tesis - uabcsbiblio.uabcs.mx/tesis/te3168.pdf · órgano en que ocurre la síntesis de enzimas...
TRANSCRIPT

UNIVERSIDAD AUTÓNOMA DE BAJA
CALIFORNIA SUR
ÁREA DE CONOCIMIENTO DE CIENCIAS DEL MAR
DEPARTAMENTO ACADÉMICO DE BIOLOGÍA
MARINA
TESIS
ESTUDIO DEL PROCESO DE PROTEÓLISIS EN EL JUGO
GÁSTRICO DE LANGOSTA AMERICANA (Homarus
americanus), POR LA ACCIÓN DE PROTEINASAS
DIGESTIVAS.
QUE PARA OBTENER EL TÍTULO DE:
BIÓLOGO MARINO
PRESENTA
IVÁN RODRÍGUEZ SIORDIA
DIRECTOR:
Betsaida Bibo Verdugo
LA PAZ, B. C. S. AGOSTO DE 2014


Resumen
La proteólisis es un proceso de hidrólisis de proteínas llevado a cabo por medio de
enzimas llamadas proteinasas, la cual se efectúa por la ruptura de enlaces peptídicos
que unen a los aminoácidos. Este proceso puede llevarse a cabo de manera
secuencial en la que una o varias proteinasas son necesarias para que otras
proteinasas actúen. Efecto que se ha observado en el tracto digestivo de
invertebrados como platelmintos e insectos. Este trabajo demuestra un mecanismo
similar en el jugo gástrico de langosta americana, Homarus americanus. El jugo
gástrico de langosta americana es ácido, pH ~4.7, y presenta cisteíno, aspártico y
serino proteinasas. En el presente estudio se evaluó la hidrólisis de los sustratos
protéicos: albúmina sérica bovina, hemoglobina y actina del músculo de Agropecten
circularis, por acción de las proteinasas del jugo gástrico de langosta americana y se
evaluó el efecto de los inhibidores específicos: Pepstatin A, E-64 y Pefabloc,
inhibidores de proteinasas aspártico, cisteíno y serino respectivamente. La hidrólisis
se cuantificó por el porcentaje de proteína remanente en SDS-PAGE y se analizó por
densitometría de imagen. El tiempo en el que ocurre la hidrólisis máxima de cada
sustrato, dependió de la naturaleza del mismo. Las proteínas comerciales BSA y
hemoglobina se hidrolizaron entre 6 y 8 h, mientras que la hidrólisis de actina,
utilizada para analizar la hidrólisis de una proteína homóloga a las hidrolizadas en el
jugo gástrico de langosta americana en el medio natural, finalizó en 1 h. El ensayo de
inhibición señala que si se inhiben las aspártico y cisteíno proteinasas la hidrólisis no
se detiene, por otro lado, al inhibir las cisteíno la hidrólisis se detiene
considerablemente, sin embargo, al inhibir tanto aspártico y cisteíno proteinasas la
hidrólisis se detiene. Estos resultados indican que las proteinasas que
sinérgicamente están participando en el proceso de proteólisis, son las de tipo
cisteíno y aspártico, lo cual se convalida dado que ambas proteinasas presentan su
máxima actividad a pH ácido. Los resultados también sugieren que las aspártico y
cisteíno proteinasas están involucrados en un proceso secuencial en la hidrólisis de
BSA y Actina, en el que se requiere la participación inicial de las cisteíno proteinasas,
para que las de tipo aspártico puedan intervenir, es decir, las cisteíno proteinasas
constituyen un paso limitante en la hidrólisis de estos sustratos. Por otra parte, la
hidrólisis de la hemoglobina puede completarse por las aspártico y cisteíno
proteinasas de forma individual y por lo tanto, ninguna de ellas constituye un paso
limitante en el proceso.

AGRADECIMIENTOS
En primer lugar quiero agradecer a mis padres, a ellos les debo todo lo que soy. Mi
madre, mi heroína, mi pilar en esta vida, por darme todas las fuerzas y las ganas de
salir adelante. Mi padre, mi ejemplo a seguir, por todos los concejos que me ha dado,
por el apoyo para llegar a cumplir todas mis metas.
Agradezco a mi directora de Tesis Betsaida Bibo, por dirigir éste trabajo, por
contagiarme su entusiasmo y ganas de trabajar, además por todos los consejos y
ánimos, pero sobre todo, por brindarme su amistad.
Agradezco al Dr. Fernando García Carreño, por abrirme las puertas del laboratorio
de BQ, por ayudarme a ampliar mi mente y formular nuevas metas, por involucrarme
en el mundo de la bioquímica y hacer que me guste, como me gusta ahora.
Agradezco a mi abuelo Pancho, mi inspiración, por inculcarme el valor de la
responsabilidad y las ganas de superarse, por enseñarme que la vida no es fácil,
pero es bonita.
Agradezco a mi familia, principalmente a mis tías: Marta, Nena y Yaya, por brindarme
todo su amor y cariño y estar apoyándome siempre.
Agradezco a la Técnico María de Los Ángeles Navarrete del Grupo de BQ por el
apoyo técnico y el entrenamiento en el manejo de técnicas, equipo y organización del
trabajo de laboratorio, además por los consejos y críticas y, no menos importante,
por todo el cariño y muestras de afecto que ha dado.
Agradezco a mi amigo Micro, porque te fuiste de esta vida, cuando apenas
empezaba, pero tu alegría y cariño permanece en cada día de mi vida.
Agradezco a Juan Ángel, porque sin ti, yo ya estuviera loco (más), por apoyarme
siempre, ayudarme a levantarme cuando me caía, ser mi sostén y mi mano derecha.
Agradezco a mis amigos: Diego, Ely, Cari, Rodrigo Edgar y Kalid por ser mi apoyo
durante todos estos años, por todo el cariño, afecto y ánimos que han dado desde

que nos conocimos, tanto en los buenos momentos como en los no tan buenos. Son
mi familia, lo sabe, ocupan un espacio importante en mi vida y en mi corazón.
Agradezco a mis amigos de Las Palma: Patricia, Josef, Evita y Michele, por ser parte
de mi vida, porque estando del otro lado del mundo, representan para mí una familia.
Gracias por ayudarme a abrir las puertas de mi mente y enseñarme a ver las cosas
desde una perspectiva más amplia. Los quiero con toda mi alma.
Agradezco a Tania, Bere y Cristi, por permanecer en mi vida a pesar de la distancia y
por ser unas verdaderas amigas.
Agradezco a todos mis compañeros de la carrera, que de mayor o menor medida,
influyeron en lo que soy ahora, principalmente a Lupita, Janna, Kenia, Zuri, Airy,
Yors, Vale, Nico, Memo. ¡GRACIAS POR TODO!
Agradezco a Erika Torres Ochoa y Sergio Flores Ramírez, por ser parte de mi
comité, por todas sus observaciones y críticas y además por ser parte importante de
mi formación académica.

ÍNDICE
1. INTRODUCCIÓN ........................................................................................................................ 1
2. ANTECEDENTES ....................................................................................................................... 4
2.1 Las proteinasas ..................................................................................................................... 4
2.2 Inhibidores de proteinasas .................................................................................................. 5
2.3 Hidrólisis de proteína por proteinasas digestivas en invertebrados ............................. 7
2.4 Proteinasas del sistema digestivo de crustáceos ............................................................ 8
2.5 El organismo de estudio: langosta americana (Homarus americanus) ..................... 10
2.6 Las proteinasas digestivas de langosta americana ...................................................... 10
3. JUSTIFICACIÓN ....................................................................................................................... 13
4. PREGUNTA CIENTÍFICA ........................................................................................................ 14
5. OBJETIVOS ............................................................................................................................... 15
5.1 Objetivo general ............................................................................................................... 15
5.2 Objetivos específicos ...................................................................................................... 15
6. HIPÓTESIS ................................................................................................................................ 16
7. MATERIAL Y MÉTODO ........................................................................................................... 17
7.1 Obtención y procesamiento de jugo gástrico de langosta americana ........................ 17
7.2 Cuantificación de concentración de proteína y actividad proteolítica total ................ 17
7.3 Identificación de proteinasas activas a pH fisiológico ................................................... 17
7.4 Extracción de Actina de Argopecten circularis. ............................................................. 19
7.5 Ensayo de hidrólisis de proteínas .................................................................................... 20
7.6 Ensayo de inhibición de proteinasas ............................................................................... 21
7.7 Análisis estadístico ............................................................................................................. 23
8. RESULTADOS .......................................................................................................................... 24
8.1 Las proteinasas del jugo gástrico de langosta americana. .......................................... 24
8.2 Proteínas del músculo de Actinopecten circularis. ........................................................ 26

8.3 Monitoreo temporal del proceso de poroteólisis ............................................................ 26
8.4 Ensayo de inhibición de proteinasas ............................................................................... 30
9. DISCUSIÓN ............................................................................................................................... 34
9.1 Identificación de proteinasas en el jugo gástrico de langosta americana. ................ 34
9.2 Monitoreo de la hidrólisis de proteína. ............................................................................ 35
9.3 Participación de las proteinasas en la hidrólisis. ........................................................... 36
10. CONCLUSIONES...................................................................................................................... 40
11. RECOMENDACIONES ............................................................................................................ 41
12. REFERENCIAS ......................................................................................................................... 42
13. LISTA DE TABLAS Y FIGURAS ............................................................................................. 48

1
1. INTRODUCCIÓN
La proteólisis es un proceso de hidrólisis llevado a cabo por las enzimas
llamadas proteinasas, la cual se efectúa por la ruptura de enlaces peptídicos que
unen a los residuos de aminoácidos de las proteínas (Kato, 1999). Las proteinasas, o
enzimas proteolíticas, son las responsables de llevar a cabo esta función. Estas
enzimas se clasifican en exoproteinasas y endoproteinasas, de acuerdo a la
localización del enlace peptídico que hidrolizan dentro de una proteína, los primeros
cortan el enlace peptídico en un amino acido no terminal y las endoproteinasas
únicamente en aminoácidos terminales. Las endoproteinasas, a su vez, se clasifican
en cuatro grupos, de acuerdo a su mecanismo de catálisis en: serino, cisteíno,
aspártico y metalo proteinasas (Rawlings y Barrett, 1993). Estas enzimas operan en
muchos procesos biológicos, tales como la digestión de proteínas provenientes del
alimento, coagulación de la sangre, síntesis de hormonas, apoptosis, entre otros
procesos celulares (Berg, et al. 2002).
La participación de proteinasas en distintos procesos fisiológicos, es a menudo
organizada en cascadas proteolíticas y/o su partición es orquestada de forma
secuencial. Los procesos enzimáticos que ocurren en cascada, son empleados por
los sistemas para alcanzar una respuesta rápida por efecto de una señal; como
ocurre en el proceso de coagulación de la sangre que donde se presenta una
cascada de activación de zimógenos (Krem y Di Cera, 2002). En el sistema digestivo
de mamíferos, la cascada de activación de enzimas digestiva inicia con la secreción
de éstas en el duodeno, en forma de zimógenos; las enteroquinasas, que son
secretadas en el duodeno al entrar la proteína del alimento, activan a las tripsinas,
las cuales subsecuentemente activan otras enzimas como tripsinógeno,
quimotripsinógeno, proelastasa, procarboxipeptidasa y prolipasa. A su vez, las
proteinasas digestivas actúan de manera secuencial para producir péptidos
pequeños y aminoácidos libres que pueden ser absorbidos por las células epiteliales

