universidad de costa ricarepositorio.sibdi.ucr.ac.cr:8080/jspui/bitstream/123456789/6565/1/... ·...
TRANSCRIPT

UNIVERSIDAD DE COSTA RICA
SISTEMA DE ESTUDIOS DE POSGRADO
Caracterización, Composición, Abundancia y Biomasa del Zooplancton en el Golfo Dulce durante
el período 1997-1998
Tésis sometida a la consideración de la Comisión del Programa de Estudios de Posgrado en Biología para optar por el grado
de Magister Scientae
Marco Antonio Quesada Alpízar
Ciudad Universitaria " Rodrigo Facio " Costa Rica
2001

DEDICATORIAS
A mi padre, quien siempre ha estado a mi lado apoyándome todos estos años. A
mi padre y mi madre, por enseñarme la importancia y el valor del esfuerzo y la
honestidad ... gracias.
ii

AGRADECIMIENTOS
A mi director de tesis, Dr. Alvaro Morales, por su permanente interés, consejos y guía académica. Al comité integrado por el M.Sc. Gerardo Umaña y el Dr. José Antonio Vargas, cuyos consejos mejoraron de manera significativa el presente trabajo.
A los profesores Jenaro Acuña, Ornar Lizano y Juan Bautista Chavarría por su disponibilidad y acertados comentarios y recomendaciones
Un especial agradecimiento al Dr. Alvaro Morales y al Lic. Eleazar Ruiz por el enorme apoyo en las giras de campo. Sin su dedicación e interés este trabajo no se hubiera realizado.
A Jollyana Malavasi, una excelente profesora y amiga que me enseño a ir siempre hacia adelante.
Al personal administrativo del CIMAR, en especial a Carmen Durán, quienes siempre brindaron su ayuda desinteresada, más allá de su deber, para la realización de este proyecto.
A Laura May y Arturo Dominici por su ayuda en las giras de campo. A Laura May, Edgardo Díaz, Ernesto Brugnoli, Arturo Dominici y Esteban Estrada por sus consejos y críticas y por ser los amigos que siempre estuvieron ahí.
A Laura, por siempre empujarme cuando dudé, darme fuerzas cuando flaqueé y levantarme cuando caí. Sin vos simplemente no se hubiera podido. Gracias china.
iii

"Esta tesis fue aceptada por la comisión del Programa de Estudios de Posgrado en Biología de la Universidad de Costa Rica, como requisito parcial para optar al grado de Magister Scientae."
Daisy Arroyo Mora, M.Sc. Representante de la Decana del Sistema de Estudios de Posgrado
Alvaro Morales Ramírez, Ph.D. Director de Tesis
Gerardo Umaña Villalobos, M.Sc. Asesor
José Antonio Vargas Zamora, Ph. D. Asesor
Virginia Salís Alvarado, Ph.D. Directora del Programa de Posgrado en Biología
Marco Antonio Quesada Alpízar, B.Sc. Candidato
IV

INDICE GENERAL
PORTADA s/n
DEDICATORIAS ii
AGRADECIMIENTOS iii
HOJA DE APROBACION iv
INDICE GENERAL V
LISTA DE CUADROS viii
LISTA DE FIGURAS ix
Capítulo 1
Distribución Vertical y Variación Temporal y Espacial de la Biomasa del Zooplancton en el Golfo Dulce durante el fenómeno El Niño 1997-1998.
Resumen
Introducción
Objetivos
Material y Métodos
l. Sitio de Estudio
11. Factores Climáticos
111. Hidrografía
IV. Colecta y Procesamiento de Muestras
V. Análisis Estadísticos
Resultados
l. Parámetros Meteorológicos
11. Parámetros Físico-Químicos
i. Temperatura
ii. Oxígeno Disuelto
iii. Salinidad
iv. Transparencia del Agua
111. Aspectos Biológicos: Biomasa del Zooplancton
Discusión
Consideraciones Finales
Referencias
1
2
7
8
8
8
9
9
14
15
15
15
24
30
31
34
48
61
63

Capítulo 11
Caracterización, Composición y Abundancia del Zooplancton en el Golfo Dulce durante el período 1997-1998
Resumen
Introducción
Objetivos
Material y Métodos
l. Sitio de Estudio
11. Factores Climáticos
111. Hidrografía
IV. Colecta y Procesamiento de Muestras
V. Análisis Estadísticos
Resultados
1.1. Abundancia y Composición: Aspectos Generales
1.11. Abundancia y Composición: Distribución por Capa
Capa Superficial (P1)
Capa Intermedia (P2)
Capa Profunda (P3)
1.111. Abundancia y Composición: Variación Temporal
Capa Superficial (P1)
Capa Intermedia (P2)
Capa Profunda (P3)
11.1. Abundancia y Composición: Copépodos
11.11. Abundancia y Composición: Distribución por Capa
Capa Superficial (P1)
Capa Intermedia (P2)
Capa Profunda (P3)
11.111. Abundancia y Composición: Variación Temporal
Capa Superficial (P1)
Capa Intermedia (P2)
Capa Profunda (P3)
Discusión
Composición del Zooplancton
Abundancia y Distribución de Copépodos
vi
72
73
78
79
79
79
80
80
83
85
85
92
92
93
95
96
102
103
104
108
115
116
117
118
120
121
132
134
138
146
151

Consideraciones Finales
Referencias
Apéndice 1
Especies de copépodos reportados para el Golfo Dulce
Apéndice 2
160
162
173
Matriz de datos de biomasa (mg/m 3)de zooplancton, temperatura(ºC), oxígeno disuelto (mg/I) y salinidad (PSU) para cada una de las estaciones, profundidades, tamaños y fechas analizadas 175
Apéndice 3
Matriz de datos de abundancia (indiv/m3) de zooplancton para cada una de las
estaciones, fechas y profundidades analizadas 189
Vll

LISTA DE CUADROS
Cuadro 1.1. Ubicación geográfica de las estaciones de muestreo de zooplancton establecidas en el Golfo Dulce, Costa Rica ..................... .. .... ............ 11
Cuadro 1.2. Ambito de variación, promedio y desviación estándar de los valores de biomasa total, temperatura, oxígeno disuelto y salinidad para cada una de las estaciones muestreadas durante el período 1997-98, Golfo Dulce ..... 23
Cuadro 1.3. Valores de biomasa de zooplancton (mg/m3) reportados para diversas
regiones del planeta ................. .... ......................... ......... ............. 57
Cuadro 2.1. Ubicación geográfica de las estaciones de muestreo de zooplancton establecidas en el Golfo Dulce, Costa Rica ..................... ....... ........... 81
Cuadro 2.2. Abundancia (indiv./m3) de los distintos grupos de zooplancton identificados
en la capa superficial (P1) para cada una de las estaciones durante el período 1997 -1998, Golfo Dulce ................................................... 88
Cuadro 2.3. Abundancia (indiv./m3) de los distintos grupos de zooplancton identificados
en la capa intermedia (P2) para cada una de las estaciones durante el período 1997 -1998, Golfo Dulce ................................................... 89
Cuadro 2.4. Abundancia (indiv./m3) de los distintos grupos de zooplancton identificados
en la capa profunda (P3) para cada una de las estaciones durante el período 1997 - 1998, Golfo Dulce ................................................... 90
Cuadro 2.5. Abundancia (indiv./m3) de los distintos géneros de copépodos identificados
en la capa superficial (P1) para cada una de las estaciones durante el período 1997 - 1998, Golfo Dulce .................................................. 112
Cuadro 2.6. Abundancia (indiv./m3) de los distintos géneros de copépodos identificados
en la capa intermedia (P2) para cada una de las estaciones durante el período 1997 - 1998, Golfo Dulce ... .. ............................................. 113
Cuadro 2.7. Abundancia (indiv./m3) de los distintos géneros de copépodos identificados
en la capa profunda (P3) para cada una de las estaciones durante el período 1997 - 1998, Golfo Dulce .................................... .............. 114
Cuadro 2.8. Valores de abundancia de zooplancton (indiv./m3) reportados para diversas
regiones del planeta .................................................................... 140
viii

LISTA DE FIGURAS
Figura 1.1. Ubicación geográfica de los puntos de muestreo, Golfo Dulce, Costa Rica.1 O
Figura 1.2. Criterio de muestreo utilizado para la colecta de zooplancton, basándose en la posición de la termoclina en la columna de agua y categorías de tamaño utilizadas para fraccionar las muestras ................................................ 13
Figura 1.3. Parámetros meteorológicos durante el período 1997 - 98 para la zona del Golfo Dulce: a) promedios de temperatura (ºC) máxima y mínima, b) promedios mensuales de precipitación . ............................... .. .............. 16
Figura 1.4.Variación temporal en las isoclinas de a) temperatura (ºC), b) oxígeno disuelto (mg/I) y e) salinidad (PSU) correspondiente a la estación 01, Golfo Dulce, durante el período 1997 - 1998 .................................. ... ........... 17
Figura 1.5.Variación temporal en las isoclinas de a) temperatura (ºC), b) oxígeno disuelto (mg/I) y e) salinidad (PSU) correspondiente a la estación 02, Golfo Dulce, durante el período 1997-1998 .... ... ........ ..... .. .. . .............. ... .. .. .. 18
Figura 1.6.Variación temporal en las isoclinas de a) temperatura (ºC), b) oxígeno disuelto (mg/I) y e) salinidad (PSU) correspondiente a la estación 03, Golfo Dulce, durante el período 1997 - 1998 ................................................ 19
Figura 1.7.Variación temporal en las isoclinas de a) temperatura (ºC), b) oxígeno disuelto (mg/I) y e) salinidad (PSU) correspondiente a la estación 04, Golfo Dulce, durante el período 1997 -1998 ................................................ 20
Figura 1.8.Variación temporal en las isoclinas de a) temperatura (ºC), b) oxígeno disuelto (mg/I) y e) salinidad (PSU) correspondiente a la estación 11, Golfo Dulce, durante el período 1997 - 1998 ................................................ 21
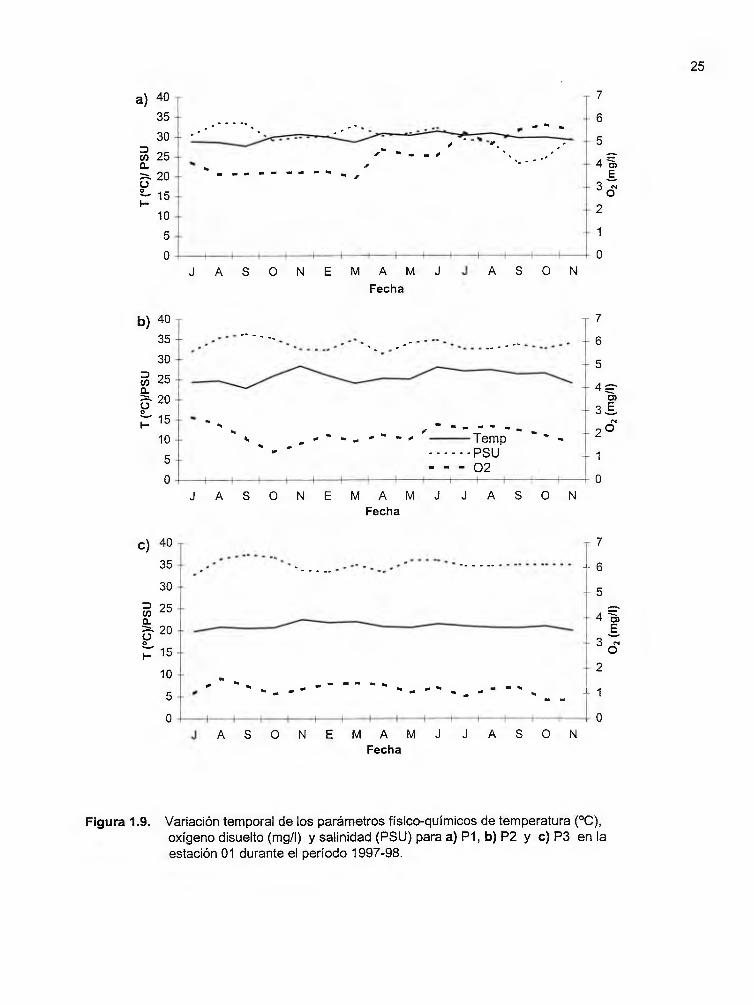
Figura 1.9. Variación temporal en los parámetros fisico-químicos de temperatura (ºC), oxígeno disuelto (mg/I) y salinidad (PSU) para a) P1, b) P2 y e) P3 en la estación 01 durante el período 1997-98 .............................................. 25
Figura 1.1 O. Variación temporal en los parámetros fisico-químicos de temperatura (ºC), oxígeno disuelto (mg/I) y salinidad (PSU) para a) P1, b) P2 y e) P3 en la estación 02 durante el período 1997-98 .............................................. 26
Figura 1.11. Variación temporal en los parámetros fisico-químicos de temperatura (ºC), oxígeno disuelto (mg/I) y salinidad (PSU) para a) P1, b) P2 y e) P3 en la estación 03 durante el período 1997-98 .............................................. 27
Figura 1.12. Variación temporal en los parámetros fisico-químicos de temperatura (ºC), oxígeno disuelto (mg/I) y salinidad (PSU) para a) P1, b) P2 y e) P3 en la estación 04 durante el período 1997-98 .............................................. 28
ix

Figura 1.13. Variación temporal en los parámetros fisico-químicos de temperatura (ºC), oxígeno disuelto (mg/I) y salinidad (PSU) para a) P1, b) P2 y e) P3 en la estación 11 durante el período 1997-98 ......................... . ... .. ............... 29
Figura 1.14. Variación temporal en la transparencia del agua (m) para las estaciones a) 01, b) 02, e) 03, d) 04 y e) 11 durante el período 1997-98, Golfo Dulce ..... 32
Figura 1.15. Promedio y desviación estándar de la transparencia del agua en cada estación de muestreo durante el período 1997-1998 .............................. 33
Figura 1.16. Valores promedio y desviación estándar de la biomasa total por capa (P1, P2, P3) para cada estación: a) 01, b) 02, e) 03, d) 04 y e) 11, durante el período 1997-98 .......................... ..... . ........................................ . .... 35
Figura 1.17. Variación temporal en la biomasa total (mg/m3) del zooplancton en cada
capa (P1, P2 y P3) para las estaciones a) 01, b) 02, e) 03, d) 04 y e) 11, durante el período 1997-98 ........................................ . .... . ...... .......... 36
Figura 1.18. Variación temporal y aporte por clase de tamaño (x) (T1: x > 1000µm; T2: 1000µm > x > 500µm; T3: 500µm > x > 150µm) de la biomasa (mg/m3
) del zooplancton en las capas P1 (a), P2 (b) y P3 (e), Golfo Dulce, estación 01 .......... . .. .. . . ............................. ... .. .... . ............. . .......... .. ............. 38
Figura 1.19. Variación temporal y aporte por clase de tamaño (x) (T1: x > 1000µm; T2: 1000µm > x > 500µm; T3: 500µm > x > 150µm) de la biomasa (mg/m3
) del zooplancton en las capas P1 (a), P2 (b) y P3 (e), Golfo Dulce, estación 02 ............. . ................................. .. ... ....................... . ... .... ............ 39
Figura 1.20. Variación temporal y aporte por clase de tamaño (x) (T1: x > 1 OOOµm; T2: 1000µm > x > 500µm; T3: 500µm > x > 150µm) de la biomasa (mg/m3
) del zooplancton en las capas P1 (a), P2 (b) y P3 (e), Golfo Dulce, estación 03 ............. . .................. .. ......... . .. .... ............................................. 40
Figura 1.21. Variación temporal y aporte por clase de tamaño (x) (T1 : x > 1 OOOµm; T2: 1000µm > x > 500µm; T3: 500µm > x > 150µm) de la biomasa (mg/m3
) del zooplancton en las capas P1 (a), P2 (b) y P3 (e), Golfo Dulce, estación 04 ........ ... ............................ ........ .............................. ..... ............. 41
Figura 1.22. Variación temporal y aporte por clase de tamaño (x) (T1: x > 1 OOOµm; T2: 1000µm > x > 500µm; T3: 500µm > x > 150µm) de la biomasa (mg/m3
) del zooplancton en las capas P1 (a), P2 (b) y P3 (e), Golfo Dulce, estación 11 ........ ... . .. ........................ .. .... .... ............................. .... .............. 42
Figura 1.23. Promedio y desviación estándar de la biomasa del zooplancton (mg/m3)
para cada clase de tamaño (T1, T2, T3), en cada profundidad y para cada estación a) 01, b) 02, e) 03, d) 04 y e) 11, Golfo Dulce .......................... .43
Figura 1.24. Análisis de conglomerado para las estaciones de muestreo respecto a la biomasa del zooplancton en la capa superficial (P1 ), aplicando el coeficiente de similitud Bray-Curtis y uniones de promedio por grupo ..... .45
X

Figura 1.25. Regresión lineal entre la biomasa del zooplancton en la capa P1 y a) la profundidad del disco de Sechii, b) la profundidad de la capa de mezcla ....................................................................................... 47
Figura 2.1. Criterio de muestreo utilizado para la colecta de zooplancton, basándose en la posición de la termoclina en la columna de agua ................................ 82
Figura 2.2. Valores promedio y desviación estándar de la abundancia (indiv./m3) general
de zooplancton y copépodos en las capas superficial (P1 ), intermedia (P2) y profunda (P3) en el Golfo Dulce, 1997-1998 ..................................... 85
Figura 2.3. Valores promedio y desviación estándar de la abundancia de zooplancton (indiv./m3
) para cada estación y capa de muestreo, Golfo Dulce, 1997-1998 ........................................................................................... 86
Figura 2.4. Valores promedio y desviación estándar de la abundancia de zooplancton (indiv./m3
) para cada una de las fechas y capas analizadas, Golfo Dulce, 1997- 1998 .................................................................................. 87
Figura 2.5. Composición del zooplancton y porcentaje de abundancia de los principales grupos taxonómicos en el Golfo Dulce, 1997-1998 ................................ 91
Figura 2.6.Análisis de conglomerado para los grupos de zooplancton respecto a su abundancia en la capa P1, aplicando el coeficiente de similitud Bray-Curtis y uniones de promedio por grupo. A, B y C representan los conglomerados resultantes .................................................................................... 93
Figura 2. 7 .Análisis de conglomerado para los grupos de zooplancton respecto a su abundancia en la capa P2, aplicando el coeficiente de similitud Bray-Curtis y uniones de promedio por grupo. A, B y C representan los conglomerados resultantes .................................................................................... 94
Figura 2.8.Análisis de conglomerado para los grupos de zooplancton respecto a su abundancia en la capa P3, aplicando el coeficiente de similitud Bray-Curtis y uniones de promedio por grupo. A, B y C representan los conglomerados resultantes .................................................................................... 95
Figura 2.9. Variación temporal en la abundancia (indiv./m3) de tres estadías de
desarrollo de copépodos en las capas a) P1, b) P2 y e) P3 en la estación 01, Golfo Dulce ............................................................................ 97
Figura 2.1 O. Variación temporal en la abundancia (indiv./m3) de tres estad íos de
desarrollo de copépodos en las capas a) P1, b) P2 y e) P3 en la estación 02, Golfo Dulce .......................................................................... 98
Figura 2.11. Variación temporal en la abundancia (indiv./m3) de tres estadías de
desarrollo de copépodos en las capas a) P1, b) P2 y e) P3 en la estación 03, Golfo Dulce ............................................................................. 99
xi

Figura 2.12. Variación temporal en la abundancia (indiv./m3) de tres estadías de
desarrollo de copépodos en las capas a) P1, b) P2 y e) P3 en la estación 04, Golfo Dulce ........................................................................... 100
Figura 2.13. Variación temporal en la abundancia (indiv./m3) de tres estadías de
desarrollo de copépodos en las capas a) P1, b) P2 y e) P3 en la estación 11, Golfo Dulce ........................................................................... 101
Figura 2.14. Análisis de conglomerado para las estaciones muestreadas respecto a la abundancia de zooplancton en las capas a) superficial (P1 ), b) intermedia (P2) y e) profunda (P3), aplicando el coeficiente de similitud Bray-Curtis y uniones de promedio por grupo. Notar diferentes escalas de distancia en cada caso ................................................................................. 106
Figura 2.15. Regresión lineal entre la abundancia general de zooplancton (indiv/m3) y
a) la profundidad de la capa de mezcla (m) y b) la biomasa del zooplancton (mg/m3
) ................................................................... 107
Figura 2.16. Abundancia promedio (indiv/m3) y desviación estándar de copépodos,
copepoditos y nauplius para cada una de las capas muestreadas durante el período de estudio, Golfo Dulce ................................................ 108
Figura 2.17. Valores promedio y desviación estándar de la abundancia de copépodos (indiv./m3
) para cada estación y capa de muestreo, Golfo Dulce, 1997-1998 ......................................................................................... 109
Figura 2.18. Valores promedio y desviación estándar de la abundancia de copépodos (indiv./m3
) para cada una de las fechas y capas analizadas, Golfo Dulce, 1997-1998 ................................................................................. 110
Figura 2.19. Abundancia relativa de los principales géneros de copépodos en el Golfo Dulce, 1997-98 ........................................................................... 115
Figura 2.20. Análisis de conglomerado para los géneros de copépodos respecto a su abundancia en la capa P1, aplicando el coeficiente de similitud Bray-Curtis y uniones de promedio por grupo. A, B y C representan los conglomerados resultantes .................................................................................. 117
Figura 2.21. Análisis de conglomerado para los géneros de copépodos respecto a su abundancia en la capa P2, aplicando el coeficiente de similitud Bray-Curtis y uniones de promedio por grupo. A, B, C y D representan los conglomerados resultantes .................................................................................. 119
Figura 2.22. Análisis de conglomerado para los géneros de copépodos respecto a su abundancia en la capa P3, aplicando el coeficiente de similitud Bray-Curtis y uniones de promedio por grupo. A, B, C y D representan los conglomerados resultantes .................................................................................. 120
Figura 2.23. Variación temporal en la abundancia (indiv./m3) de copépodos, copepoditos
y nauplius en las capas a) P1, b) P2 y e) P3 en la estación 01, Golfo Dulce ........................................................................................ 122
xii

Figura 2.24. Variación temporal en la abundancia (indiv./m3) de copépodos, copepoditos
y nauplius en las capas a) P1, b) P2 y e) P3 en la estación 02, Golfo Dulce ..... ..... ......................................................... .... ................. 123
Figura 2.25. Variación temporal en la abundancia (indiv./m3) de copépodos, copepoditos
y nauplius en las capas a) P1, b) P2 y e) P3 en la estación 03, Golfo Dulce ........ .. ............................... .......... ................. ........ ... .. . ...... 124
Figura 2.26. Variación temporal en la abundancia (indiv./m3) de copépodos, copepoditos
y nauplius en las capas a) P1, b) P2 y e) P3 en la estación 04, Golfo Dulce ...... .. ................................. ......... ................. ....... .............. 125
Figura 2.27. Variación temporal en la abundancia (indiv./m3) de copépodos, copepoditos
y nauplius en las capas a) P1, b) P2 y e) P3 en la estación 11, Golfo Dulce ....... .. ............................................................................... 126
Figura 2.28. Variación temporal en la abundancia (indiv./m3) de los principales géneros
de copépodos en las capas a) P1, b) P2 y e) P3 en la estación 01, Golfo Dulce ..... ..... ............................. ......... .. .............. .. ........ .. ............ 127
Figura 2.29. Variación temporal en la abundancia (indiv./m3) de los principales géneros
de copépodos en las capas a) P1, b) P2 y e) P3 en la estación 02, Golfo Dulce ......................................... ................................. ... ........... 128
Figura 2.30. Variación temporal en la abundancia (indiv./m3) de los principales géneros
de copépodos en las capas a) P1, b) P2 y e) P3 en la estación 03, Golfo Dulce ....... ... ................. ... ........... ....... .. ...................................... 129
Figura 2.31. Variación temporal en la abundancia (indiv./m3) de los principales géneros
de copépodos en las capas a) P1, b) P2 y e) P3 en la estación 04, Golfo Dulce ........ ... ............................................................... .............. 130
Figura 2.32. Variación temporal en la abundancia (indiv./m3) de los principales géneros
de copépodos en las capas a) P1, b) P2 y e) P3 en la estación 11, Golfo Dulce .......................................................................... .. ............ 131
Figura 2.33. Análisis de conglomerado para las estaciones muestreadas respecto a la abundancia de géneros de copépodos en las capas a) superficial (P1 ), b) intermedia (P2) y e) profunda (P3), aplicando el coeficiente de similitud Bray-Curtis y uniones de promedio por grupo. Notar las diferentes escalas en cada caso ................................................................. ............ 136
Figura 2.34. Regresión lineal entre la abundancia de copépodos (indiv/m3) y la biomasa
general del zooplancton (mg/m3) .... • . ....... .....•...•..•............ . ....•......• 137
xm


2
INTRODUCCIÓN
El término plancton fue acuñado por el alemán Victor Hensen ( 1887) y proviene
del griego 'planao' que significa derivar. Esta definición comprende a todos aquellos
organismos que están a la deriva en el agua y cuyas habilidades de locomoción son
insuficientes para soportar corrientes, por lo que se incluye a gran número de taxones,
como bacterias, microalgas y hongos, huevos, larvas y adultos de un gran número de
invertebrados, así como huevos y larvas de algunos peces (Bougis 1976; Sommer 1989;
Lenz 2000). El fitoplancton (autótrofos) puede ser separado del zooplancton
(heterótrnfos) tanto sobre la base de su tipo de nutrición como en cuanto a su
morfología, fisiología y la presencia de ambos grnpos hace que l1as comunidades
plláncticas sean autosuficientes (Zeitzshel 1978; Smetacek 1988; Kennish 1994 ).
El zooplancton juega un papel clave en las cadenas alimentarias pelágicas,
donde actúa como agente activo en la transferencia de energía de los productores
primarios a los consumidores secundarios (crustáceos, peces), así como en la
regeneración de nutrimentos a través de la excreción (lkeda 1985; Kane 1993; Skjoldal
et al. 2000). De éste modo, los estudios sobre zooplancton son utilizados para elaborar
modelos de flujo de carbono y nitrógeno, predicción de productividad de pesquerías a
partir de observaciones directas, para estimar la producción (o las pérdidas) de los
principales grupos de organismos oceánicos y como indicadores de calidad de agua
entre otras cosas (Razouls et al. 1994 ).
El medio en ell que las comunidades pláncticas se desarroll'an es muy complejo,
tanto desde el punto de vista de las interacciones biológicas como del punto de vista del
ambiente físico. Así, por ejemplo, las condiciones oceanográficas y climáticas llegan a
afectar el nivel y los tiempos de la producción primaria del fitoplancton, lo cual trae
consecuencias sobre el zooplancton y peces. Las condiciones oceanográficas, a su
vez, afectan de una manera directa al' zooplancton, al influenciar su metabolismo,
comportamiento y distribución (Zeitschel 1978; Lenz 2000; Skjoldal et al. 2000).
Entre los factores más importantes que controlan la distribución del zoop'lancton
se encuentran la profundidad, el estado trófico del área y la temperatura. La
profundidad, por ejemplo, es un factor determinante que permite diferenciar al plancton

3
nerítico del oceánico. El primero se caracteriza por una alta proporción de
merozooplancton, debido a la proximidad del fondo marino. Caso contrario es el del
zooplancton oceánico, que más bien se caracteriza por una ausencia general de
meroplancton y por la presencia de organismos que presentan migración vertical (Davis
1984; Harris 1987; Malina 1996; Van Wangelin & Wolf 1996; Skjoldal et al. 2000).
La temperatura es considerada como uno de los principales factores que
determinan la diversidad de especies en los océanos, además de ser un factor que
afecta directamente la tasa de crecimiento de los organismos. Así, por ejemplo, en
ecosistemas donde las aguas se caracterizan por sus bajas temperaturas, las especies
dominantes necesitan mantener una biomasa relativamente alta para asegurar su
supervivencia debido a que la productividad es baja. Mientras tanto, en ecosistemas de
aguas cálidas, es posible que se dé un rápido crecimiento de las poblaciones debido a
la alta tasa de crecimiento individual (Longhurst & Pauly 1987b; Lenz 2000). Otros
factores, cuyas fluctuaciones pueden representar un estrés físico para los organismos
pláncticos son la salinidad y el oxígeno disuelto. Estos llegan a limitar la distribución de
las especies y a disminuir el crecimiento y el tamaño corporal de algunos grupos
(Zeitzschel 1978; Dessier & Donguy 1985; Harris 1987; Longhurst & Pauly 1987b; Wiafe
& Frid 1996; Lenz 2000).
Las termoclinas se dan a menudo en los océanos, especialmente en zonas
tropicales y subtropicales, y juegan un papel fundamental en la distribución vertical del
zooplancton. Por ejemplo, se ha determinado que las concentraciones de fitoplancton
asociadas a la termoclina están acompañadas de agregaciones de especies de niveles
tróficos superiores. De hecho, la alta biomasa de autótrofos y heterótrofos en esta zona
hace de la termoclina un lugar de intensa herbivoría, depredación y competencia (Ortner
et al. 1978; Zeitzschel 1978; Longhurst 1985; Harris 1987; Longhurst & Pauly 1987;
Wiafe & Frid 1996). Al respecto, Longhurst (1985) hace énfasis en la importancia que
tiene una estructura física estable en la termoclina para mantener la gran diversidad de
especies de zooplancton que se observa en esta parte de la columna de agua.
La región del Pacífico Oriental Tropical (POT) se caracteriza por ser una zona de
alta producción biológica debido a la presencia de una zona de afloramiento ecuatorial,
la cual presenta una significativa variabilidad intra- e interanual. La variabilidad
intraanual es causada por cambios en la intensidad de los vientos, lo cual resulta en

4
máximos de clorofila y biomasa durante el invierno en el hemisferio Sur. La variación
interanual, por su parte, se relaciona con el fenómeno El Niño (ENSO) (White et al.
1995). Este consiste de un calentamiento anómalo de las corrientes limítrofes orientales
del Pacífico inducidas por anomalías en el campo de vientos del Pacífico Oriental. Esta
oscilación irregular se propaga hacia el Pacífico oriental con una ola interna (olas
Kelvin). Como resultado del acoplamiento atmósfera-océano se observan efectos
físicos a gran escala cerca de las costas, como por ejemplo, un calentamiento de la
capa de mezcla y el hundimiento de la termoclina y la nutriclina. Entre los efectos
biológicos de ENSO, se espera una disminución en la productividad primaria debido a la
baja entrada de nutrimentos (disminuye o cesa el afloramiento en las regiones del POT)
lo cual afecta de manera directa e indirecta al zooplancton (Dietrich 1963; Bougis 1976;
Margalef 1985; White et al. 1995; Philander 1996; Thurman 1996; Lavaniegos &
Gonzalez-Navarro 1999; NOAA 2000).
Todos los aspectos mencionados con anterioridad contribuyen con que el
zooplancton no se encuentre distribuido homogéneamente en el plano horizontal ni en el
vertical. Al contrario, el zooplancton más bien forma agregaciones que pueden ser el
resultado de respuestas de los organismos a gradientes de temperatura, salinidad,
movimiento del agua, intensidad de luz, depredadores y alimento, entre otros, así como
a comportamientos sociales complejos. Por otra parte, el tamaño y forma de las
agregaciones pueden diferir entre localidades, especies e inclusive entre estadías de
desarrollo de una especie (Ortner et al. 1978; Omori & Hammer 1982; Wiafe & Frid
1996). La organización espacial y temporal de las comunidades pláncticas en el
ambiente pelágico de los océanos es, por lo tanto, muy dinámica. Lo anterior es
producto de una combinación de efectos de los factores físicos sobre las especies, así
como de los procesos biológicos característicos de cada especie (ej. reproducción)
(Harris 1987; Longhurst & Pauly 1987).
Dentro de la evidente complejidad que muestran los ambientes marinos y en
especial las comunidades pláncticas, Buskey (1993) sugiere que para lograr
comprender las redes tróficas marinas se requiere de información sobre abundancia,
biomasa y tasa de interacción entre todos los niveles tróficos. En lo que a la biomasa
del zooplancton se refiere, el peso seco y/o el peso orgánico (peso libre de ceniza) son
los principales métodos recomendados como medida estándar para su estimación

5
(Omori 1978). La cuantificación de la biomasa del zooplancton se reconoce como un
índice importante para estimar las tasas de variabilidad estacionales y anuales en la
producción secundaria de los ecosistemas marinos. Más aún, el peso es considerado
como una medida confiable del tamaño de un animal y a menudo es utilizado para
cuantificar el flujo de energía en comunidades acuáticas (Giguere et al. 1989; Kane
1'993).
La existencia de metodologías tales como la caracterización del tamaño del
zooplancton sobre la base de la talla (o en combinación con otros factores ecológicos)
es de gran utilidad, especialmente al rel'acionarse con cuantificaciones de biomasa.
Esto se debe a que la talla de los organismos tiene un gran valor ecológico y está
fuertemente l1igada a las características hidrográficas y espacio-temporales de una
reg1ión. La distribución de tallas está asociada a la estructura trófica de la comunidad, lo
que lleva implícita la idea de que la eficiencia en la alimentación y la vulnerabilidad a la
depredación son determinadas por los tamaños corporales de los organismos
zoopláncticos (Valdés et al. 1990). Además, el uso de categorías de tamaño en el
zooplancton aporta información que permite comparar el plancton de distintas regiones y
refleja de manera precisa las características de la comunidad que están rel1acionadas
con el flu~o de energía (Valdés et al. 1990). De hecho, estudios en esta área han sido
de gran valor en l'a elaboración de modelos de simulación de la estructura de las
comunidades pláncticas (Harris 1987).
Respecto a los estudios acerca de las comunidades pláncticas que se han
realizado hasta la fecha en la costa Pacífica de Costa Rica, estos se pueden considerar
como escasos y concentrados sobre todo en el Golfo de Nicoya, el principal sistema
estuarino del país además de que han estado restringidos principalmente al componente
merozooplánctico (Epifanio & Dittel 1984; Hossfeld 1996; Melina 1996). De manera más
reciente, se han llevado a cabo otros estudios en diversas áireas de l1as ciencias
marinas, incluidos varios trabajos sobre el holozooplancton, en varios puntos de la costa
Pacífica del país. Entre estos, destacan los llevados a cabo por Morales & Jakob (en
prep.); quienes estudiaron l1a abundancia, distribución vertical, biomasa y biometría de
los ostrácodos en el Golfo Dulce; Morales & Vargas ( 1995); quienes describieron las
especies comunes de copépodos en el Golfo de Nicoya; Morales (1996); quien elaboró
la lista de especies de copépodos para el Pacífico costarricense con notas sobre su

6
distribución y Morales & Nowazyck (en prep.), quienes estudiaron la distribución,
abundancia y biomasa del zooplancton gelatinoso en el Golfo Dulce. Por otra parte,
Morales (en prensa) discute aspectos sobre la biodiversidad de copépodos en las cos1as
de Costa Rica.
La información generada hasta la fecha ha sido la suficiente como para plantear
los primeros modelos tróficos para el Golfo Dulce (Wolff, Hartmann & Koch 1996) y el
Golfo de Nicoya (Wolf et al. 1998). Estos modelos representan un primer paso y una
herramienta muy práctica para la comprensión del funcionamiento de dos de los
principales sistemas marino-costeros del país. Sin embargo, aún es mucho lo que resta
por hacer, en especial en lo que se refiere al comportamiento y descripción detallada de
las comunidades pláncticas a través de períodos de tiempo prolongados. Esta
necesidad es aún mayor en aquellas zonas del país donde se ha generado
relativamente poca información hasta la fecha . Tal es el caso del Golfo Dulce, ubicado
en la costa Pacífica sur del país y que posee características morfológicas y
oceanográficas similares a las esperadas en un fiordo, lo cuál resulta poco común en
zonas tropicales (Richards et al. 1971; Thamdrup et al. 1996; Umaña 1998) y le hacen
más atractivo para futuras investigaciones. Es una zona que, además, recibe un
creciente impacto humano producto de la deforestación, las zonas agrícolas y el
crecimiento demográfico (Spongberg & Davis 1998, Umaña1998). Por esto, resulta
particularmente importante generar información que sirva como base para el desarrollo
de futuras estrategias de manejo tanto del Golfo Dulce como de los ecosistemas
terrestres que lo afectan de manera directa.
Con base a lo anterior y a la importancia que los estudios a largo plazo revisten
para lograr una mayor comprensión de los sistemas marino-costeros del país y su futuro
manejo, se plantea la siguiente investigación. La información generada a partir de éste
trabajo será además muy valiosa para la actualización del modelo trófico del Golfo Dulce
y servirá de base para el planteamiento de futuras investigaciones en la zona.

7
OBJETIVOS GENERALES
A. Describir la distribución temporal y espacial de la biomasa de zooplancton en el
Golfo Dulce durante un período de dos años (1997-1998).
B. Describir los gradientes físicos del agua en la zona de estudio durante el mismo
período.
C. Evaluar el posible efecto del fenómeno El Niño sobre el desarrollo de la comunidad
de zooplancton en el Golfo Dulce durante 1997 y 1998.
OBJETIVOS ESPECIFICOS
A. Determinar si existe variación espacial y temporal en la biomasa del zooplancton en
el Golfo Dulce.
B. Estudiar la distribución vertical de la biomasa de zooplancton en cada una de las
estaciones de muestreo y determinar si existen diferencias en la biomasa de las
distintas capas muestreadas.
B. Determinar si existen diferencias en el aporte de biomasa de tres clases de tamaño
de zooplancton y si existen diferencias al comparar las capas muestreadas, las
estaciones y las fechas de muestreo.
D. Comparar las distribuciones de biomasa en las estaciones con variables físicas
como temperatura, salinidad, oxígeno disuelto, profundidad del disco de Sechii y
profundidad de la capa de mezcla con el fin de establecer las posibles relaciones
entre dichas variables.
E. Comparar los datos obtenidos durante esta investigación con estudios previos
llevados a cabo en períodos anteriores al fenómeno El Niño con el fin de determinar
el posible efecto de dicho fenómeno sobre la distribución de la biomasa del
zooplancton en el Golfo Dulce.

8
MATERIAL Y METODOS
l. Sitio de Estudio
El Golfo Dulce es una bahía semicerrada, tectónicamente formada, del Pacífico
tropical oriental, localizado en la costa sur de Costa Rica, entre los 8º 27' y 8º 45' latitud
norte y los 83º 07' y 83º 30' longitud oeste (Fig. 1.1 ). Está separado del océano Pacífico
por la Península de Osa, un área montañosa y cubierta de bosque. El Golfo tiene cerca
de 50 km de longitud y de 1 O a 15 km de ancho, con un área superficial de
aproximadamente 680 km2 y se caracteriza por poseer una cuenca interna de pendiente
fuerte con una profundidad máxima de 215 m y una cuenca externa poco profunda de
aproximadamente 70 m (Richards et al. 1971; Hartmann 1994 ).
Estas características tienden a restringir la circulación de agua en la cuenca
interna del Golfo y a favorecer la formación de aguas profundas anóxicas. Su
circulación asemeja la de los fiordos de altas latitudes y es uno de tan solo cuatro
sistemas de éste tipo conocidos en los trópicos (Richards et al. 1971 ).
11. Factores Climáticos
El clima de la zona puede definirse como muy húmedo y caliente. En general,
llueve durante todo el año, pero se puede distinguir una estación lluviosa determinada
por el movimiento de la zona de convergencia ínter-tropical. La cuenca del Golfo recibe
de 4000-5000 mm de lluvia por año y la estación lluviosa se extiende desde abril hasta
diciembre, alcanzando su pico en los meses de agosto y octubre, cuando el promedio
mensual de lluvia varía entre los 520 mm y los 860 mm. La precipitación por lo general
excede la evaporación durante al menos 8 meses al año, razón por la cual las tierras se
mantienen saturadas de agua. Esto favorece la rápida erosión de los terrenos
expuestos y da cabida al desbordamiento de ríos, lo que incrementa el ingreso de
nutrimentos y materia orgánica a las aguas del Golfo. Las máximas temperaturas
atmosféricas se registran durante la estación seca, época durante la cuál también se da
un incremento en el número de horas diarias de brillo solar (Hartmann 1994; Wolf et al.
1996).

9
111. Hidrografía
Las características de fiordo del Golfo Dulce se reflejan en la composición de sus
masas de agua. Dos capas son evidentes: una capa superficial cálida (19 - 30 ºC) y
diluida (< 34 PSU) que alcanza los 50 - 60m de profundidad, justo sobre la profundidad
del umbral ubicado en la parte externa del Golfo. La otra capa la componen las aguas
profundas y físicamente homogéneas, con un mínimo de temperatura de 15.4 ºC y
salinidades que alcanzan los 34.8 PSU. La mezcla y circulación de estas aguas es
influenciada por las fuerzas de mareas, el viento, el ingreso de agua al sistema, el
afloramiento de agua subsuperficial y la topografía de la cuenca (Richards et al. 1971;
Córdoba & Vargas 1996).
El río más grande, el Coto-Colorado, tiene un transporte medio anual de 177
m3/s. El flujo total de agua dulce al Golfo no excede los 1000 m3/s por lo que
probablemente no afecta significativamente los patrones de circulación dentro del Golfo.
Sin embargo, las fuertes lluvias durante los meses críticos de la estación lluviosa
pueden afectar significativamente el flujo de agua dulce sobre períodos cortos de
tiempo, especialmente cuando los suelos se saturan de agua (Hartmann 1994; Umaña
1998).
La renovación de las aguas profundas en el Golfo Dulce es lenta y restringida a
dos períodos de entrada de agua subsuperficial (Hartmann 1994 ). Se ha identificado
masas densas de agua que pasan sobre el umbral externo y entran a la cuenca, donde
se hunden rápidamente y desplazan las aguas profundas hacia arriba (Richards et al.
1971)
IV. Colecta y Procesamiento de Muestras
Se seleccionó un total de cinco estaciones de muestreo (Cuadro 1.1) distribuidas
en la cuenca del Golfo Dulce con el fin de obtener muestras de las partes interna
(estación 01), media (estación 03) y externa (estación 11) (Fig. 1.1). Las estaciones 02
y 04 se ubicaron cerca de la desembocadura de los ríos Esquinas y Coto-Colorado,
respectivamente.

a3·20'
10"001
_ ¡
Figura 1.1. Ubicación geográfica de los puntos de muestreo, Golfo Dulce, Costa Rica. llapm Dnlldo de M«*ia 1'98
10

11
Es importante señalar que las estaciones 02 y 03 del presente estudio se ubican
muy cerca de las estaciones 05 y 03 (respectivamente) usadas por Richards et al.
( 1971 ). Así mismo, la estación 11 corresponde a la que fue previamente muestreada
entre los años 1993 y 1994 durante la campaña de investigación del buque
oceanográfico B.O. Victor Hensen, coordinada por el Centro de Investigación en
Ciencias del Mar y Limnología (CIMAR) de la Universidad de Costa Rica en conjunto
con el Centro de Ecología Marina Tropical (ZMT) de la Universidad de Bremen,
Alemania (Wolff & Vargas 1994)
El programa de muestreo comprendió un total de 17 visitas al área, realizadas
una vez al mes a lo largo de un período de dos años consecutivos, entre abril de 1997 y
noviembre de 1998. Dicho programa incluyó muestreos tanto durante la época seca
como la lluviosa, con el fin de asegurarse que los patrones encontrados correspondan a
patrones estacionales y no a problemas metodológicos.
Cuadro 1.1. Ubicación geográfica de las estaciones de muestreo de zooplancton establecidas en el Golfo Dulce, Costa Rica.
Estación Coordenadas Descripción Prof.(m)
GD-01 08º 42' N; 83º 27' O Frente a Rincón de Osa 200
GD-02 08º 40' N; 83º 22' O Frente al Río Esquinas 200
GD-03 08º 36' N; 83º 12' O Frente a Bahía Golfito 150
GD-04 08º 33' N; 83º 11' O Frente a río Coto-Colorado 70
GD-11 08º 29' N; 83º 12' O Desembocadura del Golfo 70
Los muestreos se realizaron en un bote Zodiac equipado con una estructura
metálica con un sistema de poleas con el fin de facilitar la extracción de las muestras de
agua y de plancton. Previo a los muestreos de zooplancton en cada estación, se
colectó una serie de muestras de agua usando una botella Niskin de 2L a las siguientes
profundidades: O, 10, 20, 30, 40, 50, 60, 70 y 100 m. Durante 1998 se tomó una
muestra adicional de agua a 150 m de profundidad en las estaciones 01, 02 y 03. Para
cada una de las muestras, se determinó la salinidad (PSU) con un refractómetro de

12
mano, marca ATAGO S/Mill-E, así como la temperatura (ºC) y oxígeno disuelto (mg
0 2/1), con un medidor de oxígeno YSI 52. Además, en cada estación se midió la
profundidad del disco de Sechii y las condiciones climáticas como nubosidad, condición
del viento y oleaje.
Una vez determinada la posición de la termoclina en la columna de agua, se
procedió a realizar tres arrastres verticales en cada estación con una red cónica para
zooplancton de 50 cm de diámetro de boca y 153 µm de poro de red. Con ayuda de un
mecanismo de cierre remoto para la red, se realizó un arrastre por encima, uno por
debajo y otro a través de la termoclina (Fig.1.2). Los volúmenes de agua filtrados fueron
Z(m)
-
T (ºC)
Termoclina
11100um
500·um
150 um
D ·· -·~'"" .. _ .........
""""""- --·
T1
T2
T:i
P1
P2
P3
Figura 1.2. Criterio de muestreo utilizado para la colecta de zooplancton, basándose en la posición de la termoclina en la columna de agua y categorías de tamaño utilizadas para fraccionar las muestras.

13
calculados matemáticamente conociendo el área de la boca de la red y la distancia de
arrastre en cada caso. Además, se calculó un promedio de temperatura, oxígeno y
salinidad para cada capa muestreada. Una vez colectadas, las muestras fueron fijadas
en formalina al 4 % (Omori & Fleminger 1976) y llevadas a los laboratorios del CIMAR
para ser procesadas. Se debe aclarar que se realizó solamente un arrastre a cada
profundidad debido a razones logísticas. Además, resulta importante recalcar que se
tomó en cuenta la estructura física de la columna de agua (en lugar de intervalos de
profundidad predeterminados) como criterio para realizar los muestreos de zooplancton.
La campaña de muestreo generó un total de 246 muestras (17 fechas x 5
estaciones x tres profundidades), considerando que en la gira de abril de 1997 no se
muestreo la estación 02 debido a problemas logísticos y que durante la gira de junio del
mismo año no se muestreo las estaciones 04 y 11 debido a problemas climatológicos.
Por esta razón, todas las muestras colectadas en dichas fechas se excluyeron de los
análisis de biomasa, los cuales por lo tanto, se realizaron sobre una base de 15 fechas
de muestreo (225 muestras).
Una vez en los laboratorios del CIMAR, las muestras fueron fraccionadas,
utilizando un separador de plancton Folsom (WILDCO, modelo 1831F10), para obtener
alícuotas de no menos de 1/16ªvº del volumen total. Seguidamente, cada alícuota (x) fue
fraccionada en tres clases de tamaño: 1) x > 1000 µm; 2) 500 µm < x < 1000 µm; 3) 150
µm < x < 500 µm. Los volúmenes restantes de cada muestra fueron lavados con agua
destilada para eliminar la formalina y transferidos a alcohol al 70% para su preservación.
Las fracciones de tamaño obtenidas fueron utilizadas para la determinación de
biomasa, para lo cuál se siguió el método de medida de peso seco según Beers (1976).
Para la filtración se utilizó filtros de fibra de vidrio (GF/C Whatmann) que fueron
previamente secados y pesados en una balanza analítica. Seguidamente, cada fracción
de tamaño fue filtrada con ayuda de una bomba al vacío y un sistema de filtrado
MILLIPORE. Así mismo, una vez filtrada cada fracción, se procedió a lavar el filtro
correspondiente con agua destilada con el fin de remover las sales y evitar sobre
estimaciones (Omori 1978). Posteriormente, los filtros fueron colocados en una estufa a
60 ºC por espacio de 8-10 horas, siguiendo las recomendaciones de Lenz (1971).
Finalmente, los filtros se dejaron aclimatar a temperatura ambiente dentro de un

14
desecador por espacio de 3-4 horas, al cabo de las cuales fueron pesados de nuevo
hasta peso constante. Los datos de biomasa fueron calculados en mg/m3, tomando en
cuenta el volumen de agua filtrado en cada caso.
V. Análisis Estadísticos
Las pruebas estadísticas se llevaron a cabo con el programa SYSTAT 8.0. Se
aplicaron pruebas de estadística descriptiva (promedio, desviación estándar, ámbito)
para las variables de biomasa, temperatura, salinidad y oxígeno disuelto para cada
estación y para cada capa de agua muestreada (P1 , P2, P3) por estación. Además se
llevó a cabo un análisis de conglomerado (aplicando el coeficiente de similitud Bray
Curtis y uniones de promedio por grupo) con el fin de agrupar a las estaciones de
muestreo con respecto a la biomasa del zooplancton. Así mismo, se realizaron pruebas
de ANDEVA (prueba post-hoc: Tukey) con un nivel de confianza del 95% (Bailey 1995),
con el fin de evaluar las posibles diferencias de biomasa entre estaciones, fechas,
profundidades y tallas. Finalmente, se aplicó un análisis de regresión lineal con el fin de
cuantificar la posible relación funcional entre la biomasa y las variables de profundidad
del disco de Sechii, profundidad de la capa de mezcla, temperatura, salinidad y oxígeno
disuelto.

15
RESULTADOS
l. Parámetros Meteorológicos
Al observar los valores de temperatura atmosférica (Figura 1.3a) resulta claro
como durante 1997 y 1998 las mayores temperaturas se dieron durante el período de
febrero a mayo. En el caso del primer año, el mes con mayores temperaturas fue abril,
con un promedio de 35.5ºC, mientras que para 1998, es el mes de marzo, con un
promedio de 35ºC. Resulta importante aclarar que para 1997 no se obtuvieron datos de
temperatura para los meses de marzo, agosto y septiembre. Con respecto a las
temperaturas mínimas registradas durante el período de estudio, estas correspondieron
a los meses de diciembre, enero y febrero. En ambos años, los valores más bajos de
temperatura se dieron durante el mes de enero, con 20.2ºC y 21.1 ºC respectivamente.
Con respecto a la precipitación, durante 1997 se tuvo un total de 3008. 7 mm de
lluvia, para un promedio mensual de 250. 73 mm, mientras que para 1998 el total fue
mayor, con 4776 mm de lluvia para un promedio mensual de 398 mm. Los meses más
lluviosos durante 1997 fueron mayo y junio, con 591.1 mm y 376.6 mm respectivamente.
En el caso de 1998, los meses con mayor precipitación fueron junio y julio, con 854.8
mm y 665.3 mm respectivamente (Figura 1.3b).
11. Parámetros Físico-Químicos
• Temperatura
Los gráficos de isolíneas correspondientes a los valores de temperatura (ºC),
salinidad (PSU) y oxígeno disuelto (mg/I) para cada estación (Figuras 1.4 - 1.8) ofrecen
información muy interesante respecto al comportamiento de las aguas del Golfo Dulce
durante el período de estudio. Para el caso de la temperatura, en las estaciones 01 y 03
(Figura 1.4a y 1.6a) resulta claro un calentamiento abrupto de las aguas de abril a junio
de 1997. El mismo calentamiento de observa en las estaciones 04 y 11 (Figuras 1.7a y
1.8a) entre los meses de abril a julio. Para el caso de la estación 02 (Figura 1.5), no se
cuenta con datos físicos para el mes de abril, sin embargo, el calentamiento de las
aguas parece haberse dado entre abril y junio, al igual que en las estaciones 01 y 03, lo

a)
35 + 1
:: 1 §: 1 ...
.. . -................................. .. ... . -·· - ...... .. ... -....... . . . .. .. .. . .. ....... ··-
.... .. .. .. .... . 1- 20 -' •
15
10
5
o E-97 F A M J J o
b) 900 t 800
700 ' 1
1 I 600 t :~ 500 1
l 400 í .... 300 ~ c.. 1
200 t 100 1
T
o E- F M A M J J A 97
N D E-98 F M A
s o N D E· F M 98
M J
A M
---T max · · · · · · T min
J A s
J J A
o
s
Figura 1.3. Parámetros Meteorológicos durante el período 1997-98 para la zo1 Golfo Dulce: a)promedios de temperatura(ºC) máxima y mínima, b) pror mensuales de precipitación. Fuente: Instituto Meteorológico Nacional. Estación Meterológica Coto 47: 08' 82° 59' O; 8 m.s.n.m.
16
.. ·-... .. ..
1--l
N D
o N D

a)
z (m)
b)
z (m)
e)
17
Ab-97 J A S O N E-98 M All M J J A S O N
Figura 1.4. Variación temporal en las isoclinas de a) temperatura (ºC), b) oxígeno disuelto (mg/I) y e)
salinidad (PSU) correspondiente a la estación O 1, Golfo Dulce, durante el período 1997-1998.

a)
z (m)
b)
z (m)
e)
z (m)
{)
a
20
30
40
50
60
70
100
150
o
10
20
30
40
50
60
70
100
150
18
J.97 J A :s o N ~~81 ,.. A M ~ ~ s o N
Figura 1.5. Variación temporal en las isoclinas de a) temperatura (ºC), b) oxígeno disuelto (mg/I) y e)
salinidad (°loo) correspondiente a la estación 02, Golfo Dulce, durante el periodo 1997-1998.

19
a) o
10
20
30
z (m) 40
50
60
10
100
100
()
b) 10
20
30 z
(m) 40
50
60
70
100
150
o e)
10
.20
30
z 40
(m) 50
Ab-97 J J A S O N E-98 M A M J J A S O N
Figura 1.6. Variación temporal en las isoclinas de a) temperatura (ºC), b) oxígeno disuelto (mg/I) y e)
salinidad (PSU) correspondiente a la estación 03, Golfo Dulce, durante e/ período 1997-1998.

20
o a)
10
20
z 30
(m) 40
50
60
b) 10
20
30 z
(m) 40
50
e) o
10
20-z
(m) 30 -
40 -
Ab-97 J J A S O N E-96 M A M J A S O N
Figura 1.7. Variación temporal en las isoclinas de a) temperatura (ºC), b) oxígeno disuelto (mg/I) y e)
salinidad (PSU) correspondiente a la estación 04, Golfo Dulce, durante el período 1997-1998.

a)
b)
10
20
z (m)3º
40
o
10
20
z 30
(m) 40
50
60
o e)
10
20
z 30
(m) 40
50
60
\
Ab-97
21
J A s o N E-98 M A M .J J A s o N
Figura 1.8. Variación temporal en las isoclinas de a) temperatura (ºC), b) oxígeno disuelto (mg/I) y e)
salinidad (PSU) correspondiente a la estación 11 , Golfo Dulce, durante el período 1997-1998.

