porqué un mono no puede jugar baseball
DESCRIPTION
Paper que enseña porque el mono no puede jugar baseballTRANSCRIPT

Elastic energy storage in the shoulder and the evolution of high-speed throwing in Homo
(Article begins on next page)
The Harvard community has made this article openly available.Please share how this access benefits you. Your story matters.
Citation Roach, Neil T., Madhusudhan Venkadesan, Michael J. Rainbow,and Daniel E. Lieberman. 2013. “Elastic energy storage in theshoulder and the evolution of high-speed throwing in Homo.”Nature 498 (7455): 483-486. doi:10.1038/nature12267.http://dx.doi.org/10.1038/nature12267.
Published Version doi:10.1038/nature12267
Accessed March 9, 2016 12:21:26 PM EST
Citable Link http://nrs.harvard.edu/urn-3:HUL.InstRepos:11879202
Terms of Use This article was downloaded from Harvard University's DASHrepository, and is made available under the terms and conditionsapplicable to Other Posted Material, as set forth athttp://nrs.harvard.edu/urn-3:HUL.InstRepos:dash.current.terms-of-use#LAA

Elastic energy storage in the shoulder and the evolution of high-speed throwing in Homo
Neil T. Roach1,2, Madhusudhan Venkadesan3, Michael J. Rainbow4, and Daniel E.Lieberman1
1Department of Human Evolutionary Biology, Harvard University, Cambridge, Massachusetts02138, USA2Center for the Advanced Study of Hominid Paleobiology, Department of Anthropology, TheGeorge Washington University, Washington, District of Columbia 20052, USA3National Centre for Biological Sciences, Tata Institute of Fundamental Research, Bangalore,Karnataka 560065, India4Spaulding National Running Center, Department of Physical Medicine and Rehabilitation,Harvard Medical School, Cambridge, Massachusetts 02138, USA
AbstractAlthough some primates, including chimpanzees, throw objects occasionally1,2, only humansregularly throw projectiles with high speed and great accuracy. Darwin noted that humans’ uniquethrowing abilities, made possible when bipedalism emancipated the arms, enabled foragers toeffectively hunt using projectiles3. However, there has been little consideration of the evolution ofthrowing in the years since Darwin made his observations, in part because of a lack of evidence onwhen, how, and why hominins evolved the ability to generate high-speed throws4-8. Here, weshow using experimental studies of throwers that human throwing capabilities largely result fromseveral derived anatomical features that enable elastic energy storage and release at the shoulder.These features first appear together approximately two million years ago in the species Homoerectus. Given archaeological evidence that suggests hunting activity intensified around this time9,we conclude that selection for throwing in order to hunt likely played an important role in theevolution of the human genus.
Compared with other carnivores, hominins are slow, weak, and lack natural weapons such asfangs and claws. Yet, hominins were eating meat by at least 2.6 Ma, and likely hunting largeprey by 1.9 Ma (Supplementary Note 1). Although contemporary hunter-gatherers rarelyrely on throwing to kill prey, earlier hominins probably needed to throw projectilesfrequently in order to acquire and defend carcasses before the relatively recent inventions ofthe atlatl and bow10. We can therefore surmise that the ability to throw well would confer astrong selective benefit to early hunters. However, in order to test when and how homininsevolved the ability to throw projectiles effectively, it is necessary to understand boththrowing biomechanics and how changes in hominin anatomy affect throwing performance.
Correspondence and requests for material should be addressed to NTR ([email protected])..Author Contributions NTR designed the study, collected and analyzed the data, and wrote the paper in conjunction with DEL. MVcontributed to the experimental design, power calculations, and to editing the paper. MJR helped analyze the data and edit the paper.
Supplementary Information is linked to the online version of the paper at www.nature.com/nature.
Author Information Reprints and permissions information is available at www.nature.com/reprints. The authors declare nocompeting or financial interests.
Europe PMC Funders GroupAuthor ManuscriptNature. Author manuscript; available in PMC 2013 December 27.
