universidad autónoma metropolitana iztapalapa148.206.53.84/tesiuami/uami10822.pdf · etiología y...
TRANSCRIPT

1
Universidad Autónoma Metropolitana
Iztapalapa
Ciencias Biológicas y de la Salud
“Evaluación del efecto causado por la Desnutrición de segundo y tercer grado enla serie leucocitaria y eritrocitaria de sangre periférica de ratas desnutridas”.
Proyecto de Investigación
Que para obtener la Licenciatura en
Biología Experimental
P R E S E N T A
Elsa Cervantes Ríos
Con la asesoría:
Dra. Alda Rocío Ortiz Muñiz
M. en C. Leonor Rodríguez Cruz
Diciembre, 2003.

2
I. Antecedentes
Desnutrición.
Conceptualmente, nutrición debe entenderse como el conjunto de funciones deasimilación y desasimilación de nutrientes que mantiene la vida, el crecimientocorporal y el desarrollo de las funciones orgánicas, (Cooper, 1966).
Por otro lado, la desnutrición es un estado patológico caracterizado por la faltade aporte adecuado de proteínas, calorías y/o de nutrientes acordes con lasnecesidades biológicas del organismo, que produce un estado catabólico ysistémico alterados pero potencialmente reversibles (Brunner, 1988).
El cuerpo humano se compone de moléculas en cambio constante, las cualesposeen un patrón característico de organización estructural y funcional cuyoequilibrio resulta de la relación entre la velocidad de síntesis (dependiente delaporte y utilización de los nutrientes) y la velocidad de degradación (Calzada,1998).
Durante cada momento de la existencia, la nutrición está íntimamente ligadacon el fenómeno biológico del crecimiento en un equilibrio que puede manifestarsepor el aumento, mantenimiento o disminución de la masa y del volumen, queconforman al organismo en relación con el momento previo, así como por laadecuación a las necesidades del cambio de forma, función y composicióncorporal.
Cuando la velocidad de síntesis es menor que la de degradación, la masacorporal disminuye, situación que no puede mantenerse por tiempo prolongado, yaque las disfunciones orgánicas que la acompañan son incompatibles con la vida.Por ello, es necesario comprender que la desnutrición daña las funciones celularesde manera progresiva, afectándose primero el depósito de nutrientes yposteriormente la reproducción, el crecimiento, la capacidad de respuesta alestrés, el metabolismo energético, los mecanismos de comunicación y deregulación intra e intercelular, y finalmente la generación de la temperatura, por loque de no resolverse a tiempo conduciría a la muerte del organismo (Calzada,1998).
Etiología y clasificación de la desnutrición.
La desnutrición puede ser definida como un estado de desequilibrio en el que elorganismo requiere una mayor cantidad de nutrientes de los que recibe. Ladesnutrición puede clasificarse en base a los factores que la causan, comoprimaria, secundaria o mixta (Gómez y col., 1956).

3
v Desnutrición primaria: se debe a la ingesta insuficiente de alimento, ya seaporque éste no se encuentre disponible o porque aunque existe no se consume.Por lo general tiene origen socioeconómico y cultural, así mismo se relaciona conel poder adquisitivo insuficiente.
v Desnutrición secundaria: se produce cuando el alimento no es consumido ono es debidamente utilizado por el organismo, porque existen condiciones que:
a) Interfieren con la ingestión: enfermedades neurológicas, motoras,psiquiátricas, estomatológicas, infecciosas y/o digestivas que producen anorexia ovómito.
b) Aumentan los requerimientos energéticos y/o las necesidades deregeneración tisular: infecciones crónicas, quemaduras, traumatismo múltiple,hipertiroidismo, insuficiencia cardiaca, etc.
c) Interfieren con la digestión y absorción: deficiencias enzimáticas digestivascongénitas o adquiridas, enfermedad celiaca, fibrosis quística del páncreas,insuficiencia hepática, alteraciones de vías biliares, pancreatitis, procesosinflamatorios crónicos de tubo digestivo como enfermedad de Crohn o colitisulcerativa crónica inespecífica, infecciones o infestaciones enterales, etc.
d) Dificultan la utilización: Diabetes Mellitus, neoplasias, enfermedadescongénitas del metabolismo de carbohidratos, proteínas o grasas; enfermedadespulmonares crónicas, cardiopatías congénitas, etc.
e) Aumentan la excreción: diarrea crónica, fístulas enterales o urinarias,problemas renales que afectan al glomérulo (síndrome nefrótico), a la funcióntubular o ambos (insuficiencia renal crónica), quemaduras extensas, eczemacrónico, etc.
Se ha determinado que la desnutrición se presenta con mayor frecuencia en losniños, ya que sus requerimientos de proteínas son cinco veces mayores que lasde los individuos adultos (Waterlow y col., 1960).
Tomando como referencia otros parámetros, el Doctor Federico Gómezpropuso, en 1956, una clasificación de la severidad de la desnutrición basada enel déficit de peso para la edad, a la cual se han realizado modificacionesposteriores, de tal manera que actualmente la intensidad puede clasificarse en:
a) Desnutrición de primer grado: el peso corporal corresponde de 76 al 90% delesperado para la edad y la talla.
b) Desnutrición de segundo grado: el peso corporal corresponde de 61 al 75%del esperado para la edad y la talla.

4
c) Desnutrición de tercer grado: el peso corporal es menor al 60% del esperadopara la edad y la talla (Gómez y col., 1956).
Dentro de este grado de severidad se encuentran dos tipos de desnutricióngrave: El Marasmo y Kwashiorkor.
La desnutrición tipo marasmo, se presenta cuando existen deficiencias en elaporte de proteínas y calorías, principalmente se presenta en niños menores de unaño, aún cuando también puede observarse en la edad adulta. Es producto de lafalta de alimentos y de los medios para adquirirlos, sus causas son:
1. Destete. El cese de la lactancia materna antes de que el niño sea capaz derecibir una dieta balanceada
2. Pobreza. La falta de alimento es sin duda un factor importante
Los síntomas dependen de las causas, de la gravedad y de la duración de ladesnutrición y pueden ser pérdida de la grasa corporal, estómago hinchado, ojoshundidos, cara de viejito, pérdida de peso, cambios en el color del cabello,diarreas frecuentes y una apariencia física de inanición severa (Vega,1999).
Por otro lado, kwashiorkor es un término aplicado a un niño que comesuficientes carbohidratos pero no tiene proteínas en su dieta, generalmente sepresenta en niños de 1 a 5 años (Behrman y col., 1986). La característica principales el edema, además se. presentan otros síntomas como son: que el cabellocambia de color y cae con facilidad al tocarlo, la piel siempre está reseca y serasga con facilidad, el niño no crece, tiene antebrazos muy delgados, se muestratriste, tiene anemia, presenta diarrea semilíquida, no tiene ganas de comer, tieneel estómago hinchado y apatía mental pronunciada (Sauerwein y col., 1997). Elanálisis clínico revela que se retiene gran cantidad de agua, así como pérdida depotasio y retención de sodio. Existe una reducción general de proteínasplasmáticas, sobre todo en la albúmina. (Manocha, 1972).
Hematopoyesis.
La hematopoyesis se define como la serie de elementos concatenados que seinician a nivel unicelular con autoduplicación, seguidos de diferenciación ymaduración, culminando con la producción de células sanguíneas funcionales. Losprincipales tipos de células sanguíneas proceden de una célula tallo pluripotencial,en el individuo adulto las células madre se localizan principalmente en la médulaósea roja (Sans y col., 2001).
Se considera a la diferenciación como la secuencia de hechos genéticos quepermiten a una célula sintetizar productos específicos, los que le confierenpotencialidad para determinada función. La maduración es la secuencia defenómenos bioquímicos y morfológicos iniciados por la diferenciación y que

5
confieren capacidad funcional a la célula. Tanto las células estromales como lashematopoyéticas tienen un precursor común, la célula totipotencialhematopoyética (CTH) (Fig.1). Las propiedades que definen a la CTH son sucapacidad de autoduplicación, la que resulta en progenies con las mismascaracterísticas de la CTH primitiva (unidad formadora de colonias de blastos UFC-B1), y la de dar origen a todas las células sanguíneas: eritrocitos, granulocitos(neutrófilos, eosinófilos y basófilos), monocitos/macrófagos y plaquetas, así comolos linfocitos T y B (Dzierzak, y col., 1995).
Hematopoyesis
CHT = célula totipotencial hematopoyética. UFC= unidad formadora de colonias. UFB = unidadformadora de brotes. BI= blastos. LM = linfoide mieloide. GEMM = granulocitos, eritrocitos, monocitos,megacariocitos. L = linfoide. EG = eritrocitos y granulocitos, megacariocitos. M = monocitos. G =granulocitos. Meg = megacariocitos. Bas = basófilos. Eo = eosinófilos. LB = linfocitos B. LT = linfocitosT. LGG = linfocitos grandes granulares.
Figura.1 La hematopoyesis es la serie de elementos concatenados que se inician a nivelunicelular con autoduplicación, seguidos de diferenciación y maduración, culminando con laproducción de elementos formes sanguíneos funcionales.
Los factores de crecimiento hemolinfopoyético son indispensables en el procesode formación de células sanguíneas y se dividen en interleucinas (IL) y factoresestimulantes de colonias (FEC). Las características generales de estas citocinas

