posibilidades de regeneración del manglar en la zona de...
TRANSCRIPT

Ecología de los bosques tropicales
Posibilidades deregeneración delmanglar en la zona dela Ciénaga Grandede Santa Marta(Colombia)

Ecología de los bosques tropicales
Posibilidades deregeneración delmanglar en la zona dela Ciénaga Grandede Santa Marta(Colombia)
Carola Elster
Eschborn 1998

Número de la serie: TÖB F-II/6s
Publicado por la: Deutsche Gesellschaft fürTechnische Zusammenarbeit (GTZ) GmbHPostfach 518065726 Eschborn (Alemania)
Responsable: Tropenökologisches Begleitprogramm (TÖB)Dr. Claus Bätke
Autora: Carola ElsterJustus-Liebig-Universität Gießen, Institut fürBotanik I, Senckenbergstr. 17-21, 35390 Gießen(Alemania)
Redacción: Richard Haep, Dr. Gudrun Bruzak
Producción: TZ Verlagsgesellschaft mbH, 64380 Roßdorf(Alemania)
Precio simbólico: DM 10,-
ISBN: 3-933984-15-7
© 1998 Reservados todos los derechos

PrefacioLos ecosistemas tropicales son la base para la vida de una inmensa mayoría dela población mundial. Sin embargo, la progresiva destrucción y degradación delos recursos naturales en los países en desarrollo amenazan con hacer vanoslos esfuerzos dirigidos a lograr un desarrollo sostenible y a combatir de maneraeficaz la pobreza.Dentro del marco de la Cooperación al Desarrollo, el Programa de ApoyoEcológico (TÖB) tiene el propósito de contribuir a tratar, evaluar eimplementar con eficiencia las informaciones y las experiencias adquiridas eneste ámbito.El Programa de Apoyo Ecológico es un proyecto de carácter suprarregionalrealizado por la Gesellschaft für Technische Zusammenarbeit (GTZ) GmbHpor encargo del Ministerio Federal de Cooperación Económica y Desarrolloalemán (BMZ).Previa solicitud, el programa fomenta estudios complementarios sobre temasde relevancia ecológica para las regiones tropicales. La idea es contribuir alperfeccionamiento de estrategias para la protección y el uso sostenible de losecosistemas tropicales y desarrollar, a partir de ello, instrumentos innovadorespara una cooperación al desarrollo que tenga debidamente en cuenta losaspectos del medio ambiente.La integración de los conocimientos científicos en las actividades deasesoramiento ayuda a los respectivos proyectos a llevar a la práctica losacuerdos internacionales pertinentes, en particular la Agenda 21 y laConvención sobre la Biodiversidad, a los que el BMZ atribuye una especialrelevancia.Un elemento importante del concepto en que se basa el programa es el hechode que científicos alemanes y locales abordan conjuntamente temas orientadosa la aplicación. Con ello, el programa significa también un aporte importante alperfeccionamiento de los expertos nacionales conforme a las exigencias de lapráctica, así como a la ampliación de los conocimientos específicos de estetema de la ecología tropical en los países en desarrollo.A través de unos textos inteligibles y sucintos, la serie de publicaciones delPrograma de Apoyo Ecológico pone los resultados de los estudioscomplementarios y las recomendaciones de actuación de ellos derivadas alalcance de las organizaciones e instituciones que actúan en el campo de laCooperación al Desarrollo así como de todos aquellos que se interesan portemas propios de la política de protección del medio ambiente y del desarrollo.
Dr. H. P. SchipulleJefe de la Sección Protección delAmbiente y los Recursos Naturales,Gestión de Recursos Forestales
Dr. J. FriedrichsenJefe de la División Producción y ProtecciónVegetales, Investigaciones Agronómicas, Sistemasde Explotación Agropecuaria
Ministerio Federal de CooperaciónEconómica y Desarrollo (BMZ)
Deutsche Gesellschaft für TechnischeZusammenarbeit (GTZ) GmbH

Indice
GLOSARIO .....................................................................................III
RESUMEN ......................................................................................IV
ABSTRACT..................................................................................VIII
1 INTRODUCCIÓN........................................................................ 11.1 La zona objeto del estudio y sus particularidades ............................ 11.2 Análisis del problema...................................................................... 31.3 Entorno del proyecto....................................................................... 61.4 Objetivos ........................................................................................ 8
2 RESULTADOS.......................................................................... 102.1 Las zonas estudiadas..................................................................... 102.2 Pruebas de establecimiento de plántulas en condiciones
controladas ................................................................................... 112.2.1 Tasas de germinación de diásporas de diferentes
orígenes ............................................................................ 122.2.2 Influencia de ligeras diferencias en el nivel del
agua y en la concentración de la sal en elestablecimiento y el crecimiento en altura de lasplántulas............................................................................ 13
2.2.3 Influencia de ligeras diferencias del nivel del aguay de la concentración de la sal en la mortalidad delas diásporas ..................................................................... 15
2.2.4 Pérdida de vitalidad de las diásporas porinundación ........................................................................ 16
2.2.5 Pérdida de vitalidad de las diásporas enconcentraciones de sal elevadas ........................................ 17
2.2.6 Influencia de la sequedad y de la irradiación solaren la vitalidad de las diásporas .......................................... 18
2.2.7 Influencia de las condiciones de iluminación conun buen suministro de agua ............................................... 20

Indice
II
2.2.8 Relación entre el peso y el desarrollo de lasdiásporas........................................................................... 20
2.2.9 Mortalidad de plántulas en caso de inundacióntotal .................................................................................. 21
2.2.10Resistencia de diásporas y plántulas a latemperatura ....................................................................... 22
2.2.11Plagas fitófagas de la simiente........................................... 242.3 Desarrollo de la regeneración natural y evolución de los
factores edáficos a lo largo del año en el campo............................ 282.3.1 Factores ambientales ......................................................... 282.3.2 Desarrollo de las plántulas ................................................ 32
2.4 Biología de las inflorescencias y difusión de las diásporasde Laguncularia ........................................................................... 37
2.5 Pruebas de plantación ................................................................... 402.5.1 Plantación de diásporas..................................................... 412.5.2 Cultivo de plántulas .......................................................... 422.5.3 Trasplante de plantas jóvenes............................................ 452.5.4 Multiplicación por estacas................................................. 462.5.5 Destrucción total de plántulas de Avicennia por
orugas de lepidópteros ...................................................... 48
3 RELEVANCIA PARA LA APLICACIÓN YRECOMENDACIONES .............................................................. 533.1 La regeneración natural del manglar.............................................. 533.2 La regeneración artificial del manglar............................................ 55
3.2.1 La siembra de diásporas.................................................... 563.2.2 El establecimiento de plántulas.......................................... 573.2.3 El establecimiento de plantas jóvenes................................ 583.2.4 El establecimiento de estacas ............................................ 593.2.5 Simiente y cultivo de las plantas........................................ 603.2.6 Elección de las parcelas .................................................... 623.2.7 La época de plantación...................................................... 633.2.8 Perspectivas ...................................................................... 63
4 BIBLIOGRAFÍA ....................................................................... 66

Glosario
Tajeas: (en inglés: box culverts) pasajes construidos bajo la carretera
que permiten el intercambio de agua entre diversas zonas de
la Ciénaga Grande
Ciénaga: pantano, laguna pantanosa
CORPAMAG: Corporación Autónoma Regional del Magdalena y de la
Sierra Nevada de Santa Marta; organismo regional de medio
ambiente encargado, entre otras cosas, de la dirección de la
construcción de canales en la zona estudiada así como de la
coordinación de proyectos científicos y sociales
Diáspora: unidad de difusión; dado que los frutos del mangle pueden
germinar ya antes de su difusión o durante la misma, se
observa con frecuencia una difusión de plántulas provistas ya
de raíces. En el informe presente se habla de diásporas hasta
la formación de las hojas, hablándose luego de plántulas
DNP: Departamento Nacional de Planeación
INDERENA: Instituto Nacional de los Recursos Naturales Renovables y
del Ambiente
INVEMAR: Instituto de Investigaciones Marinas y Costeras ”José Benito
Vives De Andreis”, Punta de Betín.
n: número total de valores medidos
PRO-CIÉNAGA: proyecto colombiano-alemán en el que cooperan la
CORPAMAG y la GTZ; dirección de la construcción de
canales en la zona estudiada así como coordinación de
proyectos científicos y sociales
Calderas de sal: zonas llanas, muy salinizadas y desprovistas de vegetación
que se forman con frecuencia en lagos y lagunas en proceso
de desecación; en parte son aprovechadas por los pescadores
locales para la obtención de sal

Resumen
IV
Resumen
La Ciénaga Grande de Santa Marta es una laguna salobre ubicada en la
costa colombiana del Caribe. Es el principal centro pesquero del país, siendo
de gran importancia, tanto ecológica como económica, para las tierras
limítrofes. Intervenciones humanas en el sistema hidrológico de las lagunas
provocaron sin embargo la salinización extensiva y una desecación parcial
de los suelos en el noroeste de la zona así como la muerte de un 60% de los
manglares que pueblan las áreas aluviales de la laguna. Debido a esto ha
disminuido notablemente la productividad del ecosistema, y las bases vitales
para animales y seres humanos están amenazadas o incluso han sido ya
destruidas.
En la actualidad se trabaja en la restauración de las condiciones hidrológicas
originales, a fin de hacer posible una regeneración del ecosistema. Objetivo
del presente proyecto es descubrir qué factores ambientales son importantes
para el establecimiento del manglar y si es posible la reforestación de las
grandes áreas en la actualidad desprovistas de vegetación.. A tal fin se
realizaron experimentos en condiciones controladas y ensayos de plantación
con Avicennia germinans, Laguncularia racemosa y Rhizophora mangle,
las tres especies principales de mangle que crecen en la zona estudiada.
Además se estudió la influencia de los factores ecológicos en el
establecimiento y el desarrollo de las plántulas de mangle en áreas pobladas
y despobladas de bosque. Los resultados muestran que en principio es
posible una regeneración, a condición de que disminuya suficientemente la
salinidad. Sin embargo, dicha regeneración será lenta, ya que la superficie
despoblada de bosques, de 30.000 ha, es muy grande y el aporte natural de
diásporas es muy escaso, siendo en algunos lugares inexistente. Por esta

Resumen
V
razón debería considerarse la posibilidad de una forestación que acelerara el
proceso de regeneración.
En las zonas pobladas de bosque, el crecimiento de las plántulas está
sometido ante todo a la influencia de la salinidad y el establecimiento y la
mortalidad de las plantas jóvenes se ven afectados principalmente por las
inundaciones estacionales. La polinización y la producción de frutos de
Laguncularia así como la regeneración natural del bosque con las tres
especies de mangle están garantizadas a pesar de la alta tasa de mortalidad
de las plántulas.
En las zonas despobladas de bosque, la regeneración está dificultada
adicionalmente por las elevadas temperaturas del suelo y por la desecación
de las capas superiores del suelo. En algunos lugares, la concentración de
sal aumenta durante la época seca a valores superiores al 150% debido a la
elevada evaporación, muriendo a causa de ello todas las plántulas. Por su
tamaño relativamente pequeño, las diásporas de Avicennia y Laguncularia
tienen dificultades extremadamente grandes para establecerse. Una
inundación de sólo 5 cm impide ya su establecimiento, siendo entonces
arrastradas por las aguas. Los rayos solares directos las resecan en apenas
uno a tres días. También las plántulas de estas dos especies son muy
sensibles. La inundación, el viento, las olas, temperatura elevadas y
sequedad del suelo son los mayores obstáculos para un establecimiento
persistente. Las diásporas y plántulas de Avicennia son destruidas además
con frecuencia por plagas fitófagas.
No obstante, la mayor parte de las dificultades que se plantean a la
forestación puede soslayarse tomando medidas apropiadas. Muros de adobe
o piedra protegen del viento y de las olas, impidiendo el arrastre de las

Resumen
VI
diásporas. Zanjas llenas de agua y sombra protegen las diásporas y las
plántulas del resecamiento y el sobrecalentamiento, mientras que una
irradiación solar intensa combinada con un abastecimiento suficiente de
agua fomenta el desarrollo y la vitalidad de las tres especies. Con redes
contra insectos pueden retenerse las diásporas al mismo tiempo que se corta
el paso a los insectos fitófagos. Mientras las concentraciones de sal sean
bajas, el establecimiento de Rhizophora es, por la forma y el tamaño de las
diásporas, considerablemente más fácil que el de las otras dos especies,
siendo además menor la mortalidad. Esto hace que con frecuencia no sean
necesarias medidas adicionales para la plantación de Rhizophora.
En general, la siembra de diásporas es el método más sencillo y económico
de forestación, seguido del establecimiento de plántulas. Sin embargo, la
mortalidad de Avicennia y Laguncularia es muy alta utilizando estos dos
procedimientos. En el caso de la Rhizophora, el establecimiento de
diásporas proporciona resultados mucho mejores que la plantación de
plántulas, teniéndose en condiciones favorables una mortalidad reducida.
Plantas jóvenes de mayor tamaño de las tres especies estudiadas así como
estacas de Laguncularia resultan ser notablemente más robustas frente a
inundaciones, viento, olas y sequía, así como frente al ataque de fitófagos. Si
bien el cultivo de plantas y estacas es más laborioso y costoso, para algunas
zonas es recomendable la plantación de plantas grandes. Otro aspecto
positivo es que pueden florecer y fructificar en corto tiempo, iniciando así
una regeneración natural.
Como simiente es aconsejable recurrir a diásporas grandes, intactas y en lo
posible frescas, procedentes de árboles vigorosos. Todas las plantas
deberían cultivarse en condiciones análogas a las de las áreas a forestar, o al

Resumen
VII
menos deberían aclimatarse, antes de su plantación en el campo, a las
condiciones extremas que puedan darse (altas temperaturas e irradiación
solar intensa). Además, al seleccionar el lugar de plantación ha de atenderse
a que las condiciones ambientales sean lo más favorables posibles en cada
caso. El final de la época de lluvias principal ha resultado ser el período más
conveniente para la plantación.

Abstract
The Ciénaga Grande de Santa Marta, a brackish lagoon system on the
Caribbean coast of Colombia, is the main fishing ground of the country.
Locally, it is of great ecological and economical importance. Human induced
changes in the hydrological system caused hypersalinisation of the soils. As
a result, about 60% of the mangroves growing in the lagoon’s flood plain
died, thereby decreasing the productivity of the ecosystem and endangering
or destroying the food source for both animals and humans in the area.
Presently, efforts are beeing made to reestablish the original hydrological
system. The objective of the present project is to identify the regeneration
patterns in the forests, to define the conditions for mangrove regeneration in
the Ciénaga Grande, and to ascertain whether regeneration of the large
unforested area is possible.
Experiments under controlled conditions, measurements in bare and forested
areas, as well as reforestation experiments showed, that regeneration is
possible. As the bare area is large (30,000 ha) and presently very few or no
propagules arrive naturally, the regeneration will be slow. Reforestation with
Avicennia germinans, Laguncularia racemosa and Rhizophora mangle, the
most important mangrove species of the area, can accelerate the process.
In the forests, the growth rate is mainly determined by salinity and the
mortality by seasonal flooding. Pollination and fruit production of
Laguncularia are ensured as well as the natural regeneration in the forests in
spite of high seedling mortalities.
In the bare areas, the regeneration is aggravated by high salinities and
temperatures as well as by the dessication of the top soil layers. The

Abstract
IX
establishment of the small Avicennia and Laguncularia propagules is
especially difficult. They cannot establish themselves in flooded soils, they
are carried away by the current, and in direct sunlight they dry out rapidly.
The seedlings of these two species are also very sensitive. Flooding, wind,
waves, high temperatures and dry soils all impact their survival. The
propagules and seedlings of Avicennia are frequently destroyed by
phytophagous insects. Several precautionary measures can alleviate these
problems. Clay or stone walls protect against wind, waves and secondary
dispersal; trenches and shade against drought and overheating. The
propagules can be held together and protected against phytophagous insects
by nets. However, mortality will remain high. Shape and size of the
propagules make the planting of Rhizophora easy. They can survive without
precautionary measures and the mortality is low under favorable
circumstances. The planting of propagules is much more successful than that
of seedlings. The main problem is high salinity. High solar radiation supports
growth and viability of all three species when sufficient water is present.
Sowing of propagules is the easiest and most economical way of
reforestation, followed by planting of seedlings. In both cases high
mortalities are to be expected. Bigger saplings of the three species and shoot
cuttings of Laguncularia proved to be less sensitive to flooding, wind,
waves, drought and phytophagous attack. Although it takes more effort and
is more expensive to grow saplings and shoot cuttings, it is nevertheless
recommended to plant them in some areas. They can flower and fruit within
a few months and then start a new natural regeneration.
Large, undamaged, fresh propagules from vital trees should be used as
seeds. All seedlings and saplings should be grown in conditions similar to
those in the area they will be planted in. At a minimum they must be adapted

Abstract
X
to high temperatures and solar radiation before transplanting to areas
suitable for mangrove growth. The most advantageous time for planting is
the end of the main rainy season.

1 Introducción
1.1 La zona objeto del estudio y sus particularidades
La Ciénaga Grande de Santa Marta, ubicada en la costa colombiana del
Caribe (Ilustración 1), es no sólo la laguna más grande del país, con una
extensión de 450 km², sino también la más productiva (Garcés & De La
Zerda 1994, PRO-CIÉNAGA 1993). Se encuentra en la parte oriental del
delta del Río Magdalena, atravesado por lagunas y canales o "caños". En la
zona aluvial de esta laguna de agua salobre crecían hasta hace unos 40 años
extensos manglares que cubrían una superficie superior a las 51.000 ha
(González 1991). En estos bosques se presentan en conjunto cinco especies
de mangle: Avicennia germinans (L.) Stearn, Laguncularia racemosa (L.)
Gaertn. f. y Rhizophora mangle L., que constituyen la práctica totalidad de
la población arbórea, mientras que de Conocarpus erectus L. y Acrostichum
aureum L. existen sólo pocos ejemplares. Originalmente, el ecosistema
proporcionaba hábitat y alimento a numerosas especies animales de peces,
reptiles y mamíferos, y su particular relevancia como lugar de estadía para
aves nativas y migratorias hizo que en 1964 se declarara una extensión de
21.000 ha "Parque Nacional Natural Isla de Salamanca" (Sánchez Páez
1988). Siguió en 1977 la creación del "Santuario de Fauna y Flora Ciénaga
Grande de Santa Marta" con 23.000 ha. La riqueza piscícola hizo del
sistema de lagunas el centro pesquero más importante de Colombia, y
25.000 familias viven en la actualidad de la pesca en aldeas ribereñas y
lacustres (Garcés & De La Zerda 1994).
La exportación de nutrientes desde la zona de manglares influye además de
modo importante en la riqueza piscícola de las zonas marítimas limítrofes.
Muchas especies piscícolas, tanto de agua dulce como de agua salada,
necesitan las áreas de agua salobre rodeadas de manglares como lugares de
Introducción
2
incubación o como hábitat transitorio. En consecuencia, también la pesca
marítima, económicamente importante, está íntimamente relacionada con la
Ciénaga Grande (Garcés & De La Zerda 1994, Sánchez Páez 1988).
Además de los peces, otro factor económico de gran importancia para la
población de los alrededores (Garcés & De La Zerda 1994) es la abundancia
de ostras (Crassostrea rhizophorae) y crustáceos de la Ciénaga Grande. La
madera de los mangles se corta también (con frecuencia ilegalmente en el
interior del parque natural) a fin de utilizarla para la construcción de casas,
como leña, etc.
Ilustración 1: La zona del proyecto en la costa colombiana del Caribe.