22
cual es de esperarse dado que la estación 02 se encuentra en un punto intermedio entre
estas. Resulta interesante además, observar que en todas las estaciones la columna de
agua presentó los valores más bajos de temperatura durante abril de 1997 y que dichos
valores no se volvieron a repetir durante el período de estudio.
Un aspecto muy importante es el de los patrones de enfriamiento de la columna
de agua que se dan durante los meses de marzo - abril y agosto - septiembre de cada
año. Estos son evidentes por la elevación de las isotermas durante el período citado
para su posterior descenso en los dos meses siguientes. Este patrón se repite en cada
una de las estaciones muestreadas (Figuras 1.4a - 1.8a) y es producto de períodos de
entrada de agua (subsuperficial) oceánica fría a la cuenca del Golfo Dulce.
En lo que se refiere a los valores promedio de temperatura por capa muestreada
(P1, P2, P3) (Cuadro 1.2), el valor mínimo en todo el período de estudio fue de 18.3 ±
1.5ºC, en la capa profunda de la estación 03, mientras que el máximo (31.5 ±O. ?°C) se
dio en la capa superficial de la estación 11 . Al observar con más detalle el
comportamiento de las temperaturas promedio por año para cada una de las estaciones
(Figuras 1.9 -1.13), resulta evidente la manera en la que varían las condiciones físicas
con la profundidad en el Golfo Dulce.
En general, la capa superficial (P1) de todas las estaciones se caracterizan por
tener promedios anuales de temperatura que varían entre 29.2 y 29. ?°C. Para el caso
de las estaciones con- mayor profundidad (01, 02 y 03), las capas intermedia (P2) y
profunda (P3) tienen temperaturas promedio anuales de 24.6 - 25. ?°C y de 20.4 - 21 ºC
respectivamente. Mientras tanto, en las estaciones con menor profundidad (04 y 11 ),
los promedios anuales de temperatura en las capas P2 y P3 varían entre 26.9 - 27.3ºC y
23.4 - 23.?°C respectivamente. El caso de las estaciones 04 y 11 se considera por
separado debido a que su menor profundidad de fondo viene a influenciar las
temperaturas mínimas esperadas para las capas intermedia y profunda. En todas las
estaciones es posible observar que la temperatura promedio de las capas P1 y P3 se
mantiene relativamente estable durante todo el período de estudio (Figuras 1.9a -
1.13a). Como se aprecia en las figuras 1.4 - 1.8, las condiciones de la columna de agua
cambian muy poco después de los 100 m de profundidad. En la capa P2, los valores de
temperatura para todas las estaciones descienden durante el mes de

23
Cuadro 1.2. Ambito de variación, promedio y desviación estándar de los valores de biomasa total, temperatura, oxígeno disuelto y salinidad para cada una de las estaciones muestreadas durante el período 1997-98, Golfo Dulce.
Estación Biomasa {mg/m3} Tem~ {ºC} Oxig {mg/I} Sal (PSU}
Est. 01 N 45 45 45 45 Min. 1.08 19.83 0.73 23 Max. 32.42 31.13 5.73 37.33 Prom. 7.01 25.43 2.51 32.77 O.E. ± 7.15 ± 3.73 ± 1.5 ± 2.98
Est. 02 N 45 45 45 45 Min. 0.34 19.33 0.8 26.33 Max. 29.80 30.88 5.23 37.25 Prom. 6.49 24.83 2.71 33.02 O.E. ± 7.16 ± 3.65 ± 1.44 ± 2.34
Est. 03 N 45 45 45 45 Min. 0.91 18.33 0.7 26.37 Max. 25.23 31 5.65 38 Prom. 6.79 24.79 2.88 32.98 O.E. ± 5.98 ± 3.9 ± 1.56 ± 2.8
Est. 04 N 45 45 45 45 Min. 0.83 20.5 1.1 22 Max. 54.51 31 6.3 37 Prom. 12.16 26.61 3.46 31.93 O.E. ± 9.64 ± 2.84 ± 1.32 ± 3.29
Est. 11 N 45 45 45 45 Min. 1.76 21 1.25 23 Max. 47.84 31.5 6.2 37 Prom. 12.76 26.88 3.61 32.54 O.E. ± 9.82 ± 2.77 ± 1.21 ± 2.74

24
septiembre de ambos años así como en abril de 1998 (de manera más leve) (Figuras
1.9b - 1.13b ). Esto se refleja en un ascenso de las isotermas y, por lo tanto, de la
termoclina en tales fechas.
• Oxígeno Disuelto
Se pueden apreciar, aunque de manera menos clara, patrones similares en el
comportamiento de las isolíneas de oxígeno disuelto para cada estación (Figuras 1.4b -
1.8b ). En éste caso, al observar la isolínea correspondiente a 1 mg02/I, por ejemplo, se
aprecia como esta se hace más profunda aproximadamente durante los mismos
períodos en los que se observa el enfriamiento de las aguas. Lo anterior es más claro
para el caso de las estaciones 01, 02 y 03 (Figuras 1.4b, 1.5b, 1.6b), las cuales además
tienen mayor profundidad. En estas estaciones, resalta el hecho de que se den
concentraciones relatiivamente altas de oxígeno (hasta 1 mg02/I) a profundidades
mayores a los 100 m, especialmente en los períodos correspondientes a la entrada de
agua oceánica. Durante el primer año de estudio, en las estaciones 01, 02 y 03 se
aprecia como las isolíneas son relativamente más profundas en abril, (la estación 02 no
cuenta con datos de oxígeno disuelto para abril y junio de 1997) y durante agosto
septiembre, para volver a subir en octubre. Al, año siguiente el aumento en la
concentración de oxígeno disuelto en aguas profundas se da durante los meses de
marzo y septiembre, lo que coincide con los períodos de entrada de agua oceánica.
En el caso de las estaciones 04 y 11 (Figuras 1. 7b, 1.8b ), el patrón de
movimiento de las isolíneas de oxígeno disuelto no resulta tan claro. En ambos casos,
durante el mes de abril (1997) la columna de agua presenta niveles al,tos de oxígeno
hasta el fondo. Un aspecto en el que coinciden todas las estaciones es que durante los
meses en los que se da un calentamiento de las aguas se observa una disminución en
los niveles de oxígeno disuelto, especialmente en las capas superficiales (P1 ).
Con respecto a los valores promedio de oxígeno disuelto por capa (Cuadro 1.2),
el mínimo corresponde a 0.7 ± 0.6 mg/I en la capa profunda de la estación 03, mientras
que el máximo es de 6.3 ± 1.3 mg/I, en la capa superficíal de la estación 04. Al observar
los valores correspondientes a la concentración de oxígeno disuelto por capa, la P1 en
todas las estaciones se caracterizó por tener promedios anuales de entre 4.4 y 5 mg/I.
a) 40 T 7
35 6 .... -- .... -.. .. ... ,. ... .. 30 •' .. ... ~ - -. ,. ... 5
::::> .. ,,. en 25 ,, .. -- ,,. c.. .. ,, ........... - 4'§, .. :::::.. 20 ---- - .. - E ,, (.) 3-:; !!- 15 o .... 2 10
5
o o J A s o N E M A M J J A s o N
Fecha
b) 40 7
35 - ... .... 6 .. , ... .. ... .... . . . . .... .. ..... .. .... .. • P"
30 5 ::::> 25 en c.. 4::? rr 20 C>
o 3.§.. ;: 15 .. .. -- "' .. , -- .. 2º
10 ... .. .. .... .... .. ,,. Temp .... .. .. 5 · - - · - - PSU - 02 o o
J A s o N E M A M J J A s o N Fecha
e) 40 7
35 ... ..... .. ........ .. ..... -- ~ ......... .. .. .. ... ........... ....... 1 6 ... . '
30 5 ::::> 25 ........ en 4 "§, c.. ~- ..___ rr 20 - - E o 3--;::; ;: 15 o
10 2 .. ,,. .. .. -- -- -.. .. .. --- .. --.. .. .. - -.. 1 5 .. .. o o
J A s o N E M A M J J A s o N Fecha
Figura 1.9. Variación temporal de los parámetros físico-químicos de temperatura (ºC), oxígeno disuelto (mg/I) y salinidad (PSU) para a) P1, b) P2 y e) P3 en la estación O 1 durante el período 1997-98.
25

a) 40 7
35 6 ........ . .. 30 ·- - ... . .... ... ....
• - - # -- - ..... : 5 ~ 25
.. .... --,. . ... ... .. ,. ,. -.. ,. 4"§, c.. --- - ..
:::...20 E (.)
l ~ª ~15 1-
10
5
oL, J A s o N E M A M J ,J A s o N
Fecha
b) 40 T f: 35 ... .... - .. -. . -- ... - ........ -........ ... .. .. .. .. . -· . .. 30 5
::::> en 25 4E:" c.. ::::=.. 20 , .. en (.)
' .. 3.§.
~ 15 - ' \ .. N - ... o 1- .. -.. .. .. .. ... .. .... 10
... .. Temp .. 2
"' .... ..
5 - - - - - - PSU r: - 02 o
J A s o N E M A M J J A s o N Fecha
e) 40 - 7
35 1 . . . .... .. ....
.............. .. .. -........ --- .. ...... -...... - ............. 6 .............. 30 5
::::> 25 -en 4 =a, c.. ---------::::=.. 20 ~ E
(.) 3-;:: ~ 15 .. o 1- I .. 2 10 ..
I .. .. .. .. - ,. .. 5 I - ,. ----.. .... -... -- - 1 ..... o o
J A s o N E M A M J J A s o N Fecha
Figura 1.1 O. Variación temporal de los parámetros físico-químicos de temperatura (ºC), oxígeno disuelto (mg/I) y salinidad (PSU) para a) P1, b) P2 y e) P3 en la estación 02 durante el período 1997-98.
26

a) 40 T? 35 ... - 6
••• -¡.. • ......... .. - '~ ,. .. 30
::l ' ~ .... .. .. " ~- -- ... '.:'~ 5 .. .. .. .. .
en 25 .. .. .. .. .. .... ... --...... ~
e: , .... 4 Ci u 20
, .s ~ 3 N
1- 15 o 10 2
5 1
o o J A s o N E M A M J J A s o N
Fecha
b) 40 T - 7 .. ... ..... ... ...
35 t .... ................... 6 . . . 30 5
~ 25 42 ~ 20 , .. Cl
u .. 3É.
~:n I " .... .. .. ,. .. .. -.. .. N
I - - .. " .. o Temp "
.. .. 2
· · · · · · PSU 1 - 02 o o
J A s o N E M A M J J A s o N Fecha
e) 40 T 7 .. .... .. ... 35 .. .... .. .................. -- ...... 6 30
.. 5
::l 25 ~
en 4 '§, a. ;:::.. 20 E u 1 3 ';:; ~ 15 + o 1- !
10 i 2 ,. .. .. - .. ' .. .. .. -.. .. .. .. " .. 1
~ I " .. ... -..
o J A s o N E M A M J J A s o N
Fecha
Figura 1.11. Variación temporal de los parámetros físico-químicos de temperatura (ºC), oxígeno disuelto (mg/I) y salinidad (PSU) para a) P1, b) P2 y e) P3 en la estación 03 durante el período 1997-98.
27