6
incluyen: estructura glicoproteíca, actividad in vitro e in vivo a bajasconcentraciones, son producidas por diferentes tipos de células, generalmenteregulan más de una línea celular y muestran efecto aditivo o sinérgico con otrosfactores de crecimiento, modulan la expresión de genes que regulan la producciónde citocinas (Lee y col., 1993).
Sistema Inmunológico.
El término sistema inmunológico se refiere a aquella porción del tejido somáticoencargado de mantener la integridad del organismo por medio de la defensa delmismo. El sistema inmunológico evolucionó en el sentido de proteger a losvertebrados contra la infección por parte de microorganismos y parásitos mayores,de hecho casi cualquier macromolécula, siempre que sea extraña al receptor,puede inducir una respuesta inmunitaria; toda sustancia que pueda desencadenaruna respuesta inmunológica recibe el nombre de antígeno. Este sistema incluyecélulas fijas y móviles con capacidad de fagocitosis (fagocitos: granulocitos ymonocitos) o con propiedades inmunológicas (linfocitos), y órganos linfoides comoel timo, el bazo, nódulos linfáticos, amígdalas, placas de Peyer y acumulacionesde linfocitos y macrófagos (Alberts y col., 1998).
Existen dos clases principales de respuestas inmunológicas: (1) Inmunidadinnata (no específica), compuesta por los macrófagos y sus precursores,monocitos y neutrófilos (polimorfonucleares) y (2) Inmunidad adquirida (adaptativao específica) que incluye respuestas por anticuerpos (humoral) y respuestasmediadas por células (Field, 1996).
Inmunidad innata
Los granulocitos abarcan a los neutrófilos, basófilos y eosinófilos. Losneutrófilos provienen de la médula ósea, tienen un núcleo denso que tiene de dosa cinco lóbulos y un citoplasma que contiene gránulos azul-rosados (azurofílicos) overde-azules (Fig. 2); los gránulos se dividen en primarios (los cuales aparecen enlos promielocitos) y secundarios (que predominan en el neutrófilo maduro). Ambostipos de gránulos tienen un origen lisosomal, los primarios contienenmieloperoxidasa, fosfatasas ácidas e hidrolasas, mientras que los secundarioscontienen colagenasa y lactoferrina. Este tipo de células responden ante unaamplia variedad de situaciones inflamatorias. Su respuesta es frecuentemente noespecífica y se activan ante cualquier ruptura de las superficies endotelialesmigrando desde la sangre hasta el sitio dañado, esto es seguido por unadegranulación y liberación de su contenido lisosomal, estos materiales crean unambiente hostil para la viabilidad y replicación microbiana, su vida media es dealrededor de 10 horas (Hoffbraud y col., 1997).
Los eosinófilos y basófilos son subcategorías especializadas de granulocitos,los basófilos (Fig. 3) tienen una función antimicrobiana similar a la de losneutrófilos, además producen sustancias como la heparina y la histamina lascuales participan en procesos fisiológicos del organismo, este tipo celular se

7
encuentra raramente en la sangre periférica. Los eosinófilos (Fig. 4) son similaresa los neutrófilos, excepto porque sus gránulos tienen una coloración roja intensamuy característica (debido a que contienen proteínas básicas) y raramente tienenmas de dos lóbulos, participan en reacciones alérgicas, contra invasionesparasitarias y en la remoción de fibrina formada durante la inflamación (Hoffbraudy col., 1997).
Los monocitos tienen un núcleo no segmentado de perfil variable, puede seroval, redondo y/o con incisuras. El citoplasma es abundante de un color azul-gris ycontiene una fina granulación azurófila. El monocito abandona la sangre periféricay finalmente se instala en los tejidos en forma de histiocito y macrófago.Citoquímicamente los monocitos se caracterizan por su riqueza en esterasasinespecíficas, fosfatasa ácida, lisozima, beta glucuronidasa, catepsina yarilsulfatasa. (Fig. 5) (Sans y col., 2001). Los macrófagos son células quecontienen en su interior restos de material fagocitado, son muy ricos en hidrolasasácidas y esterasas inespecíficas en adaptación a su intensa capacidad digestiva,también tienen muramidasa, proteasas neutras, inhibidores enzimáticos y factorquimiotáctico de los neutrófilos; habitualmente no contienen peroxidasa (Sans ycol., 2001).Los monocitos permanecen muy poco tiempo en la médula ósea, salende ella para circular en sangre de 20 a 40 horas, después de este tiempo
Figura 2. Morfología deneutrófilos circulantes ensangre periférica.
Figura 3. Morfología de eosinófiloscirculantes en sangre periférica.
Figura 4. Característicasmorfológicas de basófilos desangre periférica.

8
abandonan la sangre para entrar a los tejidos en donde llevan a cabo su función(Sans y col., 2001).
La función de los neutrófilos y monocitos se puede dividir en tres fases:quimiotaxis, fagocitosis y digestión.
La quimiotaxis consiste en la movilización y migración de las células, el fagocitoes atraído hacia la bacteria o al sitio de la inflamación gracias a las sustanciasliberadas por los tejidos dañados. En la fagocitosis el reconocimiento del materialextraño al organismo (bacterias, células muertas, etc.) es debido a la opsonizaciónde éste con inmunoglobulinas. Los macrófagos también tienen un papel en lapresentación del antígeno al sistema inmune y secretan IL-1, TNF, IL-3, GM-CSF,G-CSF, M-CSF, IL-4 e IL-6. La destrucción y digestión ocurre por dos vías: En lavía oxígeno dependiente el superóxido, el peróxido de hidrógeno y otras especiesactivadas del oxígeno se generan partir de éste y NADPH. En los neutrófilos elperóxido de hidrógeno reacciona con mieloperoxidasas y haluro intracelular paramatar a las bacterias. En el mecanismo no oxidativo se genera una disminucióndel pH dentro de las vacuolas fagocíticas dentro de las cuales se liberan lasenzimas lisosomales, en adición la lactoferrina que se encuentra en los gránulosde los neutrófilos es liberada y actúa como un poderoso bacteriostático (Hoffbraudy col., 1997 ).
Inmunidad adquirida
Esta respuesta inmune depende principalmente de dos tipos de linfocitos: Lascélulas B y las células T. En los mamíferos, las células B derivan de la médulaósea, los linfocitos T también se generan en la médula ósea pero experimentan unproceso de maduración en el timo.
Las respuestas por anticuerpos implican la producción de proteínas que recibenel nombre de inmunoglobulinas. Los anticuerpos circulan por la sangre y penetranen otros fluidos corporales donde se unen específicamente al antígeno extrañoque ha desencadenado su producción. La unión del anticuerpo inactiva a virus ytoxinas bacterianas bloqueando su capacidad de unirse a los receptores de lascélulas diana, la unión de los anticuerpos también marca a los microorganismos
Figura 5. Morfología demonocitos de sangre periférica.

9
invasores para su destrucción, bien facilitando su ingestión por una célulafagocítica o mediante la activación del complemento.
La respuesta mediada por células, implica la producción de célulasespecializadas que reaccionan con los antígenos situados sobre la superficie deotras células del huésped que se encuentran infectadas. La célula reactiva puedematar estas células infectadas, que presenten proteínas de reconocimiento en susuperficie, en otros casos, la célula reactiva secreta señales químicas que activana los macrófagos para que destruyan a los organismos invasores. Los linfocitos seencuentran en grandes cantidades en la sangre, en la linfa y en órganos linfoidescomo el timo, los nódulos linfáticos y el apéndice (Alberts y col., 1998).
Morfológicamente los linfocitos de sangre periférica son una subpoblaciónheteromorfa. Se distinguen dos tipos: uno mayoritario formado por células depequeño tamaño, de núcleo con contorno algo irregular; cromatina condensada,una relación núcleo-citoplasma elevada y habitualmente agranulares. El segundotipo tiene una relación núcleo-citoplasma más baja, contiene varios gránulosazurófilos en su citoplasma y se le conoce como linfocito grande granular (LGG).La población de LGG constituye entre el 10 y el 15% de las célulasmononucleadas, son ricos en fosfatasa ácida y otras hidrolasas ácidas (Fig. 6)(Sans y col., 2001).
Los linfocitos B constituyen la minoría de la población linfocitaria en sangreperiférica (entre un 5 y 15%), tienen un núcleo redondo con cromatina condensadasin nucléolo aparente, citoplasma escaso, gránulos citoplasmáticos ocasionales yuna basofilia citoplasmática menor; posee numerosos monoribosomas ypoliribosomas dispersos a través del citoplasma, un aparato de Golgi pequeño,varias mitocondrias y un retículo endoplásmico escaso (Sans y col., 2001).
Una de la más importantes características del sistema inmunológico es sucapacidad para producir una respuesta altamente específica, la cual es unaconsecuencia de la presencia de un receptor particular en la superficie de loslinfocitos T y B, el sistema inmunológico contiene muchas clonas de linfocitos,cada una de estas clonas tiene un receptor que muestra una pequeña diferenciaen su estructura con respecto a las otras, y como consecuencia, solamente seunirá a un antígeno específico.
Figura 6. Morfología de linfocitoscirculantes en sangre periférica.

