redalyc.proteÍnas antifÚngicas contra p. infestans en los espacios intercelulares de ... ·...
TRANSCRIPT

Revista Colombiana de Química
ISSN: 0120-2804
Universidad Nacional de Colombia
Colombia
Riaño A., Luz Dary; Zamora E., Humberto Miguel
PROTEÍNAS ANTIFÚNGICAS CONTRA P. INFESTANS EN LOS ESPACIOS INTERCELULARES DE
HOJAS DE TOMATE (Lycopersicon esculentum cerasiforme) POSIBLES DEFENSINAS DE
PLANTAS
Revista Colombiana de Química, vol. 34, núm. 1, enero, 2005, pp. 7-23
Universidad Nacional de Colombia
Bogotá, Colombia
Disponible en: http://www.redalyc.org/articulo.oa?id=309026662007
Cómo citar el artículo
Número completo
Más información del artículo
Página de la revista en redalyc.org
Sistema de Información Científica
Red de Revistas Científicas de América Latina, el Caribe, España y Portugal
Proyecto académico sin fines de lucro, desarrollado bajo la iniciativa de acceso abierto

PROTEÍNAS ANTIFÚNGICAS CONTRA P. INFESTANS EN LOSESPACIOS INTERCELULARES DE HOJAS DE TOMATE (Lycopersiconesculentum cerasiforme) POSIBLES DEFENSINAS DE PLANTAS
ANTIFUNGAL PROTEINS AGAINST P. INFESTANS FROMINTRACELLULAR SPACE OF TOMATO LEAVES (Lycopersicon
esculentum cerasiforme) POSSIBLE DEFENSINS OF PLANTS
Luz Dary Riaño A.* y Humberto Miguel Zamora E.**
Recibido: 27/09/04 – Aceptado: 20/05/05
RESUMEN
Se aislaron y caracterizaron parcialmenteproteínas antifúngicas de los espacios in-tercelulares de hojas de tomate Lycopersi-
con esculentum cerasiforme, variedadque ha mostrado resistencia en campo aPhytophthora Infestans; se observó quedespués de inoculación con el patógenodichas proteínas se acumularon sistémi-camente en la planta. Las proteínas iden-tificadas mostraron características de De-fensinas de plantas, una nueva familia deproteínas con bajo peso molecular, cargapositiva a pH fisiológico y actividad anti-fúngica evaluada in vitro contra P. infes-
tans. Los análisis electroforéticos en ge-les de poliacrilamida con SDS-Tricina encondiciones reductoras y no reductoras,sugirieron que están asociadas en tríme-ros y tetrámeros y poseen pesos molecu-lares de 5,2 kDa.
Palabras clave: espacios intercelula-res, hojas, tomate, proteínas PR (relacio-nadas con patogenicidad), Defensinas deplantas, actividad antifúngica.
ABSTRACT
Antifungal Proteins from intercellularspace of Tomato leaves Lycopersicon es-
culentum cerasiforme were isolated andpartially characterized; this variety hadshown resistance against Phytophthora
infestans, after inoculating plants with thepathogen it was observed that these pro-teins were accumulated systemically. Theisolated proteins be haved as plant defen-sins, a novel protein family, with low mo-lecular weight, cationics at physiologicalpH and Antifungal Activity in vitro eva-luated against P. infestans. Electrophore-tic analyses in SDS Tricine-PAGE, underreducing and non-reducing conditions
7
REVISTA COLOMBIANA DE QUÍMICA, VOLUMEN 34, No. 1 DE 2005
* Departamento de Química, Facultad de Ciencias, Universidad Nacional de Colombia, Sede Bogotá.
** Departamento de Química, Facultad de Ciencias, Universidad Nacional de Colombia, Bogotá, Colombia. Correo elec-trónico: [email protected]

suggested that, they were associated astrimers and tetramers; and have molecu-lar weights of 5,2 kDa.
Key words: Intercellular space, lea-ves, tomato, PR (pathogenesis related)proteins, Plant Defensins, Antifungal ac-tivity.
INTRODUCCIÓN
En la periferia de la membrana plasmáti-ca ocurren eventos moleculares útilespara estudiar las interacciones plan-ta-patógeno, de allí que se considere el es-pacio intercelular (1, 2) como el primersitio de respuesta de la planta al ataque demicroorganismos, además de construiruna fuente importante de proteínas PR (3,4, 5). Se han aislado diversas proteínasPR de plantas de tomate, tabaco y papaencontrando en algunas de ellas actividadantifúngica (6) contra microorganismoscomo Phytophthora infestans (7), Clavi-
bacter michiganensi, Botrytis cinerea yFusarium oxisporum, los cuales atacangeneralmente estos cultivos (8, 9).
Aunque las plantas no poseen un siste-ma de defensa como el nuestro (10), encondiciones de estrés y en presencia deagentes patógenos producen péptidos ca-tiónicos (11), llamados defensinas con ca-racterísticas tan similares a los que posee-mos, que se han considerado como siformasen parte de un sistema inmune enella (10, 12). Las defensinas son péptidosricos en cisteína, los cuales difieren enlongitud, número de cisteínas, enlaces ypatrón de plegamiento (13); se han consi-derado como elementos ancestrales yesenciales de defensa en vertebrados(13), insectos (14) y plantas (15-17).
En plantas las defensinas (18) son pép-tidos pequeños (5kDa), básicos, con unpatrón de plegamiento 3D característicoestabilizado por 8 cisteínas enlazadas pordisulfuros; su estructura, determinada porRMN, comprende una triple hebra �-lami-nar con una �-hélice en paralelo (19). Eneste trabajo se identificó la presencia deposibles defensinas en los espacios inter-celulares de hojas de tomate luego de la in-ducción de resistencia sistémica en lasplantas, y se determinó actividad antifún-gica contra Phytophthora infestans.
La mayoría de las defensinas de plan-tas que han sido aisladas provienen de se-millas, principalmente de rábano, de teji-dos de flores y otras especies, siendo éstala primera vez que se identifican en hojasde tomate (20-23). En nuestro laboratoriose han identificado y caracterizado paraesta variedad de tomate enzimas como�1,3-glucanasas y quitinasas, las cualesfueron consideradas como parte del siste-ma de defensa de la planta al ataque de P.
infestans (24-26).
MATERIALES Y MÉTODOS
Material vegetal
Las semillas de L. esculentum cerasifor-
me, variedad resistente, fueron suminis-tradas por Corpoica del Banco de Germo-plasma Nacional, número de accesoL1568. Las plantas crecieron en condi-ciones de invernadero; al cabo de 18 se-manas se obtuvieron 220 plantas adecua-das para inducir resistencia sistémica en110 de ellas empleando Phytophthora in-
festans, las 110 restantes se utilizaroncomo control.
8
REVISTA COLOMBIANA DE QUÍMICA, VOLUMEN 34, No. 1 DE 2005