Published in final edited form as:Nature. 2013 June 27; 498(7455): 483–486. doi:10.1038/nature12267.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

Throws are powered by rapid, sequential activation of many muscles, starting in the legs andprogressing through the hips, torso, shoulder, elbow and wrist11-14. Torques generated ateach joint accelerate segmental masses, creating rapid angular movements that accumulatekinetic energy in the projectile until its release. It has been shown that internal (medial)rotation around the long axis of the humerus is the largest contributor to projectilevelocity15. This rotation, which occurs in a few milliseconds and can exceed 9,000°/sec13, isthe fastest motion the human body produces. Although previous research has focused on theinternal rotator muscles of the shoulder11,16,17, these muscles alone cannot explain howhumans generate so much internal rotational power. For one, calculations of the maximumpower production capacity of all the shoulder’s internal rotator muscles indicate that thesemuscles can contribute, at most, half of the shoulder rotation power generated during thethrowing motion (Supplementary Notes 2,3). Peak internal rotation torque also occurs wellbefore the humerus starts to rotate internally12. Furthermore, variation in muscle fiberorientation in these muscles produce actions other than internal humeral rotation that reducepower output for this action.
Elastic energy storage has been shown to be an important source of power amplification formany high-powered movements18,19. We propose that several evolutionarily novel featuresin the human shoulder help store and release elastic energy to generate much of the powerneeded for rapid humeral rotation during human throwing. According to this model, energystorage occurs during the arm-cocking phase (Fig. 1A), which begins with completion of alarge step towards the target. As the foot hits the ground, the arm is already externallyrotated, horizontally extended, and abducted nearly 90° at the shoulder, with forearm flexionapproaching 90° at the elbow13. As the cocking phase begins, large torques are generated byrapid rotation of the torso towards the target and by the activation of the major shoulderhorizontal flexor, Pectoralis major11,16. The positioning of the shoulder and elbow at thistime increase the mass moment of inertia around the long axis of the humerus, causing theforearm and hand to lag behind the accelerating torso. Further, a flexed elbow during thecocking phase allows passive inertial forces to externally counter rotate the arm, stretchingthe short, parallel tendons, ligaments, and elastic components of muscles that cross theshoulder, potentially storing elastic energy in the large aggregate cross-sectional area ofthese structures (Supplementary Note 4). Then, when the biceps deactivate and elbowextension begins, the arm’s moment of inertia is reduced, allowing these stretched elementsto recoil, releasing energy, and helping to power the extremely rapid internal rotation of thehumerus (Supplementary Note 5).
Three derived morphological features of humans not present in chimpanzees, our closestextant relatives, play a major role in storing and releasing elastic energy during throwing(Supplementary Note 6). First, humans’ tall, mobile waists decouple the hips and thorax,permitting more torso rotation20, in turn enabling high torque production over a large range-of-motion (ROM), needed to load the shoulder’s elastic elements. Second, humeral torsion,the angle between humeral head orientation and the axis of the elbow, is 10-20° lower inhuman throwers’ dominant arms compared to chimpanzee humeri5. Decreased torsionextends the rotational ROM at the shoulder externally21,22, potentially enabling more elasticenergy storage during the cocking phase. Finally, humans have a more laterally orientedglenohumeral joint, which aligns the P. major flexion moment around the same axis as thetorso rotation moment. This orientation allows humans to increase the arm’s moment ofinertia by abducting the humerus in line with the torso rotation and shoulder flexion torques,maximizing resistance to both (Fig. 1B/C/D). In contrast, chimpanzees’ more craniallyoriented glenohumeral joint and limited ability to produce torso rotation torque requiresthem to maximize inertial loading by abducting their humeri more than humans to bringtheir arm in line with the P. major flexion moment. This increased abduction, however,would force chimpanzees to position their elbow in a more extended posture to maximize
Roach et al. Page 2
Nature. Author manuscript; available in PMC 2013 December 27.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

the arm’s moment of inertia, resulting in a costly reduction in elbow extension during thethrow.