10
Después de la activación por ciertos antígenos, particularmente polisacáridos, lacélulas B proliferan y maduran para transformarse en células plasmáticas, lascuales secretarán inmunoglobulinas específicas, por otro lado, los linfocitos T nopueden unirse al antígeno libre en solución y requieren de células presentadorasdel antígeno, el cual es presentado en asociación con moléculas del complejomayor de histocompatibilidad. Aunque la primera interacción entre células T, B ypresentadoras del antígeno depende de la presencia del antígeno adecuado, unavez que se ha dado las células activadas liberan factores amplificadores, IL-1, IL-2y otros factores de crecimiento y proliferación, los cuales inducen proliferación ydiferenciación de las clonas activadas.
Desnutrición e inmunosupresión.
A pesar de los grandes avances que se han logrado en la prevención ytratamiento de la desnutrición calórico-proteica (DCP), ésta sigue constituyendo unproblema de salud en el mundo y en particular en América Latina. La OrganizaciónPanamericana de la Salud estima que entre el 10 y el 20% de los niños sufrendesnutrición moderada o severa (Nájera y col., 2001).
La relación entre el estado nutricional y el sistema inmune ha cobrado unaimportancia relevante desde hace muchos años, teniendo en cuenta que unaamplia variedad de nutrientes son esenciales para garantizar una adecuada salud,ya que tienen un impacto sobre la inmunocompetencia del huésped (Good yFernandes., 1980).
Por otro lado, se han hecho numerosos estudios respecto a este problema, y seha reportado que la desnutrición conlleva a una profunda inmunodeficiencia, lacual se ve reflejada en el decremento de la inmunidad celular, la humoral y la noespecífica (Lesourd y Mazari, 1997).
Desnutrición e inmunidad innata.
Monocitos/ Macrófagos y polimorfonucleares.
La desnutrición provoca alteraciones en la función de los monocitos, estudioshechos con ratas han demostrado que induce una baja secreción de IL-1 yconsecuentemente se reduce la respuesta febril a endotoxinas. De hecho lasfunciones de los monocitos están alteradas en animales con desnutrición,incluyendo la fagocitosis, la secreción de interleucinas y la producción del factor denecrosis tumoral (Lesourd y Mazari, 1997).
Por otro lado, la respuesta fagocítica se ve reducida en animales y humanos pordeficiencia de vitamina A vitamina C, vitamina E y Zinc (Scrimshaw y SanGiovanni,1997). Las funciones de los polimorfonucleares están alteradas en animalesexperimentales, especialmente la actividad antibacteriana, que se ve disminuidadebido a un decremento en la producción y liberación de la enzima superoxidasa(Lesourd, 1997). También se ha reportado una disminución en la función y número

11
de los neutrófilos por deficiencia de vitamina C en animales y humanos(Scrimshaw y SanGiovanni, 1997).
El proceso de fagocitosis se ve desfavorecido en la DCP, aunque la ingestiónde partículas por los fagocitos se muestra intacta de forma general, la destrucciónde éstas no es eficiente debido a una actividad metabólica reducida de las víasintracelulares involucradas en el proceso de fagocitosis. Con respecto a lainmunidad no específica, la desnutrición calórico-proteica tiene una fuerteinfluencia sobre ésta, pues se han detectado diversas alteraciones, entre lascuales se encuentran las siguientes:
a)La secreción de algunos componentes del complemento estaseveramente deprimida, pues C3, C4 y el factor B se encuentran en bajasconcentraciones.
b)Las funciones de los polimorfonucleares se ven alteradas,principalmente su actividad bactericida, la cual está relacionada con labaja producción y secreción de enzimas y radicales libres como elsuperóxido.
c)Las funciones de los monocitos se ven afectadas porque hay unabaja producción de IL-1.
Inmunidad específica.
La desnutrición actúa directamente sobre los linfocitos. En sangre periférica,los linfocitos T se encuentran poco diferenciados, hecho que puede ser medidopor el decremento de linfocitos maduros CD3+ y el incremento de linfocitosinmaduros CD2+ CD3- en niños y ancianos. (Good y Fernandes 1980).
Por otro lado, estudios con animales desnutridos han mostrado que loscambios en los linfocitos T son mayores o mas graves en sangre periférica queen otros tejidos linfoides, estos cambios se reflejan en el hecho de que lasfunciones de los linfocitos T se ven afectadas, se ha observado:
-Disminución en la proliferación celular-Reducción en la secreción y función de citocinas-El poder de respuesta de los linfocitos NK a agentes patógenos está
retardado y-La citotoxicidad celular es poco efectiva (Good y Fernandes, 1980).
Inmunidad humoral y producción de anticuerpos.
Aunque está menos alterada, la inmunidad humoral también se ve afectada, losefectos adversos de la DCP incluyen una baja producción de anticuerpos,decremento en la producción de IgA secretora (Reddy y col., 1976). Por otro lado,la respuesta a las vacunas también se ve afectada, la desnutrición está asociadacon una baja respuesta de los anticuerpos así como una baja afinidad de losanticuerpos secretados al antígeno.

12
Timo.
La inmunidad es en parte un producto del tejido linfoide del organismo, éste seencuentra en el timo, los ganglios linfáticos, las amígdalas, partes del bazo, deltracto gastrointestinal y la médula ósea.
La DCP causa una atrofia generalizada de los tejidos linfoides,fundamentalmente en niños, el timo (Chandra, 1982), el bazo, las amígdalas, lasplacas de Peyer y los nódulos linfáticos se ven seriamente afectados conevidencias histológicas de atrofia significativa en las áreas de linfocitos T de estostejidos (Good, 1981). La DCP provoca cambios morfológicos evidentes a nivel detejidos en órganos linfoides primarios como el timo con alteraciones en la síntesisde hormonas tímicas (Chandra, 1983), que muestran una atrofia significativa y conello, afectaciones en las regiones T de otros órganos linfoides periféricos. Estasmodificaciones morfológicas inciden en el proceso de maduración de linfocitos Tmaduros, aspecto que se ha asociado además con altos niveles de la enzimadesoxinucleotidil transferasa, considerada como un indicador del estado demaduración de las células T y con ello a un estado de inmunocompetenciadeficiente (Lesourd. y Mazari, 1997).
Además, Chandra reportó que una atrofia del timo reduce significativamente elporcentaje de linfocitos circulantes (Chandra, 1991). El decremento en lasfunciones del timo juega un papel muy importante en disfunción de la respuestainmune celular, esto se ve respaldado porque la incubación de linfocitos de niñoscon DCP con timulina reestablece la cantidad o el balance en el número de Tcirculantes (los linfocitos T inmaduros disminuyen y los linfocitos T madurosincrementan su número) (Lesourd. y Mazari, 1997).
Posiblemente el deterioro de la respuesta inmune mediada por células en losniños con DCP se deba a que las cuentas totales de linfocitos están disminuidas ymuy especialmente la proporción y número de linfocitos T circulantes (Chandra,1979), por el contrario Gorodezky y col. (1986) mostraron que el porcentaje delinfocitos presentes en los desnutridos tiende a ser ligeramente mayor que enniños eutróficos.
Desnutrición-mecanismos de defensa inmunológica
A la desnutrición se le relaciona con alteraciones en diferentes mecanismos dedefensa inmunológica, principalmente con aquellos que están mediados porcélulas La respuesta inmunológica humoral está menos afectada en losdesnutridos. Se ha observado una evidente asociación entre las infecciones y ladesnutrición (Woodward, 1998). Diversos autores han señalado que ladesnutrición es la principal causa de inmunodeficiencia en todo el mundo (Dai yMcMurray, 1998; Chandra, 1999).