Los análisis se realizaron en el líquidode lavado intercelular (LLI) obtenido apartir de las hojas.
Propagación del patógeno
El Oomycete P. infestans aislamientoM49, fue suministrado por la profesoraCelsa García, directora del Laboratoriode Fitopatología (Universidad Nacionalde Colombia). Éste se cultivó en medioagar, con arveja y sacarosa 20, 60, 20g/litro (24-26) y se mantuvo a 20 ºC en laoscuridad. Los esporangios de P. infes-
tans se suspendieron en agua destilada de-sionizada a temperatura ambiente y seajustó la concentración a 1x104 zoospo-ras/mL, y así se utilizó para la inocula-ción de las plantas y para la determina-ción de actividad antifúngica.
Inducción de resistencia sistémica
Para la inducción (27, 28) de proteínasPR se utilizaron plantas de 4 meses lascuales se inocularon con la suspensión deesporangios de P. infestans aplicándolassobre algunas de las hojas en forma de go-teo y aspersión, finalmente se dejaron in-cubando por 3 días en oscuridad con unahumedad relativa entre 80-90%, y des-pués de 15 días aparecieron los primerossíntomas (28).
Obtención del LLI
Se examinó el líquido de lavado intercelu-lar (29, 30) utilizando dos sistemas de in-filtración diferentes: agua destilada a 4 ºCy buffer fosfato pH 7 10 mM NaHPO4, 15mM NaH2PO4, 100 mM KCl, 2 mMEDTA, 0,15% PVP, 5 mM ácido ascórbi-
co a 4 ºC (31-33) tanto para hojas de plan-tas sanas como para inoculadas.
Se utilizó la siguiente nomenclatura parala identificación de los extractos: 11A=LLI planta sana extraído con agua, 11B=LLI planta sana extraído con buffer, 21A=LLI planta inoculada extraído con agua,21B= LLI planta inoculada extraído conbúfer.
Determinación de proteína
La proteína se determinó según el métodode Bradford modificado por Zor y Selin-ger (34).
Aislamiento de proteínas de los LLI(35, 36)
La proteína de los extractos (11A, 11B,21A, 21B) se precipitó con sulfato deamonio hasta un 30% de saturación, a 4ºC, se centrifugó a 7000 g durante 30 mi-nutos a 4 ºC; el sobrenadante obtenido sellevó hasta un 75% de saturación y se dejó2 horas a 4 ºC, se centrifugó nuevamentea 7000 g durante 30 minutos a 4 ºC, elprecipitado se resuspendió con 5 o 6 mLde buffer fosfato de sodio 50 mM, pH 6 sedializó contra éste durante 2 días a 4 ºCcon cambios cada 8 horas; finalmente secuantificó la proteína, se obtuvo un pa-trón electroforético, se realizó el ensayode actividad antifúngica, se fraccionó enalícuotas y se almacenó a –20 ºC.
Purificación y caracterización parcialde las proteínas en 21A y 21B
Cromatografía de tamiz molecular
Las proteínas de los extractos obtenidos apartir de las hojas, y que mostraron en el
9
REVISTA COLOMBIANA DE QUÍMICA, VOLUMEN 34, No. 1 DE 2005

perfil electroforético la presencia de pro-teínas con pesos moleculares inferiores a6,5 kDa (último marcador patrón), se se-pararon por cromatografía de tamiz mole-cular empleando una columna de 120 x1,5 cm empacada con Sephadex G25, uti-lizando como buffer de equilibrio y deelución fosfato de sodio 50 mM, pH 6; laseparación se realizó con un flujo de 18mL/h, siguiendo la proteína con lecturasde absorbancia a �=280 nm, �=250 nmy �=214 nm, longitudes de onda a las quetambién absorben cisteínas y los enlacesdisulfuros (32). Se determinó proteínapara cada “pool”, así como un perfil elec-troforético para identificar las fraccionescon proteínas de bajo peso molecular, yse evaluó la actividad antifúngica en cadacaso.
Cromatografía de intercambio
aniónico (38)
Las proteínas de las fracciones obtenidasdespúes de cromatografia de tamiz mole-cular (CTM) de los extractos 21A y 21B,que en el perfil electroforético presenta-ban pesos moleculares bajos, se separa-ron de acuerdo con su carga empleandouna columna de intercambio aniónicoPharmacia 10/16, con un lecho de 10 mlde DEAE Fraktogel TSK 650M ®, equili-brada en buffer fosfato de sodio 50 mMpH 6, y las proteínas retenidas se eluye-ron con un gradiente lineal NaCl 0-1M auna velocidad de flujo de 16 mL/h.
Las fracciones catiónicas de cada cro-matografía se concentraron mediante pre-cipitación con sulfato de amonio hasta75% de saturación, luego de centrifuga-ción a 7000 g durante 30 minutos a 4 ºC,se resuspendieron y dializaron contrabuffer fosfato de sodio 50 mM pH 6 a 4
ºC durante 2 días cambiándola cada 8 ho-ras, utilizando membranas de 3500 PM;se cuantificó proteína total, se determinóel peso molecular mediante electroforesisSDS-tricina-PAGE en condiciones re-ductoras y no reductoras, y se evaluó laactividad antifúngica. Las proteínas anió-nicas que se eluyeron con el gradiente sa-lino se dejaron para análisis posteriores.
Electroforesis SDS-Tricina-PAGE (38)
La purificación parcial de las proteínas secontroló mediante la determinación de lospesos moleculares relativos, empleandoun sistema de electroforesis discontinuocon geles de poliacrilamida, en condicio-nes denaturantes adecuado para péptidosy proteínas en el rango entre 1-100kDa.
Fijación, tinción y decoloración
de proteínas
Las bandas de proteínas se fijaron conuna solución de ácido tricloroacético 5%por 30 min; la tinción de las bandas serealizó con azul de comassie R en eta-nol-ácido acético 2:1 por 2 horas. Los ge-les se decoloraron utilizando etanol: áci-do acético: agua 5:5:2 (38).
Determinación de peso molecular
Se empleó el sistema de electroforesisSDS-Tricina-PAGE en geles T 10% C3%. Las muestras se prepararon en condi-ciones reductoras empleando �-mercap-toetanol para estimar el peso molecular re-lativo; las electroforesis en condiciones noreductoras se realizaron para establecer laposible formación de trímeros y tetráme-ros (38).
10
REVISTA COLOMBIANA DE QUÍMICA, VOLUMEN 34, No. 1 DE 2005

