spinnentiere, arachnida - museu nacional -...
TRANSCRIPT

r·,
(
Spinnentiere, Arachnida
Weher knechte, 'Opiliones
Von Professor Dr. Jochen Martens, Mainz
unter Mitarbeit von
Albert Ausobsky, Bischofshafen Professor Dr. Paolo Marcello Brignoli, Roma Dr. Jürgen Gruber, Wien Dr. Vlaclimlr Silhavy OSc, Tfebic Dr. Wojciech Star~ga, Warszawa Dr. Konrad Thaler, Innsbruck
Mit 815 Abbildungen
VEB GUSTAV FISCHER VERLAG JENA· 1978
...

I,
Vorwort
Mit dieser Lieferung des "DAHL" wird erBtmals eine Tiergruppe in z\l·eiter Bearbeitung vorgelegt. Die erste hat 1928 Alfred KAES'l'NER durchgeführt, und seine knappe Darstellung erfaßte alle wesentlichen Daten, die damals über die einheimischen Arten der Opilim1es bekannt waren. In den folgenden 50 .fahren entwickelte sich die Kenntnis mitteleuropäischer W'eberlmechte unterschiedlich. Hinsichtlich des systematisch-taxonomischen Aspektes - Grundlage für alle anderen zoologischen Teildisziplinen - lutt. sich vor allem Carl-Friedrich RoEWER Verdienste erworben. Er erweiterte die Zahl der Taxa in fünf Jahrzehnten publikatarischer Tätigkeit erheblich, wirkte aber auf andere "Wissensgebiete - selbRt auf das eigene - wenig befruchtend. Die Biologie· der Weberknechte wurde in diesem Zeitraum nur beiläufig untersucht und fand über den spärlichen Wissensstand hinaus, den ebenfalls KAESTNER 1926 in der Reihe "Biologie der Tiere Deutschlands" und 1935-1937 im "Ha-ndlmch der Zoologie" dokumentiert hatte, nur in wenigen Abhandlungen wissenschaft.Iiches Interesse. Erst 1956 hat VladimJr SrLH.-I.VY im Weberknecht-Band der "Famut ÖSR" viele neue biologische und anatomische Einzelheiten mitgeteilt; <tber sein Wissen ist im deutschen Sprachraum leider weitgehend ungenutzt geblieben. In jüngster Zeit nimmt sich ei11e französische Schule um Christiau ,JuBER'rHTE intensiv der Biologie und Ökologie der Opiliones an.
Alle neuen Fakten, die in diesen J"ahrzelmt.en iiher äußere Morphologie, Taxonomie, Verbreitung und Biologio der mittolt'uropäischert "Weberknechte erarheitet \\'Orden sind, vpr~ucht. der vorliPgenrln Band zus<tmmen%ufassen. dariiber hinnu8 a.ber <tuch möglichst viele noch unpublir.iert.e Daten ein%ubeziehen. Bei diP~l'm arheitsa.ufwendig<•n Vorlmben hnk ich mn.nnigfaJt.ige Unterstützung erfahren -und \'Or <1llen anderen sind meine Freunde und l\Htarheiter A. Au::;onSKY, Prof. Dr. P. M. BRIONOLL Dr. J. GRUBER, Dr. V. SrLH'AVY CSc, Dr. W. il'l'ARI)G~ und Dr. K. THALER ~r.u nennen, die dem gemoinHamen 'Verk zum Rang einer bpachtliehen Informa timmquelle verholfen hahen. Ihnen gilt mein besonderer und sehr herzlicher Danlc Daneben lutben viele weitere Freunde und Kollegen des In- und Anslandes sowohl ihr Wi~sen ab ().nch }bterial nuH eigener oder von ihnen ,-erwalteter Sammlung zur Verfiigm{g gt>stPllt. Sie hier allo zn erwähnen ist nicht möglich; für viele stellvertretend seien genannt:
:Fra.u Dr. St. AvRAni (Bulmn,st}, Prof. Dr. R. BRAUN (l\fainz). Dr. E. DREsco (Pa-ris), Dr. D. DUMITRESCU (BukareHt), Prof. Dr, H. FRANZ (Wien), .Dr. lli. GRASSHOFF (Frankfurtfl\1.), Dres. D. und 0. VON HELVERSEN (Freiburg). Prof. Dr. Ch . . JunERTH'IE (l\1oulis), Prof. Dr. H. KAum (Bergen), l'rof. Dr. R. KrNZELBAOit (l\Iainz), Prof. Dr. 0. KRAUS (Frankfmt, jet-zt. Hall)lmrg), llk D. W. li'L~CKIE (Stockport), Dr.l\L .i\Ion.rTz .(ße.rliu), Prof. Dr. P. PALMliliEN (Helsinki), Dr. H. PAUI"us (Freiburg), Prof. Dr. H. PE'l'ERS (Diissel!lorf), Dr. H. PIEPER (Kiel}, Dr.

6 Vorwort
W. RAllELER, ]'rau Dr. G. RAOK (Hamburg), Frau Dr. JYI. RAMBLA (Barcelona), Mr. J. H. P. SA..NKEY (Dorking), Dipl.-Biol. W. Sa.HAWALLER (Mainz), Dr. H. SoHMALFuss (Tühingen), Prof. Dr. R. So:HusTER (Graz), Prof. Dr. S. SuzuKI (Hiroshima), Prof. Dr. M. VAoß'oN (Paris).
Die Herren Prof. Dr. F. G. BARTH' und Prof. Dr. E. W' AOHMANN stellten dankenswerterweise z. T. unpublizierte Originale ihrer Zeichnungen zur Verfügung.
Einen großen Teil der Illustrationen, hauptsächlich die auf geprägtem Papier ausgeführten Darstellungen der dorsalen Zeichnungsmuster und die Habitusbilder, verdanke ich vor allem der Sorgfalt und dem Geschick von Frau G. T:HOMAS, überdies dem von Fräulein R. BRENDEL und Fräulein G. RöHRIG. Die beiden letztgenannten wurden aus Mitteln der Deutschen Forschungsgemeinschaft bezahlt, der a.uch an dieser Stelle gedankt sei.
Dankbar anerkennend müssen auch die Herausgeher der DAHL-Reihe und die Mitarbeiter des VEB Gusttw ],ischer Verlag genannt werden, durch deren ständiges Interesse und hilfreiches Bemühen dem Werk ein guter Start zuteil wurde.
Mainz, im Januar 1977 Jochen MARTENS.
Inhaltsübersicht
I. Allgemeiner Teil
1.1. Einführung
1.2. Morphologie
I.~. I. Großgliederung eh~:-; 1\:örpers ].:?.:!. nor,;;alseito . l.:t:l. Vent••ah;eite 1.2.4. Augen . . . 1.2.5. Ht.inkdrü:-;mt 1.2.(1. Atemsystem 1.2. 7. Extremitäten . 1.2.8. Genitalorgane. 1.2.9. Sekundäre Geschlechtsinerkmale 1.2. 10. Rtridula.t.ionsorgane
1.3. Biologie . . . . .
I.:l.l. Biotope . . . . . l.:l.2. Vertikalverbreitung 1.:1.3. Nahrung . . . 1.3.4. Sozjalverhaltcn . . 1.:1.5. Fortpfl~tnztmg , . 1.:3.6. Eiablage, Eizahl, Brutfürsorge, Part·henogenese ].:{. 7. Lebensclaner, Phänologie . . .
I
1.4. Verbreitung und Zoogeographie
1.5. System. . . . . . . . . . .
1.6. Sammlungs- und Bestimmungstechni.k
I.ü.l. Xat.ionale Samn1lnngen
2. Spezieller Teil
Subordo Cyphophthalmi
'Familio. Hironidae Genus Sil'O .
Subordo Laniatores .
Suprafamilin Travunioidea
b"'amilia Tt·ayuniidae . . Gc:nn8 Peltonychüc. . .
.Familia Erebon1a.~tridae . Genus Holo~<tcotole·mm~< .
13
13
14
16 16 21 2:l 24 26 26 29 38 38
:10
39 41 41 42 43 44 45
40
51
57
59
50
60 60
68
69 69 70 76 76

8 Inhaltsübersicht
Suprafamilia Gonyleptoidea
Familia Phalangodidae
Genus 8cotolemon.
Subordo Palpatores . . . .
Suprafamilia Troguloidea
Familia Nemastmnatidae
Genus Nemastoma . Genus Paranernastonut Genus Hist1·icostoma Genus Oarin.ostoma, . Genus Oe·ntetostom.a . Genus ltfitostoma . .
li'a1nilia Dicranolasmatidae
Gonus Dicnmolasma
Familia 'rrogulidn.e . . .
Genus ~'1·ogulu.y Genus Anela::nnocephalus.
Suprafamilia Ischyropsalidoidell.
Familia Ischyro psaliclidae
Genus lschy•·opsaUs.
li"atnilia Sabaoonidae
Genus ~-.'}abac.~on .
Suprafamilia Phalangioidea
Familia Phalangiidae . .
Rtthfamilia Phalangiinae.
Geum; Phalangiwn . . G~nm; 1\-letaplwlanrtium 0NlU8 Op1:Uo . ..•. Oenus Platybu:I/US Genus .:.lfc.,ta.pltlt,ljbHJlUS Gllll UH .1li t~yalnuws Gt\llU:-:> R-ilaenu . . Gfnus Eudasylobu.'J Oenns Lophop·i.Ho . Genu~ Zaclwus . . Genus Eaacnus. .
Snbfatnilht Oligolophinae
Genu:; OUgolophu.<J . Ge.Hu:'-1 Pm·oligoloplw6 Genus Laeiwius Genus Oclicll-u-9 . . . Gf!tH1:3 JlUopw~. . .
Suhfamilia Gyautinau
Genus U.IJUS . • • • Genus Dir.ra.IIO)JillJH'·'l Gonu:; Amilrnus . ,
86
87 87
90
91 91 93
116 129 134 139 141
150 ( ( 152
162
103 175
186
180 187
225 2~i)
228
22U
230 231 23ü 239 257 270 27:1 287 29(1 297 :JOl 304
309. :wu 3lll 322 :l:J:l !34:3
n5 :!56 :J05 372
3.
4.
Subfamilia Sclerosomatinae
Genus Homalenotus • • Genus ABtrobunus . ,
Subfamilia Leiobuninae ,
Genus Leiobunum Genus Nelima
Schriftenverzeichnis
Register .
Inhaltsübersicht !)
~76
377 380
39:1 393 417
431
451
J

Verzeichnis der Abkürzungen
ab<l Abd ad. .-l.po AUS
abdomiual Abdomen adult ..\.dultu• Apopl1y•e Österreich
ba basal Bo BRD BUL
Coph CH Chel CSSR Cx
DAN DDR <lo do-cli Do Dö
FR
GR GRB
I'l'
Borste Bunclosropublik DeutsohlEmd Bnlga.rit.nl
Cepha.lot.lwrax Schweiz Chelicero '1~scltecltoslowakoi
Coxa.
Dänexnark Deutsche Demokratische Hepublik dorsal dorso-di•tal Dorn Dörnohen
Franluoiclt
Griec:lwnla.nd Britischn Insoln
Italien
iuv. Iuv. iuvenis Jungtier (für Nymphe und Larve)
1\:ö Korn I~örnchen
L Länge Ia. lateral Ii links Lam Supracholioeral-Lamelle
)fA Marokko nl,~, 1ne- n:Iedian, medio-me-t.li medio-distal 1nm 1\fillhneter Mt Metatarsus
K nördl.
Op an Op gen östl.
Norden nördlich
Operculum anale Operculum genitale östlich
Pa l'alpus l'edipalpus pa.ra-rno para-median pro-la pro-lateral Pt Patella.
re rechts Roc soxn Uoceptaculum seminis REM Hasterelektronenmikroskop RtT Humänien
f:j Süden SCH Schweden SP Spanien St Sternit subad. subadult· Subadult.us •üdl. südlich
~l'u
'l'erg thorak Ti Tu Tu oo 1'r
UdSSR UNG
\V westl.
YU
'l'ßl'::illH
Tcrgit thoralml 'l.'ibia Tubc•·kel 'l'ubet• oculorun1 Trochanter
Howjotunion Ungarn
ventral \'entro~tlistßl
\Vest.e11 wesWeh
J ugoslawicn

12 Verzeichnis der Abkürzungen
Liste der erwähnten Museen
BM(NH) MCSV MHNG MNB MNHNP NHMB NHMW PANW SMF UNMB ZMH
London, British Museum (Natural History) Verona, Museo Civico cli r-Jtoria Naturale Genf~ Museum d'Histoire NatureHe Berlin, J.\riuseum für Naturkunde, Zoologisches )luseunl Paris, Mus8un1 national cl'Historo Nattue1le Basel, Naturhistorisches I\iusoun1 Wien, Naturhistorisches l\Iusenm Warschau, Instytut. Zoologiczny, Polska Ah:ademia Nauk Frankfut·t am Main, Senckenberg-l\'[useuJn Budapest. Ungarisches Nat.urhistm·isches 1\Iuseun1 Hamburg, Zoologisches Institut und :Museum der Universit·ät
Initialen der Sammler bzw. Mitarbeiter (siehe 'l'itolhlatt):
A. A. Albe1·t AusoBSKY, I'. B. Paolo Marcello BltiGNOLI, J. G. Jiirgon GIWBEJt, J. :li. Jochon l\fARTENS, V. R. VJadimfr Sn,uAv'S'·, \\'. H. "c-ojcinnh S•rARfilOA, K. T. J(onrnd ~rHALEn..
1. Allgemeiner Teil
1.1. Einführung
Gebietsabgrenzung: Der geographische Rahmen ist weiter gezogen, als in dieser Reihe sonst üblich. Außer den Arten, die im Gebiet der beiden deutschen Staaten vorkommen, werden auch jene deB weiteren Mitteleuropa behandelt - nämlich die der Benelux-Länder, die der Britischen Inseln, der südlichen Teile von Norwegen und Schweden, der VR Polen, der ÖSSR, der VR Ungarn, die NW-Jugoslawiens (Slowenien, Istrien), die des g11nzen Alpenraumex mit Österreich und der Schweiz sowie die der italienischen und französischen Alpenzone; ferner sind die Arten der Po-Ebene bis zum N-l!'uß des Apenniu erfaßt. Dagegen ist das außeralpine Frankreich nicht einbezogen worden.
In diesem geographischen Bereich sind Artenbestand, Verbreitung der einzelnen Arten sowie Biologie und Ökologie vieler Arten soweit bekannt, daß es berechtigt ist, die Grenzen so weit zu ziehen.
Verbreitungskarten: Auf ihnen ist - mit wenigen Ausnahmen - jeweilti das gesamte Areal dargestellt. Trotz bisher noch geringer Arealkenntnis bei vielen Arten habe ich mich in den meisten Fällen für die l!'lächenkarte und nur bei Arten, die vergleichsweise kleine Areale besiedeln, für die objektivere Punktkarte entschieden. Das ist damit zu begründen, daß sich die Areale vieler A1·ten auf der Basis der bisher bekannten Verbreitung und ihrer ökologischen Präferenda durchaus verläßlich interpretieren und teilweise auch extrapolieren lasxen. Der aktuelle Wissensstand kann im Text überprüft werden. Dort sütd nur gesicherte, in einigen Fällen gesichert erscheinende Daten übemummen worden; auf ex>tkte Dolmmentation wurde Wert gelegt.
Die allermeisten der Karten werden hier erstmals publiziert; somit sind sie in vieler Hinsicht ergänzungs- und verbesserungsbedürftig. Sie sollen zur Überprüfung und Erweiterung der Kenntnisse ~tnregen.
System: Die system>ttisehe Gliederung aller im Gebiet behandelten Gruppen und·dm·über hinaus die Großgliederung des gesamten Systems der Opiliones wurden revidiert; .Änderungen gegenüber den bisherigen Meinungen sind begründend dargelegt worden. Es war Grundsatz, möglichst konservativ zu verfahren und .\nderungen nur dort zu billige!),, wo sie hinreichend fundiert erschienen. So sind z. B. - trotz manch gegenteiliger Meinung - die ehemaligen Unterfamilien der Phalangüdae als solche beibehalten worden. Dem10eh führten neue Erkenntnisse selbst auf dem Familien- und Überfamilien-Niveau zu tiefgreifenden .i\.nderungen.
Alle z,1·eifelhaften Arten wurden möglichKt an Hand von Typenmaterial revidiert. Somit konnten praktisch alle UnKicheren 'Fälle der Zuordnung, die besonders ltus Arbeiten von RoEWER und. einigen neuerett \'On HADZr entstanden waren, geklärt werden.
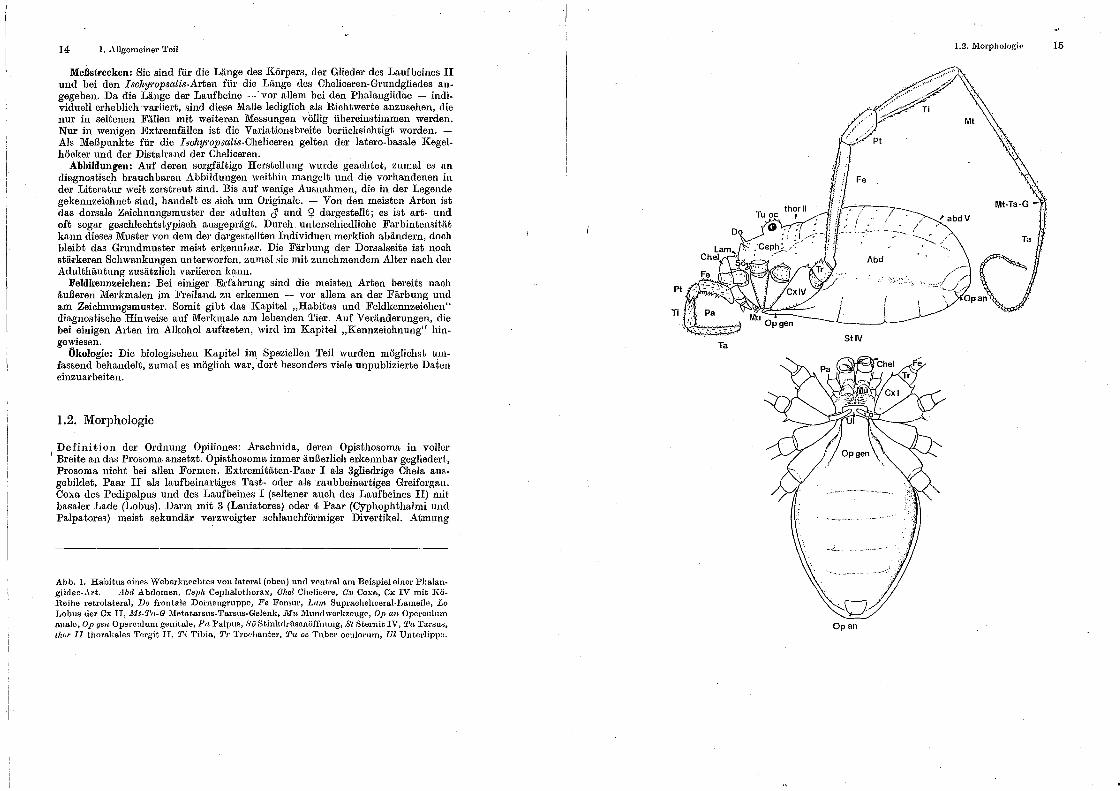
14 I. Allgemeiner Teil
Meßstrecken: Sie sind für die Länge des Körpers, der Glieder des Laufbeines II und bei den Ischyropsalis-AJ:ten für die Länge des Cheliceren-Grundgliedes angegeben. Da die Länge der Laufbeine -·vor allem bei den Phalangiidae - individuell erheblich varüert, sind diese Maße lediglich als Richtwerte anzusehen, die nur in seltenen Fällen mit weiteren Messungen völlig übereinstimmen werden. Nur in wenigen Extremfällen ist die Variationsbreite berücksichtigt worden. -Als Meßpmili:te für die Ischyropsalis-Cheliceren gelten der latero-basale Kegelhöcker und der Distalrand der Cheliceren.
Abbildungen: Auf deren sorgfältige Herstellung wurde geachtet, zun:al es an diagnostisch brauchbaren Abbildungen weithin mangelt und die vorhandenen in der Literatur weit zerstreut sind. Bis auf wenige Ausmthmen, die in der Legende gekennzeichnet sind, handelt es sich um Originale. - Von den meisten Arten ist das dorsale Zeichnungsmuster der adulten (f und ~ dargestellt; es ist art- und oft sogar geschlechtstypisch ausgeprägt. Durch. unterschiedliche Farbintensität kann dieses Muster von dem der darge~tellten Individuen merklich abändern, doch bleibt das Grundmuster meist erkennlmr. Die Färbung der Dorsalseite ist noch stärkeren Schwankungen unterworfen, zumal Hie mit zunehmendem Alter nach der Adulthäutung zusätzlich variieren kann.
Feldkennzeichen: Bei einiger Erfahrung sind die meisten Arten bereits nach äußeren Merkmalen im Freiland, zu erkennen -:- vor allem an der Färbung und am Zeichnungsmuster. Somit gibt das Kapitel "Habitus und Feldkennzeichen" diagnostische Hinweise auf Merkmale am lebenden Tier. Auf Veränderungen, die bei einigen Arten im Alkohol auftreten, wird im Kapitel "Kermzeichnung" hingewiesen.
Ökologie: Die biologischen Kapitel im Speziellen Teil wurden möglichst umfassend behandelt, zuma.I es möglich war, dort besonders viele unpublizierte Daten einzuarbeiten.
1.2. Morphologie
Definition der Ordnung Opiliones: Arachnida, deren Opisthosoma in voller ' Breite an d!t~ Prosoma anset.zt. Opisthosoma immer äußedich erkennbar gegliedert,
Prosoma nicht bei allen Formen. Extremitäten-Paar I als 3gliedrige Chela ausgebildet, Paar II als laufbeinartiges Tast- oder als raubbeinartiges Greiforgan. Coxa des Pedipalpus und des Laufbeines I (seltener auch des Laufbeines II) mit basaler Lade (Lobus). Darm mit 3 (Laniatores) oder 4 Paar (Cyphophthalmi und Palpatores) meist sekundär verzweigter schlauchförmiger Divertikel. Atmung
Abb. I. Habitus eines Weberknechtes von lateral (obon) und vent.ral am Beispiel einer Phalangiidae-Art. - Abd Abdomen, Ceph Cephalothorax, Ohel Chelicore, Gx Coxa, Cx IV mit KöReihe retrolateral, Do frontale Dornengruppe, Fe Forum·, Lam Supracheliceral-Latnelle, Lo Lobus der Cx II, JUt-'1.1a-G 1\fetatarsus-Ta.rsu~-Gelenk, lrlu J\hmdwerkzeuge, Op a.n Opercultnn anale, Op gen Operculum genitale, Pn Palpns, ,)'ö Stinlulrüsenüffnnng, St Sternit IV, Ta Tarsus, thor 11 thorakales Tergit. IT, Ti 'l'ibia, Tr Troc.-IJantor, Tu oc Tuber oculorum, Ul Unt.erlippo.
1.2. MOl'}JlJoJogiP- 15
Ti
Opan
16 I. Allgerneiner Teil
durch baumartig verästelte Tracheen, nur 1 Paar Stigmen auf dem 2. ~terni~ des Opisthosoma. Samenübertragung durch echtes Begattungsorgan (Pems; her Cyphophthalmi möglicherweise differierend).
1.2.1. Großgliederung des Körpers
Der Körper (Abb. I) ist in 2 Tagmata gegliedert, ~rosoma ( = Cephalothor_a~) u_nd Opisthosoma. Beide sind stark ineinar~der :--ersc~rankt, so daß ge~taucht-eif~~mrge bis rundliche Gestalt resultiert. Folghch smd ernzeine Segmentrmge des Korpers nur noch unvollständig erhalten oder völlig reduziert. Embryologische Untersuchungen erlauben den Schluß, daß die Höchstzahl der ursprünglich angelegten Segmente 16 beträgt (WrNKLER 1957). .
Das Prosoma besteht aus ursprünglich 6 Segmenten, von denen nur noch dre beiden thorakalen differenzierbar sind. Diesen sind die Extremitäten zugeordnet: Chcliceren: I, Pedipalpen II, Laufbeine: III- VI. Das Opisthosoma umfaßt ursprünglich 10 Segmente, von denen das Prägenitalsegment reduzie!t und nur noc~ embryonal nachweisbar ist. Die segmentale Zuordnung von Tergrten und Sternrten ist während der Embryog(lnese möglich, nicht mehr bei Jungtieren oder Er-wachsenen. '
1.2.2. Dorsalseite
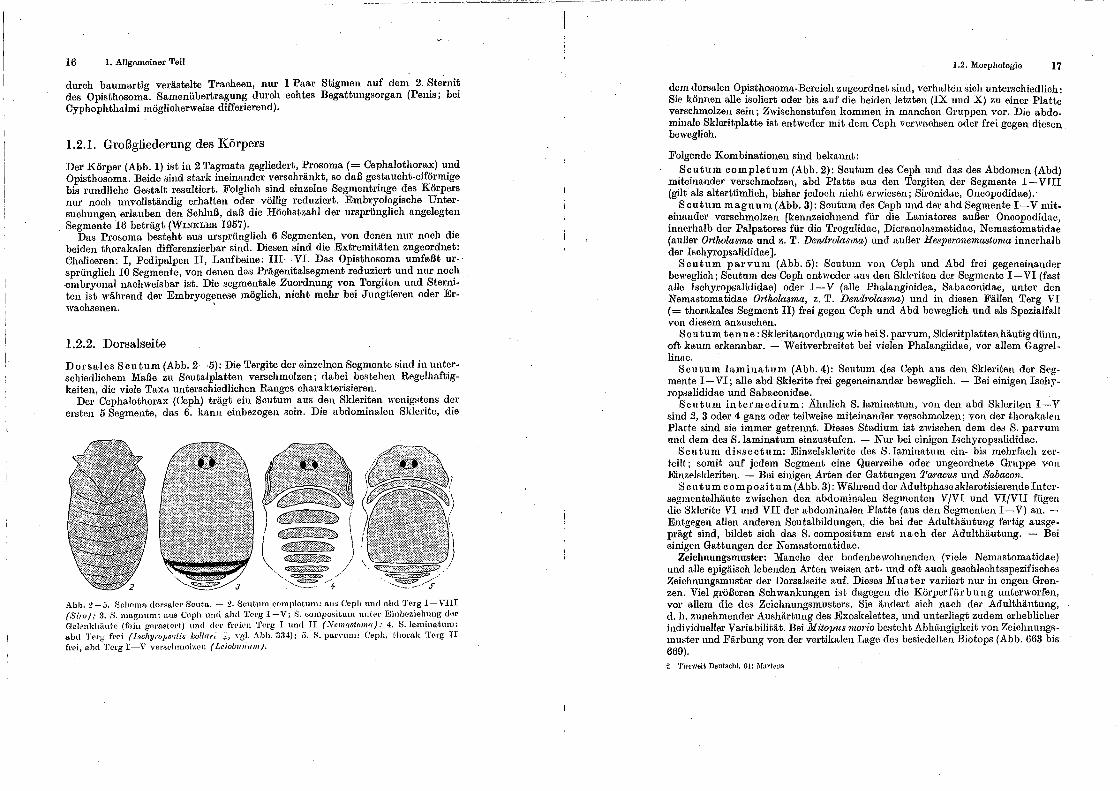
Dorsales Senturn (Abb. 2-5): Die Tergite der einzelnen Segmente sind in unterschiedlichem Maße zu Soutalplatten verschmolzen; dabei bestehen Regelhaftigkeiten die viel!" Taxa unterschiedlichen Rang!ls charakterisieren.
Der' Cophalothorax (Ceph) trägt ein Scutum. aus ~en Skleri.ten wenigHt~ns d~r ersten 5 Segmente, das 6. kann einbezogen sem. Dre abdommalen Sklente, ehe
Abb. 2 -ö. ßr:lwma dor:::aler Scut.a. - 2. Scutntn eompletun1: aJL:-:> C'epiL und o.bd Terg I- VIII (Siro); :1. H. magnum: aus Ccph uud ahd 'l'erg I-V; S. eurnpo~itum mltPr Ein~JezieiL.ung dt.'lr Gch:mkhiiutc (feiu gera.';:tert) und dPr fr<'ien Torg I unct II (~lemaBtomu); 4. S. lammat.um: abd TPJ'g frei (Jschyrop,grdi8 kollm·: :;:, Ygl. Abb. ·334); 5. K paiTum: C'epll. thorak TC\rg II frei, abcl 'fcrg I-V ver::H:Innol't.Nl (Leiolnwum).
1.2. Morphologie 17
dem dorsalen Opisthosoma-Bereich zugeordnet sind, verhalten ~ich unterschiedlich: Sie können alle isoliert oder bis auf die beiden letzten (IX und X) zu einer Platte verschmolzeu sein; Zwischenstufen kommen in manchen Gruppeu vor. Die abdominale Skleritplatte ist eutweder mit dem Ceph verwachsen oder frei gegen diesen beweglich.
Folgende Kombinationen sind bekannt: Scutum completum (Abb. 2): Scut.um des Ceph und das des Abdomen (Abd)
miteinander verschmolzen, abd Platte aus den Tergiten der Segmente I-VIII (gilt als altertümlich, bisher jedoch nicht erwiesen; Sironidae, Oncopodidae). ·
Scutum magnum (Abb. 3): Souturn des Ceph und der abd Segmente I-V miteinander verschmolzen [kennzeichnend für die Laniatores außer Oncopodidue, innerhalb der Palpatores für die Trogulidae, ;Dicranolasmatidae, Nemastomatidae (außer Ortholasma und z. T. Dendrolasma) und außer Hesperonemastoma innerhalb der· Ischyropsalididae].
Souturn parvum (Abb. 5): Scutum von Ceph und Abd frei gegeueinander beweglich; Senturn des Ceph entweder <tus den SklPriten der Segmente I-VI (fast alle Ischyropsalididae) oder I-V (alle Phalangioidea, Sabaconidae, unter den Nemastomatidae Ortholasma, z. T. Dendmlasma) und in diesen Fällen Terg VI ( = thorakales Segment II) frei gegen Ceph und Abd beweglich und als Spezialfall von diesem anzusehen.
Scu tum tenue: Sideritanordnungwie beiS. parvum, Skleritplattenhäutigdünn, oft, k1tum erkennbar. - Weitverbreitet bei vielen Pha.Jangiidae, vor allem Gagrellinae.
Senturn laminaturn (Abb. 4): Souturn des Ceph aus den Skleriten der Segmente I-VI; alle abd Sklerite frei gegeneinander beweglich. - Bei einigen Isch~'ropsalididae und Sabaconidae.
Senturn intermedium: Ahnlieh S.laminatum, von den abd Sklerittm I-V sind 2, 3 oder 4 ganz oder teilweise miteinander verschmolzen; von der thorakalen Plat.te sind sie immer getrennt. Dieses Stadium ist zwischen dem de~ S. parvum tmd dem des S.laminatum einzustufen. - Nur bei einigen Ischyrops!tlididae.
Souturn dissectum: Einzelsiderite des S.laminatum ein- bis mehrfach zerteilt.; somit auf jedem Segment eine Querreihe oder ungeordnete Gruppe VOlt
Einzelslderiten. - Bei einigen Arten der Gattungen Paramts und Sabacon. S cutum composi tum (Abb. 3): Während der Adultphasesklerotisierendeiuter
segmentalhäute zwischen den abdominalen Segmenten VJVI und VI/VII fügen die Siderite VI und VII der abdominalen Platte (aus den Segmenten I-V) an. -Entgegen allen anderen Scutalbildungen, die bei der Adulthäutung fertig ansgeprägt sind, bildet sich das S. compositum erst nach der Adulthäutung. - Bei einigen Gattungen der Nemastomatidae.
Zeichnungsmuster: Manche der bodenbewohnenden (viele Nemastomatidae) und alle epigäisch lebenden Arten weisen art- und oft auch geschlechtsspezifisches Zeichnungsmuster der Dorsalseite auf. Dieses Muster variiert nur in engen Grenzen. Viel größeren Schwankungen ist dagegen die Körperfärbung unterworfen, vor allem die des Zeichnungsmusters. Sie ändert sich nach der Adulthäutung, d. h. zunehmender Aushärtung des Exoskelettes, und unterliegt zudem erheblicher individueller Variabilität. BeiMitopus mon:o besteht Abhängigkeit von ZeichnungRmust.er und Färbung von der vertikalen Lage des besiedelten Biotops (Abb. 663 bis 669). 2 'J'it>rwelt Deutschl. 64: 1\Inrtr.ns

18 I. Allgemeiner Teil
Abb. 6. Echte Haare· (Haarsensillen) und unechte Haare auf dem d'-Pedipalpen-TarsuH. a) Astrobunus laevipea (CANESTRINI). 81, 82, 83 3 verschiedene Haarsensillen-Typen unbekann· ter Modalität, (j Ausfuhröffnungen von Hautdrüsen; unechte Haare sind nicht bezeichuot. - b) Opilio saxatilis C. L. KocH. S Haai·sensillum, Ou massive Cuticular-Dornen (als Reihe auf der Ventralseite); unechte Haare sind nicht bezeichnet.
Abb. 7. Einzelner Zahn aus der Cheliceren-Schneide von Histricostoma dentlpalpe (CANESTRINI), - Alle Zeichnungen nach REM-Aufnahmen.
Integument: Neben der vielfältigen Gliederung des Integuments in Sklerite und Scuta ist es zusätzlich stark strukturiert - mehr als bei den meisten anderen Arachniden. In vielen Gruppen treten als Makroskulptur reliefartige Strukturen in Form von Höckern (Brückendornen z. B. der Nemastomatidae, z. B. Abb. 199, 221), Wülsten und großen Zapfen, Dornen und Tuberkeln auf dem dorsalen Scntum (z. B. Gagrellinae) oder auf den Pedipalpen (z. B. Laniatores, Abb. 85, 96) und Cheliceren (Iscfl.yropsalis, Abb. vgl. Spez. Teil) auf. Viele dieser Kutikularbildungen tragen ihrerseits Haare oder kleine Tuberkel. Als Mikroskulptur können kleinere, meist solide Reliefbildungen der Kutikula bezeichnet werden. Dazu gehören (innervierte) Haare bzw. massive Mikrotriellien (unechte Haare), Bezähnelung und feine Granulation, überdies Stridulationsorgane (Abb. 192). Di.e echten (=innervierten) Haare (=Sinneshaare) können auch als eigene Gruppe angesehen werden.
Abb. 8. Kugelhaar vom Pedipalpus einer Nemastomatidae-Art. - A Ausbuchtung des Haarhohlraumes, B Binnenkanal, Oh Chaetoiden anf dem Schirm, E End faden, H" Haare unbekannter Bedeutung, K 500-700 Abreite Kanäle in der Haarwand, Kh Kugelhaare auf der Palpent.ibia, L Lamellen des Schirmstiels, St Schirmstreben, Wv Wandverdickung. Nach ·WACH
MANN 1970.
a)
I I
b)
I. 2. Morphologie 19
E
H L ..ffJ\1\..\.t.lz.,:
l.(;;ü
B
dl
20 I. Allgemeiner Teil
Die Behaarung (im weiteren Sinne) des Körpers spielt bei der Gestaltung der Körperoberfläche bei den Opiliones nur eine untergeordnete Rolle und fällt lediglich ·bei den Sinneshaaren der Extremitäten stärker ins Gewicht. Auch deren äufere Struktur ist weniger vielgestaltig als die anderer Arthropoden-Gruppen, und manche Typen, z. B. Trichobothrien, fehlen ganz (HANSEN 1917). Unechte Haare (Mikrotrichien) treten dagegen häufiger und in großer Vielgestaltigkeit auf, vor allem an den Extremitäten.
Auf typische SpezialisierungL•ll sei hingewiesen: Drüse1.haare (= Kugelhaare) der Nemastomatidae und Dicranolasmatidae -innervierte Haare mit haareigenen Drüsenzellen, die ihr Sekret durch das Haar nach außen abgeben ( Abb. 8, RrMSKYKoRSAKOW 1924, WACHMANN 1970). Hierher gehören auch Fiederhaare auf den Pedipalpen der Ischyropsalidioidae und vieler Phalangioidea - z. T. ebenfalls mit Drüsenfunktion. Fiederhaare auf dem Ovipositor vieler Sironidae (Abb. 38 bi,; 39) und Caddidae (Abb. 41) arbeiten wahrscheinlich rein sensorisch.
Die meisten Mikrotrichien stellen auf den distalen Extremitätenpartien (Palpen, Laufbeine) die Hauptmasse des dort oft diohten Haarkleides dar. Oft sind sie distad geneigt und liegen dem Integument dicht an. Spezialisierte Formen unechter Haare stellen die ventralen Körnchenfelder auf den Pedipalpen-Tarsen vieler Phalangiidae-cl' dar (Abb. 6b). ,
Für Arachniden kennzeichnende Sensillen sind die Spaltsinnesorgane. Sie treten oberflächlich als schmale Spalten auf und sind in wechselnder Anordnung iiber die Körperoberfläche und über die Extremitäten verteilt. Bei Echten Spinnen (Araneae) kommen sie einzeln vor, in lockeren Gruppen oder dicht gedrängt als "Lyraförmige Organe". Auf Körper und Extremitäten der Spinne Oupiennius salei KEYSERLING wurden insgesamt 3300 Einzelspalten gezählt (BARTH & LIBERA 1970). Bei Weberknechten sind die Spaltsinnesorgane spärlicher verteilt, als Einzelorgane (einzelne Schlitze) oder in lockeren Gruppen, denen nach der Definition von BARTH & STAGL (1976) nur bedingt der Terminus "Lyraförmiges Organ'' zukommt, da die Einzelspalten zu jenen anderer Arachniden vergleichsweise weit voneinander getrennt sind, z. T. über 10 p,m. Auch die Orientierung der Schlitze ist anders als bei Arachniden sonst iiblich: Die zu Gruppen angeordneten Schlitze stehen mehr oder weniger senkrecht zur Längsachse des Beines, nicht parallel. Auf dem Laufbein I von Amilenus aumntiacus (Srl'IION) fanden BARTH & STAGL (1976) Spaltsinnesorgane nur auf Coxa, Trochanter und Femur; J. G. (in litt.) entdeckte einen einzelnen Schlitz auf dem letzten Tarsalglied anderer PhalangiidenArten. BARTH & STAGL teilen die Schlitze bei Amilenus 2 Lyraförmigen Organen. 6 Gruppen von Einzelschlitzen und 6 großen Einzelschlitzen zu; insgesamt 45 Einzelorgane (Abb. 15); vgl. auch EDGAR (1963).
Die Spaltsinnesorgane sind in unterschiedlicher Weiije dem mechanischen Sinn zugeordnet. Nach den bisherigen Untersuchungen bei den Araneae nehmen sie· Verbiegungen des Exosklettes wahr, sie können Krallenbewegungen registrieren und spielen in vielfältiger Weise bei der Orientierung eine Rolle. Es gibt überdies Belege für Luftschall-Empfindlichkeit; möglicherweise perzipieren sie die höherfrequenten Anteile von Fluggeräuschen, die die Trichobothrien (der Araneae) nicht wahrnehmen. Somit können sie eine Rolle beim Beutefang spielen. - Eingehende Untersuchungen der Spaltsinnesorgane von Weberknechten stehen bisher aus.
Das Integument der Trogulidae, Dicranolasmatidae und ScleroHomatinae ist auffällig drüsig-papillös. Mit seinem Sekret. werden MinemllJestandteile des Bodens
I
1.2. Morphologie 21
a~f die Körperoberfläche aufgeklebt und verhelfen diesen Gruppen in Verbindung 1mt langsamer Bewegungsweise zu einer "Erdmimikry". Auf den Pedipalpen der Sclerosomatinae erhärtet das Sekret - meist ohne Inkrustierung von Fremdkörper~ - zu einem glasig spröden Überzug. Farbige Hautsekrete bei Gagrellinen beschreibt LoMAN (1905). Bei Arten der Nemastomatidae, Dicranolasmatidae und Ischyropsalididae sind Hautdrüsen in den Cheliceren der d' konzentriert und zu auffälligen Drüsenorganen entwickelt (vgl. Kap. 1.2.9.).
1.2.3. Ventralseite
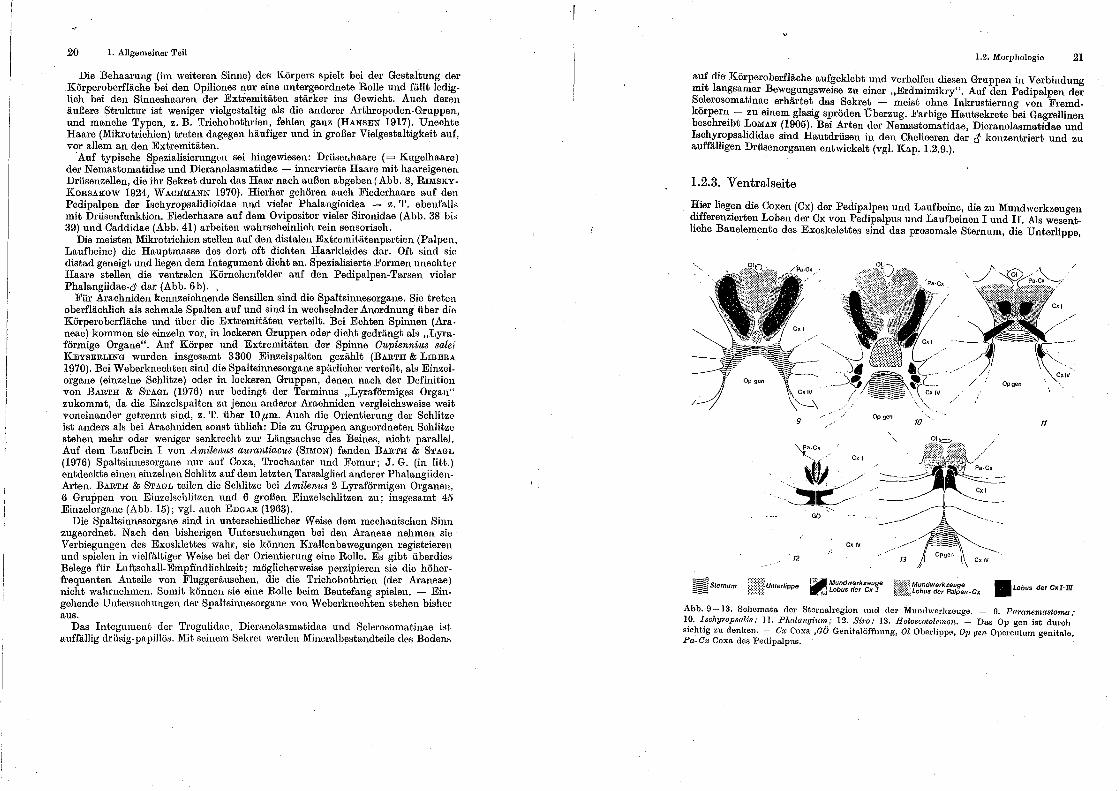
Hier liegen die Coxen (Cx) der Pedipalpeu und Laufbeine, die zu Mundwerkzeugen differenzierten Loben der Cx von Pedipalpus und Laufbeinen I und II. Als wesentliche Bauelemente des Exoskelettes sind das prosamale Sternum, die Unterlippe,
==" =:::::::::::l'Sternum
9
12
mmmm Unterlippe :·:·:·:·:-:.·
Op gen
Cx IV
Ii!'?,~ ··. Mundwerkzeuge .. , Lobus der Cx I
.... o.i..:.l
10 11
.Lobus der Cxi-Ill
Abb. 9-13. Schemata der Sternalregion und der 1\fundwerkzeuge. - 9. Paranemastoma;· 10. Ischyropsalis; II. Plwlangium; 12. Siro; 13. Holoscotolemon. - Das Op gen ist dUl•ch sichtig zu denken. - Cx Coxa ,aö Genitalöffnung, Ol Oberlippe, 07J gen Operculum genitale, Pa-C.v Coxa des Pedipalpus.
22 I. Allgemeiner Teil
das Operculum genitale, die opisthosomalen Sternite und das Operculuni anale zu nennen (Abb. 1). Durch die starke Verschränkung von Pro- und Opisthosoma sind vor allem prosamale Exoskelett-Teile oft stark reduziert, oder sie fehlen ganz. Das ist besonders dann der Fall, wenn sich die Cx der LaufbeiM beider Körperhälften median berühren, wie bei den Cyphophthalmi und den Laniatores (Abb. 12, 13). Der Bau des ventralen Exoskelettes ist für das Niveau der Überfamilien, bisweilen für das der Familie und selten sogar der Gattung (z. B. bei den Sironidae) typisch. Dieser Merkmalskomplex ist geeignet, Gattungsgruppen strittiger Zuordnung auf ihre Stellung zu überprüfen.
Mundwerkzeuge: z.T. häutige, z.T. skierotisierte Teile der Cx-Loben von Pedipalpen und Laufbein I, selten auch von Laufbein II (Erebomastridae, Abb. 13). Die großen Loben der Cx II der Phalangiidae (Abb. 11) gehören zwar zum weiteren Mundbereich, fungieren aber nicht als Mundwerkzeuge. Bei einigen Gruppen können wenig· abgegliederte Loben des Laufbeines II in das Mundfeld einbezogen werden: in geringem Maße bei den Phalangodidae, markant ausgeprägt bei den Erebomastridae (Abb. 13). Dort begrenzen die median zusammenstoßenden (nicht verwachsenen) Loben pyramidenartig den caudalen Teil des Mundfeldes - also die weichhäutigen Loben der Laufbein-Cx I. - Die Mundöffnung liegt zwischen den Loben der .Cx.
Am Freßakt beteiligen sich überdies die Cheliceren; Tastfunktion bei der Beute~ aufbereitung im Nahbereich übernehmen die Tarsen der Pedipalpen.
Unterlippe: Hohle dünnhäutige Platte, die von der Bauchwand hinter Beinpaar I aufgefaltet ist (KAESTNER 1935); sie deckt von caudal die Mundwerkzeuge ab. Die Unterlippe ist bei den Phalangioidea und Ischyropsalidoidea der Palpatores meist kräftig entwickelt, klein bei den Troguloidea, offenbar fehlend bei den Laniatores und Cyphophthalmi.
Sternum: zwischen den Cx der Laufbeine als einheitliche Platte, entweder mit den Cx zu fester Platte verwachsen (Troguloidea; viele, vielleicht alle Gruppen der Laniatores) oder lose im Membranfeld gelagert (Phalangioidea, Ischyropsalidoidea), bei den Cyphophthalmi vielleicht ganz fehlend.
Operculum genitale: Falte des 2. opisthosomalen Sternits; es bildet mit der gegenüberliegenden Bauchdecke die quergestellt-schlitzförmige tertiäre Geschlechtsöffnung. Die Falte fehlt allen Arten der Cyphophthalmi; deren Geschlechtsöffnung ist offen (Abb. 12). Das Operculum der Laniatores ist klein; es liegt im Bereich der Cx IV (Abb. 13); bei den Phalangioidea erstreckt es sich (meist) über das Gebiet aller Cx, und der Vorderrand liegt unmittelbar hinter den Mundwerkzeugen (Abb. 11). Die übrigen Gruppen der Palpatores nehmen Mittelstellungen zwischen diesen Extremen ein (Abb. 9-10).
Opisthosomale Ster1dte: z. T. miteinander verschmolzen (2. und 3. Sternit), z. T. als selbständige Sternit-Spangen erhalten, die gegeneinander gelenkig beweglich sind (im allgemeinen 4.-7. Sternit). Bei den Sironidae bleiben z. T. nur die Sternite VIII und IX frei, und bei den Sironidae, Oncopodidae und Trogulidae bilden die flächig angelegten Sternite II-VIII bzw. II-VII ein einheitliches Ventralscutum.
Opercul um anale: Es entspricht dem Tergit X, umgeben wird es meist von der Cormut ana.lis - rudimentären Tergiten und Sterniten, die bei den einzelnen Gruppen unterschiedlich ausgeprägt sind. Bei den Cyphophthalmi können St VIII und IX und T IX zu einem Ring verschmelzen, der das Operculum umgibt (z. B. Si1·o). Bei den Laniatores ist. die Corona. reduziert: St VIII und IX sind ver-
1.2. Morphologie 23
schmolzen, T IX fehlt. Bei den Palpatores finden sich Überg~nge zwischen fe~lender (manche Phalangiidae) bis vollständiger Corona analis (z. B. Troguhdae, Nemastomatidae).
1.2.4. Augen
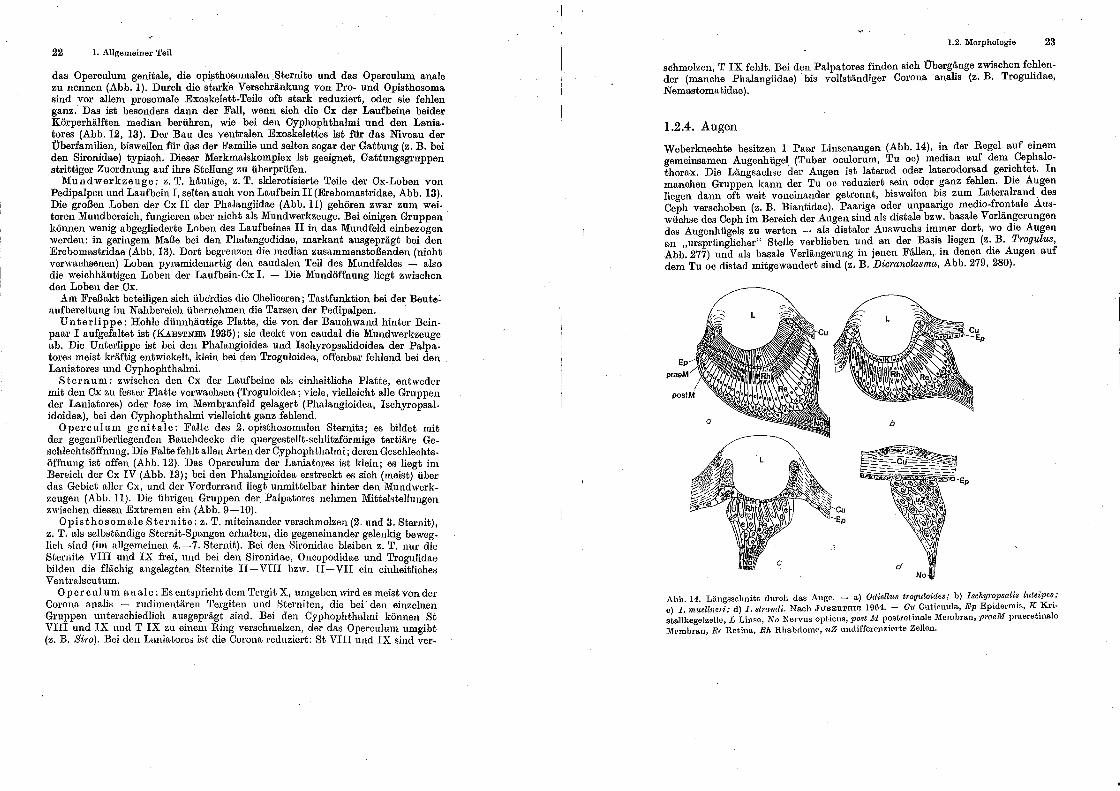
Weberknechte besitzen 1 Paar Linsenaugen (Abb. 14), in der Regel auf einem gemeinsamen Augenhügel (Tuber oculorum, Tu oc) median auf dem. Cephalothorax. Die Längsachse der Augen ist laterad oder laterodorsad gert?htet. In manchen Gruppen kann der Tu oc reduziert ~ein. oder ~anz fehlen. Dte Augen liegen dann oft weit voneinander getrennt, btsweilen b~s zum ~ateralrand. des Ceph verschoben (z. B. Biantidae). Paa:rige ode! unpaar1ge medw-fr~_ntale Auswüchse des Ceph im Bereich der Aug~n smd als dtstale ~zw. basale Verlan?erungen des Augenhügels zu werten - als dtstaler Auswuchs n~me_r dort, wo dte Augen an "ursprünglicher" Stelle verblieben und an der Basts hegen (z. _B. Trogulus, Abb. 277) und als basale Verlängerung in jenen Fällen, in denen dte Augen auf dem Tu oc distad mitgewandert sind (z. B. Dioranolasma, Abb. 279, 280).
Abb. 14. Längsschnitt duroh das Auge. - a) Od·iell«s t•·og«loides; b) Ischyl'opsali~ l«teipes.; c) I. muellncri; d) J. stmndi. Nach JUBERTHIE 1964. - a,: Cuticuula, Ep Eptdermts, K .Krt· st.allkege1zelle, L Linse, ];.;r 0 Nervus opticus, po.r;t lti postret1nale Membran, praelJ!l praeretu1a.le Mf'rnhra.n~ Re Ret.ina, Rh Rhahclome, 'uZ undifferenzie.rte Zellen.

24 I. Allgemeiner Teil
Derart verlängerte Augenhügel sind mehrfach unabhängig voneinander entstan, den; bekannt sind sie von den Troguloidea (Nemastomatidae-Ortholasmatinae, Trogulidae, Dicranolasmatidae) und von den Ischyropsalidoidea (Ischyropsalididae: Geratolasma), ebenso von den Caddidae (Austropsopilio) und einigen Triaenonychidae.
Bei hochspezialisierten troglobionten Arten aus ganz verschiedenen Verwandtschaftsgruppensind Tu oc und Augen bis auf geringe Reste oder sogar völlig reduziert. Im hier behandelten Gebiet sind das Ischy1'0psalis stmnili (Augen bei einigen Populationen ganz fehl'3nd) und Mitostoma anophthalmum (Augen und Tu oc völlig reduziert), ferner die zum Euedaphon zählenden ßiro itU?·icorius und 8. carpaticus. (Unter den Sironidae besitzen überhaupt nur ßtylocellus-Arten rudimentär entwickelte Augen.)
Zwischen normaler Ausprägung der Augen und ihrer völligen Reduktion bes~ehen vielfältige Übergangsstadien. Allgemein läßt sich eine Korrelation zwischen Ökologie und Entwicklungsstand der Augen erkennen. Über "normal" ausgebildete Augen verfügen jene Gruppen, deren Arten an die Perzeption von Tageslicht adaptiert sind (z. B. Phalangiidae). Linse, Glaskörper und Retina sind vollständig und funktionstüchtig.
Bei einigen Gattungen, deren Arten z. T. ver~teckt in den obersten Bodenschichten, z. T. in Höhlen leben, lassen sich Stadien der Augenreduktion verfolgen, z. B. bei Ischyropsalis (Abb. 14 b-d).
Augenreduktion der kavernikolen A1·ten verläuft nach dem Prinzip der Differenzierungsverzögerung. Danach werden die unterschiedlichen Reduktionsstadien bereits während der Embryogenese angelegt (JUBERTHIE & Muiiioz-CUEVAS 1973).
Viele Arten mit besonders großem Tuber oculorum und großen Augen sind tagaktiv (z. B . .IYiegabunus), andere zeichnen sich zudem durch überaus schnelle und geschickte J!'ortbewegung aus (z. B. Gaildo agiUs, BANKS 1892, SHEAR 1975; Sclulitt durch die Augen in JUBERTHIE & Mu5wz-CuEvAs 1973). Inwieweit bei diesen Arten das Sehvermögen beim Beuteerwerb eine Rolle spielt, ist im einzelnen nicht geklärt.
1.2.5. Stinkdrüsen
Sie sind ausnahmslos bei allen Opiliones vorhanden, doch differieren innerer B!tU und die Lage der Austrittsöffnung bei den einzelnen Gruppen: meist auf der Dorsalseite des Ceph lateral nahe dem Frontalrand - bei den Palpatores auf der Höhe der Cx I, bei den Laniatores auf der Höhe der Cx II, bei den Cyphophthahni zwischen den Cx II und III auf der Spitze eines großen lateralen bzw. darsolateralen Kegels (Ahb. 57). Bei den Trogulidae, Dicranolasmatidae, Nemastonm- . tidae und Phalangiidae-Solerosomatinae, ferner bei den Ischyropsalididen Geratolasma und Hesperonemastoma sind die Öffnungen late.ral unter den Rand des Prosoma versehohen und von oben nicht sichtba.r.
Soweit das Sekret nicht sofort an Ort und Stelle weiterverwendet wird, fließt es in Ka.nälen der Körperoberfläche zu anderen Körperteilen. Diese Kanäle -jederseits einer - liegen entweder lateral aur dem Scutum und auf den freien Tergiten (8cotolernon lespesi: Phalangodidae) oder unterhalb des Souturn lateral und somit. seitlich geöffnet (Oncopus: Oncopodidae) oder auf der Oberseite der Laufbein-Coxen (Gonyleptidae). Das Sekret fließt in den Kanälen caudad, wo es
1.2. Morphologie 25
sich entweder als Tropfen sammelt oder auf der· Hüfte bzw. auf der Körperwand verteilt (JUBERTHIE 1961 b, 1976).
Funktion: Das Sekret dient als Wehrsekret und wird unterschledlich eingesetzt. Bei den Phalangiidae tritt es als Tropfen bei stärkerem Druck auf den Körper aus oder bei Angriff räuberischer Insekten, z. B. Ameisen; im Normalfall scheinen in dieser Gruppe überwiegend gasförmige bzw. flüchtige Komponenten zu wirken, worauf der starke Geruch schließen läßt, den es bei vielen Arten ausströmt. Lebende in einem Gefäß eingeschlossene Individuen betäubt das eigene Sekret (SAVORY 1962). Bei allen anderen Gruppen wird es - soweit bis jetzt bekannt - in Überwiegend flüssiger Form appliziert.
ßiro rubens (Sironidae) tupft das Sekret mit einem Laufbein-Tarsus vom Drüsenkonus ab und versucht, es am Angreifer abzustreifen. Es kommt vor, daß sich die sezernierte Flüssigkeit über eirten Teil des Prosomas verteilt oder sich auf der behinderten Extremität ausbreitet (JuBERTHIE 1961 a). Im Versuch mit Sekret kontaminierte Milaoarthropoden (Collembolen, Spinnen, terrestrische Isopoden, 8cutigerella) starben sofort, von Dämpfen erfaßte Individuen wurden unterschiedlich lange Zeit betäubt (JUBERTHIE 1976).
Vonones say·i (Cosmetidae) produziert ein Chinon, das (im Versuch) bei Druck auf den Körper abgegeben wird; zu diesem Tropfen fließt Verdauungsflüssigkeit aus der Mundöffnung. Die vorderen Laufbein-Tarsen beschmierenmit diesem Gemisch den Angreifer und wehren ihn erfolgreich ab (ErsNER, KLUGE, CARRELL & MEINWALD 1971, EISNER 1972).
Heteropachyloiilellus robustus (Gonyleptidae): Beobachtungen über den Einsatz des Sekretes liegen nicht vor. Im Versuch hat sich das Sekret, ein Gemisch von Chinonen (vgl. unten), als hochwirksames flüchtiges Antibiotikum erwiesen. Es schädigte Kulturen von Arten aus wenigstens 18 Bakterien- und Protozoen-Gattungen, z. B. 6 Stämme von 8taphylococcus au1·eus (EsTABLE, ARDAO, BRASIL & FIESER 1955). Bakteriostatischer Einfluß auf der Cuticula-Oberfläche der sezernierenden Tiere ist somit nicht auszuschließen.
Leiobunun~ jormosurn und L. spem:osum (Phalangiidae-Leiobmlinae) benutzen das Sekret als Abwehrstoff: Inmitten einer Kolonie der Ameise Solenapsis saevissi?na wird beim Angriff einzelner Ameisen das Sekret freigesetzt und die eigenen Beine gegen den sekretfeuchten Körper gepreLlt. Ameisen, die das kontaminierte Bein ergreifen, lassen es oft sofort los und beginnen, sich zu reinigen. Sie zeigen sich irritiert und laufen unkaardiniert (BLtJM & EDGAR 1971).
Chemismus: Vonones sayi: Gemisch aus 2,3-Dimethyl-1,4-henzochinon, nach der Sekretion versetzt mit Darmflüssigkeit (EISNER, KLUGE, CARRELL & MEINWALD 1971).
Heteropachyloiilellus robustus: Gemisch aus einer Serie von 1,4-Benzoohinonen ("Gonyleptidin"), im einzelnen: 2,3-Dimethyl-1,4-Benzochinon, 2,5-Dimethyl-1,4-Benzochinon und 2,3,5-Trimethyl-1,4-Benzochinon. Diese Chinone sind als Sekrete noch nicht bei anderen Arthropoden gefunden worden (ESTABLE, ARDAO, BRASIL &
· FIESER 1955, FIESER & ARDAO 1956). Leiobunum Jormosum und L. speciosurn: Das Sekret wurde als 4-Methyl-3-
Heptanon identifiziert (BLuM: & EDGAR 1971).

26 I. AUgemeiner Teil
1.2.6. Atemsystem
Das Tracheensystem wird im allgemeinen lediglich durch 1 Paar Stigmen ventiliert, die im Bereich des 2. opisthosomalen Sternits liegen, teils frei auf der Sternalfläche (z. B. Sironidae, viele Familien der Laniatores, Ischyropsalidoidea außer Ceratolasma und Hesperonemastoma), teils tief versenkt in einem Spalt retro-lateral entlang der Cx IV (Phalangioidea, Troguloidea). .
Die Stigmen sind selten ungeschützt offen und dann durch den Hinterrand der Cx IV vollständig abgedeckt (Phalangüdae). Bei allen anderen Gruppen werden sie durch unterschiedlich gestaltete Borsten oder Chitin-Brücken bzw. -Leisten meist dicht vergittert. Der Gasaustausch erfolgt im wesentlichen durch J?üfusio~l, und bei den Phalangioidea unte):'stützen Atemmuskeln nahe dem Stigma die Ventilation.
Akzessorische Stigmen finden sich als abgeleitete Merkmale distal und basal an den Tibien der Laufbeine der Phalangioidea. Die dort mündenden Tracheen versorgen die distalen Glieder der oft extr~m langen Beine. Somit könn~n ~~e Nerven autotomierter Beine weiter mit Sauerstoff versorgt werden; das erhalt fur einige Zeit ihre Fähigkeit, zuckende Bewegungen auszuführen (LoMAN" 1896, WASGESTIAN"-SOHALLER 1968).
1.2. 7. Extremitäten
In allen postembryonalen Entwicklungsstadien treten 6 Extremitätenpaare auf; das I. Paar als 3gliedriges Greiforgan (Chelicere), das 2. Paar als Tastorgan oder/ und als Hilfsorgan beim Beuteerwerb (Pedipalpus), die 3.-6. Paare (Laufbeine) als Organe der FoFtbewegung und das 2. Paar zusätzlich als Tastorgan für den Fembereich spezialisiert.
Cheliceren: Immer 3gliedrig, das 3. Glied bildet eine Chela. In den meisten Fällen sind die Cheliceren klein - das Grundglied wesentlich kürzer als der Körper (Abb. I). Exzessive Verlängerungen - Grundglied so lang oder länger als der Körper - treten unabhängig voneinander in verschiedenen Familien auf (Ischyropsalididae, Abb. 332-334; Nipponopsalididae, Phalangüdae Megalopsalididae).
Die Chelicereu tragen oft sekundäre d'-Geschlechtsmerkmale: meist aufgeblähte Höcker und Buckel oder Apophysen auf 1. und 2. Glied. Bei den Troguloidea und bchyropsalidoidea (Palpatores) sind die Apophysen auf dem Grundglied (und z. T. auf dem 2. Glied) mit Drüsenorganen kombiniert, die im Balzverhalten eine Rolle spielen (Kapitell.3.5. ). Die Drüsen liegen wenigstens teilweise im Grundglied: Entweder sind sie in der Apophyse konzentriert (z. B. Dicranolasma, einzelne Sabacon-Arten), oder sie durchziehen das ganze (Nernastoma, Mito8torna) oder große Teile des Cheliceren-Grundgliedes (viele Isckyropsalis-Arten), oder sie reichen bis in das Prosoma (Pamnemastoma, Histricostoma). Das Sekret tritt durch eine Porenplatte (Nemastorna, Isckyropsalis, Dicranolasrna), durch teilweise gebündelte · Einzelkanäle (wenigstens 1 Sabacon-Art) oder durch einen einzigen großen Sammelkanal, der sich als weitlumiger Porus nach außen öffnet ( Paranernastorna, Carinastoma, Hist1'icostoma), auf die Oberfläche der Chelicere (MARTENS & ScKAWALLER 1977).
Die Cheliceren-Zähne sind einheitlich kräftig skierotisiert und kompakt; nur bei den Troguloidea und Ischyropsalidoidea tritt ein zweiter Typ hinzu: feine
r 1.2. Morphologie 27
hyaline und gleichmäßig serial angeordnete kleine Zähne im größeren Teil der Schneide, bei verlängerten Cheliceren ( Isckyropsalis, Nipponopsalis, Taracus) nur auf dem kleineren Teil der Schneide. Jeder dieser Zähne ist distal mit feinem Sägeblatt-Besatz versehen (Abb. 7).
Die Cheliceren inserieren unter dem Ceph-Frontalrand in den Cephalothorax. Zwischen diesem und den Cheliceren sind Gelenkhäute ausgespannt, deren proxi· maler Bereich abgeglied.ert und mit Teilen des Ceph-Frontalrandes die Supracheliceral-Lamellen (Lam, Abb. 1) bilden können: unterschiedlich stark skierotisierte Plättchen, die einzelne Dornen oder Dornenreihen tragen können.
Pedipalpus: Er gliedert sich ähnlich den Laufbeinen in Coxa (Cx), Trochanter (Tr), Femur (Fe), Tibia (Ti) und Tarsus (Ta); Metatarsus (Mt) fehlt, und der Ta ist immer eingliedrig. Die Tarsalkralle fehlt in vielen Fällen. Die Pedipalpen dienen nicht "mehr" der Lokomotion, unterstützen sie aber in vielen Fällen als Klammerorgan bei Klettermanövern (z. B. Phalangiidae). Große Bedeutung hat der Pedipalpus als Tastorgan für den Nahbereioh, z. B. bei Beutefang und Nahrungsaufnahme. Tastfunktion im Fernbereich übernehmen die 2. Laufbeine. Vor allem der Tarsus trägt zahlreiche Sinneshaare, von denen die Fiederhaare der Isohyropsalidoidea und die sezernierenden Drüsenhaare der Troguloidea zusätzlich spezialisiert sind (Abb. 6, 8). Deren Bedeutung für den Nahrungserwerb ist nicht abschließend g~klärt (RIMSKY-KORSAKOW 1924, WAOHMAN"N 1970).
Weitverbreitet ist die Funktion des Palpus als Raubbein, vor allem bei den Laniatores. Dann sind die Sinneshaare bis auf wenige lange und kräftige reduziert und stehen meist subapical auflangen Zapfen- Auswüchsen der einzelnen Palpenglieder. Beide Palpen zusammen bilden eine wirksame Einrichtung, Beutetiere wie mit einem Fangkorb zu umfassen und festzuhalten (Abb. 96).
Laufbeine: Die 4 Laufbein-Faare an den prosamalen Segmenten III-VI sind in Cx, Tr, Fe, Ti, Mt und Ta gegliedert, das letzte Tarsalglied mit Krallenorgan. Im einfachsten Falle bestehen Mt und Ta aus nur je einem Glied (die meisten Sironidae, Anartkrotarsus unter den Trogulidae, einige Oncopodidae). Sonst sind Ta und oft auch Mt vielfach sekundär unterteilt, b'-i spezialisierten Gruppen (z. B. Phalangüdae-Leiobuninae) oft in über 100 Einzelglieder (vgl. Abb. 15). Dann sind die Gelenke oft nicht voll gegeneinander beweglich, besonders im Mt-Bereich, sondern stellen nnr Zonen verdünnter Cuticula dar. Das Mt-Ta-Gelenk ist kräftig sklerotisiert, weit auslenkbar und in der Vielzahl der sekundären Gelenke an Mt und Ta leicht auffindbar (Abb. I, 15).
Die vielfach gegliederten Tarsen stellen ein innerhalb der Arthropoden einmaliges Greiforgan dar: Sie können lassoartig um Substratteile - Grashalme, Blätter, dünne Aste - geschlungen werden und ermöglichen schnell-fördernde Fortbewegung im räumlichen Maschenwerk von Büschen und Bäumen oder im oberen Bereich von Grashorsten, Stauden und dichtem Buschwerk. Auf ebenem Boden werden die Ventralseiten deJ; distalen Tarsenglieder "fußartig" aufgesetzt und mit der Kralle zusätzlich im Substrat verankert (Abb. 15, rechte Seitenfigur; vgl. KAESTN"ER 1931 a).
Form und Zahl der Tarsalkrallen der einzelnen Laufbeine sind unterschiedlich und für größere systematische Gruppen charakteristisch. Die Krallen der Beine I und II sind im allgemeinen einfach, gegenüber diesen die der Beine III und IV oft dimorph und komplizierter strukturiert.
Einfache, stark gekrümmte Kralle an allen Laufbeinen: Palpatores, Beine I und II der Phalangodidae, Erebomastridae. Doppelkralle: 2 gleich gestaltete

28
0::: 0 ~. w 1-z <1:
1. Allgemeiner Teil
E E :l..E
11
1.2. Morphologie 29
Krallenäste, die meist(? immer) einer gemeins1tmen Basis aufsitzen: BeiiY III und· IV der Phalangodidae, Erebomastridae (Abb. 100) und vieler andArer Laniatores. Sohildkralle: do-ve abgeflachte Kralle, lateral unsymmetrisch gekilrbt (Travuniidae, Abb. 76-77, und andere Travunioidea).
Jungtiere vieler Laniatores tragen im Krallenbereich ein Arolium; es fehlt den Oncopodidae und Triaenonyohidae. Krallenbau und ontogenetische Entwicklung der Krallenform sind nicht geklärt und bedürfen der Überprüfung.
Drüsenbildungen an den Laufbeinen als sekundäre d'-Geschlechtsorgane wurden erst von wenigen Gruppen beschrieben, sind aber wahrscheinlich weiter verbreitet als bisher bekannt. Bei allen Arten der Sironidae mündet auf dem d'-Ta IV eine (mei>;t spornförmige) Adenostyle, die durch einen Porns Sekret nach außen leitet (Abb. 53, 65). Vergleichbare Tarsaldrüse rnit Adenostyle ist von dem Phalangodiden Ausobskya athos MAR'l'ENS bekannt - dort auf Tarsus Il, und Holoscotolemon ·unicolor RoEWER besitzt ein Drüsenorgan im verdickten Mt III. Diese Drüsen haben wahrscheinlich Funktion im Fortpflanzungsverhalten.
Die meisten Arten der Phalangiidae besitzen die Fähigkeit, arretierte Beine, die nicht mehr bewegt werden körmen, zu autotomieren. Ausgenommen sind einige Arten mit vergleichsweise kurzen Beinen, z. B. Egaenus convexus 0. L. KocH {J. G. in litt.). Der Bruch findet an einer präformierten Stelle zwischen Trochanter und Femur statt und verheilt schnell (WASGESTIAN-SCHALLER 1968). Regeneration verlorener Beine findet nicht statt.
1.2.8. Genitalorgane
Gegenüber (fast) allen anderen Araolmiden-Gruppen zeichnen sich die Opilione> durch echte Begattungsorgane aus, einen Penis, dem die Legeröhre der <;> als homologes Organ entspricht. Die Öffnung der Geschlechtswege ist auf der Körperunterseite weit rostrad versehohen und liegt - bei den einzelnen Taxa unterschiedlich - auf der Höhe der Cx I (z. B. Phalangüdae), Cx II (z. B. Nemastomatidae, Trogulidae) oder Ox IV (viele Familien der Gonyleptoidea). Die Geschlechtsöffnung wird durch das Operculum genitale verschlossen - einer Falte des 2. opisthosomalen Segmentes. Es fehlt den Cyphophthalmi (Abb. 12).
Die Gonade ist U-förmig gebogen (Ahb. 16: G); an den Enden schließt je ein Ovidukt bzw. Vas deferens an; beide vereinigen sich zum Uterus internus. Er i~>t. meist außerordentlich lang und kann vom vorgestülpten Penis bzw. von der Legeröhre distad nachgezogen werden. Der Uterus interuns mündet in einen kompliziert strukturierte:1 Uterus externus. Er ist ektodermaler Herkunft und stellt eine Einstülpung des Integuments dar, die sichvom Rand der äußeren Geschlechtsöffnung tief in das Körperinnere erstreckt. An ~einer Basis, im Körperinnern. ändert der Uterus externus seine Lage um 180° (Abh. 16: rechts bzw. links neben "pG'') und zieht - handschuhfingerförmig eingestülpt - als meist kräftig skierotisierter Penis hzw. als Legeröhre distad. Im distalen Bereich wendet er sich nochmals nach innen um, zieht - als Samenkanal - weit proximad und schließt dort an den nicht cuticularisierten Uterus interuns an (Abb. 16: pG). Somit bestehen die ektoderm~tlen Genitalorgane aus 3 Schichten, von denen die mittlere die Außenwand von Penis bzw. I"egeröhre, die innere den Samenkanal bzw. die Wand des Lumens der Legeröhre und die äußere die Scheide von Penis bzw. Ovipositor,
.d. h. die Außenwand des Uterus externus, darstellen.

30 1. Allgemeiner Teil
lß äKö
Sk/Ue
Abb. 16. Schema des d'-Genitalapparat~s. - äKo äußere Körperwand, G Gonade, Gd Gonu dukt, Gl Glans, Op (Jen Operculum,genitale, P Propulsionsorgan, Pe Penis, pG Übergang zwi· sehen Ui und Ue, 8 Scheide, Sn, Sa.menreservoir, sG sekundäre Gesohlechtsöffnung, Skj ue Sa.meuknnal/Uterns extornus, tG tertiäre Gesohlechtsöffnung, Ui Uterus internus, T1· Truncu:-.. Nach .TuBER~'HIE 1964, verändert.
Durch den l<;omplizierten Bau der ektodermalen Geschlechtsorgane lassen sieh 3 Geschlechtsöffnungen (GÖ) unterscheiden (Abb. 16):
a) primäre GÖ: zwischen I. und 2. abd Sternit; nur bei den Cyphophthalmi noch erhalten, denen das Operculum genitale fehlt (Abb. 12); bei den übrigen Gruppen durch Falte des 2. opisthoRomalen Sternits überdeckt..
b) sekundäre GÖ: distale Öffnung von Penis (=Öffnung des SamenkamtlH, Abb. 16: sO) bzw. OvipoRitor.
c) tertiäre GÖ: Schlitz, gebildet von distaler Falte deB 2. opisthosomalen (abd) Sternits ( = Genitaldeckel) und vom proHomalen Sternum bzw. der Basis der Unterlippe (Abb. 16: tG).
Die AusHtülpung der Genitalorgane erfolgt durch Körperbinnendruck, hervorgerufen durch Rumpfmuskelkontraktion; die Ovipositorscheide wird zusätzlich durch die eigene Muskulatur bewegt. Da der Schlitz der tertiären GÖ distad geöffnet ist, werden Penis bzw. Legeröhre entsprechend distad und in der Längsachse des Körpers vorgestreckt. Die handschuhfingerförmig nach außen umgc" krempelte Scheide von Penis bzw. Legeröhre verleiht beiden Organen vergleichsweise große Reichweite und - meist unterstützt durch verschiedene Muskelsysteme - hohe Beweglichkeit.
Die Einstülpung geschieht durch 2 Muskelpaare: nämlich Längsmuskeln der Scheide, ferner durch Retraktoren von der Basis des Organs zur Hinterwand deH Körpers, die neben dem After angeheftet sind, unterstützt durch Nachlassen des Körper binnendruckes.
1.2. l\Iorphologie 31
Penis Der Penis (Abb. 16: P) ist ein meist langes und schmales Chitinrohr, das sich
in den längeren Truncus und die distale, meist abgesetzte Glans gliedert. Die Oberfläche tleJ Truncus ist unstrukturiert glatt und läßt auch bei mehrtausendfacher Vergrößerung (REM) keine oder so gut wie keine Skulpturiarung erkenneu. Die Glans kann vielfach in Membranen und Sklerite aufgegliedert und mit Sinnesborsten besetzt sein. Durch Truncus und Glans zieht der Samenkanal (Abb. 16: Sk), der auf einem Stylus mündet. Dieser schließt entweder distal an die Glans an oder wird erst während der Kopulation durch Expansion der kompliziert strukturierten Glans freigelegt (Abb. 32-35).
Der Terminus Glans (z. B. Abb. 16: Gl) kann nicht eindeutig definiert werden. Die Verhältnisse sind überall dont übersichtlich, wo Truncus und "Glans" durch ein Gelenk gegeneinander beweglich sind (z. B. Abb. 407: G). Weniger klar sind sie dort, wo in den Truncus distal expandierbare Sklerite und Membranen eingelagert sind, eine Gelenkzone zwischen Truncus und einer distalen Glans aber fehlt (z. B. Abb. 22-27). Begrifflich noch schwerer faßbar sind die Verhältnisse in solchen Fällen, in denen Gelenkzone und expandierbare Sklerite fehlen, z. B. ·bei Ptychosoma. Gar nicht anwendbar scheint die Dichotomie "Truncus-Glans" bei den Sironidae zu· sein. deren Penis kurz und stark dorso-ventral abgeplattet ist. Als Glans könnten do"rt die ventralen Greifzangen bezeichnet werden (z. B. Abb. 58-61). Es ist letztlich eine Frage der Übereinkunft, auch solche Penes als in Truncus und Glans gegliedert zu bezeichnen, die eine "typische", durch Gelenk abgesetzte Glans vermissen lassen. Da die distalen Penisteile in den allermeisten Fällen in irgendeiner Weise gegenüber dem einförmigen Truncns differenziert und mit Sinnesborsten bewehrt sind, wird vorgeschlagen, durchweg diese Terminologie anzuwenden.
Die Penes der Arten:, die zu verschiedenen höheren Taxa gehören, sind unterschiedlich strukturiert.
Weitverbreitet sind Muskeln im Truncus-Rohr, die Truncus und Glans nach dem Prinzip des Muskel-Sehnen-Zuges in funktionelle Beziehung setzen. Es lassen sieb mehrere Muskularisierungstypen unterscheiden (MARTENS 1976):
I. Drei Muskelstränge: zwei para-mediane, die zu den lateralen Loben ziehen, ferner ein medianer, der wahrscheinlich zu dem ventralen Klammerapparat führt. Dieser vergleichsweise komplizierte Typ kommt allein bei den Sironidae vor. Bei allen bisher untersuchten Arten ist das Organ kurz gestaucht, dorso-ventral abgeplattet und ventral mit komplizierter Greifzangen-Armatur versehen (HANSEN & SeERENSEN 1904, RAFALSKI 1958; Abb. 58-61, 69-70).
2. Zwei Muskelstränge: Sie liegen jeweils lateral im basalen Teil des Truncus; ihre Sehnen ziehen zur ventro-lateralen Seite der Glans. Die beiden Muskelkörper sind entweder über den größeren Teil des Truncus verteilt; dann sind die Sehnen. kurz (unspezialisierter Typ), oder sie konzentrieren sich auf eine basale TruncusErweiterung; dann sind die Sehnen lang (spezialisierter Typ). - Dieser PenisTyp ist auf die Familien Nemastomatidae, Trogulidae, Dieranolasmatidae und. Nipponopsalididae der Unterordnung Palpatores beschränkt (vgl. Abb. im Spez .. Teil).
Zwei Funktionsweisen sind denkbar: a) durch alternierenden Muskel-Sehnen-Zug kann die Glans laterad oder auch
ventrad bewegt werden. Membranöse Zonen im Übergangsbereich Truncus-Glans
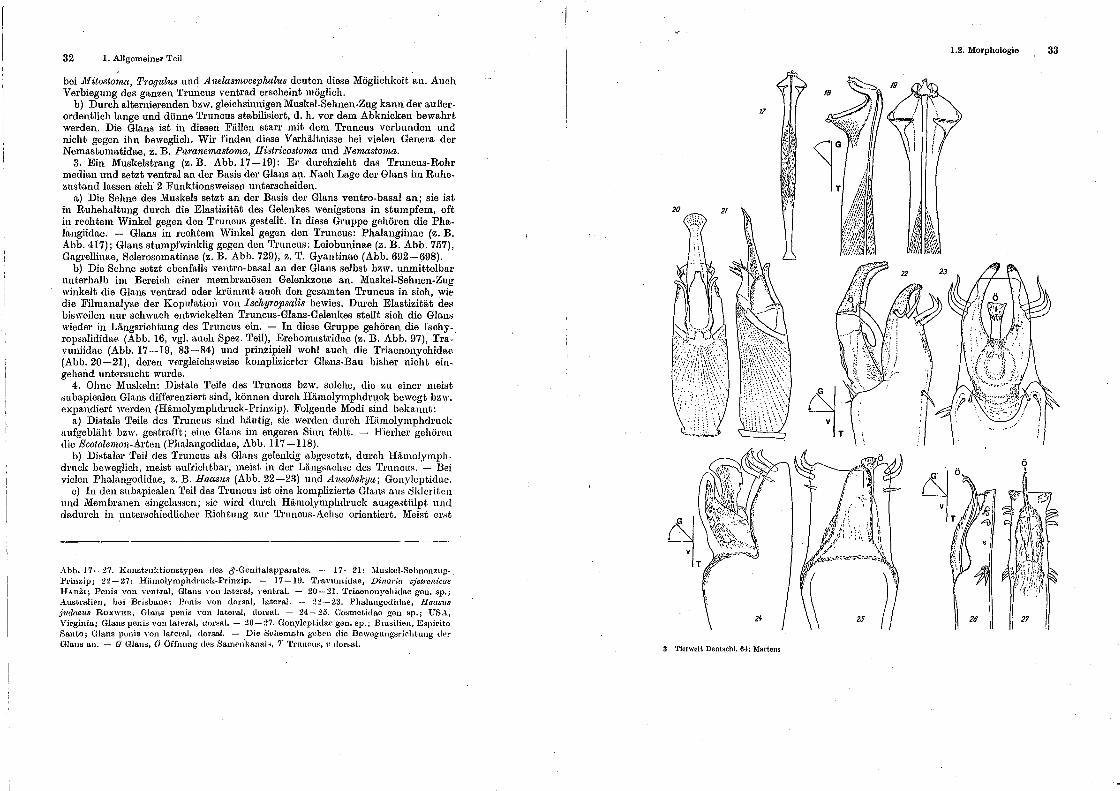
32 I. Allgemeiner Teil
bei llfitostornct, Tmgulus und Anelasrnocephalus deuten diese Möglichkeit an. Auch Verbiegung des ganzen Truncus ventrad erscheint möglich.
b) Durch alternierenden bzw. gleichsinnigen Muskel-Sehnen-Zug kann der außerordentlich lange und dünne Truncus stabilisiert, d. h. vor dem Abknicken bewahrt werden. Die Glans ist in diesen Fällen starr mit dem Truncus verbunden und nicht gegen ihn beweglich. Wir finden diese Verhältnisse bei vielen Genera der Nemastomatidae, z. B. Paranemastoma, Hist1·icostoma und Nemast01na.
3. Ein Muskelstrang (z. B. Abb. 17-19): Er durchzieht das Truncus-Rohr median und setzt ventral an der Basis der Glans a11. Nach Lage der Glans im Ruhezustand lassen sich· 2 Funktionsweisen unterscheiden.
a) Die Sehne des Muskels setzt an der Basis der Glans ventro-basal an; sie ist in Ruhehaltung durch die Elastizität des Gelenkes wenigstens in stumpfem, oft in rechtem Winkel gegen den Truncu~ gestellt. In diese Gruppe gehören die Phalangiidae. - Glans in rechtem Winkel gegen den Truncus: Phalangiinae (z. B. Abb. 417); Glans stumpfwinklig gegen den Truncus: Leiobuninae (z. B. Abb. 757), G!tgrcllinae, Sclerosomatinae (z. B. Abb. 729), z. T. Gyantinae (Abb. 692-(198).
b) Die Sehne setzt ebenfalls ventro-basal an der Glans selbst bzw. unmittelbar unterhalb im Bereich einer membranäsen Gelenkzone an. Muskel-Sehnen-Zug winkelt die Glans ventrad oder krümmt auch den gesamten Truncus in sich, wie die Filmanalyse der Kopulatim1 von Ischyropsalis bewies. Durch Elastizität des bisweilen nur schwach entwickelten Truncus-Glans-Gelenkes stellt sieh die Glans wieder in Längsrichtung des Truncus ein. - In diese Gruppe gehören die Isehyropsalididae (Abb. 16, vgl. auch Spez. Teil), Erebomastridae (z. B. Abb. 97), Travuniidae (Abb. 17-19, 83-84) und prinzipiell wohl auch die Triaenonychidae (Abb. 20-21), deren vergleichsweise komplizierter Glans-Bau bisher nicht ein-. gehend untersucht wurde.
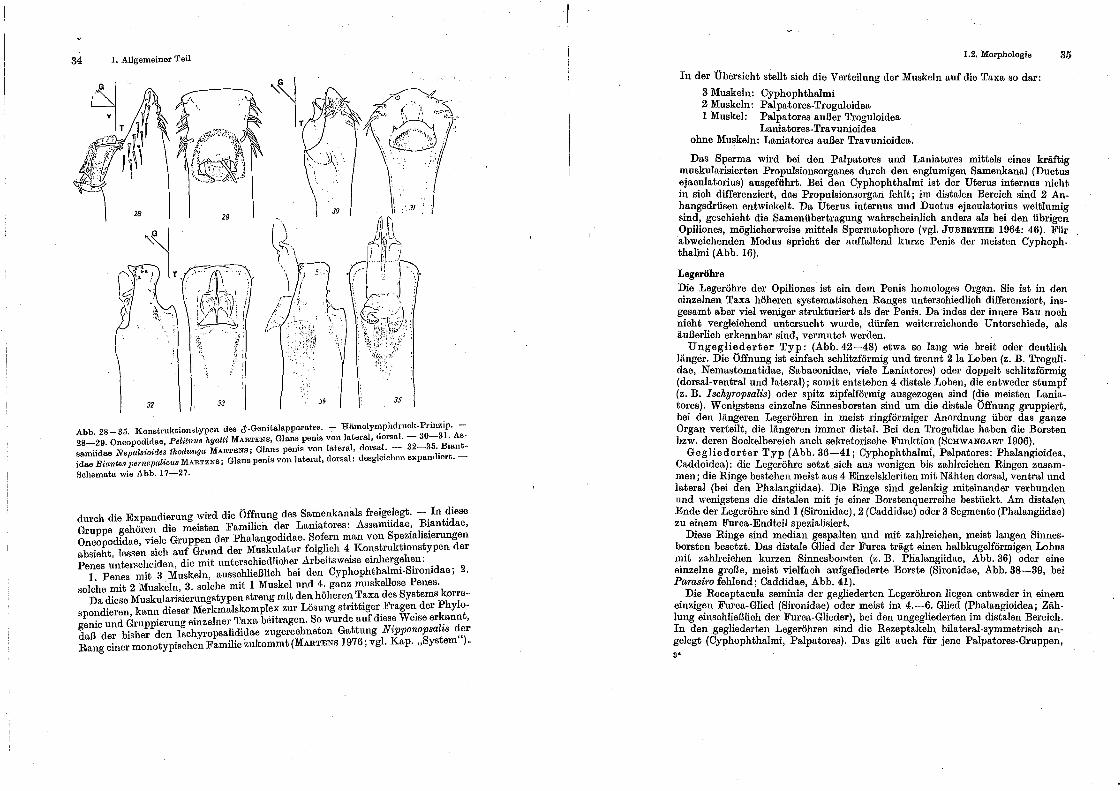
4. Ohne Muskeln: Distale Teile des 'I'runcus bzw. solche, die zu einer meist subapicalen Glans differenziert sind, können durch Hämolymphdruck bewegt bzw. expandiert werden (Hämolymphdruck-Prinzip). Folgende Modi sind bekannt:
a) Distale Teile des Truncus sind häutig, sie werden durch Hiimolymphdruck >tufgebläht bzw. gestrafft; eine Glans im engeren Sinn fehlt. - Hierher gehören die Scotolemon.-Arten (Pha.Iangodidae, Abb.ll7-ll8).
b) Distaler 'feil des Truncu~ als GhtnB gelenkig ahgeHet.zt, durch Hämolymrlhdmck beweglich, meist anfrichtbar, meist in der Läng~ach~c de~ TrunouK. - Bei vielen Phahtngodidae, z. B. Haasus (Abb. 22-23) und A~t8obskya; Gonyleptidae.
c) In den subapicalen Teil des Truncus ist eine komplizierte Ghtns au8 t:lklcritcn und Membra,nen eingelas8en; sie wird durch Hämolymphdruck autige.~tülpt und dadurch in unterschiedlicher Richtung zur Trnncus-AchBe orientiert. 1\IIeist er.,t
Abb. 17- ~7. Konstruktionstypen des d'-Genitalapparatos. - 17-21: :lluskel-Schnonzug-. Prinzip; 2~-27: Hämolyn1phclrnck-Prinzip. - 17-IU. 'rra\'uniidae, Dinaria ·v;'et1'enicae HAnZr; Penis von v<mt.ral, Glans von lateral, ventral. - 20-21. 'l'riaenonychidae gen. sp.; ..:\.ust.ralir.n, bei Brisbane~ PP-ni1:1 vou dorsal, lateral. - ~:!--23. Phalangodidae, Haasus }udaeus RoEwEn, Glans penis von lateral, dorsal. - 24-25. Cosn1etidao gen sp.; USA, Virginia; Glans penh~ von latPral, dorsal.- 20-27. Gonylept.ida<-"~ gen. sp.; Brasilien, E8pirito Santo; (Hans pt1nis von lateral, dorsal. - Die Schemata gebt-n die Bewegungsriehtung tl<."t'
GlanH au. - G Glans, Ö Öffnung des Sa:mt>nkanal~, '1' 'L'rutwu~, ·l• dor::-;a.l.
1.2. Morphologie 33
18
I?
20 21
~;;;,~lf,, ·::.·.\; •. :.· .. · .. .- .. :.·.
''.:';: -::: ,.·
~.J\~\· ..
25
3 Tierwelt Dcntschi. 64: :Martens
34 l. Allgemeiner Teil
32 33 35
Abb. 28-35. Konstruktionstypen des d'·Genitalapparate~. - Hämolymphdruck-_Pdnzip. 28-29. Oncopodidae, PeUtnus hyatti MARTENS, Glans. pems von lateral, dorsal.- 30-31: As· samiidae Nepalsioirles thodunga MARTENB; Glans pents von lateral, dorsal: - 32-35 .. Bmntidae B·ianteat;ernep(tlic·us MARTENS; Glanspenis von lateral, dorsal; desglmchen expant.hert.-
Schemata wie Abb. 17-27.
durch die Expandierung wird die Öffnung des Sa~enkanals freig~.legt. -.In ?iese Gruppe gehören die meisten Familien der Lamatores: Assamndae~ ~~ant1dae, Oncopodidae viele Gruppen der Phalangodidae. Sofern man von Spez1ahs1ernngen absieht, lass~n sich auf Grund der Musk~latur folgl~ch 4. Ko~struktionstypen der Penes unterRcheiden, die mit unt.erschiedhcher Arbeitsweise emherg~h~n: .
1. Penes mit 3 Muskeln, ausschließlich bei den Cyphophthalmi-SITonidae; 2. Holehe mit 2 Muskeln, 3. solche mit 1 Muskel und 4. ganz muskellose Penes. · Da diese Muskularisierungstypen streng mit den höheren Taxa des Systems korrespondieren, kann dieser Merkmalskoniple.x zur Lösung strittiger.F:age~ der Phylogenie und Gruppierung einzelner Taxa hertragen. So wurde auf dies~ Weise er~annt, daß der bisher den Ischyropsalididae zugerechneten Gattung Ntpponopsalts ~?r Rang einer monotypischen Familie zukommt (MARTENS 1976; vgl. Kap. "System )-
I 1.2. Morphologie
In der Übersicht stellt sich die Verteilung der Muskeln auf die Taxa so dar:
3 Muskeln: Cyphophthalmi 2 Muskeln: Palpatores.Troguloidea 1 Muskel: Palpatores außer Troguloidea
Laniatores-Travunioidea ohne Muskeln: Laniatores außer Travunioidea.
35
Das Sperma wird bei den Palpatores und Laniatores mittels eines kräftig muskularisierten Propulsionsorganes durch den englumigen Samenkanal (Ductus ejaculatorius) ausgeführt. Bei den Cyphophthalmi ist der Uterus internus nicht in sich differenziert, das Propulsion.sorgari fehlt; im distalen Bereich sind 2 Anhangsdrüsen entwickelt. Da Uterus interuns und Ductus ejaculatorius weitlumig sind, geschieht die Samenübertragung wahrscheinlich anders als bei den übrigen Opiliones, möglicherweise mittels Spermatophore (vgl. JUBERTHIE 1964: 46). Für abweichenden Modus spricht der auffallend kurze Penis der meisten Cyphophthalini (Abb. 16).
Legeröhre Die Legeröhre der Opiliones ist ein dem Penis homologes Organ. Sie ist in den einzelnen Taxa höheren systematischen Ranges unterschiedlich differenziert, insgesamt aber viel weniger strukturiert als der Penis. Da indes der innere Bau noch nicht vergleichend untersucht wurde, dürfen weiterreichende Unterschiede, als äußerlich erkennbar sind, vermutet werden.
Ungegliederter Typ: (Abb. 42-48) etwa so lang wie breit oder deutlieh länger. Die Öffnung ist einfach schlitzförmig und trennt 2 Ia Loben (z. B. Trogulidae, Nemastomatidae, Sabaconidae, viele Laniatores) oder doppelt schlitzförmig (dorsal-ventral und lateral); somit entstehen 4 distale Loben, die entweder stumpf (z. B. Ischyropsalis) oder spitz zipfeiförmig ausgezogen sind (die meisten Laniatores). Wenigstens einzelne Sinnesborsten sind um die distale Öffnung gruppiert, bei den längeren Legeröhren in meist ringförmiger Anordnung über das ganze Organ verteilt, die längeren immer distal. Bei den Trogulidac haben die Borsten bzw. deren Sockelbereich auch sekretorische Funktion (SOHWANGART 1906).
Gegliederter Typ (Abb. 36-41; Cyphopht.halmi, Palpatores: Phalangioidea, Caddoidea): die Legeröhre setzt sich aus wenigen bis zahlreichen Ringen zusammen; die Ringe bestehen meist aus 4 Einzelskleriten mit Nähten dorsal, ventral und lateral (bei den Phalangiidae). Die Ringe sind gelenkig miteinander verbunden und wenigstens die distalen mit je einer Borstenquerreihe bestückt. Am distalen Ende der Legeröhre sind 1 (Sironidae), 2 (Caddidae) oder 3 Segmente (Phalangiidae) zu einem Furca-Endteil spezialisiert.
Diese Ringe sind median gespalten und mit zahlreichen, meist langen Sinnesborsten besetzt. Das distale Glied der Furca trägt einen halbkugelförmigen Lobus mit zahlreichen kurzen Sinnesborsten (z. B. Phalangiidae, Abb. 36) oder eine einzelne große, meist vielfach aufgefiederte Borste (Sironidae, Abb. 38-39, bei Parasiro fehlend; Caddidae, Abb. 41).
Die Receptacula seminis der gegliederten Legeröhren liegen entweder in einem einzigen Furca-Glied (Sironidae) oder meist im 4.-6. Glied (Phalangioidea; Zählung einschließlich der Furca-Glieder), bei den ungegliederten im distalen Bereich. In den gegliederten Legeröhren sind die Rezeptakeln bilateral-symmetrisch angelegt (Cyphophthalmi, Palpatores). Das gilt auch für jene Palpatores-Gruppen, s•

'
I. Allgemeiner Teil
36
l . ---, ~~- -~---~rH f,-, __ -1 /-~
~ --~_: __ ._l_. ~-------- --\ r --· -· ----l r· ----- -·----...,
t-· _____ ( _ _j
J8
J9
Abb. 36-41. Gegliederte Ovipositores. - 36-37. Pha.langiidae. 36. Leiobunum •·otundum (LATREILLE), 36. a) linkes distales Furca-Gliecl von L. •·otundum; 37. Ast•·obunus helle;oi (AussERER). - 38-39. Sironidae. 38. Sh·o duricm·ius (JoSEPH), 38a. dist-ale Furca-Gliede•· von S. du•·icorius; 39. Siro crn-paticus RAFALSKI, diat.ale Glieder. Nach RAFALSKI 1956. 40-41. Caddiclae. 40. Caddo agilis BANKS: 41. Tasma.rwpiUo fuscu.s HIOKMAN, 40 und 41 nach GRUBER 1974b.
(
[FJ ( / i
\ /. .. I 'I ·.•
' \ L\':
I I :
LJ
/J
/
i I
. ·, ,, \'-~\' ~'-.
\I\
' ' i
'I ., i ~
(._ _____________ __ j
...
1.2. Morphologie
;o. ~· ! . ' .J] . /
''>1'/-v,/o {~,
_,(t : ,., :
.. , •• , L ,l't.. '.• ·...-r- --
1::-:_ .. --. '
\ I
\·.
t :/ I
~ j I
I
37
Ahb. 42-48. Ungegliederte Ovipositores. - 42. Erebom<ridae, Holoscotolemon unicolor RoEWER; 48. Sabaconidae, 8abacon simoni DREsco; 44. Ischyropsalididae, Ischyropsalis he./lwigi (PANZER); 45. 'l'ra.vuniirla.e, Pelton1Jchia clavigera. (SnroN); 46. 'l'rogulidae, T•·ogulus nr-paeformis (ScoPOLI);- 47. Dicra.nolaßma.ticlae, D·ic1·anolasma scab~·um (HERBST); 48. Nema.~tomatidae, Histricostoma dentipalpe (:4. USSERER).
die ungegliederte Legeröhren besitzen (Troguloidea., Isohyropsalidoidea). In den ungegliederten Legeröhren der Laniatores sind sie dagegen radiär-symmetrisch 4strahlig mit 8 Rezeptakeln oder Vielfachem davon angelegt. Die Form der Rezeptakeln ist bei den höher evoluierten Gruppen (soweit bisher untersucht) artspezifisoh, z. B. bei den Phalangioidea. Meist Rind die Unterschiede jedoch gering und als Identifizierungshilfe wenig geeignet.