2
del intestino (Barboza et al. 2008). Por otro lado, en invertebrados se ha descrito que
las proteinasas actúan de manera secuencial, es decir, que las proteinasa de un tipo
catalítico actúan primero sobre el sustrato, y posteriormente, los productos de la
hidrolisis, continúan siendo hidrolizados por las mismas enzimas o por proteinasas
de otro tipo catalítico (Brunelle et al. 1999; Williamnson et al. 2004; Delcrox et al.
2006; Horn et al. 2009).
Los crustáceos, son un grupo de invertebrados que pertenecen al phylum
Arthropoda, uno de los más abundantes y diversos del reino animal. Los crustáceos
presentan variaciones estructurales y fisiológicas en el sistema digestivo, las cuales
dependen de los hábitos alimenticios, la disponibilidad de alimento, las condiciones
del hábitat y el estadio de vida (Biesiot y McDowell, 1990; García-Carreño y Haard
N.F. 1993; García-Carreño et al. 1994 y Johnston y Yellowlees 1998). En los
crustáceos, las proteinasas digestivas son de mecanismos catalíticos variados,
incluyendo serino, cisteíno, metalo y aspártico proteinasas; se ha propuesto que la
transición de cisteíno/aspártico a serino proteinasas en el proceso evolutivo ocurrió
en artrópodos y/o moluscos (Delcrox et al. 2006). Las proteinasas de los crustáceos
son secretadas por la glándula digestiva, también llamada hepatopáncreas, que es el
órgano en que ocurre la síntesis de enzimas digestivas, la absorción de los nutrientes
del alimento y almacenamiento de lípidos y glucógeno (Ceccaldi, 1977). Los
principales componentes proteinolíticos de los sistemas digestivos de las especies de
crustáceos estudiadas a la fecha son serino proteinasas (tripsinas y quimotripsinas)
(Galgani et al. 1985; Tsai et al. 1991, Muhlia-Almazán y García-Carreño, 2003).
La langosta americana, Homarus americanus, es un crustáceo bentónico del
orden Decápoda, perteneciente a la familia Nephropidae. El jugo gástrico de langosta
americana es un medio ácido de pH ~4.7. Algunas proteinasas digestivas de este
organismo han sido purificadas y caracterizadas desde el punto de vista bioquímico y
molecular. Estas proteinasas pertenecen a los tipos catalíticos aspártico (Rojo et al.,

3
2010a, 2010b) y cisteíno (Layckoc et al., 1989, 1991). Sin embargo, poco se ha
ahondado en el estudio del tipo de proteinasas activas bajo las condiciones de pH del
jugo gástrico y que probablemente estén involucradas en la proteólisis que ocurre
dentro de la cavidad gástrica de langosta americana. Tampoco se ha descartado que
las serino proteinasas, que comúnmente son abundantes en otras especies de
crustáceos decápodos, sean activas en la proteólisis. Aunque se han purificado
proteinasas del jugo gástrico de langosta americana, aún no se conoce si estas
orquestan un proceso de proteólisis ordenado, en que la participación de las
proteinasas es consecutiva, o si bien es un proceso que ocurre al azar. El objetivo de
esta tesis fue estudiar las proteinasas digestivas de la langosta americana por medio
de técnicas electroforéticas y desarrollo de zimogramas. Así también, estudiar la
hidrólisis de diversas proteínas por acción de las proteinasas del jugo gástrico e
investigar el posible papel en la proteólisis mediante el uso de inhibidores de los
principales tipos catalíticos de proteinasas, más específicamente si la participación
de alguna de ellas es determinante para el inicio de la proteólisis.

4
2. ANTECEDENTES
2.1 Las proteinasas
Las proteinasas de tipo serino (EC 3.4.21) son una clase de enzimas
proteolíticas caracterizadas por la presencia de un residuo de serina en su sitio
activo. Estas pueden ser encontradas tanto en eucariotas como procariotas y están
involucradas en procesos como la digestión, la homeostasis, la apoptosis, la
reproducción y la respuesta inmune (Porgal, 2005). Las serino proteinasas más
estudiadas son las quimotripsinas las cuales presentan residuos de Ser196-Asp102-
Hist5 en su sitio activo (Hedstrom, 2002).
Las proteinasas de tipo cisteíno (EC 3.4.22), son proteínas involucradas en
muchos procesos bioquímicos que ocurren en los organismos, cuya función principal
es la degradación de péptidos y proteínas. Las proteinasas de tipo cisteíno se dividen
en dos grupos las citosólicas que conforman las calpainas y las lisosomales que
constituyen las catepsinas (catepsina B, C, H, K, L, M, N, S, T, V y W) (Grzonka et al.
2001). La actividad enzimática de las cisteíno proteinasas está relacionada con la
presencia de una diada catalítica formada por un residuo de cisteína e histidina, los
cuales se encuentran presentan actividad en un intervalo de pH de 3.5 a 8.9 (Turk et
al. 1997). La familia de la papaína ha sido mayormente estudiada de este tipo de
proteinasas, la cual tiene como residuos catalíticos a Cys25 y His159. La actividad
enzimática de las cisteíno proteinasas está relacionada con la presencia de una
diada catalítica formada por un residuo de cisteína e histidina, los cuales se
encuentran presentan actividad en un intervalo de pH de 3.5 a 8.9 (Turk et al. 1997).
Las aspártico proteinasas (EC 3.4.23) comprenden un grupo de enzimas
homólogas que incluye a pepsina, quimiosina, renina, catepsina D. Con la excepción
de la renina, su pH óptimo es ácido y todas tienen dos residuos de ácido aspártico

5
involucrados en el mecanismo catalítico (Pear y Blundell, 1984). Las aspártico
proteinasas tienen secuencias características en la región de los dos residuos
catalíticos de ácido aspártico: Asp32-Thr-Gly-Ser en el dominio terminal N y Asp215-
Thr-Gly-Ser/Thr en el dominio terminal C, ambos tienen afinidad por aminoácidos
hidrofóbicos (Davies, 1990).
2.2 Inhibidores de proteinasas
Los inhibidores de proteinasas son compuestos que interactúan con las
proteinasas de tal manera que las inhabilitan para llevar a cabo su función catalítica.
La función fisiológica de estos inhibidores es evitar que ocurra la ruptura de los
enlaces peptídicos de las proteínas, lo que los hace importantes en la iniciación,
mantenimiento y finalización de varios procesos biológicos. Los inhibidores de
proteinasas son estructuras químicas variadas y se encuentranse encuentran
presentes en múltiples formas, así como en numerosos tejidos de plantas y animales,
incluso en microorganismos (Laskowski y Kato, 1980).
Cada inhibidor presenta en su superficie un ambiente químico ideal para que,
al contacto con un tipo determinado de enzima, pueda reconocerlo como posible
sustrato (Laskowski y Kato, 1980). El Pepstatin A, el L-trans-Epoxysuccinyl-
leucylamido(4-guanidino)butano (E-64) y el Pefabloc, son algunos inhibidores que se
emplean comunmente en la caracterización de proteinasas, los cuales inhiben a las
aspártico, cisteíno y serino proteinasas, respectivamente. Estos pueden ser
empleados para procesos de extracción y purificación, análisis de procesos
secuenciales de proteinasas y estudios bioquímicos que requieren proteínas
(Hernández-Cortez et al. 1997; Navarrete del Toro et al. 2006; Rojo et al. 2010a).
El Pepstatin A es un inhibidor reversible de enzimas ácidas como Pepsina,
HIV-, Catepsina D y Renina. Este inhibidor es insoluble en agua, sin embargo, puede

6
diluirse en Sulfóxido de Dimetilo (DMSO). Para que este inhibidor pueda llevar a
cabo su efecto, deben de estar presentes tres elementos: un grupo 3 (S)-hidroxilo en
el tercer residuo de Pepstatina, un grupo isopropilo o su equivalente en el primer
residuo y una porción de grupo dipeptil Ala-Sta en C-terminal (Richt y Sun, 1980). En
trabajos previos el inhibidor Pepstatin A ha sido empleado con el fin de identificar,
purificar y caracterizar las aspártico proteinasas crustáceos (Navarrete del toro et al.
2006; Rojo et al. 2010a).
Otro inhibidor utilizado es el E-64, se ha utilizado como un potente inhibidor
irreversible para muchas cisteíno proteinasas, tales como la papaína, ficina,
actinidina, la catepsina B y L (Matsumoto et al. 1999). El E-64 reacciona con el sitio
activo de la enzima mediante la formación de un enlace tioéter con el grupo
sulfhidrilo del residuo catalítico de la sisteína, sin embargo, se ha observado que no
reacciona con las proteinasas de otras clases (Handa et al. 1978; Matsumoto et al.
1989). Anteriormente este inhibidor ha sido utilizado para inhibir la actividad de
cisteíno proteinasas en decápodos, con el fin de identificar aislar y caracterizar a este
tipo de proteinasas (Laycock et al 1989; Teschke y Saborowski, 2005; Navarrete del
Toro et al. 2006).
Finalmente el inhibidor 4-[2-aminoethyl]benzenesulfonyl fluoride (Pefabloc) es
un inhibidor irreversible específico de serino proteinasas, cuya mayor cualidad es su
solubilidad en soluciones acuosas y la ausencia de toxicidad. Es frecuentemente
utilizado en sustitución del PMSF, ya que tiene características de inhibición similares.
Éste inhibidor reaccionando rápidamente con el residuo de serina de la proteinasa,
formando así una enzima acilada estable, lo que resulta en una disminución de la
actividad enzimática (Mintz, 1993).

7
2.3 Hidrólisis de proteína por proteinasas digestivas en invertebrados
El estudio del proceso de proteólisis del alimento en invertebrados se ha
centrado principalmente en nemátodos parasitarios e insectos. En varios de estos
organismos la hidrólisis de proteína es llevada a cabo por proteinasas del mismo tipo
catalítico, mientras que en otras especies, el proceso es llevado a cabo por
proteinasas de tipos catalíticos diversos. Sin embargo, se ha observado que existen
otros mecanismos por los cuales la proteólisis se lleva a cabo, la cual involucra un
proceso secuencial (Williamnson et al.2004; Delcroix, et al. 2006; Brunelle et al.
1999; Horn et al. 2009).
Williamnson et al. (2004) describió un proceso secuencial de hidrólisis de
hemoglobina por las proteinasas de un anquilostoma (Ancylostoma caninum), un
parásito exclusivo de mamíferos. En este proceso la aspártico proteinasa, tipo
catepsina D (Ac-APR-1) y una cisteíno proteinasa tipo catepsina B (CP-2) inician el
proceso de hidrólisis, posteriormente la metalo proteinasa MEP-1 participa en la
hidrólisis.
Delcroix, et al. (2006) estudiaron la proteólisis en el parásito Schitosoma
mansoni, vector de la enfermedad Esquitosomeasis. Para la hidrólisis de albúmina se
observaron que las cisteíno proteinasas, catepsina B1 y L1, presentes en el sistema
digestivo de este organismo, son necesarias para el inicio del proceso,
posteriormente se les une la catepsina D y la parte final de la hidrólisis la llevan a
cabo la catepsina B1 y C y una metalo aminoproteinasa. Por otro lado, en la
degradación de hemoglobina, la aspártico proteinasa catepsina D es iniciadora del
proceso, posteriormente se le unen las catepsinas B1 y L1 y la parte final de la
hidrólisis ocurre como se describió en el caso de la albúmina.

