transporte de membrana -...
TRANSCRIPT

Transporte
de
Membrana

Tipos de Transporte
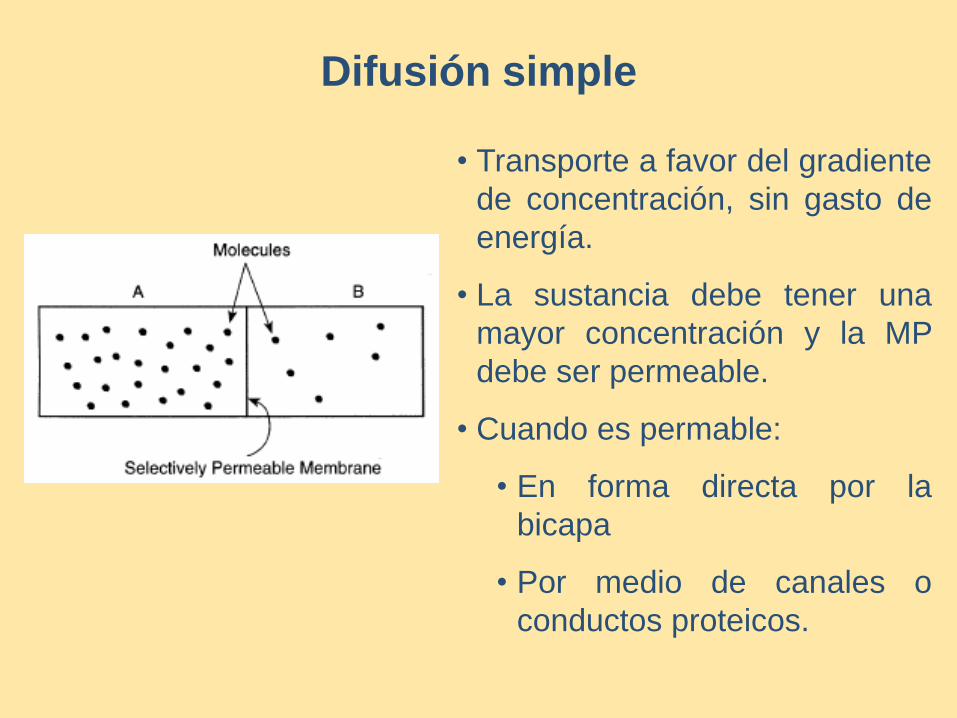
Difusión simple
• Transporte a favor del gradiente
de concentración, sin gasto de
energía.
• La sustancia debe tener una
mayor concentración y la MP
debe ser permeable.
• Cuando es permable:
• En forma directa por la
bicapa
• Por medio de canales o
conductos proteicos.
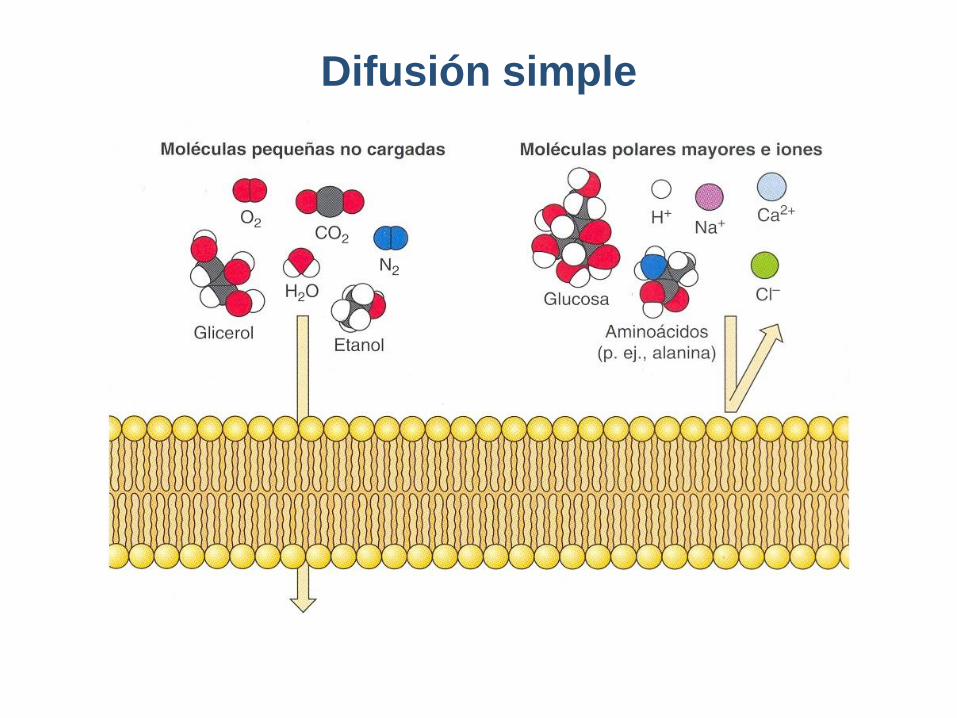
Difusión simple
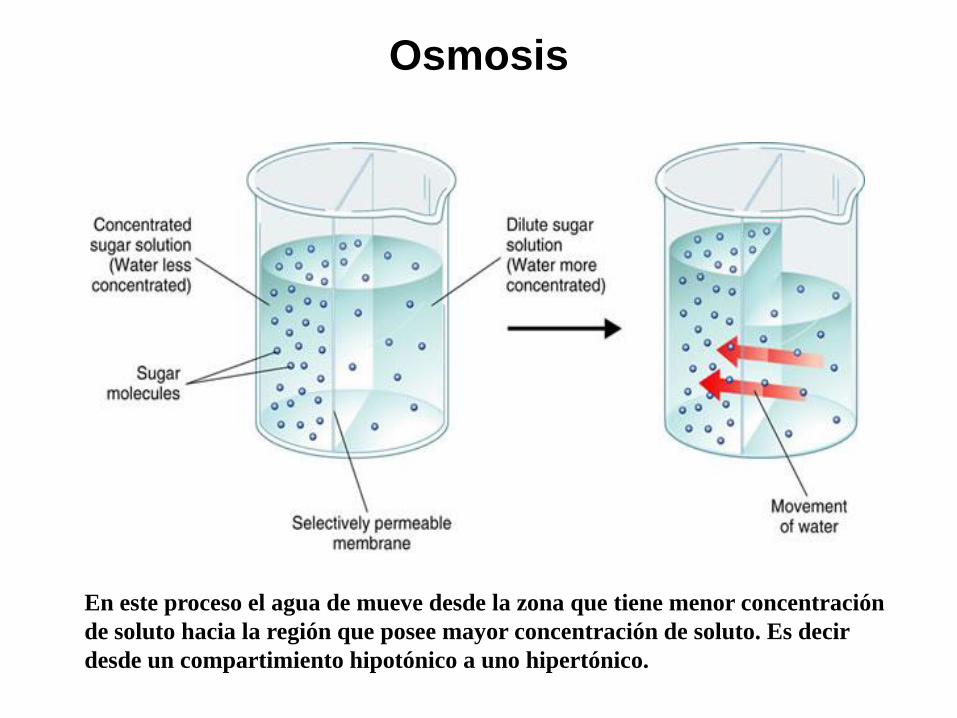
Osmosis
En este proceso el agua de mueve desde la zona que tiene menor concentración
de soluto hacia la región que posee mayor concentración de soluto. Es decir
desde un compartimiento hipotónico a uno hipertónico.