La Ciénaga Grande es una laguna costera ubicada en la desembocadura del
Río Magdalena, con una importancia ecológica y económica extraordinaria.
El funcionamiento del ecosistema depende en gran medida de la existencia
de manglares intactos.
1.2 Análisis del problema
A diferencia de los manglares costeros, el manglar de la zona objeto del
estudio no está caracterizado por los efectos de la bajamar y la pleamar, sino
por la alternancia de los periodos de lluvia y de sequía. Ni siquiera en la
zona norte de la Ciénaga Grande, donde la influencia de las mareas es más
marcada, la diferencia de nivel de las mareas supera los 30 cm, mientras que
el nivel de las aguas puede fluctuar en más de medio metro entre las épocas
de lluvia y las de sequía. En consecuencia, grandes zonas pobladas por
manglar están inundadas durante meses en época de lluvias, mientras que
más tarde están en seco durante muchas semanas. En los periodos secos, las
aguas freáticas salinas ascienden hasta la superficie del suelo debido a la
gran aridez, pues la evapotranspiración triplica aproximadamente las
precipitaciones anuales (INDERENA 1984). Debido a ello se forman
delgadas costras de sal en los charcos desecados. Las costras sólo se
disuelven de nuevo durante las épocas principales de lluvias, entre agosto y
noviembre, produciéndose una lixiviación de los suelos. El sistema de
lagunas estaría constantemente salinizado si no fuera por el agua dulce que
los afluentes aportan desde la Sierra Nevada de Santa Marta en el este y al
Río Magdalena por el oeste (PRO-CIÉNAGA 1993). Bajo estas
condiciones, la intervención humana en el sistema hidrológico de la Ciénaga
Grande acarreó una intensa salinización de amplias zonas del sistema de

Introducción
4
lagunas, debido a lo cual murieron grandes extensiones de manglar. Una de
las intervenciones más graves en el régimen hídrico la constituyó la
construcción de una carretera entre Ciénaga y Barranquilla. El trazado se
realizó en 1956 atravesando la estrecha lengua de tierra denominada "Isla de
Salamanca", que separa la Ciénaga Grande del mar y que ocho años más
tarde sería declarada Zona natural protegida (Ilustración 1). Si bien
originalmente se había proyectado la construcción de tajeas rectangulares
(box-culverts), se cegaron prácticamente todos los canales de enlace entre
las partes del sistema de lagunas situadas al norte y al sur de la carretera.
Las pocas tajeas realmente construidas no tenían dimensiones suficientes,
por lo que tanto el intercambio de agua como el drenaje del suelo quedaron
casi totalmente obstruidos (Sánchez Páez 1988). Como consecuencia de ello
se produjo muy pronto un desecamiento y una salinización local de los
suelos en las proximidades de la nueva carretera. La situación se vio
empeorada por la desecación parcial de las zonas pantanosas del noroeste,
emprendida con el propósito de utilizarlas para la agricultura y la ganadería.
Los aportes de agua dulce procedentes del Río Magdalena se vieron
interrumpidos debido a ello o a la intensa sedimentación del Río Magdalena
(Garcés & De La Zerda 1994, Sánchez Páez 1988). A partir de finales de
los años 60 comenzó así desde el noroeste la muerte de grandes extensiones
de manglares debido a la progresiva salinización, a la que entre tanto ha
sucumbido casi un 60 % del total del área boscosa (Ilustración 2).

Introducción
5
Ilustración 2: Manglares muertos en los alrededores del Caño Clarín. (Foto: J. Polanía)
Desde 1956 se perdieron así unas 580 ha anuales de bosque (entre 1956 y
1968: 175 ha/año y entre 1968 y 1987: 985 ha/año) (González 1991). Con
frecuencia se conservó como máximo una estrecha franja de manglar vivo en
las orillas de extensiones de agua importantes y canales menos salinizados
(Ilustración 2). Los árboles supervivientes son en su mayor parte Avicennia,
que es la especie de mangle más resistente a la sal de las que se presentan en
la zona objeto del estudio. También están amenazas de muerte grandes
partes de las 21.000 ha de bosque aún restantes. Esto significa la pérdida de
las bases vitales para los animales que viven en y del manglar, viéndose
afectados asimismo los ecosistemas vecinos. También la población que
habita en las aldeas de los alrededores se ve directamente afectada por la
enorme disminución de la riqueza piscícola de la Ciénaga
Grande (PRO-CIÉNAGA 1993).

Introducción
6
Intervenciones humanas en el sistema hidrológico de la Ciénaga Grande han
sido causa de una salinización extensiva de los suelos y de la muerte del
60% de los manglares. La productividad del ecosistema ha disminuido por
ello en gran medida y las bases vitales para animales y seres humanos están
amenazadas o han sido ya destruidas.
1.3 Entorno del proyecto
Ya en 1964 se detectó la incipiente muerte de manglares y pronto se
localizaron sus causas. La administración del parque INDERENA impulsó
en 1978 la limpieza y reapertura de canales y presentó el "Plan de
emergencia para la restauración del Parque Nacional Natural Isla de
Salamanca", en el que se ponían de relieve los problemas y las posibles
soluciones. Además se estimaron los costos de la construcción de canales y
se planificaron medidas de educación ambiental, investigación y protección
de la zona (INDERENA 1978). En los años siguientes, primero el
INDERENA, luego la DNP/CORPAMAG, la GTZ y otras organizaciones
comenzaron a estudiar con mayor detalle los problemas, sus causas y sus
repercusiones, así como las posibilidades de solucionarlos a través de la
construcción de canales (PRO-CIÉNAGA 1993).
Sólo diez años más tarde se iniciaron las obras hidráulicas. Una de las
primeras tajeas se construyó en 1989 en Barra Vieja (Sánchez Páez 1988),
siguiéndole después otras. En enero de 1996 quedó concluido el dragado del
caño Clarín (Ilustración 1), con lo que se restableció el primer canal entre el
Río Magdalena y la parte norte de la Ciénaga Grande. Más al sur
comenzaron los trabajos en otros dos canales de enlace, el Caño Aguas
Negras y el Caño Renegado-Condazo.

Introducción
7
El objetivo de las actividades es restablecer en la medida de lo posible las
condiciones hidrológicas originales del sistema de lagunas. A través del
incremento del aporte de agua dulce debe reducirse a largo plazo el
contenido de sal de los suelos, haciendo posible una regeneración del
manglar y, con ello, del conjunto del sistema (Elster et al., en impresión).
Colaboradores de la Corporación Autónoma Regional del Magdalena y de la
Sierra Nevada de Santa Marta (CORPAMAG) se encargan de supervisar las
obras en los canales así como de estudiar y solucionar los problemas
sociales que los daños causados al medio ambiente han originado en las
aldeas de pescadores. Son apoyados por la GTZ en el proyecto "PRO-
CIÉNAGA". Los cambios experimentados por el ecosistema son
documentados por el Instituto de Investigaciones Marinas (INVEMAR). El
Gobierno estudia, además, la realización de actividades de reforestación
extensiva a fin de repoblar las 30.000 ha afectadas hasta el momento por la
muerte de manglares.
La recopilación de datos correspondientes al presente proyecto tuvo lugar
entre julio de 1994 y julio de 1996. El INVEMAR proporcionó laboratorios
y embarcaciones para estos trabajos. Participó en el proyecto Laura Victoria
Perdomo, que aprovechó los datos por ella recopilados para redactar su tesis
de grado y que hasta hoy se ocupa de las pruebas de plantación. Una parte
de sus datos se ha utilizado en este informe.
Las causas de la muerte de los manglares se detectaron en fecha temprana,
pero sólo mucho más tarde se iniciaron medidas para su eliminación. En la
actualidad varias organizaciones cooperan en el restablecimiento de las
condiciones hidrológicas originales, y estudian los cambios ecológicos.

Introducción
8
1.4 Objetivos
Es probable que tampoco después de la apertura de los canales proyectados
se puedan restablecer por completo las condiciones hidrológicas originales
del sistema de lagunas. Pero ya una reducción de la concentración de sal a
valores aceptables para los manglares permite confiar en una regeneración
importante de la zona. El objetivo del presente trabajo es estudiar las
relaciones entre las condiciones ecológicas y el desarrollo de plántulas en la
zona de la Ciénaga Grande. Al respecto debe comprobarse si en principio es
posible una regeneración de las áreas no pobladas por el bosque. Además,
realizando pruebas de plantación debería responderse a la pregunta de si
conviene reforestar y, en caso afirmativo, de qué modo. Aparte de las
mediciones de los factores edáficos y del crecimiento y la mortalidad de
plántulas de mangle en la zona de la Ciénaga Grande, se han realizado
también estudios comparativos en el Parque Nacional Natural Tayrona,
ubicado unos 50 km al noreste de la Ciénaga Grande (Ilustración 1). En
algunas zonas costeras del Parque Tayrona se presentan grandes
concentraciones de sal, especialmente durante las épocas secas, y en las
hondonadas ubicadas detrás del dique costero se forman incluso extensas
calderas de sal, exentas de vegetación, que en parte se conservan incluso
durante los períodos de lluvias. Con frecuencia, estas calderas de sal están
rodeadas de manglares, y en las zonas vecinas crecen poblaciones vigorosas
de mangle. Por esta razón deberían realizarse pruebas de germinación y
establecimiento de plántulas en condiciones controladas, a fin de comprobar
si aquí se han originado ecotipos particularmente resistentes a la sal que
puedan tomarse en consideración para la regeneración de la Ciénaga
Grande. También se ha estudiado el comportamiento de las diásporas
germinantes frente a distintas concentraciones de sal y distintos niveles del
agua, a fin de descubrir qué especie resulta más apropiada para una

Introducción
9
repoblación en las condiciones propias de la zona objeto del estudio, a saber
la Avicennia germinans, resistente a la sal, o las otras dos especies también
frecuentes, la Laguncularia racemosa y la Rhizophora mangle.
Adicionalmente debería estudiarse con mayor detalle la biología
reproductiva de la Laguncularia, ya que hasta ahora se dispone de pocas
informaciones sobre esta especie.
Objetivo del presente proyecto era averiguar en qué condiciones puede
regenerarse el manglar y si es posible una regeneración de las grandes zonas
actualmente despobladas de bosque. Al mismo tiempo debería responderse
también a la pregunta de si es recomendable realizar actividades de
reforestación.

2 Resultados
A continuación se presentan los resultados más importantes en lo que atañe
a la regeneración del manglar. La descripción exacta de los métodos, la
evaluación estadística así como otros resultados detallados pueden
consultarse en la obra de Elster (1997).
2.1 Las zonas estudiadas
A. Bosques intactos, Ciénaga Grande: En las zonas estudiadas pobladas
por bosques (Ilustración 1), la producción de diásporas y el establecimiento
de plántulas alcanzaban índices elevados, garantizando una regeneración
natural del manglar. La diferencia principal entre las cuatro zonas está dada
por el grado de humedad del suelo, la cual depende, a su vez, de la
granulometría de los sedimentos limosos y del relieve del terreno. Las zonas
ubicadas a mayor altura sólo se inundaban durante la época de lluvias
principal, mientras que las hondonadas estaban inundadas durante casi todo
el año, al menos hasta el nivel de la superficie del suelo.
B. Zonas degradadas, Ciénaga Grande:
1. Barra Vieja: La primera zona degradada era una pequeña isla arenosa,
rodeada por una laguna poco profunda. En esta zona el manglar murió hace
unos 30 años debido a la construcción de una carretera. En 1989 se
restableció el enlace con la Ciénaga Grande al construir una tajea por debajo
de la carretera, con lo que comenzó una lenta regeneración natural. En la
época del estudio, aproximadamente un 15% de la isla estaba poblado de
nuevo por manglar, si bien los árboles más grandes alcanzaban una altura de
sólo 2 m; un 30% de la superficie de la isla estaba cubierto de plantas
halófitas de las especies Batis maritima L. y Sesuvium portulacastrum L..

Resultados
11
El resto estaba aún exento de vegetación, si bien el agua traía hasta aquí
muchas diásporas de Avicennia y, periódicamente, de Laguncularia y
Rhizophora.
2. Km 17: Los manglares ubicados en el kilómetro 17 del Caño Clarín, un
canal de enlace entre el Río Magdalena y la Ciénaga Grande, murieron hace
20 años, cuando los sedimentos acarreados por el Río Magdalena cegaron el
canal. Cuando en enero de 1996 se procedió a la reapertura del canal,
después de un largo periodo de obras, pudieron establecerse de nuevo
plántulas en estos suelos, gracias a la influencia del agua dulce. Sin
embargo, durante la época del estudio la mayor parte del área lodosa,
parcialmente inundada, presentaba sólo tocones de mangles muertos. A
orillas del canal crecían todavía árboles adultos de Avicennia y
Laguncularia, que alcanzaban una altura máxima de 4 m, y en algunos
puntos se encontraban Batis y Sesuvium.
C. Ensenadas del Parque Tayrona: En las dos zonas boscosas estudiadas
en este parque nacional (Ilustración 1), los árboles adultos producían
cantidades suficientes de diásporas, pero el desarrollo de las plántulas se
veía impedido con frecuencia por la sequía. La sequedad era intensificada
por el alto contenido de arena de los suelos, y sólo directamente en las
orillas había siempre suficiente agua para un establecimiento persistente de
las plántulas.
2.2 Pruebas de establecimiento de plántulas en
condiciones controladas
A fin de poder determinar cuantitativa y cualitativamente la influencia de
diversas condiciones ambientales, se realizaron pruebas en condiciones

Resultados
12
controladas. El objetivo de ello era comprender mejor la interacción de los
distintos factores en el complejo ecosistema de la Ciénaga Grande. Todas
las pruebas se realizaron en macetas llenas de arena, que se regaron con
agua del mar. Si no se dan datos especiales, la concentración de sal era de
entre el 30 y el 32‰ aproximadamente (agua de mar diluida), y el nivel del
agua se situaba en la superficie del sustrato o como máximo 1 cm por
encima del mismo. Se registraron el crecimiento de las plántulas, la
mortalidad y la biomasa.
2.2.1 Tasas de germinación de diásporas de diferentes orígenes
Se sembraron diásporas de diferentes zonas estudiadas del Parque Tayrona y
de la Ciénaga Grande en suelos con diferente concentración salina (0 a
150‰ ) y con distintos niveles de agua (5 cm por debajo de la superficie del
sustrato, en la superficie y 5 cm por debajo de la superficie del sustrato
(5 cm, 0 cm, +5 cm)), a fin de averiguar si en las proximidades de las
calderas de sal del Parque Tayrona se habían desarrollado ecotipos
particularmente resistentes a la sal. Si bien el desarrollo de las diásporas
procedentes de zonas distintas presentó diferencias significativas, no se
pudo comprobar la existencia de un ecotipo particularmente resistente a la
sal. En lugar de ello, y con independencia del origen geográfico, se
consiguieron las mejores tasas de germinación con diásporas grandes,
intactas, aun no germinadas y que se habían tomado directamente del árbol o
que habían estado sólo durante corto tiempo sobre el suelo. Las diásporas
que habían estado ya durante un tiempo prolongado sobre suelos secos o
que habían flotado en el agua resultaron tener menos vitalidad. Se
presentaron también elevados índices de mortalidad en el caso de diásporas
procedentes de árboles sometidos a un fuerte estrés. Posiblemente, las

Resultados
13
diferencias genéticas entre las poblaciones de manglares existentes desde el
punto de vista de su resistencia a la sal quedaron en un segundo plano en
comparación con la edad y las condiciones de almacenamiento de las
diásporas, así como con la vitalidad de los árboles madre, y son por lo tanto
irrelevantes para las medidas de regeneración.
Las tres especies investigadas presentaron diferentes resistencias a la sal. La
mayor resistencia la presentó la Avicennia. En esta especie se observó un
crecimiento normal, si bien más lento, de las plántulas incluso con un 60‰
de salinidad, mientras que en el caso de la Laguncularia y también de la
Rhizophora sólo pudo observarse crecimiento hasta un 40‰ . Con un 90‰
de salinidad ya no pudo establecerse ninguna plántula de Avicennia, lo que
en el caso de otras especies ocurría ya a partir de un 60‰ .
2.2.2 Influencia de ligeras diferencias en el nivel del agua y en la
concentración de la sal en el establecimiento y el crecimiento
en altura de las plántulas
Con independencia de su origen, las diásporas sembradas con diferentes
concentraciones de sal (16, 32, 40‰ ) y niveles del agua (-5 cm, 0 cm,
+5 cm) presentaron claros puntos óptimos de desarrollo. Todas las especies
crecieron tanto más rápidamente cuánto más baja era la concentración salina
del medio (Ilustración 3). La influencia de la salinidad en la velocidad de
crecimiento se puso de relieve con mayor intensidad en el caso de la
Rhizophora y con menor intensidad en el caso de la Avicennia.
Resultados
14
02468
101214161820
40‰-5cm
32‰-5cm
16‰-5cm
40‰0cm
32‰0cm
16‰0cm
40‰+5cm
32‰+5cm
16‰+5cm
[Cre
cim
ient
o en
altu
ra]
Laguncularia Avicennia Rhizophora
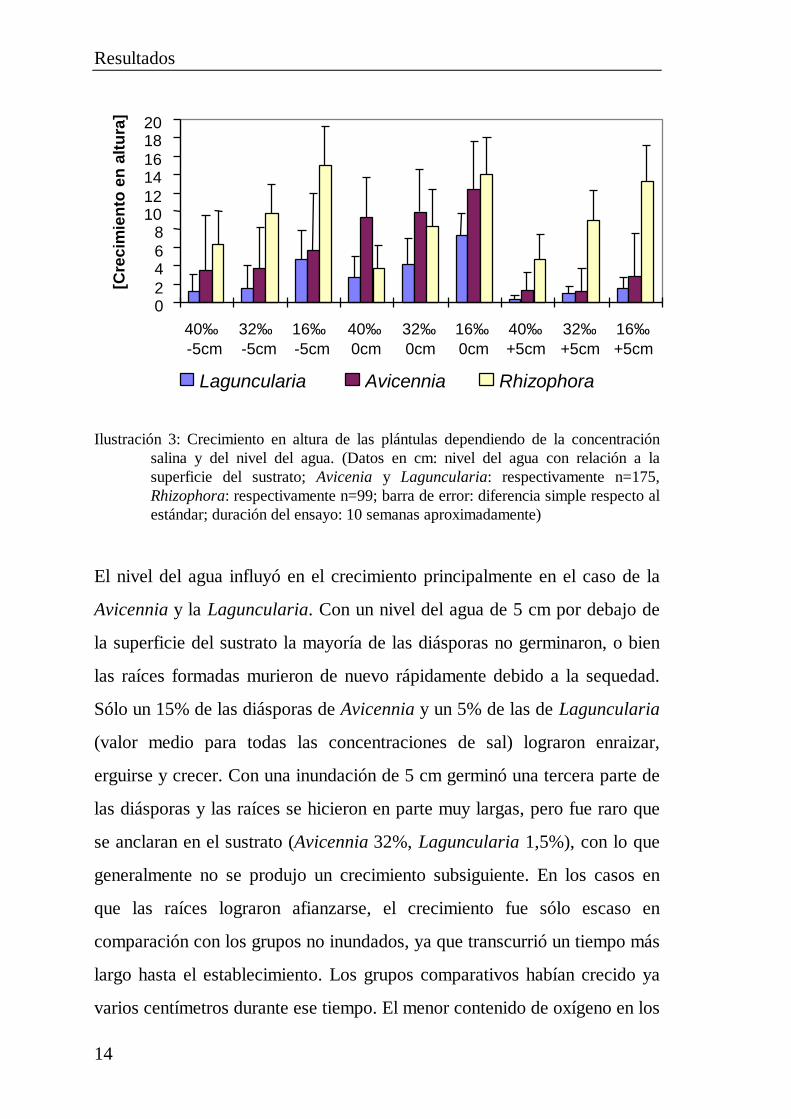
Ilustración 3: Crecimiento en altura de las plántulas dependiendo de la concentraciónsalina y del nivel del agua. (Datos en cm: nivel del agua con relación a lasuperficie del sustrato; Avicenia y Laguncularia: respectivamente n=175,Rhizophora: respectivamente n=99; barra de error: diferencia simple respecto alestándar; duración del ensayo: 10 semanas aproximadamente)
El nivel del agua influyó en el crecimiento principalmente en el caso de la
Avicennia y la Laguncularia. Con un nivel del agua de 5 cm por debajo de
la superficie del sustrato la mayoría de las diásporas no germinaron, o bien
las raíces formadas murieron de nuevo rápidamente debido a la sequedad.
Sólo un 15% de las diásporas de Avicennia y un 5% de las de Laguncularia
(valor medio para todas las concentraciones de sal) lograron enraizar,
erguirse y crecer. Con una inundación de 5 cm germinó una tercera parte de
las diásporas y las raíces se hicieron en parte muy largas, pero fue raro que
se anclaran en el sustrato (Avicennia 32%, Laguncularia 1,5%), con lo que
generalmente no se produjo un crecimiento subsiguiente. En los casos en
que las raíces lograron afianzarse, el crecimiento fue sólo escaso en
comparación con los grupos no inundados, ya que transcurrió un tiempo más
largo hasta el establecimiento. Los grupos comparativos habían crecido ya
varios centímetros durante ese tiempo. El menor contenido de oxígeno en los