8
En insectos se ha estudiado el proceso de proteólisis por las proteinasas
digestivas (Boldbaatar, 2006; Ahn y Zhu-Salzman, 2009). Sin embargo, existen
pocos estudios que señalen la presencia de un proceso secuencial de proteólisis.
Brunelle et al. (1999) identificaron el papel de la catepsina D en la digestión de
proteína del escarabajo de la papa de Colorado (Leptinotarsa decemlineata) en la
hidrólisis de diferentes proteínas, siendo esta proteinasa indispensable para iniciar el
proceso, posteriormente la quimotripsina y las catepsinas B y H participan en la
hidrólisis y finalmente las carboxilo y amino proteinasas producen los péptidos y
aminoácidos son absorbidos. Horn et al. (2009) describió un proceso secuencial en la
hidrólisis de hemoglobina, llevada a cabo por las proteinasas de la garrapata común
europea (Ixodes ricinus), en donde la catepsina D soportada por la catepsina L y
legúmina son las encargadas de iniciar la hidrólisis, la cual es continuada por la
catepsina C y B.
2.4 Proteinasas del sistema digestivo de crustáceos
Se han llevado a cabo diversos estudios para conocer la fisiología de la
digestión de proteína en decápodos, principalmente en las especies de importancia
económica de la industria acuicultora. (Lee et al. 1984; Teschke y Saborowski, 2005,
García-Carreño y Haard et al.1993). En las especies estudiadas a la fecha, las
proteinasas del tipo serino , tales como, tripsinas y quimotripsinas, se reconocen
como las más abundantes y se les considera como las responsables de la digestión
de la proteína del alimento, estas enzimas proteolíticas presentan su máxima
actividad a pH 8 (Galgani et al. 1985; Souza-Buarque et al. 2009) Tsai, et al. 1991
estudiaron y diferenciaron las quimotripsinas de tres peneidos (Penaeus monodon,
Penaeus japonicus y Penaeus penicillatus), encontrando similitudes y diferencias en
este tipo de proteinasas. Las tres especies presentaron carboxiloproteinasas A y B,
quimotripsinas y tripsinas, cada una de ellas representadas por varias isoenzimas,
con las quimotripsinas como una de las endoproteinasas digestivas más abundantes.
Hernández-Cortés et al. 1997, en un estudio de purificación y caracterización de

9
quimotripsinas del camarón Penaeus vannamei, identificaron la máxima actividad a
pHs alcalinos y su gran tolerancia a cambios de temperatura.
Se han revisado una serie de estudios en relación a la caracterización de las
enzimas digestivas presentes en el sistema digestivo de distintas especies de
langostas queladas y espinosas. En los estudios realizados, Celis-Guerrero et al.
(2004), caracterizaron las enzimas presentes en jugo gástrico, hepatopáncreas y
contendido intestinal de la langosta Panulirus interruptus, encontrando que las serino
proteinasas son las enzimas más activas, entre estas encontraron tripsinas y
quimotripsinas. Perera et al. (2008) determinaron la presencia de serino y
metaloproteinasas en un estudio de caracterización de proteinasas del jugo gástrico
de Panulirus argus.
Otros estudios han mostrado la participación de la cisteíno proteinasas en
conjunto con proteinasas de tipo serino, en el proceso de hidrólisis de proteína
(Laycock et al. 1989; Teschke y Saborowski, 2005; Vijayaraghavan et al. 2011). Un
ensayo de inhibición de proteinasas en dos especies de la familia Crangonidae
(Crangon crangon y Crangon allmani), señalaron la presencia de cisteíno
proteinasas, cuya inhibición produce una disminución del 70% en el proceso,
mientras que la inhibición de serino petidasas únicamente un 30% (Teschke y
Saborowski, 2005).
Existen pocos estudios donde se le atribuya un papel en la hidrólisis de
proteína a las proteinasas de tipo aspártico en crustáceos Glass y Stark (1994)
sugieren la presencia de proteinasas tipo-pepsina en el jugo gástrico de Homarus
gamarus; estas enzimas fueron posteriormente abordadas por Navarrete del toro et
al. (2006), en un estudio donde compararon las proteinasas de distintos decápodos,

10
bajo diferentes tratamientos de inhibición y pH, demostró la presencia de proteinasas
de tipo aspártico a pH ácido en varias especies de decápodos.
2.5 El organismo de estudio: langosta americana (Homarus americanus)
La langosta americana se distribuye en el Atlántico Norte desde Cape Hatteras
en Carolina del Norte, hasta Labrador y Newfoundland en Canadá; habita en fondos
rocosos cubiertos de algas a no más de 50 metros de profundidad, a una
temperatura que oscila entre los 4 y 26°C (Factor, 1995). Esta especie es
considerada un organismo omnívoro generalista-oportunista, es decir, que pueden
alimentarse de una gran diversidad de organismos, tales como crustáceos,
pelecípodos, gasterópodos, equinodermos, gusanos marinos, macroalgas e incluso
cadáveres; la elección del alimento se relaciona con la abundancia del mismo
(García y Kensler, 1980; Elner y Campbell, 1987). La langosta americana es de gran
importancia económica y ecológica, debido a su valor en la industria de alimentos y a
su importancia trófica en el Atlántico Norte (Brusca y Brusca, 2005; Factor, 1995).
2.6 Las proteinasas digestivas de langosta americana
Existen algunos estudios de caracterización y evaluación de proteinasas
digestivas de langosta americana. Brockerhoff et al. (1970), estudiaron las enzimas
digestivas del jugo gástrico de langosta americana y encontraron la presencia de
lipasas, proteinasas como tripsina y quimotripsina, carboxiloproteinasas A,
desoxiribunocleasas, ribunocleasas, fosfatasas y quitobiasas. En este estudio
también se midió el pH del jugo gástrico, señalando que es un medio ácido (pH ~4.7)
y se encuentra por debajo del pH encontrado regularmente en otros crustáceos
decápodos (que varía usualmente por encima de pH 5) (Ceccaldi, 1997). El pH del
jugo gástrico de langosta fue monitoreado en diversas especies por Navarrete y

11
colaboradores en 2006, donde se encontró que en especies del género Homarus es
más ácido que en especies como Panulirus interruptus, Cáncer pagurus, Calinectes
arcuatos y Calinectes belicosus. En un estudio de inhibición de la actividad
proteolítica del jugo gástrico de langosta americana, se encontró que el 80% de la
actividad es suprimida por efecto del E-64 (Layckoc et al. 1989).
En el estudio de Navarrete et al. (2006), se destaca que a diferencia de otras
especies de crustáceos decápodos, en langosta Europea (Homarus gamarus) la
proteólisis a pH ácido es mayor que la encontrada a pH alcalino; lo que sustenta que
el proceso proteólisis ocurre principalmente a pH ácido. Del análisis de inhibición, el
Pepstatin A y el E-64 son los inhibidores que tienen mayor efecto en la actividad a pH
3, lo que sugiere que además de las cisteíno proteinasas reportadas por Layckoc et
al. (1989), las aspártico proteinasas también tienen participación activa en la
proteólisis, dado que en este caso el pH del ensayo es más próximo al pH fisiológico
del jugo gástrico de la langosta.
A la fecha se han purificado y caracterizado las proteinasas digestivas de los
tipos catalíticos aspártico y cisteíno a partir del jugo gástrico de langosta americana
(Homarus americanus) (Laycock et al. 1989 y 1991; Rojo et al 2010a y 2010b). La
cisteíno proteinasa purificada por Layckoc et al (1989) es una enzima de 28 kDa de
peso molecular, con un pH óptimo de pH 4.5. Esta proteinasa fue purificada por
afinidad al sustrato y por cromatografía de intercambio iónico; en todos los
experimentos utilizaron DTT para activar a las cisteíno proteinasas y amortiguador de
acetato de sodio 100mM a pH 4.7, para nivelar el pH. Años después, se investigó la
secuencia de esta enzima y se determinó que es una catepsina L, la cual presenta 3
izoenzimas: LCP1, LCP2 Y LCP3 (Laycock et al. 1991). Rojo et al. (2010a y 2010b)
mediante técnicas de biología molecular investigaron la naturaleza de las aspártico
proteinasas de langosta americana y europea (Homarus gamarus) y se identificaron
dos isoenzimas, la catepsina D1 (~32 kDa) y catepsina D2 (~45 kDa), los transcritos

12
de la catepsina D1 se encontraron únicamente en el tejido de la glándula digestiva,
mientras que los de la catepsina D2 se encontró en todos los tejidos analizados. Esto
sugiere que la catepsina D1 es una proteinasa digestiva. La secuencia traducida del
RNAm de la catepsina D1 coincide con secuencias obtenidas por análisis de
espectrometría de masas de la aspártico proteinasa purficada del jugo gástrico de
langosta americana y europea. La catepsina D1 del jugo gástrico de langosta tiene
su máxima actividad a pH ácido (pH ~4.7), además presenta una gran especificidad
al sustrato a bajas temperaturas (Rojo et al. 2013).

13
3. JUSTIFICACIÓN
Las proteinasas del sistema digestivo de langosta americana (Homarus
americanus) han sido estudiadas por diferentes autores (Laycock et al. 1989;
Laycock et al. 1990; Navarrete del Toro et al. 2006; Rojo et al. 2010a, 2010b). Se han
identificado cisteíno y aspártico proteinasas activas en el jugo gástrico a pH ácido, a
las cuales se les atribuye que participan en la hidrólisis de proteína, sin embargo, se
desconoce si las proteinasas del tipo serino, participan en el proceso. De igual
manera se desconoce el tipo de proteinasas activas al pH fisiológico del jugo gástrico
de langosta y que por lo tanto son potenciales a participar en el proceso de hidrólisis
de proteína del alimento.
La caracterización de la actividad proteolítica llevada a cabo por las enzimas
digestivas de langosta americana constituye una parte importante en el estudio de la
fisiología digestiva de este organismo, además provee información complementaria
para el estudio de sus necesidades alimenticias y nutricionales (Holthuis, 1991). La
langosta americana es una especie de gran importancia económica, ya que
representa uno de los recursos de mayor valor monetario en algunos sitios en los
que se distribuye (Nicosia y Lavalli, 1999), dado esto, el conocimiento de la biología y
ecología de la especie en función es de gran importancia para su manejo y
conservación, así como para asegurar una explotación sustentable del mismo (Ward,
2000).
Por otro lado, debido las sus propiedades físicas, químicas y catalíticas de las
enzimas proteolíticas, pueden ser utilizadas para hidrolizar proteínas con propósitos
industriales. Por lo que el estudio de estas proteinasas podría contribuir a dar
información acerca del potencial como herramienta biotecnológica en las industrias,
alimentaria, farmacéutica y cosmética (Rasmussen y Morrissey, 2007).

14
4. PREGUNTA CIENTÍFICA
¿Las proteinasas del jugo gástrico que intervienen en la proteólisis, están
involucradas en un proceso secuencial o su participación en el proceso ocurre
meramente al azar? Si el proceso es llevado a cabo de manera secuencial, ¿cuál es
el papel que desempeña cada tipo de proteinasa?

15
5. OBJETIVOS
5.1 Objetivo general
• Describir el proceso de la proteólisis in vitro, que efectúan las proteinasas en
el jugo gástrico de langosta americana (Homarus americanus), a pH
fisiológico.
5.2 Objetivos específicos
• Identificar la composición de proteinasas presentes en el jugo gástrico de H.
americanus
• Describir el proceso de proteólisis en el jugo gástrico de H. americanus, a pH
fisiológico, utilizando extractos de proteína soluble de músculo de organismos
marinos y proteínas comerciales.
• Identificar el papel de las proteinasas en el proceso de proteólisis ocurrido en
el jugo gástrico de H. americanus, a pH fisiológico.

16
6. HIPÓTESIS
Si en otros sistemas digestivos de invertebrados, que basan la proteólisis en
aspártico y cisteíno proteinasas, la hidrólisis es un proceso secuencial que es
iniciado por alguna de ellas, permitiendo así la participación del resto, entonces las
proteinasas del jugo gástrico de langosta americana están involucradas en un
mecanismo similar.