We tested the effects of these derived features on throwing performance using high-speed,3D kinematic and kinetic data from 20 experienced human throwers to quantify powerproduction at the shoulder during overhand baseball throwing (Supplementary Note 7).During the arm-cocking phase, the throwers’ humeri externally rotate 57±15 (mean±s.d.)past the active ROM limit achieved using their own muscular power, indicating passivestretching of the ligaments, tendons, and muscles crossing the shoulder. Inverse dynamicsanalysis shows that during this period, the shoulder produces an opposing internal rotationtorque, causing a sustained period of power absorption (Fig. 2). During arm-cocking, thenegative work of shoulder rotation averages −201±70J, with an average power of−631±337W. In contrast, the total rotational work of the subsequent internal rotation motionis 346±116J, with power during acceleration averaging 3,847±1,697W. If 90% of thenegative work during arm-cocking is stored and returned elastically23, this energy canaccount for 54±15% of the internal humeral rotation work done during a typical throw.
Elastic energy storage at the shoulder also augments the generation of joint velocity andpower at the elbow. During acceleration, the elbow extends at very high angular velocities(2,434±552°/sec) despite large amounts of negative power and work (−246±63J), indicatingthat the triceps alone are not powering this rapid extension (Fig. 2). As previous studies haveshown, elbow extension is powered primarily by more proximal segments15,24, especiallythe shoulder.
An additional line of evidence for the importance of elastic energy storage comes fromexperimentally limiting shoulder rotational ROM with therapeutic braces (SupplementaryNotes 8-11), which restricted external rotation by 24±9°. During brace trials, shoulderrotation beyond the active ROM decreased by 50±36% and shoulder work during arm-cocking decreased by 39±16% (rmANOVA p < 0.001) (Fig. 3). Shoulder rotation workduring the subsequent acceleration phase was not significantly different between conditions,but average shoulder rotation power during acceleration decreased significantly (−16±35%,rmANOVA p = 0.036). Wearing a shoulder brace also decreased elbow negative workduring acceleration by 20±21% (rmANOVA p < 0.001). Overall, these work and powerreductions from less elastic energy exchange significantly reduced humeral rotation angularacceleration (−24±29%, rmANOVA p < 0.001) and elbow extension angular velocity(−21±10%, rmANOVA p < 0.001), reducing ball speed by 8±6% (MANOVA, p < 0.001).
Natural variation in humeral torsion (Supplementary Note 12) yields similar performanceeffects. It has long been known that athletes such as pitchers have lower degrees of humeraltorsion, by 10-15°, in their throwing versus non-throwing arms21,22,25. By maintainingrelatively lower, juvenile levels of torsion into adulthood26, throwing athletes increasepower generation by shifting the humerus’ rotational ROM externally21,22. This shift allowsfurther external rotation during arm-cocking and increases internal rotation duringacceleration (Fig. 4A), permitting more elastic energy storage and release (Fig. 4B/C). It isunknown whether the plasticity of humeral torsion is greater in humans than in other taxa,but we speculate that plasticity in humans may be advantageous, enabling low torsion topersist in the throwing arm, while higher torsion (useful for manipulative tasks) develops inthe non-throwing arm5,25.
When high-speed throwing first evolved is difficult to test because the first projectiles wereprobably rocks and untipped wooden spears (Supplementary Notes 7,13). However, many ofthe derived morphological features that help human throwers store elastic energy can beassessed in the fossil record (Supplementary Note 14). These features evolved in a mosaic
Roach et al. Page 3
Nature. Author manuscript; available in PMC 2013 December 27.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

fashion, some predating the evolution of Homo. Tall, decoupled waists first appear inAustralopithecus as adaptations for locomotion20. Low humeral torsion also appears inAustralopithecus, likely resulting from the release of the forelimbs from weight-bearingduring quadrupedal locomotion, and is present in early Homo5 (Fig. 4D). Although variationin glenoid orientation exists within Australopithecus27, a fully lateral glenoid position is firstdefinitively present in Homo erectus28 (Supplementary Notes 15,16). Such laterally orientedshoulders probably decreased the mechanical advantage of the scapular rotator musclesduring climbing, and probably had little or no effect on stone tool production. Throwingperformance would likely also have benefited from low, wide shoulders, long legs, andhyperextendable wrists, which are present in H. erectus20,29. Although some of thesefeatures were probably selected for functions other than throwing, their combinedconfiguration, first present in H. erectus, would have benefited throwing performance byenabling elastic energy storage in the shoulder, providing a selective advantage duringhunting (Supplementary Note 1). Furthermore, high-speed throwing was likely a criticalcomponent of a suite of hunting behaviors that allowed early members of the genus Homo tothrive in new and varied habitats both in and out of Africa.