Osmosis

Osmosis
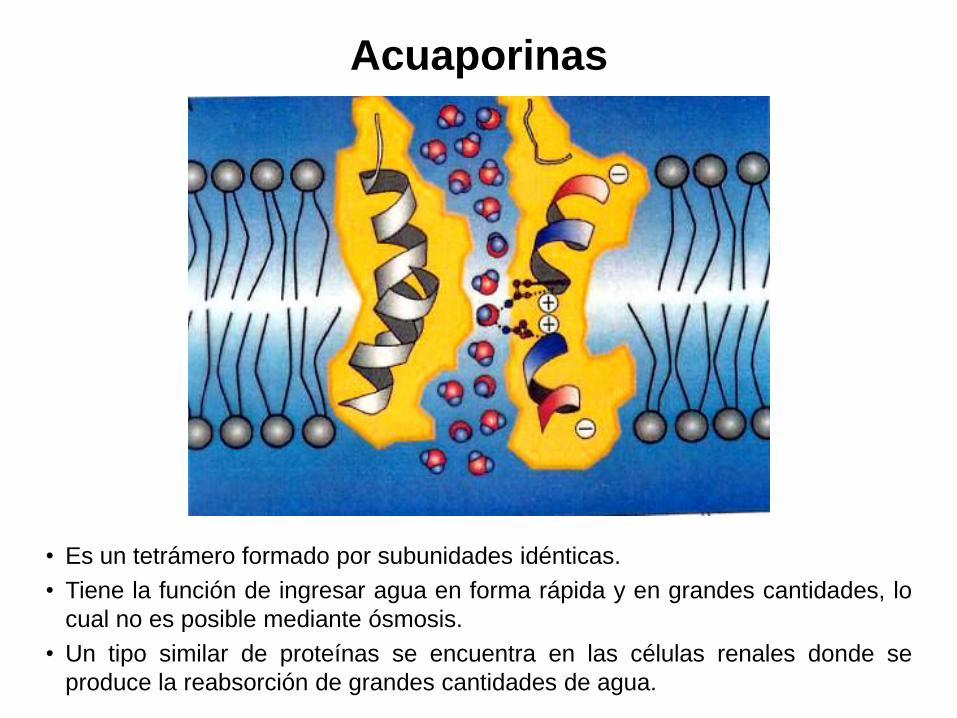
Acuaporinas
• Es un tetrámero formado por subunidades idénticas.
• Tiene la función de ingresar agua en forma rápida y en grandes cantidades, lo
cual no es posible mediante ósmosis.
• Un tipo similar de proteínas se encuentra en las células renales donde se
produce la reabsorción de grandes cantidades de agua.

