
Ácidos graxos livres corrente sanguínea – Albumina músculo, coração e fígado Taxa de hidrólise de triacilglicerídeos regula o catabolismo de ácidos graxos
95% da energia de um triacilglicerol reside nos ácidos graxos ~5% no glicerol
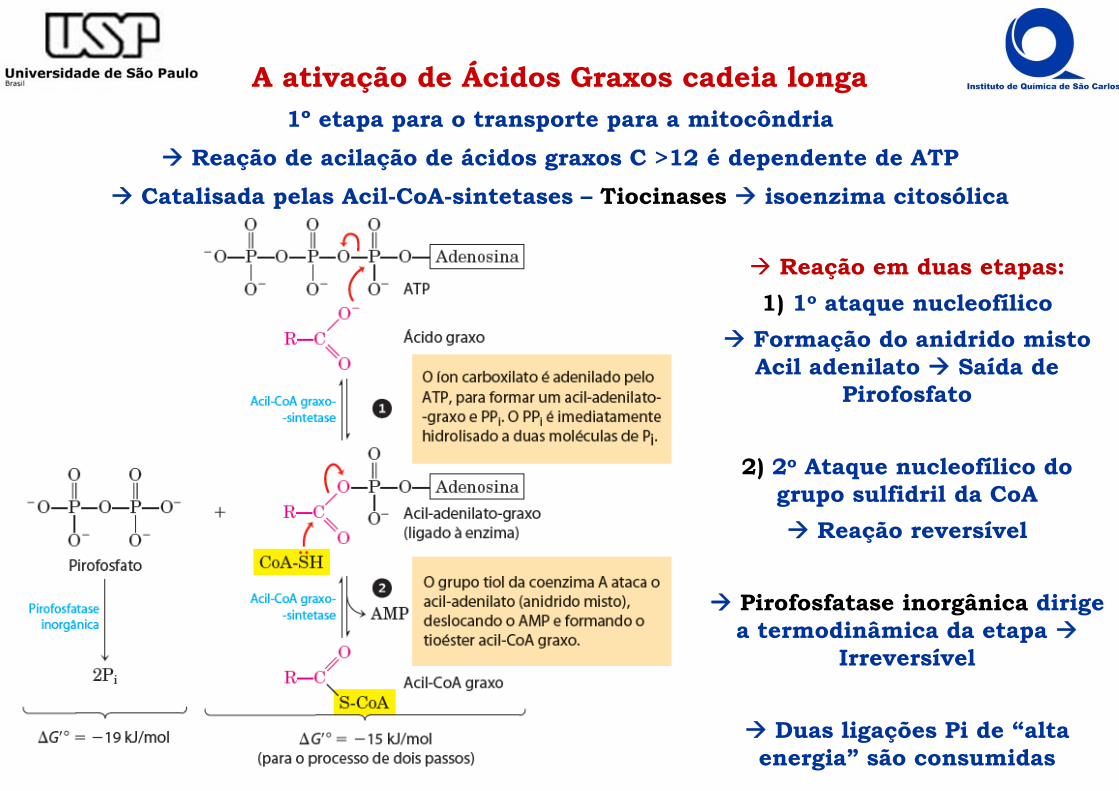
A ativação de Ácidos Graxos cadeia longa1º etapa para o transporte para a mitocôndria
Reação de acilação de ácidos graxos C >12 é dependente de ATP Catalisada pelas Acil-CoA-sintetases – Tiocinases isoenzima citosólica
Reação em duas etapas:1) 1o ataque nucleofílico
Formação do anidrido misto Acil adenilato Saída de
Pirofosfato
2) 2o Ataque nucleofílico do grupo sulfidril da CoA Reação reversível
Pirofosfatase inorgânica dirige a termodinâmica da etapa
Irreversível
Duas ligações Pi de “alta energia” são consumidas

A β-oxidação ocorre na mitocôndria! Transporte do Acil-CoA C > 12 pela membrana mitocondrial interna
Ácidos graxos com C ≤ 12 tem livre trânsito na MMI
- Transferência via Carnitina- Catalisada pelas isoenzimas Carnitina-palmitoil-transferases- A reação ocorre próxima ao equilíbrio
Ligações de energia livre de hidrólise similar- Mantém o balanço de CoA citosólica e mitocondrial- Transporte do ácido graxo para mitocôndria é ponto de regulação

A β-oxidação ocorre na mitocôndria! Uma translocase transporta a Acil-carnitina pela membrana mitocondrial interna
Proteína carreadora de carnitina
Carnitina-palmitoil-transferase I Ponto de controle por inibição
- Malonil-CoA
Carnitina-palmitoil-transferase II Transporte passivo
- Segue o gradiente de concentração

A estratégia da β-oxidação! 4 reações enzimáticas sequenciais = remoção de Acetil-CoA
1o Oxidação
3o Oxidação
2o Hidratação
4o Clivagem
Ciclos adicionais
Total: 7 Acetil-CoA
2 complexos multienzimáticostrifuncionais CANALIZAM os substratos pela via reacional
Cadeias com 12 ≥ C face interna da MMI
Cadeias com 12 ≤ C matriz mitocondrial

