tesis: inmunoexpresiÓn de la molÉcula clase ii del
TRANSCRIPT

t!)/y,21 /.,3~
UNIVERSIDAD NACIONAL AUTÓNOMA DE MÉXICO
FACULTAD DE ODONTOLOGÍA.
INMUNOEXPRESIÓN DE LA MOLÉCULA CLASE JI DEL COMPLEJO PRINCIPAL DE
HISTOCOMPATIBILIDAD Y LA CUANTIFICACIÓN DE CÉLULAS DENDRÍTICAS EN EL GRANULOMA PERIAPICAL, GRANULOMA PERIAPICAL CON
PROLIFERACIÓN EPITELIAL Y QUISTE PERIAPICAL.
TESIS.
PARA OBTENER EL TITULO DE:
CIRUJANO DENTISTA.
PRESENTA:
RODOLFo(__GóMEZ MACÍAS.
Dr. Juan Carlos ;~:::~ández Guerrero~~ DIRECTOR DE TESI~
Dr. Andrés Eliú Castel/ Rodríguez.
Asesores: M.C. Judith Álvarez Pérez. Biól. Miguel A. Herrera Enriquez.
C.D. CMF. Gabriel Loranca Fragoso.
México, D.F. 7 octubre del 2003. TESIS CON FALLA DE ORIGEN

UNAM – Dirección General de Bibliotecas
Tesis Digitales
Restricciones de uso
DERECHOS RESERVADOS ©
PROHIBIDA SU REPRODUCCIÓN TOTAL O PARCIAL
Todo el material contenido en esta tesis esta protegido por la Ley Federal del Derecho de Autor (LFDA) de los Estados Unidos Mexicanos (México).
El uso de imágenes, fragmentos de videos, y demás material que sea objeto de protección de los derechos de autor, será exclusivamente para fines educativos e informativos y deberá citar la fuente donde la obtuvo mencionando el autor o autores. Cualquier uso distinto como el lucro, reproducción, edición o modificación, será perseguido y sancionado por el respectivo titular de los Derechos de Autor.

OCTUBRE 2003 RODOLFO GOMEZ ;..·tACÍ.'\S
AGRADECIMIENTOS
A DIOS por estar a mi lado en todos los momentos de mi vida y darme
la Fé y fuerza necesaria para ver hacia el horizonte, ponerme en este camino
y guiarme con sabiduria al bien y hasta donde estoy.
A mi mamá MARIA EVA MACÍAS GÓMEZ, que siempre me ha
apoyado en miles de cosas, tu sabes que yo no estaria aqui sino fuera por
todos tus esfuerzos y sacrificios para que pudiera llegar hasta aquí. Gracias
por ser mi mejor amiga y darme ánimos para seguir adelante. TE AMO
MAMÁ.
A mi papá FIDEL ROSAS MARTINEZ, no tengo palabras con que
agradecerte todo lo que me enseñaste y lo que me diste. Gracias por
ayudarme a realizarme en todos los aspectos de mi vida, con tus consejos,
regaños y acciones que ayudaron a mi formación profesional y sentirte
conmigo en todo momento y en mi corazón. TE AMO PAPÁ
A mis hermanos MARIA DE LOS ANGELES, SILVIA, MIGUEL
ANGEL, JOSE LUIS, JUAN CARLOS Y MIS SOBRINOS PAULINA,
JESÚS, KAREN, DANIELA Y EL NUEVO INTEGRANTE DE LA FAMILIA,
por existir y estar a mi lado física y moralmente, y ser ustedes mis mejores
amigos, apoyarme y respetarme en todo momento. LOS AMO
A toda mi familia (abuelitos, tíos, primos, cuñados) por quererme,
apoyarme y ayudarme durante todo este tiempo, gracias por darme ese

OCTUBRE 2003 RODOLFO GOMEZ :0.1..\l'iAS
cariño tan-· e~pecial y enseñarme varias cosas de la vida y hacerme la
persona que ahora soy. G_RACIAS A TODOS.
A mis amigos DON MEMO, DOÑA MALENA, PEDRO, GISELA,
NORMA, MARITZA, NORA, KARINA, MARISOL, ALEJADRO, ERIQUE,
DOLORES, LETICIA, MAGALI, Lab. TEODORO PÉREZ SALAZAR, Téc
Acad. VERONICA RODRÍGUEZ _MATA Y A TODA LA BANDA DE
AMACUZAC, por apoyarme y estar conmigo en los momentos malos y
buenos durante todo este tiempo, darme consejos para ser mejor cada día.
GRACIAS A TODOS USTEDES.
A todos mii:;,R()mpañeros desde los de la facultad hasta los de la
preparato.ria, por boryipartir todos los días de escuela y tener su paciencia y
cariño conmigo. A TODOS SIN EXCEPCIÓN, GRACIAS.
A todos mis profesores, que me brindaron sus conocimientos, su
ayuda y tiempo para realizarme como persona, enseñarme a ver la
Odontología de una manera distinta, Dr. JUAN CARLOS C. HERNÁNDEZ
GUERRERO, Dr ANDRÉS E. CASTELL RODRÍGUEZ, C.D. CMF GABRIEL
LORANCA FRAGOSO Y MUY ESPECIALMENTE M.C. JUDITH ÁLVAREZ
PÉREZ. GRACIAS.
EL SIGUIENTE TRABAJO ES DEDICADO A LAS PERSONAS QUE POR
CUALQUIER COSA SON MUY IMPORTANTES EN MI VIDA, GRACIAS
POR CONFIAR EN Mi, SIN SU APOYO Y CARIÑO NO LO HUBIERA
LOGRADO.
RODOFO GÓMEZ MACÍAS.

OCTUBRE DEL 2003 GÓMEZ MACÍAS RODOLFO.
INDICE.
1.1.1. Resumen. 1 1.2.1. Introducción. 2 1.3.1. Granuloma periapical. 4 1.3.2. Etiopatogenia. 5 1 .3.3. Histología. 9 1.3.4. Características clínicas. 1 O 1.3.5. Características radiográficas. 11 1.4.1. Quiste periapical. 11 1.4.2. Quite periapical verdadero. 12 1.4.3. Quiste periapical en bolsa. 14 1.4.4. Etiología. 15 1.4.5. Histopatología. 15 1.4.6. Características clínicas. 15 1.4.7. Características radiográficas. 16 1.5.1. Etiología de la periodontitis apical crónica. 16 1.6.1. Procesos de defensa del huésped
en las lesiones periapicales. 19 1. 7.1. Células dendríticas. 24 1. 7 .2. Ultraestructura de las células de Langerhans. 26 1.7.3. Origen de las células de Langerhans. 26 1. 7.4. Funciones de las células de Langerhans. 30 1.7.5. Identificación y fenotipo de las células de Langerhans. 36
a).-lmpregnación metálica. b).-Histoquímica enzimática. C).-Marcadores inmunológicos.
1.8.1. Antecedentes. 41 1.9.1. Planteamiento del problema. 44 1.9.2. Justificación. 44 1.9.3. Hipótesis de investigación. 45 1.9.4. Hipótesis nula. 45 1.9.5. Objetivos. 45 1.9.6. Tipo de estudio. 45 1.9.7. Universo o población. 45 1.9.8. Metodología 45 1.9.9. Material y método. 45 1.10.1.Recursos financieros 49 1.10.2.Recursos humanos 49 1.10.3. Criterios de inclusión 49 1.10.4.Criterios de exclusión. 49 1.10.5.Variable dependiente 50 1.10.6. Variables independientes. 50 1.10.7. Unidad de observación. 50

OCTUBRE DEL 2003 GÓMEZ MACÍAS RODOLFO.
1.10.8. Método para procesamiento de datos. 1.11.1. Resultados. 1 .12. 1. Discusión. 1.13.1. Conclusión. 1.14.1. Producción científica 1.15.1. Bibliografía. 1.16.1. Apéndice.
50 51 62 63 64 66 86

OCTUBRE DEL 2003 GÓMEZ MACÍAS RODOLFO.
ATPasa. 87. CDs. CFD: CID.· CL,
·CV. · Fe. GM-:CSF.
GP. GPE; HIV. HLA. ICAM. lgA. lgD. lgE. lgG~ lgM. IL-1 a 15. INFy LB LP Lp LT. MHC. NK. PBS. PHA. PMN. QP. QPV. QPB. TCR. ThO. Th1. Th2. ZIO.
. S-100.
Índice de abreviaturas.
Adenosintrifosfatasa Antígeno 87. Células dendríticas. Células dendríticas foliculares. Células dendríticas interdigitantes. Células de Langerhans. Células de vela. Fracción cristalizable. Factor estimulador de colonias de Granulocito y Macrófagos. Granuloma periapical. Granuloma periapical con proliferación epitelial. Virus de inmunodeficiencia humano. Antígeno leucocitaria de histocompatibilidad. Molécula de adhesión intercelular. lnmunoglobulina A. lnmunoglobulina D. lnmunoglobulina E. lnmunoglobulina G. lnmunoglobulina M. lnterleucina 1 a 15. lnterferon y Linfocitos B. Lesiones periapicales. Ligamento periodontal. Linfocitos T. Complejo principal de hitocompatibilidad Natural Killer. Amortiguador salino de fosfato. Finohematoglutinina. Polimorfonucleares Quiste periapical. Quiste periapical verdadero. Quiste periapical en bolsa. Receptor de células T. Linfocito T auxiliares O. Linfocito T auxiliares 1. Linfocito T auxiliares 2. Zinc-Yodo-Osmio. Proteína S-1 OO .

OCTUBRE DEL 2003 GÓMEZ MACÍAS RODOLFO.
INDICE DE FOTOGRAFIAS, TABLAS Y GRAFICAS.
Figura 1. 4 Figura 2. 22 Figura 3. 29 Figura 4. 31 Figura 5. 36 Figura 6. 54 Figura 7. 54 Figura 8. 55 Figura 9. 55 Figura 10. 55 Figura 11. 56 Figura 12. 56 Figura 13. 56 Figura 14. 57 Figura 15. 57 Figura 16. 57 Figura 17. 58 Figura 18. 58 Figura 19. 58
Tabla 1. 39 Tabla 2. 59 Tabla 3. 60 Tabla 4. 61
Grafica 1. 54 Grafica 2. 59 Grafica 3. 60

OCTUBRE DEL 2003 GÓMEZ MACÍAS RODOLFO.
1.1.1. RESUMEN.
Las células dendríticas (CD) se han estudiado en diferentes procesos
infecciosos, sin embargo, existe escaso conocimiento y poca información
publicada a cerca de la relación de las CD en el granuloma periapical (GP),
granuloma con proliferación epitelial (GPE) y quiste periapical (QP), lo cual
nos brida un campo fértil para investigar y dilucidar el papel que desempeñan
las CD en el desarrollo y perpetuación de estas lesiones.
El Objetivo del presente estudio fue demostrar la expresión de la molécula
clase 11 del Complejo Principal de Histocompatibilidad (CMH) y la Proteína
S-100 de las CD en los GP, GPE y QP que se presentan en cavidad oral.
En el GPE con el marcador de HLA-DR se observo una x= 134.60 CD/ mm2,
en el GPE con el marcador S-100 se observo una x= 125.91 CD/mm2 , en el
GP con el marcador de HLA-DR se observo una x= 109.82 CD/ mm2, en el
GP con el marcador S-100 se observo una x= 106.07 CD/ mm2, en el QP con
el marcador HLA-DR se observo una x= 114.37 CD/mm2 y en el QP con el
marcador S-100 se observo una x=110.02 CD/ mm2.
El GPE tuvo una diferencia estadísticamente significativa con el GP y QP, sin
embargo, no hubo diferencia estadísticamente significativa cuando se
compararon el GP con el QP entre ellos, además las CD tuvieron una mayor
distribu_ción en el tejido epitelial que en el infiltrado inflamatorio crónico, en este
último la~ CD se encontraron cerca de los linfocitos y vasos sanguíneos.
Por.10'~~~:1.;~~:s~tros concluimos que las CD presentan los antígenos a los
linfoci~Os'i e inician y modulan la respuesta inmune primaria, siendo estas
cél_ulas las responsables de infiltrado de los linfocitos y probablemente
teniendo un papel primordial en el desarrollo del tejido epitelial aunque los
mecanismos aun no son bien entendidos.

OCTUBRE DEL 2003 GÓMEZ MACIAS RODOLFO.
1.2.1. INTRODUCCIÓN.
La organización mundial de la salud (OMS) en 1995 y otros autores han
clasificado a las lesiones periapicales de origen pulpar en tres grupos
principales, basándose en las características clínicas, radiográficas e
histopatológicas en: periodontitis apical aguda, periodontitis apical cronica
y abscesos apicales Cl-7>_
La periodontitis apical crónica es la inflamación y destrucción del
periodonto apical como consecuencia de Ja necrosis pulpar, se
caracteriza por la presencia de un infiltrado inflamatorio crónico y en
ocasiones se observa Ja proliferación de células epiteliales. Estos
elementos celulares aíslan los productos bacterianos y/o de necrosis
pulpar, que son desechados desde el conducto radicular a la zona
periapical es>, los cuales tienen la capacidad de activar Jos procesos de
defensa del huésped <9>.
Histológicamente Ja periodontitis apical crónica se clasifica como GP y QP
los cuales a su vez se subdividen en: GPE con base en la presencia de
células epiteliales dentro del infiltrado inflamatorio crónico, y en un quistes
periapical verdadero (QPV) o en un quiste periapical en bolsa (QPB) de
acuerdo a la relación quiste cavidad con el conducto radicular del diente
afectado respectivamente <1
•2
•1º·11
'.
Probablemente estas lesiones son las más frecuentemente observadas
en la practica clínica odontológica, por Jo que es importante conocer los
diversos mecanismos por Jos cuales se desarrollan y se perpetúan (1,2,10,11)
En este tipo de lesiones participan células como macrófagos, neutrofilos y
linfocitos T y B, sin embargo, aún no se reconoce el papel de las CD y/o CL
en .estas. Las CD y/o CL son reconocidas como las células presentadoras
de antígenos (CPA) más eficientes del organismo humano, estas tienen la

OCTUBRE DEL 2003 GÓMEZ MAC[AS RODOLFO.
capacidad de presentar antígenos a las células T, siendo estas la llave
para iniciar y modular la respuesta inmune, ya que puede influenciar en la
diferenciación de las ThO en Th1 y Th2; cada una de estos linfocitos
auxiliares secretan diferentes interleucinas (IL) que estimulan o deprimen la
respuesta inmune celular y/o humoral <12>.
3

OCTUBRE DEL 2003 GÓMEZ MAC[AS RODOLFO.
1.3.1. GRANULOMA PERIAPICAL.
Es uno de Jos estadios avanzados de la periodontitis apical crónica, el GP
muestran un infiltrado inflamatorio crónico como respuesta a un estímulo
(físicq, químico o biológico) este infiltrado prolifera para aislar los
productos bacterianos y/o de necrosis pulpar, que son desechados desde
el conducto radicular a la zona periapical cai, estos tienen la capacidad de
activar la respuesta inmune del huésped C9J_ El GP se desarrolla al no ser
posible Ja eliminación del estímulo proveniente del conducto radicular del
órgano dental. El GP reemplaza al ligamento periapical (lp), al hueso
apical, en ocasiones al cemento y la dentina radicular. Esta lesión puede
llegar a medir varios milímetros y para su estudio se han dividido en
varias capas concéntricas con distintas características microscópicas que
se describirán más adelante, sin embargo en algunos casos se ha
observado proliferación de células epiteliales provenientes de los restos
epiteliales de Malassez, siendo esta Ja base para clasificar a los GP en:
GP y/o GPE (1.2.>_
,,,..,,, .... "'.
TESIS CQ~T FALLA DE ORIGEN
fig.1 Granuloma periapical. Takahashi K 199a<13>

OCTUBRE DEL 2003 GÓMEZ MACIAS RODOLFO.
1.3.2. ETIOPATOGENIA.
El infiltrado inflamatorio crónico se desarrolla como un mecanismo de
aislamiento del contenido del conducto radicular, ya sean bacterias y/o
productos de la necrosis pulpar o medicamentos irritantes que se
comportan como haptenos en una reacción antígeno-anticuerpo, todos
ellos actuando de forma persistente <14-19
>_ En este caso, en la reacción
inflamatoria periapical crónica entra en juego la segunda línea de defensa
celular compuesta por los macrófagos, linfocitos, células plasmáticas, y
fibroblastos; así el proceso queda limitado por una pseudocápsula <1
·2>.
El mecanismo por el cual se establece un GP, ha sido estudiado durante
muchos años, y en la actualidad se considera que se trata de un
fenómeno de tipo inmune. Hay estudios que indican que los complejos
antígeno-anticuerpo y las reacciones mediadas por lgE <20>, podrían iniciar
el daño tisular en las lesiones periapicales (LP), de la misma forma que
los fenómenos de hipersensibilidad retardada podrían perpetuar el
procei:fo:
Otros autores describieron la secuencia inmunopatogénica que rodea la
formación de un GP <1
·2>. Así, la presencia de grandes cantidades de
bacterias provenientes del conducto radicular y sus toxinas así como los
restos de necrosis pulpar, poseen una alta capacidad antigénica que no
escapa a nuestro mecanismos inmunológico. En un primer momento se
produce un acumulo de leucocitos polimorfonucleares que fagocitan a los
antígenos <21
•22>, a su vez, las células cebadas aumentan el fenómeno
inflamatorio mediante la liberación substancias vasoactivas <23
>_ En una
segunda fase de esta secuencia aparecen monocitos y los linfocitos, que
posteriormente, se encargan de digerir todos los restos celulares y
antígenos presentes: muchos de los monocitos se transforman en
5

OCTUBRE DEL 2003 GÓMEZ MAC[AS RODOLFO.
macrófagos para cumplir con esta tarea <24
-26>. Los antígenos una vez
presentes en Ja superficie celular son transportados hasta los ganglios
linfáticos transmitiendo Ja información a Jos linfocitos T !25>.
Todos Jos trabajos recientes sobre Ja etiopatogenia de Jos GP, se han
encaminado fundamentalmente a estudiar de los componentes celulares
inflamatorios, así como Jos numerosos mediadores químicos de la
respuesta inmunológica <27·28
'.
En un estudio histológico de 12 GP se encontró que Ja células
inflamatorias crónicas más importantes son Jos linfocitos con un (44%),
seguido de Jos macrófagos con un (30%), las células plasmáticas con un
(15%) y finalmente los neutrofilos con un (12%) <28
'.
Cuando se estudiaron a las células plasmáticas que elaboran las
inmunoglobulinas, se comprobó que Ja lg más frecuente fue Ja JgG
representando el 81.9%. Otras inmunoglobulinas encontradas fueron Ja
lgA (11.4%), Ja lgM (5.4%), la JgE (1.1 %) y Ja JgD (0.2%) <25>_ La presencia
·de ÍgE ~o fue constante en todas las muestras y estos autores sugieren
quei'.además del daño tisular provocado por el depósito de complejos
inmunes;:las reacciones de hipersensibilidad inmediata mediadas por lgE
pueden jugar algún papel en la patogénesis !29>.
En 1985, un .estudio de 90 LP determinó la presencia de diversas
inmunoglobulinas en el líquido sobrenadante de cultivos de estas • . >'¡ '·
lesiones;¡L:ainmunoglobulina cuya presencia se determinó en el 100% de
cultivo ·f~~.:,ík1gG, mientras que la JgA Jo fue en el 65%. No se detectó
lgM, mi~~tµ~~· que la lgE fue positiva en 40 de los 90 casos estudiados
(45%). Es.Íos resultados confirmaron los datos anteriormente citados y
refuerzan la teoría inmune !20•30>.
Otras células relacionadas con la patogenia de los GP son las células
cebadas <23>, cuya misión es la de liberar gran cantidad de productos
mediadores de la inflamación que precipitan. acentúan. agravan o
6

OCTUBRE DEL 2003 GÓMEZ MAC[AS RODOLFO.
perpetúan Ja respuesta inmune. Se ha encontrado una alta incidencia de
estas células, especialmente asociadas a zonas con aumento de Ja
inflamación. Además existe otro tipo de células en Jos GP, cuyo papel
respondería a fenómenos de reacción a cuerpo extraño, son las células
gigantes multinucleadas. Aparecen aisladas, más frecuentemente
asociadas a cristales de colesterol y en ocasiones a fibras de celulosa en
el periápice. Al mismo tiempo, estrechamente relacionados con Jos
mecanismos de limpieza y autolisis han sido descritos Jos macrófagos,
cuyo estudio con microscopia electrónica puso de manifiesto una activa
fagocitica de partículas extrañas y eritrocitos en el área periapical <32>. Se
ha demostrado la presencia de células natural killer (NK) en las GP, lo
que indica que su papel defensivo en el control de la infección del con
ducto radicular podría ser importante, así como su participación en la
patogenia de estas lesiones <31 >.
A pesar de que existen suficientes pruebas sobre Ja participación de la
respuesta inmune humoral en la etiopatogenia de Jos GP, la
extraordinaria predominancia de los linfocitos T sobre los B en estas
lesiones indica también, una importante participación de Ja respuesta
inmune de tipo celular. Los resultados son bastante contradictorios puesto
que mientras _unos autores encuentran un elevado porcentaje de linfocitos
B, la .. mayoíra "de los trabajos que analizan el infiltrado inflamatorio .. ,, ... - ' ' ,_;.
medianl:e(a.nticuerpos monoclonales indican un predominio claro de los
linfociib~}~;¿;'¡j;J'¡6~ B. Otros estudios han determinado las proporciones
de diver~h~·tlpos celulares presentes en las LP de tal forma que el 51.7%
fueron linfódtos T, el 39. 7% neutrofilos, 3.6% macrófagos, 2.1 % células
plasmáticas, 2.1 % mastocitos y el 0.8% Eosinófilos <21·2 ª>. La presencia de
los linfocitos T es prácticamente constante en las GP y todos los autores
establecen su proporción entre el 25 y el 55% del total de células
inflamatorias aunque para algunos autores, los linfocitos T son más
frecuentes en los infiltrados difusos y los B en Jos infiltrados de tipo focal;
también se ha constatado que la proporción de linfocitos T aumenta en
las lesiones de mayor tamaño.
7

OCTUBRE DEL 2003 GÓMEZ MACIAS ROOOLFO.
Es importante el estudio de las diversas subpoblaciones de linfocitos T
que pueden aparecer en las GP. La proporción de linfocitos inductores y
supresores varia de unos trabajos a otros; mientras que algunos autores
<32
•33> han demostrado en los GP una proporción de ambos subtipos de
linfocitos T (en tejido sano la proporción inductor/supresor es 2: 1 ). parece
existir un ligero predominio de los supresores.
No obstante, las diferencias observadas entre los diversos trabajos re
visados no son concluyentes en un sentido u otro y las variaciones en el
índice : inductor/supresor, podrían determinar los diversos momentos
· ~~6iuti~os del GP. Los linfocitos inductores predominarían en las forma
· adtida .. ·:~i~ntras que los supresores predominarían en las formas
crÓhic~~~}Ee~tas variaciones podrían ser un reflejo del mecanismo
patogé~i~6i§de la transformación del GP a QP <25·26>.
' ,·._\: ·:·.:.~··:~<-~~}~;_~t!!;~~; ... : . ',_ . Otro factÓ~\'f~t~resante a considerar es la relación entre las células
presente'~t~R1i;e1}·c;8 .yJa sintomatologia <281
. En este sentido, se ha
ens~~~f~~g~~~:ff~.~.Siyor proporción de linfocitos T cuanto mayor es el
tamaficide;>la lesióri. Así, en lesiones pequeñas, la proporción fue del
3Ó"Ío,·ri-;¡~h~t~~~ c:iL~ ~n las lesiones más grandes alcanzó el 50% de todas
la~.6'éfú1~~· rriononucleares del infiltrado.
lgÜalmente se observó una proporción significativamente mayor de
mo11ocitos/macrófagos entre los casos que presentaron alguna sin
tomatologfa (dolor espontáneo, a la palpación o percusión del diente) que
en . los.• ... asintomáticos, también encontraron una proporción
signffib.ati~arneme ~ayer de leucotrieno L TB4 y de polimorfonucleares
entre el G8;;sin'torl1át16'ó·:que entre el asintomático <21
·281 Estudios más
recient~~ ikcii6~{J'ri''íi'i;;:{'g·~rtancia de las interleucinas en la patogenicidad del GPPº;:· .. ' "/.: .:,; .·. .
8

OCTUBRE DEL 2003 GÓMEZ MACIAS RODOLFO.
1.3.3. HISTOPATOLÓGIA.
A continuación, se describe el desarrollo del GP. La presencia de restos
necróticos pulpares y/o bacterianos (zona de necrosis/infección), dan
lugar a la difusión del material tóxico, hacia y ligeramente más allá de la
zona de coalescencia del tejido pulpar, hasta el tejido conjuntivo
periodontal <1
•2>. Alrededor de cada foramen se forman centros de
infiltración . celular que no se detectan radiológicamente. Se produce
dil~t:~ci~n capilar y los neutrofilos (PMN) se sitúan en la inmediata
proximidad de la zona de necrosis (zona 1) <1
·2
·21
·22 l_
Estas células están rodeadas por grandes masas de linfocitos y células
plasmáticas. La respuesta crónica leve inicia con la periodontitis apical
crónica incipiente progresa a medida que difunden los productos
necróticos y bacterianos, desde el conducto radicular hacia el periápice
<5•9> •. La toxicidad de los irritantes del conducto radicular se reduce por la
actividad del exudado líquido y celular de la zona de contaminación (zona 11) (1,2).
Esta menor toxicidad estimula a células indiferenciadas para formar
osteoclastos multinucleados, que reabsorben al hueso periapical
contaminado. En este momento, se observa en la radiografía un
ensanchamiento del espacio periodontal apical. En última instancia la
hendidura abierta en el hueso que rodea la lesión se rellena con un
infiltrado Í°nflamatorio crónico formando la zona 111 o zona de irritación <1
·2 >. . . .
El. GP contiene nuevos capilares y fibroblastos jóvenes que poseen la
capaddac:Í d~ rep~~ar la zona periapical. Asimismo tiene la función de .• -<··-_,. :..··, --, ___ ,
aislar los"· ~gentesL~ausales de las LP, su resistencia a la infección
aumenta coh;1~;pr~:7~ncia de linfocitos y células plasmáticas <20
·25>, así
corrio > dé i&~iZfa~_Y Índiferenciadas e histiocitos transformados en
ma~rófag~~:'j .. a pre'sencia de las células plasmáticas suele indicar la
síntesis acti~a 'de anticuerpos <29
·3 º)_ Los macrófagos sufren una
9