a) 40 + 7
35 •' I" .. 6
' , ..
30 .. .. . .. 5 -- .. .. "' ::::> 25 .. ... ---.. ~
CJ) ... .. 4'§,
~ 20 -. . .. .. -"
E (.) 3--;:; ~ 15 o
10 2
5
o o J A s o N E M A M J J A s o N
Fecha
b) 40 T 7
35 ............ . .. .. ...... . ,. . 6
30 .......... 5
~ 25 ~
~ 20 1 .. -- -.. 4 '§, .. .... .. E
(.) .. .. -.. .. .. - 3--;:; .. .. - -- ~ -
~~~L--- ..
Temp o 2
· · · · · · PSU
- 02 o 1- o
J A s o N E M A M J J A s o N Fecha
e) 40 T 7
35 .... .. -~ ........ -... •"" "' ... ~. .. "' . .. ... 6
30 5 ::::> 25 ~
CJ) -+- 4:::: e: 1 Cl
u 20 1 E o ... .. .. ,,... + 3--;:; ¡::- 15 ... .. .. , o .... -.. ..
' ' -- ' - -~2 10 .. .. ' ... 5 • 1
o 1- o J A s o N E M A M J J A s o N
Fecha
Figura 1.12. Variación temporal de los parámetros físico-químicos de temperatura (ºC), oxígeno disuelto (mg/1) y salinidad (PSU) para a) P1, b) P2 y e) P3 en la estación 04 durante el período 1997-98.
28

a} ;~ f 30 t
~ 25 T g,_ 20 1
~ 15 r
~ ... , .. .. ........ .. . . • • • •• ·• • • r 11,
--~-------;.....,-~. --~- ._..;.---.~~~~ .. ::--::--:.~,7·~. ~--:~~- ~· ·.;....~·~· ~-:_ _ __________ ,.
.,
7
6
5 -4 'a, E
3~ o 2 ~ '~ t
O -;-----t~-r-~r---+~-;-~1---1---1~-.--~+---+~--+-~+--+~+ o
b} 40
35
30
J A S O N E M
.................. .. . ..
A Fecha
M J
.. .
J A s o N
7
.. 6 . .. 5 i
~ 25 + .. 4~Cl a. .. .. .. ::::.. 20
1 .... ... .. .. ... .. - • • - .. ""' .._ E
u 1 11111... .. .. .. .. 3~
~ 15 + ---Temp o
'~ l+~----+-~.-------+---1r---+-~-- ~~·-~--~~-~-u--~~-+---+~-+---.----+ : e) 40 T
35 : T 30 +
J A S O N E M A M J J A S O N Fecha
.. . ........... .. .......... .. .....
T 7
6
5 -~ 25 1
~ 20 11 .. 4 'a,
¡::- 15 ..
10
,,. ~-----
.. " .. ... - - "' .. ,,. .... ... -- .. ..
E 3~ o 2
5
0 +---+~--+-~+-~~--1-~1---r~-+-~1---+-~-+---+~~~+--+ 0
J A S O N E M A M Fecha
J J A S O N
Figura 1.13. Variación temporal de los parámetros físico-químicos de temperatura (ºC), oxígeno disuelto (mg/I) y salinidad (PSU) para a) P1, b) P2 y e) P3 en la estación 11 durante el período 1997-98.
29

30
En el caso de las estaciones 01, 02 y 03, las capas P2 y P3 tienen promedios
anuales en la concentración de 0 2 de 1.9 - 2.6 mg/I y 1.1 - 1.3 mg/I respectivamente.
En las estaciones 04 y 11 los promedios anuales de las capas P2 y P3 variaron entre
3.3 - 3.5 mg/I y 2.1- 2.3 mg/I respectivamente (Figuras 1.9 -1.13). Con respecto a las
curvas correspondientes a la concentración de oxígeno disuelto para la misma capa, se
aprecia que en la estación 01 (Figura 1.9a) se da un incremento en los valores de 0 2 a
partir de abril de 1998, alcanzando los valores más altos hacia julio y octubre del mismo
año. Para el caso de la estación 02 (Figura 1.1 Oa), dichos incrementos se dan en
agosto (1997) y abril (1998) mientras que en la estación 03 (Figura 1.11 a) suceden en
agosto de 1997 y los meses de abril, julio y septiembre de 1998. Finalmente, las
estaciones 04 y 11 (Figuras 1.1'2a y 1.13a) presentan aumentos en el nivel de 0 2
disuelto en los meses de julio (1997-98) y abril (1998), as i como un descenso en los
meses de agosto y septiembre de ambos años. Las capas P2 y P3 de las estaciones
01, 02 y 03 presentan valores altos de 0 2 durante el mes de agosto (1997) que
disminuyen hacia octubre del mismo año. En el caso de las estaciones 04 y 11, el
incremento en los val'ores de 0 2 en las capas P2 y P3 se da en junio de ambos años y
en enero de 1998.
• Salinidad
En las figuras correspondientes a las curvas de salinidad para cada estación
(Figuras 1.4c-1.8c) es posible observar períodos en los que se da un aumento de la
salinidad en aguas profundas, como es de esperar en caso de entradas de masas de
agua oceánica. Para todas l'as estaciones, estos períodos corresponden a los meses de
septiembre (1997-98), marzo y mayo de 1998. Para el caso de las estaciones 01 y 02
es posible observar como la salinidad del agua superficial disminuye durante los meses
de junio (1997) y septiembre-octubre (1997-98). En las restantes estaciones los
períodos de dilución de las aguas superficiales se dan en los meses de septiembre
( 1997), j1ulio, agosto y noviembre de 1998. Algo importante que se debe considerar es
que las bajas salinidades observadas en algunos casos probablemente coinciden más
bien con períodos de lluvia que se dieron días y hasta horas antes de los muestreos, por
lo que los valores bajos observados en algunas fechas no coinciden de manera exacta
con la curva de precipitación mensual (Figura 1.3b)

31
Finalmente, el valor mínimo de salinidad promedio corresponde a 22 ± 9.9 PSU,
en la capa superficial de la estación 04, mientras que el máximo es de 38 ± 1.0 PSU, en
la capa profunda de la estación 03 (Cuadro 1.2). Los valores de salinidad promedio por
año para cada capa no presentaron fluctuaciones fuertes y sus valores se mantuvieron
dentro de los siguientes rangos para todas las estaciones: 28.9 -30.5 PSU en la P1;
32.7 - 33.7 PSU en la P2 y 34.1 - 34.9 PSU para la P3 (Figuras 1.9 -1.13).
Por último, los valores de salinidad en todas las estaciones y capas muestreadas
(Figura 1.9 - 1.13) se comportan de manera similar, con pequeños incrementos en
agosto - setiembre (1997) y marzo y mayo de 1998. Además, en todas las estaciones
se observa un descenso en la salinidad en abril de 1998, con el inicio de la estación
lluviosa (Figura 1.3b). También se observan salinidades promedio bajas en julio de
1998 (estaciones 03, 04 y 11) y septiembre (estaciones 01-02) de 1998. Los bajos
valores de salinidad en la P1 en las estaciones externas (Figuras 1.12 y 1.13)
observados en julio de 1998 evidencian sin duda el gran efecto del río Coto-Colorado
sobre esta parte de la cuenca del Golfo Dulce.
• Transparencia del Agua
La transparencia del agua (Figura 1.14) también registró importantes cambios a
lo largo del período de estudio. Todas las estaciones, excepto la estación 03, presentan
un aumento en la transparencia hacia el mes de agosto de 1997. Entre los meses de
septiembre a noviembre del mismo año la transparencia del agua disminuye en todas
las estaciones y fluctúa de manera variable. Sin embargo, se vuelve a dar un aumento
considerable en la transparencia en todas las estaciones entre enero y marzo de 1998,
alcanzando valores de entre 9.8 m (est. 03) y más de 15 m (est. 02 y 11). A partir de
éste punto, los valores empiezan a disminuir progresivamente hasta el final del estudio,
dándose solo un pequeño aumento en la visibilidad en el mes de mayo, especialmente
en las estaciones 01 y 03. A lo largo de todo el período de estudio, la menor visibilidad
se presentó en la estación 03, con un promedio de 4.1 ± 2.4 m y la estación 04, con 4.8
± 2.7 m mientras que los mayores valores se dieron en la estación 02 (7.8 ± 3.9 m), 01
(7.3 ± 4.1 m) y 11 (6.9 ± 3.9 m) (Fig. 1.15).

a) ~5 .s N
10
15
b) ~ 1 4
'E6
Ѫ 10 12 14 16
12 14 16
12 14 16
.... 9 ..,
..... CXl <O CXl CXl "' 9 9 cr» 9 Ol "' z ::::; :i .., Ji z
1 '
Figura 1.14. Variación temporal en la transparencia del agua (m) para las estaciones a) 01 , b) 02, e) 03, d) 04 y e) 11 durante el período 1997-98, Golfo Dulce.
32

12
10
8
6
4
2
o
·~
· ~
·~
,__
i :1 1
-- I¡ 1
1 .. '
1 1- '" ... .1
,i 1
Est 01 Est 02 Est03 Est04 Est 11
Figura 1.15. Promedio y desviación estandar de la transparencia del agua en cada estación de muestreo durante el período 1997-1998.
33
1--
--
--
T

34
111. Aspectos Biológicos: Biomasa del Zooplancton
Al observar la biomasa total de zooplancton por capa (Figuras 1.16, 1.17) resulta
evidente que los mayores valores así como las principales fluctuaciones en dichos
valores se dan en la capa superficial (P1) de todas las estaciones. En general, los
valores de biomasa fluctuaron entre 0.343 mg/m3 (Est. 02, P3) y 54.51 mg/m3 (Est. 04,
P1 )(Cuadro 1.2). Los valores más altos de biomasa, así como las mayores
desviaciones, siempre se dieron en la capa superficial (P1 ). En el caso de las
estaciones 11 y 04, los valores promedio correspondientes a P1 fueron 19.42 ± 12.49
mg/m3 (coeficiente de variación, CV = 64%) y 18.99 ± 11.98 mg/m3 (CV = 63%)
respectivamente. Mientras tanto, los mayores promedios de biomasa, correspondientes
a las estaciones 01, 02 y 03, fluctuaron entre 12.07 ± 6.28 mg/m3 (CV = 52%) y 14.65 ±
7.36 mg/m3 (CV = 50%) y también se dieron en la capa superficial. De este modo,
resulta evidente que las estaciones externas no solo presentan valores más altos de
biomasa sino que también presentan una mayor fluctuación en estos valores a través
del tiempo.
Como se puede observar en las Figura 1.17, los menores valores de biomasa se
dieron, la mayor parte de las veces, en la capa profunda (P3). De hecho, al observar los
promedios de biomasa por estación, es la P3 la que siempre registra los menores
valores de biomasa (Figura 1.16). En esta figura se observa que los promedios de
biomasa de las estaciones 01, 02 y 03 en P3 son similares, fluctuando entre 1.3 ± 0.45
mg/m3 (CV = 35%) y 2.3 ± 1.15 mg/m3 (CV = 50%) Sin embargo, los valores de
biomasa en P3 de las estaciones 04 y 11 son considerablemente más altos que en las
otras estaciones, con valores de 8.25 ± 4.63 mg/m3 (CV = 56%) y 9.12 ± 5.63 mg/m3
(CV = 62%) respectivamente. De nuevo, son las estaciones externas las que muestran
un mayor grado de variación en los valores de biomasa a esta profundidad. Los
mayores valores de biomasa observados en las estaciones externas con respecto a las
internas en la capa P3 se deben probablemente a que las estaciones externas son
menos profundas y presentan concentraciones relativamente altas de oxígeno disuelto
por debajo de la termoclina, por lo que la disponibilidad de oxígeno no es un factor
limitante para la distribución del zooplancton como si lo es para las estaciones internas,
donde disminuye rápidamente después de los 70 m de profundidad.

Total P3 Total P2 Total P1
e) 30 ~
25 1 20 ~
l15 j
Total P3 Total P2 Total P1
10 . T 5
o -j-----'----L---.--'---"'---,----''--I_~
25
20 "'e ~15
10
5
Total P3 Total P2 Total P1
o +---"'---'·---.---'--''---~___J'----1-,
Total P3 Total P2 Total P1
Figura 1.16. Valores promedio y desviación estandar de la biomasa total por capa (P1, P2, P3) para cada estación: a) 01, b) 02, c) 03, d) 04 y e) 11, durante el período 1997-98.
35

a)60 ,
50 j 40 ~
30 ' . \ ¡.
20 I ' d) 60 - - • Totál P1 .. '
.. "" I \. • I ., 50
10 .. ---Total P2
' \
o 40 ' 1 - ··-Total P3
"' ' 1
b)60 -§, 30 • E
50 20
40 10
o 30 '
' I \ I 60 1 20 ' I e)
I ' ,. .. ' ... ' I
... , ' I ... 50 ... 10 ' ' ' ., ... I '
I 40 •, ' ' o "' ' -§, 30 \ '
c) so l E \'
20 50 -
~ 401
... /'
10 ... ......
o E 30 -¡ <.. )> en o z m ;:: )> ;:: ~
<.. ?' en o z
20 j". ,¡, ,¡, ,¡, cO ,¡, cO cO ,¡, ,¡, cO "'
,¡, cO '° .... .... .... .... .... "' "' "' "' "' "' "' "' CD CD
i'· , ... I
' I
1: 1 -~ ';- ?' ~ o z m ;:: )> ;:: <.. ;t; en o z "' "' '° ,¡, ,¡, cO '° ,¡, ,¡, '° ,¡, ,¡, ,¡, .... .... .... .... .... CD CD CD CD "' "' CD CD CD CD
Figura 1.17. Variación temporal en la biomasa total (mg/m3) del zooplancton en cada capa (P1,
P2 y P3) para las estaciones a) 01, b) 02, e) 03, d) 04 y e) 11 durante el período 1997-98.
36

37
Para el caso de la estación 01 (Figura 1.17a), la biomasa total en la P1 muestra
incrementos en los meses de julio y octubre de 1997 así como en el período abril - julio
y en septiembre de 1998. En caso de la estación 02 (Figura 1.17b ), se observa un
patrón similar a la estación 01 en cuanto a la fluctuación de los valores de biomasa
durante 1997 acompañados por incrementos en los meses de mayo y septiembre de
1998. Tanto para la estación 01 como para la 02, los valores de biomasa en la capa P3
se mantienen relativamente estables durante todo el período de estudio. En lo que se
refiere a la capa P2, la estación 01 muestra picos de biomasa durante agosto y octubre
de ambos años, mientras que los mayores valores para la estación 02 se dan en abril y
octubre de 1998.
La biomasa de la estación 03 (Figura 1.17c), en general, se comporta de manera
distinta a las estaciones 01 y 02, presentando muchas fluctuaciones en el tiempo. En el
caso de la biomasa en P1, los principales incrementos se dan en los meses de julio y
septiembre de 1997, así como en mayo y octubre de 1998. En la P2 también se
observan muchas fluctuaciones, con un incremento progresivo en la biomasa a partir de
septiembre de 1998. De igual manera, entre los cambios de biomasa que se dan en la
P3, destaca el pico en octubre de 1997, el cuál coincide con un descenso en los valores
correspondientes a P1 y P2.
Las estaciones 04 y 11 (Figuras 1.17d,e ), por su parte, presentan un
comportamiento similar a lo largo del tiempo en cuanto a la biomasa total de sus capas.
En ambos casos se observan incrementos en los valores de todas las capas en los
meses de julio, septiembre y noviembre de 1997, así como en noviembre de 1998,
cuando superan los valores de biomasa total correspondientes de las demás estaciones.
Además, los valores de biomasa total en la capa P3 de las estaciones 04 y 11 son
superiores a los valores correspondientes de las demás estaciones durante la mayor
parte del período de estudio.
Las figuras 1.18 - 1.22 muestran la variación en el tiempo de la biomasa total por
fracción de tamaño "x" (T1: x > 1000 µm, T2: 500 µm < x < 1000 µm, T3: 150 µm < x <
500 µm) y por capa (P1, P2, P3) para las estaciones 01, 02, 03, 04 y 11
respectivamente. En general, la biomasa de las capas superficiales (P1) en todas las
estaciones (Figuras. 1.18a - 1.22a) está dominada por la fracción más pequeña de
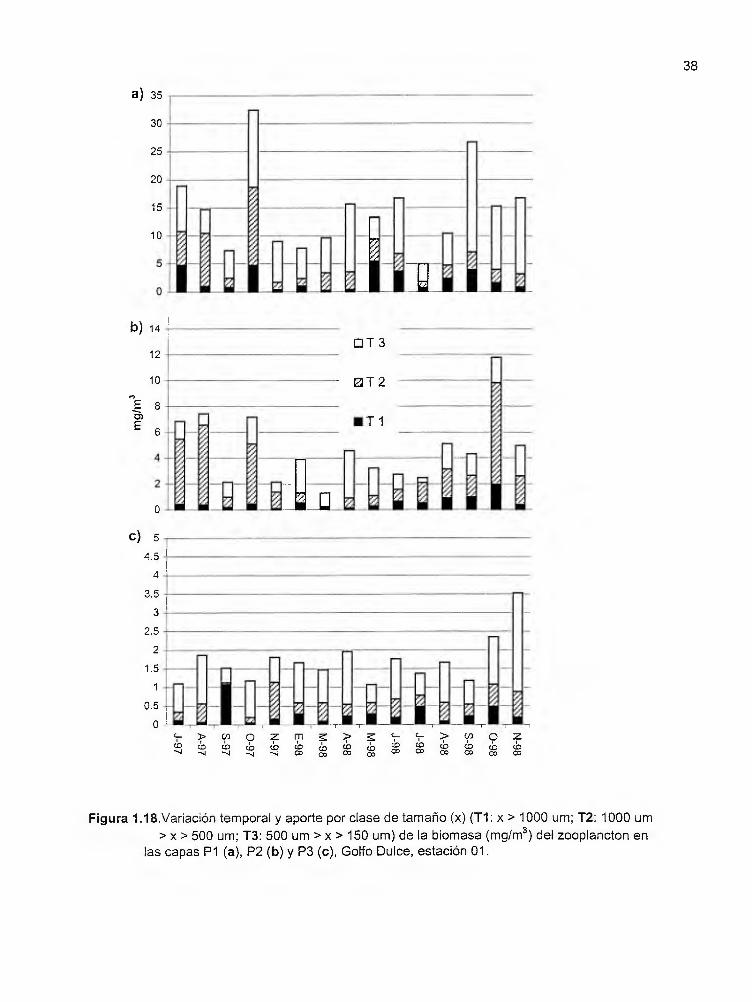
a) 35 -30
.... 25 :
20 i -· - ---- -1 15
5
n - ,_ ,_ -F"
. 1 1 - 1 -
1 ,_ ,_ 1- - I'-- 1-
,_ - 1- _][] ,= - L-- -
1
: ~ ~ 1
. ' ~ ~ :. ~ ~
10
o
b) 14
DT 3 12
10 0T2 .., E 8 e, E 6
•T 1
4
2
o
e) 5
4 .5
4
3.5 i
3
2.5
2 -1.5 ~
-- e--1
0.5 ~ - ~ -
o t• 'f ~ -i"'" c.... :¡.. (J) {o <O {o -.¡ -.¡ -.¡
Figura 1.18.Variación temporal y aporte por clase de tamaño (x) (T1 : x > 1000 um; T2: 1000 um > x > 500 um; T3: 500 um > x > 150 um) de la biomasa (mg/m3
) del zooplancton en las capas P1 (a), P2 (b) y P3 (e), Golfo Dulce, estación 01.
38

a) 30
25
20 ....
15 F-
- -10
5 ,_ -
o
b)
: : 1 10
.., E Di
8
E 6
e) 3
2.5
2
1.5 W--- -
lL 0.5
o
_, .... 1 -
' 1 ;
-,_... ,_ ' -- - -
~ lT~ -
;·
,_...
-F" 1 -
- - - 1 -1
- - - - ,_
~ ~ .....- , ~ ! í T
m
'° CD ' )>
'° CD
-
-1
~
'
' -,1 - - -
-~ -
:[ - - - - --1 1
1
' 1 - ', i~ 1-¡ - ~ -~
rLI 1 1 Í"_
OT3
0T 2
• T 1
-
tr~-~ -
-
c.... c....
'° '° CD CD
U)
'° CD
9 <O CD
z
'° CD
Figura 1.19.Variación temporal y aporte por clase de tamaño (x) (T1 : x > 1000 um; T2: 1000 um
> x > 500 um; T3: 500 um > x > 150 um) de la biomasa (mg/m3) del zooplancton en
las capas P1 (a), P2 (b) y P3 (c), Golfo Dulce, estación 02.
39

a) IO
25
-20 --
- ,--
~ ·- -15 - -5
,! 1-·m ,__ ,_ -]- ' ~ - ,, - - ,_ ~ '- ~ - 1 ~ -
~ ~ ... ., i ~ ' ' ! ~ 1 -,_ I , - ,_
10
o
b) 20
18 -DT3 -
16 -14
12
1
~ 10 E
8 -
0T2 ~ -
• T 1 v:. /
6 r '-4
2
o
e) : +-1---• i-----------3 ~: ' ¡
2 11--~~~~~~~'
1
z cb -..¡
m cb co
~ cb co
1" <O co
~ cb co
c... c... cb cb co co
1" <O co
Ul cb co
o cb co
z cb co
Figura 1.20.Variacíón temporal y aporte por clase de tamaño (x) (T1 : x > 1000 um; T2: 1000 um > x > 500 um; T3: 500 um > x > 150 um) de la biomasa (mg/m3
) del zooplancton en las capas P1 (a), P2 {b) y P3 (c), Golfo Dulce, estación 03.
40

a) 60
1
50
1 40
l 30
20
10
o
b) 30
25 1
2J
"'~ 15 E
10
e) 20
18
16
14
12
10
8
6
4
2
o
_ , 1
1
~
í -~-
-
~ ~
~ ~ -
- -
- ---,- ---¡-
t.. cb -..¡
-
(/)
cb -..¡
_ ,
i 1
1 1
1
'
1
,._ 1 -
,_
~º D ~
~
' mt ,_, --- - ¡
1 ,_ ~
'
- ·--,-
z cb -..¡
1
m S:: cb cb CD CD
- --- - ,_ -~ .,. ' ~ 1
-1,
I!
·- ! 1
n líil ~
t.. t.. cb cb CD CD
~~ ---,_ ,_
i ,-~
-~ ~
)>
cb CD
DT3
EZIT 2
.•T1
--
--¡;¡i
~
--
o cb CD
--,_
-
z cb CD
-
-
-
--
-'--;
Figura 1.21.Variación temporal y aporte por clase de tamaño (x) {T1 : x > 1000 um; T2: 1000 um > x > 500 um; T3: 500 um > x > 150 um) de la biomasa (mg/m3
) del zooplancton en las capas P1 (a), P2 (b) y P3 (c), Golfo Dulce, estación 04.
41

a) 50 -45
40 --35 --30
25
20 -15 -- ,_ -10
5
o
-- - -:-o- ~
1
b) 30
25
20
~ 15 E
10
5
o
e) 25
20
-15
-10
1
5 -- -o (, ~ 61 " .. ,-
-
-
i
- - ~ -r 1
- - 1'- -- ~ .... 1
--' 1-_í - ~ -i- iHJ- ~
L=i DT3
0T2
1
f-
f-
.._
1-
1
1
-
--
-c..
•T 1 I_
-1
J 1
fdt ~j-1
en 9 ID '° O> O>
~
z ID O>
-
-
-
Figura 1.22.Variación temporal y aporte por clase de tamaño (x) (T1 : x > 1000 um; T2: 1000 um > x > 500 um; T3: 500 um > x > 150 um) de la biomasa (mg/m3
) del zooplancton en las capas P1 (a), P2 (b} y P3 (c), Golfo Dulce, estación 11.
42

~10 E
5
b) 20
15
e)_ 20
15
'"'¡: Oi10 E
5
2
2
DT1
DT2
•T3
~ d) ~ --.,
3
OT1
CIT2
•T3
20 l 1
15 J
.., ..§ 10 CI E
5
o
e)_ 20
3 15
.., ..§ 10 CI E
5 OT1
OT2
•T3 o
3
DT1
OT2
•T3
1 2 3
DT1
CIT2
•T3
Jrl ;
1 2 3
Figura 1.23. Promedio y desviación estandar de la biomasa del zooplancton (mg/m3) para cada
clase de tamaño (T1, T2. T3), en cada profundidad y para cada estación: a) 01, b) 02, e) 03, d) 04 y e) 11, Golfo Dulce.
43

44
tamaño (T3). Así mismo, los principales incrementos observados en la biomasa total de
la capa P1 se deben principalmente a incrementos en la biomasa de T3. Cabe destacar
que en el mes de octubre de 1997 de da un pico en la biomasa de la capa superficial
donde las fracciones T2 y T3 contribuyen significativamente.
Al observar las figuras correspondientes a la capa intermedia (P2) ( 1.18b -
1.22b) se observa que la biomasa total tiene, proporcionalmente, un mayor aporte por
parte de la fracción T2. Lo anterior resulta más claro para el caso de las estaciones O 1,
02 y 03 (Figuras 1.18b - 1.20b ), en especial durante 1998. En las estaciones 04 y 11
(Figuras 1.21, 1.22) la contribución a la biomasa de las tres fracciones de tamaño es
mucho más equitativo, aunque en la capa P1 las fracciones T1 y T2 contribuyen
relativamente más con la biomasa total que en las estaciones internas. De esta manera,
en las estaciones externas no se aprecia una marcada dominancia de la fracción
pequeña sobre la biomasa, como sí sucede en las estaciones internas. Finalmente,
respecto a la capa profunda (P3), la fracción T3 recobra importancia en cuanto a su
aporte a la biomasa total, especialmente en las estaciones 01 y 02 (Figuras 1.18c,
1.19c ). En la capa profunda de las estaciones 03, 04 y 11 (Figuras 1.20c - 1.22c) la
contribución de T3 deja de ser tan importante y, al igual que en la capa intermedia, la
fracción T2 aporta un porcentaje importante a la biomasa total.
La Figura 1.23 refleja las mayores biomasas, en todas las fracciones de tamaño
y capas, que se dieron en las estaciones externas (04 y 11 ). Así mismo, permite
observar como, en la capa superficial, la fracción de tamaño menor (T3) siempre fue la
dominante, seguida por las fracciones de tamaño T2 y T1, en ese orden. Lo anterior no
se refleja claramente en las capas intermedia y profunda, donde las fracciones de
tamaño T3 y T2 contribuyen de manera muy similar a la biomasa total.
La similitud de las estaciones 01-02 y 04-11 viene a confirmarse al aplicar un
análisis de conglomerado a los datos de biomasa (Figura 1.24 ). A partir de éste análisis
resulta evidente la relación entre dichas estaciones así como el hecho de que la
estación 03, a pesar de comportarse distinto a todas las demás estaciones, está más
relacionada a las estaciones internas que a las estaciones 04 y 11. Los mismos
resultados se obtienen en la prueba de ANDEVA, la cuál demuestra la existencia de
diferencias significativas entre todas las estaciones (F=1O.70, p < 0.05). Sin embargo, la

45
EST 01
EST 02
EST 03
EST 04
EST 11
o 5 10 15
Distancias
Figura 1.24. Análisis de conglomerado para las estaciones de muestreo respecto a la biomasa del zooplancton en la capa superficial (P1 ), aplicando el coeficiente de similitud Bray-Curtis y uniones de promedio por grupo.

46
prueba post-hoc de Tukey indica que no existen diferencias entre las estaciones
internas, 01, 02 y 03 (p > 0.05), ni entre las estaciones externas, 04 y 11 (p > 0.05), pero
si existen diferencias significativas en la biomasa entre las estaciones internas y las
externas (p < 0.05). También existen diferencias significativas en la biomasa entre las
profundidades muestreadas (F= 75.38, p < 0.05). Estas diferencias se presentan al
comparar la capa P1 con P2 y P3, pero no se dan entre las dos últimas (Tukey, p >
0.05). Además, se determinó que existen diferencias significativas de biomasa entre las
fechas de muestreo (F=3.46, p < 0.05). Sin embargo, se detectó una interacción entre
los factores de fecha y estación (F=1.53, p < 0.05), lo que indica que las diferencias
observadas entre las estaciones varían con la fecha y viceversa. También existe una
interacción entre los factores de fecha y profundidad (F=1.54, p < 0.05) indicando que
las diferencias observadas entre las fechas varían también con la profundidad de
muestreo y, por lo tanto, que las diferencias existentes entre las profundidades varían
con las distintas fechas.
La prueba de ANDEVA indica que las diferencias observadas en la biomasa de
las distintas fracciones de tamaño son significativas {F=43.65, p < 0.05) y que estas
diferencias se dan entre cada una de las fracciones de tamaño consideradas (Tukey, p <
0.05). Además, como era de esperar, se determinó que existe una interacción entre los
factores de profundidad y tamaño de la fracción (F= 27.02, p > 0.05), lo que significa que
las diferencias existentes entre las distintas clases de tamaño varían con las
profundidades muestreadas y viceversa.
Finalmente, se logró determinar la existencia de una relación lineal leve pero
significativa entre la biomasa total del zooplancton en la capa superficial {P1) y a) la
profundidad del disco de Sechii (p = 0.009, R2 = 0.089) (Fig. 1.26a), b) la profundidad de
la capa de mezcla, tomada como la profundidad de la capa sobre la termoclina, P1 (p =
0.001, R2 = 0.140) (Figura 1.26b). En ambos casos, se observa una disminución en los
valores de biomasa a medida que aumenta la transparencia del agua y la profundidad
de la capa de mezcla, respectivamente. No se detectó ninguna relación significativa (p
> 0.05) entre la biomasa y las variables físicas evaluadas (oxígeno disuelto, temperatura
y salinidad).

a)
60
50
M' E40 -Cl E ';'30 U) n:I E o 20 -i:ñ
10
o
b)
60
50 M' .É40 Cl E ';'30 fil n:I
.§20 al
10
o o
o
o
o
o
o o o o o
o o
es 9 ce. :%. o ~o Cb ~~º
o
CO o 0o o o cP 0o o o 00 o o
2 4
<>
<> <> <> - o ~ <>
10
6
o
o
e
1 20
8 Sechii (m)
o
o ºº o o
o o o
10
<>
<>
3
~· 30
Prof. mezcla (m)
y= -0.7836x + 20.65,
R2 = 0.0893, p = 0.009
o O fl! B o o
12 14
8 -o
16
y = -0.4002x + 25.808,
R2 = 0.1403, p = 0.001
g $
1 40 50
Figura 1.25. Regresión lineal entre la biomasa del zooplancton en la capa P1 y a) la profundidad del disco de Sechii, b) la profundidad de la capa de mezcla.
47

48
DISCUSION
Los fenómenos biológicos y oceanográficos asociados con el zooplancton se dan
sobre enormes escalas de espacio y tiempo, por lo que no es posible conocer de
manera precisa y exacta su distribución. El conocimiento generado hasta ahora sobre
estos aspectos, es producto de lo que se ha derivado de los muestreos de campo,
modelaje y experimentación en el laboratorio (Lenz 2000).
Los estuarios son sistemas que se caracterizan por ser poco profundos, bien
mezclados verticalmente y donde el ingreso de agua dulce regula la salinidad (Riley
1967). Por el contrario, los fiordos, a los que se asemeja el Golfo Dulce, más bien se
distinguen por tener una fuerte estratificación así como aguas profundas hipóxicas o
anóxicas. Como un ambiente para el plancton, los fiordos presentan condiciones que
por lo general se consideran virtualmente oceánicas. Además, ofrecen ventajas, con
respecto a los estuarios, para el estudio de sus comunidades pláncticas debido a que el
transporte de organismos por corrientes se ve minimizado considerablemente (Aksnes &
Magnesen 1983).
En general, como se esperaba, se comprobó que las aguas del Golfo muestran
gradientes verticales pronunciados en los valores de temperatura, salinidad y oxígeno
disuelto, los cuales descienden rápidamente con la profundidad. Esta es una
característica del Golfo Dulce que ha sido reportada en investigaciones anteriores
(Richards et al. 1971; Córdoba & Vargas 1996; Thamdrup et al. 1996; Wolff et al. 1996)
y se debe en gran medida, como lo apuntan estos autores, a la morfología de su
cuenca, que presenta un umbral poco profundo en la parte externa que restringe la
circulación de agua, especialmente en la capa profunda, la cual incluso llega a
desarrollar condiciones anóxicas en ciertas épocas del año.
Por esta razón, se espera observar junto a la estratificación de las aguas una
estratificación biológica, de manera que la mayor parte de los organismos se encuentren
distribuidos en la capa superficial rica en oxígeno. Esto también concuerda con los
resultados del presente estudio, donde los valores más altos de biomasa se dan en la
capa superior de la columna de agua (P1 ), la misma donde, además, los promedios de
biomasa por estación siempre fueron superiores (Figura 1.16). De hecho, la capa P1

49
tuvo valores de biomasa significativamente distintos de las capas P2 y P3. Sin
embargo, estas últimas no se diferenciaron significativamente, lo que no resulta
sorprendente debido a que la fuerte estratificación y los fuertes gr adientes en las
variabl1es fíisicas que presentan las aguas del1 Golfo favorecen la presencia de dos capas
de agua distintas: la capa de mezcla (superficial) y la capa profunda, las cuales, como
se comprueba en las Figuras 1.9 - 1.13, tienen características físi.cas distintas ..
El anterior es un aspecto que también ha sido reportado con anterioridad en el
Golfo Dulce (Richards et al. 1971; Córdoba y Vargas 1996; Thamdrup et al. 1996).
Autores como Alcaraz (1980), Sameoto (1984), Allison & Wishner (1986) y White et al.
( 1995) han obtenido resultados similares en otras regiiones y han determinado, al igual
que aquí, que la mayor parte de la biomasa del zooplancton se haya distrfüuiída en los
estratos superiores (sobre la termoclina) de la columna de agua y que debajo de esta
profundidad los valores de biomasa caen marcadamente, mostrando un gradiente
vertical en su distribución.
Estudios previos realizados en fiordos noruegos i'nd'ican que la biomasa del
zooplancton en estos sistemas tiende a disminuir hacia su parte interna y que, por lo
general:, la biomasa está dominada por organismos pequeríos {Aksnes & Magnesen
1983). De acuerdo a Wolff et al. (1996) y a los resultados obtenidos en éste estudio, el
Golfo Dulce presenta una distribución de la biomasa similar a lo observado por Aksnes
& Magnesen, donde los valores más alitos se dan en las. estaciones ubicadas en la parte
externa del Golfo (estaciones 04 y 11) y de hecho es la fracción de tamaño pequeño
(T3) la que domina la biomasa en todas las estaciones. Entre l1os grupos que dominan
la biomasa de T3 destacan los géneros de copépodos Oithona, Paracalanus y Oncaea
junto con copepoditos y nauplius (Capítulo 2). Mientras tanto, la biomasa en T2 es
domi1nada principalmente por el grupo de los osfrácodos y algunos copépodos de mayor
tamaño pero menos abundantes como Eucalanus, Acartia y Labidocera. Finalmente, la
biomasa de la fracción de tamaño superior a las 1000 µm es dominada princiipalmen~e
por quetognatos, pequeñas hidromedusas y algunas larvas de crustáceo.
Como lo apuntan Wo'lff et al. (1996), la dominancia de la fracción de tamaño
pequeño en el Golfo Dulce indica que la comunidad de zooplancton del Golfo Dulce es,
en general, poco desarrollada, debido a que la mayor parte de l1a energía es

50
aprovechada por los organismos pequeños. Un aspecto importante de señalar es que,
en el Golfo Dulce, la dominancia que muestra la fracción de tamaño T3 sobre la
biomasa tiende a disminuir luego de los períodos de entrada de agua oceánica y
también hacia el segundo año de muestreo, cuando la intensidad de El Niño empieza a
caer. Lo anterior se podría deber a que, ante tales circunstancias, se da una mayor
entrada de nutrimentos (vía agua oceánica o debido al mayor caudal de ríos) al Golfo,
con lo que se daría un mayor desarrollo de la comunidad de plancton, reflejándose, a su
vez, en una mayor contribución de las tallas T1 y T2 a la biomasa total. Esto también
vendría a explicar el hecho de que la contribución de las tres fracciones de tamaño sea
más equitativa en las estaciones externas. Además, estas estaciones, que están
sometidas a una mayor influencia oceánica y del río Coto-Colorado, son las que en
promedio muestran los mayores valores de biomasa a todas las profundidades, así
como las mayores fluctuaciones (CV > 60%)(Figura 1.16).
La entrada de masas de agua oceánica a la cuenca del Golfo Dulce no es un
evento desconocido. Anteriormente se ha reportado que masas de agua fría,
relativamente rica en oxígeno y de salinidades altas entran al Golfo Dulce en distintos
momentos del año (Richards et al. 1971; Hartmann 1994; Thamdrup et al. 1996). Así
mismo, se sabe que estos eventos son relativamente lentos y provocan la advección de
nutrimentos a la zona fótica (Richards et al. 1971; Brown et al. 1989b; Thamdrup et al.
1996; Wolff et al 1996; León-Morales & Vargas 1998). Sin embargo, la periodicidad de
estas entradas de agua permaneció hasta ahora desconocida. Los datos físicos
obtenidos en el presente estudio no solo permiten comprobar el fenómeno de entrada
de masas de agua oceánica la cuenca del Golfo, como se observa en las figuras 1.4 -
1.8, sino que dichos eventos muestran una marcada periodicidad, al coincidir en los
meses de abril y septiembre de cada año.
El hecho de que el agua oceánica posea características distintas al agua de
zonas costeras, hace que sea relativamente fácil detectarla en las curvas
correspondientes a los factores de temperatura y salinidad (Dietrich 1963; Brown et al.
1989a, b; Thurman 1996). Al observar las isotermas y las isohalinas correspondientes a
cada estación muestreada, se puede apreciar claramente que se dan fluctuaciones en
los valores de temperatura y salinidad en los períodos mencionados, poniendo en

51
evidencia la entrada de masas de agua oceánica relativamente fría y alta salinidad al
Golfo Dulce.
Sin embargo, dicha entrada de agua no resulta tan evidente en las curvas de
oxígeno disuelto. A pesar de que es posible observar incrementos en los valores de
oxígeno en las aguas profundas y superficiales que coinciden con los períodos
mencionados, dicho patrón no siempre es claro ni consistente con las fechas de entrada
de agua, lo que probablemente se debe a que dicha variable es afectada por factores
como la mezcla vertical, la productividad primaria, la tasa respiratoria de organismos
heterótrofos y la acción de organismos descomponedores (Richards et al. 1971; Tilman
et al.1982; lkeda 1985; Reynolds 1989; Thurman 1996). Estos factores vendrían a
enmascarar la entrada de agua rica en oxígeno disuelto a la cuenca del Golfo Dulce, si
es que su detección no se da a corto plazo.
Con respecto a la fuerza que mueve la entrada de masas de agua oceánica al
Golfo Dulce, Richards et al. (1971) sugirieron originalmente que fenómenos de
afloramiento costero estacional son los responsables de dichos eventos. Un
afloramiento es aquel proceso mediante el cuál agua profunda, fría y rica en nutrimentos
es transportada hacia la superficie, usualmente debido a corrientes ecuatoriales
divergentes o a corrientes costeras que arrastran agua superficial lejos de la costa
(Zeitzschel 1978; Brink 1985; Longhurst & Pauly 1987a; Brown et al. 1989a; Thurman
1996; NOAA 2000). El transporte de nutrimentos a la superficie resulta muy importante
para los ecosistemas marinos debido a que es una de las variables que controlan la
biomasa y estructura de comunidades del fitoplancton (Tilman et al. 1982).
El Pacífico Oriental Tropical (POT) se caracteriza por tener aguas relativamente
muy productivas, lo que se debe precisamente a los sistemas de afloramiento presentes
en dicha zona, tales como el de Perú (uno de los sistemas más productivos del planeta)
el domo de Costa Rica, el afloramiento de Papagayo y el de Panamá. Los dos últimos
ocasionados por vientos que se mueven en dirección fuera . de la costa debido,
principalmente, a la topografía de cada sitio (Fielder et al. 1991; White et al. 1995).
Se ha observado que en el POT se dan típicamente dos máximos en la
producción primaria y que, aparentemente, el zooplancton varía de forma sincrónica con

52
la clorofila (Longhurst & Pauly 1987). Al respecto, Dessier & Donguy (1985) encontraron
que en el Pacífico ecuatorial, entre los 4°$ y 'los 4ºN, los máximos anual'es en clorofila
se dan en los meses de agosto (cuando el afloramiento ecuatorial es intenso) y entre
marzo y mayo (afloramiento de menor intensidad). Estos picos en clorofila, fueron
observados por los autores precisamente en zonas de muestreo ubicadas frente a las
costas de Panamá. Al mismo tiempo, observaron incrementos en la abundancia de
copépodos que coinciden con las fechas anteriormente señaladas.
Los resultados anteriores vienen a apoyar lo ya afirmado en la presente
investigación respecto a la entrada de agua oceánica a la cuenca del Golfo. De hecho,
la periodicidad observada en las entradas de agua (en los meses de abril y agosto
septiembre) coincide con las fechas de afloramiento observadas por Dessier & Donguy
( 1985), por lo que es razonable afirmar que el sistema de afloramiento frente a las
costas de Panamá es la principal fuerza que impulsa la entrada de agua oceánica a la
cuenca del Golfo. Más aún, resulta interesante observar que, de acuerdo a los datos
obtenidos, los principales picos en la biomasa del zooplancton que se observan en cada
una de las estaciones (Figuras 1.17-1.22) coinciden o se dan poco después (un mes) de
los períodos en los que se da la entrada de agua oceánica a la cuenca del Golfo Dulce
(Figuras 1.4, 1.5, 1.6, 1.7 y 1.8).
El ingreso de aguas ricas en nutrimentos a la capa de mezcla (P1 ), que ha sido
reportada anteriormente como pobre en nutrimentos (Wolff et al. 1996), vendría a
provocar un impulso en el desarrollo del fitoplancton, de herbívoros, de depredadores
pláncticos y finalmente un incremento en el micronecton (Viinogradov et ali. 1973, en
Longhurst & Pauly 1987; Bougis 1976; Dessier & Donguy 1985; Smetacek & Pollehne
1986; Benovic et al. 1994; Ortner et al. 1995; White et al. 1995; Gasol et al. 1997;
Micheli 1999). De esta manera, el incremento en la abundancia de fitoplancton
representa un cambio en las condiciones alimentarias que vienen a afectar, directa e
indirectamente, la abundancia, composición y estructura de la comunidad de
zooplancton (Lampert & Schober 1980; Allison & Wishner 1986; Arcos & Fleminger
1986; Smetacek & Pollehne 1986; Reynolds 1989; Kane 1993; Ortner et al. 1995; Wiafe
& Frid 1996; Brett & Golman 1997; Harris 1987). Esto, como se mencionó antes, se ve
reflejado en los datos de biomasa del zooplancton obtenidos (Figura 1.17), donde se
observan picos en la biomasa que se dan en los períodos posteriores a la entrada de

53
agua al Golfo. Estos incrementos en la biomasa del zooplancton son la consecuencia
directa de los posibles cambios en la productividad del fitoplancton que se dan luego de
la entrada de nutrimentos al sistema del Golfo Dulce.
Con respecto a los altos valores de biomasa observados en las estaciones
externas, estos podrían deberse a que la parte externa de la cuenca del Golfo, a parte
de ser poco profunda, esta más expuesta a recibir pequeñas pulsaciones de agua
oceánica que vendrían a aportar nutrimentos a la capa de mezcla y por lo tanto, a
incrementar la productividad en esta zona. Esta explicación coincide con lo sugerido por
Richards et al. (1971), Thamdrup et al. (1996), y von Wangelin & Wolf (1996) con
respecto a la gran influencia que ejerce la entrada de aguas oceánicas a la cuenca del
Golfo Dulce sobre su hidrografía y biología.
Otro aspecto importante es que, debido a que las estaciones externas reciben
una mayor incidencia de viento (mayor oleaje, obs. pers.), la mezcla vertical de la
columna de agua puede jugar un papel importante en cuanto a la importación de
nutrimentos de aguas profundas, así como en el transporte aguas ricas en oxígeno
disuelto hasta el fondo (Figuras 1. 7b, 1.8b ). Además, la cercanía del Río Coto-Colorado
a la estación 04 implica una entrada constante de nutrimentos (Umaña 1998), lo que
tiene efectos sobre las estaciones adyacentes. Lo anterior se refleja, por ejemplo, en
las bajas salinidades observadas en la capa superficial en el mes de julio de 1998 en las
estaciones externas (Figuras 1.12-1.13) así como en los amplios frentes de agua dulce
(obs. pers) asociados a este río luego de períodos de fuertes lluvias.
La entrada de nutrimentos resulta importante debido a que en zonas tropicales,
donde típicamente se dan termoclinas poco profundas y de carácter permanente,
nutrimentos como los fosfatos, nitratos y silicatos son a menudo limitantes para los
productores primarios. Por esta razón, su ingreso al sistema provenientes de fuentes
externas (ríos), así como su regeneración por medio de bacterias y excresiones de
diversos organismos, tiene una gran importancia para el desarrollo de las comunidades
de fitoplancton y zooplancton (Dietrich 1963; Zeitzschel 1978; Longhurst 1985; Dortch &
Packard 1989; Vanni & Findlay 1990; Fielder et al. 1991; Wylie & Curie 1991; Thurman
1996; Wolff et al. 1996; Harris 1997).

54
Los ríos que drenan en el Golfo Dulce, por su parte, actúan como vehículos de
transporte de materiales y nutrimentos hacia las aguas del Golfo por lo que el impacto
de las actividades humanas en la cuenca del Golfo Dulce, como agricultura,
deforestación, urbanización, etc., se viene a reflejar en el estado de sus ríos (Umaña
1998). El enriquecimiento de las aguas debido a nutrimentos antropogénicos viene a
alterar la disponibilidad de recursos y representa una perturbación de tipo "bottom-up"
en los ecosistemas marinos (Benovic et al. 1994; Micheli 1999). De hecho, Umaña
(1998) observó que la variable que mostró mayor variabilidad durante su estudio fue la
de sólidos en suspensión, especialmente en los ríos Coto-Colorado y Rincón, donde se
da un mayor impacto de actividades agrícolas, deforestación y minería.
Benovic et al. ( 1994 ), por ejemplo, determinaron en su estudio en el Adriático,
que los valores más altos en la biomasa del zooplancton se dieron en regiones poco
profundas cercanas a la costa, donde había altos niveles en la entrada de nutrimentos
(descarga de ríos, contaminación, etc.). Además, aquellos disturbios que se dan en el
ecosistema y que resultan en un ingreso de nutrimentos provocan explosiones de
fitoplancton y, por lo tanto, un dominio temporal de especies oportunistas que tienen
tamaños corporales menores (Cowles et al. 1987). De hecho, en regiones en las que se
dan valores altos de producción primaria llegan a ser dominadas en abundancia por
organismos zoopláncticos con rangos de tamaño pequeño (Piontkovski et al. 1995),
como es el caso del Golfo Dulce.
Con base a lo anterior, resulta razonable suponer que la alta productividad en la
parte externa del Golfo Dulce es controlada en gran medida por la entrada de
nutrimentos vía el Río Coto-Colorado y la influencia oceánica (pulsaciones de
nutrimentos). La factibilidad de que se den pulsaciones de agua oceánica (ricas en
nutrimentos) que afecten la parte externa del Golfo Dulce se evidencia en observaciones
de Richards et al. (1971) y Thamdrup et al. (1996), donde se reportan cambios drásticos
en la columna de agua debidos a la entrada de agua oceánica. Sin embargo, la época
del año en la que se dan estas observaciones no coincide con los importantes períodos
de entrada de agua observados en esta investigación (se trata de estudios a corto
plazo).

55
De acuelido a Wolff et al. (1996), la productividad y la clorofila de la columna de
agua del Golfo Dulce es mucho más baja que en el Golfo de Nicoya y coincide con los
valores promedio esperados para aguas tropicales costeras de carácter mesotrófico.
Sin embargo, considerando que el Golfo Dulce es un sistema costero, muestra valores
promedio de biomasa bajos (Cuadro 1.2), más bajos que los observados en otros
sistemas costeros (Cuadro 1.3), como el Golfo de Nicoya (Mornles-Ramírez & Brugnoli
Olivera, en rev.), donde la sola biomasa de copépodos herbívoros supera la biomasa del
zooplancton en el Golfo Dulce. En este caso, hay que hacer la salvedad de que el Golfo
de Nicoya es un sistema de carácter estuarino, muy distinto al Golfo Dulce (Wolff et al.
1998). Ante esto, los datos obtenidos en la presente investigación coinciden con Wolff
et aL ( 1996), quienes caracterizan al Golfo Dulce como un sistema con una muy fuerte
influencia oceánica, con una productividad primaria baja y cuya productividad general
(10 g/m2) es una de las más bajas al ser comparada con más de cuarenta sistemas
simi1lares que cuentan con modelos tróficos equivalentes.
Se debe considerar, además, que el presente estudio estuvo influenciado por el
fenómeno El Niño. Este no solo reduciría la intensidad de l'os fenómenos de
afloramiento costero que afectan al Golfo Dulce sino que, como se observa en los datos,
generó una marcada reducción en la precipitación durante 1997 (Figura 1.3b) y
probablemente una reducción en el caudal de los ríos y su aporte de nutrimentos al
Golfo. Todo lo anterior implica que el ingreso de nutrimentos al Golfo Dulce se ve
severamente limitado durante períodos de El Nliño y esto se vendría a reflejar en la
productividad del sistema. A pesar de que no se cuente con datos sobre productividad
primaria o secundaria, el hecho de que los datos de biomasa reflejen importantes
incrementos luego de la entrada de masas de agua oceánica (Figura 1.17) viene a
apoyar el planteamiento anterior y a sustentar la hipótesis de que el sistema del Golfo
Dulce se asemeja más a un sistema olig:o-mesotrófico cuando se ve afectado por el
fenómeno El Niño debido, como se mencionó, a la limitada entrada de nutrimentos al
sistema.
A pesar de lo anterior, estas observaciones son valiosas ya que demuestran que
se dan pequeñas pulsaciones en l'a entrada de agua en otras épocas del año,
influenciadas, a su vez, por pulsaciones en sistemas de afloramiento en zonas zonas
adyacentes al Golfo Dulce. Al respecto, se ha reportado que los procesos de

56
afloramiento varían considerablemente a lo largo de escalas temporales de 2 a 10 días
que, a su vez, se ven determinadas, en gran medida, por la meteorología (Brink 1985;
Margal'ef 1985; White et ali. 1995).
Entre las posibles razones por las que estas pulsaciones pequeñas en la entrada
de agua al Golfo Dulce no hayan sido detectadas en la presente investigación se
encuentra el efecto negativo sobre los sistemas de afloramiento que tiene el fenómeno
de El Niño (Dietrich 1963; Bougis 1976; Margalef 1985; White et al. 1995; Philander
1996; Thurman 1996; Lavaniegos & Gonzalez-Navarro 1999; NOAA 2000). Esto pudo
haber Inhibido dichas pulsaciones durante el período de estudio. Otra explicación radica
en el hecho de que se tomaran muestras solamente una vez al mes, con lo que se
pierde resolución en los datos y aumentan las probabilidades de que dichas
pulsaciones, de haberse presentado, no hayan sido detectadas.
Otro aspecto importante es que los picos observados en la biomasa por lo
general coinciden con períodos de alta turbidez en las aguas del Golfo (Figura 1.14 ). De
ahí que se 'haya encontrado una relación lineal entre la biomasa y la visibilidad (Figura
1.24a). La poca visibilidad observada en junio de 1997 en todas las estaciones, por
ejemplo, se debe probablemente tanto a la entrada de agua oceánica que se da en abril
como al hecho de que el pico de precipitación de 1997 es en mayo. Factores como l1a
precipitación afectan, tanto directa como indirectamente (descarga de ríos), los valores
de salinidad en la capa superficial durante ciertos períodos del año, lo que sin duda
tiene consecuencias sobre las comunidades pláncticas (estímulo sobre la productividad
primaria)(Harris 1987; Wylie & Curie 1991; Thurman 1996; Wolff et al. 1996). La
descarga de sedimento de los ríos, por su parte, puede tener el efecto inmediato de
reducir la transparencia del1 agua así como de enriquecer la concentración de
nutrimentos (Riley 1967). Por consiguiente, se puede suponer que los cambios
observados a lo largo del período de estudio en la turbidez del agua se deben a una
combinación de efectos de ,Ja entrada de agua oceánica en la cuenca del Golfo y a
períodos lluviosos que conllevan al ingreso de gran cantidad de materia orgánica y
sedimentos en suspensión a la cuenca del Golfo.

57
Cuadro 1.3. Valores de biomasa de zooplancton (mg/m3) reportados para diversas
regiones del planeta.
Autor Sitio de Estudio
Zenkevitch (1963) *
Gapishko (1971) *
Philippon ( 1972) *
Mar de Barents
Océano Indico
Antártico
Mar de Bering
Mediterraneo
Golfo de Aden Lenz(1973)*
Tranter (1973) * Oceánico
Costero
Barkhatov et al. (1973) *
Antártico
Sargasso
Pacífico Oriental
Long lsland
Irlanda
Sydney, Australia
India
Sub-Antártico
Nueva Zelanda
Nagasawa & Marumo (1975)* B. Suruga, Japón
Vladimirskaya (1976) * Escocia
Buskey (1993) B. Nueces, EE.UU.
Morales-Ramírez &
Brugnoli-Olivera Golfo de Nicoya, CRC
Quesada-Alpízar Golfo Dulce, CRC
* Tomado de Raymont 1983
** Biomasa de copépodos herbívoros (artículo en revisión)
***Biomasa de zooplancton en la capa superficial (sobre la termoclina)
Biomasa (mg/m3)
40
500-1000
(región de afloramiento)
46-362
250-1000
9-30
2-51
41
87
14
66
83
104
57
20
>500
25-63
>100
25-46
55.4**
19.4 - 12***

58
Sin embargo, en las estaciones 01, 02 y 03 se observa una mayor visibilidad en
el mes de mayo de 1997 acompañada de un aumento en la biomasa . Lo anterior
también puede ser producto de la entrada de agua oceánica que se observó en abril de
ese año, lo cual, como se mencionó antes, ocasiona una cascada de respuestas en las
comunidades de fitoplancton y zooplancton. En este caso, es probable que los altos
valores de biomasa se deban a la existencia de una alta proporción de herbívoros que
están consumiendo a los productores primarios, por lo que aumenta la transparencia del
agua. De hecho, en la zona ecuatorial se han registrado altos valores en fa abundancia
de zooplancton al mismo tiempo en el que la clorofila desciende a valores mínimos, lo
que indica que el zooplancton podría ser uno de los principales factores limitantes para
e~ desarrollo de la producción de fitoplancton (Bougis 1976; Alcaraz 1985; Dossier &
Donguy 1985; Fielder et al. 1991; Brett & Goldman 1997).
En el caso de la estación 03, fa biomasa a lo largo del período de estudio
presenta un comportamiento similar al observado en las estaciones 01 y 02 aunque
resulta más fluctuante a lo largo del1 tiempo. De hecho, algunas de estas fluctuaciones
coinciden más bien con fas que se dan en las estaciones 04 y 11 (Figura 1.17). Lo
anterior refleja el carácter de transición de la estación 03 entre las partes interna y
externa del Golfo Dulce, mostrando sin embargo, mayor similitud a fas estaciones
internas, como se observa en el análisis de conglomerado (Figura 1.24). Un aspecto
interesante, por ejemplo, es el hecho de que se den picos en la biomasa en las
estaciones internas (01 y 02) en el mes de octubre de 1997, mientras que para la misma
fecha se dé una caída en los valores de biomasa de las estaciones 03, 04 y 11. Sin
embargo, el descenso en los valores de biomasa que se observa en la estación 03 no
es tan grande como el que se da en las estaciones externas. Esto podría estar
relacionado con la entrada de nutrimentos arrastrados por ríos en la parte interna del
Golfo. En éste caso, en octubre de 1997 se observa una dilución en fa salinidad en las
estaciones internas, especialmente en la estación 01 (Figuras 1.18c, 1.19c) lo que no
sucede en las restantes estaciones (Figuras 1.20c, 1.21 c, 1.22c). Esto parece indicar
que se dio una entrada de agua dulce y nutrimentos en la parte interna del Golfo (Río
Rincón, R. Esquinas) que pudo provocar un aumento en la productividad en ese sector
del Golfo Dulce, no llegando a afectar las estaciones externas.

59
Un aspecto más a considerar para el caso de la estación 03 es que su
comportamiento también se ve afectado por el hecho de encontrarse muy cerca de
Golfito, uno de los principales centros de población en el Golfo Dulce. Esta cercan[a
hace que esta estación esté más expuesta a la descarga de materia orgánica asociada
a los centros de población humana (León-Morales 1998; Sponberg & Davis 1998) y por
lo tanto, presente una mayor productividad que las estaciones 01 y 02. De hecho, la·
estación 03 es la que presentó las menores transparencias del agua en el presente
estudio (Figura 1.15).
Por otra parte, la baja precipitación que se dio durante 1997, con respecto a los
valores esperados para la zona (Caen 1983; Van Wangelin & Wolff 1996), pone en
evidencia los efectos del fenómeno de El Niño, que se presentó justo a partir de ese año
en el Pacífico ecuatorial. Otro aspecto que soporta la anterior afirmación es el hecho de
que las temperaturas más bajas que registró la columna de agua durante el período de
estudio se dieron en el mes de abril de 1997, justo antes de que se reportaran los
primeros incrementos en la temperatura del agua (3-5°C) en las estaciones
oceanográficas de las islas Galápagos, en el Pacífico Oriental (NOAA 2000). De hecho,
en el mes de junio del mismo año, el Golfo Dulce mostró un incremento significativo en
la temperatura del agua con respecto a abril, como se evidencia en la profundización de
las isotermas (Figuras 1.4, 1.5, 1.6, 1.7 y 1.8).
En zonas tropicales , a menudo se dan variaciones en la productividad primaria
asociadas a variaciones en las condiciones hidrográficas (corrientes, afloramientos, etc.)
o dimáticas (viento, lluvia , entre otros, Bougis 1976). De este modo, pequeños cambios
en una variable ambiental, tal como la temperatura, pueden provocar grandes cambios
biológicos al alterar las ventajas competitivas de una especie sobre otra (Harris 1987).
Así, por ejemplo, se han reportado menores concentraciones de clorofila y nutrimentos
así como menores abundancias de diatomeas y dinoflagelados durante períodos de El
Niño en bahía de San Quintín (Baja California) y la zona oceánica adyacente (Galindo
Bect et al. 1999).
También se ha observado que, en las aguas ricas en nitratos del POT, los
nutrimentos son agotados cuando se alcanzan temperaturas superiores a los 25°C,
como las que se dan con la entrada del fenómeno El Niño (ENSO) (Fielder et al. 1991 ).

60
Dichas disminuciones en la entrada de nutrimentos a las capas superiores y por lo tanto
en la productividad primaria implican una disminución general en la productividad de
niveles tróficos superiores con lo que es posible observar un descenso en la biomasa
del zooplancton así como un cambio en 11a composición de especies. Desde éste punto
de vista, resulta razonable la relación lineal obtenida entre la biomasa y la profundidad
de la capa de mezcla, ya que con la entrada de ENSO se da un hundimiento de la
termoc'lina y una caída en la productívídad debido a la carencia de nutrimentos. Por
supuesto, se espera que estos efectos se vean revertidos tan pronto el fenómeno
disminuye en intensidad (Dietrich 1963; Bougis 1976; White et ali. 1995; Philander 1996;
Thurman 1996; lavaniegos & Gonzalez-Navarro 1999).
Otro aspecto a resaltar es el hecho de que durante la época seca (enero-marzo)
se da un descenso general en la biomasa de todas las estaciones, excepto la 03, donde
la biomasa muestra un ligero incremento. Este descenso en los valores de biomasa del
zooplancton está asociado a lo encontrado por Morales-Ramírez & Nowaczyk (en prep.),
quienes reportan un desarrollo temprano en la comunidad de zooplancton gelatinoso del
Golfo Dulce, producto del fenómeno El Niño en el verano de 1997. Estas condiciones
favorecieron que el carácter oceánico de las estaciones externas se extendiera hacia la
parte ,interna del Golfo Dulce, con lo que se observó un incremento en la biomasa del
zooplancton gelatinoso. Dichos resultados hacen suponer que el descenso en la
biomasa del zooplancton se deba precisamente al incremento en la biornasa de los
depredadores gelatinosos, los cuales pueden llegar a controlar la biomasa de
organismos más pequeños (Raymont 1983; Purcell 1992; Lenz 2000).
Los resul,tados de esta investigación, con una clara dominancia de la fracción T3
sobre las fracciones de tamaño más grandes, no concuerdan con lo determinado por
van Wangelin & Wolff (1996), quienes encontraron mayores valores de biomasa en la