17
7. MATERIAL Y MÉTODO
7.1 Obtención y procesamiento de jugo gástrico de langosta americana
Los ejemplares adultos de langosta americana fueron comprados en un
mercado local de la ciudad de San Francisco, California, EU. El jugo gástrico de
éstos se extrajo de los especímenes vivos y en ayuno, expuestos a condiciones
controladas de salinidad, temperatura y suministro de alimento; por succión,
utilizando una jeringa equipada con tu tubo de diámetro pequeño que se insertó por
la cavidad oral hasta la cavidad gástrica. Inmediatamente después del muestreo se
determinó el pH de las muestras y se transfirieron a tubos de 1.5 mL, los cuales
fueron centrifugados por 10 min a 10 000 g a 4 °C, con el fin de separar los sólidos.
El sobrenadante fue liofilizado y posteriormente almacenado a -20 °C. El extracto se
resuspendió en agua destilada a una concentración de 20 mg/mL, esta preparación
se empleó en los análisis posteriores.
7.2 Cuantificación de concentración de proteína y actividad proteolítica total
La concentración de proteína del jugo gástrico se cuantificó por el método de
Bradford (1976) utilizando BSA como proteína estándar. La actividad proteolítica total
se cuantificó empleando el método reportado por Díaz-López, et al. (1998), con la
modificación de que se empleó amortiguador de acetato de sodio 100mM a pH 4.7.
7.3 Identificación de proteinasas activas a pH fisiológico
Para determinar los tipos de proteinasas que se encuentran activas en el jugo
gástrico de langosta americana se evaluó la actividad enzimática mediante la técnica
de zimograma en geles de acrilamida 12% al pH fisiológico del jugo gástrico (pH 4.7)

18
y a pH alcalino (pH 8.0). Antes de la electroforesis se mezclaron 5 L de jugo
gástrico con 5 L de los inhibidores 1 mM Pefabloc, 1 mM E-64 y 1 mM Pepstatin A,
se incluyó un control que contenía DMSO, el solvente del Pepstatin A. El jugo
gástrico se incubó con los inhibidores durante una hora a temperatura ambiente.
Para contrastar los resultados, se repitió el ensayo ahora incubando con jugo
gástrico previamente tratado con ditrioteitol (DTT) 1.25 mM, con la finalidad de
observar el efecto de este sobre las proteinasas en los zimogramas, posterior al
tratamiento con DTT se siguió la misma metodología que se indica para las muestras
no tratadas con este agente.
Después de la incubación, la reacción fue frenada por la incorporación de
amortiguador de muestra 4X para SDS-PAGE. Las proteínas del jugo gástrico se
separaron en SDS-PAGE 12%. Para la evaluación de los zimogramas a pH ácido,
después de la electroforesis el gel se sometió a tres baños de 10 minutos con
amortiguador de acetato de sodio (100Mm, pH 4.7), con la finalidad de ajustar el pH
del gel. Inmediatamente después, los geles se incubaron a 4°C por 30 min con el
sustrato hemoglobina al 0.25% disuelta en amortiguador de acetato de sodio
(100mM, pH 4.7) y posteriormente a temperatura ambiente por 90 min. Para evaluar
la actividad a pH alcalino (pH 8), se empleó el procedimiento anterior, omitiendo los
baños de ajuste de pH, dado que éste ya presenta un pH alcalino tras la
electroforesis, además se utilizó caseína al 3% como sustrato, disuelta en
amortiguador Tris (50mM a pH 8.0). Finalizado el tiempo de incubación los geles
lavados y teñidos con la solución de Coomassie (0.05% azul de Coomassie R, 40%
Metanol, 7% ácido acético). Bandas claras en un fondo azul indica la presencia de
actividad proteolítica. El efecto de los inhibidores se medirá por comparación de la
intensidad de banda comparando los tratamientos con el control.

19
7.4 Extracción de Actina de Argopecten circularis.
Se realizó la extracción de proteína de músculo aductor de almeja (Argopecten
circularis), utilizando la metodología propuesta por Rodrigáñes, et al. (2011), con la
finalidad de emplearlo como sustrato en los experimentos posteriores a la par de
proteínas comerciales seleccionadas. A. circularis presenta una estrecha relación
evolutiva con los organismos de los cuales se alimenta esta langosta (Elner y
Campbell, 1987).
Los especímenes de A. circularis fueron adquiridos en un mercado local y
trasportados al laboratorio. El músculo aductor se disectó y se cortó en trozos de
aproximadamente un gramo, se añadió amortiguador (0.1M Tris-HCl, 0.25 sucrosa,
1Mm EDTA) a razón de 3:1. La extracción de proteína soluble se realizó por
maceración en un mortero y posteriormente se centrifugó a 10 000g por 10 min a 4
°C en tubos de plástico de 2mL. Después de la centrifugación, se recuperó el
sobrenadante y la proteína soluble se extraída se concentró por ultrafiltración en
membrana Amicon de 10 000 kDa de peso molecular de corte a 4 000 g por una hora
a 4 °C. Se cuantificó proteína del extracto de proteína soluble del extracto y se ajustó
la concentración a 6 mg/mL. El extracto de proteína soluble del músculo de A
circularis se almacenó a -20 ° C hasta su uso.
Se utilizó SDS-PAGE 8% para identificar la composición y masa molecular de
proteína en el músculo de A. circularis (Yamada et al. 2000). Se identificó a actina,
una proteína representativa de músculo, tanto de vertebrados como de
invertebrados, la que fue utilizada como sustrato.

20
7.5 Ensayo de hidrólisis de proteínas
Para investigar el proceso de proteólisis por las proteinasas digestivas de
langosta americana, se hicieron ensayos con tres sustratos: albúmina sérica bovina,
hemoglobina y extracto de proteína soluble del músculo de A. circularis. Un primer
ensayo de actividad con cada sustrato se realizó para obtener una aproximación al
tiempo en que se agota el sustrato en la reacción, para luego hacer reacciones en las
que se incluyeron inhibidores de aspártico, cisteíno y serino proteinasas.
Todas las reacciones se llevaron a cabo en tubos de plástico de 0.5 mL y a
temperatura ambiente. Para investigar el tiempo en que el sustrato se ha agotado en
la reacción, se incubó jugo gástrico (0.15 U/mL de actividad proteolítica a pH ácido)
con 1.25 mM DTT por 10 minutos. Las mezclas de reacción consistieron en: 20 L de
jugo gástrico tratado con DTT, 20 L de DMSO (que se emplearía en los siguientes
ensayos debido a que es el solvente de uno de los inhibidores empleados), 20 L de
amortiguador de acetato de sodio (350 mM, pH 4.7) y 40 L de agua destilada, las
reacciones se iniciaron con la adición de sustrato, todos los sustratos se emplearon a
una concentración de 6 mg/mL.
Las mezclas fueron incubadas por 1200 min, durante los cuales se llevaron a
cabo muestreos a los 0, 15, 60, 240 y 1200 min después de haber comenzado la
incubación, en cada uno de ellos se extrajeron 15 L de la muestra y se les adicionó
5 L de amortiguador de muestra 4X para DTT-SDS-PAGE (Laemmli, 1970). Se
colocaron los tubos en baño de agua hirviendo por 10 minutos para detener la
reacción. Posteriormente se refrigeraron hasta condensar y se centrifugaron a 10 000
g por 5 min a temperatura ambiente, para obtener el líquido condensado en las
paredes del tubo. Los productos de la hidrólisis en la mezcla de reacción de BSA y
de Actina se separaron por SDS-PAGE 12% y los productos de hidrólisis de

21
hemoglobina por SDS-PAGE 15%. Los geles se tiñeron para revelar proteína con
solución de Coomassie.
Los geles se documentaron en un equipo de análisis de imagen (GEL-DOCTM
IMAGE; Bio-Rad). Para el análisis se identificó la banda que corresponde a la
proteína comercial y a la actina en el extracto de proteína soluble del músculo de A.
circularis. Se comparó la cantidad de proteína remanente en cada tratamiento con la
resolución de la proteína utilizada como sustrato, mediante la técnica de
densitometría de imagen, utilizando el software (ImageLab 4.0.1). Con este
experimento se determinó el tiempo en que ocurre la hidrólisis completa del sustrato.
Luego se repitió el mismo experimento realizando los muestreos en el periodo de
tiempo identificado, con la finalidad de tener una aproximación más exacta a este.
7.6 Ensayo de inhibición de proteinasas
Con el objetivo de estudiar de qué manera están actuando las proteinasas del
jugo gástrico de langosta americana en la hidrólisis de los diferentes sustratos y de
esta manera poder identificar la presencia de proceso sucesivo en la hidrólisis, se
llevó a cabo un ensayo de inhibición, utilizando los inhibidores específicos de
proteinasas Pepstatin A (100 M), E-64 (100 M) y Pefabloc (100 M), los cuales
tienen la función de inhibir a las proteinasas de tipo aspártico, cisteíno y serino
respectivamente.
De igual manera que en el ensayo de hidrólisis de proteína, todas las
reacciones se llevaron a cabo en tubos de 0.5 mL a temperatura ambiente. Se inició
el ensayo con la activación de las cisteíno proteinasas del jugo gástrico (0.15 U/mL
de actividad proteolítica a pH ácido) incorporando 1.25 Mm de DTT al extracto y
dejando incubar por 10 min. Posteriormente se incorporaron los inhibidores, el
22
amortiguador de acetato de sodio (350 mM, pH 4.7) y el DMSO, como se muestra en
la tabla I y se dejaron en incubación por 30 min, finalmente se adicionaron 20 L de
sustrato (BSA, Hemoglobina y extracto de proteína soluble del musculo de A.
circularis) y se incubaron hasta el tiempo de hidrólisis máxima de cada uno,
identificado en el experimento anterior. Al finalizarla incubación se extrajeron 15 L
de cada una de las muestras y se les adicionó 5 L de amortiguador de muestra 4X
para DTT-SDS-PAGE. Se colocaron los tubos en baño de agua hirviendo por 10
minutos para detener la reacción. Posteriormente se refrigeraron hasta condensar y
se centrifugaron a 10 000 g por 5 min a temperatura ambiente, para obtener el
líquido condensado en las paredes del tubo. Los productos de la hidrólisis en la
mezcla de reacción de BSA y de Actina se separaron por SDS-PAGE 12% y los
productos de hidrólisis de hemoglobina por SDS-PAGE 15%. Los geles se tiñeron
para revelar proteína con solución de Coomassie. Este experimento se llevó a cabo
por triplicado para cada una de los sustratos experimentales.
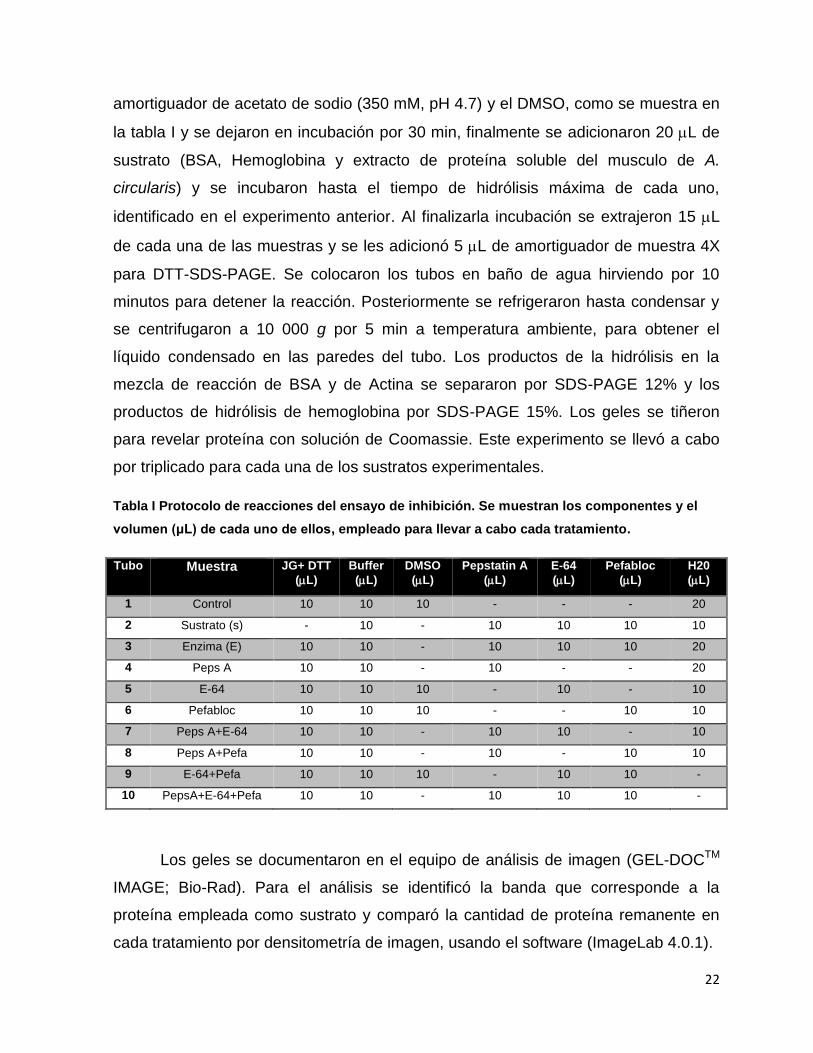
Tabla I Protocolo de reacciones del ensayo de inhibición. Se muestran los componentes y el
volumen (μL) de cada uno de ellos, empleado para llevar a cabo cada tratamiento.
Tubo Muestra JG+ DTT
(L)
Buffer
(L)
DMSO
(L)
Pepstatin A
(L)
E-64
(L)
Pefabloc
(L)
H20
(L)
1 Control 10 10 10 - - - 20
2 Sustrato (s) - 10 - 10 10 10 10
3 Enzima (E) 10 10 - 10 10 10 20
4 Peps A 10 10 - 10 - - 20
5 E-64 10 10 10 - 10 - 10
6 Pefabloc 10 10 10 - - 10 10
7 Peps A+E-64 10 10 - 10 10 - 10
8 Peps A+Pefa 10 10 - 10 - 10 10
9 E-64+Pefa 10 10 10 - 10 10 -
10 PepsA+E-64+Pefa 10 10 - 10 10 10 -
Los geles se documentaron en el equipo de análisis de imagen (GEL-DOCTM
IMAGE; Bio-Rad). Para el análisis se identificó la banda que corresponde a la
proteína empleada como sustrato y comparó la cantidad de proteína remanente en
cada tratamiento por densitometría de imagen, usando el software (ImageLab 4.0.1).

