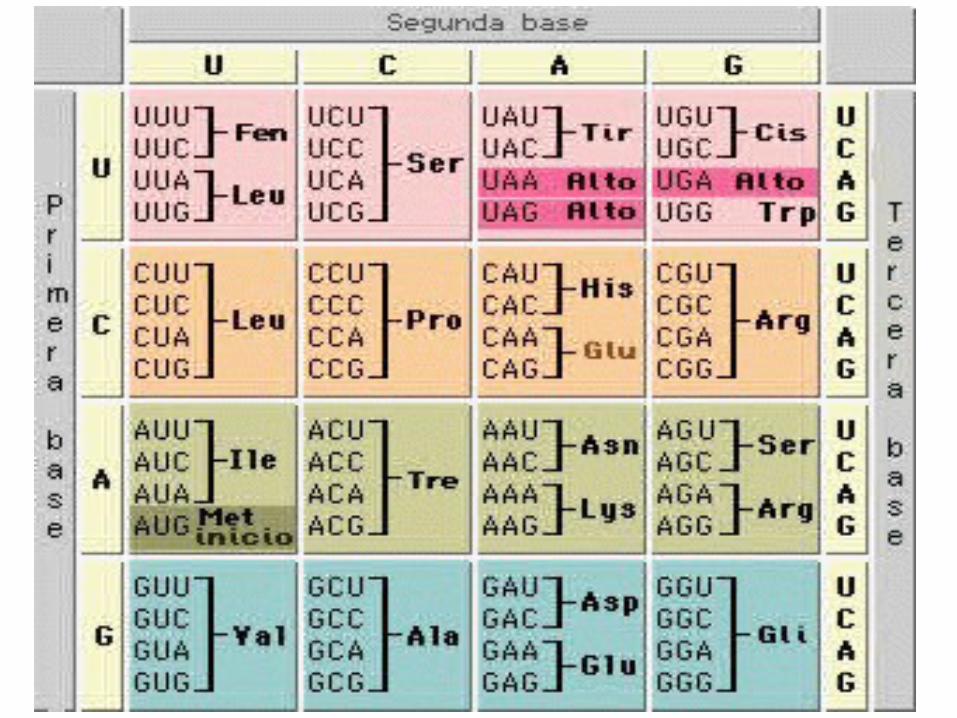
transcripción sólo
TRANSCRIPT

LA TRANSCRIPCIÓNEs una síntesis de ARN
Copia complementaria de un segmento de ADN.
Sigue la complementaridad de bases
El ARN sintetizado o transcrito porta la información contenida en el ADN