Repetição da mesma estratégia!Outras vias usam a mesma estratégia de oxidação
- Desestabilizar sequencialmente a ligação C—C e depois quebrar

A rota da β-oxidação! 4 Isoenzimas acil-CoA-Desidrogenase todas com FAD como coenzima
- Diferentes enzimas para tamanho de ácidos graxos diferentes- Acil-CoA-Desidrogenase de cadeias muito longas: Ácidos graxos com 12-18 C
- Acil-CoA-Desidrogenase de cadeias longas: Ácidos graxos com 10-16 C- Acil-CoA-Desidrogenase de cadeias médias: Ácidos graxos com 6-10 C- Acil-CoA-Desidrogenase de cadeias curtas: Ácidos graxos com 4-6 C
Todas estão ligadas à cadeia de transporte de elétrons- Transferência direta de e’ pela Flavoproteína de transferência de elétrons – FTE
Sem fluxo de H+ pela MMI!
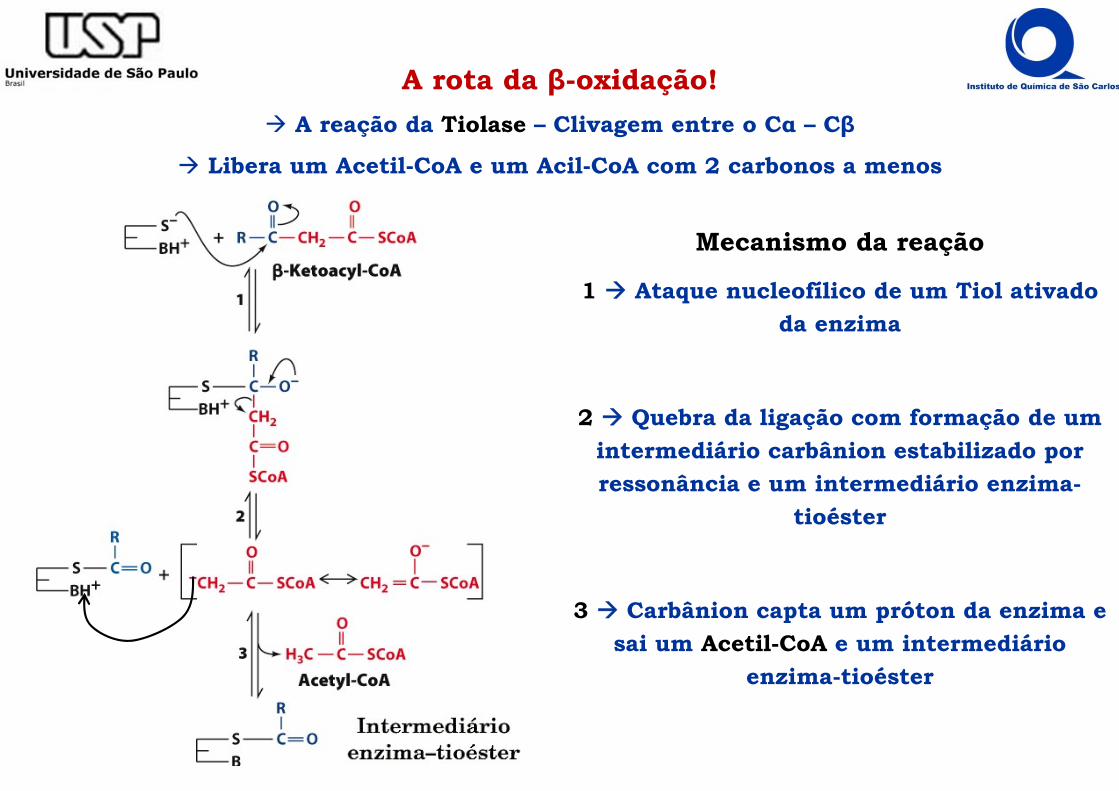
A rota da β-oxidação! A reação da Tiolase – Clivagem entre o Cα – Cβ
Libera um Acetil-CoA e um Acil-CoA com 2 carbonos a menos
Mecanismo da reação
1 Ataque nucleofílico de um Tiol ativado da enzima
2 Quebra da ligação com formação de um intermediário carbânion estabilizado por ressonância e um intermediário enzima-
tioéster
3 Carbânion capta um próton da enzima e sai um Acetil-CoA e um intermediário
enzima-tioéster