13
No obstante, existen informes controversiales con relación a los elementosinvolucrados en la inmunosupresión. Un ejemplo representativo de lo anterior esque, en diversos trabajos, como se mencionó anteriormente, se ha observado unamarcada reducción en la proporción y número de linfocitos T circulantes en niñoscon desnutrición, principalmente los CD4 (Chandra, 1991). Al contrario, en otrosestudios se ha encontrado que en los desnutridos, la proporción de estos linfocitostiende a ser similar o incluso mayor que en los niños bien nutridos, esto pareceindicar que la alteración inmunológica está relacionada con una deficientefuncionalidad y no con una alteración en el número de células (Gorodezky y colet., 1986; González y col., 1997; Nájera y col., 2001). Lo anterior refleja que serequieren estudios adicionales para el mejor conocimiento de los cambiosinmunológicos asociados con la desnutrición.
II. Justificación.
La desnutrición es uno de los principales problemas de salud en el país ya queafecta a un porcentaje importante de la población y es, asociada con otrosfactores, responsable de la mayor parte de las muertes por causas evitables y deconsiderables daños a la salud de la niñez.
Para el estudio de las alteraciones asociadas con la desnutrición, los modelosen animales de laboratorio han sido de gran utilidad. Estos estudios tienendiversas ventajas.
El período de lactancia de una rata corresponde al período de infancia de unindividuo, los estudios realizados durante este período pueden aportar datos útilespara dilucidar los efectos de la desnutrición sobre los valores normales deleucocitos y eritrocitos durante esta etapa de la vida.
III. Objetivo.
Evaluar el efecto de la desnutrición de segundo y tercer grado sobre los valoresnormales de células leucocitarias y eritrocitarias en la sangre periférica de rataslactantes.
IV. Hipótesis.
Dado que la desnutrición produce un estado metabólico y sistémico alterado enlos organismos, los valores normales de las células leucocitarias y eritrocitarias ensangre periférica de ratas lactantes de la cepa Wistar registrarán alteraciones ensus valores normales.

14
V. Metodología
1. Inducción de desnutrición.
El método utilizado para inducir la desnutrición fue por competencia dealimento, el cual consistió en formar camadas de 15 a 17 ratas recién nacidas pornodriza, mientras que para obtener el grupo de ratas bien nutridas (BN), secolocaron de 6 a 7 crías por nodriza. A partir de ese momento el peso de las ratasse registró cada tercer día por un período de 21 días (hasta el destete de lascrías).
Concluido este tiempo, los datos registrados al día 21, fueron utilizados paracalcular el déficit de peso de cada individuo, y así determinar el grado dedesnutrición en el que las crías se encontraban. Bajo este criterio, se consideróque las ratas con un déficit de peso mayor de 10, pero menor de 25% sufríandesnutrición de primer grado, por otro lado, si las crías registraban un déficit mayorde 25 pero menor de 40% padecían una desnutrición moderada o de segundogrado (DN2), finalmente si las crías presentaban un déficit de peso mayor de 40%mostraban una desnutrición grave o de tercer grado (DN3) (Ortiz et. al., 1999).
Una vez hechos los cálculos, se procedió a la selección y marcaje de losindividuos de estudio, para pasar a la siguiente etapa.
2. Obtención de muestras sanguíneas de los tres grupos de estudio.
Para obtener sangre periférica de las crías, se le colocaba a cada una en unacámara de éter, antes de que la cría muriera, se le retiraba de la cámara y seprocedía a extraer la sangre por punción cardiaca con jeringas previamenteheparinizadas. Enseguida se retiraba la aguja de la jeringa y la sangre eracolocada en tubos de ensaye previamente marcados cuidando que ésta resbalepor las paredes del tubo sin hacer burbujas.
3. Procesamiento de las muestras obtenidas.
Las muestras de sangre periférica obtenidas de los tres grupos de estudio sesometían a los siguientes procedimientos:
a) Cuenta de eritrocitos.
Se hizo una dilución de las muestras de sangre 1:200, colocando en un tubo deensaye 199 µl de líquido de Dacie (Formaldehído al 40%, Citrato de sodio al 3%) +1µl de sangre periférica, después se procedió a tomar una alícuota de 100 µl quefue colocada en la cámara de Neubauer, la cual era colocada en una cámara

15
húmeda durante 10 minutos. Transcurrido este tiempo, se procedió a contar almicroscopio con el objetivo 40x a todas las células que se encontraran en loscinco cuadros pequeños de la cámara que están marcados con una R (Fig. 6).
Una vez contados los eritrocitos, se procedió a hacer los siguientes cálculospara obtener el número de células por mL:
No. de eritrocitos = No. de eritrocitos contados ÷5 X 200 X250
b) Cuenta de leucocitos.
Se hizo una dilución 1:20 de las muestras de sangre obtenidas de la siguientemanera: en un tubo de ensaye previamente marcado se agregaron 190 desolución de Turk (Ácido acético al 2%) + 10 µl de sangre periférica. Se tomó unaalícuota de 100 µl de la muestra y se colocó en la cámara de Neubauer a la que sele colocó en una cámara húmeda. Pasado este período de tiempo se procedió acontar al microscopio con el objetivo 40x a los leucocitos presentes en los cuadrosgrandes marcados con una W (Fig. 6).
Después se procedió a hacer los siguientes cálculos para determinar el númerode leucocitos por mL:
No. de leucocitos = No. de leucocitos contados ÷4 X 20 X 104
c) Cuenta diferencial de leucocitos.
El primer paso consistió en hacer un frotis de la muestra de sangre obtenidapara lo cual se requieren portaobjetos limpios, desengrasados y secos, de estosportaobjetos se eligió uno con bordes lisos que fue utilizado como extensor. En elextremo derecho de un portaobjetos se colocó una gota de sangre deaproximadamente 2 mm, con el extensor aplicado sobre el portaobjetos en unángulo de aproximadamente 450 se esperó a que la gota se extendiera a lo largodel borde de contacto del portaobjetos, que después se deslizó en sentidocontrario rápida y uniformemente.
Los frotis se dejaron secar a temperatura ambiente y posteriormente se tiñeroncon colorante de Wright (0.24g de colorante de Wright en 100 mL de metanol).
Este método de tinción se hizo de la siguiente manera: Se colocaron los frotissobre una gradilla de tinción y se cubrieron con colorante de Wright durante 15minutos, después sin eliminar el colorante, se adicionó con una pipeta pasteur 2-3volúmenes de Buffer salino de fosfato pH 6.8 dejando actuar la mezcla durante 10