23
7.7 Análisis estadístico
Para cada tratamiento de sustratos, se probó normalidad y homocedasticidad
de varianzas y se realizaron análisis estadísticos en el software STATISTICA 7, para
determinar diferencias entre cada tratamiento de inhibidores. Mediante un análisis de
varianza de una vía (ANOVA) se determinó la presencia de diferencias significativas
(p≤0.05) entre cada tratamiento de cada uno de los sustratos y posteriormente se
utilizó la prueba de diferencia de medias Tukey (Zar, 1984) para determinar grupos
sin diferencias significativas.

24
8. RESULTADOS
8.1 Las proteinasas del jugo gástrico de langosta americana.
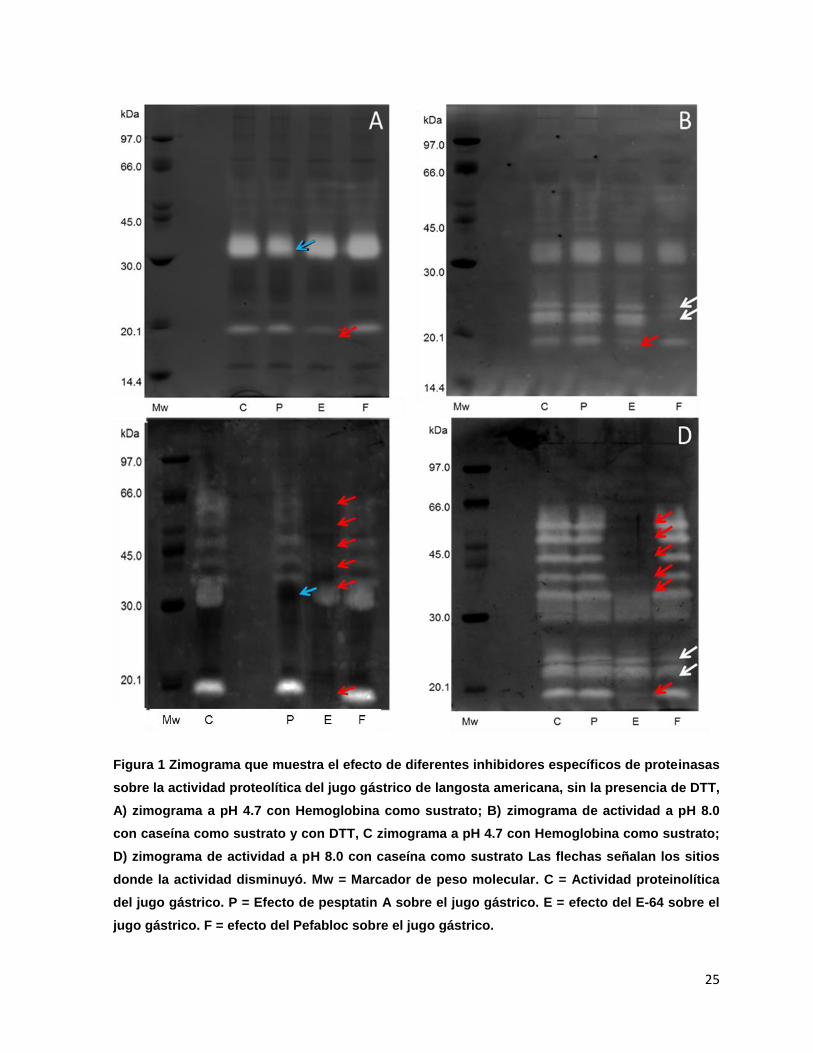
La actividad de las proteinasas del jugo gástrico de langosta americana se
identificó mediante un análisis del efecto de diferentes inhibidores, utilizando como
base las bandas en los zimogramas realizados. Se observó que a pH 8.0 hay nueve
bandas de actividad, de las cuales seis de ellas no son visibles en los carriles donde
se colocaron las muestras con E-64 (Figura 1-C), lo cual indica que pertenecen a la
clase cisteíno (18.5, 37, 42, 48, 54 y 60 kDa), otras dos bandas disminuyen
su actividad en las muestras que contenían Pefabloc, por lo que se puede considerar
una proteinasa de la clase serino (Figura 1-D); la banda restante pertenece a la clase
aspártico (37 kDa) dado que se vio reducida su actividad en las muestras que
fueron incubadas con Pepstatin A como se observa en la figura 1-A.
Por otro lado, en el zimograma a pH 4.7 incubado con DTT se identifican seis
bandas de actividad, todas ellas observadas en el zimograma a pH 8.0, sin embargo,
presentan una mayor intensidad (Figura 1-A,B), lo que hace que la cisteíno petidasa
de 37kDa no se pueda identificar. Las muestras que contenían Pefabloc como
inhibidor no vieron afectada su actividad, lo cual señala que las serino proteinasas no
están activas a pH 4.7 (Figura 1-A,C).
25
Figura 1 Zimograma que muestra el efecto de diferentes inhibidores específicos de proteinasas
sobre la actividad proteolítica del jugo gástrico de langosta americana, sin la presencia de DTT,
A) zimograma a pH 4.7 con Hemoglobina como sustrato; B) zimograma de actividad a pH 8.0
con caseína como sustrato y con DTT, C zimograma a pH 4.7 con Hemoglobina como sustrato;
D) zimograma de actividad a pH 8.0 con caseína como sustrato Las flechas señalan los sitios
donde la actividad disminuyó. Mw = Marcador de peso molecular. C = Actividad proteinolítica
del jugo gástrico. P = Efecto de pesptatin A sobre el jugo gástrico. E = efecto del E-64 sobre el
jugo gástrico. F = efecto del Pefabloc sobre el jugo gástrico.

26
8.2 Proteínas del músculo de Actinopecten circularis.
Por medio de electroforesis se identificó las proteínas presentes en el músculo
de Argopecten circularis, Miosina (220 kDa), Paramiosina (111 kDa) y Actina (42
kDa) (Figura 2). Dado la abundancia de la Actina, bajo las condiciones
experimentales de este trabajo, se tomó como proteína referente en el resto de los
experimentos.
Figura 2 SDS-PAGE de las principales proteínas presentes en el músculo de A. circularis. LMH
y HMW son marcadores de peso molecular, alto y bajo respectivamente.
8.3 Monitoreo temporal del proceso de poroteólisis
Mediante un análisis de intensidad de banda por densitometría, se realizó un
monitoreo de los productos de hidrólisis de los sustratos a diferentes tiempos de
reacción por las proteinasas del jugo gástrico de langosta americana, para evaluar el
tiempo en que el sustrato se agota en la reacción.
27
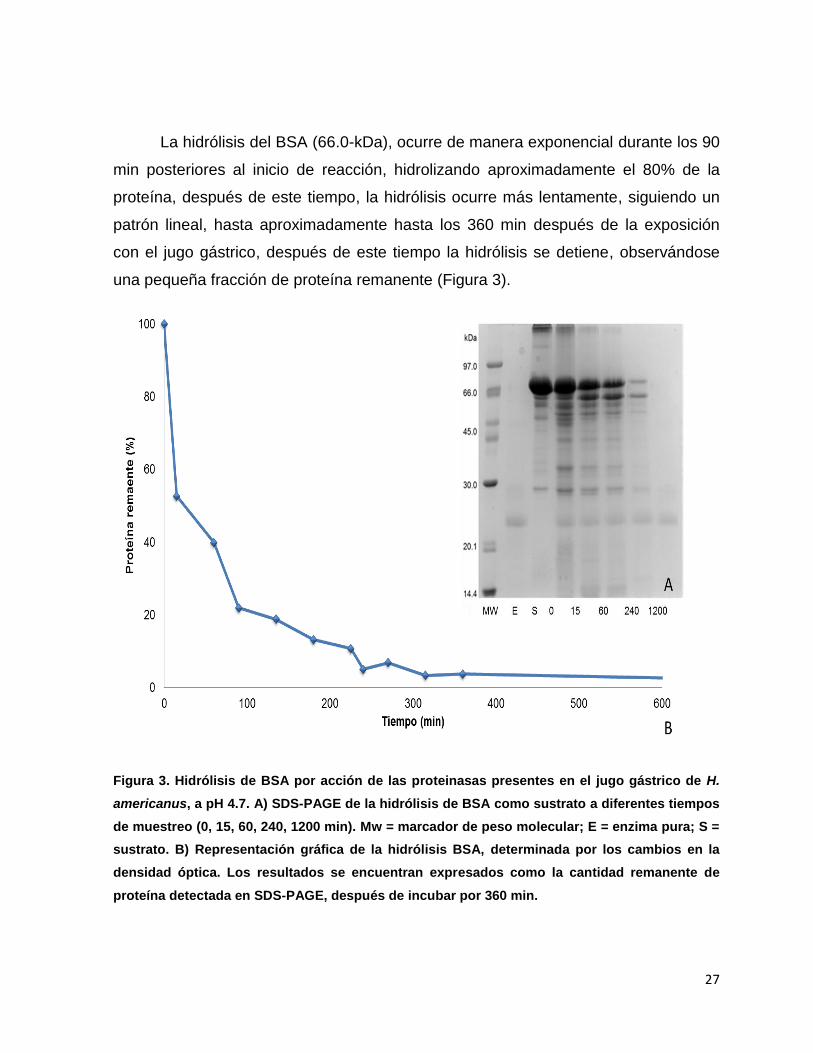
La hidrólisis del BSA (66.0-kDa), ocurre de manera exponencial durante los 90
min posteriores al inicio de reacción, hidrolizando aproximadamente el 80% de la
proteína, después de este tiempo, la hidrólisis ocurre más lentamente, siguiendo un
patrón lineal, hasta aproximadamente hasta los 360 min después de la exposición
con el jugo gástrico, después de este tiempo la hidrólisis se detiene, observándose
una pequeña fracción de proteína remanente (Figura 3).
Figura 3. Hidrólisis de BSA por acción de las proteinasas presentes en el jugo gástrico de H.
americanus, a pH 4.7. A) SDS-PAGE de la hidrólisis de BSA como sustrato a diferentes tiempos
de muestreo (0, 15, 60, 240, 1200 min). Mw = marcador de peso molecular; E = enzima pura; S =
sustrato. B) Representación gráfica de la hidrólisis BSA, determinada por los cambios en la
densidad óptica. Los resultados se encuentran expresados como la cantidad remanente de
proteína detectada en SDS-PAGE, después de incubar por 360 min.