A rota da β-oxidação A reação da Tiolase – Clivagem entre o Cα – Cβ
Libera um Acetil-CoA e um Acil-CoA com 2 carbonos a menos
Mecanismo da reação
4 Ataque nucleofílico de um Tiol da CoAlivre e protonação de um grupo básico na
enzima
5 Reorganização eletrônica e liberação da Acil-CoA encurtada em 2 carbonos
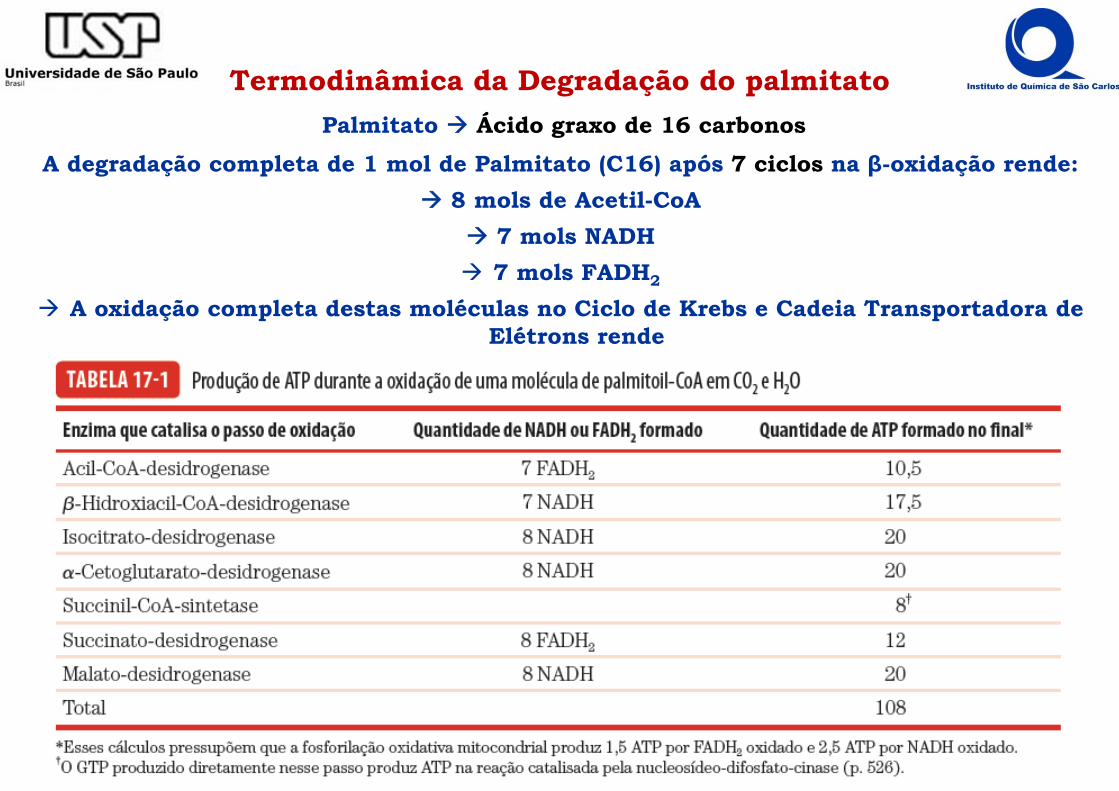
Termodinâmica da Degradação do palmitatoPalmitato Ácido graxo de 16 carbonos
A degradação completa de 1 mol de Palmitato (C16) após 7 ciclos na β-oxidação rende: 8 mols de Acetil-CoA
7 mols NADH 7 mols FADH2
A oxidação completa destas moléculas no Ciclo de Krebs e Cadeia Transportadora de Elétrons rende

Termodinâmica da Degradação do Palmitato! A degradação de 8 mols de Acetil-CoA no ciclo do ácido cítrico rende:
8 mols GTP = 8 mols ATP 24 mols NADH 8 mols FADH2
Somando ao rendimento na β-oxidação:
8 mols ATP 31 mols NADH 15 mols FADH2
Na cadeia transportadora de elétrons e fosforilação oxidativa:
1 mol NADH = 2,5 mol ATP 1 mol FADH2 = 1,5 mol ATP
Rendimento Total:8 mols ATP +
31 mols NADH x 2,5 77,5 mols ATP + 15 mols FADH2 x 1,5 22,5 mols ATP =
108 mols ATP
Duas ligações fosfato foram consumidas na Ativação do palmitato! RENDIMENTO final = 106 mols ATP x 30,5 kJ/mol = 3230 kJ/mol
33% do valor teórico em condições padrão ~60% da energia em condições fisiológicas
Aproximadamente 3,5 vezes ao rendimento da oxidação da glicose 1 mol de Glicose = 30-32 mols ATP
NADH + H+ + ½ O2 NAD+ + 1 H2OFADH2 + ½ O2 FAD + 1 H2O
H2O metabólica
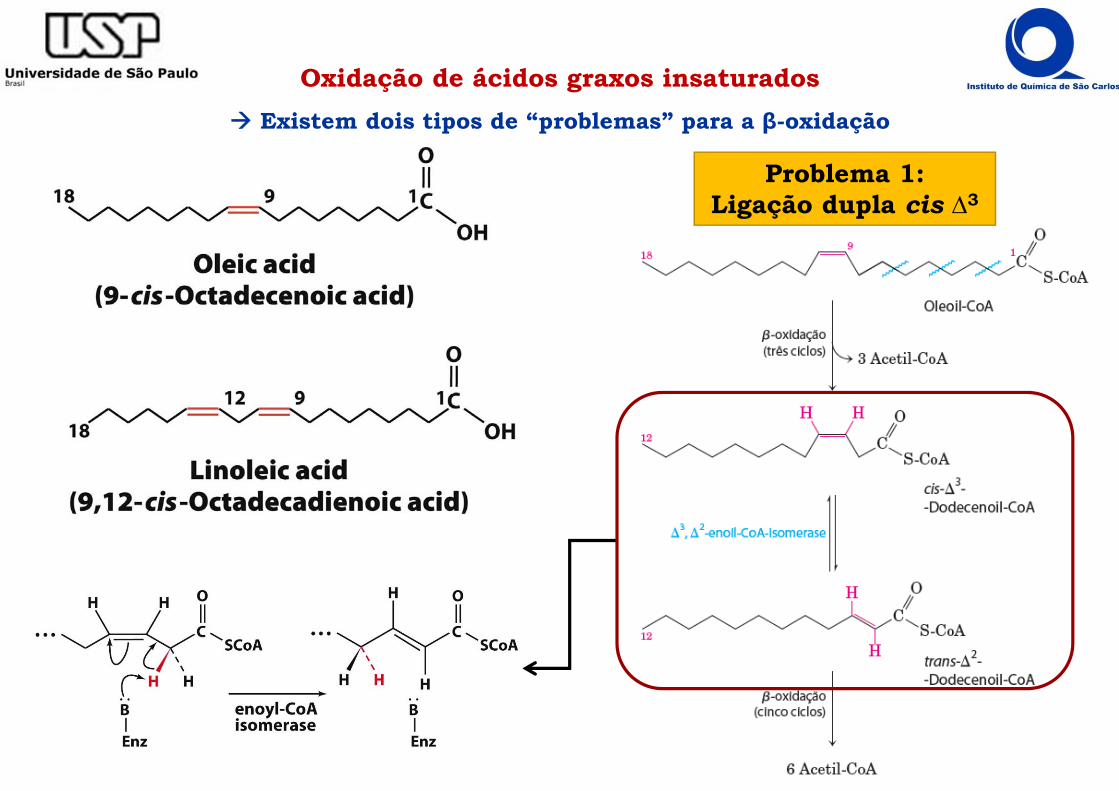
Oxidação de ácidos graxos insaturados Existem dois tipos de “problemas” para a β-oxidação
Problema 1:Ligação dupla cis ∆3

