universidad de chile facultad de ciencias químicas y...
TRANSCRIPT

UNIVERSIDAD DE CHILE Facultad de Ciencias Químicas y Farmacéuticas
PARTICIPACIÓN DEL FACTOR DE CRECIMIENTO NERVIOSO EN LA REGULACIÓN DE LA ANGIOGÉNESIS OVÁRICA
Tesis presentada a la Universidad de Chile para optar al Grado Académico de Doctor en Bioquímica
POR
MARCELA JULIO PIEPER
Directores de Tesis
Dra. Carmen Romero Osses Dr. Hernán Lara Peñaloza
Santiago- Chile 2006

ii

iii
UNIVERSIDAD DE CHILE Facultad de Ciencias Químicas y Farmacéuticas
Informe de Aprobación de Tesis de Doctorado Se informa a la Comisión de Grados Académicos de la Facultad de Ciencias Químicas y Farmacéuticas, que la tesis de Doctorado en Bioquímica presentada por el candidato
MARCELA JULIO PIEPER Ha sido aprobada por la Comisión informante de tesis como requisito para optar al Grado de Doctor en Bioquímica en Examen de Defensa de Tesis rendido el día _____________________________________________________________ Directora de Tesis Dra. Carmen Romero Osses ____________________ Co-director de Tesis Dr. Hernán Lara Peñaloza ____________________ Comisión informante de Tesis Dr. Sergio Lavandero (Presidente) ____________________ Dr. Jorge Garrido ____________________ Dr. Mario Herrera-Marschitz ____________________ Dr. Luis Sobrevía ____________________ Dr. Luis Valladares ____________________

iv
Agradecimientos
A mis directores de Tesis, Dra. Carmen Romero y Dr. Hernán Lara, por haberme ayudado a crecer y por enseñarme que la recompensa más valiosa es la satisfacción del trabajo bien realizado. A mi colega y compañero de vida, Javier, por su apoyo incondicional, por compartir conmigo su sabiduría y fortaleza, y por su enorme paciencia en cada etapa de esta travesía. A mi amiga Ximena, por su sabio consejo, su cariño y lealtad a toda prueba. A mis compañeros de laboratorio Patty y Enrique, quienes pasaron a ser parte de mi familia durante estos años. Al personal del Laboratorio de Endocrinología: Jaime, Egardo, Verónica, Salomé, Erminia, Carlos, Daniela, Filomena y Soledad, por su amistad, ayuda y colaboración, la cual fue indispensable en el desarrollo de este trabajo. A Alfonso Paredes, Mauricio Venegas, Sergio Andrés y Donald Brown, por su generosa ayuda. A Sergio Ojeda y sus colaboradores: María Eugenia Costa, Bredford Kerr, Greg Dissen, Claudio Mastronardi, por recibirme generosamente en su laboratorio y por prestarme una valiosa ayuda durante estos años. A toda la gente del Programa de Fertilización Asistida del Hospital Clínico Universidad de Chile, al personal del Programa de Genética y a la gente del pabellón de Maternidad de la misma institución, por su colaboración y paciencia. Al personal de la Oficina de Apoyo a la Investigación Clínica del Hospital Clínico Universidad de Chile, por su permanente colaboración. Al Dr. Luis Sobrevía y Paola Casanello, por su valiosa ayuda con las técnicas de cultivo celular. A Gabriel Aravena y Fidel Albornoz, por su importante apoyo. Finalmente, quiero agradecer a mis padres, por su generosidad y su amor, por entregarme la posibilidad de estudiar y por haberme apoyado en cada objetivo. Mi fortaleza e inspiración es el inmenso cariño de mi familia.

v
Publicaciones y presentaciones a congresos
Publicaciones:
1. Julio-Pieper M, Lara HE, Bravo JA, Romero C (2006). Effects of nerve growth factor (NGF) on blood vessels area and expression of the angiogenic factors VEGF and TGFbeta1 in the rat ovary. Reprod Biol Endocrinol 4:57-67.
2. Salas C, Julio-Pieper M, Valladares M, Pommer R, Vega M, Mastronardi C, Kerr B,
Ojeda SR, Lara HE, Romero C (2006). Nerve growth factor-dependent activation of trka receptors in the human ovary results in synthesis of FSH receptors and estrogen secretion. J Clin Endocrinol Metab 91:2396-2403.
En preparación: Julio-Pieper M, Lozada P, Muñoz Y, Miranda C, Vantman D, Cortínez A, Alba F, Ojeda SR, Kerr B, Mastronardi C, Dissen GA, Lara HE, Romero C. Nerve growth factor-induced increase on vascular endothelial growth factor expression is dependent of trka receptor activation and ERK1/2 phosphorylation in human granulosa cells.
Presentaciones a congresos nacionales: 1. Valladares M, Muñoz Y, Julio M, Pömmer R, Vega M, Lara HE, Romero C. El factor
de crecimiento nervioso (NGF) induce la expresión de receptores de FSH funcionales, a través de su receptor de alta afinidad TrkA. XIV Reunión Anual de la Sociedad Chilena de Reproducción y Desarrollo, La Serena, Chile, 2003.
2. Muñoz Y, Valladares M, Julio M, Campos X, Selman A, Vantman D, Miranda C, Cortínez A, Alba F, Lara HE, Romero C. Acción del factor de crecimiento nervioso (NGF) en cáncer ovárico epitelial. XIV Congreso Chileno de Endocrinología y Metabolismo, Arica, Chile, 2003.
3. Campos X, Oddo D, Selman A, Martinez L, Moyano L, Yazigi R, Julio M,
Mastronardi C, Kerr B, Dissen G, Ojeda S, Lara H, Romero C. Expresión del receptor TrkA y VEGF y su relación con el grado de diferenciación del cáncer ovárico epitelial. XV Reunión Anual Sociedad Chilena de Reproducción y Desarrollo, El Quisco, Chile, 2004.
4. Julio-Pieper M, Lara HE, Bravo JA, Vantman D, Miranda C, Cortínez A, Alba F,
Ojeda SR, Paredes A, Kerr B, Mastronardi C, Dissen G, Romero C. El factor de crecimiento nervioso regula la expresión de VEGF en el ovario. XVI Reunión Anual de la Sociedad Chilena de Reproducción y Desarrollo, Reñaca, Chile, 2005.

vi
5. Lozada P, Julio-Pieper M, Miranda C, Vantman D, Cortínez A, Alba F, Lara HE,
Romero C. NGF aumenta el mRNA de TrkA y c-Myc en células de granulosa humana. XVI Reunión Anual de la Sociedad Chilena de Reproducción y Desarrollo, Reñaca, Chile, 2005.
6. Tapia V, Julio-Pieper M, Campos X, Candia E, Selman A, Miranda C, Vantman D,
Cortínez A, Carvajal A, Alba F, Lara HE, Romero C. AKT está involucrada en el aumento de la expresión del factor de crecimiento de endotelio vascular (VEGF) por efecto del factor de crecimiento nervioso (NGF) en ovario humano. XVI Reunión Anual de la Sociedad Chilena de Reproducción y Desarrollo, Reñaca, Chile, 2005.
7. Julio-Pieper M, Lara HE, Bravo JA, Dissen G, Ojeda SR, Romero C.
“Vascularización ovárica: rol del factor de crecimiento nervioso”. XVII Reunión Anual de la Sociedad Chilena de Reproducción y Desarrollo, Reñaca, Chile, 2006.
8. Tapia V, Julio-Pieper M, Lozada P, Miranda C, Vantman D, Cortínez A, Carvajal A,
Alba F, Lara H, Romero C. “Rol indirecto del factor de crecimiento nervioso en la angiogénesis normal del ovario humano”. 2006. XVII Reunión Anual de la Sociedad Chilena de Reproducción y Desarrollo, Reñaca, Chile, 2006.
Presentaciones a congresos internacionales: 1. Julio M, Valladares M, Salas C, Muñoz Y, Campos X, Pommer R, Vega M, Ojeda
SR, Lara HE, Romero C. Role of nerve growth factor (NGF) in human granulosa cells. Endocrine Society´s 86th Annual Meeting, New Orleans, USA, 2004.
2. Julio-Pieper M, Lara HE, Romero, C. VEGF but not TGFbeta1 expression is
increased by NGF in neonatal rat ovaries. X Panamerican Association for Biochemistry and Molecular Biology Congress, Pinamar, Argentina, 2005.

vii
Becas y Premios Becas
1. 2002 a 2005: Beca Conicyt de doctorado. 2. 2004 a 2005: Beca Conicyt de apoyo a la realización de tesis doctoral
3. 2004: Beca del Departamento de Postgrado de la Universidad de Chile para
financiamiento parcial de tesis doctoral.
4. 2006: Beca del Departamento de Postgrado de la Universidad de Chile para finalización de tesis doctoral.
Premios 2006: Premio Oriana Josseau a la estudiante destacada del Doctorado en Bioquímica.

viii
Financiamiento
1. Beca Conicyt de Apoyo a la realización de tesis doctoral AT-4040059. Investigador responsable: Marcela Julio.
2. Beca del Departamento de Postgrado de la Universidad de Chile para
financiamiento parcial de tesis doctoral PG/90/2003. Investigador responsable: Marcela Julio.
3. Beca del Departamento de Postgrado de la Universidad de Chile para
finalización de tesis doctoral PG/67/2005. Investigador responsable: Marcela Julio.
4. Proyecto Fondecyt 1030661. Investigador responsable: Dra. Carmen Romero
5. Proyecto DI SAL 02/2-2. Investigador responsable: Dra. Carmen Romero

ix
Tabla de contenido Página1. Introducción 1
1.1. Angiogénesis 11.2. Angiogénesis en el ovario 21.3. Factor de crecimiento nervioso (NGF) en el ovario 51.4. NGF como probable factor angiogénico en el ovario 61.5. Factor de crecimiento de endotelio vascular (VEGF) 61.6. Factor de crecimiento transformante β (TGFβ) 81.7. Relación de NGF, TGFβ y VEGF 91.8. Hipótesis 111.9. Objetivos específicos 11
2. Materiales y métodos 142.1. Técnicas de cultivo celular 14
2.1.1. Aislación y cultivo de células de granulosa humana 142.1.2. Aislación y cultivo de células de endotelio de vena umbilical humana
15
2.2. Experimentos en animales 162.2.1. Ratas 162.2.2. Ratones mutantes 17
2.2.2.1. Ratones NGF knockout 172.2.2.2. Ratones trkA knockout 172.2.2.3. Ratones transgénicos 17
2.2.3. Cultivo de ovarios 182.2.4. Denervación ovárica 19
2.3. Análisis cuantitativos y semicuantitativos 192.3.1. ELISA 192.3.2. Western blot 192.3.3. Inmunohistoquímica 202.3.4. Extracción de RNA y reacción de transcripción reversa 222.3.5. Reacción en cadena de la polimerasa (PCR) 222.3.6. PCR de tiempo real 232.3.7. Cell-Titer 25
2.4. Análisis estadístico 253. Resultados 26
3.1. Efecto de NGF sobre VEGF y TGFβ1 en ovario de roedores 263.1.1. Efecto de NGF sobre los factores VEGF y TGFβ1 en ovario de ratas neonatas en cultivo
26
3.1.2. Efecto de la ausencia de NGF sobre los factores VEGF y TGFβ1 en ratones neonatos
31
3.1.3. Efecto de NGF sobre los factores VEGF y TGFβ1 en ratas sometidas a denervación ovárica
32
3.1.4. Efecto de la sobreexpresión ovárica de NGF sobre los factores VEGF y TGFβ1 en ratones transgénicos
36

x
Página
3.1.5. Resumen de los resultados: Efecto de NGF sobre los factores VEGF y TGFβ1 en ovario de roedores
37
3.2. Efecto de NGF sobre los factores VEGF y TGFβ1 en células de la granulosa humana
38
3.2.1. Resumen de los resultados: Efecto de NGF sobre los factores VEGF y TGFβ1 en células de la granulosa humana
40
3.3. Evaluación de un posible mecanismo de acción de NGF sobre el incremento de VEGF y TGFβ1 en modelos de ovario de roedor y de humano
41
3.3.1. Participación del receptor trkA 413.3.2. Señalización intracelular vía ERK 453.3.3. Factor transcripcional inducible por hipoxia (HIF1α) 483.3.4. Resumen de resultados: Evaluación de un posible mecanismo de acción de NGF sobre el incremento de VEGF y TGFβ1 en modelos de ovario de roedor y de humano
49
3.4. Efecto de NGF en la vascularización ovárica 503.4.1. Efecto de NGF en un modelo de angiogénesis in vitro 503.4.2. Efecto de NGF en la vascularización ovárica in vivo, en ratas sometidas a denervación ovárica
52
3.4.3. Efecto de la sobreexpresión ovárica de NGF sobre la vascularización en ratones transgénicos
54
3.4.4. Resumen de resultados: Efecto de NGF sobre la vascularización ovárica
55
4. Discusión 564.1. Nuevos roles de NGF en la función ovárica 564.2. NGF, VEGF y la angiogénesis del ovario 614.3. NGF no modifica los niveles de TGFβ1 en el ovario 654.4. Mecanismos del aumento de VEGF inducido por NGF en el ovario 664.5. Comentarios finales y proyecciones 69
5. Conclusiones 716. Referencias 727. Anexos 83
7.1. Anexo 1. Resumen de los modelos usados en este estudio 837.2. Anexo 2. Carta del Comité de Etica de la Facultad de Ciencias Químicas y Farmacéuticas
84
7.3. Anexo 3. Carta del Comité de Etica del Hospital Clínico Universidad de Chile
85
7.4. Anexo 4. Consentimiento Informado para Investigación Clínica 867.5. Anexo 5. Información sobre los cultivos celulares 877.6. Anexo 6. Marcadores de la presencia de células de granulosa 88

xi
Indice de figuras PáginaFigura 1.1 Efecto de la administración de VEGF trap sobre el ovario 3Figura 1.2. Mecanismos fisiológicos de angiogénesis 4Figura 2.1. Mapa del transgén de NGF 18Figura 2.2. Evaluación de CD-31 21Figura 3.1. Análisis del contenido de mRNA de VEGF y TGFβ1 en
ovarios de rata cultivados por 2 horas con NGF 27
Figura 3.2. Análisis del contenido de mRNA de VEGF y TGFβ1 en ovarios de rata cultivados por 8 horas con NGF
27
Figura 3.3. Análisis del contenido de mRNA de VEGF y TGFβ1 en ovarios de rata cultivados por 24 horas con NGF
28
Figura 3.4. Ensayo de inmuno-preadsorción de los anticuerpos utilizados para la evaluación de VEGF y TGFβ1
28
Figura 3.5. Inmunohistoquímica de VEGF en ovarios de rata cultivados por 24 horas con NGF
29
Figura 3.6. Análisis de la inmunohistoquímica de VEGF en ovarios de rata cultivados por 4 y 24 horas en presencia de NGF
30
Figura 3.7. Inmunohistoquímica de TGFβ1 en ovarios de rata neonata cultivados por 24 horas con NGF
30
Figura 3.8. Análisis de la inmunohistoquímica de TGFβ1 en ovarios de rata cultivados por 4 y 24 horas con NGF
31
Figura 3.9. Análisis del contenido de mRNA de VEGF y TGFβ1 en ovarios de ratones NGF knockout y silvestres
32
Figura 3.10. Análisis densitométrico de la marca de NGF en ovarios de ratas sometidas a la sección del nervio ovárico superior durante seis días, y ratas controles
33
Figura 3.11. Análisis semicuantitativo del contenido de mRNA de VEGF (isoformas 120 y 164) y TGFβ1 en ovarios de ratas sometidas a la sección del nervio ovárico superior durante seis días, y ratas controles
34
Figura 3.12. Análisis semicuantitativo del contenido de mRNA de VEGF y TGFβ1 en ovarios de ratas sometidas a la sección del nervio ovárico superior durante tres días y ratas controles
34
Figura 3.13. Inmunohistoquímica de VEGF en ratas sometidas a la denervación de los ovarios por 6 días
35
Figura 3.14. Cuantificación de la inmunorreactividad de TGFβ1 en ratas sometidas a la denervación ovárica por 3 ó 6 días
36
Figura 3.15. Análisis del contenido de mRNA de VEGF y TGFβ1 en ovarios de ratones que sobreexpresan NGF en el ovario
37
Figura 3.16. Inmunohistoquímica para trkA, en células de granulosa humana recién aisladas
39
Figura 3.17. Análisis del contenido de mRNA de VEGF y TGFβ1 en células de granulosa estimuladas con NGF
39

xii
PáginaFigura 3.18. Análisis de la secreción de VEGF al medio en células de
granulosa tratadas con NGF 40
Figura 3.19. Análisis del contenido de mRNA de VEGF y TGFβ1 en ovarios de ratones trkA knockout y silvestre
42
Figura 3.20. Análisis de los niveles de mRNA para las isoformas de p75 en ovarios de ratones trkA knockout y silvestres
43
Figura 3.21. Contenido de mRNA para VEGF 165 en células de granulosa tratadas con NGF y un inhibidor del receptor trkA (K252a)
44
Figura 3.22. Niveles de VEGF en el medio de células de granulosa tratadas con NGF y un inhibidor del receptor trkA (K252a)
44
Figura 3.23. Western blot de ERK en células de granulosa incubadas con NGF
45
Figura 3.24. mRNA de VEGF 121 en células de granulosa tratadas con NGF y un inhibidor de la fosforilación de Erk 1/2 (U0126)
46
Figura 3.25. mRNA de VEGF 165 en células de granulosa tratadas con NGF y un inhibidor de la fosforilación de Erk 1/2 (U0126)
46
Figura 3.26. Niveles de VEGF en el medio de células de granulosa tratadas con NGF y el inhibidor 1/2 (U0126)
47
Figura 3.27. mRNA de TGFβ1 en células de granulosa tratadas con NGF y un inhibidor de la fosforilación de Erk 1/2 (U0126)
47
Figura 3.28 mRNA para HIF1α en células de granulosa tratadas por 8 horas con NGF
48
Figura 3.29. mRNA para HIF1α en ovarios de rata de 2 días tratados con NGF por 2 horas
49
Figura 3.30. Proliferación de células HUVEC 51Figura 3.31. Tinción de CD-31 en ratas denervadas 52Figura 3.32. Área porcentual de marca positiva para vasos en ovario
de ratas sometidas a la sección del nervio ovárico superior durante 6 días
53
Figura 3.33. Área porcentual de marca positiva para vasos en ovario de ratas sometidas a la sección del nervio ovárico superior durante 3 días
53
Figura 3.34. Marca de CD-31 en ovarios de ratones que sobreexpresan NGF
54
Figura 3.35. Área porcentual de marca positiva para vasos en ovarios de ratones transgénicos que sobreexpresan NGF
55
Figura 4.1. Modelo propuesto 62Figura 4.2. Secuencia de eventos para el efecto de NGF en el ovario 64Figura 4.3. Mecanismos de señalización involucrados 68Figura 7.1. Marcadores positivos de células de granulosa 88

xiii
Indice de tablas y figuras
Página Tabla 1.1 Información de los partidores usados en este estudio 24Tabla 7.1. Resumen de los modelos evaluados durante esta tesis 83Tabla 7.2. Información general del origen de las células de granulosa
utilizadas en este estudio 87
Tabla 7.3. Datos clínicos de las pacientes de las cuales se obtuvieron las muestras de células de granulosa
87
Tabla 7.4. Información general de las células HUVEC utilizadas en este estudio
87

xiv
Abreviaturas
DNA: Acido desoxirribonucléico
cDNA: Acido desoxirribonucléico complementario
RNA: Ácido ribonucléico
mRNA: Ácido ribonucléico mensajero
Pb: Pares de bases
FSH: Hormona folículoestimulante
GnRH: Hormona liberadora de gonadotrofinas
hCG: Gonadotrofina coriónica humana
hMG: Gonadotrofina de la menopausia humana
HIF1α: Factor inducible por hipoxia 1 alfa
HUVEC: Células de endotelio de vena umbilical humana
i.m.: Intramuscular
LH: Hormona luteinizante
NGF: Factor de crecimiento nervioso
PCR: Reacción en cadena de la polimerasa
s.c.: Subcutáneo
TGFβ: Factor de crecimiento transformante beta
TβR: Receptor de TGFβ
VEGF: Factor de crecimiento de endotelio vascular

xv
Resumen
Participación del factor de crecimiento nervioso en la regulación de la angiogénesis ovárica
La formación de vasos sanguíneos a partir de otros preexistentes, o angiogénesis,
es un proceso esencial en el desarrollo del tejido normal. La mayor parte de la
angiogénesis fisiológica ocurre en la etapa embrionaria, y en el adulto, el endotelio
vascular es un tejido con un bajo índice de proliferación. Sin embargo, los ovarios
presentan cíclicamente procesos asociados a angiogénesis. En el ovario, la
angiogénesis acompaña al desarrollo folicular y a la formación del cuerpo lúteo, y de no
existir concepción, esta red vascular involuciona. La extraordinaria rapidez con que los
vasos sanguíneos aparecen y son reabsorbidos en cada ciclo ovárico, hace de este
sistema un modelo ideal para el estudio de los elementos regulatorios de la
angiogénesis, por lo que su mejor comprensión podría significar un gran avance para la
investigación de la angiogénesis en eventos fisiológicos y patológicos.
Distintos elementos regulan la angiogénesis en el ovario y otros tejidos. Entre ellos,
el factor de crecimiento de endotelio vascular (VEGF) y el factor de crecimiento
transformante beta (TGFβ) han sido asociados a eventos importantes en la
angiogénesis ovárica fisiológica y patológica. El factor de crecimiento nervioso (NGF)
también se ha relacionado con angiogénesis en otros tejidos, como el ganglio cervical
superior en ratas neonatas. Además este factor gatilla la proliferación de células
endoteliales humanas. NGF, a través de sus receptores trkA y p75, ejerce acciones
esenciales tanto en el desarrollo ovárico como en la función del ovario. De hecho, NGF
está presente en el ovario humano desde la etapa fetal. Sin embargo, se desconoce si
NGF participa en la angiogénesis ovárica. Por otra parte, NGF modula la expresión de
TGFβ1 y VEGF en distintos tipos celulares, pero tampoco se sabe si en el ovario dicha
expresión es regulada por esta neurotrofina. En esta Tesis se propuso estudiar la participación de NGF como factor angiogénico en el ovario, y su acción en los
niveles de proteína y mRNA de los factores VEGF y TGFβ1.

xvi
Para cumplir con este objetivo, se trabajó con ovarios de ratas neonatas cultivados
con NGF, ovarios de ratones neonatos NGF-deficientes (NGF KO), ratas prepúberes
con niveles ováricos elevados de NGF, inducidos por denervación, y ratones
transgénicos prepúberes que sobreexpresan NGF en el ovario. Se analizaron los
niveles de mRNA y/o proteína de VEGF y TGFβ1 en los ovarios de estos animales.
También se cultivaron células de granulosa humana y se estimularon con NGF, para
luego determinar los contenidos de mRNA de VEGF y TGFβ1 y los niveles de proteína
secretada en el medio de cultivo. Los resultados obtenidos en ovario de roedores y en
el cultivo de células de granulosa indican que NGF incrementa tanto el mRNA como la
proteína de VEGF. Los niveles de TGFβ1 no se modificaron por NGF en los modelos
estudiados.
Con el objetivo de conocer el mecanismo mediante el cual NGF induce los cambios
observados en el ovario, se estudió la participación de trkA, el receptor de alta afinidad
para NGF. Se analizaron ovarios de ratones neonatos trkA-deficientes (trkA KO). Se
determinó que en estos animales el contenido de mRNA de VEGF y TGFβ1 es similar
al encontrado en ratones silvestres, probablemente por un sistema de compensación
que involucra el aumento de la expresión p75, el receptor de baja afinidad para NGF.
También se analizó el efecto de K252a, inhibidor del receptor trkA, en células de
granulosa humana tratadas con NGF. La preincubación con el inhibidor revirtió en
forma significativa el aumento de mRNA y proteína de VEGF inducido por NGF.
A continuación, se quiso saber si NGF inducía la activación de MAPK asociadas a la
fosforilación del receptor trkA en el ovario, y si la producción del mensajero o proteína
de VEGF y TGFβ1 dependía de esta vía de señalización. Para estudiar esto se utilizó
el modelo de células de granulosa humana en cultivo. Análisis de western blot
mostraron que hubo un incremento significativo en la proporción de proteínas ERK 2
fosforiladas respecto de ERK 2 totales, luego de 5 minutos de estimulación con NGF.
Además, el incremento en la expresión de mRNA y proteína de VEGF fue prevenido
por la incubación previa con U0126, un inhibidor de la proteína MEK, responsable de la
fosforilación de ERK 1/2.

xvii
El factor de transcripción HIF1 es uno de los responsables de la activación
transcripcional de VEGF como respuesta a la hipoxia, pero también frente al estímulo
con factores de crecimiento. No se produjo un cambio en el contenido de mRNA de la
isoforma inducible de HIF1, (HIF1α) en células de granulosa humana estimuladas con
NGF 50 ng/ml por 8 horas. Sin embargo, en ovario de rata neonata en cultivo, el
contenido de mRNA de HIF1α aumenta en los tejidos estimulados con NGF por 2
horas.
Finalmente, se quiso verificar si existían acciones indirectas de NGF sobre la
proliferación de células endoteliales, a través del aumento en los niveles de VEGF.
Para esto se realizó un ensayo de angiogénesis in vitro, en el cual el medio
condicionado de células de granulosa tratadas con NGF se utilizó para estimular
células endoteliales provenientes de cordón umbilical humano (células HUVEC). El
medio condicionado no indujo mayor proliferación de células endoteliales,
probablemente debido a que la concentración de VEGF en éste era muy baja para
estimular la proliferación en las células HUVEC. Sin embargo, es muy importante
destacar que estudios in vivo realizados en ratas con niveles ováricos elevados de
NGF, inducidos por denervación, y en ratones transgénicos que sobreexpresan NGF
en el ovario, demostraron que esta neurotrofina es capaz de incrementar la
vascularización en ovario de roedores prepúberes. Los resultados aquí reportados no
permiten establecer si el efecto de NGF sobre la vascularización ovárica es una
consecuencia del aumento de VEGF.
La señalización de NGF es esencial para la ovulación. Por otra parte, se produce
un marcado aumento de la expresión ovárica de VEGF en la etapa periovulatoria. Los
resultados presentados indican que NGF es capaz de aumentar la expresión de VEGF
y la vascularización ovárica. De acuerdo con lo anterior, existiría una asociación entre
el alza periovulatoria de NGF, y el aumento de VEGF y de los capilares ováricos. Esto
podría tener importantes aplicaciones en el área de la medicina reproductiva y las
terapias contra el cáncer, ya que amplía la información acerca de los elementos
reguladores de la angiogénesis en el ovario.

xviii
Summary
Role of nerve growth factor in the regulation of ovarian angiogenesis
The formation of blood vessels that arise from the preexisting vasculature, a process
also known as angiogenesis, is essential for normal tissue development. Physiological
angiogenesis occurs mainly in the embryo and fetus, whereas in the adult, the vascular
endothelium is a tissue with low proliferation rate. However, cyclical changes
associated to the angiogenic process are produced in the ovaries. Angiogenesis
supports follicular and luteal development, and in the non-conceptive cycle, this
vascular network is reabsorbed. During each ovarian cycle, blood vessels rapidly
appear and regress, being an ideal model to study the regulatory elements of
angiogenesis. The understanding of the mechanisms underlying ovarian angiogenesis
could be of great importance for investigating of this process both in physiological and
pathological events.
Ovarian angiogenesis is regulated by different elements. Among them, vascular
endothelial growth factor (VEGF) and transforming growth factor beta (TGFβ) have
been associated to physiological and pathological ovarian angiogenesis. Nerve growth
factor (NGF) has also been associated with angiogenesis in tissues, such as superior
cervical ganglia in neonatal rats. It is also able to increase proliferation of human
endothelial cells. NGF, by activating trkA and p75 receptors, induces essential
processes in ovarian development and function. In fact, NGF is present in the human
ovary from fetal stage. However, it is not known if NGF has a role in ovarian
angiogenesis. On the other hand, NGF modulates TGFβ1 and VEGF expression in
various cell types, but whether it does so in the ovary, is unknown. The aim of this Thesis was to investigate the role of NGF as an ovarian angiogenic factor, and its
effect on protein and mRNA levels for VEGF and TGFβ1.
To this aim, the following models were used: neonatal rat ovaries cultured in the
presence of NGF, ovaries from neonatal NGF-knockout mice (NGF KO), prepuberal
rats with high levels of ovarian NGF, induced by denervation and prepuberal transgenic

xix
mice, designed to overexpress NGF in the ovary. The levels of mRNA and/or protein for
VEGF and TGFβ1 were analyzed in these animals ovaries. Also, human granulosa cells
were cultured with NGF to determine the levels of mRNA for VEGF and TGFβ1, and the
concentration of the respective proteins in the culture media. The results obtained with
rodent ovaries as well as with cultured granulosa cells indicate that NGF is able to
increase both VEGF mRNA and protein. TGFβ1 expression was not modified by NGF in
the models studied.
To know about the mechanism involved in the effects of NGF in the ovary, the role of
trkA receptor (the high-affinity receptor for NGF) was evaluated. Neonatal trk-A
knockout mice were analysed. mRNA levels for VEGF and TGFβ1 were found to be the
same as those present in wild-type mice ovaries. There is the possibility that the lack of
trkA was compensated by an increase of p75, the low affinity NGF receptor. The effect
of K252a, a trkA inhibitor, was studied in granulosa cells. We found that the pe-
treatment with this inhibitor significantly reverted the NGF-induced increase of VEGF
mRNA and protein levels.
We were interested to elucidate if NGF could induce the activation of MAPK that are
associated to the phosphorylation of trkA receptor in the ovary, and if the expression of
VEGF and TGFβ1 was dependent on this signaling pathway. To evaluate these issues,
cultured granulosa cells were used. Western blot analysis showed that a significant
increase of the phosphorylated ERK 2/total ERK 2 ratio was induced after 5 minutes of
NGF stimulation. Besides, the NGF-induced increase of VEGF mRNA and protein
levels was prevented after using U0126, an inhibitor of MEK, the kinase responsible for
Erk 1/2 phosphorylation.
Transcription factor HIF1 is important for transcriptional activation of VEGF in
response to hypoxia, but also after stimulation by other growth factors, such as IGF-1.
No change in inducible subunit of HIF1 (HIF1α) mRNA was found in human granulosa
cells cultured with NGF by eight hours. Still, HIF1α mRNA increases significantly in
cultured neonatal rat ovaries after 2 hours stimulation with NGF.

xx
Finally, we wanted to verify if there was an indirect stimulation of endothelial cells
proliferation driven by NGF. To this aim, an in vitro angiogenesis assay was made, by
using conditioned media obtained from NGF-treated granulosa cells, to stimulate
human umbilical cord endothelial cells (HUVEC). We found that conditioned media did
not induce a greater proliferation on endothelial cells. A probable reason is that VEGF
concentration was too low to stimulate HUVEC proliferation in those conditioned
medium. However, a remarkable observation is that in vivo studies performed in
prepuberal rats with high levels of ovarian NGF, induced by denervation and prepuberal
transgenic mice, designed to overexpress NGF in the ovary, showed that this
neurotrophin is able to increase vascularization in prepubertal rat ovaries. The results
here presented don’t explain if the effect of NGF on ovarian vascularization is a
consequence of the increase of VEGF expression.
NGF signaling is essential to the ovulatory cascade. On the other hand, a marked
increase of ovarian expression of VEGF is produced in the periovulatory stage. The
data reported here indicate that NGF is able to increase both ovarian VEGF expression
and vascularization. According to this, an association can be established between the
periovulatory elevation of ovarian NGF, VEGF and capillaries. These results expand the
actual information about regulatory elements of ovarian angiogenesis, so they could
have important applications on reproductive and anti-cancer medicine.