OCTUBRE DEL 2003 GÓMEZ MACIAS RODOLFO.
morfodiferenciación transformándose en células epiteloídes, con una
coloración ácidofila (color rosado) con la tinción de hematoxilina y eosina,
también se ha observado la presencia de células gigantes multinucledas y
algunos macróf~gos parecen células espumosas porque su citoplasma
está vacuolado, ellas ingieren material procedente de las células locales
degeneradas. Finalmente, las células espumosas liberan el material
lipídico, apareciendo agujas o cristales de colesterol. Muchos GP también
contienen franjas o islotes de epitelio <32>. Hay que recordar que esta zona
de irritación no contiene microorganismos viables.
El GP se puede comparar con el granuloma pulpar; sin embargo, a
diferencia de la respuesta crónica del tejido conjuntivo pulpar, se
desarrolla una cuarta zona (zona de estimulación). La toxicidad de los
agentes irritantes del conducto radicular se diluyen y reducen en la
periferia de la zona de granulación, tales agentes irritantes producen la
estimulación de los fibroblastos y osteoblastos locales. Los fibroblastos
forman un muro de fibras de colágena con el fin de encapsular todo el
complejo inflamatorio, separando el infiltrado inflamatorio crónico del
hueso. Los osteoblastos actúan óptimamente en esta zona y sintetizan
matriz ósea adicional sobre la superficie del hueso viejo y reabsorbido
durante los períodos de regresión inflamatoria. Los defectos óseos
reabsorbidos del extremo radicular suelen ser secuela del infiltrado
inflamatorio crónico <1-2>.
1.3.4. CARACTERÍSTICAS CLÍNICAS.
El GP es habitualmente asintomático, no hay dolor a la percusión ni el
diente presenta movilidad, los tejidos blandos periapicales pueden o no
estar sensibles a la palpación. El diente afectado no responde a las
pruebas térmicas ni eléctrica. Es importante resaltar que si los irritantes
pulpares invaden este tejido periapical, se forma un absceso agudo o fénix (1.2)
10

OCTUBRE DEL 2003 GÓMEZ MACIAS RODOLFO.
1.3.5. CARACTERÍSTICAS RADIOGRÁFICAS.
La imagen radiolúcida del GP esta bien definida en comparación con el
absceso apical crónico. El GP y el QP son las lesiones radiolúcidas más
frecuentes <1
·2>.
De hecho se ha reportado que aproximadamente el 95% de las imágenes
radiolúcidas apicales son GP. Las describen como lesiones relativamente
pequeña, de bordes bien definidos, de forma redonda alrededor del ápice
del diente afectado <1
·2 >.
1.4.1. QUISTE PERIAPICAL.
En general se considera que Jos QP constituyen una secuela directa
GPE, pero no en todos las ocasiones se transforman en QP.
El QP se definen como una cavidad patológica revestida de epitelio
escamoso estratificado este tejido deriva de los restos epiteliales de
Malassez, conteniendo material semisólido o restos celulares. El infiltrado
inflamatorio crónico que rodea al tejido epitelial contiene las mismas
caract~~l~tlt~s celulares que un GP. ' ' :- : ; ~ :: ·,
Existen, d~s categorías distintas de QP: (1) los que contienen cavidades
completárnente encerradas dentro de la mucosa epitelial llamados QP
verdaderos, y (2) los que contienen cavidades revestidas por epitelio,
pero abiertas a los conductos radiculares. Estos últimos se describieron al
principio como QP en bahía, pero recientemente se les ha llamado QP en
bolsa. Más de la mitad de las lesiones quísticas son QP verdaderos, y el
resto son QP en bolsa. Si se tiene en cuenta la diferencia estructural
entre las dos categorías de quistes, es posible que las vías patogénicas
que conducen a su formación varíen en ciertos aspectos <1
·1º·11 >.
11

OCTUBRE DEL 2003 GÓMEZ MACIAS RODOLFO.
1.4.2. QUISTE PERIAPICAL VERDADERO.
Se han realizado intentos para explicar la patogenia de los quistes
periapicales verdaderos. Se ha dicho que el proceso que conduce a su
formación pasa por tres fases. Se cree que durante la primera fase
proliferan los restos epiteliales de Malassez en letargo, probablemente
bajo la influencia de factores de crecimiento liberados por diversas células
residentes en la lesión. Durante la segunda fase se forma una cavidad
revestida de epitelio <1•1º· 11 >.
Se han propuesto dos hipótesis para explicar la formación de la cavidad
quística:
1. La teoría de la deficiencia nutricional se basa en la asunción de
que las células centrales de las hebras epiteliales perdían su
fuente de nutrición, y experimentaban necrosis por degeneración
licuefactiva. Los productos acumulados, a su vez, atraerían
granulocitos neutrofilos hacia el área necrótica. Tales
mícrocavidades, que contienen células epiteliales degeneradas,
infiltrados leucocitos y exudado tisular, confluyen para formar la
cavidad quística, revestida por epitelio escamoso estratificado.
2. La teoría del absceso postula que el epitelio en proliferación rodea
al absceso formado por necrosis y lisis del tejido, dada la tendencia
intrínseca de las células epiteliales a cubrir la superficie de tejido
conectivo expuesto.
3. Durante Ja tercera fase, el quiste crece, aunque todavía no se ha
aclarado de forma adecuada la causa de tal crecimiento.
Las teorías basadas en la presión osmótica han perdido importancia
durante Jos últimos años, conforme el interés de los investigadores se
centró. en el estudio de las bases moleculares de la quistogénesis. El
hecho de que el QP en bolsa cuya luz comunica con el conducto radicular
12

OCTUBRE DEL 2003 GÓMEZ MACIAS RODOLFO.
necrótico pueda crecer, elimina a la pres1on osmótica como un factor
potencial en el desarrollo del QP. Aunque todavía no se dispone de
evidencia, la dinámica tisular y los componentes celulares del QP
sugieren posibles vías moleculares para la expansión del quiste. Los
neutrofilos persistentes en la luz del quiste proporcionando una fuente
continua de prostaglandinas, que pueden extenderse a través de la pared
epitelial porosa hacia los tejidos adyacentes. La población celular
residente en el área extraepitelial contiene numerosos linfocitos T y
macrófagos, de los que se sabe que son capaces de producir una
variedad de citocinas, en particular IL-1 B. Las prostaglandinas y las
citocinas inflamatorias pueden activar a los osteoclastos, desarrollando
una reabsorción ósea, También se ha descrito la presencia de moléculas
efectoras (MMP-2) en QP <1
•1º· 11 >.
Desde el punto de vista histopatológico, el QPV tiene cuatro componentes
fundamentales: (1) cavidad quítica; (2) pared epitelial; (3) tejido
extraepitelial, y (4) cápsula de colágeno. La cavidad, completamente
encerrada dentro de la mucosa epitelial, contiene generalmente tejido
necrótico y, en ocasiones, restos de colesterol y eritrocitos. El grosor del
epitelio escamoso estratificado puede variar desde unas pocas hasta
muchas capas de células. La microscopia electrónica de barrido de la
superficie interna del quiste revela la presencia de células epiteliales
planas y globulares (es decir, evidencian la superficie del epitelio y los
neutrofilos que sobresalen a través de los espacios Intercelulares). El
tejido existente entre la mucosa epitelial y la cápsula fibrosa suele
contener números vasos sanguíneos y células infiltrantes, entre las que
prédominan los linfocitos T y B, las células plasmáticas y los macrófagos.
Los neútrofll~s. que son numerosos en la mucosa epitelial, se encuentran
rara vez en el área extraepitelial <1
•1º·11 >.
13

OCTUBRE DEL 2003 GÓMEZ MACÍAS RODOLFO.
1.4.3. QUISTE PERIAPICAL EN BOLSA.
El QPBse desarrolla probablemente por la acumulación de neutrofilos
alrededor del orificio apical, en respuesta de la presencia de bacterias
dentro del conducto radicular apical. Los microabscesos así formados
pueden quedar rodeados por epitelio en proliferación que, al entrar en
contacto con la punta radicular forman un collar epitelial con inserción
epitelial. Este collar sella el conducto radicular infectado y los
microabscesos, separándolos del medio periapical. Cuando los neutrofilos
más externos se destruyen y se desintegran, el espacio ocupado por ellos
se convierte en un saco microquístico. La presencia en el conducto
radicular apical de microbios, sus productos y células necrosadas
procedentes de la luz del quiste, atrae más granulocitos neutrofilos al
crear un gradiente quimiotáctico. Sin embargo, la luz en forma de saco,
biológicamente situada fuera del medio periapical, actúa como una
trampa para los neutrofilos en fase de migración. Conforme se acumulan
las células necróticas, el saco se agranda para acomodar los residuos, y
se puede formar un divertículo voluminoso comunicado con el espacio del
conducto radicular, que se extiende en el área periapical. Se produce
reabsorción ósea y degradación de la matriz, relacionadas con el
agrandamiento de la bolsa quítica, probablemente por mecanismos
moleculares similares a los operantes en el QPV. Desde el punto de vista
de la patogenia, la estructura, la dinámica tisular y el beneficio del
huésped, la extensión en forma de bolsa del espacio canalicular radicular
tiene mucho en común con una bolsa periodontal, a lo que se debe el
nombre de QPB. Desde el punto de vista histológico, la mucosa epitelial
escamosa estratificada y el resto de la pared quística son similares a los
de un Qpv<1.10.11¡.
14

OCTUBRE DEL 2003 GÓMEZ MACIAS RODOLFO.
1.4.4. ETIOLOGÍA.
Se forma a partir de una irritación previa de la pulpa, por agentes físicos,
químicos o bacterianos, que han causado necrosis pulpar y una
periodontitis apical crónica o granuloma pero no en todos las ocasiones,
donde se estimulan Jos restos epiteliales de Malassez, que se encuentran
en el periodonto de letargo '1•1º·11>.
1.4.5. HISTOPATOLOGÍA.
Se observa una capa de epitelio escamoso estratificado, conteniendo
restos necróticos, células inflamatorias y restos de colesterol, que son
observados como numerosas hendiduras aciculares en Ja zona central del
quiste. La literatura refiere que un 29% a 43% de los quistes presentan
estos cristales de colesterol, y adjudican su presencia a Ja desintegración
de los eritrocitos, linfocitos, células plasmáticas y/o macrófagos.
Se ha descrito que generalmente el revestimiento suele ser incompleto, al
estar ulcerado presentando células inflamatorias; el líquido contenido es
pálido y eosinófilo; se ha reportado que el GP y el QP son muy similares,
pero Ja diferencia radica en Ja presencia de una cavidad recubierta de
epitelio rellena de un material líquido o semi-sólido <1
•1º·11 >.
1.4.6. CARACTERÍSTICAS CLÍNICAS.
Siempre estará asociado a un diente con pulpa necrótica o en su defecto a
un diente tratado endodoncicamente, por Jo que las pruebas de vitalidad
serán negativas. La palpación en la zona apical puede ser negativa, pero
en ocasiones se puede sentir una crepitación similar a cuando se aprieta
una pelota de ping-pong.
El QP no presenta síntomas vinculados con su desarrollo, excepto cuando
aparece una infección de tipo aguda. Sin embargo el QP puede crecer
hasta llegar a ser una tumefacción evidente para el paciente. En algunas
15

OCTUBRE DEL 2003 GÓMEZ MACÍAS RODOLFO.
ocasiones la presión del quiste puede provocar el desplazamiento de los
dientes vecinos, movilidad, y clínicamente se puede observar como las
coronas se proyectan fuera de su posición normal en boca,
radiográficamente los ápices se separan.
Es importante saber que los QP, pueden infectarse y presentar una
sintomatología similar a un absceso apical agudo, y posteriormente
fistulizarse y supurar '1·1º· 11 >.
1.4.7. CARACTERÍSTICAS RADIOGRÁFICAS.
Se observa una amplia imagen radiolúcida de contornos precisos, rodeada
de una línea radiopaca nítida de mayor densidad, asociada a un diente con
necrosis pulpar. Pero es casi imposible diferenciar a un GP de un QP. Se
ha reportado que si la lesión es muy grande, es más probable que sea un
QP y no un GP ' 1·1º· 11 >.
1.5.1. ETIOLOGÍA DE LA PERIODOTITIS APICAL CRÓNICA.
Las bacterias son la causa principal y más común de inflamación pulpar y
periapical <33>. De hecho, la relación entre la inflamación periapical y la
infección bacteriana quedó muy bien establecida en el estudio realizado por
Kakehashi <34>· él realizó exposiciones pulpares en ausencia de
microorganismos, él no observó lesiones periapicales, y por el contrario se
observaba reparación tisular; en cambio en las pulpas expuestas en
presencia de microorganismos se producia necrosis y LP. Estudios
subsecuentes confirman la relación causa-efecto entre la infección pulpar y
la periodontitis apical crónica C35 >.
Por consiguiente el punto de partida más usual en la inflamación pulpar y
periapical es la caries. No es necesario que se establezca una
comunicación directa con el tejido pulpar, ya que diversas especies
bacterianas, así como sus componentes (Lipopolisacáridos LPS), pueden
16

OCTUBRE DEL 2003 GÓMEZ MACIAS RODOLFO.
alcanzar la pulpa a través de los túbulos dentinarios. Así mismo, otros
procesos destructivos dentales como las abrasiones, erosiones o
traumatismos que expongan la dentina, posibilitan la llegada de las
bacterias a la pulpa de igual forma a través de los túbulos dentinarios
permeables. También las baderias pueden penetrar los túbulos dentinarios
o los conductos accesorios a través de un saco periodontal; o por la
microfiltración de una restauración defectuosa. Adicionalmente la literatura
refiere un fenómeno denominado "anacoresis" el cual consiste en la
migración de bacterias desde un foco infeccioso a través del torrente
sanguíneo hacia la pulpa de un diente que no está infectado pero que
presenta una inflamación pulpar, por ejemplo, producto de un traumatismo.
Por último, la generación de calor y la desecación de los túbulos dentinarios
en procedimientos restauradores pueden lesionar el tejido pulpar,
produciendo alteraciones vasculares e iniciando una inflamación por la
liberación de neuropéptidos y citoquinas <33
>_
Cuando las bacterias desarrollan una inflamación en la pulpa y no se
efectúa un tratamiento precoz, en un período variable, la inflamación se
extiende y puede llegar a la necrosis. Las bacterias y sus componentes
alcanzarán el periodonto a través del orificio apical o de los conductos
accesorios produciendo una periodontitis <33
>_
Estudios microbiológicos en conductos infectados crónicamente coinciden
en que los microorganismos mayormente aislados son anaerobios y Gram
negativos. Como conclusión podemos decir que las principales bacterias
involucradas en la infección pulpar y periapical son las siguientes <35>:
• Prevotellas - Profiromonas
• Peptostreptococos - Streptococos
• Enterococos - Campilobacter
• Fusobacterium - Eubacterium
• Propionibacterium
17

OCTUBRE DEL 2003 GÓMEZ MACIAS RODOLFO.
Cabe destacar que diferentes estudios en animales demuestran que a
medida que los anaerobios estrictos aumentan, los anaerobios facultativos
disminuyen <35>_
La severidad de la inflamación periapical ha sido directamente relacionada
con microorganismos dentro de los conductos radiculares y a su exposición
prolongada. En contraste, no hay evidencia de que la pulpa necrótica
pueda producir inflamación periapical <35>.
También un estudio demostró que los microorganismos interactúan entre si
haciéndose más virulentos; dentro de ellos podemos destacar a Prevotellas
y Porfiromonas, Fusobacterium y Peptostreptococos los cuales mostraron
incremento en la destrucción ósea. De igual forma, hay estudios que
demuestran una relación directa entre los síntomas agudos y la presencia
de infección por microorganismos como Prevotellas y Porfiromonas <35>.
Por otro lado, se han diseñado diversos métodos para identificar las
bacterias causantes del daño pulpar y periapical, en conducto radiculares
infectados de órganos dentales humanos. Entre las técnicas de
identificación de los microorganismos se encuentra: el cultivo de bacterias,
métodos inmunológicos y la biología molecular.
a).· La técnica de cultivo de bacterias no es muy confiable porque tiene
algunos errores, ya que el crecimiento de los microorganismos patógenos,
varia en el tipo y el número de bacterias debido al medio de cultivo
empleado para identificar bacterias en los conductos radiculares infectados (36-40)
b). Losmétodos inmunológicos necesitan de anticuerpos específicos contra
bacterias .. Los resultados pueden dar falsos positivos por las reacciones
cruzadas con anticuerpos que no son específicos para los microorganismos
utilizados ..
c). Lastécnicas de biología molecular incluyen a la reacción en cadena de
Ja polimerasa del ADN (PCR) de las colonias bacterianas obtenidas de los
conductos radiculares infectados <41 >. Esta técnica se usa para detectar
18

OCTUBRE DEL 2003 GÓMEZ MAC[AS RODOLFO.
bacterias, sin embargo, las pruebas de algunos microorganismos tienen
problemas P.ºr su baja especificidad y los clones de ADN resultantes
pueden presentar baja especificidad <41 >.
Además de lo mencionado anterior se aúnan los traumatismos dentales,
los fracasos endodóncicos (sobreinstrumentación, sobreirrigación y
sobreobturación), al mal uso de productos químicos en la desinfección del
conducto radicular y a los materiales utilizados en la obturación <1
·2>.
1.6.1. PROCESOS DE DEFENSA DEL HUESPED EN LAS LESIONES
PERIAPICALES.
La histopatología de la periodontitis periapical crónica es la misma que se
observa en otros tejidos de granulación que se desarrollan en el tejido
conjuntivo que rodea el área dañada. En la periodontitis periapical crónica
se encuentra un gran número de células inmunocompetentes como
neutrofilos (PMN) <25
•29>. macrófagos, linfocitos, células plasmáticas,
células gigantes, células NK y células cebadas <21
·28
·23>. Los PMN <
21•22>, y
los macrófagos son importantes en la respuesta inmune mediada por
células, en la opsonización y la fagocitosis.
Las células T y B son componentes celulares de la periodontitis periapical
crónica. Estas células juegan un papel central en la respuesta inmune
celular y humoral respectivamente teniendo un papel importante en la
patogenicidad de estas lesiones <13>.
La diferencia en la proporción de las células inmunocompetentes entre los
GP, .. GPE .: y QP está estudiado y se reportó que las células T
coop~raclÓra~/inductoras (Th/i) existen en mayor proporción, que las
célul~~ ..f. i'Jpr~soras/citotoxicas (Ts/c). Por otro lado los macrófagos,
aumentan e~. los QP en comparación con los GP. Otros investigadores
reportaron que no hay diferencias significativas en la proporción de las
subclases de las células T <13>.
Se ha desarrollado experimentalmente GP para demostrar la presencia
de las CPA (CD): Estas células son capaces de capturar el antígeno en
19

OCTUBRE DEL 2003 GÓMEZ MAC[AS RODOLFO.
los órganos periféricos y migrar a los órganos linfoides, donde presentan
el antígeno a las células T.
Los linfocitos T que reconocen los antígenos son las células que van a
dar origen a las clonas de linfocitos T colaboradores (Th) y T citotóxicos
. (Te) (13>.
Los linfocitos Th son células que después de haber reconocido un
antígeno expresan una respuesta dirigida a modular la reactividad que
tienen otros linfocitos sensibilizados por los mismos determinantes. Esta
modulación la llevan a cabo liberando substancias (citocinas) que actúan
estimulando o suprimiendo la proliferación y la diferenciación de otros
linfocitos <13>.
A medida que se multiplican los linfocitos T sensibilizados por un
antígeno, se forma sub-poblaciones encargadas de mantener la memoria
inmunológica. Estas últimas células tienen una vida media mucho más
prolongada que la de las células efectoras de la respuesta colaboradora o
citotóxica <13>.
Los linfocitos T colaboradores se caracterizan porque expresan sobre su
membrana _el antígeno CD4. En los ratones y en varias otras especies se
han ericoñtrado dos subpoblaciones de linfocitos colaboradores, las
cu~les t1'~~'~íc:lo denominadas Th1 y Th2. Los linfocitos Te se caracterizan
p6rq~'~y~-~~ie~~~ sobre su membrana el antígeno CDS.
Las··~;~~5ij"gp.oblaciones de linfocitos Th expresan el mismo marcador de
menibrari-~;;'.'co4, pero se pueden diferenciar entre sí porque las . ' --. . ~ ' . ~ . ( .
interleuCinás que producen están dirigidas a la inducción de respuestas r '' ·,,:
inmunolÓgiéas diferentes.
Los jinf~cltos Th2 producen citocinas que estimulan las células
encargadas de la respuesta humoral mediada por anticuerpos y los
linfocitos Th1 producen citocinas que estimulan las células encargadas de
la respuesta celular.
Probablemente, a las dos subpoblaciones proceden de células que
expresan un fenotipo común, ThO, capaz de sintetizar todas las diferentes
20

OCTUBRE DEL 2003 GÓMEZ MACIAS RODOLFO.
citocinas que más adelante van a ser producidas solamente por uno u
otro tipo de células colaboradoras <13>.
La clasificación de las dos subpoblaciones de linfocitos T colaboradores
se ha basado d acuerdo con su capacidad para producir citocinas.
Por ejemplo, la IL-2 producida por los Th1 es también un factor
estimulante de la proliferación y diferenciación de los linfocitos B. aunque
generalmente esa actividad inhibe la presencia del IFNy.
Al estudiar la colaboración de las células Th1 y Th2 para estimular la
proliferación de los linfocitos Te, tampoco se han encontrado diferencias
absolutas, aparentemente, las dos sub-poblaciones de células Th pueden
colaborar tanto, para estimular como para inhibir la proliferación de los
linfocitos Te <13>.
Algunas de las citocinas producidas por la sub-poblaciones de linfocitos
colaboradores tienen un efecto supresor sobre la proliferación y las
actividades biológicas de algunos linfocitos.
El IFNy producidos por Th1 (y también por las NK) inhibe la proliferación
de los linfocitos Th2, mientras que IL-10 producida por Th2 (y también por
los monocitos y los macrófagos) inhiben la proliferación de los Th1, así
como la síntesis de.las citoci.nás IFNy e IL-2 .
. •. ··.· << > .... :.·:.~5; .• Algunos experiril'~~-iB~1,'.iwJ;.~itro ~ugieren que la IL-4 puede estimular la
' •e ,:• '. ·< ..... • :.:·~·,,':'_ .;:.i·:·f .. ~'f~.;~\•.:\:·~·;'.,~i~:~'.:-.··. •;e
diferenciaciór¡'de;~hO ~n::rh2, mientras que el IFNy y la IL-2 promueven la
difere~~c;¡~;~I~~~·~~:i-tiri·:~~· +ti 1.
· ·· ; :·§L.1!;~~°'r;;r·F .: LaPGE2·1.nhiÍJe'.1a·proliferación de citocinas por íos linfocitos Th1, pero no
modifica la~~ti~i~lá~:;d~ l~s linfocitos Th2 <13>.
21
TESIS CON FALLA DE OfilGEN

OCTUBRE DEL 2003
11 ·I!
rnt
\
GÓMEZ MACÍAS RODOLFO.
::. ~ .• -~.~ r ·,, \
,..~·· ••· r11.:··i •. · ( ·, ' ' i. Thl ,,.rl (~ { 11 \·... j /'
··~ ~· fl):ll(. ., '-,,: .. 2-
1 "- 11·· , ... , ., • ....... "7 ... ··· ..... • .
l 1L·1 .. Í {'J r 11 .. 111 \ \'':$ ,/ IL·I , .. /
11. 13 IL"' -- r:'r .. ' 11.: "·" ,,--.......
(
-.._,_/ 11,m ~ \ 11.·13 _(--"'
rh2C:<¡..i u) JI 1\ /' IÁG
'-..._/ cmoL <"0111'--" °';. 1,;.~
l¡;E
Fig.2. Proceso de diferenciación de lifocitos Th1 y Th2 con algunas
citocinas involucradas en su desarrollo. Takahashi K 1998 <13l
En Jos últimos años se ha demostrado que las citocinas clasificadas en el
cuadro anterior como características de una u otra sub-población de
linfocitos colaboradores pueden ser producidas por varias células
diferentes, como parte de una respuesta que, en algunos casos, facilita el
desarrollo de lesiones tisulares, citotóxicas o de hipersensibilidad y que en
otros casos, estimula fundamentalmente la producción de anticuerpos. Por
esa razón, actualmente se observa una tendencia a referirse a ellas como
las citocinas que son características de una respuesta celular o humoral
(tipo Th1 o tipo Th2) respegtivamente (13>_
A pesar de las diferencias mencionadas, tanto las células Th1 como Th2 de
ratón tienen la misma capacidad para producir IL-3 y los factores
estimulantes de la formación de colonias de monocitos, macrófagos y
granulocitos. En los humanos también ha sido posible diferenciar las dos
subpoblaciones de linfocitos T colaboradores Th 1 y Th2. No obstante, la
separación funcional de estas dos sub-poblaciones humanas no es tan
22 TESIS CON FALLA DE ORIGEN

