repÚblica bolivariana de venezuela...
TRANSCRIPT

REPÚBLICA BOLIVARIANA DE VENEZUELA
UNIVERSIDAD DEL ZULIA
FACULTAD EXPERIMENTAL DE CIENCIAS
DIVISIÓN DE ESTUDIOS PARA GRADUADOS
MAESTRÍA EN CIENCIAS BIOLÓGICAS
MENCIÓN ECOLOGÍA ACUÁTICA
ABUNDANCIA Y BIOMASA DEL BACTERIOPLANCTON EN AGUAS
SUPERFICIALES EN LA BAHIA EL TABLAZO Y ESTRECHO DEL LAGO DE
MARACAIBO Y SU RELACIÓN CON FACTORES BIÓTICOS Y ABIÓTICOS
Trabajo de Grado presentado como requisito para optar al Grado de Magíster Scientiarium en
Ciencias Biológicas. Mención Ecología Acuática
Autor: Lcda. María Caicedo Mora
Tutora: Dra. Luz Marina Soto
Co-Tutor: Dr. Carlos López
´
Maracaibo, Octubre 2012

ABUNDANCIA Y BIOMASA DEL BACTERIOPLANCTON EN AGUAS
SUPERFICIALES EN LA BAHIA EL TABLAZO Y ESTRECHO DEL LAGO DE
MARACAIBO Y SU RELACIÓN CON FACTORES BIÓTICOS Y ABIÓTICOS
--------------------------------------------------------
Lcda. María de los Ángeles Caicedo Mora
C.I. No.: 15.195.685
Teléfono: (0412) 0786347
B/12 de Octubre calle 93
Correo electrónico: [email protected]
-----------------------------------------------
Dra. Luz Marina Soto


DEDICATORIA
A DIOS todopoderoso, por su infinita presencia en mi vida, a la memoria de mi padre, a
mi familia por su apoyo y paciencia y especialmente a mi hijo Carlos Eduardo, que es el motor
que me impulso a seguir adelante, te dedico mi nuevo triunfo esta nueva etapa de mi vida. Te
Amo.

AGRADECIMIENTOS
A Dios todopoderoso, por darme la vida, por sostenerme, guiarme y permitirme seguir a tu
lado a pesar de mis debilidades. Además por darme la sabiduría para enfrentar las dificultades
que se presentaban en el día a día.
A la memoria de mi padre José Francisco Caicedo Córdoba, por darme odas las herramientas
necesarias para caminar en esta vida, y por impulsarme seguir escalando peldaños para continuar
mejorando en cada aspecto de mi vida.
A mi mama Nelly Mora y mis hermanos, por su apoyo, paciencia en acompañarme alcanzar
esta meta, y sobretodo por cuidar de mi hijo mientras me dedicaba a estudiar.
A mi hijo Carlos Eduardo, por su paciencia y tolerancia mientras estaba fuera de casa ocupada
en mis obligaciones académicas, por llenar mi vida de ternura y alentar mi corazón con su
sonrisa.
A la Dra. Luz Marina Soto, tutora de este trabajo por su confianza depositada en mí, por su
constante apoyo, por compartir sus conocimientos, sus sabios consejos y su material
bibliográfico.
Al Dr. Carlos López, cotutor de este trabajo por todo su apoyo y ayuda incondicional, por
compartir sus conocimientos, sus consejos y toda su confianza.
A Mayré Prieto, mi mejor amiga y madrina de mi hijo, por todos los momentos compartidos a
lo largo d esta maestría, los recuerdos con mucho cariño, buenos y malos, pues todos ellos nos
dejaron una enseñanza. Gracias por tu amistad y permanecer siempre allí. Te Quiero mucho
A Luis Soto y Adanoris Camargo, por su apoyo incondicional, por los momentos compartidos
en laboratorio de Zooplancton y sus buenos consejos. Así mismo, a Mario Nava, Nínive Espinosa
y Néstor Pereira, A TODOS MIL GRACIAS…

Caicedo, María. Abundancia y Biomasa del Bacterioplancton en aguas superficiales en La
Bahía El Tablazo y el Estrecho del Lago de Maracaibo y su relación con factores bióticos y
abióticos. Universidad del Zulia. Facultad Experimental de Ciencias. Departamento de Biología.
Maestría en Ciencias Biológicas - Mención Ecología Acuática. Venezuela. 2012. 72 p.
RESUMEN
Se estimó la abundancia y biomasa del bacterioplancton en aguas superficiales en Bahía El
Tablazo y Estrecho del Lago de Maracaibo y su relación con factores bióticos y abióticos,
durante doce meses de muestreo para el periodo 2009. La abundancia bacteriana varió entre
7,51E+08 - 3,22E+09 cel/l en la Bahía el Tablazo y 5,09E+08 - 2,15E+09 cel/l en el Estrecho del
Lago. Las bacterias libres y adheridas presentaron valores promedio de 1,29E+09 cel/l y
4,81E+08 cel/l respectivamente en Bahía y en Estrecho 9,05E+08 cel/l y 4,24E+08 cel/l
respectivamente; representando los cocos el principal morfotipo encontrado. Para los parámetros
fisicoquímicos solo se encontró correlación para abundancia bacteriana total con temperatura (r=
0,305 p=0,018) para la bahía y para el estrecho con la salinidad (r= 0,039 p=0,008) y
transparencia (r= -0,291 p=0,024). Para nutrientes y abundancia de ciliados, rotíferos y
nanoflagelados no se encontró correlación en ambas zonas. Durante el muestreo la biomasa
bacteriana total promedio fue de 8,09E+01 µg C/l en Bahía y 7,58E+01 µg C/l en el Estrecho. La
biomasa total presento una correlación negativa con la transparencia (r= -0,316 p=0,014) para
bahía y Estrecho, mientras que en el resto de las variables fisicoquímicas, nutrientes y factores
bióticos no se encontró correlación. No obstante, los morfotipos cocos adheridos presentaron una
correlación negativa con la abundancia de ciliados (r= -0,384 p=0,028) en Bahía y cocos y
bastones adheridos (r= -0,384 p=0,028) en Estrecho. Por otro lado, la biomasa del morfotipo coco
presento correlación con la abundancia de nanoflagelados (r= -0,315 p=0,014). Los resultados
sugiere que los valores de abundancia y biomasa de bacterioplancton incrementan con las
variables de temperatura y salinidad, mientras que disminuyen con transparencia y por pastoreo
de ciliados y nanoflagelados, donde las bacterias adheridas son las principalmente pastoreadas.
Palabras claves: Bacterioplancton, Cadena trófica, Bucle microbiano
E-mail: [email protected]

Caicedo, María. Abundancia y Biomasa del Bacterioplancton en aguas superficiales en La
Bahía El Tablazo y el Estrecho del Lago de Maracaibo y su relación con factores bióticos y
abióticos. Universidad del Zulia. Facultad Experimental de Ciencias. Departamento de Biología.
Maestría en Ciencias Biológicas - Mención Ecología Acuática. Venezuela. 2012. 72 p.
ABSTRACT
The abundance and biomass of bacterioplankton in surface waters in the Bay and Strait Tablazo
the Maracaibo of Lake and its relation to biotic and non-biotic factors, for twelve months of data
for the period 2009. Bacterial abundance ranged from 7.51 E 08 to 3.22 E 09 cells / l in the Bay
Tablazo and 5.09 E 08 to 2.15 E 09 cells / l in the Straits Lake. Free and attached bacteria showed
average values of 1.29 E 09 cells / l and 4.81 E 08 cells / l respectively in Bay and Strait 9.05 E
08 cells / l and 4.24 E 08 cells / l respectively, representing the major coconut morphotype found.
Physicochemical parameters for correlation was found only for total bacterial abundance with
temperature (r = 0.305 p = 0.018) for the Bay and Strait with salinity (r = 0.039 p = 0.008) and
transparency (r = -0.291 p = 0.024). For nutrients and abundance of ciliates, rotifers and
nanoflagellates no correlation was found in both areas. During sampling the total bacterial
biomass averaged 8.09 g E 01 C / l in Bahia and 7.58 E 01 mg C / l in the Strait. The total
biomass presented a negative correlation with transparency (r = -0.316 p = 0.014) for Bay and
Strait, while the rest of the physicochemical variables, nutrients and biotic factors found no
correlation. However, adhering cocci morphotypes showed a negative correlation with the
abundance of ciliates (r = -0.384 p = 0.028) in Bahia and cocci and rods attached (r = -0.384 p =
0.028) in the Strait. On the other hand, the biomass of coconut morphotype correlated with the
abundance of nanoflagellates (r = -0.315 p = 0.014). The results suggest that the values of
abundance and biomass of bacterioplankton variables increase with temperature and salinity,
while decrease with transparency and grazing ciliates and nanoflagellates, where bacteria are
attached mainly grazed.
Keywords: bacterioplankton, trophic chain, microbial loop
E-mail: [email protected]

INDICE GENERAL
Pág.
Dedicatoria
Agradecimientos
Índice de Ilustraciones
Resumen
Abstract
Introducción 13
Marco Teórico 15
1. Hidrodinámica del Sistema del Lago de Maracaibo 15
2. Bacterioplancton 16
3. Cadena trófica microbiana o Bucle microbiano 17
4. Distribución del Bacterioplancton 18
5. Cuantificación del Bacterioplancton 19
5.1. Abundancia Bacteriana 19
5.2. Biomasa Bacteriana 20
6. Factores Abióticos del Bacterioplancton 22
6.1. Materia Orgánica 22
6.2. Nutrientes 23
7. Factores Bióticos del Bacterioplancton 25
Objetivos 27
Objetivos General 27
Objetivos Específicos 27
Marco Metodológico 28
1. Área de Estudio 28
2. Recolección de las Muestras Abióticas 30
2.1. Parámetros Fisicoquímicos 30
2.2. Nutrientes 30
3. Recolección de las Muestras Bióticas 31
3.1 Bacterioplancton 31
3.2. Nanoflagelados Heterotróficos 31

3.3. Ciliados y Rotíferos 31
4.- Análisis de Factores Bióticos 32
4.1. Abundancia de Bacterioplancton 32
4.2. Biovolumen y Biomasa del Bacterioplancton 33
4.3. Abundancia Nanoflagelados 34
4.4.- Abundancia de Ciliados y Rotíferos 35
5. Análisis de los datos 35
Resultados y Discusión 36
1.- Variables Fisicoquímicos 36
1.1.-Temperatura 36
1.2.- Salinidad 37
1.3.- Transparencia 39
1.4.- Oxígeno Disuelto 40
1.5.- Profundidad 41
2.- Nutrientes 42
2.1.- Nitrito 42
2.2.- Nitrato 44
2.3.- Fósforo Total 45
2.4.- Ortofosfatos 46
3.- Abundancia y Biomasa del Bacterioplancton 47
3.1.- Abundancia 47
3.2.- Biomasa 52
4.- Abundancia de Nanoflagelados, Ciliados y Rotíferos 55
4.1.- Nanoflagelados 55
4.2.- Ciliados 57
4.3- Rotíferos 58
5.- Relación de abundancia y biomasa bacterioplancton con factores abióticos 60
5.1- Bahía El Tablazo 60
5.2.- Estrecho del Lago 61
6.- Relación de abundancia y biomasa bacterioplancton con factores bióticos 62
Conclusiones 64
Recomendaciones 65
Literatura Citada 66

ÍNDICE DE ILUSTRACIONES
FIGURAS TITULOS Pág. 1
Bucle microbiano
14
2
El Sistema del Lago de Maracaibo, Estado Zulia
27
3 Imagen Satelital de Bahía El Tablazo y Estrecho del Lago de
Maracaibo
28
4a Variaciones de Temperatura Mensual en Bahía El Tablazo y El
Estrecho de Lago Maracaibo.
36
4b Valores de Temperatura por Estación en Bahía El Tablazo y El
Estrecho de Lago de Maracaibo.
36
5a Variaciones de Salinidad Mensual en Bahía El Tablazo y El Estrecho
de Lago de Maracaibo.
38
5b Valores de Salinidad por Estación en Bahía El Tablazo y El Estrecho
de Lago de Maracaibo.
38
6a Variaciones de Transparencia Mensual en Bahía El Tablazo y
Estrecho del Lago de Maracaibo.
39
6b Valores de Transparencia por Estación en Bahía El Tablazo y
Estrecho del Lago de Maracaibo.
39
7a Variaciones de Oxígeno disuelto Mensual en Bahía El Tablazo y El
Estrecho de Lago de Maracaibo.
40
7b Valores de Oxígeno disuelto por Estación en Bahía El Tablazo y El
Estrecho de Lago de Maracaibo.
41
8a Variaciones de Profundidad Mensual en Bahía El Tablazo y Estrecho
del Lago de Maracaibo.
42
8b Valores de Profundidad por Estación en Bahía El Tablazo y Estrecho
del Lago de Maracaibo.
42
9a Variaciones de Nitrito Mensual en Bahía El Tablazo y El Estrecho
de Lago de Maracaibo.
43
9b Valores de Nitrito por Estación en Bahía El Tablazo y El Estrecho de
Lago de Maracaibo.
43
10a Variaciones de Nitrato Mensual en Bahía El Tablazo y Estrecho del
Lago de Maracaibo.
44
10b
Valores de Nitrato por Estación en Bahía El Tablazo y Estrecho del
Lago de Maracaibo.
44
11a Variaciones de Fósforo Total Mensual en Bahía El Tablazo y El
Estrecho de Lago de Maracaibo.
45
11b Valores de Fósforo Total por Estación en Bahía El Tablazo y Estrecho 46

del Lago de Maracaibo.
12a Variaciones de Ortofosfato Mensual en Bahía El Tablazo y Estrecho
del Lago de Maracaibo.
46
12b Valores de Ortofosfato por Estación en Bahía El Tablazo y Estrecho
del Lago de Maracaibo.
47
13a Variaciones de Bacterioplancton Mensual en Bahía El Tablazo y
Estrecho del Lago de Maracaibo.
48
13b Valores de Bacterioplancton por Estación en Bahía El Tablazo y El
Estrecho de Lago de Maracaibo.
48
13c Variaciones de Morfotipos Cocos y Bastones Mensual en Bahía El
Tablazo.
50
13d Variaciones de Morfotipos Cocos y Bastones Mensual en Estrecho del
Lago de Maracaibo.
50
13e Variaciones de Bacterias Libres y Adheridas Mensual en Bahía El
Tablazo.
51
13f Variaciones de Bacterias Libres y Adheridas Mensual en Estrecho del
Lago de Maracaibo.
52
14a Variaciones de Biomasa de Bacterioplancton Mensual en Bahía El
Tablazo y Estrecho del Lago de Maracaibo.
53
14b Valores de Biomasa de Bacterioplancton Por Estación en Bahía El
Tablazo y Estrecho del Lago de Maracaibo.
53
15a Variaciones de abundancia de Nanoflagelados Mensual en Bahía El
Tablazo y El Estrecho de Lago de Maracaibo.
55
15b Valores de Abundancia de Nanoflagelados por Estación en Bahía El
Tablazo y El Estrecho de Lago de Maracaibo.
56
16a Variaciones de Abundancia de Ciliados Mensual en Bahía El Tablazo
y El Estrecho de Lago de Maracaibo.
57
16b Valores de Abundancia de Ciliados por Estación en Bahía El Tablazo
y El Estrecho de Lago de Maracaibo.
58
17a Variaciones de Abundancia de Rotíferos Mensual en Bahía El
Tablazo y El Estrecho de Lago de Maracaibo.
59
17b Valores de Abundancia de Rotíferos por Estación en Bahía El
Tablazo y El Estrecho del Lago de Maracaibo.
59

INTRODUCCIÓN
En los ecosistemas acuáticos el papel de las bacterias es conocido desde hace mucho ya que
son descomponedoras de la materia orgánica y agentes que participan en el reciclado de
minerales indispensables para las comunidades autótrofas, pero solo recientemente biólogos y
ecólogos acuáticos han empezado profundizar en los estudios sobre las funciones que las
bacterias y sus depredadores desempeñan en la cadena tróficas. El concepto de “bucle
microbiano”, que implica a las bacterias como competidoras tanto por los nutrientes como por
otras moléculas orgánicas con organismos que pertenecen al mismo compartimiento trófico y
dimensional, ha impulsado los estudios sobre estos organismos, contribuyendo a formular nuevas
hipótesis e interpretaciones sobre red microbiana y sobre los factores que influyen en ellos
(Cognetti y Magazzu, 2001).
El bucle microbiano, se refiere a un modelo de transferencia de energía simple, mediante la
vía del carbono y el ciclo de nutrientes a través de los componentes de las comunidades acuáticas
pelágicas e incluyen procariotas, y algunos eucariotas, organismos unicelulares, tanto autótrofos
como heterótrofos, bacterias pelágicas, pico y nanoplancton, nanoflagelados heterotróficos, virus,
ciliados y zooplancton; y todos están integrados en una red planctónica (Petrúcio, 2003; Álvarez,
2005). Así, la transferencia de energía existente en la cadena trófica microbiana compone la base
de la gran cadena trófica de organismos superiores (Weinbauer y Hofle, 1998; González, 2006).
(Fig.1)
Figura 1. Bucle microbiano (tomado de: Azam y col.,1998)