1
1. Introducción
1.1. Angiogénesis La formación del sistema vascular comienza tempranamente en el desarrollo de los
mamíferos, con la formación y agregación de células precursoras (angioblastos) en el
embrión. En el saco vitelino se produce una estrecha asociación entre las células
hematopoyéticas primitivas y el endotelio en desarrollo, lo que sugiere la existencia de
un precursor común, denominado hemangioblasto (Rossant y cols. 2002). En cambio,
en el embrión, la formación de los primeros vasos sanguíneos se produce en ausencia
de hematopoyesis, lo que sugiere que los precursores celulares son solamente
angioblastos (Rossant y cols. 2002). La formación de vasos sanguíneos en el embrión
y en el saco vitelino a partir de la agregación de angioblastos formados de novo, y su
incorporación en una red vascular primitiva de vasos endoteliales simples es conocida
como vasculogénesis. El desarrollo de un sistema de vasos maduro que ocurre con
posteriorioridad, involucra un proceso mucho más complejo de remodelamiento y
refinamiento del patrón inicial, con la proliferación y ramificación de vasos a partir de
otros vasos existentes. Este segundo proceso es conocido como angiogénesis (Papetti
y cols. 2002, Rossant y cols. 2002).
La angiogénesis es un proceso esencial en el desarrollo del tejido normal. Sin
embargo, varias etapas son necesarias para el establecimiento de la red capilar. En
primer lugar, el tejido que requiere neovascularización emite una señal angiogénica,
que fragmenta y remodela la membrana basal del vaso sanguíneo preexistente,
ocurriendo posteriormente la migración de células endoteliales hacia la zona de
estimulación, y la proliferación de las células endoteliales para formar una monocapa
que adquiere una estructura tubular. Finalmente, los nuevos capilares se diferencian en
arteriolas o vénulas (Redmer y cols. 1996, Fraser y cols. 2001, Papetti y cols. 2002,
Rossant y cols. 2002).

2
La médula ósea postnatal contiene un subtipo de células progenitoras que tienen la
capacidad de migrar a la circulación periférica para diferenciarse hacia células
endoteliales maduras (Hristov y cols. 2003). La mayor parte de la angiogénesis
fisiológica ocurre en la etapa embrionaria. En el adulto, el endotelio vascular es un
tejido con un bajo índice de proliferación, y sólo eventos puntuales, como la
cicatrización de heridas, hacen que se active la neovascularización en forma transitoria
(Papetti y cols. 2002). Estudios recientes indican que células precursoras endoteliales,
derivadas de la médula ósea, ayudan al reestablecimiento de la función de órganos
isquémicos, posiblemente por medio de la inducción de angiogénesis en zonas con un
aporte de oxígeno disminuido, o a través de la reendotelización de vasos dañados
(Hristov y cols. 2003). La proliferación vascular incontrolada o persistente se asocia a
numerosas condiciones patológicas, como el crecimiento tumoral (Papetti y cols. 2002).
Sin embargo, la angiogénesis es un proceso normal en los tejidos reproductores
femeninos. Los ovarios, el útero y la placenta están sometidos a cambios cíclicos
asociados a angiogénesis. En el ovario, la angiogénesis es necesaria para el
desarrollo folicular y la formación del cuerpo lúteo, y los vasos sanguíneos formados en
un ciclo en que no existe concepción son reabsorbidos, lo que refleja que la
neovascularización en este tejido se encuentra muy regulada fisiológicamente (Fraser y
cols. 2001).
1.2. Angiogénesis en el ovario En la mayoría de los tejidos, la disminución en la presión parcial de oxígeno
(hipoxia) es el principal estímulo para la síntesis de factores angiogénicos. En cambio,
en el ovario, la expresión de estos factores durante etapas como el desarrollo folicular
y la formación del cuerpo lúteo, parece estar regulada en forma predominante por las
hormonas luteinizante (LH) y foliculoestimulante (FSH) (Fraser y cols. 2001).
La angiogénesis comienza tempranamente durante el desarrollo del folículo, y
mientras éste sigue creciendo, se limita a la capa tecal. Luego del alza de LH, el
folículo experimenta profundos cambios que permiten la formación del cuerpo lúteo:
después de la ovulación, sobreviene una rápida neovascularización que atraviesa la
membrana basal, alcanzando las células de granulosa luteinizadas (Redmer y cols.

3
1996). Desórdenes en este proceso de vascularización parecen ser incompatibles con
la función ovárica ya que se ha demostrado en primates que la inhibición de la
angiogénesis ovárica suprime el desarrollo folicular (Figura 1.1; Wulff y cols. 2002).
Figura 1.1. Efecto de la administración de VEGF Trap (receptor truncado del factor de crecimiento del endotelio vascular) sobre el desarrollo de los folículos ováricos de primates. Se muestran secciones de ovarios de animales que recibieron una inyección subcutánea con el vehículo (control) o con VEGF Trap durante toda la fase folicular del ciclo ovárico. Las secciones de tejido fueron teñidas con hematoxilina-eosina. En el ovario control se observan dos folículos dominantes saludables, mientras que en el animal tratado con VEGF Trap existe una supresión del desarrollo folicular (adaptado de Wulff y cols. 2002). La estrecha asociación que existe entre la angiogénesis y los procesos de
foliculogénesis y formación del cuerpo lúteo, implican que el control de la angiogénesis
sea un elemento crucial en la función ovárica normal (Otani y cols. 1999). Más aún, la
extraordinaria rapidez con que se suceden la neovascularización y la reabsorción de
los vasos sanguíneos en cada ciclo ovárico, hace de este sistema un modelo ideal para
el estudio de los elementos regulatorios de la angiogénesis, por lo que su mejor
comprensión puede significar un gran avance hacia la aplicación de este modelo en la
investigación de la angiogénesis en eventos fisiológicos y patológicos.
Una variedad de factores regulan positiva y negativamente la angiogénesis en el
ovario y otros tejidos: polipéptidos solubles, interacciones célula-célula o célula-matriz
extracelular y efectos hemodinámicos coordinan este proceso. Las células endoteliales,
los pericitos, los fibroblastos, y algunos integrantes del sistema inmune expresan

4
diferentes citoquinas y factores de crecimiento que encuentran su blanco en células o
en componentes de la matriz extracelular y afectan la migración de las células
endoteliales, su proliferación, la formación de tubos y la estabilización de los vasos
sanguíneos (Figura 1.2; Fraser y cols. 2001, Papetti y cols. 2002). Durante las últimas
décadas, varios factores de crecimiento y citoquinas se han encontrado en tejidos
lúteos, células lúteas, o células de granulosa luteinizadas en cultivo. Entre ellos se
encuentran los factores de crecimiento: epidermal, análogos a insulina, nervioso,
transformantes, de necrosis tumoral α, de fibroblastos y del endotelio vascular (Redmer
y cols. 1996).
Figura 1.2. Mecanismos fisiológicos de angiogénesis. La angiogénesis depende de la coordinación de varios procesos independientes, cada uno de los cuales es dirigido por uno o más efectores específicos (Papetti y cols. 2002).
1. Desestabilización de los vasos -Ang2/Tie2
2. Hiperpermeabilidad de los vasos -VEGF, VE caderina Remodelamiento de la matriz -TGFβ
3. Prolif. endotelial -VEGF -FGF -EGF
4. Migración endotelial -Integrina αvβ3 -VEGF -FGF
5. Contacto cél.-cél. -VE caderina -ephrin B2/eph B4
6. Formación del tubo -FGF -PDGF -TNFα -Eph 2A
7. Migrac. y prolif. endotelial -PDGF -Ang1/Tie2
8. Diferenciación pericitos -TGFβ
9. Estabilización de los vasos -Ang1/Tie2 -PDGF -VE caderina -TGFβ

5
1.3. Factor de crecimiento nervioso (NGF) en el ovario
NGF forma parte de un grupo de neurotrofinas esenciales para la sobrevida,
diferenciación y mantención de las células nerviosas. Sin embargo, en la actualidad se
ha convenido que NGF es un factor con acciones pleiotrópicas, involucrado en el
desarrollo de tejidos no nerviosos en el sistema inmune, cardiovascular y endocrino
(Sofroniew y cols. 2001). En algunos tejidos, el dímero de NGF es secretado en
conjunto con otras 2 proteínas también diméricas, denominadas NGFα y NGFχ. Estas
proteínas son serín proteasas de la familia de la calicreina, y su función más probable
sería procesar al pre-NGF para dar la forma madura (Wiesmann y cols. 2001). Por este
motivo, a NGF con frecuencia se le denomina NGFβ, ya que la actividad trófica ha sido
asignada a esa parte del complejo secretado (Wiesmann y cols. 2001). NGF se une a
dos receptores de membrana: trkA, de alta afinidad y con actividad tirosina kinasa, y
p75, de baja afinidad, sin actividad kinasa. En el sistema nervioso, la unión de NGF a
trkA activa la vía Ras/Erk y PI3K, que llevan a la diferenciación y sobrevida neuronal.
Por otra parte, la unión de NGF a p75 activa las vías de JNK y la generación de
ceramidas, que promueven la apoptosis (Sofroniew y cols. 2001, Kishibe y cols. 2002). A través de sus receptores trkA y p75, NGF ejerce acciones esenciales en el
desarrollo y la función del ovario, y junto con otras neurotrofinas y sus receptores, está
presente en el ovario humano desde la etapa fetal (Anderson y cols. 2002). En
roedores, NGF también es crítico para el desarrollo folicular temprano y la ovulación.
La inmunoneutralización de NGF durante la vida postnatal temprana de la rata provoca
un crecimiento folicular pobre, retrasa la primera ovulación, altera la ciclicidad y la
fertilidad (Lara y cols. 1990a). Ratones knock out para NGF muestran un retraso en el
desarrollo folicular, lo que podría deberse a la pérdida de la señal proliferativa de NGF
(Dissen y cols. 2001). Además, a los siete días, estos animales muestran niveles
ováricos reducidos del RNA mensajero (mRNA) para el receptor de FSH, lo que indica
que NGF participa también en la diferenciación del ovario en desarrollo (Romero y
cols. 2002).

6
Durante el desarrollo folicular, NGF y trkA se expresan principalmente en la teca y
las células intersticiales del ovario de rata, con un máximo luego del alza de LH (Dissen
y cols. 1996). Sin embargo, en ovarios de ratón de siete días se ha encontrado tanto el
mRNA como la proteína de trkA en las células de granulosa (Dissen y cols. 2001). En
ovario de bovino, las células de granulosa expresan el mRNA de trkA, sin importar el
tamaño del folículo (Dissen y cols. 2000). Asimismo, las células de granulosa humana
son capaces de responder a NGF, induciendo la proliferación celular, la
esteroidogénesis (Salas 2002) y la expresión de receptores para FSH (Salas y cols.
2006).
1.4. NGF como probable factor angiogénico en el ovario NGF también ha sido asociado a la actividad angiogénica. En células endoteliales
humanas derivadas de cordón umbilical, NGF gatilla la proliferación celular y aumenta
la fosforilación de trkA y de ERK. NGF también induce la proliferación de células de la
microvasculatura dérmica humanas, en las que además aumenta la expresión de
moléculas de adhesión (Cantarella y cols. 2002). En embriones de pollo, NGF induce la
angiogénesis de la membrana corio-alantoidea de manera independiente de VEGF
(Cantarella y cols. 2002). Sin embargo, en cerebro de ratas neonatas, y en músculo
esquelético de ratón NGF induce angiogénesis, probablemente a través del incremento
en la expresión de VEGF (Calzà y cols. 2001, Emanueli y cols. 2002). En ovario
humano y de rata, sin embargo, no está claro si el efecto de NGF sobre la proliferación
celular es directo o mediado por otro factor de crecimiento. Además, se desconoce si
esta neurotrofina cumple algún rol en el proceso de angiogénesis ovárica.
1.5. Factor de crecimiento de endotelio vascular (VEGF) VEGF es uno de los principales factores que regulan la angiogénesis del ovario, y
su expresión es diferencial de acuerdo al tamaño y estado funcional del folículo. En el
ovario humano, la síntesis del mRNA para VEGF comienza en las células de granulosa
y teca de los folículos secundarios tardíos. En la granulosa de los folículos terciarios se
produce un aumento en esta expresión, mientras que en folículos atrésicos se reduce.
Luego de la ovulación, el mRNA de VEGF es producido por las células de granulosa
luteinizadas (Fraser y cols. 2001). VEGF ejerce varios efectos en las células

7
endoteliales: incrementa la permeabilidad de los vasos sanguíneos, regula el balance
entre la producción de activadores del plasminógeno e inhibidores de este proceso.
Estudios in vitro describen que VEGF estimula específicamente la proliferación y
migración del endotelio vascular, e inhibe la apoptosis de este tipo de células (Papetti y
cols. 2002).
VEGF ejerce sus efectos a través de, al menos, dos receptores con actividad
tirosina kinasa: Flt-1 (VEGFR1) y KDR/Flk-1 (VEGFR2) (Shibuya 2001, Dvorak 2002),
presentes en las células endoteliales de los vasos sanguíneos que irrigan al folículo o
al cuerpo lúteo (Fraser y cols. 2001). Al unirse a sus receptores, VEGF inicia una
cascada de eventos de señalización que comienza con la autofosforilación de ambos
receptores, seguida de la activación de numerosas proteínas río abajo, como la
fosfolipasa Cγ, PI3-K, ERK y otras. El receptor Flk-1 mediaría la permeabilidad
microvascular y la subsecuente proliferación y migración de las células endoteliales.
Las señales mediadas por Flt-1 no han sido bien caracterizadas, pero se ha descrito
que Flt-1 regularía negativamente algunas vías de señalización y efectos biológicos
mediados por Flk-1 (Shibuya 2001, Dvorak 2002). Moléculas relacionadas con VEGF
también se unen a estos receptores: el factor de crecimiento placentario (PlGF) y
factores con un grado de homología a VEGF, como VEGF-B, pueden formar
heterodímeros con VEGF y potenciar su señalización (Papetti y cols. 2002).
El gen de VEGF posee varias isoformas generadas por splicing alternativo, siendo
las más frecuentes los polipéptidos de 206, 189, 165 y 121 aminoácidos en el humano
(en la rata y el ratón, las correspondientes isoformas son un aminoácido más cortas).
De acuerdo con su afinidad por componentes de la matriz extracelular, las isoformas
de VEGF tienen distinta capacidad de difusión: VEGF121 es altamente soluble;
VEGF165 también es soluble pero una parte permanece unida a la matriz extracelular.
Por otra parte, VEGF189 y VEGF206 quedan casi completamente retenidas en la
matriz, pudiendo ser liberadas por proteólisis (Ferrara 2001). Estudios de actividad
mitogénica sobre células endoteliales han indicado que la isoforma 164/165 combina
características óptimas de biodisponibilidad con una alta actividad biológica (Ferrara
2001, Dvorak 2002).

8
1.6. Factor de crecimiento transformante β (TGFβ)
El TGFβ es el prototipo de los factores de crecimiento multifuncionales que regulan
eventos claves en el desarrollo. En la célula blanco, TGFβ se une a un complejo
heterodimérico de receptores serina/treonina kinasa tipo I y tipo II (Böttner y cols.
2000). El receptor tipo I actúa río abajo del receptor tipo II y propaga la señal al núcleo,
al fosforilar miembros específicos de la familia Smad, regulados por receptor (R-
Smads). Las R-Smads fosforiladas pueden formar complejos con otros componentes
de la familia, y acumularse en el núcleo, donde participan en la regulación
transcripcional de genes blanco (Böttner y cols. 2000).
Hasta el momento, se han aislado cinco isoformas de TGFβ, de las cuales TGFβ1,
TGFβ2 y TGFβ3 se expresan en tejido de mamíferos (Böttner y cols. 2000). Sólo
recientemente se está haciendo evidente la importancia del TGFβ en la morfogénesis
capilar y en la mantención de la integridad de las paredes de los vasos sanguíneos,
tanto en el desarrollo embrionario como en la vida postnatal (Pepper 1997). Ratones
knock out para TGFβ1, mueren durante la gestación, o bien en la etapa postnatal, y
sus tejidos extraembrionarios presentan vasos sanguíneos dilatados y frágiles, con
uniones intercelulares interrumpidas. En el caso de los ratones knock out para el
receptor II de TGFβ, este fenotipo se produce en los tejidos intraembrionarios (Pepper
1997, Papetti y cols. 2002), reafirmando que TGFβ es un importante promotor de la
angiogénesis.
TGFβ1 está presente en el ovario humano y se expresa por la mayoría de los tipos
celulares durante la etapa folicular y lútea del ciclo ovárico (Dissen y cols. 1994,
Chegini y cols. 1992a, Chegini y cols. 1992b). En ovario de hámster, la expresión del
mRNA para los receptores I y II de TGFβ (TβRI y II) está regulada diferencialmente por
gonadotrofinas y esteroides ováricos y su incremento se correlaciona con el desarrollo
folicular durante el ciclo estral (Roy 2000). La revascularización de autotransplantes de
ovario en rata está asociada a un incremento en la expresión de TGFβ1 y VEGF
(Dissen y cols. 1994). Por otra parte, en el cáncer ovárico, TGFβ estimula el
crecimiento tumoral en forma directa al promover la proliferación e invasión celular, y

9
en forma indirecta, al inducir la expresión de VEGF y metaloproteinasas, favoreciendo
la angiogénesis (Brown y cols. 2000). En esta acción, TGFβ1 sería la forma más activa
(Poon y cols. 2001). Sin embargo, el papel de TGFβ en la angiogénesis del cáncer
ovárico es un tema que genera controversia, y al igual que en otros tipos de carcinoma,
parece depender del estado de avance del tumor, entre otros factores (Saito y cols.
1999, Akhurst 2002).
1.7. Relación de NGF, TGFβ y VEGF
La conexión que existe entre NGF y los factores TGFβ y VEGF ha sido ampliamente
demostrada in vitro, utilizando células PC12. El tratamiento de células PC12 con NGF
incrementa significativamente los niveles de mRNA para TGFβ1, y se acompaña de un
aumento en la secreción de TGFβ1. Este efecto es mediado por un elemento
específico en el promotor del gen de TGFβ1 (Kim y cols. 1994). En este mismo tipo
celular, NGF incrementa la expresión del mRNA de VEGF y la acumulación de la
proteína. Además, el medio condicionado de células PC12 estimuladas con NGF
induce la proliferación de células endoteliales humanas, respuesta que es prevenida
cuando se agrega un anticuerpo neutralizante anti-VEGF (Middeke y cols. 2002).
Los factores de crecimiento NGF, TGFβ y VEGF, todos ellos importantes en el
desarrollo ovárico, parecen estar interrelacionados en el ovario y otros tejidos. En la
rata, transplantes intraoculares de células cromafines aumentan la producción del
mRNA para TGFβ1 al ser tratados previamente con NGF (Förander y cols. 2000).
Durante el proceso de neovascularización del ganglio cervical superior en ratas
neonatas, se observó un aumento de la densidad de vasos sanguíneos cuando los
animales se trataron con NGF. Este efecto se correlaciona directamente con un
aumento en la expresión de VEGF, lo que sugiere que la angiogénesis puede ser
regulada a través de la activación de agentes angiogénicos producidos directamente
por las neuronas, y que NGF estimula indirectamente este proceso (Calzà y cols.
2001). Un incremento en la expresión de VEGF y TGFβ1, dependiente de
gonadotrofinas, está asociado a la revascularización de autotransplantes de ovario en
rata (Dissen y cols. 1994), y NGF parece estar involucrado en ciertas patologías

10
ováricas, tales como el ovario poliquístico (Lara 2000) al igual que VEGF (Agrawal y
cols. 1998). La expresión del receptor trkA, de los receptores TβRI y II y de VEGF se
incrementa previo a la ovulación (Dissen y cols. 1996, Hazelton y cols. 1999, Roy
2000).
En resumen, el factor de crecimiento nervioso tiene un rol de gran importancia en el
desarrollo y la función ovárica. Sin embargo, aunque en otros tejidos se ha demostrado
que NGF induce la angiogénesis, se desconoce si NGF participa en la angiogénesis
ovárica. Por otra parte, NGF modula la expresión de TGFβ1 y VEGF en distintos tipos
celulares, pero tampoco se sabe si dicha expresión es regulada por esta neurotrofina
en el ovario.

11
1.8. Hipótesis
NGF estimula la angiogénesis ovárica, regulando los niveles de mRNA y/o proteína de
los factores de crecimiento VEGF y/o TGFβ1.
1.9. Objetivos específicos Para estudiar el efecto de NGF en la angiogénesis del ovario, se decidió trabajar
con diferentes modelos de estudio, como son ovarios de ratas y ratones de distintas
edades y células de granulosa humana porque:
a) existe suficiente evidencia en roedores donde se demuestra que NGF está
involucrado en el desarrollo del ovario (antes que éste responda a gonadotropinas) y
en la ovulación (cuando el ovario comienza a ciclar y responde a gonadotropinas)
(Dissen y cols. 1996, Ojeda y cols. 2000, Dissen y cols. 2001, Romero y cols. 2002).
b) nuestro grupo de trabajo había demostrado por primera vez que en ovario humano
adulto, NGF y su receptor de alta afinidad trkA se expresaban en las células de
granulosa y responden a esta neurotrofina (Salas y cols. 2006).
Además, este último modelo tiene la ventaja de que es fácil de obtener a través de
los programa de fertilización asistida, por ser un producto de desecho. Habiendo
antecedentes en la literatura que indicaban que VEGF y TGFβ1 se expresan en estas
células al igual que NGF (Mulheron y cols. 1992, May y cols. 1995, Lee y cols. 1997,
Martinez-Chequer y cols. 2003, Salas y cols. 2006), se hacía un muy buen modelo de
estudio, ya que permitiría conocer en parte la señalización de NGF en la expresión de
los factores angiogénicos a estudiar.
Las ventajas y características de cada modelo utilizado se detallan en la Tabla 7.1,
incluída en la sección Anexos.

12
Objetivo específico 1: analizar el efecto de NGF sobre los niveles de mRNA y
proteína de los factores VEGF y TGFβ1 en diferentes modelos de ovario.
Como las gonadotrofinas inducen la expresión del factor angiogénico VEGF en este
órgano (Dissen y cols. 1994, Christenson y cols. 1997, Wulff y cols. 2002), el trabajo
con animales de una edad inferior a los 35 días permitió independizarse de la influencia
de las gonadotrofinas.
Los modelos estudiados se detallan a continuación:
a) Ovarios de roedores en período de desarrollo ovárico (previo a la aparición de
receptores a gonadotrofinas)
i) Ratones NGF knock-out de siete días
ii) Ratones trkA knock-out en el día del nacimiento
iii) Ovario de rata de dos días, cultivado en presencia de NGF.
b) Ovarios de roedores durante el período de 25 a 30 días, etapa previa a la aparición
de la respuesta cíclica estral
i) Ratas con ovario denervado, que presentan niveles ováricos elevados de NGF
(Lara y cols. 1990b).
ii) Ratones transgénicos que sobreexpresan NGF en el ovario.
c) Células de granulosa provenientes de ovarios humanos adultos.
Objetivo específico 2: evaluar el mecanismo de señalización asociado a los
cambios en la expresión de TGFβ1 y/o VEGF en células de granulosa humana
estimuladas con NGF. Como se indicó antes, las células de granulosa como modelo de ovario humano se
eligieron porque responden a NGF in vitro (Salas 2002, Valladares 2003, Salas y cols.
2006), y expresan in vivo tanto VEGF como TGFβ1 (Chegini y cols. 1992a, Fraser y
cols. 2001). Además, las células de granulosa humanas y de otros mamíferos expresan

13
y/o secretan VEGF y TGFβ1 in vitro en respuesta a estímulos con gonadotrofinas o
factores de crecimiento (Mulheron y cols. 1992, May y cols. 1995, Lee y cols. 1997,
Martinez-Chequer y cols. 2003). Además, nuestro grupo de trabajo demostró por
primera vez que las células de la granulosa humana recién aisladas expresan el
receptor de alta afinidad de NGF, trkA (Salas y cols. 2006).
En distintos sistemas se ha demostrado que el efecto mitogénico de NGF requiere
de la activación de su receptor trkA, y de la vía de las ERK (Descamps y cols. 2001,
Cantarella y cols. 2002). Por lo tanto, se propuso utilizar el modelo de células de
granulosa para saber si la acción de NGF en este proceso induce la activación de ERK
1/2 asociadas a la fosforilación del receptor trkA, y si la producción del mRNA y/o
proteína de VEGF y TGFβ1 depende de esta vía de señalización.
Objetivo específico 3: determinar si NGF modula la vascularización ovárica, y si
esta acción es directa, o mediada por VEGF/TGFβ1.
Para la primera parte del objetivo se realizó una evaluación de la presencia de
vasos sanguíneos en ovario, utilizando como modelo los roedores durante el período
de 25 a 30 días, descritos de manera más extensa en el objetivo 1:
i) Ratas con ovario denervado, que presentan niveles ováricos elevados de NGF.
ii) Ratones transgénicos que sobreexpresan NGF en el ovario.
Finalmente, para saber si la acción de NGF sobre la vascularización ovárica era
mediada por cambios en la secreción de VEGF/TGFβ1, se usaron células de granulosa
estimuladas con NGF. El medio condicionado se transfirió a un cultivo de células
endoteliales (células HUVEC, provenientes de vena umbilical humana), en las cuales
se determinó la proliferación celular, en ausencia y presencia de anticuerpos
neutralizantes anti-TGFβ1, anti-VEGF, para saber si TGFβ1 y VEGF actúan como
factores angiogénicos en este sistema. También se neutralizó NGF, para descartar que
los cambos observados fueran producto del NGF utilizado para estimular a las células
de granulosa.