• AQP0. Se expresa en las células fibrilares del cristalino, en las cuales cumple un papel
primariamente estructural, aunque su función aún está lejos de ser comprendida
completamente; fue llamada LMIP (Lens Major Intrinsec Protein), debido a que es una de
los péptidos más abundantes en estas células, constituyendo la mitad de todas sus
proteínas.
• AQP1. Es la acuaporina más abundante en las membranas animales y posiblemente la de
expresión menos selectiva. Ha sido la acuaporina prototipo pues fue la primera en ser
descrita y por tanto es la más estudiada hasta el momento. Fue descubierta inicialmente en
eritrocitos, pero su presencia se ha demostrado en la mayor parte de epitelios, sobre todo
abundante en túbulo proximal renal (TPR) y segmento descendente delgado (SDD) del asa
de Henle en el riñón, en todos los tipos de endotelio, en los epitelios de cristalino y córnea.
• AQP 2. Es expresada exclusivamente en membranas apicales en los túbulos distales y
colectores renales y es la responsable de la permeabilidad apical al agua de este segmento
del nefrón; su actividad es dependiente de hormona antidiurética (ADH).
• AQP 3. Es expresada en membranas basolaterales de túbulos renales, coexistiendo con
AQP2 en el mismo tipo de células; la función de ambas acuaporinas está acoplada, pues
AQP3 es la responsable de la permeabilidad al agua de la membrana basolateral.
• AQP4. Es la acuaporina más abundante en el cerebro, donde fue aislada por primera vez;
es activada por fosforilación mediada por diversos sistemas.
• AQP 5. Está localizada en la membrana apical de células epiteliales en múltiples glándulas,
tales como las sudoríparas, lacrimales, salivares y submucosas respiratorias; su principal
papel fisiológico consiste en regular el flujo de agua hacia la luz glandular.

AQP7. Fue identificada inicialmente en tejido adiposo donde es expresada ampliamente,
aunque también parece estar presente en muchos otros tejidos como espermatocitos y
túbulo proximal renal. Es permeable a glicerol y parece ser una ruta alterna para la salida
del glicerol producido durante la lipólisis, sin embargo su papel fisiológico aún está siendo
evaluado.
AQP8. Está presente exclusivamente en membranas intracelulares; ha sido hallada en
células epiteliales de túbulo proximal renal, túbulo colector renal, yeyuno, íleon, colon,
bronquios y glándulas salivales; además parece estar presente en hepatocitos y testículos.
Es la única acuaporina que exhibe permeabilidad a la úrea, pero su función específica aún
permanece en estudio.
AQP9. Fue identificada en hepatocitos, células en las cuales parece cumplir su principal
papel. Es también permeable a otros solutos de bajo peso molecular y puede funcionar
como una ruta de entrada para glicerol durante la gluconeogénesis. Su función en
situaciones de control metabólico extremo es aumentar la fuente de glicerol en el
hepatocito, probablemente funcionando en concierto con la AQP7 en tejido adiposo.
También es expresada en leucocitos, donde se ha encontrado que es permeable a arsenita,
un agente usado en quimioterapia para el tratamiento de ciertos tipos de leucemias
mielocíticas, por lo cual su expresión en células tumorales podría tener cierta significancia
terapeútica.
AQP10. Es la más recientemente informada, fue hallada en duodeno y yeyuno, pero al
parecer se encuentra en epitelios en forma inespecífica, pero aún no hay datos acerca de
su significancia funcional.
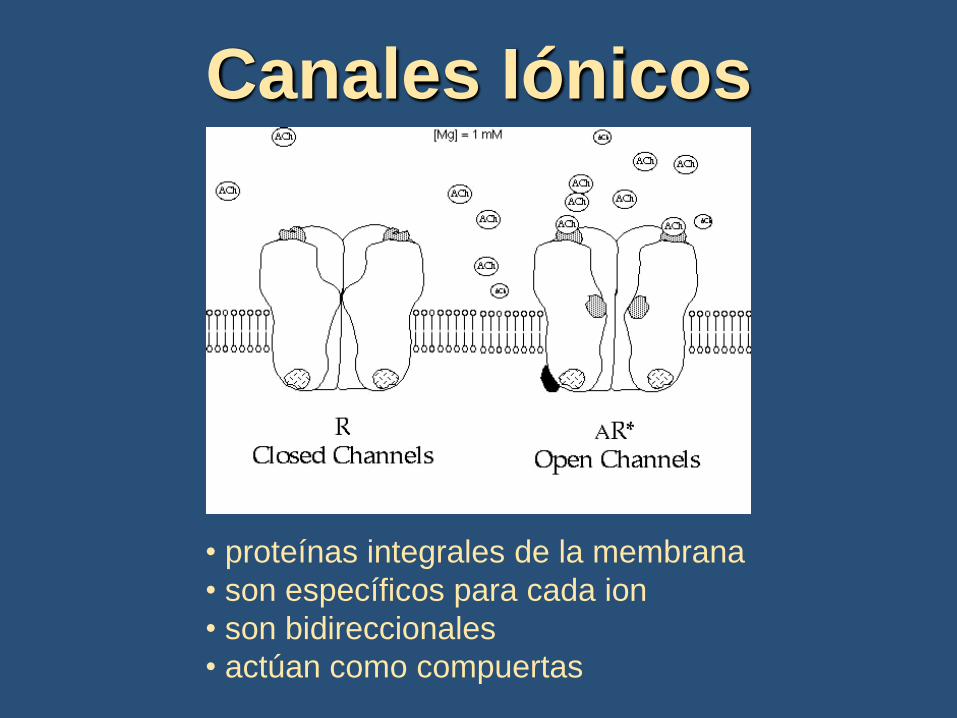
Canales Iónicos
• proteínas integrales de la membrana
• son específicos para cada ion
• son bidireccionales
• actúan como compuertas

Mecanismo voltaje
la apertura o cierre depende de la diferencia
de carga iónica a ambos lados de la MP.
sustancias químicas
son regulados por el enlace de un ligando
particular a la proteína del canal.
mecánicamente?
se abren mediante el estiramiento de la MP a
través de las fibras del citoesqueleto.