14
De esta forma, el crecimiento bacteriano en el medio acuático puede estar limitado por
factores, entre los que se puede destacar: la calidad de la materia orgánica disuelta (Carlson y
Ducklow, 1996; Eiler y col., 2003), los nutrientes inorgánicos (Thingstad y col., 1999), las
infecciones virales (Middelboe y col., 1996), las concentraciones de micronutrientes (Church y
col., 2000) y pastoreo por depredadores (Godinho y col., 2007).
La eutrofización de los sistemas lacustres mediante el incremento de la carga de nutrientes, es
uno de los cambios que más afectan a estos ecosistemas. Cambios en la abundancia y estructura
de la red trófica microbiana tienen efectos muy importantes en el flujo de nutrientes y energía que
se da en la columna de agua. El conocimiento de estos flujos que se dan a través del
compartimiento microbiano es un requisito indispensable para predecir cómo los ecosistemas
acuáticos pueden verse afectados por estas perturbaciones (Cognetti y Magazzu, 2001; Lucinda,
2007). Otro factor, que afecta el crecimiento bacteriano es la depredación o pastoreo por parte
principalmente por protistas. Algunas investigaciones han revelado que la depredación por
protistas tiene un gran impacto en la disminución del número de bacterias, la morfología,
taxonomía y estructura de sus comunidades (Masín y col., 2003). La biomasa bacteriana es
directamente depredada por protozoarios que más tarde son consumidos por metazoos dentro de
los niveles tróficos más altos en los ecosistemas acuáticos (Tzaras y col., 1999).
En general, la mayoría de los trabajos publicados sobre redes tróficas acuáticas proviene en
gran parte de la investigación realizada en sistemas templados y es poco lo que se conoce al
respecto para sistemas tropicales. En Venezuela y especialmente en el estado Zulia, son escasos
los estudios en los que se integren a los microrganismos como parte del ecosistema marino. El
presente trabajo es el primer acercamiento desde el punto de vista ecológico al bacterioplancton
en el sistema del lago de Maracaibo, permitiendo ofrecer información sobre la abundancia y
biomasa del bacterioplancton en aguas superficiales en la Bahía el Tablazo y Estrecho del Lago
de Maracaibo, y las relaciones con algunos factores bióticos y abióticos; a partir del cual se
generaran las primeras informaciones sobre bacterioplancton en esta importante área Lacustre.

OBJETIVOS
Objetivos General
Establecer la relación entre la abundancia y biomasa del bacterioplancton en aguas
superficiales en la Bahía el Tablazo y Estrecho del Lago de Maracaibo con factores bióticos y
abióticos.
Objetivos Específicos
1.- Estimar la abundancia y biomasa del bacterioplancton en aguas superficiales en la Bahía el
Tablazo y Estrecho del Lago de Maracaibo.
2.- Estimar la abundancia de nanoflagelados, ciliados y rotíferos en aguas superficiales de la
Bahía del Tablazo y Estrecho del Lago de Maracaibo.
3.- Correlacionar la abundancia y biomasa del bacterioplancton en la Bahía del Tablazo y
Estrecho del Lago de Maracaibo con factores bióticos y abióticos.

MARCO TEÓRICO
1.- Hidrodinámica del Sistema Lago de Maracaibo
El Sistema del Lago de Maracaibo está afectada por: el régimen de mareas del Mar Caribe que
a su vez, se ven afectadas por la forma del Golfo de Venezuela que actúa como una caja de
resonancia, produciendo mayor amplitud en la marea que se produce a la entrada de la Bahía del
Tablazo; el régimen de caudales de los 135 ríos que escurren al lago producto de los períodos de
distribución bi-modal de las lluvias (Abril- Mayo y Septiembre – Octubre); el régimen de viento
predominante de dirección Nor-este. (Alisios) y la Fuerza de Coriolis (Herman, 2006).
Las variaciones en los niveles de agua en el extremo abierto del Golfo (mar abierto) son
determinadas por la atracción del sol, la luna y la posición relativa de la tierra con respecto a
ellos. Son estas variaciones la que inducen la entrada y salida del agua al Lago a través de las
bocas de San Carlos, Cañonera y Cañonerita, en un proceso periódico (Rincón y Godoy, 2006).
La marea en el Golfo es derivada de la onda del Caribe; en su parte noreste conserva sus mismas
características (mareas mixtas predominantemente diurnas). Al penetrar al Golfo y debido a sus
dimensiones, los componentes semidiurnos son aumentados notablemente por resonancia,
mientras que los diurnos permanecen relativamente inalterados. Las mareas que son semidiurnas,
se propagan desde el Golfo hacia el Lago y disminuyen progresivamente a todo lo largo del
estuario, desde un rango promedio de 110 cm. En el mareógrafo del Malecón (en el Golfo a corta
distancia de la boca principal del estuario en San Carlos-Zapara) hasta 18-20 cms.
En la Ensenada en el Estrecho cerca de la entrada del Lago, y 7,8-14 cms. En la Salina, Costa
Nororiental del Lago (Rodríguez, 2000). Aunque las mareas del Lago son muy leves puede
tomarse como un rango promedio de 12,5cm en la Salina y 6 cm en Bobures, en la Costa
Sudoriental del Lago. Existe un desfasamiento de 2,5 hasta 3,5h entre las mareas en la boca
principal del estuario en San Carlos-Zapara y los caudales medidos en el estrecho, a la altura de
Maracaibo, que trae como consecuencia que con el cambio de marea entrante a saliente y
viceversa, las corrientes se invierten pero lo hacen con un retardo del orden antes indicado y de
tal manera que cuando las mareas bajan, los flujos en el estrecho no son hacia el norte (salientes)
como cabría esperarse normalmente, sino que lo hacen hacia el sur y cuando las mareas suben las
corrientes en vez de ser hacia el sur, son hacia el norte (Molines y col, 1989).

17
Los vientos predominantes sobre el sistema son los llamados vientos Alisios que circulan en
dirección noreste de noviembre hasta abril (época de sequía) y disminuyen lo que resta del año
(época de lluvia). La circulación de estos vientos predominantes en el Lago originan un
desbalance en el sistema de viento, que contribuye a producir el movimiento de las masas de agua
del epilimnio del lago en sentido contrario a las agujas del reloj (Herman, 1997). La circulación
en el Lago de Maracaibo es en el sentido contrario a las agujas del reloj con velocidades típicas
de 0,2 m/s. En la bahía del Tablazo es un poco mas compleja y fuertemente afectada por las
mareas.
La salinidad del Lago de Maracaibo proviene del aporte de las aguas del Golfo de Venezuela
en forma de una intrusión estacional, que genera conjuntamente con el comportamiento
hidrodinámico un cono hipolimnético anóxico, donde ocurre la mayor acumulación de materia
orgánica, es decir, actúa como una fuente de nutrientes (Herman, 1997). El incremento de la
salinidad en el lago se debe al proceso de profundización del canal de navegación y a la
reducción del escurrimiento en las cuencas aportadoras de agua dulce. Antes de dicha
profundización, la intrusión de agua salina al Lago solo se producía durante la estación seca
(Rincón y Godoy, 2006).
El agua salina procedente del Golfo de Venezuela se mezcla con el agua del Lago en la Bahía
del Tablazo debido a la acción de las mareas y los vientos (Harlemann y col., 1967) y fluye por el
fondo del Estrecho de Maracaibo hasta depositarse en el fondo del Lago; generando el aspecto
más característico del Lago, el cual es la estratificación salina, mientras que la temperatura del
agua es prácticamente constante durante todo el año (Rincón y Godoy, 2006).
2.- Bacterioplancton
El término bacterioplancton se refiere básicamente a la fracción picoplanctónica del plancton,
es decir, los microorganismos con tamaños entre 0,2 y 2,0 µm. Estos microorganismos
suspendidos en el agua se pueden clasificar como autóctonos y alóctonos. Los primeros serían
originarios del agua y, por lo tanto, estarían adaptados a las condiciones del hábitat, en tanto que
los alóctonos permanecerían transitoriamente en el ecosistema, en un corto o largo tiempo

18
dependiendo de las características del medio. Sin embargo, no existe una delimitación clara entre
uno y otro (Fogg, 1995).
Las bacterias planctónicas son principalmente Gram negativas (80% a 95%) y a menudo son
móviles, pleomórficas y pigmentadas. Abundan en la zona litoral de los cuerpos de agua, la
mayoría no están libres en el agua, sino adheridos a partículas, especialmente materia orgánica.
Su biomasa es pequeña en comparación con los productores primarios, pero su actividad es más
importante en el reciclaje de nutrientes. El número de bacterias varía mucho en relación con la
cantidad de materia orgánica, la cual puede ser alóctona o autóctona. Por lo general el recuento
de organismos viables varía de 100 cel/mL a 1000 cel/mL
para los lagos templados (Campbell,
1993).
Las actividades de los microorganismos en el agua son múltiples e incluyen muchos procesos
relacionados con las cadenas alimentarias y flujo de energía. En primer lugar, las bacterias
reciclan y disuelven las partículas orgánicas y liberan nutrientes inorgánicos que pueden ser
asimilado por el fitoplancton. En segundo lugar, toman carbono disuelto del medioy lo
incorporan al carbono orgánico particulado, formando de esta manera una reserva disponible y
fuente directa de carbono y energía para niveles tróficos superiores (Elser y col., 1995; Petrúcio
2003).
3.- Cadena trófica microbiana o Bucle microbiano
Las ideas sobre la estructura de las redes tróficas pelágicas han venido cambiando desde 1980.
El concepto tradicional de la cadena planctónica microbiana “fitoplancton-zooplancton” se ha
complementado con el concepto de bucle microbiano (Azam y col., 1983). El bucle microbiano,
se refiere a un modelo de transferencia de energía simple, mediante la vía del carbono y el ciclo
de nutrientes a través de los componentes de las comunidades acuáticas pelágicas, e incluyen
procariotas y algunos eucariotas, organismos unicelulares, tanto autótrofos como heterótrofos,
bacterias pelágicas, pico y nanoplancton, nanoflagelados heterotróficos, virus, ciliados y
zooplancton; y todos están integrados en una red planctónica (Weisse, 1991). La transferencia de
energía existente en la cadena trófica microbiana constituye la base de la gran cadena trófica de

19
organismos superiores (Jürgens, 1994; Weinbauer y Hofle, 1998; Petrúcio, 2003).
Azam y col., (1983) resumieron la información disponible y actualizaron el modelo de
Pomeroy (1974). Ellos propusieron el termino “microbial loop” (bucle o red microbiano ) para
describir como una fracción significativa de la producción primaria se desvía fuera de la
rutaalimenticia tradicional, hacia un trama trófica microbiana en donde las bacterias crecen sobre
materia orgánica disuelta y particulada, y son consumidas por pequeños flagelados no
pigmentados, los cuales a su vez son consumidos por protozoarios grandes.
El bucle microbiano consta de un complejo sistema de diferentes microorganismos, los
procesos responsables de la utilización y transferencia de la materia orgánica y de su producción
a través de la fotosíntesis, la utilización de bacterias heterótrofas y la producción de biomasa
bacteriana (Azam y col., 1983). También se ha asumido que los virus causan la muerte de las
bacterias acuáticas, a través de lisis viral. De esta manera los nutrientes que liberan las bacterias
al morir son aprovechados por otros microorganismos. Así los virus aceleran el flujo de los
nutrientes, como el carbono y el nitrógeno en el ecosistema (Middelboe y col., 2001).
El concepto de bucle microbiano ha cambiado la tradicional transferencia de los nutrientes y
los niveles de energía de bacterias a los metazoos a una compleja red trófica. De este modo se
han venido realizando grandes esfuerzos en investigaciones dedicadas al estudio de las bacterias
en una gran variedad de los ecosistemas acuáticos (Weisse, 1990). Este nuevo concepto, ha
permitido nuevos avances para cuantificar la biomasa y la actividad de las bacterias heterotróficas
en el sistema acuático, confirmando que la actividad metabólica bacteriana es altamente
significativa y se ha cambiado el concepto sobre el papel de las bacterias en la transformación y
flujos de carbono y nitrógeno en los ambientes acuáticos, pasando de ser simplemente
mineralizadoras de partículas de detritus a recicladoras y competidoras por nutrientes orgánicos e
inorgánicos junto con el fitoplancton (Zehr y Ward, 2002; González y col., 2006).
4.- Distribución del bacterioplancton
El bacterioplancton es limitado por la disponibilidad de sustratos orgánicos y nutrientes,

20
especialmente del fósforo y nitrógeno. En consecuencia, cabría esperar un aumento general en la
biomasa bacteriana en un sistema acuático que este cargado de materia orgánica y nutrientes
(Wetzel, 2001).En general, el número y la biomasa de bacterias aumentan con el incremento de la
productividad y de las concentraciones de elementos inorgánicos y compuestos orgánicos en los
lagos. A pesar de las grandes variaciones estacionales, el número y biomasa de bacterias
aumenta en las aguas continentales de oligotróficas a eutróficos. Los niveles más altos de
bacterias se han observado en los lagos tropicales alcalinos y en los embalses eutróficos,
probablemente debido que los grandes embalses estudiados tienen afluencia de los ríos que
reciben los desechos industriales y municipales. El número de bacterias es notablemente menor
en los lagos ácidos distróficos, que contienen altas concentraciones de material húmico (Wetzel,
2001). La biomasa y la producción bacteriana planctónicas suele es menor durante el invierno
que en verano en lagos y embalses templados; esta condición se correlaciona con una
disminución de las temperaturas en el invierno y la reducción de la carga de partículas disueltas
y de sustratos orgánicos autóctonos (fitoplancton y plantas de litoral) y alóctonos cuando las
fuentes de los suelos están congelados (Wetzel, 2001).
5.- Cuantificación del Bacterioplancton
5.1.- Abundancia Bacteriana
La mayoría de las bacterias han sido tradicionalmente clasificadas en base a su sensibilidad a
reacciones bioquímicas específicas de interés clínico. Actualmente se considera que para la
caracterización de la diversidad bacteriana se deben incluir aspectos fenotípicos, genéticos e
información acerca del hábitat y la comunidad, lo que permite distinguir nuevas categorías de
microorganismos (Arocena y col., 1999).
Uno de los métodos para la cuantificación del bacterioplancton se efectúa por microscopía de
epifluorescencia, mediante la cual se ha observado que la abundancia bacteriana en el medio
acuático es dos a tres órdenes de magnitud mayor que la obtenida con las técnicas de cultivo
tradicional en placa o usando microscopía óptica de contraste de fases. La técnica más utilizada
para determinar abundancia total de bacterias, fue la descrita por Porter y Feig (1980), que

21
consiste en teñir el DNA bacteriano de doble cadena con un fluorocromo específico (DAPI),
complejo que excitado con luz de 365 nm que fluorece azul brillante, indicando la presencia de
células bacterianas, aunque no necesariamente activas.
Para la identificación taxonómica individual de células bacterianas, los métodos más
aceptados son: secuenciación de ARN ribosómico mediante detectores específicos de
oligonucleótidos (De Long, 1993), la identificación de anticuerpos mediante inmunofluorescencia
(Campbell, 1993), el registro de secuencias genéticas mediante PCR (Atlas, 1993), homología de
DNA-DNA (Allsopp y col., 1995), hibridación de DNA-ARN y las técnicas de secuenciación de
ARN (5S, 16S y 23S) (Montoya y col., 1996), y la hibridización fluorescente “in situ” (FISH)
empleando microscopía de epifluorescencia, microscopía focal laser o citometría de flujo
(Pernthaler y col., 2003).
La incorporación de timidina y leucina durante la síntesis de DNA y de proteínas,
respectivamente, son los métodos más comunes para estimar la producción bacteriana en el
medio acuático. El principal problema radica en relacionar en forma precisa las tasas de
incorporación de ambos marcadores con las tasas reales de producción bacteriana, lo que depende
del conocimiento exacto de varios componentes celulares (concentración de DNA y proteína por
célula, relación timidina/ADN y dilución intracelular del marcador) (Arocena y Conde, 1999).
Sin embargo, el empleo de las técnicas moleculares aún es limitado, ye que necesitan del
empleo de reactivos y equipamientos costosos, por lo que muchos investigadores siguen usando
los esquemas de clasificación apoyados en los métodos tradicionales, con el auxilio de la
microscopía electrónica (Wistreich, 1998).
5.2.- Biomasa Bacteriana
La biomasa es un parámetro básico considerado en ecología microbiana. Conociendo la
biomasa se puede evaluar la actividad energética de la misma y su potencial como fuente de
alimento para los niveles tróficos superiores. Puede ser definido como la masa de bacterias vivas
(ya sea peso seco, contenido de carbono o proteínas) en un hábitat dado. Sin embargo, para fines
ecológicos es más útil determinar la biomasa como carbono utilizando un apropiado factor de