14
2. Materiales y métodos
2.1. Técnicas de cultivo celular 2.1.1. Aislación y cultivo de células de granulosa humana
Las células se obtuvieron mediante aspiración folicular guiada por ultrasonido,
procedimiento que se realizó en pacientes que participaron en el programa de
fertilización asistida del Hospital Clínico Universidad de Chile. Todas las pacientes
entregaron su consentimiento informado (se adjunta una copia del consentimiento en la
sección Anexos) y el protocolo de trabajo fue aprobado por el Comité de Ética
Institucional (se adjunta carta en la sección Anexos).
La sincronización de los ciclos ováricos se indujo mediante la administración de un
agonista de GnRH (Lupron 14 UI s.c., Abbott, Abbott Park IL, USA, por siete días
previos a la menstruación, seguidos de 7 UI s.c. adicionales entre los días 10 y 12 del
ciclo menstrual). La ovulación se indujo mediante la administración secuencial de FSH
recombinante (Puregon Pen 200 UI s.c., Organon, Roseland NJ, USA), en forma diaria
por tres días, seguida de la administración diaria de hMG (Pergonal 150 UI i.m.,
Organon, Roseland NJ, USA). El progreso del desarrollo folicular se siguió por
ultrasonido, y cuando al menos tres folículos alcanzaron un diámetro de 18 mm, se
administró una dosis única de hCG (Pregnyl 10.000 UI i.m., Organon, Roseland NJ,
USA). Las células de granulosa se colectaron 34 horas más tarde, en el momento de la
aspiración de los ovocitos. Algunos datos clínicos de las pacientes de las cuales se
obtuvieron las muestras de células de granulosa utilizadas en este estudio se
resumieron en las Tablas 7.2 y 7.3, en la sección Anexos.
Las células se recuperaron de los aspirados foliculares por centrifugación (290 g por
10 minutos), se resuspendieron en medio DMEM/F12 (Sigma Chemicals, St Louis MO,
USA), suplementado con bicarbonato de sodio (600 mg/l, Sigma Chemicals, St Louis
MO, USA), penicilina (50 mg/l, Laboratorio Chile, Santiago, Chile), gentamicina (80
mg/l, Laboratorio Chile, Santiago, Chile), estreptomicina (50 mg/l, Laboratorio Chile,
Santiago, Chile) y ketoconazol (5 mg/l, Laboratorio Chile, Santiago, Chile) y se
depositaron sobre una solución compuesta por un 50% de Percoll (Sigma Chemicals,

15
St Louis, MO, USA) y un 50% de medio DMEM/F12. Se formó una gradiente al
centrifugar por 40 minutos a 480 g, y las células recuperadas de la interfase se lavaron
dos veces con medio para eliminar el remanente de Percoll. El pellet de células se
resuspendió en 1 ml de medio y se realizó el recuento celular mediante el test de
exclusión de azul tripán (Sigma Chemicals, St Louis, MO, USA) en un hemocitómetro.
Parámetros como el número de folículos aspirados, número y vitalidad de las células
aisladas, se informan en la Tabla 7.2 (ver Anexos).
La presencia de células de granulosa fue verificada mediante análisis funcionales y
de expresión, como se indica en la Figura 7.1, en la sección Anexos. Las células se
sembraron a una densidad de 5 × 105 por pocillo en placas de cultivo (6-pocillos,
Becton Dickinson, USA) recubiertas con gelatina, con 3 ml de medio DMEM/ F12
preparado como se describió anteriormente, a 37°C en un ambiente húmedo con 5%
CO2 / 95% aire. 18 horas más tarde, el medio se reemplazó y las células se incubaron
con NGF (50 ng/ml) (Sigma Chemicals, St Louis MO, USA) por 0 a 10 min para los
experimentos de señalización intracelular, o bien por 8 h, para los análisis de RNA
mensajero y proteínas secretadas al medio de cultivo. La concentración de NGF a
utilizar se estableció en estudios previos de nuestro grupo de trabajo (Lozada 2006).
Los inhibidores utilizados en este estudio se aplicaron 30 minutos antes de la adición
de NGF (K252a 100 nM, Calbiochem, Darmstadt, Alemania; U0126 10 µM o LY294002
30 µM, ambos de Cell Signaling Technologies, Beverly MA, USA). El medio de cultivo
se recolectó y almacenó a –80°C hasta su utilización y las células se procesaron para
la extracción de RNA total o proteínas. En el caso de los medios condicionados que se
utilizaron para estimular células HUVEC, éstos se concentraron tres veces mediante
columnas Microcon (Millipore, Billerica MA, USA), y almacenados a –80°C hasta su
utilización. En cada experimento se utilizó una preparación celular proveniente del
conjunto de folículos obtenidos de una paciente individual.
2.1.2. Aislación y cultivo de células de endotelio de vena umbilical humana Los cordones umbilicales se obtuvieron en el Departamento de Obstetricia y
Ginecología del Hospital Clínico Universidad de Chile. Se recibieron en PBS frío, y y se
mantuvieron a 4 °C hasta ser procesados dentro de las 48 horas siguientes al parto.

16
Para aislar las células endoteliales, se canuló la vena umbilical y se incubó con
colagenasa por 30 min a 37 °C. Por cada 20 cm de cordón se aplicaron 10 ml de una
solución de colagenasa (1 mg/ml, Roche, Mannheim, Alemania) en medio de cultivo
DMEM/ F12 preparado como se describió anteriormente Los glóbulos rojos presentes
en la suspensión celular se eliminaron con una gradiente de Percoll al 50% (Sigma
Chemicals, St Louis, MO, USA). La gradiente se centrifugó 30 minutos a 480 g, y las
células recuperadas de la interfase se lavaron con medio para eliminar el remanente de
Percoll. El pellet de células se resuspendió en 1 ml de medio, y se realizó el recuento
celular mediante el test de exclusión de azul tripán (Sigma Chemicals, St Louis, MO,
USA) en un hemocitómetro. Parámetros como el número y vitalidad de las células
aisladas se informan en la Tabla 7.4 (ver Anexos). Se sembraron 5 × 104 células por
pocillo en placas de cultivo recubiertas con gelatina (96-pocillos; Becton Dickinson,
Franklin Lakes NJ, USA) y en 200 µl de medio con un 20% de suero fetal bovino
(Hyclone, Logan UT, USA). Se incubaron a 37°C en un ambiente húmedo con 5% CO2
/ 95% aire. El medio fue reemplazado por medio DMEM/ F12 sin suero 24 horas antes
de cada experimento.
Las células endoteliales se estimularon con medios condicionados de células de
granulosa controles o tratadas con NGF. En cada caso se aplicaron 100 µl de medio
condicionado por 48 h. En los tratamientos con anticuerpos anti-VEGF (anti-VEGF [C-
1], sc-7269 Santa Cruz Biotechnology, Santa Cruz CA, USA) o anti-NGF (antisuero K-
595-2, proporcionado por el Dr. Urbansky, Oregon Regional Primate Research Institute,
Portland, Oregon, USA), se trabajó con una dilución 1:500 y el medio condicionado se
preincubó con cada anticuerpo por 30 min a 37°C, antes de aplicar esta mezcla al
cultivo celular. El ensayo de proliferación celular se explica más adelante.
2.2. Experimentos en animales 2.2.1. Ratas Todos los procedimientos realizados con animales fueron aprobados por el Comité
de Ética de la Facultad de Ciencias Químicas y Farmacéuticas de la Universidad de
Chile (se adjunta carta en la sección Anexos). Ratas Sprague Dawley se obtuvieron del

17
bioterio de la Facultad de Ciencias Químicas y Farmacéuticas de la Universidad de
Chile. Los animales se mantuvieron bajo condiciones controladas de temperatura (21
°C) y luz (12 horas de luz, desde las 08:00 a las 20:00 horas, y 12 horas de oscuridad).
2.2.2. Ratones mutantes 2.2.2.1. Ratones NGF knockout Ratones de la cepa C57BL/6-AB1, con una mutación NGF +/- (The Jackson
Laboratory, Bar Harbor ME, USA) fueron cruzados con ratones B6D2F1/J (The
Jackson Laboratory, Bar Harbor ME, USA). Los heterocigotos obtenidos se cruzaron
entre sí para generar los ratones NGF -/- usados en este estudio. Las crías
sobrevivieron hasta dos semanas, manteniendo el fenotipo mutante original (Crowley
1994). Los ovarios fueron retirados el día 7 postnatal y el genotipo de los animales se
confirmó por análisis de PCR del DNA de la cola. Los ovarios de estos animales fueron
donados gentilmente por el Dr. Sergio Ojeda (Oregon Regional Primate Research
Institute, Portland, Oregon, USA).
2.2.2.2. Ratones trkA knockout Los ovarios se retiraron el día del nacimiento, y el genotipo de los animales se
confirmó por análisis de PCR del DNA de la cola. Los ovarios de estos animales fueron
donados gentilmente por el Dr. Sergio Ojeda.
2.2.2.3. Ratones transgénicos Los ratones transgénicos usados en este estudio son de la cepa B6D2 y fueron
diseñados para sobreexpresar NGF en el ovario, puesto que contienen un transgén
formado por un fragmento del gen de NGF humano, que se encuentra bajo el control
transcripcional del promotor de la 17α-hidroxilasa (Figura 2.1). Esta enzima se expresa
principalmente en las células de la teca, presentes en el ovario y productoras de
andrógenos.
Los ovarios se obtuvieron el día 32 postnatal. Los tejidos destinados para los
estudios de mRNA se almacenaron a –80 °C. Los ovarios utilizados para
inmunohistoquímica se fijaron con Bouin acuoso e incluyeron en parafina. Los ovarios

18
de estos animales fueron donados gentilmente por el Dr. Sergio Ojeda (Oregon
Regional Primate Research Institute, Portland, Oregon, USA).
Figura 2.1. Mapa del transgén de NGF. La secuencia promotora (17α-hidroxilasa) es derivada de ratón (Youngblood y cols. 1992), el intrón A de la insulina II fue sacado de rata, el minigén de NGF se obtuvo de humano y la señal de poliadenilación provino de rata. Todos los componentes, a excepción del promotor, provienen de un casete de expresión de NGF (Hoyle y cols. 1998).
2.2.3. Cultivo de ovarios Ovarios de ratas de 2 días de edad se disecaron en asepsia y cultivados a 37 °C en
una interfase de medio de cultivo y aire (George y cols. 1987), en una atmósfera
húmeda de 5% CO2 / 95% aire. Los ovarios se cultivaron en placas de 24 pocillos, y
cada pocillo contenía 1 ml de medio DMEM/ F12 preparado como se describió
anteriormente. Los tejidos se estimularon por 2, 4, 8 y 24 h con NGF 100 ng/ml (Sigma
Chemicals, St Louis, MO, USA), y los ovarios contralaterales, cultivados con medio
solo, se usaron como controles. La concentración de NGF se estableció en estudios
previos (Romero y cols. 2002). En el caso de los estudios de mRNA, los ovarios se
almacenaron a –80 °C al finalizar el cultivo. Los tejidos utilizados para
inmunohistoquímica se fijaron con Bouin acuoso e incluyeron en parafina.
m17α Hidroxilasa hNGF intrónregión de
poliadenilación
promotor región codificante

19
2.2.4. Denervación ovárica Ratas hembra de 23 a 24 días se anestesiaron con isoflurano 1% v/v, 2,5 l/min
(Baxter Healthcare Co, Guayama, Puerto Rico). La disección del nervio ovárico
superior se realizó en forma aséptica en ambos ovarios, a través de una incisión en la
línea media dorsal. Este procedimiento ha mostrado inducir un incremento en NGF
dentro de la glándula (Lara y cols. 1990b). Debido a que la denervación induce la
hipertrofia del ovario contralateral (Gerendai y cols. 1978), los controles consistieron en
animales sometidos a una operación simulada. Tres o seis días después, los ovarios
se retiraron y almacenaron a –80 °C para estudios de mRNA. Este tiempo se
estableció en un trabajo previo (Lara y cols. 1990b). Los tejidos utilizados para
inmunohistoquímica se fijaron con Bouin acuoso e incluyeron en parafina.
2.3. Análisis cuantitativos y semicuantitativos 2.3.1. ELISA La concentración de VEGF en el medio de cultivo de células de granulosa humana
se midió usando un ELISA (Quantikine Human VEGF Immunoassay; R&D Systems
Inc., Minneapolis, MN, USA). Este ensayo altamente específico reconoce VEGF 165 y
121, y no tiene reacción cruzada con otros factores de crecimiento o citoquinas. Para
cuantificar los resultados se utilizó un lector de EIA (Sigma, St. Louis, MO, USA),
programado para leer a una emisión de 450 nm. Los coeficientes de variación intra- e
inter- ensayo fueron 6,9 y 8,5%, respectivamente. En el caso de TGFβ1, se usó un
ELISA de características similares al de VEGF (Quantikine Human TGFβ1
Immunoassay; R&D Systems Inc., Minneapolis, MN, USA).
2.3.2. Western blot
Las células de granulosa cultivadas (1 X 106) se lisaron con 75 µl de buffer de lisis,
que contenía NaCl 150 mM, Tris-HCl 50 mM, Tritón X-100 1%, EDTA 1mM, EGTA 1
mM, PMSF 1 mM, ortovanadato de sodio 1 mM, leupeptina 10 mg/ml, aprotinina 1,8
mg/ml, fluoruro de sodio 2 mM, pirofosfato de sodio 2 mM y ditiotreitol 1 M. Todos estos
reactivos eran de Sigma, St. Louis, MO, USA. De cada muestra, 50 microgramos de
proteína se fraccionaron por electroforesis en gel de poliacrilamida al 12% (Sigma, St.
Louis, MO, USA), en condiciones denaturantes, y luego transferidos a una membrana

20
de nitrocelulosa (0,2 µm, BioRad, CA, USA). La unión no específica se bloqueó por
incubación en PBS (pH 7,4) con 2% de leche descremada y 0,2% v/v de Tween-20
(Sigma, St. Louis, MO, USA) durante 1 hora. Luego, las membranas se incubaron con
un anticuerpo policlonal de conejo anti fosfo-p44/42 o anti-p44/42 (1:1.000, 4º C, por
toda la noche, Cell Signaling Technologies, Beverly, MA, USA), y posteriormente con
un anticuerpo secundario conjugado con peroxidasa (1:15.000, temperatura ambiente,
2 h, Cell Signaling Technologies, Beverly, MA, USA). Las proteínas se detectaron
usando un sistema quimioluminiscente (PerkinElmer Life Sciences, MA, USA). Los
films autorradiográficos se escanearon y analizaron con un programa de densitometría
(UN-SCAN-IT gel 4.1, Silk Scientific Corporation, Orem UT, USA).
2.3.3. Inmunohistoquímica Los ovarios se fijaron por inmersión en solución de Bouin y embebidos en parafina.
Secciones seriadas de 4 µm se colocaron sobre portaobjetos silanizados. Se realizó
una recuperación antigénica de las muestras en un recipiente con una solución
tamponada de citrato de sodio 10 mM, pH 6 (Merck, Darmstadt, Alemania), el cual se
mantuvo en una olla vaporera (Oster, Boca Raton FL, USA) en ebullición por 40 min.
Las muestras se dejaron reposar por 20 min adicionales. Las peroxidasas endógenas
se inhibieron mediante la incubación por 30 min con una solución de perhidrol (Merck,
Darmstadt, Alemania) al 2% en metanol (Merck, Darmstadt, Alemania). La unión no
específica se bloqueó por incubación con PBS (pH 7.4) con 5% de leche descremada
durante 30 min. Para la detección inmunohistoquímica de VEGF y TGFβ1 se utilizaron
anticuerpos preparados en ratón y conejo, respectivamente (anti-VEGF [C-1], sc-7269
and anti-TGFβ1 [V] sc-146, ambos de Santa Cruz Biotechnology, Santa Cruz CA, USA,
en una dilución 1:50). Las secciones de tejido se incubaron toda la noche a 4 ºC, y
luego con el anticuerpo secundario apropiado, en una dilución 1:300, durante media
hora. Se utilizó DAB como cromógeno (LabVision Co, Freemont CA, USA), a excepción
de la detección de VEGF en los ovarios de rata neonata, para la cual se utilizó NBT-
BCIP (Sigma, St. Louis, MO, USA). Como controles negativos se utilizaron secciones
adyacentes incubadas en ausencia del anticuerpo primario, o con anticuerpo pre-
adsorbido con el respectivo antígeno purificado. En estos controles no hubo desarrollo
de color. Además, se probó la especificidad de los anticuerpos anti-VEGF y anti-TGFβ1

21
realizando inmuno western-blots de extractos de proteínas provenientes de células de
granulosa humana y de cáncer ovárico. En el caso de VEGF se observó la presencia
de una banda única, lo cual ha sido descrito para este anticuerpo en la literatura (Shao
y cols. 2005), y se trataría de la isoforma de 165 aminoácidos (Corne y cols. 2000).
Para TGFβ1 no se pudo detectar la presencia de bandas en el blot.
En los casos en que se obtuvo una tinción bien definida, se realizó un conteo celular
y los datos se expresaron como H-Score, el que se calculó de la siguiente manera: el
porcentaje de células con intensidad de tinción equivalente a 1+, más el porcentaje de
células con intensidad de tinción 2+, más el porcentaje de células con intensidad de
tinción equivalente a 3+ (Anaf y cols. 2002). En el caso de una tinción difusa, la
intensidad de la marca se evaluó usando un programa de digitalización automatizado,
(UN-SCAN-IT gel 4.1, Silk Scientific Corporation, Orem UT, USA) y los datos se
expresaron como número de píxeles.
Figura 2.2. Evaluación del área de tinción positiva para el marcador de vasos sanguíneos CD31 en ovario de rata. El programa Image J permite delinear los vasos sanguíneos de acuerdo al criterio del observador, y cuantifica el área delineada, entregando los resultados expresados en píxeles. En A se muestra una foto de la señal de CD31 obtenida en un ovario de rata. En B se muestra la sobreposición del delineado de los vasos y la misma foto. Magnificación original 100X. Barra = 100 µm.
La detección inmunohistoquímica del marcador de vasos sanguíneos CD31 se
realizó con un anticuerpo preparado en cabra (anti-PECAM-1 [M-20], de Santa Cruz
Biotechnology, Santa Cruz CA, USA, en una dilución 1:100). Las secciones de tejido se
incubaron toda la noche a 4 ºC, y luego se incubaron con el anticuerpo secundario
A B

22
apropiado, en una dilución 1:300, durante 30 min. Se utilizó DAB como cromógeno. El
área de vasos sanguíneos se evaluó con un programa automatizado, como se indica
en la Figura 2.2 (Image J 1.36b, NIH, USA) y los datos se expresaron como porcentaje
del número de píxeles del área total de la sección.
2.3.4. Extracción de RNA y reacción de transcripción reversa El RNA total se extrajo de los tejidos o células en estudio mediante el método de
isotiocianato de guanidinio y fenol cloroformo (Trizol Reagent, Invitrogen, Foster City
CA, USA), siguiendo las instrucciones del fabricante. La concentración y pureza del
RNA se midió en un espectrofotómetro a 260 y 280 nm. El cDNA se sintetizó en un
volumen final de 20 µl, usando 1 µg de RNA total. La mezcla de reacción contenía 0,5
µl de random hexamers (500 ng/µl), 1 µl de dNTPs (10 mM), 4 µl de buffer de reacción
5x (Tris-HCl 250 mM pH 8,3, KCl 375 mM, MgCl2 15 mM), 2 µl de DTT (0,1 M), 1 µl de
inhibidor de ribonucleaseas (10 U/µl) y 1 µl de transcriptasa reversa M-MLV (200 U/µl).
Todos estos reactivos eran de Invitrogen, Foster City CA, USA. La reacción se incubó a
37º C por 60 min e inactivó por congelación.
2.3.5. Reacción en cadena de la polimerasa (PCR) Las secuencias de los partidores específicos para los genes examinados, y el
número de ciclos de amplificación se muestran en la Tabla 1.1. Previamente, nuestro
grupo de trabajo evaluó la expresión del mRNA de β-actina (en muestras humanas) y
ciclofilina (en muestras de roedores), y no se encontró un efecto regulatorio inducido
por NGF en las condiciones de trabajo descritas, permitiendo su uso como un control
interno confiable.
El cDNA se amplificó en una mezcla de reacción de 25 µl, usando 1 µl de DNAc de
hebra simple. Las condiciones para la PCR fueron las siguientes: 2,5 µl de buffer de
reación 10 x (Tris-HCl 200 mM pH 8,4, KCl 500 mM), 1 µl de MgCl2 (50 mM), 0,5 µl de
dNTPs (10 mM each), 1,25 µl de cada partidor (10 µM), 0,15 µl de DNA polimerasa (5
U/µl). Todos estos reactivos eran de Biotools, Madrid, España. Las mezclas de
reacción se incubaron en un termociclador (Eppendorf, Foster City CA, USA) por 2 min

23
a 94 °C. Luego se realizó el siguiente programa de ciclos: denaturación a 94 °C por 45
seg, alineamiento a 62 °C por 1 min y extensión a 72 °C por 1 min. El número óptimo
de ciclos se determinó experimentalmente analizando la fase exponencial de la
reacción de amplificación. Los productos de PCR se separaron en un gel de agarosa al
2%, con bromuro de etidio (200 ng/ml, Invitrogen, Foster City CA, USA). Estándares de
peso molecular (100 bp Ladder, Biotools, Madrid, España) se corrieron en paralelo,
para validar el tamaño predicho de las bandas de amplificado. Los geles se
visualizaron bajo luz ultravioleta, se fotografiaron y analizaron con un programa de
densitometría (UN-SCAN-IT gel 4.1, Silk Scientific Corporation, Orem UT, USA).
En el caso de los partidores de VEGF, tanto de humano como de rata y ratón, estos
fueron diseñados para amplificar diferentes isoformas de splicing, correspondientes a
189, 165, 145 y 121 aminoácidos. Esto permitió evaluar todas las isoformas
detectables en cada determinación, y lo que se reporta en los resultados corresponde a
las isoformas que fueron detectables en cada caso.
2.3.6. PCR de tiempo real 500 ng de RNA total se sometieron a una transcripción reversa, como se indicó
anteriormente, y el cDNA resultante se amplificó por PCR de tiempo real (Salas 2006).
Los partidores y la sonda interna (Tabla 1.1) se diseñaron con el programa Primer
Express (Perkin Elmer Applied Biosystems, Wellesley MA, USA), para alinear entre los
exones 5 y 7 del mRNA de VEGF humano, de manera de amplificar sólo la isoforma
165. La sonda utilizada tiene la marca fluorescente 6-carboxifluoresceína en el extremo
5’ y el apagador tetrametilrodamina en el extremo 3’. Para normalizar los resultados se
utilizó el RNA ribosomal 18s, usando partidores comerciales (TaqMan Ribosomal RNA
Control Reagents Kit, Perkin Elmer Applied Biosystems, Wellesley MA, USA). La
reacción se realizó en un volumen de 10 µl, utilizando 2 µl de la mezcla de cDNA
(obtenido luego de la transcripción reversa), 5 µl de TaqMan Universal PCR Master Mix
(PerkinElmer Applied Biosystems, Wellesley MA, USA), 250 nM de cada sonda (VEGF
o 18S) y 300 nM de cada partidor para VEGF ó 10 nM de cada partidor para 18S. El
PCR consistió en un alineamiento inicial de 2 min a 50 ºC, seguido de 10 min de
denaturación a 95 ºC y luego 40 ciclos de una denaturación por 15 seg a 95 ºC y un
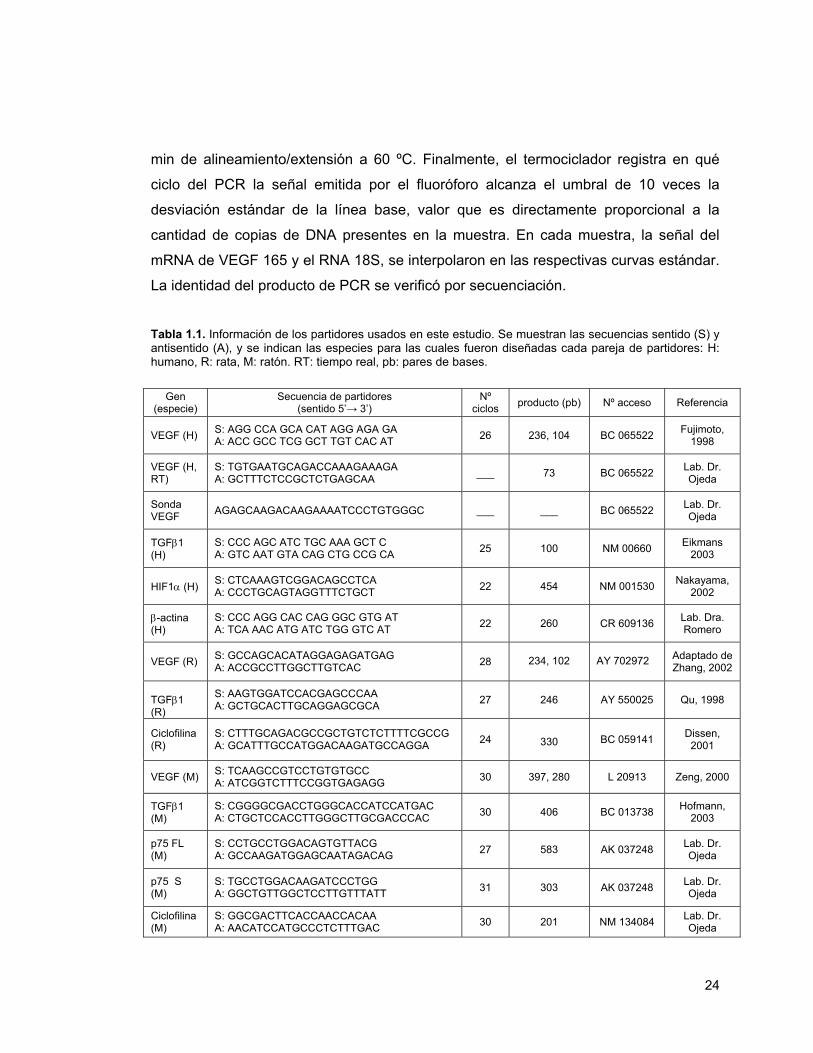
24
min de alineamiento/extensión a 60 ºC. Finalmente, el termociclador registra en qué
ciclo del PCR la señal emitida por el fluoróforo alcanza el umbral de 10 veces la
desviación estándar de la línea base, valor que es directamente proporcional a la
cantidad de copias de DNA presentes en la muestra. En cada muestra, la señal del
mRNA de VEGF 165 y el RNA 18S, se interpolaron en las respectivas curvas estándar.
La identidad del producto de PCR se verificó por secuenciación.
Tabla 1.1. Información de los partidores usados en este estudio. Se muestran las secuencias sentido (S) y antisentido (A), y se indican las especies para las cuales fueron diseñadas cada pareja de partidores: H: humano, R: rata, M: ratón. RT: tiempo real, pb: pares de bases.
Gen (especie)
Secuencia de partidores (sentido 5’→ 3’)
Nº ciclos producto (pb) Nº acceso Referencia
VEGF (H) S: AGG CCA GCA CAT AGG AGA GA A: ACC GCC TCG GCT TGT CAC AT 26 236, 104
BC 065522
Fujimoto, 1998
VEGF (H, RT)
S: TGTGAATGCAGACCAAAGAAAGA A: GCTTTCTCCGCTCTGAGCAA ___ 73
BC 065522
Lab. Dr. Ojeda
Sonda VEGF AGAGCAAGACAAGAAAATCCCTGTGGGC ___ ___
BC 065522
Lab. Dr. Ojeda
TGFβ1 (H)
S: CCC AGC ATC TGC AAA GCT C A: GTC AAT GTA CAG CTG CCG CA 25 100 NM 00660 Eikmans
2003
HIF1α (H) S: CTCAAAGTCGGACAGCCTCA A: CCCTGCAGTAGGTTTCTGCT 22 454 NM 001530 Nakayama,
2002
β-actina (H)
S: CCC AGG CAC CAG GGC GTG AT A: TCA AAC ATG ATC TGG GTC AT 22
260
CR 609136 Lab. Dra.
Romero
VEGF (R) S: GCCAGCACATAGGAGAGATGAG A: ACCGCCTTGGCTTGTCAC 28
234, 102
AY 702972 Adaptado de
Zhang, 2002
TGFβ1 (R)
S: AAGTGGATCCACGAGCCCAA A: GCTGCACTTGCAGGAGCGCA 27 246
AY 550025 Qu, 1998
Ciclofilina (R)
S: CTTTGCAGACGCCGCTGTCTCTTTTCGCCG A: GCATTTGCCATGGACAAGATGCCAGGA 24 330
BC 059141 Dissen,
2001
VEGF (M) S: TCAAGCCGTCCTGTGTGCC A: ATCGGTCTTTCCGGTGAGAGG 30 397, 280 L 20913 Zeng, 2000
TGFβ1 (M)
S: CGGGGCGACCTGGGCACCATCCATGAC A: CTGCTCCACCTTGGGCTTGCGACCCAC 30 406 BC 013738 Hofmann,
2003
p75 FL (M)
S: CCTGCCTGGACAGTGTTACG A: GCCAAGATGGAGCAATAGACAG 27 583 AK 037248 Lab. Dr.
Ojeda
p75 S (M)
S: TGCCTGGACAAGATCCCTGG A: GGCTGTTGGCTCCTTGTTTATT 31 303 AK 037248 Lab. Dr.
Ojeda
Ciclofilina (M)
S: GGCGACTTCACCAACCACAA A: AACATCCATGCCCTCTTTGAC 30 201 NM 134084 Lab. Dr.
Ojeda

25
2.3.7. Ensayo de proliferación de células HUVEC mediante Cell Titer El ensayo Cell Titer (Promega, Madison WI, USA) es un método colorimétrico que
permite determinar el número de células viables en estudios de proliferación. El
compuesto MTS tetrazolium es reducido por las células, con lo que se forma un
compuesto coloreado (formazán), que es soluble en el medio de cultivo. La cantidad de
formazán se mide como la absorbancia a 492 nm y es directamente proporcional al
número de células vivas en cultivo. Luego de la incubación con medios condicionados
por 48 h, las células HUVEC se trataron con 20 µl de reactivo Cell-Titer e incubaron por
1-4 h a 37 °C. La absorbancia a 492 nm se determinó en un equipo Multiskan Ex. En
todos los casos se restó la señal basal (100 µl de medio de cultivo y 20 µl de reactivo
Cell-Titer, sin células).
2.4. Análisis estadístico El análisis estadístico de los datos se realizó con la asesoría de la Oficina de Apoyo
a la Investigación Clínica del Hospital Clínico Universidad de Chile. Se utilizó el test no
paramétrico de Mann-Whitney, debido a que los números de muestras evaluadas en
este trabajo eran muy bajos para asegurar la normalidad de las poblaciones
estudiadas. Un p < 0.05 se consideró estadísticamente significativo. Cuando se
compararon más de dos grupos, se realizó un test de Mann-Whitney mejorado, el cual
incluye un análisis de varianzas, y luego como post-test, una prueba de Mann-Whitney
para evaluar las diferencias entre los distintos grupos.