Oxidação de ácidos graxos insaturados Os problemas são contornados por enzimas adicionais
Problema 2: Ligação dupla cis ∆4
Não é substrato para a β-oxidação
A ligação cis ∆4 inibe a Enoil-CoA hidrataseque atua somente em substratos trans

Oxidação de ácidos graxos de cadeia impar
Intermediário do ciclo do ácido
cítrico
Produto da oxidação da Met, Val e Ile
Reação radicalar incomum dependente de Cobalamina =
Vitamina B12
- Comuns em plantas dieta de mamíferos
Produto final do último ciclo da β-oxidação é Acetil-CoA e Proprionil-CoA

Regulação da oxidação de ácidos graxos- Estritamente regulada
A entrada na mitocôndria determina o destino do ácido graxo - Mitocôndria β-oxidação
- Citoplasma Conversão em triacilglicerídeos Principal ponto de regulação Carnitina-acil-transferase I
Inibição por Malonil-CoA sinaliza disponibilidade de Acetil-CoA via Glicólise
Razão alta NADH/NAD+ inibe a β-hidroxiacil-CoA-
desidrogenase

Regulação da oxidação de ácidos graxos Glucacon dispara a fosforilação, via PKA e AMPK, da Acetil-CoA-carboxilase (ACC)
= inibição da síntese de malonil-CoA no citoplasma Glucacon ativa fator de transcrição CREB no músculo, fígado e tecido adiposo
Fatores de transcrição PPARα ativam a síntese - Síntese de proteínas envolvidas na β-oxidação ativada, incluindo a Carnitina-acil
transferase I e II

Outros sítios de oxidação de ácidos graxosPeroxissomos/glioxissomos organela especializada em oxidar ácidos graxos
Animais e vegetaisDiferencia da via mitocondrial
em 3 aspectos principais
1) Ácidos graxos de cadeia > 20 C
2) 1º passo oxidativo Acil-CoA oxidade geração de
H2O2 a partir de FADH2 e O2- Não é gera ATP
- Catalase consome o H2O2
3) 2º passo oxidativo NADH deve ser exportado para ser
reoxidado
Acetil-CoA formada é exportada
Em vegetais: glioxissomos
Ciclo do glioxilatopermite uso de Acetil-CoA
como percussor Biosintético na
germinação

A ω-oxidação Ácidos graxos de 10-12C são oxidados em ambas as extremidades no retículo
endoplasmático- Fígado e rins
- Via minoritária em mamíferos importante se a β-oxidação tem falhas Envolve a oxidação do C ω usando O2 oxidase de função mista
Conta com a participação da álcool desidrogenase e aldeído desidrogenase
Oxidação em terminação dupla
Gera ácidos dicarboxilicos = succinato
Entra diretamente no ciclo de Krebs

Formação de Corpos cetônicos Ácidos graxos são os combustíveis no estado de jejum A cetogênese ocorre no fígado durante a gliconeogênese
No hepatócito Acetil-CoA sofre conversão em “corpos
cetônicos” solúveis para transporte para outros tecidos
- Permite depleção de oxaloacetato para a gliconeogênese