Ensayo de antifungicidad
Ensayo microespectrofotométrico
La actividad antifúngica se midió espec-trofotométricamente de acuerdo con tra-bajos anteriores (39) empleando cajaspara microtitulación de 96 pozos y un lec-tor de micro placas Biorad modelo 680.
El método consiste básicamente en to-mar medidas de absorbancia a 595 nm demicrocultivos del organismo que se va aevaluar y una dosis de la proteína de laque se presuma posee actividad inhibido-ra de crecimiento para el microorganis-mo. Los microcultivos de control conte-nían 20 �L de la muestra protéica paraensayo en condiciones estériles y 80 �Lde la suspensión de esporangios, inactiva-dos por el calentamiento durante 10 mi-nutos en baño de agua en ebullición. Lasplacas de microcultivos se incubaron atemperatura ambiente durante 48 horasen la oscuridad. El porcentaje de inhibi-ción de crecimiento se definió así (39):Ec. 1
Bioensayo con azul de Lactofenol (40)
La actividad de los extractos crudos y delas fracciones proteicas obtenidas duranteel crecimiento de P. infestans se evaluóempleando placas de microtitulación de96 pozos con 20 �L de muestra y 80 �L deuna suspensión de esporangios del mi-croorganismo, en medio de cultivo líqui-do con arveja y sacarosa; luego de 5 díasde incubación se tiñeron los micelios conazul de lactofenol y se observaron al mi-croscopio de luz.
RESULTADOS Y DISCUSIÓN
Inducción de resistencia sistémica
Del número de plantas inoculadas, el93% presentó sintomatología de infec-ción en hojas, no se observaron daños niseñales de infección en flores, tallo ni fru-tos en ninguna de las plantas tratadas, evi-denciándose la tolerancia de la variedadde tomate al ataque del patógeno, tenien-do en cuenta que no hubo destrucción delcultivo.
Extracción del LLI
Los dos tipos de extractos del LLI obteni-dos, uno empleando agua destilada a 4 ºCy otro con buffer fosfato de sodio pH 7�,se realizaron teniendo en cuenta las ca-racterísticas catiónicas y altamente bási-cas de las defensinas a pH fisiológico, seutilizó una fuerza iónica alta para solubi-lizarlas.
Las condiciones iniciales de los cuatroLLI obtenidos se resumieron en la Tabla1, donde se aprecia que la cantidad deLLI de plantas inoculadas extraído conbuffer es mayor que aquel de plantas sa-nas, lo mismo sucedió con los extraídoscon agua; de igual forma la cantidad pro-teína total fue mayor en plantas inocula-das que sanas.
En los LLI obtenidos con buffer el por-centaje de inhibición fue mayor en 21B(49%) que en 11B (39%), y en los extraí-dos con agua fue mayor en 21A (45%) queen 11A (25%), sugiriendo que en las plan-tas inoculadas la actividad aumentó luego
11
REVISTA COLOMBIANA DE QUÍMICA, VOLUMEN 34, No. 1 DE 2005
controlAbs
ensayoAbscontrolAbsxInhibición
nm
nmnm
595
595595
*
**100%
� Buffer fosfato de sodio pH 7 = 10mMNaHPO4, 15mM NaH2PO4, 100mM KCl,2mM EDTA, 0,15% PVP y 5mM Ácidoascórbico a 4 ºC

de la infección, comparada con la de lasplantas sanas; se presentó mayor actividaden los extractos con buffer, indicando quecon este sistema se logró extraer mayorcantidad de “compuestos antifúngicos”que con agua (Figura 1).
Aislamiento de proteína
La cantidad de proteína en los LLI, du-rante la precipitación con sulfato de amo-nio hasta un 30% de saturación no fueconsiderable, comparada con la del 70%;tampoco hubo actividad antifúngica yaque los porcentajes de inhibición presen-taron valores negativos en todos los ca-
sos, indicando que no hubo inhibición decrecimiento y que por el contrario, P. in-
festans continuó creciendo en el medio;éstos resultados permitieron asegurar lacontinuación de las proteínas antifúngicasen la fracción del 70%, como se indica enla Tabla 2.
Los porcentajes de inhibición de creci-miento más altos correspondieron a losextractos 21A y 21B, confirmando unainducción de respuesta de defensa de lasplantas al ataque de Phytophthora infes-
tans luego de inoculadas.
La actividad antifúngica que presenta-ron los extractos 11A y 11B, correspon-
12
REVISTA COLOMBIANA DE QUÍMICA, VOLUMEN 34, No. 1 DE 2005
Figura 1. Evaluación de actividad antifúngica en LLI. Los controles en la fila A. En la fila B muestras 11A 20�L (5 �g proteína), 11B 20 �L (5 �g), 21A 20 �L (10 �g), 21B 20 �L (13 �g). En todos los casos se utilizó 80�L (1x104 zoosporas/mL) de suspensión de esporangios de P. infestans.
Tabla 1. Condiciones iniciales de extractos de líquido de lavado intercelular de hojas deplantas de tomate (Lycopersicon esculentum cerasiforme)