22
conversión. En este sentido, una gran cantidad de factores han sido estimulados utilizando
diferentes métodos, técnicas de preparación, organismos y condiciones de crecimiento y más que
dar claridad a la medición, el problema se ha incrementado en los últimos años (Wetzel, 2001).
Zingel (1999), afirma que las variaciones de la biomasa son atribuibles a los diferentes
estados nutricionales de las bacterias en sus hábitats. Sin embargo, Norland (1993) concluye que
las diferentes relaciones de carbono-volumen se deben básicamente a variaciones en la
calibración y la escala, por lo que se haría necesaria la homogenización de criterios que conllevan
a la definición de un procedimiento estándar.
La biomasa, en la práctica es muy difícil de determinar, pero muchos métodos se han
desarrollado con el propósito de determinar algún parámetro que pueda ser correlacionado con la
biomasa. Estos parámetros incluyen conteos totales celulares, así como varios constituyentes
químicos diferentes tales como: lipopolisacáridos, ácido murámico, bacterioclorofila, lípidos
específicos y otros. No obstante, uno de los métodos más populares es la estimación de la
biomasa bacteriana a partir del volumen celular, en combinación con los conteos celulares totales
(Kemp y col., 1993; Arocena y Conde, 1999).
La biomasa puede ser calculada a partir de los conteos por microscopio de epifluorescencia y
las medidas de las células para el volumen celular. Este último se puede realizar a través de la
técnica digital que es ampliamente usado en combinación con la microscopía de epifluorescencia,
para determinar el tamaño y la forma de las bacterias. El procedimiento consiste en la obtención
de una imagen mediante un microscopio, el mejoramiento digital y la medición propiamente
dicha. El equipo más básico consiste en un microscopio de epifluorescencia, una cámara de video
para trasmitir la imagen al procesador, un digitalizador para convertir la imagen análoga, una
computadora y un programa digital para almacenar y procesar la información digital, y dos
monitores para observar la imagen y la operación con los datos (Arocena y Conde, 1999).
Para obtener el volumen de cada morfotipo de bacteria, estas pueden ser asemejadas a esferas
para los cocos y cilindros para los bacilos, utilizando la fórmula del volumen de estas
figurasgeométricas (Bratback y Dundas, 1984; Di Siervi, 2002). El biovolumen se calcula

23
multiplicando los números de células contadas por microscopia por el volumen de las células
(Bratback y Dundas, 1984; Di Siervi, 2002).Con el fin de expresar la biomasa bacteriana en
gramos de carbono por mililitro, es necesario transformar el volumen obtenido en los conteos
microscópico en contenido celular.
6.- Factores Abióticos que afectan el bacterioplancton
El crecimiento y el desarrollo de bacterias heterotróficas normalmente están limitados por la
disponibilidad de algunos recursos tales como sustratos orgánicos de carbono, nutrientes,
especialmente fósforo y nitrógeno e infecciones virales. Cada uno de estos factores puede limitar
el crecimiento bacteriano de diferentes formas en escala espacio-temporal (Ducklow y Carlson,
1992). Así mismo, Pomery y Wiebe (2001) reportan que la cantidad y calidad de materia
orgánica, concentración de nutrientes, temperatura, salinidad, concentración de oxígeno, entre
otros, ejercen un control en la abundancia y crecimiento del bacterioplancton.
6.1-Materia orgánica
El flujo de materia orgánica a través de las bacterias es una amplia vía donde en promedio la
mitad de la producción primaria en los sistemas acuáticos, es llevado a cabo por las bacterias por
la red microbiano. Todo esta red microbiano involucra distintos procesos, como por ejemplo: lisis
celular (Middelboe y col., 1996; Agustí y col., 1998), consumo de células bacterianas (Nagata,
2000) transferencia de sustrato (Kirchman, 1990; Blight., 1995). Sin embargo, el efecto principal
del consumo del carbono orgánico por el bacterioplancton es la producción de biomasa y dióxido
de carbono (CO2). La producción de cualquiera de estas dos variables tiene una profunda
influencia en el ciclo del carbono.En medios oceánicos el carbono orgánico disuelto (COD) es
predominantemente consumido por las células bacterianas heterotróficas, que en última instancia
son las responsables del consumo de alrededor del 50% de la producción primaria total en estos
ecosistemas (Furhman y Azam, 1982; Azam y col., 1983).
El bacterioplancton tiene la capacidad de tomar carbono disuelto del medio, el cual puede
obtener de alimentación negligente del zooplancton, exudación directa del fitoplancton (o

24
productores primarios en general), excreción, acción exoenzimática de bacterias y hongos sobre
detritus, y por la lisis de bacterias y algas provocada por virus. Una vez asimilado por las
bacterias, el carbono orgánico disuelto se convierte en particulado y pasa a constituir la
comunidad bacteriana. Al ser incorporado permanece disponible y puede ser transferido por
consumo a los bacteríovoros y a partir de ellos ser exportado del sistema (Thingstad y col., 1993;
Bratbak y col., 1998; Thingstad, 2000).
La despolimerización de la materia orgánica tiene lugar, principalmente, mediante la actividad
de las enzimas hidrolíticas segregadas al medio por las bacterias heterótrofas y los hongos
quienes juegan un papel importante en este proceso. Los biopolímeros, como las proteínas, los
polisacáridos (celulosa, quitina y almidón, entre otras) y los ésteres organofosfórico (ácidos
nucleicos y fosfolípidos, entre otras) son los componentes predominantes en los ambientes
acuáticos no contaminados, por lo tanto son la fuentes nutricionales mas importantes de las
bacterias heterótrofas (Chróst y Rai., 1992).
El estado nutricional del ambiente regula la velocidad y actividad de la exoenzimas
microbianas. La síntesis de exoenzimas es regulada por las concentraciones de materia orgánica
disuelta utilizable, que es la requerida por las bacterias para su metabolismo y crecimiento. Por
otro lado, la actividad de las exoenzimas también muestra una marcada variación temporal en las
capas superficiales y profundas de la columna de agua. En la capa superficiales, la máxima
actividad enzimático ocurre cuando el fitoplancton se encuentra en un estado avanzado de
desarrollo (Chróst, 1991).
6.2.- Nutrientes
El fósforo es comúnmente el nutriente limitante del bacterioplancton. Por ejemplo la adición
fósforo inorgánico mostró estimular la producción de bacterioplancton en agua del lago diluida
(Toolan y col, 1991).Las adiciones de nutrientes orgánicos mostró que los sustratos orgánicos por
sí mismos, no incrementan el índice de crecimiento del bacterioplancton en un lago oligotrófico,
pero la adición fósforo únicamente incrementa el crecimiento (Coveney y Wetzel, 1992).

25
Rejas y col., (2005) reportaron que el fósforo resultó ser un fuerte limitante del crecimiento
bacteriano, pero no se detectó efecto del enriquecimiento con nitrógeno ni con carbono orgánico
disuelto (DOC), en estudios realizados mediante seis experimentos in situ sobre limitación de
nutrientes en un sistema oligotróficos (Laguna bufeos, Bolivia).
En estudio realizado por Farjalla y col., (2006) señalan que la disponibilidad de fósforo y
nitrógeno son los principales factores responsables de la baja utilización de carbono orgánico
disuelto (DOC) por las bacterias en la laguna Imboassica, Brasil.
Muchos estudios de los ecosistemas marinos y de agua dulce han señalado que la
abundancia de células o la producción bacterioplancton se correlaciona positivamente con la
biomasa o la producción del fitoplancton (Azam y col., 1983; Fuhrman y col., 1991; Pace, 1993;
Robarts y col., 1996). Varios estudios han interpretado que esto significa que el crecimiento de
bacterioplancton se suma a del fitoplancton (Currie, 1990 y Pace, 1993), es decir, el crecimiento
del bacterioplancton es directamente estimulado por el fitoplancton. Se presume que el
acoplamiento ocurre porque las bacterias heterótrofas derivan su energía a partir del metabolismo
de fuentes externas de carbono orgánico (Currie, 1990 y Pace, 1993).
El crecimiento del bacterioplancton y fitoplancton es estimulado por nitrógeno inorgánico (N)
y fósforo (P). En algunos casos, esto también ha sido interpretado como prueba de acoplamiento
(Pace, 1993). De acuerdo con esta interpretación, los nutrientes inorgánicos estimulan el
crecimiento del fitoplancton y la mayor disponibilidad de exudados de algas estimulan el
crecimiento del bacterioplancton. Sin embargo, también es posible que el fitoplancton y
bacterioplancton sean estimulados directamente por adiciones de nutrientes (Brett y col., 1999).
Los altos requerimientos de nitrógeno y fósforo de las bacterias se debe a su alto contenido
de ácidos nucleicos y proteínas (Church y col., 2000), así como a la alta relación
superficie/volumen (Elser y col., 1995), que las convierte en consumidoras potenciales
competidoras con el fitoplancton (Elser y col., 1995; Church y col., 2000; Joint y col., 2002).
7.- Factores Bióticos que afectan el bacterioplancton

26
Entre los principales consumidores de bacterias están los protozoarios que son organismos
microscópicos y crecen en diferentes ambientes acuáticos, ya sean marinos o de agua dulce. Los
nanoflagelados heterotróficos (HNF), son generalmente los protozoarios más abundantes de la
zona eufótica de los sistemas acuáticos, y considerados como un componente importante en la red
alimenticia pelágica (Azam y col., 1983; Tzaras y col., 1999; Godinho y col., 2007). Varios
estudios han reconocido que estos protozoarios, son los principales consumidores de bacterias
(Simek y col., 1990; Godinho y col., 2007). Ellos controlan las poblaciones bacterianas a través
del pastoreo, convirtiéndolas en grandes partículas, las cuales son utilizadas como alimentos para
grandes protozoarios y metazooarios (Tzaras y col., 1999; Zhao y col., 2003).
La depredación por protistas sobre las poblaciones bacterianas es selectiva, la eficacia de la
depredación y la vulnerabilidad de la presa dependen de una gran variedad de factores (Simek., y
col., 1999). La mayoría prefieren células bacterianas de tamaños intermedios, ya que si depredan
células muy grandes o muy pequeñas es menos eficiente (Chrzanowski y Simek, 1990). Además
del tamaño, la forma es otra característica que puede influenciar la selectividad por parte de los
protistas (Hahn y Hofle 2001).
En estudios realizados en la laguna Bufeos en Bolivia (Rejas y col., 2005), se evidenció que
los HNF fueron los principales depredadores de bacterias, y que el coeficiente de pastoreo
aumentó en el experimento, cuando se agregaron ciliados. Adicionalmente, se conoce que estas
poblaciones de protistas son reguladas por los rotíferos, especialmente los ciliados que debido a
su tamaño pueden ser fácilmente pastoreados por ellos.Las comunidades zooplanctónicas
dulceacuícolas están constituidas esencialmente por rotíferos y crustáceos (los ciliados y
flagelados heterotróficos pueden ser incluidos dentro del zooplancton pero, generalmente, se
estudian aparte). Ellos son consumidores de bacterias en poblaciones marinas y de agua dulce,
encargándose de controlar las densidades bacterianas (Jiménez-Gómez y col., 1994).
Por otra parte, los rotíferos juegan un papel fundamental en las cadenas tróficas pelágicas (la
mayoría de ellos se alimentan de pequeños microorganismos, tales como microalgas, bacterias,
levaduras y protozoos). Son un eslabón entre el fitoplancton y los consumidores secundarios,
pero su importancia se acrecienta porque pueden transferir materia y energía desde bacterias y

27
partículas detríticas de pequeño tamaño, que son recursos no utilizables por otros organismos
planctónicos (Conde y col., 2004).
Oom (1997) y Rejas y col (2005) enfatizan la importancia de las bacterias como recursos
alimenticios para rotíferos, y destacó la relación entre el tamaño de la bacteria y las especies de
rotíferos que la consumen. Así los rotíferos juegan un papel importante en la transferencia de
energía en la red trófica y el reciclaje de nutrientes.

MARCO METODOLÓGICO
1.- Área de Estudio
El sistema del Lago de Maracaibo está ubicado en la zona nor-occidental de Venezuela, en el
Estado Zulia, aproximadamente entre 8º22’y 11º51’de altitud norte y entre los 70º30’y 73º24’de
longitud oeste. Posee cerca de 13.820 Km2 y una gran riqueza petrolífera en donde se extrae la
mayor parte de la producción de crudo del país (González, 2007). El sistema se ha dividido en
cuatro zonas bien delimitadas y diferenciadas, debido a sus características físico-geográficas,
estas son: El Lago, El Estrecho, La Bahía el Tablazo y El Golfo de Venezuela (Rodríguez, 2000).
(Fig. 2)
Figura 2. El Sistema del Lago de Maracaibo, estado
Zulia, Venezuela. (Villasmil; Mendoza, 2001)
La Bahía el Tablazo presenta una marcada forma rectangular con un largo de Oeste a este de
30 km y un ancho de Norte a Sur de 20 Km. Se encuentra ubicado entre los 10º 48' y los 11º el
Golfo de Venezuela, por lo cual es una típica área estuarina de aguas salobres, en la cual se
mezclan las aguas del golfo que penetran al lago con las dulces que salen de éste y del río Limón.
Situada entre el lago de Maracaibo y Es considerablemente más somera que el Estrecho, está
recorrida por un canal natural que forma un arco al este de la Isla de Pescadores, con profundidad
mínima de 5 m (Rodríguez, 2000)

30
El Estrecho, une al lago con la Bahía de El Tablazo, tiene un área de 480 Km2, con una
longitud máxima de 40 km de norte a sur y un ancho que varía entre los 6 y los 17 km. Está
situado entre los 10º y 27’ y los 10º y 48' de latitud norte y entre el 71º 29’ y los 71º 41’ de lon-
gitud oeste. La zona sublitoral está caracterizada por presentar dos franjas sublitorales de poca
profundidad, que bordean a una franja más angosta de 10 m de profundidad al oeste de la cual
corre el canal de navegación. La circulación de las aguas de esta área se ve influenciada
principalmente por el agua que entra o sale del lago, aunque los vientos también intervienen pero
debido a lo angosto de la zona su acción se ve limitada (Herman,1997).
El presente estudio se llevó a cabo en dos zonas diferentes del Sistema del Lago de Maracaibo
y son: La Bahía El Tablazo y El Estrecho del lago de Maracaibo. El muestreo de campo se realizó
horizontalmente en 5 estaciones enumeradas para cada zona de estudio. Para la Bahía el Tablazo
correspondió las estaciones del 0 al 4 y para el Estrecho del Lago de Maracaibo las estaciones del
5 al 9. Estás fueron previamente establecidas con ayuda de un Sistema de Ubicación Geográfica
(GPS) y tomada como punto de referencia algunos lugares específicos, los cuales se indican en la
figura 3. Se realizaron muestreos mensuales, durante el periodo correspondiente de Enero a
Diciembre del año 2009.
Figura 3. Imagen satelital de la Bahía El Tablazo y Estrecho del Lago de
Maracaibo- estado Zulia. Distribución horizontal de las zonas de muestreos
y estaciones. Tomado de Google Eart. http//www.googleearth.com
2.- Recolección de las muestras abióticas

31
2.1.- Parámetros fisicoquímicos
Se determinó los parámetros temperatura, salinidad, oxígeno disuelto y transparencia en las
muestras de aguas, con el fin de conocer las condiciones ambientales donde se desarrollan las
bacterias. Las mediciones de estos parámetros se realizó in situ con ayuda de una sonda
multiparamétrica sumergible marca Horiba para el caso de temperatura, salinidad y oxígeno
disuelto.
Para los parámetros como transparencia y profundidad fueron medidos directamente al agua,
donde se midió la transparencia con un disco de Secchi de 30cm2 de diámetro con dos colores
pintados e intercalados, sujetos a una cuerda aproximadamente de 50 m.
Para la profundidad se utilizó un ancla de hierro y una cuerda con una medida aproximada de
30 m. Estas longitudes fueron medidas con una cinta plástica, los datos fueron anotados en
libretas de campo con un lápiz grafito y fueron enumerados de acuerdo a cada estación.
2.2.- Nutrientes
Se tomaron submuestras de 500 ml sin filtrar, para el análisis de los nutrientes: nitritos,
nitratos, Ortofosfatos y fósforo total. Se almacenaron en una cava con hielo hasta su traslado al
laboratorio Ambiental en el Departamento de Ingeniería Sanitaria y Ambiental (DISA), donde el
análisis se realizó, a través del método estándar APHA y col, (1998).
La determinación de cada nutriente se realizaron a través de los siguientes procedimientos:
método colorimétrico reducción de Cadmio para nitrito, método colorimétrico NED-
Dihidrocloruro y Sulfanilamida para el nitrato, método colorimétrico ácido ascórbico con previa
digestión con persulfato para fósforo total y el método colorimétrico ácido ascórbico para
ortofosfato.
3.-Recolección de las Muestras Bióticas

32
3.1.-Bacterioplancton
Para la recolección de bacterias se tomaron muestras de agua de la Bahía y el Estrecho en las
10estaciones (repetido) a nivel de superficie, mediante un envase plástico de aproximadamente 1
L de capacidad, directamente del agua sin filtrar.
A partir de esta muestra se tomaron submuestras de 10 ml en un vial para el contaje
bacteriano la cual inmediatamente se fijó con formaldehído hasta obtener una concentración final
de 2%.
3.2.- Nanoflagelados Heterótrofos
Para la recolección de nanoflagelados heterótrofos, se tomaron muestras de agua de la Bahía
y el Estrecho en las 10estaciones (repetido) a nivel de superficie, mediante un envase plástico de
aproximadamente 1 L de capacidad, directamente del agua sin filtrar.
Las muestras fueron tomadas del recipiente que contenía 1 litros de agua inicial, a través de
viales de 25 ml de capacidad con tapas. Seguidamente se tomó una submuestras 10 ml de agua y
se fijó con 10 ml de formalina, obteniéndose así 20 ml de muestra a una concentración final de
2%.
3.3.- Ciliados y rotíferos
Se tomaron 15 litros de agua en recipientes plásticos de 18 litros de capacidad directamente de
la superficie (0 m) y sin filtrar. Seguidamente se recolecto una submuestra de 500 ml para el
estudio de los protozoarios (ciliados) y rotíferos en envases plásticos de polietileno con tapa de
rosca (previamente lavados y rotulados con fecha y número de la estación) agregando
inmediatamente 8 ml de lugol y 12 ml aprox. de formalina al 2%.. Las muestras fueron colocadas
cava plástica con hielo para su traslado al laboratorio donde se refrigeraron a 4ºC por 48 horas o
hasta el momento de su análisis.