Today, technological advances such as the bow and arrow, nets, and firearms have reducedcontemporary hunter-gatherers’ reliance on thrown projectiles, but the human ability andproclivity to throw persists in many sports, where athletes rely on the same mechanics(Supplementary Note 7). In this modern context, the evolution of adaptations for elasticenergy storage during human throwing has implications for the high prevalence of injuries inthrowing athletes. Paleolithic hunters almost certainly threw less frequently than modernathletes, who often deliver more than 100 high-speed throws in the span of a couple ofhours. Unfortunately, the ligaments and tendons in the human shoulder and elbow are notwell adapted to withstanding such repeated stretching from the high torques generated bythrowing, and frequently suffer from laxity and tearing12,30. While humans’ unique ability topower high-speed throws using elastic energy may have been critical in enabling earlyhunting, repeated overuse of this motion can result in serious injuries in modern throwers.
METHODSSubjects
Data were collected from 20 male subjects (ages 19-23). Nineteen of the subjects werecollegiate athletes (16 baseball players, 3 non-throwing athletes). Prior to enrollment in thestudy, all participants were required to pass a throwing performance task (SupplementaryNote 17) in order to exclude poor throwers. For all subjects, we collected weight,information on relevant injury/medical history, and basic anthropometric data (height,segment lengths and circumferences, joint range of motion). Humeral torsion was estimatedusing range of motion measures31. All subjects provided informed written consent inaccordance with the Harvard Committee on the Use of Human Subjects.
KinematicsKinematic data were collected at 1000 Hz using an eight-camera Vicon T10s 3D infraredmotion capture system (Vicon Inc, Centennial CO, USA). Each subject had twenty-onepassive reflective markers taped on the throwing arm and torso (Supplementary Note 18).Subjects were given approximately 5 minutes to stretch and warm up before recording. Afterthe warm up period, subjects were tasked to throw a 144g baseball at a 1m-radius targetfrom 10m away. The subject then threw 8-10 normal pitches and 8-20 pitches using aDonjoy Shoulder Stabilizer (Donjoy Inc, Vista, CA) brace that restricts external rotationalrange of motion at the shoulder (Supplementary Note 19). As a sham, data from anintermediate condition in which the brace was applied but not tightened were also collected
Roach et al. Page 4
Nature. Author manuscript; available in PMC 2013 December 27.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

(Supplementary Note 8). Ball speed was measured using a Sports Radar Model 3600 radargun. Ball release was timed using a synched FlexiForce A201 force sensor (Tekscan Inc,Boston MA, USA) collected at 1000 Hz taped to the palmar side of the distal phalanx of thethird digit and synched with a 30 Hz Canon Vixia HV30 digital video camera (Canon Inc,Tokyo, Japan). In order to filter the kinematic data, a residual analysis32 of the entirethrowing trial and the critical period during the humeral internal rotation motion wascalculated in MATLAB (version R2010b) (Supplementary Note 20). A Butterworth 2nd
order low-pass filter (cutoff 25Hz) was applied and marker gaps up to 100 frames wereinterpolated using C-Motion Visual3D software (v4). For analysis, each motion was thensubdivided into five standard phases of the throw: windup/stride, arm cocking, armacceleration, arm deceleration, and follow-through14.
KineticsJoint Euler angles were calculated and inverse dynamics analyses were performed usingmass distribution data from Dempster33 in Visual3D. Joint angular velocities, moments, andpower were calculated using each joint’s instantaneous axis of rotation (Supplementary Note23). The sequence of rotations at each joint is described in Supplementary Note 22. Jointwork was calculated in MATLAB using the trapz function.