dientes a las plantas sanas, sugirió que laplanta posee su propio mecanismo de de-fensa y que en condiciones de patogenici-dad éste se refuerza, confiriéndole mayorresistencia a la enfermedad (resistenciasistémica).
Los porcentajes de inhibición provie-nen de la aplicación de la ecuación 1 conlas lecturas realizadas a 595 nm, de loscontroles y las muestras a los 30 minutosy a las 48 horas después de iniciado el en-sayo, así: los valores negativos y menoresde 10% indican no antifungicidad, y losvalores inferiores a 20% y superiores a10% indican actividad deficiente.
El perfil electroforético en la Figura 2mostró zonas enriquecidas de proteína entodos los casos; los carriles correspon-dientes a las plantas inoculadas, 21A y21B presentaron proteínas de peso mole-cular inferiores a 6,5 kDa (asignado parael patrón de PM más bajo en el marca-dor), mientras que en 11A y 11B no seapreciaron; del mismo modo el número yla intensidad de las bandas fue menor, aun
cuando la cantidad de proteína dispuestaen todos los pozos fue similar. (Pie de Fi-gura 2). Teniendo en cuenta que se busca-ban proteínas de bajo PM se decidió puri-ficar las que estaban presentes en losextractos 21A y 21B.
13
REVISTA COLOMBIANA DE QUÍMICA, VOLUMEN 34, No. 1 DE 2005
Tabla 2. Actividad antifúngica de proteínas en LLI durante la precipitación fraccionadahasta 30 y 70% de saturación de sulfato de amonio
Figura 2. Perfil electroforético SDS-TRICINA-PAGE en condiciones reductoras de proteínas en LLIconcentradas y dializadas. Muestras 11A 20 L (46 gproteína), 11B 20 �L (66 �g), 21A 10 �L (64 �g),21B 10 �L (81 �g). PM. Marcadores Sigma de bajopeso molecular (2 �g/2 �L). BSA patrón (1 �g/5 �L).Geles espaciador y separación T 10% C 3%.
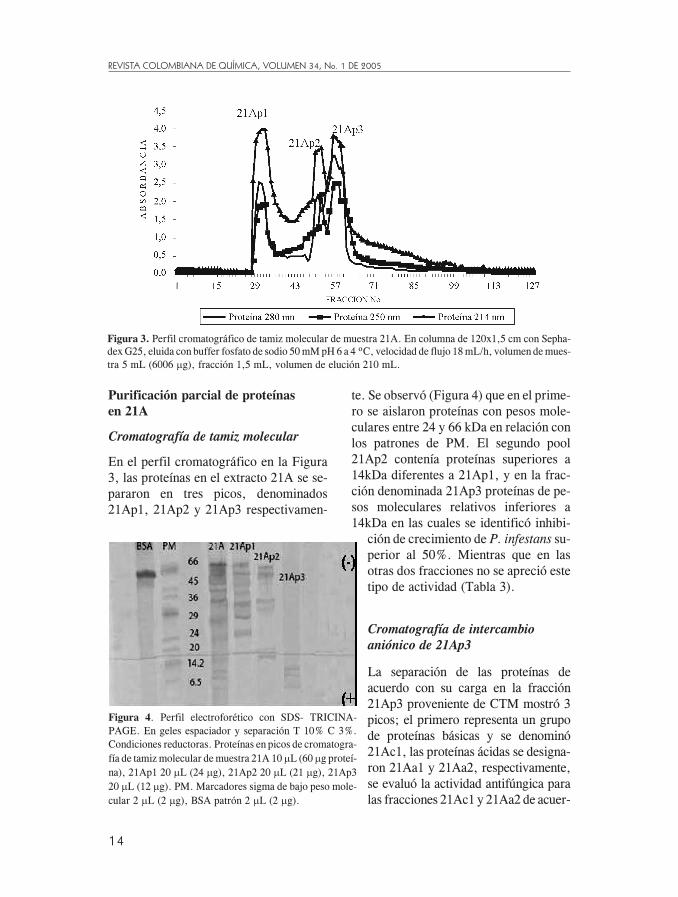
Purificación parcial de proteínasen 21A
Cromatografía de tamiz molecular
En el perfil cromatográfico en la Figura3, las proteínas en el extracto 21A se se-pararon en tres picos, denominados21Ap1, 21Ap2 y 21Ap3 respectivamen-
te. Se observó (Figura 4) que en el prime-ro se aislaron proteínas con pesos mole-culares entre 24 y 66 kDa en relación conlos patrones de PM. El segundo pool21Ap2 contenía proteínas superiores a14kDa diferentes a 21Ap1, y en la frac-ción denominada 21Ap3 proteínas de pe-sos moleculares relativos inferiores a14kDa en las cuales se identificó inhibi-
ción de crecimiento de P. infestans su-perior al 50%. Mientras que en lasotras dos fracciones no se apreció estetipo de actividad (Tabla 3).
Cromatografía de intercambio
aniónico de 21Ap3
La separación de las proteínas deacuerdo con su carga en la fracción21Ap3 proveniente de CTM mostró 3picos; el primero representa un grupode proteínas básicas y se denominó21Ac1, las proteínas ácidas se designa-ron 21Aa1 y 21Aa2, respectivamente,se evaluó la actividad antifúngica paralas fracciones 21Ac1 y 21Aa2 de acuer-
14
REVISTA COLOMBIANA DE QUÍMICA, VOLUMEN 34, No. 1 DE 2005
Figura 3. Perfil cromatográfico de tamiz molecular de muestra 21A. En columna de 120x1,5 cm con Sepha-dex G25, eluida con buffer fosfato de sodio 50 mM pH 6 a 4 ºC, velocidad de flujo 18 mL/h, volumen de mues-tra 5 mL (6006 �g), fracción 1,5 mL, volumen de elución 210 mL.
Figura 4. Perfil electroforético con SDS- TRICINA-PAGE. En geles espaciador y separación T 10% C 3%.Condiciones reductoras. Proteínas en picos de cromatogra-fía de tamiz molecular de muestra 21A 10 �L (60 �g proteí-na), 21Ap1 20 �L (24 �g), 21Ap2 20 �L (21 �g), 21Ap320 �L (12 �g). PM. Marcadores sigma de bajo peso mole-cular 2 �L (2 �g), BSA patrón 2 �L (2 �g).