26
3. Resultados
3.1. Efecto del NGF sobre los factores VEGF y TGFβ1 en ovario de roedores
A fin de simplificar los modelos de estudio, se trabajó con animales que aún no
habían comenzado a ciclar. Las gonadotrofinas incrementan la expresión del factor
angiogénico VEGF en el ovario (Dissen y cols. 1994, Christenson y cols. 1997, Wulff y
cols. 2002), por lo tanto, para no apantallar el efecto de NGF se decidió usar como
modelo, ovarios de roedores en período de desarrollo ovárico (entre el día del
nacimiento y los 7 días post-natal, previo a la aparición de receptores a gonadotrofinas)
y durante la etapa entre la adquisición de receptores a gonadotrofinas y la aparición de
la respuesta cíclica estral, que ocurre aproximadamente a los 35 días post-natal (Mc
Gee y cols. 2000). Las ventajas y características de cada modelo utilizado se resumen
en la Tabla 7.1, en la sección Anexos.
3.1.1. Efecto in vitro del NGF sobre los factores VEGF y TGFβ1 en ovario de ratas
de 2 días de vida
Se trabajó con ovarios de rata de 2 días de vida, cultivados por distintos tiempos en
medio HAM-F12, en presencia de NGF (100 ng/mL). La edad de los animales, así
como las condiciones de cultivo se seleccionaron en base al trabajo de Romero y cols.
(2002), donde se demostró que en esa etapa del desarrollo ovárico y con esa
concentración de NGF los ovarios de ratas de 2 días de vida responden incrementando
los niveles de mRNA del receptor de FSH. Se utilizó NGF humano, que tiene
aproximadamente un 90% de homología con el de rata. Las Figuras 3.1, 3.2 y 3.3 muestran los resultados del efecto de NGF (100 ng/ml) al
incubar los ovarios de rata de dos días de vida por 2, 8 y 24 horas respectivamente.
Solamente a las 2 h de cultivo se encontró un incremento significativo en el contenido
de mRNA de las isoformas de VEGF120 y VEGF164. Sin embargo, no se encontraron
cambios en los niveles de mRNA de TGFβ1 a los distintos tiempos estudiados.

27
Figura 3.1. Análisis semicuantitativo del contenido de mRNA de VEGF (isoformas 120 y 164) y TGFβ1 en ovarios de rata cultivados por 2 horas en presencia de NGF (100 ng/ml). Los niveles de mRNA se expresaron como el número de pixeles de la banda del transcrito en estudio, dividido por el número de pixeles de la banda de Ciclofilina (en unidades arbitrarias, o UA). Los datos mostrados corresponden al promedio ± error estándar de un n=5 para VEGF y 3 para TGFβ1. NGF indujo un incremento estadísticamente significativo del contenido de mRNA de las isoformas 120 y 164 de VEGF (p < 0,05).
Figura 3.2. Análisis semicuantitativo del contenido de mRNA de VEGF (isoformas 120 y 164) y TGFβ1 en ovarios de rata cultivados por 8 horas en presencia de NGF (100 ng/ml). Los niveles de mRNA se expresaron como en la Figura 3.1. Los datos mostrados corresponden al promedio ± error estándar de un n=3. NGF no indujo cambios significativos del contenido de mRNA de los genes estudiados.
VEGF 120 VEGF 164 TGFβ10.0
0.5
ControlNGF (100 ng/ml)
**
Con
teni
do d
e A
RN
m (U
A)
VEGF 120 VEGF 164 TGFβ10.0
0.5
1.0
1.5ControlNGF (100 ng/ml)
Con
teni
do d
e A
RN
m (U
A)

28
Figura 3.3. Análisis semicuantitativo del contenido de mRNA de VEGF (isoformas 120 y 164) y TGFβ1 en ovarios de rata cultivados por 24 horas en presencia de NGF (100 ng/ml). Los niveles de mRNA se expresaron como en la Figura 3.1. Los datos mostrados corresponden al promedio ± error estándar de un n=3 para VEGF y un n=5 para TGFβ1. NGF no indujo cambios significativos del contenido de mRNA de los genes estudiados.
Figura 3.4. Ensayo de inmuno-preadsorción de los anticuerpos utilizados para la evaluación de VEGF y TGFβ1 por inmunohistoquímica en ovario de rata. Antes de tratar los tejidos con el anticuerpo anti-VEGF ó anti-TGFβ1, el anticuerpo se incubó con el péptido correspondiente, a 37 ºC por 15 min. En la figura se muestra el resultado de las inmunohistoquímicas realizadas con anticuerpos sin preadsorber y preadsorbidos (magnificación original 10X, recuadro interior 40X).
VEGF 120 VEGF 164 TGFβ10.0
0.5
ControlNGF (100 ng/ml)
Con
teni
do d
e A
RN
m (U
A)

29
La evaluación de las proteínas VEGF y TGFβ1 en ovario de rata se hizo por
inmunohistoquímica. La especificidad de los anticuerpos utilizados se verificó mediante
un ensayo de inmuno-preadsorción (Figura 3.4).
Cuando se evaluó la proteína VEGF en ovarios cultivados con NGF por 24 h, se
observó la presencia de marca en todo el tejido ovárico, especialmente en los folículos
primarios, estructuras en las cuales se detectó VEGF en la capa de células de
granulosa (Figura 3.5). NGF incrementó significativamente la marca de VEGF con
respecto al control, tanto para la tinción general del tejido como para la intensidad de
marca localizada en los folículos ováricos (Figura 3.6 B). No se encontró una diferencia
significativa en la proteína VEGF cuando los ovarios se estimularon con NGF por un
tiempo más corto (4 h), como se indica en la Figura 3.6 A.
Figura 3.5. Inmunohistoquímica de VEGF en ovarios de rata cultivados por 24 h con NGF (100 ng/ml). La proteína VEGF fue detectada específicamente en la capa de células de granulosa de folículos primarios (ver flechas en A, B y D) pero también en otros tipos celulares en el resto del tejido (ver cabezas de flecha en C y D). A y C muestran la marca encontrada en ovarios control, y B y D los respectivos tejidos tratados con NGF, en los que se aprecia una mayor intensidad de marca. El control negativo de la inmunohistoquímica se muestra en E. Magnificación original 100X. Barra = 50 µm.

30
Figura 3.6. Análisis semicuantitativo de la inmunohistoquímica de VEGF en ovarios de rata cultivados por 4 (A) y 24 h (B) en presencia de NGF (100 ng/ml). Los datos mostrados corresponden al promedio ± error estándar de un n=6 para las 4 h y un n=5 para las 24 h. El estímulo de NGF por 24 h indujo un incremento estadísticamente significativo de la intensidad de marca de VEGF (* p < 0,05; ** p < 0,01) tanto para la tinción general del ovario como para la tinción folicular.
En el caso de TGFβ1, el análisis de la inmunohistoquímica permitió observar que la
marca se encuentra específicamente en los ovocitos de folículos de distinto tamaño
(Figura 3.7). Sin embargo, luego de hacer una evaluación por H-Score, no se
encontraron diferencias en la intensidad de tinción de TGFβ1 entre el grupo de ovarios
estimulado con NGF y el grupo control en los tiempos de cultivo estudiados (Figura
3.8).
Figura 3.7. Inmunohistoquímica de TGFβ1 en ovarios de rata cultivados por 24 h con NGF (100 ng/ml). La marca se detectó específicamente en los ovocitos. A y B muestran resultados representativos de la marca encontrada en un ovario control y uno estimulado con NGF, respectivamente. El control negativo de la inmunohistoquímica se muestra en C. Magnificación original 100X. Barra = 50 µm.
Marca general Marca folicular
0
1000
2000
ControlNGF 100 ng/ml
5000
6500*
**
Inm
unor
reac
tivid
ad d
e VE
GF
(pix
eles
)
Marca general Marca folicular
0
1000
2000
3000ControlNGF 100 ng/ml
Inm
unor
reac
tivid
ad d
eVEG
F (p
ixel
es)
A B
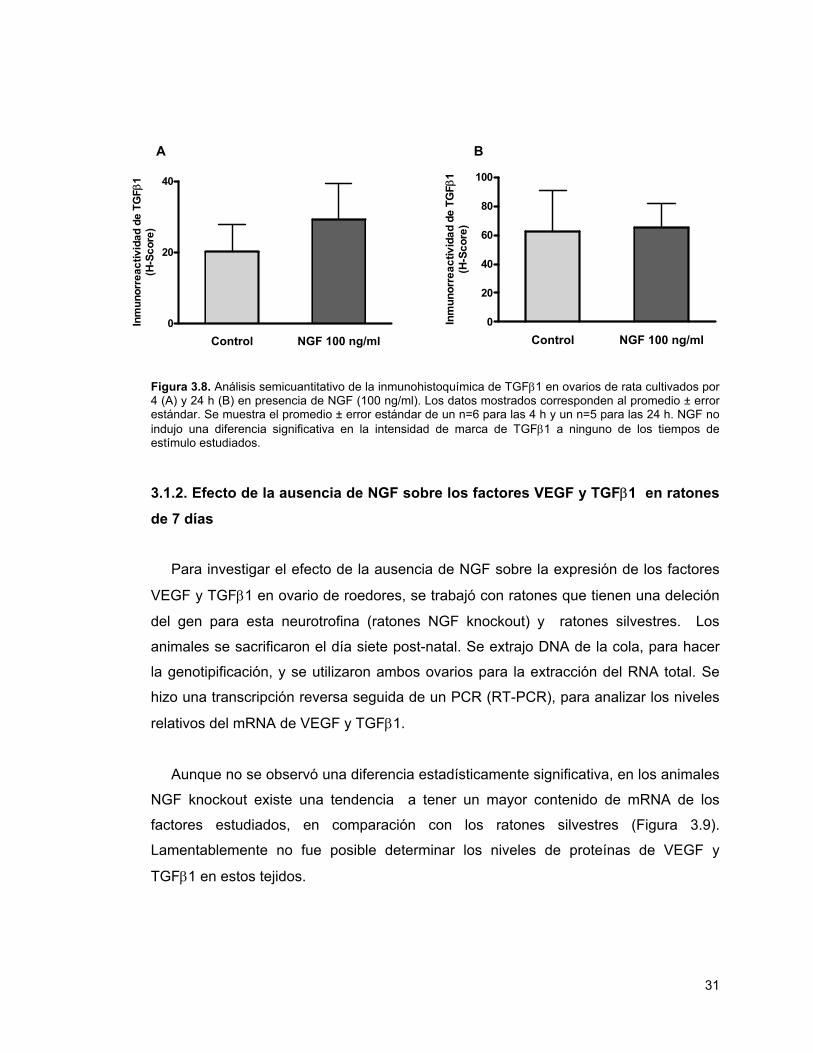
31
Figura 3.8. Análisis semicuantitativo de la inmunohistoquímica de TGFβ1 en ovarios de rata cultivados por 4 (A) y 24 h (B) en presencia de NGF (100 ng/ml). Los datos mostrados corresponden al promedio ± error estándar. Se muestra el promedio ± error estándar de un n=6 para las 4 h y un n=5 para las 24 h. NGF no indujo una diferencia significativa en la intensidad de marca de TGFβ1 a ninguno de los tiempos de estímulo estudiados. 3.1.2. Efecto de la ausencia de NGF sobre los factores VEGF y TGFβ1 en ratones
de 7 días Para investigar el efecto de la ausencia de NGF sobre la expresión de los factores
VEGF y TGFβ1 en ovario de roedores, se trabajó con ratones que tienen una deleción
del gen para esta neurotrofina (ratones NGF knockout) y ratones silvestres. Los
animales se sacrificaron el día siete post-natal. Se extrajo DNA de la cola, para hacer
la genotipificación, y se utilizaron ambos ovarios para la extracción del RNA total. Se
hizo una transcripción reversa seguida de un PCR (RT-PCR), para analizar los niveles
relativos del mRNA de VEGF y TGFβ1.
Aunque no se observó una diferencia estadísticamente significativa, en los animales
NGF knockout existe una tendencia a tener un mayor contenido de mRNA de los
factores estudiados, en comparación con los ratones silvestres (Figura 3.9).
Lamentablemente no fue posible determinar los niveles de proteínas de VEGF y
TGFβ1 en estos tejidos.
Control NGF 100 ng/ml0
20
40
60
80
100
Inm
unor
reac
tivid
ad d
e TG
Fβ1
(H-S
core
)
Control NGF 100 ng/ml0
20
40
Inm
unor
reac
tivid
ad d
e TG
Fβ1
(H-S
core
)A B

32
Figura 3.9. Análisis semicuantitativo del contenido de mRNA de VEGF (isoformas 120 y 164) y TGFβ1 en ovarios de ratones NGF knockout y silvestres de 7 días de edad. Los niveles de mRNA se expresaron como en la Figura 3.1. Los datos mostrados corresponden al promedio ± error estándar de un n=4 para los animales silvestres y un n=5 para los knockout. La ausencia de NGF indujo un incremento del contenido de mRNA de los genes estudiados, que no fue estadísticamente significativo. 3.1.3. Efecto del NGF sobre los factores VEGF y TGFβ1 en ratas sometidas a
denervación ovárica
Se ha descrito que, en la rata, la denervación quirúrgica del ovario induce un alza
local en los niveles de NGF (Lara y cols. 1990b). La neurotrofina normalmente es
transportada en forma retrógrada desde el terminal nervioso hacia el cuerpo neuronal,
por lo que la denervación impide que sea retirada del ovario, induciendo la
acumulación de la proteína. Esto se demostró en un estudio de denervación por seis
días (Lara y cols. 1990b). En base a estos hallazgos, ratas de 23-24 días se
sometieron a la disección del nervio ovárico superior, y seis días después se extrajeron
los ovarios, para determinar los niveles de mRNA de VEGF y TGFβ1 mediante PCR y
para localizar y semicuantificar las proteínas por inmunohistoquímica. Como control, se
utilizaron ratas a las que se les practicó una operación simulada.
VEGF 120 VEGF 164 TGFβ10.0
0.5
1.0SilvestreNGF knockout
Con
teni
do d
e A
RN
m (U
A)

33
El incremento local de NGF en los ovarios denervados se verificó por
inmunohistoquímica. Se decidió usar esta técnica porque, si bien no posee una alta
sensibilidad, estaba disponible en nuestro laboratorio en ese momento.
El estudio densitométrico mostró un incremento en la marca para NGF en ovario de
ratas denervadas con respecto a las controles (Figura 3.10). Este aumento fue
significativo para la señal encontrada en la teca, concordando estos resultados con lo
encontrado por Lara y cols. (1990b), mediante una técnica de EIA en ovario total de
ratas denervadas.
Figura 3.10. Análisis densitométrico de la marca de NGF en ovarios de ratas sometidas a la sección del nervio ovárico superior durante seis días, y ratas controles. Los datos mostrados corresponden al promedio ± error estándar de un n=5 para los animales controles y un n=6 para los denervados. La denervación ovárica indujo un incremento en la señal de NGF evaluada por inmunohistoquímica, que fue estadísticamente significativo en la teca (p < 0,05). Los niveles de mRNA de VEGF y TGFβ1 no se modificaron por efecto de la
denervación ovárica, al menos en el grupo de animales analizados (Figura 3.11). Ante
la posibilidad de que el cambio en el mRNA de VEGF y TGFβ1 ocurriera más
tempranamente y fuera indetectable a los 6 días de denervación, se determinó el
contenido de estos mRNA a los 3 días de denervación (Figura 3.12). Nuevamente, no
hubo diferencias significativas entre ratas controles y denervadas.
Granulosa Teca0
1000
2000
ControlesDenervadas (6 días)
*
Pixe
les
-bkg
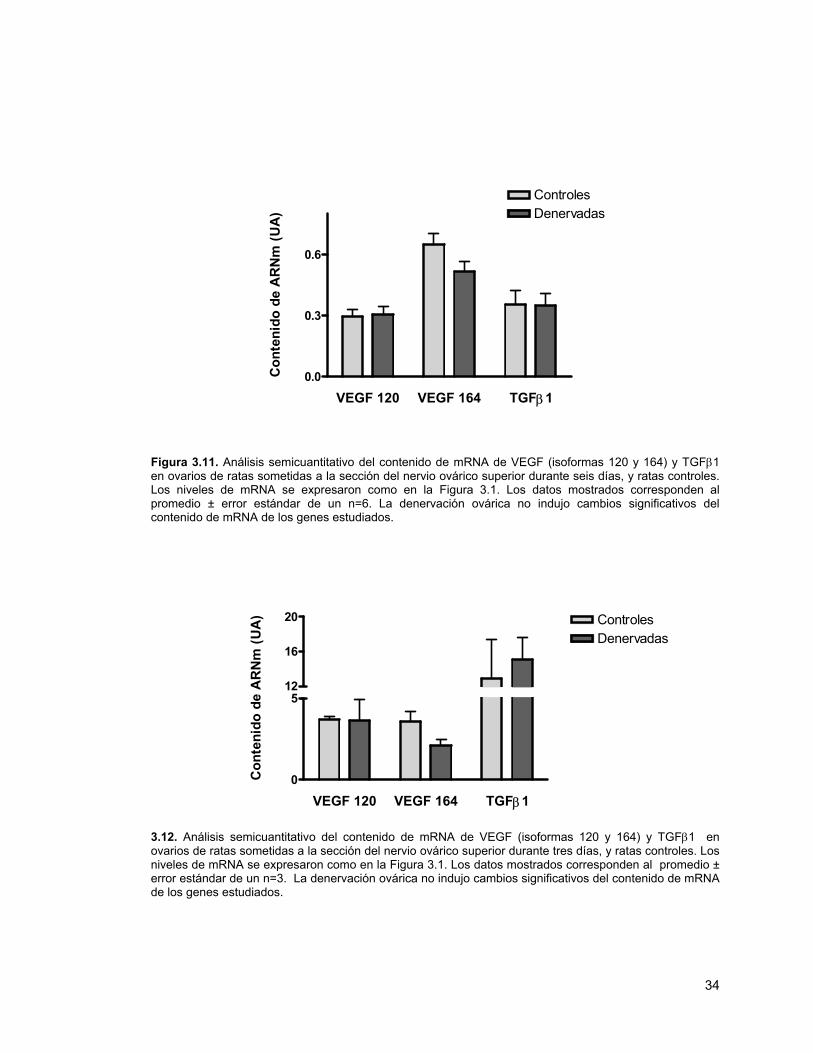
34
Figura 3.11. Análisis semicuantitativo del contenido de mRNA de VEGF (isoformas 120 y 164) y TGFβ1 en ovarios de ratas sometidas a la sección del nervio ovárico superior durante seis días, y ratas controles. Los niveles de mRNA se expresaron como en la Figura 3.1. Los datos mostrados corresponden al promedio ± error estándar de un n=6. La denervación ovárica no indujo cambios significativos del contenido de mRNA de los genes estudiados.
3.12. Análisis semicuantitativo del contenido de mRNA de VEGF (isoformas 120 y 164) y TGFβ1 en ovarios de ratas sometidas a la sección del nervio ovárico superior durante tres días, y ratas controles. Los niveles de mRNA se expresaron como en la Figura 3.1. Los datos mostrados corresponden al promedio ± error estándar de un n=3. La denervación ovárica no indujo cambios significativos del contenido de mRNA de los genes estudiados.
VEGF 120 VEGF 164 TGFβ10.0
0.3
0.6
ControlesDenervadas
Con
teni
do d
e A
RN
m (U
A)
VEGF 120 VEGF 164 TGFβ10
5
ControlesDenervadas
12
16
20
Con
teni
do d
e A
RN
m (U
A)

35
Los efectos de la denervación en los niveles de la proteína VEGF en ovario de rata
se analizaron por inmunohistoquímica. La inmunorreactividad para VEGF se
incrementó en los ovarios de ratas sometidas a la denervación por seis días (Figura
3.13). La marca se presentó solamente en folículos secundarios pequeños,
principalmente en la capa de células de granulosa adyacente al ovocito, y en algunos
casos se observó marca en la membrana plasmática del ovocito. La gráfica que se
muestra en la Figura 3.13 da cuenta de la inmunorreactividad presente en células de
granulosa.
Figura 3.13. A y B: Fotografías representativas de la inmunohistoquímica de VEGF en ratas control y ratas sometidas a la denervación de los ovarios por 6 días, respectivamente. C: Control negativo de la inmunohistoquímica. D: Cuantificación de la inmunorreactividad de VEGF por el método de H-Score. Los datos mostrados corresponden al promedio ± error estándar de un n=3. En las ratas denervadas se encontró un incremento significativo del H-Score (p < 0,01).
Controles Denervadas0
10
20
30
40 *
VEGF
Inm
unor
reac
tivid
ad d
e VE
GF
(H-S
core
)
D

36
En el caso de la evaluación inmunohistoquímica de la proteína de TGFβ1, no se
encontró diferencias entre ratas controles y tratadas, tanto para 6 como para 3 días de
denervación ovárica (Figura 3.14).
Figura 3.14. Cuantificación de la inmunorreactividad de TGFβ1 en ratas sometidas a la denervación de los ovarios por 3 (A) ó 6 días (B), y sus respectivos controles. La inmunorreactividad se evaluó usando el método de H-Score. Los datos mostrados corresponden al promedio ± error estándar de un n=3 para los 3 días y un n=6 para los 6 días de denervación. No se encontraron diferencias significativas entre los grupos. 3.1.4. Efecto de la sobreexpresión ovárica de NGF sobre los factores VEGF y
TGFβ1 en ratones transgénicos
Se trabajó con ovarios de ratones transgénicos, diseñados para que
sobreexpresen NGF específicamente en el ovario (Dissen 2005, trabajo no
publicado). Se utilizaron animales de 30 días de vida. En este caso fue posible
detectar mediante PCR la isoforma 188 de VEGF, además de las isoformas 120 y
164. Se encontró que los ratones que sobre expresan NGF en el ovario poseen
niveles de mRNA de VEGF 120 y 188 significativamente superiores que los
detectados en ratones silvestres (Figura 3.15). VEGF 164 solamente muestra una
tendencia al aumento.
Controles Denervadas (6d)0
50
100
Inm
unor
reac
tivid
ad d
e TG
Fb1
(H-S
core
)Controles Denervadas (3d)
0
50
100
150
Inm
unor
reac
tivid
ad d
e TG
Fb1
(H-S
core
)
A B

37
Por otra parte, los niveles de mRNA de TGFβ1 encontrados en los ovarios de
ratones transgénicos no fueron diferentes de aquellos detectados en animales
silvestres.
Figura 3.15. Análisis semicuantitativo del contenido de mRNA de VEGF (isoformas 120, 164 y 188) y TGFβ1 en ovarios de ratones silvestres y ratones que sobreexpresan NGF en el ovario. Los niveles de mRNA se expresaron como en la Figura 3.1. Los datos mostrados corresponden al promedio ± error estándar de un n=5 para cada grupo de animales. Los ratones que sobreexpresan NGF en el ovario poseen niveles de mRNA de VEGF 120 y 188 significativamente superiores que los ratones silvestres (* p < 0,05; ** p < 0,01)
3.1.5. Resumen de los resultados: Efecto de NGF sobre los factores VEGF y
TGFβ1 en ovario de roedores
NGF incrementa el contenido de mRNA y de proteína de VEGF en ovarios de rata
de 2 días de vida. En el caso de TGFβ1, NGF no indujo cambios en los niveles de
mRNA ni de proteína en las condiciones de cultivo estudiadas.
En ratones carentes de NGF no se observó una diferencia significativa en los
niveles de mRNA de VEGF y TGFβ1, en comparación con animales control,
posiblemente debido a un efecto compensatorio a la falta de NGF.
VEGF 120 VEGF 164 VEGF 188 TGFβ1
0.0
0.5
1.0
1.5 Ratones silvestres
Ratones quesobreexpresan NGF
**
*
Con
teni
do d
e A
RN
m (U
A)

38
En el modelo de rata con denervación ovárica, procedimiento que induce un
incremento local de NGF in vivo, se encontraron niveles de proteína de VEGF
significativamente superiores a los de animales controles. En el caso de TGFβ1, no
hubo modificaciones en el mRNA ni en la proteína producto de la denervación, para los
tiempos estudiados.
En ovarios de ratones que sobre expresan NGF, hubo mayores niveles de mRNA
para VEGF 120 y 188 que en ovarios de ratones silvestres. No se detectaron
diferencias para TGFβ1.
3.2. Efecto de NGF sobre los factores VEGF y TGFβ1 en células de la granulosa
humana
Las células de granulosa se eligieron como modelo de ovario humano porque
responden a NGF in vitro (Salas 2002, Valladares 2003, Salas y cols. 2006), y
expresan tanto VEGF como TGFβ1 in vivo (Chegini y cols. 1992a, Fraser y cols. 2001).
Además, las células de granulosa humanas y de otros mamíferos expresan y/o
secretan VEGF y TGFβ1 in vitro en respuesta a estímulos con gonadotrofinas o
factores de crecimiento (Mulheron y cols. 1992, May y cols. 1995, Lee y cols. 1997,
Martinez-Chequer y cols. 2003).
Nuestro grupo de trabajo demostró por primera vez que las células de la granulosa
humana recién aisladas expresan trkA, el receptor de alta afinidad de NGF. Se detectó
tanto el mRNA (resultados no mostrados) como la proteína en células recién aisladas
(Figura 3.16; Salas y cols. 2006). Esto permite justificar que las células en cultivo sean
capaces de responder a un estímulo con NGF.