38 I. Allgemeiner Teil
1.2.9. Sekundäre Geschlechtsmerkmale
Sie betreffen fast immer die 3 und treten als Proportionsänderung der Gliedmaßen in Erscheinung (meist einzelner Glieder der Cheliceren, Palpen und Laufbeine), als exzessive Änderung der Bewehrung der Gliedmaßen (vor allem Palpen und Laufbeine) und in einigen Gruppen sogar als gänzlich neue Organe, die den ~ fehlen. Als sekundäre Geschlechtsmerkmale der Cf können Reduktionen des dorsalen Scutum angesehen werden (Phalangiidae, einzelne Ischyropsalis-Arten); das verschafft ihnen "Erleichterung" während der Gravidität. Die folgenden Beispiele beschränken sich im wesentlichen auf die europäische Fauna; sie betreffen 3-Merkmale.
I. Größen- und Proportionsänderung von Extremitäten: Exzessiv au~gebildete Cheliceren: Phalangimn opilio (Abb. 408-409), Zachezts C?'i8ta (Abb. 561). - Sonderbildungen am Pedipalpus: Apophysen und Dorne an l?emur und Patella: viele Nemastomatidae (Abb. 189) und Sabaconidae. Verdickung bzw. Aufblähung einiger Glieder der La.ufbeine, meist I uild II: z. B. Metaphalanghtm. prop1:nq1tum, Opilio .saxatilis, Zadwu8 c1·ista. - Diese Merkmale sind selbst innerhalb von Pop,ulationen recht. variabel und können sogar nahezu auf die schlichten Cf-Ausprägungen reduziert sein. Die 3-Laufbeine sind bei den allE>rmeisten der hier belumdelten Gruppen absolut und relativ länger als bei den ~ (vgl. :Maße im Spez. Teil).
2. Verstärkte Bewehrung der Extremit.äten: Körnchen.reil1e bzw. Körnchenfeld auf dem Palpen-TarstiB der meiste11 Phalangiidae; Dorn-Konzentration auf den bas,tlen Gliedern der Ltmfbeit1e, mei~t .J!'emora, z. B. bei JJ1etaphalang-i1un propinqmtm. und Opilio sa.mtilis.
3. Neue, nur den 3 eigene Organe: Epidermale Drüsen zur Pheromon-Produktion. - An den Cheliceren: bei den meisten Nemastomatidae (l'lfARTENS 196i, SILHAVY 1967a), bei vielen Ischyropsalididae (Ischy·rop~a/is. Cera.tolas-ma), Dicranolasmatidae (Dicranolasma) und Sabaconidae (Abb. im Spez. Teil.). - Am Pedip'nlpu~: Euda8ylobu-s nicaeensi..; (.Femur, Patella); Le·iobumnn 1'UJJe8tre (Tibia); Dicranolasma scabntm und D. opiliolloirles, De1idmlasrnct mimb·ile (Patella), einige Ortholasrna-Arten, Ilespemnemastomrt (Femur bis Tibia; .T. G. in litt.); Hinwlphaldngiurn {MARTENS 19i3b); Nelima sem.proni (Tarsus). - An den Laufbeinen: an Ta IV bei allen Sironidae, an Ta II bei Au.sob.skya atho8 (iVIAR'rENs l972b). an Ta IH bei Holoscofolemon ·uuicolor.
1.2.10. Stridulationsorgane
Kommunikation mitteb Stridulation tritt Yergleieltslreise selten auf, hat sich aber in mehreren Gruppen unabhängig voneinander entwickelt; im hier beh>tndelten Gebiet sind Stridulationsorgane auf Histricostoma dentipalpe und H. argenteolww./cttum beschränkt. Erzeugt und perzipiert wird 1mhrscheinlich in allen Fällen Substratschall. Untersuchungen~ ur Funktion gibt es bi~her nicht.
1.3. Biologie 39
Folgende St.ridulationsorgane sind bisher bekannt geworden(GRUBER l969a): I. Organ symmetrisch; 2 gleichgestaltete Oberflächen arbeiten gegeneinander,
hier die Medialseite der Cheliceren: a) am Cheliceren-Grundglied der Nemastomatiden Histricostoma dentipalpe
und H. argenteolunulatum (JUBERTIDE 1957b; Abb. 185, 191-192), Mediostoma Jmmerale, bei dem Travuniiden Peltonychia sat·ea (JUBERTHIE 1968, Abasola s.).
b) am 2. Cheliceren-Glied: S-afrikanische Arten der Roeweriana-Gruppe der Triaenonychidae (LAWRENCE 1938), bei dem mittelamerikanischen Phalangodiden Panzosus hispidulus (RoEWER l949a: 15).
2. Organ asymmetrisch; 2 verschieden gestaltete Organteile arbeiten gegeneinander.
c) Femur des Pedipalpus medio-dorsal gegen Zähnchen des Tuber oculorum: Giljarovia st·ridula, G. rossica, G. redikorzevi und Mediostoma ceratocephalum (KRATocnvfL 1958b, ÜHARITONOY 1946, GRUBER 1976).
d) Medial:;eite des Pedipalpen-Femur gegen das Cheliceren-Grundglied laterodorsal: bei dem Ischyropsalididen Ceratoln.sma tt·icantha.
Die Aufstellung läßt erkennen, daß Stridulationsorgane ganz ähnlicher Lage, Beschaffenheit und ArbeitHweise (z. B. am Cheliceren-Grundglied) in nicht näher miteinander' verwandten Familien vorkommen, andererseits in ein und derselben Gattung ganz verschiedene Organe vorkommen können ( 1]!J ediostoma). Das mindert entscheidend den Wert dieses Merkmalskomplexes für taxonomische Erwägungen, wie ihn KRATOCHVfL noch 1959 bei der Aufst·ellung der Nemastomatiden-Unterfltmilie Giljaroviinae benutzte.
1.3. Biologie
1.3.1. Biotope
Trotz vergleichsweise geringer Artenzahl haben die Weberknechte eine Fülle verschiedener Habitustypen hen·orgebracht (Abb. 1, 57, 96, 213, 277-279, 332 bis 334). Folglich sind Lebensweise und bevorzugte Biot.ope ganz unterschiedlich, und unter den terrestrischeil Biotopen gibt es nur wenige, die sie nicht besiedeln.
Die größte Artenzahllebt in der Bodenauflage und dort vorzugs\l'eise in Fallaubund Humuslagen, ebenso unter morschem Holz und tief in den Boden eingebetteten Steinen (Förna, A-Horizont des Bodens).
Dieser großen Gruppe i~t hoher Feuchtigkeitsanspruch gemeinsam, das PräferPndum lieot zwischen 80 und 100% RL. Bei einigen Arten ist die Bindung an hohe Luft.f~uchtigkeit (und dan{it indirekt an ausgeglichenen Temperaturgang) so groß, daß sie fast nur in umnitt.elbarer Nähe fließenden Wassers vorkommen, z. B. die C/ya.s-lu:ten, Ischyropsalis cadi und Para-nemastoma bicuspidatu.rn. Während Trockenperioden dringen kleine Arten in tiefere Bodenschichten ein, bisweilen bis 1m unter die Oberfläche, nachgewiesen für Si1'0 rubens und Scotolemon doriae; Siro legt in diesen tieferen Bodenschichten auch Häütungskammern an.
Zu diesen rein terrikol lebenden Gruppen gehören in der europäischen J!,auna die Sironidae. Nemastomatidae, Trogulidae, Ischyrops>tlididae, Dicr>tnolasmatid>te,

40 I. Allgemeiner Teil
Sclerosomatinae und alle Laniatores. Dieser Gruppe nahe stehen einige. Gattungen der Phalangiidae, die überwiegend die Bodenschicht ~ewohnen und.n~r geleg?ntlich auch höhere Strata aufsuchen können (z. B. Ol~golopkus, Lacmms korrtdus und L. epkippiatus). . .
Zu den Terrikolen haben auch einige Arten zu zählen, dre nicht rm humosen und feuchten Oberboden leben, sondern die Initialstadien von Böden besiedeln, nämlich Geröllschutt im Gebirge, meist in den Alpen und dort vielfach ü~er der Baumgrenze. Hierher gehören z. B. Mitostoma alpinum, M. ckr~somelas~ JJ_zcrar:opalpus gasteinensis und Mitopus glacialis. Besonders extreme B~o.tope hrnswht.hch täglichen und jahreszeitlichen Temperaturganges, Sonnen~xposrtron .un_d re~atrv~r Luftfeuchtigkeit nehmen die Megabunus-Arten, .auch M~top':'s glaczalM, ~m. dre in den Alpen Felswände über der Baumgrenze besredeln. Nur eme Art, M. dwdema, lebt im Flachland ersatzweise auch auf Baumstämmen.
In der einheimischen Fauna ist die Zahl der Arten viel geringer, die Bich in höheren Strata aufhalten, z. B. in der Krautschicht oder noch höheren Strata der vertikalen Zonierung. Meist werden - wahrscheinlich wiederum aus Gründen des Wasserhaushaltes - bodennahe Strata bevorzugt, und nur nachts finden bei re-lativ hoher Luftfeuchtigkeit Vertikalwanderungen statt, die bis in die Kronen yon hohen Bäumen führen können. Paroligolophus agrestis wurde bis 9 m hoch gefunden. Diese Arten gehören in Europa ausschließlich den Phalangiidae. an, in _den Tropen aber auch manchen Laniatores-Gruppen, z. B. den Gonyleptrdae in S-Amerika.
Die allermeisten Arten leben vor Insolation und somit zugleich vor hohen Temperaturen geschützt. Folglich ist die .A..rten~ahl in ariden Gebieten .reduziert. ,und die wenigen an diese Bedingungen adaptrerten ~rten haben Re~fe- ~nd Eortpflanzungszeit ttuf die feuchtesten Monate verlegt (m Europa z. B. rm Mrttelmeer-
gebiet). . . .. . . , Bei mittäglicher Sonnenbestrahlung vermag m ~er _nutteleuropmschen E ~tt~ua
nur Phalangimn opilio tnlf' Vegetation ttktiv zu sem; .m den Alpen leb~n e1n~ge Arten (auch) tagsüber an Felswänden aktiv und merden dabe~ auch uüt:ni<!Ve Sonnenbestra.hlung nicht, z. B. Leiolmmtm. !'Ose·um, 11fegabunus !'hznocer:o8, Mttopus qlacialis (dieser meist mwhtaktiv) und weniger markant auch D~m·anopalzms yasteinensis. Obwohl bei weitem nic~t alle fels?ewohnende~l Arten d~r ~pen t~gaktiv sind (z. B. die Gyas-Arten, Lewbun·um l~mbat·um, M~topus glac~alzs), schemt dieser Aktivitätsrhythmus bei den ungünstigen Klima-Verhältnissen der Alpen Vorteile zu bieten.
Wenige Arten dringen auf spärlich bewachsene, .s~eppena~~ige Bi~t~pe vor, _darunter auch solche mit hoher Insolation, z. B. Op~lw sa.xatzl~s, Laczn~lts kOI'?'t.rlus, Trogulus ti·icarinatlts, Odiellus .spinosus, Nelim.a silvatica ~nd Ane~asmocep~alus ca-mbridge·i (vg!. ökophysiologische Daten, so wert ~ekannt,. rm Spe~rell?n Terlj. .
Gegen Feuchtigkeitsschwankungen und InsolatiOn wemg emJ?fmdhche Arten ye1·mögen auch das Innere von Städten zu besie~e.ln; da_d~ch smd manche :on ihnen passiv weit verschleppt worden, z. B. Opdw panet·mu~ und Phalangt-·~"!1' opilio in die Nearktis, 0. pa~·ietinus zusätzlich n~ch Tasmamen und P_h. opzlw nachNeuseeland, ferner Nelima. doriae nach Austrahen und Neuseeland, Lewb·u-nltm lim.batmn von den Alpen und manchen Mittelgebirgen in viele Städte nördlich der Alpen bis Skandinavien (vgl. S. 50-51). ~ach jetziger Kenntnis legen die meisten Ar~en die Eier i~ den ~oden r'd"'r
in vorjährige Pflanzenteile ab, wie 1liitopns -morw - drc verglewhsw<1ISe trocken-
1.3. Biologie 41
adaptierten auch in Felsritzen ( Le-iohunum roseurn) oder gar in 11-fauerwerk ( L. limbatum.J. In besonders feuchten Biotopen werden die Eier einfach in einem Schleimpaket an die Unterseite von Steinen oder altem Holz angeklebt (lscky. ropsalis, Paranemastoma).
Da die Jugendstadien wenig skierotisiert und in hohem Maße gegen Feuchtigkeit~verlust empfindlich sind, leben sie fast ausschließlioh am und im Boden. Erst die Subadult-Stadien beginnen, höhere Strata einzunehmen.
Obligatorisch troglobionte Arten leben im weiteren Mitteleuropa nur in Gebieten, die glazial nicht devastiert waren, z. B. in den S-Alpen und im jugoslawischen Karst. Im hier behandelten Raum gehören dazu Arten der Gattungen Mitostoma, Isckyropsalis und Peltonych-ia (vgl. dazu Kap. 1.4. "Verbreitung und Zoogeographie").
1.3.2. Vertikalverbreitung
.-\.llgemeingültiges kann nur in gan;r, beNchränktem Umfange ausgesagt 11·erden, da sich die einzelnen Arten in den verschiedenen Teilen ihrer Areale nicht einheitlich verhalten. Viele Arten überwiegend mitteleuropäisch-atlantischer Zugehörigkeit, die im engeren Mitteleuropa sowohl das Flachland, die Mittelgebirge als <tnch noch die mittleren Lagen der Alpen besiedeln, leben in den S-europäischen, medit.erran beeinflußten Arealausläufern nur noch in den hohen und höchsten Gebirgslagen, oft weit über der Baumgrenze, z. B. Phalangi1tm opilio auf dem thessnli;;chen Olymp und Oligolophlts t1·ülens in den bulgarischen Hochgebirgen. Selhxt in den Alpen sind die oberen Verbreitungsgrenzen nicht einheitlich, sond<'l' t wechseln z. B. nach Pflat1zenbedeckung und Exposition. Einzelheiten vergleir•h(• man im Speziellen Teil.
In Mitteleuropa einschließlich der N-Abdachung der Alpen haben die meiste:1 Arten vergleic.hsweise große Vertikalverbreitung, die von (nahe) Meo1·eshöhe hi~ in die mittleren Lagen der Alpen reichen kann. Für die meisten dieser euryöken Arten bildet die Baumgrenze die obere Arcalgrenze.
Nur wenige Arten dringen über diesen Bereich in die alpine Matten- und G-eröllzone ein. Zu ihnen gehört Mitopus morio, der mit fa-st 3 000 m breitem Arenlgiirtel den ausgedehntest;en Höhenbereich aller europäischen Weberknechte einnimmt.
Die in den oberen Lagen des Areals sied-elnden Individuen von .Mitopus morio und Platybumts bucephal·us sind kurzbeiniger als Tiere aus Tieflandspopulationea, nicht dagegen solche von 111-itostomrt chrysomelas.
Ausschließlich im oberen Bereich der Waldgrenze, in der Matten- und Gletscherzone der Alpen, leben einige spezialisierte Felswand- bzw. Geröll-Bewohner (vgl. Kap. 1.3.1. Biotope).
1.3.3. Nahrung
\:Veberkncchte leben vor allen Dingen räuberisch. Sie erbeuten Arthropoden, mit denen sie den Lebensmum teilen. Im Falle der Förna- und Humu:~sohicht-Bcwohner sind das hauptRächlieh Milben, Collembolen und andere hodenlebende Arthropoden und deren Larvenstadien, soweit sie iiberwültigt werden können. Jmw Arten, die

42 l. Allgemeiner Teil
in die Kraut- und Strauchschicht aufsteigen oder Felswände besiedeln, erbeuten dort auch die Nal!l'ungstiere. Rilaena t?·iangularis (IMMEL 1955, Platybunus bucephalus) vermag (nach Gefangenschaftsbeobachtungen) selbst fliegende Insekten zu fangen; sie werden mit den tasterartig verlängerten Tarsen und Metat.ar~en vor allem des 2. Laufbeinpaares aus der Luft herabgeschlagen. Wahrschemhch vermögen die meisten plantikolen Phalangiiden auf diese Weise ~u jage~, do?h fehlen darüber unmittelbare FreilandbeobachtungeiL IschyropsalM kollan greift (in Gefangenschaft) mit Untemtützung der vorderen Laufbeine und der Cheliceren fliegimde Drosophila, in der Lurgrotte bei Graz, seinem natürlichen Lebensraum, erbeutet er die Assel llfesoniscus alpicola (NEUHERZ mündl., vgl. auch NEUHERZ 1975, MARTENS 1975c).
Als Nalll'ungsspezialigten haben die meisten, wenn nicht gar alle Troguliden und Ischympsalis hellwigi zu gelten, die sich überwiegend von Mollusken ernähren. D<'mentsprechend sind Troguliden auf kalkarmen und somit molluskenarmen Böden selten oder fehlen ganz.
Tmgulus (nepaeformi8 und t1·icarinat1ts) und Anela.smocephalus camb1·idg~i verfolgen Solmocken sellmt biK in die hinteren Umgänge des Gehäuses, und Sie vermögen die Weichkörper völlig daraus zu entfernen.
Ischyropsalis hellwigi bricht dagegen die Schalen von Gehäuseschnecken auf, bis die Scheren den Wcichkörp~r fassen und Teile von ihm abschneiden können (MARTENS 1965b, 1975b). Gelegentlich werden von ihr in Gefangenschaft auch Arthropoden angenommen, z. B. Dipteren, und die Subspezies I. hellwigi lucantei scheint - möglicherweise populationsgebunden - Schnecken überhaupt nicht als Nahrungstiere zu kennen.
Neben den tierischen selbstgefangenen Beuteobjekten nehmen Phalangiiden auch frisches Aas an, z. B. zerquetschte Schnecken. Daneben werden pflanzliche St.offe gefressen, z. B. faulendes Obst. In Gefa,ngenschaft lassen sich viele Arten mit. eingeweichtem Brot längere Zeit am Leben erhalten (KAESTNER 1931a, SANKEY 1949a, RüHM 1926).
Bei der Nahrungsaufnahme werden in der Regel vom Beutetier Teile abgeschnitten. mit den Cheliceren und von den Mundwerkzeugen durchgeknetet und in den Ö~ophaguH aufgenommen. Chitinreste sind im Kot nachweisbar. Die Verdmmng erfolgt somit W>thrscheinlich mir zum kleinen Teil extraintestinaL
Fast alle Weberknechte sind stark wasserabhängig und trinken regelmäUig. J'ndiYiduen, die längere Zeit gedurstet haben, können innerhalb weniger Minuten HO viel Wasser aufnehmen, daß ihr Abdomen prall gefüllt ist.
1.3.4. Sozialverhalten
Die meisten Weberknechte leben solitär und treffen sich nur kurzfristig während der Paarungszeit. Viele Arten der Phalangiidae, vor allem der Leiobuninae und Gagrellinae, kommen jedoch an geschiitzten Stellen zu Tagesruhe· und zu Überwint.erungsgesellschaften zusammen. Dort ~itzen Rie dicht gedrängt neben- und übei·einander und berühren sich gegenseitig, meist mit den taktil besonders empfindlichen Tarsen der L;tufbeinP. Einzelne gestörte Individuen alarmieren durch i!ll'e Bewegung die ganze GeRellschaft., die sich darauf in wenigen Sekunden aufzulösen vermag. In Mexiko wurde eine Ruhegesellsclmft von Leiobumtm cactontm auf 70000 Individuen geschätzt (WAGNER 1954). Überwinterungsgesellschaften
1.3. Biologie 43
von Am.ilenus a?t1Ytntiacus in Höhlen können Hunderte oder gar Tausende von Indh·i~luen umfassen. Versch~edene mitteleuropii:ische Leiob·ttn·um-Arten, vor allem L. hmbatum., versammeln stch an den Tagesruheplätzen höchstens zu mehreren Dutzend Individuen.
1.3.5. Fortpflanzung
Die ,Kopulation erfolgt bei den meisten der bisher untersuchten Arten schnell, ohne Vorspiel oder ausgeprägtes Balzverhalten. Die Partner stehen sich frontal gegenüber, und der Penis dringt zwischen den Cheliceren des ~ in dessen Ge. !lchlechtsöffrmng und in die Legeröhre ein.
Dit>ser Modus wurde schon 1850 von lVIENGE und 1882 von DE GRAAF für Pha. lau-f!illm opilio beschrieben und seither vielfach für Phalangiidae-Arten bestätigt (Zu~a.nunenfassung bei lVIARTENs 1969 b).
Während der Kopulation umfassP.n dio d'-Pedipalpen im allgemeinen die Basis dt"H :3. Beinpam·es des <;' nn.he deKsen Körper. Das 2. d'-Beinpaar wird auch in die>!<'lll Falle als Tastorgan für den Fernbereich eirtgesetzt. Apophysen an Chelicc·n·n und Pedipalpen des d' dienen in mar10hen (allen n Fällen dazu, die ~-Chelice. n•rt oder andere Extremitäten zu arretieren bzw. in eine bestimmte Lage zu drängt•n und dann zu fixieren. :Mit dem I. Beinpaar wird regelmäßig der Körper de:; ~ bt•t.astet. - Das <;' von Leiobun·Mm vittatnm (SAY) soll mit Palpus und Cheliceren dc<n Penis zur Geschlechtsöffnung leiten.
.... -
.\lob. 40. IschyropsaliB hellwigi (PANZ~tt). -- Kopulation, link• 0 . Unmittelbar vor der Kopu· laf.jl)n berührC'n die ~·?I!Inndwerl\:zeugo (Pfeil) das Drüst'!nfL"ld ( = Bürstenregion) des J·Chelic·eJ'~"ngrmldgliedes; das ~ kla.Intnert sioh rnit seinen CJu"lieeren· Klatwn an Pectipttlpns und Olmlir:t>re des 0 ft""~st-. Aus I\ I ARTEXS 1969 b.

44 I. Allgemeiner Teil
In allen beschriebenen Fällen sind die arterkennenden Mechanismen bisher nicht aufgeklärt worden.
Ein viel komplizierteres Verhalten mit ausgeprägter Balz zeigen jene Ischyropsalis-Arten, deren ~ ein Drüsenorgan im Cheliceren-Grundglied besitzen. Wähl'end der Werbung bietet das es dem 'j' ein Sekret dar, das auf einem Porenfeld des Cheliceren-Grundgliedes austritt; Das 'j' hebt das Prosoma an und richtet die Cheliceren auf, so daß seine Mundwerkzeuge freiliegen. Mit ihnen bringt das es da~ auf dem Porenfeld austretende Sekret in Berührung. Erst dann findet die Kopulation statt (Abb. 49; MARTENS 1969b, 1975a; für Ischyropsalis hellwigi). Diesen Kopulationstyp mit vorgeschalteter Geschmacksbalz (gustatorische Balz) darf man bei vielen Arten vermuten, deren 3 ebenfalls Drüsenorgane auf den Cheliceren tragen; das sind im hier behandelten Gebiet alle Arten der Nemastomatidae, Dicranolasmatidae, ferner die Sabaconidae (vielleicht nicht alle Arten) und unter den Nmnerikanischen Ischyropsalididae Oeratolasma und einige andere Gattungen.
Inwieweit Drüsenorgane am es-Pedipalpus eine Rolle im Kopulationsablauf spielen, ist bisher ungeklärt (z. B. Euda8ylobus nicaeensis, Leiobunum rupest1·e, Himalphalangium-Arten; vgl. Kap. 1.2.9. "Sekundäre Geschlechtsmerkmale).
Die Partner der T1'ogul1t8-Arten tricarinatus und nepaefo1'mis berühren sich bei der Kopulation mit den Stern!!>lflächen; dabei weist die des es nach oben, die des ~nach unten. Die Penisscheide dieser (der 1) Tmgulus-Arten besitzt keine lateralen Versteifungen, so daß sie mit dem Penis während der Kopulation um 180° verdreht werden kann (PABST 1953).
Die weichhäutigen, sackartig eingestülptenlateralen Flügelungen am Penis vil'ler Leiohunum und Nelima-luten haben möglicherweise eine Funktion als Leitstruktur für die Furca der Legeröhre, in die der Stylus des Penis (und weiter in die Receptacula seminis) eindringt (vgl. SILHAVY 1949, T. 8, Abb. 9, ferner die PetliHAbbildungen im Speziellen Teil). Dasselbe mag für Rillen im Operculum genitalo \'On Paroligolophus agrestis gelten, der einen extrem dünnen Penis besitzt (Ahh. 594 und 598).
Bei manchen Phalangiiden finden zwischen den es oft Kämpfe um die 'i' Htatt.; dabei versuchen die es von Phalangium opilio, sich mit den langen Chelicen·nHörnern gegenseitig abzudrängen. Das war schon MENGE (1850) hekannt. ·
1.3.6. Eiablage, Eizahl, Brutfürsorge, Parthenogenese
Die Eier werden meist in Erdspalten der obersten Bodenschicht a.hgelegt, die die langen und beweglich gegliederten Legeröhren der Phalangiidae mit tastenden Bewegungen aufsuchen; auch Hohlräume in morschen Pflanzenteilen werden mit Eiern beschickt, z. B. von Mitopus morio. Die Trogulus-Arten (ob alle 1) und Anelasmocephal1t-Y camb1'idgei belegen leere Schneckenschalen - meist die ihrer Beutetiere - mit ihren nur a.us wenigen Eiern bestehenden Gelegen. Das Gelege wird durch ein feines Gespinst von der Außenwelt isoliert. 8im rubens legt in Höhlungen der oberen Bodensc:licht ab, und nur in Mikrobiotopen mit hoher relativer Luftfeuchtigkeit deponieren einige Arten die Eier auch frei hängend unter hohl liegendem Holz, unter Steinen oder Pflanzenteilen, z. B. Ischyropsa.lis hellwigi, I. wrli und Pamnemastoma quadripunctat·um. In Höhlen werden die Gelege oft an den \Vänden hoch über dem Boden angehängt, z. B. von den G-ylr8-Arten, von J.schympsali8 strwuli und von Pamnemastoma silli.
1.3. Biologie 4:i
Die Eier werden in größeren oder kleineren Gruppen abgesetzt - das schwankt individuell und· auch nach der systematischen Stellung der einzelnen Arten. Gelege von über 300 Eiern (Odiellus spinosus bis 200, Platybunus pinetorum bis 275) sind bisher von keiner Art bekannt geworden. Das 2. und 3. Gelege sind im allgemeinen kleiner als das erste. Siro 1'1tbens und Scotolemon dm·iae legen die Eier einzeln ab. Die Tendeonz zur Einzelablage scheint auch (oder besonders n bei troglobionten Arten zu bestehen und ist von Ischympsalis strandi beschrieben worden (JUBERTHIE 1973). Bei solchen Arten ist die Eizahl insgesamt gering, das einzelne Ei aber groß und nährstoffreich (vgl. bei I. strand·i).
Die Eier eines Geleges haften entweder locker aneinander (z. B. Phalangiidae; RüFFER 1966), oder sie sind von einer glasklaren, zähen Gallerte umhüllt, die nach der Eiablage durch Wasseraufnahme quillt (z. B. Nemastomatidae, IschyropsalisArten).
Die allermeisten Arten kennen weder Brutfürsorge noch Brutpflege. Unter den Laniatores wurden jedoch einige Arten entdeckt, die einfache Form der BrutfiirKorge treiben, und möglicherweise ist diese Verhaltensweise viel weiter verbreitet als bisher bekannt. Die 'j' dieser Arten verlassen die Eier nach der Ablage nicht. sondern bewachen sie während der Embryonalentwicklung. Erst wenn sich die geschlüpften Jungtiere zerstreuen, löst sich der "Familienverband" auf. Die ersten Hinweise auf Brutfürsorge innerhalb der Opiliones lieferte STOLL (1897, vgl. dazu LoMAN 1900); erst in neuerer Zeit gibt es entsprechende Mitteilungen von CAPOCASALE & BRUNA TREZZA (1964), MITCHELL (1971), ,JUBERTHIE & MUNOZ-CUEVAS (1971) und von 0. v. HELVERSEN (in litt.). Alle Fälle betreffen - soweit nähere Angaben vorliegen - S-amerikanische Gonyleptidae. FüRSTER (1954) beschreibt entsprechende Verhaltensweisen von neuseeländischen Triaenonychidae.
Weberknechte sind im Normalfall zweigeschlechtig; bei einigen Arten sind es indes selten oder überhaupt noch nicht nachgewiesen worden. Im hier behandelten Gebiet gilt das für Platybunus pinetorum (einzelne Populationen im südl. Schwarzwald und am Stadtrand von Stuttgart ohne es), Megabumts lesserti (aus Salzburg keine es bekannt), Megabunus d1:adema (erst 1 es beschrieben). Ferner gehört in diese Gruppe Oacldo agilis (Caddidae, N-Amerika und Japan; erst 1 es bekannt, GRUBER 1974) und Orosbycus dasycnemus (N-Amerika und Japan, bisher keine es beschrieben). Diese Arten bzw. einzelne Populationen dieser Arten vermehren sich parthenogenetisch. Gründe für diese "lokale" bzw. "obligatorische" Parthenogenese sind nicht bekannt.
Auf den Testes mancher Phalangiidae entwickeln sich vergleichsweise häufig auch Eistadien; wahrscheinlich ist die es-Geschlechtstätigkeit dieser Intersexe dadurch nicht eingeschränkt. Die meisten Fälle beziehen sich aus Phalangium opilio. Eine Zusammenfassung hat BLASZAK (1968) geliefert; siehe auch BLANC (1881).
1.3.7. Lebensdauer, Phänologie
Die individuelle Lebensdauer scheint für einige größere Taxa typisch zu sein wenngleich hinsichtlich der tropischen Formen noch fast keine Daten vorliegen. Individuell am langlebigsten sind offensichtlich die phylogenetisch ältesten Gruppen. 8·im 1'Ubens soll bis 6-9 Jahre alt werden, wobei 2-4 Jahre auf die Zeit der Postembryonalentwicklung fallen (JUBERTHIE 1964). Möglicherweise haben auch die Oncopodidae ähnlich hohe Lebensdauer, doch fehlen bisher Daten. Die meisten

46 I. Allgemeiner Teil
Laniatores werden wahrscheinlich 2-3 Jahre alt ·:- soweit lediglich die Formen gemäßigter Breiten berücksichtigt werden. Die Arten der höchst evaluierten, "modernen" Gruppen leben in Mitteleuropa etwa I ,Jahr, wobei sich der Zyklus oft über 2 Jahre verteilt. Unter den mitteleuropäischen Artenlassen sich folgende Zyklen abgrenzen (Einzelheiten im Speziellen Teil):
Etwa einjährig: a) Eiablage im Herbst; die Eier überwintern., Postembryonal-Entwicklung im
Frühjahr, Reifehäutung im Sommer oder Herbst nach dem Jahr der Eiablage. -Hierher die meisten Phalangiidae und Nemastomatidae.
b) Eiablage und Beginn der Postembryonal-Entwicklung im Sommer des Jahre:> der Eiablage, Reifehäutung im nächsten Frühjahr. - Hierher die Arten der Gattungen Platybunus, Megabunus, Rilaena; Amilenus aurantiacus durchläuft die Reifehäutung im unterirdischen Winterquartier.
c) Reifezeit ganzjährig, ohne deutlich abgesetzte Zyklen. - Hierher DicTanopalpus gasteinensis, Leiobunum lirnbatum in den Alpen (zwei ineinander verHchränkte Zyklen), Gyas.
Etwa zweijährig: a) deutlich abgegrenzter Zyklus: Eiablage im Sommer/Herbst, Ende der Poxtembryonal-Entwicklung im nächsten Frühjahr, Reifehäutung im Sommer, Überdauerung als ad. etwa bis zum Frühjahr des 3. Lebensjahres. - Hierher Nernustoma bimac·ulatum.
b) kein deutlich abgegrenzter Zyklus: Eiablage in der wärmeren Jahreszeit ohne deutliche Präferenda; vom Zeitpunkt der Eiablage abhängig die Spanne bis zur Adulthäutung. - Hierher die Arten der Trogulidae und Dicranolasmatidae.
Mehr als zweijährig: Junge und reife Tiere während des ganzen Jahres, Fortpflanzung wahrscheinlieh auf die warme Jahreszeit beschränkt. - Hierher die Sim-Arten. Als ad. in Gefa.ngenschaft genommeneS. duricori1tslebten dort noch nach 3 Jahrert und 8 Monaten (J.M.).
In der Postembryonal-Phase werden ein Larven- und mehrerf' Nymphenstadien durchschritten. Die Larve sprengt mit einem (oder zwei) Eizähnen die Eil1iillc. Sie ist wenig beweglich und häutet sich nach höchstens wenigen Stunden z1mt ersten Nymphen-Stadium. Die Zahl der Nymphenstadien schwankt zwixclum 4 und 8, ist a.ber intraspezifisch weitgehend konstant (allenfalls 4 ansta.t.t 5, odet· 5 ansta.tt 6). Am häufigsten treten 4 und 6 Nymphen auf (JuBERTJIIE 1964).
Im speziellen Teil werden alle Jugendstadien mit "iuv." bezeichnet, das letzte soweit notwendig als "subad." differenziert. Nach der Adulthäutung finden keine weiteren Häutungen statt. Autotomiarte Extremitäten werden während der Poxt- · embryonal-Entwicklung nicht regeneriert.
1.4. Verbreitung und Zoogeographie
Das weitere Mitteleuropa ist Einzugsgebiet von Arten ganz unterschiedlicher Herkunft hinsichtlich des Htammesgeschichtlichen Alters, des Entstehungsgebietes der einzelnen Gruppen und ihrer glazialbedingten Refugialgebiete. Allein diese
1.4. Verbreitung und Zoogeographie 47
Gegebenheiten machen Mitteleuropa bereits zu einem zooge()graphisch bemerkenswerten Gebiet. Es kommt hinzu, daß von den 110 behandeiten Arten viele als End?miten des Gebietes gelten müssen, von denen z. B. 28 Arten (26 %) wiederum nur m den Alpen vorkommen. Mehrere Gruppea haben sich im engerea europäischen Bereich eiaschließlich des M;ttelmeergebiete~ entwickelt und sind auch hen te auf diesen Teil der Paläarktis beschränkt Typische Arealbilder und junge Speziationsabläufe - bedingt durch Klimaschwankungen des Pleistozäns - )a.,;sen ~ich vor allem in vielen tcrdkolen Gruppen nachweisen.
Im Baltischen Bernstein wurden Vertreter der Familien Phalangodidae, Nemastom!ttidae, Sabaconidae, Caddidae, Phalangiidae (Phalangiinae, Gyantinae, Gagrellinae) nachgewiesen (KooH & BERND't: 1854, BISHOP & ÜROSBY I924, RoEWER 1939j, ferner ein Vertreter der Dicranolasmat.idae (J. M. rev.).
Überregionale Beziehungen:
Beziehungen zur Nearktis:
Die Übereinstimmungen sind gering; sie betreffen das Familienniveau bei alten, das Gattungs- und Artniveau bei wenigen jungen Gruppen:
Sironidae: Die Gattung Siro ist formenreich vor allem in SO-Europa und Vorderasien vertreten, 2 (genitalmorphologisch bisher nicht untersuchte) Arten in NAmerika.
Nemastumatidae: Die Nemastomatinae sind eine btark aufgespaltene Gruppe allein der W-Paläarktis, die offenbar hier entstan4en ist. Die wenigen 0-paliiarktischen und nearktischen Arten der Unterfamilie Ortholasmatinae weichen in vielen Merkma-len stark ab; sie haben eine seit langem eigenstänr.lige Entwicklung durchlaufen.
Erebomnstridae: Eine artenarme Reliktgruppe in Europa (I Gattung) und NAmerika ( 4 Gattungen); _es besteht keine generische Übereinstimmung.
Travuniidae: Eine ebenfalls artenarme Reliktgruppe in Europa (in glazial unbeeinflußten Gebieten, oft in Höhlen), Japan und Korea (I Gattung) und N-Amerika (I Art). Alle Teilpopulationen sind auf dem Gattung~niveau differenziert.
Sabaconidae: Eine monotypische Familie mit holarktisch er, meist weit disjunkter Verbreitung; in Europa überwiegend in südwestl. Gebirgen; auch aus dem Baltischen Bernstein bekannt. Die Verwandtschaftsbeziehungen innerhalb der Familie b~w. Ga.ttung sind nicht geklärt.
Ischyropsalididae: l'ypenreiche Familie in der ganzen Paläarktis; die einzige europäische Gat.tung lschyropsalis ist endemisch. Engere Beziehungen haben 0-Paläarktis und Nearktis mit der Gattung Taracus. Mit dem habituell stark abweichenden Gemtolasma tricantha (westl. N-Amerika) besteht eine holarktische-Westseitendisjunktion. .
Phalangiidae: Gruppen- und 1artenreiche Familie in der ganzen Holarktis. -
Phalangiinae: Schwerpunkt in der W-J?.aläarktis, offenbar weniger artenreich in der 0-Paläarktis und spärlich in der Athiopis; in der Nearktis fehlend (Opilio parietinus und Phalangium op-iiio offensichtlich eingeschleppt). - Oligolophinae: sowohl in der Palä- wie in der Nearktis; Übereinstimmungen im Gattungs. (Odiellu8) wie Art-Niveau ( Mitopus morio). - Leiobuninae: die zahlreichen Arten aus der W-Paläarktis und Nearktis stehen trotz generi~cher Überein~timmung ("Leiobunum'' nach jetziger AuffasHung) nicht in enger Beziehung,

48 I. Allgemeiner 'l'eil
Beziehungen innerhalb der Palä,arktiR: Sie sind bisher ka11m zu überBehen, da vor allem die 0-paläarktisch-asiatische
Fauna nicht genügend bekannt ist; so ist z. B. völlig offen, wie weit in Europa weitverbreitete arboreale Arten nach 0 reichen und den Ural überschreiten. Als sibirische Waldart ist bisher nur Mitopus morio erkennbar, und entsprechende Beziehungen scheinen bei Oli{!olophus zu bestehen (mit 0. aspersus in Japan und 0. tienmushanensis in Korea), ebenso möglicherwei~e in der heterogenen Gattung Opilio.
Am S-Rand des paläarktischen Asien, z. B. im Himalaya, der von der übrigen 0-Paläarkt.is durch Trockengebiete isoliert ist (Mghanistan und W-Himalaya, Hochsteppen Tibets) leben überwiegend Gruppen indisch-orientalischer Zugehörigkeit (z. B. Biantidae, Assamiidae, viele Leiobuninae), alte paläarktische Reliktgruppen (Sabacon) oder junge am Ort dilferenr.ierte Gattungen mit paläarktischer Beziehung (Himalpl!alangium, z. T. Opilio). Somit sind die Beziehungen zur europäischen Fauna minimal.
Beziehungen innerhalb der W-Ptdäarktis: Endemit.en: Trogulidae, östl. bis in den Elburs, im gesamten Mittelmeergebiet und dort der größte Formenreichtum; möglicherweise ausstrahlend bis in die .Äthiopis. Beziehungen zu habituell ähnlichen Formen 0-Asiens und N-Amerikas (Ortholasma. Dendmlasrna, Ceratolasma) bestehen entgegen vielfacher Annahme nicht.
Nemastomatidae: Nemastomatinae: östl. bis in das Elburs-Gebirge und in den K!tuknsus; lokal in N-Mrika.
Dicranolasmatidae: Überwiegend in der Mediterraneis östl. bis in die Gebirge des N-Irak. Die monotypische Familie hat keine (engen) Beziehungen zu den Trogu.Iidae, wie vielfach angenommen, wohl aber zu den Nemastomatidae.
Beziehungen innerhalb Mitteleuropas: Die mitteleuropäische Weberknecht-Fauna gehört zum ganz überwiegenden Teil dem Arboreal a,n. Ihre Geschichte und heutige Verbreitung sind damit in hohem Maße mit jener der Waldbedeckung in diesem Gebiet verknüpft. In vielen J!'äi!en iBt aus dem rezenten Arealbild und den ökologischen Präferenda der einzelnen Arten dtts glaziale Überdauerungsgebiet auf den südlichen europäischen HalbinsBill mühelos erkennbar. Unter diesen Refugialgebieteu, von denen. postglazinlo Bt•siedlungsschübe ausgingen, nehmen die Bnllmnhalbinsel und (weniger) di& Iberische Halbinsel die erote Stelle ein, während der Apenninen-Halbinsol nur untergeordnete Bedeutung zukommt. Das Karpaten-Gebiet ist ebenfalls als Refugium erkeimbar, doch scheinen von dort keine Vorstöße in das engere Mitteleuropa erfolgt• zu sein.
Balkanrefugium: DPs8e.n arboreale Arten haben z T. mediterran bzll'. sub-. mediterran, z. T. montan geprägte klimatische Präferenda. Die Artengruppe ist somit recht heterogen und untfaßt verschiedene Komponenten. Die einzelnen Arten haben z. T. ganz Mitteleuropa erneut besiedelt - montane Arten vorwiegend die Alpen oder nur deren südl. Massive, vielfach auch die Mittdgebirge. In diese Gruppe gehören:
Holoscotolemon un·icolor (Abb. 107}, Siro dw·icorb1s (ALb. 68), 1-'•·ogulus t•·icm·inatus (Abb 267}, T. tingifo•·mis (Abb. 267}, Anelasmocephalus hadzi.i (Abb. 290}, Histricostoma de"#JJalp (.Abb. 193}, Ccwinostoma <•cwi>wtum, Nemastoma lug<~bre (1; Abb. 139}, Dicmnolasma scab,·u' (Abb. 245), A.sii'Ob<~mts diuaricus (Abb. 756), A.lwlleri (Abb. 756), A. lacvipes (Abb. 747}
1.4. Verbreitung und Zoogeographie 49
Eyae-nus conve:vu.s-(Abb. 576), Zacheus c1·ista (wahrsc1u~inlich keine arboreale, sundnrn moditonanenmlisohe Art.; Abb. 566}, Opilio •·uzickai (Abb. 452), 0. dina.·icus (Abb. 445), Laci11ius de"ti· ge,., L. hon·idus (wahrscheinlich mediterran-oren1isoh), Aniilenus au1-antiacus (Ahb. 728) .. Ndhna sempmni (Abb. 799).
Iberisches Refugium: Dessen Arten nelunetl heute atlantisch bzw. atlantomediterran geprägte Areale oin.
A t.lantisoh: Nemastoma bimaculatum (Abb. 139), Homalenotus q"adridentat"s (Abb. 747), 01-igolophus hanseni, Pa.roligolophua ag>·estis, P. meadii, Leiobunum rotundum, L. blackwalli.
At.lanto-mediterran: Anelasmocephalus cambl'idgei (Abb. 290}, Odiellus spi"osus (Abb. 630), 0. t•·oguloides, Nelima gotMca (Abb. 815}.
Apenninen-Refugium: Von dort Ausbreitung in den Alpenraum; rezentes Areal meiRt apenninisch-alpin. Astrobunus bemardinus (Abb. 747}, Bcotolemon doria •.
Karpaten-Refugium: Dessen Arten auch heut·e noch im wesentlichen auf den Karpaten-Bagen beschränkt.
S.i1'o carpaticus, Pm·anemastomct lJilU, P. ko<:h7:, Ischyropsa.lis manicata (Abb. 350), Platybu11118
pallidus (Abb. 467).
Vorläufig nicht sicher einzuordnen sind t•inige Arten mit "mitteleuropäischem" Art>a.l:
Pamnemastoma qu<~dripunctatum (Abb. 161}, T•·ogulus nepaejormis (Abb. 276), Nem<~stoma lugubre (Ahb. 139), Oligolophus tridens.
Endemiten der Alpen: Ihre Zahl ist mit 28 Arten (26% des gesamten Alteubestandes des hier behandelten Gebietes) vergleichsweise hoch. Sie werden hier mit Hinweis auf geographische Beziehungen der nächst verwandten Arten bzw. Artengruppen genannt.
Holoscotolemon. leasiniense, H. oreophilum: beide nahe verwandt~ sonst innerhalb der Gat,. tung isoliert. - PcltoniJchia leprieut·i: epigäische Arten in den Pyrenäen .. }{avernikole iJn l(arst. - Auelasmocephalus rujitm·sis (Abb. 290}: nächste Verwandte unbekannt. - Dicranolasma tHt1l}J<·I' (Ahb. 245}: im Apennin D. cristatmn. - Nemastoma bidentatmn relictum (Ahb. 150): die beiden anderen Subspezies von N. b. im Alpenbereich und auf der BalkanhalbinseL -N. scltuelle1·i (Abb. 134}: nächste Verwandte unbekannt. - PManemastoma bicuspidatum (Ahb. 108): Gattung l'aranemastonw auf der Balkanhalbinsel in starker Aufspaltung begriffeu; niiehstverwandte Art nicht bekannt. - 11>1itostom<~ alphmm (Abh. 134}: nächst verwandt. 11-1. cl!.rysomelas. - M. anophthalmum: nächste Verwandte unbekannt.. - Gentetastoma centetes (Abh. 134}: Gattung G. in den Pyreniien und auf der Iberischen Halbinsel stark aufspaltend. - 1schympsalis ccwli (Abb. 350): im Apennin I. adamii, in den Pyrenäen 1. lute-ipes. - 1. dentiJ.WltJis: verwandt mit I. !.~Oum··i, z. T. stark abgeleitete Höhlenfm•men aus dieser Verwandt,. :sl'l1aft ::.ind die ebenfaHs alpin-endemischen 1. hadz#, I. strand·i und I. ravasinii. - I. helt•et.icct (Abb. 394}: nächste Verwandte ')!'bekannt. - Sabacon sinwm:: Gattung 8. in Europa nur im Zentraln1assiv, in den Pyreniien und in clen Kantabrischen Gebirgen; hier insgesan1t wa1lr::<C'heinlich naliverwanllte Artengruppc. - Platyb'unus alpinoreUctu.s: Vül'\\-·an<lte Formen wahr•oheinlich auf d<•r BalkanhalbinseL - Dicra"O)Jalpus gasto·inensis (Abb. 710}: 'Gattung D. aufspaltend vor allem in den Pyreiläen uud J{antabl'ischen Gebirgen. - Gyas anrvulatus (Abb. 691, 701): Gat.tnng 0. nur mit. einer weiteren nahe verwandten Art. - lvlltopus ylaciali.s (Ahb. 677}: •lark •pezialisierte Form innerhalb cler Gattung. - lflegabunus losse•'li, M.,·kinoce•·os, 1J;1. arma.t,.s, M. vignai (alle Abb. 517): dio Gattung 111. ist mit Platybunus, l!Ietc;platybu·ll?ts und R-llaena na]w ,·erwandt; innerhalb der Gattung stehen sich armatus und {e,r;setti besontler~ nahP. - Le·iohunum 1'<~lig·iosurn (Abb. 765): nHchSt.c Verwandte unhekn.nnt-.
-4 Tierwelt. Dc-nt!\t'hl. 64: JI.Inrtrns

50 I. Allgemeiner 'l'cil
Von dieNen Alpen-Endemiten sind ohligntoriHche, z. T. stark nbgcleitete 'l'roglobionten:
.tl1itostoma anophtltalmum, I.schyroJJsaUs clentipalpls, I. tJYl'etwect alpi-nulct, I. harlzii, I. st1·ancli, I. 1'avasinil.
Eng lokalisierte Areale in vielen S-Alpen-Massiven geben Hinweise auf wenig· stem partielle Glazialüberdauerung in diesen "Massifs de Refuge". Spätere Arealltusweitung Iutt nicht oder nur in geringem Maße stattgefunden. Hierher gehören:
Ho!oscotolemon lessi,.·ien.•e und H. oreophilum (Abb. 107), Ce11tetostoma centetes (Abb. 134), Dicranolasma 1"'"1'"'' (Ahb. 245), Anelasmoce.phalus ntjitm·sis (Abb. 290), Iscltyropsalis lwZ...r:tica (.:\hb. :394, troglobionto Ischyropsalis-Arten :::~iohe oben), Sabacon simon·i, Platybunus a.ljJino-1'elictu..'J; .~.11ega.ln1n.·us armat·us, .tll. ·vz'gnwi, 111. 1'kinocr.ros (alle .. ..\hb. 517).
Als orcoalpine Nunatnk-ÜLerdauerer können die über der Baumgrenze siedellidnn J<'elxxchutt- und Fel~wand-Be,mlmer gmYertet \\·erden:
Dicranopalpu8 ga8teinen8i8 (Abh. 710) und Jiitopn8 glaciaUs (.Abh. 677); huide xind Ho mit nnch zentral-alpin v•whrcitct, und ill. glaciali8 ist nahezu auf diexe Alpentdie hexchränkt. 'ßei den 111e(lrtbu.J!?l8-Arton le88e1·ti, rhinoceros, a1'1natus und vigna.-i, die ebenfnlJs (auch) über der Baumgrenze Iebon (aber nur lokal in den ZentralAlpen vorkommen), ist zentr>,J.I-alpine Glaziltl-Überrlnuerung nuf Nunatalmm weniger wahrscheinlich. Bei diesen Arten muß eher mit postglazialer Riiok\mnderung von mndalpinen Maxxif.~ de refuge in devastierte Alpenteile gerechnet \\'erden (vgl. oben).- Bei Nem.a8toma bidenta.hvrn relictzm~, dtt~ die Subalpin-Stufe !Jesiedelt. ist Nunatnk-Überdauerung wenig wahrscheinlich.
Addonlpine Arten scheinen nicht zu existieren. Ob das Aren! von 1llegabu.m'" rlictdemn (Abh. 530) entsprechend gedeutet werden darf, ist unsicher.
Re~ente Area.Ie sind bei vielen Arten disjunkt; diese Disjunktionen xind nnte.rschiedlich ltlt und hisweilen erst postglazial entstanden.
Anclererxeits weisen aiiopatrische Arealenahverwandter Arten auflänger dauerndt> Disjunktion eines ursprünglich kontinuierlichen Areals hin.
Poxtgl az in! cli~jungiert sind die Areale folgender Arten: Holoscotolemonjaqueti. Nema81oma. tri.ste (Abb. 130), Trorrulu-8 tingiformi.s (Abh. 267), Oyas titamts (Abb. ti91), Platyb·unus pinetorwn (Abb. 467). J>. bucephalus (Abb. 480).
Wenig~tens inttll'glnzial clixjungiert. sind die Arenie folgender Arten, dPren Teilpopulationen bisher ent.weder gnr nicht oder subspezifiHCh und sognr ~pezifixeh differenziert sind: [s,·h.yroJMalis he!l11:i(fi he!lwigi - I. h.lucaute.i (Ahh. :!08); 1. pyremtea. jJyrena.r.a - I. p. alpin:ultt {Ahh. :J50); H·istr·icostom.a t/e·uUpalpe - H. t:trgenteolwwlatum. (~-\bb. 19:1); D·icnmolasma pcn.tper - D. cl"istatum (Ahb. 245); Eudasylobus uicaeeusis (Abb. :34:l: gehört mögJic:Jwrwei::ie in die vorhergehende Gruppe); Ocliellus coroJtatus - 0. ::e('ar-iensls; Astrobu·wlls heller{ - A. kochi ( Ahb. 756); A6"lrob·unus lat.~t:it>es - .-1. bt!I'Urtrrtinus (Abh. 747); .:.Ye.limu doriae - ~V. ,.Jn'sar.IJ/.'i.f;;. (Ahh. 815; gehöt·t mijgJitdiL'l'wei~ß in die n.n·lierg'f!IH.•nde
Gruppe).
Sekundäre Arealerweit.erungen Mehrere europäixche Arten xind offen~ichtlich durch menschlielw Ein\\'irlmugen weit über ihre nr~prünglichen \'erhreitungsgebietn .. hinan~ ver~chleppt \Wrden: ~ie haben zum Teil innerhalb Europax, zum Teil iu UIJerxee Jehonsfähige Kolonien
1.5. Systom 51
gegt;ündet, ihr SiedlungHgebiet dort aber nur in anthropogen degradierten Habitaten aktiv erweitern können. AndererReits sind keine außereuropäischen Arten in Europa heimisch geworden, möglicherweise mit der Ausnahme von Opilio pa1'ietinus. Einzelheiten zu der nachstehenden Liste vergleiche man im Speziellen Teil.
Tfogulus tricarinatua: USA, St.aat N<"w Yorli", bei Roehoster. Phalangium opilio: In vielen Staat.en der USA, Neuseelancl. Opilio pariet·in.us: Ver1nutlich ausgehend von SO-Europa (Kaukasus-Gebiet) und Vorclerasion
heute in engem Anschluß an menschliche Siedlungen über große Teile Europas verbreitet, überdies naeh USA, l{anada und Tasmanien verHc!hleppt.
Opilio sa.xatil-ls: Große Teile cles mitteleuropiiif.;chen Arl'als nach \Valdvernichtung und enge Bindung a.n H.udm•alhiotopo ersehlossen; möglicher\'v'(liso oingosclllcppt nach IsraeL
Opilio 'l'uz·ickwi: Htndtgobict uncl Umgebung von 'Vien. Rilae1ut tdangulari.s: UR.-'\, Staat \Vashington, Hnatt.lc~; Kanada, Brit,ish Columhia. Oligolophus t1·i<lens: URA, Staat. Vormont .. Pm·oligolotJhus ct(JJ'estz'.s: UHA; Ktaat Washington, H('lat.tlc; Ka.nada, Brit.ish Columhia. Lfliobunutn Umbotum: ViPifiwl1 in HUidtc nöJ·dlü·h dt>J' Alpnn vcl'~C!hleppt, vereinzelt bis Sücl-
Hchwednn. Eeiobunnrn ti.sclae: Rüci-Englawl. ]\'elim((. rlorirte: Kreta, for·ner A11:=;trulicn 11ncl Ncu::;Ae]aru1.
1.5. System
Die klassische Gliederung der Opilionc~ in die 8 Unterordnungen Cyphopbthalmi La.nintores und Pnlpatores geht auf THORELJ, (1876) und SnvroN (1879) zurück; Rie wurde von HANSEN & SOERENSEN (1904) weitergefiihrt. ROE\VER (1923) lutt diese Einteilung übernommen, aber während 50jiihriger Tätigkeit nicht modifiziert !Jzw. weiterentwickelt. Erst in jüngerer Zeit xind einzelne Gruppen unter großRystematischem Aspekt revidiert \\'Orden; eine Gesmntwertuug unter einheitlich vergleichenden GesichtRpunkteu steht. LiHher <WR.
Die wichtigsten neuerenAnRichten iiber den Bereieh von der Fnmilienlmtegoric aufwärts sollen hier xki~ziert werden.
Suborclo Cyphophthalmi
Der Rang der Gruppe als Unterorclnüng mit der einzigen Familie Sironidae ist unverändert geblieben. Ob\\'ohl manche Merkma.Ie der äußeren Morphologie auf eine altertümliche Gruppe schließen laxHell (z. B. Tarsa.I-Gliederung, fehlende.-< Operculum genitale), erscheinen andere Merkmale modernen Gruppen ähnliclt und müssen möglicherweiHe nls tu;~prllngliclw AuRprägung angexehen werden (z. B. die vielfach gegliederte Legeröhre). Ob diu komplizierte .Pcnisarmntur im Rahmen aller Opiliones als urspriiuglich gewertet wNden muß. i~t ninht. geklärt ..
Die Gnt.tungen der Sironinac hat. .JurmR1'll'IE ( 1!)70) revidiert.
Gliederung: Hubordo Cyphophthaln1i Rnwx 187!} mit. dc.w roin7.igrn l•~arnilie Rironidao (Riro~ ninaß, Sty1ocr~llinao).

52 I. Allgemeiner Teil
Subordo Laniatores
Zweimal wurden Versuche unternommen, die von THORELL (1870) hegründeto Unterordnung aufzugliedern - im Hinblick auf die "aberranten" Familien Triaenonychidae und Oncopodidae. . . .
I. LollfAN (1900, 1903b) wertete die Triaenonychidae nls eigene Unterordnung Insidiatores und stellte sie neben die LaniatoreH. LollfAN fand größere Differenzen zwischen den Laniatores alter Fassung und den Triaenonychidae nls zwiHchen den Laniatores und den merklich abweichenden Oncopodidae. HANSEN & SoERENSEX (1904) und später RoEWER (1923) übernahmen diese Gliederung nicht, zmnnl RoEWER Merkmale der inneren Anatomie gänzlich unberücksichtigt ließ.
Sinngemäß, wenn auch teilweise mit anderen Merkmalen, f~~rte K_RATOCHViL {1958a) die Dichotomie der Laniatores in Form von 2 Überfaunben weiter: On?opodoidea und Travunioidea - mit Merkmalen d~r Krallenfor~ der Laufhem-1'm·Ken UI und IV. DicKe beiden Gruppen werden Im selben Umfange noch besS<'I' - und in phylogenetischem Sinne wahrscheinlich zuverlitssiger - durch clie Peni~Konstruktion umgrenzt: Travunioidea mit Muskel-Sehnen-System, das die Glanx bewegt; Oncopodoidea mit mu,skellosen Penes, deren Glans durch Hämolymph-druck bewegt wird (MARTENS 1976). . . . , .
2. SrLHAVY (1961a) löste aus der Überfanuhe Oncopodmdea KRATOOHVIL dit• Oncovodidae und erhob sie in den Rang einer eigenen Unterordnung Oncopodomorphi mit der einzigen Familie Oncopodidae. Daneben be~eß er ~ie weit.aus größere Restgruppe der Laniatores als Unterordnung Gonyleptimorplu. Damit waren die Laniatores erneut aufgelöst und in 2 Teilgruppen derselben Ranghöhe aufgewertet worden.
Schon LOMAN (l903b) trennt·f\ innerhalb useiner" Laniatorns (\'ermindert mn die lnsidiat·Ol'P~ = 'I'riaenonychidae) die Oncopoclidae als Sterrhonoti von den übrigen Familien, den Can1ptu· noti, ab unci wies damit. ebenfalls auf die Sonderstellung del' Oncopodidae hin.
Äußere Morphologie und die Genitalmorphologie (möglichel'}miso Klammel'mcchaniHmus, Anklänge an die Sironicbe) weisen den Oncopodidae Z\\'aJ' isoliertt· StPllung zu, rechtfertigen aher kaum den R>tng einer eigenen Unterordnung. Somit. werden sie hier aiK eigene Überfamilie Oncopodoidea. n. comb. aufgefaßt. Sie entxprieht. nicht den Oncopodoidett KnATOCHVfL; er bezog n.uch die Gon~'-leptoidea im hier vcrt.retenen Sinne e.in. . _ . ~ _ .
Die Restgruppe der La.niatores, dw Unterordnung Gonyleptunorplu SILHAVY. aliedert Sn.HAVY ähnlich KRATOOHVIL in die Überfmnilien Travunioidea und Gonvleptoiden (diese ohne die Oncopodidae; vgl. oben). Diese Einteilung erscheint Rinn~oll. Beieie Uberfamilien sind verläßlich abgrenzbar und bilden sehr wahrschein- . lieh monophyletische Einheiten. .. . . .
Die Travunioidea sind in vielen Merkmalen nrspnmglwh (z. B. mit PetusmuskPl. mannigfache AuHprägung der Beinklauen III und IV, bipolar disjunkte Verbrei-
h~). . . Für die Gonyleptoidea iKt· Monophylic Lisln.ng mcht nach we1sbar, aber nach allen .
Vmständen wahrHcheinlioh. Immerhin iHt. denkbar. daß der Penismuskel, der allP Trnvunioidea. chara.kteriKiert, mehrfaeh reduziert wurdP. Folglich könnten "Gonyleptoidea."-Gruppen (ohne Penismuskel!) auf ganz \'erschiedenen Nivmws de:; SyKtems ontst>tnden 8ein. Die Erebomn-xtl'idn.e mit. PPJÜKmuskel (Merknml de~·
1.5. System 53
Trav~mioidea) und Doppelkrallen an den Tarsen Hf und IV (Merkmal der Gonyleptmdea) deuten diese Möglichkeit an. In diesem Falle wären die Erebomastridae eine konservative Schwestergruppe aller Gonyleptoidea (MARTENS 1976).
Der Aufwertung einzelner Teilgruppen der Laniatores zu eigenständigen Unterordnungen (LOMAN, SILHAVY) ist die Einheitlichkeit dieser Gruppe in vielen Merkmalen entgegenzuhalten - dies trotz Formenmannigfaltigkeit und Notwendigkeit der Aufteilung in viele Fainilien. Umfassende Merkmale sind: do;.•sales Scutum als~· magnum (Ausnahme Oncopodidae), Oorona analis fehlend; Ooxa I beweglich; Pedipalpus als Raubbein mit unterschiedlicher Spezialisierung, oft SubchelaBildung; Beinklauen trotz Unterschieden in den einzelnen Gruppen im Dimorphismus an Bein III und IV übereinstimmend; grobmorphologisch einheitlicher Ovipositor-Bau.
Es wird deshalb vorgeschlagen - unter Betonung der umfassenden Gemeinsamkeiten - die Unterordnung LaniatoreR bestehen zu lassen und den Unterschieden ihrer Teilgruppen durch Untergliederung in Überfamilien Rechnung zu trageiL
Nach den vorstehend dargelegten Begründungen lautet die Gliederung:
Subordo Laniatores THORELL, 1876
8uprafamilia Travunioiclea ABSOLON & K.n.ATOOHViL 1932 Travuniidae, Triaenonychirlae, Erebomastrido.e; Pentanychidae, ? Synthetonychidae.
Suprafamilia Oncopocloidea THORELL, 1876; nov. comb. (neo Oncopodoidea l{RATOCHViL, diese = Gonyleptoidea und Oncopocloidea im hier vert.retenen Sinne). Oncopoclidae.
Suprafamilia Gonyleptoidea SUNDEVALJ., 1833; nov. comb. (neo Gonyleptimorphi i='hLHAVY, diese= Travunioidea und Gonyleptoidea in1 hier ver•trete .. nen Sinne). Gonylepticlae, Phalan~odidae, Cosmeticlae, Assamiidae, Podoctidae, Stygnicla.e, Biantidae, Agoristenidae.
Subordo Palpatores
Die Unterordnung PalpatoreR wurde von THORELJ. (1876) aufgestellt; HANSEN & SoERENSEN (1904) untergliederten sie in die "Triben" Eupnoi und Dyspnoi, u. a. nach Merkmalen der Tracheen-Morphologie und der Stigmenverteilung an den Beinen, des Baues von Ovipositor und des Operculum genitale.
SILHAVY (196la) erhebt die Triben in den Rang eigener Unterordnungen. Die Famili~ngliederun~ der Dyspnoi behält er entsprechend RoEWER (1923) bei; den Eupnm ordnet er Jedoch 3 allstatt bisher 1 Familie zu: Leiobuninae und Sclerosomatinae der Phalangiidae erhalten FamilielU'aug.
Gegen die Aufwertung der Eupnoi und Dyspnoi zu Unterordnungen wendet. ;;ich GRUBER (1974b) mit dem Argunient. vergleichsweise hoher Eigenständigkeit der Ca.ddidae, die aber zugleich Merkmale beider Gruppen miteinander vereinigen. Bei RoEWER (1923) Rt.ehen die Arten dieser Gruppe a.Js Oligolophinae-Phalangiidae und Am·opsopilionidae bei den Eupnoi hzw. Dyspnoi, hei SILHAVY (196la) als Acropsopilionidae und Oaddinae-Phalangiidae bei den Dyspnoi.
Wahrscheinlich. haben die Oaddidae als altertümliche Ba-sisgruppe der Palpa.tores zu g<'ltcn, ehe den Eupnoi näher steht als den Dyspnoi: Merkmale des Yentralen Exoskelette8, der LegeröhrP u. n. (vgl. GRUBER 1974b). SHEAR (1975)

G+ 1. Allgemeiner Teil
räumt- dt·n Caddidae zu Recht den Rang einer Überfamilie Gaddoitbi ein. Somit läßt sich der hohe Rang der Dyspnoi und Eupnoi al~ Unterordnungen (SILHAVY) wegen dt1r vermit-t-elnden Stellung der Caddidae kaum aufrechterhalten.
Rücken nun die Eupnoi und Dyspnoi durch die vermittelnden Caddidae in unerwartet enge Beziehungen, so bleibt die Monoplwlie von deren Teilgruppen zu bestätio·en; sie wurde bisher vorausgesetzt.
Die Eu~noi (sensu HANSEN & SoERENSEN, i. e._ Ph_al~ngiidae s. I.) :;ind als vei•o-leichHweise junge und in hoher Blüte stehende emhe1thche Gruppe zu \\'<'rten. Du;ch die svnapomorphen Merkmale dm· akzesKorischen St-igmen an den LaufbeinTibien und. die freien Loben der Coxa li sind sie kla.r umrissen. Die wichtigste Familie bilden die Phalangiidne mit zahlreichen Unterfamilien z. T. ganz untersdliedlicher Habitustypen. Trotz gegenteiliger Meinung von SrL:HAVY (1961) und 1\L-!.RTENS (1973 b) mlissen auch die Sclero:-:om•lt-in.>te - im Ra-hmen der \Velt.fanna - ihren Unterfamilien-Rang behalten. In 0-Asien gehen sie offenbar st-ufenlos in rlio Leiohuninae-Gagrcllinae iihe1·.
Die D_vspnoi htthen Hich d_ngcfelt a-1~ het<•ro?en e!'ll'iese~t; H_i<> t~~nfns~~n Tt'~lgruppen mit recht- unterscluedhclwr xystenwhxcher \Vert1gke!t .. S1e musxen m eine viel größere Zahl von l!',tmilicn aufgeteilt werden, al~ RoEWEH (1923) und SrLHAVY (1961 a) ammhm<'n. J?ttx läßt sich mit genitalmorphologischen Merkmalen henTiindcn.
Bei allen Palpatores wird die GlanH Iutch dem !VIu:-:kel-Sehnenzug-Prinzip htnn·gt (für die Caddiclae nicht durchgehend geklärt, vgl. Abb. fiir C'adell(b in KAURI 1961 und SHEAR 1075). Ein grundlegender Unterschied teilt die Arten der Dyspnoi in 2 Gruppen: Bei den Arten der einPn ixt. im Pcnis-Rolu· lediglich ein Muxkel bzw. eine Sehne vorhanden, bei den anderen d>tgegen zwei (Einzelheiten zur Konxt-l'llktion und Funktion vgl. Iütp. 1.2.8. "Genita.!organe").
Zur er.<ten Familiengruppe gehören die Ixchyropsalididae und Sahaconidtte, möglicherweixe eine weitere monot-ypische l!'amilie Croxb,vcidae.
Die >tndcre Gruppe iRt. umfangreicher; zu ihr zählen die Trogulidae, Nemaxtomatidae und die Dicranolasmatidae. Ihre Zugehörigkeit zu den Trogulidac, die hi~her vermutet- wurde und <tlx derPn Ui!terfamilie sie bixl>tng gefiihrt wmdon, i~t sicher nicht richtig. Die Beziehungen zu den .N't•nutstomatidae xind viel enger (Einzolheiten im Spezidien Teil).
ÜIJerrascllenderll'eise gel1ört in dieHe Gruppe - Penis mit- z"·ei Muskeln- auch die .0-asiatische Gattung N1:pponops(tlis. Sie kann HO!llit nicht zu den hoh,rropxalididao gehö1·en (vgl. llfAR'l'ENS & SuzuKI 1966) - nicht einmal in deren l<'nmiliengruppe. Auch den Familien der anderen Familiengruppe läßt xic xich nicht zu~n-dn.en: MAH~rENS (1976) erkannte ilu Rlx .N'ipponop~alididae Familienrang zu.
Die Pa.Ipatores lassen sich somit- zu 4- Familiengruppen bündeln, die mit der hi~hcrigen Zweiteilung iu Eupnoi und Dyspnoi nicht- in Einklang gebracht 11'(11'
den können. Als notwendige l!'olge miisxpn die Kategot·ien Eupnoi und Dyxpnoi aufo·eo-eb<·n werden; an ihrt> Stelle treten + Überfamilien. Damit wird schiirft•re Bii~d:lung und bessere, ohjektin• 'übersieht der Vennmdtschaftsverhältnisse eneicht. Dann lautet die GliNlerung rlt'r Palpat-orc~:
Nuhurdo J>aipatore~ THOHELL~ lH76
8upr·afatnilin. Caddoidoa ßAXKs, lSH~
(Eupnoi pal't., U~' .... pnoi part.: aw:t.) C'addicia<'
1.6. Sammlungs- und Bestimmungstechnik 55
Snprafanüli~ l)halangioi(lea SUNDEVALL, 18:l:~
(Eupnoi auct.) Phalangiidae, Megalopsalididae, Noopilionidae.
8uprafa1nilia Ischyropsalidoiclea SoERENSEN, 1884 (Dyspnoi auct.; part.) Ischyropsaliclidae, Snbaconidae, ? Croshycidae.
Suprafamilia 'l'roguloidea SuNDEVALL, 18:l3 (Dysp••oi auct .. ; part.) -Trogui.idae, Nema.c.:;tonlatidae, ~leranola.smat.idao, Nippon.opsnlididae.
1.6. Sammlungs~ und Bestimmungstechnik
Für die Arten mit rein terrestrixcher Lebenxweixe sind alle Methoden anwendbar, die Nich für andere tenikole Arthroporf<,n bewährt haben.
F <t n.g: Viele der ganz kleinen Formen (Sironidae, NemaHtomatidae, 'froguliclae, Laniatores) erbeutet man durch Aus~iehen der oberen Bodenschichten in Wäldern und Gebüschen oder am Spülsaum vou B>tchufern. Arten- und individuenreich ,;incl tiefgründige, alte Fprna- bzw. Humusablagerungen. Als Sieb verwen~et man die handelsüblichen Käfersiebe; die Maschen aus Drahtgeflecht sollen eme lVIaschenweite von 5-7 mm haben. Selbst große Troguliden-Arten, z. B. T. t·ingiform.i-s, fallen durch Mascl1en von 6 mm Seitenlänge. Viele auch der kleinen Arten findet man bereits durch Wenden von altem Holz und Steinen oder durch Ausschütteln von Moos- und Farnbewuchs auf einem weißen Tuch.
Bodenfallen ("Barberfallen") sind nützlich, vor allem, wenn sie längere Zeit an einer Stelle stehen können. Sie werden zu einem Drittel oder bis zur Hälfte mit Äthvlenglykol. ers!ttzweise mit 4%igem Formol, gefüllt und am Fangplatz so eingegt:·a.ben, daß. der Rand zu ebener Erde >tbschließt, und mit einem Stück Rinde oder einem flachen Stein hohl abgedeckt.
.Automatische Bodenstreu-Eklektoren, die meist- zur Extraktion besondei'H kleiner Boden-Arthropoden benutzt 11·erden, eignen sich fiir Weberknechte weniger, rht "ie nur die g>tnz kleinen Arten erfitssen. z. B. Siro und die kleineil Arten der Nema.stomatidae ..
Alle in höheren Sti-ata. lebenden Arten sucht man in ihren TageNverst-ecken >tuf, a.lso ttuf Busclnn•rlc an Ba-umstämmen, Felswänden, Hauxwänden und an Bretterzäunen. Immer gtil~stig und meist von artenreichen Populat-ionen besiedelt sind die Puterseiten von kleinen Brücken aus Mauerwerk.
Von Stauden können Weberknechte gestreift, von höhe1'em Gebüsch auf ein mJißes Tuch geklopft oder geschüttelt- werden. Die meist-en Arten werden beim f'lt.reifen allerdings stark beschädigt.
Kleine auf Holz oder Steinen!~ ufencle Individuen IasRen sich mit angefeuchteter Fingerkuppe aufnehmen: feuchter Pinsel oder (.Federstahl-) Pinzette sind meist zu umxti\ndlich.
:Flüchtende Individuen langbe;niger Arten fiingt man, indem man mit 3 oder 4 Fingem einer H>tnd zugleich möglichst viele Beine zu faKxen Htwht. Nur ~o ist rlie Ge.falu gering, daß Beine autot.omiert \\'erden.
DR ~tlle Arten (auch) nachtaktiv sind, können vor allem die hmgheinigen Arten (Phalangiidae) nachts mit kräftiger Lnmpe (Taschenla-mpe) gesucht w<>rden.

56 I. Allgemeiner Teil
S >t m ml u ng: Individuen, die konservi.ert we~·don sollen, w~rde~1 in 70%i~em · Alkohol abgetötet und nach wenigstens einmaligem Wechsel m diesem Medmm
aufbewahrt. Wer eine Sammlung anlegt, soll sie von Anfang an so aufbauen. daß sie bei ständigem Materialzuwachs übersioh~lieh un~ jede ~erie mit. we1~igen Hnn~griffen auffindbar bleibt. Das ist leicht errerchba~, mdem Jede Serre, dre nur IJ~dividuen einer Art, von einem Fundort und von emem Datum enthalten soll, eme fortlaufende Nummer erhält die auf dem Etikett mit den Funddaten (Art, Datum, Ort Höhenlage Sammler ökologisehe Notizen, ggf. Koordinaten) vermerkt wird. Zusätzlich wird' auf dem Plan-Boden des Sa: umlungsröh,.chens ein kleines Papier· stück, das dieselbe Nummer trägt, mit der Beschriftung nach unten mit einem 'Vattebausch festgedrückt. Die Röhrchen werden unter Ausschluß von Luftbittsen ebenfalls mit Watte verschlossen und aufrecht stehend - mit der Mündung nach unten - in einem gut schließenden Glas mit weiter Öffnung in Alkohol aufbewahrt .. :Mit doppelter Buchführung bleibt die Sammlu.ng übersichtlich: in.einem Einwmgshuch werden die Serien mit allen Angaben, dre auch auf dem Etrkett verzewhnet sind, fortlaufend eingetragen. Eine n[l,ch Arten geordnete Kartei, in der wiederum alle Etikett-Angaben vermerkt sincl, ermöglicht es, den Bestand an Arten und Gattungen leicht zu überblicken und einzelne Serien sohneil aufzufinden.
Bestimmungstechnik - Präparation: Vor jedem Bestimmungsvei•such überzeuge man sich davon, daB tttt.sächlich adulte ~xe~pla~~ vorliegen. N~r ·~uf solche sind die Schlüssel zugeschmtten, und nur sie smd uherhaupt verlaßhell bestimmbar.
Das Operculum genitale adulte!' Individuen ist distal, also auf der Höhe der Coxa I (!I-IV bei Laniatores) zu einem schmalen Schlitz geöffnet und meist etwas lippenartig aufgewölbt. Bei Jungtier~n aller Altcrsstufen fehlt di~ser Schlitz inu~wr, zudem ist deren Sternalhereich zwrschen den Coxen I-IV cmgcsenkt, schlwßt also nicht in gleicher Höhe mit dem Ventralbe.reich der Coxen ab.
Bei den Ph[l,langioidea ist das Operculum breit geöffnet., ebenso bei den Nemn~to. matidae und Ischyropsalididae, vergleichswei~e schmal bei den Sabaconidae, Png bei den Trogulidae; bei ihnen ist besonders sorgfältig der Alterszustand zu prüfeiL Sironidae haben kein Operculum genitale; die Genitalöffnung der Adulten lit>gt sichtbar offen (Abb. 12).
:Merkmale der äußeren Morphologie sind bei schwacher Binokular-Vergrößenmg leicht erkennbar. Davon ausgenommen können Merkmale an der Basis der Extre· mitäten sein, die durch andere Körperteile verdeckt sind. Dazu gehören die Cht~li· ceren, der Pedipalpus und die Coxen der Laufbeine. Die Cheliceren sollen mit feiner Pinzette ausgezupft, die Pedipalpen zwischen Coxa und Troch>tnter ahgctrennt werden. Es empfiehlt sich, jeweils die rechten Glieder abzulösen; diese ;;ind im Speziellen Teil abgebildet. Die Coxen der LM~fbeine ( Höckerreihen bei den Lt>io-. buninae!) lassen sich mit 2 Pinzet.ten auseina.nderdri.icken.
Für die Bestimmung sind die d'-Genitalorgane von größter Bedeutung. Sie liegen unter der Körperdecke und lassen Rich meist nur unter Beschädigung des kon~l'l'· Yierten Exemplares freilegen. Dazu wird mit einem feinen und spitzen Skalpt'll der Genitaldeckel lateral aufgeschnitten und die diHtale Ansatzstelle der Peni$scheide abD"etrennt. Der Genitaldeckel läßt sich dann zurückklappen und Penis hzw, Lege1~m·e aus dem Körper und aux der Scheide lösen. Bei den Troguli•kn wird der Schnitt am besten durch die weichhäutigen Pleuren geführt und tla~ Ventralscutum abgeklappt.
r
1.6. Sammlungs. und Bestimmungstechnik 57
Schwierig sind Individuen der stark skierotisierten Laniatores zu handlutbon. Bei ihnen werden am besten bereits beim Sammeln unmittelbar vor der Konscrvier~mg durch leichten D_ruck auf das Abdomen Penis bzw. Legeröhre ausgestülpt. _ Dre harten und zuglerch kleinen Sironiden müssP•l mit spitzer Pinzette bzw. Skalpell zwischen Coxen und.Sternalplatte vorsichtig aufgemeißelt werden.
Die freigelegten bzw. abgelösten Organteile können unmittelbar in Alkohol betrachtet werdeu. Besonders kleine Objekte (Penes der Sironidae und kleiner Nemastomatidae-Arten) sind auf Objektträgern mit Hohlschliff abgedeckt unter dem Mikroskop zu untersuchen. Die Gestalt der Legeröhren ist für die Spezios. Diagnostik unwesentlich, wichtig dagegen die der Receptacula seminis. D& Kie meist unter Lagen von Muskeln und Bindegewebe verdeckt sind, müssen sie &ufgehellt werden. Das geschieht mit Nelkenöl, Glycerin, Terpineol oder K>trbo!Xylol. Die Legeröhren können unmittelbar aus dem Alkohol in diese Flüssigkeiten überführt werden. Bei Terpineol-Beh>tndlung empfiehlt es sich, den Alkohol etwas abtrocknen zu lassen. Nur bei den sehr massigen Legeröhren der Laniatores und Ischyropsalidoidea kann es uotwendig sei11, das Bindegewebe mit Kalilauge oder Natriumsulfid zu mazerieren.
Pigmentierte Cuticula läßt sich !Jp,i einigem Geschick mit Skalpell und feiner Pinzette abpräparieren.
Dauerpräparate sollten nur von Legeröhren und stark dorso-ventral abgephttteten Penes angefertigt werden ( Siro, Oarino.stoma, H1:st1·icostO?na, Paranemastoma). Die Penes der Arten aller anderen Gattungen sind stark dreidimensional struktu. riert, und in Dauerpräparatenlassen sich die räumlichen Beziehungen nur unvol!. kommen erfassen. Abgelöste Organe sollen in kleinen Röhrchen - mit Watte verschlossen - bei den zugehörigen Tieren aufbewahrt werden.
Zur mikroskopischen Untersuchung, &uch zum Zeichnen, ist es oft notwendig, das Objekt exakt auszurichten und zu fixieren. Es kann an ein Quaderglas, rhtK auf ei11em Objektträger angeklebt ist, mit etwas Glycerin angeheftet und zu jed!n" beliebigen Lage ausgerichtet werden. Nicht be11etzte Penes verziehen und krümmen sich. Fixierung in Alkohol auf einem Wattebausch, einem Stück Fließpapier odnr unter einem Deckglas ist ebenso möglich (vgl. SrLI!AVY 1969a).
1.6.1. Nationale Sammlungen
\Yer über VVeberknechte publiziert~ solJte nic.:ht versäumen, der jeweils zustii.urligen Zentl·n.Jsamm!ung Belegexemplare zur Verfügung zu stellen; das gilt besonders für Typenexemplai'O bei Neubeschreibungen.
Die arachnologischen Sammlungen mit überregiona]er Becleut.ung im deut·schsprad1igen Ranmsincl:
Bundesrepublik Deutschland.: Natur-:Nluseum Senckenberg der Senckenbergis<'hou Kat.urforschenden Gesellschaft, 6 Fr Iinkfurt am Main I, Senckenberg-Anlage 25; Evert.o.. brat.en-Abteilung.
Deutsche Demokratische Republik: i\[usemn für Naturkuncle, Zoologisches Museum, 104 Ber!in, Invalidenst•·· 43; Arachnologischo Abt.eilung.
Ost.erreich: Naturhistorisches Museum, 1014 Wien, Burgring 7; 3. Zoologisehe Abteilung.
Schweiz: ~at.urhistorisches 1\-Inseuin, Augustinergasse 2, 40.11 Basel. - l\'Iuscum d'Histoire No.turelle, Route de l\falagnou1 1211 Genßve.

2. Spezieller Teil
Bestimmungstabelle für d1e Unterordnungen der Opiliones
Laufbein I a1n längsten; oluw rru ot~ und ohuo Augen; Öffnungen der Stinkdt·üsen auf :! l·wnischen, Ia vom Cr.ph abstehondcn Höc.•l,:C'rl'i. (Ozophoron; Abb. 57). - Im hior bohantl~lten Gebiet nur lokal im 0 und SO . . . . . . . . . . . . . Cyphophthalmi, S. 69 IJaufbein II am Hingsh•n; tnit Tu oe und mit Augen (Ausnahme: fohlend bei einigen troglubionte.n .Artfln der Alpen); ÖffnnngPn der Htinkdrilsr\ß nicht auf konis(·hen HöC'kern, sondern alf:> oYalo bis ~:whlitz.förmigc Vf'rtieftmg auf rlen1 Coph, do oder Ja. . . . . . . . 2
2 Pa mit kloin<1r Kralle odor Krallefehlewl (z. B. Abh. 411, 12:l), Kralle nie als Gmiforgan (Subclwla) gl'gen den Ta anlegbar; falls Ti und Ta dos Pa mit konischen Za.pfen· besetzt, rlann nie 2reihig anguorrhwt (z. B. Ahb. 483, 554); J(örpol' ni€! gelb, bernsteinfarben. -hn hier be-hanrlelton Gebiet woitverhroitet. . . . . . . . . . . . . Palpatores, H. 90 Pedipalpus mit großer. Kralle, als Rubc•lwlla gegen don Ta einschlagbar (Abh. 96, 101); Ti und Ta d0s Pa immer mit 2 Roillen (jcw(\il::; Jä) lnugor Zapfen und aufgl'Se:t.ztct• Bo bo~ wehrt (z. B. Abb. 101); :Körper gell>-bernst<•infarbcn. - Im hier behanclolton Gebiet nur im Be•·eich der Alpen uncl sücll. der Alpen. . . . . . . . . . . . . Laniatores, R. 68
Subordo Cyphophthalmi
1879 Cyphophthalmi RBION, Arachn. d" France, 7: 142. 1!l~:J Cyphophthalmi - RoEWER, 'Yelwrlmochto der Erclo: 42.
Körpergliederung: Dorsum mit Scutum completum: Opisthosoma mit 8 vorwach:>eiten unrl 2 freien Tergiten, von diesen daR hintero aiK Operculum anale. Corona analis kombinimt aus dem (freien) Tergit IX und den Stemiten Vlli und IX, die~e in unterschiedlichem M>tße verschmolzen; ent1reder völlig frei (P.u.rcellia H.HiSEN & SoERENSEN) oder ein e!nheit.liche.r den Analdeckel umfassender Rii1g. Prosomal-Sternum reduziert. Operculum genitale fehlend; Genitalöffnung unbedeckt.
Genitalmorphologie: Truncus penis kurz-gedrungen, ohne Glans und Stylus, ~tark do-ve abgeflacht, do l oder 2 Paar rlurch Muskeln bewegliche Greifzangen; Be~rehrung mit langen Borsten in ,\'rt- und gattun.gst,vpi~char Anordnung. - Ovipu~itor: 2-3fach so htng 11·ie der Körper, an~ gelenkig miteinander V6rbundenen Ringen zu~ammenge~etzt; die Furca eingliedrig, 2-:Jfach so lang wie die übrigen Ovipositor-Ringe. Ringe mit. je einer Borstem:eihe, Furca mit Rpeziali~iertem Seusillum. Inneret Genitalapparat mit 2 Anhangsdrü:<en. ohne Propul:<ion~orgau (möglicherweise Spermatophoren-Besamting).
Pedipalpus kurz, laufl.l('införmig, TarsuK mit Ht.iftförmigt>r Klaut'. Laufbeine kurz, höchstens körperlang. Bein l immer am li\ngstl'Jl. Tarsus meist ungegliedert, selten 2glit'drig, Q'-Ta lV mit Dri\Henorgan (Adenost~·le), Ausfuhrporus für da~

60 Familia Sironidae
Sekret >tuf abgesetztem Kegelhöoker. - Unguigrnd mit Muskeln wen.igsten~ im letzten Tarsalglied (bei Siro duricorius [.JoSEPH]; J. M.). Öffnungen der Stinkdrüsen (Ozophore) auf je einem stumpfen Kegel para-me bis Ia. - Augen meist fehlend; wenn vorhanden, dann weit voneinander getrennt am Lateral-Rand des c~ph.
Gliederung: 1 Familie, Sironidae.
Familia Sironidae
1879 Sironidae SIMDN, Arachn. de France, '1: 143. 1923 Sironidae - ROEWER, Weberknechte d. Errle: 42.
Einzige Familie der Unterordnung Cyphophthalmi, gekennzeichnet durch deren. ~ferkmale. ·
Verbreitung: weltweit, mit Ausnahme der Türkei bisher 1ücht auf dem a~iatischen Festland nachgewiesen. Die beiden Unterfamilien mit unterschiedlichen Verbreitungsschwerpunkten; Sironinae im westl. und östl. N-Amerika, im iiber"·iegend mediterranen Europa, Kloinasien, Sri Lanka (Ceylon), Japan, S-Chile, S-Afrika, N-Australien, Nense~land, Neukaledonien. - Stylooellinae: lokal im nördl. S-Amerika, auf der Iberischen Halbinsel, westl. Zentralafrika, Malayi,;cher Archipel.
Subfamilia Sironinae
1904 Sironini IIA.NSEN & SoERENSEN, On two Orclo•·s of Araohnida, 88.
Eine Unterfamilie der Sironidae, gekennzeichnet durch: Cx I und II der Laufbeine untereinander und gegen die fest miteinander verwachsenen Cx III und IV beweglich. Sternum !dein oder fehlend.
Verbreitung: 15 Gattungen (JuBERTHIE & MA.ssoun 1976) mit weltweiter \'prbreitung, wahrscheinlich bipolar mit extratropischem Schwerpunkt. JuBER'rlHE (1970) unterscheidet eine Gattungsgruppe S-hemisphärischer (Gond\\'ana-) Verbreitung in Chile, S-Mrika, Sri Lanka, Neuseeland (Chelioereu-Ii'inger mit 2 ZahnTypen, Ozophoren auf dem Ceph do) und eine solche mit N-hemi~phärisoher Verbreitung (Cheliceren-Finger nur 1nit einem Typ von Zähnen, Ozophoren auf dem Ceph Ia) in N-Amerika, Europa mit Vorder-Asien, Japan und auffälligerweise auch S-Mrika (Parapu1·celUa RosAs CosTA.), was möglicherweise auf KonYergenz zu dieser Gattungsgruppe beruht (.TuBERTHIE 1970, JuBERTHIE & MA.ssoun 1976).
Größter Formenreichtum - soweit bisher bekannt - im östl. S-Europa und in Neuseeland.
Genus Siro LA.TREILLE, 1796
1796 Siro LATREILLE, Precis. C'aract. Ins.: 185. 1868 CyphojJh.thaln..us JosEPH, Berlin. ent. Z., 12: 241-250. 1923 Sim - RoEWER, \V eberknechte d. Erde: 52. 1958 Sü·o (Oeosz~1·o) RAFALSKI, Acta Zoo]. Cracov., 2: 5:15.
~ominat-Gattung der Sironidae-Sironinao, gekennzeichnet durch: Q'-Genitalmorphologie: Trunous penisgestaucht unterxetzt, ve Greifapparat in der di Hälfte;
Genus Siro 61
diei<er immer aus 1 Paar großer, mediad oder latemd gerichteter, in sielt gekrünunter Zapfen, ferner meist kleinen häutigen Loben me oder para-me, diese selten längcl' als die Zangen-Zapfen (duricorius). Sinneshorsten bzw. Cutioula-Emergenzen in distinkten Gruppen di, do ve, und beidseitig Ia. - Muskulatur als kräftiger Strtmg me zu den Zangen-Zapfen ziehend, ferner je 1 kleiner Ia Strang zu den In Loben- hzw. Ba-Gruppen (vgl. Abb. 69). Cormut analis: gEschlossener Ring aus den Stenüten VIH und IX und dem Tcrgit IX, Op anale do-di abgeflacht rundlich (Abb. 62-64). Mt und Ta der Laufbeine jeweils 1gliedrig, nicht tuberkuliert. .Adenostyle auf Q'-Ta IV zunächst stumpf konisch, distad dünn röhrenförmig und oberhalb der Öffnung des Sekretkanals fein spitz bzw. breit gerundet verliingert oder 1 in sich gekrümmter Haken seitlich auf den stumpfen Konus aufgesetzt (Abb. 53-56, 65-67). Bezahnung der Cheliceren nur aus 1 Zahntyp. Kegel der Stinkdrüsen Ia etwa um den Kegel-Durehmesser vom Ceph-Lateralrand entfl'rnt. Sekundäre Geschlechtsmerkmale: Drüse im Q'-Ta IV mit arttypisch Kt.rnkturiertem Ausfuhrkanal; Kiel auf dem c3'-0p anale.
Verbreitung: llnominelle Arten in Europa (S-Frankreich, N-Karpaten, 0-Alpen. auf der Balkanhalbinsel in Jugoslawien, Bulgarien, Albanien, Griechenland südl. bis Euböa, auf den In~eln Levkas und Korfu), in Kleinasien auf dem Bithynischen Olymp und bei Yalova (GRUBER 1969b); inN-Amerika in den Staaten Oregon und Virginia (HOFFMA.N 1963).
Die Aufteilung des Formenbestandes in Arten stößt auf erhebliche Schwierigkeiten. Da alle Formen allopatrisch leben, lällt sich oft nicht entscheiden, welche Merkmale alK artsignalisierend angesehen werden dürfen. W nhrscheiulich kommt der Q'-Genitalmorphologie sowie der Lage und Gestalt der Adenostyle (Ausfuhrkanal der Tarsaldrüse) das größte Gewicht zu. Nach dem jetzigen Kenntnisstand erscheint es nicht sinnvoll, Subspezies zu benennen.
Typus·Art (rluroh Monotypie): Si1·o •··ubens LATREILLE, 1804.
Bestimmungstabelle für die Arten A<lenostyle a11f d'-'l'a IV gerade ge•treckt, die beiden di Drittol kanülonförrnig(Abh. 65-67); in <len SO-Alpen und in NW-Jugosla.wion . . . . . . . . . . . . . . duricorius, S. 61 Adl~nostyle auf cr-Ta IV niedrig und stumpf-ltonisch, seit.linh llliinnnr, in Mich gekrüJmntrr Haken ansetzend, spitz und dünn auslaufend (Ahb. 7:l- 74); Iolm1 in den N-Karpaten . .
Siro durieorius (JosEPH, 1868) Ahb. 50-68
1868 Cyphophthalm"s dw·icorius JosEPii, Berlin. ont. z.; 12: 241. 1923 Siro d"ricolius - ROEWER, w~herknechto d. Erde: 5:l.
carpaticus, S. 66
Habitus und Feldkennzeichen (AJ;>,h. 57): auffallend klein und kur:>.beinig, milbenähnlich, rostbraun; in dicke11 Fallaubsohiohten: SO-Alpen.
Kennzeichnung: Körpermalle (in mm): L c3' 1,70-1,85; L Cj? 1,82-1,95. Laufbein I (Q', in Klammern Cj?): Fe 0,50 (0,53) Pt 0,25 (0,25) Ti 0,35 (0,35) Mt 0,2 (0,2) Ta. 0,50 (0,50).
Genitalmorphologie (Abb. 58-61): 'l.'runcus pe1üs breit, gedrungen, distad genmdet zugespitzt, di Hälfte stark do-ve abgeplattet.; dort in eine!' ve Kavität der Greifapparat: nach me weisend 2 massige, konisch verjüngte zangenförmigt· .Zapfen, deren Innenseite heza.hnt, do da\'on ( = da.hiltter gl'legen), 2 lange und

62 Fami1ia Sironidae
Abb. ;30-57. 8iJ'O duricorÜ/.'J (.JosEPH). - no. 1'-Lanfbein 1 von Io.tera]; ;}}. 6-l)edipalpus \'(111 1nedial; 52. ö-CiteJioero von 1nedial; ;)!L 6-Tarsus IV von JatoJ'al; AUS Tröpolach/Gil.iftal; 64-50. 0'-'.rarsus IV von <lorsal: 54. fT Tagliamonto-Tal; ,;,;, YU Kroatien, Borg Z0ljak; 56. YU Zavala. - 57. Habit-us von <lorsa.l.
schlanke, glatte, ehenfalls konisch verjüngte, >1ber nicht gekrümmte Zapfen, diPse die o-ekrümmten Zapfen regelmäßig ühergipfelnd. An der Basis der Zapfen kkitw, una~ffällige und schwer zu sehende häutige Lohen. - Chaotataxie de~ Truucus in engen Gn\nzen variabel: do 3-4 (meist 4), di 4, ve 3-4, Ia .ie. 3-4 lange Bo, bisweilen beidseitig unterHchiedlich. - Legeröhre (Ahb. 40-41): nut 22 Segment<·n (außer den Furka-Zttpfen), Bo des letzten Segmentringes heRanders lang.
I
_ ___)
J/( ____ "'~>. 62
I>" __ '·--·/ i.. / .. ;.·-- ··--'·
\ } 65
/f; </ ri
~IL
Genus Siro
61 i/ / ''I l p 1/ \
. \ I I 11 "I • I' !lt
/·lifrY.l\"·:'-'1 t:. I •· ; . z~~-~~--t--.._, ,
j~~~?f'(.\ ··.\': • I ' "II "" I . ·, : ./• ,, 'I '• (Ii\
'Jn· !\,· . ..,, u 'I \ ;) . ' ! .·· . . ~r~ . I
..____,:__ ____ _
67
) i
)
Ahb. 58-67. Siro dul'icorh.ts (JoSEPH). - 58-61. Penis von Yentra.l mit· Chaotohtxie-Dio.~ramm (Unter8eit.e entspl'ioht- \'cntral): 58. IT 'l.'a.gliamento-Tal; 59.1'1' Ploso/Brbmn; 60. AUS rj•I'Öpola.chfGailtal; 61. YU Zavala. - 02-64. Ca.ucla.lende des K.örpors, ~bon J, unten ~: (12. YU '.raglia.monto-Ta.l; 6:l. AUH Tröpolaoh/GCiiltal; 04. YU Zava.la. - ß.o-67. AdonM!yle d•'S ,;-.Tarsus TV: Oii. TröpoJa.,h; 66. Taglia.mcnto·'ra.l; 67. YU Berg Zeljak.