Formação de Corpos cetônicos Acetil-CoA é convertida em acetoacetato, D-β-
hidroxibutirato e acetona1) 1o Condensação de 2 Acetil-CoA Tiolase2) 2o Condensação de + 1 Acetil-CoA HMG-CoA-sintase3) Clivagem de 1 Acetil-CoA HMG-CoA-liase
Destinos do acetoacetato1) Redução a D-β-hidroxibutirato2) Descaboxilação espontânea para acetona
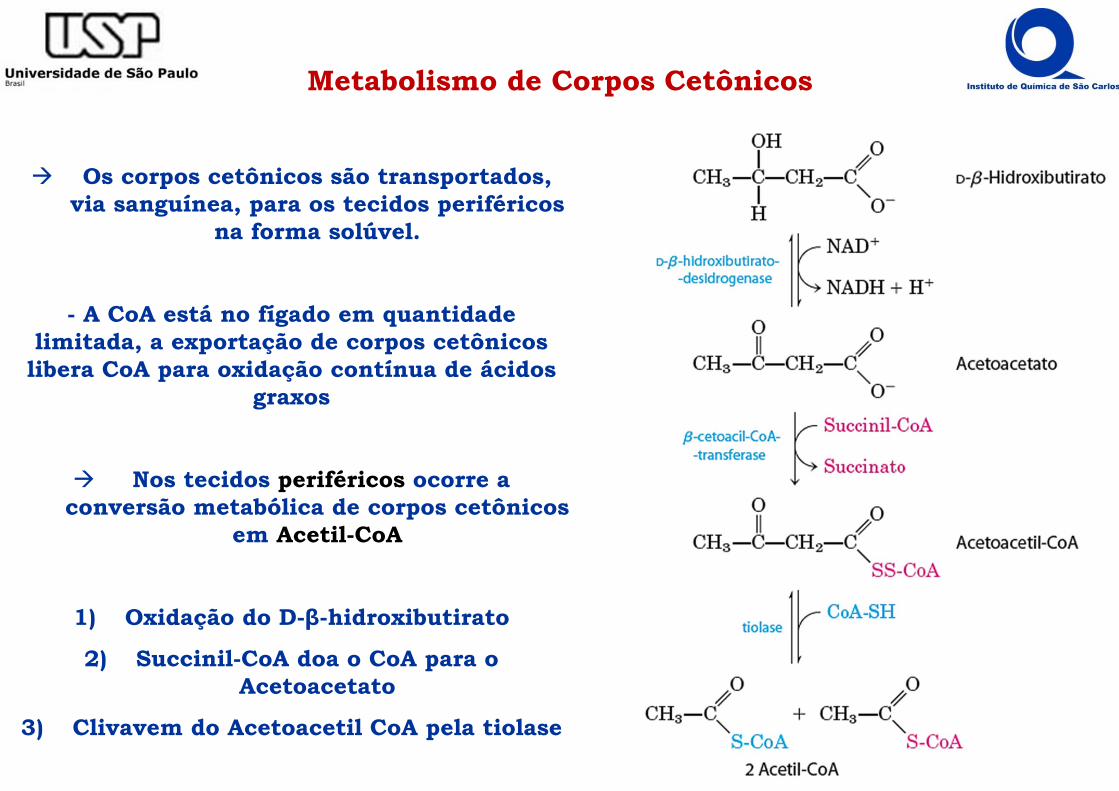
Metabolismo de Corpos Cetônicos
Os corpos cetônicos são transportados, via sanguínea, para os tecidos periféricos
na forma solúvel.
- A CoA está no fígado em quantidade limitada, a exportação de corpos cetônicos
libera CoA para oxidação contínua de ácidos graxos
Nos tecidos periféricos ocorre a conversão metabólica de corpos cetônicos
em Acetil-CoA
1) Oxidação do D-β-hidroxibutirato
2) Succinil-CoA doa o CoA para o Acetoacetato
3) Clivavem do Acetoacetil CoA pela tiolase
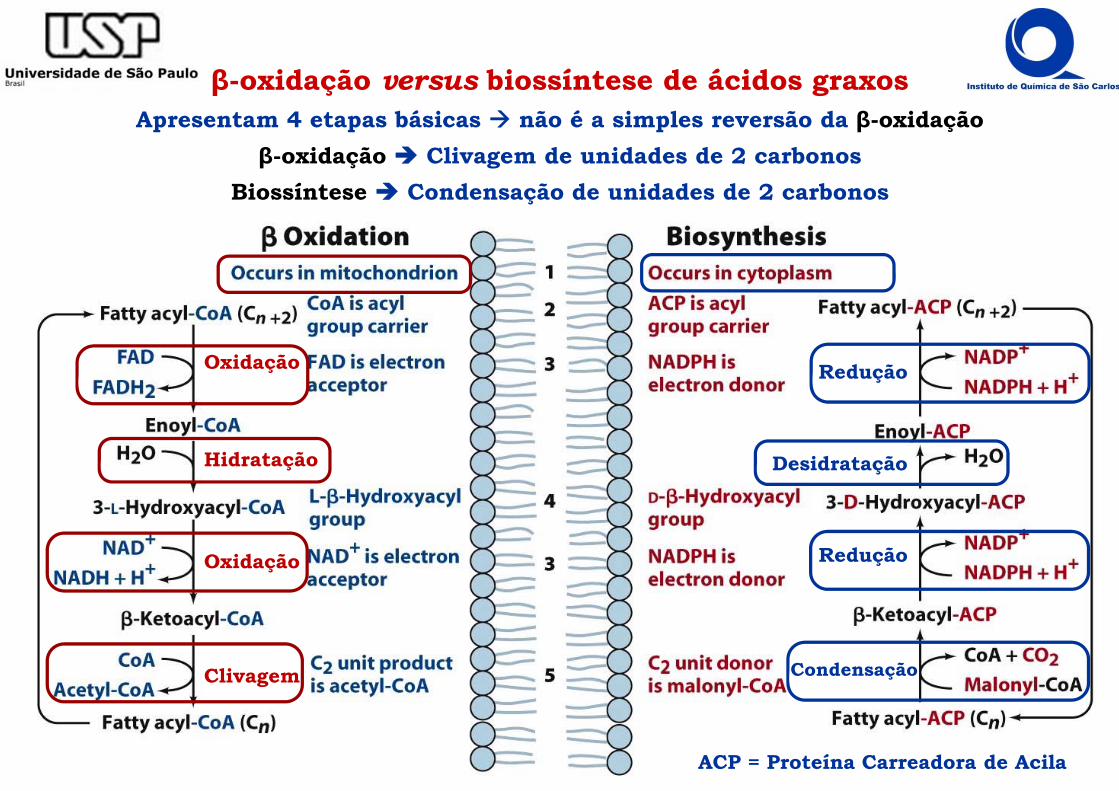
β-oxidação versus biossíntese de ácidos graxosApresentam 4 etapas básicas não é a simples reversão da β-oxidação