Resultados
15
suelos constantemente inundados puede considerarse otro factor que impide
el crecimiento. Los efectos negativos de los niveles de agua bajos o altos se
vieron potenciados por concentraciones salinas elevadas (Ilustración 3). Por
el contrario, el crecimiento de las plántulas de Rhizophora estuvo poco
influenciado por el nivel del agua. Suelos relativamente secos y bien
ventilados resultaron ser los más favorables para esta especie. En las
primeras cuatro a seis semanas, la germinación y el crecimiento se
desarrollaron con mayor rapidez en el caso de la Avicennia, mientras que la
Laguncularia se estableció con la mayor lentitud. En las semanas siguientes,
así como con niveles de agua desfavorables, el crecimiento de la Avicennia
se retrasó con respecto al de la Rhizophora.
2.2.3 Influencia de ligeras diferencias del nivel del agua y de la
concentración de la sal en la mortalidad de las diásporas
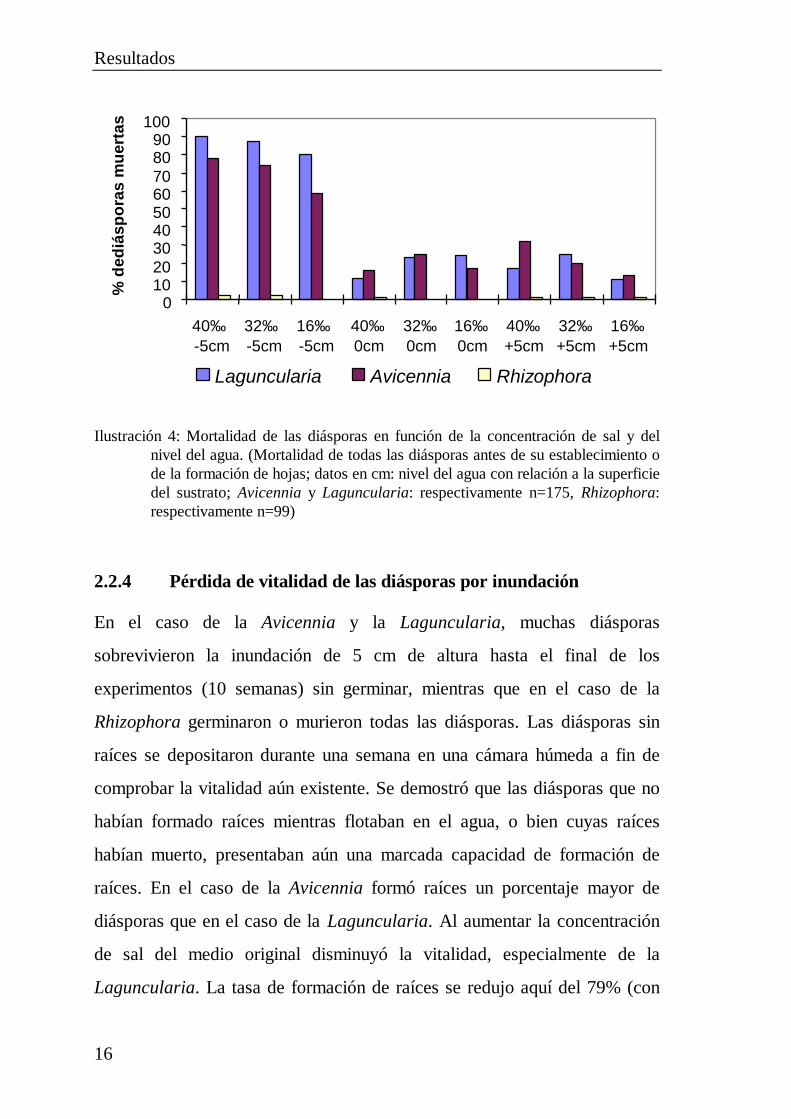
En el caso de la Rhizophora murieron pocas diásporas. La mortalidad
aumentó ligeramente al incrementarse las concentraciones de sal (del 16 al
40‰ ), no viéndose influenciada apenas por el nivel del agua. La mortalidad
fue considerablemente superior en el caso de la Laguncularia y la
Avicennia. La tasa de mortalidad dependió visiblemente tanto de la
concentración de sal como del nivel del agua. Una baja concentración de sal
y un nivel suficiente de agua resultaron favorables para la supervivencia. Las
mayores tasas de mortalidad de ambas especies se observaron con un nivel
del agua situado 5 cm por debajo de la superficie del sustrato. En este caso,
el nivel del agua resultó ser más importante que la concentración de sal para
la supervivencia (Ilustración 4).
Resultados
16
0102030405060708090
100
40‰-5cm
32‰-5cm
16‰-5cm
40‰0cm
32‰0cm
16‰0cm
40‰+5cm
32‰+5cm
16‰+5cm
% d
ediá
spor
as m
uert
as
Laguncularia Avicennia Rhizophora
Ilustración 4: Mortalidad de las diásporas en función de la concentración de sal y delnivel del agua. (Mortalidad de todas las diásporas antes de su establecimiento ode la formación de hojas; datos en cm: nivel del agua con relación a la superficiedel sustrato; Avicennia y Laguncularia: respectivamente n=175, Rhizophora:respectivamente n=99)
2.2.4 Pérdida de vitalidad de las diásporas por inundación
En el caso de la Avicennia y la Laguncularia, muchas diásporas
sobrevivieron la inundación de 5 cm de altura hasta el final de los
experimentos (10 semanas) sin germinar, mientras que en el caso de la
Rhizophora germinaron o murieron todas las diásporas. Las diásporas sin
raíces se depositaron durante una semana en una cámara húmeda a fin de
comprobar la vitalidad aún existente. Se demostró que las diásporas que no
habían formado raíces mientras flotaban en el agua, o bien cuyas raíces
habían muerto, presentaban aún una marcada capacidad de formación de
raíces. En el caso de la Avicennia formó raíces un porcentaje mayor de
diásporas que en el caso de la Laguncularia. Al aumentar la concentración
de sal del medio original disminuyó la vitalidad, especialmente de la
Laguncularia. La tasa de formación de raíces se redujo aquí del 79% (con

Resultados
17
una concentración de sal del 16‰ ) al 26% (con una concentración de sal del
40‰ ). En el caso de la Avicennia, la diferencia fue sólo del 6% para una
tasa de formación de raíces del 89% en una concentración de sal del 16‰ .
2.2.5 Pérdida de vitalidad de las diásporas en concentraciones de
sal elevadas
Si bien en el caso de concentraciones de sal muy elevadas (90‰ , 120‰ y
150‰ ) se produjo con frecuencia una formación de raíces de las tres
especies de mangles o un desarrollo de los hipocotiledones de las diásporas,
no llegaron a formarse plántulas. En lugar de ello murió más tarde la
práctica totalidad de la siembra. Este proceso se desarrolló más rápidamente
con salinidades elevadas que con salinidades bajas. Las diásporas que aún
no habían muerto inequívocamente después de dos meses y medio se
volvieron a sembrar con una concentración de sal del 10%. Mientras que en
el caso de la Laguncularia no pudo iniciarse con ello el desarrollo y las
diásporas habían muerto ya ostensiblemente en un 100%, en el caso de la
Avicennia y la Rhizophora pudieron desarrollarse algunas diásporas hasta el
estado de plántulas normales, o al menos se formaron aún raíces antes de su
muerte definitiva. Un 3% de las diásporas de Rhizophora e incluso hasta un
12% de las diásporas de Avicennia alcanzaron el estado de plántulas (tabla.
1).
Laguncularia Avicennia RhizophoraSalinidad alcomienzo
Mortalidadal comienzo
Establecidasmás tarde
Mortalidadal comienzo
Establecidasmás tarde
Mortalidadal comienzo
Establecidasmás tarde

Resultados
18
90‰ 100%? 0% 73% 9% 17% 3%120‰ 100%? 0% 81% 12% 21% 0,5%150‰ 100%? 0% 97% 0% 26% 0%
Tab. 1: Mortalidad de diásporas con salinidades elevadas y establecimiento posteriorcon salinidad del 10‰ . (Plazo: diez semanas; Avicennia y Laguncularia:respectivamente n=264, Rhizophora: respectivamente n=176; ?= la mortalidadno se pudo comprobar con seguridad).
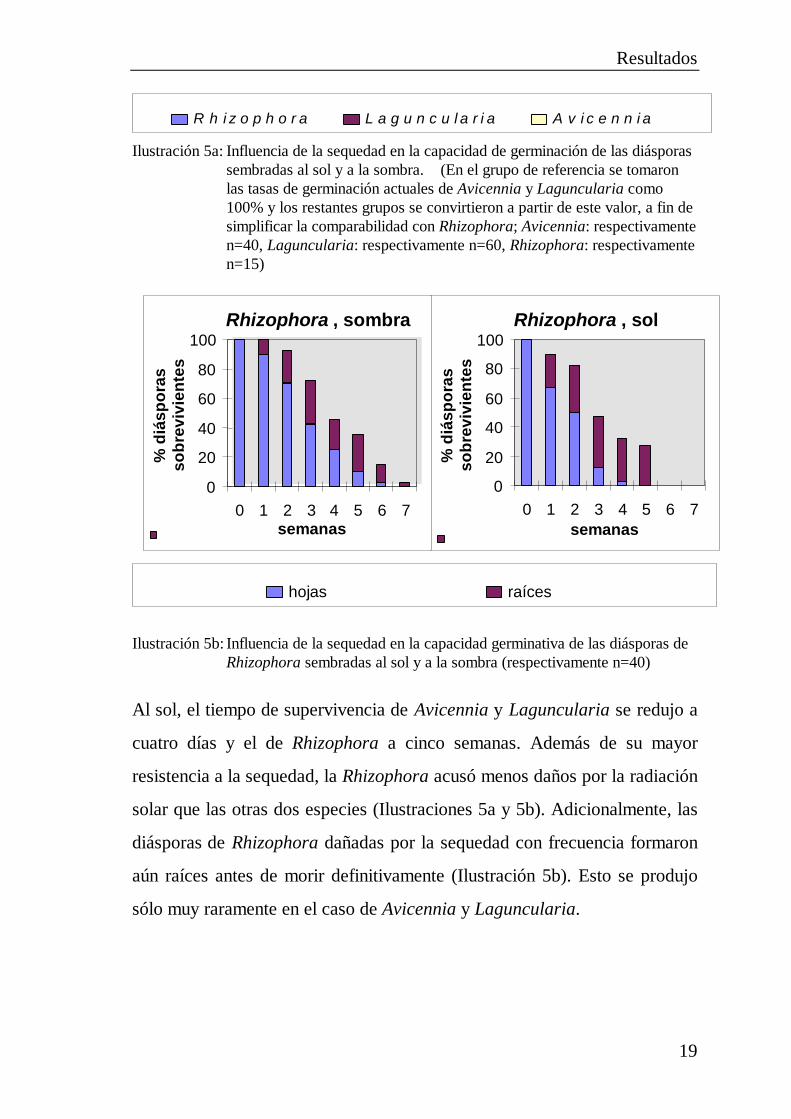
2.2.6 Influencia de la sequedad y de la irradiación solar en la
vitalidad de las diásporas
A fin de comprobar la influencia de la sequedad y la irradiación solar en la
vitalidad, se sembraron diásporas sobre arena seca, protegiendo una parte de
ellas con sombra y dejando las restantes expuestas a los rayos solares
directos. A continuación se regaron las diásporas. La capacidad de
germinación de las diásporas de Laguncularia y Avicennia sembradas sobre
sustrato seco con sombra se había perdido totalmente después de siete a
ocho días. En el caso de las diásporas de Rhizophora, de tamaño mucho
mayor, la capacidad de germinación no se perdió por completo hasta
después de pasar siete semanas (Ilustraciones 5a y 5b).
sol
0102030405060708090
100
0 1 2 3 4 5 6 7días
% d
iásp
oras
ger
min
adas
sombra
0102030405060708090
100
0 1 2 3 4 5 6 7días
% d
iásp
oras
ger
min
adas
Resultados
19
R h i z o p h o r a L a g u n c u l a r i a A v i c e n n i a
Ilustración 5a: Influencia de la sequedad en la capacidad de germinación de las diásporassembradas al sol y a la sombra. (En el grupo de referencia se tomaronlas tasas de germinación actuales de Avicennia y Laguncularia como100% y los restantes grupos se convirtieron a partir de este valor, a fin desimplificar la comparabilidad con Rhizophora; Avicennia: respectivamenten=40, Laguncularia: respectivamente n=60, Rhizophora: respectivamenten=15)
hojas raíces
Ilustración 5b: Influencia de la sequedad en la capacidad germinativa de las diásporas deRhizophora sembradas al sol y a la sombra (respectivamente n=40)
Al sol, el tiempo de supervivencia de Avicennia y Laguncularia se redujo a
cuatro días y el de Rhizophora a cinco semanas. Además de su mayor
resistencia a la sequedad, la Rhizophora acusó menos daños por la radiación
solar que las otras dos especies (Ilustraciones 5a y 5b). Adicionalmente, las
diásporas de Rhizophora dañadas por la sequedad con frecuencia formaron
aún raíces antes de morir definitivamente (Ilustración 5b). Esto se produjo
sólo muy raramente en el caso de Avicennia y Laguncularia.
Rhizophora , sol
0
20
40
60
80
100
0 1 2 3 4 5 6 7semanas
% d
iásp
oras
sobr
eviv
ient
es
Rhizophora , sombra
0
20
40
60
80
100
0 1 2 3 4 5 6 7semanas
% d
iásp
oras
sobr
eviv
ient
es

Resultados
20
2.2.7 Influencia de las condiciones de iluminación con un buen
suministro de agua
Disponiendo de un buen suministro de agua, la irradiación solar directa tuvo
repercusiones positivas en el desarrollo de las plántulas. Las diásporas de
todas las especies germinaron mejor y las plántulas se desarrollaron más
rápidamente y se hicieron más vigorosas. Además, en el caso de Avicennia y
Laguncularia disminuyó el índice de mortalidad. La influencia del sol
resultó ser más marcada en el caso de la Laguncularia. Ninguna de las
diásporas de esta especie protegidas por sombra (5 -10% de la intensidad
solar) pudo establecerse y formar hojas en el plazo de las primeras doce
semanas. La tabla 2 ofrece una visión de conjunto de las tasas de
establecimiento al final del experimento.
Laguncularia Avicennia Rhizophora
Sol 57% 71% 100%Sombra 0% 63% 21%
Tab. 2: Formación porcentual de hojas de diásporas después de doce semanas a lasombra (5-10% de intensidad solar) y expuestas a los rayos solares directos.(Laguncularia y Avicennia: respectivamente n=150, Rhizophora:respectivamente n=50)
2.2.8 Relación entre el peso y el desarrollo de las diásporas
Las diásporas pesadas de Avicennia y Rhizophora germinaron y crecieron
más rápidamente y murieron en un porcentaje menor que las livianas. La
correlación existente entre el peso y la mortalidad de las diásporas presentó
un carácter muy marcado en ambas especies, pero en conjunto la mortalidad
de la Avicennia fue considerablemente superior al de Rhizophora: en el caso
de Avicennia murió un 45% del grupo más liviano (Rhizophora: 3%) y un
29% del grupo más pesado (Rhizophora: 0%). En el caso de la
Laguncularia no se detectó ninguna correlación entre el peso y la
Resultados
21
mortalidad. Aquí se dieron las condiciones más favorables en el segundo
grupo más pesado (36% de mortalidad), mientras que la máxima mortalidad
se alcanzó en el grupo más pesado (45%).
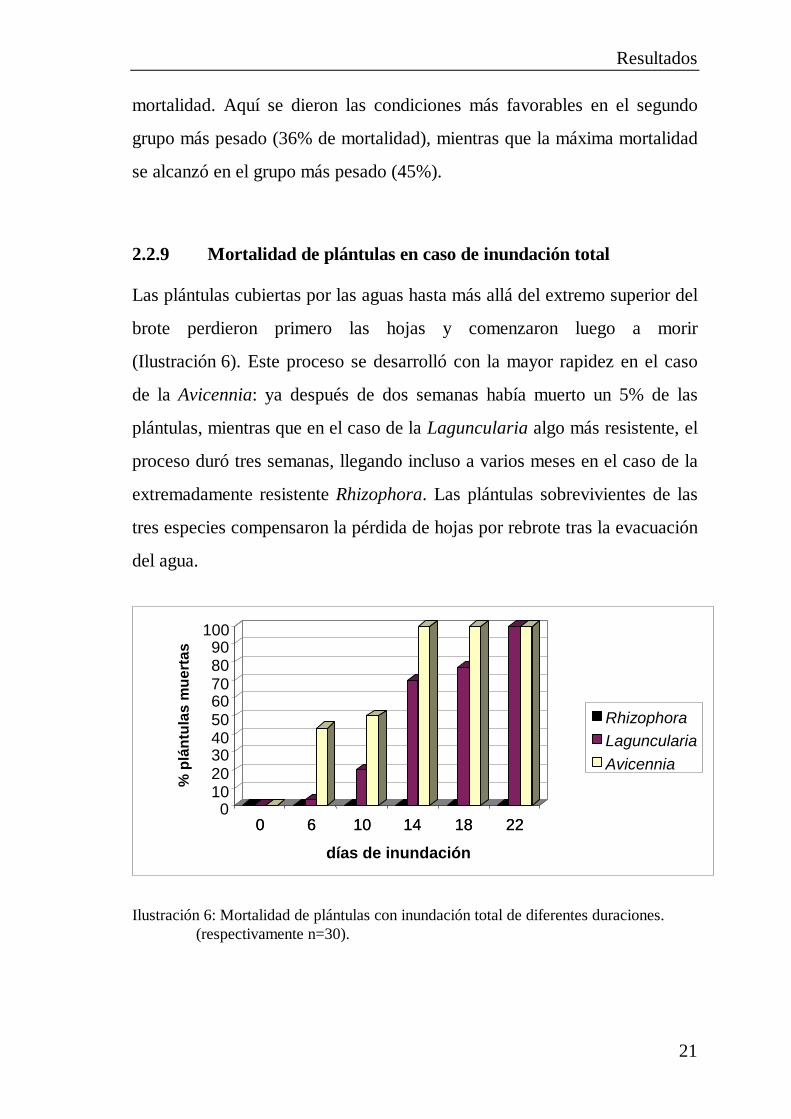
2.2.9 Mortalidad de plántulas en caso de inundación total
Las plántulas cubiertas por las aguas hasta más allá del extremo superior del
brote perdieron primero las hojas y comenzaron luego a morir
(Ilustración 6). Este proceso se desarrolló con la mayor rapidez en el caso
de la Avicennia: ya después de dos semanas había muerto un 5% de las
plántulas, mientras que en el caso de la Laguncularia algo más resistente, el
proceso duró tres semanas, llegando incluso a varios meses en el caso de la
extremadamente resistente Rhizophora. Las plántulas sobrevivientes de las
tres especies compensaron la pérdida de hojas por rebrote tras la evacuación
del agua.
0 6 10 14 18 220
102030405060708090
100
% p
lánt
ulas
mue
rtas
0 6 10 14 18 22
días de inundación
RhizophoraLagunculariaAvicennia
Ilustración 6: Mortalidad de plántulas con inundación total de diferentes duraciones. (respectivamente n=30).