33
4.- Análisis de Factores Bióticos
4.1.- Abundancia de bacterioplancton
Se tomó una alícuota de 1ml a partir de las muestras de 10 ml, el cual fue filtrado con ayuda
de un filtro nucleopore de policarbonato negro (diámetro: 25 mm, tamaño de poro: 0,2μm), al
vacío sin exceder los 5 KPa (Kepner y Pratt, 1984). Debajo del filtro de policarbonato negro se
colocó un filtro de Backup (diámetro: 25 mm, tamaño de poro: 2 1/4μm) para promover la
dispersión homogénea de la muestra sobre el filtro de policarbonato negro (Kepner y Pratt,
1994). La muestra se lavó con 2 ml de agua estéril aproximadamente.
Posteriormente, las muestras fueron coloreadas con un 1 ml Naranja de Acridina (5 μg/ml) y
se dejó reposar por 2 min, pasado el tiempo se filtró completamente al vacío, se lavó con 2 ml de
agua estéril (2 veces).El filtro se removió con ayuda de una pinza de punta fina para así no dañar
la membrana y se colocó en un portaobjeto humedecido con aceite de inmersión y, para finalizar,
se agregó una gota de aceite de inmersión entre la membrana y el cubre objetos para observarse
inmediatamente al microscopio.
Las muestras se observaron, bajo inmersión utilizando un microscopio (Olympus CH2) con un
sistema de epifluorescencia y cámara digital incorporada, bajo luz azul, se identificaron y
contaron las bacterias. Para ello, se consideró el número de bacterias presentes en la rejilla
(campo) hasta enumerarse un mínimo de 20 campos por muestra (Kirchman, 1993).
Cálculos del número de bacterias en una muestra
Donde:
N= Número de bacterias totales en la muestra
n= Número promedio de bacterias contadas en una unidad (cuadrito, línea, rejilla, campo)
S= Superfície de filtro
P= Área contada
filtradovolumen
dilucionx
p
sxpromedion )(

34
4.2.- Biovolumen y Biomasa del Bacterioplancton
Para la obtención del volumen de cada morfotipo de bacteria, éstas fueron asemejadas a
esferas (coco) y cilindros (bastones), utilizando la fórmula del volumen de estas figuras
geométricas (Bratback y Dundas, 1984). De esta manera, se tiene que:
Biovolumen de los cocos: V= 1/6. π . d3
Donde:
d= Diámetro
π= 3,1416
Biovolumen de los Bastones: V= 1/4. π . d2 . h + 1/6.π.d³
Donde:
π= 3,1416
d= Diámetro
h= Altura
Para estimar el biovolumen se medieron 20 organismos por muestras empleando un
micrómetro del ocular y la platina. Se consideraron las medidas de diámetro y longitud para los
morfotipo bastones y solo diámetro para los morfotipos de cocos. Finalmente, el biovolumen se
calculó multiplicando los números de células contadas por microscopía por el volumen de las
células (Bratback y Dundas, 1984).
La biomasa se determinó en forma separada para cada grupo, expresándolo en gramos de
carbono por mililitro, por lo cual fue necesario transformar los biovolúmenes (μm3
cel-1
)
obtenido, en carbono celular. Basándose en el uso del factor de conversión de carbono que indica
que 1 μm3 equivale a 2,2.10
-10 mgC μm
3 (Bratback y Dundas, 1984)
.4.3.- Abundancia de Nanoflagelados

35
Para determinar la abundancia de los nanoflagelados, se tomó una alícuota de 5ml de la
muestra, éstas a su vez fueron filtradas con un filtro nucleopore de policarbonato negro
(diámetro: 25 mm, tamaño de poro: 1μm), al vacío sin exceder los 5 KPa (Blöem y col., 1986).
Debajo del filtro de policarbonato negro se colocó un filtro Millipore (diámetro: 25 mm, tamaño
de poro: 0,45μm) para promover la dispersión uniforme de la muestra sobre el filtro de
policarbonato negro (Caron, 1983).
El filtro se lavó tres veces con 2 ml de una solución buffer de Trizma 0,1M (pH 4.0). Para la
preparación del buffer se diluyó 15,76 g de Trizma en 1L de agua destilada estéril. Luego se tiñó
con 2 gotas de solución de Primulina (63 mg/L) y se dejó reposar por 5 minutos. Posteriormente,
la muestra fue filtrada en su totalidad por medio de un sistema de filtración al vacío, luego el
filtro fue lavado tres veces con 2 ml del buffer Trizma (Blöem y col., 1986), la membrana fue
removida y colocada sobre un portaobjeto con una fina capa de aceite de inmersión (Cargille
tipoB), se agregó una gota de aceite de inmersión entre el filtro y el cubreobjeto.
Se examinaron las muestras, bajo inmersión utilizando un microscopio (Olympus CH2) con un
sistema de epifluorescencia y cámara digital incorporada al microscopio, a través del objetivo
100x y bajo luz azul se contaron los nanoflagelados. Para esto, fueron contados 25 campos por
filtros (Blöem y col., 1986). Posteriormente, el número promedio de células por campo observado
fue convertido a organismos por mililitros por conocimiento de volumen de la muestra (x 0,5
debido a la dilución durante la preservación), el área del campo de observación y el área del filtro
cubierta por la muestra, utilizando la siguiente ecuación (Caron, 1983):
# de células x ml= [(# de células/campos) (área analizada/área de campos)]
[(0,5) (volumen de la muestra)]
4.4.- Abundancia de Ciliados y Rotíferos
Las muestras fueron agregadas en cilindros graduados de 1000 ml manteniéndose estables por
48 horas o hasta que se completó la sedimentación el microzooplancton y el mesozooplancton se
asentó en el fondo y se descartó el sobrenadante (400ml) con ayuda de una manguera y un envase
plástico, utilizando el resto (100ml) para la estimación de ciliados y rotíferos. Seguidamente, se
extrajo una submuestras de 1 ml, con ayuda de una pipeta de Hensel–Stempel (1ml) y ésta se

36
trasfirió inmediatamente a la cámara de conteo de (Sedgwick-Rafter), donde se contaron 3
cámaras de 1 ml por cada muestra, con ayuda de un microscopio óptico Motic (BA400) a (40x y
100x) (Bloem y col., 1986). La abundancia de las especies se expresó como número de
organismos por litros.
5.- Análisis de los datos
Los datos obtenidos de abundancia y biomasa del bacterioplancton, la abundancia de
nanoflagelados, ciliados y rotíferos (factores bióticos), así como los datos fisicoquímicos y de
nutrientes (factores abióticos) recolectados en in situ, fueron analizados estadísticamente y
graficados mediante software SPSS 19.
Se realizó unos análisis de Correlación Simple (Pearson) entre los factores bióticos y
abióticos, mencionados anteriormente; para establecerse la relación con respecto a la abundancia
y biomasa del bacterioplancton en muestras de agua de la Bahía El Tablazo y estrecho del Lago
de Maracaibo. Finalmente se realizaron comparaciones mediante la Prueba T Student para los
factores bióticos y abióticos de las muestras de agua de la Bahía El Tablazo y Estrecho del Lago
de bióticos y abióticos de las muestras de agua de la Bahía El Tablazo y Estrecho del Lago de
Maracaibo.

RESULTADOS Y DISCUSIÓN
1.- Variables Fisicoquímicos
1.1.-Temperatura
En la Bahía El Tablazo la temperatura del agua varió entre 22 y 32 °C, con una temperatura
promedio de 28,47±2,31 °C durante los doce meses de muestreo (Fig.4a). Los valores mayores de
temperatura se registraron durante el estudio en la estación 4 y el menor valor en la estación 0
(Fig.4b). Para el Estrecho de Maracaibo la temperatura del osciló entre 26 °C y 34 °C, con una
temperatura promedio de 29,69±1,80°C durante los doce (12) meses de muestreo (Fig.4a). Los
valores mayores de temperatura se registraron durante el estudio en la estación 9 y el menor
valor en la estación 5 (Fig.4b).
Figura. 4a. Variaciones de Temperatura mensual en Bahía El
Tablazo y Estrecho del Lago de Maracaibo. (Caicedo, 2009)
Figura. 4b. Valores de Temperatura por estación en Bahía El Tablazo y
Estrecho del Lago de Maracaibo. (Caicedo, 2009)

37
En ambas zonas de estudio se encontraron diferencias significativas (t= -7,125, P= 0,002). Los
valores de temperatura fueron altos en las estaciones pertenecientes al Estrecho, que en Bahía; es
posibles que las variaciones de temperatura entre las estaciones este asociadas a la diferencia de
hora en que fueron recolectadas las muestras. No obstante, las variaciones observadas en
temperatura durante todo el período de muestreo en ambas zonas de estudio, fueron mínimas, lo
que se atribuye a que el sistema del Lago de Maracaibo se encuentra ubicado en una zona tropical
donde la temperatura se mantiene relativamente constante en el curso del año (Roldán y Ramírez,
2008).
Wetzel (2001) señala que los ecosistemas acuáticos tropicales, las temperaturas tienden a
sufrir pocas variaciones y, debido a que la temperatura se mantiene relativamente alta durante
todo el año, la columna de agua tiende a ser muy estable. Según Siqueira, y col, (2003) esas
oscilaciones observadas en aguas superficiales están sujetas a las influencia de las condiciones
climáticas de la región, así como acción de las corrientes y vientos.
Rodríguez (2002) presentó valores de temperatura similares a los obtenidos en este estudio,
donde observó que la temperatura del agua en el Sistema del lago de Maracaibo ocurre pequeña
variaciones diarias, estacionales y verticales.
1.2.- Salinidad
En la Bahía El Tablazo la salinidad del agua fluctuó entre 4 UPS y 12 UPS, con una salinidad
promedio de 8,40 ± 2,31 UPS durante los doce meses de muestreo (Fig.5a). Los valores mayores
de salinidad se registraron durante el estudio se registraron en la estación 3 y el menor valor en la
estación 4 (Fig.5b). Para el Estrecho de Maracaibo la salinidad del agua fluctuó la entre 3 UPS y
12 UPS, con una salinidad promedio de 5,49 UPS y una desviacion estándar 2,16 durante los
doce (12) meses de muestreo (Fig.5a). Los valores mayores de salinidad se registraron durante el
estudio se registraron en la estación 5 y el menor valor en la estación 8 (Fig.5b).
Los valores de salinidad en las estaciones pertenecientes a la Bahía el Tablazo mostraron ser
mayores que en el Estrecho del Lago de Maracaibo, encontrándose diferencias significativas

38
entre ambas zonas (t= -3,045, P=0,002). Esto puede atribuirse a que la salinidad en la Bahía
muestra cambios causados por el intercambio de la salinidad del golfo de Venezuela, producidos
por las mareas dominantes, y además, debido a las variaciones anuales de precipitación y
escorrentía terrestre, afectando sensiblemente la salinidad de esta agua. Para el Estrecho del Lago
de Maracaibo la salinidad fluctúa de forma marcada debido a las precipitaciones sobre la cuenca
del lago (Rodríguez, 2002). Adicionalmente, Esclapés y col., (1997) y Febres y Masciangioli
(2000), demostraron la tendencia de la distribución de la salinidad en las zonas del Estrecho y
Bahía, siendo la dinámica de las mareas la responsable del intercambio de agua dulce y salada en
dichas zonas.
Figura. 5a. Variaciones de Salinidad mensual en Bahía El Tablazo
y Estrecho del Lago de Maracaibo. (Caicedo, 2009)
Figura. 5b. Valores de Salinidad estacional en Bahía El Tablazo y Estrecho
del Lago de Maracaibo. (Caicedo, 2009)

39
1.3.- Transparencia
En la Bahía El Tablazo los valores de transparencia varió entre 0,26 m y 1,31 m, con un
promedio de 0,71 m y una desviacion estándar 0,24 durante los doce meses de muestreo (Fig. 6a)
Los valores mayores de transparencias se registraron durante el estudio en la estación 4 y el
menor valor en la estación 0 (Fig.6b). Para el Estrecho de Maracaibo la transparencia osciló
entre0,7 m y 1,82 m, con un promedio de 1,11 ± 0,19 m durante los doce meses de muestreo
(Fig.6a).Los valores mayores de transparencia durante el estudio se registraron en la estación 9 y
el menor valor en la estación 5 (Fig.6b)
Figura. 6a. Variaciones de Transparencia mensual en Bahía El Tablazo
y Estrecho del Lago de Maracaibo. (Caicedo, 2009)
Figura. 6b. Valores de Transparencia estacional en Bahía El Tablazo y Estrecho
del Lago de Maracaibo. (Caicedo, 2009)

40
Los valores de transparencia para las estaciones del Estrecho del Lago de Maracaibo
mostraron ser mayor que en la Bahía el tablazo durante todo el estudio, encontrándose diferencias
significativas entre ambas zonas (t=-10,057, P= 0,002). Esto pude deberse a que las estaciones
pertenecientes a la Bahía presentan menor profundidad y, consecuentemente, mayor turbidez
causada por la resuspensión de sedimentos; mientras que en el Estrecho la profundidad es mayor
y por ende la turbidez es menor (Wetzel, 2001). El oleaje generado por los vientos alisios que
soplan del noreste, puede provocar la turbidez del agua en las zonas de menor profundad,
generando mayor cantidad de sólidos suspendidos y, por lo tanto, la disminución de la
transparencia (Rodríguez, 2000).
1.4.- Oxígeno disuelto
En la Bahía El Tablazo los valores de oxígeno disuelto oscilaron entre 4,42 y 6,58 mg/l, con
un promedio de 5,48 mg/l durante los doce meses de muestreo (Fig.7a). Los valores mayores de
oxígeno disuelto se registraron en la estación 0 y el menor valor en la estación 4 (Fig.7b). Para el
Estrecho del Lago de Maracaibo el oxígeno disuelto oscilaron la entre 5,3 y 7,3 mg/l, con un
promedio de 6,34 mg/l durante los doce (12) meses de muestreo (Fig.7a). Los valores mayores de
oxígeno disuelto durante el estudio se registraron en la estación 6 y el menor valor en la estación
5 (Fig.7b).
Figura. 7a. Variaciones de Oxígeno disuelto mensual en Bahía El Tablazo
y Estrecho del Lago de Maracaibo. (Caicedo, 2009)

41
Figura. 7b. Valores de Oxígeno disuelto estacional en Bahía El Tablazo y
Estrecho del Lago de Maracaibo. (Caicedo, 2009)
Los valores de oxígeno disuelto para las estaciones del Estrecho del Lago de Maracaibo y
Bahía El Tablazo no mostraron diferencias significativas (t= -1,3476 P= 0,180). Las
concentraciones de oxígeno disuelto presentaron valores por encima del limite de 4,0 mg/l,
establecido para el desarrollo adecuado de diversos organismos para la vida acuática y para la
oxidación química y bioquímica del material orgánico e inorgánico (Avila,2009).
1.5.- Profundidad
En la Bahía El Tablazo los valores de profundidad oscilaron entre 1,08 m y 4,22 m, con un
promedio de 2,32 m y una desviacion estándar 0,56 durante los doce (12) meses de muestreo
(Fig.8a). Los valores altos de profundidad se registraron durante el estudio en la estación 3 y el
menor valor en la estación 0 (Fig8b). El Estrecho del lago de Maracaibo los valores de
profundidad oscilaron la entre 1,5 m y 13,15 m, con un promedio de 8,64 m durante los doce (12)
meses de muestreo (Fig.8a). Los valores altos de profundidad durante el estudio se registraron en
la estación 9 y el menor valor en la estación 5 (Fig.8b).
Según lo obtenido por la prueba T-student existe diferencias significativas entre ambas zonas
(t= -22,199 p = 0,002). La Bahía El Tablazo es considerablemente más somera que el Estrecho,
con profundidades mínimas de 5m de acuerdo a lo indicado por Rodríguez (2002).