39
Figura 3.16. Inmunohistoquímica para trkA, en células de granulosa humana recién aisladas. Luego de la fijación de las células, se utilizó un anticuerpo anti-trkA (Santacruz) en una dilución 1:100. La detección se realizó con diaminobenzidina, y se hizo una contratinción con hematoxilina. En el control negativo no se agregó anticuerpo primario (Salas y cols. 2006).
El estímulo de NGF en una concentración de 50 ng/mL produjo un incremento
significativo de los niveles de mRNA de las isoformas 121 y 165 de VEGF. En el caso
de TGFβ1, el incremento detectado no alcanzó la significancia estadística (Figura
3.17).
Figura 3.17. Análisis semicuantitativo del contenido de mRNA de VEGF (isoformas 121 y 165) y TGFβ1 en células de granulosa estimuladas con NGF (50 ng/mL) por 8 horas. Los niveles de mRNA se expresaron como en la Figura 3.1. Los datos mostrados corresponden al promedio ± error estándar de un n=9. Las células cultivadas con NGF poseen niveles de mRNA de VEGF 121 y 165 significativamente superiores que las controles (* p < 0,05).
Cuando las células fueron tratadas con NGF, se encontraron dos tipos de
poblaciones celulares, aquellas que responden a NGF y las que no responden;
posiblemente por diferencias en la maduración de ellas, principalmente por su grado de
VEGF 121 VEGF 165 TGF β10.0
0.5
1.0ControlNGF (50 ng/mL)
**
Niv
eles
de
AR
Nm
(UA
)
Anti-trkA Control negativo

40
luteinización (producción y secreción de progesterona). En las células que responden a
NGF, se produce un incremento de un 20% en la secreción de VEGF al medio de
cultivo con respecto al control. (Figura 3.18).
En el caso de TGFβ1, la mayoría de las muestras de medio de cultivo se encontró
bajo el límite de detección para la técnica de ELISA. Dentro de las muestras que
tuvieron niveles detectables de TGFβ1, se observó una ligera tendencia al aumento por
efecto de NGF, pero al ser un número muy reducido de muestras, no se pudo hacer
ningún análisis estadístico (resultados no mostrados).
Figura 3.18. Niveles de VEGF en el medio de cultivo de células de granulosa tratadas con NGF (50 ng/mL) por 8 horas. La proteína VEGF se evaluó por ELISA. Los resultados fueron agrupados de acuerdo a la respuesta observada: cuando hubo más de un 10% de cambio (aumento o disminución respecto al control), se clasificó como “con respuesta”, y cuando se produjo menos de un 10% de cambio, se clasificó como “sin respuesta”. Se indica el promedio ± SEM y el número de experimentos de un n=12 para el grupo con respuesta y un n=12 para el grupo sin respuesta. En la población con respuesta, NGF indujo un incremento significativo en los niveles de VEGF secretado al medio de cultivo (*** p < 0,005).
3.2.1. Resumen de los resultados: Efecto de NGF sobre los factores VEGF y
TGFβ1 en células de granulosa humana
En células de granulosa humana aisladas y en cultivo, i) NGF incrementó
significativamente los niveles de mRNA de VEGF y la secreción de VEGF. Estos
resultados concuerdan con lo encontrado en ovario de rata, donde también se observó
un aumento en VEGF en células de granulosa, lo que valida el modelo in vitro de
Con respuesta Sin respuesta 0
50
100
150 ControlNGF (50ng/ml)***
VEG
F en
el m
edio
de
culti
vo(r
elat
ivo
al c
ontro
l)

41
células de granulosa humana, para posteriores estudios acerca del mecanismo de
acción de NGF sobre la angiogénesis ovárica. ii) la presencia de un grupo de muestras
que no responde a NGF aumentando la secreción de VEGF puede ser explicada por
una heterogeneidad en la capacidad de traducción o de secreción, ya que cada
muestra de células granulosa proviene de un conjunto de folículos aspirados, que no
se encuentran en el mismo grado de maduración en todas las pacientes. Sin embargo,
dicha heterogeneidad no se manifiestó cuando se analizaron los efectos de NGF sobre
los niveles de mRNA de las distintas isoformas de VEGF.
En el caso de TGFβ1, NGF no indujo cambios en los niveles de mRNA. No se
encontaron niveles detectables de la proteína de TGFβ1 en los medios de cultivo
analizados.
3.3. Evaluación de un posible mecanismo de acción de NGF sobre el incremento
de VEGF y TGFβ1 en modelos de ovario de roedor y de humano
3.3.1. Participación del receptor trkA NGF tiene dos receptores conocidos: trkA y p75. La señalización vía trkA induce la
mayoría de los efectos neurotróficos clásicos del NGF (Sofroniew y cols. 2001).
Además, este receptor es de gran importancia en el ovario, siendo esencial en la
ovulación (Dissen y cols. 1996).
Con el objetivo de conocer su importancia en la expresión de los factores VEGF y
TGFβ1 en ovario de roedores, se trabajó con ratones que tienen una deleción del gen
para trkA (ratones trkA knockout) y con ratones silvestres. Los animales se sacrificaron
el mismo día del nacimiento, se extrajo DNA de la cola para hacer la genotipificación, y
se utilizaron ambos ovarios para la extracción del RNA total. Se hizo una transcripción
reversa seguida de un PCR (RT-PCR), para analizar los niveles relativos del mRNA de
VEGF y TGFβ1.
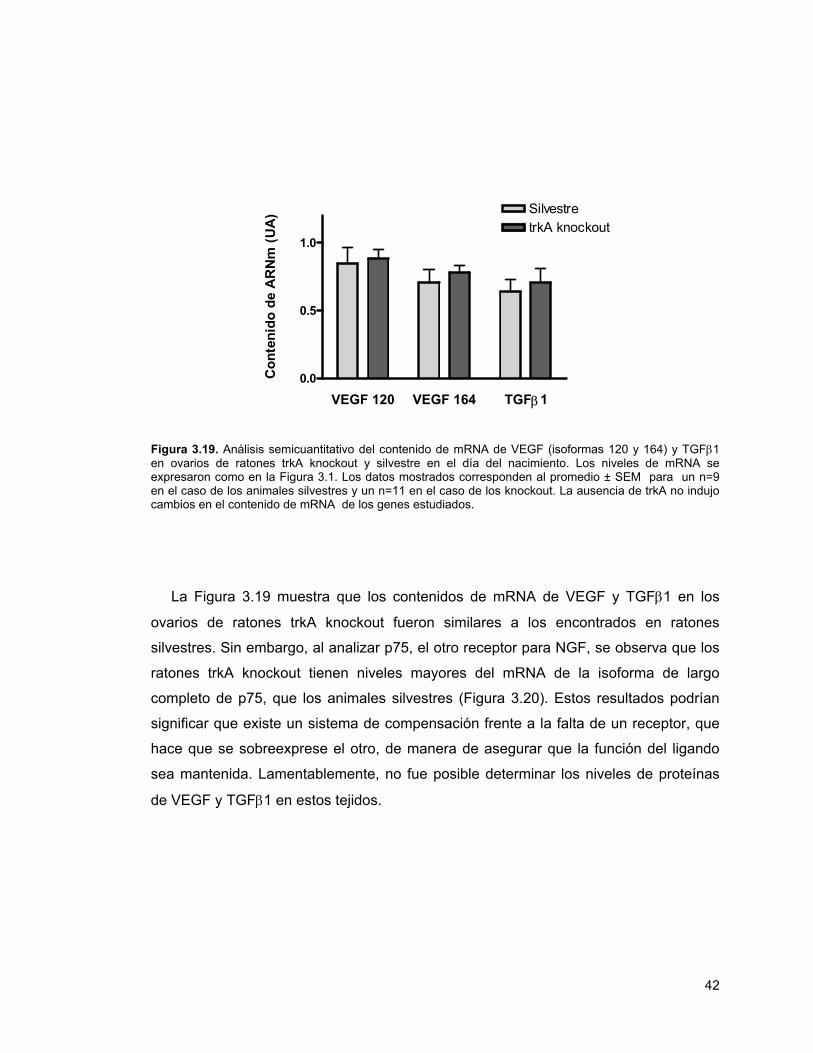
42
Figura 3.19. Análisis semicuantitativo del contenido de mRNA de VEGF (isoformas 120 y 164) y TGFβ1 en ovarios de ratones trkA knockout y silvestre en el día del nacimiento. Los niveles de mRNA se expresaron como en la Figura 3.1. Los datos mostrados corresponden al promedio ± SEM para un n=9 en el caso de los animales silvestres y un n=11 en el caso de los knockout. La ausencia de trkA no indujo cambios en el contenido de mRNA de los genes estudiados.
La Figura 3.19 muestra que los contenidos de mRNA de VEGF y TGFβ1 en los
ovarios de ratones trkA knockout fueron similares a los encontrados en ratones
silvestres. Sin embargo, al analizar p75, el otro receptor para NGF, se observa que los
ratones trkA knockout tienen niveles mayores del mRNA de la isoforma de largo
completo de p75, que los animales silvestres (Figura 3.20). Estos resultados podrían
significar que existe un sistema de compensación frente a la falta de un receptor, que
hace que se sobreexprese el otro, de manera de asegurar que la función del ligando
sea mantenida. Lamentablemente, no fue posible determinar los niveles de proteínas
de VEGF y TGFβ1 en estos tejidos.
VEGF 120 VEGF 164 TGFβ10.0
0.5
1.0
SilvestretrkA knockout
Con
teni
do d
e A
RN
m (U
A)

43
Figura 3.20. Niveles de mRNA para las isoformas de largo completo y corta de p75 en ovarios de ratones trkA knockout y silvestres. Los niveles de mRNA se calcularon como se indica en la Figura 3.1. Los datos mostrados corresponden al promedio ± error estándar de un n=5 para los animales silvestres y un n=5 para los knockout. Los ratones trkA knockout tenían un mayor contenido de mRNA de la isoformas de largo completo de p75, aunque la diferencia no alcanzó a ser estadísticamente significativa.
También se analizó el efecto de un inhibidor del receptor trkA en células de
granulosa humana tratadas con NGF. Se utilizaron partidores específicos para la
detección del mRNA de VEGF 165 por PCR de tiempo real.
La Figura 3.21 muestra que la preincubación con K252a (100 nM) revirtió en forma
significativa el aumento en el contenido de mRNA de VEGF 165 inducido por NGF.
Cuando se analizó VEGF en el medio de cultivo, se encontró que la preincubación con
K252a (100 nM) también revirtió en forma significativa el aumento en la secreción de
VEGF inducido por NGF (Figura 3.22).
0.0
0.5
trkA silvestretrkA knockout
p=0.057
p75 (isoforma delargo completo)
p75 (isoforma corta)
Niv
eles
de
AR
Nm
(UA
)

44
Figura 3.21. Contenido de mRNA para VEGF 165 en células de granulosa tratadas con NGF y un inhibidor del receptor trkA (K252a). El mRNA de VEGF 165 se midió por PCR de tiempo real. Se utilizó el RNA 18S para normalizar los resultados. NGF indujo un incremento significativo en el contenido de mRNA de VEGF 165, el que fue revertido por la preincubación con K252a. Los datos mostrados corresponden al promedio ± SEM de un n=4. * p < 0,05 NGF vs Control y ≠ p < 0,05 NGF vs K252a + NGF, respectivamente.
Figura 3.22. Niveles de VEGF en el medio de cultivo de células de granulosa tratadas con NGF y un inhibidor del receptor trkA (K252a). Los niveles de VEGF presentes en el medio de cultivo se midieron por ELISA. Los datos mostrados corresponden al promedio ± SEM (n=4). * p < 0,05 vs Control y ≠ p < 0,05 NGF vs K252a + NGF, respectivamente.
0
1
2
Condición de cultivo
*
≠
ControlK252a (100 nM)NGF (50 ng/ml)NGF + K252a
AR
Nm
VEG
F 16
5/A
RN
18S
(UA
)
0
50
100
150ControlK252a (100 nM)NGF (50 ng/ml)K252a + NGF
*≠
Condición de cultivo
VEG
F en
el m
edio
de
culti
vo(R
elat
ivo
al c
ontro
l)

45
3.3.2. Señalización intracelular vía ERK
En distintos sistemas se ha demostrado que el efecto mitogénico de NGF requiere
de la activación de su receptor trkA, y de la cascada de ERK (Descamps y cols. 2001,
Cantarella y cols. 2002). En este objetivo se utilizó un modelo de angiogénesis ovárica
para saber si la acción de NGF en este proceso requiere de la activación de las ERK a
nivel de la fosforilación del receptor trkA para la producción del mRNA y/o proteína de
VEGF y TGFβ1.
Para ello se determinó en células de granulosa humana si la incubación con NGF
por 0, 5 y 10 min incrementaba la fosforilación de ERK 1/2. La Figura 3.23 indica que
hubo un incremento significativo en la proporción de proteínas ERK 2 fosforiladas
respecto de ERK 2 totales, luego de 5 min de estimulación con NGF. Sin embargo, no
se encontró un aumento significativo para la razón ERK 1 fosforilada / ERK 1 total. Lo
anterior es un reflejo de la alta dispersión en los resultados obtenidos para esta
isoforma de ERK, puesto que en algunos casos la señal de la proteína fosforilada en el
tiempo 0 era muy débil (ver Figura 3.23 A).
Figura 3.23. Inmunowestern blot de ERK fosforiladas y ERK totales en células de granulosa incubadas por 0, 5 ó 10 min con NGF (50 ng/ml). En A se muestra una fotografía de un western blot representativo. En B se muestra el resultado de la densitometría de los western bots. Los datos mostrados corresponden al promedio ± SEM de un n=8. El estímulo de NGF indujo un rápido incremento en la cantidad relativa de ERK 2 fosforilada (* p < 0,01), efecto que es revertido a los 10 minutos de iniciado el estímulo (≠ p < 0,05).
NGF 50 ng/ml
0 min 5 min 10 min
ERK 1/2 fosforilada
ERK 1/2 total
A B ERK fosforilada / ERK total
P-ERK1/ERK1 P-ERK2/ERK20
1
2 t = 0t = 5 mint = 10 min
*
≠
ERK
fosf
orila
da /
ERK
tota
lre
lativ
o al
con
trol
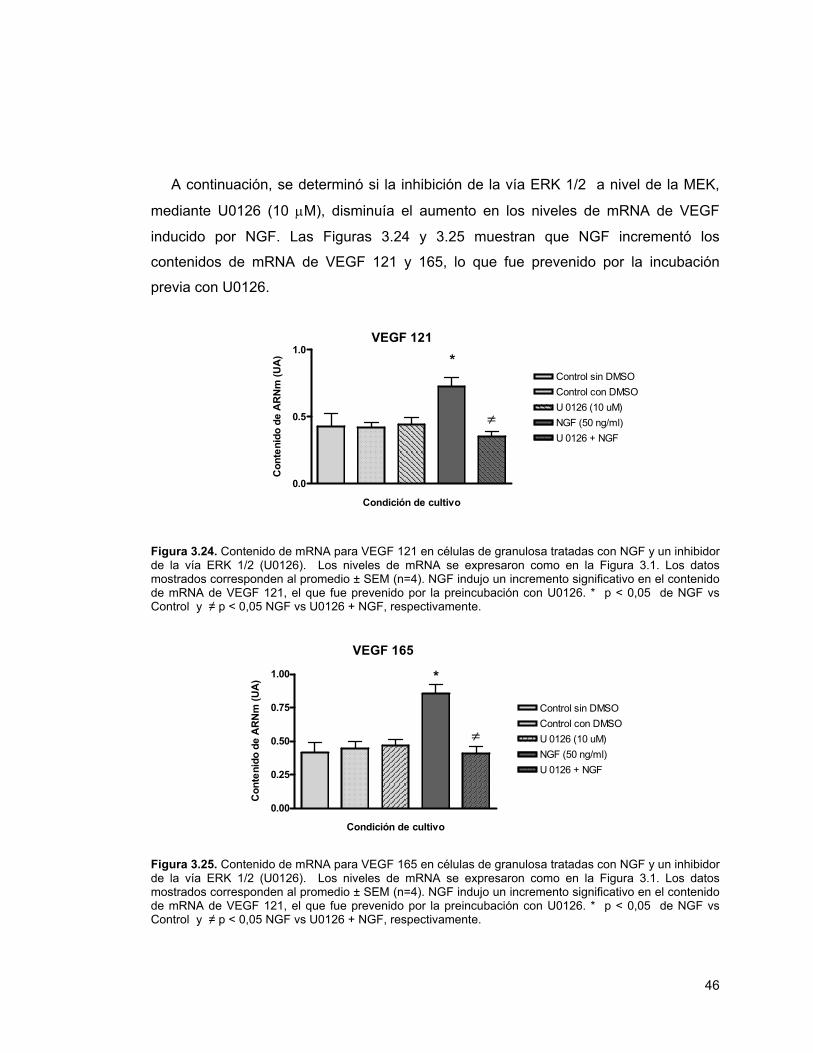
46
A continuación, se determinó si la inhibición de la vía ERK 1/2 a nivel de la MEK,
mediante U0126 (10 µM), disminuía el aumento en los niveles de mRNA de VEGF
inducido por NGF. Las Figuras 3.24 y 3.25 muestran que NGF incrementó los
contenidos de mRNA de VEGF 121 y 165, lo que fue prevenido por la incubación
previa con U0126.
Figura 3.24. Contenido de mRNA para VEGF 121 en células de granulosa tratadas con NGF y un inhibidor de la vía ERK 1/2 (U0126). Los niveles de mRNA se expresaron como en la Figura 3.1. Los datos mostrados corresponden al promedio ± SEM (n=4). NGF indujo un incremento significativo en el contenido de mRNA de VEGF 121, el que fue prevenido por la preincubación con U0126. * p < 0,05 de NGF vs Control y ≠ p < 0,05 NGF vs U0126 + NGF, respectivamente.
Figura 3.25. Contenido de mRNA para VEGF 165 en células de granulosa tratadas con NGF y un inhibidor de la vía ERK 1/2 (U0126). Los niveles de mRNA se expresaron como en la Figura 3.1. Los datos mostrados corresponden al promedio ± SEM (n=4). NGF indujo un incremento significativo en el contenido de mRNA de VEGF 121, el que fue prevenido por la preincubación con U0126. * p < 0,05 de NGF vs Control y ≠ p < 0,05 NGF vs U0126 + NGF, respectivamente.
VEGF 165
0.00
0.25
0.50
0.75
1.00
Control sin DMSOControl con DMSOU 0126 (10 uM)NGF (50 ng/ml)U 0126 + NGF
*
≠
Condición de cultivo
Con
teni
do d
e A
RN
m (U
A)
VEGF 121
0.0
0.5
1.0
Control sin DMSOControl con DMSOU 0126 (10 uM)NGF (50 ng/ml)U 0126 + NGF
≠
*
Condición de cultivo
Con
teni
do d
e A
RN
m (U
A)

47
Figura 3.26. Niveles de VEGF en el medio de cultivo de células de granulosa tratadas con NGF y un inhibidor de MEK (U0126). La proteína se cuantificó por ELISA, y los datos mostrados corresponden al promedio ± SEM de un n=4. * p < 0,05 de NGF vs Control y ≠ p < 0,01 NGF vs U0126 + NGF, respectivamente. Al analizar el efecto del inhibidor de MEK en los niveles de proteína de VEGF
secretados al medio de cultivo luego de un estímulo con NGF, encontramos que se
repite lo observado para el caso del mRNA de VEGF: NGF incrementó los niveles de
VEGF secretado al medio, lo que fue prevenido por la incubación previa con U0126
En el caso de TGFβ1, NGF indujo un incremento que no alcanzó a ser significativo.
Sin embargo, la preincubación con U0126 disminuyó significativamente el contenido de
mRNA de este factor de crecimiento (Figura 3.27).
Figura 3.27. Contenido de mRNA para TGFβ1 en células de granulosa tratadas con NGF y un inhibidor de la MEK (U0126). Los niveles de mRNA se expresaron como en la Figura 3.1. Los datos mostrados corresponden al promedio ± SEM de un n=4. NGF no indujo un incremento significativo en el contenido de mRNA de TGFβ1, sin embargo, este disminuye significativamente cuando se preincuba con U0126. * p < 0,05.
TGF β1
0.0
0.5
1.0
1.5
Control sin DMSOControl con DMSOU 0126 (10 uM)NGF (50 ng/ml)U 0126 + NGF
*
Condición de cultivo
Con
teni
do d
e A
RN
m (U
A)
Proteína VEGF
0
50
100
150
200Control sin DMSOControl con DMSOU 0126 (10 uM)NGF (50 ng/ml)U 0126 + NGF
*
≠
Condición de cultivo
VEG
F en
el m
edio
de
culti
vo(r
elat
ivo
al c
ontro
l)
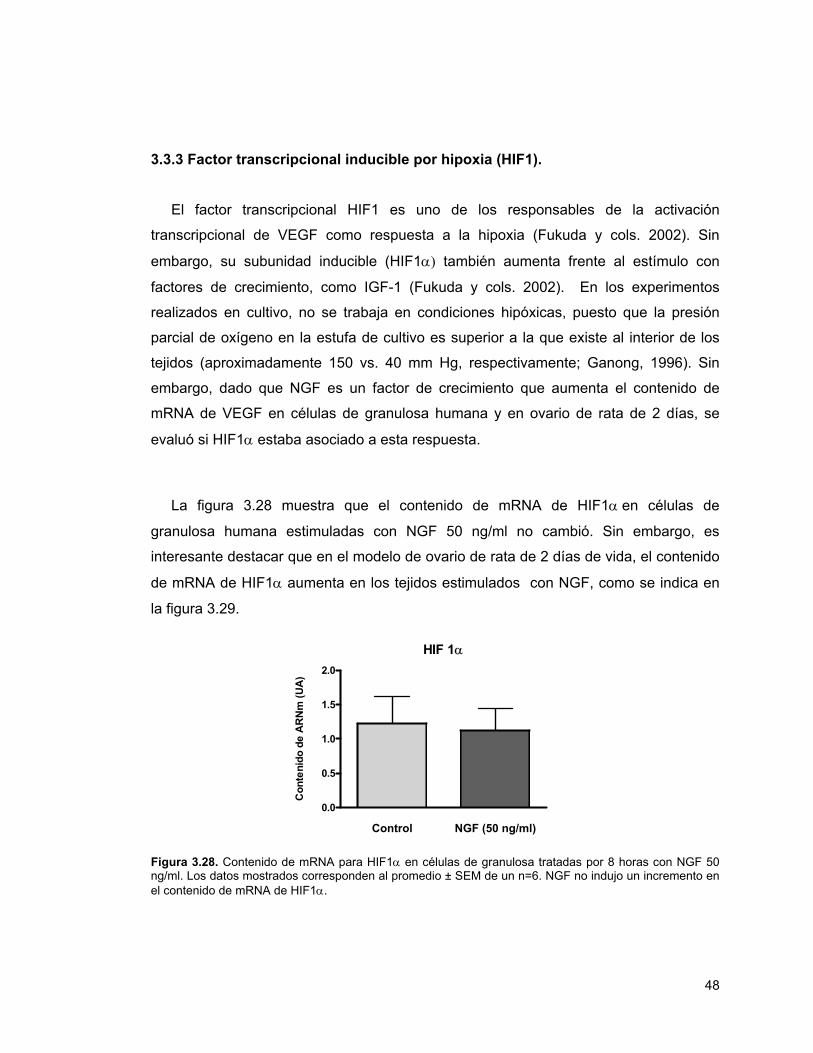
48
3.3.3 Factor transcripcional inducible por hipoxia (HIF1). El factor transcripcional HIF1 es uno de los responsables de la activación
transcripcional de VEGF como respuesta a la hipoxia (Fukuda y cols. 2002). Sin
embargo, su subunidad inducible (HIF1α) también aumenta frente al estímulo con
factores de crecimiento, como IGF-1 (Fukuda y cols. 2002). En los experimentos
realizados en cultivo, no se trabaja en condiciones hipóxicas, puesto que la presión
parcial de oxígeno en la estufa de cultivo es superior a la que existe al interior de los
tejidos (aproximadamente 150 vs. 40 mm Hg, respectivamente; Ganong, 1996). Sin
embargo, dado que NGF es un factor de crecimiento que aumenta el contenido de
mRNA de VEGF en células de granulosa humana y en ovario de rata de 2 días, se
evaluó si HIF1α estaba asociado a esta respuesta.
La figura 3.28 muestra que el contenido de mRNA de HIF1α en células de
granulosa humana estimuladas con NGF 50 ng/ml no cambió. Sin embargo, es
interesante destacar que en el modelo de ovario de rata de 2 días de vida, el contenido
de mRNA de HIF1α aumenta en los tejidos estimulados con NGF, como se indica en
la figura 3.29.
Figura 3.28. Contenido de mRNA para HIF1α en células de granulosa tratadas por 8 horas con NGF 50 ng/ml. Los datos mostrados corresponden al promedio ± SEM de un n=6. NGF no indujo un incremento en el contenido de mRNA de HIF1α.
HIF 1α
Control NGF (50 ng/ml)
0.0
0.5
1.0
1.5
2.0
Con
teni
do d
e A
RN
m (U
A)
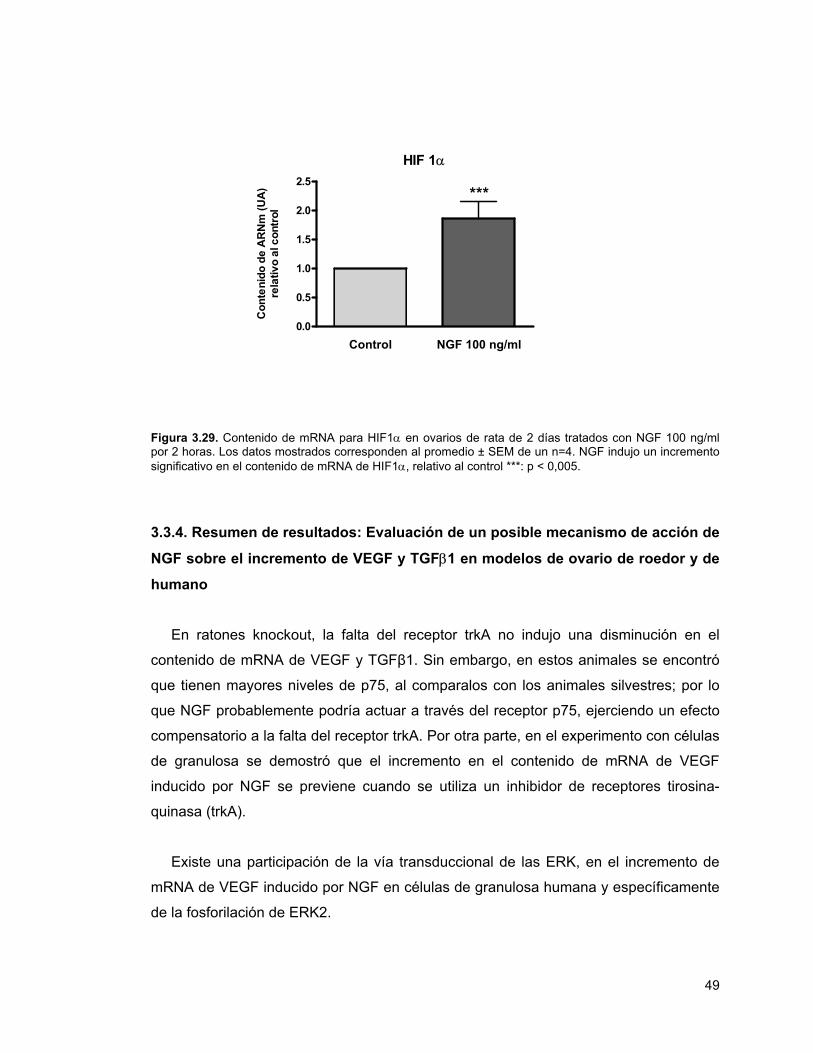
49
Figura 3.29. Contenido de mRNA para HIF1α en ovarios de rata de 2 días tratados con NGF 100 ng/ml por 2 horas. Los datos mostrados corresponden al promedio ± SEM de un n=4. NGF indujo un incremento significativo en el contenido de mRNA de HIF1α, relativo al control ***: p < 0,005. 3.3.4. Resumen de resultados: Evaluación de un posible mecanismo de acción de
NGF sobre el incremento de VEGF y TGFβ1 en modelos de ovario de roedor y de
humano
En ratones knockout, la falta del receptor trkA no indujo una disminución en el
contenido de mRNA de VEGF y TGFβ1. Sin embargo, en estos animales se encontró
que tienen mayores niveles de p75, al comparalos con los animales silvestres; por lo
que NGF probablemente podría actuar a través del receptor p75, ejerciendo un efecto
compensatorio a la falta del receptor trkA. Por otra parte, en el experimento con células
de granulosa se demostró que el incremento en el contenido de mRNA de VEGF
inducido por NGF se previene cuando se utiliza un inhibidor de receptores tirosina-
quinasa (trkA).
Existe una participación de la vía transduccional de las ERK, en el incremento de
mRNA de VEGF inducido por NGF en células de granulosa humana y específicamente
de la fosforilación de ERK2.
HIF 1α
Control NGF 100 ng/ml0.0
0.5
1.0
1.5
2.0
2.5***
Con
teni
do d
e A
RN
m (U
A)
rela
tivo
al c
ontr
ol