mayoría de los grupos de tamaño durante el verano, cuando además se observó un
mayor y más continuo espectro de biomasa. Lo anterior puede ser la consecuencia de
que el Golfo Dulce haya sido afectado por el fenómeno El Niño desde principios de
1997, por lo que el desarrollo de sus comunidades pláncticas no se ajustó al modelo
propuesto por van Wangelín & Wolf y no se llegó a dar el' desarrollo esperado durante la
época seca.

61
Consideraciones Finales
De acuerdo a lo anterior y a lo determinado en la presente investigación, la
productividad del Golfo Dulce parece estar gobernada por el ingreso periódico de
nutrimentos de origen oceánico, así como de nutrimentos que ingresan como parte de la
descarga de los ríos de la zona. Como se mencionó con anterioridad, los picos en
biomasa que presenta el sistema parecen estar directamente relacionados a estos dos
factores, siendo ese efecto más dramático durante períodos de El Niño, cuando los
afloramientos se debilitan y la precipitación disminuye. Es posible que, bajo condiciones
normales, cuando ,los afloramientos son intensos (Philander 1996; Thurman 1996;
Lavaniegos & Gonzalez-Navarro 1999; NOAA 2000), se den entradas de agua más
frecuentes en la cuenca del Golfo Dulce, como lo observaron Richards et al. ( 1971) y
Thamdrup et al. (1996). Sin embargo, el patrón de entrada de agua observado en esta
investigación, por presentar la misma periodicidad que el afloramiento en Panamá,
probablemente represente los períodos más importantes y estables en el fenómeno de
entrada de agua al Golfo Dulce. Estas entradas de agua, junto con la mayor descarga
de agua (y por lo tanto, nutrimentos) por parte de los ríos que se espera durante años
en que no se presente ENSO, ponen a disposición de las comunidades pláncticas una
mayor cantidad de nutrimentos, con lo que se observan valores de biomasa
relativamente superiores a los observados en esta investigación (von Wangelin & Wolff
1996).
Del mismo modo, este trabajo comprueba que la influencia de aguas oceánicas
es, como se esperaba, más significativa en la parte externa del Golfo Dulce, que viene a
representar un sistema de transición entre las condiciones oceánicas de mar abierto y
las condiciones costeras en la parte interna de la cuenca. Lo anterior, se viene a
reflejar, por supuesto, en los valores de biomasa obtenidos en el presente trabajo e
incluso en la composición de la infauna en la zona externa del umbral (León-Morales &
Vargas 1998).
Un aspecto importante que se debe de tener siempre presente es que, al medir
la biomasa del zooplancton y estimar su producción, a menudo hay que lidiar con toda la
comunidad. El emplear tamices con distintos tamaños de poro permite una clasificación
por tamaño así como separar los componentes principales. Este es un método que ha

62
probado ser muy exitoso, dependiendo del espectro de tamaños y de la diversidad de
especies encontrados. Sin embargo, se debe considerar que dichos datos no son otra
cosa que resultados metodológicos y que la realidad del sistema estudiado es mucho
más compleja (Pastel et al. 2000). No existe a la fecha una teoría lo suficientemente
desarrollada y aceptada que explique las grandes variaciones espacio-temporales de la
biomasa y la productividad de los organismos marinos (Piontkovski et al. 1995).
El Golfo Dulce sigue considerándose como un sistema relativamente poco
productivo (comparado con el Golfo de Nicoya, por ejemplo) y que, en general, presenta
pocas señales de contaminación o deterioro (Umaña 1998; Sponberg & Davis 1998).
Sin embargo, se debe prestar atención al continuo crecimiento urbano y agrícola, a la
actividad minera y a la alta y creciente tasa de deforestación que se da en la cuenca del
Golfo Dulce (Umaña 1998; Sponberg & Davis 1998; Lobo 2000, com. pers.). Si tales
actividades continúan creciendo sin mayor regulación, se podrían dar serias
consecuencias sobre las comunidades biológicas del Golfo Dulce.

63
REFERENCIAS
Aksnes, D. & T. Magnesen.1983. Distribution, development and production of Calanus
finmarchicus (Gunneros) in Lindaspollene, western Norway, 1979. Sarsia 68:195-
208.
Alcaraz, M. 1980. Evolución y distribución vertical de la biomasa de zooplancton
expresada en carbono y nitrógeno orgánicos, relación C/N y carbono detrítico en
una zona marina afectada por aguas residuales procedentes de la ciudad de
Barcelona. lnv. Pesq. 44(2): 265-274.
Allison, S. & K. Wishner. 1986. Spatial and temporal patterns of zooplankton biomass
across the Gulf stream. Mar. Ecol. Prog. Ser. 31: 233-44.
Arcos, F. & A. Fleminger. 1986. Distribution of filter-feeding calanoid copepods in the
eastern equatorial Pacific. CalCOFI Rep., Vol XXVII, pp.170-187.
Bakun, A. 1990. Golbal climate change and intensification of coastal ocean upwelling.
Science 24 7: 198-201.
Beers, J.R. 1976. Determination of zooplankton biomass: Gravimetric methods. pp.35-
83. In H.F. Steedman (ed.). The UNESCO Press, Paris.
Benovic, A., S. Fonda-Umani, A. Malej & Speecchi. 1994. Net zoopplankton biomass in
the Adriatic sae. Mar. Biol. 79: 209-18.
Bougis, P. 1976. Marine Plankton Ecology. American Elsevier Publishing Company, lnc,
N.Y.pp. 355.
Brett, M.T. & C.R. Goldman. 1997. Consumer versus resource control in freshwater
pelagic food webs. Science 275: 384-386.
Brink, K. 1985. Sorne aspects of physical processes in coastal upwelling. lnt. Symp.
Upw. W. Afr. 1: 5-14.

64
Brown, J., A. Colling, D. Park, J. Phillips, D. Rothery & J. Wright.1989a. Seawater: its
composition, properties and behaviour, pp., G. Bearman (ed.). Pergamon Press.
Brown, J., A. Colling, D. Park, J. Phillips, D. Rothery & J. Wright.1989b. Ocean
Circulation, pp., G. Bearman (ed.). Pergamon Press.
Buskey, E.J.1993. Annual pattern of micro- and mesozooplankton abundance and
biomass in a subtropical estuary. J.Plank.Res. 15(8):907-24.
Coen, E.1983. Climate, p. 35-46, In D. Janzen Costa Rican Natural History, University of
Chicago Press.
Córdoba, R. & J.A. Vargas.1996. Temperature, salinity, oxygen and nutrient profiles ata
200 m station in Golfo Dulce, Pacific coast of Costa Rica. Rev. Bici. Trap. 44(3):
233-236.
Cowles, T, M. Reman, A. Gauzens & N. Copley.1987. Short term changes in the biology
of a warm-core ring: zooplankton biomass and grazing 32(3):653-64.
Davis, C.C. 1984. Planktonic Copepoda (lncluding Monstrilloida). pp 67-92. In K.A.
Steidinger & L.M. Walker (eds.), Marine Plankton Life Cycle Strategies. CRC
Press, lnc.
Dessier, A & J.R. Donguy.1985. Planktonic copepods and environmental properties of
the eastern equatorial Pacific: seasonal an d spatial variations. Deep sea Res.
32(9):1117-1133.
Dietrich, G. 1963. General oceanography: an lntroduction. lnterscience Publishers.
Dortch, Q. & T. Packard.1989. Differences in biomass structure between oligotrophic and
eutrophic marine ecosystems. Deep Sea Res. 36(2):223-40.
Epifanio, C & A. Dittel.1984. Seasonal abundance of brachyuran crab larvae in a tropical
estuary: Gulf of Nicoya, Costa Rica, Central America. Estuaries 7:501-505.

65
Fiedler, P.C., Philbrick, V. & F.P. Chavez.1991. Oceanic upwelling and productivity in the
eastern tropical Pacific. Limnol. Oceanogr. 36(8):1834-1850.
Galindo-Bect, M., LA. Galindo-Bect, J. Hernández-Ayón, F. Ley-Lou & S. Alvarez
Borrego.1999. Efecto de El Niño en los nutrimentos y el carbón orgánico total de
una laguna costera del noroeste de Baja California. Ciencias Mar. 25(2):225-37.
Gasol, J.M., del Giorgio, P.A. & C.M. Duarte.1997. Biomass distribution in marine
planktonic communities. Limnol. Oceanogr. 42(6):1353-1363.
Giguére, L., J. St-Pierre, B. Bernier, A. Vézina & J. Rondeas.1989. Can we estimate the
true weight of zooplankton samples after chemical preservation?. Can. J. Fish.
Aquat. Sci., vol. 46.
Harris, R. 1987. Spatial and Temporal organization in marine plankton communities.
Organization of communities. Ed. Blackwell Sci. Pub.
Harris, R., P. Wiebe, J. Lenz, H. Skjodal & M. Huntley. 2000. Zooplankton Methodology
Manual. Academic Press.
Hartmann, H.J. 1994. The Golfo Dulce Marine Environment. Review and Critica!
Assesment. pp. 34. Department of Biology of La Rochelle, France. Unpublished.
Hossfeld, B.1996. Distribution and biomass of arrow worms (Chaetognatha) in Golfo de
Nicoya and Golfo Dulce, Costa Rica. Rev. Biol. Trap. 44(3): 157-172.
lkeda, T. 1985. Metabolic rates of epipelagic marine zooplankton as a function of body
mass and temperature. Mar. Biol. 85:1-11.
Kane, J. 1993. Variability of zooplankton biomass and dominant species abundance on
Georges bank, 1977-86. Fish. Bull. 91:464-74.
Kennish, M.J.1994.Practical Handbook of Marine Science. 2nd ed. CRC Press.

66
Lampert, W. & U. Shober.1980. The importance of "threshold" food concentrations.
pp.264-267. In W.C. Kerfoot (ed.), Evolution and Ecology of Zooplankton
communities. Special Symposium Vol 3. American Society of Limnology and
Oceanography. Univ. Press of New England.
Lavaniegos, B. & E. Gonzalez-Navarro.1999. Cambios en la comunidad de copépodos
durante el ENSO de 1992-93 en el canal de San Lorenzo, Golfo de California.
Ciencias Mar. 25(2): 239-65.
Lenz, J. 1971. Zur methode der Sestonbestimmung. Kieler Meersforsch. 27: 180-193.
Lenz, J. 2000. lntroduction. In Harris, R., P. Wiebe, J. Lenz, H. Skjodal & M. Huntley.
( eds.) Zooplankton Methodology Manual. Academic Press.
León-Morales, R. & J. Vargas.1998. Macroinfauna of a tropical fjord-like embayment:
Golfo Dulce, Costa Rica. Rev. Biol. Trap. 46(6):81-90.
Longhurst, A. 1985. Relationships between diversity and the vertical structure of the
upper ocean. Deep Sea Res. 32(12):1535-1570.
Longhurst, A. & D. Pauly.1987a. Circulation of tropical seas and oceans.pp. 27-62. In
Ecology of tropical Oceans. Academic Press, lnc.
Longhurst, A. & D. Pauly.1987b. Biological Communities of tropical Oceans.pp. 63-105.
In Ecology of tropical Oceans. Academic Press, lnc.
Margalef, R. 1985. Primary pruduction in upwelling areas: energy, global ecology and
resources. lnt. Symp. Upw. W. Afr. 5(1 ): 225-332.
Micheli, F. 1999. Eutrophication, fisheries and consumer-resource dynamics in marine
pelagic ecosystems. Science 285: 1396-98.
Molina-Ureña, H.1996. lchthyoplankton assemblages in the Gulf of Nicoya and Golfo
Dulce. Rev. Biol. Trap. 44(3): 173-182.

67
Morales, A. & J. Vargas.1995. Especies comunes de copépodos (Crustacea:Copepoda)
pelágicos del Golfo de Nicoya, Costa Rica. Rev. Biol. Trap. 43(1-3):207-218.
Morales, A. 1996. Checklist of copepods from Gulf of Nicoya, Coronado Bay and Gulf
Dulce, Pacific coast of Costa Rica, with comments of their distribution. Rev. Biol.
Trap. 44(3): 103-113.
Morales-Ramírez, A & Brugnoli-Olivera, E. El Niño 1997-1998 impact on the plankton
dynamics in the Gulf of Nicoya, Pacific coast of Costa Rica. En revisión, Rev.
Biol. Trap.
Morales-Ramírez, A & J. Jakob. Seasonal ·vertical distribution, abundance, biomass and
biometrical relationship of ostracods in Golfo Dulce, Pacific coast of Costa Rica.
En prep.
Morales-Ramírez, A & J. Nowaczyk. Distribution, abundance, biomass and composition
of gelatinous zooplankton in Golfo Dulce, Pacific coast of Costa Rica, during the
transition wet-dry season 1997-98: possible impact of El Niño. En prep.
NOAA. 2000. National Oceanographic and Atmospheric Agency home page. In
http://www.noaa.gov/
Omori, M. & A. Fleminger. 1976. Fixation and preservation of various marine taxa:
Laboratory methods far processing crustacean zooplankton, pp. 282-286. In
H.F.Steedman (ed.). The UNESCO Press, Paris.
Omori, M.1978. Sorne factors affecting on dry weight, organic weight and concentrations
of carbon and nitrogen in freshly preserved zooplankton. lnt. Revue ges
Hydrobiol. 63(2):261-69.
Omori, M. & W. Hammer.1982. Patchy distribution of zooplankton: behavior, population
assessment and sampling problems. Mar. Biol. 72: 193-200.

68
Ortner, P., P. Weibe, L. Haury & S. Boyd.1978. Variability in zooplankton biomass
distribution in the northern Sargasso Sea: the contribution of Gulf stream cold
core rings. Fish. Bull. 76(2):323-34.
Philander, G.1996. El Niño and La Niña, pp. 72-87, In R.G. Pirie (ed.), Oceanography:
contemporary readings in ocean science, 3rd ed. Oxford Univ. Press.
Piontkovski, S., R. Williams & T. Melnik.1995. Spatial heterogenity, biomass and size
structure of plankton of the lndian ocean: sorne general trends. Mar. Ecol. Prog.
Ser 117:219-27.
Postel, L., H. Fock & W. Hagen.2000. Biomass and Abundance. In Harris, R., P. Wiebe,
J. Lenz, H. Skjodal & M. Huntley (eds.) Zooplankton Methodology Manual.
Academic Press.
Price, N.M., Ahner, B.A. & M.M. Morel.1994. The equatorial Pacific Ocean: grazer
controlled phytoplankton populations in an iron-limited ecosystem. Limnol.
Oceanogr. 39(3):520-534.
Purcell, J.E.1992. Effects of predation by the scyphomedusae Chrysaora quinquecirrha
on zooplankton populations in Chesapeake Bay, USA. Mar. Ecol. Prog. Ser.
87:65-76.
Raymont, J.E.G.1983. Plankton and productivity in the oceans, 2no ed., vol. 2
Zooplankton. Pergamon Press, pp.24-52; 333-46.
Razouls, S., F de Boveé, C. Razouls & M. Panouse.1994. Are size spectra of
mesozooplankton a good tag for characterizing pelagic ecosystems? Vie Milieu
44( 1 ): 59-68.
Reynolds, C.S. 1989. Physical determinants of phytoplankton succession. pp. 9- 56. In
U. Sommer (ed.), Plankton Ecology: Succesion in plankton communities.
Springer-Verlag.

69
Richards, F.A. , Anderson, J.J. & J.D. Cline. 1971. Chemical and Physical observations
in Golfo Dulce, an anoxic basin on the Pacific coast of Costa Rica. Limnol.
Oceanogr. 16( 1 ):43-50.
Riley, G.A.1967. The Plankton of Estuaries. pp. 316-326. In G.H. Lauff (ed.). Estuaries.
American Association for the Advancement of Science, Washington D.C.
Sameoto, D. 1984. Vertical distribution of zooplankton biomass and species in
northeastern Baffin bay related to temperature and salinity. Polar biol. 2:213-24.
Skjodal, H.,P.H. Wiebe & K. G. Foote.2000. In Harris, R., P. Wiebe, J. Lenz, H. Skjodal &
M. Huntley (eds.) Zooplankton Methodology Manual. Academic Press.
Smetacek, V. 1988. Plankton characteristics.pp.93-130. Ecosistems of the world .
Elsevier, N. Y.
Smetacek, V. & Pollehne.1986. Nutrient cycling in pelagic systems: a reappraisal of the
conceptual framework. Ophelia 26: 401-28.
Sommer, U. 1989. Toward a Darwinian ecology of plankton. pp.1-8. In U. Sommer (ed .),
Plankton Ecology: Succesion in plankton communities. Springer-Verlag.
Sponberg, A. & P. Davis. 1998. Organochlorinated pesticide contaminants in Golfo
Dulce, Costa Rica. Rev. Biol. Trop. 46(6): 111-24.
Thamdrup, B. , Canfield , O.E. , Ferdelman, T.G., Glud, R.N. & J.K. Gundersen.1996. A
biogeochemical survey of the anoxic basin Golfo Dulce, Costa Rica. Rev. Biol.
Trop. 44(3):19-33.
Thurman, H.V. 1996. Essentials of oceanography, pp. Prentice Hall, lnc.
Tilman, D., S. Kilham & P. Kilham. 1982. Phytoplankton community ecology: The role of
limiting nutrients. Ann. Rev. Ecol. Syst. 13:349-72.

70
Umaña, G. 1998. Characterization of sorne Golfo Dulce drainage basin rivers (Costa
Rica). Rev. Biol. Trop. 46(6):125-35.
Valdés,L., M.T. Alvarez-Ossorio & M. Varela.1990. Relación entre la estructura de tallas
y la biomasa del zooplancton nerítico en la plataforma de La Coruna. Bol. lnst.
Esp. Oceanogr. 6(2): 97-106.
Vanni, M.J. & D.L. Findlay. 1990. Trophic cascades and phytoplankton community
structure. Ecology 71(3):921-937.
von Wangelin, M. & M. Wolff.1996. Comparative biomass spectra and species
composition of the zooplankton communities in Golfo Dulce and Golfo de Nicoya,
Pacific coast of Costa Rica. Rev. Biol. Trop. 44(3): 135-155.
White, J., X. Zang, L. Welling, M. Roman & H.Dam.1995. Latitudinal gradients in
zooplancton biomass in the tropical Pacific at 140°W during the JGOFS EqPac
study: effects of El Niño. Deep Sea Res. 11, 42(2-3): 715-33.
Wiafe, G. & C.L. Frid .1996. Short-term temporal variation in coastal zooplankton
communities:the relative importance of physical and biological mechanisms.
J.Plank. Res. 18(8):1485-1501.
Wolff, M. & J.A. Vargas (Eds.).1994. R.V. Victor Hensen Costa Rica Expedition
1993/1994. Cruise Report. Center for Tropical Marine Ecology (ZMT).
Bremen. 109p.
Wolff, M., Hartmann, H.J. & V. Koch. 1996. A pilot trophic model for Golfo Dulce, a fjord
like tropical embayment, Costa Rica. Rev. Biol. Trop. 44(3):215-231 .
Wolff, M., V. Koch,J.B. Chavarría & J.A. Vargas.1998. A trophic flow model of the Golfo
de Nicoya, Costa Rica. Rev. Biol. Trop. 46(6): 63-80.
Wylie , J.L. & D.J. Currie.1991. The relative importance of bacteria and algae as food
sources for crustacean zooplankton. Limnol. Oceanogr. 36(4):708-728.

71
Zeitzschel, B. 1978. Oceanographic factors influencing the distribution of plankton in
space and time. Micropaleontology. 24(2): 139-159.

Capítulo 11
Caracterización, Composición y Abundancia del Zooplancton en el Golfo Dulce durante el período 1997-1998
Resumen
72
Se analizó un total de 86 muestras de las cuales se cuantificó un total de 75 225 organismos. A partir de esto, se estimó una abundancia promedio de zooplancton de 3793.35 ± 4867.25 indiv./m3
. En general, la comunidad de zooplancton del Golfo Dulce es dominada por el grupo de los copépodos (71.47%), seguidos de apendicularias (8.25%), ostrácodos (7.99%) y huevos de invertebrados (5.43%), entre otros. De los 29 géneros de copépodos identificados, tan solo cinco resultaron responsables del 91.5% de la abundancia total: Oithona (46.9%), Paracalanus (21.4%), Oncaea (15.1%), Euterpina (4.5%) y Corycaeus (3.6%). Al igual que para el caso de la biomasa, se determino que la abundancia del zooplancton (y de los copépodos) es significativamente mayor en la capa superficial (P1) de la columna de agua, lo que confirma la presencia de una fuerte estratificación en la actividad biológica. Además, las estaciones internas (01 y 02) mostraron valores ligeramente menores en cuanto a la abundancia de zooplancton que las estaciones externas. Los dos resultados anteriores coinciden con la distribución de la biomasa del zooplancton. Además, y al igual que para el caso de la biomasa, la entrada de masas de agua oceánica parece tener un importante efecto sobre la comunidad de zooplancton. El hecho de que el sistema fuera dominado por géneros como Oithona, Paracalanus y Oncaea durante el período de estudio no resulta sorprendente debido a que son grupos típicos de sistemas estables y bajos en nutrimentos. Esto último pudo haberse visto favorecido por el hecho de que el fenómeno El Niño afectara el Pacífico Tropical durante el período de estudio. Es posible que la dominancia de Oithona se vea disminuida durante períodos en los que El Niño no está presente.

73
INTRODUCCION
El plancton es el más abundante y ampliamente distribuido grupo ecológico del
planeta y se encuentra tanto en hábitats marinos como dulce acuícolas a todas
profundidades (Perkins 1974; Zeitzschel 1978, Raymont 1983). Factores físicos tales como
la temperatura, salinidad, oxígeno disuelto, turbulencia y la disponibilidad de nutrimentos,
entre otros, tienen una enorme influencia sobre el plancton (Margalef 1978; Zeitzschel 1978).
En este sentido, el éxito de una población depende no sólo de la disponibilidad de alimento y
la abundancia de depredadores sino también de las dinámicas de su ambiente físico, el cual
afecta, entre otras cosas, la eficiencia en la alimentación, la susceptibilidad a la depredación,
el transporte y el éxito de reclutamiento (Dietrich 1963; Raymont 1983; Harris 1987; Kane
1993; White et al. 1995; Wiafe & Frid 1996; NOAA 2000).
De esta manera, los distintos patrones de respuesta al ambiente físico desarrollados
por el plancton se reflejan tanto en las diferentes estructuras de la red alimentaria como en la
composición de especies y en la biomasa de los distintos componentes (Tilman et al. 1982;
Smetacek 1988; Fieldler et al. 1991 ). Por lo tanto, la variabilidad estacional del plancton
depende de la historia de vida de los organismos y de su interacción con los factores
ambientales. Debido a lo anterior, la medición de los factores ambientales que se presentan
en la zona de estudio resulta muy importante para poder explicar las diferencias espaciales y
temporales de la productividad biológica (Harris 1987). Además es importante señalar que
las fluctuaciones que se dan en dichos factores a lo largo del tiempo son, por lo general, más
complejas y significativas en áreas costeras, debido a la combinación de influencias
continentales y oceánicas que se presentan en dichas áreas (Dietrich 1963; Thurman 1996).
Por lo tanto, los estudios que se concentran en analizar la estructura de las
comunidades pláncticas se han enfocado principalmente en considerar los gradientes físicos
como áreas donde los patrones son más fácilmente detectables y donde el papel de las
interacciones bióticas que condicionan la composición de las comunidades son mejor
evaluadas. Esto se debe al hecho de que el zooplancton, micronecton y animales
epipelágicos no se encuentran distribuidos homogéneamente en el plano horizontal ni el
vertical (Omori 1978; Omori 1982; Raymont 1983; Davis 1984; Harris 1987; White et al. 1995;
Wiafe & Frid 1996).

74
El zooplancton es considerado como un agente activo en la transferencia de energía
de los productores primarios {fitoplancton) a carnívoros grandes (peces y crustáceos, entre
otros), así como en la regeneración de nutrimentos a través de la excreción (Raymont 1983;
lkeda 1985; Harris 1987; Bottger-Schnack 1995). Dentro del zooplancton, hay gran
diversidad de tamaños y hábitos alimentarios, y se distinguen tres grupos principales: el
merozooplancton, al cuál pertenecen los estadías larvales de invertebrados, como por
ejemplo larvas de crustáceos y cordadas bénticos así como larvas de peces; el
holozooplancton, compuesto por aquellos organismos que pasan todo su ciclo de vida en
asociaciones pláncticas, como copépodos, cladóceros, ostrácodos y rotíferos, y el
ticoplancton, el cual usualmente resulta ser de menor importancia y consiste primariamente
de organismos que accidentalmente son arrastrados desde el fondo, como por ejemplo
isópodos, cumáceos y mísidos. Esta última categoría representa una potencial fuente de
alimento para el zooplancton omnívoro y carnívoro, así como para peces planctívoros
( Jeffries 1967; Perkins 197 4; Raymont 1983; Smetacek 1988; Kennish 1994 ).
La importancia del zooplancton y particularmente de los copépodos para el estudio de
la eficiencia en la transferencia de materia orgánica, ha sido evidente tras numerosos
trabajos en este campo (Longhurst 1985, Sterner 1986, Kleppel 1993, Hopcroft & Roff 1996).
Tradicionalmente se le ha dado mucho énfasis al flujo de materia y energía en los
ecosistemas acuáticos. Sin embargo, la tendencia actual apunta a prestarle una mayor
atención a las especies individuales de zooplancton y sus dinámicas poblacionales en la
ecología marina. Esto se debe a que se ha reconocido la gran influencia de los procesos
físicos sobre el plancton y que esta influencia debe tratarse a nivel de especie (Lenz 2000).
Los copépodos, son los organismos holozoopláncticos "suspensívoros" más diversos
del ambiente marino (de 55% a 95% de todo el plancton) (Raymont 1983; Longhurst 1985;
Longhurst & Pauly 1987; Braun et al. 1990; Madhupratap et al. 1990; Bottger-Schnack 1995).
Representan un enlace entre los procesos de transferencia energética del fitoplancton a
organismos de niveles superiores (Petipa 1978 en Morales y Vargas 1995; lkeda 1985) y son
considerados como los productores secundarios metazoarios más importantes en los
ecosistemas marinos pelágicos, tanto en términos de abundancia como de biomasa
(Smetacek 1988; Hopcroft & Roff 1998a).

75
Sin embargo, la mayor parte de la investigación sobre este grupo ha sido desarrollada
en zonas templadas y se ha enfocado sobre especies grandes y sus estadías tardíos de
desarrollo. En zonas tropicales el espectro de tamaño de la comunidad de copépodos se
desplaza hacia las especies más pequeñas y estadías tempranos de desarrollo, donde los
nauplius, por ejemplo, son relativamente más abundantes debido a que la reproducción es
continua (Hopcroft & Roff 1998a y b ). El estudio de los sistemas tropicales oceánicos y
costeros es de gran importancia por las diferentes condiciones bióticas y abióticas que se
presentan y así ha quedado demostrado tras estudios realizados por investigadores como
Zeitzschel (1978), Alcaraz (1985), Cheng (1986), Bollens & Frost (1991), Hartmann (1994),
Morales & Vargas (1995), Heywood (1996) y Morales (1996). En años recientes, se le ha
prestado especial atención a los copépodos ciclopoideos y poecilostomatoideos marinos
debido a su gran abundancia en la mayoría de los ambientes oceánicos y su frecuente
dominio numérico entre el zooplancton metazoario. Además, entre los ciclopoideos y
poecilostomatoideos, las familias Oithonidae y Oncaeidae (respectivamente) usualmente
constituyen una fracción significativa de la biomasa de copépodos. En el Mar del Norte, por
ejemplo, Oithonidae contribuye con el 25% de la biomasa de copépodos en ciertas épocas
del año (Sabatini & Ki0rboe 1994 ), mientras que ambos grupos llegan a constituir hasta el
70% de los copépodos en el Mar Rojo (Bottger-Schnack 1988). No obstante, es
relativamente poco lo que se conoce sobre la abundancia de copépodos en estuarios
tropicales.
Los estuarios se encuentran entre los sistemas costeros más complejos e
importantes, tanto en zonas templadas como tropicales. El papel que juegan los estuarios
como áreas naturales para deshove, crianza de juveniles y como hábitat de numerosas
especies marinas es ampliamente conocido (Caspers 1967). Estos sistemas, además, se
caracterizan por presentar una alta productividad primaria y secundaria, por lo que son
considerados entre los ecosistemas más productivos del planeta (Caspers 1967; Riley 1967;
Perkins 1974; Thurman 1996). Sin embargo, a pesar de que la fertilidad de las aguas
costeras y estuarinas es generalmente alta, el balance de elementos esenciales no es
regulado de una manera simple. Durante gran parte del año los nutrimentos son usados tan
rápido como son devueltos al agua, y la tasa de reciclaje llega a ser crucial en determinar
qué factor será más importante en determinar la producción (Riley 1967).

76
El Pacífico de Costa Rica cuenta con dos importantes ambientes estuarinos
protegidos, de baja energía, con tamaños, orientaciones y delineaciones geográficas
similares: el Golfo de Nicoya y el Golfo Dulce (Morales 1996). Las investigaciones hasta la
fecha se han concentrado en el primero, sin embargo, aún resta mucho por hacer en ambas
zonas, especialmente en el Golfo Dulce. Este golfo posee características que lo hacen un
sistema particularmente interesante tanto a nivel tropical como mundial. Lo anterior por
cuanto se puede catalogar también como un fiordo tropical debido a su morfología. Un
fiordo, en general, se caracteriza por ser un sistema bien estratificado y por tener aguas
profundas hipóxicas o anóxicas. Además, como ambiente para el plancton presenta
condiciones a menudo virtualmente oceánicas. Los estuarios, por su parte, son por lo
general poco profundos y bien mezclados, además de que se da un significativo ingreso de
agua dulce que regula la salinidad del sistema (Richards 1965; Caspers 1967; Riley 1967;
Thurman 1996). El trabajo en fiordos representa una buena oportunidad para estudiar la
dinámica poblacional del zooplancton debido a que el transporte de organismos por
corrientes, que frecuentemente representa un problema cuando se realizan series
temporales de muestreo en sistemas pelágicos, se ve minimizado (Aksnes & Magnesen
1983).
Richards et al. ( 1971) fueron los primeros en describir efectivamente las
características físicas del Golfo Dulce. Más tarde, Yukihara (1991 en Hartmann 1994) realizó
un breve estudio en la zona del Golfo Dulce, donde se determinó que el volumen total de
plancton es más bajo durante la época seca y que la composición del zooplancton consiste
principalmente de copépodos y quetognatos, así como larvaceos, ostrácodos y larvas de
bivalvos. Más tarde, entre los años de 1993 y 1994, el Centro de Investigaciones en
Ciencias del Mar y Limnología (CIMAR) de la Universidad de Costa Rica en conjunto con el
Centro de Ecología Marina Tropical (ZMT) de la Universidad de Bremen, Alemania, unieron
esfuerzos para llevar a cabo una campaña oceanográfica que incluyó el Golfo de Nicoya
además de otros puntos de la costa Pacífica de Costa Rica, como el Golfo Dulce y Bahía
Coronado (Wolff & Vargas 1994 ). Esta campaña generó una gran cantidad de información
científica que resulta especialmente importante para el Golfo Dulce, dado lo poco que se
conoce acerca de la biología de éste sistema (Rev. Biol. Trop. 44(3), 46(6)).
Recientemente se han realizado otras investigaciones en el Golfo Dulce, como la
determinación de la abundancia, distribución vertical, biomasa y biometría de los ostrácodos
(Morales & Jakob, en rev.) y un estudio sobre la distribución, abundancia y biomasa del

77
zooplancton gelatinoso (Morales & Nowazyck, en prep.). Resulta trascendental darle
seguimiento a la investigación científica en el Golfo Dulce debido a que es una zona en la
que, en los últimos años, se ha dado un crecimiento demográfico acelerado, deforestación
desmedida, incremento de las zonas agrícolas, así como un aumento en la actividad turística
(Spongberg & Davis 1998, Umaña 1998; Obs.pers. 1997-2000),
Con base a lo anterior, la importancia que reviste el estudio de la variabilidad espacial
y temporal de las comunidades de zooplancton se basa en que la información generada es
clave para la comprensión del funcionamiento de los ecosistemas costeros, lo cual, en el
caso del Golfo Dulce resulta ser un factor clave para el desarrollo de futuras campañas de
investigación y el planteamiento de estrategias de conservación. Por lo tanto, los objetivos
de la presente investigación se plantean de la siguiente manera:

78
OBJETIVOS GENERALES
A. Describir la abundancia y composición del zooplancton en el Golfo Dulce con base en
muestras recolectadas en seis períodos de muestreo durante un lapso de dos años
(1997-1998).
B. Medir los gradientes de los parámetros físicos del agua asociados a la composición de
especies observada.
C. Evaluar el posible efecto del fenómeno El Niño sobre el desarrollo de la comunidad de
zooplancton en el Golfo Dulce durante 1997 y 1998.
D. Determinar si existe variación espacial y temporal en la composición del zooplancton en
el Golfo Dulce.
OBJETIVOS ESPECIFICOS
A. Estudiar si existen diferencias en la composición (grupos) y abundancia del zooplancton
en cada una de las estaciones y al comparar las distintas capas muestreadas.
B. Comparar la composición y abundancia de zooplancton en las estaciones con variables
físicas como temperatura, salinidad, oxígeno disuelto y profundidad del disco de Sechii
con el fin de establecer las posibles relaciones entre dichas variables.
C. Aprovechando la presencia del fenómeno El Niño durante el período de estudio, se
compararán los datos obtenidos durante esta investigación con estudios previos con el fin
de determinar el posible efecto de dicho fenómeno sobre la abundancia y composición
del zooplancton en el Golfo Dulce.
D. Estudiar si existen diferencias en la composición (géneros) y abundancia de los
copépodos en cada una de las estaciones y al comparar las distintas capas
muestreadas.

79
MATERIAL Y METODOS
l. Sitio de Estudio
El Golfo Dulce es una bahía semicerrada, tectónicamente formada, del Pacífico
tropical oriental, localizado en la costa sur de Costa Rica, entre los 8º 27' y 8º 45' latitud
Norte y los 83º 07' y 83º 30' longitud Oeste (Fig 1.1 ). Está separado del océano Pacífico por
la Península de Osa, un área montañosa y cubierta de bosque. El Golfo tiene cerca de 50
km de longitud y de 1 O a 15 km de ancho, con un área superficial de aproximadamente 680
km2 y se caracteriza por poseer una cuenca interna de pendiente fuerte con una profundidad
máxima de 215 m y una cuenca externa poco profunda de aproximadamente 70 m (Richards
et al. 1971; Hartmann 1994 ).
Estas características tienden a restringir la circulación de agua en la cuenca interna
del Golfo y a favorecer la formación de aguas profundas anóxicas. Su circulación asemeja la
de los fiordos de altas latitudes y es uno de tan solo cuatro sistemas de éste tipo conocidos
en los trópicos (Richards et al. 1971 ).
11. Factores Climáticos
El clima de la zona puede definirse como muy húmedo y caliente. En general, llueve
durante todo el año, pero se puede distinguir una estación lluviosa determinada por el
movimiento de la zona de convergencia inter-tropical. La cuenca del Golfo recibe de 4000-
5000 mm de lluvia por año y la estación lluviosa se extiende desde abril hasta diciembre,
alcanzando su pico en los meses de agosto y octubre, cuando el promedio mensual de lluvia
varía entre los 520 mm y los 860 mm. La precipitación por lo general excede la evaporación
durante al menos 8 meses al año, razón por la cual las tierras se mantienen saturadas de
agua. Esto favorece la rápida erosión de los terrenos expuestos y da cabida al
desbordamiento de ríos, lo que incrementa el ingreso de nutrimentos y materia orgánica a las
aguas del Golfo. Las máximas temperaturas atmosféricas se registran durante la estación
seca, época durante la cuál también se da un incremento en el número de horas diarias de
brillo solar (Hartmann 1994; Wolff et al. 1996).

80
111. Hidrografía
Las características de fiordo del Golfo Dulce se reflejan en la composición de sus
masas de agua. Dos capas son evidentes: una capa superficial cálida ( 19 - 30 ºC) y diluida
(< 34 PSU) que alcanza los 50 - 60m de profundidad, justo sobre la profundidad del umbral
ubicado en la parte externa del Golfo. La otra capa la componen las aguas profundas y
físicamente homogéneas, con un mínimo de temperatura de 15.4 ºC y salinidades que
alcanzan los 34.8 PSU. La mezcla y circulación de estas aguas es influenciada por las
fuerzas de mareas, el viento, el ingreso de agua al sistema, el afloramiento de agua
subsuperficial y la topografía de la cuenca (Richards et al. 1971; Córdoba & Vargas 1996).
El río más grande, el Coto-Colorado, tiene un transporte medio anual de 177 m3/s. El
flujo total de agua dulce al Golfo no excede los 1000 m3/s por lo que probablemente no
afecta significativamente los patrones de circulación dentro del Golfo. Sin embargo, las
fuertes lluvias durante los meses críticos de la estación lluviosa pueden afectar
significativamente el flujo de agua dulce sobre períodos cortos de tiempo, especialmente
cuando los suelos se saturan de agua (Hartmann 1994; Umaña 1998).
La renovación de las aguas profundas en el Golfo Dulce es lenta y restringida a dos
períodos de entrada de agua subsuperficial (Hartmann 1994 ). Se ha identificado masas
densas de agua que pasan sobre el umbral externo y entran a la cuenca, donde se hunden
rápidamente y desplazan las aguas profundas hacia arriba (Richards et al. 1971)
IV. Colecta y Procesamiento de Muestras
Se seleccionó un total de cinco estaciones de muestreo (Cuadro 2.1) distribuidas en
la cuenca del Golfo Dulce con el fin de obtener muestras de las partes interna (estación 01 ),
media (estación 03) y externa (estación 11) (Fig. 2.1 ). Las estaciones 02 y 04 se ubicaron
frente a la desembocadura de los ríos Esquinas y Coto-Colorado, respectivamente.
Es importante señalar que las estaciones 02 y 03 del presente estudio se ubican muy
cerca de las estaciones 05 y 03 (respectivamente) usadas por Richards et al. (1971). Así
mismo, la estación 11 corresponde a la que fue previamente muestreada entre los años 1993

81
y 1994 durante la campaña de investigación del buque oceanográfico B.O. Victor Hensen,
coordinada por el Centro de Investigación en Ciencias del Mar y Limnología (CIMAR) de la
Universidad de Costa Rica en conjunto con el Centro de Ecología Marina Tropical (ZMT) de
la Universidad de Bremen, Alemania (Wolff & Vargas 1994)
El programa de muestreo comprendio un total de 17 visitas al área, realizadas una
vez al mes a lo largo de un período de dos años consecutivos, entre abril de 1997 y
noviembre de 1998. Dicho programa incluyó muestreos tanto durante la época seca como la
lluviosa, con el fin de asegurarse que los patrones encontrados correspondan a patrones
estacionales y no a problemas metodológicos.
Cuadro 2.1. Ubicación geográfica de las estaciones de muestreo de zooplancton establecidas en el Golfo Dulce, Costa Rica.
Estación Coordenadas Descripción Prof. (m)
GD-01 08º 42' N; 83º 27' O Frente a Rincón de Osa 200
GD-02 08º 40' N; 83º 22' O Frente al Río Esquinas 200
GD-03 08º 36' N; 83º 12' O Frente a Bahía Golfito 150
GD-04 08º 33' N; 83º 11' O Frente a río Coto-Colorado 70
GD-11 08º 29' N; 83º 12' O Desembocadura del Golfo 70
Los muestreos se realizaron en una embarcación tipo Zodiac, equipada con un
cabrestante portátil con el fin de facilitar la extracción de las muestras de agua y de plancton.
Previo a los muestreos de zooplancton en cada estación, se colectó una serie de muestras
de agua usando una botella Niskin de 2L a las siguientes profundidades: O, 1 O, 20, 30, 40,
50, 60, 70 y 100 m. Durante 1998 se tomó una muestra adicional de agua a 150 m de
profundidad en las estaciones 01, 02 y 03. Para cada una de las muestras, se determinó la
salinidad (PSU) con un refractómetro de mano, marca ATAGO S/Mill-E, así como la
temperatura (ºC) y oxígeno disuelto (mg 0 2/1), con un medidor de oxígeno YSI 52. Además,
en cada estación se midio la profundidad del disco de Sechii y las condiciones climáticas
como nubosidad, condición del viento y oleaje.

82
Una vez determinada la posición de la termoclina en la columna de agua, se procedio
a realizar tres arrastres verticales en cada estación con una red cónica para zooplancton de
50 cm de diámetro de boca y 153 µm de poro de red . Con ayuda de un mecanismo de cierre
remoto para la red, se realizó un arrastre por encima, uno por debajo y otro a través de la
termoclina (Fig.2.1 ). Los volúmenes de agua filtrados fueron calculados matemáticamente
conociendo el área de la boca de la red y la distancia de arrastre en cada caso. Además, se
T (ºC)
P1
Z(m) Termoclina P2
P3
Figura 2.1. Criterio de muestreo utilizado para la colecta de zooplancton, basándose en la posición de la termoclina en la columna de agua.
calculó un promedio de temperatura, oxígeno y salinidad para cada capa muestreada. Una
vez colectadas, las muestras fueron fijadas en formalina al 4 % (Omori & Fleminger 1976) y
llevadas a los laboratorios del CIMAR para ser procesadas. Se debe aclarar que se realizó

83
solamente un arrastre a cada profundidad debido a razones logísticas. Resulta importante
señalar que se tomó en cuenta la estructura física de la columna de agua (en lugar de
intervalos de profundidad predeterminados) como criterio para realizar los muestreos de
zooplancton.
La campaña de muestreo comprendía las siguientes fechas: abril, agosto y noviembre
de 1997 y marzo, abril y setiembre de 1998 que juntas suman un total de 86 muestras
considerando que en la gira de abril de 1997 no se muestreó la estación 02 debido a
problemas. Una vez en los laboratorios del CIMAR, las muestras fueron fraccionadas,
utilizando un separador de plancton Folsom (WILDCO, modelo 1831 F10), para obtener
alicuotas de no menos de 1/32ªvº del volumen total. El análisis cuantitativo se llevó a cabo en
una cámara Bogorov modificada. Haciendo uso de un estereoscopio LEICA MZ 8 (6.3 - 50
x), todos los organismos en cada alícuota fueron identificados y contados. Los copépodos
fueron el grupo a analizar, aunque también se determinó la abundancia de todos los grupos
taxonómicos presentes. Para el caso de los copépodos, la identificación se llevó hasta el
nivel de género e incluso al nivel de especie cuando fue posible. Los volúmenes restantes
de cada muestra fueron lavados con agua destilada para eliminar la formalina y transferidos
a alcohol al 70% para su preservación.
V. Análisis Estadístico
Las pruebas estadísticas se llevaron a cabo con el programa SYSTAT 8.0. Se
aplicaron pruebas de estadística descriptiva (promedio, desviación estándar, ámbito) para las
variables de biomasa, temperatura, salinidad y oxígeno disuelto para cada estación y para
cada capa de agua muestreada (P1, P2, P3) por estación. Los datos de abundancia fueron
transformados con la función lag (x + 1) para mejorar la distribución de los datos causada por
un alto número de ceros y valores cercanos a cero en las matrices de abundancia (Taggart &
Frank 1987). Además se llevó a cabo varios análisis de conglomerado (aplicando el
coeficiente de similitud Bray-Curtis y uniones de promedio por grupo) con el fin de agrupar a
las estaciones y los grupos taxonómicos con respecto a su abundancia relativa. Los análisis
de conglomerado usando índices de similitud derivados de Bray-Curtis resultan apropiados
debido a que no se ven afectados por las ausencias típicas en las listas de fauna marina
(Field et al. 1982, en Longhurst 1985; Clarke & Ainsworth 1993). Así mismo, se realizaron
pruebas de ANDEVA (prueba post-hoc: Tukey) con un nivel de confianza del 95% (Bailey

84
1995), con el fin de evaluar las posibles diferencias de abundancia entre estaciones, fechas,
profundidades y grupos taxonómicos. Finalmente, se aplicó un análisis de regresión lineal
con el fin de cuantificar la posible relación funcional entre las abundancias de zooplancton y
copépodos con las variables de profundidad del disco de Sechii, profundidad de la capa de
mezcla, temperatura, salinidad y oxígeno disuelto.

85
RESULTADOS
1.1 Abundancia y Composición: Aspectos Generales
Del total de 86 muestras analizadas se cuantificó e identificó un total de 75 225
organismos, para un promedio de 874.7 ± 526.4 organismos por muestra. Con respecto al
valor promedio de organismos por capa, la capa superficial {P1) contó con una abundancia
significativamente mayor de organismos (ANDEVA, F= 66.7, p < 0.05), con un promedio de
7979.46 ± 5845.25 indiv./m3 (CV=73.25%), seguida por las capas P2, con 1667.39 ± 1610.08
indiv./m3 (CV=96.56%) y P3 con 1431.85 ± 2271.12 indiv./m3 (CV=158.6%)(Figura 2.2).
Nótese que los valores de abundancia de ·zooplancton presentaron una mayor variación en
las capas intermedia y profunda. A pesar de que existen diferencias en la abundancia de
zooplancton entre las tres capas, al observar los resultados de la prueba post-hoc de Tukey,
se determinó que existen diferencias entre la capa superficial y las profundidades P2 y P3 (p
< 0.05), pero no entre estas últimas (p > 0.05), que presentan valores de abundancia
similares (Figura 2.2).
..,
15000 l 12500
1 10000 E ~ "O 7500 e:
5000
2500
o
1
DZ
.-...
.. , :r., ..
ol 1
1
P1
oop anc on EIC o pepo d os
I T I Ir·
T 11 l 1 1 1
1 1
P2 P3
Figura 2.2. Valores promedio y desviación estándar de la abundancia (indiv./m3) general de
zooplancton y copépodos en las capas superficial (P1 ), intermedia {P2) y
profunda (P3) en el Golfo Dulce, 1997-1998.

86
Las diferencias en cuanto a la abundancia del zooplancton se dieron también a nivel
de estaciones (F= 2.97, p < 0.05), sin embargo, la prueba Tukey no logró demostrar la
existencia de diferencias entre las estaciones al compararlas individualmente (p > 0.05). No
obstante, como se puede apreciar en la Figura 2.3, los valores de abundancia en las
estaciones O 1 y 02 (internas) fueron ligeramente menores para cada una de las tres capas
muestreadas, al compararlos con las demás estaciones de muestreo.
20000 1
15000 .
("')
E :? 10000 "C e:
1
OP1 OP2 •P3
2 3 4 11 Estación
Figura 2.3. Valores promedio y desviación estándar de la abundancia de zooplancton
(indiv./m3) para cada estación y capa de muestreo, Golfo Dulce, 1997-1998.
Además, resalta el hecho de que el valor más alto en los promedios de abundancia
se dio en la estación 03, tanto en la capa superficial (9921,6 ± 8334,3 indiv./m3) como en la
intermedia (2626,7 ± 3102,2 indiv./m3). Las estaciones 01 y 02, por su parte, presentaron
valores de abundancia en la capa superficial de 6976,2 ± 2559, 1 indiv./m3 y 5892, 7 ± 4239, 7
indiv./m3 respectivamente, los más bajos de las cinco estaciones. Con respecto a la capa
intermedia (P2) las abundancias más altas se dieron en las estaciones 03 , 04 y 11, con
valores que fluctúan entre 1585,2 ± 759,9 indiv./m3 (Estación 11) y 2626,7 ± 3102,2 indiv./m 3
(Estación 03). Aquí, las estaciones internas (01 y 02) también presentaron las abundancias
más bajas, con valores de 946,4 ± 1069,5 indiv./m3 y 1307,8 ± 1270,9 indiv./m3
respectivamente. Finalmente, la capa profunda mostró una tendencia similar a las capas
anteriores. Los mayores valores fluctuaron entre 1662,5 ± 849,2 indiv./m 3 (Estación 11) y

87
2491,7 ± 3191,9 indiv./m3 (Estación 04), mientras que los menores valores de 426,5 ± 274,4
indiv./m3 y 419,8 ± 364,7 indiv./m3 se dieron en las estaciones 01 y 02 respectivamente.
Al analizar los valores de abundancia correspondientes a cada una de las fechas
muestreadas, la prueba de ANDEVA demuestró la existencia de diferencias significativas
entre estas (F= 8.83, p < 0.05). La prueba de Tukey, sin embargo, mostró que dichas
diferencias se dieron entre la fecha 1 (abril 1997) y las fechas 2, 3, 4, 5 y 6 (p < 0.05) pero no
entre estas últimas (p > 0.05). Lo anterior también resulta evidente al observar la Figura 2.4,
donde los valores promedio de abundancia de zooplancton en la primera fecha (18 700.9 ±
4208.8 indiv./m3, en P1) son los mayores que se dan durante el estudio para las tres capas
muestreadas. Los valores en P1 de las demás fechas fluctuaron entre 3820.8 ± 1057.3
indiv./m3 en agosto, y 8042.1 ± 3404.9 indiv./m3 en abril de 1998. Además se aprecia una
tendencia hacia el incremento en la abundancia promedio en la P1 en las dos últimas fechas
analizadas para 1998 (con respecto a los meses anteriores), sin llegar a aproximarse, sin
embargo, a los valores de abundancia observados en abril de 1997. En vista de lo anterior,
resulta razonable suponer que la alta variación que se dio en los valores promedio de
abundancia de zooplancton por estación (Figura 2.3) se debe principalmente a las altas
abundancias observados en la primera fecha. Los valores completos de abundancia para
todas las profundidades se pueden apreciar en los cuadros 2.2, 2.3 y 2.4.
30000 DP1 DP2 •P3
25000
"' 20000
~ =g 15000 -
Ab-97 Ag-97 Nov-97 Mar-98 Ab-98 S et-98
Figura 2.4. Valores promedio y desviación estándar de la abundancia de zooplancton
(indiv./m3) para cada una de las fechas y capas analizadas, Golfo Dulce, 1997-
1998.

Cuadro 2.2. Abundancia (indiv/m3) de los distintos grupos de zooplancton identificados en la capa superficial (P1) para cada
una de las estaciones durante el período (1997-1998), Golfo Dulce. Los cinco grupos más importantes en cada estación se indican entre paréntesis.
Gru~o Estación 01 Estación 02 Estación 03 Estación 04 Estación 11
Copépodos 3750,35 ± 1048,04(1} 3589,80 ± 2886,28(1} 4621,87 ± 3423,82(1} 4153,72 ± 2364,08(1} 4256,02 ± 3824,61 (1} Copepoditos 958,06 ± 565,62(2} 650,91 ± 672,88(3} 1659, 70 ± 1278, 79(2} 1201,09 ± 827,61(2} 1127,66 ± 719,54(2} Nauplius 283,75 ± 329,98 287,43 ± 233,69(5} 873,53 ± 822,45(4} 1106,97 ± 1162, 16(3} 918,96 ± 1045,34(4} Hidromedusas 46,63 ± 54,24 20,99 ± 22,47 45,26 ± 71,27 17,00 ± 24,98 37,56 ± 54,22 Ctenoforos 2,45 ± 3,40 1,87 ± 4,17 4,35 ± 7,29 0,73 ± 1,78 2, 18 ± 3,64 Pteropodos 59,59 ± 69, 73 39,23 ± 43,90 157, 12 ± 259,48 134,24 ± 233, 78 160,77 ± 327,20 Gastropodos 42,38 ± 41,39 13,07 ± 8,58 50,32 ± 31,76 44,68 ± 29,20 147,09 ± 291,25 L. Molusco 42,41±46,79 16,07 ± 20,91 40,30 ± 29,86 15,24 ± 12,24 72,91 ± 122,88 L.Poliqueto 42,85 ± 44,01 25,35 ± 24,80 38,97 ± 21,13 37,21±26,19 45,92 ± 63,39 L. Equinodermo 32,05 ± 78,50 0,44 ± 0,97 0,45 ± 1,11 0,52 ± 1,27 6,29 ± 11,34 L.Crustaceo 14,39 ± 27,06 6,98 ± 6,82 42,44 ± 73,21 46,02 ± 84, 73 34, 73 ± 68,44 Cladoceros 445,04 ± 966,39(4} 136,51 ± 273,76 24,61 ± 33, 17 7,05 ± 8,21 5,80 ± 10,18 Ostracodos 322,62 ± 210,23(5} 322,57 ± 62,76(4} 346,58 ± 364,56 333,47 ± 198,39(5} 580, 11 ± 1073,61 (5) Apendicularias 798,25 ± 392,53(3} 657,37 ± 470,96(2} 1127,84 ± 840,67(3} 795,78 ± 438,80(4} 919,48 ± 584,02(3} Salpas 9, 19 ± 14,96 15,48 ± 15,95 10,16 ± 13,71 6,53 ± 8,60 10,54 ± 11,43 Anfipodos 2,42 ± 5,92 0,73 ± 1,78 4,84±11,85 Quetognatos 47,23 ± 33,25 47,66 ± 26,79 112,29 ± 117,14 123,04 ± 24,41 145,95 ± 128,99 L. Braquiópodo 0,60 ± 1,48 2,30 ± 4,04 0,91±2,22 1,45 ± 3,55 1,24±1,97 Actinotrocas 1,81 ± 4,44 Larvas de pez 3,02 ± 3,68 6,39 ± 11,70 2, 18 ± 3,64 3,87 ± 5,63 Huevos de lnvert. 73,06 ± 51,92 58,65 ± 42,69 756,09 ± 1762,81 (5) 47,48 ± 34, 15 201,96 ± 400,38 Anfioxo 0,48 ± 1, 18 TOTAL 6976,22 ± 2559, 12 5892,67 ± 4239,68 9921,60 ± 8334,32 8075,13 ± 4623,80 8683,88 ± 8298,81
88

Cuadro 2.3. Abundancia (indiv/m3) de los distintos grupos de zooplancton identificados en la capa intermedia (P2) para cada
una de las estaciones durante el período (1997-1998), Golfo Dulce. Los cinco grupos más importantes en cada estación se indican entre paréntesis.
Grupo Estación 01 Estación 02 Estación 03 Estación 04 Estación 11
Copépodos 416,92 ± 501,26(1) 666,32 ± 697,32(1) 1340,60 ± 1458,76(1) 1193,06 ± 501,31(1) 971,35 ± 469,64(1) Copepoditos 115,18 ± 166,95(3) 222,05 ± 135,64(2) 353,97 ± 417,29(3) 277,72 ± 142,14(3) 263,71±166,43(2) Nauplius 49,77 ± 102,31(5) 74,81 ± 95,27(5) 407,06 ± 849,84(2) 65,31 ± 42,97(4) 83,13 ± 60,15(4) Hidromedusas 11,99 ± 14,50 9,45 ± 11,76 22, 19 ± 25,29 9,44±4,12 3,71±1,52 Pteropodos 3,30 ± 7,54 0,47 ± 1,04 5,51 ± 9,43 4,66 ± 8,08 5,71 ± 5,74 Gastropodos 3,55 ± 2,32 18,66 ± 18,65 20,36 ± 21,40 13,51 ± 9,34 25,86 ± 26,94 L. Molusco 6,48 ± 6,48 20,47 ± 18,80 12,95 ± 13,02 11,99±10,76 16,01 ± 16,20 L.Poliqueto 22,42 ± 13,29 9,22 ± 8,00 11,93 ± 12,68 4,55±5,19 8,74 ± 10,90 L. Equinodermo 0,19 ± 0,48 L.Crustaceo 1,94 ± 3,02 4,47 ± 3,44 14,42 ± 22,92 0,47 ± 1,04 1,54±1,99 Cladoceros 28,46 ± 68,33 3,52 ± 6,29 0,58 ± 1,30 0,47 ± 1,04 Ostracodos 188,42 ± 183,22(2) 149,80 ± 231,53(3) 216,99 ± 169,54(4) 347,90 ± 337,78(2) 100,84 ± 81,28(3) Apendicularias 72,34 ± 111, 77(4) 85,00 ± 106,26(4) 138,75±263,18(5) 43,45 ± 27,58(5) 59,49 ± 48,74(5) Salpas 1,55 ± 3,81 13,93±31,15 2, 18 ± 4,87 1,16 ± 2,60 0,93 ± 2,09 Anfipodos 0,52 ± 1,27 0,00 ± 0,00 0,82 ± 1,83 0,82 ± 1,83 0,23 ± 0,52 Quetognatos 7,17 ± 8,66 7,06 ± 8,03 25,07 ± 40,80 15,73 ± 4,98 19, 16 ± 6,54 L. Braquiópodo 0,23 ± 0,52 0,00 ± 0,00 1,52 ± 1,73 0,27 ± 0,61 Larvas de pez 0,58 ± 1,30 Huevos de lnvert. 16,20 ± 14,23 21,80 ± 17,53 53,33 ± 58, 70 23,55 ± 24,97 23,88±16,10 L Calamar 0,14 ± 0,30 Anfioxos 0,23 ± 0,52 TOTAL 946,40 ± 1069,53 1307,84 ± 1270,89 2626,72 ± 3102,24 2015,06 ± 839,36 1585, 16 ± 759,91
89

Cuadro 2.4. Abundancia {indiv/m3) de los distintos grupos de zooplancton identificados en la capa profunda {P3) para cada
una de las estaciones durante el período (1997-1998), Golfo Dulce. Los cinco grupos más importantes en cada estación se indican entre paréntesis.
Grupo Estación 01 Estación 02 Estación 03 Estación 04 Estación 11
Copépodos 163,07 ± 121,11(1) 90, 18 ± 50,79(2) 903,57 ± 1770, 18(1) 1380,05 ± 1906,15(1) 827, 11 ± 484,03(1) Copepoditos 44,67 ± 37,71(3) 55,41 ± 56,69(4) 239,91 ± 413, 12(3) 439,56 ± 630,87(2) 356, 13 ± 245,95(2) Nauplius 15,21 ± 19,62 0,95 ± 1,65 241,96 ± 533,30(2) 33,80 ± 34,53(5) 55,71 ± 56,29(4) Hidromedusas 6,41 ± 5,92 2,54 ± 1,68 5,26 ± 8,13 6,95±8,10 9,88 ± 5,93 Ctenoforos 20,41 ± 49,99 Pteropodos 0,19 ± 0,48 11,20 ± 26,52 12,44 ± 26,00 1,31±1,95 Gastropodos 1,86 ± 1,21 5,03 ± 5,02 24,09 ± 45,62 25,62 ± 25,13 33,26 ± 41,92(5) L. Molusco 2,12 ± 2,85 0,92 ± 0,90 27,34 ± 57,79 9,90 ± 7,75 12,66 ± 12,91 L.Poliqueto 37,57 ± 31,72(4) 60,05 ± 59,70(3) 47,99 ± 34,88 7,14 ± 6,21 18,02 ± 22,77 L. Equinodermo 20,41 ±49,99 L.Crustaceo 0,97 ± 1,52 0,23 ± 0,32 20,03 ± 47,07 8,28 ± 19,94 1,30 ± 2,04 Cladoceros 3,34 ± 6,63 0,37 ± 0,83 O, 19 ± 0,48 125,56 ± 306,41(4) 1,77 ± 1,83 Ostracodos 17,70 ± 22,77(5) 5,53 ± 3,93(5) 142,30 ± 296,79(5) 328,10 ± 272,73(3) 309,22 ± 126,69(3) Apendicularias 12,02 ± 11,89 5,56 ± 5,01 136,00 ± 313,80 31,29 ± 51,54 13,81±11,02 Salpas 0,31 ± 0,76 1,21 ± 2,96 0,78 ± 1,90 Anfipodos 13,61 ± 33,33 0,44 ± 0,97 Quetognatos 1,78 ± 2,45 0,47 ± 1,04 23,56 ± 52,55 7,48 ± 6,79 6,97 ± 5,37 L. Braquiópodo 0,31 ± 0,76 0,09 ± 0,21 0,93 ± 2,09 Actinotrocas 1,21 ± 2,96 Larvas de pez 0,36 ± 0,81 Huevos de lnvert. 118,64 ± 202, 15(2) 192,45 ± 249,56(1) 202,69 ± 314,0 (4) 20,31 ± 29,04 13,68 ± 15,80 Anfioxos 0,31 ± 0,76 TOTAL 426,50 ± 274,38 419,80 ± 364,72 2028,49 ± 3583,44 2491,68 ± 3191,91 1662,54 ± 849, 15