StatisticsKinetic data were standardized to phase length, interpolated, and resampled using customMATLAB code to yield comparable data across all trials and subjects (Supplementary Note21). Individual subject means were compared across experimental conditions using repeatedmeasures ANOVA or MANOVA where appropriate. All statistical analyses were conductedusing JMP software (v5). Differences were considered to be significant at alpha < 0.05.
Supplementary MaterialRefer to Web version on PubMed Central for supplementary material.
AcknowledgmentsWe would like to thank the Wyss Institute, Leia Stirling, Andy Biewener, Richard Wrangham, Susan Larson, BrianRoach, Laszlo Meszoly, Amanda Lobell, and many undergraduate research assistants for their feedback, help, andsupport.
Funding was provided by the National Science Foundation (BCS-0961943 to NTR/DEL), the American School forPrehistoric Research (to NTR/DEL), and the Wellcome Trust/DBT India Alliance (500158/Z/09/Z to MV).
References1. Goodall, J. The chimpanzees of Gombe: Patterns of behavior. Harvard University Press; 1986.
2. Westergaard GC, Liv C, Haynie MK, Suomi SJ. A comparative study of aimed throwing bymonkeys and humans. Neuropsychologia. 2000; 38:1511–1517. [PubMed: 10906376]
3. Darwin, C. The descent of man, and selection in relation to sex. John Murray; 1871.
4. Churchill, SE.; Rhodes, JA. The evolution of hominin diets: Integrating approaches to the study ofPaleolithic subsistence. Hublin, JJ.; Richards, MP., editors. Springer; 2009.
5. Larson SG. Evolutionary transformation of the hominin shoulder. Evol Anthropol. 2007; 16:172–187.
6. Calvin, WH. The throwing madonna: Essays on the brain. Bantam; 1983.
7. Fifer F. The adoption of bipedalism by the hominids: A new hypothesis. Hum Evol. 1987; 2:135–147.
Roach et al. Page 5
Nature. Author manuscript; available in PMC 2013 December 27.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

8. Bingham PM. Human evolution and human history: A complete theory. Evol Anthropol. 2000;9:248–257.
9. Stanford, CB.; Bunn, HT. Human Evolution Series. Ciochon, RL.; Wood, B., editors. OxfordUniversity Press; New York: 2001.
10. Shea JJ. The origins of lithic projectile point technology: evidence from Africa, the Levant, andEurope. J Archaeol Sci. 2006; 33:823–846.
11. Hirashima M, Kadota H, Sakurai S, Kudo K, Ohtsuki T. Sequential muscle activity and itsfunctional role in the upper extremity and trunk during overarm throwing. J Sports Sci. 2002;20:301–310. [PubMed: 12003275]
12. Fleisig GS, Andrews JR, Dillman CJ, Escamilla RF. Kinetics of baseball pitching withimplications about injury mechanisms. Am J Sports Med. 1995; 23:233–239. [PubMed: 7778711]
13. Pappas AM, Zawacki RM, Sullivan TJ. Biomechanics of baseball pitching. A preliminary report.Am J Sports Med. 1985; 13:216–222. [PubMed: 4025673]
14. Dillman CJ, Fleisig GS, Andrews JR. Biomechanics of pitching with emphasis upon shoulderkinematics. J Orthop Sports Phys Ther. 1993; 18:402–408. [PubMed: 8364594]
15. Hirashima M, Kudo K, Watarai K, Ohtsuki T. Control of 3D limb dynamics in unconstrainedoverarm throws of different speeds performed by skilled baseball players. J Neurophysiol. 2007;97:680–691. [PubMed: 17079349]
16. DiGiovine NM, Jobe FW, Pink M, Perry J. An electromyography analysis of the upper extremityin pitching. J Shoulder Elbow Surg. 1992; 1:15–25. [PubMed: 22958966]
17. Gowan ID, Jobe FW, Tibone JE, Perry J, Moynes DR. A comparative electromyographic analysisof the shoulder during pitching: Professional versus amateur pitchers. Am J Sports Med. 1987;15:586–590. [PubMed: 3425786]
18. Astley HC, Roberts TJ. Evidence for a vertebrate catapult: elastic energy storage in the plantaristendon during frog jumping. Biol Lett. 2012; 8:386–389. [PubMed: 22090204]
19. Patek SN, Nowroozi BN, Baio JE, Caldwell RL, Summers AP. Linkage mechanics and poweramplification of the mantis shrimp’s strike. J Exp Biol. 2007; 210:3677–3688. [PubMed:17921168]
20. Bramble DM, Lieberman DE. Endurance running and the evolution of Homo. Nature. 2004;432:345–352. [PubMed: 15549097]
21. Osbahr DC, Cannon DL, Speer KP. Retroversion of the humerus in the throwing shoulder ofcollege baseball pitchers. Am J Sports Med. 2002; 30:347–353. [PubMed: 12016074]
22. Reagan KM, et al. Humeral retroversion and its relationship to glenohumeral rotation in theshoulder of college baseball players. Am J Sports Med. 2002; 30:354–360. [PubMed: 12016075]
23. Bennett MB, Ker RF, Dimery NJ, Alexander RM. Mechanical properties of various mammaliantendons. Zool Lond (A). 1986; 209:537–548.