OCTUBRE DEL 2003 GÓMEZ MACIAS RODOLFO.
clara como la que ha sido observada en los ratones. Actualmente, la
principal dificultad para identificar las sub-poblaciones Th 1 y Th2 consiste
en que no se conocen todavía marcadores fenotipicos de cada sub
población. Por otra parte, a medida que se realizan más estudios, se
identifican nuevas poblaciones de células que producen las mismas
cit~C:i~.~~ qLJe los linfocitos Th 1 o Th2. Como ejemplo se puede mencionar
que losH~focitos B, los monocitos y Jos macrófagos producen las citocinas
IL-10 ~ 1C12 y que las células NK producen IFN gama como lo hacen los
· nñfocita~·fííf<13>. • ~ ' !,,;; ;. ,,.~:·~··
Vari6~:,~'Jtg·r~s· han propuesto la aplicación de criterios diferentes para
dividir •. i§~ni6fó~itos Th humanos en células Th1 y Th2. Uno de ellos es la
medida.de·la respuesta proliferativa al estimularlos con mitogenos. Esta
otra clasificación sólo toma en cuenta la respuesta de los linfocitos
obtenidos de los tejidos y la sangre circulante de pacientes con diferentes
enfermedades. Así por ejemplo, cuando se colocan en un medio de
cultivo, la mayoría de las células T co4<+> que infiltran la glándula tiroides
de pacientes con tiroiditis autoinmune, ellos responden al estimulo de
mitógenos como la fitohematoglutinina (PHA) formando clonas que
pueden producir IFNy, pero no IL-4. Al contrario, las células T co4<-> que
tienen receptores específicos para alergenos y que se obtuvieron de
pacientes alérgicos, casi no producen IFNy y tienen aumentada la síntesis
de IL-4 e IL-5 cuando se cultivan en las mismas condiciones y bajo
estimulación con PHA <13>.
La condición de reposo y/o actividad en la que se pueden encontrar los
linfocitos T para expresar el receptor para la IL-2 ( IL-2R) o CD25 sobre su
membrana, fue conocido inicialmente como Tac, se observo que los
linfocitos T activados aumentan la síntesis de este receptor y su
expresión sobre la superficie de la membrana.
Todos los linfocitos de las sub-poblaciones que se mencionaron tienen el
receptor de membrana RCT que les permite reconocer antígenos. Los
linfocitos T también tienen sobre su membrana correceptores como el
ligando del antígeno CD40 (CD40L) y varias moléculas de adhesión
23

OCTUBRE DEL 2003 GÓMEZ MACIAS RODOLFO.
intercelular que son necesarias para su interacción con otras células. Por
ejemplo, la falta del antígeno CD40L, impide que los linfocitos Th
estimulan a los linfocitos B y como una consecuencia de este defecto de
los linfocitos Th, se provoca una disminución en la producción de los
anticuerpos <13J.
Recientemente se ha reconocido que en los GP la respuesta inmune
mediada por anticuerpos predomina más que la respuesta inmune
mediada por células. Además, un estudio en animales sustenta que las
células T tienen un papel menor en la patogenicidad de estas lesiones.
Está claro que la evaluación simple del marcador fenotípico para las
subclases de células T no refleja adecuadamente el proceso inmune que
puede estar ocurriendo en las lesiones, por lo tanto, los efectos
funcionales de las células T ya activadas en las lesiones, podrían
evaluarse con técnicas inmunohistoquímicas y de biología molecular, y
con ello discutir las funciones de las células T <20J.
1.7.1. CÉLULAS DENDRÍTICAS.
Las OC son una familia de células originadas en la medula ósea, estas
localizan en diferentes tejidos (piel, ganglios linfáticos, timo), o formando
redes en las áreas que ocupan los linfocitos T o cercanas a éstos. Las
funciones más importante de estas células; son endocitar el material
antigénico, digerirlo enzimáticamente, unir los determinantes antigénicos
resultantes a las moléculas de histocompatíbilidad propias y finalmente
transportar los antígenos extraños hasta los ganglios linfáticos de drenaje,
a través de los vasos linfáticos aferentes. Durante su paso por los vasos
linfáticos, estas células se adaptan y cambian su morfología
denominándoseles células veladas. Una vez dentro de los ganglios
linfáticos <5 ºl, las células veladas se localizan en la paracorteza donde se
encuentran las células T. Una vez localizadas en este sitio las células
veladas extienden sus proyecciones citoplasmáticas que se interdigitan
entre los linfocitos T para presentar los antígenos. Ahora estas CPA

OCTUBRE DEL 2003 GÓMEZ MACIAS RODOLFO.
reciben la denominación de CD interdigitantes por su particular
disposición en los ganglios linfáticos <43
>_ En general las CD se
caracterizan porque expresan sobre su membrana una cantidad
abundante de moléculas de histocompatibilidad Clases 1 y 11 !44.4
5>_ así
como antígenos CD1 <45
-47
>_ Las CD también expresan sobre su
membrana una gran cantidad de moléculas de adhesión, tales como LFA-
1 (COiia), ICAM-1 (CD54), LFA-3 (CDSS) y también poseen moléculas co
estimulatorias como 87.1 (COSO) y 87.2 (CD86) <4 ª>. Todas estas
moléculas son necesarias para las interacciones entre las CD y los
linfocitos T. A diferencia de los macrófagos, las CD no producen IL-1. Sin
embargo, sintetizan IL-2 y a través de esta interleucina, contribuyen a la
co-estimulatorias de los linfocitos T. Por último en su citoplasma
contienen también una proteína denominada S-100 <49
•50>.
Las CD pueden clasificarse en varios grupos según su localización. En la
piel se les denomina CL y estas se localizan entre los queratinocitos de la
epidermis. Las CL son sensibles a la radiación ultravioleta, de modo que
su número disminuye después de la exposición prologada a la luz del sol
y se caracterizan porque contienen abundantes granulaciones
citoplásmicas, llamadas gránulos de Birbeck. Son células móviles que,
después de endocitar los antígenos que penetran al cuerpo por la piel,
pueden movilizarse a través de la epidermis y la dermis, atraviesan la
membrana basal, alcanzan los vasos linfáticos y llegan hasta los ganglios
linfáticos regionales. A medida que avanzan por los ganglios linfáticos,
presentan un proceso de maduración mediante el cual reducen su
capacidad para atrapar y procesar enzimáticamente los antígenos,
mientras que aumenta su capacidad de presentación antigenica. Cuando
llegan a los ganglios, se localizan en la región paracortical que contiene
linfocitos T, a los cuales les presentan los determinantes antigénicos de
las moléculas que endocitaron cuando estaban en la piel <50>.
Recientemente se ha caracterizada las CD del timo, llamadas también
células estrelladas. Son CPA que se caracterizan porque tener un
abundante cantidad de moléculas de histocompatibilidad clase 11 sobre la
25

OCTUBRE DEL 2003 GÓMEZ MACÍAS RODOLFO.
superficie de su membrana. Como un órgano linfoide primario, el timo no
participa activamente en el inicio de la respuesta inmune humoral o
celular. Sin embargo, las CD del timo son importantes para la selección
(positiva o negativa) de los timocitos y para la eliminación de los linfocitos
autorreactivos que maduran en esta glándula. Lo mismo que CD
expresan sobre su membrana el antígeno 87 que es el ligando del
antígeno CD28 en la membrana de los linfocitos T 51.
También hay otras CD tal como las CD de la medula, del tejido intersticial,
de los ganglios y de la sangre <52
.53
>_
1.7.2 ULTRAESTRUCTURA DE LAS CL.
La microscópica electrónica le dio a las CL una identidad peculiar
diferenciándola de las otras células epidérmicas, además de que
incrementó de manera considerable el conocimiento acerca de ellas. En un
artículo clásico, Birbeck, Breathnatch y Everall (1961) describieron el patrón
estructural básico de las CL: Un núcleo celular con una apariencia
marcadamente indentada debido a la presencia de profundas hendiduras,
un aparato de Golgi bien desarrollado, un citoplasma claro en comparación
con los queratinocitos adyacentes, la ausencia de filamentos intermedios
de prequeratina, escaso lisosomas, Ja carencia de desmosomas en su
membrana plasmática y la presencia de un organelo citoplásmico
característico. Este organelo, al cual se le denominó gránulo de Birbeck,
puede tener forma de bastón o de raqueta de tenis <54>_
1.7.3. ORIGEN DE LAS CÉLULAS DE LANGERHANS.
Por muchos años, las CL fueron consideradas erróneamente como un
componente del sistema nervioso periférico derivado del neuroectodermo.
Lo anterior se debió a dos razones: la técnica de cloruro de oro utilizada
26

OCTUBRE DEL 2003 GÓMEZ MACIAS RODOLFO.
para evidenciar a las CL también impregna al tejido nervioso y Langerhans
e investigadores posteriores consideraron que los procesos dendriticos de
las CL recordaban a las fibras nerviosas. Otra teoría ampliamente discutida
a mediados del siglo, fue la posible relación de las CL con los melanocitos.
Las CL fueron consideradas como melanocitos desgastados <55>, estados
previos a melanocitos melanogénicos <55
> o posiblemente melanocitos
inmaduros o en reposo C57J. Sin embargo, al examinar la epidermis y los
primordios de extremidades de ratones deprivados experimentalmente de
crestas neurales, con microscopia fotónica C55> y electrónica C
59>, se
demostró la presencia de CL pero no de melanocitos, evidenciando con
esto que estas dos células no estaban relacionadas entre sí.
En 1979, dos grupos de investigadores dieron a conocer al mismo tiempo,
que las CL derivaban de las células de la medula ósea. Katz y cols C50>,
realizaron dos series de experimentos: El primero consistió en transplantar
piel de ratón A/J a ratones híbridos F1 (A/J x Balb/c). Después de tres
semanas, con antisueros aloespecíficos se determinó que las CL dentro de
la piel transplantada no retenían la antigenicidad de las moléculas clase 11
del complejo principal de histocompatibilidad (CMH) del donador sino que
expresaban la del híbrido, mientras que los queratinocitos retenían la
antigenicidad de las moléculas clase 1 del CMH del donador, indicando que
las CL habían derivado de un grupo de células móviles. En el segundo
grupo de experimentos, se crearon ratones quiméricos al inyectar
intravenosamente células de médula ósea alogénica o semialogénicas
(híbrido F1) a un ratón irradiado letalmente. En los ratones transplantados
alogenicamente, a los 32 días, se encontró que el 33% de las CL
epidérmicas del receptor, expresaban las moléculas clase 11 del donador y
en los rat~~,es transplantados semialogénicamente, a los 85 días se
encontró qUe más del 80% de las CL epidérmicas se habían originado en el
tejido donado. Ellos concluyeron que las CL eran derivadas de, y
continuamente repobladas por células precursoras móviles que se
originaban en la médula ósea. Al mismo tiempo, Frelinger y Cols '61•62>,
investigaron el origen de las moléculas clase 11 epidérmicas en la piel de
27

OCTUBRE DEL 2003 GÓMEZ MACIAS RODOLFO.
ratones quimencos, mediante la inmunoprecipitación indirecta de estas
moléculas con aloantisueros de ratón. Ellos encontraron que el fenotipo de
las moléculas clase 11 epidérmicas en el ratón quimérico coincidía con el de
la medula ósea del donador. Diez semanas después del transplante, las
células· que sintetizaban las moléculas clase 11 eran originadas
exclusivamente por el tejido donado. Ya que las CL son las únicas células
en la epidermis normal que expresan las moléculas clase 11 ellos
concluyeron que las CL habían derivado de la médula ósea. Estas
observaciones en el ratón se confirmaron posteriormente en el humano, al
observar que en la piel de una mujer de 19 años que padecía aplasia
medular, las CL mostraban fluorescencia nuclear en el cromosoma Y, ya
que se le había realizado un transplante alogénico de médula ósea de un
varón <53>.
En la actualidad, los estudios acerca del linaje de las CL están
encaminados a identificar a la células madre de médula ósea que les da
origen. En 1985, Goordyal y cols <54>, demostraron en la médula ósea del
humano la presencia de células con el fenotipo de CL (CD1 y HLA-DR) y
macrófagos. Estudios más detallados en estos progenitores, revelaron que
las células CD1a+ de la médula ósea también eran positivas a la lectina
aglutinina de cacahuate (PNA +) y que expresaban además las moléculas
CD4, CD14, CD33, HLA-DR, HLA-DP y receptores para el C3b y para el FC
de la lgG <55
•55>. Además, se observó una aparente transformación de
macrófagos a CL en la piel de pacientes con diferentes leucemias en los
cuales se había realizado transplantes de médula ósea <57>. Estos estudios
sugirieron que las CL y los macrófagos podían tener un antecesor común.
Recientemente, el cultivo de células de la médula ósea y de las células de
la sangre periférica en presencia del factor estimulador de colonias de
granulo y macrófagos (GM-CSF) ha permitido la generación de
progenitores de células dendríticas <5ª·59>, algunas de las cuales tienen la
morfología y el inmunofenotipo de CL aunada a la capacidad de inducir
una gran respuesta proliferativa de células T alogénicas <70
>_
28

OCTUBRE DEL 2003 GÓMEZ MACIAS RODOLFO.
Además, Ja adición de TNFcc y de GM-CSF a un cultivo de células de
sangre de cordón umbilical de humano, permitió el aislamiento de un
precursor que expresaba CD34 <71
> y que originó tanto monocitos como
CD/CL que tenían las siguientes características.
a) marcadores de membrana CD1, CD4 y moléculas clase 11 MHC, b)
presencia de gránulos de Birbeck y c) capacidad de interrelacionar
antígenos por endocitosis mediada por receptores y una gran capacidad
estimular a Jos linfocitos T alogénicos <12>. En 1996, Rozanzwanjg, Canque y
Gluckman, en unos experimentos muy elegantes, establecieron la vía de
diferenciación de las células dendríticas que van desde un precursor CD34+
que después de algunos días en cultivos se torna en CD34- y al mismo
tiempo aparecen los antígenos CD13, CD4 y HLA- DR. Posteriormente, las
células expresaron el antígeno CD1 y moléculas de adhesión y co
estimulatorias. Se puede considerar que los precursores de las células
dendríticas se diferencian in vitro, primero en células CD1 a- que recuerdan
a las células dendríticas sanguíneas y después en CD1 a+, que son
similares a las CL y células dendríticas de los órganos linfoides <73>.
~ .. * ~¡ ~;· ~ :,
. ·.~-•.;t .. ·::...:·
f.:6•L.o.":I'
Cf•M<J11! t:.'t • •. ' "'
'"''U~:.r1~.:~:~.~. ', .·: ,: ··-- ... ·
;_~: ........ - ''..~ ;.~.~:~:.' :;f'hll" tr-on<"•-• '"'r•l"•iro;¡or µl~1llflC>le-flCl~I .,,•f'l<.Udft ., :
fig.3 Origen de las CL. Tomado de Rozenzwajg et al., 1996<74>
29
TESIS CON FALLA DE ORIGEN

OCTUBRE DEL 2003 GÓMEZ MACIAS RODOLFO.
1.7.4. FUNCIONES DE LAS CL.
Tan sólo siete años después de la descripción de las CL realizada por
Langerhans en el año 1868 y Ranvier <75
>_ se consideró que las CL eran
linfocitos, posteriormente, Arnstein <75
> y Herxheimer <77>, las consideraron
como leucocitos pasajeros (es decir, en transito en la epidermis para
dirigirse a otro lugar). En este sentido es pertinente preguntarnos ¿cuáles
hubieran sido las consecuencias para la investigación acerca de las CL si
se hubiera proseguido por ese camino en esos años? La
inmunocompetencia en el contexto de las CL fue especulada primero por
Billingham y Silvers <16
> y por Pruneiras <79>, quienes hipotetizaron que las CL
podían capturar material antigénico y desempeñar una función en la
respuesta inmunológica primaria. Sin embargo, el concepto de que las CL
son CPA se afirmó por las relaciones estructurales que tenían las CL con
otras células epidérmicas. En el curso de estudios ultraestructurales de los
eventos celulares que ocurren en las reacciones de hipersensibilidad de
contacto en humanos y cobayos <60
•61 >, observaron la estrecha aposición
entre CL y linfocitos, así como la presencia de CL con antígeno en su
superficie, dentro de · los vasos linfáticos dérmicos y en los ganglios
linfáticos regionales.'
Las CL funcionan C::omo células cutáneas presentadoras de antígenos que
inducen potentes repuestas inmunológicas primarias dependientes de
linfocitos T CD4+/CD8-. Para disparar estas respuestas inmunológicas es
necesario que los receptores de los linfocitos T (RCT) reconozcan a los
antígenos en asociación con las moléculas clase 11 del CMH. Estos
complejos antígeno/moléculas clase 11 están presentes en la superficie
celular de las CPA. Sin embargo, para que los linfocitos T puedan
reconocer antígenos protéicos, se necesita que éstos sean procesados <62>.
30

OCTUBRE DEL 2003 GÓMEZ MACIAS RODOLFO.
Lo anterior involucra tres pasos (fig 4).
~ La captación de antígenos por las CPA.
~ El procesamiento de los antígenos mediante proteolisis en
vesículas intracelulares.
~ La presentación de un péptido conteniendo el determinante
antigénico
Aunque la secuencia anterior es cierta, el reconocimiento de haptenos
puede ser por conjugación directa de éste con moléculas clase 11 del CMH
en la superficie de las células presentadoras de antígenos <53
.54>.
fig.4. Captación, procesamiento y presentación del antígeno
Chu AC,et al., 1997(53·54>.
Además, para completar la activación de las células T CD4+/CDS-, las
células presentadoras de antígenos están equipadas con otras moléculas
de adhesión ca-estimuladoras. Estas moléculas presentes en la superficie
de las CPA interactúan con sus ligandos en los linfocitos T (55>. Estas
interacciones no sólo estabilizan la unión del RCT con el complejo
antígeno/moléculas clase 11, sino que también proveen señales de
activación adicionales <85·86>.
Algunas interacciones receptor/ligando juegan un papel funcional
importante en la activación de linfocitos T, por ejemplo: ICAM-1/LFA-1,
31 TESIS CON
FALLA DE ORIGEN

OCTUBRE DEL 2003 GÓMEZ MACIAS RODOLFO.
ICAM-3/LFA-1, LFA-1/ICAM-1, LFA-3/CD2, B7-1/CD28, 87-1/CTLA4, B7-
2/CD28, B7-2/CTLA4, y CD40/CD40L. La ausencia de señales co
estimulatorias como 87 pueden conducir a una pérdida de respuesta de los
linfocitos T a estimulación antígeno-específica subsecuente (alergia) <57>.
Por otro lado, las células presentadoras de antígenos pueden producir
señales ca-estimuladoras a través de la producción de citoquinas como las
IL-1, IL-6, IL-12, e IL15. La IL-12. Atrae mucha la atención, ya que esta
citoquina derivada de las CPA es esencial para la producción de IFN y y
para la generación de linfocitos T ayudadores del tipo 1 (Th1) <55>, los cuales
están asociados con la inmunidad mediada por células, incluyendo la
hipersensibilidad por contacto.
Las CL poseen todas las características necesarias de las CPA para la
activación de linfocitos T. Las CL aisladas en fresco (son el equivalente in
vitro de las CL intraepidérmicas) pueden en forma eficiente captar y
degradar intracelularmente antígenos complejos <59
•90>. Durante el complejo
in vitro, las CL adquieren una fuerte expresión de moléculas clase 11 del
CMH y de moléculas ca-estimuladoras como ICAM-1, ICAM-3, LFA-3, 87-
1, 87-2 y CD40, las cuales juegan un papel importante en la activación de
linfocitos T <91 -95>. Además, las CL son capaces de producir IL-1 <95>, IL-6 <97>, IL-12 <95> e IL-15 <99>,
Por otro lado, estudios in vitro utilizando ca-cultivos de linfocitos T con
suspensiones celulares enriquecidas en CL o células epidérmicas totales
han demostrado que las CL pueden funcionar como CPA en respuestas
antígeno específicas de linfocitos T <10º· 101 >, como células estimuladoras en
reacciones leucocíticas mixtas alogénicas y autólogas <100
·1º2
·103
•104>, como
células accesorias en respuesta de linfocitos T inducidas por lectinas <103
·105>
y como cel.ulas inductoras en la generación de linfocitos T citotóxicos
alorre.a6Uvos y haptenos específicos <106
·1º1>. Pruebas de estas capacidades
funcionales demostraron las siguientes observaciones, las CL son capaces
de reemplazar las funciones estimuladoras de monocitos/macrófagos en
respuestas dependientes de CPA en linfocitos T, las respuestas de
linfocitos T fueron abrogadas cuando las CL fueron selectivamente
32

OCTUBRE DEL 2003 GÓMEZ MAC[AS RODOLFO.
eliminadas de suspensiones celulares epidérmicas previo al ca-cultivo y
finalmente el enriquecimiento de CL provocó un aumento paralelo de las
respuestas de linfocitos T.
Las respuestas proliferativas antígeno específico inducidas por CL en los
linfocitos están restringidas por las moléculas clase 11 del CMH, ya que los
anticuerpos monoclonales anti-HLA-DR específicamente pueden bloquear
esta clase de respuestas <1º4
·10ª> Célula a célula, las CL son
significativamente más potentes, como CPA, que los monocitos de Ja
sangre periférica en la inducción de la respuesta contra antígeno y
aloantígenos <109
•111 >. Esta superioridad de las CL fue recalcada por la
observación de que clonas níquel específicas de linfocitos T humanos
derivados de lesiones cutáneas inducidas experimentalmente, fueron
activadas cuando el níquel fue presentado por las CL, mientras que las
mismas clonas no respondieron cuando el níquel fue presentado por
monocitos sanguíneos <112>. Además, las CL no sólo presentan haptenos
como el cloruro de níquel o proteínas solubles como el toxoide tetánico, el
derivado proteico purificado (PPD) del Mycobacterium o proteínas de
Candida Albicans, sino que también son capaces de presentar antígenos
celulares complejos tales como proteínas de eritrocitos de borrego <113>.
Ya que las CL son fuertemente estimuladoras de respuestas inmunes
alogénicas, se considera que tienen un papel muy importante en el rechazo
de injertos cutáneos <114
·115>. En este sentido, la capacidad de
alosensibilización de las CL es muy alta, ya que se ha observado que con
tan sólo 1 O de estas células un ratón puede ser sensibilizado a moléculas
clase 1 y clase JI del MHC <116>.
Mucho de Jo que se conoce a cerca de la función de las CL proviene de
estudios de sensibilización en reacciones de hipersensibilidad por contacto
(respuesta inmunológica tipo IV). Los sensibilizadores son substancias
químicas relativamente pequeñas (haptenos) que al estar unidas a un
acarreador proteico llegan a ser inmunogénicos.
Frecuentemente el acarreador es una proteína epidérmica. Algunos de
estos sensibilizadores por contacto como, el TNBS o el cloruro de níquel,
33

OCTUBRE DEL 2003 GÓMEZ MAC[AS RODOLFO.
pueden unirse de manera directa y con gran afinidad a las moléculas clase
11 del CMH de células presentadoras de antígenos. Por otro lado, la
inducción de hipersensibilidad por contacto en piel parece depender de la
densidad de las CL.
En este sentido, no se logra una inducción de hipersensibilidad por
contacto en zonas de piel carentes naturalmente de CL como la cola del
ratón o áreas de piel carentes experimentalmente por radiación ultravioleta
<117•118
·119> o por substancias quimicas. El papel crucial de las CL como
células presentadoras de antigenos fue establecido por Shelley y Juhlin
<120>, quienes demostraron una acumulación específica de sensibilizadores
de contacto en las CL. El papel selectivo de las CL en este proceso lo
sugirieron los resultados de los experimentos en los que se demostró que
los sensibilizadores por contacto se acumulan en la superficie celular de las
CL, mientras que otro tipo de substancias no lo hacen (121 >. Otros trabajos
revelaron que las CL poseen un alta tasa de endocitosis por medio de la
cual captan. moléculas extracelulares <122> y que son capaces de unir
antígenos porrne~io de receptores de manosa <123>.
Algunos estL~ici;s d~~~straron que los sensibilizad ores de contacto afectan
::~zw~~~~¡~~i~ittr;d~ª:i:~ ":~" ;::~::":"~:~·::os~:~::~ sensibi1Ízaéforas5 Cós 'efectos obser-Vados en las CL incluyen un aumento en
la expreiiÓ~\je las molÉlculas clase 11 del CMH <124>, pérdida de actividad de
ATPa~á <"125>~'.áurnento del retículo endoplasmico rugoso (124>, aumento en el
número de los gránulos de Birbeck .<125> y aumento en la endocitosis
mediada por receptores evidenciada por aumento en el número de
vesículas recubiertas, endosomas y lisosomas P25·126>. Sin embargo,
también se pueden observar <124
·121
> en Experimentos en los que se
realizaron aplicaciones de sensibilizadores por contacto, demostraron una
caída de más del 40% en el número de CL a las 24 hrs siguientes (128>.
Muchos estudios (129·130
> apoyan el concepto generalmente aceptado de
que estas CL "cargadas de antígenos", migran a través de los vasos
linfáticos aferentes, hacia los ganglios linfáticos de drenaje, donde activan a
3-J.