Hay que tener en cuenta que el ARN tiene uracilo en lugar de timina y es de cadena simple
(monocatenario).TIPOS DE ARN:
FUNCIONALES: No se traducen en proteínas: ARNr; ARNt o ARNs; ARN np y ARNcp.
INFORMATIVOS: Se van a traducir en proteínas: ARNm .
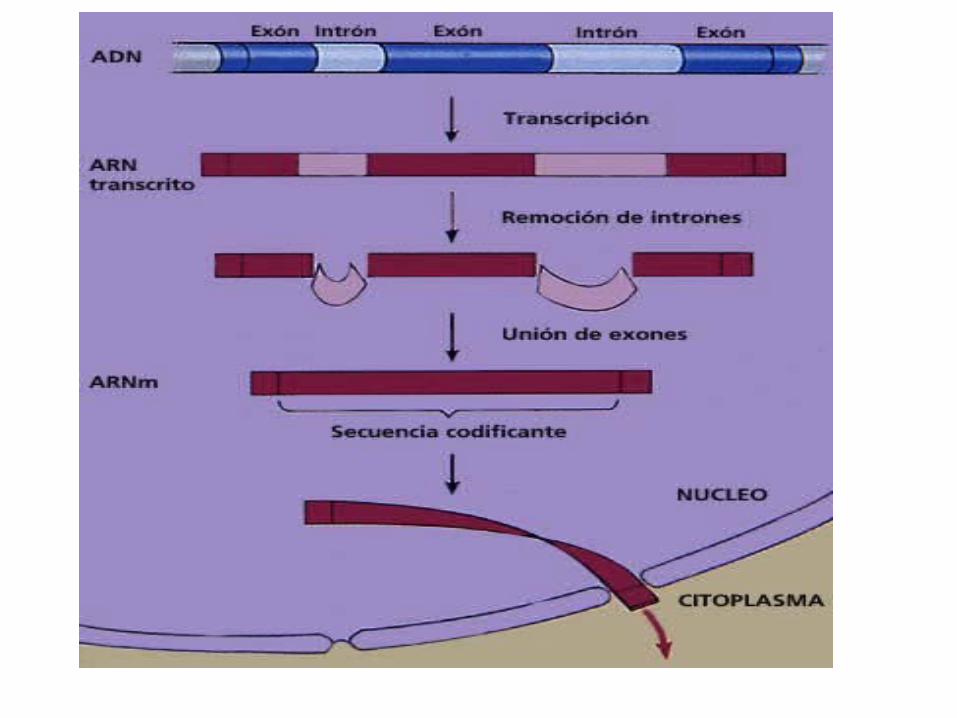
En bacterias este transcrito primario ya es ARN maduro; en eucariotas el transcrito primario es ARN heterogéneo nuclear (ARNhn); éste se procesa y se convierte en ARN maduro y después se traduce a
proteína
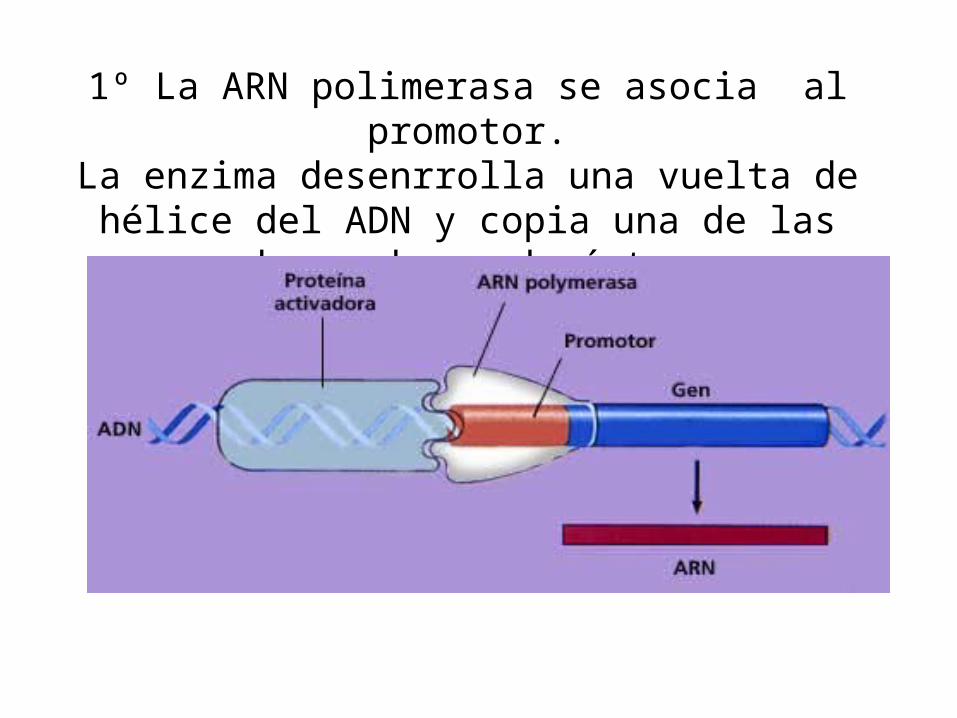