Tipos de Canales Iónicos
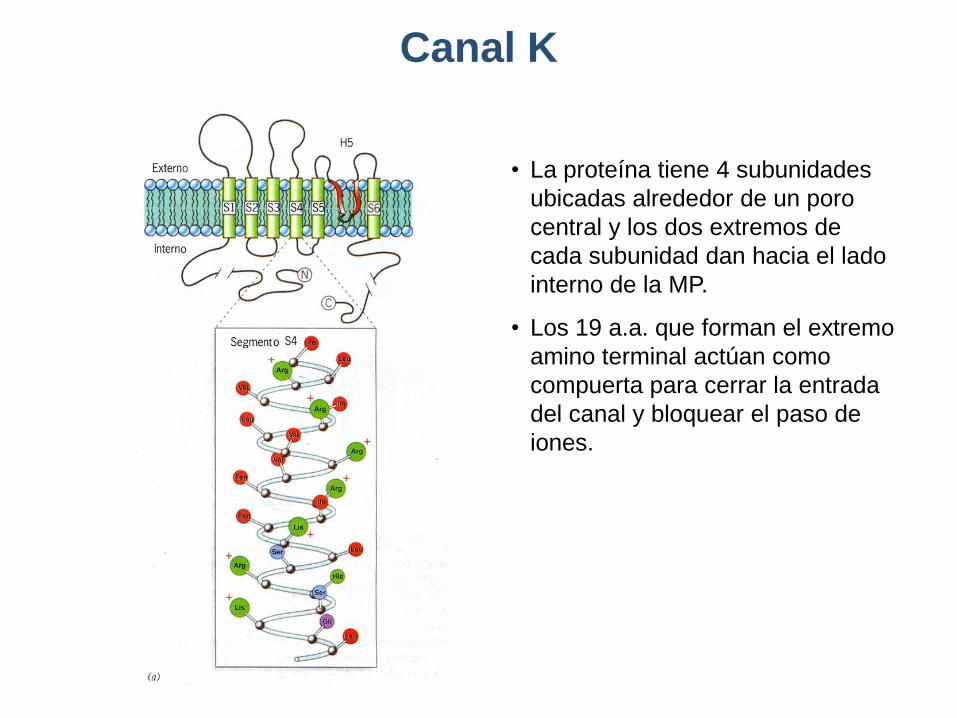
Canal K
• La proteína tiene 4 subunidades
ubicadas alrededor de un poro
central y los dos extremos de
cada subunidad dan hacia el lado
interno de la MP.
• Los 19 a.a. que forman el extremo
amino terminal actúan como
compuerta para cerrar la entrada
del canal y bloquear el paso de
iones.

Canal K
• Los cambios de carga iónica alteran las interacciones entre los a.a. Con
carga + con los a.a. cargados negativamente de otra parte de la proteína.
• Se produce un cambio en la conformación de la proteína, que hace que el
extremo de la proteína salga de su lugar y libere el poro.
• Una vez abierto, pueden entrar más de 1 millón de iones K por segundo,
pero pasa 1 ion por vez (poro= 0,3 nm diám.; K mide 0,2 nm).
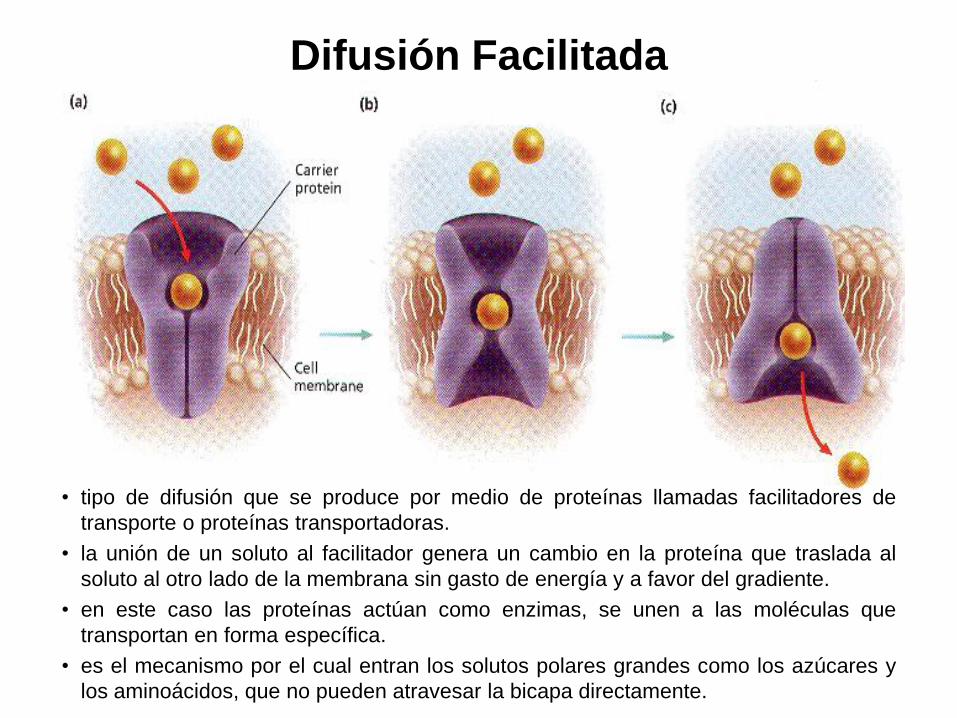
Difusión Facilitada
• tipo de difusión que se produce por medio de proteínas llamadas facilitadores de
transporte o proteínas transportadoras.
• la unión de un soluto al facilitador genera un cambio en la proteína que traslada al
soluto al otro lado de la membrana sin gasto de energía y a favor del gradiente.
• en este caso las proteínas actúan como enzimas, se unen a las moléculas que
transportan en forma específica.
• es el mecanismo por el cual entran los solutos polares grandes como los azúcares y
los aminoácidos, que no pueden atravesar la bicapa directamente.