50
En ovarios de rata de 2 días de vida, NGF incrementó los niveles de mRNA de la
subunidad inducible del factor transcripcional HIF a las 2 h de cultivo, lo que podría ser
un componente responsable del aumento del contenido de mRNA de VEGF. El hecho
de que en células de granulosa humana estimuladas con NGF por 8 horas no se
observara un incremento en el mRNA de HIF, probablemente podría deberse a que
este factor transcripcional se active a tiempos cortos de estímulo con NGF.
3.4. Efecto de NGF en la vascularización ovárica 3.4.1. Efecto de NGF en un modelo de angiogénesis in vitro.
La proliferación de las células endoteliales es un componente importante en el
proceso de neovascularización. En el estudio de la angiogénesis, uno de los modelos
más ampliamente utilizados es el de proliferación de células endoteliales provenientes
de vena de cordón umbilical humano, también conocidas como HUVEC (Verheul y
cols. 2000, Middeke y cols. 2002, Cantarella y cols. 2002, Papetti y cols. 2002). Se
aprovechó entonces la capacidad que tiene la célula de granulosa de responder a NGF
y de secretar factores angiogénicos, para crear un modelo de angiogénesis ovárica in
vitro que permita verificar acciones indirectas de NGF sobre la proliferación de células
endoteliales. Para cumplir con este objetivo se cultivaron HUVEC en presencia de
medios condicionados, provenientes de células de granulosa estimuladas con NGF y
de las respectivas células control. Para discriminar si los efectos observados eran
generados por el VEGF presente en el medio condicionado, se trató este medio con un
anticuerpo anti-VEGF. También se usó un anticuerpo anti-NGF, para descartar el
efecto de la neurotrofina que se usó para estimular las células de granulosa.
La proliferación celular de las HUVEC se evaluó por la técnica de Cell Titer. Esta
técnica consiste en la conversión de un compuesto (MTS tetrazolium) en otro de
diferente color (formazán), proceso que es realizado por las células en cultivo. La
cantidad de formazán generado se detecta por espectrometría y es proporcional al
número de células metabólicamente activas presentes.
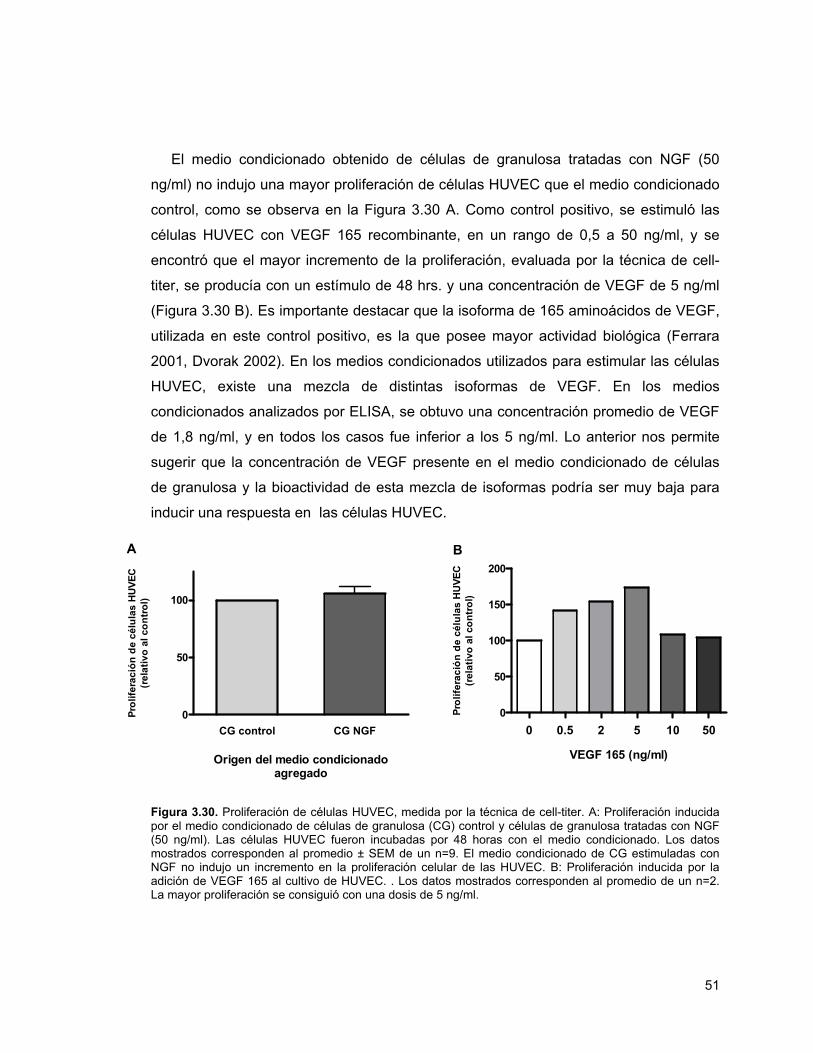
51
El medio condicionado obtenido de células de granulosa tratadas con NGF (50
ng/ml) no indujo una mayor proliferación de células HUVEC que el medio condicionado
control, como se observa en la Figura 3.30 A. Como control positivo, se estimuló las
células HUVEC con VEGF 165 recombinante, en un rango de 0,5 a 50 ng/ml, y se
encontró que el mayor incremento de la proliferación, evaluada por la técnica de cell-
titer, se producía con un estímulo de 48 hrs. y una concentración de VEGF de 5 ng/ml
(Figura 3.30 B). Es importante destacar que la isoforma de 165 aminoácidos de VEGF,
utilizada en este control positivo, es la que posee mayor actividad biológica (Ferrara
2001, Dvorak 2002). En los medios condicionados utilizados para estimular las células
HUVEC, existe una mezcla de distintas isoformas de VEGF. En los medios
condicionados analizados por ELISA, se obtuvo una concentración promedio de VEGF
de 1,8 ng/ml, y en todos los casos fue inferior a los 5 ng/ml. Lo anterior nos permite
sugerir que la concentración de VEGF presente en el medio condicionado de células
de granulosa y la bioactividad de esta mezcla de isoformas podría ser muy baja para
inducir una respuesta en las células HUVEC.
Figura 3.30. Proliferación de células HUVEC, medida por la técnica de cell-titer. A: Proliferación inducida por el medio condicionado de células de granulosa (CG) control y células de granulosa tratadas con NGF (50 ng/ml). Las células HUVEC fueron incubadas por 48 horas con el medio condicionado. Los datos mostrados corresponden al promedio ± SEM de un n=9. El medio condicionado de CG estimuladas con NGF no indujo un incremento en la proliferación celular de las HUVEC. B: Proliferación inducida por la adición de VEGF 165 al cultivo de HUVEC. . Los datos mostrados corresponden al promedio de un n=2. La mayor proliferación se consiguió con una dosis de 5 ng/ml.
CG control CG NGF0
50
100
Origen del medio condicionadoagregado
Prol
ifera
ción
de
célu
las
HU
VEC
(rel
ativ
o al
con
trol)
0 0.5 2 5 10 500
50
100
150
200
VEGF 165 (ng/ml)
Prol
ifera
ción
de
célu
las
HU
VEC
(rel
ativ
o al
con
trol
)
A B

52
3.4.2. Efecto de NGF en la vascularización ovárica in vivo en ratas con denervación ovárica.
Se analizó el efecto del incremento de NGF intraovárico (inducido por sección del
nervio ovárico superior, como se indicó en el punto 3.1.3) en la vascularización del
tejido ovárico. Para ésto, en secciones de ovario de ratas controles y denervadas se
realizó una inmunohistoquímica contra CD31, un marcador específico de vasos
sanguíneos, y se cuantificó el área marcada. Se normalizó el área de vasos por el área
de la sección del ovario.
Figura 3.31. Tinción de CD-31 en ratas con niveles elevados de NGF, a consecuencia de la disección del nervio ovárico superior. Se observan vasos en el área que rodea a los folículos ováricos. A y B indican la marca encontrada en ovarios de ratas control, en donde los capilares se indican con una flecha y los vasos pequeños con una cabeza de flecha. En C y D se muestran los ovarios de ratas denervadas por 6 días, en los que se encontró una mayor área de marca para vasos sanguíneos. El control negativo de la inmunohistoquímica se muestra en E. Magnificación original 100X. Barra = 100 µm.
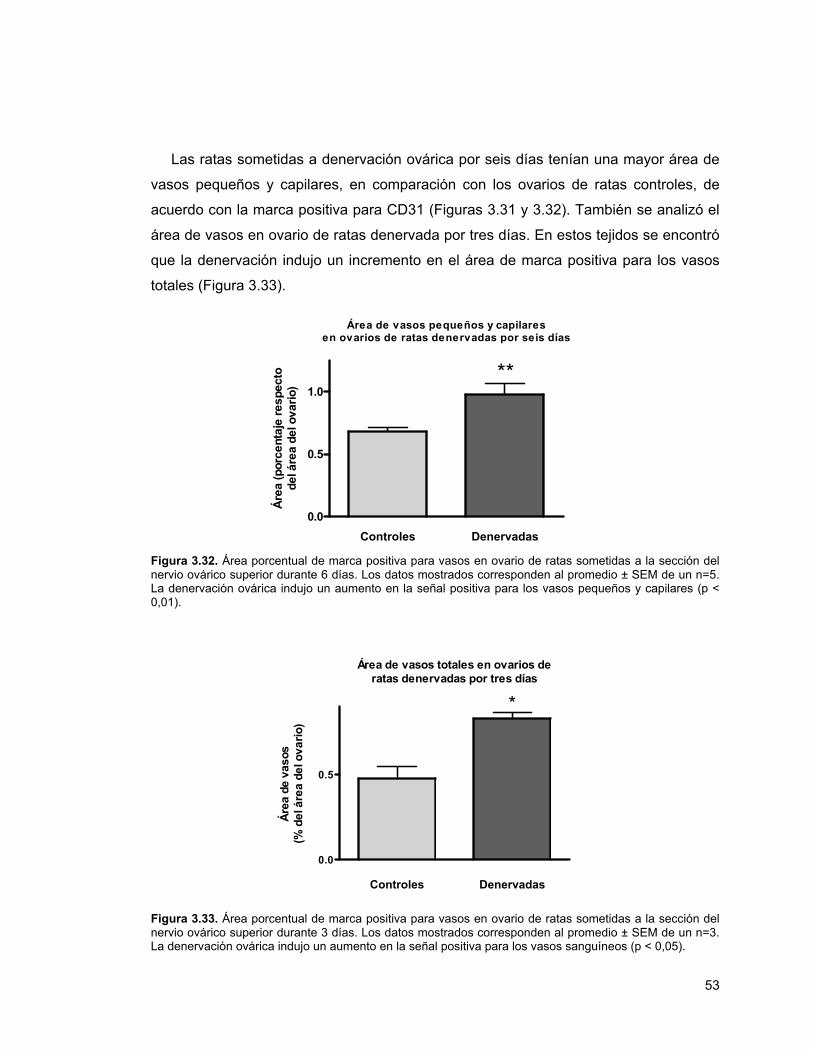
53
Las ratas sometidas a denervación ovárica por seis días tenían una mayor área de
vasos pequeños y capilares, en comparación con los ovarios de ratas controles, de
acuerdo con la marca positiva para CD31 (Figuras 3.31 y 3.32). También se analizó el
área de vasos en ovario de ratas denervada por tres días. En estos tejidos se encontró
que la denervación indujo un incremento en el área de marca positiva para los vasos
totales (Figura 3.33).
Figura 3.32. Área porcentual de marca positiva para vasos en ovario de ratas sometidas a la sección del nervio ovárico superior durante 6 días. Los datos mostrados corresponden al promedio ± SEM de un n=5. La denervación ovárica indujo un aumento en la señal positiva para los vasos pequeños y capilares (p < 0,01).
Figura 3.33. Área porcentual de marca positiva para vasos en ovario de ratas sometidas a la sección del nervio ovárico superior durante 3 días. Los datos mostrados corresponden al promedio ± SEM de un n=3. La denervación ovárica indujo un aumento en la señal positiva para los vasos sanguíneos (p < 0,05).
Área de vasos totales en ovarios deratas denervadas por tres días
Controles Denervadas
0.0
0.5
*
Área
de
vaso
s(%
del
áre
a de
l ova
rio)
Área de vasos pequeños y capilaresen ovarios de ratas denervadas por seis días
Controles Denervadas0.0
0.5
1.0**
Área
(por
cent
aje
resp
ecto
del á
rea
del o
vario
)

54
3.4.3. Efecto de la sobreexpresión ovárica de NGF sobre la vascularización en ratones transgénicos
De la misma manera que en el punto 3.4.2, se analizó el efecto de la
sobreexpresión intraovárica de NGF sobre la vascularización del ovario en ratones
transgénicos. Se realizó un estudio de inmunohistoquímica, pero en este caso, al
tratarse de otra especie, el anticuerpo utilizado marcó específicamente los capilares y
no los vasos sanguíneos de mayor calibre.
Como se muestra en las Figuras 3.34 y 3.35, los ratones transgénicos diseñados
para sobreexpresar NGF en el ovario tuvieron una mayor área de capilares en este
tejido que los ratones silvestres.
Figura 3.34. Tinción de CD-31 en ovarios de ratones que sobreexpresan NGF. Se observan capilares en el área que rodea a los folículos ováricos. En A se muestra la marca encontrada en ovario de ratón silvestre, en donde los capilares se indican con una flecha. En B se muestra el ovario de un ratón transgénico, donde se encontró una mayor área de marca para vasos sanguíneos. El control negativo de la inmunohistoquímica se muestra en C. Magnificación original 40X. Barra = 200 µm.

55
Figura 3.35. Área porcentual de marca positiva para capilares en ovario de ratones que sobreexpresan NGF en este tejido y en ovario de ratones silvestres. . Los datos mostrados corresponden al promedio ± SEM de un n=6. NGF indujo un aumento en la señal positiva para los capilares (p < 0,05).
3.4.4. Resumen de resultados: Efecto de NGF sobre la vascularización ovárica
En un ensayo de angiogénesis in vitro, el medio condicionado de células de
granulosa tratadas con NGF no indujo mayor proliferación de células endoteliales. Una
explicación a lo observado es que la concentración de VEGF secretado en respuesta a
NGF en el medio condicionado de células de granulosa, fue muy baja para estimular la
proliferación en las células HUVEC.
En el modelo de rata con denervación ovárica, que induce un incremento local de
NGF in vivo, se encontró una mayor vascularización del tejido, tanto a los tres como a
los seis días post-cirugía, en comparación con los controles.
En ovarios de ratones que sobreexpresan NGF también se observa una mayor área
de marca positiva para capilares que en ovarios de animales silvestres.
Capilares en ovario de ratón
Controles Transgénicos (NGF)
0.0
0.5
1.0
1.5
2.0
*
Área
de
capi
lare
s(%
del
áre
a de
l ova
rio)

56
4. Discusión
4.1. Nuevos roles de NGF en la función ovárica Diversos eventos de la fisiología ovárica son controlados por factores de
crecimiento. Entre ellos, el factor de crecimiento nervioso (NGF) y su receptor de alta
afinidad, trkA, parecen tener roles importantes en, al menos, dos etapas esenciales
para la función reproductiva:
1) durante el desarrollo ovárico temprano, proceso en el cual NGF induce:
a) proliferación celular, promoviendo el paso de folículos primordiales a primarios,
y de primarios a secundarios (Ojeda y cols. 2000, Dissen y cols. 2001).
b) diferenciación, induciendo la adquisición de receptores para FSH (Romero y
cols. 2002).
2) durante los ciclos ováricos, en los cuales NGF induciría la expresión de
receptores para FSH y la secreción de estrógenos (Salas y cols. 2006), proceso
indispensable para que ocurra la ovulación (Dissen y cols. 1996).
En el período periovulatorio, y especialmente luego de la expulsión del óvulo, se
produce una etapa de intensa vascularización del cuerpo lúteo (Fraser y cols. 2001).
En este momento se van a sobre expresar distintos factores de crecimiento
angiogénicos, entre los que se puede destacar el factor de crecimiento del endotelio
vascular (VEGF), el cual es esencial para la proliferación endotelial (Koos 1995, Fraser
y cols. 2001). Este estudio muestra que NGF incrementa tanto la expresión de VEGF
como el área de vasos sanguíneos en el ovario, lo que permite conectar dos eventos
de gran importancia para la función ovárica. Además, como se ha demostrado que
VEGF se expresa en células de granulosa de folículos pre-antrales y antrales (Fraser y
cols. 2001), al igual que NGF y trkA (Salas et al, 2006), estos resultados indican que
NGF podría ejercer una acción autocrina en el incremento de la expresión de VEGF en
el ovario.

57
En la mayoría de los tejidos, la hipoxia es el principal estímulo para la síntesis de
factores angiogénicos. En cambio, en el ovario, la expresión de estos factores durante
etapas como el desarrollo folicular y la formación del cuerpo lúteo, parece estar
regulada en forma predominante por las hormonas luteinizante (LH) y
foliculoestimulante (FSH) (Fraser y cols. 2001). Para determinar distintos parámetros
asociados a la angiogénesis en este tejido, evitando la influencia de las gonadotrofinas,
decidimos trabajar con roedores en etapas tempranas de la vida ovárica, es decir,
previo a la aparición de los ciclos estrales (McGee y cols. 2000). La Tabla 7.1, incluída
en la sección Anexos, indica las características de cada modelo evaluado en este
estudio.
Este trabajo reporta por primera vez que VEGF se expresa en ovarios de roedores
de 0 a 7 días de vida. Lo anterior sugiere que este factor de crecimiento podría tener
funciones distintas a la inducción de la vascularización en etapas tempranas del
desarrollo ovárico, ya que en este momento no ocurren los eventos angiogénicos
cíclicos que se manifiestan en el ovario adulto. Posibles funciones de VEGF en esta
etapa serían promover la proliferación de células somáticas o incrementar la
permeabilidad vascular para permitir la extravasación de nutrientes y hormonas. Se
sabe que VEGF incrementa la proliferación de poblaciones celulares, específicamente
de las células del endotelio vascular (Papetti y cols. 2002) y que aumenta el número de
fenestraciones en los capilares (Papetti y cols. 2002), por lo que no sería extraño que
este factor de crecimiento cumpliera funciones similares en el ovario en desarrollo. En
este estudio se demostró que NGF induce un incremento temprano en la expresión de
VEGF en ovarios de rata de 2 días. Es sabido que trkA, el receptor de alta afinidad
para NGF, se encuentra expresado en el ovario de los roedores luego del nacimiento
(Dissen y cols. 1995, Dissen y cols. 2001). Esta información es clave para explicar los
resultados obtenidos, ya que aunque se ha descrito que NGF es capaz de regular de
manera positiva la expresión de sus propios receptores (Sofroniew y cols. 2001), si los
niveles de receptor trkA en el ovario en el momento del nacimiento hubieran sido muy
bajos, probablemente no habríamos tenido una respuesta tan rápida como la
observada, ya que habría sido necesaria una síntesis de novo de trkA, para poder
señalizar de manera eficiente e incrementar la expresión de VEGF.

58
Para el caso del mRNA de VEGF en ovarios de rata de 2 días, se observó un
aumento luego de 2 horas de estímulo con NGF. En cambio, la proteína mostró un
incremento significativo luego de 24 horas de estimulación. Este retraso en el aumento
de proteína con respecto al aumento del mRNA podría estar reflejando una mayor
estabilidad del mensajero y/o proteína, lo que permitiría la acumulación de VEGF, y
que el aumento de esta proteína con respecto al control fuera evidente recién varias
horas más tarde. Acerca de la vida media de VEGF, se ha reportado que en la
circulación ésta corresponde a 3 min (George y cols. 2000). Suponemos que en los
tejidos VEGF podría tener una mayor estabilidad, pero lo más probable es que una
combinación en el aumento de estabilidad tanto del mRNA como de la proteína de
VEGF sea responsable del resultado observado luego de 24 horas de estímulo con
NGF. Por otra parte, la vida media de NGF en cultivo tampoco se ha descrito, pero en
el plasma corresponde a 7,2 min (Poduslo y cols. 1996). Esto hace suponer que en el
medio de cultivo, la vida media de NGF no debe ser tan larga como para explicar
efectos de NGF sobre su receptor durante varias horas luego de iniciado el cultivo. En
conjunto, estos datos apuntan a que NGF podría actuar de manera rápida sobre sus
receptores en el tejido ovárico en cultivo, mientras que los efectos de esta acción se
observarían horas más tarde.
Una observación interesante con respecto a la proteína de VEGF detectada en
ovarios de rata de 2 días por inmunohistoquímica, es que se encontró marca
específicamente en las células de granulosa, pero también se pudo detectar una señal
difusa en el resto del tejido ovárico. De acuerdo con las indicaciones del fabricante, el
anticuerpo utilizado se obtuvo utilizando como inmunógeno un fragmento de VEGF que
contenía los aminoácidos 1 al 145. Lo anterior permite suponer que el anticuerpo es
capaz de reconocer tanto las isoformas solubles de VEGF (como VEGF 165 y 121)
como las isoformas que poseen una alta afinidad por los componentes de la matriz
extracelular, quedando retenidas en ella (como VEGF 189 y 206). La señal difusa de
VEGF encontrada en el tejido ovárico de rata de 2 días podría corresponder a las
isoformas solubles, que tienen una mayor capacidad de distribuirse luego de ser
secretadas. En cambio, la marca detectada en el interior de las células de granulosa, o

59
en las cercanías de los folículos, podría corresponder a una mezcla de las distintas
isoformas de VEGF producidas.
El complejo NGF/trkA podría estar actuando como un regulador de la expresión
ovárica de VEGF en los primeros días de vida postnatal de los roedores. Sin embargo,
en este estudio no se encontraron diferencias en la expresión de VEGF en ovarios de
ratones NGF o trkA knockout con respecto a ratones silvestres. Se sabe que en el
ovario de roedores también existe expresión de otras neurotrofinas luego del
nacimiento (Ojeda y cols. 2000, Dissen y cols. 2001), pero esto no necesariamente
quiere decir que la acción de las neurotrofinas durante el desarrollo ovárico temprano
sea redundante: en los ovarios de ratones knockout para NGF hay menos folículos
primarios y secundarios, existe una mayor cantidad de ovocitos que no quedan
incorporados dentro de una estructura folicular, y la proliferación celular se encuentra
reducida (Dissen y cols. 2001). Además, la inmunoneutralización de NGF en la etapa
postnatal temprana en la rata interfiere con el crecimiento de folículos antrales y retrasa
la pubertad (Lara y cols. 1990b). Lo anterior indica que la presencia de otras
neurotrofinas no evita trastornos a nivel del desarrollo ovárico en los ratones NGF
knockout. Pero a la luz de los resultados presentados en esta tesis, suponemos que
en el caso de la expresión de VEGF, otras neurotrofinas sí podrían estar compensando
la falta de NGF en los animales NGF knockout. Se piensa que podría existir una cierta
promiscuidad en la unión a receptores de distintos miembros de la familia de
neurotrofinas. Por ejemplo, la neurotrofina 4 (NT4) es capaz de unirse a trkA e inducir
la autofosforilación de este receptor (Ip y cols. 1992). De acuerdo con esto, al no estar
presente NGF, podría existir un incremento en los niveles de otras neurotrofinas, como
NT-4, lo cual favorecería algún tipo de interacción con trkA.
En el caso de los ratones trkA knockout, podrían ocurrir al menos dos efectos: 1)
que se incrementara la expresión de NGF, y éste actuara sobre otros receptores, por
ejemplo, receptores para otras neurotrofinas. Lo anterior se basa en reportes que
describen efectos neuroprotectores de NGF en poblaciones neuronales que no
expresan los receptores para NGF (Sofroniew y cols. 2001). 2) Otra opción es que
existiera una elevación de p75 (el receptor de baja afinidad para NGF), lo cual

60
permitiría que NGF señalizara por esta vía. En esta Tesis se exploró la posibilidad de
un efecto compensatorio vía el incremento de p75. Se evaluaron los niveles de mRNA
de p75 (el receptor de baja afinidad para NGF) en los ovarios de ratones trkA knockout,
y se encontró que el contenido de mRNA de la isoforma de largo completo de p75
estaba aumentado (aunque no de manera estadísticamente significativa) en los ratones
trkA knockout, en comparación con los silvestres. Aunque inicialmente se pensaba que
los efectos de la señalización NGF/p75 eran antagónicos a los de NGF/trkA, en la
actualidad existe información que apoya que la señalización vía p75 también puede
contribuír a la proliferación y diferenciación celular (Botchkareva y cols. 1999, Giuliani y
cols. 2004), de modo que en los ratones trkA knockout podría producirse una
compensación de esta naturaleza. Es importante dejar claro que, la mayor parte de los
ratones trkA knockout mueren antes de llegar a la madurez, y ninguno logra sobrevivir
más de 2 meses (Smeyne y cols. 1994), lo que indica que, aunque en estos roedores
se desarrollen sistemas que les permitan resistir algunos días, la falta del receptor trkA
es finalmente incompatible con la vida.
Recientemente se descubrió una nueva variante de splicing de trkA en tumores
cerebrales humanos. Esta nueva isoforma promueve la señalización intracelular en
forma independiente de NGF, y en este tipo de lesiones antagoniza los efectos de la
activación de los receptores trkA conocidos originalmente (Tacconelli y cols. 2004). Si
bien no se ha descrito la presencia de esta isoforma de trkA en otras especies, lo
anterior nos hace reflexionar acerca de la complejidad de respuestas que podrían
esperarse en animales knockout que solamente carecen de las isoformas conocidas
anteriormente.
Para concluir con este punto, habría sido importante realizar el estudio de los
ratones knockout a distintas edades en el período comprendido entre el día del
nacimiento y los 7 días post-natal, ya que se ha descrito que tanto NGF como trkA son
importantes en distintos procesos a lo largo de esta etapa (Ojeda y cols. 2000, Dissen
y cols. 2004). Lamentablemente, no existió la posibilidad de seleccionar la edad de los
ratones knockout utilizados, y se tuvo que trabajar con los tejidos que estaban
disponibles y que pertenecían a animales de edades ya establecidas.