42
Figura. 8a. Variaciones de Profundidad mensual en Bahía El Tablazo
y Estrecho del Lago de Maracaibo. (Caicedo, 2009)
Figura. 8b. Valores de Profundidad estacional en Bahía El Tablazo y
Estrecho del Lago de Maracaibo. (Caicedo, 2009)
2.- Nutirentes
2.1.- Nitrito
En la Bahía El Tablazo el nitrito del agua se encontró entre 0,008 y 0,047 mg/l, con un
promedio de 0,029 mg/l durante los doce meses de muestreos (Fig. 9a). Los valores mayores de
nitrito se registraron durante el estudio en la estación 3 y el menor valor para la estación 0 (Fig.
9b). El Estrecho de Maracaibo el nitrito del agua osciló entre 0,022 y 0,049 mg/l, con un

43
promedio de 0,032 mg/l durante los doce (12) meses de muestreo (Fig.9a). Los valores mayores
de nitrito se registraron durante el estudio en la estación 7 y el menor valor en la estación 6
(Fig.9b).
Figura. 9a. Variaciones de Nitrito mensual en Bahía El Tablazo y
Estrecho del Lago de Maracaibo. (Caicedo, 2009)
Figura. 9b. Valores de Nitrito estacional en Bahía El Tablazo y
Estrecho del Lago de Maracaibo. (Caicedo, 2009)
Entre ambas zonas de estudio no se encontró diferencias significativas (t = -0,620, P = 0,536)
con respecto al nitrito. Rodríguez (2000), señala que los valores de nitrito en aguas superficiales
para Estrecho y Bahía son relativamente bajos y muy uniformes; los valores altos se registraron
durante la estación de lluviosa.
Los nitritos en el agua se producen por la actividad bacteriana ejercida sobre el amonio y
nitrógeno orgánico, pero usualmente no se encuentran en concentraciones altas debido a su rápida
oxidación a nitrato (Roldan y Ramírez, 2008).

44
2.2.- Nitrato
En la Bahía El Tablazo el nitrato del agua varió entre 0,001 y 0,165 mg/l, con un promedio de
0,042 mg/l durante los doce meses de muestreo (Fig.10a). Los valores mayores de nitrato se
registraron durante el estudio en la estación 0 y el menor valor en la estación 1 (Fig.10b). El
Estrecho de Maracaibo el nitrato del agua osciló entre 0,003 y 0,132 mg/l, con un promedio de
0,037 mg/l durante los doce (12) meses de muestreo (Fig.10a). Los valores mayores de nitrato se
registraron durante el estudio en la estación 7 y el menor valor en la estación 9 (Fig.10b).
Figura. 10a.Variaciones de Nitrato Mensual en Bahía El Tablazo y
Estrecho del Lago de Maracaibo. (Caicedo, 2009)
Figura. 10b. Valores de Nitrato por Estación en Bahía El Tablazo y
Estrecho del Lago de Maracaibo. (Caicedo, 2009)
Entre ambas zonas de estudio no se encontraron diferencias significativas (t = 0,584, P =
0,560) con respecto al nitrato. El incremento de las precipitaciones determina el transporte de
45
nutrientes, especialmente los fosfatos y compuestos de nitrógeno en forma de nitratos en grandes
cantidades, desde las cuencas de los ríos afluentes del lago, producto de las actividades
agropecuarias, camaroneras, industriales y urbanas (fuentes puntuales) que se desarrollan en ellas
(Herrera y col. 2004).
La concentración de nitratos puede aumentar por el vertimiento de contaminantes domésticos
e industriales y de la actividad agrícola (Roldán y Ramírez, 2008).
2.3.- Fósforo Total
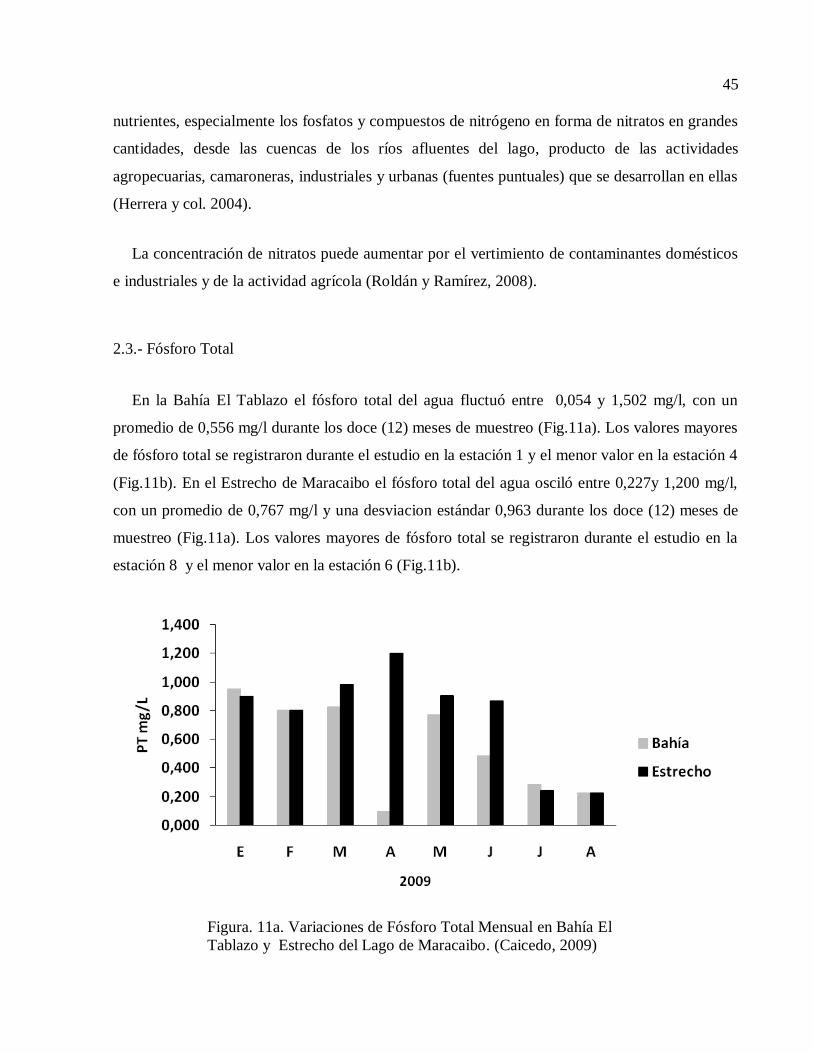
En la Bahía El Tablazo el fósforo total del agua fluctuó entre 0,054 y 1,502 mg/l, con un
promedio de 0,556 mg/l durante los doce (12) meses de muestreo (Fig.11a). Los valores mayores
de fósforo total se registraron durante el estudio en la estación 1 y el menor valor en la estación 4
(Fig.11b). En el Estrecho de Maracaibo el fósforo total del agua osciló entre 0,227y 1,200 mg/l,
con un promedio de 0,767 mg/l y una desviacion estándar 0,963 durante los doce (12) meses de
muestreo (Fig.11a). Los valores mayores de fósforo total se registraron durante el estudio en la
estación 8 y el menor valor en la estación 6 (Fig.11b).
Figura. 11a. Variaciones de Fósforo Total Mensual en Bahía El
Tablazo y Estrecho del Lago de Maracaibo. (Caicedo, 2009)

46
Figura.11b. Valores de Fósforo Total por Estación en Bahía El Tablazo y
Estrecho del Lago de Maracaibo. (Caicedo, 2009)
Entre ambas zonas de estudio no se encontraron diferencias significativas (t = -1,308, P=
0,194) con respecto al fósforo total. El fósforo es el elemento que desempeña el papel más
importante en el metabolismo biológico. En comparación con los demás nutrientes, es el menos
abundante y al mismo tiempo, es el factor más limitante en la productividad primaria (Roldán y
Ramírez, 2008).
2.4.- Ortofosfatos
En la Bahía El Tablazo el ortofosfato del agua osciló entre 0,004 y 0,312 mg/l, con un
promedio de 0,041 mg/l durante los doce meses de muestreo (Fig.12a). Los valores mayores de
Ortofosfato se registraron durante el estudio en la estación 3 y el menor valor en la estación 2
(Fig.12b).
Figura. 12a. Variaciones de Ortofosfato Mensual en Bahía El Tablazo
y Estrecho del Lago de Maracaibo. (Caicedo, 2009)

47
Para el Estrecho de Maracaibo el ortofosfato del agua osciló entre 0,011 y 0,711 mg/l, con un
promedio de 0,056 mg/l y una desviacion estándar 0,121 durante los doce (12) meses de muestreo
(Fig.12a). Los valores mayores de Ortofosfato nitrato se registraron durante el estudio en la
estación 6 y el menor valor en la estación 5 (Fig.12b).
Figura.12b. Valores de Ortofosfato por Estación en Bahía El Tablazo y
Estrecho del Lago de Maracaibo. (Caicedo, 2009)
Entre ambas zonas de estudio no se encontró diferencias significativas (t = -0,680, P = 0,498)
con respecto al ortofosfato. La presencia de ortofosfato, está relacionada el metabolismo de las
plantas y fitoplancton, por lo que el ortofosfato se consume más rápidamente; esto trae como
consecuencia que su concentración sea muy baja en lagos tropicales (Roldán y Ramírez, 2008).
3.- Abundancia y Biomasa del Bacterioplancton
3.1.- Abundancia
La abundancia de bacterioplancton en las aguas superficiales de Bahía el Tablazo presentó un
promedio de 1,77x109 cel/l, con un máximo 3,22x10
9 cel/l y un mínimo de 7,51 x10
8 cel/l durante
los doce meses de muestreo. En los meses de Junio y Septiembre se observaron los valores
máximos de bacterioplancton 2,62 x109 cel/l y 3,22 x10
9 cel/l; mientras que los recuentos más
bajos se observaron durante el mes de Enero y Octubre 7,51 x108 cel/l y 8,70 x108 cel/l
respectivamente (Fig.13a). Para la estación 0 se encontraron los valores más altos de

48
bacterioplancton y los más bajos en la estación 4 (Fig. 13b).
Para el Estrecho de l Lago de Maracaibo, la abundancia de bacterioplancton presentó un
promedio de 1,33x109 cel/l, con un máximo de 2,15x10
9 cel/l y un mínimo 5,09x10
9 cel/l,
durante los doce meses de muestreo (Fig. 13a). En los meses de marzo y junio se observaron los
valores máximos de bacterioplancton 2,01 x109 y 2,15x10
9 cel/l respectivamente, mientras que
los recuentos más bajos se observaron durante el mes de octubre y diciembre 5,09x108 cel/l y
8,08 x108 cel/l. Para la estación 7 se encontró los valores altos de bacterioplancton y los bajos en
la estación 9(Fig.13b)
Figura. 13a. Variaciones de Abundancia de Bacterioplancton mensual
en Bahía El Tablazo y Estrecho del Lago de Maracaibo. (Caicedo, 2009)
Figura. 13b. Valores de Abundancia de Bacterioplancton por Estación en
Bahía El Tablazo y Estrecho del Lago de Maracaibo. (Caicedo, 2009)

49
Los valores de abundancia de ambas zonas de estudio fueron similares a los encontrados en El
Amazonas Brasilero por Wissmar y col. (1991), donde registraron abundancias bacterianas en
diferentes ecosistemas del Amazonas que oscilan entre 9 x108cel/l y 1,1 x10
10cél/L; Sommaruga
(1995) en el lago Rodo de Uruguay, encontró una abundancia bacteriana que fluctuó entre
1,5X109
cel/l y 2,0X1010
cel/l y Pinilla (2007) en el lago Boa (Colombia) indica promedios que
fluctuaron entre 4,1 x 109
y 1,4 x 1010
cel/l. En otros sistemas acuáticos de diferente condición
trófica, se encontró valores de bacterias alrededor de 6 x108a 1 x10
10cel/l (Cole y Pace, 1995;
Mariazzi y col., 1998; Zingel, 1999 y Castillo y col., 2004), semejantes a los obtenidos en este
estudio.
Los recuentos de bacterias resultaron ser elevados en comparación a los obtenidos por Dupontt
(2003) y Rincón y col. (2007) para la zona litoral del Lago de Maracaibo (entre 6,5 - 6,7 x 105
cel x mL-1
). Por otra parte, Abarzúa y col., (1995) y Miranda (2009) reportan también valores
bajos de bacterias en el Golfo de Arauco, Chile (3,4 x 107 cel/l) y en el río Amazonas, Brasil
(9,18 x 105cel/l a 1,85x 10
6cel/l) respectivamente. Los promedios de abundancia de bacterias
presentaron diferencias significativas (t = -18,524, P˂ 0,05), prevaleciendo los valores altos en la
Bahía.
Los morfotipos de bacterias observados en la Bahía durante la investigación corresponden a la
forma cocos con un promedio de 1,11 x109 cel/l y bastones con un promedio 6,67 x10
8 cel/l,
predominando los cocos durante el estudio (Fig. 13c). Para el Estrecho del Lago de Maracaibo los
morfotipos de bacterias están representados también por las formas cocos y bastones con un
promedio de 8,32 x108 cel/l y 4,97 x10
8 cel/l respectivamente; encontrándose un predominio del
morfotipo coco, coincidiendo a lo obtenido en el área Bahía el Tablazo (Fig.14d).
Esto valores obtenidos este estudio concuerda con los resultados encontrados en Bahía de
Biscay por Artigas (1998), en un estuario del Amazonas, por Chicheportiche (2007) y Rincón y
col., (2007) en la zona litoral y pelágica de la Costa Oriental del Lago de Maracaibo. Es probable
que con los cambios fisicoquímicas que ha experimentado el Lago de Maracaibo en los últimos
años, haya condicionado a la aparición dominante de un morfotipo bacteriano en particular. Es
conocido que los cocos tienden a ser más resistentes a ciertas condiciones como la presión
50
osmótica elevada y la desecación, que las formas bacilares (Rincon y col., 2007; Madigan y col.
2004).
Figura. 13c. Variaciones de Morfotipos Cocos y Bastones Mensual
en Bahía El Tablazo. (Caicedo, 2009)
Figura. 13d. Variaciones de Bacterias Libres y Adheridas Mensual
en Estrecho del Lago de Maracaibo. (Caicedo, 2009)
Barragán y col. (2009) indicaron una dominancia de células bacterianas pequeñas en la Bahía
de Gaira, especialmente durante el período seco, que pudo ser consecuencia de la limitación por
nutrientes orgánicos (COD), argumento que se apoya en la observación frecuente de células de
forma esférica durante todo el período de muestreo. Cuando los procariotas crecen en ambientes
sin limitación por nutrientes orgánicos, aparecen células de mayor volumen y si están limitadas
por nutrientes inorgánicos aparecen formas alargadas, pero cuando están limitadas por carbono,