Regulación de la Captación de Glucosa
• Los transportadores de glucosa se almacenan en las paredes de vesículas formadas
por invaginación de la membrana plasmática (endocitosis).
• Cuando aumenta la concentración de insulina las vesículas se mueven hacia la
periferia de la célula y se fusionan con la membrana para liberar los transportadores.

Transporte Activo
Primario
ATP
ATPasas o Bombas
Secundario
Gradientes de
Concentración Iones
Proteínas
Cotransportadoras
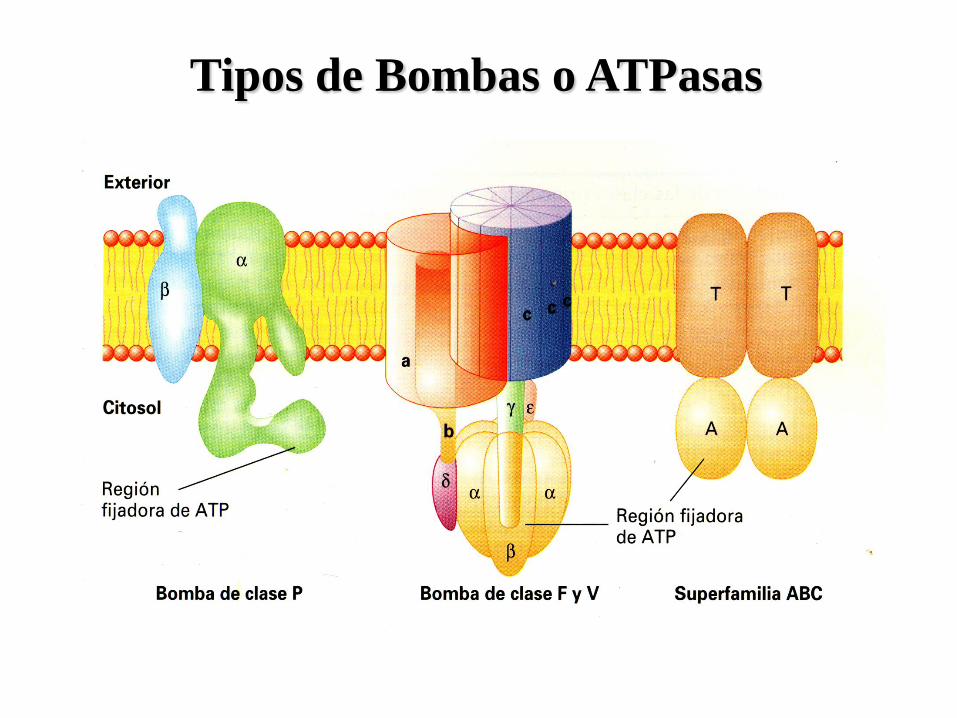
Tipos de Bombas o ATPasas

Bombas Tipo P
• son proteínas formadas por una subunidad α y una subunidad β, que en la mayoría
de los casos forman dímeros, pero a veces también tetrámeros.
• la α tiene el sitio de unión al ATP y la β desempeña funciones reguladoras.
• durante el transporte la subunidad α se fosforila, de ahí el nombre de tipo P.
• los iones transportados se mueven a través de la subunidad α fosforilada.
• esta clase incluye a las bombas de Na/K, bombas de Ca y bombas de H.

Bomba de Ca
• La subunidad alfa de la proteína tiene sitios de fijación para 2 iones Ca.
• Luego de unirse el Ca se fija el ATP a un sitio específico y se produce la hidrólisis del
ATP liberándose ADP + P.
• El fosfato liberado se une a la proteína produciendo un cambio de conformación que
traslada los dos iones Ca al otro lado de la membrana.
• Una vez realizado el transporte, el fosfato se libera y cambia la conformación de la
proteína para poder transportar nuevamente dos iones.