Resultados
22
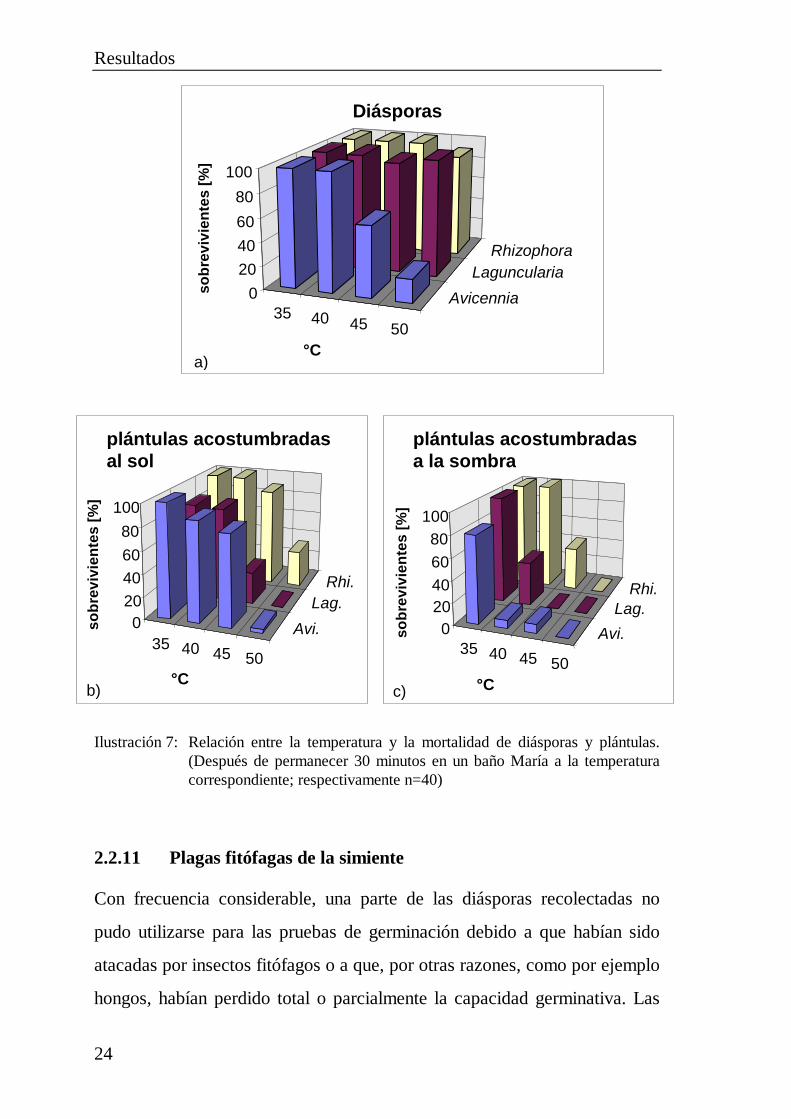
2.2.10 Resistencia de diásporas y plántulas a la temperatura
Para experimentar la resistencia a la temperatura se depositaron plántulas
cultivadas al sol o a la sombra durante 30 minutos con sus raíces en un baño
María a diferentes temperaturas, mientras que también se pusieron al baño
María diásporas durante el mismo tiempo. Luego se plantaron en macetas
llenas de arena y protegidas por sombra. Las diásporas demostraron ser muy
insensibles a las altas temperaturas. Ni siquiera una temperatura de 50°C
produjo una pérdida apreciable de vitalidad en el caso de la Laguncularia y
la Rhizophora, mientras que las diásporas de Avicennia germinaron aún en
un 20% a esa temperatura (Ilustración 7a). Dado que los suelos de las áreas
despobladas de bosque se calientan como máximo a un poco más de 50°C,
es posible una regeneración de estas superficies con diásporas, a condición
de que el sustrato no se reseque adicionalmente.
Las plántulas cultivadas al sol y acostumbradas a temperaturas elevadas
reaccionaron en forma algo más sensible al calor que las diásporas
(Ilustración 7b). Hasta 45°C, la tasa de supervivencia de las plántulas sólo
se vio claramente limitada en el caso de la Laguncularia (28%). Sin
embargo, la temperatura de 45°C no se alcanzó en el campo a la
profundidad normal de las raíces (aprox. 5 cm) ni siquiera en el caso de
suelos desnudos de vegetación. En consecuencia puede decirse que si bien
la regeneración natural se ve dificultada por temperaturas elevadas, sigue
siendo posible mientras se disponga de agua suficiente.
Por el contrario, las plántulas cultivadas a la sombra reaccionaron en forma
relativamente sensible frente al calor (Ilustración 7c). Ya a una temperatura
de 40°C se vio muy limitada la vitalidad de las plántulas de Laguncularia y
Avicennia. La Rhizophora presentó una menor sensibilidad, pero también en
el caso de esta especie sobrevivió a la temperatura de 45°C únicamente el

Resultados
23
40% de las plántulas cultivadas a la sombra. Por lo tanto, las plántulas
cultivadas a la sombra no son apropiadas para la plantación en los suelos
desnudos de vegetación y muy recalentados de la Ciénaga Grande (cap.
2.3.1 Factores ambientales).
Resultados
24
Ilustración 7: Relación entre la temperatura y la mortalidad de diásporas y plántulas.(Después de permanecer 30 minutos en un baño María a la temperaturacorrespondiente; respectivamente n=40)
2.2.11 Plagas fitófagas de la simiente
Con frecuencia considerable, una parte de las diásporas recolectadas no
pudo utilizarse para las pruebas de germinación debido a que habían sido
atacadas por insectos fitófagos o a que, por otras razones, como por ejemplo
hongos, habían perdido total o parcialmente la capacidad germinativa. Las
35 40 45 50
Avicennia
LagunculariaRhizophora
020406080
100
sobr
eviv
ient
es [%
]
°C
Diásporas
a)
35 40 45 50
Avi.Lag.
Rhi.
020406080
100
°Cc)
plántulas acostumbradasa la sombra
sobr
eviv
ient
es [%
]
35 40 45 50
Avi.
Lag.Rhi.
020406080
100
°C
plántulas acostumbradasal sol
b)
sobr
eviv
ient
es [%
]

Resultados
25
diásporas atacadas se separaron y se guardaron. Una parte de las diásporas
atacadas por insectos se sembró sobre arena y se regó con agua de mar, a fin
de comprobar la tasa de germinación.
En todas las parcelas estudiadas se detectó una gama de especies fitófagas
similar para las diásporas de todas las especies de mangle, pero se
apreciaron fluctuaciones en la tasa de infestación (véase la Ilustración 8).
Generalmente, el ataque de fitófagos fue mayor en el caso de la Avicennia
(16%). Las diásporas de esta especie fueron atacadas principalmente por
larvas de pirálidas, si bien con frecuencia conservaron la capacidad
germinativa. Por el contrario, en el caso de la Laguncularia, el ataque de
larvas de gelechiidas (12%, véase el cap. 2.4) produjo casi siempre la
muerte. Las diásporas de Rhizophora apenas resultaron atacadas por
fitófagos (4%) y su vitalidad se vio limitada escasamente por ello. Las
diásporas con una capacidad germinativa reducida por otros factores fueron
muy raras en el caso de la Laguncularia y Rhizophora, mientras que en el
caso de la Avicennia alcanzaron un 8%.
Ilustración 8: Ataque de fitófagos y diásporas con capacidad germinativa reducida porotras razones. (Area estudiada: La Rinconada (1994); Resto: diásporascon capacidad germinativa reducida; Avicennia: n= 1924, Laguncularia:n= 3447 y Rhizophora: n= 279)
Rhizophoramangle
fit.4%resto
1%
bien95%
Lagunculariaracemosa
fit.12%resto
0%
bien88%
Avicenniagerminans
resto8%
fit.16%
bien76%

Resultados
26
La disminución, con frecuencia considerable, de la tasa de germinación
debida al ataque de fitófagos en el caso de Laguncularia y Avicennia se vio
compensada, en condiciones naturales, por la elevada producción de
diásporas. Sin embargo, para la producción de plántulas en condiciones
controladas deberían desecharse las diásporas atacadas, ya que las larvas
atacan muchas diásporas consecutivamente, pudiendo causar así graves
daños.

Resultados
27
Ecotipos con diferentes resistencias a la sal no son de importancia decisiva
para la regeneración en la zona estudiada. En lugar de ello, en lo que se
refiere a las plantas debería atenderse a que se utilicen diásporas grandes e
intactas tomadas directamente de árboles vigorosos o que hayan
permanecido sobre el suelo sólo durante corto tiempo y en condiciones
favorables. La vitalidad de las diásporas de Avicennia y Laguncularia se
extingue, en caso de sequedad, en el plazo de pocos días, mientras que si
disponen de suficiente humedad pueden resistir durante semanas a pesar de
concentraciones salinas elevadas. La capacidad de germinación de ambas
especies disminuye en gran medida por el ataque de fitófagos. En el caso de
la Rhizophora son menos importantes las condiciones de almacenamiento.
Las diásporas resisten a la sequedad hasta siete semanas y son dañadas sólo
raramente por fitófagos. Las diásporas y plántulas acostumbradas al sol de
las tres especies de mangles son sorprendentemente resistentes a las altas
temperaturas.
En general, las condiciones más favorables para la germinación son
concentraciones bajas de sal y un nivel del agua a la altura de la superficie
del sustrato. El asentamiento de la Rhizophora depende en mayor medida de
la concentración de sal y el de Avicennia y Laguncularia del nivel del agua.
Las tasas de establecimiento de la Rhizophora son muy superiores a las de
las otras dos especies. Además, las plántulas son claramente más resistentes
a la inundación total. Las intensidades luminosas elevadas repercuten
positivamente en las plántulas de mangle si el suministro de agua es bueno.

Resultados
28
2.3 Desarrollo de la regeneración natural y evolución de
los factores edáficos a lo largo del año en el campo
Durante dos años se examinaron mensualmente los factores ecológicos y el
crecimiento en longitud y la mortalidad de plántulas recién restablecidas.
Los factores edáficos se midieron en lo posible a una profundidad de 5 cm
en el suelo, en las aguas subterráneas así como en las aguas superficiales,
mientras que la intensidad luminosa se midió directamente sobre las
plántulas.
En las parcelas estudiadas se documentaron generalmente los factores
siguientes entre las 9°° y las 11°° horas:
- Concentración de sal - Potencial Redox - pH
- Temperatura - Humedad del suelo - Nivel del agua
- Contenido de oxígeno - Intensidad luminosa
2.3.1 Factores ambientales
Nivel del agua y humedad del suelo
En la Ciénaga Grande de Santa Marta, el nivel del agua osciló en el
transcurso del año generalmente alrededor de aprox. 50 cm (Ilustración 9).
En dos parcelas estudiadas se midió incluso una diferencia de 80 cm entre la
época de lluvias y la época seca. El aumento del nivel del agua en el sistema
de lagunas fue causado no sólo por las precipitaciones locales, sino también
por la influencia del nivel del agua en los afluentes (lluvias en la Sierra
Nevada de Santa Marta y en el interior del país), y durante la temporada de
lluvias principal se produjeron con frecuencia inundaciones que duraron
cuatro o más meses. Las tierras más bajas se vieron inundadas también en

Resultados
29
parte dos veces al año, habiéndose medido niveles máximos del agua de 35
cm, mientras que las zonas situadas a mayor altura no resultaron inundadas,
o sólo en escasa medida.
La humedad se vio muy influenciada por el nivel del agua en suelos limosos,
oscilando generalmente entre el 40% y el 80%. Debido a la escasa
capacidad de retención de agua de los suelos arenosos, la humedad en las
zonas costeras, caracterizadas generalmente por altas concentraciones de
sal, se situó en valores claramente inferiores, del 15% al 35%, y las
fluctuaciones del nivel del agua repercutieron menos en la humedad del
suelo. Adicionalmente, la capacidad de retención de agua se vio disminuida
aquí también por el escaso contenido de humus, de manera que las capas
superiores del suelo de las zonas sin vegetación se resecaron rápida y casi
totalmente debido a la exposición total a los rayos solares. Las zonas sin
vegetación con sedimentos muy finos y con un alto porcentaje de material
orgánico se localizaron generalmente más hacia el interior. También aquí
disminuyó mucho el contenido de agua durante las épocas secas. A 5 cm de
profundidad alcanzó únicamente un 20% aproximadamente. Las capas
superiores del suelo se secaron por completo y las partículas especialmente
pequeñas fueron arrastradas por el viento, depositándose en forma de polvo
fino en las zonas protegidas del viento.
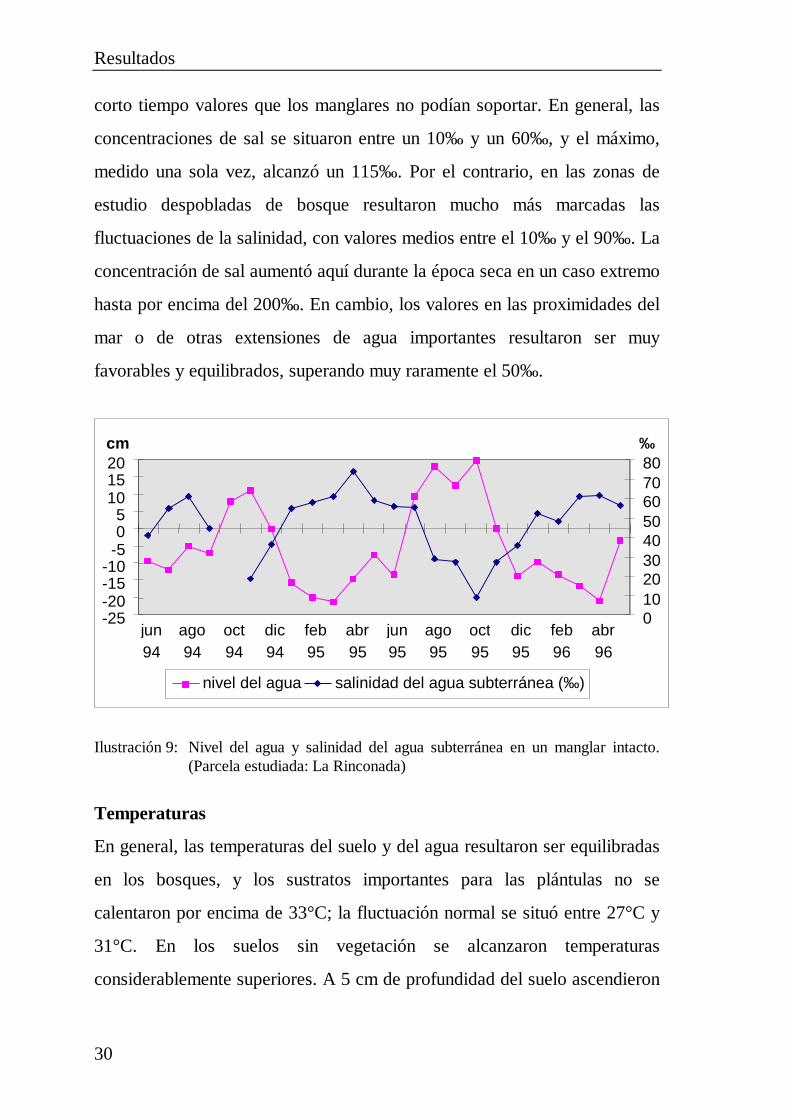
Concentraciones de sal
La disminución de las concentraciones de sal durante la temporada de
lluvias dependió principalmente de la inundación de los suelos, con la
consiguiente lixiviación de sales, y no del nivel de las aguas subterráneas. La
salinidad osciló relativamente poco en las áreas estudiadas pobladas por
bosques (Ilustración 9). Si bien aumentó especialmente en el agua del suelo
durante las épocas secas, incluso entonces alcanzó raramente y durante
Resultados
30
corto tiempo valores que los manglares no podían soportar. En general, las
concentraciones de sal se situaron entre un 10‰ y un 60‰ , y el máximo,
medido una sola vez, alcanzó un 115‰ . Por el contrario, en las zonas de
estudio despobladas de bosque resultaron mucho más marcadas las
fluctuaciones de la salinidad, con valores medios entre el 10‰ y el 90‰ . La
concentración de sal aumentó aquí durante la época seca en un caso extremo
hasta por encima del 200‰ . En cambio, los valores en las proximidades del
mar o de otras extensiones de agua importantes resultaron ser muy
favorables y equilibrados, superando muy raramente el 50‰ .
-25-20-15-10-505
101520
jun94 94
oct94
dic94
feb95
abr95
jun95 95
oct95
dic95
feb96
abr96
cm
01020304050607080‰
nivel del agua salinidad del agua subterránea (‰ )
ago ago
Ilustración 9: Nivel del agua y salinidad del agua subterránea en un manglar intacto.(Parcela estudiada: La Rinconada)
Temperaturas
En general, las temperaturas del suelo y del agua resultaron ser equilibradas
en los bosques, y los sustratos importantes para las plántulas no se
calentaron por encima de 33°C; la fluctuación normal se situó entre 27°C y
31°C. En los suelos sin vegetación se alcanzaron temperaturas
considerablemente superiores. A 5 cm de profundidad del suelo ascendieron

Resultados
31
hasta por encima de 41°C, y a 1 cm de profundidad se midieron incluso
temperaturas superiores a 50°C.
pH
Los suelos poblados de bosque presentaron generalmente un pH entre 6 y 7.
El agua subterránea fue normalmente algo más ácida, y el agua superficial
algo menos. Sin embargo se detectaron, especialmente en el suelo,
fluctuaciones muy irregulares, alcanzándose valores de pH desde 4 hasta 8.
En general, los valores de pH fueron más altos durante la temporada húmeda
del año que durante la temporada seca. Los suelos sin bosque presentaron
una reacción entre neutra y ligeramente alcalina, con pH entre 6,5 y 8.
Potenciales Redox
Los potenciales Redox medidos fluctuaron mucho en todas las parcelas
estudiadas. En general tenían valores negativos (entre -200 mV y -400 mV),
alcanzando valores extremos de -500 mV. Sólo como excepción se
registraron potenciales Redox superiores a 100 mV, y tan sólo en suelos no
inundados o en el agua superficial. Las diferencias entre los terrenos
poblados y despoblados de bosque fueron escasas.
Luz
La intensidad luminosa en el interior de los bosques fue en término medio de
sólo 11 Klux. Osciló entre valores extremos de 1,6 y 57 Klux, alcanzando
las manchas de luz un 99% de los valores que se presentaron a campo
abierto. Fuera del bosque, la intensidad luminosa media resultó ser unas
cinco veces superior, situándose en valores de 50 a 60 Klux.

Resultados
32
2.3.2 Desarrollo de las plántulas
Los factores ambientales influyeron mucho en el desarrollo de las plántulas,
sin bien debe decirse que ni en las zonas pobladas de bosque ni en las
despobladas pudo demostrarse una influencia de los valores pH y de los
potenciales Redox en las plántulas, no existiendo tampoco una relación entre
el contenido de oxígeno y el desarrollo de las plántulas.
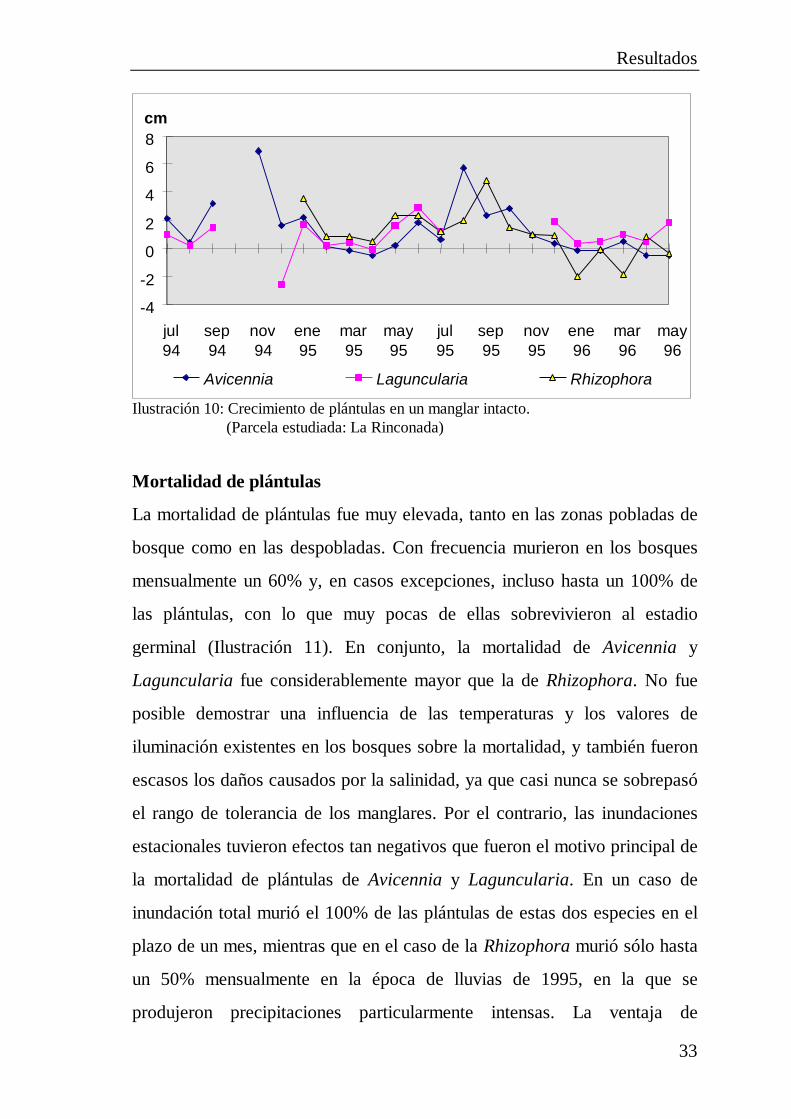
Crecimiento de las plántulas
El crecimiento de los mangles se ve favorecido por salinidades bajas, con lo
que las plántulas crecieron muy lentamente al comienzo de la temporada
seca y con la mayor rapidez al comienzo de la época de lluvias principal. En
los bosques, la altura media de las plántulas varió con ello entre -1 y
4 cm/mes (Ilustración 10).
Los impulsos de crecimiento más intensos, si bien sólo muy breves, se
registraron en el caso de la Laguncularia, con 12 cm por mes, mientras que
el máximo crecimiento en longitud de la Avicennia y la Rhizophora resultó
ser considerablemente menor (6 y respectivamente 5 cm/mes).
Especialmente en el caso de la Avicennia se observaron incluso valores de
crecimiento negativos, que alcanzaron hasta los -3 cm. Son atribuibles a la
muerte de las puntas de los brotes, p. ej. por resecamiento o al ser comidas
por insectos, y se presentaron ante todo hacia el final de la época de lluvias
o al comienzo de la temporada seca. Las muy breves fluctuaciones de la
intensidad luminosa causadas con frecuencia por manchas de luz tuvieron en
los bosques poca influencia en el crecimiento de las plántulas. Por el
contrario, en los terrenos despejados el crecimiento de las plántulas éste se
vio muy favorecido por la luz, de modo que en el caso de la Laguncularia y
la Avicennia se alcanzaron hasta 10 cm/mes a lo largo de cinco meses.
Resultados
33
-4
-2
0
2
4
6
8
jul94
sep94
nov94
ene95
mar95
may95
jul95
sep95
nov95
ene96
mar96
may96
cm
Avicennia Laguncularia Rhizophora
Ilustración 10: Crecimiento de plántulas en un manglar intacto.(Parcela estudiada: La Rinconada)
Mortalidad de plántulas
La mortalidad de plántulas fue muy elevada, tanto en las zonas pobladas de
bosque como en las despobladas. Con frecuencia murieron en los bosques
mensualmente un 60% y, en casos excepciones, incluso hasta un 100% de
las plántulas, con lo que muy pocas de ellas sobrevivieron al estadio
germinal (Ilustración 11). En conjunto, la mortalidad de Avicennia y
Laguncularia fue considerablemente mayor que la de Rhizophora. No fue
posible demostrar una influencia de las temperaturas y los valores de
iluminación existentes en los bosques sobre la mortalidad, y también fueron
escasos los daños causados por la salinidad, ya que casi nunca se sobrepasó
el rango de tolerancia de los manglares. Por el contrario, las inundaciones
estacionales tuvieron efectos tan negativos que fueron el motivo principal de
la mortalidad de plántulas de Avicennia y Laguncularia. En un caso de
inundación total murió el 100% de las plántulas de estas dos especies en el
plazo de un mes, mientras que en el caso de la Rhizophora murió sólo hasta
un 50% mensualmente en la época de lluvias de 1995, en la que se
produjeron precipitaciones particularmente intensas. La ventaja de