OCTUBRE DEL 2003 GÓMEZ MACIAS RODOLFO.
linfocitos CD4+. Durante su migración, las CL viajan a través de los vasos
linfáticos diferenciándose en células de veladas. Una vez que llegan a los
ganglios linfáticos regionales de drenaje, las células veladas adquieren un
fenotipo de células interdigitales. De hecho, los datos experimentales
indican que las CL originan tanto a las células veladas como a las
interdigitantes, ya que estas últimas pueden mostrar la presencia de
gránulos de CL en un lapso de 6 a 72 hrs después de Ja sensibilización con
un antígeno en la piel. Además, el número de células veladas en la linfa
puede ser hasta 1 O veces mayor poco después de la sensibilización con un
antígeno; y cuando utilizan antígenos marcados, se ha demostrado que las
únicas células que Jos muestran son las células en veladas y las
interdigitnates.
Los mecanismos por los cuales las CL emigran de la epidermis hacia los
ganglios linfáticos todavía están claros del todo. Sin embargo, se relacionan
a un aumento en la expresión de moléculas clase 11 <131 >, cambian en el
patrón de secreción de citoquinas por parte de las células epidérmicas <132>,
secreción de interleucinas IL-113 (IL- I 13) <133•134
> y factor de necrosis tumoral
o. (FNT-o.) <135
> y cambios en la expresión de moléculas de adhesión que
disminuyan las interacciones adhesivas entre las CL y los queratinocitos y/o
la matriz extracelular <13ª·1137>. Una vez en los ganglios linfáticos, estas
células se localizan en las zonas T-dependientes, donde activan en forma
antígeno-específica, a linfocitos en reposo (CD45RO+). lo anterior conduce
al desarrollo y expansión clonal de linfocitos T antígeno específicos de
memoria (CD45RO+) los cuales pueden regresar a la piel por circulación
sanguínea, si muestran el antígeno asociado a los linfocitos cutáneos
(CLA). Este ciclo funcional de las CL está resumido en la Fig.5.
35

OCTUBRE DEL 2003
Ag
GÓMEZ MACIAS RODOLFO.
DER...'\llS
~ ---~
fig.5. Origen, captación, endocitosis y procesamiento del antígeno,
migración y presentación del mismo a Jos linfocitos T. Modificado de
Teunissen: Histochem J 24:697-716,1992.
1.7.5. IDENTIFICACIÓN Y FENOTIPO DE LAS CELULAS DE
LANGERHANS.
Las CL no se observan con Ja fijación y tincíón rutinaria de cortes en
parafina de piel, ya que con la tinción de hematoxilina-eosina aparecen
irregularmente como células claras en los estratos superficiales de la
epidermis <13ª>. Si.n embargo, las CL muestran un patrón característico de
tinción y diversas técnicas se han empleado para Ja identificación de las
CL, entre las que se encuentran las impregnaciones metálicas,
histoquímícas enzimáticas, inmunohistoquimocas y microscopia electrónica
de transmisión.
36
TESIS CON FALLA DE ORIGEN

OCTUBRE DEL 2003 GÓMEZ MACIAS RODOLFO.
a) Impregnaciones metálicas.
Las CL se identificaron originalmente con impregnación en una solución
ácido de cloruro de oro, con la cual se observan como "elementos
dendríticos intraepidérmicos cuyas prolongaciones alcanzan Ja capa córnea
y las capas inferiores de Ja epidermis". Aunque las CL se impregnan
selectiycimente, Ja técnica es caprichosa y los resultados son difíciles de
reproducir <139>_
Otra impregnación metálica empleada para identificas a las CL es el
método de zinc-yodo-osmio, el cual se aplicó con éxito para la
demostración de las CL en piel humana normal y patológica, tanto con
microscopia fotónica como electrónica <140
.141
>_
b) Histoquímica enzimática.
Está demostrada la presencia de varias enzimas en las CL, como a.-D-
manosidasa <142> fosfatasa alcalina <
143> aminopeptidasa (144) ¡3-
glucoronidasa <145>, colinesterasa <
145> y peroxidasa endógena <
145> Sin
embargo, los procedimientos enzimáticos más útiles para Ja observación de
las CL son las histoquímicas para demostrar la actividad de ATPasa y de
esterasa inespecífica.
A principios de los años 60, varios grupos de investigadores informaron de
la actividad de ATPasa en las CL, lo cual fue un hallazgo circunstancial ya
que el propósito de las investigaciones era otro. Mustakallio <147>, trato de
demostrar que los elementos neurales epidérmicos podían teñidos por la
reacción histoquímica para ATPasa, describió la tinción de nervios
papilares y sus extensiones intraepidérmicas y la red dendritica de CL.
Desgraciadamente Mustakallio <147
> intentó relacionar a Ja ATPasa de las
células dendríticas con funciones neurales.
37

OCTUBRE DEL 2003 GÓMEZ MACIAS RODOLFO.
c) Marcadores inmunológicos.
Una manera más precisa de identificar a las CL es emplearon anticuerpos
para la identificación de marcadores citoplásmicos y de superficie. En este
sentido, han sido identificadas una gran variedad de moléculas, a nivel de
microscopia de luz y electrónica, usando diferentes técnicas de tinción
(inmunoperoxidasa, inmunofluorescencia o marcaje con inmune-oro) con
distintas formas de procesamiento del tejido (cortes en frío, suspensiones
celulares epidérmicas). Por otro lado, la identificación de estos marcadores
le han dado a las CL una identidad característica y han proporcionado una
mejor comprensión de su papel funcional en la presentación de antígenos.
Dentro del perfil fenotipo, las moléculas del CMH son esenciales para que
las CL desempeñen su función ya que están asociadas a la presentación
de antígenos. Estás moléculas se dividen en dos grupos principales:
moléculas clase 1 y clase 11 los cuales son necesarias para la presentación
de péptidos antigénicos <155J_
Estos complejos antígeno-molécula del CMH, expresados en la superficie
de las células presentadoras de antígenos, pueden ser reconocidos
específicamente por linfocitos T a través de sus receptores. Además, la
interacción entre el CMH-antígeno y el receptor de células T está
acompañada por la unión de moléculas de superficie en los linfocitos con
las moléculas del CMH. Esto es, las moléculas clase 11 y clase 1 de las CL
se unen a los antígenos CD4 y CDS, respectivamente, en los linfocitos
<149•15ºl. Las moléculas clase 1 están involucradas en la presentación de
antígenos .endógenos (como los antígenos tumorales), mientras que las
moléculas clase U lo están en la presentación de antígenos exógenos <151
J.
38

OCTUBRE DEL 2003 GÓMEZ MACIAS RODOLFO.
Tabla 1: Perfil fenotípico de las CL epidérmicas humanas.
Marcador Presencia Marcador Presencia
Específicos de CL Marcadores de Mo/Mq
Ag-Lag + C015 (LeuM1) -(+) (leuM2)
Moléculas MHC C014 (LeuM3) Clase 1 (HLA-ABC) + CD68 Clase 11 (HLA-OR) ++ OKM5 Clase 11 (HLA-OP) ++ Clase 11 (HLA-OQ) ++ Recept y Mol. de Adhesión
CD64 (Fc-1R1) Marcadores de células T COW32 (FcgR11) + CD la ++ C016 (Fcg R111) CD1b C035 (CR1) CD1c + C021 (CR2) CD2 CD11b (CR,; C,biR; gp155/95) + CD3 C011c (gp 150195) + CD4 + co1a (cadena p C011a,b,c) + CDS C011a (LFA-1; gp 180/95) CD7 cosa (LFA-3! +(-) coa C054 (ICAM-1) +(·) Receptor de anlicuerpos CD29 (cadena b de VLA) + Receptor yo -(+) CDw49b (VLA-2) +
VLA-3 + Marcadores de células B CDw49d (VLA-4) ++ CDIO CDw49f (VLA.Q) ++ CD19 C025 (IL·2R) CD20 CD22 Otros Marcadores CD24 C045(T200) + CD40 -(+) 8100 +
Modificado de Teunissen: Histochem J 24: 697-716, 1992
Al igual qu~)odas las.células nucleadas, las CL expresan moléculas clase 1
del CMH <152•153> y usando anticuerpos monoclonales contra un
determinante monomórfico clase 1 (W6/32) se encontró que los niveles de
estas· moléculas eran parecidas a queratinocitos y las CL por
inmunofluorescencia y citometria de flujo mientras que otros describieron
niveles mayores de las moléculas clase 1 en los queratinocitos <154>.
Rowden <155> y Klareskog <
155> fueron los primeros en describir la expresión
de moléculas clase 11 en las CL del humano y desde entonces está bien
39

OCTUBRE DEL 2003 GÓMEZ MACIAS RODOLFO.
establecido que las CL muestran en su superficie moléculas codificadas por
los genes HLA-DR (análogos a los 1-E murinos) C157>,- DQ (análogos a los 1-
A murinos) <157> y -DP <
158>. En la epidermis normal parece existir un
traslapamiento completo de estos antígenos en las CL <158>. Con dos
excepciones, las CL son las únicas células en la epidermis que expresan
estas moléculas. Las células acrosiríngeas en la epidermis humana normal
forman la porción intraepidérmica de los conductos de glándulas
sudoríparas, son fuertemente positivas para HLA-DR pero no para el -DQ y
-DP <159>. Los queratinocitos en piel con procesos patológicos también
pueden expresar moléculas clase 11 <150>.
Las CL son fuertemente positivas para el HLA-DR, sin embargo, se pueden
distinguir dos sub-poblaciones por la intensidad de tinción con un anticlase
11. Así, encontró que el 25% de las CL en suspensiones epidérmicas <151> y
del 5-10% en láminas epidérmicas <152
> se tiñen más brillantemente con
fluorescencia. Lo anterior parece indicar que las CL más brillantes son más
capaces en la presentación de los antígenos. También está calculado que
la cantidad de determinantes DR en las CL es de 50 a 100 veces mayor
que la de los monocitos/macrófagos <153>. Esta característica coloca a las CL
como las CPA por antonomasia.
En Fithian <154> y Murphy <155> observaron que las células de Langerhans del
humano se teñían fuertemente con el anticuerpo OKT6. Esta reactividad
llamó fa atención ya que con anterioridad se había descrito que el antígeno
T6 era un antígeno de diferenciación de timocitos <156>. En la actualidad, a
este antígeno se le conoce como CD1 y está dividido en los subgrupos de
diferenciación CD1 a (glucoproteína de 49-kDa), CD1 b (glucoproteína de
45-KDA) y CD1 c (glucoproteina de 43-kDa) <151>. Estas glucoproteinas, al
igual que las moléculas clase 1, están asociadas con la [32-microglobulina
<168> y se sugiere que las moléculas CD1 pueden ser la contraparte humana
de los antígenos leucémicos del timo murino <169·170>.
Todas las CL epidérmicas expresan los antígenos CD1 a, expresan
débilmente los antígenos CD1 c y no expresan las CD1 b <171 >. Por otro lado,
-+O

OCTUBRE DEL 2003 GÓMEZ MACIAS RODOLFO.
los queratinocitos no expresan ningún antígeno CD1 a, estás pruebas son
muy útiles para la identificación de las CL <112>.
Las CL humano expresa también un marcador citoplasmático característico
llamada proteína S-100 <173
·174>. Esta proteína también están presente en
melanocitos <175>, células dendríticas de la dermis <
174· 1176>, células
interdigitantes de órganos linfoides <175
·174
•177>, células dendríticas epiteliales
de las amígdalas <175> y células gliales del cerebro <174>. En el ratón, las
células dendríticas del bazo también son S-100+ <174>, La familia de
proteínas S-100 esta muy conservada a través de la escala filogenética y
comprenden un pequeño grupo de proteínas unidoras de Ca++. Entre otras
funciones está comprobado que participan en el crecimiento de las neuritas
<179>. Ésta puede ser una explicación del porqué de su presencia en los tipos
celulares que tienen forma dendrítica (la única excepción parecen ser las
células dendríticas foliculares) <174>. Este marcador distingue a las CL de los
monoctitos/macrófagos, ya que estos últimos no la expresan. Por otro lado,
hasta el momento, sólo existe un anticuerpo monoclonal específico para las
CL, el NLDC-145 <150>. Este anticuerpo monoclonal reconoce únicamente a
las CL de ratón, pero no a la de otras especies incluido el hombre. La
proteína reconocida por el anticuerpo NLDC-145 es un receptor llamado
DEC-205 <181•182
•183>, el cual aparentemente está involucrado en la captación
de antígenos. Es de notar que los macrófagos son negativos para el NLDC-
145 (180).
1.8.1. ANTECEDENTES.
A pesar de que se ha investigado el papel que desempeñan las CD y/o
CL en diferentes procesos infecciosos, existe poca información publicada
en relación a las CD y/o CL en las LP.
En un modelo experimental. en ratas 1994 se expuso la pulpa dental del
primer molar inferior para producir una LP y se observo los cambios en el
numero y la distribuciÓ~ d~ los macrófagos y las CD durante 8 semanas
.. J.1

OCTUBRE DEL 2003 GÓMEZ MACÍAS RODOLFO.
utilizando un anticuerpo monoclonal OX6, Okiji reporto el incremento de
estas células a medida que los días pasaban, las CD en las LP mostraron
diversas morfologías. Él concluyo que las CD están involucrados en el
desarrollo y la perpétLiación de las LP, además menciono que estas
células podían. actuarcor.no CPA, siendo esenciales para la iniciación de
la respuesta inmun·a1Ógica primaria <184>.
Otros autores han utilizado anticuerpos monoclonales para demostrar la
presencia y la distribución de Jos linfocitos, macrófagos y las CL en el
ligamento periodontal (lp), GP y QP <185>.
En el 2001 otros autores desarrollaron un modelo experimental para
observar la distribución y el número CD en diversas regiones del lp, (zona
apical, mesial, distal y furcal) del primer molar inferior de una rata, por
medio de los anticuerpos monoclonales OX6, ED-1, ED2, OX62, ED9
(CD14), OX42 (CD1 :ll:i), 8A2 (CD11c) y WT.1 (CD11a), los resultados . -.,- ... . :,
refieren .una variáción en la densidad de las CD por mm2 en las diversas ' . ·. .' .
reglones del lp, las CD se localizaron alrededor de los vasos sanguíneos,
aunque no fueron bien caracterizadas debido a que los marcadores
utilizados en este estudio los comparten diferentes tipos celulares
residentes del lp, sin embargo concluyo que las CD pueden funcionar como
CPA(186l.
En otro modelo experimental en el 2001, se indujo quirúrgicamente LP en
el 1er molar inferior de una rata durante 8 semanas, para observar en el
microscopio electrónico la proporción relativa de los macrófagos y las CD,
la hipótesis fue que estas células están involucradas en la patogenicidad
de las LP. Los resultados apoyaron que las CD expresaban una alta
cantidad de moléculas clase 11 CMH que juegan un papel primordial en la
presentación de antígenos a las células T y sugieren que las CD pueden
ser la llave para iniciar la respuesta inmune primaria 1187>.

OCTUBRE DEL 2003 GÓMEZ MACIAS RODOLFO.
Las CD expresan grandes cantidades moléculas clase 11 MHC tienen una
gran capacidad para presentar antígenos a las células T y activan la
respuesta inmune primaria, teniendo un papel principal en la
patogenicidad de las LP como ya se había reportado en otros estudio <188>.
A los GP, GPE, QP y quistes residuales humanos se les aplico una técnica
de inmunohistoquímica para determinar el nivel de CD1 a con el objetivo de
caracterizar a las CD en estas lesiones, estas fueron positivas en todas las
lesiones concluyendo que las CD están involucradas en el desarrollo del
tejido epitelial y responsable del infiltrado inflamatorio en estas LP <189>.
-1-3

OC:TUBRR DF.T. 2001 GÓl\fEZ Mc\c:fr\S RODOT.Fll.
1.9.1. PLANTEAMIENTO DEL PROBLEMA.
Los GP, GPE y QP tienen una alta prevalencia e incidencia en la cavidad
bucal, esto nos obliga a conocer los diversos mecanismos por lo cuales
se establecen estas lesiones.
Las ultimas investigaciones se han enfocado fundamentalmente al estudio
de los componentes celulares y extracelulares de la respuesta inflamación
y/o inmunológica de los GP, GPE y QP, para dilucidar el papel que juegan
en estas lesiones.
No obstante que en la década pasada se identificaron CD, en estas
lesiones al momento actual se desconoce el papel que juegan en los GP,
GPE y QP humanos.
1.9.2 JUSTIFICACIÓN.
El escaso conocimiento y la poca información publicada acerca de las CD
en los GP,GPE y QP humanos, nos brida un campo fértil para investigar y
dilucidar el papel que desempeñan estas células en el desarrollo del estas
lesiones.
Seria interesante determinar la expresión de la moléculas clase 11 del
CMH y proteína S-100 debido a que las variaciones en la expresión
podría tener una injerencia directa el incremento o disminución en el
número de las CD, además en el incremento o disminución del infiltrado
inflamatorio y en el desarrolla del tejido epitelial en estas lesiones.

OC:TIJRRF. OF.I. 21101 GÓJ'\IF.7. Mr\C"'.ÍAS ROOOT.FQ
1.9.3. HIPOTESIS DE INVESTIGACIÓN.
H 1 .• El número de CD se incrementa en los GP, GPE y QP humanos.
1.9.4. HIPOTESIS NULA.
H 1 .• H 1 .• El número de CD se incrementa en los GP, GPE y QP humanos.
1.9.5. OBJETIVOS .
1.- Cuantificar el número de CD en los GP, GPE y QP humanos mediante
la expresión de a molécula clase li del CMH y la proteína S-100.
2.- Determinar la distribución de las CD en los GP, GPE y QP humanos.
1.9.6. TIPO DE ESTUDIO.
Descriptivo, observacional y prospectivo.
1.9.7. UNIVERSO O POBLACIÓN.
Para el presente estudio se seleccionaron al azar 11 individuos de ambos
sexos que presentaran las características clínicas y radiográficas de las
LP determinadas por la OMS en 1995 y 1 espécimen de encía sana para
el caso control.
1.9.8. METODOLOGÍA.
1.9.9. MATERIAL Y MÉTODO.
Las muestras para el presente estudio se obtuvieron en el Departamento
de Exodoncia y Cirugía Oral de la Facultad de Odontología de la UNAM,

OíTlJBRF, DF.L 200.1 GÓJ\fF.7. l\L-\l.TAS RODOLFO
Campus e.u. La recolección de las muestras estuvo basada en las
características radiográficas y sintomáticas de Ja periodontitis periapical
crónica preestablecidas por OMS de 1995, estas características estuvieron
presentes en todos Jos pacientes seleccionados independiente de la causa
etiológica.
Una vez seleccionado el paciente, se aisló y desinfectó el área de trabajo,
posteriormente se realizo Ja técnica de anestesia local y/o regional con
dentocaína al 2% con una aguja corta, se debridó con una legra y se realizó
Ja extracción con un elevador recto del numero 2 ó 3, si era necesario se
utilizó un Fórceps (el número dependió del diente afectado). Una vez
extraído el diente afectado se diseco el tejido blando del tejido duro con una
hoja de bisturí del número 15, todo el material utilizado para la obtención de
Ja muestra fue de Ja marca HU-FRIDAY.
Antes de fijar Ja muestra se lavó en suero fisiológico por unos segundos
para eliminar Jo más posible de tejido hemático, para Ja técnica de cortes en
parafina se fijo en formalina amortiguada al 10% durante 24 horas, al final
de este tiempo se lavaron en agua corriente durante 2 hrs y se colocaron
en el Histoquinet Tissue-Tek 11 Processo Modelo 4640-B Sakura
FJNETECHNICAL CO., L TO Tokio Japan MR para posteriormente
embeberlo en parafina con Ja ayuda Tissue-Tek 11 Modelo Tissue
Embedding Center MR.
Para la técnica de cortes en congelación las muestras se fijaron
inmediatamente en paraformaldéhido al 4% al 2M con un pH 7.2 a 7.4
durante dos horas, después se lavaron tres veces en Buffer de fosfato al
0.2 M con un pH de 7.2 a 7.4 durante 15 minutos cada uno. Posteriormente
se realizaron dos lavados con suerosa, el primero se realizara al 14%
durante 24 hrs y el segundo al 16% por 1 hr al final las muestras se
colocaron criomoldes de papel de estaño de 25x20x5 mm, y se embebieron
en un medio criosolidificable (OCT-Compound, Tissue-Tek 4583,
46

OC:TlJRRR DRT, '>Om GÓl\fF.7. l\f:\dAS RODOT.FO.
Distribuidor BayerMR S.A) mediante se colocó en un medio de inmersión
altamente criogénico que permitió la congelación rápida a 70°C (Reico).
Una vez realizado estos pasos independientemente de la técnica se
procedió a etiquetar y enumerar las muestras. Una vez realizad esto se
procedió a realizar cortes seriados, para la técnica de cortes por parafina
nos auxiliamos de un micrótomo LKB 8ROMMA retracting microtome
ROTARY ONE MR y para la técnica de cortes en congelación nos
auxiliamos de un criostato American Optical corporation Cryo-Cut
microtome MR, los cortes se realizaron a un espesor de 6 a 7 micras y se
colocaron cada una de las muestras en laminillas marcadas y enumeradas
en orden de secuencial del 1 al 11 en porta objetos seleccionados para
microscopio, manufactura de espejos s.a Mandesa 26 X 76 m.m, Las
laminillas fueron preparadas previamente con poli-L-lisina y enumeradas
(Sigma DiagnosticsMR, INC, San Louis, U.S.A). Al primer corte realizado, se
le· hizo una tinción de H-E para realizarle un estudio histopatológico para
incluir o excluir las muestras en nuestro estudio, la técnica H-E utilizada fue
tomada del Teory and Practic of Histotechnology, Sheehan D.C. Hrapchak.
8.8. The C.V. mosby Company, second, edition, 1980, St Louis. .. . . -
. . .
Los derrlá~ cortes se lavaron en PBS 3 veces por 5 minutos e/u, una vez
hidratad~ el tejido:se procedió a bloquear la peroxidasa endógena con
H202 81.3% .1: 1·0 por 1 O minutos, posteriormente se lavó en PBS/albumina
(albu;nÚ1~ 6~~i,~ots1~maMR) 3 veces por 5 minutos . . , •.. ,"··; ·. ->·'i~:<s--'.'
.,~ ~,·,._:_:::~r~~;:~-;z:.~:-,1c::,:-·-·. -
PosterÍorf11~g~~V~·~t<'cblocaron los anticuerpos primarios, primero un anti
H LÁ~DR '.cb'~~·~,~~:;dilúC::ión de 1:100 (anti-mouse, ZymedMR, San Francisco,
Califórni~.\:T.~'.A) para identificar la molécula clase 11 del CMH de las CD en
las . mu~stras seleccionadas para el presente estudio en las laminillas
enumeradas con número pares, el segundo anticuerpo primario utilizado en
este estudio fue un anti-proteína S-100 con una diluci'on de 1 :40 (rabbit,
ICN, BiomedicalsMR, 688121) que se coloco en las laminilla enumeradas
.J-7

OC:TUBRF. DEL 2001 GÓMF.7. M-\l,ÍAS RODOT.170
con los números nones durante toda noche a una temperatura de 4ºC en
un refrigerador Samsung SR368 (No Frost - Clean Back MR).
Al día siguiente se lavaron en PBS/albumina 3 veces por 5 minutos cada
laminilla. Después de transcurrido el tiempo indicado se procedió a colocar
los anticuerpos secundarios anti-mouse (coat anti-mouse lgG, Biotinilado,
INC, BiomedicalsMR) y anti-rabbit (coat anti-rabbit lgG, Biotinilado, INC,
BiomedicalsMR) respectivamente por una hora a 37°C, en una incubadora
(Modelo feliza 343, Equipo S:A MR).
Después se lavaron con PBS/albúmina 3 veces por 5 minutos, al termino
de este paso se coloco el complejo (Biotina Avidina Peroxidasa) por una
hora a 37°C en una incubadora.
Y se volvió a lavar con PBS/albúmina 3 veces por 5 minutos. Para revelar
nos auxiliamos de la DAS Diamino Bencidina (SigmaMR) la técnica de
inmunohistoquímica fue tomada del In situ Gene Expression Analysis
Industrias Bioselec, catalog 2001-2002 se realizaron modificaciones de
acuerdo al fabricante.
Al termino de esto se contratiñó con hematoxilina, se coloco resina, se puso
un cubreobjeto de la misma marca de las laminillas, se observaron y
cuantificaron las CD en el microscopio óptico (ZEISS México 1,25X) con un
objetivo de 40X, una cámara lúcida (ZEISS474623-9902 West Germany),
una lámpara de luz amarrilla y una hojas prefabricadas que contenían uno
cuadro un tamaño de 25 x 25 micras en el microscopio óptico, en cada
laminilla se cuantificaron diez campos por laminilla posteriormente se sacó
la media y se multiplicó por un millón de micras dividiéndose por 225 x 225
micras, dando el número de CD por milímetro cuadrado después se hizo la
la estadística y se tomaron fotografías con el microscopio óptico olimpos y
la cámara de la misma marcar.
.J.8

OC:TIJRRF. DF.T. 2001 GÓl\fF.7. Mr\C':ÍAS RODQT .FO.
1.10.1. RECURSOS FINANCIEROS.
Facultad de Medicina Departamento de Biología Celular y Tisular
(Laboratorio de lnmunohistoquímica). y la División de Estudios de Posgrado
e investigación de la Facultad de Odontología (Laboratorio de Inmunología)
de la Universidad Nacional Autónoma de México financio el presente
estudio.
1.10.2. HUMANOS.
M.C. Judith Alvarez Pérez.
CD. CMF. Gabriel Loranca Fragoso.
Dr. Juan Carlos C. Hernández Guerrero, inmunólogo.
Dr. Andrés Eliú Castel! Rodríguez, especialista en el tema.
Biól. Miguel A Herrera Enriquez, especialista en estadística.
1.10.3. CRITERIOS DE INCLUSIÓN.
Granuloma periapicales, granulomas con proliferación epitelial, quistes
periapicales.
1.10.4. CRITERIO DE EXCLUSIÓN.
Lesiones mal fijadas, mal cortadas ó insuficientes.
49
TESIS CON FALLA DE ORIGEN

OC:TIJBRF, DRT. "'001 GÓMR7. j\LAC:Ír\S RODOT.FO.
1.10.5. LAS VARIABLES DEPENDIENTES.
Granulomas periapicales.
Granulomas periapicales con proliferación epitelial.
Quiste periapical.
1.10.6. LAS VARIABLE INDEPENDIENTES.
Molécula clase 11 del CMH.
Proteína S-100.
1.10.7. UNIDAD DE OBSERVACIÓN.
En el estudio histopatológico fue Ja distribución de las células
inflamatorias dentro de las lesiones incluidas en el presente estudio, Ja
presencia o ausencia de células epiteliales, Ja transformación quística de
las lesiones y Ja relación quiste cavidad con el conducto radicular del
diente afectado.
En el estudio lnmunohistoquímico fue la expresión de la molécula clase JI
del CMH y Ja proteína S-100 de las CD en Jos GP, GPE y QP humanos
por su densidad/mm 2•
1.10.8. MÉTODO PARA PROCESAMIENTO DE DATOS.
Los datos se capturaron en tablas especialmente diseñadas (Tabla 1,2,3,4)
se vaciarán en el programa Microsoft Excel, y el estudio estadístico se
realizó en el paquete S.P.S.S. (versión 2000). se calcularán medidas de
tendencia central. La significancía estadística se obtuvo del 95% por medio
de la T de Student se realizó las pruebas de ANOVA.
50