64 Familia Sironidae
Cheliceren (Abb. 52): schlank, im ba Drittel durch do und .ve Konkavität jener Teil abgescluiürt, der unter den Ceph-Frontalrand und in den Ceph ragt; do, Ia und ve kräftig granuliert, medial glatt; 2. Glied glatt, schlank, unbewehrt bis auf I Borste auf der Ba des festen Fingers. Schneiden beider Finger mit je 8 stumpfen Zähnen.
Pedipalpus (Abb. 51): untersetzt; d'-Tr do und ve mit spitzen Do, übrige Glieder rauh, ohne auffällige Do, Pt di etwas erweitert, Ta distad wenig tropfenförmig; alle Glieder fein beborstet, Ta besonders dicht. - ~ wie d', doch Ta nur unwesentIich verdickt.
Laufbeine (Abb. 50): auffallend kurz, plump, rauh bekörnelt von Tr bis Ti, schütterer Besatz langer Bo, auffallend kurze und dicht stehende nur auf Ta I ve. -Drüse des d'-Ta IV: in der ba Hälfte des Ta, dort Exoskelett nach do und pro-Ja aufgewölbt; Ausfuhrkanal (Adenostyle) zunächst breit konisch, dann als schmaler und langer röhrenförmiger Kanal, dessen Öffnung sub-apical, Kanal über der Öffnung nur auf I Seite fein und spitz ausgezogen; Adenostyle schräg 'über dem Ta modiad geneigt (Abb. 53-56, 65-67).
Dorsalseite (Abb. 57): Zeichnung und Färbung: einfarbig rötlich-braun, einheitlich flach granuliert, schütterer Besatz feiner Haare frontal am Ceph bz'r. auf den hinteren abd Areae. Segmentgrenzen durch geringfügig dunklere Färbung und reihenweise angeordnete Granula markiert. - Ventralseite (Abb. 62-64): Op anale unregelmäßig rundlich oval, me Kiel das Op caudad etwas überragend; Kiel variierend: in den Extremen entweder breit und flach und dann über das ganze Op. laufend oder schmal und dann oft nicht den di Rand erreichend, caudal immer am breitesten. Kiel innerhalb engumschriebener Populationen einheitlich. - Cfl ohne Kiel, Op meist stärker gerundet.
Verbreitung (Abb. 68): Typ: SO-alpin-dinarisch. - Nur die N-Grenze dc~ Areals im Bereich der S-Alpen läßt sich mit hinreichender Genauigkeit überschauen. Auf der Ballranhalbinsel sind weitere Arten bekannt, deren Abgrenzung gegen du1'icm·ius oft nicht eindeutig möglich ist, zumalnur in ganz wenigen Fällen die genitalmorphologischen Verhältnisse dargestellt worden sind. Auch die Verbreitungsmuster (Sympatrie-Allopatrie) sind hisher nicht beachtet worden und harren der Abklärung. - Locus typicus: Sloll'enien, Luegger Grotte ( = Predjama, nordwestl. Postojna).
Italien: Plose bei Brixen (HOLDHAUS leg., Beleg NHMW; Bestätigung Nwünscht); "iiclöstl. des Passes Ma~uia (STROFHAL leg.); CariliaJ '.roimezzo und VerzegniH (CooREAtAK HIGO). - ös terreioh: Die bisher btlh:annte N·Grenze des Areals wird durch foJgende Fundort~.~
bolegt (vgl. Abb. 68): Kärnten: Doberbaoh-Graben bei Ratton<lorf/Gailtal (SCHUSTER 1975), südl. Tröpolach (J. G.leg.), zwischen Vorderberg und Feistritz/Gailtal (SCHUSTER 1975), Bll·ibei·g (SCHUSTER in litt.), Grat-schitzen-Graben/Karawanken, sücll. und südwestl. <!es Reutschacher Sees (hoide f:icHUSTER 1975), Ebenthai bei Klagenfurth (J"ANCZYK 1956), Kurathöhle bei Grafenstein (HöLZEL 1958) und sücll. Sand, Griffenor Berg (beide STROUHAL leg.), Johanno•burg südöstl. St. PaulfLavanttal (FRANZ leg.); - Steiermark: J(lause bei Deut-Rehlandsborg (SCHUSTER 1960); Malt-schach n<;rdl. des Pösnit-zbacltes; N-Hänge des Sausa!; Leibnitz (alle SCHUSTER in lit·t.). - Südl. dieser Orte ist. du.-rico1'itts in lGirnten und Steiermark weitYet·breitet. Nennungen fü.I' den Ossiach~r Tauern (I{ ÜHNELT 1943; auf der linken Dran-Seite nördl. Punkt i in Abb. 68) beruhen auf Irrtum bei der Drucldegt\)lg (SCHUSTER in lit-t.). - Jugoslawien: Im Anschluß-an den Österreichischen Arealteil in Slowenien häufig und kontinuierlich verbreitet, ebenso in den gebirgigen Teilen Kroatiens (z. B. Berg Ui'ka bei Opatija, J. ~1. leg.), :-;üd1. bis Bosnien-Herzl"gowina (Za,·alaJ J. G. leg.); woitere Einzelheiten nicht. bekannt. Nennungen fü.r clic> Jn~Pl Korfti/Grif'(·]wnlaucl (/'·". d1.tr·.fco1·lu.9 cmjurmus KRATOOHVh,) und fiit•
Genus Siro 65
·· ....
""-------~ , .. Siro duricorius 0 JO 100
kff;
Abb. 68. N-Grenze des Areals \'On 8-il'o d111'icnl'ius (J"oSEPH). - IT a PlosefBri~en, b 'l'agliaJUento4Tal, Mauria, r. Ca.rnia, Tolmezzo und Vf'rzegnis; AUS d-e Karnische Alp~n: d Dober4 bach-Graben, e südl. Tröpolac.J1, .f Gailtal,. zwischen Vorderberg und Feistrit1., u Bleibu1·g. k Villaeher Alpe, i Gmtschitzen-Graben, j -l: Hattnitz: j Umgebung Kautschacher See, k Ebnnt-hal. l Kurathöhle nnd südl. St>nrl, m Griff~ner Berg, n JohannesbergfSt. Paul, o Klause/ Deutschlandsberg, p Maltschaeh, nördl. des Pösnitzbaches, q N-Hänge des Sau"al, ,. Leibnit7 - Siidl. dieser Grenzorte i"t die Art m•1t Yerbreitet, vgl. Punkte.
die '.rürkei (S. cl. yaloven<Jis GRUBER, S. d. bithynicus GRUBER) sind eher auf eigenständige Arten zu beziehen; mit Bestimmtheit gilt das für yalovensis und bitkynicus (J. M. rev., J. G. in litt.).
Ökologie: Biotopansprüche: S. du1·icm·ius ist ein stenöker Bewohner bodenfeuchter Wälder unterschiedlicher Zusammensetzung, gern in illyrischen Laub:Mischwäldern, in Buchenwald-Gesellschaften und in Slowenien besonders in BuchenFichten-Mischwäldern, Diese Waldtypen besitzen dichten Kronenschluß und meist dicke Fallaub- oder Humusdecke; dadurch sind die oberen Bodenschichten isoliert und ständig gleichmäßig durchfeuchtet. Diese Biotop-Eigenschaften gelten auch für lichtere Wälder, etwa im Bereich der Baumgrenze an der oberen vertikalen Arealgrenze, z. B. aufgelockerter Lärchenwald-Rand (Karawanken) oder Latschenzonen im außeralpinen slowenischen Areal. In allen Waldtypen lebt dun'corius in dicken, vermodernden Fallaublagen in der Humusschicht oder - während längerer Trockenperioden - in Ritzen und Spalten der oberen Bodenschichten, in· detritusarmen, lichten Wäldern auch auf der Unterseite tief in den Boden eingewachsener Steine. 5 Tierwelt Deutschl. 6-!: Mm·teus
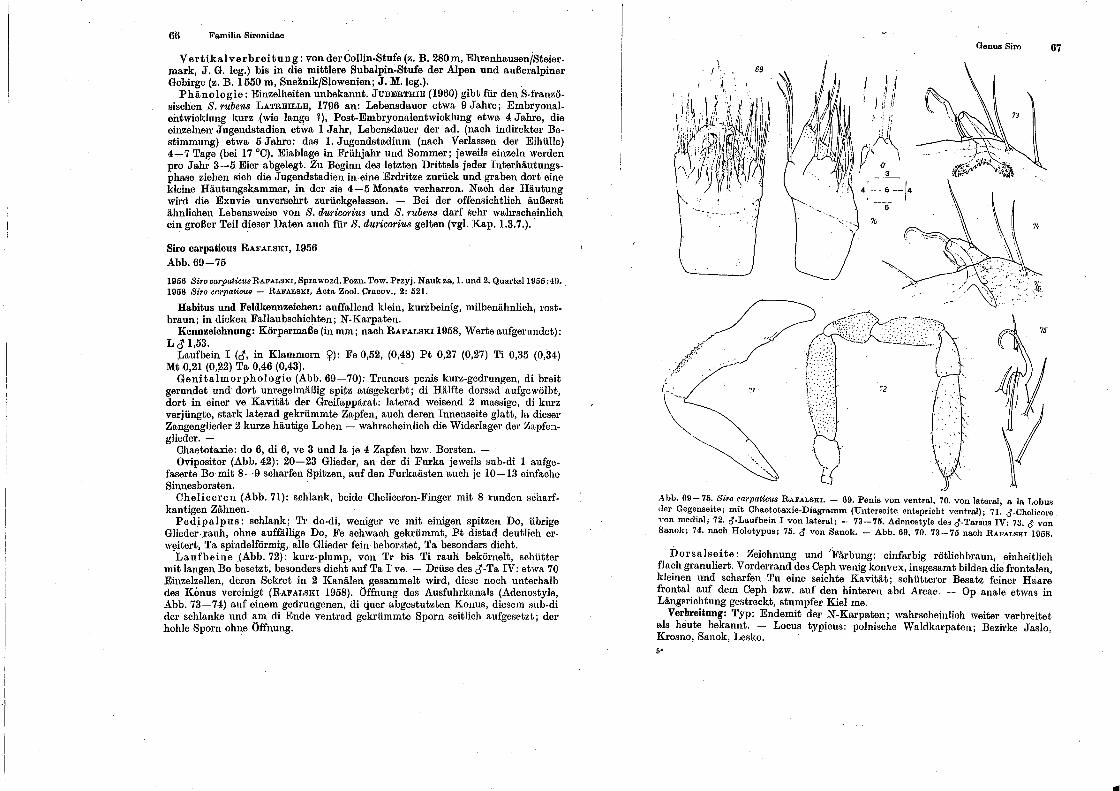
66 Familia Sironidae
Vertikalverbreitung: von derCollin-Stufe (z. B. 280m, EhrenhausenfSteiermark, J. G. leg.) bis in die mittlere Subalpin-Stufe der Alpen und außeralpiner Gebirge (z. B. 1550 m, SneznikfSlowenien; J. M.leg.). .
Phänologie: Einzelheiten unbekannt. JuBERTmE (1960) gibt für den S-französischen S. rubens LATREILLE, 1796 an: Lebensdauer etwa 9 Jahre; Embryonalentwicklung kurz .(wie lange 1), Post-Embryonalentwicklung etwa 4 Jahre, die einzelnen· Jugendstadien etwa 1 Jahr, Lebensdauer der ad. (nach indirekter Bestimmung) etwa 5 Jahre: das 1. Jugendstadium (nach Verlassen der Eihülle) 4~7 Tage (bei 17 °0). Eiablage in Frühjahr und Sommer; jeweils einzeln werden pro Jahr 3~5. Eier abgelegt. Zu Beginn des letzten Drittels jeder Interhäutungsphase ziehen sich die Jugendstadien in eine Erdritze zurück und graben dort eine kleine Häutungskammer, in der sie 4~5 Monate verharren. Nach der Häutung wird die Exuvie unversehrt zurückgelassen. ~ Bei der offensichtlich äußerst ähnlichen Lebensweise von S. duricorius und S. rubens darf sehr wahrscheinlich ein großer Teil dieser Daten auch fürS. duricorius gelten (vgl. Kap. 1.3.7.). ·
Siro carpaticus RAFALSKI, 1956 Abb. 69~75
1956 Siro carpaticusRAFALSKI, Sprawozd. Pozn. Tow. Przyj. Nauk za, 1. und 2. Quartal1955:49. 1958 Siro carpaticus - RAFALSKI, Acta Zoo!. Cracov., 2: 521.
Habitus und Feldkennzeichen: auffallend klein, kurzbeinig, rnilbenähnlich, !'Ostbraun; in dicken Fallaubschichten; N-Karpaten.
Kennzeichnung: Körpermaße (in mm; nach RAFALSKI 1958, Werte aufgerundet): L .j" 1,53.
Laufbein I (.j", in Klammern ~): Fe 0,52, (0,48) Pt 0,27 (0,27) Ti 0,35 (0,34) Mt. 0,21 (0,22) Ta 0,46 (0,43).
Genjtalmorphologie (Abb. 69-70): Truncus penis kurz-gedrungen, di breit gerundet und dort unregelmäßig spitz alisgekerbt; di Hälfte dorsad aufgewölbt, dort in einer ve Kavität der Greifapparat: laterad weisend 2 massige, di kurz verjüngte, stark laterad gekrümmte Zapfen, auch deren Innenseite glatt, ht dieser Zangenglieder 2 kurze häutige Loben- wahrscheinlich die Widerlager der Z<tpfe:1-glieder.-
Chaetotaxie: do 6, di 6, ve 3 und Ia je 4 Zapfen bzw. Borsten. -Ovipositor (Abb. 42): 20-23 Glieder, an der di Furka jeweils sub-di 1 aufge
faserte Bo mit 8-9 scharfen Spitzen, auf den Furkaästen auch je 10-13 einfache Sinnes borsten.
Cheliceren (Abb. 71): schlank, beide Cheliceren-Finger mit 8 runden scha.rfkantigen Zähnen.
Pedipalpus: schlank; Tr do-di, weniger ve mit einigen spitzen Do, übrige Glieder·rauh, ohne auffällige Do, Fe schwach gekrümmt, Pt distad deutlinh erWt:Jitert, Ta spiudelförmig, alle Glieder fein beborstet, Ta besonders dicht.
Laufbeine (Abb. 72): kurz-plump, vön Tr bis Ti rauh bekörnelt, schütter mit langen Bo besetzt, besonders dicht auf Ta 1 ve. - Drüse des Ö"-Ta IV: etwa 70 Einzelzellen, deren Sekret .in 2 Kanälen gesammelt wird, diese noch unterhalb des Konus vereinigt (RAFALSKI 1958). Öffnung des Ausfuhrkanals (AdenoHtyle, Abb. 73-74) auf einem gedrungenen, di quer abgestutzten Konus, diesem sub-di der schlanke und am di Ende ventrad gekrümmte Sporn seitlich aufgesetzt; der hohle Sporn ohne Öffnung.
Genus Siro 67
\ ( . 'l
"".'
··.· .~.
..\bb. 69-75. Si•·o carpaticus RAFALSKI. - 69. Penis von ventral, 70. von lateral, a Ia Lobus der Gegenseite; mit Chaetotaxie-Diagramm (Unterseite entspricht ventral); 71. d'-Chelicere von medial; 72. d'-Laufbein I von lateral; - 73-75. Adenostyle des d'-Tarsus IV: 73. d' von Sanok; 74. nach Holotypus; 75. d' von Sanok. - Ahb. 69, 70, 73-75 nach RAFALSKI 1958.
Dorsalseite: Zeichnung und 'Färbung: einfarbig rötlich braun, einheitlich fla?h granuliert. Vorderrand des Ceph wenig konvex, insgesamt bilden die frontalen, klemen und scharfen Tu eine seichte Kavität; schütterer Besatz feiner Haare frontal auf dem Ceph bzw. auf den hinteren abd Areae. - Op anale etwas in Längsrichtung gestreckt, stumpfer Kiel me.
Verbreitung: Typ: Endemit der N-Karpaten; wahrscheinlich weiter verbreitet als heute bekannt. - Locus typicus: polnische Waldlmrpaten; Bezirke Jaslo. Krosno, Sanok, Lesko. · 6•

I I
68 Familia Siro,;idao
. ~'o~en: ~al~k~rpaten: Boz. Jaslo, südl .. Zmigrod Nowy; Bez. Kt·o•tw; Jl,.,, Hnnok, Bnl'g Oil1 l\.anueu be1 Sanok; Bo?.. I.Jesh:o, 'Vothna.Kett.c, s·iidwe::;tl. HüngP dC\H HPrJte~ .Hnatowe Borclo (RAFALSKI 1958). · ·
Dies_e Fundorte liegen in unmittelbarer Grenznähe zur ÖSSR und UdSSR."" daß ca~-pat1-cu.9 w~hrs~heinlich auch dort vorkommt. - 8. ca1-paticu8 hat die Glazialzeiten offenswhtl_wh an seinem heutigen Verbreitungsgebiet verbracht. Das heut., hnkannte Areal hegt nur wenige Kilometer vom Rand des InlandeisschildeN wäiiIei~d seiner größten Ausdehnung entfernt. . Ö~ologie: Biotopa.~sp_rüche: Die vier hisher bekannten J<'undpunkte liegen m_ dtchten_ und ursprunghohen Laub- und Mischwäldern, meist Buchen. fern .. r· Etchen, Bn·ken, _Tannen und Kiefern, mit Unterwuchs aus Haselnuß-~büsch. !>ort lebt_ carz~at!cus in dicken und dichten Fallaublagen oder unter großen, tief ms Erdreich eingebetteten und von Moos überwucherten Steinen. ~ Vertikalverbreitung: Die Fundorte liegen in 300 (.hwlo), 440 (i':lanok),
4o0 (Kro::;no) und 750 m (Lesko; RAFA.LSKI 1961).
Subordo Lamatores
1870 Laniatores •.raoRELL, Ann. 1\Ius. Genova, 8: 469. 187!l Mecostet-hi SrMON, Ann. Soc. ent. Beige, 22: 183. HJ23 Laniatores - RoEWER, Weberknechte d. Erde: 55.
K~rpergli~derung: Dorsum im typischen Falle mit Scutum magnum, selten ( Oncopodmdea) nn~ S. completum; Segmente des Ceph nicht abgesetzt bzw. erkennbar; ~ymp_h;n nnt !lOhen Verscl~mel~ungsgra~en der Scutum-Sklerite (S. magmmL Immei : )._ - Op1sthosoma meist mit 8 Tergiten und Operculum anale, selten (einige Travummde~, BRIGGS 1971) Tergit IX frei, quergestellt vor dem Op an; 9 Sternite Corona anahs fehlend. ' . Prosmnal-~ternun~ (soweit v?rhanden): lang und schmal, bei einzelnen Gruppen
c,l-Udad erweitert (Tr1aenonycludae); Op genfastausnahmslos klein und weit dixtad vorgeschoben, Prosomal-Sternum teilweise bedeckend.
Genitalmorphologie: Penis meist in Truncus und Glans geglieclf•rt; Glans entweder durch Muskel-Sehnen-Zug (dann immer 1 Muskelstrang) oder durch Hämolymphdruck (Muskel fehle»;d) bewegbar. Innerer Genitalapparat mit Propulsion~Ol:~an (Sper~abesa~ung ~uttels Kopulation). - Ovipositor immer kurz-ungeglied~rt, nur ~ptcal mit we~ugen Borsten, Apex 2-'-4lappig, innerer Bau (? meist) nut 4strahhger Symmetne: 8 Rec sem oder Vielfaches davon. _Pedipa~pus: kräftig, Klauen immer entwickelt, als Subch~la dnschlagba.r und
nut kraftiger Bedornung Raubbeine bildend. · .. ~aufbeine: 2. länger als 1., z. T., auch 4. Bein verlängert, Cx I beweglich, die ulmge1~ ~tar~·; Lohns der Cx I unbeiYeglich, ohne freies Sklerit. - KIHnen: an Bein T ~nd. II Je 1 emfac~1e Klaue, an B~in. III und IV r~gelmäßig komplexer:Doppelklaw,, ~cluldl~laue, Dreizacl~klaue ~ ~ratarsuK Ill und IV der Nymphen meist mit Arohm.n. (mcht: On.~opodHlae, 'Inaenonychidae). - Plantigrad.
1-Jt~nkdrüsen-OffJ!.ung am Scutum-Rand oberhalb des Vordorrand<!x \·on C!x 11. , ~hederung: 3 Überfamilien: Oncopodoidea (nur OncopodidaP.). Tra 1•1w.ioidea
(C.Jn~derung vgl. 8. 52, 69), Gonyleptoidca (1·gl. H. 52, 86).
Genuß Siro ß9,
Bestimmungstabelle für die Überfamilien und die Familien der Laniatores
1 Kralle von Laufbein III und IV als do-ve abgeflachte einästige Schilcll<ralle mit beidseitiger grober Bezahnung (Abb. 76, 77). - Freilebend in den S-Alpen, troglobiont im jugoslawischen Karst. . . . . . . . . . . . • . Travunioidea, T1·avuniidae, S. 69 Kralle von Laufbein Ill und IV zweiästig, die beiden Einzelkrallen la glatt (Abb. 100) 2
2 Fe des Pa do mit 3-4 auffälligen stumpf-kegeligen Do (z. B. Abb. 101). - Weitverbreitet in den O·Alpen •.•...•.... , • . . . Travunioidea, Erebomastridae, 1
1•. 76
l<'e des Pa do glatt, allenfalls mit einzelnen niedrigen 'l'u oder Borsten (Abb. II9). - LGkal in N-Italien und Istrien. • • • . . • . . • . . Gonyleptoidea, Phalangodidae, 8. 87
Suprafarnilia Travunioidea
1958 Travunioi<)ea KRATOCHVlL, Pr!i<Je Brn. zakl. lJesk. Al<ad. ved, 30: 380.
Eine Überfamilie der Unterordnung Laniatores, gekennzeichnet durch: 0'-Genitalmorphologie: Penis deutlich in Truncus und apikale Glans gegliedert, diese durch Muskel-Sehnen-Zug gegen die Längsachse des Truncus beweglich. Der einzige Muskelstrang entweder fast auf den ganzen Truncus verteilt oder in bulbusförmiger Basis konzentriert.
Ovipositor: kurz und ungegliedert, etwa so lang wie breit_ oder wenig länger als breit, di zu 4 stumpfen Loben ausgezogen; Bewehrung mit Borsten spärlich.
Sternum meist vorhanden, oft nur rudimentär keilförmig. Krallen der Tarsen der Laufbeine III und IV: eine Hakenkialle mit Zahnung
la (selten zusätzlich ve) oder als ungezähnte Doppelkralle auf gemeinsamer Ba~k Gliederung: Die Überfamilie umfaßt die Travunüdae (disjunkt und wahr
scheinlich reliktär holarktisch), Triaenonychidae (vorwiegend disjunkt süd-hemisphärisch) (S-Afrika mit Madagaskar, Australien, Neukaledonien, Neuseeland, SAmerika, subantarktische Inseln, reliktär im nordwestl. N-Amerika), Erebomastridae (reliktär holarktisch), Synthetonychidae (Neuseeland), Pentanychidae (westl. N-Amerika).
Familia Travuniidae
1923 Pelta.eonychidae ABSOLON & KRATOCHViL, Pfiroda, 25: 15:l. 1932 Travuniidae AsscLON & KRATOOHViL, ;\iit.t. Höhlen- u. 1\:arstforschung. Berlin 1932
• (3): 9 (nom. nov.). 1933 Tria.enonychidae-Travuniinae - HADZI, Bull. Aoad. Scieno. Math. Natm. Bcograd,
B 1:68.
Eine Familie der Laniatores-Travunioidea, gekennzeichnet dm<ch: Q'-Genital: morphologie: Truncus entweder extrem schmal und Glans gelenkig abgesetzt (z. B. Peltonychia) und Muskel in bulbusförmiger Basis konzentriert oder TruncuR untersetzt, Muskulatur fast übe1>"den ganzen Truncus verteilt, Glans nicht gelenkig abgesetzt (z. B. Dind1'ia).
Tarsalkrallen der Laufbeine III und IV zu Schildkralle (Peltonychium) differenziert: do-ve abgeflacht, seitlich mehrfach gezähnt, oft unsymmetrisch.
Sternum zwischen den Cx III und IV ausgespannt, keil- bis stäbchenförmig. Verbreitung: disjunkt und offen::;ichtlich reliktär in S-Europa, 0-Asien und N
Amerika. - Europa: genitalmorphologisch geprüft und sicher abgrenzbar bisher nur 2 Gattungen: Peltonychia RoEWER und Dina1·ia RoEWER. Bisher nicht genital-

I
70 li'amilia. Travuniidae
morphologisch differenzierbar sind die Gattungßn Kr.t;ttochvi~iol(t_R,Q.~.\YI!JR,, Arhasu.s· RoEWER, Abaaola· STRAND, Trav·unia Aßso:Lo:N''& 'k:RA.To<iH:VfJ:, 'uiid 'lJdema?'i.noa Roil\v:E&: Ehi'Teii der betroffenen Arteil ist·iiuf P~ltonychlli 1 1Hiri'Diriaria zü h<'-
zie!r~h; ~i'Ä'k~~ i~·,f~l>~~~ uh4 ~~r~.~'~ dj:, Mr~OSHI ( ~~~,;,): ;~q~Uz;';l~~ ·.; J 9~6, 1972) zu Pelton,ychta stellen,,di!l;!llbe;r,()In \)Igenes, G:en)lpl;lprä~()ntw~·e,n vw:t Y~rza pulcra SuzJJia .. nah!lst.l;lb,en.:,:Nenerd.ings, ;stellt SuzuiU (I9~5a, b) alle jap~niti,chea und koreanischen: Arten d!ln ;Travuuioidea.zu den Tr.iaenonychidae (8 Arte.n i.' •nur Yuria pulcra• bleibt· bei' den .!I'ra vunüdae. Die genitalmorphologischen Ktutei·ieu s.cheint> n diese Aufteilung nicht zu rechtfertigen.
N-Amerika: I troglobionte Art im Staat Washington, Spelr:onychia aengeri BRIGGS. · !.L_ , . . . .,., .. 'i
Einen Überblick über die Eutdeckungsgeschichte der Formen deR .iugoshtl~iHehen Karstes gibt HAnzr (1933). · • ..
f I! .,0, ~ • " ... f ,·;,_,"
GBausiPeltonychia RoEWER,' 1935 ... ,
102ii Ai;~.lota·~".ll~~~; i~~i. A~z. 7'{~5 'c/.r~m., n. fiir,Abs~lonia Hdi!ii\:~rt; pm·r.: A. sin'ea RoE-' '\~~k''i9!i5):· .. '"'' ·•·;< .. · ,,,,,::·. · ....... ;;,·• ..•. , .. , ., ·, .. , .• ," ;, ;.! . "·' . "
1935 Pelt~nycMa RoEw~a, Arch. Zoo!. exper. gen., 78: 55. . . . ""· 1935 1Krato6~i;iliolti· Roi;rwfua/ Aroh': 'ZboE'expe~. ·gen'.; 78': ·64''/part:;' 'nhr 'd ;,; 'Mt.eii i;itroli'llht
RoEwER·und· caverrdcola· ·RÜEWEit;'tStB.tus vön-·navarica.: (SIM6N] bisiltw··.liilgelirüft). 1935 Hadziana ROEWER, Arcl! .. Zoi>I..exper. gen.,•7.S:.69. ,,.,,., '· · .:,.·;
Begriirfd~hg d~r:s;Yn~Jynii~·: ·~o!;,o/:Em (l9:i5) 't.rennte die ~Gattung~h d\Ji· Tranmiida•• auRschließiich 'mittels Merkriuileh d~r"Mßiir~n''lHorplü'>logi&, voi allen't• n~Clt d~r Glitederung dP!' I.aufbeincTatisen iiiitl. d.er,Bestacheluiig des Pedipa1pus: Die~e -Metkniale, die: bereit" beidsnitig an einem ]ndividllull?- •V.a.rliereQ. könnehJ 'llnlreißen 'keine·sfß.Us:Artengruppen natüi·Jir·lu·{J· Ver. wandtschaft. Verl]fßli~he;I;Iiinveise gibt .a110li·.an dieser Gruppe:aUe:hl'der Bau 'des·ö'·Gimitnlappara,t.es, ·Demnach< sind ,cli~·'<ht~ndeJC. in der ,:J;.is~e.:genannteÄ .. Gattung~n·al•.·!Iii"hst .. veqv>!n<lt. anz_98Ahon u~.d. _ill. ßine~ .e:i~zig~n GattQ~lg --~?-s~m~~~J.~uf~~.sen_." Solltß ~~_ich .;l.ba~spla,; ·/.'tnq{nd?t.'···'i (Hoiiw.l&, i9i5) ',;, Ab;ol~n·ia. C .. :·die 2. A;t ~~~r Gatt~ng' Abds~ia :.:,"~,;;;.h ;,.·~,: f>.~(i~;,u,;hw gel1örig orweisen~ so müßte diese Gattung aus Prioritätsgründen Abasola 8Tu\:~~~J·T928 Jw.lf.ll'!L Da. aber Abasola nur von HoEWER (1935) und JUBERTHIE (1968, 1972) lwuurzt worden i•t, kann nicht von allgemeinem Gebrauch dieses Nan1ens gesprochen worclel!.j-:~.~:~P.-l!tr(·:'•titerhin der eingebürgerte Name Peltonyokia RoEWElt, 1935 verwendet werden.
Eine Gattung der Travuniidae;"gekennieichnet durch: '"" d'-Gimitaln'wrphologie: Truricus '):ienis 'lliüg nnd. dürti\., par;ilie1i'lmdig·, iiul· cli~
Basis. ~rw«lit~~t !J,bgf!R~~zt .\ll1d, d9,1;t die .:(v,(u*ql~~urlw:q~l)p.tr,i\lF~i· (),i. 1lnt~;r}l~tl!J dPI' Glans knopfförmig angeschwollen. Glans in gerader Verlängerung deR Trunom•. d~r?h. )~i;us}~?lzu~. ~1w~g!)c,h:. Gl,~ps, g~rup.g~t}.ä!;t~]ioh! g~; !;t)~, I, grc~ßer·: L~~u~, an f die~ell:l; .. 2. Smzws~()r~ten,;._pifnung de,s S!tmeqkanal~. 1/-,U( eiTlem Ztpfel di;nlf. -~: ]:;eg_llr?lir~:i~ur~,)~ { Z'ipfehi ,vedangert. iitit; mft elnzel~eii. 13;~; }:aum 'Iang~>t· als (lie ,Lqpen. ~arsalgliederung der. Laufbeine artverschieden; 'he.i trnglobiontPil Arten oft beidseitig variierend. Tu oc groß, et1i'as vom Cepli~Froi1takand 'a bgtTilokt. bei Troglobionten reduziert. Kleine Formenvon I bis'fast 2 mm" . . . ·~ .. ·,
Ver~reitu~g: ~t\y(t.,l? Ar,t~ß I~JI!gqsll\.~':i~o\~~~ ·~cil'r~:i; 'h{,~e~~ ~~AJ!le1~; {h d{·n Pyrenat)n und Ifftp.tabri~clH;nyeR)fg':Jl.;J?Ie ~'Pl!l'll,I~.cl!J~ben~ttA ~~:t~n 1~e.hnwn rlen n:eitaus g~·ößten' Tei\. C~flS. ~~al~, eiP, aJl~I:i~che ,l:J:al!JjnseJ),. ()QligatoriKche TroglooJnonpen ko";ll.men nur lokal Im Karst und m den Pyrenäen vor.
Typus-'A;·t (Festlegung <Iurch RoEWER l93;i): Sco!olemo" lepr·ieuri I.uc.". ISMI.
Bestimmungstabelle für die Arten (Unberücksichtigt bleibt P. !enius RoEWER)
Genus Peltonychia 71
1 t:lcutum do grob granuliert, Hinterrand <!er freien Tergite gesägt (Abb. 78)i H.öhl~n S u~~ l<'reiland der S-Alpen . . . . . . · · · • • • • • • • •. · · • : · epr~eur~, · Scutum do matt, nicht grob granuliert, Hinterrand dar freten Tergite glatt (~~b. s: • 94~
postumicola, s. n 2 Glans pen~s länger als ?reit (~hb. 8
9) · . · :. ' · j1s~ ~ u 'rr:es~e~ ~0~ ·Giansgelenk bis zur
Glans pems so lang Wie breit (Abb. 82' Langsac gabria, S. 73 Ansatzetelle des me Stylus) . · · · • · · · • · · · · • ' ' '
Peltonyehia Jeprieuri (LucAs, 1860) Abb. 76-80
1860 Sco!ole.moll lepriewii LucAs, Ann. Soc. ent. France (3) 8: 974. 1923 Scotolemon leprieuri - RoEWER, Weberknechte d. Erde: 100. l!l28 8colo/e.mon helveticus SCHENKEL, Zoo!. Anz., 78: 4~. . 1935 Peltunychia leprieu1·i - RoEWER, Arch. Zoo!. exper. gen,, 78: 56. l9:l5 Kra!ochviliola parolinia RoEWER, Arch. Zoo!. exper. gen., 7'8: ~7. . .
Begründung der Synonymie: Von allen genannten Arte~ Ia? typisch~s bGz,~·ß. topBotyp:ches d 1 \? A ffällige Überemstimmung m ro e, ewe rung
(leprieuri) Material vor'. je oc I nu: ,·. ;I m des Tu oc lassen kaum Zweifel, daß lediglich des Souturn und der Pedipalpen sowie < te or eine einzige Art anzuei•kennen ist. Die Bestätigung durch J steht aus. . .
Habitus und Feldkennzeiehen: klein und kurzbeinig, rostgelb; m Freiland und
Höhlen der S-Alpen. S t )·I 55 b' Kennzeichnung: Körpermaße (in mm): L ~ (c3' unbelmnnt; nur cu um · , I:;
I,~~ufbein n (~, c3' unbekannt): Fe I,I5 Pr ~,4 Ti I,O ~t I,O Ta 1,15. Genitalmor hologie: unbekannt, da keme c3' ~orhegen. . ... Cheliceren {Abb. 79): Basalglied schlank, do-ch Buokel schwach ausgepragt,
2 Glied massig, mit kräftigen Bo vor allem frontaL . .. . · Pedipalpus (Abb. 80): mäßig schlank; Fe do nur werug ~ewo~bt, dor~. ~~:
zahlreichen kleinen Do. - Übrige Bedornung: F~. ve .4-5, ~edm~ 2 '.Pt me Ia s 1 weiz) Ia 3 · Ta medial und la je 2, Ia zusat,zhch I nel klemerer Do. .
( -t~u~~eine': alle' Glieder rund, fein bekörnelt un~ behaart, vor.allem Pt und T~ kräftig behöckert, im Profil "gesägt", vergleichsweise kurz .. ~. Ghed~ru~g derira.
· · B d II' Orso)· I 3 II - III 4, IV 4; "helvetzca (Tessm). I 3, 5, ii{~e~:"~ 3u~o r~ 4· parolinid": I 3,'n 6/7, III 4, IV 4. (Unterschiede an ?eide~ l::ieiten ein~s /Tieres si~d durch SchrägRtrich angezeigt). - Kralle de.s Laufbems I\
vgL Abb. 76-77. I... d k I rostgelb Seg Dorsalseite (Abb. 78): Zeichnung und Färbung: \.orper un e ' . -t auf dem Souturn hell abgesetzt. - Bewahrung: Souturn und freie
mAn .gre;~en kt' t f den thorak und abd Areae Felder grober Körnchen, T~rgite eidndpunf .Ier T' au sa"g,t - Tu oc· groß hochgewölbt, kräftig bekörnelt, Hmterran er rc.Ien erg ge · · ' .. II · b frontal steil abfallend, vom Ceph-~ronta~rand etwas abgeruckt; Augen { em, a er deutlich ausgeprägt und k.räftig pigmentiert. C
Verbreitung: Typ: Endemit der zentralen S-Alpen.·- Locus typicus: omer See Höhle Buco dell' Orso. ..
.' , · . Tcssin Val Bedrett.o (SOHE><KEL 1928); Tessin, Höhle Ca' di Vecc, Frasco (DR~eco Sch" ~IIZt ·,.... 'comer See Buco clell' Orso (LucAs 1860); Trient.inei' Alpen, Brenta-fal .. 1959). - a 1en: . , · .
Höhle Paroliui bei Oliero (RoEWER 19:l5).

72 Fatuilia 'l'ravuniidae Uenus Peltonychia 7H ·
Ökologie: Biotopansprüche: P.leprieuri lebt im Freiland wie in Höhlen, doch werden Höhlenbiotope offeilSichtlich nur zufällig besiedelt. Die morphologischen Verhältnisse deuten auf eine Freilandform.
Vertikalverbreitung: die beiden datierten Funde in der Schweiz bei 1400 m (Val Bedretto) und 1215 m (Ca' di Veco); die Höhle Buco dell' Orso liegt auf 615m.
Peltonychia gabria RoEWER, 1935 Abb. 81-87
1935 Peltonychia g<1b1'ia RoEwER, Arch. Zoo!. exper. gen., 78: 60.
Habitus und Feldkennzeichen: auffallend klein und langbeinig, hellgelb. Troglobiont des jugoslawischen Karstes.
Kennzeichnung: Körpermaße (in mm): L ~ 1,1; L ~ 1,25. Laufbein II (~, in Klammern ~): Fe 1,15 (1,05) Pt 0,40 (0,35) Ti 1,0 (0,9) Mt
1,0 (0,9) Ta 1,25 (1,15). Genitalmorphologie (Abb. 81-84): Truncus penis lang, dünn und parallel
randig, Basis abgesetzt; Glans etwa so lang wie b:ceit, la-di in 2 stumpfe und kurze Loben verlängert, dort 2 unterschiedlich lange Sinnesborsten, die beiden la lang und rechtwinklig abstehend; Samenkanal auf einem di-me Stylus mündend.
Cheliceren (Abb. 86): di Höcker des Grundgliedes stark entwickelt, 2. Glied klobig, frontal mit einigen Höckern und Bo.
Pedipalpus (Abb. 85): schlank; Fe ve 4 (~: 3), medio-di 1, Pt: medial2, la I; Ti und Ta medial und la je 3 große Do; Tarsalkralle vergleichsweise klein.
Laufbeine: kurz (II: 4,7 mm), alle Glieder rund, fein beborstet; Tarsalgliederung: I 3; II 5/6; III 4; IV 4.
Dorsalseite (Abb. 87): Zeichnung und Färbung: Körper zeichnungslos einfarbig rostgelb. - Bewehrung: außer feiner Granulierung glatt, Segmentgrenzen meist deutlich abgesetzt. - Tu oc: kräftig entwickelt, breit glatt, zum Rand des Ceph sanft abfallend. Linsen der· Augen kräftig entwickelt, diese stark pigmentiert; sie wirken nicht wie die Augen eines Troglobionten.
Verbreitung: Typ: Endemit des jugoslawischen Karstes. Locus typicus: ~iehe "Jugoslawien".
Jugoslawien: Slowenien1 Gabria jama bei Triest. - Nur vom Locus typicus bekannt.
Ökologie: Biotopansprüche: Die Typenserie stammt aus einer Höhle. Die äußeren Merkmale sind z. T. aber nicht die eines e:x:tremen Troglobionten (z. B .. Pigmentierung der Augen). Es ist trotzdem unwahrscheinlich, daß P. gabria auch epigäisch lebt.
Peltonychia postumicola (RoEWER, ·1935) Abb. 88-94
193ö Hadziana postumicola RoEWER, Arch. Zoo!. exper. gen., 78: 69. 1935 Kmtoohviliola cavemioola RoEWER, Arch. Zoo!. exper. gen., 78: 68.
Begründung der Synonymie: Von K. oavemicola sind nur 'j! bekannt, so daß ein schlüssigeL' Vergleich mit H. poBtumicola vorerst nicht möglich ist. Weitgehende Übereinstimmung in der Proportion und Bewahrung der Peclipalpen und der Cheliceren und in der Körpergröße deuten an, daß beide Arten synonym sind. Die Bestät.igung durch eS bleibt abzuwarten.

74 Familia Travuniidae Genus Peltonychia 75
Habitus und Feldkennzeichen: auffallend klein und langbeinig, hellgelb. Troglabiont der SO-Alpen und des-Karstes.
Kennzeichnung: Körpermaße (in mm): L Cf 0,95-1,1; L ~ 1,1-1,3. Laufbein II (Cf, in Klammern ~): Fe 2,40 (2,55) Pt 0,45 (0,55) Ti 2,05 (2,05)
Mt 1,75 m Ta 1,5 (1). Genitalmorphologie (Abb. 88-89): Truncus penislang und schmal, Musku
latur in der wenig erweiterten Basis konzentriert. Glans länger als breit, unterhalb zweier di-la Loben eingeschnürt, auf diesen Loben je 2 Sinnesborsten. Mündung des Samenkanals auf quer abgestutztem Stylus di-me.
Cheliceren (Abb. 91): Grundglied di mit do Buckel, Basis nur schwach erweitert, 2. Glied frontal mit wenigen Tu und aufgesetzter Borste.
Pedipalpus (Abb. 90): auffallend schlank. Fe ve 3, me-di 2 Do, Pt medial 2, ve-Ia 1; Ti und Ta medial und Ia mit je 3 großen Do; alle mit langer, di säbelartig aufgesetzter Borste.
Laufbeine: zur Körpergröße vergleichsweise lang (II: über 8 mm), dünn, glatt, nur schütter behaart. - Gliederung der Ta nur geringfügig variierend: Ta I: 3/4, II: 5-7, III: 4, IV: 4. Bisweilen rechts und links an einem Individuum variierend (durch Schrägstrich angedeutet). Kralle von Laufbein IV vgl. Abb. 92-93. .
Dorsalseite (Abb. 94): Zeichnung und Färbung: einfarbig rostgelblich ohne deutliche Zeichnung. - Bewehrung: bis auf feine und gleichmäßige Bekörnelung glatt, verstreut winzige Bo; Segmentierung in Areae deutlich. - Tu oc: kräftig entwickelt und hochgewölbt, Linse der Augen winzig, bei den meisten Individuen mit kräftigem Pigmentrand.
Ökologie: Biotopansprüche: Die morphologischen Verhältnisse deuten auf eine obligatorisch troglobiont lebende Art; Freilandfunde sind nicht bekannt und auch nicht zu erwarten.
Verbreitung: Typ: troglobionter Endemit der SO-Alpen und des jugoslawischen Karstes._ - Locus typicus: Adelsherger Grotte.
Italien: Grotta del Puint bei Pradis/Friaul ("cavernicola"). - Jugoslawien: Slowenien, Po•tojnska jamafAdelsberger Grotte bei Postojna (postumicola).
Peltonychia tenuis RoEWER, 1935
1935 Peltonychia tenuis ~OEWER, Arch. Zoo!. exper. gen., 78: 59.
Der Halotypus ist ein Jungtier, dem jegliche artspezifischen Merkmale fehlen; er kann deHhalb nicht zu den bekannten Arten in Beziehung gesetzt werden. Neues Material vom Locus t.ypicus, der Martinova jama bei Matteria/Istrien, ist dazu notwendig.
Abi>. 81-87. Peltonychia gabria RoEWER. - 81. Glanspenis von lateral, 82. von ventral; 8:J. Penis von lateral; 84. von ventral; 85." Q'-Pedipalpus von medial; 86. Q'-Chelicere von meclial; 87. Q'-Habitus von dorsal. - Holotypus; YU Gabria Jama.
Abb. 88-94. Peltonychia postumicola ROEWER. - 88. Glanspenis von lateral; 89. von ventral; 90 .. :)-Pedipalpus von medial; 91. Q'-Chelicere von medial; 92. Laufbein IV, letztes Tarsalglie.rl mit Klaue von lateral; 93. Tarsalklaue in Aufsicht; 94. 6'-Habitus von dorsal. - Holot.yJ.mS; YU Adelsherger Grotte.
I

76 Familia Erebornastridae
Familia Erebomastridae
1935 Oladonychiinae ILmzr, Bio!. Generalis, 11: 69. 1969 Erebomastridae BRIGGS, Pan-Pac: Entomologist, 45: 37.
Eine Familie der Laniatores-Travunioidea, gekennzeichnet durch: Q'-Genitalmorphologie: Penis in Truncus und gelenkig abgesetzte Glans geglie
dert, Muskulatur entweder in kleiner bulbusförmig abgesetzter Basis konzentriert (Holoscotolemon) oder auf den größten Teil des Truncus verteilt (amerikanische Gattungen). Glans durch Muskel-Sehnen-Zug beweglich.
Tarsalkrallen der Laufbeine III und IV mit Doppelkralle auf gemeinsamp.r Basis; Krallen selten mit einigen Zähnchen.
Sternum: schmal stabförmig zwischen den Cx II und III, weitgehend mit ihnen verwachsen und oft kaum differenzierbar.
Coxae der Laufbeine II mit großem me Lobus disto-ventrad gerichtet, dM Mundfeld caudad abschließend.
Verbreitung: holarktisch; HoloscotolBmon ROEWER (einzige europäische Gattunl!"lErebomaster CoPE, Oryptomaster BRIGGS, Theromaster BRIGGS, Speleoma8ter BRIGns (N-Amerika).
Genus Holoscotolemon RoEWER, 1915
1915 Holoscotolemon RoEWER, Arch. Naturgesch., 81: 5. 1915 111etascotolemon RoEWER, Arch. Naturgesch., 81: 6. 1935 Querilhacia RoEWER, Arch. Zoo!. exper. gen., 78: 37. 1935 Scotolemops RoEWER, Arch. Zoo!. exper. gen., 78: 40. 1935 Scotolemoniscus RoEWER, Arch. Zoo!. exper. gen., '18: 42. 1935 Brigestus RoEWER, Arch. Zoo!. exper. gen., 78: 45. 1935 Lucassa RoEWER, Arch. Zoo!. exper. gen., '18: 48. 1935 Bukowir1a RoEwER, Arch. Zoo!. exper. gen., '18: 51. 1973 Poleneciana HAnzr, Razprave, Dissertat. Ljubljana, 16: ll. 1973 Trentania HAnzr, Razprave, Dissertat. Ljubljana, 16: 14. 1973 Sketia HAnzr, Razprave, Dissertat. Ljubljana, 16: 16. 1973 Euscotolemon HADZr, Razprave, Dissertat. Ljubijana, 16: 18.
Begründung der Synonymie: Alle europäischen Arten der Erebomastriclae stimmen in der Genitalrnorphologie, in Gestalt und Lage der Ox II und in Gestalt und Bewehrung der PNlipalpen prinzipiell üb~rein; es ist berechtigt, sie in einer einzigen Gattung zusammenzufa..,~en. Früher benutzte Merkmale, vor allem die Zahl und die Verteihmg der Tarsalglieder der Lmtfbeine, erscheinen für die Gattungsgliederung nur von geringem Gewicht, wenn nicht gar b(\· cleutungslos.
Eine Gattung der Erebomastridae, gekennzeichnet durch: Q'-Genitalmorphologie: Trunous penislang und auffallend schlank, Basi.~ schwach
erweitert, dort die Muskulatur konzent'riert, Truncus sonst meist parallelranclig, nur unterhalb des Glansgelenkes wenig angeschwollen. Glans in gerader Verlängerung des Truncus, durch Muskel-Sehnenzug beweglich, do-ve abgeplattet, di mit 2 Paar Sinnesborsten. - ~: Legeröhre in 4 Zipfel ausgezogen; der ve Zipfel kürzer alR die übrigen; nur ausnahmsweise spärlich beborstet. - Tu oc niedrig, um seinen LängsmesHer vom Ceph-Frontalrand entfernt. - Pedipalpus: massig-gedrungen. - Ta der Laufbeine I und II mit lästiger, III und IV mit 2ästiger Kralle, heide Äste auf kurzem, ·gemeinsamem Stiel. - Cx II ba zu 2 langen, me-di Apodemen
Genus Holoscotolemon 77
ausgezogen zur Abdeckung des ve-caudalen Teiles des Mundvorraumes. Chcliccren-Grundglied mit ve-ba Apodem.
Verbreitung: 5 Arten allopatrisch in den Gebirgen 0-, Mittel- und W-Europas; jaqueti in den Karpaten und S-Jugoslawien, unicolorin den 0-Alpen und NW-Jugoslawien, lessiniense in den zentralen S-Alpen, o1·eophilum in den SW-Alpen und q·uerilhaci im Zentralmassiv. - Holoscotolemon hat in der Paläarktis keine Vervmndten; weitere Gattungen sind in N-Amerika verbreitet.
Typus-Art (durch Monotypie): Holoscotolemon unicolor ROEWER, 1915
Bestimmungstabelle für die Arten (Gilt nur für ~. ohne H. querilhaci, siehe S. 86)
1 Glanspenis pantoffeiförmig ausgehöhlt; Sinnesborsten unauffällig klein (Abb. 104-105); im Karpaten-Bagen und in S-Jugoslawien . . • . . . . . . . . . . . jaquetii, S. 80 Glans penis nicht pantoffeiförmig ausgehöhlt, sonelern plan (z. B. Abb. 98); Sinnesborsten groß um! zapfenförmig; in den Alpen und in NW-Jugoslawien . . . . . . . . . . 2
2 Truncus penis an der Basis und in der Mitto schwach bauchig erweitert; Glans außer clen zapfenförmigen Sinnesborsten Ia mit je 1 basad weisenden Haken (Abb. 98) . . . . . •
· unicolor, S. 77 Trunous penis nur an der Basis bauchig oder sackeiförmig erweitert, in der Mitte parallelranclig oder schwach konvex . . . . . . . . . . . . . . . . . . . . . . . . . 3
3 Glans penis Ia (von clo oder ve betrachten) konvex, breiteste Stelle so breit oder geringfügig breiter als das cli Ende des •rruncus (Abb. 109). . . . . . . . . oreophilum, S. 85 Glans penis Ia (von do oder ve betrachten) geradlinig oder geringfügig konkav, wesentlich schmaler als das di Ende des Truncus (Abb. II3) . . . . . . . . . . Jessiniense, S. 82
Holoscotolemon unicolor RoEWER, 1915 Abb. 95-102, 107 191ö Holoscotolemon unicolor RoEWER, Arch. Naturgesch., 81 A: 5. 1923 Holoscotolemon unicolor - RoEWER, Weberknechte d. Erde: 102. 193ö Lucassajerruginea RoEWER, Arch. Zoo!. exper. gen., '18: 52. 1949 Holoscotolemon jusitarsus RoEWER, Senckenberglana, 30: 289. 1973 a Poleneciana terricola HAnzr, Razprave, Dissertat. Ljubljana, 16: 11. l97:l a Trentania antoniana HADZI, Razprave, Dissertat. Ljubljana, 16: 14. 19i:J a Sketia borisi HADZI, Razprave, Dissertat. Ljubljana, 16: 18. 1973 a Euscotolemon novaki HAnzr, R;.zpravo, Dissertat. Ljubljana, 16: 19.
Begründung der Synonymie: Die von HADzr anfgestellten Taxa sind auf unicol01· zu beziehen; das belegen die Abbildungen der Peclipalpen und des ~-Gen~talapparates. Von Lucassa ferruginea, Holoscotolemon fusitarsus und 8ketia borisi lag Typenmaterial vor.
Habitus und Feldkennzeichen: (Abb. 96): bernsteingelb, klein und kurzbeinig. - In den 0-Alpen spärlich, aber weitverbreitet.
Kennzeichnung: Körpermaße (in mm): L (nur Scutum magnum): d' 2,0-2,4; ~ 2,2-.2,4; L gesamt: d' 2,7-4,~; ~ 2,6-3,1.
Laufbein II (Q', in Klammern ~):Fe 1,0 (1,0) Pt 0,7 (0,7) Ti 1,8 (1,8) Mt 2,4 (2,3) Ta 2,0 (2,2).
Tarsalgliederung der Beine I-IV:~ I 4 + 3 oder 5 + 3, ~ 3 + 3 oder 4 + 3; II d' 16-17, ~ 14,16 oder 17; III d' 4, ~5-6, IV~ 5-6, ~ 7. - Zahl oft rechts und links verschieden.
Genitalmorphologie (Abb. 97-99): 1'runcus penis schlank; Basis klein, distacl kontinuierlich bi~ etwa zur Mitte erweitert, dann verschmälert, unterhalb

78 Familia Erebomastridae
Abb. 95-102. Holoscotolemon unicolor RoEWER. - 95. d'-Habitus von dorsal; 96. von lateral; 97. Penis von ventral (links) und iateral; 98. Glans penis von ventral (links) und lateral; 99. Gl~ns penisvon distal (nach REM-Aufnahme; Pfeil: Öffnung des Samenkanals); 100. Laufbein IV, letztes Tarsalglied mit Klaue von lateral; 101. cl'-Pedipalpus von medial; 102. d'-Clmlicere von medial. - 95-96: Holotypus; sonst: AUS WeißbachJLofer.
Genus Holoscotolemon 79
der Glans knopfförmig erweiterb. "Glans parallelrandig, distad in 2 basad weisende hakenartige Flügel.ausgezogen. 4 Sinnesborsten: I Paar plump-hornförmige rechtwinklig zur Glans ventrad gerichtet, I Paar pilzförmige in der Längsachse der Glans. Öffnung des Samenkanals als feiner Stylus nahe der Pilzhaare (Abb. 99, Pfeil). - Legeröhre vgl. Abb. 43.
Cheliceren(Abb. I02): schlank, do-baeingeschnürt, spärlich beborstet; 2.Glied frontal mit kräftigen Höckern, davon do-frontal meist ein großer abgesetzt.
Pedipalpus (Abb. IOI): Fe do kräftig gewölbt, dort 4-5 große Do, ve 5; Pt nur medial 1 großer und 1 kleiner Do; Ti und Ta mit je 1 ve-medialen und ve-la Reihe von Do: Ti medial 3, Ia 4, Ta medial 2-3, Ia 3-4, jeweils unterschiedlich groß; ~ Ti und Ta beiderseits 4.
Laufbeine: rund und kurz, alle Glieder bis auf Ta und Calcaneus des Mt unregelmäßig granuliert und mit kurzen Borsten besetzt; Glieder bis Mt-Astragalus schuppig gra!luliert, Calcaneus und Ta-Glieder nur mit Mikrotrichien-Besatz. Kralle des Laufbeins IV vgl. Abb. 100.
Dorsalseite (Abb. 95-96): Zeichnung und Färbung: bernsteingelb, nur Augen schwarz pigmentiert; bisweilen deutet graue Chagrinierung undeutliches Muster an, sonst zeichnungslos. - Bewahrung: je eine Körnchen-Querreihe auf den abd Areae I-V, den freien Terg I-III, auf Area III oft mehr als nur 1 Reihe; 2 BoPaare seitlich am Ceph-Frontalrand.
Verbreitung: (Abb. 107): Typ: alpin-dinarisch; Endemit der 0-Alpen und des südl. Vorlandes, wahrscheinlich nur teilweise geschlossenes Areal in den nördl. und südl. Kalkalpen und im außeralpinen Slowenien. - Locus typicus: Osttirol, Lienz. ·
Österreich: N-Tirol: Kaisergebirge, Kufsteiner Raum (THALER 1966b); Salzburg: Hohe Tauern, N-Abdachung: isoliert im Kaprunerund Gasteiner Tal (Gasteiner Klamm, Naßfeld/ Böckstein); Niedere Tauern: Taurachtal/Untertauern; in der Salzbw·ger Kalkalpenzone weitverbreitet, z. B. im Einzugsgebiet von Saalach· und Salzach-Quertal: südl. Weißbach bei Lofer (GRUBER 1966), Schwarzbergklamm, Unken, Hochkönig, Hagen- und Tennengebirge, Roher Göll, Osterhorngruppe (alle A. A. leg.), Untersberg bei Salzburg (SCHÜLLER & SPERLING leg., GuuBER 1966), in der voralpinen Flyschzone isoliert auf dem Kolomansberg (A. A. leg.). Ober-Österreich: Sengsengebirge, SW-Seite (RoEWER 1949, fusitarsus); Nieder-Österreich: südl. Wang, Boz. Scheibbs (RESSL leg., GUUBER 1974a); - Steiermark: Deutschlandsberg (SCHUSTER 1960); südl. Koralpe, zwischen Mauthnereck und St. Lorenzen (GRUBER 1974a); Windische Büheln, südöstl. Spielfeld; Grazer Bergland, westl. St. Radegund (alle GRUBER 1974a); Stadtrand von Graz, Labgraben bei Gösting (SCHUSTER 1972);- Kärnten: St. Oswald ob Eibiswald (beide FuANZ leg.); Villaeher Alpe, 0-Hang; Ossiacher Tauern, N-Hang; Arria~her Klamm (alle KÜHNELT 1942); Garnitzen Klamm bei Hermagor (A. A.leg.); - 0-Tirol: Lienz (RoEWER 1915). -Jugoslawien: Slowenien, Golnik (Poleneciana terricola), Trenta (Trentania antoniana), Pilenca Höhle bei Slovenj Gradec (Euscotolemon novaki), Brezno Höhle/Mokrecu (Sketia borisi; alle HAnzr 1973a, b); Doline an der Krizna jama bei Grahovo (.T. M.leg.). Italien: Cadore, Villanova di Borca (i:!rLHAVY 1969b), Parco di Fusine bei Tarvisio (A. A. & .T. M. leg.). - BRD: bisher nicht nachgewiesen, aber in den Berchtesgadener Alpen mit Sicherheit zu erwarten.
Zur Verbreitungsgeschichte: Große Teile des Areals, vor allem jene nörrll. der AlpenHauptkette, hat unicolor postglazial besiedelt, denn diese Gebiete waren während der Glazialzeiten vom Eisstromnetz bedeckt (Klammkalkzone im Taurach-, Gasteiner-und Kaprunertal, Goldberg-Gruppe südl. Gastein). Von einem Präglazialrelikt, wie bisweilen vermutet und bereits von THALER (1966b) angezweifelt, kann in diesem Gebiet keine Rede sein.
Ökologie: Biotopansprüche: Wo unicolor in hoher Populationsdichte vorkommt, besiedelt es vorzugsweise Buchenwälder auf Kalkuntergrund, der durch
..

80 Faroilia Erebomastridae
Qe.;;teinstrümmer zerklüftet und mit einer dicken Schicht alten LaubeK bedeckt ist. Die Mikrohabitate in den Geröllspalten sind ganzjährig gegen Temperaturschwankungen isoliert, und selbst im Winter sinken die Temperaturen unter einer mehr als 50 cm dicken Schnee- und Laubschicht nicht unter den Gefrierpunkt: in I wurden ad. und iuv. einschließlich der Begleitfauna aktiv gefunden (A. A. in litt.).
Vertikalverbreitung: niedrigste Funde nm 270m (Windische Büheln, GRuBER I974a), dichtbesiedelter Arealgürtel in der Montan-Stufe von 500 bis etwa IIOOm, oberhalb llüü'm nur ausnahmsweise lmal auf I660 m (Naßfeld südl. Gastein, alle A. A.leg.).
Phänologie: ad. und iuv. eurychron; in Salzburg Nachweise von ad. in allen schneefreien Monaten (IV -X), zusätzlich in I (alle A. A. leg.), iuv. auch in XI. In weniger gegen Außeneinflüsse isolierten Biotopen scheint etwa ab A XI Rückzug in tiefere, geschützte Bodenschichten stattzufinden (GRUBER I974a).
Holoscotolemon jaqueti (CORTI, I905) Abb. 103-106
19il5 Phalangodes jaqueti CORTI, Bull. Soc. Sei. Bucarest, 14: 224. 1915 Metqscotolemon granulatus ROEWER, Arch. Naturgesch., 81: 7. 1935 Brigestus grarfulatus - RoEWER, Arch. Zoo!. expt\r. gen., 7'8: 47. 1935 Br·igestus robustus RoEWER, Arch. Zoo!. exper. gen., 7'8: 46. 1935 Bukowina monticola RoEWER, Arch. Zoo!. exper. gen., 78: 46. 1935 Scotolemops dacicus RoEWER, Arch. Zoo!. exper. gen., 78: 40. 1957 Bukowina hormuzachii ClnnEI, Stlll;l. Cerc. ~ti., Iasi, 8: 59.
Begründung der Synonymie: Die Identität von B. granulatus, B. rob!l8!us, M, granula!?IH. B. mon!icola undS. dacicus konstatierteAvRAM(1968 a). Die zahlreichenneuenFunde vonAvRAM deuten darauf hin, daß im Karpaten-Bagen lediglich eine Laniatores-Art vm•kommt; auch das Material der Sammlung RoEWER gehört nur einer Art an. Es wird vorgeschlagen, den ält.esten verfügbare:p. 'Namen zu verwenden, zumal keiner der jüngeren allgemein bt'uutzt, wurde und nicht als schut~bedürftig gelten lmnn.
Habitus und Feldkennzeichen: bernsteingelb, klein, kurzbeinig. - Im KarpatenBagen und in S-Jugoslawien.
Kennzeichnung: Körpermaße (in mm): L (nur Souturn magnum): <3 2,4-2,9; ~ 2,I-2,2. L gesamt: <3 2,6-3,2; ~ 2,7-2,9.
Laufbein II (($,in Klammern ~):Fe 2,I (I,9) Pt 0,8 (0,6) Ti I,8 (I,6) Mt 2,0 (I,8) Ta 2,I (1,8).
Tarsalgliederung-der Laufbeine: I: r3 5 bis 6 + 3, ~ 3 + I, 4 +I, 4 + 2, 5 + 3. - II: r3 6 bis ll + 4, 6 bis ll + 5; ~ 6 bis 10 + 3, 6 bis 10 + 4, 6 bis 10 + 5. - III: r3 und ~ 4. -IV: <3 und ~ 4.
Genitalmorphologie (Abb. 103-105): Truncus penis schlank, paralleh,andig, Basis erweitert, kontinuier-lich in den übrigen Truncus-Teifübergehend, unterhalb der Glans knopfartig. Glans do pantoffelartig ausgehöhlt, di 2 Paar kurz-plumper Borsten.
Cheliceren: I. Glied schlank, Basis kaum abgesetzt, do 3 große, distad geneigte Do; alll 2. Glied 2 kräftige Höcker do-frontal, ba kleinere jeweils mit Borste.
Pedipalpus (Abb. 106): Fe do gewölbt, mit 5 kräftigen gekrümmten Do, ve meist 4 große borstentragende, kleine medial; an Pt nur medial 2 kräftige Do; Ti und Ta mit je einer ve-medialen und ve-lateralen Reihe; Ti media14, la 4 (unterschiedlich groß), Ta medial und la je 3 große Do.
Genus Holoscotolemon
103 i I
Abb. 103-106. Hol~scotolemonjaqu;<ti (ConTI).- 103. Penis von ventral (links) und lateral; 104-105. ~!ans pems von ventral (hnks) und lateral: RU Grotte Topolni~a bzw. YU bei Foca· 106. d'-Ped1palpus von media.I. '
La ufbein_e: alle .Glieder rund, einheitlich fein granuliert und beborstet. Do~salselte: Zewhnung und Färbung: einheitlich rost- bis bernsteingelb stel
le~weise g~aue Soh~eier, ohne distinkte Zeichnung. - Bewahrung: gleich~äßig f~m gra~uliert, veremzelt winzige .Borsten. - Tu oc: niedrig, Augen klein schwarz pigmentiert. '
Ver~reitung: Typ: SO-europäisch-karpatisch. Nach jetziger Kenntnis di.sjunktes Areal Im .. Karpaten-Bogen und in S-Jugoslawien. Locus typicus: Umgebung von Azuga, sudl. Kronstadt. ;;
.Rum~nien: wah~scheinlich in allen Gebirgs-Gegenden verbreitet; in den 0-Kar aten be1 Brodm~ und Slatw&ra, im S bis in die Nähe der Donau (Bäile Herculane und Ba:aterBerge; sudhchs~e Fundorte); im N bis in die Apusenischen Gebirge (pestera de Ia Galasenil n~he der un.gansc~en Grenze. In den S-Karpaten fehlen bislang Funde zwischen den Flüssen Jml und Dimbov1ta ~vgl. AVRA~ 1968a). - UdSSR: Karpato-Ukraine: Bogdan (BARTOS 1939). - Jugoslaw1en: Bosmen-Herzegowina: 20-30km südöstl. Foca, Suha-Cemerno· Montenegro: Taratal, bei Djur~e':iea-Tara (beide A. A. leg.). Nachweise aus der CSSR{Siowaket und aus Ungarn stehen aus; Sie smd aber zu erwarten, da jeweils grenznahe Funde bekannt sind . .ß Tiel'welt Deutschl. 64: Martens
81