β-oxidação Clivagem de unidades de 2 carbonosBiossíntese Condensação de unidades de 2 carbonos
ACP = Proteína Carreadora de Acila
Oxidação
Oxidação
Hidratação Desidratação
Redução
Redução
CondensaçãoClivagem
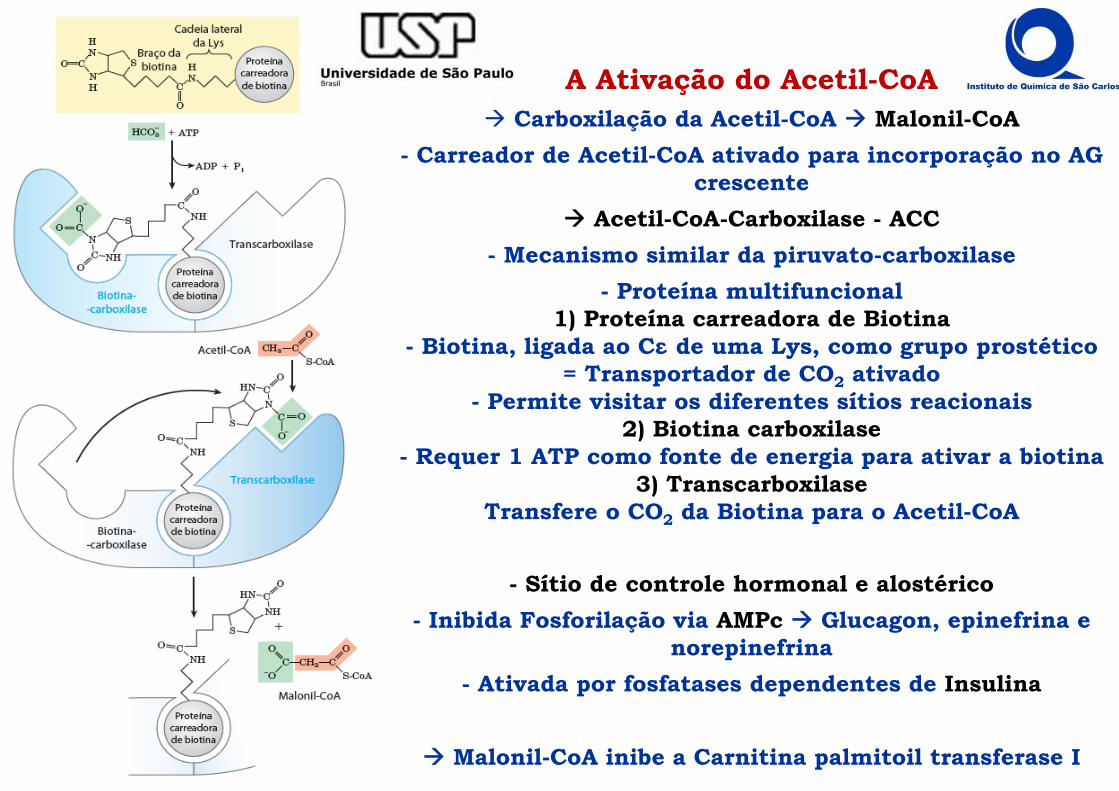
A Ativação do Acetil-CoA Carboxilação da Acetil-CoA Malonil-CoA
- Carreador de Acetil-CoA ativado para incorporação no AG crescente
Acetil-CoA-Carboxilase - ACC- Mecanismo similar da piruvato-carboxilase
- Proteína multifuncional1) Proteína carreadora de Biotina
- Biotina, ligada ao Cε de uma Lys, como grupo prostético= Transportador de CO2 ativado
- Permite visitar os diferentes sítios reacionais2) Biotina carboxilase
- Requer 1 ATP como fonte de energia para ativar a biotina3) Transcarboxilase
Transfere o CO2 da Biotina para o Acetil-CoA
- Sítio de controle hormonal e alostérico- Inibida Fosforilação via AMPc Glucagon, epinefrina e
norepinefrina- Ativada por fosfatases dependentes de Insulina
Malonil-CoA inibe a Carnitina palmitoil transferase I