16
minutos. Enseguida se lavó el portaobjetos bajo agua corriente y se dejó secar atemperatura ambiente.
Una vez seco el portaobjetos, se procedió a contar al microscopio con elobjetivo 40x un total de 200 leucocitos recorriendo la laminilla en zig-zag. Elnúmero de linfocitos, monocitos, neutrófilos, eosinófilos y basófilos se registró conayuda de un contador manual, se calcularon los valores porcentuales.
Una vez obtenidos los valores porcentuales, se procedió a encontrar losnúmeros absolutos por medio de la siguiente relación:
Números absolutos = (% subpoblación ÷100) X (Leucocitos totales)
44.. AAnnáálliissiiss eessttaaddííssttiiccoo..
Los datos obtenidos se analizaron mediante la prueba estadística de U-MannWhitney.
VI. Resultados.
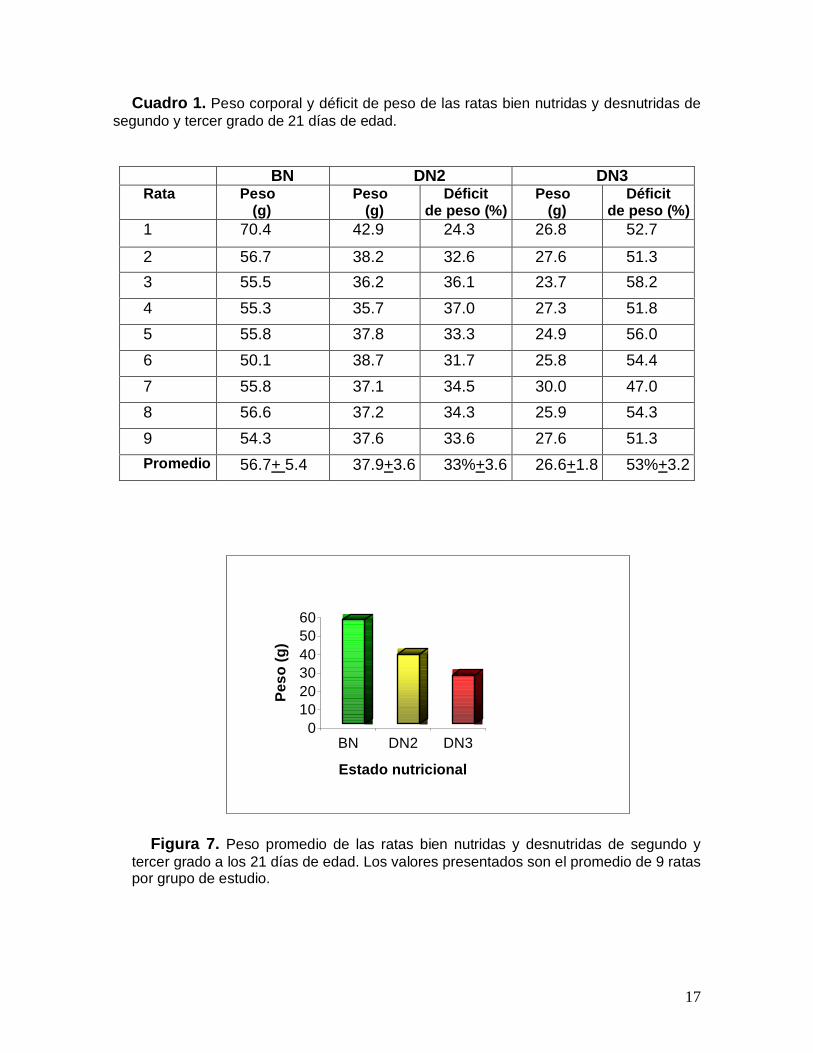
Se estudiaron un total de 27 ratas, 9 ratas de cada uno de los grupos deestudio. El grupo de las ratas BN mostró un peso promedio de 56.7 g + 5.4 sindéficit de peso, el grupo de ratas DN2 registró un peso promedio de 37.9 g + 3.6con un déficit de peso de 33%+ 3.6 y el grupo de ratas DN3 tuvo un pesopromedio de 26.6 g +1.8 con un déficit de peso de 53%+3.2 (Cuadro 1, Fig. 7).
Figura 6. Cuadrícula de la Cámara de Neubauer
17
Cuadro 1. Peso corporal y déficit de peso de las ratas bien nutridas y desnutridas desegundo y tercer grado de 21 días de edad.
Figura 7. Peso promedio de las ratas bien nutridas y desnutridas de segundo ytercer grado a los 21 días de edad. Los valores presentados son el promedio de 9 rataspor grupo de estudio.
BN DN2 DN3Rata Peso
(g)Peso (g)
Déficitde peso (%)
Peso (g)
Déficitde peso (%)
1 70.4 42.9 24.3 26.8 52.7
2 56.7 38.2 32.6 27.6 51.33 55.5 36.2 36.1 23.7 58.24 55.3 35.7 37.0 27.3 51.85 55.8 37.8 33.3 24.9 56.06 50.1 38.7 31.7 25.8 54.47 55.8 37.1 34.5 30.0 47.08 56.6 37.2 34.3 25.9 54.39 54.3 37.6 33.6 27.6 51.3Promedio 56.7+ 5.4 37.9+3.6 33%+3.6 26.6+1.8 53%+3.2
0102030405060
Peso
(g)
BN DN2 DN3
Estado nutricional
18
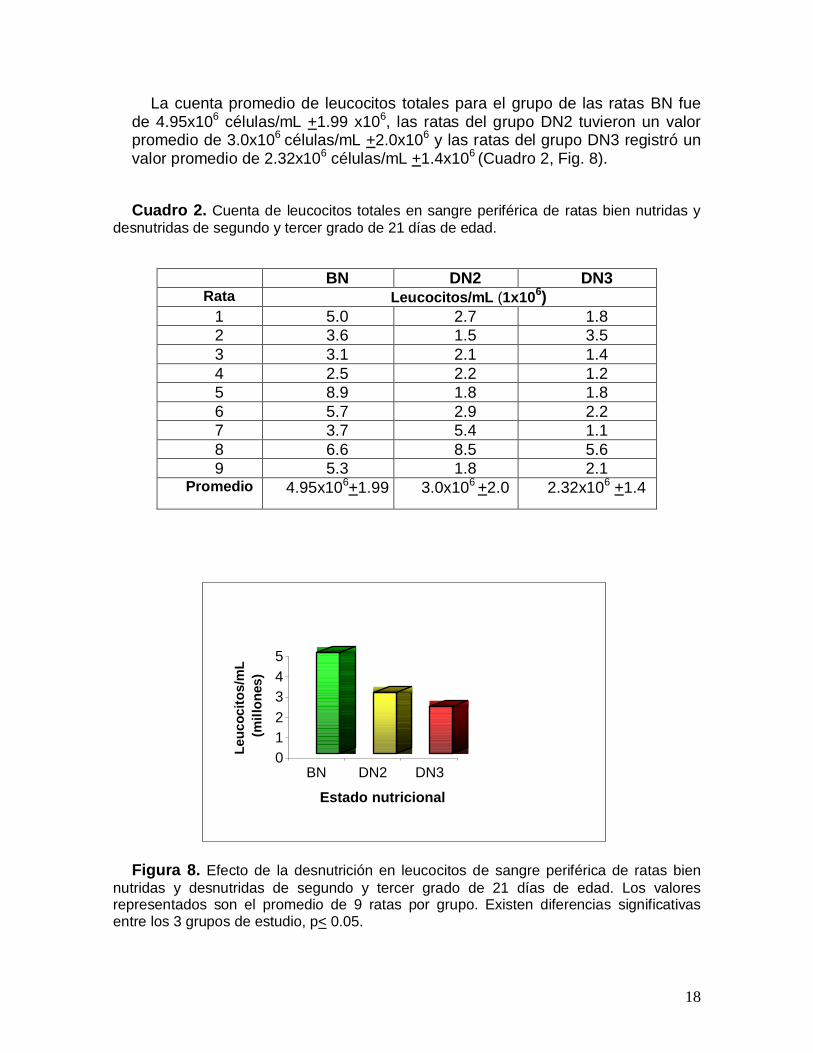
La cuenta promedio de leucocitos totales para el grupo de las ratas BN fuede 4.95x106 células/mL +1.99 x106, las ratas del grupo DN2 tuvieron un valorpromedio de 3.0x106 células/mL +2.0x106 y las ratas del grupo DN3 registró unvalor promedio de 2.32x106 células/mL +1.4x106 (Cuadro 2, Fig. 8).
Cuadro 2. Cuenta de leucocitos totales en sangre periférica de ratas bien nutridas ydesnutridas de segundo y tercer grado de 21 días de edad.
BN DN2 DN3Rata Leucocitos/mL (1x106)
1 5.0 2.7 1.82 3.6 1.5 3.53 3.1 2.1 1.44 2.5 2.2 1.25 8.9 1.8 1.86 5.7 2.9 2.27 3.7 5.4 1.18 6.6 8.5 5.69 5.3 1.8 2.1
Promedio 4.95x106+1.99 3.0x106 +2.0 2.32x106 +1.4
Figura 8. Efecto de la desnutrición en leucocitos de sangre periférica de ratas biennutridas y desnutridas de segundo y tercer grado de 21 días de edad. Los valoresrepresentados son el promedio de 9 ratas por grupo. Existen diferencias significativasentre los 3 grupos de estudio, p< 0.05.
012345
Leuc
ocito
s/m
L (m
illon
es)
BN DN2 DN3
Estado nutricional
19
Con respecto a la cuenta diferencial, para el grupo de ratas BN fue de: linfocitos82.9%+8.8, neutrófilos 8.5%+5.2, monocitos 7.6%+4.6, basófilos 0.3%+0.1 yeosinófilos 0.05%+0.02 (Cuadro 3). El grupo de las ratas DN2 registró lossiguientes valores: linfocitos: 84.9%+5.7, neutrófilos 8.1%+4.5, monocitos7.2%+2.0, basófilos 0.1%+0.2 y eosinófilos 0.1%+0.1 (Cuadro 4). Las ratas delgrupo DN3 mostraron los siguientes valores: linfocitos 84.5%+5.0, neutrófilos6.7%+3.0, monocitos 7.7%+3.3, basófilos 0.16%+0.2 y eosinófilos 0.1%+0.2(Cuadro 5). Mediante la prueba estadística de U- Mann Whitney se determinó queno existe diferencia significativa entre el porcentaje de los diferentes tipos deleucocitos entre los 3 grupos de estudio, p< 0.05.
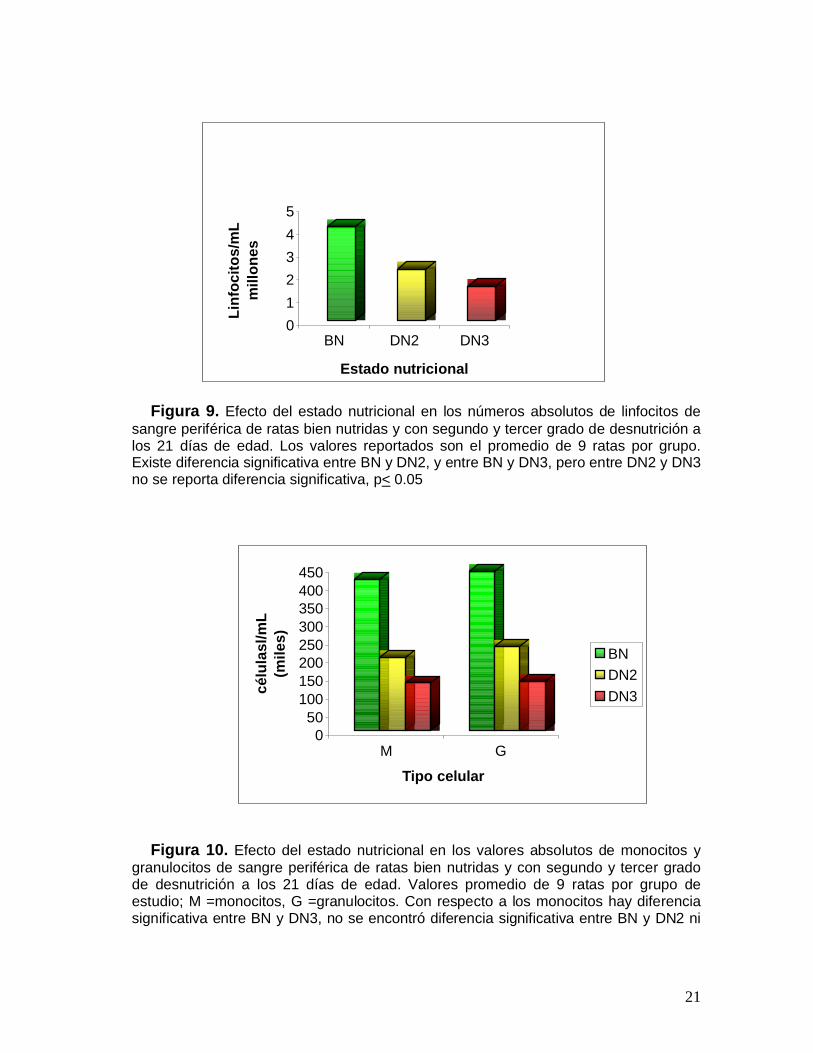
Al calcularse los números absolutos promedio para cada grupo se encontró quepara el grupo BN: linfocitos 4.95x106 células/mL, monocitos 0.42x106 células/mL ygranulocitos 0.44x106células/mL. Del mismo modo, para el grupo de las ratasDN2, los valores registrados fueron: linfocitos 2.24x106 células/mL, monocitos0.22x106 células/mL y granulocitos 0.23x106 células/mL. El grupo de las ratas DN3mostró los siguientes valores: linfocitos 1.48x106 células/mL, monocitos 0.13x106
células/mL y granulocitos 0.13x106 células/mL (Fig.9, 10,11).
Cuadro 3. Porcentaje de cada uno de los tipos de leucocitos de sangre periférica deratas bien nutridas a los 21 días de edad.
Rata Linfocitos(%)
Neutrófilos(%)
Monocitos(%)
Basófilos(%)
Eosinófilos(%)
1 79.5 12.0 6.5 2.0 02 77.5 9.0 13.0 0.5 03 93.5 1.5 0.5 0 04 71.0 20.0 9.0 0 05 73.5 14.5 11.5 0.5 06 92.5 2.0 5.5 0 07 84.5 8.0 7.0 0 0.58 80.5 7.5 12.0 0 09 94.5 2.0 3.5 0 0
Promedio 82.9+8.8 8.5+5.2 7.6+4.6 0.3+0.1 0.05+0.02