OC:TI TBRF. DF.T. 2001 GÓl\fF.7. MAC:fAS RODOT.FO.
1.11.1. RESULTADOS.
En el estudio histopatológico, observamos dos GP, se determinó la
presencia de neutrofilos, macrófagos, linfocitos y células plasmáticas (fig 6)
y números vasos sanguíneos en algunas zonas de la lesión (fig. 14), siete
GPE en los cuales observamos las mismas características celulares que
los GP, en los GPE se encontraron células epiteliales con diferentes grados
de proliferación (fig.6) y 2 QP en los cuales observamos una cavidad
epitelial y un material semisólido (fig. 7) (graf 1 ).
En los grupos experimentales y el caso control fueron positivas para los
marcadores HLA-DR y S-100 de las CD (fig 9,17 y 18.), teniendo una
mayor densidad las CD en el tejido epitelial (membrana basal) (fig. 10, 11 y
12) que en el infiltrado inflamatorio crónico (fig.13, 17 y 18), en este
infiltrado observamos a las CD en la proximidad con vasos sanguíneos (fig.
15 y 16.) y los linfocitos (fig.17 y 18). Las CD presentaron distintas
características morfológicas esto pudo ser a causa de como se realizaron
los cortes (fig. 19).
La ES con el marcador de superficie HLA-DR tuvo una X= 74.66 CD por
mm2 y una DE = 32.24 CD por mm2 en 50 campos, la ES con el marcador
citoplasmáticc;i S-100 tuvo una X= 67.16 CD por mm2 y una DE = 30.19 CD
por mm2 ~n'sbcampos, en los GPE con el marcador de superficie HLA-DR
tuvo úna ~.;~{i~4.6o CD por mm2 y una DE = 69. 11 CD por mm2 en 350
campos; en'lósiÚPE.con .el marcador citoplasmático S-100 tuvo una X= 125.9~1 CD e~iJh~~f 1:1na DE = 53.54 CD por mm2 en 350 campos, en los
GP con el''rija~~~"tó'f}'cie superficie HLA-DR tuvo una X= 109.82 CD por
mm2 ·y 'uri~"'[)E.:,:,;~'g~~64 CD por mm2 en 100 campos, en los GP con el
marcad~r,ciitbt1~~hí~~j(;~:.s~1oortuvo unaX= 106.07 CD por mm2 y una DE
= 45.41 ¿·i:)"¡f6~·~~2 ~n 100 '6am~0
Ós,, en los QP con el marcador de
~up~rll~i~·HLÁ-DR tuvo una x = 114:37co por mm2 y una DE= 53.88 CD
por mrn2 en 100 campos y en los QP con el marcador citoplasmático S-100
51

OCTIJJ3RE DET. 20111 GÓMEZ MAC:Ír\S RODOT.FO.
tuvo una X= 110.02 CD por mm2 y una DE= 51.63 CD por mm2 en 100
campos (graf. 2 y Tabla 2).
En este estudio realizamos la prueba de Kruskal-Wallis para determinar las
diferencias estadísticamente significativas entre los grupos experimentales
(GP, GPE y QP) y el caso control (ES). esto sucedió cuando el valor de la
media de Z>1.96.
En el caso control con el marcador de superficie HLA-DR exhibió
diferencias estadísticamente significativas cuando se comparó con los
grupos experimentales, tanto para HLA-DR como para la proteína S-100.
GPE con HLA-DR Z = 6.92; p<0.05, GPE con S-100 Z = 6.61; p<0.05, GP
con HLA-DR Z = 3.64; p<0.05, GP con S-100 Z = 3.86; p<0.05, QP con
HLA-DR Z = 4.24; p<0.05, QP con S-100 Z = 3.92; p<0.05, pero cuando se
comparó el caso control con el marcador citoplasmático S-100 no hubo
diferencias estadísticamente significativas.
El caso control con el marcador citoplasmático S-100 exhibió diferencias
estadísticamente significativas cuando se comparó con los grupos
experimentales, tanto para el marcador HLA-DR como para la proteína S-
100. GPE con HLA-DR Z = 8.30; p<0.05, GPE con S-100 Z = 7.99;
p<0.05, GP con HLA-DR Z = 4.85; p<0.05, GP con S-100 Z = 5.07;
p<0.05, QP con .HLA-DR Z = 5.44; p<0.05, QP con S-100 Z = 5.13;
p<0.05, pe;b -~¿~licio se comparó el caso control con el marcador de
superficie lionul:>o diferencias estadísticamente significativas. ,. . " . " ..
El GPE co11 el marcador de superficie HLA-DR exhibió diferencias
estadístiC:arl"lénte significativas cuando se comparó con los GP, QP y con
el casocontrol, tanto para el marcador HLA-DR como con proteína S-100.
GP con .HLA-DR Z = 3.65; p<0.05, GP con S-100 Z = 3.32; p<0.05, QP
con HLA-DR Z = 2.74; p<0.05, QP con S-100 Z = 3.22; p<0.05, pero
cuando se comparó el GPE con el marcador citoplasmático S-100 no
hubo diferencias estadísticamente significativas.
52 TESIS CO~T
FALLA DE ORIGEN

OC:TI!Bl~F. DEI, :wrn GÓl\rEZ MAc:f r\S RODOT .FO.
El GPE con el marcador citoplasmático S-100 exhibió diferencias
estadísticamente significativas cuando se comparó con los grupos
experimentales y con el caso control, tanto para HLA-DR como para la
proteína S-1 OO. GP con HLA-DR Z = 3.24; p<0.05, GP con S-100 Z = 2.90;
p<0.05, QP con HLA-DR Z = 2.33; p<0.05, QP con S-100 Z = 2.81; p<0.05,
pero cuando se comparó con el GPE con el marcador de superficie HLA
DR no hubo diferencias estadísticamente significativas.Entre los GP y QP
ya sea con el marcadores HLA-DR y/o S-100 no hubo diferencias
estadísticamente significativos ya que el valor Z=-1.96; p>0.05 por lo cual
no las reportamos en este estudio (tabla. 3).
53
TESIS CON FALLA DE ORIGEN

fig. 6 (A y 8) H-E 40 X. Se observa un granu)oma periapica) y un granuJoma periapicaJ con proliferación epitelial. respectivamente .
fig. 7 (C) Se observa un quiste periapicaJ con un materia) semisólido.
Graf. 1 Diagnostico histopatologico de las lesiones periapicales.
63.63%
54
•GP. •GPE CQP
TESIS CO~T FALLA DE ORIGEN
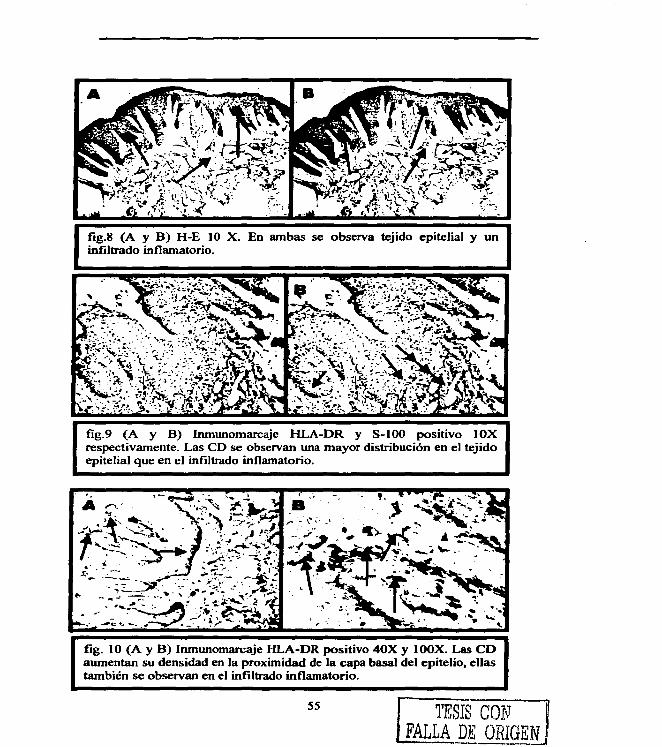
fig.8 (A y B) H-E 10 X. En ambas se observa tejido epitelial y un infiltrado inflamatorio.
fig.9 (A y B) Irununomarcaje HLA-DR y S-100 positivo IOX respectivamente. Las CD se observan una mayor distribución en el tejido epitelial que en el infiltrado inflamatorio.
~ -
fig. 10 (A y B) Inmunomarcaje 1-Il.A-DR positivo 40X y IOOX. Las CD aumentan su densidad en la proximidad de la capa basal del epitelio, ellas también se observan en el infiltrado inflamatorio.
SS TESIS CON FALLA ~-ORIGEN

fig.11 (A y B) lnmunomarcaje S-100 positivo 40X y lOOX. Las CD aumentan su densidad en la proximidad de la capa basal del epitelio, ellas también se observan en el infiltrado inflamatorio.
fig.12 (A y B) Inmunomarcaje HLA-DR y S-100 positivo lOOX respectivamente. Las CD en el tejido epitelio, ellas también se observan en el infiltrado inflamatorio (B).
B
fig.13 (A y B) Inmunomarcaje HLA-DR y S-100 positivo lOOX respectivamente. Las CD se observan en el infiltrado inflamatorio.
56 TESlS CON FALLA DE ORIGEN
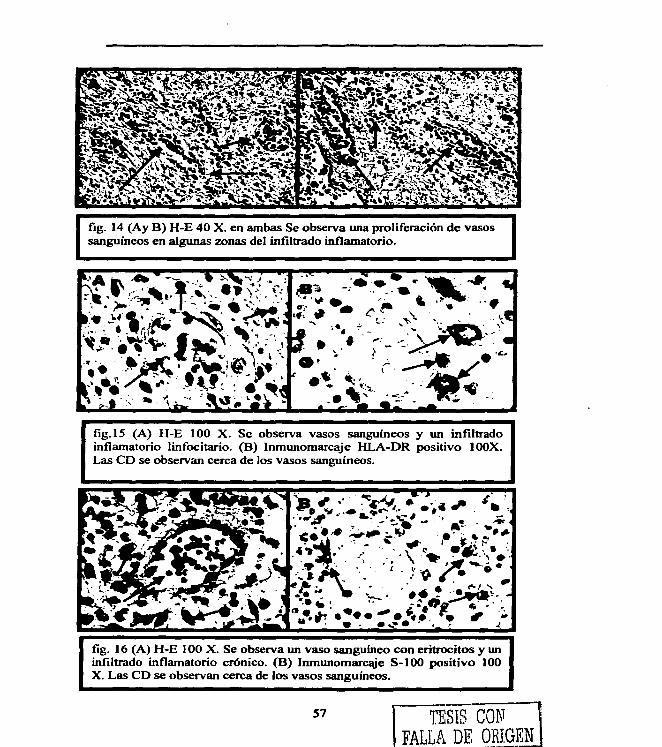
fig. 14 (Ay B) H-E 40 X. en ambas Se observa una proliferación de vasos sanguíneos en algunas zonas del infiltrado inflamatorio.
fig.15 (A) H-E 100 X. Se observa vasos sanguíneos y un infiltrado inflamatorio línfocitario. (B) lnmunomarcaje Ill..A-DR positivo lOOX. Las CD se observan cerca de los vasos sanguíneos.
fig. 16 (A) H-E 100 X. Se observa un vaso sanguíneo con eritrocitos y un ínfiltrado inflamatorio crónico. {B) lnmunomarcaje S-100 positivo 100 X. Las CD se observan cerca de los vasos sanguíneos.
51 TESIS CQ~T " FALLA DE ORIGEN l
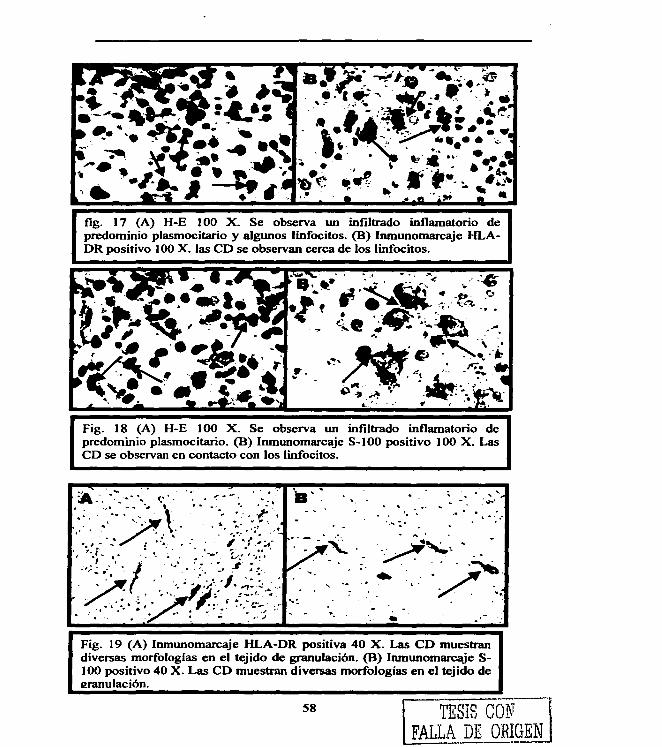
•• '!Ji.• ~·1:·· ~ ... ~ ..• ~ J#: .:.:.- . . . -· .. •· 1 .p-. .e-a~ ',_) .t
. '.- -~ -~. :~~·¡ ~ 1 ·(~, -~ . .. l"~J:• Cj ~. • • .... ,. """5'""!~ .. ;· • -~- . _'-: .•. <· ~-::.·.:,·
,,. ~ ~ •• ..,. --a~ • •· . . .... e ... • .. • • ... • .. ....... • .• ~ .. ·.· .• •4
11 ~ • ~ , • ·~ . • _,, ... •• .. • - . • •. ..._A '& (~ ~...,, ~ • .. - ~ - .. ~ .. ·• .¡t._ • ~ ....... . . . . ' •· . . .,,. fig. 17 (A) H-E 100 X. Se observa un infiltrado inflamatorio de predominio plasmocitario y algunos linfocitos. (B) Inmunomarcaje lfi.ADR positivo 100 X. las CD se observan cerca de los linfocitos.
Fig. 18 (A) H-E 100 X. Se observa un infiltrado inflamatorio de predominio plasmocitario. (B) Inmuoomarcaje S-100 positivo 100 X. Las CD se observan en contacto con los linfocitos.
- ..... ·. ' · .... :.
.. ..... . .
A O O \ ... -
.. ~, .... -~:· .- .. ~ ... "/ ·.~ •
Fig. 19 (A) Inmuoomarcaje lfi.A-DR positiva 40 X. Las CD muestran diversas moñologías en el tejido de granulación. (B) Irununomarcaje SI 00 positivo 40 X. Las CD muestran diversas moñologías en el tejido de eranulación.
58 TESIS CQ~T -;] FALLA DE OR!9EN

Gnf. 2 Densidad de CD en GP, GPE y QP.
i E el ~
"CI
e ~
E ·= z
Lesiones periapicales.
Tabla 2.
S-100
lhHcl ~
Media de los grupos experimentales y el caso control.
Lesiones Campos Medias periapicales ES CMH 11 50 74.66 ES S-100 50 67.16 GPE CMH 11 350 134.60 GPE S-100 350 125.91 GP CMH 11 100 109.82 GP S-100 100 106.07 QP CMH 11 100 114.37 QP S-100 100 110.02
Desviación estándar 32.24 30.91 69.11 53.54 58.64 45.41 53.88 51.63
59
Valor Valor minimo máximo 19.75 138.27 19.75 177.77 19.75 493.82 39.50 316.04 19.75 256.79 19.75 217.28 39.50 316.04 39.50 316.04
TESIS COl\T FALLA DE ORIGEN
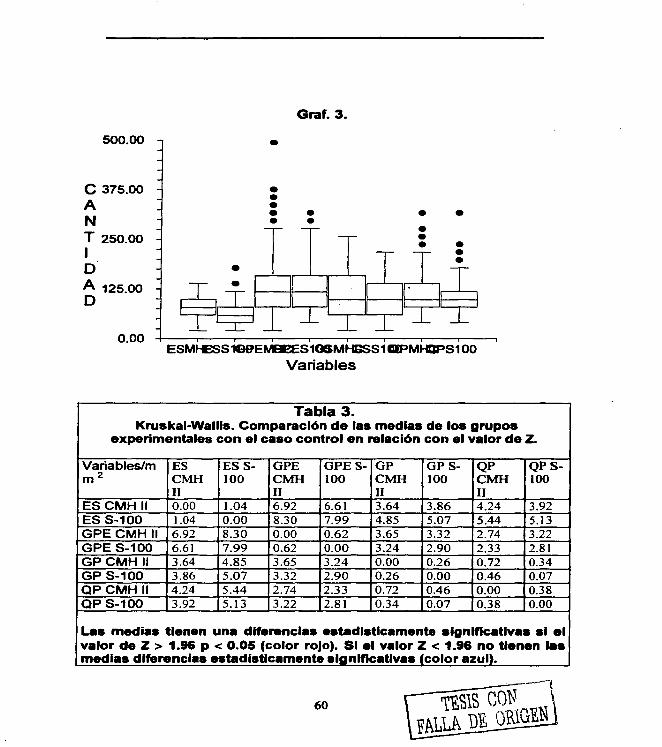
500.00
e 375.oo A N T 250.00 1 o A 125.00 o
0.00
•
• • • • •
Graf. 3.
• • • •
ESMl-ESS 1GIPEIVIEES10SMHGSS1GIPMHJPS100
Variables
Tabla 3. Kruskal-Wallis. Comparación de las medias de loa grupos
experimentales con el caso control en relación con el valor de z.
Variables/m ES ES S- GPE GPES- GP GPS- QP QPS-m2 CMH 100 CMH 100 CMH 100 CMH 100
11 11 11 11 ES CMH 11 O.DO 1.04 6.92 6.61 3.64 3.86 4.24 3.92 ES S-100 1.04 0.00 8.30 7.99 4.85 5.07 5.44 5.13 GPE CMH 11 6.92 8.30 O.DO 0.62 3.65 3.32 2.74 3.22 GPE S-100 6.61 7.99 0.62 0.00 3.24 2.90 2.33 2.81 GP CMH 11 3.64 4.85 3.65 3.24 0.00 0.26 0.72 0.34 GP S-100 3.86 5.07 3.32 2.90 0.26 0.00 0.46 0.07 QP CMH 11 4.24 5.44 2.74 2.33 0.72 0.46 0.00 0.38 QP S-100 3.92 5.13 3.22 2.81 0.34 0.07 0.38 0.00
U.a medias tienen una diferencias .. tadlatlcamente algnmcatlvaa el el valor de Z > 1.96 p < 0.05 (color rojo). SI el valor Z < 1.96 no tienen ... medias diferencias eabldlsttcamente elanlficatlvaa (color azul).
60
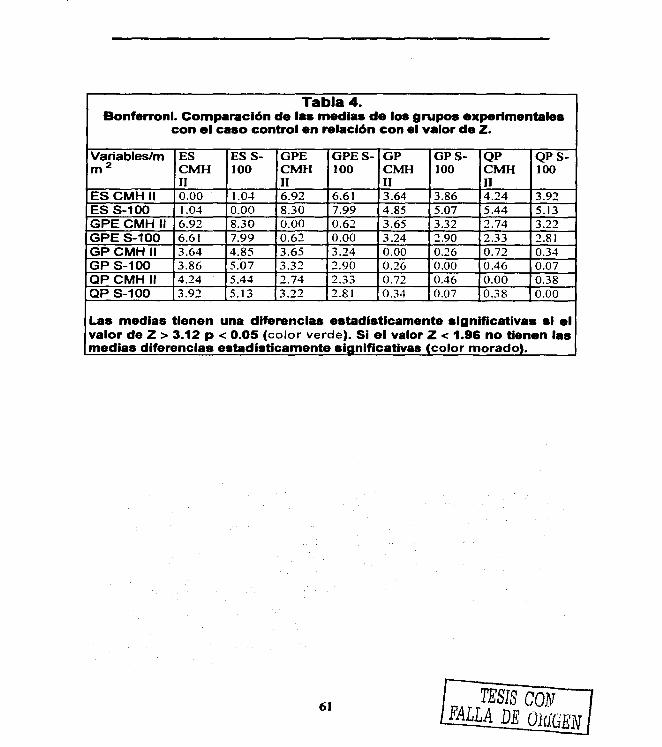
Tabla 4. Bonferronl. Comparación de las medias de los grupos experimentales
con el caso control en relación con el valor de Z.
Variables/m ES ES S- GPE GPES- GP GPS- QP QPS-m2 CMH 100 CMH 100 CMH 100 CMH 100
11 11 11 11 ES CMH 11 0.00 1.04 6.92 6.61 3.64 3.86 4.24 3.92 ES S-100 1.04 0.00 8.30 7.99 4.85 5.07 5.44 5.13 GPE CMH 11 6.92 8.30 0.00 0.62 3.65 3.32 2.74 3.22 GPE S-100 6.61 7.99 0.62 0.00 3.24 2.90 2.33 2.81 GP CMH 11 3.64 4.85 3.65 3.24 0.00 0.26 0.72 0.34 GP S-100 3.86 5.07 3.32 2.90 0.26 0.00 0.46 0.07 QP CMH 11 4.24 5.44 2.74 2.33 0.72 0.46 0.00 0.38 QP S-100 3.92 5.13 3.22 2.81 0.34 0.07 0.38 0.00
Las medias tienen una diferencias estadistlcamente significativas al el valor de Z > 3.12 p < 0.05 (color verde). Si el valor Z < 1.96 no tienen las medias diferencias estadisticamente slanlflcativas (color morado).
I
61 . TESIS CON i FALLA DE Olíf GEN_J

Ol.TI JBRF. DEL 2ílíl1 GÓJ\fEZ MAl.Ír\S RODOT.FO.
1.12.1. DISCUSIÓN.
Las CD no se observan con fijación y tinción rutinaria de cortes en
parafina, ya que con la tinción de H-E aparecen como "células claras", sin
embargo; las CD muestran un patrón característico de tinción y diversas
t'técnicas se han empleado para su identificación, entre las que se
encuentran la impregnaciones metálicas, histoquímica enzimáticas,
inmunohistoquímica y microscopia electrónica de transmisión.
Diversos autores han utilizado anticuerpos monoclonales en varios
estudios experimentales para demostrar la expresión de la molécula clase
11 del CMH de las CD en las LP, ya que estas molécula tiene un papel
preponderante en la presentación de antígenos (Okiji et al., 1994;
Tomoatsu K et al., 2001; Kaneko et al., 2001; L. Kan et al., 2001).
En este estudio se demuestra la expresión de la molécula clase 11 del
CMH de las CD en los GP, GPE y QP humanos, nosotros sugerimos que
esta molécula participa activamente en la presentación de antígenos
bacterianos y/o de necrosis pulpar que provienen del conducto radicular
para posteriormente presentarlos a las células T siendo esta la llave para
iniciar y regular la respuesta imnune primaria, además tiene un papel
primordial en el desarrollo y perpetuación de los GP, GPE y QP, pero los
mecanismo todavía no son bien entendidos (Okiji et al., 1994; Kaneko et
al., 2001).
La molécula, Clase 11 del CMH de las CD tuvo una mayor expresión cuando
en··ei~stÜdid.histopatológico observamos células epiteliales en diferentes
grados de proliferación tal fue el caso del GPE en donde la densidad de
las CD se incremento, pero cuando en el estudio histopatológico
observamos un QP o un GP el numero de CD presentes en estas lesiones
no tuvieron diferencias estadísticamente significativas, esto corrobora el
estudios realizado por T. Suzuki et al., 2001, por lo que también,
sugerimos que probablemente las CD tienen un papel primordial en el
desarrollo del tejido epitelial.
La molécula clase 11 de CMH no sólo es expresada en las CD si no
también, en los macrófagos y linfocitos B (Okiji et al., 1994; Tomuatsu K et
62

OCTITBRF. DF.T. :wrn GÓi\fF.Z i\fr\r.Ír\S RODOLFO.
al., 2001; Kaneko et al., 2001 ), Sin embargo las CD presentaron una
morfología dendrítica con numerosas y largas proyecciones
citoplasmáticas características distintivas de las CD (fig. ), estas fueron
observadas con mayor claridad en el tejido epitelial de los GPE y QP.
Sin embargo las CD presentaron diferentes morfologías en el infiltrado
inflamatorio crónico (Okiji et al., 1994; L. Kan et al., 2001) esto
probablemente fue a causa de como se realizaron los cortes.
En presente estudio también se demostró la expresión de la proteína S-
100 en estas lesiones. La proteína S-100 participa en el crecimiento de
las neuritas, esta puede ser una explicación del porque de su presencia
en tipos celulares que exhiben una forma dendrítica (la única excepción
parecen ser las CD foliculares, las cuales no expresan la proteína S-100).
Este marcador citoplasmático diferencio a las CD de Jos
monocitos/macrófagos en el infiltrado inflamatorio crónico de estas
lesiones ya que los macrófagos no expresa la proteína S-1 OO.
Los resultados obtenidos mostraron una mayor densidad de CD en el
tejido epitelial, sin embargo en el infiltrado inflamatorio crónico las CD
fueron observadas cerca de Jos vasos sanguíneos y además tenían una
estrecha relación con las linfocitos como lo demostró (L. Kan et al., 2001;
T. Suzukí et al., 2001).
La proteína S-100 exhibió una menor expresión que la molécula clase 11
del CMH en las CD en las lesiones incluidas en este estudio, sin embargo
no hubo diferencias significativas entre estos dos marcadores en Jos GP,
GPE y QP lo cual determino que las células cuantificadas en estas
lesiones eran CD.
1.13.1. CONCLUSIÓN.
Se ha reportado que el tejido epitelial funciona como una barrera física
para que Jos producto bacterianos y/o de necrosis pulpar no se diseminen,
pero no solo tienen esta función si no estas células epiteliales secretan el
factor de crecimiento (Epidermal Thymocyte Activador factor-1) el cual
63 r TES!S CO!il ~ FALLA DE ütüliEt;