61
El ovario humano adulto expresa NGF y trkA (Abir y cols. 2005, Salas y cols. 2006).
Como se dijo antes, NGF incrementa la expresión del receptor de FSH y la secreción
de estrógeno en células de granulosa humana (Salas y cols. 2006). En los ovarios de
oveja, cerdo, vaca y cabra también se expresa NGF y tanto el receptor trkA como p75
(Mattioli y cols. 1999, Ren y cols. 2005, Levanti y cols. 2005). En roedores, NGF se
encuentra expresado en la pubertad y la adultez (Dissen y cols. 1995, Dissen y cols.
2000, Lara y cols. 2000). En este estudio reportamos que NGF es capaz de
incrementar el mRNA y/o proteína de VEGF en ovario de rata y ratón, y en células de
granulosa humana. Con respecto a este último modelo, se observó que todas las
muestras de granulosa respondieron al estímulo con NGF, con mayores niveles de
mRNA de VEGF. En cambio, para el caso de la proteína de VEGF cuantificada en el
medio de cultivo, existía un grupo de muestras sin respuesta. Al reunir todas las
muestras (con y sin respuesta) en un mismo grupo, de todas maneras se producía un
aumento significativo de VEGF por efecto de NGF (p < 0,05). Sin embargo, se decidió
que era más ventajoso separar las muestras en dos grupos, ya que era probable que el
grupo clasificado como “sin respuesta” efectivamente correspondiera a una población
de células con un menor grado de maduración y que requiería de más tiempo para
montar la maquinaria de traducción y/o secreción de VEGF.
4.2. NGF, VEGF y la angiogénesis del ovario Los resultados presentados en esta tesis indican que NGF aumenta la
vascularización del ovario de roedores, proceso que podría ser consecuencia directa
de la unión de NGF al receptor trkA presente en las células endoteliales, o bien podría
ser una acción indirecta mediada por el aumento de VEGF.
En la Figura 4.1 se muestra un resumen del modelo propuesto para la cadena de
eventos mediante los cuales NGF estaría regulando la angiogénesis ovárica. Se sabe
que, luego del alza de LH que antecede a la ovulación, hay un aumento agudo en la
expresión de NGF y trkA en el ovario de rata (Dissen y cols. 1996). Si la acción de NGF
o el receptor trkA son bloqueados en uno de los ovarios, se produce una inhibición
ipsilateral de la ovulación (Dissen y cols. 1996). Lo anterior permite concluír que la

62
señalización de NGF es esencial para la ovulación. Por otra parte, se produce un
marcado aumento de la expresión ovárica de VEGF en la etapa periovulatoria (Shweiki
y cols. 1993, Berisha y cols. 2000). La inhibición de la unión de VEGF a sus receptores
suprime el desarrollo folicular y la ovulación en primates (Wulff y cols. 2002). Los
resultados presentados en esta Tesis indican que NGF es capaz de aumentar la
expresión de VEGF y la vascularización ovárica. De acuerdo con lo anterior, se
propone una asociación entre el alza periovulatoria de NGF, y el aumento de VEGF y
de los capilares ováricos (Figura 4.1).
Figura 4.1. Modelo propuesto para el orden de eventos luego del alza de LH que antecede a la ovulación. Los eventos 1 y 2 fueron descritos por Dissen y cols. en 1996, y lo indicado en 3a y 3b ha sido reportado por Shweiki y cols. (1993), Berisha y cols. (2000) y Fraser y cols. (2006), entre otros. Este trabajo plantea una probable asociación entre ambos eventos, dada por el incremento de VEGF inducido por NGF.
Para determinar si los efectos de NGF sobre la vascularización ovárica fueron una
consecuencia del aumento de VEGF (ver Figura 4.2), se realizó un estudio in vitro
utilizando células HUVEC. La red vascular presente en el ovario está formada por
microvasos, por lo que el modelo de HUVEC no corresponde al mismo tipo de vasos
que están presentes en el ovario. Sin embargo, en el estudio de la angiogénesis, este
un modelo ampliamente utilizado. Por ejemplo, Middeke y cols. (2002) evaluaron la
Ovulación
1. Alza de LH preovulatoria 2. Incremento ovárico
de mRNA de NGF
Aumento periovulatorio de la expresión de VEGF
3b. Aumento de capilares
Fase folicular Fase lútea
3a. Aumento post-ovulatorio de la
expresión de VEGF

63
participación de NGF en la angiogénesis del feocromocitoma, y utilizaron como modelo
in vitro células PC12 estimuladas con NGF, para posteriormente transferir el medio a
un cultivo de HUVEC. Finalmente, en ese estudio se determinó que al neutralizar
VEGF en ese medio condicionado, se inhibía la proliferación de las HUVEC. Este
antecedente nos hizo pensar que el modelo de HUVEC tendría aplicación en el objetivo
de determinar si los medios condicionados de células de granulosa incubadas con
NGF, tenían la capacidad de estimular la proliferación de células del endotelio vascular,
y si VEGF estaba involucrado en este proceso.
Los resultados aquí reportados no nos permitieron establecer si los efectos de NGF
sobre la vascularización ovárica fueron una consecuencia del aumento de VEGF (ver
Figura 4.2). En el ensayo de angiogénesis in vitro, el medio condicionado de células de
granulosa tratadas con NGF no indujo mayor proliferación de células endoteliales que
el medio control. Suponemos que la concentración de VEGF secretado en respuesta a
NGF en el medio condicionado de células de granulosa fue muy baja para inducir una
respuesta en las células HUVEC, por el siguiente motivo: las concentración de VEGF
en el medio condicionado de células de granulosa se encontraba en el orden de 1
ng/ml, y correspondía a una mezcla de isoformas con mayor o menor actividad
biológica. Se probó que la mayor respuesta de proliferación se alcanzaba cuando las
células HUVEC eran estimuladas exclusivamente con VEGF 165 (la isoforma más
activa), en una dosis de 5 ng/ml. De lo anterior se infiere que los medios condicionados
deberían haber estado mucho mas concentrados para poder inducir la proliferación de
las células HUVEC. De cualquier modo, si nos situamos en el contexto de la fisiología
del ovario, el estrecho contacto que se produce entre la célula de granulosa y la célula
endotelial luego de la ovulación, permite que localmente se alcancen concentraciones
muy elevadas de las distintas moléculas secretadas por la granulosa. Esto significa que
puede haber un efecto paracrino que no se reprodujo en el experimento realizado.
Se podría pensar que la condición de hiperoxia que se produce en la cámara de
cultivo, en comparación con la disponibilidad de oxígeno en los tejidos
(aproximadamente 150 vs. 40 mm Hg, respectivamente; Ganong, 1996) podría haber
afectado la capacidad de proliferación de las HUVEC. Hubiera sido ideal trabajar en

64
condiciones que se asemejaran más al microambiente ovárico, pero lamentablemente
no se disponía de los medios para generar dichas condiciones. Por otra parte, hay que
considerar que el experimento realizado se diseñó en base al trabajo de Middeke
(2002), en el cual se utilizaron las mismas condiciones de cultivo aquí reportadas.
Además, nosotros encontramos que VEGF 165 de todas maneras estimuló la
proliferación de las células endoteliales in vitro, lo que permite suponer que la
condición de hiperoxia de nuestros cultivos no influyó de forma importante en los
resultados obtenidos.
Figura 4.2. Secuencia de eventos que podrían explicar el efecto de NGF sobre la angiogénesis del ovario. Los resultados presentados en esta tesis indican que NGF es capaz de aumentar la expresión de VEGF y la vascularización ovárica. No fue posible demostrar si los efectos de NGF eran mediados por VEGF, pero los datos reportados en sistemas como feocromocitoma y cerebro de rata indican que sería una vía probable (Calzà y cols. 2001, Middeke y cols. 2002).

65
Otro parámetro que habría sido interesante evaluar, era la formación de estructuras
tipo tubo (tubulogénesis), en los cultivos de HUVEC tratados con los medios
condicionados. Esto no se pudo realizar debido a que, para poder evaluar la
tubulogénesis, habría sido necesario sembrar las células endoteliales en placas de
mayor dimensión, lo que hubiera requerido volúmenes importantes de medio
condicionado. El sistema de trabajo con células de granulosa humana no genera
grandes cantidades de células, por lo que no fue posible producir un volumen de
medio condicionado que permitiera dicho experimento.
Además del efecto conocido que tienen factores de crecimiento como VEGF sobre
el endotelio vascular (fragmentación y remodelamiento de la membrana basal del vaso,
migración y proliferación de células endoteliales), se ha propuesto que también podrían
actuar sobre los precursores de células endoteliales que existen en la médula ósea,
induciendo su movilización hacia la circulación para alcanzar el tejido que emite
señales angiogénicas (Hristov y cols. 2003). En estos momentos existe poca
información sobre los elementos que regulan la maduración in vivo de estos
precursores hacia células endoteliales completamente diferenciadas.
También existe la posibilidad de que el aumento de VEGF por efecto de NGF,
específicamente la isoforma 165 en humano y 164 en roedores, de la cual una parte
permanece retenida en la matriz extracelular, podría ser una de las señales para que
las células endoteliales migraran hacia las células de granulosa como ocurre
inmediatamente después de la ovulación cuando se forma el cuerpo lúteo (Fraser y
cols. 2006).
4.3. NGF no modifica los niveles de TGFβ1 en el ovario
TGFβ1, el otro factor angiogénico evaluado en este estudio, no mostró variaciones
inducidas por NGF en los modelos de ovario analizados. En el ensayo de ELISA
realizado en el medio de células de granulosa tratadas con NGF, TGFβ1 ni siquiera
pudo ser detectado. Lo más probable es que esto se haya debido a la vida media de 3
minutos del factor de crecimiento. (Wakefield y cols. 1990) Sin embargo, se sabe que
TGFβ1 está presente en el ovario humano, y es expresado por la mayoría de los tipos

66
celulares durante la etapa folicular y lútea del ciclo ovárico (Dissen y cols. 1994,
Chegini y cols. 1992a, Chegini y cols. 1992b). Los resultados aquí presentados, en
conjunto con la información de que en el ovario de hámster, la expresión de los
receptores de TGFβ tiene un comportamiento cíclico que parece estar regulado por las
gonadotrofinas (Roy 2000), nos llevan a pensar que la acción de TGFβ1 en el ciclo
ovárico podría estar controlada a nivel del receptor en lugar del ligando. Sería muy
interesante evaluar si NGF induce cambios en el receptor de TGFβ, ya que esto sería
concordante con los efectos angiogénicos atribuidos a TGFβ1 (Pepper y cols. 1997,
Brown y cols. 2000).
4.4. Mecanismos del aumento de VEGF inducido por NGF en el ovario Con respecto al mecanismo por el cual NGF estaría incrementando la expresión de
VEGF en los modelos estudiados, los resultados obtenidos indican que esta
neurotrofina estaría actuando sobre el receptor trkA. En este estudio se utilizó el
inhibidor K252a, el cual corresponde a un alcaloide cuya estructura es análoga al ATP
(Kase y cols. 1987), lo que le permite actuar como inhibidor competitivo de proteínas
kinasas. Pese a su acción inespecífica, el inhibidor K252a ha sido ampliamente
utilizado para estudiar la señalización vía trkA y para bloquear la unión de NGF a este
receptor (Knusel y cols. 1992, Dissen y cols. 1996, Sofroniew y cols. 2001). En el
estudio aquí presentado, se decidió que K252a era una buena opción para discriminar
si en el modelo de células de granulosa, el incremento de VEGF inducido por NGF era
vía trkA, un receptor con actividad kinasa, o vía p75, que no posee actividad kinasa.
Con posterioridad al diseño de los experimentos aquí reportados, apareció una
publicación que describe que el receptor p75 no se encuentra presente en ovario
humano adulto (Abir 2005). Por otra parte, el ovario de roedores expresa el mRNA de
p75 luego del nacimiento, pero este receptor no está presente en las células de
granulosa ni en los ovocitos (Hsueh y cols. 1994). Además, ovarios de ratones
deficientes para p75 tienen una apariencia normal (Ojeda y cols. 2000), por lo que el rol
de este receptor en el desarrollo ovárico temprano no está del todo claro, y resulta
difícil hacer un análisis acerca de la participación que podría tener p75 en los efectos
de NGF sobre VEGF en el ovario de roedores de 0 a 7 días.

67
Una amplia variedad de vías de señalización intracelular han sido descritas para
NGF (Sofroniew y cols. 2001), pero la mayor parte de los estudios se han realizado en
la línea celular PC-12, derivada de feocromocitoma de rata (Sofroniew y cols. 2001), y
existe muy poca información acerca de las vías de señalización activadas por NGF en
el ovario. Con respecto a esto, un estudio en células de ovario de hámster chino
(células CHO), indica que NGF induce la activación de ERK1/2 y de PLC-gamma1
(Zapf-Colby y cols. 1999). En esta tesis se encontró que NGF induce una rápida
activación de ERK2 en células de granulosa humana, y que al inhibir la vía de las ERK,
se prevenía el incremento de VEGF inducido por NGF. No se encontró un aumento
significativo para la razón ERK 1 fosforilada / ERK 1 total, lo que podría ser un reflejo
de la alta dispersión en los resultados para esta isoforma de ERK, puesto que en
algunos casos la señal de la proteína fosforilada en el tiempo 0 era muy débil. No se
debe descartar otras vías de señalización en este proceso, como la de PI3 kinasa, que
también puede ser gatillada por NGF luego de unirse a trkA (Sofroniew y cols. 2001), y
que ha sido asociada al aumento en la expresión de VEGF (Wang y cols. 1999).
La disponibilidad de oxígeno es crucial para el metabolismo celular. El factor
inducible por hipoxia (HIF1) es un factor de transcripción que regula la homeostasis
celular en condiciones hipóxicas, permitiendo la activación de genes esenciales para la
adaptación de las células a situaciones de bajos niveles de oxígeno (Déry y cols.
2005). HIF1 puede aumentar la actividad del promotor de VEGF en respuesta a un
estímulo de hipoxia, pero también frente a las señales de otros factores de crecimiento.
Asimismo, NGF es capaz de regular los niveles de la subunidad inducible de HIF1
(HIF1α) (Naranjo-Suárez y cols. 2003). Una observación interesante es que en el
trabajo de Naranjo-Suárez y cols. (2003), se encontró que NGF disminuía los niveles
de HIF1α. Sin embargo, recientemente se publicó que otra neurotrofina, BDNF, induce
la expresión de VEGF a través del aumento de HIF1α (Nakamura y cols. 2006).
Nosotros encontramos que en ovarios de rata de 2 días cultivados con NGF por 2
horas se producía un incremento del mRNA de HIF1α, lo que permite sugerir que
podría ser un componente involucrado en el aumento de mRNA de VEGF. En células

68
de granulosa el mRNA de HIF1α no se modificó por efecto de NGF en las condiciones
de cultivo analizadas, pero resultaría interesante estudiar el mRNA de este factor a
tiempos cortos de estímulo con NGF, puesto que los cambios en la expresión de HIF1α
podrían ser transitorios. Por otra parte, es probable que en el tejido se logre mantener
una mayor hipoxia con respecto a las células, pero descartamos que el resultado
obtenido (aumento de mRNA de HIF1α en tejido y no en células) sea consecuencia de
esto, ya que se ha descrito que la hipoxia contribuye a la estabilidad de la proteína de
HIF1α, pero no a la de su mRNA (Dèry y cols. 2005).
Figura 4.3. Mecanismos de señalización involucrados en el aumento de VEGF inducido por NGF. En células de granulosa se demostró que es necesaria la señalización de trkA así como la vía de ERK 1/2 para que se produzca el incremento en el mRNA y la proteína de VEGF inducidos por NGF. En el caso de HIF1α, los datos obtenidos en el modelo de células de granulosa no indican que sea un componente involucrado en este proceso. Sin embargo, los resultados de cultivo de ovario de rata de 2 días indican que podrían existir cambios tempranos en el mRNA de HIF1α asociados al estímulo de NGF. En esta tesis no se estudió si existía una asociación entre trkA, ERK 1/2 y HIF1α luego del estímulo con NGF. Los datos de la literatura indican que esta sería una vía de señalización posible (Sofroniew y cols. 2001, Qian y cols. 2004, Jeong y cols. 2005).
Resumiendo los mecanismos de señalización explorados en esta tesis, en células
de granulosa se demostró que la activación de trkA así como la vía de ERK 1/2 son
necesarias para que se produzca el incremento en el mRNA y la proteína de VEGF
inducidos por NGF (ver Figura 4.3). En el caso de HIF1α, los datos obtenidos en el
modelo de células de granulosa no indican que sea un componente involucrado en
este proceso. Sin embargo, de acuerdo con los resultados de cultivo de ovario de rata
de 2 días, podrían existir cambios tempranos en el mRNA de HIF1α asociados al

69
estímulo de NGF (ver Figura 4.3). En esta tesis no se estudió si los eventos de
activación de trkA, ERK 1/2 y la producción de HIF1α luego del estímulo con NGF se
encontraban asociados, pero los antecedentes de la literatura indican que esta sería
una vía de señalización posible (Sofroniew y cols. 2001, Qian y cols. 2004, Jeong y
cols. 2005). Sería interesante evaluar el efecto de la combinación de NGF e hipoxia, lo cual
podría simular la condición que se puede producir en el caso del transplante de tejidos,
donde podríamos tener niveles altos de NGF, como resultado de la denervación, y una
situación de hipoxia, dada por la vascularización disminuída luego de la cirugía. Se ha
reportado que NGF prolonga los efectos neuroprotectores que se producen como
respuesta a un episodio de hipoxia en células PC12 (Gross y cols. 2000). Si
trasladamos estos datos al transplante de tejidos, se podría pensar que, luego de la
cirugía se produce una condición de hipoxia que elevaría los niveles de HIF1α. A
continuación, los niveles aumentados de NGF (generados por la denervación del tejido
transplantado), podrían potenciar aún más la acción de HIF1α, lo cual tendría como
resultado una mejor respuesta de vascularización del tejido, y además una mejor
adaptación a la hipoxia.
4.5. Comentarios finales y proyecciones Patologías diversas como el hemangioma, la psoriasis y la mayoría de las
condiciones neoplásicas han sido relacionadas con defectos en la angiogénesis (Yu y
cols. 2004, Veale y cols. 2005, Ferrara y cols. 1997). Pacientes con ovario poliquístico
tienen un mayor flujo sanguíneo ovárico, lo que probablemente está relacionado con
los elevados niveles de VEGF plasmático que presentan estas mujeres (Agrawal y
cols. 1998). En ratas a las que se les ha inducido ovario poliquístico mediante la
administración de esteroides, se produce un aumento de NGF y p75 ováricos, y la
administración de un antisuero anti-NGF en conjunto con un RNA antisentido para p75
normaliza los ciclos ováricos y la capacidad ovulatoria en la mayoría de estos animales
(Lara y cols. 2000). Lo anterior indica que desórdenes de la fertilidad, como el ovario
poliquístico u otros asociados a problemas en la angiogénesis, podrían ser generados

70
por una desregulación en la expresión o función de NGF, que finalmente resulte en una
producción aberrante de VEGF.
Una proyección interesante de los resultados aquí presentados apunta a la
preservación de la función ovárica en mujeres que deben recibir tratamientos con
radiación para erradicar tumores de la región abdominal. Con respecto a esto, se ha
descrito el caso de una mujer que sufría de cáncer cérvico uterino, cuyo ovario fue
removido y transplantado en el brazo. Luego de la intervención, el tejido fue capaz de
re-vascularizarse e innervarse, y finalmente recuperó la ciclicidad y la capacidad
ovulatoria (Hilders y cols. 2004). Esta observación es congruente con los resultados
presentados en esta Tesis, ya que podría existir un incremento de NGF inducido por el
corte de los nervios del ovario, lo cual a su vez podría aumentar la expresión de VEGF,
favoreciendo así la re-vascularización del tejido. Por lo tanto, los datos presentados
abren un nuevo campo en el cual se podría mejorar la posibilidad de éxito en
procedimientos de esta naturaleza. La terapia con NGF para promover la cicatrización
de lesiones ha tenido resultados prometedores (Chiaretti y cls. 2002), por lo que resulta
interesante sugerir que esta neurotrofina podría tener un efecto beneficioso en el
transplante del ovario u otros órganos.
Por otra parte, también se ha estudiado el rol que juega NGF en una importante
patología ovárica: el cáncer epitelial (Davidson y cols. 2001, Davidson y cols. 2003,
Campos y cols. 2006). Esta condición por lo general tiene un mal pronóstico, ya que se
trata de tumores silentes, es decir, no dan síntomas hasta que la enfermedad se
encuentra muy avanzada (Davidson y cols. 2001). Los tumores ováricos son altamente
angiogénicos, y se demostró que NGF es capaz de aumentar la expresión de VEGF en
el tejido ovárico transformado, efecto que sería mediado por el receptor trkA (Campos y
cols. 2006). De acuerdo con lo anterior, una posible terapia para reducir la
angiogénesis y el tamaño de este tipo de tumores podría ser el uso local de
anticuerpos anti-NGF. Similares estrategias se encuentran bajo estudio, mediante el
uso del inhibidor de receptores tipo Trk (K252a) para regular el crecimiento de células
derivadas de tumores pulmonares (Pérez-Piñera y cols. 2006).

71
5. Conclusiones
• Los resultados obtenidos en distintos modelos de ovario de roedores tanto in vitro
como in vivo, así como en un modelo de ovario humano in vitro, indican que NGF
actúa sobre las células ováricas incrementando el mRNA de distintas isoformas del
factor angiogénico VEGF. Además NGF aumenta los niveles de la proteína de
VEGF, tanto la que está presente en el tejido ovárico, como la secretada al medio
de cultivo. TGFβ1, el otro factor de crecimiento evaluado en esta tesis, no fue
modificado por NGF en los modelos estudiados.
• El incremento de VEGF inducido por el estímulo de NGF en el ovario, sería un
efecto dependiente del receptor de alta afinidad trkA, ya que la inhibición de esta
vía lleva los niveles de VEGF a valores similares al control. Resultados preliminares
sugieren que en ausencia de trkA, el receptor de baja afinidad, p75, podría tener
alguna participación en el incremento de VEGF en el ovario en desarrollo.
• La activación de la vía transduccional de las ERK también participa en el
incremento de VEGF inducido por el estímulo de NGF. En cuanto al factor
transcripcional HIF los resultados aquí presentados no son suficientes para sugerir
que podría tener un rol en el proceso descrito.
• En un ensayo de angiogénesis in vitro, el medio condicionado de células de
granulosa tratadas con NGF no indujo mayor proliferación de células endoteliales.
Sin embargo, en modelos de angiogénesis in vivo, NGF indujo la vascularización
del ovario de roedores de 25 a 30 días, aumentando el área de marca positiva para
vasos sanguíneos.
• Los resultados de esta tesis sugieren que NGF puede actuar sobre la angiogénesis
ovárica en forma indirecta, aumentando los niveles de VEGF en el ovario.

72
6. Referencias
Abir R, Fisch B, Jin S, Barnnet M, Ben-Haroush A, Felz C, Kessler-Icekson G, Feldberg
D, Nitke S, Ao A (2005). Presence of NGF and its receptors in ovaries from human
fetuses and adults. Mol Hum Reprod 11:229-236.
Agrawal R, Sladkevicius P, Engmann L, Conway GS, Payne NN, Bekis J, Tan SL,
Campbell S, Jacobs HS (1998). Serum vascular endothelial growth factor
concentrations and ovarian stromal blood flow are increased in women with
polycystic ovaries. Hum Reprod 13:651-655.
Akhurst RJ (2002). TGF-β antagonists: Why suppress a tumor suppressor? J Clin
Invest 109:1533-1536.
Anaf V, Simon P, El Nakadi I, Fayt I, Simonart T, Buxant F, Noel JC (2002)
Hyperalgesia, nerve infiltration and nerve growth factor expression in deep
adenomyotic nodules, peritoneal and ovarian endometriosis. Hum Reprod
17:1895-1900. Anderson RA, Robinson LLL, Brooks J, Spears N (2002). Neurotropins and their
receptors are expressed in the human fetal ovary. J Clin Endocrinol Metab 87:890-
897.
Berisha B, Schams D, Kosmann M, Amselgruber W, Einspanier R (2000). Expression
and tissue concentration of vascular endothelial growth factor, its receptors, and
localization in the bovine corpus luteum during estrous cycle and pregnancy. Biol
Reprod 63:1106-1114.
Botchkareva NV, Botchkarev VA, Chen LH, Lindner G, Paus R (1999). A role for p75
neurotrophin receptor in the control of hair follicle morphogenesis. Dev Biol
216:135-153.
Böttner M, Krieglstein K, Unsicker K (2000). The transforming growth factor-βs:
structure, signaling, and roles in nervous system development and functions. J
Neurochem 75:2227-2240.
Brown MR, Blanchette JO, Kohn EC (2000). Angiogenesis in ovarian cancer. Baillieres
Best Pract Res Clin Obstet Gynaecol 14:901-918.

73
Calzà L, Giardino L, Giuliani A, Aloe L, Levi-Montalcini R (2001). Nerve growth factor
control of neuronal expression of angiogenetic and vasoactive factors. Proc Natl
Acad Sci USA 98:4160-4165.
Campos X, Munoz Y, Selman A, Yazigi R, Moyano L, Weinstein-Oppenheimer C, Lara
HE, Romero C (2006). Nerve growth factor and its high-affinity receptor trkA
participate in the control of vascular endothelial growth factor expression in
epithelial ovarian cancer. Gynecol Oncol 2006, Aug 24; [Epub ahead of print].
Cantarella G, Lempereur L, Presta M Ribatti R, Lombardo G, Lazarovici P, Zappala G
, Pafumi C, Bernardini R. (2002). Nerve growth factor– endothelial cell interaction
leads to angiogenesis in vitro and in vivo FASEB J 16:1307-1309.
Chegini N, Flanders KC (1992a). Presence of transforming growth factor-β and their
selective cellular localization in human ovarian tissue of various reproductive
stages. Endocrinology 130:1707-1715.
Chegini N, Williams RS (1992b). Immunocytochemical localization of transforming
growth factors (TGFs) TGF-α and TGF-β in human ovarian tissues. J Clin
Endocrinol Metab 74:973-980.
Chiaretti A, Piastra M, Caresta E, Nanni L, Aloe L (2002). Improving ischaemic skin
revascularisation by nerve growth factor in a child with crush syndrome. Arch Dis
Child 87:446-448.
Christenson LK, Stouffer RL (1997). Follicle-stimulating hormone and luteinizing
hormone/chorionic gonadotropin stimulation of vascular endothelial growth factor
production by macaque granulosa cells from pre- and periovulatory follicles. J Clin
Endocrinol Metab 82:2135-2142.
Corne J, Chupp G, Lee CG, Homer RJ, Zhu Z, Chen Q, Ma B, Du Y, Roux F, McArdie
J, Waxman AB, Elias JA (2000). IL-3 stimulates VEGF and protects against
hyperoxic acute lung injury. J Clin Invest 106:783-791.
Crowley C, Spencer SD, Nishimura MC, Chen KS, Pitts-Meek S, Armanini MP, Ling LH,
McMahon SB, Shelton DL, Levinson AD, Phillips HS (1994). Mice lacking nerve
growth factor display perinatal loss of sensory and sympathetic neurons yet
develop basal forebrain cholinergic neurons. Cell 76:1001-1011.
Davidson B, Lazarovici P, Ezersky A, Nesland JM, Berner A, Risberg B, Tropé CG,
Kristensen GB, Goscinski M, van de Putte G, Reich R (2001). Expression levels of

74
the nerve growth factor receptors trka and p75 in effusions and solid tumors of
serous ovarian carcinoma patients. Clin Cancer Res 7:3457-3464.
Davidson B, Reich R, Lazarovici P, Nesland JM, Skrede M, Risberg B, Tropé CG,
Flørenes VA (2003). Expression and activation of the nerve growth factor receptor
trka in serous ovarian carcinoma. Clin Cancer Res 9:2248-2259.
Déry MA, Michaud MD, Richard DE (2005). Hypoxia inducible factor-1: regulation by
hypoxic and non-hypoxic activators. Int J Biochem Cell Biol 37:535-540.
Descamps S, Toillon RA, Adriaenssens E, Pawlowski V, Cool SM, Nurcombe V, Le
Bourhis X, Boilly B, Peyrati JP, Hondermarck H (2001). Nerve growth factor
stimulates proliferation and survival of human breast cancer cells through two
distinct signaling pathways. J Biol Chem 276:17864-17870.
Dissen GA, Hill DF, Costa ME, Dees WL, Lara HE, Ojeda SR (1996). A role for trka
nerve growth factor receptors in mammalian ovulation. Endocrinology 137:198-
209.
Dissen GA, Hirshfield AN, Malamed S, Ojeda SR (1995). Expression of neurotrophins
and their receptors in the mammalian ovary is developmentally regulated: changes
at the time of folliculogenesis. Endocrinology 136:4681-4692.
Dissen GA, Lara HE, Fahrenbach WH, Costa ME, Ojeda SR (1994). Immature rat
ovaries become revascularized rapidly after autotransplantation and show a
gonadotropin-dependent increase in angiogenic factor gene expression.
Endocrinology 134:1146-1154.
Dissen GA, Paredes A, Romero C, Dees WL, Ojeda SR. Neural and neurotrophic
control of ovarian development. En: Leung P, Adashi EY. The ovary 2a ed. EEUU,
Elsevier Academic Press, 2004. pp 11-18.
Dissen GA, Parrott JA, Skinner MK, Hill DF, Costa ME, Ojeda SR (2000). Direct effects
of nerve growth factor on thecal cells from antral ovarian follicles. Endocrinology
141:4736-4750.
Dissen GA, Romero C, Newman Hirshfield A, Ojeda SR (2001). Nerve growth factor is
required for early follicular development in the mammalian ovary. Endocrinology
142: 2078-2086.

75
Dvorak HF (2002). Vascular Permeability Factor/Vascular Endothelial Growth Factor: A
critical cytokine in tumor angiogenesis and a potential target for diagnosis and
therapy. J Clin Oncol 20:4368-4380. Eikmans M, Baelde HJ, Hagen EC, Paul, LC, Eilers PHC, De Heer E, Bruijn JA (2003).
Renal mRNA levels as prognostic tools in kidney diseases. J Am Soc Nephrol
14:899-907.
Emanueli C, Salis MB, Pinna A, Graiani G, Manni L, Madeddu P (2002). Nerve
growth factor promotes angiogenesis and arteriogenesis in ischemic hindlimbs.
Circulation 106:2257-2262.
Ferrara N (2001). Role of vascular endothelial growth factor in regulation of
physiological angiogenesis. Am J Physiol 280: C1358-C1366.
Ferrara N, Davys-Smith T (1997) The biology of vascular endothelial growth factor.
Endocrine Rev 18:4-25.
Förander P, Krieglstein K, Söderström S, Strömberg I (2000). Mutual induction of
TGFβ1 and NGF after treatment with NGF or TGFβ1 in grafted chromaffin cells of
the adrenal medulla. Exp Neurol 164:303-313.
Fraser HM, Wulff C (2001). Angiogenesis in the primate ovary. Reprod Fertil Dev
13:557-566.
Fraser HM (2006). Regulation of the ovarian follicular vasculature. Reprod Biol
Endocrinol 4:18-26.
Fujimoto J, Ichigo S, Hirose R, Sakaguchi H, Tamaya T (1998). Expressions of vascular
endothelial growth factor (VEGF) and its mRNA in uterine endometrial cancers.
Cancer Lett 134:15-22.
Fukuda R, Hirota K, Fan F, Jung YD, Ellis LM, Semenza GL (2002). Insulin-like growth
factor 1 induces hypoxia-inducible factor 1-mediated vascular endothelial growth
factor expression, which is dependent on MAP kinase and phosphatidylinositol 3-
kinase signaling in colon cancer cells. J Biol Chem 277:38205-38211.
Ganong WF. Funciones pulmonares. En su: Fisiología Médica 15ª ed. México, Editorial
El Manual Moderno, 1996. pp. 715-733.
Gerendai I, Marchetti B, Maugeri S, Roxas MA, Scapagnini U (1978). Prevention of
compensatory ovarian hypertrophy by local treatment of the ovary with 6-OHDA.
Neuroendocrinology 27:272-278.