28
En el caso de la hemoglobina (14.4-kDa), la degradación ocurre rápidamente
durante los primeros 15 minutos, luego disminuye la cantidad de proteína cerca del
50%. Posterior a esto, el potencial de hidrólisis ocurre de una manera más lenta, en
donde se presentó patrón lineal, hasta degradarse completamente pasados 480 min
desde el inicio de la reacción (Figura 4).
Figura 4. Hidrólisis de Hemoglobina por acción de las proteinasas presentes en el jugo gástrico
de H. americanus, a pH 4.7. A) SDS-PAGE de la hidrólisis de Hemoglobina como sustrato a
diferentes tiempos de muestreo (0, 15, 60, 240, 480, 600, 1200 min). Mw = marcador de peso
molecular; E = enzima pura; S = sustrato. B) Representación gráfica de la hidrólisis de
hemoglobina, determinada por los cambios en la densidad óptica. Los resultados se
encuentran expresados como la cantidad residual de proteína detectada en SDS-PAGE,
después de incubar por 480 min.
A

29
El efecto de la hidrólisis de la actina (42 kDa) ocurre de una manera similar a
la hidrólisis del BSA y la hemoglobina, es decir, se presenta un decremento
exponencial durante el inicio de la reacción, hasta disminuir a aproximadamente un
80% de la cantidad de proteína original durante los primeros 15 min; además
presenta un decremento lineal que se prolonga hasta los primeros 60 min de la
reacción, posterior a este tiempo, la degradación de proteína ha ocurrido casi en su
totalidad (Figura 5).
Figura 5. Hidrólisis de Actina de A. circularis por acción de las proteinasas presentes en el jugo
gástrico de H. americanus, a pH 4.7. A) SDS-PAGE de la hidrólisis de Actina de extracto
muscular de A. circularis, a diferentes tiempos de muestreo (0, 15, 60, 240, 360, 1200 min). Mw =
marcador de peso molecular; E = enzima pura; S = sustrato. B) Representación gráfica de la
hidrólisis Actina, determinada por los cambios en la densidad óptica. Los resultados se
encuentran expresados como la cantidad residual de proteína detectada en SDS-PAGE,
después de incubar por 240 min.

30
8.4 Ensayo de inhibición de proteinasas
La hidrólisis de BSA, en las muestras que fueron incubadas con los inhibidores
Pepstatin A, E-64 y Pefabloc por separado, muestra variaciones significativas en la
cantidad de proteína remanente. Las muestras incubadas con Pepstatin A y Pefabloc
presentan una cantidad de proteína cercana al 0%, lo que indica que la hidrólisis se
llevó a cabo en su totalidad, en cambio las muestras incubadas con E-64 muestran
cerca del 60% de proteína remanente, lo cual señala que la hidrólisis de proteína se
llevó a cabo parcialmente. Por otro lado, el resultado cambia al incubar las muestras
con mezclas de los distintos inhibidores. Las muestras incubadas con Pepstatin A y
E-64, al igual que las incubadas con los tres inhibidores presentaron valores
estadísticamente iguales a la cantidad de proteína pura utilizada como sustrato, lo
que indica que la hidrólisis se detuvo. Sin embargo, no ocurre lo mismo al incubar las
muestras con Pepstatin A y Pefabloc, ya que la hidrólisis no se detiene. Al incubar E-
64 y Pefabloc, se presenta el mismo efecto que incubar únicamente con E-64. Como
se muestra en la figura 6-B y corroborado con el análisis de varianza y la prueba
Tukey (P < 0.05) los tratamientos que contenían Pepstatin A+E-64 y Pepstatin A y la
combinación de los tres inhibidores son significativamente iguales al control negativo;
de igual manera, las muestras que contenían E-64 y E-64+Pefabloc son
estadísticamente iguales entre sí; así mismo, las muestras que contenían Pepstatin
A, Pefabloc y la combinación de ambos son significativamente iguales (Figura 6).

31
Figura 6 Representación gráfica (A) de la hidrólisis de BSA por acción de las proteinasas
presentes en el jugo gástrico de H. americanus, en ausencia o presencia de inhibidores
específicos. La imagen muestra la cantidad de proteína remanente, observada en SDS-PAGE
(B), después del tiempo de hidrólisis máxima (360 min), comparada con la cantidad de proteína
del sustrato puro (S). Cada barra representa la media de las tres réplicas. Barras con la misma
letra señalan que no existen diferencia significativas (P < 0.05). (MW = Marcador de peso
molecular; C = control; JG = jugo gástrico; P = Pepstatin A; E = E-64; F = Pefabloc).
La hidrólisis de hemoglobina, en las muestras que fueron incubadas con
Pepstatin A, E-64, Pepstatin A + Pefabloc y E-64 + Pefabloc mostraron muy poca
variación en el grado de hidrólisis, mostrándose valores que van desde el 15% al
30% de cantidad de proteína remanente, lo cual, en base a análisis estadísticos, los
hace iguales (P < 0.05). Por otro lado, en las muestras incubadas con los inhibidores
Pepstatin A + E-64 y con la mezcla de los tres inhibidores, se observa una cantidad
de proteína remanente que va del 85% al 90% respectivamente, lo cual, junto con el
control, los hace estadísticamente iguales (Figura 7).

32
Figura 7 Representación gráfica (A) de la hidrólisis de BSA por acción de las proteinasas
presentes en el jugo gástrico de H. americanus, en ausencia o presencia de inhibidores
específicos. La imagen muestra la cantidad de proteína remanente, observada en SDS-PAGE
(B), después del tiempo de hidrólisis máxima (480 min), comparada con la cantidad de proteína
del sustrato puro (S). Cada barra representa la media de las tres réplicas. Barras con la misma
letra señalan que no existen diferencia significativas (P < 0.05). (MW = Marcador de peso
molecular; C = control; JG = juago gástrico; P = Pepstatin A; E = E-64; F = Pefabloc).
La hidrolisis de la Actina extraída de Argopecten circularis, presenta gran
semejanza con la hidrólisis de BSA, en relación a la manera en que los inhibidores
actúan sobre las proteinasas, ya que el resultado al utilizar Pepstatin A y Pefabloc
por separado y en una mezcla de ambos, no producen ningún efecto sobre la
hidrólisis, consumiéndose casi el 100% de la proteína. Al incubar con E-64 de
manera individual y en combinación con Pefabloc, los efectos sobre la hidrólisis son
muy similares, observándose cantidades de proteína remanente de 27%. Los efectos
al incubar las reacciones con Pesptatina A y E-64 en conjunto, muestran los mismos
resultados que se observan con BSA y Hemoglobina, ya que frenan casi en su
totalidad la hidrólisis, de igual manera ocurre al incubar con la mezcla de los tres

33
inhibidores. Los análisis estadísticos muestran resultados muy similares al ensayo
realizado con BSA (Figura 8).
Figura 8. Representación gráfica (A) de la hidrólisis de BSA por acción de las proteinasas
presentes en el jugo gástrico de H. americanus, en ausencia o presencia de inhibidores
específicos. La imagen muestra la cantidad de proteína remanente, observada en SDS-PAGE
(B), después del tiempo de hidrólisis máxima (240 min), comparada con la cantidad de proteína
del sustrato puro (S). Cada barra representa la media de las tres réplicas. Barras con la misma
letra señalan que no existen diferencia significativas (P < 0.05). (MW = Marcador de peso
molecular; C = control; JG = jugo gástrico; P = Pepstatin A; E = E-64; F = Pefabloc).

34
9. DISCUSIÓN
9.1 Identificación de proteinasas en el jugo gástrico de langosta americana.
Las serino proteinasas tripsina y quimotripsina, son los componentes
proteinolíticos más comunes en el tracto digestivo de crustáceos decápodos
estudiados a la fecha (Tsai et al. 1991; Celis-Guerrero et al. 2004; Navarrete del Toro
et al. 2011). Dado el mecanismo de catálisis de estas proteinasas, su pH óptimo es
alcalino (pH ~8.0). Brockerhoff et al. (1970) determinó la presencia de serino
proteinasas en Homarus americanus a pH alcalino; sin embargo, Rojo et al. (2010a)
señalan que no presentan actividad al pH fisiológico de la langosta, lo cual concuerda
con los resultados obtenidos en este estudio. Las serino proteinasas se encontraron
activas en los zimogrmas a pH alcalino pero no así al pH fisiológico de la langosta, lo
cual sugiere que estas no participan en el proceso de hidrólisis de proteína al pH
fisiológico.
En el presente trabajo se destaca la presencia de una aspártico proteinasa,
que corresponde a la catepsina D1 purificada y caracterizada por Rojo et al. (2010a;
2010b; 2013). Sin embargo, la actividad más conspicua identificada en los
zimogramas corresponde a cisteíno proteinasas, tipo catalítico del que se
identificaron múltiples bandas de actividad. De las cisteíno proteinasas identificadas
en el presente trabajo, al menos una de ellas corresponde a una Catepsina L
(Laycock et al. 1991).
Por otro lado, es evidente la presencia de una proteinasa de tipo aspártico y
varias del tipo cisteíno activas tanto a pH fisiológico como a pH alcalino; sin embargo,
a pH alcalino observa disminuida su actividad, ya que, tanto las aspártico como
cisteíno proteinasas se caracterizan por tener un pH óptimo y estabilidad a pH ácido

35
y son inestables a pH alcalino (Mihalyi, 1978; Whitaker, 1994). De las proteinasas
que se encontraron activas en los zimogramas, a pH ácido y alcalino, se puede
identificar la catepsina L reportada por Laycock et al. (1991) y la catepsina D1
reportada por Rojo et al. (2010a,b). La actividad mostrada por las cisteíno
proteinasas a pH 8.0, a pesar de saber que su actividad es muy baja a este pH
(Whitaker, 1994) se debe posiblemente a la presencia de ditioterietol (DTT) en las
muestras, ya que este compuesto protege a las cisteíno proteinasas del contacto con
el oxígeno, manteniendo un ambiente reductor (-SH) y de esta manera pueden
mantener su actividad (Price, 1991).
9.2 Monitoreo de la hidrólisis de proteína.
El objetivo del presente estudio fue conocer el mecanismo de hidrólisis de
proteína por acción de las proteinasas del jugo gástrico de H. americanus, y así
observar el comportamiento de la hidrólisis de tres distintas proteínas a través del
tiempo. Dos de los sustratos empleados son proteínas comerciales (BSA y
Hemoglobina), mientras que la actina, se extrajo del músculo abductor de la almeja
catarina (A. circularis) con el fin de representar de una manera más cercana, el
proceso de proteólisis en el jugo gástrico, en condiciones naturales (Figura 5).
El presente estudio muestra que BSA, Hemoglobina y Actina, son proteínas
que son hidrolizadas por las enzimas del jugo gástrico, sin embargo, la eficiencia de
las proteinasas para llevar a cabo la hidrólisis, depende del sustrato. En todos los
experimentos se empleó la misma cantidad de unidades de actividad proteolítica
total, sin embargo, el tiempo en que se registró el agotamiento del sustrato varió.
Mientras que la hidrólisis de la Actina es muy eficiente (~60 min) (Figura 5), la
hidrólisis del BSA y la hemoglobina se lleva a cabo muy lentamente (~360 min y ~
480 min, figura 3 y 4 respectivamente). La variación en la duración y la manera en
que se lleva a cabo la hidrólisis de las proteínas depende principalmente de la

36
solubilidad, la complejidad estructural y la composición de aminoácidos de las
proteínas (Rodrigañez, et al. 2011).
9.3 Participación de las proteinasas en la hidrólisis.
Los resultados obtenidos del ensayo de inhibición, apuntan a que las serino
proteinasas no representan un papel importante en la hidrólisis de proteína, ya que a
este tipo de proteínas están generalmente inactivas, de tal manera que al incluir
inhibidores de serino proteinasas, la hidrólisis de los sustratos proteicos puede
continuar (Rawlings y Salvesen, 2003). Esto sugiere que las proteinasas
involucradas en esta función son las de tipo cisteíno y aspártico.
Los resultados arrojados en los ensayos de inhibición específica de
proteinasas señalan que al inhibir las proteinasas de tipo cisteíno, la hidrólisis de
proteína se detiene un 65% en el caso del BSA y un 50% en el caso de la actina, sin
embargo, al inhibir las proteinasas de tipo aspártico la hidrólisis no se detiene, ya que
la hidrólisis se lleva a cabo en su totalidad (Figura 6 y 8 respectivamente). Al inhibir
tanto aspártico como cisteíno proteinasas, la hidrólisis se detiene casi en su totalidad,
observando una cantidad de proteína remanente muy similar a la de la proteína pura
utilizada como sustrato. Este mismo fenómeno ocurre al inhibir los tres tipos de
proteinasas, y dado que las serino proteinasas no están participando, se sugiere que
la hidrólisis de la proteína por parte de las cisteíno proteinasas es necesaria para que
las de tipo aspártico puedan participar en la hidrólisis del sustrato. Este fenómeno
apunta a un posible proceso secuencial de proteólisis, en donde, una vez que las
cisteíno proteinasas hayan hidrolizado a la proteína, las aspártico proteinasas en
sinergismo con las cisteíno van a hidrolizar los péptidos restantes hasta degradarlos
a aminoácidos o péptidos más pequeños (Figura 9). Un proceso secuencial en el
proceso de proteólisis ha sido identificado en el tracto digestivo de vertebrados
(Lenaric et al. 1991) donde la catepsina D se encarga de fragmentar a la proteína y