Resultados
34
Rhizophora residió por una parte en su mayor resistencia absoluta (cap.
2.2.9) y por otra parte en el tamaño de las diásporas, que ya al comienzo de
la germinación levantan la punta del brote por encima del nivel normal de las
aguas (cap. 2.5.1). Dado que las plántulas de Avicennia y Laguncularia
reaccionaron de forma tan sensible a las inundaciones, las zonas situadas a
mayor altura resultaron ser particularmente apropiadas para su
establecimiento, siempre y cuando no bajara excesivamente el nivel de las
aguas subterráneas. En el bosque, un descenso del nivel del agua superior a
63 cm por debajo de la superficie del suelo no incrementó sin embargo la
mortalidad de plántulas.
La muerte extensiva de todas las plántulas de Avicennia y Laguncularia así
como de muchas de Rhizophora, que se produjo periódicamente durante
épocas de fuertes precipitaciones, fue un proceso natural que se vio
compensado por una elevada producción de frutos. Al escurrirse las aguas al
comienzo de la época seca permanecieron sobre el terreno nuevas diásporas,
que germinaron. En muchas orillas se formaron entonces densos tapices de
plántulas, que sin embargo se aclararon ya después de corto tiempo debido
al ataque de insectos (cap. 2.5.5) o al oleaje. Sin embargo, la regeneración
natural del bosque deja de producirse con frecuencia durante varios años.
Por ejemplo, en manglares inundados estacionalmente, la regeneración está
garantizada sólo en años secos, mientras que en suelos secos sólo es posible
en años húmedos. Debido a esto se forman grupos de plantas de la misma
edad, faltando las de los años intermedios.
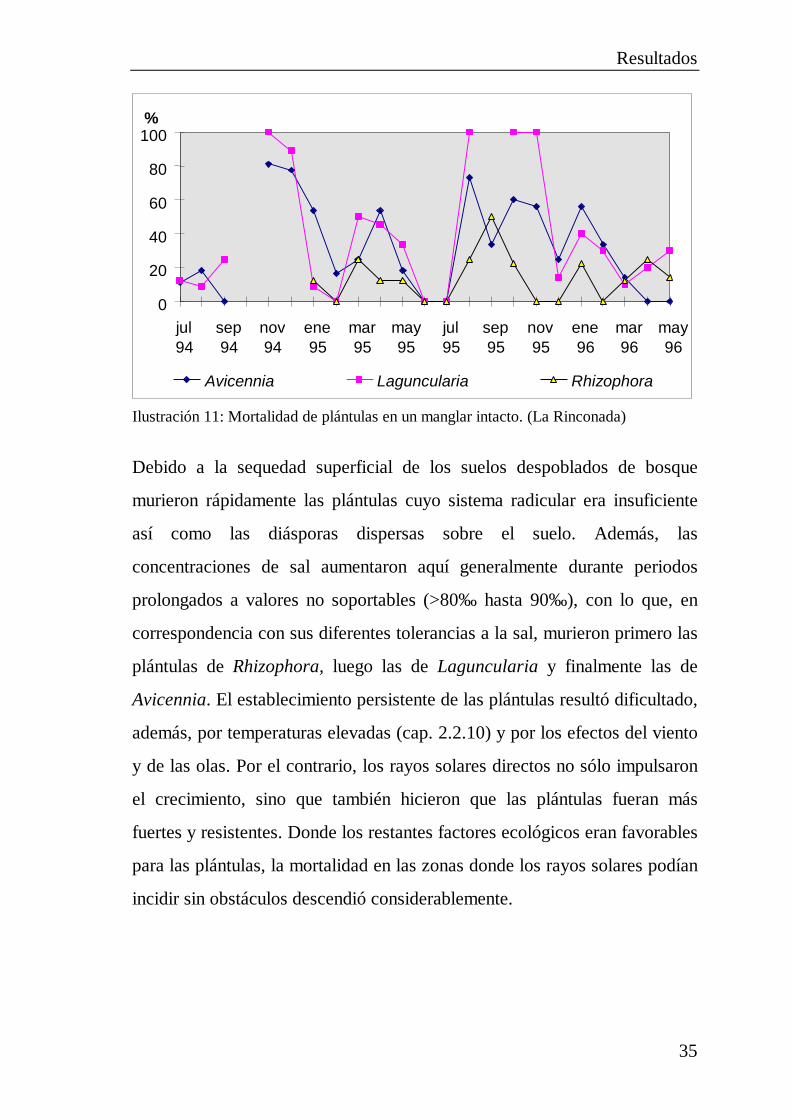
Resultados
35
0
20
40
60
80
100
jul94
sep94
nov94
ene95
mar95
may95
jul95
sep95
nov95
ene96
mar96
may96
%
Avicennia Laguncularia Rhizophora
Ilustración 11: Mortalidad de plántulas en un manglar intacto. (La Rinconada)
Debido a la sequedad superficial de los suelos despoblados de bosque
murieron rápidamente las plántulas cuyo sistema radicular era insuficiente
así como las diásporas dispersas sobre el suelo. Además, las
concentraciones de sal aumentaron aquí generalmente durante periodos
prolongados a valores no soportables (>80‰ hasta 90‰ ), con lo que, en
correspondencia con sus diferentes tolerancias a la sal, murieron primero las
plántulas de Rhizophora, luego las de Laguncularia y finalmente las de
Avicennia. El establecimiento persistente de las plántulas resultó dificultado,
además, por temperaturas elevadas (cap. 2.2.10) y por los efectos del viento
y de las olas. Por el contrario, los rayos solares directos no sólo impulsaron
el crecimiento, sino que también hicieron que las plántulas fueran más
fuertes y resistentes. Donde los restantes factores ecológicos eran favorables
para las plántulas, la mortalidad en las zonas donde los rayos solares podían
incidir sin obstáculos descendió considerablemente.
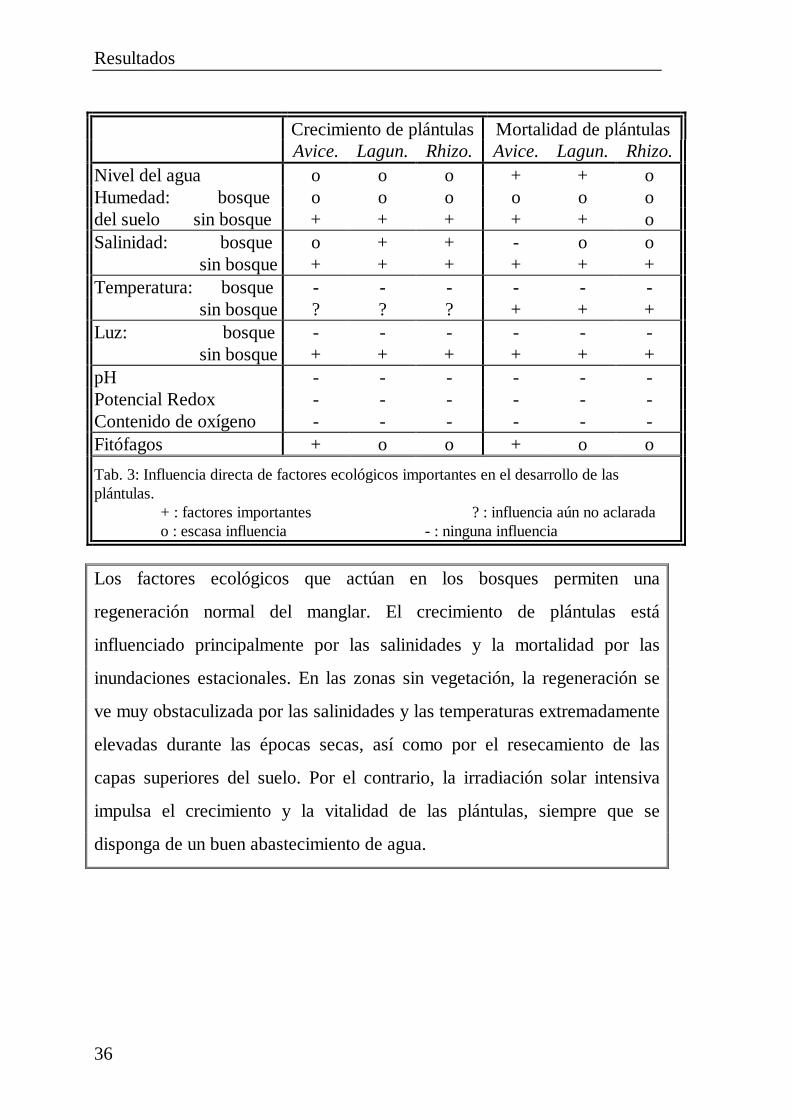
Resultados
36
Crecimiento de plántulas Mortalidad de plántulasAvice. Lagun. Rhizo. Avice. Lagun. Rhizo.
Nivel del agua o o o + + oHumedad: bosque o o o o o odel suelo sin bosque + + + + + oSalinidad: bosque o + + - o o sin bosque + + + + + +Temperatura: bosque - - - - - - sin bosque ? ? ? + + +Luz: bosque - - - - - - sin bosque + + + + + +pH - - - - - -Potencial Redox - - - - - -Contenido de oxígeno - - - - - -Fitófagos + o o + o o
Tab. 3: Influencia directa de factores ecológicos importantes en el desarrollo de lasplántulas. + : factores importantes ? : influencia aún no aclarada o : escasa influencia - : ninguna influencia
Los factores ecológicos que actúan en los bosques permiten una
regeneración normal del manglar. El crecimiento de plántulas está
influenciado principalmente por las salinidades y la mortalidad por las
inundaciones estacionales. En las zonas sin vegetación, la regeneración se
ve muy obstaculizada por las salinidades y las temperaturas extremadamente
elevadas durante las épocas secas, así como por el resecamiento de las
capas superiores del suelo. Por el contrario, la irradiación solar intensiva
impulsa el crecimiento y la vitalidad de las plántulas, siempre que se
disponga de un buen abastecimiento de agua.
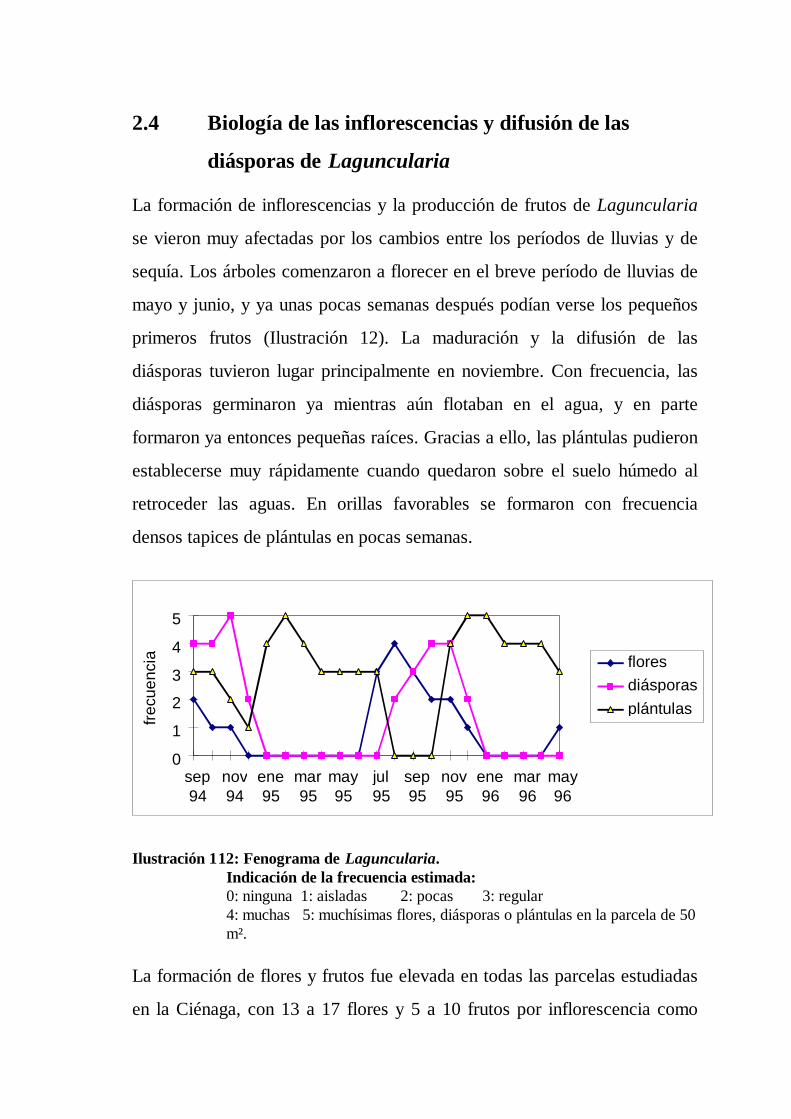
2.4 Biología de las inflorescencias y difusión de las
diásporas de Laguncularia
La formación de inflorescencias y la producción de frutos de Laguncularia
se vieron muy afectadas por los cambios entre los períodos de lluvias y de
sequía. Los árboles comenzaron a florecer en el breve período de lluvias de
mayo y junio, y ya unas pocas semanas después podían verse los pequeños
primeros frutos (Ilustración 12). La maduración y la difusión de las
diásporas tuvieron lugar principalmente en noviembre. Con frecuencia, las
diásporas germinaron ya mientras aún flotaban en el agua, y en parte
formaron ya entonces pequeñas raíces. Gracias a ello, las plántulas pudieron
establecerse muy rápidamente cuando quedaron sobre el suelo húmedo al
retroceder las aguas. En orillas favorables se formaron con frecuencia
densos tapices de plántulas en pocas semanas.
0
1
2
3
4
5
sep94
nov94
ene95
mar95
may95
jul95
sep95
nov95
ene96
mar96
may96
floresdiásporasplántulas
frecu
enci
a
Ilustración 112: Fenograma de Laguncularia.Indicación de la frecuencia estimada:0: ninguna 1: aisladas 2: pocas 3: regular4: muchas 5: muchísimas flores, diásporas o plántulas en la parcela de 50m².
La formación de flores y frutos fue elevada en todas las parcelas estudiadas
en la Ciénaga, con 13 a 17 flores y 5 a 10 frutos por inflorescencia como

Resultados
38
término medio. No se pudo demostrar una relación entre la formación de
flores y/o frutos y la concentración de sal en los suelos. Sólo en condiciones
extremas, como las que se dan en una zona muy seca y elevada del Parque
Tayrona, descendió notablemente el éxito de la reproducción. Aquí se
contaron por término medio sólo ocho flores y dos frutos por cada
inflorescencia, lo que es atribuible posiblemente al gran estrés causado por
la sequedad.
En conjunto, sólo un 37% de las flores llegó a formar diásporas maduras, un
30% cayó aún antes de la formación del fruto y otro 33% cayó durante la
fase de maduración. La razón principal de que abortaran los frutos
inmaduros pudo ser el ataque de larvas de polillas de la familia de las
gelechiidae. También un porcentaje muy elevado de las diásporas maduras
fue atacado con frecuencia por larvas de polillas, lo que impidió su
germinación (cap. 2.2.11). El intenso ataque de los insectos no es atribuible
probablemente al elevado grado de degradación de la Ciénaga Grande, ya
que también en otras zonas de manglares se describe el ataque de insectos
como una de las principales razones de la destrucción de diásporas (Clarke
1992, Robertson et al. 1990, Robertson 1991).
Si bien en la zona estudiada la producción de frutos, gracias a la elevada
intensidad de floración, fue suficiente para la regeneración natural del
bosque, no pudo aclararse, sin embargo, si permitirá también la repoblación
de las zonas exentas de bosques en un tiempo previsible. Además, la
difusión de las diásporas sólo estaba asegurada en las proximidades de
árboles con frutos y en zonas ribereñas favorables o en zonas de inundación,
mientras que a las zonas apartadas y sin vegetación no llegaba con
frecuencia ninguna diáspora. El principal motivo de ello fue que las
corrientes procedentes de bosques aptos para la reproducción aparecen muy
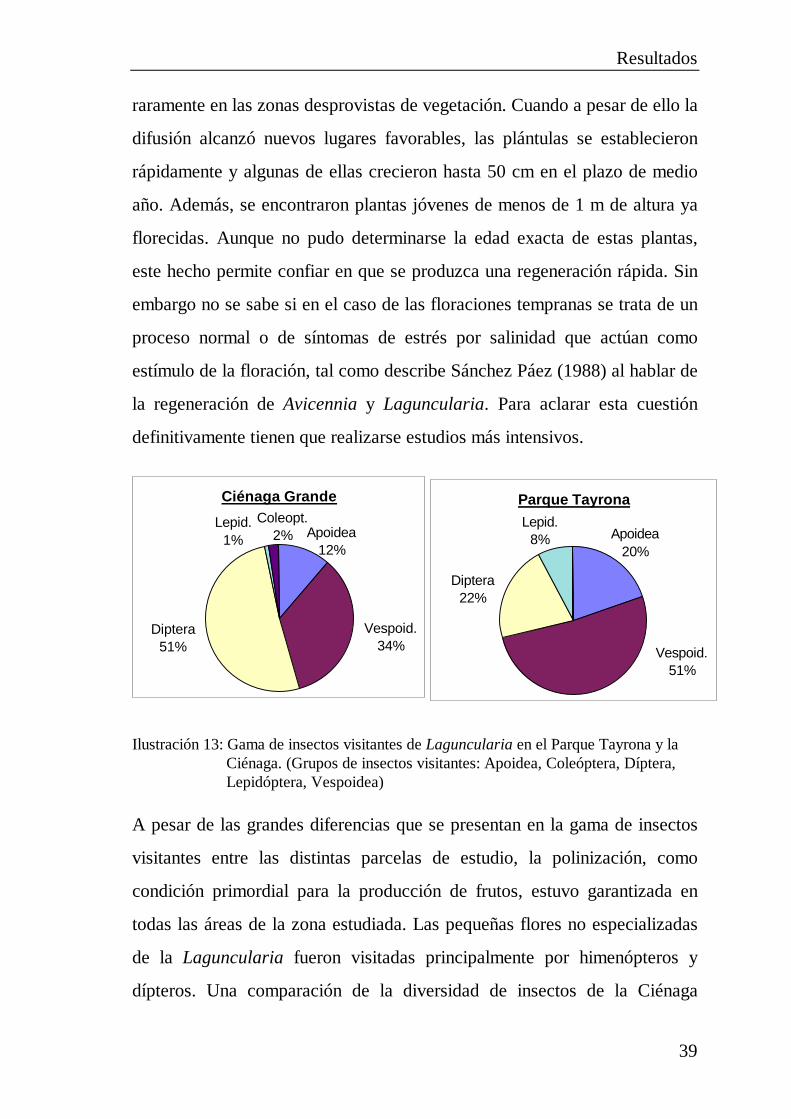
Resultados
39
raramente en las zonas desprovistas de vegetación. Cuando a pesar de ello la
difusión alcanzó nuevos lugares favorables, las plántulas se establecieron
rápidamente y algunas de ellas crecieron hasta 50 cm en el plazo de medio
año. Además, se encontraron plantas jóvenes de menos de 1 m de altura ya
florecidas. Aunque no pudo determinarse la edad exacta de estas plantas,
este hecho permite confiar en que se produzca una regeneración rápida. Sin
embargo no se sabe si en el caso de las floraciones tempranas se trata de un
proceso normal o de síntomas de estrés por salinidad que actúan como
estímulo de la floración, tal como describe Sánchez Páez (1988) al hablar de
la regeneración de Avicennia y Laguncularia. Para aclarar esta cuestión
definitivamente tienen que realizarse estudios más intensivos.
Ilustración 13: Gama de insectos visitantes de Laguncularia en el Parque Tayrona y laCiénaga. (Grupos de insectos visitantes: Apoidea, Coleóptera, Díptera,Lepidóptera, Vespoidea)
A pesar de las grandes diferencias que se presentan en la gama de insectos
visitantes entre las distintas parcelas de estudio, la polinización, como
condición primordial para la producción de frutos, estuvo garantizada en
todas las áreas de la zona estudiada. Las pequeñas flores no especializadas
de la Laguncularia fueron visitadas principalmente por himenópteros y
dípteros. Una comparación de la diversidad de insectos de la Ciénaga
Ciénaga Grande
Vespoid.34%
Diptera51%
Lepid.1%
Coleopt.2% Apoidea
12%
Parque Tayrona
Diptera22%
Apoidea20%
Lepid.8%
Vespoid.51%