90

91
El grupo más abundante fue el de los copépodos, con un 71.47% del promedio de
abundancia total (adultos = 51.28%; copepoditos = 13.82%; nauplius = 6.37%), seguidos de
apendicularias (8.25%), ostrácodos (7.99%), huevos de invertebrados (5.43%), larvas de
poliquetos (1.8%) y quetognatos (1.03%) (Figura 2.5). Estos seis grupos representaron el
95.97% de la abundancia total en las muestras analizadas. El restante 4.03% de los
organismos se distribuye en un total de 15 grupos taxonómicos, como cladóceros,
pterópodos, larvas de moluscos, salpas y larvas de pez, entre otros. Las cifras completas de
abundancia por grupos taxonómicos se presentan en los cuadros 2.2, 2.3 y 2.4 para las
capas de muestreo P1, P2 y P3 respectivamente. También es de esperar que, ante la
presencia de tan solo seis grupos dominando prácticamente un 96% de la abundancia, la
prueba de ANDEVA demuestre la existencia de diferencias significativas en la abundancia de
los distintos grupos (F= 165.13, p < 0.05).
8.2% O Copépodos
O Copepoditos
O Apendicularia 1 .·
1.8% s . /
O Ostracodos / 4.5% 1.03% El Nauplius
'\ 0.9%
D H. de 1 nvert '<..,
[[) L.Poliqueto
' 0.8% ~ Quetognatos
51.3% ~ Gastropodos
Figura 2.5. Composición del zooplancton y porcentaje de abundancia de los principales
grupos taxonómicos en el Golfo Dulce, 1997-1998.
Se detectó, además, una interacción entre los factores de fecha y grupos (F= 1.838, p
< 0.05), lo que indica que las diferencias observadas entre las fechas de muestreo variaron
de acuerdo a los distintos grupos taxonómicos, así como que las diferencias encontradas
entre los grupos variaron de acuerdo a la fecha. Lo anterior refleja que la abundancia de los
grupos fluctuó de manera distinta en el tiempo, lo cual indica que los diferentes grupos
responden de manera distinta a los factores bióticos y abióticos que se dan en la columna de

92
agua. También existió una interacción, como es de esperar, entre los factores de
profundidad y grupo (F= 9.429, p < 0.05), con lo que se confirma que las diferencias
existentes en la abundancia de los grupos varían con la profundidad y viceversa.
1.11. Abundancia y Composición: Distribución por Capa
La abundancia relativa de los principales grupos de zooplancton (aquellos que,
juntos, dominan más del 85% de la abundancia en cada sitio) para cada capa de agua
muestreada varió de la siguiente manera:
Capa superficial (P1)
Como se deduce de los datos, en el cuadro 2.2, el grupo de zooplancton que dominó
en todas las estaciones en la P1 fue el de los copépodos (adultos), con abundancias entre el
46.6% (Est. 03) y el 60.9% (Est.02). Los copepoditos son el siguiente grupo en importancia
en todas las estaciones excepto en la 02. Su abundancia relativa fluctuó entre 11.1 % (Est.
02) y 16. 7% (Est. 03). En el caso de la estación 02, la abundancia de copepoditos es apenas
superada por la de apendicularias ( 11.2% ), grupo que, en general es el tercero en
importancia en la P1 (9.9-11.4%). Los nauplius (4.9-13.7%) y ostrácodos (4.1 -6.7%) son
otros de los principales grupos en la P1. Cabe destacar que en la estación 01 aparecen los
cladóceros (6.4%), siendo la única estación adonde los nauplius fueron desplazados de los
cinco grupos más importantes. Así mismo, en la estación 03, los huevos de invertebrados se
constituyeron en el quinto grupo en importancia, con un 7.6% de la abundancia.
Lo anterior se refleja claramente en el análisis de conglomerado efectuado sobre la
abundancia de grupos en la capa P1 (Figura 2.6), donde, como se puede apreciar, los
copépodos se separaron como un grupo aparte debido a su alta abundancia y junto a estos
se agruparon los taxones anteriormente señalados: ostrácodos, nauplius, copepoditos y
apendicularias (conglomerado A), estos dos últimos con abundancias similares. Se destaca
luego una segunda agrupación con los taxones que presentaron abundancias medias a
bajas, como los huevos de invertebrado, quetognatos, hidromedusas y larvas de poliqueto
(conglomerado B). De este grupo, se separan primero los cladóceros y salpas. Los
primeros, como se mencionó antes, son un grupo importante en la estación 01. Finalmente,
se separó un tercer conglomerado con aquellos taxones con abundancias muy bajas y que
se pueden catalogar como grupos raros en las muestras analizadas (conglomerado C).
Ejemplos son las larvas de equinodermos y de pez, anfípodos y ctenóforos.

A
B
e
Copép. Copdit.
Apendic . Nauplius---'
Os trác . H . invert. ----,. Pterop .
Quetog . Gastrop .---. L. molus - . Hidrom . L. poliqu . L. e rus ta """.----' Salpas Cladóc. L.equ inoct--------..__ _____ _, L. pez Ctenof. Lingula Anfípodo--~ L.calam Anfioxo Actinotro .., . ._ ___ ..,_----...---------..
0,0 0,5 1,0 Distancia
1,5 2,0
93
Figura 2.6.Análisis de conglomerado para los grupos de zooplancton respecto a su
abundancia en la capa P1 , aplicando el coeficiente de similitud Bray-Curtis y
uniones de promedio por grupo. A, B y C representan los conglomerados
resultantes.
Capa intermedia (P2)
Al igual que en la capa superficial, los copépodos adultos fueron el grupo dominante
en la P2 , con entre 44% (Est. 01) y 61.3% (Est. 11) de la abundancia relativa (Cuadro 2.3).
La abundancia general del zooplancton y de éste grupo, sin embargo, disminuyó
considerablemente con respecto a la P1 (Figura 2.2). Además, la abundancia relativa de los
ostrácodos (6.4 - 19.9%) aumentó con respecto a la capa P1, mientras que en caso de las
apendicularias, su abundancia relativa decreció en P1 (2.2 - 7.6%), pero se mantuvo dentro
de los cinco grupos más importantes. Los copepoditos también siguen siendo un grupo
importante en esta capa, con abundancias relativas de entre 12.2% y 17%. Finalmente, el

A
B
e
Copép. Os trae. Copdit. N auplius
Apendic. H. invert. -----. Quetog. Gastrop. L.molus · _ Hidrom. L. poliqu.
Cladóc. ---- ---. Salpas L. crusta .r-_----' Pterop.
Anfípodo Lingula Anfioxo L.calam Ctenof.
Actinotro L.equino L. pez
0,0 0,5 1,0
Distancia 1,5
94
2,0
Figura 2.7.Análisis de conglomerado para los grupos de zooplancton respecto a su
abundancia en la capa P2, aplicando el coeficiente de similitud Bray-Curtis y
uniones de promedio por grupo. A, B y C representan los conglomerados
resultantes.
grupo de los nauplius se mantuvo con abundancias moderadas (3.2 - 15.5%), aunque en
general su abundancia relativa con respecto a los demás grupos pareció disminuir en la
mayoría de las estaciones. Es importante destacar que los cinco grupos anteriormente
señalados dominaron más del 89% de la abundancia en la P2 y son los mismos en todas las
estaciones. Al observar el análisis de conglomerado para la capa intermedia (Figura 2. 7), los
copépodos, al igual que en la capa superficial, se separaron de primeros entre los grupos
más abundantes. Luego, se separaron juntos los ostrácodos y copepoditos por un lado y los
nauplius y apendicularias por el otro. Estos grupos conforman el conglomerado A El
conglomerado de los taxones con abundancias medias (conglomerado B) es más reducido
que en la capa superficial y destacan los huevos de invertebrados, quetognatos,
hidromedusas y larvas de poliqueto. Finalmente el grupo de los taxones raros

95
(conglomerado C) fue más amplio que en la superficie, conteniendo doce taxones, entre los
que destacan cladóceros, salpas, larvas de crustáceo y pterópodos
Capa profunda (P3)
Como se aprecia en la Figura 2.2 y como se mencionó anteriormente, la abundancia
de zooplancton en la capa profunda es ligeramente (mas no significativamente) menor a la
que se observa en la capa intermedia. Los copépodos continuaron dominando la comunidad
de zooplancton (38.2 - 55.4%) en todas las estaciones excepto en la 02, donde los huevos
de invertebrados (45.9%) desplazaron a los copépodos a un segundo plano (21.5%) (Cuadro
2.4).
A
8
e
Los huevos de invertebrado, por su parte también aparecieron como un grupo
Copép. Copdit.
Ostrác. L. poliqu .. ----,_ ____ _JI
H. invert. ----N au pi ius -_:::::::-_-J----,
Apendic. Gas trap.----.... Hidrom. -----. L. molus . ,
Quetog. L. e rus ta . Pterop.
Ciado c. L equinod Ctenof.
Anfípodo Salpa Actinotro Anfioxo L. calam L. pez Lingula
0,0 0,5 1,0
Distancia 1,5 2,0
Figura 2.8.Análisis de conglomerado para los grupos de zooplancton respecto a su
abundancia en la capa P3, aplicando el coeficiente de similitud Bray-Curtis y
uniones de promedio por grupo. A, 8 y C representan los conglomerados
resultantes.

96
importante en las estaciones 01 (27.8%) y 03 (10%), pero sin llegar a ser el grupo principal
en abundancia y no estuvo dentro de los cinco grupos principales que dominaron la
abundancia en las estaciones 04 y 11. Los copepoditos siguen siendo importantes en las
estaciones 01, 02 y 03 (10.5-13.2%) y en especial en las estaciones 04 y 11 (17.6% y
21.4% respectivamente). Los ostrácodos, por su parte, también se mantuvieron como uno
de los grupos importantes en todas las estaciones. Sin embargo, son relativamente más
abundantes en las estaciones 03, 04 y 11 (7%, 13.2% y 18.6% respectivamente) que en la
01 y 02 (4.1% y 1.3% respectivamente). Del mismo modo, los nauplius no aparecieron
dentro de los cinco principales grupos en las estaciones internas (01 y 02), pero si lo hicieron
en las restantes estaciones (Est. 03=11.9%; Est. 04=1.4%; Est. 11=3.4%). A pesar de esto,
los nauplius en general disminuyeron en importancia en la P3 con respecto a la capa P2.
Otros grupos importantes que aparecieron en la capa profunda son las larvas de poliqueto,
en las estaciones 01 y 02 (8.8% y 14.3%), los cladóceros, en la estación 04 (5%) y los
gastrópodos holopláncticos en la estación 11 (2% ).
Al igual que en las capas anteriores, los cinco grupos dominantes se separaron
claramente en el análisis de conglomerado (Figura 2.8), con los copépodos, copepoditos y
ostrácodos como los grupos más importantes, seguidos de huevos de invertebrado y larvas
de poliqueto, dos grupos que adquieren importancia en esta profundidad (conglomerado A).
Nauplius y apendicularias pasaron a formar parte del conglomerado de especies con
abundancia media (conglomerado B), junto con hidromedusas, quetognatos y pterópodos,
entre otros. Finalmente, está el conglomerado de los grupos raros (conglomerado C), de
donde el grupo principal es el de los cladóceros, que se separó primero de los grupos menos
abundantes.
1.111. Abundancia y Composición: Variación Temporal
Las Figuras 2.9 - 2.13 describen la variación temporal en la abundancia de los tres
grupos más importantes en la composición general del zooplancton en cada una de las
capas muestreadas en el Golfo Dulce: copépodos, ostrácodos y apendicularias, los cuales
contribuyeron con más del 67% a la abundancia total. A continuación se detallan las
fluctuaciones temporales en la abundancia de cada grupo para las tres capas y para cada
estación:

12000 ~~~~~~~~~~~~~~~-
a) DCop
10000 +-------------• Ostrac OApendic
.., 8000 +----------------~ E > ~6000 +----------------c:
"'e ~2000 +----------------"'C e:
Ab Ag Nov Mar Ab Set Fecha
Figura 2.9. Variación temporal en la abundancia (indiv/m3) de tres estadías de desarrollo de
copépodos en las capas a) P1, b) P2 y e) P3 en la estación 01, Golfo Dulce.
97

12000 ..------~---------
a) 10000 +-----------
DCop
•Ostrac
ClApendic
~8000 +----~---------~-=a ~6000 +------~--~--' ~ _ ____,,....___
..,E ~2000 +----------------"'C e:
O +----'-'----~·º1-.J.....--1.D..1-_ _.o.......__ , =---,
Ab Ag Nov Mar Ab Set Fecha
Figura 2.10. Variación temporal en la abundancia (indiv/m3) de tres estadios de desarrollo de
copépodos en las capas a} P1, b) P2 y e} P3 en la estación 02, Golfo Dulce.
98

a)12000
10000
8000 "'e > :e 6000 e:
4000
2000
o
3000
"'e ~ 2000 "'C e:
e)
..,
1000
o
4500
4000
3500
3000
..§ 2500 > ] 2000
1500
1000
500
o
- DCop 1
• Ostrac -' DApend
'
, ~~-=1L:fk Q r ._ 1 ~ 1 ,
-
--
_e ·- - rt ......... o 1 1
Ab Ag Nov Mar Ab Set Fecha
Figura 2.11. Variación temporal en la abundancia (indiv/m3) de tres estadías de desarrollo de
copépodos en las capas a) P1, b) P2 y e) P3 en la estación 03, Golfo Dulce.
99

a)12000 OCop
10000 • Ostrac
DApendíc
8000 .., E > :e 6000 .::
4000
2000
o
b) 4000
3000
.., E ~ 2000 "O
·= 1000
JL, o
e) 1500
1000
Ab Nov Mar Fecha
Ab Set Ag
Figura 2.12. Variación temporal en la abundancia (indiv/m3) de tres estadías de desarrollo de
copépodos en las capas a) P1, b) P2 y e) P3 en la estación 04, Golfo Dulce.
100

12000 ~---------------~
a) 10000
"'e sooo > ] .6000
"'e >
4000
DCop
• Ostrac
DApendic
~ 2000 ..¡_~~~------------~
500 +----------!
Ab Ag Nov Mar Ab Set Fecha
Figura 2.13. Variación temporal en la abundancia (indiv/m3) de tres estadías de desarrollo de
copépodos en las capas a) P1, b) P2 y e) P3 en la estación 11, Golfo Dulce.
101

102
Capa Superficial (P1)
En este caso, se puede observar que en todas las estaciones y todas las fechas los
copépodos fueron el grupo dominante. En la estación 01 (Figura 2.9a) los copépodos
presentan un pico en agosto de 1997, una pequeña caída hacia el mes de marzo de 1998 y
un aumento en la abundancia en los meses de abril y setiembre de 1998, lo que coincide con
los picos de biomasa observados en la misma capa (capítulo 1 ). Las apendicularias
mostraron una mayor abundancia que los ostrácodos e incrementan su número a lo largo de
los últimos cuatro muestreos. Los ostrácodos, por su parte, presentan pequeños
incrementos en agosto y marzo. Para el caso de la estación 02 (Figura 2.1 Oa) destaca el
gran incremento en la abundancia de copépodos que se dio de marzo a abril de 1998,
también observado en la estación 01, pero no tan marcadamente. Las apendicularias, al
igual que en la estación 01, dominan sobre los ostrácodos a lo largo de la mayor parte de las
fechas y presentan un leve repunte hacia el mes de setiembre de 1998.
La estación 03 (Figura 2.11 a), por su parte, al igual que las estaciones 04 y 11
(Figuras 2.12a y 2.13a), mostró una muy alta abundancia de copépodos, apendicularias y
ostrácodos en el mes de abril de 1997. El dominio de los copépodos disminuye
marcadamente para el mes de agosto de 1997, cuando los números de organismos en los
tres grupos (y la biomasa general del zooplancton) bajan marcadamente. Así mismo, los tres
grupos mostraron un incremento en el mes de noviembre de 1997. Al igual que en las
estaciones internas, las apendicularias dominan sobre los ostrácodos durante todas las
fechas analizadas, especialmente durante 1998.
El comportamiento de los copépodos a lo largo de las fechas en la estación 04
(Figura 2.12a) es el mismo que en la estación 03, lo que refleja la estrecha relación entre
ambos sitios. Una vez más, las apendicularias dominan sobre los ostrácodos durante la
mayoría de las fechas a excepción del mes de agosto de 1997, cuando las apendicularias
tienen una leve caída en su abundancia. Resulta importante observar que los aumentos que
se dan en la abundancia de apendicularias, en los meses de noviembre de 1997 y abril de
1998, coinciden con los picos en la abundancia de copépodos y en la biomasa total de
zooplancton
El caso de la estación 11 (Figura 2.13a), finalmente, también es similar al de las
anteriores estaciones, con altas abundancias de copépodos en los meses de abril y
noviembre de 1997 (pico de biomasa) y con las apendicularias dominando sobre los
ostrácodos la mayor parte del año, excepto por el mes de abril de 1997, cuando el número

103
de ostrácodos es superior. Al igual que en la estación 04, las apendicularias mostraron un
incremento en su abundancia al mismo tiempo que los copépodos, en noviembre de 1997.
Sin embargo, a diferencia de la estación 04, en la estación 11 no se obseliVa un aumento en
la abundancia de copépodos ni de apendicularias en el mes de abril' de 1998.
Capa Intermedia (P2)
La estación 01 (Figura 2.9b) mostró, en el mes de abril de 1997, una capa intermedia
con relativamente pocos organismos y donde los tres grupos de interés tienen abundancias
bastante bajas. Sin embargo, en agosto se observa un importante incremento en la
abundancia de los ostrácodos que los lleva a dominar la comunidad, aún sobre los
copépodos. A partir de éste muestreo, los ostrácodos disminuyen en su abundancia y los
copépodos pasan a dominar. Los tres grnpos mostraron un i>ncremento en el mes de abri1I de
1998, sin embargo, la abundancia de copépodos y apendicularias decrece para setiembre de
1998.
Para todas las demás estaciones, la capa intermedia fue dominada en itodas las
fechas por los copépodos. El comportamiento de los valores de abundancia de los tres
grupos en la estación 02 (Figura 2.10b) es muy similar a la estación 01, con valores bajos
entre las fechas de agosto y marzo, para un posterior incremento, especia1lmente en
copépodos y apendicularias, en el mes de abril de 1998 (incremento en la biomasa del
zoopancton).
La capa P2 en la estación 03 (Figura 2.11 b ), al igual que la capa superficial, se
comporta de manera distinta a las estaciones internas. En primer lugar, mostró valores muy
altos en la abundancia de copépodos durante la primera fecha (abri ll 199'7) que cayeron
marcadamente en agosto de 1997. Posteriormente, l·a abundancia de copépodos solo
mostró un pequeño pico en marzo de 1998 y un descenso en los dos muestreos siguientes.
Las apendicularias so·lo superan a los ostrácodos durante abril1 de 1997 para luego caer a
valores muy bajos que se mantienen durante las restantes fechas. Los ostrácodos, por su
parte, mostraron un incremento importante en el mes de agosto de 1997, que coincide con el
observado en la estación 01. Dichos valores luego descienden y se mantienen relativamente
constantes.

lü4
Con respecto a la estación 04 (Figura 2.12b ), los copépodos mostraron valores
moderados en agosto de 1997, un descenso en noviembre y u1n aumento en marzo y abril de
1'998, luego de lo cual los valores descienden marcadamente. Los ostrácodos presentaron
dos incrementos en su abundancia, en las fechas de agosto de 1997 y abril de 1998,
mientras que la apendicularias mostraron valores bajos durante todo el período de estudio.
Finalmente, la estación 11 (Figura 2. 13b) presenta abundancias muy bajas de los tres grupos
en agosto. Los ostrácodos y apendicularias mantuvieron abundancias bajas con pocas
fluctuaciones a lo largo del perfodo dé estudio. Los copépodos, mientras tanto, mostraron un
incremento para noviembre de 1997 y un segundo pico para abril de 1998.
Capa Profunda (P3)
Al igual que en las capas anteriores, los copépodos fueron el grupo dominante en la
capa profunda de todas las estaciones. Las estaciones 01, 02 y 03, sin embargo, se
comportan de manera muy distinta en la capa profunda. La estación 01!, (Figrna 2.9c) mostró
valores mínimos de abundancia en el mes de abril de 1997 así como un 1incremento en
agosto y luego un incremento más importante entre marzo y abril de 1998. Su abundancia
cae luego para la última fecha. Los ostrácodos por su parte, mostraron tan solo un pequeño
pico en noviembre de 1997, justo cuando se da una pequeña caída en la abundancia de
copépodos. Las apendicularias mostraron un incremento en su abundancia hasta en el mes
de marzo de 1998
La estación 02 (Figura 2.1 Oc) mostró su máxima abundancia de copépodos en
noviembre de 1997, al mismo tiempo que en la estaci.ón 01 se da un descenso en dicho
grupo. l a abundancia de copépodos luego desciende en cada fecha de muestreo hasta
alcanzar valores mínimos en la última fecha. Con respecto a los otros dos grupos, los
ostrácodos parecen dominar levemente a las apendicularias durante 1997 mientras que lo
opuesto ocurre al' año sigu iente. En general, ambos grupos mostraron abundancias muy
bajas en esta capa. En la estación 03 (Figura 2.11 c), resalta el hecho de las muy altas
abundancias que se dan, especialmente de copépodos, en la primera fecha. Dichos valores
presentan un descenso en agosto de 1997 y se mantienen así en las restantes fechas.
Solamente los copépodos mostraron un leve repunte en setiembre de 1998.

105
Para el caso de las estaciones externas, copépodos y ostrácodos mostraron valores
altos en los meses de abril y agosto de 1997, que disminuyen en la tercera fecha.
Posteriormente, los tres grupos mostraron un importante incremento en sus abundancias en
el mes de abril de 1998. En 1.a estación 11' (Figura 2.13c), a diferencia de la 04 (Figura
2.12c), se observan valores bajos en agosto de 1997 que luego, para el caso de copépodos
y ostrácodos, aumentan en el mes de noviembre. Los copépodos también mostraron un
incremento muy importante en abril y setiembre de 1998. Los ostrácodos mantienen una
abundancia relativamente constante en todas las fechas a partir de marzo. Las
apendicularias mostraron valores muy bajos sin cambios importantes a lo largo de todas las
fechas.
Los patrones anteriormente descritos para cada una de las estaciones se reflejan en
los análisis de cong!lomerado efectuados para cada una de las capas muestreadas (Figura
2.14 ). Como se puede ver, en las capas P1 y P3 se agrupan las estaciones in,ternas (01 y
02) aparte de las demás estaciones y esta separación es mayor en el caso de la capa
profunda. Resulta 1interesante el hecho de que 1los análisi1s de conglomerado realizados
sobre los valores de biomasa (capítulo 1) dieron como resultado una representación simi11ar.
La única diferencia se dio en que la estación 03 resultó estar más relacionada a las
estaciones internas al analizar la matriz de biomasa. Sin embargo, al considerar los datos de
abundancia por grupos, esta situación cambi'8. En la capa superficial (Figura 2.14a) se
separan las estaciones internas (01 y 02) de las restantes. Las estaciones internas, a pesar
de su similitud, se separan claramente una de la otra, a diferencia de la capa P3, donde son
más similares. En el caso de las restantes tres estaciones en P1, llama la atención el hecho
de que las estaciones 03 y 11 sean muy similares y que la estación 04 se separe ligeramente
de estas.
En el caso de la capa intermedia (Figura 2.14b), la única estación que se separa
claramente en cuanto a la composición del zooplancton es la 01. Por otra parte, las
estaciones más similares en cuanto a su composición son la 11 I y la 04, lo cual coincide con
el analisis efectuado sobre los valores de biomasa. Resulta interesante que la estación que
más se les parece es la 02 y que la estación 03 guarda muy poca similitud con estas. Lo
anterior ,indica que las estaciones 01' y 03 tienen comunidades de zooplancton con
composiciones muy características y que l1as diferencian de las demás estaciones en la capa
intermedia.

106
Finalmente, en la capa profunda (Figura 2.14c) se agrupan claramente las estaciones
internas, con composiciones muy similares y las estaciones 11 y 03, relativamente con un
menor grado de similitud. La estación 04 se separa de las restantes y se constituye en un
caso a parte en cuanto a la composición del zooplancton a esta profundidad.
a) o 1
o 2
o 4
1 1
o 3
b) o 1
o 2
1 1
o 4
o 3
e) o 4
o 3
1 1
o 1
o 2
o ' o o '1
o , o a .1
' o ' 2 o ' 3
1
1
Distanc ia
1
1
o ' 2 Dis tancia
1 1
o ,4 o ,5
1) , 3 o , 4
o.o 0,1 0,2 0,3 0,4 0,5 0,6 0 ,7 0,8 D is t a nc i a
Figura 2.14. Análisis de conglomerado para las estaciones muestreadas respecto a la
abundancia de zooplancton en las capas a) superficial (P1 ), b) intermedia (P2)
y e) profunda (P3), aplicando el coeficiente de similitud Bray-Curtis y uniones
de promedio por grupo. Notar diferentes escalas de distancia en cada caso.

107
Como último punto, se logró determinar la existencia de una relación lineal leve pero
significativa entre la abundancia del zooplancton en la capa superficial (P1) y: a) la
profundidad de la capa de mezcla (p = 0.002, R2 = 0.347) y b) la biomasa del zooplancton en
la capa P1 (p = 0.024, R2 = 0.2026) (Figuras 2.15a y b, respectivamente). En el primer caso,
se observa una disminución en la abundancia de zooplancton al aumentar la profundidad de
la capa de mezcla la mientras que en el segundo caso se da, como es de esperar, un
aumento en la biomasa a medida que aumenta la abundancia del zooplancton. En este
último caso no se espera una relación importante debido a que un aumento en la abundancia
puede deberse a organismos pequeños, por lo que no se esperará un aumento lineal ni
proporcional en los valores de biomasa. No se detectó ninguna relación significativa (p >
0.05) entre la abundancia del zooplancton y las demás variables físicas evaluadas
(profundidad del disco de Sechii, oxígeno disuelto, temperatura y salinidad).
12000 l M'
E 10000 > 'O ~ 8000
ci.. o
600) o N (1)
·¡:; 4000 e
(1) "O e ::i 2000 ..e <t:
Q
o
• • • •
y =-151.83x +9968.6
ff = 0.3473, p = 0.002
Figura 2.15. Regresión lineal entre la abundancia general de zooplancton (indiv/m3) y a) la
profundidad de la capa de mezcla (m) y b) la biomasa del zooplancton (mg/m3).

108
11.1 Abundancia y Composición: Copépodos
Como se mencionó con anterioridad, los copépodos constituyeron el grupo más
abundante dentro del zooplancton (capturado con una red de 153 µm) en el Golfo Dulce. En
las 86 muestras analizadas, se cuantificó un total de 38 576 copépodos adultos
pertenecientes a un total de 29 géneros distintos (Apéndice 1 ), para un promedio de 448.6 ±
331.3 copépodos por muestra. Al mismo tiempo, se cuantificó un total de 1 O 394
copepoditos y 4 794 nauplius, para un promedio de 120.9 ± 84.7 copepoditos y 55.7 ± 64.7
nauplius por muestra. Al calcular las abundancias promedio para cada caso, se obtiene en
primer lugar a los copépodos, con 1913.3 ± 2511.8 indiv/m3 (CV=131%) seguidos por
copepoditos (557.9 ± 725.1 indiv/m3, CV=130%) y nauplius (354.3 ± 709.1 indiv/m3
,
CV=200% ). Como se puede apreciar, los coeficientes de variación en la abundancia de cada
grupo son bastante altos, especialmente en el caso de los nauplius. Como se observa en las
figuras 2.2 y 2.16, la mayoría de los copépodos se concentra en la capa superficial (4091.1 ±
2688.2 indiv/m3, CV=65.7%), mientras que las capas intermedia (898.4 ± 817.3 indiv/m3
,
CV=91 %) y profunda (545.5 ± 880.9 indiv/m3, CV=161%) presentan abundancias mucho
menores. Es importante resaltar el hecho de que la variabilidad relativa de los datos
aumenta con la profundidad.
6000
.., 4500 E ~ "t:I ·= 3000
1500
L
o ...____._ _ __._ __ P1
O Copépodos O Copepoditos • Nauplius
P2 P3
Figura 2.16. Abundancia promedio (indiv/m3) y desviación estándar de copépodos,
copepoditos y nauplius para cada una de las capas muestreadas durante el
período de estudio, Golfo Dulce.

109
Al analizar los datos mediante una prueba ANDEVA, se comprobó que existen
diferencias significativas en los valores de abundancia a las distintas profundidades (F =
43.8, p < 0.05). Dichas diferencias se dieron, al igual que cuando se consideró la
abundancia de todos los grupos, entre la capa superficial y las restantes dos capas (p <
0.05), pero no entre estas últimas ( P2 y P3) (p > 0.05), de acuerdo a la prueba post-hoc de
Tukey.
Así mismo, la prueba de ANDEVA demostró la existencia de diferencias significativas
entre las estaciones de muestreo (F=1O.7, p < 0.05). Dichas diferencias se dieron al
comparar las estaciones internas (01 y 02) contra las restantes (03, 04 y 11) (p < 0.05), pero
no entre la 01 y 02 ó entre las estaciones 03, 04 y 11. Lo anterior indica que las estaciones
internas son claramente distintas a las restantes estaciones en cuanto a la composición de la
comunidad de copépodos. Como se puede ver en la Figura 2.17, estas diferencias son muy
7500 1
OP1
OP2
.., 5000 J •P3 E -.:!:
"O .5
2500
2 3 4 11
Figura 2.17. Valores promedio y desviación estándar de la abundancia de copépodos
(indiv./m3) para cada estación y capa de muestreo, Golfo Dulce, 1997-1998.
bajas en la P1, donde la abundancia de copépodos es ligeramente menor en las estaciones
internas. La Figura 2.17 también ilustra claramente la existencia de diferencias significativas
en la abundancia de copépodos entre las tres capas de muestreo.
En general, la abundancia de copépodos en la capa superficial varió poco entre
estaciones, con valores que fluctuaron entre 3589.8 ± 2886.3 cop./m3 (CV=80.4%) en la
estación 02, y 4621.9 ± 3423.8 cop./m 3 (CV=74%) en la estación 03. Al igual que para el

110
caso de la abundancia general de zooplancton, la estación 03 es la que presentó los
mayores valores de abundancia de copépodos en las capas superficial e intermedia. En el
caso de esta última, los valores en las estaciones 01 y 02 variaron entre 416.9 ± 501.3
cop./m3 (CV=120%) y 666.3 ± 697.3 cop./m3 (CV=105%), respectivamente. Mientras tanto,
los valores de abundancia en la P2 para las restantes estaciones resultaron un poco más
altos, fluctuando entre 971.4 ± 469.6 cop./m3 (CV=48%) en la estación 11y1340.6 ± 1458.8
cop./m3 (CV=109%) en la estación 03. Finalmente, las diferencias entre las estaciones se
hacen más obvias al observar los valores de abundancia correspondientes a la capa
profunda. En este caso, las estaciones 01 y 02 tienen valores de 136.1 ± 121.1 cop./m3
(CV=89%) y 90.2 ± 50.8 cop./m3 (CV=56%), respectivamente. Estos, contrastan con los
valores más altos en las restantes estaciones, que fluctúan entre 827.1 ± 484 cop./m 3
(CV=59%) en la estación 11y1380±1906.1 cop./m3 (CV=138%) en la estación 04.
Con respecto a la abundancia de copépodos por fecha (Figura 2.18), se observó una
gran similitud con los patrones de abundancia de zooplancton en general (Figura 2.4). La
prueba de ANDEVA demostró que existen diferencias significativas en la abundancia de
copépodos entre las fechas de muestreo (F=6.5, p < 0.05). Las mayores abundancias de
copépodos se observan en el mes de abril de 1997 en todas las capas muestreadas y dichos
15000 l DP1 DP2 • P3
i i
12500 ~
10000 1
.., 1 E -.? 7500
"C e:
5000
2500
o Ab-97 Ag-97 Nov-97 Mar-98 Ab-98 Set-98
Figura 2.18. Valores promedio y desviación estándar de la abundancia de copépodos
(indiv./m3) para cada una de las fechas y capas analizadas, Golfo Dulce, 1997-
1998.

111
valores no se vuelven a repeti r durante el período de estudio. La prueba post-hoc de Tukey
mostró, al igual que con las abundancias totales de zooplancton, que las diferencias entre
fechas se dan solamente entre la fecha 1 (abril de 1997) y las restantes (p < 0.05) y no entre
estas últimas, lo cual se debe, como ya se mencionó, a los (considera,blemente) mayores
valores de abundancia que se dan en la primera fecha con respecto a las demás. Un
aspecto interesante es que los valores de abundancia en la capa profunda (P3) superan a los
de la P2 durante esta primera fecha. Al igual que para las abundancias de zooplancton, la
de copépodos también presentó un leve incremento en los dos últimos meses del período de
estudio (con respecto a marzo de 1998). Además, cabe señal8!f que los altos valores en la
abundancia que se observan en el mes de abril de 1997, son probablemente responsables,
en gran medida, de la alta variabilidad en los valores de abundancia por estación (Figura
2.17).
Del total de copépodos contados, 18 11 O fueron del orden Cyclopoida (46.9%), 11
138 del orden Calanoida (28.9%), 7204 del orden Poecilostomatoida (18.7%) y 2 124 del
orden Harpacticoida (5.5% ). Como se mencionó antes, se identificó un total de 29 géneros
de copépodos, del total de 30 que hasta la fecha han sido reportados para la zona del Golfo
Dulce (Morales 1996). La especie que no fue detectada en el presente estudio es
Subeucalanus subcrassus (O. Calanoida ), reportada para la zona por M!ora1les ( 1996 ). El
orden Calanoida es el que presentó mayor riqueza, con 16 familias y 20 géneros, seguido de
los ordenes Harpacticoida y Poecilostomatoida, con 3 familias y 4 géneros cada uno y
finalmente el orden Cyclopoida, con una familia y un género identi,ficado ( Oithona spp).
A pesar de que se identificó un total de 29 géneros en el1 presente estUJ dio, tan solo
cinco resultaron responsables del 91.5% de la abundancia total de copépodos en el Golfo
Dulce. Se trata de Oithona spp. (46.9%), Paracalanus spp. (21.4%), Oncaea spp. (15.1 %),
Euterpina sp. (4.5%) y Corycaeus spp. (3.6%)(Figura 2.19). El1 restante 8.5% de la
abundancia de copépodos se distribuye en los 24 géneros restantes. Los resultados
completos de abundancia se presentan en los cuadros 2.5, 2.6 y 2.7, correspondientes a las
capas P1, P2 y P3 respectivamente.
El hecho de que tan sólo cinco géneros concentren más del1 90% de la abundancia
total de copépodos, explica la existencia de diferencias significativas al comparar los valores

Cuadro 2.5. Abundancia (indiv/m3) de los distintos géneros de copépodos identificados en la capa superficial (P1) para
cada una de las estaciones durante el período (1997-1998), Golfo Dulce. Los cinco grupos más importantes en cada estación se indican entre paréntesis.
Género Estación 01 Estación 02 Estación 03 Estación 04 Estación 11
Canthocalanus 5.93 ± 10.18 2.40 ± 4.67 14.51 ± 35.55 1.24 ± 1.97
Undinula 0.84 ± 1.31 1.04 ± 2.54 4.15±10.16
Acrocalanus 138.73 ± 153.34(5) 46.92 ± 24. 76 95.58 ± 105.10 70.90 ± 88.32 72.15 ± 73.61
Paracalanus 964.19 ± 575.67(2) 872.81 ± 865.34(2) 1045.48 ± 739.81(2) 948.48 ± 626.73(2) 1075.7 ± 1163.42(2)
Eucalanus 2.42 ± 3.86 5.80 ± 12.98 17.87 ± 22.83 26.43 ± 51.44 42.60 ± 86.02
Euchaeta 2.42 ± 5.92
Temora 1.45 ± 3.55 6.05 ± 8.66 2.87 ± 3.67 3.94 ± 3.44 12.34 ± 16.62
Centropages 24.27 ± 24.57 27.48 ± 47.52 115.66 ± 242.15 61.47 ± 100.01 74.43 ± 155.55
lucicutia 1.93 ± 3.52 1.93 ± 4.74 0.73 ± 1.78 0.73 ± 1.78
Pseudodiaptomus 3.39 ± 5.33 3.63 ± 8.89 2.18 ± 3.64
Candacia 6.87 ± 7.69 5.71 ± 7.07 2.49 ± 2.81 6.56 ± 4.34
labidocera 3.61 ± 4.94 2.90 ± 4.50 1.24 ± 1.97 5.32 ± 8.29
Acartia 94.66 ± 87.32 27.04 ± 20.97 422.26 ± 910.03(3) 154.25 ± 278.1 o 105.04 ± 150.65
Oithona 2013.99 ± 1028.2(1) 1803.55 ± 1260.4(1) 1856.83 ± 557.26(1) 1827.7 ± 637.13(1) 1768.7 ± 1464.54(1)
Euterpina 174.53 ± 206.58(4) 213.74 ± 345.82(4) 369.03 ± 630.58(4) 245.36 ± 354.23(5) 241.45 ±319.30(5)
Clytemnestra 1.45 ± 3.55 0.45 ± 1.11 1.45 ± 3.55 1.45 ± 3.55
Macrosetella 3.41±7.28 1.74 ± 3.89 10.49 ± 10.92 3.21 ± 5.43 16.97 ± 27.48
Microsetella 1.97 ± 3.53 1.16 ± 2.60 5.23 ± 7.48 9.43 ± 12.44 13.06 ± 24.00
Oncaea 216.39 ± 277.45(3) 405.93 ± 379.28(3) 343.14 ± 467.97(5) 347.26 ± 576.88(4) 489.23 ± 718.02(3)
Corycaeus 88.56 ± 69.38 144.18 ± 151.32(5) 300.37 ± 409.14 404.27 ± 542.63(3) 286.62 ± 395.99(4)
Copilia 2.42 ± 5.92 7.77 ± 17.56 3.14 ± 5.84
Saphirina 0.48 ± 1.18 1.45 ± 3.55
Caloca/anus 2.90 ± 7.11 12.58 ± 7.53 10.37 ± 20.53 14.93 ± 12.19 23.12 ± 14.39
Clausocalanus 13.10 ± 16.67 10.31 ± 9.78 6.65 ± 13.86 3.21 ± 3.78 6.01 ± 7.00
TOTAL 3750.35 ± 1048.04 3589.80 ± 2886.3 4621.87 ± 3423.82 4153.7 ± 2364.08 4256.0 ± 3824.61
112
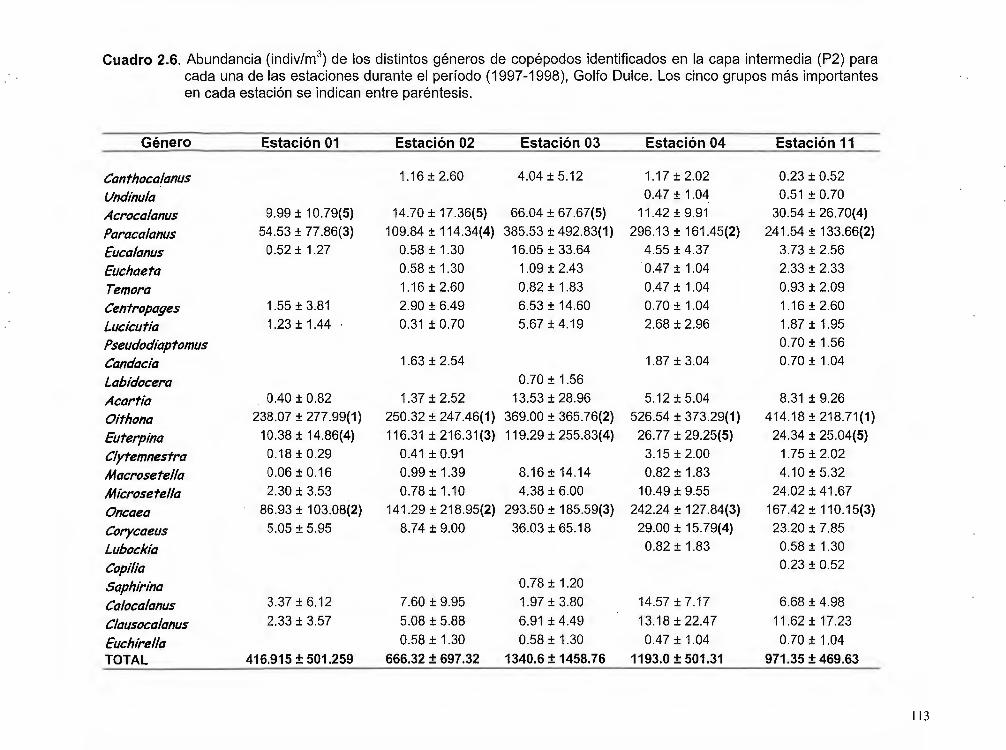
Cuadro 2.6. Abundancia (indiv/m3) de los distintos géneros de copépodos identificados en la capa intermedia (P2) para
cada una de las estaciones durante el período (1997-1998), Golfo Dulce. Los cinco grupos más importantes en cada estación se indican entre paréntesis.
Género Estación 01 Estación 02 Estación 03 Estación 04 Estación 11
Canthoca/anus 1.16 ± 2.60 4.04 ± 5.12 1.17 ± 2.02 0.23 ± 0.52
Undinula 0.47 ± 1.04 0.51 ± 0.70
Acrocalanus 9.99 ± 10.79(5) 14.70 ± 17.36(5) 66.04 ± 67.67(5) 11.42 ± 9.91 30.54 ± 26.70(4)
Paracalanus 54.53 ± 77.86(3) 109.84 ± 114.34(4) 385.53 ± 492.83(1) 296.13 ± 161.45(2) 241.54 ± 133.66(2)
Euca/anus 0.52 ± 1.27 0.58 ± 1.30 16.05 ± 33.64 4.55 ± 4.37 3.73 ± 2.56
Euchaeta 0.58 ± 1.30 1.09 ± 2.43 0.47 ± 1.04 2.33 ± 2.33
Temora 1.16 ± 2.60 0.82 ± 1.83 0.47 ± 1.04 0.93 ± 2.09
Centropages 1.55 ± 3.81 2.90 ± 6.49 6.53 ± 14.60 0.70 ± 1.04 1.16 ± 2.60
Lucicutia 1.23 ± 1.44 0.31 ± 0.70 5.67 ± 4.19 2.68 ± 2.96 1.87 ± 1.95
Pseudodiaptomus 0.70 ± 1.56
Candada 1.63 ± 2.54 1.87 ± 3.04 0.70 ± 1.04
Labidocera 0.70 ± 1.56
Acartia 0.40 ± 0.82 1.37 ± 2.52 13.53 ± 28.96 5.12 ± 5.04 8.31 ± 9.26
Oithona 238.07 ± 277.99(1) 250.32 ± 247.46(1) 369.00 ± 365.76(2) 526.54 ± 373.29(1) 414.18 ± 218.71(1)
Euterpina 10.38 ± 14.86(4) 116.31±216.31(3) 119.29 ± 255.83(4) 26.77 ± 29.25(5) 24.34 ± 25.04(5)
C!ytemnestra 0.18 ± 0.29 0.41 ± 0.91 3.15 ± 2.00 1.75 ± 2.02
Macrosetel/a 0.06 ± 0.16 0.99 ± 1.39 8.16 ± 14.14 0.82 ± 1.83 4.10 ± 5.32
Microsetel/a 2.30 ± 3.53 0.78±1.10 4.38 ± 6.00 10.49 ± 9.55 24.02 ± 41.67
Oncaea 86.93 ± 103.08(2) 141.29 ± 218.95(2) 293.50 ± 185.59(3) 242.24 ± 127.84(3) 167.42±110.15(3)
Corycaeus 5.05 ± 5.95 8.74 ± 9.00 36.03 ± 65.18 29.00 ± 15.79(4) 23.20 ± 7.85
Lubockia 0.82 ± 1.83 0.58 ± 1.30
Copilia 0.23 ± 0.52
Saphirina 0.78 ± 1.20
Ca!ocalanus 3.37 ± 6.12 7.60 ± 9.95 1.97 ± 3.80 14.57 ± 7.17 6.68 ± 4.98
C!ausoca!anus 2.33 ± 3.57 5.08 ± 5.88 6.91 ± 4.49 13.18 ± 22.47 11.62 ± 17.23
Euchirella 0.58 ± 1.30 0.58 ± 1.30 0.47 ± 1.04 0.70 ± 1.04
TOTAL 416.915 ± 501.259 666.32 ± 697 .32 1340.6 ± 1458.76 1193.0 ± 501.31 971.35 ± 469.63
113

Cuadro 2.7. Abundancia (indiv/m3) de los distintos géneros de copépodos identificados en la capa profunda (P3) para
cada una de las estaciones durante el período (1997-1998), Golfo Dulce. Los cinco grupos más importantes en cada estación se indican entre paréntesis.
Género_ Estación 01 Estación 02 Estación 03 Estación 04 Estación 11
Canthocalanus 0.05 ± 0.12 2.92 ± 5.73 3.11 ± 6.54 3.08 ± 5.67
Acrocalanus 1.08 ± 1.31 1.55 ± 2.34 53.11 ± 123.3(5) 41.27 ± 56.84 29.31±16.1(4)
Paracalanus 26.42 ± 23.93(3) 6.02 ± 7.57(3) 175.21 ± 359.7(3) 423.0 ±718.9(1) 168.87 ± 109.0(3)
Eucalanus 0.05 ± 0.12 0.14 ± 0.30 1.21 ± 2.96 14.38 ± 21.19 3.19 ± 4.40
Euchaeta 3.63 ± 8.89 2.07 ± 2.49 1.40 ± 2.09
Temora 0.41 ±0.75 0.09 ± 0.21 0.35 ± 0.55 2.72 ± 6.67 0.90 ± 1.24
Temoropia 0.18 ± 0.44 0.92 ± 1.43 1.15 ± 1.15 0.34 ± 0.83
Centropages 1.06 ± 1.68 0.33 ± 0.51 40.18 ± 97.63 26.63 ± 62.97 1.31 ± 2.92
Lucicutia 0.10 ± 0.24 0.23 ± 0.52 0.59 ± 0.47 2.83 ± 4.20 1.74 ± 2.38
Pseudodiaptomus 0.09 ± 0.21 0.15 ± 0.21 1.36 ± 3.33 0.93 ± 2.09
Candacia 0.28 ± 0.56 0.05 ± 0.12 0.34 ± 0.83
Labidocera 2.57 ± 5.86
Acartia 0.81 ± 0.95 43.29 ± 103.22 20.80 ± 49.81 4.69 ± 5.99
Oithona 61.39 ± 51.33(1) 17.28 ± 12.33(2) 192.7 ± 321.9(2) 303.6 ± 411.9(2) 287.9 ± 179.6(1)
Euterpina 7.74 ± 8.24 (4) 2.83 ± 1.26(5) 108.21 ± 249.2(4) 113.7 ± 244.7(4) 11.15 ± 7.58
C/ytemnestra 0.36 ± 0.89 0.14 ± 0.30 0.53 ± 0.87 2.43 ± 4.25 0.83 ± 1.15
Macrosetella 0.44 ± 0.82 0.46 ± 0.89 11.71 ± 26.27 23.32 ± 35.78 1.96 ± 2.71
Microsetella 1.63 ± 2.18 1.03 ± 2.04 4.70 ± 6.72 13.35 ± 15.74 10.08 ± 13.70
Oncaea 56.93 ± 44.57(2) 54.08 ± 35.22(1) 208.67 ± 371.8(1) 294.9 ±213.3(3) 263.55 ± 154.5(2)
Corycaeus 3.06 ± 3.37(5) 0.94 ± 0.98 40.68 ± 86.74 68.77 ± 150.5(5) 18.44 ± 16.14(5)
Lubockia 0.19 ± 0.48
Saphirina 0.16±0.38
Ca/ocalanus 0.13 ± 0.20 3.66 ± 5.38 0.78 ± 1.90 4.40 ± 3.33
C/ausocalanus 0.87 ± 1.23 3.95 ± 8.45(4) 3.85 ± 3.01 20.73 ± 38.13 12.96 ± 24.22
Spinocalanus 3.63 ± 8.89
Euchirella 0.31 ± 0.48 0.44 ± 0.97
TOTAL 163.07±121.11 90.18 ± 50.79 903.57 ± 1770.18 1380.05±1906.15 . 827.11±484.03
114

115
de abundancia de los distintos grupos (F=160.8, p < 0.05). También resulta importante
señalar que, en concordancia con los resultados correspondientes al análisis de grupos de
15.1% 4.5%
46.9% O Oithona O Paracalanus O Oncaea
2.6%
- 1.9%
· 0.9%
- 3.2%
O Euterpina llJ Corycaeus IIlJ Acroca/anus
E3 Acama ~ Centropages ll3 Otros
Figura 2.19. Abundancia relativa de los principales géneros de copépodos en el Golfo Dulce ,
1997-98.
zooplancton , al analizar los resultados de abundancia de copépodos también se detectó una
interacción entre los factores de grupo (género) y profundidad (F=11.7, p< 0.05), lo que
indica que los distintos géneros de copépodos varían en forma diferente con la profundidad y
viceversa. En otras palabras, la composición de la comunidad de copépodos en el Golfo
Dulce, al igual que la de zooplancton, varía con la profundidad. También se detectó
interacción entre los factores de grupo (género) y fecha (F=2.9, p< 0.05), por lo que, al igual
que con el zooplancton en general, los distintos géneros de copépodos varían en forma
diferente con la fecha, así como las diferencias que se dan entre las fechas varían de
acuerdo a los distintos géneros.
11.11. Abundancia y Composición: Distribución por Capa
Como se acaba de mencionar, existen diferencias tanto en la abundancia como en la
composición de la comunidad de copépodos al comparar las distintas capas de agua
muestreadas, por lo que a continuación se detallan los distintos patrones de abundancia
relativa de los principales grupos de copépodos en cada una de las tres capas.

116
Capa superficial (P1)
En esta capa, todas las estaciones fueron dominadas claramente por el género
Oithona, sin embargo, las estaciones internas mostraron mayores porcentajes de
abundancia, con 53.7% en la estación 01 y 50.2% en la 02. Las estaciones restantes, por su
parte, presentaron abundancias de Oithona que varían entre 40.2% (Est. 03) y 44% (Est. 11 ).
El segundo género en importancia en todas las estaciones fue Paracalanus, con
abundancias que fluctuaron entre 22.6% (Est. 03) y 25.3% (Est. 11 ). El tercer género en
importancia en las estaciones internas es Oncaea, con un 5.8% en la estación 01 y un 11.3%
de abundancia en la estación 02. En la estación 03, el género Acartia fue el tercero en
importancia, con un 9.1 % de abundancia. Lo anterior solamente se dio en esta estación, ya
que Acartia no vuelve a entrar dentro de los cinco géneros más importantes en ninguna otra
estación, a ninguna otra profundidad. Las estaciones 04 y 11, por su parte, tuvieron a
Corycaeus (9.7%) y Oncaea (11.5%) como tercer grupo en importancia, respectivamente. El
género Euterpina, también se encontró dentro de los géneros más abundantes, con un
cuarto lugar en las estaciones 01 (4.7%), 02 (5.9%) y 03 (8%), mientras que en las
estaciones externas se colocó en quinto lugar con un 5.9 % en la estación 04 y 5. 7% en la
11. En esta última, el cuarto lugar en abundancia fue ocupado por Corycaeus, con un 6. 7%.
Oncaea también resultó ser un grupo importante en las estaciones 03 y 04, donde tiene
porcentajes de 7.4% y 8.4% respectivamente. El género Acrocalanus sólo apareció dentro
de los principales grupos en la estación 01, con un 3. 7% de abundancia. El detalle de las
abundancias de todos los grupos presentes en la capa superficial se puede observar en el
cuadro 2.5.
El análisis de conglomerado para la capa superficial (Figura 2.20), permitió
comprobar los patrones antes descritos, donde los géneros más abundantes se agruparon
en el conglomerado A. Aquí se distinguen los dos géneros dominantes en la capa superficial
del Golfo Dulce durante el período de estudio: Oithona y Paracalanus. Junto a estos están
los géneros Euterpina, Corycaeus y Oncaea, también abundantes en la superficie, además
de otros como Acartia, Centropages y Acrocalanus. Estos son los géneros de copépodos
que dominan más del 90% de la abundancia de éste grupo en el Golfo Dulce. En el
conglomerado B, se agruparon los géneros con abundancias de medias a bajas, como por
ejemplo Eucalanus, Ca/ocalanus, C/ausoca/anus y Macrosetella. Finalmente, en el
conglomerado C se agruparon los géneros menos abundantes, que se presentaron solo
ocasionalmente en las muestras, como por ejemplo Temoropia, Ponte/fina, Sapphirina,
Lucicutia y Copilia, entre otros.

e
B
A
Canthocalanus ---~ Copilia
Pseudodiaptomus Ludcutia
C/ytemnestra Undinula E u cha e
Sapphirina Euchirella Lubockia
Pleuromam11 Temoropia Pon tellina
Spinoca/anu Labidoce Candacia Te mora
Microsetel/a Macrosetella
C/ausocalanus -----L _ _J Calocalanus - --i_ _ __J
Eucalanus Centropages --~ Acrocalanus ---' 1
Acartia Corycaeu Euterpina Oncaea
Paracalanus Oithona
0,0 0,5
117
1,0 1,5 2,0 2,5 Distancia
Figura 2.20. Análisis de conglomerado para los géneros de copépodos respecto a su
abundancia en la capa P1, aplicando el coeficiente de similitud Bray-Curtis y
uniones de promedio por grupo. A, B y C representan los conglomerados
resultantes.
Capa intermedia (P2)
En esta capa el género Oithona volvió a ser el más importante tanto en las estaciones
internas (Est. 01 = 57.1 %, Est. 02 = 37.6%) como en las externas (Est. 04 = 44%, Est. 11 =
42.6%). La estación 03 sin embargo, fue dominada, a diferencia de las demás, por el género
Paracalanus (28.8%) mientras que el género Oithona se ubica en un cercano segundo lugar
(27.5%). El segundo género en importancia en las estaciones internas fue Oncaea, con un
20.9% y un 21.2% respectivamente. En las demás estaciones, Oncaea se ubicó en el tercer
lugar de abundancia, con entre 17.2% (Est. 11) y 21.9% (Est. 03). Resulta importante
destacar que la abundancia relativa del género Oncaea en la P2 aumentó con respecto a la

118
capa superficial. El segundo lugar de abundancia en las estaciones externas fue
Paracalanus, con 24.8% (Est. 04) y 24.9%(Est. 11 ). Este mismo género ocupó un tercer
lugar en la estación 01 ( 13.1 % ) y un cuarto lugar en la 02 ( 16.5% ). Finalmente, Euterpina se
constituyó en un género importante al ser el tercero en importancia en la estación 02
(17.5%), el cuarto en las estaciones 01 (2.5%) y 03 (8.9%) y el quinto en las estaciones
externas (Est. 04 = 2.2%, Est. 11 = 2.5%). El género Acrocalanus apareció también como
grupo importante en todas las estaciones excepto la 04 y presentó abundancias entre 2.2%
(Est. 02) y 4.9% (Est. 03). Los datos completos de abundancia de copépodos en la capa
intermedia se pueden observar en el Cuadro 2.6.
Al observar el análisis de conglomerado correspondiente a la capa intermedia (Figura
2.21), se distingue que existen tres géneros claramente dominantes (conglomerado A):
Oithona, Oncaea y Paracalanus. A parte, se separa otro grupo de géneros (conglomerado
B) importantes en cuanto a su abundancia en la capa intermedia: Euterpina, Acrocalanus,
Corycaeus, Microsetella, C/ausocalanus y Ca/ocalanus. El conglomerado C, muestra a
aquellos géneros con abundancias medias o bajas, como Acartia, Eucalanus, Centropages y
Canthocalanus. Finalmente, los grupos raros se separan en un último conglomerado (D),
con géneros como Sapphirina, Labidocera, Undinula, Temora y Euchaeta, entre otros.
Capa profunda (P3)
Finalmente, en la capa profunda (Cuadro 2.7), Oithona solo llegó a dominar, y de
manera menos clara, en las estaciones 01 (37.6%) y 11 (34.8%). El género Oncaea adquirió
aún más importancia en esta profundidad y dominó las estaciones 02 (60%) y 03 (23.1 % ),
mientras que en las estaciones 01 (34.9%) y 11 (31.9%) ocupó un segundo lugar. Caso
aparte se da en la estación 04, que fue dominada por Paracalanus (30.7%) y Oithona (22%).
Aquí, Oncaea ocupó el tercer lugar, con 21.4%. Paracalanus, por su parte, fue el tercer
género en importancia en las restantes estaciones con abundancias que variaron entre 6. 7%
(Est. 02) y 20.4% (Est.11 ). Finalmente, Euterpina se mantuvo como uno de los cinco
géneros más importantes con abundancias de 4.8% en la estación 01 y 3.1 % en la 02. Las
estaciones 03 y 04 presentaron abundancias un poco más altas, con 12% y 8.2%
respectivamente. Sin embargo, el género Euterpina no entró dentro de los grupos más
importantes en la estación 11, donde Acrocalanus ocupó su lugar con un 3.5% de la
abundancia. Este género también apareció en la estación 03, donde presentó un 5.9% de
abundancia.

A
B
e
D
1 Oithona ==--""'!---------------. Oncaea -Paracalanus _ _,- -
Euterpina------, Acroca/anus ---, Corycaeus Microsetel/a - --,___ __ _¡
Clausocalanus Ca/ocalanus
Acartia Eucalanus Ludcuüa -- -~ ,
Macrosetel/a Centropages
Canthocalanus Sapphirina Lab1docera
Spinocalanus Pleuromamma
Temoropia Pon tellina
Copilia Pseudod1aptom
Undinula Lubockia Euchire/la Te mora Euchae
Clytemnestra Candacia~---~~--------..,...._ ___ _
0,0 0,5 1,0 Distancias
1,5 2,0
119
Figura 2.21 . Análisis de conglomerado para los géneros de copépodos respecto a su
abundancia en la capa P2, aplicando el coeficiente de similitud Bray-Curtis y
uniones de promedio por grupo. A, B, C y D representan los conglomerados
resultantes.
El análisis de conglomerado correspondiente a la capa profunda (Figura 2.22)
también viene a corroborar lo anteriormente explicado. El conglomerado A, de nuevo,
muestra tres géneros muy abundantes y que por lo tanto dominaron a esta profundidad:
Paracalanus, Oithona y Oncaea. El conglomerado 8, por su parte, separó los géneros que, a
pesar de no ser dominantes, tienen un aporte importante a la abundancia total. Se trata de
géneros como Euterpina, Corycaeus y Acrocalanus, por un lado, junto con otros géneros un
poco menos representados, como Acartia, Centropages, Eucalanus y Clausocalanus. El
conglomerado C, en éste caso, esta compuesto por géneros cuya abundancia en la capa
profunda fue baja, como por ejemplo Ca/ocalanus, Canthocalanus, Euchaeta y Temora.

120
Finalmente, los grupos con abundancias muy bajas, se separaron en un conglomerado
distinto (D), que está compuesto por grupos como Pontellina, Undinula, Pleuromamma, y
Copilia, entre otros. Un aspecto interesante es que los grupos que presentaron abundancias
consideradas como muy bajas son prácticamente los mismos en todas las profundidades.
D
e
8
A
Spinocalanu Labidoce . Temoropia Euchirel/a Can dacia Lubockia
Sapphirina Copi/ia
Pleuromamma Undinula
Pon tellina Pseudodiaptomus
Te mora C/ytemnestra
Ludcutia Euchaeta
Canthocalanus Calocalanus
Clausocalanus Microsetella
Eucalanus----' Macrosetella - - --,__ _ _J Gen tropa ges
Acartia Acrocalanus - ---, __
Corycaeu Euterpina ·---J:--~
1 Paracalanus
Oithona Oncaea ;;;=='"---.....,.----.....---- --.---------.
0,0 0,5 1,0 Distancia
1,5 2,0
Figura 2.22. Análisis de conglomerado para los géneros de copépodos respecto a su
abundancia en la capa P3, aplicando el coeficiente de similitud Bray-Curtis y
uniones de promedio por grupo. A, 8 , C y D representan los conglomerados
resultantes.
11.111. Abundancia y Composición: Variación Temporal
Las variaciones temporales en la abundancia del grupo de los copépodos resultan de
gran importancia en la comunidad de zooplancton debido a que éste es, por amplio margen,
el grupo más abundante en el Golfo Dulce.

121
Capa Superficial (P1)
Las Figuras 2.23a -2.27a describen la variación en la abundancia de copépodos
copepoditos y nauplius en la capa superficial para cada una de las estaciones de muestreo.
Como se puede observar, en la estación 01 (Figura 2.23a) se dio un alto número de
copepoditos en abril de 1997, número que disminuyó en la siguiente fecha de muestreo. En
cambio, los copépodos empezaron con una abundancia moderada que mostró un incrementó
para la fecha 2. Resulta importante señalar el incremento que se observó en la abundancia
de los tres grupos en abril de 1998, lo que también se refleja en un aumento en la biomasa.
Lo mismo se observó en la estación 02 (Figura 2.24a), donde los tres grupos mostraron
abundancias de moderadas a bajas en las primeras fechas y un fuerte incremento en abril de
1998. En ambas estaciones, la abundancia de nauplius en 1998 resultó superior a la
observada en 1997.
En las estaciones 03, 04 y 11 (Figuras 2.25a - 2.27a), se observaron altos valores de
abundancia en copépodos, copepoditos y nauplius en la primera fecha, cuando los valores
de biomasa también fueron altos. En las tres estaciones, la abundancia de los tres grupos
disminuyó para agosto de 1997 mientras que los copépodos presentaron un incremento
importante en noviembre de 1997,cuando la biomasa del zooplancton también incrementa.
En el caso de copepoditos y nauplius, no se observó un pico tan claro en noviembre, sin
embargo, sus abundancias presentaron un leve crecimiento a partir de esta fecha. Las
estaciones 03 y 04 también presentaron un leve incremento en la abundancia de copépodos
en el mes de abril de 1998, caso que no se da en la estación 11.
Por otra parte, las figuras 2.28a - 2.32a muestran la variación temporal de cuatro de
los principales géneros de copépodos en el Golfo Dulce. En la estación 01 (Figura 2.28a),
resulta interesante que la capa superficial en la primera fecha fue dominada por Paracalanus
solo durante abril de 1997, ya que Oithona pasó a dominar por el resto del período de
estudio. Lo mismo se observó en las estaciones 03, 04 y 11 (Figuras 2.30a - 2.32a). De
hecho, en la estación 01, Oíthona mostró su máximo pico de abundancia en el mes de
agosto de 1997, al mismo tiempo que la concentración de Paracalanus presentó un
descenso. Para el mes de marzo de 1998, las abundancias de Paracalanus y Oncaea
aumentaron, mientras que la de Oíthona bajó considerablemente. Finalmente, en el mes de
abril de 1998, se observó un incremento en la abundancia de los cuatro géneros. En el caso

a) 12000 OCop