Bomba Na/K

Bomba Na/K
• es similar a la de Ca, pero esta posee dos subunidades alfa y dos subunidades beta.
• la subunidad beta es esencial para el plegamiento de la subunidad alfa cuando son
sintetizadas, pero no participa directamente en el transporte.
• en el proceso, la célula saca tres iones Na e ingresa dos iones K por cada molécula
de ATP hidrolizada.
• el mecanismo de acción es similar a la bomba de Ca, pero en este caso la bomba
transporta iones en ambas direcciones.

Bomba Na/K
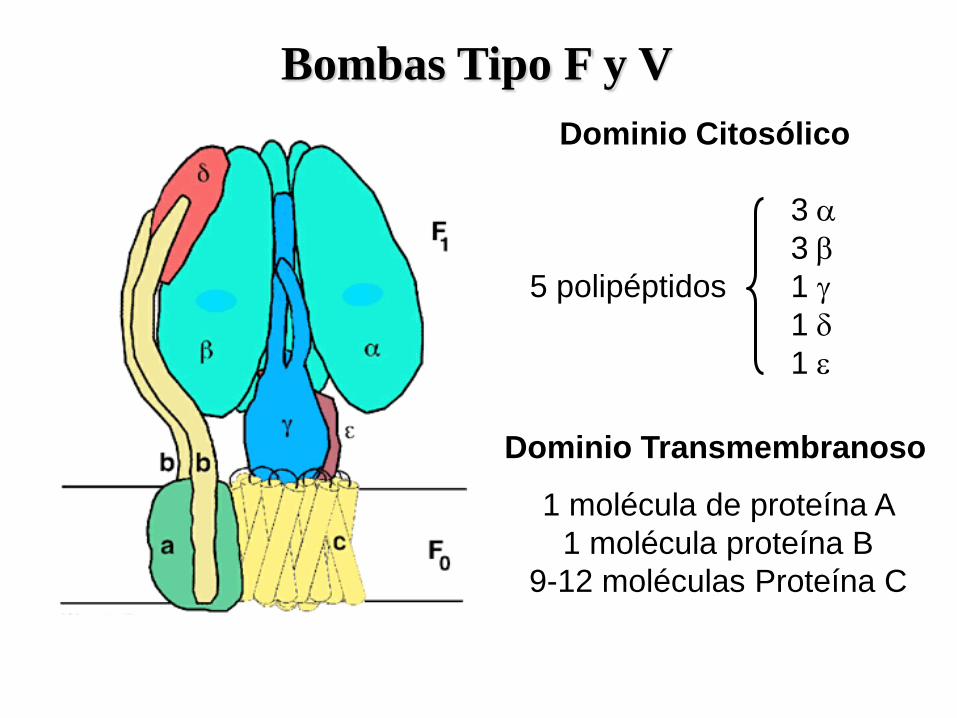
Bombas Tipo F y V
ggdfss
3 a
3 b
5 polipéptidos 1 g
1 d
1 e
Dominio Citosólico
Dominio Transmembranoso
1 molécula de proteína A
1 molécula proteína B
9-12 moléculas Proteína C

Bombas Tipo ABC
• Su nombre deriva del inglés ATP binding cassette o sea
casete de fijación al ATP.
• Esta familia incluye un grupo de 100 proteínas
diferentes que se encuentran en diferentes organismos.
• Cada proteína ABC es específica para una sustancia
particular o un grupo de sustancias relacionadas.
• Transportan diversos tipos de sustancias tales como:
iones, monosacáridos, péptidos, polisacáridos e incluso
proteínas completas.
• Tienen 4 polipéptidos, dos insertados en la membrana
(T) que forman el canal y dos en la cara intracelular
que tienen los sitios el ATP (A).
• Los 4 polipéptidos pueden estar fusionados en 1 o 2
polipéptidos con dominios múltiples.
• La secuencia de los dominios A es 30 a 40% homóloga
en todos los miembros de esta superfamilia, lo que
indica un origen evolutivo común.