Resultados
40
Grande con la del Parque Tayrona puso de relieve que el Parque Tayrona
presenta una mayor riqueza de especies, especialmente de himenópteros
(Ilustración 13) (Ciénaga: 14 especies visitantes, Parque Tayrona: 18
especies). Sin embargo, las flores fueron visitadas en la Ciénaga Grande con
mayor frecuencia que en el Parque Tayrona. Así, en la parcela con mayor
afluencia de insectos de la Ciénaga Grande, cada flor fue visitada por
término medio una vez cada horas y en el Parque Tayrona sólo cada 20
horas. Dado que no se detectaron diferencias significativa en la formación
de frutos de los arbustos de Laguncularia del Parque Tayrona y de la
Ciénaga Grande, la polinización estuvo garantizada en ambas zonas a pesar
de las grandes diferencias existentes en la variedad y en la frecuencia de los
insectos visitantes.
La Laguncularia florece con gran intensidad en todas las zonas estudiadas
de la Ciénaga Grande y la polinización, como condición primordial para la
- también elevada - producción de frutos, está garantizada en toda la zona
estudiada. A pesar de las elevadas pérdidas, el número de frutos formados
es suficiente para una regeneración natural del bosque, si bien debido a las
corrientes no se produce la difusión de diásporas a todas las áreas
despobladas de bosque.
2.5 Pruebas de plantación
Las pruebas de plantación se realizaron durante dos años en la Barra Vieja y
en el Km 17. Se establecieron cuatro grupos distintos de las tres especies de
mangles, y en cada grupo se estudiaron la mortalidad y el crecimiento
durante al menos un año. A continuación se tratan con detalle los distintos

Resultados
41
resultados sólo en casos de excepción, a fin de conservar la claridad del
estudio. Los resultados detallados los describe Elster (1997).
2.5.1 Plantación de diásporas
Sólo diásporas cuya capacidad de germinación pareció estar intacta (sin
ataque de fitófagos, sin infestación de hongos ni zonas dañadas, etc.) se
utilizaron como simiente. De las diásporas de Avicennia y Laguncularia
sembradas en 1994 en la Barra (160 de cada especie), sobrevivieron sin
embargo el primer mes sólo una plántula de Laguncularia y cinco de
Avicennia, debido a que las diásporas fueron arrastradas a las zonas de
plantación apenas inundadas y al margen de la laguna, y el sol las quemó en
zonas de plantación secas (Ilustración 14). Debido al tamaño y a la forma,
las diásporas de Rhizophora (Ilustración 14) pueden introducirse en la tierra,
lo que evita que se pierdan al ser arrastradas por las aguas. Gracias a ello se
establecieron sin problemas casi en un 100%. En 1995 se alcanzaron
mejores resultados con Avicennia y Laguncularia. En este año, el arrastre
de las diásporas se evitó formando pequeños diques y construyendo
armazones de tablas provistos de redes para insectos. Además pudo evitarse
el resecamiento de las diásporas sembrándolas en zanjas de plantación de
profundidad suficiente. En condiciones favorables se estableció así más del
60% de las diásporas de Laguncularia, mientras que en el caso de la
Avicennia se estableció más del 50%.

Resultados
42
Ilustración 14: Diásporas de Laguncularia, Avicennia (a, resecadas) y Rhizophora (b).
Algunas diásporas de las tres especies de mangles fueron dañadas por insectos
no identificados después de la siembra, siendo las pérdidas de Avicennia y
Laguncularia mucho más elevadas que las de Rhizophora. Además, las
diásporas y plántulas de Avicennia fueron destruidas con frecuencias por
orugas de lepidópteros (cap. 2.5.5), y los hipocotiledones de Rhizophora, con
frecuencia ya formados, fueron atacados en las proximidades del suelo, con
formación de agujeros. El tamaño de tales agujeros fue incluso tan grande, que
las plantas se tumbaron y murieron. Es posible que los ataques se debieran a
unas avispas de gran tamaño existente en la zona estudiada.
2.5.2 Cultivo de plántulas
Las plántulas destinadas a pruebas de plantación se cultivaron en parte al sol y
en parte a la sombra, con concentraciones de sal del 36‰ , hasta que
desarrollaron de una a tres parejas de hojas. Al plantar luego las plántulas en

Resultados
43
la época de lluvias se obtuvieron en un principio mejores resultados con
Rhizophora (del 40% al 70% sobrevivió el primer mes) que con las otras
dos especies. Las plántulas de Rhizophora demostraron ser menos sensibles
a las fluctuaciones del nivel del agua, al oleaje, al viento y a temperaturas
elevadas. Además no se observó que las hojas fueran comidas por insectos
fitófagos. Hacia el final de la época seca de 1995 se produjo sin embargo
una mortalidad masiva debido al aumento de la salinidad del suelo.
Las plántulas de Avicennia y Laguncularia plantadas también en la época de
lluvias reaccionaron a la plantación con un sensibilidad tal, que muchas de
las plántulas murieron ya en el primer mes. Las altas concentraciones de sal
relacionadas con la tardanza del inicio de la época de lluvias a finales de
1995 y la sequedad acarrearon más tarde la muerte de casi todas las
plántulas sobrevivientes. También las plántulas procedentes de la
regeneración natural en las proximidades de los lugares de plantación
murieron en ese período casi por completo. Sólo sobrevivieron al período
seco inusualmente prolongado plantas de más edad, con sistemas radiculares
extensos, así como plántulas ubicadas en las orillas de las lagunas, donde
disponían constantemente de agua fresca y no hipersalina. La muerte de una
gran parte de las plántulas durante y, especialmente, hacia el final de la
época seca fue un proceso natural en los suelos despoblados de bosque, ya
que aquí la regeneración se ve dificultada por los rayos solares directos y los
suelos estaban muy resecos y altamente salinizados. Por el contrario, en los
bosques los períodos secos prolongados no representaron ningún problema
importante.
Otras razones de la mortalidad fueron el estrés del trasplante, que se vio
intensificado por las elevadas temperaturas reinantes durante la plantación
(cap. 2.2.10), la inundación (cap. 2.2.9 y 2.3.2), suelos blandos en los que
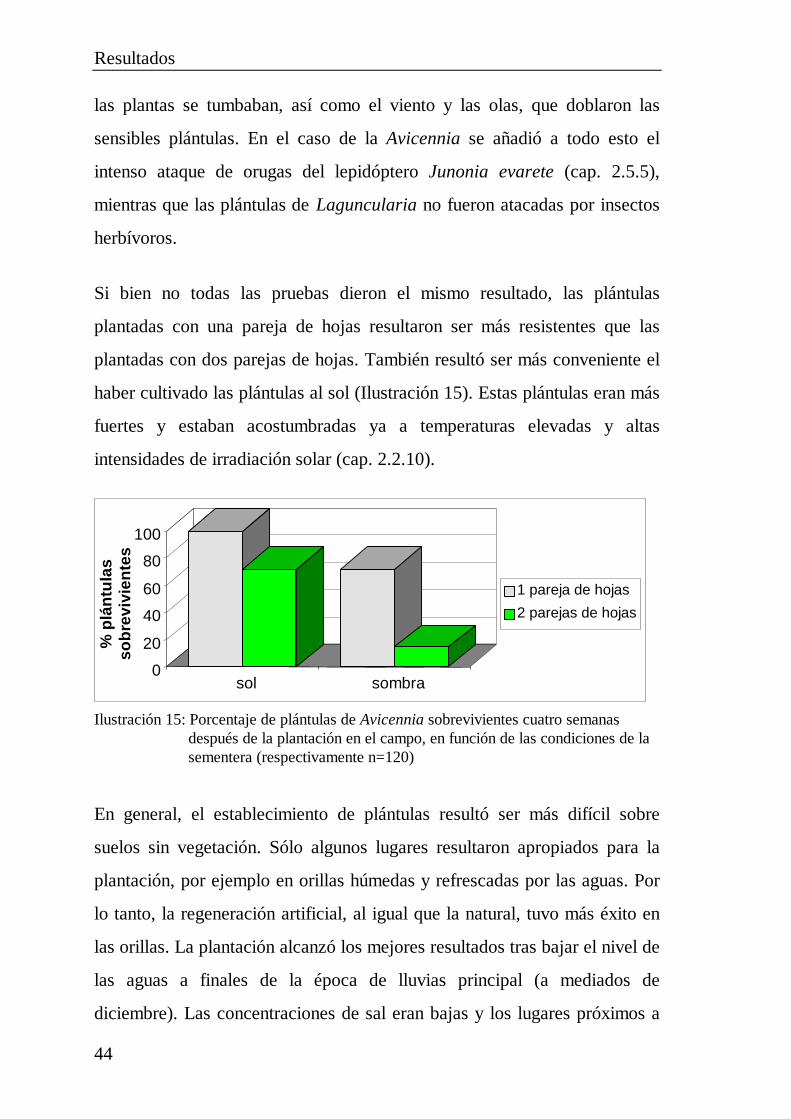
Resultados
44
las plantas se tumbaban, así como el viento y las olas, que doblaron las
sensibles plántulas. En el caso de la Avicennia se añadió a todo esto el
intenso ataque de orugas del lepidóptero Junonia evarete (cap. 2.5.5),
mientras que las plántulas de Laguncularia no fueron atacadas por insectos
herbívoros.
Si bien no todas las pruebas dieron el mismo resultado, las plántulas
plantadas con una pareja de hojas resultaron ser más resistentes que las
plantadas con dos parejas de hojas. También resultó ser más conveniente el
haber cultivado las plántulas al sol (Ilustración 15). Estas plántulas eran más
fuertes y estaban acostumbradas ya a temperaturas elevadas y altas
intensidades de irradiación solar (cap. 2.2.10).
sol sombra0
20
40
60
80
100
% p
lánt
ulas
sobr
eviv
ient
es
1 pareja de hojas2 parejas de hojas
Ilustración 15: Porcentaje de plántulas de Avicennia sobrevivientes cuatro semanasdespués de la plantación en el campo, en función de las condiciones de lasementera (respectivamente n=120)
En general, el establecimiento de plántulas resultó ser más difícil sobre
suelos sin vegetación. Sólo algunos lugares resultaron apropiados para la
plantación, por ejemplo en orillas húmedas y refrescadas por las aguas. Por
lo tanto, la regeneración artificial, al igual que la natural, tuvo más éxito en
las orillas. La plantación alcanzó los mejores resultados tras bajar el nivel de
las aguas a finales de la época de lluvias principal (a mediados de
diciembre). Las concentraciones de sal eran bajas y los lugares próximos a

Resultados
45
las orillas, húmedos en aquel momento, no volvieron a ser inundados hasta
el siguiente período de lluvias. En otros lugares con salinidades bajas fue
posible plantar Rhizophora debido a su robustez. Sin embargo, la siembra
de diásporas de Rhizophora dio siempre resultados más favorables que el
establecimiento de plántulas de Rhizophora.
2.5.3 Trasplante de plantas jóvenes
Las plantas jóvenes trasplantadas en agosto y diciembre de 1995 tenían una
altura de 30 a 150 cm y se habían tomado de manglares de regeneración aún
natural. La mortalidad de Laguncularia, de sólo un 11%, fue
considerablemente más baja que la de Avicennia y Rhizophora. Igual que en
el caso de las plántulas, también en el caso de las plantas jóvenes resultó ser
más desfavorable para el establecimiento el comienzo de la época de lluvias
(agosto) que el final del período de lluvias (diciembre). En el caso de la
Avicennia y la Rhizophora murió más de un 60% de las plantas del grupo
temprano, mientras que del grupo plantado más tarde murió sólo un 40%
aproximadamente (Ilustración 16). Probablemente, esto fue atribuible al
descenso de las salinidades causado por la inundación. La mortalidad fue
siempre máxima en los primeros tres meses tras la plantación, debido al
estrés del trasplante. Una vez aclimatadas, murieron sólo estacas aisladas
((Ilustración 16). En conjunto, las plantas jóvenes resultaron ser
considerablemente más resistentes que las plántulas a inundaciones, viento y
olas, sequedad, suelos blandos así como al ataque de fitófagos.
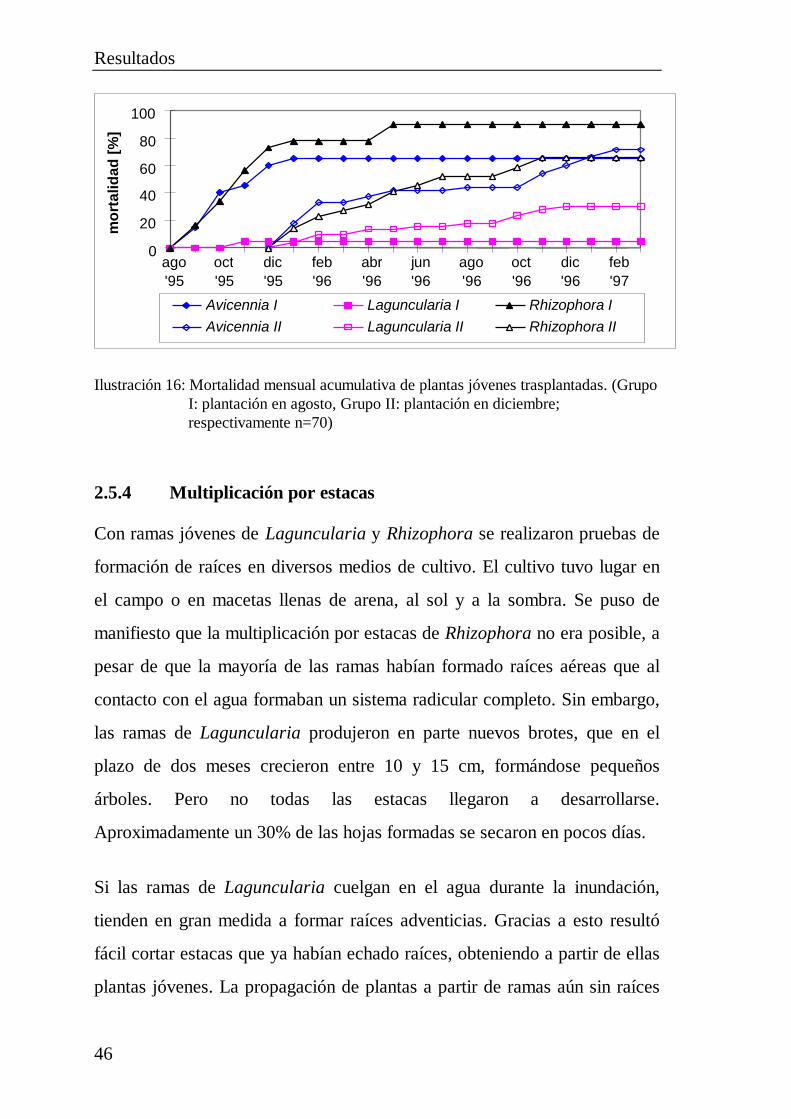
Resultados
46
0
20
40
60
80
100
ago'95
oct'95
dic'95
feb'96
abr'96
jun'96
ago'96
oct'96
dic'96
feb'97
mor
talid
ad [%
]
Avicennia I Laguncularia I Rhizophora IAvicennia II Laguncularia II Rhizophora II
Ilustración 16: Mortalidad mensual acumulativa de plantas jóvenes trasplantadas. (GrupoI: plantación en agosto, Grupo II: plantación en diciembre;respectivamente n=70)
2.5.4 Multiplicación por estacas
Con ramas jóvenes de Laguncularia y Rhizophora se realizaron pruebas de
formación de raíces en diversos medios de cultivo. El cultivo tuvo lugar en
el campo o en macetas llenas de arena, al sol y a la sombra. Se puso de
manifiesto que la multiplicación por estacas de Rhizophora no era posible, a
pesar de que la mayoría de las ramas habían formado raíces aéreas que al
contacto con el agua formaban un sistema radicular completo. Sin embargo,
las ramas de Laguncularia produjeron en parte nuevos brotes, que en el
plazo de dos meses crecieron entre 10 y 15 cm, formándose pequeños
árboles. Pero no todas las estacas llegaron a desarrollarse.
Aproximadamente un 30% de las hojas formadas se secaron en pocos días.
Si las ramas de Laguncularia cuelgan en el agua durante la inundación,
tienden en gran medida a formar raíces adventicias. Gracias a esto resultó
fácil cortar estacas que ya habían echado raíces, obteniendo a partir de ellas
plantas jóvenes. La propagación de plantas a partir de ramas aún sin raíces
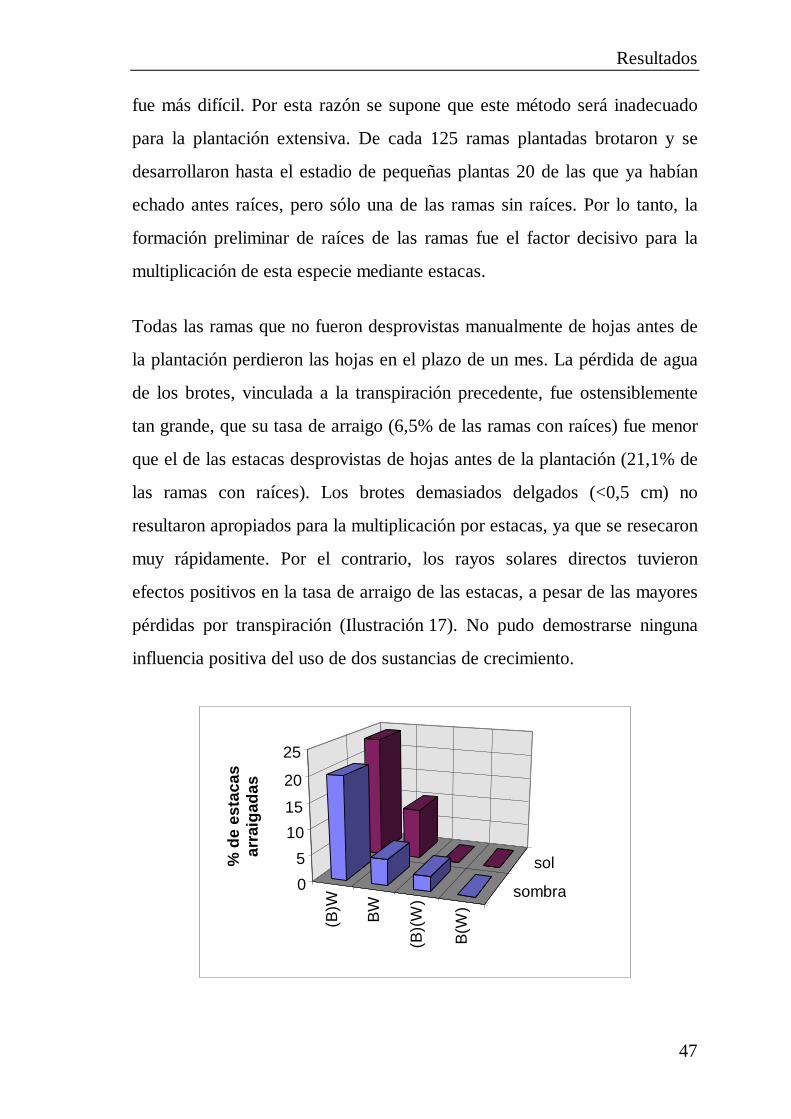
Resultados
47
fue más difícil. Por esta razón se supone que este método será inadecuado
para la plantación extensiva. De cada 125 ramas plantadas brotaron y se
desarrollaron hasta el estadio de pequeñas plantas 20 de las que ya habían
echado antes raíces, pero sólo una de las ramas sin raíces. Por lo tanto, la
formación preliminar de raíces de las ramas fue el factor decisivo para la
multiplicación de esta especie mediante estacas.
Todas las ramas que no fueron desprovistas manualmente de hojas antes de
la plantación perdieron las hojas en el plazo de un mes. La pérdida de agua
de los brotes, vinculada a la transpiración precedente, fue ostensiblemente
tan grande, que su tasa de arraigo (6,5% de las ramas con raíces) fue menor
que el de las estacas desprovistas de hojas antes de la plantación (21,1% de
las ramas con raíces). Los brotes demasiados delgados (<0,5 cm) no
resultaron apropiados para la multiplicación por estacas, ya que se resecaron
muy rápidamente. Por el contrario, los rayos solares directos tuvieron
efectos positivos en la tasa de arraigo de las estacas, a pesar de las mayores
pérdidas por transpiración (Ilustración 17). No pudo demostrarse ninguna
influencia positiva del uso de dos sustancias de crecimiento.
(B)W
BW
(B)(
W)
B(W
)
sombra
sol0
5
10
15
20
25
% d
e es
taca
sar
raig
adas