20
Cuadro 4. Porcentaje de cada uno de los tipos de leucocitos de sangre periféricade ratas con segundo grado de desnutrición a los 21 días de edad.
Rata Linfocitos(%)
Neutrófilos(%)
Monocitos(%)
Basófilos(%)
Eosinófilos(%)
1 72.5 18.5 8 0.5 02 89.5 4.5 6.0 0 03 85.0 9.0 6.0 0 04 84.5 7.5 7.5 0.5 05 83.0 10.0 6.5 0 1.06 83.5 6.0 10.5 0 07 81.5 9.0 10.0 0 08 93.0 3.0 4.0 0 09 87.5 5.5 7.0 0 0
Promedio 84.9+5.7 8.1+4.5 7.2+2.0 0.1+0.2 0.1+0.1
Cuadro 5. Porcentaje de cada uno de los tipos de leucocitos de sangre periférica deratas con tercer grado de desnutrición a los 21 días de edad.
Rata Linfocitos(%)
Neutrófilos(%)
Monocitos(%)
Basófilos(%)
Eosinófilos(%)
1 88.0 6.5 4.5 0.5 0.52 75.5 11.5 13.0 0 0.53 90.0 3.0 6.5 0.5 04 82.5 8.0 9.5 0 05 87.5 7.5 4.5 0.5 06 86.0 8.5 5.5 0 07 86.0 5.5 8.5 0 08 91.0 4.0 5.0 0 09 74.5 12.5 12.5 0 0
Promedio 84.5+5.0 6.7+3.0 7.7+3.3 0.16+0.2 0.1+0.2
Cuadro 6. Promedio de los números absolutos para los 3 grupos de estudioconsiderando a los neutrófilos, basófilos y eosinófilos como un solo tipo celular(granulocitos).
Estadonutricional
Linfocitos Monocitos Granulocitos
Células/mL (1x106)BN 4.59 0.42 0.44
DN2 2.24 0.22 0.23DN3 1.48 0.13 0.13
21
Figura 9. Efecto del estado nutricional en los números absolutos de linfocitos desangre periférica de ratas bien nutridas y con segundo y tercer grado de desnutrición alos 21 días de edad. Los valores reportados son el promedio de 9 ratas por grupo.Existe diferencia significativa entre BN y DN2, y entre BN y DN3, pero entre DN2 y DN3no se reporta diferencia significativa, p< 0.05
Figura 10. Efecto del estado nutricional en los valores absolutos de monocitos ygranulocitos de sangre periférica de ratas bien nutridas y con segundo y tercer gradode desnutrición a los 21 días de edad. Valores promedio de 9 ratas por grupo deestudio; M =monocitos, G =granulocitos. Con respecto a los monocitos hay diferenciasignificativa entre BN y DN3, no se encontró diferencia significativa entre BN y DN2 ni
050
100150200250300350400450
célu
lasl
/mL
(mile
s)
M G
Tipo celular
BNDN2DN3
012345
Linf
ocito
s/m
Lm
illon
es
BN DN2 DN3
Estado nutricional

22
entre DN2 y DN3. Los valores para granulocitos no muestran diferencia significativaentre ninguno de los 3 grupos de estudio, p< 0.05.
El recuento promedio de eritrocitos para el grupo BN fue de 65.15x106
células/mL + 18.15x106, las ratas del grupo DN2 mostraron un valor promediode 27.55x106 células/mL +12.42x106 y el grupo de ratas DN3 tuvo un valorpromedio de 28.20x106 células/mL +18.50x106 (Cuadro 7).
Cuadro 7. Cuenta de eritrocitos en sangre periférica de ratas bien nutridas ydesnutridas de segundo y tercer grado a los 21 días de edad.
BN DN2 DN3Rata Eritrocitos/mL (1x106)
1 87.5 16.3 30.82 85.6 35.6 28.73 59.2 31.0 33.54 73.5 17.5 35.15 53.0 52.2 69.06 82.4 26.2 8.27 58.4 31.1 17.78 52.8 28.1 24.29 34.0 10.0 6.6
Promedio 65.15+18.15 27.55+12.42 28.20+18.50
Figura 11. Efecto de la desnutrición en eritrocitos de sangre periférica de ratas biennutridas y desnutridas de segundo y tercer grado de 21 días de edad. Los valoresrepresentados son el promedio de 9 ratas para cada grupo de estudio. Se encontródiferencia significativa entre BN y DN2 y entre BN y DN3, para los grupos DN2 y DN3 nose registró diferencia significativa .p< 0.05
010203040506070
Eritr
ocito
s/m
L(m
illon
es)
BN DN2 DN3
Estado nutricional

23
VII. Discusión.
En el presente trabajo se realizó el recuento de leucocitos y eritrocitos desangre periférica de ratas bien nutridas (BN) y desnutridas (DN) de segundo ytercer grado. También se analizaron las proporciones de los diferentes tipos deleucocitos circulantes de sangre periférica de ratas BN y DN para determinaralguna posible alteración en ellas. Con el recuento diferencial se evalúa ladistribución y morfología de los glóbulos blancos, lo que proporciona informaciónmás específica respecto al estado inmunológico, que la obtenida utilizandoúnicamente el recuento de leucocitos totales (Hamilton y Rose, 1987).
Es importante hacer hincapié en que los estudios previamente realizados en loscuales se hace recuento diferencial de leucocitos, las ratas utilizadas son ratasadultas e incluso de algunas otras cepas, mientras que en este estudio seutilizaron ratas lactantes (21 días de edad) de la cepa Wistar.
Leucocitos totales
De acuerdo con los datos obtenidos, el número de leucocitos en el grupo deratas bien nutridas fue en promedio de 4.95x106, el cual concuerda con datosreportados previamente en un estudio en el que se evaluaron los valores debiometría hemática en ratas de la cepa Wistar de entre dos y tres meses de edad,donde se señala que el contenido de leucocitos en sangre, oscila entre 4x106 y12x106 células/mL (Ruiz y Fuentes, 1996). Con respecto a los valores obtenidosen este trabajo en ratas DN2 y DN3 éstos fueron de 3.24x106 y 2.32x106
células/mL respectivamente; es decir, se observó una disminución en el númerode leucocitos en las ratas desnutridas con respecto a las bien nutridas. Estosvalores son más bajos que los previamente reportados en rata (Ruiz y Fuentes,1996). Además están de acuerdo con estudios previos realizados con niños condesnutrición calórico-proteica (DCP) en los que las cuentas de leucocitos totalesson mas bajas que en niños sanos (Sauerwein y col., 1997).
En la sangre de las ratas desnutridas de tercer grado se presentaron los valoresmás bajos de leucocitos, lo que pudiera ser debido a posibles alteraciones en lamédula ósea que es donde son producidos los leucocitos; con respecto a estoBetancourt y col. (1992) observaron en ratas severamente desnutridasalteraciones en la producción de células en la médula ósea.
Por el contrario, un estudio realizado con niños con desnutrición calórico-proteica (DCP), se observó incremento en el número de leucocitos en estos niñoscon respecto a niños bien nutridos, sin embargo, este aumento se encontró dentrode los intervalos normales reportados para niños (González, 1997). Esta diferenciapodría deberse a que en el caso de los niños éstos se encuentran infectados porlo que existe un incremento en el número de leucocitos, lo que no sucede con lasratas de este estudio. Con relación a lo anterior resulta importante mencionar que