51
se presentan tamaños mas pequeños (Vrede y col., 2002; Rancy, 2004).
No obstante, la bacteriovoría es otro factor que afecta la distribución de tamaños y biomasa de
la comunidad bacteriana, cuando el control por depredación es más importante que el sustrato
disponible, la comunidad bacteriana puede favorecer la presencia de una mayor abundancia de
células más pequeñas y activas (Psenner y Sommaruga, 1992; Hahn y Hofle, 2001). Algunas
bacterias pueden mostrar una absoluta resistencia contra los depredadores, por ejemplo, las
grandes bacterias filamentosas, y las que se agrupan en microcolonias. Mientras que otras pueden
ser susceptibles, tales como las unicelulares o de tamaño reducido, obteniéndose así una relativa
resistencia (Hahn y Hofle, 2001). De esta manera, el consumo de bacterias por depredadores
puede cambiar el tamaño de las células y la composición de la comunidad bacteriana.
Por otra parte, durante el estudio se encontraron un promedio de bacterias libres de 1,29 x109
cel/l y 4,81 x108 cel/l para bacterias adheridas donde predominaron las bacterias libres sobre las
adheridas (Fig.13e) para La Bahía El Tablazo; mientras que para el Estrecho del Lago de
Maracaibo el número de bacterias libres promedio fue de 9,05 x108 cel/l y 4,24 x10
8 cel/l para
bacterias adheridas donde prevalecieron de nuevo las bacterias libres sobre las adheridas (Fig.
13f). Cabe destacar que los morfotipos cocos fueron los más abundantes en la abundancia de
bacterias libres y adheridas para ambas zonas de estudio.
Figura. 13e. Variaciones de Bacterias Libres y Adheridas
Mensual en Bahía El Tablazo. (Caicedo, 2009)

52
Figura. 13f. Variaciones de Bacterias Libres y Adheridas
Mensual en Estrecho del Lago de Maracaibo. (Caicedo, 2009)
El porcentaje de bacterias adheridas fue menor (27 a 32%) que las bacterias libres (62 a 63%),
resultados semejantes a los encontrados por Artigas (1998) en un estuario de Gironde y Miranda
(2009) en estuario del Amazonas, donde aproximadamente el 30% de las bacterias eran adheridas
y más del 60% eran libres. Las bacterias adheridas no pueden ser consideradas ecológicamente
insignificantes en el bacterioplancton pues cumplen diversas funciones: tienen una contribución
relevante para la biomasa total de bacteria; funcionabilidad en ambientes diferentes a las bacterias
libres; aumenta la calidad nutricional de las partículas y son importantes fuentes alimenticias para
otros organismos mayores del plancton, cuando son consumidas directamente.
3.2.- Biomasa
En la Bahía El Tablazo, la biomasa bacteriana varió entre 1,48x101 a 1,73 x10
2 µg C/ l, con
promedio de 8,09 x101 µg C/l durante todo el estudio. Durante los meses julio y septiembre se
registraron los valores máximos de biomasa bacteriana 1,45x102µg C/l y 1,73x10
2µg C/l
respectivamente; mientras que los valores mínimos se observaron durante los meses de marzo y
abril con 1,48x101 µg C/l y 2,01x10
1µg C/l. (Fig. 14a). Para la estación 0 se encontraron los
valores más alto de biomasa de bacterioplancton y los más bajos en la estación 3 (Fig. 14b). En el
Estrecho del lago la Biomasa bacteriana total presentó un promedio de 7,58 x101 µg C/l, con
máximo de 1,54x102 µg C/l y un mínimo de 1,32x10
1 µg C/l, durante los eses de muestreo.

53
En los meses de junio y julio se observaron los valores más altos de biomasa bacteriana
1,54x102 µg C/l y 1,36x10
2 µg C/l respectivamente, mientras que los valores de biomasa más
bajos se observaron en el mes de marzo con 1,32x101 µg C/l (Fig. 14a). Para la estación 7 se
encontraron los valores alto de recuento de bacterioplancton y los bajos en la estación 9 (Fig.
14b).
Figura. 14a. Variaciones de Biomasa de Bacterioplancton Mensual en Bahía
El Tablazo y El Estrecho del Lago de Maracaibo. (Caicedo, 2009)
Figura.14b. Valores de Biomasa de Bacterioplancton Por Estación en Bahía
El Tablazo y El Estrecho del lago de Maracaibo. (Caicedo, 2009)

54
Los valores de biomasa son similares a los señalados por Araújo (2004) en un sistema lagunar
en Brasil donde fluctuó entre 123 a 269 μg C/l y Bojanic y col., (2006), en la zona eutrófica de la
bahía de Kastela (21,04 a 78,93 μg C/l). Sin embargo, Barragán y col (2009) reportaron que en
aguas superficiales de la Bahía de Gaira en Colombia, la biomasa bacteriana varió entre 0,88 y
3,21 μg C/l, valores bajos en comparación al de este estudio. Los valores de biomasa se vieron
afectados por la dinámica hidro-climática, donde la disponibilidad de nutrientes orgánicos,
limitan la comunidad bacteriana en la bahía de Gaira. De esta manera, variación hidro-climática,
podría afectarlos aportes de nutrientes al cambiar su cantidad, su disponibilidad y la capacidad de
las células para asimilarlos, una situación que finalmente generaría cambios en la producción
bacteriana (Church y col., 2000; La Ferla y col., 2005).
Las diferencias en las variaciones de abundancia y biomasa bacteriana entre ambas zonas de
muestreo fueron significativas (t = -18,524, P ˂ 0,001 y t = -13,942, P ˂ 0,001 respectivamente).
De esta manera, La Bahía El Tablazo presentó los valores más altos de abundancia y biomasa de
bacterias con respecto al Estrecho del Lago, lo que puede atribuirse a las variaciones en los
factores fisicoquímicos como salinidad, temperatura y transparencia que presentaron diferencias
significativas entre ambas zonas de muestreo, como se comentó anteriormente.
No todos los microorganismos pueden crecer y reproducirse bajo idénticas condiciones, cada
uno tiene su intervalo de tolerancia a las diferentes variables en que puede crecer y reproducirse.
Fuera de este intervalo, es posible que sobreviva, pero en condiciones estáticas relativas
conocidas como estado latente (Atlas y Bartha, 2002). Sin embargo, en la naturaleza las
condiciones ambientales que predominan son el resultado de la interacción de muchos factores, lo
que permite la coexistencia de diferentes comunidades al mismo tiempo y el predominio de un
grupo u otro, de acuerdo a las condiciones que prevalezcan en cada momento (Kirchman y
William, 2000).
No obstante, la limitación por nutrientes orgánica (COD) es otro factor que puede influir en
estas variaciones, debido a que las bacterias depende de la disponibilidad de sustratos orgánicos,
siendo un factor limitante y uno de los mecanismos de control más importantes de la producción
bacteriana (Eiler y col., 2003). Al ser incorporado permanece disponible y puede ser transferido
por consumo a los bacteríovoros y a partir de ellos ser exportado del sistema (Bratbak y col.,

55
1998; Thingstad, 2000). Además, la bacteriovoría (Pereira y col., 2005; Eskinazi-Santanna y
Björnberg, 2006) y la lisis viral (Fuhrman y Schwalbach., 2003; Lymer y col., 2008), han sido
sugeridas por varios estudios como factores importantes para la estructuración de las
comunidades de bacterioplancton.
4.- Abundancia de Nanoflagelados, Ciliados y Rotíferos
4.1- Nanoflagelados
La abundancia de nanoflagelados presentó un promedio de 8,65 x105 orgl/l en las aguas
superficiales de Bahía el Tablazo, durante el periodo de estudio. Los valores máximos de
nanoflagelados se encontraron en el mes de abril con 1,91 x106 orgl/l; mientras que los valores
mínimos se observaron durante el mes de Febrero 3,20 x105 orgl/l (Fig.15a). Para la estación 2 se
encontraron los valores más altos de nanoflagelados y en la estación 3 los más bajos (Fig. 15b).
Figura. 15a. Variaciones de abundancia de Nanoflagelados mensual en
Bahía El Tablazo y El Estrecho del Lago de Maracaibo. (Caicedo, 2009)
En el Estrecho del Lago de Maracaibo, la abundancia de nanoflagelados mostró un promedio
de 7,29x105 orgl/l, durante el periodo de estudio. Los valores máximos de nanoflagelados se
encontraron para el período de Marzo con 2,00x106 orgl/l; mientras que los valores mínimos se
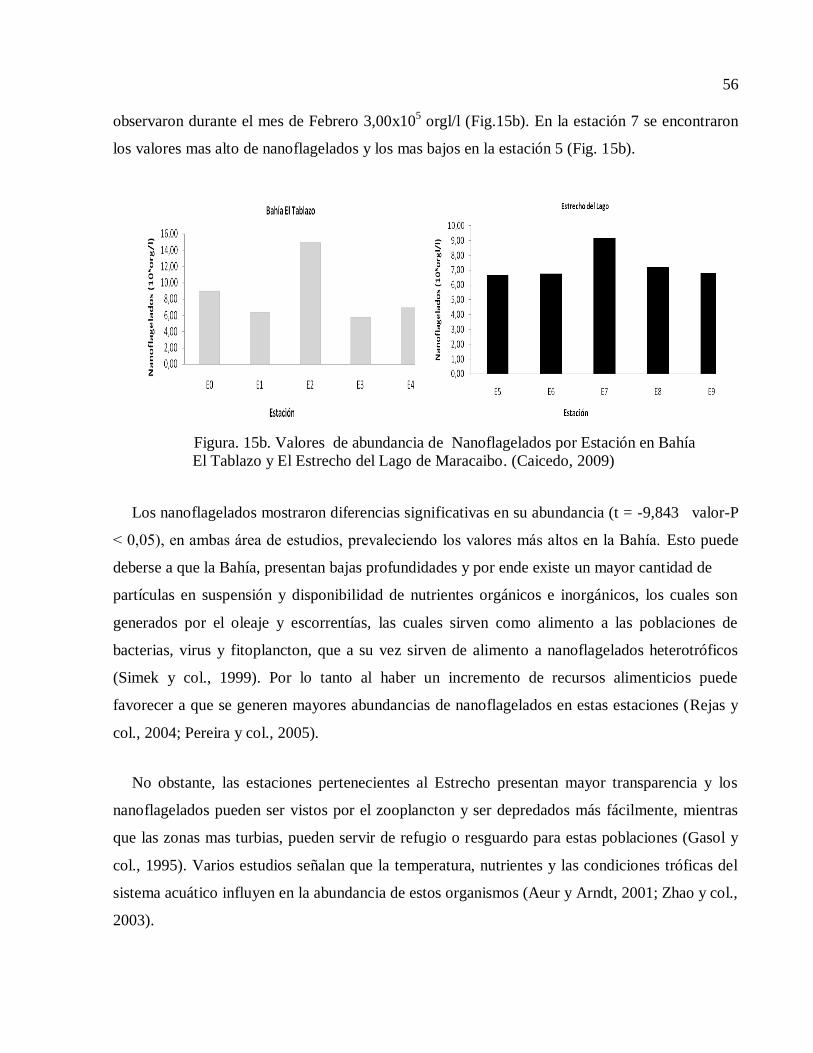
56
observaron durante el mes de Febrero 3,00x105 orgl/l (Fig.15b). En la estación 7 se encontraron
los valores mas alto de nanoflagelados y los mas bajos en la estación 5 (Fig. 15b).
Figura. 15b. Valores de abundancia de Nanoflagelados por Estación en Bahía
El Tablazo y El Estrecho del Lago de Maracaibo. (Caicedo, 2009)
Los nanoflagelados mostraron diferencias significativas en su abundancia (t = -9,843 valor-P
˂ 0,05), en ambas área de estudios, prevaleciendo los valores más altos en la Bahía. Esto puede
deberse a que la Bahía, presentan bajas profundidades y por ende existe un mayor cantidad de
partículas en suspensión y disponibilidad de nutrientes orgánicos e inorgánicos, los cuales son
generados por el oleaje y escorrentías, las cuales sirven como alimento a las poblaciones de
bacterias, virus y fitoplancton, que a su vez sirven de alimento a nanoflagelados heterotróficos
(Simek y col., 1999). Por lo tanto al haber un incremento de recursos alimenticios puede
favorecer a que se generen mayores abundancias de nanoflagelados en estas estaciones (Rejas y
col., 2004; Pereira y col., 2005).
No obstante, las estaciones pertenecientes al Estrecho presentan mayor transparencia y los
nanoflagelados pueden ser vistos por el zooplancton y ser depredados más fácilmente, mientras
que las zonas mas turbias, pueden servir de refugio o resguardo para estas poblaciones (Gasol y
col., 1995). Varios estudios señalan que la temperatura, nutrientes y las condiciones tróficas del
sistema acuático influyen en la abundancia de estos organismos (Aeur y Arndt, 2001; Zhao y col.,
2003).

57
De esta forma, se obtuvo valores de nanoflagelados mayores a los indicados por Hall y col.
(2004) en aguas subtropicales en Nueva Zelanda (910x103 a 180x10
3 cel/l) y Eissler y col. (2010)
en la columna de agua de la Costa del Pacífico Sudamericano (0,02x103 a 0,6x10
3 cel/l).
Mientras que Dupuy y col. (2007) señalan valores mayores de abundancia de nanoflagelados a a
los obtenidos en este estudio (3,2x108 a 2,5x10
8 cel/l).
3.2.- Ciliados
En Bahía el Tablazo la abundancia de ciliados presentó un promedio de 6,55 x104 org/l,
convalores máximos para Febrero de 2,58 x104 orgl/l; mientras que los mínimos se observaron
durante los meses de Abril, Mayo y Septiembre 0,00 orgl/l (Fig.16a). En la estación 1 se
encontraron los valores más alto ciliados y en la estación 4 los más bajos (Fig. 16b). Para las
aguas superficiales del Estrecho del Lago de Maracaibo, la abundancia de ciliados mostró un
promedio de 1,37x104orgl/l, con valores máximos en Febrero con 3,96x10
6orgl/l; mientras que
los mínimos se observaron durante los meses Abril, Mayo y Septiembre 0,00 orgl/l (Fig.17a).
Para la estación 6, se encontraron los valores más alto de ciliados y los más bajos en la estación 9
(Fig. 17b).
Figura. 16a. Variaciones de Abundancia de Ciliados Mensual en Bahía
El Tablazo y El Estrecho del Lago de Maracaibo. (Caicedo, 2009)

58
Figura. 16b. Valores de Abundancia Ciliados por Estación en Bahía El
Tablazo y El Estrecho del lago de Maracaibo . (Caicedo, 2009)
Los ciliados mostraron diferencias significativas entre ambas zonas de estudio (t = -5,183
valor-P ˂ 0,05), prevaleciendo mas en la Bahía El Tablazo que en el Estrecho del Lago. Esto
puede ser debido a que la Bahía El Tablazo son las que presentan menor profundidad y una gran
cantidad de partículas en suspensión, siendo la zona donde existen los requerimientos adecuados
para el desarrollo de bacterias, fitoplancton y nanoflagelados, los cuales sirven como fuente de
alimentación de los ciliados (García y Lara, 2001; Eskinazi-Santanna y Björnberg, 2006; Pereira
y col., 2005) lo cual favorece el crecimiento y reproducción de los ciliados y por ende un
aumento de su abundancia.
Los promedios de ciliados son bajos a lo señalado por Sommaruga (1995) y Hwang y Heath
(1997) para lagos eutróficos (5,5 y 4,52 x 105 cel/l) e hipereutróficos (9 a 2,15x105 cel /l)
tropicales y templados y Dupontt (2003) para la zona litoral del Lago de Maracaibo (2,44 a
2,76x106 cel/l). Sin embargo, los promedios de ciliados son similares a la obtenida por Rincón y
col. (2007) en la zona litoral de la costa oriental del Lago (2,58x105 cel/l).
3.3.- Rotíferos
Los rotíferos en la Bahía el Tablazo presentaron una abundancia promedio de 5,14E+00org/l, con
valores que se mantuvieron por debajo 1,00x101 orgl/l casi durante el periodo de muestreo
excepto para Mayo y Septiembre (Fig.17a). En la estación 3 se encontraron los valores más alto

59
rotíferos y los más bajos en la estación 0 (Fig. 17b). Con respecto al Estrecho del Lago, la
abundancia de rotíferos mostró un promedio de 1,02x101orgl/l, con valores máximos para el mes
de Septiembre con 2,00x106orgl/l; mientras que durante los de más meses no se encontraron estos
organismos (Fig.17a). Para la estación 7 se encontraron los valores más alto y los más bajos en la
estación 8 (Fig. 17b).
Figura. 17a. Variaciones de Abundancia de Rotíferos Mensual en Bahía
El Tablazo y El Estrecho del Lago de Maracaibo. (Caicedo, 2009)
Figura. 17b. Valores de Abundancia Rotíferos por Estación en Bahía El
Tablazo y El Estrecho del lago de Maracaibo. (Caicedo, 2009)

60
La abundancia de rotíferos en ambas zonas de estudios fue relativamente baja, sin embargo
predominaron más en el Estrecho que la Bahía El Tablazo. Estas especies aunque pueden ser
encontradas en aguas estuarina o marinas (Theis, 2000), exhiben gran diversificación en agua
dulce pues su origen es dulceacuícolas (Roldan y Ramírez, 2008).
Estudios realizados han mostrado que existen pocas especies de rotíferos capaces de
toleraraguas salobres (Fontaneto y col., 2006, Zakaria y col., 2007; Medeiros y col., (2010), la
salinidad puede influenciar en la producción de huevos, en la eclosión y en la madurez de las
poblaciones de rotíferos (Sarma y col., 2005).No obstante, Ávila (2009) indicó que en las zonas
costeras del sistema de Lago de Maracaibo, los rotíferos representan el segundo grupo
zooplanctónicos más abundante ( 29,4%), siendo la costa de Mara y Maracaibo las zonas donde
mas predominan estos organismo.
5.- Relación de abundancia y biomasa bacterioplancton con factores abióticos
5.1- Bahía El Tablazo
Se encontró una correlación positiva y significativa entre la abundancia de bacterias con la
temperatura (r = 0,305 p=0,018), por lo que a medida que aumenta la temperatura aumenta la
abundancia de bacterioplancton en esta zona de muestreo. Los morfotipos bastones tanto libres
como adheridos mostraron esta correlación positiva con la temperatura (r = 0,316 p=0,014; r =
0,289 p=0,025), indicando que ellas se benefician con los aumentos de temperatura en
comparación a los morfotipos cocos. Dentro de los factores reguladores del crecimiento y la
fisiología de las bacterias marinas, el factor temperatura en particular ha sido estudiado por
seruna variable que determina potencialmente el crecimiento bacteriano (Pomeroy y Wiebe,
2001).
Existe evidencia de la interacción entre temperatura y la utilización del sustrato orgánico por
las bacterias heterotróficas, de acuerdo a los datos de campo de Pomeroy y Wiebe (2001), donde
se ha observado en el medio marino que generalmente las bacterias heterotróficas de ambientes
tropicales funcionan cerca de la temperatura óptima de asimilación y crecimiento y también en

61
ambientes templados por lo menos durante el verano. Sin embargo, para ambientes templados
durante invierno operan cerca de la temperatura de crecimiento nulo con excepción de episodios
con alta disponibilidad de sustrato (Pomeroy y Wiebe, 2001).
La temperatura de crecimiento puede afectar profundamente el estado fisiológico de las
bacterias y en respuesta a la variación de la temperatura, éstas alteran la síntesis de proteínas,
membranas y una variedad de otras estructuras celulares como resultado de adaptarse a las
nuevas condiciones ambientales (Bayles y col., 2000). Esto también ha sido reportado por otros
autores (Wouters y col., 1999; O’Connell y Thomashow, 2000, O’Connell y col., 2000), quiénes
concuerdan que las bacterias realizan complejas modificaciones estructurales y fisiológicas que
les favorezca mantener la fluidez de la membrana celular.
La biomasa presentó una correlación negativa con la transparencia (r = -0,316 p=0,014), a
medida que aumenta la transparencia disminuye la biomasa del bacterioplancton. Considerando
que esta zona presenta menor profundidad y, consecuentemente, mayor turbidez causada por la
resuspensión de sedimentos, es posible que sean poco visibles las bacterias por los
depredadores.Por otro lado, los resultados estadísticos señalaron que no hay relación de la
abundancia y biomasa del bacterioplancton en la Bahía El Tablazo con el resto de las variables
fisicoquímicas y nutrientes.
5.2.- Estrecho del Lago
Los análisis estadísticos mostraron una correlación positiva y significativa entre la abundancia
de bacterias con la salinidad (r = 0,039 p=0,008), por lo que a medida que aumenta la salinidad
aumenta la abundancia de bacterioplancton en el estrecho del Lago de Maracaibo. La salinidad en
esta zona de muestreo presenta cambios causados por las dinámicas de las mareas (responsables
del intercambio de agua dulce y salado en dicha zona), variaciones anuales de precipitaciones y
escorrentía terrestre, afectando sensiblemente la salinidad de estas aguas. (Esclapés y col., 2000).
Se encontró una correlación negativa entre la abundancia de bacterias con la transparencia

62
(r = -0,291 p=0,024). La biomasa también presento una correlación negativa con la transparencia
(r = -0,294 p=0,023). Dado a que esta zona de muestreo es mas profunda hay mayor
transparencia, y las bacterias son fáciles presa para los depredadores. Con respecto al resto de las
variables fisicoquímicas y nutrientes no presentaron relación para la abundancia y biomasa de
bacterioplancton.
A pesar que comúnmente se ha citado el fósforo y el nitrógeno como determinantes en la
dinámica del bacterioplancton en diferentes sistemas acuáticos (Farjalla, 2001; Muylaert y col.,
2002; Castillo y col., 2003), no se observó correlación significativa en este estudio. Posiblemente
los aportes de los ríos y las actividades antropogénicas pueden ser importantes, no limitando al
crecimiento bacteriano. Por ello, seria necesario considerar otros factores abióticos no estudiados
aquí para una mejor caracterización de la dinámica del bacterioplancton.
En ambas zonas de estudio, se evidencia que las variables ambientales influyen en la dinámica
del bacterioplancton, y a su vez que cada uno de estos sistemas acuáticos posee característicos
específicas (hidrodinámica, variables bióticas y abióticas) que afectan a dicha comunidad. Sin
embargo, existen otros factores abióticos no analizado en este estudio, como es la materia
orgánica disuelta, la cual es un factor limitante y uno de los mecanismos de control más
importantes de la comunidad bacteriana (Eiler y col., 2003).
6.- Relación de abundancia y biomasa bacterioplancton con factores bióticos
En la Bahía El Tablazo, la abundancia y biomasa de bacterioplancton no presentaron
correlación con la abundancia de ciliados, nanoflagelados y rotíferos. No obstante, los morfotipos
cocos adheridos presentaron una correlación negativa con la abundancia de ciliados (r = -0,384
p=0,028), a medida que aumenta la abundancia de éstos disminuye la abundancias de cocos
adheridos. Para el Estrecho del Lago, la abundancia y biomasa de bacterioplancton no
presentaron correlación con la abundancia de ciliados, nanoflagelados y rotíferos. Sin embargo se
encontró que los morfotipos cocos y bastones adheridos presentaron una correlación negativa con
la abundancia de ciliados (r = -0,276 p=0,032), lo también obtenido en las aguas de Bahía el
Tablazo.

63
Adicionalmente, los resultados estadísticos señalan que la biomasa del morfotipo coco
presentó una correlación negativa con la abundancia de nanoflagelados (r = -0,315 p=0,014) en el
Estrecho. Por lo tanto, los nanoflagelados evidencian una selección por tamaño, concordando a
lo señalado por Posch y col. (1999), donde el patrón de tamaño es constante al momento de la
selección de presas (bacterias) por parte de los nanoflagelados. Adicionalmente, los resultados
indican que para el periodo septiembre a febrero los morfotipos cocos disminuyeron sus valores
de abundancia y biomasa de bacterioplancton, este hecho podría sugerir que la depredación por
parte de los nanoflagelados podría tener influencia en ello.
En general, ambas aéreas de estudio, se observa que las bacterias parecen ser afectadas por
depredación de los ciliados principalmente las bacterias adheridas en comparación a las libres,lo
cual podría explicar los bajos índices de éstas durante todo el estudio. Los ciliados se alimentan
de poblaciones bacterianas, pero también de algas y detritos particulado (Corliss, 2002;
Fernández, 2004). Precisamente por ser bacterias adheridas se podría suponer por tanto, que los
morfotipos cocos y bastones pudieran ser depredados por los ciliados cuando éstos consumen
partículas en suspensión (Artigas, 1998).
La abundancia y biomasa de bacterioplancton podrían ser controladas por factores abióticos,
sin embargo, es difícil concluir satisfactoriamente, pues es necesario considerar ciertos aspectos.
Por ejemplo, ver el efecto de la depredación sobre la comunidad bacteriana, teniendo en cuenta
no solamente tamaño y forma, sino también otras propiedades de los procariotas planctónicos que
los hacen ser consumidos por los depredadores (Matz y Jurgens, 2001). En cuanto a la parte
abiótica considerar nutrientes orgánicos (COD) e incluso acoplamiento con el fitoplancton
(Gutiérrez 1997; Joint y col., 2002 y Barragán, 2009).
Por lo tanto, es de vital importancia para lograr comprender la dinámica del bacterioplancton
considerar en forma conjunta todos las factores ambientales e interacciones que puedan
presentarse dentro de estas zona de estudio y el cómo afecta a dicha comunidad.

CONCLUSIONES
1.- La abundancia y la biomasa del bacterioplancton mostró mayores valores en la Bahía el
Tablazo con respecto al Estrecho del Lago, sugiriendo que existe ciertos factores en el agua que
favorecen mejor una zona que otra. La comunidad bacteriana resulto relacionada en la Bahía El
Tablazo por la temperatura, mientras que en el Estrecho con la salinidad y transparencia.
2.- La abundancia de nanoflagelados, ciliados y rotíferos mostró valores elevados de recuentos en
la Bahía El Tablazo, concordando a lo obtenido en el bacterioplancton, por lo que los factores
abióticos sugieren son los responsables de estas variaciones en las áreas de estudio.
3.- Los factores abióticos, fundamentalmente la salinidad, temperatura y transparencia sugieren
ser los controladores del crecimiento y desarrollo de la comunidad bacteriana en ambas áreas de
estudio.
4.- Los morfotipos presenten en ambas zonas de muestreo correspondieron a las formas cocos y
bastones, encontrándose un predomino de cocos en todo el estudio, es posible que los cambios
que ha presentado el sistema del Lago de Maracaibo en los últimos años, haya condicionado la
presencia de un morfotipo en particular de mayor resistencia y adaptabilidad.
5.- Las bacterias libres predominaron sobre las adheridas, y estas últimas mostraron una
correlación con la abundancia de ciliados, lo que podría explicar los bajos valores de bacterias
adheridas.

RECOMENDACIONES
1.- Estudiar la abundancia y biomasa de bacterioplancton en relación a otros factores abióticos
que no se analizaron en este trabajo como materia orgánica disuelta, amonio, material en
suspensión y turbidez.
2.- Un mayor número de muestreos en escala vertical y horizontal, así como en frecuencia
temporal, que involucren un mayor grupo de microorganismos participantes en las cadenas
tróficas acuáticas, permitirán visualizar mejor cómo se dan las interacciones entre el
bacterioplancton y el resto del plancton.
3.- Considerar dentro de los factores bióticos a la comunidad fitoplanctónico, pues se han
reportados interacción entre microalgas y bacterias apuntándolas como mecanismo de control
para estas.

LITERATURA CITADA
Agustí, S; Satta, M.; Mura M; Benavent, E. 1998. Dissolved esterase activity as a tracer of
phytoplankton lyses: evidence of high phytoplankton lyses in the NW Mediterranean. Limnol.
Oceanogr. 43:1832-1849.
Allsopp, D.; colwell,R. R. y Hawksworth, D. L. 1995. Microbial diversity and ecosystem
function. University Press, Cambridge. 482p.
Araújo, F. 2004. Ecologia do protozooplancton, bacterioplancton e virioplancton em um sistema
Fluvial-Lagunar do nordeste Brasileiro. Teses Doutorado. Unversidade do são Carlos. Pág. 26-97
Arocena, R. y Conde, D. 1999. Métodos en ecología de aguas continentales. Con ejemplos de
limnología en Uruguay. Edición D.I.R.A.C. Montevideo – Uruguay. Pág. 97
Artigas, L. 1998. Seasonal variability in microplanctonic biomasses in the Gironde dilution
plume (Bay of Biscay); relative importance of bacteria. Oceanológica Acta. Vol. 21. No. 4
Atlas, R. y Bartha, R. 2002. Ecología microbiana y microbiología ambiental (4 ed.). Editorial
Pearson Educación, S.A., Madrid, España, 677 pp.
Atlas, R. 1993. Detecting gene sequences using the polymerase chain reaction. En: Kemp P y col.
(eds.) Handbook of methods in aquatic microbial ecology. Lewis Publ. London: 267-271
Auer, B. y Elzer, U. 2004. Comparison of pelagic food webs in lakes along a trophic gradient and
with seasonal aspects: influence of resource and predation. JournaL of Plankton Research Vol. 26
N° 6 Pág. 697–709
Apha. 1999. Standard methods for the examination of water and wastewater.
APHA/AWWA/WPCF. Washington
Azam, F.; Fenchel, T.; Field, J.; Gray, J.; Meyer-Reil, L. y Thingstad, F. 1983. The ecological
role of waters-column microbes in the sea. Mar. Ecol. Prog., Ser. 10: 257-263.
Azam F. 1998. Microbial control of oceanic carbon flux: the plot thickens. Science 208:694-696.
Barragán, R.; Canosal, A. y Niñoa, J. 2009. Bacterioplancton en Bahía Gaira, Mar Caribe
(Colombia): comparación de la variabilidad en abundancia y biomasa bacteriana durante
diferentes períodos. Bol. Invest. Mar. Cost. 38 (2). Pág. 75-90
Biddanda, B. y Cotner J. 2003. Love handles in aquatic ecosystems: the role of dissolved organic
carbon drawdonwn, resuspended sediments, and terrigenous inputs in the carbon balance of Lake
Michigan. Ecosystems 5:431-445.
Blight, S.; Bentley, T.; Lefevre, D.; Robinson, C.; Rodriguez, R.; Rowlands, J.; Williams, P.
1995. Phasing of autotrophic and heterotrophic plankton metabolism in a temperate coastal
ecosystem. Mar. Ecol. Prog. Ser. 209:1-17.
Bloem, J.; Bar-Gillison, M.; Cappenberg, T. 1986. Fixation, counting and manipulating of
heterotrophic nanoflagellates. Applied environmental microbiology 52: 1266—1272.
Bratbak G. 1985."Bacterial biovolume and biomass estimations." Applied and Environmental
Microbiology 49:1488-1493
Bratbak, G. 1993. Microscope methods for measuring bacterial biovolume: epifluorescence
microscopy, scanning electron microscopy, and transmission electron microscopy. En Handbook

of Methods in Aquatic Microbial Ecology, Kemp, P, Sherr B. F., Sherr, E. B., Cole, J., (eds) lewi
Lewis Publishers:309-318.
Brett, M.; Fred, S.; Lubnow, M.; Müller, A. y Goldman C.1999. Nutrient control of
bacterioplankton and phytoplankton dynamics. Aquatic Ecology 33: 135–145.
Bojanic, N.; Solic, M.; Krstulovic, N.; Sestanovic,S.; Gladan,Z.; Marasovic, I. y Brautovic, I.
2006. The role ciliates within the microbial food web in the eutrophicated part of Kastela Bay
(middle Adrict Sea). España. Scietia marina 70(3)
Brussaard, C. 2004. Viral control of phytoplankton populations - a review. Journal of Eukaryotic
Microbiology, 51, 125–138.
Campbell, L. 1993. Inmunofluorescence method for the detection and characterization of marine
microbes. En Kemp P y col. (eds). Handbook of methods in aquatic microbial ecology. Lewis
Publ. London: 295-302
Carlson, C. y Ducklow, H. 1996. Growth of bacterioplankton and consumption of dissolved
organic carbon in the Sargasso Sea. Aquat. Microb. Ecol. 10: 69-85.
Castillo, M.; Kling, G.; Allan, J. 2003. Bottom- up controls on bacterial production in tropical
lowland river. Limnol. Oceanog. 48; 1466-1475
Chicheportiche, J. 2007. Approche du compartiment microbien et phytoplanctonique do la zone
estuarinne amazonienne. 2 eme. année option Génie. Enverinnment.
Chrzanowski, T.; Sterner, R. y Elser, J. 1995. Nutrient enrichment and nutrient regeneration
stimulate bacterioplancton growth. Microb Ecol 29: 221–230
Cognetti, S. y Magazzu, G. 2001. Biología Marina. Ariel Ciencia. España. Pag 327-328
Conde, J.; Ramos, E. y Morales, R. 2004. El zooplancton como integrante de la estructura trófica
de los ecosistemas lénticos. Ecosistemas 13 (2): 23-29. Instituto del Agua, Universidad de
Granada, Ramón y Cajal 4, 18071 Granada, España.
Coveney, M. y Wetzel, R.1992. Effects of nutrients on specific growth rate of bacterioplankton in
oligotrophic lake water cultures. Appl Envir Microbiol 58: 150–156
Chrost, R. 1991. Ectpenzymes in aquatic environment: microbial strategy for substrate supply.
Verh. Internat. Verein. Theor. Angew. Limnol. 24: 3926-3942
Chrost, R. 1992. Ectpenzymes activity and bacterial secondary production in nutrient
improverished and enriched freshwater mesocosms. Microb. Ecol. 3:123-127
Church, J.; Hutchins, D. y Ducklow, H. 2000. ―Limitation of bacterial growth by dissolved
organic matter and iron in the Southern Ocean‖. Appl. Environ. Microbiol. 66(2):455-466
Chrzanowski, T. and Simek, K. (1990) Prey-size selection by freshwater fagellated protozoa.
Limnol. Oceanogr. 35, 1424-1436.
Currie, D. 1990. Large-scale variability and interactions among phytoplankton, bacterioplankton
and phosphorus. Limnol. Oceanogr., 35: 1437-1455.
Del Giorgio y Cole J. 2000. In microbial ecology of the oceans. (Ed.) David. L. Kirchman.
Wiley-Liss y Sons. Inc. Publication 289-235pp.
De Long, E. 1993. Single cell identification using fluorescently labeled, ribosomal RNA-specific
probes. En Kemp P y col. (eds.) Handbook of methods in aquatic microbial ecology. Lewis Publ.
London: 285-294