Ol.TUBRF. DF.T. 2003 GÓJ\fF.Z l\f.·\l.Ír\S RODOT.FO.
posiblemente puede atraer a las CD ha este tejido y aumentar su número
de las mismas, las cuales actúan como inmunovigilantes.
Durante mucho anos sea tratado de entender como se lleva acabo la
proliferación del tejido epitelial en las LP para esto se han surgido
diferentes hipótesis, pero no fue hasta 1996 cuando LM. Lin propone que
las IL-1 y la IL-6 modifican el receptor membrana! de los restos epiteliales
de Malassez que están en el lp en el targo y junto con el factor de
crecimiento epidérmico (EGF) estimulan la multiplicación de las células
epiteliales en las LP, el aumento de las CD en las lesiones incluidas en este
estudio pueden estimular la secreción de estas interleucinas, por diferentes
tipos celulares residentes de estas lesiones. Sin embargo estas IL también
son quimiotacticas para las CD.
Aunque sea investigado si las CD presentan receptores membranales
para la histamina y las prostaglandinas estas ultimas facilita la migración y
maduración de las CD in vitro, nosotros sugerimos que se deben realizar
más investigación sobre los receptores superficiales de las CD para
verificar si estas células expresan receptores membranas para las
diferentes sustancias quimiotacticas secretadas por diferentes tipos
celulares presentes en las LP y saber con certeza si tienen alguna
influencia sobre el infiltrado de las CD,
Las CD pueden posiblemente ser las responsables del infiltrado de las
células T en las LP, ya que cuando aumenta el número de las CD
aumentan las células T para afirmar esto se necesitan entender los
mecanismo por los cuales sucede estos.
1.14.1. PRODUCCIÓN CIENTÍFICA.
Este trabajo fue presentado en el congreso lnternational Workshop on
Langherhans Cells 2003 in Tokyo Japan
6-l-
TESIS CO.N FALLA DE ORIGEN
QUANTIFICATION OF DENDRITIC CELLS IN HUMAN CHRONIC APICAL PERIODONTITIS.
C.D. Rodolfo Gómez Macias•. M.C. Judith Alvarez Perez .... C.D. Dolores Jiménez Farfan·. C.D. Gabriel Loranca Fragoso-. Dr. Juan Carlos C. Hernández Guerrero·. Dr. Andrés E. Castell Rodriguez .....
División de Estudios de Posgrado e Investigación (Laboratorio de lnmunologla) de la Facultad de Odontologla•. Departamento de Biologia Celular y Tisular (Laboratorio de lnmunohistiquimica) de la Facultad
de Medicina .. de la Universidad Nacional Autónoma de México.
l:"liTHODl'CTIOS.
1 hC' \\"orlJ lk.:ihl• Or;anu;iu.,11 tWHOI m l'J'J~. d.:iuolkJ 1h.: ro.:ri;ip1.:.:1h:• lnu,l11J uf pulr.ir on¡¡m 111 11111.:c 111.un 11ruup• bu.:<! 011.:luu.:.:il r.ul1u11r.1pl1t.: .1n¡J hn1u11.11lmlo¡¡u;.1I .:h.:11.1.:t..~rut•.:• .i.:u1.: ;ip1.:.1I p.:riuJcm11111 .:hnm1.: .:1pro;.1I p.:rwd"nt1111 .:inJ .:ip1.:;il .:ibJ.:.:u.:1 Cl110111.: .:ip1.:.1l p.;11uJ1.11111t1• " .:l•.ir.i..1.:1u.:J l" Lh.: 11111.:imm:il1011 .:uod dC'11ru.:uun uf .ip1.:.:1I p.:ru,,J.in10 .:u ;a 1~·1ult uf p1.Jp.11 n..:'11»11 1hm\11111 11r:111ul.ir .mJ cpulu:h.JI prohli:r.mon 1'1-ol1f.:r~uon of 1hc1.: 1111111:1 11r.Jl;uc h;i.:1e11;1I p1r.JJu.:11 ;inJ or ueerou.: ro;i11 .. 111.:h .ne rc¡ce1eJ from 1he rJJ1.:ul.:1r .;:o,.Ju..:1 10 lhe p.:n;ip1.;:;il 1one ;mJ h.nc 1hc .;:.:ip,1.;:11• 10 ;¡.;hui.: 1mmunolo11c.:1I re1pon1e1 H111ol11i:1..:.:11l~ 1hc .:hromc: 3p1..:;il pc11oJon11t1t c.:1n be .;:l.1u11icJ ;11 .:1p1.:;1I 1r;1nulon1.:1 (AGll. cp1Lhch.1I prohf.:r.111on ¡,:r;111ulon1.11EG11 or .J r;1J1cul.Jr c~u 1RC11. bcmw tlu:sc l.:110111 fri:querul~ ob•.:ncd 111 !111: chni.:.:il ptJcHcc Thcrc .1tC J1tli:u:n1 .;:clluJ.u 1~ pc1 1h.u p.:11Uc•p.:11.: m thc mdu.;:uon of 1hc 1111n1uni: rc1J'(Jn1e. 1ud1 ;is 1he ;1nt11en Je11Jr111e prncn1m1 .;:elh In 1;111 Jec;iJc th.:rc 1>1;rc 1ome tlu\111:1 rdJ1m11hc dc1clopmcn1 oflhc pcn;ipic;il le11on1 .md dcndntl.: ecfü!DCI ;mJ.'or L1r¡erhJn1 edh ILCI Thc ;i1n1 oflhc prc•cnl 1tuJ~ "ª'lo J.;:tcmun.: 1hc J11t11but1on ;1nd quw11fic;111011 of1he OC 111 AG1 EG1 .:1nJ RC1 b~ mc:an1 of thi: e•prc:nton f>Í the el:lu 11 molecule1 of lhc ,\IJJOI lh.iocomp.ihh1h11 Compl..:• t\lllCJ.ind thi: c~1opl;111m.: pro1c111 S-ltltl
'1ATl::Hl.\LS \ .... D :\IETllODS.
El.:•cn urnpl.:1 ol .:hromc .irn:;;il pcn0<..lon111 .. ;inJ '"º of h..:;ihh~ 11111!!1\.1 tGH¡ .u .:onuol ¡¡roup \lcrc 111.:luJcd rn our 1tuJ1 .S.u11pl.:1"ere1cle1;tcJ 3nJ oh1;imcJ 111 thc ~l;i .. llo F.:1.:1.iJ Ocp.:inmcnl f;icuh~ oí Odotllolo¡¡• L '.'<.iA \I ;1,;eord1n¡: to thc rulc1 c11:ibli1hcJ b~ \\.'110 lo chmc:.:11 3nJ r.:1Jm¡¡r.:>ph1c: J1;1¡r.110t1t .:>.:,;ordu11110 1hc pcn;1p1c;il le11mu S.:implc1 "ere rnx:cncJ 10 obt.:111111111.:tomcrcu 1cdmn1oí1•.:1r.:illi11 .rnJ frcc11n!j. bl0<:l..1 fnm• c:.i;;h 1.1mplc "" ubt.:>lnc:J 11 1c11.1l 1i;c1101u ;ind trc.:11cJ thclll ;i, follou1 onc \\.JI u•eJ lur lus10p.11holo~1.:.:il .!1.J!j.110111 !he olhcr lll 1.ec110111 \\C'fC proceneJ for 1m111unou.i1111n¡¡ to i.lcmon1tr;i1i; c/;i11 I/ nmlcculc1 c•pri:u1on 311J pm1c1n S·llHJ 1'01111\c cclh \\ere ob1c:'"'cJ ;inJ counlcd 11111111.1 1111..:ro1copc i:qu1ppcJ "llh .:i lm:1J c:.:in1cra ,1nJ .:> -40."\ nb¡ccll•c Tcu nucmu;op1c licld1 \\ere ob1cnc:J and thc po•Ul\C ,;cll1 for bolh m;irl..c1t \\C'IC counti;J .:iml thc numbcr "<11 c•prc11c:J u1 111111:
HESL'LTS.
A1.a:ordm!i to \\.'110 1 i:111en;i rcbtcd to thc lu11op:a1!mloy1c.:il J1.1¡110111 of pcn.Jp1c:il l.:110111 \\C fouuJ u follo\\0:1 T"o AG1 .. 1111h prctcnce oí neulroplult m;icroph.1!;CI, J, mpJm,;~tcs <111d plum;itic ccllt 1>11h
•.11;;ulJr prol1!cr;111on 1c•cn ep11hch3I gr.3111110111:1 tEG1) "h1!h cpuh.:h.:ll ccll1 or1¡;rn3l111~ 1hc cp11hch.:1I 1ctt of ~l.ilJuc:.t .:11 J1tl'.:rc111 Jci;rec1 of .:c:Jlular rroldcramm .inJ ""' RC1 '"th p.11holo111c31 c:l\lllct. C<l\CIO:J '"'" •<Jt1J11101H llral1lied ,;p11hc:J1;¡ Jlld J •cm .. ohJ 111.11.;rul H..ir;iph1c h
Mostop~ttoiolog><:.al Da9no111 ol P•ri•p>e~I L•''º"'
AGi. EG1. RC1 .ind UH li.id f'ltl'l\I 1c ~di~ lo 1hc c'IJ" JI mok"1lc1 ut 1hc .\lll(' 3ni.I 10 1111: c~toploHmlC
ptot.:m 5.100 Thcsc po11tl\C cdlt \\CIC touml 111 !h..: q111hdt.il .inJ i;1.i11ulo11 t1nuc .ind 111 thc UC1!:hbmhood ol thc blooi.I •c,•.:I• ¡¡¡~ l ·'11
ITJ .. '~· -· ·:·.
:) : . t 1~ 1 l.HJHul.ir [ 1"'"' .. 11 11 .IJ" hi,; ~ tn.i11uL¡r [1nuc "11h ll m<>lc·culc .,b¡ct1•.: I•"''\ l'rutcm \.11>11 ''htcll\..: ltu1\
DD h.: ; t,rJnulJr l1.•uc"11h.J."' 11\; ·ILi1.mul.11 lhu1..: ""h H ;,,.,¡""''k uhicll\C -4u\ í'l\>lcm "·llll!ul•1c!l'c ¡u\
65
EJF.,10c,, Ep11hch;il T1uuc .. 11h cl;inll molcc11lc ObJCll\C lfHI;\'. D F11llDC1nEp1thch:al
T111uc \Ulh Protc1n SltHlob¡c1nc IUll:\
\\111:11 lh<: numbcr ofpo11111c cclls for bu1h cl;111 11 mokculc1 :111J S-IUll protcm n1:ul.crt \\ere comp;iu:d \\1lh AG• 1111J RC1 1h...rc: \\ere not 1111n11ic;int d11Tercn~c~ In co111r;i1t, \\C founJ 111nilic:ant 1.hfTCJcncc bch•ccn AG1 :and RC1 1uth rc1pcct lo EG1 ;mJ of;il/!luce ¡:roup1 "nh conllol ¡¡roup{T.:lblc l 11nd 11r11phic 11)
T.\BLE 1
Cnmpos ti.kdml/mm· Standard Standard Minimun Mn.ximun Range De"iation Error
-GH•MHC 50 74.66 32.24 4.56 19.75 138.27 118.51 G!-tS-100 so 67.16 30.91 4.37 19.75 177.77 158.02 AGsiMHC IOO 10982 58.6-1 5.86 19.75 256.79 237.03 AGsiS-100 100 106.07 45.41 4.54 19.75 217.28 197.53 EGs!MHC 3:50 134.60 69.11 3.69 19.75 493.82 475.04 EGsiS-100 350 125.91 53.54 2.86 39.50 316.04 276.54 RCs!MHC 100 114.37 53.88 5.38 39.50 316.04 276.54 RCsiS-100 IOO 110.02 51.63 5.16 39.50 316.04 276.54
GRAPHICll
·--·------ -·----------- -- J OC Density on Dlrfrrenl Pulnplcal Lesio111
·~··· Lrsiuns
UISCUSSJO!'li,
R.:1ult1obt11111c:J1n lhu s1ud~ iho" 1h:i11l11: 1 .. i1on1 ,;om;i1n cctl c•P'"'"'5111g d11H 11MUC11101.:cuk• ••••h 11 Jcntrm.: p1ulilc Tl11111i,:rcc1\\1th:a prc•1ou11111J• IL.:incl..u,;I ;il !IHllJ Al lhi; momcnl. thCJ.: u fcu 111form;111011 r..:l.JtcJ of thc role of O\ 111 1hc p:atho¡:cn..:i11 uf 111.., J><.!ri.ip1c:il h,:11ont 111 111111111111 Cc:n;;iml~ thcrc "·11110 "•mlkJlll i.l1rkrcncc bct\\cc111hi: uumh..:r of OC ;imung 1h.: c;i1c1 of ,\Gi ;¡nJ RC• ~om;1hck1~. lhc J1ffcn:ii.:.: ".:11 r.:m.11l..11bk 1>11h r.:¡¡a1J lo 1lu; .::n,.·1 of EG• Ho\\ 10 C•pl;irn 1hu" Tnn pou1b1hl1ct 1h.11 h3•c 11111pon. t:\t\I F1111 1ha1 thc cp11Ju;;hJI cclh 1c1:1ch; c~wL.1nc1 ;i1 1hc ET..\f-J tC:p1Jcr111;il Tl1•111rn.;\I..: ·\cu .. uur Factor·l 1 th.Jt cou!J •ni.Ju~;: ;i pa•hfrr.iuun ol OC. •..:~,,,,J ba.:1 .. ·11.:d produ,.11or11ccro11c c.:lh 11111~ mJucc .m a.:11•311011 ;inJ prol1fcr<1t1011 of OC 1h31. 111 rc111111. 11131 111111111.ilc 1hc :1.:1111111011 uf Thl .:cll, Lh;it 1.:~1c1.: IL-1 311J IL-•• 1h;1\. 111 rrc1i:n~c or EGF 1Epnh,;l1,ll Grn .. ,h f.i..:1011 "'ª' c.i11•.: ;i prolth;ratmn of c¡"1l11;h,1I cclh ¡p• "'!C pl11cc 10 t/1.: l•Hlll.llU>R uf1hc EG• lh11h h•¡>o1h.:ws :irc bcmi; c•aoum..:J 111 om l.1hu1aioo
TESI8 CON FALLA DE ORIGEN

DR. RODOLFO GOMEZ MACIAS
Agradecemos nos haya enviado su trabajo titulado: INMUNOEXPRESIÓN DE
LA MOLÉCULA CLASE 11 DEL COMPLEJO PRINCIPAL DE
HISTOCOMPATIBILIDAD Y CUANTIFICAIÓN DE CÉLULAS DENDRÍTICAS EN
LA PERIODONTITIS Api.
Para ser considerado por el Comité Científico del /1 Encuentro
Iberoamericano y XI Encuentro Nacional de Investigación en Odontología.
Nos es muy grato informarle que este trabajo fue evaluado
satisfactoriamente para ser incluido en el programa de dicho evento en la
categoría de: ESPECIALIDAD en la Modalidad de ORAL con la clave OE04
Siendo el día de su presentación el día JUEVES 6 a las 18:50p.m. en el
SALÓN EL AL TO del Centro de Convenciones William O. Jenkins de la
Ciudad de Puebla.
Las presentaciones en Modalidad Oral, se llevarán a cabo con los siguientes
lineamientos:
1- La duración de su exposición será de 15 minutos y 5 minutos más
para preguntas y comentarios.
2- Durante la exposición en el podium existirá un semáforo con luz
verde, faltando 3 minutos se encenderá la luz amarilla y terminado el
tiempo de exposición se encenderá la luz roja, automáticamente se
apagará el proyector y se encenderá la luz del auditorio, dando por
terminada la exposición.
3- La decisión del Comité Científico fue: el trabajo que rebase el tiempo
permitido no tendrá derecho a preguntas y comentarios y quedará
eliminado automáticamente del concurso.

OCTUBRE DEL ?003 GÓMEZ MACÍAS RODOLFO.
1.15.1. BIBLIOGRAFÍA.
1. Torabinejad M, Walton R. "Lesiones perirradiculares" en: Ingle J
Bakland, L. Me. Graw-Hill lnteramericana. 4ª edición. Capítulo 8.
1996.
2. Marshall H, Smulson y James C. Hagen. "Patología pulpoperiapical y
consideraciones inmunológicas en: Franklin S. Weine. Terapéutica en
endodoncia. Editorial Salvat Editores,S.A. 2ª edición. Capítulo 4. 1995.
3. Lasala A. "Patología pulpar y periapical" en: Endodoncia. Editorial
Ediciones Científicas y Técnicas.4ª Edición. Capítulo 4. 1992.
4. Ingle J, Walton R." Lesiones perirradiculares" en: Ingle, J., Bakland, L.
Endodoncia. Editorial McGraw-Hill lnteramericana. 4ª Edición. Capítulo
8. 1996.
5. Smulson M, Hagen J, Ellenz S. "Patolog'ia pulpoperiapical y
consideraciones inmunológicas" en: Wiene.F. Tratamiento
Endódontico. Editorial Harcourt Brace. 5ª Edición. Capítulo 4. 1997.
6. Nair P. "Pathology of apical periodontitis" en: Piitt Ford T. R., Orstavik
D. Essential endodotology. Editorial Blackwell Science Ud. Capítulo 4.
1998.
7. Pumarola J, Canalda C. "Patología de la pulpa y del periapice" en:
Canalda C. Brau E. Endodoncia. Editorial Masson. Capítulo 6. 2001.
8. Nair SP, Meghji S. Wilson M, Reddik, White P, Henderson B.
Bacterially induced bon e destruction: mechanisms and
misconceptions. lnfection and lmmunity 1996; 64: 2371-80.
9. Meghji s, Qureshi W. Henderson B, harris M. The role of endotoxin and
cytokine in the pathogenesis of Odontogenic cysts. Archives of Oral
Biology 1996; 41: 523-21.
1 O. Simon J. "Patología periapice" en: Cohen S. Vías de la pulpa. Editorial.
ELSEVIER SCIENCE. 7ª Edición. Capítulo 11. 2000.
66-'

OCTUBRE DEL 2003 GÓMEZ MACÍAS RODOLFO.
11. Nair P. "Fisiopatologia del periapice" en: Cohen S. Vías de la pulpa.
Editorial. ELSEVIER SCIENCE. 8ª Edición. Capítulo 12. 2002.
12. Reid SD, Penna G and Adorini L. The control of T cell responses by
dendritic cell subsets. Current Opinion in lmmunology 2000; 12: 114-
121.
13. TAKAHASHI K. Microbiological, pathological, lnflammatory,
immunological and molecular biological aspect of periradicular
disease. lnternational Endondontic Journal 1998; 31 : 311-325.
14. Kakehashi s, Stanley HR, Fitzgerald RJ. The effects of surgical
exposures of dental pulps in germ- free conventional laboratory rats.
Oral surgery, Oral Medicine and Oral pathology 1965; 20: 340-9.
15. lwu C, Macfarlene TW. Mackenzle D. Stenhouse D. The microbiology
of periapical granuloma. Oral surgery, Oral Medicine and Oral
pathology 1990; 69: 502-5.
16. Barnett f, Stevens R and Tronstad L. Demostration of Bateroides
intermedius in periapical tissue using indirect immunofluorescen
microscopy. Endodontics and Dental Traumatology 1990; 6: 153-6.
17. Wayman BE, Murata SM, Almeida RJ, Fowler CB. A bacteriologica
and histological evaluation of 58 periapical lesions. Journal of
Endodontics 1992; 18: 152-5.
18. Cross DL, Ellender JA, Smith GI. Simultaneus hybridization and
subsequent colour detection of subgingival bacteria! DNA on colony
lifts. Archives of Oral Biology 1993; 38: 931-5.
19. Melvin WL, Assad DA, Miller GA Gher ME, Simonson L, York Ak.
Comparison of DNA probe and ELISA Analysis methods and their
association with adult periodontitis. Journal or periodontology 1994; 5:
576-82.
20. Nevins ';\J, levine S, Faitlowicz Gayer Y, Svetcous. Sensitization via
lgE-mediated mechanism in patients wich chronic periapical lesions.
Journal of Endodontias 1985; 11: 228-30.
67

OCTUBRE DEL 2003 GÓMEZ MACÍAS RODOLFO.
21. Cassatella MA. The production of cytokines by polymorphonuclear
neutrophils. lmmunology today 1995; 21: 1292-7.
22. Miller GA, Demayo T, Hutter JW. Production of interleukin-1 by
polimorphonuclear leukocytes resident in perirradicular tissue. Journal
ofEndodontics 1996;22:346-51.
23. Ledesma Montes C, Garcés OM. Células cebadas en granulomas
periapicales humanos. Patología 1996; 34: 107-111.
24. Nilsen R. Johannesen AC. Skaug N. Matre in situ characterization of
mononuclear cells in human dental periapical lnflammatory lesions
using monoclonal anbodies. Oral surgery, Oral Medicine and Oral
Pathology 1984; 58: 160-5.
25. Akamine A, Hashiguchi 1, Toriya Y Maeda K. lmmunohistochemical
examination on the localization of macrophages and plasma cells in
induced rat periapical lesions. Endodontics and Dental traumatology
1994a; 10: 121-8.
26. Barkhordar RA, Desouza YG, Francisco S. Human T- Lymphocyte
subpopulations in periapical lesions. Oral surgery, Oral Medicine and
Oral Pathology 1958; 65: 763-6.
27. Pulven WH, Taubman MA, Smith DJ. lmmunocompetents in human
dental periapical lesions. Archives of Oral Biology 1978; 23: 435-43.
28. Matsuo T, Ebisu S, Shimabukuro Y, Ohtake T, Okada H. Quantitative
analysis of immunocompetents cell in human periapical: correlations
with the clinical findings of the involved teeth. Journal of Endodontics
1992; 18: 497-500.
29. Mattews JB, Masan GI. Jmmunoglobulin producing cells in human
periapical granuloma. British of oral surgery 1983; 21: 192-7.
30. Kettering JO, Torabinejad M. Presence of natural Killer cells in human
chonic periapical Jesions. lnternational Endodontic Journal 1993; 26:
344-7.
68

OCTUBRE DEL 2003 GÓMEZ MACÍAS RODOLFO.
31. Matsuk Y, Yamamoto T, Hara K. Detection of inflammatory cytokine
messenger RNA ( mRNA ) expressing cells in human inflamed gingiva
by combined in situ hibrydization and immunohístochemistry.
lmmunology 1992; 76: 42-7.
32. Lin MI, Wang SL, Wu Wang C, Chang KM and Leung. Detection of
epidermal growh factor receptor in inflammatory periapical lesions.
lnternational Endodontic Journal 1996; 29: 179 -184.
33. Pumarola J, Canalda C. "Etiopatogenia de la enfermedad pulpar y
periapical" en: Canalda C. Brau E. Endodoncia. Editorial Masson.
Capitulo 5. 2001.
34. Kakehashi S, Stanley H.R., Fitzgeral R.J. The effects of surgical
exposures of dental pulps in germ-free and conventional laboratory
rats. Oral surgery, Oral Medicine, Oral Pathology, Oral Radiology and
Endodontics. 1995; 20; 340-349.
35. Stashenko P. "Etiology and pathogenesis of pulpitis and apical
periodontitis" en: Pitt Ford T.R., Orstavik D. Essential endodontology.
Editorial Blackwell Science Ud. Capitulo 3. 1998.
36. Baumgartner JC: Microbiology and pathologic aspects of endodontics.
Current Opinion in Dentistry 1991 a; 1: 737-43.
37. Trowbrige HO, Stevens BH. Microbiologic and pathologic aspects of
puf paf and periapical disease. Current Opinion in Dentistry 1992; 2: 85-
92.
38. Simon JHS. Periapical pathology. In: Cohen S and Burns RC, eds.
Pathways of the pulp 6 1h edn, pp 337-62 1994. Baltimore, MA: Mosby.
39. Kettering JO. Trabinejab M. Microbiology and immunology, In : Cohen
S. Burns RC, eds. Pathways of the pulp 6 1h edn, pp 363-76, 1994.
Baftimore, MA: Mosby.
40. Brook 1 , Grimm S, Kielich RB. Bacteriology of acute periapical
abscess in children. Journal of endodontics 1981; 7: 378-80.
69

OCTUBRE DEL 2003 GÓMEZ MACÍAS RODOLFO.
41. Brook 1, Frazier EH. Gher ME. Aerobic and anaerobic microbiology of
periapical abcess. Oral Microbiology and lmmunology 1991; 6: 123-5.
42. Sundquist G. Johansson E, Sjogren V. Prevalence of black-pigmonted
Bacteroides species in root canal infections. Journal of
Endodontics 1989; 15: 13-8.
43. Teunnissen MBM. Dynamic nature and function on epidermal
Langherhans cells in vivo emphasis on human Langerhans cells.
Histochem J 1992; 24: 697-716.
44. Bjorkman PJ, Saper MA; Samraour B, Bennet WS, Strominger JL,
Wiley OC. the foreign antigen binding site and T cell recognition
regions of class 1 histocompatibility complex. Nature 1987; 329: 512-
518.
45. Ooyle C, Strominger JL. lnteration between CD4 and class 11 MHC
molecules mediates cell adhesion. Nature 1987; 330: 256- 259.
46. Norment AM, Salter RO. Parham P, Engelhard VH, Littman DR. Cell
cell adhesion mediated by CD 8 and class 1 molecules. Nature 1988;
336: 79- 81.
47. Calabi F, Jarvis JM, Martin L, Milstein C. Two clases of CD1 genes.
Eur J lmmunol 1989; 19: 285-292.
48. Amiot M, Bernard A, raymund 8, Knapp W, Desclidre C, Boumsell L.
Heterogeneity of the first cluster of differentation: characterization and
epitopic mapping of three Cd1 molecules on normal human thymus
cells .. lnmunol 1986; 36:1752-1768.
49. ChÚ Ac. Morris J: The Keratinocyte. In Skin Inmune System Ed: Jan D.
Sos. 2~d Edition .. CRC. Press, Boca Raton, 1997
50. C~c~hia >.o;< Michetti F, Donato R. lmmunochemical and
immu~~cytochemical localization of S-1 oo antigen in normal human
skin: N~tqf~'1981; 294: 85-93. -•, .. - .. , -·
51.- L.otzaºTM~. Thomson W.A. Dendritic Cells. Editorial Hardcover 1999; ...
6: 1-1043.
70