Proteína carreadora de acila = ACP
Acyl carrier protein
- Proteína de 10 kDa
- Grupo fosforil da fosfopanteteína é esterificada a uma Ser
- Ancora o ácido graxo crescente para a reação com o Malonil-CoA

Síntese sequencial de AGÁcido Graxo Sintase I (AGS I) vertebrados e fungos
Polipeptídeo multifuncional 240 kDa é um homodímero 7 domínios funcionais canalização dos substratos ligados covalentemente
6 atividades catalíticas (Tioesterase não é mostrada)
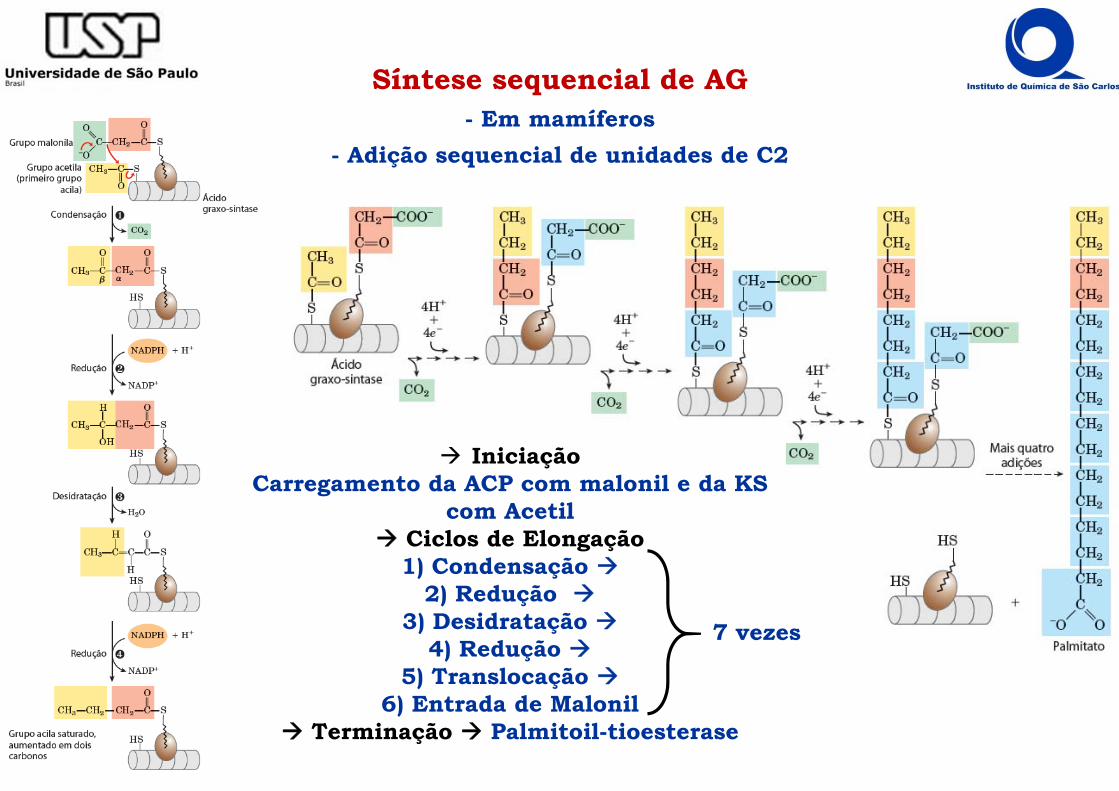
Síntese sequencial de AG- Em mamíferos
- Adição sequencial de unidades de C2
IniciaçãoCarregamento da ACP com malonil e da KS
com Acetil Ciclos de Elongação
1) Condensação 2) Redução
3) Desidratação 4) Redução
5) Translocação 6) Entrada de Malonil
Terminação Palmitoil-tioesterase
7 vezes

Síntese sequencial de AGÁcido Graxo Sintase I
Ciclos de Elongação1) Condensação β-cetoacil-ACP-sintase
2) 1º redução β-cetoacil-ACP-reductase
3) Desidratação β-cetoacil-ACP-desidratase
4) 2º redução Enoil-ACP- redutase
5) Translocação malonil/acetil-CoA-ACP-transferase
6) Entrada de Malonil malonil/acetil-CoA-ACP-transferase
IniciaçãoCarregamento da ACP com malonil
e da KS com Acetil