do con el interés del estudio hacia las pro-teínas catiónicas de bajo peso molecular, yse establecieron diferencias en cuanto a ac-tividad antifúngica entre los dos tipos deproteína (21Ac1 y 21Aa2) de diferentecarga (Figura 5 y Tabla 3).
Hubo inhibición de crecimiento del31% en la fracción 21Ac1, en la aniónica(21Aa2) se obtuvo un porcentaje negativo(-1%) indicando actividad nula. De aquí seconcluyó que en el extracto 21A existíanproteínas catiónicas, de pesos molecularesrelativos inferiores a 14kDa y con activi-dad antifúngica in vitro contra Phytoph-
thora infestans del orden de 31% de inhi-bición con cerca de 5 g de proteína.
Purificación parcial de proteínasen 21B
Cromatografía de tamiz molecular
Luego de la separación por tamiz molecu-lar (Figura 6) se encontraron proteínascon pesos moleculares inferiores a 14kDa en 21 Bp3; en las fracciones 21Bp2 y21Bp1 se separaron moléculas con PMsuperiores a 20000 Da, presentándose elmayor número de éstas en 21Bp1 (Figura7). Nuevamente, dado que se buscabanproteínas de bajo peso molecular, se tomóel pool 21 Bp3 para realizar la separaciónde proteínas por carga y su posterior eva-luación de actividad antifúngica.
15
REVISTA COLOMBIANA DE QUÍMICA, VOLUMEN 34, No. 1 DE 2005
Tabla 3. Actividad antifúngica de proteínas en “pooles” de cromatografía de tamiz mole-cular de las muestras 21A y 21B
Figura 5. Perfil cromatográfico de intercambio aniónico de 21Ap3 de plantas de tomate resistente (Lycopersicon
esculentum cerasiforme). Columna 10/16 de Pharmacia con DEAE Fraktogel TSK 650M® eluida con Buffer fos-fato de sodio pH 6 a 4 ºC, gradiente lineal NaCl 0-1 M, velocidad de flujo 16 mL/h. Cantidad de muestra 2 mL(1203 �g), volumen de fracción 1,5 mL, volumen de elución 200 mL

En todos los “pooles” hubo inhibiciónde crecimiento visto como una disminu-ción en la intensidad de color de miceliosde P. infestans teñidos con el azul de lacto-fenol; el porcentaje de inhibición fue ma-yor para 21Bp3 (Tabla 3), por lo tanto, sepudo confirmar que la extracción de lasproteínas con actividad antifúngica y bajospesos moleculares, en el LLI de hojas detomate resistente, en las que se había indu-cido resistencia sistémica, fue mejor conbuffer fosfato pH 7 que aquella realizadacon agua destilada únicamente.
Cromatografía de intercambio
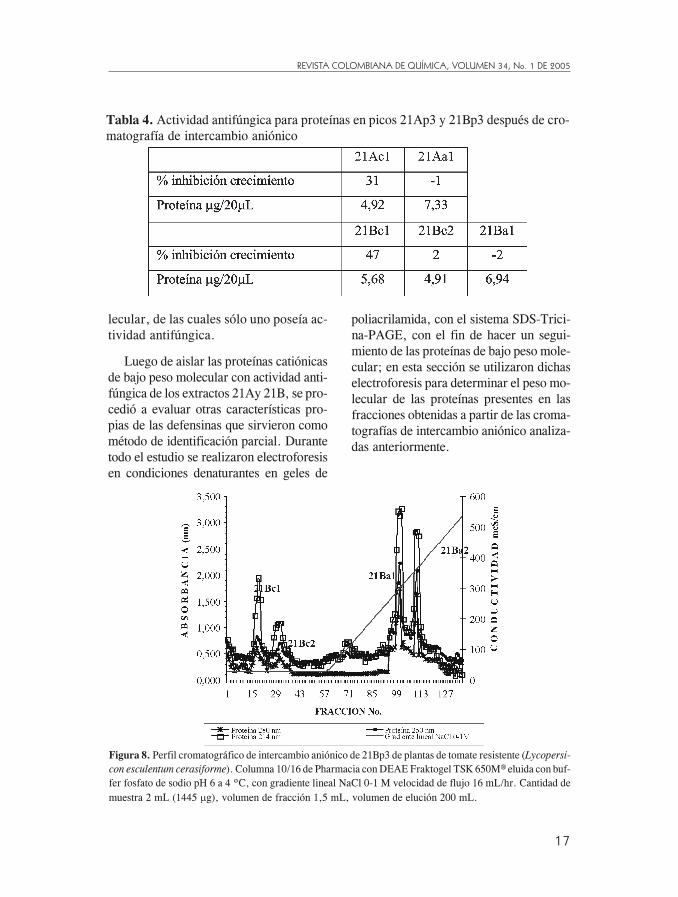
aniónico de 21Bp3
Se observaron 4 picos correspondientesa cuatro tipos de proteínas separadas deacuerdo con su carga; en ellos se en-cuentran 2 grupos básicos, 21Bc1,21Bc2, y dos ácidos 21Ba1, 21Ba2.
Al evaluar la actividad se encontróque 21Bc1 inhibió el crecimiento in vi-
tro de P. infestans, con un 47,2%, en21Bc2 se esperaba un comportamientosimilar, pero no presentó actividad anti-fúngica (Tabla 4). Con la fracción 21Ba1se obtuvo un valor negativo en porcenta-je, por lo que se pudo afirmar que en elLLI de hojas de tomate inoculadas ex-traído con buffer fosfa0to pH 7 (21B), seencontraron 2 grupos de proteínas catió-nicas (21Bc1 y 21Bc2) de bajo peso mo-
16
REVISTA COLOMBIANA DE QUÍMICA, VOLUMEN 34, No. 1 DE 2005
Figura 6. Perfil cromatográfico de tamiz molecular de muestra 21B. En columna de 120x1,5 cm con SephadexG25, eluida con buffer fosfato de sodio 50 mM pH 6 a 4 ºC, velocidad de flujo 18 mL/h, volumen de muestra 6mL (7636 �g), fracción 1,5 mL, volumen de elución 180 mL.
Figura 7. Perfil electroforético con SDS-TRICINA- PAGE,condiciones reductoras. Picos después de cromatografíade tamiz molecular de muestra 21B 10 �L (80 �g), 21Bp120 �L (30 �g), 21Bp2 20 �L (28 �g), 21Bp3 20 �L (14�g). PM. Marcadores sigma de bajo peso molecular 2 �L(2 �g), BSA patrón 2 �L (2 �g). Geles espaciador y sepa-ración T 10% C 3%.
lecular, de las cuales sólo uno poseía ac-tividad antifúngica.
Luego de aislar las proteínas catiónicasde bajo peso molecular con actividad anti-fúngica de los extractos 21Ay 21B, se pro-cedió a evaluar otras características pro-pias de las defensinas que sirvieron comométodo de identificación parcial. Durantetodo el estudio se realizaron electroforesisen condiciones denaturantes en geles de
poliacrilamida, con el sistema SDS-Trici-na-PAGE, con el fin de hacer un segui-miento de las proteínas de bajo peso mole-cular; en esta sección se utilizaron dichaselectroforesis para determinar el peso mo-lecular de las proteínas presentes en lasfracciones obtenidas a partir de las croma-tografías de intercambio aniónico analiza-das anteriormente.
17
REVISTA COLOMBIANA DE QUÍMICA, VOLUMEN 34, No. 1 DE 2005
Figura 8. Perfil cromatográfico de intercambio aniónico de 21Bp3 de plantas de tomate resistente (Lycopersi-
con esculentum cerasiforme). Columna 10/16 de Pharmacia con DEAE Fraktogel TSK 650M® eluida con buf-fer fosfato de sodio pH 6 a 4 ºC, con gradiente lineal NaCl 0-1 M velocidad de flujo 16 mL/hr. Cantidad demuestra 2 mL (1445 �g), volumen de fracción 1,5 mL, volumen de elución 200 mL.
Tabla 4. Actividad antifúngica para proteínas en picos 21Ap3 y 21Bp3 después de cro-matografía de intercambio aniónico