Resultados
48
Ilustración 17: Arraigo de estacas de Laguncularia al sol y a la sombra(B= estacas con hojas, (B)= hojas arrancadas antes de la plantación,W= con enraizamiento en el árbol por inundación, (W)= sin raíces)
Con estacas se consiguieron en pocos meses arbolillos de 1 a 1,5 m de
altura. Su plantación ahorró por lo tanto mucho tiempo en comparación con
la siembra de diásporas o con el cultivo y plantación de plántulas. Además,
con la multiplicación por estacas pudo soslayarse el muy crítico estadio de
las plántulas, en el que se produce una elevada mortalidad (cap. 2.5.1 y
2.5.2). También fue posible colonizar zonas inundadas, ya que las estacas no
fueron tumbadas por las olas ni cubiertas por el agua, no produciéndose en
consecuencia la muerte por tales factores.
2.5.5 Destrucción total de plántulas de Avicennia por orugas de
lepidópteros
Muchas plántulas de Avicennia fueron comidas por orugas herbívoras del
lepidóptero Junonia evarete Cramer. La incidencia del ataque fue mucho
mayor en las zonas despobladas de bosque (con frecuencia hasta el 100%)
que en las boscosas (generalmente, por debajo del 50%). Además, en los
bosques sólo se observó con frecuencia la ingestión de hojas aisladas o
partes de las hojas, mientras que en campo abierto las orugas devoraron
frecuentemente la plántula completa, dejando sólo un pequeño resto del
brote. Después de un ataque intenso las plántulas en general murieron,
mientras que las plantas menos atacadas brotaron de nuevo.
La Junonia evarete se presentó en la Ciénaga Grande casi exclusivamente
durante la época de lluvias principal, desapareciendo durante el período seco
(Ilustración 18), lo que puede estar causado por una diapausa o por
migraciones (Elster et al., enviado). Debido a la aparición estacional del
insecto, las diásporas que germinaron al comienzo de la época seca tuvieron
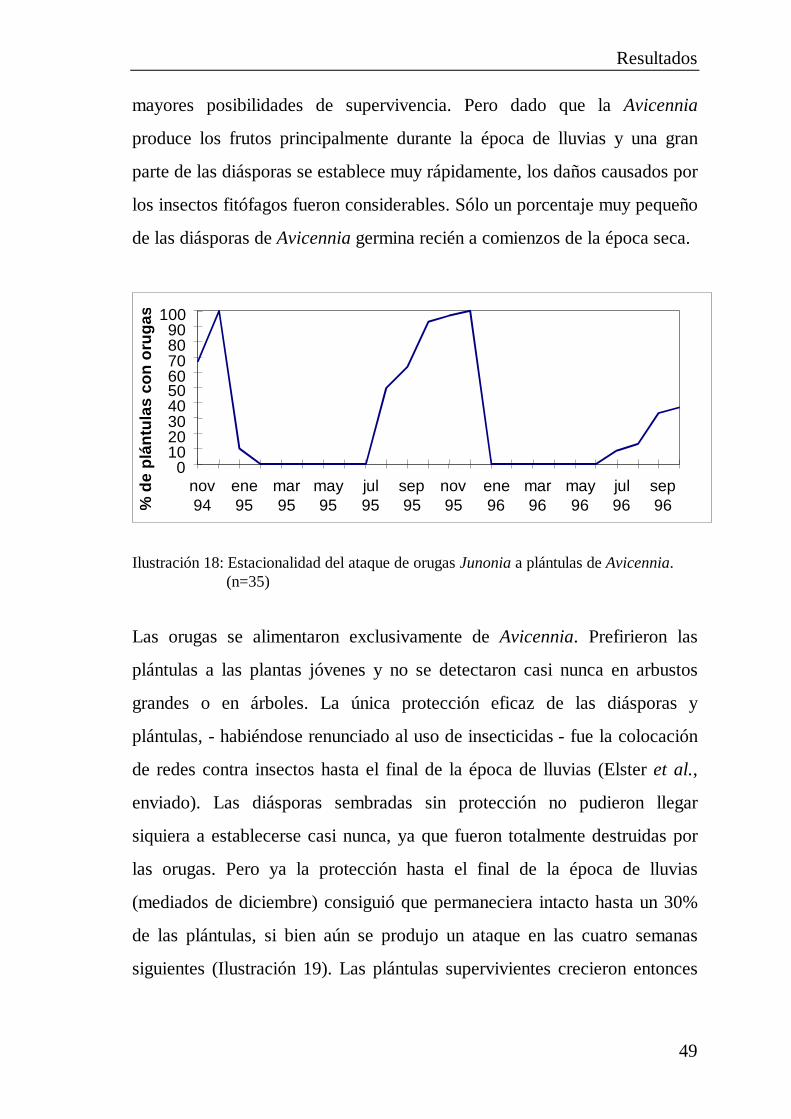
Resultados
49
mayores posibilidades de supervivencia. Pero dado que la Avicennia
produce los frutos principalmente durante la época de lluvias y una gran
parte de las diásporas se establece muy rápidamente, los daños causados por
los insectos fitófagos fueron considerables. Sólo un porcentaje muy pequeño
de las diásporas de Avicennia germina recién a comienzos de la época seca.
0102030405060708090
100
nov94
ene95
mar95
may95
jul95
sep95
nov95
ene96
mar96
may96
jul96
sep96%
de
plán
tula
s co
n or
ugas
Ilustración 18: Estacionalidad del ataque de orugas Junonia a plántulas de Avicennia.(n=35)
Las orugas se alimentaron exclusivamente de Avicennia. Prefirieron las
plántulas a las plantas jóvenes y no se detectaron casi nunca en arbustos
grandes o en árboles. La única protección eficaz de las diásporas y
plántulas, - habiéndose renunciado al uso de insecticidas - fue la colocación
de redes contra insectos hasta el final de la época de lluvias (Elster et al.,
enviado). Las diásporas sembradas sin protección no pudieron llegar
siquiera a establecerse casi nunca, ya que fueron totalmente destruidas por
las orugas. Pero ya la protección hasta el final de la época de lluvias
(mediados de diciembre) consiguió que permaneciera intacto hasta un 30%
de las plántulas, si bien aún se produjo un ataque en las cuatro semanas
siguientes (Ilustración 19). Las plántulas supervivientes crecieron entonces
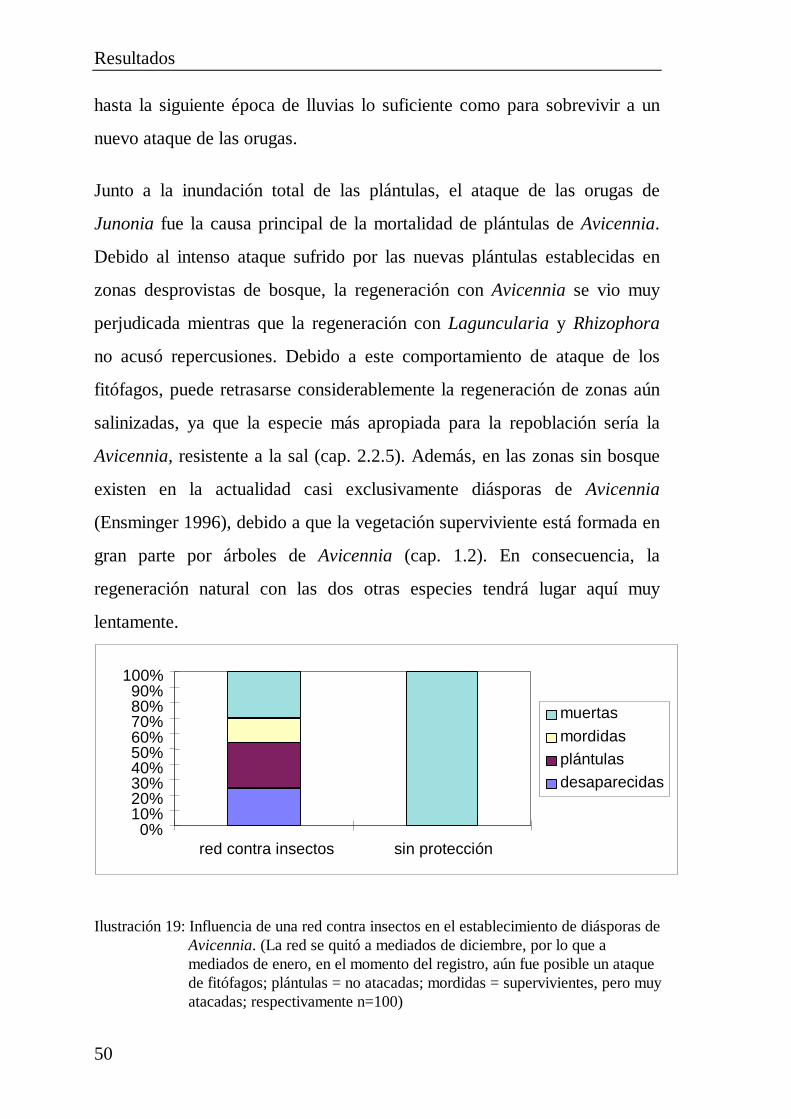
Resultados
50
hasta la siguiente época de lluvias lo suficiente como para sobrevivir a un
nuevo ataque de las orugas.
Junto a la inundación total de las plántulas, el ataque de las orugas de
Junonia fue la causa principal de la mortalidad de plántulas de Avicennia.
Debido al intenso ataque sufrido por las nuevas plántulas establecidas en
zonas desprovistas de bosque, la regeneración con Avicennia se vio muy
perjudicada mientras que la regeneración con Laguncularia y Rhizophora
no acusó repercusiones. Debido a este comportamiento de ataque de los
fitófagos, puede retrasarse considerablemente la regeneración de zonas aún
salinizadas, ya que la especie más apropiada para la repoblación sería la
Avicennia, resistente a la sal (cap. 2.2.5). Además, en las zonas sin bosque
existen en la actualidad casi exclusivamente diásporas de Avicennia
(Ensminger 1996), debido a que la vegetación superviviente está formada en
gran parte por árboles de Avicennia (cap. 1.2). En consecuencia, la
regeneración natural con las dos otras especies tendrá lugar aquí muy
lentamente.
Ilustración 19: Influencia de una red contra insectos en el establecimiento de diásporas deAvicennia. (La red se quitó a mediados de diciembre, por lo que amediados de enero, en el momento del registro, aún fue posible un ataquede fitófagos; plántulas = no atacadas; mordidas = supervivientes, pero muyatacadas; respectivamente n=100)
0%10%20%30%40%50%60%70%80%90%
100%
red contra insectos sin protección
muertasmordidasplántulasdesaparecidas

Resultados
51
La presencia de orugas con una frecuencia superior al promedio en las zonas
despobladas de bosque permite suponer que el ataque masivo de Junonia
está causado por el alto grado de degradación de la Ciénaga Grande. El
ataque intenso de insectos fitófagos en ecosistemas degradados es un
fenómeno que se observa con frecuencia en las zonas templadas. Si bien el
proceso de regeneración se ve claramente dificultado por el ataque masivo
de Junonia, a largo plazo, tras la disminución de las salinidades, es posible
que el establecimiento de nuevas plantas de Rhizophora y Laguncularia
conlleve una disminución de la población de Junonia. Además, es posible
que en el futuro las orugas de este lepidóptero sean diezmadas por enemigos
naturales, como son las avispas icneumónidas.
Por su tamaño comparativamente pequeño, las diásporas de Avicennia y
Laguncularia se enfrentan en parte a dificultades extremas para el
establecimiento. En caso de inundación son arrastradas, mientras que los
rayos solares directos las resecan rápidamente. También las plántulas de
ambas especies son muy sensibles. Inundaciones, viento, olas, temperaturas
elevadas y sequedad del suelo son los problemas principales para su
establecimiento. Especialmente en el caso de la Avicennia, las diásporas y
las plántulas son destruidas además por fitófagos. Sin embargo, si se toman
medidas apropiadas pueden soslayarse la mayoría de las dificultades en las
forestaciones. Muros de adobe o piedra protegen del viento, de las olas y del
arrastre de las diásporas; zanjas llenas de agua y sombra protegen de
resecamiento y sobrecalentamiento. Mediante redes contra insectos pueden
retenerse las diásporas y al mismo tiempo se corta el paso a los fitófagos.
Pero la mortalidad es elevada a pesar de todo. Por su forma y su tamaño, el
establecimiento de la Rhizophora es más sencillo incluso sin tomar medidas

Resultados
52
adicionales, y la mortalidad es escasa en condiciones favorables. El
establecimiento de diásporas de esta especie da unos resultados
considerablemente mejores que la plantación de plántulas; el principal
problema para el establecimiento son las salinidades elevadas.
Las plantas jóvenes de mayor tamaño de las tres especies estudiadas así
como las estacas de Laguncularia son mucho más robustas frente a
inundaciones, viento, olas, sequedad y ataque de fitófagos. En consecuencia,
en algunas zonas es recomendable la plantación de plantas grandes, a pesar
de que la obtención de plantas y estacas resulta más laboriosa y cara.
Además, dichas plantas grandes pueden florecer y fructificar en corto
tiempo, iniciando así una regeneración natural.
Todas las plántulas y plantas tienen que acostumbrarse antes del transplante
a las extremas intensidades de irradiación solar y las temperaturas elevadas,
plantándolas luego en lugares favorables.

3 Relevancia para la aplicación y recomendaciones
3.1 La regeneración natural del manglar
Si bien las zonas de la Ciénaga Grande y del Parque Tayrona pobladas de
manglar están aún relativamente intactas, la mortalidad de plántulas es muy
elevada. En los bosques, la causa principal de la muerte de plántulas es la
inundación durante la época de lluvias principal, que se cuenta entre las
particularidades naturales de la zona estudiada. En esto no ha cambiado
nada el hecho de que el nivel de las inundaciones se haya reducido un poco
debido a los cambios hidrológicos experimentados durante los últimos años.
Dado que las plántulas de Laguncularia y Avicennia reaccionan con gran
sensibilidad a la inundación total prolongada, mientras que superan bien
fluctuaciones breves del nivel de las aguas (cap. 2.2.9), el establecimiento
de plántulas en la Ciénaga Grande resulta naturalmente más difícil que en
los manglares costeros, que están primariamente bajo la influencia de la
marea baja y la alta. Las inundaciones persistentes que se producen en la
zona estudiada pueden significar localmente la muerte de todas las plántulas,
mientras que en zonas situadas a mayor altura o en años secos sobreviven
siempre suficientes plántulas para garantizar una regeneración natural del
manglar. Las plantas jóvenes de mayor tamaño y los árboles no sufren sin
embargo efectos persistentes.
La repoblación de las tierras antaño cubiertas de manglar resulta difícil, ya
que aquí el crecimiento de las plantas está obstaculizado por otros factores.
Entre las condiciones negativas cuentan los vientos alisios racheados, que
inciden sin obstáculos por la falta de vegetación, y el oleaje por ellos
provocado. El viento dobla especialmente las sensibles plántulas de
Laguncularia y Avicennia con frecuencia, produciendo su muerte. En

Relevancia para la aplicación y recomendaciones
54
ocasiones, los alisios arrastran también desde el mar nubes de polvo fino y
algas secas que se sedimentan sobre las plántulas, asfixiándolas o
doblándolas. El establecimiento de plántulas de Avicennia se ve
obstaculizado adicionalmente por un ataque masivo de orugas de
lepidóptero durante la época de lluvias, haciéndose casi imposible en
determinados lugares (cap. 2.5.5).
La frecuente presencia de suelos arenosos relativamente secos en las zonas
sin vegetación de las proximidades de la costa, dificultan adicionalmente el
establecimiento y el crecimiento de las plántulas, ya que tales suelos no
pueden retener la humedad aportada por inundaciones ligeras y frecuentes.
Las diásporas se resecan antes de poder germinar (cap. 2.2.3 y 2.2.6), y las
plántulas están expuestas a un gran riesgo de desecación (cap. 2.3.2). El
problema es agudizado por los rayos solares, que inciden sin obstáculos, y
por el consiguiente resecamiento y gran calentamiento de las capas
superiores del suelo (cap. 2.2.10). Además, en las zonas despobladas de
bosque y muy degradadas, la regeneración es obstaculizada por la alta
salinidad, que durante las épocas secas con frecuencia llega a matar todas
las nuevas plantas jóvenes establecidas (cap. 2.3.2)
La concentración de sal es el factor sobre el que más puede influir el
hombre. Es posible reducirla abriendo más canales entre la Ciénaga Grande
y el Río Magdalena, hasta el punto que vuelva a ser posible una
regeneración. Debido a los factores ambientales negativos antes
mencionados, a la extensión de las superficies sin vegetación y a la escasa
presencia de diásporas, la regeneración resulta muy difícil y se retrasará
mucho. A fin de acelerarla y de garantizar una composición equilibrada de
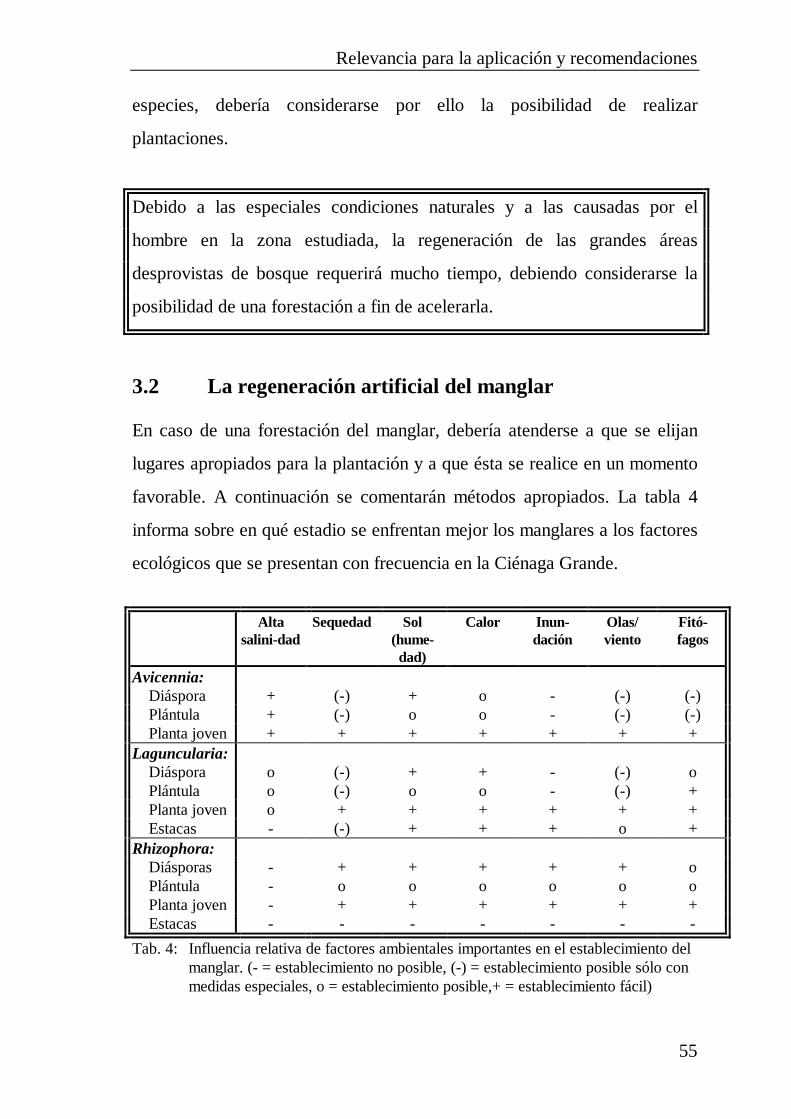
Relevancia para la aplicación y recomendaciones
55
especies, debería considerarse por ello la posibilidad de realizar
plantaciones.
Debido a las especiales condiciones naturales y a las causadas por el
hombre en la zona estudiada, la regeneración de las grandes áreas
desprovistas de bosque requerirá mucho tiempo, debiendo considerarse la
posibilidad de una forestación a fin de acelerarla.
3.2 La regeneración artificial del manglar
En caso de una forestación del manglar, debería atenderse a que se elijan
lugares apropiados para la plantación y a que ésta se realice en un momento
favorable. A continuación se comentarán métodos apropiados. La tabla 4
informa sobre en qué estadio se enfrentan mejor los manglares a los factores
ecológicos que se presentan con frecuencia en la Ciénaga Grande.
Altasalini-dad
Sequedad Sol(hume-dad)
Calor Inun-dación
Olas/viento
Fitó-fagos
Avicennia: Diáspora + (-) + o - (-) (-) Plántula + (-) o o - (-) (-) Planta joven + + + + + + +Laguncularia: Diáspora o (-) + + - (-) o Plántula o (-) o o - (-) + Planta joven o + + + + + + Estacas - (-) + + + o +Rhizophora: Diásporas - + + + + + o Plántula - o o o o o o Planta joven - + + + + + + Estacas - - - - - - -Tab. 4: Influencia relativa de factores ambientales importantes en el establecimiento del
manglar. (- = establecimiento no posible, (-) = establecimiento posible sólo conmedidas especiales, o = establecimiento posible,+ = establecimiento fácil)