Devetter, M. y Sed’a, J. 2003. Rotifer fecundity in relation to components of microbial food web
in a eutrophic reservoir. Hydrobiologia 504: 167–175
Di Siervi, M. 2002. Bacterias. Aquate. Instituto de Limnología ― Dr. Raul Ringulet‖ Vol. 7 . Pag.
1-53
Ducklow, H y Carlson, C. 1992. Oceanic Bacterial Production. Adv. Microbiol. Ecol. 12:113-
181.
Dupontt, J. 2003. Depredación bacteriana por protozoarios presentes en muestras de agua
procedentes del Lago de Maracaibo. Tesis de Maestría, Facultad Experimental de Ciencias,
Universidad del Zulia, 80 pp.
Eiler, A.; Langenheder, S.; Bertilsson, S. y Tranvik, L. 2003. Heterotrophic bacterial growth
eficiency and community structure at different natural organic carbon concentrations. Appl.
Environ. Microbiol., 69 (7): 3701-3709.
Elser, J.; Stabler L. y Hassett, R. 1995. Nutrient limitation of bacterial growth and rates of
bacterivory in lakes and oceans: a comparative study. Aquat. Microb. Ecol. 9: 105-110.
Esclapés, G. 2000. El Sistema de Maracaibo. Instituto Venezolano de investigaciones científicas.
Segundo edición. Pág. 125-135
Fajarlla, V. 2001. Bacterial density and biomass, and relations with abiotic factors, in 14 coast
Lagoons of Rio Janeiro State. Oecologia brasiliensisvol IX, 65-76
Farjalla, V.; Enrich-Prast, A.; Esteves, F. y Cimbleris, A. 2006. Bacterial Growth and DOC in a
Tropical Coastal Lagoon. Braz. J. Biol., 66(2A): 383-392
Fenchel, T. 1982. Ecology of heterotrophic microflagellates IV. Quantitative occurrence and
importance as bacterial consumers. Marine ecology progress series 9: 352.
Fischer, U. y Velimirov, B. 2002. High control of bacterial production by viruses in a eutrophic
oxbow lake. Aquatic Microbial Ecology, 27, 1–12
Fogg, G. 1995. Some comments on picoplankton and it importante in the pelagic ecosystem.
Aquatic. Microbaol ecology.Vol 9 pp 33-39
Fuhrman, J. 1991. Bacterioplankton roles in cycling of organic matter: a microbial food web
uptake. In: Brookhaven National Laboratory Symposium, Plenum Press, 1: 1-22
Fuhrman, J. y Noble, R. 1995. ―Viruses and protists cause similar bacterial mortality in coastal
seawater‖. Limnol. Oceanogr. 40(7): 1236-1242.
Fuhrman, J. 1999. ―Marine viruses and their biogeochemical and ecological effects‖.Nature. 399:
541-548.
Fuhrman, J. y Schwalbach, M. 2003.Viral Influence on Aquatic Bacterial Communities. Marine
Biological Laboratory.April 204: 192–195.
González, A.; Paranhos, R.; Lutterbach, M. 2006. Heterotrophic Bacteria abundances in Rodrígo
de Freitas lagoon (Río de Janeiro, Brazil). Brazilian Journal of micribiology, 37:428-433.
González, L. 2007. El sistema del lago de Maracaibo. Ecología Social Ambiental,Zulia.
Disponible en: hptt//socioambiente.blospot.com/2007/04/el-sistema-del-lago-de maracaibo-html.
[Fecha de consulta 23 septiembre 2011]
Gude, H. 1979. Grazing by protozoa as selection factor for activated sludge bacteria. Microb.
Ecol. 5, 225-237.

Gude, H. 1988. The role of grazing on bacteria in plankton succession, (in:) Plankton Ecology:
Succession in Plankton Communities., red. Sommer U., Brock/Springer, Berlin, pp:337-364.
Ngando, T. 2005. Estimates of protozoan- and viral-mediated mortality of bacterioplankton in
Lake Bourget (France). Freshwater Biology, 50, 627–645.
Hahn, M. y Höfle, M. 2001. Grazing of protozoa and its effect on populations of aquatic bacteria.
FEMSMicrob. Ecol. 35: 113–121
Hwang, S. y Heath, R. 1997. The distribution of protozoans across trophic gradient, factors
controlling their abundance and importance in the plankton food web. J. Plank. Research 19(4):
491–518.
Hofmann, W. y Höfle, M. 1993. Rotifer population dynamics in response to increased bacterial
biomass and nutrients: a mesocosm experiment. Hydrobiologia 255/256: 171–175.
Jiménez-Gómez, F.; Rodríguez, V. y B. Bautista. 1994. Trophic interactions in the microbial food
webat a coastal station in the Alboran Sea (Western Mediterranean) in winter. (II). Size selective
flagellate feeding on bacteria and its implication on the microbial loop size-structure. Sci. Mar.
58: 153-159.
Joint, I.; Henriksen, P.; Fonnes G.; Bourne, D.; Thingstad, T. y Riemann, B. 2002.―Competition
for inorganic nutrients between phytoplankton and bacterioplankton in nutrient manipulated
mesocosms‖. Aquat. Microb. Ecol. 29: 145-159
Jürgens, K. 1994. Impact of Daphnia on planktonic microbial food wes. A Review. Mar.
Microbial. Food Webs. 8: 295-324
Kemp, P; Sherr, B.; Sherr, E. y Cole; J. 1993.Handbook of Methods in Aquatic Microbial
Ecology. Lewis Publishers, Estados Unidos. Pp 776
Kirchman, D. 1990. Limitation of bacterial growth by dissolved organic matter in the sub arctic
Pacific. Mar. Ecol. Prog. Ser. 62:47-54.
Landry,M. y Hassett, R. 1982. Estimating the grazing impact of marine micro-zooplankton. Mar.
Biol., 67, 283–288.
La Ferla, R. y Leonardi, M. 2005. Ecological implications of biomass and morphotype variations
of bacterioplankton: an example in a coastal zone of the Northern Adriatic Sea (Mediterranean).
Marine Ecology (26) 82–88
Lymer, D.; Lindstro¨ E. y Vrede, K. 2008. Variable importance of viral-induced bacterial
mortality along gradients of trophic status and humic content in lakes. Freshwater Biology (2008)
53, 1101–1113
Madigan, M.; Martinko, J. y J. Parker. 2004. Biología de los microorganismos (9 ed.). Ediciones
Prentice Hall. Inc., Madrid, España, 986 pp.
Miranda, K. 2007. Caracterização dinamica do comunidade do bacterioplancton no Estuario do
Rio Amazonas(canal do Norte). Teseapresentad ao programa de pósgraduacao em
biodiversidade tropical de UniversiidadeFederal de AMapa. Pág. 17-23;56-79
Middelboe, M.; Borch, N. y Kirchman, D. 1995. ―Bacterial utilization of dissolved free amino
acids dissolved combined amino acids and ammonium in the Delaware Bay estuary: effects of
carbon and nitrogen limitation‖. Mar. Ecol. Prog. Ser. 128: 109-120.
Middelboe, M.; Jorgensen, N. y Kroer, K. 1996. ―Effects of viruses on nutrient turnover and
growth efficiency of noninfected marine bacterioplankton‖. Appl. Environ. Microbiol. 62(6):

1991-1997.
Middelboe, M.; Hagstro¨m, A.; Blackburn, N.; Sinn, B.; Fischer, U. ; Borch, N.H.; Pinhassi,
J.;Simu, K. y Lorenz, M.G. 2001. Effects of Bacteriophages on the Population Dynamics of Four
Strains of Pelagic Marine Bacteria. Marine Mikrobiologie, Fachbereich 2: Biologie/Chemie,
Zentrum fu¨r Umweltforschung und Umwelttechnologie, Universita¨t Bremen, D-28359 Bremen,
Germany
Montoya, R.; Gómez, P.; Gonzalez M.; Henriquez, M. y Candia, A. 1996. Amplificación de
genes por PCR y su aplicación al estudio de la biodiversidad.BOL. Soc. Biof. 67:43.51.
Noble, R. y Fuhrman, J. 1998. Use of SYBR Green I for rapid epifluorescence counts of marine
viruses and bacteria. Aq. Microb. Ecol. 14:113-118.
Norlan, S. 1993. The relationship between biomass and volume of bacteria. En Kemp P y col.
(eds.) Handbook of methods in aquatic microbial ecology. Lewis Publ. London: 303-307
Ovreas, L.; Bourne, D.; Sandaa R.; Casamayor, E.; S. Benlloch, V.; Goddard, G.; Smerdon,M.;
Heldal y Thingstad T. 2003. ―Response of bacteria and viral communities to nutrient
manipulations in seawater mesocosms‖. Aquat. Microb. Ecol. 31: 109-121.
Pace ML. 1993. Heterotrophic microbial processes. In Carpenter SR and Kitchell JF (eds). The
trophic cascade in lakes. Cambridge University Press, Cambridge, pp. 252–277.
Pernthaler, J.; Pernthaler, A. y Amann, R. 2001. Fluorescence in situ hybridization (FISH) with
Rrna- targeteg oligonucleotide. Probes. En: Methodsin microbiology. Vol. 30 Marine
Microbiology. Ed. Jhon H. Paul. Academic Press, 543p.
Pernthaler, J.; Pernthaler, A. y Amann, R. 2003. Automated enumeration of groups of marine
picoplankton after fluorescence in situ hybridization. Applied and Environmental Microbiology
69:2631-2637.
Petrúcio, M. 2003. Produtividade Bacterioplâtonica e fitoplâtonica mo ecosistemas Aquáticos do
Trecho Médio da Bacia do Rio Doce. MG. Sâo Carlos : UFSCAr, Tese de Doutorado.
Pomeroy, L. 1974. The ocean’s food web, a changing paradigm. BioScience, 24, 499–504.
Pomeroy, L. y Weibe, W. 2001. Temperature and substrates as interactive timiting factors for
marine heterotrophic bacteria. Aquatic. Microb. Ecol. 23:187-204
Porter, K. y Feig, S. 1980. The use of DAPI for identifying and counting aquatic microbiota.
Limnol. Ocenagr.25:943-948
Proctor, L. y Fuhrman, J. 1990. Viral mortality of marine bacteria and cyanobacteria. Nature
343:60-62.
Psenner, R. y Sommaruga, R. 1992. Are rapid changes in bacterial biomass caused by shifts from
top-down to bottom-up control? Limnol. Oceanogr., 37 (5): 1092-1100
Oom- Wilms A. 1997. Are bacteria an important food source for rotifers in eutrophic lake?
Racy, F. 2004. Aspectos numéricos, morfológicos e morfometricos das comunidades bacterianas
eem diferentes escalas trófica e temporal, em reservatórios. Dissertacao (Mestrado em Ecologica
e Recursos Naturais) Universidae Federal do São Carlos
Ram, A.; Chandramohan, D. 2007. Bacterial growth efficiency in tropical estuary; seasonal
variability subsidized by allocthonous carbon. Microbial ecology, 53:591-599
Reche, I.; Pace, M. y Cole, J. 1998. Interactions of photobleaching and inorganic nutrients in

determining bacterial growth on colored dissolved organic carbon. Microb Ecol 36: 270–280
Rejas, D.; Muylaert, K. y Luc De M. 2005. Trophic interactions within the microbial food web in
a tropical floodplain lake (Laguna Bufeos, Bolivia). Rev. Biol. Trop. (Int. J. Trop. Biol. ISSN-
0034-7744) Vol. 53 (1-2): 85-96, March-June 2005 (www.tropiweb.com)
Rincón, N.; Dupontt, J. y Díaz, L. 2007. Bacterias y protozoarios ciliados de muestras de agua de
la Costa Oriental del Lago de Maracaibo. Bol. Centro Invest. Biol. Volumen 41, No. 3, pp. 309–
322 Universidad del Zulia, Maracaibo, Venezuela
Robarts, R.; Zohary, T.; Waiser, M. y Jacobi, Y. 1996. Bacterial abundance, biomass, and
production in relation to phytoplankton biomass in the Levantine Basin of the southeastern
Mediterranean Sea. Mar Ecol Prog Ser 137:273-281
Rodríguez, G. 2000. Fisiograf{ia del sistema de Maracaibo. En Rodríguez, G. El sistema de
Maracaibo. 2da Edición.Instituto Venezolano de Investigaciones Científicas. Pág. 7-19
Sommaruga, R. 1995. Microbial and classical food webs: A visit to a hypertrophic lake. FEMS
Microbiol Ecol. 17: 257–270.
Steward, J.; Smith D. y Azam, F. 1996. Abundance y production of bacteria and viruses in the
Bering and Chukchi Sea. Mar. Ecol. Prog. Ser.131:287-300.
Suttle, C. y Chen, F. 1992. Mechanisms and rates of decay or marine virases in seawater. Appl.
Environ. Microbiol. 58:3721-3729
Thingstad, T.; Pérez, M.; Pelegri S.; Dolan S. y Rassoulzadegan F. 1999. Trophic control of
bacterial growth in microcosms containing a natural community from northwest Mediterranean
surface waters. Aquat Microb. Ecol. 18: 145-156.
Tijdens, M.; Van de Waal, D.; Slovackova, H.; Hoogveld, H. y Gons, H. 2008. Estimates of
bacterial and phytoplankton mortality caused by viral lysis and microzooplankton grazing in a
shallow eutrophic lake. Freshwater Biology, 53, 1126–1141
Toolan, T.; Wehr, J. and Findlay, S. 1991. Inorganic phosphorus stimulation of bacterioplankton
production in a meso-eutrophic lake.
Vrede, K.; Heldal, M; Norland, S y Bratbak, G. 2002. Elemental composition (C,N,P) and cell
volume of exponentially growing and nutrient-limited bacterioplankton. Appl.Environ.
Microbiol., 68 (6): 2965-2971.
Watson, S.; Novistky, T. ; Quinby,H.; y Valois, F. 1997. Determination of bacterial number y
biomasa in the marine. Enviroment. Appl. Envioron. Microbiol. 33: 940-947
Weinbauer, M. y Hofle , M. 1998. Significance ofvirallysis and flagellate grazing as factors
controlling bacterioplanckton productionin a eutrophic lake. Appl. Environ. Microbiol. 64: 431-
438
Weisse, T.; Mu¨ ller, H., Pinto-Coelho, R. M., Schweizer, A.,Springmann, D. y Baldringer, G.
1990. Response of the microbial loop to the phytoplankton spring bloom in a large prealpine lake.
Limnol. Oceanogr., 35, 781–794.
Weisse, T. 1991. The microbial food web and its sensitivity to eutrophication and contaminant
enrichment: a cross-system overview. Internat. Rev. ges. Hydrobiol., 76, 327–337.
Wetzel, R. 2001. Limnology. Lake and rivers ecosystems. Third edition. Pag. 489-506
Wilhelm, S. y Suttle, C. 1999.Viruses and nutrient cycles in the sea. Biosciences 49:781-788.

Wistreich, G. A. 1998. Microbiology perspectives a photographic Survey of the microbial world.
Ed. Prestice Hall. A. Viasom Co. upper Saddle River, New Jersey 189p
Zhao, Y.; Yu, Y.;Yeng W. y Shen, Y. 2003. Growth and production of free-living heterotrophic
nanoflagellates in a eutrophic lake – lake. Donghu,Wuhan,China. Hidrobiología 498:85-95
Zehr, J.; Ward, B. 2002. Nitrogen cycling in this ocean: new perspectives on processes and
paradigms. Appl. Environ. Microbiol. 68(3): 1015-1024.
Zingel, P. 1999. Pelagic ciliated in a shallow eutrophic lake: community structure and seasonal
dynamics. Arch. Hydrobiol. 146. (4): 495-511