Determinación de peso molecular
en condiciones reductoras
Se encontraron proteínas catiónicas depeso molecular 5,2 kDa en los extractos21A y 21B, algunas otras con masas de7,7 kDa de las cuales estas últimas tienenmenor posibilidad de ser defensinas (pro-teínas de 5 kDa).
De acuerdo con los resultados que semuestran en la Tabla 5, en la fracciónproteíca denominada 21Ac1 del LLI deplantas inoculadas extraído con agua(21A), se encontró una proteína catiónicade peso molecular relativo de 5,2 kDa, lacual posee actividad antifúngica in vitro
contra P. infestans. Por otro lado, en lafracción 21Aa1 del mismo LLI, se encon-traron 3 proteínas aniónicas con pesosmoleculares diferentes; 13,8, 15,2 y 16,8
kDa, las cuales no poseen actividad an-tifúngica y su porcentaje de inhibiciónde crecimiento fue negativo.
En las fracciones 21Bc1 y 21Bc2 delLLI de plantas inoculadas extraído conbufffer (21B), se encontraron dos pro-teínas de pesos 5,2 y 7,7 kDa, sugirien-do que éstas pertenecen a ambas frac-ciones; es probable que cada fracciónsea una proteína y contenga trazas de laotra, ya que de acuerdo con la actividadantifúngica evaluada, 21Bc2 no presen-tó dicha actividad mientras que 21Bc1,
sí. ¿Cuál de las dos proteínas confiere laactividad? Éste es un cuestionamiento porresolver, habría que separarlas totalmen-te para evaluar su actividad individual yestablecer la diferencia.
Determinación de peso molecular
en condiciones no reductoras
En cuanto a la fracción 21Ba1 pertene-ciente al mismo extracto anterior se en-contraron 2 proteínas aniónicas con pe-sos moleculares de 7,7 y 15,2 kDa; deacuerdo con las observaciones anterio-res dicha fracción no presentó actividadantifúngica.
Las proteínas no reducidas proporcio-naron bandas finas; para 21Ac1 y 21Bc2(Tabla 5) se apreciaron dos bandas conuna masa molecular aparente de 14,2 y19,8 kDa en ambos casos. En 21Bc1 y en21Bc2 se encontró una banda con pesomolecular estimado de 14,2 y otra de 19,8kDa respectivamente. Los pesos molecu-lares de las proteínas evaluadas antes encondiciones reductoras sugirieron que es-tas proteínas surgieron como oligómerosconstituidos por protómeros de aproxi-madamente 5 kDa; de ser así estas estruc-
18
REVISTA COLOMBIANA DE QUÍMICA, VOLUMEN 34, No. 1 DE 2005
Tabla 5. Pesos moleculares de proteínas catió-nicas estimados en condiciones reductoras
Tabla 6. Pesos moleculares de proteínascatiónicas estimados en condiciones no re-ductoras

turas involucraron los puentes disulfuroentre cisteínas intactas por la ausencia delagente reductor. Las bandas de 14,5 kDarepresentarían un trímero (en 21Ac1,
21Bc1), mientras que las de 19,8 kDa (en21Ac1, 21Bc2) un tetrámero.
Morfología de inhibición
de crecimiento de P. infestans
Se tomaron microfotografías de algunasde las pruebas realizadas en las placas demicrotitulación empleando un microsco-pio de luz con objetivo 100X y se observóel deterioro del saco esporángico de P. in-
festans; éstas fueron tomadas 96 horasdespués de la incubación de la suspensiónde esporangios de Phytophthora infestans
de 1x104 células/ml en medio líquido dearveja y sacarosa, en presencia de 20l21Ac1 (4,92g), 20l 21Bc1 (5,68g), 20l21Bc2 (4,91 g), 10 minutos antes de to-mar la microfotografía, se adicionó azulde metileno 0,01% a los microcultivospara hacer contraste. Se observaron frag-mentos de micelio cenocítico (no septa-do) característico de P. infestans luego deuna degradación (Figura 11).
Las observaciones anteriores permi-tieron establecer el efecto antifúngico
19
REVISTA COLOMBIANA DE QUÍMICA, VOLUMEN 34, No. 1 DE 2005
Figura 9. Perfil electroforético con SDS-TRICINA-PAGE. Condiciones reductoras. Proteínas en frac-ciones básicas de cromatografía de intercambio ióni-co. 21Ac1 10 �L (29 �g), 21Bc1 10 �L (23 �g),21Bc2 10 �L (22 �g). PM. Marcadores sigma debajo peso molecular 2 �L (2 �g), BSA patrón 2 �L(2 �g). Geles espaciador y separación T 10% C 3%.
Figura 10. Perfil electroforético de proteínas conSDS-TRICINA-PAGE. Condiciones no reducto-ras. Proteínas de fracciones básicas después de cro-matografía de intercambio iónico. 21Ac1 15 �L (36�g), 21Bc1 15 �L (35 �g), 21Bc2 15 �L (34 �g).PM. Marcadores sigma de bajo peso molecular 2�L (2 �g), BSA patrón 2 �L (2 �g). Geles espacia-dor y separación T 10% C 3%.
Figura 11. Morfología de inhibición de crecimientode P. infestans luego de acción de proteínas catióni-cas de bajo peso molecular extraídas de plantas de to-mate resistente (Lycopersicon esculentum cerasifor-
me). A. zoosporangio intacto. B. 21Ac1, C. 21Bc1,D. 21Bc2.