OCTUBRE DEL 2003 GÓMEZ MACÍAS RODOLFO.
52. Pérez TA, Castel! A, Farfan CJ. La Célula de Langerhans. l. Origen y
morfología. LABORAT acta 1989; Y:.: 28-33.
53. Castel! A, Pérez TA, Farfan CJ. La Célula de Langerhans. 11. Funcuón.
LABORAT acta 1989; Y:.: 23-26-
54. Pérez TA, Castel! A, Farfan CJ. La Célula de Langerhans. 111.EI
sistema de las Célula de Langerhans. LABORAT acta 1989; 'Y:.: 25-29.
55. Masson P. Pigment cells in man. In: the Biology of the melanosome
(M. Gordon, ed). Spec Publ NY Acad Sci 1948; 4: 15-51.
56. Zelickson AS. The Langerhans cell. J lnvest Dermatol 1965; 44: 201-
212.
57. Zelickson AS. Granule formation in the Langerhans cell. J lnvest
Dermatol 1959; 47: 498-503.
58. Silvers WK. An histogical and experimental approach to determine the
relationship betweet gold impregnated dendritic Cell and melanocytes.
Am J Anat 1957; 100: 225-240.
59. Breathnach AS. Silver WK, Smith S, Heyner S. Langerhans cell in
mouse skin experimentally deprived of its neural crest component. J
lnvest Dermatol 1968; 50: 147-160.
60. Katz SI, Tamaki K, Sachs D. Epidermal Langerhans cells are derived
from cells originating in gane marrow. Natura 1979; 282: 324-326.
61. Frelinger GJ, Hood L, Hill S, Frelinger JA. Mouse epidermal la
molecules have a bone marrow origin. Nature 1979; 282: 321-324.
62. Frelinger JA, Frelinger GJ.Bone marrow origin of la molecules purified
from epidermal cells. J invest Dermatol 1980; 75: 68-75.
63. Volc-Platzer 8, Stingl G, Wolff K, Hinterberger W, Schnedl W.
Cytogenetic identification of epidermal Langerhans cell in a bone
marrow-graft recipient. New Engl J Med 1984; 31 O: 1123-1124.
64. Goordyal P, lssacson PG. lmmunocytochemical characterization of
monocyte colonies of human bone-marrow: a clue to the origin of
71

OCTUBRE DEL ?003 GÓMEZ MACÍAS RODOLFO.
Langerhans cells and interdigitating reticulum cells. J. Pathol 1985;
146: 189-195.
65. Gothelf Y, Sharon N, Gazit E. Fractionation of human bone marrow
mononuclear cells with peanut agglutinin: phenotypic characterization
with monoclonal antibodies. Hum lnmunol 1986; 17: 37-44.
66. De Fraissinette A, Schmitt D, Dezutter-Dambuyant C, Guyotat D,
Zabot MY. Thivolet J: Culture of putative Langerhans cell bone marrow
precursors: Characterization of their phenotype. Exp Hemotol 1988;
16: 764-768,
67. Murphy G, Messadi D, Fonferko E, Hancock W. Phenotypic
transformation of macrophages to Langerhans cells in skin. Am J
Pathol 1986; 123: 401-406.
68. !naba K, lnaba M, Romani N, Aya H, Deguchi M, lkchara S.
Generation of large numbers of dendritic cells from mouse bone
marrow cultures supplemented with granulocyte/macrophage colony
stimulating factor. J Exp Med 1992; 176: 1693-1702.
69. Reíd C, et al. ldentification of hematopoietic progenitors of
macrophages and dendritic Langerhans cells (DL-CFU) in human bone
marrow and peripheral blood. Blood 1990; 76: 1139-149.
70. Egner W, Mckenzie JL, Smith SM, Beard MEJ, Hart DNJ. ldentification
of potent mixed leukocyte reaction-stimulatory cells in human bone
marrow. Putative differentioation stage of human blood dendritic cells.
J lmmunol 1993; 150: 3043-3053.
71. Santiago-Schwarz F, Belilos E, Diamond B, Carsons SE. TNF in
combination with GM-CSF enhances the differentiation of neonatal
cord blood stem cells and macrophages. J Leuk Biol 1992; 52: 27 4-
281.
72. Caux C, Dezutter- Dambuyant C, Schmitt D, Banchereau J. GM-CSF
and TNF a cooperate in generation of dendritic Langerhans cells.
Nature 1992; 360: 258-261.
72

OCTUBRE DEL 2003 GÓMEZ MACÍAS RODOLFO.
73. Rosenzwaijg M, Canque B, Gluckman JC. Human dendritic cell
diferentiation pathway from CD34+ hemopoietic precusor cells. Blood
1996; 87: 535-544.
74. Ranvier L. Traité technique d'histologie. Savy ed., Paris 1875.
75. Arnstein SB. Die nerven bei behaarten haut. Acad Wiss Wein Math
Nat KI 1876; 74: 203-07.
76. Herxheimer K. Uber pemphigus vegetans nebst bemerkungen uber die
natur de Langerhansschen zellen. Arch Derm Syph (Berlín) 1896; 36:
140-149.
77. Billingham RE, Silvers WK. Sorne unsolved problems in the biology of
skin. In Biology of the skin and hair growth. Lyne AG and Short Bf eds.
Angus & Robertson, Sydney, 1965.
78. Prunieras M. lnteractions between keratinocytes and dendritic cells. J
lnvest Dermatol 1969; 52: 1-17.
79. Silberberg. Ultrastrucutral studies of Langerhans cells in contact
sensitive and primary irritant reactions to mercuric chloride. Clin Res
1971; 19: 715 ( Abst ).
80. Silberberg l. Apposition of the mononuclear cells to Langerhans cells
in contact allergic reactions. An ultrastructural study. Acta
Dermatovener (Stockh) 1973; 53: 1-12.
81. Unanue ER, Allan PM. The basis far the immunoregulatory role of
macrophages and other accesory cells. Science 1987; 236: 551-557.
82. Clement L T, Shevach EM. Characterization of MHC antigens on TNP
modified cells. Mol lmmunol 1979; 16: 67-73.
83. Sinigaglia F. The molecular basis of metal recognition by T cells. J
lnvest Dermatol 1994; 102: 398-406.
84. Springer TA. Adhesion receptors of the immune system. Nature 1990;
346-355: 425.
85. Geppert TD, et al. Accesory cells singnals involved in T -cell activation.
lmmunol Rev 1990; 117: 5-9.
73

OCTUBRE DEL 2003 GÓMEZ MACÍAS RODOLFO.
86. Jenkins MK. The role of cell division in the induction of clona! anergy.
lmmunol Today 1992; 13: 69-74.
87. Trinchieri G. lnterleukin-12 and its role in the generation of Th1 cells.
lmmunol Today 1993; 14: 335-342.
88. Romani N, Koi,de S, Crowley M, Witmer-Pack M, Livinstone AM,
Fathman CG, !naba K, Steinman RM. Presentation of exogenous
protein antigens by dendritic cells to T cell clones: lntact protein is
presented best by immature epidermal Langerhans cells. J Exp Med:
1989; 169: 1169-1777.
89. Streilein JW, et al. Functional dichotomy between Langerhans cells
that present antigen to naive and to memory/effector T lymphocytes.
lmmunol Rev 1990; 117:159-165.
90. Teunissen MBM, Rongen HAH, Bos JD. Function of adhesion
molecules lymphocyte function-associated antigen-3 and intercellular
adhesion molecule-1 on human epidermar Langerhans cells in antigen
specific T cell activation. J lmmunol 1994; 152: 3400-3406.
91. Teunissen MBM, Koomen CW, Bos JD. lntercellular adhesion
molecule-3 (COSO) on human epidermal Langerhans cells participates
in T-cell activation. J lnvest Dermatol 1995; 104: 995-998.
92. Girolomoni G, Zambruno G, Manfredini; R, Zacchi V, Ferrari S,
" ccissarizia A 1 Gianetti A. Expression of 87 costimulatory molecule in
cÜl~~~~·ctf1~man epidermal Langerhans cells is regulated at the mRNA
leve!. J lnvest Dermatol 1994; 103: 54-59.
93. Staquet MJ, et al. A surface glycoprotein complex related to the
adhesive receptors of the VLA family, shared by epidermal Langerhans
cells and basal keratinocytes. J lnvest Dermatol 1989; 92: 739-745.
94. Péguet-Navarro J. Funcional expresion of CD40 antigen on human
epidermal Langerhans cells. J lmmunol 1995; 155, 4241-4248.
95. Sauder DN, Dinarello CA, Morhenn VB. Langerhans cell production of
interleukin-1. J lmmunol 1984; 82: 605-612.
74

OCTUBRE DEL ?003 GÓMEZ MACÍAS RODOLFO.
96. Schreiber S. Cytokyne patterns of Langerhans cells isolated from
murine epidermal cell cultures. J. lmmunol 1992; 149: 3525-29.
97. Kang K. IL-12 synthesis by human Langerhans cells. J lmmunol 1996;
156: 1402-1411.
98. Blauvelt A. lnterleukin-15 mRNA is expressed by human keratinocytes,
Langerhans cells, and blood-derived dendritic cells and its down
regulated by ultraviolet B radiation. J lnvest Dermatol 1996; 106: 1047-
1056.
99. Stingl G, Katz SI, Clement L, Green 1, Shevach E. lmmunologic
functions of labearing epidermal Langerhans cells. J lmmunol 1978;
121 :2005-2013.
1 OO. Hauser C, Katz SI. Activation and expansion of hapten and protein
specific T helper cells from nonsensitized mice. Proc Natl Acud Sci
USA 1988; 85: 5625-5628.
101. Braathen LR, Thorsby E. Studies on human epidermal Langerhans
cells. l. Allo-activating and antigen presenting capacity. Scand J
lmmunol 1980; 11: 401-408.
102. Teunissen MBM, Wormmester J, Kapsenberg ML, Bos JD: Enrichment
of unlabeled human Langerhans cells from epidermal cell suspensions
by discontinuous density gradient centrifugation. J lnvest Dermatol
1988; 91: 358-362.
103. Pollack MS, Goldenhersh M, Chin-Luois J, Safai B. The function study
of DR-positive human epidermal Langerhans cells in mixed cell
cultures with allogeneic lymphocytes. Clin lmmumol lmmumopathol
1982;;24: 15-25,. •J , •. • ~·- T· "
104~ Sheyniu~ A, Grovnik Ko, Anderson J. Epidermal Langerhans cells as
~~ces~~ry cells in Con A stimulation of thymocytes in guinea pig.
Scand J lmmunol 1983; 17: 283-290.
105. Pehamberger H, et al. Epidermal cell-induced generation of cytotoxic
T-lymphocyte responses against alloantigens or TNP-modifed
75

OCTUBRE DEL ?003 GÓMEZ MACÍAS RODOLFO.
syngeneic cells. Requirement far la-positive Langerhans cells. J lnvest
Dermatol 1983; 81: 208-211.
106. Steiner G, et al. Epidermal cells as accessory cells in the generation of
allo-reactive and hapten-specific cytotoxic T lymphocyte (CTL)
responses. J lmmunol 1985; 134: 736-7 41.
107. Braathen LR, Thorsby E. Studies on human epidermal Langerhans
cells. IV: HLA-DR restriction of Langerhans cell dependent antigen
activation of T lymphocytes. Scand J lmmunol 1982; 15: 55-61.
108. Bjercke S, Elgo J, Braathen L, Thorsby E. Enriched epidemal
Langerhans cells are potent antigen-presenting cells far T cells. J.
lnvest Dermatol 1984; 83: 286- 289.
109. Sontheimer RO. The mixed epidemal cell-lymphocyte reaction l.
Human epidermal cells elicit a greater allogeneic lymphocyte responce
than de autologous peripheral blood lymphoid cells. J lmmunol 1983;
130: 2612-2614.
11 O. Res P, et al. The crucial role of human dendritic antigen-presenting cell
subsets in nickel- specific T-cell proliferation. J lnvest Dermatol 1987;
88: 550-554.
111. Kapsenberg ML, et al. Nickel-specific T lymphocyte clones derived
from allergic nickel-contact dermatitis lesions in man: heterogeneity
based on requirement of dendritic antigen-presenting cell subsets. Eur
J lmmunol 1987; 17: 861-865.
112. Halliday GM, Muller HK. Langerhans cell presentation of sheep red
blood cells induces antibody production. lmmunol cell Bici 1987; 65:
71-76.
113. Streilein JW, Longsberry LW, Bergstresser PR. Depletion of epidermal
Langerhans cells and la immunogenicity from tapestripped mouse skin.
J Exp Med 1982; 155: 863-871.
76

OCTUBRE DEL 2003 GÓMEZ MACÍAS RODOLFO.
114. Rico MJ, Streilein J.: Comparison of alloimmunogenicity of Langerhans
cells and keratinocytes from mouse epidermis. J lnvest Dermatol 1987;
89: 607-610.
115. Mckinney EC, Streilein JW. ON the extraordinary capacity of allogeneic
epidermal Langerhans cells to prime cytotoxic T cells in vivo. J
lmmunol 1989; 143: 1560-1564.
116. Toews GB, Bergstresser PR, Streilein JW. Langerhans cells: sentinels
of skin-associated lymphoid tissue. J lnvest Dermatol 1993; 75: 78- 82.
117. Stingl G, Gazze-Stingl LA, Aberer W, Wolff K. Antigen-presentation by
murine epidermal Langerhans cells and its alteration by ultraviolet B
light. J lmmunol 1981; 127: 1707-1711.
118. Rheins LA, Nordlund JJ. Modulation of the population density of
identifiable epidermal Langerhans cells associated with enhancement
or suppression of cutaneous immune reactivity. J lmmunol 1986; 136,
867-876.
119. Shelley WB, Juhlin L. Selective uptake of contact allergens by the
Langerhans cell. Arch Dermatol 1977; 113: 187-189.
120. Botham PA, Rattray NJ Walsh ST Riley EJ. Control of the immune
response to contact sensitizing chemicals by cutaneous antigen
presenting cell. Brit J Dermatol 1987; 117- 1-9.
121. Jiang W, Swiggard WJ, Heutler C, Peng M, Mirza A, Steinman RM,
Nussenzweig MC. The receptor DEC-205 expressed by dendritic cells
and thymic epithelial cells in involved in antigen processing. Nature
.1995; 375: 151-157.
122. sa'llú~tci~/ F, Cella M, Lanzavecchia A. Dendritic cells use • . ,,,. "·-· - . ~-,;:·-.,; . .:<o:: '·
rri~~;Ópik~cytosis and the mannose receptor to concentrate
macromolecules in the major histocompatibility complex class 11
compartment: downregulation by cytokines and bacteria! products. J.
Exp Med 1995182: 289,.
77

OCTUBRE DEL 2003 GÓMEZ MACÍAS RODOLFO.
123. Picut CA, Lee CS, Lewis RM. Ultrastructural and phenotypic changes
in Langerhas cells induced in vitro by contact allergens Br. J Dermatol
1987; 116: 773-784.
124. Hanau D, Fabre M, Schmitt DA, Lepoittevin JP, Stampt JL, Grosshans
E, Benezra C, Cazenave JP. ATPasa and morphologic changes in
Langerhans cells induced by epicutaneous applications of a sensitizing
dose of DNFB. J invest Dermatol 1989; 92: 689-694.
125. Kolde G. Knop J. Different cellular reaction patterns of epidermal
Langerhans cells after application of contact sensitizing. toxic, and
tolerogenic compounds. A comparative ultrastrutural and morphometric
time-course analysis. J. lnvest Dermatol 1987; 89: 19-26.
126. Zhao B, Wang BH: Cytochemical and ultrastructural studies of the
Langerhans cells. Sequential observations in experimental contact
allergy reaction: lnt J Dermatol 1985; 24: 653- 659.
127. Weinlich G, Sepp N, Koch F, Schuller G. Romani N: Evidence that
Langerhans cells rapidly disappear from the epidermis in response to
contact sensitizers but not to tolerogens/nonsensitizers. Arch Dermatol
Res 1989; 281: 556A- 559.
128. Hoefsmid ECM, Duijveestijn AM, Kamperdijk EWA. Relation between
Langerhan cells, veiled cells, and interdigitating cells. lmmunobiol
1982; 161: 255-262.
129. Knight KL, et al. The role of dendritic cells in the initiation of immune
responses to contact sensitizers. l. in vivo exposure to antigen. Cell ., __ ·. ~· ..
lmmunor1985; 94: 427-433.
130. Yamashita K. Yana A. Migration of murine epidermal Langerhans cells
to ·regional lymph nades : engagement of majar histocompatibility
complex class 11 antigens induces migration of Langerhans cells.
Microbio! lmmunol 1994; 38: 567-578.
78

OCTUBRE DEL 2003 GÓMEZ MACÍAS RODOLFO.
131. ·· Erík AH;·~Kati~~l·~~rly molecular events in the induction phase of ' - < ':--:·/"''>" ;:--.-::,.:,;,,~·.::r;:::_··-·'::;_·,;:.:,,.,
contáct hypersensitivity. Proc Natl Acad Sci USA 1992; 89: 1398-
.1412.;:.,{·.(;:,f¡~W~~Jr"".: Enk 'A.H;.'eU=:íbAn essential role far Langerhans cell-derived IL-113. in . ·. - . ,., . - "::: ~ . - . . . " -
132.
the i~iiiation~ófprirnary immune responces in the skin. J lmmunol .--- -···· .,.-_·-o-;-; .,,e·- •
.19~3; 150: 3698-3704.
133. Rambukkana A, et al. Effects of contact allergens on human
Langerhans cells in skin organ culture: migration modulation of cell
surface molecules and early expression of interleukin- 1 B Protein. Lab
lnvest 1996; 74: 422-427.
134. Cumberbatch M, Kimber l. Dermal tumor necrosis factor-alpha induces
dendritic cells migration to lymph nades and possibly provides one
stimulus far Langerhans celll migration. lmmunol 1992; 75: 257-266.
135. Kobayashi Y, et al. Development of motility of Langerhans cell through
extracellular matrix by in vitre hapten contact. Eur J lmmunol 1994; 24:
2254-2263.
136. Staquet M, et al. Role of specific succesive contacts between
extracellular matrix by in vitre hapten contact. Eur J Cell Biol 1995; 66:
342-347.
137. Breathnach AS; The cell of Langerhans. lnt Rev Cytel 1975;18: 1-28.
138. Zelickson A. .Mottaz J: Localization of gold chloride and . -·. '~.'
adénosinfriphosphatase in human Langerhans cells. J lnvest Dermatol
196a:· 51: ~65;:372. 139. Niebave(·G; et al. Osmium zinc iodide reactive sites in the epidermal
Langerhah~ cell'. J Cell Biol 1969; 43: 80-89.
140. Rodríguez EM, Caorsi l. A second look at the ultrastructure of the
Langerhans cell of the human epidermis. J Ultrastruc Res 1978; 65:
279-295.
141. Elleder M. Actiivity of a-D-mannosidase in human Langerhans
epidermal cells. Virchows arch 8 Cell Patl 1975; 19: 93-96.
79 -~ . . .
: ~ :,

OCTUBRE DEL 2003 GÓMEZ MACÍAS RODOLFO.
142. Khalil HM, Nitiuthai S, Allen J. Alkaline phosphatase-positive
Lahgerhans cells in the epidermis of cattle. J lnvest Dermatol 1982; 79:
47-51.
143. Gschnait F, Brenner W. kinetics of epidermal Langerhans cells. J lnest
· Dermatol 1979; 73: 566-569.
144. Mackenzie IC. Bickenbach JR, Rittman BR. Reactivity of epidermal
Langerhans cells to a histochemical method for demonstration of beta
glucuronidase. J lnvest Dermatol 1982; 78: 239-242.
145. Lyne AC. Chase HB. Branched cells in the epidermis of the sheep.
Nature 1966; 209: 1357-1358.
146. Dubertret L, Breton-Gorius J, Fosse M, Touraine R. Peroxidatic activity
in endoplasmic reticulum of normal human Langerhans cells J.
Reticuloendothel Soc. 30 1981; 30: 313-316.
147. Mustakallio K: Adenosine triphosphatase activity in neural elements of
human epidermis. Exp. Cell Res 1962; 28: 449-451.
148. Bjorkman PJ, Saper MA; Samraouri 8, Bennet WS, Strominger JL,
Wiley DC. The foreign antigen binding site and T cell recognition
regions of class 1 histocompatibility complex. Nature 1987; 329: 512-
518.
149. Doyl~ C, Strominger JL. lnteration between CD4 and class JI MHC
molecules mediates cell adhesion. Nature 1987; 330: 256-259.
150. ~Ór~~~t AM, Salter RO. Parham P, Engelhard VH, Littman DR. Cell
ceU adhesion mediated by CDS and class 1 molecules. Nature 1988;
336: 79-81.
151. Acolla RS, Adorini L, Sartoris S, Sinigaglia F, Guardiola J. MHC
orchestrating the immune response. Jmmunol Today 1995; 16: 8-11.
152. Bronstein Br, Murphy GF, Harrist TJ, Ruiter DJ, Mihm MC, Bhan AK.
Location of HLA A,B,C, antigens in dendritic cells of normal human
skin: an immunoelectron microscopic study. J lnvest Dermatol 1983;
80: 481.
80

OCTUBRE DEL 2003 GÓMEZ MACÍAS RODOLFO.
153. Staquet MJ, Dezutter- Dambuyant C. Schmitt D. Reactrivity patterns of
the panel of antibodies far clustering in the T cell section in normal
human skin (keratinocytes and Langerhans cells) Tissue Antigens
1989; 33: 121-125.
154. Gielen V, Schmitt D. Thivolet J: HLA class 1 antigen (heavy and light
chain) expression by Langerhans cells and keratinocytes of the normal
human epidermis: ultrastructural quantification using immunogold
labeling procedure. Arch Dermatol Res 1988; 280: 131-137.
155. Rowden G, Lewis MG, Sullivan AK. la antigen expression on human
epidermal Langerhans cells. Nature 1977; 268: 247-248.
156. Klareskog L, Malmnas-Tjerlund U, Forsum U, Peterson PA. Epidermal
Langerhans cells express Ja antigens Nature 1977; 268: 248-249.
157. Kaufman JF. Aufray C, Korman AJ, Shakleford DA, Strominger J. The
class JI molecules of the human and murine majar histocompatibility
complex cell. Natura 1984; 36: 1-9.
158. Sontheimer RO, Stastny P, Nunez G. HLA-DR region antigen
expression by human epidermal Langerhans cells. J lnvest Dermatol
1986; 87: 707-710.
159. Murphy .GF, et at. Ultrastructural documentation of HLA-DR antigen
reácti~ity.in normal human acrosyringeal epithelium. J lnvest Dermatol - - .. ,,, ,-''
1983; 81: 181-186.
160. AúbockJ, et al. HLA-DR expression on keratinocytes is a common
featÍ.ire of diseased skin. Br J Dermatol 1986; 114: 465-472.
161. [)'e.iutter-Dambuyant C, Cordier G, Schmith D, Faure M, Jaquoi C,
· Thivoíet · J. Quantitative evaluation of two distinct cell populations
'3~,p~essing HLA-DR antigens in normal human epidermis. Br J
Dernatol 1984; 111: 1-11.
162. Romani N. Lenz A, Glassel H, Stossel H, Stanzl U, Majdic O, Fritsch
P, Schuller G. Cultured human Langerhans cells resemble lymphoid
81

OCTUBRE DEL 2003 GÓMEZ MACÍAS RODOLFO.
dendritic cells in phenotype and function. J lnvet Dermatol 1989; 93:
600-609.
163. Bjercke S, Gaudernack G, Braathem LR. Enriched Langerhans cells
express more HLA-DR determinants than blood-derived adherend cells
(monocytes and macrophages). Sean J lmmunol 1985; 21: 489- 492.
164. Fithian E, Kung P, Goldstein G, Rubenfeld M, Fenoglio, Edelson R.
Reactivity of Langerhans cells with hibridoma antibody. Proc Natl Acad
Sci 198178: 2541-2544.
165. Murphy GF, Bahn AK, Sato S, Mihm MC, Harrist TJ. A new
immunologic marker for human Langerhans cells. New England J Med
1981; 304: 791-792.
166. Reinherz EL, Sclossman SF. The differentiation and function of human
Tlymphocytes. Cell 1980; 19: 821-829.
167. Calabi F, Jarvis JM, Martín L, Milstein C. Two classes of CD1 genes.
Eur J lmmunol 1989;19: 285-292.
168. Amiot M, Bernard A, Raynal B, Knapp W, Desclidre C, Boumsel L.
Heterogeneity of the first cluster of differentiation: Chacterization and
epitopic mapping of trhee CD1 molecules on normal human thymus
cells J lmmunol 1986; 136: 1752-1258.
169. Knowles RW, Bodmer WF. A munoclonal antibody recognizing a
human thymus leukemia-like antigen associated with b2-microblobulin,
Eur J lmmunol 1982; 12: 676-684.
170. Lerch .PG, Vande Rijin M, Schrier P, Terhosrt C. Blochemical
có"mparision of the T6 antigen and HLA-A,B, antigens. Human immunol
1983; 6: 13-30.
171. Schmitt O, Dezutter- Dambuyant C, Brochier J, Thivolet J.
Subclastering of CD1 monoclonal antibodies based on the reactivity
on human Langerhans cells. lmmunol Lett 1986; 12: 231-235.
82

OCTUBRE DEL 2003 GÓMEZ MACÍAS RODOLFO.
172. Ashworth J,. Kahan MC Breatnach SM. Flow cytometric analysis and
sortingofHLA-DR+ CD1+ Langerhans cells. BrJ Dermatol, 1989; 114:
173. Coé'chi~ : D, . Michetti F, Donato R. lmmunochemical and
iinmunocytochemical localization of S-100 antigen in normal human
skin. Nature1981; 294: 85-91.
174. Rowden G, Boudreau S. Higley H. Langerhans cells and extra
epidermal dendritic cells, an investigation in laboratory animals and
man with lmmunomorphological methods. Scant J lmmunol 1985; 21:
471-476.
175. Wen DR, Bhuta S, Herschman HR, Gaynor RB, Cochran AJ: S-100
protein: a marker far melanocytic tumors. Ann NY Acad Sci 1990; 487:
261.
176. Wood GS, Tumer RR, Shiurba HR, Eng L, Warnke RH. Human
dendritic cells and macrophages, In situ immunophenotype definition
of subset that exhibit specific morphologic and microenviromental
characteristic. Am J Pathol 1985; 119: 73-78.
177. Takahashi K, et al. lmmunohistochemical localization and distribution
of S-100 proteins in the human lymphoreticular system. Am. J Pathol
1984; 116:497:-513.
178. Weinb_e.rg[)§;;pinkus GS, Murphy GF. Tosillar epithelial dendritic cells: - ':.' ,_-.. ~:'~;;:?~!~'i:.::l;f.:.:l~}~;.';)~:.,~-.~·> demonstr.a
OCTUBRE DEL 2003 GÓMEZ MACf AS RODOLFO.
NLDC-145: purification, characterization, and N-terminal amino acld
sequence. Cell lmmunol 1995; 165: 302-309.
182. lnaba K, et al. Tissue distribution of the DEC-205 protein that is
detected by the monoclonal antibody NLDC-145. 1e. Expression on
dendritric cells and other subsets of mouse lymphocytes. Cell immunol
1995; 163: 148-153.
183. Witmer-Pack MD, et al. Tissue distribution of the DEC-205 protein that
is detected by the monoclonal antibody NLDC-145. 11 Expression in
situ in lymphoid and nonlymphoid tissues. Cell immunol 1995; 163:
167-183.
184. Okiji T, Kawashima N, Kosaka T, Kobayashi and suda H. Distribution
of la Antigen-Expressing Nonlymphoid cells in Varius Stages of
lnduced Periapical Lesions in Rat Molars. Journal of Endodontics
1994; 20 (1): 27-31.
185. Gao Z, Mackenzie IC, Rittman Br, korszun AK, William DM and
Cruchley AT. lmmunocytochemical examination of immune cells in
periapical granulamata and odontogenic cysts. J. Oral Pathol 1998; 17
(2): 84-90.
186. Kaneko T, Okiji T, kan L, Takagi M and Suda H. Ultrastructural
Analysis of MHC Class 11 Molecule Expressing Cells in Experimentally
lnduced periapical lesions in the Rat. Journal of Endodontics 2001; 27
(5): 337-342.
187. Kaneko T, Okiji T, Kan L, Suda H. Takagi. An immunoelectron
microscopic study of class 11 majar histocompatibility complex
molecule- expressing macrophages and dendritric cells in experimental
rat periapical lesions. Archives of oral Biology 2001; 46: 713-720.
188. Kan L, Okiji T, Kaneko T and Suda H. Localization and density of
myeloid leucocytes in the periodontal ligament of normal rat molar.
Archives of Oral Biology 2001; 46: 509-520.
84