37
de esta manera las cisteíno en sinergia con la catepsina D, actúan rompiendo los
enlaces peptídicos de los fragmentos más pequeños. De igual manera, dentro del
grupo de los artrópodos se ha observado un mecanismo similar. Se ha demostrado
que en el proceso de degradación del alimento del escarabajo de la papa de
Colorado (Leptinotarsa decemlineata), la catepsina D inicia la hidrólisis de la
proteína, rompiendo los enlaces peptídicos de esta, produciendo fragmentos, las
cisteíno participar en la hidrólisis de proteína (Brunelle et al. 1991), de igual manera
se ha identificado este mismo proceso en el sistema digestivo del escarabajo de la
semilla de frijol (Callosobruchus maculatus) (Ahn y Zhu-Salzman, 2009).
La catepsina D y la catepsina L son las únicas proteinasas que han sido
identificadas en el jugo gástrico de langosta americana a pH ácido (Laycock, et al.
1998; Rojo et al. 2010a, 2010b). Se sabe que la catepsina D tiene afinidad por lo
aminoácidos hidrofóbicos (leucina y fenilalanina principalmente) (Sun, et al. 2013), de
igual manera la catepsina L muestra afinidad por este mismo tipo de aminoácidos
(Rawlings y Barrett, 1993). Al poseer afinidad por el mismo tipo de aminoácidos, se
sugiere que otros factores, independientes a la composición de aminoácidos, están
siendo determinantes para que se efectúe la proteólisis.
De manera general, las proteínas en su estructura terciaria, presentan la
mayor parte de sus aminoácidos hidrofóbicos orientados hacia el interior de la
molécula (Bajaj y Blundell, 1984), lo que haría imposible la participación tanto de la
catepsina D como de la catepsina L en la proteólisis. Sin embargo, la catepsina L de
langosta americana podría presentar características únicas de especificidad,
reconociendo además residuos hidrofílicos, tal como se ha reportado para catepsinas
L de otros invertebrados (Horn et al. 2009). Esto facilitaría a la catepsina L participar
inicialmente en la hidrólisis y, posteriormente, permitir a la catepsina D participar en
el proceso. Otro posible escenario, que sustenta la participación de las cisteíno
proteinasas en el inicio de la hidrólisis de proteína, dada la presencia de varias

38
bandas de actividad observadas en los zimogramas a pH ácido, sugiere que el jugo
gástrico de langosta americana podría presentar otros tipos de cisteíno proteinasas,
que están participando en la hidrólisis, por ejemplo catepsina B, la cual ha sido
identificada recientemente en otros sistemas de decápodos (Aoki et al. 2004;
Stephens et al. 2012). La catepsina B tiene afinidad por aminoácidos hidrófobicos
grandes y además tiene la particularidad de reconocer residuos básicos,
principalmente arginina (Hasnain et al. 1993), los cuales, al ser hidrofílicos, suelen
encontrarse en la periferia de las proteínas (Berg et al. 2002). Facilitando así, que las
proteinasas puedan hidrolizar el sustrato.
Figura 9 Diagrama de la degradación de BSA y Hemoglobina por acción de las péptidas del
jugo gástrico de H. americanus. La proteína ingerida que llega al jugo gástrica es fragmentada
por acción de las cisteíno proteinasas, lo cual expone los sitios catalíticos para que las
aspártico proteinasas puedan actuar; ambas enzimas degradan los fragmentos a péptidos para
que, posteriormente, puedan ser absorbidos por el epitelio.

39
Por otro lado la manera en que actúan de las proteinasas en la proteólisis, va
a estar determinada por la naturaleza del sustrato (Sun et al. 2013). Al incubar la
Hemoglobina con las enzimas del jugo gástrico, se observa que tanto las proteinasas
de tipo cisteíno como las de tipo aspártico pueden llevar a cabo la hidrólisis de
proteína, ya que independientemente de la inhibición de alguna de ellas, la
hemoglobina es hidrolizada, esto señala que ambas proteinasas participan en la
proteólisis, posiblemente de manera sinérgica y, por lo tanto, no existe un paso
limitante para que se lleve a cabo la hidrólisis. Un estudio donde llevaron a cabo
reacciones de inhibición de cisteíno y aspártico proteinasas de Plasmodium
falciparum, vector de la malaria, para hidrolizar hemoglobina, demostró la
inespecificidad de ambas proteinasas al sustrato, de igual manera demostraron la
necesidad de ambos tipos para llevar a cabo la hidrólisis de hemoglobina (Semenov
et al. 1989). Algo similar ha sido observado en las enzimas proteolíticas de tipo
aspártico CmCatD y cisteíno CmCPS del escarabajo de la semilla del frijol
(Callosobruchus maculatus), de igual manera actúan en conjunto, sin embargo,
cuando las aspártico proteinasa está ausente, la cisteíno aumenta su actividad (Ahn
y Zhu-Salzman, 2009); en estos trabajos la catepsina D siempre es más eficiente,
que otros tipos de proteinasas en hidrolizar a la hemoglobina. Por otro lado, se ha
sugerido que la catepsina D, apoyada en menor medida por la catepsina L, están
involucradas en el primer paso del proceso de hidrólisis de hemoglobina y
posteriormente el resto de las proteinasas continúan con la hidrólisis (Horn et al.
2009). Dado que estas dos proteinasas, son las únicas que se han identificado bajo
las condiciones de estudio, se podría sugerir que éstas den inicio a la hidrólisis,
permitiendo a los demás tipos de proteinasas, aún no identificados, participar en la
hidrólisis.

40
10. CONCLUSIONES
En el jugo gástrico de Homarus americanus se presentan proteinasas de tipo
aspártico, cisteíno activas a pH fisiológico (pH 4.7) y con poca actividad a pH
alcalino, por el contrario, se encuentran serino proteinasas activas a pH alcalino y
ausentes de actividad a pH ácido.
La Albúmina sérica bovina, la hemoglobina y la actina, son proteínas que son
hidrolizadas por las proteinasas del jugo gástrico.
El proceso de hidrólisis de las proteínas va a depender del sustrato sobre el que
actúan las proteinasas.
El proceso de proteólisis del jugo gástrico de Homarus americanus sobre algunas
proteínas se lleva a cabo de forma secuencial, ya que la intervención de la
proteinasas de tipo cisteíno, en la hidrólisis de algunas proteínas, representan un
paso limitante para que las proteinasas de tipo aspártico puedan involucrarse en
el proceso.

41
11. RECOMENDACIONES
Se recomienda llevar a cabo investigaciones posteriores, con el fin de
identificar los tipos de cisteíno proteinasas, aún desconocidas, observadas en los
zimogramas a pH fisiológico y llevar a cabo estudios utilizando inhibidores de cada
una de ellas para, de esta manera, describir con más detalle el procesos de hidrólisis
de proteína en el jugo gástrico de langosta americana.

42
12. REFERENCIAS
Ahn, J-E. K. Zhu-Salzman. 2009. CmCatD, a cathepsin D-like protease has a
potential role in insect defense against a phytocystatin. J. Insect Physiol.55:
678-685.
Barboza, P. S., Parker, K. L., Hum, I. D. 2008. Integrative wildlife nutrition. Ed.
Springer. 360 pp.
Berg, J. M., Tymoczko, J. L. y Stryer, L. 2002. Biochemistry. 5th edition.WH Freeman.
Nueva York. 1515 pp.
Biesiot, P.M., McDowell Capuzzo, J., 1990. Changes in digestive enzyme activities
during early development of the American lobster Homarus americanus Milne
Edwards. J. Exp. Mar. Biol. Ecol. 136, 107–122.
Bradford, M. 1976. A rapid and sensitive method for the quantitation of microgram
quantities of protein utilizing the principle of protein-dye binding. Anal Biochem
72:248–254.
Brockerhoff, R., Hoyle, R. J., and Hwang, P. C. 1970. Digestive enzymes of the
American lobster (Homarus americanus).J Fish Res Bd Can 27:1357–1370.
Brunelle, F., Nguyen-Quoc, B., Cloutier, C. and Michaud, D.1999.Protein hydrolysis
by Colorado potato beetle, Leptinotarsa decemlineata, digestive proteinases:
the catalytic role of cathepsin D. Arch. Insect Biochem. Physiol. 42, 88–98.
Bajaj, M. and Blundell, T. 1984. Evolution and the tertiary structure of proteins. Ann.
Rev. Biophys. Bioeng. 13: 453-92
Brusca, R. C. y Brusca, G. J. 2005. Invertebrados. 2da edición. McGraw
Hill/Interamericana de España, S. A. 1005 pp.
Ceccaldi, H. J. 1997. Anatomy and physiology of the digestive tract of Crustaceans
Decapods reared in aquaculture. Advances in Tropical Aquaculture. AQUA-
COP. IFREMER Actes de Colloque, 243-259.

43
Celis-Gerrero, L. E., García-Carreño, F. L., Navarrete del Toro, M. A., 2004.
Characterization of proteases in the digestive system of spiny lobster
(Panulirus interruptus). Mar.Biotechnol. 6, 262–269.
Davies, D. R. The structure and function of aspartic proteinases. Annu. Rev. Biophys.
Biophys. Chem. 1990.19:189-215
Delcroix, M., Sajid, M., Caffrey, C. R., Lim, K. C., Dvorák, J., Hsieh, I., Bahgat, M.,
Dissous, C., McKerrow, J. H. 2006. A multienzyme network functions in
intestinal protein digestion by a platyhelminth parasite. J Biol Chem 281:
39316-39329.
Díaz- López, M. and García-Carreño, F. L. 2000 Applications of Fish and Shellfish
Enzymes in Food and Feed Products. In: Seafood Enzymes. Marcel Dekker,
Inc. 571-618.
Elner, R. W., Campbell, A. 1987. Natural diets of lobster Homarus americanus from
barren ground and macroalgal habitats of southwestern Nova Scotia, Canada.
Mar. Ecol. Prog. Ser. 37, 131–140.
Factor, J. R. 1995. Biology of the Lobster: Homarus americanus. Academic Press.
528pp.
García, A., Kensler, C. B. 1980. Las langostas de México: su biología y pesquería.
An. Inst. Cienc. del Mar y Limnol. Univ. Nal. Autón. México. 7(2): pp. 111-128.
García-Carreño, F. L., Haard, N. F. 1993. Characterization of proteinase classes in
langostilla (Pleuroncodes planipes) and crayfish (Pacifastacus astacus)
extracts. J Food Biochem 17: 97–113.
García-Carreño, F. L., Hernández-Cortés, M. P., and Haard, N. 1994. Enzymes with
peptidase and proteinase activity from the digestive system of a fresh water
and a marine decapod. J AgricFoodChem 42:145–146.
García-Carreño, F. L., Navarrete-del Toro, M. A. 1997. Clasification of proteases
without tears. Biochemical Education 25(3):161-167.

44
Glass, H., y Stark, J. 1994. Protein digestion in the European lobster Homarus
gammarus (L). Comp BiochemPhysiol108B(2): 225–235.
Hanada, K., Tamai, M., Ohmura, S., Sawada, J., Seki, T., Tanaka, I. 1978. Structure
and synthesis of E-64, a new thiol protease inhibitor. Agric. Biol. Chem. 42,
529-536
Hasnain, S., Hirama, T., Huber, C. P., Mason, P., Mort J. S. 1993. Characterization of
cathepsin B specificity by site-directed mutagenesis. The importance of Glu245
in the S2-P2 specificity for arginine and its role in transition state stabilization. J.
Biol. Chem, 268, 235-240.
Hedstrom, L. 2002. Serine protease mechanism and specificity. Chem. Rev., 102:
4501-4524.
Hernández-Cortés, P. J., Whitaker, R., García-Carreño, F. L. 1997. Purification and
characterization of chymotrypsin from Penaeus vannamei, Journal of Food
Biochesmestry, 21: 497-514.
Horn, M., M. Nussbamerova, M. Sanda, Z. Kuvarova, J. Srba, Z. Franta, D. Sojka, M.
Bogyo, C. R. Caffrey, P. Kopacek, et al. 2009. Hemoglobin digestion in blood-
feeding ticks: mapping a multipeptidase pathway by functional
proteomics. Chem. Biol. 16:1053–1063.
Holthuis, L. B. 1991. Marine lobsters of the world. FAO species catalogue. 13 (125):
292 p.
Johnston D. J. and Yellowlees D. 1998. Relationship between dietary preferences
and digestive enzyme complement of the slipper lobster Thenus orientalis
(Decapoda: Scyllaridae). J. Crustac. Biol. 18: 126 – 135.
Laskowski, M. Jr., Kato, I. 1980. Protein inhibitors of proteinases. Annual Review of
Biochemistry. 49, 593-626.
Laemmli, U. K. 1970. Cleavage of structural proteins during the assembly of the head
of bacteriophage T4.Nature 227, 680 – 685.