10000 El Copdts
• Nauplius
8000 .., E ~ 6000 "'C .s
4000
2000
o
b) 4000
"'E ~ 2000 +-~~~~~~~~~~~~~~~~~~~~~~~
.E
1500
.., E ~ 1000 -e e:
500
o [L °=' 1
[6.. rk.. • 1 o.._,_
Ab Ag Nov Mar Ab Set Fecha
Figura 2.23. Variación temporal en la abundancia (indiv/m3) de copépodos, copepoditos y nauplius
en las capas a) P1 , b) P2 y e) P3, estación 01, Golfo Dulce.
122

a)12000 .-------------------- - ----0 Cop
10000 +--------------------º Copdts • Nauplius
M 8000 +---------------------~~-E > ~6000 +----------~~~~~~~1
ME ~ 2000 -1---~~~~-----------------"0 .!:
Ab Ag Nov Mar Ab Set fecha
Figura 2.24. Variación temporal en la abundancia (indiv/m3) de copépodos, copepoditos y nauplius
en las capas a) P1, b) P2 y e) P3, estación 02, Golfo Dulce.
123

a) 12000 O Cap
10000 OCopdts
• Nauplius
8000 .., E ~ 6000 "'C .::
4000
2000
o
b) 4000
3000
.., E ~ 2000 "'C
.E
1000
o
e)
4000
3000 .., E ~ "'C e: 2000
1000
o Ab Ag Nov Mar Ab Set
Fecha
Figura 2.25. Variación temporal en la abundancia (indiv/m3) de copépodos, copepoditos y nauplius
en las capas a) P1, b) P2 y e) P3, estación 03, Golfo Dulce.
124

aJ2000 -i---------------------~~-~ DCop
10000 +--------------------O Copdts •Nauplius
8000
"'E
~ººº e:
4000
2000
o _ _..._
"'E z2000 +------------------.......---------~ "'C .!:
1000 +----- ·
c)2000 ~-----------------------~
1500
"'E z1000 _L "'C .!:
Ab Ag Nov Mar Ab Set Fecha
Figura 2.26. Variación temporal en la abundancia (indiv/m3) de copépodos, copepoditos y nauplius
en las capas a) P1, b) P2 y e) P3, estación 04, Golfo Dulce.
125

a) 12000 ¡ DCop 1
1
10000 1 DCopdts
• Nauplius
8000 M
_§ -~ 6000 "'C .E
4000
2000
o -r-[h. -,
b) 4000
3000 1
.., E ~ 2000 'O .!:
1ººº r 1
o [l
e) 2000
1500
.., E :? 1000 'O .!:
500 ·--
lli. -.,--
Ag Nov Mar Ab Set Fecha
Ab
Figura 2.27. Variación temporal en la abundancia (indiv/m3) de copépodos, copepoditos y nauplius
en las capas a) P1, b) P2 y e) P3, estación 11, Golfo Dulce.
126

a)
4000 .+--~--------
OOithona
• Paracafanus
D Oncaea
Q Corycaeus ..,3000 '-----' 1----------- - --
~ :e e 2000 -+------ · r--
1000 -i-1
c)1000 ~-----------------
750 +--------------~--~
"'E > :0500 +-----------------c
250 +-----------------~
.JJ _fh Jh a .Jb... o+-- -. 1 • 1 - , •
Ab Ag Nov Mar Fecha
Ab Set
Figura 2.28. Variación temporal en la abundancia (indiv/m3) de los principales géneros de
copépodos en las capas a) P1, b) P2 y e) P3 en la estación 01, Golfo Dulce.
127

a)
4000
3000 "'e > '5 .: 2000
1000
o
b)
1000
"'e > '5 r::
500
()
e) 1000
750 '
500
250
o
t
Ab Ag Nov Mar
OOithona • Paracalanus O Oncaea lll Corycaeus
Ab Set
...,
Figura 2.29. Variación temporal en la abundancia (indiv/m3) de los principales géneros de
copépodos en las capas a) P1, b) P2 y e) P3 en la estación 02, Golfo Dulce.
128

a) 1
4000 _i ---------
2000
1000
o+
b)
500
o u e) 1000
750
500
Ab Ag Nov Mar
O Oithona • Paracalanus Cl Oncaea ra Corycaeus
Ab Set
Figura 2.30. Variación temporal en la abundancia (indiv/m3) de los principales géneros de
copépodos en las capas a) P1, b) P2 y e) P3 en la estación 03, Golfo Dulce.
129

a} D Oithona
4000 • Paracalanus EJ Oncaea
li!!I Corycaeus 3000
2000 1
1000
o
b}
1000
500 +--------1
o
e} 2000
1750
1500
1250
1000
750
500
250
o Ab Ag Nov Mar Ab Set
Figura 2.31 . Variación temporal en la abundancia (indiv/m3) de los principales géneros de
copépodos en las capas a) P1, b} P2 y e) P3 en la estación 04, Golfo Dulce.
130

a) D Oithona 4000 • Paracalanus
D Oncaea 21 Corycaeus
3000
2000
1000
o ___ Jl:b b)
1000
1 1
e) 1000
750
500
-1
h ·- ¡, -
,.,
mil 1·0 · ~
250
o Ab Ag Nov Mar Ab Set
Figura 2.32. Variación temporal en la abundancia (indiv/m3) de los principales géneros de
copépodos en las capas a) P1, b) P2 y e) P3 en la estación 11, Golfo Dulce.
131

132
de la estación 02 (Figura 2.29a), se observó un claro dominio de Oíthona durante todo el
período de estudio. No se cuenta, sin embargo, con datos para el mes de abril de 1997,
cuando Paracalanus dominó en todas las demás estaciones. Resalta, también, el
incremento que se observó en la abundancia de los cuatro géneros durante el mes de abril
de 1998.
La estación 03 (Figura 2.30a) mostró un comportamiento similar a lo que ocurre en
las estaciones externas. La primera fecha mostró a Paracalanus como el género dominante,
seguido de Oíthona, Oncaea y Corycaeus, estos dos últimos con abundancias bastante altas.
La abundancia de todos los géneros, sin embargo, disminuyó para agosto. Paraca/anus,
Oncaea y Corycaeus no volvieron a mostrar aumentos en su abundancia en el resto del
período de estudio. Oíthona, por su parte, mostró dos picos importantes en los meses de
noviembre y abril de 1998.
En la estación 04 (Figura 2.31 a), por su parte, llama la atención el hecho de que
Oíthona fue el género menos abundante en la primera fecha. Además, Oncaea y Corycaeus
disminuyeron en número, mostrando valores bajos por el resto de las fechas. Oíthona
dominó la comunidad y, junto con Paracalanus, experimentó picos en los meses de
noviembre de 1997 y abril de 1998. Por último, la estación 11 (Figura 2.32a) mostró, como la
estación anterior, altas abundancias en la primera fecha. Oíthona mostró un muy importante
incremento en su abundancia en noviembre, acompañado por un incremento de menor
proporción de Paracalanus. Para el final del período de estudio, todos los géneros mostraron
abundancias relativamente bajas.
Capa Intermedia (P2)
En la capa intermedia (P2), como se mencionó antes, y al igual que en la capa
superficial, los copépodos fueron el grupo dominante, como se puede observar en las figuras
2.23b -2.27b. En el caso de las estaciones 01 y 02 (Figuras 2.23b y 2.24b), se aprecia que
las abundancias de copépodos, copepoditos y nauplius en las primeras cuatro fechas fueron
bajas y que se dio un muy importante incremento en los valores de abundancia de los tres
grupos en el mes de abril de 1998, cuando se da un aumento en la biomasa en esta capa.
La estación 03 (Figura 2.25b) solo mostró altas abundancias de los tres grupos en el mes de
abril de 1997. Estos valores disminuyeron considerablemente en la siguiente fecha y se

133
mantuvieron así a lo largo del período de estudio, excepto por un pequeño incremento en la
abundancia de copépodos que se observó en el mes de marzo. Finalmente, las estaciones
04 y 11 (Figuras 2.26b y 2.27b) mostraron un incremento en las abundancias de los tres
grupos a partir de marzo de 1998 para alcanzar valores máximos en abril del mismo año.
Al observar las abundancias de los géneros Oithona, Paracalanus, Oncaea y
Corycaeus en la estación 01 (Figura 2.28b), destaca que el primero dominó durante todo el
período de estudio, seguido por Oncaea, que como se mencionó antes, mostró mayores
abundancias relativas a esta profundidad. Resulta interesante, también el aumento en los
valores de abundancia de Oithona, Paracalanus y Oncaea que se observó en el mes de abril
de 1998. La estación 02 (Figura 2.29b) mostró un patrón similar, con bajos valores de
abundancia hasta marzo de 1998. En abril de 1998, se dió un incremento en la abundancia
de Oncaea, que pasó a dominar la comunidad sobre Oithona y Paracalanus. Sin embargo,
la abundancia de Oncaea cayó en la última fecha, mientras que la de Oithona y Paracalanus
aumentó.
La estación 03 (Figura 2.30b), al contrario de las estaciones internas, mostró altos
valores de abundancia en los cuatro géneros durante la primera fecha. En este caso, la
comunidad se vio dominada por Paracalanus, seguida de Oithona y Oncaea. Sin embargo,
todas las abundancias disminuyeron en agosto. Oncaea y Oithona mostraron un incremento
en su concentración en marzo (mientras Paracalanus tiene valores mínimos), pero sus
valores bajaron en las últimas fechas.
La estación 04 (Figura 2.31b) fue dominada en agosto por Oncaea y Paracalanus, sin
embargo sus abundancias cayeron y pasó a ser dominada por Oíthona a partir de noviembre.
Los tres géneros tienen un importante incremento en el mes de abril de 1998. Luego de
esto, las concentraciones de Oithona y Paracalanus descendieron mientras que la de
Oncaea continuó subiendo de fecha a fecha. La estación 11 (Figura 2.32b ), mostró un
patrón muy similar a la estación 04, con un dominio de Oithona y dos importantes aumentos
en noviembre de 1997 y abril de 1998, cuando se observaron incrementos en la abundancia
del zoolancton. Al igual que en la estación anterior, la concentración de Oncaea mostró una
curva ascendente hacia el final del período de estudio, mientras Ja de Oíthona y Paracalanus
disminuyó.

134
Capa Profunda (P3)
En este caso, como se mencionó con anterioridad, las abundancias de copépodos,
copepoditos y nauplius fueron considerablemente más bajas que en las capas superiores.
En la estación 01 (Figura 2.23c), los copepoditos y nauplius presentaron valores muy bajos
durante todo el período de estudio, pero con una tendencia al incremento en las fechas de
1998. Los copépodos, por su parte, presentaron un incremento moderado que se dio a partir
de marzo de 1998 y incrementó al siguiente mes (al igual que en P1 y P2). La estación 02
(Figura 2.24c), presentó un comportamiento hasta ahora no observado en las demás
estaciones, debido a que se observó un aumento en la abundancia de copépodos y
copepoditos en el mes de noviembre: A la fecha siguiente (marzo de 1998), sin embargo, la
abundancia de copépodos cayó drásticamente, mientras que la de copepoditos mostró un
aumento, superando incluso a los copépodos adultos. En esta estación la abundancia de
nauplius resultó muy baja a lo largo de todo el año.
La estación 03, por su parte (Figura 2.25c) mostró el mismo patrón que en la capa
P2, donde se dieron altas abundancias de los tres grupos en la primera fecha. En la estación
04 (Figura 2.26c) se observó una alta concentración de copepoditos en la primera fecha para
descender en las siguientes dos fechas de muestreo. El mismo decrecimiento se observó en
el caso de los copépodos. Destaca que, al igual que en P2, se dio un pico en las
abundancias de los tres grupos durante abril de 1998. Finalmente, la estación 11 (Figura
2.27c), presentó muy bajas abundancias en el mes de agosto así como un aumento en
copépodos y copepoditos en el mes de noviembre de 1997, cuando se da un importante pico
en la biomasa del zooplancton. Marzo de 1998 representó un decrecimiento en las
abundancias de estos dos grupos para incrementar a partir del siguiente mes (abril 1998).
En la capa profunda (P3), la estación 01 presentó abundancias de copépodos
relativamente bajas en sus principales géneros (Figura 2.28c). En las primeras dos fechas
Oncaea fue el grupo dominante. Sin embargo, la concentración de Oithona aumentó en las
restantes fechas, llegando a dominar sobre los demás géneros, mientras que Oncaea se
mantuvo en un segundo lugar de abundancia. En general, Oithona, Paracalanus y Oncaea
mostraron un incremento progresivo en sus abundancias a partir del mes de noviembre de
1997 hasta el mes de abril de 1998. La estación 02 (Figura 2.29c), presentó bajas
abundancias en todos sus géneros y se vio dominada claramente por Oncaea. Es

135
únicamente en este último género donde se notaron dos pequeños incrementos en
noviembre de 1997 y abril de 1998.
La estación 03 (Figura 2.30c), al igual que en sus capas superiores, mostró altos
valores de abundancia en los cuatro principales géneros de copépodos durante el mes de
abril de 1997. En esta primera fecha, la comunidad fue dominada por Oncaea, Paracalanus
y Oithona, en ese orden. Las siguientes cuatro fechas, se caracterizaron por
concentraciones muy bajas en todos los grupos y no se dieron fluctuaciones importantes, a
excepción de un pequeño incremento en Oncaea en marzo de 1998. En la última fecha, sin
embargo, se da un aumento en los tres principales géneros y la comunidad pasó a ser
dominada por Oithona.
Al igual que la estación 03, la 04 (Figura 2.31c) mostró altas abundancias en los
principales géneros de copépodos durante la primera fecha y la comunidad se vio dominada
por Paracalanus. Los valores de abundancia bajaron drásticamente para la siguiente fecha y
la comunidad pasó a ser dominada por Oncaea en noviembre y marzo. En el mes de abril de
1998, Oithona y Oncaea presentaron un aumento en la abundancia, mientras que
Paracalanus y Corycaeus mostraron valores bajos por el resto del período de estudio.
Finalmente, la estación 11 (Figura 2.32c) fue claramente dominada por Oithona y Oncaea
durante todo el período de estudio, dándose incrementos en la abundancia en noviembre de
1997 y abril - setiembre de 1998.
Los patrones anteriormente descritos sobre la abundancia de los distintos grupos en
cada una de las capas muestreadas en la columna de agua sirven como base para explicar
los análisis de conglomerado que se observan en la figura 2.33. En las tres capas de
muestreo, las estaciones internas (01 y02) se separaron claramente del resto y mostraron
mayor similitud entre sí (en cuanto a su composición y abundancia) en la capa profunda. Las
demás estaciones se agrupan juntas pero su grado de similitud varió en cada capa. Así, por
ejemplo, las estaciones 04 y 03 se mostraron más próximas en las capas superficial y
profunda. Mientras tanto, en la capa intermedia la estación 04 parece estar más relacionada
a la estación 11 y la estación 03 se mostró más separada de éstas que en P1 y P3.

136
a) o 2
o 1
o 4
o 3
1 1
O ,O o '1 o ,2 o ,3 o ,4 o ,5 Distancia
b) o 1
o 2
o 4
o 3
O ,O o '1 o ,2 o ,3 o ,4 D is ta n c ia
e) o 3
~ o 4
-o 1
o 2 J
O ,O o ' 1 o ,2 o ' 3 o ,4 o '5 o ,6 o ' 7 Distancia
Figura 2.33. Análisis de conglomerado para las estaciones muestreadas respecto a la
abundancia de géneros de copépodos en las capas a) superficial (P1 ), b)
intermedia (P2) y e) profunda (P3), aplicando el coeficiente de similitud Bray
Curtis y uniones de promedio por grupo. Notar las diferentes escalas en cada
caso.

137
Finalmente, se determinó la existencia de una relación lineal leve entre la abundancia
de copépodos en la capa superficial (P1) y la biomasa del zooplancton en la capa P1 (Figura
2.34 ). En éste caso, se observó un aumento en la biomasa a medida que aumenta la
abundancia de copépodos. Al igual que en el caso de la relación entre la abundancia de
zooplancton y la biomasa, no se espera que esta relación sea muy fuerte debido a que los
copépodos, a pesar de ser dominantes, solo representan una fracción del zooplancton y, al
presentar gran variedad de tamaños, no guardan una relación más fuerte ni proporcional con
los cambios en biomasa. Por último, no fue posible detectar ninguna relación significativa (p
> 0.05) entre la abundancia del zooplancton y las otras variables físicas, como profundidad
del disco de Sechii, profundidad de la capa de mezcla, oxígeno disuelto, temperatura y
salinidad.
filX)
~?OCO "' _§ filX) > '6 :§, 9:m
ui 1
.á} 400) ' o.. 8 3Xl)
-a •
•
§ :mJ •• .o •
••• •
•
y=58.!nlx +2A01.6 R =Q2118, p=O.CY.21
<( 1cro •
º L~ ~---,....--------.. o 10 3)
Bcrnasa (rrg'm'}
40
Figura 2.34. Regresión lineal entre la abundancia de copépodos (indiv/m3) y la biomasa
general del zooplancton (mg/m3).

138
DISCUSION
El ambiente marino no sólo es muy variable en espacio y tiempo sino que es
dominado por gradientes verticales que se dan en los parámetros fisico-químicas
(temperatura, nutrimentos, presión hidrostática) y biológicos (producción primaria, presión de
depredación y disponibilidad de alimento) (Harris 1987). Estos gradientes ocasionan
distribuciones verticales específicas de fitoplancton y bacterioplancton que, a su vez,
representan gradientes en términos de cantidad y calidad de alimento, para el zooplancton.
Así, por ejemplo, se han reportado altas concentraciones de zooplancton asociadas con la
placa bacteriana en la oxiclina en hábitats estratificados (De Meester & Vyverman 1997).
El Golfo Dulce presenta un comportamiento distinto de la mayoría de los ecosistemas
costeros tropicales y está dominado por el flujo de energía y biomasa dentro del grupo
pelágico, con lo que se asemeja más a un sistema oceánico abierto que a uno estuarino
(Wolff et al. 1996). Las características y dinámicas del plancton en la zona del Golfo Dulce
son en gran parte, desconocidas. Estudios previos parecen mostrar una fuerte estratificación
de la actividad biológica junto con una estratificación física y una limitada zona fótica
(Richards et al. 1971; Córdoba & Vargas 1996; Thamdrup et al. 1996; von Wangelin & Wolff
1996; Wolff et al. 1996).
En primer lugar, el zooplancton en la cuenca del Golfo Dulce presentó
concentraciones significativamente mayores en la capa superficial, lo cual coincide además,
con la distribución de biomasa de zooplancton determinada en el mismo estudio (capítulo 1 ).
Este mismo patrón de distribución de zooplancton ha sido reportado en otras zonas del
mundo por autores como Nishida & Marumo (1982), Longhurst (1985), Bottger-Schnack
(1988, 1990, 1994, 1995), Madhupratap et al. (1990) y Saltzman & Wishner (1997a y b).
Tanto Ja distribución de biomasa como la distribución vertical del zooplancton, concentradas
en la capa superficial, se justifican desde el punto de vista de los fuertes gradientes verticales
en las variables físicas que presentan las aguas de la zona de estudio (Richards et al. 1971;
Córdoba & Vargas 1996; Thamdrup et al. 1996; von Wangelin & Wolff 1996; Wolff et al.
1996), los cuales, a su vez, se deben a la morfología de la cuenca del Golfo Dulce. Como se
mencionó en el capítulo 1, la capa superficial se distingue por una relativamente alta
temperatura y concentración de oxígeno disuelto, lo cual favorece la alta concentración y
biomasa de zooplancton observadas (Raymont 1983; Davis 1984; White et al. 1995; Wiafe &

139
Frid 1996). Al respecto, Madhupratap et al. (1990), por ejemplo, en su estudio en el Indo
Pacífico, encontraron que el zooplancton se concentra en la capa sobre la termoclina,
presentando biomasas y densidades hasta cinco veces superiores a las encontradas por
debajo de la termoclina. Longhurst (1985), además, en su estudio en el Pacífico Tropical
Oriental, determinó la existencia de una fuerte planctoclina, separando el epizooplancton en
las capas superiores, del mesozooplancton en las capas profundas. Lo anterior es un
aspecto esperado en sitios que presentan una fuerte termoclina.
Con respecto a la productividad primaria, solamente existe un conjunto de datos
tomado hace más de 30 años. Los resultados obtenidos, de < 720 mgCm2d-1, son
relativamente bajos si se comparan con los valores de productividad del Golfo de Nicoya
(690-2530 mgCm2d-1). Basándose en lo anterior, las aguas del Golfo Dulce se pueden
catalogar como moderadamente productivas (Hartmann 1994 ). Este es un factor que
favorece la concentración del zooplancton en la capa superficial (P1) del Golfo Dulce, ya que
se ha observado una concentración del fitoplancton en la superficie y hasta una profundidad
de 40 m (Kuever et al.1996). Por debajo de esta profundidad el fitoplancton es escaso y en
su mayor parte corresponde a células muertas. Otras formas de detritus vegetal, con
estructuras celulares de origen terrestre (manglares y palmeras) constituyen una fracción
significativa de la materia orgánica particulada en la parte superior del sedimento, pero es
raro en la columna de agua (Kuever et al.1996).
En relación con lo anterior, la abundancia de zooplancton en el Golfo Dulce mostró
valores que se pueden considerar entre bajos y moderados al compararlos con los valores
reportados para otros sistemas (Cuadro 2.8). Resulta importante mencionar, sin embargo,
que los valores de biomasa de zooplancton encontrados en el Golfo Dulce son bastante
bajos al compararlos con otros sistemas, incluido el Golfo de Nicoya (Cuadro 1.2), por lo que
se le puede considerar como un sistema oligo-mesotrófico. Así mismo, la biomasa del
zooplancton en el Golfo Dulce está dominada por la fracción de tamaño pequeña (500 µm >
x > 150 µm) y al observar la composición del zooplancton, destaca el fuerte dominio de los
copépodos (adultos, copepoditos y nauplius), dominados a su vez por grupos pequeños,
como Oithona, Paracalanus y Oncaea. La composición, distribución y abundancia del grupo
de los copépodos serán abordadas con mayor detalle más adelante.
También de manera coincidente con la distribución de biomasa, se determinó que no
existen diferencias significativas en la abundancia del zooplancton al comparar las capas P2

140
Cuadro 2.8. Valores de abundancia de zooplancton (indiv/m3) reportados para diversas
regiones del planeta.
Autor
Zooplancton
Bottger-Schnack ( 1994)
Bottger-Schnack (1995)
Razouls et al. ( 1994)
Brugnoli-Olivera
Quesada-Alpízar (2001)
Copépodos
Brugnoli-Olivera
Hopkins ( 1977)*
Hopcroft et al. {1998a)
Géneros
Uye & Sano (1995)
Carlsson et al. (1995)
Laundry et al. ( 1985)
*Tomado de Raymont (1983)
Sitio de Estudio
Mar Arábigo
Mar Rojo
Golfo Aden
Indico (oceánico)
Indico (costero)
Golfo de Nicoya, CRC
Golfo Dulce (superficie)
Golfo de Nicoya, CRC
Florida
Jamaica
Japón
Mar Barents
Océano Pacífico
Japón
Columbia Británica
Abundancia (indiv/m3)
6000-9000
2000
1 X 1 04 - 1 X 1 05
0.5 X 105
2 X 105
6636 (estac. seca)
9822 (estac. transición)
9941 (estac. lluviosa)
7979.46 ± 5845.25
2192 (estac. seca)
6858 (estac. transición)
6564 (estac. lluviosa)
93 100 -108 600
5000
5.98 x 105 Oithona sp.
500 - 2000 Oithona sp.
O - 40 000 Oithona sp.
1 x 106 Oithona sp.
40 000 Corycaeus sp.

141
y P3. Este aspecto se ajusta a lo esperado, puesto que, el Golfo Dulce presenta una
estructura de corrientes de tres capas en la parte externa y una circulación estuarina de dos
capas en la parte interna (Svendsen et al. 2001 ). Además de dichos patrones de circulación,
al observar las características físicas de la columna de agua en el Golfo Dulce, se distinguen
dos capas de agua principales, una superficial (O - 50/60m), cálida con salinidades bajas y
altas concentraciones de oxígeno y una profunda, fría, con salinidades altas y poco oxígeno
disuelto (Richards et al. 1971; Córdoba & Vargas 1996; Thamdrup et al. 1996). Estas
características de la columna de agua en el Golfo Dulce son las que, en conjunto, definen la
distribución de la biomasa y de los organismos, con altos valores en la capa superficial y
valores relativamente bajos por debajo de esta.
Otro patrón interesante es que las estaciones externas mostraron mayores valores de
abundancia de zooplancton, en todas las capas, con respecto a las estaciones internas (01 y
02). Lo anterior, nuevamente, coincide con las distribuciones de biomasa del zooplancton
(capítulo 1) y con lo reportado por Wolff et al. (1996) y Aksnes & Magnesen (1983). Como se
indicó en el capítulo 1, las partes interna y externa de la cuenca se diferencian tanto biológica
(en términos de biomasa) como físicamente. Esto se puede explicar sobre la base de que
las estaciones externas parecen ser afectadas con mayor frecuencia por pulsaciones de
agua oceánica profunda y rica en nutrimentos (Richards et al. 1971; Thamdrup et al. 1996;
von Wangelin & Wolff 1996), una mayor incidencia de viento (obs. pers.), lo cual significa una
mayor mezcla vertical en la columna de agua, y la cercanía del principal río de la cuenca del
Golfo Dulce, el Coto-Colorado, que representa también una entrada importante de
nutrimentos al sistema (Benovic et al. 1994; Umaña 1998). Es la combinación de estos
factores lo que favorece que existan altas concentraciones de oxígeno en el fondo (P3)
(Figuras 1. 7 y 1.8). Esto viene a favorecer, por ejemplo, las (relativamente) altas
concentraciones de zooplancton que se dan en la P3 de la parte externa del Golfo, en
contraposición con las estaciones internas, que presentan bajas concentraciones de oxígeno
y zooplancton en la P3.
Sin duda, el ingreso de materia orgánica ha aumentado en los últimos años, debido
principalmente a la deforestación y la expansión de la agricultura en partes de la cuenca del
golfo. Prueba de ello es la gran cantidad de materia vegetal que se observa flotando en las
aguas del Golfo Dulce, especialmente luego de períodos de fuertes lluvias (obs.pers). A
pesar de esto, la profundidad de la oxiclina no parece haber cambiado dramáticamente en

142
los últimos años, probablemente debido a los períodos de renovación de las aguas profundas
(Richards et al. 1971; Hartmann 1994; Córdoba & Vargas 1996; Thamdrup et al. 1996).
La entrada de nutrimentos de origen antropogénico es, también, un posible factor
responsable de que la estación 03, en la parte media de la cuenca, sea la que presente las
mayores concentraciones de zooplancton en las capas P1 y P2, así como la menor
visibilidad del agua (Figura 1.15). Como se aprecia en la figura 2.1, la estación 03 se ubica
frente a la entrada del principal puerto y centro de población del Golfo Dulce: Golfito. Esto
favorece que la estación 03 esté más expuesta a la entrada de nutrimentos debido a la
descarga de aguas residuales y materia orgánica asociadas a poblaciones humanas (León
Morales & Vargas 1998; Sponberg & Davis 1998; Acuña en prep.). De hecho, los
nutrimentos son una de las variables más importantes que controlan las comunidades de
fitoplancton (Tilman et al. 1982) y por lo tanto, de zooplancton.
Además, las estaciones 03 y 04 están expuestas a ser afectadas por la entrada de
agua dulce, dada su cercanía a la desembocadura del río Coto-Colorado. Se ha
comprobado que la entrada de ríos puede acarrear grandes cantidades de nutrimentos a un
sistema marino (Benovic et al. 1994; Umaña 1998). Así, por ejemplo, una gran parte del
nitrógeno orgánico disuelto en las áreas costeras se da en la forma de sustancias húmicas.
Estas se desprenden de los sólidos y son transportadas por ríos a las aguas costeras. La
cantidad de sustancias húmicas que entran a áreas costeras podría ser significativa en
regiones donde las cuencas hidrográficas están dominadas por áreas boscosas, como el
Golfo Dulce. De hecho, las sustancias húmicas constituyen la mayor fracción de la materia
orgánica disuelta (MOD) en la mayoría de los cuerpos de agua (Carlsson et al. 1995).
Carlsson et al. (1995) también reportan que la presencia de sustancias húmicas
representa un estímulo pequeño pero significativo en el crecimiento del fitoplancton. De este
modo, la entrada de nutrimentos a la capa superficial favorece cambios en la dominancia de
productores primarios, herbívoros y carnívoros de la comunidad plánctica (Bougis 1976;
Ortner et al. 1978; Dessier & Donguy 1985; Smetacek & Pollehne 1986; Benovic et al. 1994;
White et al. 1995; Gasol et al. 1997). Es precisamente la combinación de estos factores lo
que podría favorecer la presencia de una alta concentración de organismos así como la
mayor turbidez del agua en la estación 03. El efecto de la entrada de agua dulce sobre la
composición del zooplancton en las estaciones 03 y 04 será tratado más adelante.

143
Otro aspecto muy importante es la variación temporal observada en los valores de
abundancia del zooplancton. De acuerdo a los resultados obtenidos, los promedios más
altos en la abundancia del zooplancton en todas las profundidades se presentan en abril de
1997 (Fig. 2.4). Luego de esta fecha, la concentración de zooplancton en todas las capas
muestreadas disminuye significativamente y se mantiene de esa manera hasta el final del
período de estudio. Como se mencionó en el capítulo anterior, el mes de abril de 1997 es el
que presenta las menores temperaturas en la columna de agua en todo el período de estudio
y coincide con las fechas previas a la llegada del fenómeno El Niño (ENSO) al Pacífico
Ecuatorial Tropical (NOAA 2000). Además, el mes de abril coincide con las fechas
esperadas, para que se dé la entrada de agua oceánica a la cuenca del Golfo Dulce, en
sincronía con el fenómeno de afloramiento y los máximos de clorofila que se observan en las
costas de Panamá en esta época del año (Dessier & Donguy 1985).
Estas entradas de agua oceánica, rica en nutrimentos, a la cuenca del Golfo Dulce
(Richards et al. 1971; Thamdrup et al. 1996; Wolff et al. 1996; León-Morales & Vargas 1998;
Acuña en prep.) vendrían, como se mencionó antes, a estimular la productividad primaria y
por lo tanto, a afectar de manera directa e indirecta la abundancia, composición y estructura
de la comunidad de zooplancton (Lampert & Schober 1980; Allison & Wishner 1986; Arcos &
Fleminger 1986; Smetacek & Pollehne 1986; Harris 1987; Reynolds 1989; Wiafe & Frid
1996). Con base a lo anterior, se justifica que en la primera fecha del presente estudio (abril
1997) se hayan dado valores altos en la abundancia de zooplancton, ya que las
características físicas de la columna de agua correspondientes a esta fecha indican que se
pudo haber dado una entrada de agua oceánica.
De la misma manera, se logra explicar la relación inversa encontrada entre la
abundancia del zooplancton y la profundidad de la capa de mezcla, ya que los mayores
valores de abundancia de zooplancton coinciden con las fechas de entrada de agua
oceánica al Golfo Dulce (abril de 1997-98 y setiembre 1998) (termoclina sube) y por lo tanto,
con los picos de biomasa observados. Esto implica que los altos valores de abundancia y
biomasa, por lo general, coinciden con épocas en que la termoclina se encontraba a menor
profundidad debido a la entrada de masas de agua oceánica. Lo anterior, a su vez, vendría
a justificar la relación positiva detectada entre las variables de biomasa y abundancia de
zooplancton.

144
Mientras que hasta hace algunos años se consideraba a la termoclina como una
barrera física y ecológica (Angel 1985, Bainbridge 1972 en Longhurst 1985), Longhurst
( 1985) encontró más bien una alta concentración de especies en esta zona. Esto se debe a
que la termoclina representa un gradiente de luz, temperatura, nutrimentos y densidad. La
alta diversidad observada en la termoclina se debe a que la picnoclina favorece una
estructura de multicapas con gradientes ambientales y discontinuidades que representa una
serie de microhabitats que son ocupados por distintas especies (Longhurst 1985). En el
Golfo Dulce, las constantes fluctuaciones que se dan en la abundancia de los distintos
grupos de zooplancton y copépodos se podrían deber a los fenómenos de entrada de agua y
el consiguiente movimiento de la termoclina, lo que provoca que no se den condiciones
estables en la columna de agua durante largos períodos de tiempo.
Las fluctuaciones físicas que se dan en la columna de agua debido a la entrada de
masas de agua oceánica, tienen un considerable efecto sobre la comunidad de zooplancton.
Lo anterior se refleja con mayor claridad al observar que la abundancia de zooplancton en la
P3 tiene un coeficiente de variación muy alto (CV = 158.6%). Como se menciona con
anterioridad, la entrada periódica de una masa de agua rica en nutrimentos y oxígeno, que
renueva las aguas profundas de la cuenca, viene a estimular incrementos en la abundancia
del zooplancton en toda la columna de agua, que más tarde caen de nuevo al disminuir los
niveles de oxígeno en la capa profunda (Vidal 1980; Raymont 1983; Smetacek 1988; Roman
et al. 1993 Saltzman & Wishner 1997 a y b ).
Al respecto, Madhupratap et al. (1990) encontraron que la baja abundancia de
zooplancton por debajo de la termoclina en el Indo-Pacífico se debe, al menos en parte, a la
baja concentración de oxígeno disuelto en dichas aguas. Las zonas con mínimos de oxígeno
existen en todos los océanos del mundo y son el resultado de la degradación de los
productores primarios, el alto consumo de oxígeno por parte del zooplancton y bacterias, así
como de una pobre mezcla vertical por debajo de la termoclina. En estas regiones, la
distribución del zooplancton sigue el patrón de distribución del oxígeno y mostró un descenso
general de la biomasa con la profundidad (Richards 1965; Thurman 1996; Saltzman &
Wishner 1997a). Asociado a esto, Saltzman & Wishner (1997a) encontraron que más del
50% de la biomasa y del 80% de la abundancia de zooplancton se distribuyen en los 100 m
superiores del Pacífico Oriental Tropical (POT). Otro aspecto que demuestra la importancia
del oxígeno disuelto para la distribución del zooplancton es lo encontrado por Roman et al.

145
(1993), quienes observaron que al darse un aumento en la concentración de oxígeno en las
aguas del fondo de la Bahía de Chesapeake se dio un incremento en el número de
copépodos, mientras que en las épocas en las que el agua profunda era anóxica, cesó el
comportamiento de migración vertical en algunos grupos.
La caída que se da en los valores de abundancia en las fechas siguientes a abril de
1997 se debe probablemente al efecto del fenómeno El Niño. Este no solo reduce la
intensidad de los fenómenos de afloramiento costero en el Pacífico Ecuatorial (Dietrich 1963;
Philander 1996; Thurman 1996; NOAA 2000), sino que provoca un hundimiento en la
termoclina (Figuras 1.4 - 1.8) e, indirectamente, un agotamiento de los nutrimentos en la
capa de mezcla, con lo que se da uña disminución en la productividad primaria y por lo tanto,
en la abundancia y biomasa del zooplancton (Bougis 1976; Margalef 1985; Longhurst &
Pauly 1987; Fielder et al. 1991; White et al. 1995). Además, cabe mencionar que los dos
eventos más intensos de El Niño en más de 100 años se dieron en las pasadas dos
décadas, en los años 1982 y 1997. En ambos casos el fenómeno ENSO fue de
relativamente corta duración, prolongándose por alrededor de un año (Federov & Philander
2000). En el caso de El Niño 1982-83, Rojas de Mendiola et al. (1985) determinaron que
produjo un desequilibrio de los diferentes componentes del ecosistema en la costa peruana,
afectando en primera instancia al fitoplancton. Además, se observó que los cambios en la
abundancia y distribución y composición de las comunidades de fitoplancton guardan una
relación directa con la intensidad del fenómeno. Estos cambios en el fitoplancton por efecto
de El Niño afectan necesariamente a los niveles tróficos superiores. Al respecto, Morales &
Brugnoli (en prensa) encontraron diferencias en la estructura del fitoplancton en la zona de
Punta Morales ( 1997), que se vieron reflejadas en los valores de biomasa del zooplancton y
que fueron atribuidas al fenómeno El Niño.
El fenómeno ENSO, además, provocó una disminución en la precipitación observada
en la zona del Golfo Dulce durante 1997 (Figura 1.3b). La consecuencia directa de las bajas
precipitaciones es, por supuesto, una disminución en el caudal de los ríos y en la entrada de
nutrimentos a la cuenca del Golfo Dulce. Debido a lo anterior, resulta razonable que se
observara una disminución en la biomasa y la abundancia del zooplancton en el área de
estudio una vez entrado el fenómeno El Niño, aproximadamente en el mes de mayo de 1997
(NOAA 2000).

146
Composición del Zooplancton
Al igual que la mayoría de los sistemas marinos del planeta (Raymont 1983;
Longhurst 1985; Longhurst & Pauly 1987; Braun et al 1990; Madhupratap et al. 1990;
Bottger-Schnak 1995; Saltzman & Wishner 1997a, b) el Golfo Dulce es dominado por los
copépodos (71.4 7% ), tanto sobre como por debajo de la termoclina. Los siguientes dos
grupos en importancia son las apendicularias y ostrácodos, los cuales, junto con los
copépodos, representaron el 87. 7% de la abundancia total del zooplancton en el Golfo Dulce.
Lo anterior ratifica que el Golfo Dulce, a diferencia del Golfo de Nicoya (Brugnoli 1998; Wolff
et al. 1998; Morales, com. pers.) es un sistema dominado claramente por el holozooplancton
y donde el merozooplancton representa solamente una pequeña fracción de la abundancia
total. Además, la presente investigación detectó que la composición de la comunidad de
zooplancton en el Golfo Dulce varía con la profundidad.
Longhurst ( 1985), observó que los principales grupos que componen el zooplancton
en el Pacífico Oriental Tropical son los copépodos (65%), tunicados (7.9%), quetognatos
(6.5%), ostrácodos (4.3%) y eufausidos (3.4%). Además determinó que del total de 158
especies de distintos grupos, tan solo 17 de ellas dominaban un 75.4% de la abundancia
total en las muestras. En el caso del Golfo Dulce, los grupos dominantes en la P1 de todas
las estaciones son, invariablemente, copépodos, apendicularias y ostrácodos.
Destaca que en la estación 01, a diferencia de las demás estaciones, los cladóceros
son uno de los cinco principales grupos en la composición del zooplancton. Los cladóceros
son organismos típicos del plancton nerítico, ya que todos sus miembros tienen estadías de
reposo bénticos, por lo que su distribución está restringida a áreas poco profundas. A pesar
de su importancia como productores secundarios y su capacidad de alimentarse de
partículas muy pequeñas, tienden a ser extremadamente estacionales, alcanzando
abundancias muy altas durante períodos cortos y usualmente se dan durante las épocas
cálidas del año (Raymont 1983; Longhurst 1985; Smetacek 1988; Braun et al. 1990).
Especies como Penilia avirostris, por ejemplo, se reproducen rápidamente cuando las
condiciones son favorables (Smetacek 1988). Evadne y Podon, por otra parte, son dos
géneros de cladóceros raptoriales (selectivos) que, al igual que Penilia, producen huevos que
se mantienen en reposo durante condiciones desfavorables (Longhurst 1985; Smetacek
1988). La presencia de cladóceros en la estación 01 se podría estar viendo favorecida por
su circulación de carácter estuarino, ya que recibe el caudal de varios ríos cercanos, como el

147
Rincón y Esquinas y es una estación próxima a la costa y algunos parches de manglar, lo
que favorecería el ciclo reproductivo de este grupo.
Con respecto a las apendicularias, sus mayores abundancias siempre se encontraron
en la capa superficial de todas las estaciones. Algunos picos en la abundancia de este grupo
en la capa superficial coinciden con picos en la abundancia de copépodos (noviembre de
1997, abril de 1998), lo que indica que probablemente durante esas fechas se dieron
condiciones favorables en la cantidad y/o calidad del alimento que, a su vez favorecieron el
crecimiento de dichos grupos. La importancia de las apendicularias radica en que se les
considera entre los organismos más abundantes y omnipresentes dentro del
mesozooplancton no crustáceo. Su corto tiempo de generación y el hecho de tengan la
capacidad de alimentarse de partículas muy pequeñas favorece rápidos aumentos en sus
tamaños poblacionales (Acuña & Anadón 1992). Se ha observado, por ejemplo, que la
comunidad de apendicularias en el mar Cantábrico exhibe una sucesión anual en la
abundancia relativa de especies y que la estructura de su comunidad está acoplada a las
variaciones en temperatura. Las apendicularias tienen una baja regulación metabólica, por lo
que son muy sensibles a la temperatura del agua. De este modo, la abundancia de
apendicularias en cierta estación depende principalmente de la productividad primaria. Sin
embargo, los patrones de abundancia relativa dependen de la temperatura a la cual ocurre la
producción y que el tiempo de generación y la fecundidad en apendicularias están
inversamente relacionados a la temperatura (Taggart & Frank 1987; Acuña & Anadón 1992).
Todos estos aspectos fortalecen la presencia de apendicularias en el Golfo Dulce asociada a
la entrada de masas de agua oceánica.
Otro aspecto interesante se da en la P1 de la estación 03, donde los huevos de
invertebrados se constituyen en un componente importante. Esto se debe probablemente a
que dicha estación está cerca de la bahía de Golfito, la cuál se caracteriza por ser poco
profunda y con abundantes parches de manglar. Lo anterior favorece la presencia de gran
número de invertebrados bénticos que se están reproduciendo en la zona, por lo que es más
probable encontrar un mayor número de huevos en dicha zona. Además, las playas rocosas
en punta Gallardo y punta Mula, cercanas a la estación 03, podrían favorecer la presencia de
crustáceos y moluscos que contribuirían con las altas concentraciones de huevos
observadas en ésta estación (756 ± 1762 huevos/m3).

148
Al observar la capa intermedia (P2), destaca, al igual que en la capa superior, el
dominio del grupo de los copépodos. Sin embargo, es importante recalcar que, como se
esperaba, la abundancia de zooplancton (al igual que la biomasa) en esta capa es
significativamente menor que en la capa P1. Como se mencionó antes, los fuertes
gradientes verticales en las condiciones físicas del Golfo Dulce (Richards et al. 1971;
Córdoba & Vargas 1996; Thamdrup et aL 1996) provocan un fuerte gradiente vertical , tanto
en la biomasa (capítulo 1) como en la abundancia del zooplancton .
También destaca que la abundancia de ostrácodos en la capa intermedia sea
superior a 11a encontrada en la capa superficial , lo que coincide con lo reportado por
Longhurst ( 1985) respecto a que los ostrácodos son un grupo especializado en
profundidades subtermoclina. Para el caso de las apendicularias y nauplius, se observa un
patrón opuesto al antenior, con una mayor abundancia en la capa superficial que en llas
capas intermedia y profunda. Una vez más, dicha distribución coincide con lo reportado por
la literatura respecto a que la máxima abundancia de estos grupos se espera en la capa de
mezcla y está asociada , sobretodo, a altas concentraciones de fitoplancton (Esnal 1986;
Lagadeuc et al. 1997; Saltzman & Wishner 1997a).
Un aspecto que destaca en la capa profunda es que los huevos de invertebrado se
constituyen en un grupo importante en las estaciones 011, 02 y 03, las cuales son las tres
estaciones más profundas en el presente estudio (se muestrearon hasta los cien metros).
Por esta razón, resulta razonable suponer que la relativamente alta concentración de huevos
observada en dichas estaciones se debe a los procesos de liberación de huevos en las
capas superiores de la columna de agua y su consiguiente precipitación hacia mayores
profundidades donde, dados los bajos niveles de oxígeno , los huevos mueren . Lutz et al.
( 1992) y Ro man et al. ( 1993 ), por ejemplo, han determinado que la incubación de huevos de
Acartia tonsa, Labidocera sp . Oithona y Centropages es suprimida cuando la concentración
de oxígeno es menor a los 0.02 mi 0 2/I o al menos retardada cuando hay bajos niveles de
oxígeno disuelto. En las estaciones externas no solo hay buenos niveles de oxígeno en la
capa profunda, sino que la columna de agua no es tan profunda, por lo que no se esperaría
que precipitara tan alto número de huevos. Las bajas concentraciones de oxígeno también
justifican el hecho de que se hayan encontrado relativamente pocos nauplius en la capa
profunda de las estaciones O 1 y 02.

149
Otra característica interesante es que los ostrácodos continúan siendo un grupo
importante en la P3, sobretodo en las estaciones 03, 04 y 11. La razón de dicha distribución
recae nuevamente en la distribución del oxígeno disuelto en la columna de agua. Las
estaciones internas, a'I tener una capa P3 más profunda presentan menores concentraciones
de oxígeno (ver capítulo 1 ), por lo que la distribución de los organismos en esta capa se ve
seriamente limitada. En el mar Arábigo por ejemplo, se ha determinado que la deficiencia de
oxígeno en sus aguas profundas limita drásticamente la distribución del zooplancton
(Bóttger-Schnack 1994 ). Las estaciones externas, por su parte, presentan niveles más altos
de oxígeno disuelto, mientras que la 03, por ser una estación de carácter intermedio,
presenta niveles más altos de oxígeno que lo observado en las estaciones 1internas. Otro
argumento importante sobre el mayor número de ostrácodos en las estaciones media y
externas se debe a que son estaciones más productivas, por lo que además de más
organismos, se encontrará una mayor cantidad de detritus que representan una potencial
fuente de alimento a un grupo como los ostrácodos.
Los anteriores no son los únicos aspectos respecto a la abundancia y composición
del zooplancton que coinciden con la distribución de oxígeno disuelto y con las fechas de
entrada de agua oceánica en el Golfo Dulce. Así, por ejemplo, la concentración de
copépodos en todas las capas de 1las estaciones internas aumenta en el mes de agosto de
1997, coincidiendo con mayores valores de oxígeno disuelto para esa capa, así como en
abril de 1998, coincidiendo con las fechas de entrada de agua oceánica. De hecho, en la
capa intermedia de todas las estaciones se presentan incrementos en la abundancia, en
especial de copépodos, que coinciden en abril de 1998 y las estaciones 03, 04 y 11
presentan importantes incrementos en la abundancia de distintos grupos en abril de 1997.
El que las estaciones internas no presenten altos valores en la abundancia de
organismos en fa primera fecha de muestreo se puede deber a varias razones: en primer
lugar, la parte interna del Golfo presenta un patrón de circulación de sus aguas distinto al que
se presenta en las estaciones externas (Svendsen et al. 2001 ). Otro posible factor se
relaciona con el hecho de que la entrada de agua oceánica al Golfo Dul'ce es un fenómeno
gradual. Por esta razón, existe la posibilidad de que el poco desarrollo mostrado por la
comunidad de zooplancton en abril de 1997 en la parte interna de la cuenca se deba a que la
masa de agua oceánica tenía un menor tiempo de residencia en la parte interna que en la
parte externa de la cuenca, la cual pudo responder más temprano a dicho fenómeno.

150
Resulta muy interesante observar que los principales incrementos en la abundancia de
zooplancton observados, se deben principalmente a grupos como copépodos y
apendicularias, los cuales, como se mencionó con anterioridad, responden rápidamente a
incrementos en l'a productividad primaria provocados por fenómenos de afloramiento e
intrusión de nutrimentos en general.
Las asociaciones anter1iores se reflejan claramente en los análisis de conglomerado
respectivos (Figura 2.14), donde se puede observar que las estaciones internas se agrupan
en las capas P1 y P3 de muestreo. En estas capas se puede observar que la estación 04 se
separa de las demás, lo que se debe probablemente a la influencia que ejerce sobre la
comunidad de zooplancton el río Coto-Co,lorado, el más importante en la cuenca del Golfo
Dulce. En la capa P2 destaca principalmente el que la única estación que se separa
claramente es la estación 01 y que las estaciones más similares son las externas (04 y 11 ),
las cuales además, presentan una columna de agua con características físicas muy
similares. La capa P2 presenta mayor variación en la comunidad de zooplancton que P1
debido a que es la capa que comprende 'la termoclina, donde se dan fuertes gradientes en
temperatura, densidad, concentración de oxígeno y de nutrimentos, entre otros (Longhurst
1985; Longhurst & Pauly 1987; Philander 1996; Thurman 1996; Lavaniegos & Gonzalez
Navarro 1999).
Los efectos del fenómeno ENSO, el cual afecta la profundidad de la termoclina en la
columna de agua (Dietrich 1963; Philander 1996; Thurman 1996; NOAA 2000), así como los
eventos de entrada de agua oceánica, representan disturbios a nivel' de la termoclina, que se
reflejan en las fluctuaciones observadas en la comunidad de zooplancton y en el hecho de
que las estaciones 02 y 03 no se agruparan claramente en el análisis de conglomerado
correspondiente a P2. Así mismo, las variaciones por períodos cortos que se dan en las
abundancias de los distintos grupos del zooplancton pueden ser resultado de disturbios en la
organización espacial del sistema, provocados por turbulencia, advección de marea,
afloramiento de aguas, etc. (Madhupratap et al. 1990). En el caso del Golfo Dulce, Svendsen
et al. (2001) determinaron que el viento, la entrada de agua dulce, las mareas y la topografía
de la cuenca son factores que tienen una influencia significativa sobre la circulación en el
Golfo Dulce. Además, determinaron que existen diferencias en la estructura de corriientes al
comparar las partes interna y externa del Golfo Dulce.

151
En general, resulta claro que existe una variación significativa, tanto en la abundancia
como en la composición de fa comunidad de zooplancton, al comparar la capa superficial con
las capas P2 y P3, así como al comparar las partes interna y externa de la cuenca. Lo
anterior se debe principalmente, como se explicó, a los fuertes gradientes verticales,
espaciales y temporales en las variables físicas que se dan en el Golfo Dulce.
Abundancia y Distribución de Copépodos
Los patrones de distribución vertical específicos de las especies y su relación a las
condiciones ambientales son aspectos importantes de la estructura y función de las
comunidades pláncticas marinas (Bottger-Schnack 1990). De esta manera, los copépodos
herbívoros, carnívoros y omnívoros tienen distintas distribuciones verticales en relación con
la termoclina (Longhurst 1985; Longhurst & Pauly 1987).
Como se mencionó antes, los copépodos fueron el grupo más abundante en el Golfo
Dulce a todas las profundidades. Dentro de este grupo, el estadía que más variación mostró
a través del tiempo fue el de los nauplius, con un coeficiente de variación del 200%. Al
respecto, Lagadeuc et al. (1997) determinaron en su estudio que en una columna de agua
estratificada, la mayor biomasa de fitoplancton se encontró cerca de la superficie y que los
nauplius estuvieron concentrados sobre la picnoclina, en aguas ricas en fitoplancton. Los
resultados de la presente investigación indican que la reproducción de los copépodos (y por
lo tanto la abundancia de nauplius) durante el período de estudio fluctuó de manera
importante, probablemente como resultado de la inestabilidad que el sistema mostró debido
a los efectos combinados del fenómeno El Niño y los eventos de entrada de masas de agua
oceánica.Se ha determinado que eventos como las tormentas, por ejemplo, pueden provocar
cambios dramáticos en la salinidad de un sistema (hasta 20 PSU en seis horas). Los
estadías larvales de organismos son particularmente vulnerables a estos cambios rápidos en
las condiciones ambientales, ya que se ve afectada su supervivencia, crecimiento, actividad
y tasa de desarrollo, entre otros factores (Richmond & Woodin 1999).
La importancia de los nauplius recae en el hecho de que es un grupo que a menudo
domina numéricamente la comunidad de copépodos. Sin embargo, su contribución a la
biomasa es mucho menor que la de copepoditos y adultos (Roff et al. 1995 en Hopcroft &
Roff 1998b ). Además, son intermediarios críticos entre las redes alimentarias clásica y
microbiana ya que debido a su pequeño tamaño y alta abundancia tienen la habilidad de

152
alimentarse de partículas en el rango de tamaño picoplánctico, las cuales generalmente no
están a disposición de copepoditos y adultos.
La influencia del fenómeno El Niño resulta evidente al observar que los valores de
abundancia y composición de la comunidad de zooplancton en el Golfo Dulce presentan sus
valores más altos en abril de 1997, antes de la entrada de ENSO (NOAA 2000) y dichos
valores caen significativamente durante las restantes fechas. Esto podría representar una
consecuencia indirecta de la disminución en la cantidad de nutrimentos en la capa de mezcla
que provoca ENSO (Fielder et al. 1991 ). Así mismo, la entrada de masas de agua oceánica
(abril de 1998, por ejemplo) podría provocar un aumento en la abundancia de grupos como
apendicularias y copépodos, incluso en la capa profunda que por lo general mostró bajos
valores de abundancia en todos los grupos. También se puede observar la presencia de un
mayor número de copépodos de mayor tamaño, de géneros típicamente oceánicos, como
Eucalanus, Candacia, Euchaeta, Labidocera y Lucicutia (Raymont 1983, Longhurst 1985;
Saltzman & Wishner 1997b). Estos géneros presentan, en general , bajas abundancias
durante el período de estudio, sin embargo se les encuentra con mayor frecuencia en las
fechas en las que se observa la entrada de masas de agua oceánica. Además, en abril de
1998 se observa un incremento en los cuatro principales géneros de copépodos en todas las
capas. Uno de estos géneros, Eucalanus, es considerado como un buen indicador de la
entrada de masas de agua oceánica al mar Rojo (Bottger-Schnack 1990). De hecho,
muchos grupos de plancton son muy útiles como indicadores de masas de agua en particular
y de su movimiento (Perkins 1974).
Al igual que para el caso del zooplancton, al analizar la abundancia y composición de
los copépodos se encontraron diferencias al comparar las profundidades de muestreo, lo que
evidencia una vez más la existencia de una fuerte planctoclina en el Golfo Dulce así como un