contra el crecimiento de Phytophthora in-
festans, producido por las proteínas catió-nicas de bajo peso molecular, aisladas delos espacios intercelulares, de hojas ino-culadas de tomate resistente (Lycopersi-
con esculentum cerasiforme) encontradasen este trabajo.
CONCLUSIONES
Se obtuvieron tres fracciones con cuatroproteínas catiónicas provenientes de losespacios intercelulares de hojas de plantasde tomate resistente con peso molecularbajo y actividad antifúngica con un altoporcentaje de inhibición in vitro frente a P.
infestans, características que permiten de-nominarlas como defensinas de plantas.
Se establecieron las condiciones paraaislar y purificar parcialmente posiblesdefensinas de los espacios intercelularesde plantas de tomate, ya que las investiga-ciones realizadas hasta este momento ha-bían involucrado tejidos como semillas yel endospermo de diversas especies, y noexistía ningún trabajo publicado que mos-trara la presencia de tales proteínas en ho-jas de tomate.
Las proteínas localizadas en los espa-cios intercelulares –y caracterizadascomo posibles defensinas en este estudio,mostraron actividad antifúngica frente aP infestans, lo cual muestra que dichasproteínas están implicadas en el mecanis-mo de defensa de la planta y en la resis-tencia en campo que se ha observado enesta variedad de tomate. Es la primeravez que luego de inducción de resistenciasistémica se tienen indicios de la presen-cia de defensinas en plantas de tomate, es-pecialmente en las hojas y aún más en una
variedad perteneciente a nuestro Germo-plasma Nacional.
AGRADECIMIENTOS
Los autores agradecen a la UniversidadNacional de Colombia – Facultad deCiencias por facilitar las instalacionespara el desarrollo de este trabajo, a laDeutschen Gesellschaf für TechnischeZusammenarbeit (GTZ) y a la WorldUniversity Service (WUS) por la dona-ción de algunos de los equipos y reacti-vos, y a la División de InvestigacionesBásicas (DIB) por la financiación esteproyecto.
REFERENCIAS BIBLIOGRÁFICAS
1. Cervone, F.; Hahn, M. G.; De Lo-renzo, G.; Albersheim, P. (1989).Host-Pathogen Interactions. Plant
Physiol. 90, 542-548.
2. Barón, C.; Zambryski C. (1995).The Plant response in Pathogenesis,symbiosis and wounding variationson a common theme. Ann. Rev. Gen.29, 107-129.
3. Bowles, D. J. (1990). “Defense-re-lated proteins in higher plants”. Ann.
Rev. Biochem. 59, 873-907.
4. Ryals, J. A.; Uknes, S.; Ward, E.(1994). Systemic Acquired Resistan-ce. Plant Physiol. 104, 1109-1112.
5. Kessmann, H.; Staub, T.; Hofmann,C.; Maetske, T.; Herzog, J. (1994).Induction of Systemic acquired di-sease resistance in plants by chemi-cals. Ann. Rev. Phytopathol. 32,439-459.
20
REVISTA COLOMBIANA DE QUÍMICA, VOLUMEN 34, No. 1 DE 2005

6. Woloshuk, C. P.; Meulenhoff, J. S.;Sela-Buurlage, M.; Cornelissen, B.J. C. (1991). Pathogen-Induced Pro-teins with Inhibitory Activity towardPhytophthora infestans. Plant Cell.
3, 619-628.
7. Niderman, T.; Genett, I.; Bruyere,T. (1995). Pathogenesis-relatedPR-1 proteins are antifungal (isola-tion and characterization of three 14kDa proteins of tomato and of a basicPR-1 of tobacco with inhibitory acti-vity against Phytophthora infestans).Plant. Physiol. 108, 17-27.
8. Nielsen, K. K.; Nielsen, J. E.; Ma-drid, S. M.; Mikkelsen, J. A.(1990). New antifungal proteinsfrom sugar beet (Beta vulgaris L.)
showing homology to non- specificlipid transfer proteins. Plant Mol.
Biol. 31, 539-552.
9. Kristensen, A. K.; Brusted, J.; Niel-sen, J. E.; Mikkelsen, J. D.; Niel-sen, K. K. (2000). Partial characteri-zation and localization of a noveltype of antifungal proteins isolatedfrom sugar beet Leaves. Plant Sci.159, 29-38.
10. Salisbury, F. B.; Ross, C. W.(1994). Fisiología Vegetal. Grupoeditorial Iberoamericana, pp.395-449.
11. Selitrennikoff, C. P. (2001). Anti-fungal Proteins. Appl. Env. Microb.67, 2883-2894.
12. Van Loon, L. C.; Strien Van, E. A.(1999). The families of pathogene-sis-related proteins, their activities,and comparative analysis of PR-1
type proteins. Plant. Pathol. 55,85-97.
13. Ganz, T.; Lehrer, R. I. (1994). De-fensins. Curr. Opin. Immunol. 6,584-589.
14. Lamberty, M.; Ades, S.; Uttenwei-ler, S.; Brookhart, G.; Bushey, D.;Hoffmann, A. (1999). Insect immu-nity. Isolation from the LepidopteranHeliothis virescens a novel insect de-fensin with potent antifungal acti-vity. J. Biol. Chem. 274, 9320-9326.
15. Broekaert, W. F.; Terras, F. R. G.;Cammue, B. P. A.; Osborn, R.W.(1995). Plant defensins: Novel anti-microbial peptides as components ofthe Host defense system. Plant
Physiol. 108, 1353-1358.
16. Thomma, B. P. H. J.; Cammune, B.P. A.; Thevissen, K. (2002). PlantDefensins. Planta. 216, 193-202.
17. Terras, F. R. G.; Torrekens, S.; VanLeuven, F.; Osborn, R. W.; Cam-mune, B. P. A.; Broekaert, W. F.(1993). A new family of basic cystei-ne-rich plant antifungal proteinsfrom Brassicaceae species. FEBS
Letter 316, 233-240.
18. Terras, F. R. G.; Schoofs, H. M. E.;De Bollet, M. F. C.; Van Leuven,F.; Cammune, B. P. A.; Broekaert,W. F. (1992). “Analysis of two no-vel classes of Plant Antifungal pro-teins from Radish (Raphanus sativus
L.) seeds. J. Biol. Chem. 267,15301-15309.
19. Almeida, M. S.; Cabral, K. M.;Kurtenbach, E. (2002). Solutionstructure of Pisum sativum defensin
21
REVISTA COLOMBIANA DE QUÍMICA, VOLUMEN 34, No. 1 DE 2005