82 Familia Erebomastridae
Ökologie: Biotopans prüohe: H. jaqueti besiedelt die Bodenstreuschicht in Gebirgswäldern, vor allem in Moos und Fallaub, unter morschem Holz und Steinen. Diese Pritferenz für feucht-kühle Biotope präadaptiert jaqueti zum Leben in Höhlen, in denen gebietsweise (z. B. Karst der Mehedinti Berge; AVRAM 1968) große Populationen vorkommen. Die besiedelten Höhlen wiesen hohe relative Luftfeuchtigkeit auf (nahe 100%), sie sind ohne Luftzug und nahrungsrdoh. In Höhlen, deren Temperatur während des ganzen Jahres nicht unter 9 °0 absinkt, sind die Tiere zu jeder Jahreszeit aktiv. Zu morphologischen Umbildungen hat das Leben in Höhlen bisher nicht geführt.
Vertikalverbreitung: AVRAM & DUMITRESOU (1969) geben für Rumänien die Montan- und Submontan-Stufe an.
Phänologie: ad. in Höhlen eurychron, wahrscheinlich ebenso im Freiland; ~aus Höhlen mit legereifen Eiern in VIJVII und XII; 1 ~mit 16 Eiern im Uterus, Durohmesser 0,75 mm (AVR.A.M 1968). Lebensdauer der ad. unbekannt, wahrscheinlich aber länger als 1 Jahr.
Nahrung: In Gefangenschaft fraß jaqueti gern den troglobionten Käfer Sophrochaeta sp., mit dem er in vielen Höhlen zusammen vorkommt; im Darmkanal wurde der Palpus einer Nemastomatidae-Art und die Tarsalklaue einer Linyphüde gefunden (AVRAM 1968).
Holosootolemon lessiniense n. sp. Abb. 107, 112-116
Diagnose: Eine Art der Gattung Holoscotolemon, gekennzeichnet durch die d'-Genitalmorphologi.e und die Verbreitung.
Habitus und Feldkennzeichen: bernsteingelb, kurzbeinig; Pedipalpen massig und kräftig bestachalt; in den Lessinischen Bergen.
Kennzeichnung: Körpermaße (in mm): L (nur Souturn magnum) d' 2,2; ~ 2,1. - L gesamt: d' 2,9; ~ 2,7.
Laufbein II (d', in Klammern ~):Fe 1,6 (1,5) Pt 0,6 (0,5) Ti 1,4 (1,3) Mt 1,5 (1,4) Ta 1,5 (1,4). - Tarsalgliederung der Laufbeine: I: d' 4; ~ 4. - II: d' 10; ~ 9. -III: d' 4, ~ 4. -IV: d' 4, ~ 4.
Genitalmorphologie (Abb. 112-113): Truncus sehr schlank, parallelrandig, Basis sockelförmig abgesetzt und dort die Muskulatur konzentriert, unterhalb des Glans-Gelenkes lo).otig aufgetrieben. Glans kurz, von do bzw. ve nahezu parallelrandig, allenfalls geringfügig konkav, wesentlich schmaler als das knotig erweiterte Distal-Ende der Glans; 4 große Sinnesborsten distal.
Cheliceren (Abb. 115): Grundglied schlank, deutlich vom Basisteil abgesetzt, do nur 2 kräftige :ijaken-Do. 2. Glied do-di 2 kräftige Do mit Borste, ba unregelmäßige Tuberkulierung, vor allem nahe der Schere mit langen Bo.
Pedipalpus (Abb. 114): Fe: do Reihe aus 6 Do, ve mit 4 großen und beigestellten kleinen Do; Pt: 2 mediale Do; Ti und Ta: jeweils 1 va-medialer und ve-la Do; Ti: medial 4, la 5; Ta medial 3, la 3, wenige kleine und unscheinbare Tu eingestreut.
Laufbeine: kurz, alle Glieder rund, durch Tuberkel-Besatz und feine Haare rauh wirkend.
Dorsalseite: Zeichnung und Färbung: Körper und Beine bernsteingelb, graue Chagrinierung auf Teilen des Ceph. - Bewahrung: Areae IV und V des Souturn nnd die freien Terg mit mehreren Querreihen niedriger Höcker, meist beborstet,
Genus Ho!oscotolemon
so 100 km
Abb, 107d. HPunktkarte der Verbreitung von Holosootolemon unicolor ROEWER H l .. n. sp. un . oreophilum n. sp. , ' . esstn-z,ense
Kö-Gruppen nahe den Frontalecken des Ce h T · we;gbtu?terkuliAert, Linse der Augen klein, du~k~l ;gm~n~i:~t~1~b ~~~~gewölbt,
er re1 ung ( bb. 107): Typ: wahrscheinlich E d 't d Le .' · · und benachbarter Gebir t" k L . n e~ er ss1mschen Berge
. gss oc e. - oous typ10us: Lassmische Berge. Mater.al: Italien: Gardasee-Gebiet: Monti Lessini· Cerro Va'o ·
typus SMF 29179, VIGNA leg ll VII 1968 p d t '. 0
• J di Squaranto, I ß Holo-1968. . . . . - o es erm, I + Paratypoid, VIGNA leg. 17. VII.
Ar~e~:~:~;:n~!t d~~8~~igen. bish~r aus den Alpen bekannten Holoscotolemon-
~~~::h::e;' :iftä~~=~z~:~~=i:~~P::~i~.c~:;d:!~Ar:~ e~~~ei~r~~:;l:~r&~~~ do v b fl ht d' b · · el e1 en ten 1st.d1e Glans penis
- ~ a ge ac ; 1e e1den Sinnesborsten-Paare stehen in der Längsriebt von uncus und Glans, und die einzelnen Borsten sind kräfti · ung ~=rGt_cht tspezialisier.t (vgl. die V~~hältnisse bei unicolor). Grö~ee~~~c;:~~o~~~: weise k::n,r~~:~ub;!;a~i:a:~a:~:\v!:~!~t:~n::~~:~:r d!~s GJ::~~:::ll-~chs-~~~:~~~:; !;~~)eTh~um dagegdenb ~erklich größer (bei etwa gleicher Trunc~~~ li . . • a onvex un re1ter als das Distal-Ende des Truncus Zwar
Beglt votnllesszmArense nur 1 d' vor, doch erlauben die Verhältnisse bei allen an. deren o osco o emon- ten den Schi ß d ß · h
di b 'd Arte . h u • a es S1c um konstante Merkmale handelt e e1 e n sw er trennen. · ' 6*
83
) I
I
84 Fa.milia Erebomastridae
Abb. 108-111. Holoscotolemon oreophilum n. sp.- 108. Penis von ventral (links) und lateral; 109. Glans penis von ventral (links) und lateral; 110. d'-Pedipalpus von medial; 111. 6-Chelicere von medial. - IT Col d i Tenda..
Abb. 112-116. Holoscotolemon lessiniense n. sp. - 112. Penis von ventral (links) und lateral; 113. Glans peu"is von ventral (!jnks) und lateral; 114. d'-Pedipalpus von medial; 115. d'-Cheliceru von medial; 116. Tuber oculorum von lateral (Pfeil weist frontad). - IT Monti Lessini.
J

Genus Holoscotolemon 85
Wahrscheinlich stehen lessiniense und oreophil"rn in einem SchwestergruppenVerhältnis. Beide besiedeln die Massifs de Refuge der S-Alpen, wo sie möglicherweise die gesamten Glazialzeiten überdauert und sich in der Isolation aus der gemeinsamen Stammform differenziert haben.
Ökologie: keine Daten verfügbar; wahrscheinlich ähnlich oreophil"rn sowohl im Waldgürtel als auch über der Baumgrenze.
Holoscotolemon oreophilum n. sp. Abb. 107-111
1908 Scotolemon querilhaci - Gozo, Boll. Soc. ent. ItaJ., 38: 134.
Diagnose: Eine Art der Gattung Holoscotolemon, gakeunezeichnet durch die Genitalmorphologie und die Verbreitung.
Habitus und Feldkennzeichen: bernsteingelb, kurzbeinig; vergleichsweise groß (größte Art der Gattung), Pedipalpen massig und kräftig bestacheil; in den SW-Aipen.
Kennzeichnung: Körpermaße (in mm): L (nur Senturn magnum) c1 2,4-2,8; ~ 2,3-2,5. - L gesamt: c1 3,4-4,1; ~ 3,0-3,5.
Laufbein II (<1, in Klammern~): Fe 2,1 (1,8) Pt 0,7 (0,6) Ti 1,7 (1,5) Mt 2,0 (1,8) Ta 2,0 (1,9).
Genitalmorphologie (Abb. 108-109): Truncus sehr schlank, parallelrandig, Basis sockelförmig abgesetzt und dort die Muskulatur konzentriert, unterhalb des Glans-Gelenkes knotig aufgetrieben. Glans lang, von do bzw. ve deutlich konvex, breiter als das knotig erweiterte Distal-Ende der Glans; 4 große Sinnesborsten di.
Cheliceren (Abb. 111): wie bei lessiniense; do 2 kräftige Haken-Do. 2. Glied do-di mit 2 kräftigen Do mit Borste; ba unregelmäßig tuberkuliert, vor allem nahe der Schere mit langen Borsten.
Pedipalpus (Abb. 110): Fe: do Reihe aus 5-6 Do, ve mit 4 großen und beigextellten kleinen Do; Pt: 2 mediale Do; Ti und Ta: jeweils 1 ve-mediale und ve-la Do-Reihe; Ti medial4, Ia 3-4; Ta medial 3, Ia 3 zapfenartige Do.
Laufbeine: kurz, alle Glieder rund, durch Tuberkel-Besatz und feine Haare rauh wirkend.
Dorsalseite: wie bei les.9iniense. Varia bil i tä t: Die kennzeichnenden genitalmorphologischen Merkmale sind
bei allen vorliegenden c1 einheitlich. Verbreitung (Abb. 107): Typ: Endemit der SW-Alpen. - Locus typicus: Co! di
Tenda. Material: Italien: 1\feeralpen: Col di Tenda (~ Halotypus SMF 29180, VIGNA leg. 16. VI.
1968); - Ligurische Alpen: Serra. di Pamparato, Tana. dell' Orso (I(! 1 ~ Paratypen; MoRISI leg. 23.1II. 1969 bzw. MoROANTINI 13.1I. 1972); Pamparato, Tana delle Turbiglie (I cJ Para· typus, CASALE leg. 11. X. 1972); Rossa.na, Grottä. dei Partigiani (I(! Paratypus, MORISI leg. 20.1I. 1972); Alta Val Pesio (2 !f Paratypen Monrsr leg. 22. VI. 1972).
Holoscotolemon-Funde aus den Ligurischen Alpen sind bereits seit längerem bekannt; Gozo (1908, Scotolemon querilhaci) nennt als Fundorte: Grotta sul Monte Penna und Grotta di Val di Pesio.
Beziehungen: siehe bei lessiniense. Ökologie: Biotopansprüche: im einzelnen nicht bekannt, scheint aber in der
Regel höher im Gebirge zu leben als das 0-alpine unicolor und in den ökologischen Präferenda eher lessirii.ense vergleichbar; häufig in Höhlen.

86 Fe.milia. Erebomastridae
Vertikalverbreitung: die einzigenAngaben sind 1600m (Co! diTenda) Ullll 1700-2000 m (Alta Val Pesio).
Phänologie: wahrscheinlich eurychron; ad. liegen vor aus li, III, VI und X.
Holoscotolemon querilhaci (LuoAs, 1864)
1864 Scotolemon querilhaci LuoAS, Amt. Soc. Ent. France, (4) 6: 216. 1879 Phalangodes querilhaci - SrMoN, Arachn. de France, 7: 153. 1923 Scotolemon querilhaci - RoEWER, Weberlmechte d, Erde: 101. 1935 Querilhacia que1-ilhaci - RoEWEU, Arch. Zool. exp8r. gßn. 78: 38. 1935 Scotolemoniscua austriacus RoEWER, Arch. Zool. expBr. gßn. 78: 42. 1935 Scotol'emoniscus styriacus RoEWER, Arch. Zool. exper. glm. 78: 43.
In der Sammlung RoEWER hefi11den sich 3 Serien einer Holoscotolemon-Art, nach denen RoEWER die beiden Arten Scotolemoniscus austriacus und Sc. stydacus beschrieben hat. Alle Individuen dieser Serien stimmen mit H. querilhaci überein (Areal in S-Frankreich); beide nominellen Arten sind somit als Synonyme von querilhaci zu betrachten. Die Nennungen für Österreich beruhen zweifellos auf Fundort-Verwechslungen; im hier behandelten Gebiet ist querilhaci als nicht nachgewiesen zu betrachten.
Suprafamilia Gonyleptoidea
1968 Oncopodoidea KBATOCHViL, PrSce Brn. zßkl. Öesk. Akad. v~d. 80: aao (part.: Oncopodoidea. und Gonyleptoidea. im hier vertretenen Sinno).
1961 Gonyleptimorphi SrLnAvY, Verh. Ent. Congr. Wien, 2: 261 (part.: Gonyleptoidea und Truvwüoidea. in hier vertretenen Sinne).
Eine Überfamilie der Unterordnung Laniatores gelrennzeichnet durch: d'-Genitalmorphologie: Penis ohne Muskel, meist deutlich in Trunous und sub
apicale (selten apicale) Glans gegliedert, Glans durch Hämolymphdruck beweglich und/oder deren Sklerite expandierbar, selten ohne bewegliche Teile.
Ovipositor: meist kurz und ungegliedert, di zu 2 stumpfen Loben ausgezogen; Bewehrung mit Borsten spärlich, die Loben oft beträchtlich überragend.
Krallen der Laufbeine 111 und IV als zwei (immer isolierte 1) Einzelkrallen. Gliederung: Mit zahlreichen Familien bzw. Unterfamilien weltweit verbreitet;
Status und Abgrenzung der einzelnen Gruppen weitgehend ungeklärt. - Gonyleptidae (neotropisch), Cosmetidae (neotropisch, siidl. nearktisch), Phalaugodidae (Verbreitung schwanke:1d mit der Abgrenzung der Familie: Holarktis, IndoAustralis und Neotropis); früher Unterfamilien der Phalangodidae, jetzt als eigene Familien anerkannt: Podoctidae (Orientalis, Japan, Australien, tropisches Afrika mit Madagaskar, Kuba), Biantidae (Äthiopis, Indo-Orientalis), Assamiidae (Äthiopis, Indo-Orientalis), Agoristenidae (Karibische Inseln), Trionyxellidae (OrientaIis, Äthiopis), Stygnidae. - Im hier behandelten Gebiet nur 1 Art der Phalallgodidae.
Genus Scotolemon
Familia Phalangodidae
)879 Phalangodidae SIMON, Arachn. de }1,ranne, 1: 148. 1!123 Phalangodida.e - ROEWER, Weberknechte der Erde: 64 (pa.rt.). 1958 Paralolida.e KRAToonvfr., PrBce Brn. ztikl. Öeak. Aka.d. v~d. 30: 394.
87
Eine ~amilie der Laniatores-Gonyleptoidea, gekennzeichnet durch d'-?erutalmo:phologie: vgl. Definition der Gonyleptoidea. Ber der germgen Kenntnis der einzelnen Teil-Gruppen kann augenblicklich
noch kei':'e Definition für die ganze Familie gegeben werden. V~rbrertung_: Zahlreiche Unterfamilien (deren Status meist ungeklärt) im
t_ropmchen Asw~ (wenige Arten im Himalaya), auf den pazifischen Inseln, Austrahen, ganz Amerrka, N- und S-Mrika. .. In J!!uropa; Scotolemon LUOAB, im westlichen und zentralen Mittelmeergebiet ostl. brs lstnen, Lola KRATOOHVfL auf der dalmatischen Insel H var Paralola KRATOOHVfL im Balkan-Gebirge, Ptychosoma SoERENSEN im westl. 's-Europa, Ausobskya MARTENS in N-Griechenland.
Genus Scotolemon LuoAs, 1860
1860 Scotolemon LucAs, Ann. Soc. ent. J.i'ranco, (3) 8: 973. 1879 Phalangodea - SrM:ON, Arachn. do Francs, 7: 149. 1923 8cotolemon - ROEWEU~ Weberknechte d. Erde: 97. 1935 Scotolemon - RoEwEn.~ Arch. Zool. exp8r. g8n., 18: 27.
Eine Gattung der Phalaugodidae, gekennzeichnet durch: d'-Genitalmorphologie: Truncus penis plump untersetzt bis langgestreckt
P."~allelrandig oder distad wenig erweitert, do-di mit einem häutigen, teils sklero: trsrer.ten Sack, durch Hämolymphdruck schwell- und aufrichtbar, di Ende im BereiCh _des Sackes stark d?-ve a~g.eplattet, vcntrad gekrümmt, bisweilen zweigeJappt. Smnesborsten groß, m spezifisch wechselnder Zahl di-ve und Ja z. T. sicheiförmig gekrümmt. - Legeröhre kurz, di 2 kurze Loben, von Borsten umstellt. Dorsal~ freie Te:g 1-III ~n unmittelb~rer Verlängerung an das Scutum magnum anschließend, nwht auf dre Ventralseite umgeklappt. Sternum zwischen Cx III und IV ausgespannt. - Tu oc deutlich und groß, unmittelbar am Ceph-Frontalrand anschließend.
Tarsalgliederung der Laufbeine offenbar regelmäßig 3-5-4-5. Sekundäre Geschlechtsmerkmale: bei 2 Arten d'-Tr IV mit einem caurlal weisenden, medialen Dorn.
Verbreitung: 7 nominelle Arten im westl. Mediterran-Gebiet: Iberische Halbinsel (Kantabrische_s _Gebirge, Pyrenäen, S-Spanien). Balearen, Apenninen-Halbinsel, Korsika, Sardiruen, S-:tl.lpen. Der Status der einzelnen Arten ist nicht abgeklärt.
Typus-Art (durch Monotypie): Scotolemon leap88ii LuoAs, 1860.
Scotolemon doriae PAVESI, 1878 Ahh. 117-122
1878 Scotolemon doriae PAVESI, Ann. Mus. Civ. Genua, 12: 565. 1923 Scotolemon terricola - ROEWER, '\Veber1mechte d. Erde: 98 (part.). 1935 Scotole.mon doriae - RqEWER, Arch. Zool. expßr. gßn., 18: 33.

88 Familia Phalangodidae
Habitus uud Feldkennzeichen: klein, kurzbeinig, hellgelb. - Im behandelten Gebiet in den S-Alpen.
Kennzeichnung: Körpermaße (in mm): L (nur do Soutum): c11,0-1,1; ~ 0,9 bis 1,1. .
Laufbein II (c1, in Klammern~): Fe 1,1 (0,9) Pt 0,4 (0,3) Ti 0,9 (0,7) Mt 0,8 (0,6) Ta 0,9 (0,8).- Gliederung der Ta I-IV bei c1 und~ regelmäßig; I 3; II 5; Ill 4; IV 5.
Genitalmorphologie (Abb. 117 -118): Trunous penisplump untersetzt, distad wenig erweitert, in 2 breiten, ventrad umgebogenen Zipfeln endend. Ohne diffe-
Abb. 117 -122.· Scotolemon doriae PAVESI. - 117. Penis von ventral {links) und lateral; 118. Glanspenis von ventral {links) und lateral; 119. O'·Pedipa.lpus von medial; 120. Q'-Laufbein IV, Tr und Fe von lateral; 121. Q'-Chelicere von medial; 122. Q'-Habitus von dorsal. - IT Riva, Mt.Brione.
Genus Sootolemon 89
renzierte Glans, doch di Ende im Bereich der Sinnesborsten do abgeflacht und dort ein häutiger, durch Hämolymphdruck schwollbarer Sack; di-la je 4 kräftige basad gerichtete gekrümmte Do; Gestalt und Lage der übrigen Sinnesborsten vgl. Abb. 118.
Cheliceren (Abb. 121): untersetzt, l. Glied do eingeschnürt, di bucklig, ohne ve Apodem.
Pe.dipalpus (Abb. 119): Fe mäßig gewölbt; Zapfen-Do-Bewehrung: Fe ve 3, me-d1 1, Pt je 1 medial und ve, Ti und Ta je 2 medial und la; Klaue klein.
Laufbeine: kurz, alle Glieder rund, fein bekörnelt und schütter mit Borsten besetzt, c1-Tr mit kräftigem, ve Sporn, leicht geschwungen und rauh bekörnelt, di glatt und knopfartig verdickt, me-ventrad gerichtet (Abb. 120).
Dorsalseite (Abb. 122): Zeichnung und Färbung: einfarbig bernsteingelb und zeichnungslos.
Bewahrung: außer feiner Bekörnelung und einigen Höckern auf den freien Terg III und IV unbewehrt. - Tu oc: groß und hoohgewölbt, über den Ceph-Frontalrand vorgezogen, etwas distad weisend. Linse mit Durchmesser von 30 p,m, nicht bikonvex (wie bei lespesi LuoAs), sondern von Uhrglasform, mit nur 16 Sahzellen (JUBERTHIE 1957b, 1964).
Beziehungen: Nächst verwandt ist wahrscheinlich S. terricola SIMON, ebenfalls mit Apophyse an c1-Tr IV. Es bestehen Unterschiede in der Genitalmorphologie, in den Proportionen des Sporns an Tr IV und in der Pigmentierung der Augen.
Verbreitung: Typ: N- und zentral-mediterran. - Weitverbreitet auf der Apenninen-Halbinsel, in den Alpen und in S-Frankreich; möglicherweise existieren nur lokale Einzelvorkommen ohne zusammenhängendes Areal. - Locus typicus: Latium, Albaner Berge. ·
Frankreich: vereinzelt in den Dep. Haute-Garonne und Herault; Meeralpen, Baume ,]o ColombierfRoquefort {JuBERTHIE 1957a). - Italien: San Remo {RoEWER 1935); GardaHueGebiet: Riva, MonteBrione (K. T.leg.); Ligurien, Boccadasse {Gozo 1908), weit.ere Fundorte bei BRIGNOLI {1968) und MAROELLINO (1970, 1973b); Apoanisehe Alpen, Üarrara {ROEWER 1935). Mte. Gabberi; PratolinofF!orenz, Arezzo {MARCELLINO 1970); Latium, Albaner Berge {PAVESI 1878), Umgebung von Rom, Monti Ernici, Pantinische Inseln (BRIGNOLI 1968); Sardinien, Insel Asinara {BRIGNOLI 1968); Korsika, Ponte Nuevo {JuBRETIDE 1957a); Sizilien, Palermo, Grotta Conza {MAROELLINO 1970). - Jugoslawien: Istrien, Rovinj, S. Giovanni (K.T.leg.), .
Ökologie: Biotopansprüche: S. doriae lebt je nach temporären Feuchtigkeitsverhältnissen in der Laubschicht des Bodens, unter Holz und tief eingebetteten Steinen, während Trockenperioden tiefer in Rissen und Spalten, in Regenwurmgängen und Bauten von Kleinsäugern bis l m unter der Oberfläche (JUBERTHIE l957a, 1964); Waldbedeckung des Bodens scheint nicht notwendig zu sein.
Auf derart versteckte, zeitweilig subterrane Lebensweise deuten die pigmentlosen und stark reduzierten Augen, die manche Autoren ganz übersehen haben (z. B. ROEWER 1935). ,;
Vertikalverbreitung: keine Daten, jedoch ist doriae keinesfalls ein Gebirgstier.
Phänologie: wahrscheinlich eurychron.
..

90 Familia Phalangc;>didae~
Subordo Palpatores
'1876 Palpatores THORELL, Ann. Miia. Genova: 462. 1879 Plagiostethi SIMON, Faunade France, '1: 157. 1904 Palpatores HANSEN & SOERENSEN, On two Orders of Arachnida: 79. 1923 Palpatores - ROEWER, Weberknechte d. Erde: 632.
Körpe~gliederung: Dors~~ mit Souturn parvum (meiste Gruppen der Isohyr~psalidmdea und Phalangioidea) oder S. magnum (meiste Gruppen der Trogulmdea), Auflösungstypen nur bei wenigen Gruppen, dann oft hoohvariabel; S. mag~um erst bei. der ~dulthäutung angelegt. - Opisthosoma mit 8-9 Tergiten, Tergit IX als Seitenteil~ der Corona analis entwickelt oder fehlend, Tergit X = ~peroulum anale; Sterrute VIII-IX kleiner, die Corona ana]is bildend oder reduZiert.
Prosom~l-Sternum:_ ~YP .kurz-breit, frei oder mit d~n Cx verschmolzen; Operoulum.gerutale unterschiedlich groß, meist das Prosomal-Sternum ganz bedeckend.
Gerutalmorpholog.~e: Penis meist ~entlieh in Truncus und Glans gegliedert, mit 1 .oder 2 M~skelstrangen, durch diese Glans beweglich. Innerer Genitalapparat mit. Propuls10nsorg~n (Spermabes~mung mittels Kopulation). - Ovipositor: variabel, konstant Innerhalb der eiUZelnen Fari:rilien: überkörperlang und vielgliede~g (~haHtngioidea}.bis ~ngegliedert und kurz (übrige Überfamilien, Caddoidea nur ~eilweise). Bes~tz mit Smnesborsten z. T. komplex (reihenweise, komplizierte Sensillen), z. T. einfach oder fast fehlend. Innerer Bau bilateralsymmetrisch · 2 Rec sem oder Vielfaches davon. ·
Pedipalpus: ~laue klein, ru~imentär ?der ganz fehlend, nie als wirkungsvolle Subch~la ausgeb!l~et, Palpus rue.Raubbem, sondern Tastorgan für den Nahbereich und Hilfsorgan bei. d~r LokomotiOn. - Laufbeine: 2. meist beträchtlich länger als 1., al~e Cx starr mitemander v~rbunden, Lobus der Cx I (als ?) mit be'weglichem S.klent. - Klauen an allen Bemen einfach, einschließlich•der Nymphen. - Plantigrad,
Gliederung: ~ ~berfamilien: Caddoidea (nur Caddidae), Troguloidea (vgl. S. 91), Ischyropsalidmdea (vgl. S. 186), Phalangioidea (vgl. S. 228).
Besiimmungstabelle für die tJberfainilien und Fainilien der Palpatores
Pa von do frei sichtbar, ohne mo am Ceph-Frontalrand entspringende Kappe . . . 2
- Pa von do nicht sichtbar, von einer me am Ceph-Frontalrand entspringenden Kappe verd~ckt (Abb. 277- 280) . . . . . · . . . . . . . . . . . . . . . . . . . . . 5
2 Ta des Pa mit di Krallo, meist klein aber immer deutlich erkennbar und schwarz abgesetzt (z. B. Abb. 411) · · · · . . . . . . . . . . . Phalangioidea, Phalangiidae, S. 229 Ta des Pa ohne di Kralle . . . . . . . . . . . . . . . . . . ; . . . . . . 3
3 Chel "normal" proportioniert, d. h. I. Glied wesentlich kürzer als der Körper . . . . 4
Chel überproportioniert lang, I. Glied etwa so lang oder länger als der Körper (z. B. Abb. 332-334) · . . . . . . . . . . . Ischyropsalidoidea, Jschyropsalididae, S. 186
4 kein Glied des Pa aufgebläht verdickt; besonders Ti und Ta des Pa mit zahlreichen an der Spitze kugeligen Haaren besetzt (z. B. Abb. 160). - Weitverbreitet in Mitteleu:cpa
Trcguloidea, Nemastomatidae, S. 91 vor allem Ti und Ta des Pa keulig verdickt (Abb. 404e); Haare an der Spitze nicht kugelig verdickt; Ta gegen Ti geklappt. - Lokal in den SW-Aipen ............. .
Ischyropsalidoidea, Sabaconldae, S. 225
Genus Scotolemon 91
5 Augen an der Basis der Kappenäste auf dem Ceph (Abb. 277, 278), Ta der Laufbeine höchstens 4gliedrig, Sternite zu einheitlichem ve Souturn verwachsen und nicht gegeneinander beweglich, durch feine helle Mediane 2geteilt. - Weitverbreitet in Mitteleuropa
Troguloidea, Trogulidae, S. 162 Augeü etwa in der Mitte der Kappenäste, nie auf dem Ceph (Abb. 279, 280); Ta der Laufbeine mehr als 4gliedrig; Sternite nicht zu ve Scutum verwachsen, immer gegeneinander beweglich, ohne hellen Medianstreif. - Im hier behandelten Gebiet nur lokal in den S-Alpen und im südöstl. Mitteleuropa Troguloidea, Dicranolasmatidae, S. 150
Suprafamilia Troguloidea
1833 Trogulides SUNDEVALL, Consp. Arachn.: 35 (part.).
Eine Überfa~nilie der Unterordnung Palpatores, gekennzeichnet durch: d'-Genitalmorphologie: Penis mit zwei Muskeln, deren Sehnen an den Ventro
lateral-Seiten der Glans ansetzend; Glans in gerader Verlängerung des Truncus und la-ba oft mit "Fenstern" weicher Cuticula, me-do und me-ve Verstrebungen zwischen Truncus und Glans. Bewegungsmöglichkeit allenfalls geringfügig, möglicherweise (auch ~) laterad.
Ovipositor länger als breit, ungegliedert, di Furoa zweigliedrig mit ba 4 (ovalen) und di 2 (halbmond-förmigen) Skleriten, schütterer Besatz mitBorsten auf kleinen Warzen, die längsten Borsten ;tuf der Furca. . Sternum zwischen den Cx ausgespannt und mit diesen in Form einer festen
Brücke verwachsen. Tracheenstigmen vergittert; an den Tibien der Laufbeine keine ·akzessorischen
Stigmen .. Pedipalpus laufbeinartig groß bis rudimentär reduziert, Tarsalklaue fehlend. Zähne der Cheliceren z. T. massig-massiv, z. T. zart-hyalin. Dorsales Souturn als
S. parvum (Nipponopsalididae, Ortholasma) bzw. als S. magnum (Trogulidae, Dioranolasmatidae, Nemastomatidae ohne Ortholasma und Denrlrolasma), selten Reduktionsformen ( N ipponopsalis).
Corona analis vollständig entwickelt. Gliederung: Die Überfamilie umfaßt die Nemastomatidae (disjunkt holarktisch
verbreitet), Trogulidae (W-paläarktisch), Dioranolasmatidae (W-paläarktisch) und Nipponopsalididae (mono~ypisc~, Nipponopsali~ MARTENS ~ SuzuKI; O:paläar~tisch). Nahe verwandt sind die Nemastomatidae und Dwranolasmatidae; sie stehen wahrscheinlich in einem Schwestergruppenverhältnis.
Familia Nemastomatidae
187!i- Nemastomatidae SrMON, Arachn. de France, '1: 277. 1923 Nemastomatidae - RoEWER, Weherknechte d. Erde, 650 (part.).
Eine Familie der Palpatores-Troguloidea, gekennzeichnet durch: d'-Genitalmorphologie: Trunous penismeist lang und schmal, der Basisteil zwie
belförmig erweitert in gerader Verlängerung des übrigen Trunous (einige Nemastomatinae Ortholasmatinae) oder in spitzem Winkel gegen den Truncus gekrümmt (die me~ten Nemastomatinae). Basisteil selten ~latt ge~an~et, meist ~wischen den beiden M11skeln tief gespalten. - Muskulatur Im BasiSteil konzentriert, durch 2

92 Familia Nemastomatidae
meist lange Sehnen mit der Glans Ia verbunden. Glans in gerader Verlängerung des Truncus, nicht oder nur unwesentlich breiter als dieser; Stylus meist kurz, wenn lang und schmal, meist asymmetrisch geschwungen; Besatz mit Sinnesborsten variierend. - Legeröhre: die beiden di Furka-Äste kurz gerundet, mit wenigen langen Sinnesborsten.
Cheliceren: d'-Grundglied meist mit Drüsenorga.il, Drüsenzellen teilweise tief in den Ceph versenkt; Ausfuhröffnung für das Sekret als Einzelporus oder als Porenplatte auf oder an der Basis einer Apo.
Pedipalpus: alle Glieder mit Ausnahme von Cx und Tr mit Kugelhaaren, konzentriert auf Pt, Ti und Ta, selten reduziert (Dendrolasma BANKS); Ta-Klaue fehlend.
Ta der Laufbeine vielgliedrig, ohne abgegliederten Distalteil des Mt (Calca-neus). ,
Souturn als S. magnum (Nemastomatinae) oder S. parvum mit Tendenz zur Verschmelzung zum S. magnum (Ortholasmatinae) ausgebildet. Cuticularstruktur des Souturn nicht warzig, oft mit Amboßzähnchen in girlandenförmiger Reihung, solche Strukturen regelmäßig auf den Rändern der Cx (reduziert bei Troglohionten).
Sternum deutlich entwickelt, zwischen den Cx 1-III ausgespannt und mit diesen fest verwachsen, caudal tief ausgekehlt.
Verbreitung: disjunkt holarktisch, größte Formenfülle in der W-Paläarktis (Nemastomatinae), wenige Formen in der Q.Paläarktis und Nearktis (Ortholasmatinae). - W-Paläarktis: von der Iberischen Halbinsel und den Atlas-Ländern N-Afrikas durch ganz Europa (ob im Ural ?), in Vol.'der-Asien bis zum Kaukasus und Elburs.Gebirge.
Nennungen von "Nemastoma" für N-Amerika sind auf Hesperonemastoma GRUBER zu beziehen (lschyropsalididae, vgl. dort).
Noch 1951 ordnete RoEWER über 90% aller Arten nur einer Gattung, "Nemastoma", zu. Erst später wurden·Versuche unternommen, diese heterogene Großgattung in natürliche Einheiten aufzugliedern und sie mit genitalmorphologischen Kriterien zu charakterisieren (SILHAVY 1966b). Eine angemessene Einteilung ist bisher nur für den hier ·behandelten mittel-europäischen Bereich gefunden worden.
0-Paläarktis und Nearktis: DieN-amerikanischen (Ortholasma BANKS, DendrolasmaBANKS) und japanischen (Dendrolasma) Gattungen, die bisher zu den Trogulidae gerechnet wurden (z. B. RoEWER 1923, SuzUiu 1974), stehen weder mit diesen noch mit ~en Dicranolasmatidae in engen verwandtschaftlichen Beziehungen. So kann z. B. der unpaare me-di Ceph-Auswuchs (ein Derivat des Tu oc) nur als Konvergenz zu dem paarigen Auswuchs der Arten dieser beiden Familien angesehen werden. Die genitalmorphologischen Verhältnisse deuten auf enge Beziehungen zu den Nemastomatidae: die beiden Muskelstränge in erweiterter Basis konzentriert, diese me tief gespalten; ferner Brückendornreihen auf dem Souturn wie bei vielen Gattungen der Nemastomatidae; Metatarsen ohne Calcaneus.
Auf Vorschlag von J. GRUBER (in litt.) sollen beide Gattungen als Unterfamilie Ortholasmatinae zu den Nemastomatidae gezählt werden.
.Subfamilia Nemastomatinae
1879 Nemastomatidae SrMoN, Arachn. de France, 7: 277. 1959 Giljaroviinae KRATOCHviL, Zoo!. Zur., 38: 1344.
Genus Nemastoma 93
Mit den Merkmalen der Nemastomatidae, gekennzeichnet durch: d'-Genitalmorphologie: Truncus peni~ gerade gestreck.~, ~asis markant a.ufge
bläht und abgesetzt, oft in spitzem Winkel gegen den ubrigen Trun~us-Te!l gebogen. Glans auch nie bulbus-förmig, oft do-ve abgeflacht, Stylus deuth?h e~kenn· bar, meist lang pfriemförlnig, aber von Gattm~g zu Ga~~ung unterschiedlich. -·Ceph-Frop.talrand nie Init me Auswuchs bzw. distad verlangertem Tu oc.
Cheliceren: 2. Glied der d' glatt, ohne frontale Apo. Typus-Gattung: Nemastoma C. L. KooH.
Bestimmungstabelle für die Gattungen
do Souturn mit Girlanden aus Brückendorn-Reihen, wenigstens als Querreihen zwischen den thorak Areae, meist ebenso auch auf den abd Areae (Abb. 198, 199), oft das ganze Senturn Ja umrandend (Abb. 220, 221) . . . . . . . . · · · · · · · · · · • · • 2 do Souturn ohne Girlanden aus Brüokendorn-Reihen .. · . · · · · · · · · · · • 4
·2 do Souturn ohne Stabdorneu-Bewahrung (vgl. Abb. 199, 220) · · · · · · · · · · · 3. do Souturn mit langen Stabdornen auf den abd Areae und den freien Tergiten (vgl. Abb. 213) ·· Centetostoma, S. 139
:3 Brückendorn-Reihe umrahmt das ganze do Scutum, frei lediglich der Frontalrand me (Abb. 220); Beine fadenförmig dünn und lang; Färbung des Körpers bra~n, oft mit unauffälligen Goldflecken do . . . . . . . . . . . . · · · · · · · · .MJtostom~, S. ~41 Brüokendorn-Reihe umrahmt nie das ganze do Scutum, caudaler Rand Immer frei; Beme kurz. Färbung des Körpers tiefschwarz, 2 auffällige Silberflecken auf dem Ceph . . . .
' Carinostoma, S. 13•1
4 do Souturn nie mit Stabdornen; leicht gewellt glatt oder auf den abd Areae I und II auf 2 para-me breiten Sockeln je 1 sjmnipf-konischer Tu (vgl. Abb. 159, 166); d'-Pa-Pt immer
unbewehrt . . . . . . . . . . . · · · · · · · · · · · · · · · · · · · · · · 5 do Souturn mit Stabdornen auf den abd Areae I-: V (bei !f bisweilen m~au~fällig kurz, von Ja betrachten! Abb. 188); d'-Pa-Pt mit Dorn mediO·<h ...... HistriCostoma, S. 1.29
5 do Souturn glatt oder die einzelnen abd Areae durch seichte Furchen angedeutet; kleme Arten bis höchstens 2,5 mm . . . . . . . . · · · · · · · · · · · Nemastoma, S. 93 do Souturn auf den abd Areae I (und II) mit je 2 para-me breiten Sockeln und aufgesetztem stumpf-konischem Tu (d', bei !f Ullauffällig oder ganz fehlend; von Ia betrachten); große Arten von etwa 4 bis über 5 mm . . . . . . . . . . . . . • . Paranemastoma, S. 116
Genus Nemastoma. C. L. KooH, 1836
1836 Nomastoma C. L. Koon, in HAHN & C. L. KooH, Die Arachniden, 3: 71 (part.). 1923 Nomastoma - RoEWER, Weberknjlchte d. Erde: 651 (part.). 1951 Nomastoma - RoEWER, Senckenbergiana, 32 (1/4): 96 (part.). 1968 Nomastoma - GRUBER & MARTENS, Senckenbergiana bio!., 49 (2): 139.
Nominat-Gattung der Nemastomatidae, gekennzeichnet durch: d'-Genitalmorphologie: Truncus penis lang und. schmal, meist nah~zu recht
winklig gegen die erweiterte und tiefgespa~tene Bas_rn abgebogen; Glans m gerader Verlängerung des Truncus und k~um breiter a~s dJeser; .~o:ve abgeplatte~. Glans symmetrisch oder leicht asymmetrisch, lateral m1t ~ -8 kra~ti~en, basad gerwhteten Haken, ausnahmsweise fehlend. Stylus der Glans Immer 1astig. '

94 Familia Nemastomatidae
Soutum: primär S. magnum, alte und stark skierotisierte Individuen mit S. compositum, d. h. abd Terg VI, VII und selten VIII mit dem S. magnum verschmolzen. Glatt-glänzend bis rauh bekörnelt, ohne Kegeldorn- oder BrückendornBewehrung; tiefschwarz, oft mit thorak Gold- oder Silberfleckenzeichnung.
Chelicerendrüse: .zahlreiche Einzeldrüsen mit getrennten Ausfuhröffnungen (Porenfeld) auf deutlich abgesetzter Apo des Grundgliedes, medial oder frontal.
Pedipalpus bei einigen Arten mit Do- oder Kö-Bewehrung an Fe oderfund Pt di-medial. - Laufbeine kurz und robust. - Kleine Arten bis höchstens 2,5 mm. -Sek. Geschlechtsmerkmale: Apo auf d'-Chel-Grundglied, Do- oder Kö-Bewehrung auf d'-Pa. . .
Verbreitung: 7 Arten im größten Teil Europas; von den Kantabrisohe!l· Gebirgen über ganz Mitteleuropa einschließlich der Alpenländer, Italien, Britische Inseln mit Färöer und Orl!:neys, S-Island, Skandinavien mit Ausnahme des höchsten Nordens, Finnisch-Lappland bis 66 °N, in 0-Europa wenigstens bis Moskau, nördl. Teil der Balkanhalbinsel (nicht Griechenland), entlang der Schwarzmeerküste bis wenig östl. des Bosporus. - Außerhalb des hier behandelten Gebietes N. transsylvanicum GRUBER & MARTENS 1968.
Nach genitalmorphologischen Elfwägungen bilden bimaculatum, lugubre, schuelleri, triste und transsylvanicum eine Gruppe engverwandter Arten; von ihr isoliert stehen dentigerum und bidentatum.
Typus-Art (Festlegung durch THORELL I876): Phalangium bimaculalum FABRroxus, I775).
Bestimmungstabelle der Arten (Gilt nur für (!, Pedipalpus und Chelicere zur Betrachtung abtrennen)
Pt des Pa. mit medialer Apophyse (z. B. Abb. 140). . . . . . . . . . . . . . 5 Pt des Pa. ohne mediale Apophyse (z. B. Abb. I23) . . . . . . . . . . . . . 2
2 Apo des Cheliceren-Grundgliedes deutlich zweizipflig (Abb. I25), Glans penis Ia. mit je 9 kleinen Haken • . . . . . . . . . . . . . . . . . . . . . . bimaculatum, S. 95 Apo des Cheliceren-Grundgliedes do gerundet, ohne deutlichen Zipfel (z. B. Abb. I28), Glans penis Ia. mit I oder 3 großen Haken . . . . . . . . . . . . . . . . . . . 3
3 Fe des Pa in der di Hälfte medial und ve aufgetrieben (Abb. I3I), Glans penis Ia mit 3 kräftigen, sich überdeckenden Haken; nur 0-Alpen . . . . . . . . schuelleri, S. 100 Fe des Pa nicht auffällig aufgetrieben, Ja Haken der Glans penis (1 oder 3) überdecken sich nicht; auch außerhalb der Alpen . . . . . . . . . . . . . . . . . . . . . 4
4 Glanspenis mit je I großen ve weisenden Haken (Abb. 136), Ceph meist mit 2 Silberflecken, selten schwarz . . . . . . . . . . . . . . . . . . . . . . . . . . lugubre, S. 103 Glans penismit je 3 kleineren ve weisenden Haken (Abb. I27), nie mit Silberflecken . . .
triste, S. 97 5 Glans penis Ja ohne Haken, ve und do mit je 1 Bo-Feld (Abb. I53a-c), Apo des Chel-Grund·
gliedes trapezoid (Abb. I52a-o) . . . . . . . . . . . . . . . . dentigerum, S. 114 Glanspenis Ja mit je 2 kräftigen, gelenkig inserierten, lanzettförmigen Stacheln (Abb. I47), Apo des Chel-Grundgliedes nicht trapezoid (z. B. Abb. 145) . . . ·. . . . . . . . 6
6 Fe des Pa medial mit spitzen Tu (Abb. 140, 143), Apo des Chel-Grundgliedes mehr oder wenig spitz ausgezogen (Abb. I41, 144-146), Sekretionsfeld (helle Chitinstrukturen) mediad gerichtet . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 7 Fe des Pa medial ohne Tu (Abb. 148), Apo des Chel-Grundgliedes knopfförmig, Sekretions· feld (helle Chitinstrukturen) frontad gerichtet (Abb. 149) . . . . . . . . . . . . . .
bidentatum relictum, S. 112 7 Fe des Pa stark keulenförmig, me-di mit 2-3 kräftigen spitzen Tu (Abb. 140), Apo des
Chel-Grundgliedes auffällig groß, halbmondförmig (Abb. 141) ............ . bidentatum bidentatum, S. 107
Genus Nemastoma 95
Fe des Pa schwach keulenförmig (Abb. I43), me-di mit meist 2 kleinen Tu, Apo des ChelGrup.dgliedes nahezu rechteckig: Oberseite wenig konkav, Vordel'lll'ite wenig konvex (Abb. 144, 145). ·. : . • .. · . . . . . . . . . . . . . . .. . bidentatum sparsum, S. 111
Nemastoma bimaculatum (FARRroms, 1775) Abb. 123-125, 139
1775 Phalangium bimaculatum FABRIOIUS, Systema Entomologiae: 440. 1879 Nemastoma lugubre - SIMON, Arachn. de France, 7: 28I (part.). 1923 Nemastoma lugubre-bimaculalum - ROEWER, Weberknechte d. Erde: 654 (part.). 1951 Nemastoma lugubre-bimaculalum - ROEWER, Senckenbergiana 32: I26 (part.). 1968 Nemastoina bimaculalum - GRUBER · & MAR;ENS, Senckenbergiana bio!. 49: 149 (dort
ausführliche Synonymie). ·
Begründung der Synonymie (vgl. GRUBER & MARTENS 1968): Locus typicus von Phalangi"m bimaculat"m FABRICIUS - einer genitalmorphologisch klar gekennzeichneten, habituell aber N. lugubre außerordentlich ähnlichen Art - ist "Anglia". Die Namen wurden unterschiedlich für beide Arten gebraucht; viele Autoren verwandten durchgehend den ältereln, und 'in Mitteleuropa wurde nur "lugubre" benutzt. RoEWER (1914) koppelte beide zu l!tgubre-bimaculalum. Einfarbig schwarze Individuen nannte er lugubre-unicolor, und stellte sie in den Rang von Unterarten. Da sich lugubre-unicolor aber auf N. triste bezieht, ist Subspezies-Rang nicht vertretbar (vgl. "Variabilität"). N. lugubre und bimaculatum vikariieren weitgehend; so ist die Zuordnung der meisten Nennungen jetzt noch möglich.
Habitus und Feldkennzeiohen: klein, ·tiefschwarz, mit 2 großen Silberflecken auf dem Ceph; in W-und NW-Europa, lokal syntop mit N.lugubre.
Kennzeichnung: Körpermaße in mm: L d' 2,0-2,2; L ~ 2,3-2,5. Laufbein II (d', in l~lammern ~):Fe 1,6 (1,7) Pt 0,5 (0,5) Ti 1,3 (1,3) Mt 2,2 (2;2)
Ta 1,9 (1,9). Pseudogelenke der Fe: I 0, II 1, III 1, IV 2-3.
Abb. I23-125. Nemastoma bimaculatum (FABRICIUS). - 123. [!-Pedipalpus von meclial;. 124. Glans ·penis von ventral (links), dorsal und lateral; 125. (!-Cheliceren-Grundglied von medial (oben) und lateral. - SP Prov. Pamplona.

96 Familia Nemastomatidae
Genitalmorphologie (Abb. 124): Truncus penisohne Absatz in die symmetrische Glans übergehend, diese lanzettförmig zugespitzt, Ia mit je 1 Reihe von 8 kleinen basad gerichteten Dornen; 2 Paar kleine Sinnesborsten an der Basis des Stylus. Glans in sich geschwungen und dorsad gerichtet.
Cheliceren (Abb. 125): d'-Basalglied mit großer distad gerichteter Apo, di breit gerundet me-di in einen spitzen Zipfel ausgezogen, medial konkav und mit Haaren besetzt, dort das Sekretionsfeld.
Pedipalpus (Abb. 123): Ti do-ba mit stark granulierter Beule; übrige Glieder normal proportioniert.
Laufbeine: Fe, Pt, Ti I schwach keulig verdickt, Fe besonders do-proximal mit zerstreuten, etwa in 2 Längsreihen stehenden Kö, besonders auffallend an der Basis des Fe IV; bei lugubre (auch ~ !) fehlend.
Dorsalseite: Souturn und freie Tergite im Vergleich zu lugubre glatt, niedrige weit auseinanderstehende Körnchen nur auf den thorak und abd Areae gehäuft und deutlich hervortretend. Färbung tiefschwarz; auf dem Ceph I Paar große Silberflecken mit unregelmäßigem Umriß. Souturn compositum bei d' nur schwach ausgebildet, selbst bei ~bleibt Terg VII durchunverhärtete Membran vom Sou-turn getrennt. ·
Variabilität: Schwärzlinge ohne thorak Fleckenzeichnung kommen nur selten vor, gelegent· lieh lokal gehäuft. Einzelne sind aus England bekannt (Bakewell, Shoreham, Cantonbury Ring, SANKEY 1953a; Cheshire, MAOKIE 1968). - Frankreich: Tal des Gave d'Arrena bei Lour. des/Hautes-Pyr., 1500 m: von 20 ~ u. 10 !? nur 3 mit Fleckenresten, alle anderen völlig schwarz. 2 ~' 3 !? aus demselben Tal aus etwa I 000 m waren dagegen normal gefleckt; ferner Dep. Cantal (SIMON 1879), Belgien (BEOKER 1896).
Verbreitung (Abb. 139): Typ: atlantisch. -Geschlossenes Areal von den Kautabrisahen Gebirgen und Pyrenäen, ganz Frankreich, Britische Inseln, lokal in Skandinavien und BRD. - Locus typicus: Britische Inseln.
Spanien: Kautabrisehe Gebirge, Picos de Etll'opa, Espinama (J. M.leg.). -Frankreich: kontinuierlich verbreitet einschließlich der Pyrenäen, lokal oder völlig fehlend im mediterranen Küstenatreifen, östl. der Rhöne und an der 0-Grenze zur BRD, Grenzverlauf dort nicht bekannt. - Belgien (BECKER 1896; J. G. rev.) .. - BRD: Rheinland linksrheiniach: Venusberg und Marieuforater Tal bei Bonn (J. M.leg.), rechtsrheinisch Siebengebirge (VERHOEFF leg.). - Gesamte Britische Inseln einschließlich der Shet.lands, Orlmeys und Färöer (BRISTOWE 1949, N. lugubre). - Island (HENRIKSEN 1932, N. lugub•·e; J. G. rev.). - Norwegen: Skjolden am Joatedalsbre (WuNDERLICH 1973, in litt.; J. M. rev., Pfeil in Abb. 139), darüber hinaus in der westl. Küstenzone von Kristiansand (58° N) nordwärts weit verbreitet (KAURI in litt.) und erreicht die Lofoten (HAtrGE in litt.). Bisher keine Nachweise aus den Niederlanden, dort aber zu erwarten. In Belgien, im Rheinland und wahrscheinlich lokal in Norwegen leben bimaeulatum und lugubre syntop. Die weitgehende Vikarianz zwischen beiden ist offensichtlich nicht durch Konklll'renz bedingt.
Ökologie: Biotopansprüche: N. bimaculatum lebt hauptsächlich in Wäldern mit hoher Bodenfeuohtigkeit, meist geschützt vor Insolation; im extrem atlantischen Gebiet auch in offenem Gelände, in Wiesen, in Feuchtgelände mit Carex und Equisetum. Hohe Luftfeuchtigkeit und ausgeglichener Temperaturgang müssen gewährleistet sein. Mit diesen Präferenda übereinstimmend die ökophysiologischen Daten: bevorzugte relative Luftfeuchtigkeit 69-79% (Vertrauensgrenzen), bevorzugte Temperatur bei 100% RL 9,6 °0 (Vertrauensgrenzen 7,3 und 11,9 °0; ToDD 1949). Der Temperaturwert liegt, verglichen mit anderen terrikolen Arten (z. B. Anelasmocephalus cambridgei) auffallend niedrig.
i. \;. .. ~.
Genua Nemastoma 97
Vertikalverbreitung: Von den Küstenlandschaften der Nordsee und des Atlantik wahrscheinlich bis an die Waldgrenze in den Pyrenäen (südl. Lourdes 1500 m) und in den Kautabrisehen Gebirgen (Picos de Europa 1250 m, beide J. M. leg.).
Phänologie: ad. eurychron; für die Britischen Inseln PlllLLIPSON (1959), W!LLIAMS (1962), SANKEY & SAVORY (1974); ad. der neuen Generation ab IV (TODD 1949), V (WILLIAMS 1962) bzw. A VIII (PHILLil'SON 1959), Hauptaktivitätsperiode V-VIII. Eiablage E VIII - A XII (PHILLIPSON 1959); Zyklus von Eiablage bis Tod der ad. erstreckt sich somit über 3 Jahre.
Nemastoma triste (0. L. KooH, 1835) Abb. 126-130
I83ö Phalangium triste C. L. KocH; in HERRICH-SCHÄFFER, Fauna Ins. Germ., 34: 128. 1923 Nemastoma triste - RoEWER, Weberknechte d. Erde, : 655. 1968 N~mastoma t·riste - GRUBER & MARTENS, Senckenbergiana bio!., 49: 167 (dort die übrige
Synonymie).
Habitus und Feldkennzeichen: klein, einfarbig, tiefschwarz. ~Im Gelände meist nicht sicher bestimmbar, da gebietsweise mit anderen, ebenfalls schwarzen Arten sympatrisch (Sohwärzlinge von N. lugubre und N. schuelleri, alle Subspezies von 1{. bidentatum).
Kennzeichnung: Körpermaße (in mm) L d' 1,5-1,7; L ~ 1,7-1,9. Laufbein II (Q', in Klammern ~):Fe 1,4 (1,5) Pt 0,5 (0,4) Ti 1,1 (1,2) Mt 1,6 (2,0)
Ta 1,5 (1,6). Pseudogelenke der Femora I 0, ausnahmsweise 1; II 1-3, meist 2; III 1; IV 1-4, meist 2-3; oft beidseitig verschieden.
1 .].\",.. -:~'· ~-
. '. /, '•.' ·'· y;
Abb. 126-129. Nemastoma t1-iste (C. L. KocH). - 126. ~-Pedipalpus von medial; 127. Glans penis. von ventral (links);· dorsal und lateral; 128. ~-Cheliceren-Grundglied von medial (oben) und lateral; 129. Penis. - BRD Böhmerwald, Luaen.
· 7 Tierwelt Deutsch!. 64: ~fartens

i I 98 Familia Nemastomatidae
Genitalmorphologie (Abb.l27): Truncus penis vor allem in der di Hälfte nahezu parallelrandig, stark do-ve abgeplattet; Glans nicht vom Truncus abgesetzt, diese· nahezu symmetrisch, lediglich der Stylus etwas in sich gekrümmt. Glans la mit je 3 kräftigen, basad gerichteten Hakendornen, davon apikal jeweils ein weiterer kleinerer. Wenige Bo am Stylus. ··
Cheliceren (Abb. 128): d'-Basalglied mit nahezu quadratischer; vom Grundglied nicht abgesetzter do-di Apo; diese do-di wenig konkav und do etwas nach medial umgebogen, also undeutlich zweizipflig; geringer Bo-Besatz medial im Bereich des Sekretionsfeldes.
Pedipalpus (Abb. 126): ohne sek. d'-Geschlechtsmerkmale, Ti do undeutlich konkav.
Laufbeine: Fe bis Ti I leicht keulig verdickt, alle übrigen Glieder normal schlank.
Dorsalseite: Souturn und freie Terg dicht granuliert und fein punktiert; auf den Areae I-V nur andeutungsweise Verdichtungen größerer Granula, jedoch ohne markante Höcker; Souturn von la nur schwach wellig; tiefschwarz ohne Gold- oder Silberflecken-Zeichnung. - Bei alten !j2 Terg VI in das Souturn compositum einbezogen; vor Terg VII immer ein heller Querstreif nicht verdunkelter Membran. ·
I
Variabilität: Im gesamten Areal bemerkenswert einheitlich: Gestalt von Oheliceren und Pedi-palpen aus den deutschen Mittelgebirgen und den Österreichischen Alpen gleich, doch kann. bei Individuen alpiner Herkunft die Granulierung des Scutum· reduziert sein, so daß fast glatte Partien vorkommen. Die Zahl der femoralen Pseudogelenke schwankt dagegen erheblich.
Verbreitung (Abb. 130): Typ: postglazial reliktärer .Endemit der O~Alpen und einiger Mittelgebirge. - Locus typicus: Regensburg.
Alpenländer: Österreich: Vorarlberg: Großvermunt, Flexenbachtal nördl. Zürs (westlichste alpine Fundorte, J. G. leg.); - in Tirol, Salzburg und Ober-Österreich in allen Hauptlandschaften: Lungauer Becken, Zentral-Alpen, Grauwackenzone, Kalkalpen, Salzburger Becken, in der Flyschzone, im unteren Salzachtal (eiszeitliche Endmoränen), im Oberen Weilhartforst und im südöstl. Hausruck (etwa auf Linie Burghausen-Wels); - weiter östl. in OberÖsterreich in den Kalkhoch- und Kalkvoralpen, Sauwald (J. G. in litt.), nördl. der Donau im Mühtviertel nördl. Linz (A. A.leg.); - Nieder-Österreich: wohl allgemein verbreitet in den Kalkhoch- und Kalkvoralpen, belegt sincl das Lunzer Gebiet, Ötscher, Raxalpe, Schneeberg und Wechsel (St. Oorona, östlichster Fundort in den Alpen, J. G. in litt.); fehlt im Wienarwald und der Buckligen Welt nordöstl. vom Wechsel, nächstgelegener Fund zum Wiener Becken: Sierningbach-Tal nordwestl. Ternitz (490 m), im westl. Nieder-Österreich bis ins Alpenvorland (Purgstall, 300m, RESSL leg.), fernerins Donautal (Strudengau) und in das nördl. anschließende Waldviertel (bis 220m herab); nicht in den Donau-Auen und in der Ebenenund Hügelstufe des östl. Nieder-Österreich mit der auffälligen Ausnahme des Bisamberges an der Donau nördl. Wien (J. G. leg.); - Steiermark: überall in den alpinen Landesteilen, jedoch keine Nachweise aus dem 0-steierischen Hügelland und der Grazer Bucht; - Kärnten: weitverbreitet in der S-Abdachung der Hohen Tauern und in den Gurktaler Alpen, aus den Karswanken (Bärental, A. A. leg.) und Karnischen Alpen erst wenige Funde in der SubalpinStufe, wo N. bidentatum vorherrscht; fehlt in den 0-Karawanken (z. B. um Eisenkappel) und in der Sattnitz (J. G. in litt.). - Italien: S-Tirol, Sexten (A. A.leg.); fehlt wahrscheinlich in großen Teilen der S-Abdachung der Zentral-Alpen, z. B. nicht bei DeutschnofenfBozen, Sarntheim, Meran und im Vintschgau (A. A. in litt.). - BRD: im Anschluß an die Österreichischen Alpenteile weit verbreitet; bisher keine Nachweise aus dem Allgäu.
Mittelgebirge: BRD: Böhmerwald und Bayerisoher Wald: Rache!, Lusen, Arber (J. M. leg., KIRCHNER & al. 1967), Regensburg (0. L. KocH 1835), Happurg bei Nürnberg (L. KocH.
Genus Nemastoma
Abb. 130. Gesamtareal von Nemastoma triste (0. L. KocH).
1877); Vogelsberg, Taufstein; Fichtelgebirge (beide J. M. leg.). _ DDR. Erz ebir e (SoH · ~47, N: lugubre unicolor). - ÖSSR: Böhmerwald/Bubin, Marianske fazneg (Mari~~~~ ~ I~HAVY 1956, 1966). - Polen: Riesengebirge, Schneekoppenkegel (BRUNN leg. Beleg ZMH
· · rev., vgl. FrcKERT 187?)- - Belgien: Hautes-Fagnes: Hockai, Moup.; (CooREMA~ 1949, J. G: rev.) .. - Außeralpmes Österreich vgl. unter "Alpenländer, Österreich".
Ökologte: Bwtop~_nsprüche: N. triste besiedelt bodenfeuchte Waldgesellschafte~, z. B. Laubwalder unterschiedlicher Zusammensetzung ebenso die ur. :'egetatwn :'"on _;'3äch~n und Flüssen und angrenzender Hochst~udenfluren, oh~~ ~ndes auf die N~he fheße_nden _Wassers angewiesen zu sein; in den Alpen auch in . wergs~rauchhe~den und m alpmen Rasen über der Baumgrenze, und seltener auch m Habitaten mit trocke_ner_ Bodendecke, z. B. am Bisamberg nördl. Wien (J. G leg.). Aufenthalt_ ausschließlich am Boden in der Detritussohicht, unter Holzteile~ un~ locker gesch_whteten Blockhalden, sofern sie beschattet sind. _ In den Mittel gebrrgen _lebt tnste syntop mit N. lugubre und in den 0-Alpen mit N schuell ~ und N. bzdentatum. · en
Vertikal ver brei tung: Alpen: In allen Höhenstufen; in Salzburg in der Montan~ (50_0-1;oo m) und Subalpin~Stufe (1200-2000 m) etwa gleichmäßi verbreitet (Je 46 Y~ der Serie~ von_n ~·'380, A. A. in litt.), in der Alpin-Stufe (20~0 bis 2400 ~)nur 5 Yo der Senen; die hochsten Funde in Salzburg in den Zentral-Alpen und nordl._ Kalkalpe~. (2270 _und 2300 m), niedrige Funde in den Salzachauen (380m), Bisa~berg nordl. Wien 235-350 m (J. G. leg.), nördl. Purgstall 280 m (RESSL leg.); lll· den Berchtesgadener Alpen (BRD) bis 2320 m (Schneiber östl. des Gr. ~undstod, H~MS leg.). - In den Norischen Alpen, dem Arealschwer unkt von-N. bzdentr;-tum r~lzctum, fehlt triste in der Subalpin-Stufe: Die obere Ve~breitungsgrenze liegt be1 etwa I 600 m, als::~ 700 m tiefer als in den Hohen Tauern. 7•
99

100 Familia Nemastomatidae
Bei postglazialer Neubesiedlung hat triste nicht (oder ?-ur lokal mi~ geringem Erfolg) in die relictum-Populationen eindringen können, die dort wemgstens das letzte Glazial überdauert haben. Umgekehrt gelang es b. relictum (fast) nicht, postglazial triste-Siedlungsgebiet zu erreichen, z. B. in den Hohen und Niederen Tauern. Dieser Befund läßt auf sehr ähnliche, wenn nicht gar gleiche Einnischung beider Arten schließen.
Mittelgebirge: Bevorzugt in den höheren Lagen bzw. in niedrigen Gebirgsstöcken allein in den Gipfelbereichen, z. B. Vogelsberg/Taufstein (770 m). Arealteile außerhalb von Gebirgen sind Ausnahmen: nördl. Linz (um 280m, A. A. leg.); Alpenvorland in Nieder-Österreich: Pnrgstall (300m), Waldviertel (220m), Bisamberg nördl. Wien (235-350 m; alle J. G. in litt.).
Phänologie: ad. eurychron; Nachweise in Salzbnrg von III-I (A. A. in litt.); bei PurgstallfNieder-Österreich in allen Monaten außer I. - Generationenfolge: von M VII - E IX alte, ausgefärbte neben jungen, hellbraunen ad. der neuen Generation (Fallenfänge, Bisamberg, J. G.) somit Überschneidungzweier Generationen. - Sex ratio: Handfänge 733 ~: 737 ~ (Salzburg, A. A. in litt.), 155 ~ : 106 ~ (Purgstall, Nieder-Österreich, RESSL leg.), Fallenfänge 85 ~: 101 ~ (Bisamberg, J. G. leg.).
Nemasto~a sohuelleri GRUBER & MARTENS, 1968 Abb. 131-134
1968 Nemastoma schuell.,·i GRUBER & MARTENS, Sencl<enbergiana bio!., 49: 160.
Habitus und Feldkennzei~hen: klein, tiefschwarz, mit 2 großen Silber: oder GoldFlecken auf dem 'ceph, ~-Pa-Fe sub-di angeschwollen; nnr in den 0-Alpen.
Kennzeichnung: Körpermaße (in mm): L 1,8-1,9; L ~ 2,0-2,2.
r~ ~-/~;·~ 132 ~\~
j:, ~ b ~ t\ j4 1: \I ;f4 '\ (' / \\\ j/ ~; ! .. '_i\
''.-l' 1 l I . \ i } "
.·~ I 133
-~-
/
- -.__ ._· _._<·~_> . .,.....
\. -.: ,/. --~-:__.
Abb. 131-133. ·NemaBtoma. schuelle1'i GRUBER & MARTENS, - 131. d'-Pedipalpus von medial; 132. Glanspenis von ventral (links), dorsal und lateral; 133. d'·Oheliceren-Grundglied von medial (oben). und lateral. - Nach Holotypus.
Genus Nemastoma 101
Laufbein II (~,in Klammern ~):Fe 1,5 (1,4) Pt 0,5 (0,5) Ti 1,1 (1,1) Mt 2,1 (2.1) Ta 1,8 (1,9).
Pseudogelenke der Femora: I 0, II 1, III 1, IV 3. Genitalmorphologie (Abb. 132): Truncus penis im di Drittel bauchig er
weitert, znr Glans versohmälert, im Querschnitt angenähert gleichseitig dreieckig; eine Flachseite weist ventrad. Glans Ia ausgeschwungen und einschließlich des Stylus etwas unsymmetrisch; Ia mit je 3 kräftigen, basad gerichteten Haken, sich do dachziegelartig überdeckend. Glans ve stark sklerotisiert; Stylus ventrad gekrümmt, mit einigen knrzen Bo besetzt.
Cheliceren (Abb. 133): ~-Grundglied do-di mit schräg nach vorn gerichteter Apo, von la und me nahezu quadratisch, Ecken leicht gerundet; das mediale Sekretionsfeld etwas vertieft und mit feinen Haaren umstellt.
Pedipalpus (Abb. 131): ~-Fe an der di Hälfte mit me und ve Auftreibung, Pt di wenig nach ve abgebeugt, Ti do wenig konkav.
Laufbeine: ~-Fe I schwach keulig verdickt, alle anderen Glieder dieses und der übrigen Beine normal.
Dorsalseite: Souturn und freie Tergite stark granuliert, vor allem Ceph und Tu oc; thorak Terg II und abd Areae I-IV mit je 1 Paar para-me Tu, auf dem thorak Terg und den Areae I-II zu einem breiten Höcker verschmolzen. Ceph mit 2 Ia Gold- oder Silber-Flecken, bisweilen in je 2 Einzelflecke aufgelöst oder gänzlich reduziert. ~: do Höckerbildung weniger markant, ähnlich bei anderen Arten der Gattung.
Variabilität: Die thorak Silberflecken können denselben Reduktionsvorgängen wie bei N. lugubre unterliegen, so daß auch bei schuelleri alle Übergänge von gezeichneten bis (selten) einfarbig schwarzen Individuen vorkommen. Abhängigkeit von der vertikalen Lage des Siedlungsgebietes wie bei lugubre ließ sich bisher aber nicht nachweisen.
Beziehungen: Sie bestehen - unter genitalmorphologischem Aspekt - vor allem zu N. triste. Nahe Verwandtschaft beider Arten scheinen aber manche Besonderheiten auszuschließen.
Verbreitung (Abb. 134): Typ: Endeinit der 0-Alpen. - Locus typicus: Mur-Tal, Lungau.
Österreich: N-Tirol: Oberperfuß-Rögerl (K T. leg., westlichster Fundort); Salzburg, N-Abdachung der Hohen Tauern: Untersulzbach-, Habach-, Hollersbach-, Stubach-, Rapruner-, Rauriser TalfHüttwinkeltal, Gasteinertal, Großarltal; Lungau: beim :Kraftwerk Rotgülden; Nockberge und GurktaJer Alpen: BundschuhfFeldseite; Rendlbruckgraben; Turrach (alle A. A.leg.); Steiermark: Ennstal westl. Adrnont (FRANZ leg.), Fischbacher Alpen: östl. Stanz; Feistritz-Tal westl. Wechsel, öst.l. Rettenegg (beide STROUHAL leg.; bisher östlichster Fundort); Kärnten: Mallnitz, Mirnock bei Radenthein, Taohiernock bei Gmünd, Afritz, Amberger Alm (bisher südlichster Fundort; alle A. A. leg.). Außer den Funden bei Innsbruck, arn Wechsel und in c:len Fischbacher Alpen liegen alle schuelleri-Nachweise in den Hohen Tauern und in den Gurktaler Alpen; nördl. der Längstäler von Inn, Salzach und Enns scheint schuelleri zu fehlen, praktisch gesichert ist das ffu• die (von A. A.) gründlich untersuchten Gebiete nördl. des Salz· ach-Längstales .
Ökologie: Biotopansprüche: N. sckuelleri lebt in bodenfeuchten Waldgesellschaften und dort bevorzugt an besonders feuchten Stellen wie Bachufern, Gräben und Gerinnen in Ainus-Gesellschaften )lud Hochstaudenfluren, ferner in und an oberflächlich rinnendem Wasser und in dessen unmittelbarer Umgebung, dort gern unter locker eingebetteten und lose aufliegenden Steinen und Holzteilen. Verglichen mit N. triste und dem extrem hygrophilen Pamnemastoma bicuspi-
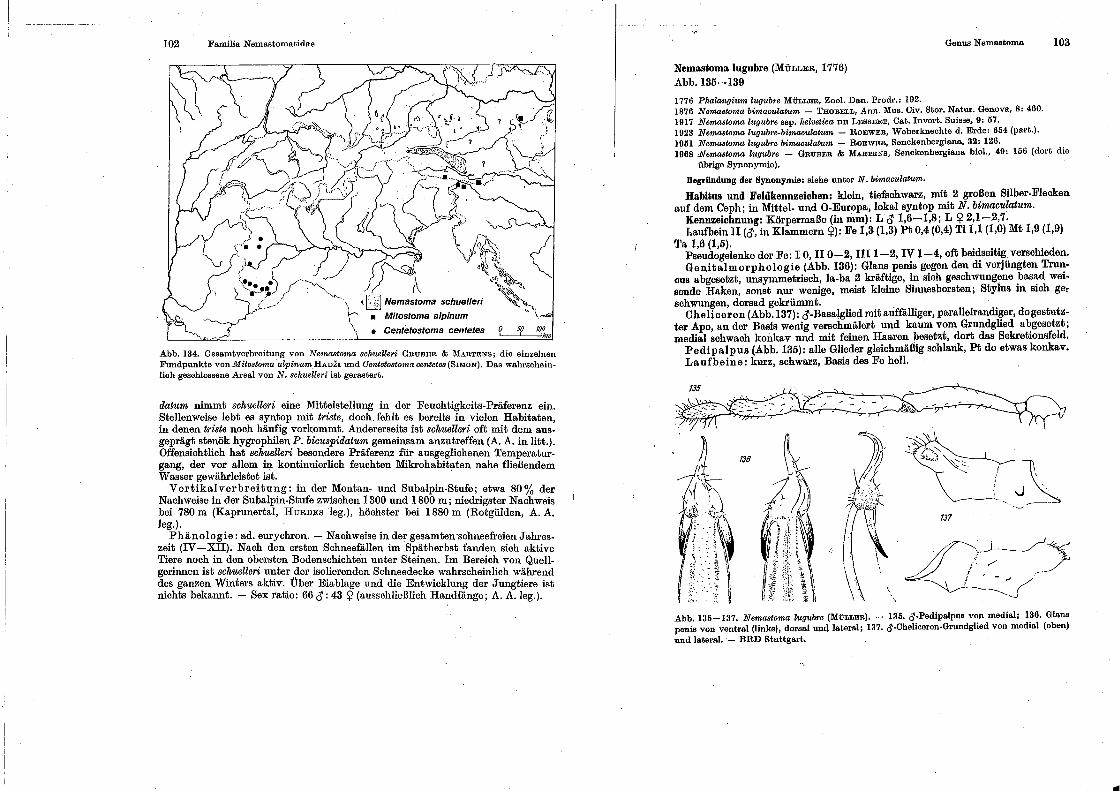
102 Familia Nemastomatidae
• Centetastoma centetes o so
Abb. 134. Gesamtverbreitnng von Nemastoma schuelleri GRUBER & MARTENS; die einzelnen Fundpunkte von Mitostoma alpin um HADZI nnd Oentetostomn centetes (SrMoN). Das wahrscheinlich geschlossene Areal von N. schuelleri ist gerastert.
datum nimmt schuelleri eine Mittelstellung in der Feuchtigkeits-Präferenz ein. Stellenweise lebt es syntop mit triste, doch. fehlt es bereits in vielen Habitaten, in denen triste noch häufig vorkommt. Andererseits ist schuelleri oft mit dem ausgeprägt stenök hygrophilen P. bicuspidatum gemeinsam anzutreffen (A. A. in litt.). Offensichtlich hat schuelleri besondere Präferenz für ausgeglichenen Temperaturgang, der vor allem in kontinuierlich feuchten Mikrohabitaten nahe fließendem Wasser gewährleistet ist.
Vertikalverbreitung: in der Montan- und Subalpin-Stufe; etwa 80% der Nachweise in der Subalpin-Stufe zwischen 1300 und 1800 m; niedrigster Nachweis bei 780 m (Kaprunertal, HuRDES leg.), höchster bei 1880 m (Rotgülden, A. A. leg.).
Phänologie: ad. eurychron. -Nachweise in der gesamten·schneefreien Jahreszeit (IV -XII). Nach den ersten Schneefällen im Spätherbst fanden sich aktive Tiere noch in den obersten Bodenschichten unter Steinen. Im Bereich von Quellgerinnen ist schuelleri unter der isolierenden Schneedecke wahrscheinlich während des ganzen Winters aktiv. Über Eiablage und die Entwicklung der Jungtiere ist nichts bekannt. - Sex ratio: 66 c1: 43 ~(ausschließlich Handfänge; A. A. leg.).
Nemasioma Iugubre (MüLLER, 1776) Abb. 135-139
Genus Nemastoma
1776 Phalangium lugubre MÜLLER, Zoo!. Dan. Prodr.: 192. 1876 Nemastoma bimaculatum - THORELL, Ann. Mus. Civ. Stor. Natur. Genova, 8: 460. 1917 Nemastoma lugubre ssp. helvetica DE LESSERT, Cat. lnvert. Suisse, 9: 57.
103
1923 Nemaetoma lugubre-bimaoulatum - RoEWER, Weberknechte d. Erde: 654 (part.). 1951 Nemastoma lugubre bimaculatum - RoEWER, Senckenbergiana, 32: 126. 1068 Nemastoma lugubre - GRUBER & MARTENS, Senokenbergiana bio!., 49: 156 (dort die
übrige Synonymie).
Begründung der Synonymie: siehe nnter N. bimaculatum.
Habitus und Feldkennzeichen: klein, tiefschwarz, mit 2 großen Silber-Flecken auf dem Ceph; in Mittel- und 0-Europa, lokal syntop mit N. bimaculatum.
Kennzeichnung: Körpermaße (in mm): L ö-1,6-1,8; L ~ 2,1-2,7. Laufbein II (d", in Klammern~): Fe 1,3 (1,3) Pt 0,4 (0,4) Ti 1,1 (1,0) Mt 1,9 (1,9)
Ta 1,6 (1,5). Pseudogelenke der Fe: I 0, II 0-2, III 1-2, IV l-4, oft beidseitig verschieden. Genitalmorphologie (Abb. 136): Glanspenis gegen den di verjüngten Trun
cus abgesetzt, unsymmetrisch, la-ba 2 kräftige, in sich geschwungene basad weisende Haken, sonst nur wenige, meist kleine Sinnesborsten; Stylus in sich gec schwungen, dorsad gekrümmt. .
Cheliceren (Abb.137): ö--Basalglied mit auffälliger, parallelrandiger, dogestutzter Apo, an der Basis wenig verschmälert und kaum vom Grundglied abgesetzt; medial schwach konkav und mit feinen Haaren besetzt, dort das Sekretionsfeld.
Pedipalpus (Abb. 135): alle Glieder gleichmäßig schlank, Pt do etwas konkav. Laufbeine: kurz, schwarz, Basis des Fe hell.
)A. 11 i#
((\ \\··. I.
137
Abi>. 135-137. Nemastoma lugubre (MÜLLER). - 135. d'·Pedipalpus von medial; 136. Glans penisvon ventral (links), dorsal und lateral; 137. d'·Che!ioeren-Grundg!ied von medial (oben) und lateral. - BRD Stuttgart.
..

104 Familia Nemastomatidae
Abb. 138. Nemaseoma lugubre (MÜLLER). - Variabilität des dorsa.Ien Zeichnungsmusters (dunkel angelegt). - normale Zeichnung: a) AUS Wien; reduzierte Zeichnung: b) SCH Hailand; c) DÄNinsei Bornholm; d) SCH Halland, e) DÄN Insel Bornholm; exzessive Zeichnung: f) AUS Wien; g) UNG oberes Theiß-Tal.
Dorsals&ite: Souturn und freie Tergite fein gepunktet und - segmentweise konzentriert - stark granuliert, aber dennoch schwach glänzend; abd Areae erhaben; auf Areae I bis höchstens IV para-me je 1 besonders intensiv bekörneltes Tuberkel paar. Auf den thorak Terg la je 1 großer Silberfleck, bisweilen auf abd Areae übergreifend. Mehrwöchige adulte und somit ausgehärtete Individuen immer tiefschwarz, sonst bräunlich. Souturn compositum bei alten Tieren, vor allem <f'; meist jedoch zwischen Tergit VI und VII ein unsklerotisierter Me:q~branstreif.
Variabilität (Abb. 138): Individuen einzelner Populationen neigen dazu, die großen Silberflecke auf dem Ceph zurückzubilden oder sogar völlig zu reduzieren. Die Ausdehnung der Fleckung liegt bereits bei der Imaginalhäutung fest; Schrumpfungen durch fortschreitende Sklerotisierung des Exoskelettes kommen vor.
Die Verkleinerung der Silberflecken läßt sich daran erkennen, daß ihnen vom Rand her gelbes Pigment eingelagert wird, das mehr und mehr verdunkelt und schließlich schwarz wird. Das hohe Alter dieser Individuen ist auch am Souturn compositum erkennbar (vgl. Gattungsdiagnose). - Die Verteilung der Schwärzlingspopulationen innerhalb des Areals is6 unterschiedlich, und ihre Abhängigkeit von äußeren Faktoren nicht immer klar ersichtlich. In den mitteleuropäisohen Mittelgebirgen lösen sich beide Formen in einer bestimmten Höhenstufe ab: in der Gipfelregion des Taufsteins (Vogelsberg, 770 m) finden sich Individuen mit reduzierter Fleckung neben (selten) ganz schwarzen. In den polnischen Waldkarpaten (Bieszczady) leben bis 700-750 m (auch) normal gefleckte Tiere, zwischen 550-600 m bis auf die höchsten Gipfel (z. B. Polonina Wetlinska, 1253 mundauf Halicz und Krzemien über 1300 m) meist einfarbig schwarze, nur zwischen 600 bis 800 m eine Obergangsform mit reduzierten .Flecken (STARI!GA 1966a). Die einzige Österreichische Schwärzlings-Population liegt im Gegensatz zu bisherigen Erfahrungen im Tiefland: Grub an der March (170·m, J. G. leg.). Knapp die Hälfte der hier gesammelten Tiere sind einfarbig schwarz. Gelegentlich kommen Individuen mit exzessiver Zeichnung vor (Abb. 138).
Verbreitung (Abb. 139): Typ: subatlantisch (-mitteleuropäisch), in auffallender Vikarianz zu N. bimaculatum. - Geschlossenes Areal von den Benelux-Ländern, 0-Frankreich, das engere Mitteleuropa und Skandinavien über die europäische UdSSR bis auf die Balkanhalbinsel südl. bis Bulgarien. - Locus typicus: Däne-mark. ·
Genus Nemastoma 105
Abb. 139. Mittel- und Teile des 0-europäischen Areals von Nemasloma lugubre (MüLLER) und Gesamtareal (außer Shetland-Inseln, Island und W-Küste von Norwegen) von N. bimaculalum (FABRICIUS).
Arealgrenzen: W-Grenze westl. des Rheins (in Belgien und Teilen der nor<lwestl. BRD syntop mit N. bimaculaeum) und entlang des Unterlaufs der Rhöne, jedoch im einzelnen nicht belegt. - S-Grenze: im engeren Mitteleuropa in der S eh weiz, dort außeralpin woitverbreitet; in den alpinen Landesteilen: Berner Oberland: Adelboden, EngstligenfaJI, in Nidwalden südl. des Vierwaldstättersees (SCHENKEL 1923), Merligen nm·döstl. des Thuner Sec" (RCHENKEL 1936), Axalp südl. Brienz (LESSERT 1917); Grindelwald südl. des Faulhorns und Obbürgon in Nidwalden (ScHENKEL 1918); - Oberes Rbeintal: Chur (Beleg SMF), oberstes Tessin: Bedret.I.n. Ronco (SCHENKEL 1929). Im Ober-Wallis wahrscheinlich zum großen Teil fehlend, ebermo wohl im Tessin. - Weiter östl. umfaßt das Areal in Österreich einen Großtei(Vorarlbergs (J. n. in litt.), ferner in der BRD das Allgäu (z. B. Balderschwang/Oberstdorf, J. M.leg.); aus dnm östl. anschließenden bayerischen Alpenvorland in der Partnachklamm südl. Garmisoh-Partenkirchen (VERHOEFF leg., Beleg ZMB); ,die Grenze ist sonst in Bayern nicht belegt. - Öste•·reioh: im östl. Landesteil nördl. der ri'onau: Böhmische Masse und Weinviertel mit Marchfeld sowie im Donautal und dort Charakterart in den Auwäldern, sürll. der Donau in Nieder-Österreich nur aus den Hainburger Bergen bekannt (J. G. in litt.). - Ungarn: im W entlang der Donau, aber nördl. davon in Ö S SR /S-Siowakei bis zur Donau nachgewiesen: große Schüttinsel (GULIÖKA 1957), !ltilrovo (!lrLHAvY 1972); weiter im 0" im Cserhat-Gebirge (SzALAY 1949), an der Theiß erst im Oberlauf(aufwärts etwa von Toka.j, KOLOSVARY 1969); fehlt somit w<>itgehend im pannonischen Tiefland bis auf seine nörcll. Gebirge und Hügelländer. - SO-Grenze: Rumänien: für die 0-, S- und W-Karpaten vielfach genannt. (AVRAM & DHrrJITREROl' 196!1, GRrBER & MARTENS 1968), SW-Karpaten nahe dem Eisernen Tor, Almajului-Gohirge (Dn-

106 Familia Nemastomatidae
MITRESCU 1972); außerhalb der Karpaten: Moldau, Bro~teni bei Iasi (BECKER 1879) und nördl. Dobrtidscha, Slava Rusa (AVRAM & DUMITRESOU 1969); - {J dS SR /Karpato-Ukraine: Rachov (BARTOS 1949). - Bulgarien: vereinzelt im W (W. S. in litt.), - 0-Grenze: im europäischen Teil der UdSSR, bisher erst mangelhaft bekannt: SchodnjafMo~kau (GRESE 1911); Podolien: Djakivci westl. Vinnica(MonrN 1934); PetrodvorecfLeningrad(ROEWER 1951); Lettland, mehrere Fundstellen (TUMSS 1963); ehemaliges Ostpreußen: bei Königsberg (Kaliningrad) (LE Ror 1913).
Ökologie: Biotopansprüche: N. lugubre ist als euryöker und hygrophiler Bodenbewohner weitgehend auf beschattetes Gelände beschränkt: vorzugsweise in Laubwald, auch in aufgelockerten Gesellschaften, z. B. Feldgehölzen, Buschreihen, Flußauen, soweit nur ein Mindestmaß an Bodenfeuchtigkeit gegeben ist. .Auch MEINERTZ (1964a) bezeichnet lugubre als Laubwaldform. Im pannonischen Raum Österreichs in .Auwäldern, in collinen Laubmischwäldern, z. B. im Hügelland des Weinviertels nördl. der Donau (J. G. in litt.) . .Aufenthalt in der Bodenstreu, unter Laub, Steinen und altem Holz; mit .Ausnahme der unmittelbaren Arealgrenzen ist lugubre häufig und kommt meist in hoher Siedlungsdichte vor.
Vertikalverbreitung: Von den Nord- und~Ostseeküsten bis in mittlere und höhere Lagen der Mittelgebirge, dort bisher keine Nachweise über 1300 m. In diesem Höhenbereich ist lugubre - soweit es dort noch vorkommt - spärlich und wird 1\'eitgehend von N. triste ersetzt. In den schweizerischen W-.Alpen, wo potentiell konkurrierende .Arten fehlen, steigt lugubre bis in die obere SubalpinStufe: BedrettoJRonco: 1500-1600 m (ScHENKEL 1929), .Axalp: 1790 m (LEs-SERT 1917). .
Phänologie: ad. eurychron, möglicherweise lokal stenochron sommerreif ohne Überwinterung. BRD: Raum Erlangen, Fallenfänge: ad. von VII-XII, .AktivitätsMaximum X-XI (PFEIFER 1956); Mainz: VI-II (HöREGOTT 1962); Vogelsberg, Taufstein: ad. neuer Generation neben ad. der Vorjahresgeneration in VIII (J. M.leg.); DDR: Greifswald: I-XII, .Abundanz-Maximum IX-XII, .AktivitätsMaximum E X-.A XI (BROEN & MoRITZ 1963). - Niederlande: ad. I-XII, .Aktivitäts-Maximum IX-XII (SPOEK 1963); Lauwerszeepolder, Fallenfänge VI-XII, .Aktivitäts-Maximum IX-X, iuv. VI-VII, dort möglicherweise keine Überwinterung der ad. (MEIJER 1972). - Dänemark: Fänge ganzjährig außer I (MEINERTZ 1964c). - Österreich: ad. neuer Generation mit denen der VorjahresGeneration ab VI, ad. überwintern, große iuv. in V-VI (J. G. in litt.).
Die meisten Daten deuten auf Eurychronie mit ad. der neuen Generation ab VI und zeitlicher Überschneidung alter und neuer Generation; .Aktivitäts-Maximum von IX-XII mit lqkal unterschiedlichen Spitzen; strenge Stenochronie glaubt nur MEIJER (I972) für ein erst kürzlich von lugubre besiedeltes Polder-Gebiet gefunden zu haben. Der Befund allein mittels Fallen ist indes nicht zweifelsfrei. - Gelege überwintern. Sex ratio: 1220d': 846 ~ (Fallenfänge, MEIJER 1972 für niederländische Polder); 966 d': 1229 ~ (Handfänge, MEiNERTZ 1964o für Dänemark).
Nemastoma bidentatum RoEWER, 19i4 .Abb. 140-150
1914 Nemastoma bidentatum RoEWER, Arch. Naturgesch., 80 A: 141 (part..: Typenserie en.thält auch N. triste),
Vorbemerkung: Das Basalglied der d'-Chelicere, deren Apo und der d'-Pedipalpus weisen bei Individuen verschiedener geographischer Herkunft markante Unterschiede auf, sind aber regional einheitlich. Die einzelnen Formen sind überwiegend nach äußeren l'llerkmalen zu
Genus Nemastoma 107
trennen· genital h I · h · d · . . h ib d morp o ogrsc . sm sre -.., gemessen an den großen Spezies-Unterschieden Inner a. . er ~attung - wertgehend einheitlich. Diese (3) Formen vikariieren, so daß es b~re;htrgt. rst, ?'e als Subspezies anzusehen. Trotzdem darf nicht übersehen werden, daß zumm est eme ~res~r Fo~men (relictttm) gegenüber den beiden anderen (bidentatum und
8 ar·
Btt~} unte~scluedlwhe .. ökologische Präferenda zeigt, die auf mehr als nur subspezifische ~erschredenhert deuten konnten. Allen gemeinsam sind folgende diagnostisphen Merkmale:
G~ni~al.morphologie: Truncus penisvon der sockelförmigen Basis zur Glans kont~nmer!wh. verbreitert und ohne .Absatz in die Glans übergehend. Glans symmetrisch, In einen schmalen und spitzen Stylus verlängert, nur ve mit je einem para-m~ Feld langer Borsten, la je I Paar lanzettlich-breiter Stacheln. Chehce_re~: d'-Grundglied mit meist' großer, do zipfeiförmig verlängerter und
an der ;sasiS emgesohnürter .Apo; Einzelheiten vgl. unten. . Pedipa~pus :d'-P~ me-di zu einer markanten .Apo verlängert; vgl. unten.
. L.aufbeme: relativ lang, kein Glied merklich verdickt, Fe I-IV do-di mit Je .eine~ hellen F!eck ;. Fe IV. außen mit .2-3 Längsreihen .spitzer Körner, somit bei geeignete~ BlickWinkel mit gesägter Kontur. Maße vgl. ·unten.
Dorsalseite: Souturn und freie Terg lackschwarz bis matt mit feiner 't zerstreuter Granu!ier~ng, oh~e Silberflecken-Zeiohnung; auf thorak Terg II ::~ abd .Areae I-Iy Je em deutliches flaches Höokerpaar, caudad divergierend;.Scutum von la somit markant wellenförmig.
yerbreitung (.Abb. 150): 0-.Alpen, Karpaten-Länder, Balkanhalbinsel ohne Gneohenland, NW-Türkei. Einzelheiten vgl. unten.
Bestimmungstabelle für die Subspezies (Gilt nur für .d'; vgl. auch Schlüssel für die Nemastoma-Arten, S. 94)
Fe des Pedipalpus me-d! mit d?utlichen Kö oder Tu bewehrt (Abb. 140, 143), Sekretionsfeld der Chel-Apo m~drad gerrohtat (helle Vertiefilllg, mit Borsten umstellt, Abb. 141, 144). . . . . . . . . • . . . . . . . • . . . 2 Fe des Pedipalpus unbewehrt (Abb. 148), Sekretion~f~ld. d~r. Chei-Ap~ fro~t~d· g~riohtet (Abb. 149) · · · · · · · · · ,' · · . · · . . · . . bidentatum reliotum, S. 112
2 Chel-Apo auffallend groß und drstad vorgewölbt, halbmondförmig· Grundglied m k t bucklig auf~ewölbt (Abb. 141) . . . . . . . . . . . . bidentat~m bidentatum, ";, ~;7 Chel-AP.o mcht auffallend groß, eher klein-knopfförmig; Grundglied do nicht markant aufgswolbt, aUenfalls sanft gerundet (Abb. 144, 145) . . . . bidentatnm sparsum, s. 1ll
Nemastoma bideniatum bidentatum RoEWER, 1914 .Abb. 140-142, 150
1914 Nemastoma bidentatum ROEWER, Arch. Naturgesch., SO A: 141, Abb. 20a-d 1923 Nemastoma bidentatttm - RoEWER, Weberknechte d. Erde: 685, Abb. 817a.-d (part) 1968 Nemastoma. bi.~en~atttm bidentatum -- GRUBER & MARTENS, Senckenbergiana bio!., 4D;
143 (dort dre ubrrge Synonymie). 1973a Nemastoma (Lttgub•·ostoma) trist~plttridentatum HADZI Razprave Dissertat LJ'ubl'
16: 43. ' , . Jana,
1973 a Nemastoma (Stridulostoma) seliskari HADZI, Razprave, Dissertat. Ljubljana, 16: 48,
Habitus und Feldkennzeichen: klein, tiefschwarz, d'-Chelicere mit auffallend großer, h~lbmondför~ger .Apo. - Nur in den SO-Alpen. ·
Kennze.whnung:. Korpermaße (in mm): L d': 1,5-I,6; ~: I,6-1,8. Laufbem II (d', m Klammern ~): Fe 1,2 (0 8) Pt 0 4 (0 4) Ti I I (I 1) Mt 1 9 (2 0)
Ta 1,6 (1,6). ' ' ' ' ' ' '

108 Familia Nemastomatidae
(
.. 'i-1_"./
Genus Nemastoma 109
. Genitalmorphologie (.Abb. 142): vgl. unter N. bidentatum. Stylus schulterförmig von der Glans abgesetzt.
Cheliceren (.Abb. 141): di Hälfte des d'-Grundgliedes mit do buckelförmiger Aufwölbung; do-di .Apo auffällig groß, vom Basalglied deutlich abgesetzt •und annähernd halbmondförmig; Sekretionsfeld medial, gekennzeichnet durch helle, seichte Vertiefung, von feinen Haaren umstellt. - ~-Chelicere glatt, ohne .Apo.
Pedipalpus (.Abb. 140): Fe keulenförmig aufgebläht und eingekrümmt, me-di 3 kräftige Do, bisweilen 2 verschmolzen; Pt gekrümmt, di mit stumpfer .Apo, Ti ba mit kleinem Höcker bzw. Beule me-do, doch nicht immer deutlich ausgeprägt.
Variabilität: Bewahrung des Pa-Fe: meist sind 3 Do vorhanden, der mittlere am längsten; einzelne Do aber gelegentlich verwachsen, zahlenmäßig reduziert oder rechts m1d links verschieden ausgeprägt. Im slowenisch-kroatischen Hybridgebiet (mit N. b. sparsum, siehe unten) unterliegt die Form der Chelioeren-Apo gl'Oßen populationsgebrmdenen (nicht individuellen!) Schwankungen, sie tendiert dann je nach Lokalität mehr zu b. bidentatum oder b. sparsum siehe Abb. 146). '
Verbreitung (.Abb. 150): Typ: Endemit der SO,.Alpen und Sloweniens. - Locus typicus: Herkunftsangabe für das Originalmaterial unrichtig; hier festgelegt auf Österreich, Karawanken, Feistritz.
Ö st-erreioh: S-Kärnten: Gailtal, Karnische Alpen, Villaeher Alpe, Sattnitz, Karawanken, Julisehe Alpen· (GRUBER & MARTENB 1968), GerlitzenfViUach (A. A.leg.); - S-Steiermarl<: Possruok. - In Österreich verläuft die bisher bekaruite N -Grenze entlang der Orte Tröpolach, Möltschaoh, Sekirn, Sattnitz/Ebenthal, südöstl. Griffen (von dort nur ~I) und 5 km südl. Leutsohaoh und Pößnitz (Abb. 150B). - Italien: Friaul, Laghi di Fusine bei 'l'arvisio (A. A, & J. ;\!.leg.). - Jugoslawien: Slowenien: im Anschluß an das alpine Areal (z. B. Trigla\·. HADZI 1931) weit verbreitet: bei Bled, Berg Nanos, Postojna, Ljubljana (GRUBER & MARTENS 1968), Umgebung von Cerknica (J. M. leg.). - Öst!. reicht das b. bidentatum-Areal (inselartigi) bis in das nördl. Kroatien: bei Zagreh (VERHOEFF leg.), Dugo Selo/Zagreb (J. M. leg.). - Populationen von Zupanja, Novo Mesto und vom Sneznik (Sohneeberg) im b. bidentatumfb. sparsum - Hybridgebiet (viele Lokalitäten in Slowenien und Kroatien) lassen in unterschiedlicher Ausprägung Merkmale von b. sparsum erkennen. - Über Areal-Beziehungen zu b. Bparsu?/1 und b. reliotum siehe dort, zur Verbreitrmgsgesohiohte siehe bei b. reliotum.
Ökologie: Biotopansprüche: N. b. bidentatum ist bekannt aus Flußauen mit Grauerlen, Weiden und Fichten, aus unterschiedlich strukturierten Laubmisch-
Abb. 140-149. Nemastoma bidentatum RoEWER.
Abb. 140-142. N. bidentatum bidentatum RoEWER. - 140. cl'-Pedipalpus von medial; 141. cl'-Cheliceren-Grundglied von medial (oben) rmd lateral; 142. Stylus penis. AUS Karawanken, Feist-ritz.
Abb. 143-147. N. bidentatum sparsum"GRUBER & MARTENS, - 143. cl'-Pedipalpus von medial. AUS Leithagebirge; Holohypus; 144-146. cl'-Chelioeren-Grrmdglied ·iron medial (oben) rmd lateral; 144. AUS Leithagebirge; 145. BUL Sozopol; 146. YU Kroatien, Zupanja (Hybridform zwischen N. b. sparsum· undN. b. bidentatum); 147. Glanspenis von ventral (links) und lateral.
Abb. 148-149. N. bidentatum relictum GRUBER & MARTENS. - 148. cl'-Pedipalpus von medial; 149. cl'-Cheliceren-Grundglied von medial (oben) und lateral. AUS ·salzburg, Tappenkarsee; Holotypus.

HO Familia Nemastomatidae
50 km
•• ••
..... ..... bidentatum
• • • •
.,,
•
••• •
••
•
•
• sparsum
••
B Abb. 150. A. Gesamtverbreitung von Nemastoma bidentatum RoEWER, aufgegliedert naeh den Subspezies. - B. Kontaktgebiet der 3 Subspezies in SO-Österreich. Von b. relictum sind a.lle bekannten Fundorte dargestellt, von b. sparsum nur die grenznahen Fundpunkte numeriert und von b. bidentatum nur die grenznahen eingezeichnet. Fundorte Nr. 1-17. N. b. relictum: I. Schuhflicker, 2. Tappenkarsee, 3. Lignitztal, 4. Weißpriachtal, 5. Anlauftal, 6. GroßarltaJ, 7. Rotgüldensee, 8. Gößgraben, 9. Tschiernook, 10. Katschbergpaß, 11. Lungauer Nockberge, 12. Turracher Höhe, 13. Flattnitz, . 14. Murau, 15. Zirbitzkogel, 16. Stubalpe, 17. Koralpe. Fundorte Nr. 18-29. N. b. bid~ntatum: 18. Naßfeld, 19. Tröpolaoh, 20. Möltschach, 21. Sekirn 22. Maria Rain, 23. Sattnitz, Ebenthal, 24. Eisenkappel, 25. süd-südöstl. Griffen (nur '?!), 26. I km südl. Leutschaoh, 27. 5 km südl. Leutschach, 28. Pößnitz, äst!. vom Kreuzwirt, 29. 2 km westl. Leutschach. Fundorte Nr. 30-40. N. b. sparsum: 30. Windische Büheln, 2,5 km nördl. Bahnstation Pesnies, 31. Langegg, 32. Leutschach, 33. Griffener Berg, 34. Köflach, 35. Pernegg, 36. Stanz im Mürztal, 37. äst!. Rettenegg, 38. Semmering, 39. Grünbachsattel, 40. Ruine Starhemberg/ Oberpiesting.
wäldern in der Collin- und Montan-Stufe, in der Subalpin-Stufe (ab 1500 m) in Fichten-Lärchen-Wäldern, im Bereich der Baumgrenze in Wacholder- und Grünerlen-Beständen. Im Bereich der Waldgrenze dringt b. bidentatum allenfalls in Zwergstrauchgesellschaften ein, nie in die alpine Rasen- und PolsterpflanzenStufe.
Vertikalverbreitung: In Kärnten von 420 bis ausnahmsweise 1750 m (Gerlitzen; A. A.leg.) belegt. In der S-Steiermark abwärts bis350m und330m (südl. bzw. westl. von Leutschach; J. G. in litt.).
Genus Nemaston1a lll
Phänologie: ad. eurychron, iuv. und frisch gehäutete ad. (Exoskelett hellbraun, ohne Senturn compositum) M VII zusammen mit vorjährigen ad. (Slowenien, bei Divaca, um 400 m), zur seihen Zeit in 600-700 m (z. B. bei Cerknica) nur iuv. neben vorjährigen ad. (J. M. leg.) .
Nemastoma bidentatum sparsum GRUBER & MARTENS 1968 (Abb. 143-147, 150)
1956 Nemastoma bidentatum - i'lrLHAVY, Fauna ÖSR. 7: 132, Abb. 243 -246 . 1968 Nemastoma bidentatum Bpar•um GRUBER & MARTENS, Senckenbergiana bio!., 49: 146
(dort die übrige Synonymie) .
Habitus und Feldkennzeichen: klein, tiefschwarz, d'-Chelicere mit markanter, do leicht zipflig ausgezogener Apo; Verwechslungsmöglichkeit nur in den 0-Alpen mit N. b. bidentatum, N. b. relictum und N. triste.
Kennzeichnung: Körpermaße (in mm): L d': 1,5-1,6; ~: 1,6-1,7. Laufbein II (d', in Klammern !j?): Fe 1,3 (1,3) Pt 0,4 (0,4) Ti 1,0 (1,0) Mt 2,0 (2,0)
Ta 1,5 (1,5) . Genitalmorphologie (Abb. 147): vgl. unter N. bidentatum; Styl1,1s ohne
schulterartigen Absatz in die Glans übergehend. Cheliceren (Abb. 144-146): di Hälfte des d'-Grundgliedes do ohne buckelför
mige Aufwölbung, sondern in gerader Linie verlaufend; do-di Apo weniger groß als bei b. bidentatum, von la oder medial fast rechteckig, ve Seite konvex, do in kleinen Zipfeln ausgezogen, dadurch frontal etwas konkav. Sekretionsfeld me, wenig vertieft und mit feinen Haaren umstellt.
Pedipalpus (Abb. 143): auffallend schlanker als bei der Nominatform, Fe nur angedeutet keulenförmig, me-di nur 2 (selten 3) kleine Do, Pt me-di mit schlanker Apo; Ti ba ohne Höcker.
Variabilität: Pa und Cheliceren-Apo in 0-alpinen {Leithagebirge) und bulgarischen Populationen {Sozopol) bemerkenswert einheitlich, doch zeigen die bulgarischen Individuen do stärker aufgewölbtes Chel-Grundglied. Bewahrung des Pa-Fe bisweilen bis auf einen Do reduziert. Über Verhältnisse im slowenisch-kroatischen Hybridgebiet vgl. bei b. bidentatum.
Verbreitung (Abb. 150): Typ: SO-europäisch-karpatisch. - Geschlossenes Areal von den 0-Alpen über große Teile der nördl. Balkanhalbinsel bis in die NW-Türkei. - Locus typicus: Österreich, Leithagebirge, Sonnenberg.
Österreich: 0-Alpen und Randlandschaften des pannonischen Beckens {Einzelheiten siehe unten). - ÖSSR: nordöstl. bis Bardejov {Bartfeld) {SOERENSEN 1894; W. S. rev.). -Ungarn und Rumänien: offenbar weit verbreitet, möglicherweise lokal fehlend: südlichste Nachweise in Rumänien: Bäile Herculane (Herkulesbad) und Comana (J. G. in litt.) Dobrudscha und W-Karpaten {DUMITRESOU 1972). - Bulgarien: Osogovogebirge und Sozopol (W. S. in litt.). - Jugoslawien: südl. bis Bosnien-Herzegowina: Plasa {Beleg SMF) und Split {Beleg NHMW), -Montenegro: Moraca-Tal, A. A.leg.); -in Slowenien siedelt die Nominatform {siehe dort). - Türkei: Bithynien, 10 km östl. Izmit (J. G. leg.).
In Nieder-Österreich und Steiermark wird die westl. Arealgrenze markiert {vgl. Abb. 150B): Windische Büheln, 2,5 km nördl. Bahnstation Pesnica, Langegg, Leutschach, Köflach, Pernegg, Stanz im Mürztal, östl. Rettenegg, Semmering, Grünbachsattel, .Ruine StarhembergfOberpiesting, ferner {auf der Karte Abb. 150B nicht mehr erfaßt) Neulengbach {48°l2'N, l5°55'E) am NW-Rand des Wienarwaldes und Großweikersdorf (nörrll. der Donau, 48°28'N, l5°59'E). Der S-Teil der durch diese Orte bestimmten Grenzlinie nähert sich streckenweise bis aufmehrere Hundert{!) m der N-Grenze der Noihinatform: Lautschach (sparsum), 1 km südl. Lautschach (bidentatum); Langegg (sparsum), 5 km südl. Leutschach; Pößnitz, äst!. vom Kreuzwirt

112 •· Familia Nemastomatidae
(bidentatum). Die Fundpunkte "Leutschach" und "1 km südl. Leutschach" sind etwa 700 m voneinander entfernt, "Pößnitz" und "Langegg" 1.250 m.-- In diesem Bereich parapatrische:J;l Vorkommens liegt kein Hinweis auf eine Hybridzone vor. Intermediäre Individuen sind von dort unbekannt und beide Formen verhalten sich hier wie Arten.
Ökologie: Biotopansprüche: Größte Siedlungsdichte besteht wahrscheinlich in Laubmischwäldern der Collin-Stufe, z. B. in Querco-CaJ"pinetalil!-, auch in Quercus pubescens-Wäldern; fehlt zwar den Donau-Auen, doch werden Au-Gese11-schaften, wie Erlen- und Weidengebüsche an Bachrändern, nicht gemieden; b. sparsurn lebt auch in hoher S~edlungsdichte in den feuchten Eichen-Mischwäldern der Save-Niederung (bei Zupanja: J. G. leg.).
Vertikalverbreitung: vom Tiefland (Save-Tal: 90 m, J. G. leg.) bis in die montane (Bosnien-Herzegowina: Plasa; 1400 m) und selten in die Subalpin-Stufe (Steiermark, südwestl. unter dem Hochwechsel: 1650 m; A. A. leg.). Eine spatsumähnliche Population lebt in der Latschenzone des slowenischen SneznikfSchneeberg bis in mindestens 1650 m Höhe.
Phänologie: wahrscheinlich eurychron; in VIII-IX frisch gehäutete ad. der neuen Generation, ad. der Vorjahresgeneration mindestens bis VII, halbwüchsige bis subad. iuv. von V-A VIII (Material aus Nieder-Österreich, J. G. leg.), somit Verhältnisse ganz ähnlich den anderen Arten der Gattung.
Nemastoiha bidentatum relictum GRUBER & MARTENS, 1968 Abb. 148-150 -
1968 NemaBtoma bidentat.um reliotum GRUBER & MARTENS, Senckenbergiana bio!., 49: 147. ·
Habitus und Feldkennzeichen: klein, tiefschwarz, Q'-Chelicere mit kleiner, gerundet-knopfförmiger Apo. - Im 0-alpinen Areal Verwechslungsmöglichkeiten mit dem syntop auftretenden N. triste gegeben. ·
Kennzeichnung: Körpermaße (in mm): L d': 1,6-1,9; L Sj!: 1,9-2,2. Laufbein II (d', in Klammern Sj!): Fe 1,1 (1,2) Pt 0,4 (0,4) Ti 1,0 (0,9) Mt 1,7 (1,7)
Ta 1,3 (1, 3). · Genitalmorphologie: vgl. unter N. bidentatum. Stylus ohne schulterartigen
Absatz in die Glans übergehend; Truncus extrem dünn, im Vergleich auffallend schmaler als b~i b. bidentatum und b. sparsum. ·
Cheliceren (Abb. 149): di Hälfte des Grundgliedes do leicht konvex, Apo knopfförmig, do gerundet, frontal glatt senkrecht abgeschnitten, mit der ve Fläche rechten Winkel bildend; Sekretionsfeld der DrÜse auf der Frontalfl~che der Apo, dort mit feinen Haaren besetzt (bei allen anderen bidentatum-Formen liegt die Sekretionsfläche medial).
Pedipalpus (Abb. 148): d'-Fe nur schwach keulig verdickt, ohne me-di Do, me-di Pt-Apo schlank und spitz ausgezogen ..
Variabilität: ist in dem vergleichsweise kleinen Verbreitungsgebiet nicht festste11-bar, vor a11em nicht solche, die auf Hybridbildung mit den beiden anderen bidentatum-Subspezies schließen ließe.·
Verbreitung (Abb .. 150): Typ: Endemit der SO-Alpen. - Locus typicus: Österreich, Kleinarltal, Tappenkarsee.
Österreich: (aJle Angaben A. A. in litt.): östl. Hohe Tauern.(Höhenzug zwischen Großarltal und Gasteinertal, Rotgüldensee), westl. Niedere Tauern (Kleinarltal, Weißpriaohtal, Lignitztal) über die Norischen Alpen östl. bisStubalpe und Koralpe. In den Norischen Alpen sind belegt: Lungauel' und Kärntner Nockberge, zwischen Katschberg, Tschiernoci< bei MiHer-
Genus Nemastoma 1.13
haii, Flattnitz und Murau, Zirbitzkogel, Stub- und Koralpe. Reliotum fehlt offensichtlich bereits östl. der Mur in den Niederen Tauern (z. B. häufig bei Murau südl. der Mur, fehlend aber auf
. der Stolzalpe unmittelbar nördl. der Mur). Diese Einschränkung gilt nicht für die westlichsten Niederen Tauern, wo b. reliotum die Mur-Quelle nach N und 0 postglazial geringfügig umgangen hat. Im Mürztal und am Semmering lebt bereits b. BparBum, und im W fehlt reliotum mit Sicherheit an der N-Abdachung der mittleren und westl. Hohen Tauern, besiedelt zwar N- und 0-Flanke der Ankogel- und Reißeckgruppe (Hänge über Malta-Tal und Gößgraben, Anlauf- und inneres Großarltal). Im W der Reißeck- und SW der Allkogelgruppe fehlt relictum gleichfalls (keine Funde mehr im Seebach-, Dösen-, Rieckenbach-Tal und am Schoberboden). Ebensowenig gelangen Nachweise in der Kreuzeck-Gruppe (Teuchlgraben) und in der Goldberg-Gruppe (Böseck, N aßfeldsattel),
An den Abhängen von Glein-, Stub-, Kor- und Saualpe ist eine Kontaktzone zwischen b. reliot"m und b. BparBum zu erwarten. Die Entfernung zwischen einander nächstgelegenen l>is jetzt bekannten Fundorten beträgt nur 14 km. Möglicherweise besteht zwischen beiden formen bereits Fortpflanzungsisolation; das lassen die großen morphologischen Unterschiede an den d'-Cheliceren-Apo und die differierende ökologische Einnischung vermuten.
Vertikalverbreitung: Relicturn lebt ausschließlich in der Subalpin-Sttd'e; die bisher bekannten Fundorte liegen zwischen 1360 und 1880 m mit größter Siedlungsdichte zwischen 1550 und 1750 m; 70% der Individuen (n = 250) stammen aus diesem Bereich (A. A. in litt.). Zur Phänologie sind keine Daten bekannt.
Zur Verbreitungsgeschichte des bidentatum-Komplexes: Die Entste· hung der morphologisch und teilweise auch ökologisch auffallend differierenden Subspezies läßt sich mit der Glazialgeschichte des Gebietes in Einklang bringen. Die Gebiete mit hoher relictum-Siedlungsdichte (Nockberge der Gurktaler Alpen, die Sau-, Stub- und Koralpe) gehören zu den Massifs de refuge, die während der (aller 1) Glazialzeiten immer unvergletschert waren. Dort sind prä- oder interglaziale Faunenanteile erhalten geblieben. Sie haben unter den extremen Bedingungen der Glazialzeiten eine eigene Entwicklung durchlaufen; b. relictum, das sich in dieser Zeit differenziert hat, ist zu ihnen zu zählen. Postglazial hat es sein Areal nur wenig nach N und NW erweitern können, denn in den selbst während der Würm-Vereisung hochgradig devastierten östl. und mittlerenHohen Tauernfehlt relictum (über Arealbeziehungen zu N. triste siehe dort). Die beiden anderen Subspezies b. bidentatum und b. sparsum werden in 2 getrennten Refugien die Glazialzeiten verbracht haben, b. bidentatum möglicherweise in den Karnischen und Julisehen Alpen sowie in den Karawanken, wo es noch heute den Schwerpunkt seiner Verbreitung hat. Sparsum (bzw. seine Ausgangsform) war . wahrscheinlich von jeher im Karpatenbecken und auf der Balkanhalbinsel weitverbreitet. Postglaziale Arealerweiterungen brachten alle 3 Formen miteinander in Ko11takt: Gebietsweise scheinen sie ·sich zu vermischen (b. sparsum und b. bidentatum in Slowenien und Kroatien, wohl lokal in Serbien) teilweise aber auch - bei gleicher ökologischer: Valenz -unmittelbar nebeneinander zu bestehen (b. sparsum und b. bidentatum in Kärnten und Steiermark). Die Beziehungen zwischen b. relictum und b. sparsum bzw. b. bidentatum sind noch unbekannt. Möglicherweise- können beide, da sie ökologisch differieren, unvermischt nebeneinander leben (sympatrisch und syntop). Das wiirde b. relictum in den Rang einer eigenständigen Art erheben.
8 Tierwelt Deutschl. 64: Marians

114 Familia Nemastomatidae
Nemastoma dentigerum .CANESTRINI, 1873 Abb. 151-153
1873 Nemastoma dentigerum CANESTRINI, Atti Soo. Veneto·Trent., 2: 51. 1923 Nemastoma deritigerum - RoEWER, Weberknechte d. Erde: 657.· 1968 Nemastoma dentigerum - GRUBER & MARTENB, Senckenbergiana bio!., 49: ·153 (dort
die übrige Synonymie).
Habit~s und Feldkennzeichen: klein, tiefschw~rz ohne Silberfleckenzeichntmg, d'-Cheliceren-Grundglied mit knopfartig abgesetztel.' Apo. - In' den S-Alpen weitverbreitet, ·nur lokal nördl. der Alpen.
Kennzeichnung: Körpermaße (in mm): L d': 1,5-1,7; L ~: 1,6-1,9. Laufbein II (d', in Klammern~): Fe 1,2 (1,2) Pt0,5 (0,4) Ti 1,0 (1,0) Mt 1,8 (1,7)
Ta 1,4 (1,4). Pseudogelenke der Femora: I 0; II l-2; III I; IV 2'-3; an Fe IV oft beidseitig
verschieden. · Genitalmorphologie (Abb. 153): Der schmale Truncus ohne Absatz in. die
symmetrische Glans und die~e in einen langen, schmalen und manchmal in sich geschwungenen Stylus übergehend. Glans ve mit langen, in 2 Ia ;Längsgruppen geordnetrm Bo, do kürzere Bo; an der Basis des Stylus wenige seht kurze Bo, do bisweilen zu spindeiförmigen Stacheln verbreitert.
Chelice,ren (Abb. 152): d'-Basalglied mit auffällig großer, ba eingeschnürter Apo, von Ia und me .trapezoid, do etwas konvex, und dort proximal in einen kleinen Zipfel ausgezogen. Längsachse der Apo gegen die des Grundgliedes verwinkelt, somit deren Medialseite etwas nach außen gedreht. - Sekretionsfeld der Drüse medial; feinstruktureHer Bau vgl. MARTENS (1973a). .
Pedipalpus (Abb. 151): d'-Fe di keulenförmig, sub,di mit. einer kräftigen, medial weisenden stumpfen Apo ausgezogen. ~-Fe weniger keulig, Fe und Pt ohne Apo.
Laufbeine: kurz und robust, Fe und Ti I keulig verdickt. Beinglieder mit · bezeichnender Mikroskulptur; fein und regelmäßig punktiert und granuliert,
schwarz. Dorsalseite: Souturn und freie Tergite durch feiueiJ und unauffällige Granulie
rung mit mattem Glanz, auch Tu oc nur schwach bekörnelt; ohne paramediane Höckerreihe; tiefschwarz, ohne Fleckenzeichnung.
Variabilität: (J aus den Gottischen und Penninischen Alpen weichen durch zipflig ausgezogene Chel-Apo deutlich von allen anderen bisher bekannten Pop ab und ähneln einzelnen Pop von N. bidentatum bidentatum außerordentlich (vgl. Abb. 152c und 145). Da diese Chel-F.orm auf einige Gebirgsstöcke der SW-Alpen beschränkt zu sein scheint, kommt diesen Pop wahrscheinlich Subspezies-Rang zu; Benennung schon jetzt verbreitet die geringe Arealkenntnis. - Einzelne Individuen aus Pop der mittleren S-Alpen können durch spindeiförmige Stacheln. der Glans (vg!. Abb. 1.53c) differieren. · '
Verbreitung: Typ: adiiato-mediterran. Geschlossenes Areal im zentralen Mittelmeer-Gebiet; S-Alpen und Apenninen-Halbinsel; nördl. der Alpen nur (reliktäre) Arealsplitter. __; Locus typicus: Italien, Altichiero bei Padua.
Alpenländer: Schweiz: Mt. Generose/Lugano (VERHOEFF leg.); Mt. Ceneri zwischen Bellinzona und Lugano (STROUHAL leg.). - Italien: wahrscheinlich in den gesamten randliehexponierten Alpenteilen, aber möglicherweise an vielen Stellen nur lokal. - Ligurische Alpen: zwischen Pontinvrea und Giusvalla; Meeralpen: Umgebung von Viozene, Pian Rosso; südwestl. Cuneo, Co! di Tenda, Val Grande; Cottische Alpen: Giaveno nördl. Pinerolo, Alpe Colombini
Genus Nemastoma 115
!SI
IS2a,
'S3~· a b.
\.~ .
Abb. 151-153~ Nemastoma dentigerum CANESTRINI. - 151, (J-Pedipalpus von medial; 152. (J-Cheliceren-Grm:idglied von lateral (links) und medial: a) BRD Stuttgart, b) IT Gardasee, Rocca di Manerba; c) IT Oropa nördl, Biella; 153. Glans penis von lateral (links) und dorsal: a) und b) BRD Stuttgart, c) IT Brunate.
und westl. Forno (bisher westlichster 'Fundorte in den Alpen); Co! Lis zwischen Almesi und Viu; Penniniache Alpen: Santuario di Oropa nördl. Biella (alle K. T.leg.); Tessiner Alpen: Varzo (DRmsoo & DRESOO·DEROUE!r 1960); Lago Maggiore, Stresa (R. v. HELVERBEN leg,); Brunate/Comer See; Veltlin/Valtellino, Tirano und Bondrio (J. M.leg.); Gardasee-Gebiet, Mte. Baldo-Massiv: Mte. Altissimo (PAULUS leg., SOH.A.WALLER leg., K. T. leg.), Val Cerdro (K. T. leg.); Dolomiten, Spluga del Romit, Miane; Mt, Pizzoo (beide PAOLETTI leg.). Die Nennung für Sestro ( = Sestre/Sexten, Dolomiten; GnuBEB & MARTENB 1968) hat sich trotz intensiver Nachsuche nicht bsstätigen lassen (A. A. in litt.), Außerhalb der Alpen in Italien wahrscheinlich weiter verbreitet, als die bisherigen Nennungen erkennen lassen: AltichierofPadua (CANE· 8•

116 Familia Nemastomatidae
STRINI 1873), Euganeisohe Hügel (!irLHAVY 1969), Apuanisohe Alpen (MAROELLINO 1970), Florenz (Beleg SMF), Schlucht von Tivoli (VERHOEFF leg:), Asl'romonte (Beleg SMF) \ffid Gargano (Foresta umbra; ARGANO leg.). - Jugoslawien: nur in Slowenien und Kroatien im Anschluß an das alpine Areal: Divaöa/Triest (VERHOEFF leg.), BukovjefSelva cli Piro (SBORDONI leg.); Kroatien: Berg Uöka (J. M.leg.), OrahovioafRijeka (Beleg PAN Warszawa). - BRD: Filder Ebene südl. Stuttgart, zwischen den Orten Rohr, Mähringen und Vaihingen (J. M.leg.), Rhein-Main-Gebiet: Mainzer Universitäts-Wäldchen, Gonsenheimer WaldfMainz (J. M. leg.), Mainz-Gustavsburg (SOHAWALLER leg.), Eiebrieher Schloßpark/Wiesbaden, <;!eisen-heim/Rheingau .(beide v. HELVERSEN leg.); Sieg-Mündung bei Bonn (Beleg SMF). ·
Ökologie: Biotopansprüche: N. dentigerum lebt in der Humusschicht bodenfeuchter Laubwälder, in den Arealsplittern nördl. der Alpen vorzugsweise in offenem, parkartigem Gelände: lichter Wald, Gehölze, Buschreihen und Obstpflanzungen mit Grasunterwuchs, bisweilen sogar in gänzlich unbeschattetem Gelände (Stuttgart). Im Stuttgarter Areal-Splitter vikariiert dentigerum kleinräumig mit lugubre: dentigerum im offenen Gelände mit Buschreihen und Obstpflanzungen, lugubre in angrenzendem Laubmischwald (GRUBER & MARTENS 1968). Einzelheiten über die S-alpinen und italienischen Populationen sind nicht bekannt.
Vertikalverbreitung: In Abhängigkeit von beschatteten, bodenfeuchten Biotopen wahrscheinlich von nahe Meereshöhe (bei Divaca etwa 400 m) bis etwa 2000 m (Mte. Baldo, K. T.leg.).
Phänolo.gie: ad. eurychron; nur aus Populationen nördl. der Alpen bekannt: ad. werden etwa 1 Jahr alt, vorjährige und neue Generation - kenntlich an unterschiedlicher Ausfärbung und Härtung des Exoskelettes - überschneiden sich in VJVI. Die neue Generation lebt somit bis VJVI des folgenden Jahres. Ob in beiden Jahren Eier abgelegt werden, ist ungewiß, aber möglich (vgl. bei N. bimaculatum). Iuv. schlüpfen wahrscheinlich erst nach Überwinterung der Eier im folgenden Jahr; die letzte Reifehäutung findet etwa VIJVII statt (J. M. nach Befunden im Rhein-Main-Gebiet).
Genus Paranemastoma REDIKORZEW, 1936
!923 Nemastoma - RoEWER, Weberknechte d. Erde: 651 (part.).
1936 Pa.-anemastoma REDIKORZEW, Trav. Inst. Zoo!. Acad. Sc. URSS, 3: 54. 1958 Nemastoma (Dromedostoma) KRATOCHViL, Praoe Brn. zakl. Ces. Akad.-ved., 30:529.
Eine Gattung der Nemastomatidae-Nemastomatinae, gekennzeichnet durch: d'-Genitalmorphologie: Truncus penislang und extrem schmal, Basis erweitert,
vom übrigen Truncus markant abgesetzt, aber nicht abgewinkelt; tiefgespalte'n und in den beiden Hälften die beiden Muskelstränge konzentriert, Glans in gerader Verlängerung des Truncus, kaum breiter als dieser, differenziert nur durch stärkere Sklerotisation und winzige Sinnesborsten. Spitze der Glans in 2 Äste gespalten, .der rechte schmalere (von ve) den kürzeren und breiten linken wenig übergipfelnd. - Souturn: para-me Kegeldorn-Paare, am stärksten entwickelt auf abd Area II, bei ~ schwächer ausgeprägt. ·
Cheliceren-Drüse: Drüsenzellen von der Basis des Cheliceren-Grundgliedes proximal bis etwa zur Höhe des Tu oc um einen gemeinsamen Ausfuhrkanal verteilt, dessen Öffnung als Porus do-di auf dem Chel-Grundglied an der proximalen Basis der Apo.
Körper immer tiefschwarz, meist mit Gold- oder Silberflecken-Zeichnung. - d'und ~-Pedipalpus ohne Do-Bewehrung.
Genus Paranemastoma 117
Große Arten bis fast 5 mm; die Gattung umfaßt, die größten Nemastomatiden überhaupt.
Verbreitung: etwa 20 Arten in MitteJ-Europa mit den Alpenländern, östl. bis in die Karpaten und auf die südl. Balkan-Halbinsel und auf einige KykladenInseln; im Nahen Osten bis in die Pontischen Gebirge, in den Kaukasus und in das Elburs-Gebirge. - Nennungen für die Iberische Halbinsel bedürfen der Bestätigung; die Gattung fehlt allen Gebieten nördl. der Mittelgebirge. - Im hier behandelten Gebiet 4 Arten, größte Formenvielfalt mit lebhafter Aufspaltung in Subspezies und Lokalformen in den Gebirgen der Balkanhalbinsel. Nemastoma polonicum RoEWER, 1951 (= Paranemastoma p.), das angeblich aus dem oberen San-Tal Galiziens stammt, wurde dort nicht wiedergefunden; offensichtlich liegt eine Fundortverwechselung vor (ST.ARF;GA 1965b).
Typus-Art (durch Monotypie): Paranemastoma superbum REDIKORZEW, 1936.
Besfunmungsta.belle für die Arten
1 Souturn tiefschwarz, ohne Silber- oder Goldfleckenzeichnung (Abb. 166); nur in den Alpen bicuspidatum, S. 121
Souturn tiefschwarz, mit markant abgesetzter Silber- bzw. Goldfleckenzeichnung . , 2 2 Goldfleckenzeichnung auf dem Ceph Ia in Form einer schrägstehenden Acht (Abb. HiS).
d'-Cheliceren-Grundglied (von do und von Ia) mit deutlich abgesetzter, konusförmiger Apo (Abb. 156, 157), in Mitteleuropa weit verbreitet . . . . . . . quadripunctatum, S. 117 Goldfleckenzeichnung auf dem Ceph bis auf einige winzige Flecken fehlend oder dort als große und auffällige Girlanden (Abb. 173, 180), d'-Cheliceren-Grundglied (von do und Ia) nur mit undeutlicher, knotiger Apo (Abb. 171, 178) .............. , 3
3 auf dem Ceph Ia immer große Girlanden aus Goldflecken (Abb. 180), d'-Cheliceren-Grundglied mit stumpf-konischer Apo, hinter dem Sekretionsporns flach abfallend (Abb. 178), in den Karpaten-Ländern . . . • . . . . . . . . . • . . . . . . . . silli, S. 126 Ceph Ia schwarz oder dort höchstens einige helle Flecken, nie ausgedehnte Girlanden-Zeichnung (Abb. 173), d'-Cheliceren-Grundglied mit stumpf-konischer Apo, hinter dem Sekretionsporus nochmals hoch aufgewölbt (Abb. 171); in den Beskiden und Karpaten . , . .
Paranemastoma quadripunctatum (PERTY, 1833) Abb. 154-161
1833 Phalangium quadripunotatum PERTY, Delect. An. art., 3: ·204. 1879 Nemastoma quadripunotatum - SIMON, Arachn. de France, '1: 279.
koohi, S. 124
1923 Nemastoma quadripunotatum-quadripunolatum - RoEWER, Weberknechte d. Erde: 660. 1940 Nemastoma quadripunotatum var. leasinensis CAPORIACCo, Fol. Zoo!. hydro bio!., 10: 34. 1951 Nemastoma quadripunotatum quadripunotatum - RoEWER, Senckenbergiana, 32: 105. 1951 Nemastoma ohauoioum ROEWER, Senckenbergiana, 32: 105. 1951 Nemastoma alfken·i RoEWER, Senckenbergiana, 32: 106. 1951 Nemastoma gerljohi RoEWER, Senckenbergiana, 32: 107 . 1951 Nemastoma auatriaoum RoEWER, Senckenbergiana, 32: 108. 1951 Nemastoma nervosum RaEWER,'·Senckenbergiana, 32: 119. 1951 Orosbyouslengeradorfi RoEWER, Senckenbergiana, 32: 152. 1956 Nemastoma quadripunotatum - !irLHAVY, Fauna CSR, 7: 132. 1973 Nemastoma (Nemastoma) mediosignaturn HADZr, Razprave Dissertat., Ljubljana, 16: 24. 1973 Nemastoma (Nemastoma) slovenioum HADZI, Razprave Dissertat., Ljubljana, 16: 26. 1973 Nemastoma (Nemastoma) triglavense HADZI, Razprave, Dissertat., Ljubljana, 16: 27. 1973 Nemastoma (Nemastoma) quadripunotatum oarniolioum HADZI, Razprave, Dissertat.,
Ljubljana, 16: 28. 1973 Nemastoma (Nemastoma) emonenae HADZI, Razprave, Dissertat., Ljubljana, 16: 29.
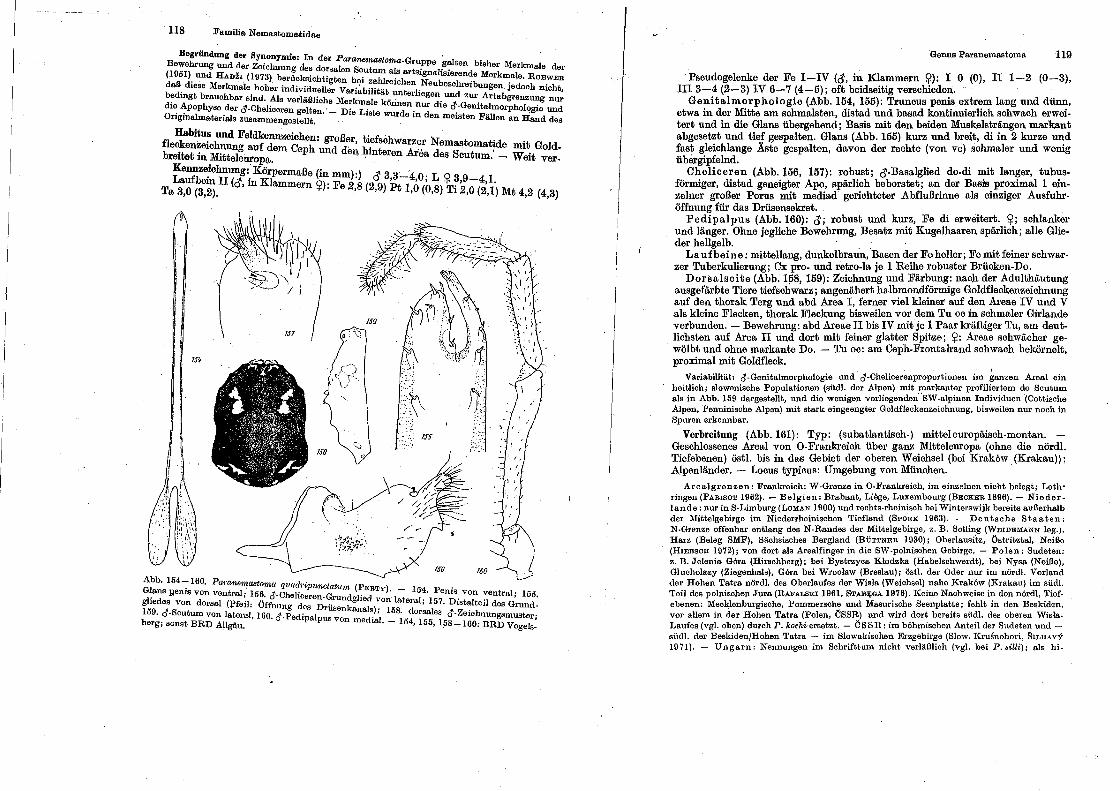
· 118 Familia Nemaatomatidae
Begründung der Synonymie: In der Paran•mastom , . Bewahrung und der Zeichnung des dorsalen Souturn a a-Gru!'pe .g~lten bisher Merkmale der (1951)_ und lliDzr (1973) berücksichtigten bei zahlre~ :rtsrgnalrsreren~e Merkmale. RoEWEB daß drese Merkmale hoher individueller Vari~biJit··t IC tenl'Neubesohrerbungen jedoch nicht, b~dingt brauchbar sind. Als verläßliche Merkmale ~ö un er regen. und zur. Artabgrenzung nur dre Apophyse der d'-Cheliceren gelten. D' L' t nn;n nur dre d'-Gerutalmorphologie und Originalmaterials zusammengestellt. . - re rs e wur e in den meisten Fällen an Hand des
Habitus und Feldkennzeichen· gr ß t' :E h · 'n
fleckenzeichnungauf dem Ceph 'un: der, 1t :c war;"'e'r .~.~emastoma~de mit Goldbreitet in Mittelenropa. en · · n eren .tUea des Scutum. - Weit ver-
~::;b~:~ungt~permaße (in mm):) (J 3,3-4,0; L ~ 3,9 _ 4,1. Ta 3,0 (3,2). ((J, ammern ~):Fe 2,8 (2,9) Pt 1,0 (0,8) Ti 2,0 (2,1) Mt 4,2 (4,3)
160 f Abb. 154-;-160, PamnemaBtoma qu.ad•·ipunctatum (PERT ) . , Glans Ilems von ventral· 156 "' Ch I' G Y • - 154. Pems von ventral· 155 . ' • o- e ICeren- rundgl 'ed 1 ' • ghedes von dorsal (Pfeil· Öffnung de D .. k ' von ateral; 157. Distalteil des Grund. 1"9 "' S . • s rusen anals). 158 I I "' . o · o· cutum von lateral 160 d' p d' 1 ' · 'orsa es o·Zewhnungsmuster· berg; sonst BRD Allgäu. ' ' . 8 rpa pus von medial. - 154, 155, 158-160: BRD Vogels:
IS6
Genus Paranemastoma 119
· Pseudogelenke der Fe I-IV ((J, in Klammern ~}: I 0 (0}, II 1-2 (0-3}, III 3-4 (2-3} IV 6-7 (4-5}; oft beidseitig verschieden.
Genitalmorphologie (Abb. 154, 155}: Truncus penisextrem lang und dünn, etwa in der Mitte am schmalsten, distad und basad kontinuierlich schwach erweitert und in die Glans übergehend; Basis mit den beiden Muskelsträngen markant abgesetzt und tief gespalten. Glans (Abb. 155} kurz und breit, di in 2 kurze und fast gleichlange Äste gespalten, davon der rechte (von ve} schmaler und weuig übergipfelnd.
Cheliceren (Abb. 156, 157): robust; (J-Basalglied do-di mit langer, tubusförmiger, distad geneigter Apo, spärlich beborstet; an der Basis proximal 1 einzelner großer Porus mit mediad gerichteter Abflußrinne als einziger Ausfuhröffnung für das Drüsensekret.
Pedipalpus (Abb. 160}: (J; robust und kurz, Fe di erweitert. ~; schlanker und länger. Ohne jegliche Bewahrung, Besatz mit Kugelhaaren spärlich; alle Glie-der hellgelb. . .
La. ufbeine: mittella.ng, dunkelbraun, Basen der Fe heller; Fe mit feiner schwarzer Tuberkulierung; Cx pro- und retro-la. je 1 :Reihe robuster Brücken-Do.
Dorsalseite (Abb. 1!)8, 159}: Zeichnung und Färbung: nach der Adulthäutung ausgefärbte Tiere tiefschwarz; angenähert halbmondförmige Goldfleckenzeichnung auf den thorak Terg und a.bd Area. I, ferner viel kleiner auf den Areae IV und V als kleine Flecken, thorak Fleckung bisweilen vor dem Tu oc in schmaler Girlande verbunden. - Bewehrung: a.bd Areae II bis IV mit je 1 Paar kräftiger Tu, am deutlichsten auf Area II und dort mit feiner glatter Spitze; ~: Areae schwächer gewölbt und ohne markante Do. - Tu oc: am Ceph-Frontalrand schwach bekörnelt, proximal mit Goldfleck.
Variabilität: d'·Genitalmorphologie und d'-Chelicerenprop~rtionen im ganzen Areal ein haitiich; slowenische Populationen (südl. der Alpen) mit markanter profilier teil\ do Scutum als in Abb. 159 dargestellt, und die wenigen vorliegenden .. SW-alpinen Individue:O. ·(Cottische Alpen, Penninische Alpen) mit stark eingeengter Goldfleckenzeiclmung, bisweilen nur noch in Spuren erkennbar.
Verbreitung (Abb.161}: Typ: (subatla.ntisch-} mitteleuropäisch-montan. -Geschlossenes Areal von 0-Frankreich über ganz Mitteleuropa (ohne die nördl. Tiefebenen} östl. bis in das Gebiet der oberen Weichsel (bei Krak6w (Krakau)); Alpenländer. - Locus typicus: Umgebung von München. ·
Arealgrenzen: Frankreich: W-Grenze in 0-Frankreich, im einzelnen nicht belegt; Loth· ringen (PARISOT 1962). -Belgien: Brabant, Liege, Luxembourg (BECKEB 1896). - Niederlande: nur in S-Limburg (LOMAN 1900) und rechts-rheinisch bei Winterswijk bereits außerhalb der :Viittelgebirge im Niederrheinischen Tiefland (SPOEK 1963). - Deutsche Staaten: N-Grenze offenbar entlang des N-Randes der Mittelgebirge, z. B. Solling (WEIDEMANN leg.), Harz (Beleg SMF), Sächsisches Bergland (BÜTTNER 1930); OberlausÜ,z, Östritztal, Neiße (HIEBSOH 1972); von dort als Arealfinger in die SW-polnischen Gebirge.- Polen: Sudeten: z. B. Jelenia. G6ra (Hirschberg); bei ·ßystrzyca Klodzka (Habelschwerdt), bei Nysa (Neiße), Glucholazy (Ziegenhals), G6ra. bei Wroclaw (Breslau); östl. der Oder nur im nördl. Vorland der Hohen Tatra nördl. des Oberlaufes der Wisla (Weichsel) nahe Kra.k6w (Krakau) im südl. Teil des polnischen Jura (RAFALSKI 1961, STARlj)GA 1976). Keine Nachweise in den nördl, Tiefebenen: Mecldenburgische, Pommersehe und Masurische Seenplatte; fehlt in den Beskid0n, vor• allem in der Hohen Tatra (Polen, ÖSSR) und wird dort bereits südl. des oberen '\\'isla.Laufes (vgl. oben) durch P. kochiersetzt.- ÖSSR: im böhmischen Anteil der Sudeten undsüdl. derBeskiden/Hohen Tatra. - im Slowakischen Erzgebirge (Slow. Iüusnohori, SILHAVY 1971). - Ungarn: Nennungen im Schrifttum nicht verläßlich (vgl. bei P. •illi); als hi-

120' Familia Nemastomatidae
Mitostoma chrysomelas
Abb. 161. Gesamtareal von Paranemastoma quadripunctatum (PERTY) und Mitostoma chrysomelas (HERMANN).
reichend wahrscheinlich· können gelten: Sopron (KoLOSVARY 1929, Nemastoma nervosum) Köszeg,_ Zircf:~akony-_Gebirge (beide KoLOSVARY 1965, N. nervosu";). - Alpen1änder; Sohwe1z: we1tverbrmtet, doch nur wenige Nachweise aus den südl. Alpenmassiven (vgl. LEBBEBT 1917). - Österreich: 0-Grenze entlang des 0-Alpenrandes und seiner Vorketten unter Ausschluß der pannonisch beeinflußten Ebenen und Hügelländer im NO des Landes: fehlt im N-Burgenland mit Leithagebirge (GRUBER 1960), im Wiener Becken, im Viertel unter dem Manh~rtsberg (= Wei:~J.Viertel) ~nd im anschließenden außeralpinen ÖSSRJS-Mähren. _ J ugoslaw1en: Bachargebuge (HAnzr 1973 a, Nemastoma qu. carnio!ioum); Karawanken, z. B. Wurzen-Paß (J. M.leg.); Julisehe Alpen, z. B. Triglav (HAnzr 1931) bis in das außeralpine Slowenien, z. B. Cerknica und Sneznik (Sclmeeberg) (J. M. leg.); wahrscheinlich südl. bis Kroatien, ~och S-Grenze nicht belegt. - S-Grenze in den S-Alpen, doch in den mediterran-exponierten sudl. Kalkalpen nur spärlich und meist lokal verbreitet. - Frankreich: DauphineAlpen; Isere, Sappey (SrMON 1879). - Italien: Cottische Alpen: C. Lis zwischen Almesi und Viu; Penninische Alpen: Prov. Novara; Valsesia, Alpi Pile (beide K. T.leg.); Aosta-Tal (MAROELLINO 1973b); Vall. Tonente Dolca bei Biella; Val Sessera/Bocchetto Sessera; Lago di l'rfucro'.'e bei Oropa; - Lein~ be~ Torino (alle MAROELLINO 1975d); Prov. Varese, Mt. Colonna; Dolom1ten: z. B. Bozen; Trtentmer Alpen: Valsugana (alle K. T.leg.), Cima Galbana (ÜAPO-RIACCO 1940). .
Genus Para.nemastoma 121
Ökologie: Biotopansprüche: P. quadripunctatum lebt in bodenfeuchten Wäldern unterschiedlicher Zusammensetzung, auch an Bestandsrändern und selbst in Feldgehölzen und Gebüschen, soweit die Bodendecke durch Fallaub, Detritus, morsches Holz oder Gesteinsschutt hinreichend strukturiert ist. Unter günstigen Bedingungen wird sogar Freiland besiedelt, z. B. Latschen-Hochmoore, Verlandungszonen von Seen, selbst an Gerinnen auf xerothermen Hängen (A. A. in litt.) und gelegentlich selbst auf Schuttplätzen unter Gesteinsschutt (J. M. leg.), Aufenthalt in der Bodendecke, soweit sie hinreichend durchfeuchtet ist. Insgesamt ist die ökologische Valenz von quadripunctatum größer als die von bicuspidatum, mit dem es in den Alpen an extrem staunassen Stellen zusammen vorkommt (A. A. in litt.); dringt auch gern in Stollen und Höhlen ein. - Als Mindesterfordernis ermittelte IMMEL (1954) 75% relative Luftfeuchtigkeit; Präferenz bei nahe 100%.
Vertikalverbreitung: vom Flachland (Niederlande, Belgien, N-Rand der Mittelgebirge) und alle Höhenstufen der Mittelgebirge z. B. Böhmerwald, Lusen 1200 m, J. M. leg.) bis in die Su balpiri-Stufe der Alpen. - Österreich, Salzburg: gleichmäßig in der. Collin- (400-500 m) und Montan-Stufe (500-1200 m), in beiden Stufen 77% der Serien (n = 304); in der Subalpin-Stufe (1200-2000 m), Siedlungsdichte viel geringer (23 %) ; steigt nicht über die natürliche Waldgrenze. Höchster Fund in den Hohen Tauern: 1 780 m, Grauwackenzone 1750 m, nördl. Kalkalpen 1700 m (A. A. in litt.). Mit der Angabe von 2300 m im Dachstein erscheinen FRANz & GUNHOLD (1954) nicht glaubhaft. - Italien: Penninische Alpen, Mte. Rosa: 1870 m (MAROELLINO 1973b), Karnische AlpenfCarnia, F.lla Foredar 1850 m (CAPORIAOOO 1927).
Phänologie: ad. und iuv. eurychron, Hauptreifezeit im Frühsommer. Da ad. wahrscheinlich 1 Jahr oder sogar wenig länger leben (in Gefangenschaft 14 Monate, IMMEL 1954), ist die Gene1ationenfolge schwierig zu überblicken. - Alpen, Salzburg: Hauptreifezeit V -IX, in der Subalpin-Stufe (1200-1400 m) unter Steinen größere Ansammlungen gemeinsam überwinternder ad. (HURDES leg.). Iuv. von I-XII; im Winter aktiv unter hohem Schnee (A. A. in litt.), - Außeralpines Mitteleuropa: Niederlande ad. und iuv. I-XII (SPOEOK 1963); BRD: III-X (IMMEL 1954, J. M.leg.); Hauptreifezeit von V-VIII (J. M. leg.) bzw. VI-VIII (PFEIFFER 1956). - Eiablage E III-M VIII, meist V-VI; pro Gelege 6-35 Eier, selten weniger als 10, im Durchschnitt 22,5 (~ach 28 Gelegen aus Freiland und Labor, davon 5 ~ mit je 2 und 3 ~ mit je 3 Gelegen); 7 Jugendstadien; Postembryonalentwicklung in 20 Wochen, soweit sie im Sommer/Frühherbst abgeschlossen werden kann (IMMEL 1954). Sex ratio: 435 Ö': 429 ~ (A. A. in litt. für Salzburg).
Paranemastoma bicuspidatum (C. L. KooH, 1835)
Abb. 162-168
1835 Nemastoma bicuspidatum 0. L. KocH in HERRICH-SOHÄFFER, Fauna Ins. Germ., 124, no. 16.
1923 Nemastoma quadripunctatum-bicuspidatum - RoEWER, Weberknechte rl. Erde: 663.
Habitus und Feldkennzeichen: großer, tiefschwarzer Nemastomatide ohne Goldoder SHberfleckenzeichnung, 1 do Höckerpaar. -Nur in den Alpen.
Kennzeichnung: Körpermaße (in mm): L Ö' 2,9-3,1; L ~ 3,2-3,7.

122 Familia Nemastomatidae
Laufbein II (rf, in Klammern~): Fe 3,3 (3,5) Pt 0,9 (0,9) Ti 2,3 (2,5) Mt 5,1 (5,4)
l'a 3,7 (3,9).1 nk d F I IV(:!- in Klammern 0): I-.(-), II 2-4 (1-4), Pseudoge e e er e - o ' '~' . ·
III 0-4 (2-4) IV 4-6 (4-6). d. h I · ' h 1 · (Abb 162 163): Truncus penisextrem lang un sc ma
Gemtalmorp o ogie ·. b kontinuierlich verbreitert, Basis markant u:terh:l~ ~~:inGI::r ;e~~jt::.ruJia~s (~bb. 163) schmal, von den beiden Ästen der
:a!::: s~chmale {von ve) wenig i~b~$pfel~~i~~dd~~;e:~ ~!:~:=~~~::~pf koni-
hCheAlicerenn ~~:~ lp6!x~!~l~r B:a~: eYn großer mediad geöffneter Porus als
so er po, a .. r h Austrittsstelle des Drüsensekretes, Beborstung spar IC •
Pedipalpus (Abb. ~67): robust, auffallend kurz; hellgelb.
Genus Paranemastoma 123
Abb. 168. Gesamtareal von Par,anema8toma bicuspidatum (C. L. KooH).
Laufbeine: vergleichsweise lang, Fe I und III di schwach keulig verdickt; je nach Alter gelb bis dunkelbraun; Cx mit je 1 Reihe Brückendornen pro- und retro-la.
Dorsalseite (Abb. 166): Körper bis auf die Gelenkhäute tiefschwarz und zeiohnungslos. - Bewahrung: rauh, aber nicht auffällig gekörnt, die einzelnen abd Areae aufgewölbt, Area II mit 1 kräftigen para-me Röckenpaar und aufgesetzter feiner Spitze; bei ~ niedriger. - Tu oc.: kräftig entwickelt, rauh tuberkuliert.
Verbreitung (Abb.168): Typ: Endemit hauptsächlich der 0-Alpen; Areal nach jetziger Kenntnis fast ganz auf die Österreichischen Alpenteile beschränkt. - Locus typicus: Hohe Taueru,. Gastein.
Schweiz: WaiJis, Berisal (LEssERT 1917); Montreux (RoEWER 1914; diese Nennung scheint ·nicht hinreichend gesichert). - Österreich: N-Tiro1: Obergurgl, Oberperfuß, Umgebung von Innsbruck: Patsch, Axams; Arzt.al (alle K. T.leg.); - Salzburg (alle Angaben A. A. in litt.): auf der N-Abdachung der Hohen Tauern.in allen Tälern vom KrimmlerAchental bis zum Großarltal; Niedere Tauern: Kleinii.rl-, Taurach-, Forstau- und Weißpriachtal, PreberGebiet; Lungauer Nockberge; im Salzburger Teil der Kitzbüheler.Aipen nach 0 bis Zell am See; in der Kontaktzone zwischen Grauwacke und nördl. Kalkalpen am S- und 0-Fuß des Hochkönig-Massivs; nördl. Kalkalpen: N-Fuß des Tennengebirges; - Ober-Österreich: Kalkalpen, Pyhrnpaß (KRITSOHER leg.); - Steiermark: Kontaktzone zwischen Grauwacke und nördl. Kalkalpen: Ramsau am Dachstein (FRANZ & GUNHOLD 1954; A. A. leg.), Niedere Tauern: Stolzalpe (A. A.leg.), Koralpe: St. Oswald/Eibiswald (FRANZ leg.), Fischbacher Alpen: nordwestl. Wenigzell, östl. Rettenegg (beide STROUHAL leg.); Kärnten: Mallnitz, Kolbnitz, Taohiernock bei Gmünd, Schlatzing/Maltatal, St. Oswald bei Radenthein, Mirnock bei
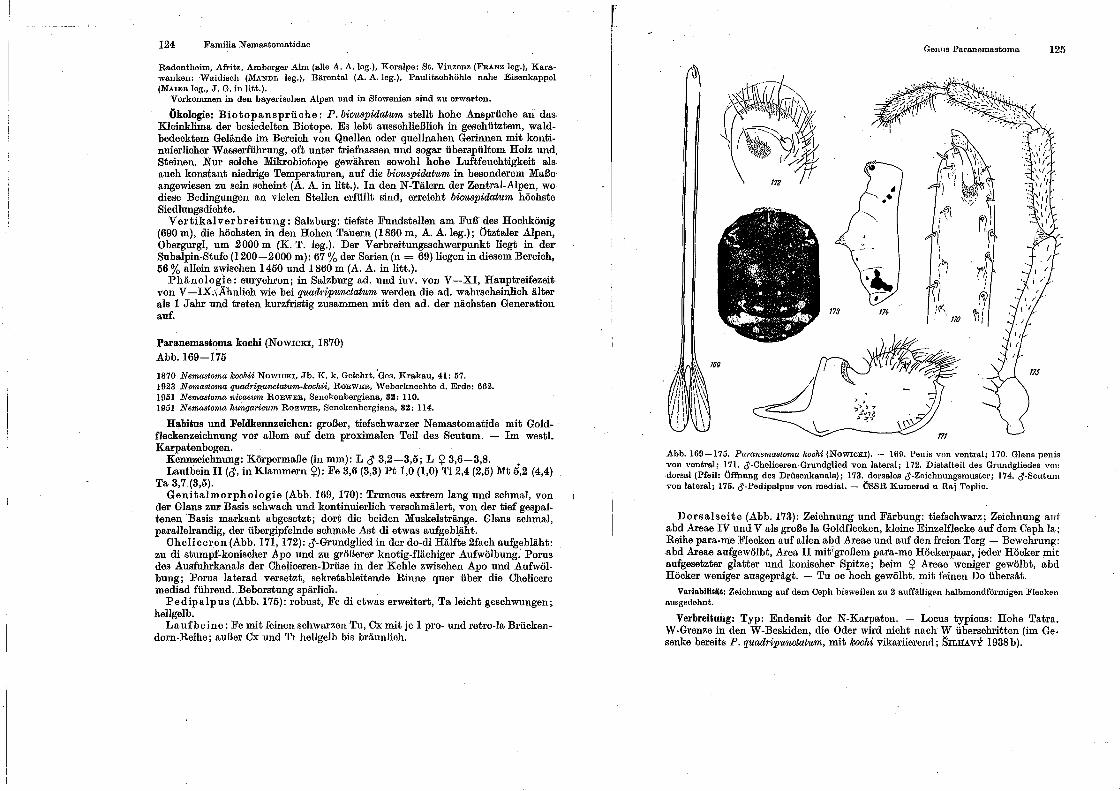
124 Familia Nemastomatidae
Radentheim, Afritz, Amberger Alm (alle A. A. leg.), Koralpe.: St. Vinzenz (FRANZ leg.), Karswanken: Waidisch (MANDL leg.), Bärental (A. A.leg.), Paulitschhöhle nahe Eisenkappel (MAlER leg., J. G. in litt.).
Vorkommen in den bayerischenAlpen und in Slowenien sind zu erwarten.
Ökologie: Biotopansprüche: P. bicuspidatum stellt hohe Ansprüche an dasKleinklima der besiedelten Biotope. Es lebt ausschließlich in geschütztem, waldbedecktem Gelände im Bereich von Quellen oder quellnahen Gerinnen mit kontinuierlicher Wasserführung, oft unter triefnassen und sogar überspültem Holz und. Steinen. Nur solche Mikrobiotope gewähren sowohl hohe Luftfeuchtigkeit als auch konstant niedrige Temperaturen, auf die bicuspidatum in besonderem Maße angewiesen zu sein scheint (A. A. in litt.). In denN-Tälernder Zentral-Alpen, wo diese Bedingungen an vielen Stellen erfüilt sind, erreicht bicuspidatum höchste Siedlungsdichte.
Vertikal ver breitung: Salzburg: tiefste Fundstellen am Fuß' des Hochkönig (690 m), die höchsten in den Hohen Tauern (1860 m, A. A.leg.); Ötztaler Alpen, Obergurgl, um 2000 m (K. T. leg.). Der Verbreitungsschwerpunkt liegt in der Subalpin-Stufe (1200-2000 m): 67% der Serien (n = 69) liegen in diesem Bereich, 56% allein zwischen 1450 und 1860 m (A. A. in litt.).
Phänologie: eurychron; in Salzburg ad. und iuv. von V -XI, Hauptreifezeit von V -IX:iÄhnlich wie bei quad1·ipunctatum werden die ad. wahrscheinlich älter als 1 Jahr und treten kurzfristig zusammen mit den ad. der nächsten Generation auf.
Paranemasioma kochi (NowiOKI, 1870) Abb. 169-175
1870 Nemastoma kochii NowiCKI, Jb. K. k. Gelehrt. 'Ges. Krakau, 41: 57. 1923 Nemastoma quadripunctatum-kochii, RoEWER, Weberknechte d. Erde: 662. 1951 Nemastoma nicawm RoEWER, Senckenbergiana, 32: llO. 1951 Nemastoma hungaricum RoEWER, Senckenbergiana, 32: 114.
Habitus und Feldkennzeichen: großer, tiefschwarzer Nemastomatide mit Goldfleckenzeichnung vor allem auf dem proximalen Teil des Scutum. - Im westl. Karpaten bogen.
Kennzeichnung: Körpermaße (in mm): L iJ 3,2-3,5; L ~ 3,6-3,8. • Laufbein II ((],in Klammern~): Fe 3,6 (3,3) Pt 1,0 (1,0) Ti 2,4 (2,5) Mt 5,2 (4,4)
Ta 3,7 (3,5). Genitalmorphologie (Abb. 169, 170): Truncus extrem lang und schmal, von
der Glans zur Basis schwach und kontinuierlich verschmälert, von der tief gespaltenen 'Basis markant abgesetzt; dort die beiden Muskelstränge. Glans schmal, parallelrandig, der übergipfelnde schmale Ast di etwas aufgebl.äht.
Cheliceren (Abb. 171, 172): iJ-Grundglied in der do-di Hälfte 2fach aufgebläht: zu di stumpf-konischer Apo und zu größerer knotig-flächiger Aufwölbung. Porus des Ausfuhrkanals der Cheliceren-Drüse in der Kehle zwischen Apo und Aufwölbung; Porus laterad versetzt, sekretableitende Rinne quer über die Chelicere mediad führend .. Beborstung spärlich.
Pedipalpus (Abb. 175): robust, Fe di etwas erweitert, Ta leicht geschwungen;. hellgelb.
Laufbeine: Fe mit feinen schwarzen Tu, Cx mit je 1 pro- undretro-la Brückendorn-Reihe; außer Cx und Tt hellgelb bis bräunlich,
Gen11s Paranemastoma 125
,_:: .. ----~ --
Abb. 169-175. Pm-anemastoma kochi (NoWIOKI). -- 169. Penis vom ventral; 170. Glanspenis von ventral; 171. Q'-Cheliceren-Grundglied von lateral; 172. Distalteil des Grundgliedes vo" -dorsal (Pfeil: Öffnung des Drüsenkanals); 173. dorsales Q'-Zeichnungsmuster; 174. Q'-Scutum von lateral; 175. Q'-Pedipalpus von medial. - ÖSSR Kumerad u Raj Teplic.
Dorsalseite (Abb. 173): Zeichnung und Färbung: tiefschwarz; Zeichnung auf abd Areae IV und V als große la Goldflecken, kleine Einzelflecke auf dem Ceph la; Reihe para-me Flecken auf allen abd Areae und auf den freien Terg - Bewahrung: .abd Areae aufgewölbt, Area II Init'großem para-me Höckerpaar, jeder Höcker mit aufgesetzter glatter und konischer Spitze; beim ~ Areae weniger gewölbt, abd Höcker weniger ausgeprägt. - Tu oc hoch gewölbt, mit feinen Do übersät.
Variabilität: Zeichmmg auf dem Ceph bisweilen zu 2 auffälligon halbmondförmigen Flecken ausgedehnt.
Verbreitung: Typ: Endeinit der N-Karpaten. - Locus typicus: Hohe Tatra. W-Grenze in den W-Beskiden, die Oder wird nicht nach W überschritten (im Gesenke bereits P. quadripunctatum, mit kochi vikariierend; SrLHA.VY l938b).

126 Familia Nemastomatidae
Polen: Barania G6ra bei Oieszyn (KuLCZYNBXI 1876).; Babia G6ra; Hohe Tatra (beirle RAFALS:KI 1961), W-Teil der Waldkarpaten; Einzugsbereich des oberen San, zahlreiche · Fundorte (STARJlGA 1966). - ÖS SR: Im Bereich der Hohen und Niederen Tatra weitverbreitet (EliLHAVY 1956, 1972) und nach Westen die Oder nicht überschreitend. Für das slowakische Erzgebirge, wo quadripunctatum vorkommt (s. d.), bisher nicht belegt; Vihorlat-Gebirge (!;;rLHAVY 1950).- UdSSR: Karpato-Ukraine: Oberlauf der Theiß, im Gebiet um Rachov, dort angeblich zusammen mit P. sil!i (BARTO§ 1939b). - Rumänien~ Spärlich und Areal wohl diskontinuierlich; AVRAM (1973) nennt neben I 056 Exemplaren silli nur 18 kachi ohne Angabe des Fundortes; S-Karpaten: Oiucas-Massiv und Oberlauf des Teleajen: Oulmea Buzaianuiui, Mt. Ro~u, Mt. Babe~ (DUMITRESCU I 973); Funde aus den 0-, W- und S-Karpaten nenne~> auch AVRAM & DUMITRESOU (1959). Nennungen (und besonders Wiederholungen) vieler Autoren für Gebiete außerhalb des Karpatenbogens haben sich in keinem Falle bestätigen lassen.
Ökologie: Biotopansprüche: In Gebirgswäldern auf gut durchfeuchtetem Boden; Einzelheiten sind nicht bekannt. ·
Vertikalverbreitung: offenbar nicht im Vorland der Karpaten, sondern nur in mittleren und höheren Lagen des Gebirges. Babia G6ra: 1000-1200 m (RAFALSKI 1961), W-Teil der Wald-Karpaten: 1000-1150 m (S:J'ARI>GA 19~g). - Keim~ Daten zur Phänologie.
Paranemastoma silli (HERMAN, 1871) Abb. 176-182
1871 Nemastama sillii HERMAN, Abh. Siebenbürg. Ver. Nattll'wiss. Hermannstadt, 21: 28. 1894 Nemastomct lirzeatum SoERENSEN in LENDL, Tern1. Füzetek, 17: 20. 1923 Nemastama quadripunctatum-sillii, RoEWER, Weberknechte d. Erde: 662. 1951 Nemastama bishapi RoEWER, Senckenbergiana, 32: 106. 1951 Nernastoma zilchi RoEWER, Senckenbergiana, 32: 120 1951 Nemastoma wiehlei RoEWER, Senckenbergiana, 32: JJ4. 1951 Nemastoma ru.manicum RoEWER, Senckenbergiana, 32: 125. 1951 Nemastama rnahilevum RoEWmR, Senckenbergiana, 32: 125.
Begründung der Synonymie: siehe unt.er P. quarlritJunctatum.
Habitus und Feldkennzeichen: großer tiefschwarzer und auffallend langbeiniger Nemastomatide mit reicher Goldfleckenzeichnung. - Nur im Karpatenbogen.
Kennzeichnung: Körpermaße (in mm): L Cf gesamt: 3,2-5,0; nur Scutum: 3,0-3,7. ~gesamt: 4,0-5,6; nur Scutum: 5,5-4,2.
Laufbein II (Cf, in Klammern.~): Die beiden (f:Maße beziehen sich auf ein kurzbeiniges und ein langbeiniges Individuum; langbeiniges Cf und das ~ stammen aus einer Population (Pe~tera Bohoui). Fe 3,6/7,7 (7,0) Pt 1,1/1,3 (1,3) Ti 2,5/4,6 (4,3) Mt 6,3/11,6 (ll,2) Ta 3,9/6,0 (6,2). - Extrem-Maße de~ Cf-Fe II: 4,0-8,0 mm (Av&AM 1973). Pseudogelenke der Fe I-IV: I 0-4 (meist P-2), II 2-9 (meist 3-6), III 1-8 (meist 3-6), IV 4-ll (meist 5-9); oft rechts und links unterschiedlich (AVRAM 1973a).
Genitalmorphologie (Abb. 176, 177): Weist unter den 4 ParanemastomaArten des Gebietes einen besonders schmalen Truncus penis auf; Basis wie bei den übrigen Arten markant abgesetzt und tief gespalten. Glans unterhalb der Gabelung etwas erweitert, der schmalere der beiden Glans-Äste vergleichsweise dünn, di unverdickt und den stumpfen Ast deutlich übergipfelnd; in diesen beiden Iderkmalen bicuspidatum ähnlich. Glans von kocki dagegen parallelrandig und mit anderer Kontur der Innenseite des stumpfen Glans-Astes.
Genus Paranemastoma 127
~\
I,
'I
I ~i
l', ~.
('JJ'\·.··. :t:\ . ~ i . .
176
'· .182 !78
0 Abb. 176-182. Puranernastama silli (HERMAN). - 176. Penis von ventral; In. Glanspenis von ventral; I 78. Q'-Oheliceren-Grundglied von lateral; 179. Distalteil des Grundgliedes von dorsal (Pfeil: Öffnung des Drüsenkanals); 180. dorsales d'-Zeichnungsmuster; 181. d'-Scntnm von lateral; 182. Q'-Pedipalpus von medial. - RU Pe~tera Bohni.
Cheliceren (Abb. 178, 179): robust; Cf-Grnndglied do-di mit 1 frontal-geneigten, stumpf-konischen und wenig erhabenen Apo. Ausfuhrperus der G~elic~renDrüse proximal an der Basis der Apo, etwas mediad versetzt und von Ia mcht swhtbar (bei kocki von la sichtbar), ,f\bflußrinne mediad gerichtet. - ~-Chel ähnlich, da Cf-Apo nicht deutlich abgesetzt, ohne Porus.
Pedipalpus (Abb. 182): auffallend schlank und (variierend) rela.tiv lang, wesentlich graziler als der von kocki. Kugelhaare vor allem an TI und Ta: nur v~reinzelt an Pt ve. - Durch diese Merkmale von den 3 anderen Arten Im Gebiet scharf getrennt.
Laufbeine: Länge (siehe oben) von Population zu Population stark schwankend, weniger individuell innerhalb geschlossener Populationen (z. B. in Höhlen). Bei Individuen aus Populationen mit langen Beinen Fe unverdickt, bei kurzbeinigen

128 Familia Nemastolllatidae
Individuen Fe I und II bisweilen di leicht verdickt. ~ Mittelbraun, Basen der Fe hellgelb, Fe und Pt dünn schwarz bekörnelt.
Dorsalseite (Abb. 180, 181): Zeichnung und Färbung: Souturn tiefschwarz mit hochvariabler Gold- und Silberflecken-Zeichnung. Ceph mit in sich gegliedertem fronto-la Band, di durch den Tu oc unterbrochen; auf denthorakund abd Areae in den Extremen entweder bis auf einige la Flecken völlig schwarz oder (mit zahlreichen Übergängen) auf den thorak und abd Areae eine para-me in sich gegliederte Girlande, an die des Ceph anschließend. - Bewehrung: die abd Areae, vor allem d'-Area Il, wulstig aufgewölbt, je 1 para-me Höckerpaar auf den Areae !IIV, auf Area II mit schlankem und glänzendem Konus, seltener auch auf Area III und IV. - Tu oc: groß, hochgewölbt und stark bekörnelt.
Variabilität: P. silli ist eine äußerst plastische Art (Av:aAM 1973a); Länge und ·Proportion der Laufbeine, Zahl der Pseudogelenke der Fe, die do Fleckenzeichnung und Bewahrung variieren in weitem Spektrum, lediglich die do-cli Apo der d'-Cheliceren scheint weitgehend konstan.t zu sein. - Die extrem langbeinigen Individuen gehören meist zu Höhlenpopulationen, auch Individuen mit intermediären Beinmaßen loben meist unterirdisch, während kurzbeinige regelmüßig Freiland-Biotope besiedeln. Bei troglobionten Individuen ist die Fleckenzeichnung rnicher, und die do Tuberkel desAhd tendieren zur Reduktion. Von einer festen Regelläßt sich aber offenbar nicht sprechen (AVRAM 197:Ja). - Siehe auch "Kennzeichnung" und "Dorsal.seite".
VerbreituJg: Typ: Endemit der. Karpaten. - Locus typicus: Hermannstadt (= Sibiu). · · ·
Häufigste und weitverbreitete Pamnem.aBtorna-Art des rumänischeil ~nd ukrainischen Teiles des Karpaten-Bogens; Grenzverlaufinden NW-~arpaten unbekannt, zumal ältere Nennungen nur bedingt verwendbar sind.
UdSSRfKarpato-Ukraine: Munkacz (ROEWER 1951; NemaBtoma mohilevum,. N. hungm-icum). -Ungarn: Angaben im Schrifttum durchweg unsicher, da mehrere Arten (silli, kochi, quadripunctatum) ständig verwechselt ww·den; hier erfolgt Zuordnung nach geographischer Evidenz: Tiszakarad (KoLOSVARY 1965, N. nervosum), Simontomya (ROEWER 1951, N. wiehlei; Zuordnung unsicher). - Funde in W- Ungarn (Köszeg, Sopron, ZircfBakony-Gebirge) betreffen wahrscheinlich P. quadripunctatum.
Bisher keine Nachweise aus dem polnischen und slowenischen Teil der Karpaten (RAFALSKI 1961, W. S. in litt., tlrLHAVY 1956), und selbst aus dem Vihorlat--Gebirge (Grenze ÖSSRfUdSSR) kennt tliLHAVY (1950) nur P. kochi. '
Ökologie: Biotopansprüche: P. Billi lebt vorzugsweise in Gebirgswälde1·n der Karpaten, meidet aber thermisch begünstigte Gebiete, z. B. xerotherme waldbedeckte Standorte im südsiebenbürgischen Hügelland, nicht (WEIBS 1975). Aus Rumänien sind zahlreiche Höhlenpopulationen bekannt, die morphologisch differieren, wohl aber mit epigäisch lebenden Populationen in genetischem Austausch -stehen.
Ver.tikalverbreitung: keine Daten. . :P)lä'~10logie: ad. eurychron; wahrscheinlich überlagern sich mehrere Genera
tionen, die Mehrzahl der ad. Individuen scheint in VII-IX aufzutreten (WEIBS 1975) .. In günstig gelegenen Höhlen. sind alle Individuen ganzjährig aktiv mit kontinuierlichem Zyklus (AvRAM & DUMITRESCU 1969). - Entwicklung: optimale Tßmperatur für die Embryonalentwicklung zwischen 16 und 17 °C (27 Tage, 100% der iuv. schlüpfen), zwischen 12 und 16 °C 32 Tage (ebenfalls 100 %), bei höheren Temperaturen zwar schnellere Entwicklung doch stark abnehmende Schlüpfrate (bei 20-20,5 °C noch 10 %) ; 7 Jugendstadien. Einzelheiten - auch die der Embryonal,Ex~twicklung ~.bei AvRA.M (1973b).
Genus Histricostonla 129
Genus Histricostoma KRATOCHvfL, 1958
192:l Remastoma - ROEWER, Weberknenhte d. Erd~: flli1 (part.). 11lii8 a Hist•·icostoma KRATOCHviL, Prace Brne zakl. Ces. Akad. ved, 30: 520. 1958a Genteto8/oma (Gretostoma) KRATOCHviL, Prace Brne zakl. ÖeA. Akad. vM, 30: 530.
Eine Gattung der Nemastomatidae, gekennzeichnet durch: d'-Genitalmorphologie: Truncus penis lang und dünn, Basis erweitert, gegf'n
den Truncus abgesetzt, me nicht gespalten. Glans in gerader Verlängerung des Truncus und so breit wie dieser. Spitze der Glans 2ästig; der rechte schmale (von do) übergipfelt den linken breiteren und kürzeren; bisweilen 1 kräftig-spitzer Zapfen an der Basis des schmalen Astes entspringend. - Dorsales Scutum: auf den Areae I-IV oder I-V je 1 para-me Paar dünner, konisch verjüngter StabDornen; selten mit Girlanden von Brücken-Dornen. - Cheliceren-Drüse: Ausfuhröffnung( eil) medio- bis do-di auf stumpfkegelförruiger Apo, bisweilen kaum vom Grundglied abgesetzt, dann ~-ähnlich. - Stridulationsorgane bei einigen Arten auf der Medial-Seite beider Cheliceren-Grundglieder in gleicher Ausprägung (Abb. 19~)- Sekundäre Geschlechtsmerkmale: d'-Pa-Pt me-di mit großem, gelenkig ins~rxertemKegel-Dorn; doStabdornen bei d' länger als bei ~- -Körperfarbe der ad. tiefschwarz, nur wenige Arten mit Silberfleckenzeichnung. Kleine bis mittelgroße Arten von 1,5 bis 3 mm .
Verbreiiung: 5 Arten in Mittel- und S-Europa sowie in Vorder-Asien: dentipalpe (AUSSERER) hauptsächlich in den Alpen, argenteolunulaturn (CANESTRINI) auf der Apenninen-Halbinsel, drenBkii KRATOOJIV:iL in Bulgarien, creticurn (ROEWER) im westl. Kleinasien und in der S-Ägäis, caucaBicurn (REDIKORZEW) im Kaukasus. Sympatrie zweie.r Arten ist nicht bekannt. - Im hier behandelten Gebiet kommen 1, vielleicht 2 Arten vor. ·
HistricoBtoma ist mit CarinoBtorna nächst verwandt im Sinne eines Schwestergruppenverhältnisses (synapomorpher Bau der Cheliceren-Drüse, vgl. MARTENS & SeRAWALLER 1977), und auch zu ParanernaBtorna bestehen enge Beziehungen. _Trenn~nde Merkmale (bezogen auf CarinoBtorna und ParanernaBtorna) im Bau des d'-Gemtalapparates, in der d'-Palpenbewehrung und (meist) in der Scutalbewehrung, ferner (bezogen auf ParanernaBtorna) im Bau der d'-Chelicerendrüse.
Typus-Art (l!'estlegung dm·ch KRATOOHViL 1958): Histricostoma d•·enskii KRATOmrviL, 1958.
Bestimmungstabelle der Arten
Dorsalseite tiefschwarz (nur in den S-Alpen ausnahmsweise mit kleinen Silberflecken auf dem Oeph); St.-idulationsorgan auf der Innenseite des Chel-Grundgliedes mit- et.wa 20 engstehenden Rippen (Abb. 185); in den· Alpen und in W-Jugoslawien .•.......
dentipalpe, S. 13(• Dorsalseite mit 2 großen Silberflecken .~uf dem Ceph, 2 kleine auf dem Souturn oaudal; Stridulationsorgan auf der Innenseite des Chel-Grundgliedes mit etwa 70 engstehenden Rippen (Abb. 191); auf der Apenninen-Halbinsel ...... argenteolunulatum, S. 133
9 'l'ierwelt Deutsc:hl. 64: i\Iarteus

130 Familia Nemastomatidae
Histricostoma dentipalpe (AussERER, 1867) Abb. 183-'189,-193
1867 Nemastom·a dentipalpis AusSEREJR, Verb. Zoo!. Gas. Wien, 17: 168. 1923 Nemastoma dentipalpe - RoEJWER, Weberlmechte d. Erde: 667. 1973 Histricostoma (Histricostoma) slovenicum HAnzr, Razprave, Dissertat., Ljubljana, 16: 49.
Habitus und Feldkennzeichen: mittelgroß, tiefschwarz mit relativ langen Beinen; auf dem abd Souturn 5 Dq-Paare (Lupe!; Abb. 187, 188). - In den Alpen und Dinarischen Gebirgen.
183
I. rf i I
I J
I~ 18~
191
/,,;/ '@' i
~~~
tl), }Vt
/89
/ /
.. " ··v
V . ·v
·''.'.,
' '\( V
,>····'~·1i:······V····:··
Abb. 183-189. Hist•·icostoma dentipalpe (AusSERER). 183. Penis von ventral; 184. Glans penis von ventral. BRD Allgäu; 185. Q'·Cheliceren-Grundglied von medial. CH Davos; 186. Distalteil des Grundgliedes von dorsal (Pfeil: Öffnung des Drüsenlmnals); 187. Bewahrung des dorsalen Scutum; 188. d'·Scutum von lateral; 189. d'-Pedipalpus von medial.
Abb. 190-191. Histricostoma argenteolunulatum (CANEJSTRINI). - 190. Glanspenis von ventral. IT Rom, Palatin; 191. Q'-Cheliceren-Grunclgliecl von medial. - IT Mocliso, Trivento.
Genus Histricostoma 131
Kennzeichnung: Körpermaße: L Ö" 1,9-2,3; L ~j:· 2,2 -2,5. Laufbein II (Ö", in Klammern ~)Fe 2,8 (2,8) Pt 0,6 (0,6) Ti 1,9 (1,9) Mt 3,9 (4.1)
Ta 2,7 (2,9). · Pseudogelenke der Femora (in Klammern ~): I: 0 (0); II: 6-7 (6-7); III:
2-4 (3); IV: 5-8 (5-7); oft rechts und links unterschiedlich. Genitalmorphologie (Abb. 183-184): Trunous penis schlank und parallel
randig, Basis deutlich bulbusförmig aufgebläht, me nicht gespalten; Glans nicht vom Trunous abgesetzt, durch unterschiedliche Sklerotisation kenntlich, do-ve abgeplattet; Spitze der Glans 2ästig: 1 kurz-stumpfer und 1 übergipfelnder schlanker Ast. Besatz mit Sinnesborsten spärlich.
Cheliceren (Abb. 185-186): d"-Grundglied mit stumpfer do-di Apo, mit medialer Kavität und dort Austrittstelle für das Sekret in großem Einzelporus und gering entwickeltem Porenfeld; Apo do-di und medial beborstet. -Auf ö- und ~!Grundglied me-di unterhalb der Apo ein beidseitig gleich strukturiertes Stridulationsorgan (Abb. 192): unteres Feld aus 20 schmalen und ongstehenden, oberes Feld
Abb. 192. Stridulationsorgan der linken Chelicere von Histricostoma. dent·iprtlpe (AU•J"ERElll). Nach REM-Foto.
,iiPP§NM•

132 Familia Nemastomatidae
6-10 breiteren und we~tstehenden Leisten. Ais Plectrum und Pars stridens aus . f b 'd Cl I' . dien~n wechselseitig die 'feilorgane au eJ en 1e 1ceren. . . .
P d . 1 8
(Abb 189)· kurz und gedrungen; nur Q'-Pt me-di mit emer ge-e Ipa pu · · F. b' T ·
1 nki inserierten hakenförmigen Apo; Kugelhaare von < e I~ a. . .. . eLagufbeine: kräftig, dabei auffällig lang, d' und '12-Fe I schwach keubg verdwkt; Cx und Tr schwarz, übrige Glieder braun. . . d .. 1 Dorsalseite (:Abb. 187, 188): Färbung und Zewhnm_1g; tiefschwarz. un ~e10 1-
1 · den Alpen treten lokal Individuen mit emem Paar klemer Silber-nungs os; m I V '"lbt d "t je flecken auf dem Ceph auf. - Bewehrung: abd Areae - · gewo. un nn . 1 para-me caudad geneigtem Stabdorn~n-Paar, bei d' länger als .~ei J; ~~~t:rer:J~ VI bisweilen 1 Paar kürzerer und spitzerer Do; Ceph-~ronta af a ~ e höckert, ebenso Hinterrand von Area V, Senturn sonst fem granu Iert .. - u oc di mit kräftigen Tuberkeln.
· · · l l t · b · de stehen wahrscheinlich in Beziehungen: Nächst verwandt Ist H. m-genteo unu a um, e> St "dul t'onsorgan
einem Schwestergruppen-Verhältnis. Konstant trennende Merkmale: am • r1 a 1
und an der Glans penis.
Verbreitung (Abb. 193): Typ: alpin-dinarisch. - Area:I-kontinuierlich von de.n französischen W-Alpen über den gesamten Alpenzug mit Ausnahme des NO b1s · d' d' · h Geb;~ge Locus typicus· Gebiet um Innsbruck, Husslhof. 1n 1e ~nansc en . ~. . - ·
H. argenteolunutatum '----·"-~-~so~m
Hl·8e1-,·costoma denti~al"'e (AussERER) und H, a.rgenteo/1t1t~~laturn Abb. 193. Gesamtareal von .- .-(CANESTRINI).
Genus Histricostoma 133
Frankreich: Hautes·Alpes, Monestier, Bosodon; Isert-~; lA Aappey; Jura: I 10p. Ain (alle SrMON 1879); zweifellos weiter verbreitet· als bisher beka.mtt. - Schweiz:NW-Grenze wahrscheiulich im Jura (auch französischer Anteil); Kanton Vaud (LESSERT 1917), Umgebung rnn Basel (ScHENKEL leg.; Coll. EIIL.HAvY). - BRD: in den Allgäuer (z. B. Umgebunr: von Oberstdorf, J. M. leg.) und Bayerischen Alpen (KAESTNER 1927) weitverbreitet; möglicherweise lokal im Alpenvorland; Schwäbische Alb, A)theimfRiedlingen (WuNDERLICH leg.; nach jetziger Kenntnis isolierter Fundort). - Österreich: in den nördlichen Kalkalpen östl. et.wa bis zur Saalach (belegt bei Unken, A. A. leg~). aber fehlend davon südl. in den Kitzbüheler Alpen und auf der N-Abdachung der Hohen Tauern (A. A. in litt,). fnrnnr fehlend in allen übrigen Teilen der NO-Alpen. von den Niederen Tauern ostwärts. Dort verläuft dieN-Grenze am S-Fuß rler Hohen Tauern und der Norischen Alpen, über die Kamwauiren und Julisehen Alpen. ··· Italien: S-Grenze auf der S-Abdachung der italienischen S-Alperi, z. B. Mte. Rosa (CANESTRINI 1872, MARcELLrNO 1972), Mte. Bre bei Lugano (R. v. HELVERSEN leg.), nördl. GardaRee-Gebiet; Valsugana öst.l. Trient (beide K. T.leg.); Bosco del Cansiglio (PAOLETTI leg.). Der bisher am meisten südöstl. im westl. Alpenbogen exponierte Fundort (in Nachbarschaft zum argenteoltn>ll.· latum-Areai) isti Meeralpen, Umgebungvon Viozene, PianRosso(K. T.leg.).- Jugoslawien: Areal erst mangelhaft bekannt, zumal Siedlungsdichte im südl. Teil gering; Slowenien: häufig auch in den außeralpinen Teilen, z. B. Umgebung von Cerknica (J: M. leg.); Kroatien: z. B. Plitvitzer Seen (WUNDERLICH leg.); Bosnien und Herzegowina: bei Sarajevo (KRATOCHViL 1946), Suha bei Foca; Montenegro: Durmitor-Massiv, Komov"i-Gebirge (alle A. A. leg.). -Albanien: Vermo~a (PENTHER leg., Beleg NHMW).
Ökologie: Biotopansprüche: H. de.ntipalpe ist eine euryöke Art, die eine Vielzahl montaner und alpiner Biotope besiedelt. Alle Mikrohabitate sind von Wäldern unterschiedlicher Zusammensetzung beschattet und somit meist vor direktet· Insolation geschützt. Sonnige Waldränder und Gebüsch - selbst isolierte Gebüschgruppen - werden an ·vielen Stellen jedoch nicht gemieden. Möglicherweise besiedelt de.-ntipalpe auch offene Wiesen, soweit der Boden auch bei längeren Trockenperioden ausreichend feucht bleibt. Die große Vertikalverbreitung unterstreicht. das breite Biotopspektrum; die Waldgrenze scheint aber an keiner Stelle des Areals nach oben überschritten zu werden. Aufenthalt in feuchten Bodenstrauund Fallaubschichten, unter Steinen und Holzteilen . . Vertikal ver brei tung: im ganzen Areal von der Collin-. bis in die ohere
Subalpin-Stufe belegt; Nachweise von 490 (nördl. Alpenrand) bis 1840 n1 (S~Tirol, Meran; HuRDES leg,), im .Veltlin bis 2000 m (Val di Rezzalo, J. M. leg.) Lokal werden wahrscheinlich beide Extremwerte unter- bzw. überschritten.
Phänologie: ad . .euryohron. - Lebensdauer je Generation einschließlich der Jugendstadien wahrscheinlich zwischen.l5 )lnd 18 Monaten; 2 Generationen überschneiden sich jeweils im Sommer, z. B. A-M VII in Slowenien (600-700 m) nehen alten vorjährigen ad. auch halbwüchsige iuv., aber noch keine frischgehäuteten jungen ad. (die an hellem, noch nicht ausgehärtetem Exoskelett leicht kenntlich sind), ad. der neuen Generation somit etwa A VIII (;T. M.leg.). In anderen Höhenlagen sind geringe jahreszeitliche Verschiebungen zu erwarten.
Histricostoma argenteolunulatum (CANESTRINI, 1875) Abb. 190-191, 193
1875 Nemastoma dentipalpe var. argenteo-lunulatum ÜANESTRINI, Atti Sec. Ven.-Trent. Sei. Natur. Padova, 1875: 4.
1923. Yemastoma argenteolunulatum - RoEWER, Weberknechte d. Erde: 668.
Habitus und Feldkennzeiohen: mittelgroß, tiefschwarz mit 2 großen halbmon.dförmigen Flecken auf dem Ceph und 2 kleine am Hinterrand des Senturn; auf dem Senturn 5 Do-Paare (Lupe!). -Auf der Apenninen-Halbinsel.

134 Familia Nemastomatidae
Kennzeichnung: Körpermaße (in mm): K (J 1,9-2,~; L ~ 2,2-2,6. Laufbein II ((],in Klammern ~): Fe 2,4 (2,1) Pt 0,6 (0,5) Ti 1,7 (1,5) Mt 3,4 (2,8)
Ta 2,5 (2,1). Pseudogelenke der Femora (in Klammern ~): I: 0 (0); II: 4-7 (4--;_6); III:
1-2 (1-3); IV: 3-5 (4-5); oft rechts und links unterschiedlich. Genitalmorphologie (Abb. 190): Truncus und Glans wie dentipalpe, ab
weichend Feinbau der Glans: stumpfer Ast der Glans flach und ohne distad weisende, auf Sockel stehende Bo, übergipfelnder Ast der Glans schlanker als bei dentipalpe, an seiner Basis ein langer und spitzer Zapfen, mit dem schlanken GlansAst nahezu parallelverlaufend, etwas mehr als die Hälfte seiner Länge erreichend.
Chelioeren (Abb. 191): wie bei dentipalpe; Stridulationsorgan abweichend: unteres Feld aus etwa 70 schmalen und engstehenden Leisten, oberes Feld aus 5-8 breiten und weitstehenden Leisten.
Laufbeine und Pedipalpus: wie bei dentipalpe; (]-Fe I kräftig verdickt. Dorsalseite: Zeichnung und Färbung: ausgehärtete Tiere tiefschwarz, auf
dem Ceph mit 2 großen halbmondförmig nach außen geöffneten weißen Flecken, 2 viel kleinere am Caudalende des Souturn; - Bewahrung wie bei dentipalpe; 'j?-Stabdornen kurz, oft nur im Profil erkennbar.
Beziehungen: vgl. bei dentipalpe. •\
Verbreitung (Abb. 193): Typ: adriato-mediterran, Endemit der Apenninen-Halbinsel. - Loous typicus: Calabrien.
Italien: geschlossenes Areal auf der gesamten Apenninen-Halbinsel mit Korsika;· Sardinien und Sizilien sowie zahlreichen kleinen Randinseln, z. B. Elba und Ischia. Die nördl. Arealgrenze ist bisher nicht belegt; MAROELLINO (1972, 1973b) nennt argenteolunulatum aus den Apuanischen Alpen bzw. aus der Umgebung von Genua. Ob argenteolunulatum und dentipalpe in den SW-Alpen hybridisieren, bleibt zu prüfen; möglicherweise schließen sich beide bei engster geographischer Nachbarschaft aus. In der Po-Ebene scheint keine HistricostomaArt vorzukommen.
Ökologie: Biotopansprüche: euryök und weitgehend an die mediterranen Klima-Verhältnisse augepaßt; lebt außer in verschiedenen Waldformationen auch in wenig oder gar nicht beschattetem Gelände, soweit die Luftfeuchtigkeit ausreichend hoch ist, z. B. auf Wiesen mit Gebüsch, in Straßengräben, eto.
Vertikal ver brei tung: von nahe Meereshöhe (Elba, Ischia) bis in höhere Lagen des Apennin, z. B. Velino, I 700 m (VIGNA leg.).
Phänologie: ad. eurychron; Generationenfolge unbekannt.
Genus Carinastoma KRATOCHViL, 1958
1923 Nemastoma - ROEWER, Weberknechte d. Erde: 671 (part.). 1958a Mitostoma (Oarinostoma) KnATOOHViL, Prace Brn. zakl. Ces. Akad. vM. 30 (12): 530.
Begründung des Gattungsranges von Oa.-inostoma: KnATOCHVIL hat jene Arten, die mit Nemastoma carinatum; nahe verwandt sind~ als Untergattung Oa·rinoatoma zu Mitostoma ge. stellt. Er zog dabei Ähnlichkeiten in der dorsalen Brückendorn-Bewahrung als Gliederungskriterium heran. Diese recht variablen äußeren Strukturen können innerhalb der Nemastoma· tidae jedoch kaum subgenerische Verhältnisse ausdrücken; viehnehr sprechen die markanten genitalmorphologischen Unterschiede zwischen den Mitostorna- und den Oarin.ostoma-Arten, die nicht durch Übergänge miteinander verbunden sind, für den Gattungsrang beider Gruppen.
Genus Oarinostoma 135
·Eine Grtttung der Nemastomatidae, gekennzeichnet durch: (J-Genitalmo~phologie: Trunous penis lang und vergleichsweise breit, stark
do-ve abgeplattet; Basis erweitert, aber nicht scharf vom 'i'runcus abgesetzt, me nicht gespalten. Glans in gerader Verlängerung des Truncus, nicht oder nur unwesentlich breiter als dieser; Spitze der Glans 2ästig, der rechte schmalere ~von do) den "linken breiteren übergipfelnd; Stylus gerade gestreckt. Glans Ia nnt 7-10 kräftigen, basad gerichteten hyalinen Haken. - Dorsales Souturn mit Girlanden von Brückendorn-Reihen in spezifisch wechselnder Anordnung auf dem Ceph und den vorderen abd Areae; ohne Stab- oder Kegeldornen.
Öheliceren-Drüse: Öffnung des Ausfuhrkanals als einzelner Porus me-d~ auf dem Basalglied und zu einer flachen, mit Borsten umstellten W a:nne erweitert. (J-Grundglied ohne Apo, sondern do lediglich angesoh~ollen. - Pedipalpus: ~~tersetzt, ohne Do- oder Apo-Bewahrung. Färbung des Korpers und der Extrem1taten tiefschwarz, 1 Paar großer Silberflecken auf dem Ceph. - Kleine Arten von 1,5 bis knapp 2 mm. .
Verbreitung: Die scharf umrissene Gattung ~mfaßt 3 Arten m den ~lpen und auf der Balkan-Halbinsel: carinatum RoEWER 1n den SO-Alpen und aut der westl. :Balkan-Halbinsel; elegans SoERENSEN auf der südöstl. Balkan-Halbinsel, ornatum JlADzi in der südwestl. Balkan-Halbinsel; Sympatrie zweierArten ist aus Montenegro bekannt.
Typus-Art (Festlegung durch KRATOCHviL 1958): Nema.~toma carinatum RoEWER, 1914.
Bestimmungstabelle der Arten
I caudal der beiden Silberflecke auf dem Ceph 3 Querreihen von Brückendornen (Abb. 198, 199); Glans penis breit 1anzettlich, Stylus stumpf gerundet (Abb. 194, 195); in den SOAlpen, W-und S-Jugoslawien . . . . . . . • . • . . . . . . . carinatum, S. 135 caudal der beiden Silberflecke auf dem Ceph 2 Querreihen von Brückendornen (Abb. 205, 206), Glans penis schmal lanzettlich, Stylus lang-ausgezogen (Abb. 201, 202); KarpatenLänder nnd Bulgarien. . . . . . . . . . . . . . . . . . . . . . . elegans, S. 137
Carinastoma carinaturn (RoEWER, 1914) Abb. 194-200 1914 Nemastoma carinaturn RoEWER, Arch. Naturgesch., 80: 165. 1923 Nemastoma carinaturn - RoEWER, Weberknechte d. Erde: 671.
Habitus und Feldkennzeichen: klein, schwarz, mit 2 großen Silberflecken auf dem Ceph und zwei kleinen auf dem Hinterende des Scutum. - SO-Alpen lind W .,Jugoslawien. ·
Kennzeichnung: Körpermaße (in mm): L (J 1,5-1,6; L ~ 1,5-1,8. Laufbein II ((], in Klammern ~): Fe 1,3 (1,2) Pt 0,35 (0,4) Ti 0,8 (1,0) Mt 1,5
(1,5) Ta 1,0 (1,5). " Pseudogelenke der Femora der Laufbeine: I 2. - II 2-5. - III 2-3. - IV
2-3· oft rechts und links unterschiedlich. G~nitalmorphologie (Abb. 194-195): Glans an der Basis wenig breiter als
der Trunous, distad breit konisch zulaufend, Stylus-Ast der Glans kurz und stumpf. di wenig angeschwolli.m, den anderen kürzeren Ast übergipfelnd; Glans stark do-ve abgeplattet, an ihren Seitenrändern je 1 Reihe 8-9 breit konisohet, basad gerichteter hyaliner Haken.
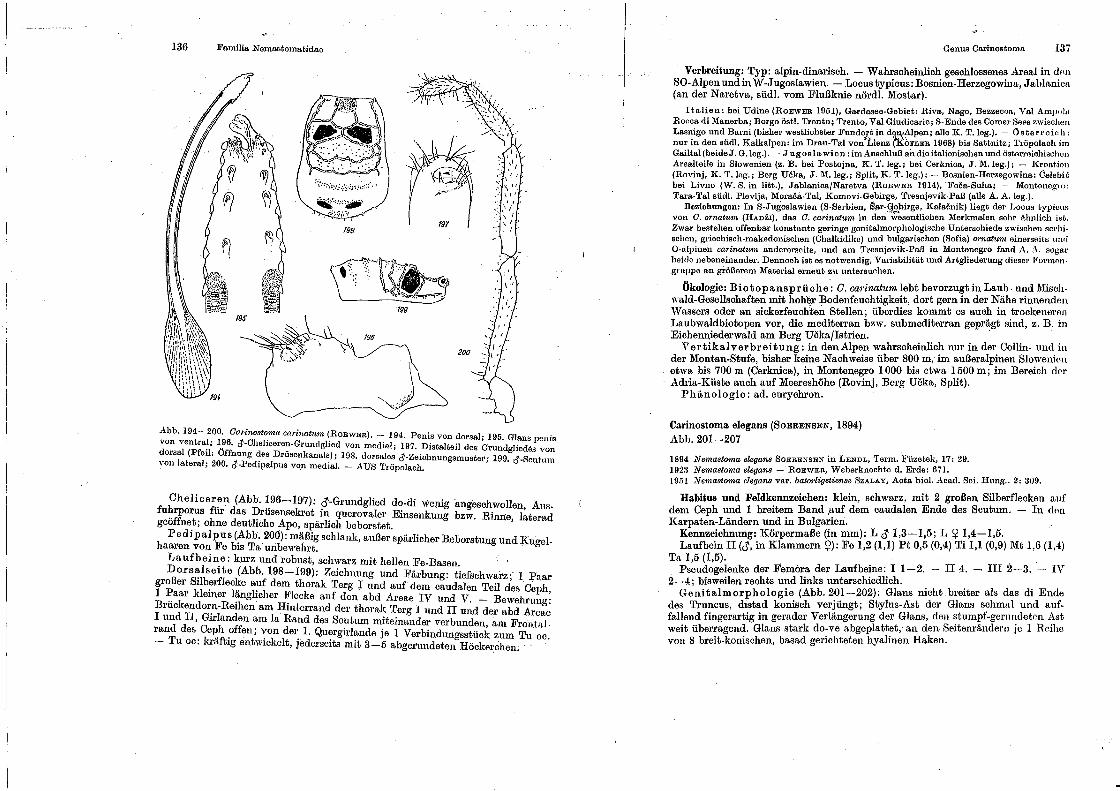
136 Familia Nemastomatidae
''\ 1-k "(/--~~,;J~ ,, ... ~~·
197
199
19(f
200
::•\j I
.~ \ ''. _; \r'.
''' \'.f 'I
. \ I I
Abb. 194-200. Carinastoma carinatum (ROEWER}.- 194. Penis von dorsal· 195 GI · von ventral; 196. d'-Cheliceren-Grundglied von medial. 197 DI"staltei"l le G, .d l~nds penis d 1 (Pü ·1 Öff ' · < s run g Ie es von
orsa ei: nung des Drüsenkanals}; 198. dorsales d'-Zeichnungsmuster• 199 d' s t von lateral; 200. d'-Pedipa.lpus vo.n medial. - AUS Tröpolach. , , - cu mu
Chelicer~n (Abb. ~96-197): d'-Grundglied do-di wenig ·angeschwollen, Ausfu~rporus für das Drusensekret in querovaler Einsenkung bzw. Rinne 1 t d geoffne.t; ohne deutliche Apo, spärlich beborstet. ' a era
Ped1palpus (:Abb._200): mäßig schlank, außer spärlicher Beborstungund Ku-gelhaaren von Fe b1s Ta unbewehrt.
Laufbein.e: kurz und robust, schwarz mit hellen Fe-Basen. · Dors~lse.Ite (Abb. 198-199): Zeichnung und Färbung: tiefschwarz;· I Paar
großer Sllbeiflecke auf dem thorak Terg I und auf" dem caudale T ·1 d ·c· 1 I Pa kl · 1-- 1. I Fl · n e1 es eiJ 1 ar emer ang 10 1er ecke auf den abd Areae IV n11d V B hr ' B .. k d R ih . · - ewe ung:
ruc en o~n- e en am Hmterrand der thorak Terg I und II und der abd Areae I und II, Grrlanden am la Rand des Souturn miteinander verbunden, am Fro"ntalrand des Cel?,h ?ffen; :'on de1: I. Quergirlande je I Verbindungsstück zum Tu 00•
- Tu oc: kraft1g entwwkelt, Jederseits mit 3-5 abgerundeten Höckenihen; · ·
Genus Carinastoma 137
Verbreitung: Typ: alpin-dinarisch. - Wahrscheinlich geschlossenes Areal in dHn SO-Alpen und in W-J ugoslawien. - Locus typicus: Bosnien-Herzegowina, ,J ablanica (an der Naretva, südl. vom Flußknie nördl. Mostar).
Italien: bei Udine (ROEWER 1951), Gardasee-Gebiet: Riva, Nago, Bezzeccn, Val Amp"l" Roc~ca di Manerba; Borgo östl. Trento; Trento, Val Giudicarie; S~Ende des Corner ~eAs zwischen Las~igo und __ Barni (bisher w?stlichster Fund~ft.in d~"(Alpen; alle K. _T.leg.). - Österreieh: nur m den sudl. Kalkalpen: Im Drau-Tal von Lienz ('KoFLER 1968} bis Sattnitz; Tröpolach im Gailtal (beideJ. G. leg.). -Jugoslawien: imAnschluß an dieitalienischen und österrniohischon Arealteile in Slowenien (z. B. bei Postojna, K .. T. leg.; bei Cerlmica, J. M.!eg.); - Kroat.ien (Rovinj, K. T. log.; Berg Uöka, J. M. leg.; Split, K. T. leg.); - Bosnien-Herzegowina: Ce!ebic bei Livno (W. S. in litt.), Jablanica/Naretva (RoEWER 1914}, ·Foila-Suha; -· Mont-enegro: Tara-Tal südl. Plev!ja, M?raila-Tal, Komovi-Gebirge, Tresnjevik-Paß (alle A. A. leg.).
Beziehungen: In S-J ugoslawien (S-Serbien, iil~>r;9,~.birge, Kalacnik} liegt der J,ocus typicus von 0. ornaturn (HAnzr), das 0. carinatum in den wesentlichen Merkmalen sehr ii-lmlich i<t• . Zwar bestehen offenbar konstante geringe genitalmorphologische Unterschiede zwisch~n serbisclJen, gdechisch-makedonischen (Chalkidikc} und bulgarischen (Sofia} ornatum einerseit" "'"' 0-nlpinen ca1·inatum anderers~its, und am Tresnjevik-Paß in Montenegro fand A. A. soga.r hei<le nebeneinander. Dennoch ist es notwendig, Variabilität und Artgliederung dieser T<'urmnngmppe an größerem Material erneut zn untersuchen.
Ökologie: Biotopansprüohe.: C. ca1·inatum lebt bevorzugt in Laub- und .M:isohwald-Gesellsohaften mit holi~r Bodenfeuchtigkeit, dort gern in der Nähe rinnenden Wassers oder an sickerfeuchten Stellen; überdies kommt es auoh in trockenerm1 Laubwaldbiotopen vor, die mediterran bzw. submediterran geprägt sind, z. B. in Eichenniederwald am Berg Ucka{Istrien. ·
Vertikalverbreitung: in denAlpen wahrscheinlich nur in der Collin- und in der Montan-Stufe, bisher keine Nachweise über 800 m,· im außeralpinen Slowenien etwa bis 700m (Cerknica), in Montenegro 1000 bis etwa 1500m; im Bereich der Adria-Küste auch auf Meereshöhe (Rovinj, Berg Ucka, Split).
Phänologie: ad. eurychron.
Carinostoma elegans (SOERENSEN, 1894) Abb. 201-207
1894 Nemastoma elegans SOERENSEN in LENDL, Term. lfiizetek, 17: 29. 1923 Nemastoma elegana - RoEWER, Weberknechte d. Erde: 671. 19.51 Nem.astoma clegans var. batorligetiense SzALAY, Acta bio!. Acad. Sei. Hung .. 2: 309.
Habitus und Feldkennzeichen: klein, schwarz, mit 2 großen Silberflecken auf dem Ceph und I breitem Band ,auf dem caudalen Ende des Scutum. In don Karpaten-Ländern und in Bulgarie1i.
Kennzeichnung: Körpermaße (in mm): L d' 1,3-1,5; L ~ 1,4--,-1,5. Laufbein II (d', in Klammern~): Fe 1,2 (1,1) Pt 0,5 (0,4) Ti 1,1 (0,9) Mt 1,6 (1,4)
Ta 1,5 (1,5). Pseudogelenke der Fernara der Laufbeine: I 1-2. - II 4. - III 2-3. ·_ IV
2-4: bisweilen rechts und links unterschiedlich. Genitalmorphologie (Abb. 201-202): Glans nicht breiter als das di Ende
des Truncus, d1stad konisch verjüngt; Stylus-Ast der Glans schmal und auffallend fingerartig in gerader Verlängerung der Glans, den stumpf-gernncletl'n Ast weit überragend. Glans stark do-ve abgeplattet, an den Seitenrändern ,je I Reihe von 8 breit-konischen, basad gerichteten hyalinen Haken.

138
~ I;
r I ~
Familia Nemastomatidae
(1 ~
i·
11 I'
~ \"' ,)
202
201
Abb. 201-207. Carinastoma elegans (SOERENSEN).- 201. Penis von ventral; 202. Glanspenis von ventral; 203. cl'·Cheliceren-Grundglied von medial; 204. Distalteil des Grundgliedes von dorsal (Pfeil: Öffnung des Drüsenkanals); 205. dorsales cl'-Zeiohnungsmuster; 206. cl'·Soutum von lateral; 207. cl'-Pedipalpus von medial. - BUL Pisaneo bei Ruse.
Cheliceren (Abb. 203-204): d'-Grundglied do-di etwas angeschwollen, Ausfuhrporus für das Drüsensekret in querovaler Einsenkung bzw. Rinne, mediad geöffnet; ohne deutliche Apo, spärlich beborstet.
Pedipalpus (Abb. 207): wie bei carinatum mäßig schl!ink, außer spärlicher Beborstung und Kugelhaaren von Fe bis Ta ohne Bewehrung.
Laufbeine: kurz und robust, vor allem Fe bis Ti I schwach, aber nicht keulig verdickt; schwarz, mit hellen Fe-Basen.
Dorsalseite (Abb. 205-206): Zeichnung und Färbung: tiefschwarz; 1 Paar großer Silberflecke auf dem thorak Terg I und auf dem caudalen Teil des Ceph, Querstreif auf abd Area V. - Bewehrung: Brückendorn-Reihen am Hinterrand der thorak Areae I und II und der abd Area I, Girlanden am Rand des Ceph miteinander verbunden und den ganzen Frontalrand umgreifend, von der 1. Quer-
Genus Centetastoma 139
gixlande je 1 Verbindungsstück zum.Tu oo. - Tu oc kräftig entwickelt, jederseits mit 3 Do.
Verbreitung: T;yp: SO-europäisch-montan, karpatisch. - Wahrscheinlich ge~chlossene_s :Areal m de~ Karpa_ten-Ländern und in 0-Bulgarien. - Locus typicus: ~ der Originalbeschreibung mcht bestimmt, hier festgelegt auf Ungarn, Batorliget.
· Ungarn: In den heutigen Grenzen nur Lelesz bei Satoraljaujhely (8. W. in litt.) und Batorliget _(8ZALAY 1952); allenfalls lokal in der Pannonischen Tiefebene. - UdSSRJKarpatoUkrame:Munkaoz(RoEWER 1923), OberlaufderTheiß, Rachov(BARTOS 1939b). -Rumänien: wahrscheinlich kontinuierlich im ganzen Karpatenbogen einschließlich des Banater- und BibarGebirges (DuMITRESOU 1972, W. S. in litt.), östlichste Funde Bretcu bei Tirgu Sacuiso (W. S. in litt.) und im Gebiet von Vranoea (ÜLTEAN & DUMITRESCU 1973; vgl. auch AVRAM & DumTREs.ou 1969); bisher keine Nachweise aus den rumänischen NO-Karpaten. - Bulgarien: nur tm NO, hauptsächlich im Balkan-Gebirge und in den Rhodopen, west!. bis zum 9ipkaPaß bei Gabrovo, südl. bis Stara Zagara (Zmejovo und Starazagorski bani; alle W. S.leg.). -Ö S S _R : bisher kein Nachweis; kommt aber wahrscheinlich in der S-Slowakei vor, da Funde unmtttelbar an der Grenze auf ungarischer Seite. - Jugoslawien: bisher nicht nachgewiesen, aber im serbischen Golobinje-Gebirge zu erwarten, da häufig nördl. der Donau im BanaterGebirge.
Arealbeziehungen : Nach jetziger Kenntnis vikariiert elegans mit ornatum auf der BalkanHalbinsel. Das ornatum-Araal reicht in Bulgarien östl. bis Gara Koprivlltica und Kalofer (beide Balkan-Gebirge) und Asenova krepost bei Asenovgrad (Rhodopen; alle W. S. leg.). Beide Arten sind morphologisch und wahrscheinlich auch ökologisch so ähnlich, daß sie konkurrieren und somit allenfalls lokal sympl>trisch bzw. syntop vorkommen (Über carinaturn und ornatum in 8-Jugoslawien vgl. in der Gattungs-Diagnose).
Ökologie: Biotopansprüche: Vorwiegend in gleichmäßig durchfeuchteten Humusschichten von Laub- und Mischwäldern; Einzelheiten nicht bekannt.
Vertikalverbreitung: vom Flachland bis in mittlere Gebirgslagen. In Bulgarien, wo das Areal am besten bekannt ist, von 100-1200 m (W. S. in litt.).
Phänologie: ad. eurychron; Einzelheiten unbekannt.
Genus Centetostoma KRAToo:HviL, 1958
1923 Nemastoma - ROEWER, Weberknechte d. Erde: 651 (part.). l958a Gentetastoma (Oentetostoma) KRATOCHViL, Prace Brn. zakl. Ces. Akad. ved, 30 (12):
530.
Eine Gattung der Nemastomatidae, gekennzeichnet durch: 0'-Genitalmorphologie: Truncus penis langgestreckt-schlank, bulbusförmig er
weiterte Basis klein, vom Truncus scharf abgesetzt, me gespalten. Glans geringfügig breiter als Truncus und di zu meist langem, gerade gestrecktem Stylus ausgezogen, immer lästig; als Bewahrung neben langen Stacheln (bei einigen Arten fehlend) kleine Sinnesborsten. - 'Dorsales Souturn mit auffallender Bewahrung langer Stab-Dornen unterschiedlicher Anordnung; Anlage auf den abd Areae immer para-me, auf Area V und auf den freien Terg oft als Querreihe · kleine akzessorische _Do gelegentlich auf den Areae II-IV; Do di stumpf und' körnig rauh. - Cheliceren-Drüse: große Apo auf dem Chel-Grundglied di-frontal, etwas m~diad eingekrümmt und in dieser medialen Kavität das Sekretionsfeld; Apo bei allen Arten der Gattung vergleichsweise einheitlich. - Farbe des Körpers und de~ Extr~mitäten gelb bis dunkelbraun, vielfach mit Goldflecken-Zeichnung; Laufbeme meiSt fadenartig dünn, vergleichsweise lang. Pedipalpus bei c3' und ~
i+WW

140 Familia Nemastomat.idae
kurz 1;nd schlank, nur ausnahmsweise mit medio-di Pt-Apo· (bei . dentipateU.ae DRESOo). - Kleine Arten von 1,2 bis etwa 2 mm.
Verbreitung: Etwa 10 nominelle Arten in den Pyrenäen und in den Gebirgen der nördl. Iberischen Halbinsel, 1 Art in den W -Alpen (und somit !im hier behandelten Gebiet). Viele Arten besiedeln lediglich isolierte Klein-Areale in einzelnen Gebirgsstöcken; 1 Art lebt troglobiont (sexmucronatum SIMoN). Die meisten Arten wurden von DRESOo (1967a) und RAMBLA (1968b) revidiert ("N. bacillijerumGruppe"),
Typus-Art (Festlegung durch I(RATOOHViL 19/l8a): Nernastoma centetes SrMoN, 1881.
Contetostoma centetes (SIMoN, 1881) Abb. 134, 208-"--213
1881 Nemastoma centetes 13rMON, Bull. Zoo!. Franca, 6: 89. 1923 Nemastoma centetes - RoEWER, Weberkn<'cl1te d. Erde: 674.
Habitus und Feldkennzeich~n: klein, tiefschwarz, mit reicher do Stabdorn- 1 Bewehrung (Lupe!); lokal in den SW-Alpen.
Kennzeichnung: Körpermaße (in mm): L c! 1,3-1,4; L ~ 1,2-1,4. Laufb!),in II (r!, in Klammern~): Fe 1,5 (1,5) Pt 0,3 (0,3) Ti 1,3 (1,4) Mt 1,5 (1,4)
Ta 1,3 (1,2) .. Pseudogelenke der Fe(()', in Klammern~; 6 c!, 3 ~):I 0 (0). - II 6-8 (4."--8).
- III .2-3 (2-3). IV 4-5 (4-5); Anzahl oft beidseitig verschieden. Genitalmorphologie (Abb. 208-209): Truncus· penis schlank und geradc
gestreckt, Basis klein, bulbusartig erweitert; Truncus unterhalb der Glans wehig erweitert und .dort zu 2 9-orsad gerichteten schmalen para-me Tascllen ausgehöl1lt; Glans symmetrisch, schmallanzettlich.
Oheliceren (Abb. 210): r!-Grundglied mit massiger. do, distad geneigter Apo, di in stumpfen Zipfel verlängert, medial tief konkav und dort Austrittstelle für das Drüsensekret. ·
Pedipalpus (Abb. 212): robust und vergleichs~eise kurz, Kugelhaare an Ti und Ta.
Laufbeine (Abb. 211): robust und kurz, r!· und ~:Fei distad keU:!ig verdickt, weniger auffallend auch Fe III und IV, ebenso Ti III und IV; Fe I kräftig tuherkuliert.
Dorsalseite (Abb. 213): Zeichnung und Färbung: zeichnungslos tiefschwarz. ·Bewehrung: lange und dünne di etwas angeschwollene Stab-Do: Area I-IV. mit je 2 para-me,.Area V mi~ 8, davon die 4 mittleren am längsten, auf den freien Terg I und II 6-8. - Brückendorn-Reihen am proximalen Rand von thorak Terg I und II und entlang des Oeph-Frontalrandes und des Lateralrandes.
Variabilitäi': GeringfÜgig in der Zahl der Stab-Do auf Al:ea V und auf den freien Terg; siehe auch Pseudogelenke der Fe. · ·
Verbreitung: (Abb. 134): Typ; Endemit der SW-Alpen. - Locus typicnR: J.Vteer-alpen: St. Martin Lantosque. ·
Frankreich: Meera.lpen, SL Ma.rtin La.ntosque. - Italien: Gra.jische Alpen: Gran Pal'a· tliso-Massiv, mehrere Fundorte (MARCELLINO 1975d; K. T. leg.); Gottische Alpen: Colle della. Finestia. (VIGNA leg.), Pian de1 Re (K. T.leg.), Val Chisone/Rifugio Jervis, Val Varaita/S. Anno. di Bellino (beide MAROELLINO 1975d); l'ifeeralpen: S. Anno. Yina.dio, V alle MaleAic'io (VIGNA leg.), Val <:;esso, Terme di Valdieri (K. T. leg.), Limonetto ('VIGNA leg.); Vallone S. AnnafCuneo; Val Pesio/Rifugio Garelli (beide ll.fAROELLINO 1975d); Ligurische Alpen:'Cirna. :IIargua.reis, Sella di Carnina. (VIGNA leg.), oberhalb 1\Jiroglio; westl. Orrna. (beide K. '1':. leg.).
GAnus Mitostoma 141
Abb. 208-213. Gontelastoma cen!e!es (SrMON). _ 208. Penis . , , von ventral; 2HI. J·Cheliceren-Grundglied von medial· 2ll von v_entra.J, JO!J. lxians P""'" cl'·PPrlipalpus von medial· 213 J H b 't I ' . La.nfbem I, Fo von la!.et·al: 212. nestra. ' ' . - a I us von at.eral. - TT Cot.t.ische Alpen, C:niJp, fieliRr Fi-
Ökologie: Bioptopansprüche und Vertikalverb · t . o!fenbar sowohl in der subalpinen Waldzone 'Wie an de:~ ~~g. 0. centßt~s lebt Krmnmho]z. und Zwergstrauchzone; Einzelheiten sind nich~ ~:~:ze tU!~d ~~ rl~r al);~:.15~0-.I ~00, 2000 ~; .Ligurische Alpen: 1900 m; Gottische A~~~: 2IOOe:-
ano og1e. wahrsohemlwh euryohron; Generationenfolge unbekannt. '
GPnus Mitostoma RoEWER, 1951
19~1 jf~tostomn RoEWER, Senckenbergli.na, 32: 140 (patt.). 19o8. M>tostoma. ·- KRATOCHVIL, Pr8.ce Ern. zakl. Öes. Akad. v<ld, 30: 53.0 (part.).
Eme Gattung der Nemastomatidae gekennzeichnet durch: d'·Genitalmorphologie: Trunous penis plump untersetzt b d · ·
in die erweiterte und me gespaltene Basis übergehend Basis' nic'i:: kontdinuiTei'Iwh aus gewinkelt GI · d V 1 ' gegen en run-
.. . · .. ans m gera er er ängerung des Truncus. wenig breiter als d' . bulhusform1g mit Skleriten und (möglicherweiR<' Hchwellha.ren) Mem,brane~~s=~~

I42 Familia Nemastomatidae
k~einer Sporn wahrscheinlich ausklappbar (Sp in Abh. 2I5); nur wenige kurze Smnesborsten. Scutum: ausgeprägte Bewahrung aus Girlanden von Brückendorn~ Re~hen; be~ maxiinaler Ausprägung am Außenrand des ganzen Souturn und jeweils am Hinterrand derthorakund abd Areae, ebenso auf dem Tu oc; Brückendorn-Bewehrung und Tu oc ausnahmsweise fehlend. Cheliceren-Drüse: Sekrßti~nsfeld an! einer m~ist langen und fast aufrechtstehenden Apo immer auf der Proximal-Serte, d~rt eme dichte B~ste aus feinen und langen Haaren; Apo (und Drüse 1) ausnahmswerse fehlend (grac~le REDIKORZEW). 2. d'-Glied do-frontal mit mediad gekrümmter und spitzer Apo. Pedipalpus; auffallend lang und schlank, außer Kugel~aar- ~nd Borsten-Bewehrung glatt. Laufbeine: meist fadenartig dünn und verglerchswerse lang .. Farbe des Körpers und der Extremitäten honiggelb bis dunkelbraun, oft wemg abgesetzte Goldfleckenzeichnung.
Verbreitung: Die scharf umrissene Gattung umfaßt etwa I5 nominelle Arten mit ko~~inuierlichem Areal von den Kautabrisehen Gebirgen, den Pyrenäen und den Brrtrschen _Inseln über gan~ Mittele~ropa einschließlich S-Skandinavien (Insel Gotland, nwht Nor~egen), dre Alpenlander, 0-Europa mit UdSSR/Lettland, den Karpaten-Bogen, die Balkanhalbmsel mit N-Griechenland Apenninen-Halbinsel mit Sizilien und Sardinien; Kaukasus. '
Typus-Art (festlegung durch RoEWER 1951): Phalangium chrysomelas HERMANN, 1804.
Bestimmungstabelle für die Arten
Dorsalseite mit Girlanden aus Brücken dornen; Tu oc und Augen deutlich entwickelt . . 2 Dorsalseite ohne Girlanden aus Brückendornen; Tu oo und Augen fehlend; in Höhlen der Bergam~ker Alpen ·.. . . . . . . . . . . . . . . . . . . . anophthalmum, s. 149
2 Dorsalseite rau~ bekor":elt, Ceph um den Tu oo und das abd Souturn braun, thorak Segmente und breite Mediane auf dem Abd hellgelb; weitverbreitet in l\Iitteleuropa ein. sohließli?h der Alpen . . . . . . . . . . . . . . . . . . . . . ohrysomelas, s. 142 Dorsalseite glatt g~änzend, nahezu einfarbig braun, nur undeutliches para-me Fleckenpaar auf Area V; lokalmden SO-Alpen ................. alpinum, s. 147
Mitostoma chrysomclas (HERMANN, I804) Abb. I6I, 2I4-22I
1804 Phalangium chl-ysomelas HERMANN, :Wem. aptorologique, 108. 1879 Nemastoma chrysomelas - ßiMON, Amchn. d. France, 7: 285. 1923 Nemastoma chrysomelas - ROEWER, Weberiowehte d. Erde: 669. 1929 Nemastoma spinosum HNATEWYTSCH, Zoo!. ,Tb. (Sysl.), 29: 231. 1949 Nemastoma onjbicum ÜAPORIAcco, Atf.i Mus. Civ. Stor. Natur. Trieste, 17: 121. 1950 Nemastoma janet..•cheki ScHENKEL, Rev. Suisse Zoo!. 57: 766. 1951 Mitostoma cMysomelas chrysomelas - ROEWER, ßenckenbergiana, 32: 142. 1951 l.iitostoma silhavyi ROEWEI<, Senckenbergiana, 32: 143. . 1951 l.iitostoma hadzii ROEWER, Senckenbergiana, 32: 146. 1962 Orosbycus biilckensis LoKsA, Zoo!. Anz., 168: 265. 1962 Orosbycus t1·ansdanubicus LÜKSA, Zoo!. Anz., 168: 267. 1963 Mitostoma ch7"1JBOmelas co11jusum SPOEK, Zoo!. Verh., Leiden, 63: 21. 1965 Mitostoma vosag01·um RA>IBLA, Pub!. Inst. Bio!. Apl., 24: 152. 1973 Mitostoma { Mitostoma) sketi HAn:lh, Razprave, Dissertat.; Ljubljana, 16: 50. 1973 Mitostoma ( Mitostoma) ch>-ysomelasjrancochorion·iHADZI, Razprave, Dissertat., Ljubljana,
16: 52.
1973 M?:tostoma (Mitostoma) ch>-ysomclas poleneci ffAnzi, Razprave, Dissertat., Ljubljana, 16: 56.
Genus Mitostoma 143
Begründung der Synonymie: Innerhalb der Formenvielfalt der Gattung Mito.Ytoma ·.vurden meist individuell"n Ausprägungen der do Brückendorn-Bewahrung und der Körperfärh1mg "Jleziesanzeigender Charakter zugeschrieben. Das hat sich in praktisch allen Fällen als irrig
·erwiesen, da diese äußeren Merkmale z. B. in dem vergleichsweise großen Areal von M. chryao· melas stark variieren können und da sich die Farbmerkmale mit fortschreitendem Alter nach der Adulthäutung bei jedem Individuum verändern. Vielmehr ist den genitalmorphologischen Verhältnissen und noch mehr den sekundären ö'-Geschlechtsmerkma1en am I. nnrl 2. Chelicerenglied besondere Bedeutung zuzumessen. Legt man diese Merkmale zugrunde, dann erweisen sich alle in der Liste ternär benannten Formen nur als individuelle Varianten, die nebenHinander vorkommen können (vgl. SPOE.K 1963), Homit also gar keine Subspezins darstellen. Dagegen hat sich das als Subspezies beschriebene a./pinum H ADZI als gut abgrenzbare eigene Art erwiesen (s. d.). Nemastoma jrmetscheki, Orosbycus hu•kkensis und 0. transdanubicus wurden Hach Jungtieren von chrysomelas heHr,hrieben; der Typus von N. orobicum, ein subad. Q', gehört zu einer unbestimmbaren Mitostoma-Art - nach geographisoher Evidenz zweifellos zu clwysomelas (J. G. rev.).
Habitus und Feldkennzeichen: klein, dunkelbraun-hellgelb gemustert, Beine fadenartig lang; unter Steinen und in Detritus; träge Bewegungen. -' Weitver breitAt in Mitteleuropa.
Kennzeichnung: Körpermaße (in mm): L d' I,4-1,6; L ~ 1,7-2,0. Laufbein II (d', in Klammern~): Wien, Praterauen; I60 m: Fe 3,6 (3,4) Pt 0,.15
(0,5) Ti 2,65 (2,9) Mt 5,4 (4,7) Ta 2,5 (2,2); Hohe Tauern, Stubachtrt!, 2000 m: Fe 3,9 (3,9) Pt 0,4 (0,4) Ti 3,2 (3,2) Mt 5,4 (5,1) Ta 2,4 (2,3).
PRaudogelenke der Fe: I 5-8, II 9-15, III 3-7, IV 0-10; oft beidseitig Vf·l'·
schieden. Genitalmorphologie (Abb. 214-2I6): Truncus penis untersetzt, nahezu die
ganze ba Hälfte keulig erweitert und me tief gespalten, dort die MuRkulatur konZAutriert. Glans bulbusförmig vom Truncus abgesetzt und wenig breiter alR dieser; F~inzelheiten der Sklerit-Bewehrung vgl. Abh. 2I5-216.
Che!iceren (Abb. 217 -2I8): gedrungen untersetzt; d' do-di mit laugAr und gerundeter, proximallöffelförrnig ausgehöhlter Apo, dort dichte Bürste ferner und langer Haare und die Sekretionsfläche der Drüse; Apo etwa so lang wie der di Durchmesser des Basalgliedes. Am 2. Glied do-frontal I hakig mediad gekrümmte Apo. ~ ohne Apo an beiden Chel-Gliedern; sonst wie d' spärlich behorstet. ·
Pedipalpus (Abb. 2I9): bei d' und ~lang und schlank, fast laufbeinartig, ohnn Haken- odm· Apo-Bewehrung; Kugelhaare von Fe bis Ta.
Laufhein e : ill). Verhältnis zum Körper außerordentlich lang, dünn fadenförmig, braun, Fe bis Ti di aufgehellt, CJr di mit dunkler Fleckung.
Dorsalseite (Abb. 220-22I): Zeichnung und Färbung: variabel, entsprechend dem Aushärtungsgrad nach der Adulthäutung; Region um den Tu oc und das abd Scutum meist braun, thorak Segmente und breite Mediane auf dem A.bd hellgelb, oft mit silbrigen Guanin-Ablagerungen, bisweilen auch la; auf den freien Terg 2 para-me Flecken.- Bewehrung: ganzes Scutum mit Brücken-Do umrandet, ferner je 1 Reihe am Hinterrand 1aller thorakund abd Areae; je I Querverbindung zum Tu oc zur 1. Reihe und von dieser zur 2. Besonders in diesem Bereich hohe Variabilität durch Ausfall einzelner Brücken-Do. Areae rauh bekörnelt, mit verstreuten größeren Tuberkeln. - Tu oc.: Linse groß, para-me je 1 Reihe kräftiger Brücken-Do.
Variabilität: Sie betrifft die do Zeichnung, bedingt durch unterschiedlich<• Sklerotisation des Exn•kelet,tes, die die Zeichnung verschieden stark hervortreten läßt; besonders alte Individuen sind auffallend konti·astreich gezeichnet. Einzelne Brücken·Do fehlen oft im Median-Teil
...
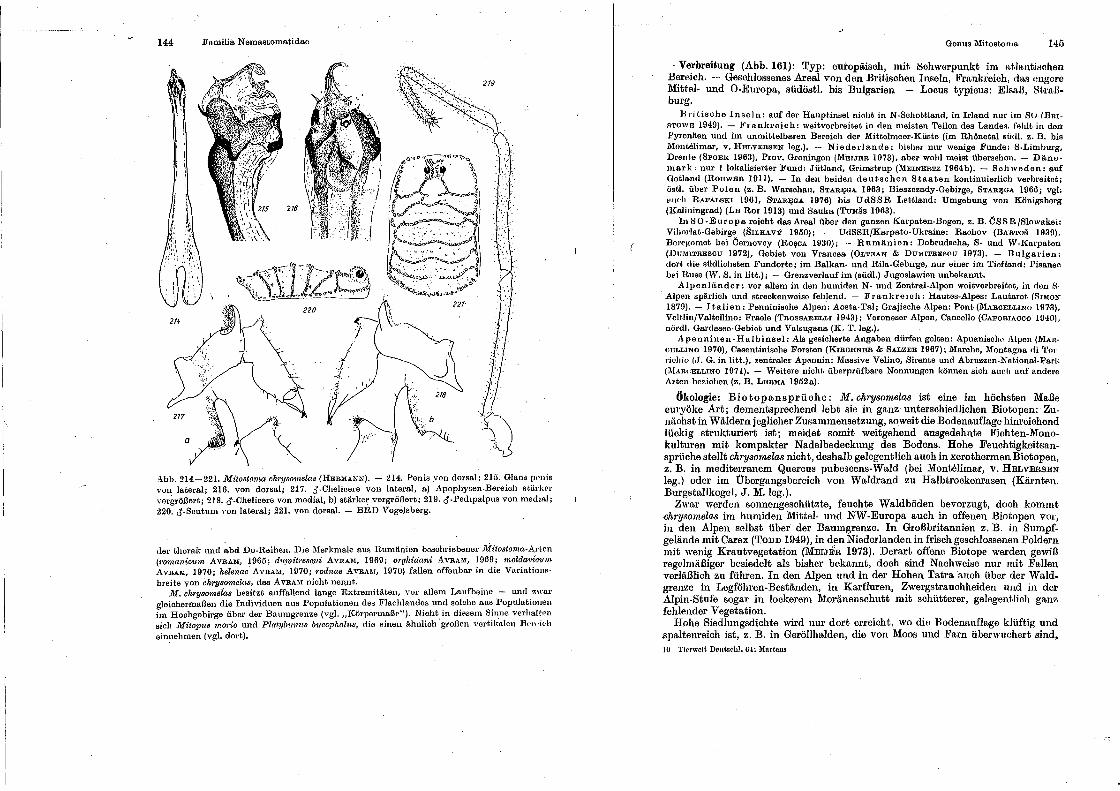
144 Familia Nemastomatidae
Abb. 214-221. Mitostoma chi'ysomelas (HERMANN). - 214. Penis von dorsal; 215. Glans pt•nis von lateral; 216. von dorsal; 217. J-Chelicere von lateral, a) Apophysen-Bereich städ<t•t· vergrößert; 218. d'-Chelicere \'On medial, b) stärke~ vergrößert; 219. d'-Pedipalpus von medial; 220. d'-Scutum von lateral; 221. von dorsal. - BRD Voge)sherg.
der t.Iwrak und abd D.o~Reihen. Die 1\ferkmale aus Rumänien beschriebener Mitostoma. 8 Arten (1'omanicu.Jn AVRAI\f, 1965; d·up1#resc:u.i AVRAM, 1969; orghidani AvRA,M, 1969; moldavicum AvRAM, 1970; helenae AvRAM, 1970; rodnae AVRAM, 1970) fallen offenbar in die Variationsbreite von chrysomelas, das AvRAM nicht nennt~
M. ch•·ysomelas besitzt auffallend lange Extremitäten, vor allem Laufbeine - und Z\rar gleichermaßen die Individuen aus Populationen des Flachlandes und solche aus Populationen im Hochgebirge über der Batm1grenze (vgl. ,,I{örpertnaß~"). Nicht. in diesein Sh~ne verhalten sich Mitopus morio und Platybu·nus bucephalus, die einen ähnlich großen vertikalen RPrt·ieh einnehmen (vgl. dort),
Genua Mitostoma 145
-Verbreitung {Abb. 161): Typ: europäisch, mit t:!chwerpunk.t im at.lantiHchen Bereich. --- Geschlossenes Areal von den Britischen Inseln, Frankreich, das •mgore Mittel- und 0-Europa, südöstl. bis Bulgarien - Locus typicus: ElsaU, Straß· burg.
H ri tische Inseln: auf der Haupt.insel ninht in N-Schottlanrl, in Irland nur im SO (.BRIBTOWE 1949). - Frankreich: weitverbreitet in den meisten Teilen des Landes. fehlt in den Pyrenäen und im unmittelbaren Bereich der Mittelmeer-Küste (im Rhönetal südl. z. B. bis Montelimar, v. HELVERBEN leg.). - Niederlande: bisltAr nur wenige Funde: S-Limburg, Drente (SPOEK 1963), Prov. Groningen (MEIJER 1973), aber wohl meist übersehen. - Dänemark: nur I lokalisierter Fund: Jütland, Grimstrup (MEINERTZ 1964b). - Schweden: auf Gotlan<l (ROEWER 1911). - In den beiden deutschen Staaten kontinuierlich verbreitet; östl. über Polen (z. B. Warsohan, STAR'i]GA 1963; Bieszczady-Gebirge, STAR'i]GA 1966; vgl: Hu<·h RAFALSKI 1961, STAR'i]GA 1976) bis UdSSR Lettland: Umgebung von Königsberg (Kaliningrad) (LE Ror 1913) und Sauka (Tu>tss 1963).
InSO-Europa reicht das Areal über den ganzen Karpaten-Bogen, z. B. ÖSSR/Slowakei: Vihorlat.-Gebirge (ilrLHAVY 1950); - UdSSR/Karpato-Ukraine: Rachov (BARTml Hl39), Bnrel(omet bei Öernovcy (Ro~oA 1930); -- Rumänien: Dobrudsoha, S- und W-Karpaten (DUMITRESCU 1972), Gebiet von Vranoea (ÜLTEAN & DUMITRER!HJ 1973), - Bulgarien: dort. die südlichsten Fundorte; im Balkan- und Rila-Gebir-ge, nur einer im Tiefland: Pisanec bei Ruse (W. S. in litt.); - Grenzverlauf im (südl.) Jugoslawien unbekannt.
Alpenländer: vor allem in den humiden N- und Zentral-Alpen weitverbreitet, in den S· Alpen spärlich und streckenweise fehlend. - Frankreich: Hautes-Alpes: Lautaret IRrMON 1879). - Italien: Penninische Alpen: Aosta-Tal; Grajische Alpen: Pont (lliAROELLINo 197·3), Veltlin/Valtellino: Fraeie (TROSSARELLI 1943); Veroneser Alpen, Cancello (CAPORIAOCO 1940), nördl. Gardasee-Gebiet und Valsugana (K. T. leg.).
A penninen-Halbinsel: Als gesicherte Angaben dürfen gelten: Apuanische Alpen (MAROEJ,LINO 1970), Gasentinische Forsten (KIRCHNER & SALZER 1967); Marche, Montagna cti Tm· richio (,J. G. in Jit.t.), zentraler Apennin: Massive Velino, Sirente und Abruzzen-National-Park (MAR"ELLINO 1971). - Weitere nicht überprüfbare Nennungen können sich aueh auf andere .At•ten beziehen (z. B. LERMA 1952a).
Ölwlogie: Biotopansprüche: M. chrysomelas ist eine im höchsten Maße euryöke Art; dementsprechend lebt sie in ganz unterschiedlichen Biotopen: Zunächst in Wäldern jeglicher Zusammensetzung, soweit die Bodenauflage hinreichend lückig strukturiert ist; meidet somit weitgehend ausgedehnte Fichten-Monokulturen mit kompakter Nadelbedeckung des Bodens. Hohe Feuchtigkeitsansprüche stellt chrysomelas nicht, deshalb gelegentlich auch in xerothermen Biotopen, z. B. in mediterranem Quercus pubescens-Wald (bei Montelimar, v. HELVERSEN leg.) oder im Übergangsbereich von Waldrand zu Halbtrockenrasen (Kärnten. Burgstallkogel, J. M. leg.).
Zwar werden sonnengeschützte, feuchte Waldböden bevorzugt, doch kommt chry8omelas im humiden Mittel- und NW-Europa auch in offenen Biotopen vor, in den Alpen selbst über der Baumgrenze. In GroUbritannien z. B. in Sumpfgelände mit Carex (Tonn 1949), in den Niederlanden in frisch geschlossenen Poldern mit wenig Krautvegetation {MEIJER 1973). Derart offene Biotope werden gewiß regelmäßiger besiedelt als bisher bekannt, doch sind Nachweise nur mit Fallen verläUlich zu führen. In den Alpen und in der Hohen Tatra auch über der Waldgrenze in Legföhren-Beständen, in Karfluren, Zwergstrauchheiden und in der Alpin-Stufe sogar in lockerem Moränenschutt mit schütterer, gelegentlich ganz fehlender Vegetation.
Hohe Siedlungsdichte wird nur dort erreicht, wo dit• Bodenauflage klüftig und .spaltenreich ist, z. B. in Geröllhalden, die von Moos und Farn überwuchert sind, lU Tierwelt DflntschJ. 04! Martens

146 Familia Nemastomatidae
in übereinandergeschichtetem alten Holz oder in lockerer Auflage alten Laubes.· Die auffällige Langbeinigkeit von chrysornelas (und fast aller anderen 111-itostoma-
Arten) steht damit im Einklang. Gelegentlich steigt chrysornelas von den Spaltensystemen der Bodenauflage in
höhere Strata auf: in der Montan- und unteren Subalpin-Stufe Salzburgsan Baumstämmen und Felswänden bis 1,5 m hoch (A. A. in litt.).
Vertikalverbreitung: Von der Küste der Nord- und Ostsee bis in die AlpinStufe der Alpen. Funde in den W- und mitteleuropäischen Niederungsgebieten sind zwar spärlich, do"ch der Nachweis (außerhalb von Waldgesellschaften) auch schwierig. In. den. Mittelgebirgen weitverbreitet: z. B. Vogelsberg, 770 m (J. M. leg.); Böhmerwald, Arber, 1100 ll1 (KIRCHNER & al. 196~); BieszczadyJWaldkarpaten, Krzemien, 1335 m (ST.ARiiGA 1966), polnische Hohe Tatra, 2000 m (STARF;G.A 1976a); Apennin: hisetwa 1900 m (KIRCHNER & SA.LZER 1967 a, MARCELLINO 1970, 1971); in Bulgarien selten ab 100m, normalerweise von 800-1700 m (W. S.
in litt.). Die Besiedlung der Alpen scheint nicht überall kontinuierlich an die der Alpon-
vorländer anzuschließen: aus der Collin-Stufe (400-500 m) des Salzburger Beckens erst 1 Fund, dort sonst nur auf jenen Bergen, die bereits in die Montan-Stufe aufragen (Möp.chsberg, Rainberg), regelmäßig danri in der Montan- und SubalpinStufe (500''-1200 bzw. 1200-2000 m; dort 51% aller Funde, n = 83 Serien; A.A. in litt.). Im nieder-östßrreichischen Erlaf- und Ybhs-Gebiet besteht dagegen durch· gehende, wenn auch spärliche Verbre.itung von280m (um Purgstall) bis über 1800 m (Gipfeh;egionen der Kalkalpen; RESSL leg., J. G. in litt.).
In den Österreichischen Zentral-Alpen steigt chrysornelas bis 2200 und 2280 m (A. A. für Salzburg). - Die höchsten Funde in den Alpen: 2780 m (Colle Pinter, Aosta; Gozo 1908), 2600 m (Aosta-Tal, Penninjsche Alpen, MARCELLINO 1972), bei 2820 m am Aletschgletscher (LESSERT 1917), bei 2700 m in N-Tirol (bei Ober-
gurgl, A. A. leg.). Phänologie: ad. eurychron; Generationen-Abfolge bisher nicht völlig über-
schaubar, zumal offenbar lokal variierend. Ad. wahrscheinlich während des ganzen Jahres; bestätigt bisher nur für die Niederlande, denn in Gebieten mit langer Schneebedeckung nur schwer nachweisbar.
Ad.: BRD und außeralpines Österreich (Salzburg): V-X (J.M., A.A.); Ober-Österreich: V-XII (REsSL leg.); alpines Salzburg: V-X (A. A. in litt.); Polen: E 111-M XII (STAMGA 1976); ÖSSR: VIII-E XI (i:JILH.AVY 1956); Niederlande: I-XI (SPOEK 1963), VI-XII (MEIJER 1973); Britische Inseln: Surrey: A V -Xll
(S.ANKEY 1949). Iuv.: BRD und außeralpines Österreich: V-A X; alpines Salzburg: IV-X;
Polen: M IV -E XII; Niederlande: VI-XII. Wahrscheinlich überwintern ad., iuv. und Gelege. Überwinterte ad. sterben
Frühjahr/Sommer und können in dieser Zeit mit den ersten ad. der neuen Generation zusammen vorkommen. Ob sie in beiden Jahren Gelege produzieren, ist nicht
bekannt.
Genus Mitostoma 147
Mitostoma alpinum {HADZI, 1931) Abb. 134, 222-228
1199~11 MNe!"astoma chrysomelas a!pinum HADZI, Prirod. Razprave 1: 109 u ttostoma chry la l · ' · 1973a Mitostoma so!"e s apmum - RoEWER, SenckAnhergiana, 32: 143. .
LJ"ublJ"a 16~~4ttostoma) chrysomelas multidenticulatum HAnzr, Razpra.v~, Dissertat. na, . u . :' .\. . •
1973a Mitostoma (Mito to ) h ia · · · 16: 55. . s . ma c rysome s mwhteh HA:ilzr, Razpfave, Dissertat .. Ljuhljana,
Begründung der Syno y • . •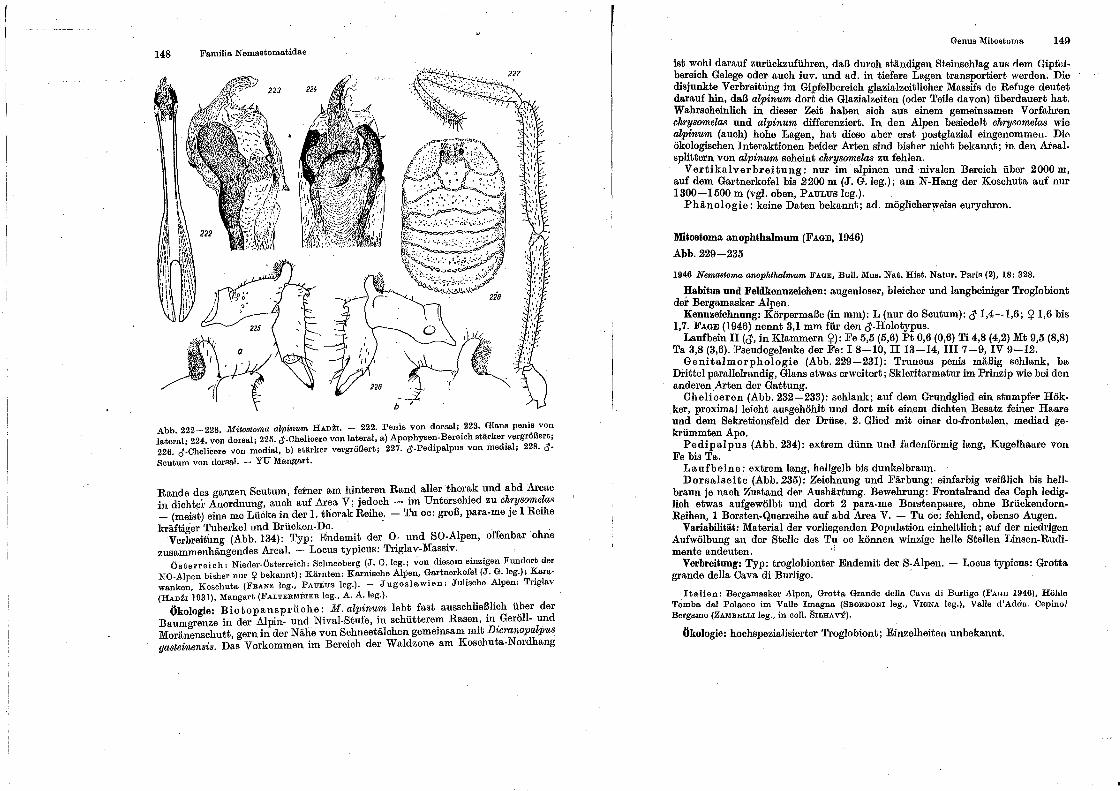
148 Familia Nemnstomatidae
Abb. 222-228. Mitostoma alpinum HADZI. - 222. Penis von dorsal; _223. _Glans peni.~ von lateral; 224. von dorsal; 225. d'-Chelicere von lateral, a) Apophysen-Berewh starker_vergroßert; 226. d'-Chelicere von medial, b) stärker vergrößert; 227. d'-Pedipalpus von med18l; 228. d'Soutum von dorsal. - YU Mangart.
Rande des ganzen Scutum, ferner am hi~teren Ra~d aller thor~k und abd Areae in dichtßr Anordnung, auch auf Area V; Jedoch - 1m Unterschied zu ch:ysome_las _ (meist) eine me Lücke in der I. thorak Reihe_. - Tu oc: groß, para-me Je 1 Reihe kräftiger Tuberkel und Brücken-Do. .
Verbreitimg (Abb. 134): Typ: Endemi~ der 0_- und SO:Alpen, offenbar ohne zusammenhängendes Areal. - Locus typwus: Triglav-Massiv.
Österreich: Nieclet-o"sterreich: Schneeberg (J. G.leg.; von diesem einzigen Fundort der NO-Alpen bisher nur ~ bekannt); Kärnten: Karnische Alpen, ~artnerlw.fel (J. G. leg.); ~ara: wanken, Koschuta (FRANZ leg., PAULUS leg.). - Jugoslawien: Julisehe Alpen: Tr1gla' (HADZI 1931), Mangart (FALTERMIÜER leg., A. A. leg.).
Ökologie: Biotopansprüche: M. alpinu;n leb~ fast ausschlie~lich ~?er der Baumgrenze in der Alpin- und Nival-Stufe, m schuttere~ Rase~, In _Geroll- und Moränenschutt, g~rn in der Nähe von Schneetälchen gememsam mit D~cranopalpus gasteinensis. Das Vorkommen im Bereich der Waldzone am Koschuta-Nordhang
Genus Mitostoma 149
ist wohl darauf zurückzuführen, daß durch ständigen Steinschlag auH <lern Gipfelbereich Gelege oder auch iuv. und ad. in tiefere Lagen transportiert werden. Die disjunkte Verbreitung im Gipfelbereich glazialzeitlicher Massifs de Refuge deutet darauf hin, daß alpinumdort die Glazialzeiten (oder Teile davon) überdauert hat. Wahrscheinlich in dieser Zeit haben sich aus einem gemeinsamen Vorfahren chrysomelas und alpinum differenziert. In den Alpen besiedelt chrysomelas wie alpinum (auch) hohe Lagen, hat diese aber erst postglazial eingenommen. Di11 ökologischen Interaktionen beider Arten sind bisher nicht bekannt; in den Arealsplittern von alpinum scheint chrysomelas zu fehlen.
Vertikalverbreitung: nur im alpinen und·nivalen Bereich über 2000m, auf dem Gartnerkofel bis 2200 m (J. G. leg.); am N-Hang der Koschuta auf nur 1300-1500 m (vgl. oben, PAULUS leg.).
Phänologie: keine Daten bekannt; ad. möglicherrveise eurychron.
Miiosioma anophthalmum (FAGE, 1946)
Abb. 229-235
1946 Nemastoma anophthalmum FAGE, Bull. Mus. Nat. Hiat. Natur. Paris (2), 18: 328.
Habit1111 und Feldkennzeichen: augenloser, bleicher und langbeiniger Troglobiont der Bergamasker Alpen.
Kennzeichnung: Körpermaße (in mm): L (nur do Soutum): if 1,4-1,6; ~ 1,6 bis 1,7. FAGE (1946) nennt 3,1 mm für den {f-Holotypus.
Laufbein II ((f, in Klammern~): Fe 5,5 (5,6) Pt 0,6 (0,6) Ti 4,8 (4,2) Mt 9,5 (8,8) Ta 3,8 (3,6). Pseudogelenke der Fe: I 8-10, II 13-14, III 7-9, IV 9-12.
Genitalmorphologie (Abb. 229-231): Truncus penis mäßig schlank, ba Drittel parallelrandig, Glans etwas erweitert; Skleritarmatur im Prinzip wie bei den anderenArten der Gattung.
Cheliceren (Abb. 232-233): schlank; auf dem Grundglied ein stumpfer Hökker, proximalleicht ausgehöhlt und dort mit einem dichten Besatz feiner Haare und dem Sekretionsfeld der Drüse. 2. Glied mit einer do-frontalen, mediad gekrümmten Apo.
Pedipalpus (Abb. 234): extrem dünn und fadenförmig lang, Kugelhaare von Fe bis Ta.
Laufbeine: extrem lang, hellgelb bis dunkelbraun. Dorsalseite (Abb. 235): Zeichnung und Färbung: einfarbig weißlich bis hell
braun je nach Zustand der Aushärtung. Bewahrung: Frontalrand des Ceph lediglich etwas aufgewölbt und dort 2 para-me Borstenpaare, ohne BrückendornReihen, 1 Borsten-Querreihe auf abd Area V. - Tu oo: fehlend, ebenso Augen.
Variabilität: Material der vorliegenden Population einheitlich; auf der niedrigen Aufwölbung an der Stelle des Tu oc können winzige helle Stellen Linsen-Rudi-mente andeuten. ''
Verbreiiung: Typ: troglobionter Endemit der S-Alpen. - Locus typicus: Grotta grande della Cava di Burligo. ··
Italien: Bergamasker Alpen, Grotta Grande della Cava di Burligo (FACIE 1946), Höhle Tomba da! Polacco im Valle Imagna (SBORDONI leg., VIGNA leg.), Valle cl'Addu. Cepino/ Bergamo (ZAMBELLI leg., in coll. f:!rLHAVY).
Ökologie: hochspezialisierter Troglobiont; Einzelheiten unbekannt.
..
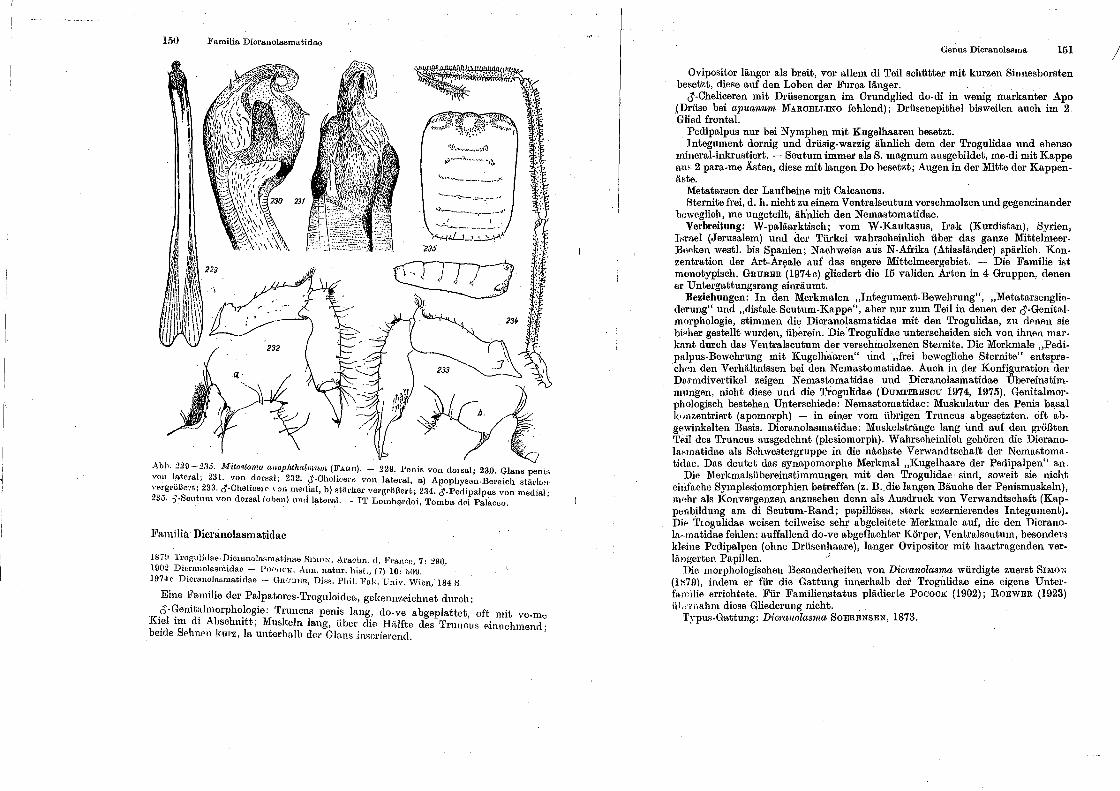
150 Familia Dicranolasmatidae
II •. ·
~..". ...... _______ ,t'~
~--·-.-----<~'
--~-- ----~--~.J..Lj_,~,....\,1\
"235
~yt· ~-"")· . ~t"j;,, . ,.
Abh. ~29-2~l~; Mitostoma anophtlwtmum (FA<>E). - 22H. Penis von dorsal; 23.0. Glans peni" von lateral; c31. von dor"'•l; 232. ß'-Chelicere von lateral, a) Apophysen-Bereich stärk.;,· ~-e:gr~ßert; 233. c3'-ChelicerP 1·on merlial, b) stiirker vergrößert.; 234. c3'-Pedipalpus von medial; 23a. ::>:Scutum von dorsal (oben) uncllateral. -- IT Lomh~rdei, Tomba del Palitcco.
l!'amilia Dicninolasmatidac
l87tt Trogulitlae-Dicranolasmatinae SIMox~ Arachn. d. Ji'ra.ur:f:J 7: ~90. 190.:? Dic:ranolasn1idae - Pru•ocn. Ann. natur. hi::;t., (7) 10: 509. . 197-h· Dicrunolasmatidae -- Gn.r;DER, Diss. PJ.il. Val.:. Univ. \VümJ 184 :-:;.
Eine Familie der Palpatores-Troguloidea, gekennzeichnet durch: . . 0' -t?enit:thnorpho_Iogie: Truncus penis htng, do-ve abgeplattet, oft mit ve-me K1~l nn d1 Ahschmtt; Muskeln lang, über die H;i.lfte des Trunnus f'innehmend: be1de Sehnc:n kU!'z, Ia unterhalb de1: Glans inserierend. ·
I . I
Genus Dicranolasma 151
Ovipositor länger als breit, vor allem di Teil schütter mit kurzen Sinnesborsten besetzt, diese auf den Loben der Furca länger.
d'-Cheliceren mit Drüsenorgan im Grundglied do-di in wenig markanter Apo .(Drüse bei apuanum MAROELLINO fehlend); Drüsenepithel bisweilen auch im 2. Glied frontal.
Pedipalpus nur bei Nymphen mit Kugelhaaren besetzt. Integument dornig und drüsig-warzig ähnlich dem der Trogulidae und ebenso
mineral-inkrustiert. - Souturn immer als S. magnum ausgebildet, me-di mit Kappe an~ 2 para-me .Ästen, diese mit langen Do besetzt; Augen in der Mitte der Kappenäste.
Metatarsen der Laufbeine mit Calcaneus. Sternite frei, d. h. nicht zu einem Ventralsouturn verschmolzen und gegeneinander
l•eweglich, me ungeteilt, ähnlich den Nemastomatidae. Verbreitung: W-paläarktisch; vom W-Kaukasus, Irak (Kurdistan), Syrien,
I."mel (Jerusalem) und der Türkei wahrscheinlich über das ganze MittelmeerBc11:ken westl. bis Spanien; Nachweise aus N-Afrika (Atlasländer) spärlich. Konzentration der Art-Ar~Jale auf das engere Mittelmeergebiet. - Die Familie ist monotypisch. GRUBER (1974c) gliedert die 15 validen Arten in 4 Gruppen, denen er Untergattungsrang einräumt. · .
Beziehungen: In den M;erkmalen "Integument-Bewehrung", "Metatarsengliederung" und "distale. Scutum-Kappe", aber nur zum Teil in denen der d'-Genitltlmorphologie, stimmen. die Dicranolasmatidae mit den Trogulidae, zu df'nen sie bi~her gestellt wurden, überein. Die'Trogulidae unterscheiden sich von ihnen markant durch das Ventralsouturn der verschmolzenen Sternite. Die Merkmale "Pedipalpus-Bewehrung mit Kugelliaaren" und "frei bewegliche Sternite" entsprech(•n den Verhältnissen bei den Nemastomatidae. Auch in !ler Konfiguration der Darmdivertikel zeigen Neniastomatidae und Dicranolasinatidae Übereinstinlnnmgen, nicht diese und die Trogulidae (DuMITREsou 1974, 1975). Genitalmorphologisch bestehen Unterschiede: Nemastomatidae: Muskulatur des Penis basal k· .nzentriert (apomorph) - in einer vom übrigen Truncus abgesetzten.. oft abgf'win.kelten Basis. Dioranolasmatidae: Muskelstränge lang und auf den größten Tf\il des Truncus ausgedehnt. (plesiomorph). Wahrscheinlich gehören die DicranolaHmatidae als Schwestergruppe in die nächste Verwandtschaft der Nemastoma. tidae. Das deutet das synapomorphe Merkmal "Kugelhaare der Pedipalpen" an.
Die Merkmalsübereinstimmungen mit den Trogulidae · sind, soweit sie nicht eiri.fache Symplesiomorphien betreffen (z. B..die langen Bäuche der Penismuskeln), m"hr als Konvergenzen anzusehen denn als Ausdruck von Verwandtschaft (Kappenbildung am di Scutum-Rand; papillöses, stark sezernierendes Integument). Di,. Trogulidae weisen teilwei.~e sehr abgeleitete Merkmale auf, die den Dicranola.--matidae fehlen: auffallend do-ve abgeflachter Körper, Ventralscutum, besonders kleine Pedipalpen (ohne Drüsenhaare), langer Ovipositor mit haartragenden ver-län4erten Papillen. ·
Die morphologischen Besonderheiten von Dicranolasma würdigte zuerst SIMON (lh7!1), indem er für die Gattung innerhalb der Trogulidae eine eigene Unterfan;ilie errichtete. Für Familienstatus plädierte PocooK (1902); ROEWER (1923) iiln•-.·r.ahm diese Gliederung nicht .
Typus-Chtttun.g: Dicmuolasma SoERENSEN, 1873.
I

-· . 152 Familia Dicranolasmatidae
Genus Dicranolasma SoERENSEN, 1873
1873 Dicranolasma SOERENSEN, Naturhist. 'L'idsskr., (3) 8: 516. 1873 Amopaum.SOERENSEN, Naturhist; Tidsskr., (3) 8: 517. 1876 Dicranolasma - THORELL, Ann. Mus. Genova, 8: 467. 1879 Dicranolasma - SIMON, Arachn. de France, 7: 293. 1879 Amopaum - SrMON, Arachn. de France, 7: 290. 1903 Dicranolasma - DAHL, SB Ges. naturforsch. Freunde, Berlin: 278. 1903 Amopaum - DAHL, SB. Ges. naturforsch. Freunde, Berlin: 289. 1923 Dicranolasma - RoEWER, Weberknechte d. Erde: 641.
Nominatgattung der Dicranolasmatidae, gekennzeichnet durch: d'-Genitalmorphologie: Penis etwa halb so lang wie der Körper, Truncus do-ve
abgeplattet, di im Querschnitt etwa 3eckig; Glans iri gerader Verlängerung des Truncus, ohne äußerlich erkennbare Gelenkzone in den Truncus übergehend; Glans rri.eist wenig breiter als das di Ende des Truncus, mit 25-60 stachelartigen Sinnesborsten besetzt. Legeröhre weichhäutig, ungegliedert, mit schütterer, di dichterer Beborstung, Furca 2gliedrig; Rec sem als 2 Ia Säckchen oder Gruppen solcher. - Körper oval, etwa halb so breit wie lang, di in Verlängerung des Tu oc bzw. des Ceph kopfartige Kappe von etwa I Viertel Körperlänge aus 2 para-me Ästen, A~gen auf halber Länge der Kappenäste. - Dorsalseite mit Souturn magnum ;\Corona analis voll entwickelt; abd Sterrute IV-VII frei beweglich, me nicht geteilt. l-aufbein-Cx unbeweglich, ohne 'Randhöcker-Reihen. - Cheliceren und Pedipalpus kurz, in Ruhe unter der di Kappe verborgen. Basalglied der d'Cheliceren mit kegelförmigem Höcker do-di, auf dessen Spitze 1 Borstenschopf;. Höcker meist mit Drü.senorgan. - Pedipalpus ohne strukturelle Besonderheiten; bei iuv. ruft zapfenförmiger Bewehrung auf Fe und weniger Pt und Kugelhaare auf Ti und Ta, diese den ad fehlend. Laufbeine kurz, 1-3fache Körperlänge.
Ad. meist stark erdinkrustiert, frei davonlediglich Calcaneus, Tarsen, Pedipalpen, Cheliceren und Mundteile.
Verbreitung: 15 valide Arten in der mediterran geprägten W-Paläarktis (J. G. in litt.); in S-Europa von der südl. Iberischen Halbinsel nördl. bis S-Frankreich, Meeralpen, südl. 0-Alpen, Karpaten-Länder, Apenninen- und Balkan,Halbinsel, Atlas-Länder, Kleinasien östl. bis Irakisch-Kurdistan, westl. Kaukasus, im SO am 0-Rand des Mittelmeeres.
Typus-Art (durch Monotypie): Opilio scabe1· HERBST, 1799. SoEBENSEN (1873) ordnete dem neueingeführten Gattungsnamen Dicranolasma die Spezies Opilio scaber als einzige sichere Art zu; tatsächlich lag ihm jedoch die Art D. opilionoides (L. KocH, 1867) vor und somit eine "falsch bestimmte Typus-Art" (J. G. in litt.). Somit kann nur durch eine Entscheidung der Nomenklatur-Kommission die Typus-Art für die Gattung Dicranolasma endgültig festgelegt werden. Da sich. aus der Entscheidung keine nomenklatorischen Konsequenzen ergeben werden, soll hier dem ersten revidierenden Autor, (THORELL 1876),. gefolgt werden, der D. scabrum als Typus-Art nannte.
Bestimmungstabelle für die Arten (Chelicere und Palpus abtrennen; Palpus zwischen Coxa und Trochanter)
I Tr des Palpus ve mit langem Zapfen und aufgesetzter Borste (z. B. Abb. 239) . Z Tr des Palpus ohne Zapfen, Borsten höchstens auf niedrigem Sockel (Abb. 236), weitverbreitet in SO-Europa . . . . . . scabrum, S. !53
2 Zahl der Glieder an Tarsus II: 5-6 . . . . . . . . . . . . . . . . . . . . . . 3
Oenus Dicranolasma
Zahl der Glieder an Tarsus II: 8-18; im Gebiet mu• in den SW-Alpen soet•enseni, S. 157
3 Zahl der GHeder an Tarsus II: 5 (selten 6), kleine Art. (3,3-4,0 mm); nur im nardaseoGebiet. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . pauper, S. 16! Zahl der Glieder an Tarsus II: 6; größere Art (4,0··-4,6 mm); nur in den Me~ralpen und in den Apenninen . . . . . . . . . . . . . . . . . . . . . . . . cristatum, S. lHO
Dicranolasma scabrum (HERBST, 1799) Abb. 236-238, 245-247, 279-'--280
1799 OpiÜo scaber HERBST, Natursyst. ungefl. Insekten, 3: 15. 1903 Dicranolasma scabrum - DAHL, SB. Ges. naturforsch. Freunde, Berlin, 1903: 291. 1923 Dicranolasma scabrum - ROEWER, Weberknechte d. Erde: 643 (part.: nicht die itali<'
nischen und französischen Funde). 1927 Dicranolasma schmidti HADZI, Bull. Ass. Mus. Slov., 1/8 B: I I.
Begründung der Synonymie: Der von HERBST begründete Name wurde oft auf ancloJ'f' Arten bezogen, vor allem auf das W-mediterrane soerenseni (vgl. dort). Erat DAHL (1903) f~harakterisierte Bcabrum eindeutig; er schloß aber soerenBeni in die Synonymie ßln und trug damit zu fortgesetzter Verwirrung der Benennung bei. J. G. (in litt.) hat den größten Teil des publizierten Materials revidiert.
Habitus und Feldkennzeichen: große Art mit hohen Tarsalzahlen ( d' Ta II: 10-16) do Erdbekrustung immer flach (nie als hoher Buckel); nur im SO des hier behandelten Gebietes (Abb. 279-280).
Kennzeichnung: Körpermaße (in mm): L gesamt d' 4,6-5,9; ~ 4,8-6,2. L Souturn (mit frontaler Kappe und Do der Area V) q 4,1-5,9; ~ 4,5-5,6_. -Laufbein II (q, in Klammern ~; Leitha-Gebirge): Fe 3,5 (3,1) Pt 0,9 (0,9) Tx 2,9 (2,5) Mt 3,1 (2,9) Ta 2,1 (1,8). .
Genitalmorphologie (Abb. 246-247): Truncus perus do-:re ab.geplattet, von der Basis distad kontinuierlich verschmälert, schmalster Bereich dicht unterhalb des Glansansatzes. Glans länglich schmal, in sich geschwungen; Bewehrung au~> etwa 45 geraden bis leicht gebogenen stachelartigen Do auf allen E)eitcm der Glans; Stylus nicht deutlich abgesetzt, (von dof.la) in tric?terförmig erweiterten transparenten Mündungslippensaum endend; links-do mit transparenter Lamelle.
Cheliceren (Abb. 238): d'-Grundglied do-di mit breitem, do abgeflachtem Höcker, zum 2. Glied steil abfallend; auf der Spitze des Höckers mit kleinem Haar-büschel. - ~-Chelicere ohne Höcker, flach.. . .
Pedipalpus (Abb. 236-237): Tr ve mit 2 langen Bo auf_medrxgem S~~kel, Fe schlank do schwach erweitert, 3+6 ve-la Bo; Pt etwa 2 Dnttel der Fe-Lange, nur bei d' kräftig konvex, mit do-di Haarschopf und kleinem Drüsenfeld. ~-Pt nicht verdickt und normal behaart.
Laufbeine: Fe bis Ti I, III und IV keulig verdickt, bei Ö' stärker als bei~; Fe I und IV mit medio-do Do-Kiel, Fe bis Mt nur bedornt, Mt IV mit 2-4 Pseudogelenken di, Calcaneus II doppelt so lang . wie dick, bei den übrigen Beinen fast so lang wie dick. - Tarsal-Gliederung: I: d' 6, selten 5 oder 7; ~ 5, selten 4 oder 6. _ II: q 10-16, meist 11-13; ~ 6-9, meist 8. - III: q 6, selten 7; ~ 6, selten 5. - IV: q 6, selten 5; ~ 6, selten 5. .
Dorsalseite (Abb. 279-270) di Kappe (von do) gerundet, Augen etwa m deren Mitte, von frontal ohne me herabgezogenen Zipfel. - Senturn-Querfurchen auf dem Souturn nur angedeutet: hinter dem thorak Terg II und vor_ abd Area V. Area I mit breitem rundlichen Hügel me; Scutum bedornt, Do dicht stehend,
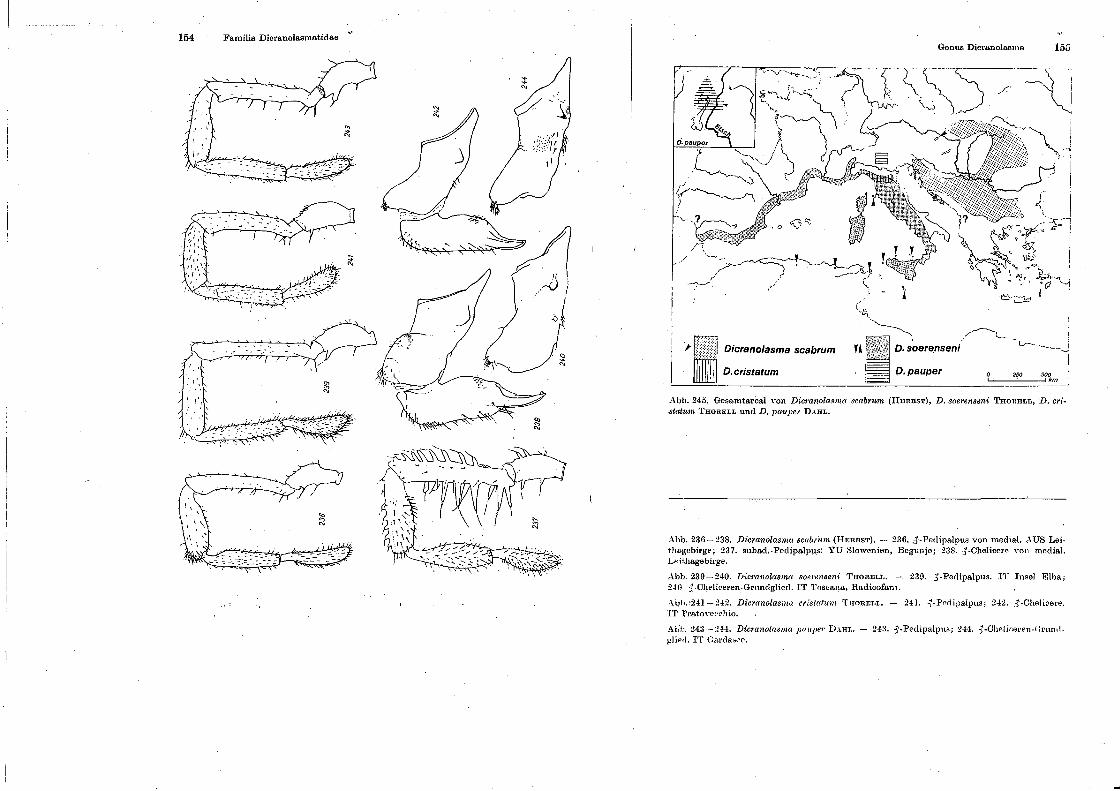
154 Familia Dicranolasmatidae Genus Dicranolasma 15G
/
(:.",,
L_~_l~:!:!~:!!:~!:::~~1_:_._;:_:1_:_:a_o,_/:_:_m_a_s_c_a_b_ru_m ___ '_' .§~~'--~~1_;;_/)_·. s_P~_ae_:_:_:_,s_e_n_/_~ _ _'::==25':::::=,_=··::'~~-~ Abb. 245. Gesan1ta.r8al von Dim·anolcumw .gcabrum (HERBST}, D. soerenseni THOliELL, D. crf ... st<~t.um THORELL und D. paupe.;• DAHL.
Abb. 236-238. Dicmnolasnw SC((/Ji"Uin (HERBST), - 2~6. 3'-Pe<lipalpus von medtal. AUS Leithagcbirge; 237. subad.-Pedipalp\t.s; YU Slowenien, Begunje; :238 . .:)'-Chelicere YOn medial. L1·1·jt,hagebirge .
.-\bh. 239-241). lJicranolasma. soeJ'Mtseni THORELT.. _, 2a9. j'-Peclipalpus. I'f Insel Elba; 24il 1-0heliceren-Grtmdglied. IT Toscana, Radicofant.
\.üh. ~241- 242. Dicranolasma cristatum THORELL. - :241. -)-Pr>clipalpus; 242. -~-Chelicere.
IT :wrato,·er-c•hio.
Ab'b. 243 -:!44. Dicwanolrc.:ww ptntper DAHI., - 24~. j'-Pedipalpu~; 244. :)-Ch~linere.n-Cirundg]if.HI. IT Ga.rda.-;t"'P.

156 Familia Dicranolasmatidae
nicht auffallend lang, Anordnung segmental (Einzelheiten vgl. Abb. 280); auf Area I zu einem auffallenden Büschel vereinigt, weniger deutlich auf den anderen Areae. Am Ceph-Frontalrand neben der Kappe je 2 lange horizontale Zapfen,. der äußere gegabelt.
Verbreitung (Abb. 245): Typ: europäisch-montan; karpatisch-dinarisch: - Geschlossenes Areal hauptsächlich in den Karpaten-Ländern nnd Jugoslawien unter Ausschluß des pannonischen Beckens, Ausläufer bis Bulgarien un<} N-Griechenland.- Locus typicus: "Ungarn" in den damaligen Grenzen, somit Typenmaterial wahrscheinlich aus dem Karpaten-Gebiet stammend; hier einengend festgelegt. auf Sibiu, Rumänien.
Osterreich: Isolierte Splitterareale im Leithagebirge und in den Leitha-Auen; sie sind wahrscheinlich durch Einschleppung bedingt (KRITSOHER 1959, GRUBER 1960, 1974c). -ÖSSR: Slowakei: auf der S-Abdachung derW-und Waldkarpaten, bisher nicht in den Außenketten wie Kleine und Weiße Karpaten und nicht in der Hohen Tatra; westl. bis zum NitraFluß (Öakajovce bei Nitra, L.{o 1957); in der östlichsten Slowakei liegt der nördlichste Ftmdpunkt des Artareals (dolina Roztol<a, STARF,)GA 1966). - Ungarn: anschließend an den slowaldschen Arealteil an der nördl. Landesgrenze; Cserhat-Gebirge: Als6peteny und Csorhatszentivan (SzALAY 1949); nicht in den pannonisch beeinflußten Tiefebenen. -Rumänien: in allen gebirgigen L.;_ndesteilen weit verbrei"tet; Siebenbürgen, Bihar-Gebirge, Banster Gebirge etc.; östlichster Fund in den 0-Karpat.en (CiRDEI 1955). - Bulgarien: im W-Balkan, im Osogolvo-Gebirge und in den W-Rhodopen, östl. etwa bis Asenovgrad (W. S. leg.). -Griechenland: 0-Makedonien: Kerkini-Gebirgejl!'[omos Serrai und Koula-GebirgejNomos. Xanthi (beide A. A. leg.). - Jugoslawien: Makedonien: Rasce (10 km westl. Skopje, HADZI 1942); - Kosmet: Bar-Gebirge (A. A. leg.); - Montenegro: Tresnjevik (A. A.leg.); - Bosnien-Herzegowina: Sarajevo (Belog ZMB); Celebic, Jaice (Belege ZMB und PANW); - Serbien: von den Inselbergen des pannonischen Beckens nur von der Fruska Gora nachgewiesen (Beleg NHMW); - über Kroatien und Slowenien wird die nörcll. Adria z. B. boi Rijelm erreicht; W-und N-Grenze bei Divaca (J. G. in litt.) und Kobarid (A. A. leg.); der nördlichste Fund am S-Rand der SO-Alpen (Jesenice/Assling, am S-Fuß der Karawanken; Beleg ZMB), deren N-Hang nicht erreicht wird.
Ökologie: Biotopansprüche: Vorwiegend in Laubmischwäldern der Collinund Montan-Stufe, z. B. in Eichenmischwäldern oder Buchenwäldern in Istrien, Slowenien, in den Karpaten und in N-Griechenland; gelegentlich auch in trockenen Flaumeichen-Buschwäldern und sogar in offenem, nahezu unbeschattetem Gelände (S-Siebenbürgen, WEIBS 1975) z. T. hoher Insolation. Fehlt im allgemeinen in Auwäldern und feuchten Pflanzengesellschaften an Bachufern; das Vorkommen in den schottergründigen Leitha-Auen dagegen beruht wahrscheinlich auf Einschleppung. Nur bei Rijeka-Opatija liegen Biotope auch in unmittelbarem Einflußbereich der Mittelmeerküste; sonst siedelt scabrum nur im mitteleuropäisch und SO-europäisch gemäßigten Bereich. - Nur im und auf dem Boden, gern in Steinhalden, in morschem Holz und unter Steinen, dringt auch in die Eingangsregion von Höhlen ein.
Vertikalverbreitung: Slowakei: von etwa 200 bis wenig über 600m (Uo I957, SILHAVY 1950, STARF;GA 1966), die obere Arealgrenze fällt etwa mit der unteren Grenze der geschlossenen Nadelwaldzone zusammen (Uo 1957); in Rumänien dementsprechend in der "montanen und submontanen" Zone; in Bulgarien obere Grenze deutlich höher: 1 Fund zwischen 1450 und I500 m (W. S. leg,); in Jugoslawien (Montenegro, Kosmet, Makedonien) zwischen I 000 und 1400 m; die wenigen Fundorte in (N-) Griechenland zwischen 500 und 850 m (A. A.leg.); im nördl. Adria-Küstengebiet von nahe Meereshöhe (Rijeka) bis etwa I 000 m
Genus Dioranolasma 157
(Berg Ucka! 950 ~), im _benachbarten Slowenien bis etwa 650 in (MAirNERT leg., J. M. ~-eg.); I~ Leithagebll'ge von 130 bis 480 m (J. G. leg.). . Ph~_nologie: Ad. _und iuv. eurychron. - Eiablage im späten Frühjahr und 1m Fruhsom~er, wemger auch im Herbst (trächtige ~ in Fallen, J. G. leg.), in Kul~ur- vorWiegend in VII und VIII, aber auch zu anderen Jahreszeiten; wahrs~hemlich mehrere. Gelege jährlich, Eizahlen klein, meist 1-3, möglicherweise bis 10; ad. leb.ten _m Kultur. über 3 Jahre. Hauptaktivitätszeit in IV/V und im Herb~t (J. G. m htt., Material-Herkunft: Leithagebirge); Eurychronie von ad. u~d ~uv. auch in Rumänien (S-Siebenbürgen, WEIBS 1975). - 7 Jugendstadien, die siCh unter Gefangenschaftsbedingungen nur langsam entwickeln; Dauer der Pos~embryonalentwicklung ungeklärt. - Sex ratio: Handaufsammlung LeithagebJrge: 181 c!: .223 ~ (55,8%: 44,2 %), Fallenfänge Leithagebirge: 36 c!: 95 ~ (~7,5: 72,5%); Museumssammlungen (nicht Leithagebirge): 57 c!: 75 ~ (J. G. in htt.).
Dicranolasma soerenseni THORELL, 1876 Abb. 239-240, 245, 248-249
1876 Dicranolasma Soerensenii THORELL, Ann. Mus. Genova, 8: 505. 1870 Dicranolasma scabrum - SrMON, Arachn. de France,.7: 294. 1879 Dicranolasma lalifrons SIMON, Araohn. de Franoe, 7: 295. 1879 Amopaum Soorensani - SrMON, Arachn. de France, 7: 292. 1003 Dicranolasma scabrum - DAHL, SB. Ges. naturforsoh. Freunde, Berlin: (7) 292 (part.). 1923 Dicranolasma opilionoides - RoEWER, Weberknechte d. Erde: 643 (p~trt.: das meiste für
lta!ien und Sizilien genannte M~tterial). 1923 Dicranolasma scabrum - ROEWER, Weberl<nechte d. Erde: 643 (part.: Nennungen für
Oberitalien, Riviera, Korsika). 1950 Dicranolasma wiehlei KRAus, Mitt. Zoo!. Mus. Ber)in, 85: 295.
BegrUndung der Synonymie: Diese in der ganzen W-Mediterrane.is weitverbreiteten Art wurde unter vielen verschiedenen Namen im Schrifttum gen~trmt; die Bel~ge der mejsten Nennungen hat J. G. ·(in litt.). revidiert. Hier wird der älteste verfügb~tre Name benutzt, da. keiner der jüngeren Namen fest eingeführt und die Stabilität der Namengehring nicht gefährdet ist.
Habitus und Feldkennzeichen: große Art mit hohen Tarsalzahlen (Ta II: 11-18) und vergleichsweise langen Beinen; do Erdbekrustung oft als deutlicher Höcker ausgeprägt; im hier behandelten Gebiet nur in den äußersten SW-Alpen.
Kennzeichnung: KörpermaUe (in mm): L gesamt c! 4,6-6,4; ~ 4,3-6,3. L Souturn (mit frontaler Kappe und Do der Area IV) c! 'l,5-6,0; ~ 4,1-5,9.- Laufbein II (c!, in Klammern ~): Fe 4,2 (4,0) Pt 1,0 (1,0) Ti 3,4 (3,3) Mt 3,5 (3,5) Ta 2,2 (2,1).
Genitalmorphologie (Abb. 248-249): Truncus penis wenig über dem ba Ende distad kontinuierlich verjüngt, am schmalsten dicht unterhalb der Glans und kräftig eingeschnürt unmittelbar an der Ansatzstelle der Glans. Glans (Abb. 249) langgestreckt konisch; di E;p.de in kurzen s-förmig geschwungenen und in sich gedrehten Stylus auslaufend; am Stylus (von do) links abstehender 3eckiger Zipfel herablaufend. Glans mit etwa 50 schlanken, meisl< geraden stachelartigen Do .besetzt.
Cheliceren (Abb. 240): c!-Grundglied mit breit kegelförmigem, distad sanft .ansteigendem, frontal steil abfallendem Apo-artigem Höcker, do-frontalauf dem Höcker ein Haarbüschel, darunter Drüsenorga.n. - ~-Chelicere ohne Höcker, :flach.

I
I
158 Farnilia Dicranolasrnatidae
2~6 2~8 / 250
i
I! I' i
Abb. 246-247. Dicranolasma scabtam (HERBST). - 246. Penis von dorsal (links) und latet·al; 247. Glanspenis von dorsal. AUS Leithagebirge.
Abb. 248-249. 'Dicranolasma. soet·enseni THORELL. - 24R. Penis von dorsal; 249. Glanspenis von dorsal. IT Toscana.
Genus Dicranolasma
Pedipalpus (Abb. 239): Tr ve mit 2 Bo, di1• rli davon auf hohem Hocke!, Fe schlank, ve-la mit 2+9 {meist 7 j-8) geraden und k;rii.ftigen Bo; Pt etwa 2 Drittel rler F!>-Länge, bei Cf nur unwesentlich dicker als bei :;?.
Laufbeine: Fe bis Mt nur bedornt, ohne·Papillen-Besatz; Fe bis Ti I, III und IV deutlich, aber nicht auffallend spindeiförmig verdickt; bei ~weniger stark ausgeprägt. - Pseudogelenke: Fe II mit wenigen hellen Ringen, Ti II ebenso, dort oft fehlend; Mt II mit 3-12 deutlichen, die di davon beweglich; Mt IV: 2-6 di. Calcaneus II mehr als doppelt so lang wie hn'it·: Honst kürzer als· breit. - Tarsalmiederung: I: Cf 6-8, meist 6 oder 7; ~ 5-·7, meist 6.- II: Cf Il-18, meist. 12-13; ~ 8-13, meist 9-11. -- IIJ: Cf 6-8, meist 7; '2 6-7, meiRt 7. -- IV: 3 6-8, meist 7; ~ 6-7, meist 7.
Dorsalseite: di Kappe vergleichsweise· breit und Ia winklig konvex, Augen vor der Mitte der Kappenäste gelegen: von. frontal ohne medio-ventra.d gebogmwn Zipfel. Ceph me gegen die Kappe ansteigend, aber in sich flach,· ohne Höcker, mit. 2 Längsgruppen z. T. langer Do hinter der Kappe; Area I ohne me Erhöhung, Areae I-IV mit lockereil Do-Querreihen, auf I-III para-me stark hervortretend.
Verbreitung (Abb. 245):' Typ: W-meiliterran. - Wahrscheinlich (nahezu) geschlossenes Areal in den küstennahen Gebieten des gesamten westl. Mittelmeergehiet.es einschließlich der Apenninen-Halbinsel. - Locus typicus: Meeralp•·•• hei Nizza.
Italien: Wahrscheinlich auf der ganzen Apenninen-Halbinsel mit Schwerpunkt in den .,.,.,tf. Gebirgsabdachungen {vgl. BRIGNOLI 1968, wiehlei), Ioim! auch bis in Küstenlagen; Piemonte, Borgo südl. Dalmazzo, Grotta di Tetto Rafel {BRIGNOLI 1968); ·wenig$!Pns in ·den italienischen Alpen reicht ein Arealfinger bis in die C!ott.ischen nnd Grajischen Alpen: norrlwest,l. Saluzzo, Bagnolo Piemonte nördl. Barge; Umgebung \'Oll GJa\'(~no nördl. Pinerolo; Va1 Ln~
oa.na bei Cuorgno westl. Ivrea, T. Soana (alle K. T. leg.); - Ins.,ln: ?vfalta {ZILCH leg.), Sizihr·" mit Aalischen und Agadisehen Inseln {MAROELLINO 1973a, wiehl•·i), Sardinien, Korsika (J. U. in litt .. ); Elba {J. M, leg.). - Frankreich: 0-;E'yrenäen, küstennahe Gebiet.e der Meeralpen und lokal in rlen Cevennen (J. G. in litt.). - Spanien: kiist.Pnnabe Gebiets rm Sund 0, Granarla-Katalonien, Areal wahrscheinlich erst zum kleinen Teil erfaßt. - N. A f r 1 k /'1.:
Algeden und Tnne,ien; die Nennungen sind nicht, an Hanrl der Belege überprüft, (J. U. '" litt.).
In Italien kommt soeren.seni sympa.tiisch mit. cl'istat?un THORELL, hil·tum L"· MAN, opiz.iouoides (L. Koch) und apuanum MAROELLINO vor·; mit c>·istaturn und a.puanu.rn Ü;t Syntopie belegt (,J. G. in litt.).
Ökologie: Biotopansprüche: meist in beschattetem, vor unmittelbarer ln'" Iation geschütztem Gelände, meist in Laubwäldern nnterschiedlich~'r Prägung, z. T. auch nur Gebüsche und ganz selten in offenem Gelände. Aufenthalt unter Steinen, in Geröllhalden, unter morschem Holz und in der Humusschicht des Bodens; besiedelt auch gern Höhlen und pflanzt sich dort wahrscheinlich auch fort. BRIGNOT.I (1968) nennt soerenseni troglophil. Höhlenpopulationen sollen geringfügig längere Beine und höhere ~arsalzahlen auf;reisen IMAROELLINO 1965).
Abb. :!5U --:!51. Du·ranolasma cJ'i8f.at?.tnt T:a:oRELL. - 250. Penis von dorsal (links) und latc·ral; :!5 L (}lau~ ponis von dorsal. rr Praton~nchio.
Ah!J. 252-25::1. Dicranolasma ]Utnper DAJIL. - 252. Penis von dorsal; 253. Glans rmlis \'Oll
dor;;;aJ. IT Gardas:~eP.

160 Familia Dioranolasmatidae
Vertikal ver breitung: von Tieflagen nahe Meereshöhe bis 800 m (Bronte am Ätna, BJotiGNOLI 1968), 700 m (Elba, J. M.leg.), 600-700 (Höhlen in Piemont), 650 m (Apuanische Alpen, MAROELLINO 1971), bis 1000 m (Korsika, DRESOO 1950); soerenseni steigt weniger hoch ins Gebirge und erscheint im öko1omsehen Praeferendum stärker mediterran geprägt als andere Arten mit ähnlicher Verbreitung, z. B. cristatum (HERBST) und apuanum MAROELLINO.
Phänologie: ad. und iuv. wahrscheinlich eurychron. - JUBERTmE (1964, scabrum) nennt 1-3 Eier je Gelege; Jugendstadien sin4 nicht vollständig bekannt, 6 lage!J. J. G. (in litt.) vor, doch treten wahrscheinlich wie bei scabrum 7 auf. Sex ratio: aus Sammlungen sind 51 Q'und 67 ~bekannt.
Dicranolasma cristatum THORELL, 1876 Abb. 241-24.2, 245, .250-251
1876 Dicranolasma cristatum THORELL, Ann. Mus. Genova, 8: 506. 1879 Dicranolasma cristatum - SIMON, Arachn. de France, 7: 295. 1903 Dic1·anolasma cristatum - DAHL, SB. Ges. naturforsch. F'reunde, Berlin: 290. 1923 Dim·anolasma cristatum - ROEWER, Weberknechte d. Erde: 642.
Habitus und Feldkennzeichen: Vergleichsweise kleine und kurzbeinige Art mit geringer Tar~enzahl (Ta II: 6) ; do Erdbekrustung oft in Fo11m eines deutlichen Höckers; nur in der nördl. Apenninen-Halbinsel. ·
Kennzeichnung: Körpermaße (in mm): L gesamt d' 4,4-4,5; ~ 4,0-4,6. Laufbein II (Q', in Klammern~): Fe 2,8 (2,7) Pt 0,8 (0,8) Ti 2,3 (2,.2) Mt 2,3 (.2,3)
Ta 1,1 (1,.2). Genitalmorphologie (Abb. 250-251): Truncus penis von der Basis distad
kontinuierlich verjüngt; Glans breit löffelförmig, deutlich breiter als das di Ende des Schaftes, stark do-ve abgeplattet, mit stark konvexen Seitenrändern asymmetrisch gegen den Stylus verjüngt; Stylus kurz schlauchförmig und (von do) nach links abgebogen. Glans mit etwa 60 meist basad gekrümmten stachelartigen Do, kleinere nahe der Basis des Stylus.
Cheliceren (Abb. 242): Q'-Grundglied do-cli mit breit-kegelförmigem, distad sanft ansteigendem und frontal steil abfallendem Apo-artigem Höcker, etwas 'mediad geneigt, frontal auf dem Höcker ein Haarbüschel, darunter Drüsenorgan. - ~Chelicere ohne Höcker, flach.
Pedipalpus (Abb. 241): Tr ve mit 3 kräftigen Bo, die di auf Sockel, Fe gedrungen mit meist 3-6·ve-la Bo; Pt kurz, etwa .2 Drittel Fe-Länge erreichend.
Laufbeine: Fe bis Ti I, III und IV deutlich verdickt, Fe I und IV mit DoKiel, bis zum Mt auch Kugel-Papillen; nur ausnahmsweise an Mt IV Pseudogelenke; Calcaneus II 'etwas länger als breit; an den übrigen Beinen kürzer. -Tarsal-Gliederung: bei d' und ~I: 5, II: 6, III: 6, IV: 6. ·
Dorsalseite: di Kappe in Aufsicht gerundet, Augen etwa in der Mitte der Kappenäste, Äste distad konvergierend ventrad gebogen, dadurch von frontal ventrad gerichteter Zipfel erkennbar. Ceph zur Kappe ansteigend, auf abd Area 1 me Höcker; Bewahrung des Souturn schütter, nur mit wenigen, auffallend langen Do. Ceph hinter der Kappe mit 2 Längsgruppen solcher; thorak Terg II mit schütterer Querreihe; Area I mit Gruppe divergierend stehender längerer Do (5-12), Areae II-IV mit para-me Gruppen aus je 1-31angen Do, auf Atea V am Hinterrand schräg abstehend in Querreihe unterschiedlich lange Do.
Gon11A Dicranolat=:~Jilt1 l6l
·VI."rbreitunp; (Abh. 245): Typ: l<Jnrlemit der nördl. Apenninen. - Lor·11s typicn;;: bei Florenz.
Iialien \rom nördl. Apennin (Tosco-Emilianischer Ap1,3nnin~ Ligl~risnher Apouni11 und Apnanischen Alpen) his zum S~Fuß der Meera.lpen; südlichster :B'undort bei FJorr:nz, nördlif·h ster hei BuAaHa (Ligurisoher Apennin), westlichster bei Finale (J. G. in litt.).
In <len Apoanisehen Alpen lebt cristatum sympatrisClh mit soerenseni~ lokalsyntop m1t mpua~ ·IIUn! )IAROELLINO {JioiARCELLINO 1971).
Ökologie: Biotopansprüche: D. cristaturn ist nur aus Wald oder waldähnlichen Vegetationsformen bekannt: Buchenwald (M.AROELLTNO 1971), lichter Buschwald (KIRCHNER & SALZER 1967), "castagni" (ZANGHERI 1966); Funde in Höhlen existieren nicht.
Vertikalverbreitung: Die Funde vonM.AROELLINo(l97l) in denAlpiApuani liegPn zwischen 600 und mindestens 1250 m; in diesen Bereich ordnen sich auch die 'renigen anderen bekannten Daten ein.
Phänologie: ad. wahrscheinlich eurychron; Sex ratio: nach vorliegendem l\1atpJial und Literaturangaben etwa 1 : 1.
Dicranolasma pauper DA.HL, 1903 Abh. 243-245, 25.2-253
lOfJ:l Dicrmwlasma pa,.per DAHL, SB. Ges. naturforsch. Freunde, Berlin {7): 290. 192:l Dicranolasrna pauper - RoEWER, Weberknechte rl. Erde: 642.
Habitus und Feldkennzeichen: kleine und kurzbeinige Art mit geringer 1';tr~enzahl (Ta II: 5); im nördl. Gardasee-Gebiet.
Kennzeichnung: Körpermaße (in mm): L gesamt d' 3,3-3,8; ~ 3,5-4,0. La.ufbein II (Q', in Klammern~): Fe 2,4 (2,2) Pt 0,7 (0,6) Ti .2,0 (1,8) Mt 2,0 (1,8)
Ta 1.0 (1,0). Genitalmorphologie (Abb. 252--253): Truncus penis distad kontinuierlich
n'rjüngt, Glans breit löffelförmig und nur wenig breiter als das di Ende des Truncus. rlie konvexen Seitenränder gegen den Stylus verjüngt, insgesamt weniger ausladend gerundet als bei cristatum THORELL, Stylus kurz schlauchförmig und (von do) nach links abgebogen.
Cheliceren (Abb. 244): Q'-Grundglied do-di mit breit-kegelförmigem, distarl geneigtf'm Apo-artigem Höcker, frontal steil abfallend; Höcker mit di Haarbüschel !Proportionen ganz ähnlich wie bei cristatum).
Pedipalpus (Abb. 243): Tr ve mit 2 steifen Bo, die di auf hohem Kegelhöcker; Fe gedrungen, mit 3+5 ve-la steifen Borsten; Pt beim d' ve-di schwach kom·ex; Ti et\m 1 Viertel länger als Pt; Ta schlank.
Laufbeine: Fe bis Ti I, III und IV deutlich keulig verdickt, bedmnt.. bis Mt übf'\rrl.ies mit Kugelpapillen besetzt, Mt ohne Pseudogelenkl}. Calcaneus von Bein II deutlich länger als breit, an den übrigen Beinen kürzer. -Tarsal-Gliederung: bei 0 und ~I: 5, ll: 5, III: 6, IV: 6; ·Ta II ausnahmsweise (bisher bei 2 c)) 6gliedrig.
Dorsalseite: ähnlich cristatu.rn; schütter bedornt, z. T. mit auffallend langen, wenigen Do; auf abd Area I kleiner Höcker mit 4-6 langen para-me Do, AreaP IT-IY mit 1-3, e beiL~O Hinterrand von Area V mit schräger Querreihe yor 11.llem pa.ra-me la.nger Do.
Beziehungen: D. paupe·r bildet mit cristat.um. und dem S-italienischen Mrtum Lmu~ eine Gruppe nahverwandter Arten, die sich in der Genitalmorphologie und den Proportionen der Q'-Ohei-Apo ähneln. Vermutlich steht paUJIM' mit cn·n Tit>I'Wf!lt Dentsch1. 64: l\furtt>US

162 Familia Trogulidae
statum in einem Sohwe&tergruppen-Verhältnis; die rezenten Areale beider Arten sind nur durch die Po-Ebene getrennt. · : ·
Verbreitung (Abb. 245): Typ: Ende~it des n?rd~. Gardasee-Gebretes und nahegelegener Gebirgsstöcke. - Locus typwus: Gehret rm NW des Gardasees.
Italien: Das bis jetzt bekannte Areal besteht aus einem kleinen Gebiet im NW und 0 des Gardasees: Mt. Tremalzo (SCHAWALLER leg.); Riva, Fonale-Schlucht. (VERH?EFF leg.), Val d'Ampola, Bezze~cca Mt. Vies, Val Conci, Corno, Val Scalglia; Val Gmchcart~, Ponte ~rche oberhalb Comano; ebendort bei Ragoli (alle K. T.leg.); Brenta-Gruppe, Val d Algone (SOHA· WALLER leg.); Lessinische Berge: Pasubio, östl. Pianodelle Fugazze (K. T.leg.).
Die geringe Arealgröße und die Konzentration ~~f Gebirgsstöcke nahe bekannten Massifs de Refuge (Brescianer Alpen und Lessuusche Berge). lassen das rez~nte Verbreitungsgebiet als wahrscheinlich glazialbedingtes Reliktareal ers~h~men (vgl. die Verhältnisse bei J!latybumts alpin~relictu~ und Holoscotolemon less~mense, die ebenfalls Reliktareale un Gardasee-Gebret besredeln). .
Ökologie: Biotopansprüche: bisher erst ungenüg~n~. bekannt; Materral de~ Sammlung VERHOEFF (Belege MNB) aus "feuchtem Gelande _am Abha;ng un~er Oliven", bei Riva in "Laubgebiiseh" der Ponale-Schlucht, Im Pasubro-~cbi~t 1
in lichtem Buchenwald mit Kalksteinblöcken, im nordwestl. Gard~see-0ebiet m Buchen- und Fichten-Buchen-Mischwald, in krautiger UfervegetatiOn ernes Ba-ches, meis~ unter Steinen; bisher keine Funde in Höhlen. . ..
Vertikalverbreitung: von Tieflagen um 100m (Gardasee) bis uber 1000 m (Pasubio-Gebiet: 1000 m; nordwestl. Gardasee: 700-900 m, dort vielleicht bis 1300 m; K. T.leg.).
Phänologie: ad. wahrsnheinlich eurychron.
Familia Trogulidae
1833 Trogulides SUNDEVALL, Consp. Ara"lm.: 35._ . 1872 'l'rogulidae - SrMON, Ann. Soc. ent. Frmwe, 2: 2H\l (part.). !"879 Trogulidae - SrMON, Araehn. de Franco, 7: 28!J (part.). 1923 Trogulidae - RoEWEit, Weborlmechte d. Erde: ß:J:l (part.).
Eine Familie der Palpatores-Troguloidea, gekennzeichnet durch: . . d'-Genitalmorphologie: Truncus penis do-ve abgeflacht, vergleiChsweise k~rz
und zart schlank bis untersetzt, Muskulatur in über der Hälfte des TruneuH, Ihn bisweile~ ganz ausfüllend; mit 2 kurzen zur. Basi~ der Glans z_iehenden Sehnen. Glans in gerader Verlängerung des Truncus, m meist asymmetrischen Stylus vor-längert; Bewahrung mit Sinnesborsten spärlich. . . . .
1
Ovipositor: ungegliedert kurz, nur wenig länger als breit, vor allen: di T~Ii _nut Querreihen dichtstehender Wt1rzen und ttufgesetzter kurzer Borste; d1e zweighedrige Furca wenig differenziert, bis auf wenige kurze. Borsten glatt.
Cheliceren klein, ohne DrüRenorgane; unter JTIIl-di Ceph-Kappe verborg~n. . . Pedipalpus: auffallend klein; bei Anelnsm.ocephal~ts Ta etwa so lang wie Tibm,
spärlich beborstet, immer ohne Kugelhaare. . . Met<ttarsen der Laufbeine mit Calcaneus; Laufbeme kurz, d. h. Bem II nur
wenig länger als der Körper. Zahl der Tarsenglieder meist gattungsspez!fisch. Die S~ernite II-VII zu flachem und starrem Scutum verschmolzen, mcht gegen
einander b~weg!ich; die ursprünglichen Gelenkhäute erkennbar, zusätzlich 1 Mediannaht.
1 Genus Trogulna
· Souturn me-di zu einer Kappe._aus 2 Fortsätzen des Tu oc verlängert, und mit langen :r<;egelpapillen besetzt;. Augen auf Hügel an der Basis der Kappenäste; z. T. unterschiedliche Ausprägung bei den einzelnen Gattungen, Integumflnt rauhwarzig und drüsig, mineralinkrustiert.
_Verb~eitung: W-paläarktisch, von den Kaukasus-Ländern über ganz Europa (mcht m den kaltgemäßigten nördl. Gebieten, nördl. bis Schonen), das Mittelmeergebiet mit N-Mrika, östl. bis N-Iran (Elburs); mit größter Formvielfalt im östl. und zentralen Mittelmeergebiet: Trogulus LATREILLE (ganze W-Paläarktis), Anelasmocephalus SIMON (atlanto-mediterran), Galathocratus SIMON mit Trogulocrat~s .R?EWER (zentrales und östl. Mittelmeergebiet, Vorder-Asienund N-Afrika), Kojmwt~s RoEWER (Griechenland und Kreta), Anarthrotarsus SILHA.VY (Kreta, Korfu), Platybessobius RoEWER (Kaukasus und Kreta).
Weitere Gattungen, die meist zu den Trogulidae gerechnet werden - DendrolaBma BANKS (N-Amerika und J"apan), Ortholasma BANKB (N-Amerika), OeratolaBma GooDNIGRT & GoonNIGRT (N-Amerika) und DicranolaBma SeERENSEN (W-Paläarktis) - gehören zu anderen Familien: NemaHtomatidae: Dend•·olasma und Ortholasma; Ischyropsalididae: Oe-ratolasma; ])iera.nolasmatidae: Dicranolasma.
Bestimmungstabelle für die Gattungen
I Körper extrem do-ve abgeflacht; große Arten über 5 mm (selten ab 4,5 mm), Giiederung der Laufbein-Tarsen I-IV immer 2-2-3-3 . . . . . . . . . • . Trogulus, S. 163 Körper bolzenförmig, nicht extrem do-ve abgeflacht; kleine Arten bis 4 mm (in den SWAlpen selten bis 5,4 mm), Gliederung der Laufbein-Tarsen I- IV (im hier behandelten Gebiet.) 2-3-3-3, 3-3-4-4 oder 4-4-4-4 . . . • • . . . . Anelasmocephalus, S. 175
Genus Trogulus LATREILLE, 1802
1802 Trogulus LATREILLE, Rist. Natur. Crust. Ins., 3: 61. 1879 Trogulus - SrMoN, Arachn. de France, 7: 302. 1879 Metopoctea SIMON; Arachn. de France, "/: 309. 1923 Trogulus - RoEWER, Weberknechte d. Erde: 635.
Nominat-Gattung der Trogulidae, gekennzeichnet durch: d'-Genitalmorphologie: Truncus penis do-ve abgeplattet, Glans in gerader Ver
längerung des Truncus, Stylus dorsad abgewinkelt oder in Verlängerung von Truncus und Glans. - Körper langgestreckt oval, do-ve a,uffallend brettartig abgeflacht; Ceph-Frontalrand zu einer runden Kappe aus 2 halbkreisförmigen .Ästen verlängert, an deren Basis der niedrige Tu oc, Augen klein. Sterrute zu einheitlichem und gut kenntlichem Ventralsouturn verschmolzen. - Pedipalpus kurz und gedrungen, Sinnesborsten vor allem auf Ti und Ta. Gliederung der Laufbein-Ta I-IV regelmäßig 2-2-3-3. - Ohne sekundäre Geschlechtsmerkmale.
Verbreitung: etwa 20 nominelle Arten in Europa, Vorder-Asien (Türkei, LevanteBecken, Kaukasus), N-Mrika (Algerien bis .Ägypten). Im hier behandelten Gebiet kommen 4 Arten vor.
Typmi-Art (Festlegung durch THORELL 1876): Acarus nepaiformis ScoPOLI, 1763.
Bestimmungstabelle für die Arten ( Größenangaben für Dorsalsouturn einschließlich Kappe, für.;! und 1'l 1 kleine Art (4,6-5,8 mm), Endglied des Ta. li wesentlich länger (1,5-2,2mal) als das
Grundglied. . . . . . . . . . . . . . . . . . . . . . . . . tricarinatus, S. 164

164 Familia Trogulidae
größere Art (über 7 mm), Endglied des Ta II nicht wesentlich länger (höchstens 1,5mal) ·oder ebenso lang wie das Grundglied . . . . . . . . . . . . . . , , . . . . . . 2
2 Glieder des Ta II genau oder nahezu gl~ich lang, große Art (8,0_:ll,O mm); 0-Alpen u~cl N-Jugoslawien . . . . . . . . . . . . . . . . . . . . . . . . tingiformis, S. 172 Endglied des Ta II länger als das Grundglied . . . . . . . . . . . . . . . • . 3
3 mittelgroße Art (6,1-8,3 mm), me Rückenkiel schwach entwickelt oder fehlend; c!: Stylus gegen Glans gewinkelt; weitverbreitet in Mitteleuropa . . . . . • . nepaeformis, S. 168 große Art (9,0.:_13,1 mm), me Rückenkiel markant; (S: Stylus in gerader Verlängerung der Glans; im l>ehandelten Gebiet nur in den SW-Aipen und in Istrien ..... , •. , ..
Trogulus .tricarinatus (LINNAEUS, 1767) Abb. 263-267, 273-274
1767 Phalangium t1·icarinatum LrNNAEus, Syst. Naturae, 12. ed.: 1029. 1879 TI'Dgulus asperatus - SrMON, Araclm. de France, 7: 304. 1923 T1·ogulus tricarinatus - ROEWER, Weberknechte d. Erde: 604.
coriziformis, S, 1 7 4
1971 T1·ogulus oltenicus AVItAM, Trav. Iust. Speol. Emile Racovitza, 10: 242.
Begründung der Synonymie: T. olteniws muß nach den Abbildungen des Autors ((S-Genitalmorphologie, Ta-Proportion des Laufbein II) tricm·inatus zugerechnet werden; auch . die Körperlänge Ijegt im Variationsbereich von tricarinatus. ·
Habitus und Feldkennzeichen: kleinste Tl"ogulus-Art im Gebiet, bis 6 mm lang; Ta des Laufbein II: Endglied fast doppelt so lang wie Grundglied (gegen hellen Hintergrund betrachten); weitverbreitet in Mitteleuropa.
Kennzeichnung: Körpermaße (in mm): L ({ 4,6-6,5 (Griechenland: 4,4-4,9); L ~ 5,3-5,8 (selten 4,7).
Laufbein II (({,in Klammern~): Fe 2,0 (2,0) Pt 0,8 (0,8) Ti 1,1 (1,0) Mt 1,4 (1,4) Ta 1,2 (1,0). Längenverhältnis von Grundglied zu Endglied des Laufbeines II: 1: 1,55-2,25, d. h. Endglied anderthalb bis mehr als doppelt so lang wie das Grundglied (Abb. 263-264).
Genitalmorphologie (Abb. 273-274): Truncus penis bandartig, vergleichsweise schlank, im ba Drittel schwach verengt. Glans groß, von la auffallend schmal, mit langem, vom Glans-Körper nicht abgesetztem und nur wenig nach links dorsad gebogenem Stylus, Iinks-la (von do) mit häutiger Membran; nur wenig variabel.
Laufbeine: robust, Bein II länger als der Körper, rund, Fe I mit kräftigem do Kiel, auf den übrigen Fe ein solcher nur angedeutet, alle mit je 1 ba Pseudogelenk. Proportion der Tarsen von Bein II vergleichsweise variabel (siehe unter "Keimzeichnung").
Pedipalpus (Abb. 26ß): ohne spezifische Besonderheiten. Dorsalseite: Zeichnung und Färbung: einheitlich erdbraun. - Bewahrung:
me Kiel auf den abd Arcae nur angedeutet; ohne Ia Wulst; T~J oc niedrig, an der Basis der Kappe. .
Variabilität: Proportionen der Ta von Laufbein II vergleichsweise variabel (Abb. 263-264), aber regional einheitlich; die Länge beider Glieder differiert nach jetziger Kenntnis am meisten(= Index-Werteam höchsten, über 2) in slowenischen Populationen, hei allen untersuchten Populationen des engeren Mittaleuropa nördl. der Alpen liegt er etwa zwischen 1,6 und 1,8. N-griechische tt·icm·inatus (Chalkidike) sind auffallend schmal und klein: 5 ({von 4,4-4,9 mm; es ist fraglich, ob sie tatsächlich zu dieser Art gehören. Größenunterschiede zwischen if und ~ gering entwiclmlt.
. I
~~'( ..... -t·r ' ~~------y---- --- . --. r-
- - . _ ____.J---___ ----------------251;
255 •. ~"~ ~ •
·\·~-~ ~"" ----- ____..L___. ---.. __ j~
~;~ ~- - ----r-- - --- ·-----.,1"'
··~ ---~~-- ---·-----
256
257
\·
/
258 ,.;.-,
"':~·~_-:=--._]:------ --) 259 . ''>··-- -·.. ·-----·
260' .: ~~-: y _______ ~~~ r,,. :~--- .. ----y--··~
. '~----~ ----· .'
261
1:.;.7:: ~:·;- --~-- --------__) 't. _____ f .. __ _
Genus Tt·ognJus Hii)
Abb. 254- ~5.1. '.P1·ogulus coriz2jm·mis C. L. KocH. -- Laufbein TI, 'l,arsalghf d(:.t· .. ~54. '2, ~ ).:'}, rJ ·-- V R· Meera.lpen.
Abh 256-257. Trogulus tingiformis C. L. KooH. - Laufbein II, Tarsalglieder .. ~iif;. ,;, ,-\[),-.,
Villaober Alpe; 257. c!, IT Garchtsee.
Abb. 258-262. Trogidus nepaeformis (ScoPoLr). - 258-2(11. Laufbein II, Tarsalglieder, 258. ;·, FR Cerdon; 259. ~. IT Dolomiten, Sei"; 260. <f, BRD St.uttgart; 261. (S, FR 11nntelimar; ~6~. :)'-0belicPren-Grundglied von lateral.
-'.hh. 263-266, Tmg11lus tricar.:11atuk ILnu;:E). - 263-264. Laufbein II, TarsalgJieckr. 2fi>l ~ ~TS Karawanlu:n, Boclental; 264. -~, YTJ Slowenien, Triglav; 265. (S-ChelioPren-Grundgl wd \'On latera-l: 266. t Peclipalpus von lateral. - 'l\Jaßstab (nur für Tarsen): 1 mn'
-

166 Familia Trogulidae
T. lingiformis
Abb. 267. Gesamtareale von T1·ogulus tricarinatus (LrNNE) und T. tingiformis C. L. KocH.
Ve~br~itung (Ab.b. 267): Typ: europäisch (-subatlantisch bis kontinental). _ T. tncannatus besiedelt neben T. nepaejormis das größte geschlossene Areal aller Trogulus-Arten. - Locus typicus: Dresden.
Spa.nien: möglicherwei~e spärlich in den Kautabrisehen Gebirgen (Galicien, Prov. Lugo; KRAUS 1959) .. - Frankr?Ich: Fundorte in den Departements Villers-sur-Mer, Seine-et-Oise und Aube (Sr>roN 1879), mit Ausnahme der Mediterran-Gebiete aber wahrscheinlich viel weiter verbreitet. - Britische Inseln: nur in den Grafschaften an oder nahe der S-Küste: CornwaU, Su~seJO, Kent, Dorset, l3erkshire, Hampshire (B:arsTOWE 1949, SANKEY & SAVORY 1974). --: Belgien: Luxembourg, Redu (BEOKER 1896). - Niederlande: nur in den süd); Teilen Limburgs, dort auf Kalkuntergrund (SPOEK 1963). - BRD und DDR: weitverbreitet einschließlich der nördlichen Tiefebenen, lokal spärlich oder auch ganz fehlen,d (vgl. LE Ror i913 KR":EPELIN 1896). - ~änemark: l.?kal im S des Landes: Kolding Fjord, Vejle Fjord, auf Aero und La~geland, sudl. von Paesto auf 0- und W-1\Iöen und aufBornholm; aUeOrte liegen a~ der 0-Kuste bzw. auf Inseln (MEINERTZ l964b). - Schweden: im östl. Schonen bei VIttaby, Ryadal und MeUb!. (LOHMA,:;rDER 1950). - Polen: hauptsächlich in den Karpaten und Sudeten, entlang der großeren Flusse und erreicht bei Gdansk (Danzig) die Ostsee (LE Ror
I
i I
Genus Trogulna 167
1913, RAFALSKI 1961, STAR!i!GA 1966 und in litt.). - ÖSSR: gleichmäßig und weitverbreitet (EirLHAVY 1956). -Rumänien: vereinzelt Funde in den 0- und S-Karpaten unu in den Banater Bergen (AVRAM 1971). - Bulgarien: spärlich, aber in aUenLandesteilen (W. S. in litt. - Griechenland: Chalkidike, Cholomon (A. A., J. M. leg.); - südöstl. bis nach Türkisch'l'hralden (GRUBER 1969).
Alpenländer: oft nur lokal, und in besonderem Maße von Kalkgestein abhängig. -Schweiz: alte und bisher nicht bestätigte Funde aus den Kantonen Genf, Vaud, Wallis und Basel {LESSERT 1917). - Österreich: in allen Hanptlandschaften, spärlich in den Zentralalpen (J. G., A. A. in litt.). - Jugoslawien: im Anschluß an die alpinen Arealteile (z. B. 'l'riglav, HAnZl: 1931) weitverbreitet, südl. wenigstens bis Montenegro (z. B. Moraca-Tal, A. A.leg.). - Italien: keine gesicherten Daten aus den Alpen; für die Apenninen-Halbinsel nennt MAROELLINO {l970a) Funde in den Apuanischen Alpen; Glans-Abbildungen erlauben indes kaum zweifelsfreie Zuordnung. - N -Amerika: USA, eingeschleppt nach Rochester, New York (MuCHMORE 1963).
Ökologie: Biotopansprüche: T. tricarinatus besiedelt vorzugsweise Waldformationen ganz unterschiedlicher Zusammensetzung, von bodenfeuchten Auwaldgesellschaften über mesophile Wälder bis zu vergleichsweise trockenen Wäldern auf Sandboden (z. B. Kiefernwälder bei Mainz, HöREGOTT 1963, J. M. leg.), ebenso in trockenen Gebüsohen (S-exponierte Hänge im Kaiserstuhl, J. M.leg.) und sogar auf Trockenrasen (Eichberg, S-Schwarzwald; HELVERSEN & MARTENS 1971). In den Alpen in Auwäldern und bodenfeuchten Laubmischwäldern, gelegentIich in baumlosem Gelände (A. A. in litt.); häufig in den Donau-Auen der Umgebung Wien.s an Stellen, die nicht (oder nur wenig) überschwemmungsgefährdet sind.
Gegenüber den mitteleuropäischen Arten nepaejormis und tingijormis erscheint tr-icarinatus eurytoper; er lebt auch in den Präferenda dieser Arten, überschreitet sie aber in deren Areal sowohl zur trockeneren wie zur (wechsel-) feuchten Seite.
Auch bei tricarinatus besteht über die Nahrungstiere Bindung an Kalkböden, die besonders im Bereich der Arealgrenzen deutlich wird. So wird für England ausdrücklich die Vorliebe für Buchenwälder auf Kalkuntergrund betont (BRISTOWE 1949, SANKEY & SAVORY 1974), und auch in Mitteleuropa ist die Siedlungsdichte auf nicht zu feuchten Kalkböden besonders hoch. Auf solchem Untergrund sind die Nahrungstiere, vor allem Gehäuseschnecken, zahlreich. In ebenfalls molluskenreichen, aber viel feuchteren Auwaldbiotopen lebt tricarinatus meist seltener. -In Mitteleuropa Vorkommen oft syntop mit nepaejormis, in den Alpen und im außeralpinen Slowenien zusätzlich mit tingiformis.
Vertikal ver breitung: Von nahe Meereshöhe bis in die Collin- und MontanStufe der Alpen gleichmäßig verbreitet; die Verhältnisse in den höheren Lagen der Alpen weitgehend ungeklärt: in Salzburg Funde bisher nur bis 1200 m (A. A. in litt.); nach FRANZ & GUNHOLD (1954) in den Haller MauernfAdmaut in 1800 bis 1900 m; im nieder-österreichischen Ötscher-Gebiet bis an die Waldgrenze ("Krummholzzone", Höhe unbekannt), bei Lunz bis 1200 m (REsSL leg.). Am Muttler/ Schweiz, Graubünden angeblich sogar bis 2000 m (1; LESSERT 1917); in Bulgarien von nahe Meereshöhe bis 1500 m (W. S. in litt.).
Phänologie: Ad. und iuv. eurychron; ad. mehrjährig. - In Gefangenschaft wurden folgende Daten ermittelt (PAJ!ST 1953): pro Jahr 6-12 Gelege aus 1-8 Eiern (meist 2-5), im Durchschnitt 3,0; Eier 1,1-1,5 mm lang, 0,4-0,8 mm breit. Eiablage während der ganzen Aktivitätsperiode, mit Gipfeln in IV/V und IX. Ablage der Eier wie bei nepaeformis in leere Schneckengehäuse, mit derbem Gespinst nach außen isoliert, - Entwicklungsdauer abhängig von der jahreszeitlichen Verteilung der Eiablage: 42 bis (bei Überwinterung) 207 Tage, Postem-
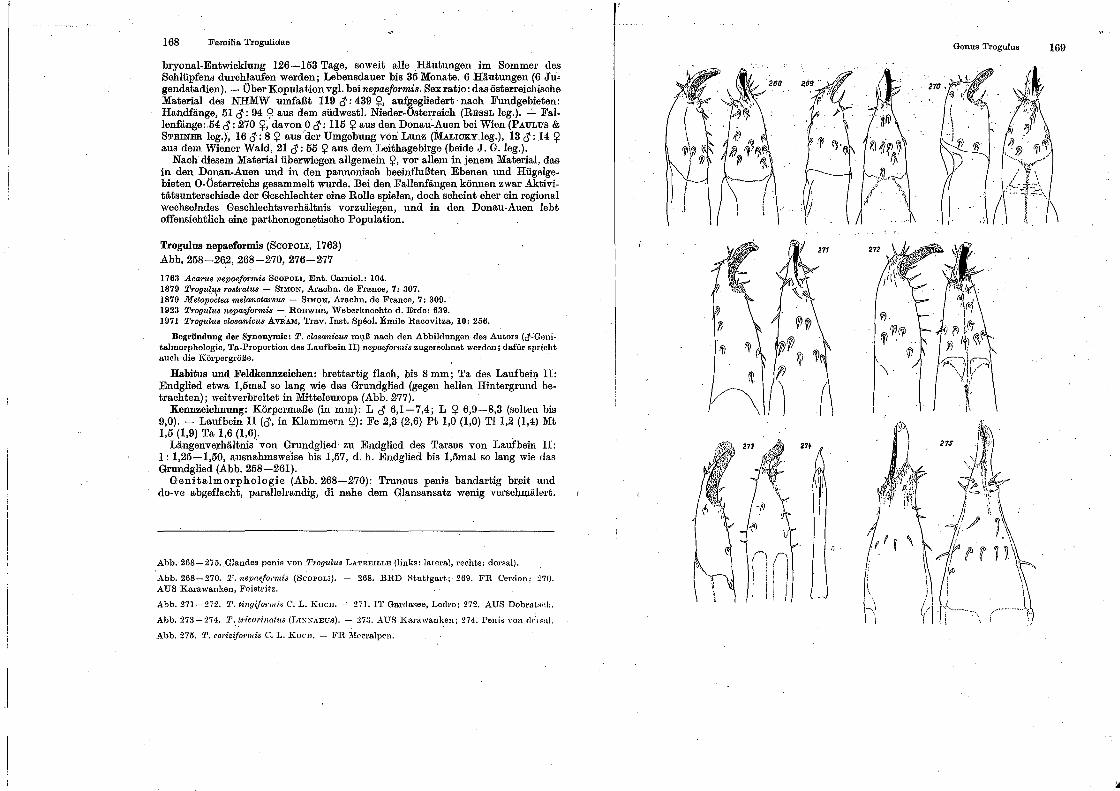
168 Familia Trogulidae
bryonal,Entwicidung 126-153 Tage, soweit alle Häutungen .im Sommer des Schlüpfens durchlaufen werden; Lebensdauer bis 35 Monate. 6 Häutungen (6 Ju• gendstadien). - Über Kopulation vgl. bei nepaejormis. Sex ratio: das Österreichische Material des NHMW umfaßt 119 Cf: 439 ~. aufgegliedert· nach Fundgebieten: Handfänge, 51 Cf: 94 ~ aus dem südwestl. Nieder-Osterreich (R:EssL leg.) . ...:. Fallenfänge:.54 Cf: 270 ~.davon 0 Cf: ll5 ~aus den Donau-Auen bei Wien (PAuLus & STEINER leg.), 16 Cf: 8 ~ aus der Umgebung vöri Lunz (MALIOKY.leg.), 13 Cf: 14 ~ aus dem Wiener Wald, 21 Cf: 55 ~ aus dem Leithagebirge (beide J. G. leg.).
Nach diesem Material überwiegen allgemein~. vor allem in jenem Material, das in den Donau-Auen und in den pannonisch beeinflußten Ebenen und Hügelgebieten 0-Österreichs gesammelt wurde. Bei den Fallenfängen können zwar Aktivitätsunterschiede der Geschlechter eine Rolle spielen, doch scheint eher ein regional wechselndes Geschlechtsverhältnis vorzuliegen, und in den Donau-Auen lebt offensichtlich eine parthenogenetische Population.
Trogulus nepaeformis (SooPOLI, 1763) Abb. 258-26~, 268-270, 276-277
I 763 Acarus nepaeformis SooPOLI, Ent. Carniol.: 104. 1879 Trogulu.,• rostratus - SIMON, Arachn. de France, "1: 307. 1879 Metopoctea melanotm·sus - SIMON, Arachn. de France, 7: 309. 1923 Trogulus nepaeformis - ROEWER, Weberknechte d. Erde: 639. 1971 Trog1tlus closanicus AvRAM, Trav. Inst. Speo!. Emile Racovitza, 10: 256.
Begründung der Synonymie: T. closanicus m~ß nach den Abbildungen des Autors (d';Genitalmorphologie, Ta-Proportion des Laufbein li) nepaeformis zugerechnet werden; dafür sprieht auch die Körpergröße.
Habitus und Feldkennzeichen: brettartig flach, bis 8 mm; Ta des Laufbein II: Endglied etwa 1,5mal so lang wie das Grundglied (gegen hellen Hintergrund bet.rachten); weitverbreite.t in Mitteleuropa (Abb. 277).
Kennzeichnung: Körpermaße (in mm): L Cf 6,1-7,4; L ~ 6,9-8,3 (selten bis 9,0). - Laufbein II {Cf, in Klammern ~): Fe 2,3 (2,6) Pt 1,0 {1,0) Ti 1,2 (1;t) Mt 1,5 {1,9) Ta 1,6 (1,6). .
Längenverhältnis ·von Grundglied zu Endglied des Tarsus von Laufbein ll: 1: 1,25-1,50, ausnahmsweise bis 1,57, d. h. Endglied bis 1,5mal so lang wie das Grundglied (Abb. 258-261).
Genitalmorphologie (Abb. 268-270): Truncus penis bandartig breit und do-ve abgeflacht, parallelrandig, di nahe dem Glansansatz wenig verschmälert.
Abb. 268-275. Glandespenis von T1·ogulus LATREILLE (links: lateral, rechts: dor•al) ..
Abb. 268-270. T. nepneformis (SCOPOLI), - 268. BRD Stuttgart.;· 269. FR Cerdon; 270. AUS Karawanken, Feist.ritz.
Abb. 271-272. T. tir~gifo•·mis C'. L. KocH. --'- 271. IT Gar<la,ee, Ledro; 272. AUS Dobrat.sd1.
Abb. 273-274. T .. tricarinatus (LIXXAEl.'S). - 27:l. AUS Karawanken; 274. Penis \'On dn''"''l.
Abb. 275. T. col'iziformls C. L. KocH. - FR )[l'eralpl'n.
Genus Trogulus 169

170 Familia Trogulidae
Glans variabel: von do schlank bis bauchig zum Stylus verschmälert; Stylus (von la betrachten) in kontinuierlicher Verlängerung der Glans wenig bis stark dorsad gekrümmt oder deutlich abgewinkelt (Abb. 268 bzw. 270). Stylus Ia (Glans von do betrachten) unterschiedlich ausgeschwungen.
Cheliceren (Abb. 262): alle Glieder schlank, ohne spezifisch Besonderheiten. Pedipalpus: klein, die einzelnen Glieder wenig differenziert, spärliche Bebor-
stung vor allem an Ti und Ta. · Laufbeine: robust, 2. Paar länger als der Körper, alle Glieder rund, Fe I mit
kräftigem do Kiel aus verklebten und mit Erdpartikeln durchsetzten Tubenhaaren; dieser Kiel auf Fe !I-IV ebenfalls aber viel schwächer vorhanden. Glieder der Ta II siehe oben.
Dorsalseite (Abb. 277): Zeichnung und Färbung: zeichnungslos erdbraun, mit feinen Erdpartikeln inkrustiert. - Bewahrung: niedriger me Kiel, auf dem Ceph undeutlich distad gegabelt, Ia Rand des abd Souturn schwach wulstartig gewölbt, die einzelnen Areae durch niedrige Kiele kenntlich.
Variabilität: Sie betrifft vor allem das Längenverhältnis der Tarsalglieder von ( Laufbein II (siehe "Kennzeichnung", Abb. 258-261) und die Gestalt der Glans penis. Als weitverbreitet und für nepaeformis allgemein zutreffend muß die Proporti~>n der Glans des d' von Stuttgart (Abb. 268) gelten; 1 Ö' aus denKarawanken (Abb! 270) weicht davon besonders stark ab; es lebte in einer Population "normaler" nepaeformis. - Vgl. Bemerkungen bei der Gattungsdiagnose.
Verbreitung (Abb. 276): Typ: europäisch (atlantisch bis kontinental). - Vergleichsweise großes Areal in vielen Teilen des überwiegend atlantisch beeinflußten W- und MitteJ-Europa und der mehr kontinentalen Klimabereiche SO-Europas;· extrem atlantische Gebiete (z. B. Britische Inseln) werden ebenso gemieden wie die besonders kontinentalen 0-Europas, Arealschwerpunkt im gemäßigten Mitteleuropa. - Locus typicus: Herzogtum Krain, entspricht etwa dem heutigen Slowenien.
Iberische Halbinsel: Portugal: Monte Pedral bei Porto, Leiria Pombal, Sorra de Sico (RAMBLA 1967). - Spanien: Kantabrische Gebirge, Baskische Provinzen einschließlich Logronjo (KRAUS 1959, 1961; ferner J. M.leg.). - Frankreich: ih den gesamten Pyrenäen (zahlreiche Lokalitäten; J. M. leg.); ferner wahrscheinlich in den meisten übrigen Landesteilen mit Ausnahme des mediterranen Küstenstreifens, wohl auch in Teilen der Atlantik-Küste fehlend (zahlreiche Fundorte bei SrMON 1879).
Engeres Mitteleuropa: kontinuierlich verbreitet in beiden deutschen Staaten mit Ausnahme der nördl. Tiefebenen. - Polen: nur in den südöstl. Landesteilen an der Grenze zur ÖSSR; Kreise Gorlice, Sanok, Przemysl, Ustrzyki Dolne (RAFALSKI 1961, STARljlGA 1966, I 1976), bisher nicht östl. der Oder. - ÖSSR: in Mähren und in der.Slowakei verbreitet, Nachweise aus Böhmen scheinen zu fehlen (EliLHAVY 1956). Nordöstl. Arealgrenze somit offensichtlich in der östl. DDR, am N-Fuß der polni•chen Beskiden und in den N-Karpaten.
SO-Europa und Balkan-Länder: Ungarn: bisher nicht sicher belegt (SZALAY 1968). -Rumänien: erst wenige Funde (AVRAM 1971, z.T. closanicus; WErss 1975);- ebenso in Bulgarien (W. S. in litt.). - Jugoslawien: wahrscheinlich überall, doch in Küstennähe wohl nur in waldbestandenen Gebirgen (z. B. Berg Uöka bei Rijeka, J. M.leg.); südl. nachgewiesen bis Bosnien-Herzegowina (z. B. Gacko und Foöa, A. A. leg.). - UdSSR: nördl. Küste des Schwarzen Meeres, Soci (STARljlGA 1966); dieser Fundort wahrscheinlich nicht isoliert. -
Alpenländer: kontinuierliche Verbreitung, auch in den südl. italienischen Alpen. -Apenninen-Halbinsel: wohl nur in Gebirgen. südl. bis zum Mateso-Massiv (MARCELLINO 1971).
Genus Trogulus 171
Abli. 276. Gesamtareal von Trogulus nepaeformis (SooPOLI).
Ökologie: Biotopansprüche: T. nepaeformis lebt in der Humusschicht und in der Bodenauflage, unter gefallenem Holz, soweit es Kontakt mit dem Untergrund hat, und unter Steinen. Da gleichmäßige und hohe Luftfeuchtigkeit gefordert wird (Grenzwerte unbekannt), ist der Biotop fast immer beschattet, meist durch verschiedene Waldtypen, bisweilen nur von Gebüsch, z. B. Ainus an der Waldgrenze; nur gelegentlich in unbeschattetem Ruderalgelände, dann unter isolierenden dicken Schutt- und Gesteinspackungen. In den Alpen (Salzburg, A. A. in litt.) in Auwäldern, in bodenfeuchten Mischwäldern und in deren Mantelgesellschaften (Corylus), an der oberen Verbreitungsgrenze auch in lichten Beständen von Larix und Ainus und selbst in Lägerflora.
Gegenüber tricarinatus bevorzugt nepaeformis (in Mitteleuropa) mehr Biotope der Montanstufe mit ausgeglichenem, feucht kühlem Mikroklima, und dort tritt er häufiger auf als tricarinatus. T. nepaeformis fehlt z. B. in den niedrig gelegenen und überschwemmungsgefährdeten Donau-Auen bei Wien (J. G. in litt.).
Vertikal ver brei tung: In Mitteleuropa außerhalb der Alpen in allen Höhenstufen, doch bisher offensichtlich nicht aus dem unmittelbaren Küstenbereich vorliegend. Im nordöstl. Österreich in der Ebenen-Stufe spärlich, nicht in den Donau-Auen; Nachweise aus dem Wiener Becken, Laxenburg-Schloßpark, 175m {J. G. leg.); 0-Rand des Leithagebirges um 150m: Schützeuer Tiergarten bei Eisenstadt, Martal-Wald bei Neusiedl (J. G.leg.); im Hügelland ab 200 m dann überall regelmäßig. In den Alpen in der Collin-, Montan- und Subalpin-Stufe; in Salzburg die höchsten Funde bei 1450, 1580 und 1760 m (A. A. in litt.), in Nieder-Österreich

172 Familia Trogulidae
bis 1500 m (Hochkar, RESSL leg.); in den Berchtesgadener Alpen bis 1800 m (HARMS leg.); im Tessin bis 1500 m (LESSERT 1917), im Veltlin bis 2000 m (Val di Rezzalo, J. M. leg.). An den südl. Arealgrenzen vorzugsweise in oberen Gebirgslagen: Apenirinen-Halbinsel, Matese-Massiv, 1700 und 2000 m, Gran Sasso, 1500 m (MAROELLINO 1971); Bulgarien: von Tieflagen um 100m bis 1800 man der Waldgrenze (W. S. leg.); Spanien: Picos de Europa bis mind. 1300 m; Frankreich: Ariege, bis 1400 m (beide J. M.leg.).
Biologie: in der lokalen Siedlungsdichte besteht deutliche Abhängigkeit von kalkführenden Bodenschichten; auf diesen erreicht nepaejormis höchsten Individuenbestand; er beruht wahrscheinlich auf den dort zahlreich vorkommenden Mollusken, die als Nahrungstiere dienen (PABST 1953). Er fällt Schnecken bis etwa eigener Körpergröße an; dabei hält er mit den Vorderbeinen das Gehäuse, schneidet mit den Cheliceren Teile aus dem Schneckenkörper und schiebt sich in die vorderen Umgänge des Gehäuses, wenn sich die Schnecke in die hinteren Gewfudeumgänge zurückzieht. Das Gehäuse selbst wird nicht aufgebrochen. SILI!A.VY (1957) stellte lediglich die Aufnahme von Schneckenschleim fest. In ausgefressene 1 Gehäuse werden die Eier abgelegt und nach außen mit einem luftdurchlässigen, festen Gespinst abgeschlossen (PABST 1953). Kopulationsverhalten vgl. Kap. 1.3., s. 44. Phä~ologie: ad. und iuv. eurychron; ad. mehrjährig. - In Gefangenschaft
wurde ermittelt (PABST 1953): Gelegegröße schwankend zwischen 1 und 8 Eiern (meist 2-4), im Mitte laus 862 Gelegen 3,3; 1,1-1,5 mm lang, 0,4-0,8 mm breit, Eiablage während der ganzen Aktivitätsperiode,. (lie durch die Temperatur-Verhältnisse bestimmt wird. - Entwicklungsdauer abhängig von der jahreszeitlichen Verteilung der Eiablage: 44 bis (bei Überwinterung) 226 Tage; im Frühjahr abgelegte Eier entwickeln sich in 11/. Monaten, im Herbst abgelegte benötigen gegen 9 Monate; davon abhängig die Postembryonal-Entwicklung in 3-9 Monaten. Sex ratio: österreichisches Material des NHMW 395 (j': 244 ~; aufgegliedert nach der Fangmethode: Handfänge 95 Ö': 68 ~(davon 33 (j': 11 ~aus dem südwestl; NiederÖsterreich, RESSL leg.); - Fallenfänge 300 (j': 176 ~ (davon 34 (j': 16 ~ aus der Umgebung von Lunz, MALIOKY leg.). Bei nepaeformis überwiegen (im Gegensatz zu tricarinatus) die (j'.
Trogulus tingiformis C; L. KocH, 1848 Abb. 256-257, 267,.271~272
1848 Trogu!us tingiformis C. L. KooH, in HAHN & KocH, Die Arachniden, 16: iO. 1923 T•·ogu!us tingijormis - RoEWER, ·Weberknechte d. Erde: 636. 1971 Trogu!us grubari AvRAM, Trav. Inst. Speol. Emile Racovitza,.IO: 259.
Begründung der Synonymie: T. grubari AVRAM ist, nach den Abbildungen des Autors !e[eht kenntlich, auf tingijorm.is zu beziehen. Die Abbildung der Glans penis von tingijomds in· tliLHAVY ( 1968 c; Steiermark, SMF 6206), auf die sich AvRAM zum Vergleich stützt, ist irrefühmnd, da sie ein verletztes oder mißgebildetes Organ darstellt .. Der Stylus der Glans Yon .tingifonn•:., bricht leicht ab; dabei entstehen Verhältnisse, wie sie iliLHAVY (I. c.) abbildet.
Habitus und Feldkennzeichen: brettartig flach; große Art bis über 10 mm; kräftiger me Kiel do; Ta des Laufbein II: Endglied und Grundglied etwa, gleichlang (vor hellem Hintergrund betrachten). -- In den 0-Alpen.
Kennzeichnung: Körpermaße (in mm): L d' 8,0-9,7; L ~ 9,7-11,1. · Laufbein II ((j', in Klammern<;?): Fe 3,3 (4,3) Pt 1,3 (1,5) Ti 1,8 (2,3) :m 2,3(:t0)
Ta 2,1 (2,1).
Genus Trogulna 173
l,ängenverhältnis von Grundglied zu Endglied des Tarsus von Laufbein II: 1: ~,93;--1,10; beide Gli~der also nahezu gleich lang (Abb. 256-257). . .•
Gemtalmorpholog1e (Abb .. 271-272): Truncus penisvergleichsweise schlank dis~ad ~ontinuierlic~ verjüngt; Glans nur undeutlich abgesetzt, Stylus lang, unter~ schiedhch stark - Im Extremfall nahezu rechtwinklig - dorsad gegen die Glans abgebogen; Glans kräftig bedornt, vor allem do.
Pedipalpus und Cheliceren: ohne spezifische Eigenschaften. · Laufbeine: robust; Paar II länger als der Körper; alle Glieder rund, Fe I mit
kräftigem do Kiel aus verklebten Tubenhaaren, auf den übrigen Fe Kiel kaum angedeutet. Glieder des Ta II auffallend lang, Proportionen vgl. unter "Kennzeich-nung". .
Dorsalseite: Zeichnung und Färbung: einfarbig dunkelbraun bis schwarzbraun, erdinkrustiert. - Bewahrung: auf dem abd Souturn me meist scharf gekielt, cauda,J besonders auffallend, Querwülste auf den einzelnen Areae angedeutet; rauh bekömelt. - Tu oo: an der Basis der Kappe kräftig entwickelt, mit seichter Läng~furche me.
Variabilität: nicht auffallend; Körpergröße stark geschlechtsdimorph; Proportionen der Ta von Laufbein II·- im Gegensatz zu vielen anderen Arten - wenig variierend.
Verbreitung (Abb. 267): Typ: disjunkt alpin-dinarisch und karpatisch. - Geschlossenes Areal in den 0-Alpen und W-jugoslawischen Gebirgen, ferner - bisher erst mangelhaft bekannt - in den Karpaten. - Locus typicus: Oberbayern.
Alpen-Areal: BRD: Schneizelreuth, Saalachtal (ZILOH leg., Beleg SMF), sonst nur die allgemeine Angabe "Oberbayern" iri der Originalbeschreibung (0. L. KocH 1848). - Österreich: westlichste Funde im Land Salzburg (Raurisertal in den Hohen Tauern, Mühlbach am Hochkönig, Untersberg/Salzburg), also in den Kalkalpen und. auf Kalkböden in den Zentralalpen; ferner (? isoliert) auf dem Rainberg, einem Inselberg im Salzburger Becken (alle A. A. in litt.); östl. in den nörcll. Kalkalpen bis zum Wienerwald (Kahlenberg und Leopoldsl;lerg, J. G. in litt.), in Steiermark und Kärnten weitverbreitet (z. B. Villaeher Alpe, J. M. leg.). -Italien: bisher erst 3 Funde: Gardasee-Gebiet, V. Ledro, Leano Mt. Guil (K. T. leg.), das 3 weü•ht. von N-alpinen Populationen geringfügig ab (vgl. Abb. 271); Karnische Alpen, Arta (ScHWEIGER leg.). - Fehlt in den W-Alpen.
Balkan-Areal: Jugoslawien: im Anschluß an das alpine Areal weitverbreitet; z. B. Slowenien: Grahovo bei Cerknica, SneznikJSohneeberg (beide J. M. leg.), Gottseitee (J. G. leg.): Kroatien: Plitvice (KNUSE leg.), südl. bis Bosnien-Herzegowina: 10 km sücll. Banja Lulm, Foca (beide A. A. leg.).
Karpaten-Areal: Rumänien: 0-Karpaten: Schlucht von Virghis, Bil•gau Berge (AvRAui 1971, qruberi); Gebiet um Vrancea (OLTEAN & DuMITRESOU 1973). - S-Karpaten: Retezat Ma•sh·, Berge von Sebes, Plateau von Meheclintzi. Das Karpaten-Areal ist wahrscheinlich (viel) ausgedehnter als bisher bekannt.
Ökologie: Biotopansprüche: In den Salzburger Alpen lebt tingiformis vorwiegend in Buchenwald-Gesellschaften (Fagetum silvatica, Abieti;Fagetum, AceriFagetum), gelegentlich in Hainbuchen-Bergulmen-Holunder- oder in GrauerlenBeständen (A. A. in litt.); am nordwestl. Stadtrand von Wien (Ausläufer des Wienerwaldes) vereinzelt auch in vergleichsweise trockenen Biotopen, z. B. in xerothermem Buschwald am SW-Hang des Leopoldsberges (J. G.leg.). Die generelle Bindung an gleichmäßig durchfeuchtete Bodendecke in geschlossenen Wäldern gilt ebenso für den slowenischen außeralpinen ArealteiL An der oberen Verbreitungsgrenze dringt tingijol"fl!'is in die Latschenzone ein (z. B. Villaeher Alpe tmd SneznikfSchnee berg, beide .T. M. leg.).
..

174 Familia Trogulidae
Vertikal ver breitung: Ini westl. Nieder-Österreich von etwa 270m (Erlafsohluoht bei Purgstall) bis um 1200 m (RESSL leg.); in den Salzburger Alpen bisher nur in der Collin- und Montan-Stufe zwischen 480 und 1400 m (A. A. in litt.); die Subalpin-Stufe wird wenigstens in den südl. Kalkalpen erreicht (Villaoher Alpe, 1900 m), lokal möglicherweise bis 2000 m (vgl. FRANZ & GuN.Iro~:p 1954); in Slowenien (außeralpin) Nachweise um 600 m und 1550-1600 m (J. M.leg.).
Phänologie: Ad. und iuv. wahrscheinlich euryohron und ad. mehrjährig. - Ad. nachgewiesen zwischen A V und "X, iuv. von IV -XI (A. A., J. G., J. M., RESSL leg.). ·
Trogulus ooriziformis C. L. KocH, 1839 Abb. 254-255, 275
1839 Trogulus- ooriziformis C. L. KocH, in HAHN & KocH, Die Arachniden, 5: 128. 1879 Trogulus oristatus SrMON, Arachn. de France, 7: 308. 1023 Trogulus oristatus - RcEWER, Weberknechte d. Erde: 636. 1923 Trogulus ooriziformis - ROEWER, Weberknechte d. Erde: 639. 1968 Trogulus ooriziformis - BRIGNOLI, Frag. entomol., Roma, 5: 274.
Begründung der Synonymie: Die Zuordnung des Namens coriziformis C. L. KocH zu einer bestimmten Art ist nicht zweifelsfrei, zumal KocH die Herkunft seines Typenmaterials nicht angibt. RoEWER (1923) bezieht Material aus großen Teilen des westl. Mittelmeergebietes einschließlich N-Afrikas auf diesen Namen; seine Unterlagen sind jedoch, wie die teilweise Überprüfung ergab, heterogen. Es erscheint zweckmäßig, der Deutung von BRIGNOLI zu folgen, der die große, auf der Aperminen-Halbinsel weitverbreitete Art als ooriziformis C. L. KooH bezeichnet. Alte, vermutlich z. T. syntypische Tiere von oristatus SrMON (MNHNP) stimmen mit italienischen Individuen völlig überein (J. M. rev.).
Habitus und Feldkennzeiohen: besonders brettartig flach und auffallend groß, bis 13 mm, scharfer me Kiel do; Ta des Laufbeines II: Endglied geringfügig länger als Grundglied. - In den SW-Alpen und in Istrien.
Kennzeichnung: Körpermaße (in mm): L (] 9,0-10,1; L 5j! (alpine und außeralpine Populationen) (9,3) 10,4-13,1.
Laufbein II ((],in Klammern 5j!): Fe 4,0 (4,6) Pt 1,3 (1,5) Ti 2,0 (2,4) Mt 2,7 (2,9) Ta 1,8 (2,0).
Längenverhältnis von Grundglied zu Endglied des Tarsus von Laufbein Il: 1: 1,20-1, 56; d. h. Endglied ein Fünftel bis etwa die Hälfte länger als das Grundglied (Abb. 254-255).
Genitalmorphologie (Abb. 275): Trunous peni~ im Verhältnis zur Körpergröße klein, von ba kontinuierlich distad verschmälert; Glans nicht gegen den Trunous abgesetzt bzw. abgewinkelt, stumpf konisch zum Stylus versohmälert; Stylus ebenfalls gestreckt in gerader Verlängerung der Glans, an diesem (von do) linksseitig ·hyaline Membran ausgespannt, von Ia stärker hervortretend; Bebor-stung unauffällig kurz, spärlich. .
Cheliceren: schlank-glatt, ohne spezifische Besonderheiten. Pedipalpus: kurz, merklich schlanker als bei nepaeformiB, Ta etwas aufge
bläht, nur spärlich beborstet. Laufbeine: robust, vergleichsweise kurz (vgl. tingijormiB); alle Glieder rund,
Fe I auf der ba Hälfte mit einem streng ausgerichteten palisadenartigen Kamm langer, konischer Zapfen mit aufgesetzter kurzer Bo, die einzelnen Bo kürzer als der Fe-Durchmesser; Kamm bei 5j! schwächer entwickelt. Ta von Laufbein II vergleichsweise kurz; Proportionen vgl. oben.
Genus Anelasmocephalus 175
Dorsalseite: Zeichnung und Färbung: erdbraun und mit Milleralpartikeln inkrustiert. - Bewahrung: kräftiger me Kiel auf den abd Areae I-V, doch hei den alpinen Populationen weniger markant als bei jenen der Apenninen-Ha.binsel. - Tu oc: niedrig, Augen weit voneinander getrennt.
Verbreitung: Typ: W-mediterran (bis iberisch-subatlautisch). - Locus ty pious: Herkunft des Typenmaterials nicht bekannt; L. t. hier festgelegt auf Meeralpen, Saint-Martin de Lautosque, dem Fundgebiet von T. criBtatuB SrMoN, einem Synonym von T. corizijormiB.
Italien: Schwerpunkt des Areals wahrscheinlich auf der Apenninen-Halbinsel, dort offensichtlich kontinuierlich verbreitet: Latium, Lucanien, Calabrien (BRIGNOLI 1968), Gebirgsmassive Sirente, Matese und Abruzzen-Nationalpark (MARCELLINO 1971), Toskana, Umbrien, Kampanien, Calabrien und Insel Capri (ROEWER 1923). - Frankreich: Meeralpen: SaintMartin de Lantosque, Montboron (SIMON 1879, oristatus); in der Sammlung SIMON (MNHNP) überdies Belege aus Cagnis, Cannes, La Roquette und von einigen weit<Jren, nicht näher benannten Orten
1 alle in den Meeralpen.- Jugoslawien: Istrien, Rovinj; erster Nachweis
(v. HELVERSEN leg.). -Iberische Halbinsel: Spanien: Pyrenäen, Massiv des Mt. Perdu (NIETHAMMER leg.), ferner Nachweise aus den Provinzen Pontevedra (Galicien, Sierra de Gondomar), Jaen (Andalusien, Cazorla; alle KRAUS 1959, FRANZ leg.). - Portugal: Porto (ROEWER 1923).
Ökologie: Biotopansp~üohe: T. corizijonniBiebt auf der Apenninen-Halbinsel in Vielzahl (auch) offener und hoher Insolation ausgesetzter Biotope sowohl im Flachland wie im Gebirge; somit offensichtlich im weitesten Sinne euryök mit
·Vorliebe für mediterrane Klimabedingungen. J. M. vermißte corizijormi8 in den regenreichen, N -exponierten Waldgebieten der französischen Pyrenäen und der nördl. Kautabrisehen Gebirge; dort kommt (1 allein) nepaejor-miB häufig vor.
Vertikalverbreitung: von nahe Meereshöhe (z. B. Rovinj) bis in höhere Lagen der Apenninen-Halbinsel (z. B. 800 m in Latium, BRIGNOLI 1968).
Genus Anelasmocephalus SrMON, 1879 1873 Anelasma SoERENSEN, Naturhist. Tidsskr., (3) 8: 519 (präocc.). 1879 Anelasmooephalus SrMON, Arachn. de France, '/: 297. 1923 Anelasmooephalus - RoEWER, Weberlmechte d. Erde: 644.
Eine Gattung der Trogulidae, gekennzeichnet durch: (J-Genitalmorphologie: Truncus penis do-ve bandförmig abgeflacht; Glans ver
gleichsweise lang, schmal, deutlich gegen den Truncus abgesetzt, di hakig gebogen (bei 1 Art plan-tellerartig verbreitert). Körper tropfenförmig, nur mäßig do-ve abgeplattet. Tu oo am Ceph-Frontalraud in 2 stumpfe oben verlängert, von diesen distad und laterad weisend Schirm aus langen, konisch verjüngtem Tu. Pa und Chel unter diesem Tu-Schirm fast völlig verborgen. Laufbeine kurz und robust, mit drüsigen Tu und langen Bo dicht besetzt, alle Glieder mit Ausnahme des Caloaneus und der Ta dicht mit Erdreich verklebt. - Gliederung der Ta der Laufbeine I-IV: intraspezifisch Jr.onstant, interspezifisch variierend; bisher bekannte Kombinationen: 2-2-3-'3 (lycoBinuB), 2-3-3-3 (hadzii), 3-3-4-4 (cambridgei, puBillus, pyrenaicus), 4-4-4-4 (rujitarBis). - Ohne sekundäre Geschlechtsmerkmale.
Verbreitung: etwa 8 nominelle Arten im überwiegend atlantisch und mediterran beeinflußten Klimabereich W- und Mitteleuropas. Von S-Spanien (Andalusien), den Kautabrisehen Gebirgen und Pyrenäen und S-England über ganz Mitteleuropa einschließlich der Alpenläuder, Mittel- und N-Italien, Sizilien, Korsika, Sardinien,

176 Familia Tcrogulida<-
Balearen; N-if'rika (Algerien). - Anelasmocephalus fehlt in Skal).dinavien (mit Dänemark) und 0-Europa (Polen, ÖSSR, Ungarn, Rumänien). Syrtopie zweier Arteri ist aus den Pyrenäen und Kautabrisehen Gebirgen bekannt.
Typus·Art (Festlegung durch TnoRELL 1876): AnelaBma ZycoBinum SoERENSc:N, 1873.
Besiimmungstabelle für die Arten {A. pyrenaicUB ist nicht berücksichtigt, vgl. S. 184)
1 Gliederung der Laufbein-Tarsen I-IV: 3-3-4-4, Körperlänge 2,6-3,~ mm; weitverbreitet in Mittel-Europa. . . . . • . . . . . . . . . . . . oambridgei, S. 176 Laufl.•in-Tarsen I-IV anders gegliedert. . . . . . . . . . . . • . . . . . . . 2
2 Glieuerung der Laufbein-Tarsen I-IV: 2-3-3-3, Körperlänge 2,2-~,5 mm; in den Ü·Alpen und NW.Jugoslawien .................•.. hadzii, S. 181 Gliederung der Laufbein·Tarsen I-IV: 4-4-4-4, Körperlänge 3,6-5,4 mm; in den SW ·Alpen . . . • . . . . . . . . . . . . . . . . . . . . • . rufitarsis, S. 180
Anelasmocephalus cambridgei (WESTWOOD, 1874) Abb. 278, 283-285, 290
1874 TroguZus cambridgii WESTWOOD, Thesaur. ent. Oxon.: 202. 1923 Ane7,a
1smocepha!UB cambridgei- ROEWER, Weberknechte d. Erde: 646.
Habitus und Feldkennzeichen: klein, Körper schlank tropfenförmig, struppig wirkend, erdfarben; weitverbreitet in Mitteleuropa (Abb. 278).
Kennzeichnung: Körpermaße (in mm): L ö- 2,6-3,5; L ~ 3,1-3,9. Laufbein II (Ö", in Klammern ~):Fe 1,0 (1,0) Pt 0,5 (0,4) Ti 0,8 (0,9) Mt 1,0 (1,0)
Ta 0,8 (0,7). Genitalmorphologie (Abb. 284, 296): Truncus penis (von dofve) breit, par
allelrandig, nur im di Viertel wenig verjüngt. Glans(Abb. 296) untersetztund kurz, gegen den Truncus verschmälert abgesetzt, di Ende zu breit löffelförmigem Haken halbkreisförmig dorsad umgebogen; mit wenigen kurzen Sinnesborsten.
Cheliceren: schlank, ohne Besonderheiten. Pedipalpus: kurz und untersetzt, ö--Pt und ö--Ta etwas aufgebläht. Laufbeine: kräftig-robust; alle Glieder außer Calcaneus des Mt und Ta mit
langen Warzen und aufgesetzter Bo von bis doppelter Warzenlänge; di oft gekrümmt, am längsten auf Fe I; do oft zu Kiel konzentriert. - Tarsalgliederung der Laufbeine I-IV: 3-3-4-4; das ba Glied vom 2. Glied nur durch Ringfurche getrennt und dort wenig gelenkig (Abb. 283).
Dorsalseite (Abb. 278): Zeichnung und Färbung: durch ·angeklebte Mineralpartikel des Bodens erdbraun, Gliederung in abd Areae nicht erkennbar. - Be-wahrung: nur fein tuberkuliert. .
Variabilität: Genitalmorphologisch ist cambridyei offensichtlich im ganzen Areal außerordentlich einheitlich (vgl. Abb. 296 und 297, Individuen aus Stuttgart und 8-Frankreich); auch die Tarsalgliederung ist konstant. Die Körpergröße variiert sehr, regional aber nur in engen Grenzen; die kleinsten Individuen sind aus den 0-Pyrenäen bekannt.
Verbreitung (Abb. 290): Typ: atlantisch-subrnediterran. - Geschlossenes Areal von den Kautabrisehen Gebirgen und Pyrenäen über fast ganz Frankreich, 8-England und das engere Mitteleuropa östl. etwa bis zur Eibe. Locus typicus: Britische Inseln, Bloxworth bei Blandford, Dorset.
Genus Anelasmocephalus
Abb. 277-280: H~bitus von dorsal; - 277. Trogulus nepaejormiB {Scop . ·Cephaluscambndge•{WESTWOOD)· ''79-280 n· l OLI), 278. AnelasmoIateral. ' • . wrano asma scabrum {HERBST); 280. Scut-um von
12 1.'1erwelt Deutsch!. 64: Mnrtens
177

178 Familia Trogulidae
Abb. 281. Anelasmocephalus cf. rufitarsis SIMON. - Laufbein Il, Tarsalglieder: eS, IT Lom•
bardei, Cepino Imagna. Abb. 282. Anelasmocephalus rufitarsis SrMON. - Laufbein Il, Tarsa.Jglieder: eS· IT Meeralpel),
S. Anna Vinadio. Abb. 283-285. Anelasmocephalus cambridgei (WEsTWOOD). - 283. Laufbein _n, Tarsalglieder: eS, BRD Stuttgart; 284. Penis von dorsal (links} und lateral; 285. cS-Pedtpalpus von
lateral. Abb. 286-287. Anelasmocephalus hadzii n. sp. - 286. Laufbein Il, Tarsalglieder: eS, AUS Übelbach/Gieinalpe; 287. Penis von dorsal (links} und lateral.
Abb. 288-289. Anelasmocephalus pyrenaicus n. sp. - 288. Laufbein Il, 'Darsalglie~er: eS, SP Sierra de Aralar; 289. Penis von dorsal (links) und lateral. - Homologe Organe m ver·
gleichbaren Maßstäben dargestellt.
Arealgrenzen: Spanien: Kantabrische Gebirge, z. ~· Pic~s de Europa und Sierra de Aralar (J. M. Jeg.); weitere Nennungen für die nördl. Gebtrge bet KRAUS (1_961). - Frankreich: z. B. Hautes- Pyrenees (Lourdes), Ariege (bei St. Gi;ons), P_yrenees-Orrenta.Jes (Ba~~sur-Mer; alhi J. M. leg.), ferner im ganzen übrigen Frankrerch (vgl. ~IMON 1879) wa~schet~rch mit Ausnahme der Alpen (vgl. unten). -Britische Inseln: nur 1m S d~r :S:a':'ptr_nsel~ nor~I. bis in die Grafschaften Flint, Anglesy und Derby, nördl. davon ~ahrscheinhch tsohert m Mrd. W Yorkshire (BRISTOWE 1949, SANKEY & SAVORY 1974). - Beigten: Brabant,_z. B. Umg~bung vo~ Brüssel, Namur und Luxembourg (BECKER 1896). - Niederlande: btshe~ nur m der Provinz Limburg (SPOEK 1963). - BRD und DDR: in der Nordde~ts?hen Trafebene nur
I U b H ornburg (KRAEPELIN 1896). 0-Grenze wahrschemhch westl. der Eibe; Joka : mge ung von ~ • . K" · I
· t Fundorte haben zu gelten· Simmelsberg/Rhön (PIEPER leg.), bet Bad rssmgen a s expomer e · B R b t · (WEIDNER 1954), Hartmannshof und Happurg bei Nürnberg (L. KocH 1877), urg a ens em
im Sächsischen Bergland (BÜTTNER 1930).
Genus Anelasmocephalus 179
Anelasmocepha/us cambridgei
. 1]'{{:::? ~ ~J • J ::::::::::::::: A.ruf1tars1s .·.·.·.·.·.·.·.
Abb. 290. Gesamtareale von Anelasmocephalus cambridgei (WESTWOon), A. rufitarsis SrMON und A. hadzii n. sp.
Alpenländer: Frankreich: SrMON (1879) nennt für die Hochalpen Sappey und Bourg d'Oisans; die Zuordnung zu cambridgei erscheint nicht gesichert (vgl. bei rufitarsis). - Schweiz Becken des Genfer Sees (LESSERT 1917; Monthey, J. M. leg.); Jura: St. Ursanne (MARTI leg.); Delemont (v. HELVERSEN leg.), Gilgenberg, Oberdorf, Schwarzwaldalp bei Altzellen (ScHENKEL 1923), Gadmental (SCHENKEL 1936), MuttlerfGraubünden (LEBSERT 1905; ob diese Art ?), bei Basel {LESSERT 1917). - Öster_reich: Vorarlberg: PfänderfBregenz (JANETSOHEX 1961), Götzig' und Klaus (Belege NHMW).
Keine Nachweise aus den mittleren und östl. Alpenteilen (siehe bei A. hadzii); fehlt in Skandinavien und östl. der Eibe.
Ökologie: Biotopansprüche: A. cambridgei lebt in der obersten Boden- und Fallaubschicht von Laub- und Mischwald-Formationen mit unterschiedlichen Feuchtigkeitsverhältnissen - im atlantischen Bereich ebenso wie an der Mittalmeerküste; z. B. in dünnem Moosrasen in feuchtem Laubwald in Bachnähe (bei Stuttgart), in dicken, feuchten Humusschichten auf Kalkuntergrund (Pyrenäen), in GebüschanS-exponiertem Trockenhang (Kaiserstuhl; alle J. M.leg.), Flaumeichenwald bei MontelimarJRhöne-Tal (v. HELVERSEN leg.), S-exponiertes Buchen-12•

180 Familia Trogulidae
1ltangenholz bei MontheyfWallis (J. M.leg.). Auch in unbeschattetem_ Ge!ände, selbst unmittelbar an der Mittelmeer-Küste - soweit die Substratfeuchtigkeit ausreicht, z. B. Wiesen und Weiden (S-England, ToDD 1949), fallaubbedeckter Abflußgraben in Weinberg (S-Frankreich, Banyuls-sur-Mer, J. M. leg.)._ - Wahr• scheinlich hat als begrenzender Faktor primär die Substratfeuchtigkeit zu gelten, weniger der Temperaturgang in den Mikrobiotopen. . .
Die öko-physiologischen Daten bestätigen die Toleranz warm-expomerter BIOtope: Temperatur-Präferenz bei _16,3 °0 (Vertraue_nsgrenzen 15,~ un_d 17,5 °01-Diese Werte entsprechen jenen, die an dem xerophilen und auch ahnhch verbreiteten Odiellus spinosus gewonnen wurden (vgl. bei diesem). - Vertrauensgrenzen für die Feuchtigkeits-Präferenz bei 61 und 67 %; sie liegt deutlic~ unter d~r des ebenfalls terrikolen Nemastoma bimaculatum (69 und 79 %) und ISt verglewhbar mit der des euryöken~ in höheren Strata lebenden Leiobunum rotundum (61 und 69 %) (alle Angaben nach ToDD 1949). Da A. cambridgei von Mollusken !eh~, ist er in der Verbreitung weitgehend von Kalkböden abhängig; auf Urgestem 1st er meist selten oder fehlt völlig.
Vertikalverbreitung: von Meereshöhe (S-England, S-Frankreich) über die Mittelgebirge (z. B. Rhön, Bieberstein, 500 m) bis in höhere Lagen der Pyrenäen
. (Col de la CorefAriege, 1400 m, J. M. _leg.). - Fu~~e aus _noch größere1_1: ~öhen in den Alpen (z. B. LESSERT 1917) bleiben zu bestätigen; s1e betreffen moghcher-weise andere Arten. '
Phänologie: ad. und iuv. eurychron; Fortpflanzung wahrscheinlich nur während des Sommerhalbjahres; als Ad. in Gefangenschaft genommene Individuen wurden 23 bzw. 34 Monate alt, also fast 3 Jahre. - 5; produzieren pro Jahr 20 bis 25 Gelege (x = 2 Eier, n = 543 Gelege, Streuung 1-6). Eier schlank kommaförmig, L 0,8-1,1, B 0,3-0,4 mm; Eiablage immer in leere Sch~10c~ens~halen, meist solche von Beutetieren. - Embryonal-Entwicklung unterschiedlich Je nach Ablagezeit der Eier, in Gefangenschaft 47-206 Tage, am kürzesten während der Sommermonate. - Postembryonal-Entwicklung ebenfalls abhängig von der Jahreszeit; im Sommer 1-2, im Winter fast 6 Monate (31-191 Tage); iuv., die in: IX schlüpfen, werden noch im seihen Jahr reif. - 5 Häutungen bis zum ad., also nur 4 Jugendstadien (alle Angaben nach P.ABST 1953). .
Anelasniocephalus rufitarsis SnmN, 1879 Abb.281-282,290, 295
1879 Anelasmocephalus rufitarsis SrMON, Arachn. de France, 7: 298. 1923 Anelasmocephalus rufitarsis - RoEWER, Weberknechte d. Erde: 645.
Habitus und Feldkennzeichen: größte Anelasmocephalus-Art, kurzbeinig, strup-pig-wirkend, erdbraun; nur in den S(W)-Alpen. . ..
Kennzeichnung: Körpermaße (in mm): L 0' 3,6-4,2; L 5;3,8-5,4; aufgeschlusselt nach Fundorten: 0' 3,6 (Meeralpen); 4,2 (Lombardei); 3,8 (Brenta-Gruppe); 5; 3,8, 3,9 (Ligurische Alpen); 4,2, 4,4 (Lombardei); 5,~ (Bas~es-Alpes, MNHNP 2.317). - Laufbein II (0', in Klammern 5;): Lombardei, Cepmo Imagna: F~ 1,4 (1,4) Pt 0,6 (0,6) Ti 1,3 (1,3) Mt 1,4 (1,4) Ta 1,2 (1,2). Meeralpen (0'), Ligunsche Alpen (5;): Fe 1,2 (1,3) Pt 0,5 (0,4) Ti 1,1 (1,1) Mt 1,~ (1,1) Ta 0,9 (0,8). ..
Genitalmorphologie (Abb. 295): Truncus pems entsprechend der Korpergröße lang, nur schwach do-ve abgeplattet, von der Basis zur Glans kontinuierlich verjüngt, ve durch Krümmung konkav. Glans markant gegen den Truncus ab-
Genus Anelasmocephalus 181
gesetzt, deutlich schmaler als dieser und in den Proportionen ähnlich ca, 1bridgei; insgesamt schlanker, von do stärker gestreckt, di Krümmung der Glans (von Ia) weniger stark gerundet, flacher; Bewahrung mit Sinnesborsten spärlich.
Cheliceren und Pedipalpus: ähnlich cambridgei. Laufbeine: kräftig-robust; alle Glieder außer Calcaneus des Mt und Ta mit
langen Warzen und aufgesetzter Borste von bis doppelter Warzenlänge; struppig wirkend. - Tarsalgliederung der Laufbeine I-IV: 4-4-4-4; Unteneilung der beiden großen gelenkig verbundenen Glieder jeweils ba als feine Einschnürung (Abb. 281-282).
Dorsalseite: Ceph me meist mit "Kiel" aus engstehenden Tubenwarzen; Abd schwach gewölbt, nur vereinzelt mit Tubenwarzen, konzentriert am Hinterrand von Area V und auf den freien Terg; ganzer Körper mit Erdpartikeln inkrustiert.
Variabilität: Die wenigen zur Verfügung stehenden Individuen dieser wenig bekannten Art (darunter typisches bzw. syntypisches Material der Sammlung SIMON) weichen in der Körpergröße und in den Proportionen und Maßen der Laufbein-Ta voneinander ab, so daß sich unter "rufitarsis" möglicherweise mehrere Arten verbergen. - Körpermaße vgl. oben. - Ta der Laufbeine: Größen- und Proportionsunterschiede zwischen Individuen aus der Lombardei und aus den Meeralpen (vgl. Abb. 281-282). - Behaarung der Laufbeine: Haare auf den Tubenwarzen entweder dicht stehend und di etwas gekrümmt (Lombardei) oder schütter verteilt und gerade gestreckt (j.\1eeralpen und Ligurische Alpen). Genitalmorphologisch besteht - bis auf geringe Größenunterschiede - Einheitlichkeit der beiden verfügbaren 0' aus den (disjunkten ~ vgl. unten) Teilarealen.
Verbreitung (Abb. 290): Typ: Endemit der SW-Alpen. - Weit verbreitet in den südwestl. Alpenmassiven, doch offensichtlich überall spärlich und nur lokal auftretend. - Locus typicus: Basses-Alpes, Digne.
Frankreich: Basses-Alpes, Digne (SIMON 1879). - Italien: Ligurische Alpen: Colle della Pala (COLONELLI leg.); Meeralpen: S. Anna Vinadio, Valle Maledicia (VIGNA leg.); Cottische Alpen: Chianale, Val Vareita (MARCELLINO 1972, cambridgei), Val Chisone, Rifugio Jervis; Grajische Alpen: LeinifTorino; LombardorefPiemonte; Gran Paracliso, Val Soana/Campiglia (alle MARCELLINO 1975d, camb,.idgei; nicht rev., Zuordnung nicht völlig gesichert); Bergamasker Alpen: Cepino Imagna (BrNI leg.), I Moi, Val Cavallina östl. Bergamo (V ALLE leg.); Brenta-Gruppe: Val Rendena, Fisto (bisher östlichster l!'undort, SeRAWALLER leg.).
Ökologie: Biotopansprüche und Vertikalverbreitung: In den Meeralpen bis 1 700 m, in den Ligurischen Alpen bis 2000 m nachgewiesen und somit wahrscheinlich über der Waldgrenze; tiefste Funde bei 520 m (Va.] Cavallina) und 600 m (Brenta, Val Rendena). - Einzelheiten über die Biotopbindung sind nicht bekannt.
Anelas~ocephalus hadzii n. sp. Abb. 286-287, 290-292
1942 Anelasmocephaltts cambridge·i - HAnzr, Razprave, Dissertat., Ljubljana, 2: 58. 1968 Anelasmocephalus lycosinus - AfARCELLI:XO, Ann. Mus. Civ. Stor. Natur. Genova, 77:
ll8 (part.: Alto Isonzo).
Diagnose: Eine Art der Gattung Anelasrnocephalus der 0-Alpen und der westl. Balkanhalbinsel, gekennzeichnet durch die 0'-Genitalmorphologie, die Körpergröße und die Tarsalgliederung der Laufbeine.

182 Familie. Trogulidae Genus Anele.smocephalus 183
Habitus und Feldkeunzeichen: klein, Körper gedrungen tropfenförmig, struppig wirkend, erdfarben; in den 0-Alpen und in NW-Jugoslawien.
Kennzeichnung: Körpermaße (in mm): L c12,1-2,2; ~ L 2,3-2,5. Laufbein II (c1, in Klammern~): Fe 0,7 (0,8) Pt 0,3 {0,4) Ti 0,5 (0,6) Mt 0,7 (0,8) ·
Ta 0,7 (0,7). Genitalmorphologie (Abb. 287, 291): Truncus penis lang, von der Basis
distad kontinuierlich verschmälert, stark do-ve abgeplattet, ··ba stärker als di; Glans (Abb. 291) deutlich gegen den Truncus abgesetzt, absolut und relativ lang und schmal, vom Gelenk distad stetig verschmälert, di Ende (von la) zu kleinem, dorsad gebogenen löffelartigem Haken gekrümmt; dieser Teil deutlich kleiner als bei eambridgei und übrige Glans dadurch länger erscheinend. Besatz mit Sinnesborsten spärlich.
Cheliceren (Abb. 292) und Pedipalpus: ähnlich cambridgei. Laufbeine: kräftig-robust; alle Glieder außer Calcaneus der Mt und Ta mit
langen Warzen und aufgesetzter Bo; Warzen meist dicht mit Erdreich beklebt, · Beine dadurch einförmig dick und rund erscheinend. - Tarsalgliederung der
Laufbeine: 2-3-3-3; Gelenk zwischen dem 1. und 2. Glied an Ta I und II nur als feine Einschnürung (Abb. 286).
Dorsalseite: Zeichnung und Färbung: durch Inkrustierung von Erdpartikeln schwärzlich-braun. - Bewahrung: über und über mit flachen Warzen bedeckt, diese jeweils mit kurzer, aufgesetzter Borste.
Beziehungen: Es ist seit langem bekannt, daß in den 0-Alpen eine Anelasmocephalus-Art vorkommt. Sie wurde fälschlich entweder für cambridgei (HADZI 1942, FRANZ & GUNHOLD 1954) oder für lycosinus gehalten (MAROELLINO 1968, 1970). Die ,S der Typenserie von lycosinus SOEBENSEN (2 ,S, Mittel-Italien, Gennazzano., J. G. rev.) besitzen die Tarsalgliederung 2-2-3-3 und andere Proportionen der Glanspenis als die 0-alpinen Populationen und können deshalb nicht mit diesen als artgleich angesehen werden. MAROELLINO ( 1968, lycosinus) nennt von der nördl. Aperminen-Halbinsel (O-Ligurien, Ligurisoher Apennin) Individuen mit der Tarsal-Formel 2-3-3-3, die mit der von hadzii aus den 0-Alpen übereinstimmt. Eine Serie aus diesem Gebiet (Etruskischer Apennin, Abetone; 2 ,S SMF 11751) weicht von hadzii genitalmorphologisch stark ab (Abb. 293), so daß in N-Italien außerhalb der Alpen offenbar eine weitere, bisher nicht beschriebene Art vorkommt. Die Beziehung~n dieser mit hadzii möglicherweise allopatrischen Form bleiben vorerst offen.
Verbreitung (Abb. 290): Typ: alpin-illyrisch. - Geschlossenes Areal in den S-, 0-, und NO-Alpen (unter Aussparung der Zentralalpen) und im angrenzenden alpinen und außeralpinen Jugoslawien. - Locus typicus: Österreich, Karawanken, Eisenkappet - Alle mit Stern (~) markierten Serien gelten als Paratypen.
Abb. 291-297. Glandes penis von An.~lasmocephalus SrMON (links: lateral, rechts: dorsal).
Abb. 291. A. hadzii n. sp. - AUS Übelbach/Gleinalpe.·Abb. 292. ,S-Ohelicere von lateral.
Abb. 293. Anelasmocephalus sp. - IT N-Apennin, Abetoml (SMF 11 751).
Abb. 294. A. pyrenaicus n. sp. - SP Sierra de Araler.
Abb. 295. A. cf. rufitarsis SnroN - IT Lombardei, Oepino Imagna.
Abb. 296-297. A. cambridgei (WESTWoon). - 296. BRD Stuttgart; 297. FR 0-Pyrenäen, FontfrMe.

184. Familia Trogulidae
Schweiz: * Monte Bre bei Lugano {FBANZ leg., bisher westlichster Fundort; vgl, auch LEBBEBT [1917] für MuttlerfGraubünden angeblich auf.2000m, ob diese Art?)- Italien:* Corner See nördl. Gravedo~a {FBANZ leg.); • Veltlin, bei Sandrio (J. M. leg.); Gardasee-Gebiet: Riva, Val Ampola (K. T. leg.); • Monti Lessini; Cerro {VIGNA leg.); S-Tirol, * Meran {HUBDES ieg.). ~ Österreich: Hohe Tauern, Hochalm-Gruppe, Gössgraben {HöLZEL leg.); Gailtaler Alpen: z. B. * MöltschachfVillach {STBOUHAL leg., NHMW); Karnische Alpen: z . .ß. • südl. Tröpolach; Karawanken: z. B. Obir-Massiv, Jovanberg/Eisenkappel, I c! Halotypus {SOHWEI· GER leg., NHMW), Golica-Massiv, Turmalpe {SOHEEBPELTZ leg., NHMW), Lavanttaler Alpen: z. B. • JohannisbergfSt. Paul; Steiermark: Grazer Bergland: z. B. * Rein {beide F:aANZ leg., NHMW); * Raab-KlammfWeiz, bisher östlichster Fund in den Alpen {STBOUHAL leg., NHMW); * Gleinalpe, Übelbach (J. M. leg.); Hoch!antsch-Gebiet, * Bärenschütz-KlammfMixnitz; Eiseuerzer Alpen: z. B. Gößeck, Kaisertal; Eri.nstaler Alpen/Gesäuse: z. B.: * Hartelsgraben; nördl. Kalkvoralpen des östl. Ober-Ö.: z. B. *Klein Reifling {alle FBANZ leg., NHMW) bis zum westl. Nieder-Ö.: * Lunza. See {MALIOKY leg.). - Jugoslawien: Triglav, Vrata Tal {HADZI 1928, cambridgei), oberer Isonzo, Mt. Kuck {MABOELLINO 1968, lycosinus); im außeralpinen Slowenien weitverbreitet, doch spärlich: z. B. Umgebung von * Cerknica; • SneznikfSchneeberg {beide J. M. leg.), * Kocevje fGottschee {GANGLBAUER leg., NHMW); Kroatien: * Plitvitzer Seen {STUBANY leg., NHMW); Bosnien:* Nemila, nördl. Zenica {REITTEB leg., NHMW).
Ökologie: Biotopansprüche: A. hadzii ist ein ausgeprägter Waldbewohne~, vor allem von verschiedenen Buchenwald-Gesellschaften auf Kalkuntergrund, die in der Montan-Stufe der Aipen, die er überwiegend besiedelt, und im illyrischen Arealteil vorherrschen. Meist in tiefgründigen Humusablagerungen, die konstante Luftfe~Öhtigkeit und ausgeglichimen Temperaturgang gewährleisten.
Der weitaus größte Teil des Areals ist ozeanisch geprägt; kontinental beeinflußte Alpenteile werden nur ausnahmsweise besiedelt, und die dort bekann~e~ Fund?rte sind durch feuchtigkeitsliebende Pflanzengesellschaften charakteriSiert. Nwht waldbedeckte Biotope werden nur selten eingenommen, z. B. Trockenrasen ·bei Lunz (dort aber 2000 mm Jahresniederschlag), Magerwiese bei Bämdorf; möglicherweise über der Baumgrenze im Engadin (J. G. in litt.).
Vertikalverbreitung: Das 0-alpine Areal liegt zwischen 400 und 1300 m, das slowenisch-außeralpine zwischen 600 (z. B. bei Postojna) und I200 m (Berg Sneznik; beide J. M. leg.). Die Mehrzahl der alpinen Funde zwischen 500 und I 000 m, also in der Montan-Stufe; im Engadin möglicherweise bis 2000 m, doch Bestätigung erwünscht (LESSERT I 9 I 7, cambridgei).
Phänologie: ad. und iuv. eurychron.
Deri vatio nominis: Die Art trägt ihren Namen zu Ehren von Prof. Dr. Jovan HADZI (t 1972), der sich u~ die Erforschung der Arachnidenfauna Jugoslawiens Verdienste erworben hat.
Anelasmocephalus pyrenaicus n. sp.
Abb. 288-289, 294
19ö9 Anelasmocephalus !ycosinus - KRAUS, :\litt. Zoo!. I'llus. Berlin, 35: (2}: 296. 1961 Anelasmocephalus lycosinus - KRAUS, Senckenbergiana bio!., 42 {4): 339. 1968 Anelasmocepha!us lycosinus - MARCELLINO, Ann. l\Ius. Civ. Ster. Natur. Genova, 77:
ll9, Abb. 8.
Diagnose: Eine Art der Gattung Anel~moceP_haltts der P~enä~n u~d Kar~~abrischen Gebirge, gekennzeichnet durch dre Gemtalmorphologie, dw Korpergroße und die Tarsalgliederung der Laufbeine.
Genus Anelasmocephalus ,185
Habitus und Feldkennzeichen: klein, Körper gedrungen, caudal auffallend breit, erdfarben braun; unter Feldbedingungen meist nicht mit Sicherheit von dem syn.top lebenden A. camb1·idgei zu trennen; in den Pyrenäen und Kautabrisehen Gebirgen.
Kennzeichnung: Körpermaße (in mm): L (J2,I-2,5; L <j! 2,4-2,6. Laufbein II ((J, in Klammern Cj!): Fe 0,7 (0,8) Pt 0,3 (0,3) Ti 0,6 (0,6) Mt 0,6 (0,7)
Ta 0,6 (0,6). Genitalmorphologie (Abb. 289, 294): Truncus peniskurz gestaucht, in der
ba Hälfte bandartig do-ve abgeplattet, distad verjüngt und kräftig rund aufgewölbt; Truricus in sich stark ventrad gekrümmt. Glans (Abb. 294) in gerader Verlängerung des Truncus, doch deutlich llom Truncus abgesetzt, Gelenkzone wenig entwickelt; Glans - im Gegensatz zu allen anderen Anelasmocephalus-Arten - kurz, bulbig gestaucht, di Ende zu schraubigem Unigang verdreht und fronto-di '"tellerartig abgeflacht. Besatz mit kurzen Sinnesborsten spärlich.
Cheliceren und Pedipalpus: ähnlich jenen der anderen Arten, ohne strukturelle Besonderheiten.
Laufbeine: kräftig-robust, kurz, meist dicht mit Erdreich beklebt, reicher Besatz mit tubusartigen Warzen und aufgesetzter Bo, diese entweder zur Oberfläche senkrecht abstehend oder eingekrümmt und in die Erdkruste einbezogen. - Tarsalgliederung der J"aufbeine: 3-3-4-4, Gelenk zwischen I. und 2. Glied an Fe I und II nur als feine Einschnürung markiert (Abb. 288).
Dorsalseite: zeichnungslos erdfarben, mit lockerem Besatz von tubusartigen Warzen und aufgesetzter, caudad gekrümmter Borste.
. Beziehungen: Unter allen bisher bekannten Anelasmocephalus-Arten fällt pyrenaicus durn], die besonderen genitalmorphologischen Verhältnisse auf; sie machen es augenblicklich unmöglich, ihn mit anderen Arten in nähere Beziehung zu bringen. Die äußeren morphologiscl16n Churaktere sind bei pyrenaictts ebenso gleichförmig ausgeprägt wie bei allen anderen Arten. Dio Tarsalgliederung 3-3-4-4 teilt er mit cambridgei und pusillus, doch ist dieses llferkmal zur Beurteilung von Verwandtschaftsverhältnissen wenig aussagekräftig.
Verbreitung: Typ: atlantisch, Endemit der Pyrenäen und Kautabrisehen Gebirge. - Locus typicus: spanische Zentralpyrenäen, V alle de Ordesa ((J HolotypnH SMF 29I86, 4 (J4 <j! Paratypaide SMF 11727; alle FRANZ leg. 12. VIII. 1955).
Spanien: Nachweise in den spanischen N-Provinzen Asturien, Santander, Pamplona; in den Zentralpyrenäen und {spärlich} in Catalonien z. B. 11ft. Serrat {KRAUS 1959, 1961, lycusinus; dort Liste zahlreicher Fundorte; alles FRANZ leg., später auch J. M. leg.). - Frank. reich: wahrscheinlich in den ganzen Pyrenäen weitverbreitet, dech bisher nur Naehweise aus dem Dep. Ariege: Pie de l'Estelas, Lac de Bethmale, llfines du Bocart (J. 1\L leg.).
Ökologie: Biotopansprüche: Wie die meisten Anelasmocephalus-Arten ein Bewohner der oberen dauerfeuchten Bodenschichten - vor allem solcher montaner Laubwälder mit besonders ausgeglichenen Feuchtigkeitsverhältnissen. Durch diese spezialisierten Ansprüche scheint das Areal weniger kontinuierlich zu sein als das des syntop lebenden cambridgei, der wesentlich größere ökologische Baudbreite besitzt.
Vertikal ver breitung: bisher kaum bekannt, doch wahrscheinlich hauptsiichIich im Montan-Bereich der besiedelten Gebirge. - Spanien: Sierra de Aralar: 450-650 m, Picos de Europa: 1250-I 300m; Frankreich: Dep. Ariege: 600 bis IOOO m (alle J. M.leg.).

186 Familia Isohyropsalididae
Suprafamilia Ischyropsalidoidea
1879 ·rsohyropsalidae SrMoN, Arachn. de France, 7: 265. 1923 Ischyropsalidae - RoEWER, Weberknechte d. Erde: 680. 11!76 Ischyropsalidoidea ~ MARTENS, Entomol. germ., 3 (1/2): 64.
· Eine Überfamilie der Unterordnung Palpatores, gekennzeichnet durch: d'-Genitalmorphologie: Penis mit 1 Muskel, Sehne an der Ventral-Seite der
Glans ansetzend, diese durch Muskelzug ventrad abwinkelbar. Legeröhre entweder kurz, ungegliedert und spärlich beborstet, zu 4 stumpfen
Zipfeln ausgezogen: Ischyropsalididae; mit 2 gerundeten Loben: Sabaconidae, oder Legeröhre lang, dicht und annähernd in Querreihen beborstet (Sabaconidae in 0-Asien). Sternum frei, an den Seitenrändern nicht mit den Cx verwachsen. Tracheenstigmen vergittert; Tibien der Laufbeine ohne akzessorische Stigmen.
Pedipalpen groß, kräftig laufbeinartig entwickelt; Tarsalklaue reduziert oder ganz fehlend (Orosbycus, vgl. unten). ~Spezialisierte Borsten auf dem Pedipalpus -soweit vorhanden -als "Fiederborsten". (
Zähne der Cheliceren z. T. massig-massiv, z. T. zart-hyalin. Dorsales Scutum meist als S. parvum ausgeprägt, bisweilen weiter aufgegliedert als S. laminaturn und s. dissectum; ausnahmsweise als s. magnum (Hesperonemastoma GRUBER).
Gliederung: Sicher abgrenzbar sind die Ischyropsalididae und die Sabaconidae (beide disjunkt holarktisch verbreitet). In die Überfamilie Ischyropsalidoidea gehört auch die Gattung Orosbycus RoEWER (in NO-Amerika und Japan); möglicherweise repräsentiert sie eine monotypische Familie (d' unbekannt).
Bestimmungstabelle für die Familien: S. 90.
Familia Ischyropsalididae
1879 Ischyropsalidae SIMON, Arachn. de France, 7: 265. 1904 Ischyropsalidoidae - HANSEN & SoERENSEN, On two Orders 6f Arachn.: 82. 1923 Ischyropsalidae - ROEWER, Weberknechte d. Erde: 680 (part.). 1969a Ischyropsalididae - MARTENS, Zoo!. Jb. Syst., 96: 133.
Eine Familie der Palpatores-Ischyropsalidoidea, gekennzeichnet durch: d'-Genitalmorphologie: Truncus penis gerade gestreckt oder wenig in sich ge
krümmt, Muskulatur wenigstens die Hälfte des Truncus einnehmend, bisweilen ihn ganz ausfüllend; die Sehne medioventral an der Glans ansetzend, -durch Muskelzug ventrad abwinkelbar. Glans in gerader Verlängerung der Truncus, nicht I von ihm abgesetzt, Gelenkzone meist undeutlich und nur ve entwickelt.
Ovipositor: etwa so lang wie breit, ungegliedert, ohne deutlich abgesetzten FurcaTeil, di meist in 4 stumpfe Zipfel ausgezogen, Bewahrung mit Sinnesborsten spärlich.
Cheliceren entweder exzessiv verlängert (bei 2 Gattungen) oder normal proportioniert, oft mit Drüsenorgan.
Pedipalpus meist schlank-laufbeinartig, Tarsalklaue rudimentär entwickelt oder völlig reduziert. Spezialisierte sekretorische Haare auf den Palpengliedern -soweit vorhanden - "gefiedert".
Dorsales Souturn variierend; entweder als S. magnum (Hesperonemastoma GRU' BER}, S. parvum (Ischyropsali8 0. L. KocH, Taracus SIMON) oder S.laminatum
Genus Ischyropsalis 187
(einzelne I~chyropsalis-Arten); variable Reduktionsformen zwischen S. parvum und S. lammatum (S. dissecturn und S. intermedium) bei troglobionten Ischyropsalis-Arten. ·
Sternum deutlich entwickelt, zwischen Cx I und II ausgespannt, nicht mit ihnen verwachsen.
Verbreitung: disjunkt holarktisch. - In der Palaarktis mit Schwerpunkt in Mittel- und W-Europa ( Ischyropsalis); Sibirien (Taracus); in N -Amerika die Gattungen Taracus, Hesperonemastoma und Oeratolasma GooDNIGHT & GooDNIGltr (Orosbycus ROEWER stellt möglicherweise eine monotypische Familie dar).
Entgegen früherer. Annahme gehört Nipponopsalis (Japan und Korea; vgl. MARTENS & StrzUKI 1966) nicht zu den Ischyropsalididae, sondern repräsentiert innerhalb der Troguloidea (vgl. dort) eine eigene Familie.
Genus Ischyropsalis C. L. KocH, 1839
1839 Ischyropsalis C. L. KooH, Übersicht d. Arachnidensystems, 2: 24. 1923 lschyropsalis - RoEWER, Weberknechte d. Erde: 680. 1924 Ischyropsalis - MÜLLER, Zoo!. Jb. Anat;, 411: 405. 1954 Ischyropsalis (Odontopalpa) HAnzr, Razpra.ve, Dissertat., Ljubljana, 1: 107. 1969a Ischyropsalis - "MARTENS, Zoo!. Jb. Syst., 96: 133.
Nominatgattung der Ischyropsalididae, gekennzeichnet durch: d'-Genitalmorphologie: Trunous penis untersetzt bis lang gestreckt, ba Hälfte
schwach erweitert, do-ba und ve-ba tief ausgekehlt; Glans in gerader Verlängerung des Truncus, ve :init schwärzlichem Sklerit, auf diesem 2 para-me BorstenFelder, solche a-uf der Glans auch do und - viel kleiner - la. Glans zu feinem Stylus ausgezogen, do-laterad gekrümmt. Gelenkzone zwischen Glans und Truncus. Cheliceren: Grundglied nahezu so lang oder deutlich länger als der Körper, mit 2. Glied zusammen immer länger als der Körper. d'-Grundglied me-do mit buckliger Aufwölbung oder kurzer Apo, dort meist dichtstehender Besatz feiner Bo, die "Bürste", darunter Drüsenorgan. - Dorsales Scutum: c3' mit S. parvum, S. intermedium oder S. laminaturn; ~ mit S. parvum oder S. laminatum.
Sternum frei, d. h. nicht mit denCx der Laufbeine bzw. derenLoben verschmolzen; Unterlippe groß. - Sekundäre Geschlechtsmerkmale: d'-Chel-Grundglied meist mit Drüsenorgan; Austrittstelle des Sekrets in einem Porenfeld im Bereich der "Bürste", dort oft do-di bzw. da-medialer Höcker bzw. Apo. ·
Verbreitung: Die klar umgrenzte Gattung ist auf Europa beschränkt. 17 Arten in Gebirgen von N-Portugal, in den Kautabrisehen Gebirgen und den Pyrenäen, im Zentral-Massiv, wahrscheinlich in den gesamten Alpen, nördl. der Alpen nur rechts-rheinisch bis zum N-Rand der Mittelgebirge, Apenninen-Halbinsel, W-Jugoslawien, Karpaten-Bogen. Größte Arten-Konzentration auf der nördl. PyrenäenHalbinsel (mind. 5 Arten) und in den Alpen (9 Arten). 9 Arten leben in glazialen Refugialgebieten obligatorisch troglobiont und haben entsprechende Umwandlungen erfahren (Augenverlust, Scutum-Variabilität, Verlängerung der Cheliceren und Laufbeine). Das Siedlungsgebiet dieser Arten ist meist klein und reliktär auf die Kautabrisehen Gebirge, die Pyrenäen, die S-Alpen und die Dinarischen Gebirge beschränkt. 11 Arten besiedeln das hier behandelte Gebiet.
Gliederung: 3 Artengruppen lassen sich unterscheiden: 1. Sklerit der Glans klein, spärlich beborstet, Cheliceren-Drüse stark entwickelt (1 Art, Abb. 319); 2. Sklerit der Glans groß, basad zugespitzt, mit me Kiel, Cheliceren-Drüse stark

ISS Familia Isohyropsalididae
entwickelt (8 Arten, Abb. 379~382); 3. Sklerit der Glans groß, basad breit 2lap. pig, ohne Kiel, Cheliceren-Drüse fehlend. oder undeutlich entwic~elt· (8 Arten,
Abb. 316-318, 351-353). Die Arten der emzelnen Gruppen stehen m engem verwandtschaftlichen Verhältnis; wahrscheinlich kommt ihnen Untergattungs-Rang zu.
Beziehungen: Gattungen mit habituell ähnlichen Arten, d. h. mit stark verlängerten Cheliceren, leben in Japan und Korea (Nipponopsalis ~T1' 'TS -~ Su~UKI) und in 0-Sibirien und N-Amerika (Taracus SrMON 1879). Von d1esen gehort N~ppondpsalis mit 2 Penis-Muskeln nicht zu den Ischyropsalididae. Mit Ischyropsalis nahe verwandt ist die N-amerikanische Gattung Ceratolasma GooDNIGHT & GoonNIGHT 1942; die Cheliceren von 0. tricant'ßu sind "normal" proportioniert. ·
Typus-Art (Festlegung dw·ch THORELL 1876): Ischy>·opsalis kollari C. L. KocH, 1839.
Besiimmungstabelle für die Arten c! (Glans penis, z. B. Abb. 316, unter dem Operculum genitale) 2 ~ (Ovipositor, Abb. 44, unter dem Operculum genitale) . . . . . . . . 12
2 Abd do mit Souturn parvum (Sklerite 1-V versohmolzen,z. B. Abb. 361) .'1 Abd do mit Souturn laminaturn (Sklerite isoliert, Abb. 334) oder Souturn intermedium (2-4 Sklerite verschmolzen, übrige isoliert, Abb. 332). . . . . . . . . . . . . . . . . 10
3 Chel-Grundglied gedrungen massig bis schlank, Bewahrung mit zahlreichen, untersohiedliph großen Do, davon auf der do Seite entweder 3-5 in Längsreihe markant hervortretend oder alle ungeordnet gruppiert . . . . . . . . . . . . . . • . . . · · · · · · 4 Chel-Grundglied auffallend lang und dünn, ·völlig glatt oder allenfalls mit wenigen nied-rigen Tu ................... · · .. · · · · · · · · · · · 9
4 Chel-Grundglied massig gedrungen, Bewahrung do mit vielen unterschiedlich großen Do, nie zu markanter Reihe ausgerichtet (Abb. 298-300, 335-337) . . . . . . . . . lf Chel-Grundglied schlank, Bewahrung do ntlt' spärlich, markant 1 Reihe aus 3-5 großen, zu I Reihe ausgerichteten Do (z. B. Abb. 320, 364, 371, 383) . . . . . . . . . . . 6
5 di Drittel des Chel-Grundgliedes do mit dichtem Besatz feiner Haare ("Bürste"), dort olmH Do-Bewehrung (Abb. 298-300), abd Souturn gewellt; weitverbreitet in Mitteleuropa mit O-Alpen . . . . . . . . . . . . . . . . . . . . . . . . . hellwigi hellwigi, S. 190 di Drittel des Chel-Grundgliedes do ohne, allenfalls mit spärlichem und schütterem Besatz feiner Haare (keine "Bürste"), dort mit. Do-Bewehrung (Abb. 337), abrl Sout-urn plan; 0-Alpen . . . . . . . . . . . . . . . . . . . . . . . . . . . kollari, S. 203
6 Chel-Grundglied do mit Reihe aus 3 großen Do (Abh. 364, 383) . . . . . . . . . . 7 Chel-Grundglied do mit mehr als 3 (höchstens 6) großen Do (Ahb. 320, 371) J
7 Che!-Grundglied do-di schwach bucklig aufgewölbt (von Ja betrachten), punktförmige Bürste am medio-di Rand (Ahb. 364-366), in den W-Alpen zwischen Bodensee undGenfer-See . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . oarli, S. 213 Chel-Grundglied do-di halbkugelförmig aufgewölbt, schwarzes Exoskelet.t im Bereich r!er ( schütteren Bürste gelblich (Abb. 383); in Höhlen der Schweiz. . . . . nelvetica, S. 219
8 Pa des Pedlpalpus ve-di mit spitzer Apo; in den Penninischen und Bergamasker Alpen · dentipalpis, S. 197
Pa des Pedipalpus ve-di ohne spitze Apo; im Karpatenbogen . . . . . . .nanicata, S. 216 9 Chel-Grundglied do-di knotig erweitert (Ahb. 400); in Höhlen der Cottischen Alpen . . .
pyrenaea alpinula, S. 224 Chel-Grundglied do-di nicht knotig erweitert (Abb. 324-326), Höhlen der SO-Alpen und NW-Jugoslawiens. . . . . . . . . . . . . . . . . . . . . . . muellneri, S. 200
10 Chel-Grundglied lang, bis 5,5 mrn; do Do groß, länger als der ba Durchmesser der Chel, distarl gekrümmt (Abb. 309, 354) . . . . . . . . . . . . . . . . . . . . . . . 11 Chel-Grundglied kürzer, bis 4,2 mm, do Do kürzer als der Durchmesser der Chel, in rechtem Winkel abstehend (Abb. 344- 345); in Höhlen der Karawanken unrl Steiner Alpen . . .
· hadzii, S. 207
Genus Ischyropsalis 189
11 Che!-Grundglied do-di knotig aufgebläht; in Höhlen der Venetianer Alpen (Abb. 354-355) ravasinii, S. 210
Chel-Grundglied do-di nicht knotig aufge-bläht (Abb. 309- 311) in Höhlen des Monte Baldo und der Monti Lessini . . . . . : . . . . . . . . . . . . . . . . . strandi, S. 195
12 Abd do mit Souturn parvum (Sklerite I-V verschmolzen, z. B. Abb. 361) . . . . . 13 Abd do mit S. laminaturn (Sklerite isoliert, Abb. 334) . • . . . . . . . . 14
13 Chel-Grundglied massig gedrungen, do zahlreiche unterschiedlich große Do ohne distinktes Lagemuster (Abb. 301); weitverbreitet in Mittel-Etlt'opa mit 0-Alpen . . . . . . . •
hellwigi hellwigi, S. 190 Chel-Grundglied schlank, do mit I Reihe aus 3 großen Do (Abb. 367); W-Alpen zwischen Bodensee und Genfer See . . . . . . . . . . . . . . . . . . . . . . carli, S. 213
14 Chel-Grundglied massig-gedrungen oder lang-grazil, Bewahrung mit zahlreichen unterschiedlich großen Do, davon do entweder 3-5 in Längsreihe markant hervortretend, selten ungeordnet verteilt, länger oder kürzer als der Dtlt'ohmesser des Grundgliedes . . . 15 Chel-Grundglied auffallend lang und dünn, do ntlt' spärliche Bedornung, die einzelnen Do immer wesentlich kleiner als der Dtlt'chmesser des Chel-Grundgliedes. . . . . . • . 20
15 Chel-Grundglied massig-gedrungen, Do auf dem Chel-Grundglied do klein, Do nie länger als dessen halber Dtlt'ohmesser im mittleren Bereich (Abb. 338, 374) 16 Chel-Grundglied lang-grazil, do Do auf dem Chel-Grundglied groß, wenigstens einzelne länger als dessen Dtlt'chmesser an der Basis (Abb. 312, 321, 356-358) . . . . . . . 21
16 do Do des Chel-Grundgliedes nicht zu markanter Reihe geordnet, dicht stehend (Abb. 338-339), in den 0-Alpen weitverbreitet . . . . . . . . . . . . . . . • kollari, S. 203
~ we!lig:stens die größten Do des Chel-Grundgliedes zu I Längsreihe ausgerichtet (z. B. Abb. 321, 346, 374) . . . . . . . . . . . . . . . . . . . . . . . . 17
17. d.o Do des Chel-Grundgliedes in rechtem Winkel abstehend (Abb, 346); in Höhlen der Karawanken und Steiner Alpen. . . . . . . . . . . . . . . . . . . . hadzii, S. 207 do Do des Chel-Grundgliedes distad geneigt . . . . . . . . . . . . . 18
18 do Do dicht beieinander stehend, breit konisch (Abb. 374-375), weitverbreitet im Karpa-ten-Bagen . . . . . . . . . . . . . . . . . . . . . . . . . . . manioata, S. 216
· do Do dtlt'ch breite Lücken getrennt, schmal konisch (Abb. 322, 401) . . . . . . . 19 19 'abd Sklerite jeweils mit Querreihe kleiner Borsten, in Höhlen der Penninischen und Berga
'masker Alpen . . . . . . . . . . . . . . . . . . . . . . . . dentipalpis, S. 197 abd Sklerite ohne Querreihe kleiner Borsten; in (? Höhlen) der S-Schweiz helvetica, S. 219
20 cli Hälfte des Chel-Grundgliedes do glatt, allenfalls mit kleinen Tuberkeln (Abb. 401), in . Höhlen der Cottischen Alpen . . . . . . . . .- . . . . . . pyrenaea alpinula, S. 224 di Hälfte des Chel-Grundgliedes do so bewehrt wie an der ba Hälfte (Abb. 327-328), in Höhlen der SO-Alpen und NW-Jugoslawiens . . . . . . . . . . . mqellneri, S. 200
21 nur in Höhlen des Monte Baldo und der Monti Lessini . . . . . . . . . strandi, S. 195 nur in Höhlen der Venetianer Voralpen . . . . . . . . ravasinii, S. 210 nur in Höhlen der Penninischen und Bergamasker Alpen. . . . . . dentipalpis, S. 197
Ischyropsalis hellwigi (PANZER, 1794) Abb.44, 49,298--308,319,362
Bestimmungstabelle für die Subspezies
Area des abd Souturn dtlt'ch seichte Furchen deutlich gegeneinander abgesetzt, Truncus penisunterhalb der Glans nur schwach geflügelt; weitverbreitet in Mitteletlt'opa .
hellwigi hellwigi, S. 190 Area des abd Souturn nahezu ftlt'chenlos ineinander übergehend, Truncus penis unterhalb
· cier Glans stark geflügelt; lokal in den Pyrenäen und Kantabrischen Gebirgen . . . . . hellwigi Jucantei, S. 194
.-.
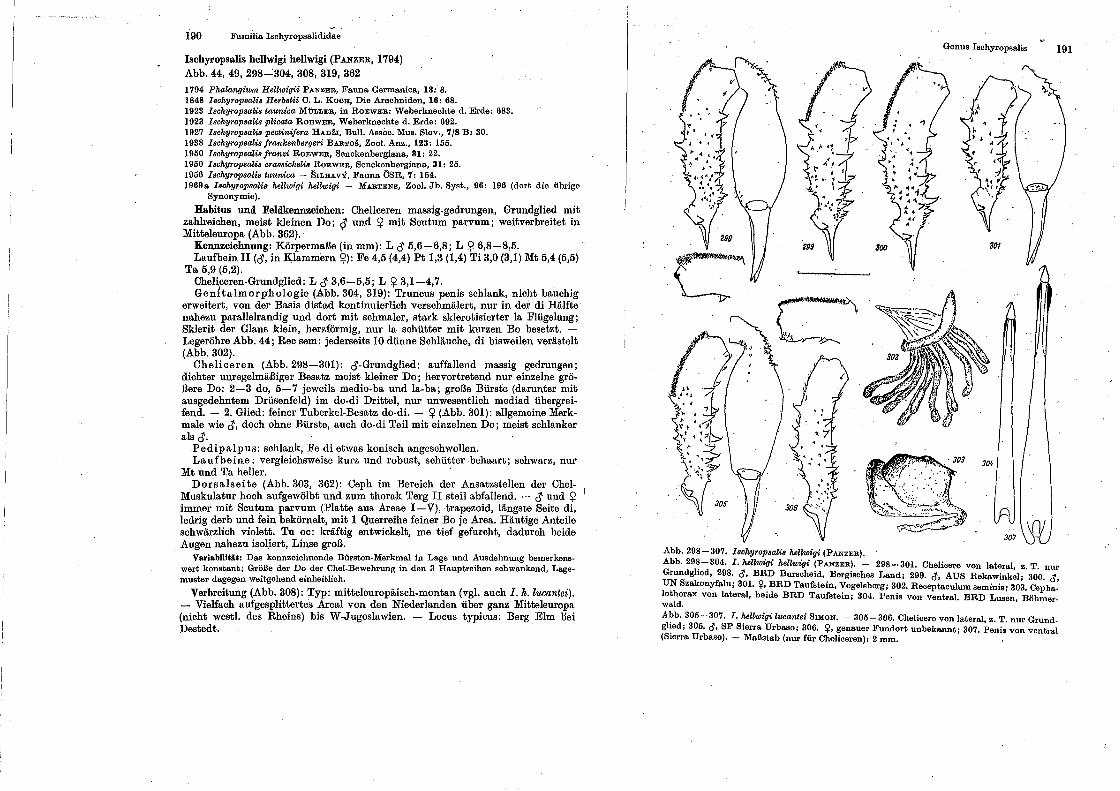
190 Familia lschyropsalididae
Ischyropsalis hellwigi hellwigi (PANZER, 1794) Abb.44, 49,298~304,308, 319,362
1794 Phalangium Hellwigii PANZER, Fauna Germanica, 13: 8. 1848 Iavhyropaalis Herbstii C. L. KooH, Die Arachniden, 16: 68. 1923 Isohyropsalis taunica MÜLLER, in ROEWER: Weberknechte d. Erde: 683. 1923 Isvhyropsalis plicata ROEWER, Weberknechte d. Erde: 692. 1927 Isvhyropsalis pectinijera HADZI, Bull. Assoc. Mus. Slov., 't/8 B: 30. 1938 Isohyropsalis jrankenbergeri BARTOS, Zoo!. Anz., 123: 155. 1950 Isohyropsalisjranzi RoEWER, Senckenbergiana, 31: 22. 1950 Isohyropsalis orassiohelis RoEWER, Senckenbergiana, 31: 25. 1956 Isohyropsalis tauniva - !lrLHAVY, Fauna ÖSR, 1: 154. 1969a Isohyropsalis hellwigi hellwigi - MARTENS, Zool. Jb. Syst., 96: 196 (dort die übrig<>
Synonymie).
Habitus und Feldkennzeichen: Cheliceren massig.gedrungen, Grundglied mit zahh·eichen, meist kleinen Do; iJ un.d ~ mit Souturn parvum; weitverbreitet in Mitteleuropa (Abb. 362).
Kennzeichnung: Körpermaße (in mm): L (J 5,6-6,8; L ~ 6,8-8,5. · Laufbein II ((J, in K;lammern ~):Fe 4,5 (4,4) Pt 1,3 (1,4) Ti 3,0 (3,1) Mt 5,4 (5,5)
Ta 5,9 (5,2). Cheiiceren-Grundglied: L (J 3,6-5,5; L ~ 3,1-4,7. Genftalmorphologie (Abb. 304, 319): Truncus penis schlank, nicht bauchig
erweitert, von der Basis distad kontinuierlich verschmälert, nur in der·di Hälfte nahezu parallelrandig und dort mit schmaler, stark skierotisierter la Flügelung; Sklerit der Glans klein, herzförmig, nur la schütter mit kurzen Bo besetzt. -Legeröhre Abb. 44; Rec sem: jederseits 10 dünne Schläuche, di bisweilen verästelt (Abb. 302). .
Cheliceren (Abb. 298-301): (J-Grundglied: auffallend massig gedrungen; dichter unregelmäßiger Besatz meist kleiner Do; hervortretend nur einzelne größere Do: 2-3 do, 5-7 jeweils medio-ba und la-ba; große Bürste (darunter mit ausgedehntem Drüsenfeld) im do-di Drittel, nur unwesentlich mediad übergreifend. - 2. Glied: feiner Tuberkel-Besatz do-di. - ~ (Abb. 301): allgemeine Merkmale wie (J, doch ohne Bürste, auch do-diTeil mit einzelnen Do; meist schlanker als (J.
Pedipalpus: schlank, Fe di etwas konisch angeschwollen. Laufbeine: vergleichsweise kurz und robust, schütter behaart; schwarz, nur
Mt und Ta heller. · Dorsalseite (Abb. 303, 362): Ceph im Bereich der Ansatzstellen der Chel
Muskulatur hoch aufgewölbt und zum thorak Terg II steil abfallend. - (J und ~ immer mit Souturn parvum (Platte aus Areae 1-V), trapezoid, längste Seite di, ledrig derb und fein bekörnelt, mit 1 Querreihe feiner Bo je Area. Häutige Anteile schwärzlich violett. Tu oc: kräftig entwickelt, me tief gefurcht, dadurch beide Augen nahezu isol.iert, Linse groß.
Variabilität: Da>! kennzeichnende Bürsten-Merkmal in Lage und Ausdehnung bemerkenswert konstant; Größe der Do der Chel-Bewehrung in den 3 Hauptreihen schwankend, Lagemuster dagegen weitgehend einheitlich.
Verbreitung (Abb. 308): Typ: mitteleuropäisch-montan (vgi. auch I. h. lucantei). - Vielfach aufgesplittertes Areal von den Niederlanden über ganz Mitteleuropa (nicht westi. des Rheins) bis W-Jugoslawien. - Loous typicus: Berg Elm liei Destedt.
Genus Ischyropsalis 191
Abb. 298-307. lavhyropsalis hellwigi (PANZER). Abb. 298-304. I. hellwigi hellwigi (PANZER). - 298-301. Chelicere von lateral, z. T. nur Grundglied, 298. ~. BRD Burscheid, Bergisches Land; 299. ~. A(TS Rekawinkel; 300. ~. UN Szakonyfalu; 301. ~. BRD Taufstein, Vogelsberg; 302. Receptaculum seminis; 303. Cephalothora.x von lateral, beide BRD Taufstein; 304. Penis von ventral. BRD Lusen, Böhmerwa.ld.
Abb. 305-307. I. hellwigi lucantei SalON, - 305-306. Chelicere von lateral, z. T. nur Grundgl~ed; 305. ~. SP Sierra Urbaso; 306. ~. genauer Fundort unbekannt; 307. Penis von ventral (Sterra Urba>!o). - Maßstab (nur für Cheliceren): 2 mm. .

192 Familia Ischyropsalididae
""' . ~· "'-T T'' t IIT • /schyro;;,aiis hellwlgi hellwigi
T /, hettwigi M:an~
Abb. 308. Punktkarte der 'Verbreitung von lsohyropBaliB hellwigi hellwigi (PANZER) und I. hellwigi lucantei SIMON.
Niederlande: Meerdink bei Winterswijk(SPOEK\n litt.). - BRD und DDR: wahrschein· !ich in den Mittelgebirgen allgemein verbreitet, lokal auch im Flachland; zwischen Main und Inn über weite Strecken spärlich oder völlig fehlend; nicht in den links-rheinischen Mittelgebirgen. -Nordwest!. Mittelgebirge und Vorland: Wuppertal-Beyenburg; ~affrat~1, Wüstenhammer, Barmer Talsperre (alle LösER leg.); Sauerland, Naturschutzgebiet Stimmstamm (RENSOH 1955); Münsterland, Wolbeck (FARWIOK & RADE 1878), Herford (B~NER 1~55); Elm bei Destedt (PANZER 1794); bei Göttingen (BELLMANN 1975); - westl. Mlttelgebuge: Siebengebirge (VERHOEFF 1900); Taunus: Schlangenbad, Altkönig, Hohemark, Münster, Feldberg (HESSE, v. HEYDEN, A. MüLLEP., ScHAWALLER, ZUR STRASSEN leg .. 1820-19?6); zwischen Frauenstein und Geergenborn (HAACKER leg.); Vogelsberg, Taufstern und Geiselstein (J. M. leg., PIEPER leg.); Rhön: Schafstein, Lettengraben, Milseb~_g, Ste.llberg (_alle PIEPER in litt.); Rupboden bei Bad Brückenau (NIETHAMMER leg.); - nordostl. Mitteigebuge und Vorland: Thüringer Wald, Inselsberg; Bürge! bei Jena (beide HoFFMANN in litt.); Naunhof südöstl. Leipzig (HIEBSCH in Iist.); Erligt bei Frohburg (MARTIN 1971); Erzgebirge, Fichtelberg-Gebiet (BÜTTNER 1930); - südl. und südöstl. Mitt~lgebirge: ?den~ald: Hirschhorn (WIDER, unveröff. MS); Waldmichelbach (HEINZ leg.); Bärental bei Hatdeiberg (JANDER mdl.); Spessart (HAHN H\34); Franken: Umgebung von Erlangen (PFEIFER 1956); Pretzfeld (MATTHES & GLANDT 1971); Schwarzwald und Stuttgart: Neuenbürg bei Karlsruhe (Wt;,NDER· LICH leg.); Herrenalb (v. HAGEN 1973); Stuttgart-Rotenberg (WEHRMAKER leg.); - Bohmerwald: Berge Lusen, Rache! und Arber (alle J. M. leg.); Dreisesselberg und Dreitannenriegel (GAUCKLER in litt.); - Alpen und Alp.envorland: Spitzingsee bei Schliersee. (FALTERMEIER leg.); Berchtesgaden, oberhalb der Wimbachklamm (PIEPER leg.). - Polen: m den Sudeten,
Genus IschY,.opsalis 193
m E!Je:!y-Massiv und, weit östlich vorgeschoben, in der Umgebung von Ojc6w im Jura zwischen Czestochowa. und Kra.k6w (Kraka.u) (STAR~GA 1976 a); im einzelnen: RiesengebirgeJKrkonolle, Snezne Jamy (BARTOs 1938), Bystrzyca Klodzka, Mieldtygorze (Beleg PANW); Landeck/ Glatz (Beleg MNB), G6ra Sob6tka. (RAFALSKI 1961). - ÖSSR: Böhmen: 1\farianskß Lazne (Marienba.d) (1liLHAVY 1966), Mittelgrund/Eibe (BARTes 1938), Altvater/PradM, Schlucht des Oppafalles (ZAVREL 1930) ;_Mähren: Rimov bei Ti'ebiö (1liLHAVY 1958), Höhle Byci Skala bei Brne/Brünn; Morava-Be~kydy, Lysa hora (beide Belege Coll. 1liLHAVY); Schutzgebiet Kluöi bei Ti'est/Jihlava (ROHACEK 1971).
Österreich: weitverbreitet in der Montan- und lokal in der Subalpin-Stufe der Alpen, seltener im Mittelgebirge (Böhmische Masse); fehlt im pannonischen Gebiet und westl. der Umgebung von Innsbruck: ""'": Tirol: ·"Innsbruck" (Beleg NHMW); Sellraintal, Egghöfe bei St· Quirin (HAUSER leg.); Oberperfuß-R"angger Köpfl (THALER 1966 b); ValserTal, Tscheischbaoh (KRAUS leg.); '- Salzburg: Trockene Klammen/Elsbethen (ScHÜLLEB leg., A. A.leg.; Belege SMF); Saalfelden, Kaltenbachtal (HuRDES leg., Beleg SMF); Untertauern bei Radstadt; Lung_a,u, Rotgülden; Stubaal,ttal (alle A. A. leg.); - Ober-.Österreich: Sauwald, Kesselbachtal (J. G.leg.); - Nieder-österreich: Kalkvoralpen, Gscheid südl. der Reisalpe; Edlitz (PAULus leg.) 1 Wienerwald, R~kawink~l; mittlerer Rotgrabe~ westl. Klosterneuburg (alle J,. (}. leg.); ...,.-Wien: Wienerwald, am Kalbeterberg und am Heuberg (beide J. G.leg.); - Steiermark: ;De.utschlandsberg (STROUHAL leg.);' Poßruck' südl. Schloßberg '(J. G. leg.); Windische Büheln; 2km westl. Leutschach (KREISSL & J. G.leg.); Lichtmaßgraben bei Admont (Beleg SMF)r - Kärnten: Karawanken, Obir und Lobniggraben (ScHEERPELTZ leg.); Bärental (A. A . .Jeg.); Drauta.I' oberhalb Villaoli (Beleg SMF); Vellachta.l, Seifengraben (STROUHAL leg.); Koralpe, St. Lorenzen und bei St. Vinzenz (beide FRANZ leg.); Viktring bei Klagenfurt (PAULUS leg.);. Villaeher Alpe (J. M. leg,); Wörther See (Beleg SMF); Windisch-La,ndsberg (HAnl!I 1954, taunica). - Unga~n: 0-Rand der Alpen, Güns~r Gebirge, Köszeg/GiiJ1s (KOLOSVABY 1965); Höhle bei Szakonyfalu (LOKSA leg., Beleg UNMB). - Jugoslawien: Slowenien: Friedrichsteiner Wald südl. Gottschee (J. G.leg.); Umgebung von Cerknica (J. M. leg.); -. Kroatien: Zagreh (Beleg SMF); Brod Moravice im Gorski Kotar (Beleg PANW); Berg Kum, Zla.tica (HADZr 1942, t~>unica); -Bosnien-Herzegowina: Trabevia planina bei Sarajevo (HADZI 1954; südlichster Ftindort, taunica). · . -.
Ökologie: Biotopansprüche: Unter den hier behandelten Ischyropsalis-Arten ist hellwigi am wenigsten spezialisiert, stellt aber dennoch so hohe Ansprüche hinsichtlich Feuchtigkeit und Temperaturgang, daß sie meist nur lokal vorkommt. Sie lebt überwiegend in geschlossenen Laub- und Nadelwäldern der Mittelgebirge und der Alpen, nur selten in solchen des Flachlandes. Die oberen Bodenschichten müssen ständig durchfeuchtet und kleinräumig strukturiert sein ~ meist durch Fallaubsohichten, überwuchertes morsches Holz und Gesteinstrümmer. Die Nähe fließ~nden Wassers benötigt hellwigi viel weniger als z. B. kollari und carli; ausnahmsweise auch in offenem Gelände, meist nur an der oberen vertikalen Arealgr~nze (z. B. Villaeher Alpe, 1900 m). In forstlich intensiv bewirtschafteten Wäldern ist hellwigi selten oder fehlt ganz. Vgl. auch LÖSER (1977). ·
•Vertikalverbreitung: vom Flachland nördl. der Mittelgebirge (Niederlande,. .50 ·m) bis in die untere und ausnahmsweise auch obere Subalpin·Stufe der Alpen. -Öst~rreioh; in Salzburg von 640 bis 1530 bzw. 1550 m in den Zentral-Alpen (A. A. 1eg.); in N-Tirol etwa bis 1600 m (K. T. leg.); in Kärnten, Villaeher Alpe, bis in .die.J.atschel).zone bei etwa 1900 m (J. M, leg.); tiefstgelegene Fundorte im Wienarwald ,um 300m, in der Steiermark 330m, im Sauwald (außeralpin) 300-m (alle ,J. G. in litt.).
In den Hohen Tauern überschneiden sich die Arealgürtel von hellwigi (obere Grenze) und von kollari (untere Grenze) auf etwa 100-250 m; beide Arten leben in diesem Bereich syntop. Ob dieser auffallend schmale Gürtel gemeinsamen Vor-13 Tierwelt Deutsch]; 64: Martens
•

·---·-·-··--·---194 Familia Ischyropsalididae
kommans durch bestimmte Biotoppräferenzen oder durch Konkurrenz bedingt ist, läßt sich vorerst nicht entscheiden.
Phänologie: stenochron sommer-und herbstreif. - BRD: VogelsbergfTaufstein: ad. AVII-E X; iuv. bis M-E IX; Gelege überwintern. Ob iuv. aus IX zu überwintern vermögen, ist bisher nicht belegt und kaum wahrscheinlich (MARTENS 1969a). - Österreich: iuv. ausnahmsweise bis EX undankleinklimatisch günstigen Biotopen vielleicht überwinternd (A. A. in litt.). -Eiablage an die Unterseite feuchter Holzteile, an.Moos und Steine. Die Eier sind in Gallerte eingehüllt die durch Wasseraufnahme quillt und die Eier voneinander trennt. - Eizahl (nach Laboruntersuchungen): je ~ 3-5 Gelege mit je 3-39 Eiern, insgesamt mind. 60 Eier. Embryonalentwicklung bei 10-11 °0 70 und 77 Tage (2 Fälle); bis zur Adulthäutung 6 Jugendstadien, Lebensdauer in Gefangenschaft etwa 7 Monate (bis I des folgenden Jahres; alle Daten J. M.).
Biologie: Die Cheliceren-Drüse hat Funktion im Fortpflanzungs-Verhalten, besonders in der Werbung, die nach dem Typ der gustatorischen Balz verläuft: Unmittelbar vor der Kopulation bringt das er die Bürsten-Region (und somit, das zugleich austretende Sekret) seiner Chel-Grundglieder an die Mundregion des~. das den Vorderkörper und die ersten beiden Beinpaare vom Substrat abhebt. Mit seinen Cheliceren verklammert es sich an Cheliceren und/oder Pedipalpus des ($. Erst .,d,ann findet die Kopulation statt (Abb. 49; MARTENS 1969b; M.!BTENS 1975a). -I. hellwigi ist ein Nahrungsspezialist, der sich vorzugsweise von Gehäuse, und Nacktschnecken ernährt. Auch hartschalige Gehäuse vermag er mit seinen gedrungen-massigen Chel leicht aufzubrechen (VERliOEFF 1900, MARTENS 1965; 1975b).
Ischyropsalis hellwigi lucantei SrMoN, 1879 Abb. 305-308
1879 Ischyropsalis lucantei SrMON, Arachn. d. France, 7: 273. 1881 Ischyropsalis superbus SrMON, Ann. Soo. espan. Hiat. Nat., 10: 129. 1969a Ischyropsalis hellwigi lucantei - MARTENS, Zoo!. Jb. Syst., 96: 200 (dort die übrige
SY!J.onymie). ,
Kennzeichnung: Mit den Maßen und Merkmalen der Nominatform; abweichend: Areae des abd Scutum nahezu furchenlos ineinander übergehend; Truncus penis unterhalb der Glans stark geflügelt (Abb. 307).
Verbreitung (Abb. 308): Typ: Endemit der Kautabrisehen Gebirge und der ·Pyrenäen (vgl. auch I. h. hellwigi). - Locus typicus: Grotte de Betharram heil Bagneres de Bigorre.
Frankreich: Fundorte in den DepartementsHautes-und Basses-Pyrenees. -Spanien: östl. Kautabrisehe Gebirge: Sierra Urbaso, Sierra de Ara!ar (Einzelheiten in MARTENB 1969a).
Ökologie: Biotopansprüche: In Höhlen und Freiland, von nahe Meereshöhe bis mindestens 650 m (Sierra de Aralar).
Phänologie: wahrscheinlich stenochron sommerreif in V -X; im Gegensatz zur Nominatform überwintern iuv. (1 und Gelege) (MARTENS 1969a: 145).
. lschyropsalis strandi KRATooliViL, 1936 Abb.309-315, 318,332,394
Genus Isohyropsalis 195
1936 Ischyropsal~s strandi KRA.Toonvfr., Festschrift Strand, 1: 249. 1947 IschyropsaZ.s ruffoi CA.PORIAOOO, Mem Mus Civ Verona 1• 139 1950 lechyropsalis knirschi ROEWER, Senok~nber~an~, 31: 27: ' ' 1,969a Ischyropsalis strandi - MARTENB Zoo! Jb Syst 96· 239 (do t d' "b · s ·
• 1 • • ., • r 1e u .r1ge ynonym.1e).
_Habitus und Feldkennzeichen: Laufbeine und Cheliceren auffallend lang diese mitldBewehrung aus !p'Oßen und dicht stehenden Zapfen; nur in Höhlen des Monte Ba o un~ der Monti Lessini (Abb. 332).
Kennzeichnung: Körpermaße (in mm): L er 4 6-6 2· L ~ 5 4-6 9 Laufbein II (er, in Klammern ~; Spluga Car~ene): Fe 7 7 (7 5) Pt 15 (15) T'
6,5 (6,2) Mt 10,5 (10,1) Ta 9,8 (9,2). ' ' ' ' 1
Cheliceren-Grundglied: L er 5,3-5,5; L ~ 4,7-5,8.
313
Abb. 309-3~5. Ischyropsalis strandi KRAToOHVfL. - 309-312. Chelioere von lateral, z. T. nur Grun.dghed; 309. (S; 310. (S; 311. !?; 312. !?, alle IT Monte Baldo; 312. Buoo della Preta· 313. Pems von ventral; 314. Reoeptaculum seminis; 315. Cephalothorax von lateral (all~ · Monte Baldo). - Maßstab (nur für Cheliceren): 2 mm. 18*

196 Familia Ischyropsalididae
Geni talmorpha"logie (Abb. 313, 318): Truncus penisnicht .baucuig, ... von d!lr Basis distad kontinuierlich verschmälert, etwa im letzten Drittel nahezu parallelrandig; Sklerit der Glans breit und lang, Ia wenig gerundet, basad undeutllch 2zipflig, somitmedio-basad nur schwach konkav, Beborstung spärlich in 2 achmalert para-me Reihen, Ia markanter Kranz kurzer basad an ·Länge zunehmenderJ3o; aiie Bo derb. - Rec sem: Schläuche· dick, in der ba Hälfte. mihezu rechtwinklig abge-bogen; beidseitig 3 oder 4 Schläuche (Abb~ 314). · ··· ·
Chelice·ren (Abb. 309-312): · &; Grrindglied:· .. schlank ·und ·auffallen(l lang (strandi ·hat neben ravasinii die: längsten Cheliceren · aller Ischyropsalis-Arten) ;. do 5-10 unterschiedlich große distad geneigte Do vor aiiem in ·der ba Hälfte;' ve-la 7-10, ve-me 9~11 Do; oft an beiden Gliedern unterschiedlich ausgeprägt; kleinere Tu la ·und spärlich medial, nilr teilweise in die Hauptreihen einbezogen; Grundglied do-di etwas angeschwollen, doch ohne Apo i:tnd ohne Bürsten-Feld. 2. Glied: an der Basis frontal niedrige Tu und einzelne Bo. - ~: ohne distinkte Unterschiede zum a. . .
Pedipalpus: &-Pt me-di bisweilen mit Apo-artiger Vorwölbung, alle Glieder fein beborstet. · · 1
Laufbeine: Auffallend lang, dunkelbraun, nur Mt und Ta heiigelb; schütter mit feinen Haaren besetzt. Dors~lseite (Abb. 315, 332): Ansatzstellen der Chel-Mus~ulatur hinter der
Augenregion (soweit vorhanden!) deutlich erhaben, zum thorak Terg Il steil abfallend, dieses mit 8-14 kräftigen, di gerichteten Do; Ceph kräftig granuliert und mit einzelnen größeren borstentragenden Tu. - Tu oc: variabel: entweder stark reduziert oder einschließlich der Augen völlig fehlend. ·- Scutum: & mit S. intermedium, ~ mit S. laminaturn; &-Scutum oft fast bis zum S. laminaturn zergliedert,
· doch wenigstens 2 Areae immer. in losem Zusammenhang, entweder I/II oder II/III; meist auch I und IV mit kleinen Kontaktstellen zu den benachbarten Areae, V fast immer isoliert (Abb. 332). Areae rauh, mit je 1 Querreihe niedriger Tu und jeweils aufgesetzter langer Bo. Cx, vor allem Cx I, und Op gen warzig i:Jehöckert und mit langen Bo.
Variabilität: Sie betrifft die Ausprägung von Tu oc und Augen, das &-Scutum und die Chel-Bewehrung. - Tu oc: nach Populationen unterschiedlich; entweder einschließlich der Augen fehlend; auch Retinazonen rückgebildet· (Abb.14d; JUBERTmE 1964: 104), doch soll nach MUNOZ-CUEVAS (1976) eine rudimentäre Retina vorhanden sein. Der Cornea-Bereich ist in diesem Falle degeneriert, und an seine Stelle tritt eine Kavität in der Kutikula (MUNoz-CuEVAS 1976, Populationen der Höhlen Buso dell'Arena und Buso Rena). In anderen Populationen kann der Tu oc angedeutet sain und winzige pigmentlose Augen tragen. Bisweilen sind diese nur einseitig ausgebildet; ·W!lnn beide vorhanden, dann meist unterscJ.!.iedlich stark ausgeprägt (Abb. 315, 332). - Vor allem die Augenreduktion und d,ie geringe Sklerotisation des Souturn weisen strandi als extrem adaptierte troglobionte Art aus.
Verbreitung (Abb. 394): Typ: ·Endemit des Monte Baldo und der Monti Lessini. - Locus typicus: Grotta dei Cervi bei Malga Prato.
Italien: Monte Baldo, Grotta dei Cervi bei Malga Prato (KRATOOHV:iL 1936, JUBE:&THIE 1963, 1964); .Belege vom Monte Baldo, wahrscheinlich alle aus derselben Höhle, in den Sammlungen NHMW·und SMF (sub knirschi). - Monti Lessini: Nachweise stammen aus folgenden Höhlen: .Cogolo de Zavatin; Bus· de Ia Fanta; Spluga de Ia Preta (auch FOLLIS leg.); Grotta del Monte Caobo; Covolo di Velo, Buso dell' :Arena, Busode Ia Neve de Ia Gaibana, Busode
Genus Ischyropsalis 197
la.Nef de Ia Coe (alle nach·.:!"uBERTHIE 1964); Spluga Carpene, S. Mauro ~i Saline (CASALE leg., VIGNA leg.), Buso Vaio Spiazzoletti, Selva di Progno (CoRltADI leg.);"Buso della Rana bei Malo (CASALE leg.), wahrscheinlich identisch mit Buso Rena bei Mu:Noz-CuEVAS (1976).
Nennungen von strandi für die Bergamasker Alpen (Val Seril),na, Grotte Bus del Tacoi) und in den Venezianer Alpen (Cansiglio-Massiv; MARTENS 1969a) haben sich an Hand umfangreicheren un:d neu gesammelten Materials nicht bestätigen lassen (vgl. unter dentipalpis und ravasinii). · 1
Ökologie: Biotopansprüche: I. strandi ist in seinem engbegrenzten ~eal ausschließlich troglobiont und wurde noch nie im Freiland gefunden.· Alle b1sher näher untersuchten Höhlen liegen hoch im Gebirge (zwischen 875 und 1691 m, die meisten über 1400 m) und zeichnen sich durch besonders niedrige Temperaturen aus: JuBERTmE (1974) gibt Werte zwischen 1,5 °0 (Buso de la Neve de la Gaibana, 1691 m, VII. 1962) und 8 °0 an (Grotta del Monte Caobo1 Covolo di Velo, 875 m, XI. 1971). . ·
Phänologie: In den Höhlen sind ad. und iuv. während des ganzen Jahres anzutreffen. Die Eier werden einzeln meist an feuchte Felspartien der Höhlenwände abgelegt -eine Ausnahme bei Ischyropsalis, denn sonst immer in größeren_Gruppen in Schleimpaket. Eiablage einzeln im Abstand von mehreren ~agen (b1s Wochen ~). 2 ~ legten 45 Eier in 9 Monaten (bi~ zum ~ode) bzw. 59 m 1 ~ahr; umgerechnet auf die Lebensdauer im Adultstadmn;t erg1bt ~as etwa 120 ~1er. Dauer der· Embryonalentwicklung 142 (± 4) Tage be1 5-6,5 C. Das geschatzte Temperatur-Optimum der Entwicklung liegt bei etwa 10 °0, bei 1_2 °0 setzt die Embryonalentwicklung nicht ein. Dauer der Postembryonal-EntwiCklung unbekannt, Lebensdauer der ad. etwa 2,5 Jahre (alle Angaben nach JUBERTHIE 1974).
Vertikalverbreitung: Die von strandi besiedelten Höhlen liegen zwischen 875 und 1691 m · diese Beschränkung auf hoch gelegene und somit kalte Höhlen läßt sich mit de~ Anpassung der Embryonal-Entwicklung an niedrige.,_T~mp?raturen begründen. Diese Adaptation hat möglicherweise während der Glaz1alze1ten in den Refugialgebieten des Baldo-Lessini-Massivs stattgefunden, also im rezenten Areal. Zu aktiver Arealerweiterung ist strandi un~r jetzigen Klimabedingungen nicht fähig.
Ischyropsalis dentipalpis CANESTRINI, 1872 Abb. 317, 320-323, 394
1872 Ischyropsalis dentipa!pis CANESTRINI, Ann. Mus. Civ. Stor. natur. Genova, 2: 9. 1923 Ischyropsalis dentipa!pis - ROEWE:&, Weberknechte d. Erde: 686.
Begründung der Namenszuordnung: Der Locus typicus von dentipa!pis ist das Aosta-Tal in den Penninischen Alpen; MARTENS ( 1969 a) lag kein topatypisches Material vor; "dentipa!pis" aus dem Tessin (DRESOO 1959), das li<!A.RTENS (1. c.) zur Kennzeichnung dieser Art heranzog, gehört trotz äußerer .Ähnlichkeiten in eine ganz andere Artengruppe (siehe unter he!vetica). Ob die beiden weit disjunkten Populationen aus den Penninischen und Bergamasker Alpen (vgl. unten), die hier zu dentipa!pis gestellt werden, tatsächlich unter einem Namen zusammengefaßt werden dürfen, muß an Hand neuen Materials aus dem Aosta-Tal überprüft werden.
Habitus und Feldkennzeiohen: Chel-Grundglied do-diknotig aufgetrieben, Souturn parvum (&) bzw. S.laminatum (~)- & unter Feldbedingungen nicht von helvetica zu trennen, ~ ununterscheidbar. Endemit der Penninischen und Bergamasker Alpen.
\
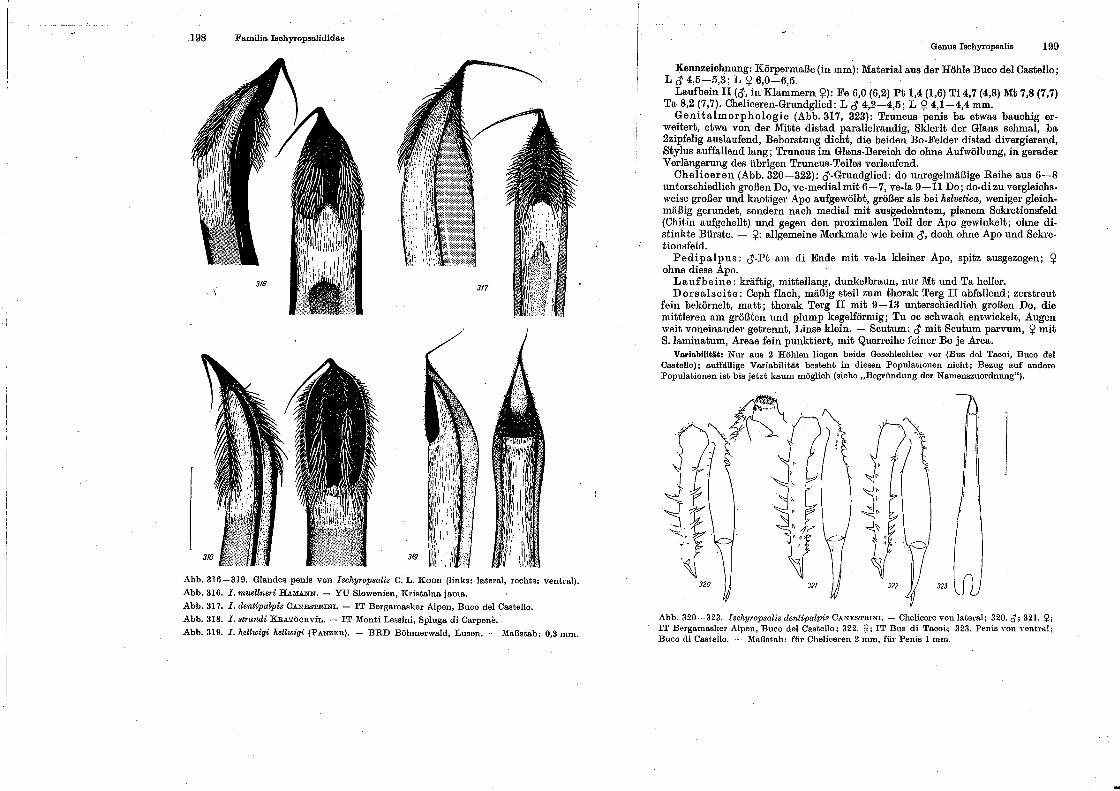
.198 Familia. Iachyropaalidida.e
319
Abb. 316-319. Glandes penia von IBchyropBalis C. L. KocH (links: lateral, rechts: ventral).
Abb. 316. I. muellne•·i IIAMANN. - YU Slowenien, Kristalna jama.
Abb. 317. I. den!ipalpiB ÜANESTlllNI. - IT Berga.masker Alpen, Buco del Ca.stello.
Abb. 318. I. stmndi KRATOOHViL. - IT Monti Lessini, Spluga di Carpene.
Abb. 319. I. hellwigi hellwigi (PANZER). - BRD Böhmerwald, Lusen. - Maßstab: 0,3 mm.
Genus Iachyropsalis 199
Kennzeichnung: Körpermaße (in mm): Material aus der Höhle Buco del Castello; L ~ 4,5-5,3; L ~ 6,0-6,5.
Laufbein II (~.in Klammern~): Fe 6,0 (6,2) Pt 1,4 (1,6) Ti 4,7 (4,8) Mt 7,8 (7,7) Ta 8,2 (7,7). Cheliceren-Grundglied: L ~ 4,2-4,5; L ~ 4,1-4,4 mm.
Genitalmorphologie (Abb. 317, 323): Truncus penis ba etwas bauchig erweitert, etwa von der Mitte distad parallelrandig, Sklerit der Glans schmal, ba 2zipfelig auslaufend, Beborstung dicht, die beiden Ba-Felder distad divergierend, Stylus auffallend lang; Truncus im Glans-Bereich do ohne Aufwölbung, in gerader Verlängerung des übrigen Truncus-Teiles verlaufend.
Cheliceren (Abb. 320-322): ~-Grundglied: do unregelmäßige Reihe aus 6-8 unterschiedlich großen Do, va-medial mit 6-7, ve-la 9-11 Do; do-di zu vergleichsweise großer und knotiger Apo aufgewölbt, größer als bei kelvetica, weniger gleichmäßig gerundet, sondern nach medial mit ausgedehntem, planem Sekretionsfeld (Chitin aufgehellt) und gegen den proximalen Teil der Apo gewinkelt; ohne distinkte Bürste. - ~: allgemeine Merkmale wie beim~. doch ohne Apo und Sekretionsfeld.
Pedipalpus: ~-Pt am di Ende mit ve-la kleiner Apo, spitz ausgezogen; ~ ohne diese Apo.
Laufbeine: kräftig, mittellang, dunkelbraun, nur Mt und Ta heller. Dorsalseite: Ceph flach, mäßig steil zum thorak Terg II abfallend; zerstreut
fein bekörnelt, matt; thorak Terg II mit 9-13 unterschiedlich großen Do, die mittleren am größten und plump kegelförmig; Tu oc schwach entwickelt, Augen weit voneinander getrennt, Linse klein. - Scutum: ~ mit Scutum parvum, ~ mit S. laminatum, Areae fein punktiert, mit Querreihe feiner Bo je Area.
Variabilität: Nur aus 2 Höhlen liegen beide Geschlechter vor (Bus del Tacoi, Buco del Castello); auffällige Variabilität besteht in diesen Populationen nicht; Bezug auf andere Populationen ist bis jetzt kaum möglich (siehe "Begründung der Namenszuordnung").
I I
323 ul Abb. 320-323. Ischyropsalis dentipalpis ÜANESTRINI.- Chelicere von lateral; 320. 6; 321. <;?; IT Bergamasker Alpen, Buco del Castello; 322. ~; IT Bus di Tacoi> 323. Penis von ventral; Bueo di Castello. - Maßstab: für Cheliceren 2 mm, für Penis I mm.

200 Familia Isohyropsalididae
Verbreitung (Abb. 394): Typ: (überwiegend) troglobionter Endemit der.SWAlpen. - Locus typicus: Aosta-Tal, Gressoney St. Jean.
Italien: Penninische Alpen, Aosta-Tal, Gressoney St. Jean (CANEBT.&INI 1872); Bornade Ia Glace, oberhalb von· Chabaudey (Gozo 1908); Brusson, in künstlichem Stollen (nur iuv., GoBETTI leg.). - Bergamasker Alpen: Val Seriana, Boario di Grono, Höhle·"Bus di Tacoi (HADZI 1954, I. aff. strasseri; ferner Brni leg.); Roncobello, Höhle Buco del Castello (BINI leg.).
Die Zuordnung der Höhlenpopulationen aus den Bergamasker Alpen zu dentipalpis ist nicht zweifelsfrei, da keine adulten Individuen vom Loous typious von dentipalpis vorliegen.
Ökolog!e: Biotopansprüche: Alle Belege aus den Bergamasker Alpen wurden in Höhlen gesammelt, doch weist Gozo (1908) ausdrücklich darauf hin, daß diese Art im Aosta-Tal nicht troglobiont sei.
Vertikalverbreitung: Die Höhle Buco del Castello liegt 700 m hoch (BRIG· l'(OLI 1972).
Ischyropsalis muellneri HAMAJ.I1'N, 1898 Abb.316,324-331, 333,350
18981 lschyropsaUs Müllneri HAMANN, Zoo!. Anz., 21 (570): 535. 1914 Ischyropsalis corcyraea RoEWER, Aroh. Naturgesoh., 80 A (3): 108. 1923 Ischyropsalis helwigii - RoEWER, Weberknechte d. Erde: 683 (part.). 1950 lsohyropsalis alfkeni RoEWER, Senokenbergiana 31: 19. 1956 Isohyropsalis helwigii - tliLHAVY, Fauna ÖSR, 7: 151. 1969a Isohyropsalis muellneri- MARTENB, Zoo!. Jb. Syst., 96: 245 (dort die übrige Synonymie).
Habitus und Feldkennzeichen: sehr langbeinig, Cheliceren lang und auffallend schlank, fast glatt (Ö') oder geringfügig bedornt (~);fast nur in Karsthöhlen Jugoslawiens (Abb. 333).
Kennzeichnung: Körpermaße (in mm): L Ö' 4,8-5,5; L ~ 4,8-5,8. Laufbein II (Ö', in Klammern ~): Fe 7,4 (8,0) Pt 1,5 (1,6) Ti 6,6 (6,5) Mt 10,8
(10,3) Ta 9,4 (9,7). - Cheliceren-Grundglied: L Ö' 4,9-5,4; L ~ 4,5-6,0. Genitalmorphologie (Abb. 316, 329): Truncus penis kräftig-gedrungen, ba,
Hälfte erweitert, aber nicht bauchig, die di stark skierotisierte Ventral-Seite schwach geflügelt; Sklerit der Glans breit und kurz, basad deutlich 2zipflig, Sklerit überwiegend auf dem ba Teil in 2 para-me Feldern beborstet. Rec sem: Schläuche kurz und dick, mäßig bis stark gekrümmt (Abb. 331).
Cheliceren (Abb. 224-228): J, Grundglied lang und schlank, distad kontinuierlich schwach erweitert und etwas mediad geschwungen, medio-do in winziger, spitz auslaufender Apo endend; Bewahrung fast fehlend; do und ve-la Do-Reihe lediglich durch wenige stumpfe Tu angedeutet; ohne Bürste bzw. Drüsenfeld. .2. Glied: auf dem di Drittel frontal Reihen kleiner Höcker und Bo. - ~. Proportionen wie Ö', Bewahrung ausgeprägter: markante Reihen kleiner Do ve-la, ve-me und do.
Pedipalpus: robust; bei Ö' und ~Ta di schwach tropfenförmig erweitert, mit dichtem Besatz feiner und anliegender bzw. längerer schütter verteilter Haare.
Laufbeine: auffallend lang, Basis der Fe hellgelb geringelt, Mt und Ta hellgelb, übrige Glieder mittel- bis dunkelbraun, matt.
Dorsalseite (Abb. 330, 333): Ceph nur schwach aufgewölbt, fein granuliert, 'thorak Terg II mit 7-9 großen, di gerichteten Do; ohne Tu oc im eigentlichen
Genus Isohyropsalis 201
Abb. 324-331. Ischyropsalis muellneri HAMANN. -'- 324-328. Chelicer~ vo 1 t · 1 T nur Grundglied; 324. ($, YU Slowenien~ Öastitljiva jama; 325. ($, Fundort w~e ~2":~ a2~·. ·: g~nauer Fun~ort unbekannt, angeblich Insel Korfu; 327. \?, Castitljiva jama; 328. \?. o:vfe ~27;. 31~.9. ~ems von ventral; 330. Cephalothorax von lateral; 331. Receptaculum seminis(~lln
ast1t JIVa Jama). - Maßstab (nur für Cheliceren): 2 mm.
Si~ne, Aug~n isoliert und ';eit voneinander entfernt, Linse klein. Scutum: Ö' mit S._mterme~mm oder S.lammatum; ~immer mit S.laminatum. J-Scuta variabel· alle. ü?ergange. von abd Platte aus den Areae I-III (folgende Sklerite frei) bi~ zu Isoharten Emzel-Areae. Die Ablösung der einzelnen Areae (vom rezent nicht m_ehr auftretend~n) S.J?~rvum (aus Areae I-V) unterliegt folgender Regel: Area I Wird durc~ la Einsch~urung undfoder me Perforation von der Platte aus Areae I-III. gelost: dann ~rd auch die Restplatte aus Areae II und III zerteilt, so daß S: la'?ma~um .resultiert. - Jedes ad. Individuum repräsentiert natürlich nur 1 S~admm m dieser Evolutionsreihe. ''- Scuta bzw. Areae uud Ceph ausgefärbter Tiere braun-schwarz, Membranen weißlich. Cx ve auffällig spitz warzig· und lang beborstet. ·
Variabi~itäi: Bewahrung des J-Chel-Grundgliedes bisweilen geringfügig stärker als abge~Jldet; Scutum vgl. unter "Dorsalseite".
.Verbreitung (Abb. 35~): Endemit ~er Julisehen Alpen und des jugoslawischen Karstes. - Locus typ10us: Slowemen, Castitljiva luknia, bei RadovlJ'icafRadman:Jsdorf.
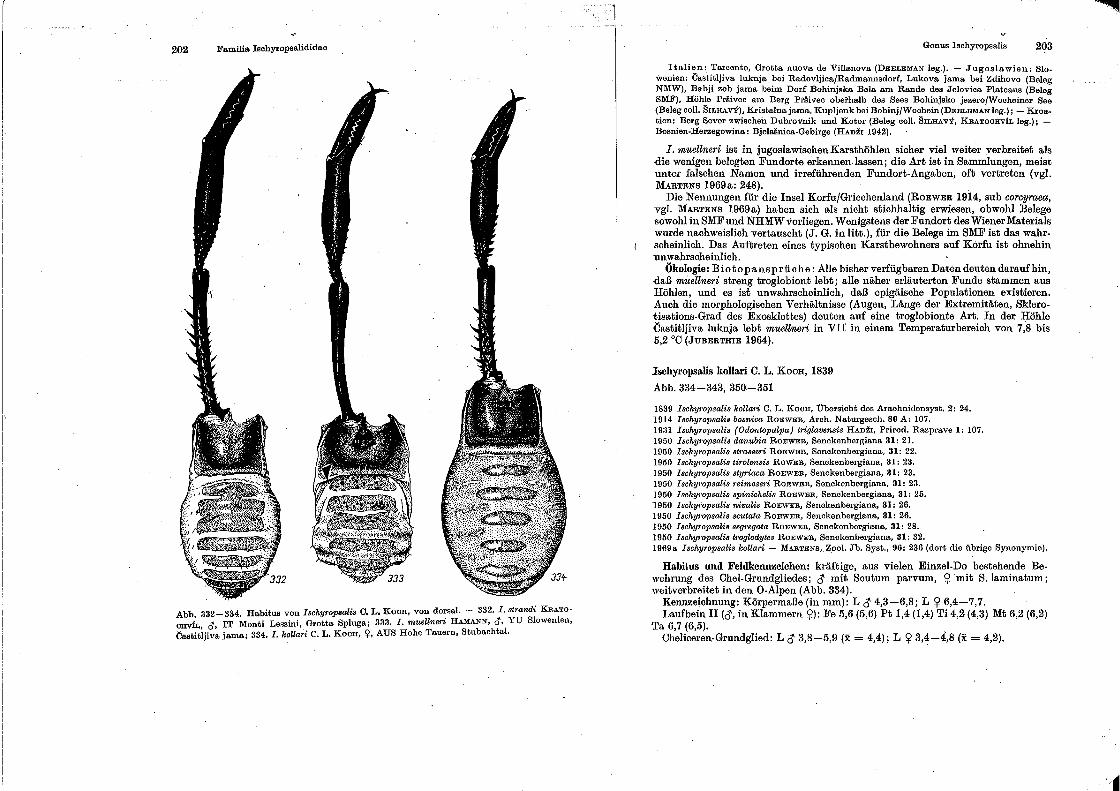
202 Familia Ischyropsalididae
Abb. 332-334. Habitus von IBchyropsalis 0, L, KocH, von dorsal. - 332. I. •~randi KR":TO· oHviL, es, IT Monti Lessini,. Grotta Spluga; 333. I. muellneri !IAMANN, es. y u Slowemen, öastit.ljiva jama; 334. I. kollari C. L. KooH, Cf, AUS Hohe Tauern, Stubachtal.
Genus Ischyropsalis 203
Italien: Tarcento, Grotta nuova de Villanova (DEELEMAN leg.). - Jugoslawien: Slowenien: Castitljiva luknja bei RadovljicafRadmannsdorf, Lukova jama bei Zdibovo (Beleg NMW), Babji zob jama beim Dorf Bohinjska Bela am Rande des Jelovica Plateaus (Beleg SMF), Höhle Pr1iivec am Berg Pr1iivec oberhalb des Sees Bohinjsko jezerofWocheiner See (Beleg coll. EirLHAVY), Kristalna jama, Kupljenk bei BohinjfWoohein (DEELEMAN leg.); - Kroatien: Berg Sever zwischen Dubrovnik und Kotor (Beleg coll. EirLHAVY, KRATOOHViL leg.); -Bosnien-Herzegowina: Bjela1inica-Gebirge (HAnzr 1942).
I. muellneri iso in jugoslawischenKarsthöhlen sicher viel weiter verbreitet als die wenigen belegten· Fundorte erkennen. lassen; die Art ist in Sammlungen, meiso unter falschen Namen und irreführ(mden Fundort-Angaben, oft vertreten (vgl. MARTENS 1969a: 248).
Die Nennungen für die Insel Korfu/Griechenland (RoEWER 1914, sub corcyraea, -vgl. MARTENS 1969a) haben sich als nicht stichhaltig erwiesen, obwohl Belege sowohl in SMF und NHMW vorliegen. Wenigstens der Fundort des Wiener Materials wurde nachweislich vertauscht (J. G. in litt.), für die Belege im SMF ist das wahrscheinlich. Das Auftreten eines typischen Karstbewohners auf Korfu ist ohnehin unwahrscheinlich.
Ökologie: Biotopansprüche: Alle bisherverfügbaren Datendeuten daraufhin, -daß muellneri streng troglobiont lebt; alle näher erläuterten Funde stammen aus Höhlen, und es ist unwahrscheinlich, daß epigäische Populationen existieren. Auch die morphologischen Verhältnisse (Augen, Länge der Extremitäten, Sklerotisations-Grad des Exosklettes) deuten auf eine troglobionte Art. In der Höhle ·Castitljiva luknja lebt muellneri in VII in einem Temperaturbereich von 7,8 bis 5,2 °0 (JUBERTHIE 1964).
1schyropsalis kollari C. L. KooH, 1839
Abb. 334-343, 350-351
1839 Ischyropsalis kollari C. L. KooH, Übersicht des Arachnidensyst. 2: 24. 1914 Ischyl'OJJsalis bosnica RoEWER, Arch. Naturgesch. 80 A: 107. 19:H Ischyropsalis (Odontopalpa) triglavenBis HAnzr, Prirod. Razprave I: 107. 1950 Ischy•·opsalis danubia RoEWER, Senckenbergiana 31: 21. 1950 Ischy•·opsalis strasseri RaEWER, Senckenbergiana, 31: 22. 1950 Ischy•·opsalis tirolensis Row'ER, Senckenbergiana, 31: 23. 1950 Ischyropsalis styriaca RaEWER, Senckenbergiana, 31: 23. 1950 Ischyropsalis reimoseri RaEWER, Senckenbergiana, 31: 23. 1950 Ischyropsalis spinichelis RaEWER, Senckenbergiana, 31: 25. 1950 I schy.-opsalis nivalis Ra EWER, Senckenbergiana, 31: 26. 1950 Ischy•·opsalis scutata RaEWER, Senckenbergiana, 31: 26. 1950 Ischy•·opsali• segregata RoEWER, Senckenbergiana, 31: 28. 1950 Ischy.-opsalis troglodytes RaEWljlR, Senckenbergiana, 31: 32. 1969a Ischy•·opsalis kollari - MARTENB,)!'!ool. Jb. Syst., 96: 236 (dort die übrige Synonymie).
Habitus und Feldkennzeichen: kräftige, aus vielen Einzel-Do bestehende Bewahrung des Chel-Grundgliedes; rJ mit Scutum parvum, ~ mit S.laminatum; weitverbreitet in den 0-Alpen (Abb. 334).
Kennzeichnung: Körpermaße (in mm): L rJ 4,3-6,8; L ~ 6,4-7,7. Laufbein II (rJ, in Klammern ~):Fe 5,6 (5,6) Pt 1,4 (1,4) Ti 4,2 (4,3) Mt 6,2 (6,2)
Ta 6,7 (6,5). Cheliceren-Grundglied: L rJ 3,8-5,9 (X. = 4,4); L ~ 3,4-4,8 (X. = 4,2).

.204 Familia Ischyropsalididae
. Genitalmorphologie (Abb. 340, 351): Truncus penis schlank; ba verbreitert, aber nicht bauchig, di Drittel nahezu parallelrandig, Ia skierotisierte Randleiste zur Glans bisweilen wenig erweitert; Sklerit der Glans rundlich bis länglich, ba~ad .2zipflig ausgezogen, Beborstung dicht, Bo lang. - Rec sem: 4-5 lange und dünne· Schläuche, meist stark in sich gekrümmt, bisweilen .2zipfelig (Abb. 34.2). ·
Chelioeren (Abb. 335-339): d'-Chel-Grundglied: kräftig-untersetzt und dicht. bedornt; do Reihe aus 8-11 Do, nicht zu gerader Linie ausgerichtet, sondern gegeneinander versetzt, ba Do am größten; ve-la streng ausgerichtete Reihe ans. 9-11 Do unterschiedlicher Größe, ve-medial 6-8 Do; do-di ohne 'deutliche Aufwölbung oder Apo und ohne Bürste, oft mit schütter stehenden Haaren; Drüsenfeld schwach entwickelt oder fehlend. - .2. Glied: do-ba stark bewehrt mit Do-Feld. ~:ohne deutliche Unterschiede zum d'· ·;
Pedipalpus (Abb. 343): d'-Fe und Pt dimeist etwas angeschwollen, in manoben Populationen d'-Pt ve-la mit kleiner und spitz ausgezogener Apo. - meist in Abhängigkeit von der Höhenlage des besiedelten Biotops (vgl. Kap. "Variabilität").
Abb. 335-342. IschyropsaliB kollari C. L. KooH. - 335-339. Chelicere von lateral, z. T. nur Grundglied; 335. ~.AUS Gastein, Syntypus; 336. ~.AUS Grödnertsl; 337. ~. IT Cima Undici Val Sugana; 338. !f, AUS Seeriegelhöhle; 339. !f, AUS Ramsau; 340. Penis von ventral, IT PlosefBrixen; 341. Cephalothorax von lateral; 342. Receptaculum seminis (beide Stubachtal). -Maßstab (nur für Cheliceren): 2 mm.
Genus IschY,.opsalis .205
Laufbeine: mittellang, robust; bräunlich schwarz, Mt und Ta heller. · Dorsalseite (Abb. 334, 341):· Ceph hinter der Augenregion steil zum thorak
Terg II ?'bfallend, dieses mit 8-12 kleinen Do; Scutum: d' mit S. parvum, nahezu quadratisch, Ecken gerundet, einzelne Areae fein bekörnelt mit je I Bo-Querreihe · sel~eri Areae IV und V, gelegentlich auch III, nicht völlig mit dem übrigen 'Sou~ tm:q. verschmolzen; g: S.laminatum (Abb. 334), Sklerite der Areae III-VI bei -einigen Populationen me halbiert (z. B. Lurgrotte). . ...
:var.iabilität:' Anlage der Chel-Be.wehrung immer einheitlich, ·individuell aber hoohvariabel; die 3'Hauptreihen (do, ve-la, va-medial) deutlich, Do aber nie so lang wie das Grundglied breit und nie zu I Linie ausgerichtet. - ~-Chel-Grundglied bei kräftigen Individuen do,di flachknotig aufgetrieben, dort schütter behaart. - Die ~ aus Populationen des inittleren und oberen Teiles, des vertikalen Areals meist.mit deutlicher. Apo an Pa-Pt (Abb. 343): Im Stubachtalf
Abb. 343. IschyropsaliB kollari C. L. KocH. Variabilität der lateralen Patellar-Apophyse des Pedipalpus in Abhängigkeit der vertikalen Lage des Siedlungsgebietes. - a-f: AUS Hohe 'Tauern, Stubachtal; a) und b) 1640 m;' c) 1640-1660 m; d) 1740 m; e) 1865 m, f) 2020 m; :g) IT PlosefBrixen, etwa'2300 m; h) IT Mühi.walder Joch, 2360 m; i) IT Cimi> Undici, Val Sugana, 2050 m; k) AUS Tirol, Wolfendorn-Mäuerlscharte, 2200-2600 m.

206 Familia Ischyropsalididae
Hohe Tauern in 1640 m ohne Apo,. bei Individuen von 1740 und 1865 m· Apo deutlich und bei 2 020 m auffallend stark ausgeprägt. Dieselbe Korrelation besteht bei den meisten anderen Populationen, andererseits fehlt die Apo bei einigen sehr hoch siedelnden Populationen (Brenner, Wolfel).dorn-Mäuerlscharte, 2200-2600 m).
Beziehungen: I. kollari steht in nahem verwandtschaftlichen Verhältnis zu englokalisierten troglobionten Arten der S-Alpen, nämlich hadzii, strandi, ravasinii und dentipalpis (d. non sensu MAl!.TENS 1969a). Diese Arten haben offensichtlich in kleinen und reliktären Einzelpopulationen die GlazialzeiteninS-alpinen Refugialgebieten überdauert; wahrscheinlich gehen sie auf einen gemeinsamen präglazialen Vorfahren zurück. Die nur ausnahmsweise troglobiont lebende kollari hat ihr heutiges Areal erst postglazial eingenommen (vgl. Kap. Biotopan• sprüche). ·
Verbreitung (Abb. 350): Typ: Endemit der 0-Alpen. - Gesamt-Areal etwa vom Brenner undSchlern östl. bis zum Hochschneeberg und Wechsel.- Locus typicus: Hohe Ta.uern, Gastein.
Österreich: N-Tirol: Brenneralpen, Wolfendorn-Mäuerlscharte (K. T.leg.); Windlöcher, Seeriegelhöhle (Belege SMF); - 0-Tirol: Lienz, Peba-Klamm (KOFLER leg.); - Salzburg: Verbreitungsschwerpunkt auf der N-Abdachung der Hohen Tauern, vereinzelt in den Niederen Tauern, bisher keine Funde in den nördl. Kalkalpen westl. der Salzach und in der Grauwackenzone (A. A. in .litt.) - im einzelnen: Krimmler Achental (A. A. leg.), Habachtal südwestl. Fazeneck (HuTz leg.), Stubachtal (A. A. & al. leg.), Kaprunertal südl. der Salzburger Hütte (HURDES leg.), Käfertal, Stollenfenster (WÖRGÖTTER leg. fide K. T.), Hüttwinkeltal (A. A. & al. leg.)'\ Böckstein, Thermalstollen (A. A. leg.), Gasteiner Tal (KoLLA.R leg. 1855, Belege NHMW), Großarltal, Kleinarltal, Weißpriachtal .und Lignitztal (alle A. A.leg.), Tennengebirge, Sommereckkessel-Höhle (SoHÜLLER 1933, hellwigi); Osterhorngruppe, KühllochTrattberg (LOVOIK leg.); Schmittenstein, Gugelau-Alm (A. A. leg.); Ober-Österreich: Gosau (Beleg SMF), Traunstein bei Ebensee, Gipfelregion; Kasberg, Doline am Kamm gegen Schwalbenmauer (beide ROEWER 1950); - Nieder-Österreich: Hochkar bei Göstling (RESSL leg.); Hochkarschacht bei Göstling (STROUHAL & VoRNATSCHER 1975); Lunz, Herdengelhöhle (Vo:RNATSCHER leg.); Lunz, zwischen Obersee und Pauschenalm, Dürrenstein und Ybbstaler Hütte (alle K. T.leg.); Lunz, Schwabenreithhöhle und Poschenreitherschacht (beide HAUSER leg.); Gipfel des Ötsoher bei Gaming (RESSL leg.); Hochschneeberg (Beleg SMF); Mira[uoke nordwestl. Pernitz (POSPICHAL leg.); Taubenloch (= Hermannshöhle) bei KirchbergfWechsel (Beleg NHMW);- Steiermark: Dachstein-Massiv: GjaidsteinfGuttenberg-Haus (Beleg SMF); RamsaufSchladming (Beleg SMF); Totes Gebirge, Sinterhans/Brettstein (Beleg NHMW); Almherger Eis- und Tropfsteinhöhle bei Grundlsee (Beleg NHMW); Admont (Beleg SMF); Hoohschwab, Gipfel des Ebenstein und Mitteralm bei Aflenz (beide FRANZ & GuNHOLD 1954); · Weinbergerloch in Triebeinalpe bei Gußwerk (MAYER leg.); Veitsch (Beleg NHMW); Raxalpe (Beleg SMF); Eiseuerzer Alpen, Gipfel des Zeyritzkampel (FRANZ & GuNHOLD 1954); Lurgrotte bei Graz (VORNATSOHER & al. leg., J. M.leg., Belege SMF, NHWM); Seckauer Zinken, Kar der Goldlacke; Gleinalpe, Speikkogel; Totes Gebirge, Umgebung Steyrer See (FRANZ & GUN· HOLD 1954); - Kärnten: Mallnitz: Seebachtal oberhalb Sohwussner Hütte; GutenbrunnWolligeralm; Gmünd, Taohiernock (alle A. A.leg.). - Italien: Pustertal, Gruppechtspitze über Terenten (PEEZ leg.); Mühlwalder Joch nördl. Bruneck (PESKOLLER leg.); Weißenbach nördl. Bruneck (SCHULZE leg.); Plose bei Brixen (HOLDHAUS leg., K. T.leg.); Sohlern (K. T. leg.); Grödner Tal (Beleg SMF); Dolomiten, Setla-Joch (STRASSER leg.); Rolle-Paß (Beleg MNB); Cima Undici, Valsugana, östl. Trient (K. T.leg.); Monti Lessini, Cima Pasta, unter Stein in Doline (CoRNACOHIA leg., Beleg MNHNP). - Jugoslawien: Slowenien: TriglavMassiv, Kredarico (HAnzr 1931, triglavensis); Triglav-Massiv, Pod Sternarjem am Berg Skrlatica; Bachergebirge ( = Pohorje), Ribniska koca (beide HADZI 1954, triglavensis).
Ökologie: Biotopansprüche: I. kollari lebt auf der N-Abdachung der Hohen Tauern - dem ·Gebiet größter Siedlungsdichte - in subalpinen Blockwäldern (Rhodoreto-Vaccinietum cembretosum), deren Böden von Gesteinstrümmern be-
Genus Isohyropsalis 207
deckt sind und die ein dichter Teppich von Moosen, Vaccinium und Rhododendron überzieht. Diese bodennahe isolierende Vegetationsschicht bewirkt zusammen mit;
· Quellgerinnen konstante Temperatur- und Feuchtigkeitsverhältnisse, auf die unter den freilebenden lschyropsalis-Arten kollari in hohem Maße angewiesen ist. Im Winter bietet hohe Schneebedeckung zusätzliche Isolation; die auch in der kältesten Jahreszeit wahrscheinlich fast normale Aktivität erlaubt (A. A. in litt.).
Viel geringer ist die Siedlungsdichte oberhalb der Waldgrenze, und nur in den S-Alpen scheint kollari in der Alpin-Stufe weit verbreitet zu sein. Sie lebt dort in Karfluren und Schneetälchen-Gesellschaften, unter Holz und tief eingebetteten Steinen, meist in Wassernähe. Gemäß diesen ökologischen Präferenzen werden Höhlen gern besiedelt, aber nur in der niedrig gelegenen· LurgrottejSteiermark besteht eine feste Abhängigkeit vom Höhlenmilieu (MARTENS 1969a, NEUHERZ 1975). -Über Nahrung uridNahrungsaufnahme vgl. Kap. 1.3.3.
Vertikal ver brei tung: im Freiland von der Montan- bis in höhere Lagen der Alpin-Stufe, lokal mit unterschiedlichem Schwerpunkt. In den Hohen Tauern v:on 1460 m (obere Montan-Stufe) bis 2150 m (oberste Subalpin-Stufe) noch unterhalb der klimatischen Baumgrenze; 80% der Individuen (n = 184) stammen aus 15-1800 m (A. A. in litt.), im südwestl. Nieder-Österreich (Lassingalpen) Freilandfänge in subalpinen Wäldern (um 1100 bis 1300 m) bis in Gipfellagen (fast 1900 m), Höhlenfunde wenig tiefer (etwa 900 m; RESSL leg., MAL:roKY leg., KünNELT 1'948, THALER 1963). In den S-Alpen ist die Verbreitungsgrenze weiter nach oben verschoben; z. B. 2360 m am Mühlwalder Joch, zwischen 2200 und 2600 m •am Wolfendorn-Mäuerlscharte. Die von kollari besiedelte LurgrottejGraz liegt in nur 350 m Höhe. Die dort lebende Population ist offenbar von epigäischen über größere Entfernung separiert.
Phänologie: Bisher nur von den Populationen der Hohen Tauern hinreichend genau bekannt; dort stenochron mit 2 Gipfeln in VI und E VIII-X. Die Gipfel stammen offenbar von 2 verschiedenen Generationen, deren Reifezeiten ineinander verschachtelt sind:
Generation I: Eiablage in IX/X, Überwinterung der Eier, Schlüpfen im Frühjahr, wohl IVJV, Reifehäutung ab VIII, Tod der ad. ab XI. - Generation II: Eiablage in VI, Schlüpfen der iuv. ab VIII, Überwinterung halbwüchsiger iuv. und möglicherweise Reifehäutung vor oder während der Schneeschmelze in IVJV, Tod der ad. in VI (Q'), vereinzelt erst in VII(~)-
In den Hohen Tauern sind beide Generationen durch ein Populationstief in VII scharf voneinander getrennt. Das braucht nicht auszuschließen, daß sich "alte" und "junge" ad. in VII treffen und möglicherweise auch miteinander kopulieren können. Die Reifezeit in anderen Populationen, vor allem aus solchen in Gebieten über der Baumgrenze, läßt sich bisher nicht überblicken.
Ischyropsa.lis hadzii RoEWER, 1950:
Abb. 344-349, 352, 394
1851 Pha!angium cancroides ScHMIDT, Ber. Mitth. Freund. Naturwissensch. Wien, 7: 61 (prä-occ. durch MüLLER 1776).
192.6/27 I. cancroides - HADZI, Bull. Mus. Slov. '1/8: 26. 1950 Ischyropsalis hadzii RoEWER, Senokenbergiana 31: 52 (Nom. novum). 1969a Ischyropsalis hadzii - MARTENS, Zoo]. Jb. Syst., 96: 242. (dort die übrige Synonymie).
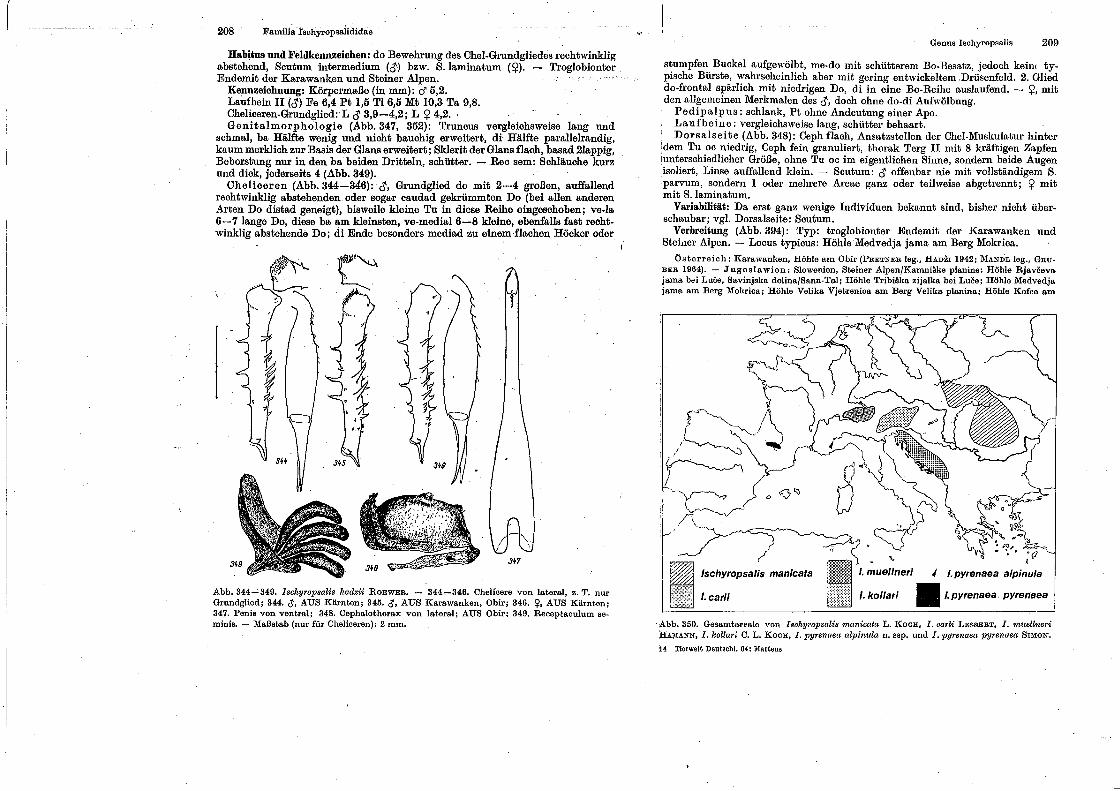
208 Famili~ IsohyropsaÜdidae
Hab~tus und Feldkennzeichen: do Bewahrung des Chel-Grundgliedes rechtwinklig abstehend, Souturn intermedium (d') bzw. S.laminatum (~). - Troglobionter Endemit der Karawanken und Steiner Alpen.
Kennzeichnung: Körpermaße (in mm): d 5,2. Laufbein II (d') Fe 6,4 Pt 1,5 Ti 6,5 Mt 10,3 Ta 9,8. Cheliceren-Grundglied: L d' 3,9-4,2; L ~ 4,2. Genitalmorphologie (Abb. 347, 352): Truncus vergleichsweise lang und
schmal, ba Hälfte wenig und nicht bauchig erweitert, di- Hälfte parallelrandig, kaum merklich zur Basis der Glans erweitert; Sklerit der Glans flach, basad 21appig, Beborstung nur in den ba beiden Dritteln, schütter. - Rec sem: Schläuche kurz und dick, jederseits 4 (Abb. 349).
Cheliceren (Abb. 344-346): d', Grundglied do mit 2-4 großen, auffallend rechtwinklig abstehenden oder sogar caudad gekrümmten Do (bei allen anderen Arten Do distad geneigt), bisweile kleine Tu in diese Reihe eingeschoben; ve-la 6-7 lange Do, diese ba am kleinsten, va-medial 6-8 kleine, ebenfalls fast rechtwinklig abstehende Do; di Ende besonders mediad zu einem ·flachen Höcker oder
3+7
Abb. 344-'349. Ischyropsa!is hadzii RoEWER. - 344-346. Chelfoere von lateral, z. T. nur Grundglied; 344. <J, AUS Kärnten; 345. <J, AUS Karawanken, Obir; 346. !j?, AUS Kärnten; 347. Penis von ventral; 348. Cephalothorax von lateral; AUS Obir; 349. Reoeptaculum seminis. - Maßstab (nur für Chelioeren): 2 mm.
Genus Isohyropsalis 209
stumpfen Buckel aufgewölbt, me-do mit schütterem Ba-Besatz, jedoch kein< typische Bürste, wahrscheinlich aber mit gering entwickeltem .Drüsenfeld. 2. Glied da-frontal spärlich mit niedrigen Do, di in eine Ba-Reihe auslaufend. - ~. mit den allgemeinen Merkmalen des d', doch ohne do-di Aufwölbung.
Pedipalpus: schlank, Pt ohne Andeutung einer Apo. Laufbeine: vergleichsweise lang, schütter behaart. Dorsalseite (Abb. 348): Ceph flach, Ansatzstellen der Chel-Muskulatur hinter
:dem Tu oc niedrig, Ceph fein granuliert, thorak Terg II mit 8 kräftigen Zapfen iunterschiedlicher Größe, ohne Tu oc im eigentlichen Sinne, sondern beide Augen isoliert, Linse auffallend klein. - Scutum: d' offenbar nie mit vollständigem S. parvurn, sondern 1 oder mehrere Areae ganz oder teilweise abgetrennt; ~ mit mit S.larninaturn.
Variabilität: Da erst ganz wenige Individuen bekannt sind, bisher nicht überschaubar; vgl. Dorsalseite: Scutum.
Verbreiiung (Abb. 394): Typ: troglobionter Endemit d.er Karawanken und Stei.1er Alpen. - Locus typicus: Höhle Medvedja jarna am Berg Mokrica.
Österreich: Karawanken, Höhle am Obir (PRETNER leg., HADZI 1942; MANDL leg., GRU· BER 1964). - Jugoslawien: Slowenien, Steiner AlpenfKamniske planine: Höhle Rjavceva ;jama bei Luöe, Savinjska dolina/Sann·Tal; Höhle Tribiska zijalka bei Luce; Höhle Medvedja jarna am Berg Mokrica; Höhle Velika Vjetrenica am Berg Velika planina; Höhle Kofce am
I
I. muellneri J t.pyrenaea alpinula
f. kollari llt.pyrenaea. pyrenaea
· Abb. 350. Gesamtareale von Ischyropsalis manicata L. KOCH, I. car!i LESSERT, I. muellneri HAMANN, I. kollmi C. L. KocH, I. pyrenaea alpinula n. ssp. und I. pyrenaea pyrena.ea SrMoN.
14 Tierwelt Deutsch!. 64: nrartens

210 Familia Ischyropsalididae
Berg Velika planina (alle HAnzr 1942). Julisehe Alpen: Höhle Visevnik jama auf der Alm Viäevnik planhm, nördl. Bohinjsko jezerojWocheiner See (Bestimmung nicht gesichert, da nur iuv. vorliegend ; MARTENS 1969a).
Ökologie: Biotopansprüche und Vertikalverbreitung: I. kadzii ist bisher nur in Höhlen gefunden worden; und die rudimentären Augen dieser Art deuten an, daß sie obligatorisch troglobiont lebt. Einzelheiten über die ökologischen Bedingungen der besiedelten Höhlen sind nicht bekannt. Eine Höhle im Obir-Massiv (auf dem Wege zur Reiner Hütte) liegt 1600 m hoch (HADZI 1942; ~ETNER in litt.).
Ischyropsalis ravasinii HADZI, 1942
Abb. 353-359, 394
1942 Ischy•·opsa!is ravasinii HAnzr, Razprave, Dissertat. Ljubljana, 2: 11.
Habitus und Feldkennzeichen: Chel-Grundglied auffallend lang und stark ]Jewehrt, bei c'{ do-di knotig erweitert; nur in Höhlen des S-alpinen Cansiglio-Massivs.
Kennzeichnung: Körpermaße (in mm): nach Material aus der Höhle Bus de la Genziq,na: L c'{ 5,7-6,0; L ~ 7,0-7,8.
Laufbein II (({,in Klammern ~):Fe 7,3 (7,2) Pt 1,6 (1,7) Ti 5,7 (5,9) Mt 9,3 (9,3) Ta 9,4 (9,2).
Cheliceren-Grundglied: L c'{ 5,0-5,4; L ~ 5,0-5,4; ~ aus der Höhle Bus del Pal (Holotypus): 5,7.
Genitalmorphologie (Abb. 353, 359): Truncus penis schlank, nicht bauchig erweitert, in der di Hälfte nahezu parallelrandig, Glans schmal, ve Sklerit ba zweizipfelig, die beiden Bo-Felder di divergierend, Glans do in gerader Verlängerung des Truncus, nicht bucklig abgesetzt.
Cheliceren (Abb. 354-358): Unter allen Arten der Gattung am längsten, bei einzelnen Populationen (Bus de Ia Genziana) aber nicht länger als bei stmnd-i. ({, Grundglied: do unregelmäßige Reihe aus 7-9 unterschiedlich langen Do, nicht Rtreng ausgerichtet, sondern Einzel-Do gegeneinander versetzt, kleinere Do rechts und links beigestellt; ve-la etwa 10 verschieden lange Dp, ve-medial 7-9 auffallend lange ~nd schlanke; di zu einem medial geneigten auffälligen buckelförmigen Knoten vergrößert, mit flacher, nach links oben (von do) geneigter Sekretionsfläche (Chitin bräunlich aufgehellt), aber ohne distinkte Bürste. 2. Glied do-ba mit dichtem Feld großer und stumpfer Höcker. - ~:allgemeine Merkmale wie ({, doch ohne di Apo und Sekretionsfeld.
Pedipalpus: laufbeinförmig lang, ({-Pt ve mit kleiner, spitz ausgezogener Apo, dem ~fehlend; glatt glänzend, mit schütterer, unterschi!)dlich langer Behaarung.
Laufbeine: lang und robust, schütter behaart, bräunlich schwarz, Mt und Ta heller.
Dorsalseite: Ceph hinter der Augenregion zum thorak Terg II steil abfallend, matt, zerstreut fein bekörnelt, einzelne größere Tu mit aufgesetzter Bo; thorak Terg II mit 9-13 zapfenförmigen Do, die mittleren am größten. -Tu oc niedrig, breit, median seicht gefurcht, Linse der Augen klein. - Scutum: c'{ mit S. parvum, doch bei den Individuenzweier Höhlen, aus denen Material vorliegt, Area V' teilweise oder sogar ganz vom übrigen Schild abgetrennt; ~. S. laminaturn; zerstreut granuliert, mit unterbrochener Querreihe kleiner Bo je Area.
Genus Ischyropsalis
352
Abb. 351-353. Glandes penis von Ischyrof,sa!is C. I •. Komr (links: lateral, rechts: ventral);
Abb. 351. I. koll,.,·i C. L. KocH. - AUS Stubachtal.
Abb. 352. I. hadzii RoEWER. - AUS Obir.
Abb. 353. I. ravasinii HAnzr. - IT Cansiglio, Bus de Ia Genziana. - Maßstab: 0,3 mm.
u•
211
•
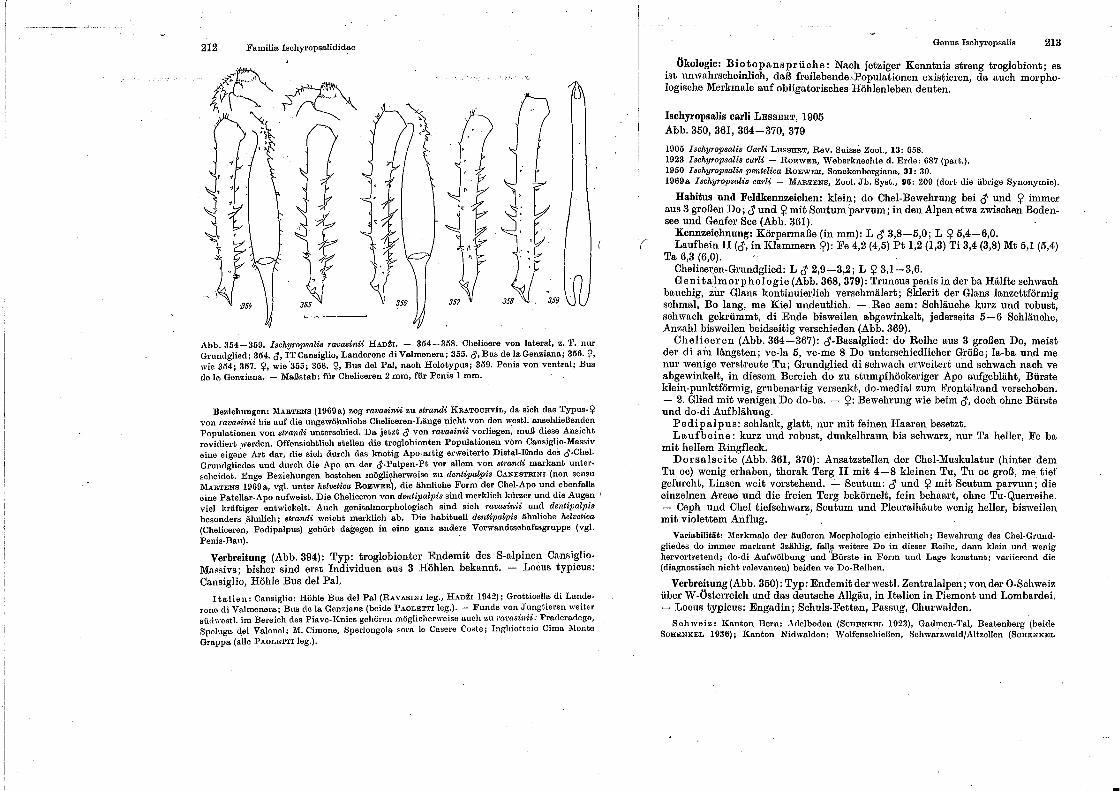
212 Familia Ischyropsalididao
Abb. 354-359. Ischyropsalis ravasinii HAnzr. - 354-358. Chelicere von lateral, z. 1'. nur Grundglied; 354. <J, IT Cansiglio, Landerene di Valmenera; 355. <J, Bus dela Genziana; 356. !f, wie 354; 357. !f, wie '355; 358. !f, Bus del Pa!, nach Holotypus; 359. Penis von ventral; Bus de Ia Genziana. - Maßstab: für Cheliceren 2 mm, für Penis 1 mm.
Beziehungen: MARTENS (1969a) zog ravasinii zu strandi KRATOOHVIL, da sich das Typus·!? von ra·vasinii bis auf die ungewöhnliche Cheliceren-Länge nicht von den ":estl. ·anschließenden Populationen von st.-andi unterschied. Da jetzt iJ von ravasinii vorliegen; muß diese Ansicht revidiert werden. Offensichtlich stellen die troglobionten Populationen vom Cansiglio-Massiv eine eige1~e Art dar, die sich durch das knotig Apo-artig erweiterte Distal-Ende des <J·ChelGrundgliedes und durch die Apo an der (]-Palpen-Pt vor allem von strandi markant unterscheidet. Enge Beziehungen bestehen möglicherweise zu dentipalpis CANESTRINI (non sensu MARTENS 1969a, vgl. unter helvetica ROEWER), die ähnliche Form der Chel-Apo und ebenfalls eine Patellar-Apo aufweist. Die Cheliceren von dentipalpis sincl merklich kürzer und die Augen viel kräftiger entwickelt. Auch genitalmorphologisch sind sich ravasinii uncl dentipalpis besonders ähulich; strandi weicht merklich ab. Die habituell dentipalpis ähnliche helt•etica (Cheliceren, Pedipalpus) gehört dagegev. in eine ganz andere Verwandtschaftsgruppe (vgl. Penis-Bau). ·
Verbreitung (Abb. 394): Typ: troglobionter Endemit des S-alpinen CansiglioMassivs; bisher sind erst Individuen aus 3 Höhlen bekannt. - Locus typicus: Cansiglio, Höhle Bus del Pal.
Italien: Cansiglio: Höhle Bus del Pa! (RAYASINI leg., HADZI 1942); Grotticella di Landerono cli Valmenera; Bns de Ia Genziana (beide PAOLETTI leg.). - Funde von Jungt.ieren weiter süclwestl. im Bereich des Pia.ve-I\:nies gehöron möglicherweise auch zu '1'cn·a.sinii: Praderadego .. Speluga 4e1 Valonel; l\'1. Cimone .. Sperlongola. sora le Casere Coste; Inghiottoio Cima :J\[onte Grappa (alle PAOLETTI leg.).
(
Genus Ischyropsalis 213
. Ökologie: Bi;>t?pansprüche: Nach jetziger Kenntnis streng troglobiont; es 1st .unwahrschemhch, daß freilebende·.Populationen existieren, da auch morphologische Merkmale auf obligatorisches Höhlenleben deuten.
Ischyropsalis carli LESSERT, 1905 Abb.350, 361,364--370,379
1905 lschy•·opsalis Oarli LESSERT, Rev. Suisse Zoo!., 13: 658. 1923 lschyropsalis carli - RoEWER, Weberknechte d. Erde: 687 (part.). 1950 lschyropaalia pentelica RoEWER, Senckenbergiana, 31: 30. 1969a Ischyropsalis carli - MARTENS, Zoo!. Jb, Syst., 96: 209 (dort die übrige Synonymie).
Habitus und Feldkennzeichen: klein; do Chel-Bewehrung bei($' und ~ immer aus 3 großen Do; ($'und ~ mit Senturn parvum; in den Alpen etwa zwischen Bodensee und Genfer See (Abb. 361).
Kennzeichnung: Körpermaße (in mm): L ($' 3,8-5,0; L ~ 5,4-6,0. Laufbein li (($',in Klammern ~): Fe 4,2 (4,5) Pt 1,2 (1,3) Ti 3,4 (3,8) Mt 5,1 (5,4)
Ta 6,3 (6,0). Cheliceren-Grundglied: L ($' 2,9-3,2; L ~ 3,1-3,6. Genitalmorphologie (Abb. 368, 379): Truncus penis in der ba Hälfte schwach
bauchig, zur Glans kontinuierlich verschmälert; Sklerit der Glans lanzettförmig schmal, Bo lang, me Kiel undeutlich. - Rec sem: Schläuche kurz und robust, schwach gekrümmt, di Ende bisweilen abgewinkelt, jederseits 5-6 Schläuche, Anzahl bisweilen beidseitig verschieden (Abb. 369).
Cheliceren (Abb. 364-367): ($'-Basalglied: do Reihe aus 3 großen Do, meist der di am längsten; ve-la 5, ve-me 8 Do unterschiedlicher Größe; la-ba und me nur wenige verstreute Tu; Grundglied di schwach erweitert und schwach nach ve abgewinkelt, in diesem Bereich do zu stumpfhöckeriger Apo aufgebläht, Bürste klein-punktförmig, grubenartig versenkt, do-medial zum Frontalrand verschoben. - 2. Glied mit wenigen Do do-ba. - ~: Bewehrung wie beim($', doch ohne Bürste und do-di Aufblähung.
Pedipalpus: schlank, glatt, nur mit feinen Haaren besetzt. Laufbeine: kurz und robust, dunkelbraun bis schwarz, nur Ta heller, Fe ba
mit hellem Ringfleck. Dorsalseite (Abb. 361, 370): Ansatzstellen der Chcl-Muskulatur (hinter dem
Tu oc) wenig erhaben, thorak Terg li mit 4-8 kleinen Tu, 'i'u oc groß, me tief gefurcht, Linsen weit vorstehend . ..:.... Scutum: ($'und ~mit Souturn parvum; die einzelnen Areae und die freien Terg bekörnelt, fein behaart, ohne Tu-Querreihe. - Ceph und Chel tiefschwarz, Souturn und Pleuralhäute wenig heller, bisweilen mit violettem Anflug. ·
Variabilität: Merkmale der äußeren Morphologie einheitlich; Bewahrung des Chei-Grundgliedes do immer markant 3zählig, falw weitere Do in dieser Reihe, dann klein und wenig hervortretend; do-di Aufwölbung und Bürste in Form und Lage konstant; variierend die (diagnostisch nicht relevanten) beiden ve Do-Reihen.
Verbreitung (Abb. 350): Typ: Endemit der westl. Zentralalpen; von der 0-Schweiz über W-Österreich und das deutsche Allgäu, in Italien in Piemont und Lombardei. - Locus typicus: Engadin; Schuls-Fettan, Passug, Churwalden.
Schweiz: Kanton Bern: Adelboden (SCHENKEL 1923), Gadmen-'l'al, Beatenberg (beide ScHENKEL 1936); Kanton Niclwalclen: ''Volfenschießen, Schwarzwalcl/Altzellen (SCHENKEL

214 Familia Ischyropsalididae Genus Ischyropsalis 215
368.
Abb. 364-370. Ischyropsa!is cm.zi LESSERT. - S64-367. Chelicere von lateral, z. T. nur Grmulglied; 364 . .;!, CH Wolfenschießen; 365. eS, IT Porlezza; 366. eS, CH Adelboden; 367. ~, AUS Sünser See; 368. Penis von ventral; 369. Receptaculum seminis; 370. Cephalothorax von lateral, AUS Groß-Vermunt. - Maßstab (nur für Cheliceren): 2 mm.
1923); Kanton Graubünden: Schuls-Fettan, PassugfChur, Churwalden (alle LESSERT 1905); Tessin: Mte. Ceneri (STROUHAL leg.), Castel di Chiasso (Beleg NHMB), Grotta del Mago (PAVESI 1875, manicata). Cuvel La s-Chala (STRINATI 1966); Castel San Pietro (LESSERT 1917); Kanton St. Gallen: Wildenmannlislöchi, Höhle bei Freienbach (beide STRINATI 1966). - Italien: Piemont, Varzo (DiEsoo & DRESCO·DEROUET 1960); Lombardei: Corno (Beleg SMF), Grotta Tre Crocette im Berg Campo dei Fiori (CASALEI eg.), Grotta di Cunardo (Beleg NHMW); Veltlin: östl. Borrnio, Valfurva; bei Sandrio (beide J. M. leg.). - Österreich: Vorarlberg: Damüls, Sünser See (K. T. leg.), Groß-Vermunt (J. G.leg.), Mittagsflul1 zwischen Au und Seimepfau (STROUHAL leg.); N-Tirol: Ötztal, Obergurgl (K. T. in litt.); UrnhausenfStuibenfall (KULL· MAN" leg.).- BRD: Allgäu, Umgebung von Balderschwang; Oberstdorf, Faltenbach-Wasser· fälle (beide J. M.leg.), FischenfHirschspl'Ung (BÜTTNER 1966). ·
Ökologie: Biotopansprüche: I. carli besiedelt Wald-Biotope, dort gern an besonders feuchten Stellen, an rinnendem Wasser oder an Quellhorizonten mit
Abb. 360-363. Habitus von Ischy1·opsalis C. L. KocH, von dorsal. - 360. I. pyrenae<t alpinul<t n. ssp., eS, IT Crissolo; 361. I. car!i LESSERT, eS, BRD Allgäu, Baldersch'jVang; 362. I. hellwigi hellwigi (PANZER), eS, BRD Wupperthal-Beyenburg; 363. I. manicata L. KocH, eS, CSSR Zilina.

216 Familia Ischyropsalididat>
Staunässe, vielfach ttn Wasserfällen. Bei ausreichend hoher Luftfeuchtigkeit ist Beschattung des Biotops nicht notwendig; dann auch iu Almgelände oder in niedrigem Gebüsch in der Nähe der Baumgrenze. Kavemikole Populationen .;ind ttus der Lombardei und ttus Piemont bekannt, doch besteht keine obligatorische Bindung an Höhlen.
Vertikalverbreitung: Von der Montan-Stufe (800 m, MittagsfluL; Oberstdorf) bis in die ohere Subalpin-Stufe (1960 m, Groß-Vermunt); Schwe1·punkt der vertikalen Verbreitung bisher nicht bekannt.
P hä n o I o g i e : stenochron sommer- und herbstreif mit unterschiedlichen Maxima in den einzelnen Höhenstufen. ---' Allgäu und Vorarlberg: ad. A VI-A VIII belegt, Reifezeit aber wahrscheinlich weiter in den Hei.·bst verlängert, da kleine iuv. noch E VII. - Schweiz: ad. bis E IX (MARTENS 1969a). - Ob ad. und iuv. aus VIfVII zur seihen Generation gehören, ist nicht sicher, da ad. vielleicht überwintern. - Gelege mit 13 Eiern A VII im Veltlin, 1550 m.
Ischyropsalis manicata L. Koch, 1865 Abb. 350, 363, 371-378, 380.
1865 Ischyropsalis manicata L. KocH, Z. Ferdinandeum 1'il'Dl Vorarlberg, (3) 12: 164. 1916 lschyropsalis rlacica RoEWER, Arch. Naturgosch., 82 A: 153. 1934 Ischi/ropsalis helvctica milleri KRATOOHViL, Acta Soc. nat. morav., 9: 13. 1950 Ischyropsalis käst1>eri ROEWER, Senckenbergiana, 31: 19. 1950 Ischyropsalis austriaca ROEWER, Scnckonbergiana, 31: 22. 1950 Ischyropsalis balcanica RoEWER, Senckonhorgiana, 31: 30. 1969a Ischyropsalismanicata - MARTENS, Zool.Jb. Syst., 96: 216 (dort die übrige Synonymie).
Habitus und Feldkennzeichen: klein und kurzheinig, ~- und 5j2-Cheliceren-Grundglied kräftig bedornt, mit Souturn parvum (~) bzw. S. laminaturn (5j2) - Einzige Ischyropsalis-Art im Karpaten-Bogen (Abb. 363).
Kennzeichnung: Körpermaße (in mm): L ~ 4,6-5,5; L 5j2 6,3-7,7. Laufbein II (~,in Klammern 5j2): Fe 4,0 (3,7) Pt 1,2 (1,3) Ti 2,9 (2,9) Mt 4,5 (4,2)
Ta 5,6 (5,2). Cheliceren-Grundglied: L ~ 2,8-3,4; L 5j2 3,1-3,9. Genitalmorphologie (Abb. 376, 380): Truncus penis untersetzt,' in det' ba
Hälfte schwach bauchig, di nahezu parallelrandig, doch unterhalb der GlanR wenig erweitert; Sklerit .der Glans schmallanzettförmig, schütter mit langen Bo besetzt. - Rec sem: jederseits 4 kurze, in sich gekrümmte Schläuche.
Cheliceren (Abb. 371-375): ~, Grundglied do mit Längsreihe von 3-5 unterschiedlich großen und meist wenig gegeneinander versetzten Do, ve-la mit 3 großen, ve-medial mit 5 Do, ba am kleinsten; zwischen diesen auffälligen DoElementen Ia und medial kleine Höcker und tuberkelartige Erhebungen eingeschoben; di Viertel des Grundgliedes keulenartig aufgetrieben und etwas ventmd abgebogen, do-medial in stumpf-auslaufende, wulstige Apo verlängert. Bürste großflächig di des I. Do, von dort mediad übergreifend und bis zum di Ende des Grundgliedes ausgedehnt. 2. Glied nur do-frontal mit einigen Tu; 5j2: allgemeine Merkmale wie bei ~, doch ohne Bürste und ohne do-di Auftreibung des Grundgliede~ und somit viel schlanker erscheinend.
Pedipalpus: glatt und schlank, mit feiner Beborstung. Laufbeine: kurz und kräftig, bis auf Mt und Ta dunkelbraun-schwärzlich,
Fe ba mit gelblichem Ringfleck.
Genus Isohyropsalis 217
1' ..
:l{J ' ~,, ; '
375
:m 37/J
Abb. 371-378. lschy•·opsalis manicata L. KooH. - 371-375. Chelicere von lateral, z. T. nur Grumlglied; 371. cJ, RU Batrina Bilea; 372. cJ, "Tatra"; 373. cJ, ÖSSR Zilina; 374. !?, richtiger Fundort unbekannt; 375. !?, "Tatra"; 376. Penis von ventral; 377. Cephalothorax von lateral, RU Kronstadt; 378. Receptaculum seminis. - Maßstab (nur für Cheliceren): 2 mm.
Dorsalseite (Abb. 363, 377): Ansatzstellen der Chel-Muskulatur hinter dem Tu oc kräftig gewölbt, Ceph fein granuliert, thorak Terg II mit 6 zapfenartigen Tu unterschiedlicher Größe; Tu oc groß, me stark gefurcht, Linse groß und· auffallend vorgewölbt. - Scutum: ~mit S. parvum, 5j2 mit S. laminatum, die einzelnen Areae fein granuliert, mit je I Querreihe feiner Tu.
Variabilität: Ihr unterliegt vor allem die Ausprägung der Chel-Bewehrung; die markante Do-Reihe des Grundgliedes do durch eingeschobene Tu von Ia aufgelockert und dadurch "unübersichtlich". Im ganzen Areal·sind konstant Lage und Form der Bürste und der CheJ. Apo und die Form ?er Glans penis.
VerbreUung (Abb. 350): Typ: karpatisch; Areal von den polnischen und tschechoslowakischen Beskiden bis in die rumänischen S-Karpaten. - Locus typicus: Siebenbürgen.
Polen: in den gesamten Beskiclen einschließlich der Tatra, wahrscheinlich nicht in den Pieninen, nicht westlich der oberen Oder (STARJ!GA 1976); - im einzelnen: Si'lsk (Schlesien), Rogozna/Kt•eis Rybnik (Guzy leg. 1884, Beleg PANW); Barania Gora /Kreis Cieszyn; Babia G6rafKreis Wadowice; Hohe Tatra (alle RAFALSKI 1961); BieszczadyfW-Teil der Wald· karpaten: zahlreiche Fundorte in den Kreisen Ustrzyki Dolne (z. B. Berg Tarnica), Lesko (Ustrzyki G6rne, Wolosatld Tal; Borg Rozki, Berg Polonina Wetlinska, Berg Jaslo); Kreis Sanok (STARJ!GA 1966), - ÖSSR: von den mährischen Beskiden und vom mittelslowakischen Gebirge über Niedere und Hohe Tatra weitverbreitet; - Mähren: FrydlandfOstrasice (iliL-

2JS Fan1ilia Isrhrrop$n.lit1idiw
AhO. :J7H-:t82. Glnudos pPni::; ,-ou /xclturojuutli~ C. L. 1\.ocu (linl\.R lntl't'nl~ I'Prhtt;, yontral).
Abb. 3iU. l. curli ]JHKStcnT. - BRD Allgiiu~ Bu.lclors\~hwn..ng-.
Ahh. :lSO. /. muuicatn IJ, 1\.ocu. - RlT Tran:o~t4ylntnist·lw Alpl'H.
1\hh. :l~H. [. Jl,IJJ'CIHtCa alpinula u. ssp. - IT Crissolo.
Ahb. :l8:?. I. hrli.'Cticrr H(H~\ri•~U. ··- l'H 1'P:'-I::tin. 14'ru.st•o.
Genus Ischyropsalis 219
HAVY 1973); - Slowakei: z. B. 2ilina (Sillein) (Belege Colt. E!ILHAVY); bei Kremnies (Krein· nitz) (Belege UNMB); Hohe Tatra (E!ILHAVY leg.); 0-Slowakei, Kamenica im Vihorlat-Gebirge (E!ILH.WY 1950); Zadieske doline (EliLHAVY 1968). - UdSSR/Karpato-Ukraine: Bogdan' (K:&ATOOHviL 1934, helltvigi); Rachov (B.A:RTOs 1939, dacica). - Rumänien: offenbar in allen gebirgigen Landesteilen weitverbreitet und lokal häufig; Zusammenstellung der bisher bekannten Fundorte bei AVRAM (1964a) und M.A:RTENS (1969a), Ergänzungen bei AVRAM & DUMITRESOU (1969), DUMITRESOU· (1972, 1973) und ÜLTEAN & DUMITRESOU (1973). Nennungen z. T. unter verschiedenen Namen)· für Gebiete außerhalb des Karpaten-Bogens haben sich in Iminern Falle bestätigen lassen (vgl. MARTENS 1969a);
Ökologie: Biotopansprüche: I. manicata besiedelt schattige, feuchu-küble Waldbiotope; sie lebt dort unter Holzueilen, unter Steinen, in Blockhalden oder in der Bodenauflage in Fallaub und Moos, oft in der Nähe fließenden Wassers. Aus Rumänien sind viele höhlenlebende Populationen bekannt, doch weisen diese Individuen keine Spezialisierungen auf, die sie von freilebtmden unterscheiden würden; sie leben in einem Temperaturbereich von 4-12 °C (AVRAM 1964).
Vertikalverbreitung: offenbar nur in mittleren und höheren Gebirgslagen. -Polnische Waldkarpaten: Nachweise zwischen 600 und 1350 m (W. S. in litt.); Rumänien: zwischen 700 und 1300 m {DUMITRESCU 1973, ÜLTEAN & DUMITRESCU 1973); von manicata besiedelte Höhlen liegen zwischen 305 und 1370 m (beide im Bezirk Crisana, AVRAM 1964a).
Phänologie: keine Einzelheiten bekannt; möglicherweise überwintern ad. (AVRAM 1964a fide SILHAVY); Höhlenpopulationen eurychron.
Ischyropsalis helvetica ROEWER, 1916 Abb. 382-394
1916 Ischyropsalis helvetica RoEWER, Arch. Naturgesch., 82 A: 152. 1923 Isohyropsalis helvetica - ROEWER, Weberknechte d. Erde: 691 (paro.: nur ~-Diagnose). 1959 IschyrojJBalis dentipalpis - D&Esco, Ann. Speleol., 14: 383. 1969& Ischyropsalis dentipalpis- MARTENS, Zoo]. Jb. Syst., 96: 201.
Begründung der Synonymie: Der Typus von helvetica ist ein ~ mit der Fundort-Angabe "Tessin". D&Esoo (1959) bezeichnet neues Material aus dem Tessin als dentipalpis, da das ihm vorliegende (] wie dentipalpis aus dem Aasta-Tal (dem Locus typicus von dentipalpis) eine Patellar-Apo am Palpus aufweist - ein Merkmal, das nur bei wenigen IschyropsalisArten auftritt.
Die verwandtschaftlichen Beziehungen der Tessin-Population zu dentipalpis aus dem Aosta-Tal/Penninische Alpen ist bisher nicht zweifelsfrei geklärt, da kein topatypisches Material von dentipalpis vorliegt und nur die genitalmorphologischen Verhältnisse Klärung bringen können. Da die geographisch dem Tessin nächst benachbarte Pppulation in den Bergam&>~ker Alpen in eine ganz andere Artengruppe gehört (siehe unter dentipalpis, Abb. 320- 323), wird Yorgezogen, für die Tessin-Population(en) den ältesten verfügbaren Namen zu verwenden.
Habitus und Feldkennzeichen: do. Bewehrun.g des Chel-Grundgliedes auffallend lang und dünn; c3' mit do-di halbkugelförmiger Apo, Senturn magnum, ~: S.laminatum. c3' unter Feldbedingungen nicht von deritipalpis zu trennen; ~ ununterscheidbar.
Kennzeichnung: Körpermaße (in mm): L c3' 5,2; L ~ 7,2. Laufbein II {c3', in Klammern~): Fe 7,0 (6,9) Pt 1,6 (1,7) Ti 5,6 (5,7) Mt 8,7 {8,6)
Ta 7,8 {7,7). Cheliceren-Grundglied: c3': 3,9; L ~: 4,3.
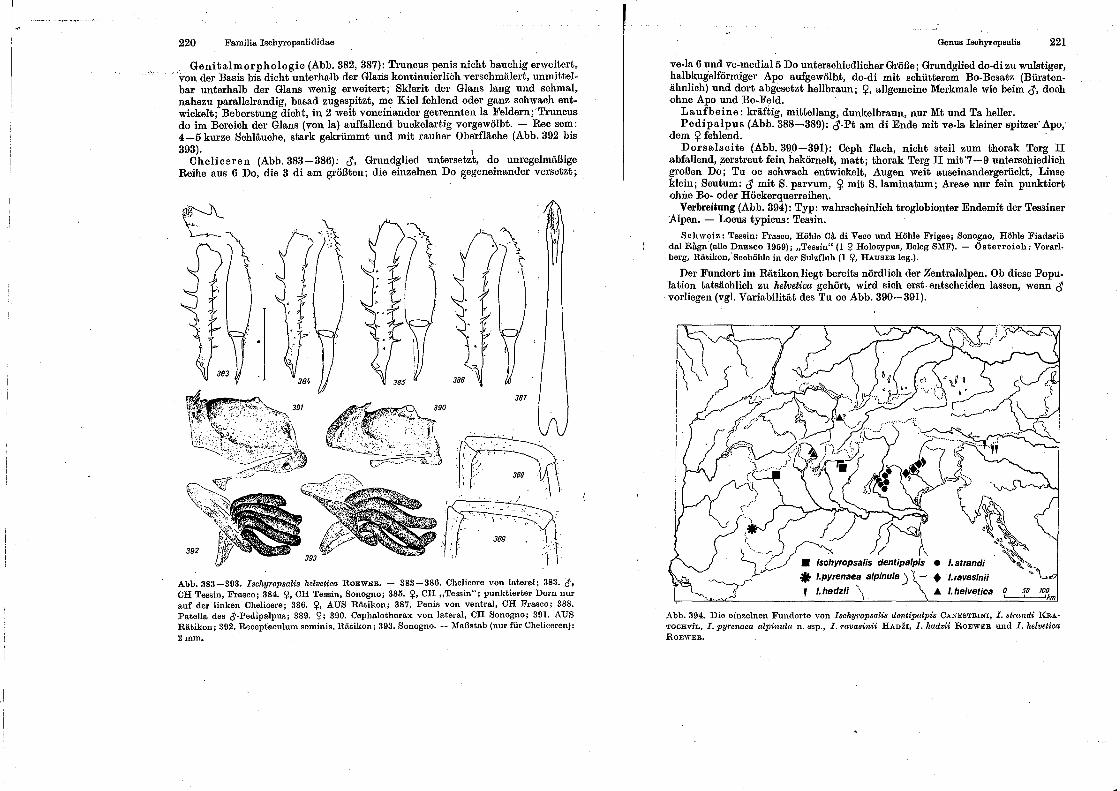
220 Familia Ischyropsalididae
. Genitalmorphologie (Abb. 382, 387): Trunc]ls penisnicht bauchig erweitert, . von der Basis bis dicht unterhalb der Glans kontinuierlich verschmälert, unmittel
bar unterhalb der Glans wenig erweitert; Sklerit der Glans lang und schmal, nahezu paral!elrandig, basad zugespitzt, me Kiel fehlend oder ganz schwach entwickelt; Beborstung dicht, in 2 weit voneinander getrennten la Feldern;"Truncus do im Bereich der Glans (von Ia) aUffallend buckelartig vorgewölbt. - Rec sem: 4-5 kurze Schläuche, stark gekrümmt und mit rauher Oberfläche (Abb. 392 bis 393). 1
Cheliceren (Abb. 383-386): rJ, Grundglied untersetzt, do unregelmäßige Reihe aus 6 Do, die 3 di am größten; die einzelnen Do &:egeneinander versetzt;
Abb. 383-393. Ischyropsalis helvetica RoEWER. 383-386. Chelicere von lateral; 383. c!, CH Tessin, Frasco; 384. !j?, CH Tessin, Sonogno; 385. !j?, CH "Tessin"; punktierter Dorn nur auf der linken Chelicere; 386. !j?, AUS Rätikon; 387. Penis von ventral, CH Frasco; 388. Patella des c!-Pedipalpus; 389. !j?; 390. Cephalothorax von lateral, CH Sonogno; 391. AUS Rätikon; 392. Receptaculum seminis, Rätikon; 393. Sonogno. -Maßstab (nur für Chelieeren): 2mm.
Genus Isohyropsalia 221
ve-Ja 6 und ve-medial5 Do unterschiedlicher Größe; Grundglied do-di zu wulstiger, halbkrtgelförmiger Apo aufgewölbt, do-di mit schütterem Bo-Besatz (Bürstenähnlich) und dort abgesetzt hellbraun; ~. allgemeine Merkmale wie beim rJ, doch ohne Apo und Bo-Feld.
Laufbeine: kräftig, mittellang, dunkelbraun, nur Mt und Ta heller. Pedipalpus (Abb. 388-389): d'-Pt am di Ende mit ve-Ia kleiner spitzer· Apo;
dem ~ fehlend. Dorsalseite (Abb. 390-391): Ceph flach, nicht steil zum thorak Terg II
abfallend, .zerstreut fein bekörnelt, matt; thorak Terg II mit"7-9 unterschiedlich großen Do; Tu oc schwach entwickelt, Augen weit auseinandergerückt, Linse klein; Scutum: d' mit S. parvum, ~mit S.laminatum; Areae nur fein punktiert ohne Bo- oder Höckerquerreihen.
Verbreitung (Abb. 394): Typ: wahrscheinlich troglobionter Endemit der Tessiner :Alpen. - Locus typicus: Tessin.
Seit weiz: Tessin: Frasco, Höhle Ca di Veco und Höhle Frigee; Sonogno, Höhle Fiadariö da! Ragn (alle DRESCO 1959); "Tessin" (1 !j? Holotypus, Beleg SMF). - Österreich: Vorarlberg, Rätikon,' Seehöhle in der Sulzfluh (1 !j?, HAusER leg.).
Der Fundort im Rätikon liegt bereits nördlich der Zentralalpen. Ob diese Population tatsächlich zu helvetica gehört, wird sich erst. entscheiden lassen, wenn rJ vorliegen (vgl. Variabilität des Tu oc Abb. 390-391).
+ ·t.ravaslnii
.&. I. he/vetica o so
Abb. 394. Die einzelnen Fundorte von Ischyropsalis dentipalpis CANESTRINI, I; strandi KRATOCHviL, I. pyrenaea alpinul<t n. ssp., I. ravasinii HAnzr, I. hadzii RoEWER und I. helvetica ROEWEB.
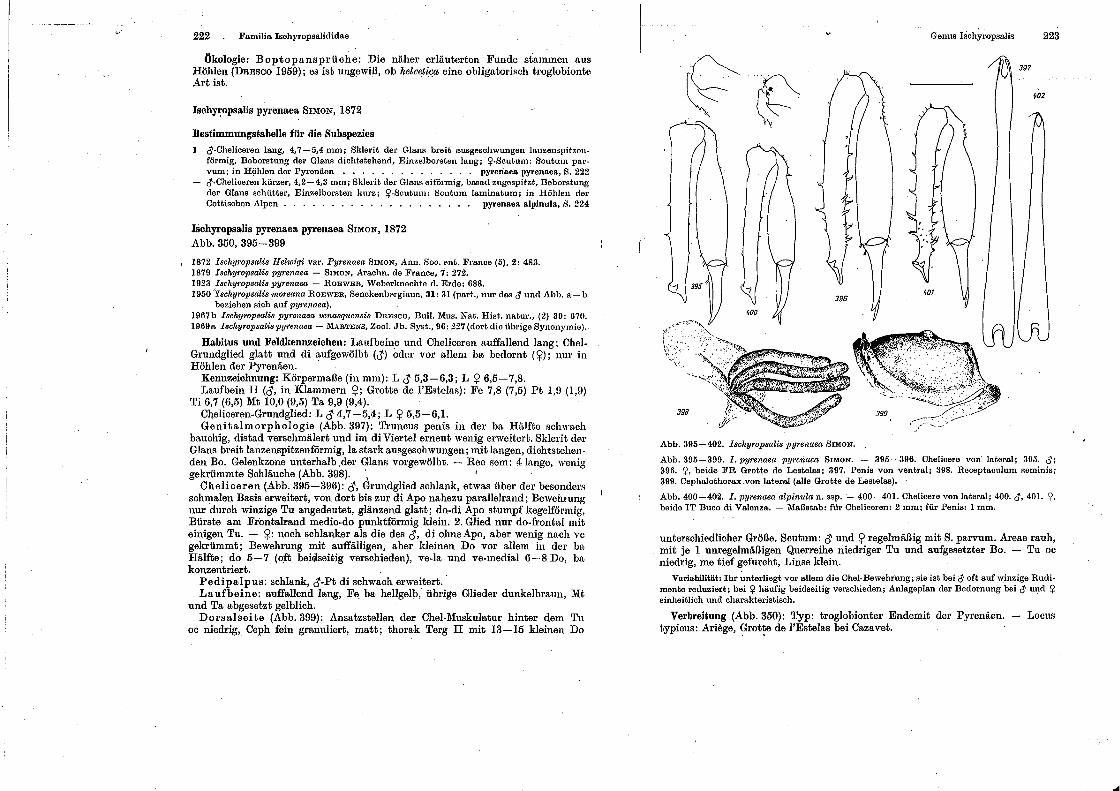
-·· 222 Familia Ischyropsalididae
Ökologie: Boptopansprüche: Die näher erläuterten Funde stammen aus Höhlen (DRESCo 1959); es ist ungewiß, ob helt•etica eine obligatorisch troglobionte Art ist.
IschYJ;opsalis pyrenaea SIMON, 1872
Bestimmungstabelle für die Subspezies
d'-Cheliceren lang, 4, 7-5,4 mm; Siderit der Glans breit ausgeschwungen lanzenspit.zonförmig, Beborstung der Glans dichtstehenrl, Einzelborsten lang; 'f-Scutum: Scutum parvum; in H~Hllen der Pyronäen . . . . . . . . . . . . . . pyrenaea pyrenaea, S. 222 d'-Cheliceren kürzer, 4,2-4,3 mm; Siderit der Glans eiförmig, basacl zugospit.zt., Beborstung der Glans schütter, Einzelborsten kurz; 'f-Scut.um: Souturn laminaturn; in Höhlen <'I er Cottischen Alpen . . . . . . . . . . . . . . . . . . . . pyrenaea. alpinula, S. 224
Ischyropsalis pyrenaea pyrenaea SIMON, 1872 Abb. 350, 395-399
, 1872 Isohy•·opsa!is Helwigi var. Pyrenaea SrMON, Ann. Soc. ent. France (5), 2: 483. 1879 Isohyropsa!is py•·enaea - SrMON, Arachn. de France, 7: 272. 1923 Ischy••opsa!is pyrenaea - RoEWER, Weberknechte <'1. Erde: 688. 1950 ·''Isohyropsa!is moreana RoEWER, Senckenbergiana, 31: 31 (part.., nur das d' und Abb. a- h
beziehen sich auf pyrenaea). I967b Isohyropsalis pyrenaea venasquensis DRESCO, Bull. Mus. Nat. Hist. natur., (2) 39: 070. 1969a Isohyropsalis py1·enaea - MARTENS, Zoo!. Jb. Syst., 96: 227 (dort die übrige Synonymie) ..
Habitus und Feldkennzeichen: Laufbeine und Cheliceren auffallend lang; ChelGrundglied glatt und .di aufgewölbt (d') oder vor allem ba bedornt (~); nur in Höhlen der Pyrenäen.
Kennzeichnung: Körpermaße (in mm): L d' 5,3-6,3; L ~ 6,5-7,8. Laufbein II (d', in Klammern ~; Grotte de l'Estelas): Fe 7,8 (7,5) Pt 1,9 (1,9)
Ti 6,7 (6,5) Mt 10,0 (9,5) Ta 9,9 (9,4). C4eliceren-Grundglied: L d' 4,7-5,4; L ~ 5,5-6,1. Genitalmorphologie (Abb. 397): Truncus penis in der ba Hälfte schwach
bauchig, distad verschmälert und im di Viertel erneut wenig erweitert. Sklerit der Glans breit lanzenspitzen.förmig, la stark ausgeschwungen; mit langen, dichtstehenden Bo. Gelenkzone unterhalb.der Glans vorgewölbt.- Rec sem: 4lange, wenig gekrümmte Schläuche (Abb. 398). ', 1
Cheliceren (Abb. 395-396): d', Grundglied schlank, etwas über der besonders schmalen Basis erweitert, von dort bis zur di Apo nahezu parallelrand; Bewehrung nur durch winzige Tu angedeutet, glänzend glatt; do-di Apo stumpf kegelförmig, Bürste am Frontalrand medio-do punktförmig klein. 2. Glied nur do-frontal mit einigen Tu. - ~:noch schlanker als die des d', di ohne Apo, aber wenig nach ve gekrümmt; Bewehrung mit auffälligen, aber kleinen Do vor allem in der ba Hälfte; do 5-7 (o.ft beidseitig verschieden), ve-la und ve-medial 6-8 Do, ba konzentriert.
Pedipalpus: schlank, d'-Pt di schwach erweitert. Laufbeine: auffallend lang, Fe ba hellgelb,' übrige Glieder dunkelbraun, Mt
und Ta abgesetzt gelblich. Dorsalseite (Abb. 399): Ansatzstellen der Chel-Muskulatur hinter dem Tu
oc niedrig, Ceph fein granuliert, matt; thorak Terg II mit 13-15 kleinen Do
I
Genus Ischyropsalis 223
397
W2
Abb. 395-402. Isohyropsalis pyrenaea SIMON.
Abb. 395-399. I. pyrenaect py•·enae<! SrMoN. - 395-396. Chelicere von· lateral; 395. Q'; 396. 'f, beide FR Grotte de Lestelas; 397. Penis von ventral; 398. Receptaculum Aeminis; 399. Cephalothorax .von lateral.(alle Grotte de Lestelas).
Abb. 400-402. I. pyrenaea alpinula n. ssp. '-- 400-401. Chelicere von lateral; 400. Q', 401. 'f, beide IT Buco di Valenza. - Maßstab: für Cheliceren: 2 mm; für Penis: I mm.
unterschiedlicher Größe. Scutum: d' und ~regelmäßig mit S. parvum. Areae raub, mit je 1 unregelmäßigen Querreihe niedriger Tu und aufgesetzter Bo. - Tu oc niedrig, me tief gefurcht, Linse klein.
Variabilität; Ihr unterliegt vor allem die Chel-Bewehrung; sie ist bei d' oft auf winzige Rudimente reduziert; hei 'f häufig beidseitig verschieden; Anlageplan der Bedornung bei d' U\ld 'f einheitlich und charakteristisch.
Verbreitung (Abb. 350): Typ: troglobionter Endemit der Pyrenäen. - Locus typicus: Ariege, Grot~ de l'Estelas bei Cazavet.

224 Familia Ischyropsalididae
Frankreich: Nachweise in den Departements Ariege, Hante-Garonne, Haute-Pyr6nees {DRESCO I967b; MARTENS I969a, dort Liste aller Eilizelfunde).
Ökologie: Biotopansprüche: I. p. pyrenaea scheint fast ausschließlich troglobiont zu leben; die besiedelten Höhlen liegen zwischen 500 und 1500 m (JuJlERTmE 1961 c). Offenbar werden nur die dunklen hinteren Höhlenteile eingenommen, wo ausgeglichener Temperaturgang herrscht; die Extreme lieg~n bei 7 und 11,5 °C. - DRESCO (1967b) meldet 2 Freiland-Funde am Port de Venasque (Dep. Haute-Garonne, 2400 m). Diese freilebenden (individuenarmen) Populationen der höheren Lagen der Pyrcnäen sind bisher übersehen worden.
Vertikal ver brei tung: In Höhlen zwischen 500 und 1500 m, einzelne Freifunde bei 2400 m (vgl. oben).
Phänologie: troglobionte Populationen eurychron. - Die ~ legen erstmals etwa einen Monat nach der Reifehäutung Eier, bis zum Tode etwa 3-6 Gelege, die zusammen 30-60 Eier enthalten (im Mittel10; unter natürlichen Bedingungen wurde ein Gelege mit 8 Eiern gefunden); Embryonalentwicklung 58 Tage (bei 11,5 °C), doch schlüpfen nur etwa 50% der Eier; pyrenaea durchläufu. meist 7 Jugendstadien, Postembryonalentwicklung in 6-7 Monaten, Lebenszeit der ad. 6-8 Monate (alle Angaben nach JullERTillE 1964).
Ischyr~psalis pyrenaea. a.lpinula. n. ssp.
Abb.350, 360,381,394,400-402
Diagnose: Eine Subspezies von I. pyrenaea, gekennzeichnet durch die Körpergröße, die d'-Genhalmorphologie, das abd ~-Scutum und die Verbreitung (Abb. 360).
Habiius und Feldkennzeichen: Wie die der Nominatform, doch wesentlich kleiner; ~mit Souturn laminatum; nur in den Cottischen Alpen.
Kennzeichnung: Köl.\permaße (in mm): L d' 4,6-5,1; L ~ 5,9. Laufbein II (d', in Klammern~); d' Grotta di Rio Martino, ~ Buco di V~lenza. Fe 5,3 (6,1) Pt 1,7 (1,6) Ti 3,9 (4,6) Mt 6,3 (7,3) Ta 7,1 (7,5). - Chehceren-
Grundglied: L d' 4,2-4,3; L ~ 4,8. . , Genitalmorphologie (Abb. 381, 402): Truncus penis schlank, breiteste Stelle
mehr basal als bei der Nominatform; Glans eiförmig, ba spitz ausgezogen, Beborstung schütter, Einzelborsten kurz. - ?elenkzo~e nicht wulstig vorge~ölbt:.
Cheliceren (Abb. 400-401): wie be1 der Nommatform, doch wesentlich kurzer; in Form der do-di Apo und in Lage der Bürste besteht volle Übereinstimmung.
Dorsalseite (Abb. 360): wie bei der Nominatform, doch aufdem thorak Terg II nur 4-5 kleine Do. - Scutum: d' mit S. parvum, ~ mit S. laminatum.
Beziehungen: I. p. alpinula ist durch geringe Körpergröße, abweichende Genitalmorphologie vor allem im Glans-Bereich und Souturn laminaturn des ~recht auffällig von der Nominatform verschieden. Beieie Formen st.immen in der Chelicerim-Morphologie (Bewahrung, Proportionen, do-di Apo) weitgehend überein, doch deuten einige Einzelheiten auf der Medialseite der ChelApo (Abb. 395, 400) auch dort auf geringe Unterschiede. Es ist somit nicht auszuschließen, daß alpinula bereits Artstatus erreicht hat (vgl. auch die viel geringeren Unterschiede zwischen den Subspezies von I. hellwigi, Abb. 298-307).
Verbreitung (Abb. 350, 394): Typ: Endemit der SW-Alpen. - Locus typicus: Cottische Alpen, Crissolo, Grotta di Rio Martino.
Genus Sabaoon 225
Italien: Cottische Alpen: Piemonte, Crissolo, Buco di Valenza (I <!, I~ Paratypoide, AcTIS leg, 7.Y. 1959; VIGNA leg. 24. VIII. I967); Crissolo, Grotta di Rio ~artino (I<! Holetypus SMF 29 I81, CAsALE leg. I2. III. 1972; 2 iuv. MORISI leg. I6. III. 1969; I iuv. LoNGHETTO leg. I5. III. I970); Massicchio del Monviso, Colle dell'Agnello (I ~ Paratypeid ÜSELLA leg. VII. I967).
Ökologie: Biotopansprüche: Es liegen Individuen aus 2 Höhlen und 1 Einzeltier aus dem Freiland vor, so daß I. p. alpinula zu der großen Gxuppe der sowohl troglobiont wie epigäisch lebenden Arten zu zählen ist. Der Freilandfund liegt weit über der Baumgrenze in der Alpin-Stufe.
Vertikalverbreitung: Im einzelnen nicht bekannt; die Höhle Buco di Valenza liegt 1500 m hoch {:ßRIGNOLI 1972), der eine epigäische Fundort auf 2700 m.
Familia Sabaconidae
I923 Ischyropsalidae - RoEWER, Weberknechte d. Erde: 680 (part.). 1970 Sabaconidae DRESOO, Bull. Mus. Nat. Hist. Natur., (2) 41: 1210.
Eine Familie der Palpatores-Ischyropsalidoidea, gekennzeichnet durch: d'-Genitalmorphologie: Truncus meist extrem do-ve abgeflacht, abgesetzter
Basis-Teil dorsad umgebogen und dort die Muskulatur konzentriert; die zur Glans ziehende Sehne somit lang. Glans in gerader Verlängerung des Truncus.
Ovipositor: in den Proportionen variierend: entweder kurz und ungegliedert (nur wenig länger als breit) oder wesentlich länger als breit und dann durch ringförmig angeordnete Reihen kleiner Borsten gegliedert wirkend (vgl. SuzuKI 1974). Loben der di Furca halbkugelförmig und dicht beborstet.
Cheliceren: d'-Grundglied do mit Apo-artiger Anschwellung, dort meist ein Drüsenorgan lokalisiert.
Pedipalpus: Ti und Ta keulig bzw. stumpf spindeiförmig aufgebläht, mit dichtem Besatz sekretorischer Borsten vom ·"gefiederten" Typ. Tarsalkralle rudimentär oder bisweilen fehlend.
Dorsales Souturn variierend; entweder als S. parvum, S.laminatum oder S. dissecturn ausgebildet oder {nur bei ~) ganz fehlend.
Sternum undeutlich entwickelt und schwach sklerotisiert; zwischen den Cx I-IV ausgespannt und nicht mit diesen verwachsen.
Verbreitung: disjunkt holarktisch; die Familie ist monotypisch (vgl. unter Gattung Sabacon).
Genus Sabacon SrMoN, 1879
I879 Sabacon SIMON, Arachn. de France, 7: 266. I923 Sabacon - ROEWER, Weberknechte .d. Erde: 694. I972a Sqbacon - MARTENS, Senckenberglana bio!., 53: 307.
Nominat-Gattung der Sabaconidae, gekennzeichnet durch: d'-Genitalmorphologie: Truncus penis schlank, do-ve abgeplattet, kurzer Basis
teil markant abgesetzt, tief gespalten und dorsad umgeklappt, zwischen diesem Gabel-förmigen Abschnitt die Muskulatur ausgespannt. Glans meist in gerader Verlängerung des Truncus, entweder zu feinem Stylus ausgezogen oder mit komplizierter Axmatur versehen. - Pedipalpus kurz und untersetzt, vor allem Ti und Ta 15 Tierwelt Deutsch!. 64: Martens

. 226 Familia Sabaconiclae
unterschiedlich aufgebläht; Ta gegen die Medial-Seite der Ti geklappt und dort in eine haarfreie Zone eingepaßt. Ti und Ta mit dichtem Besatz gefiederter Haare. - Cheliceren klein, Q'-Grundglied meist (1 immer) mit do-di Drüsenfeld, oft in auffälligem do Höcker oder Apo konzentriert.
Dorsales Scutum: Q', S. parvum oder S.laminatum; ~. S.laminatum, S. dissecturn oder Senturn völlig reduziert.
Sekundäre Geschlechtsmerkmale: Q'-Chel-Grundglied mit Drüsenfeld, meist in do Höcker bzw. Apo lokalisiert; Q'-Pa-Pt meist mit medio-di spitzem Do bzw. mit Do- oder Kö-Reihe; abd Sentalplatten der Q' (soweit ausgebildet) größer als die der ~. d. h. aus den Sideriten einer größeren Zahl von Segmenten verschmolzen; ~-Scuta unterliegen stärkerer Reduktion.
Verbreitung: Areal relilüär holarktisch; in Europa 4 Arten in den Kautabrisehen Gebirgen, Pyrenäen, dem Zentral-Massiv, in den SW-Alpen und N-Apennin; im Himalaya bisher 8 Arten aus Nepal und Bhutan, in Japan 9 Arten, davon ·1 auch in Korea; in China (Mandschurei) 1 Art; in Sibirien (Tunguska-Jenisei) 1 Art; in N-Amerika 4 Arten (östl. und westl. USA und Kanada). Die größte Formenfülle besteht wahrscheinlich in den glazial nicht oder nur geringfügig devastierten Gebieten am S-Rand der Paläarktis: S-Abdachung des Himalaya und Japan, wahrscheinlich auch - bisher unentdeckt - in vielen Teilen Chii1as. - .:ijie Gattung Sabacon ist aus dem Baltischen Bernstein bekaunt (ROEWER 1939). - 1 Art erreicht das hier behandelte Gebiet.
Typus-Art (durch Monotypie): Sabacon pm·acloxum Sr.ro;:;, 1879.
Sabacon simoni DRESOO, 1952 Abb. 43, 403-405
1952 Sabacon simoni DRESCO, Ann. Soc. ent. France, 121: 123. 1976 Sabacon simoni - THALER, Bull. Brit. arachn. Soc. 3: 205.
Habitus und Feldkennzeichen: Kleine bodenlebende, gelbgraue Art ohne distinkte Zeichnung, Pedipalpen auffallend aufgebläht; nur in den Meeralpen, Ligurischen Alpen und im nördl. Apennin.
Kennzeichnung: Körpermaße (in mm): L Q' 2,8-3,0; L ~ 3,5-4,0. Laufbein II (Q', in Klammern ~; 2. ~-Maß nach Syntypus): ·Fe 2,4 (2,4/1,9)
Pt 0,9 (0,9/0,7) Ti 2;6 (2,5/2,0) Mt 3,8 (3,4/2,5) Ta 4,4 (3,9/3,7). Genitalmorphologie (Abb. 403a-c): Truncus penis kurz gestaucht und
bandartig flach, di schwach erweitert; Glans als kurzes gegen den Truncus rechtwinklig abgebogenes kurzes Stück, Stylus stilettartig lang ausgezogen, gegen die ( Glans wiederum rechtwinklig abgesetzt. Truncus ve rillenförmig - di divergierend - vertieft. Im Winkel zwischen Truncus und Glans großer, häutiger und durch Hämolymphdruck offenbar schwallbarer Sack; dort auf der do Seite des Truncus je 1 para-me Feld langer und feiner Borsten; ~. Ovipositor kurz, kaum länger als breit, nicht in Segmente gegliedert, sondern di-la neben dem me Spalt 2 bräunliche fast halbkugelförmige Loben, mit kurzen Sinnesborsten besetzt. Weitere, kttum größere Sinnesborsten in lockerem und breitem Kranz im mittleren Teil der Logeröhre.
Cheliceren (Abb. 404a-b): Q', schlank-schmächtig, vor allem das Grundglied, dieses do-proximal mit länglichem, do planem zylindrischem Höcker, do mit dichtem Besatz feiner Haare. 2. Glied frontal schütter mit derben Borsten, wenige auch auf Grundglied di. - ~. ohne do Höcker des Grundgliedes.
Genus Sabacon 227
~OS
Abb. 403-405. Sabacon ~imoni DRESCO, - 493a-c) Penis von ventral, lateral und dorsal; 404a-b) ~- u~d ~-Chehcere von medial; 404c und e) c!· und ·~-Pedipalpus von medial; 404d) mediO-distale Bewahrung der Patplla des Q'-Pedipalpus; 405. dorsales Zeichnungsmuster, ~. - 403a-c, 404a, 404c-d: IT Passo di Cerreto; 404b 404e 405· nach Syntypus FR Foret de Turini. ' ' · •
15:.1<

228 Familia Phalangiidae
Pedipalpus (Abb. 404c-e): (f, die einzelnen Glieder sc~wa~ll, ke~Ii[i aufgebläht, vor allem Ti und diese dorsad geschwungen, Pt me-d1 m1t Reihe großer, proximad kleinerer massiver Tu (Abb. 40~d)! ~. ~chla~er, vgl. Abb. 404e_: .
Laufbeine: alle Glieder rund, nur fem m Langsreihen beborstet.; braunhch, Fe und Ti di etwas aufgehellt. · ·
Dorsalseite (Abb. 405): Zeichnung und Färbung: bräunlich bis hellviolettgrau, ohne deutliches Zeichnungsmuster. - Bewahrung: winzige Bo am Ceph-Frontalrand me Bo-Paar auf dem thorak Terg II; Querreihe kaum erkennbarer Bo auf den abd Areae. Scutum: Cf und ~auf den abd Area nur je 1 kleines me Skleritplättchen als S. laminatum. - Tu oc: niedrig gewölbt, me tief gefurcht, wegen des durchscheinenden Pigments der Augen tiefschwarz, me etwas aufgehellt.
Variabilität: Die !j! der Typenserie haben maridich kürzere Beine als die Individuen aus dom
Ap~jl,nin (vgl. "Kennzeichnung").
Verbreitung: Typ: Endemit der SW-Alpen und des nördl. Apennin. - Locus typicus: französische Meeralpen, Foret de Turini.
Frankreich: Meeralpen, Foret de Turini (leg. 1. IX. 1915, 2 !j! Holo- und !'arat~pus_coll. SilliON MNHNP 23863, DRESOO 1952; ferner VIGNA leg. 18. VIII. 1972). - I tahen: L1gur1SChe Alpen, Serra di Pamparato, Tana dell' Orso (MoRISI leg., MoRGANTINI leg.); Pamparato, ~ana delle'lrurbiglie (CASALE leg.); Passo Fascette, Arma d'la Ciosa (BoLoGNA leg.); Ap~amsche Alpen: Turrite Secca bei Castelnuovo; Toseaniseher Apennin: Abetone und Passo d1 Cerreto
(alle THALER 1976).
Ökologie: Biotopansprüche: THALER (1976) fand simoni in fe~~hten Waldschluchten. Damit scheinen die Präferenda denen der anderen europaischen Sabacon-Arten zu entsprechen und auf eine besonders hygrophile Art hinzudeuten. Aufenthalt in der Bodenauflage, unter Holz und Steinen, gern in der Nähe von rinnendem Wasser.
Vertikal ver brei tung: Im Foret de Turini (Meeralpen) auf 1500 m, in den Apuanischen Alpen und im Apennin zwischen 800 un~ 11?0 m. .
Phänologie: möglicherweise stenoehren herbstreif; ~Ie Typenserie_ (nu_r ad.) stammt aus IX, MX in der Toscana nur ad. (K. T. leg.), mv. aus !fen Ligurischen Alpen in VIII und M X.
.Suprafamilia Phalangioidea
1833 Phalangides SUNDEVALL, Consp. Arachn.: 35. 1876 Phalangioidae - THORELL, Arm. Mus. Cir. Stor. Genova, 8: 462. 1879 Phalangiidae - Sr:MoN, Arachn. de Fran<le, 7: 158. 1923 Phalangiidae -· RoEWER, Weberknechte d. Erde: 697. ·
Eine Überfamilie der Unterordnung Palpatores, gekennzeichnet durch: d'-Genitalmorphologie: Penis mit 1 Muskel, Sehne an der Ventralseit~ der Glans
.ansetzend· diese durch Sehnenzug in Längsrichtung des Truncus ausrwhtbar. -Legeröhre' lang und in meist zahlreiche, gelenkig miteina.?-de: ve:bund~ne Ringe gegliedert; di Furca 3gliedrig; Sternum scl:mal s~ai~genformig bis breit plattenförmig, zwischen den Cx III ausgespannt, mch~ mit Ilu;en _verwachsen.
Tracheenstigmen offen, mit Entapophyse fur Ventilatwnsmuskulatur; akzessorische Stigmen an den Tibien der Laufbeine.
Familia Phalangiidae 229
Loben der Cx II frei, nicht mit anderen Teilen der Sternalfläche bzw. Unterseite verwachsen.
Pedipalpen groß, kräftig-laufbeinartig entwickelt mit deutlicher und funktionstüchtiger Tarsalklaue (fehlt einem Teil der Neopilionidae: Vibone KAURI).- Spezialisierte Borsten auf dem Pedipalpus - soweit vorhanden - "gefiedert".
Zähne der Cheliceren einheitlich-massiv strukturiert; ohne Gruppe hyaliner Zähne (Zähnchen bei Neopilio australis LAWRENCE verdoppelt).
Gliederung: Die Überfamilie umfaßt die Phalangiidae (weltweit außer Madagaskar, Neuseeland und andere Inseln, südl. S-Amerika), Neopilionidae und Megalopsalididae (Abgrenzung und Verbreitung bisher nicht abschließend geklärt).
Bestimmungstabelle für die Familien bzw. Unterfamilien S. 229.
Familia Phalangiidae
1879 Phalangiidae - SI:MoN, Araclm. de France, 7: 158 (part.). 1923 Phalangiidae - RoEWER, Weberknechte d. Erde: 697 (part.).
Eine Familie der Palpatores-Phalangioidea, gekennzeichnet durch: (f-Genitalmorphologie: Truncus meist langgestreckt und schmal; Glans in deut
licher Gelenkzone dorsad in rechtem oder stumpfem Winkel gegen den Truncus abgewinkelt; Glans di mit(in unterschiedlichem Maße) gelenkig inseriertem Stylus. Glans (meist) mit 2 Paar Sinnesborsten. Muskulatur als einzelner Strang über nahezu den ganzen Truncus verteilt oder seltener in bauchig abgesetzter Basis konzentriert. Sehne des Muskels ve an der Basis der Glans ansetzend, diese durch Muskel-Sehnen-Zug in Längsrichtung des Trunous ausrichtbar.
Ovipositor: bis mehrfach körperlang, in zahlreiche gegeneinander gelenkig bewegliche Ringe gegliedert, diese mit je 1 Borsten-Querreihe; di Furca aus jeweils 3 Gliedern, auf dem Endglied je 1 kleiner halbkugelförmiger Lobus mit zahlreichen Sinnesborsten.
Pedipalpus laufbeinförmig, mit kräftig entwickelter Tarsa'lkralle, diese glattrandig oder bezähnelt.
Cheliceren normal proportioniert, bei d' oft mit auffälligen sekundären Geschlechtsmerkmalen; Grundglied ve glatt oder mit kräftigem, distad gerichtetem Dorn.
Dorsales Souturn meist weichhäutig und schwach entwickelt; wenn deutlich erkennbar als S. parvum ausgeprägt .
Loben der Cx II groß und frei beweglich; unterschiedlich, aber immer schwach gegeneinander gewinkelt.
Verbreitung: paläarktisch (Phalangiinae, Leiobuninae, Oligolophinae,Gyantinae, Solerosomatinae), nearktisch ,(einige Oligolophinae), äthiopisch (Phalangiinae), indo-orientalisch, indo-malayisch und neotropisch (Gagrellinae), wenige Sclerosomatinae auch indo-orientalisch. ;;
Bestimmungstabelle für die Unterfamilien der Phalangiidae (Chelicere zur Betrachtung abtrennen)
Kralle des Pa-Ta glatt, nicht gezähnt (etwa 30fache Vergrößerung notwendig) 2 Kralle des Pa-Ta kammartig gezähnt (z. B. Abb. 745) . . . . . . . . . . . 3
2 Chel-Grundglied ve glatt, ohne distad geneigten Zahn (z. B. Abh. 408- 410), bisweilen ve dichtes Feld kleiner Zähne (z. B. Abh. 573) . . . . . . . . . . Phalangiinae. S. 230