cambio en la composición de la comunidad de copépodos con la profundidad. Asociado a lo
anterior, se ha observado que en el Mediterráneo, al igual que en el Golfo Dulce y otros
sistemas marinos tropicales, la abundancia de copépodos disminuye con la profundidad
(Longhurst 1985; Longhurst & Pauly 1987; Saltzman & Wishner 1997a, b). Además, en la
presente investigación se determinó que existen diferencias entre las estaciones, que se
pueden agrupar en dos grupos: las estaciones internas (01 y 02) y las estaciones externas
(04 y 11 ), lo que es un reflejo de los distintos patrones de circulación así como de las
diferencias que existen en la descarga de agua dulce, nutrimentos e influencia oceánica que

153
se dan en ambas partes de la cuenca del golfo. La estación 03, por poseer un carácter
intermedio no se llega a relacionar claramente con ninguno de los dos grupos.
Lo anterior se confirma en los resultados de un estudio previo realizado por Morales
(1996), donde el autor determinó que existen diferencias en los grupos de copépodos en el
Golfo Dulce. Según el autor, la parte externa del Golfo se caracteriza por la presencia de
especies oceánicas como Paracalanus aculeatus, Pleuromamma gracilis, Lucicutia ova/is,
Candacia catula, Euchaeta wolffendeni y Oncaea mediterranea. Además, Morales identificó
las especies Coryceaus flaccus y C. speciosus, lo que refleja el carácter oceánico de las
aguas en la parte externa del Golfo. Al contrario, la parte interna del Golfo se caracteriza
más bien por un grupo heterogéneo de copépodos, con especies tanto neríticas, como
Pseudodiaptomus wrigthi, Acartia danae, A. c/ausi y Canthocalanus pauper, como con
especies oceánicas, como Scolicithricella marginata, Saphirina nicromaculata y Oncaea
conifera. Actualmente Morales (en prep.) describe las especies más comunes para el Golfo
con notas diagnósticas para su identificación.
Como se mencionó, la estación 03 resulta particularmente interesante debido, no solo
a su carácter intermedio entre las partes interna y externa de la cuenca, sino a la influencia
humana (posible eutroficación) debido a la cercanía de la ciudad de Golfito (León-Morales &
Vargas 1998; Sponberg & Davis 1998). Lo anterior se plantea basándose en el hecho de
que dicha estación presenta los mayores valores de abundancia de zooplancton en general y
de copépodos así como los menores valores de visibilidad del agua. Las estaciónes 03, 04 y
11 además, son las que presentan un mayor número de copépodos en la capa profunda
(P3). En el caso de la primera, esto está asociado, como ya se explicó, con la alta
productividad (alta biomasa y abundancia de zooplancton) asociada a su cercanía a Golfito,
así como a la influencia de la parte externa de la cuenca. Las estaciones externas, por su
parte, presentan mayores concentraciones de oxígeno en la capa profunda y una mayor
influencia oceánica, lo que favorece la presencia de un mayor número de organismos en ese
estrato, al contrario de lo que sucede en las estaciones internas, que presentan bajas
abundancias de organismos en la P3. La influencia oceánica se refleja también en la
composición del zooplancton gelatinoso en el golfo, la cual indica una importante influencia
sobre la parte externa del Golfo (Morales-Ramírez y Nowaczyk, en prep.).

154
Al observar la composición de la comunidad de copépodos, destaca que el grupo
dominante, Cyclopoida, es a la vez el que menor riqueza presenta, con solamente un género:
Oithona. Lo anterior no resulta sorprendente en vista de que los oithonidos son los
ciclopoideos de vida libre más exitosos del ecosistema pelágico marino. A pesar de su
tamaño (< 1.00 mm) se encuentran en todos los océanos del mundo a densidades
relativamente altas. En el Mar Rojo, por ejemplo, los ciclopoideos llegan a constituir
alrededor del 50-70% de los copépodos (Bottger-Schnack 1988). A pesar de esto, son pocos
los estudios que se han concentrado sobre este grupo (Nishida & Marumo 1982; Sabatini &
Ki0rboe 1994; Bottger-Schnack 1995; Uye & Sano 1995). Entre los ciclopoideos, la familia
Oithonidae usualmente constituye una fracción significativa de la biomasa en zonas
templadas. En el mar del norte, por ejemplo, Oithonidae contribuye con el 25% de la
biomasa de copépodos en ciertas épocas del año (Sabatini & Ki0rboe 1994). En un estudio
realizado en el Pacífico Occidental e Indico Oriental (Nishida & Marumo 1982), se determinó
que la mayoría de los oithonidos se distribuyen en la zona epipelágica, especialmente en la
capa de mezcla, lo que coincide con los resultados del presente estudio. El hecho de que
pocas especies dominen algunas comunidades de plancton significa que las especies raras
intervienen en solo una pequeña parte del flujo de energía de la comunidad (Longhurst
1985). Paffenhofer (1993) afirma que, en general se ha descuidado el estudio de los taxones
más pequeños en tamaño, como por ejemplo los estadíos juveniles de muchos taxones y
grupos como Oithonidae u Oncaeidae, los cuales podrían ser los metazoarios más
abundantes de los océanos.
De acuerdo a los resultados obtenidos por Vidal (1980), para optimizar las tasas de
crecimiento en ambientes con recursos alimentarios limitados, las especies pequeñas y
estadíos tempranos de desarrollo de especies grandes deben vivir en la capa superficial
cálida. Sin embargo, una vez que las especies grandes alcanzan un tamaño corporal crítico,
solamente pueden alcanzar su crecimiento óptimo en aguas más frías. Lo anterior viene a
justificar el hecho de que zonas tropicales y subtropicales con aguas oligotróficas y
termoclinas profundas, se caracterizan por la dominancia de especies pequeñas (Vidal
1980), como se observó en el Golfo Dulce durante el período de estudio.
Los mayores porcentajes de abundancia de Oithona se dan en las estaciones
internas, lo que probablemente se debe a la menor influencia oceánica en la parte interna del
Golfo Dulce. Al igual que en la presente investigación, Fosshagen (1980) observó

155
diferencias en la composición de especies de copépodos al comparar las partes interna y
externa de un fiordo. Así mismo, dicho autor reporta un incremento en las especies típicas
de aguas oceánicas al darse fenómenos de entrada de agua al fiordo estudiado. Además,
asume que la baja biomasa de zooplancton en la parte interna del fiordo, al igual que lo que
se presenta en el Golfo Dulce, se puede deber a la entrada de agua dulce. Fosshagen
(1980) describe a los géneros Oíthona, Paracalanus y Oncaea como típicos de sistemas
estables y bajos en nutrimentos. Precisamente estos tres géneros en ese orden, junto con
Euterpína y Corycaeus son los grupos que dominan el 91.5% de la abundancia de
copépodos en el Golfo Dulce. Destaca que en la estación 03 el género Acartía ocupa un
tercer lugar en importancia en la capa P1, sin embargo no aparece dentro de los cinco
principales géneros en ninguna ofra estación a ninguna otra profundidad.
Los copépodos del género Acartía se caracterizan por ser un grupo típicamente
eurihalino (Cervetto et al. 1999) por lo que frecuentemente se les encuentra en ambientes
estuarinos (como el Golfo de Nicoya) (Brugnoli-Olivera 1998), ya que pueden resistir cambios
bruscos en salinidad. Es probable que éste género haya tenido una mayor importancia en la
estación 03 debido a que esta estación, como se dijo, se encuentra cerca de uno de los
principales centros de población en el Golfo Dulce, por lo que la consiguiente eutroficación
de las aguas podría favorecer a Acartía sobre otros géneros. Sin embargo, el aspecto más
importante probablemente sea la proximidad del río Coto-Colorado, el cual, al provocar
frecuentes fluctuaciones en la salinidad, lo que favorece la presencia de Acartía en esta
estación. La baja abundancia de Acartía en la estación 04, mucho más cerca del río Coto
Colorado, se debe probablemente a la fuerte influencia oceánica a la que también está
expuesta la parte externa del Golfo Dulce. Otra razón por la que Acartía no es un género
importante en el Golfo Dulce como lo es en el Golfo de Nicoya radica en que Oíthona
presenta un metabolismo más bajo, lo que le permite ser más exitoso que Acartía en
ambientes con bajos niveles de oxígeno (Longhurst 1985), como el Golfo Dulce.
A pesar de que las tasas de depredación individuales de los géneros canívoros
Corycaeus y Oncaea son relativamente bajas, su alta abundancia en las comunidades
pláncticas favorece el que puedan llegar a tener un impacto significativo sobre las
poblaciones de presas (Landry et al. 1985). En el Golfo Dulce, Oncaea es uno de los
principales grupos de copépodos y su alta abundancia se podría ver favorecida por los altos

156
números de herbívoros pequeños (Oíthona y Paracalanus) que dominan el sistema y les
pueden servir de alimento (obs. pers.).
Al respecto, resulta interesante que en algunos meses (marzo 1998, capa P1,
estaciones 01, 02 y 03, por ejemplo), se observan descensos en la abundancia de Oíthona
que coinciden con incrementos en la abundancia del género Oncaea. Incluso, el género
Oncaea llega a dominar la comunidad de copépodos en algunos meses del año,
especialmente en P2 y P3, lo que podría estar relacionado, al menos en parte, con el efecto
de la depredación de Oncaea sobre Oíthona (Raymont 1983; Lenz 2000).
En el Golfo Dulce, Oncaea tarnbién mostró otros aspectos muy interesantes, como el
hecho de que su abundancia relativa aumenta con la profundidad, lo que coincide con lo
observado por Bottger-Schnack (1994) en el mar Arábigo y mar Rojo respecto a que la
abundancia relativa de poecilostomatoideos aumenta con la profundidad. Bottger-Schnack
(1995) también reporta un aumento en la abundancia de Oncaea en el mar Rojo y el Golfo de
Aden al aumentar la profundidad. De igual manera, se observa que en la capa P2 del Golfo
Dulce Oncaea pasa a ocupar un segundo lugar en importancia, superando al género
Paracalanus que ocupa dicho puesto en la capa P1. Así mismo, como se mencionó, Oncaea
adquiere mayor importancia en las capas P2 y P3 profunda llegando a dominar en algunas
fechas en las estaciones 02 y 03. Lo anterior coincide con la distribución observada para
éste género en aguas del POT y se debe principalmente a la naturaleza depredadora de los
oncaeidos, ubicándose en capas subtermoclina como estrategia de forrageo (Longhurst
1985).
A pesar de las variaciones en la dominancia de los géneros a las distintas
profundidades, Oithona, Paracalanus y Oncaea son invariablemente los géneros que
dominan la columna de agua en el Golfo Dulce. Lo anterior resulta particularmente
interesante si se considera la biología de dichos grupos y su comportamiento en ambientes
como el Golfo Dulce. Paffenhofer (1993), por ejemplo, determinó que el género Oncaea
tiene un patrón de reproducción muy variable, llegando a producir en promedio de 9 a 30
huevos por hembra por día, mientras que el período de vida se estima que varía entre los
29.1 y 40.7 días. En el mismo estudio, el autor estimó que cada hembra del género Oíthona
produce aproximadamente entre 1.6 y 3 huevos por hembra por día, con un período de vida
de hasta 71.5 días. Como se puede ver, Oithona produce mucho menos huevos por día que

157
Oncaea, sin embargo, si se considera todo el período reproductivo de cada especie, las
diferencias son en realidad bajas. En general, la producción de huevos durante un mayor
tiempo de vida de los géneros Oithona y Oncaea podría contribuir a sus altas
concentraciones, lo que convierte a las especies de estos géneros en verdaderos
estabilizadores ecológicos (Paffenhofer 1993)
Estos géneros, además, no producen una corriente de alimentación y su capacidad
de nado es limitada, por lo tanto no consumen mucha energía en estos aspectos. Además,
ambos géneros son morfológica y anatómicamente menos derivados que los calanoideos,
por lo que son, a su vez, poco especializados, lo cual favorece su sobrevivencia en un amplio
rango de ambientes y condiciones, con lo que podrían lograr mantener sus poblaciones bajo
condiciones poco ventajosas (Paffenhofer 1993). El ser generalista le permite a un animal
modificar su dieta con la variación en el ambiente alimentario. Además, una dieta diversa
puede aumentar la probabilidad de que el copépodo obtenga una ración nutricionalmente
completa (Kleppel 1993).
En este sentido, resulta interesante el hecho de que todas las estaciones fueron
dominadas por Paracalanus en el mes de abril de 1997, antes de que entrara el fenómeno El
Niño. Lo anterior coincide con lo reportado por van Wangelin y Wolff (1996), quienes
observaron que el Golfo Dulce estaba dominado por calanoideos pequeños y no por
ciclopoideos. Vale aclarar que dicho estudio se realizó entre diciembre de 1993 y febrero de
1994, cuando el POT no estaba afectado por un fenómeno ENSO. Lo anterior indica que la
entrada del fenómeno ENSO favoreció la dominancia observada del género Oithona durante
todo el período de estudio. Además, al observar la variación temporal en los principales
géneros de copépodos, Oithona parece responder más rápida y ampliamente a los
fenómenos de entrada de agua oceánica.
Por lo tanto, el crecimiento y el tiempo de desarrollo a menudo están asociados con la
temperatura ambiental, concentración de recursos, estadía de desarrollo y tamaño corporal,
así como combinaciones de estas variables (Vidal 1980; Escribano et al. 1997; Hopcroft et al.
1998b). Al respecto, Vidal (1980) afirma que el tamaño corporal final en especies pequeñas,
debido a que requieren relativamente menos alimento para alcanzar su máximo crecimiento,
depende más bien de la temperatura ambiental. Para el caso de especies más grandes, la
concentración de alimento puede llegar a jugar un papel más importante. En este caso, la

158
limitación de alimento es un factor que a menudo se ha sugerido como un elemento que
restringe el crecimiento somático de los copépodos (Jónasdóttir 1994; Hopcroft et al. 1998b;).
De hecho, existe evidencia de que la concentración de fitoplancton en el mar puede llegar a
ser tan importante como la temperatura en determinar el tamaño corporal de herbívoros
pláncticos (Vidal 1980). Cada especie responde de manera distinta a las concentraciones de
alimento disponible, de manera que las diferencias entre las especies son amplias bajo
condiciones de poco alimento (Boyd et al. 1980).
Con respecto a la dominancia de Oithona spp. durante la mayor parte del período de
estudio, se ha observado que bajo condiciones fluctuantes, Oithona spp. mantiene una
biomasa casi constante, en contraposición con especies pequeñas de calanidos, como
Paracalanus parvus y Pseudocalanus spp. (PaffenhOfer 1993). Debido a lo anterior, Oithona
spp. contribuye relativamente más a la biomasa de copépodos durante épocas
"desfavorables" (alta depredación, poco alimento, etc.), que precisamente parece ser lo que
sucede en el Golfo Dulce. De hecho, se ha observado que cuando hay poco alimento, los
flagelados llegan a constituir una importante fuente de alimento para Oithona spp., pero no
para otros copépodos calanoideos (PaffenhOfer 1993; Sabatini & Ki0rboe 1994 ). De hecho,
de acuerdo a Estrada (2000 en prep.), en el mes de noviembre de 1997 se observó una baja
biomasa de flagelados en la zona de Golfito, lo que se podría deber en parte a la presión de
forrageo por parte del zooplancton. Como se puede apreciar, el hecho de que Oithona
presente bajas tasas de alimentación, crecimiento, reproducción y mortalidad, así como su
habilidad para soportar condiciones adversas, favorece el que sus poblaciones se puedan
mantener estables en el tiempo (Paffenh6fer 1993).
Rao & Urosa (1974 en Raymont 1983) estudiaron el Golfo de Cariaco, en Venezuela,
el cual al igual que el Golfo Dulce, es una cuenca anóxica tropical que presenta fenómenos
de entrada de agua oceánica profunda. Sin embargo, este no es el único aspecto que
comparten los dos sistemas. En este lugar también se ha determinado que se da una
entrada de agua oceánica como respuesta a vientos que mueven las aguas superficiales
fuera del Golfo (Richards 1965). Además, la entrada de agua al Golfo promueve la
producción primaria y llega a causar fluctuaciones importantes en el plancton. El grupo
dominante es el de los copépodos (65% ), seguido de cladóceros, apendicularias y
quetognatos. Los principales géneros de copépodos son Temora, Paracalanus,
Clausoca/anus, Oithona, Oncaea y Microsetella (Zoppi 1964 en Raymont 1983; Rao & Urosa

159
1974 en Raymont 1983). Finalmente, Zoppi (1964 en Raymont 1983) describió que la fauna
en el Golfo de Cariaco es una mezcla de especies neríticas y oceánicas, al igual que lo
observado en el Golfo Dulce (Morales 1996).
La variación temporal de los principales géneros de copépodos en el Golfo Dulce
coincide bastante bien tanto con los principales incrementos en biomasa, como con las
fechas en las que se detectó la entrada de masas de agua oceánica. De hecho, se
determinó la existencia de una relación significativa entre las variables de abundancia de
copépodos y la biomasa del zooplancton. Como se discute en el capítulo 1 y en la presente
discusión, la entrada de masas de agua oceánica probablemente representa un ingreso de
nutrimentos a la cuenca del Golfo Dulce (Acuña en prep.), lo cual a su vez tiene
consecuencias directas sobre las comunidades biológicas. En una serie de experimentos,
Hopcroft et al. (1998b) determinaron que el incremento de recursos mediante la adición de
nutrimentos causa un incremento de un 32% en las tasas de crecimiento en sitios
mesotróficos (como el Golfo Dulce), pero sólo un 17% de incremento en un sitio eutrófico.
En ambos sitios los copepoditos mostraron un desarrollo más rápido y mayores tamaños
como respuesta a la adición de nutrimentos (Hopcroft et al. 1998b ). Por lo tanto, no resulta
sorprendente el observar incrementos en la biomasa y abundancia del zooplancton luego de
la entrada de masas de agua oceánica.
Finalmente, al observar los análisis de conglomerado, resalta que las estaciones
internas (01 y 02) se agrupan en las tres capas de muestreo, lo cual indica que comparten
una comunidad de copépodos con abundancias y composiciones similares. Así mismo, la
influencia del río Coto-Colorado se manifiesta en la similitud que presentan las estaciones 03
y 04 en las capas P1 y P2. Las estaciones externas (04 y 11) se agrupan en P3 debido a
que comparten una columna de agua con características similares, como la fuerte influencia
oceánica, los niveles de oxígeno disuelto y la profundidad máxima, lo que determina las
características de la comunidad de copépodos en la P3. La estación 11 sin embargo, se
separa de las demás en las capas P1 y P2 producto de la fuerte influencia oceánica a la que
está sujeta la comunidad de zooplancton en dicha estación y que la hacen distinta de las
demás.
Consideraciones Finales

160
Al estudiar la distribución y ciclos estacionales del zooplancton marino, el investigador
se enfrenta al problema de que el medio marino es altamente dinámico, siempre en
movimiento y donde se mezclan constantemente distintos tipos de agua. Debido a lo
anterior, por lo general resulta virtualmente imposible darle seguimiento a la misma población
de organismos. Por lo tanto, las observaciones usando estaciones fijas a menudo
representan una mezcla de secuencias de distintas poblaciones de plancton y de cambios
estacionales de las mismas estaciones. Con el fin de determinar el impacto de los
fenómenos anteriormente señalados, los programas de muestreo se acompañan de una
detallada documentación oceanográfica.
No obstante, se debe tener en cuenta que las fluctuaciones en cada una de las
variables afectan de manera distinta a los diferentes taxones y estadías de desarrollo en el
zooplancton. Además de las fluctuaciones en las variables físicas, debe considerarse que
los distintos grupos de zooplancton interactuan entre sí (depredación, competencia, etc.) de
maneras muy complejas, por lo que resulta muy difícil, sino imposible explicar todas las
variaciones temporales que se observan en los distintos grupos de zooplancton. A menudo
se da por un hecho que los factores abióticos son determinantes en la distribución de los
organismos y no se toma en cuenta que muchos de los aspectos del comportamiento animal
están regidos por las relaciones tróficas (Esnal 1986).
Además, debe de considerarse las limitaciones a las que se ven sometidos los
estudios zooplancticos debido a las imprecisiones de muestreo asociadas al tamaño de poro
de red utilizado. Las comparaciones de distintos estudios, a menudo dan impresiones
erróneas de las comunidades en estudio si no se considera el hecho de que existe una gran
variación en el tamaño del poro de red utilizado por los diferentes autores (Hopcroft et al.
1998a; Pastel et al. 2000; Sameoto et al. 2000). Hopcroft et al (1998a), por ejemplo,
compararon los resultados al usar dos redes para zooplancton, de 64 µm y 200 µm, en la
bahía de Kingston. Cada red dio impresiones muy distintas de la estructura de la comunidad
de copépodos, tanto en términos de biomasa como de abundancia. Los muestreos con la
red de poro más pequeño evidenciaron una mayor contribución relativa de los grupos
pequeños, como nauplius y el género Oithona, a la biomasa total. Sin embargo, las
abundancias de Acartia, Centropages, Paracalanus, C/ausocalanus, Oithona, Corycaeus y
Euterpina fueron mayores cuando se usó la red de 200 µm. La comunidad de copépodos en

161
Kingston, al igual que en el Golfo Dulce, es dominada por especies pequeñas y estadías
tempranos de desarrollo. Según Hopcroft et al. (1998a), lo anterior se podría deber a que la
predominancia de nanofitoplancton favorezca a copépodos pequeños.
Todos los aspectos anteriormente señalados limitan en alguna medida los alcances
de los estudios zooplancticos. A pesar de lo anterior, la presente investigación logró
alcanzar todos los objetivos planteados en torno a la descripción de la comunidad de
zooplancton en el Golfo Dulce y arroja luz alrededor de los posibles efectos del fenómeno
ENSO sobre las comunidades pláncticas en la zona de estudio. En general el sistema del
Golfo Dulce muestra un fuerte dominio por parte de los copépodos, en especial del género
Oithona. Las fluctuaciones que se observan en la composición de las comunidades de
zooplancton y copépodos podrían reflejar el ingreso de nutrimentos al sistema provenientes
de la descarga de ríos y la entrada de masas de agua oceánica. Lo anterior se apoya en la
similitud temporal que presentan los incrementos que se dan en la abundancia de algunos
grupos y los incrementos observados en la biomasa del zooplancton durante el mismo
período. A pesar de las posibles limitaciones metodológicas del presente estudio y de los
trabajos en zooplancton en general, la presente investigación señala claramente las
tendencias que se dan en cuanto al comportamiento de la comunidad de zooplancton del
Golfo Dulce. Además, al describir su composición y las fluctuaciones temporales observadas
durante dos años, se crea una referencia importante sobre el sistema que permitirá plantear
futuras investigaciones.

162
REFERENCIAS
Acuña, J. L. & R. Anadón.1992. Apendicularian assemblages in a shelf area and their
relationship with temperature. J. Plank. Res. 14(9): 1233-50.
Adrian, R. & T.M. Frost. 1993. Omnivory of cyclopoid copepods: comparisons of algae and
invertebrates as food far three differently sized species. J. Plank. Res.15(6): 643- 58.
Aksnes, D. & T. Magnesen.1983. Distribution, development and production of Calanus
finmarchicus (Gunneros) in Lindaspollene, western Norway, 1979. Sarsia 68:195-208.
Alcaraz, M. 1985. Vertical distribution of zooplankton biomass during summer stratification in
the western Mediterranean. Proceedings of the 191h European Ma. Biol. Symp.
Cambridge Univ. Press, pp.135-43.
Allison, S. & K. Wishner. 1986. Spatial and temporal patterns of zooplankton biomass across
the Gulf stream. Mar. Ecol. Prog. Ser. 31: 233-44.
Angel, M.V. 1985. Vertical Migrations in the oceanic realm: possible causes and probable
effects, pp. 45- 70. In Rankin, M.A. (ed.) Contributions in Marine Science, vol. 27.
Marine Science lnst., Univ. of Texas, Austin.
Arcos, F. & A. Fleminger. 1986. Distribution of filter-feeding calanoid copepods in the eastern
equatorial Pacific. CalCOFI Rep., Vol XXVII, pp.170-187.
Bailey, N.T. 1995. Statistical Methods in Biology. Cambridge Univ. Press.
Benovic, A., S. Fonda-Umani, A. Malej & Speecchi. 1994. Net zoopplankton biomass in the
Adriatic sea. Mar. Biol. 79: 209-18.
Bollens, S. & B. Frost. 1991. Diel Vertical migration in zooplankton rapid individual response
to predators. J. Plank. Res. 13 (6): 1359-65.

163
Bottger-Schnack, R. 1988. Observations on the taxonomic composition and vertical
distribution of cyclopoid copepods in the Central Red Sea. Hydrobiol. 167/168: 311-
18.
____ _____ 1990. Community structure and vertical distribution of ciclopoid
copepods in the Red Sea. Mar. Biol. 106:473-85.
___ _ _____ 1994. The microcopepod fauna in the Eastern Mediterranean and
Arabian seas: a comparison with the Red Sea fauna. Hydrobiol.292/293: 271-82.
_ ________ 1995. Summer distribution of micro and small mesoplankton in the Red
sea and Gulf of Aden , with special reference to non-calanoid copepods. Mar. Ecol.
Prog. Ser. 118: 81-102.
Bougis, P. 1976. Marine Plankton Ecology. American Elsevier Publishing Company, lnc,
N.Y.pp. 355.
Boyd, C. , S. Smith & T.J . Cowles.1980. Grazing patterns of copepods in the upwelling system
of Perú. Limnol. Oceanogr. 25(4):583-96.
Braun, J., M.T. Alvarez-Osorio, M.L. Fernandez-Puelles & J.A. Camiñas. 1990. Estudio sobre
la estructura, biomasa y producción del meso- y microzooplancton en la plataforma de
Galicia en abril de 1982.Bol. lnst. Esp. Oceanogr. 6(2):81-96.
Brugnoli-Olivera, E.1998. Factores abióticos y bióticos que regulan la estructura y dinámica
de la comunidad fitoplánctica en la zona de Punta Morales, Golfo de Nicoya. Tesis de
Maestría, Universidad de Costa Rica, San José, Costa Rica.
Calbet, A & M. Alcaraz1996. Effects of constant and fluctuating food supply on egg
production rates of Acartia grani (Copepoda:Calanoida). Mar. Biol. Prog . Ser. 140:33-
39.

164
Carlsson, P., E. Granéli, P. Tester & L. Boni.1995. lnfluence of riverine humic substances on
bacteria, protozoa, phytoplankton and copepods in a coastal plankton community.
Mar. Ecol. Prog. Ser. 127: 213- 21.
Caspers, H. 1967. Estuaries: analysis of definitions and biological considerations, pp. 6-8. In
G.H. Lauff (ed.) Estuaries. American Association for the Advancement of Science,
Washington D.C.
Cervetto, G., R. Gaudy & M. Pagano.1999. lnfluence of salinity on the distribution of Acartia
tonsa (Copepoda, Calanoida). J. Exp. Mar. Biol. and Ecol. 239:33-45.
Chen, Y.Q. 1986. The vertical distribution of sorne pelagic copepods in the Eastern Tropical
Pacific. CalCOFI Rep., vol. XXVII, pp.205-227.
Clarke, K.R. & M. Ainsworth. 1993. A Method of linking multivariate community structure to
environmental variables. Mar. Ecol. Prog. Ser. 92: 205-19.
Córdoba, R. & J.A. Vargas.1996. Temperature, salinity, oxygen and nutrient pretiles ata 200
m station in Golfo Dulce, Pacific coast of Costa Rica. Rev. Biol. Trop. 44(3): 233-236.
Davis, C.C. 1984. Planktonic Copepoda (lncluding Monstrilloida). pp 67-92. In K.A. Steidinger
& L.M. Walker (eds.), Marine Plankton Life Cycle Strategies. CRC Press, lnc.
Deibel, D. 1988. Filter feeding by Oikopleura vanhoeffeni: grazing impact on suspended
particles in cold ocean waters. Mar. Biol. 99:177-86.
DeMeester, L. & Vyverman.1997. Diurnal residence of the larger stages of the calanoid
copepod Acartia tensa in the anoxic monimolimnion of a tropical meromictic lake in
New Guinea. J. Plank. Res. 19(4): 425-34.
Dessier, A & J.R. Donguy.1985. Planktonic copepods and environmental properties of the
eastern equatorial Pacific: seasonal an d spatial variations. Deep sea Res. 32(9):
1117-1133.

165
Dietrich, G. 1963. General oceanography: an lntroduction. lnterscience Publishers.
Escribano, R., C. lrribarren & L. Rodríguez.1997. lnfluence of food quantity and temperature
on development and growth of the marine copepod Ca/anus chi/ensis from northern
Chile. Mar. Biol. 128: 281-88
Esnal, G. 1986. Tunicados planctónicos (Chordata, Tunicata), distribución y relaciones
tróficas. PHYSIS (Buenos Aires), Secc. A, 44(106):51-57.
Federov, A. V. & S. G. Philander. 2000. Is El Niño changing? Science, vol. 288, pp. 1997-
2001.
Fielder, P.C., V. Philbrick & F.P. Chavez.1991. Oceanic upwelling and productivity in the
Eastern Tropical Pacific. Limnol. Oceanogr. 36(8): 1834- 50.
Fosshagen, A. 1980. How the zooplankton community may vary within a single fjord system,
pp. 399- 405. In H.J. Freeland, D.M. Farmer & C.D. Levings (eds.), Fjord
Oceanography. Plenum Press. N.Y. & London.
Gasol, J.M., del Giorgio, P.A. & C.M. Duarte.1997. Biomass distribution in marine planktonic
communities. Limnol. Oceanogr. 42(6): 1353-1363.
Hansen, P.J., P.K. Bj0rnsen & B. Hansen. 1997. Zooplankton grazing and growth: scaling
within the 2-2000µm body size range. Limnol. Oceanogr. 42(4): 687-704.
Harris, R. 1987. Spatial and Temporal organization in marine plankton communities.
Organization of communities. Ed. Blackwell Sci. Pub.
Hartmann, H.J. 1994. The Golfo Dulce Marine Environment. Review and Critica! Assesment.
pp. 34. Department of Biology of La Rochelle, France. Unpublished.
Heywood, K. 1996. Diel vertical migration of zooplankton in the Northeast Atlantic. J. Plank.
Res. 18(2): 163.

166
Hopcroft, R., J. Roff, & D. Lombard.1998a. Production of tropical copepods in Kingston
Harbour, Jamaica: the importance of small species. Mar. Biol. 130: 593- 604.
Hopcroft, R., J. Roff, M. Webber & J. D. Witt. 1998b. Zooplankton growth rates: the influence
of female size and resources on egg production of tropical marine copepods. Mar.
Biol. 132: 79-86.
Hopcroft, R.& J. Roff. 1996. Zooplankton growth rates: diel egg production in the copepods
Oithona, Euterpina and Corycaeus from tropical waters. J. Plank. Res .. 18(5):789- 803.
________ .1998a. Zooplankton growth rates: the influence of size and resources in
tropical marine copepodites. Mar. Biol. 132: 67-77.
______ __ 1998b. Zooplankton growth rates: the influence of size in nauplii of
tropical marine copepods. Mar. Biol. 132:87-96.
lkeda, T. 1985. Metabolic rates of epipelagic marine zooplankton as a function of body mass
and temperature. Mar. Biol. 85: 1-11.
Jeffries, H.P. 1967. Saturation of estuarine zooplankton by congeneric associates, pp. 501-
08. In G.H. Lauff (ed.) Estuaries: American Association of the Advancement of
science, Washington D.C.
Jónasdóttir, S. H. 1994. Effects of food quality on the reproductive success of Acartia tonsa
and Acartia hudsonica: laboratory observations. Mar. Biol. 121 :67- 81.
Kane, J. 1993. Variability of zooplankton biomass and dominant species abundance on
Georg es Bank, 1977-86. Fish. Bull. 91: 464-7 4.
Kennish, M.J.1994.Practical Handbook of Marine Science. 2nd ed. CRC Press.
Kleppel, G. S.1993. On the diets of calanoid copepods. Mar. Ecol. Prog. Ser. 99: 183- 95.

167
Kuever, J., Wawer, C. & R. Lillebrek. 1996. Microbiological observations in the anoxic basin
Golfo Dulce, Costa Rica. Rev. Biol. Trop. 44 (3): 49-57.
Lagadeuc, Y, M. Boulé & J. Dodson. 1997. Effect of vertical mixing on the vertical distribution
of copepods in coastal waters. J. Plank. Res. 19(9): 1183- 1204.
Lampert, W. & U. Shober.1980. The importance of "threshold" food concentrations. pp.264-
267. In W.C. Kerfoot (ed.), Evolution and Ecology of Zooplankton communities.
Special Symposium Vol 3. American Society of Limnology and Oceanography. Univ.
Press of New England.
Landry, M., J.M. Lehner-Founier & V. Fagerness.1985. Predatory feeding behavior of the
marine cyclopoid copepod Corycaeus anglicus. Mar. Biol. 85:163-9.
Lavaniegos, B. & E. Gonzalez-Navarro.1999. Cambios en la comunidad de copépodos
durante el ENSO de 1992-93 en el canal de San Lorenzo, Golfo de California.
Ciencias Mar. 25(2): 239-65.
Lenz, J. 2000. lntroduction. In Harris, R., P. Wiebe, J. Lenz, H. Skjodal & M. Huntley. (eds.)
Zooplankton Methodology Manual. Academic Press.
León-Morales, R. & J. Vargas.1998. Macroinfauna of a tropical fjord-like embayment: Golfo
Dulce, Costa Rica. Rev. Biol. Trop. 46(6):81-90.
Longhurst, A. R. 1985. Relationship between diversity and the vertical structure of the upper
ocean. Deep-Sea Res. Vol. 32 (12): 1535-70.
Longhurst, A. & D. Pauly.1987a. Circulation of tropical seas and oceans.pp. 27-62. In Ecology
of tropical Oceans. Academic Press, lnc.
Lutz, R.V., N. H. Marcus & J. P. Chanton.1992. Effects of low oxygen concentrations on the
hatching and viability of eggs of marine calanoid copepods. Mar. Biol. 114:241- 4 7.

168
Madhupratap, M., S.R. Sreckumaran, P. Haridas & G. Padmavati.1990. Response of
zooplankton to physical changes in the enviranment: coastal upwelling along the
central west coast of India. J. Coastal Res. 6(2): 413-26.
Margalef, R. 1978. Life forms of phytoplankton as survival alternatives in an unstable
enviranment. Oceanologica Acta 1(4): 493.
_ ____ 1985. Primary pruduction in upwelling areas: energy, global ecology and
resources. lnt. Symp. Upw. W. Afr. 5(1 ): 225-332.
Morales, A. & J. Vargas.1995. Especies comunes de copépodos (Crustacea:Copepoda)
pelágicos del Golfo de Nicoya, Costa Rica. Rev. Biol. Trap. 43(1-3): 207-218.
Morales, A. 1996. Checklist of copepods from Gulf of Nicoya, Coronado Bay and Gulf Dulce,
Pacific coast of Costa Rica, with comments of their distribution. Rev. Biol. Trap. 44(3):
103-113.
Morales-Ramírez, A & J. Jakob. Seasonal vertical distribution, abundance, biomass and
biometrical relationship of ostracods in Golfo Dulce, Pacific coast of Costa Rica. En
pre p.
Morales-Ramírez, A & J. Nowaczyk. Distribution, abundance, biomass and composition of
gelatinous zooplankton in Golfo Dulce, Pacific coast of Costa Rica, during the
transition wet-dry season 1997-98: possible impact of El Niño. En prep.
Nishida, S. & R. Marumo.1982. Vertical distribution of cyclopoid copepods of the family
Oithonidae in the Western Pacific and Eastern lndian Oceans. Bull. Plank. Soc. Japan
29(2): 99-118.
NOAA. 2000. National Oceanographic and Atmospheric Agency home page. In
http://www. noaa. gov /

169
Omori, M. & A. Fleminger. 1976. Fixation and preservation of various marine taxa: Laboratory
methods for processing crustacean zooplankton, pp. 282-286. In H.F.Steedman (ed.).
The UNESCO Press, Paris.
Omori, M .1978. Sorne factors affecting on dry weight, organic weight and concentrations of
carbon and nitrogen in freshly preserved zooplankton. lnt. Revue ges Hydrobiol.
63(2):261-69.
Omori, M. & W. Hammer.1982. Patchy distribution of zooplankton: behavior, population
assessment and sampling problems. Mar. Biol. 72: 193-200.
Ortner, P., P. Weibe, L. Haury & S. Boyd.1978. Variability in zooplankton biomass distribution
in the northern Sargasso Sea: the contribution of Gulf Stream cold-core rings. Fish.
Bull. 76(2): 323-34.
Paffenhofer, G. A. 1993. On the ecology of cyclopoid copepods (Crustacea; Copepoda).J.
Plank. Res. 15(1): 37- 55.
Perkins, E.J.1974. Plankton.pp. 71-104. In The Biology of Estuaries and Coastal Waters.
Academic Press, lnc.
Philander, G.1996. El Niño and La Niña, pp. 72-87, In R.G. Pirie (ed.), Oceanography:
contemporary readings in ocean science, 3rd ed. Oxford Univ. Press.
Postel, L. H. Fock & W Hagen.2000. Biomass and Abundance. In Harris, R., P. Wiebe, J.
Lenz, H. Skjodal & M. Huntley. (eds.) Zooplankton Methodology Manual. Academic
Press.
Raymont, J.E.G.1983. Plankton and productivity in the Oceans. Vol. 2. Zooplankton. 2nd ed.
Pergamon Press,.pp.
Razouls, S., F. DeBoveé, C. Razouls & M. Panouse.1994. Are size spectra of
mesozooplankton a good tag for characterizing pelagic ecosystems? Vie Milieu 44(1 ):
59-68.

170
Reynolds, C.S. 1989. Physical determinants of phytoplankton succession. pp. 9- 56. In U.
Sommer (ed.), Plankton Eoo 1logy: Succesion in plankton communities. Springer
Verlag.
Richards, F.A. 1965. Anoxic basins and fjords, pp. 611-645. In J.P. Riley & G. Skirrow (eds.),
Chemical Oceanography, Vol. 1, Academic Press, London & NY.
Richards, F.A., Anderson, J .J. & J.D. Cl1ine. 1971. Chemical and Physi'cal observations in
Golfo Dulce, an anoxic basin on the Pacific coast of Costa Rica. Limnol. Oceanogr.
16(1 }: 43-50.
Richmond, C. E. & S.A. Woodin. 1999. Effect of salinity reduction on oxygen consumption by
larval estuarine invertebrates. Mar. Biol. 134: 259-67.
Riley, G.A.1967. The Plankton of Estuaries. pp. 316-326. In G.H. Lauff (ed.). Estuaries.
American Association for the Advancement of Science, Washington D.C.
Rojas de Mendiola, B., O. Gomez & N. Ochoa.1985. Efectos del fenómeno El Niño 1982 - 83
sobre el fitoplancton de la costa peruana. Simp. lnt. Afl. O Afr., l1nst. lnv. Pesq., v.I:
417- 33.
Roman, M.R., A.L. Gauzens, W.K. Rhinehart & J.R. White.1993. Effects of low oxygen waters
in the Chesapeake Bay zooplancton. Limnol. Oceanogr. 38(8): 1:603- 114.
Sabatini, M. & T. Ki0rboe. 1994. Egg production, growth and development of the cyclopoid
copepod Oíthona símílís. J. Plank. Res. 16(10): 1329-51.
Saltzman, J. & K.F. Wishner. 1997a. Zooplankton ecologiy in the eastern tropical Pacific
oxygen minimum zone above a sea mount: 1. General trends. Deep-Sea Res. 1,
vol.44 (6): 907-30
_ ___________ 1997b. Zooplankton ecology in the eastern tropical Pacific
oxygen minimum zone above a sea mount: 2. Vertical distribution of copepods. Deep
Sea Res. 1, vol.44 (6):931-54.

171
Sameoto, D. , P.Wiebe, J. Runge, L. Poste!, J. Dunn, C. Miller & S. Coombs. 2000. Collecting
Zooplankton. In Harris, R., P. Wiebe, J. Lenz, H. Skjodal & M. Huntley. (eds.)
Zooplankton Methodology Manual. Academic Press.
Smetacek, V. & Pollehne.1986. Nutrient cycling in pelagic systems: a reappraisal of the
conceptual framework. Ophelia 26: 401-28.
Smetacek, V.1988. Plankton characteristics. In Ecosystems of the World 27. Continental
Shelves. Elsevier, N.Y.
Sponberg, A. & P. Davis. 1998. Organochlorinated pesticide contaminants in Golfo
Dulce, Costa Rica. Rev. Biol. Trap. 46(6):111-24.
Sterner, R. 1986. Herbivores direct and indirect effects on algal populations. Science vol.
231: 605.
Svendsen, H., R. Rosseland, S. Myking, J.A. Vargas, O.G. Lizano & E.J. Alfara. 2001. A
physical-oceanographical study of Golfo Dulce. Preliminary Report. Geophysical
lnstitute, Univ. Bergen and CIMAR, Univ. De Costa Rica.
Taggart, C. & K.T. Frank.1987. Coastal upwelling and Oikopleura ocurrence: a model and
potential application to inshore fisheries. Can J. Fish. Aquat. Sci., vol.44:1729 - 36.
Thamdrup, B., Canfield, O.E., Ferdelman, T.G., Glud, R.N. & J.K. Gundersen.1996. A
biogeochemical survey of the anoxic basin Golfo Dulce, Costa Rica. Rev. Biol. Trap.
44(3):19-33.
Thurman, H.V. 1996. Essentials of Oceanography, pp. Prentice Hall, lnc.
Tilman, D., S.S.Kilham & P. Kilham.1982. Phytoplankton community ecology: the role of
limiting nutrients. Ann. Rev. Ecol. Syst. 13:349- 72.
Umaña, G. 1998. Characterization of sorne Golfo Dulce drainage basin rivers (Costa Rica).
Rev. Biol. Trap. 46(6):125-35.

172
Uye, S. & K. Sano.1995. Seasonal reproductive biology of the small cyclopoid copepod
Oithona davisae in a temperate eutrophic inlet. Mar. Ecol. Prog. Ser. 118: 121-128
Vidal, J. 1980. Physioecology · of zooplankton.1. Effects of phytoplankton concentration,
temperature and body size on the growth rate of Calanus pacificus and
Pseudocalanus sp. Mar. Biol. 56: 111-134.
van Wangelin, M. & M. Wolff.1996. Comparative biomass spectra and species composition of
the zooplankton communities in Golfo Dulce and Golfo de Nicoya, Pacific coast of
Costa Rica. Rev. Biol. Trap. 44(3): 135-155.
White, J., X. Zang, L. Welling, M. Reman & H.Dam.1995. Latitudinal gradients in zooplancton
biomass in the tropical Pacific at 140ºW during the JGOFS EqPac study: effects of El
Niño. Deep Sea Res. 11, 42(2-3): 715-33.
Wiafe, G. & C.L. Frid.1996. Short-term temporal variation in coastal zooplankton
communities: the relative importance of physical and biological mechanisms. J.Plank.
Res. 18(8): 1485-1501.
Wolff, M. & J.A. Vargas (Eds.).1994. R.V. Victor Hensen Costa Rica Expedition 1993/1994.
Cruise Report. Center far Tropical Marine Ecology (ZMT).Bremen. 109p.
Wolff, M., Hartmann, H.J. & V. Koch. 1996. A pilot trophic model far Golfo Dulce, a fjord-like
tropical embayment, Costa Rica. Rev. Biol. Trap. 44(3): 215-231.
Wolff, M., V. Koch,J.B. Chavarría & J.A. Vargas.1998. A trophic flow model of the Golfo de
Nicoya, Costa Rica. Rev. Biol. Trap. 46(6): 63-80.
Zeitzschel, B. 1978. Oceanographic factors influencing the distribution of plankton in space
and time. Micropaleontology. 24(2):139-159.

Orden
Calanoida
APENDICE 1
Especies de copépodos reportados para el Golfo Dulce 1
Familia
Aetideidae
Calanidae
Clausocalanidae
Calocalanidae
Paracalanidae
Spinocalanidae
Eucalanidae
Euchaetidae
Temoridae
Metridinidae
Centropagidae
Lucicutiidae
Especie
Euchirel/a sp. Giesbrecht 1888
Canthocalanus pauper Giesbrecht 1888 Undinula vulgaris Dana 1852
C/ausocalanus sp. Giesbrecht 1888
Ca/ocalanus sp. Giesbrecht 1888
Acrocalanus gibber Giesbrecht 1888
Paracalanus aculeatus Giesbrecht 1888 P. parvus Claus 1863
Spinocalanus sp.
Eucalanus elongatus Dana 1849 t Subeucalanus subcrassus Giesbrecht
1888
Euchaeta Wolffendeni Scott A. 1909 Euchaeta sp. Philippi 1843
Temora discaudata Giesbrecht 1889 Temoropia mayumbaensis Scott 1894
Pleuromamma gracilis Claus 1863
Centropages furcatus Dana 1849
Lucicutia ova/is Wolffenden 1911
Pseudodiaptomidae Pseudodiaptomus wrigthi Johnson 1964
Candaciidae
Pontellidae
Acartiidae
Candacia catula Giesbrecht 1889
Labidocera acuta Dana 1849 L. /ubbocki Giesbrecht 1892
Acartia clausi Giesbrecht 1889 A. danae Giesbrecht 1889 A. /i/ljeborgii Giesbrecht 1889 A. tonsa Dana 1848
173

Cyclopoida Oithonidae
Harpacticoida Tachidiidae
Clytemnestridae
Miraciidae
Poecilostomatoida Oncaeidae
Corycaeidae
Sapphirinidae
1 Morales (1996) t Especie no detectada en el presente estudio.
Oithona plumifera Baird 1843
O. similis Claus 1863
Euterpina acutifrons Dana 1852
C/ytemnestra rostrata Brady 1883
Macrosetella gracilis Dana 1848 Microsetella sp.
Oncaea conifera O. mediterrranea
Giesbrecht 1891 Claus 1863
Corycaeus flacus Giesbrecht 1891 C. speciosus Dana 1849 Copilia sp. Dana 1849
Sapphirina nigromacu/ata Claus 1863
174

175
APENDICE 2
Matriz de datos de biomasa (mg/m3)de zooplancton, temperatura(ºC), oxígeno disuelto (mg/I)
y salinidad (PSU) para cada una de las estaciones, profundidades, tamaños y fechas
analizadas
Estación Fecha Profundidad Tamaño Biomasa Temp Oxigeno Salinidad
1 J-97 1 1 4.706 28.800 4.120 30.400
1 A-97 1 1 0.980 28.600 3.580 33.400 1 S-97 1 1 0.784 27.600 3.615 33.400 1 0-97 1 1 4.706 29.875 3.650 29.000 1 N-97 1 1 0.392 30.500 3.675 29.750 1 E-98 1 1 1.046 29.875 3.700 29.875 1 M-98 1 1 0.261 28.500 3.425 32.750 1 A-98 1 1 0.392 30.667 4.667 30.000 1 M-98 1 1 5.490 30.167 4.400 30.667 1 J-98 1 1 3.660 31.125 4.350 32.000 1 J-98 1 1 0.784 30.125 5.400 29.125 1 A-98 1 1 2.353 30.625 4.875 28.500 1 S-98 1 1 3.922 29.500 5.500 23.000 1 0-98 1 1 1.569 29.667 5.733 24.333 1 N-98 1 1 0.784 29.000 5.500 29.750 1 J-97 1 2 6.078 28.800 4.120 30.400 1 A-97 1 2 9.510 28.600 3.580 33.400 1 S-97 1 2 1.667 27.600 3.615 33.400 1 0-97 1 2 13.987 29.875 3.650 29.000
N-97 1 2 1.307 30.500 3.675 29.750 E-98 1 2 1.307 29.875 3.700 29.875
1 M-98 1 2 3.137 28.500 3.425 32.750 1 A-98 1 2 3.137 30.667 4.667 30.000 1 M-98 1 2 3.922 30.167 4.400 30.667 1 J-98 1 2 3.137 31.125 4.350 32.000 1 J-98 1 2 1.046 30.125 5.400 29.125 1 A-98 1 2 2.353 30.625 4.875 28.500 1 S-98 2 3.137 29.500 5.500 23.000 1 0-98 2 2.353 29.667 5.733 24.333 1 N-98 1 2 2.353 29.000 5.500 29.750 1 J-97 1 3 8.137 28.800 4.120 30.400 1 A-97 1 3 4.216 28.600 3.580 33.400 1 S-97 1 3 4.902 27.600 3.615 33.400 1 0-97 1 3 13.725 29.875 3.650 29.000 1 N-97 1 3 7.320 30.500 3.675 29.750 1 E-98 1 3 5.490 29.875 3.700 29.875 1 M-98 1 3 6.275 28.500 3.425 32.750 1 A-98 1 3 12.157 30.667 4.667 30.000 1 M-98 1 3 3.922 30.167 4.400 30.667 1 J-98 1 3 9.935 31.125 4.350 32.000 1 J-98 1 3 3.137 30.125 5.400 29.125

176
1 A-98 1 3 5.752 30.625 4.875 28.500 1 S-98 1 3 19.608 29.500 5.500 23.000 1 0-98 1 3 11.373 29.667 5.733 24.333 1 N-98 1 3 13.595 29.000 5.500 29.750 1 J-97 2 1 0.441 24.333 2.733 32.000 1 A-97 2 1 0.392 24.667 2.433 35.333 1 S-97 2 1 0.196 22.833 1.800 36.333 1 0-97 2 1 0.458 25.833 1.200 35.250 1 N-97 2 1 0.098 28.333 1.550 32.500 1 E-98 2 1 0.523 26.000 1.950 32.250 1 M-98 2 1 0.131 24.000 1.650 35.000 1 A-98 2 1 0.131 25.167 1.950 31.250 1 M-98 2 1 0.294 25.000 1.740 34.000 1 J-98 2 1 0.654 28.000 2.413 34.750 1 J-98 2 1 0.523 27.000 2.225 32.500 1 A-98 2 1 0.915 27.333 2.325 32.500 1 S-98 2 1 0.980 26.250 2.175 33.625 1 0-98 2 1 1.961 26.500 1.940 32.600 1 N-98 2 1 0.392 24.000 1.600 34.000 1 J-97 2 2 5.049 24.333 2.733 32.000 1 A-97 2 2 6.176 24.667 2.433 35.333 1 S-97 2 2 0.784 22.833 1.800 36.333 1 0-97 2 2 4.641 25.833 1.200 35.250 1 N-97 2 2 1.275 28.333 1.550 32.500 1 E-98 2 2 0.784 26.000 1.950 32.250 1 M-98 2 2 0.131 24.000 1.650 35.000 1 A-98 2 2 0.784 25.167 1.950 31.250 1 M-98 2 2 0.784 25.000 1.740 34.000 1 J-98 2 2 0.915 28.000 2.413 34.750 1 J-98 2 2 1.569 27.000 2.225 32.500 1 A-98 2 2 2.222 27.333 2.325 32.500 1 S-98 2 2 1.667 26.250 2.175 33.625 1 0-98 2 2 7.843 26.500 1.940 32.600 1 N-98 2 2 2.222 24.000 1.600 34.000 1 J-97 2 3 1.373 24.333 2.733 32.000 1 A-97 2 3 0.882 24.667 2.433 35.333 1 S-97 2 3 1.176 22.833 1.800 36.333 1 0-97 2 3 2.092 25.833 1.200 35.250 1 N-97 2 3 0.784 28.333 1.550 32.500 1 E-98 2 3 2.614 26.000 1.950 32.250 1 M-98 2 3 1.046 24.000 1.650 35.000 1 A-98 2 3 3.660 25.167 1.950 31.250 1 M-98 2 3 2.157 25.000 1.740 34.000 1 J-98 2 3 1.176 28.000 2.413 34.750 1 J-98 2 3 0.392 27.000 2.225 32.500 1 A-98 2 3 1.961 27.333 2.325 32.500 1 S-98 2 3 1.667 26.250 2.175 33.625 1 0-98 2 3 1.961 26.500 1.940 32.600
N-98 2 3 2.353 24.000 1.600 34.000 J-97 3 1 0.123 19.833 1.000 32.667
1 A-97 3 1 0.074 20.833 1.600 36.333 1 S-97 3 1 1.078 20.500 0.980 37.333

177
0-97 3 1 0.049 20.667 0.967 36.667 N-97 3 0.157 22.500 1.167 33.750
1 E-98 3 1 0.294 21.833 1.367 33.333
1 M-98 3 1 0.098 22.000 1.400 35.000
1 A-98 3 1 0.235 20.875 1.350 33.250
1 M-98 3 1 0.294 20.667 1.033 36.000 1 J-98 3 1 0.196 21.500 1.233 36.000 1 J-98 3 1 0.490 21.000 0.867 34.667
1 A-98 3 1 0.098 20.667 1.167 34.667
1 S-98 3 1 0.235 20.625 1.250 35.000 1 0-98 3 1 0.490 21.000 0.733 35.000
1 N-98 3 1 0.196 20.000 0.733 35.000 1 J-97 3 2 0.221 19.833 1.000 32.667 1 A-97 3 2 0.490 20.833 1.600 36.333
1 S-97 3 2 0.049 20.500 0.980 37.333 1 0-97 3 2 0.147 20.667 0.967 36.667 1 N-97 3 2 0.980 22.500 1.167 33.750 1 E-98 3 2 0.294 21.833 1.367 33.333 1 M-98 3 2 0.490 22.000 1.400 35.000 1 A-98 3 2 0.314 20.875 1.350 33.250 1 M-98 3 2 0.294 20.667 1.033 36.000 1 J-98 3 2 0.490 21.500 1.233 36.000 1 J-98 3 2 0.294 21.000 0.867 34.667 1 A-98 3 2 0.490 20.667 1.167 34.667 1 S-98 3 2 0.314 20.625 1.250 35.000 1 0-98 3 2 0.588 21.000 0.733 35.000 1 N-98 3 2 0.686 20.000 0.733 35.000 1 J-97 3 3 0.760 19.833 1.000 32.667 1 A-97 3 3 1.299 20.833 1.600 36.333 1 S-97 3 3 0.392 20.500 0.980 37.333 1 0-97 3 3 0.980 20.667 0.967 36.667 1 N-97 3 3 0.667 22.500 1.167 33.750 1 E-98 3 3 1.078 21.833 1.367 33.333 1 M-98 3 3 0.882 22.000 1.400 35.000 1 A-98 3 3 1.412 20.875 1.350 33.250 1 M-98 3 3 0.490 20.667 1.033 36.000 1 J-98 3 3 1.078 21.500 1.233 36.000 1 J-98 3 3 0.588 21.000 0.867 34.667 1 A-98 3 3 1.078 20.667 1.167 34.667 1 S-98 3 3 0.627 20.625 1.250 35.000 1 0-98 3 3 1.275 21.000 0.733 35.000 1 N-98 3 3 2.647 20.000 0.733 35.000 2 J-97 1 1 3.627 28.400 4.800 30.800 2 A-97 1 1 1.373 28.200 5.040 33.600 2 S-97 1 1 0.490 27.500 4.450 33.000 2 0-97 1 1 1.699 29.000 3.875 31.250 2 N-97 1 1 0.196 29.800 3.875 30.000 2 E-98 1 1 0.784 28.600 3.880 30.400 2 M-98 1 1 0.261 28.875 4.400 33.250 2 A-98 1 1 0.980 30.333 4.600 29.667 2 M-98 1 1 5.490 29.833 4.200 32.000 2 J-98 1 1 1.569 30.875 4.675 31.250

178
2 J-98 1 1 1.046 30.000 4.550 29.000
2 A-98 1 1 2.876 30.000 5.225 28.500
2 S-98 1 1 8.627 29.250 4.850 28.000
2 0-98 1 1 2.353 29.333 5.133 26.333
2 N-98 1 1 1.765 28.200 4.740 30.200
2 J-97 1 2 6.765 28.400 4.800 30.800
2 A-97 1 2 6.569 28.200 5.040 33.600
2 S-97 1 2 2.647 27.500 4.450 33.000
2 0-97 1 2 5.621 29.000 3.875 31.250
2 N-97 1 2 6.667 29.800 3.875 30.000
2 E-98 1 2 1.569 28.600 3.880 30.400
2 M-98 1 2 0.523 28.875 4.400 33.250
2 A-98 1 2 2.745 30.333 4.600 29.667
2 M-98 1 2 5.098 29.833 4.200 32.000
2 J-98 1 2 1.046 30.875 4.675 31.250
2 J-98 1 2 1.307 30.000 4.550 29.000
2 A-98 1 2 2.353 30.000 5.225 28.500
2 S-98 1 2 3.137 29.250 4.850 28.000
2 0-98 1 2 4.706 29.333 5.133 26.333
2 N-98 1 2 3.137 28.200 4.740 30.200
2 J-97 1 3 7.059 28.400 4.800 30.800 2 A-97 1 3 2.451 28.200 5.040 33.600
2 S-97 1 3 5.490 27.500 4.450 33.000
2 0-97 1 3 11.503 29.000 3.875 31.250
2 N-97 1 3 12.353 29.800 3.875 30.000
2 E-98 1 3 3.725 28.600 3.880 30.400
2 M-98 1 3 3.922 28.875 4.400 33.250
2 A-98 1 3 5.490 30.333 4.600 29.667
2 M-98 1 3 16.471 29.833 4.200 32.000
2 J-98 1 3 9.935 30.875 4.675 31 .250
2 J-98 1 3 2.353 30.000 4.550 29.000 2 A-98 1 3 5.490 30.000 5.225 28.500
2 S-98 1 3 18.039 29.250 4.850 28.000
2 0-98 1 3 11.373 29.333 5.133 26.333
2 N-98 1 3 5.490 28.200 4.740 30.200
2 J-97 2 1 0.637 23.833 2.517 32.333
2 A-97 2 1 0.392 24.500 3.667 34.667
2 S-97 2 1 0.588 22.333 2.580 36.333
2 0-97 2 1 0.196 24.000 1.500 35.500
2 N-97 2 1 0.784 26.667 1.750 32.667 2 E-98 2 1 0.098 24.333 2.000 32.500 2 M-98 2 1 0.131 24.075 2.500 34.750 2 A-98 2 1 1.569 24.875 2.375 33.000 2 M-98 2 1 1.176 24.000 2.020 33.800 2 J-98 2 1 1.699 26.875 2.650 34.250 2 J-98 2 1 0.654 25.375 2.050 32.000 2 A-98 2 1 1.307 25.500 2.475 32.750
2 S-98 2 1 1.667 24.625 2.175 33.500 2 0-98 2 1 1.961 26.000 2.425 32.000 2 N-98 2 1 0.392 22.000 1.600 34.000
2 J-97 2 2 1.373 23.833 2.517 32.333
2 A-97 2 2 0.833 24.500 3.667 34.667

179
2 S-97 2 2 0.392 22.333 2.580 36.333 2 0-97 2 2 0.588 24.000 1.500 35.500 2 N-97 2 2 0.588 26.667 1.750 32.667 2 E-98 2 2 0.196 24.333 2.000 32.500 2 M-98 2 2 0.523 24.075 2.500 34.750 2 A-98 2 2 3.399 24.875 2.375 33.000 2 M-98 2 2 2.549 24.000 2.020 33.800 2 J-98 2 2 1.046 26.875 2.650 34.250 2 J-98 2 2 1.046 25.375 2.050 32.000 2 A-98 2 2 0.784 25.500 2.475 32.750 2 S-98 2 2 2.843 24.625 2.175 33.500 2 0-98 2 2 5.882 26.000 2.425 32.000 2 N-98 2 2 3.529 22.000 1.600 34.000 2 J-97 2 3 1.912 23.833 2.517 32.333 2 A-97 2 3 0.882 24.500 3.667 34.667 2 S-97 2 3 0.882 22.333 2.580 36.333 2 0-97 2 3 1.046 24.000 1.500 35.500 2 N-97 2 3 1.569 26.667 1.750 32.667 2 E-98 2 3 0.980 24.333 2.000 32.500 2 M-98 2 3 1.046 24.075 2.500 34.750 2 A-98 2 3 4.967 24.875 2.375 33.000 2 M-98 2 3 2.255 24.000 2.020 33.800 2 J-98 2 3 0.392 26.875 2.650 34.250 2 J-98 2 3 0.392 25.375 2.050 32.000 2 A-98 2 3 0.523 25.500 2.475 32.750 2 S-98 2 3 2.451 24.625 2.175 33.500 2 0-98 2 3 2.092 26.000 2.425 32.000 2 N-98 2 3 4.314 22.000 1.600 34.000 2 J-97 3 1 0.147 20.000 1.050 33.250 2 A-97 3 1 0.196 20.333 2.533 35.750 2 S-97 3 1 0.490 19.333 2.000 37.250 2 0-97 3 1 0.147 19.500 1.467 36.500 2 N-97 3 1 0.147 21.333 1.200 34.000 2 E-98 3 1 0.098 21.333 0.933 33.375 2 M-98 3 1 0.294 21.667 1.733 35.000 2 A-98 3 1 0.078 21.500 1.100 34.500 2 M-98 3 1 0.392 20.500 1.133 35.250 2 J-98 3 1 0.294 21.833 1.200 35.750 2 J-98 3 1 0.147 20.500 0.800 34.000 2 A-98 3 1 0.392 21.000 1.033 34.750 2 S-98 3 1 0.196 20.625 1.225 35.000 2 0-98 3 1 0.078 20.500 1.050 35.000 2 N-98 3 1 0.098 20.000 1.067 35.000 2 J-97 3 2 0.441 20.000 1.050 33.250 2 A-97 3 2 0.490 20.333 2.533 35.750 2 S-97 3 2 0.294 19.333 2.000 37.250 2 0-97 3 2 0.245 19.500 1.467 36.500 2 N-97 3 2 0.245 21.333 1.200 34.000 2 E-98 3 2 0.196 21.333 0.933 33.375 2 M-98 3 2 0.098 21.667 1.733 35.000 2 A-98 3 2 0.235 21.500 1.100 34.500 2 M-98 3 2 0.441 20.500 1.133 35.250

180
2 J-98 3 2 0.294 21.833 1.200 35.750 2 J-98 3 2 0.049 20.500 0.800 34.000
2 A-98 3 2 0.686 21.000 1.033 34.750 2 S-98 3 2 0.157 20.625 1.225 35.000 2 0-98 3 2 0.314 20.500 1.050 35.000 2 N-98 3 2 0.686 20.000 1.067 35.000 2 J-97 3 3 1.176 20.000 1.050 33.250 2 A-97 3 3 0.980 20.333 2.533 35.750 2 S-97 3 3 0.686 19.333 2.000 37.250 2 0-97 3 3 0.980 19.500 1.467 36.500
2 N-97 3 3 1.373 21.333 1.200 34.000
2 E-98 3 3 0.980 21.333 0.933 33.375 2 M-98 3 3 0.588 21.667 1.733 35.000 2 A-98 3 3 0.863 21.500 1.100 34.500