76
George FW, Ojeda SR (1987). Vasoactive intestinal peptide enhances aromatase
activity in the neonatal rat ovary before development of primary follicles or
responsiveness to follicle-stimulating hormone. Proc Natl Acad Sci USA, 84:5803-
5807.
George ML, Eccles SA, Tutton MG, Abulafi M, Swift RI (2000). Correlation of plasma
and serum vascular endothelial growth factor levels with platelet count in colorectal
cancer: clinical evidence of platelet scavenging. Clin Cancer Res 6:3147-3152.
Giuliani A, D'Intino G, Paradisi M, Giardino L, Calza L (2004). p75 (NTR)-
immunoreactivity in the subventricular zone of adult male rats: expression by
cycling cells. J Mol Histol 35:749-758.
Gross J, Leclere N, Heldt J, Muller I, Kietzmann T, Andreeva N (2000). Differential
effects of hypoxia on untreated and NGF treated PC12 cells. Life Sci 67:2595-
2603.
Hazelton DA, Hamilton TC (1999). Vascular endothelial growth factor in ovarian cancer.
Curr Oncol Rep 1:59-63.
Hilders CG, Baranski AG, Peters L, Ramkhelawan A, Trimbos JB (2004). Successful
human ovarian autotransplantation to the upper arm. Cancer 101:2771-2778.
Hofmann M, Van der Wee KS, Dargart J, Dirami G, Dettin L, Dym M (2003).
Establishment and characterization of neonatal mouse sertoli cell lines. J Androl
24:120-130.
Hoyle GW, Graham RM, Finkelstein JB, Nguyen KP, Gozal D, Friedman M (1998).
Hyperinnervation of the airways in transgenic mice overexpressing nerve growth
factor. Am J Respir Cell Mol Biol 18:149-157.
Hristov M, Erl W, Weber PC (2003). Endotelial progenitor cells: mobilization,
differentiation and homing. Arterioscler Thromb Vasc Biol 23:1185-1189.
Hsueh AJ, Billig H, Tsafriri A (1994). Ovarian follicle atresia: a hormonally controlled
apoptotic process. Endocr Rev 15:707-724.
Ip NY, Ibañez CF, Nye SH, McClain J, Jones PF, Gies DR, Belluscio L, Beau MM,
Espinosa R, Squinto SP, Persson H, Yancopoulos GD (1992). Mammalian
neurotrophin-4: structure, chromosomal localization, tissue distribution, and
receptor specificity. Proc Natl Acad Sci USA 89:3060-3064.

77
Jeong HJ, Kim JB, Hong SH, An NH, Kim MS, Park BR, Park RK, Kim HM (2005).
Vascular endothelial growth factor is regulated by hypoxic stress via MAPK and
HIF1α in the inner ear. J Neuroimmunol 163:84-91.
Kase H, Iwahashi K, Nakanishi S, Matsuda Y, Yamada K, Takahashi M, Murakata C,
Sato A, Kaneko M (1987). K-252 compounds, novel and potent inhibitors of protein
kinase C and cyclic nucleotide-dependent protein kinases. Biochem Biophys Res
Commun 142:436-440.
Kim SJ, Park K, Rudkin BB, Dey BR, Sporn MB, Roberts AB (1994). Nerve growth
factor induces transcription of transforming growth factor-β1 through a specific
promoter element in PC12 cells. J Biol Chem 269:3739-3744.
Kishibe K, Yamada Y, Ogawa K (2002). Production of nerve growth factor by mouse
hepatocellular carcinoma cells and expression of trka in tumor-associated arteries
in mice. Gastroenterology 122:1978-1986.
Knusel B, Hefti F (1992). K-252 compounds: modulators of neurotrophin signal
transduction.
J Neurochem 59:1987-1996.
Koos RD (1995). Increased expression of vascular endothelial growth/permeability
factor in the rat ovary following an ovulatory gonadotropins stimulus. Biol Reprod
52:1426-1435.
Laitinen M, Ristimäki A, Honkasalo M, Narko K, Paavonen K, Ritvos O (1997).
Differential hormonal regulation of vascular endothelial growth factors VEGF,
VEGF-B, and VEGF-C messenger ribonucleic acid levels in cultured human
granulosa-luteal cells. Endocrinology 138:4748-4756.
Lara HE, Dissen GA, Leyton V, Paredes A, Fuenzalida H, Fiedler JL, Ojeda SR (2000).
An increased intraovarian synthesis of nerve growth factor and its low affinity
receptor is a principal component of steroid-induced polycystic ovary in the rat.
Endocrinology 141:1059-1072.
Lara HE, Hill DF, Katz KH, Ojeda SR (1990b). The gene encoding nerve growth factor
is expressed in the immature rat ovary: effect of denervation and hormonal
treatment. Endocrinology 126:357-363.
Lara HE, Mc Donald JK, Ojeda SR (1990a). Involvement of nerve growth factor in the
female sexual development. Endocrinology 126:364-375.

78
Lee A, Christenson LK, Patton PE, Burry KA and Stouffer RL (1997). Vascular
endothelial growth factor production by human luteinized granulosa cells in vitro.
Hum Reprod 12:2756-2761.
Levanti MB, Germana A, Abbate F, Montalbano G, Vega JA, Germana G (2005). TrkA
and p75NTR in the ovary of adult cow and pig. J Anat 207:93-96.
Lozada P (2006). Rol del factor de crecimiento nervioso en la regulación de la
proliferación y la angiogénesis del ovario. Tesis para optar al Título de Bioquímico.
Anales de la Facultad de Ciencias Químicas y Farmacéuticas, Universidad de
Chile.
Martinez-Chequer JC, Stouffer RL, Hazzard TM, Patton PE, Molskness TA (2003).
Insulin-like growth factors-1 and -2, but not hypoxia, synergize with gonadotropin
hormone to promote vascular endothelial growth factor-a secretion by monkey
granulosa cells from preovulatory follicles. Biol Reprod 68:1112-1118.
Mattioli M, Barboni B, Gioia L, Lucidi P (1999). Nerve growth factor production in sheep
antral follicles. Domest Anim Endocrinol 17:361-371.
May JV, Turzcynski CJ, Ramos L, Mau Y (1995). Differential involvement of protein
kinase C in the regulation of transforming growth factor-β (TGF-β) secretion by
porcine theca and granulosa cells in vitro. Endocrinology 136:1319-1322.
McGee EA and AJW Hsueh (2000). Initial and cyclic recruitment of ovarian follicles.
Endocrine Rev 21:200-214.
Middeke M, Hoffmann S, Hassan I, Wulffnderlich A, Hofbauer LC, Zielke A (2002). In
vitro and in vivo angiogenesis in PC12 pheochromocytoma cells is mediated by
vascular endothelial growth factor. Exp Clin Endocrinol Diabetes 110:386-392.
Mulheron GW, Bossert NL, Lapp JA, Walmer DK, Schomberg DW (1992). Human
granulosa-luteal and cumulus cells express transforming growth factors-beta type 1
and type 2 mRNA. J Clin Endocrinol Metab 74:458-460.
Nakamura K, Jackson JK, Martin KC, Kiichiro B, Woo C, Thiele CJ (2006). Brain-
derived neurotrophic factor activation of TrkB induces VEGF expression via HIF-1α
in neuroblastoma cells. Cancer Res 66:4249-4255.
Nakayama K, Kanzaki A, Hata K, Katabuchi H, Okamura H, Miyazaki K, Fukumoto M,
Takebayashi Y (2002). Hypoxia-inducible factor 1 alpha (HIF-1 alpha) gene
expression in human ovarian carcinoma. Cancer Lett 176:215-223.

79
Naranjo-Suárez S, Castellanos MC, Álvarez-Tejado M, Vara A, Landázuri MO, del Peso
L (2003). Down-regulation of hypoxia-inducible factor-2 in PC12 cells by nerve
growth factor stimulation. J Biol Chem 278:31895-31901.
Ojeda SR, Romero C, Tapia V, Dissen GA (2000). Neurotrophic and cell-cell dependent
control of early follicular development. Mol Cell Endocrinol 163:67-71.
Otani N, Minami S, Yamoto M, Shikone T, Otani H, Nishiyama R, Otani T, Nakano R
(1999). The vascular endothelial growth factor/fms-like tyrosine kinase system in
human ovary during the menstrual cycle and early pregnancy. J Clin Endocrinol
Metab 84:3845-3851.
Papetti M, Herman IM (2002). Mechanisms of normal and tumor-derived angiogenesis.
Am J Physiol 282:C947-C970.
Pepper MS (1997). Transforming growth factor-beta: vasculogenesis, angiogenesis, an
vessel wall integrity. Cyitokine Growth Factor Rev 8:21-43.
Pérez-Piñera P, Hernández T, García-Suárez O, de Carlos F, Germana A, del Valle M
Astudillo A, Vega JA (2006). The Trk tyrosine kinase inhibitor K252a regulates
growth of lung adenocarcinmas. Mol Cell Biochem, Jul 22; [Epub ahead of print].
Poduslo JF, Curran GL (1996). Permeability at the blood-brain and blood-nerve barriers
of the neurotropic factors: NGF, CNTF, NT-3, BDNF. Brain Res Mol Brain Res
36:280-286.
Poon RT, Fan ST, Wong J (2001). Clinical implications of circulating angiogenic factors
in cancer patients. J Clin Oncol 19:1207-1225.
Qu Q, Perälä-Heape M, Kapanen A, Dahllund J, Salo J, Väänänen HK, Härkönen P
(1998). Estrogen enhances differentiation of osteoblasts in mice bone marrow
culture. Bone 22:201-209.
Quian D, Lin HY, Wang HM, Zhang X, Liu DL, Li QL, Zhu C (2004). Normoxic induction
of the hypoxic-inducible-factor-1α by interleukin-1β involves the extracellular
signal-regulated kinase 1/2 pathway in normal human cytotrophoblast cells. Biol
Reprod 70:1822-1827.
Redmer DA, Reynolds LP (1996). Angiogenesis in the ovary. Rev Reprod 1:182-192.
Ren L, Medan MS, Weng Q, Jin W, Li C, Watanabe G, Taya K (2005).
Immunolocalization of nerve growth factor (NGF) and its receptors (trkA and
p75LNGFR) in the reproductive organs of Shiba goats. J Reprod Dev 51:399-404.

80
Romero C, Paredes A, Dissen GA, Ojeda SR (2002). Nerve growth factor induces the
expression of functional FSH receptors in newly formed follicles of the rat ovary.
Endocrinology 143:1485-1494.
Rossant J, Howard L (2002). Signaling pathways in vascular development. Ann Rev
Cell Dev Biol 18:541-573.
Roy SK (2000). Regulation of transforming growth factor-β-receptor type I and type II
messenger ribonucleic acid expression in the hamster ovary by gonadotropins and
steroid hormones. Biol Reprod 62:1858-1865.
Saito H, Tsujitani S, Oka S, Kondo K, Ikeguchi M, Maeta M, Kaibara N (1999). The
expression of transforming growth factor-β1 is significantly correlated with the
expression of vascular endothelial growth factor and poor prognosis of patients
with advanced gastric carcinoma. Cancer 86:1455-1462.
Salas C (2002). Efecto del factor de crecimiento nervioso (NGF) en la proliferación y
secreción de esteroides en células de granulosa humana. Tesis para optar al
Título de Químico Farmacéutico. Anales de la Facultad de Ciencias Químicas y
Farmacéuticas, Universidad de Chile.
Salas C, Julio-Pieper M, Valladares M, Pommer R, Vega M, Mastronardi C, Kerr B,
Ojeda SR, Lara HE, Romero C (2006). Nerve growth factor-dependent activation of
trkA receptors in the human ovary results in synthesis of FSH receptors and
estrogen secretion. J Clin Endocrinol Metab 91:2396-2403.
Shao J, Jung C, Sheng H (2005). Prostaglandin E2 stimulates the beta-catenin/T cell-
factor dependent transcription in colon cancer. J Biol Chem 28:26565-26572.
Shibuya M (2001). Structure and function of VEGF/VEGF receptor system involved in
angiogenesis. Cell Struct Funct 26:25-35.
Shweiki D, Itin A, Neufeld G, Gitay-Goren H, Keshet E (1993). Patterns of expression of
vascular endothelial growth factor (VEGF) and VEGF receptors in mice suggest a
role in hormonally regulated angiogenesis. J Clin Invest 91:2235-2243.
Shih SC, Claffey KP (2001). Role of AP-1 and HIF-1 transcription factors in TGF-beta
activation of VEGF expression. Growth Factors 19:19-34.
Smeyne RJ, Klein R, Schnapp A, Long LK, Bryant S, Lewin A, Lira SA, Barbacid M
(1994). Severe sensory and sympathetic neuropathies in mice carrying a disrupted
Trk/NGF receptor gene. Nature 368:246-249.

81
Sofroniew MV, Howe CL, Mobley WC (2001). Nerve growth factor signaling,
neuroprotection, and neural repair. Annu Rev Neurosci 24:1217-1281.
Tacconelli A, Farina AR, Cappabianca L, Desantis G, Tessitore A, Vetuschi A, Sferra R,
Rucci N, Argenti B, Screpanti I, Gulino A, Mackay AR (2004). TrkA alternative
splicing: a regulated tumor-promoting switch in human neuroblastoma. Cancer Cell
6:347-360.
Valladares M (2003). Rol del factor de crecimiento nervioso (NGF) en la regulación de
la expresión del receptor de FSH en células de granulosa humana. Tesis para
optar al Título de Bioquímico. Anales de la Facultad de Química y Biología
Universidad de Santiago de Chile.
Verheul HMW, Jorna AS, Hoekman K, Broxterman HJ, Gebbink MFBJ, Pinedo HM (2000). Vascular endothelial growth factor–stimulated endothelial cells promote
adhesion and activation of platelets. Blood 96:4216-4221.
Wakefield LM, Winokur TS, Hollands RS, Christopherson K, Levinson AD, Sporn MB
(1990). Recombinant latent transforming growth factor beta 1 has a longer plasma
half-life in rats than active transforming growth factor beta 1, and a different tissue
distribution. J Clin Invest 86:1976-1984.
Wang T, Horng S, Chang C, Wu H, Tsai Y, Wang H, Soong Y (2002). Human chorionic
gonadotropin-induced ovarian hyperstimulation syndrome is associated with up-
regulation of vascular endothelial growth factor. J Clin Endocrinol Metab 87:3300-
3308.
Wang D, Huang HJ, Kazlauskas A, Cavenee WK (1999). Induction of vascular
endothelial growth factor expression in endothelial cells by platelet-derived growth
factor through the activation of phosphatidylinositol 3-kinase. Cancer Res 59:1464-
1464.
Wiesmann C, de Vos AM (2001). Nerve growth factor: structure and function. Cell Mol
Life Sci 58:748-759.
Wulff C, Wilson H, Wiegand SJ, Rudge JS, Fraser HM (2002). Prevention of thecal
angiogenesis, antral follicular growth, and ovulation in the primate by treatment
with vascular endothelial growth factor trap R1R2. Endocrinology 143:2797-2807.
Youngblood GL, Payne AH (1992). Isolation and characterization of the mouse P450 17
alpha- hydroxylase/C17-20-lyase gene (Cyp17): transcriptional regulation of the

82
gene by cyclic adenosine 3',5'-monophosphate in MA-10 Leydig cells. Mol
Endocrinol 6:927-934.
Yu Y, Flint AF, Mulliken JB, Wu JK, Bischoff J (2004). Endothelial progenitor cells in
infantile hemangioma. Blood 103:1373-1375.
Zapf-Colby A, Eichhorn J, Webster NJ, Olefsky JM (1999). Inhibition of PLC-gamma1
activity converts nerve growth factor from an anti-mitogenic to a mitogenic signal in
CHO cells. Oncogene 18:4908-4919.
Zeng G, Gao L, Birklé S, Yu RK (2000). Supression of ganglioside GD3 expression in a
rat F-11 tumor cell line reduces tumor growth, angiogenesis and vascular
endotelial growth factor production. Cancer Res 60: 6670-6676.
Zhang L, Yang N, Conejo Garcia JR, Mohamed A, Benencia F, Rubin SC, Allman D,
Coukos G (2002). Generation of a syngeneic mouse model to study the effects of
vascular endothelial growth factor in ovarian carcinoma. Am J Pathol 161:2295-
2309.

83
7. Anexos
Anexo 1: Resumen de los modelos estudiados en esta tesis Tabla 7.1. Resumen de los modelos evaluados durante esta tesis. Para cada uno de los modelos estudiados, se indican los antecedentes que apoyan su uso, y el aporte de este modelo al estudio. Cabe hacer notar que existe similitud en los eventos más importantes en el desarrollo ovárico en roedores y humanos (McGee cols. 2000), así como también en los principales cambios en la vascularización y la expresión de factores angiogénicos en este órgano durante el ciclo ovárico (Redmer y cols. 1996). Lo anterior hace compatible el trabajo con estas distintas especies, así como las conclusiones y/o proyecciones de los datos obtenidos.
Modelo Antecedentes que apoyan el uso de este modelo Aporte de este modelo
Cultivo de ovario de rata de 2 días
Este modelo responde al estímulo con NGF, produciéndose la diferenciación del ovario (expresión de receptores para FSH; Romero y cols. 2002)
Permite el estudio temporal del efecto de NGF en el tejido ovárico
Ratón NGF knockout
La ausencia de NGF en ratones afecta el desarrollo folicular temprano (Ojeda y cols. 2000)
Permite estudiar in vivo el efecto de la ausencia de NGF sobre los factores VEGF y TGFβ1
Ratón trkA knockout
La señalización vía trkA es necesaria para que NGF pueda participar en el proceso de la ovulación (Dissen y cols. 1996)
Permite estudiar in vivo el efecto de la carencia del receptor trkA sobre los factores VEGF y TGFβ1
Rata denervada
La denervación del ovario induce una acumulación local de NGF, con un 80% de incremento respecto a ovarios controles (Lara y cols. 1990b)
Permite un estudio del aumento local de NGF en el tejido ovárico in vivo, y los efectos de esta condición sobre los factores VEGF y TGFβ1 pero también sobre la vascularización ovárica.
Ratón que sobre expresa NGF
Los ovarios de estos ratones responden al aumento de NGF, el cual genera un mayor número de folículos antrales y una disminución de la fertilidad (Dissen y cols. 2004, resultados no publicados)
Permite el estudio de los efectos crónicos del aumento de NGF en el ovario sobre los factores VEGF y TGFβ1 y la vascularización ovárica.
Células de granulosa humana
Estas células responden a NGF in vitro (Salas 2002, Valladares 2003, Salas y cols. 2006), expresan y secretan VEGF y TGFβ1 al ser incubadas con factores de crecimiento o gonadotrofinas (Mulheron y cols. 1992, May y cols. 1995, Lee y cols. 1997, Martinez-Chequer y cols. 2003)
Facilita el estudio mecanístico, es decir, la señalización a través de receptores y vías transduccionales luego del estímulo con NGF
Células HUVEC
Para estudiar el rol de NGF en la angiogénesis del feocromocitoma, se estimularon células PC12 con NGF. Se transfirió este medio condicionado de PC12 a un cultivo de HUVEC, y se determinó que al neutralizar VEGF en ese medio, se inhibía la proliferación de HUVEC (Middeke y cols. 2000)
Permite determinar si los efectos de NGF sobre la vascularización ovárica fueron una consecuencia del aumento de VEGF

84
Anexo 2. Carta del Comité de Etica de la Facultad de Ciencias Químicas y Farmacéuticas

85
Anexo 3. Carta del Comité de Etica del Hospital Clínico Universidad de Chile

86
Anexo 4: Consentimiento informado para investigacion clinica
NOMBRE DEL PROYECTO: PARTICIPACIÓN DEL FACTOR DE CRECIMIENTO NERVIOSO EN LA REGULACIÓN DE LA ANGIOGÉNESIS OVÁRICA. INVESTIGADOR RESPONSABLE:
Bq. Marcela Julio Pieper. Tesista de doctorado. Laboratorio de Endocrinología y Biología de la Reproducción Hospital Clínico Universidad de Chile SUPERVISOR:
Dra. Carmen Romero Osses. Bioquímico. Laboratorio de Endocrinología y Biología de la Reproducción Hospital Clínico Universidad de Chile Mi médico tratante me ha explicado que, como parte de mi tratamiento, debo someterme a un procedimiento de aspiración folicular. Se me ha solicitado donar una parte del líquido folicular sobrante después de la aspiración, para un proyecto de investigación donde se está estudiando el mecanismo de la angiogénesis en el ovario normal y en cáncer ovárico epitelial. Estoy de acuerdo en participar en este estudio, y doy mi consentimiento para que utilicen este material, ya que de esta manera ayudo en el conocimiento científico, lo que a su vez en el futuro puede ser de gran ayuda para las mujeres con cáncer ovárico. Yo entiendo que: a) Cuando me practiquen la aspiración folicular, procedimiento que ha indicado el médico, como parte de
mi tratamiento, se obtendrán ovocitos y células de granulosa. Las células de granulosa, que normalmente son desechadas, se utilizarán para los análisis de este estudio, lo que no interfiere en nada con mi tratamiento. El que se utilice este material en este estudio no implica ningún riesgo para mí.
b) Cualquier pregunta que yo quiera hacer con relación a mi participación en este estudio deberá ser contestada por la Bioquímico Marcela Julio Pieper, tesista de Doctorado e Investigadora Responsable del Proyecto, o la Dra. Carmen Romero Osses, supervisora del proyecto.
c) Yo podré negarme a participar de este estudio en cualquier momento sin ser obligada a dar razones y sin que esto me perjudique en mi calidad de paciente o usuaria del Hospital Clínico de la Universidad de Chile.
d) Los resultados de este estudio pueden ser publicados, pero mi nombre o identidad no será revelado y mis datos clínicos y experimentales permanecerán en forma confidencial, a menos que mi identidad sea solicitada por ley.
e) Mi líquido folicular se utilizará única y exclusivamente en los estudios que estoy autorizando a través de este documento.
f) Mi consentimiento está dado voluntariamente sin que haya sido forzada u obligada. Si usted desea hacer alguna consulta acerca de este estudio, puede llamar a la Bq. Marcela Julio P., o a la Dra. Bq. Carmen Romero O. a los teléfonos 6788304-6788305. FIRMA DEL INVESTIGADOR PRINCIPAL FIRMA DEL PACIENTE O O MEDICO RESPONSABLE REPRESENTANTE LEGAL FECHA ………………………………………………………
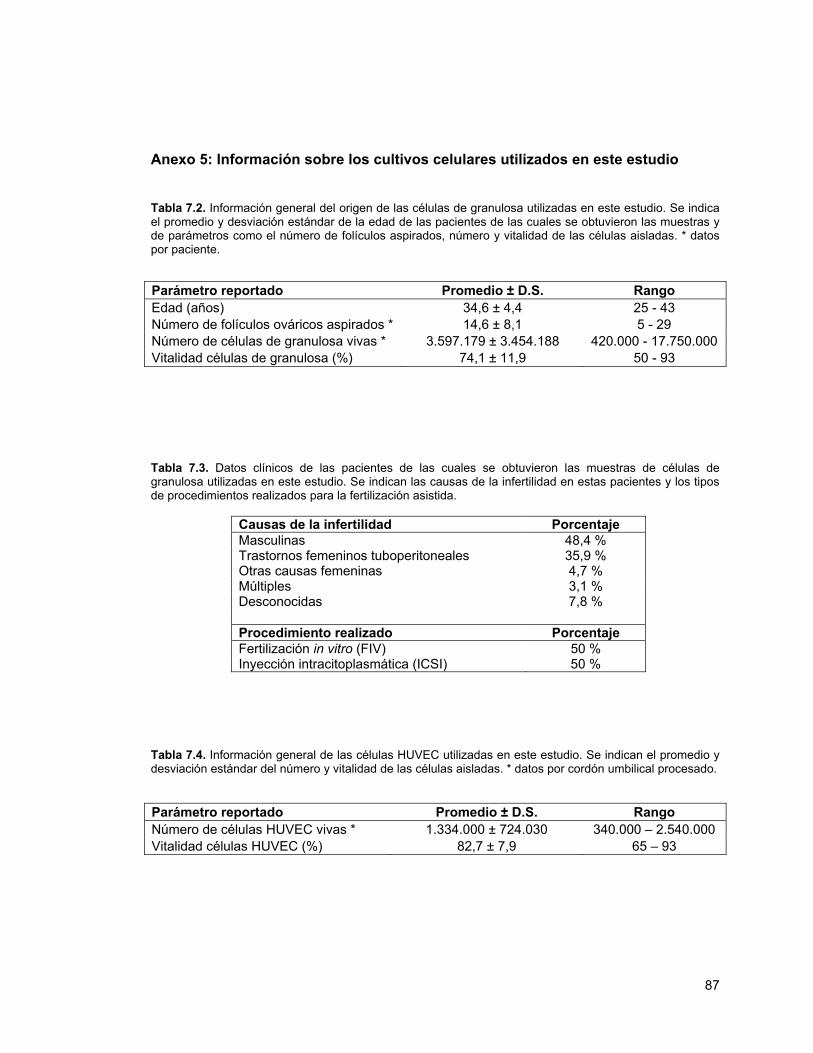
87
Anexo 5: Información sobre los cultivos celulares utilizados en este estudio Tabla 7.2. Información general del origen de las células de granulosa utilizadas en este estudio. Se indica el promedio y desviación estándar de la edad de las pacientes de las cuales se obtuvieron las muestras y de parámetros como el número de folículos aspirados, número y vitalidad de las células aisladas. * datos por paciente. Parámetro reportado Promedio ± D.S. Rango Edad (años) 34,6 ± 4,4 25 - 43 Número de folículos ováricos aspirados * 14,6 ± 8,1 5 - 29 Número de células de granulosa vivas * 3.597.179 ± 3.454.188 420.000 - 17.750.000 Vitalidad células de granulosa (%) 74,1 ± 11,9 50 - 93 Tabla 7.3. Datos clínicos de las pacientes de las cuales se obtuvieron las muestras de células de granulosa utilizadas en este estudio. Se indican las causas de la infertilidad en estas pacientes y los tipos de procedimientos realizados para la fertilización asistida.
Causas de la infertilidad Porcentaje Masculinas 48,4 % Trastornos femeninos tuboperitoneales 35,9 % Otras causas femeninas 4,7 % Múltiples 3,1 % Desconocidas 7,8 % Procedimiento realizado Porcentaje Fertilización in vitro (FIV) 50 % Inyección intracitoplasmática (ICSI) 50 %
Tabla 7.4. Información general de las células HUVEC utilizadas en este estudio. Se indican el promedio y desviación estándar del número y vitalidad de las células aisladas. * datos por cordón umbilical procesado. Parámetro reportado Promedio ± D.S. Rango Número de células HUVEC vivas * 1.334.000 ± 724.030 340.000 – 2.540.000 Vitalidad células HUVEC (%) 82,7 ± 7,9 65 – 93

88
Anexo 6: Marcadores de la presencia de células de granulosa en los cultivos estudiados
Figura 7.1. Marcadores positivos de células de granulosa en los cultivos estudiados. En A se muestra una inmunofluorescencia del receptor de FSH realizada en células de granulosa en cultivo. Se observa el núcleo en rojo, y el receptor de FSH, con una localización citoplasmática, en verde. También se detectó el mRNA para el receptor de FSH en los cultivos de células de granulosa (datos no mostrados, Valladares, 2003). Este receptor se expresa exclusivamente en las células de granulosa. Magnificación original 40X. Barra = 10 µm. En B se muestra el resultado de la determinación de estradiol en células de granulosa estimuladas con NGF. Los resultados se expresaron como porcentaje de cambio, ya que la secreción basal de estradiol estaba en un rango de 10 a 400 pg/ml. Nuevamente, las células de granulosa son las únicas células ováricas que tienen la capacidad de secretar estradiol. Los datos mostrados corresponden al promedio ± SEM de un n=12. * p < 0,01 de NGF 10 ng/ml vs Control y ≠ p < 0,01 NGF 100 ng/ml vs Control, respectivamente. Los datos presentados en A y B fueron adaptados de Valladares (2003) y Salas (2002), respectivamente.
A Estradiol secretado
0
50
100
150
200
250ControlNGF (10 ng/ml)NGF (100 ng/ml)
*≠
Condición de cultivo
Secr
eció
n de
Est
radi
ol(r
elat
ivo
al c
ontro
l)
B