Histidina Permeasa
• existe una proteína soluble fijadora de histidina ubicada en el espacio periplasmático que
asiste en el transporte.
• esta proteína fija la histidina y la dirige a las subunidades T, a través de las cuales el
aminoácido atraviesa la membrana impulsado por la hidrólisis de ATP.
• las células de E. coli mutantes para las subunidades de la permeasa o la proteína
fijadora son incapaces de transportar histidina hacia el interior celular.
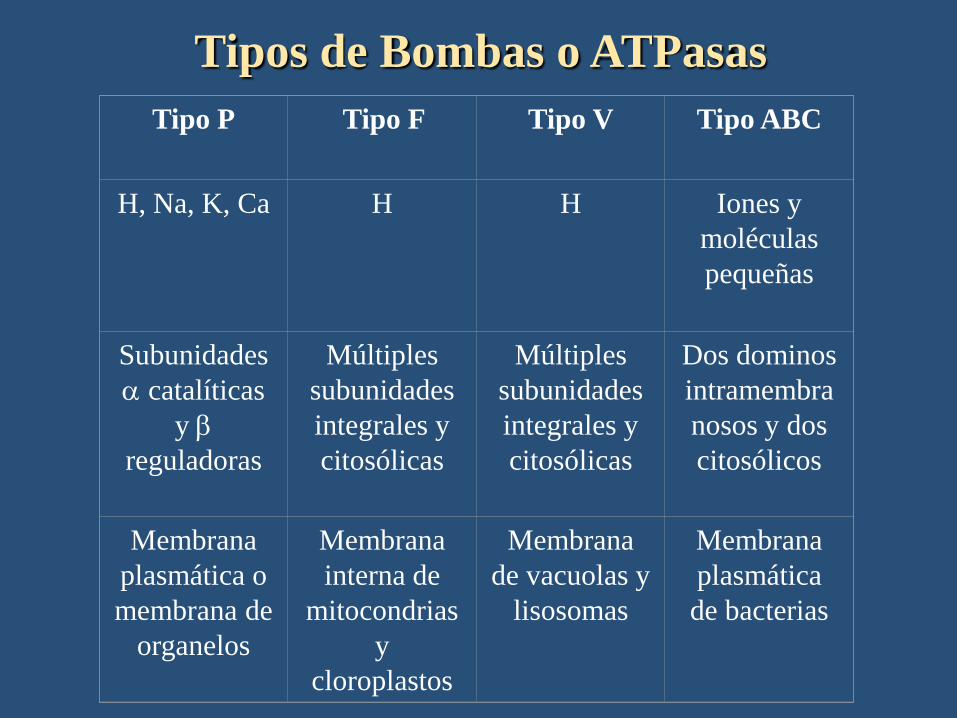
Tipo P Tipo F Tipo V Tipo ABC
H, Na, K, Ca H H Iones y
moléculas
pequeñas
Subunidades
a catalíticas
y b
reguladoras
Múltiples
subunidades
integrales y
citosólicas
Múltiples
subunidades
integrales y
citosólicas
Dos dominos
intramembra
nosos y dos
citosólicos
Membrana
plasmática o
membrana de
organelos
Membrana
interna de
mitocondrias
y
cloroplastos
Membrana
de vacuolas y
lisosomas
Membrana
plasmática
de bacterias
Tipos de Bombas o ATPasas

Transporte Activo Secundario
• las proteínas no utilizan directamente ATP como fuente de energía
• emplean los gradientes de concentración de iones Na o H para mover
sustancias en contra del gradiente
• el proceso se denomina
cotransporte y las proteínas
que participan se llaman
proteínas cotransportadoras
• mediante este tipo de
mecanismo son transportados
algunos iones, aminoácidos y la
glucosa.

Simporte Na-Glucosa
• Como el sodio tiene una
concentración baja en la célula
siempre tiende a entrar.
• La célula utiliza este gradiente de
concentración para acoplar la
entrada del Na con la entrada de
glucosa en contra del gradiente de
concentración.
• Mediante este mecanismo las
proteínas incorporan dos iones Na
junto con una molécula de glucosa.
• O sea que este tipo de transporte
secundario es un simporte, ambas
sustancias van en el mismo sentido.

• En las células de músculo
cardíaco hay una proteína que
transporta Na a favor de su
concentración y saca Ca en
contra del gradiente de
concentración.
• Este antiporte es el responsable
de mantener baja la
concentración de Ca en el
citosol.
• Cada 3 iones Na que entran a la
célula, se extrae un ion Ca.
Ca2
Na+
Antiporte Ca-Na