2 M-98 3 3 1.029 20.500 1.133 35.250 2 J-98 3 3 0.392 21.833 1.200 35.750 2 J-98 3 3 0.147 20.500 0.800 34.000 2 A-98 3 3 0.686 21.000 1.033 34.750 2 S-98 3 3 0.392 20.625 1.225 35.000 2 0-98 3 3 0.471 20.500 1.050 35.000 2 N-98 3 3 0.686 20.000 1.067 35.000 3 J-97 1 1 1.961 28.750 3.775 31.250 3 A-97 1 1 1.176 29.100 5.640 33.200 3 S-97 1 1 8.235 28.200 5.150 30.800 3 0-97 1 1 0.588 29.500 4.675 30.500 3 N-97 1 1 0.196 30.000 4.500 30.000 3 E-98 1 1 0.784 29.625 4.325 30.250 3 M-98 1 1 4.444 29.250 4.775 33.500 3 A-98 1 1 2.745 31.000 5.100 29.667 3 M-98 1 1 1.176 30.333 4.600 35.000 3 J-98 1 1 1.307 30.250 4.450 31.250 3 J-98 1 1 2.092 30.125 5.175 26.375 3 A-98 1 1 1.046 29.125 4.800 26.500 3 S-98 1 1 3.137 29.250 5.650 28.000 3 0-98 1 1 3.529 29.167 5.267 28.333 3 N-98 1 1 3.137 28.125 4.850 29.250 3 J-97 1 2 9.020 28.750 3.775 31.250 3 A-97 1 2 1.912 29.100 5.640 33.200 3 S-97 1 2 4.510 28.200 5.150 30.800 3 0-97 1 2 1.634 29.500 4.675 30.500 3 N-97 1 2 0.980 30.000 4.500 30.000 3 E-98 1 2 2.353 29.625 4.325 30.250 3 M-98 1 2 2.876 29.250 4.775 33.500 3 A-98 1 2 1.176 31.000 5.100 29.667 3 M-98 1 2 3.529 30.333 4.600 35.000 3 J-98 1 2 1.307 30.250 4.450 31.250 3 J-98 1 2 1.569 30.125 5.175 26.375 3 A-98 1 2 2.353 29.125 4.800 26.500 3 S-98 1 2 3.137 29.250 5.650 28.000 3 0-98 1 2 5.294 29.167 5.267 28.333 3 N-98 1 2 5.752 28.125 4.850 29.250 3 J-97 1 3 14.248 28.750 3.775 31.250

181
3 A-97 1 3 2.941 29.100 5.640 33.200 3 S-97 1 3 0.784 28.200 5.150 30.800 3 0-97 1 3 1.765 29.500 4.675 30.500 3 N-97 1 3 8.039 30.000 4.500 30.000 3 E-98 1 3 5.490 29.625 4.325 30.250 3 M-98 1 3 8.105 29.250 4.775 33.500 3 A-98 1 3 5.098 31.000 5.100 29.667 3 M-98 1 3 7.843 30.333 4.600 35.000 3 J-98 1 3 3.137 30.250 4.450 31.250 3 J-98 1 3 6.013 30.125 5.175 26.375 3 A-98 1 3 2.614 29.125 4.800 26.500 3 S-98 1 3 10.980 29.250 5.650 28.000 3 0-98 1 3 13.529 29.167 5.267 28.333 3 N-98 1 3 7.582 28.125 4.850 29.250 3 J-97 2 1 0.490 24.500 2.400 33.000 3 A-97 2 1 0.637 23.667 3.667 35.333 3 S-97 2 1 1.176 22.167 2.930 36.333 3 0-97 2 1 0.065 24.750 2.200 34.500 3 N-97 2 1 0.784 25.667 2.525 32.333 3 E-98 2 1 0.523 25.625 2.850 31 .000 3 M-98 2 1 0.523 24.500 2.500 35.000 3 A-98 2 1 25.000 2.633 31 .333 3 M-98 2 1 0.392 24.625 2.375 36.750 3 J-98 2 1 0.654 27.250 2.900 34.250 3 J-98 2 1 0.261 26.625 2.725 31.125 3 A-98 2 1 0.131 23.625 1.975 33.000 3 S-98 2 1 0.882 24.000 2.800 33.667 3 0-98 2 1 1.275 23.250 2.125 33.750 3 N-98 2 1 3.660 23.250 1.950 34.000 3 J-97 2 2 4.052 24.500 2.400 33.000 3 A-97 2 2 3.382 23.667 3.667 35.333 3 S-97 2 2 0.196 22.167 2.930 36.333 3 0-97 2 2 0.523 24.750 2.200 34.500 3 N-97 2 2 2.451 25.667 2.525 32.333 3 E-98 2 2 1.046 25.625 2.850 31.000 3 M-98 2 2 1.046 24.500 2.500 35.000 3 A-98 2 2 25.000 2.633 31.333 3 M-98 2 2 3.725 24.625 2.375 36.750 3 J-98 2 2 1.699 27.250 2.900 34.250 3 J-98 2 2 2.353 26.625 2.725 31 .125 3 A-98 2 2 1.046 23.625 1.975 33.000 3 S-98 2 2 1.471 24.000 2.800 33.667 3 0-98 2 2 3.627 23.250 2.125 33.750 3 N-98 2 2 11 .242 23.250 1.950 34.000 3 J-97 2 3 1.667 24.500 2.400 33.000 3 A-97 2 3 3.676 23.667 3.667 35.333 3 S-97 2 3 2.941 22.167 2.930 36.333 3 0-97 2 3 1.242 24.750 2.200 34.500 3 N-97 2 3 1.569 25.667 2.525 32.333 3 E-98 2 3 3.660 25.625 2.850 31 .000 3 M-98 2 3 2.614 24.500 2.500 35.000 3 A-98 2 3 25.000 2.633 31 .333

182
3 M-98 2 3 2.745 24.625 2.375 36.750 3 J-98 2 3 1.830 27.250 2.900 34.250 3 J-98 2 3 2.745 26.625 2.725 31.125 3 A-98 2 3 0.784 23.625 1.975 33.000 3 S-98 2 3 1.373 24.000 2.800 33.667 3 0-98 2 3 2.549 23.250 2.125 33.750 3 N-98 2 3 4.575 23.250 1.950 34.000 3 J-97 3 1 0.686 19.167 0.767 34.333 3 A-97 3 1 0.074 18.333 1.667 36.333 3 S-97 3 1 0.294 18.333 1.200 38.000 3 0-97 3 1 0.196 19.167 0.700 36.000 3 N-97 3 1 0.147 20.333 0.980 34.000 3 E-98 3 1 0.196 21.500 1.267 31.333 3 M-98 3 1 0.098 21.833 1.667 35.000 3 A-98 3 1 0.392 21.750 1.400 32.750 3 M-98 3 1 1.176 20.833 1.333 38.000 3 J-98 3 1 0.588 22.833 1.733 35.333 3 J-98 3 1 0.196 20.833 0.867 33.833 3 A-98 3 1 0.882 19.833 0.933 35.000 3 S-98 3 1 0.471 20.375 1.600 35.000 3 0-98 3 1 0.392 20.333 1.033 35.000 3 N-98 3 1 0.196 19.833 1.000 35.000 3 J-97 3 2 0.686 19.167 0.767 34.333 3 A-97 3 2 0.172 18.333 1.667 36.333 3 S-97 3 2 0.098 18.333 1.200 38.000 3 0-97 3 2 1.373 19.167 0.700 36.000 3 N-97 3 2 0.294 20.333 0.980 34.000 3 E-98 3 2 0.196 21.500 1.267 31.333 3 M-98 3 2 0.392 21.833 1.667 35.000 3 A-98 3 2 0.392 21.750 1.400 32.750 3 M-98 3 2 11.078 20.833 1.333 38.000 3 J-98 3 2 1.078 22.833 1.733 35.333 3 J-98 3 2 0.196 20.833 0.867 33.833 3 A-98 3 2 1.275 19.833 0.933 35.000 3 S-98 3 2 0.941 20.375 1.600 35.000 3 0-98 3 2 0.784 20.333 1.033 35.000 3 N-98 3 2 0.588 19.833 1.000 35.000 3 J-97 3 3 1.422 19.167 0.767 34.333 3 A-97 3 3 0.662 18.333 1.667 36.333 3 S-97 3 3 1.765 18.333 1.200 38.000 3 0-97 3 3 3.824 19.167 0.700 36.000 3 N-97 3 3 0.784 20.333 0.980 34.000 3 E-98 3 3 0.980 21.500 1.267 31.333 3 M-98 3 3 0.686 21.833 1.667 35.000 3 A-98 3 3 1.020 21.750 1.400 32.750 3 M-98 3 3 0.980 20.833 1.333 38.000 3 J-98 3 3 0.882 22.833 1.733 35.333 3 J-98 3 3 0.980 20.833 0.867 33.833 3 A-98 3 3 0.980 19.833 0.933 35.000 3 S-98 3 3 1.098 20.375 1.600 35.000 3 0-98 3 3 0.784 20.333 1.033 35.000 3 N-98 3 3 1.961 19.833 1.000 35.000

183
4 J-97 1 1 2.745 29.333 5.067 30.333 4 A-97 1 1 1.307 29.500 4.125 32.000
4 S-97 1 1 4.183 27.167 4.545 27.667
4 0-97 1 1 0.196 29.667 4.967 30.333
4 N-97 1 1 6.667 30.333 5.185 29.000
4 E-98 1 0.392 29.167 4.533 31.500
4 M-98 1 1 4.314 30.167 4.633 33.333 4 A-98 1 1 2.353 31.000 5.150 29.000
4 M-98 1 1 1.569 29.667 4.733 34.333 4 J-98 1 1 3.922 30.333 5.300 27.667 4 J-98 1 1 3.137 29.500 6.300 22.000
4 A-98 1 1 5.490 29.750 5.800 23.000 4 S-98 1 1 2.745 28.333 5.067 29.333 4 0-98 1 1 1.569 28.833 4.800 28.333 4 N-98 1 1 3.137 28.333 5.433 25.000 4 J-97 1 2 7.843 29.333 5.067 30.333 4 A-97 1 2 0.654 29.500 4.125 32.000
4 S-97 1 2 9.412 27.167 4.545 27.667 4 0-97 1 2 1.176 29.667 4.967 30.333 4 N-97 1 2 11.373 30.333 5.185 29.000
4 E-98 1 2 1.176 29.167 4.533 31.500 4 M-98 1 2 3.529 30.167 4.633 33.333 4 A-98 1 2 3.137 31.000 5.150 29.000 4 M-98 1 2 1.569 29.667 4.733 34.333 4 J-98 1 2 4.706 30.333 5.300 27.667 4 J-98 1 2 3.922 29.500 6.300 22.000 4 A-98 1 2 3.137 29.750 5.800 23.000 4 S-98 1 2 3.137 28.333 5.067 29.333 4 0-98 1 2 4.706 28.833 4.800 28.333 4 N-98 1 2 4.706 28.333 5.433 25.000 4 J-97 1 3 17.059 29.333 5.067 30.333 4 A-97 1 3 2.614 29.500 4.125 32.000 4 S-97 1 3 17.255 27.167 4.545 27.667 4 0-97 1 3 5.882 29.667 4.967 30.333 4 N-97 1 3 36.471 30.333 5.185 29.000 4 E-98 1 3 8.235 29.167 4.533 31.500 4 M-98 1 3 6.667 30.167 4.633 33.333 4 A-98 3 12.549 31.000 5.150 29.000 4 M-98 3 9.804 29.667 4.733 34.333 4 J-98 3 10.980 30.333 5.300 27.667 4 J-98 3 9.412 29.500 6.300 22.000 4 A-98 3 5.490 29.750 5.800 23.000 4 S-98 3 9.804 28.333 5.067 29.333 4 0-98 3 12.157 28.833 4.800 28.333 4 N-98 1 3 12.549 28.333 5.433 25.000 4 J-97 2 1 0.980 28.000 3.667 31.667 4 A-97 2 1 0.098 27.000 3.067 34.333 4 S-97 2 1 3.137 24.750 2.900 35.000 4 0-97 2 1 0.098 27.333 2.733 34.000 4 N-97 2 1 1.765 29.333 3.130 30.667 4 E-98 2 1 0.784 26.833 3.533 33.500 4 M-98 2 1 0.000 25.833 3.067 34.667

184
4 A-98 2 0.784 27.375 3.050 31.250 4 M-98 2 1 4.706 26.833 3.233 36.000 4 J-98 2 1 1.569 28.667 3.867 33.000 4 J-98 2 1 3.529 28.200 3.800 30.000 4 A-98 2 1 1.046 28.500 3.775 30.500 4 S-98 2 1 2.157 24.333 2.933 33.667 4 0-98 2 1 0.588 26.333 3.333 31.333 4 N-98 2 1 5.490 25.375 3.000 31.500 4 J-97 2 2 1.520 28.000 3.667 31.667 4 A-97 2 2 0.392 27.000 3.067 34.333 4 S-97 2 2 11.503 24.750 2.900 35.000 4 0-97 2 2 1.275 27.333 2.733 34.000 4 N-97 2 2 3.529 29.333 3.130 30.667 4 E-98 ·2 2 1.176 26.833 3.533 33.500 4 M-98 2 2 1.373 25.833 3.067 34.667 4 A-98 2 2 7.582 27.375 3.050 31.250 4 M-98 2 2 7.451 26.833 3.233 36.000 4 J-98 2 2 1.961 28.667 3.867 33.000 4 J-98 2 2 1.569 28.200 3.800 30.000 4 A-98 2 2 1.830 28.500 3.775 30.500 4 S-98 2 2 2.745 24.333 2.933 33.667 4 0-98 2 2 3.922 26.333 3.333 31.333 4 N-98 2 2 6.667 25.375 3.000 31.500 4 J-97 2 3 1.814 28.000 3.667 31.667 4 A-97 2 3 0.343 27.000 3.067 34.333 4 S-97 2 3 13.595 24.750 2.900 35.000 4 0-97 2 3 2.549 27.333 2.733 34.000 4 N-97 2 3 5.294 29.333 3.130 30.667 4 E-98 2 3 2.353 26.833 3.533 33.500 4 M-98 2 3 2.745 25.833 3.067 34.667 4 A-98 2 3 4.967 27.375 3.050 31.250 4 M-98 2 3 5.490 26.833 3.233 36.000 4 J-98 2 3 3.922 28.667 3.867 33.000 4 J-98 2 3 2.549 28.200 3.800 30.000 4 A-98 2 3 2.353 28.500 3.775 30.500 4 S-98 2 3 1.961 24.333 2.933 33.667 4 0-98 2 3 2.157 26.333 3.333 31.333 4 N-98 2 3 5.490 25.375 3.000 31.500 4 J-97 3 1 1.765 25.500 3.100 32.000 4 A-97 3 1 5.948 23.500 2.100 35.500 4 S-97 3 1 2.745 22.833 2.133 35.667 4 0-97 3 1 0.980 23.500 2.167 35.667 4 N-97 3 1 0.392 27.667 2.550 31.667 4 E-98 3 1 0.392 24.667 2.933 34.000 4 M-98 3 1 2.157 23.000 2.167 35.000 4 A-98 3 1 0.980 23.000 1.400 32.667 4 M-98 3 1 0.784 23.333 1.967 37.000 4 J-98 3 1 2.353 25.667 2.900 34.667 4 J-98 3 1 1.176 22.750 1.450 32.500 4 A-98 3 1 0.196 22.500 1.700 34.333 4 S-98 3 1 0.392 21.000 1.733 35.000 4 0-98 3 1 0.784 22.167 1.667 34.667
185
4 N-98 3 1 3.137 20.500 1.100 32.500
4 J-97 3 2 9.314 25.500 3.100 32.000
4 A-97 3 2 2.222 23.500 2.100 35.500
4 S-97 3 2 4.510 22.833 2.133 35.667
4 0-97 3 2 3.725 23.500 2.167 35.667
4 N-97 3 2 3.333 27.667 2.550 31.667 4 E-98 3 2 0.784 24.667 2.933 34.000 4 M-98 3 2 3.725 23.000 2.167 35.000 4 A-98 3 2 3.529 23.000 1.400 32.667 4 M-98 3 2 3.725 23.333 1.967 37.000 4 J-98 3 2 4.510 25.667 2.900 34.667 4 J-98 3 2 0.784 22.750 1.450 32.500 4 A-98 3 2 3.137 22.500 1.700 34.333 4 S-98 3 2 0.294 21.000 1.733 35.000 4 0-98 3 2 3.725 22.167 1.667 34.667 4 N-98 3 2 3.922 20.500 1.100 32.500 4 J-97 3 3 6.078 25.500 3.100 32.000 4 A-97 3 3 1.699 23.500 2.100 35.500 4 S-97 3 3 7.059 22.833 2.133 35.667 4 0-97 3 3 1.176 23.500 2.167 35.667 4 N-97 3 3 3.137 27.667 2.550 31.667 4 E-98 3 3 2.353 24.667 2.933 34.000 4 M-98 3 3 1.569 23.000 2.167 35.000 4 A-98 3 3 3.529 23.000 1.400 32.667 4 M-98 3 3 3.529 23.333 1.967 37.000 4 J-98 3 3 4.118 25.667 2.900 34.667 4 J-98 3 3 1.176 22.750 1.450 32.500 4 A-98 3 3 1.569 22.500 1.700 34.333 4 S-98 3 3 0.588 21.000 1.733 35.000 4 0-98 3 3 2.157 22.167 1.667 34.667 4 N-98 3 3 8.627 20.500 1.100 32.500 11 J-97 1 1 4.052 28.833 5.000 31.333 11 A-97 1 0.915 29.250 4.500 33.500 11 S-97 1 12.026 28.500 4.650 31.333 11 0-97 1 0.784 29.500 4.800 32.000 11 N-97 1 3.922 30.667 4.735 29.333 11 E-98 1 2.745 30.167 4.667 30.000 11 M-98 1 3.529 30.167 4.733 33.667 11 A-98 1 6.275 31.500 5.350 30.000 11 M-98 1 1 0.392 29.833 4.867 34.667 11 J-98 1 1 3.529 30.500 5.000 31.000 11 J-98 1 1 2.353 30.000 6.200 23.000 11 A-98 1 1 0.392 29.833 5.533 29.333 11 S-98 1 1 5.490 28.333 4.667 30.333 11 0-98 1 1.961 29.333 5.267 27.333 11 N-98 1 7.451 28.667 5.433 27.667 11 J-97 2 5.490 28.833 5.000 31.333 11 A-97, 2 0.784 29.250 4.500 33.500 11 S-97 2 13.595 28.500 4.650 31.333 11 0-97 2 8.235 29.500 4.800 32.000 11 N-97 2 10.980 30.667 4.735 29.333 11 E-98 2 3.529 30.167 4.667 30.000

186
11 M-98 1 2 2.353 30.167 4.733 33.667 11 A-98 1 2 1.569 31.500 5.350 30.000
11 M-98 1 2 3.137 29.833 4.867 34.667 11 J-98 1 2 3.529 30.500 5.000 31.000 11 J-98 1 2 0.784 30.000 6.200 23.000 11 A-98 1 2 0.392 29.833 5.533 29.333 11 S-98 1 2 2.353 28.333 4.667 30.333 11 0-98 1 2 2.353 29.333 5.267 27.333 11 N-98 1 2 5.098 28.667 5.433 27.667
11 J-97 1 3 11.373 28.833 5.000 31.333 11 A-97 1 3 5.229 29.250 4.500 33.500 11 S-97 1 3 20.915 28.500 4.650 31.333 11 0-97 1 3 10.980 29.500 4.800 32.000 11 N-97 1 3 32.941 30.667 4.735 29.333
11 E-98 1 3 9.020 30.167 4.667 30.000 11 M-98 1 3 5.882 30.167 4.733 33.667 11 A-98 1 3 7.059 31.500 5.350 30.000 11 M-98 1 3 7.451 29.833 4.867 34.667 11 J-98 1 3 9.020 30.500 5.000 31.000 11 J-98 1 3 7.843 30.000 6.200 23.000 11 A-98 3 7.451 29.833 5.533 29.333 11 S-98 3 9.020 28.333 4.667 30.333 11 0-98 3 10.980 29.333 5.267 27.333 11 N-98 1 3 16.078 28.667 5.433 27.667 11 J-97 2 1 2.810 27.750 4.200 32.000 11 A-97 2 1 1.176 27.000 3.800 35.000 11 S-97 2 1 9.150 27.000 3.300 34.500 11 0-97 2 1 0.196 27.000 2.867 34.667 11 N-97 2 1 1.961 29.333 3.200 30.333 11 E-98 2 1 1.176 27.833 3.533 31.667 11 M-98 2 1 1.176 26.833 3.800 35.000 11 A-98 2 1 1.307 27.500 3.425 30.500 11 M-98 2 1 1.569 28.167 3.767 35.667 11 J-98 2 1 1.569 28.667 3.667 33.000 11 J-98 2 1 5.098 28.100 3.680 29.200 11 A-98 2 1 1.176 27.833 3.667 31.333 11 S-98 2 1 2.941 24.833 3.033 34.333 11 0-98 2 1 3.725 25.833 3.333 31.667 11 N-98 2 1 8.105 25.875 3.000 34.750 11 J-97 2 2 1.895 27.750 4.200 32.000 11 A-97 2 2 3.039 27.000 3.800 35.000 11 S-97 2 2 4.706 27.000 3.300 34.500 11 0-97 2 2 5.294 27.000 2.867 34.667 11 N-97 2 2 4.314 29.333 3.200 30.333 11 E-98 2 2 0.784 27.833 3.533 31.667 11 M-98 2 2 1.373 26.833 3.800 35.000 11 A-98 2 2 1.569 27.500 3.425 30.500 11 M-98 2 2 1.569 28.167 3.767 35.667 11 J-98 2 2 0.980 28.667 3.667 33.000 11 J-98 2 2 1.961 28.100 3.680 29.200 11 A-98 2 2 0.980 27.833 3.667 31.333 11 S-98 2 2 1.569 24.833 3.033 34.333

187
11 0 -98 2 2 1.961 25.833 3.333 31.667 11 N-98 2 2 5.882 25.875 3.000 34.750 11 J-97 2 3 2.026 27.750 4.200 32.000 11 A-97 2 3 4.020 27.000 3.800 35.000 11 S-97 2 3 9.935 27.000 3.300 34.500 11 0-97 2 3 2.157 27.000 2.867 34.667 11 N-97 2 3 4.118 29.333 3.200 30.333 11 E-98 2 3 3.529 27.833 3.533 31 .667 11 M-98 2 3 2.353 26.833 3.800 35.000 11 A-98 2 3 3.660 27.500 3.425 30.500 11 M-98 2 3 2.157 28.167 3.767 35.667 11 J-98 2 3 3.333 28.667 3.667 33.000 11 J-98 2 3 2.549 28.100 3.680 29.200 11 A-98 2 3 1.176 27.833 3.667 31 .333 11 S-98 2 3 7.059 24.833 3.033 34.333 11 0-98 2 3 4.902 25.833 3.333 31 .667 11 N-98 2 3 12.026 25.875 3.000 34.750 11 J-97 3 1 0.490 25.833 3.333 32.667 11 A-97 3 1 0.196 23.000 2.500 35.500 11 S-97 3 1 2.941 24.167 2.450 35.667 11 0-97 3 1 0.588 23.667 2.400 35.667 11 N-97 3 1 3.529 27.000 2.800 31.667 11 E-98 3 0.196 25.000 3.200 33.333 11 M-98 3 1.569 23.167 2.367 35.000 11 A-98 3 1.176 23.000 1.967 33.000 11 M-98 3 1 1.569 24.000 2.033 37.000 11 J-98 3 1 1.569 26.000 2.667 34.333 11 J-98 3 1 2.353 23.500 1.750 32.500 11 A-98 3 1 0.392 22.500 1.867 34.667 11 S-98 3 1 0.588 21.667 2.233 35.000 11 0-98 3 1 1.569 21 .833 1.733 35.000 11 N-98 3 1 3.529 21 .000 1.250 35.000 11 J-97 3 2 2.059 25.833 3.333 32.667 11 A-97 3 2 1.863 23.000 2.500 35.500 11 S-97 3 2 9.020 24.167 2.450 35.667 11 0-97 3 2 0.392 23.667 2.400 35.667 11 N-97 3 2 8.235 27.000 2.800 31.667 11 E-98 3 2 2.745 25.000 3.200 33.333 11 M-98 3 2 7.059 23.167 2.367 35.000 11 A-98 3 2 3.529 23.000 1.967 33.000 11 M-98 3 2 2.157 24.000 2.033 37.000 11 J-98 3 2 4.510 26.000 2.667 34.333 11 J-98 3 2 1.176 23.500 1.750 32.500 11 A-98 3 2 0.588 22.500 1.867 34.667 11 S-98 3 2 2.353 21.667 2.233 35.000 11 0-98 3 2 2.745 21.833 1.733 35.000 11 N-98 3 2 9.020 21.000 1.250 35.000 11 J-97 3 3 9.020 25.833 3.333 32.667 11 A-97 3 3 1.765 23.000 2.500 35.500 11 S-97 3 3 4.706 24.167 2.450 35.667 11 0-97 3 3 0.784 23.667 2.400 35.667 11 N-97 3 3 5.294 27.000 2.800 31.667
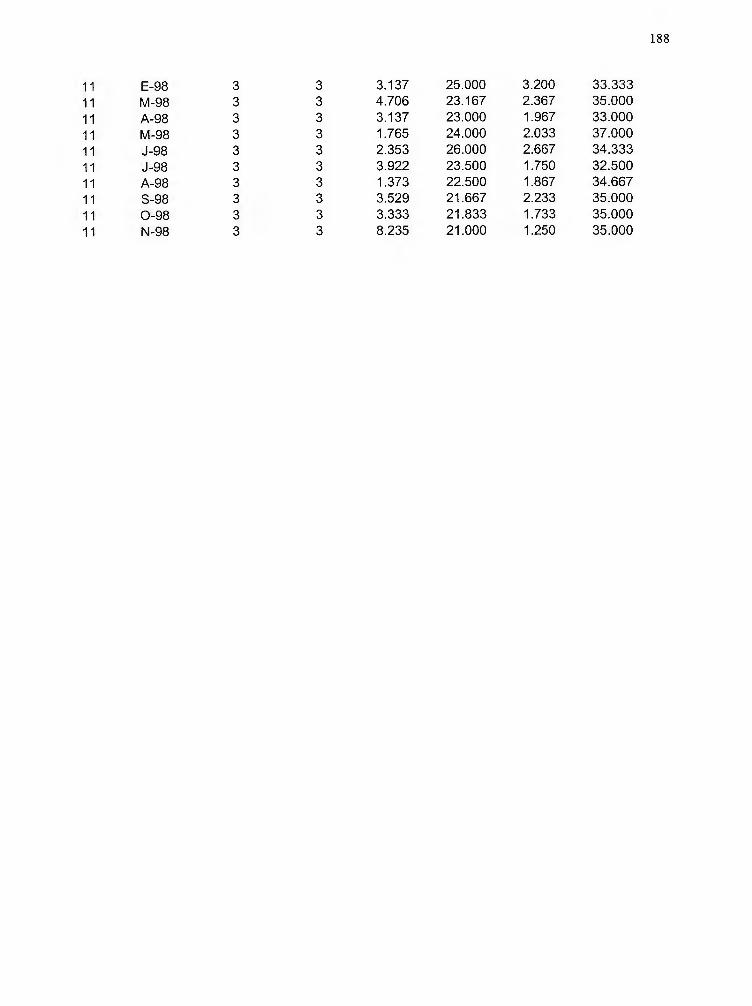
188
11 E-98 3 3 3.137 25.000 3.200 33.333 11 M-98 3 3 4.706 23.167 2.367 35.000
11 A-98 3 3 3.137 23.000 1.967 33.000
11 M-98 3 3 1.765 24.000 2.033 37.000 11 J-98 3 3 2.353 26.000 2.667 34.333
11 J-98 3 3 3.922 23.500 1.750 32.500 11 A-98 3 3 1.373 22.500 1.867 34.667 11 S-98 3 3 3.529 21 .667 2.233 35.000 11 0-98 3 3 3.333 21.833 1.733 35.000 11 N-98 3 3 8.235 21.000 1.250 35.000

189
APENDICE 3
Matriz de datos de abundancia (indiv/m3) de zooplancton para cada una de las estaciones,
fechas y profundidades analizadas
Estacion 1 Prof. 1 Fecha Abr-97 Ago-97 Nov-97 Mar-98 Abr-98 Set-98 Grupo-Canthoca/anus 3.62806122 o o 5.8047619 26.1221939 o Undinula o o o o o o Acroca/anus 438.995408 60.6423469 27.9887755 60.95 87.0739796 156.728571 Paracalanus 1248.05306 417.499235 363.854082 690.766667 1219.03571 1845.91429 Euca/anus o o o 5.8047619 8.70739796 o Euchaeta o o o o o o Temora o o o o 8.70739796 o Temoropia o o o o o o Pleuromamma o o o o o o Centropages 50.7928571 4.66479592 3.10986395 34.8285714 52.2443878 o Lucicutia o o o 2.90238095 8.70739796 o Pseudodiaptomus o o o o o o Candacia o o o o o o Labidocera o o o o o o Pontellina o o o o o o Acartia 246.708163 4.66479592 133.72415 52.2428571 95.7813776 34.8285714 Oithona 366.434184 3181.39082 2612.28571 1256.73095 2142.0199 2525.07143 Euterpina 228.567857 4.66479592 18.6591837 29.0238095 226.392347 539.842857 C/ytemnestra o o o 8.70714286 o o Macrosetel/a 18.1403061 2.33239796 o o o o Microsetella o o 3.10986395 o 8.70739796 o Oncaea 25.3964286 9.32959184 15.5493197 577.57381 565.980867 104.485714 Corycaeus 79.8173469 18.6591837 6.21972789 113.192857 191.562755 121.9 Lubockia o o o o o o Copilia o o o o o o Saphirina o o o 2.90238095 o o Calocalanus o o o 17.4142857 o o C/ausocalanus o o 3.10986395 40.6333333 26.1221939 8.70714286 Spinocalanus o o o o o o Euchirella o o o o o o Copépodos 2706.53367 3703.84796 3187.61054 2899.47857 4667.16531 5337.47857 Copepoditos 1701.56071 137.61148 603.313605 824.27619 1428.01327 1053.56429 Nauplius 105.213776 23.3239796 99.5156463 307.652381 922.984184 243.8 Hidromedusas 152.378571 13.9943878 3.10986395 49.3404762 34.8295918 26.1214286 Ctenoforos o o 3.10986395 2.90238095 o 8.70714286 Pteropodos 185.031122 18.6591837 o 58.047619 87.0739796 8.70714286 Gastropodos 94.3295918 23.3239796 3.10986395 20.3166667 17.4147959 95.7785714 L. Molusco 61.6770408 18.6591837 o 52.2428571 121.903571 o L.Poliqueto 97.9576531 97.9607143 3.10986395 14.5119048 8.70739796 34.8285714 L. Equinodermo 192.287245 o o o o o

190
L.Crustaceo 68.9331633 o o 8.70714286 8.70739796 o Cladoceros 188.659184 o o 69.6571 429 2411.94923 o Ostracodos 156.006633 592.429082 46.6479592 531.135714 296.051531 313.457143 Apendicularias 493.416327 468.81199 463.369728 899.738095 1044.88776 1419.26429 Salpas o o o 20.316666.? o 34.8285714 Anfipodos o o o o o o Quetognatos 65.305102 25.6563776 12.4394558 31.9261905 104.488776 43.5357143 Lingula 3.62806122 o o o o o Actinotroca 10.8841837 o o o o o Larvas de pez 3.62806122 o o 5.8047619 8.70739796 o Huevos de lnvert 105.213776 o 34.2085034 116.095238 130.610969 52.2428571 Calamar o o o o o o Anfioxo o o o 2.9023809'5 o o
Estacion 2 Prof. 1 Fecha Abr-97 Ago-97 Nov-97 Mar-98 Abr~98 Set-98
Canthocalanus o o o o o Undinula o o o o o Acroca/anus 27.9887755 32.6517857 29.0246599 83.96403G6 60.95 Paraca/anus 338.197704 100.132143 348.295918 2071.11276 1506.33571 Euca/anus o o 29.0246599 o o Euchaeta o o o o o Temora o o 11.6098639 1'8.6586735 o Temoropia o o o o o Pleuromamma o o o o o Centropages 11.6619898 2.17678571 111.6098639 11 1, .952041 o Lucicutia o o o o o Pseudodiaptomus o o o o o Candacia 6.99719388 o o 18.6586735 8.70714286 Labidocera o o o 9.32933673 8.7G714286 Pontellina o o o o o Acartia 2.33239796 30.475 11.6098639 55.9760204 34.8285714
Oithona 1170.86378 814.117857 743.031293 2789.47168 3500.27143 Euterpina 4.66479592 28.29821143 40.6345238 820.981 '633 174.142857 Clytemnestra o o o o o Macrosetella o o o o 8.70714286 Microsetella o o 5.80493197 o o Oncaea 60.6423469 19.5910714 493.419218 942.26301 513.721429 Corycaeus 58.309949 4.353571143 75.4641156 382.502806 200.264286 Lubockia o o o o o Copilia o o o o o Saphirina o o o o o Ca/ocalanus o 15.2375 11.6098639 18.6586735 17.4142857 C/ausocalanus 2.33239796 2.17678571 11.6098639 9.32933673 26.1214286 Spinoca/anus o o o o o Euchire/la o o o o o Copépodos 1683.99133 1049.21071 1822.74864 7332.85867 6060. H 143 Copepoditos 83.9663265 185.026786 388.930442 1725.9273 870.714286 Nauplius 20.9915816 56.5964286 417.955102 541.101531 400.528571 Hidromedusas 2.33239796 4.35357143 52.2443878 37.3173469 8.70714286

191
Ctenoforos o o o 9.32933673 o Pteropodos 11 .6619898 o 29.0246599 111 .952041 43.53571 43 Gastropodos 11.6619898 13.0607143 23.2197279 o 17.4142857 L. Molusco 4.66479592 o 29.0246599 46.6466837 o L.Poliqueto 41.9831633 8.70714286 5.80493197 9.32933673 60.95 L. Equinodermo o 2.17678571 o o o L.Crustaceo 2.33239796 o 5.80493197 9.32933673 17.4142857 Cladoceros 2.33239796 o 46.4394558 625.065561 8.70714286 Ostracodos 275.222959 322.164286 307.661395 429.14949 278.628571 Apendicularias 305.544133 533.3125 557.273469 410.490816 1480.21429 Salpas 2.33239796 o 11 .6098639 37.3173469 26.1214286 Anfípodos o o o o o Quetognatos 13.9943878 26.1214286 63.8542517 55.9760204 78.3642857 Lingula o 2.17678571 o 9.32933673 o Actinotroca o o o o o Larvas de pez o o o o o Huevos de lnvert 18.6591837 124.076786 34.8295918 37.3173469 78.3642857 Calamar o o o o o Anfioxo o o o o o
Estacion 3 Prof. 1 Fecha Abr-97 Ago-97 Nov-97 Mar-98 Abr-98 Set-98 Especie Canthoca/anus o 2.72104592 11.6619898 o o o Undinula o 2.72104592 2.33239796 o o o Acrocalanus 275.732653 10.8841837 156.270663 87.0739796 43.5369898 o Paracalanus 2554.1551 775.498087 688.057398 748.836224 731.421429 774.935714 Eucalanus 58.0489796 2.72104592 o 29.0246599 o 17.4142857 Euchaeta o o o o o o Temora o 2.72104592 o 5.80493197 8.70739796 o Temoropia o o o o o o Pleuromamma o o o o o o Centropages 609.514286 5.44209184 9.32959184 34.8295918 17.4147959 17.4142857 Lucicutia o o o 11.6098639 o o Pseudodiaptomus o o o 11.6098639 o 8.70714286 Candacia o 8.16313776 o 17.4147959 o 8.70714286 Labidocera o o o o 8.70739796 8.70714286 Ponte/Ji na o o o o o o Acartia 2278.42245 40.8156888 25.6563776 110.293707 8.70739796 69.6571429 Oithona 1639.88367 1357.80191 2640.27449 1219.03571 2333.58265 1950.4 Euterpina 1654.39592 48.9788265 118.952296 139.318367 139.318367 113.192857 Clytemnestra o 2.72104592 o o o o Macrosetel/a 29.0244898 5.44209184 2.33239796 17.4147959 8.70739796 o Microsetel/a o o 2.33239796 11.6098639 o 17.4142857 Oncaea 1277.07755 40.8156888 186.591837 284.441667 43.5369898 226.385714 Corycaeus 1131.9551 78.9103316 97.9607143 162.538095 165.440561 165.435714 Lubockia o o o o o o Copi/ia 14.5122449 o o o o o Saphirina o o o o o o Caloca/anus o 10.8841837 51.3127551 o o o Clausocalanus o 2.72104592 2.33239796 34.8295918 o o

192
Spinocalanus o o o o o o Euchirella o o o o o o Copépodos 111 522.7224 2399.9625 3995.3977 2925.68571 3509.08138 3378.37143 Copepoditos 3816.72041 370.062245 706.716582 1346.74422 1236.45051 2481.53571 Nauplius 1204.51633 117.004974 184.259439 644.347449 748.836224 2342.22143 Hidromedusas 188.659184 10.8841837 2.33239796 34.8295918 8.70739796 26.1214286 Ctenoforos o o o 17.4147959 o 8.70714286 Pteropodos 682.07551 19.0473214 20.9915816 81.2690476 104.488776 34.8285714 Gastropodos 87.0734694 43.5367347 11.6619898 81,.2690476 60.9517857 17.4142857 L. Molusco 87.0734694 21.7683673 2.33239796 34.829591,8 60.9517857 34.8285714 l.Poliqueto 72.5612245 46.2577806 16.3267857 46.4394558 17.4147959 34.8285714 L. Equinodermo o 2.72104592 o o o o L.Crustaceo 188.659184 38.0946429 4.66479592 23.2197279 o o Cladoceros 29.0244898 5.44209184 o 87.0739796 26.1221939 o Ostracodos 1059.39388 119.72602 389.510459 179.952891 95.7813776 235.092857 Apendicularias 2670.25306 187.752168 786.0181112 806.885544 1001.35077 1314.77857 Salpas 29.0244898 o o 5.804931:97 26.1221939 o Anfipodos 14.5122449 o o o o o Quetognatos 348.293878 68.026148 62.9747449 98.6838435 52.2443878 43.5357143 Lingula o 5.44209184 o o o o Actinotroca o o o o o o Larvas de pez 29.0244898 o 9.32959184 o o o Huevos de lnvert 4353.67347 o o 34.8295918 52.2443878 95.7785'714 Calamar o o o o o o Anfioxo o o o o o o
Estacion 4 Prof. 1 Fecha Abr-97 Ago-97 Nov-97 Mar-98 Abr-98 Set-98 Especie Canthocalanus 87.0748299 o o o o o Undinula o 6.21972789 o o o o Acrocalanus 239.455782 68.4170068 30.475 8.70714286 78.3642857 o Paracalanus 2111.56463 488.248639 1097.1 383.114286 713.985714 896.86199 Eucalanus 130.612245 6.21972789 o 4.35357143 o 17.4147959 Euchaeta o o o o o o Temora o 6.21972789 4.35357143 4.35357143 8.70714286 o Temoropia o o o o o o Pleuromamma o o o o o o Centropages 261.22449 3.10986395 13.0607143 21.7678571 60.95 8.70739796 Lucicutia o o o 4.35357143 o o Pseudodiaptomus 21.7687075 o o o o o Candacia o 6.21972789 4.35357143 4.35357143 o o Labidocera o 3.10986395 4.35357143 o o o Ponte/Ji na o o o o o o Acartia 718.367347 15.5493197 100.132143 30.475 17.4142857 43.5369898 Oithona 1131.97279 2015.191,84 2869.00357 1240.76786 1654.35714 2054.94592 Euterpina 957.823129 31.0986395 100.132143 113.192857 208.971429 60.9517857 Clytemnestra o o o o o 8.70739796 Macrosetella o 6.21972789 o 13.0607143 o o Microsetella o o o 30.475 8.70714286 17.4147959 O ne a ea 1523.80952 1'02.62551 78.3642857 152.375 113.192857 113.196173

193
Corycaeus 1502.04082 118.17483 161.082143 261.214286 278.628571 104.488776 Lubockia o o o o o o Copi/ia 43.537415 3.10986395 o o o o Saphirina o o o o o o Caloca/anus 21.7687075 15.5493197 8.70714286 o 8.70714286 34.8295918 C/ausocalanus o 6.21972789 o 4.35357143 o 8.70739796 Spinocalanus o o o o o o Euchirel/a o o o o o o Copépodos 8751.02041 2901.50306 4471.11786 2276.91786 3151.98571 3369.76301 Copepoditos 2764.62585 279.887755 944.725 988.260714 1062.27143 1166.79133 Nauplius 3308.84354 146.163605 235.092857 1192.87857 1114.51429 644.347449 Hidromedusas 65.3061224 6.21972789 o 21.7678571 o 8.70739796 Ctenoforos o o o 4.35357143 o o Pteropodos 609.52381 21.7690476 60.95 43.5357143 60.95 8.70739796 Gastropodos 87.0748299 15.5493197 26.1214286 17.4142857 60.95 60.9517857 L. Molusco 21.7687075 o 8.70714286 8.70714286 17.4142857 34.8295918 L.Poliqueto 87.0748299 18.6591837 34.8285714 13.0607143 34.8285714 34.8295918 L. Equinodermo o 3.10986395 o o o o L.Crustaceo 217.687075 6.21972789 30.475 13.0607143 o 8.70739796 Cladoceros o 3.10986395 4.35357143 17.4142857 17.4142857 o Ostracodos 391.836735 668.620748 404.882143 187.203571 130.607143 217.684949 Apendicularias 783.673469 373.183673 1284.30357 592.085714 1367.02143 374.418112 Salpas 21.7687075 o o 8.70714286 8.70714286 o Anfipodos o o o 4.35357143 o o Quetognatos 108.843537 102.62551 139.314286 108.839286 113.192857 165.440561 Lingula o o o o 8.70714286 o Actinotroca o o o o o o Larvas de pez o o 4.35357143 o 8.70714286 o Huevos de lnvert 21.7687075 93.2959184 34.8285714 13.0607143 34.8285714 87.0739796 Calamar o o o o o o Anfioxo o o o o o o
Estacion 11 Prof. 1 Fecha Abr-97 Ago-97 Nov-97 Mar-98 Abr-98 Set-98 Especie Canthocalanus o 3.10986395 o 4.35357143 o o Undinula o 24.8789116 o o o o Acroca/anus 217.683673 71.5268707 56.5964286 34.8285714 21.7678571 30.475 Paracalanus 3410.37755 575.32483 949.078571 304.75 513.721429 700.925 Euca/anus 217.683673 3.10986395 4.35357143 13.0607143 o 17.4142857 Euchaeta 14.5122449 o o o o o Temora 43.5367347 o o 8.70714286 17.4142857 4.35357143 Temoropia o o o o o o P/euromamma o o o o o o Centropages 391.830612 15.5493197 4.35357143 8.70714286 13.0607143 13.0607143 Lucicutia o o 4.35357143 o o o Pseudodiaptomus o o o 8.70714286 o 4.35357143 Candacia 14.5122449 3.10986395 4.35357143 8.70714286 4.35357143 4.35357143 Labidocera 14.5122449 o o o 17.4142857 o Pontellina o o o o o o Acartia 406.342857 27.9887755 95.7785714 47.8892857 4.35357143 47.8892857

194
Oithona 2554.1551 1013.81565 4366.63214 1397.49643 413.589286 866.360714 Euterpina 885.246939 71.5268707 204.617857 108.839286 65.3035714 113.192857 Clytemnestra o o o o o 8.70714286 Macrosetella 72.5612245 3.10986395 4.35357143 4.35357143 13.0607143 4.35357143 Microsetel/a o o o 13.0607143 4.35357143 60.95 Oncaea 1944.64082 80.8564626 126.253571 213.325 256.860714 313.457143 Corycaeus 1088.41837 87.0761905 148.021429 208.971429 65.3035714 121.9 Lubockia o o o o o o Copilia 14.5122449 o o o 4.35357143 o Saphirina o o o o o 8.70714286 Caloca/anus 43.5367347 34.2085034 17.4142857 13.0607143 4.35357143 26.1214286 C/ausocalanus o 18.6591837 4.35357143 4.35357143 o 8.70714286 Spinoca/anus o o o o o o Euchirel/a o o o o o o Copépodos 11334.0633 2033.85102 5990.51429 2403.17143 1419.26429 2355.28214 Copepoditos 2292.93469 267.448299 1628.23571 766.228571 831 .532143 979.553571 Nauplius 2975.0102 161.712925 330.871429 979.553571 444.064286 622.560714 Hidromedusas 145.122449 6.21972789 13.0607143 34.8285714 o 26.1214286 Ctenoforos o o o 8.70714286 o 4.35357143 Pteropodos 827.197959 37.3183673 4.35357143 65.3035714 13.0607143 17.4142857 Gastropodos 740.12449 3.10986395 13.0607143 21 .7678571 47.8892857 56.5964286 L. Molusco 319.269388 9.32959184 13.0607143 o 30.475 65.3035714 L.Poliqueto 174.146939 18.6591837 17.4142857 21 .7678571 8.70714286 34.8285714 L. Equinodermo 29.0244898 o 4.35357143 4.35357143 o o L.Crustaceo 174.146939 12.4394558 4.35357143 o 8.70714286 8.70714286 Cladoceros o o o 26.1214286 4.35357143 4.35357143
Ostracodos 2757.32653 118.17483 370.053571 60.95 17.4142857 156.728571 Apendicularias 1538.29796 643.741837 1776.25714 666.096429 435.357143 457.125
Salpas 29.0244898 12.4394558 o 17.4142857 o 4.35357143
Anfipodos 29.0244898 o o o o o Quetognatos 391 .830612 74.6367347 104.485714 126.253571 21 .7678571 156.728571
Lingula o 3.10986395 4.35357143 o o o Actinotroca o o o o o o Larvas de pez 14.5122449 o o o 4.35357143 4.35357143
Huevos de lnvert 1015.85714 o 87.0714286 21.7678571 8.70714286 78.3642857
Calamar o o o o o o Anfioxo o o o o o o
Estacion 1 Prof. 2 Fecha Abr-97 Ago-97 Nov-97 Mar-98 Abr-98 Set-98
Grupo Canthoca/anus o o o o o o Undinula o o o o o o Acrocalanus 4.76190476 o 8.16122449 1.55510204 27.9887755 17.4910714
Paracalanus 4.08163265 21.7673469 15.6423469 8.16428571 205.25102 72.2964286
Euca/anus o o o o 3.10986395 o Euchaeta o o o o o o Temora o o o o o o Temoropia o o o o o o Pleuromamma o o o o o o Centropages o o o o 9.32959184 o

195
Lucicutia o 2.72091837 o 1.55510204 3.10986395 o Pseudodiaptomus o o o o o o Candacia o o o o o o Labidocera o o o o o o Pontel/ina o o o o o o Acartia 2.04081633 o o 0.38877551 o o Oithona 70.7482993 136.045918 158.463776 68.8132653 796.12517 198.232143 Euterpina 0.68027211 5.44183673 6.80102041 4.27653061 40.4282313 4.66428571 Clytemnestra o o 0.68010204 0.38877551 o o Macrosetel/a o o o 0.38877551 o o Microsetella o o 1.36020408 1.94387755 9.32959184 1.16607143 Oncaea 57.1428571 29.930102 49.647449 61.4265306 295.437075 27.9857143 Corycaeus o 8.1627551 o 3.11020408 15.5493197 3.49821429 Lubockia o o o o o o Copilia o o o o o o Saphirina o o o o o o Calocalanus o o o 1.16632653 15.5493197 3.49821429 Clausoca/anus o 2.72091837 o 0.77755102 9.32959184 1.16607143 Spinoca/anus o o o o o o Euchirella o o o o o o Copépodos 139.455782 206.789796 240.756122 153.955102 1430.53741 329.998214 Copepoditos 59.8639456 27.2091837 25.1637755 28.7693878 450.930272 99.1160714 Nauplius o 5.44183673 4.76071429 9.33061224 258.118707 20.9892857 Hidromedusas 4.08163265 21.7673469 4.08061224 1.16632653 37.3183673 3.49821429 Ctenoforos o o o o o o Pteropodos o o o o 18.6591837 1.16607143 Gastropodos o 5.44183673 6.12091837 1.94387755 3.10986395 4.66428571 L. Molusco 3.40136054 o 7.48112245 5.83163265 18.6591837 3.49821429 L.Poliqueto 21.0884354 24.4882653 25.1637755 17.8836735 43.5380952 2.33214286 L. Equinodermo o o o o o 1.16607143 L.Crustaceo 5.44217687 o o o 6.21972789 o Cladoceros 2.04081633 o o 0.77755102 167.932653 o Ostracodos 0.68027211 473.439796 71.4107143 36.9336735 279.887755 268.196429 Apendicularias 22.4489796 13.6045918 8.84132653 20.2163265 295.437075 73.4625 Salpas o o o o 9.32959184 o Anfipodos o o o o 3.10986395 o Quetognatos 0.68027211 o 13.6020408 3.49897959 21.7690476 3.49821429 Lingula o o o o o o Actinotroca o o o o o o Larvas de pez o o o o o o Huevos de lnvert 6.12244898 40.8137755 12.9219388 3.11020408 24.8789116 9.32857143 Calamar o o o o o o Anfioxo o o o o o o
Estacion 2 Prof. 2 Fecha Abr-97 Ago-97 Nov-97 Mar-98 Abr-98 Set-98
Canthocalanus o o o 5.8047619 o Undinula o o o o o Acroca/anus 3.06122449 39.6464286 o 26.1214286 4.66479592 Paracalanus 45.9183673 69.9642857 13.9928571 116.095238 303.211735

196
Eucalanus o o o 2.90238095 o Euchaeta o o o 2.90238095 o Temora o o o 5.8047619 o Temoropia o o o o o Pleuromamma o o o o o Centropages o o o 14.5119048 o Lucicutia o o 1.5547619 o o Pseudodiaptomus o o o o o Candacia o o o 5.8047619 2.33239796 Labidocera o o o o o Pontellina o o o o o Acartia 1.02040816 o o 5.8047619 o Oithona 65.3061224 73.4625 79.2928571 452.771429 580.767092 Euterpina 3.06122449 o 4.66428571 499.209524 74.6367347 C/ytemnestra 2.04081633 o o o o Macrosetella 2.04081633 o o 2.90238095 o Microsetella o o 1.5547619 o 2.33239796 Oncaea 8.16326531 55.9714286 62.1904762 531.135714 48.9803571 Corycaeus 8.16326531 o 9.32857143 2.90238095 23.3239796 Lubockia o o o o o Copilia o o o o o Saphirina o o o o o Ca/ocalanus o o 3.10952381 23.2190476 11.6619898 C/ausocalanus o 6.99642857 1.5547619 14.5119048 2.33239796 Spinocalanus o o o o o Euchirella o o o 2.90238095 o Copépodos 138.77551 246.041071 177.242857 1715.30714 1054.24388 Copepoditos 118.367347 271.694643 46.6428571 377.309524 296.214541 Nauplius 2.04081633 5.83035714 20.2119048 217.678571 128.281888 Hidromedusas 3.06122449 3.49821429 o 29.0238095 11.6619898 Ctenoforos o o o o o Pteropodos o o o o 2.33239796 Gastropodos 4.08163265 3.49821429 7.77380952 40.6333333 37.3183673 L. Molusco 3.06122449 9.32857143 34.2047619 46.4380952 9.32959184 L.Poliqueto 7.14285714 4.66428571 o 20.3166667 113.9943878 L. Equinodermo o o o o o L.Crustaceo 4.08163265 o 3.10952381 5.8047619 9.32959184 Cladoceros o o 3.10952381 14.5119048 o Ostracodos 10.2040816 23.3214286 17.102381 551.452381 146.941071 Apendicularias 3.06122449 12.8267857 49.752381 95.7785714 263.560969 Salpas o o o 69.6571429 o Anfipodos o o o o o Quetognatos o 2.33214286 1.5547619 17.4142857 13.9943878 Lingula o 1.16607143 o o o Actinotroca o o o o o Larvas de pez o o o 2.90238095 o Huevos de lnvert 20.4081633 45.4767857 4.66428571 5.8047619 32.6535714 Calamar o o o o o Anfioxo o o o o o

197
Estacion 3 Prof. 2 Fecha Abr-97 Ago-97 Nov-97 Mar-98 Abr-98 Set-98 Especie Canthoca/anus 10.8843537 o 8.1627551 o 1.16607143 Undinula o o o o o Acroca/anus 174.14966 77.5510204 59.8602041 5.8047619 12.8267857 Paracalanus 1229.93197 416.326531 114.278571 55.1452381 111.942857 Eucalanus 76.1904762 o o 2.90238095 1.16607143 Euchaeta o o 5.44183673 o o Temora o 4.08163265 o o o Temoropia o o o o o Pleuromamma o o o o o Centropages 32.6530612 o o o o Lucicutia 10.8843537 o 8.1627551 5.8047619 3.49821429 Pseudodiaptomus o o o o o Candacia o o o o o Labidocera o o o o 3.49821429 Pontellina o o o o o Acartia 65.3061224 o o o 2.33214286 Oithona 1001.36054 106.122449 157.813265 356.992857 222.719643 Euterpina 576.870748 4.08163265 2.72091837 11.6095238 1.16607143 Clytemnestra o o o o o Macrosetella 32.6530612 8.16326531 o o o Microsetel/a o o 2.72091837 14.5119048 4.66428571 O nea ea 565.986395 130.612245 285.696429 368.602381 116.607143 Corycaeus 152.380952 o 10.8836735 8.70714286 8.1625 Lubockia o o o o o Copilia o o o o o Saphirina o o 2.72091837 o 1.16607143 Ca/oca/anus o o o 8.70714286 1.16607143 C/ausocalanus 10.8843537 o 10.8836735 5.8047619 6.99642857 Spinocalanus o o o o o Euchirella o o o 2.90238095 o Copépodos 3940.13605 746.938776 669.345918 847.495238 499.078571 Copepoditos 1099.31973 200 136.045918 171.240476 163.25 Nauplius 1926.53061 o o 55.1452381 53.6392857 Hidromedusas 65.3061224 24.4897959 5.44183673 8.70714286 6.99642857 Ctenoforos o o o o o Pteropodos 21 .7687075 o o 5.8047619 o Gastropodos 54.4217687 28.5714286 5.44183673 8.70714286 4.66428571 L. Molusco 10.8843537 8.16326531 10.8836735 34.8285714 o L.Poliqueto 32.6530612 4.08163265 13.6045918 o 9.32857143 L. Equinodermo o o o o o L.Crustaceo 54.4217687 12.244898 5.44183673 o o Cladoceros o o o 2.90238095 o Ostracodos 87.0748299 510.204082 138.766837 208.971429 139.928571 Apendicularias 609.52381 24.4897959 19.0464286 23.2190476 17.4910714 Salpas 10.8843537 o o o o

198
Anfipodos o 4.08163265 o o o Quetognatos 97.9591837 4.08163265 8.1627551 5.8047619 9.32857143 Lingula o o o o o Actinotroca o o o o o Larvas de pez o o o o o Huevos de lnvert 141.496599 81.6326531 35.3719388 o 8.1625 Calamar o o o o o Anfioxo o o o o o
Estacion 4 Prof. 2 Fecha Abr-97 Ago-97 Nov-97 Mar-98 Abr-98 Set-98 Especie Canthoca/anus o 1.16607143 4.66428571 o o Undinula o 2.33214286 o o o Acrocalanus o 25.6535714 9.32857143 5.80493197 16.325 Paracalanus 420.408163 195.9 317.171429 470.19949 76.9607143 Eucalanus 4.08163265 2.33214286 11.6607143 o 4.66428571 Euchaeta o o o o 2.33214286 Temora o o 2.33214286 o o Temoropia o o o o o P/euromamma o o o o o Centropages o 1.16607143 2.33214286 o o Lucicutia 4.08163265 o 6.99642857 o 2.33214286 Pseudodiaptomus o o o o o Candacia o 2.33214286 6.99642857 o o Labidocera o o o o o Ponte/Ji na o o o o o Acartia o 6.99642857 6.99642857 11 .6098639 o Oithona 261 .22449 535.226786 524.732143 1131 .96173 179.575 Euterpina o 15.1589286 58.3035714 58.0493197 2.3321 4286 Clytemnestra 4.08163265 2.33214286 4.66428571 o 4.66428571 Macrosetella 4.08163265 o o o o Microsetella o 9.32857143 11.6607143 5.80493197 25.6535714 Oncaea 444.897959 99.1160714 184.239286 226.392347 256.535714 Corycaeus 44.8979592 23.3214286 18.6571429 46.4394558 11.6607143 Lubockia 4.08163265 o o o o Copilia o o o o o Saphirina o o o o o Ca/oca/anus 12.244898 24.4875 18.6571429 5.80493197 11.6607143 C/ausocalanus 53.0612245 1.16607143 4.66428571 o 6.99642857 Spinoca/anus o o o o o Euchirella o o 2.33214286 o o Copépodos 1257.14286 948.016071 1196.38929 1962.06701 601.692857 Copepoditos 183.673469 188.903571 417.453571 446.979762 151.589286 Nauplius 53.0612245 25.6535714 97.95 121.903571 27.9857143 Hidromedusas 12.244898 2.33214286 11.6607143 11 .6098639 9.32857143 Ctenoforos o o o o o Pteropodos o 18.6571429 4.66428571 o o Gastropodos 16.3265306 3.49821429 4.66428571 17.4147959 25.6535714 L. Molusco 4.08163265 5.83035714 4.66428571 29.0246599 16.325 L.Poliqueto 12.244898 3.49821429 o o 6.99642857

199
L. Equinodermo o o o o o L.Crustaceo o o 2.33214286 o o Cladoceros o o o o o Ostracodos 877.55102 51.3071429 128.267857 481 .809354 200.564286 Apendicularias 20.4081633 85.1232143 48.975 46.4394558 16.325 Salpas o o o 5.80493197 o Anfipodos 4.08163265 o o o o Quetognatos 20.4081633 19.8232143 9.32857143 17.4147959 11.6607143 Lingula 4.08163265 1.16607143 o o 2.33214286 Actinotroca o o o o o Larvas de pez o o o o o Huevos de lnvert 65.3061224 6.99642857 11 .6607143 5.80493197 27.9857143 Calamar o o o o o Anfioxo o 1.16607143 o o o
Estacion 11 Prof. 2 Fecha Abr-97 Ago-97 Nov-97 Mar-98 Abr-98 Set-98 Especie Canthocalanus o 1.16607143 o o o Undinula 1.36020408 1.16607143 o o o Acroca/anus 8.16122449 36.1482143 16.325 17.4142857 74.6367347 Paraca/anus 29.9244898 248.373214 242.542857 397.62619 289.217347 Eucalanus 4.08061224 o 6.99642857 2.90238095 4.66479592 Euchaeta o 2.33214286 4.66428571 o 4.66479592 Temora o o 4.66428571 o o Temoropia o o o o o Pleuromamma o o o o o Centropages o o o 5.8047619 o Lucicutia o 2.33214286 2.33214286 o 4.66479592 Pseudodiaptomus o 3.49821429 o o o Candacia o 1.16607143 2.33214286 o o Labidocera o o o o o Ponte/lina o o o o o Acartia 1.36020408 8 .1625 23.3214286 8.70714286 o Oithona 90.4535714 479.255357 436.110714 696.571429 368.518878 Euterpina o 18.6571429 60.6357143 37.7309524 4.66479592 Clytemnestra o 1.16607143 o 2.90238095 4.66479592 Macrosetel/a 12.9219388 o 4.66428571 2.90238095 o Microsetella o 2.33214286 13.9928571 5.8047619 97.9607143 Oncaea 25.8438776 143.426786 128.267857 217.678571 321.870918 Corycaeus 17.002551 13.9928571 32.65 29.0238095 23.3239796 Lubockia o o o 2.90238095 o Copilia o 1.16607143 o o o Saphirina o o o o o Calocalanus 5.44081633 11.6607143 4.66428571 11 .6095238 o C/ausocalanus 6.80102041 2.33214286 6.99642857 o 41.9831633 Spinoca/anus o o o o o Euchirella o 1.16607143 2.33214286 o o Copépodos 203.35051 979.5 993.492857 1439.58095 1240.83571 Copepoditos 25.1637755 222.719643 307.842857 487.6 275.222959 Nauplius 16.322449 34.9821429 144.592857 145.119048 74.6367347

200
Hidromedusas 3.4005102 2.33214286 2.33214286 5.8047619 4.66479592 Ctenoforos o o o o o Pteropodos o 1.16607143 13.9928571 8.70714286 4.66479592 Gastropodos 4.08061224 6.99642857 16.325 31.9261905 69.9719388 L. Molusco 2.04030612 2.33214286 9.32857143 29.0238095 37.3183673 L.Poliqueto 4.08061224 1.16607143 4.66428571 5.8047619 27.9887755 L. Equinodermo o o o o o L.Crustaceo 0.68010204 2.33214286 o o 4.66479592 Cladoceros o o 2.33214286 o o Ostracodos 37.4056122 144.592857 48.975 49.3404762 223.910204 Apendicularias 12.9219388 45.4767857 74.6285714 136.411905 27.9887755 Salpas o o 4.66428571 o o Anfipodos o 1.16607143 o o o Quetognatos 10.2015306 16.325 20.9892857 20.3166667 27.9887755 Lingula 1.36020408 o o o o Actinotroca o o o o o Larvas de pez o o o o o Huevos de lnvert 48.2872449 27.9857143 13.9928571 5.8047619 23.3239796 Calamar 0.68010204 o o o o Anfioxo o o o o o
Estacion 1 Prof. 3 Fecha Abr-97 Ago-97 Nov-97 Mar-98 Abr-98 Set-98 Grupo Canthocalanus o o o 0.29158163 o o Undinula o o o o o o Acroca/anus 0.51020408 o 1.08836735 3.49897959 o 1.39928571 Paraca/anus 1.53061224 36.7346939 5.44183673 27.1170918 67.1730612 20.5228571 Euca/anus o o o 0.29158163 o o Euchaeta o o o o o o Temora o o o 0.58316327 1.86591837 o Temoropia o o 1.08836735 o o o Pleuromamma o o o o o o Centropages o o o 2.62423469 3.73183673 o Lucicutia o o o 0.58316327 o o Pseudodiaptomus 0.51020408 o o o o o Candacia o o o 0.29158163 o 1.39928571 Labidocera o o o o o o Pontellina o o o o o o Acartia 1.53061224 o 2.17673469 1.16632653 o o Oithona 2.04081633 11.2244898 62.0369388 106.427296 132.480204 54.1057143 Euterpina o 1.02040816 2.17673469 8.74744898 20.525102 13.9928571 Clytemnestra o o 2.17673469 o o o Macrosetella o 2.04081633 o 0.58316327 o o Microsetella o o 1.08836735 4.95688776 3.73183673 o Oncaea o 87.755102 27.2091837 78.7270408 117.552857 30.3178571 Corycaeus o o o 6.70637755 5.5977551 6.06357143 Lubockia o o o o o o Copi/ia o o o o o o Saphirina o o o o o o Ca/ocalanus o o o 0.29158163 o 0.46642857

201
C/ausoca/anus o o o 2.91581633 1.86591837 0.46642857 Spinoca/anus o o o o o o Euchirel/a o o o o o o Copépodos 6.12244898 138.77551 104.483265 245.803316 354.52449 128.734286 Copepoditos 5. 10204082 17.3469388 41, .3579592 67.0637755 108.223265 28.9185714 Nauplius 1.53061224 o 1.08836735 34.9897959 44.7820408 8.86214286 Hidromedusas 5.10204082 14.2857143 4.35346939 1.16632653 13.0614286 0.46642857 Ctenoforos o o o o o o Pteropodos o o o 1.16632653 o o Gastropodos 0.51020408 4.08163265 1.08836735 1.7494898 1.86591837 1.86571429 L. Molusco 0.51020408 o 1.08836735 3.20739796 7.46367347 0.46642857 L.Poliqueto 2.55102041 79.5918367 56.595102 17.2033163 59.7093878 9.795 L. Equinodermo o o o o o o L.Crustaceo 2.55102041 o 3.26510204 o o o Cladoceros 1.02040816 o o 1.7494898 16.7932653 0.46642857 Ostra codos o 5.10204082 60.9485714 24.7844388 7.46367347 7.92928571 Apendicularias 1.02040816 5.10204082 6.53020408 33.8234694 16.7932653 8.86214286 Salpas o o o o 1.86591837 o Anfipodos o o o o o o Quetognatos 1.02040816 o o 4.08214286 5.5977551 o Lingula o o o o 1.86591837 o Actinotroca o o o o o o larvas de pez o o o o o o Huevos de lnvert 2.04081633 510.204082 167.608571 7.28954082 9.32959184 15.3921429 Calamar o o o o o o Anfioxo o o o o 1.86591837 o
Estacion 2 Prof. 3 Fecha Abr-97 Ago-97 Nov-97 Mar-98 Abr-98 Set-98
Canthoca/anus o o o o o Undinula o o o o o Acrocalanus 2.04068878 5.44183673 o o 0.27204082 Paracalanus 14.965051 13.6045918 o 0.46642857 1.08816327 Euca/anus 0.68022959 o o o o Euchaeta o o o o o Temora o o o 0.46642857 o Temoropia o 1.36045918 o o 3.2644898 Pleuromamma o o o o o Centropages o o 1.16607143 0.46642857 o Lucicutia o o 1.16607143 o o Pseudodiaptomus o o o 0.46642857 0.27204082 Candacia o o o o 0.27204082 Labidocera o o o o o Pontel/ina o o o o o Acartia o o o o o Oithona 37.4126276 12.2441327 19.8232143 11.1942857 5.71285714 Euterpina 2.04068878 1.36045918 4.66428571 2.79857143 3.2644898 Clytemnestra 0.68022959 o o o o Macrosetella 2.04068878 o o o 0.27204082 Microsetella o o 4.66428571 0.46642857 o

202
Oncaea 53.0579082 111.557653 33.8160714 53.1728571 18.7708163 Corycaeus 2.04068878 o o 1.86571429 0.81612245 Lubockia o o o o o Copilia o o o o o Saphirina o o o o o Calocalanus o o o o o Clausocalanus 0.68022959 19.0464286 o o o Spinoca/anus o o o o o Euchirella o o o o o Copépodos 115.639031 164.615561 65.3 71.3635714 34.005102 Copepoditos 17.6859694 88.4298469 139.928571 13.06 17.9546939 Nauplius o o o 0.93285714 3.80857143 Hidromedusas o 2.72091837 4.66428571 2.33214286 2.99244898 Ctenoforos o o o o o Pteropodos o o o o o Gastropodos 13.6045918 5.44183673 2.33214286 1.86571429 1.90428571 L. Molusco 2.04068878 o 1.166071'43 1'. 399285 71 o L.Poliqueto 85.7089286 153.731888 19.8232143 26.5864286 14.4181633 L. Equinodermo o o o o o L.Crustaceo 0.68022959 o o 0.46642857 o Cladocer;os o o o 1.86571429 o Ostracodos 10.2034439 8.1627551 1.16607143 1.86571429 6.25693878 Apendicularias o 4.08137755 2.33214286 12.1271429 9.24938776 Salpas o o o o o Anfipodos o o o o o Quetognatos o o 2.33214286 o o Lingula o o o 0.46642857 o Actinotroca o o o o o Larvas de pez o o o o o Huevos de lnvert 408.137755 516.97449 9.32857143 12.5935714 15.2342857 Calamar o o o o o Anfioxo o o o o o
Estacion 3 Prof. 3 Fecna Abr-97 Ago-97 Nov-97 Mar-98 Abr-98 Set~98
Especie Canthocalanus 14.5122449 o o 1.16607143 1.86571429 o Undinula o o o o o o Acrocalanus 304.757143 o 3.401 114796 5.8303571 4 2.79857143 1.86571429 Paracalanus 907.015306 24.4897959 12.9243622 11.6607143 8.39571429 86.7557143 Eucalanus 7.25612245 o o o o o Euchaeta 21.7683673 o o o o o Temora o o o 1.16607143 o 0.93285714 Temoropia o o 2.72091837 2.33214286 0.93285714 0.93285714 P/euromamma o o o o o o Centropages 239.452041 o 0.68022959 o 0.93285714 o Lucicutia o 1.02040816 0.68022959 o 0.9'3285714 0.932857114 Pseudodiaptomus o o o o o o Candacia o o 2.04068878 o o o Labidocera 14.51 22449 o o o o 0.93285714 Pontellina o o o o o o

203
Acartia 253.964286 o 2.04068878 o o 3.73142857 Oithona 834.454082 26.5306122 27.2091837 34.9821429 29.8514286 203.362857 Euterpina 616.770408 2.04081633 4.08137755 5.83035714 8.39571429 12.1271429 C/ytemnestra o o 2.04068878 1.16607143 o o Macrosetella 65.305102 1.02040816 2.04068878 o 0.93285714 0.93285714 Microsetella o o 0.68022959 2.33214286 8.39571429 1A.7914286 Oncaea 965.064286 35.7142857 22.4475765 80.4589286 42.9114286 105.412857 Corycaeus 217.683673 4.08163265 2.72091837 4.66428571 6.53 8.39571429 Lubockia o o o 1.16607143 o o Copilia o o o o o o Saphirina o o o o o 0.93285714 Ca/oca/anus 14.5122449 o 1.36045918 2.33214286 1.86571429 1.86571429 Clausoca/anus 7.25612245 o 1.36045918 2.33214286 5.59714286 6.53 Spinoca/anus 21.7683673 o o o o o Euchirella o o o o 0.93285714 0.93285714 Copépodos 4506.05204 94.8979592 88.4298469 157.419643 121.271429 453.368571 Copepoditos 1073.90612 16.3265306 37.4126276 52.4732143 69.0314286 190.302857 Nauplius 1327.87041 1.02040816 1.36045918 15.1589286 7.46285714 98.8828571 Hidromedusas 21.7683673 1.02040816 2.72091837 2.33214286 2.79857143 0.93285714 Ctenoforos o o o o o o Pteropodos 65.305102 o o o o 1.86571429 Gastropodos 116.097959 o o 2.33214286 7.46285714 18.6571429 L. Molusco 145.122449 o o 5.83035714 5.59714286 7.46285714 L.Poliqueto 87.0734694 85.7142857 59.1799745 9.32857143 35.4485714 11.1942857 L. Equinodermo o o o o o o L.Crustaceo 116.097959 1.02040816 o 1.16607143 0.93285714 0.93285714 Cladoceros o o o 1.16607143 o o Ostracodos 747.380612 3.06122449 7.48252551 31.4839286 41.9785714 22.3885714 Apendicularias 776.405102 1.02040816 5.44183673 6.99642857 5.59714286 20.5228571 Salpas 7.25612245 o o o o o Anfipodos o o o o o o Quetognatos 130.610204 o o 2.33214286 o 8.39571429 Lingula o o o o o o Actinotroca 7.25612245 o o o o o Larvas de pez o o o o o o Huevos de lnvert 188.659184 816.326531 193.185204 5.83035714 o 12.1271429 Calamar o o o o o o Anfioxo o o o o o o
Estacion 4 Prof. 3 Fecha Abr-97 Ago-97 Nov-97 Mar-98 Abr-98 Set-98 Especie Canthocalanus 16.3265306 o o 2.33214286 o o Undinula o o o o o o Acroca/anus 146.938776 65.3061224 20.9892857 6.99642857 4.66479592 2.72040816 Paraca/anus 1844.89796 489.795918 88.6214286 32.65 74.6367347 7.48112245 Euca/anus 48.9795918 32.6530612 o 4.66428571 o o Euchaeta o o o 2.33214286 4.66479592 5.44081633 Temora 16.3265306 o o o o o Temoropia o o o o o 2.04030612 Pleuromamma o o o o o o

204
Centropages 155.102041 o o 4.66428571 o o Lucicutia o o o 6.99642857 9.32959184 0.68010204 Pseudodiaptomus 8.16326531 o o o o o Candacia o o o o o o Labidocera o o o o o o Ponte/Ji na o o o o o o Acartia 122.44898 o 2.33214286 o o o Oithona 1110.20408 87.0748299 156.253571 86.2892857 359.189286 22.4433673 Euterpina 612.244898 o 3.49821429 25.6535714 37.3183673 3.4005102 Clytemnestra o 10.8843537 o 2.33214286 o 1.36020408 Macrosetella 73.4693878 65.3061224 1.16607143 o o o Microsetel/a o o o 20.9892857 37.3183673 21.7632653 Oncaea 661.22449 337.414966 163.25 240.210714 335.865306 31.2846939 Corycaeus 375.510204 21.7687075 2.33214286 6.99642857 4.66479592 1.36020408 Lubockia o o o o o o Copilia o o o o o o Saphirina o o o o o o Ca/ocalanus o o o o 4.66479592 o Clausoca/anus o 97.9591837 9.32857143 11.6607143 o 5.44081633 Spinoca/anus o o o o o o Euchire/la o o o o o o Copépodos 51 91.83673 1208.16327 447.771429 454.767857 872.316837 105.415816 Copepoditos 1706.12245 370.068027 117.773214 139.928571 256.563776 46.9270408 Nauplius 24.4897959 21.7687075 9.32857143 30.3178571 102.62551 14.2821429 Hidromedusas o 21.7687075 5.83035714 9.32857143 o 4.76071429 Ctenoforos 122.44898 o o o o o Pteropodos 65.3061224 o o 4.66428571 4.66479592 o Gastropodos 16.3265306 54.4217687 3.49821429 9.32857143 60.6423469 9.52142857 L. Molusco 16.3265306 21.7687075 1.16607143 9.32857143 4.66479592 6.12091837 L.Poliqueto o 10.8843537 o 6.99642857 9.32959184 15.6423469 L. Equinodermo 122.44898 o o o o o L.Crustaceo 48.9795918 o o o o 0.68010204 Cladoceros 751.020408 o o 2.33214286 o o Ostracodos 661.22449 663.945578 152.755357 219.221429 261.228571 10.2015306 Apendicularias o 10.8843537 12.8267857 23.3214286 135.279082 5.44081633 Salpas o o o o 4.66479592 o Anfipodos 81.6326531 o o o o o Quetognatos o 10.8843537 6.99642857 6.99642857 18.6591837 1.36020408 Lingula o o o o o o Actinotroca o o o o o o Larvas de pez o o o o o o Huevos de lnvert o 65.3061224 3.49821429 48.975 o 4.08061224 Calamar o o o o o o Anfioxo o o o o o o
Estacion 11 Prof. 3 Fecha Abr-97 Ago-97 Nov-97 Mar-98 Abr-98 Set-98 Especie Canthoca/anus o 2.33214286 13.0607143 o o Undinula o o o o o Acrocalanus 39.9068027 51.3071429 15.2375 26.1214286 13.9943878

205
Paraca/anus 52.6044218 177.242857 67.4803571 243.8 303.211735 Eucalanus 7.25578231 o o 8.70714286 o Euchaeta o 2.33214286 o o 4.66479592 Temora o 2.33214286 2.17678571 o o Temoropia o o o o o Pleuromamma o o o o o Centropages o o 6.53035714 o o Lucicutia o o 4.35357143 4.35357143 o Pseudodiaptomus o 4.66428571 o o o Candacia o o o o o Labidocera o o o o o Pontellina o o o o o Acartia 7.25578231 o 2.17678571 o 13.9943878 Oithona 76.1857143 307.842857 145.844643 391.821429 517.792347 Euterpina 1.81394558 11.6607143 6.53035714 21.7678571 13.9943878 C/ytemnestra 1.81394558 2.33214286 o o o Macrosetel/a 5.44183673 o 4.35357143 o o Microsetella o 4.66428571 o 13.0607143 32.6535714 Oncaea 70.7438776 298.514286 143.667857 356.992857 447.820408 Corycaeus 5.44183673 13.9928571 13.0607143 13.0607143 46.6479592 Lubockia o o o o o Copilia o o o o o Saphirina o o o o o Caloca/anus 3.62789116 4.66428571 o 4.35357143 9.32959184 Clausocalanus 1.81394558 6.99642857 o o 55.977551 Spinoca/anus o o o o o Euchirel/a o o 2.17678571 o o Copépodos 273.905782 890.878571 426.65 1084.03929 1460.08112 Copepoditos 43.5346939 307.842857 235.092857 522.428571 671.730612 Nauplius 3.62789116 23.3214286 58.7732143 43.5357143 149.273469 Hidromedusas 12.697619 13.9928571 o 8.70714286 13.9943878 Ctenoforos o o o o o Pteropodos o o 2.17678571 4.35357143 o Gastropodos 12.697619 9.32857143 10.8839286 26.1214286 107.290306 L. Molusco 5.44183673 18.6571429 2.17678571 4.35357143 32.6535714 L.Poliqueto 3.62789116 o 21.7678571 8.70714286 55.977551 L. Equinodermo o o o o o L.Crustaceo 1.81394558 o o o 4.66479592 Cladoceros o 2.33214286 2.17678571 4.35357143 o Ostra codos 195.906122 527.064286 287.335714 269.921429 265.893367 Apendicularias 5.44183673 30.3178571 19.5910714 4.35357143 9.32959184 Salpas o o o o o Anfipodos o o 2.17678571 o o Quetognatos 5.44183673 11.6607143 13.0607143 o 4.66479592 Lingula o o o o 4.66479592 Actinotroca o o o o o Larvas de pez 1.81394558 o o o o Huevos de lnvert 38.0928571 11.6607143 o o 18.6591837 Calamar o o o o o Anfioxo o o o o o