Relevancia para la aplicación y recomendaciones
56
3.2.1 La siembra de diásporas
La siembra de diásporas sólo resulta prometedora si las aguas no las
arrastran o impiden su establecimiento (cap. 2.2.2), y si las diásporas no son
resecadas por los rayos solares directos y el calor (cap. 2.2.6 y 2.2.10). Por
su tamaño, las diásporas de Rhizophora son mucho menos sensibles a estos
factores ambientales que las de las otras dos especies. Las diásporas de
Rhizophora pueden afianzarse en el suelo y germinar incluso con una ligera
inundación (cap. 2.2.3). Ni siquiera un ligero oleaje o una corriente
moderada representan un gran problema para ellas. Además, la Rhizophora
se establece en un porcentaje mucho mayor que las otras dos especies. Por
lo tanto es recomendable plantar Rhizophora en suelos ligeramente
inundados o menos húmedos. Dado que el mayor problema para esta especie
lo representan las salinidades altas, en los lugares muy salinizados debería
procederse a una forestación con Avicennia o Laguncularia, a pesar de los
mayores problemas de establecimiento. En tal caso deben disponerse
obstáculos que impidan que las diásporas puedan alejarse a la deriva en caso
de inundación. Para ello pueden prestar buenos servicios pequeños
armazones de tablas con redes contra insectos así como diques de fango que
no sean cubiertos por las aguas. Después de la siembra en suelos húmedos
es recomendable una protección temporal con sombra, para que no se
sequen demasiadas diásporas por los efectos de los rayos solares directos. Si
se dispone de la humedad suficiente y tras el establecimiento completo de
las plántulas, debería renunciarse a la sombra, ya que en estas condiciones la
irradiación solar intensa tiene efectos positivos en el desarrollo de las
plántulas.
Si las condiciones son favorables, la siembra de diásporas es apropiada para
la recolonización, ya que las plántulas así producidas son más resistentes

Relevancia para la aplicación y recomendaciones
57
que las trasplantadas. Además, ésta es la forma más sencilla y barata de
forestación, pues no es necesario cultivar plantas y la siembra de diásporas
requiere menos tiempo que la plantación de plantas jóvenes. Sin embargo,
las pérdidas que se producen con este método son extremadamente
elevadas.
3.2.2 El establecimiento de plántulas
La plantación de plántulas previamente cultivadas resulta difícil, ya que
dichas plántulas (especialmente las de Laguncularia, pero también las de
Avicennia) son muy delicadas. Al estrés del trasplante se debe en gran
medida la alta mortalidad que se produce. Por esta razón, el estrés debería
mantenerse al nivel más bajo posible (cap. 3.2.5).
Si bien las plántulas de Rhizophora son mucho más resistentes que las de las
otras dos especies en lo que atañe a casi todos los factores estudiados, hay
que decir que la tasa de mortalidad es aún menor en caso de siembra directa
de las diásporas en el campo, desarrollándose mejor las plantas jóvenes así
obtenidas. Para esta especie debería renunciarse por lo tanto totalmente al
cultivo previo de las plántulas, plantando directamente las diásporas en el
campo.
En comparación con las diásporas, las plántulas de Avicennia y
Laguncularia tienen la ventaja de que pueden crecer sin problemas en el
caso de ligera inundación, mientras que las diásporas se establecen
entonces sólo con dificultad o son arrastradas por las aguas (cap. 2.2.2).
Bajo determinadas circunstancias puede ser necesario proteger las plántulas
del viento y las olas. Pequeños muros de adobe o piedra pueden prestar
buenos servicios para ello. Si el suelo está demasiado seco, puede

Relevancia para la aplicación y recomendaciones
58
procederse a un establecimiento en pequeñas zanjas que conduzcan agua.
Para evitar una muerte extensiva de plántulas de Avicennia debida al ataque
masivo de orugas de Junonia, es posible proteger las plántulas con redes
contra insectos durante la primera época de lluvias.
En conjunto, la plantación de las plántulas es más laboriosa que la siembra
de diásporas. Además, las plántulas tienen que cultivarse primero, lo que
encarece este método de forestación.
3.2.3 El establecimiento de plantas jóvenes
La plantación de plantas jóvenes de 1 m de altura aproximadamente resulta
muy prometedora, especialmente en el caso de la Laguncularia. La
mortalidad suele ser más baja que la de las plántulas (cap. 2.5.3). Otra
ventaja es el tamaño y la robustez de las plantas jóvenes. En lugares de
plantación poco apropiados para la regeneración con diásporas o plántulas
(inundación permanente o importante, sequedad o calor excesivos que
impidan la germinación o produzcan la muerte de plantitas pequeñas), en
general es aún viable la plantación de plantas más grandes.
Otra ventaja consiste en que las plantas jóvenes pueden florecer y fructificar
pronto. Dan sombra y con sus ramas y neumatóforos retienen las diásporas y
plántulas que flotan a la deriva, pudiendo acelerar así la regeneración natural
en sus proximidades. Sin embargo debe prestarse atención a que las plantas
sean plantadas en lugares apropiados o debidamente preparados. La
sequedad excesiva puede evitarse, por ejemplo, excavando zanjas para
plantación, siendo así posible forestar incluso zonas situadas a mayor altura
o superficies secas. Estas zonas serían muy difíciles de repoblar con
diásporas o plántulas, y ello sólo en años muy húmedos. Si las orillas están

Relevancia para la aplicación y recomendaciones
59
expuestas a un fuerte oleaje, deberían protegerse con muros de piedra antes
de la plantación, mientras que deberían evitarse zonas con inundaciones
demasiado altas (>30 cm) y de duración excesiva, ya que aquí la mortalidad
es muy elevada, no estando asegurado un establecimiento persistente en el
caso de la Avicennia y la Laguncularia.
Para una repoblación extensiva no es conveniente utilizar las plantas jóvenes
de los bosques, ya que con ello se deterioran manglares aún intactos. Por lo
tanto, las plantas jóvenes han de ser cultivadas. Dado que la plantación de
plantas de mayor tamaño es, además, más difícil y exige más tiempo que la
siembra de diásporas o plántulas, ésta es la alternativa de forestación más
cara. Por ello es recomendable aprovechar las grandes ventajas de las
plantas jóvenes de mayor edad para establecer islas de vegetación en zonas
a las que en la actualidad no sea arrastrada ninguna diáspora por las
corrientes. Se convertirán muy rápidamente en centros de una nueva
regeneración natural. De este modo procurarán una reforestación natural de
las zonas pantanosas, inaccesibles para el hombre y que, en consecuencia,
no pueden forestarse.
3.2.4 El establecimiento de estacas
La multiplicación por medio de estacas sólo es posible sin costos y trabajos
excesivos en el caso de la Laguncularia. Las estacas deberían haber echado
raíces ya en el árbol, y pueden precultivarse en macetas llenas de arena o
bien pueden plantarse directamente en el campo (cap. 2.5.4). Para reducir a
un mínimo las pérdidas por transpiración, es recomendable deshojar al
menos en parte las estacas después de cortarlas, y conservarlas en bolsas de
plástico hasta la plantación.

Relevancia para la aplicación y recomendaciones
60
La multiplicación por estacas tiene así las mismas ventajas que la plantación
de plantas jóvenes grandes (cap. 3.5.3), pero resulta considerablemente más
barata al evitarse fases de almáciga prolongadas. Por lo tanto, en el caso de
la Laguncularia puede reemplazarse la forestación con plantas jóvenes por
la forestación con estacas.
3.2.5 Simiente y cultivo de las plantas
No se pudo comprobar un comportamiento de germinación diferente en
función del origen geográfico. Por lo tanto, para forestar las zonas
actualmente despobladas de bosque de la Ciénaga Grande, la procedencia
geográfica de la simiente es irrelevante. Sin embargo, es importante prestar
atención a que las diásporas estén maduras y a que antes de su recolección
hayan estado expuestas a condiciones ambientales favorables. Diásporas
viejas, ya germinadas o dañadas tienen menos vitalidad, al igual que
aquellas que se han encontrado en un ambiente demasiado seco. Tales
diásporas generalmente no son apropiadas para una regeneración. Sólo las
que han estado protegidas del resecamiento por el pericarpio pierden su
vitalidad con menor rapidez. Una estadía prolongada en el agua tiene
también repercusiones negativas en la supervivencia, especialmente en el
caso de salinidades elevadas (cap. 2.2.4, 2.2.5 y Rabinowitz 1978). A este
respecto, la Rhizophora es mucho más resistente que las otras dos especies.
La recolección del árbol (Rhizophora y Laguncularia) así como la recogida
de diásporas totalmente frescas (Avicennia) han resultado ser
particularmente convenientes. También las condiciones reinantes durante el
transporte y el eventual almacenamiento pueden ejercer una gran influencia
en la disposición a la germinación. Ha resultado ser conveniente guardar las
diásporas de Avicennia y Laguncularia en cajas de cartón o de un material
similar. De este modo, las diásporas están protegidas de daños mecánicos y

Relevancia para la aplicación y recomendaciones
61
no se resecan. Si el almacenamiento se va a prolongar dos o más días, es
recomendable abrir las cajas, ya que de lo contrario las diásporas,
especialmente las de Avicennia, comienzan a enmohecerse o a pudrirse. Sin
embargo, las diásporas no deberían extenderse sin protección, a fin de evitar
su resecamiento. Dado que en el caso de la Avicennia se pudren
rápidamente en especial los pericarpios, éstos deberían desprenderse antes
de un almacenamiento prolongado de las diásporas. Tal operación resulta
sencilla después de haber tenido las diásporas a remojo en agua. De todo
modos deberá evitarse un almacenamiento demasiado prolongado, pues la
vitalidad de las diásporas disminuye rápidamente. Además, los fitófagos
presentes en las simientes, comienzan a atacar las diásporas aún intactas.
Las diásporas de Rhizophora son menos delicadas y, en casos
excepcionales, pueden conservarse durante semanas en bolsas de plástico
con pequeños orificios de ventilación.
Para el cultivo de plántulas y plantas jóvenes deberían tenerse en cuenta las
condiciones a que se enfrentarán en las zonas a forestar. Las plantas tienen
que aclimatarse a posibles condiciones extremas, a fin de que el estrés
propio del trasplante sea lo menor posible. Particularmente importante es
que las plantas se cultiven al sol. Si esto no es posible, deberían
acostumbrarse al menos algunas semanas antes de su plantación a los rayos
solares directos y a las consiguientes altas temperaturas. También se
recomienda mantener la salinidad durante el cultivo a un nivel similar al que
existirá en las zonas a forestar en el momento de la plantación.
Es conveniente mantener las plantas en la fase de almácigo (o cultivo
previo) durante el mayor tiempo posible, a fin de evitar trasplantarlas en el
sensible estadio de plántula. En opinión de mucho autores es recomendable

Relevancia para la aplicación y recomendaciones
62
un período de seis meses, o mejor aún, de un año (Hoffmann et al. 1985,
Saenger & Siddiqi 1993, Siddiqi et al. 1993).
3.2.6 Elección de las parcelas
Los resultados ponen de manifiesto que la plantación con fines de
regeneración rápida de manglares muertos en forma extensiva se desarrolla
más positivamente allí donde también es de esperar una regeneración natural
rápida en el caso de que lleguen diásporas a dichas zonas. Se trata de
lugares de baja salinidad, con agua suficiente, protegidos del viento y de las
olas y, en lo posible, refrescados por agua. Se tratará por lo tanto
principalmente de zonas ribereñas sometidas a los efectos compensadores de
extensiones importantes de agua, pero que no sean inundadas en exceso.
Puede resultar muy positiva la plantación a las orillas de lagos o lagunas
grandes que no se calienten excesivamente durante el día. La elección de las
parcelas apropiadas es uno de los factores más importantes para el éxito de
la forestación. Sin embargo debería forestarse primariamente donde no sea
de esperar una regeneración natural por diásporas acarreadas por las aguas.
No obstante, aún se sabe muy poco sobre los mecanismos de difusión
reinantes en la Ciénaga Grande (corrientes y las direcciones de difusión a
ellas vinculadas así como puntos de varado principales y disponibilidad de
las diásporas). Estos factores deberían investigarse antes de proceder a una
forestación. De este modo podrían obtenerse también informaciones más
exactas sobre la velocidad de regeneración del conjunto del sistema.
En parcelas de salinidad más elevada debería realizarse la forestación con
Avicennia, ya que esta especie es más insensible a las altas concentraciones
de sal.

Relevancia para la aplicación y recomendaciones
63
3.2.7 La época de plantación
Es recomendable realizar la plantación hacia el final de la principal época de
lluvias, ya que entonces las probabilidades de supervivencia son más altas,
tanto para las diásporas como para las plantas. Las concentraciones de sal
son bajas en ese período. Además, los suelos aún están lo suficientemente
húmedos para un establecimiento del mangle y no volverán a ser inundados
hasta después de siete u ocho meses. Hasta entonces, y si las condiciones
son favorables, las plántulas habrán alcanzado un tamaño suficiente como
para no ser cubiertas por las aguas. Al final de la época de lluvias hay
también grandes cantidades de diásporas maduras de las tres especies de
mangle investigadas en la zona objeto del estudio. Si en el caso de la
Avicennia se quiere evitar el uso de redes contra insectos para proteger las
plántulas de las orugas de Junonia, en el caso de esta especie no debería
comenzar el establecimiento de diásporas y plántulas hasta el comienzo de
la época seca.
3.2.8 Perspectivas
La alta mortalidad de plántulas de mangle es un fenómeno natural, por lo
que en toda forestación hay que contar con grandes pérdidas. A pesar de
estas grandes pérdidas y de que la regeneración natural se desarrollará
previsiblemente sólo con lentitud (Ilustración 20), las zonas actualmente
despobladas de bosque de la Ciénaga Grande volverán a ser colonizadas en
cuanto las salinidades puedan reducirse a una medida soportable por el
manglar. La condición realmente primordial para el éxito de todas las
medidas de forestación es por lo tanto el restablecimiento de las
características hidrológicas originales del sistema de lagunas.

Relevancia para la aplicación y recomendaciones
64
La posibilidad más sencilla y barata de forestación es la siembra de
diásporas. Sin embargo, las pérdidas son muy elevadas, especialmente en el
caso de la Avicennia y la Laguncularia. Incluso zonas ligeramente
inundadas son difíciles de repoblar con las diásporas de estas dos especies,
por lo que aquí se recomienda la plantación de plántulas o la siembra de
diásporas de Rhizophora. Las zonas más inundadas, al igual que las zonas
secas, pueden forestarse con estacas de Laguncularia o con plantas jóvenes
grandes de las tres especies. Este es sin embargo el método más caro y el
que más tiempo requiere. Es recomendable especialmente en las
proximidades de zonas pantanosas, no accesibles para el hombre, ya que las
plantas relativamente grandes pueden convertirse en un plazo muy corto en
centros de regeneración natural para las tierras del entorno.
La simiente debería estar madura y ser en lo posible fresca, y las plantas
jóvenes deberían cultivarse en condiciones equivalentes a las que se darán
en las zonas a forestar. De este modo, las plantas se aclimatan a las
condiciones extremas a las que posiblemente tendrán que enfrentarse,
reduciéndose así el estrés del trasplante. Además debería plantarse en lo
posible hacia el final de la época de lluvias. Muy importante es también que
la forestación se realice sólo en zonas apropiadas para la regeneración del
mangle.

Relevancia para la aplicación y recomendaciones
65
Ilustración20: Regeneración natural en Barra Vieja. (Con la construcción de una tajeacomenzó aquí hace unos nueve años la regeneración del manglartotalmente muerto)

4 Bibliografía
CLARKE, P.J. 1992. Predispersal mortality and fecundity in the greymangrove (Avicennia marina) in southeastern Australia. AustralianJournal of Ecology 17: 161-168.
ELSTER, C. 1997. Beziehung zwischen ökologischen Faktoren und derRegeneration dreier Mangrovenarten im Gebiet der Ciénaga Grandede Santa Marta, Kolumbien. Dissertation, 220 págs., Justus-LiebigUniversität Gießen.
ELSTER, C., PERDOMO, L., POLANÍA, J. & SCHNETTER, M.-L. Junoniaevarete larvae control Avicennia germinans recruitment and survivalin a disturbed mangrove forest in Colombia. Ecology, enviado.
ELSTER, C., PERDOMO, L. & SCHNETTER, M.-L. Posibilidades de larecuperación del manglar en la Ciénaga Grande de Santa Marta,(Colombia). La Tadeo, en imprenta.
ENSMINGER, I. 1996. Hydrologische Veränderungen am Canal Clarín undihre Bedeutung für die Regeneration salzgeschädigter Mangrove.Diplomarbeit, 114 págs., Justus-Liebig Universität Gießen.
GARCÉS G., D.M. & DE LA ZERDA L., S. 1994. Gran Libro de los ParquesNacionales de Colombia. Círculo de lectores S.A., 230 págs., Santaféde Bogotá.
GONZÁLEZ, E. 1991. El manglar de la Ciénaga Grande de Santa Marta:Ecosistema en peligro de extinción. Colombia, sus gentes y regiones.IGAC. 21:2-21.
HOFFMANN, W.E., DURAKO, M.J. & LEWIS, R.R. 1985. Habitat restorationin Tampa Bay. Tampa Bay area scientific information symposium,Proceedings: 636-657, Florida Sea Grant College, Report Number 65.

Bibliografía
67
INDERENA - Divisiones fauna terrestre y parques nacionales. 1978. Plande emergencia para la restauración del Parque Nacional Natural Islade Salamanca. Mimeografiado, 41 págs., INDERENA, Santafé deBogotá.
INDERENA - Divisiones fauna terrestres y parques nacionales. 1984.Colombia Parques Nacionales. 264 págs., INDERENA, Santafé deBogotá.
PRO-CIÉNAGA. 1993. Plan de recuperación del complejo lagunar de laCiénaga Grande de Santa Marta. DNP, CORPAMAG, GTZ. Informefinal, 59 págs., ejemplar no publicado, Santa Marta.
SAENGER, P. & SIDDIQI, N.A. 1993. Land from the sea: The mangroveafforestation program of Bangladesh. Ocean & Coastal Management20: 23-39.
SÁNCHEZ Páez, H. 1988. Hacia la salvación del Parque Nacional NaturalIsla de Salamanca. TRINEA (Act. Cient. Tecn. INDERENA) 2: 505-527.
SIDDIQI, N.A., ISLAM, M.R., KHAN, M.A.M. & SHAHIDULLAH, M. 1993.Mangrove nurseries in Bangladesh. Mangrove Ecosystems OccasionalPapers no 1, págs. 14, International Society of Mangrove Ecosystems,Okinawa.
RABINOWITZ, D. 1978. Dispersal properties of mangrove propagules.Biotropica 10(1): 47-57.
ROBERTSON, A.I., GIDDINS, R. & SMITH, T.J. 1990. Seed predation byinsects in a tropical mangrove forest: extent and effects on seedviability and the growth of seedlings. Oecologia 83: 213-219.
ROBERTSON, A.I. 1991. Plant-animal interactions and the structure andfunction of mangrove forest ecosystems. Australian Journal ofEcology 16: 433-443.

Bibliografía
68
TOMLINSON, P.B. 1986. The botany of mangroves. 419 págs., CambridgeUniversity Press, Cambridge.

Deutsche Gesellschaft fürTechnische Zusammenarbeit (GTZ) GmbH
Tropenökologisches Begleitprogramm (TÖB)Programa de apoyo ecológicoPostfach 5180D-65726 EschbornRepública Federal de Alemania
Fax +49-(0)6196-79-7413E-Mail: [email protected]: http://www.gtz.de/toeb

Im Auftrag des Bundesministeriums für wirtschaftlicheZusammenarbeit und Entwicklung (BMZ)