Antiporte HCO3-Cl
• permite la eliminación del CO2 de los tejidos
durante la respiración
• el CO2 pasa a la sangre y de allí entra a los
eritrocitos, donde es convertido en
bicarbonato (HCO3) por una enzima
• Los iones bicarbonato son transportados
hacia afuera de los eritrocitos en un antiporte
con Cl
• ello permite que casi todo el HCO3 sea
transportado por el plasma sanguíneo fuera
de las células
• lo cual aumenta la cantidad de CO2 que
puede transportarse desde los tejidos hasta
los pulmones.
• si no hubiera intercambio aniónico, el
aumento de HCO3 en el eritrocito haría que
su citosol se tornase alcalino
• el intercambio de HCO3 por Cl- hace que el
pH citosólico permanezca cercano a la
neutralidad.
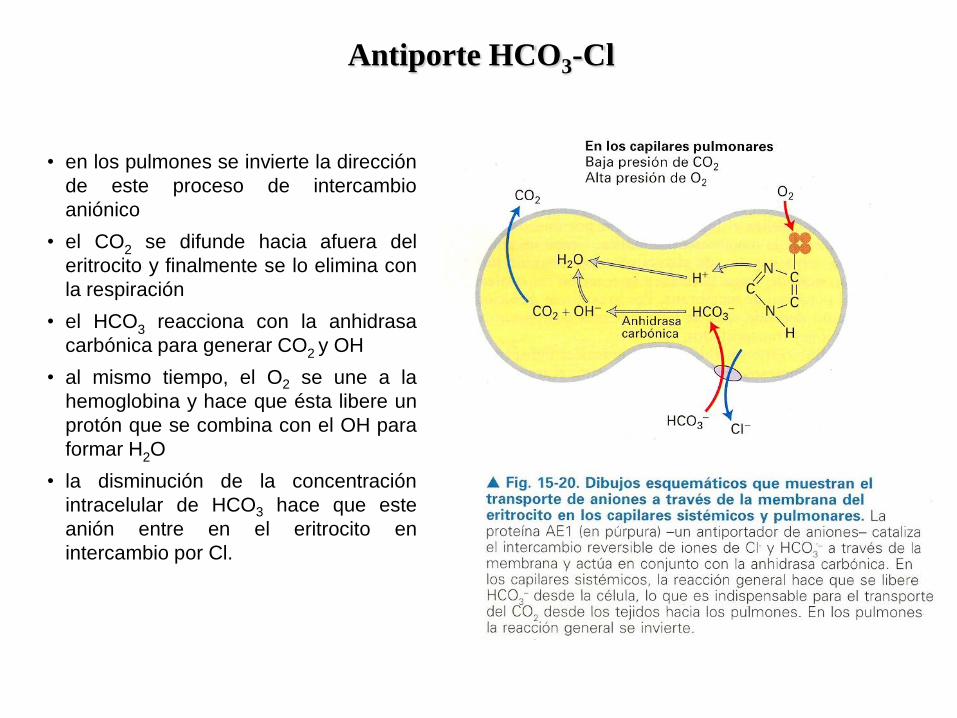
Antiporte HCO3-Cl
• en los pulmones se invierte la dirección
de este proceso de intercambio
aniónico
• el CO2 se difunde hacia afuera del
eritrocito y finalmente se lo elimina con
la respiración
• el HCO3 reacciona con la anhidrasa
carbónica para generar CO2 y OH
• al mismo tiempo, el O2 se une a la
hemoglobina y hace que ésta libere un
protón que se combina con el OH para
formar H2O
• la disminución de la concentración
intracelular de HCO3 hace que este
anión entre en el eritrocito en
intercambio por Cl.
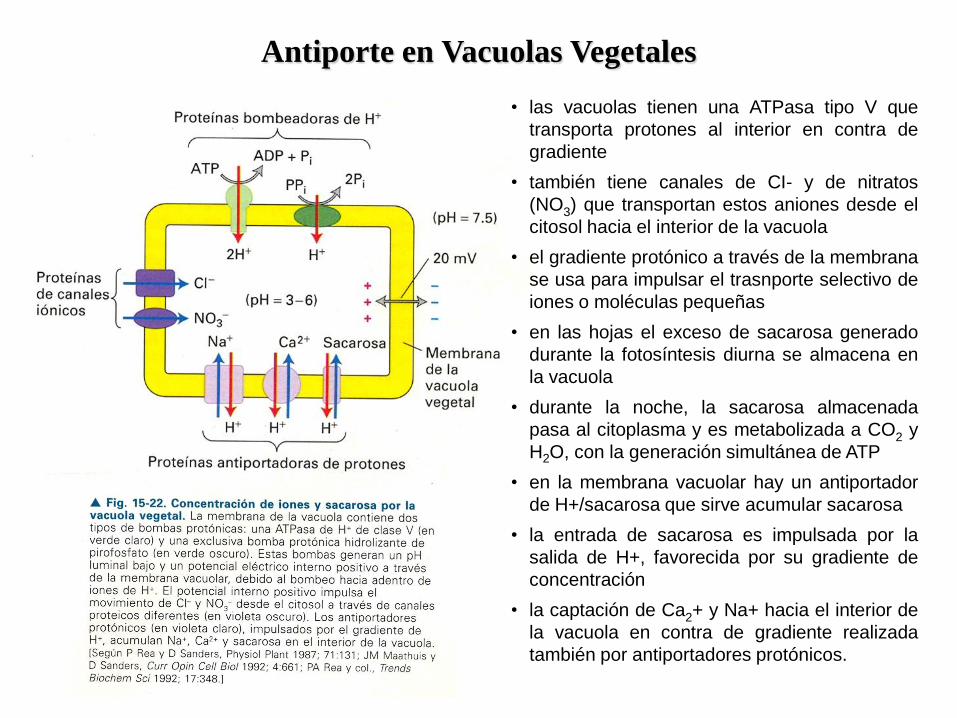
Antiporte en Vacuolas Vegetales
• las vacuolas tienen una ATPasa tipo V que
transporta protones al interior en contra de
gradiente
• también tiene canales de CI- y de nitratos
(NO3) que transportan estos aniones desde el
citosol hacia el interior de la vacuola
• el gradiente protónico a través de la membrana
se usa para impulsar el trasnporte selectivo de
iones o moléculas pequeñas
• en las hojas el exceso de sacarosa generado
durante la fotosíntesis diurna se almacena en
la vacuola
• durante la noche, la sacarosa almacenada
pasa al citoplasma y es metabolizada a CO2 y
H2O, con la generación simultánea de ATP
• en la membrana vacuolar hay un antiportador
de H+/sacarosa que sirve acumular sacarosa
• la entrada de sacarosa es impulsada por la
salida de H+, favorecida por su gradiente de
concentración
• la captación de Ca2+ y Na+ hacia el interior de
la vacuola en contra de gradiente realizada
también por antiportadores protónicos.