24
una de las ventajas de los estudios realizados con animales de experimentaciónes la facilidad de controlar factores extranutricionales, como las infecciones queson muy frecuentes en los niños.
Cuenta diferencial (Porcentajes)
Los datos de la cuenta diferencial de leucocitos: linfocitos, monocitos,neutrófilos, basófilos y eosinófilos, mostraron valores similares entre las ratas biennutridas y las ratas con desnutrición moderada y severa. Estos datos no muestrandiferencia entre cada uno de los grupos de ratas estudiados y concuerdan con unestudio previo realizado con ratas de la cepa Lewis (250 – 350 g), en el cualreportan que la proporción de linfocitos: 72-93% con una media de 84%,monocitos: de 1.0-3% con una media de 3%, neutrófilos: 6% a 20% con una mediade 11%, eosinófilos 1%-4% con una media de 2% y basófilos de 0-1% (Ruiz yFuentes,1996). También otro estudio realizado con ratas adultas de la cepa Wistaren el cual incluyen a los neutrófilos, basófilos y eosinófilos como un solo tipocelular (granulocitos), reportan valores para estas ratas de: granulocitos 13.6%,monocitos: 5.8% y linfocitos 80.6% (Peakman y col.,1993).
El porcentaje de monocitos determinado en este estudio fue de 7.6%, el cual seencuentra dentro del intervalo reportado para ratas (1-8%) (Ruiz y Fuentes,1996);sin embargo si se comparan valores promedio reportados previamente de 3% y de5.8%, con un intervalo de 5.2%-7.1% encontrado en un estudio realizado porcitometría de flujo en ratas adultas de la cepa Lewis (Pekman y col,1993), seobserva una disminución en el porcentaje de monocitos, lo que podría deberse aque en este caso las ratas estudiadas son ratas lactantes de 21 días, a diferenciade Ruiz y Fuentes (1996) que utilizaron ratas adultas de 2 a 3 meses. También enhumanos se ha encontrado que el promedio de monocitos es mayor en neonatos ydisminuye al pasar a la etapa adulta (Billove y col., 1997).
Con respecto a los linfocitos en las ratas BN se observó 82.9% de las células deeste tipo celular, este valor se encuentra dentro del intervalo previamentereportado para ratas adultas que es entre 73% y 92% con una media de 84%(Ruíz,1996). Sin embargo, el porcentaje de linfocitos reportado en el presenteestudio es ligeramente mayor al encontrado por Peakman y col. (1993) en el cualel valor promedio es de 80.6% con un intervalo de 79.1 a 82.0%, esta pequeñadiferencia podría deberse a que estos datos fueron obtenidos mediante uncitómetro de flujo a diferencia del presente estudio y del de Ruíz (1996) en loscuales se realizó el recuento mediante tinción con colorante de Wright y conmicroscopio óptico.
Al analizar la proporción de linfocitos presentes en ratas desnutridas se observóque las DN2 registraron 84.9% y en las DN3 fue de 84.5%, estos valores coincidencon el 84% encontrado por Ruiz (1996), por el contrario se encuentra ligeramentemayor a los valores proporcionados por Peakman y col. (1993) de 79.1-82.0% delinfocitos en ratas adultas.

25
Con respecto al porcentaje de neutrófilos en las ratas BN se observó un 8.7%de este tipo celular, estos datos se encuentran dentro del intervalo (6-20%)observado por Ruiz (1996); de la misma manera los datos observados para DN2 yDN3 (8.1% y 6.7% respectivamente) entran dentro del intervalo de valoresnormales reportados por Ruiz (1996).
Cuenta diferencial (Números absolutos)
El recuento diferencial es el número relativo de leucocitos de cada tipo en lasangre. Al multiplicar la cifra porcentual de cada tipo por el recuento total deleucocitos de cada especie celular, se obtienen datos que proveen un diagnósticomás exacto respecto a la presencia de alteraciones en la proporción de célulasblancas (Hamilton y Rose, 1987). Por lo anterior, se calculó el número absoluto decada uno de los tipos de células blancas en los tres grupos de estudio.
El número absoluto de linfocitos en las ratas bien nutridas fue de 4.09x106
células/mL y se observó disminución significativa (P<0.05) en las ratas desnutridasde segundo y tercer grado, 2.24x106 células/mL y 1.48x106 células/mL,respectivamente. Este descenso en el contenido de linfocitos en las ratasdesnutridas concuerda con datos previos los que mostraron que niños condesnutrición calórico-proteica presentaron disminución en las cuentas totales delinfocitos, muy especialmente la proporción y número de linfocitos T circulantes(Chandra, 1979).
Con respecto al número absoluto de granulocitos (neutrófilos, eosinófilos ybasófilos) el valor para las ratas BN fue de 4.4 X 105 y se observó una disminuciónen el grupo de ratas DN2 de 2.3 X 105 y las DN3 de 1.3 X 105, sin que existadiferencia estadísticamente significativa. En un estudio previo, realizado con ratasdesnutridas, se analizó sangre periférica mediante citometría de flujo y sedeterminó que no existió alteración en la proporción de granulocitos encomparación con las ratas bien nutridas (Ortiz y col., 1999).
Al analizar el número absoluto de monocitos en los tres grupos de ratas seobservó que existe disminución en las ratas DN2 (0.02x106 células/mL) y en lasratas DN3 (0.13x106 células/mL) con respecto a las BN (4.17x106 células/mL) ysolamente se observó diferencia significativa entre las ratas BN y las DN3.
Eritrocitos.
El recuento promedio de eritrocitos para el grupo BN fue de 65.15x106
células/mL + 18.15x106, las ratas del grupo DN2 mostraron un valor promedio de27.55x106 células/mL +12.42x106 y el grupo de ratas DN3 tuvo un valor promediode 28.20x106 células/mL +18.50x106. En estudios previos se ha demostrado que

26
las cuentas de eritrocitos son mas bajas en niños desnutridos que en niños sanos(Sauerwein y col.1997).
La cuantificación del número de leucocitos totales y de eritrocitos totalesevidenció una disminución en estos valores en ratas desnutridas con respecto alas bien nutridas. Es importante señalar que en las ratas con desnutrición desegundo grado o moderada (DN2) se observaron cambios similares a losencontrados en las ratas con desnutrición severa (DN3). Se considera que estasalteraciones se pueden deber a diversos factores, ya que la desnutrición es unsíndrome capaz de generar diversos problemas.
La hematopoyesis se encuentra regulada y, en una persona sana, por logeneral las cantidades de células nuevas que se forman son proporcionales a lascantidades de células viejas que se pierden o mueren. Es conveniente anotar queel tiempo de vida de las diferentes células sanguíneas es muy variable y puede irdesde horas (neutrófilos) o días (eritrocitos) hasta 20 a 30 años para algunoslinfocitos. Los mecanismos que regulan la hematopoyesis dependen de variosfactores. El primero es el control en la producción de las diferentes citocinas queestimulan la médula y que son producidas por las células del estroma. El segundoes el control simultáneo en la producción de las citocinas estimulantes de lamédula que son de origen exógeno y derivan de macrófagos y linfocitos Tactivados. El tercero consiste en la regulación en la expresión de los receptorespara las citocinas hematopoyéticas. Finalmente, el cuarto es el control de laspoblaciones de células maduras mediante la estimulación de la muerte celularprogramada y el inicio de la apoptosis (Dzierzak y Medvinsky, 1995).
En diversas circunstancias fisiológicas y/o patológicas, la hematopoyesis puededisminuir o bien incrementarse para aumentar el número de algunas poblacionesde células sanguíneas. La desnutrición puede modificar la producción de lascélulas sanguíneas y, al variar alguno de los cuatro valores mencionados,provocan cambios en el contenido de tejido linfoide disponible. Un aspectoadicional por estudiar es definir si todos los mecanismos que regulan lahematopoyesis están alterados de igual forma o si alguno de ellos es de mayorrelevancia con relación a la deficiencia nutricional.
Estudios previos han demostrado que la desnutrición calórico-proteica (DCP)provoca alteraciones en la respuesta inmune en niños (González y col., 1997), yque ésta es la mayor causa de inmunosupresión en el mundo (Field, 1996). Sinembargo, existen informes controversiales con relación a los elementosinvolucrados en la inmunosupresión Un ejemplo representativo de lo anterior esque, en diversos trabajos se ha observado una marcada reducción en laproporción y número de linfocitos T circulantes en niños con desnutrición,principalmente los CD4 (Chandra, 1991). Al contrario, en otros estudios se haencontrado que en los desnutridos, la proporción de estos linfocitos tiende a sersimilar o incluso mayor que en los niños bien nutridos, esto parece indicar que laalteración inmunológica está relacionada con una deficiente funcionalidad y no conuna alteración en el número de células (Gorodezky y col., 1986; González y col.,