OCTUBRE DEL 2003 GÓMEZ MACÍAS RODOLFO.
189. Susuki T, Kumamot H, Ooya K and Motegi K. lmmunohistochemical
analysis of CDla-labeled Langerhans cells in human dental periapical
inflammatory lesions-correlation with inflammatory cells and epithelial
cells. Oral Diseases 2001; 7: 336-343.
85
OCTUBRE DEL ?003
1.16.1. APENDICE.
CAMPOS µ'
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
20.
21.
22.
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
39.
40.
41.
42.
43.
44.
45.
ES HLA·DR
5
2
4
5
4
4
5
4
7
6 2
4
2
3 2
4
5
3
5 4
3 5 4
3
3
7
5
7
6
3
1. 3
1 3
2
4
3
3
2
6
4
7
3
5
ES S-100
4
3
3
2
3 2
4
2
4
1
4
7
3
6 3
2
2 3
2
2
3
2
6
5 5
6
3
3
4
3
4 2 3
2 3 4
3 4
3
3 2
9
3
6
GPE HLA·DR
4
2 3 5
12
12
10 9
7
9
3
6
5
5
2
3
6
4
11 8
9
8
8
8
11 13
10 10
8
11
10 12
10 10
6
8
10 11
9
9
5
2 4
5
4
GPE s.100
4
6
11 7
10
11 4
7
7
9
7
4
8
9 6
8
3
3
8 6
4 5 3
4
9
6 7
7 3
5 5 3
2
4
9
5
7
7
7
6
10 5
4
6
8
86
GÓMEZ MACÍAS RODOLFO.
GP HLA·DR
8
9
5 10
5
7
7
7
3 3
11 4
6
5 6 4
5 4
6 5 4
4
4
3
5
5
5
4
7 10
2
5
7 6 5 3
5
3
8
6
9
9
6
10 6
GP S-100
8
10 6
4
5
5
4
3
3
11
4
11
4
7 5
7
6
3
4
4
7
4
5
2
3
12
6
4
5 2
4
6
6
8 4
7
5
6
6
6
6
3
4
3
9
QP HLA-DR
3
5
4
3
9
4
4
3
5 3
4
3
9
4
7
9
6 9
5 9
11
5 16
9
4
4
4
3
14
7
9
2
7
10
8 5
4
4
6
4
5
5
8
13
5
QP S-100
8
9
10
5
5
7
7
7
3 3
11
4
6
5
6 4
5
4
6
5 4
4 4
3
5 5
5 4
7
10 2 5
7
6
5 3 5 3
16
6
9
9
6
10 6
1000000 50625
19.7530864

OCTUBRE DEL 2003
46.
47.
48.
49.
50.
51.
52.
53.
54.
55.
56.
57.
58.
59.
60.
61.
62.
63.
64.
65.
66.
67.
68.
69.
70.
71.
72.
73.
74.
75.
76.
77.
78.
79.
80.
81. 82.
83.
84.
85.
86.
87.
88.
89.
90.
91.
92.
93.
4
5
3 2
3
3
3
5 2
3
2
3
4
6
5
3
2
2
4
1
7
4
2
5
3
6
4
4
5
8
3 4
4
2
3
3
3 3
2
4
3
3
3
5
5
6
3
5 5
4
4
4
6
3
3
4
3
4
6
8
8
5 3
3
2
2
5
3
7
3
7 2
5
4
2
5
6
7
7
5 7 3
7
3
6 5
5 3
9
10
5
7 3 6
3
2 2
6
5 5
6
5
5
87
GÓMEZ MACÍAS RODOLFO.
6
8
6 5
5
4
7
7
7
3
4
5
5 3 2 6
5 4
8
3
4
7
5
5 4
5
7 4
3
3 4
5
7
7
5
7
7 3
5
4
8 4
5 4
3
6
4
5
5
3
5
.6
6
6
5
10
9
4
5
5
5
5
4
5
5
6
4
9
3 5
7
3
7
2 3 7
7
6
5 5
3
5 5
7
8
5 3
6
9
4
6
5
8
8
12
4
10
11
10
8 4
3
2
5
4
5 4
8 5
4
4
8
4
3
6
5
7
5
4
5
4
3
4
5
5
3
6
4
5
3
6
6
5
5
7
4
6
4 6
12
4
5
6
8
6 3
16
5
3
3
2
5
4
12
5 3
6
4
5
5
5
5
4
2
3
3
4
6
10
8
5
5
4
5
5
5
4
7
8 3
3
5
7
7
5
6
11
2
6
5

OCTUBRE DEL ?QQ3 GÓMEZ MACÍAS RODOLFO.
94. 6 3 3 5 5 2 95. 6 6 3 5 6 4 96. 4 5 3 3 5 97. 7 5 5 5 8 4 98. 3 4 2 6 6 3 99. 5 6 4 5 4 6 100. 7 2 4 7 5 5 101. 13 6 102. 25 4 103. 14 11 104. 10 8 105. 13 8 106. 10 6 107. 13 10 108. 15 8 109. 8 9 110. 10 8 111. 15 8 112. 11 6 113. 10 6 114. 6 5 115. 5 3 116. 7 8 117. 11 5 118. 9 6 119. 12 5 120. 8 5 121. 10 8 122. 8 12 123. 13 4 124. 10 9 125. 15 9 126. 16 10 127. 10 8 128. 17 8 129. 9 5 130. 14 7 131. 8 4 132. 7 6 133. 3 7 134. 4 5 135. 3 6 136. 3 7 137. 12 12 138. 10 6 139. 3 7 140. 12 8 141. 19 9
88

OCTUBRE DEL 2003 GÓMEZ MACÍAS RODOLFO.
142. 16 11 143. 10 15 144. 17 11 145. 10 14 146. 12 16 147. 16 12 148. 16 9 149. 17- 9 150. 13 16 151. 4 13 152.
153. ..__ 4 11
3 6 154. 4 4 155. 3 6 156. 3 5 157. 8 5 158. 2 4 159. 18 5 160. 6 2 161. 4 9 162. 6 6 163. 4 8 164. 7 9 165. 5 9 166. 4 11 167. 2 11 168 .. 9 14 169. 4 4 170. 4 2 171. 10 4 172. 5 7 173. 10 3 174. 7 8 175. 4 5 176. 5 7 177. 12 3 178. 6 5 179. 13 3 180. 16 5 181. 3 6 182. 6 6 183. 8 2 184. 8 5 185. 5 6 186. 4 7 187. 3 8 188. 3 10 189. 4 9
89

OCTUBRE DEL 2003
190.
191.
192.
193.
194.
195.
196.
197.
198.
199.
200.
201.
202.
203.
204.
205.
206.
207.
208.
209.
210.
211.
212.
213.
214.
215.
216.
217.
218.
219.
220.
221.
222.
223.
224.
225.
226.
227.
228.
229.
230.
231.
232.
233.
234.
235.
236.
237.
90
GÓMEZ MACÍAS RODOLFO.
6 7
7 12
6 14
4 11
3 6
6 4
8 4
3 4
8 8
5 5
10 6
7 4
9 7
8 4
11 10
6 5
5 5
9 8
7 7
8 9
8 8
5 6
5 5
10 4
4 7
4 8
7 8
8 6
6 5
7 6
8 3
6 4
5 5
8 4
6 6
5 7
6 6
3 5
4 3
7 9
6 6
5 8
3 4
9 7
6 4
6 10
9 7
4 9

OCTUBRE DEL 2003
238.
239.
240.
241.
242.
243.
244.
245.
246.
247.
248.
249.
250.
251.
252.
253.
254.
255.
256.
257.
258.
259.
260.
261.
262.
263.
264.
265.
266.
267.
268.
269.
270.
271.
272.
273.
274.
275.
276.
277.
278.
279.
280.
281.
282.
283;
284.
285.
91
GÓMEZ MACÍAS RODOLFO.
3 8 4 11
5 5
2 6
6 8 5 8 7 7
6 9
8 6
5 4
6 4
4 10
7 6
13 6
4 5
7 6
7 4
8 5
9 6
6 7
5 5
8 6
2 6
3 11
4 4
6 2 6 5
17 2
8 5
5 4
5 5
4 2
6 11
3 5
4 3
7 6
8 7
2 8
5 3
4 4
4 5
2 6
3 7
5 3
4 4
8 3
8 6
9 3

OCTUBRE DEL 2003
286.
287.
288.
289.
290.
291.
292.
293.
294.
295.
296.
297.
298.
299.
300.
301.
302.
303.
304.
305.
306.
307.
308.
309.
310.
311.
312.
313.
314.
315.
316.
317.
318.
319.
320.
321.
322.
323.
324.
325.
326.
327.
328.
329.
330.
331.
332.
333.
92
GÓMEZ MACÍAS RODOLFO.
8 4
5 4
5 4 4 6
6 6 2 5 8 7
5 6 6 5 9 4 8 6
7 5
7 6
4 11
13 4
8 9 10 7
3 8 4 10
7 9
11 9
7 10
7 11
11 8 9 5
9 3 11 3
10 2 8 2
7 9
10 7
8 10 6 12
6 11
4 5
3 2
12 3
4 7
10 7
7 9
4 6
6 8
6 8
3 4
6 7
7 8
8 11
9 5

OCTUBRE DEL 2003
334.
335.
336.
337.
338.
339.
340.
341.
342.
343.
344.
345.
346.
347.
348.
349.
350.
93
GÓMEZ MACÍAS RODOLFO.
8 4 3 2 6 5 7 2 7 6 9 11
12 6 12 12
11 13
12 9 11 7
8 5
12 13 10 6 10 12
3 7
6 7

OCTUBRE DEL 2003
CAMPOS mm'
1.
2.
3.
4.
5.
6.
7.
8.
9. 10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
20.
21.
22.
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
39.
40.
41.
42.
43.
44.
45.
46.
47.
ES HLADR
98.765
39.506
79.012
98.765
79.012
79.012
98.765
79.012
ES 5-100
79.012
59.259
59.259
39.506
59.259
39.506
79.012
39.506
79.012
GPE HLA-DR
79.012
39.506
59.259
98.765
237.037
237.037
GPE S-100
79.012
118.519
217.284
138.272
197.531
217.284
197.531 '79.012 177.778. 138.272
138.272 . 138.272 138.272
118.519
39.506
79.012
39.506
59.259
39.506
79.012
98.765
59.259
98.765
79.012
59.259
98.765
79.012
59.259
59.259
19.1s3 _~1iy.118 .m.778
79.012 59.259 138.272
138.272
98.765
138.272
118.519
59.259
19.753
59.259
19.753
59.259
39.506
79.012
19.753
59.259
59.259
39.506
118.519
79.012
138.272
59.259
118.519
59.259
39.506
39.506
59.259
39.506
39.506
59.259
39.506
118.519
98.765
98.765
118.519
59.259
59.259
79.012
59.259
79.012
39.506
59.259
19.753
39.506
59.259
79.012
59.259
79.012
59.259
59.259
39.506
138.272 177.778
59.259 59.259
98.765 118.519
79.012 59.259
98.765 59.259
118.519. 79.012
98. 765 158.025
98.765 177.778
39.506 118.519
59.259 158.025
118.519 59.259
79.012 59.259
217.284 158.025
158.025
177.778
158.025
158.025
158.025
217.284
256.790
197.531
197.531
158.025
217.284
197.531
237.037
197.531
197.531
118.519
-158.025
197.531
217.284
177.778
177.778
98.765
39.506
79.012
118.519
79.012
98.765
59.259
79.012
177.778
118.519
138.272
138.272
59.259
98.765
98.765
59.259
39.506
79.012
177.778
98.765
· 138.272
138.272
138.272
.118.519
197.531
98.765
~79.012
98.765_ . 1°18.519
79.012 158.025 59.259 · 5!Í.259
39.506 79.012
94
GÓMEZ MACÍAS RODOLFO.
GP HLA-DR
158.025
177.778
98.765
197.531
98.765
138.272
138.272
138.272
59.259
59.259
217.284
79.012
118.519
98.765
118.519
79.012
98.765
79.012
118.519
98.765
79.012
79.012
79.012
59.259
98.765
98.765
98.765
79.012
138.272
197.531
39.506
98.765
138.272
118.519
98.765
59.259
98.765
59.259
158.025
118.519
177.778
177.778
.118.519
197.531
118.519
118.519
158.025
GP 5-100
158.025
197.531
118.519
79.012
98.765
98.765
79.012
59.259
59.259
217.284
79.012
217.284
79.012
138.272
98.765
138.272
118.519
59.259
79.012
79.012
138.272
79.012
98.765
39.506
59.259
237.037
118.519
79.012
98.765
39.506
79.012
118.519
118.519
158.025
79.012
138.272
98.765
118.519
118.519
118.519
118.519
59.259
79.012
59.259
177.778
79.012
98.765
QP HLA-DR
59.259
98.765
79.012
59.259
177.778
79.012
79.012
59.259
98.765
59.259
79.012
59.259
177.778
79.012
138.272
177.778
118.519
177.778
98.765
177.778
217.284
98.765
316.049
177.778
79.012
79.012
79.012
59.259
276.543
138.272
177.778
39.506
138.272
197.531
158.025
98.765
79.012
79.012
118.519
79.012
98.765
98.765
158.025
256.790
98.765
237.037
79.012
QP 5-100
158.025
177.778
197.531
98.765
98.765
138.272
138.272
138.272
59.259
59.259
217.284
79.012
118.519
98.765
118.519
79.012
98.765
79.012
118.519
98.765
79.012
79.012
79.012
59.259
98.765
98.765
98.765
79.012
138.272
197.531
39.506
98.765
138.272
118.519
98.765
59.259
98.765
59.259
316.049
118.519
177.778
177.778
118.519
197.531
118.519
118.519
158.025
OCTUBRE DEL 2003
48.
49.
50.
51.
52.
53. 54.
55.
56.
57.
58.
59.
60.
61.
62.
63.
64.
65.
66.
67.
68.
69.
70.
71.
72.
73.
74.
75.
76.
77.
78.
79.
80.
81.
82.
83.
84.
85.
86.
87.
88.
89.
90.
91.
92.
93.
94.
95.
59.259
39.506
19.753
59.259
98.765
39.506
59.259
79.012
118.519
98.765
59.259
39.506
39.506
79.012
19.753
138.272
79.012
39.506
98.765
59.259
118.519
79.012
79.012
98.765
158.025
59.259
79.012
79.012
39.506
59.259
59.259
59.259
59.259
39.506
79.012
59.259
19.753
59.259
59.259
98.765
98.765
118.519
. 59.259
98.765
118.519
158.025
158.025
98.765
59.259
59.259
39.506
39.506
98.765
59.259
138.272
59.259
138.272 39.506
98.765
79.012
39.506
98.765
118.519
138.272
138.272
98.765
138.272
59.259
138.272
19.753
59.259
118.519
98.765
98.765
59.259
177.778
197.531
98.765
138.272
59.259
118.519
59.259 98.765 ·.· 39.506
79.012 .· . 39.506
79,012': 118.519
79.012
118.519
.59.259 .•
59.259
·79.012
118.519
118.519
98.765
.. 98.765
1,18.519
98.765
98.765
59.259
118.519
95
GÓMEZ MACÍAS RODOLFO.
118.519
98.765
98.765
79.012
138.272
138.272
138.272
59.259
79.012
98.765
98.765
59.259
39.506
118.519
98.765
79.012
158.025
59.259
79.012
138.272
98.765
98.765
79.012
98.765
138.272
79.012
59.259
59.259
79.012
98.765
138.272
138.272
98.765
138.272
138.272
59.259
19.753
98.765
79.012
158.025
79.012
98.765
79.012
59.259
118.519
19.753
59.259
59.259
98.765
59.259
98.765
118.519
118.519
118.519
98.765
197.531
177.778
79.012
98.765
98.765
98.765
98.765
79.012
98.765
98.765
118.519
79.012
177.778
59.259
98.765
138.272
59.259
138.272
39.506
59.259
138.272
138.272
118.519
98.765
98.765
59.259
98.765
98.765
138.272
158.025
98.765
59.259
118.519
177.778
79.012
118.519
98.765
158.025
158.025
98.765
98.765
197.531
217.284
197.531
158.025
79.012
59.259
39.506
98.765
79.012
98.765
79.012
158.025
98.765
79.012
79.012
158.025
79.012
59.259
118.519
98.765
138.272
98.765
79.012
98.765
79.012
59.259
79.012
98.765
98.765
59.259
118.519
79.012
98.765
59.259
118.519
118.519
98.765
98.765
138.272
79.012
118.519
79.012
118.519
237.037
79.012
98.765
98.765
118.519
118.519
59.259
316.049
98.765
59.259
59.259
39.506
98.765
79.012
237.037
98.765
59.259
118.519
79.012
98.765
98.765
98.765
98.765
79.012
39.506
59.259
59.259
79.012
118.519
197.531
158.025
98.765
98.765
79.012
98.765
98.765
98.765
79.012
138.272
158.025
59.259
59.259
98.765
138.272
138.272
98.765
118.519
217.284
39.506
118.519
98.765
39.506
79.012

OCTUBRE DEL 2003 GÓMEZ MACÍAS RODOLFO.
96. 79.012 19.753 98.765 59.259 59.259 98.765
97. 138.272 98.765 98.765 98.765 158.025 79.012
98. 59.259 79.012 39.506 118.519 118.519 59.259
99. 98.765 118.519 79.012 98.765 79.012 118.519
100. 138.272 39.506 79.012 138.272 98.765 98.765
101. 256.790 118.519 102. 493.827 79.012
103. 276.543 217.284
104. 197.531 158.025
105. 256.790 158.025
106. 197.531 118.519
107. 256.790 197.531
108. 296.296 158.025
109. 158.025 177.778
110. 197.531 158.025
111. 296.296 158.025
112. 217.284 118.519
113. 197.531 118.519
114. 118.519 98.765 115 •. 98.765 59.259
116. 138.272 158.025
117. 217.284 98.765
118. 177.778 118.519
119. 237.037 98.765
120. 158.025 98.765
121. 197.531 158.025
122. 158.025 237.037
'123. 256.790 79.012
124. 197.531 177.778
125. 296.296 177.778 Ú6. 316.049 197.531
127. 197.531 158.025
128. 335.802 158.025
129. 177.778 98.765 130. 276.543 138.272
131. 158.025 79.012
132. 138.272 118.519
133. 59.259 138.272
134. 79.012 98.765
135. 59.259 118.519
136. 59.259 138.272
137. 237.037 237.037
138. 197.531 118.519
139. 59.259 138.272
140. 237.037 158.025
141. 375.309 177.778
142. 316.049 217.284
143. 197.531 296.296
96

OCTUBRE DEL ?QQ3
144.
145.
146. 147. -
148.
149.
150.
151. 152.
153.
154.
155.
.156.
157.
158.
159.
160.
161.
162.
163.
164.
165.
166.
167.
168.
169.
170.
171.
172.
173.
174.
175.
176.
177.
178.
179.
180.
181.
182.
183.
184.
185.
186.
187.
188.
189.
190.
191.
97
GÓMEZ MACÍAS RODOLFO.
335.802 217.284
197.531 276.543
237.037 316.049
316.049 237.037
316.049 177.778 335.802. 177.778
256.790 316.049
79.012 256.790
79.012 217.284
59.259 118.519
79.012 79.012
59.259 118.519
59.259 98.765
158.025 96.765
39.506 79.012
355.556 98.765
118.519 39.506
79.012 177.778
118.519 118.519
79.012 158.025
136.272 177.778
98.765 177.778
79.012 217.284
39.506 217.284
177.778 276.543
79.012 79.012
79.012 39.506
197.531 79.012
98.765 138.272
197.531 59.259
138.272 156.025
79.012 96.765
98.765 138.272
237.037 59.259
118.519 96.765
256.790 59.259
316.049 98.765
59.259 116.519
118.519 118.519
158.025 39.506
158.025 98.765
98.765 116.519
79.012 136.272
59.259 156.025
59.259 197.531
79.012 177.778
118.519 136.272
138.272 237.037

OCTUBRE DEL ?003 GÓMEZ MACÍAS RODOLFO. ·'
192. 118.519 276.543
193. 79.012 217.284
194. 59.259 118.519
.... ___
195. 118.519 79.012
196. 158.025 79.012
197. 59.259 79.012
198. 158.025 158.025
199. 98.765 98.765
200. 197.531 118.519
201. 138.272 79.012
202. 177.778 138.272
203. 158.025 79.012
204. 217.284 197.531
205. 118.519 98.765
206. 98.765 98.765
207. 177.778 158.025
208. 138.272 138.272
209. 158.025 177.778
210. 158.025 158.025
211. 98.765 118.519
212. 98.765 98.765
213. 197.531 79.012
214. 79.012 138.272
215. 79.012 158.025 216 .. 138.272 158.025
217. 158.025 118.519
218. 118.519 98.765
219. 138.272 118.519
220. 158.025 59.259
221. 118.519 79.012
222. 98.765 98.765
223. 158.025 79.012
224. 118.519 118.519
225. 98.765 138.272
226. 118.519 118.519
227. 59.259 98.765
228. 79.012 59.259
229. 138.272 177.778
230. 118.519 118.519
231. 98.765 158.025.
232. 59.259 79.012
233. 177.778 138.272
234. 118.519 79.012
235. 118.519 197.531
236. 177.778 138.272
237. 79.012 177.778
238. 59.259 158.025
239. 79.012 217.284
98

OCTUBRE DEL ?003
240.
241.
242.
243.
244.
245.
246.
247.
248.
249.
250.
251.
252.
253.
254.
255.
256.
257.
258.
259.
260.
261.
262.
263.
264.
265.
266.
267.
268.
269.
270.
271.
272.
273.
274.
275.
276.
277.
278.
279.
280.
281.
282.
283.
284.
285.
286.
287.
99
GÓMEZ MACÍAS RODOLFO.
98.765 98.765
39.506 118.519
118.519 158.025
98.765 158.025
138.272 138.272
118.519 177.778
158.025 118.519
98.765 79:012
118.519 79.012
79.012 197.531 . 138:212· 118.519
256.790 118.519
79.012 98.765
138.272 118.519
138.272 79.012
158.025 98.765
177.778 118.519
118.519 138.272
98.765 98.765
158.025 118.519
39.506 118.519
59.259 217.284
79.012 79.012
118.519 39.506
118.519 98.765
335.802. 39.506
158.025 98.765
98.765 79.012
98.765 98.765
79.012 39.506
118.519 217.284
59.259 98.765
79.012 59.259
138.272 118.519
158.025 138.272
39.506 158.025
98.765 59.259
79.012 79.012
79.012 98.765
39.506 118.519
59.259 138.272
98.765 59.259
79.012 79.012
158.025 59.259
158.025 118.519
177.778 59.259
158.025 79.012
98.765 79.012

OCTUBRE DEL ?003
288.
289. 290.
291.
. 292.
293.
294.
295.
296.
297.
298.
299.
300.
301.
302.
303.
304.
305.
306.
307.
308.
309.
310.
311.
312.
313.
314.
315.
316.
317.
318.
319.
320;
321.
322.
323.
324.
325.
326.
327.
328.
329.
330.
331.
332.
333.
334.
335.
100
GÓMEZ MACÍAS RODOLFO.
98.765 79.012
79.012 118.519
118.519 118.519
39.506 98.765
158.025 138.272
98.765 118.519
118.519 98.765
177.778 79.012
158.025 118.519
138.272 98.765
138.272 118.51'9
79.012 217.284
256.790 79.012
158.025 177.778
197.531 138.272
59.259 158.025
79.012 197.531
138.272 177.778
217.284 177.778
138.272 197.531
138.272 217.284
217.284 158.025
177.778 98.765
177.778 59.259
217.284 59.259
197.531 39.506
158.025 39.506
138.272 177.778
197.531 138.272
158.025 197.531
118.519 237.037
118.519 217.284
79.012 98.765
59.259 39.506
237.037 59.259
79.012 138.272
197.531 138.272
138.272 177.778
79.012 118.519
118.519 158.025
118.519 158.025
59.259 79.012
118.519 138.272
138.272 158.025
158.025 217.284
177.778 98.765
158.025 79.012
59.259 39.506

OCTUBRE DEL ?003
336. 337 •.
338.
339. 340.
341.
342.
343.
344.
345.
346.
347.
348.
349.
350.
101
GÓMEZ MACÍAS RODOLFO.
118.519 98.765
138.272 39.506
138.272 118.519
177.778 217.284
237.037 118.519
237.037 237.037
217.284 256,790
237.037 177.778
217.284 138.272
158.025 98.765
237.037° 256.790
197.531 118.519
197.531 237.037
59.259 138.272
118.519 138.272