Síntese sequencial de AG!Balanço energético para a síntese de palmitoil.
Custo da ativação: 7 Acetil-CoA + 7 CO2 + 7 ATP 7 Malonil-CoA + 7 ADP + 7 Pi
7 Ciclos de condensação, desidratação e dupla redução:1 Acetil-CoA + 7 Malonil-CoA + 14 NADPH + 14 H+ 1 Palmitoil + 7 CO2 + 8 CoA + 14 NADP+ + 6 H2O
Balanço global8 Acetil-CoA + 7 ATP + 14 NADPH + 14 H+ 1 Palmitoil + 8 CoA + 7ADP + 7 Pi + 14 NADP+ + 6 H2O
Potencial redutorEnergia Química para
ativação do Malonil-CoA
A Biossíntese de AG ocorre no citoplasma1) Presença de todas as enzimas
2) Alta razão NADPH/NADP+ 75 em hepatócitos via pentose-fosfato e enzima málica- NADH/NAD+ 0,0008 permite glicólise ocorrer concomitantemente

A Biossíntese de AG ocorre no citosol!
Piruvato sofre descarboxilação
oxidativa na mitocôndria
- Ação da piruvato-desidrogenase
Transporte do acetil-CoA da
mitocôndria para o Citosol
Via Sistema de transporte de
tricarboxílico: Citrato:Malato
- Transfere unidades Acetil- da
mitocôndria para o citoplasma
- Sistema pode transferir força redutora
da mitocôndria para o citoplasma

Regulação da Biossíntese de ácidos graxos Citrato sinaliza combustível disponível na mitocôndria ativador alostérico da
Acetil-CoA-carboxilase (ACC) - Inibe a fosfofrutoquinase I na via glicolítica
Palmitoil, produto final da biossíntese, inibe a ACC Insulina ativa fosfatase que desfosforila a ACC ativando-a formação de malonil-CoA
= inibição da Carnitina-acil-transferase I Glucacon dispara a fosforilação, via PKA e AMPK, da ACC
= inibição da síntese de malonil-CoA no citoplasma

Os destinos do Palmitoil
Precursor para a biossíntese de outros AG- estearato
- Sistema de alongamento de ácidos graxos- Envolve enzimas específicas do RE- Acetil-CoA na forma de Malonil-CoA
InsaturaçõesPalmitato e estearato são precursores para os
equivalentes monoinsaturados
Poli-insaturações plantas e bactérias- AG essenciais
- Precursores dos eicosanoides
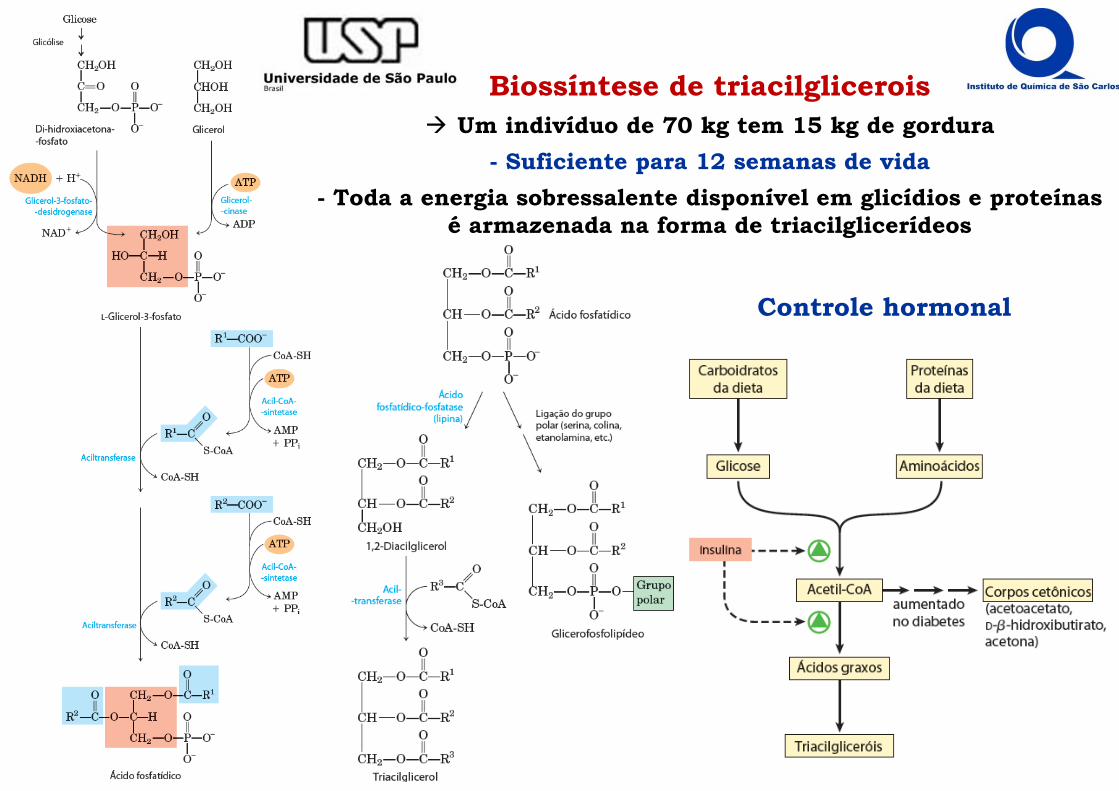
Biossíntese de triacilglicerois Um indivíduo de 70 kg tem 15 kg de gordura
- Suficiente para 12 semanas de vida- Toda a energia sobressalente disponível em glicídios e proteínas
é armazenada na forma de triacilglicerídeos
Controle hormonal