27
1997; Nájera y col., 2001). Lo anterior refleja que se requieren estudiosadicionales para el mejor conocimiento de los cambios inmunológicos asociadoscon la desnutrición.
Los datos obtenidos en el presente estudio indican una disminución en losvalores de leucocitos totales en los grupos experimentales, lo que sugiere que laalteración en la respuesta inmunológica debida al estado de desnutrición en quese encuentran las ratas, está relacionada con la deficiencia en número de células,resulta interesante estudiar en un futuro si hay alteraciones en la funcionalidad deestas células. Aquí también es esencial mencionar que la mayoría de los trabajosrealizados se han abocado a estudiar la desnutrición grave o de tercer grado. Sinembargo, se considera importante evaluar adicionalmente la desnutriciónmoderada, ya que su frecuencia es alta y los probables cambios que se presentenpodrían contribuir al agravamiento de esta condición. Los datos obtenidos en estetrabajo indican que es esencial profundizar en el estudio de las alteraciones quese presentan en este grado de desnutrición.
VIII. Conclusiones.
Con base en los resultados obtenidos, se concluye que:
a) Los números totales de leucocitos y eritrocitos en las ratas bien nutridasestán dentro del intervalo de valores normales reportados para ratas adultas de lacepa Wistar.
b) Las ratas desnutridas experimentalmente durante la lactancia (DN2 y DN3),registran valores más bajos, lo cual podría estar relacionado directamente con unadeficiencia en la respuesta inmunológica.
c) Los valores obtenidos de leucocitos y eritrocitos en los organismos condesnutrición de segundo grado y tercer grado, descienden conforme seincrementa el déficit de peso.
d) El valor absoluto de linfocitos, monocitos y granulocitos fue mayor en lasratas con nutrición adecuada y descendió conforme aumentó el déficit de peso.
e) Los resultados del presente trabajo reafirman que para el análisis de lasprobables alteraciones en las células sanguíneas, es necesario registrar ademásde los porcentajes los números absolutos de cada tipo celular.

28
IX. Bibliografía.
z Alberts, B. y Col. (1998). Biología molecular de la célula. Segunda edición.Ed. Omega, S.A.pp 1066-1072.
z Berhman R., Vaughm V. y Nelson, W. (1986). Tratado de pediatría. 9aedición. Editorial Iberoamericana. México, pp 139-190.
z Betancourt, M., Ortiz, R. Y González, C. (1992). Proliferation index in bonemarrow cells from severely malnourihed rats during lactation. Mutat. Res.283:173-177.
z Brunner, L. S. (1988). Enfermería Médico Quirúrgica. Sexta Edición, EdInteramericana. pp 71-73.
z Calzada,L.R.(1998). Fundamentos Fisiopatológicos. Primera Edición, Ed.McGraw-Hill Interamericana.
z Cooper,B.(1966). Nutrición y dieta. Décimo cuarta edición, Ed.Interamericana. pp 600.
z Chandra, R.K. (1991). Nutrition and Inmunity: lesson from the past and newinsights into the future. Am. J Clin Nutr 53: 1087-101.
z Chandra, R. K. (1979). T y B lymphocyte subpopulation and leukocyteterminal deoxynucleotidyl transferase malnutrition. Acta Pediat, Scad. 68:841-845.
z Chandra, R. K. (1982). Inmunología de los transtornos nutricionales.Editorial El Manual Modeno. México, D. F. Pp 18-48.
z Chandra, R. K. (1983). Numerical and functional deficiency in T helper cellsin protein energy malnutrition. Clin. Exp. Immunol. 51: 126-132.
z Chandra, R. K. (1991). 1990 Mc Collum Award Lecture. Nutrition andimmunity: lessons from past and new insights into the future. Am. J. Clin.Nutr. 53: 1087-1101.
z Chandra R.K. 1999. Nutrition and immunology: from the clinic to cellularbiology and back again. Proc Nutr Soc. 58: 681-683.
z Dai G. y McMurray D. 1998. Altered cytokyne production and impairedantimycobacterial immunity in protein-malnourished guinea pigs. Infectionand Immunity. 66: 3562-3568.

29
z Dzierzak, E. and Medvinsky, A. (1995). Mouse embrionic hematopoiesis.TIG 11(9).
z Field, C. (1996). Using inmunological techniques to determine the effect ofnutrition on T-cell function. Can J. Physiol. Pharmacol. 74:769-777.
z Good R., West and Fernandes G. (1980). Nutritional modulation of inmuneresponses. Federation Proc. 39:3098-3104.
z Good, R. A. (1981). Nutrition and immunity. Clin. Immunol. 1: 3-8.
z Gomez, F., R. R. Galván, S. Frerenk, and J. Vazquez. (1956). Mortality insecond and third degree malnutrition. J. Trop. Pediatr. 2:77-83.
z González C., Rodríguez L. Bonilla E. Betancourt M., Siller N., Zumano E.And Ortiz R. (1997). Electrophoretic analysis of plasmatic and lymphocytessecreted proteins in malnourished children. Med. Sci. Res. 25: 643-646.
z Gorodezky C., Betancourt M., Salazar-Mallén M., Amezcua M. And CraviotoJ. (1986). Blast tranformation inmexican malnourished children. Rev. LatAmer. Microbiol. 28: 217-220.
z Hamilton, H. K. y M. B. Rose. (1987). Diagnóstico Clínico. Ed.Interamericana. México.
z Hoffbraud, A. L.,Pettit, J. E. (1997). Essential haematology. Ed. Wolfe. pp141-149,161-165,174.
z Lee, G. R., et. al. (1993). Clinical hematology. Novena edición. Ed. Ley &Febiger. pp 48-50, 135, 248.
z Lesourd B. y Mazari L. (1997). Inmune responses during recovery fromprotein-energy malnutrition. Clinical Nutrition. 16:37-44.
z López L. C. (1992). Enfermería Sanitaria. Primera Edición Ed.Interamericana. pp 204-206.
z Manocha. (1972). Malnutrition and retarded human development. Publish.C. C.Thomas. N. Y. pp 350.
z Nájera O., González C., Toledo G., Cortés E., López L., Betancourt M. yOrtiz, R. 2001 (a). Early activation of T, B and NK lymphocytes in infectedmalnourished and infected well-nourished children. J. Nutr. Immunol. 5: 85-97.

30
z Nájera, O., et. al. (2001). CD45RA and CD45RO isoforms in infectedmalnourished and infected well-norished children. Clin Exp Immunol;126:461-465.
z Ortiz, R., Cortés, E., Pérez, L. (1999). Desnutrición experimental porcompetencia de alimento durante la lactancia y su efecto sobre la fórmulaleucocitaria en sangre periférica. La revista hispanoamericana 4: 33-39.
z Peakman M. Tredger J.M. et. al. (1993). Analysis of peripheral bloodmononuclear cells in rodents by three-colour flow cytometry using a smallvolume lysed whole blood technique. Journal of Immunological Methods.158:87-94.
z Reddy, V, Raghuramulu N, Bhaskaram C. (1976). Secretory IgA in proteincalorie malnutrition. Arch Dis Child; 51:871-874.
z Ruiz, E., Fuentes A. (1996). Evaluación de la biometría hemática en la ratade laboratorio. Rev. Méx. Patol. Clin. 43:80-85.
z Sánchez, A. V. (2000). Inmunocompetencia en la desnutrición proteico-energética
z Sans-Sabrafen, J. y Col. (2001). Hematología clínica. Cuarta edición. Ed.Hartcourt. Madrid, España. pp 18-33.
z Sauerwein,. R., Mulder J. y Col. (1997). Inflammatory mediators in childrenwith protein energy malnutrition. Am. J. Clin. Nutr. 65: 1534-1539.
z Scrimshaw, N. y SanGiovanni, P. (1997). Synergism of nutrition, infectionand inmunity; an overview. Am. J. Clin. Nutr. 66:4645-4775.
z Vega, F. L. (1999). Hitos conceptuales en la historia de la desnutriciónproteico-energética. Salud pública de México 41:326-333.
z Woodward B. 1998. Protein, calories, and immune defenses. Nutr. Rev. 56:S84-S92.