by high resolution 1NMR: plant de-fensins, identical backbone with dif-ferent mechanisms of action. J. Mol.
Biol. 315, 749-757.
20. Terras, F. R. G.; Eggermont, K.;Kovalera, V.; Ralkhel, N. V.;Osborn, R. W.; Kester, A.; Torre-kens, S. (1995). Small Cystei-ne-Rich antifungal proteins from Ra-dish: Their role in host defense.Plant Cell. 7, 573-588.
21. Osborn, R. W.: De Samblanx, G.W.; Thevissen, K.; Goderis, I.; VanLeuven, F.; Broekaert, W. F.(1995). Isolation and characterisa-tion of plant defensins from seeds ofAsteraceae, Fabaceae, Hipp.ocasta-naceae and Saxifragaceae. FEBS Let-
ters 368, 257-262.
22. Cammune, B. P. A.; De Bollet, M.F. C.; Terras, F. R. G.; Proost, P.;Van Damme, J.; Broekmert, W. F.(1992). Isolation and characteriza-tion of a Novel Class of Plant Anti-microbial Peptides from Mirabilis
jalapa L. Seeds. J. Biol. Chem. 267,2228-2233.
23. Van den Bergh, K. P. B.; Proost, P.;Van Damme, J.; Cooseman, J.(2002). Five disulfide bridges stabi-lize a hevein-type antimicrobial pep-tide from the bark of spindle tree(Euonymus europeus L.). FEBS let-
ter. 530, 181-185.
24. Moreno, Y. (2000). Análisis diferen-cial de las 1,3-glucanasas presentes enplantas de tomate Lycopersicon escu-
lentum cerasiforme y su patógenoPhytophthora infestans. Trabajo de
grado. Químico. Universidad Nacio-nal de Colombia - Bogotá.
25. Ceron, L. (2000). Determinación yanálisis comparativo de proteínas re-lacionadas con la patogénesis (PR)de los espacios intercelulares deplantas de tomate resistente (Lyco-
persicon esculentum cerasiforme) ysusceptible (Lycopersicon esculen-
tum Mill) antes y después de la infec-ción con Phytophthora infestans
(Mont de Bary. Trabajo de grado.Químico. Universidad Nacional deColombia- Bogotá.
26. Velásquez, D. E. R. (2003). Carac-terización parcial de Quitinasas enlos espacios intercelulares de plantasde tomates resistentes (Lycopersicon
esculentum cerasiforme) y suscepti-bles (Lycopersicon esculentum Mill)a Phytophthora infestans”. Trabajode grado. Químico Universidad Na-cional de Colombia - Bogotá.
27. Youn Yi, S.; Kook, H. B. (1996).Diferenctial induction and accumu-lation of -1,3-glucanase and chitina-se isoforms in soybean hypocotylsand leaves after compatible and in-compatible infection with Phytoph-
thora megasperma f.sp. glycinea.
Physiol. Mol. Plant. Pathol. 48,179-192.
28. Anfoka, G.; Buchenauer, H. (1997).Systemic acquired resistance in to-mato against Phytophthora infestans
by pre-inoculation with tobacco ne-
crosis virus Physiol. Mol. Plant.
Pathol. 50, 85-101.
29. Rohringer, R.; Ebrahim-Nesbat, F.(1983). Proteins in intercellular
22
REVISTA COLOMBIANA DE QUÍMICA, VOLUMEN 34, No. 1 DE 2005

Washing Fluids from Leaves of Bar-ley (Hordeum vulgare L.). J. Exp.
Bot. 34, 1589-1605.
30. Hammond-Kosack, K. E. (1983).Preparation and analysis of interce-llular fluid. Plant Physiol. 72,326-332.
31. Kroll, K. A.; Brunstedt, J.; Nielsen,J. E.; Nielsen, K. K. (2000). Partialcharacterization and localization of anovel type of antifungal protein inIWF from sugar beet leaves. Plant
Sci. 159, 29-38.
32. Cammue, B. P. A.; De Bolle, M. F.C.; Terras, F. R. G.; Proost, P.; VanDamme, J.; Broekaert, W. F.(1992). Isolation and characteriza-tion of a novel class of Plant antimi-crobial Peptides from Mirabilis jala-
pa L. seeds. J. Biol. Chem. 267,2238-2233.
33. Terras, F. R. G.; Torrekens, S.; VanLeuven, F.; Osborn, R. W.; Vander-leyden, J.; Cammue, B. P. A.; Broe-kaert, W. F. (1993). A new family ofbasic cysteine-rich plant antifungalproteins from Brassicaceae species.
FEBS letter 316, 223-240.
34. Zor, T.; Selinger, Z. (1996). Linea-rization of the Bradford Protein as-say increases its sensitivity: Theore-tical and experimental Studies. Anal.
Biochem. 236, 302-308.
35. Van Jagow, G.; Schägger, H.(1994). A practical guide to mem-brane protein purification. Acade-mic Press, San Diego, USA, pp.3-21.
36. Linn, S. (1990). Strategies and con-siderations for Protein Purifications,in Methods in Enzimology, vol. 142,Academic Press, pp. 9-83.
37. Morris, C. J. O. R.; Morris, P.(1976). Ion-Exchange chromato-graphy, in Separation Methods inBiochemistry, Pitman Publishing,pp. 251-357.
38. Schagger, H.; Von Jagow, G.(1987). Tricine-Sodium dodecyl Sul-fate- Polyacrylamide Gel Electrop-horesis for the separation of proteinsin the Range from 1 to 100 kDa.Anal. Biochem. 166, 368-379.
39. Terras, F. R. G.; Schoofs, H. M. E.;De Bollet, M. F. C., Van Leuven,F.; Vanderleyden, J.; Cammue, B.P. A.; Broekaert, W. F. (1992).Analysis of two novel classes of plantantifungal proteins from radish(Raphanus sativus L.) Seeds. The J.
Biol. Chem. 267, 15301-15309.
40. Woloshuk, C. P.; Meulenhoff, J. S.;Sela-Buurlage, M.; Van den Elzen,P. J.; Cornelissen, B. J. C. (1991).“Pathogen-induced proteins withInhibitory Activity toward Phytoph-thora infestans” Plant cell. 3:619-628.
23
REVISTA COLOMBIANA DE QUÍMICA, VOLUMEN 34, No. 1 DE 2005