24. Werner SL, Fleisig GS, Dillman CJ, Andrews JR. Biomechanics of the elbow during baseballpitching. J Orthop Sports Phys Ther. 1993; 17:274–278. [PubMed: 8343786]
25. Pieper HG. Humeral torsion in the throwing arm of handball players. Am J Sports Med. 1998;26:247–253. [PubMed: 9548119]
26. Cowgill LW. Humeral torsion revisited: a functional and ontogenetic model for populationalvariation. Am J Phys Anthropol. 2007; 134:472–480. [PubMed: 17657784]
27. Haile-Selassie Y, et al. An early Australopithecus afarensis postcranium from Woranso-Mille,Ethiopia. Proc Natl Acad Sci U S A. 2010; 107:12121–12126. [PubMed: 20566837]
28. Walker, A.; Leakey, RE. The Nariokotome Homo erectus skeleton. Harvard University Press;1993.
29. Richmond BG, Begun DR, Strait DS. Origin of human bipedalism: The knuckle-walkinghypothesis revisited. Yearb Phys Anthropol. 2001; 44:70–105.
30. Snyder SJ, Karzel RP, Del Pizzo W, Ferkel RD, Friedman MJ. SLAP lesions of the shoulder.Arthroscopy. 1990; 6:274–279. [PubMed: 2264894]
31. Roach NT, Lieberman DE, Gill TJ IV, Palmer WE, Gill TJ III. The effect of humeral torsion onrotational range of motion in the shoulder and throwing performance. J Anat. 2012; 220:293–301.[PubMed: 22257273]
Roach et al. Page 6
Nature. Author manuscript; available in PMC 2013 December 27.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

32. Winter, DA. Biomechanics and motor control of human movement. 2nd edn. Wiley-Interscience;1990.
33. Dempster, WT. Space requirements for the seated operator. Wright Air Development Center; 1955.
Roach et al. Page 7
Nature. Author manuscript; available in PMC 2013 December 27.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

Figure 1. Model of elastic energy storageArm-cocking and acceleration phases of the overhand throw (A). Humans (left) andchimpanzees (right) differ in arm abduction and elbow flexion during throwing (B) becauseof differences in shoulder orientation, which alters the major line of action of the Pectoralismajor (C). Aligning the long axis of the humerus with the major axis of P. major and flexingthe elbow maximizes inertia to shoulder flexion torque and loads the elastic ligaments in theshoulder. However, chimpanzee morphology is compromised between maximizing humeralrotation or elbow extension. Signatures of shoulder orientation found in the scapula (D) canbe used to reconstruct hominin shoulder orientation.
Roach et al. Page 8
Nature. Author manuscript; available in PMC 2013 December 27.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

Figure 2. Shoulder rotation and elbow flexion/extension powerMean shoulder rotational power (with 95% confidence intervals) shows a sustained periodof negative power and work during arm-cocking, between stride (STR) and maximumexternal rotation (MER) - white. This negative work is recovered during acceleration,between MER and release (REL) - gray. Recovered work powers both internal rotation atthe shoulder and extension of the elbow.