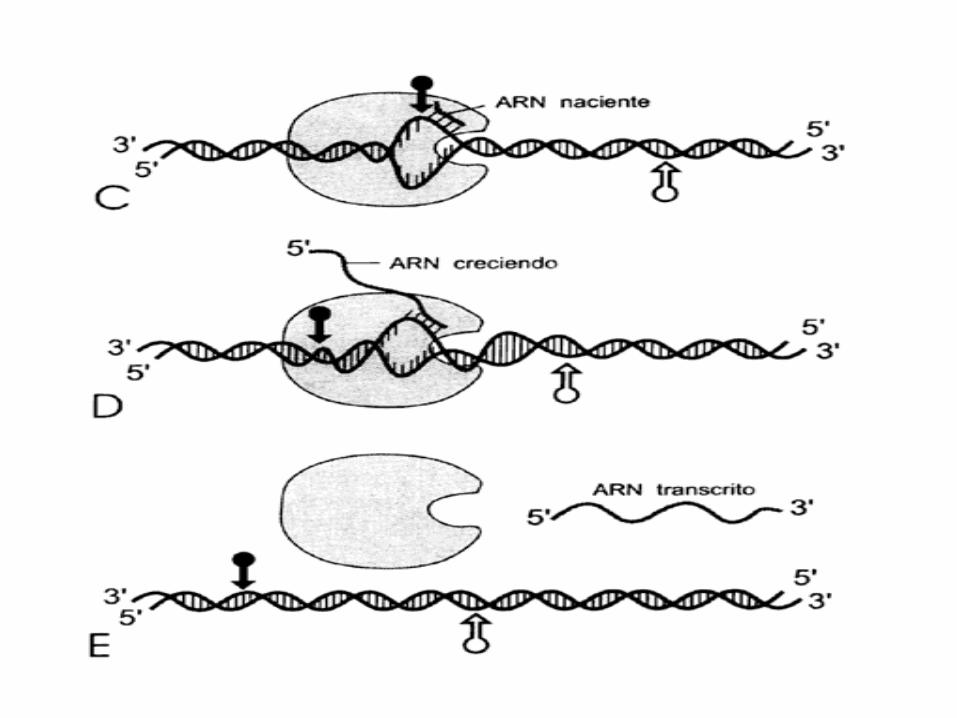
1º La ARN polimerasa se asocia al promotor.La enzima desenrrolla una vuelta de hélice del ADN y
copia una de las dos cadenas de éste.
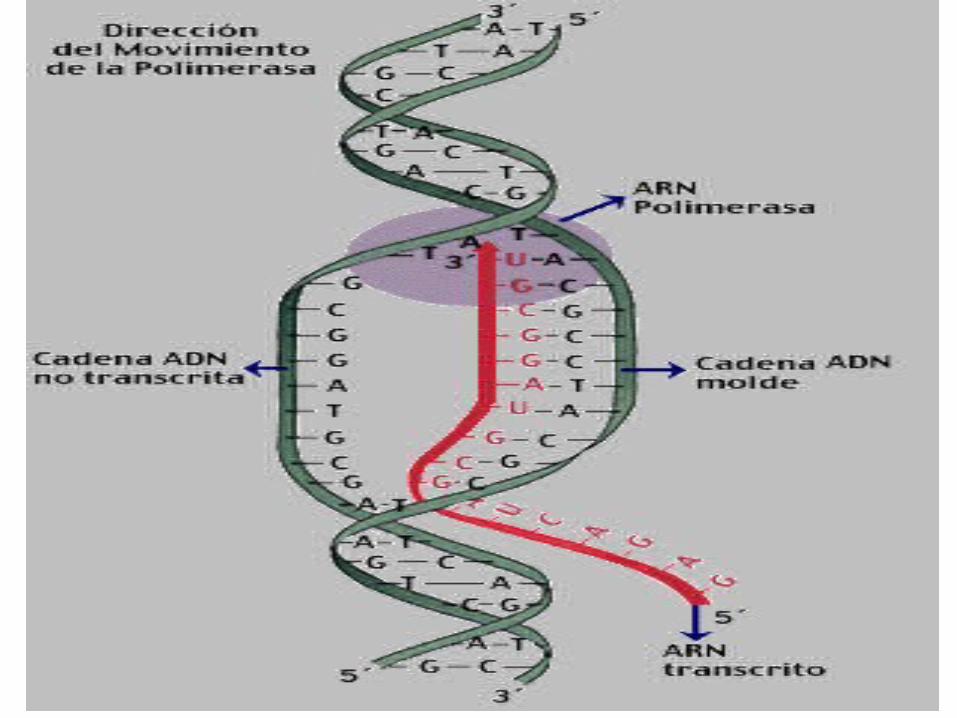
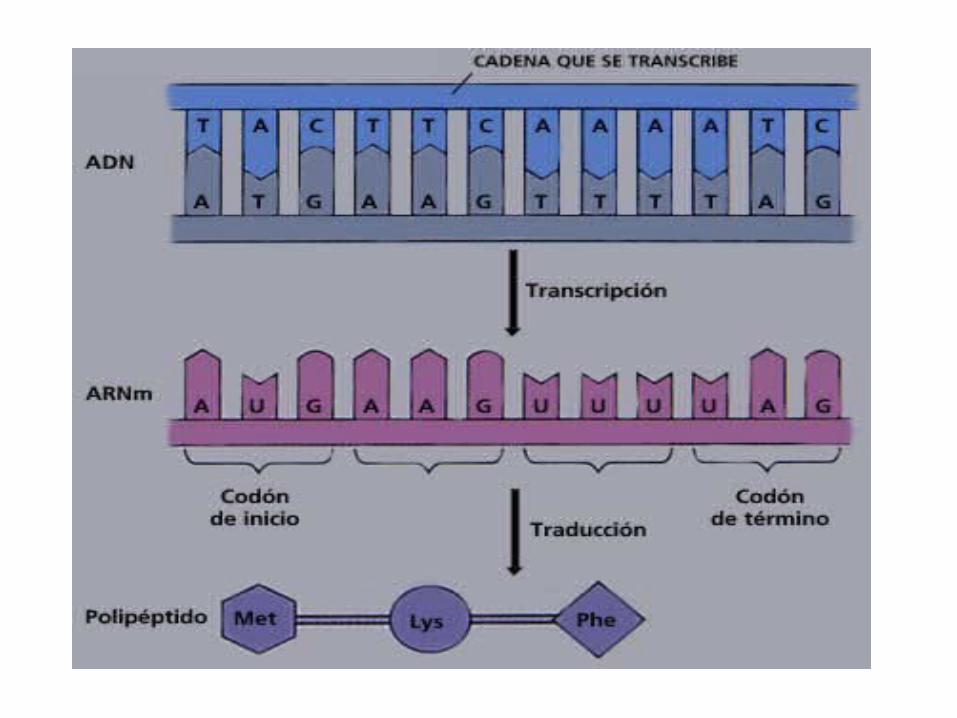
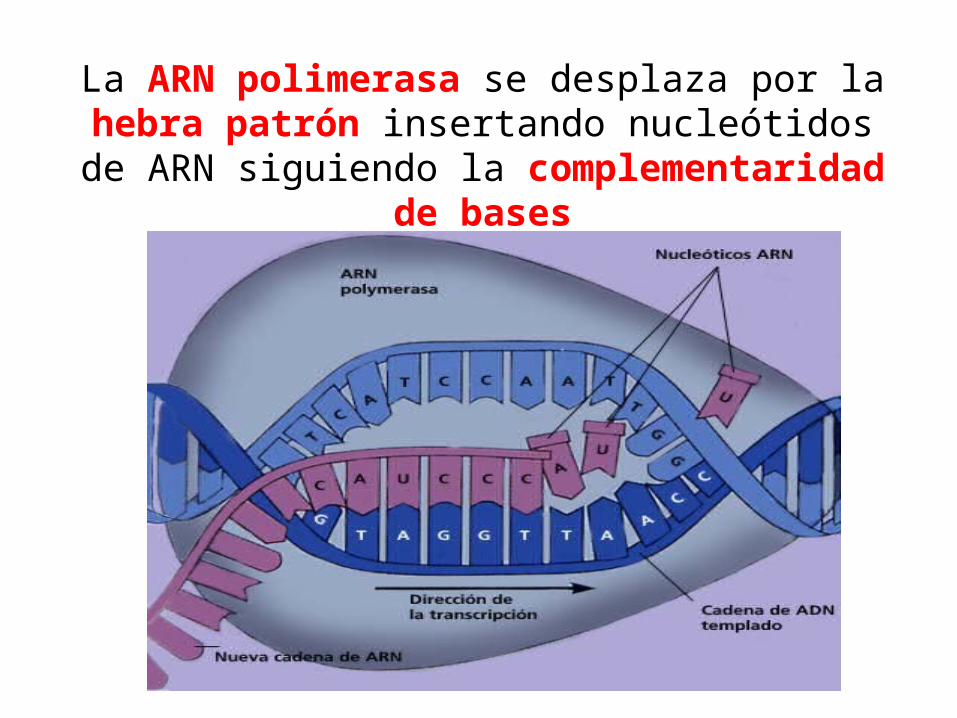
La ARN polimerasa se desplaza por la hebra patrón insertando nucleótidos de ARN siguiendo la
complementaridad de bases
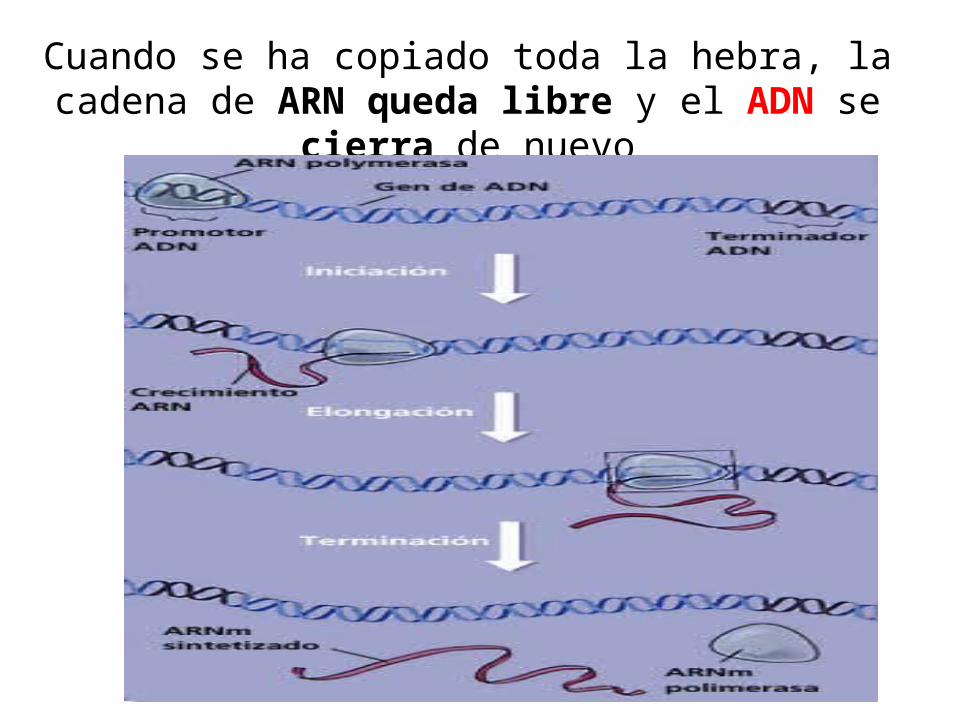
Cuando se ha copiado toda la hebra, la cadena de ARN queda libre y el ADN se cierra de nuevo

De esta forma las instrucciones genéticas copiadas o transcritas al ARN quedan listas
para salir al citoplasma.Por lo tanto el ADN es la copia maestra de la
información genética que permanece en “reserva” dentro del núcleo y el ARN es la
copia de trabajo.Este ARN que lleva las instrucciones para la
síntesis de proteínas se denomina ARN mensajero (ARNm)
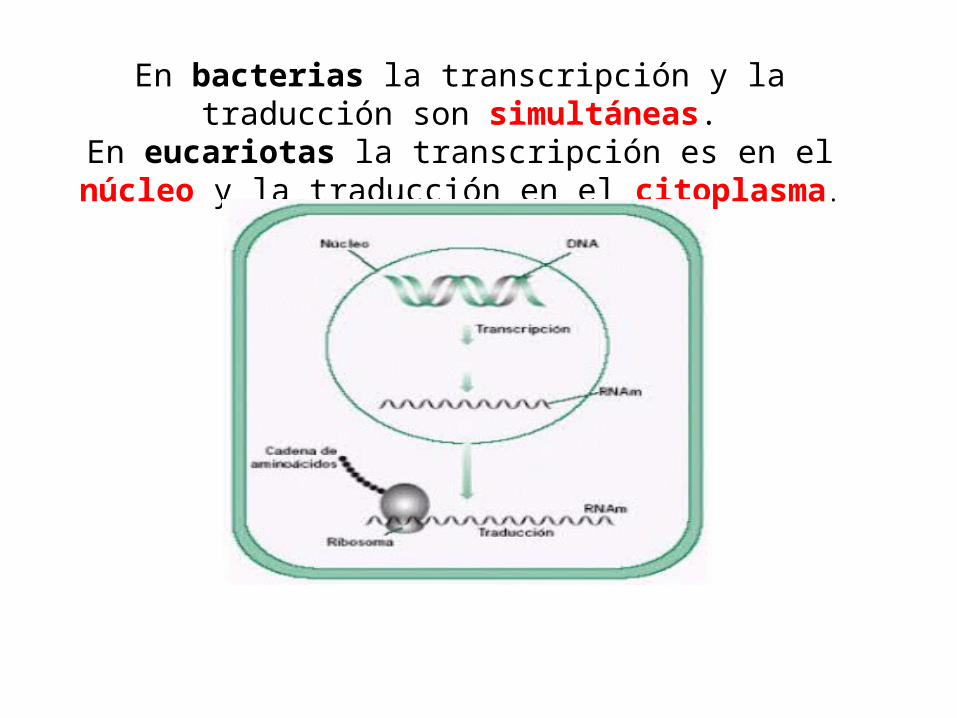
En bacterias la transcripción y la traducción son simultáneas.En eucariotas la transcripción es en el núcleo y la traducción
en el citoplasma.

En procariontes hay una sola polimerasa y esta sintetiza todos los ARN.
En eucariotas existen varias polimerasas que sintetizan los distintos tipos de ARN:
ARN pol I: precursosres de ARNrARN pol II: ARNhn, que tras procesarse se
transforma en ARNm maduroARN pol III: ARN precursores de ARNt, ARNnp
y ARN cp.A las ARN polimerasas se les conoce como
TRANSCRIPTASAS.Carecen de actividad correctora
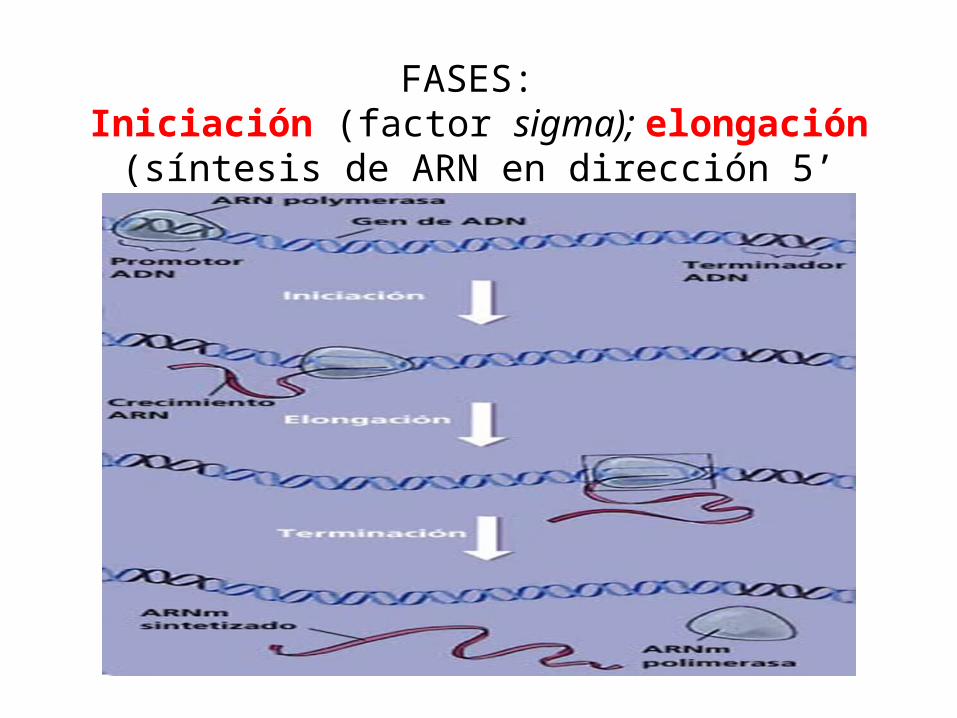
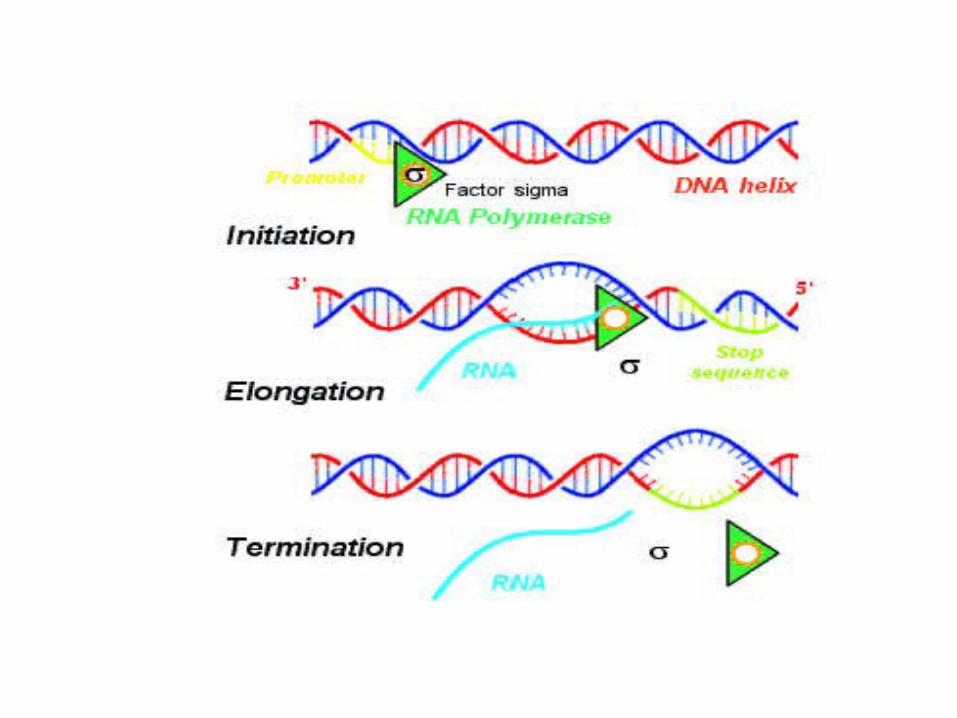
FASES: Iniciación (factor sigma); elongación (síntesis de ARN en
dirección 5’ -------- 3’), terminación

TRANSCRIPCIÓN EN EUCARIOTAS
Es bastante más compleja que en procariotas, debido principalmente a que:
1. Existen varias RNA polimerasas (RNA pol). Se conocen tres que participan en la transcripción del DNA nuclear y tanto la mitocondria como el cloroplasto poseen sus
RNA pol particulares.2. La formación del complejo de pre-iniciación en el que
participan un gran número de factores y cofactores y difiere para cada tipo de RNA pol, lo que da origen a
tres sistemas de transcripción.3. La necesidad de procesar el RNA sintetizado.
4. El RNA se sintetiza en el núcleo y debe ser llevado al citoplasma donde realizará sus funciones, mientras que
los factores de transcripción se sintetizan en el citoplasma y deben ser llevados al núcleo.

RNA POLIMERASAS EUCARIOTAS
El núcleo eucariota contiene tres tipos de RNA polque difieren en los RNAs que sintetizan:
RNA pol I, está en el nucléolo, sintetiza los precursores de los RNAs ribosómicos, 28, 5.8 y 18 S.
RNA polimerasa II, situada en el nucleoplasma, sintetiza los precursores del RNAm y de ciertos RNAs
pequeños nucleares estables. Es la más estudiada de las tres. RNA polimerasa III, presente en el nucleoplasma, sintetiza los
precursores del RNA ribosómico de 5S, de los RNAts y de una variedad de otros RNAs pequeños nucleares y citosólicos.
Además de estas enzimas nucleares, las células eucariotas contienen RNA polimerasas mitocondriales y
de cloroplastos que son independientes

FACTORES DE TRANSCRIPCIÓN
Los factores generales de transcripción (GTFs) , se fijan a los promotores, son funcionalmente semejantes al factor σ de los
procariotes:a. La proteína de unión a TATA (TBP), reconoce la caja TATA del promotor en los genes de tipo II (los que son transcritos por la RNA pol II) y se fija a ella en un proceso de gran especificidad.
Recluta después otros factores de transcripción para formar un complejo (TFIID).
b. La RNA pol II es incorporada a este complejo para formar el complejo de pre-iniciación
c. Además del TBP, otros factores de transcripción son más selectivos y específicos y se incorporan al complejo
dependiendo del gene de tipo II que será transcrito, lo cual regula la transcripción de dichos genes.

PREINICIACIÓNAl contrario de la replicación de ADN, durante el inicio de la
transcripción no se requiere la presencia de un cebador. Antes del inicio de la transcripción se necesita toda una serie
de factores de transcripción (TF) que se unen a secuencias específicas de ADN para reconocer el sitio donde la
transcripción ha de comenzar, el promotor, que se localizan en los extremos 5'-terminales de los genes, antes del comienzo del
gen, y a ellos se unen los factores de transcripción mediante fuerzas no covalentes (Van der Waals y puentes de hidrógeno. Los promotores tienen secuencias reguladoras definidas, muy conservadas en cada especie, donde las más conocidas son la
caja TATA y la caja TTGACA.

La formación del complejo de transcripción se realiza sobre el promotor TATA, allí se forma el núcleo del
complejo de iniciación. Sobre la caja TATA se fija una proteína de unión (TBP) junto con otros factores de transcripción . Todo ello forma un complejo que se llama «complejo de preiniciación cerrado» o PIC.
Cuando la estructura se abre por mediación del factor de transcripción TFII H, comienza la iniciación y al
«complejo abierto» (por su acción helicasa dependiente de ATP).

INICIACIÓN Una Helicasa separa las hebras de ADN en las TATA, ( A = T); después se unen los factores y las proteínas de transcripción (TBP, TF2D, TF2B) permitiendo, el acceso de la ARN polimerasa al molde de ADN de
cadena simple formando la «burbuja de transcripción» o apertura del ADN, de unos 18 pb,
donde empieza a sintetizarse el ARN cebador a partir del nucleótido número 10 del ADN molde de la
burbuja de transcripción. La burbuja de transcripción se llama «complejo abierto».

La ARN polimerasa es una enzima formada por 5 subunidades: 2 subunidades α, 1 subunidad β, 1 subunidad β' y 1 subunidad ω que tiene como función la unión de ribonucleótidos trifosfato. Cuando se forma el complejo abierto, la ARN polimerasa comienza a unir ribonucleótidos
mediante enlaces fosfodiéster, y una vez que se forma el primer enlace fosfodiéster, acaba la etapa de iniciación y comienza así la siguiente etapa (la
disgregación del promotor).

DISGREGACIÓN DEL PROMOTORUna vez sintetizado el primer enlace fosfodiéster, se debe deshacer el complejo del promotor para que
pueda volver a funcionar de nuevo. Una vez que la cadena transcrita alcanza una longitud de unos 23 nucleótidos, el complejo ya no se desliza y
da lugar a la siguiente fase, la elongación.La disgregación del promotor coincide con una
fosforilación de la serina 5 del dominio carboxilo terminal de la ARN polimerasa.

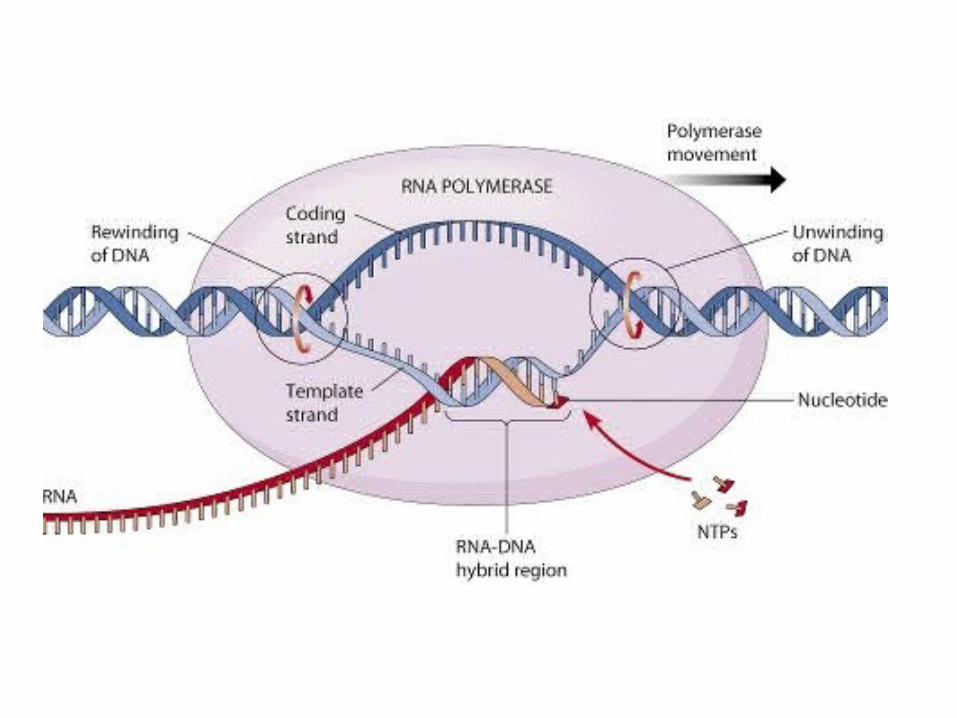
ELONGACIÓNLa ARN polimerasa cataliza la elongación de cadena
del ARN. Una cadena de ARN se une por apareamiento de bases a la cadena de ADN, y para
que se formen correctamente los enlaces de hidrógeno que determina el siguiente nucleótido del molde de ADN, el centro activo de la ARN polimerasa reconoce a los ribonucleótidos trifosfato entrantes. Cuando el nucleótido entrante forma los enlaces de
hidrógeno idóneos, entonces la ARN polimerasa cataliza la formación del enlace fosfodiéster que
corresponde.

TERMINACIÓNAl finalizar la síntesis de ARNm, esta molécula ya se ha separado
completamente del ADN (que recupera su forma original) y también de la ARN polimerasa.
La terminación está señalizada por la información contenida en sitios de la secuencia del ADN que se está transcribiendo, por lo
que la ARN polimerasa se detiene al transcribir algunas secuencias especiales del ADN ricas en guanina y citosina,
situadas en el extremo de los genes, seguidas de secuencias ricas en timina, formando secuencias palindrómicas, que cuando se transcriben el ARN recién sintetizado adopta una «estructura en horquilla» que desestabiliza el complejo ARN-ADN, obligando a separarse de la ARN polimerasa, renaturalizándose la burbuja
de transcripción.

ARN POLIMERASA EN BACTERIASEn procariotas, la misma enzima cataliza la síntesis de
todos los tipos de ARN. Está formada por cinco subunidades de aproximadamente 410 kilodaltons :
α2ββ'ω.Para unirse al promotor específico, la holoenzima
requiere un factor σ (sigma) con el que se reduce la afinidad con regiones de ADN inespecíficas,
aumentando la especificidad por regiones promotoras para formar la holoenzima de cinco subunidades α2ββ'σω (~480 kDa).

Su estructura de la ARN pol presenta una ranura de 55 Å de longitud y una anchura 25 Å que permite el
paso de la doble hélice de ADN (20 Å).La longitud de 55 Å puede aceptar la secuencia de
18 nucleótidos.Todas las unidades funcionan conjuntamente para
llevar a cabo la transcripción. La subunidad β' participa en la unión del ADN, la subunidad β contiene parte del centro activo y la subunidad σ está implicada en la iniciación de la transcripción, disociándose del resto de la enzima
una vez iniciada la transcripción.

ARN POLIMERASA EN EUCARIOTASEn eucariotas hay 3 tipos de ARN pol, una por cada tipo de ARN:
ARN polimerasa I: síntesis, reparación y revisión. Sintetiza precursores de ARNr.
ARN polimerasa II: reparación, síntesis de precursores de ARNm, microARNs y otros tipos de ácido ribonucleico. Es la más
estudiada, y requieren de factores de transcripción para que se una a los promotores del ADN.
Su estructura tridimensional ha sido dilucidada por Roger Kornberg de la Universidad de Stanford (Premio
Nobel de Química en 2006) , hijo de otro Premio Nobel, Arthur Kornberg, que recibió el galardón en 1959 por el descubrimiento
de , laADN polimerasa.ARN polimerasa III: sintetiza ARNt, ARNr de 5S y otros pequeños
ARN (ARNpequeños) encontrados en el núcleo celular (ARNp nucleares) y en el citoplasma (ARNp citoplasmáticos).

La polimerasa IV y polimerasa V: reparación en condiciones únicas.
Otros tipos de ARN polimerasa se encuentran en la mitocondria y en cloroplasto y en core del ribosoma.
ARN polimerasa en arqueasLas arqueas tienen un solo tipo de ARNPol, que
sinrtetiza todos los ARN. El ARNP arqueano es similar estructural y
mecánicamente al ARNPol bacteriano y al eucariota nuclear RNAP I al V, pero especialmente relacionado
con el ARNP II eucariota.