45
Laycock, M. V., Hirama, T., Hasnain, S., Watson, D., Storer, A. 1989, Purification and
characterization of a digestive cysteine proteinase from the American lobster
(Homarus americanus). Biochem J 263:439–444.
Lenarcic, B., Krasovec, M., Ritonja, A., Olafsson, I., Turk, V., 1991. Inactivation of
human cystatin C and kininogen by human cathepsin D. FEBS Letters 280,
211–215.
Mintz, G. R. 1993. An irreversible serine protease inhibitor. BioPharm, 6, 34.
Kato, G. J. 1999. Human genetic diseases of proteolysis. Human Mutation. 13: 87-98.
Krem, M. M. Di Cera-Enrico. 2002. Evolution of enzyme cascades from embryonic
development to blood coagulation. Trends in biochemical sciences. 27(2):67-
74.
Navarrete del Toro, M. A., García-Carreño, F. L., Díaz, L. M., Celis-Guerrero, L.,
Saborowski, R. 2006. Asparticproteinases in thedigestivetract of marine
decapodcrustaceans. J ExpZool 305A:645–654.
Navarrete del Toro, M. A., García-Carreño, F. L. y Córdova-Murueta, J. H. 2011.
Comparison of digestive proteinases in three penaeids. Aquaculture 317: 99-
106.
Nicosia, F. y Lavalli, K. 1999 Homarid lobster hatcheries: their history and role in
research, management, and aquaculture. Mar. Fish. Rev., 61, 1–57.
Pearl, L. y Blundell, T. (1984). The active site of aspartic proteinases. FEBS. Lett.
174: 96-101.
Perera, E., Moyano, F. J., Díaz, M., Perdomo-Morales, R., V. Montero,
E. Alonso, O. Carrillo y G. Galich. 2008. Polymorphism and partial
characterization of digestive enzymes in the spiny lobster Panulirus
argus.Comp. Biochem. Physiol. B 150: 247– 254.

46
Price, N. C. 1991. En: Enzyme Assay, a Practical Approach. Rickwood, D. y Hames,
B. D. Ed: The Practical Approach Series. 265-266 pp.
Polgár, L. 2005. The catalytic triad of serine peptidases. CMLS, Cell. Mol. Life Sci. 62:
2161–2172
Rasmussen, R. S. y Morrissey, M.T. Marine biotechnology for production of food
ingredients. Adv. Food Nutr. Res. 2007, 52, 237–292.
Rawlings, N. D. y Barrett, A.J. 1993. Evolutionary families of peptidases.Biochem. J.
290, 205-218.
Rich, D. H. y E. T. O. Sun. 1980. Mechanism of inhibition of pepsin by Pepstatin.
Effect of inhibitor structure on dissociation constant and time-dependent
inhibition. Biochem. Pharmacol. 29:2205–2212.
Sáenz de Rodrigánez, M. A., Medina, E., Moyano, F. J., Alarcón, F. J., 2011.
Evaluation of protein hydrolysis in raw sources by digestive proteases of
Senegalese sole (Solea senegalensis, Kaup 1958) using a combination of an
in vitro assay and sodium dodecyl sulphate polyacrylamide gel electrophoresis
analysis of products. Aquacult. Res. 42, 1639–1652
Rojo, L., Muhlia-Almazán, A., Saborowski, R., García Carreño, F. L. 2010a. Aspartic
cathepsin D endopeptidase contributes to extracellular digestion in clawed
lobsters Homarus americanus and Homarus gammarus. Mar Biotechnol
12:696–707
Rojo, L., Sotelo-Mundo, R., García-Carreño, F., Gráf, L. 2010b.Isolation,biochemical
characterization, and molecular modeling of American lobster digestive
cathepsin D1. Comp Biochem Physiol B 157:394–400.
Semenov, A. Olson, J. E. Rosenthal, P. J. 1998. Antimalarial Synergy of Cysteine and
Aspartic Protease Inhibitors. Antimicrob. Agents Chemother. 42: 2254–2258
Souza-Buarque, D., Fernandes-Castro, P., Silva-Santos, F. M., Lemos, D., Carvalho-
Junior, L. B. y Souza-Bezerra, R. 2009. Digestive peptidases and proteinases

47
in the midgut glan of the pink shrimp Farfantepenaeus paulensis (Crustacea,
Decapoda, Penaeidae) Aquaculture research, 2009, 40:861-870.
Sun, H., Lou, X., Shan, Q., Zhang, J., Zhu, X. 2013. Proteolytic Characteristics of
Cathepsin D Related to the Recognition and Cleavage of Its Target Proteins.
DOI: 10.1371/journal.pone.0065733.
Teschke, M. and Saborowski, R. 2005. Cysteine proteinases substitute for serine
proteinases in the midgut glands of Crangon crangon and Crangon allmani
(Decapoda: Caridea). J Exp Mar Biol Ecol 315:213–299.
Tsai, I. H., Lu, P. J. and Chuang, J. L. 1991. The midgut chymotrypsins of shrimps
(Penaeus monodon, Penaeus japonicus and Penaeus penicillatus), Biochimica
et Biophysica Acta -Protein Structure and Molecular Enzymology, 1080 59-67.
Vijayaraghavan, P., Flanetraj, S. R., Vincent, S. G. P. 2011. Purification and
Characterization of a Cysteine Proteinase for the Mid-gut Gland of Indian
Lobster, Panulirus homarus. World J Chem 6(1): 44-48.
Ward, R. D. 2000. Genetics in fisheries management. Hydrobiologia, 420: 191–201.
Whitaker, J.R. 1994. Principles of enzymology for the food sciences. New York,
Marcel Dekker, 2nd ed.
Williamson, A.L., Lecchi, P., Turk, B.E., Choe, Y., Hotez, P.J., McKerrow,
J.H., Cantley, L.C., Sajid, M., Craik, C.S., Loukas, A. 2004. A multi-enzyme
cascade of hemoglobin proteolysis in the intestine of blood-feeding
hookworms. J. Biol. Chem. 279,35950–35957.
Yamada, A., Yoshio, M., Oiwa, K., Nyitray, L. 2000. Catchin, a novel protein in
molluscan catch muscles, is produced by alternative splicing from the myosin
heavy chain gene. J. Mol. Biol. 295, 169–178.

48
13. LISTA DE TABLAS Y FIGURAS
Tabla II. Protocolo de reacciones del ensayo de inhibición. Se muestran los
componentes y el volumen (μL) de cada uno de ellos, empleado para llevar a cabo
cada tratamiento.
Figura 9. Zimograma que muestra el efecto de diferentes inhibidores específicos de
proteinasas sobre la actividad proteolítica del jugo gástrico de langosta americana,
sin la presencia de DTT, A) zimograma a pH 4.7 con Hemoglobina como sustrato; B)
zimograma de actividad a pH 8.0 con caseína como sustrato y con DTT, C
zimograma a pH 4.7 con Hemoglobina como sustrato; D) zimograma de actividad a
pH 8.0 con caseína como sustrato Las flechas señalan los sitios donde la actividad
disminuyó. Mw = Marcador de peso molecular. C = Actividad proteinolítica del jugo
gástrico. P = Efecto de pesptatin A sobre el jugo gástrico. E = efecto del E-64 sobre
el jugo gástrico. F = efecto del Pefabloc sobre el jugo gástrico.
Figura 10. SDS-PAGE de las principales proteínas presentes en el músculo de A.
circularis. LMH y HMW son marcadores de peso molecular, alto y bajo
respectivamente.
Figura 11. Hidrólisis de BSA por acción de las proteinasas presentes en el jugo
gástrico de H. americanus, a pH 4.7. A) SDS-PAGE de la hidrólisis de BSA como
sustrato a diferentes tiempos de muestreo (0, 15, 60, 240, 1200 min). Mw = marcador
de peso molecular; E = enzima pura; S = sustrato. B) Representación gráfica de la
hidrólisis BSA, determinada por los cambios en la densidad óptica. Los resultados se
encuentran expresados como la cantidad remanente de proteína detectada en SDS-
PAGE, después de incubar por 360 min.
Figura 12. Hidrólisis de Hemoglobina por acción de las proteinasas presentes en el
jugo gástrico de H. americanus, a pH 4.7. A) SDS-PAGE de la hidrólisis de
Hemoglobina como sustrato a diferentes tiempos de muestreo (0, 15, 60, 240, 480,
600, 1200 min). Mw = marcador de peso molecular; E = enzima pura; S = sustrato. B)
Representación gráfica de la hidrólisis de hemoglobina, determinada por los cambios

49
en la densidad óptica. Los resultados se encuentran expresados como la cantidad
residual de proteína detectada en SDS-PAGE, después de incubar por 480 min.
Figura 13. Hidrólisis de Actina de A. circularis por acción de las proteinasas
presentes en el jugo gástrico de H. americanus, a pH 4.7. A) SDS-PAGE de la
hidrólisis de Actina de extracto muscular de A. circularis, a diferentes tiempos de
muestreo (0, 15, 60, 240, 360, 1200 min). Mw = marcador de peso molecular; E =
enzima pura; S = sustrato. B) Representación gráfica de la hidrólisis Actina,
determinada por los cambios en la densidad óptica. Los resultados se encuentran
expresados como la cantidad residual de proteína detectada en SDS-PAGE, después
de incubar por 240 min.
Figura 14 Representación gráfica (A) de la hidrólisis de BSA por acción de las
proteinasas presentes en el jugo gástrico de H. americanus, en ausencia o presencia
de inhibidores específicos. La imagen muestra la cantidad de proteína remanente,
observada en SDS-PAGE (B), después del tiempo de hidrólisis máxima (360 min),
comparada con la cantidad de proteína del sustrato puro (S). Cada barra representa
la media de las tres réplicas. Barras con la misma letra señalan que no existen
diferencia significativas (P < 0.05). (MW = Marcador de peso molecular; C = control;
JG = jugo gástrico; P = Pepstatin A; E = E-64; F = Pefabloc).
Figura 15 Representación gráfica (A) de la hidrólisis de BSA por acción de las
proteinasas presentes en el jugo gástrico de H. americanus, en ausencia o presencia
de inhibidores específicos. La imagen muestra la cantidad de proteína remanente,
observada en SDS-PAGE (B), después del tiempo de hidrólisis máxima (480 min),
comparada con la cantidad de proteína del sustrato puro (S). Cada barra representa
la media de las tres réplicas. Barras con la misma letra señalan que no existen
diferencia significativas (P < 0.05). (MW = Marcador de peso molecular; C = control;
JG = juago gástrico; P = Pepstatin A; E = E-64; F = Pefabloc).
Figura 16. Representación gráfica (A) de la hidrólisis de BSA por acción de las
proteinasas presentes en el jugo gástrico de H. americanus, en ausencia o presencia
de inhibidores específicos. La imagen muestra la cantidad de proteína remanente,

50
observada en SDS-PAGE (B), después del tiempo de hidrólisis máxima (240 min),
comparada con la cantidad de proteína del sustrato puro (S). Cada barra representa
la media de las tres réplicas. Barras con la misma letra señalan que no existen
diferencia significativas (P < 0.05). (MW = Marcador de peso molecular; C = control;
JG = jugo gástrico; P = Pepstatin A; E = E-64; F = Pefabloc).