Roach et al. Page 9
Nature. Author manuscript; available in PMC 2013 December 27.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts
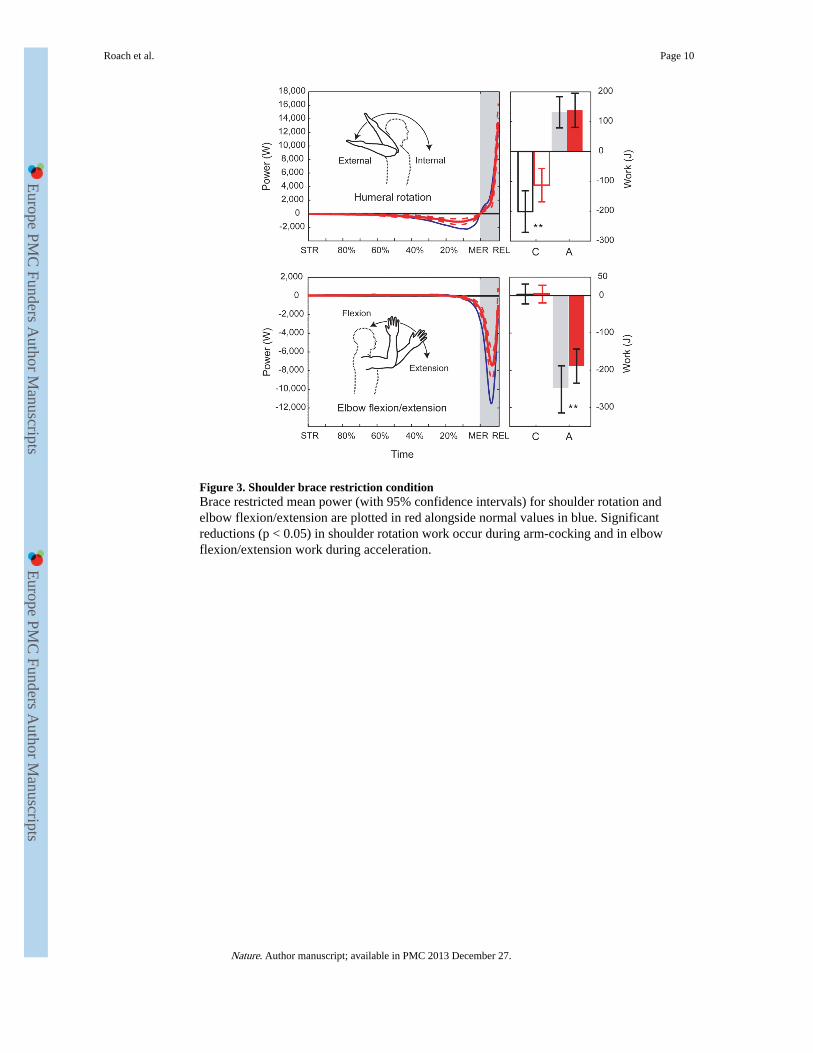
Figure 3. Shoulder brace restriction conditionBrace restricted mean power (with 95% confidence intervals) for shoulder rotation andelbow flexion/extension are plotted in red alongside normal values in blue. Significantreductions (p < 0.05) in shoulder rotation work occur during arm-cocking and in elbowflexion/extension work during acceleration.
Roach et al. Page 10
Nature. Author manuscript; available in PMC 2013 December 27.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

Figure 4. Humeral torsion and throwing performanceLow humeral torsion shifts the shoulder rotational ROM externally (A), allowing increasednegative work during arm-cocking to be stored as elastic energy (B), resulting in fasterprojectile speed (C). Humans and chimpanzees show comparable degrees of torsion5,although throwing athletes have reduced dominant arm torsion25 consistent with low torsionin Australopithecus and Homo erectus (D)5.
Roach et al. Page 11
Nature. Author manuscript; available in PMC 2013 December 27.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts